 国家高新企业 | ISO9001认证
国家高新企业 | ISO9001认证 二级病原微生物安全实验室
二级病原微生物安全实验室- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
国家高新企业 | ISO9001认证 二级病原微生物安全实验室


小编最近仔细阅读了一本书,书名是《战斗细胞:人体免疫系统奇妙之旅》(海南出版社2022年10月出版的),该书是德国科学家菲利普·德特玛(Philipp Dettmer)撰写的一本书,作者是一名信息设计师。在32岁被确诊癌症并接受了一系列手术和化疗后,他逐渐痴迷于研究免疫学,目前已成为一名科学传播者和免疫研究爱好者。
译者李超群,英国利兹大学应用翻译专业硕士,译有《饮食的迷思》《显微镜下的室友》等。
在书中,作者以亲切易懂的语言、切入场景确切的比喻,生活化的语言描绘出免疫细胞世界的模样、运行的方式、如何使我们生病或保护健康,帮助读者轻松理解复杂的免疫相关知识。
免疫系统本来是对普通人比较陌生的领域,专业名词也多,但是作者通过生动的比喻帮我们克服了概念上的障碍,例如,他将普通体细胞比作“平民”,免疫细胞则是“各种专门士兵”——肠道有益菌是可爱小兔子、抗体是小龙虾等。看书的过程就好像在看影视剧一样。就算你从未接触过医学知识,也丝毫不影响阅读和理解。
回顾我们以往的科普文章,虽然前期选定主题,查阅大量研究文献和资料,再整理撰写花费时间很多,但是没有将科普文章写得如《战斗细胞》一书一样让不同知识面的客户都可以轻而易举读懂和受益。
在此有机会阅读此书并分享书中部分关于免疫系统和肠道黏膜和肠道菌群的内容,帮助读者增进对人体免疫系统的了解,从而更好地守护自身健康。
前序:在我们生活的环境中有着不计其数的病原体,不仅有病毒,还有细菌、寄生虫等等。人类几乎每时每刻都在和环境中的危险因素接触、斗争,只不过,我们常常感知不到这个“斗争”的过程。
免疫系统的构成:一支精密完备的“老牌”军队
用作者的话说,要回答什么是人类的免疫系统,还真有点不容易,但既然叫做“系统”,也就表明它不是一个单独的东西,而是一整套完整的体系。
作者把人类免疫系统比作是“一套军队系统”,说它“是由成百上千的营地和征兵处组成的遍布全身的复杂网络”,这说的就是免疫网络,是免疫系统的“基础设施”。
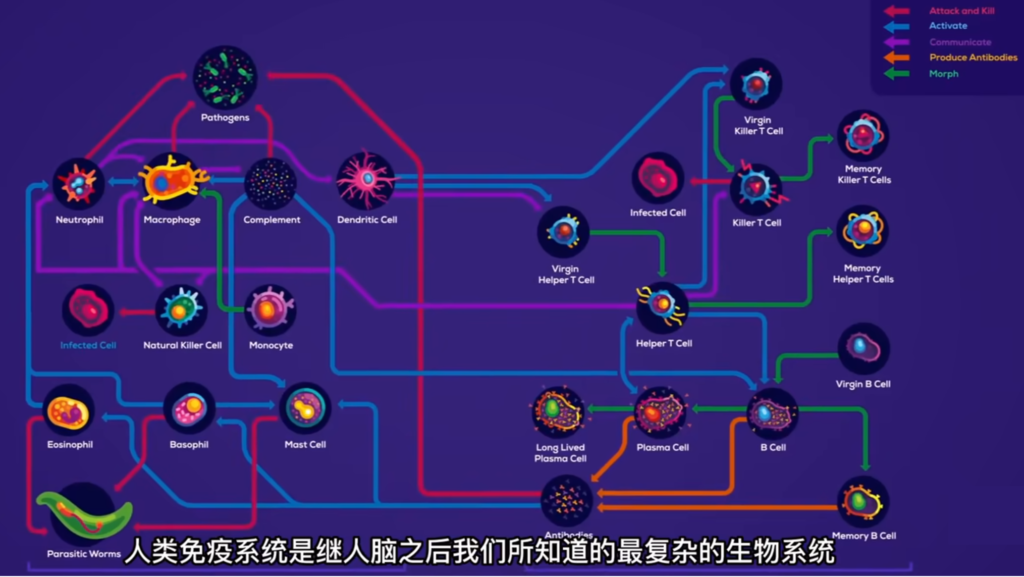
而所谓的“营地”,其实就是指免疫系统中专门的免疫器官,比如骨髓、胸腺、盲肠、扁桃体等等,这些免疫器官有些也是免疫细胞的诞生地;此外还有淋巴结。另外,免疫细胞内部还会释放出各种免疫活性物质,或者称为“细胞因子”,例如常见的有抗体、补体、免疫球蛋白等等,它们也是整个免疫系统重要的组成部分。
按照不同的免疫功能,免疫系统由两大王国组成,即先天性免疫(或说非特异性免疫),以及后天的适应性免疫(或说特异性免疫)。从名称就可以看出,两者的区别在于,先天性免疫是我们一出生就具有的,或者说不是针对特定的、单一的病原体的。
➤ 初代生命的进化与互相攻击:35亿年前,泥坑中的奇迹诞生
要是你因为每天早起上学,工作,路上堵车就觉得生活不顺,那原始的单细胞生命可以给你好好上一课。
35亿年前,地球还是一片蛮荒之地,在某个神奇的泥坑里,生命诞生了,而免疫系统要也随之出现了。我们不知道这些初代生命做过什么,又怎么活下来,但知道它们很快就开始了互相攻击。
➤ 生命的合作进化:从单细胞到多细胞
它们必须想办法把周围的化学物质转化为能为己用的东西,同时获取生成必需的能量,而其中几种细胞就盯上了“捷径”。
能偷别人干的,干嘛要自己干呢?
有几种办法实现了这一目标,比如:
可以整个同事其他细胞,或是在别的细胞上打洞并从其内部吸食营养……不过这样做颇有风险,成功了能饱餐一顿,失败了则会惨遭反噬,特别是在对手更大,更强的情况下。
风险更小的办法是潜入对手细胞内部并在此安家:分享宿主的粮食,享受它的呵护。这个办法相当成功,当然对宿主来说就很可怕了。
既然擅长“吸血”成了有效的生存技能,那么保护自己不被“吸”就成了演化上的必需。接下来的29亿年里,微生物凭借相似的招数,互相竞争厮杀。
假如有个时光机,能回到过去的微观世界一睹此类战争的宏大场面,你多半会失望,除了潮湿的石头上有一些纤薄的细胞膜之外,什么都没有。
在最初的几十亿年里,地球是个相当乏味的地方,直到生命形式的出现和飞越。
我们现在并不特别清楚,到底是什么让单打独斗的细胞开始向着密切合作并有专门分工的庞大细胞群发展的。
➤ 多细胞生物的兴起与免疫系统的重要性
大约在5亿4100万年前,多细胞动物出现了爆发性增长,体型变得肉眼可见,多样性也以极快的速度大大增加。
这当然就给刚演化出来的古代微生物们提出了一个问题:几十亿年来,生活在微型世界中的它们,无论进入任何生态系统,都要为其中的空间和资源争斗,而对这些细菌及其他微小生物来说,还有比多细胞动物更好的生态系统吗?这个系统里,从头到脚都是免费的营养!所以从一开始,多细胞生命就面临着细胞入侵者和机体寄生者的威胁。
只有能应付这种威胁的多细胞生物才能存活下来并有机会演化得越来越复杂。可惜,经过数亿年的时间,细胞和组织都没有得到好好的保存,我们看不到免疫系统的化石遗迹。但通过科技的魔法,我们可以看到生命树的各个分叉,并观察现存动物,研究它们的免疫系统。如果有些免疫特征,为生命树上差距越大的物种共同拥有,那么一般而言,这些特征也就越久远。
那么重要的问题就变成了:
➤ 免疫系统会在哪里产生区别,不同动物的免疫系统有哪些共性?
几乎所有当今的生物都有某种形式的内部防御机制,而生物越复杂,免疫系统也越复杂。选取相差悬殊的动物,比较它们的防御机制,我们就可以了解很多免疫系统的历史。
从最微小的层面来说,细菌也有抵御病毒的方法,它们不可能乖乖就范。而在动物世界中,海绵这种已经存在了5亿多年的最古老、最简单的动物,就拥有可能是最原始的动物免疫反应:“体液免疫”(“体液/humor”一词来自古希腊语,这里可不是“幽默”)。
体液免疫主要通过游离在细胞外体液中的微小蛋白来实现功能,杀伤、歼灭不该出现的外来微生物。这类防御非常有用,也非常成功,几乎所有现存的动物,包括我们人类,都有体液免疫。它没有被演化淘汰,而是变得举足轻重。原则上说,体液免疫在5亿年里都没怎么变。
但体液免疫只是个开端。多细胞动物的优势之一是就发展出了专业的防御细胞,相应的免疫就叫“细胞[介导]免疫”。
这种新型的免疫一开始就大获成功。连在蠕虫和昆虫的小小身体里,我们都发现了专门的免疫细胞卫士,它们能在小虫体内自由巡行,与入侵者正面交锋。
沿着演化之树,越是向枝头探寻,免疫系统就变得越是精妙。而在最早的脊椎动物身上,我们就已经看到了重大的创新:出现了专门的免疫器官暨细胞的“训练场”,以及最为强大的免疫原理之一——能够识别特定的敌人,并快速制造大量的针对性武器实现精准打击,还要牢牢记住它们!
即使是最原始的脊椎动物,比如样子诡异的无颌鱼,体内也有同样的免疫机制。

图源:dashangu
在数亿年的岁月里,这些防御机制变得越发复杂和精密。不过,简而言之,上面都是基本的免疫原理,它们有着出色的防御效果,所以或许在5亿年前的某些生物体内就已经存在了。因此,虽然你我的免疫系统很复杂、很强大,但其底层机制在动物界却很普遍,它擅长调动多种多样的特化细胞。
因此,从演化上来看,动物很可能没用多久其起源可以追溯回几亿年前。演化之手无须一次次地重新发明免疫系统——它发现了一套了不起的系统,并将其不断完善。
好,话题终于要回到人类,回到你我身上了。我们都在享受着数亿年来免疫系统不断演进的丰硕果实。不过,免疫系统并不只是存在于你的体内,它就是你本身。“免疫”只是对守护自身并维系生命的所有生物机制的一种表达方式。所以讨论免疫系统就是在讨论人本身。
免疫系统也不是一个单独的东西。它像一套军队系统,是由成百上千的营地和征兵处组成的遍布全身的复杂网络。它们和心血管系统一样,分布广泛,无处不在,由像高速路网一样的管道网连接起来。另外,人体还有专门的免疫器官,位于胸腔内,大小和鸡翅差不多,会随着人的衰老而逐渐退化。
基于免疫器官和免疫“基础设施”,有几百亿免疫细胞在免疫路网和血流中巡逻,可以随时应召和敌人作战。此外,人体外部组织表面也有几十上百亿的免疫细胞形成的屏障,准备抵挡入侵。

这还只是积极防御,还有1018数量级的蛋白质武器组成的防御系统,它们就像能自动组装、四处散布的地雷。免疫系统中还有专门的“大学”,供细胞学习如何识别敌人及怎样作战。这所大学有着世界上最大的生物学图书馆,可以教你识别一生中可能碰到的所有敌人并记住它们。
在最核心的意义上,免疫系统就是区分自身和异物的工具
无论对方是否恶意,并不重要,只要不是被授予自由通行证的贵宾,免疫系统都会发动攻击与歼灭,因为这些被视为潜在的威胁。对免疫系统来说,为任何“他者”冒险都不值得。没有它们的此种忠诚,我们可能活不了几天。可惜,免疫系统如果忠诚不足或忠诚过头,也会造成困扰甚至引发严重后果致人死亡,后面我们会谈到这些情况。
免疫系统的首要目标——维持稳态
虽然说区分自身和异物是免疫功能的核心,但并不是它的目标。免疫系统的首要目标是维持“稳态”,即维持体内全部细胞及物质的平衡。免疫系统真是投入了最大的努力保持平衡,也在尽心安抚自己免于反应过激,这一点我们怎么强调也不为过。
你也可以理解成,它在追求和平,追求让生存变得愉悦和轻松的稳定秩序,也就是我们所说的“健康”。健康是轻松自由地生活的基础,拥有健康,我们才能从心所欲,不受病痛的阻挡。
只有当我们失去健康时,才会真正意识到它的宝贵。健康其实是一个抽象的概念,因为它描述的并不是一种实质的存在,而是没有病痛、不受限制的状态,不是“有”而是“无”。拥有健康的时候,你觉得一切正常,感觉良好。而一旦健康离你而去,哪怕只有几天,都很难忽视人的脆弱和生命的稍纵即逝。疾病是生命中不可避免的事情。对于美好生活而言,没有什么比健康的身体更重要。
对免疫系统来说,健康就是稳态
尽管这场守护健康的战争终会不可避免地输掉,变成徒劳,但我们仍在努力夺取多一些时间,哪怕只是几个小时。总体而言,生而为人是幸福的,我们都希望自己可以活得久一点。
但维持健康殊非易事,因为我们每天都会遇到上亿的细菌和病毒,它们巴不得占领你的身体,就像几十亿年前那些单细胞生物之间发生的战争那样。
对微生物来说,你就是一套有待征服的生态系统。这片广袤无垠的土地资源丰富,适合繁殖,充满了机遇,是理想的栖居之地。当然,等我们最终离开这个世界时,它们还是会胜利的;没有了免疫系统的保护,大量疯狂的微生物组成的军团会大大加速尸体的分解过程。
预防癌症是免疫系统的头等大事之一
要担心的还不止这一大堆想要入侵的微生物,还包括可能误入歧途的身体本身,就是癌症。事实上,在你读这几页书的时候,身体里可能就有早期癌细胞正在被免疫系统悄无声息地清除掉。
免疫系统也可能出错——自身免疫性疾病、过敏
旨在保护你的免疫系统也可能出错,可能堕落。免疫系统一旦受了蒙骗,可能就会助长疾病的蔓延,或是保护癌细胞不被发现。免疫系统也可能失衡或受损,从而糊涂地攻击自身。它会把“我”误判为“他”,实实在在地攻击起它本该保护的那些身体细胞,导致一系列自身免疫性疾病,而病人不得不长期服用可能伴有严重副作用的免疫抑制药物。
过敏反应是指免疫系统对无害的物质产生了过激反应。“过敏性休克”就是一种严重的过敏反应,它凸显了免疫系统有多强大,一旦出错又多么可怕:疾病未必会马上置人于死地,而免疫系统带来的过敏性休克却能在几分钟内夺人性命。

免疫系统在帮助你的同时也会带给你痛苦
生病时的很多糟糕症状都是被激活的免疫系统引发的后果——对某些疾病来说,最严重的损伤甚至死亡都是免疫系统对入侵事件的失控反应造成的。比如,COVID-19导致的许多死亡病例都是免疫过激的结果。
伤害的累积是严重疾病的基础
免疫系统发挥防御功能的同时造成的损害会不断累积,于是今天有人认为,免疫系统的正常运转正是一些致命疾病的发展基础。因此,尽管拥有能对外界威胁做出快速有力反应的免疫系统对健康很是关键。
但同时保证它不会失控和搞破坏也很重要。就和人类社会一样,如果你的身体不得不打仗,那它至少想要快速地结束战争,赢得干净利落,而不想经历长期的被占领或冲突,那只会耗尽资源,毁坏基础设施。
人不管怎样生存和行动,都离不开世界本身及世上的一切。温馨私密的城堡,偏僻的森林木屋,遥远的社交距离,都无法让你免于和世界打交道,哪怕你是最叛逆的网瘾少年-最起码你需要稳定的食物来源,因此总要和外界有最基本的互动。

人体面临着同样的问题,因为细胞需要氧气和营养来维持生存和运转,并排出代谢产生的有害废物。就是说,资源要从外面进来,废物要从里面出去。因此,人体不可能是封闭的系统,内部总要有和外界直接接触的地方。
绝大部分病原体入侵都发生在这些内外交界之处
而这样的地方也就是人体的薄弱环节,从这里,不速之客能偷偷入侵血肉大陆。事实上,绝大部分的病原体入侵都发生在这些内外交界之处,包括从口腔一直到肛门的消化道,还有通向和外界进行交换的“洞穴系统”的多分支管道。
开头我们就说过,人体的肺和呼吸道、口腔和肠道,以及生殖道,其实是身体的外部,只是被卷到了里面。按理说,这些“管道”里铺的东西应该叫“内皮”,可惜,正确的名字是黏膜。为了听起来更酷一些,我们就叫它“沼泽之国”。
沼泽之国要解决的一大难题,方便营养进来、废物出去,同时又要让病原体难以逾越
这意味着,沼泽之国的免疫系统要和身体其他地方的不一样。肠道黏膜人体大部分都是无菌的,没有微生物,没有异物,而沼泽之国却在持续接触着各种“异物”:有待吸收的食物碎块,由此通过的可消化物质,可以自由进入并在肠道安居的有益菌,还有从空气中吸入的各种颗粒物,如污染物、灰尘等等。
这样一来,当然会有数不清的不速之客想要偷偷潜入,越过防线。其中一些是不小心闯入的无辜过客,另一些则是专门攻击人体的危险病原体。

这使得这些部位的免疫系统有着格外艰巨的职责,要维持平衡更是难上加难——因为沼泽之国的免疫系统要有一定的宽容度。
与此相反,身体多数部位的免疫系统对异物是毫不留情的。划伤之后,细菌侵入软组织,免疫系统就会发动猛烈的还击。皮下或者肌肉组织是容不得细菌的,会不惜一切代价把它们消灭掉。而这在黏膜附近就不可能了。

黏膜处的免疫系统小心处事
黏膜处的免疫系统不能像其他部位一样有攻击性,否则这些负责气体和营养交换的部位会遭到严重破坏,从而影响正常生活甚至造成死亡(许多患有自身免疫性疾病或过敏的人,就会遇到这种情况,不过我们后面再详细讲)。黏膜的免疫系统必须小心行事,被激活后要尽量把免疫反应限制在最必要的范围内。但同时,黏膜又是身体最薄弱的地方,免疫系统必须要能发挥保护作用,不能太过散漫,非常棘手。
针对入侵的第一项反制措施,就是把黏膜变成不适合危险微生物生存的死亡之地。为此,黏膜动用了一系列不同的防御系统。
如果说皮肤像广袤的荒漠,又像一道无法逾越的边境长城,黏膜就像一大片沼泽地,遍布着陷阱和巡逻的卫兵队伍。黏膜比皮肤容易入侵,但这也并不轻松。那么黏膜究竟是什么,又是怎样保护你的呢?
沼泽之国动用的第一道防线就是它本身的黏液层。黏液是细胞分泌的黏滑的物质,有点像水凝胶。你可能知道鼻涕,鼻子里面黏糊糊的东西,感冒时会流很多,看起来有点恶心。实际上黏液遍布身体里的各个地方:口腔、肠道、呼吸系统(包括肺部)、眼睑内侧等等。
与外界进行物质交换的通道表面上都有黏液。持续生产黏液的是杯状细胞,它们的样子很好玩,但我们主要讲免疫系统,就不过多介绍它们了。你可以把它们想象成被踩扁的怪虫子,不停地吐出黏液,形成了黏液层。
湿滑的黏液发挥着几方面的作用
它是一道阻止外敌入侵的物理屏障
假设你在满是污泥的泳池里游泳,还要潜到池底,而污泥有近百米深。黏液不仅是一层黏糊糊的屏障,还和皮肤荒漠一样充满了陷阱:有盐,有可以溶解微生物外部结构的酶,还有一些特殊的物质能耗尽细菌赖以为生的关键营养,从而饿死细菌。
大多数部位的黏液富含杀伤力强大的IgA抗体
所以黏液层本身就不是宜居之地。而且,黏液层不仅能为你抵御外敌,还能让你免受自身的伤害。你是否想过,胃里有那么多胃酸,你为什么没事?因为胃黏膜就是一层屏障,能保护胃壁细胞,令其不与胃酸接触。
黏液可以移动
黏膜表面是一层名为“上皮细胞”的特殊细胞,你可以把它们看成“内皮的皮肤细胞”。它的细胞膜上覆盖着形如发丝的纤毛,这种微小的细胞器聚在一起,连成大片的网。上皮细胞位于黏膜的最外层,就像“身体里的皮肤”,表面只有一层黏液。
有些地方黏液和身体内部之间只隔着薄薄的一层,一个上皮细胞的厚度。上皮可没有皮肤那么奢侈,能让好几百个细胞一层层摞在一起。也正因为这样,上皮细胞可不好对付。它们尽管不是真正的免疫细胞,但也发挥着重要的防御作用:它们特别擅长激活免疫系统,以及释放特殊的细胞因子以请求支援。它们就像民兵组织,虽不能与敌军旗鼓相当,但在身体遭遇入侵时,是非常有用的补充性防御力量。
上皮细胞的任务之一,就是借助细胞膜上的纤毛让黏液流动
有些微生物会借助纤毛移动,而上皮细胞则是摆动纤毛让表面的黏液一起有节律地流动。摆动的方向取决于上皮细胞的位置。呼吸道、鼻腔、肺里的黏液,要么从口鼻排出体外,要么绕个弯被吞进胃里。
我们在一生中会吞下相当多的黏液,虽然听起来可能有点恶心,但这种机制很是有用。毕竟,胃里充满了强酸性的胃液,绝大部分病原体都无法幸存。而肠道上皮细胞纤毛运动的方向也很清楚:从胃向肛门——吃进来的东西都从这里要排出去。
沼泽之国——合作联盟
不过黏膜形成的沼泽之国不是单一国家,而更像彼此迥然不同的多个国家,为了共同目标而结成的合作联盟。
比如在荒漠之国皮肤的情况中,脚底和腰部的皮肤厚度就有很大差异。相应的,肺部黏膜和肠道黏膜有着完全不同的功能,而女性生殖道黏膜的功能又大不一样。根据不同部位黏膜特化情况的不同,相应的免疫系统也各有特点。
在讲病毒之前,我们先看看肠道这个神奇的国度,看看它是怎么和足足几十万亿细对免疫系统来说,肠道是个非常特别的地方,因为在这里,免疫系统要应对许多复杂的挑战,才能保持身体健康和正常运转。
我们还是来把肠道想象成贯穿人体的长管道,它把一点“体外”裹进了“体内”。在这些“外”表面,即肠道黏膜上,生活着1000多种细菌,数量多达三四十万亿,还有几千种病毒,它们共同组成了“肠道微生物群”(绝大多数肠道病毒攻击的是肠道细菌而非人体)。
关于免疫系统和肠道微生物组各自的功能及其相互作用,我们还很缺乏了解。许多疾病和失调都跟两者间的失衡有关,不过要完全理解两者间的各种关系,还需要大量的研究。未来几年,我们很可能会有重大突破。
在本章中,我们会探讨免疫系统是怎样和如此众多的微生物和平共存的。
首先,肠道免疫系统是一个半封闭系统,它会试图和身体其他部位免疫系统保持一定的界线。
肠道黏膜遇到的最大挑战,就是它的防线一直在被攻破。对肠道来说,外界的侵扰永不停歇,肠道免疫系统要持续响应,分辨敌友,这和身体其他任何地方都不一样。肠道是个繁忙的地方,除了生活着组成肠道微生物组的几十万亿有机体之外,还有吃进来的所有东西。
食物消化
食物的旅程始于口腔。食物要被消化吸收,变成身体和细胞养分,第一步就是被牙齿磨碎,并被唾液浸润、炮制。唾液中含有一些可以分解食物的化学物质,从你吃第一口开始,整个消化过程就拉开了序幕。
从进食到把废物排出体外,吸收营养的时间是有限的,所以越早开始越好。嚼碎的食物接着会被吞下去,在胃酸之海里停留片刻。胃酸不仅可以帮助消化,分解结实的肉类和植物纤维,它还能淹没并杀死大量微生物,从而大大减轻免疫系统的负担。
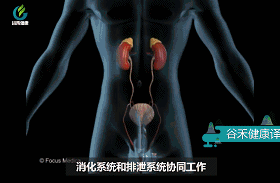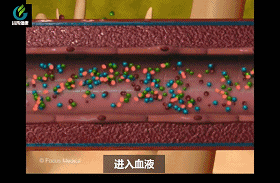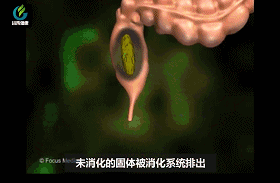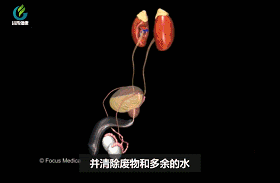
经过胃部后,食物就到了肠道。肠道长约3—7米,是最长的一段消化道。90%以上人体所需的营养都是在肠道吸收的。这里生活着大量人体必需的细菌伙伴,它们进一步分解食物,便于身体吸收营养。但这些可不是随便什么细菌。
几百万年以前,人类祖先和某些微生物群做了个小约定:
肠道细菌——租客;生成的营养物质——租金
人允许它们住在温暖的肠道里,并提供源源不断的食物;作为交换,这些微生物负责分解人类自己无法分解的碳水化合物,并生成人体自身不能合成的维生素。
肠道细菌:亦友亦敌
这些细菌就叫“[偏利]共生细菌”,“共生”(commensal)一词来自拉丁文,意为“同在一张桌前”。就和生活在皮肤表面的那些细菌“部落”一样,肠道的共生细菌也是我们的朋友。这些细菌只要不危害人体健康,免疫系统就不会攻击它们,这就是双方守约定的最理想情况。
为了维持秩序与和平,肠道细菌也像皮肤表面的细菌那样,生活在肠道黏膜的表面之上。只要肠道细菌不越线,不侵入上皮层,双方就相安无事。当然,事情不会这么简单。
细菌可不是人类真正的朋友,它们不知道什么约定,也不会去遵守任何东西。而肠道面积辽阔,细菌多得惊人,时刻都有一些共生细菌侵入肠道内壁。这就不太妙了。要是这些细菌进入血流,即进入真正的“体内”,会带来可怕的后果,甚至危及生命。而黏膜的一大目标就是防止这种情况。
肠道黏膜有三层
1
先是黏液层,内含大量抗体、防御素(前面讲皮肤时提过,它们就像微型针头,能杀死微生物)和其他一些能杀伤或杀灭细菌的蛋白质。肠道的黏液层很薄,且要有一定的通透性,好让食物中的营养物质通过,要是第一层保护太厚,你可能就要饿死了。
2
黏液层之下是肠道上皮细胞,它们是人体内外之间的真正屏障。和肺部相似,肠道的上皮层也只有一个细胞那么厚。为了更好地起保护人体内部的作用,肠道上皮细胞彼此连接得非常紧密,被特殊的蛋白紧紧粘在一起,就像真正的墙一样牢固。免疫系统严密监控着这里的情况,对于一切企图黏附到上皮细胞上的微生物,它都会毫不客气。
3
共生细菌突破防护墙的情况每时每刻都在发生,所以上皮细胞下面还有第三层,“固有层”,这里是肠道免疫系统的大本营。

在紧贴着黏膜表面的固有层之内,有特殊的巨噬细胞、B细胞和树突状细胞随时待命,等着收拾那些不速之客。
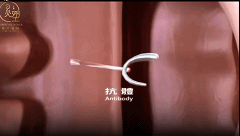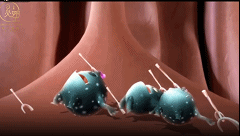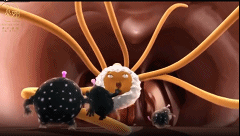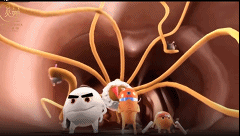
免疫系统会极力避免引发炎症
因为炎症会让肠道产生大量多余的液体,造成腹泻。腹泻不仅会使大便呈水样,还会损伤负责从食物中吸收营养的、敏感纤薄的上皮层,并迅速导致重度脱水。
很多人不知道,腹泻现在仍是重要的死因,每年约有50万儿童因此死去。所以,从几百万年前,人类这一物种登上演化的舞台开始,人体和人体免疫系统就明白要谨慎对待肠道炎症。
守卫肠道的巨噬细胞有两个特点:
一、很擅长吞噬细菌;
二、不会释放召集中性粒细胞进而引发炎症的细胞因子。
这里的巨噬细胞更像是无声的杀手,不慌不忙地吃掉越界的细菌,不会大呼小叫。

编辑
肠道的树突状细胞也很特别
大量树突状细胞直接待在上皮细胞下面,将长触手从上皮细胞之间挤过去,直接伸入肠道黏液。这样,它们就能对不安分的调皮鬼,即想侵入体内的细菌,不断进行采样。
这里就埋藏着免疫学中的一大未解之谜,将来解开谜题的人定能获得诺贝尔奖:
树突状细胞怎么知道它采样的细菌是危险的病原体,还是无害的共生细菌?
目前我们还不知道答案;我们知道的是,采样是共生细菌时,树突状细胞会让肠道免疫系统保持冷静,不要为这些细菌携带的抗原而烦恼。
肠道还有特殊的B细胞,只生成大量的IgA
这些抗体特别适合在黏液中工作,简直就是为肠道的环境而设的:
这两点对肠道来说都很重要。
IgA还有别的长处:
它们有四只钳子,伸向两个相反的方向,这让它很擅长抓住两种不同的细菌,并把它们粘在一起。于是,大量IgA可以把无法反抗的细菌黏成大团,这些菌团会成为粪便的一部分,被排出体外。毕竟,粪便有三成多都是细菌,其中相当多的细菌是被IgA粘在一起的(让人担心的是,其中约一半的细菌在被排出时仍然是活菌)。
探索肠道免疫系统的守护之路
肠道免疫系统默默地守护着你,确保内部和外界的细菌不会作乱。有了这些机制和特殊细胞,免疫系统就保证了肠道黏液不会被野心太大的共生细菌所侵扰,同时自己也不会因过激而损伤身体,如此看来,肠道免疫系统可真是一支维和部队。
但要是有真正的敌人入侵,比如有致病菌经受住了胃酸的洗礼,存活到了肠道,这些机制就成了可怕的漏洞。此时,为了尽快捕获敌人,肠道有一种名为“派尔集合淋巴结”的特殊淋巴结,它们直接长在肠道上。

会有“微皱褶细胞”直接伸入肠壁,对免疫系统可能感兴趣的对象进行采样,某种意义上它们有点像电梯,装上乘客后会直接把它们送进派尔集合淋巴结,让适应性免疫细胞能检查肠道的各种情况。这样一来,肠道就相当于拥有了超快速的免疫筛查,可以持续地密切监测肠道的细菌组成。
本文选取了《战斗细胞:人体免疫系统奇妙之旅》该书中,与肠道免疫相关的几个章节作为分享,希望大家能更轻松地了解关于肠道免疫相关的知识,进一步引发对肠道免疫的思考和探索。让我们共同努力,推动肠道免疫健康知识的普及与实践,走向更健康美好的生活。
本文用于分享和交流,部分图片如有侵权请联系我们删除。

