 国家高新企业 | ISO9001认证
国家高新企业 | ISO9001认证 二级病原微生物安全实验室
二级病原微生物安全实验室- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
国家高新企业 | ISO9001认证 二级病原微生物安全实验室

谷禾健康

人类微生物组研究正在从描述关联发展到了解整个微生态对人类的影响。虽然存在挑战,但在应用数据驱动的微生物组诊断和干预方面正在取得进展,这可能会在未来十年内带来精准医学的突破。
本文我们来探讨关于微生物组的研究进展及其对人类健康的影响及应用。
随着对微生物组的理解深入,发现巨大的潜力
过去 20 年,我们对微生物组在人类宿主生理学中作用的理解发生了重大转变。
开创性研究表明,与人类细胞和基因相比,原核生物为人类“全生物”贡献了相同数量的细胞和多倍的基因,同时影响哺乳动物从消化和新陈代谢过程到免疫甚至大脑功能。
微生物组的变化与越来越多的人类疾病有关,包括癌症。这些发现使人们普遍希望,积累的关于宿主-微生物组相互作用的知识,将迅速转化为人类疾病的治疗方法。
对微生物组的认识开始客观,同时意识到复杂性
随着该领域的稳步成熟,科学家们开始更好地了解整个微生物组。他们意识到这并不像他们最初想的那么简单。随着他们开始了解微生物之间的相互作用是多么复杂,以及将微生物组用于人类治疗有多么大的挑战,最初围绕它的兴奋和炒作开始平息。
面临挑战,需要综合多方面的努力
要克服这些挑战,许多研究者需要进行长期、数据驱动的研究。他们要综合各种看起来不相干的科学和医学学科,包括进化论、人类学和营养学等等,还要结合新技术的开发和优化,以及注意事项和混杂因素的识别和最小化,共同推动了这一探索。他们的目标是要弄清楚宿主和微生物组之间的互动关系和分子机制,以期找到因果关系。虽然充满了挑战,但只要大家协作一致,一定可以获得成果。这些成果将在未来十年内实现对可利用的治疗性微生物组目标进行更广泛和可重复的识别和调节。
微生物组研究中的挑战和新兴方法
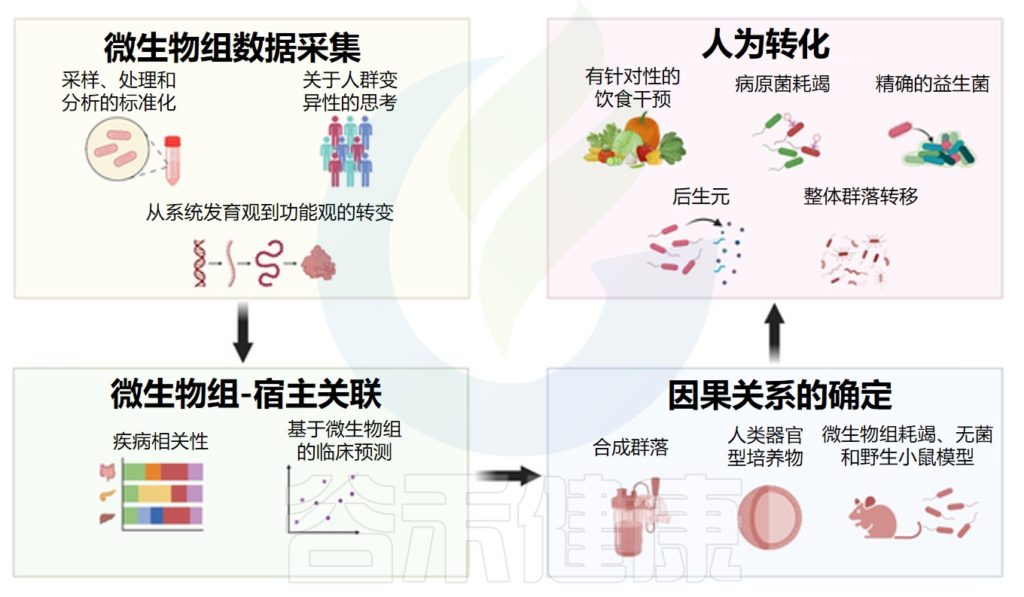
doi: 10.1371/journal.pbio.3002053
本世纪第一个十年,研究微生物组结构表征、对生理功能的重要性,由此衍生到各个领域
事实上,本世纪微生物组研究的第一个十年,集中于证明肠道微生物组对生理功能的重要性,以及在各种生理和疾病背景下基于下一代测序的微生物组群落结构表征。
这带来了微生物组与各种健康结果之间的关联、相关性和预测的识别,报告规模和细节的不断提升。与此同时,该领域广泛描述了微生物特征响应多种因素的动态调节,例如饮食、宿主免疫 、基因组学和昼夜振荡。
▼
随着这些重大进展,人们也认识到了主要的技术和概念障碍,这些障碍对微生物组发现的解释、概括和转化到临床具有挑战性。
使微生物组相关观察结果的解释复杂化的技术挑战之一是:不同研究之间样本收集、处理和分析方法的广泛差异。
此外,在微生物组处理和分析级联的每个步骤中发生的污染和注释错误可能会引入伪影和偏差,这些伪影和偏差难以解决并使真实和虚假生物信号之间的区别复杂化。
解决方案:将真实信号与混杂因素分开
了解与这些步骤中的每一个相关的范围、限制和混杂因素,对于将真实信号与混杂因素分开至关重要,特别是在微生物丰度低或无微生物丰度的样本。更好地协调这些方法,加上多种技术和生物控制,可以更准确地解释可以在人群、地域、性别和种族之间推广和复制。
然而,如果不冒技术可变性和多样性减少的风险,微生物组处理和分析标准化就不能过度执行,而技术可变性和多样性构成了推动研究创新的引擎。
▼
大多数人类微生物组研究面临的一个概念挑战是,微生物群落结构中内在的和生理上重要的个体间变异性。
如何在大样本人群数据上去识别和消除这些差异,应该是目前微生物组特征分析面临的巨大分析挑战,因为它们代表了一种固有的、难以解决的“噪音”。
解决方案:基于深度人工智能的方法等创新方法
可以识别和利用不同的微生物组配置来探索微生物组对个体人类表型的贡献,范围从血糖反应到补充剂和食物携带相同疾病易感位点的个体之间的不同疾病表现。
总而言之,使用基于深度人工智能的方法等创新方法应对技术和生物变异性的挑战,将加深微生物组知识与精准医学的合理整合。
▼
一个同样艰巨的挑战涉及从微生物组-人类特征关联到因果关系证明的进展。事实上,在建议与临床结果相关、相互关联和预测的许多微生物群中,只有少数基因产物和代谢物可以证明对生理和疾病状态有因果影响,而我们预计一部分微生物特征是先发于疾病病理改变的特征,而有一部分特征是继发于以疾病状态为特征的环境变化。
越来越多的工具正在开发中,通过揭示因果关系和机制洞察力,将前者“驱动”微生物组改变与后者“乘客”改变区分开来。
解决方案:无菌或使用抗生素的动物模型或其他改进的技术,测试单个细菌菌株或群落的影响
一种经常使用的方法依赖于微生物群耗尽(通过抗生素治疗)或微生物群缺乏(无菌)小鼠模型,这些模型允许引入单个细菌菌株或群落以测试它们对宿主生理学的影响。
抗生素杀菌模型局限性:多因素混杂,难以预测
抗生素是现代医学的基石,因为它们能够在感染期间抑制病原体的生长。抗生素的广泛特征是它们的抑制机制是抑菌(生长停止)或杀菌(致死),并且它们的活性范围是宽的或窄的。大多数抗生素的作用主要在物种单一培养中进行了研究,尽管临床环境中的治疗不可避免地会对人类肠道上的多物种群落产生意想不到的后果。抗生素会对肠道细菌造成附带损害,并降低肠道微生物群的多样性,由此可能出现艰难梭菌感染倾向增加和癌症治疗后死亡率增加。
抗生素扰动期间的群落动态很难预测,因为环境可以通过种间相互作用、pH 调节和代谢转化改变抗生素的作用。使用多种治疗方案,例如可变剂量、顺序安排多种化合物和同时使用鸡尾酒治疗使预测进一步复杂化。
同时多种药物可产生协同或拮抗作用, 由此杀死的程度分别大于或小于单独药物的总和,但是他们对肠道菌群的扰动和影响研究还甚少,检测中面临这些用药信息准确跟踪记录也较困难。而且抗生素导致的多重耐药细菌的定植也会导致抗生素耐药基因转移到微生物群中,最终会导致对各种有效抗生素的耐药性增加,并增加致命感染的风险。
其他新技术:改进的体内模型、类器官和片上器官平台
随着更多具有人类代表性的实验平台的出现,野鼠体内的微生物群与圈养外的家鼠相似,正迅速受到关注。这些改进的体内模型得到了合成人类共生群落的发展、以前“无法培养”微生物在菌株水平分辨率下的改进培养和表征,以及通过非靶向代谢组学和蛋白质组学对生物活性微生物效应因子或代谢物的无偏见阐明的补充。
类器官和片上器官平台能够研究人体组织环境中的单个细菌、细菌群落及其生物活性产物。
注:
类器官(organoid)是指由人体组织或干细胞等体外培养出的三维结构,能够模拟某些人体器官的结构和功能。类器官的制备通常是通过将干细胞或组织细胞在特定的培养条件下进行分化和自组装,形成类似于人体器官的结构。类器官的制备可以为疾病研究和药物筛选提供更真实、更可靠的模型。例如,肝脏类器官可以用于测试药物代谢和毒性,肠道类器官可以用于研究肠道微生物和肠道疾病,而大脑类器官可以用于研究神经退行性疾病等。
片上器官平台(organ-on-a-chip platform)是一种仿生学技术,它是通过微流控技术、微加工技术和3D打印技术等手段,将人体组织和器官的细胞培养在微型芯片上,模拟人体器官的生理、病理过程,以实现高通量、高精度的药物筛选、毒性测试和疾病研究。这种技术可以模拟人体器官的微环境,如细胞间相互作用、细胞信号传递、细胞分化、细胞迁移、细胞凋亡等生理和病理过程,从而更好地研究疾病的发生和发展,以及评价药物的安全性和有效性。
凭借这些新技术获得的能力,该领域正在揭示基因组测序之外的功能读数,解码难以培养和低生物量微生物组的贡献,并评估重要但研究不足的非细菌共生王国及其错综复杂的相互作用组网络。
事实上,对共生和机会病毒(包括噬菌体)、真菌和寄生虫的潜在功能及其对细菌共生生态系统和人类宿主的影响的不断探索,构成了正在推进的研究领域,当然肠道细菌还是最主要的。对这些特征不明确的共生微生物的研究需要进一步开发各种工具,包括改进的计算参考数据集、分子开发工具和体内定植模型。
通过对微生物分子和功能见解的不断扩展,可能会在微生物组研究的下一个时代开发广泛的诊断、预后和治疗人类应用。
治疗性微生物组使用的一个有前景的例子:
粪便微生物组移植 (FMT)
除了治疗常见的艰难梭菌感染之外,显示出对肝病、炎症和免疫相关疾病的积极改善效果
促进微生物组研究领域的蓬勃发展
失败的试验和风险强调了对机制的了解的重要性
为了避免人体微生物研究领域中早期技术失误:
为了辅助人类疾病的诊断和治疗需要:
作为向前迈出的积极一步,微生物组研究能够做到以下:
早期检测
识别和利用微生物组粘膜,免疫和全身信号,以便通过离散的疾病特征进行早期检测和患者分层。
治疗层面
“一刀切”的传统观念正在发生改变,人们不再简单地认为所有人的微生物组都可以通过相同的方式进行调节,而是转向将微生物组视为“信号中心”的观点,也就是说更加注重研究和借鉴微生物群落的信号传递机制,以更好地治疗相关疾病。
微生物组被视为特定环境的中继或缓冲器,整合环境和内源性信号,影响携带健康或疾病易感遗传特征的个体的生理和疾病表现。
一个备受关注的话题是“益生菌干预法”。证据表明益生菌可能有助于治疗和预防传染病,改善过敏,降低血糖,联合免疫治疗提高治疗效果。益生菌治疗会改变腔内微生物群,这种改变进一步会显着影响全身代谢,包括胰岛素抵抗,炎症,代谢等。
将微生物组作为一个高度模块化的“指纹”,有助于开发合理的、针对具体情况的和数据驱动的干预措施,例如个性化营养、精准益生菌、代谢物补充(“后生元”疗法)和定向病原体抑制模式。
这些方法的验证和整合,加上对大量微生物群及其相关生物活性产品对人类宿主的影响的持续探索,将有助于实现该领域改变人类健康的巨大前景。
参考文献
Puschhof J, Elinav E. Human microbiome research: Growing pains and future promises. PLoS Biol. 2023 Mar 17;21(3):e3002053.

谷禾健康

我们知道,口服抗糖尿病药是治疗糖尿病的有效方式之一。然而,患者对抗糖尿病药的反应程度各不相同,例如,有些患者在长期使用阿卡波糖后会产生耐药性。
阿卡波糖通常在饭前口服。它抑制人α-葡萄糖苷酶达到降血糖作用,包括唾液和胰淀粉酶,从而减缓复杂碳水化合物的消化,并降低餐后血糖水平的升高。
大部分阿卡波糖剂量在肠道中仍未被吸收,导致与肠道微生物组的强烈相互作用。
近期发表在《自然新陈代谢》杂志上的一项研究揭示了阿卡波糖“诱导降解失活”的新机制:肠道菌群可以将阿卡波糖降解,从而破坏阿卡波糖的药效。

首先鉴定肠道细菌K. grimontii TD1,在阿卡波糖失活中起重要作用。
进一步在K. grimontii TD1 中鉴定出一种阿卡波糖首选葡萄糖苷酶 Apg ,它可以将阿卡波糖降解为小分子,并失去抑制剂功能。
由于阿卡波糖被肠道细菌降解,相对较大的人群可能存在阿卡波糖耐药性风险,这也是非抗生素耐药性的一个临床相关例子。
研究过程
研究人员招募了 393 名2型糖尿病患者,发现:
阿卡波糖的治疗效果存在显著的个体差异。
在接受α-葡萄糖苷酶抑制剂阿卡波糖治疗的2型糖尿病患者队列中,无应答者的粪便样本显示出明显高于有应答者的药物降解。
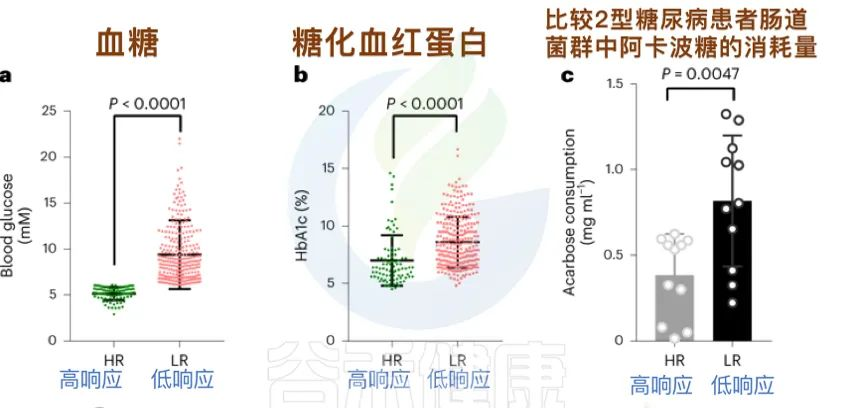
Tian J, et al., Nat Metab. 2023
这指向了微生物介导的药物降解途径。
为了分离阿卡波糖降解细菌种类,作者在逐步增加阿卡波糖浓度的条件下,对药物没反应的人的粪便样本进行培养,以富集细菌。
Tian J, et al., Nat Metab. 2023
这种方法导致分离出18种阿卡波糖降解剂,所有这些降解剂都属于Klebsiella grimontii。
Klebsiella grimontii 是兼性厌氧菌,属于变形菌门,肠杆菌科,产酸克雷伯菌复合体的一种,主要分布于小肠。
产酸克雷伯菌是人类肠道中的共生菌群,但也是一种机会致病菌,在近年来的临床分离的微生物样本中呈逐年上升的趋势。产酸克雷伯菌复合体可产生许多毒素,因其携带较多的毒力因子,并且具备携带和获得耐药基因的能力,故未来可能成为威胁人类健康的潜在病原菌,需要更多研究。
K. grimontii 这种兼性厌氧细菌主要定植于小肠,巧合的是,阿卡波糖主要靶向小肠上皮细胞中的多种淀粉酶和葡萄糖苷酶,这在空间上有利于其与K. grimontii TD1 的相互作用以及该菌株的代谢降解。
▶▷▶▷
事实上,在糖尿病小鼠模型中:
K. grimontii TD1菌株削弱了阿卡波糖的降血糖作用。
作者制作了一个T2DM小鼠模型。将小鼠分为四组,并用磷酸盐缓冲盐水、阿卡波糖、K. grimontii TD1或阿卡波糖和K. grimontii TD1处理。
阿卡波糖单独可以降低淀粉负荷后的血清葡萄糖水平;然而,这种作用在接受阿卡波糖和K. grimontii TD1的组中不太明显。这些发现表明,K. grimontii TD1降低了阿卡波糖的抗糖尿病作用。
由此,确定了降解阿卡波糖的细菌。
利用转录组学,研究人员在以淀粉为唯一碳源的最低培养基中培养了K. grimontii TD1,使细菌生长依赖于淀粉酶。通过添加阿卡波糖来抑制淀粉酶活性,迫使K. grimontii TD1上调有助于降解阿卡波糖的基因,这可以通过RNA测序进行量化。
同时,利用蛋白质组学,作者分离出细胞裂解物和具有高阿卡波糖降解活性的分离部分。这些组分中的蛋白质通过质谱法鉴定。
转录组学和蛋白质组学的发现都指向一个共同的候选者:一种新的葡萄糖苷酶,作者将其命名为阿卡波糖首选葡萄糖苷酶(简称Apg)。这是由K. grimontii TD1引起的阿卡波糖失活的关键。
在体外测定中或当在大肠杆菌中异源表达时,该酶能够将阿卡波糖水解为阿卡波糖葡萄糖和阿卡波糖。这两种产物在很大程度上都不能抑制猪α-淀粉酶。
很有意思的是,该酶在pH为7.5和温度为37°C的厌氧条件下表现最好,这些条件与人类肠道的生理学相似。

Tian J, et al., Nat Metab. 2023
那么这就引发了一个问题:
阿卡波糖如何成为细菌葡萄糖苷酶的底物,但又是人类葡萄糖苷酶(如α-淀粉酶)的抑制剂?
通过3D分子建模,作者发现葡萄糖苷酶Apg催化位点的残基与人类α-淀粉酶中的残基位置不同,这使得阿卡波糖能够水解裂解。
阿卡波糖的降解产物太小,无法有效地占据人类α-淀粉酶的配体结合口袋,导致酶的抑制作用降低。
这种独特的蛋白质结构不仅有助于理解这种酶的独特催化机制,而且为设计能够在体内阻断这种酶的抑制剂提供了潜在的靶点。
在上述2型糖尿病患者队列中,超过98%的阿卡波糖治疗患者携带具有Apg样酶的细菌,相比之下,约14.2%的健康个体携带Apg+细菌。
地理位置、性别、BMI等因素决定了其患病率和丰度的个体间差异,在分析中,Apg的丰度在不同人群之间有所不同,在中国人中最为普遍。
这些发现表明,肠道微生物群中的Apg和Apg样酶可能会影响许多患者的药物疗效和治疗成功率。
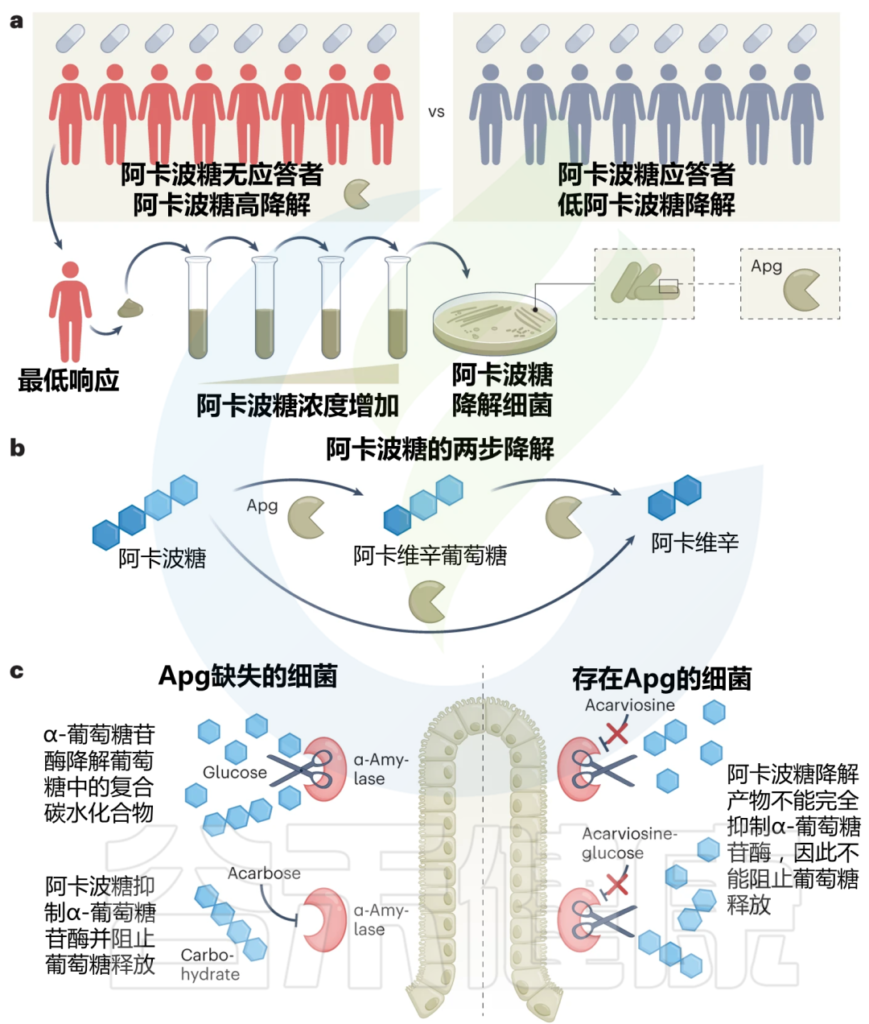
Brauny MM, et al., Nat Metab. 2023
▶ 01
这与已经描述的涉及阿卡波糖磷酸化的途径不同。阿卡波糖激酶在人类微生物组中广泛存在。这表明阿卡波糖抗性是一种常见的防御策略,在不同的肠道微生物中独立出现。为什么阿卡波糖抗性是肠道中的一个有益特征尚不清楚;也许是为了解毒相关的内源性化合物,或者成功地确保复合碳水化合物作为食物来源。
▶ 02
其他阿卡波糖降解葡萄糖苷酶以前也有描述,但从未与处理结果直接相关。由于碳水化合物代谢的基因经常过度表达,并且在肠道细菌的基因组中表现出相当大的差异,可以想象,很快就会发现更多这样的酶。
▶ 03
预计其他α-葡萄糖苷酶抑制剂以及其他基于碳水化合物的药物和食品添加剂,都可能会经历类似的过程。因此,这项研究可能只是冰山一角,很快可能会有更多这样的观测结果。
▶ 04
这种方法只需要关键细菌能够被培养。在目前的研究中,作者在纯培养中工作,但这种方法在细菌群落的培养中也应该是可行的。
总的来说,很大一部分2型糖尿病患者可能由于高水平的K. grimontii TD1而对阿卡波糖产生耐药性。对于阿卡波糖反应低的患者,可以考虑调节肠道微生态或靶向代谢酶的策略,以减少耐药性的发生。肠道微生物群的分析可用于辅助指导未来的治疗。
当然,K. grimontii TD1不太可能是肠道微生物群中唯一可以降解阿卡波糖的成员,可以对阿卡波糖降解肠道细菌以及相关代谢机制进行持续研究,从而更全面地了解微生物组与阿卡波糖的相互作用。
在一定条件下,肠道菌群可能利用一种主要机制,或几种共存方式来支持其生存,这需要更多的临床病例分析和深入研究来阐明其潜在机制,从而指导精准医疗。
微生物药物代谢很重要。更好地了解微生物组编码的药物降解途径,对于药物开发、个性化治疗及有效的治疗策略等方面具有巨大潜力。
本文提供了一种研究思路,微生物研究人员可以从中学习如何通过实验设计鉴定药物降解酶,这有助于开发个性化及有效的治疗策略。
药物开发相关人员可以考虑微生物代谢途径对药物吸收、代谢和治疗效果的影响,并在此基础上加强药物设计和开发过程中的微生物学考虑。
一些具体实验细节设计,详见原文:
Tian J, Li C, Dong Z, et al., Inactivation of the antidiabetic drug acarbose by human intestinal microbial-mediated degradation. Nat Metab. 2023 May;5(5):896-909.
谷禾健康
假单胞菌属(Pseudomonas)是最多样化和普遍存在的细菌属之一,其物种存在于沉积物、临床样本、植物(或植物根际)、患病动物、水、土壤、海洋、沙漠等,这反映在它们多变的代谢能力和广泛的适应环境的潜力上。
假单胞菌属是一类革兰氏阴性的好氧或微需氧的细菌,属于 Proteobacteria(变形菌门),代表种包括:
Pseudomonas aeruginosa(铜绿假单胞菌)
Pseudomonas putida (恶臭假单胞菌)
图源:CDC
假单胞菌属是机会性病原体,是全世界医院获得性感染的主要原因。它可以侵入多个器官系统,包括血液、肺、软组织(皮肤、肌肉和肌腱)、身体的其他部位。症状取决于感染部位。
与假单胞菌属感染相关包括免疫系统受损患者的感染和医疗器械相关感染等,如刚接受过手术的人和免疫功能低下的人,艾滋病或糖尿病患者。
假单胞菌属与人体健康相关,其中一些菌株可以引起感染和疾病,如铜绿假单胞菌(Pseudomonas aeruginosa),可以引起呼吸道、肠道、泌尿道和皮肤感染。
免疫抑制个体和住院患者的铜绿假单胞菌肠道携带率显着升高,因此感染和抗生素相关性腹泻的风险增加。
假单胞菌属与其他菌属如葡萄球菌、普雷沃氏菌属、不动杆菌属、芽孢杆菌属、肠杆菌属、肠球菌属等菌属有互作关系。
假单胞菌不被认为是肠道微生物组的典型成员,并且假单胞菌在肠道定植与发生肺部感染和死亡率的风险增加有关。
研究表明,假单胞菌与肠道屏障功能障碍和感染有关,可能导致败血症和多器官功能障碍综合征。此外,假单胞菌与炎症性肠病的发生也有关系,它在IBD患者的肠道中检测到丰度更高,而且它参与引起其他菌群的失衡会影响疾病的进展。
细菌有可能在人体内的不同部位之间转移,防止肠道定植或肠道到其它器官尤其肺部的传播,可能是预防重症患者假单胞菌感染的有效策略。
本文主要分享假单胞菌属(Pseudomonas)的生态,分类,致病因素以及与人体相关健康特性和可能的生活干预措施。
假单胞菌属(Pseudomonas),是革兰氏阴性、好氧菌属,于 1894 年被描述,是最多样化和普遍存在的细菌属之一。有些物种对人类、动物或植物具有致病性。
▼
假单胞菌属(Pseudomonas)属于假单胞菌科,包含200多个有效描述的物种。该属的成员表现出大量的代谢多样性,因此能够在广泛的生态位中定居。
它们易于体外培养,并且越来越多的假单胞菌菌株基因组序列可供使用,这使得该属成为科学研究的绝佳焦点。
Photograph by Kateryna Kon
研究最好的物种包括铜绿假单胞菌。直的或略微弯曲的棒状但非螺旋状,0.5–1.0 × 1.5–5.0 µm。由一根或数根极鞭毛运动;很少不动。在某些物种中,也可能形成短波长的侧鞭毛。有氧,具有严格的呼吸型代谢,以氧为末端电子受体;在某些情况下,硝酸盐可用作替代电子受体,从而允许厌氧生长。
大多数物种(不是全部)无法在酸性条件(pH 4.5 或更低)下生长。大多数物种不需要有机生长因子。氧化酶阳性或阴性。过氧化氢酶阳性。化学有机营养的。
▼
——分布广
假单胞菌属是变形菌门的一个庞大而多样的属。从北极苔原到热带雨林,从干旱的土壤到雨云,几乎在地球的每个角落都可以找到该属的成员。这种令人难以置信的环境适应性归功于假单胞菌非凡的新陈代谢多样性。
——野蛮生长
假单胞菌可以在 0 至 42 °C 的温度范围内生长,甚至可以在更极端的温度下存活。它们对营养的需求很少,可以利用多种碳源。尽管假单胞菌在有氧环境中生长最佳,但它们也可以利用氮进行厌氧呼吸。
它们可以作为自由生活的浮游细胞或作为生物膜群落的成员生活,并且具有将微生物信号和环境线索转化为特定生态位过程的非凡能力。
基因组分析表明,许多其他因素有助于假单胞菌属的多样性和适应性。除了赋予调节和代谢灵活性的常见基因的大量等位基因差异外,水平基因转移影响了致病性假单胞菌属的能力。
▼
第一个假单胞菌基因组测序于 2000 年,有 630 万个碱基对,是当时最大的细菌基因组测序。基因组包含大量参与分解代谢、运输、流出、运动和信号响应调节的基因。
事实上,铜绿假单胞菌基因组中超过 8% 的基因被认为参与了调控,这远远超过了在任何其他细菌基因组中观察到的百分比。很明显,假单胞菌成功的关键在于它可以表达其基因的可塑性,这是由多层监管复杂性提供的。

Sem Photograph by David M. Phillips
自 2000 年以来,已测序的 1000 多个假单胞菌基因组中的绝大多数都是铜绿假单胞菌的临床菌株。
总的来说,铜绿假单胞菌基因组的主要部分(约 4000 个基因)在所有菌株中都是保守的,代表了“核心基因组”。多达另外 20% 的基因驻留在共同代表“附属基因组”的基因组岛上。正是这个附属基因组赋予了铜绿假单胞菌的可塑性,并包含许多参与代谢、毒力和抗生素抗性的基因。
由于已在测序分离株的附属区域鉴定出大约 10 000 个独特基因,因此估计铜绿假单胞菌泛基因组可以接近甚至超过 100 000 个基因,这意味着假单胞菌这一物种的遗传库将远远超过人类。
该属的分类学多年来一直存在争议,因为许多最初包含在假单胞菌属中的细菌类群已被重新归类为其他属或来自不同类别的变形杆菌的物种。
可以根据16S rRNA、gyrB、rpoB和rpoD基因的序列分为三个谱系和13个群。
临床常见菌种主要包括:
• 铜绿假单胞菌(Pseudomonas aeruginosa)
• 斯氏假单胞菌(Pseudomonas stutzeri)
• 恶臭假单胞菌(Pseudomonas putida)
• 荧光假单胞菌(Pseudomonas fluorescens)
• 门多萨假单胞菌(Pseudomonas mendocina)
• 产碱假单胞菌(Pseudomonas alcaligenes)
• 假产碱假单胞菌(Pseudomonas pseudoalcaligenes)
其它菌种见附录一
致病性菌株见附录二
▼
详见附录三
此外,铁是细菌必需且稀缺的营养素,也是重要的信号因子。铜绿假单胞菌已经开发出多种铁吸收系统来隔离足够的铁以维持其生存,在毒力因子的释放和生物膜的形成中具有重要的调节作用。
▼
假单胞菌适应进化的特性对人类既有益又有害。
——有益
一方面,我们利用假单胞菌的力量进行生物修复和生物防治。例如,荧光假单胞菌和P. protegens已被证明在害虫控制和作物保护方面特别成功,它们被认为可以战胜和/或对抗植物病原体。
假单胞菌的分解代谢能力也被用于杀虫剂、重金属和碳氢化合物(例如石油泄漏)以及许多其他污染物的生物降解和/或解毒。
——有害
另一方面,某些种类的假单胞菌对植物和动物具有致病性,导致极难根除的感染。例如,铜绿假单胞菌是世界范围内医院获得性感染最常见的原因之一,并在使用消毒剂和抗生素后存活下来。
它可以在许多解剖部位引起感染,包括皮肤、大脑、眼睛、耳朵、泌尿道、肺部等。免疫抑制的个体,尤其是那些有过度烧伤、囊性纤维化或中性粒细胞减少症的个体,尤其处于危险之中。(这在后一章节会详细介绍)
▼
一般非特异性症状(许多疾病中常见的症状)包括:
假单胞菌感染的症状取决于它们发生在身体的哪个部位。
假单胞菌(Pseudomonas)可引起以下器官感染:
耳朵(外耳炎或游泳者的耳朵):
疼痛、水肿(肿胀)、压痛和分泌物
皮肤:
假单胞菌通过压疮、烧伤和手术伤口或从血流进入菌血症患者的皮肤,还可能引起毛囊炎等。
具体可表现为:脓肿(感染肿块)、皮疹、脓液、皮肤溃疡、坏死组织(皮肤变黑,表明组织正在死亡)。
骨骼或关节:
受影响区域行动不便、关节或背部疼痛、肿胀、疼痛或发热。
这些通常是由于静脉内 (IV) 药物使用或感染的手术伤口或受伤造成的伤口细菌传播到皮肤、骨骼和关节的结果。
伤口:
伤口有分泌物或结痂(通常是黄绿色),可能有甜味或难闻的气味。
消化道:
阑尾炎(由于治疗导致白细胞低的人发生肠道感染)腹痛、腹胀和腹泻的症状。
肺:
假单胞菌可能会导致严重的肺炎,通常发生在住院、已经生病和免疫功能低下的人身上。它可引起囊性纤维化患者的慢性肺部感染(遗传病),支气管扩张(气道永久性扩张),或慢性阻塞性肺病。
具体可表现为:呼吸急促、胸痛、咳痰(伴有白色、黄色或绿色粘液)和充血。
泌尿道:
排尿疼痛、尿液难闻或有甜味、尿急、尿液浑浊。
因此,假单胞菌感染的特征在于广泛的症状。
▼
紧急情况下:呼吸急促、心率加快、脉搏微弱、意识模糊、发烧、发抖或剧烈疼痛是败血性休克的症状,是对血流感染的全身反应。
在极少数情况下,心脏瓣膜可能会受到感染,尤其是在静脉注射药物的人群中。
脏东西的细菌可以进入血液,会传播到心脏并感染心脏瓣膜,这种情况称为心内膜炎。
使用静脉注射药物的人出现新的杂音(心音异常)可能会怀疑与假单胞菌相关的心内膜炎。
▼
未经治疗、治疗不足或复发的假单胞菌感染会导致更广泛的感染,例如败血症,这可能是致命的。
更可能出现严重并发症的高危人群包括:
值得注意的是,假单胞菌属内的抗生素耐药性正在增长。这些细菌正在寻找新的方法来避免死于以往最强大有效的抗生素。
假单胞菌属与人体健康相关,其中一些菌株可以引起感染和疾病,如Pseudomonas aeruginosa 绿脓杆菌,又称铜绿假单胞菌,可以引起呼吸道、泌尿道、皮肤感染。虽然不是正常口腔菌群的成员,但在口腔疾病和牙齿矫正治疗中可能会富集。
▼
假单胞菌属不被认为是肠道微生物群落的典型成员,肠道定植假单胞菌属与发展肺部感染和死亡风险增加有关。
假单胞菌属的过度生长可能会导致肠道炎症和系统性炎症反应。
假单胞菌属感染会引起机体的Th17细胞反应和系统性炎症反应,因此,干预Th17细胞免疫反应可能是缓解肺炎的有效方法。此外,假单胞菌属也可以通过产生GABA来影响蛋白质代谢和细胞稳态,从而影响宿主的摄食行为。
▼
假单胞菌属与其他菌属如大肠杆菌、沙门氏菌等有关,与肠道微生物群的平衡和健康密切相关。
假单胞菌属与其他菌属如葡萄球菌属和普雷沃氏菌属等,共同存在于囊性纤维化患者的肺部感染中。
假单胞菌属与其他菌属如不动杆菌属(Acinetobacter)等一起被发现在冷鲜乳中,产生脂肪酶和蛋白酶,参与乳制品和肉类的变质。
假单胞菌属与其他菌属如Bacillus、Enterobacter、Enterococcus等一起参与了肠-肺轴的微生物和免疫调节,影响呼吸道疾病的发生和发展。
▼
与假单胞菌属相关的疾病包括:免疫系统受损患者的感染和医疗器械相关感染等。免疫抑制个体和住院患者的铜绿假单胞菌肠道携带率显著升高,因此感染和抗生素相关性腹泻的风险增加。
研究表明,假单胞菌属与肠道屏障功能障碍和感染有关,可能导致败血症和多器官功能障碍综合征。假单胞菌属与肺部疾病也有关联,如支气管扩张症。
肺部疾病
研究发现,肺部假单胞菌属感染会影响肠道菌群,而通过调节肠道菌群可以影响肺部疾病的治疗效果。
肠道定植通常先于肺部感染,并且相同菌株经常在肠道和肺部中发现,表明肠道充当了假单胞菌属的储存库,可以传播到肺部和其他感染部位。
肠道微生物群的免疫调节对呼吸道疾病的发生有影响,肠道共生菌通过屏障效应提供菌群抵抗力,从而保护肠道生态环境不被改变。
以肺部假单胞菌、肠道拟杆菌和肠道酵母菌为特征的高肠-肺相互作用集群,与加重的恶化,整体支气管扩张严重相关。
炎症性肠病(IBD)
假单胞菌属与炎症性肠病的发生也有关系。
研究表明,IBD患者肠道中假单胞菌属的种类和数量与非IBD患者存在显著差异。
其中,克罗恩患者中假单胞菌属的检出率明显高于非IBD患者。此外,假单胞菌属的多样性在活动性IBD患者中也明显降低。这些结果表明假单胞菌属可能与IBD的发病机制有关。具体而言,假单胞菌属可能通过影响肠道菌群的平衡和多样性,导致肠道免疫系统的异常反应,从而引发IBD的发生。
因此,调节肠道菌群可能是预防和治疗肠道疾病的一种新策略。
▼
铜绿假单胞菌(Pseudomonas aeruginosa)是一种普遍存在的革兰氏阴性细菌,存在于许多不同的环境中,包括不同的生命来源,如动物和人类。
铜绿假单胞菌能够以最低的营养需求生存并耐受各种身体条件,这使得该生物体能够在社区和医院环境中持续存在。
图源:EYE OF SCIENCE / SCIENCE PHOTO LIBRARY
引发感染
铜绿假单胞菌是一种机会性病原体,这意味着它利用宿主防御系统中的一些漏洞来引发感染。
铜绿假单胞菌是世界范围内医院感染的罪魁祸首,其医学和兽医重要性日益增加,并且主要在免疫系统受损的患者中引起感染。
这种细菌几乎从不感染未受损的组织,但如果组织防御以某种方式受损,它就会发起攻击。
铜绿假单胞菌可能引起尿路感染、呼吸系统感染、皮炎、软组织感染、菌血症、骨和关节感染、胃肠道感染和各种全身感染,特别是在严重烧伤患者和免疫抑制的癌症和艾滋病患者中。
在免疫缺陷的人群中,如严重烧伤患者、人工呼吸机使用者和支气管扩张症患者。
在囊性纤维化患者中,铜绿假单胞菌是慢性肺部感染的主要来源,导致患者的发病率和死亡率增加。
铜绿假单胞菌感染会导致肺部炎症和肺损伤,引起细胞浸润和肺泡水肿。在肺移植患者中,铜绿假单胞菌的分离率更高,但与非囊性纤维化肺移植患者相比,死亡率没有增加。铜绿假单胞菌感染会导致肺血管通透性增加,从而引起肺水肿。
耐药性
铜绿假单胞菌显示出对抗生素的高内在耐药性,并具有获得新耐药机制的非凡能力。由于临床治疗选择有限,多重耐药 (MDR) 铜绿假单胞菌分离株的流行率不断上升是一个全球性健康问题。
对来自西班牙两个不同地区的儿童粪便样本中铜绿假单胞菌的发生率、抗菌素耐药性表型、毒力基因型和遗传谱系表征数据显示:从1,443 个粪便样本中,有72份分离分离出来铜绿假单胞菌,并检测到低水平的抗菌素耐药性:头孢他啶 (8%)、头孢吡肟 (7%)、氨曲南 (7%)、庆大霉素 (3%)、环丙沙星 (1%) 和亚胺培南 (1%);对美罗培南、阿米卡星、妥布霉素、左氧氟沙星和粘菌素敏感,此外还发现了四种多重耐药菌株。
定植
人类粪便样本中的定植率范围为 2.6% 至 24 %。关于铜绿假单胞菌分离株在健康人粪便中定殖的数据很少。
其他研究表明,肠易激综合征患者的小肠和粪便样本中铜绿假单胞菌的定植率很高。迄今为止,关于铜绿假单胞菌分离株在健康人粪便中的定植及其抗菌素耐药性和毒力特征的数据相对较少。
尽管肠道微生物群作为抵御肠道病原体的屏障,但铜绿假单胞菌通过产生一系列令人印象深刻的毒力因子,克服了肠道微生物群和先天免疫系统介导的定植抵抗。
铜绿假单胞菌凝集素 LecA 和 LecB 以及细胞外附属物,如鞭毛和菌毛,在细菌附着到宿主上起着重要作用,并与粘附有关。
此外,铜绿假单胞菌携带属于双伙伴分泌(TPS)家族(也称为Vb型分泌系统,T5bSS)的大型蛋白质系统,TPS系统主要由分泌的TpsA效应蛋白及其TpsB伙伴转运蛋白组成。TpsA1 (CdiA2, 573 kDa) 和 TpsA2 (CdiA1, 361 kDa) 通过接触依赖性生长抑制系统 (CDI) 以及粘附和生物膜形成促进细菌竞争。
因此 TPS 系统效应子被指定为对革兰氏菌有益的主要毒力决定因素阴性病原体。
▼
铜绿假单胞菌等的特殊能力引起如此多样化感染的假单胞菌属是因为它们能够产生名副其实的毒力因子库,包括毒素、生物膜、蛋白酶和溶血素。
考虑到铜绿假单胞菌的医学重要性,假单胞菌领域的大部分研究工作都致力于了解影响这些毒力因子释放的调节、生物合成和环境因素。
此外,它们还可以获得赋予进一步抗菌素耐药性的移动遗传元件;因此,可以鉴定出泛耐药菌株。
▼
假单胞菌感知和适应环境的能力导致了一系列活动,例如许多酶和其他生物分子的分泌。虽然这些毒素、降解酶和抗菌剂可以将假单胞菌的环境转变为理想的微生物生态位,但它们可能对植物和动物宿主造成严重后果。例如,绿脓素是一种蓝/绿色色素毒素,它赋予铜绿假单胞菌培养物特有的颜色,并作为一种抗菌剂可以杀死竞争性微生物。然而,它也会破坏真核细胞过程,这会对人类细胞产生不利影响。
▼
过去几年的工作提供的证据表明,群体感应是一种通用的调节机制,它允许细菌以种群密度依赖的方式发起统一、协调的反应,以完成即使不是不可能,也很难完成的任务。
群体感应系统广泛存在于假单胞菌中,人类机会性病原体铜绿假单胞菌,属于研究最广泛的细胞间通讯系统。在这种生物体中,群体感应非常复杂,由两个相互关联的 N-酰基高丝氨酸内酯 (AHL) 依赖性调节回路组成,这些调节回路由非 AHL 相关信号分子和许多调节剂进一步调制,这些调节剂在转录过程中起作用和转录后水平。这种遗传复杂性可能是导致铜绿假单胞菌具有巨大环境多样性的关键因素之一。
过去几年的工作表明,群体感应对于一系列毒力因子的表达以及铜绿假单胞菌中生物膜的形成至关重要,因此代表了设计用于治疗铜绿假单胞菌的新型药物的有吸引力的目标感染。
此外,细胞间通讯能力也在许多额外的假单胞菌中得到证实。铜绿假单胞菌,因此可以用于设计新型药物来治疗铜绿假单胞菌感染。
长时间住院、创伤ICU住院、消毒干预等与肠道假单胞菌过度生长有关,而TAP暴露与抗菌剂干预则与肠道假单胞菌过度生长有负相关。
注:TAP的暴露是指肠道上皮细胞上的TAP暴露在肠道腔中,从而使肠道内的微生物可以通过TAP进入肠道上皮细胞内部。一些研究表明,TAP的暴露与肠道微生物群失调和某些疾病(如炎症性肠病)有关。而抗菌剂干预可以抑制肠道内细菌的生长,从而减少TAP的暴露,因此与假单胞菌肠道过度生长有负相关。
对于免疫功能低下的患者,应特别注意水的纯度,有些水中可能含有假单胞菌等细菌。
研究发现用海洋益生元岩藻依聚糖补充饮食可以通过抑制分泌性毒力因子 (TpsA/CdiA) 与粘蛋白的相互作用并促进有益种群的生长,从而介导铜绿假单胞菌从肠道菌群失调和去定植中的早期恢复。
芪归饮是一种用于治疗铜绿假单胞菌感染的中药。在门和属水平上显著抑制脱铁杆菌和粘孢子菌的过度积累。鉴定了11种潜在的代谢产物,这些代谢产物在铜绿假单胞菌感染中异常表达,服用芪归饮显著逆转。
此外,部分研究表明以下物质也可以抑制肠道内的假单胞菌属。
假单胞菌属相比于其它细菌引起人们广泛的研究兴趣,该属不仅包括人类病原体铜绿假单胞菌,还包括一系列与植物致病性、生物修复和环境微生物学相关的其他重要物种。
假单胞菌被广泛用作模型原型细菌,用于研究宿主-病原体相互作用、细胞-细胞通讯系统、进化生物学、基因调控和代谢网络、分泌系统、抗生素(耐药性)、生物修复、生物膜、细菌基因组学以及与微生物学和分子生物学广泛相关的许多其他主题。
假单胞菌属因其复杂的信号和调节系统,特别是细胞密度依赖性群体感应而被广泛研究。未来只有了解复杂调节回路中涉及的机制,才能确定新型抗菌方法的可能目标。铜绿假单胞菌中的锌、铁稳态是替代抗菌策略的一个有希望的目标。
由于其复杂性和适应性,它存在于无数的环境和临床生态位中,以及它在不同层面(种内、种间和与其宿主)相互作用的能力,假单胞菌属很可能在未来许多年内吸引更多研究人员。我们期待未来有更好的解决假单胞菌属,尤其人类病原体铜绿假单胞菌的抗菌策略。
附录一:
其他菌种
附录二:
致病菌菌株
附录三:
可能参与的代谢