谷禾健康

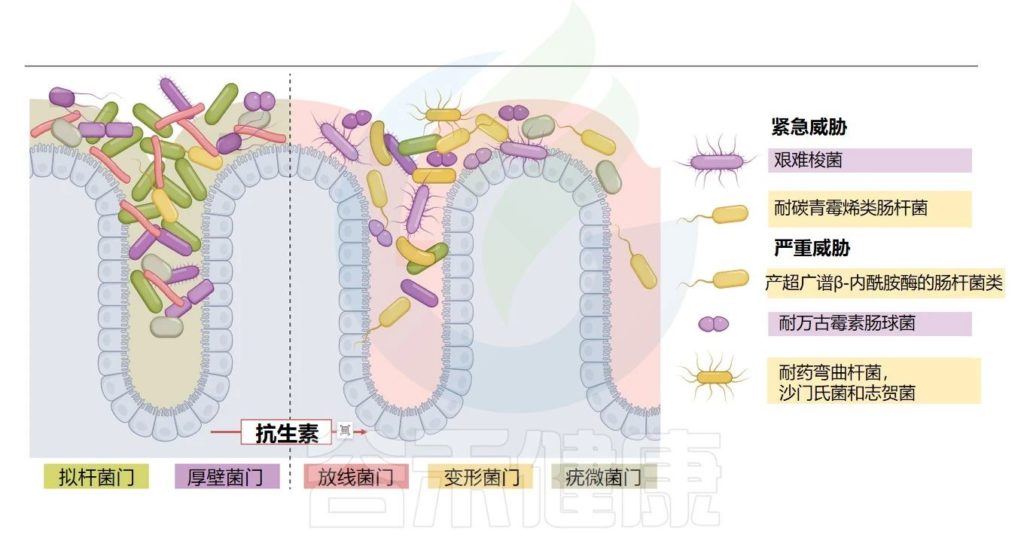
抗生素耐药性是当下和未来几十年人类健康将面临的重要挑战之一。从2000年到2015年,抗菌药物的普及导致人类抗生素消耗量增加了约65%,2015年后这个数据还在增加。最明显的后果是抗生素耐药性的增多,这使得近年来常见的感染难以治疗。
最近,由于担心出现更难治疗的肠道感染和其他感染,以及耐药病原体可能传播给更脆弱的人群(如小孩和老人),人们对使用抗生素进行严格管理和开展精准用药算是迈开了第一步。
抗生素的使用经常产生脱靶效应,这意味着抗生素的使用浓度所引发的抗菌作用超出了消除影响病原菌的作用。下一代靶向测序已证明抗生素会导致肠道细菌生物量、分类多样性和功能多样性减少。
除了耐药性之外,抗生素未能清除感染的原因还有多种。感染灶中药物的药代动力学较差或生物膜(下期专门讲)等受保护的生态位中细菌的持续存在会导致治疗失败。
此外,在感染过程中,一些细菌(持续存在)可能会由于进入抗生素持久性状态而在药物治疗后达到致命的浓度。然后会引起长期和反复感染,导致最终治疗失败。
感染期间抗生素的持久性涉及持久性在宿主免疫反应和药物的联合作用下生存的能力。病原体已经进化出多种方式来击败宿主的免疫反应,并且它们的毒力因子已经被仔细研究了几十年。然而,细菌通过持久性抵抗抗生素暴露的能力仍然更加难以捉摸。
虽然抗生素暴露对病原体清除的正向影响有利于宿主健康,但抗生素暴露对肠道微生物组组成和功能的脱靶影响可能会对宿主健康产生有害或有益的影响,具体取决于初始成分和原生微生物群落的功能。
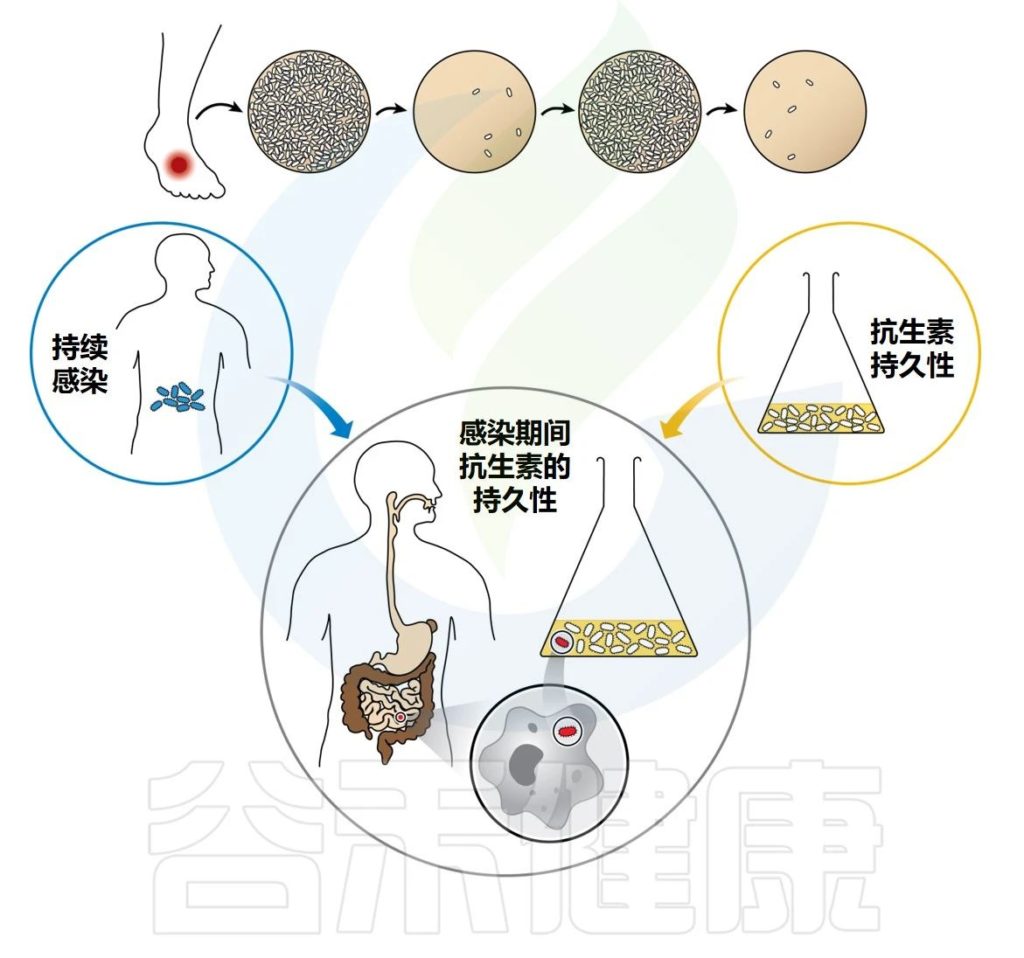
预测抗生素耐药性进化轨迹以及耐药风险的能力对于调整抗生素的给药方案以最大限度地延长其使用寿命具有重要价值。
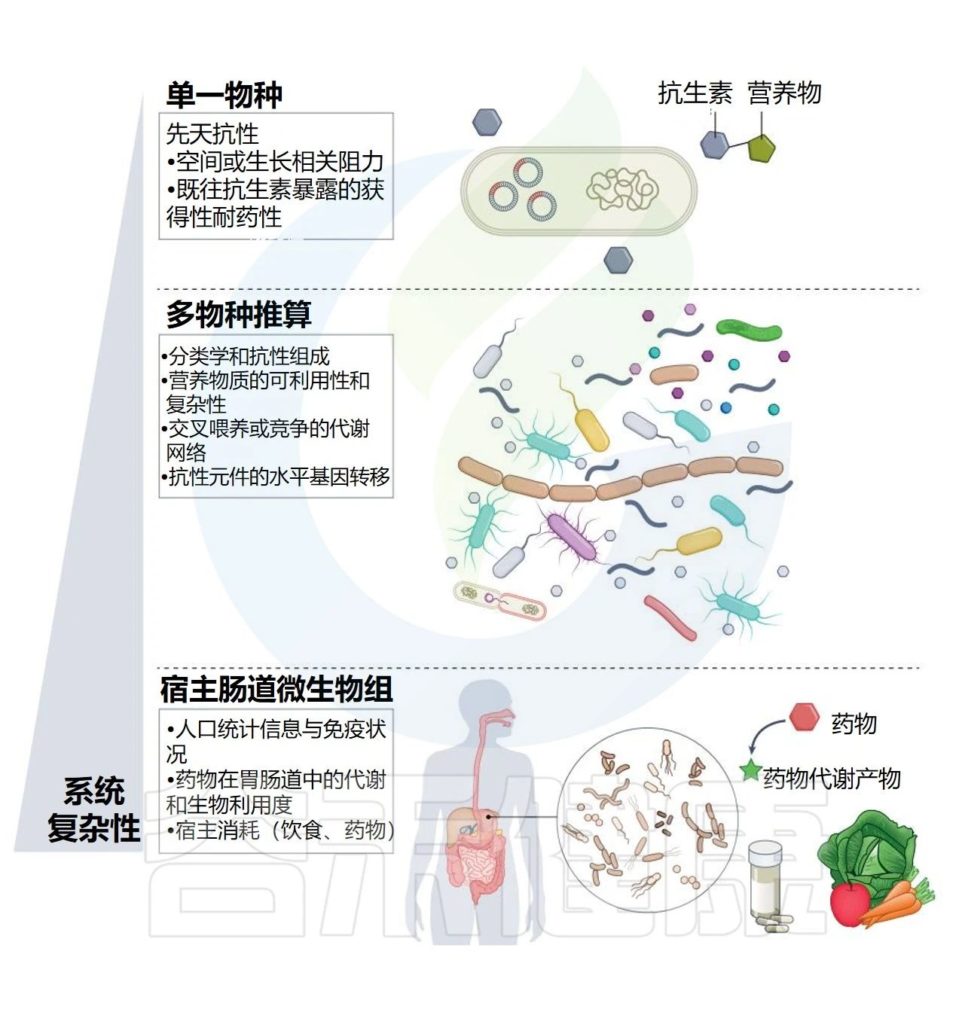
本文将讨论抗生素持久性以及微生物持久性形成在感染中的重要性,了解肠道微生物群如何对抗生素扰动做出反应以及对特别人群的影响、影响抗生素暴露对微生物群影响的因素、抗生素引起的微生物群改变的潜在健康结果以及有可能改善这些问题的策略。
抗生素持久性领域仍处于起步阶段;然而,关于定义的争议和讨论已经模糊了关键概念及其各自的重要性。在这里,采用的定义和说明与最近几十个研究小组的文章一致。
Gollan B,et al.Annu Rev Microbiol.2019
确定的定义主要集中在体外研究的细菌上,并以最小抑制浓度(MIC——明显抑制细菌生长的药物最低浓度)和杀死99%数量细菌所需的最少时间(MDK99)的测量为中心。
尽管这些测量对于区分允许细菌在实验室培养物中暴露于抗生素后存活的不同现象非常有价值,但它们在感染模型中的工作中难以进行。
因此,研究者们更多地关注不同概念的描述性定义。重要的是,一些导致抗生素暴露存活率增加的细菌行为(即抗生素持久性和表型耐药性)涉及复制群体内的表型异质性,因此最好通过群体和单细胞特征的组合来描述。
▼
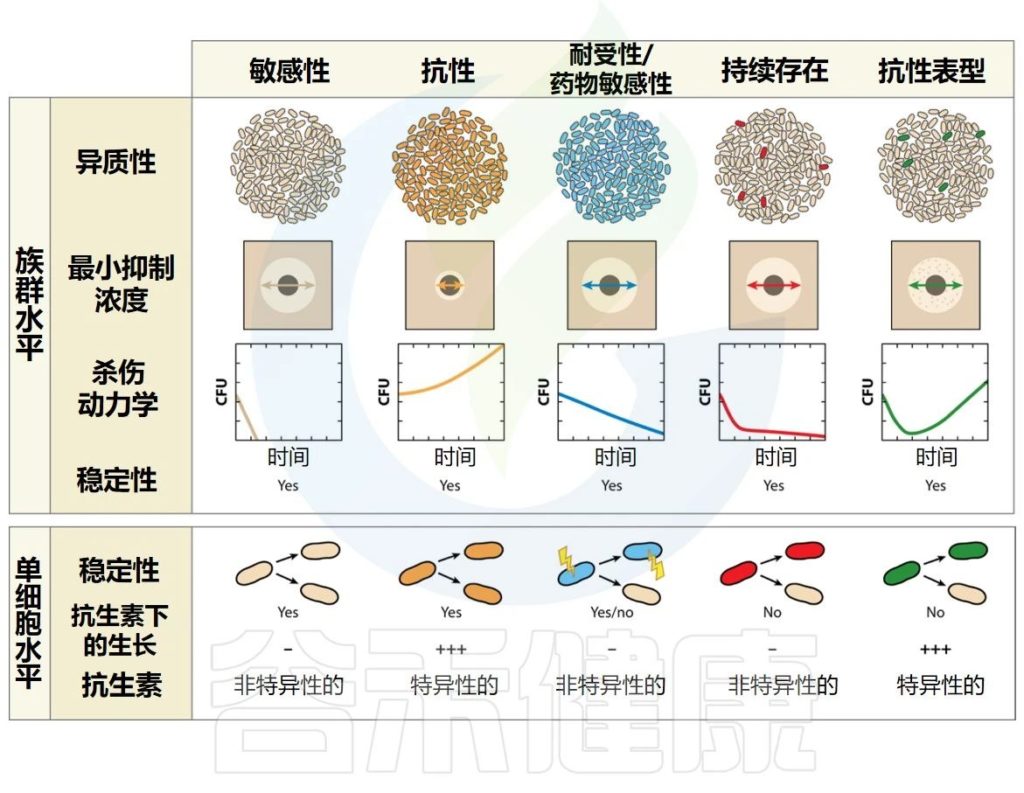
▸ 细菌的耐药性
当细菌能够在对同一物种的参考菌株致死的药物浓度下增殖时,细菌被认为对抗生素具有耐药性。
耐药性通常(但并非总是)特定于一种药物或一类药物,并且是由于非耐药亲本菌株的基因修饰(从头突变或通过水平基因转移获得)造成的。
遗传性状会代代相传
一旦获得,遗传性状就会代代相传。产生耐药性的主要原因是药物或药物靶点的输出或修改。在体外,可以通过测量增加的明显抑制细菌生长的药物的最低浓度。
▸ 群体的耐受性
当暴露于杀菌药物时,整个细菌群体可能表现出较低的杀灭率,因为群体的增长率因遗传或环境因素而降低。因此,与普通菌株相比,生长缓慢的细菌表现出相对的药物冷漠,这增加了种群在抗生素暴露下存活的时间。
从理论上讲,足够长的时间接触抗生素应该可以清除整个人群。营养缺陷型突变代表了这种耐受现象的一个极端例子。当环境条件发生变化允许恢复快速生长时,种群就会恢复到易感表型。在体外,可以通过测量杀死一定比例的群体的持续时间来量化耐受性。
▼
抗生素持久性和表型耐药性代表了抗生素存活方面的异质行为。“持久性”与“耐药性”一词指的是两个不同的概念,它们被认为是有联系的,但又是不同的。因此,它们不能互换使用。
▸ 感染持续存在
当病原体有能力在宿主体内停留较长时间时,无论宿主免疫反应如何或长期抗生素治疗如何,感染都会持续存在。
据信,持续感染至少部分依赖于感染人群的耐受性(或药物冷漠)以及抗生素的持久性,但这种联系仍然很脆弱。迄今为止,抗生素持久性领域主要忽视了持久性细菌在宿主体内生存的能力。
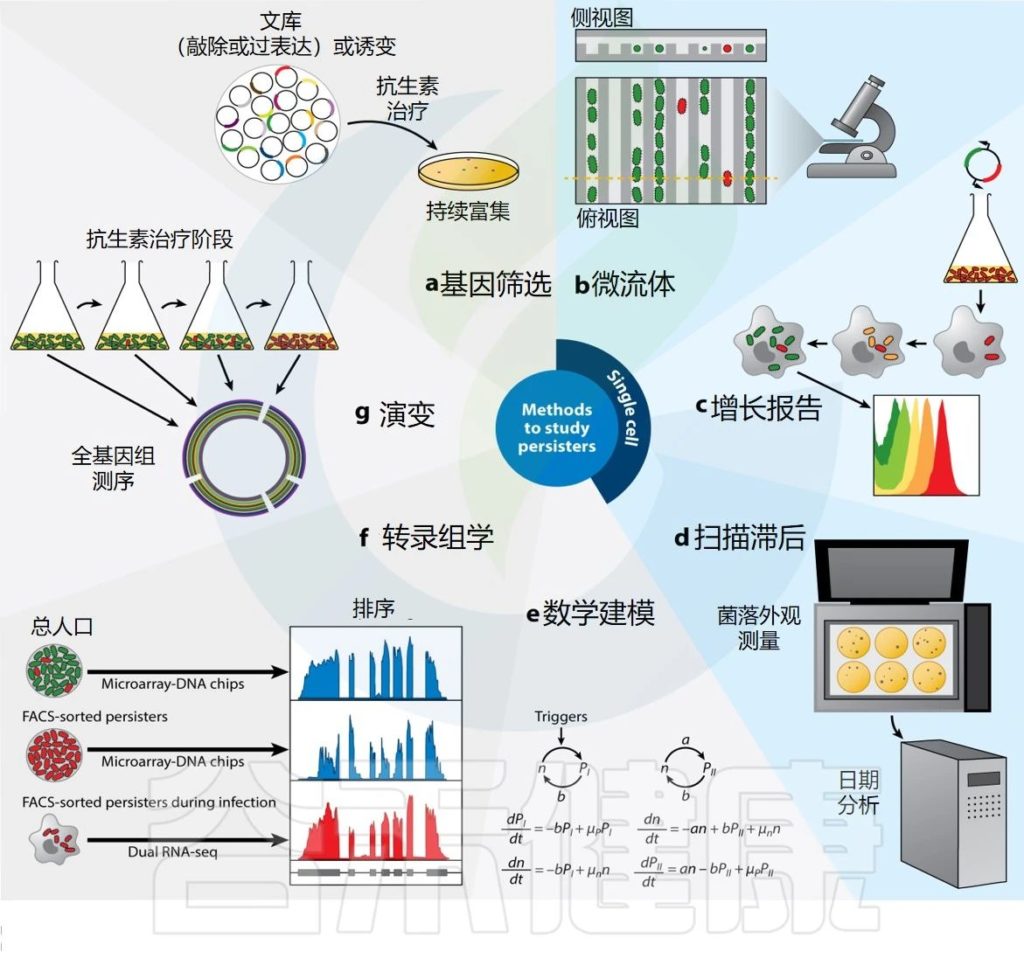
Gollan B,et al.Annu Rev Microbiol.2019
▸ 抗生素持久性
药物暴露后的这种异质群体行为是由于细菌亚群(持续存在)通过生长停滞、暴露于对其余群体致命的浓度的抗生素而得以生存的能力。
由于持续者是非定植者,因此他们的存在不会影响整个种群的MIC。在种群水平上,这种现象在几代群体中都是稳定的,这表明种群内的所有细菌都具有持续存在的遗传装备。
MIC——明显抑制细菌生长的药物最低浓度
抗生素持久性与压力有关
抗生素持久性很大程度上是由 “压力” 引发的,最好在压力缓解后进行评估,以免与耐受现象混淆。
事实上,与耐受人群相反,持久人群携带着压力暴露的记忆。由于现象的异质性,通常可能需要允许单细胞分析的工具来获取有意义的信息。
▸ 持续者
持续者是抗生素持久性种群中的个体,当暴露于对其他种群中的亲属致命的浓度的抗生素时,它们能够通过生长停滞而生存。
持久菌在药物中存活的能力是短暂的,并且持久菌后代是异质的,由易感细菌和持久菌组成。在体外,群体中存留菌的存在可以通过双相杀伤曲线来揭示,该曲线显示存活率与时间的关系,其中易感细菌被快速杀死,存留菌存活时间更长。
▼
表型耐药性是在药物暴露时可能发生的另一种异质行为,这是由于细菌亚群(耐药性)在抗生素浓度达到对其余菌群致命的情况下能够增殖的能力。
整个群体的最小抑制浓度与易感菌株相当
由于群体主要由易感细菌组成,因此整个群体的MIC(明显抑制细菌生长的药物最低浓度)与易感菌株的MIC相当。在种群水平上,这种现象在几代菌中都是稳定的,这表明种群中的所有细菌都具有抵抗力的遗传装备。
▸ 休眠
休眠是一个用于定义一系列特征的术语,从缺乏任何可测量的细胞活性到生长停滞。在这篇文章中,我们用它来表示缺乏活性。
从体内感染模型中积累的新证据强调了持续感染者在持续感染中的重要性。持续感染者在感染复发中的潜在作用是对其进行研究的重要原因。
★ 开发研究持久性的技术
由于这种生理状态的短暂性和可逆性,以及相对稀有的残留物可以产生少量的生物材料,研究抗生素残留物具有挑战性。
长期以来,最常见的方法是从基因或环境角度操纵细菌种群,并测量其对持久性水平的影响。虽然这使得能够识别涉及调节群体中持续者水平的压力和途径,但近年来通过建立能够跟踪、收集和分析持续者的方法,取得了重要突破(下图)。
Gollan B,et al.Annu Rev Microbiol.2019
很快,人们就清楚地意识到重点应该放在单细胞研究上,因为持久性细菌只是细菌种群的一小部分。微流体装置与显微镜和流式细胞术相结合的使用成为一种选择技术。微流体背后的基本原理是分离单个细菌,以便使用延时显微镜来跟踪个体随时间的行为,以检查持久细胞的历史。
然而,微流体带来的对单个细菌的直接可视化和跟踪的能力受到可同时评估的细胞数量有限的阻碍。因此,流式细胞术提供了一种替代方法,允许以高通量方式进行单细胞分析,并通过荧光报告基因的发展,增强对感兴趣细胞进行分选的能力。
因此提出休眠(代谢活动的缺乏)对于进入持久性既不是必要的,也不是充分的。第一个开发荧光稀释技术来在单细胞水平上追踪细菌分裂的人。这种方法类似于免疫学家用来追踪T细胞增殖的羧基荧光素琥珀酰亚胺酯 (CFSE) 染色。
高通量下一代测序技术的兴起,随着生物信息学分析的改进和此类平台的可访问性的增加,也为持久化领域带来了视野。
最后,除了用于跟踪持久者的实验室技术之外,数学模型还允许对持久者行为进行计算机预测。例如,已经建立了一些模型来重点了解单细胞水平上持久物形成的分子机制。
▼
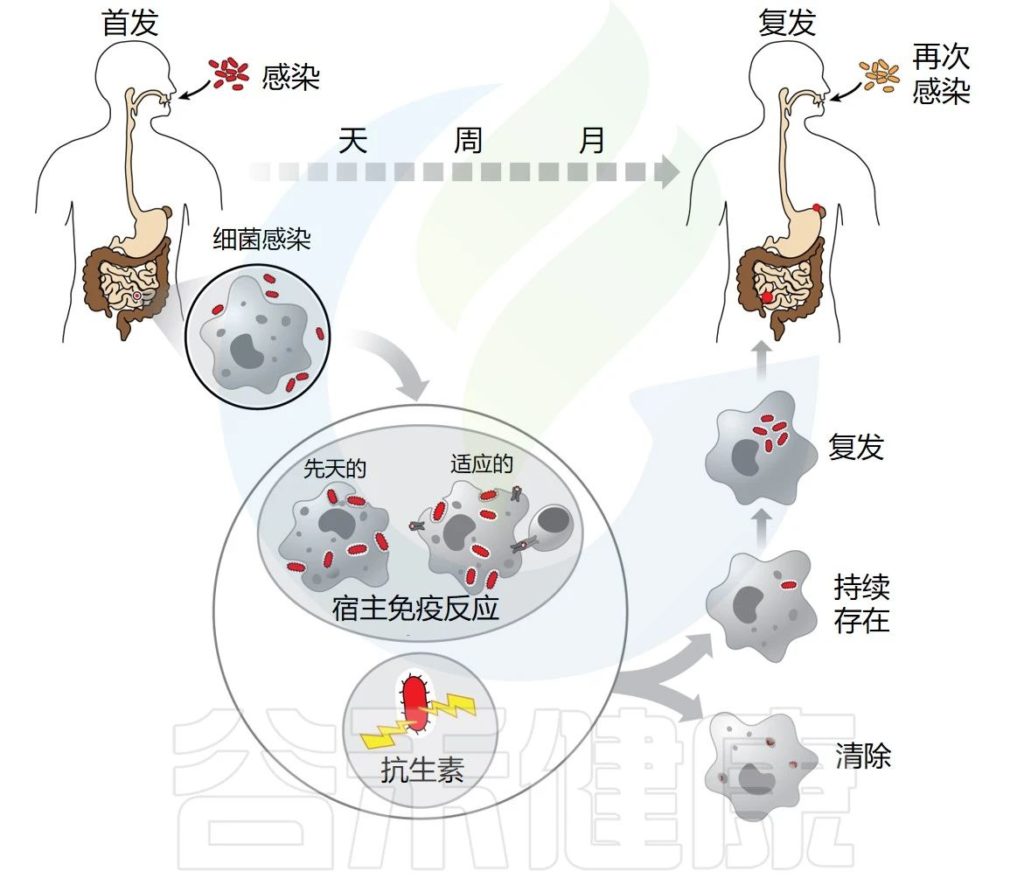
许多病原菌在宿主组织内建立持续感染,避免被免疫系统和抗生素清除。细菌感染通常会导致宿主产生先天免疫反应,然后产生适应性免疫反应,这通常足以清除感染。然而,许多病原体已经进化出复杂的生存机制来逃避免疫反应并建立持续感染。
// 急性结核感染
急性结核感染是细菌躲避免疫系统且相对难以治疗的典型例子,导致需要长时间的抗生素治疗才能治愈感染。
然而,分枝杆菌感染有时只能部分清除,并且10%的患者在其一生中会出现复发。复发不仅出现在结核病中。5%至10%的患者会出现伤寒复发。
// 其他感染
尿路致病性大肠杆菌引起的尿路感染(UTI)后,20%至30%的女性会再次出现症状;许多其他细菌感染在治疗后往往会复发。
✦许多感染会反复发作
“复发”一词用于描述这样一种现象:最初感染的症状暂时消退,直到首次感染时未清除的细菌库重新激活。
有多种原因会导致感染反复发作,例如宿主易感性增加、其他慢性合并感染以及环境和生活方式因素。
较难确定是再次感染还是复发
长期以来,无法确定感染复发是再次感染还是复发。当初次感染完全清除后,个体出现新的感染时,就会发生再感染。
▼
抗生素残留物研究中投入的许多努力都围绕着调查残留物形成的触发因素和途径。
✦持久性的形成是一种细菌应激反应
但一个总体观察结果是,持久性的形成是一种普遍的细菌应激反应。多种环境因素充当压力信号,促进持久者的产生。
此外,大多数已被确定参与持久物形成的分子途径都是应激反应元件。我们首先回顾了持续细胞形成的触发因素,然后回顾了持续细胞形成所涉及的分子途径和效应器(下图)。
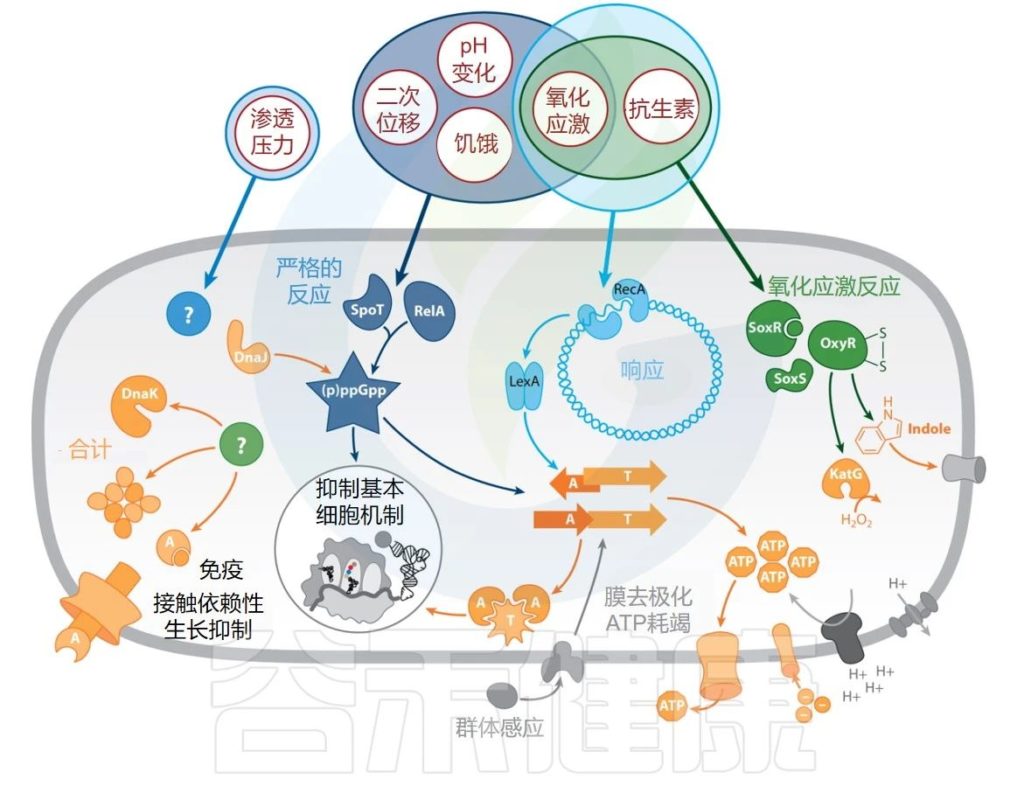
Gollan B,et al.Annu Rev Microbiol.2019
抗生素持久性的触发因素、途径和效应器。持久性的外部触发因素以红色圆圈显示。应激反应途径为蓝色或绿色(已识别的效应器为橙色,受效应器影响的细胞成分为灰色)。
缩写:A—抗毒素;T—毒素。
▼
选择压力可能是最难确定和纳入模型的因素。因此,在人类、动物和其他环境中,细菌病原体暴露于各种选择剂,这些选择剂通常存在于复杂的混合物中(例如抗生素、杀生物剂),并且其浓度可能随着时间的推移而发生很大变化。因此,在实验室环境之外通常很难确定选择的强度。
✦抗药性的选择具有较大差异
此外,抗药性的选择过程将有很大差异,具体取决于药物浓度是否足够高以(a)阻止病原体生长(如果> MIC则致命选择,如果< MIC则非致命选择)和(b)允许敏感和耐药菌同时生长。
在前一种情况下,在应用选择之前需要存在抗性突变体,富集率由群体中突变体的数量及其在特定抗生素浓度下的适应性决定。
然而,在最小抑制浓度选择过程中,可能会出现突变体,其富集率由群体中突变体的数量以及易感细胞和耐药细胞之间的适应度差异决定。因此,致死和非致死选择压力的出现率和选择的突变体类型有所不同。
高致死抗生素浓度
在高致死抗生素浓度下,在一个遗传事件中提供高水平耐药性的罕见的预先存在的大效应突变往往会被选择性富集(下图)。
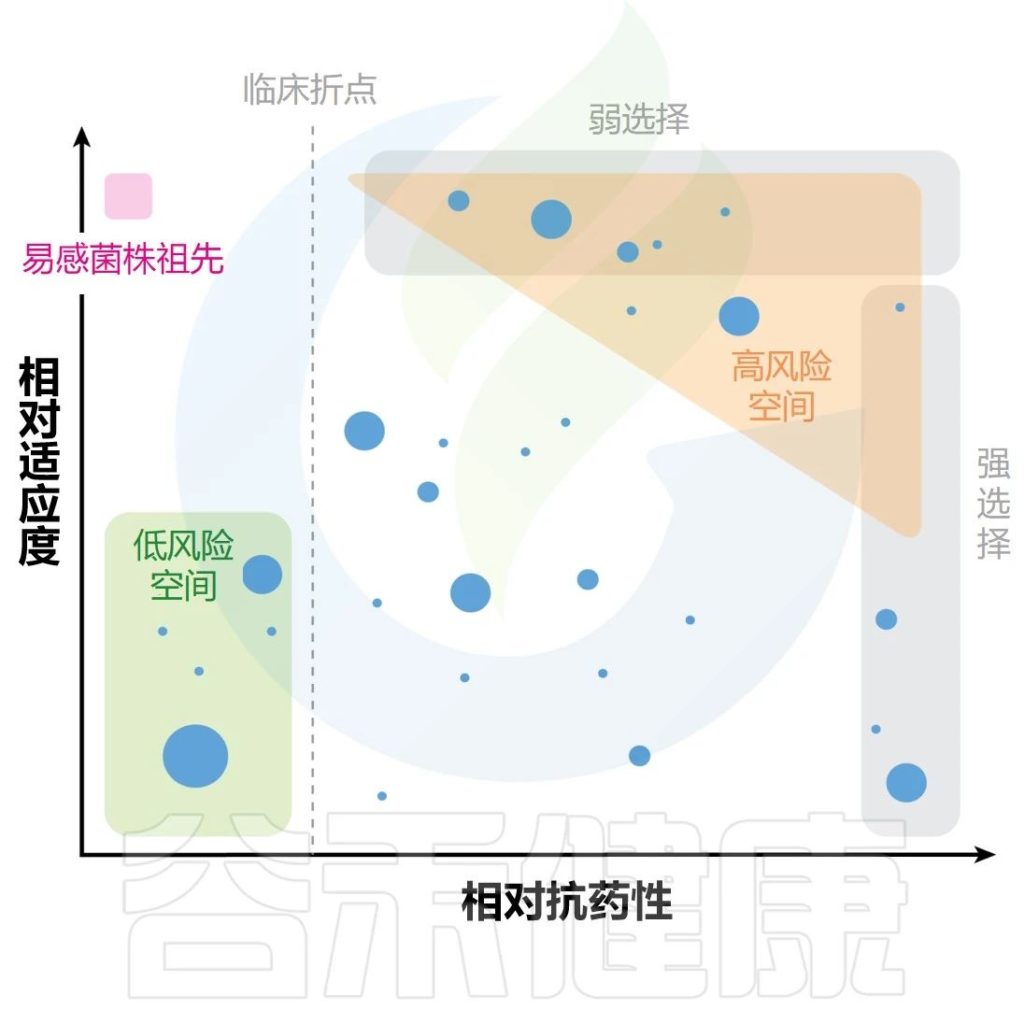
Gollan B,et al.Annu Rev Microbiol.2019
非致死性抗生素浓度
相反,在非致死性抗生素浓度下,富集的突变体通常是由许多对耐药性影响较小的突变引起的(但它们组合起来可能会产生高水平的耐药性)。
重要的是,选择越弱(即易感细菌和耐药细菌之间的适应度差异越小),对于低适应度成本的突变体(上半部分)的富集就越强。
此外,由于在弱非致死选择压力下逐步选择连续的小效应突变,通过突变或水平基因转移获得抗性的概率增加的突变菌株得到富集。
注:矛盾的是,较弱的非致死选择可能比致死选择更富集有问题的突变体(高适应度菌株和突变体)。
许多触发因素似乎都会刺激持久性的形成,从饥饿或氧化应激等营养应激再到细胞外代谢信号。
几十年来,人们都知道抗生素的杀灭率取决于细菌的生长速度,而细菌的生长速度又取决于碳源的可用性,因此控制整个细菌群体的抗生素耐受性。
✦营养缺乏是持久性形成的诱因
此外,营养缺乏似乎是持久性形成的主要诱因之一。例如,氨基酸或氮饥饿会增加铜绿假单胞菌(Pseudomonas aeruginosa)、变形链球菌(Streptococcus mutans)和大肠杆菌(Escherichia coli)等不同物种中经抗生素治疗后存活下来的细菌数量。
! 手术或抗生素治疗后患者的营养非常重要
这就涉及另外一个主题,就是手术或抗生素治疗后,患者的营养,尤其肠内营养的监测或补充非常重要。
还有充分的证据表明,细菌培养物在进入稳定期及以后往往会开始积累持续细胞,然后持续细胞的比例达到与生物膜中所见的水平一样高。事实上,研究表明,营养和氧气有限的生物膜中含有非常高比例的持久物。
除了饥饿造成的剧烈压力外,碳源的变化会破坏代谢通量(可被视为暂时饥饿),也会导致大肠杆菌的持续存在增加。
这种碳源转变伴随着严格反应关键介体四磷酸或五磷酸鸟苷 [ (p)ppGpp ] 水平的大幅增加,这可能是饮食模式改变或节食诱发的。
重要的是,通过营养转换形成的持续细胞似乎维持高 ATP 水平,从而允许非生长相关过程的继续,例如维持膜电位。
除了营养变化之外,氧化、酸性和渗透应激也被认为是持久物形成的触发因素。例如,用过氧化氢处理大肠杆菌培养物或通过向培养基中添加水杨酸盐产生活性氧会导致持久性细菌的增加。
✦吲哚促进持久细胞的形成
同样在大肠杆菌中,吲哚(一种色氨酸生物合成中间体,其产量因氧化应激而增加)被证明可以促进持久细胞的形成。
此外,无法合成海藻糖的大肠杆菌突变体产生的氧化应激与吲哚分泌增加和持久物形成同时发生。
暴露于吲哚的细菌上调了它们的oxyR调节子和噬菌体休克pspBC途径,并且这些途径的缺失降低了吲哚促进持久细胞形成的能力。
沙门氏菌也观察到对吲哚的类似反应,伴随着katG 过氧化氢酶表达的增加,有趣的是,沙门氏菌不产生吲哚。然而,这可能有利于肠道中的沙门氏菌,肠道中的沙门氏菌在感知来自其他微生物群的吲哚时升高的氧化防御反应可能会保护其免受伴随局部炎症的氧化应激。
✦pH值变化和高渗透压也促进持久性形成
pH值变化和高渗透压是促进持久物形成的其他压力。此外,抗生素本身也可能引发持久性细菌的形成,例如在大肠杆菌或金黄色葡萄球菌中,其中用亚抑制浓度的各种抗生素进行预处理显著增加了残留物水平。
✦氧化应激和抗生素诱导细菌群体的持续存在
重要的是,暴露于逐渐增加的抗生素浓度的肺炎克雷伯菌(Klebsiella Pneumoniae)临床菌株显示出持续增加的水平,以及随后出现的耐药变异体。
这一观察结果可能特别相关,因为在临床中抗生素输送和药代动力学通常会导致血液中抗生素浓度逐渐增加。这可能会增加持久菌群的规模,从而增加产生耐药性的细菌库。
同样,氧化应激和抗生素诱导的DNA损伤的结合已被证明可以诱导细菌群体的持久存在。总体而言,许多非生物胁迫导致持久细胞形成增加,大量研究充分支持了这一点,表明主要的细菌胁迫反应是这种表型的强有力的协调者。
当暴露于细菌分泌到周围环境中的细胞外代谢物时,也可以刺激持久细胞的形成。
✦细胞外代谢物也刺激持久细胞的形成
除了上面引用的吲哚的例子之外,能力刺激肽CSP(一种群体感应分子)被证明可以促进变形链球菌(Streptococcus mutans)持久细胞的形成。
这种群体感应分子似乎是一种中间体,向其他群体发出压力信号,以便其做出适当的压力反应,包括持久性形成。同样,一些假单胞菌群体感应分子不仅可以增加假单胞菌的存留细胞分数,还可以增加通常从囊性纤维化患者中分离出来的不动杆菌和伯克霍尔德氏菌的存留细胞分数。
因此,似乎不仅种内信号传导而且种间信号传导也可能促进持久物的形成。
在感染过程中,细菌会经历上述压力的组合,其强度和时间各异,可能有利于持久细胞的形成。
这提出了一种可能性,即在受感染宿主中形成的细菌持续存在可能独特地适应了它们最初形成的触发因素,并且与从生长的细菌中分离出的细菌不同,后者通常使用单一应激因子来诱导细菌持久化状态。
✦宿主巨噬细胞诱导持久化
就沙门氏菌而言,它在宿主免疫骨髓细胞中存活,其持续水平在被小鼠巨噬细胞内化后急剧增加。
结核分枝杆菌持续细胞的形成也是由与宿主巨噬细胞的相互作用触发的。
▼
鉴于刺激存留细胞形成的不同应激,看到相应的应激反应途径也在存留细胞形成中发挥作用也就不足为奇了,这可能会在细菌暴露于应激和最终导致长期生长停滞的分子效应物之间架起桥梁。
▸ 严格响应
由于饥饿是持久细胞形成的重要触发因素,因此对于严格反应和持久细胞之间的联系已经进行了大量的工作。
严格反应的关键介质是警报素 (p)ppGpp,在营养饥饿时被激活。(p)ppGpp 水平升高,与DksA协同作用,导致转录和翻译重排,使细菌能够适应稀缺资源。
当暴露于低pH值或氧化条件时,DksA会发生影响其调节功能的构象变化,因此,严格的反应不仅与饥饿有关,而且与氧化和pH应激因素有关。
影响 (p)ppGpp 合成或 DksA 的突变会显著降低革兰氏阴性和革兰氏阳性细菌中的存留细胞分数。
▸ SOS反应
SOS反应是一种DNA修复机制,在暴露于各种环境应激因素(氧化应激和抗生素暴露)时被激活,这些都是持久性的一般触发因素。
这些压力会造成损伤,产生单链DNA,而单链DNA 又会被RecA蛋白识别。RecA 随后激活LexA阻遏物的蛋白水解活性,最终导致细胞内的全局转录重排。去除这些主调节因子中的任何一个都会导致持续细胞的大量损失。
▸ ATP水平下降
在大肠杆菌和金黄色葡萄球菌中,持久性与细胞内ATP下降有关。
在这两种生物体中,处于指数期但表现出稳定期标记表达的细胞富含持久细胞。此外,用砷酸盐(一种已知能降低ATP浓度的化合物)处理呈指数增长的细菌,会导致持续细胞的大幅增加。
因此,ATP水平的降低可能是大多数细菌过程(例如翻译)关闭的普遍模式,从而导致抗生素在休眠期间持续存在。
一般来说,暴露于抗生素通常会导致微生物群落结构、物种组成和代谢能力发生重要变化。
▼
阿奇霉素是世界卫生组织消除沙眼和雅司病战略的重要组成部分。
雅司病是由雅司螺旋体引起的慢性接触性传染病,病原为雅司螺旋体,亦称纤细螺旋体。
五项随机对照试验研究了阿奇霉素预防性使用对普通人群肠道微生物群的影响。肠道微生物组的特征主要在于α多样性,它量化了单个微生物群中不同细菌类群的数量及其丰度的均匀性。
•阿奇霉素治疗后微生物群α多样性下降
据报道,12个月后微生物群α多样性下降了16%–22%。24个月的长期随访发现,阿奇霉素治疗后,乌普萨拉弯曲菌(Campylobacter upsaliensis)和人弯曲杆菌(campylobacter hominis)丰度降低,并且主要与微生物存活、生长和炎症相关的细菌代谢途径的丰度降低,这可能部分解释了所报道的对儿童死亡率的降低。
乌普萨拉弯曲菌(Campylobacter upsaliensis)在高收入环境中,特别是在农村儿童中,它是一种公认的食源性肠道病原体;而人弯曲杆菌(campylobacter hominis)与肠道炎症和菌血症有关。
三个国家的四项随机对照试验报告了短期使用的影响。与安慰剂相比,阿奇霉素治疗使微生物群α多样性降低高达32%。据报道,Akkermansia muciniphila和Proteobacteria也有所减少。
注:后一组细菌包括常见的病原体,例如大肠杆菌和弯曲杆菌属。
Fishbein SRS,et al.Nat Rev Microbiol.2023
总之,这些研究表明阿奇霉素引起的α多样性明显减少,同时肠杆菌科病原体也减少。
▼
抗生素与代谢或饮食元素之间的协同作用可以进一步塑造微生物组动力学并改变对抗生素的易感性,从而导致疾病的临床相关差异。
•高脂饮食下进行抗生素治疗会引起病理性炎症
对高脂肪饮食的小鼠进行抗生素治疗会引起肠道黏膜的病理性炎症,并增加肠杆菌(Enterobacteriaceae)的水平。这些小鼠表现出炎症性肠病的免疫症状。
Fishbein SRS,et al.Nat Rev Microbiol.2023
•饮食和抗生素结合改变了微生物和宿主的代谢
从其他几项使用饮食和化学干预来模拟西式饮食影响的小鼠研究来看,很明显,饮食和抗生素的结合改变了微生物和宿主的代谢。
具体来说,在一个模型中,这种协同干预通过改变脂肪酸代谢释放了单糖和乙醇胺,最终增加了肠道沙门氏菌感染的易感性。
另外,饮食中多糖或微生物可及的碳水化合物水平的调节对抗生素治疗结果有很大影响。在没有纤维的情况下,使用传统微生物组或人源化微生物组的小鼠在环丙沙星治疗后α多样性恢复延迟。
•含纤维饮食促进了丁酸水平的恢复
在人类实验中,在联合抗生素治疗期间,以杂食和纯素饮食喂养的志愿者,相对于那些不含纤维的液体饮食的志愿者,表现出了更快的微生物群恢复(通过α和β多样性来衡量)。
具体来说,含纤维饮食促进了粪便丁酸水平的快速恢复和具有独特氨基酸代谢的厚壁菌门的恢复。
▼
抵抗组(resistome)是指存在于肠道微生物群中的抗生素抵抗基因的集合。抗生素对肠道抵抗组具有显著但高度可变的影响。
•抗生素通常会增强肠道抵抗组
通过抗生素暴露对肠道抵抗组的改变可能引起微生物组中耐药病原体的病理性繁殖。值得注意的是,抗生素治疗最常见的结果是肠道微生物组中抵抗组负荷急剧增加(通过相对丰度的变化来衡量)。
这些研究揭示了抗生素诱导的抵抗组变化的一些一般模式。尽管增加了总抗性组负担,但抗生素可同时导致特异性的大量丧失和抗性组多样性的减少,这可能是由于分类损失。
•水平基因转移促进抗性组富集
抗生素诱导的肠道抵抗组扩张也可能由ARG编码的MGEs驱动。
在短期内,抗生素治疗后抵抗组负担的增加主要是由染色体编码ARGs的富集(即耐药细菌谱系的扩大)驱动的。
然而,此后不久(约1个月),染色体ARGs的微生物丰度急剧下降。相反,mge编码的ARGs在治疗停止后持续更长的时间,这可能是由于mge的寄生性质。(尽管缺乏抗生素介导的生长优势或细菌宿主存在适应性缺陷,它们仍在细菌群体中持续存在)
抗生素可以增加肠道中mge编码的ARGs的丰度,并导致ARGs和MGEs在患者中更频繁和更广泛(涉及更多样化的分类群)的传播。
总体而言,在临床使用和环境抗生素暴露较高的人群中,ARGs水平转移率较高。
▼
在全球范围内,使用产时抗生素预防(IAP)是预防围产期感染和相关不良妊娠结局的最常见策略。
产时抗生素预防涉及使用广谱抗生素,这些抗生素可有效对抗最有可能引起高危母亲感染的微生物,主要是B族链球菌。
建议在剖腹产手术切开前进行抗生素预防;分娩期间,母亲B族链球菌培养呈阳性或有侵袭性GBS感染风险;或出生后立即进行,以减少与手动去除胎盘、过度宫内操作或生殖道撕裂相关的感染风险。
•经过产时抗生素预防的婴儿微生物多样性降低
七项观察性研究和一项随机对照研究调查了抗生素预防与肠道微生物群之间的关联。经过产时抗生素预防治疗的母亲所生的婴儿在6-7天时的微生物群α多样性有所降低。
暴露于产时抗生素预防还与微生物群组成的更大相似性(即β多样性减少)相关,这表明母体抗生素暴露的选择性压力对婴儿的早期婴儿微生物群组成具有一致的影响。
•产时抗生素预防组以肠杆菌为主,双歧杆菌较少
一项前瞻性队列研究发现,产时抗生素预防组10日龄时放线菌门(包括双歧杆菌)的相对丰度显著减少,厚壁菌门(包括一些值得注意的致病菌群,如葡萄球菌、李斯特菌和链球菌)显著增加。
双歧杆菌是母乳喂养的婴儿肠道微生物群的重要成员,可以通过与其他重要肠道细菌(如乳杆菌)的共生交叉喂养相互作用来驱动微生物群的持续发育。
总体而言,这些研究表明,婴儿微生物群的α多样性降低与母亲产生接触抗生素相关,包括肠杆菌科相对丰度较高和拟杆菌和双歧杆菌丰度较低。
以下讨论抗生素类别、细菌进化枝、起始微生物群组成和长期微生物群组成。
▼
√不同抗生素会影响对应的菌属丰度
最近对55项人类研究的回顾总结了微生物群组成的变化,这些变化与分为22个抗生素类别的68种不同抗生素显著相关。研究表明,每种抗生素对属于一组有限属的细菌影响最强烈。
此外,受影响的细菌属的数量在抗生素类别之间和内部都存在差异。
阿奇霉素与3个属(拟杆菌、双歧杆菌、梭菌)的丰度变化相关,而其他大环内酯类药物则与2-6个属的丰度变化相关(包括前三个属,以及粪杆菌属、普雷沃菌属、瘤胃球菌、链球菌、肠球菌、嗜血杆菌或假单胞菌)。
β-内酰胺类药物,例如氨苄西林和头孢菌素,分别与8和12个属的丰度变化相关,这些属与其他β-内酰胺类药物不同,涵盖系统发育上不同的门(放线菌门、厚壁菌门、变形菌门、疣微菌门)。
√抗生素的不同分类群具有特异性作用
培养实验证实了不同抗生素的这些分类群特异性作用。例如,甲硝唑强烈抑制拟杆菌属和双歧杆菌属的生长。在厌氧培养中,而氨苄青霉素显著减少拟杆菌属和产气荚膜梭菌的生长,但其对双歧杆菌属的影响取决于细菌的生长速度。
此外,氨基糖苷类和磺胺类药物对测试的细菌几乎没有影响。大环内酯类药物对所有测试的细菌种类均具有抑菌作用(但不会杀死),但对艰难梭菌没有影响。
注意:β-内酰胺的作用具有菌株特异性,并且因所使用的特定抗生素而异。
因此,抗生素对微生物群的影响取决于所使用的抗生素,以及细菌分支的反应以及反应方式。
然而,抑制对抗生素敏感的微生物可能会破坏定植抵抗力,并为机会致病菌打开生态位,增加宿主对抗生素后感染的易感性。另外,抗生素的使用也可以增加耐抗生素的共生细菌,这对宿主有潜在的好处。
影响这些效应的因素包括不同抗生素的药代动力学、不同细菌对每种抗生素的降解和耐药机制,这些降解和耐药机制对不主动执行这些功能的其他细菌的间接影响,以及特定耐药决定因素的局部或区域流行率。
▼
考虑到抗生素作用因抗生素类别和细菌进化枝而异,微生物群的初始组成也可能决定抗生素的短期和长期作用。
√微生物群的初始组成会影响抗生素的作用
对五个队列的数据分析确定,具有增强的降解复杂碳水化合物(特别是宿主粘蛋白)的功能能力的特定物种的丰度,准确地预测了微生物群从抗生素相关的α多样性减少中恢复。
这些细菌的基线丰度可以识别使用抗生素后α多样性减少的参与者,准确度为70%。
√粘蛋白降解能力促进细菌在肠道繁殖
研究人员提出粘蛋白降解能力可能为这些细菌在肠道中重新繁殖提供了优势。此外,这些复杂碳水化合物的降解可能会产生次生代谢物,这些代谢物反过来又可以被其他细菌用作营养源以支持其再生。
注:与恢复相关的细菌包括拟杆菌属的几种。
无菌小鼠的证据支持了这一在人类身上的发现。两组均被富含普雷沃氏菌和粪杆菌或富含拟杆菌和副拟杆菌的人类粪便微生物群定植,并且两组均接受为期7天的阿莫西林克拉维酸治疗。
18天后,拟杆菌组的α-多样性对抗生素暴露更具弹性。然而,拟杆菌属的粘蛋白消化降解可以促进肠道感染,这已在艰难梭菌、啮齿类梭菌和鼠伤寒沙门氏菌中得到证实。
▼
两项分析均使用了10个月期间纵向收集的成人数据,其中包括两个为期5天的环丙沙星疗程。
√不同菌群达到新的平衡所需时间不等
一项分析确定,对于不同的细菌亚群,微生物群达到新的稳定配置所需的时间从2到42天不等。
拟杆菌属中的一个细菌亚群在两周内以相同或更高的相对丰度建立了新的平衡。然而,其他细菌如毛螺菌科(Lachnospiraceae)和一部分细菌在治疗期间有所增加,并需要42天才能恢复到使用抗生素前的水平。
相比之下,瘤胃球菌科的一组细菌在一次治疗后减少到检测不到的水平,十个月后仍未恢复。该综述发现细菌进化枝的特异性决定了抗生素对微生物群的影响。
√更短的恢复时间可能由于暴露于环境来源的细菌
对相同数据的第二次分析表明,更短的恢复时间可能是由于更多地暴露于环境来源的细菌,这些细菌可以重新定殖受干扰的微生物组。
尽管这些研究涉及的个体很少,并且最多只能代表个体间的异质性,但他们探索了微生物群丰度数据的采样时间序列,并且他们的发现得到了实验工作的支持。
√更复杂的微生物群可以减轻抗生素暴露的影响
在一项这样的实验中,两组小鼠被分配到无特定病原体的条件或暴露于土壤的一般动物条件下,并且均接受为期两周的万古霉素和链霉素治疗。
暴露于土壤的小鼠在治疗后表现出更大的α多样性和稳定性,并且在停用抗生素后更快地恢复治疗前的α多样性。这些研究表明,更受控制的外源细菌,例如复杂的益生菌混合物,可以减轻抗生素暴露对微生物组的影响。
√抗生素暴露使婴儿α多样性降低
抗生素暴露的长期影响也在婴儿中进行了研究。α-多样性降低与出生后立即治疗有关,但在产后第一年恢复。α-多样性的减少是由于毛螺菌科和肠杆菌科被抑制所致。
尽管微生物群α多样性在一岁时基本上恢复,但由于新生儿接触抗生素而导致这些早期定植菌减少对宿主健康的影响需要仔细调查。
在另一项研究中,与在那段时间没有接触过抗生素的婴儿相比,从出生到三岁接受抗生素治疗的婴儿的物种α多样性较低,但α多样性的减少幅度不大,仅在第一年明显。然而,在细菌菌株水平上,暴露于抗生素的婴儿和未暴露于抗生素的婴儿之间的差异更大。
此外,还发现部分细菌菌株只在肠道定殖一次。这些菌株通过接触抗生素而被消除,但在未经治疗的婴儿中仍然存在。相比之下,其他菌株在随访期间多次在肠道重新定殖。这些菌株更有可能在暴露和未暴露的婴儿中持续存在,这为以下观点提供了进一步的支持:再定殖是使用抗生素后微生物群恢复的驱动力。
★ 评估菌群经过抗生素治疗恢复原有结构的能力
根据菌群的基本构成,尤其是底层代谢黏蛋白以及短链脂肪酸的菌构成情况,可以评估菌群在经过抗生素治疗后恢复原有菌群结构的能力。
该恢复能力越强菌群结构较难通过抗生素改变,越弱则易受抗生素影响,导致后续菌群构成发生较大变化。该指标可以为菌群的抗生素干预选择和菌群重建提供参考。
抗生素耐药是指病原体(细菌、病毒、真菌等)不能被抗生素及其他抗感染药物有效杀死的抵御能力。由于存在抗生素滥用的情况,导致环境中许多细菌尤其是病原菌产生对特定抗生素类型的耐药性。通过对大量抗生素耐药菌基因组和耐药基因的分析,结合肠道菌群测序数据,报告中评估了肠道菌群对主要抗生素类型可能存在的耐药风险。
注意:仅作为风险提示和参考,并不直接对应抗生素耐药,具体用药和耐药检测请谨遵医嘱和进行耐药检测。

<来自:谷禾健康肠道菌群检测数据库>
总而言之,这些分析表明,微生物群从抗生素暴露中恢复可能取决于所使用的抗生素、初始微生物群组成以及受药物影响的特定细菌分类群。
此外,纵向观察研究提供了额外的证据,表明肠道的重新定植可能在抗生素暴露后微生物组的恢复中发挥关键作用。然而,还需要更多的研究来检验这几项研究提出的具体假设。
值得注意的是,仅恢复可能无法保护宿主免受抗生素扰动对微生物组的潜在长期影响。
流行病学研究已确定抗生素使用与肥胖、哮喘,过敏特应性和结直肠癌以及其他健康疾病之间存在关联。抗生素对肠道微生物组的破坏已被认为是这些关联背后的重要机制。
▼
50%或更多中低收入国家的婴儿在3个月大时携带至少一种肠道病原体,并在24个月大时经历至少一次腹泻发作。
中度至重度腹泻的最常见原因是病毒(轮状病毒、沙波病毒、诺如病毒)或细菌(大肠杆菌致病型、志贺氏菌、弯曲杆菌属)感染。
特定病原体的流行情况因国家而异。死亡率和严重腹泻发作也因国家和病原体而异。
▷抗生素降低了腹泻和痢疾导致的死亡率
抗生素的广泛目标是减轻病原菌对宿主健康的负担和影响。使用阿奇霉素进行的随机对照试验结果表明,对弯曲杆菌属具有抑制作用。促炎微生物组代谢途径可能部分解释了在这些环境中观察到的腹泻和痢疾导致的儿童死亡率持续下降。
在另一项研究中,长双歧杆菌(Bifidobacterium longum)抑制了儿科病例中分离的轮状病毒,并缩短了病毒性胃肠炎的持续时间。
▷抗生素也会扰乱肠道微生物群促进肠道感染
抗生素扰乱肠道微生物组后可促进肠道感染,这一点已在艰难梭菌、啮齿类梭菌和鼠伤寒沙门氏菌中得到证实。
或者,用抗生素扰乱肠道微生物群已被证明可以延迟轮状病毒感染并预防小鼠的诺如病毒感染。
相比之下,在艾滋病毒感染儿童中长期使用复方新诺明,4年后仍可抑制炎性草绿色链球菌以及相关的全身性炎症,从而增加该人群的防御风险。
▼
抗生素的使用与肥胖风险增加有关。接触时间越早(6个月之前)和接触频率越高,这种关联就越大。较低的肠道微生物群α多样性也与肥胖有关。
▷抗生素促进营养不良儿童的体重增加
相比之下,抗生素可以促进营养不良儿童的体重增加,尽管其影响是异质的,并且可能是由抗生素类别以及不同人群中生长限制性合并症(例如艾滋病毒和严重急性营养不良)的不同患病率驱动的。
微生物组组成对体重增加的因果影响也已在无菌小鼠中得到证实。
如果不清楚微生物群组成变化与健康结果之间的因果机制,微生物群组成通常被定义为“生态失调”,这是基于与疾病相对于健康对照的关联。
在高收入成年人群中,α多样性下降是“生态失调”的一个共同特征,“生态失调”是根据与疾病的关联来定义的。在这种情况下,α多样性减少表明肥胖和相关代谢紊乱(例如I型和II型糖尿病)的风险增加。
▷抗生素导致α多样性下降增加肥胖风险
相比之下,在纯母乳喂养的婴儿中,肠道微生物组以双歧杆菌为主。由于母乳中存在高浓度的寡糖,导致与混合或配方奶喂养的婴儿相比,α多样性较低。
更大的α多样性与较早引入补充食品和青春期超重风险增加有关。因此,婴儿早期微生物群α多样性的增加可能表明由于婴儿早期营养不足而导致“生态失调”,抗生素相关的α多样性减少对健康的影响也可能是因情况而异的。
▷新生儿抗生素暴露于儿童生长有关
抗生素暴露的时间可能有助于进一步了解具体情况的影响。在芬兰的一个由12422名足月新生儿组成的队列中,新生儿抗生素暴露(主要是静脉注射苄青霉素和庆大霉素)与男孩生命前6年生长受损有关。
一项由1707名德国新生儿组成的独立队列证实了与男孩生长的相关性。相反,新生儿期后接受抗生素疗程的次数与男女6岁时体重指数的增加有关 。
最近的一项针对小鼠的研究表明,出生时注射的低剂量青霉素会导致肥胖增加,并加剧高脂肪饮食对体重增加的影响。生长表型是在无菌接种抗生素处理小鼠的粪便微生物群时诱导的,表明改变的微生物群起着因果作用。
然而,还需要更多的研究来表征和确认促进从抗生素暴露中恢复的微生物组的成分,以及健康新陈代谢可能需要的成分,并将这些转化为人类健康。
▼
较早和更频繁的抗生素处方也与儿童哮喘风险增加有关。较低的肠道微生物群α多样性与婴儿期和儿童期的湿疹和过敏有关。
▷婴儿抗生素使用与儿童哮喘风险存在联系
最近证明婴儿抗生素使用引起的微生物群改变与5岁时哮喘风险之间存在直接联系。在出生第一年接触抗生素的婴儿到5岁时患哮喘的可能性是正常婴儿的两倍。估计这种关联的25%可归因于抗生素引起的α多样性减少或抗生素相关的特定细菌丰度变化。
▷母亲使用抗生素会影响婴儿微生物的早期定植
尽管整个婴儿期的肠道微生物组存在巨大的变异性,但婴儿期微生物组的有意义的一部分是在出生后立即从母体肠道获得的,并保留长达3年。因此,母亲接触抗生素也可能会改变分娩过程中微生物物种向婴儿的传播,并影响早期定植。
影响新生儿微生物组组成的抗生素暴露可能会对儿童发育产生长期影响。几项使用动物模型的研究表明,新生儿期对于免疫系统的成熟至关重要,这一过程依赖于新生儿肠道微生物组的定植,并且会受到抗生素消融的干扰。
小结
抗生素在治疗细菌感染和减少疾病传播方面起着积极的作用。然而,滥用和不适当使用抗生素可能导致细菌耐药性的增加,并对肠道微生物群产生负面影响。
因此,我们应该合理使用抗生素,遵循医生的建议,并在使用抗生素期间维护肠道健康。谨慎使用抗生素有助于最大限度地减少负面影响,确保抗生素继续为我们的健康服务。
人们提出了一些有前景的方法来减轻抗生素使用对微生物组的影响。
▼
共同施用益生元以促进共生细菌的生长就是这样的策略之一。在婴儿中,母乳富含复杂的低聚糖,可作为双歧杆菌生长的底物,并促进其他物种的交叉喂养。
在一个观察队列中,母乳喂养与产时抗生素预防暴露婴儿微生物组α多样性的更快恢复有关。母乳喂养的好处也可能超出其益生元含量。
•儿童益生元和抗生素联合给药使有益菌丰度升高
在儿童中,乳果糖与阿奇霉素的联合给药有助于恢复乳果糖的相对丰度。治疗后18天内,乳杆菌属(Lactobacillus)、肠球菌(Enterococcus)、Anaerostipes、经黏液真杆菌属(Blautia)和罗氏菌属(Roseburia)出现增加,而单独使用阿奇霉素可导致治疗后60天内促炎性链球菌的丰度增加。
•成人联合使用益生菌和抗生素可减轻耐药性
在成人中联合使用益生菌和抗生素也显示出一定的潜力,可以减轻耐药性遗传决定因素的抗生素选择。
然而,口服益生菌在肠道的成功定植表现出因人、肠道区域和益生菌菌株而异的相当大的变异性,这可以通过宿主和本土微生物组特征来解释。
此外,益生菌菌株中缺乏对所施用抗生素的耐药性可能会加剧微生物组中抗生素耐药性的选择,进一步限制了益生菌的潜在保护作用。
注:使用抗生素给药前收集的自备健康粪便样本进行粪便微生物群移植可能是恢复微生物组的更有效方法,尽管这对于中低收入国家来说可能不太实用。
▼
另一种策略涉及使用抗生素药物组合来实现比单一抗生素治疗更有针对性的物种特异性效果。
例如,一项调查筛选了超过1000种药物,以确定可降低抗生素广谱活性而不损害其对抗相关病原体活性的候选药物。
•红霉素对肠道共生细菌具有强抑制作用
抗凝药物双香豆素和两种非甾体类抗炎药物托芬那酸和二氟尼柳成为红霉素对肠道共生细菌(例如普通拟杆菌和均匀拟杆菌)作用的强抑制剂。
然而,红霉素对致病菌(例如金黄色葡萄球菌、肺炎链球菌和屎肠球菌)没有显著影响。
•药物组合具有物种特异性效应
另一项研究分析了近3000种抗生素、药物和食品添加剂的组合,以确定可以减轻对微生物群的附带影响而不减少对病原体影响的候选化合物。超过70%的药物组合具有狭窄的物种特异性效应,20%显示出菌株特异性效应。
▼
另一种方法是确定窄谱抗生素替代品,尽可能选择狭谱抗生素,这些抗生素只对特定类型的细菌有效,而不是广谱抗生素,后者会杀死更多的细菌,包括有益菌。
微生物组本身可能是此类窄谱化合物的来源。例如,对人类微生物组计划中的752个细菌基因组的研究发现了共生细菌携带的一组编码硫肽的基因。
•对病原菌有很强的抑制作用,对共生菌没有抑制
乳青霉素是一种由加氏乳杆菌编码的硫肽,对金黄色葡萄球菌和阴道加德纳菌等常见病原体具有很强的抑制活性,但对共生菌没有抑制作用。
苏云金素是另一种已被鉴定的抗菌素,由苏云金芽孢杆菌(Bacillus thuringiensis)产生。它已被证明可以有效对抗艰难梭菌,而不影响人类结肠的粪便培养系统中的微生物群组成。
开发此类干预措施需要更深入地了解与抗生素结合产生健康益处或不良副作用的机制,以便窄谱替代品能够实现预期结果,同时最大限度地减少不必要的风险。
最后,环境中可能还存在重要的微生物成分,有助于防止生态失调或在关键发育时期恢复肠道微生物组的生理重要子集。识别这些成分可以指导缓解干预措施的开发和测试。
有令人信服的证据表明,即使不存在抗生素耐药性,感染也很难治疗。几项依赖于对患者分离株进行深度基因组测序的纵向研究表明,这些持续性感染的复发与宿主体内细菌库的维持有关,这些细菌在免疫防御和抗生素的联合作用下幸存下来。
用于追踪动物和细胞感染模型中细菌的细菌生长停滞报告者已经证明,尽管接受抗生素治疗,但在持续感染部位仍存在不增殖或缓慢增殖的细菌。过去几年,追踪和分析稀有非生长细菌的尖端技术的发展极大地促进了抗生素持久性的研究。在这方面,这个领域正在不断发展。
总结如下:
1.与普通菌株相比,生长缓慢的细菌表现出相对的药物冷漠,这增加了种群在抗生素暴露下存活的时间。
2.药物暴露后的这种异质群体行为是由于细菌亚群(持续存在)通过生长停滞、暴露于对其余群体致命的浓度的抗生素而得以生存的能力。
3.许多病原菌在宿主组织内建立持续感染,避免被免疫系统和抗生素清除。细菌感染通常会导致宿主产生先天免疫反应,然后产生适应性免疫反应,这通常足以清除感染。然而,许多病原体已经进化出复杂的生存机制来逃避免疫反应并建立持续感染。
4.有多种原因会导致感染反复发作,例如宿主易感性增加、其他慢性合并感染以及环境和生活方式因素。长期以来,无法确定感染复发是再次感染还是复发。
5.持久性的形成是一种普遍的细菌应激反应。多种环境因素充当压力信号,促进持久者的产生。许多触发因素似乎都会刺激持久细胞的形成,从饥饿或氧化应激等营养应激到细胞外代谢信号。
6.婴儿微生物群的α多样性降低与母亲接触抗生素相关,包括肠杆菌科相对丰度较高和拟杆菌和双歧杆菌丰度较低。
7.不同的抗生素对不同细菌菌属影响不同,肠道内菌群结构的恢复与抗生素的使用和患者基线菌群有关。
8.超重,肥胖,哮喘,过敏等人群与早期和长时间抗生素的使用引起的菌群失调以及部分关键菌属的消失或下降有关。
9.未来可以发展更综合的预防措施,例如,乳青霉素是一种由加氏乳杆菌编码的硫肽,对金黄色葡萄球菌和阴道加德纳菌等常见病原体具有很强的抑制活性,但对共生菌没有抑制作用。
相关阅读:
主要参考文献
Gollan B, Grabe G, Michaux C, Helaine S. Bacterial Persisters and Infection: Past, Present, and Progressing. Annu Rev Microbiol. 2019 Sep 8;73:359-385.
Gough EK. The impact of mass drug administration of antibiotics on the gut microbiota of target populations. Infect Dis Poverty. 2022 Jun 30;11(1):76.
Shah T, Baloch Z, Shah Z, Cui X, Xia X. The Intestinal Microbiota: Impacts of Antibiotics Therapy, Colonization Resistance, and Diseases. Int J Mol Sci. 2021 Jun 20;22(12):6597.
Vuong HE, Pronovost GN, Williams DW, Coley EJL, Siegler EL, Qiu A, Kazantsev M, Wilson CJ, Rendon T, Hsiao EY. The maternal microbiome modulates fetal neurodevelopment in mice. Nature. 2020 Oct;586(7828):281-286.
Fishbein SRS, Mahmud B, Dantas G. Antibiotic perturbations to the gut microbiome. Nat Rev Microbiol. 2023 Jul 25.
Koh E, Hwang IY, Lee HL, De Sotto R, Lee JWJ, Lee YS, March JC, Chang MW. Engineering probiotics to inhibit Clostridioides difficile infection by dynamic regulation of intestinal metabolism. Nat Commun. 2022 Jul 4;13(1):3834.
Gomaa EZ. Human gut microbiota/microbiome in health and diseases: a review. Antonie Van Leeuwenhoek. 2020 Dec;113(12):2019-2040.
谷禾健康

肠道微生物群与人类宿主共同进化,对于我们的消化、代谢、神经、免疫等系统具有重要贡献,影响着人类身体和心理健康。
肠道微生物群从出生时就开始形成。生命早期的多种因素,如分娩方式、婴儿喂养方式以及母体微生物群等,都会影响肠道微生物群的初期定植。早期微生物群的组成和功能可能对宿主长期健康产生影响。
人类肠道微生物群的变化与人类生存策略的演变密切相关,随着人类从狩猎采集社群向农耕与城市生活方式的转变,肠道微生物群的多样性经历了一系列的变化。而种族、地理、文化和行为特征等宿主因素,也会对全球范围内人类肠道微生物群的多样性和组成产生影响。
近日,来自《Trends in Molecular Medicine》杂志发表的文章,从生态学的角度探讨影响不同人类种群肠道微生物群的基本驱动因素,并考察生活方式、社会经济地位和人类迁移等因素,如何通过影响肠道微生物群而对不同人群的健康产生差异。