谷禾健康
我们的身体中有很多不同的器官,组织,腺体等会产生许多信号分子来精确控制和影响身体的反应和活动,这些信号分子包括激素、神经递质、生长因子、细胞因子等。它们可以促进或抑制细胞的生长和分化,调节细胞间的相互作用和通讯,影响身体的代谢和能量平衡,参与免疫系统的细胞和分子相互作用,调节身体的内环境和外环境的适应性等。
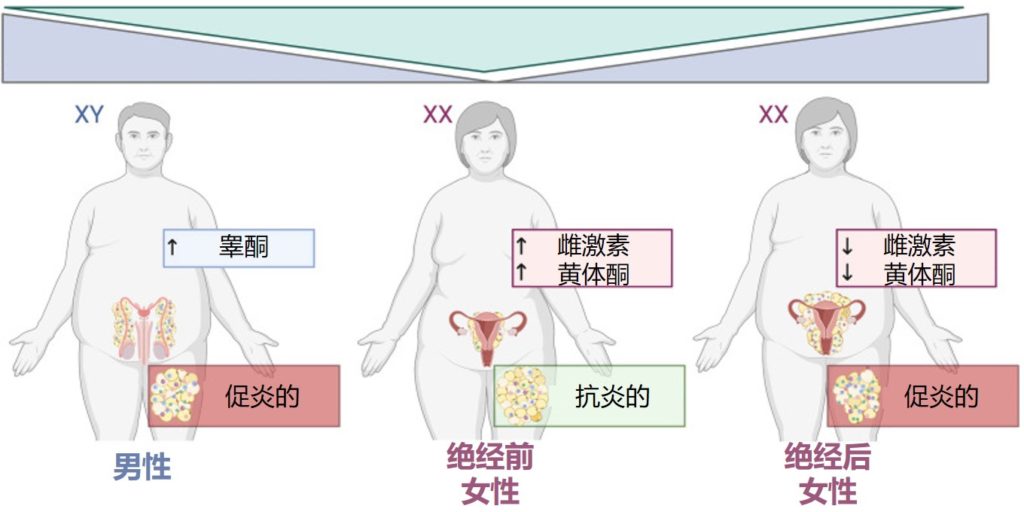
其中,激素是一类重要的生物活性物质,也可以说是信号分子,一般由特定的内分泌细胞,腺体或器官所产生,包括性激素、脂质激素、蛋白质激素、胰岛素、甲状腺激素、肾上腺素和去甲肾上腺素等。
这其中性激素是一类重要的激素,主要由生殖腺(如卵巢、睾丸)和肾上腺产生,包括雌激素、孕激素和雄激素等。它们在生殖系统中发挥重要作用,对女性月经周期、妊娠、分娩、哺乳等过程起调节作用,对男性性发育、精液生成、性欲等方面也具有重要影响。此外,性激素还与骨密度、心血管健康、认知能力等方面有关。
临床上性激素六项是生殖内分泌门诊常用的检查。性激素六项检查主要用于评估男女生殖内分泌功能,对月经失调、性功能障碍、不孕不育、生殖系统肿瘤、性染色体病等的鉴别和诊断及内分泌治疗的效果检测都具有重要的临床意义和参考价值。
近年来,越来越多的科学证据表明,肠道菌群与性激素(宿主的内源激素或外源补充激素)存在直接或间接的相互作用。性激素对于塑造肠道微生物群组成具有一定影响,与此同时,肠道微生物群本身也会影响性激素水平。
目前的研究主要基于横断面研究,基于纵向的检测肠道菌群发育以及激素水平变化的研究队列还较少,目前无法区分是性激素变化在前还是菌群变化在前,但是肠道菌群的失调和性激素的异常水平都与多种疾病的发生和发展有关,如肥胖症、炎症性肠病、乳腺增生、甲状腺疾病,妇科疾病、自身免疫疾病等。因此,保持肠道菌群和激素水平的平衡,对于维持人体的健康十分重要。
本文旨在阐述激素与肠道微生物群的相互作用,主要着重讲述性激素影响下的人体一些疾病和菌群的差异。
这些发现可能会在未来用于开发治疗激素紊乱相关疾病或与性别和激素活动相关的自身免疫性疾病、甲状腺疾病、代谢疾病甚至是压力等情绪状态的新疗法。
目录/contents
Part1:肠道微生物与激素的关联
Part2:肠道微生物通过激素对宿主的影响
Part3:性别差异下的肠道微生物群
Part4:肠道菌群与性激素的相互作用
Part5:性激素影响下的相关疾病
Part6:结语
肠道微生物群以多种方式影响动物和人类的行为,甚至可能影响人类的情绪状态和疾病状态。最近揭示了菌群的一个关键作用:调节激素分泌。
微生物和激素之间的相互作用会影响宿主的新陈代谢、免疫力和行为。这种相互作用是双向的,因为微生物群已被证明既受宿主激素影响又影响宿主激素。
肠道微生物群通过激素影响宿主情绪、免疫及代谢等
微生物群对宿主激素水平的影响可能是直接的,即微生物群产生激素,也可能是间接的,即微生物可能调节肾上腺皮质(控制焦虑和压力反应)的功能,或调节炎症和免疫反应。
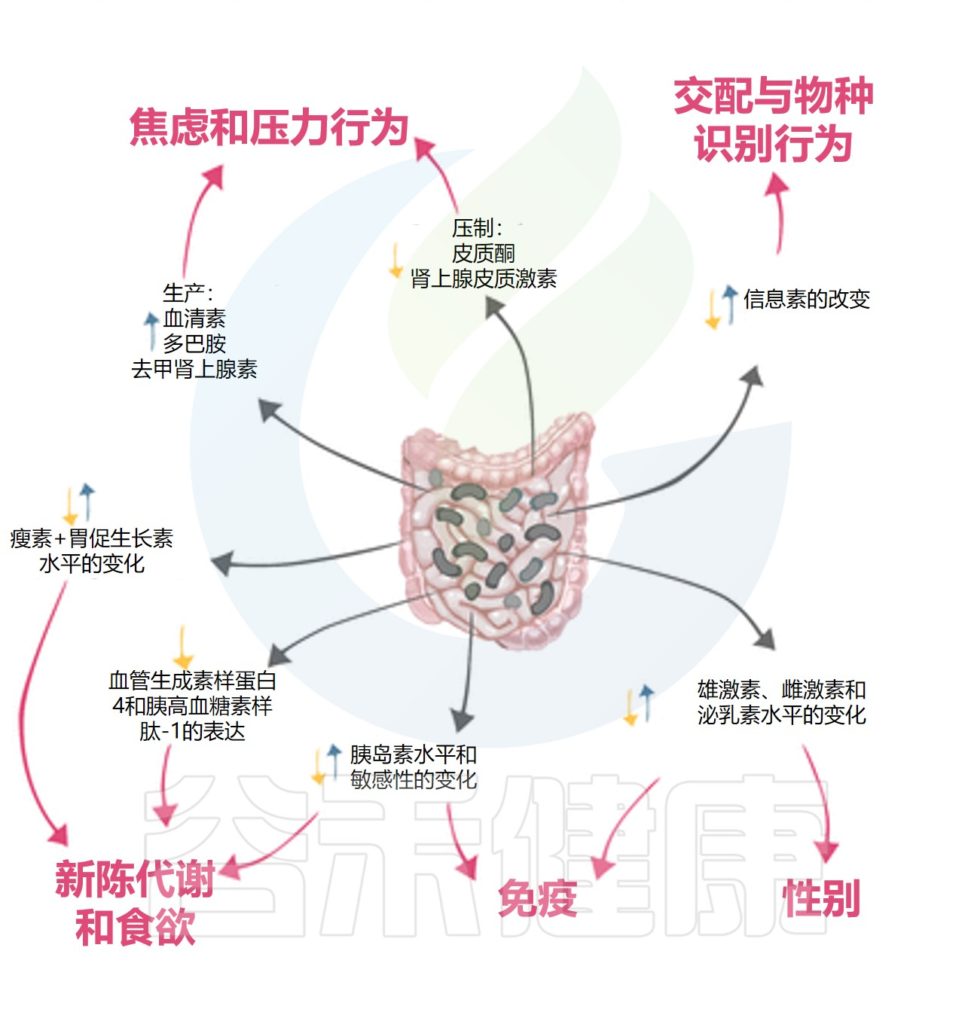
Neuman H,et al.FEMS Microbiol Rev.2015
激素水平调节微生物群的生长、毒性
多种宿主因素(如饮食、运动、情绪、健康状况、压力和性别)会导致激素水平发生变化,进而对微生物群产生多种影响(包括生长、毒力和抵抗力)。
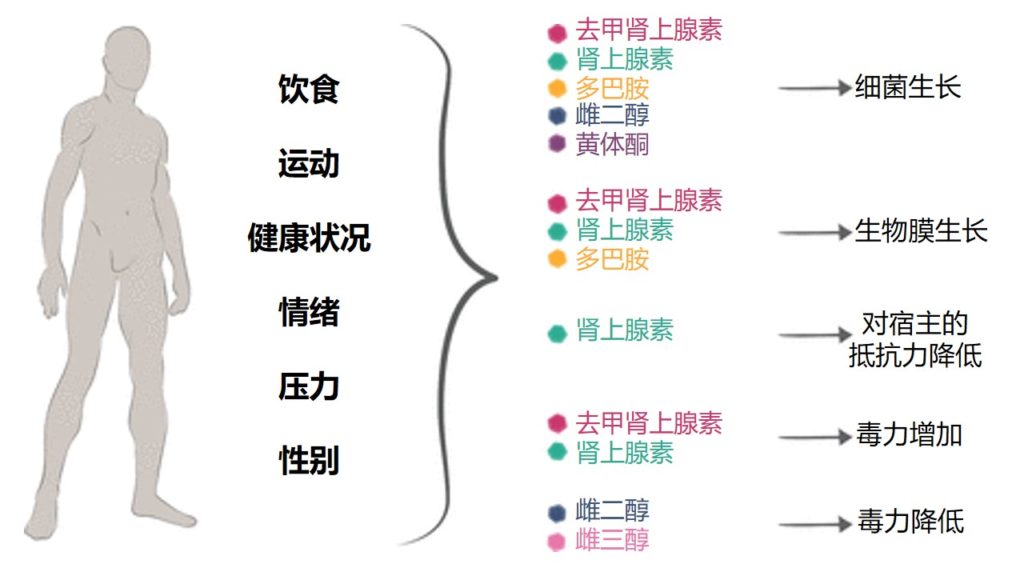
Neuman H,et al.FEMS Microbiol Rev.2015
文献中已记载激素和微生物群之间的相互作用




Neuman H,et al.FEMS Microbiol Rev.2015
▸ 菌群通过影响神经激素前体调节肠道运动、情绪、食欲等
神经内分泌细胞响应神经元输入而分泌神经激素。尽管它们被分泌到血液中以发挥全身作用,但它们也可以充当神经递质。微生物群对行为的调节(例如小鼠的焦虑)被认为是通过神经激素前体(例如血清素、多巴胺)发生的。
最近,肠道细菌被证明可以产生和响应神经激素,例如血清素、多巴胺和去甲肾上腺素。
这些机制对于研究病原体的研究人员来说很有趣,因为它们可能会影响病原体对宿主防御反应的敏感性。例如,为了响应宿主肾上腺素,沙门氏菌会下调其对宿主抗菌肽的抵抗力并诱导关键的金属转运系统,从而影响细胞中的氧化应激平衡。
// 血清素
血清素,也称为 5-羟色胺 (5-HT),是大脑中的主要神经递质之一。然而,超过90%的哺乳动物宿主血清素都存在于肠道中。
肠道血清素的分泌受饮食影响,调节肠道运动、情绪、食欲、睡眠和认知功能。这种双重作用表明血清素可能将肠道(包括其微生物群)与宿主行为联系起来。
血清素与肠易激综合征和克罗恩病有关
大脑血清素可以通过血清素转运蛋白穿过血脑屏障进入血液,这表明肠-脑轴中存在另一个联系。血清素与肠易激综合征和克罗恩病等胃肠道疾病有关,这些疾病也与微生物群的差异有关。
// 多巴胺
多巴胺由芽孢杆菌(Bacillus)和沙雷氏菌(Serratia)等细菌产生 。
无菌小鼠中的游离管腔多巴胺水平显著低于常规小鼠,并且在接种表达β-葡萄糖醛酸酶的细菌后再次升高。
多巴胺不足与帕金森病有关
这些结果表明,在以多巴胺形成不足为特征的帕金森病等疾病中,肠道细菌与多巴胺水平之间可能存在相关性。
//γ-氨基丁酸
γ-氨基丁酸 (GABA) 是哺乳动物中枢神经系统中的主要抑制性神经递质,它也由微生物群产生,并可能影响宿主行为。这很有趣,因为中枢γ-氨基丁酸受体表达的改变与焦虑和抑郁的发病机制有关。
γ-氨基丁酸影响焦虑和抑郁
已经研究了由乳杆菌产生的γ-氨基丁酸,以试图大规模发酵安全的γ-氨基丁酸。因此,向小鼠施用鼠李糖乳杆菌会改变不同中枢神经系统区域中γ-氨基丁酸受体的表达,从而减少与焦虑和抑郁相关的行为。
▸ 菌群通过调节应激激素降低宿主焦虑行为
微生物群可以通过改变应激激素水平来帮助我们保持冷静和平衡。无菌小鼠具有升高的应激激素皮质酮和促肾上腺皮质激素 (ACTH) 血浆水平以响应轻度压力,增加与焦虑和压力相关的行为。
促肾上腺皮质激素通过进一步产生皮质类固醇在下丘脑-垂体-肾上腺轴中发挥重要作用。因此,两种特定物种,瑞士乳杆菌(Lactobacillus helveticus)和长双歧杆菌,降低了大鼠和健康人类的应激激素皮质醇水平和焦虑样行为。
此外,与对照组相比,长期接受益生菌鼠李糖乳杆菌治疗的小鼠在强迫游泳测试中皮质酮水平较低,抑郁行为也较少。
▸ 肠道菌群影响甲状腺激素的转化影响代谢
肠道菌群产生的代谢物对甲状腺和内分泌功能也是必不可少的。它影响硒、铁、锌和碘的吸收,这些都是甲状腺激素产生所需的重要矿物质。它制造短链脂肪酸(SCFAs)与甲状腺激素协同工作,以改善肠道屏障功能和加强细胞间紧密连接。
肠道菌群失衡是影响甲状腺激素转化的主要障碍。事实上,20%的T4(甲状腺素)在肠道中转化为活性的T3(三碘甲状腺原氨酸)形式,肠道菌群不健康,肠道内存在过多的病原体,肠道中甲状腺激素的转化也会减少。
此外当肠道屏障功能减弱时,毒素和未完全消化的食物残渣等颗粒会逃逸,导致肠胃不适和全身炎症。炎症会在组织水平损害甲状腺激素的转化。受体受损,身体也不能将T4激素转化为活性的T3形式。
▸ 细菌通过信息素影响宿主行为
信息素是在性识别、吸引力和交配行为以及攻击行为和支配行为中发挥重要作用的激素。
信息素也称为外激素,一种分泌到一个人体外并影响他人行为的化学物质。
共生细菌和信息素之间的这种联系也发生在哺乳动物身上。两种鬣狗的气味腺中有不同的细菌群落,这与气味分泌物中不同的挥发性脂肪酸特征有关。作者推测,共生细菌产生的代谢产物提供了物种特有的气味。
这进一步支持了细菌可以产生影响宿主行为的气味特征的观点。细菌也可以在配偶选择中发挥作用:雌性小鼠不会被感染沙门氏菌的雄性小鼠的尿液所吸引。
化学信号和嗅觉刺激也在人类行为中发挥作用,未来的研究将帮助我们了解细菌产生的气味是否会影响我们自身的相互作用,甚至可能影响进化。
▸ 肠道菌群代谢性激素影响健康
肠道菌群也影响雌激素,它们通过特定的酶对雌激素进行循环利用并产生雌激素代谢物。当肠道菌群失衡或不健康时,这些过程就会被破坏,从而导致健康问题。
例如,Prevotella intermedius吸收雌二醇和黄体酮,从而促进其生长。雌激素受体ER-β表达的变化也会影响肠道微生物群的组成。
雌二醇在卵巢中合成,是雌激素中含量最高的,活性最强的激素,经肝脏代谢后,通过胆道排泄到胃肠道,部分雌二醇被胃肠道重新吸收进入血液。肠道菌群可以通过改变雌二醇的活性和非活性状态,影响血清中类固醇激素的水平。
武汉大学人民医院转化医学研究所的李艳教授团队在cell metabolism上发表研究论文,发现绝经期前抑郁症患者肠道细菌Klebsiella aerogenes(产气克雷伯氏菌)的3β-羟基类固醇脱氢酶(3β-HSD)通过降解雌二醇诱导绝经期前女性出现抑郁症。
肠道微生物群作为人体最大的菌群库,动物和人类研究发现,肠道微生物群存在性别相关差异。
影响微生物群的重要因素(例如环境或激素)取决于不同的生命周期,并且在男性和女性中会有所不同。
Kim YS,et al.World J Mens Health.2020
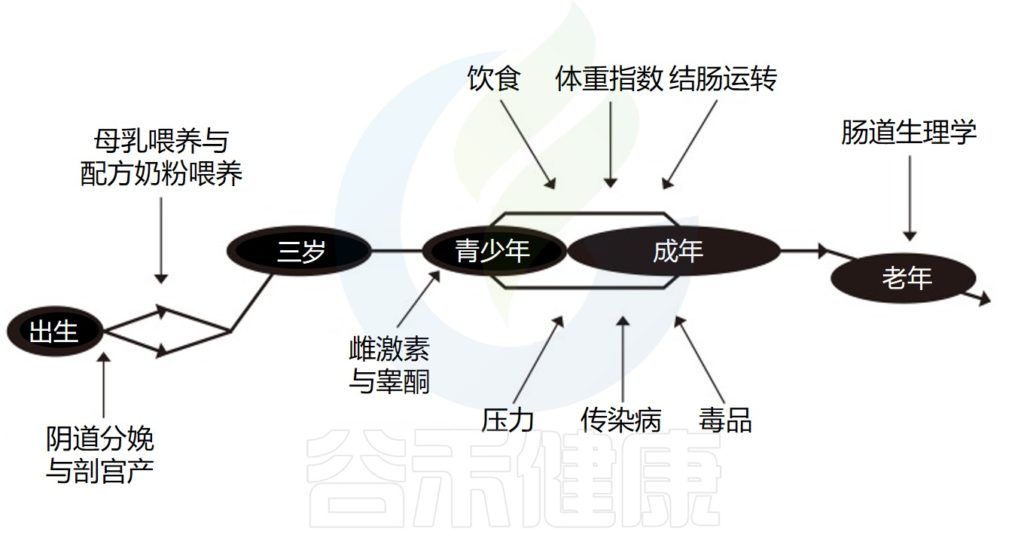
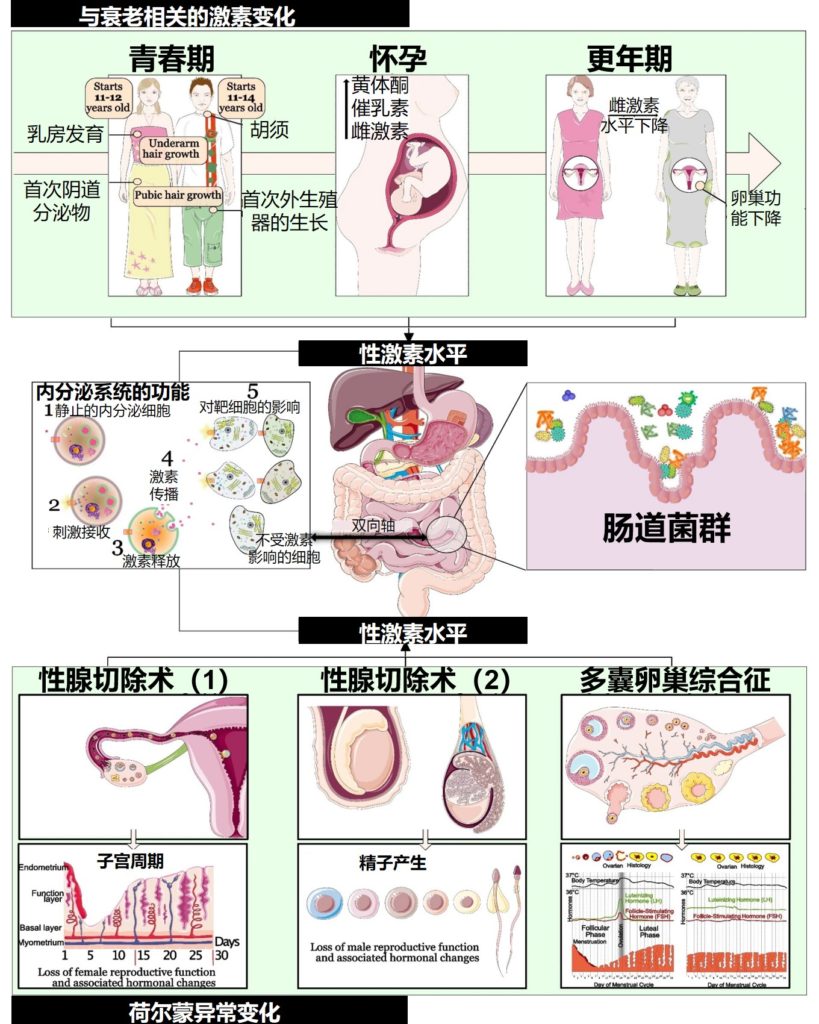
需要注意的是,性激素对肠道微生物具有显著的影响,青春期、怀孕、更年期、多囊卵巢综合征和性腺切除术等各种因素会导致性激素水平(睾酮和雌二醇)发生变化,进而导致肠道菌群组成发生变化。
肠道微生物群与性激素之间的相互作用
Santos-Marcos JA,et al.Biol Sex Differ.2023
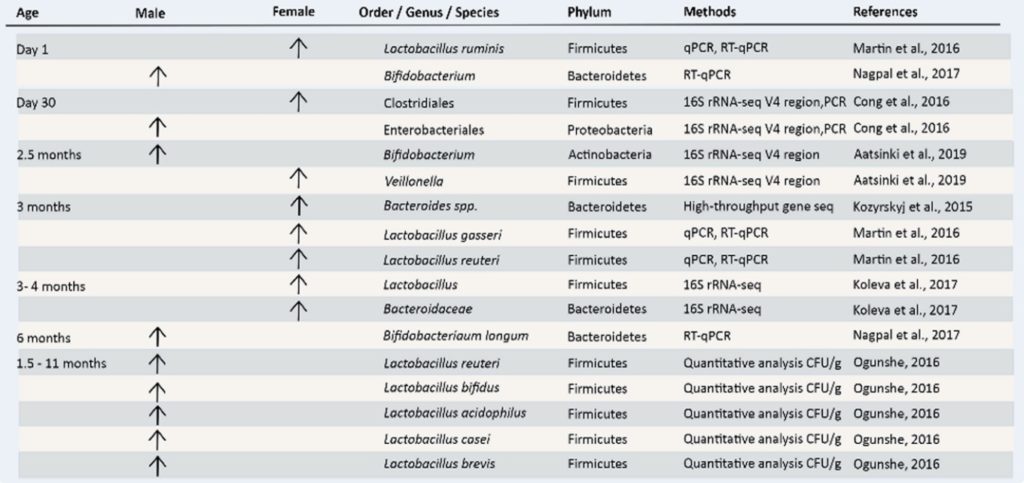
▷婴儿时期肠道菌群组成与性别相关的差异
婴儿期的特点是参与人乳低聚糖代谢的细菌种类占主导地位(据估计,婴儿细菌微生物群的25-30%来自母乳)并且肠道微生物群多样性的差异减少。
特别是,这个年龄段的特点是被属于双歧杆菌(Bifidobacterium)和链球菌(Streptococcus)的几种物种定殖,例如长双歧杆菌和嗜热链球菌。
6个月至2岁之间的年龄段的特点是将固体食物引入婴儿的饮食中,并伴随着肠道微生物群结构和功能多样性的快速增加。这是婴儿生长的关键时期,肠道微生物群开始接近成人特征。
最近对人类婴儿进行的几项研究发现了他们出生后第一年不同时间点肠道微生物群落的潜在性别依赖差异。
男婴α多样性较低
例如,对出生后前30天新生儿粪便样本的分析显示,与女性相比,男婴的α多样性较低,梭状芽孢杆菌的丰度也较高,而肠杆菌的丰度较低。
男婴双歧杆菌更丰富
在新生儿肠道中最早和最丰富的细菌定植者中,已知双歧杆菌可为宿主肠道提供健康益处。改变在从出生后一天到六个月大,在男性和女性阴道分娩的婴儿中观察到双歧杆菌丰度。发现与女孩相比,男孩的双歧杆菌水平更高。
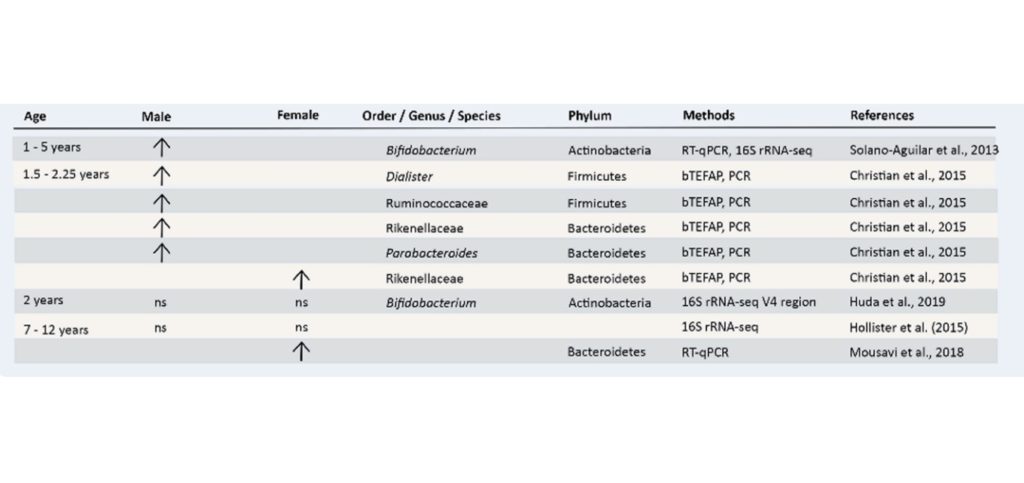
▷儿童时期肠道菌群组成与性别相关的差异
在出生后的前12个月,婴儿的肠道因饮食中引入固体食物而发生变化,随之而来的是微生物群落的变化。
特别是,在引入固体食物后,婴儿大肠中厌氧菌种群的初始定植在数量和组成上开始与成人相似。几项研究表明,在没有饮食变化或抗生素治疗等外部压力因素的情况下,与儿童相比,成年期的肠道微生物群更加稳定和有弹性。
这表明尽管出现了部分趋同,但仍存在差异,因此分析这一生命阶段与性别差异的关系很有价值。
男孩双歧杆菌与乳酸杆菌之间的正相关性比女孩更强
对来自哥伦比亚两个不同地点的277名1至5岁儿童(154名女孩和123名男孩)的评估(研究临床腹泻的存在)表明,粪便微生物群受到多种宿主因素的影响,包括年龄、健康状况、地点和性别。
有趣的是,与女孩相比,健康男孩的双歧杆菌和乳酸杆菌之间存在显著更强的正相关性,而在受腹泻影响的儿童中未观察到显着相关性。
▷青春期与性别相关的肠道菌群差异最明显!
青春期是发展的关键时期,性激素,如雌激素和睾酮,在这个年龄段人群的肠道微生物群中发挥重要作用。
在性激素的影响下异性间微生物群差距变大
一项针对青春期肠道微生物群组成的人类研究表明,与同性双胞胎相比,青少年(13-17岁)异性双胞胎的粪便微生物群差异更大。
最近的一项研究得出结论,与青春期受试者(5-15岁)相比,非青春期的α和β多样性没有差异。随着青春期的开始,梭状芽孢杆菌(Clostridium)的水平降低,而变形杆菌(Proteus)的丰度增加。
在青春期受试者中,发现Adlercreutzia、Ruminococcus、Dorea、Clostridium和Parabacteroides与睾酮水平有关。
考虑到人类这一生命阶段的可用数据有限——必须使用啮齿动物模型的研究作为替代。在小鼠身上进行的实验表明,肠道微生物群的性别差异出现在与性激素水平相关的青春期开始时。
动物实验中青春期的菌群性别差异
特别是,仅在10至13周龄的非肥胖糖尿病青春期后小鼠中检测到α多样性的差异,与雄性相比,雌性的水平显著更高。
此外,雄性小鼠表现出更高丰度的卟啉单胞菌科(porphyromonas)、韦荣球菌科、消化球菌科、乳杆菌科和肠杆菌科与女性相比。而当雄性小鼠被阉割后,这些差异就消失了。
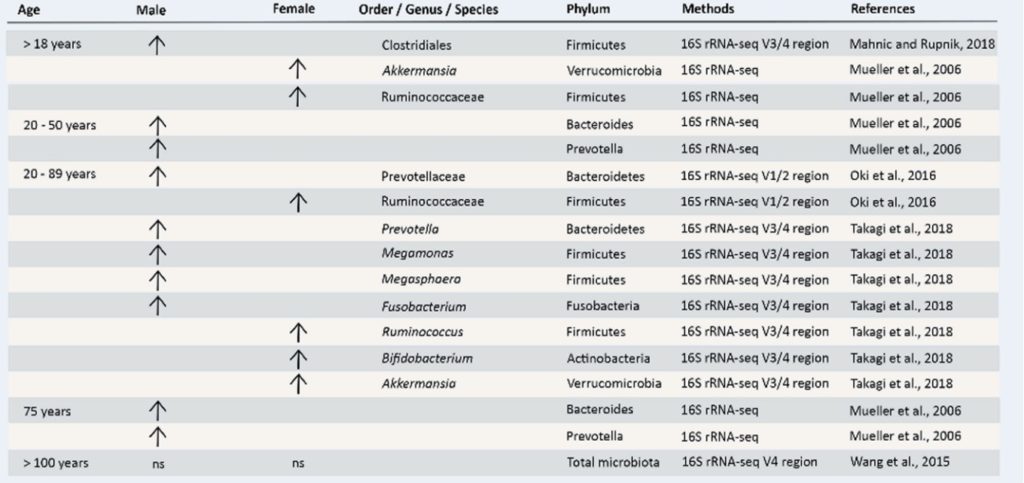
▷成年后肠道菌群组成与性别相关的差异
肠道微生物组在生命的最初几年大部分时间都不稳定,但在成年期变得更加稳定,之后细菌丰富度和组成可能伴随着特定疾病会发生变化。
在整个生命周期中,随着免疫和神经系统的成熟,肠道微生物群的伴随发育是性别二态性的,导致成年男性和女性的不同微生物群落以及免疫和神经炎症通路。
注:年龄、环境(饮食和身体活动)和地理位置在内的多种因素似乎在这些性别依赖性肠道菌群差异中发挥了作用。
分析来自结肠癌研究对照组的82名受试者得出结论,性别与肠道微生物组的整体组成显著相关。然而,这项研究包括的参与者数量相对较少,年龄范围相当广泛(30-83 岁),这可能混淆了研究的结果。
年轻女性肠道微生物α多样性比同年龄男性更高
在包括美国、英国在内的四个地理区域以及来自哥伦比亚和中国的两个队列中评估了年龄在20至69岁之间的三个大型队列中年龄、性别和肠道微生物群α多样性之间的关系。
有趣的是,与同年龄段的男性相比,年轻的成年女性(20-45 岁)表现出更高的α多样性。此外,性别和α多样性之间的关联在年轻人中比在中年人中更为明显,而当参与者的平均年龄为60岁时,男女之间的α多样性没有差异。这可以解释为更年期女性的雌激素水平下降。而更年期之后男性与女性的激素水平差异变小,肠道微生物群的相似程度更接近。
总之,对来自不同国家的男性和女性进行的人口研究发现人体肠道微生物群组成存在性别相关差异。
这意味着,性激素的动态变化会影响肠道共生,而青春期时激素的变化最大,此时男性与女性的肠道微生物群之间的差异非常明显。
注:由于该研究所覆盖的样本量还较小,年龄之间的跨度比较大,仅作为研究参考,还不能用于诊断标准。
整个生命周期中男性和女性肠道微生物群组成的差异
Valeri F,et al.Front Neuroendocrinol.2021
不仅肠道微生物群受性激素的影响,而且肠道微生物群本身也会影响激素水平。
▷ 微生物代谢雌激素进而影响多个器官
生殖内分泌功能涉及由复杂反馈机制控制的多种激素。卵巢、肾上腺和脂肪组织会产生雌激素。体内产生的或作为食物摄入的雌激素可以被肠道微生物代谢。
产生的代谢物再次影响宿主。性激素通过类固醇受体(包括雌激素受体β)直接调节细菌的新陈代谢。
具有雌激素代谢能力的肠道微生物群基因库
Yoon K,et al.J Neurogastroenterol Motil.2021
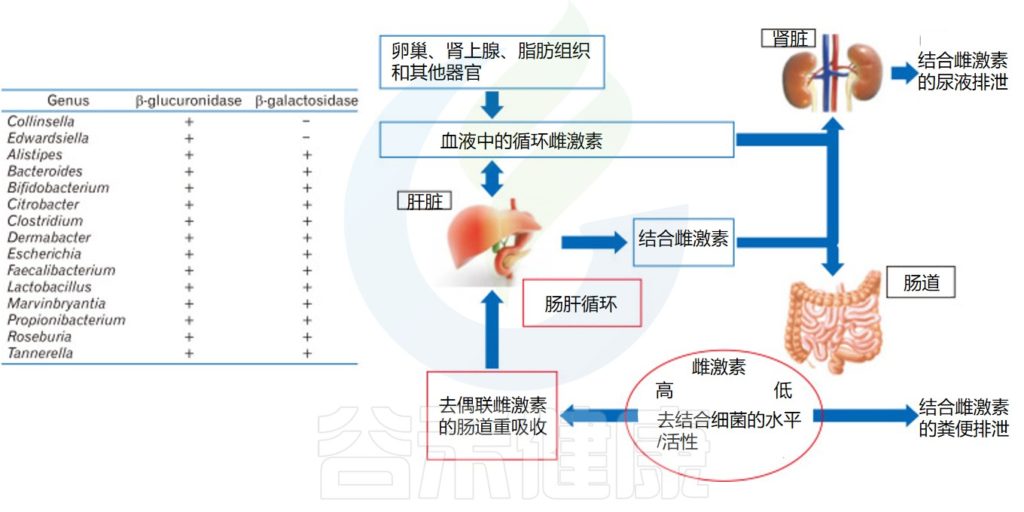
同时,具有β-葡萄糖醛酸酶活性的肠道微生物群解离胆汁中排出的结合循环雌激素。去结合使雌激素对系统的重吸收过程成为可能。去结合的雌激素通过雌激素受体循环并影响多个器官,不仅影响生殖器官,还影响骨骼系统、心血管系统和中枢神经系统。
雌激素循环途径
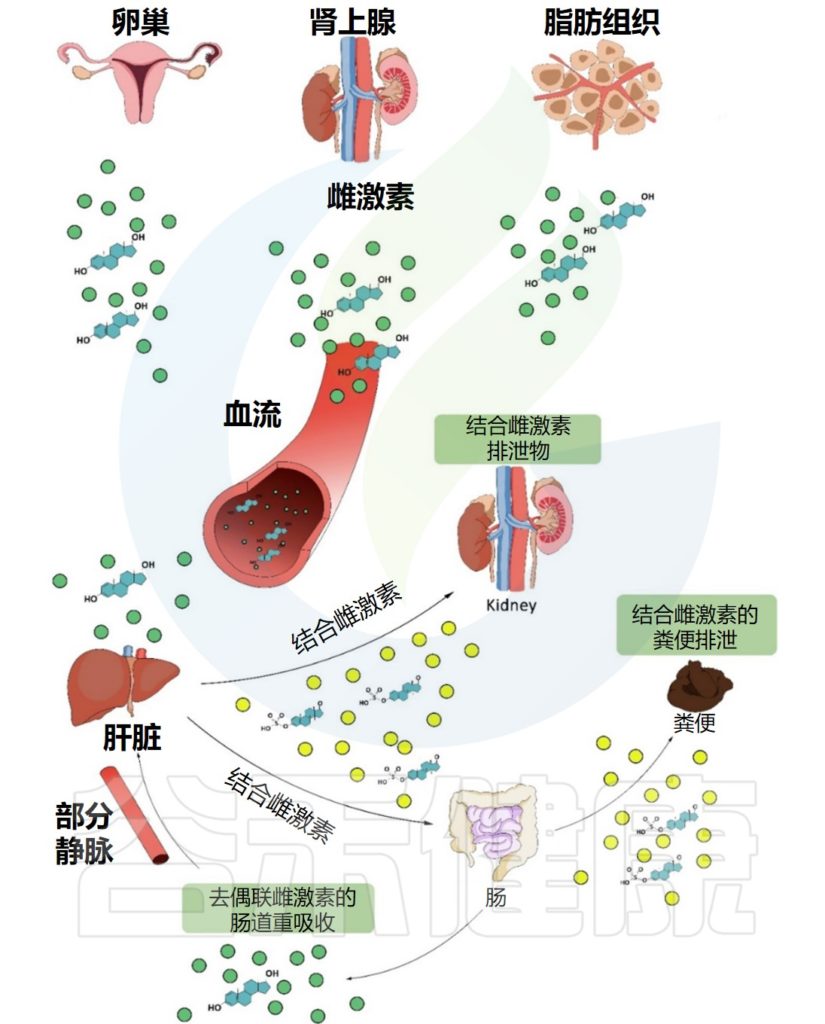
Valeri F,et al.Front Neuroendocrinol.2021
雌激素在几个组织中局部产生(主要是卵巢,但也有肾上腺和脂肪组织)。它们在血液中循环,到达多个靶器官,如肝脏和肾脏。一旦到达肝脏,它们就会通过葡萄糖醛酸化或磺化反应进行结合。
此外,结合雌激素被转化为水溶性分子,随后可以通过胆汁、尿液和粪便排出体外。某些部分的结合雌激素可以在肠道中被肠道微生物分解,并通过门静脉被重新吸收到肝脏中。
雌激素对发病机制具有影响
通常,雌激素与核受体结合,引起构象变化。通过肠道屏障通透性和免疫系统调节,中枢和外周改变与肠易激综合征 (IBS) 相关联。流行病学研究表明女性在肠易激综合征中比例更大,暗示性激素对其发病机制的影响。
雌激素受体作用的分子途径
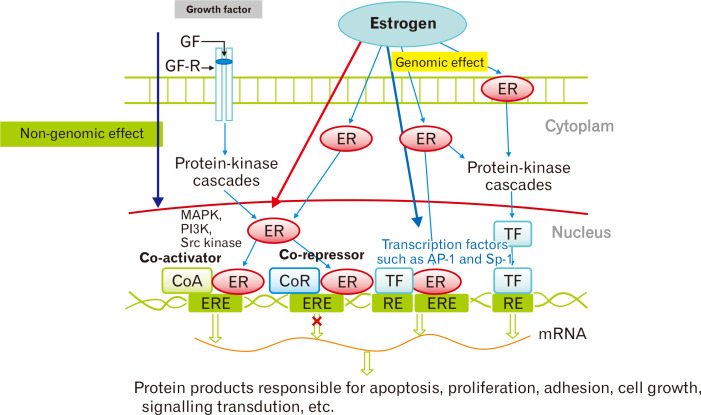
Yoon K,et al.J Neurogastroenterol Motil.2021
▷ 微生物群影响雄激素的代谢
据报道,在雄激素代谢中,肠道微生物群是一个重要的调节因子。在小鼠的小肠内容物中,葡萄糖醛酸化睾酮和双氢睾酮含量很高。远端肠道具有高水平的游离双氢睾酮。
在年轻成年男性的粪便中观察到明显高水平的未结合双氢睾酮。在无菌小鼠中,葡萄糖醛酸化睾酮和双氢睾酮较高;然而,在远端肠道中发现游离水平非常低。这意味着肠道微生物群会影响双氢睾酮和睾酮的肠道代谢。
▷ 性激素高的人拥有更多样化的肠道微生物
血清睾酮升高的男性和雌二醇水平升高的女性分别拥有更多样化的肠道微生物群,许多细菌属与睾酮(Acinetobacter, Dorea, Ruminococcus和Megamonas)和雌二醇(Slackia和Butyricimonas)水平相关。
高雄激素的女性微生物群明显改变
在这方面,患有多囊卵巢综合征(高雄激素)的女性在妊娠早期到晚期的变化中,表现出明显的微生物群改变,变形菌(Proteobacteria)和放线菌(Actinobacteria)总体增加,丰富度降低。
肠道微生物群组成的性别差异在青春期增加,随着青春期的进展,女孩的肠道微生物群变得与成年人更相似。这些结果可能还表明肠道微生物群可能会影响青春期的时间,可能是通过调节宿主性激素水平。
雌激素水平与肠道微生物多样性密切相关
在男性和绝经后女性中,尿液中的雌激素水平与肠道微生物群丰富度和α多样性密切相关,而绝经前女性雌激素水平在月经周期中收集时变化很大,并未显示出这种关联。
▷ 微生物群可以预测粪菌移植后睾酮水平
最近报道,绝经后妇女的肠道菌群与男性比绝经前妇女更相似,绝经后妇女与同龄男性之间实际上没有显著差异。
这项研究还表明,性腺类固醇与微生物群差异之间存在关联,绝经前妇女的类固醇生物合成和降解途径丰富,并且与血浆睾酮水平显著相关。
此外,微生物群可以预测人类粪便转移后人类和(经抗生素处理的)雄性小鼠的循环睾酮水平。
综上所述,这些结果表明,肠道微生物群组成的差异与性别以及不同激素状态有关。
既然知道了肠道微生物群与性激素之间存在相互作用,那么它们具体是通过什么机制影响的呢?
性激素和肠道微生物组之间相互作用机制
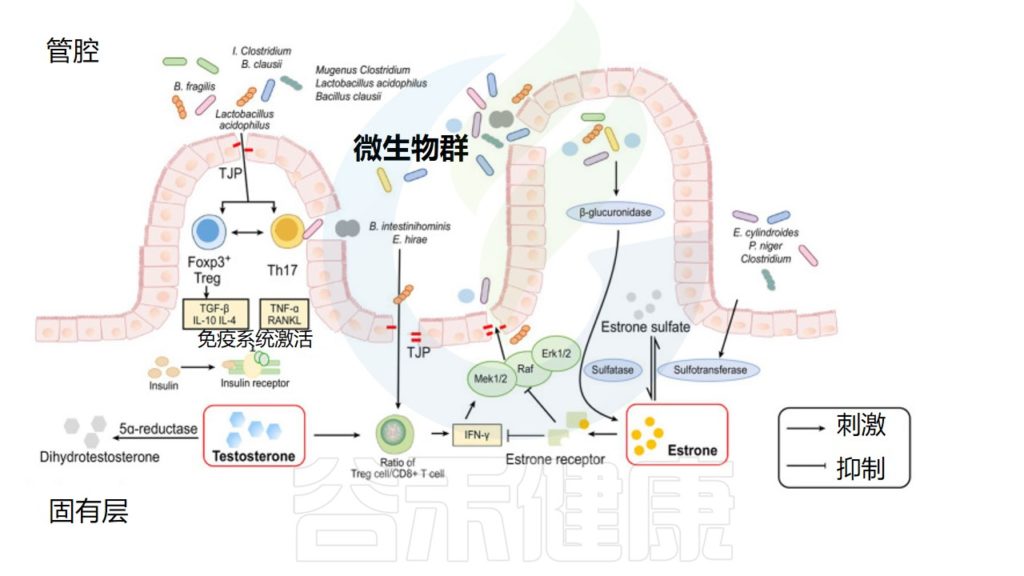
He S,et al.Front Microbiol.2021
1
微生物群通过改变胆汁酸影响性激素水平
最近有人提出,肠道微生物群的部分性别差异可能取决于胆汁酸,因为男性的胆汁酸库比女性大。在肝脏中由胆固醇合成后,它们被肠道微生物群代谢为次级胆汁酸,从而改变微生物群的结构并导致各种病理。
因此,肠道微生物群通过依赖于法尼醇X受体(FXR)的机制调节回肠中成纤维细胞生长因子15(FGF15)和肝脏中胆固醇7α-羟化酶(CYP7A1)的表达,从而调节胆汁酸的二次代谢,并抑制胆汁酸在肝脏中的合成。
FGF15抑制肝脏中CYP7A1的表达,CYP7A1是一种催化和调节胆汁酸合成第一步的酶。
胆汁酸同时也能影响微生物群的结构和丰度
已经观察到胆汁酸的减少导致细菌增殖,并且法尼醇X受体抑制细菌过度生长。
几项研究证实了胆汁酸、性激素和肠道菌群组成之间的关系。通过这种方式,给大鼠服用胆汁酸引起的微生物群变化类似于高脂肪饮食引起的变化,以牺牲拟杆菌属为代价增加厚壁菌门的水平。
此外,粪便微生物群(来自瘦供体)的移植产生了与瘦供体相似的肠道微生物群和胆汁酸库的变化,而小鼠的性腺切除术也改变了胆汁酸模式。
由于睾酮是由胆汁酸衍生的,并且如上所述,胆汁酸水平会被微生物群改变,因此微生物群可能间接影响睾酮水平的说法是成立的。
2
微生物群通过酶促激活性激素
共生微生物群落可以通过其酶的活性影响性激素水平。通过这种方式,术语“strobolome”被创造出来,定义为肠道微生物群中能够从其无活性葡萄糖醛酸苷中激活雌激素的一组基因,这主要归功于β-葡萄糖醛酸苷酶,它将雌激素解离为活性形式。这些活性雌激素进入血液并作用于雌激素受体α(ERα) 和雌激素受体β(ERβ)。
同样,最近的一项研究得出结论,肠道微生物群参与双氢睾酮 (DHT) 和睾酮的代谢和肠道去葡萄糖化,从而导致最高水平的雄激素。
肠道微生物群在性别差异中的另一种可能作用机制可以在羟基类固醇脱氢酶 (HSD) 中找到,这些酶参与类固醇激素的代谢并控制类固醇与其核受体的结合,使它们充当激活剂或抑制剂。
✦ 肠道微生物还通过植物雌激素影响代谢
除了三种主要形式的雌激素(胆固醇衍生的类固醇激素)、雌二醇(E2,在绝经前的非孕妇中占主导地位)、雌酮(E1,在绝经后占主导地位)和雌三醇(E3,在怀孕期间占主导地位),还有一种称为植物雌激素的植物化合物,其结构和功能与雌激素相似。
植物雌激素是天然存在于植物中的化合物,植物雌激素包括异黄酮,如染料木黄酮和黄豆苷原,它们主要存在于大豆中,并在被肠道微生物群代谢后被激活。从这个意义上说,肠道微生物群允许从黄豆苷元中获得O-Desmethylangolensin (ODMA) 和雌马酚,这两种物质都具有雌激素活性。
与雌激素类似,植物雌激素通过影响细胞信号传导引起生理效应,因为它们可以通过激活或抑制雌激素受体α或雌激素受体β来诱导或抑制雌激素作用,并且还可能引发表观遗传效应和细胞内信号级联反应。
植物雌激素可以调节内分泌系统改善病症
与此相关的是,几项人类研究表明,植物雌激素可以通过调节内分泌系统来改善各种病症,包括更年期症状,并且可以逆转代谢性内毒素血症的症状。
在这方面,植物雌激素代谢物雌马酚通过促进雌激素的尿液排泄和改变女性的血液水平,与降低女性激素相关疾病的风险有关,而不产生O-Desmethylangolensin与肥胖有关。
植物雌激素在饮食中被消耗,因为它们出现在水果、蔬菜、豆类和一些谷物中。事实上,饮食成分对肠道微生物群生态系统具有急性影响。通过促进更多样化和更稳定的微生物系统的发展,以植物为基础的饮食似乎对人类健康更有益。
!
药物和益生菌治疗的性别差异
已有一些关于药物治疗和微生物群性别差异的报道。
▷男女服用同一种抗炎药后微生物变化有差异
一项人体研究调查了23名健康男性和女性在非甾体抗炎药诱导的肠道屏障功能和微生物组方面的差异。使用 16S 核糖体 RNA 测序确定了十二指肠和粪便微生物群组成。健康女性的肠道通透性较低,十二指肠和粪便微生物多样性高于健康男性。
两性服用吲哚美辛后肠道通透性增加。然而,只有雌性在摄入吲哚美辛后粪便微生物多样性下降,包括普氏菌丰度增加。十二指肠微生物群组成没有表现出性别特异性变化。
▷益生菌给药存在性别差异
如今,益生菌因多种原因而流行,并且有人认为其效果可能因性别而异。
一项研究报告说,将5种乳杆菌菌株的益生菌混合物给予易患狼疮的小鼠可改善肾功能,并在雌性和阉割的雄性小鼠中显示出抗炎作用,但在性腺完整的雄性小鼠中则没有,这表明益生菌给药存在性别差异。
另一项研究报告了金枪鱼油和海藻油混合物对肠道微生物群的抗衰老作用存在性别差异。雄性小鼠在特定混合油比例下表现出比雌性小鼠更好的抗衰老效果。
肠道菌群失调引发的疾病在两性之间表现不同。研究最多的疾病主要是代谢性疾病。研究发现,性激素等多种因素影响这些疾病,它们密切相关,相互作用。
▸ 全球肥胖发病率越来越高
肥胖症的体重指数(BMI)为30kg/m2或更高,在全球各国成人和儿童中的流行率都在增加。这种病理学是复杂的遗传、社会经济和文化相互作用的结果,会导致严重的健康、经济和社会问题。
科学研究表明,一些代谢紊乱的发生与身体脂肪的分布有关,而且这种分布表现出性别二态性。脂肪倾向于堆积在男性的躯干和腹部周围,而女性的脂肪一般在臀部和大腿周围。
•腹部肥胖
腹部肥胖,尤其是内脏肥胖,会导致血糖和甘油三酯升高、高密度脂蛋白胆固醇降低和低密度脂蛋白 (LDL) 颗粒的增加,以及炎症标志物的增加 。
•臀部肥胖
相反,臀部脂肪与保护性脂质和葡萄糖水平以及降低代谢风险有关,似乎通过长期脂肪酸储存和有益的脂肪因子水平(与瘦素和脂联素水平呈正相关,与炎性细胞因子水平负相关)发挥其保护作用。
肥胖中的微生物多样性和性激素
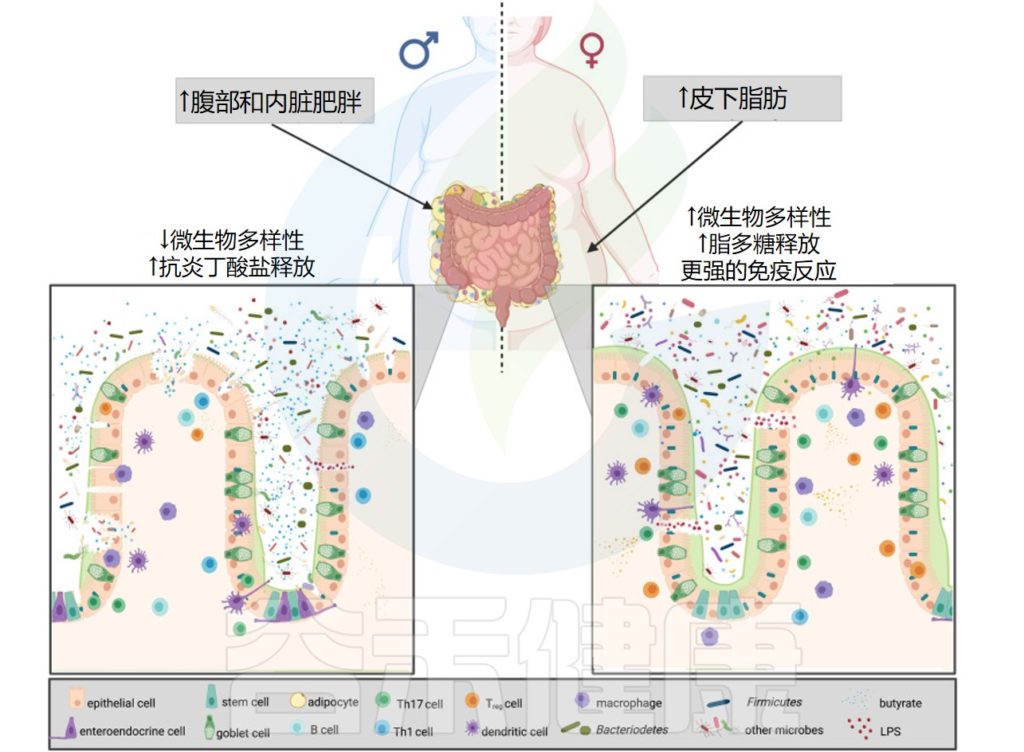
Brettle H,et al.Front Immunol.2022
在肥胖症中,拟杆菌门决定了疾病的严重程度。肥胖男性的物种丰富度较低,并且发现睾酮与厚壁菌门增加有关,因此会释放更多的抗炎丁酸盐。
另一方面,肥胖女性尽管微生物多样性更高,但雌二醇和拟杆菌增加,导致脂多糖释放更多,从而引发更强的免疫反应。
脂肪组织分布、性激素和肥胖的代谢紊乱
Brettle H,et al.Front Immunol.2022
与绝经前女性相比,男性和绝经后女性心血管风险增加,腹部/内脏肥胖和胰岛素皮下脂肪分布减少。男性和绝经后女性体内的脂肪组织比绝经前女性的脂肪组织更具促炎性。
√性激素调节身体脂肪分布
大量证据支持性类固醇调节身体脂肪分布的观点。在这方面,青春期激素的变化与不同性别之间的体重增加有关,这是由于男孩的瘦体重增加和女孩的脂肪量增加,以及男性和女性的脂肪分布。
此外,几项研究表明,一些基因参与了在体脂分布中观察到的性别二态性,以及性类固醇激素在这些基因调节中的潜在作用。
睾酮影响不同部位的脂肪堆积
在男性中,睾酮抑制腹内区域甘油三酯的摄取,并似乎促进它们在皮下区域的积累,同时导致儿茶酚胺刺激的皮下脂肪分解减少,而不是内脏脂肪分解。
这些过程似乎受到雄激素受体 (AR) 基因的影响,因为在雄激素受体敲除小鼠模型中,雄激素受体的缺失通过减少脂肪分解导致肥胖增加,尤其是晚期肥胖。
此外,caveolin-1蛋白(CAV1)似乎在脂肪堆积中起着重要作用,并且它受雌激素(雌二醇)和雄激素(双氢睾酮)的不同调节。
√性激素影响脂肪细胞功能
在细胞水平上,已经观察到性激素(雄激素和雌激素)对白色脂肪组织中脂肪细胞功能影响的差异,涉及脂肪细胞分化、脂肪分解、脂肪生成、胰岛素敏感性和脂肪因子产生/分泌等关键方面。
在这种情况下,睾酮和双氢睾酮调节小鼠间充质干细胞的分化,以雄激素依赖性方式分别促进和抑制它们向肌细胞和脂肪细胞的分化。
双氢睾酮增加脂肪分解
同样,在对人体细胞进行的体外研究中,双氢睾酮以雄激素受体依赖性方式抑制人间充质干细胞和人前脂肪细胞的脂肪形成分化,增加脂肪分解并减少脂质积累。
睾酮阻止脂肪细胞肥大和脂肪细胞生成
在阉割小鼠(雄性性腺功能减退症模型)中,脂肪量通过脂肪细胞肥大和脂肪生成增加,而当这些小鼠接受激素替代疗法时,睾酮阻止内脏和皮下脂肪量的扩张。
此外,致肥胖脂肪生成也通过抑制雄激素受体活性而升高。这项研究还显示了脂肪分布的不同调节,睾酮衍生的雌二醇和双氢睾酮分别阻止了内脏和皮下脂肪的增加。
√睾酮浓度较高利于减少内脏脂肪
睾酮浓度与向心性肥胖呈负相关,并且已发现睾酮治疗可减少具有雄激素缺乏症状和低血清睾酮水平的男性的内脏脂肪。
睾酮含量下降后脂肪更多的积累
此外,随着年龄的增长,睾丸激素的下降伴随着肥胖的增加,腹部脂肪的优先积累和内脏脂肪组织的更多积累。
还有报道称,内脏脂肪组织与生物可利用和游离睾酮呈负相关,皮下脂肪组织与性激素结合球蛋白 (SHBG) 呈负相关。
√雌二醇水平较低的女性易患向心性肥胖
在女性中,向心性肥胖与睾酮水平升高和雌二醇降低相关。更年期的荷尔蒙变化导致脂肪重新分布,与总脂肪和年龄无关,然而,一些研究表明,绝经后上半身脂肪的分布可能是由于衰老而不是绝经本身。
向心性肥胖——指患者体内脂肪沉积是以心脏、腹部为中心发展的一种肥胖类型。
最近发现,绝经前妇女的身体或躯干脂肪量与较低的总雌二醇和较高的游离雌二醇浓度有关,绝经前和绝经后妇女的总睾酮和计算游离睾酮浓度较高,性激素结合球蛋白和胰岛素样生长因子-I(IGF-I)浓度较低。
▼
代谢综合征是一种以腹部肥胖、胰岛素抵抗、高血压和高血脂为特征的病理状况,已遍布全球并导致2型糖尿病、冠心病和中风等疾病患病率上升。
性激素对肥胖、代谢综合征和2型糖尿病的影响