人类肠道中有高度多样的微生物群落,最显著的变化与工业化水平有关。
城市化、西方化的生活方式与细菌多样性减少、本地菌群(如普雷沃氏菌)和纤维降解功能丧失有关。这些差异可能导致非传染性免疫、代谢和神经系统疾病的发病率增加。
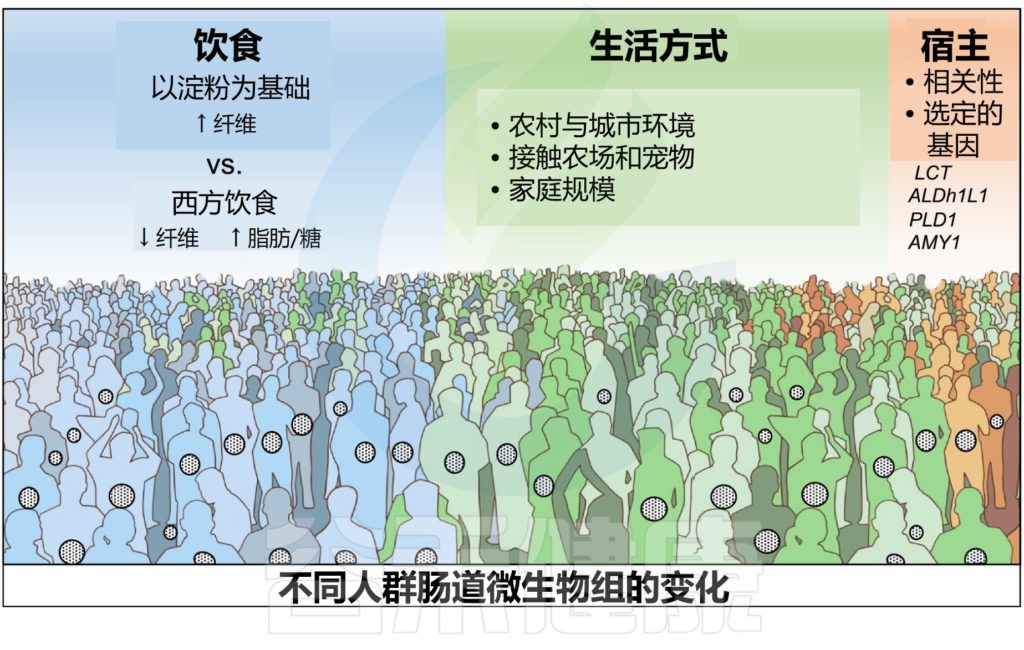
在所研究的许多生活方式和遗传因素(地理位置、遗传相关性、饮食、种族、内在环境、家庭组成等)中,饮食和生活方式实践对肠道微生物群落的形成最具影响力。
人类迁移可以揭示肠道微生物群在不同生活方式下对健康的影响。
这些信息的重要性在于,健康的或不健康的肠道微生物群不能简单适用于所有人,而应当根据特定的微生物组特征来设计适应各种人类群体的菌群失调策略。进一步了解和应对微生物组变化的探索,将成为未来疾病预防和治疗的重要方向。
通过研究微生物群落的组成,我们可以更好地了解微生物组的形成过程。这里面我们来了解几个概念:
微生物传播、环境选择、生态漂移
微生物物种在不同地点之间的传播能力,解释了微生物在人类和身体部位之间的垂直和水平传播。
另一种机制被称为环境选择,解释了微生物群落之间的适应度差异或基于生态位的相互作用,这有利于某些物种在特定环境中生存。例如,双歧杆菌属(Bifidobacterium)具有代谢母乳的能力,因此在婴儿的肠道中非常常见。
生态漂移是指人类微生物组中微生物物种丰度的随机变化。生态漂移可以减少群落内殖民化和竞争互动的作用,增加群落之间的差异。
注:扩散和环境选择在生态选择和形成微生物相互作用中发挥着更强的作用,克服了人类肠道微生物组中生态漂移的影响。
微生物之间的相互作用影响肠道微生物组的组成
有些微生物物种依赖于与其他物种的共生或互利关系。例如:
这表明在肠道微生物组中存在着合作和竞争的相互作用。
在炎症性肠病和肥胖等疾病中,已经确定了一些关键细菌物种的共存或其作用途径,这表明它们在调节疾病相关的微生物组中发挥着重要作用。
真菌在微生物组成和共生中起着重要作用
例如,肠道-真菌-细菌的相互作用有助于在生命早期形成微生物组组成,细菌和真菌α多样性之间的负相关性表明,抑制机制限制了许多真菌物种在人类肠道中的生长。
肠道中的原生动物捕食细菌
肠道中的原生动物与细菌多样性和群落组成变化有关,这表明原生动物可能通过捕食细菌产生自上而下的影响,从而增加细菌多样性。
来自不同国家的肠道微生物群研究也表明,人芽囊原虫(Blastcystis)是一种在人体肠道中常见的原生动物,与史密斯甲烷杆菌(Methanobrevibacter smithii)以及肠道优势细菌普雷沃菌(Prevotella copri)和瘤胃球菌(Ruminococcus gnavus)的丰度有关。史密斯甲烷杆菌的丰度与产短链脂肪酸菌(如Collinsella aeroffaciens、P.copri、Roseburia fadis)呈正相关。
扩展阅读:
这些研究结果强调了理解控制肠道微生物组成的生态过程和相互作用的重要性,因为我们的饮食习惯、生活方式、社会地理和其他环境因素会影响人体肠道微生物组。
人类肠道微生物组已经通过遗传相关性和同居关系的梯度进行了研究,揭示了宿主遗传和肠道微生物组成之间有趣的联系。
宿主遗传学影响特定肠道微生物类群的丰度
家庭成员拥有相似的肠道微生物组,特别是,基因相同的双胞胎表现出最高水平的相似性。为了研究宿主遗传学对肠道微生物群落的影响,研究人员分析了来自以下群体的单卵和双卵双胞胎的几个粪便样本数据集:
i)TwinsUK群体,由977个个体组成的数据集,包括1081个单卵和二卵双胞胎、具有未知结合度的双胞胎和无关个体的粪便样本
ii)Yatsunenko数据集,包括来自委内瑞拉亚马逊地区的531名美洲印第安人、马拉维农村社区居民和美国大都市地区居民的粪便样本
iii)密苏里双胞胎数据集,包括54对双胞胎
这些研究表明:宿主遗传学影响特定肠道微生物类群的丰度。
宿主遗传学的影响因类群和种群而异,例如,Christensenellaceae是TwinsUK和Yatsunenko种群中最具遗传性的细菌科,瘤胃球菌科和毛螺菌科是TwinsUK和密苏里种群中最具有遗传性的操作分类学单元。
宿主环境超过了宿主基因对肠道微生物组成的影响
相比之下,这项研究以及芬兰的一项单卵双胞胎研究都表明,拟杆菌属是一个包括主要肠道微生物物种的分类单元,是不可遗传的,其存在主要由饮食来解释。因此,虽然某些分类群受到宿主遗传学的影响,但同卵双胞胎的高度同质环境可能比遗传学更能推动其肠道微生物组成之间的相似性。这得到了几项研究的支持,这些研究表明,与非共享家庭双胞胎相比,共享家庭和饮食的非遗传相关个体之间的微生物组具有显著的相似性,这表明宿主环境超过了宿主基因对肠道微生物组成的影响。
尽管如此,人类的遗传多样性导致了与常驻微生物物种直接相关的人类种群之间的生理差异。
基因 & 肠道菌群 & 症状或饮食方式
全基因组关联研究报告了:某些宿主数量性状基因座与一些可遗传细菌类群的相对丰度和功能之间的显著联系。LCT基因座与双歧杆菌属之间的相关性在这些研究中最强,其随乳制品摄入量的变化而变化。
其他发现包括:
i)与缺血性中风相关的人类甲酸盐氧化基因乙醛脱氢酶1家族成员A1(ALDH1L1)与细菌SHA-98之间的相关性;
ii)PLD1基因和Akkermansia,其与降低肥胖风险有关;
iii)ABO血型基因和乳杆菌。
基因拷贝数的变化也与人类微生物组成有关。已知在富含淀粉的饮食人群中,负责消化过程中可消化淀粉水解的淀粉酶基因AMY1的拷贝数更高。AMY1拷贝数还与普雷沃氏菌、卟啉单胞菌、瘤胃球菌科分类群的增加有关,所有这些都可以降解耐膳食淀粉,AMY1的低拷贝数可能导致未消化的淀粉到达大肠,增加下肠道的简单淀粉发酵,并调节肠道微生物群落。
分析和制定标准化方法的重要性,尤其是在处理地理或种族分布的乳糖不耐受等特征时。值得注意的是,组成肠道微生物组的大多数微生物物种似乎不受宿主基因的影响,这表明它们大多是环境获得的。
doi.org/10.1016/j.molmed.2023.07.002
人类历史上的生活方式转变伴随着饮食、社会人口特征以及其他环境和文化因素的变化,这些因素反映在人类群体中肠道微生物多样性和组成的显著变化中。
城市环境的饮食、生活方式
如今,世界上一半以上的人口生活在城市环境中,据估计,到2050年,这一比例将增至约70%。向城市生活方式的转变带来了膳食蛋白质和脂肪摄入的增加,以及卫生条件的变化和药物负担的增加。相比之下,目前的现代狩猎采集人群以淀粉为基础,纤维摄入量要高得多,并且经常被真正的胃肠道病原体或寄生虫定植。
人类肠道微生物群随着对这些饮食变化的反应和对宿主代谢需求的适应而发生了变化。
▼
狩猎采集者的肠道菌群
例如,一组与西方生活方式接触有限的南美洲亚诺马米狩猎采集者的肠道微生物组在其他研究人群中显示出组成和功能多样性较高。
哈扎狩猎采集者的微生物多样性也高于西方城市人口。它们独特的肠道微生物组成,包括参与支链氨基酸和广谱碳水化合物代谢的基因,表明它们适应了饮食习惯和传统的觅食生活方式。狩猎采集群体表现出更高丰度的梭状芽孢杆菌、变形杆菌、螺旋体和其他与纤维降解有关的分类群,如丁酸弧菌、乳杆菌、普雷沃氏菌、瘤胃杆菌、琥珀酸弧菌和密螺旋体,以及氨基酸、毒力和维生素代谢功能。
扩展阅读:
工业化城市人群的肠道菌群
工业化城市人群的特点是以拟杆菌属、双歧杆菌属、厚壁菌门成员和将纤维发酵成短链脂肪酸的细菌为主,包括Blautia和Faecalibacterium。工业化还与更高的水平基因转移率和抗生素抗性基因的增加以及碳水化合物和外源代谢的预测代谢途径有关,这表明城市化导致了许多微生物功能和性状的消失和替代。
偏远、非工业化地区介于狩猎采集和西方生活方式之间
从生态学的角度来看,狩猎采集者的生活习惯以及这些种群与环境的接触有利于更高的微生物传播率,这是一种受西方生活方式限制的机制。微生物之间对生态位空间的竞争性相互作用也通过选择使宿主能够降解新底物的特殊类群来塑造肠道微生物多样性。其他在偏远、非工业化地区从事自给农业的非游牧传统社会,如非洲的农业学家班图人,表现出与坦桑尼亚的哈扎人相似的饮食习惯,在狩猎采集和西方生活方式之间,微生物α多样性和丰富性处于中间状态。
▼
社会经济地位显著影响健康差异,包括肠道菌群
除了与偏远或农村地区传统生活方式的差距拉开之外,人口的社会经济地位(SES)也会显著影响健康差异和不平等,推动各种因素当然也会影响微生物组的组成。其中包括食物供应和营养状况、体育活动、饮用水、空气污染、卫生和药物、寄生虫、分娩和婴儿喂养法。
影响婴儿肠道微生物组的因素
通过影响拓荒者和早期殖民者,对该生态系统产生了重要的生态影响。与母乳喂养的婴儿相比,配方奶粉喂养加速了婴儿肠道微生物组的成熟,剖腹产分娩和围产期抗生素暴露是关键因素,它们会在出生时中断初级微生物的继承和定植。
由于社会经济和文化因素,全球剖腹产出生率估计将从2018年的21.1%增加到2030年的28.5%,其中近80%的病例发生在中低收入国家。
围产期和婴儿微生物组的改变导致双歧杆菌减少、病原体增加和微生物组成熟加速,这可能导致肠道微生物失衡(微生态失调),从而导致随后的疾病,如哮喘、过敏和1型糖尿病。
扩展阅读:
社会经济地位通过饮食影响菌群
社会经济地位的影响不能与饮食实践的影响脱节,因为不良的社会经济条件与低多样性饮食有关,这可能导致肠道微生物多样性低。社会经济地位、妇女教育和产假时间也与母乳喂养的开始率和持续时间有关。有趣的是,较低社会经济地位对微生物多样性的影响在普雷沃氏菌丰度较高的个体中更为明显。普雷沃氏菌是一种工业化程度较低的细菌群。
这些发现将饮食视为人类社会中人类肠道微生物组差异的决定性媒介,与社会经济地位等社会人口学特征和健康差异密切相关。
扩展阅读:
▼
肠道真菌群的差异也与生活方式、社会人口和环境因素有关
这些差异从早期就可以识别,例如真菌属曲霉属和毕赤酵母属与念珠菌属和马拉色菌属在非工业化和工业化城市人群中的优势。先前的研究也表明,原生芽囊虫在非工业化城市人群的微生物群中更为丰富。
此外,城市生活方式与个体间和个体内古菌多样性以及古菌与细菌比率的降低有关。农村的饮食习惯与史密斯分枝杆菌数量的增加有关,史密斯分枝杆菌是一种在农村人群中更常见的古菌。
肠道病毒群之间的个体差异增加
某些肠道病毒物种与地理位置、生活方式、种族特定饮食和其他环境因素、一些药物以及传染病和慢性病有关。研究表明,随着城市化的发展,肠道病毒群之间的个体差异增加,长期城市生活与几种噬菌体(如乳酸杆菌和乳球菌噬菌体)以及一些细菌病原体相关噬菌体(包括沙门氏菌和志贺氏菌噬菌体)之间存在明显关联。
在其中一项研究中,从中国两个地区(香港和云南)的城市和农村地区收集了来自六个不同种族(白人、傣族、汉族、哈尼族、苗族和藏族)的930名健康成年人的粪便DNA病毒群。他们的研究结果表明,肠道病毒组α的多样性在不同种族之间存在显著差异。与云南居民相比,香港居民的肠道病毒组含有更多的微小病毒科噬菌体,而较少的Myoviridae、Podoviridae和Siphoviridae噬菌体。在所有城市居民中富集的病毒在汉族、苗族和藏族城市居民中也比农村居民更丰富,而在傣族和哈尼族城市居民中比农村居民不那么丰富。
对非细菌微生物在人类肠道微生物组生态学中的作用的探索才刚刚开始,但新出现的证据表明,它可以极大地塑造细菌群落。因此,非细菌微生物很可能有助于解释人类种群之间的组成和功能微生物组差异。最近的报告也将真菌在疾病发病机制中与已知的微生物组贡献联系起来,包括癌症和IBD,这表明人类社会中非细菌微生物的差异应纳入微生物组和宿主疾病范式。
人种是指定义人类群体的社会学和文化因素。微生物组成变化的种族相关模式可以用许多与宿主祖先、种族、生活方式、饮食习惯、医疗保健以及社会文化和社会经济因素相关的协变量来解释,这些协变量很难相互脱节。
种族与微生物组成的相关性
人类对几个分类群的丰度和存在与否进行了显著的选择。一项针对新加坡106名婴儿在出生前两年的多民族人群的纵向队列研究报告称,从三个月大开始,在给婴儿介绍固体食物之前,种族对个体间微生物组差异的显著影响。
在本研究的三个种族(中国人、印度人和马来人)中,印度婴儿的双歧杆菌属和乳酸杆菌属丰度较高。相比之下,中国婴儿的阿克曼菌属和拟杆菌属的丰度更高。这些发现受到母亲饮食摄入缺乏和母乳成分信息的限制,这些信息可以解释研究人群之间的种族差异。
扩展阅读:
另一项比较阿姆斯特丹六个不同种族2084人微生物组成的研究表明,某些肠道微生物类群在不同种族之间存在差异。例如,拟杆菌在苏里南更为丰富,在荷兰的梭状芽孢杆菌和在加纳人、摩洛哥人和土耳其人的普雷沃特拉属更为丰富。荷兰和南亚苏里南的参与者分别表现出最高和最低的微生物α多样性。
移民对肠道微生物组的影响与种族相关的饮食和生活方式等多因素都有关
研究的一个重要因素是移民的影响。在这项研究中,94%的非荷兰人在成年后移民到荷兰。迁移时的年龄和停留时间与它们的肠道微生物群没有显著相关性。相反,种族与所有这些因素相结合,对肠道微生物多样性和组成的影响比单独或结合但不包括种族的这些因素更强。尽管长期生活在同一环境中,但与种族相关的协变量反映了这些参与者肠道微生物组的组成,这表明移民后保持与种族有关的饮食和生活方式因素,有助于保持与荷兰居民群体的肠道微生物组差异。
种族与相似起源的人群的基因模式之间的联系
有趣的是,一些与种族密切相关的分类群也属于可遗传和/或遗传相关的微生物分类群,如Christensenellaceae、Odoribacteriaceae、Rikenellaceae,强调了种族与具有相同或类似地理和祖先起源的人群的基因模式之间的联系。微生物类群共现的模式也与人类宿主种族有关。TwinsUK群体显示,Christensenellaceae与其他可遗传细菌(Dehalobacteriaceae,RF39和SHA-98)和古菌(甲烷杆菌科)共存。
共现模式也与宿主功能调节相关
另一项针对美国1673人的多民族群体的研究也显示,Christensenellaceae12个属在不同种族(非裔美国人、亚洲或太平洋岛民、高加索人和西班牙裔)中的共现模式与人类ALDH1L1基因的遗传变异有关。Christensenellaceae相关微生物中心的共有性和可遗传性,以及它们与ALDH1L1基因(叶酸代谢和细胞增殖)表达调节的重要宿主功能的关联,可能构成了宏生物(metaorganism)水平上的共同进化策略。然而,也有可能某些分类群与宿主遗传学有关,只是因为它们与其他可遗传分类群共存,如Christensenellaceae。目前,这些有趣的发现仍然只是联系在一起,需要检验因果关系。
种族是人类社会健康差异的一个重要决定因素,可能会导致种族对肠道菌群多样性和组成的影响
例如,在一项针对80名生活在英国的黑人和白人女性的研究中,双歧杆菌在白人女性中更为丰富,这一发现与压力增加有关。相比之下,尽管饮食习惯没有显著差异,但黑人女性的微生物β多样性和拟杆菌属丰度更高。
微生物组与种族相关的结直肠癌风险
这些模式可能具有临床相关性,因为此前已经发现低丰度的双歧杆菌属以及高丰度的拟杆菌属和瘤胃球菌属与结直肠癌(CRC)有关。
在一项将该人群与美国非裔美国人进行比较的研究中,在非洲农村地区也发现了较低丰度的拟杆菌属。然而,在引入西方饮食后,该人群的结直肠癌风险才有所增加。即使将压力作为一个风险因素加以控制,非洲裔美国女性的低拟杆菌属与结直肠癌之间的关联仍然显著,这表明微生物组在种族相关因素与结直肠癌风险之间的既定联系中发挥了作用。
在非裔美国人人群中,微生物组是否可能起到中介、调节或仅仅与结直肠癌风险增加相关的作用,在人类研究中仍未得到解决。然而,脆弱拟杆菌在结肠炎相关结直肠癌小鼠模型中具有抑制肿瘤形成的作用,这表明该分类单元可能按种族依赖的方式对结直肠癌风险有因果性贡献。
扩展阅读:
从出生到成年,地理位置一直被认为是与微生物组成变化有关的因素。然而,很难将其影响与宿主的特定特征和当地环境因素脱钩。
▼
地理位置与生活方式、饮食等变量相关
一项研究比较了20个不同变量(包括人口统计、生活方式和饮食)对中国63个城市8个民族2678人肠道微生物组成的影响。这一比较表明,生活在同一地理位置的汉族和其他不同少数民族(白族、回族、苗族、蒙古族、纳西族、藏族和维吾尔族)具有相似的肠道微生物组成,这与其他城市的群体不同。虽然地理位置是个体间微生物组成变化的最强驱动因素,但作者将这一发现与生活方式和饮食等其他变量联系起来。
环境可以影响与遗传和种族有关的人类微生物群落的形成
在更精细的范围内,一项针对居住在同一省份14个区的7000多名来自同一种族(中国少数民族,包括99%的汉族)的中国人的肠道微生物组研究也显示,在共享相同地理位置的个体中,微生物组相似,这表明环境也可以塑造共享遗传和种族相关因素的人类微生物群落。需要在这些因素不同的地方进行额外的微生物组研究,以进一步了解地理位置本身的影响。
▼
地理位置对肠道微生物组组成的作用也可以从人类迁移的角度进行研究
人们永久或半永久地迁移到一个新的地方,可能会在一代或几代人的时间内对人类的生物学和健康产生深远影响。
根据《世界移民报告》,自1970年以来,移民人数增加了三倍多。2020年,全球约有2.81亿国际移民,这意味着每30人中就有一人生活在出生国以外的国家。近几十年来,由于冲突、战争和侵犯人权而被迫流离失所的人数也有所增加。截至2022年5月,联合国难民事务高级专员报告称,全球有1亿被迫旅行者和难民。
旅行和移民会改变食物供应、饮食习惯、生活方式、抗生素和抗寄生虫药物的使用、饮用水、文化和社会经济条件、心理创伤和压力、获得医疗服务的机会、气候、空气污染、环境过敏原等。
移民对藏族肠道菌群影响:微生物群的恢复力
一项研究评估了移民对30名移民咸阳的藏族妇女的影响。西藏人的微生物群与汉族人不同,这可能是由于两个群体在环境、文化、饮食和基因构成方面存在巨大差异。这项研究调查了从迁移到10个月后的微生物组,发现尽管他们的饮食发生了变化,但迁移对肠道微生物多样性、组成和基因途径没有显著影响,这表明微生物组对这些变化有较高的恢复能力。
爱尔兰旅行者饮食接近西方化,肠道菌群仍然保持着类似非工业化群体的特征
第二项研究评估了爱尔兰旅行者的微生物组,爱尔兰流浪者是爱尔兰本土的少数民族。虽然与其他欧洲游牧民族相比,他们在基因上更像爱尔兰人,但他们作为一个独特的民族生活,拥有自己的游牧文化和语言。然而,爱尔兰政府的持续压力导致2022年立法限制他们获得土地临时生活,并强制停止他们的游牧生活。
这项研究比较了118名爱尔兰旅行者的肠道微生物组,他们在游牧生活中长大,但现在永久居住在科克附近,而非爱尔兰旅行者则生活在同一地理位置。两组之间存在重要的微生物组差异,爱尔兰游民的微生物组类似于非工业化的微生物组,但主要是那些留在原地的游民(与居住在社会住房中的游民相比),有更多的兄弟姐妹,并且与动物生活在一起。
虽然是横断面的,但这项研究揭示了从小生活方式的改变,特别是与家庭条件有关的生活方式的变化,对肠道微生物组的影响。重要的是,爱尔兰旅行者的饮食转变为类似西方的饮食,但他们的微生物组仍然更接近非工业化成分。
来自爱尔兰旅行者和西藏妇女的研究表明,工业化程度较低的微生物组的保留与人口采用新生活方式的程度呈负相关,这种影响可能比饮食的变化更强烈。从生态学的角度来看,大家庭和靠近动物有助于微生物的水平扩散和增加微生物多样性。
移民的生活方式对微生物组有何影响?
专注于人类迁移模式的微生物组研究也表明,更高程度地采用新的生活方式和随后的微生物组变化可能会导致适应不良的健康后果。
例如,过渡到社会住房而不是停留在原地的爱尔兰游民群体显示出与工业化国家更常见的慢性病相关的功能性微生物组变化,尽管目前尚不清楚这些变化是否导致了爱尔兰游民与非游民爱尔兰人之间的健康差异。
移民定居后微生物多样性和功能丧失,肥胖风险增加
另一项研究检查了514名苗族和克伦族个体(包括来自泰国的第一代和第二代移民)以及19名移居美国前后的克伦族个体的肠道微生物组。移民在美国定居后,经历了微生物多样性和功能的丧失,以及微生物组成的变化。
在移民前,个体原生的普雷沃氏菌菌株以及与植物纤维降解相关的细菌酶迅速消失,与之交替出现的是拟杆菌属。同时,该研究报告称,患肥胖症的风险增加。移民时的年龄,尤其是儿童早期的年龄,以及作为第二代移民等因素增加了肥胖风险,这表明移民后代谢程序的跨代失调可能至少部分由微生物组的变化介导。
移民带来的肥胖危机:可能与饮食相关
西班牙裔/拉丁裔也描述了美国移民肥胖风险增加之间的关系,这一群体的肥胖率非常高,而且随着移民或儿童早期移民的持续时间而增加。一项针对美国西班牙裔/拉丁裔人群的肠道微生物组研究表明,与肥胖风险显著相关的细菌属也与美国接触有关,并可能与西方饮食有关。
研究还表明,一些微生物类群的共存可能解释了个体肠道微生物分类组成与环境因素的关系,如暴露于美国、饮食和肥胖。这些研究提供了证据,证明新移民获得的生活方式后肠道微生物的变化可能会加剧已知风险因素(即西方饮食)对肥胖的有害影响。
从生态学的角度来看,普雷沃氏菌等主要本土物种的丧失可能会导致人类肠道微生物群的广泛组成和功能差异,从而降低生态系统对干扰的抵御能力。当这种情况发生在生命早期,宿主发育途径更容易接受微生物组信号时,这种微生态失调可能会对宿主发育稳态造成更严重的损害,从而增加免疫和代谢性慢性疾病的风险。
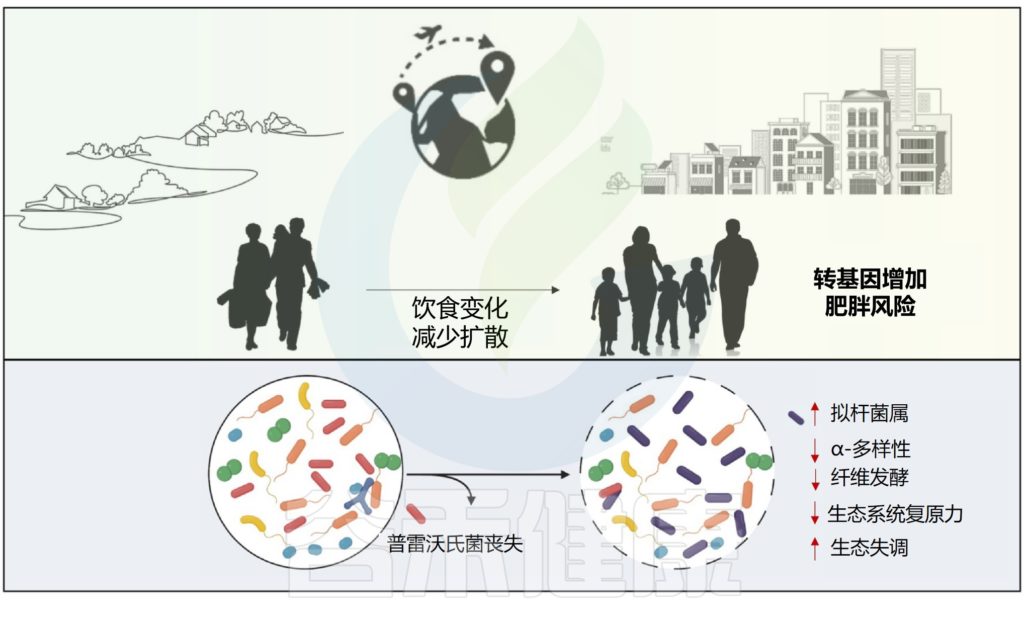
doi.org/10.1016/j.molmed.2023.07.002
基于人群的微生物组研究为微生物群落如何适应人类生活方式和遗传因素的梯度提供了一个窗口。从这些研究中,生活方式和饮食因素成为塑造肠道微生物群落的最具影响力的变量,尽管宿主基因和地理位置也发挥着重要作用。这些发现有力地证明了健康或失调肠道微生物群的普遍规模。
需要进行更多的微生物组研究,以特定人群的方式进一步调查人类微生物组组成的变化,特别是关于疾病的微生物组特征和研究不足人群中基于微生物组的治疗。这对于设计临床策略,预防或治疗世界不同地区的传染病和非传染病至关重要。
到目前为止,研究工作主要局限于更富裕的国家,忽略了社会不平等在人类微生物组中的大部分作用。将这项研究扩大到更大规模的人群的初步努力表明,工业化的城市生活方式和西方饮食与个体间细菌多样性的增加和个体内细菌多样性下降有关。换句话说,城市肠道微生物群更加个体化,每个宿主的物种数量减少。了解和解释这些差异对于定义每个群体的微生态失调和基于微生物组的诊断以及群体内的个性化药物至关重要。
将生态学理论与人类微生物组研究相结合,可以帮助人们了解更多关于早期微生物建立、对扰动的恢复力和功能多样性的生态驱动因素。通过观察早期生态系统建立过程中的微生物-微生物相互作用、人类迁徙研究中观察到的物种的损失和入侵,以及人们对非细菌微生物(古细菌组、病毒组和真菌生物组)的作用,可以更好地了解微生态失调的发展和解决。了解微生物变异的驱动因素对微生物共存的影响,可以为未来的微生物研究和治疗提供深刻的见解。
最后,对迁徙前后个体微生物组成的差异进行更多的研究,可以更深入地了解种群和人类各代肠道微生物组的稳定性或变化。移民往往发生在工业化、城市化地区,以寻求更好的生活条件。比较大型多民族和多文化社会中肠道微生物在迁移时间梯度上的变化,将有助于了解种群之间的微生物交换及其对健康或疾病轨迹的影响。
主要参考文献:
Parizadeh M, Arrieta MC. The global human gut microbiome: genes, lifestyles, and diet. Trends Mol Med. 2023 Jul 27:S1471-4914(23)00152-1. doi: 10.1016/j.molmed.2023.07.002. Epub ahead of print. PMID: 37516570.

谷禾健康

最近有客户在拿到科研报告分析结果的时候问:
“在同样的数据分析流程情况下,为什么我拿到的分析结果提取不出什么有价值的结果,而别人有着类似的项目课题,样本类型也相似,却可以拿到全面的差异分析结果,想要提取的信息一目了然?”
★项目设计和入组标准以及采样至关重要
微生物组研究涉及到生活的方方面面,从环境微生物检测,到人体疾病与微生物群的关系,再到药物与食品营养干预等,以及菌群的发酵产物等。一项好的研究一定离不开前期的项目主题优化以及合理的采样方案。
项目分析主要是依托高通量测序获取的数据和特定数据库对样本数据进行归类,识别,统计检验分析,以及差异化寻找和可视化展示,来获取全面的数据信息。
最终拿到科研分析报告结果,PDF结果报告也好,在线或平台呈现展示也好,是对结果的排列,展示和解读说明等。
个别研究者拿到结果报告后,发现结果不理想,分组没有代表性,组内差异较大,当这个时候发现前期研究不明确,或者取样不具有代表性等问题,想通过后期科研分析的手段进行补救,根据我们多年的分析经验来看,在这种情况下只能进行一定范围内的微调,差异结果大局已定。
如果想得到优质的差异分析结果靠后期数据分析筛选修改很难达到要求。这个时候如果想再次优化课题,重新取样,项目周期会延长,因此,本文就前期实验方案设计和取样环节进行详细描述和讨论,便于科研工作者获得更优质的实验结果。
一套完整的扩增子和宏基因组测序流程包括:
1. 确定实验设计与研究目的、样本规模和时间
2. 样本的采集,归纳,保存和运输
3. 预期实验结果计划,分析统计方法选择
4. 样本提取,实验,上机测序的选择
5. 数据下机,分析,与结果的呈现
一般测序公司拿到样本以后先进行样本状态查验以及样本信息核对,核对没问题后会尽快开展实验环节,包括提取,建库,上机,可以通过标准化流程减少实验误差,统一规范操作步骤;而数据分析也是通过标准的SOP分析流程拿到完整的数据分析结果,分析中分析人员可能会根据数据尝试寻找最适配的分析方法,或者增加图表展示的内容和呈现形式,以辅助客户拿到最能反映研究结果的图表。
唯有前两项主题设计与采样环节的不确定性因素较大,需要客户提前设计准备充分,同时也决定了未来数据分析结果的好坏。
实验设计就是为整个课题设定一个总的大纲,确立总的研究方向和目的;取样环节是未来能否将实验方案具体实现的关键。客户提前做好科研课题开题前这两项工作,更有利于后续测序分析的开展。
微生物组最常见的研究方法之一是高通量测序,其原理是基于微生物基因组的特异性序列(细菌16S rDNA,真菌的ITS或宏基因组DNA),进行测序和分类,分析在特定的环境下微生物群落的物种组成和功能,微生物的多样性与丰度。
根据主题的不同可以将微生物组研究主要应用到以下几个方面:
1. 将肠道微生物与疾病进行关联以探讨疾病与健康的个体之间微生物的差异性;
2. 在特定环境下找出特异性微生物,发现耐受菌群及与特定环境具有关联性的代谢通路,通过不同样本之间的比较,研究微生物群内部、微生物与环境、微生物与宿主之间的关系;
3. 通过单一因素直接作用于特定环境或宿主(例如补充维生素、给土壤施加不同的类型的肥料)研究对应的微生物群在不同时间段的变化规律。
依照不同的课题类型提前规划好实验设计方案。
▼
例如可以研究不同用药组菌群构成差异;在不同的地理环境下土壤菌群构成的变化;还可以研究不同疾病类型的病人与健康对照组的菌群做两两分组之间的差异分析。
•保证组内样本的均匀性和一致性
为了尽可能多的从不同分组当中找出差异结果,需要保证组内样本的均匀性和一致性,排除组内样本差异性过大等干扰因素。
例如研究某一种疾病的肠道菌群构成时,需要排除多种因素对菌群构成的影响:
所以在前期实验设计的时候这些细节都需要考虑到位,避免个别的干扰因素影响整体的实验效果。
不同分组直接菌属差异比较
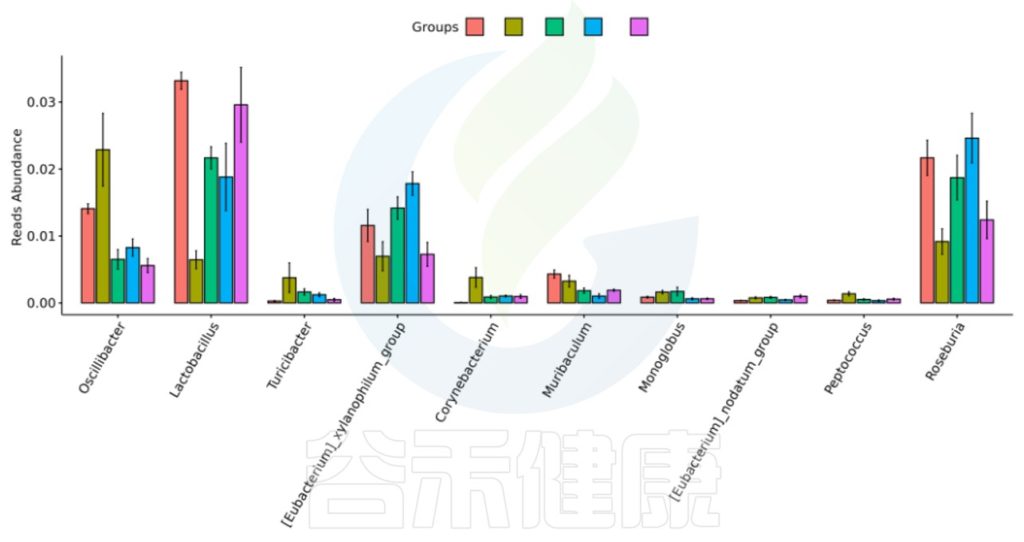
上图表明:尽管是用药组,但是不同的细分用药对菌群的差异影响不同,甚至差异还比较大。
下图可以看下每两组之间是有显著性差异的。
两两分组之间菌属差异比较
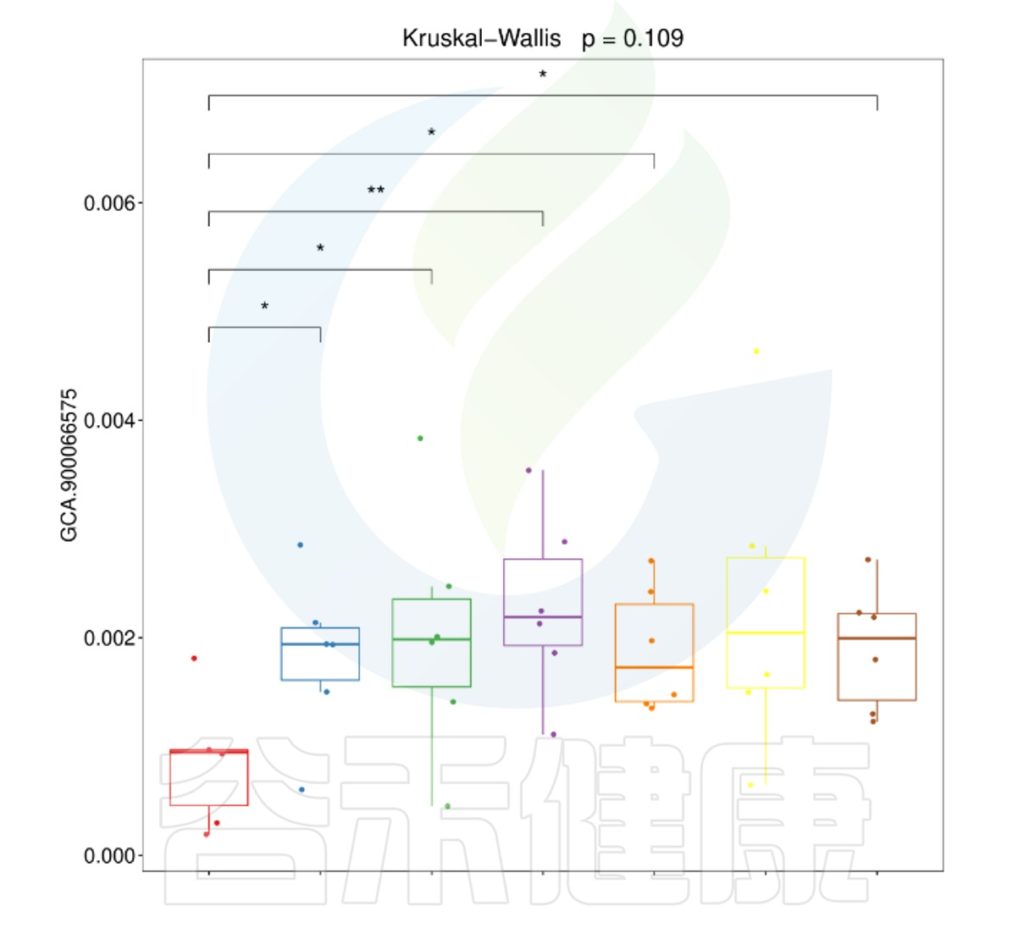
这刚好反馈在分组的时候,一定要按照研究方案不同或处理效应尽可能细分分组,或严格入组样本标准,或设置细分的干预浓度等。
合理细分分组,确保实验结果的科学性和有效性,从而推出更可靠的结论。
▼
相关性分析用于研究微生物群落内部,微生物与环境之间,微生物与宿主之间的关系。例如最常见的就是临床病人拿到的各项生理生化指标与微生物群之间做关联分析;或是环境、土壤、发酵样本的各项理化指标与微生物之间做相关性分析等。
该项目是比较自闭症儿童与正常儿童的菌群差异的项目。客户在样本信息单里详细描述了孕期各种理化指标,根据这些理化指标与菌群数据做envfit相关性分析。
从结果可以看出,自闭症谱系障碍与正常儿童之间的分组与菌群之间相关性极显著**,其次是否有先兆流产的分组与菌群之间有显著相关性*,其他的包括是否喝牛奶、孕期是否感染、妊娠高血压都与菌群有相关性。
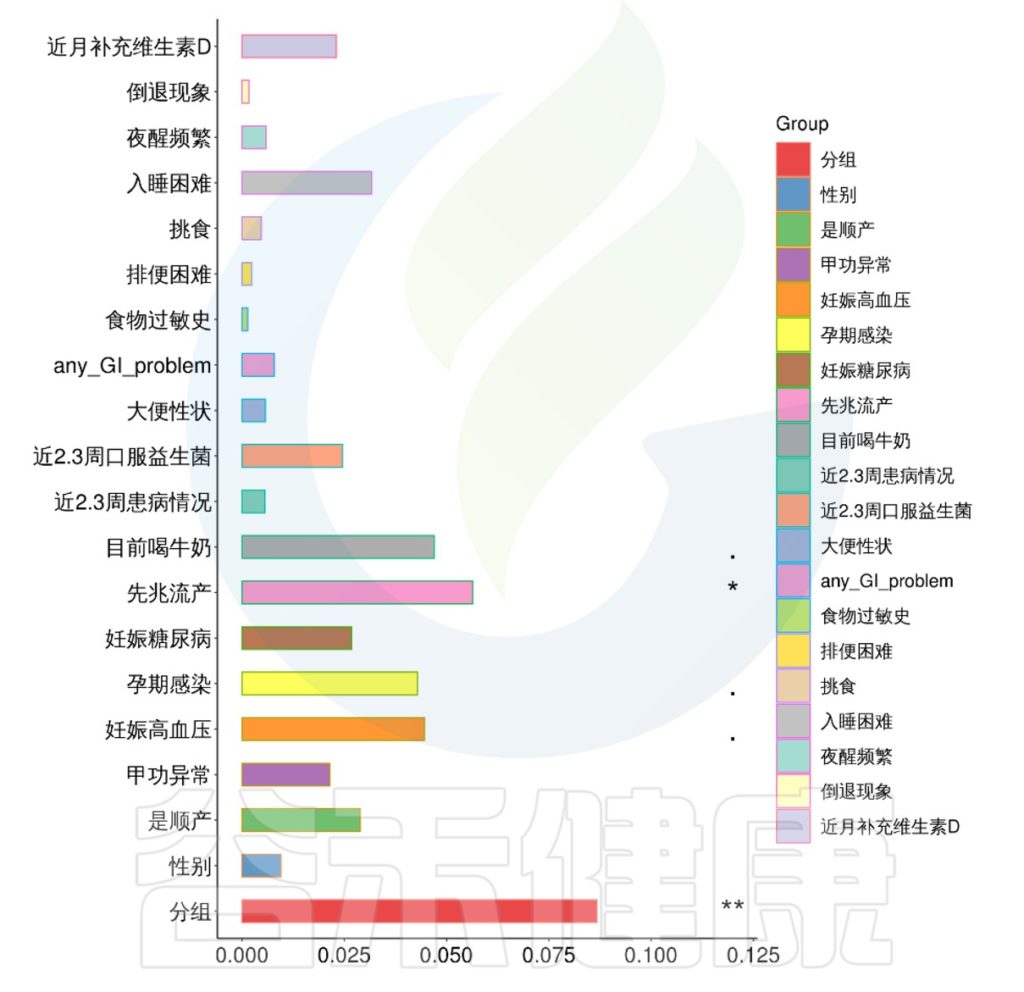
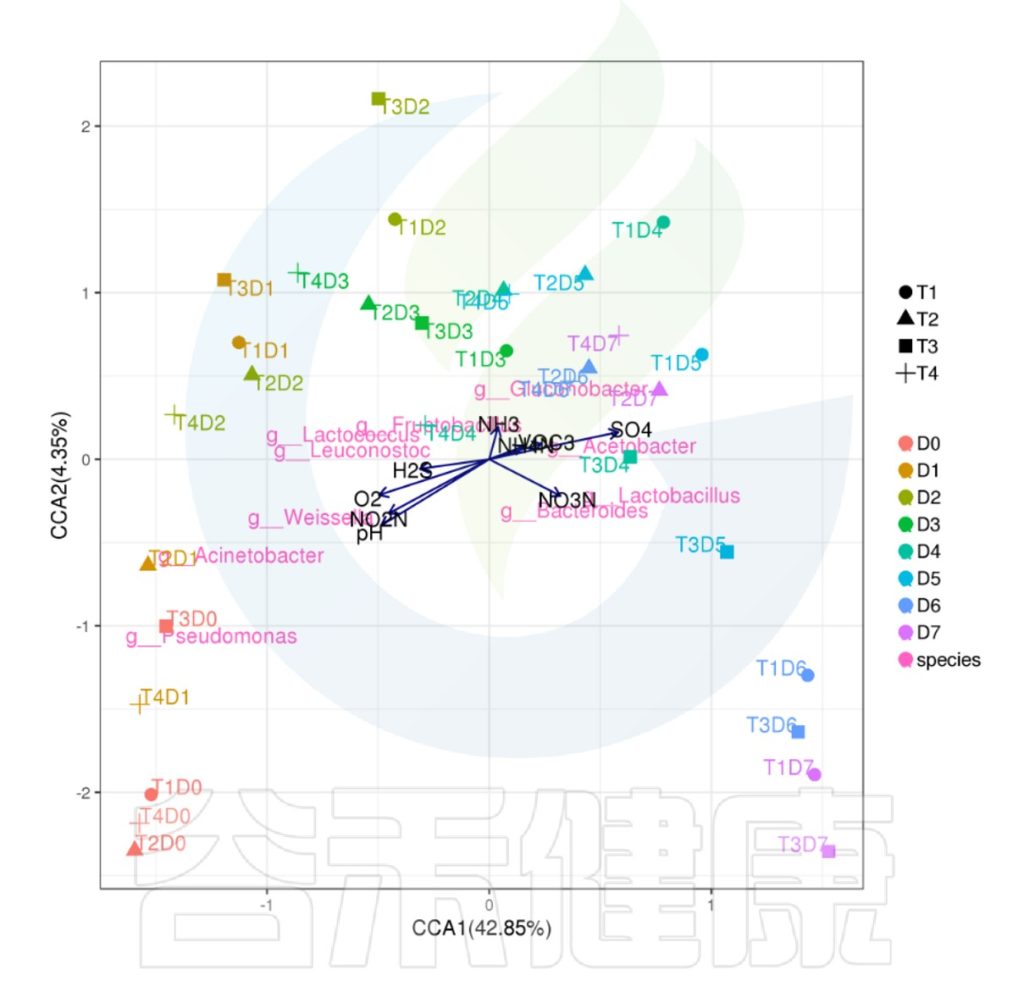
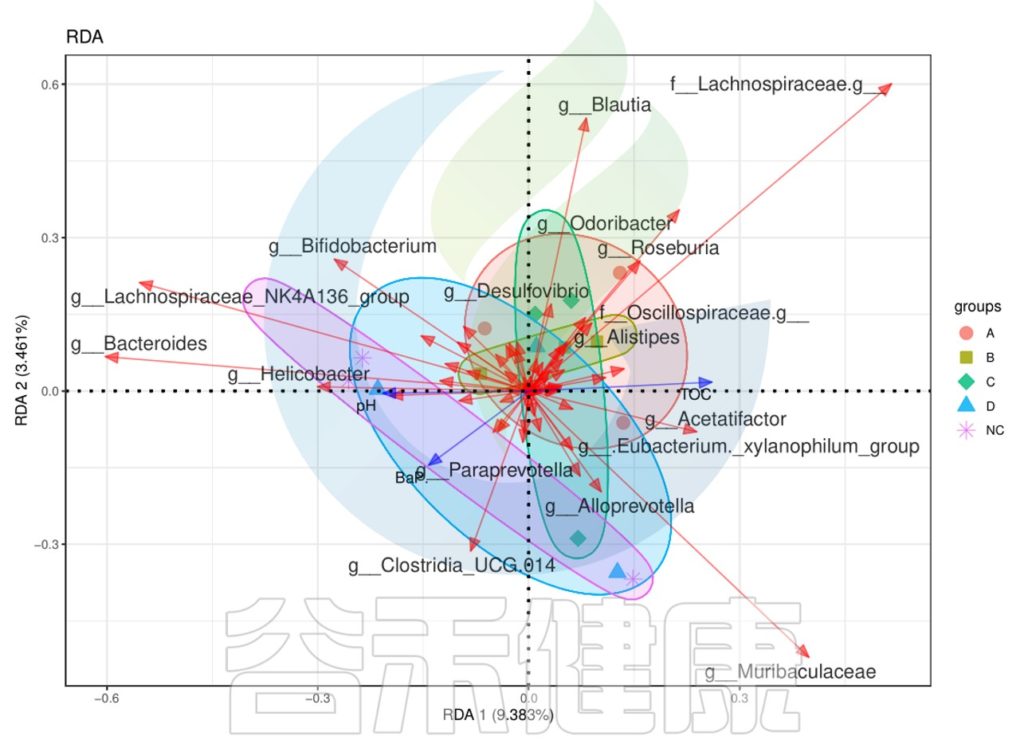
下列CCA图、RDA图可以做环境因子与菌属、样本之间的关联分析。
图中使用点代表不同的样本,从原点发出的箭头代表不同的环境因子。箭头的长度越长,表示环境因子的影响越大。夹角越小,代表相关性越高。样本点与箭头距离越近,该环境因子对样本的作用越强。
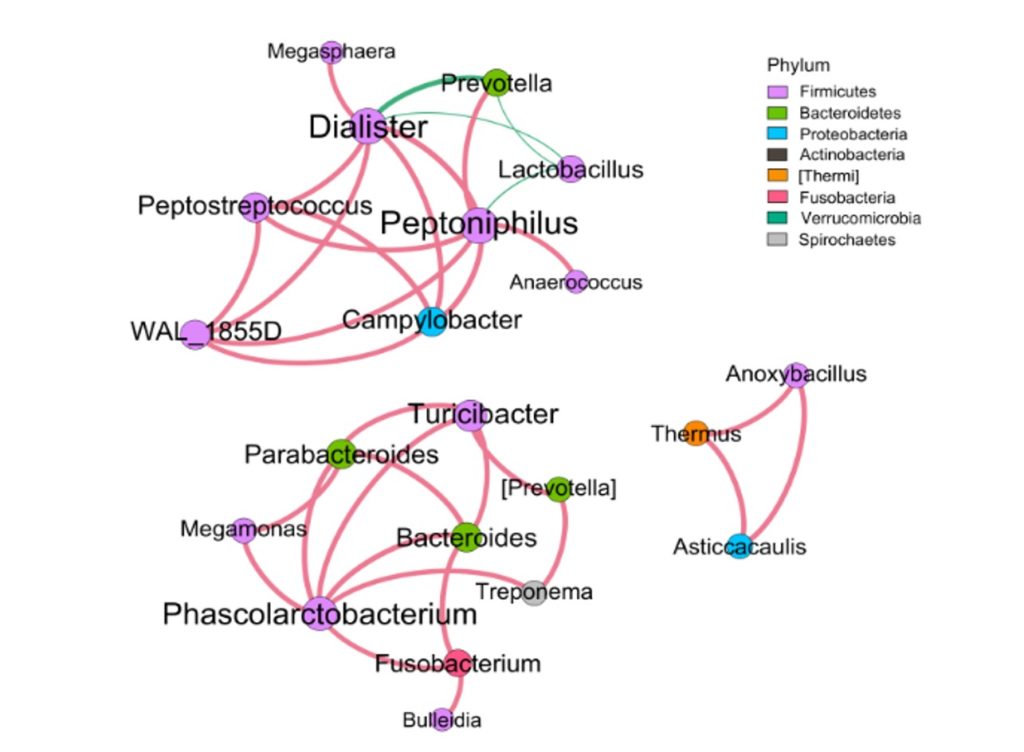
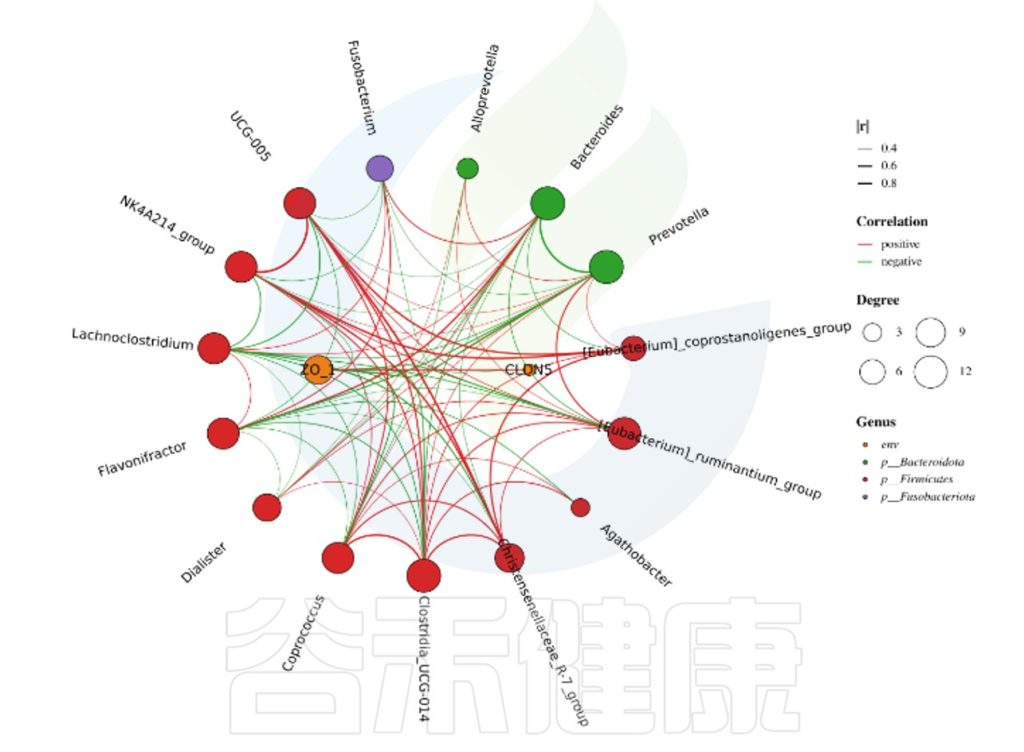
微生物相关性网络分析,可发现物种间的相互关系。图中每个节点代表属水平的物种,节点的颜色代表对应的门水平,节点越大,与之相关的属就越多,物种间的相关性用线条连接,相关性越强,线条越粗。线条红色正相关,绿色负相关。
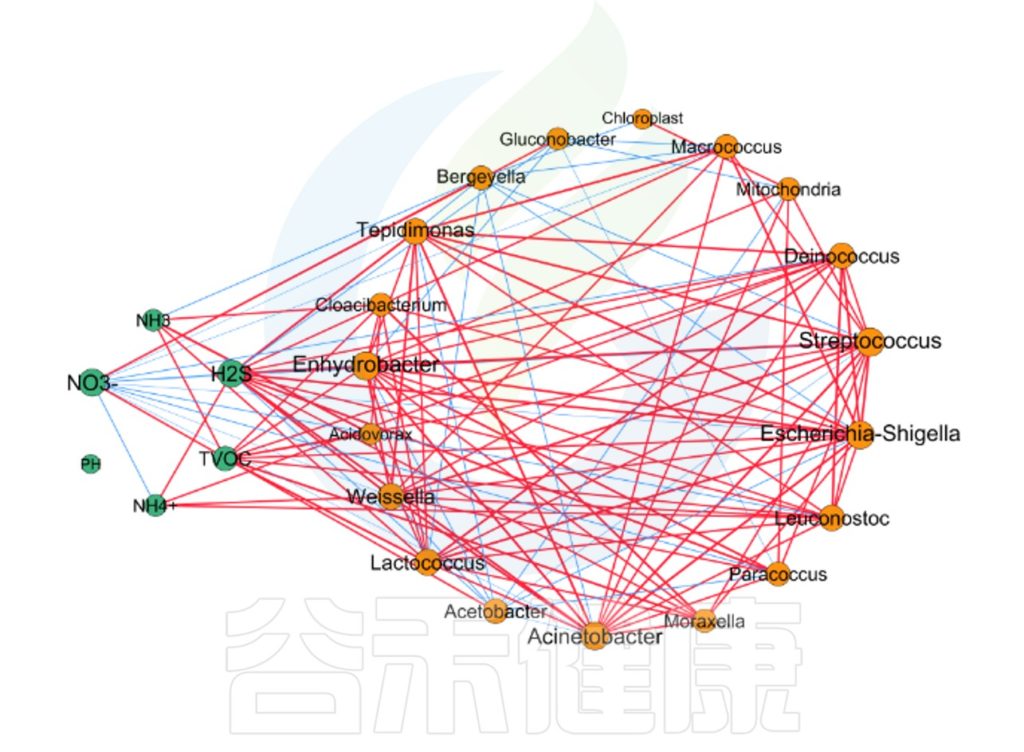
微生物与理化指标的相关性网络图
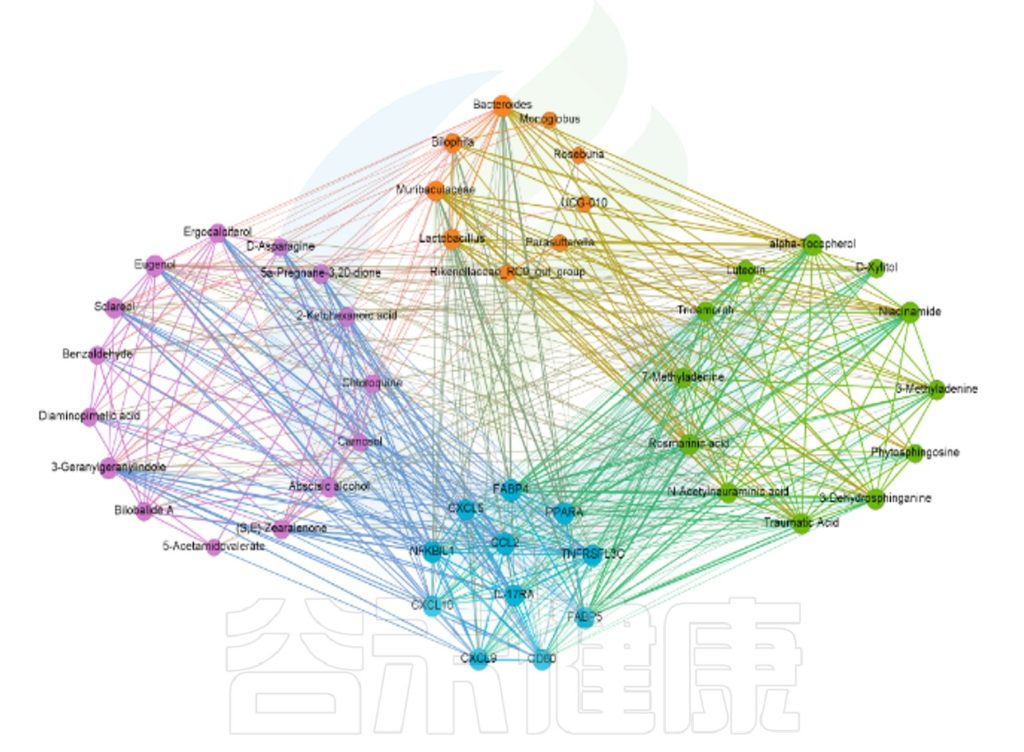
上调代谢物、下调代谢物与菌群和基因的互作关系
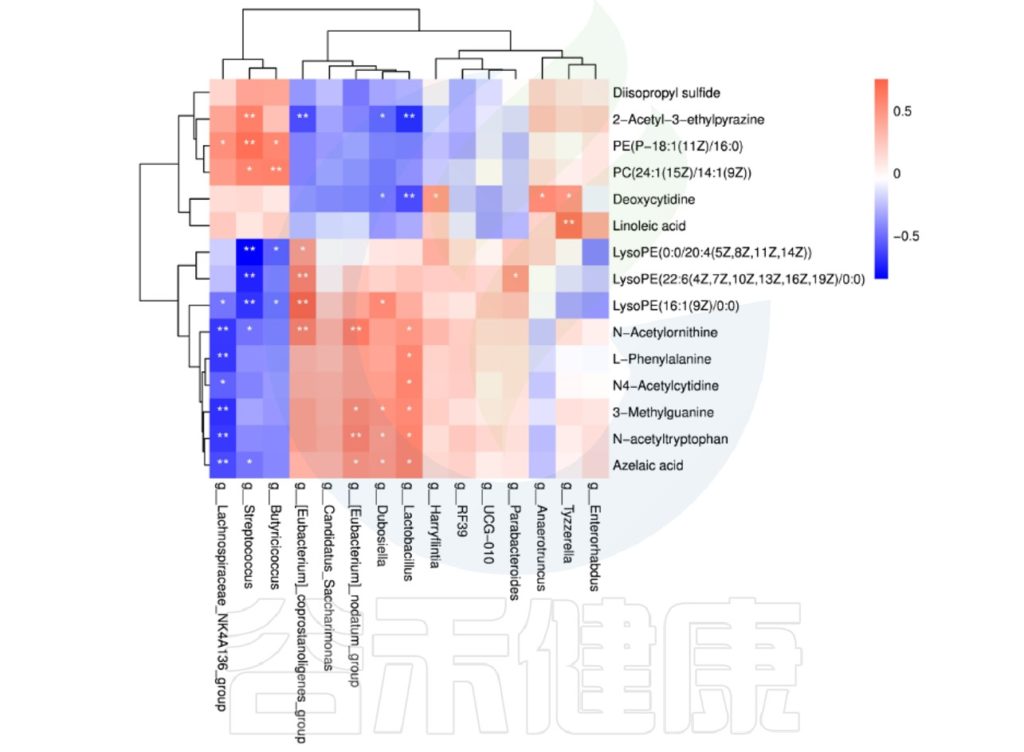
理化指标与菌群的相关性热图分析
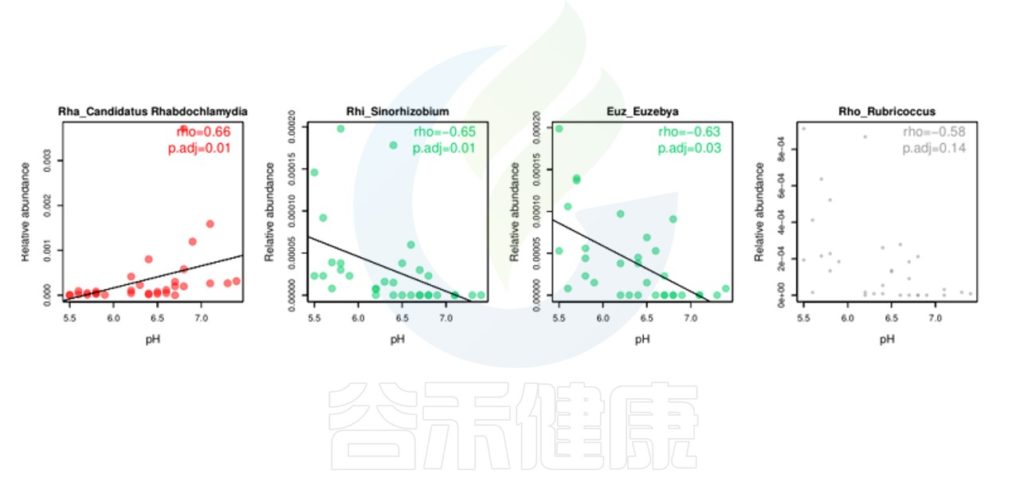
理化指标与物种的线性回归分析
•实验设计时规划好哪些指标与菌群数据之间做关联分析
在实验方案设计阶段先规划好哪些生理生化、临床指标可以跟菌群数据之间做关联分析,在取样的同时搜集好样本相关的理化指标质量和范围,有利于后期相关性分析,增强分析结果的说服力。
将详细数据填写在样本信息单里,例如:
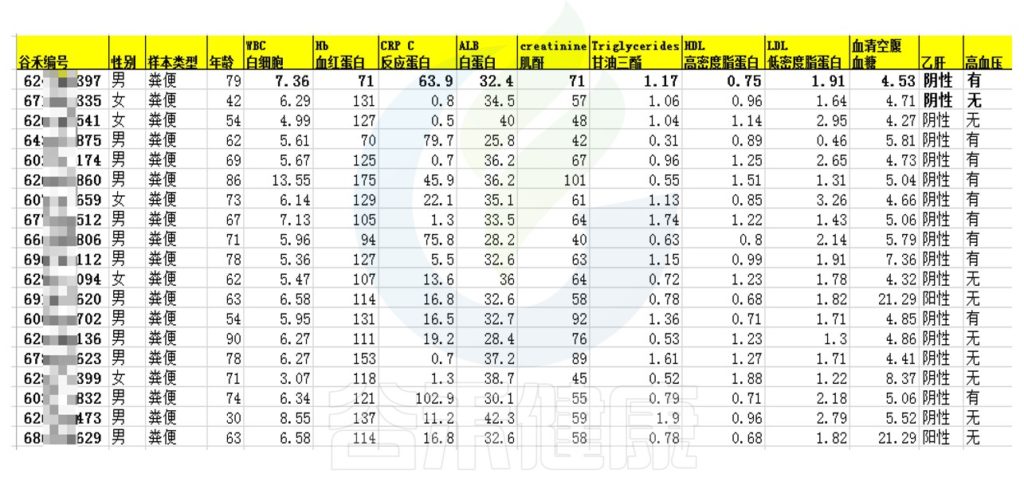
▼
微生物群是一个相对较为稳定的组合状态,在受到外部环境变化的影响下,菌群构成也会随之发生改变。在一定的时间范围内,菌群构成呈现出一定的变化规律。
•外界因素影响下肠道菌群在一定时间内变化
例如研究在抗生素药物治疗的情况下,一定时间内病人的肠道菌群变化情况。抗生素会直接影响肠道菌群的丰富度和多样性。
营养饮食的影响相对缓慢
对于益生菌,或者营养饮食等方面的摄入对肠道菌群的干预是缓慢的,被动的,需要长时间的补充摄入才能对整体的菌落构成起到影响作用,直到菌群构成趋于稳定。所以这里时间上要从几周到几个月的时间段内进行选取和采样,形成一个完整的采样周期。
婴儿的肠道菌群变化较为显著,取样间隔更短
又比如婴儿的肠道菌群在头几年变化较为显著,前期取样的时候就需要间隔更短一点,例如几天到几周采一次样,才能更细致的捕捉到婴儿肠道菌群的变化规律。
时间间隔的长短会影响微生物的时间序列分析,依据不同的样本类型,选取恰当的时间间隔周期,能观察短期或长期的菌群变化趋势,可以做个体之间的线性分析,配对T检验。
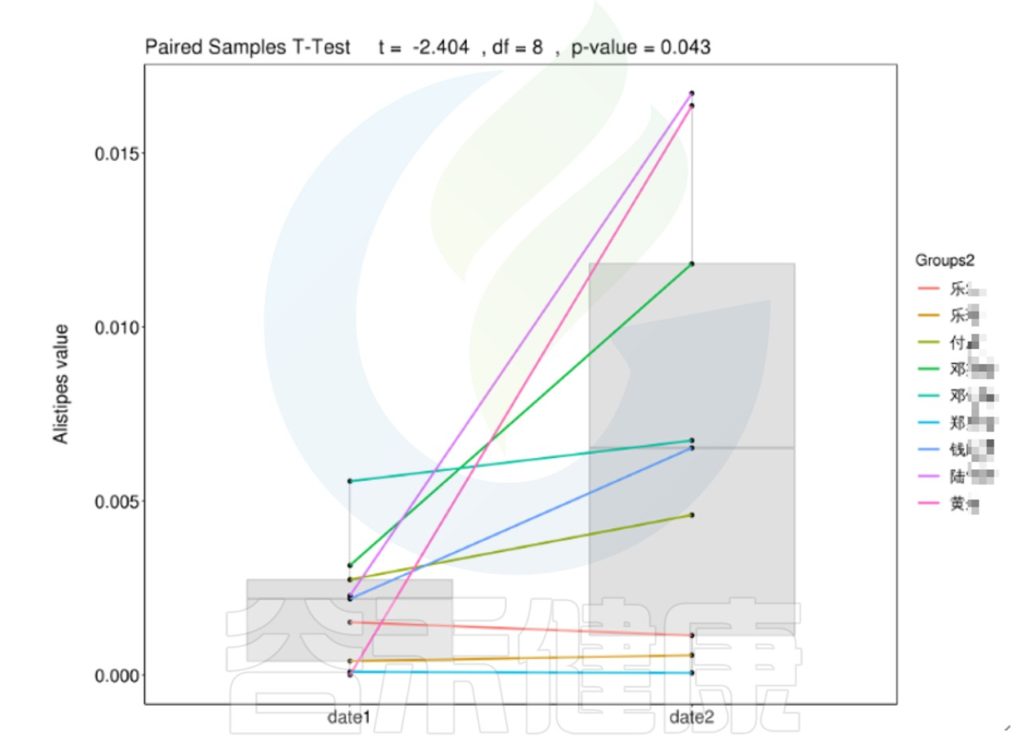
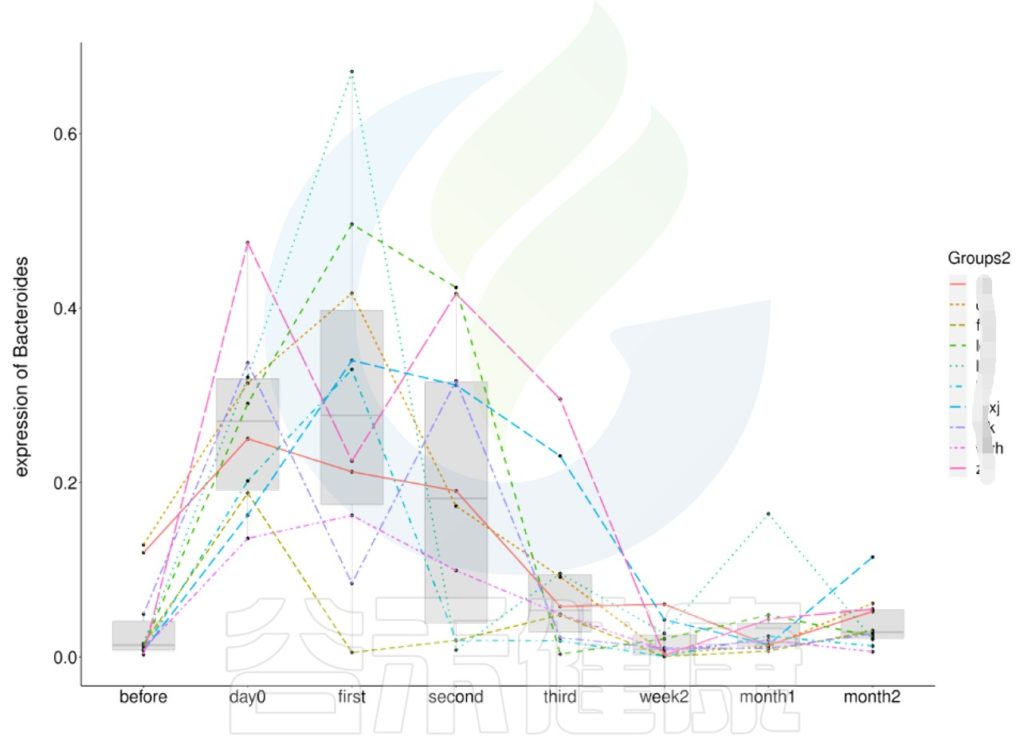
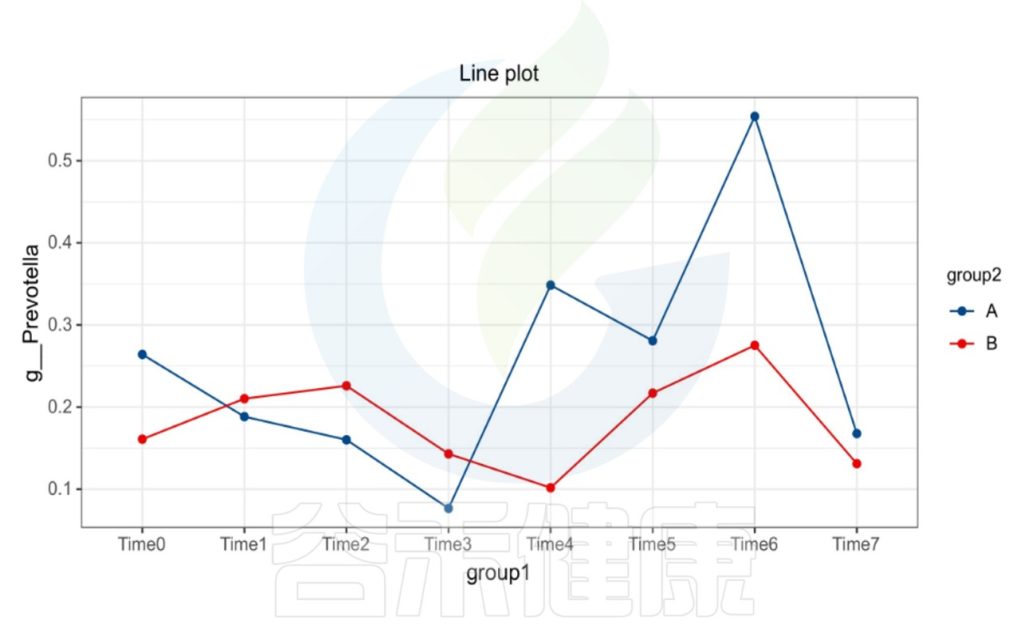
编辑
恰当的取样方法对于微生物组组学研究尤为重要,样本的收集和处理决定了后续微生物组鉴定的准确性。
前期的实验方案设计规划好以后,下一步就是落实,基于实验设计选取符合实验方案要求的高质量样本。在取样时需要注意以下几个点:
重复取样时,确定地理位置,或取样部位、取样深度、体积等都能实现标准化,以保证各个环节的一致性和可重复性。
影响肠道菌群样本的干扰因素有:年龄、地域、感染、疾病,药物治疗如抗生素、化疗、营养饮食干预等。
•收集样本时其他条件尽量保持一致
收集疾病组样本或者健康对照组样本时,尽量选择具有相同地理环境,饮食习惯,年龄段,药物治疗相关的病例作为重复样本。
采集过程的污染物也会被检测到变成样本组成的一部分,所以采样的过程中重复使用的工具,也要进行适当的消毒处理。取样的标准化和可重复性也可以避免由于不同的批次处理带来的样本菌群构成的差异性。
为了能合理的评估不同分组菌群之间的差异性或相似程度,必须要有一定的数据量来评估差异或者效应的大小。
一般来说,数据量规模越小,一组样本内的变异性越大。因此需要有更多的样本量来获得足够的统计能力。
每组至少3个重复,才能满足最低的组间差异统计检验条件,一般实验我们建议每组至少6个样本重复,临床样我们建议每组至少30个样重复。
不同的样本类型,如环境、植物、动物到人类,微生物组的来源差别较大,取样方法也有所差异。
•不同取样部位具有各自的生理特征
关于肠道菌群取样部位问题,该取粪便样本还是取肠道内容物?
根据研究发现不同的取样部位(如:粪便、粘膜活检,肠液等样本)具有各自的生理特征,肠道菌群的研究结果也有所差异。
•肠道内容物具有侵入性,用于某些特定疾病
肠道内容物由于采样方法较为复杂且具有一定的侵入性,所以在日常的应用相对较少。但是如对于一些特定的疾病或者研究问题,需要区分肠道不同部位微生物的组成或特性时,肠道组织样本或肠道内容物样本可能更加适合。
•一般情况下选择粪便样本
从实用的角度考虑,粪便样本可以自然采集、无创,且可以重复采样成为最常用的检测工具,是肠道菌群检测的最优选择。
注意
传统的粪便样本取样方法繁琐,对样本量和保存运输方式条件较为苛刻,例如单次的取样量要达到200mg以上,保存和运输过程中必须冷冻保存。一般临床上病例和健康人收集样本的配合度和接受度也不高,这也大大增加了收集样本的难度和成本。
★谷禾取样管便于采样以及储存运输
为了解决临床和实验过程中对粪便样本的取样遇到的这些实际问题和痛点,谷禾经过多年的肠道菌群检测实践,进一步对粪便样本的取样流程进行优化,开发出了专门用于肠道菌群取样的谷禾肠道菌群取样储存盒,实现了粪便样本的日常常规取样和常温储存运输。
该取样盒操作简单,取样方便,常温可以有效保存7天,日常在家就能实现常规粪便样本取样,缩短了实验人员收集样本的效率,节省了人力成本,所以该取样盒在市面上一经推出,就受到各大院校和临床机构的欢迎。同时也为谷禾做大健康方向前期收集样本阶段,提供了有力的前期技术支持。
谷禾取样管具有以下特征:
1、在家中就可轻松自行采集高质量样品
2、起始样品需要量低至0.01g左右,快速且稳定
3、样本取样7天内常温下储存和运输可保障稳定的群落结构
4、标准样品适合高通量标准化处理
5、能有效获得适用于16S 、宏基因组和qPCR等的高质量DNA
6、条形码化全样本可追溯性

取样方法也比较简单,仅需使用棉签从厕纸上沾取粪便,然后洗脱到取样管的保存液中即可,使保存液可见粪便颜色即表示取样量足够。
谷禾取样管不仅可用于人体粪便样本的采集,动物的粪便也同样适用,例如:大鼠样本用无菌棉签沾取少量粪便样洗脱到样本管的裂解液中;小鼠样本只需要放入1~2粒的粪便粒到裂解液即可。
1.如3天内使用过抗生素类,质子泵类胃药,阿片类精神药物请停药3天后进行检测;
2.感冒、腹泻或其他症状期间不影响取样,拉稀或稀便可以用棉签反复沾取粪便至取样管;
3.如长期便秘无排便可使用开塞露等辅助手段获取粪便样本;
4.取样无时间和饮食限制,取样前按照日常的用餐习惯进食即可;
5.完成取样后样本可常温有效存储一周,为保证检测时效请完成取样后尽快送检。如需长期储存,可将样本冷藏保存,-20℃冷藏最多可保存半年,-80℃可保存一年;
6.样本保存于取样管后需要拧紧瓶盖,防止取样管漏液,或者受到环境污染。

其他样本类型则需要用到传统常规的取样方法严格按照标准取样,之前我们的文章有对不同样本类型的常见取样方法作详细介绍:
微生物取样方法大全 (点击查看)
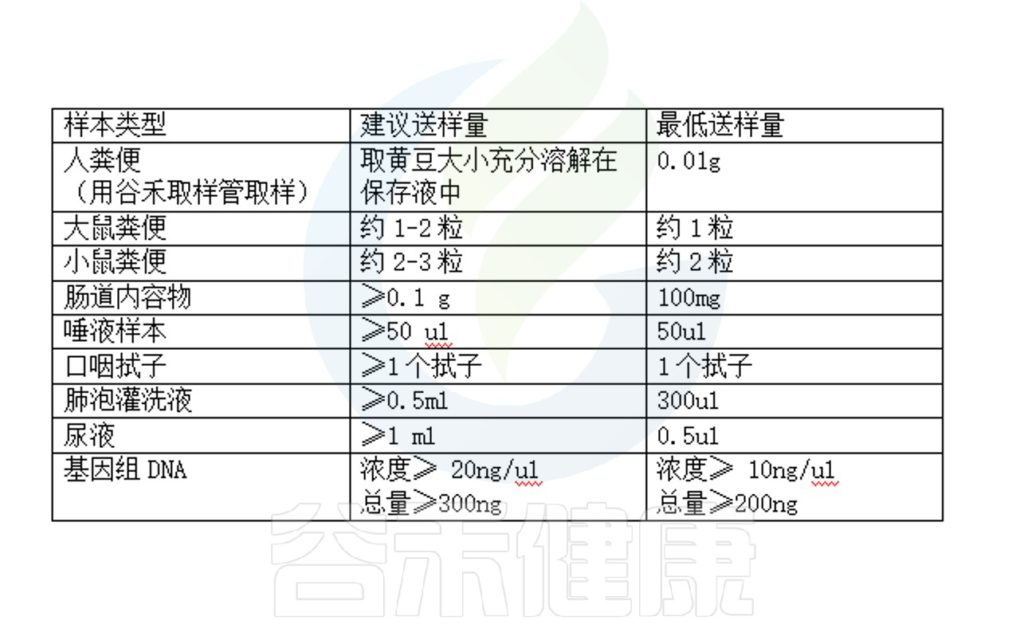
注:以上是16s测序项目送样需求,如果是宏基因组测序,需要达到常规标准,而且高宿主样本例如灌洗液,尿液,组织等样本,需要提前处理。
送样前要做好以下这些准备:
•样本信息单
样本信息单里填写好相关资料,包括谷禾编号,对应的科研分析名称,分组情况,有多少种分组情况就填几列,样本名称和分组名称由英文字母、数字、”.”组成,并以字母开头,不能以数字开头,不支持字符“- ”“_ ”、运算符、中文、空格等特殊字符。
注:名称长度最好不要超过8个字符。
如果要做相关性分析需要提前准备好样本的临床指标、生理生化指标,并将每个样本的具体测得的指标数值填写到样本信息单里。
如果对科研分析不太熟悉,不知道还能提取出哪些有用的分组信息,可以将具体的研究目的、实验方案详细写在备注里,分析人员会根据您的信息给出相应的指导意见和分析方案建议。可以提供一个纸质和电子版的样本信息单。
•样本检查
样本送样前要对样本和运输条件做检查,取样管是否拧紧,是否有漏液情况,样本不受环境污染和干扰,检查样本个数是否能核对的上,在寄样前要在快递箱上附带一张送样信息的纸条,包含以下内容:
寄样单位或联系人:xxx
寄样日期:xxx
样本个数:xxx
实验室接收到样本后会跟送样信息单进行核对,如果样本个数和信息能核对上,则继续对样本进行实验上机,如果样本个数或信息核对不上,会跟客户沟通,核对信息。
小 结
客户在做好实验方案设计和样本收集的前期准备工作以后,后续的实验和测序分析工作就交给我们来完成吧,从收到样本到科研分析报告的出具,周期一般在6~10天左右(16S v4); 15-20天(16S V3V4);15-20天(宏基因组)。
实验室在收到样本以后会先核对信息,然后进行样本DNA提取,质检,建库,测序上机。等测序数据下机以后,对原始数据拆分,质控,然后就进行科研数据分析阶段。
分析思路:
1.先了解不同分组样本都由哪些微生物构成,并计算丰富度,做α多样性分析;
2.比较不同分组微生物群落整体是否有差异,做β多样性分析;
3.进一步找出不同分组之间微生物群落的差异菌属,关键菌属;
4.根据关键菌属可以作为biomarker用于疾病诊断模型构建;
5.找出菌属与临床理化指标的相关性;
6.根据物种构成预测菌属代谢途径,基因功能预测,找出组间差异功能预测,查看菌属是怎么运行宿主的
7.将数据结果整合起来,找出不同疾病环境分组条件下,菌属的差异是否跟疾病相关,并影响宿主。
最终根据科研分析报告的研究方向和侧重科研选择不同的分析方法,并做图像化展示。
差异分析方法:
•Tukey检验
•非参数检验
•基于矩阵的Anosim检验、Adonis检验
•LEfSe分析
•随机森林和ROC曲线
相关性分析方法:
pearson相关系数
spearman相关系数
相关性网络图、相关性热图、线性回归图
功能分析:
PICRUSt2、FAPROTAX、BugBase
PCA、PCoA、NMDS
ROC、RDA
Venn图、花瓣图、Upset图
物种构成柱状图、热图、差异箱型图
GraPhlan物种进化树
metagenomeSeq
Random Forest随机森林
ROC曲线图
graphlan图
Gephi复杂网络图
三元相图
谷禾近期合作发表示例文章:
Hu Y, Li J, Ni F, et al. CAR-T cell therapy-related cytokine release syndrome and therapeutic response is modulated by the gut microbiome in hematologic malignancies[J]. Nature communications, 2022, 13(1): 1-14.
Lou M, Cao A, Jin C, et al. Deviated and early unsustainable stunted development of gut microbiota in children with autism spectrum disorder [J]. Gut, 2021 Dec 20:gutjnl-2021-325115.
Li R, Liu R, Chen L, et al. Microbiota from Exercise Mice Counteracts High-Fat High-Cholesterol Diet-Induced Cognitive Impairment in C57BL/6 Mice[J]. Oxid Med Cell Longev. 2023, 20:2766250.
Xiao W, Zhang Q, Zhao S, et al. Citric acid secretion from rice roots contributes to reduction and immobilization of Cr(VI) by driving microbial sulfur and iron cycle in paddy soil [J]. Sci Total Environ. 2023, 16:158832.
Wang X, Weng Y, Geng S, et al. Maternal procymidone exposure has lasting effects on murine gut-liver axis and glucolipid metabolism in offspring [J]. Food Chem Toxicol. 2023, 174:113657.
Liao J, Dou Y, Yang X, et al. Soil microbial community and their functional genes during grassland restoration [J]. J Environ Manage. 2023, Jan 1;325(Pt A):116488.

谷禾健康

成瘾是一种大脑疾病,受害者无法控制地对某种物质或行为产生强烈的依赖和渴求,尽管这种行为会产生有害的后果。成瘾包括一系列物质滥用障碍,例如药物、酒精、香烟,过度饮食。近年来,吸毒成瘾急剧上升,特别是阿片类药物,全世界每年有数十万人死于吸毒过量。

成瘾的特征是负面和正面情感状态的交替,这些状态可以归因于不同阶段神经递质活动的改变:
而微生物组已被证明能以多种方式影响宿主大脑,例如通过调节神经递质、通过肠脑轴与中枢神经系统相互作用,使得微生物群可以通过产生负反馈环和正反馈环来影响或参与大脑奖赏和戒断回路的神经活动,从而促进成瘾行为。
在这些回路中具有重要功能的神经递质,如多巴胺、GABA和5-羟色胺,都可以由肠道微生物群产生或调节。一些研究表明,肠道微生物可以合成苯丙氨酸和代谢左旋多巴,这两种都是多巴胺的前体,因此它们可以调节多巴胺的水平。这些过程促进了微生物影响大脑奖赏回路的途径,例如通过介导强化和奖赏的D1R和与厌恶和回避相关的D2R。
还有微生物组衍生的短链脂肪酸和色氨酸代谢,特别是它在5-羟色胺合成中的作用。总之,这些证据表明,微生物组有可能通过诱导强化积极和消极的对宿主的影响,从而影响宿主的状态和行为。
为了深入研究宿主-微生物组生态系统的生态学和进化学在成瘾机制中的影响作用,研究人员开发了一个数学框架,通过建立数学模型来模拟宿主-微生物生态系统对变化的反应动力学。利用这个模型,研究人员分析了成瘾行为的几个方面,重点关注微生物组对成瘾的起始和戒断的潜在影响。
研究人员假设,作为宿主微生物群一部分的微生物菌株已经进化出一种模式,影响宿主,从而改善了这些菌株在微生物群落中的地位。这可能会导致微生物群落在不同方向上影响宿主。比如宿主的成瘾行为改变了寄生微生物的生存环境,即使毒瘾对微生物很大程度上是有害的(例如毒素),但对一些微生物来说,它可能比对其他微生物的危害要小。这导致了微生物选择制度的转变,并扰乱了微生物组的组成。在新条件下繁殖的菌株可能会受益于宿主继续其新的行为。因此,微生物群可能在增强和维持成瘾行为方面发挥作用。
Methods
研究人员开发了一个三组分的框架来模拟宿主-微生物组相互作用及其对宿主行为的影响。
➦ 1 将宿主行为建模为空间中的连续特征。
➦ 2 宿主体内含有N个微生物菌株,随着宿主行为的改变,不同微生物菌株的丰度也会发生变化。
➦ 3 考虑微生物组对宿主行为的影响,模拟了微生物菌株分泌化合物,影响宿主奖励和戒断回路的功能,产生正向或负向的反馈(例如奖励或厌恶),从而调节宿主行为。
在模拟宿主-微生物组相互作用的模型中,其中宿主行为影响微生物组成,而微生物组则影响宿主行为。在这个模型中,研究人员使用相同的框架建立了两个模型:
具体的模型描述如下:
1 宿主的行为基线的建模描述
宿主的基线行为被表征为二维单位球体中的坐标

➨ 成瘾阶段:
即行为基线的改变,宿主的行为基线的改变表现为沿着X轴正方向的一系列移动。
在每个时间点t,步长σA(t)从均值为σ的指数分布中随机抽取。宿主行为的改变受到距离原点R的限制,代表了最大的成瘾程度。因此,宿主行为的坐标逐渐从(0, 0)移动到(R, 0),并在该阶段结束之前保持在(R, 0)位置,即从开始计算的τ个时间步长之后。
➨ 戒断阶段:
宿主通过在X轴上进行大小为 (−σA(t))的移动来逆转其行为模式,其中σA(t)从均值为σ的指数分布中随机抽取。
行为的改变下限为0。该阶段在微生物组成和宿主行为稳定之后结束,或者在该阶段开始后经过100000个时间步长之后。
2 微生物群组成的建模描述
一个由N个微生物菌株组成的群体寄居在宿主体内。每个菌株都有其独特的特征,可以用微生物组-行为空间中的坐标来建模。

考虑到微生物群中不同微生物菌株之间的资源竞争,在每次迭代中通过其总和对 xi(t+1) 的值进行归一化。由此获得微生物菌株的相对丰度,同时假设微生物群落的总丰度保持不变,类似于对整个微生物组群体设置一个承载能力。
接着,在每个菌株上设定一个恒定的低流入速率 μ / N,以避免菌株从系统中灭绝和永久消失(μ在整个过程中设置为10^(-8))。在每个时间步长中,使用上面的公式计算菌株的比例,然后计算流速和第二次归一化。
对于两菌株竞争模型(N=2),每个菌株的特征是手动定义并在结果中进行说明的。其中一种菌株会影响宿主行为,并承担反馈产生的成本,而另一种菌株不会影响宿主行为且不需要支付成本。
对于成瘾模型,在每次模拟开始时,在微生物组-行为空间内随机抽取菌株。还分析了不包括归一化的广义Lotka-Volterra模型(同样使用前向欧拉法,时间步长为1):
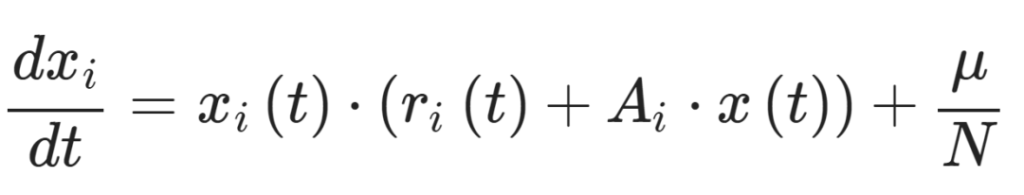
这个方程系统与上述过程类似,但有两个主要修改。
3 微生物群影响宿主行为的建模描述
假设所有或部分微生物菌株可以影响宿主的行为,且菌株可以感知它们种群中有益和有害的变化,这些变化表现为种群大小在一段时间里的增加或下降。
这些反馈将会影响宿主未来的行为轨迹。

因此,宿主轨迹远离中心、朝向中心或中性取决于过去 wh 个时间点的线性轨迹的斜率。默认设置 wh= wm -10
结合微生物代谢产物及其行为轨迹,微生物菌株 i 对宿主行为在时间 t 上的影响方向可以通过Imi(t)⋅Ib(t) 的结果来描述。
在时间t时,每个菌株的总效应强度是该菌株在微生物群落中的比例乘以( |σA(t)|⋅di)。
其中 di 表示菌株i对宿主的影响大小,|σA(t)| 是时间t的行为基线步长。微生物效应的大小 (di,i∈{1,…,N}) 是从指数分布中随机选择的,每次模拟开始时其均值为E[d],在每个时间步长中计算σA(t)。

在上述的模型中,微生物群落对宿主行为的影响是间接的,需要通过宿主的奖励回路。
为了观察微生物群落对宿主行为的直接影响,研究人员另外又建了一个模型,这个模型假设在成瘾和戒断过程中每个微生物菌株都会将宿主拉向其在微生物组-行为空间中的坐标。
首先,将宿主行为坐标减去菌株特征坐标,并对结果向量进行归一化,来确定每个微生物菌株相对于宿主行为的方向。
然后,将所有得到的向量求平均,得到整体微生物群落的影响方向。这个平均值根据每个微生物菌株的比例(xi )加权,同时每个分量也乘以菌株的影响强度(di)。
现在宿主行为的变化由宿主行为基线步长和微生物影响的总和决定,定义如下:
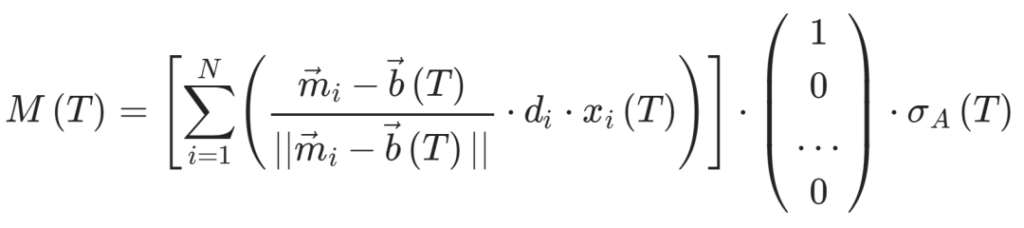
Results
基于方法中描述的模型,研究人员给出了从模型结果中观察到的宿主与微生物群的现象,如下:
1 微生物能够反应宿主行为的变化
在微生物菌株之间的竞争行为中,当其中一个菌株通过改变宿主行为而获得额外资源时,这种微生物效应就会被显现出来,反映为它在群落中的比例会增加,从而增强其继续影响宿主的能力,进一步诱使宿主继续新的行为。
如下图,模拟了两个微生物菌株之间对宿主资源的竞争,这些竞争源于宿主行为。
当微生物提供的优势超过产生这种影响的成本时,它对宿主状态的影响可能是有益的
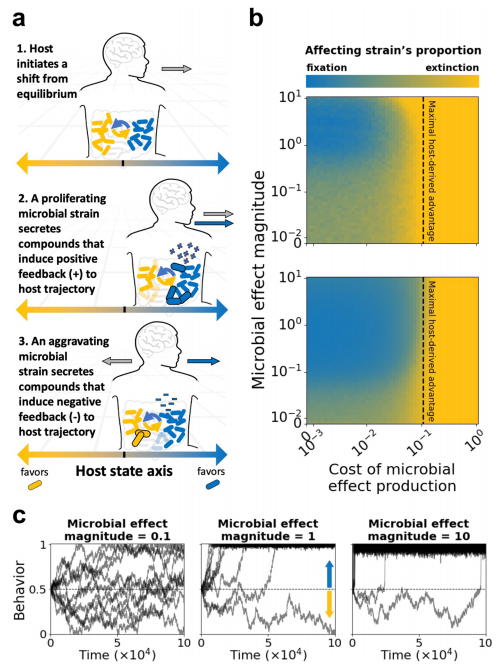
a| 模型插图。蓝色和黄色分别代表两个不同的菌株,其中蓝色的菌株可能在影响宿主的行为:它在增殖时提供正反馈,诱导宿主继续其行为趋势,在下降时提供负反馈,诱导宿主逆转其行为趋势。
b| 绘制了经过100000个时间步长后影响宿主的菌株比例的热图。宿主的行为在x轴上沿着0到1之间随机游走,代表资源的消耗。
c| 绘制了在模拟的100000个时间步长里,微生物效应的大小与宿主行为轨迹的关系。
2 宿主-微生物群的相互作用可能会加剧成瘾行为
建立了一个简单的宿主行为模式,包括三个阶段:
当宿主开始改变其行为时,有相当数量的微生物菌株支持或反对这种行为改变,如图a,这是一个微生物组-行为空间,将N个菌株随机分布在空间内,星形的移动演示了宿主行为的改变。
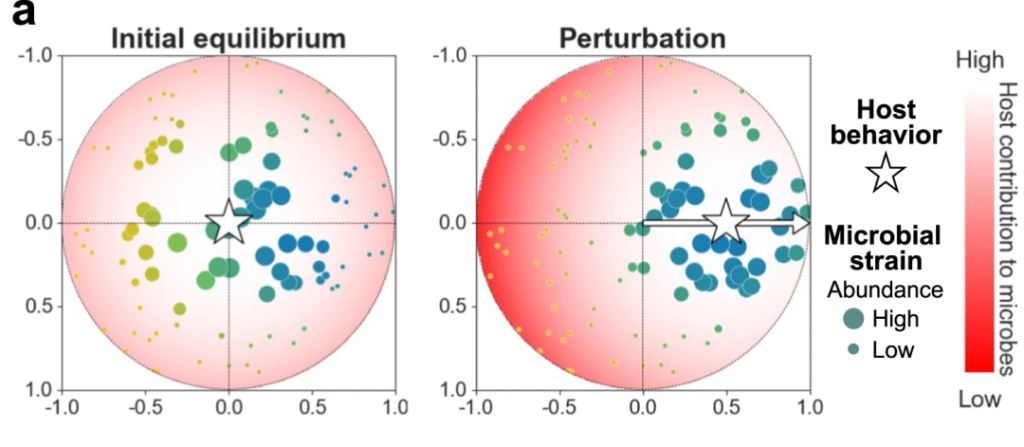
图b和c分别展示了在不同平均微生物效应量(E[d])作用下随着时间的持续,宿主行为的变化和菌株丰度及其适应度的变化。
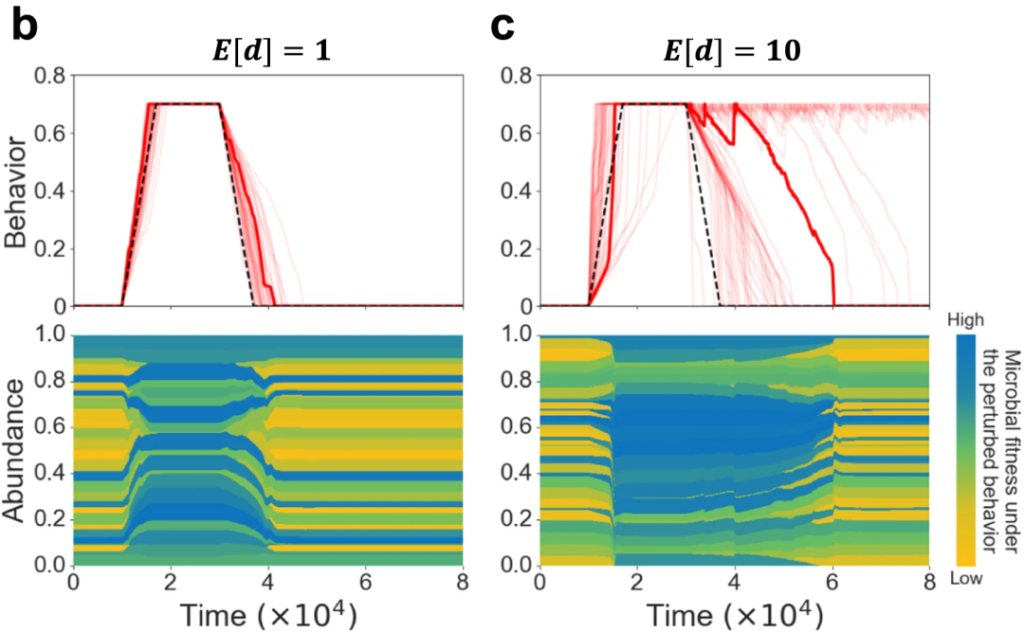
图b中的黑色虚线代表平均微生物效应量为零时的宿主行为基线。从E[d]=1到E[d]=10的宿主行为变化,可以发现微生物群的行为效应可以导致成瘾行为的加速和减速,但在大多数情况下,微生物群的行为效应减缓了戒断过程。
研究人员认为原因是在成瘾期间,微生物群转向了一个低多样性的群落,这个群落由在新的宿主行为下增殖的菌株组成。因此,在宿主开始戒断行为时,相当大一部分微生物群抵制行为变化。
3 微生物群落丰富度影响宿主成瘾行为
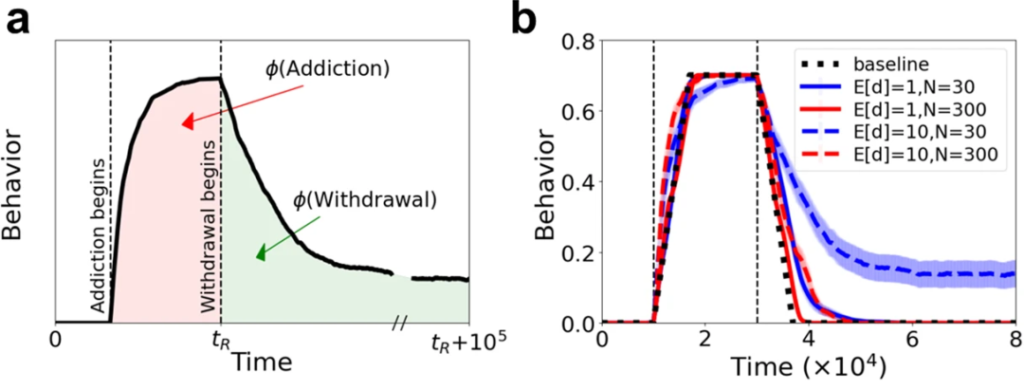
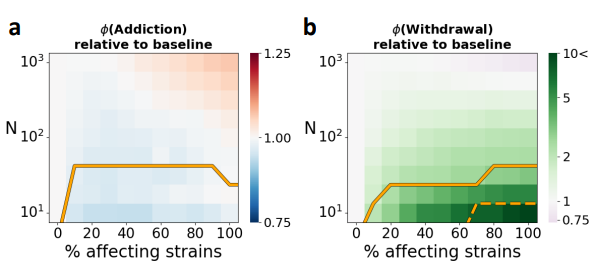
通过监测成瘾期(ϕ(Addiction))和戒断期(ϕ(Withdrawal))的微生物群落丰富度变化,发现加速成瘾或减缓戒断不仅取决于微生物效应大小,还与微生物群落丰富度有关。
如图a和b,在微生物组丰度非常高且微生物组效应强烈的情况下,微生物组实际上可能会加速成瘾和戒断过程(E[d]=10,N=300)。
在恒定微生物效应的平均大小后,然后减少影响菌株(即能够影响宿主行为的菌株)的比例,结果是总微生物组效应量的下降,并加速戒断过程。这表示即使只有少数菌株影响宿主行为,对成瘾和戒断的影响也可能是显著的。
如下图,每个像素的颜色表示相对于没有微生物组效应的基线情况下的成瘾期和戒断期的倍增或减少的情况,E[d]=5。
既然微生物群落丰富度会显著影响宿主成瘾或戒断行为,研究人员便做了一个模拟实验,在戒断期引入新的微生物菌株来增加微生物群落的丰富度和多样性,结果如下图。
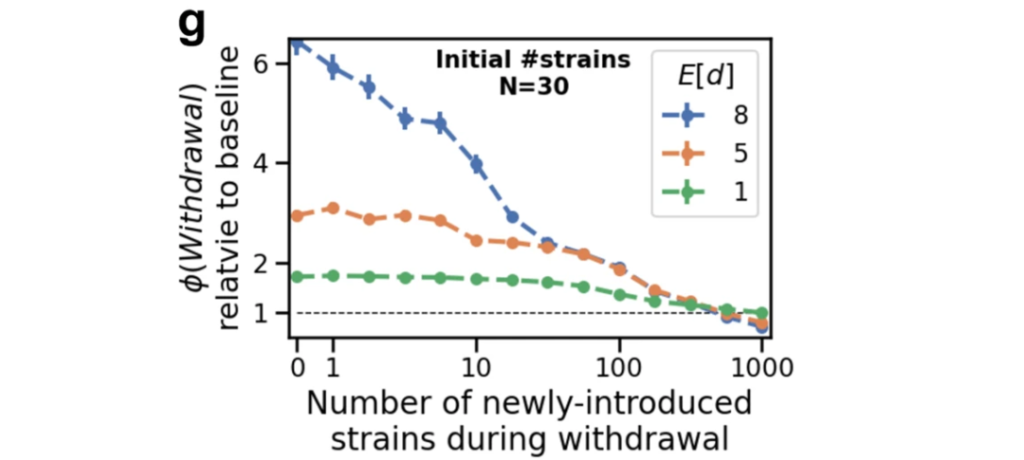
这种干预逐渐增加了微生物组内菌株之间的竞争,降低了为成瘾行为提供反馈的菌株的影响,从而促进了宿主-微生物组生态系统向其原始平衡状态的转变。
4 成瘾程度加重,微生物群越发抵制宿主戒断行为
通过研究微生物组与成瘾的最大严重程度(R)之间的相互作用,发现随着成瘾程度的加重(R值增加),宿主行为会产生一种生态制度,将微生物组引向更狭窄、多样性较低的生态位。
在新的生态建立之后,新的微生物组成可能会强烈抵制任何改变的尝试,从而减缓戒断过程。
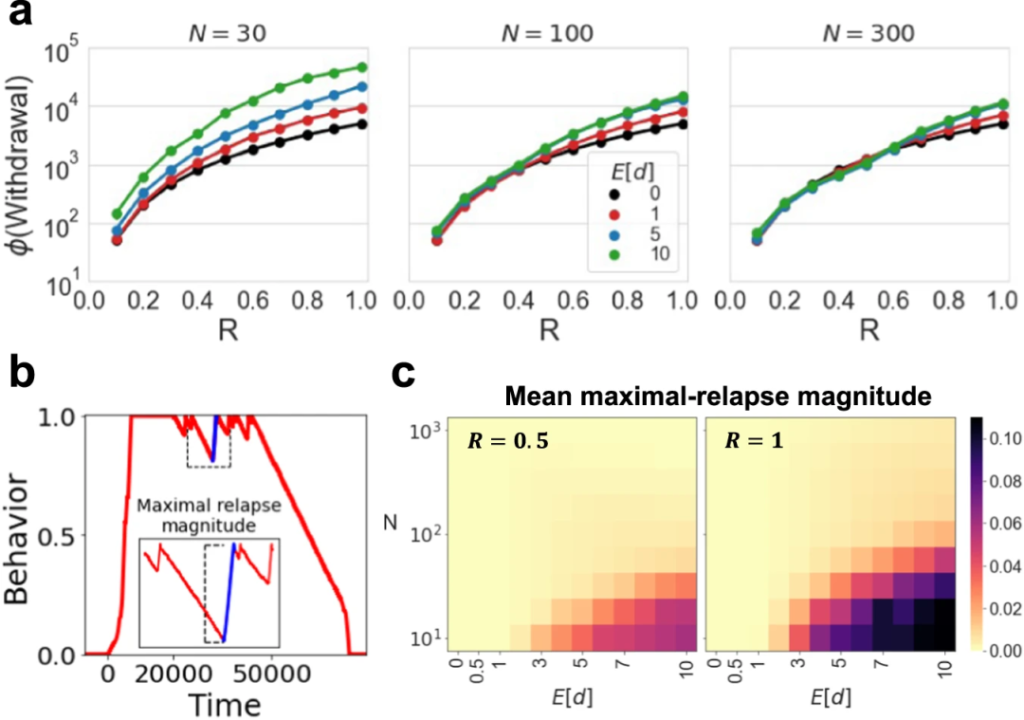
如图a,这种动态效应与微生物组对宿主行为的影响程度成正比,与微生物组的丰度成反比。当微生物组更为丰富和或其对宿主的影响相对较弱时,只有在微生物组成分发生实质性改变时,才会发生成瘾程度的加重。这意味着即使微生物组中只有少数菌株影响宿主行为,只要这些菌株发生了显著改变,成瘾问题就会加剧。
其次,还有复发的问题,即在戒断阶段发生的成瘾行为加剧。
如图b,c所示,更强的微生物效应、较低的微生物组丰富度和更剧烈的成瘾严重程度,都会导致更强烈和更频繁的复发现象。
Conclusion
模型结果表明,微生物组对宿主行为的反馈可以加重成瘾行为,使戒断更加困难,并增加复发的风险。而微生物组的丰度是这个过程的关键参数,低丰度会导致长期成瘾。
这种反馈有正向和负向,微生物组产生的代谢产物可以通过调节奖励回路对宿主行为提供正向或负向的反馈。由此,宿主-微生物群落相互作用产生了一个正反馈循环,改变了生态系统。
基于对成瘾机制的微生物组研究,也揭示了可能的成瘾治疗途径,增加微生物组的丰富度和功能多样性可能有助于成瘾的缓解和预防。而宿主的状态(包括生理和心理)与微生物群落多样性是息息相关的,比如压力和焦虑也会出现低多样性的微生物群落。
这些结论也反向验证了这个模型框架的优势:
参考文献:
Lewin-Epstein O, Jaques Y, Feldman MW, Kaufer D, Hadany L. Evolutionary modeling suggests that addictions may be driven by competition-induced microbiome dysbiosis. Commun Biol. 2023 Jul 26;6(1):782. doi: 10.1038/s42003-023-05099-0. PMID: 37495841; PMCID: PMC10372008.

谷禾健康

你是否有时会感到心悸、心慌、胸闷、气短、头晕、乏力?你是否有时感觉自己的心跳过快或过慢?
如果有上述情况,就要引起重视了,你可能存在心律失常。心律失常是最常见的心脏疾病之一,涉及到心脏的电活动节奏异常。根据发病机制和心电图特征被可以为不同类型,包括窦性心律失常、房性心律失常和室性心律失常。
长期熬夜,不健康的饮食,吸烟酗酒,压力大,过度劳累等各种因素都导致心律失常发病率越来越高和越来越年轻化。
与此同时,新冠肺炎的爆发也引起了人们对心律失常的关注。心律失常的治疗和预防,对于维持心脏健康和预防并发症的发生具有重要意义。
已有大量证据表明肠道微生物群及其代谢物对心血管健康具有重大影响,这为心律失常的治疗和预防带来了新的思路。

本文我们讨论了肠道微生物及其代谢物如何通过多种机制影响心律失常。目前公认的心律失常机制,包括结构重塑、电生理重塑、神经系统调节异常以及其他与心律失常相关的疾病。
最后介绍了与肠道微生物群相关的潜在的治疗策略,包括使用益生菌和益生元、粪便微生物移植和免疫调节剂等。
目录
1.了解心律失常
-心律失常的症状
-心律失常的分类
-发病率及高发人群
-心律失常的风险因素
2.心律失常患者的肠道微生物特征
-房性心律失常患者的肠道微生物
-室性心律失常患者的肠道微生物
-肠道菌群代谢物对心律不齐的影响
3.肠道微生物失调和心律失常的发生
-结构改造
-电生理重塑
-神经系统调节
4.其他疾病与心律失常的关系
-胃肠道疾病易诱发心律失常
-房颤患者易得胃肠道疾病
-与心律失常相关的其他疾病
5.心律失常的治疗
-饮食调整:多吃有益的脂质和膳食纤维
-补充益生菌、益生元
-药物与肠道微生物的相互作用
-粪菌移植
-免疫调节剂
-病因治疗和药物治疗
6.总结
以下是谷禾写过关于心血管疾病系列的文章,点击可查看:
-正文-
心律失常(cardiac arrhythmia)是指心脏电传导系统异常所引起的心跳不规则、过快或过慢等症状的总称。
正常心律起源于窦房结,频率60次~100次/分(成人),比较规律。窦房结冲动经正常房室传导系统顺序激动心房和心室,传导时间恒定(成人0.12~1.21秒)。
然而随着我国人口老龄化的进展和生活方式的改变,心律失常发病率快速上升,且呈年轻化和不断增长的趋势。
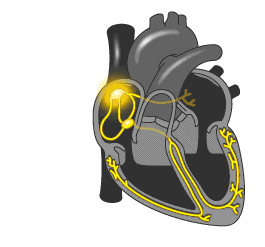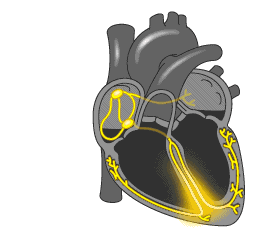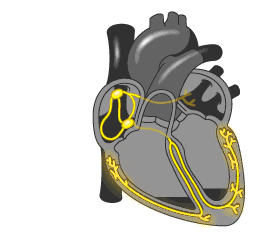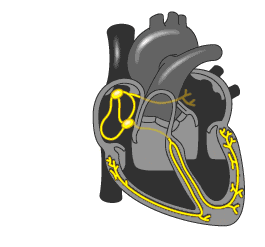
▼
心律失常的症状轻重不一,取决于发病的类型以及持续的时间,以及原发病的严重程度。
随着疾病的发生发展,患者发病早期可有心悸、出汗、乏力、透不过气等症状,此时,若心律恢复正常则无严重不适,若进一步发展可导致头晕、黑朦、晕厥,甚至猝死等。
▸ 心律失常可能伴随的症状
心律失常伴有明显的外周血流动力障碍时,还会出现相应器官受损的症状:
•脑:视力模糊、头晕、黑朦、晕厥等;
•胃肠道:腹胀、腹痛、腹泻等;
•肾:尿频、尿急、多尿等;
•肺:胸闷、气促、呼吸困难等。
▼
临床上根据心律失常的发生部位、机制及频率不同,可细分为许多种。
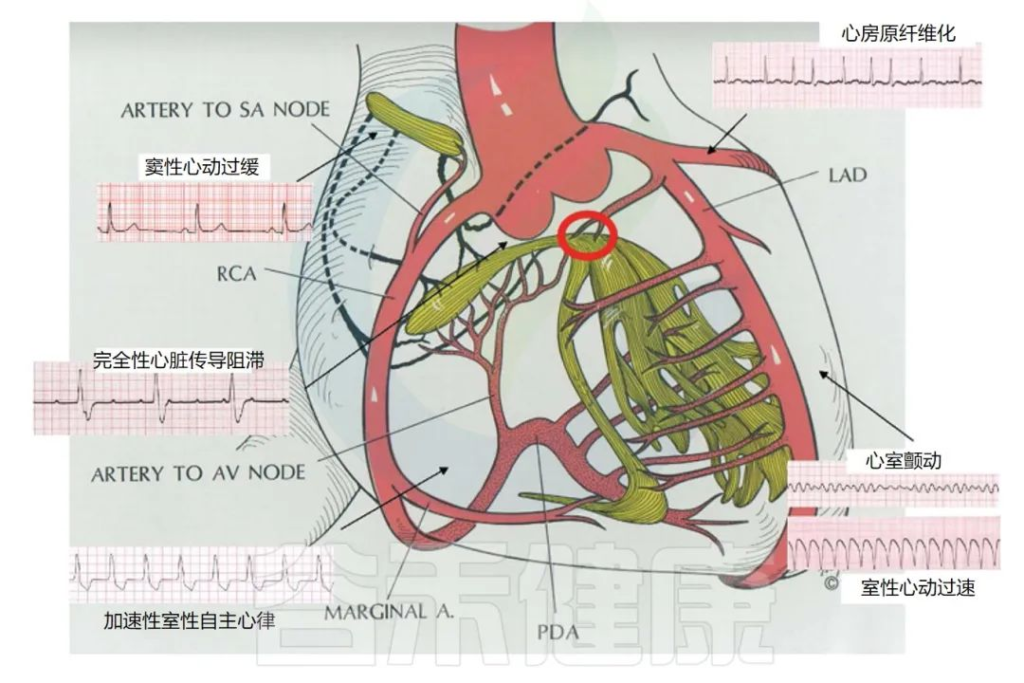
Frampton J,et al.Yale J Biol Med.2023
▸ 根据发病部位区分
按照心律失常发病部位可以分为窦性心律失常、房性心律失常、交界性心律失常、室性心律失常和束支心律失常。具体如下:
•窦性心律失常:包括窦性心动过速、窦性心动过缓、窦性停博、窦房传导阻滞、病态窦房结综合征;
•房性心律失常:包括房性早搏、房性心动过速、房室折返性心动过速、房室结折返性心动过速、房扑、房颤等;
•交界性心律失常:包括交界性早搏、交界性心动过速、房室传导阻滞等;
•室性心律失常:包括室性早搏、室性心动过速、室扑、室颤、室内传导阻滞等;
•束支心律失常:包括左束支传导阻滞、右束支传导阻滞、左前分支阻滞、左后分支阻滞等。
注:心房颤动(AF)是较为常见的心律失常。
▸ 根据发病机制区分
按发生机制可以分为冲动形成异常和冲动传导异常两大类:
•冲动形成异常:包括窦性心律失常,比如窦性心动过速、窦性心动过缓、窦性心律不齐、窦性停搏,还有异位心律失常,比如房性逸搏和逸搏心律,交界性逸搏和逸搏心律、室性逸搏和逸搏心律。
•冲动传导的异常:这种情况其实主要是包括传导阻滞,比如窦房传导阻滞、房室传导阻滞,另外还有完全性左束支传导阻滞、完全性右束支传导阻滞。
▸ 根据心律快慢区分
临床上常以心率快慢进行分类,分为快速型与缓慢型失常两大类。
•常见的缓慢型心律失常(心率<60次/分)包括:窦性心动过缓、窦性停搏、病态窭房结综合征、窦房传导阻滞。
•常见的快速型心律失常(心率>100次/分)包括:早搏、窦性心动过速、房性心动过速(心房扑动、心房颤动)、室上性心动过速、室性心动过速(心室扑动、心室颤动)等。▼
心律失常的确切发病率较难统计,根据研究数据显示,全球范围内心律失常的发病率大约在2%至4%之间,其中老年人中的患病率可能高达10%左右。
其中窦性心律不齐发病率最高,约占25%-27;窦性心动过速次之,约为20%-22%;窦性心动过缓,约13%-15%;室性早搏,约14%-16%;房颤,约11%-15%;房室传导阻滞,约5%-7%,其他各种心律失常约为5%-8%。
▸ 老年人和心血管疾病患者易发生心律失常
老年人和心血管疾病患者是心律失常的高发人群。
随着年龄的增长,心脏功能开始下降,心律失常的患病率也逐渐增加。
同时,存在高血压、冠心病、心力衰竭等心血管疾病的人群也更容易患上心律失常。
其他一些潜在因素,如家族史和遗传因素,也可能增加个体患上心律失常的风险。
▼
•年龄
随着年龄增长,心脏的功能会逐渐下降,从而增加心律不齐的风险。
•遗传
某些心律不齐类型可能与家族遗传有关,如果家族中有人患有心律不齐,那么你也可能面临较高的风险。
•心脏疾病
一些心脏疾病,如冠心病、心肌病、心脏瓣膜病等,可以增加心律不齐的风险。
•高血压
长期存在的高血压会对心脏造成损害,增加心律不齐的发生率。
•代谢紊乱
糖尿病、甲状腺功能异常等代谢紊乱疾病会对心脏功能产生影响,增加心律不齐的风险。
•药物滥用
某些药物(如某些心脏药物、精神类药物)滥用会干扰心脏的正常搏动。
•胸部手术
尤其是心脏手术、麻醉过程、心导管检查、各种心脏介入性治疗等可诱发心律失常。
此外,日常生活中也存在着很多诱发心律失常的因素,例如大量饮酒或咖啡、饮浓茶、情绪激动、剧烈运动等均可导致正常人发生快速性心律失常。
健康成年人体内存在数万亿微生物。值得注意的是,肠道微生物被称为与人类细胞基因组并列的“第二大基因组”。
几十年来,研究人员一直致力于探索心律失常的病因、机制和治疗方法。同时,不断更新的证据表明,肠道菌群不仅与人体健康和体内平衡维持密切相关,而且与包括心律失常在内的多种疾病的发生和发展密切相关。


Rashid S,et al.Ann Noninvasive Electrocardiol.2023
▼
房颤,作为心律失常的一种重要类型。肠道微生物组与房颤的关系已得到初步认识,肠道微生物组及其代谢产物在心房结构重塑和电重塑方面与房颤的发生密切相关。
宏观基因组学和代谢组学分别作为研究肠道微生物组组成及其代谢物变化的重要研究工具,在探索肠道微生物组与心律失常的关系中发挥着重要作用。
最近,一些研究报告了与健康对照组相比,房颤患者中肠道微生物群变化的特征。
▷房颤患者体内有害细菌过度生长
使用宏观基因组学和代谢组学技术分析了50名健康对照者与50名房颤患者的肠道微生物群及其代谢物的组成。
研究发现,房颤患者的肠道微生物群中基因数量显著增加,样本内多样性(香农指数)也增加,这表明他们的肠道菌群具有更高的丰度和多样性。
较多的基因和属种表明房颤患者体内可能存在多种有害细菌过度生长。瘤胃球菌(Ruminococcus)、链球菌(Streptococcus)、韦荣氏球菌(Veillonella)和肠球菌(Enterococcus)等属在房颤患者中更为富集。
瘤胃球菌具有与炎症性肠病发展相关的促炎特性;链球菌在患有高血压、慢性心力衰竭和急性心血管事件的人群中也被证明是升高的。
来自日本的一项研究表明,与对照组相比,房颤患者的肠杆菌(Enterobacter)减少,而副拟杆菌(Parabacteroides)、Lachnoclostridium、链球菌(Streptococcus)和另枝菌属(Alistipes)增加。
▷胆汁酸、油酸含量显著下降
对健康受试者和房颤患者的粪便和血清样本进行代谢组学分析,其中胆汁酸、油酸、亚油酸和α-亚麻酸的含量显著降低。
研究表明,油酸、亚油酸和α-亚麻酸具有心脏保护作用,亚油酸和α-亚麻酸可以预防和终止由溶血磷脂酰胆碱或酰基肉碱引起的心律失常。
▷丁酸弧菌等有益菌的减少
心房颤动是一种进行性疾病。根据发作持续时间可分为阵发性房颤和持续性房颤。
采用宏基因组学和代谢组学相结合的分析方法来研究房颤持续时间与肠道微生物群之间的关系。
该研究包括20名健康个体、20名持续性房颤患者、12名房颤持续时间小于12个月和8名房颤持续时间大于12个月的患者。
与对照组相比,阵发性房颤组和持续性房颤组瘤胃球菌和链球菌等的丰度增加。丁酸弧菌属随病程呈下降趋势,丁酸弧菌属可产生短链脂肪酸并对宿主有有益作用。
粪弧菌属(Vibrio faecalis spp.)随时间呈上升趋势,粪弧菌可以极大地促进慢性精神分裂症患者发生冠心病。
心律失常组和对照组之间肠道微生物群的变化
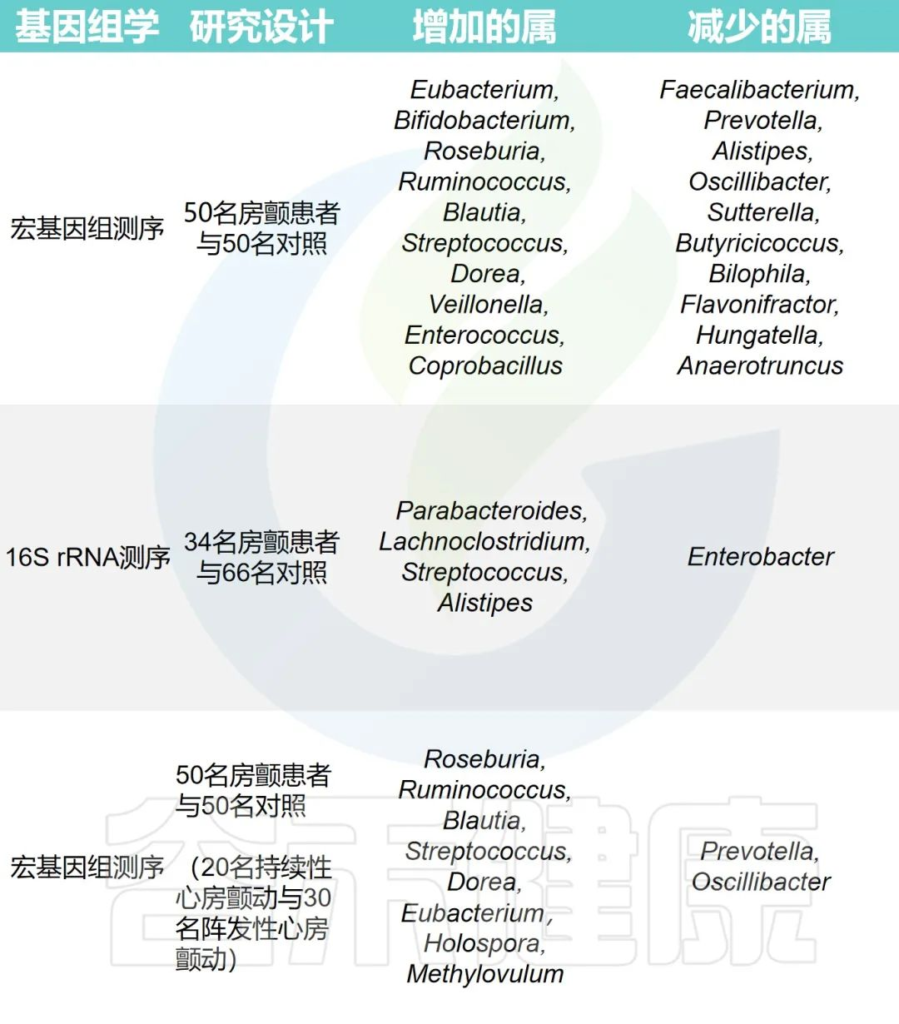