Santos-Marcos JA,et al.Biol Sex Differ.2023
√代谢综合征患者中男女的菌群差异
在代谢综合征患者中也观察到特定菌属的丰度差异。男性中韦荣氏球菌属(Veillonella)、Methanobrevibacter、Acidaminococcus、梭状芽孢杆菌(Clostridium)、罗氏菌属(Roseburia)和Faecalibacterium的丰度较高,而女性中嗜胆菌属(Bilophila)、瘤胃球菌属(Ruminococcus)和拟杆菌属(Bacteroides)的丰度更高。
√睾酮水平会影响代谢综合征
在睾酮对代谢综合征的影响中观察到的性别二态性似乎是雄激素受体依赖性的,并且已经提出了几种机制来解释睾酮水平与代谢综合征之间的关联。
男性中睾酮水平高不易患代谢综合征
在男性中,有证据表明睾酮与内脏肥胖、胰岛素抵抗和代谢综合症的发展呈负相关。沿着这些思路,在男性和啮齿动物中都报道了睾酮的雄激素受体介导的抗肥胖作用。
过量的睾酮可能导致女性胰岛素分泌受损
在女性中,据报道睾酮水平升高与胰岛素抵抗和葡萄糖耐受不良相关,因为它会降低全身葡萄糖的摄取。
关于睾酮对胰腺的作用,一项小鼠研究表明,雄激素受体调节男性胰岛β细胞生理,因此该受体的缺乏会降低葡萄糖刺激的胰岛素分泌并导致葡萄糖耐受不良。相反,有人提出,过量的睾酮可能通过雄激素受体依赖性机制导致女性胰岛β细胞功能障碍,胰岛素分泌受损。
√雌激素缺乏的女性代谢综合征患病率较高
雌激素水平似乎也影响代谢综合症的患病率。因此,大鼠卵巢切除术引起的雌激素耗竭导致大多数代谢综合征成分(脂质、葡萄糖、高密度脂蛋白和低密度脂蛋白)恶化,而在50岁以下的女性中,即经历更年期,其患病率增加。
卵巢切除的女性血糖和血压升高
此外,据报道,在接受过子宫切除术(通常伴随双侧卵巢切除术以预防后续卵巢癌)的女性中,血糖水平和高血压升高。
绝经导致性激素结合球蛋白水平下降,至少部分是由于雌激素减少,而睾酮水平在绝经期间没有改变 。从这个意义上说,绝经可以被认为是代谢综合征及其独立于年龄的所有独立组成部分的预测因素(风险因素)。
此外,已经描述了性激素结合球蛋白和代谢综合征之间的负相关,尤其是在绝经后妇女中。
▼
糖尿病包括一组疾病,根据其发展机制进行区分,这些疾病会降低调节血流中葡萄糖水平的能力,并导致长期高血糖症。
由于自身免疫和代谢过程,糖尿病有两种主要形式,即胰岛素依赖型糖尿病(1型糖尿病,T1D)和非胰岛素依赖型糖尿病(2型糖尿病,T2D)。
2型糖尿病的特征是胰岛B细胞产生的胰岛素不足和肝脏葡萄糖代谢受损,以及胰岛素抵抗,导致组织对胰岛素的反应性降低。
√糖尿病前期的症状表现出性别差异
空腹血糖受损 (IFG) 和葡萄糖耐量受损 (IGT) 作为2型糖尿病的前期阶段发生,表现出性别二态性,葡萄糖耐量受损在女性中更常见,而空腹血糖受损在男性中更常见。
有人提出,性激素可能是造成这种二态性的原因。事实上,更年期的雌激素治疗会降低空腹血糖并恶化葡萄糖耐量。
√男性的2型糖尿病发病率高于女性
此外,已证实男性的2型糖尿病发病率高于女性,这进一步支持了性激素参与这种病理学的发展。更年期2型糖尿病的风险增加,而更年期的激素治疗可能会延迟2型糖尿病的发作。
√睾酮水平影响男性2型糖尿病患病率
患有2型糖尿病的男性总睾酮和游离睾酮水平较低。与此相关的是,低水平的睾酮和性激素结合球蛋白与男性胰岛素抵抗和随后的2型糖尿病的发展有关。
低睾酮水平男性死亡率增加
此外,高水平性激素结合球蛋白和低水平睾酮的结合与2型糖尿病男性死亡率增加有关。其他研究表明,在患有2型糖尿病的男性中,低睾酮水平本身与死亡率增加相关,而睾酮替代疗法可能会提高这些男性的生存率。
同样,据报道,经过2年的睾酮治疗后,患有2型糖尿病的男性比例有所降低。此外,已发现前列腺癌的雄激素剥夺疗法会增加患糖尿病的风险。
患有2型糖尿病的男性睾酮水平往往较低
与上述一致,患有2型糖尿病的男性往往睾酮水平较低,而且他们中的大多数患有性腺功能减退症。事实上,许多研究已经证实,患有性腺功能减退症和低睾酮水平的肥胖2型糖尿病患者在接受睾酮替代疗法 (TRT) 后表现出改善的胰岛素抵抗和血糖控制。
√雌二醇水平高的男性患2型糖尿病的风险增加
关于女性荷尔蒙,雌二醇水平高的男性患2型糖尿病的风险增加,这种高雌二醇浓度与低性激素结合球蛋白浓度一起对男性患2型糖尿病的风险产生不利影响。
√女性睾酮水平较高可能易患2型糖尿病
与男性相反,女性的高睾酮水平与胰岛素抵抗和2型糖尿病相关。然而一项研究表明,中国女性性激素结合球蛋白值升高与患2型糖尿病的可能性降低相关,但雌二醇和睾酮水平与该族群中的2型糖尿病无关。
注:这些关于睾酮与2型糖尿病发病率之间关系的相互矛盾的结果可能是由于睾酮的测量,一些作者使用总睾酮而其他人使用游离睾酮,并且根据最近的一项研究,不同研究的分析方法可能不同。
此外,当性激素结合球蛋白浓度低于30nmol/L时,游离雄激素指数(FAI)不是游离睾酮的可靠指标,这将导致性激素结合球蛋白水平较低的女性可能出现研究错误。
因此,在女性中,总睾酮与2型糖尿病之间没有关联,尽管游离睾酮水平较高与2型糖尿病风险增加有关。
√雌二醇水平异常会增加2型糖尿病发病率
关于雌二醇,据报道患有2型糖尿病的绝经后女性的雌二醇水平高于健康女性。然而,基于早期初潮或绝经以及子宫切除术和卵巢切除术实践的大量证据表明,非生理性雌二醇水平(高于或低于正常值)可能是导致2型糖尿病发病率增加的原因。
月经初潮提前会增加2型糖尿病的风险
在这方面,月经初潮提前似乎会增加患2型糖尿病的风险。然而,一些研究表明,初潮早导致的2型糖尿病风险的部分原因可能是肥胖增加,因为早期月经初潮已被证明也与成年期体重指数的增加有关。
提前绝经和卵巢功能不全导致2型糖尿病风险增加
另一方面,提前绝经或卵巢功能不全会导致患2型糖尿病的风险增加。在接受双侧卵巢切除术的绝经后妇女中也观察到了类似的结果。
切除卵巢的女性2型糖尿病风险增加
最后,伴有双侧输卵管卵巢切除术(BSO)的子宫切除术显示2型糖尿病的风险高于子宫切除术本身。然而,其他研究表明子宫切除术与2型糖尿病风险增加有关,而双侧输卵管卵巢切除术本身或与子宫切除术一起并没有增加2型糖尿病的风险。
研究表明表明接受子宫切除术或卵巢切除术的女性患2型糖尿病的风险增加,但并未区分这两种情况是单独发生还是同时发生。
另一项研究表明,相对于完整女性,双侧卵巢切除的子宫切除女性的总睾酮和生物可利用睾酮水平较低,而保留卵巢的子宫切除女性的睾酮水平居中。
与此相关,绝经后妇女(子宫完整和子宫切除)的雌激素和孕激素激素疗法降低了糖尿病的发病率。
▼
肠易激综合征是一组持续或间歇发作,以腹痛、腹胀、排便习惯或大便性状改变为临床表现,胃肠道结构和生化异常的肠道功能紊乱性疾病。
√女性患肠易激综合征的比例更高
肠易激综合征 (IBS) 是一种具有性别差异的代表性疾病,通常在女性中的发生频率是男性的两倍。特别是,到三级中心就诊的患有严重症状的肠易激综合征患者的女性比例更高。
感染性结肠炎后肠易激综合征的风险增加,约10%的肠易激综合征患者在感染性结肠炎后开始出现症状。有趣的是,感染后肠易激综合征的发病率在女性中较高,这表明性别间肠道微生物群的差异可能在肠易激综合征的发病机制中发挥重要作用。
√肠易激综合征的症状存在性别差异
女性更有可能出现腹痛和便秘
肠易激综合征症状存在性别差异:具体而言,IBS-C在女性中占主导地位,而IBS-D在男性中占主导地位。
便秘型肠易激综合征(IBS-C)是肠易激综合征的一种亚型:至少25%的排便为硬粪或干球粪,且松散(糊状)粪或水样粪<25%。
腹泻型肠易激综合征(IBS-D):至少25%的排便为松散(糊状)粪或水样粪,且硬粪或干球粪<25%。
在对22项关于肠易激综合征症状性别差异的研究进行的系统回顾和荟萃分析中,发现女性比男性更可能出现腹痛和便秘相关症状。便秘型肠易激综合征在患有肠易激综合征的女性中的合并患病率为40%,而在患有肠易激综合征的男性中为21%。
男性出现腹泻症状更多
相比之下,患有肠易激综合征的男性比女性更有可能报告腹泻相关症状。腹泻型肠易激综合征(IBS-D)在患有肠易激综合征的女性中的合并患病率为 31%,而在男性中为50%。
√女性月经期间肠易激综合征症状加重
女性在月经期间报告更频繁和更严重的肠易激综合征症状,例如稀便、腹胀和腹痛加重,这些症状的增加可能与月经周期的变化有关。
在冰岛进行的一项基于人群的邮政研究表明,患有痛经的女性比没有痛经的女性更容易出现肠易激综合征症状。此外,更年期似乎与肠易激综合征症状的恶化有关。也就是说,女性在绝经后报告的腹痛比绝经前更严重。
此外,纤维肌痛、慢性疲劳综合征、慢性盆腔痛和偏头痛等经常与肠易激综合征重叠的慢性疼痛疾病的患病率在肠易激综合征女性中更高,这表明她们的症状与激素之间存在关联。
▼
甲状腺是重要的内分泌腺。甲状腺的主要功能是分泌含碘的甲状腺激素三碘甲状腺原氨酸(T3)和甲状腺素(T4)以及肽类激素降钙素,通过影响新陈代谢和组织发育来影响心血管和生殖疾病。
肠道微生物群在甲状腺疾病中起着至关重要的作用,包括桥本氏甲状腺炎 (HT) 和格雷夫斯氏病 (GD)。桥本氏甲状腺炎和格雷夫斯氏病分别是甲状腺功能减退症和甲状腺功能亢进症的主要原因。
√女性甲状腺疾病发病率高于男性
甲状腺疾病性别差异特别的显著,它的高发人群多为中年的女性,男性的患病率相对来说比较低,而之所以会这样是因为女性的内分泌活跃程度很高,没有一定的稳定性。
在平时压力大和情绪发生变化的情况下,会导致自身的免疫调节出现异常,致使甲状腺激素分泌过多,从而就会容易造成甲状腺疾病的发生。
√胆汁酸的组成可以反映甲状腺功能
胆汁酸是肠道微生物群的重要代谢产物,在甲状腺疾病中起着至关重要的作用。甲亢和甲减患者的血清胆汁酸谱不同,胆汁酸的组成可以反映甲状腺功能。甲状腺功能减退患者中最主要的胆汁酸是次级胆汁酸脱氧胆酸,而鹅去氧胆酸是甲状腺功能亢进患者中最主要的胆汁酸。
胆汁酸的组成和水平参与甲状腺激素的分泌,促甲状腺激素水平与亚临床甲减患者血清总胆汁酸呈负相关。
√短链脂肪酸与甲状腺激素相互作用影响女性情绪健康
短链脂肪酸是肠道菌群代谢的主要产物之一,短链脂肪酸与甲状腺激素三碘甲状腺原氨酸相互作用,并通过调节肠细胞基因转录来影响激素的分泌。例如,粪便短链脂肪酸酯可通过影响高脂血症和甲状腺疾病来影响女性的情绪健康。
▼
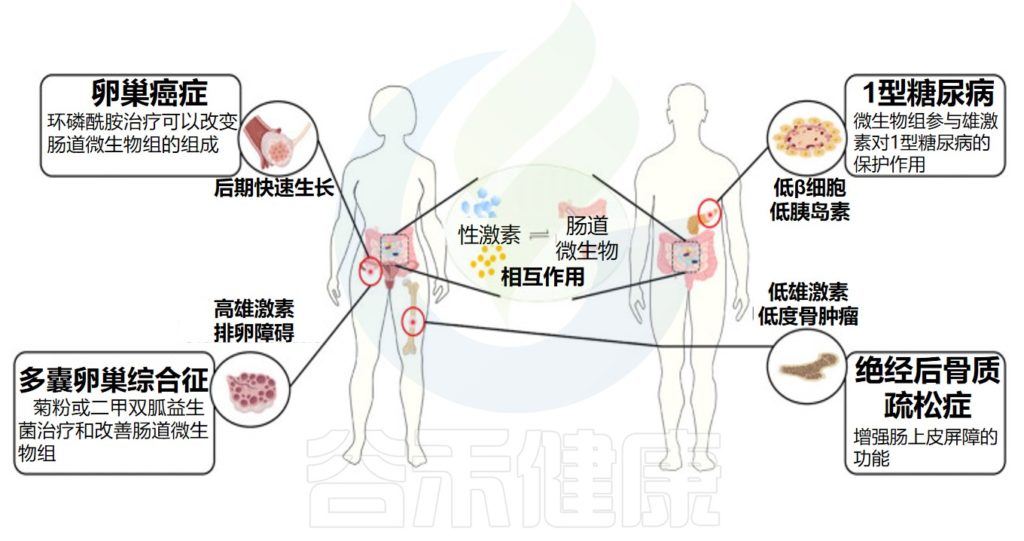
卵巢癌症的病因尚不清楚,可能与环境、生殖、行为和遗传因素有关。在这些因素中,雌激素水平的失调和雌激素的活性被认为是重要因素。
肠道微生物群可能通过影响雌激素水平参与卵巢的发育。发现17β-雌二醇治疗会改变卵巢癌小鼠模型的病理生理学,导致肿瘤提前发作、缩短总生存时间,并具有典型的乳头组织学特征。
√雌激素促进细胞粘附和迁移从而影响卵巢癌的病症
利用不同的卵巢癌症细胞系进行了一项体外研究,以确认雌激素受体在卵巢癌症病因中的需求。
他们的结果表明,雌激素治疗后,雌激素受体阳性细胞系的粘附和迁移能力增强,而雌激素受体阴性细胞系的粘附和移动能力没有显著变化。
这些结果表明,雌激素的作用与雌激素受体的表达直接相关,雌激素促进细胞粘附和迁移,从而影响卵巢癌症的转移和定植。
√微生物通过影响活性雌激素含量影响卵巢癌
如上所述,具有 β-葡萄糖醛酸酶活性的细菌可以介导雌激素的去结合,从而影响循环中活性雌激素的数量。结合雌激素水平和活性在卵巢癌病因学中所起的作用,我们可以推断肠道微生物组可能通过影响活性雌激素的量来促进雌激素驱动的疾病(如卵巢癌)的发展。
▼
绝经后骨质疏松症 (PMOP) 是一种由雌激素缺乏引起的骨质疏松症,会导致绝经后妇女骨折频率增加。
√雌激素缺乏易引起骨质疏松
目前的研究表明,肠道微生物群与骨重塑以及骨代谢疾病之间存在潜在的密切关系。
研究人员发现,在无菌小鼠中,性类固醇缺乏不能诱导破骨细胞因子表达增加、骨吸收激活,这表明肠道微生物组在性类固醇缺乏导致的骨质疏松中至关重要。
他们进一步证明,用益生菌鼠李糖乳杆菌或市售益生菌补充剂每周两次治疗性类固醇缺乏小鼠可以避免骨质疏松。这种情况的发生可能是由于肠道通透性的降低、肠道和骨髓炎症的抑制。
相比之下,补充非益生菌大肠杆菌菌株或突变鼠李糖乳杆菌并没有显示出对骨丢失的保护作用。
上述结果表明,肠道微生物群失调可能导致肠道通透性增加,并触发重要炎症通路的激活,从而导致性类固醇缺乏小鼠的骨质流失。
这些结果将雌激素缺乏引起的骨质疏松症与肠道微生物多样性、肠道通透性和炎症联系起来。
▼
√高雄激素可能导致多囊卵巢综合征
多囊卵巢综合征是育龄妇女常见的内分泌疾病,其病因可能与高雄激素血症、胰岛素抵抗和神经内分泌功能障碍有关。
一些研究关注多囊卵巢综合征患者的肠道微生物组,这些研究发现肠道微生物组与多囊卵巢综合征之间存在某种关联。然而,与多囊卵巢综合征相关的微生物的确切机制尚未确定。
√多囊卵巢综合征患者肠道微生物多样性较低
一些研究人员发现,患有多囊卵巢综合征的患者肠道微生物组的多样性明显低于健康对照组。在多囊卵巢综合征患者的肠道微生物组中,普通拟杆菌(Bacteroides vulgatus)显著升高,脱氧胆酸和熊去氧胆酸水平降低。
通过比较多囊卵巢综合征女性的粪便微生物组、肠上皮通透性和炎症状态,一些研究提出了一个假设,即肠上皮高通透性导致的毒血症与炎症、胰岛素抵抗和高雄激素血症有关。不过还需要更深入的研究来阐明内毒素血症的具体机制。
√调节肠道微生物有助于改善多囊卵巢综合征
菊粉和二甲双胍对多囊卵巢综合征的缓解作用与抗炎和肠道微生物群的调节有关,这可能有助于多囊卵巢综合症的潜在临床治疗。
环丙孕酮Diane-35(雌激素和孕激素)和益生菌可以帮助重建肠道微生物群的多样性,减少肠道菌群紊乱可以改善多囊卵巢综合征样大鼠的生殖功能。
还需要进一步的研究来确定改善肠道微生物组是否可以作为多囊卵巢综合征的有效治疗方法。
小结
根据性激素和肠道微生物组之间的相互作用,性激素相关疾病的可能治疗策略的显示在下图:
He S,et al.Front Microbiol.2021
在对性激素相关疾病治疗的过程中发现了肠道微生物群的变化,未来通过对肠道微生物的检测或许有助于判断性激素相关疾病的治疗效果。
性激素,特别是雌激素和睾酮,与肠道微生物群的结构和组成有关,性激素与肠道微生物群的相互作用对疾病的发展和性别差异具有重要影响。
肠道微生物群的特定变化可能有助于缓解激素紊乱相关的疾病,因此肠道微生物群被认为是一个重要治疗靶点。
在制定治疗策略时,确定导致疾病发病率中性别二态性的机制具有特殊的重要性,因为肠道微生物群的组成取决于与性激素的相互作用以及其他因素。治疗与这些疾病相关的肠道微生物群失调的疗法可能具有性别特异性效应。
总之,肠道微生物群与激素相互作用在疾病中的影响已经成为了一个备受关注的研究领域。肠道微生物群在激素变化及其带来的健康风险的重要作用及其检测的意义也被越来越多的人所认识。未来,随着相关研究的不断深入,肠道微生物群与激素相互作用的机制和应用价值也将会进一步得到探索和发展。
主要参考文献
Santos-Marcos JA, Mora-Ortiz M, Tena-Sempere M, Lopez-Miranda J, Camargo A. Interaction between gut microbiota and sex hormones and their relation to sexual dimorphism in metabolic diseases. Biol Sex Differ. 2023 Feb 7;14(1):4.
Li D, Sun T, Tong Y, Le J, Yao Q, Tao J, Liu H, Jiao W, Mei Y, Chen J, Liu Z, Wang G, Li Y. Gut-microbiome-expressed 3β-hydroxysteroid dehydrogenase degrades estradiol and is linked to depression in premenopausal females. Cell Metab. 2023 Apr 4;35(4):685-694.e5.
Brettle H, Tran V, Drummond GR, Franks AE, Petrovski S, Vinh A, Jelinic M. Sex hormones, intestinal inflammation, and the gut microbiome: Major influencers of the sexual dimorphisms in obesity. Front Immunol. 2022 Sep 27;13:971048.
Kautzky-Willer A, Harreiter J, Pacini G. Sex and Gender Differences in Risk, Pathophysiology and Complications of Type 2 Diabetes Mellitus. Endocr Rev. 2016 Jun;37(3):278-316.
Saklayen MG. The global epidemic of the metabolic syndrome. Curr Hypertens Rep. 2018;20(2):12.
Paschou SA, Papanas N. Type 2 diabetes mellitus and menopausal hormone therapy: an update. Diabetes Ther. 2019;10(6):2313–2320.
Valeri F, Endres K. How biological sex of the host shapes its gut microbiota. Front Neuroendocrinol. 2021 Apr;61:100912.
Kim YS, Unno T, Kim BY, Park MS. Sex Differences in Gut Microbiota. World J Mens Health. 2020 Jan;38(1):48-60.
Yoon K, Kim N. Roles of Sex Hormones and Gender in the Gut Microbiota. J Neurogastroenterol Motil. 2021 Jul 30;27(3):314-325.
He S, Li H, Yu Z, Zhang F, Liang S, Liu H, Chen H, Lü M. The Gut Microbiome and Sex Hormone-Related Diseases. Front Microbiol. 2021 Sep 28;12:711137.