Fan H,et al.Front Cell Infect Microbiol.2023
这些证据表明,有益菌的减少或致病菌的增加以及代谢物的改变可能与心房颤动密切相关,甚至可能在心脏结构重塑中发挥关键作用。
▼
室性心律失常是临床最常见的心律失常之一。之前的一项研究发现左心室间隔或后壁厚度是室性心律失常发病率和复杂性增加的重要因素。而胎儿左心室生长可能是室性心律失常的原因之一。
▷新生儿肠道微生物多样性较低影响左心室后壁厚度
研究发现胎儿左心室生长与出生时肠道微生物群的组成有关,其中新生儿左心室后壁的厚度与肠道微生物群多样性较低有关,以及抗重塑作用相关的细菌(乳杆菌等)的耗竭和与炎症功能相关的细菌的富集(肠杆菌类等)。
肠道微生物群诱导的促炎特征在胎儿发育过程中促进心室生长,孕妇早期基于微生物群的调节可能会降低室性心律失常的发生率和不良结局。
肠道可以通过促进全身炎症和感染来影响危重患者。宿主防御机制在应激和粘膜缺氧的情况下分解,导致细菌和毒素的转移,从而通过内源性和外源性炎症反应引起心肌损伤。
▷婴儿早期室性心律失常与肠道微生物相关
根据研究,室性快速心律失常与50%至75%的心源性猝死直接相关。最近发现,与健康对照婴儿相比,在婴儿猝死综合症婴儿的肠道中发现了更高比例的艰难梭菌、Cl. innocuum和多形拟杆菌(B.thetaiotaomicron)。
多形拟杆菌(B.thetaiotaomicron)似乎介导粘膜-肠道屏障的形成,并通过其对物种特异性蛋白质抗生素表达的影响来帮助抵御病原体入侵。
致病性梭菌会破坏EC紧密连接或引发炎症反应,这两者都会损害肠壁的完整性。因此,婴儿早期室性心律失常可能与异常肠道微生物的建立有关。对于婴儿的健康和早期发育,创造健康的肠道微生物被认为是至关重要的。
▼
肠道微生物代谢就会产生生物活性代谢物,这些代谢物可以直接或间接影响宿主的生理过程。
肠道微生物群衍生的代谢物可以被宿主肠道吸收,影响肠道免疫细胞,并在宿主循环中检测到。它们充当信号分子影响重要的代谢途径。一些代谢物已被证明可以调节心律不齐的促进机制。
肠道微生物群衍生代谢物对心律失常的影响
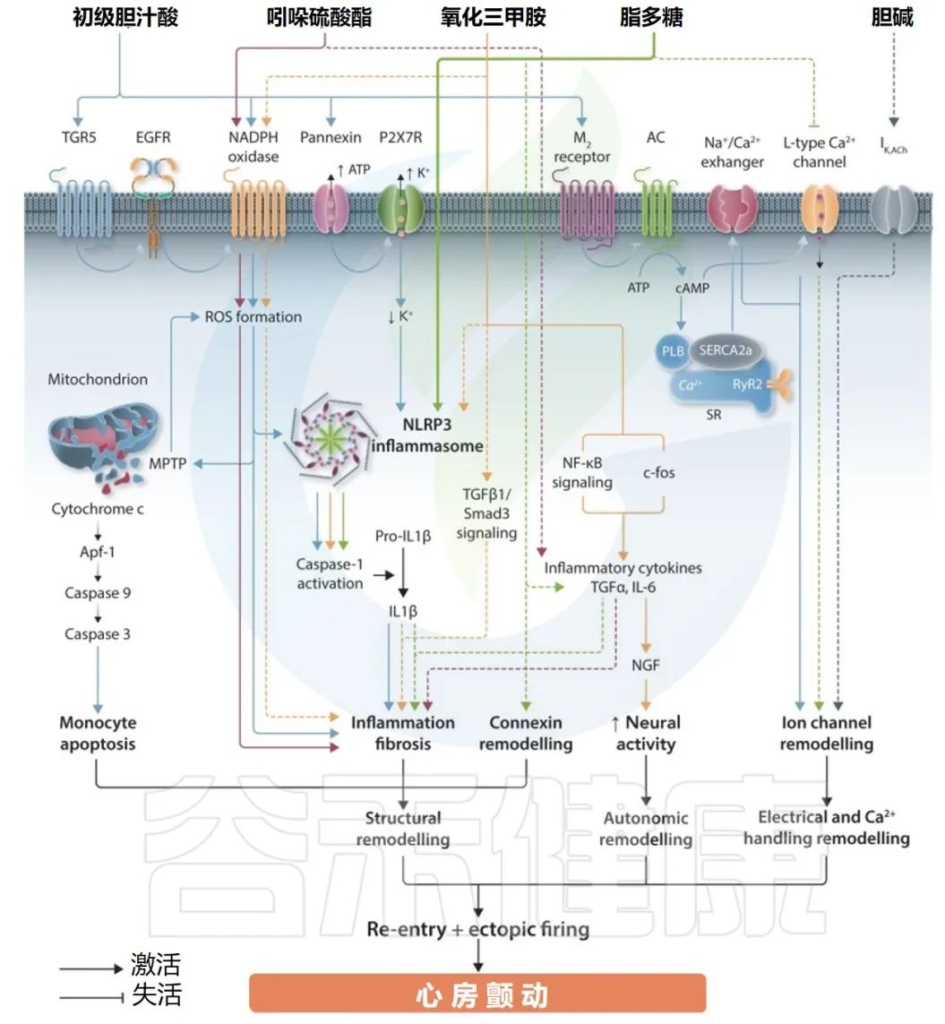
Gawałko M,et al.Cardiovasc Res.2022
1
氧化三甲胺
氧化三甲胺(TMAO)是含有肝黄素的单加氧酶氧化三甲胺的产物,也是参与房颤发病机制研究最广泛的微生物代谢产物。
氧化三甲胺过高影响房颤的发生
在犬模型中,局部注射氧化三甲胺激活心房自主神经节丛并促进心律失常,可能通过激活核因子-κB p65信号传导和增加炎症细胞因子的表达。
此外,在房颤患者的肠道中观察到三甲胺形成的微生物基因和三甲胺微生物生产者的显著增加。
尽管小型研究表明,血清氧化三甲胺水平升高可预测房颤患者的血栓栓塞事件,但这种关联的有效性和重要性需要在更多的房颤人群中进一步评估。
氧化三甲胺促进心律不齐的危险因素
此外,氧化三甲胺可能通过促进各种心律不齐危险因素(例如通过促动脉粥样硬化作用产生的代谢综合征和高血压、肾素-血管紧张素系统调节和主动脉硬化)来增强对房颤的易感性。
在蛋白质组学分析中,与非房颤受试者相比,房颤患者的心耳和血浆样本中的胆碱(TMAO前体)含量更高。
在对三个前瞻性队列进行的汇总分析中,血浆和膳食中胆碱的摄入量与后期房颤风险呈正相关。
2
硫酸吲哚酚
硫酸吲哚酚会引起氧化应激导致心律失常
在实验研究中,硫酸吲哚酚通过引起氧化应激和心肌细胞钙处理失调,增加肺静脉和左房心律失常的发生,并减少窦房结起搏器的活性。
在临床环境中,高浓度硫酸吲哚酚(≥0.65 μg/mL)可预测房颤复发风险3.7倍。相反,房颤的导管消融与硫酸吲哚酚血清浓度降低相关,这表明房颤本身可能会增加硫酸吲哚酚的产生。
在动物研究中,硫酸吲哚酚会增加促炎和促纤维化信号分子的表达,并引起氧化应激,从而可能导致房颤。
注:在这些实验研究中,所使用的硫酸吲哚酚浓度大大超出了其在血浆中的生理范围,使其在促进房颤中的潜在作用还不确定。
3
脂多糖
脂多糖是一种存在于革兰氏阴性菌(特别是埃希氏菌属)外层的内毒素。
脂多糖增加了心房促炎因子浓度
在犬模型中,给予脂多糖会增加心房促炎细胞因子的浓度,从而增加连接蛋白43的表达并导致连接蛋白偏侧化。
脂多糖会下调L型钙通道(α1C和β2亚基)的表达并缩短有效不应期。异常的钙处理和连接蛋白调节是诱导房颤的潜在机制。
L-型钙通道是一种电压依赖性钙通道。
脂多糖水平过高增加心血管疾病风险
尽管脂多糖对房颤发病机制的直接影响尚未研究,但脂多糖水平升高的房颤患者似乎有较高的不良心血管事件风险。
脂多糖可能通过加速动脉粥样硬化间接导致心律不齐,并可能诱发左心室功能障碍和心力衰竭,这两者都会增加心律不齐的风险。
4
胆汁酸
初级胆汁酸,例如鹅去氧胆酸,通过与氨基酸(牛磺酸或甘氨酸)结合形成胆汁盐,并进一步分泌到小肠中。
牛磺酸结合的胆汁酸可以通过心脏钠钙交换体刺激诱导膜电位变化并激活心肌细胞中毒蕈碱M2受体/乙酰胆碱调节的钾电流,这可能促进心房颤动。
鹅去氧胆酸等胆汁酸的水平影响心脏
鹅去氧胆酸已被证明可引起心房心肌细胞凋亡,这可能导致房颤的演变,促进结构重塑,通过法尼素X受体促进心脏损伤和纤维化,并通过NLRP3炎症小体激活加重炎症过程。
另一方面,熊去氧胆酸通过稳定细胞膜电位在预防心律失常中发挥作用。胆汁酸还可能通过参与葡萄糖和脂质代谢间接诱导房颤,并通过调节皮质醇和醛固酮水平的11β-羟基类固醇脱氢酶将血压维持在正常范围内。
5
短链脂肪酸
结肠微生物群发酵葡萄糖和膳食纤维会产生短链脂肪酸,主要是乙酸盐、丁酸盐和丙酸盐。
短链脂肪酸是房颤发病的潜在促成因素。短链脂肪酸负责促进粘液产生,从而改善肠道屏障功能,并且对于其中一些来说,还可以通过抑制组蛋白脱乙酰酶影响免疫调节。
短链脂肪酸水平过低会导致心律不齐风险升高
短链脂肪酸水平低会导致代谢物感应G蛋白偶联受体结合不良,从而损害肠道完整性并促进脂多糖等物质进入血液和组织。
丙酸盐刺激胰高血糖素样肽-1和肽YY的释放,降低肥胖风险并增加肾素释放,从而改变两个重要的心律不齐风险因素。
另外,乙酸盐会导致血脂异常,并通过大脑中副交感神经系统的激活增强胰岛素(由胰腺)和生长素释放肽(由胃粘膜)的分泌,从而促进肥胖。
心律不齐组和对照组之间的肠道微生物代谢
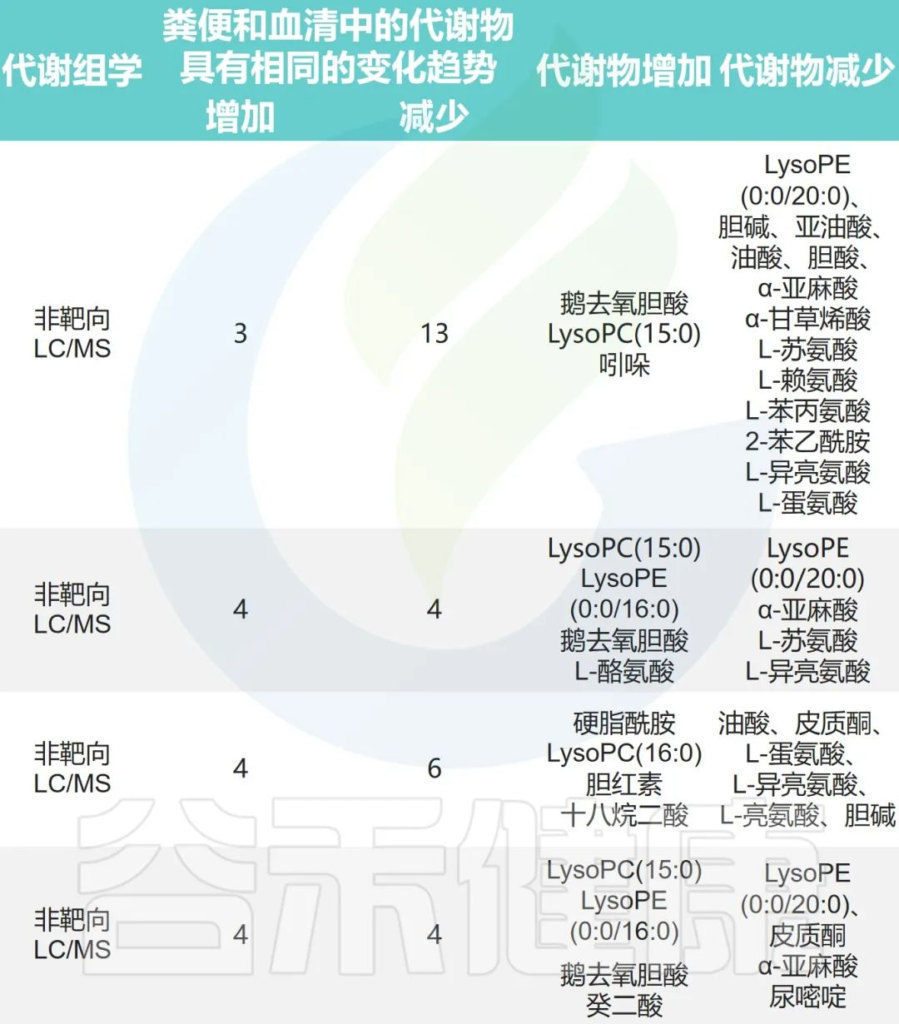
Fan H,et al.Front Cell Infect Microbiol.2023
小结
总之,这些研究表明:与健康人群相比,心律不齐人群经常伴随着肠道有益细菌的减少、心血管保护性代谢物的减少以及对健康有害的代谢物的增加。
近年来,越来越多的相关研究证明,肠道微生物紊乱与缺血性心肌病、心力衰竭、甚至心律失常有关。
下面将讲述肠道微生物如何利用代谢物、炎症因子、免疫细胞作为介质,通过神经、血液、直接作用于心肌组织,最终导致心律失常的发生。
肠道微生物群和代谢物引起心律失常的机制
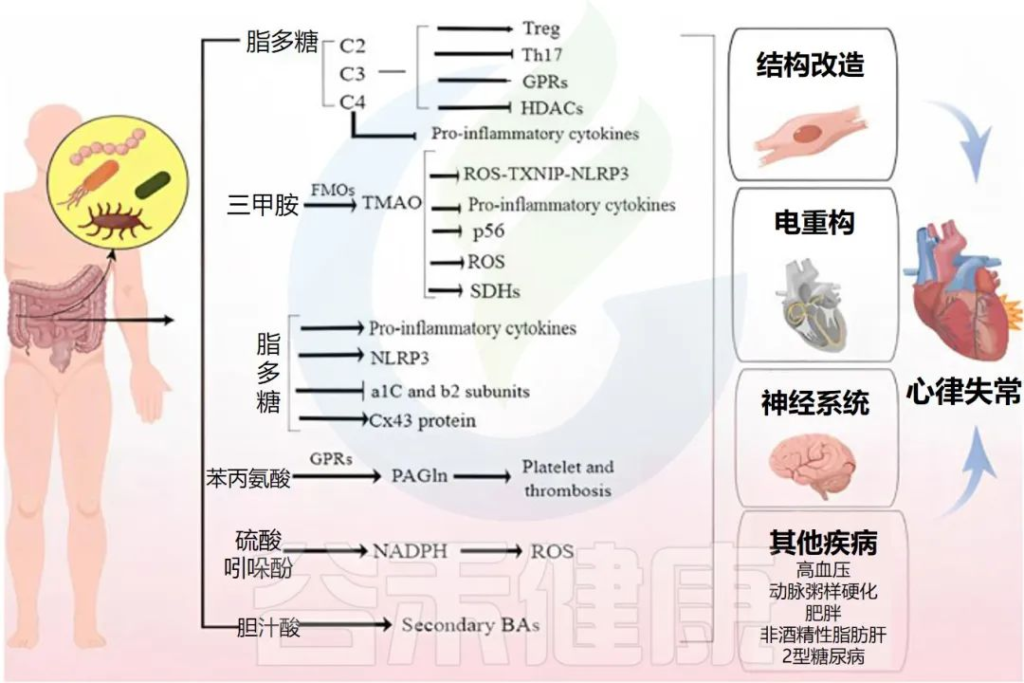
Fan H,et al.Front Cell Infect Microbiol.2023
▼
心肌重塑是指在快速心律失常的进展过程中,心肌原有的电生理和组织学特征发生了一定的变化。心肌重塑的主要表现之一是结构重塑,包括心肌细胞凋亡、心房基质纤维化等。
这主要是由于高血压、冠心病、糖尿病、肥胖等多种原因,导致心肌细胞内质网、线粒体等细胞器的结构、形态和数量发生变化,导致心肌细胞肥大、凋亡、坏死和心肌间质纤维化。
最后,肌壁不断增厚,心腔容积不断扩大,最终促进快速心律失常的发生。
短链脂肪酸通过减少炎症反应保护心血管
免疫炎症的减少可以通过肠道细菌产生的短链脂肪酸减轻心脏肥大、纤维化、血管功能障碍和高血压。
对免疫细胞的抗炎作用是通过短链脂肪酸实现的,例如丙酸可以诱导CD25+Foxp3+ Treg的分化和抑制功能。丙酸还通过抑制组蛋白脱乙酰酶或通过GPRs和嗅觉受体发挥心脏保护作用。
丙酸对心脏的保护主要取决于调节性T细胞的作用。
短链脂肪酸可以通过减少炎症反应对心血管的不利影响来预防或延缓心律失常的发生。
其他肠道代谢物通过炎症途径对心律失常的影响
肠道微生物产生的其他代谢物也可能通过炎症途径引起心律失常。
氧化三甲胺
氧化三甲胺激活NLRP3炎症小体导致人脐静脉炎症和内皮功能障碍。随着炎症标志物显著增加,活化的白细胞被招募来诱导血管炎症,这表明氧化三甲胺增强动脉粥样硬化和心血管风险的可能机制。
硫酸吲哚酚
硫酸吲哚酚可以通过氧化应激诱导肺静脉和房性心律失常。在窦性心律的患者中发现,射频电流导管消融后,患有高硫酸吲哚酚的房颤患者血清硫酸吲哚酚水平降低,这表明血清硫酸吲哚酚不仅可能诱发房颤,而且可能受到房颤的影响。
心肌肥厚和心肌纤维化影响心律失常
心肌肥厚和心肌纤维化是心律失常的重要原因。肠道微生物可以产生琥珀酸、乳酸、乙酰辅酶A,并进一步代谢成短链脂肪酸。其中一些与心肌肥大和心肌纤维化密切相关。
研究表明,丙酸可以在T细胞的帮助下减轻炎症反应,减轻小鼠心肌肥厚、纤维化、血管功能障碍和高血压。
亚油酸和α-亚麻酸抑制活性氧的产生并下调p38 MAPK途径β生长因子的激活和转化、发挥保护作用。它在心房纤维化中发挥调节作用,影响房颤的进展。
总之,有充分的证据表明,肠道微生物及其代谢物可以通过多种方式影响心肌肥大和纤维化,这很可能是心律失常发生的基础。
心肌细胞凋亡
鹅去氧胆酸已被证明可引起心房心肌细胞凋亡,这可能有助于结构重塑。鹅去氧胆酸与左心房低电压区呈正相关,可能促进心房肌细胞凋亡。
此外,丁酸通过抑制增殖和凋亡来促进血管平滑肌细胞的生长。氧化三甲胺通过琥珀酸脱氢酶复合物B亚基上调诱导活性氧促进血管内皮细胞凋亡,这可能有助于动脉粥样硬化病变的进展。
▼
心脏的功能就像机械泵一样,确保向全身和肺部持续供应血液。因此,人类心脏每天会进行100,000 次成功且协调的收缩。
由于心脏收缩受到严格的电调节,失衡会导致心律失常。这些收缩是由称为动作电位的电信号控制的。肠道微生物失衡引起的心肌重塑还反映在电生理重塑(心肌细胞膜表面离子通道的改变)。
通过钠离子通道影响心肌电活动
众所周知,钠通道是产生动作电位的关键通道,由此产生的细胞内和细胞外钠离子浓度的变化对心脏动作电位的产生很重要。
肠道微生物参与胆汁酸的代谢,胆汁酸诱导的心律失常发生的可能机制是与细胞膜和细胞膜离子通道或转运蛋白的相互作用。
研究发现牛磺胆酸通过刺激心肌中的钠钙交换诱导膜电位变化,导致内向电流密度增加和静息膜电位去极化,进而影响心肌电活动。
然而,也发现熊去氧胆酸在诱导心律失常方面无效,熊去氧胆酸偶联保护富含胆固醇的质膜免受疏水性胆汁酸的毒性作用,从而保护细胞免受心律失常的影响。
注:房颤患者的血清熊去氧胆酸偶联物水平明显较低,非熊去氧胆酸水平较高。
这表明,较高水平的毒性(致心律失常)和较低水平的保护性胆汁酸创造了一个心律失常阈值较低的环境,因此可能导致心律失常。
钾离子通道与持续性房颤相关
钾离子通道代表心脏中最多样化的离子通道,其中心脏乙酰胆碱激活的内向整流钾电流(IKACh)通道的激活是副交感神经系统对心脏功能的生理控制的重要组成部分。
目前,越来越多的研究证实,激活的内向整流钾电流被认为是持续性房颤中的背景内向整流器,因此有助于缩短动作电位持续时间并稳定高频电转子的形成,从而导致持续性房颤。
钙离子通过多种机制影响心律失常
Ca2+是一种普遍存在的第二信使,调节多种生物过程,包括激素产生、肌肉收缩、突触传递、增殖和死亡。越来越多的证据表明房颤和其他心脏疾病,与Ca2+相关。
因此,对心脏钙通道的任何影响都可能导致心律失常。在一项实验研究中,硫酸吲哚酚通过引起心肌细胞的氧化应激和钙处理失调,增加了肺静脉性心律失常和左心房心律失常的发生,并降低了窦房结起搏器的活性。
脂多糖还可以上调NLRP3系统,这除了引起结构重塑外,还可以导致心房动作电位的重新进入,促进收缩和舒张期间自发肌浆网Ca2+释放频率的增加,这可能导致去极化后延迟并触发异位活动,从而影响心肌电活动。
这些机制为钙调节促进心律失常提供了证据。
▼
人们对肠脑轴自下而上的信号传导知之甚少。然而,心脑轴的自上而下信号传导已被广泛研究。通过自主神经系统和迷走神经系统,神经系统可以控制心率。
心律失常可能是由于中枢神经系统限制副交感神经张力同时提高交感神经张力的能力而引起的。
注:神经系统调节心脏的机制非常复杂。植入的神经节丛中可能有数百到数千个自主神经元。
自主神经系统
心律失常的形成和维持在很大程度上取决于自主神经系统。
大脑和脊髓中的神经元以及连接心脏的神经组成了外源性心脏自主神经系统。在心脏本身以及沿着胸腔的大静脉中,自主神经元和神经构成了内在心脏自主神经系统的大部分。
•通过心脏自主神经系统调节房颤和室性心律失常
在犬模型中,局部注射氧化三甲胺可能通过激活 p65 核因子-κB 信号传导和增加炎症细胞因子的表达来激活心房自主神经丛并促进心律失常。
已经证明,房颤或室性心律失常的病理生理学是由心脏自主神经系统调节的。有证据表明,在发生室性心律失常之前,左心交感神经就已过度活跃,而左星状神经节的激活可大大增加室性心律失常发生的可能性。
研究结果表明,与对照组相比,静脉注射和局部氧化三甲胺治疗显著增强了左星状神经节功能和活性,从而提高了心脏交感神经张力,并恶化了缺血引起的室性心律失常。
同时,氧化三甲胺可显著促进促炎标志物的表达,如IL-1、IL-6、TNF-α,这可能进一步导致交感神经过度活跃并影响室性心律失常的发展。
迷走神经系统
调节心律受到迷走神经的负面影响。迷走神经放电增强钾通道电流,减少动作电位持续时间并稳定折叠转子。
•肠道微生物及其代谢物通过迷走神经影响心律
微生物群可以通过肠道内分泌细胞直接或间接触发迷走神经传入纤维。迷走神经传入纤维通过中枢自主神经网络刺激中枢神经系统。迷走神经传入纤维可以通过炎症反射触发传出纤维。
根据物质的不同,微生物群产生的不同代谢物可能以不同的方式激活迷走神经传入纤维。例如,油酸,一种长脂肪酸,通过胆囊收缩素介导的机制作用于迷走神经传入纤维;而丁酸,一种短脂肪酸,直接影响传入末梢。
Part 1
胃肠道疾病易诱发房颤
√较高的C反应蛋白增加心房颤动风险
炎症似乎在心律失常的发病机制中发挥着重要作用。C反应蛋白 (CRP) 升高与心房颤动相关,持续性心房颤动的平均C反应蛋白水平高于阵发性心房颤动中观察到的水平,表明炎症状态可能会促进心房颤动的持续存在。
C反应蛋白是一种非特异性炎性标志物,由肝脏合成的用于保护身体的急性时相反应蛋白。
在纵向研究中,较高的C反应蛋白会相应增加心房颤动风险。最后,在没有心血管合并症的情况下,严重败血症和急性全身炎症反应与新发心房颤动相关。
√炎症性肠病患者心房传导发生了改变
在一项研究中,观察到炎症性肠病患者的心房传导发生了改变,这是通过心电图上P波离散度的增加来衡量的。
在两项针对炎症性肠病患者的小型研究中,与参考对象相比,经胸超声心动图测量的心房机电传导时间延长。
丹麦的一项病例对照研究涉及24499例炎症性肠病病例和236275名年龄和性别匹配的对照,观察到在活动性炎症性肠病发作期间,房颤风险增加两倍,中风风险增加1.5倍。在炎症性肠病缓解期间,未观察到与房颤或中风的关联。
▼
在一些病例对照研究中,胃肠道恶性肿瘤,包括结直肠癌和食道癌,与房颤风险增加有关。
√结直肠癌患者发生房颤的风险更高
在一项研究中,与非癌症队列参与者相比,结直肠癌患者发生房颤的风险增加了一倍。
丹麦一项基于人群的病例对照研究观察到,结直肠癌患者的房颤患病率较高 [比值比为11.8],但患病率的增加仅限于癌症诊断后的前90天,这表明手术治疗癌症可能在很大程度上推动了这种关联。
√胃肠道手术后房颤风险增加
胃肠癌手术治疗后房颤的发生率很高,并且与手术结果不佳相关,包括肺部并发症、败血症、住院时间延长。
此外,在回顾性分析中,胃肠道癌症手术后发生房颤的患者在1年内发生心血管事件和60天内死亡的风险增加。
据报道,经胸食管切除术后房颤的发生率为10%至46% 。
Part 2
房颤患者易得胃肠道疾病
▼
在房颤患者中,缺血主要继发于心脏栓塞事件,而不是原位血栓栓塞。房颤患者心脏栓塞事件的主要风险因素包括既往卒中或短暂性脑缺血发作史,年龄大于75岁、高血压和结构性心脏病。
√房颤患者易发生肠系膜缺血
特别是,患有房颤的女性发生血栓栓塞的风险增加,包括肠系膜缺血。内脏-肠系膜血管系统栓塞是外周血栓栓塞疾病的第二常见部位,仅次于上肢和下肢的血管。
在一项基于房颤患者研究中,随着房颤评分的增加,发生肠系膜缺血的风险更高,评分为⩾2的患者与评分为0的患者相比,风险增加了三倍。
Part 3
与心律失常相关的其他疾病
▼
肠道微生物可以影响血压,而高血压患者通过多种作用方式参与心律失常。
√房颤、室性心律失常的发生率与高血压呈正相关
房颤、室性心律失常和猝死的发生率与以心脏重构为表现的高血压病程呈正相关。高血压患者心律失常发生的机制和表现通过心脏环境的宏观和微观变化影响心律失常的发展,导致电学和结构的改变。
高血压进一步促进左心室肥厚,这是心律失常的危险因素。长期升高的后负荷和心内压会导致心肌细胞肥大并激活心肌成纤维细胞。心肌细胞肥大和心肌成纤维细胞胶原沉积增加导致心肌质量增加,导致左心室肥厚。
已经确定,左心室肥厚可能通过诱发心肌缺血、心肌需氧量增加以及左心室舒张功能障碍而增加发生心律失常的风险。
肠道微生物群也会影响血压
据了解,缺乏肠道微生物的无菌小鼠的血压比传统小鼠的血压要低。与此同时,将人类高血压捐献者的粪便移植到无菌小鼠体内会导致这些小鼠血压升高。高血压中具有产生舒张代谢产物特性的细菌种类数量的减少也表明肠道微生物群会影响血压。
▼
动脉粥样硬化也是诱发心律失常的危险因素之一。当动脉粥样硬化影响传导系统,阻碍冲动传导时,就会引起各种心律失常。
√动脉粥样硬化会导致急性心肌梗塞与心律失常
急性心肌梗死是动脉粥样硬化的严重后果,经常与 心律失常一起发生,这会增加患者在医院接受治疗时突然死亡的风险和预后不良。
肠道微生物通过其代谢物参与动脉粥样硬化。具体而言,动脉粥样硬化患者的大肠杆菌(Escherichia coli)、克雷伯氏菌属(Klebsiella spp.)和产气肠杆菌(Enterobacter aerogenes)的丰度显著增加,而两种产生丁酸的细菌——Roseburia gutis和粪杆菌的丰度则下降。
肠道微生物的代谢产物可通过增加内皮活性氧的产生和损害内皮介导的血管舒张而参与动脉粥样硬化。
▼
肠道微生物研究表明,正常微生物群的失衡可能导致多种炎症性疾病,其中肥胖和胰岛素抵抗是肠道微生物失衡和能量失衡引起的主要疾病。
√糖耐量异常和肥胖是心律失常的重要原因
同时,糖耐量异常和肥胖也是心律失常的重要原因之一。糖尿病患者动脉粥样硬化和血栓形成的概率会增加,从而导致急性冠状动脉缺血性心脏病或心律失常。
细菌多样性的整体减少和细菌基因表达的改变被认为是影响代谢途径的主要原因,这可能与肥胖有关。
心律失常是一个缓慢发生且具有复杂机制的过程。从肠道菌群及其代谢物的角度出发,临床医生可以获得预防和治疗糖尿病/肥胖合并心律失常的新视角。尽管如此,还需要大量实验来证实。
▼
非酒精性脂肪性肝病是一种以肝脏细胞内脂肪过度沉积为主要特征的临床病理综合征,是最常见的肝脏疾病。
√非酒精性脂肪肝病提高了心律失常的易感性
非酒精性脂肪性肝病是一种多系统疾病,可能导致脂质和葡萄糖等代谢不良、炎症和氧化应激以及其他全身性有害后果。这些改变可能协同作用,增强心脏的结构、电和自主神经重塑,从而提高心律失常的易感性。
此外,心包脂肪体积或厚度与房颤的患病率和严重程度相关,大多数报告显示非酒精性脂肪肝病与心外膜脂肪体积或厚度呈正相关。
√肠道微生物通过肠-肝轴影响心律失常
肠-肝轴是肠道微生物群与肝脏之间的联系。肠道菌群功能障碍可能导致病原体相关分子模式的产生、粘膜屏障通透性增加导致肝脏炎症以及肝病的发生和进展,并且发现与健康人相比,肝病患者的微生物群多样性较低。
肠道及肝脏问题以多种方式显著影响心律失常。总之,非酒精性脂肪肝与心律失常密切相关,但现有证据并不完善,需要我们继续研究。
▼
阻塞性睡眠呼吸暂停综合征是一种在睡眠时导致呼吸暂停和通气不足的疾病,其特征是打鼾、睡眠模式紊乱、血氧饱和度频繁下降以及白天疲劳。
√睡眠呼吸暂停综合征通过多种途径诱发心律失常
阻塞性睡眠呼吸暂停综合征患者通过多种病理生理途径与心律失常的发生有关,包括呼吸暂停引起的缺氧、胸内压力变化、交感神经失衡、心房重塑、氧化应激、炎症和神经体液激活。
睡眠期间的缺氧会促进交感神经活动,这是诱发心房颤动的关键机制。因此,阻塞性睡眠呼吸暂停综合征本身可能促进心房颤动的发展。
现有的研究表明,患有心房颤动的人肠道中Ruminococcus gnavus为主的类型有所增加。因此,我们推测阻塞性睡眠呼吸暂停综合征和心房颤动病理生理过程的共同原因可能是由于肠道微生物群的改变,尤其是瘤胃球菌的增加。
▼
前不久的新冠肺炎对人类的影响是巨大的。SARS-CoV-2病毒主要影响上呼吸道,但也可能导致危及生命的肺部问题。在一些患有轻度至重度 COVID-19的人中,肠道微生物受到广泛干扰,这种干扰可能会持续长达一个月甚至更久。
√新冠肺炎是心律失常的危险因素
研究表明,新冠肺炎患者中存在心律失常的情况。这可能是由于病毒直接感染心肌细胞,或是由于免疫系统的异常反应所引起的。心律失常在新冠肺炎患者中可能表现为心率不齐、心动过速或心动过缓等症状,严重情况下甚至可能导致心脏骤停。
与其他呼吸道病毒感染一样,COVID-19可能与恶心、呕吐、腹痛和腹泻等胃肠道症状有关。由于影响肠道的途径是通过免疫和神经系统存在的,它们不仅对肠道微生物做出反应,而且还调节其组成。
循环淋巴细胞是两个器官之间的直接免疫接触,因为这些细胞不会停留在一个地方,它们在肠道和气道粘膜上巡逻。“肠肺轴”是这些器官“交流”的串扰。
综上所述,肠道微生物会通过各种分泌物引发心律失常,而SARS-CoV-2病毒由于“肠肺轴”的存在,可能是心律失常的危险因素。
心律失常作为一种心脏疾病,会给患者的健康和生活质量带来严重负面影响。因此,需要不断努力追求治疗心律失常的方法和策略。
在治疗心律失常方面,致力于整合多种治疗方法,包括药物治疗、手术治疗和其他创新疗法(例如肠道微生物干预)。需要根据患者的具体情况和心脏功能,制定最适合其需求的个性化治疗。
1
饮食调整: 多吃有益的脂质和膳食纤维
一些研究调查了不同食物成分和饮食模式对肠道菌群的影响,这可能是未来通过肠道微生物群治疗心律失常的重要目标。
•脂肪摄入过多会导致心律失常
最近的一项研究表明,房颤患者倾向于从动物脂肪中获取更多能量。至于肠道微生物群与脂肪摄入量之间的相关性,拟杆菌门与房颤患者的脂肪摄入量(特别是动物脂肪)呈弱负相关,而厚壁菌门则呈现相反的相关性。
•地中海饮食有助于降低心律不齐风险
代谢性内毒素血症和慢性炎症可能是脂肪摄入诱发房颤的机制。摄入油酸可以显著降低心血管疾病的风险,因为它可以抑制氧化应激,从而减轻心肌细胞损伤。
地中海饮食可以有益地影响肠道微生物群和相关代谢。以蔬菜为主的饮食人群具有更好的微生物代谢组。一项针对房颤患者的研究表明,地中海饮食可以降低氧化应激,从而降低房颤风险。
2
补充益生菌、益生元
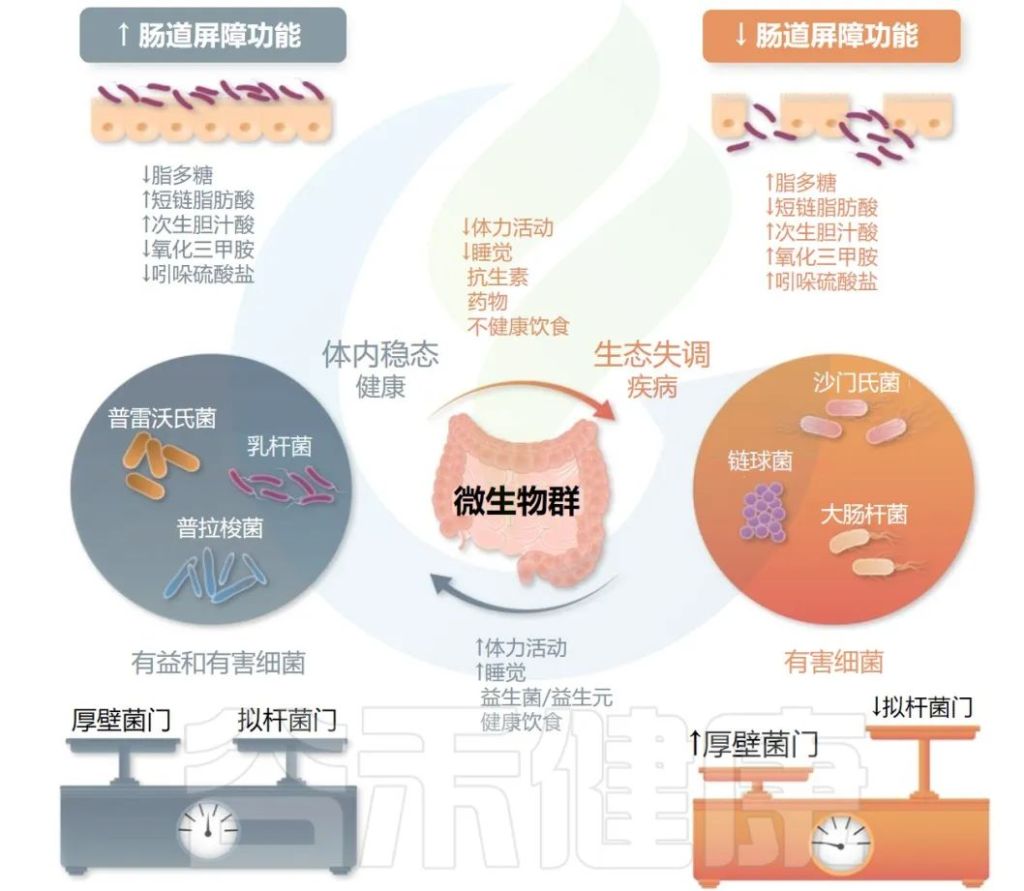
研究表明,肠道微生物的失衡与心律失常的发生有关。肠道微生物在治疗心律失常方面具有潜力。因此,调整肠道微生物的组成可能成为未来心律失常治疗的一种新方法。
编辑
Gawałko M,et al.Cardiovasc Res.2022
•服用益生菌缓解了动脉粥样硬化
几项临床研究发现,口服嗜酸乳杆菌(L.acidophilus ATCC 4356)可以通过调节小鼠的氧化应激和炎症过程来缓解动脉粥样硬化的进展。
•益生菌促进了残留药物的降解
益生菌还可以影响药物的代谢。胺碘酮是市场上主要的抗心律失常药物,具有心外毒性,因此研究人员重点寻找改善体内残留胺碘酮代谢的方法。
研究发现酿酒酵母菌株OBS2是一种具有治疗潜力的益生菌,可以在体外和体内促进残留胺碘酮的降解。
•益生元可以减少心肌损伤
益生元是一类可促进肠道益生菌生长和活性的物质,其健康益处取决于微生物的新陈代谢。目前果聚糖(低聚果糖和菊粉)和半乳聚糖(低聚半乳糖)在益生元类别中的主导地位已被数十项研究证实。
一项在大鼠缺血再灌注模型中进行的研究得出结论,阿拉伯半乳聚糖可以抑制细胞凋亡,从而减少心肌损伤。
此外,源自红酒的多酚白藜芦醇通过抑制细胞内钙的释放而具有抗心律失常特性。在小鼠模型中,白藜芦醇通过肠道微生物群重塑降低氧化三甲胺水平并增加肝脏胆汁酸合成,从而减轻动脉粥样硬化。
3
药物与肠道微生物的相互作用
大量研究表明药物和肠道微生物群之间存在双向作用,这也可能调节药物代谢。例如,迟缓埃格特菌(Eggerthella lenta)通过还原内酯环产生无活性产物来灭活地高辛。
•肠道微生物增强了心血管药物的利用度
此外,肠道微生物酶活性参与了一些心血管药物的首次通过清除,包括钙通道阻滞剂和非甾体抗炎药,当患者接受抗生素预处理时,增加了这些药物的生物利用度。
•心血管药物对肠道微生物存在显著影响
同时,许多临床使用的药物可以增强或降低肠道微生物群或肠道微生物群衍生代谢物的生长速度。对荷兰一项队列研究的1135名参与者的粪便样本进行宏基因组测序显示,使用多种心血管药物,包括他汀类药物、抗血栓药物、 受体阻滞剂和血管紧张素转换酶抑制剂,对肠道微生物组有显著影响。
4
粪菌移植
粪便微生物移植作为一种安全的治疗方法被广泛应用于胃肠道疾病及胃肠外疾病的治疗。
•粪菌移植可以有效减轻甚至治愈疾病
从健康受试者到患有菌群失调相关疾病的受试者的菌群移植可以减轻甚至治愈这种疾病。在一项研究中,将血压正常大鼠的盲肠内容物移植到自发性高血压大鼠体内可降低血压,而对自发性高血压大鼠进行粪菌移植后,血压正常的大鼠会出现高血压。
最近,将来自高脂肪饮食喂养的小鼠的粪便微生物移植到正常饮食喂养的小鼠中,他们发现对房颤的易感性显著增加。
接受高脂饮食小鼠粪菌移植的小鼠遭受由脱硫弧菌(Desulfovibrionaceae)引起的代谢内毒素血症,该血症循环脂多糖升高,破坏肠道组织结构,并增加左心房促炎因子。
5
免疫调节剂
组蛋白脱乙酰酶在钙稳态、房颤发生和心力衰竭中起着至关重要的作用,因此组蛋白脱乙酰酶抑制剂可能是潜在的治疗方法。
•组蛋白去乙酰酶抑制剂降低了心律失常的发生
组蛋白去乙酰化酶抑制剂通过拯救线粒体生物能量学来治疗心脏功能障碍,即TNF-α诱导的线粒体功能障碍,降低能量利用率。
另一项在房颤兔身上进行的研究表明,组蛋白去乙酰化酶抑制剂可以降低钙稳态诱导的房颤和肺静脉心律失常的发生,房颤持续时间缩短。
注:MPT0E014处理的心肌细胞钙瞬态振幅、钠钙交换电流和ryanodine受体表达的降低可能是组蛋白去乙酰化酶抑制剂治疗心律不齐的潜在机制。
6
病因治疗和药物治疗
•病因治疗
病因治疗包括纠正心脏病理改变、调整异常病理生理功能(如冠脉动态狭窄、泵功能不全、自主神经张力改变等),以及去除导致心律失常发作的其它诱因(如电解质失调、药物不良副作用等)。
•药物治疗
药物治疗缓慢心律失常一般选用增强心肌自律性或加速传导的药物,如拟交感神经药(异丙肾上腺素等)、迷走神经抑制药物(阿托品)或碱化剂(克分子乳酸钠或碳酸氢钠)。
治疗快速心律失常则选用减慢传导和延长不应期的药物,如迷走神经兴奋剂(新斯的明、洋地黄制剂)、拟交感神经药间接兴奋迷走神经(甲氧明、苯福林)或抗心律失常药物。
多项证据表明肠道微生物群的变化可能是心律失常的重要诱发因素。肠道微生物可以通过短链脂肪酸、胆汁酸等代谢物,改变心肌细胞的电活动,从而调节心律。
其次,肠道微生物的失衡可能导致免疫系统的异常活化,诱发心律失常的发生。某些肠道菌群的代谢产物,如脂多糖、细菌毒素等,可能引发炎症反应,损伤心肌细胞,进而导致心律失常。
然而,肠道微生物与心律失常的具体关联和机制仍需进一步的研究和探索。目前,通过调整饮食结构、补充益生菌和使用粪菌移植等策略,可以尝试改善肠道微生物组成,从而对心律失常的治疗产生积极的影响。
但需要强调的是,个体差异和复杂性限制了一刀切的推广,治疗方案应当根据具体情况进行调整和制定。
主要参考文献
Fan H, Liu X, Ren Z, Fei X, Luo J, Yang X, Xue Y, Zhang F, Liang B. Gut microbiota and cardiac arrhythmia. Front Cell Infect Microbiol. 2023 Apr 25;13:1147687.
Gawałko M, Agbaedeng TA, Saljic A, Müller DN, Wilck N, Schnabel R, Penders J, Rienstra M, van Gelder I, Jespersen T, Schotten U, Crijns HJGM, Kalman JM, Sanders P, Nattel S, Dobrev D, Linz D. Gut microbiota, dysbiosis and atrial fibrillation. Arrhythmogenic mechanisms and potential clinical implications. Cardiovasc Res. 2022 Aug 24;118(11):2415-2427.
Frampton J, Ortengren AR, Zeitler EP. Arrhythmias After Acute Myocardial Infarction. Yale J Biol Med. 2023 Mar 31;96(1):83-94.
Li W, Li C, Ren C, Zhou S, Cheng H, Chen Y, Han X, Zhong Y, Zhou L, Xie D, Liu H, Xie J. Bidirectional effects of oral anticoagulants on gut microbiota in patients with atrial fibrillation. Front Cell Infect Microbiol. 2023 Mar 24;13:1038472.
Zuo K, Yin X, Li K, Zhang J, Wang P, Jiao J, Liu Z, Liu X, Liu J, Li J, Yang X. Different Types of Atrial Fibrillation Share Patterns of Gut Microbiota Dysbiosis. mSphere. 2020 Mar 18;5(2):e00071-20.
Li N, Durgan DJ, Wehrens XHT. Gut microbiota: a key regulator of ageing-associated atrial fibrillation? Cardiovasc Res. 2022 Feb 21;118(3):657-659.
Long MT, Ko D, Arnold LM, Trinquart L, Sherer JA, Keppel SS, Benjamin EJ, Helm RH. Gastrointestinal and liver diseases and atrial fibrillation: a review of the literature. Therap Adv Gastroenterol. 2019 Apr 2;12:1756284819832237.

谷禾健康

嗜血杆菌属(Haemophilus spp) 是变形菌门,小的、多形性、不活动、无孢子的革兰氏阴性杆菌或球杆菌。它们是需氧的和兼性厌氧的。主要寄居于人和动物的咽喉和口腔黏膜,少数见于生殖道,不过很多时候在粪便中也能检出。该病原菌能引起原发性化脓性感染及严重的继发感染。
嗜血杆菌属(Haemophilus)是一组细菌的名称。它们会引起不同类型的疾病,涉及呼吸、骨骼和关节以及神经系统。嗜血杆菌属的丰度与多种疾病和健康状态有关。例如,嗜血杆菌属在多发性硬化症、结直肠癌等患者中的丰度增加。
Photograph by David M. Phillips
嗜血杆菌属的过度生长与一些炎症性肠道疾病、肠道肿瘤和代谢性疾病的发生有关。嗜血杆菌属的存在可以引起肠道炎症反应,导致肠道黏膜的损伤和疾病的发展。
其次,嗜血杆菌属与一些疾病相关的菌群存在相互作用。例如,在某些研究中发现,嗜血杆菌属与肠道菌群中的某些菌属(如Prevotella、Roseburia、Subdoligranulum和Dorea)呈负相关。此外,嗜血杆菌属可能通过影响肠道菌群的代谢产物来调节疾病的发生和发展。例如,嗜血杆菌属与肠道代谢物(如丙酸和戊酸)的浓度呈负相关。
嗜血杆菌属包括许多可引起多种感染的菌种,但它们具有共同的形态,并且在生长过程中需要血源性因子,因此该属得名。
此外,嗜血杆菌属里比较受关注的是流感嗜血杆菌(Haemophilus influenzae)。
已知荚膜类的乙型流感嗜血杆菌(或是b型流感嗜血杆菌,简称HiB)是毒性的主因之一,通常会导致 2 岁以下儿童的血流侵入和脑膜炎,偶而地它会引致蜂窝组织炎、骨髓炎及关节感染。Hib 结合疫苗是减少嗜血杆菌相关疾病发病率的重要措施。对于住院患者,联合使用氨苄西林和氯霉素或单一头孢菌素如头孢呋辛、头孢噻肟或头孢曲松是有效的治疗方法。
本文一起来了解一下嗜血杆菌属的生物学特性,代谢途径,和其他菌群相互作用,与疾病之间的关联,耐药性和抗生素治疗等,嗜血杆菌属的研究对于理解呼吸道感染的发病机制、疫苗研发和公共卫生具有重要意义。
✓
嗜血杆菌属(Haemophilus)是一种人体常见的细菌属,它可以在人体的口腔、呼吸道和消化道中寄生。属于变形菌门,巴斯德氏菌科,该科还包含另外两个属:放线杆菌属和巴斯德氏菌属。
属于该科的细菌是小型(1 × 0.3 μm)、不形成孢子的球杆菌,具有苛刻的生长要求,通常需要补充培养基才能进行体外分离。
大多数嗜血杆菌属需要两种补充剂——因子 X(血红素)和 V(烟酰胺腺嘌呤二核苷酸,NAD)——才能在人工培养基上有氧生长。
巧克力琼脂平板上生长的流感嗜血杆菌菌落
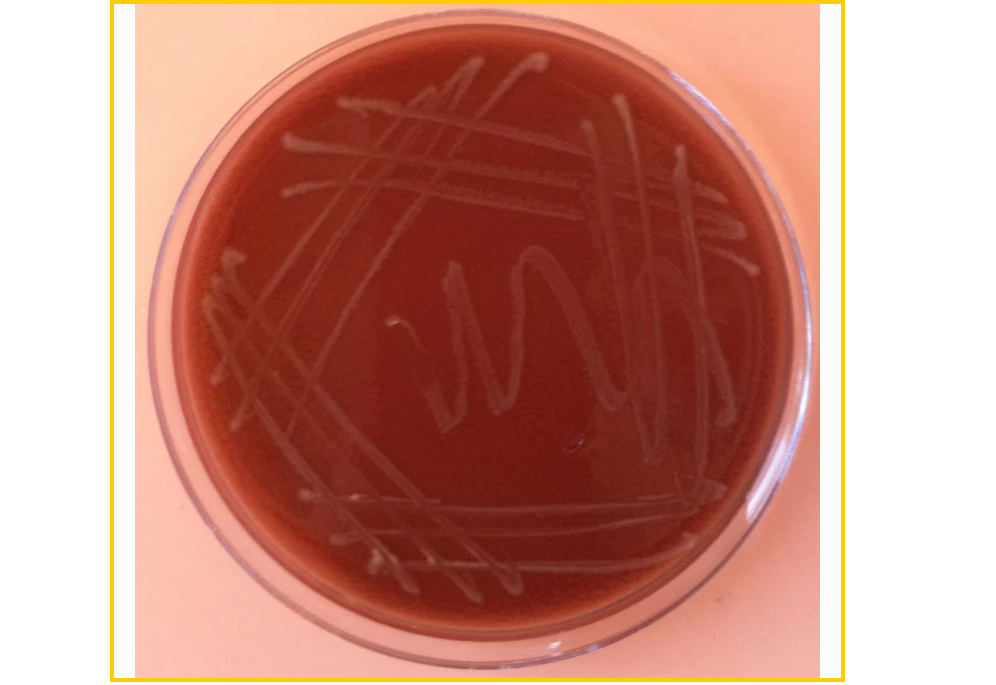
doi:10.1186/2001-1326-1-10
但是注意,当细菌在实验室培养基上生长时,流感嗜血杆菌(Haemophilus influenzae)的生存力会迅速下降,通过未知的机制在 48 至 72 小时内变得不可培养。因此,如果需要,应在嗜血杆菌菌株在固体培养基上以离散菌落形式出现后立即对其进行永久储存。
▼
– 荚膜抗原(在胶囊化菌株中):抗吞噬细胞
– 粘附素蛋白(例如HMW1、HMW2)介导与气道中的人上皮细胞的附着。
– 菌毛和主要外膜蛋白 P2:将含有唾液酸的部分结合到上皮细胞表面。
不可分型流感嗜血杆菌是定植于人类呼吸道的主要病原体。细菌在哺乳动物组织上的粘附是由菌毛介导的,被认为是定植的重要前体,而上呼吸道感染与菌毛的存在有关。这些微生物引起的呼吸道感染包括鼻窦炎、中耳炎、急性气管支气管炎和肺炎。
– IgA1 蛋白酶:在铰链区裂解免疫球蛋白 A,防止凝集并机械消除病原体
– 相变:外表面蛋白质被修饰以适应宿主环境的变化。
– 体内生物膜的产生:在急性中耳炎的发病机制中尤其重要。
急性中耳炎是最常见的儿科细菌感染,在 5 岁之前的某个时期影响多达 75% 的儿童。肺炎链球菌和流感嗜血杆菌(主要是 NTHi)造成了高达 80% 的细菌急性中耳炎。
从儿童或成人急性感染的鼻窦或婴儿和幼儿受感染的鼓膜后面吸出的化脓性物质通常含有无法分型的流感嗜血杆菌。
外膜蛋白谱研究表明,中耳和鼻咽分离株是相同的,这支持了咽鼓管定植以及随后的阻塞和感染可能是其原因的观点。中耳炎的反复发作是由不同的菌株引起的。每次感染可能与针对独特表面蛋白的抗体的出现有关。随着年龄的增长,中耳炎的发病率逐渐降低,部分原因是解剖结构的变化,部分原因是对流感嗜血杆菌的免疫力。
▼
巴斯德氏菌科的几个 16S rRNA 序列特征已得到证实,但这些特征都不是目前定义的嗜血杆菌属所特有的。DNA 的 mol% G + C 为:37–44。
模式种:
其他研究的菌种包括:
其中以下菌种有致病性,但是致病机理和其他菌种的致病性还需要更多研究确定。
其中Haemophilus ducreyi和Haemophilus aegyptius容易形成生物膜。
注:许多细菌可以形成称为生物膜的聚集结构。生物膜中的生物通常表现出与处于个体状态或浮游状态的同一生物基本不同的特性。 聚集到生物膜中的细菌可以传达有关种群大小和代谢状态的信息。 这种类型的通信称为群体感应,通过产生称为自动诱导剂或信息素的小分子来进行操作。
群体感应分子【最常见的是肽或酰化高丝氨酸内酯(AHL;特殊的信号化学物质)】的浓度与生物膜中相同或不同物种的细菌数量有关,有助于协调生物膜的行为。
扩展阅读:
▼
Haemophilus生长最佳温度为 35–37°C。氧化酶和过氧化氢酶反应因菌株而异。化学有机营养型。所有物种都可以发酵碳水化合物,葡萄糖代谢产生乙酸、乳酸和琥珀酸作为最终产物。作为专性寄生存在于人类和多种动物的粘膜上。
KEGG 途径还包括:
▼
与肺炎链球菌的相互关系
嗜血杆菌与肺炎链球菌都能在人类的上呼吸系统中发现。一项有关它们之间的竞争研究指出,在培养基内,肺炎链球菌能以过氧化氢攻击流感嗜血杆菌,及能排除在表面流感嗜血杆菌生存所需的分子。
当两种细菌一同在鼻腔内时,两星期之内,只有流感嗜血杆菌能够生存。当两种细菌分别地放在鼻腔内时,两者皆能生存。使实验鼠的上呼吸组织暴露于两种细菌时,发现有格外大量的中性粒细胞。当实验鼠只是暴露于其中一种细菌时,则不会出现这些细胞。
实验显示,接触过嗜血杆菌死菌的中性粒细胞对肺炎链球菌会强烈攻击,而未接触过流感嗜血杆菌死菌的中性粒细胞的这种攻击则较不会这么强烈。
接触流感嗜血杆菌死菌,却不会对流感嗜血杆菌活菌有所影响。这种现象有两种可能性:
(1)当嗜血杆菌被肺炎链球菌攻击时,引发了免疫系统攻击肺炎链球菌。
(2)两种细菌的结合引发了只有其中一种细菌存在所不会引发的免疫系统警报。
至于为何流感嗜血杆菌会不受免疫系统的影响则不得而知。
与其他菌的互作
在一些研究中发现,普雷沃氏菌属(Prevotella)的相对丰度与Haemophilus的相对丰度呈负相关,这种互作可能对疾病的发展和治疗产生影响。
Haemophilus与厚壁菌门中的Roseburia、Subdoligranulum和Dorea之间存在负相关关系。这意味着Haemophilus的增加可能会导致这些厚壁菌门的特定菌属减少。
而比较惊讶的是在一些研究中发现,Haemophilus的相对丰度与Akkermansia的相对丰度呈正相关,这种互作可能对肥胖和代谢性疾病的发展产生影响,但是该结论还需要在更多的研究中验证。
在一些疾病中Haemophilus和拟杆菌属的相对丰度在一些疾病中呈现出相互竞争的趋势,这种互作可能对肠道微生物群的稳态和健康产生影响。
✓
Haemophilus influenzae
听名字可能会认为该菌是引起流感的元凶,但并不是,是因为流感嗜血杆菌是1892年被德国的一位细菌学家理查德.法伊弗医生在一位流感病人身上分离得到的,因此,此后一段时间流感嗜血杆菌也被认为是引起流行性感冒的病原体,直到1933年流感病毒被分离出来。1995年,第一个活体物种嗜血流感菌(Haemophilus influenzae,1.8Mb)的基因组测序完成。
分 类
流感嗜血杆菌是嗜血杆菌属主要病原体,它可分为有荚膜或可分型菌株,根据荚膜多糖的抗原结构,有七种类型(a至f,包括e’)和无荚膜或不可分型菌株。没有荚膜的菌株则较少侵略性,但它们能诱发炎症而产生其他病症,如会厌炎(主要症状有剧烈的喉痛、吞咽困难和呼吸困难等)。
注:荚膜是细菌比较常见的结构,是位于细胞壁表面的一种粘液型物质,也是重要的致病因子。荚膜可以帮助细菌抵抗人体免疫系统的吞噬和补体介导的溶菌作用,就如细菌的“防弹衣”或“盔甲”,人体的免疫系统原本可以很轻松的击毙细菌,而一旦细菌有了“防弹装备“”,它们就更容易存活下来。
荚膜多糖进化的一个可能的原因是,与缺乏荚膜的多糖相比,它们赋予细胞了一定的抗干燥性。这可能在宿主到宿主的传播过程中提供健康益处,这是细菌生命周期的关键部分。
流感嗜血杆菌可正常寄居于我们的上呼吸道,当抵抗力下降、免疫功能较差时,才会引起呼吸道感染。因此,婴幼儿、学龄期儿童、孕妇、老人等就成为了流感嗜血杆菌的主要攻击目标。作为儿童临床中常见的条件致病菌之一,是引起儿童社区获得性肺炎仅次于肺炎链球菌的致病菌。
人体内的常居部位及携带率
Haemophilus influenzae通常可以寄居在我们的呼吸道、眼结膜和阴道黏膜上。湿润的鼻咽黏膜就是它比较喜欢的栖息场所。
人群鼻咽部的Haemophilus influenzae的携带率受到了年龄、生理因素、人群密集程度、社会经济因素以及疫苗接种率等因素的影响。在未开展Haemophilus influenzae疫苗接种之前,发达国家约有3%~5%的健康学龄儿童是无症状Haemophilus influenzae的携带者。
它也是主要在上呼吸道发现的专性人类共生菌。最近临床从血液、痰液、肺泡灌洗液及阴道分泌物等各类临床标本中均有分离到Haemophilus influenzae,而且我们在粪便高通量测序检测实践中也检出Haemophilus influenzae,表明感染明显增多。
该菌作为一种苛养菌,其生长需要V和X因子,在培养中稍不留神容易漏检。临床标本中分离出的流感嗜血杆菌形态多种多样,从球杆菌到长丝状杆菌。这种不一致的形态和经常变化的染色(尤其是革兰氏染色中的沙弗宁)可能会导致临床微生物实验室的诊断混乱。
临床感染
流感嗜血杆菌(Haemophilus influenzae)可以导致多种类型的临床感染。
该菌可在局部蔓延引起非侵袭性疾病,如无菌血症的肺炎、中耳炎、鼻窦炎和结膜炎;
还可导致更为严重的侵袭性疾病,如脑膜炎、败血症、有菌血症的肺炎、会厌炎、脓性关节炎、心包炎、骨髓炎、软组织化脓及蜂窝组织炎等。
感染率
来自我国的研究发现,1990-1992 年中国合肥地区 Hib 脑膜炎占小儿细菌性脑膜炎的 51.7%。
复旦大学附属儿科医院对上海市 2000-2005 年 266 例临床诊断为细菌性脑膜炎的患儿进行致病菌抗原检测,Haemophilus influenzae阳性率为 15.4% 。5岁以下Haemophilus influenzae脑膜炎患儿的病死率为9.7%,存活患儿中21.4%的有精神和神经系统后遗症,主要包括硬膜下积液、听力和/或智力障碍、轻度瘫痪等。
感染宿主
流感嗜血杆菌非常适合人类宿主,并且没有其他已知的自然感染宿主。
据推测,大约一万年前,人类从“狩猎采集者”的生活过渡到以农业为基础的社区生活之后,就一直接触到一系列新型细菌。那时,人类开始与牲畜近距离生活,然后可能获得了嗜血杆菌的共同祖先,它也是其他几种嗜血杆菌的祖先。宿主特异性细菌,如放线杆菌和巴斯德氏菌,现在发现于牛和猪等家畜中。
代 谢
流感嗜血杆菌Rd菌株的全基因组序列极大地增加了我们对该细菌生物学的了解。嗜血杆菌似乎在代谢上适应了富氮、微厌氧的环境。编码三羧酸 (TCA) 循环中的三种酶(乌头酸酶、柠檬酸合成酶和异柠檬酸脱氢酶)的基因缺失。这解释了在特定的实验室培养基中生长期间需要通过 α-酮戊二酸进入 TCA 循环的谷氨酸。与大肠杆菌相比,嗜血杆菌中的双组分系统较少(总共六个)(>40);双组分系统将特定的环境信号(例如 pH 值、渗透压和阳离子浓度的变化)转换为细菌细胞内程序化的协调反应。显然,嗜血杆菌适应环境变化的机制与肠杆菌科等细菌的机制不同。
判别:流感嗜血杆菌
涂片镜检:采集肺炎患者的痰,脑膜炎患者的脑脊液或者患者感染部位的分泌物等,经革兰染色后显微镜下观察,如查见上述形态图中的红色细小杆菌将有助于诊断。
细菌培养:是诊断流感嗜血杆菌感染的金标准,但对于呼吸道标本,需要医生结合临床症状、标本质量等因素综合考虑是否为感染。
还可以通过检测血清中流感嗜血杆菌抗原或抗体,咽拭子、脑脊液,粪便等标本中流感嗜血杆菌基因测序来进行快速诊断。
传染源
虽然只有少部分携带者会发展为临床病例,但携带者是流感嗜血杆菌的主要传染源。
传播途径
主要经呼吸道飞沫传播,接触患者的呼吸道分泌物也可感染。新生儿可因吸入羊水或出生时接触产道分泌物而感染。
易感人群
该细菌的目标人群是免疫抑制/免疫抑制患者、脾切除患者和未接种疫苗的患者。此外,Haemophilus influenzae引起的疾病与年龄密切相关, 在未引入疫苗的地区,有超过60%的侵袭性疾病发生在1岁之前,主要集中在6-11月月龄的婴儿。而5岁及以上的儿童就很少发生由Haemophilus influenzae引起的侵袭性疾病了(少于10%)。由此婴幼儿是我们要重点关注的人群。
没有特定症状可以将流感嗜血杆菌肺炎与其他细菌区分开来。
可以怀疑 3 个月至 3 岁的儿童患有流感嗜血杆菌肺炎;而且疫苗接种状态不存在或不完整,最近使用抗生素或使抗生素后 48-72 小时内发烧未消退。
它不仅是肺炎的常见原因,而且会引起急性中耳炎,我们还应该怀疑上述相同经历的患者,特别是那些患有中耳炎并可能影响多个家庭成员的结膜炎的患者。
治 疗
流感嗜血杆菌感染需用抗生素治疗。使用哪种抗生素取决于感染的严重程度、感染部位以及药敏结果。
如果儿童出现严重感染,应将他们收入医院并予以隔离,以防止其他人接触到他们排放至空气中的飞沫(称作呼吸道隔离),隔离应持续至开始抗生素治疗后24小时。
脑膜炎必须尽早治疗。可静脉输注头孢曲松或头孢噻肟治疗流感嗜血杆菌脑膜炎。服用皮质激素有助于预防脑部损害。
会厌炎也必须尽早治疗。患者可能需要辅助呼吸。可能需要建立人工气道,如呼吸管插管,罕见情况下可能需要开放气道(称为气管切开术)。应给予一种抗生素,如头孢曲松、头孢噻肟或头孢呋辛。
由流感嗜血杆菌引起的其它感染可使用各种口服抗生素治疗。它们包括阿莫西林/克拉维酸钾、阿奇霉素、头孢菌素类、氟喹诺酮类等。
▼
Haemophilus aegyptius
埃及嗜血杆菌(Haemophilus aegyptius)值得特别提及,因为它可以引起类似脑膜炎球菌的疾病。在南美洲,这种微生物是导致巴西紫癜的原因。这种灾难性疾病在 10 岁以下儿童中尤为突出,通常以超急性结膜炎开始,随后在 3 至 5 天内出现发烧。48 小时内会出现播散性紫癜、低血压休克和死亡。
分子研究表明,引起巴西紫癜热的所有分离株都具有遗传相关性。最初,所有报告的病例都来自巴西圣保罗和邻近的巴拉那州,但其他地区出现了新菌株,引发了人们对该疾病可能在全球范围内传播的担忧。
致病性埃及嗜血杆菌的基因组大于非致病性菌株的基因组。细菌物种之间频繁的基因交换已被证明是这种遗传添加的基础。埃及嗜血杆菌个别菌株还可引起急性结膜炎,俗称“红眼病”,有高度传染性。
▼
Haemophilus ducreyi
杜克雷嗜血杆菌(Haemophilus ducreyi)是一种高度传染性的革兰氏阴性细菌,可通过性传播。它是性病感染的原因:软下疳。杜克雷嗜血杆菌在女性中仅引起轻微症状,而在男性中,下疳却极其疼痛。它会导致快速发展的病变,以与生殖器区域剧烈瘙痒相关的深度溃疡的形式出现。显微镜检查发现杜克雷嗜血杆菌。
▼
Haemophilus parainfluenzae
副流感嗜血杆菌是一种粘膜寄生菌,特别存在于口咽、泌尿生殖系统和直肠中。这是人类最常见的嗜血杆菌属,因为它占咽部菌群的 10%,有时在唾液中大量存在。这种病原体可引起呼吸道和耳鼻喉感染,特别是在免疫功能低下的人群中。副流感嗜血杆菌还可引起突发性心内膜炎。
✓
肠道微生物群中大量存在的嗜血杆菌是否会对健康产生积极或消极影响仍然未知。据报道,多发性硬化症、结直肠癌患者的嗜血杆菌丰度增加,并且也与腺瘤患者的结直肠癌发展相关。相反,嗜血杆菌属减少与 II 型糖尿病的发展有关,并且个别 II 型糖尿病的嗜血杆菌属的完全耗尽。
▼
研究发现,类风湿性关节炎(RA)患者口腔和肠道中的Haemophilus属菌群相对丰度较低。在类风湿性关节炎患者中,Haemophilus的相对丰度降低与类风湿性关节炎特异性自身抗体(抗环瓜氨酸肽)和类风湿因子的水平降低相关。这表明Haemophilus可能在类风湿性关节炎的发病和病理过程中发挥一定的作用。
注:抗环瓜氨酸肽抗体是以合成的环化瓜氨酸多肽(CCP)为抗原的自身抗体,对类风湿关节炎(RA)具有较高的敏感性和特异性,是RA早期诊断的一个高度特异指标。
类风湿性关节炎患者的唾液、牙菌斑和粪便样本中嗜血杆菌种类减少,类风湿性关节炎治疗后部分正常化。
肠道和口腔环境极其复杂,这些环境中的微生物群起着至关重要的作用。在维持体内平衡中发挥重要作用。据报道,随着肠道微生物群与免疫系统的接触减少,过敏性以及自身免疫性和炎症性疾病的发病率和患病率显著增加。
Haemophilus与炎症反应的调节
Haemophilus可能通过调节免疫反应参与类风湿性关节炎的发展。研究发现,Haemophilus属菌群的降低与炎症标志物的水平降低相关。例如,嗜血杆菌属与 C 反应蛋白(急性炎症标志物)呈负相关。
Haemophilus与肠道菌群失调的关联
类风湿性关节炎患者肠道菌群失调可能与Haemophilus的降低有关。肠道菌群失调已被认为是类风湿性关节炎发展的一个重要因素。Haemophilus与自身抗体的水平相关:Haemophilus的降低与类风湿性关节炎患者自身抗体水平的降低相关。自身抗体在类风湿性关节炎的发病和病情进展中起着重要作用。
▼
Haemophilus是一种与结直肠癌发病和发展相关的细菌。以下是Haemophilus参与结直肠癌的发病和发展的机制和关联线索:
肠道菌群失调
Haemophilus的丰度在结直肠癌患者中较高,与肠道菌群的失调有关。肠道菌群的失调可能导致炎症反应和免疫系统的异常,从而促进结直肠癌的发展。
炎症反应
Haemophilus可能通过引起炎症反应来促进结直肠癌的发展。炎症反应可以导致细胞损伤和DNA损伤,进而增加癌症的发生风险。
免疫调节
Haemophilus可能通过影响免疫系统的功能来参与结直肠癌的发展。它可能影响肠道免疫细胞的活性和功能,从而干扰对癌细胞的清除和抑制。
其他细菌的相互作用
Haemophilus可能与其他细菌相互作用,共同参与结直肠癌的发展。例如,与口腔菌群中的Fusobacterium相互作用可能导致肠道炎症和癌症的发展。
▼
多项研究报道了肠道内Haemophilus参与了多发性硬化(MS)的发病和发展。Haemophilus可能通过分子模仿来激活自身免疫T细胞,从而导致多发性硬化的发展。研究发现,Haemophilus的蛋白质结构与多发性硬化相关的蛋白质结构相似,可能引发自身免疫反应。此外,Haemophilus的丰度变化与肠道菌群失调有关。肠道菌群失调已被证明与多发性硬化的发展和进展有关。
多发性硬化症 (MS) 是一种中枢神经系统 (CNS) 脱髓鞘疾病。遗传和环境因素,如细菌或病毒感染,涉及其多因素病因。在推定与该疾病相关的病毒因子中,EB 病毒 (EBV) 已被多次报道,流感嗜血杆菌被描述为多发性硬化症可能的细菌触发剂。尽管多发性硬化症的病因和发病机制尚未完全阐明,但人们普遍认为,针对髓磷脂和其他相关蛋白的 T 细胞介导的炎症过程起着至关重要的作用,同时 B 细胞也可能发挥作用。
在多发性硬化症中,肠道内的甲烷短杆菌属、假单胞菌属、支原菌属、嗜血杆菌属、Blautia属和 Dorea属 增加。关于肠道微生物群对炎症性中枢神经系统病理学影响的更多数据已从多发性硬化症动物模型研究中获得。这些数据支持这样的观点,即肠道微生物群失调正在积极促进多发性硬化症的发生和进展。
▼
研究发现,Haemophilus的丰度在糖尿病患者粪便中发生了改变。另人意外的是,研究显示Haemophilus的减少与2型糖尿病的发展相关。Haemophilus可能通过与免疫系统的相互作用影响糖尿病的发展。一些研究发现,Haemophilus的存在与自身抗体水平的负相关性,这可能意味着Haemophilus的减少可能导致免疫系统的异常反应,从而增加糖尿病的风险。
另一项研究发现,布劳特氏菌属(Blautia)、嗜血杆菌属(Haemophilus)丰度与TIDM(I型糖尿病)自身抗体个数相关。Haemophilus可能与肠道菌群的变化相关,从而影响糖尿病的发展。
▼
慢性心力衰竭(CHF)是指一种持续性心力衰竭状态,可以稳定、恶化或失代偿。
个别研究显示嗜血杆菌和克雷伯氏菌在CHF患者的粪便中显着富集,可能意味它们可能是CHF的粪便生物标志物。作者进一步分析了肠道微生物群与血清代谢组之间的相关性。发现志贺氏菌与生物胞素和核黄素呈负相关。而嗜血杆菌与α-乳糖、纤维二糖、异麦芽糖、乳糖、蜜二糖、蔗糖、海藻糖呈负相关。
缺血性心脏病(IHD)是全球范围内人类死亡的首要原因。近年来研究表明,人体肠道菌群通过影响氧化三甲胺(TMAO)、苯乙酰谷氨酰胺(PAGln)、三甲基-5-氨基戊酸等相关代谢产物来促进IHD的发生和发展。宁波大学附属第一医院发现IHD患者毛螺菌科(Lachnospiraceae)和瘤胃球菌科(Ruminococcaceae)显著减少。且对疾病亚组分析提示,IHD患者的肠道链球菌属(Streptococcus)、嗜血杆菌属(Haemophilus)丰度显著富集。
前面讲过链球菌属和嗜血杆菌属的互作,它们都能在人类的上呼吸系统中发现。但是这两种菌呈现竞争趋势,而在肠道中这两种菌同时富集,可能是与上呼吸道不一样的生态模式,他们致病的机理还需要进一步探究。
▼
Haemophilus参与免疫系统的互作有以下几个方面的机制和关联线索:
SLPI调节
SLPI是一种抗菌肽,它在呼吸道中的丰度受到感染的调节。研究发现,Haemophilus的存在与SLPI的丰度增加相关。SLPI的表达调节是呼吸道微生物群和呼吸道黏膜免疫系统之间维持稳态的关键机制之一。
Haemophilus与哮喘
研究发现,Haemophilus在上呼吸道中的存在与成年人免疫控制不佳相关。这表明Haemophilus诱发的免疫可能在哮喘的发病机制中起到一定的作用。
锌补充与Haemophilus
锌是胎儿免疫系统发育的重要因素。研究发现,锌补充在孕期对贫困孟加拉婴儿对结核病卡介苗和流感嗜血杆菌b型疫苗的免疫反应有影响。
Haemophilus与免疫系统防御
Haemophilus居住在上呼吸道和口腔的黏膜上。多种Haemophilus菌株,如流感嗜血杆菌和溶血性流感嗜血杆菌,可以对抗宿主免疫系统并引起人类疾病。研究发现,IgAN患者对Haemophilus parainfluenzae的IgA抗体明显增多。IgA是黏膜免疫稳态的重要介质。
▼
一项研究(n=126)显示,与健康受试者相比,处于疾病不同阶段(急性与缓解期)的参与者的肠道细菌不平衡。嗜血杆菌丰度与精神分裂症阴性症状呈正相关(r = 0.303, P = 0.021),而粪球菌丰度与精神分裂症阴性症状呈负相关(r = – 0.285, P = 0.025)。这可以作为精神分裂症不同阶段的指标,也为精神分裂症的研究提供新的视角。
▼
一项研究基于 VFDB 数据库分析了食管鳞状细胞癌患者和肺癌患者中细菌的丰度和分布,嗜血杆菌的绝对丰度可以将肺癌与健康对照区分开来(P≤0.05),该研究为食管鳞癌和肺癌的鉴别提供了可能的方法,并为后续研究物种水平上潜在的人类病原体在癌症发生中的作用提供了数据支持。
▼
癫痫
少量研究显示特发性局灶性癫痫患者肠道菌群中的弯曲菌属、代尔夫特菌属、嗜血杆菌属、奈瑟菌属丰度显著增加,并且在10.6%的癫痫患者肠道中检测出梭杆菌门,而在对照组中未检出。
强直性脊柱炎
一项来自调查强直性脊柱炎患者(包括5名同时患有IBD的AS-IBD)、59名IBD患者和105名健康对照者中获取粪便和肠道活检样本。发现强直性脊柱炎疾病活动度指数(BASDAI评分)增加的强直性脊柱炎患者和粪便钙卫蛋白>100 μg/mg的IBD患者中观察到潜在致病性嗜血杆菌属的富集。
原发性胆管炎
肠道微生物群的变化会影响肠道和肝脏,它们通过所谓的“肠-肝轴”严格连接。原发性胆汁性胆管炎,也称为“原发性胆汁性肝硬化”。该病发病机制与肠道菌群失调有关,通过引起细菌性胆管炎,进而刺激促炎症机制,促进胆管细胞的纤维化和炎症。
一些研究报告显示,原发性胆管炎患者胆汁和粪便微生物组的改变以乳酸杆菌、梭杆菌和肠球菌的高丰度和低多样性为特征。原发性胆管炎患者还表现肠道内肠球菌属,罗氏菌属,梭菌属,链球菌属,嗜血杆菌属的过度生长。
桥本甲状腺炎
福建中医药大学附属人民医院89名参与的一项研究显示,游离三碘甲状腺原氨酸(FT3)调节的Romboutsia和Haemophilus可能促进桥本甲状腺炎的发展,而游离甲状腺素(FT4)调节的Faecalibacterium和Lachnospiraceae可能保护宿主。
检出案例
以下是疾病人群粪便样本中检出的嗜血杆菌,尤其流感嗜血杆菌,该菌是病原菌,过多可导致菌群紊乱,可引发呼吸道感染,肠道检出也常见于呼吸道感染或其他感染。
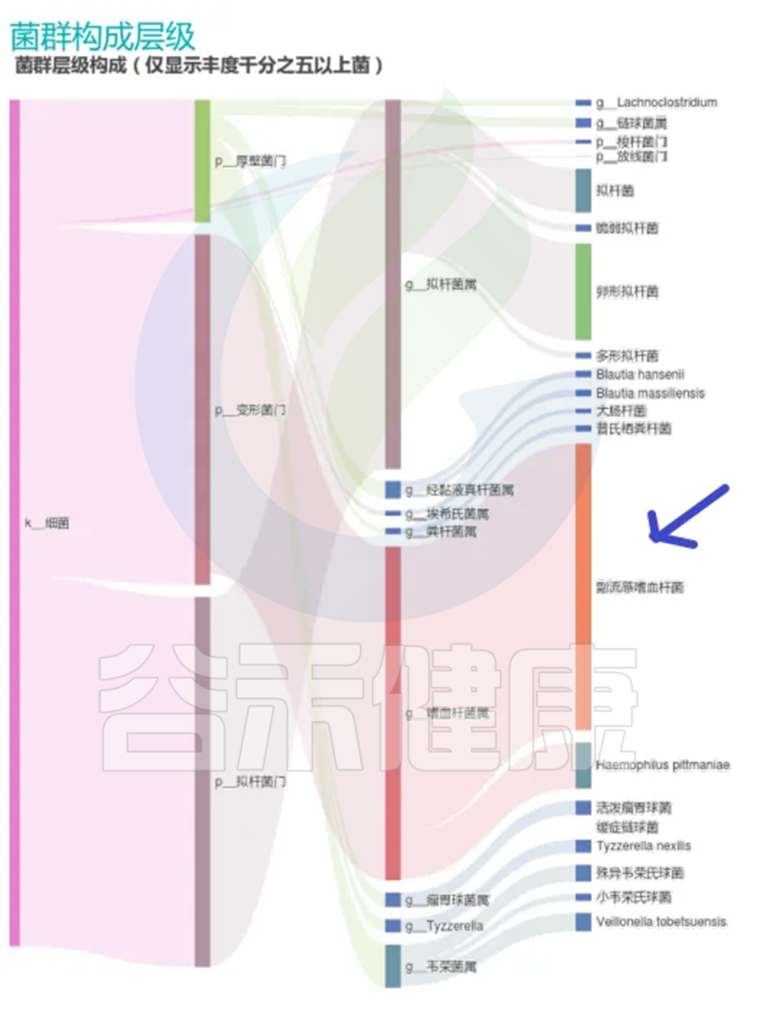
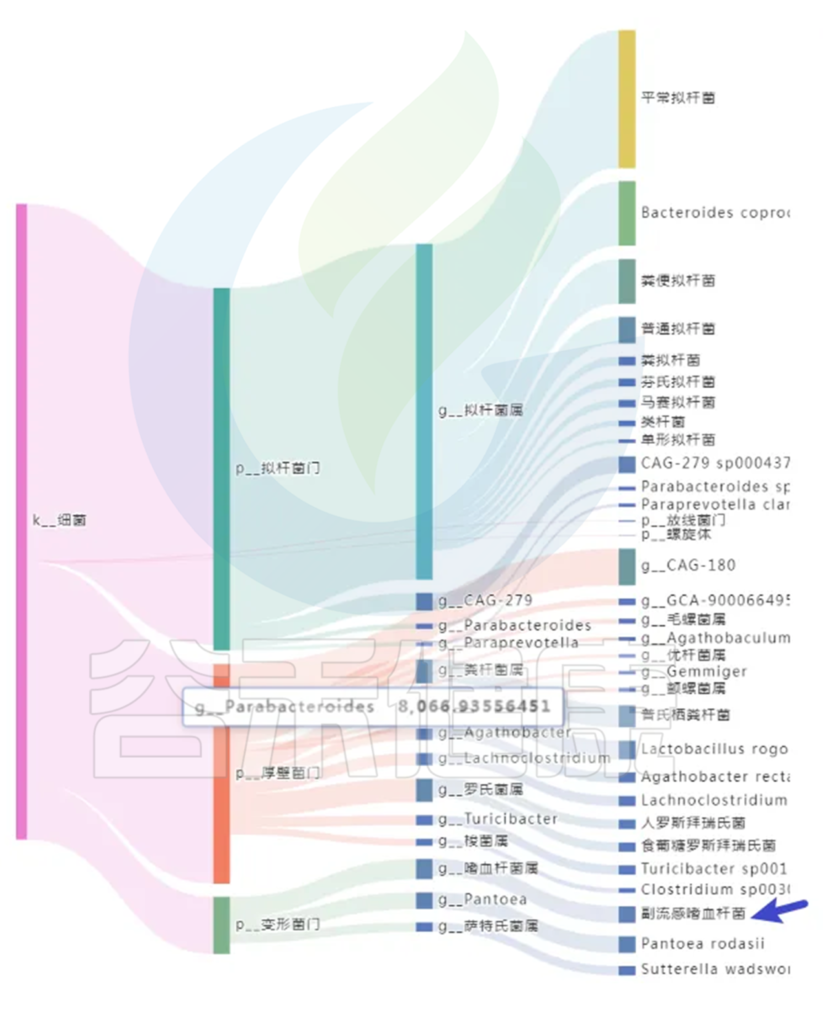
<来源:谷禾健康肠道菌群检测数据库>
(参考自:梅斯医学)
✓
氨苄西林曾经是治疗流感嗜血杆菌感染的经典药物之一,但是据2017年CHINET细菌耐药监测结果显示,流感嗜血杆菌对氨苄西林的耐药率均近45%或以上。其原因与不合理使用了3、4代头孢菌素、碳青霉烯类等广谱抗菌药物存在较大的关联。
贵州省一项研究,共收集住院患者临床分离株196株。所有研究的分离株均为不可分型流感嗜血杆菌(NTHi)。
氨苄西林、头孢克洛、阿奇霉素的耐药率分别为71.4%(140/196)、36.7%(72/196)、34.2%(67/196)。40.3% (79/196) 的菌株为 β-内酰胺酶阳性、氨苄西林耐药 (BLPAR)。β-内酰胺酶产生和 PBP3 突变的普遍存在可能导致局部氨苄青霉素耐药率较高。
临床上经验性选择氨苄西林用于临床治疗存在一定的风险;监测结果还显示,流感嗜血杆菌对除甲氧苄啶-磺胺甲唑外的其他抗菌药物敏感率均近70%或以上。因此,在未获得准确的药敏试验结果前,要根据患者病情及本地区的流调数据进行选药。比如可以考虑大环内酯类抗生素(阿奇霉素)和第1、2代菌素类抗生素为经验性治疗药物。
我们查阅其他研究资料显示,对于感染住院患者,联合使用氨苄西林和氯霉素或单一头孢菌素如头孢呋辛、头孢噻肟或头孢曲松可能是有效治疗嗜血杆菌的方法。
此外,针对儿童的b型流感嗜血杆菌(Hib),疫苗接种是预防2月龄-5周岁儿童由b型流感嗜血杆菌引起的侵袭性感染(包括脑膜炎、肺炎、败血症、蜂窝组织炎、关节炎、会厌炎等)主要的措施。
注意:以上仅供交流,不做医学建议。
主要参考文献
Chatziparasidis G, Kantar A, Grimwood K. Pathogenesis of nontypeable Haemophilus influenzae infections in chronic suppurative lung disease. Pediatr Pulmonol. 2023 Jul;58(7):1849-1860. doi: 10.1002/ppul.26446. Epub 2023 May 3. PMID: 37133207.
Shaikh N, Hoberman A, Paradise JL, Rockette HE, Kurs-Lasky M, Martin JM. Association Between Nasopharyngeal Colonization and Clinical Outcome in Children With Acute Otitis Media. Pediatr Infect Dis J. 2023 Aug 1;42(8):e274-e277. doi: 10.1097/INF.0000000000003956. Epub 2023 Apr 26. PMID: 37171965.
Zhou Y, Wang Y, Cheng J, Zhao X, Liang Y, Wu J. Molecular epidemiology and antimicrobial resistance of Haemophilus influenzae in Guiyang, Guizhou, China. Front Public Health. 2022 Dec 1;10:947051. doi: 10.3389/fpubh.2022.947051. PMID: 36530676; PMCID: PMC9751421.
Zhu C, Zheng M, Ali U, Xia Q, Wang Z, Chenlong, Yao L, Chen Y, Yan J, Wang K, Chen J, Zhang X. Association Between Abundance of Haemophilus in the Gut Microbiota and Negative Symptoms of Schizophrenia. Front Psychiatry. 2021 Jul 30;12:685910. doi: 10.3389/fpsyt.2021.685910. PMID: 34393849; PMCID: PMC8362742.
Du X, Liu J, Xue Y, Kong X, Lv C, Li Z, Huang Y, Wang B. Alteration of gut microbial profile in patients with diabetic nephropathy. Endocrine. 2021 Jul;73(1):71-84. doi: 10.1007/s12020-021-02721-1. Epub 2021 Apr 27. PMID: 33905112.
Kasai C, Sugimoto K, Moritani I, Tanaka J, Oya Y, Inoue H, Tameda M, Shiraki K, Ito M, Takei Y, Takase K. Comparison of human gut microbiota in control subjects and patients with colorectal carcinoma in adenoma: Terminal restriction fragment length polymorphism and next-generation sequencing analyses. Oncol Rep. 2016 Jan;35(1):325-33. doi: 10.3892/or.2015.4398. Epub 2015 Nov 4. PMID: 26549775.
Nocera A, Nasrallah HA. The Association of the Gut Microbiota with Clinical Features in Schizophrenia. Behav Sci (Basel). 2022 Mar 25;12(4):89. doi: 10.3390/bs12040089. PMID: 35447661; PMCID: PMC9025473.
谷禾健康
同样暴露在含有致病微生物的环境中,有的人很少生病,有的人却很容易感染并生病。这是因为每个人的机体对潜在的病原体产生的免疫应答不同。
★ 固有免疫和适应性免疫保护着人体
人体的免疫应答分为固有免疫(先天免疫)和适应性免疫(后天免疫)。
固有免疫是生物在长期进化中逐渐形成的,是机体抵御病原体入侵的第一道防线。
参与固有免疫的细胞包括白细胞、单核/巨噬细胞、树突状细胞、粒细胞、自然杀伤细胞等。
适应性免疫,是机体针对外来刺激而产生的免疫反应,包括细胞免疫和体液免疫。其中细胞免疫是指T细胞受到抗原刺激后,增殖、分化、转化为效应T细胞,当相同抗原再次进入机体的细胞中时,效应T细胞对抗原的直接杀伤作用及致敏T细胞所释放的细胞因子的协同杀伤作用,而体液免疫是指B细胞产生抗体来达到保护目的的免疫机制。
无论是先天免疫还是后天免疫,免疫细胞之间的平衡和协调互作对于每个个体应对感染和疾病的发展息息相关。
肠道微生物,病原体感染以及饮食成分在调节和诱导这些免疫细胞形成和平衡中发挥不可替代的作用。
肠道菌群主要分布在胃肠道,尤其结直肠部位,所以从上面说肠道对于免疫的作用不言而喻。而且肠道还有一个重要功能是感知外部信号并做出反应。
肠道具有大量的淋巴组织和大量分散的先天性和适应性免疫细胞。肠道及相关微生物的免疫过程也越来越多地涉及控制身体其他部位的疾病发展。这反应在肠道免疫相关的文章数量的激增。
注意,由于胃肠道沿其长度存在不同的功能和结构差异,每个部分都有不同的生理作用和免疫学成分。
小肠的免疫系统侧重于保护表面上皮细胞消化和吸收食物的能力,使其免受感染。
这些机制包括: 产生IL-17和IL-22的T细胞和先天性淋巴样细胞;抗菌肽的生产;和具有先天和细胞溶解效应功能的上皮内T细胞。调节性T细胞有助于防止对饮食抗原的超敏反应。
大肠(结肠)不参与消化,但它是大量对健康至关重要的共生微生物的储存库。结肠免疫系统将这些微生物识别为潜在的危害,并在不驱逐它们的情况下与它们“保持一定距离”。
这涉及上皮细胞更新,粘液层的产生、IgA 等抗体的产生以及大量调节性T和B细胞的存在。
免疫功能的这些不同方面由不同种类的次级淋巴器官或免疫细胞提供服务,并受到局部环境因素的影响,例如饮食成分和细菌代谢物。这其中肠上皮细胞、肠道黏膜、肠道微生物互作对于肠道免疫和全身健康的重要贡献是普遍得出的共识。
因此,进一步研究肠道微生物群与肠道免疫系统之间的相互作用机制,对于我们深入了解这种复杂关系的本质具有重要意义,并为免疫相关疾病的预防和治疗提供新的思路。
本文主要分享肠道的结构和不同部分的功能,肠道免疫涉及的主要免疫细胞及其来源和能量代谢,还有肠道微生物群在其中起到的重要作用,及其失调与免疫相关疾病的关联。
免疫中涉及的一些专业术语
Th1/Th2轴:Th1和Th2细胞是两种不同类型的辅助T细胞。Th1细胞主要分泌干扰素γ和IL-2等细胞因子,参与细胞免疫应答;而Th2细胞则主要分泌IL-4、IL-5、IL-10和IL-13等细胞因子,参与体液免疫应答。Th1/Th2轴的平衡与否,与多种疾病的发生和发展密切相关。Th2状态下容易诱发过敏性疾病;Th1状态下容易诱发自身免疫性疾病。
Th17/Treg轴:Th17和Treg细胞分别是促炎性和抗炎性的T细胞。Th17细胞主要分泌IL-17等细胞因子,参与炎症反应;而Treg细胞则主要分泌IL-10、TGF-β等细胞因子,参与免疫耐受。Th17/Treg轴的失衡,也与多种疾病的发生和发展密切相关。Th1/Th2和Th17/Treg细胞失衡与肿瘤大小、淋巴结转移和血管浸润有关。
NK/NKT轴和自然杀伤细胞:NK细胞和NKT细胞都是自然杀伤细胞,可以直接杀伤病原体和肿瘤细胞。NK细胞主要通过分泌细胞毒素杀伤靶细胞;而NKT细胞则通过分泌细胞因子和直接杀伤靶细胞等多种方式发挥作用。NK/NKT轴的失衡,也与多种肿瘤,炎症,感染等有关。
DC/T轴:DC细胞是树突状细胞,是免疫系统中最重要的抗原递呈细胞。T细胞则是免疫系统中最重要的效应细胞。DC/T轴指的是DC细胞和T细胞之间的相互作用和调节关系。DC细胞通过递呈抗原,激活和引导T细胞的免疫应答,从而对抗病原体和肿瘤细胞。
目录
第一部分:肠的解剖学和生理学
①肠道的结构
②小肠具体的结构和功能
③大肠具体的结构和功能
第二部分:肠道内丰富的免疫细胞
①肠上皮细胞——肠道免疫系统的协调员
②巨噬细胞——肠道免疫系统的关键哨兵
③先天淋巴细胞——肠道免疫的重要力量
④肠神经细胞——中心调节者
⑤肠潘氏细胞——肠道守护者,控制易感性
⑥肠杯状细胞——肠道中的前哨和护送者
⑦T细胞——免疫强弱,免疫监视
⑧B细胞——免疫耐受,产生抗体
第三部分:肠道微生物群与免疫反应
①肠是共生微生物的主要来源
②肠道微生物群——免疫系统的调控者
③肠道菌群失调与免疫相关疾病
▼
肠道是从口腔延伸至肛门的长管状结构,其主要功能是消化食物、吸收营养和水分以及排泄废物。肠道也是许多病原体的主要进入部位,并且是庞大而多样化的微生物群落家园。
小肠和大肠形成一个连续的管子。小肠始于幽门,止于回盲瓣,回盲瓣是大肠的入口。小肠分为三个主要部分,十二指肠最靠近胃,然后是空肠,最后是回肠。大肠始于盲肠,然后是升(近端)结肠、横结肠、降(远端)结肠和直肠,止于肛门。
小肠和大肠的大小明显不同,人类的小肠由多个长度为6-7米的线圈组成;结肠直径更宽且更短(大约1.5m)。
肠粘膜及其免疫装置的解剖结构
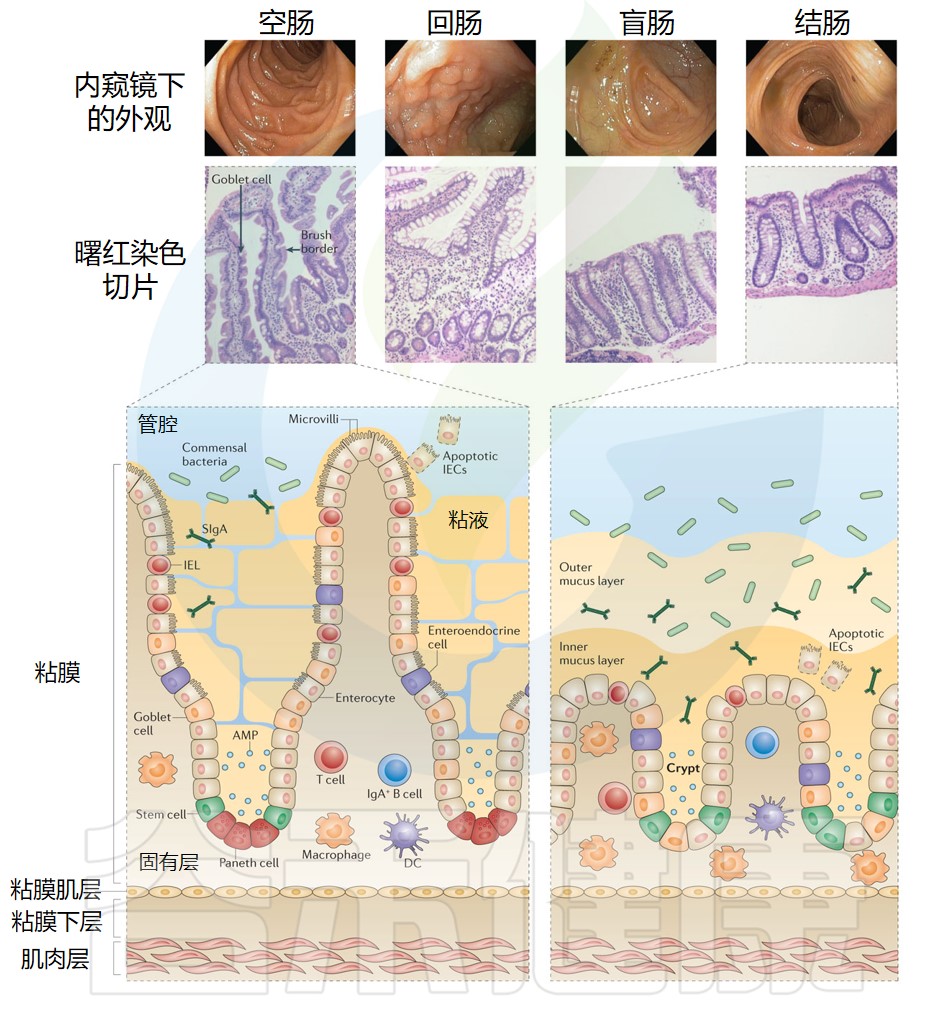
Mowat AM,et al.Nat Rev Immunol.2014
在解剖学上,肠组织通常可分为肠系膜、浆膜、肌层、粘膜下层、固有层、上皮和管腔。
▸ 固有层中有众多免疫细胞
在固有层中发现了许多适应性和先天免疫细胞,包括巨噬细胞、肥大细胞、嗜酸性粒细胞、树突状细胞、B细胞和T细胞。
此外,血管、神经、神经胶质突起延伸至整个固有层和粘膜,与这些结构相关的多种细胞和分子对于肠道功能很重要。
▸ 浆膜包裹并润滑胃肠道
肠系膜与浆膜相邻,浆膜是间皮的最外层,包裹并润滑胃肠道,使蠕动收缩不受抑制。
▸ 肌层是肠组织的最外层
肠组织的最外层统称为肌层。该区域由外部纵向肌肉层和内部圆形肌肉层组成,这些层彼此交织,拉伸。
下面详细描述的结构和细胞区室化将为胃肠道中可能发生的大量神经免疫相互作用提供解剖学参考。与这些结构相关的细胞和分子对于肠道功能非常重要。
▼
✦小肠的主要功能是消化和吸收
小肠的主要功能是营养物质的消化和吸收,起始于胃的幽门,终止于回盲瓣。在人类中,它的长度达到6-7m,按降序由十二指肠、空肠和回肠组成。
小肠的特征是绒毛的指状突起,它延伸到内腔并增加消化活性上皮的表面积,绒毛的丰富表面积,使得它们能够更充分地接触到食物中的营养分子,并将其迅速吸收进入血液循环。
相比之下,表面平坦的盲肠和结肠没有绒毛。
•十二指肠
十二指肠位于胃和空肠之间,是食物从胃进入小肠的第一段。主要功能包括:
•接受来自胃的部分消化食物,继续消化和吸收。
•分泌胰液和胆汁,帮助消化食物中的脂肪和蛋白质。
•吸收营养物质(如碳水化合物、脂肪和部分氨基酸)。
除了来自胃的管腔内容物外,十二指肠每天还通过胆总管接收0.6-1.0L胆汁。胆汁的主要成分包括胆固醇、卵磷脂、胆红素和胆汁盐,后者对食物中的脂肪颗粒具有关键的洗涤作用,并有助于吸附脂肪酸、单甘油酯、胆固醇和其他脂质。
•空肠
空肠位于十二指肠之后,是小肠的一段。主要功能包括:
•进一步消化和吸收食物中未消化的碳水化合物、脂肪和蛋白质。
•吸收维生素、矿物质和水分。
•回肠
回肠是消化系统的一部分,位于空肠之后,是小肠中最后一个部分。它具有以下主要功能:
•营养物质吸收:回肠是主要的吸收器官之一,通过其表面上的细胞吸收并转运剩余的营养物质。
•液体平衡维持:回肠通过吸收水分和电解质维持体内的液体平衡。
•消化废物储存:回肠也起到储存和排泄消化废物的作用。在回肠中,食物残渣逐渐形成粪便,然后被推进结肠,进行最终的排泄。
单糖、二糖和三糖、氨基酸、膳食脂肪、脂溶性维生素(维生素A、D、E和K)或水溶性维生素(维生素B和C)主要存在于十二指肠和空肠中,而回肠是吸收胆汁盐和维生素B12的主要场所。
小肠上段的损伤(如乳糜泻中所见)会导致严重的吸收不良、蛋白质渗漏和营养不良。
✦小肠中有众多的免疫细胞
大多数免疫过程发生在粘膜中,粘膜包括上皮细胞、下面的固有层和粘膜肌层。固有层由松散堆积的结缔组织组成,形成绒毛的支架,并包含粘膜的血液供应、淋巴引流和神经供应。它还包含许多先天性和适应性免疫系统的细胞。
淋巴细胞也存在于上皮细胞中,尽管仅被一层薄的基底膜隔开,固有层和上皮细胞形成非常不同的免疫隔室。它们的组成和功能在整个肠道中也有很大差异。
✦与小肠相关的疾病
乳糜泻
一种自身免疫性疾病,食用麸质会引发白细胞攻击小肠内壁。
克罗恩病
一种导致消化道疼痛性炎症的长期疾病。
肠易激综合症
一种与大脑和肠道之间的信号传导问题相关的常见病症,可导致胃肠道症状。
小肠细菌过度生长
小肠内菌群数量或菌群种类改变,达到一定程度并引起临床表现者。临床表现包括腹痛、腹胀、腹泻或是吸收不良。
消化性溃疡
是位于胃或十二指肠的开放性溃疡。
小肠憩室
肠腔内压力的影响或胚胎时期发育不良使小肠壁薄弱处向外膨出形成的盲囊。
▼
大肠是人体消化系统中的重要组成部分,主要分为盲肠、结肠和直肠。
•盲肠
盲肠是一个封闭的囊,是大肠的第一部分,长约6~8cm,位于腹腔右下部,与回肠交接处有回盲瓣,有孔与阑尾相连,向上续接升结肠。