谷禾健康

Dorea菌属于厚壁菌门毛螺菌科,广泛存在于人体肠道内,谷禾数据显示该菌在人群的检出率超89%。该菌最早也是从人体粪便中分离出来。
“Dorea” 目前没有一个确定的译名,Dorea是以法国微生物学家 Joel Doré 的名字命名,以表彰他对肠道微生物学的诸多贡献。
该菌是一类革兰氏阳性厌氧菌,主要存在于人类和动物的肠道中,可以利用多种底物进行发酵代谢,包括葡萄糖、果糖、乳糖和芳香族化合物等。
它可能通过诱导Treg并抑制Th17细胞的分化和功能,从而调节肠道免疫反应,维持肠道黏膜屏障的完整性和稳定性。
编辑
图源:microbiomology
Dorea菌在肠道微生态系统中的丰度和分布受到多种因素的影响,如年龄、饮食、生活方式、疾病等。在不同年龄段、不同饮食类型和不同疾病状态下存在差异,这可能与其在肠道内的营养代谢、免疫调节和菌群相互作用等方面的作用有关。
在多发性硬化症、炎症性肠病患者,甚至结直肠癌、自闭症谱系障碍以及肥胖人群中的Dorea菌高丰度富集,被认为具有促炎作用。
多数研究证实Dorea与体重指数 (BMI)、腰围和舒张压呈正相关。基线肠道内富含高丰度的Dorea菌的人群,在减重方面更困难。
然而Dorea菌在抑郁患者和患有食物过敏人群中减少,研究还表明Dorea菌可以预防或治疗过敏性鼻炎。
相较于其他的肠道重要菌属,关于Dorea 的研究资料相对有限,本文将根据该菌相关的研究结果以及检测实践数据来综合分享该菌的相关信息和健康特性及其干预措施。
该菌是不形成孢子、革兰氏染色阳性、不运动、专性厌氧的菌。葡萄糖代谢的主要终产物是乙醇、甲酸盐、乙酸盐、H2 和 CO2。不产生丁酸盐。DNA G+C 含量 (mol%):40–45.6 (Tm)。
该属的代表菌种是Dorea formicigenerans,是肠道中主要的产气菌,能够利用碳水化合物,这也是一些肠病和部分肥胖患者会伴随腹胀气的原因之一。另外还有Dorea longicatena也比较常见。
该菌属包括目前已经鉴定的常见种包括以下:
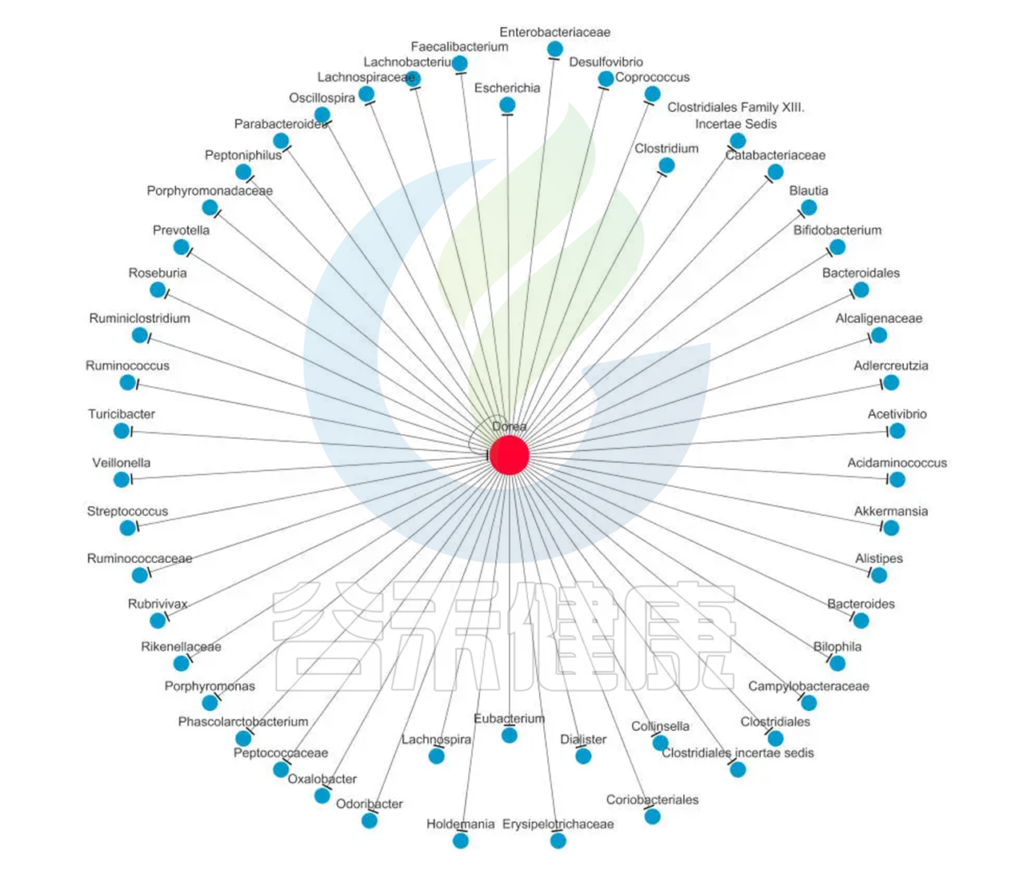
Dorea菌与肠道微生态系统中的其他菌群之间也存在着复杂的相互作用。一些研究表明,Dorea菌与Bacteroides、Faecalibacterium等菌群之间存在着协同关系,可以相互促进代谢产物的生成和利用,从而维持肠道微生态系统的平衡和稳定性。
此外,青春双歧杆菌和长双歧杆菌可以降低或抑制Dorea的丰度。
该菌作为肠道重要菌属,其与其它肠道菌群互作如下,仅供参考:
<来源:谷禾健康数据库>
➩ 非酒精性脂肪型肝炎
患有脂肪变性或非酒精性脂肪型肝炎(NASH)的儿童伴随着更高丰度的Dorea和Ruminococcus 菌属。
线粒体功能障碍,连同氧化应激和肠道菌群改变据信能够促进非酒精性脂肪型肝炎的进展。
证据表明非酒精性脂肪性肝病(NAFLD)会破坏肠-肝轴,将野生型和甲基化控制的J蛋白敲除(MCJ-KO)小鼠喂食6周的高脂饮食 (CDA-HFD),小鼠的Dorea属及Oscillospira属较WT小鼠高;研究结果阐明了MCJ-KO的特定菌群谱,表现为Dorea和Oscillopira菌属的粪便相对丰度增加,以及Allboaculum和Ruminococcus菌属的减少。
这些菌通过增强短链脂肪酸、烟酰胺腺嘌呤二核苷酸(NAD+)代谢和sirtuin(长寿蛋白)的活性,增加了无菌小鼠的脂肪酸氧化来发挥肝脏保护作用。值得注意的是,该研究强调Dorea菌属及其相关代谢产物是这种菌群依赖性保护表型的主要调节剂。
研究者进一步利用公共数据库进行分析:通过一组肝硬化患者的数据,将患者根据体重指数(BMI)分为肥胖(BMI≥30;n=71)和非肥胖(BMI<30;n=121),有趣的是,在非肥胖受试者的子集中,粪便Dorea的丰度显著降低(p=0.026),而在肥胖患者中未观察到显著差异(p=0.636)。
这些发现可能表明瘦型非酒精性脂肪型肝炎患者的肠道菌群组成发生了特定变化,特征表现为Dorea菌属的减少。
➩ 超重和肥胖
肠道菌群被认为在超重和肥胖的发展中发挥重要作用,但能量限制(特别是长期)对超重和肥胖成年人肠道微生物群的影响。发表于Genome Medicine的一项研究发现,基线时Dorea菌属丰度较高可能与随后难以减掉体重有关。
宏基因组研究也表明,Blautia,Dorea和Mediterraneibacter属的增加可能促成宿主肥胖。
Dorea formicigenans 和 Dorea longicatena 在超重/肥胖人群中的丰度更高。
基线Dorea的丰富对随后的体重减轻有适度的预测作用。
Dorea丰度与肥胖呈正相关,在糖尿病前期患者中发现其丰度增加。Dorea丰度的变化与粪中戊酸水平的变化呈负相关。Dorea高丰度可以降低产短链脂肪酸菌的丰度,导致粪便中包括戊酸在内的短链脂肪酸水平降低。
➩ 2型糖尿病
2型糖尿病患者肠道菌群中产丁酸菌减少,而潜在致病菌增多。糖尿病前期患者肠道菌群也有相应的变化:梭菌属和黏蛋白降解菌AKKermansia显著减少。此外,Dorea菌属、瘤胃球菌属和链球菌属增多。
➩ 肠易激综合征
在肠易激综合征受试者中,与健康对照组相比,观察到肠道微生物群,肠杆菌科、瘤胃球菌、梭菌、Dorea物种增加。
➩ 肿瘤息肉
肠道微生态系统组成变化与大肠肿瘤密切相关:与无腺瘤性息肉患者相比,腺瘤性息肉患者肠道微生态中拟杆菌比例相对降低,而变形菌比例相对较高,Dorea菌属及志贺菌属比例升高。
➩ 结直肠癌
结直肠癌患者中Dorea和Porphyromonas增加同时Catenibacterium和Prevotella减少的细菌属被证明产生了最准确的预测效果。Dorea在结直肠癌患者的粪便样本中比在健康对照者的粪便样本中更丰富。Dorea菌具有粘附癌细胞的能力,这可能赋予Dorea在癌性结直肠环境中的竞争优势。Dorea菌的机会主义能力带来潜在致癌作用。
➩ 牛皮癣
牛皮癣患者肠道菌群中的Ruminoccocus gnavus, Dorea formicigenerans, Collinsella aerofaciens丰度显著增加。
➩ 自闭症
与健康对照组相比,自闭症孤独症谱系障碍儿童中毛螺菌科的数量有所增加。毛螺菌科由大约几十个异质属组成。
众所周知,毛螺菌科成员中的许多属甚至可以主宰肠道微生物群,包括Dorea、Blautia、Lachnospira、Coprococcus、Roseburia和Ruminococcus。毛螺菌科成员具有广泛的代谢功能,包括合成短链脂肪酸(包括丁酸盐)、粘蛋白降解以及糖和芳香族氨基酸代谢。他们的生态失调与许多其他慢性疾病有关,例如炎症性肠病、肾脏疾病、肝脏疾病和神经行为疾病等。
➩ 过敏性鼻炎
过敏性鼻炎 (Allergic rhinitis,AR)是特应性个体接触环境中变应原后,通过IgE介导的多种细胞因子及炎性介质参与的鼻腔黏膜慢性非感染性炎症。发现过敏性鼻炎患儿的粪便Dorea菌、Dialister菌和组氨酸水平显著降低,而其差异代谢物与差异菌群无明显关联。
进一步研究发现Dorea和Ralstonia与淋巴细胞模式(Th2/Treg 比率)密切相关。有趣的是,Dorea和Ralstonia与淋巴细胞模式呈负相关:Th2/Treg 比率与Dorea呈负相关, 但与Ralstonia呈正相关。
在儿童早期,Dorea菌与屋尘螨特异性 IgE 水平呈负相关,并且似乎对过敏性鼻炎更具特异性。由于Dorea属与过敏的持续关联,因此Dorea菌丰度的改变可能通过改变对屋尘螨的过敏性致敏作用的发展,来预防或治疗过敏性鼻炎。
➩ 食物过敏
厚壁菌门的Dorea属与婴儿期的食物致敏和食物过敏呈负相关。
研究表明提出婴儿双歧杆菌(Bifidobacterium Infantis)可能通过增加Dorea和减少Ralstonia来减轻 Tm 引起的过敏反应。Dorea可能有助于诱导 Treg 并抑制 Tm 诱导的过敏反应。
注:Tm 是指过敏原(allergen)或致敏物质(sensitizing agent)。
另外一项为期 3 年的后续研究表明,Dorea在后来发生食物致敏或食物过敏的婴儿的肠道微生物群中减少,因此表明Dorea可以防止食物致敏和食物过敏。
➩ 与肠道内肠道辅酶NAD正相关
Dorea相对丰度与肠道NAD总水平呈正相关。肠道NAD指的是肠道内的辅酶NAD(nicotinamide adenine dinucleotide)分子。NAD是一种重要的辅酶,在细胞中发挥着多种重要的生物学功能,包括能量代谢、DNA修复、信号转导等等。肠道内的NAD含量和代谢状态对肠道黏膜屏障的维护、肠道免疫和菌群平衡等方面都有着重要的影响。最近的研究表明,肠道NAD水平的改变可能与多种肠道相关疾病的发生和发展有关。
➩ 心力衰竭
此外,个别研究发现直肠真杆菌和Dorea longicatena在心力衰竭患者肠道微生物群中的含量低于健康受试者。
➩ 抑郁、睡眠
重度抑郁症是一种常见的精神疾病。研究结果显示属水平上,Dorea菌与抑郁和睡眠质量同时相关。压力诱导 Dorea, Coprococcus等菌群显著变化。
Dorea在抑郁症患者中减少,患有睡眠障碍的重度抑郁症患者的Dorea formicgenerans减少。
在属水平上,摄入 3 g/d HMW β-葡聚糖会增加拟杆菌,但会减少 Dorea (P < 0.1)。
大麦干预具有更高丰度的 Akkermansia、Blautia 和 Bilophila,同时减少 Parabacteroides和 Dorea。
属于厚壁菌门的 Dialister、 Dorea 、Pseudobutyrivibrio 和 Veillonella 在使用L-谷氨酰胺后显着减少。
补充维生素D,肠道内Dorea相对丰度降低。
补充硫酸铁(口服)导致 Dorea菌物种的丰度降低。
低聚半乳糖 (GOS) 补充显示Dorea减少。
补充益生元(低聚果糖FOS 和菊粉混合物)显示 Dorea 丰度较低。
Bifidobacterium longum 增加Dorea菌。
喂食鼠李糖乳杆菌hsryfm 1301或其发酵乳28天后,大鼠的肠道微生物群和血脂得到改善。血脂与瘤胃球菌、Dorea、Enterococcus呈正相关(P<0.05)。
刺梨(RRT)的果实可食用,被认为是具有降脂作用的药用果实。12 周刺梨降低 Dorea和Coprobacillus 丰度,并促进双歧杆菌和Roseburia 丰度,来逆转高脂饮食诱导的肠道生态失调。
芒果 ( Mangifera indica L.) 果皮是工业加工获取果汁和浓缩物的副产品,富含多酚和膳食纤维。将预先消化的芒果皮提交给人类结肠的动态体外模型。Dorea属在芒果果皮发酵中的含量为 6.69%。
在抗生素(阿莫西林、四环素和甲硝唑)治疗 2 周后,Dorea formicgenerans,Eubacterium rectale的丰度下降。
白藜芦醇有效降低了Akkermansia、Dorea、Sutterella和Bilophila菌属的丰度 。
在高蛋白饮食后,Dorea会随着 BMI 的降低而降低。
Dorea菌属可以利用多种底物进行发酵代谢,包括葡萄糖、果糖、乳糖和芳香族化合物等。因此,调节饮食结构可以影响Dorea菌属的生长和代谢。
摄入动物性饮食五天后,肠球菌科、肠球菌属和 Dorea属的细菌在蠕虫感染中表现出丰度增加的趋势。
高纤维、低脂肪的饮食有利于增加Dorea菌属的丰度和多样性。
富含可发酵碳水化合物的饮食支持相对丰富的双歧杆菌、普雷沃氏菌属、瘤胃球菌属、Dorea 、 Roseburia 等。
果胶是一种水果蔬菜中发现的复杂的膳食纤维和益生元。果胶摄入增加Lachnospira、Dorea和Clostridium,其中Lachnospira的增幅最大。
中等强度的运动干预持续 3 个月后,亚阈值抑郁症的年轻青少年的肠道菌群发生变化,在属水平上增加了Coprococcus、Blautia、Dorea、Tyzzerella 、Tyzzerella nexilis 的相对丰度。
注:中等强度的运动,包括每天跑步 30 分钟,每周 4 天。
结 语
当谈到Dorea菌时,虽然仍有许多未知的领域,但我们已经取得了一些重要的研究进展。Dorea菌在肠道微生物组中扮演着重要的角色,与人类健康密切相关。它们能够帮助维持肠道菌群的平衡,支持营养物质的吸收和消化,并且可能对肥胖、炎症性肠病、自身免疫疾病等疾病的发生和发展起到重要作用。
然而,关于Dorea菌的研究仍然处于起步阶段。我们需要研究更多的样本、更多的群体,并使用更多的技术手段来深入了解这些菌属的作用机制。相信随着微生物组研究的不断深入,我们将能够更好地理解Dorea菌和其他肠道菌属对人类健康的影响,并开发更加个性化的干预管理方案。
主要参考文献:
Chaput N,Lepage P,Coutzac C et al. Baseline gut microbiota predicts clinical response and colitis in metastatic melanoma patients treated with ipilimumab.[J] .Ann Oncol, 2019, 30: 2012.
Sowah SA, Milanese A, et al., Calorie restriction improves metabolic state independently of gut microbiome composition: a randomized dietary intervention trial. Genome Med. 2022 Mar 14;14(1):30
Juárez-Fernández M, Goikoetxea-Usandizaga N, Porras D, García-Mediavilla MV, Bravo M, Serrano-Maciá M, Simón J, Delgado TC, Lachiondo-Ortega S, Martínez-Flórez S, Lorenzo Ó, Rincón M, Varela-Rey M, Abecia L, Rodríguez H, Anguita J, Nistal E, Martínez-Chantar ML, Sánchez-Campos S. Enhanced mitochondrial activity reshapes a gut microbiota profile that delays NASH progression. Hepatology. 2023 May 1;77(5):1654-1669.
Zhang Q, Yun Y, An H, Zhao W, Ma T, Wang Z, Yang F. Gut Microbiome Composition Associated With Major Depressive Disorder and Sleep Quality. Front Psychiatry. 2021 May 21;12:645045.
Dahl WJ, Rivero Mendoza D, Lambert JM. Diet, nutrients and the microbiome. Prog Mol Biol Transl Sci. 2020;171:237-263.
Chen D, Yang Z, Chen X, Huang Y, Yin B, Guo F, Zhao H, Zhao T, Qu H, Huang J, Wu Y, Gu R. The effect of Lactobacillus rhamnosus hsryfm 1301 on the intestinal microbiota of a hyperlipidemic rat model. BMC Complement Altern Med. 2014 Oct 10;14:386.
Tangestani H, Boroujeni HK, Djafarian K, Emamat H, Shab-Bidar S. Vitamin D and The Gut Microbiota: a Narrative Literature Review. Clin Nutr Res. 2021 Jul 20;10(3):181-191.
Wang R, Cai Y, Lu W, Zhang R, Shao R, Yau SY, Stubbs B, McIntyre RS, Su KP, Xu G, Qi L, So KF, Lin K. Exercise effect on the gut microbiota in young adolescents with subthreshold depression: A randomized psychoeducation-controlled Trial. Psychiatry Res. 2023 Jan;319:115005.

谷禾健康

音乐对身心的影响
近年来,环境和动物生理及心理的相关研究越来越多。环境因素的丰富性和多样性是改善动物生理和心理状态的重要研究参数。
环境因素指的是正常环境,在这种环境中,动物通过获得环境激励以做出有益的增强,使它们能够正常表达其行为和心理活动,从而改善健康状况和生长性能。
★ 音乐对人体健康具有益处

音乐这种声波刺激,影响着动物的生理和心理。以前的研究表明,伴随音乐喂养动物会影响它们的生长性能和动物生产。音乐的节奏鼓励肌肉活动并激发机体的活力。当音乐的节奏接近机体的心律时,会刺激机体分泌调节荷尔蒙。
美国开展的一项音乐疗法研究表明,它可以缓解情绪、改善消化、平衡心理状态并促进疾病康复。
★ 音乐可以辅助调节肠道菌群
中国在先秦时期就将五行音乐疗法(FEMT)引入医学领域。中国的五行音乐疗法有着悠久的历史和完整的体系。科学家观察到五行音乐疗法可以缓解焦虑症状、改善空间认知、调节肠道菌群和辅助药物治疗。
五行音乐疗法——中医理论描述了五种音乐音调、五种器官和五种元素之间的相互联系,五音分属五行,通肝、心、肺、脾、肾五脏。用角、徵、宫、商、羽五种不同的音调的音乐来治疗相对应的疾病。
虽然目前音乐疗法治疗疾病存在一定的局限性,但音乐疗法已成为临床常用的辅助治疗手段。
然而,关于喂食过程中听音乐对肠道微生物影响的报道较少。本文通过最新的研究评估喂养时伴随音乐对小鼠肠道微生物和生长性能的影响,为音乐治疗提供理论支持。
•实验选用生长状况良好、健康的小鼠
在整个研究过程中,没有动物患抑郁症、疾病或死亡,尸检和电子显微镜成像均未显示宏观和微观病理。
整个实验期间的小鼠体重
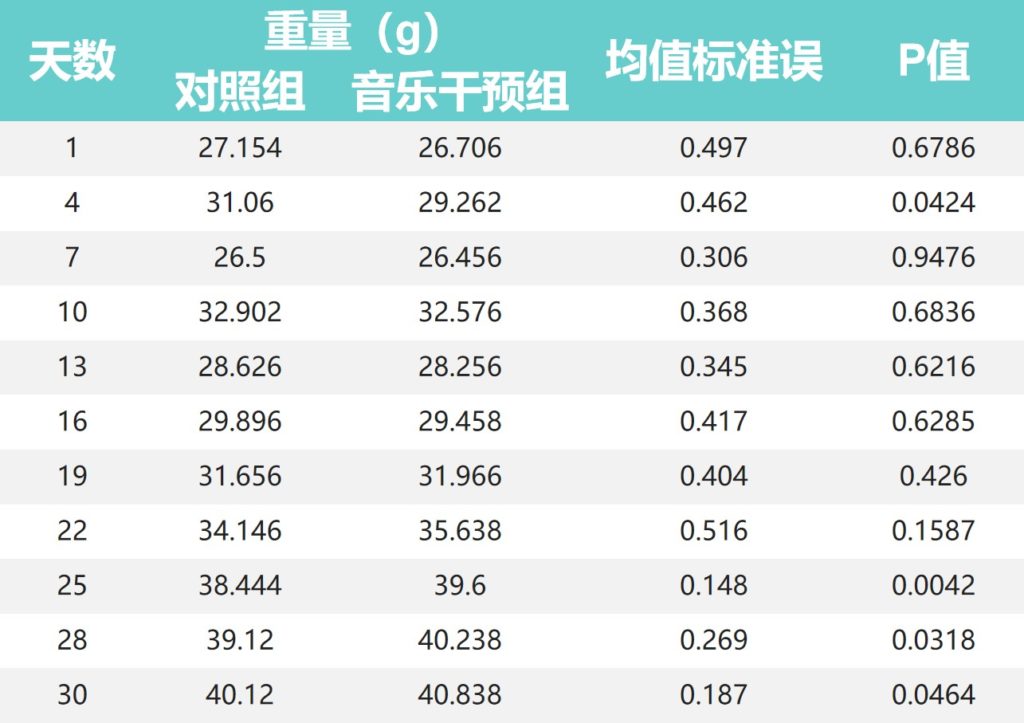
•对照组小鼠体重在前16天高于音乐干预组
对照组小鼠体重在30天喂养期的最初16天高于音乐干预组,第4天两组差异显著(P<0.05)。
•19天后音乐干预组体重开始高于对照组
然而,在第19天后,两组之间的小鼠体重发生了翻天覆地的变化,音乐干预组的体重在第19-30天高于对照组。
其中,第25天(38.444:39.600)、第28天(39.120:40.238)和第30天(40.120:40.838)组间差异有统计学意义(P<0.01;P<0.05;P < 0.05)。
•音乐干预组小鼠情绪和状态更活跃
值得注意的是,在喂食期间,我们发现音乐干预后的小鼠在情绪状况和活动状态方面比对照小鼠更活跃。
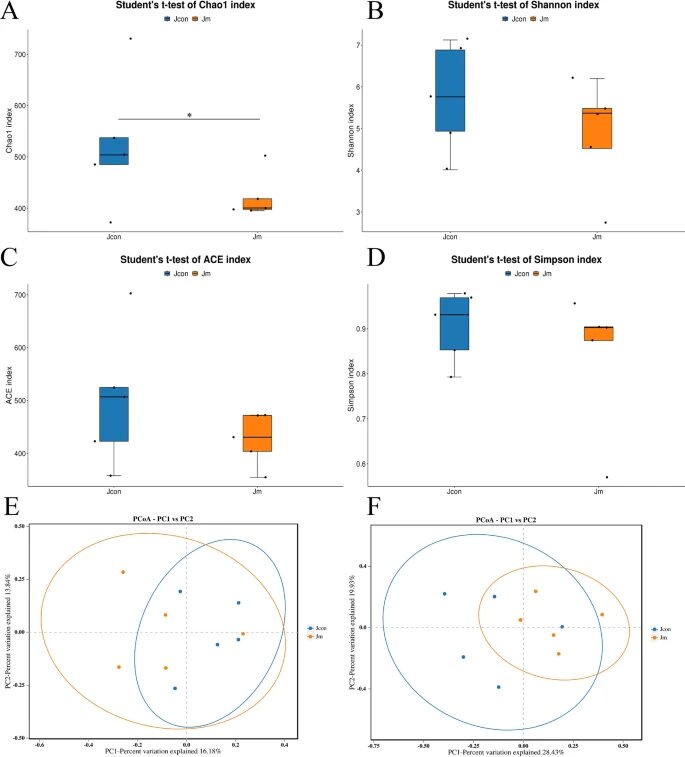
匹配了获得的合格序列,并进行了α-多样性分析,以评估对照组和音乐干预组之间肠道菌群多样性和丰度的差异。
✦伴随音乐的喂养降低了肠道菌群丰度
结果表明,对照组的微生物多样性和丰度高于音乐干预组,两组间chao1指数差异显著(P<0.05)。其他三个指数(Shannon指数、ACE指数和Simpson指数)没有显示出统计差异。
注:chao1指数——又称Chao1丰富度指数或Chao1多样性指数,是一种用于衡量群落物种多样性的指标。
综上所述,可以看出伴随音乐的喂养过程降低了肠道菌群的丰度。
✦β多样性较低
利用QIIME软件计算β多样性分析,比较样本间物种多样性的相似性。
PCoA分析基于未加权的二进制 jaccard(P>0.05)和加权的 bray curtis(P>0.05)显示两组之间没有统计学差异,表明两组之间具有高相似性和低物种多样性。
α和β多样性分析
▼
通过使用 QIIME2 软件对来自不同物种的微生物类群进行分类,评估了门和属水平上优势样本的相对比例。
✦两组中都以厚壁菌和变形菌为主
结果表明,对照组中门水平的优势菌为厚壁菌门(Firmicutes)占42.19%,变形菌门(Proteobacteria)占19.57%,蓝藻(Cyanobacteria)占13.05%和拟杆菌门(Bacteroidetes)占9.85%,占总肠道菌群的84.66%。
在音乐干预组中,门水平的优势细菌为厚壁菌门(Firmicutes)占61.42%,变形菌门(Proteobacteria)占16.00%、蓝藻占7.07%和拟杆菌门(Bacteroidetes)占4.46%,占微生物总数的88.94%。两组的微生物组成(属水平)均显示乳杆菌(Lactobacillus)为优势群落。
此外,uncultured_bacterium_f_Enterobacteriaceae(3.97%比5.83%)和uncultured_bacterium_f_Muribaculaceae(3.47%比1.62%)是两组中的优势菌株。
小结
从以上结果可以得出结论,在小鼠喂食过程中进行音乐干预可以改变肠道微生物的组成。
热图通过色带和接近程度表明多个样品的肠道微生物种群的异同。
结果表明,无论细菌门水平还是在属水平,两组样品之间的微生物丰度相似性较低,但组内样品之间的差异较小,微生物丰度相似。
小鼠肠道微生物组成分布和热图
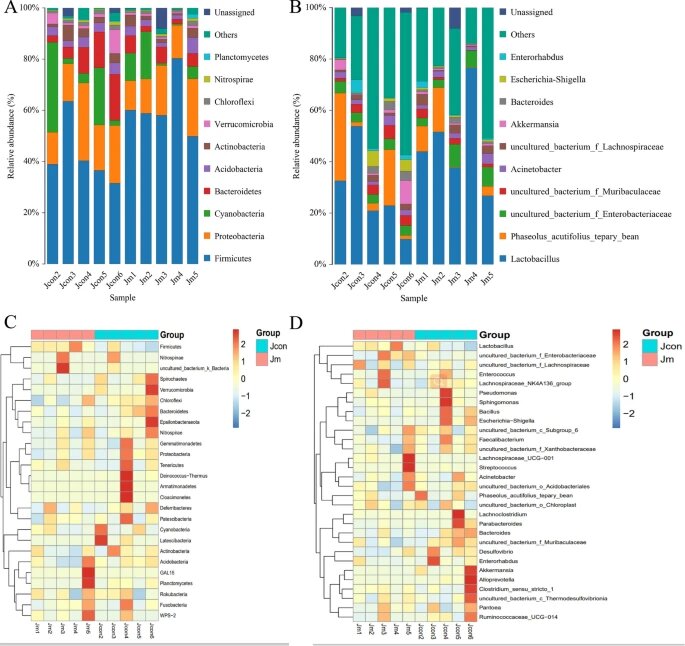
注:(A) 门级的优势细菌种类和丰度;(B) 属级的优势细菌种类和丰度;(C) 门级样品间微生物相似性热图;(D) 属级样品间微生物相似性热图。
▼
✦两组之间厚壁菌丰度存在显著差异
使用 Metastats 分析,研究了两组微生物的门和属水平组成的统计学差异。结果显示厚壁菌门的相对丰度存在显著差异(对照组0.422 ± 0.056与音乐干预组0.614 ± 0051,P < 0.05)。
Metastats分析——组间群落显著性差异分析,用于寻找组间差异物种
✦对照组丰度高于音乐干预组的菌群
在属水平上发现了11个具有统计学意义的分类群,而对照组在以下菌种丰度高于音乐干预组
uncultured_bacterium_o_Microtrichales
(P<0.0001);uncultured_bacterium_f_Micromonosporaceae
(P<0.05);
Pseudolabrys (P<0.05);
Methylobacterium (P<0.05);
uncultured_bacterium_f_Muribaculaceae
(P<0.05);
Ruminococcaceae_UCG-005 (P<0.05) 。
✦音乐干预组丰度高于对照组的菌群
与此相反,以下菌种丰度音乐干预组高于对照组:
uncultured_bacterium_f_Atopobiaceae(P<0.01);Ileibacterium(P<0.01);Lachnospiraceae_FCS020_group(P<0.05);Serratia(P<0.05);
Dietzia (P<0.05)。
门和属水平肠道细菌丰度的统计差异
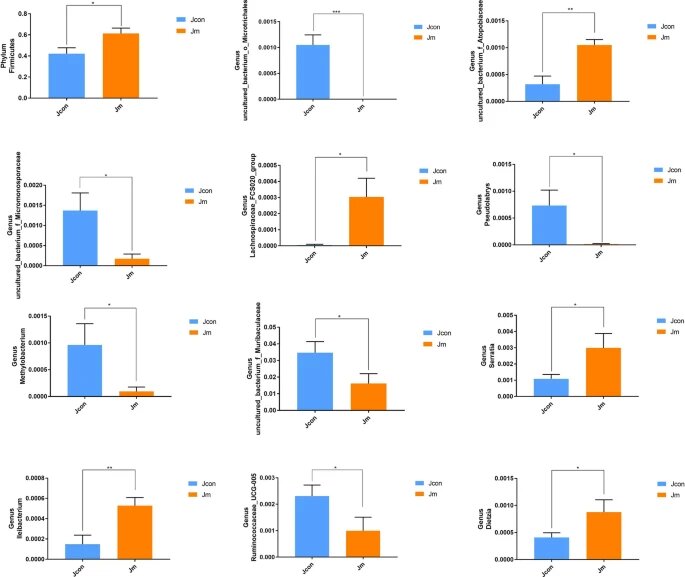
注:所有数据均表示平均值±SD(*P<0.05;**P<0.01;***P<0.001)
▼
考虑到 Metastats 分析对两组间细菌相对丰度差异的局限性,进行了 LEfSe 分析以寻找对照组和音乐干预组之间具有统计学差异的生物标志物。
LEfSe分析是一种将非参数的Kruskal-Wallis以及Wilcoxon秩和检验,与线性判别分析效应量相结合的分析手段, 能够在不同组间寻找具有统计学差异的Biomarker,其要求组内样本数≥3。
结果表明,除了上述显著差异的细菌外,还回收了15个具有统计学差异的生物标志物,对照组中最主要的群体是疣微菌门(Verrucomicrobia)、δ-变形菌(Deltaproteobacteria)和酸微菌(Acidimicrobiia),而Lleibacterium和大洋芽孢杆菌(Oceanobacillus)在音乐干预组中显著表达。
LEfSe分析和LDA评分揭示了细菌丰度的显著差异
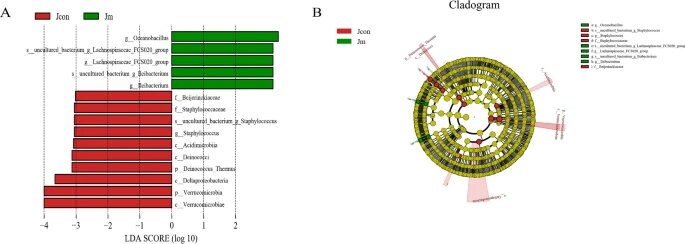
注:(A) LDA分数>3.0被认为具有统计学意义。(B) 分支图描绘了与对照组和音乐干预组相关的肠道微生物群的系统发育分布。
▼
在生态学领域,相关性通常用于构建可用于分析物种群落数据(共现模式)或组合多个数据集进行分析的网络模型。以python为基础,使用前50个属来创建基于网络的关联,该关联表示78个节点、237个边和6个社区。
✦乳杆菌是两组中最丰富的菌
我们发现,在基于网络的相关性分析中,乳杆菌(Lactobacillus)是两组中最丰富的细菌。
此外,音乐干预后小鼠(Jm组)体内乳杆菌丰度高于对照组(47.34%比28.03%),而Phaseolus_acutifolius_tepary_bean丰度低于对照组(6.24% 比 12.36%)。
✦乳杆菌与其他菌属的关联性
乳杆菌与Parabacteroides(0.6364)、Faecalibacterium(0.7455)、幽门螺杆菌(0.7720)、RB41(0.6485)呈负相关。
Phaseolus_acutifolius_tepary_bean与Candidatus_Koribacter(0.7173)和Breznakia(0.7195)呈正相关,与泛菌属(0.66060)、肠球菌属(0.6809)和Ruminococcacee_UCG-014(0.6606)呈负相关。
物种间相关性分析
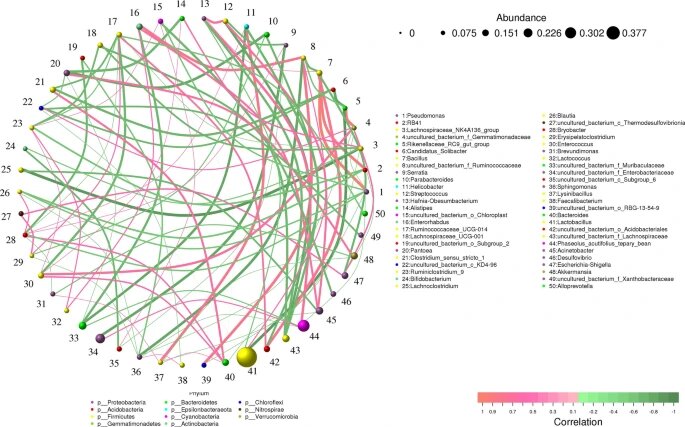
线条描绘了两个物种之间的相关性,线条粗细描绘了相关性的强度。根据线条的颜色,橙色表示正相关,绿色表示负相关。
已发现音乐对身体有积极的影响,引起协调的共振,促进器官节奏的和谐,以及一系列内分泌转变。
▼
✦提高动物的生长性能
先前的研究表明,音乐和检查声音的混合会显著提高小猪生长性能(同样的饲料量下长了更多的肉)。然而,有些音乐(例如重金属,频率<20赫兹,95–105分贝)具有相反的效果。
注:这项研究是在 El Condor Pasa 音乐上进行的,该音乐风格自然且没有过度分层。
在为期30天的喂养实验(有音乐干预)中,发现小鼠的体重在第25天时明显高于对照组。此外还显示,在音乐干预后,动物消耗了更多的食物,体重明显增加。
✦促进对营养的吸收
一些研究发现,音乐可以促进免疫系统和肠道微生物对营养的吸收,同时还可以减轻噪音的负面影响。总的来说,表明音乐可以改善体重,促进小鼠的生长和身体发育。
▼
✦增强肠道菌群的活性
先前的研究表明,音乐疗法可以减少抑郁症,它还会影响消化系统的协调共振、生长激素的分泌并增强肠道菌群的活性和多样性。
肠道微生物群是动物肠道中的微生物群落,在微生物学、医学和遗传学等肠道微生物群相关领域中观察到越来越多的研究趋势。
小鼠胃肠道α-多样性和PCoA分析表明,与对照组相比,音乐干预组肠道微生物发生了变化。有趣的是,整个多样性分析揭示了音乐组中组内样本的高度聚集和组内相似性的显著增强。
✦有助于肠道菌群的相对稳定
我们发现喂食期间的音乐干预从统计学上增加了厚壁菌和乳杆菌的丰富度,同时成倍减少了蓝藻、拟杆菌、肠道微生物中的Phaseolus_acutifolius_tepary_bean和uncultured_bacterium_f_Muribaculaceae。
厚壁菌门是数量最多的细菌,大多数为革兰氏阳性菌,呈球形或杆状,厚壁菌门的许多成员是有益菌,如乳杆菌、芽孢杆菌、双歧杆菌、丁酸梭菌等。
表明肠道微生物多样性的减少与特定细菌的减少或个别菌株的过度生长有关。这表明喂养期间的音乐干预可以减少离散的微生物群并有助于相对稳定性。
✦肠道微生物的稳态有助于防止其他疾病
肠道微生物群的稳态被发现是外部病原体入侵和定植的重要障碍,肠道微生物群的改变可能与多种疾病有关。
除了胃肠道疾病和代谢疾病,肠道菌群还与多种全身性疾病有关,例如神经系统疾病、呼吸系统疾病、心血管疾病和肿瘤疾病。
研究表明,乳杆菌等微生物可以产生乙酸盐、乳酸和抗菌物质,可以防止病原体干扰健康。 此外,它们还有助于维持肠道微生态平衡,预防和抑制肿瘤的发生,增强动物免疫力,促进消化,合成氨基酸和维生素。
总体而言,本研究描述了小鼠在进食期间听音乐情况下肠道微生物群的变化。
结果表明,音乐干预后肠道微生物群发生了显著变化,其特征是肠道细菌多样性下降和小鼠肠道微生物群组成发生变化。此外,在伴随音乐喂养小鼠期间,有益菌的数量增加,而致病菌或条件致病菌减少。
这些结果有助于理解音乐与肠道微生物群之间的关系,以及肠道微生物群会根据不同的喂养环境发生变化。本研究也为音乐疗法改善动物生长环境提高动物生理及心理状态提供了理论依据。

关于肠道菌群干预的相关措施有很多,比如说饮食、睡眠、运动、益生菌、粪菌移植、天然产物、抗生素等,这些在我们之前写过的相关主题的文章都有涉及;而关于音乐对肠道菌群的干预的研究较少,本文对此做一个详细介绍,为音乐疗法的有效性提供了理论依据。
音乐是我们日常生活中轻而易举就能接触到的,可以说是一种非常便捷的治疗方式,因为它不需要使用任何药物或器械,无需去医院等特殊的场所。通过本文的研究可以窥见音乐疗法对于干预人类肠道菌群的潜力。
对于研究人员而言,未来可以更深入了解音乐疗法的作用机制,通过研究不同类型的音乐、不同的音乐治疗对象,探索更多有效的个性化的音乐调节肠道菌群的方案,从而更好地应用于临床实践中,让大众有机会感受音乐疗法带来的不可思议的力量。
参考文献
Niu J, Xu H, Zeng G, Wang P, Raciheon B, Nawaz S, Zeng Z, Zhao J. Music-based interventions in the feeding environment on the gut microbiota of mice. Sci Rep. 2023 Apr 18;13(1):6313.

谷禾健康

目前,原核生物的系统发育分类依赖于16S rRNA基因序列,这种序列在细菌中广泛存在且高度保守,但即使16S rDNA测序的结果显示某些细菌间序列相似性≥99%,但它们仍然不能被归为同一物种。因为基因序列中的一些保守区域并不一定代表真正的进化历史,水平基因转移也可能会影响分析结果。
为此研究人员开发了一种方法,并为该方法创建了一个web应用程序Phy5,以及它的命令行版本Phy5cli。
该方法旨在对任何物种或亚种进行系统发育评估,包括与已知物种的基因序列相似性非常低的非致病菌株,可以轻松地进行系统发育树分析。
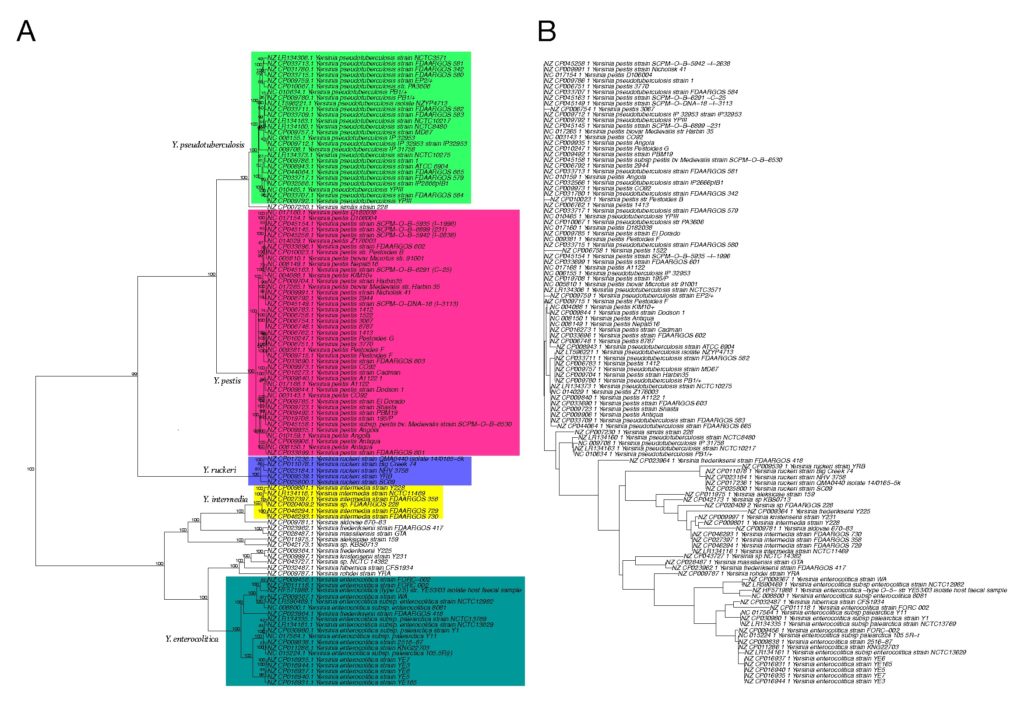
文中以E.coli、Shigella、Yersinia、Klebsiella和Neisseria spp.的系统发育关系为例进行了应用与验证。
通过分析短核苷酸的频率差异来区分高度相似物种,划分成不同的类群,从而推断它们在系统发育上的关系。
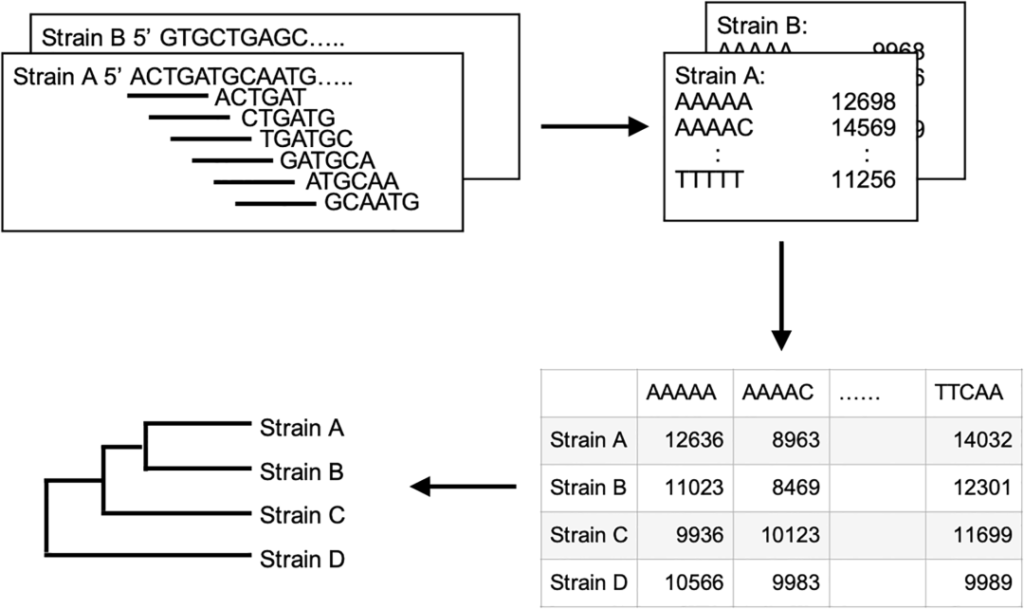
研究使用R v4.12和Biostrings软件包分别对每个细菌样品的三核苷酸、四核苷酸、五核苷酸和六核苷酸频率进行了测定。
这些K-mer(由k个连续核苷酸组成的单元)频率是构建系统发育树的重要信息。通过测定K-mer频率,可以使用层次聚类分析(hierarchical cluster analysis)将样品细菌基因组的相似性进行比较。
在此过程中,使用曼哈顿距离(Manhattan)估计样本间的差异程度,并使用沃德算法(Ward’s)来判断样本间的相似性和关联性。
在该方法中,将互补的K-mer对(例如AAA vs TTT)视为相同的核苷酸序列,以降低这种互补配对对结果的影响。如下图,就是基于五核苷酸频率的系统发育树构建方案。
1
分别基于五核苷酸频率分析和MLST构建的110个Yersinia菌株的系统发育树
如下图所示,图A为基于五核苷酸频率分析所构建的系统发育树,采用曼哈顿距离和Ward算法。
图B是采用neighbor-joining方法的多重位点序列分型(MLST) 分析所构建的系统发育树。
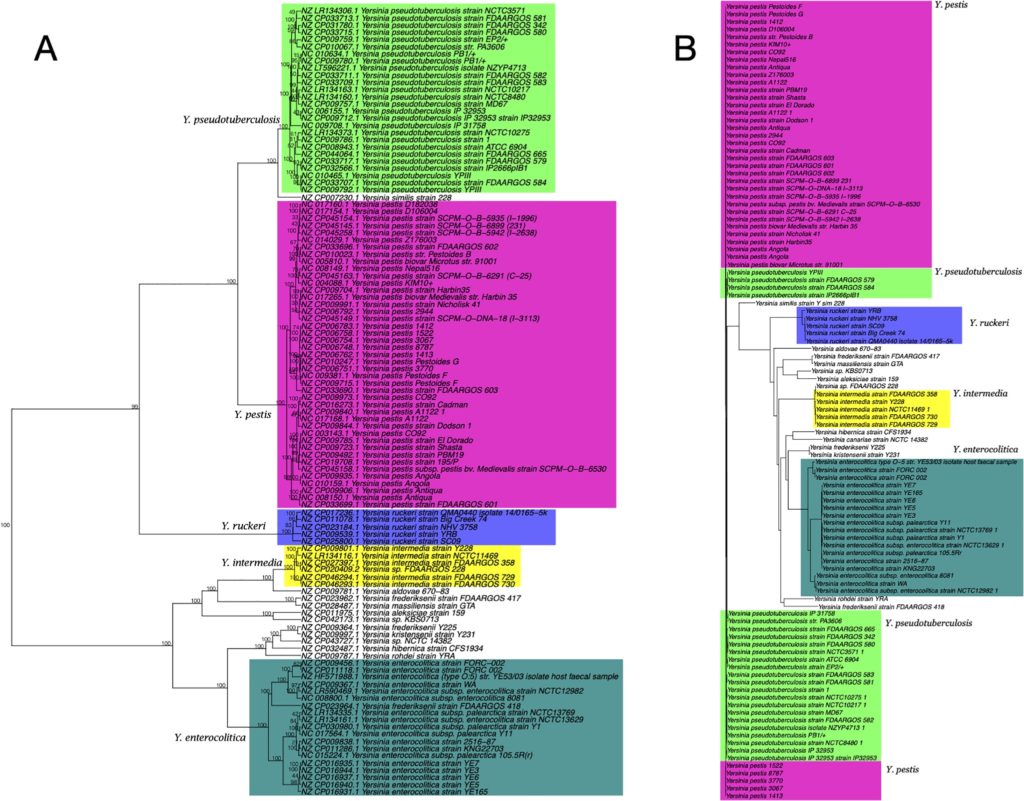
节点上的数字表示在1000个bootstrap值中出现的百分比。颜色区分不同种水平分类。
结果表示两种方法在这个特定的数据集上所产生的系统发育树是高度相似的,它们都可以很好地区分Yersinia和其他物种。
而基于16S rRNA基因序列分析所构建的系统发育树(MAFFT工具)无法区分这些物种,如下图所示,下图B为基于16S rRNA基因序列分析所构建的系统发育树。
2
基于五核苷酸频率分析方法构建系统发育树可以应用于任何物种,且仅需要通过高通量测序技术获取的短链
如下图,根据五核苷酸频率构建了E. albertii、E. coli和Shigella的系统发育树。

发现E. albertii 和致肠出血性大肠杆菌菌株(包括O157、O121和O111等)密切相关,并与Shigella和非致病性E. coli(如K-12菌株)分开排列。
Shigella菌株与肠侵袭性大肠杆菌(EIEC)可以区分开来,并与其他大肠杆菌菌株形成一个分支。
3
基于五核苷酸频率分析方法构建的系统发育树仅适用于近亲物种
研究人员从GenBank序列数据库下载了24个Ipomoea属植物的30个叶绿体基因组的核苷酸序列,并使用五核苷酸频率分析方法构建系统发育树,结果如下图,只构建出来与Ipomoea相关的物种。
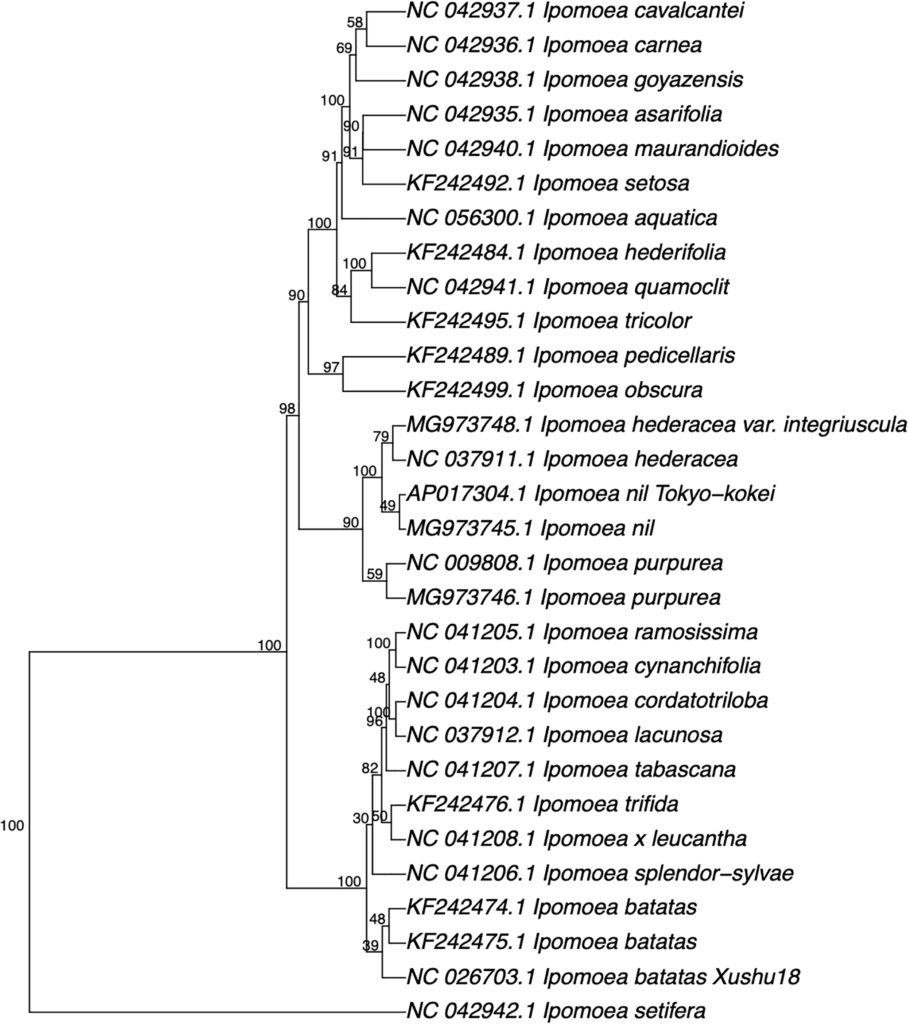
如其它隶属于Ipomoea的物种并没有被发现。
研究人员还测试了使用五核苷酸频率分析方法构建各种远亲物种的系统发育树,包括嗜热古菌和细菌。
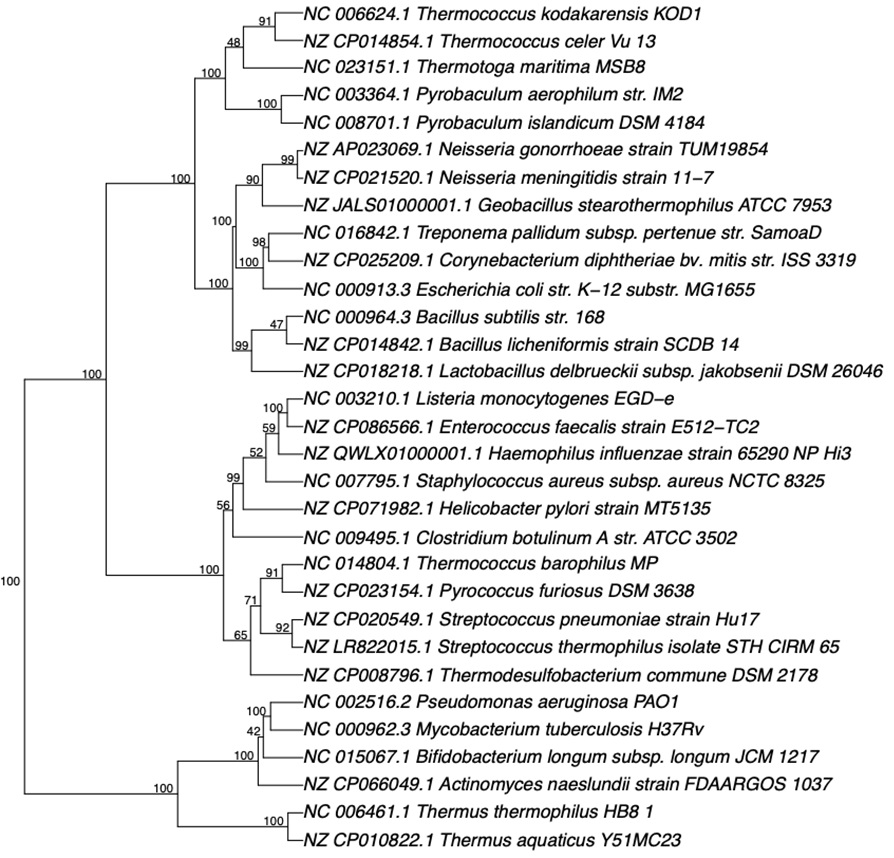
但结论与上面一致。这个方法无法准确地反映远亲物种之间的亲缘关系。
基于五核苷酸频率分析的系统发育分析是一个具有潜力的微生物系统发育分析的方法。该方法对于近亲物种表现良好,对于远亲物种,能力还不够。
研究人员将该方法制作成了Phy5的R包,工具小巧,运行速度也快,可在这里获得:
也可以访问在线版本:
参考文献:
Nakano Y, Domon Y, Yamagishi K. Phylogenetic trees of closely related bacterial species and subspecies based on frequencies of short nucleotide sequences. PLoS One. 2023 Apr 20;18(4):e0268847. doi: 10.1371/journal.pone.0268847. PMID: 37079522; PMCID: PMC10118083.
谷禾健康
粪菌移植是一项近年来备受关注的医疗技术,它涉及将健康捐赠者的粪便物质转移至患有疾病或障碍患者的胃肠道。

简单来说就是选择健康合适的人粪便,通过科学方法提取出有用的微生物,去除有害与无用的部分,然后制成制剂,给病人口服或者直接注入肠道,目的是恢复健康的肠道微生物群并改善整体健康状况。
粪菌移植的历史悠久
粪菌移植在传统中医中已有数百年的应用,粪菌移植的第一个记录可以追溯到四世纪的中国,当时人类粪便被称为黄汤,用于治疗严重腹泻的患者。
15到16世纪的中国明朝,有关于将新鲜或发酵的粪便悬浮液用于治疗腹泻、便秘和腹痛等胃肠道疾病的记载。
但直到20世纪50年代晚期,它才首次用于现代医学,以治疗由抗生素耐药菌引起的严重腹泻。
在20世纪80年代和90年代,粪菌移植主要用于治疗反复发作的艰难梭菌感染。然而,随着肠道微生物群的发现及其在健康和疾病中的作用,粪菌移植开始被探索作为治疗其他疾病的潜在方法,包括炎症性肠病、肠易激综合征甚至神经系统疾病等。

粪菌移植在未来拥有巨大前景
近年来,粪菌移植受到医学界和公众越来越多的关注,许多临床试验和研究正在进行,以评估其疗效和安全性。
粪菌移植的疗效已经在多种肠道疾病中得到证实,如艰难梭菌感染、肠炎、炎症性肠病等。粪菌移植的出现,为肠道微生物失调带来了新的治疗思路和方法,也为人类健康提供了新的希望。
随着我们对肠道微生物群研究的不断发展,粪菌移植很可能成为治疗其他疾病越来越重要的工具。谷禾在本文中将为大家介绍粪菌移植。
粪便微生物群移植 (粪菌移植) 是一种医疗程序,涉及将粪便从健康供体转移到受体的胃肠道中。已发现粪菌移植可有效治疗各种胃肠道疾病,包括艰难梭菌感染、炎症性肠病和肠易激综合征。
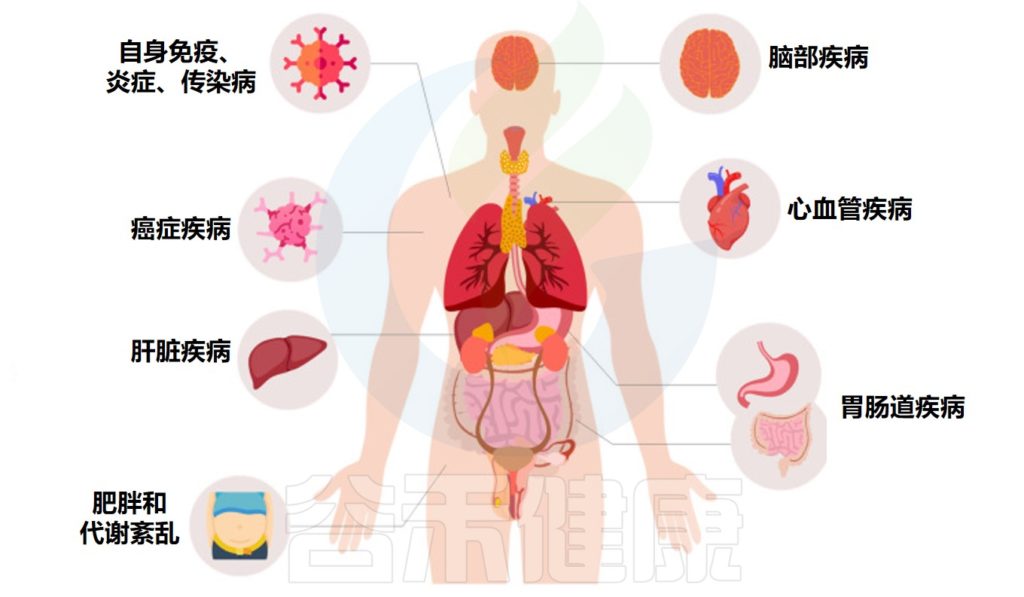
来自大量临床试验的数据表明,粪菌移植也可能对许多其他疾病具有治疗潜力,从胃肠道到肝脏疾病,神经系统疾病、自身免疫性疾病、皮肤疾病、心血管疾病、脑部疾病、肥胖和代谢综合征。
根据这些相关研究,谷禾带大家了解粪菌移植在治疗人体疾病中的作用。
人类疾病中的粪菌移植
Biazzo M,et al.J Clin Med.2022
▼
粪菌移植主要用于治疗反复发作的艰难梭菌感染(CDI),这是一种细菌感染,会引起严重的腹泻和结肠炎症。
√粪菌移植对治疗艰难梭菌感染特别有效
研究表明,粪菌移植在治疗复发性艰难梭菌感染方面特别有效,已发现粪菌移植在治疗艰难梭菌感染方面的成功率超过90%,而且几乎没有副作用。
▼
粪菌移植已被证明是治疗多种胃肠道疾病的有效方法,包括炎症性肠病(IBD)、肠易激综合征(IBS)。
炎症性肠病是一种复杂的炎症和慢性疾病,其特征是免疫失调,最终导致消化道免疫介导的损伤,包括溃疡性结肠炎(UC)和克罗恩病(CD)。肠道微生物群失调被认为是炎症性肠病发展中的关键调节事件,因此粪菌移植代表了一种可能的治疗策略。
√缓解溃疡性结肠炎患者症状
最近一项随机对照试验的荟萃分析发现,粪菌移植可有效诱导溃疡性结肠炎患者症状缓解,缓解率为36%,而对照组仅为9%。对核糖体 16S RNA 的分析表明,粪菌移植后微生物多样性增加并持续存在。
对活动期溃疡性结肠炎的儿科患者(4至17岁)进行了随机临床试验。粪菌移植组中92%的患者在第6周实现了小儿溃疡性结肠炎活动指数的改善(与安慰剂组的50%相比)。
√短期内改善了克罗恩病患者症状
评估了25名患有腹腔内炎症性肿块的克罗恩病患者使用多种新鲜粪菌移植(初始粪菌移植随后每三个月重复粪菌移植)的疗效和安全性。
超过一半的患者在第一次粪菌移植后三个月表现出临床反应的缓解,这一比例在12个月和18个月时有所下降。
注:这表明尽管粪菌移植在短期内缓解了临床症状,但未能产生持久的临床效果。
√对肠易激综合征有积极影响
六项不同的临床试验发现粪菌移植对肠易激综合征症状有积极影响:
(1) 在一项只有10名患者入组的小型临床试验中,在粪菌移植后4周,6 名患者有所改善;有趣的是,作者发现报告改善的患者接受了来自双歧杆菌含量高于无效供体的粪菌移植,这表明富含双歧杆菌的粪便供体可能是粪菌移植成功的预测因素。
(2)观察到65%的患者在通过结肠镜检查接受粪菌移植治疗三个月后症状缓解以及粪菌移植之后微生物特征向供体特征的转变,包括增加的α和β多样性。
(3)报告了腹泻型肠易激综合征患者的症状和生活质量的改善,这些患者通过胃镜接受新鲜粪便,并且(在连续分析中)增加了短链脂肪酸。
(4)报告显示,在接受来自健康供体(同种异体移植)或来自他们自己(自体移植)的粪便材料的患者中,肠易激综合征症状出现短暂改善;接受同种异体移植的患者抑郁评分有所下降。
(5)使用仅从一名健康、特征良好的供体获得的粪便样本通过胃镜粪菌移植治疗肠易激综合征患者,发现了疲劳和生活质量的改善以及细菌微生物群概况的变化、粪便短链脂肪酸的变化。
(6)在最近的一项临床试验中,招募了难治性肠易激综合征患者,主要是腹胀,并通过鼻空肠给药接受一次粪菌移植治疗;粪菌移植一年后,56%的患者报告肠易激综合征症状和生活质量有所改善。
▼
复发性肝性脑病是肝硬化的并发症(与饮酒无关),由肝功能衰竭和肠-肝-脑轴受损引起,可导致意识障碍和昏迷 。
√粪菌移植改善肝性脑病患者认知和微生态失调
粪菌移植最近被研究为一种可能的新治疗工具。第一项试验描述,通过灌肠递送的单一粪菌移植在短期内减少了肝性脑病患者的住院时间,改善了他们的认知和微生态失调。
粪菌移植治疗后一年内,与对照组相比,降低了住院率并改善了认知功能。
√粪菌移植展现出肝病的治疗潜力
广泛的研究支持粪菌移植作为一种控制肝病的疗法。粪菌移植改善了高脂肪饮食引起的肝损伤和脂质代谢,同时增加了小鼠肠道微生物群的多样性。来自对酒精性肝病有抵抗力的供体小鼠的粪菌移植可以预防酒精性肝损伤。
此外,粪菌移植已经用于慢性肝病患者。最近一项针对严重酒精性肝炎患者的初步研究表明,粪菌移植与存活率提高和腹水消退有关。
▼
多项研究探索了粪菌移植在治疗神经系统疾病方面的潜在益处,例如帕金森病、多发性硬化症和自闭症谱系障碍。
√改善帕金森病患者症状
发表在《神经病学》杂志上的一项研究发现,粪菌移植改善了帕金森病患者的症状,如便秘和睡眠障碍。
在帕金森病小鼠模型中,来自正常小鼠的粪菌移植可以减少黑质的病理特征并减轻身体损伤。
√改善多发性硬化症患者症状
发表在《神经炎症杂志》上的另一项研究发现,粪菌移植改善了多发性硬化症患者的症状,例如疲劳和抑郁。
√影响抑郁症、阿尔兹海默症状态
此外,来自重度抑郁症患者的粪菌移植在正常小鼠中诱导了抑郁症的行为/生理特征。
最后,在阿尔茨海默病小鼠模型中,源自正常对照小鼠的粪菌移植改善了认知功能。
▼
除神经系统疾病外,粪菌移植在治疗其他非胃肠道疾病(如肥胖、糖尿病和代谢综合征)方面也显示出前景。
√改善代谢综合征患者胰岛素敏感性
几项临床试验研究了粪菌移植对代谢综合征患者的的影响,显示出改善:报告说,患有代谢综合征的男性参与者接受了来自健康人群的肠道微生物群输注六周后胰岛素敏感性增加。
研究人员概括了关于胰岛素抵抗增加的发现,并补充说它取决于粪菌移植后肠道微生物群的变化。
最后,与移植正常供体粪便的受试者相比,使用代谢综合征供体粪便移植肥胖和胰岛素抵抗的男性受试者时,胰岛素敏感性降低,从而显示代谢综合征中胰岛素敏感性和微生物群之间的因果关系。
发表在《胃肠病学》杂志上的一项研究也发现,粪菌移植改善了代谢综合征患者的胰岛素敏感性。
注:代谢综合征是糖尿病的前兆。
除了这些已经确定的应用,粪菌移植还有一些新兴的用途。例如,粪菌移植被探索作为治疗肥胖和2型糖尿病的潜在方法。
▼
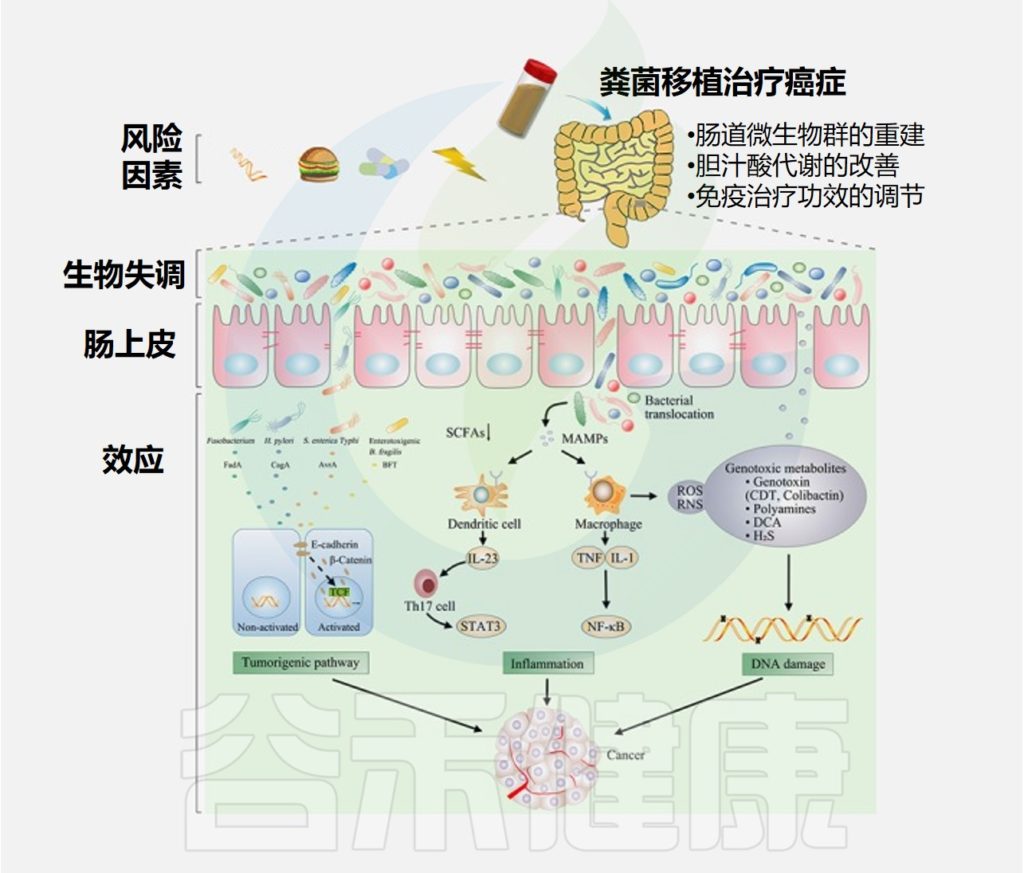
在过去的几年中,肠道微生物群在致癌过程中的作用已得到越来越多的认可。肠道微生物失调和个别细菌可以通过激活致瘤途径、诱发炎症和破坏宿主DNA来诱发癌症或影响癌症进程。
细菌影响癌症的具体机制
几种细菌拥有或产生促进β-连环蛋白与E-钙粘蛋白分离的蛋白质,激活参与癌发生的β-连环蛋白信号通路。
肠道生态失调导致细菌衍生的短链脂肪酸的产生减少。肠道生态失调通过Toll 样受体 (TLR) 的微生物相关分子模式发挥促炎作用,增加细胞产生促炎因子,从而增加致癌作用。
除了诱发炎症外,许多细菌还能够通过释放特定代谢物来破坏DNA,从而促进癌症进展。
β-连环蛋白(β-catenin)是一种细胞骨架蛋白,主要位于细胞膜,在乳腺上皮中,β-catenin游离量较少,通过介导与E-cadherin的相互作用,将细胞外黏附因子与细胞质内细胞骨架相互连接,以保持质膜的完整性,并参与细胞的黏附、迁徙与转移等过程。
β-catenin异常表达时脱离细胞间连接进入细胞质或细胞核,将失去介导细胞间链接、黏附的功能,并引起某些癌基因的表达,导致细胞癌变、转移。
β-catenin还是介导Wnt信号传导的关键分子,其在细胞核中决定着Wnt信号通路的开放或关闭,通过调节基因表达过程,在乳腺发育以及肿瘤发生的调控中起到了重要的作用。
细胞膜中β-catenin的丢失以及细胞质、细胞核中β-catenin的高表达都可以经过不同的调节途径独立发生,诱导乳腺癌的发生,并影响乳腺癌的预后。
E-钙粘蛋白(E-cadherin)是钙黏蛋白家族成员,在上皮细胞中连接细胞骨架与细胞外环境,参与细胞信号传导。在正常乳腺组织中,E-cadherin在肌上皮中呈颗粒状膜阳性,在腺上皮细胞中呈细胞膜强阳性,是区分DCIS和小叶原位癌的标志物;E-cadherin几乎在所有的DCIS细胞上呈线状膜染色,而小叶原位癌的细胞膜大多不表达E-cadherin。
令人惊讶的是,特定的微生物群物种具有调节癌症治疗的功效,显著影响癌症患者的临床预后。
√粪菌移植有助于治疗黑色素瘤
黑色素瘤是一种皮肤癌,涉及产生黑色素的黑色素细胞。最近,新工具利用免疫检查点抑制剂来增强患者对肿瘤的免疫反应(这种治疗方法称为“癌症免疫疗法”),并且有趣地发现肠道微生物组调节这种反应。
注:谷禾前面的文章中也有讲过癌症免疫疗法。
在两个具有不同肠道微生物组成的小鼠中,黑色素瘤的生长及其对抗程序性死亡配体1(PD-L1)免疫疗法的反应显著不同。
如果将来自对抗PD-1治疗有积极反应的黑色素瘤患者的粪便样本移植到无菌小鼠体内,则PD-1阻断的抗肿瘤作用会得到改善。
粪菌移植似乎有望通过转移有利的肠道微生物群来增强黑色素瘤患者的抗肿瘤免疫力。
通过粪便微生物群移植治疗癌症
Chen D,et al.Int J Cancer.2019
▼
根据最近的研究,粪菌移植可能是治疗各种皮肤病症(包括湿疹和牛皮癣)的潜在治疗选择。人体肠道微生物群在免疫调节中起着至关重要的作用,肠道微生物群的改变与各种皮肤病有关。
一些案例报告了湿疹和牛皮癣患者经粪菌移植后成功治疗。
√改善了牛皮癣患者的严重程度
在研究皮肤病学杂志上发表的一项研究中,研究人员发现粪菌移植改善了四分之三的牛皮癣患者的严重程度。
√湿疹症状有了显著改善
此外,发表在美国皮肤病学会杂志上的一项研究报告说,粪菌移植使三分之二的患者的湿疹症状有了显著改善。
总之,粪菌移植显示出作为治疗湿疹等皮肤病的巨大潜力。同样重要的是要注意粪菌移植具有潜在的风险,包括感染和不良反应。
需要更多的研究来确定粪菌移植治疗皮肤病的最佳频率和剂量。因此,在考虑将其作为皮肤病的治疗选择之前,必须彻底评估粪菌移植的潜在益处和风险。
▼
关于微生物群和粪菌移植干预在心血管疾病中的作用,只有有限的证据可用,而且大多是在动物模型中获得的。
√菌群影响房颤的易感性
最近提出了老年人肠道菌群失调在房颤发病机制中的因果作用。研究表明,将患有心房颤动的老年大鼠粪菌移植移植到年轻大鼠体内会导致更高水平的脂多糖和更高的疾病易感性。
√菌群移植改善了心肌损伤
仍然在动物模型中,实验性自身免疫性心肌炎(EAM)小鼠模型中对照小鼠的粪菌移植增加了微生物的丰富度,包括厚壁菌门/拟杆菌门的比例增加,并且由于炎症减轻而改善了心肌损伤。
尽管科学文献中很少有证据表明粪菌移植策略在心血管疾病中有治疗价值,但在动物模型和人类中获得的这些数据肯定会为进一步研究提供坚实的基础。
本章小结
未来,粪菌移植也可能用于治疗自身免疫性疾病,如类风湿性关节炎和红斑狼疮,以及精神健康状况,如抑郁症和焦虑症。
值得注意的是,这些新兴应用仍处于研究的早期阶段,需要进一步的研究来确定它们的疗效和安全性。
总的来说,目前粪菌移植主要用于治疗反复发作的艰难梭菌感染和一部分胃肠道疾病,但它在治疗其他疾病和障碍方面具有巨大的潜力,正在进行研究探索其在各个医学领域的潜在应用。
▼
移植所需的粪菌来自经过严格筛查的供体,供体捐赠样本后,用一套全自动化的机器进行分离,经过注水、搅拌、过滤、离心、沉淀等步骤,得到纯化的细菌。
粪便菌群移植过程示意图
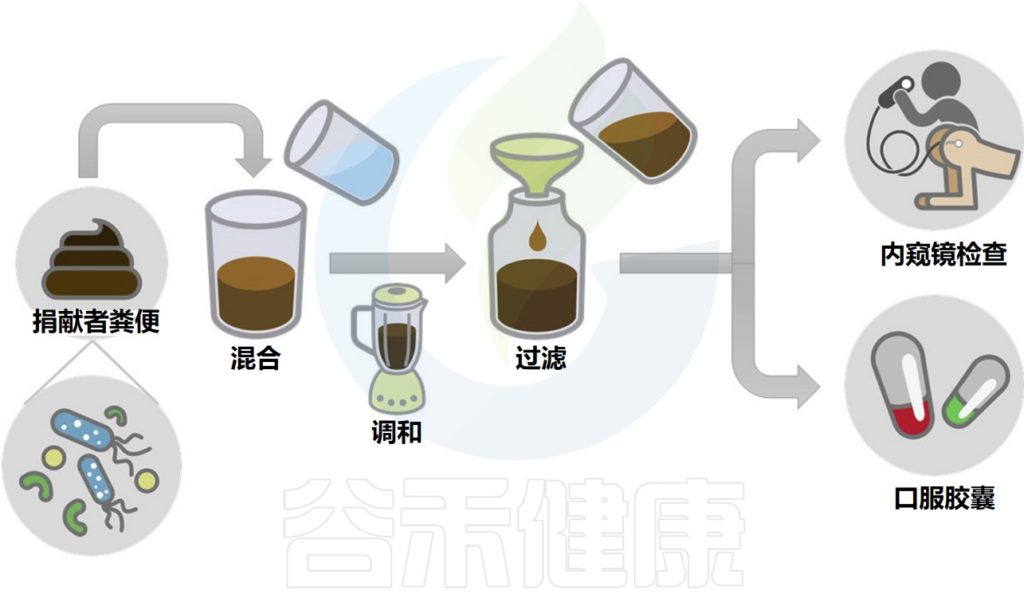
Wang JW,et al.J Formos Med Assoc.2019
其中单单过滤的步骤就要重复数次,每一道过滤用的滤网孔径都比前一道滤网的孔径更小,最后一道滤网的孔径只有0.07毫米,基本上可以把所有的杂质去除,只剩下细菌这些微生物。
分离出来的粪菌再用生理盐水制成混悬液供移植使用。
▼
“移植”过程不是简单粗暴的将一个人的粪便直接注入患者的肠道,主要是将健康供体的粪便离心纯化,获得功能菌,然后通过上消化道、中消化道和下消化道转移至受体肠道。从而重建稳定的肠道微生态环境,缓解相关症状,治疗疾病。
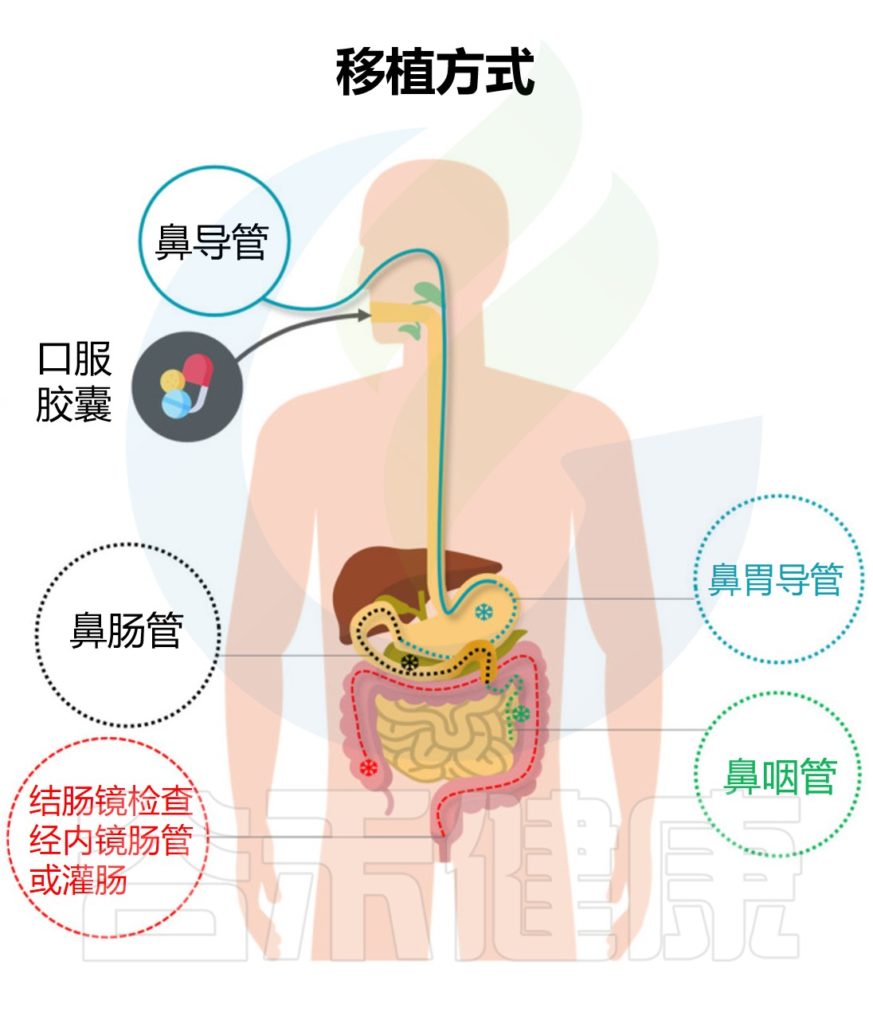
★ 粪菌移植的途径与治疗目的有相关性
例如,想要治疗溃疡性结肠炎可以采取经结肠镜灌肠。要治疗慢性乙型肝炎,则是经上消化道,可以采取:经麻醉胃镜、普通胃镜、经鼻饲管,或者用粪菌胶囊,经典方式是通过麻醉胃镜。
粪便微生物群移植的方法
Biazzo M,et al.J Clin Med.2022
不同的粪菌移植方式具有各自的优缺点,下面谷禾具体来讲述经不同部位消化道进行粪菌移植的特点。
▸ 上消化道
操作简单,总体风险较低
上消化道途径中,口服给药和鼻胃管应用广泛,操作简单,总体风险较低。但在此过程中,可能会因菌液反流或吸出而导致窒息,如果机体本身存在梗阻,则移植的菌液可能无法准确到达肠道。
其中,口服途径主要指包封粪菌材料,通过将提取的粪菌与冷冻保护剂(主要是甘油)混合制备,然后进行多重包装以保护粪菌移植材料免受胃肠道环境的影响。同时,操作人员可以根据预期的释放位置进一步选择胶囊壳的材料。目前,市售的胶囊壳通常针对胃或结肠的释放,并被配制以确保细菌的存活率和定植。
降低潜在疾病传播的概率
此外,在制作胶囊的过程中,也可以去除无症状供体中可能存在的真菌、寄生虫、病毒和部分炎症介质,以降低潜在的疾病传播概率。
总的来说,通过上消化道途径的粪菌移植具有无需昂贵器械、重复使用对患者损伤小、患者耐受性好等优点。该途径适用于不能耐受鼻肠管和胃镜移植的患者,以及需要口服定制菌的患者。
▸ 中消化道
减少移植时的细菌位移
中消化道途径主要是指鼻肠移植和经内镜肠道植管术,能够实现全肠道给药。其中鼻肠管通过导丝拉伸拉直,在胃肠蠕动正常的情况下自动通过幽门,进一步减少粪菌移植时的细菌移位。
在一项随机对照试验中报道,通过鼻肠管的粪菌移植优于单独使用万古霉素治疗艰难梭菌感染,但粪菌移植组患者也出现了腹泻、痉挛和便秘等不良事件。
什么是经内镜肠道植管术?
经内镜肠道植管术(TET)具体指的是在内镜下辅助植入固定管并固定在肠道深处,同时外端沿肠道与外部相通。TET主要包括结肠途径介导的TET和中胃肠道途径介导TET。
结肠通路介导的TET需要应用结肠镜检查,并且使用这种途径进行重复粪菌移植是昂贵的。相比之下,中胃肠道途径介导的TET更方便,更容易维持。
由于中胃肠道介导的TET不需要在内镜手术后通过X射线或其他医疗仪器进一步确认肠道位置,对于无法进行结肠镜检查肠道准备的患者,或需要重复粪菌移植的患者,中胃肠道中介导的经内镜肠道植管术也是主要选择。
▸下消化道
下消化道通路主要指灌肠、结肠镜检查、结肠通路介导的经内镜肠道植管术等操作。
灌肠侵入性较小且相对简单
其中,灌肠是一种侵入性较小且相对简单的操作。灌肠法患者耐受性好,不需要昂贵的器械,有效降低了手术风险。但这种方法需要将输注的粪悬液保留较长时间,患者保持仰卧位以减少粪便排泄,同时反复灌肠易于被患者接受。
注意:粪菌移植的细菌是否可以通过灌肠的方式保留在肠段的命题尚不明确,而且这种方式也伴随着无法到达结肠和脾曲的风险,需要多次灌注才能获得疗效和更长的手术时间,以弥补肠道菌群保留率低的缺陷。同时,由于细菌液的滞留,这种方法可能不适用于肛门括约肌松弛或尿失禁的患者。
此外,在结肠镜检查和结肠通路介导的经内镜肠道植管术方面,它具有多种优势,主要包括:
(1)能够完整显示结肠和活检可疑组织的情况,有利于疾病的诊断、识别和分期;
(2) 允许操作人员直接评估肠道炎症,并在适当的部位注入足量的供体粪便细菌;
(3) 可将肠道菌群准确移植到受影响的肠段,并进一步将细菌保留在目标肠段;
(4)可以准确地输入足量的供体粪便菌,可以在一定程度上提高治疗效率。
然而,这些手术也伴随着部分不良事件,包括加重肠道反应。
粪菌移植的主要操作方式及优缺点