它充当共生细菌的大储藏室,这些共生细菌参与无法被小肠酶处理的复杂碳水化合物,蛋白质等的发酵消化。
盲肠没有绒毛,粘膜主要由隐窝组成,只有短的平面上皮区域。杯状细胞数量众多,遍布整个隐窝。潘氏细胞很少见。盲肠是整个结肠中最宽的部分,大约20%的结肠癌发生在盲肠。
•结肠
结肠构成了大肠的大部分,包括以下四个部分:
升结肠(从盲肠向上,位于腹部右下角)
横结肠(穿过腹部)
降结肠(向下到最后的结肠部分)
乙状结肠(结肠的最后部分)
升结肠
升结肠长约10厘米,起始于盲肠的肝曲。它沿着腹部右侧延伸,连接横结肠。
横结肠
横结肠长约50厘米,靠近胃、肝脏和胆囊。它连接到降结肠,即所谓的大肠远端或左侧部分的起点。
降结肠
降结肠长约10厘米,结肠在腺曲转折向下固定在腹后壁左侧的一段为降结肠。降结肠下降至左髂嵴处与乙状结肠相连。
乙状结肠
结肠的一部分,在左髂嵴平面与降结肠相连,至第三骶椎平面续于直肠,位于左腹下部及小骨盆内,呈“乙”状或“S”形弯曲,长约40cm。
•直肠
直肠是人体消化系统中的最后一部分,位于盆腔,连接着乙状结肠和肛门。
直肠的壁内有众多的褶皱和粘膜绒毛,这增加了其表面积,有助于吸收水分和电解质。这些特点有助于保持粪便的适当湿润,以便于排泄。
!
注意
人类结肠比小肠短得多(约1.5m)且宽。尽管也被单层柱状上皮细胞覆盖,但其表面完全由散布在表面上皮平坦区域之间的隐窝组成。产生粘液的杯状细胞在小肠上皮细胞中所占比例相对较小(≤10%),但在大肠上皮细胞中所占比例≥25%。
小肠与结肠中的主要细胞与结构
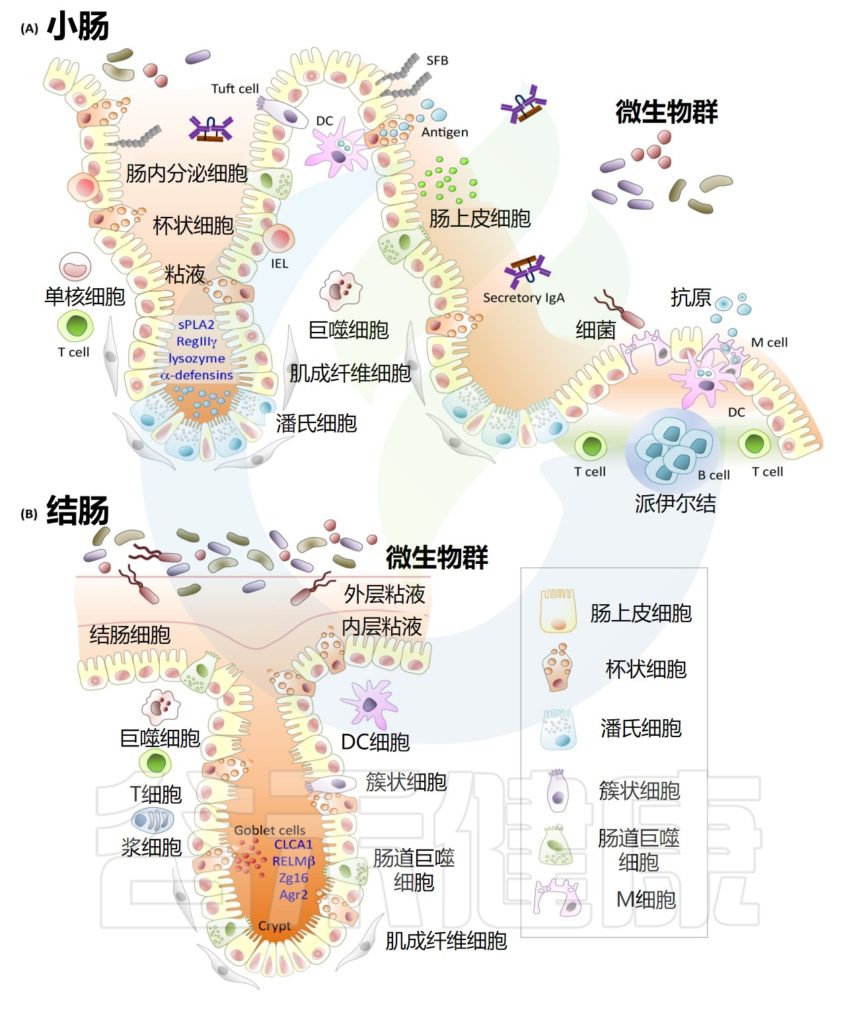
Allaire JM,et al.Trends Immunol.2018
尽管大部分膳食成分在小肠中被吸收,但每天仍有约1.5L的肠液通过回盲瓣。这种液体主要由水和电解质组成,以及半消化或未消化的食物,包括植物多糖和纤维。其中大部分被结肠上半部吸收,它们是常驻微生物群的重要营养来源。
所有结肠癌中约20%至25%起源于远端结肠,而25%至30%的癌症发生在长约15厘米的直肠。
✦与大肠相关的疾病
结肠癌
结肠癌可以始于升结肠。由于它并不总是会引起症状,因此通常直到癌症晚期才被发现。
注:当诊断出结肠癌时,病理学家将确定癌症已到达哪一层(例如最内层或最外层),以帮助确定癌症的分期。
结肠息肉
结肠息肉是发生在肠内壁上的茎状生长物。它们伸入结肠内部。如果它们变大,就会阻止废物通过。
注:息肉是癌症的先兆,在筛查结肠镜检查过程中,发现的任何息肉都将被切除,这样它们就不会继续生长并带来患癌症的风险。
憩室病
憩室病是一种在结肠内壁薄弱部分出现的向外囊状突起。它可能不会引起任何症状,除非被感染,这被称为憩室炎。憩室炎可能导致发烧、恶心、腹痛和腹泻。
注:对于40岁以上的人来说,结肠内壁出现这些小袋很常见,大多数50岁以上的人也有这种情况。
炎症性肠病
克罗恩病、溃疡性结肠炎和不确定性结肠炎是炎症性肠病的一种。炎症性肠病会引起消化道炎症和溃疡。这可能会导致消化道出血、腹痛和腹泻。
此外,炎症性肠病还可引起肠外并发症,包括疲劳、皮肤病、眼部问题和各种形式的关节炎。
▼
肠上皮细胞是位于肠道内壁的特殊细胞类型,它们构成了肠道黏膜的外层。肠上皮细胞具有多种重要功能,包括吸收营养物质、分泌消化酶、阻止有害物质进入身体内部以及促进肠道免疫响应等。
√大肠与小肠间肠上皮细胞差异较大
肠上皮是单层细胞,但是在小肠和大肠之间的结构和细胞组成上有很大差异。
在小肠中,上皮延伸到伸入管腔的结构,称为绒毛,从而增加粘膜表面积和营养吸收。结肠中没有绒毛,导致粘膜表面相对平坦,从而限制了半固体粪便通过大肠造成的潜在损害。
√肠上皮细胞内具有不同分化类型的细胞
上皮本身并不是一层直的细胞,而是由被称为“Lieberkühn 隐窝”的内陷组成。肠干细胞位于这些隐窝的底部,并产生短暂的增殖细胞,这些细胞在向上穿过过渡区时分化和成熟,肠上皮细胞最终脱落到隐窝顶端的管腔(或小肠中的绒毛)。
注:肠隐窝经历肠上皮细胞补充和更新的持续循环,在稳态条件下,估计整个隐窝每4-5天更换一次。
在肠道上皮细胞内发现了各种分化的细胞类型,每一种都执行独特和专门的功能。这些细胞类型包括:肠细胞,肠上皮细胞中负责营养和水分吸收的最主要细胞类型;各种分泌细胞,例如分泌粘蛋白的杯状细胞,分泌激素的肠内分泌细胞,以及释放抗菌因子以促进消化的潘氏细胞,保护小肠隐窝底部附近的干细胞。
还有在抵御蠕虫中起关键作用的化学感受簇细胞,以及对免疫系统摄取和最终呈递管腔抗原不可或缺的M细胞。
√肠上皮细胞在免疫中主要起屏障功能
肠上皮细胞对免疫的主要贡献是屏障功能,可防止肠腔物质与肠道内壁细胞之间的相互作用。除了不渗透性、快速更新和蠕动等因素外,上皮层首先受到杯状细胞产生的粘液的保护,其次受到无数抗菌分子的保护,例如隐菌素、溶菌酶、磷脂酶和趋化因子。
这些天然杀菌或抑菌化合物会损害多种微生物的增殖,从而阻止入侵粘膜之前大部分正在进行的定植。
尽管在大多数情况下先天免疫足以保护肠道,但适应性免疫系统效应淋巴细胞的记忆特性使病原体在再次遇到病原体时能够更特异、更有效地被消除。
因此,消化道是免疫记忆的主要部位,并且确实构成了人体B和T淋巴细胞的主要储存库。已有文献总结了在解剖学上不同的粘膜诱导位点和粘膜效应位点引发适应性免疫。抗原特异性反应的先决条件是通过粘膜上皮取样抗原并递送至下面的MALT或引流粘膜的淋巴结。
上皮细胞的能量代谢
上皮细胞的能量代谢,尤其是结肠中的能量代谢,在很大程度上取决于肠道微生物群。在生命早期,在适应性免疫系统成熟之前,未知的微生物衍生分子通过 STAT3 磷酸化以IL-23和IL-22依赖性方式激活上皮内淋巴细胞和先天淋巴细胞。
在缺乏适应性免疫的情况下,IL-23-IL-22可以控制肠道微生物群,但过度激活的IL-22产生会导致脂质代谢异常,并导致关键脂质转运蛋白的表达减少(例如, CD36、Fabp1/2),以及血清中甘油三酯和游离脂肪酸的减少。
自噬是由能量应激诱导的,以维持结肠细胞的稳态。结肠肠道微生物组产生的丁酸盐确实是结肠细胞的唯一碳源。转化为丁酰辅酶A后,它被动扩散到线粒体中,进行β-氧化,并供给三羧酸循环和氧化磷酸化以产生能量并抑制自噬激活。
Allaire JM,et al.Trends Immunol.2018
肠上皮细胞大量暴露于肠道微生物,并产生粘液和抗菌肽以保持安全距离。丁酸盐还通过下调人类肠三批细胞中的IDO1表达和犬尿氨酸途径来促进肠道稳态。
IDO1在我们前面色氨酸代谢那篇文章中讲到过:
色氨酸通过犬尿氨酸通路的分解代谢,该通路占整体色氨酸降解的约95%,形成主要最终产物NAD+。
注:犬尿氨酸通路是炎症和免疫反应的重要参与者。
首先,色氨酸被转化为N-甲酰基-L-犬尿氨酸
该反应由三种限速酶之一催化:
TDO(色氨酸 2,3-双加氧酶) -肝内
IDO1(吲哚胺 2,3-双加氧酶 1) -肝外
IDO2(吲哚胺 2,3-双加氧酶 2) -肝外
注:这三种酶都是血红蛋白,并使用分子O2作为共底物,这也使它们能够利用活性氧 (ROS) 并调节细胞内的氧化还原平衡。
IDO和TDO酶在不同的组织中表达,暴露于不同的刺激物时被诱导,表明它们在健康和疾病中具有不同的功能。
TDO在基础条件下催化色氨酸分解,而在免疫调节中具有关键作用的IDO-1受到多种刺激物的诱导和调节,例如炎症信号 。
√肠道微生物群影响肠嗜铬细胞产生血清素
在不同的肠上皮细胞类型中,肠嗜铬细胞负责产生血清素,这对免疫细胞有重大影响。结肠中血清素的产生很大程度上受到肠道微生物群,特别是孢子形成细菌代谢物的调节。
其机制尚未完全阐明,但已表明,短链脂肪酸(丁酸和丙酸)和一些次级胆汁酸(例如通过微生物生物转化产生的脱氧胆酸)可以上调TpH1表达(血清素生产中的限速酶)。
即使需要进一步研究,这些数据表明,调节肠道微生物群组成或直接施用微生物代谢物可以从治疗角度控制血清素的产生。
▼
巨噬细胞是一种具有细胞吞噬能力的细胞,源自单核细胞,而单核细胞又来源于骨髓中的前体细胞。
√巨噬细胞在肠道稳态中发挥多种作用
巨噬细胞在维持肠道稳态方面发挥多种作用。与其他组织中的巨噬细胞一样,肠壁中的巨噬细胞也具有强烈的吞噬能力。然而,虽然具有高度杀菌作用,同时还不会导致明显的炎症反应 。
吞噬
肠固有层巨噬细胞具有高度吞噬能力,负责清除凋亡和衰老的上皮细胞。通过表达组织重塑金属蛋白酶和分泌刺激上皮干细胞更新的因子,例如前列腺素E2、肝细胞生长因子和Wnt配体,它们可促进上皮完整性。
转移
巨噬细胞可以将获得的抗原转移至迁移性树突状细胞,以呈递给引流肠系膜淋巴结中的T细胞。通过产生免疫调节细胞因子,例如IL10和TGFβ,它们维持并促进局部调节性T细胞的二次扩增。
以类似的方式,巨噬细胞通过产生IL1β来支持Th17细胞和ILC3,IL1β是通过暴露于微生物群或其衍生物而诱导的。巨噬细胞也存在于肠壁的更深层,包括粘膜下层和外肌层。
促炎或抗炎
巨噬细胞是免疫系统的关键介质,几乎遍布整个胃肠道。巨噬细胞对炎症状态具有重要影响,具有促炎和抗炎功能。它们的极化状态与许多代谢紊乱有关,例如炎症性肠病。
注:炎症M1细胞和抗炎M2细胞的表型和功能之间的平衡受细胞外和细胞内刺激的调节,从而决定疾病的进展情况。
√肠道微生物诱导巨噬细胞
现在有多种证据表明微生物群在影响粘膜巨噬细胞群动态方面发挥着关键作用。
首先,微生物定植后,结肠巨噬细胞区室发生了重大变化,特别是在断奶时,单核细胞通过单核细胞“瀑布”的分化已经建立。
其次,使用广谱抗生素可以减少巨噬细胞的周转,这进一步表明共生微生物群在控制巨噬细胞周转中的作用。
此外,与正常小鼠相比,无菌小鼠肠壁中发现的巨噬细胞较少。粘膜微环境实际上可能会对巨噬细胞进行招募。
如上所述,微生物群可能构成这种分化的一种刺激因素,其他可能性可能包括饮食代谢物或蠕动产生的持续机械应力。
注:有人建议机械应力可以解释心脏中胚胎来源的巨噬细胞的替换并且除了产生低度“炎症”外,它还可以简单地阻止巨噬细胞的长期停留。
巨噬细胞的能量代谢
巨噬细胞在免疫反应过程中处于第一线,但也感知微生物群并对其做出反应以控制微生物群,而不引发有害的炎症反应。
在致病反应期间,活化巨噬细胞的代谢特征根据情况而变化。在促炎性M1巨噬细胞中,三羧酸循环被破坏,导致衣康酸和琥珀酸的积累以及糖酵解的转变。
衣康酸是免疫代谢的主要参与者,具有免疫调节和抗菌作用。它还参与琥珀酸的积累,因为它通过阻断琥珀酸脱氢酶的活性来直接抑制其氧化。
琥珀酸通过氧化产生线粒体活性氧并导致IL-1β产生,从而表现出促炎作用。
Allaire JM,et al.Trends Immunol.2018
相反,M2巨噬细胞具有完整的三羧酸循环,并且主要依赖氧化磷酸化。肠道微生物群是通过短链脂肪酸来调节这些过程。丁酸盐(但不是乙酸盐或丙酸盐)将巨噬细胞代谢重新编程为氧化磷酸化和脂质代谢,从而导致抗炎M2表型。
▼
先天性淋巴细胞家族由自然杀伤细胞、ILC1、ILC2和ILC3组成,它们参与对病毒、细菌、寄生生物和转化细胞的免疫反应。
√先天淋巴细胞对感染和损伤做出免疫反应
有不同类型的先天淋巴细胞,其特征是表达特定的膜标记、转录因子和细胞因子特征。在激活过程中,先天淋巴细胞会深刻改变其能量代谢以适应其新功能。
先天淋巴细胞,通过产生细胞因子和分泌蛋白对感染或损伤作出反应,这些细胞因子和分泌蛋白指导和增强攻击前线的免疫反应。
√肠道微生物群影响先天淋巴细胞功能
肠道微生物群影响先天淋巴细胞功能,抗生素对ILC1、ILC2和ILC3转录组程序的巨大影响证明了这一点。
ILC3是胃肠道中存在的先天淋巴细胞的主要类型。这些细胞表达RORγt,可以产生IL-17和IL-22,是炎症、感染、微生物群组成和代谢的重要调节因子。
ILC3功能例如肠上皮防御的维持,依赖于昼夜节律调节器ARNTL介导的昼夜节律信号。但众所周知,肠道微生物群是昼夜节律的影响因素。
肠道微生物群衍生的丁酸盐也可调节ILC2功能,抑制其不受控制的激活,从而抑制其在肺部炎症和哮喘中的负面作用。
▼
哺乳动物的神经系统分为中枢神经系统和周围神经系统。中枢神经系统包括大脑和脊髓,周围神经系统包括神经节,神经节是神经细胞体的聚集体,头部、颈部和内脏中的神经束从神经细胞体中产生。
注:消化道同时受自主神经和管道壁上的肠神经丛的神经支配。
√交感神经直接从大脑连接到胃肠道
从中枢神经系统到肠神经系统的外部连接由交感神经纤维和副交感神经纤维组成。离开脑后,副交感神经和交感神经可以直接突触到胃肠道。
例如,副交感迷走神经,离开后脑后,沿着食道穿过隔膜,最终突触到胃肠道。交感神经起源于脊柱,并突触到交感内脏神经节,例如腹腔神经节、肠系膜上神经节和肠系膜下神经节。副交感神经和交感神经均可直接突触至肌间神经节、平滑肌和粘膜。
√盆腔神经支配远端结肠和直肠
此外,起源于脊髓并通过骶脊神经离开的盆腔神经支配远端结肠和直肠。盆腔神经传统上被认为是副交感神经。副交感神经对交感神经节的神经支配使胃肠道的神经连接变得更加复杂。
最后,内在的肠神经系统是沿着胃肠道的神经元和神经胶质细胞的扩展网络;这些可以自主发挥作用,但也可以通过肠神经系统与外在交感神经和副交感神经的连接进行调节。因此,中枢神经系统和肠神经系统之间的通信是双向的。
√肠神经元上的受体介导重要的胃肠道功能
肠神经元上的受体介导重要的胃肠道功能。机械感受器对粘膜磨损敏感;张力感受器对拉伸敏感;化学感受器对管腔内的各种化学刺激做出反应,例如pH值、渗透压和营养物质。此外,各种受体负责调节肠道内的液体交换。
2021年美国一项研究发现肠道神经细胞可以通过分泌细胞因子IL6来调控调节性T细胞的水平。
此外,调节性细胞诱导菌很有可能是通过降低肠道神经系统密度和神经细胞IL6水平的双重作用,来诱导调节性T细胞的。
▼
潘氏细胞于19世纪末首次描述为柱状上皮细胞,胞浆内有明显的嗜酸性颗粒。它们在维持肠道稳态和调节小肠及其相关微生物菌群的生理方面发挥着不可或缺的作用。
√潘氏细胞可调节肠道稳态
潘氏细胞产生的致密颗粒含有丰富的抗菌肽和免疫调节蛋白,它们的功能是调节肠道菌群的组成。
肠道中的潘氏细胞在小肠中比较丰富,与吸收性肠细胞不同,潘氏细胞寿命长,在从干细胞分化后向下迁移到隐窝的最底部。
√潘氏细胞具有重要的抗菌作用
它们具有重要的抗菌作用,可产生溶菌酶、防御素等抗菌肽并再生胰岛衍生蛋白IIIγ,以响应白细胞介素22或刺激Toll样受体、核苷酸结合寡聚化结构域2或胆碱能神经。
潘氏细胞是专业的分泌细胞,通常通过将抗菌因子分泌到管腔中来控制肠道细菌,从而在先天免疫系统中发挥作用。潘氏细胞能够感知来自腔内细菌的信号,并通过改变这些因子的产生来做出反应,以保护上皮屏障。潘氏细胞依靠自噬来调节其分泌能力和能力。
潘氏细胞还通过产生促表皮生长因子 配体来帮助维持正常的隐窝干细胞活性,它们对于肠道稳态至关重要。
小结
总之,肠潘氏细胞在肠道内稳态的许多方面都起着关键作用,从调节与上皮密切相关的微生物区系,到维持干细胞生态位的健康,再到帮助调节炎症水平。不仅保护自己不受外来入侵,而且促进肠道的生长和发育。
这些功能在早产儿的未成熟肠道中尤其重要,早产儿的肠道发育中伴随着一个非生物菌群。因此,潘氏细胞破坏与新生儿肠道坏死的发生有机械联系是合理的。对潘氏细胞生物学的进一步了解可能会为理解新生儿肠道坏死的发展提供一条重要的途径。
▼
我们的肠道和呼吸道中布满了杯状细胞,这些细胞因其杯状外观而得名,它们能够分泌具有保护作用的凝胶状粘液,以防止肠道内细菌直接侵入肠壁组织而引发炎症反应。
这些黏液的主要成分为肠壁上杯状细胞分泌的黏蛋白。黏蛋白则肠道黏液层的重要组成部分,凝胶形成黏蛋白和跨膜黏蛋白是目前已知的两种黏蛋白。
√杯状细胞沿着胃肠道逐渐增加
与潘氏细胞相反,产生粘液的杯状细胞的频率沿着胃肠道逐渐增加。杯状细胞在远端结肠中至少占所有上皮细胞的25%,而在小肠上段则为10%或更少。
与此同时,覆盖在粘膜上的粘液层——被称为糖萼,在结肠中最厚,它由两个不同的层组成:一个附着在上皮表面的致密内层和一个外层,类似于小肠中发现的松散层。
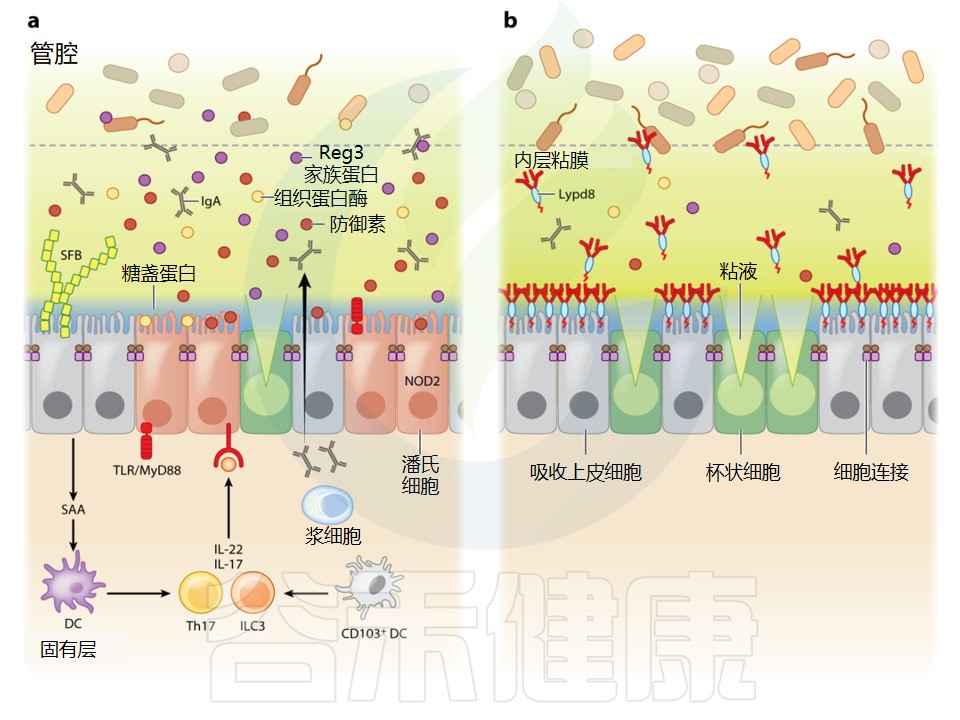
Kayama H,et al.Annu Rev Immunol.2020
√杯状细胞分泌的粘液具有抗菌作用
粘液具有抗菌作用,形成一种高度带电的凝胶,充当物理屏障,并且由对许多细菌有直接毒性的粘蛋白糖蛋白组成。
小肠较松散的糖萼也提供了抗体和抗菌肽可以粘附的基质。粘液合成的缺陷导致共生细菌更多地渗透到结肠的上皮表面,从而增加对结肠炎和结肠癌的易感性。
杯状细胞除了分泌肠道黏液层,形成肠道组织、免疫系统与肠腔微生物群、内容物分离的重要屏障,它其实还有其他意想不到的作用。
√避免一些蛋白引起免疫反应
杯状细胞还可以“护送”抗原到树突细胞。来自华盛顿大学圣路易斯医学院的研究人员在小鼠肠道中鉴定出一种“护送”食物中的抗原或蛋白到树突细胞的细胞,即杯状细胞,这样免疫系统就不会对它们发起免疫。这项发现给科学家提供一种潜在性靶标来治疗炎性肠道疾病、乳糜泻和食物过敏,这些疾病都是由于免疫系统过分活跃而导致的。
√杯状细胞与痛觉感受神经联系保护肠道
2022年,哈佛大学医学院的研究表明,肠道中的痛觉感受神经在正常情况下能够调节保护性粘液的存在,并在炎症状态下刺激肠道中的杯状细胞分泌更多粘液,从而直接发挥保护肠道的作用。
该研究详细描述了这种复杂信号级联的步骤,显示痛觉感受神经与肠道杯状细胞直接可以进行直接的串扰。
√杯状细胞增加粘液分泌阻止外来的细菌侵入
杯状细胞可以还“自杀”换取结肠的安全。
哥德堡大学的研究团队发现一类新的被称作杯状细胞的岗哨细胞能够冲洗走侵入到保护性的黏液屏障的细菌。一旦发现细菌存在于这种隐窝入口处的迹象,它们就启动一种级联反应,最终导致黏液增多,从而将细菌冲洗走。
这一发现可能为解释炎症性肠病(如溃疡性结肠炎)如何发生提供重要理论依据。
此前,该团队已证实黏液层作为第一道防线而存在。然后又证实在隐窝入口处存在一部分杯状细胞作为第二道防线阻止进入黏液层的细菌侵入结肠。
▼
T细胞在哺乳动物免疫反应中的核心作用是无可争议的。
根据所产生的细胞因子和效应细胞的生物功能特征,可将其分为辅助型T细胞1(Th1)、辅助型T细胞2(Th2)、调节性T细胞(Treg)和Th17等。
T细胞祖细胞源自骨髓中的造血干细胞。它们迁移到胸腺,在那里成熟为T细胞。随后,T细胞迁移通过次级淋巴器官,扫描呈递其同源抗原的抗原。
√肠道内含有大量的T细胞
肠道是体内最大的T细胞群之一的所在地。粘膜T细胞对于体内平衡和保护性免疫至关重要,但也在不受控制的(慢性)粘膜炎症中发挥作用,如乳糜泻或炎症性肠病,甚至可能在肿瘤的发展过程中。
胃肠道的上皮屏障是大量上皮内T细胞的家园。上皮内T细胞在功能上适应粘膜环境,是最早遇到微生物和饮食抗原的适应性免疫细胞之一。
√上皮内T细胞的免疫作用
上皮内T细胞在免疫监视中发挥重要作用,同时诱导对非病原性抗原的耐受性,从而保持单层上皮膜的完整性。
炎症性肠病和乳糜泻的特点是(复发性)胃肠道炎症占主导地位。上皮内T细胞在肠上皮屏障中的寿命长且大量存在,表明上皮内T细胞在这些疾病的复发和缓解过程以及持续的低度炎症中发挥作用。
作为组织特异性和潜在致病细胞,上皮内T细胞是治疗(非系统性)慢性组织特异性炎症性疾病(如 炎症性肠病)的理想靶点。
启动的T细胞会长期驻留在肠道中
在炎症或感染环境中,已启动的T细胞会暂时从循环中募集到肠道,但大量T细胞会永久驻留在肠道中,并且独立于淋巴和循环记忆群体而维持。
这些T细胞主要由T细胞受体αβ + CD8αβ +和TCRαβ + CD4组成+组织驻留记忆T细胞,但调节性T细胞 (TCRαβ + CD4 + FOXP3 + )、TCRγδ + T 细胞、自然杀伤T细胞和粘膜相关不变T细胞也建立长期驻留在肠道。
T细胞的能量代谢
T细胞代谢可塑性对于适应永久动态的免疫环境是必要的。肠道微生物群通过活性氧、短链脂肪酸和胆汁酸的产生以及氧化还原信号传导的修饰积极参与这一复杂过程。
效应T细胞和记忆T细胞具有不同的功能和需求,因此表现出不同的代谢。它在效应T细胞中由有氧糖酵解主导,在记忆T细胞中由脂肪酸氧化和氧化磷酸化 主导。记忆T细胞中存在融合的线粒体网络,效应T细胞中存在点状线粒体。
此外,线粒体是T细胞激活的关键组成部分,主要通过活性氧产生。通过CD3刺激T细胞会诱导钙内流,从而刺激丙酮酸脱氢酶和三羧酸循环酶的功能。
√短链脂肪酸可以增强T细胞功能
微生物群衍生的短链脂肪酸通过改变细胞代谢来增强CD8+ T细胞效应功能。肠道微生物群代谢膳食纤维产生的短链脂肪酸会刺激CD8+ T细胞中的氧化磷酸化和线粒体质量及其糖酵解能力。
此外,短链脂肪酸可以扩散到细胞质中并作为脂肪酸氧化的底物,导致乙酰辅酶A的产生,为三羧酸循环和氧化磷酸化提供燃料。
在活化的CD8+ T细胞中,短链脂肪酸,特别是丁酸,促进胆汁酸的摄取和氧化,导致三羧酸循环与糖酵解输入断开,并通过胆汁酸分解代谢和谷氨酰胺利用有利于氧化磷酸化。这种丁酸盐诱导的细胞代谢适应是分化为记忆T细胞所必需的。
短链脂肪酸对CD4+ T细胞也表现出显著影响,特别是在辅助性T(Th)17、Th1的生成方面和调节性T细胞。其机制涉及组蛋白去乙酰化酶的抑制和mTOR通路(细胞生长和代谢的主要调节因子)的调节。
最近发现了戊酸的这种联系,它可以通过为组蛋白乙酰转移酶提供额外的乙酰辅酶A并增强糖酵解和 mTOR 活性来刺激抗炎细胞因子IL-10的产生。
短链脂肪酸激活mTOR的机制
短链脂肪酸激活mTOR的机制有两种。短链脂肪酸通过对能量产生途径的作用,诱导ATP的产生和AMP的消耗,它们分别是AMP激活蛋白激酶 的抑制剂和激活剂。因此,AMPK对 mTOR的抑制剂活性被抑制,从而导致mTOR激活。
第二个潜在机制涉及短链脂肪酸的组蛋白去乙酰化酶抑制活性。短链脂肪酸与 P300/CBP(E1A 结合蛋白 p300/CREB 结合蛋白)结合,促进核糖体蛋白 S6 激酶 beta-1 (S6K1) 的乙酰化,这是mTOR 的下游靶标,从而更强有力地激活途径。
√胆汁酸对T细胞的影响
胆汁酸对T细胞也有重要影响。石胆酸的衍生物3-oxoLCA通过直接与转录因子RORγt相互作用来抑制Th17细胞的分化。
相反,石胆酸的另一种衍生物isoalloLCA促进调节性T细胞的分化。该机制涉及刺激OXPHOS和mROS的产生,从而通过增加Foxp3启动子中组蛋白 (H3K27) 乙酰化的水平来增加FOXP3的表达。
具体来说,在结肠中,胆汁酸通过胆汁酸受体 Breg 调节 RORγ+ Treg 细胞的功能,这些细胞在维持结肠稳态中发挥着重要作用。
▼
B细胞是一种在适应性免疫过程中发挥重要功能的淋巴细胞,B细胞及其所生产的抗体,对于免疫系统的正常运行是十分重要的。
√B细胞通过产生抗体阻止外来病原体
B细胞是可以产生抗体的白细胞。这些抗体或免疫球蛋白可以与有害的外来颗粒(如病毒或致病细菌)结合,阻止它们入侵和感染人体细胞。
每个B细胞携带一个单独的B细胞受体,该受体决定它可以结合哪些病原体,就像每个锁接受不同的钥匙一样。
人类肠道中B细胞谱系的两个主要区室是肠道相关淋巴组织和弥漫性淋巴组织中高度组织化的B细胞的聚集地,它们占据了大肠和小肠的广泛肠固有层 。
肠道相关淋巴组织可以以滤泡簇的形式出现,通常被称为派尔氏淋巴结或孤立的淋巴滤泡,分布在整个胃肠道中,是对从肠腔采样的抗原做出长期反应的免疫诱导位点。
研究人员使用专门设计的计算机程序来处理数百万个基因序列,这些序列会比较来自B细胞的抗体库,这取决于这些微生物是留在肠道里,还是进入血液。在这两种情况下,抗体库都改变了,但改变的方式不同,取决于暴露发生的方式。
√肠壁与血液中的抗体不同
与血液中的抗体(IgM和IgG)相比,肠壁(IgA)中存在不同种类的抗体。通过强大的基因分析,研究人员发现,肠道产生的不同抗体的范围远小于身体中央组织产生的抗体。
这意味着一旦微生物进入人体,免疫系统就有更多的可能去中和和消灭它们,而肠道中的抗体主要结合在任何时候都能看到的细菌分子。
B细胞有时会影响受损肠道的愈合
B细胞有时也会好心办坏事。来自瑞典研究组发表研究结果发现,B细胞的数量在肠道损伤后大幅增加,并且阻碍了损伤组织的愈合。这一研究结果对炎症性肠病的治疗具有重要意义。
√短链脂肪酸有助于B细胞的能量代谢
B细胞分化为浆细胞和产生抗体需要大量能量和涉及细胞代谢的整体变化。肠道微生物群衍生的短链脂肪酸有助于在不同水平上为这些过程的细胞能量引擎提供燃料,并促进抗体的产生。
短链脂肪酸转化为乙酰辅酶A,并整合到线粒体三羧酸循环循环中,从而产生ATP。短链脂肪酸还通过mTOR激活刺激B细胞中的糖酵解。短链脂肪酸衍生的乙酰辅酶A也是胆汁酸(特别是棕榈酸)合成的底物,这对于浆细胞分化并刺激抗体产生至关重要。
注:最近研究表明,异丁酸或异戊酸等分支短链脂肪酸也可以调节B细胞功能。受操纵的小鼠中缺乏分支短链脂肪酸产生导致小肠中 IgA+ 浆细胞的频率增加,以及与先天免疫细胞(如中性粒细胞、巨噬细胞和树突状细胞)表面结合的IgA水平增加。
此外,B细胞通过产生免疫球蛋白和产生IL-10的调节性B细胞的作用,在对肠道微生物群的耐受性中发挥关键作用。
肠道微生物群与免疫系统之间存在着紧密的相互作用,这种作用对于维持免疫系统的正常功能和免疫平衡至关重要。
肠道微生物群维持上皮屏障的完整性并塑造粘膜免疫系统,通过微生物代谢物、成分和对宿主细胞的附着来平衡宿主免疫防御和口服耐受性。
为了避免异常的免疫反应,上皮细胞通过构建化学和物理屏障将肠道微生物群与免疫细胞隔离开来,从而建立宿主共生互惠关系。
这是为什么人们努力表征肠道微生物群在健康和疾病中的功能。许多研究报告和我们检测实践报告了众多慢病的临床前和临床模型中微生物群的改变。尽管这些改变可能部分是由疾病状态驱动的,但最近的研究强调,改变的微生物群也可以在驱动疾病本身方面发挥核心作用。
▼
我们肠道内的良性微生物数量与我们体内的细胞数量大致相同。这些细菌大多停留在肠道内,而不是渗透到人体组织中。
不幸的是,有些渗透是不可避免的,因为肠道只有一层细胞,把我们吸收食物所需的血管和管子内部分开。
✦细菌的数量沿着胃肠道逐渐增加
细菌的数量通常会沿着胃肠道增加,从胃的高酸性环境中的每毫升100–1,000个到小肠上段的每毫升约105个,在结肠中高达每毫升1010-1011个。
然而,回肠末端可能含有比结肠中更多的细菌。这些细菌使用复杂的多糖和粘液的其他成分以及未消化的纤维作为能量来源,产生生物素、短链脂肪酸 和维生素等必需的代谢物。
Mowat AM,et al.Nat Rev Immunol.2014
由于结肠大多数物种是专性厌氧菌,难以在体外培养,还有部分种群的组成和分布尚未最终确定。然而,分子技术揭示了几个主要门类中大约200-2000 个物种;它们的相对数量因人而异,但厚壁菌门、拟杆菌门、变形菌门和放线菌门最为普遍。
也包括少部分古细菌,真菌和病毒。好氧菌在小肠上段普遍存在,而厌氧菌在结肠中占主导地位。
✦不同的疾病会导致肠道菌群分布改变
构成肠道微生物群的物种分布的改变(生态失调)与许多不同的疾病有关,从精神疾病到代谢性疾病、过敏和自身免疫病。其中盲肠中细菌密度相较于小肠较高,该部位肿瘤发生率较高的原因。
肠道不同区域接触各种饮食成分可影响免疫功能的程度也存在显著差异。例如,维生素A只存在于饮食中,也可以通过胆汁输送到小肠,类黄酮和芳基烃受体的其他配体在小肠中的含量也较高。
▼
肠道微生物通过多种机制与肠外器官中的免疫细胞进行交流。
微生物能够通过直接与免疫细胞作用,如粘附到细胞表面或被吞噬细胞摄取,来调节免疫细胞的功能和活性。
其次,微生物通过与肠道黏膜上皮细胞和巨噬细胞表面的受体结合,激活免疫细胞,使它们产生免疫应答和促炎信号,并进一步调节免疫细胞的活性。
微生物还通过产生代谢产物,如短链脂肪酸和其他微生物分子,与免疫细胞进行信号传导。这些代谢产物可以直接调节免疫细胞的活性和功能,影响它们的增殖、分化和效应物质的产生。
✦肠道微生物群影响免疫系统的发育
首先,肠道微生物群对于免疫系统的发育起着重要的影响。在婴幼儿期,肠道微生物群通过与免疫系统的交互作用,促进免疫系统的成熟和功能发展。特别是在胎儿期和早期生活中,良好的肠道微生物组成对于建立免疫耐受、预防过敏反应和自身免疫性疾病的发生非常重要。
微生物群刺激对于产生免疫球蛋白 (IgA) 的B细胞的发育和成熟、通过促进IgA类别转换的记忆浆细胞以及生发中心的发育至关重要。
此外,通过介导浆细胞样树突状细胞产生I型干扰素,微生物群增强了抗原特异性T细胞反应。
✦肠道微生物可以直接抵抗病原体
肠道微生物群赋予宿主免疫保护的最直接例子是通过对粘膜病原体(如艰难梭菌)的定植抵抗。
多样化的肠道微生物群通过各种机制帮助宿主抵抗病原体定植,包括微生物间对营养的竞争、代谢竞争和直接拮抗以及粘膜屏障完整性的强化和局部粘膜免疫防御。
通过这种方式,肠道共生微生物保护宿主免受粘膜感染,以及由于肠道病原体过度生长和转移到循环系统和远端器官中而可能发生的继发性全身感染。
此外,之前也有报道称,病原体入侵是人类代谢失调的一个特征。这些研究进一步说明了在粘膜表面需要良好的宿主/微生物群相互作用。
// 双歧杆菌对免疫细胞的影响
本文以双歧杆菌(Bifidobacterium)为例:在小鼠肥胖模型中,给予假链状双歧杆菌通过恢复调节性T细胞和B淋巴细胞的平衡状态来减少全身炎症。
类似的免疫稳态特性已归因于双歧杆菌菌株,其通过体外刺激树突状细胞,诱导Th17谱并增强调节性T细胞从幼稚淋巴细胞的分化。
双歧双歧杆菌DSM 20082裂解物刺激外周T细胞可增加CD8+ T细胞的细胞毒活性,而对CD4+ T细胞活性没有任何影响。
✦肠道微生物通过其代谢物影响免疫细胞
肠道微生物产生的代谢产物能够刺激肠道上皮细胞和免疫细胞,如树突状细胞和巨噬细胞,促进免疫细胞的活化和增殖。
肠道微生物通过膳食分子的修饰(脂肪酸、色氨酸代谢物,氧化三甲胺)、宿主胆汁酸(次级胆汁酸)以及从头细菌代谢物(多胺、维生素、支链氨基酸),可以促进全身免疫反应的调节。
肠道微生物群的免疫调节
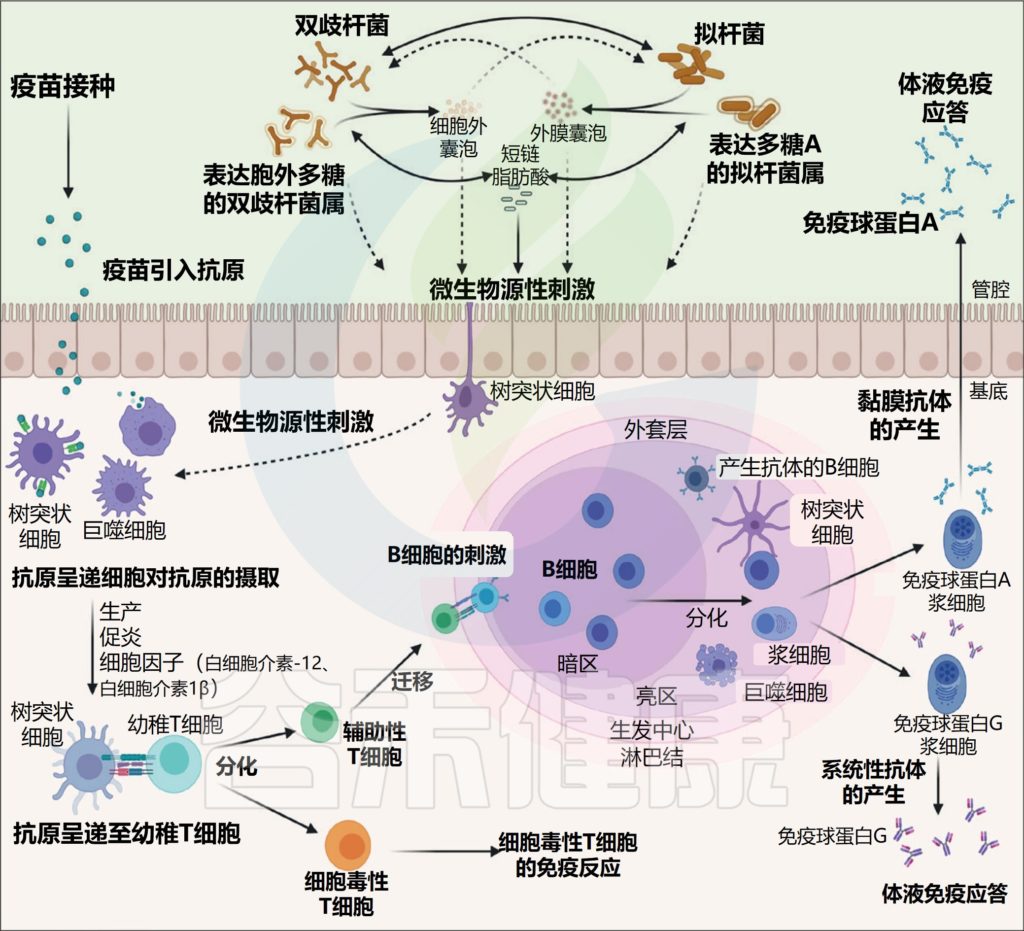
Jordan A,et al.Lancet Microbe.2022
// 乙酸盐可以激活免疫细胞
乙酸盐可以激活B细胞、T细胞亚群、中性粒细胞、巨噬细胞、树突细胞和肠上皮细胞上表达的G蛋白偶联受体43(GPR43),导致固有层调节性T细胞增殖,和调节自身抗体的产生和边缘区B细胞。
// 丙酸盐影响巨噬细胞浓度
丙酸盐诱导小鼠调节性T细胞的分化和增殖以及白细胞介素10的表达。丙酸盐激活GPR15和GPR43还通过降低组蛋白去乙酰化酶6和9的表达以及抑制 NF-κB信号传导来增加结肠调节性T细胞的浓度。
丙酸盐激活GPR41会改变骨髓造血功能,导致巨噬细胞和树突状细胞前体浓度升高,并扭曲Th2分化。
// 细菌胞外多糖影响免疫反应
脆弱拟杆菌的表面多糖A可激活巨噬细胞上的Toll样受体2,并诱导调节性T细胞的扩增和抗炎白细胞介素10的产生,从而在病毒感染期间促进强烈的抗炎反应。
来自长双歧杆菌BCRC14634的胞外多糖被证明可增强巨噬细胞产生白细胞介素,与同基因的胞外多糖阴性突变体相比,来自B.longum亚种longum 35624的胞外多糖显示通过抑制促炎细胞因子产生来抑制促炎性Th17细胞的扩增。
✦肠道微生物调节神经免疫信号
肠道和肠外器官之间的信号传导也可以通过肠道微生物调节的神经免疫机制来完成。肠道细菌可以合成γ-氨基丁酸等神经递质,并调节各种影响免疫细胞功能的神经递质。
此外,肠道细菌刺激肠内分泌细胞释放神经内分泌肽激素,这些细胞可以进入循环系统并作用于全身。
▼
肠道微生物群通过多种途径调节免疫系统的功能,包括维持免疫平衡、促进免疫耐受、保护黏膜屏障等。
➤ 自身免疫性疾病
肠道微生物群对免疫耐受的维持至关重要
自身免疫性疾病分为肠道相关疾病、非肠道相关疾病和免疫特异性疾病。
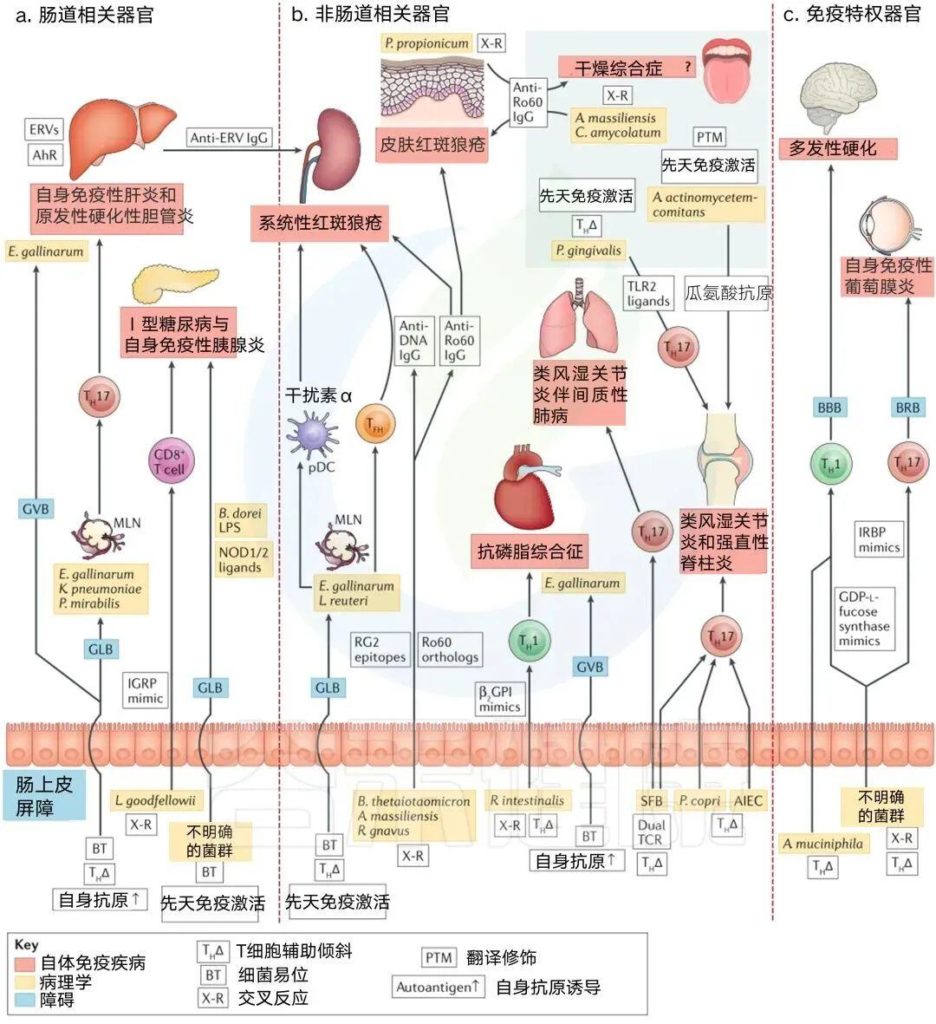
a | 肠相关器官。鸡肠球菌(Enterococcus gallinarum)向肝脏的转移促进了自身免疫性肝炎,并与肺炎克雷伯菌和奇异变形杆菌一起,通过包括TH17分化在内的多种机制导致原发性硬化性胆管炎。鸡肠球菌诱导的肝芳香烃受体(AhR)和内源性逆转录病毒(ERV)也参与了小鼠狼疮性肾炎(一种非肠道相关器官)的系统性抗ERV反应。
Leptotrichia goodfellowii 能与小鼠Ⅰ型糖尿病自身抗原胰岛特异性葡萄糖-6-磷酸酶催化亚单位相关蛋白(IGRP)发生交叉反应。在小鼠Ⅰ型糖尿病中,未定义的微生物群易位到胰腺淋巴结并激活NOD2,而来自微生物群的NOD1配体的易位驱动自身免疫性胰腺炎。
b | 非肠道相关器官。鸡肠球菌和乳杆菌在系统性红斑狼疮模型中向肠系膜淋巴结(MLNs)、肝脏和脾脏转移,从而激活先天性(浆细胞样树突状细胞(pDC))途径。
鸡肠球菌还刺激狼疮患者滤泡辅助因子(TFH)和自身抗体的产生。Ro60直系表达细菌在狼疮患者中引起Ro60自身免疫反应;肠道中的类泰奥托米克龙杆菌、口腔或肠道中的大量放线菌和阿米巴棒状杆菌,除了狼疮外,还可能导致Sjógren综合征,丙酸假丙酸杆菌与亚急性皮肤狼疮皮损通过交叉反应。
Ruminococcus gnavus在狼疮性肾炎时扩张,可与狼疮双链DNA发生交叉反应。抗磷脂综合征是一种凝血性自身免疫性疾病,鸡肠球菌传染性支气管炎也加重了这种疾病,它诱导肝脏中的自身抗原β2GPI。
口腔牙龈卟啉单胞菌、普雷沃氏菌和小肠节段丝状菌(SFB)的TH17偏斜均与类风湿关节炎的发病有关。同样,粘附性侵袭性大肠杆菌(AIEC)可导致克罗恩病相关性脊柱炎。在关节炎模型(未显示)中,SFB还使CD4+T细胞向TFH细胞倾斜,并通过携带双TCRs的TH17细胞在同一模型中引发间质性肺病。口腔中的放线菌群通过一种也能激活中性粒细胞的毒素,瓜氨酸化类风湿关节炎中的系统自身抗原。
c | 免疫特权器官。Akkermansia muciniphila在多发性硬化症中增强TH1反应,并且未定义的微生物群与GDP-l-岩藻糖合酶(一种神经炎症疾病的自身抗原)交叉反应。未定义的小鼠肠道微生物群与TH17细胞交叉反应,识别自身免疫性葡萄膜炎的自身抗原——视黄酸结合蛋白(IRBP)。
扩展阅读:
➤ 过敏性疾病
肠道微生物群可以影响免疫系统的平衡
肠道微生物群的失调可能导致免疫系统的异常激活,从而增加过敏性疾病的风险。肠道微生物群可以通过多种途径影响免疫系统,包括调节免疫应答的平衡、维持免疫耐受等。
以过敏性鼻炎为例,典型的过敏性鼻炎肠道菌群促进细胞因子的不利变化,从而促进过敏性鼻炎发展中涉及的 Th1/Th2 失衡。
过敏性鼻炎患者的肠道菌群失调及其对 Th1/Th2 平衡的影响
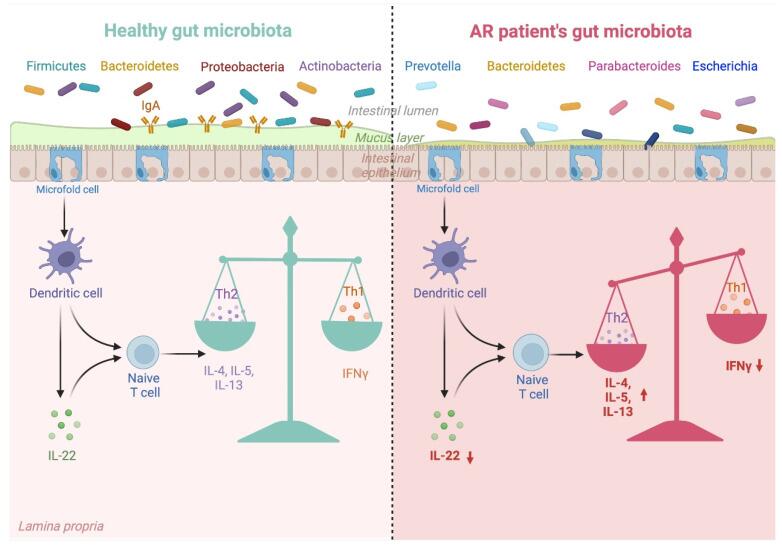
Kaczynska A, et al., Nutrients. 2022
肠道菌群的每一次紊乱都会对免疫系统产生负面影响,并破坏先天性和适应性反应。上图解释了改变的肠道微生物群组成与其对过敏性鼻炎患者免疫系统的影响之间的联系。
肠道微生物群失调影响过敏性鼻炎
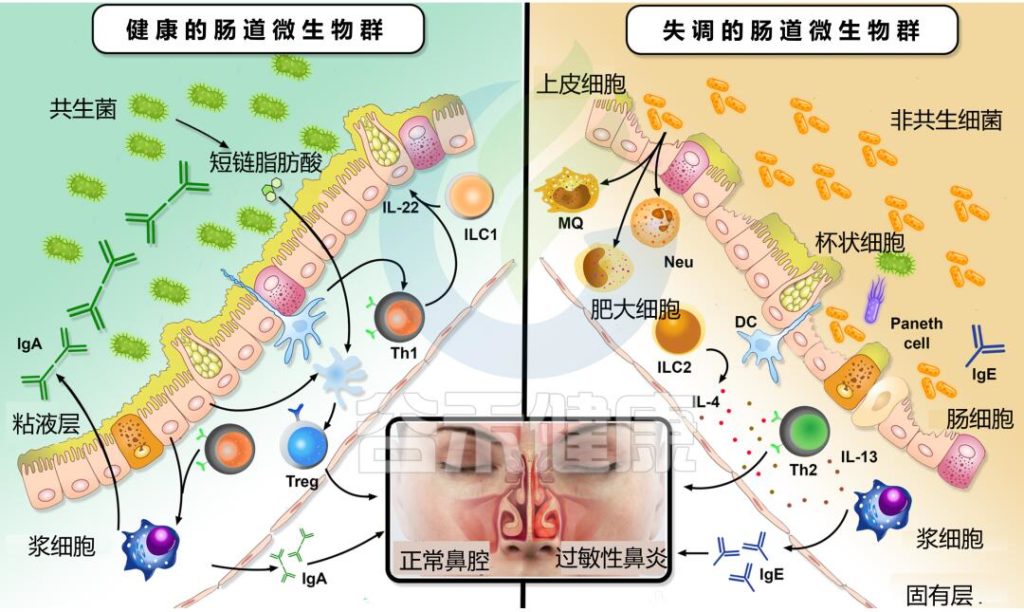
Li J, et al., Eur Arch Otorhinolaryngol. 2022
肠道微生物群影响ILC2通过肠-肺轴从肠道向肺的迁移。例如,变形菌门显著促进了上述迁移,并促进了IL-33的产生。此外,呼吸道过敏的发展与Ruminococcus gnavus的丰度增加有关联。
扩展阅读:
➤ 肠道炎症性疾病
肠道微生物群对保护黏膜屏障功能至关重要
与溃疡性结肠炎患者或健康受试者相比,克罗恩病患者的变形菌门显著增加。在炎症克罗恩病组织中,埃希氏菌(Escherichia)和志贺氏菌(Shigella)更为丰富,而在非炎症克罗恩病组织里,Pseudomonas更为广泛。
大肠杆菌和梭杆菌能够粘附粘膜,侵入肠上皮细胞,并可能加剧炎症。具核梭杆菌通过破坏上皮屏障和诱发异常炎症而加重结肠炎。
相对于炎症性溃疡性结肠炎,拟杆菌门在克罗恩病炎症粘膜中更常见。克罗恩病粘膜活检中最常见的是普通拟杆菌(Bacteroides vulgatus),克罗恩病患者中脆弱拟杆菌(Bacteroides fragilis)的数量明显高于溃疡性结肠炎患者或健康受试者。脆弱拟杆菌与产生IL-10的调节性T细胞的增加有关,这限制了促炎机制,从而有助于减少炎症。
肠道微生物群在调节宿主免疫以建立和维持肠道稳态方面起着至关重要的作用。
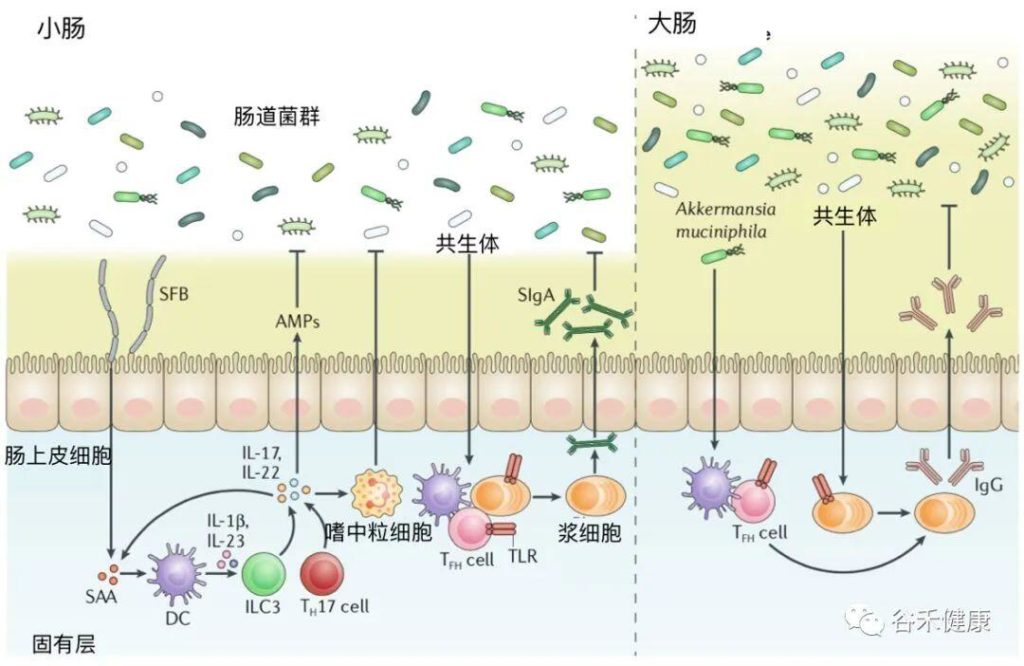
doi: 10.1038/s41577-019-0268-7
失调可能分为早期和晚期两个阶段进行。
在早期失调期间,IBD相关的遗传和环境因素可能导致致病菌的积累,这可能先于临床疾病的发展。
在晚期失调期间,肠道炎症推动了菌群的进一步变化,包括变形菌的增殖。鉴于肠道微生物群的不同菌群对宿主免疫系统和肠道屏障有有益的影响,某些菌群的缺失可能导致肠道炎症的加剧或消退。特定细菌的大量繁殖,如粘附性和侵袭性大肠杆菌,积聚在IBD患者的炎症粘膜中,可进一步促进炎症反应。
doi: 10.1038/s41577-019-0268-7
扩展阅读:
小结
综上所述,肠道微生物群与免疫系统之间的相互作用对于维持免疫系统的正常功能和免疫平衡至关重要。
良好的肠道微生物组成能够促进免疫系统的发育和成熟,调节免疫应答的程度和方向,预防免疫相关疾病的发生。
肠道内的免疫细胞在整个机体中扮演着至关重要的角色。它们作为守卫者,保护着我们免受外界病原体的入侵,并维持着肠道的免疫平衡。
肠道免疫细胞与肠道中的微生物密切合作,形成了一种复杂的共生关系。这些微生物通过调节免疫细胞的活性和功能,帮助我们抵御病原体,促进免疫平衡和整体健康。
此外,肠道免疫细胞还参与到免疫耐受的调节中,即对自身组织的免疫反应进行限制,避免自体免疫疾病的发生。肠道免疫细胞的失调可能会导致免疫炎症反应过度激活,引发炎症性肠病和其他免疫相关疾病。因此,保持肠道免疫系统的平衡和稳定对于整体健康至关重要。
了解和关注肠道免疫系统的功能及其与微生物群的相互作用对于维持整体健康具有重要意义。
主要参考文献
Mowat AM, Agace WW. Regional specialization within the intestinal immune system. Nat Rev Immunol. 2014 Oct;14(10):667-85.
Kayama H, Okumura R, Takeda K. Interaction Between the Microbiota, Epithelia, and Immune Cells in the Intestine. Annu Rev Immunol. 2020 Apr 26;38:23-48.
Allaire JM, Crowley SM, Law HT, Chang SY, Ko HJ, Vallance BA. The Intestinal Epithelium: Central Coordinator of Mucosal Immunity. Trends Immunol. 2018 Sep;39(9):677-696.
Yan Y, Ramanan D, Rozenberg M, McGovern K, Rastelli D, Vijaykumar B, Yaghi O, Voisin T, Mosaheb M, Chiu I, Itzkovitz S, Rao M, Mathis D, Benoist C. Interleukin-6 produced by enteric neurons regulates the number and phenotype of microbe-responsive regulatory T cells in the gut. Immunity. 2021 Mar 9;54(3):499-513.e5.
Cray P, Sheahan BJ, Dekaney CM. Secretory Sorcery: Paneth Cell Control of Intestinal Repair and Homeostasis. Cell Mol Gastroenterol Hepatol. 2021;12(4):1239-1250.
Yang D, Jacobson A, Meerschaert KA, Sifakis JJ, Wu M, Chen X, Yang T, Zhou Y, Anekal PV, Rucker RA, Sharma D, Sontheimer-Phelps A, Wu GS, Deng L, Anderson MD, Choi S, Neel D, Lee N, Kasper DL, Jabri B, Huh JR, Johansson M, Thiagarajah JR, Riesenfeld SJ, Chiu IM. Nociceptor neurons direct goblet cells via a CGRP-RAMP1 axis to drive mucus production and gut barrier protection. Cell. 2022 Oct 27;185(22):4190-4205.e25.