Zhang YW,et al.J Bone Miner Metab.2022
▼
粪菌移植的安全性主要与两方面因素有关,即供体的生物安全性、移植的操作安全性。
•供体的生物安全性
供体不仅要筛查排除乙肝、丙肝、梅毒、艾滋病、这些已知传染病,还要排除寄生虫、条件致病菌感染,甚至连饮食习惯、排便习惯、情绪状态、肿瘤家族史都要进行调查,不符合要求的都要排除。
供体的筛查可以说是“百里挑一”,要成为一个合格的供体是非常困难的。所有这些筛查工作都是为了保证供体的生物安全性。
•移植的操作安全性
移植的操作安全性主要与移植的方式有关。经胃镜、结肠镜的移植操作风险与常规胃镜、结肠镜检查的风险基本一致。
注:在后面的章节我们会详细讲述粪菌移植操作的具体要求。

为了评估粪菌移植的安全性,上海同济粪菌移植工作组主任等研究人员对8547名采用过粪菌移植的患者进行跟踪研究,得出以下结论:
1、回顾性分析数据表明,粪菌移植的短期和长期(从2周到5年)安全性都是比较好的,患者不要担心;
2、短期不良事件与粪菌移植给药方法有关,多为轻中度、较短暂,包括恶心、呕吐、腹泻等,只有42名(0.5%左右,很低比例)患者发生胃肠症状恶化;
3、长期随访期间的新发疾病主要是胃肠道和呼吸道疾病,发病率与一般人群无异;
4、 随访期间无粪菌移植相关感染传播事件,11例死亡与粪菌移植无关;
5、通过规范适应症和给药时机后,短期安全性和患者满意度升高。
所以,粪菌移植还是比较安全的。
粪菌移植已被证明是治疗多种胃肠道疾病的有效方法,然而,粪菌移植的潜在长期影响以及粪菌移植的最佳频率和剂量仍未得到很好的了解。
本章节旨在回顾有关粪菌移植潜在长期影响的现有文献,并为未来的研究和临床实践提供建议。
▼
虽然粪菌移植已被证明在短期内有效,但对粪菌移植长期影响的研究有限,粪菌移植对肠道微生物组和整体健康的长期影响仍不清楚。
✦可能导致抗生素抗性基因的转移
粪菌移植是一种医疗程序,它将健康捐赠者粪便中的微生物移植到受体的肠道中,以恢复或改善肠道微生物平衡。虽然粪菌移植已被证明在治疗某些疾病方面非常有效,但它的使用也存在一定争议。
一些研究表明,粪菌移植可能会产生意想不到的后果,例如抗生素抗性基因的转移或致病菌的引入。
✦病毒或其他感染因子可能通过粪菌移植传播
此外,人们还担心病毒或其他感染因子可能通过粪菌移植传播。需要更多的研究来充分了解粪菌移植的长期影响,并制定安全有效地使用该程序的指南。
▼
✦粪菌移植的监管
粪菌移植的监管因国家而异,一些国家如美国对其使用采取更为严格的方法。在美国,粪菌移植被美国食品药品监督管理局作为生物制品进行监管,只批准用于治疗未对其他治疗方法产生反应的复发性艰难梭菌感染。
在欧洲,粪菌移植通常被视为医疗程序,并由国家卫生部门进行监管。
✦粪菌移植的伦理问题
粪菌移植的伦理考虑包括知情同意、捐赠者选择以及对捐赠者和受体的潜在风险。捐赠者必须接受广泛的筛查,以确保他们健康且没有传染病,并且必须同意他们的粪便用于粪菌移植。
受体也必须提供知情同意,并且必须完全了解该程序的潜在风险和好处。粪菌移植的主要争议之一是该程序缺乏标准化。目前没有粪菌移植的标准化方案,粪便成分在捐赠者之间可能会有很大的差异。
▼
✦粪菌移植的剂量会影响其疗效
目前,粪菌移植的最佳频率和剂量仍不确定。一些研究表明,粪菌移植的剂量可能会影响其疗效,但目前还没有确定最佳剂量的标准。
✦一般需要多次粪菌移植
关于粪菌移植的频次,目前尚无明确的建议。一些研究表明,单次粪菌移植可能无法完全恢复肠道微生物群的平衡,需要多次粪菌移植才能达到最佳疗效。
单次粪菌移植可以有效治疗某些疾病,如复发性艰难梭菌感染。然而,对于其他疾病,可能需要多次粪菌移植才能获得最佳效果。
但是,多次粪菌移植也可能增加感染和其他不良反应的风险。因此,粪菌移植的频次应根据患者的具体情况和病情进行个体化的决策。
总之,粪菌移植的最佳频次和剂量应根据患者的具体情况和病情进行个体化的决策,同时应注意监测和预防不良反应的发生。
需要更多的研究来确定粪菌移植的最佳频率和剂量,以便在临床实践中更好地使用该程序。
根据《上海市菌群移植技术管理规范(2021年版)》,以下是部分粪菌移植操作要求:
▼
(一)严格遵守粪菌移植治疗操作规范和诊疗指南,严格掌握粪菌移植治疗适应证和禁忌证。
▷粪菌移植适用于治疗以下消化系统疾病
(1)艰难梭状芽胞杆菌感染:复发性/难治性艰难梭状芽胞杆菌感染。
(2)其他消化系统疾病:溃疡性结肠炎、克罗恩病、功能性便秘、肠易激综合征、菌群紊乱相关腹泻等。
▷粪菌移植治疗的其他疾病
(1)神经系统疾病:帕金森病、阿尔兹海默症、癫痫等。
(2)精神疾病:自闭症、情绪障碍、多动症、抽动症等。
(3)代谢相关疾病:代谢综合征、糖尿病等。
(4)肿瘤相关疾病:免疫治疗及放化疗所致肠炎等。
(二)除复发性/难治性艰难梭状芽胞杆菌感染外,出于其他治疗目的使用粪菌移植治疗均须以临床研究形式开展,并通过本机构伦理审查与备案。
(三)实施粪菌移植治疗前应当向患者及其家属告知治疗目的、风险、注意事项及可能发生的并发症等,并签署知情同意书。
(四)医疗机构应当建立完整的电子化临床数据库及严格的术后随访制度,在完成每例粪菌移植治疗后应当按照有关规定将治疗相关信息上报卫生健康行政部门。
▼
(1)有明确与肠道菌群紊乱有关的肠道内疾病,如肠功能障碍性疾病、肠道感染性疾病(复发性艰难梭菌感染)、肠道炎症性疾病及假性和麻痹性肠梗阻等;
(2)有明确与肠道疾病或者肠道菌群紊乱有关的肠道外疾病,如便秘合并帕金森病、便秘或腹泻合并自闭症、便秘或腹泻合并过敏性皮炎;
(3)患者需充分理解粪菌移植治疗过程及机制,并签署知情同意书。
!
出现以下情况之一不宜进行粪菌移植:
(1)肠道大面积溃疡、出血;
(2)移植通道梗阻;
(3)移植操作本身(内镜、置管、灌肠、经口饮食)的禁忌证;
(4)未通过伦理审核的临床试验或患者拒绝接受粪菌移植。
(5)生命体征(体温、心率、血压、呼吸)不稳定;
(6)严重免疫系统紊乱;
(7)孕妇或哺乳期妇女;
(8)代偿或者失代偿肝功能障碍。
▼
(1)个人情况:作息规律,饮食健康,家庭和睦,无不良性交,无吸烟、饮酒、吸毒等嗜好,无药物成瘾,近6个月未接种过疫苗或参加药物试验,近6个月未接受纹身或出现皮肤破损,近6个月无热带地区旅居史;无胃肠道病变家族史,无恶性肿瘤家族史,无传染病家族史;非孕期,非经期。
(2)开展病史和身体检查,评估供体的健康状况和是否符合粪菌移植的供体资格;选择无传染病史、无抗生素使用史、无消化系统疾病等方面的供体。
身体状态:年龄18-30周岁,体重指数(BMI)18.5-23.9 kg/m2,儿童医院开展粪菌移植,供体要求可放宽至15-30周岁。
心理状态:心理科医师或心理咨询师访谈认定心理状态良好;抑郁自评量表(SDS)、焦虑自评量表(SAS)、匹兹堡睡眠质量指数(PSQI)等评分正常。
(3)进行多项检测,如血液化验、粪便检测、病原菌检测、URTI检测等,确保找到适合的健康供体;以确保其不携带有害微生物和多重耐药菌。同时,需要考虑供体的肠道微生物组成和短链脂肪酸水平,以选择最适合的供体。
血液学检测:包括血常规、肝肾功能、电解质、C反应蛋白正常,肝炎病毒、HIV、梅毒、EB病毒、巨细胞病毒、COVID-19抗体、线虫、阿米巴等病原检测阴性;
粪便检测:粪便常规检查正常,隐血实验阴性,艰难梭菌、弯曲菌、沙门菌、志贺菌、产志贺毒素大肠杆菌及虫卵、小肠结肠耶尔森菌、致病性弧菌(副溶血弧菌、霍乱弧菌)、气单胞菌、阿米巴、孢子、诺如病毒、轮状病毒和新型冠状病毒(COVID-19)等病原学检测阴性。
(4)评估供体的肠道微生物组成,多样性和稳定性,包括菌群、真菌群和病毒群,并建立数据库;在选择供体时,除了肠道微生物群的总体多样性外,还应考虑到益生菌的丰富度,如乳酸菌、双歧杆菌等,选择粪便丁酸浓度高的供体微生物组。
(5)根据评估结果选择合适的供体,注意进行家族成员偏向,判别成熟度和完整性,同时避免与患者组群共同优势菌种重合等;一些研究表明,供体微生物组成与受体的微生物组成相似,可以提高移植成功率。
(6)移植前需对受体进行评估和准备,包括清洁肠道和停用抗生素等;为了提高移植效果,一些研究还使用抗生素预处理和肠道灌洗等方法来增强供体菌株的移植。
限食耐受性:通过限食实验完成。部分受体存在食物过敏及食物不耐受(如鸡蛋、牛奶等),根据受体要求,供体需在捐赠粪便前5天限制食物种类,如该供体不可耐受,则不可作为本次移植的捐赠者。
(7)进行移植后,对受体的微生物组成进行多次采样和分析,以评估移植效果和持续时间;
①每2个月复查以上条目,仍然符合上述要求;
②每次捐赠的粪便均留样行宏基因或者16s rDNA测序(根据各自单位检测能力),保证菌群组成及多样性稳定;
③每次捐献粪便应留样行代谢组学检测。
(8)监测受体的生理状况和不良反应,及时采取措施进行处理。
▼
(1)粪便采集:应采用无菌容器采集,粪便重量不少于100g,性状为Bristol评分标准中3-5分方为合格,立即进入菌液制作流程,或立即密封后2-8℃保存。
(2)制作流程:每次需取不少于50g粪便与无菌生理盐水以1∶3比例混合,充分搅拌混匀后过滤,从粪便排出体外至菌液制作完成应保证在2小时以内,整个处理过程应在无菌环境下操作。
注:为了避免感染窗口期的问题,菌液或胶囊制备后应待21 天后经过病毒细菌等检验合格后出库,每批产品留样至少6个月以供追溯。
(3)冻存与复融:菌液制备完成后置于-80℃保存,6个月之内使用不影响疗效;如置于-20℃保存,应在1-4周内使用。使用前置于室温复融,6小时内输注;如使用水浴锅复融,则水温不得超过37℃。
(4)质控标准:每次制作完成的菌液应随机抽样,进行致病菌监测、细胞计数及宏基因或16s rDNA菌群测序。
每50mL菌液中活细胞数应不小于2.5×10^12个,同供体同批次样本应达到菌群组成一致,6个月内同供体不同批次样本应达到菌群组成差异无统计学意义。
制作完成的菌液应有独立包装与标签。如制作胶囊,还应取胶囊内容物进行细菌活性监测与活细胞计数,其中细菌活性应不低于85%,每克内容物包含活菌数不小于10^9CFU。
▼
(1)菌液输注前需结合患者自身情况及疾病种类行肠道准备。
(2)应根据指南及操作规范选择上消化道或下消化道进行菌液输注。
(3)操作时应观察患者耐受情况,并及时处理不良反应。
▼
(1)管理原则:医疗安全(不良)事件的管理,遵循预防为主、紧急处理、合理控制、防微杜渐、持续改进的原则。
(2)上报要求:凡科室职工均有权上报医疗安全(不良)事件,可通过网络、书面、电话上报,上报时限要求在不良事件发生后24小时之内。
(3)审核、处置要求:科室负责人24小时内审核、处置科室发生的不良事件,并呈报至相关职能部门。
注意:主管职能部门自接报之日起,24小时内审核、处置不良事件;对于接报的Ⅱ级及以上不良事件,应呈报至主管院领导。
主管院领导对于接报的Ⅱ级及以上不良事件,必要时组织委员会进行讨论分析,并提交院务会。
▼
(1)每次菌液输注后密切观察患者耐受情况24小时,如有不良反应立刻处理并及时上报。
(2)1个疗程治疗结束后4周内应对患者症状学和肠道菌群改善情况进行评估,粪菌移植治疗疾病的随访及疗效判断指标主要遵循原发疾病的治疗指南。移植前后重复检测粪便及血液中传染病相关指标,有条件者可行免疫相关检测。
注意:如第1个疗程结束3周后病情无改善,可尝试更换供体,重新给予1个疗程粪菌移植,若有效可继续治疗,若仍无效则应视为粪菌移植无反应性,此时不应继续给予粪菌移植,以免耽误病情;如患者接受粪菌移植治疗后症状明显改善,可重复3-5次粪菌移植后结束治疗。
随访至末次粪菌移植结束后至少8周,有条件者应进行大于1年甚至5年以上的长期随访。
随着对微生物与宿主相互作用的了解不断加深,肠道微生物群的参与已成为一种新颖、巧妙且不可忽视的调节宿主健康的方式。
粪菌移植是一种治疗胃肠道疾病的有效方法,还有望治疗其他更多疾病。未来的研究应该重点关注粪菌移植对肠道微生物组和整体健康的长期影响,并制定安全有效地使用该程序的指南。
此外,需要进一步研究如何提高粪菌移植的改善效果,供体和受体的配型以及移植肠道菌群的定植和效果的长期性,以便在临床实践中更好地使用。
主要参考文献:
Zhang YW, Cao MM, Li YJ, Zhang RL, Wu MT, Yu Q, Rui YF. Fecal microbiota transplantation as a promising treatment option for osteoporosis. J Bone Miner Metab. 2022 Nov;40(6):874-889.
Biazzo M, Deidda G. Fecal Microbiota Transplantation as New Therapeutic Avenue for Human Diseases. J Clin Med. 2022 Jul 15;11(14):4119.
Quaranta G, Guarnaccia A, Fancello G, Agrillo C, Iannarelli F, Sanguinetti M, Masucci L. Fecal Microbiota Transplantation and Other Gut Microbiota Manipulation Strategies. Microorganisms. 2022 Dec 7;10(12):2424.
Vendrik KEW, Ooijevaar RE, de Jong PRC, Laman JD, van Oosten BW, van Hilten JJ, Ducarmon QR, Keller JJ, Kuijper EJ, Contarino MF. Fecal Microbiota Transplantation in Neurological Disorders. Front Cell Infect Microbiol. 2020 Mar 24;10:98.
Wang JW, Kuo CH, Kuo FC, Wang YK, Hsu WH, Yu FJ, Hu HM, Hsu PI, Wang JY, Wu DC. Fecal microbiota transplantation: Review and update. J Formos Med Assoc. 2019 Mar;118 Suppl 1:S23-S31.
Chen D, Wu J, Jin D, Wang B, Cao H. Fecal microbiota transplantation in cancer management: Current status and perspectives. Int J Cancer. 2019 Oct 15;145(8):2021-2031.
Stallmach A, Steube A, Stallhofer J, Grunert PC, Merkel U, Hartmann M. Fäkaler Mikrobiomtransfer – Indikationen, Risiken und Chancen [Fecal microbiota transplantation: indications, risks and opportunities]. Inn Med (Heidelb). 2022 Oct;63(10):1036-1042. German.

谷禾健康
酒精与健康

饮酒作为一种特殊的文化形式,在我们国家有其独特的地位,在几千年的发展中,酒几乎渗透到日常生活、社会经济、文化活动之中。
据2018年发表的《中国饮酒人群适量饮酒状况》白皮书数据显示,中国饮酒人群高达6亿。酒精暴露是一种环境刺激,可显著改变许多生理、心理以及周围和中枢神经系统过程。从心脏和肝功能,到对肠道微生物组和内分泌途径的影响,酒精对全身和大脑都有广泛的影响。
众多的研究表明酒精本身可以产生双重效果,当然,这取决于饮酒量和酒的生产方式。
▸ 低-中剂量
低度至中度酒精对肠道微生物组、肠道代谢物和免疫相关疾病甚至一些癌症似乎起保护和有益作用。
▸ 高剂量
高剂量酒精或长期酗酒对胃肠道、肠道微生物组、免疫细胞和营养物质起破坏和损害作用。
目前的证据还表明,不同剂量的酒精在体内不同组织部位发挥不同的作用。
总体来说,过度饮酒,尤其在青少年和部分职业人群中占比越来越高,带来的伤害和健康风险不容忽视。过度饮酒会导致许多不良的病理健康影响和沉重的医疗保健负担。
根据世界卫生组织最新的通报指出,全球每年大约有超过300万人因过量饮酒而被夺去生命,其死亡人数已经超过爱滋病、暴力和交通事故死亡人数的总和。在饮酒所造成的直接或间接的死亡事件中,男性饮酒带来的风险尤为突出。

酒精引起的胃肠道功能以及微生物群组成和代谢的变化,内毒素血症的肠道通透性与全身性炎症和组织损伤/器官病变相关。
在过去的十年中,肠道,特别是肠道微生物群与酒精性肝病之间的关联引起了研究人员的关注。酒精与消化系统密不可分。它通过肠道吸收,并在肝脏内被肝细胞代谢。过量饮酒会导致肠道微生物组和肠道上皮完整性发生改变。它会导致重要的微量营养素缺乏症,包括短链脂肪酸和微量元素,这些元素会影响免疫功能并导致肝和其它器官损伤。
而且,酒精引起的微生物组改变和肠道屏障完整性下降会对中枢神经系统产生深远影响,导致抑郁、焦虑和对酒精的渴望增加,从而增加酗酒行为,造成恶心循环。
目前,酒精的治疗干预措施主要包括药物治疗、心理治疗、社会支持等方面。然而,这些干预措施都存在着一定的局限性,例如药物治疗可能会产生副作用,心理治疗需要长时间的治疗过程,社会支持需要家庭、朋友等的积极配合。
因此,寻找一种更加有效、安全、方便的治疗方法成为了当前治疗酒精成瘾和酒精使用障碍的研究重点。近年来,越来越多的研究表明,肠道菌群可能成为新的治疗酒精成瘾和酒精使用障碍的途径。新兴研究表明,通过调节肠道菌群可以减轻酒精成瘾的症状,例如减少饮酒量、降低戒断症状等。
本文将介绍酒精摄入或饮酒与肠道菌群及其代谢,肠内外健康的关系。
↘
本文目录
01 饮酒如何影响健康
酒精进入人体过程
酒精和肠道
酒精带来的有害影响
适量酒精带来的益处
02 轻中剂量酒精对自身免疫性疾病的保护作用
短链脂肪酸
多不饱和脂肪酸
自身免疫性糖尿病
自身免疫性甲状腺疾病
系统性红斑狼疮
类风湿关节炎
多发性硬化症
03 高剂量酒精对肠道菌群及其代谢物和营养物质的促炎作用
高剂量酒精对肠道菌群组成的影响
高剂量酒精对肠道屏障功能的影响
高剂量酒精对肠道微生物代谢物的影响
长期饮酒对营养状况的影响
高剂量酒精对肠道炎症和黏膜免疫的影响
04 酒精对相关疾病的影响
酒精与肠-肝轴和肝病
-酒精代谢
-酒精相关性脂肪变性
-酒精性脂肪性肝炎
-酒精引起的纤维化和肝硬化
-肝癌
酒精与胃肠道疾病
酒精和营养不良&肥胖
-营养不良
-肥胖
酒精和心血管疾病
-血脂概括
-高血压
05 酒精与肠脑轴和酒精成瘾
06 干预措施
酒精(乙醇)是一种水溶性小分子,通过胃和近端小肠进入血流,然后分布到全身。它首先进入门静脉,门静脉直接排入肝脏,这是接触酒精最多的地方。肝脏消除了大部分酒精 (90%),而 2–5% 以原形通过尿液、汗液和呼吸排出。
身体第一个直接接触酒精的器官系统是消化系统;因此,从口腔到直肠,几乎消化系统的每个部分都会受到酒精的影响。
酒精对消化系统的病理影响部分取决于肠-肝轴。这种由肠肝循环促进的双向关系涉及消化和细菌产物从肠道到肝脏的运输,以及胆汁、抗体和细胞因子返回肠道。
过量酒精摄入已被证明会通过破坏肠道微生物组成、代谢组和肠道上皮屏障来改变这一轴。这些干扰最终会对营养吸收产生连锁反应。
酒精对肠道的影响
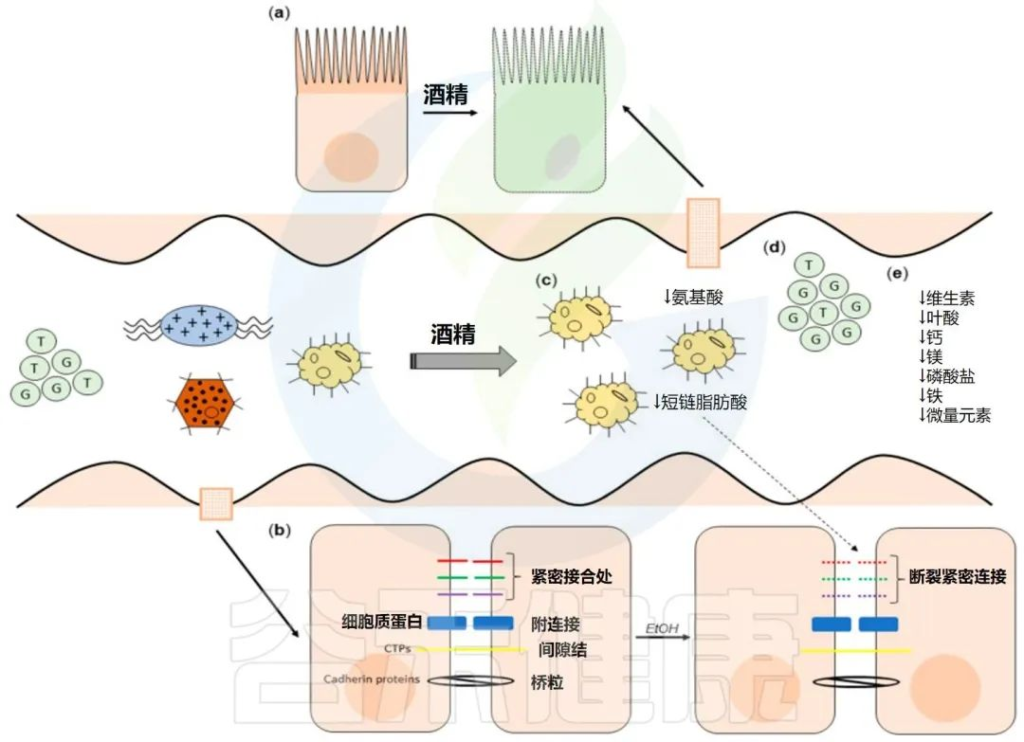
Pohl K et al. Nutrients. 2021,13(9):3170.
CTP:连接蛋白跨膜蛋白;JAM:连接粘附分子;EtOH:酒精。
( a ) 酒精对肠粘膜的组织学影响(细胞死亡、粘膜侵蚀和绒毛尖端上皮细胞丢失)。
( b ) 酒精引起的紧密连接破坏,因管腔短链脂肪酸浓度降低而加剧。
( c ) 酒精引起的生态失调导致短链脂肪酸和氨基酸浓度降低。
( d ) 次级胆汁酸浓度增加,与甘氨酸结合的比例增加。
( e ) ( a – d ) 导致的营养缺乏。
酒精对人体的影响是双重的,既有正面的作用,也有负面的作用。
▼
过量饮酒对人体各部位可能引起的有害影响
– 嘴
饮酒会刺激口腔和喉咙,这会增加患口腔癌的风险。
– 食管
酒精从口腔进入胃部时会引起刺激,从而增加患食道癌的风险。
– 胃
酒精会引起胃壁刺激和随后的炎症,这种情况也称为胃炎。胃壁过度刺激或发炎会导致受影响区域出血和溃疡。
此外,饮酒会导致营养吸收不良,从而使个人患上一系列其他疾病的风险更高。
– 大/小肠
饮酒会抑制大肠和小肠内的营养吸收。肠道蠕动也会受到影响并导致腹泻。
– 胰腺
酒精会导致胰腺发炎,也称为胰腺炎。酒精还会影响胰腺产生胰岛素的方式,如果不停止饮酒,最终会导致个体患上糖尿病。
– 肝脏
酒精是通过 ADH 和 CYP2E1 代谢的,这两种酶都是肝脏的重要功能酶。
通过干扰这些酶的正常功能并产生有害的副产物,对肝脏的影响可能包括肝脏炎症或肝炎、肝硬化、黄疸。
饮酒过多的人可能的直接有害影响
长期酗酒的危害
长期饮酒是全世界最常见的死亡原因之一。根据世界卫生组织的数据,全世界有 23 亿人饮酒,其中约 7500 万人被归类为患有酒精障碍。
酒精滥用会对多个终末器官造成损害,主要是肝脏、肠道和大脑,从而引发多系统损伤。
酒精中毒对全球发病率和死亡率有显著影响,每年约有 5.3% 的死亡是由有害饮酒造成的(世卫组织,2018 年)。一项针对印度南部农村 167,343 名成年受试者的研究发现,每天饮酒 30 年或更长时间会增加癌症相关死亡率的总体水平。
大量饮酒通常与患癌症的风险增加有关;然而,具体的剂量反应关系因癌症部位而异。
——饮酒量越多,肝癌风险越高
最近对涉及 19 组肝癌(即肝细胞癌)受试者的 16 篇文章进行的荟萃分析发现,与不饮酒者相比,饮酒量与肝癌风险之间存在线性关系。
因此,每天饮用 3 杯酒精饮料与风险增加相关,而每天饮用约 7 杯酒精饮料则风险增加高达 66%。
——不同种类乳腺癌,饮酒风险不同
乳腺癌风险也存在类似的线性关系。虽然饮酒总体上与女性患乳腺癌的风险较高有关,但这种关联并不适用于所有类型的乳腺癌。
在参加妇女健康倡议的女性中,饮酒者患雌激素阳性乳腺癌的风险增加。
而与从未饮酒的女性相比,饮酒者患三阴性乳腺癌的风险降低。
注:三阴性乳腺癌,孕激素、雌激素受体以及Her-2即表皮上皮生长因子,都是阴性,叫三阴性乳腺癌。
总之,饮酒和滥用酒精与多种癌症有关,并且这些关联的数量还在不断增加。与此同时,现在越来越清楚的是,酒精对某些慢病和癌症具有预防作用。此外,尽管酒精在预防某些癌症方面具有潜在的有益作用,但重要的是要记住,不能忽视长期酗酒的有害影响。
可能会增加以下疾病的易感性:
酒精引起的肠道菌群和代谢功能的变化可能导致
虽然酒精的新陈代谢看起来相当简单,但酒精的有害影响最常与新陈代谢过程相关,而新陈代谢过程通常会导致烟酰胺腺嘌呤二核苷酸加氢 (NADH) 的过量产生。
大量的 NADH 会导致乳酸,从而导致体内酸中毒,以及葡萄糖合成不足,从而导致低血糖。
▼
虽然大多时候我们都认为饮酒会带来许多有害影响,但过去几年进行的几项临床研究表明,适度饮酒实际上可以促进许多健康益处,例如:可能有助于降低患心脏病和死亡、缺血性中风、糖尿病的风险。这些益处的前提是要适度饮酒,那么什么是适度饮酒?
根据疾病预防控制中心的说法:
轻度饮酒:每周喝三杯或更少的酒精饮料。
适度饮酒:女性每天喝一杯酒精饮料,男性每天喝两杯。
大量饮酒:女性每天喝四杯或更多酒,男性每天喝五杯或更多酒。
注:这个定义在不同的研究中存在差异。由于参与者的主观记忆和准确的报告,准确的人类消费量很难量化。
适度饮酒具体是如何带来益处的?
适量的酒精已被证明可以提高高密度脂蛋白 (HDL) 的水平,这是一种“好”胆固醇,实际上这已经从你的身体中去除了有害的胆固醇。
当一个人的 HDL 水平较高时,它们就能更好地保护自己免受心脏病的侵害。
适度摄入不含酒精的啤酒,可以通过补充生物活性多酚和酚酸,以及通过有益菌丰富肠道微生物群多样性,对人体健康产生积极影响。
——适量饮酒与血癌发病率较低相关
有趣的是,饮酒还与几种血癌的发病率较低有关,包括非霍奇金淋巴瘤 (NHL) 和多发性骨髓瘤。对与瑞典癌症登记处相关的 420489 名被诊断患有酒精使用障碍 (AUD) 的人进行的分析还发现,他们患白血病、多发性骨髓瘤和霍奇金病的风险较低。
最近的另一项研究还表明,饮酒与白血病风险增加无关,事实上,少量饮酒(每天少于或等于一杯)与白血病发病率降低 10% 有关。
——适量饮酒与甲状腺癌和肾细胞癌风险降低有关
在肾细胞癌的案例中,男性和女性的饮酒量低至每天 1 杯,风险也会降低,而更高的酒精摄入量并没有带来进一步的好处。
——适度饮酒与直肠腺癌呈负相关
一项针对结肠和直肠腺癌的回顾性观察性研究表明,适度饮酒(每天少于 14 克)与直肠癌的发病率呈负相关。研究人员还发现,适度摄入啤酒,尤其是葡萄酒与远端结直肠癌呈负相关。
酒精在体内具有多效性
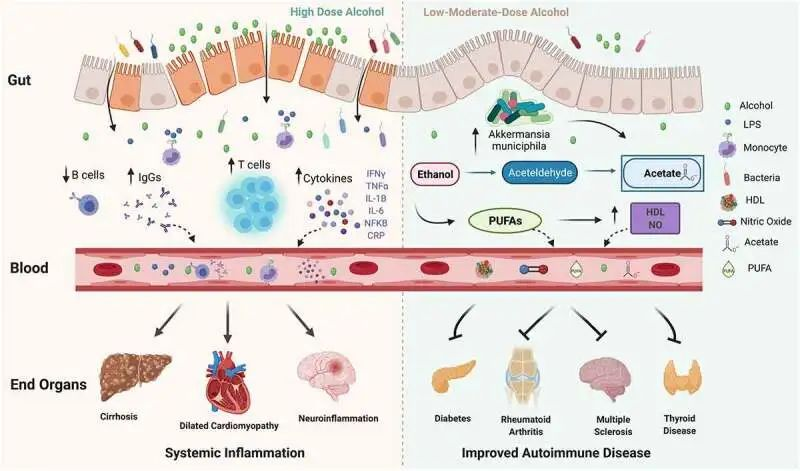
10.1080/19490976.2021.1916278
在高剂量下,酒精会破坏肠道屏障的稳定性,并可能导致肠道微生态失调、细菌壁产物、脂多糖 (LPS) 增加,后者会刺激免疫细胞上的 Toll 样受体 (TLR),并导致单核细胞、T 细胞、细胞因子和免疫球蛋白 (IgG) 水平以及 B 细胞减少。反过来,循环的炎性细胞因子、IgG 和免疫细胞会导致终末器官损伤。
在中低剂量下,酒精已被证明可以改善自身免疫性疾病的风险和进展。
低剂量酒精可能通过增加Akkermansia muciniphila对炎症产生积极影响和其他保护性肠道微生物,并有助于增加乙酸盐、多不饱和脂肪酸 (PUFA)、高密度脂蛋白 (HDL) 和一氧化氮 (NO)。
接下来展开讨论低-中剂量酒精对自身免疫性疾病的保护作用,以及高剂量酒精带来的健康危害,包括免疫系统、肠道菌群及其代谢产物、营养物质等多方面。
酒精可能对自身免疫性炎症产生有益影响的一个重要方式是通过其对肠道中脂肪酸代谢的影响。
众所周知,高剂量酒精会导致脂肪酸失调和发展为脂肪肝疾病,而在低剂量时,酒精可能有助于产生肠道衍生的抗炎脂肪酸,例如短链脂肪酸(SCFAs) 和多不饱和脂肪酸 (PUFAs)。
短链脂肪酸
低度至中度饮酒可以通过两种方式调节短链脂肪酸的产生:
1、通过改变肠道中产短链脂肪酸菌
低度至中度酒精会改变肠道中产生短链脂肪酸的微生物群,例如Akkermansia muciniphila。
短期饮酒(5 天 0.8 g/kg 胃内)会升高小鼠的Akkermansia muciniphila水平,而在消耗同等酒精强度的发酵米酒 (FRL) 的组中未观察到升高。
2、通过乙酸盐产生
酒精本身会代谢成短链脂肪酸、乙酸盐,并且喂食 Lieber-DeCarli 饮食 8 周的动物与对照组相比显示乙酸水平升高。
Lieber-DeCarli 饮食
Lieber-DeCarli 饮食是一种常用于动物实验中的饮食方案。它是由两位科学家 Lieber 和 DeCarli 在 1980 年代开发的,旨在为动物提供一种高度可控的饮食,也就是说可以根据实验需要增加或减少某种营养素的含量,以便研究不同营养素对健康和疾病的影响。
Lieber-DeCarli 饮食的特点是含有高浓度的葡萄糖和脂肪,而蛋白质含量相对较低。这种饮食的能量密度非常高,因此可以使动物快速增重。
Lieber-DeCarli 饮食在动物实验中应用广泛,特别是在研究肝脏疾病、肿瘤、代谢疾病等方面。由于该饮食的成分非常可控,可以帮助研究人员更准确地评估不同营养素对健康的影响,因此被广泛应用于基础医学和临床研究中。
需要注意的是,Lieber-DeCarli 饮食虽然在动物实验中应用广泛,但并不适合人类长期食用。这是因为该饮食的成分并不符合人类的膳食建议,长期食用可能会导致营养不良和健康问题。
多不饱和脂肪酸
低度至中度酒精保护自身免疫性疾病的另一个潜在机制,可能依赖于酒精在必需多不饱和脂肪酸代谢中的重要作用,如二十二碳六烯酸 (DHA) 和二十碳五烯酸 (EPA)。这些多不饱和脂肪酸(PUFA)可以减少活性氧的形成并充当抗炎分子。
低至中等剂量的酒精已被证明会增加多不饱和脂肪酸的产生,而在高剂量酒精下,由于脂肪酸分解代谢增加,多不饱和脂肪酸浓度会降低。
值得注意的是,多不饱和脂肪酸和多不饱和脂肪酸衍生物,如分解素、脂氧素和与自身免疫性疾病的缓解有关。
此外,多项研究还表明,多不饱和脂肪酸的增加具有心脏保护作用。由于心血管健康正在成为自身免疫性疾病结果的一个重要因素,这可能是另一种由低度至中度酒精介导的保护机制。
鉴于促炎特性,饮酒可能会导致自身免疫性疾病的风险增加或恶化。事实上,在某些炎症性疾病中,例如肠易激综合征 (IBS) 和常年性过敏,大量饮酒与疾病发作之间存在直接相关性。然而,适度饮酒似乎可以降低疾病风险、严重程度和进展。
这里介绍几种酒精相关的自身免疫性疾病,包括自身免疫性糖尿病、自身免疫性甲状腺疾病、系统性红斑狼疮、类风湿关节炎、多发性硬化症等,适度饮酒可降低其风险。
自身免疫性糖尿病
适度饮酒的有益作用已在成人非自身免疫性2 型糖尿病和自身免疫性 1 型糖尿病 (LADA) 中得到证实。
每天摄入 2–7 g 的患者风险降低了 60%
在一项自身免疫性 1 型糖尿病研究中,与每天摄入 0.01–2 g 的患者相比,每天摄入 2–7 g 的患者风险降低了 60%。这项研究还指出,与饮酒者相比,戒酒者的抗谷氨酸脱羧酶抗体 (GAD Ab) 水平更高,C 肽水平更低,对男性的影响更为明显。
注:抗谷氨酸脱羧酶抗体 (GAD-Ab) 在1型糖尿病发病前期和发病时多为阳性,而在正常人群及2型糖尿病患者中多为阴性。
葡萄酒的效果显著,可能与葡萄酒中的多酚类抗氧化物质相关
在另一项自身免疫性 1 型糖尿病研究中,发现摄入量超过 25 克/天的男性和女性的风险降低了 46%。这种效果似乎在抗 GAD 抗体水平低的患者中最强,并且与啤酒或白酒消费者相比仅限于葡萄酒饮用者。作者推测,抗 GAD 抗体水平较低的患者可能与 2 型糖尿病患者最相似,可能是葡萄酒中的多酚和羟基二苯乙烯类物质促进了酒精在自身免疫中的抗氧化或抗炎作用。
自身免疫性甲状腺疾病
与糖尿病类似,适度饮酒已被证明对自身免疫性甲状腺功能减退症和甲状腺功能亢进症均有保护作用。
例如,与对照组相比,适度饮酒与甲状腺功能减退症和格雷夫氏病的风险降低呈剂量依赖关系,无论性别或饮酒类型如何。
几项研究还发现,适度饮酒 >10 单位/周或每天至少饮酒 35 克,与自身免疫性甲状腺疾病和甲状腺过氧化物酶抗体阳性的发生概率较低有关。
系统性红斑狼疮(SLE)
多个病例对照、队列和横断面研究已经确定适度饮酒与系统性红斑狼疮风险之间存在显著的剂量依赖性关联。
在一项荟萃分析中,适度饮酒的保护作用与系统性红斑狼疮的持续时间有关,与治疗少于 5 年的患者相比,治疗少于 10 年的患者具有显著性。
另一项研究得出结论,适度饮酒可能会降低 ANA 阳性患者进展为系统性红斑狼疮的概率。
注:系统性红斑狼疮(SLE)是临床上最为常见的自身免疫性疾病之一。SLE的实验室检查包括多种项目,抗核抗体(ANA)是其中较为经常检测的项目之一。ANA阳性尤其是ANA核型为核均质性的结果往往与SLE具有较高的相关性,故临床上往往以此结果联合其他检查并结合患者症状作为诊断SLE的依据。
类风湿关节炎
与甲状腺疾病、糖尿病和系统性红斑狼疮类似,多项流行病学研究和几项机制研究支持轻度至中度酒精以 J 型或 U型剂量依赖方式在类风湿关节炎中发挥保护作用。
适度饮酒女性:较低的风险,较高的生活质量
在一项荟萃分析研究中,男性和女性的类风湿关节炎风险在10年内均有所降低,其中女性的风险降低幅度最大。
在其他研究中,与男性相比,适度饮酒的女性报告了较低的疾病活动和较高的生活质量。
然而,也有文献记载:酒精可能会阻止男性的放射学进展,并增加女性的放射学进展。与不饮酒的患者相比,中度饮酒的类风湿关节炎患者的改良健康评估问卷得分(提示功能状态改善)也显著降低。这种影响在HLA-DRB1共享表位阳性的患者中更强。
因此,类风湿关节炎患者的酒精、性别和基因构成之间可能存在有益但复杂的关系。
在类风湿关节炎中,适度饮酒与CRP 水平、IL-6水平的关联
在类风湿关节炎(RA)中,已注意到 CRP 水平呈 J 型关联:每周饮用 1-7 杯酒的患者 CRP 水平最低。
51 名适度饮酒的类风湿关节炎患者在症状出现前与 IL-6 水平呈 U 型关联,饮酒与可溶性肿瘤坏死因子受体 2 (TNFR2) 水平呈负相关。
适度饮酒:ACPA阳性患者类风湿关节炎风险降低
由于酒精会导致肝损伤,一项评估饮酒与肝脏炎症之间关系的研究报告称,每周饮酒量大于 21 单位与转氨酶相关,而每周饮酒量小于 14 单位则没有。适度饮酒还与抗瓜氨酸化蛋白抗体 (ACPA) 阳性患者的类风湿关节炎风险降低 50% 相关,并且与 ACPA 阴性类风湿关节炎患者的疾病风险降低 30% 呈反比剂量反应关系。
在类风湿关节炎小鼠模型中也注意到酒精对免疫系统的剂量依赖性影响。
适度饮酒:胶原性关节炎发病率下降
在胶原性关节炎 (CIA) 模型中,与非酒精对照组相比,适度饮酒的小鼠的 CIA 发病率降低了 40%,放射疾病严重程度降低了 50% 以上。
饮酒小鼠的 IL-21 和 IL-17A、中性粒细胞、单核细胞、浆 B 细胞和 IgG 水平也较低。酒精和乙酸盐都会影响体外和体内T 滤泡辅助 (TFH ) 细胞的功能状态,从而抑制 IL-21 的分泌。
这些发现很有趣,因为 TFH细胞通常存在于类风湿关节炎患者的滑膜关节中,也是肠道免疫的重要介质,表明肠道免疫过程与类风湿关节炎之间可能存在联系。
在另一项 CIA 研究中,适度饮酒(10% 乙醇水溶液)通过增加内源性睾酮、抑制核因子 B 活化和下调白细胞迁移来延缓 CIA 的发作并改善其进展。
多发性硬化症(MS)
同样,在多发性硬化症中,也有证据表明,适度饮酒在降低疾病风险和/或疾病进展方面具有保护作用。几项大型人口研究表明:
酒精与男女多发性硬化症风险之间存在剂量依赖性负相关
适度饮用红酒似乎与较低的扩展残疾状态量表评分相关,这表明功能有所改善,尽管适度饮酒的患者在脑部 MRI 上表现出 T2 病变体积增加。
相反,大量饮酒可能会增加患多发性硬化症的风险,尤其是男性。
也有研究认为剂量和多发性硬化症风险之间没有关联,性别可能是一个变量
一些研究指出,不同剂量的酒精与患多发性硬化症的风险之间没有关联。在这些研究中,性别可能是一个变量,可以解释酒精对多发性硬化症的影响。
例如,在一项女性护士健康研究(NHS)I和II中,不同类型的酒精与多发性麻痹症的风险没有关联。尽管这是一项针对英国国家医疗服务体系两项研究中超过90000名女性的大型研究,但258例多发性硬化症患者的队列相对较小,与男性相比,女性可能没有经历到酒精的保护作用。
例如,在多发性硬化症的动物模型,即实验性自身免疫性脑脊髓炎(EAE)中,最近的研究表明,主要是雄性小鼠在适度饮酒后疾病评分有所改善。
以上是低-中剂量酒精带来的健康益处,“凡事皆有度,过犹不及”。接下来章节来看过量饮酒可能带来哪些不利影响。
酒精与消化系统密不可分。它通过肠道吸收,并在肝脏内被肝细胞代谢。过量饮酒会导致肠道微生物组和肠道上皮完整性发生改变。它会导致重要的微量营养素缺乏症,包括短链脂肪酸和微量元素,这些元素会影响免疫功能并导致肝损伤。在某些人中,长期酗酒会导致肝病从脂肪肝发展为肝硬化和肝细胞癌。
持续酗酒会改变肠腔pH 值,促进病原体过度生长,并且还与肠道微生物群功能的改变有关,因为它会改变与肠屏障功能障碍有关的特定代谢物分泌物。酒精使用障碍患者通常表现出血浆细胞因子水平升高,例如 TNF-α、IL-10 和 CRP,这表明慢性、低度、全身性炎症。
几项研究调查了饮酒对动物和人体模型的影响,并一致表明过量饮酒与肠道微生态失调的发展有关。
简而言之,酒精已被证明可以增加变形杆菌、肠杆菌和链球菌的相对丰度,并降低拟杆菌、阿克曼氏菌和粪杆菌的丰度。其他还包括抗炎细菌(如Faecalibacterium prausnitzii和双歧杆菌等)水平下降。
在人类酒精使用障碍 (AUD) 研究中,生态失调的特征是:
扩展阅读:
肠道细菌四大“门派”——拟杆菌门,厚壁菌门,变形菌门,放线菌门
然而,并非所有研究都注意到厚壁菌门对高剂量酒精的反应有所减少。例如,在猕猴中自愿自我给予慢性高剂量酒精会导致拟杆菌减少、厚壁菌门升高,并且在饮酒期间完全没有Akkermansia muciniphila,而戒酒则恢复了基线细菌种类。
肠道微生物群稳态的破坏与这些疾病有关
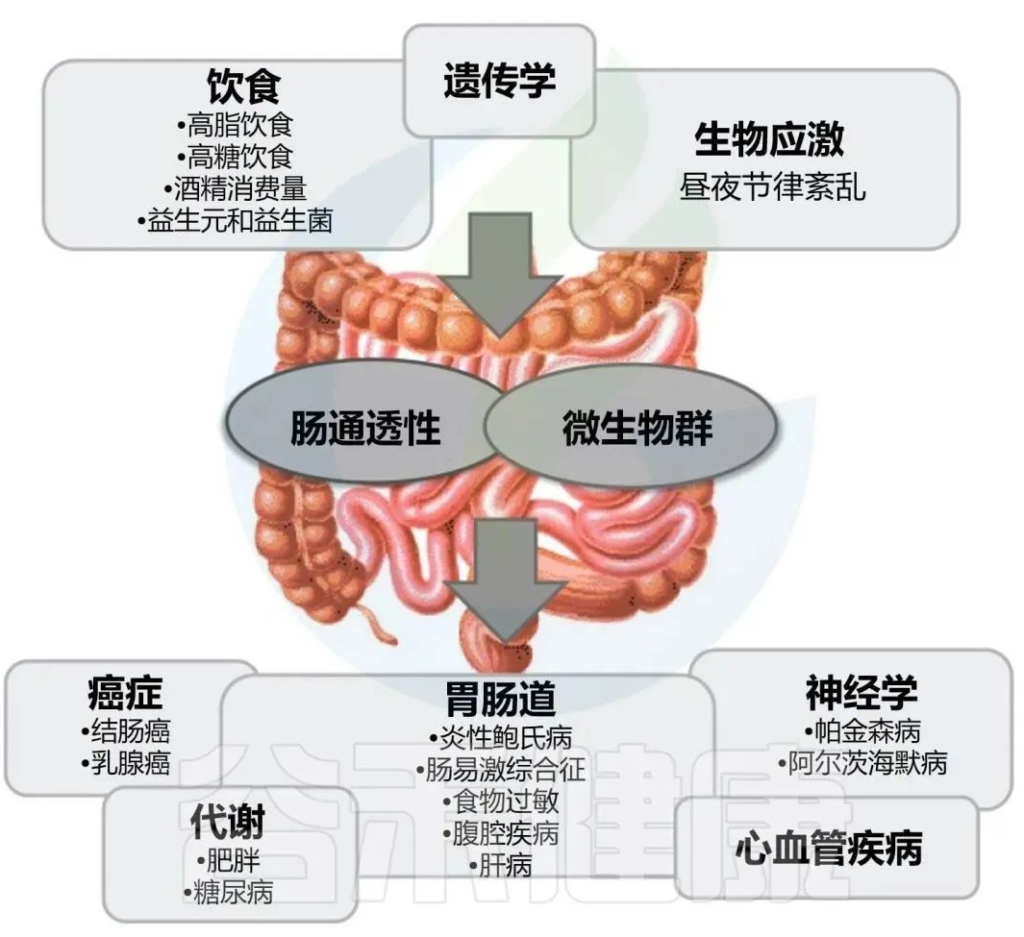
Engen PA et al., Alcohol Res. 2015;37(2):223-36.
此外,生态失调可能是由西方社会常见的环境因素引起的,包括饮食、遗传、昼夜节律紊乱和酒精饮料消费。
在酒精成瘾者中,饮酒会破坏肠道屏障功能,也称为肠漏症。肠道屏障由肠细胞、杯状细胞和影响粘液层内肠道微生物组的抗菌物质以及固有层中的众多免疫细胞组成。
酒精引起的生态失调通过其对肠道完整性的病理影响促进急性(例如,酒精性肝炎)和慢性(例如,酒精相关性肝硬化)肝病的发展。
肠粘液屏障在肠道的免疫功能中起着至关重要的作用,它的破坏会导致这些疾病状态。在这个屏障中,相邻的肠细胞被顶端“紧密连接”蛋白 claudins、occludin 和 zona occludens 结合在一起,防止病原体相关分子颗粒 (PAMP) 和细菌内毒素等管腔内容物意外转移到门脉循环中。
饮酒引起的生态失调与这些紧密连接的破坏有关。因此,随后的免疫功能障碍和循环促炎细胞因子(如肿瘤坏死因子 (TNF)-α 和IL-1β)的增加进一步破坏了肠道屏障。
扩展阅读:什么是肠漏综合征,它如何影响健康?
酒精相关的微生态失调不可避免地会影响肠道代谢,包括短链脂肪酸 (SCFA)、氨基酸和胆汁酸的显著变化。
短链脂肪酸
短链脂肪酸在维持紧密连接方面的作用越来越明显。短链脂肪酸是肠道菌群对难消化膳食纤维进行厌氧发酵的产物。
对酒精使用障碍患者的粪便代谢组分析显示,短链脂肪酸减少,这可能部分是由于生态失调对粪杆菌等产短链脂肪酸菌产生负面影响。一些小鼠模型表明,以高纤维饮食、益生菌或饮食调整形式补充短链脂肪酸,可增强肠道上皮完整性,并减少酒精模型中的肝损伤。
氨基酸
从饮食中获得的必需氨基酸(例如赖氨酸)和非必需氨基酸(例如谷氨酸)都会受到酒精影响。据推测,这是由于生态失调导致微生物-宿主共同代谢紊乱的结果。
虽然管腔氨基酸浓度会随着饮酒而下降,但一些氨基酸(如酪氨酸和苯丙氨酸)的血清水平会升高,这表明失调微生物组的代谢和吸收特征发生了改变。这种代谢失衡可能在活性氧 (ROS) 和有毒中间体水平升高的过程中发挥作用。
胆汁酸
胆汁酸已被证明在人类和大鼠饮酒后的血清和管腔内容物中都会发生变化。初级(由肝脏合成)和次级(来自细菌代谢)胆汁酸主要在小肠中发挥多种功能,并通过其类固醇结构在脂质吸收、胆固醇稳态以及激素作用中发挥关键作用。
在健康的肠肝循环中,初级胆汁酸与牛磺酸或甘氨酸结合形成分泌到肠腔中的胆汁盐。然后肠道微生物群将这些代谢为次级胆汁酸,在将它们循环回肝脏之前去除牛磺酸/甘氨酸基团。饮酒通过增加次级胆汁酸的比例和胆汁酸的总浓度,以及增加与甘氨酸而非牛磺酸结合的比例来破坏这一点。
这是由于生态失调降低了牛磺酸的生物利用度和肠肝循环率增加所致。这种破坏的后果尚不完全清楚;然而,在饮酒过程中更普遍的甘氨酸结合酸可能毒性相对更大,尽管管腔浓度高但胆汁酸的合成增加导致肝脂肪变性。
长期饮酒会减少营养吸收并导致营养不良。肠道通透性、胆汁酸概况和微生物组的改变都促成了这一点,此外,酒精代谢过程中释放的有毒代谢物和 ROS 会对肠道造成结构损伤。
特别是,长期饮酒已被证明会导致细胞死亡、粘膜侵蚀和绒毛尖端上皮细胞丢失。其后果是维生素 A、B1(硫胺素)、B2(核黄素)、B6(吡哆醇)、C、D、E 和 K 以及叶酸、钙、镁、磷酸盐、铁和微量元素锌和硒的不同程度的缺乏。
扩展阅读:如何解读肠道菌群检测报告中的维生素指标?
所有患有慢性酒精使用障碍的患者都要接受全面的营养评估,因为这些缺乏症因人而异,例如铁可能缺乏或过量。
除了上述机制外,重度饮酒者每天从营养不良的酒精饮料中获取高达 50% 的热量摄入 。此外,应该注意的是,除了慢性酒精滥用的症状影响(例如,呕吐、厌食和腹痛)之外,该群体中的社会因素(例如贫困和获得营养“完整”的饮食)也可能导致营养不良。
微生物组从高剂量酒精喂养的小鼠,转移到未接触酒精的无菌小鼠,已被证明会在受体小鼠中诱发肠道炎症。
酒精通过影响肠粘膜免疫诱发炎症
肠道炎症是由免疫系统对酒精及其代谢物产生的炎症反应引起的。酒精通过多种机制影响肠粘膜免疫。特别是,它可能首先降低粘膜中的先天免疫反应,导致对肠道病原体的易感性增加。
随后,正如在细胞培养研究中发现的那样,酒精可能会触发免疫系统反应和促进炎症反应的分子上调,包括释放炎症免疫细胞,例如白细胞和肥大细胞。
研究还表明,酒精可以直接调节先天免疫和适应性免疫,进一步促进肠道和肠道源性炎症。例如,一项针对小鼠的研究发现酒精会抑制肠道清除有害细菌的免疫反应,而其他研究发现酒精会抑制肠道粘膜免疫细胞的活性。
酒精通过引起肠道菌群失调诱发炎症
酒精相关的细菌过度生长和生态失调可能导致肠道内毒素产生增加,内毒素可与肠粘膜上的细胞结合,引起局部炎症,并易位至肠外部位,引起全身炎症。
微生物群失调可导致肠道通透性受损,并通过肠道细菌内毒素、LPS 的系统易位、免疫细胞上 TLR 和NF-kB 的激活以及炎症 iNOS 的诱导促进炎症。
酒精通过肝毒性诱发炎症
肝毒性会干扰肝脏解毒物质的能力,从而导致酒精的有毒代谢物乙醛在全身蓄积。此外,LPS 介导的肝脏驻留巨噬细胞 Kupffer 细胞的激活进一步促进了促炎细胞因子的释放和全身炎症的传播。
其他研究发现酒精影响粘膜免疫的多种方式,包括:
酒精对先天免疫系统和适应性免疫系统的影响具有剂量依赖性。
酒精对小胶质细胞(中枢神经系统的先天免疫细胞)具有显著的剂量依赖性影响
在急性酒精滥用的小鼠模型中:
酒精还以剂量依赖的方式调节适应性免疫系统
长期适度饮酒会导致 T 细胞和 B 细胞活化和增殖,而长期大量饮酒则与 T 细胞和 B 细胞耗竭和凋亡以及免疫球蛋白增加有关。
此外,长期酗酒会改变 T 细胞表型,导致幼稚 T 淋巴细胞百分比下降和记忆 T 细胞百分比升高。相反,适度饮酒与滤泡辅助性 T (T FH )细胞的调节有关。
细胞因子和炎症标志物也以剂量依赖的方式受到酒精的影响。例如,C 反应蛋白 (CRP) 和白细胞介素 6 (IL-6) 在人类重度饮酒者中升高,但与不饮酒者相比,在适度饮酒者中相对降低。
CRP 效应也可能是性别二态的,一些研究表明酒精引起的 CRP 降低是女性特有的,也有其他研究表明适度饮酒会以 U 形模式降低 CRP,而与性别无关。
以上是高剂量酒精对肠道屏障、肠道菌群及其代谢产物、营养状况、免疫系统等方面的影响。接下来章节我们具体来了解一下酒精相关疾病,包括肝病、胃肠道疾病、营养不良、肥胖、心血管疾病等。
►►►
酒精代谢
肝肠微生物轴在营养吸收和肝毒性中起着重要作用,其中肝脏是营养物质、毒素和肠道血液供应的细菌代谢产物的第一道过滤器。
酒精性肝病 (ALD) 是一种由过量饮酒引起的疾病,是全球医疗保健的负担。酒精性肝病涵盖范围广泛的肝损伤,包括无症状脂肪变性、酒精性脂肪性肝炎 (ASH)、纤维化、肝硬化和肝细胞癌 (HCC)。
酗酒患者发展为酒精性肝病的易感性是高度可变的,并且其向更晚期的进展受若干因素(即酒精滥用的量和持续时间)的强烈影响。
其中,肠道微生物群及其代谢物最近被确定为酒精性肝病病理生理学中最重要的。
酒精滥用会引发肠道菌群分类组成、粘膜炎症和肠道屏障紊乱的定性和定量改变。肠道通透性过高导致活致病菌、革兰氏阴性微生物产物和促炎性内腔代谢物转移到血液中,进一步证实了酒精引起的肝损伤。
酒精在肝脏中的代谢
酒精在肝脏中的代谢是了解其在酒精相关肝病发病机制中的作用的关键。酒精在肝细胞中主要通过乙醇脱氢酶代谢为乙醛,然后通过乙醛脱氢酶代谢为乙酸。
乙醛是一种高反应性蛋白质,会导致肝损伤。它与脂质、蛋白质和 DNA 结合形成潜在的免疫原性加合物。这些加合物可产生适应性免疫反应,导致肝细胞损伤和炎症。
线粒体结构改变可导致功能障碍,包括 ATP 生成减少、ROS 生成和乙醛脱氢酶活性降低。乙醛也是肝纤维化进程中的关键代谢物。它可以促进肝星状细胞 (hepatic stellate cells, HSCs) 中胶原 I 的合成,乙醛加合物刺激炎性细胞因子和趋化因子的释放。
乙醇脱氢酶途径可有效代谢少量酒精,但在长期接触酒精时,该途径会变得饱和,并且会显著诱导 CYP2E1。向 CYP 途径的转变导致 ROS 的产生,从而导致氧化应激。
ROS 与蛋白质结合,改变它们的结构和功能特性,并可能充当新抗原。ROS 还可以直接与 DNA 结合,造成损伤,或导致脂质过氧化产物,如 4-羟基壬烯醛 (4-HNE) 和丙二醛 (MDA),从而产生高度致癌的 DNA 加合物。
此外,在慢性重度酒精摄入中,由于乙醛介导的谷胱甘肽减少,肝脏的抗氧化清除系统受损。氧化应激的结果是诱导肝细胞凋亡和坏死。
酒精性肝损伤
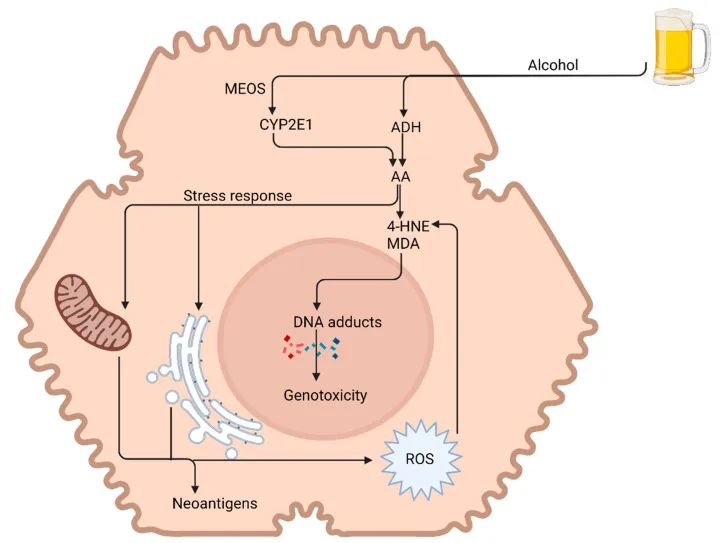
编辑
图源:biorender
乙醛 (AA) 是造成酒精对肝脏的大部分毒性作用的原因。乙醛极度亲脂,导致形成乙醛加合物——丙二醛 (MDA) 和 4-羟基壬烯醛 (4-HNE)。这与活性氧 (ROS) 一起导致 DNA 损伤和遗传毒性。
乙醛还会诱导各种细胞器(例如线粒体和内质网)的功能和结构改变。
MEOS:线粒体酶氧化系统;ADH:乙醇脱氢酶。
酒精相关性脂肪变性
脂肪变性的特征是肝细胞中脂肪(甘油三酯、磷脂和胆固醇酯)的积累,是肝脏对长期饮酒的最早反应,几乎普遍存在于慢性重度饮酒者中。
虽然它在减少饮酒后是完全可逆的,但它的存在与酒精相关肝病的进展有关,最近的一项荟萃分析发现肝硬化的年进展率为 3%。 肝脂肪变性可能通过更大的脂质过氧化和氧化应激增加肝脏炎症(脂肪性肝炎)、纤维化和肝硬化的风险。
然而,进展不仅受饮酒量的影响,还受其他因素的影响,包括性别、共存的肝病、吸烟和遗传。
慢性酒精摄入通过增加肝脂肪生成和减少肝脂肪分解导致肝脂肪变性
酒精通过多种机制诱导肝脂肪变性
图源:biorender
↑:增加;↓:减少;HSC:肝星状细胞。
酒精性脂肪性肝炎
肝脏炎症强烈影响纤维化、肝硬化和最终肝细胞癌的发展
酒精引起的肠漏症导致病原体相关分子模式(PAMP) 输送到肝脏。PAMP 与受损细胞释放的损伤相关分子模式一起激活单核细胞、巨噬细胞、枯否细胞和肝实质细胞上的先天受体【Toll 样受体 (TLR) 和 NOD 样受体 (NLR)】。
通过这些受体发出的信号导致包括 NF-κB 在内的促炎转录因子的转录增加以及促炎趋化因子和细胞因子的产生。
净效应是单核细胞、中性粒细胞和 T 细胞的流入,导致细胞死亡和肝星状细胞 (HSC) 激活的可溶性介质的释放。
除了对酒精激活的促炎性免疫反应外,酒精性肝炎患者还有免疫功能障碍的证据。肠道来源的 PAMP 对单核细胞的激活导致 T 细胞耗竭,同时产生抗炎 IL-10 的 T 细胞数量减少,单核细胞和中性粒细胞功能受损 。
酒精引起的炎症
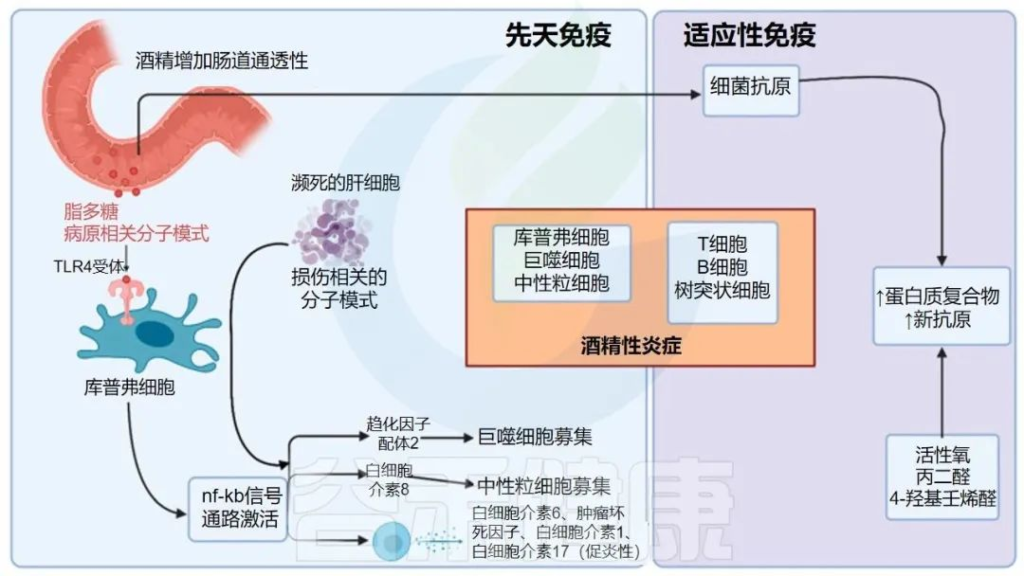
酒精对先天免疫和适应性免疫均有影响。酒精不仅会诱发肠道菌群失调,还会增加肠道通透性。病原体相关分子模式 (PAMP) 如脂多糖与 Kupffer 细胞上的 TLR4 受体相互作用,并通过 NF-κB 途径产生促炎细胞因子和趋化因子,导致肝脏炎症。
乙醛诱导各种蛋白质的结构变化并产生新抗原,从而引发适应性免疫反应并导致肝脏炎症。
CCL2:CC基序趋化因子配体2;DAMPs:损伤相关分子模式;4-HNE:4-羟基壬烯醛;IL:白细胞介素;MDA:丙二醛;NF-κB:核因子 kappa B;ROS:活性氧;TLR4:toll 样受体 4;TNFα:肿瘤坏死因子α;↑: 增加; ↓:减少。
肝细胞死亡通过多种机制发生
包括细胞凋亡、细胞焦亡、坏死和坏死性凋亡。
细胞凋亡是由直接酒精介导的肝毒性、氧化应激的诱导、存活基因 ( C-met ) 的抑制和促细胞凋亡信号分子(TNF-α 和 Fas 配体)的诱导引起的。
坏死、细胞肿胀和膜破裂也可以通过称为坏死性凋亡的程序化途径发生。
而细胞焦亡是一种依赖于 caspase-1 的程序性细胞死亡。
细胞死亡的模式可能受疾病状态的影响,早期酒精相关肝病中细胞凋亡占主导地位,但酒精性肝炎中炎性体激活驱动细胞焦亡和肝损伤扩散。
长期饮酒通过多种机制影响肝脏
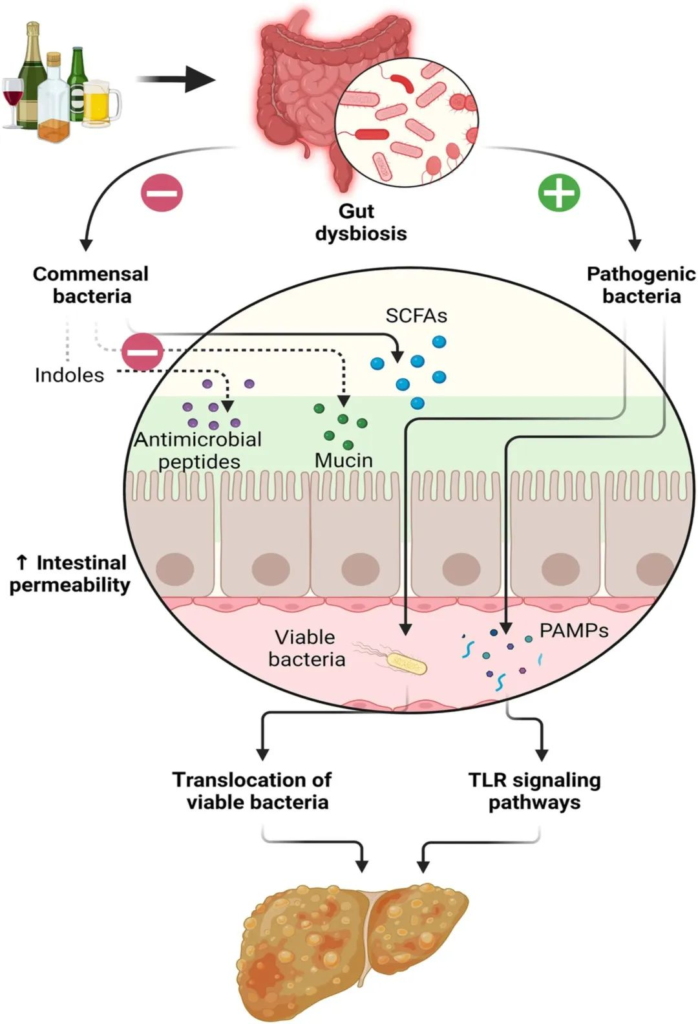
doi.org/10.1111/jgh.16199
长期饮酒会导致肠道生态失调,从而导致肠道通透性增加、肠道细菌易位和病原体相关分子模式 (PAMP),并通过 Toll 样受体 (TLR) 信号通路增加肝脏炎症和纤维化。
为什么有些人不喝酒也有可能得脂肪肝?
脂肪肝的发生与多种因素有关,包括肥胖、高血压、高血脂、糖尿病等,也与外源性酒精和内源性酒精相关。这些因素会影响肝脏的代谢功能,导致脂肪在肝脏内积累,从而引起脂肪肝。
我们一般认为的酒精摄入是指外源性酒精,是通过饮酒等途径摄入的酒精。而内源性酒精则是由人体自身产生的一种酒精,主要产生于肠道中的细菌代谢过程中。这两种酒精对人体的影响有所不同。
内源性酒精对人体的影响相对较小,因为它的产生量较少。但是,如果肠道细菌失衡,导致内源性酒精产生过多,就可能会对人体健康产生负面影响,如肝脏疾病、代谢紊乱等。
酒精引起的纤维化和肝硬化
纤维化是肝脏对破坏性刺激的伤口愈合反应,在去除刺激后可逆。在长期大量饮酒的情况下,会发生慢性炎症和纤维化,导致宽频带纤维组织沉积,扭曲肝脏结构并改变肝脏血流,导致门脉高压及其相关并发症。
当肝纤维化发展到扭曲肝脏结构并形成结节时,患者会发展为肝硬化。
活化的星形细胞 (HSC) 引起的细胞外基质沉积是肝纤维化发生和发展的关键事件。其他细胞(门静脉成纤维细胞和肌成纤维细胞)的贡献较小。HSC 被炎性细胞因子和直接被酒精及其代谢物和 ROS 激活。活化的 HSC 通过分泌趋化因子和表达粘附分子来吸引和刺激循环免疫细胞,从而使炎症反应持续存在,进而激活静止的 HSC。
肝癌
肝硬化是一种癌前状态,会增加原发性肝癌的风险,最常见的是肝细胞癌(HCC)。在全球范围内,大约 30% 的肝细胞癌是由酒精引起的。
酒精本身是一种致癌物,在肝细胞癌的背景下,通过 ROS 诱导的损伤、炎症机制及其反应性代谢物乙醛,酒精在其发展中起着特定的作用。
在大量饮酒者中,CYP 通路活性增加会产生 ROS,导致 DNA 损伤,导致细胞周期停滞和细胞凋亡并破坏基因功能,从而增加癌发生。
酒精相关性肝病患者炎症通路的激活与癌症风险增加相关
尽管机制尚未完全阐明,但可能涉及促炎细胞因子促进 ROS 积累(关于ROS平衡与健康我们准备专门搞一个主题讲下)。细胞因子的产生也与血管生成和转移发展的上调有关。此外,酒精会抑制 CD8+ T 细胞的抗肿瘤反应。
乙醛具有高反应性,并与 DNA 和蛋白质形成加合物,导致线粒体损伤和 DNA 修复机制的破坏。在具有遗传变异的人群中发现的乙醛水平升高会导致酒精脱氢酶和乙醛脱氢酶的活性改变,这与重度饮酒者患肝细胞癌的风险较高有关。
扩展阅读:深度解析 | 肠道菌群与慢性肝病,肝癌
►►►
胃肠道 (GI) 是与摄入体内任何物质的第一道接触线,特别容易受到毒素的损害。越来越多的研究表明,胃肠道健康状况不佳对身体的整体健康起着重要作用。任何可能导致胃肠道损伤的事情,其后果可能远远超出肠道。事实上,研究人员已经开始发现酒精,特别是如果长期大量饮酒,会引发肠道内启动的过程,从而促进全身炎症。
酒精使用障碍患者肠道通透性增加
对人类的研究表明,一部分酒精使用障碍 (AUD) 患者的肠道通透性实际上有所增加,这是使用称为 Cr-EDTA 的方法测量的,该方法检查口服铬的排泄。此外,那些患有酒精使用障碍且通透性增加的人更容易患肝病,表明肠道通透性可能是某些酒精使用障碍患者器官损伤的介质。
另一项研究表明,患有酒精使用障碍的人不仅肠道通透性增加,而且增加到足以让大分子通过肠道屏障。内毒素 – 也称为脂多糖 (LPS),是大分子,同一项研究发现血浆内毒素水平随着肠道通透性的增加而增加。
酒精是如何诱导肠道通透性的呢?
简短的回答是通过破坏上皮细胞本身(跨上皮通透性)和破坏上皮细胞之间的空间(细胞旁通透性),它由紧密连接、细胞骨架和一些相关蛋白组成。
跨上皮渗透性是由直接细胞损伤引起的。例如:
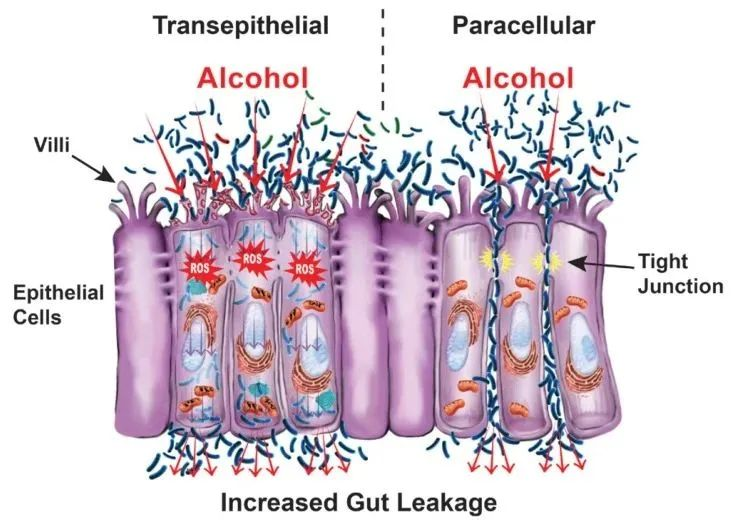
肠道屏障调节肠道内部(食物和饮料流经的地方)和肠道内部上皮细胞层另一侧的细胞和血管之间的物质通道,包括微生物产物。
破坏肠道屏障有两种方式:
肠道屏障通透性增加,使细菌及其产生的毒素离开肠道,并通过血液渗入其他器官。
酒精及代谢物如何通过触发以上两种机制?
— 酒精及其代谢物通过直接破坏细胞和通过活性氧 (ROS) 引起的氧化应激等多种机制削弱细胞膜来触发跨上皮机制。
酒精会导致细胞死亡,从而导致肠道发生变化,包括粘膜溃疡、糜烂和主要位于绒毛尖端的上皮细胞丢失;乙醛形成导致直接细胞损伤的 DNA 加合物;和酒精代谢过程中释放的活性氧 (ROS) 通过氧化应激导致直接细胞损伤。
— 酒精的代谢物通过破坏产生连接细胞和稳定细胞骨架的蛋白质的紧密连接的蛋白质,来触发细胞旁路机制。
酒精及其代谢物通过作用于将两个相邻细胞融合在一起的紧密连接复合物而引起细胞旁通透性。例如,乙醛通过重新分配蛋白质来破坏紧密连接的稳定性;酒精及其代谢物改变紧密连接蛋白的表达;和酒精的非氧化代谢物导致紧密连接再分布,破坏其屏障功能。
此外,研究表明酒精会破坏细胞的细胞骨架,即赋予它们结构的细胞边界。还有越来越多的证据表明,酒精会导致 microRNA (miRNA) 的过度表达,这些小片段的非编码 RNA 会沉默基因表达。具体而言,酒精会导致 miRNA 过度表达,从而影响与肠道屏障完整性相关的基因。
酒精和胃肠道癌症
长期饮酒会增加患主要胃肠道癌症的风险,包括食道癌、胃癌和结肠癌(结直肠癌)。这种风险通常随着饮酒量的增加以及与其他生活方式相关因素(如吸烟或代谢综合征)的增加而增加。虽然酒精最初被认为是一种直接致癌物,但研究表明酒精引起的肠道炎症可能是罪魁祸首。
代谢综合征和肥胖中出现的全身性炎症会增加多种上皮癌的风险,包括胃肠道癌,这表明酒精引起的肠道炎症造成的全身性炎症状态,也可能导致酒精引起的胃肠道和其他器官癌症发生。这个过程像滚雪球,因为随着细胞转变为癌变状态,ADH 活性会增加,而 ALDH 活性可能会降低 。
这导致氧化速率增加和清除酒精代谢物的能力降低,这反过来又可以通过对 DNA、氧化应激和肠道炎症的直接影响进一步促进癌变。
►►►
营养不良
酒精的能量价值及其对营养状况的影响
酒精是唯一提供能量 (7.1 kcal/g) 的精神药物。然而,它的卡路里被认为是“空的”,因为酒精摄入不能提供维生素和矿物质,并且它的过量摄入会导致营养状况的改变。
由于酒精的高能量值,人们普遍认为,过量饮酒会增加体重和肥胖的风险。尽管酒精密度高,但与不饮酒者相比,饮酒者的体重似乎并未增加。
当酒精摄入热量占总热量摄入的 50% 或更多时,由于微粒体乙醇氧化系统 (MEOS) 的激活,身体系统可能无法有效利用乙醇摄入提供的能量。以能量浪费为代价,MEOS 是乙醇氧化的主要肝脏途径。其诱导在戒酒后是可逆的。
饮酒与体重的关系及戒酒对营养状况的改善
在对 181 名每天消耗超过 80 克乙醇的住院男性进行观察性研究期间观察到,63.1% 的人患有厌食症,58.7% 的人体重减轻,17.8% 的人的 BMI 低于 20 kg/m2。经过三个月的戒酒期后,酗酒者的热量使用正常化,体重指数 (BMI) 和腰臀比 (WHP) 也有所增加。
作者认为,戒酒期间营养状况的改善可能是由于 MEOS 活性较低,从而减少了热量的浪费。
少量或中等程度饮酒的女性肥胖风险较低
酗酒患者因饮酒而出现代谢和营养障碍,并表现出在静息状态下大量使用能量,优先使用脂质作为能量来源,并且与对照组相比,脂肪组织比健康对照减少了 19%。
队列研究表明,与戒酒的女性相比,少量或中等程度饮酒(最多 30 克/天)的女性体重增加和肥胖的风险较低,而其他研究已经达到无论受试者的性别如何,都得出相同的结论。
研究评估了酗酒者脂肪和肌肉组织长期变化的预后价值以及戒酒对这些变化的影响。结果表明,在第一次评估后的头六个月内,原来体重较轻与较差的预后相关,无论患者是否在同一时期戒酒。然而,不禁欲与瘦体重的减少相关性更大。
低剂量饮酒是热量的重要来源
身体对酒精热量的利用可能与酒精的用量有关。酒精消耗量低于每日卡路里的 25-35% 可用作能量来源;然而,在更高的消耗量下,利用可能不完整。
在人类中,热量营养不良是世界各地数以百万计死于饥饿的人所面临的现实,并且由于价格低廉且可用性高,乙醇被摄入以代替食物。一些研究创建动物模型是为了阐明当营养不良的生物体暴露于乙醇时会发生什么。然而,结果并不是人们所期望的。
事实上,低剂量饮酒是热量的重要来源,可减轻营养不良的影响。动物研究表明,摄入低至中等剂量的酒精(最多占总热量摄入的 20%)会导致营养不良的大鼠体重增加,这表明有效利用了酒精的能量含量。然而,当酒精中的能量含量占总卡路里摄入量的 10% 或更多时,富营养化大鼠的体重就会减轻。
肥胖
个人的营养状况可能会影响酒精饮料中能量的利用。在人类中,瘦个体对酒精卡路里的利用效率可能较低,而在肥胖个体中,卡路里有助于增加体重。
酒精对体重的影响仍然是一个有争议的话题
一项长达10年的前瞻性研究表明,饮酒者的体重比不饮酒者更稳定,这表明饮酒不是肥胖的危险因素。每月长期适度饮用啤酒不会改变成年人的体重或身体成分。
一些研究表明,身体和腹部脂肪的增加与适度和频繁饮酒有关。还有证据表明,不同的饮酒方式会导致体重增加。每天摄入 ≥ 30 克酒精可能会改变能量稳态的平衡,促使食欲增加,从而导致体重增加和肥胖,无论摄入哪种类型的酒精。
为什么有可能增加肥胖的风险?
在短期内,饮酒被认为是一种食欲兴奋剂,影响用于控制食欲的神经化学和外周系统,如瘦素抑制、胰高血糖素样肽-1 和血清素,并增强γ-氨基丁酸的作用,内源性阿片类药物和神经肽 Y。因此,在没有依赖性的情况下大量饮酒以及酗酒可能会增加肥胖的风险。
饮酒与体重变化及发展为肥胖的关联因性别而异
男性将酒精添加到日常摄入的卡路里中,而女性则倾向于使用酒精替代其他能源,例如减少碳水化合物的消耗而不会增加总卡路里摄取。在考虑能量平衡时必须包括这些差异,因为它们可能会产生关于体重的不同结果。
多种肥胖风险因素与饮酒有关
例如腹部脂肪增加 、脂质氧化能力下降、高热量饮食消耗和皮质醇分泌增加。有人可能会说,与酗酒者和饮食富含脂肪的人相比,超重或肥胖的人和非日常适度饮酒者更多地利用酒精中的卡路里。
女性、饮酒者、大学生的身体、腹部和外周脂肪值也较高,表明 AUDIT 问卷中较高的分数与 BMI、体脂百分比、腰围、三头肌皮褶,手臂周长呈正相关。
喝得量少,频率高 ⇒ BMI 较低
喝得量多,频率低 ⇒ BMI 较高
一项基于 1997 – 2001 年全国健康采访调查数据的研究表明,与 BMI 相关的饮酒量和频率之间存在强烈的相反影响。喝得频率更高但喝得少(即每天喝一杯)的人 BMI 较低。另一方面,饮酒频率较低但饮酒量较大(暴饮暴食)的个体 BMI 较高。那些每天摄入 ≥ 50 g 的人的腹部肥胖风险因素增加。
与这一发现一致的是,在对英国人进行的一项研究中,有人指出,摄入 30 克或更多的酒精会增高 BMI 和体重增加的风险。
肥胖风险:当前适度饮酒者<戒酒者<酗酒者
同样,在对第三次全国健康和营养检查数据进行研究后提供的结果显示,与戒酒者相比,当前饮酒者肥胖的可能性较低,而酗酒者(一天四剂或更多剂饮酒者)肥胖的可能性更大。在报告经常饮酒但每周饮酒量少于 5 剂的人群中,肥胖风险因素显著降低。
扩展阅读:体重增长:目前为止我们所知道的一切(更新你的减肥工具箱)
►►►
血脂概况
每周饮酒三到四天与患心肌梗塞的风险较低有关
据估计,酒精的心脏保护作用可能归因于 50% 的 HDL 升高。适量饮酒(30 克乙醇/天)可使 HDL-c 浓度增加约 4 mg/dL,apoA-I 浓度增加 8.82 mg/dL,心脏病风险估计降低 24.7%。
不管饮用的饮料(葡萄酒、烈酒或啤酒)的数量或类型如何,这种改变都可能发生。此外,酒精促进 HDL (好胆固醇)的较少降解和 LDL(坏胆固醇)的更大肝脏代谢。
重度饮酒者中风风险较高
尽管饮酒会改变血脂水平并减少心血管事件,但已证明重度饮酒者中脑出血和蛛网膜下腔出血等中风的发生率高于不饮酒者 。此外,由于大量饮酒,如糖尿病、高血压性心脏病、缺血性心脏病、缺血性和出血性中风等疾病逐渐增加。
酒精的风险和益处之间的差异因人而异,科学家呼吁不应将其用作保护心血管的工具作为一项公共卫生措施加以鼓励。
在一项荟萃分析研究中描述了酒精对预防冠心病的益处的减少,其中节制和轻度和中度饮酒者显示出相同的冠心病死亡风险。
发现每天饮酒 10 克的人甘油三酯水平较高。摄入量大于 50 克/天可显著降低发生低水平 HDL-c 的风险,但会增加发生高水平胆固醇的风险。
在餐后时期,酒精会导致甘油三酯升高,并抑制游离脂肪酸 (FFA) 的氧化 。重要的是要强调,对于患有冠心病的个体,餐后高甘油三酯血症更大、持续时间更长。高甘油三酯血症或 FFA 增加与正常个体和胰岛素抵抗者的内皮血管舒张减少有关。
扩展阅读:
高血压
酒精摄入会以剂量依赖性反应升高动脉压
每天摄入超过两次的剂量是最常见的可逆性高血压原因之一 。肾素-血管紧张素系统可能参与了酒精诱发高血压的机制。
一项研究表明,急性酒精中毒改变了肾素-醛固酮系统,使人体具有正常的氢和钠平衡。血浆肾素的增加可能是由于乙醇利尿或乙醇对醛固酮分泌的抑制作用引起的脱水。渗透压、动脉压和心脏频率没有发现显著变化。后来,证实适度饮酒时肾素活性会急剧增加,从而引起液体和电解质平衡以及动脉血压的变化。
少量和适度饮酒的影响可能因性别而异
文献对这个问题的描述并不一致。在一项由女性健康研究对 28,848 名女性开展的前瞻性研究和由医生健康研究开展的另一项对 13,455 名女性开展的前瞻性研究中,少量和适度饮酒降低了女性患高血压的风险,并增加了男性患高血压的风险。
酒精成为高血压危险因素的阈值在女性中等于或大于每天 4 剂,而男性的中等水平等于或大于每天 1 剂。 然而,一些研究在考虑男性和女性的动脉血压对酒精的敏感性是否不同时存在争议。
饮酒推荐量因体重而异
由于习惯性饮酒不同,动脉血压升高在低体重个体中比在高体重个体中更为显著。因此,低体重女性预防高血压的推荐饮酒限度必须低于高体重女性。
对于重度饮酒者而言,少喝可以预防治疗高血压
在没有其他心血管疾病的健康男性和女性中调查了饮酒和当前饮酒标准与高血压风险之间的关联。与节制的人相比,那些饮酒且饮酒量很大的人患高血压的风险更高。
重度饮酒者酒精摄入量的减少显著降低了动脉收缩压和舒张压。因此,必须推荐减少饮酒作为生活方式调整措施,以预防和治疗重度饮酒者的高血压。
扩展阅读:认识肠道微生物及其与高血压的关系
以上是酒精对相关疾病的影响。长期大量饮酒后,人体对酒精容易产生依赖性,出现无法自主控制饮酒行为,也就是戒不掉、戒断症状等现象,我们通常说酒精成瘾。他们需要往往更多的酒精来获得同样的愉悦感。接下来章节,我们来看一下酒精成瘾的相关因素。
肠道和大脑是两个器官,它们在解剖学上相距甚远,但在其他方面却如此接近。我们过去的文章写过,肠道微生物组和大脑之间的代谢和神经串扰对大脑功能、情绪和行为具有重要意义。
详见:
在过去十年中,多项研究表明,全身性炎症与精神疾病(包括抑郁症和自闭症)之间存在关联。然而,全身炎症与酒精成瘾、抑郁症和自闭症等精神疾病之间相互作用的机制尚不完全清楚,肠道菌群可能是一个很好的研究目标。
全身性炎症和酒精成瘾的一个可能机制是肠道细菌产物激活外周血单核细胞,诱导细胞因子进入血流,在酒精依赖者中引起低度全身性炎症 。
肠道炎症还可以将内毒素和细胞因子传播到血液中,在那里它们可以进入中枢神经系统 (CNS),引起神经炎症。
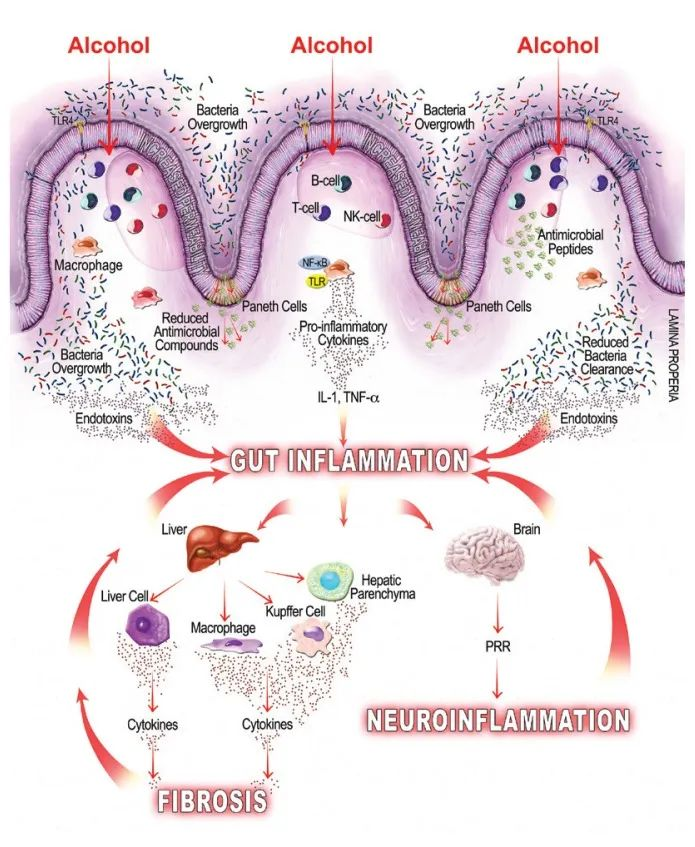
酒精成瘾与十二指肠的吸收障碍有关,导致营养不良和硫胺素的肝脏储存受损。
酒精成瘾与其他精神疾病高度相关,包括重度抑郁症、双相情感障碍以及焦虑症。重度抑郁症等情绪障碍通常先于酒瘾发作;例如,有人使用酒精来应对情绪低落。
酒精成瘾的严重程度与其渴望、认知功能障碍、焦虑和抑郁症状的强度相关
正如我们上面提到的,全身炎症可能在酒精成瘾的发展中起重要作用;肠道和肝脏的肠屏障功能障碍和炎症可能导致外周炎症并引起脑部炎症,从而诱发小胶质细胞或星形胶质细胞等脑细胞的炎症。
疾病行为理论可能将全身性炎症与酒精成瘾和情绪障碍联系起来。该理论支持外周炎症,如肠漏症,激活免疫系统并产生可到达大脑的细胞因子,导致发烧、疲劳、疲倦、无法集中注意力和退出社交;当上述行为持续存在时,可能会出现抑郁症状。
越来越多的研究将肠道微生物组的组成和功能与物质使用障碍 (SUD) 联系起来。
肠脑轴和物质使用障碍(SUD)
SUD 的特征是:尽管知道会对精神、身体和社会造成负面影响,但仍对某种物质(例如,酒精、阿片类药物和/或其他药物)长期依赖。
它们具有社会经济、生物化学、遗传以及越来越多的微生物学基础。众所周知,大脑是调节成瘾参数的重要器官,然而,通过微生物角度研究成瘾提供了一种思考:与身体相关的其他因素如何与大脑携手合作,以调解获得滥用药物的动机的新方法。
肠道和大脑通过双向、生化和神经高速公路(肠-脑轴)进行交流。肠上皮细胞下方的神经末梢接收来自肠道微生物群的代谢信号,这可能会影响行为,例如压力或焦虑。
除了与中枢神经系统发育和大脑功能有关的其他代谢物,如短链脂肪酸,肠道微生物还有助于产生一系列与情绪、认知和奖赏相关的神经递质(例如血清素和多巴胺)。
肠脑轴
图源:BioRender
这些神经递质在 SUD 的情况下特别相关;许多滥用物质通过触发大量多巴胺进入奖励途径来劫持大脑的奖励系统。这种多巴胺泛滥带来的愉悦感最终会减弱,个人可能会反复服用该物质以再次体验这些感觉。研究表明,肠道微生物参与自然(例如食物)和人工奖励(包括药物)的奖励感知,这表明 SUD 的发展/进展与肠道微生物组组成之间存在联系。
事实上,滥用酒精或药物与微生物组组成的变化有关。尽管这些改变的具体情况因物质而异,但与“健康”相关的微生物通常会减少,而具有促炎作用的微生物会增加,例如变形杆菌。这些变化伴随着关键微生物代谢物(如短链脂肪酸)的减少,具有各种全身和局部效应(例如,肠道屏障完整性的破坏)。
通常,SUD 的特征是肠道炎症增加,部分原因是这种渗漏的肠道屏障允许微生物及其产物与底层免疫细胞相互作用。激活后,这些免疫细胞会产生细胞因子,这些细胞因子不仅会引发局部炎症,还会进入循环并穿过血脑屏障。由此产生的神经炎症会改变神经元活动,包括在大脑的奖励通路内,并可能影响对物质本身的反应和耐受性。
管理物质使用障碍——肠道微生物
治疗 SUD 的策略因人和物质而异,但可能涉及药物(如阿片类拮抗剂)、咨询和行为护理。然而,这些策略可能并不总是奏效,并且可能会复发。目前药物成瘾治疗干预的成功率很低,大约40-60% 接受治疗的人最终会复发并重新使用滥用药物。
鉴于肠道微生物群和 SUD 之间出现的交叉点,发现用某些细菌补充肠道微生物群,它们的发酵产物(如短链脂肪酸)可能会减少滥用物质的影响。
例如,一项研究发现富含双歧杆菌的益生菌和Lactobacillaeae 可逆转小鼠对吗啡的耐受性。
粪便微生物群移植 (FMT) 也可能是一种选择,或许未来可以用于干预酒精成瘾或使用障碍。一项 1 期临床试验表明,与安慰剂组相比,接受富含毛螺菌科和瘤胃球菌科 FMT 的酒精使用障碍患者在 15 天后 对酒精的渴望有所减少(分别减少 90% 和 30%)。在吗啡依赖小鼠中,FMT减轻了戒断症状由阿片拮抗剂触发。由于阿片类药物耐受性容易导致剂量增加和用药过量,这些发现表明微生物可以延长药物的疗效。
以上表明肠道微生物组的未来及其对健康和福祉的影响有很大的应用,尤其是与 SUD 相关的情况。这是一个有前景的领域。
目前,酒精摄入过量的治疗干预措施主要包括药物治疗、心理治疗、社会支持等方面。以下是一些应对过度饮酒的可行性计划:
这里再介绍一些关于过量饮酒的饮食干预。
1. 豆腐
豆腐中所含的半脱氨酸是一种氨基酸,其重要性在于能解除乙醇的毒性,食用后能促进酒中乙醇的排出,从而达到解酒的目的。
2. 香蕉
香蕉还有解酒作用,因为香蕉中含有丰富的果糖,进入人体后可以稀释酒精,所以可以解酒。