谷禾健康

微量营养素缺乏或称之为“隐性饥饿”,是目前一个严重的公共卫生问题,会对儿童发育及成人健康造成严重后果。
传统的治疗和预防方法,有些时候并是很有效,并且可能产生不良副作用(例如补铁引起的消化问题)。
★ 肠道细菌与微量营养素间存在重要的相互作用
许多研究发现肠道中的共生细菌可以增加特定微量营养素的生物利用度,特别是通过去除抗营养化合物(例如植酸盐和多酚),或通过合成维生素。
肠道黏膜是抵御病原体的第一道防线。肠黏膜微生物群有助于增强肠上皮的完整性并更好地吸收微量营养素。
细菌代谢还依赖于从肠道环境中获取的微量营养素,常驻细菌可能会竞争或协作以维持微量营养素稳态。因此,肠道微生物群的组成可以通过微量营养素的可用性来调节。

本文汇集了有关微量营养素和肠道微生物群之间双向关系的最新知识,重点关注铁、锌、维生素A和叶酸(维生素B9),因为这些缺乏症是全球范围内的比较严重的公共卫生问题。
目录
1.微量营养素缺乏
2.微量营养素与宿主健康
3.营养素与细菌的相互作用:以铁为例
4.细菌在锌生物利用中的作用
5.微生物群与宿主叶酸间的关联
6.微生物群与维生素A的相互作用
7.改善人体微量营养素的建议
8.结语
微量营养素,包括矿物质和维生素,是每天只需要少量的物质,但对人体代谢至关重要。
▼
微量营养素缺乏在全世界范围内非常普遍,最新估计表明,3.72亿学龄前儿童和1.20亿育龄非孕妇缺乏一种或多种微量营养素。
维生素和矿物质缺乏,也被称为“隐性饥饿”,是一个严重的公共卫生问题,世界各个国家都存在,特别是低收入和中等收入国家的弱势群体。
在全球范围内,三分之二的妇女和儿童(主要来自经济不发达家庭)的饮食中至少缺乏一种微量营养素。全球最普遍的微量营养素缺乏症包括铁、锌、维生素A和叶酸。
★ 微量营养素缺乏会导致不良健康结果
微量营养素缺乏与多种不良健康结果相关,例如儿童发病率和死亡率增加、贫血、发育迟缓和认知缺陷。
虽然存在多种改善个人或人群微量营养素状况的策略,其中主食强化被认为是改善人群微量营养素状况最具成本效益的干预措施。
然而,由于多种原因,应对微量营养素缺乏症的策略并不总是有效。包括可接受性差(例如,孕妇铁补充剂)、强化水平差和质量控制不足(例如,维生素A强化食用油),强化食品未能惠及最贫困人群。
★ 不同营养素的吸收具有差异
微量营养素的吸收主要发生在小肠中,根据微量营养素的不同,具有不同的机制。
例如,锌和铁主要通过特定的运输载体在十二指肠和空肠中吸收,这些运输载体针对每种矿物质及其形式而异,而叶酸不仅在小肠中被吸收,而且在结肠中也被吸收,因为在这两个部位都鉴定出了转运蛋白。
微量营养素的吸收差异很大,例如,铁充足的男性成年人吸收的铁摄入量不到5%,而正常条件下摄入的锌吸收量大于30%。
注:大部分摄入的微量营养素会到达结肠。
★ 微生物群与微量营养素之间存在相互作用
在消化道中,有一个复杂的微生物群落,其浓度和多样性从口腔到结肠不断增加。尽管含有其他微生物(病毒、真菌),但大多数研究都重点关注细菌,其中厚壁菌门、拟杆菌门、放线菌门、变形菌门和疣微菌门是最重要的。
粪便细菌组成在个体之间存在很大差异,并且不仅取决于宿主的遗传学,还取决于包括饮食在内的许多环境因素。
注:大多数关于食物对肠道微生物群影响的研究都集中在大量营养素(脂肪、碳水化合物、蛋白质)上。
然而,细菌需要微量营养素才能生存。例如,细菌需要外部来源的铁,其中一些甚至开发出特定的机制,例如转铁蛋白来竞争铁源。
相比之下,许多细菌可以直接合成维生素,例如叶酸(维生素B9),因此不一定需要外部来源。
细菌影响微量营养素的生物利用度
常驻细菌可能对微量营养素产生直接作用,影响其对宿主的生物利用度,或者产生间接作用,例如通过产生能够改善上皮健康的代谢物,从而导致更好的吸收。
微量营养素影响肠道细菌的组成和功能
微量营养素的消耗反过来会影响肠道细菌的组成和功能。事实上,事实证明,补充铁可以导致潜在致病性肠杆菌科细菌的增加,以及被认为对健康具有积极影响的双歧杆菌科细菌和乳杆菌科细菌的减少。
微量营养素在控制人类健康的新陈代谢和组织功能维持中发挥着核心作用。
然而,尽管人们努力通过补充食品和饮食多样化来增加微量营养素的摄入量,全球微量营养素缺乏症的患病率仍然非常高,包括工业化国家。
推荐营养素摄入量和摄入上限
Dje Kouadio DK,et al.Crit Rev Food Sci Nutr.2023
上表提出了男性和女性一些推荐营养素摄入量和可耐受的上限摄入水平。
由于维持生长和补偿损失(例如月经)的需求更高,儿童和妇女受到的影响更大。
★ 缺乏微量营养素的症状多样且大部分在晚期
单一微量营养素缺乏的临床症状多种多样,并且通常是具有微量营养素特异性的,但一般发生在缺乏的晚期阶段。例如缺铁性贫血是缺铁的最后阶段。
▼
缺铁被认为是最常见的微量营养素缺乏症,全世界估计有超过20亿人缺铁。它会导致贫血、免疫功能受损、工作能力下降以及内分泌功能受损。
导致铁缺乏的因素包括:
(1)血红素铁摄入量低,其生物利用度高于非血红素铁
(2)富含植酸酶或酚类化合物的饮食会降低生物利用度
(3)慢性炎症,导致铁调素水平升高,下调铁吸收
▼
据估计,全球有超过10亿人缺锌,它与儿童生长迟缓相关,但也会降低免疫能力,从而增加儿童对多种疾病的易感性,包括腹泻、肺炎,也许还有疟疾。
此外,锌缺乏似乎会导致其他微量营养素缺乏。
▼
据估计,低收入和中等收入国家超过20%的育龄妇女缺乏叶酸。
怀孕期间叶酸摄入不足与胚胎最初几周的神经发育缺陷有关。在普通人群中,叶酸缺乏还会导致巨幼红细胞性贫血和神经系统症状,这些症状与维生素B12缺乏症的症状部分重叠。
▼
维生素A缺乏不仅会导致干眼症和失明,还会影响免疫功能的许多方面,导致免疫能力下降以及皮肤和上皮完整性降低。
许多低收入和中等收入国家每半年向5岁以下儿童提供高剂量维生素A补充剂,以降低儿童死亡率,但这种干预措施的有效性受到质疑。
有关铁、细菌和宿主的相互作用,参考了最近的研究,下面总结了这种复杂关系的关键要素。
铁的吸收和稳态受到严格调节
由于人类无法主动排泄铁,因此铁的吸收在十二指肠中受到严格调节。
// 血红素铁
血红素铁和非血红素铁通过两种不同的途径吸收,血红素铁被十二指肠刷膜边缘的血红素载体蛋白1(HCP1) 吸收。
// 非血红素铁
非血红素铁通常呈三价铁形式,不具有生物利用度,必须通过十二指肠细胞色素b(DCYTB) 将其还原为二价铁形式,然后再由二价金属转运蛋白 (DMT1) 转运到十二指肠刷膜上 。
注:二价金属转运蛋白的存在已被描述,铁也在回肠和结肠中被吸收,但这些吸收位点对宿主铁状态的贡献仍不清楚。
铁进入肠上皮细胞后,储存在铁蛋白中,或通过基底外侧铁转运蛋白输出到循环系统中。
铁稳态由激素铁调素调节,铁调素通过不同机制抑制铁释放到血浆中。
铁调素是由肝脏合成并分泌的富含半胱氨酸的抗菌多肽,在免疫过程中能够大量表达参与免疫反应;在机体内铁平衡的调节中起到负性调节的作用。
肠道微生物群在铁吸收中的作用示意图
Dje Kouadio DK,et al.Crit Rev Food Sci Nutr.2023
细菌可以通过不同的机制获取铁
细菌可以通过不同的机制获取铁,例如使用称为铁载体的铁特异性螯合剂,铁载体是化学性质多样的次级代谢产物,对铁具有高亲和力。
由此产生的铁-铁载体复合物通过特定的外膜蛋白(上图中的细菌A)内化。细菌还可以通过产生具有不同配体结合位点的外膜受体来利用其他细菌产生的铁载体(上图中的细菌B)。
铁载体在细菌缺铁的情况下表达
铁载体在细菌缺铁的情况下表达,因此不在铁充足的环境中表达。在厌氧条件和低pH值下,铁主要以二价铁的形式存在,可以被细菌直接转运,主要通过专用转运系统Feo,由不同的蛋白质。
细菌可以利用宿主铁化合物
细菌还可以利用宿主铁化合物,例如血红素或转铁蛋白,它们通过特定受体转运或直接吸收。此外,尽管牛奶中存在的乳铁蛋白具有抑菌功能,但它也可以被具有特定受体的细菌利用。
铁吸收的调节对于细菌非常重要,因为铁在较高浓度下有毒。在许多细菌物种中,当铁浓度过高时,铁吸收调节蛋白 (Fur) 会抑制与铁吸收相关的基因转录。
研究细菌有助于揭示新的铁吸收途径
值得注意的是,大部分关于细菌铁吸收的知识来自对病原菌的研究。长期以来,人们一直认为一些有益菌,例如乳杆菌科的成员,不需要铁来生长。然而已经在植物乳杆菌(Lactiplantibacillus plantarum)和清酒乳杆菌(Latilactobacillus sakei)中鉴定出了特定的系统。
研究这些细菌可能会揭示新的铁吸收途径。在铁吸收的主要部位十二指肠中,大约有103 个细菌/克,低于空肠(104 个细菌/克),并且远低于结肠,结肠中的细菌数量高达1012 个细菌/克。
此外,乳杆菌科属的成员在铁吸收最多的小肠中数量众多,因此这些细菌对宿主铁吸收的作用应受到更多关注,以确定它们是否在铁的生物利用度中发挥作用。
▼
铁对于许多细菌来说是必需的,它们的生长需要铁。
据计算,结肠中的铁浓度很高,并且远远超过细菌生长的最低要求。但细菌对铁的生物利用度还取决于一系列因素(铁的形式、铁形态、pH值、氧气水平等)。
不同形式的铁可用于补充或强化,例如硫酸亚铁、富马酸亚铁或乙二胺四乙酸铁钠。由于铁补充剂中含有的铁明显多于人体可以吸收的铁,因此大量未吸收的铁会留在消化道内腔中。
•铁补充剂会导致潜在致病菌水平升高
多项人类研究表明,铁补充剂或强化铁会导致潜在致病菌水平升高,并导致通常被认为是有益细菌种类减少,例如乳杆菌科成员。
最近一项针对大鼠的研究还表明,出生后补充铁会导致肠道微生物群发生改变,这种改变会持续到成年期。此外,研究表明,抗生素治疗后服用铁剂会导致肠道细菌的组成和功能发生改变。
铁对于致病细菌至关重要,哺乳动物的免疫系统已经具备了通过表达铁结合蛋白(如脂质运载蛋白-2 或乳铁蛋白)来控制细菌对铁的可用性的能力,这种现象称为“营养免疫”。
研究了补充铁对肠道微生物群的作用,总的来说,根据所使用的模型(体外、动物或人类),结果似乎有很大不同。
事实上,在补充铁期间,乳杆菌科总是减少,这在体外、小鼠和婴儿中均观察到。所用铁的化学形式似乎对细菌组成有不同的影响。此外,出乎意料的是,补充铁的效果与缺铁的效果并不直接相反。
▼
对于细菌和人体来说,非血红素铁的生物利用度低于血红素铁的生物利用度,因为它受到食物基质的成分和管腔的物理化学成分的影响。
•肠道细菌有助于更好地吸收铁
事实上,许多食物基质中的非血红素铁与抑制剂(例如多酚、纤维或植酸盐)相连,这些抑制剂可以被专用细菌酶降解,从而导致更好的铁吸收。
细菌还可以通过发酵饮食中的不可消化碳水化合物来产生短链脂肪酸,这会降低管腔pH值,从而将三价铁还原为二价铁,从而提高其被细菌和宿主的吸收。
其他一些有机酸(例如乳酸)是由许多细菌产生的,存在于整个消化道中。酸化的肠道环境还可以降解螯合微量营养素的复合物并促进铁的吸收。
扩展阅读:
▼
考虑到补铁的依从性低以及腹泻或便秘等副作用,很多研究已经测试和发现其他改善铁状态的策略。
它们包括使用益生菌、益生元和后生元等。
•使用益生菌改善铁状况
例如,将益生菌植物乳杆菌(Lactiplantibacillus plantarum 299v)与铁、抗坏血酸和叶酸一起食用,对健康、非贫血的瑞典孕妇的铁状况是安全的,可减少铁储备的损失并改善铁状况。
在另一项研究中,将果胶组装成铁纳米颗粒作为基质,用于植物乳杆菌(Lactiplantibacillus plantarum CIDCA 83114)的递送。益生菌没有毒性,生存能力没有减弱,这为用替代的铁输送策略对抗缺铁提供了新的视角。
•使用益生元帮助铁的吸收
在大鼠模型中,食用益生元寡糖通过调节肠道微生物群来增强铁吸收。摄入益生元后肠道pH值的降低可以帮助铁的吸收,此外还能以有益的方式改变肠道微生物组成。因此,益生元可以成为改善宿主铁状况的良好选择。
•食用乳铁蛋白增加铁的吸收
在肯尼亚婴儿中,研究表明,与单独服用硫酸亚铁相比,同时食用无铁乳铁蛋白(脱铁乳铁蛋白)和硫酸亚铁能够显著增加铁的吸收。
注:考虑到乳铁蛋白对肠道微生物群的影响,一些学者认为乳铁蛋白可能在婴儿铁配方中有用,因为它可以增加铁的吸收,同时可能减轻未吸收的铁对肠道微生物群的不利影响。
•使用富铁微生物
这些预防和治疗缺铁的新策略可能包括使用富铁微生物,除了从益生菌摄入中获益之外,它还可以为宿主提供大量矿物质。
在贫血小鼠中已显示,食用在铁存在下生长的酵母能够提高小鼠的血红蛋白浓度。在人类中只进行了一些尝试,其中含有富铁酵母的奶酪的吸收效率不如单独含有硫酸铁的奶酪。
注:还应该进行补充研究来评估富铁微生物在人类中的应用的全部潜力。
小结
从上面可以明显看出,肠道细菌在调节宿主铁的生物利用度方面发挥着重要作用。然而,宿主因素可以调节细菌对铁的吸收,例如,在炎症的情况下,脂质运载蛋白将与铁载体结合,从而对抗肠道细菌对铁的捕获。
最近,有人提出,共生细菌不仅使用不同的机制来释放和捕获铁,而且还在它们自己和它们的宿主之间共享铁。事实上,细菌扩散性铁载体生产的例子可以捕获铁,但不能保证这些铁将被生产细菌利用,这表明对可用铁的共同管理。
迄今为止,人类对锌、宿主和肠道微生物群之间相互作用的研究远少于铁。
★ 人体需要每日摄入锌
每日摄入锌补充剂,成人 40 毫克/天,通常被认为是安全的。
锌参与DNA合成、转录和翻译等重要过程。锌的吸收发生在十二指肠,涉及跨膜锌转运蛋白Zip4。然后,锌通过锌调节蛋白ZnT2-10转运至金属蛋白合成位点,或通过锌转运蛋白ZnT1输出至循环系统。
与铁相反,哺乳动物缺乏锌储存机制,每日摄入是必不可少的。如果过量,锌也可以主动排泄,锌通过基底外侧的 Zip5/Zip14 蛋白从循环分配到肠上皮细胞,并通过ZnT5从肠上皮细胞流出到管腔。
肠道微生物群在锌吸收中的作用示意图
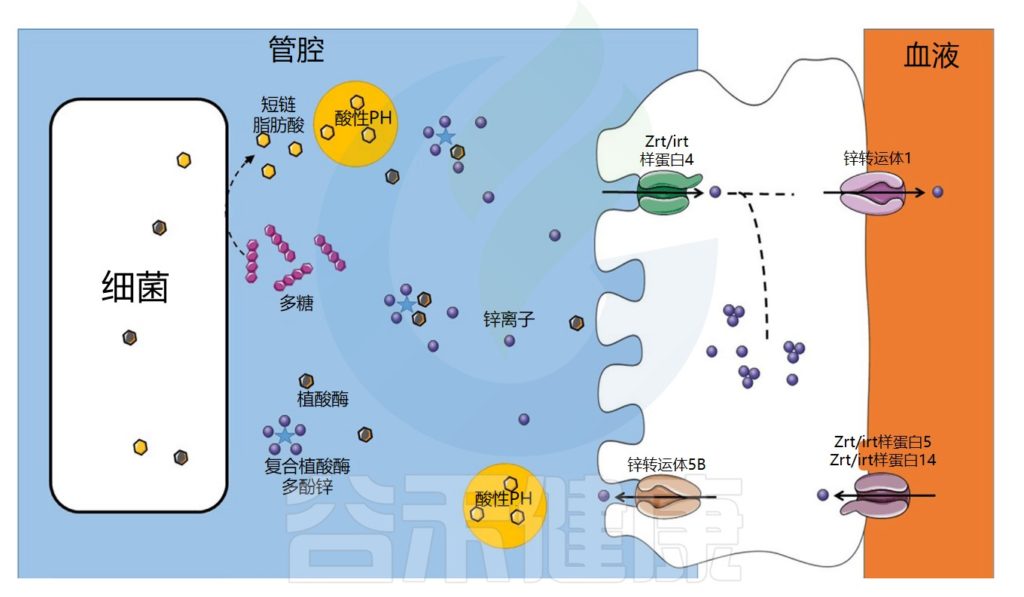
Dje Kouadio DK,et al.Crit Rev Food Sci Nutr.2023
锌的转运与吸收过程
在食物中,锌可以与抗营养因子如植酸盐或多酚络合,从而减少其吸收。未被小肠吸收的锌会到达结肠,并可能被结肠细胞吸收,因为锌转运蛋白也在结肠中表达。
在细菌中,锌通过锌伴侣蛋白转运至金属蛋白合成位点。细菌对锌的吸收受锌吸收抑制因子Zur和锌流出抑制因子CzrA的调节。一些病原菌可以通过表达高亲和力转运蛋白(ZnuABC)来克服锌限制条件。
锌浓度过高可能对某些病原菌有毒。与铁的情况一样,宿主和病原微生物之间也存在竞争。事实上,宿主还利用营养免疫来通过产生钙卫蛋白来限制锌的生物利用度。
例如,钙卫蛋白在消化道中结合锌。
▼
锌缺乏与慢性腹泻相关。补锌对肠道微生物群的影响主要在动物模型中进行研究。它已在家禽和养猪业中以药物剂量用于减少胃肠道感染和腹泻。
√锌具有抗菌活性,能够减少腹泻
锌具有抗菌活性,不同的研究强调了高剂量的锌会引起肠道不同部位(从胃到结肠)细菌组成的变化。
在断奶仔猪中,使用低剂量的锌可以通过改善小肠形态和通透性、增强屏障功能,有效促进生长性能并减少腹泻。
此外,锌还能丰富肠道微生物群中的链球菌科(Streptococcaceae)和乳杆菌科(Lactobacillaceae),并减少芽孢杆菌和肠道疾病相关病原体,包括回肠腔中的梭状芽胞杆菌和克罗诺杆菌。
注:如果缺锌小鼠补充锌是有益的,那么过量的锌饮食摄入会改变肠道微生物群并降低对艰难梭菌感染的抵抗力。
√一些菌属可以预测锌的充足性
但研究结果并不一致。事实上,一项针对鸡的研究表明,瘤胃球菌属(Ruminococcus)可以预测宿主体内锌的充足性。但在一项针对小鼠的研究中,黑色素杆菌(Melainabacteria)和脱硫弧菌属(Desulfovibrio)被确定为锌状态的潜在标志物。
▼
尽管许多代谢过程都需要锌,但关于锌状态或补充剂对肠道微生物群影响的研究却很少。
√一些细菌可以作为锌缺乏的生物标志物
最近的研究表明,尽管缺锌和锌充足的学龄儿童之间的细菌多样性相似,但缺锌儿童的个体多样性更高。特别是,粪杆菌(Coprobacter)、醋酸弧菌(Acetivibrio)、副普雷沃氏菌(Paraprevotella)和梭状芽胞杆菌在缺锌组中更为丰富。它们可以用作进一步临床诊断锌缺乏的生物标志物,但需要更多的工作来推广这一结果。
锌的摄入量也会显著增加儿童粪便细菌。在同一项研究中还鉴定出了纳米颗粒抗性细菌的一些代表。
▼
由于补锌会改变肠道微生物群的组成和功能,因此许多研究已经研究了锌与益生菌一起服用的效果。
√锌与益生菌一起补充
例如,在大鼠中,施用在富锌培养基中生长的长双歧杆菌(Bifidobacterium longum CCFM1195)比单独补充锌更能有效地增加血浆锌浓度。
此外,一些细菌群(如乳杆菌科)的丰度随着富含锌的双歧杆菌的消耗而增加。使用富含硒和锌的植物乳杆菌对小鼠进行类似的设计,与对照组或补充硒锌的组相比,能够增加循环锌浓度。这种增加伴随着肠道细菌组成的改变,乳杆菌科(Lactobacillaceae)的比例增加。
√儿童单独补充锌可能更有效
最近总结了研究中获得的数据,对于24个月以下儿童的治疗,单独补充锌可能比益生菌更有效;在学龄前儿童中同时补充锌和植物乳杆菌并不比单独补充益生菌更有效。因此,锌和益生菌联合给药的有效性需要更多的人体研究。
使用益生元似乎效率较低,因为给猪施用锌和马铃薯纤维对大多数分析指标没有显示出任何交互作用。
注:锌源和纤维补充对微生物群生态的影响是独立的。
小结
与铁的情况一样,对锌状态和宿主微生物群的研究主要集中在致病条件上,而对细菌在正常情况下的作用知之甚少。
肠道细菌、锌和宿主之间的关系值得更深入地研究,尤其是在人类中。事实上,富锌益生菌等有前途的工具正在出现,它们代表了治疗锌缺乏症的替代方案。特别是大剂量、长期补锌可能会干扰铁和铜的吸收,导致铁和铜的缺乏。
叶酸,也可以称之为维生素B9,是一种水溶性维生素。叶酸在蛋白质合成及细胞分裂与生长过程中具有重要作用,对正常红细胞的形成有促进作用。
一些共生细菌已被证明可以合成某些维生素,细菌也可以成为宿主叶酸的来源,即使整体贡献的程度不清楚。同时,叶酸的摄入会影响肠道微生物群。
叶酸的转运与吸收过程
饮食中的叶酸以单谷氨酸盐和聚谷氨酸盐形式存在,它们被肠道结合物分解为单谷氨酸盐形式,然后通过特定的转运蛋白偶联叶酸转运蛋白(PCFT)在回肠中吸收。
在肠上皮细胞中,叶酸单谷氨酸转化为活性形式5-甲基四氢叶酸(5-MTHF),它可以被运输到循环系统。
注:细胞叶酸外排可能由多个耐药蛋白(MRPs)家族介导。在结肠中,已经发现了相同的一种受体。
肠道微生物群在叶酸吸收中的作用示意图
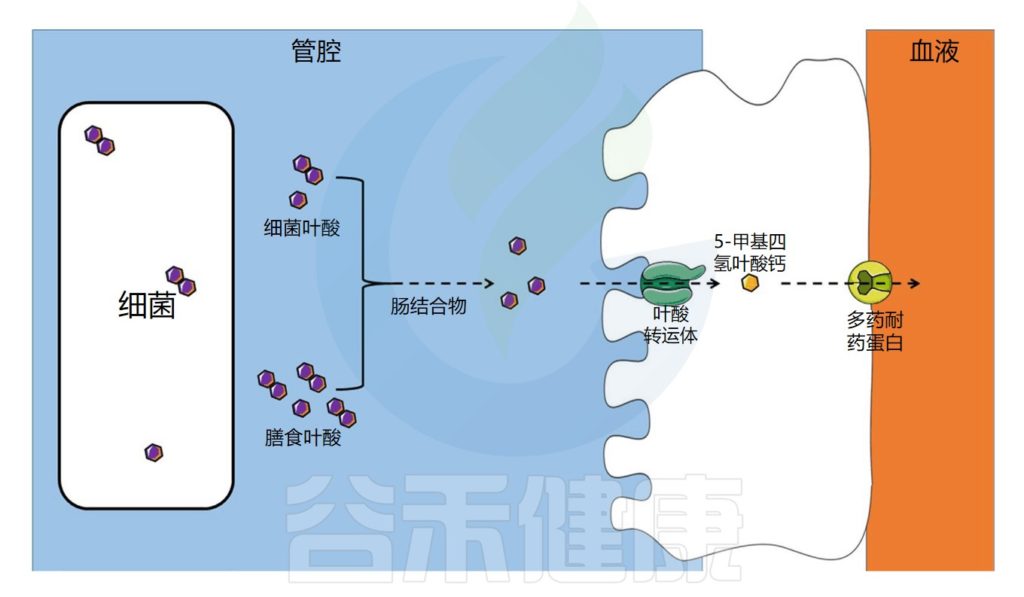
Dje Kouadio DK,et al.Crit Rev Food Sci Nutr.2023
▼
细菌的生长需要叶酸,一些细菌是原养型细菌,可以从环境中存在的前体合成叶酸,而营养缺陷型细菌必须从环境中获取叶酸。
✦肠道细菌合成叶酸
大量文献支持细菌叶酸合成对宿主叶酸状态的重要贡献。早期研究报告称,人类粪便样本中的叶酸浓度为300-500µg/天,而饮食摄入量低于100µg/天,表明肠道细菌可以合成叶酸。
事实上,512个细菌基因组的叶酸生物合成能力预测表明,其中13%的细菌基因组包含所有叶酸生物合成相关基因,而39%的细菌基因组可以在外部来源提供额外的对氨基苯甲酸的情况下合成叶酸。
总而言之,这些数据表明肠道微生物群是叶酸的重要来源,并且饮食等不同因素导致的肠道微生物群组成的变化可能会影响叶酸的需求。
✦肠道细菌可以转化叶酸以便更好地利用
由于纤维消耗而改变肠道微生物群组成可能导致结肠内容物或循环中叶酸浓度增加。此外,尽管结肠中的叶酸吸收比小肠中的叶酸吸收慢100倍,但标记的叶酸被专门包裹在结肠中并在结肠中分解,并被纳入宿主组织。
肠道细菌不仅合成可供宿主使用的叶酸,还可以将叶酸转化为更好地被宿主吸收的维生素体,正如秀丽隐杆线虫(Caenorabditis Elegans)模型所证明的那样,其中细菌可以将叶酸转化为四氢叶酸。
✦微生物代谢叶酸的能力与宿主生理状况相关
肠道微生物群的代谢能力不仅取决于其组成,还取决于宿主的生理特征。例如,与成人相比,婴儿和幼儿肠道微生物群中更频繁地发现编码参与叶酸生物合成的酶的基因。
有趣的是,受营养不良影响的儿童的微生物组显示,参与B族维生素代谢的多个途径中的基因丰度显著降低。此外,在肥胖女性中观察到叶酸水平较低,同时产生B族维生素的细菌也减少。
▼
许多产生叶酸的细菌被鉴定并成功用于增加发酵食品的叶酸含量。其中一些细菌还能够改善缺乏叶酸饮食的啮齿动物的叶酸状况。
✦产生叶酸的益生菌改善叶酸状态并调节微生物组成
使用产生叶酸的益生菌可能是改善叶酸状态并调节肠道微生物群的一种方法。在体外发现了有希望的结果,其中产生叶酸的清酒乳杆菌(Latilactobacillus sakei)被证明可以增加短链脂肪酸并改变粪便细菌组成。
在大鼠中,食用使用产生叶酸的植物乳杆菌制备的发酵奶能够恢复正常的叶酸状态,同时广泛改变肠道细菌组成。
▼
叶酸摄入量对肠道细菌的影响并不一致。
✦叶酸缺乏可能影响部分动物的肠道细菌组成
在小鼠身上发现膳食叶酸缺乏不会影响肠道细菌组成,可能是一些细菌可以在缺乏时产生叶酸,叶酸摄入量的波动对肠道微生物群的影响较小。
然而,在其他动物模型中,叶酸摄入改变了肠道微生物群的组成,例如鸡和小猪。
✦叶酸缺乏影响微生物多样性
叶酸缺乏也会影响微生物多样性,因为在限生小鼠中发现,与微量营养素充足的饮食相比,叶酸缺乏的饮食在21天后增加了β多样性,但随后的14天全饮食治疗并没有改变这一趋势。
小结
宿主的叶酸状态、食物或肠道中的叶酸产生细菌之间存在多种关联。这种关联通常是矛盾的,但有利于宿主和细菌区室之间的密切相互作用,这表明需要进行大规模人类研究,以更好地了解与叶酸可用性和改善人群叶酸状态的策略相关的宿主-微生物群相互作用。
叶酸重要天然来源有:
花生(246微克/100克)、葵花籽(238微克/100克)、芦笋(149微克/100克)、生菜(136微克/100克)、一些绿叶蔬菜和柑橘类水果等。
扩展阅读:维生素B9(叶酸)
维生素A在免疫调节中发挥着至关重要的作用,包括调节细胞因子的产生和淋巴细胞归巢至粘膜表面。它通过调节肠上皮中免疫细胞的增殖和分化,直接有助于维持肠道屏障,并在抵抗肠道病原体入侵方面发挥着至关重要的作用。
膳食维生素A以视黄醇和类胡萝卜素的形式存在,分别存在于动物和蔬菜中。
肠道微生物群在维生素A吸收中的作用示意图
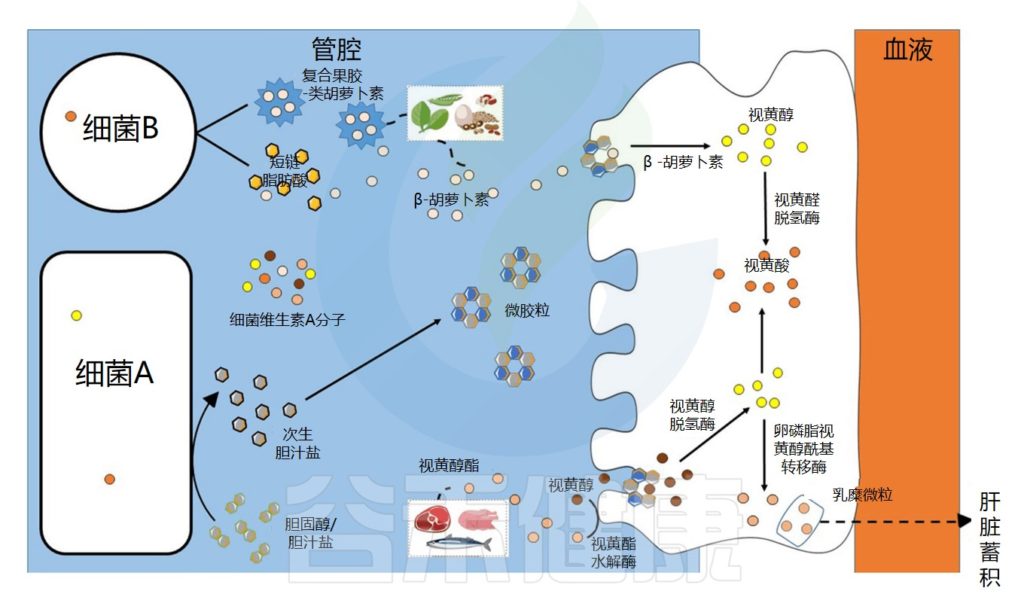
Dje Kouadio DK,et al.Crit Rev Food Sci Nutr.2023
维生素A的吸收过程
视黄醇和类胡萝卜素是膳食脂溶性成分,在被肠上皮细胞吸收之前需要通过胶束溶解 。在胃和十二指肠中发生乳化成液滴的阶段,类胡萝卜素和视黄醇的摄取发生在小肠的上半部。
在肠上皮细胞中,类胡萝卜素通过胡萝卜素-15,15′-双加氧酶直接转化为全反式视黄醛。视黄醛可被视黄醛脱氢酶不可逆地转化为具有生物活性的视黄酸分子,或被转化为视黄醇酯,储存在肝脏中。
▼
肠道微生物群在维生素A代谢中的作用可能依赖于特定酶、脂多糖的产生,脂多糖可能有助于胶束形成或直接吸收,以及胆汁稳态。
•微生物通过调节胆汁酸有利于维生素A的溶解和吸收
一项以绵羊为模型的研究表明,宿主微生物群通过调节胆汁酸分泌对维生素A吸收产生影响。在该模型中,胆汁酸水平降低与维生素A吸收减少相关,并与瘤胃球菌科丰度密切相关。
由乳杆菌(Lactobacillaceae)、双歧杆菌(Bifidobacterium)、拟杆菌(Bacteroides)或梭菌属等细菌产生的二级或三级胆汁盐的分泌可能有利于维生素A的溶解并改善其吸收,因为胆汁盐对于胶束化过程至关重要。
•肠道微生物可以直接和间接影响维生素A代谢
肠道细菌的基因组具有在动物和植物中发现的不同酶的类似物,这些酶允许乙酰辅酶A通过甲羟戊酸途径合成类胡萝卜素和类维生素A。
此外,来自拟杆菌(Bacteroides)、肠球菌(Enterococcus)和链球菌(Streptococcus)的细菌具有与宿主BCMO基因相似的brp/blh基因,可以将β-胡萝卜素转化为全反式视黄醇。
因此,肠道微生物组对维生素A代谢的影响既包括直接影响(如类胡萝卜素的合成),也包括间接影响(如调节其生物利用度)。
•肠道微生物合成相关蛋白有助于维生素A转运
类胡萝卜素的生物利用度会受到各种食物成分的影响,其中食物基质中蔬菜的细胞壁结构和组成形成物理屏障,从而限制消化酶的作用。
果胶和一些多酚(例如柚皮素)已被证明可以通过结合管腔中的胆固醇和胆汁盐来降低类胡萝卜素的体外生物利用度,从而抑制胶束的形成。
相反,果胶可以被许多共生细菌降解,并且肠道中的这种降解可能会增加类胡萝卜素的生物利用度。
此外,肠道微生物群合成酶或补体蛋白,例如细菌脂质运载蛋白,这可能有助于将类维生素A转运到肠上皮细胞中。
•肠道细菌调节宿主视黄醇的代谢
肠道微生物群可以消化膳食纤维,从而释放 β-胡萝卜素或其他结合态的微量营养素。
此外,肠道细菌群还可以调节宿主视黄醇的代谢,调节驱动免疫反应的视黄酸的产生。梭状芽胞杆菌类细菌可以通过抑制基因Rdh7的表达来调节肠道上皮中的视黄酸浓度,从而启动视黄醇向视黄酸的转化。
此外,这项研究表明,共生细菌群落降低了视黄酸的产生,视黄酸的产生通过肝脏储存形式的增加来平衡,并有利于视黄酯途径,而潜在致病菌则促进视黄酸的产生。
▼
肠道中维生素A的状态和类视黄醇的可用性可以调节肠道微生物的组成。
•维生素A调节免疫球蛋白的产生帮助保持细菌稳态
类胡萝卜素摄入、免疫功能和肠道细菌稳态之间相互作用的模型,表明维生素A可以通过调节IgA的产生来有助于保持细菌稳态。
在动物中,与未缺乏维生素A的小鼠相比,缺乏维生素A的小鼠的肠道微生物组成中,以下菌属的丰度较低:
另一项研究表明,拟杆菌在维生素A缺乏小鼠中的比例较低,维生素A缺乏组和维生素A充足组之间的几个细菌比例不同。在小鼠模型中,维生素A被证明可以调节分段丝状细菌(SFB)的丰度。
分段丝状细菌是一组能够诱导Th17细胞分化的细菌。
▼
•维生素A的摄入与微生物群多样性相关
类胡萝卜素的摄入量和状态与更大的微生物群多样性和有益细菌丰度的增加有关。
例如,在孕妇中,类胡萝卜素的摄入量和血浆浓度与肠道微生物群的α多样性呈正相关。
•维生素A影响免疫反应
维生素A对肠道微生物群组成的调节可能作为调节肠道免疫反应的间接途径。
例如,维生素A在体外和体内均被证明可以抑制诺如病毒(急性胃肠炎最常见的病毒因子)的复制,并且这种抑制似乎是通过微生物组变化直接或间接发生的,特别是肠道中的乳杆菌科。
•改善疾病患者的微生物组成
在溃疡性结肠炎患者中,摄入维生素A有益,可导致微生物群组成发生显著变化。
在自闭症儿童中,维生素A补充剂增加了拟杆菌目细菌的比例,并减少了双歧杆菌属的细菌比例。
有趣的是,一项针对306名孟加拉国新生儿的研究表明,与安慰剂相比,补充维生素A增加了男孩双歧杆菌的丰度,而这种差异在女孩中并未观察到。
对这些婴儿直至2岁的随访表明,女孩中放线菌相对丰度与血浆视黄醇呈浓度依赖性正相关,但男孩中则不然。
此外,阿克曼菌(一种与粘膜相关的细菌)的相对丰度也与女孩的血浆视黄醇呈正相关。
微生物群在肠道中参与了微量营养素的代谢和吸收,并且可以影响宿主的微量营养素状态。在这方面,给出以下一些建议:
➤ 1
摄入多样化的食物可以为肠道微生物提供更广泛的营养来源,从而促进微生物群的多样性和稳定性。
建议摄入富含膳食纤维的水果、发酵食物、蔬菜、全谷类食物以及富含微量营养素的食物,如坚果和海鲜等。
例如,香蕉钾含量高,还含有大量维生素 B6 和维生素 C。西兰花不仅富含维生素 C,还富含维生素 K、铁、钙和钾。
确保摄入足够的每种维生素和矿物质的一个简单方法就是“彩虹饮食”,彩虹饮食顾名思义就是色彩缤纷的蔬菜和水果,它们含有丰富的微量营养素,是一种健康趋势。每餐可以注意一下,是否有至少三到四种不同颜色的植物性食物,对于孩子,也可以鼓励他们去寻找不同的颜色。

下表列出了常见的维生素的食物来源,可供参考。

➤2
维生素D可以增加肠道对钙的吸收能力,并促进钙的转运到骨骼和其他组织中。
维生素 C 有助于改善植物中铁的吸收,这对于素食者来说尤其重要,可以在菠菜沙拉中添加柠檬汁。维生素C通过提高铁的溶解度或作为十二指肠细胞色素B将铁从铁还原为铁的辅助因子,有助于非血红素铁的生物利用度。
维生素B12和叶酸之间存在相互依赖的关系。维生素B12参与叶酸的代谢和激活,而叶酸则有助于维生素B12的合成和利用。
其他如,维生素E是一种脂溶性维生素,需要脂肪的存在来促进其吸收。摄入适量的脂肪可以提高维生素E的吸收率。
微量营养素之间的相互作用是复杂的,并且还受到其他因素的影响,最好的方式是通过均衡的饮食来获取各种微量营养素,因为食物中的微量营养素通常以复杂的组合形式存在,相互之间的相互作用更加协调和自然。
➤3
益生菌和益生元是一些有益的微生物和它们的营养物质,可以促进肠道的健康和微生物群的平衡。
建议摄入含有益生菌的酸奶、发酵食品、益生元的食物如洋葱、大蒜和麦麸等,以支持肠道微生物的生长和活动。
益生菌和益生元可能有助于微量营养素的最佳生物利用度,减少负面影响
例如,益生元低聚半乳糖减轻了补铁对婴儿肠道的不利影响,而罗伊氏乳杆菌与泻药氧化镁结合使用可以改善儿童的慢性便秘,而不会导致微生物组失衡。
益生元可以改变肠道微生物群,促进产短链脂肪酸的菌产生(如双歧杆菌、乳酸杆菌、真杆菌等),与钙吸收(在人类和动物模型中)、骨密度和强度(在动物模型中)增加相关。
扩展阅读:
肠道微生物群与健康:探究发酵食品、饮食方式、益生菌和后生元的影响
➤4
一般来说,在小谷物食品的自然发酵中,B族维生素的利用率可能会增加,并且可能会产生一些氨基酸,虽然说谷物含有一些维生素,但在小谷物食品发酵过程中添加乳酸菌或酵母可能会提高维生素水平。
例如,添加乳酸乳球菌N8 和布拉酵母菌Saccharomyces boulardii SAA655可将 idli 面糊中核黄素和叶酸的浓度提高 40-90%。
注:Idli 是印度的一种传统发酵面包,是一种由大米和脱壳黑豆原料的混合物,经过湿磨和传统发酵制成的糕点。
尽管小米主要用于制造面粉,但其加工(发芽和发酵)会增加谷物的铁含量。
➤5
抗生素的滥用可能破坏肠道微生物群的平衡,影响微量营养素的代谢和吸收。
抗生素还可能导致肠道黏膜的炎症反应,进一步干扰微量营养素的吸收和利用。
在使用抗生素时,建议按照医生的指导使用,并且在必要时采取预防措施,如同时摄入益生菌和益生元,以减少对肠道微生物的不良影响。
扩展阅读:
➤6
过度摄入糖或长期的压力和焦虑会对肠道微生物和宿主营养吸收产生不良后果。
深加工食品通常经过多道工序,其中许多有益的营养物质可能被去除或破坏。这些食品通常富含添加剂、防腐剂和高糖分,而这些成分可能会对微量营养素的吸收和利用产生负面影响。
通过减少摄入加工食品、管理压力等干预措施,可以帮助改善营养吸收和维持良好的微量营养素水平。
扩展阅读:
你的焦虑可能与食品添加剂有关,警惕食品添加剂引起的微生物群变化
➤7
运动可以改善血液循环,增加血液流向肠道和其他组织,从而提高微量营养素的输送和吸收。
运动还可以提高新陈代谢,提高能量消耗和细胞代谢的速度。这可能会导致对微量营养素的需求增加,从而促进其吸收和利用。
➤8
定期体检可以帮助发现潜在的营养和健康问题,及早采取措施。
定期进行体检是监测体内微量营养素的一种方法,但它可能无法提供详细的微量营养素水平信息。常规体检通常包括血液检查,可以检测一些关键的营养素水平,如铁、维生素B12、维生素D等。对于有些微量营养素,可能需要进行特殊的检测。
➤9
由于每个人的肠道微生物群和微量营养素状态可能存在差异,个体化的营养咨询和评估可以帮助确定针对个体需求的饮食和营养。
肠道菌群和微量营养素之间存在着紧密的关系和相互作用。肠道菌群中的某些菌种可以帮助合成和吸收微量营养素。这些菌种通过代谢和转化,将食物中的原始形式的微量营养素转化为人体可以吸收和利用的活性形式,提供给宿主机体所需。
同时,适当的微量营养素摄取也对肠道菌群的平衡和健康至关重要。一些微量营养素,如锌、铁等,不仅是肠道菌群的生长和代谢所必需,还能影响菌群的结构和功能。缺乏这些微量营养素可能导致菌群紊乱,而菌群紊乱又可能影响微量营养素的吸收和利用。
肠道细菌对宿主微量营养素状态的影响是一个急需关注的研究领域。同时对微量营养素代谢的研究应该扩大到包括一般肠道微生物组成,而不仅仅是病原菌。
相关阅读:
主要参考文献
Dje Kouadio DK, Wieringa F, Greffeuille V, Humblot C. Bacteria from the gut influence the host micronutrient status. Crit Rev Food Sci Nutr. 2023 Jun 27:1-16.
Amimo, J., H. Michael, J. Chepngeno, S. Raev, L. Saif, and A. Vlasova. 2022. Immune impairment associated with vitamin A deficiency: Insights from clinical studies and animal model research. Nutrients 14 (23):5038.
Baldi, A. J., L. M. Larson, and S.-R. Pasricha. 2020. Balancing safety and potential for impact in universal iron interventions. Global Landscape of Nutrition Challenges in Infants and Children 93:51–62.
Cuisiniere, T., A. Calve, G. Fragoso, M. Oliero, R. Hajjar, E. Gonzalez, and M. Santos. 2021. Oral iron supplementation after antibiotic exposure induces a deleterious recovery of the gut microbiota. BMC Microbiology 21 (1):1–15.
Kang, S., R. Li, H. Jin, H. Y ou, and G. Ji. 2020. Effects of selenium and zinc-enriched Lactobacillus plantarum SeZi on antioxidant capacities and gut microbiome in an ICR mouse model. Antioxidants 9 (10):1028.
Kramer, J., Ö. Özkaya, and R. Kümmerli. 2020. Bacterial siderophores in community and host interactions. Nature Reviews. Microbiology 18 (3):152–63.
McMillen, S., S. Thomas, E. Liang, E. Nonnecke, C. Slupsky, and B. Lonnerdal. 2022. Gut microbiome alterations following postnatal iron supplementation depend on iron form and persist into adulthood. Nutrients 14 (3):412.
Qin H, Wu H, Shen K, Liu Y, Li M, Wang H, Qiao Z, Mu Z. Fermented Minor Grain Foods: Classification, Functional Components, and Probiotic Potential. Foods. 2022 Oct 11;11(20):3155. doi: 10.3390/foods11203155. PMID: 37430904; PMCID: PMC9601907.

谷禾健康

—变形菌门
—γ变形菌纲
—肠杆菌目
—肠杆菌科
—埃希氏菌属
埃希氏菌属 (Escherichia),是一种常见的细菌。其中最著名的种是大肠杆菌(Escherichia coli),大肠杆菌是一种厌氧菌,通常生活在人和动物的肠道中。在成人的横断面研究中,大肠杆菌是超过90%个体肠道微生物组的成员,是最早在新生儿出生时定植的细菌之一。
埃希氏菌属细菌是一类适应于肠道环境的细菌,其代谢能力丰富,既可以进行好氧代谢,又可以进行发酵代谢。
大肠杆菌在正常情况下对人体无害,但某些菌株也可能引起感染和疾病。例如,某些毒力菌株的大肠杆菌可引起食物中毒,导致胃肠道症状,如腹泻、呕吐和腹痛。此外,大肠杆菌也可能引起泌尿系统感染和其他感染性疾病。
除了大肠杆菌,埃希氏菌属还包括其他一些菌株,如致病性的Escherichia albertii 和 Escherichia fergusonii。这些菌株在临床上相对较少见,但仍然可能引起感染和疾病。
本文我们来了解大肠杆菌的特性,代谢途径,和其他菌群相互作用等,可以更好地从肠道菌群的复杂性中窥探其在人体健康中的重要角色,并为预防和治疗相关疾病提供指导。
埃希氏菌属(Escherichia)是革兰氏阴性细菌中的一类,其中最为知名的是大肠杆菌(Escherichia coli)。
以下是埃希氏菌属的一些特点:
埃希氏菌属细菌主要存在于人类和动物的肠道中,是正常肠道菌群的一部分。
– 适宜温度:埃希氏菌属细菌一般生长在温度范围为20°C-45°C之间,最适生长温度约为37°C。
– 厌氧/需氧:大多数肠杆菌属细菌为好氧生物,但也有一些菌株可以在缺氧条件下生长。
– 好氧代谢:埃希氏菌属细菌能够利用氧气进行呼吸代谢,产生能量。
– 发酵代谢:埃希氏菌属细菌也可以进行发酵代谢,利用糖类等有机物产生能量。
埃希氏菌属目前包括:
序列比较表明,E. fergusonii 和 E. albertii 与大肠杆菌密切相关,而其余三个物种在进化上可能更为遥远。
志贺氏菌菌株在系统发育上与某些大肠杆菌菌株的关系比某些大肠杆菌菌株之间的关系更为密切。
– 大肠埃希氏菌属,又叫大肠杆菌(Escherichia coli):是埃希氏菌属中最为重要的菌株之一,广泛存在于人类和动物的肠道中。大肠杆菌具有多种毒力因子,包括肠毒素和肠出血性大肠杆菌毒素。
– 弗氏埃希氏菌(Escherichia fergusonii):是埃希氏菌属的另一种菌株,也存在于人类和动物的肠道中,但其致病性较低。
肠道感染的传播途径通常是粪口传播,可以通过人传人、直接接触动物或其环境,或者通过食用受污染的食物或水。
内源性感染途径也是可能的(例如细菌从肠道转移到血液),导致肠外疾病,并且更常见于免疫功能低下的人群,或患有肝硬化或接受化疗等基础疾病的人。
最重要的肠道病原体是肠沙门氏菌、大肠杆菌的某些菌株、志贺氏菌和小肠结肠炎耶尔森氏菌。在急性腹泻期间从粪便标本中分离出的肠杆菌科细菌,可能反映了粪便菌群的剧烈变化,而不是症状的原因。
代表菌大肠杆菌
大肠杆菌(Escherichia coli)是一种常见的革兰阴性细菌,存在于人类和动物的肠道中。大肠杆菌的大多数菌株对人类无害,但也有一些菌株具有致病性,可以引起不同程度的疾病。以下是一些主要的致病毒株和它们的具体信息:
大肠杆菌O157:H7
– 特点:产生肠出血性大肠杆菌毒素(Shiga毒素),引起严重的肠道感染。
– 病症:可导致急性胃肠炎、腹泻、腹痛、呕吐和血便,严重时可引起溶血性尿毒症综合征(HUS)。
– 传播途径:食物污染(尤其是未煮熟的牛肉)、接触感染(粪口传播)等。
大肠杆菌O26、O45、O103、O111、O121和O145(非O157致病毒株)
– 特点:产生肠出血性大肠杆菌毒素,与O157:H7类似,但毒力较弱。
– 病症:引起类似O157致病毒株的症状,包括腹泻、腹痛、呕吐和血便,也可引发HUS。
– 传播途径:主要通过食物污染(尤其是未煮熟的肉类、生奶和生菜)传播。
大肠杆菌O104
– 特点:产生肠出血性大肠杆菌毒素,与O157:H7类似。
– 病症:引起急性胃肠炎、腹泻、腹痛、呕吐和血便,也可导致HUS。
– 传播途径:主要通过食物污染(尤其是未煮熟的食品和生鲜蔬菜)传播。
大肠杆菌O111
– 特点:产生肠毒素,但不产生肠出血性大肠杆菌毒素。
– 病症:引起急性胃肠炎、腹泻、腹痛和呕吐。
– 传播途径:主要通过食物污染(尤其是未煮熟的肉类和生菜)传播。
需要注意的是,以上只是一些常见的致病毒株,大肠杆菌还有其他致病毒株,具体病症和传播途径可能有所不同。

总体而言,埃希氏菌属细菌是一类适应于肠道环境的细菌,其代谢能力丰富,既可以进行好氧代谢,又可以进行发酵代谢。大肠杆菌作为埃希氏菌属中最重要的代表菌株之一,具有多种致病因子,可能引起胃肠道感染和其他相关疾病。
埃希氏菌属与其他菌群共同维持肠道的稳态平衡,参与食物消化和营养吸收过程。它可以与厌氧菌共生,分解单糖、二糖和复杂多糖,为肠道提供能量和营养物质。
此外,埃希氏菌属还参与清除氧气,维持肠道的厌氧环境。埃希氏菌属在婴儿肠道中占据主导地位,可能是因为其具有上述功能,能够适应婴儿肠道的特殊环境。
然而,某些埃希氏菌属和梭状芽孢杆菌属的菌株与严重的胃肠道疾病(如炎症性肠病和腹泻)有关,这为肠道菌群成分与宿主免疫系统之间的相互作用提供了新的见解。
肠杆菌其外膜上存在一种叫做脂多糖(LPS)的分子。
LPS由三个部分组成:脂A、核心多糖和O抗原。
埃希氏菌属通过合成LPS来保护自身免受外界环境的伤害。LPS能够与宿主的免疫系统相互作用,引发炎症反应。
在健康的情况下,埃希氏菌属产生的LPS与宿主的免疫系统保持平衡,不会引发过度的炎症反应。然而,在某些情况下,如肠道菌群失调或感染,埃希氏菌属产生的LPS可能会超过宿主免疫系统的调节能力,导致炎症反应过度,对肠道和整体健康产生负面影响。
短链脂肪酸包括乙酸、丙酸和丁酸,它们是肠道内的重要能量来源,并对肠道菌群和健康产生影响。
虽然埃希氏菌通常不是短链脂肪酸的主要产生者,但它的存在和代谢活动可以对肠道微生物群落的整体代谢产生影响,并间接影响短链脂肪酸的生成。
埃希氏菌属的影响不仅局限于肠道,还可以通过肠-脑轴(MGB)对大脑功能产生影响。研究发现,埃希氏菌属的异常增加与焦虑、抑郁等精神疾病的发生有关。
大肠杆菌可以产生多种神经递质,如去甲肾上腺素、多巴胺、5-羟色胺和乙酰胆碱。这些神经递质可以通过影响神经元活动和调节情绪、压力反应等方式,对神经系统功能产生影响。
其次,大肠杆菌可以通过调节肠道微生物组的组成,影响神经递质的合成和释放。例如,它可以改变肠道微生物组中产生神经递质的菌株的丰度,从而影响神经递质的水平和功能。
此外,大肠杆菌还可以通过调节肠道微生物组与神经系统之间的相互作用,影响神经递质的信号传导和神经系统的功能。
增强:
抑制:
产生肠毒素
某些埃希氏菌株能够产生肠毒素,包括肠毒素A、肠毒素B等。这些毒素能够破坏肠道黏膜细胞,导致腹泻和肠道炎症,引起食物中毒或感染性腹泻。
破坏肠道屏障完整性
大肠杆菌可以通过多种机制破坏肠道屏障的完整性,包括改变肠上皮细胞之间的连接、破坏肠上皮细胞与肠道黏膜之间的黏附、改变肠道黏膜的结构等。这些破坏作用导致肠道屏障的通透性增加,使得细菌、毒素和其他有害物质能够穿过肠道屏障进入血液循环系统,引发炎症反应和其他肠道相关疾病。
激活肠道黏膜免疫
大肠杆菌感染会激活肠道黏膜免疫系统,导致炎症反应和免疫细胞的活化。这些炎症因子和免疫细胞的活化会进一步破坏肠道屏障的完整性,加剧肠道屏障功能的损伤。
引发肠道菌群失调
大肠杆菌感染会导致肠道菌群的失调,即有益菌的数量减少,有害菌的数量增加。这种菌群失调会进一步破坏肠道屏障的完整性,增加肠道屏障的通透性。同时,肠道菌群失调还会影响肠道黏膜免疫系统的平衡,导致炎症反应的增加。
在一些重症患者中,肠道菌群失调常常表现为埃希氏菌属的过度生长,同时伴随着其他有害菌的增加和有益菌的减少。这种失衡的菌群状态与多种疾病的发生和发展密切相关,包括肠道感染、炎症性肠病、代谢性疾病等。
▼
埃希氏菌属与炎症性肠病(IBD)之间存在着密切的关联。多项研究表明,在IBD患者的肠道中,埃希氏菌属的数量明显增加。
埃希氏菌属属于肠道菌群中的一种细菌,它可以在肠道黏膜上富集并与宿主相互作用。埃希氏菌属的增加可能与肠道黏膜的损伤和炎症反应有关。埃希氏菌属可以通过黏附和侵入肠道上皮细胞,导致炎症反应的加剧。它可以释放一些毒素和代谢产物,损伤宿主细胞膜和上皮屏障,进一步加剧肠道黏膜的损伤。
埃希氏菌属还可以释放细胞毒性扩展底物谱β-内酰胺酶(ESBL),这种酶可以降解β-内酰胺类抗生素,导致抗生素耐药性的产生。抗生素耐药性的增加可能使得治疗IBD的抗生素疗效降低,从而加剧炎症反应。
▼
大肠癌与大肠杆菌(Escherichia coli)的关联机制是多方面的。
首先,一些致病性的大肠杆菌菌株,如粘附侵袭性大肠杆菌(AIEC)和肠致病性大肠杆菌(EPEC),分泌的环调素染色质蛋白(colibactin)和效应蛋白EspF与大肠癌的发展和进展有关。
其次,大肠杆菌感染破坏了肠道微生物的微环境,从而诱导大肠癌的发生。
此外,大肠杆菌感染还与慢性炎症相关,而慢性炎症是大肠癌的一个潜在风险因素。大肠杆菌还可以通过多种策略促进结肠癌的生长和发展。
肠道微生物组的失调也被认为是大肠癌发生的因素之一。一些研究还发现,大肠癌患者的肠道微生物组成与正常人有所不同,大肠杆菌等菌株的丰度增加与大肠癌的发生相关。
▼
在摄入被致病性产志贺毒素的大肠杆菌(STEC)污染的食物或水后,Stxs可能通过M细胞摄取和胞吞作用或细胞旁转运穿过肠上皮屏障。一旦进入粘膜下层,毒素就会激活先天免疫细胞,如中性粒细胞或单核细胞,它们作为“载体”细胞在血液中输送Stxs,还可能通过局部产生促炎细胞因子进一步加剧组织损伤。
最终,毒素被转移到肾小球内皮细胞和肾小管上皮细胞,这些细胞富含毒素受体Gb3。对主要靶器官肾脏的损伤会导致腹泻相关溶血性尿毒症综合征。
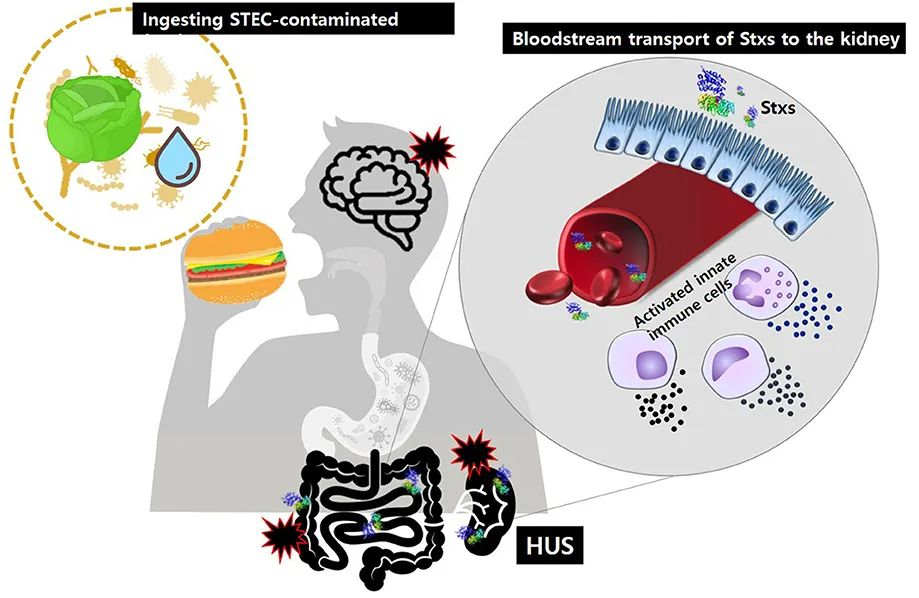
doi.org/10.3389/fcimb.2020.00273
▼
一些研究发现,在动脉粥样硬化患者中,肠杆菌属的丰度明显增加,而其他一些有益菌群的丰度明显降低。肠杆菌属的增加可能与动脉粥样硬化的发生和发展有关。
埃希氏菌属与糖尿病之间存在一定的关联。糖尿病患者往往存在高血糖状态,这为埃希氏菌属等细菌提供了良好的生长环境。研究发现,糖尿病患者的肠道菌群组成与健康人存在差异,其中埃希氏菌属的相对丰度较高。
埃希氏菌属可以产生多种促炎因子,如脂多糖和肽聚糖,从而引发宿主免疫反应,导致不同程度的肠道炎症。埃希氏菌属还可以破坏肠道上皮的完整性,导致低度炎症和自身免疫反应的发生,增加了1型糖尿病的风险。
研究还发现,糖尿病患者肠道菌群的改变与炎症、代谢紊乱等病理过程密切相关。
▼
二甲双胍的使用会导致埃希氏菌属的丰度增加
多项研究发现,二甲双胍治疗后,埃希氏菌属的数量明显增加。这种变化在2型糖尿病患者和健康人群中均有观察到。
埃希氏菌属的增加与二甲双胍的副作用有关
一些研究发现,埃希氏菌属的增加与二甲双胍治疗的胃肠道副作用有关。这可能是由于埃希氏菌属的存在导致了肠道内毒素的合成增加,从而引起胃肠道不适的症状。
埃希氏菌属的增加可能与肠道菌群的失衡有关
研究表明,二甲双胍的使用会导致肠道菌群的改变,包括埃希氏菌属的增加和其他有益菌的减少。这种菌群失衡可能与二甲双胍的疗效和副作用有关。
埃希氏菌属的增加可能与肠道炎症和代谢紊乱有关
一些研究发现,埃希氏菌属的增加与肠道炎症和代谢紊乱相关。这可能是由于埃希氏菌属的存在导致了肠道内炎症因子的释放增加,从而影响了机体的代谢功能。
▼
埃希氏菌属与非酒精性脂肪肝病(NAFLD)之间存在着密切的关联。多项研究发现,在非酒精性脂肪肝病患者中,埃希氏菌属的丰度明显增加。
埃希氏菌属可以产生乙醇,乙醇会破坏肠道屏障功能,导致肠道通透性增加。这可能是埃希氏菌属与非酒精性脂肪肝病发展相关的关键机制之一。
另外,埃希氏菌属的增加也与肝脏炎症和肝纤维化的发生相关。研究发现,在NAFLD患者中,埃希氏菌属的数量与肝纤维化的严重程度呈正相关。埃希氏菌属可能通过产生内毒素和诱导炎症反应,促进肝脏纤维化的发展。
肠道菌群失调和肠道屏障功能受损是NAFLD发展的重要因素。埃希氏菌属的增加可能导致肠道菌群失衡,进一步破坏肠道屏障功能,使内毒素进入血液循环,引发炎症反应,从而促进NAFLD的发展。
▼
埃希氏菌属与多动症存在相关性。研究发现,多动症患者的肠道菌群中有害菌如肠杆菌、大肠埃希氏菌和梭状芽孢杆菌的丰度显著增加,而有益菌如双歧杆菌、瘤胃球菌丰度显著降低。这些结果表明,某些埃希氏菌属可能与多动症的发展和症状有关。
罗伊氏乳杆菌DSM 17938已被证明具有有效的抗菌活性,调节免疫应答、减少肠道炎症。
植物乳杆菌CCFM1143对产肠毒素大肠杆菌引起的腹泻具有最明显的保护作用,通过调节炎症细胞因子、再平衡肠道菌群和调节短链脂肪酸生成的方式。
鼠李糖乳杆菌SHA113对耐药大肠杆菌QBQ009感染显示出有效的治疗效果。
短双歧杆菌FHNFQ23M3和双歧双歧杆菌FSDJN7O5可缓解产肠毒素大肠杆菌引起的腹泻症状。
荷花种子荚中的寡聚原花青素(LSPC)和益生菌(鼠李糖乳杆菌LGG和双歧杆菌Bb-12)对肠毒素性大肠杆菌(ETEC)感染的腹泻小鼠的保护作用。
阿拉伯木聚糖、菊粉和罗伊氏乳杆菌 1063 抑制粘膜中粘液的粘附侵袭性大肠杆菌。
主要参考文献:
Braz VS, Melchior K, Moreira CG. Escherichia coli as a Multifaceted Pathogenic and Versatile Bacterium. Front Cell Infect Microbiol. 2020 Dec 21;10:548492. doi: 10.3389/fcimb.2020.548492. PMID: 33409157; PMCID: PMC7779793.
Mirsepasi-Lauridsen HC, Vallance BA, Krogfelt KA, Petersen AM. Escherichia coli Pathobionts Associated with Inflammatory Bowel Disease. Clin Microbiol Rev. 2019 Jan 30;32(2):e00060-18. doi: 10.1128/CMR.00060-18. PMID: 30700431; PMCID: PMC6431131.
Sun J, Marais JP, Khoo C, LaPlante K, Vejborg RM, Givskov M, Tolker-Nielsen T, Seeram NP, Rowley DC. Cranberry (Vaccinium macrocarpon) oligosaccharides decrease biofilm formation by uropathogenic Escherichia coli. J Funct Foods. 2015 Aug;17:235-242. doi: 10.1016/j.jff.2015.05.016. PMID: 26613004; PMCID: PMC4657873.
Kim JS, Lee MS, Kim JH. Recent Updates on Outbreaks of Shiga Toxin-Producing Escherichia coli and Its Potential Reservoirs. Front Cell Infect Microbiol. 2020 Jun 4;10:273. doi: 10.3389/fcimb.2020.00273. PMID: 32582571; PMCID: PMC7287036.
Zhang S, Abbas M, Rehman MU, Wang M, Jia R, Chen S, Liu M, Zhu D, Zhao X, Gao Q, Tian B, Cheng A. Updates on the global dissemination of colistin-resistant Escherichia coli: An emerging threat to public health. Sci Total Environ. 2021 Dec 10;799:149280. doi: 10.1016/j.scitotenv.2021.149280. Epub 2021 Jul 27. PMID: 34364270.
Denamur E, Clermont O, Bonacorsi S, Gordon D. The population genetics of pathogenic Escherichia coli. Nat Rev Microbiol. 2021 Jan;19(1):37-54. doi: 10.1038/s41579-020-0416-x. Epub 2020 Aug 21. PMID: 32826992.
Leekitcharoenphon P, Johansson MHK, Munk P, Malorny B, Skarżyńska M, Wadepohl K, Moyano G, Hesp A, Veldman KT, Bossers A; EFFORT Consortium; Zając M, Wasyl D, Sanders P, Gonzalez-Zorn B, Brouwer MSM, Wagenaar JA, Heederik DJJ, Mevius D, Aarestrup FM. Genomic evolution of antimicrobial resistance in Escherichia coli. Sci Rep. 2021 Jul 23;11(1):15108. doi: 10.1038/s41598-021-93970-7. PMID: 34301966; PMCID: PMC8302606.
Khorsand B, Asadzadeh Aghdaei H, Nazemalhosseini-Mojarad E, Nadalian B, Nadalian B, Houri H. Overrepresentation of Enterobacteriaceae and Escherichia coli is the major gut microbiome signature in Crohn’s disease and ulcerative colitis; a comprehensive metagenomic analysis of IBDMDB datasets. Front Cell Infect Microbiol. 2022 Oct 4;12:1015890. doi: 10.3389/fcimb.2022.1015890. PMID: 36268225; PMCID: PMC9577114.
Wang J, Li R, Zhang M, Gu C, Wang H, Feng J, Bao L, Wu Y, Chen S, Zhang X. Influence of Huangqin Decoction on the immune function and fecal microbiome of chicks after experimental infection with Escherichia coli O78. Sci Rep. 2022 Oct 5;12(1):16632. doi: 10.1038/s41598-022-20709-3. Erratum in: Sci Rep. 2022 Nov 18;12(1):19901. PMID: 36198724; PMCID: PMC9534884.