3. 糖水
适量白糖和开水混合,甜度视醉度而定,稍凉片刻即可饮用。糖分进入人体后,会增加体内血糖浓度,降低血液中酒精浓度,加速体内酒精的排出,从而达到醒酒的目的。
4. 蜂蜜水
其实酒后喝点蜂蜜水可以有效减轻酒后头痛的症状,蜂蜜可以促进人体对酒精的吸收从而减轻酒后头痛的症状,此外,蜂蜜还有镇静催眠的作用. 多吃蜂蜜对皮肤和胃都有好处。

5.葛根水
葛根泡水喝,不管是喝前还是喝后,都有很好的效果。饮酒前先喝葛根水,可预防酒精对肝脏的损害。酒后饮用可起到醒酒作用。
6.番茄
番茄中的果糖、葡萄糖和维生素对肝脏和心脏都有保健作用,而且果糖还是解酒醒酒的好味道。酒后吃几个西红柿,可使头晕逐渐消失。
7.红薯
将生红薯磨碎,加入适量白糖,搅拌均匀后服用。
8.绿豆
绿豆适量,用温水冲净,捣烂,用沸水冲净或熬汤食用。
9.甘蔗
1 根甘蔗,去皮并榨汁。
10.盐水
在开水中加少许盐,喝下醒酒。
11.柑橘皮(陈皮)
将陈皮烘熟,研末,加食盐1.5克煮汤。
12.白萝卜
白萝卜1公斤,捣成泥取汁,分次服用。也可以在白萝卜汁中加入适量的红糖饮用,也可以生吃萝卜。

编辑
这些用食物解酒的方法虽然看似简单,但是却能够起到解酒作用。如果你也喜欢喝酒,经常喝醉,那么学习一些有效的解酒方法就显得非常重要了。对于喜欢喝酒的朋友而言,为了整体健康,请根据自身情况适度饮酒,尽可能避免过量饮酒。
总的来说,适量酒精可能给人体带来好处,尤其对自身免疫性疾病具有保护作用。低剂量酒精可能通过增加Akk菌等保护性肠道微生物,并有助于增加乙酸盐、多不饱和脂肪酸等,间接产生积极影响。
而高剂量酒精则可能带来不利影响,破坏肠道屏障的稳定性,增加肠道通透性,导致微生物成分进入循环系统,引发肝脏和其他器官的炎症反应;并可能导致肠道微生态失调、肠道菌群产物、脂多糖增加;导致单核细胞、T 细胞、细胞因子、IgG 水平以及 B 细胞减少;进而影响酒精性肝病、肥胖、胃肠道疾病、心血管疾病等发生和发展。
此外,饮酒还会影响肠道微生物组影响社交行为和精神障碍。因此,保持健康的肠道微生物组对于预防和治疗与饮酒相关的疾病具有重要意义。
未来的研究应该进一步探讨肠道菌群与酒精代谢和酒精成瘾之间的因果关系,肠道菌群的调节可能成为预防和治疗酒精成瘾的新靶点。
主要参考文献:
Jew MH, Hsu CL. Alcohol, the gut microbiome, and liver disease. J Gastroenterol Hepatol. 2023 Apr 25. doi: 10.1111/jgh.16199. Epub ahead of print. PMID: 37096652.
Meroni M, Longo M, Dongiovanni P. Alcohol or Gut Microbiota: Who Is the Guilty? Int J Mol Sci. 2019 Sep 14;20(18):4568.
Caslin B, Mohler K, Thiagarajan S, Melamed E. Alcohol as friend or foe in autoimmune diseases: a role for gut microbiome? Gut Microbes. 2021 Jan-Dec;13(1):1916278.
Zugravu CA, Medar C, Manolescu LSC, Constantin C. Beer and Microbiota: Pathways for a Positive and Healthy Interaction. Nutrients. 2023 Feb 7;15(4):844.
Engen PA, Green SJ, Voigt RM, Forsyth CB, Keshavarzian A. The Gastrointestinal Microbiome: Alcohol Effects on the Composition of Intestinal Microbiota. Alcohol Res. 2015;37(2):223-36.
González-Zancada N, Redondo-Useros N, Díaz LE, Gómez-Martínez S, Marcos A, Nova E. Association of Moderate Beer Consumption with the Gut Microbiota and SCFA of Healthy Adults. Molecules. 2020 Oct 17;25(20):4772
Crews FT, Sarkar DK, Qin L, Zou J, Boyadjieva N, Vetreno RP. Neuroimmune Function and the Consequences of Alcohol Exposure. Alcohol Res. 2015;37(2):331-41, 344-51.
González-Zancada N, Redondo-Useros N, Díaz LE, Gómez-Martínez S, Marcos A, Nova E. Association of Moderate Beer Consumption with the Gut Microbiota and SCFA of Healthy Adults. Molecules. 2020 Oct 17;25(20):4772.
Hernández-Quiroz F, Nirmalkar K, Villalobos-Flores LE, Murugesan S, Cruz-Narváez Y, Rico-Arzate E, Hoyo-Vadillo C, Chavez-Carbajal A, Pizano-Zárate ML, García-Mena J. Influence of moderate beer consumption on human gut microbiota and its impact on fasting glucose and β-cell function. Alcohol. 2020 Jun;85:77-94.
Doremus-Fitzwater TL, Deak T. Adolescent neuroimmune function and its interaction with alcohol. Int Rev Neurobiol. 2022;161:167-208.
Tirado-Kulieva VA, Hernández-Martínez E, Minchán-Velayarce HH, Pasapera-Campos SE, Luque-Vilca OM. A comprehensive review of the benefits of drinking craft beer: Role of phenolic content in health and possible potential of the alcoholic fraction. Curr Res Food Sci. 2023 Mar 4;6:100477.
Caslin B, Mohler K, Thiagarajan S, Melamed E. Alcohol as friend or foe in autoimmune diseases: a role for gut microbiome? Gut Microbes. 2021 Jan-Dec;13(1):1916278.

谷禾健康
胆汁酸是一种代谢胆固醇的物质,它具有两个亲和性不同的区域,即一部分能够与水分子相互作用(亲水),而另一部分则不能与水分子相互作用(疏水)。
由于拥有这种两亲性质,胆汁酸能够在生物体内与胆固醇等脂类物质结合并形成混合物,从而起到降低血液中胆固醇水平的作用。
胆汁酸不仅是代谢胆固醇的物质,还有其他重要的功能。胆汁酸可以通过激活肝脏和肠道中的受体,调节血糖水平、胆固醇代谢和免疫信号等。
其中,初级胆汁酸是由肝细胞合成并储存在胆囊中的,而次级胆汁酸则是由肠道中的微生物代谢初级胆汁酸形成的。
随着研究的深入,发现肠道菌群和胆汁酸在人体健康和疾病中发挥着重要的作用。肠道菌群可以代谢胆汁酸,影响其在体内的水平和作用,从而影响脂质代谢、肝胆功能和肠道健康等方面。
同时,胆汁酸也可以影响肠道菌群的组成和功能,维持肠道微生物的平衡和多样性。因此,肠道菌群和胆汁酸之间的相互作用关系对于人体健康和疾病的发生、发展和治疗具有重要的意义。
肠道微生物主要的代谢产物包括:短链脂肪酸、色氨酸、胆汁酸等。
以往我们写过短链脂肪酸(详见:你吃的膳食纤维对你有帮助吗?)、
色氨酸(详见:色氨酸代谢与肠内外健康稳态 )。
本文我们主要阐述胆汁酸及其代谢,与肠道微生物群之间的关联及其在疾病中的作用。
本文目录/contents
Part1:胆汁酸及其代谢
Part2:影响胆汁酸代谢的因素
Part3:肠道菌群与胆汁酸之间的相互作用
Part4:微生物群-胆汁酸相互作用的影响
Part5:微生物与胆汁酸在疾病中的作用
Part6:调节胆汁酸代谢的方法
Part7:结语
本文提及的专业名词缩写
FXR——法尼醇X受体,胆汁酸是FXR的天然配体,因此FXR又称为胆汁酸受体。
SHP——是LRH-1(肝脏受体同源物-1)在肝细胞激活的主要靶基因,胆汁酸通过LRH-1诱导SHP的表达,表达的SHP结合并失活LRH-1,从而抑制胆汁酸合成相关基因及SHP自身的表达。
TGR5——又称G蛋白偶联胆汁酸受体,是一种胆汁酸受体。在外周组织器官如肝、脾、肾、脂肪等多种组织中表达水平较高,能调控脂类和葡萄糖代谢。
GLP-1——胰高血糖素样肽-1是一种由肠道细胞所分泌的激素,属于一种肠促胰岛素。
FXR-Fgf15轴——回肠肠细胞中FXR的激活释放Fgf15(FGF19是人的同源基因),Fgf15通过门静脉循环到达肝细胞,与FGFR4结合并抑制CYP7A1,从而抑制肝细胞胆汁酸的合成。
当胆固醇在肝细胞内代谢时,会产生一种叫做胆汁酸的化合物。胆汁酸可以溶解脂肪,促进脂肪消化和吸收,同时还能帮助排泄体内多余的胆固醇。
胆汁酸一般可分为初级胆汁酸和次级胆汁酸
初级胆汁酸(primary bile acids):
初级胆汁酸是由肝细胞合成并储存在胆汁中的胆汁酸,它是胆汁中最主要的成分。初级胆汁酸的结构比较简单,含有羟基(-OH)和羧基(-COOH)等官能团,因此具有良好的水溶性。初级胆汁酸可以在肠道中与脂肪结合成胆汁酸盐,参与脂肪的乳化和吸收过程。
初级胆汁酸包括胆酸、鹅去氧胆酸、甘氨胆酸、牛磺胆酸、甘氨鹅去氧胆酸、牛磺鹅去氧胆酸。
次级胆汁酸(secondary bile acids):
次级胆汁酸是由肠道中的微生物代谢初级胆汁酸生成的,也称为微生物代谢产物。次级胆汁酸的结构更加复杂,它经过羟化、氧化、甲基化等反应后形成。次级胆汁酸相对于初级胆汁酸来说,具有更强的亲脂性和生物活性。
次级胆汁酸主要有去氧胆酸、石胆酸、甘氨去氧胆酸、牛磺去氧胆酸、甘氨石胆酸、牛磺石胆酸。
胆汁酸的结构有助于它们的功能。胆汁酸一般由一个甾醇核心组成,该核心由三个六元碳环和一个五元碳环组成,通常具有5β-氢和沿前两个稠合环平面的顺式结构。
一般胆汁酸的结构
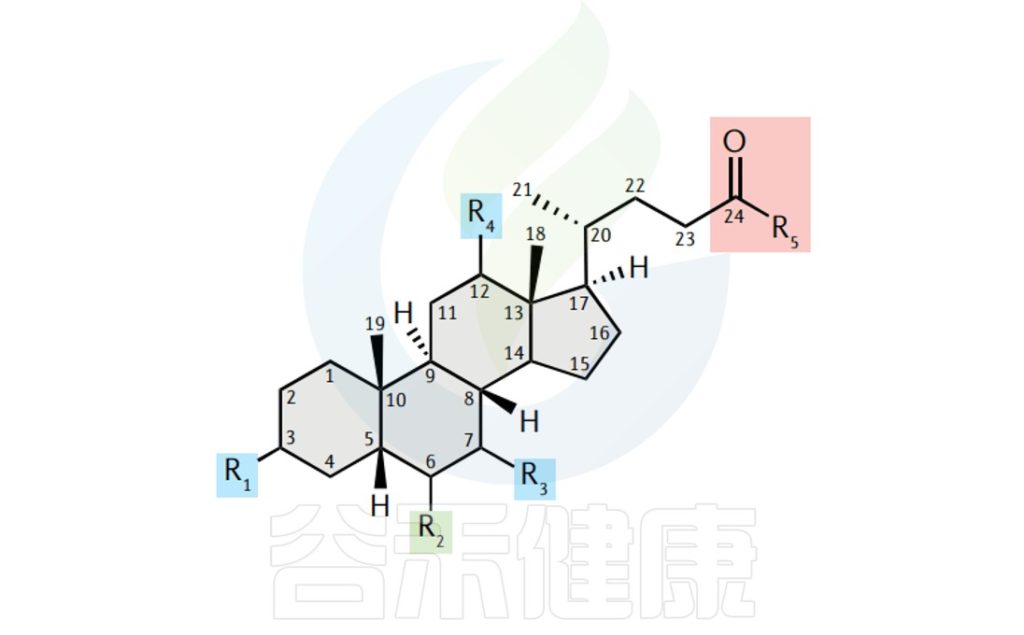
Collins SL,et al.Nat Rev Microbiol.2023
胆汁酸的种类由与其结合的羟基、羧基、硫酸根和氨基酸基团的数量和位置决定。羟基和羰基面向甾醇核心的同一侧,而甲基面向相反的一侧。这使得胆汁酸具有两亲特性,因为一侧是疏水的,而另一侧是亲水的。
胆汁酸的不同种类

Poland JC,et al.Physiology (Bethesda).2021
胆汁酸的疏水性取决于甾醇环上羟基和硫酸根的数量和位置,以及胆汁酸是否与氨基酸结合,在小鼠中主要是牛磺酸,在人类中主要是甘氨酸。
胆汁酸的肠肝循环是一个精细调节的过程。
初级胆汁酸由肝脏中的胆固醇通过胆固醇7α-羟化酶(CYP7A1)介导途径或固醇27-羟化酶(CYP27A1)介导途径合成。
然后初级胆汁酸与牛磺酸或甘氨酸结合,储存在胆囊中,然后在摄入食物后分泌到十二指肠中,以促进膳食脂质和脂溶性维生素的吸收。
95%以上的胆汁酸会被重吸收
排入肠道的各种胆汁酸约95%以上要被重吸收。回肠部的重吸收是主动重吸收,其余肠段为被动重吸收,并运回肝脏进行代谢回收。
注:虽然少量初级胆汁酸可以通过被动扩散吸收,但有效吸收需要由回肠上皮细胞中表达的顶端胆汁酸转运蛋白 (ASBT) 介导的主动转运。结合的初级胆汁酸主要由回肠胆汁酸结合蛋白 (IBABP) 通过肠细胞转运。
这个过程在人体中每天发生4到12次,并确保维持胆汁酸稳态。
人体胆汁酸的肠肝循环
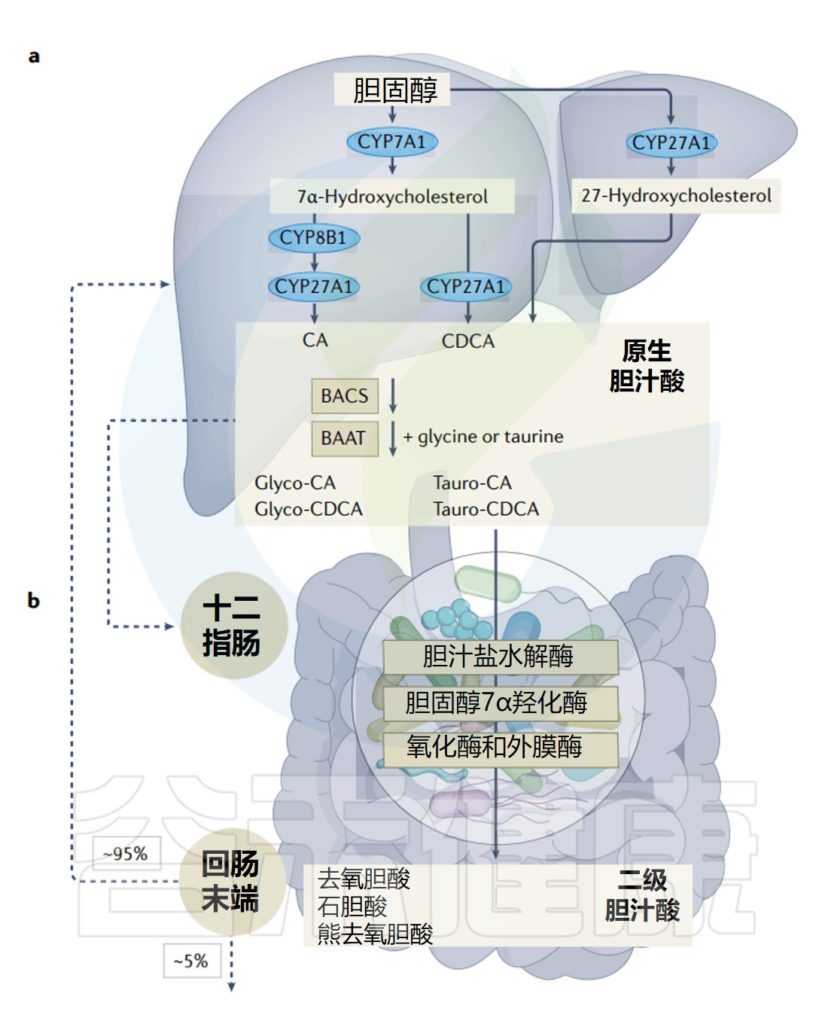
Collins SL,et al.Nat Rev Microbiol.2023
在肠道中,结合的初级胆汁酸受微生物群作用并转化为次级胆汁酸,从而进一步增加胆汁酸库的多样性和整体疏水性。
次级胆汁酸代谢的第一步是通过胆汁盐水解酶(BSH)水解氨基酸部分。胆汁盐水解酶在所有主要的肠道微生物(拟杆菌(Bacteroidetes)、厚壁菌(Firmicutes)和放线菌(Actinobacteria))中都是高度保守的,但由于它们对甘氨酸偶联或牛磺酸偶联的胆汁酸具有优先活性,因此在细菌之间是不同的。
✦细菌对胆汁酸的代谢一般是有益的
细菌胆汁酸去偶联对一般细菌有益,因为它们从氨基酸和宿主那里获得能量,它降低了胆汁酸的毒性。然而,对于胆汁盐水解酶是否对细菌有益存在一些争论。
细菌其他的作用包括氧化,脱硫,酯化和偶联。将在本文后面的章节具体展开描述。
从分类上看,多种细菌都能在体外将氨基酸与胆汁酸结合,其中双歧杆菌(Bifidobacterium)、拟杆菌(Bacteroides)和肠球菌(Enterococcus)的结合量最大。
微生物生产者的胆汁酸代谢

Cai J,et al.Cell Host Microbe.2022
药物、运动、饮食或其他不良状态导致肠道菌群组成或活性的改变都会扰乱胆汁酸代谢。
影响细菌转化胆汁酸的因素
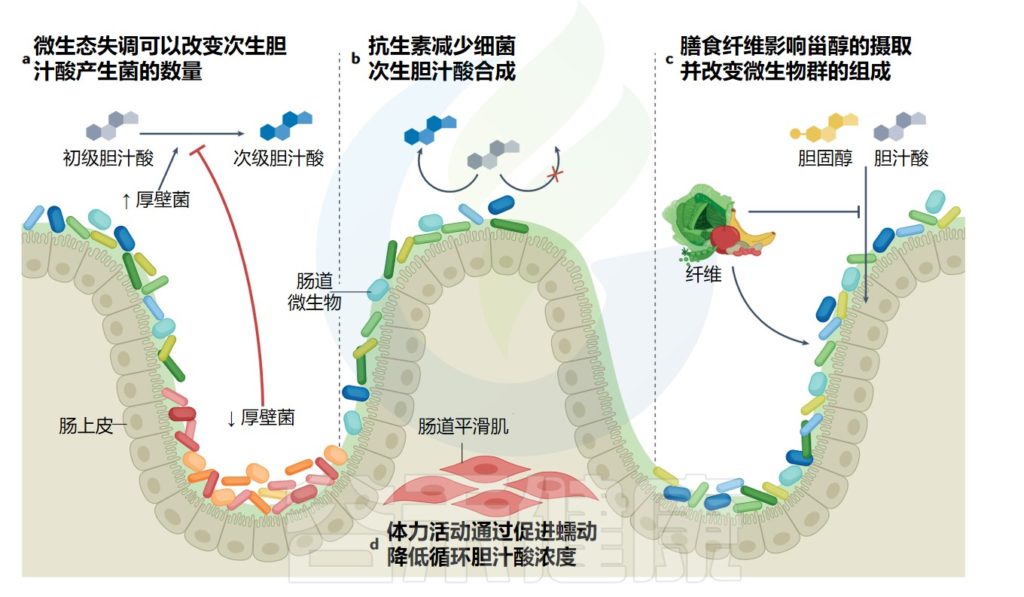
Collins SL,et al.Nat Rev Microbiol.2023
肠道微生物可以代谢胆汁酸,影响其在体内的水平和作用。肠道微生物失调可能会导致胆汁酸代谢紊乱,从而影响脂质代谢、肝胆功能和肠道健康等方面。
具体来说,肠道微生物失调可能会导致以下影响:
•胆汁酸合成减少:肠道微生物可以参与胆汁酸的合成过程,肠道微生物失调可能会导致胆汁酸合成减少,从而影响胆汁酸代谢。
•胆汁酸代谢紊乱:肠道微生物可以代谢胆汁酸,影响其在体内的水平和作用。肠道微生物失调可能会导致胆汁酸代谢紊乱,从而影响脂质代谢、肝胆功能和肠道健康等方面。
•肠道屏障功能受损:肠道微生物失调可能会导致肠道屏障功能受损,从而影响肠道对胆汁酸的吸收和代谢。
•炎症反应增加:肠道微生物失调可能会导致肠道炎症反应增加,从而影响胆汁酸代谢和肠道健康。
因此,保持肠道微生物的平衡和稳定,对于维持胆汁酸代谢的正常和健康具有重要的意义。
抗生素等药物对胆汁酸有着巨大的影响。抗生素治疗对宿主破坏最严重的代谢途径之一是胆汁酸代谢。
•影响次生胆汁酸的水平
随着细菌的耗竭,可用于解除宿主胆汁酸结合的胆汁盐水解酶减少,次级胆汁酸也不再产生。因此,使用抗生素,可观察到共轭胆汁酸和一些次生胆汁酸水平下降。
其他药物,如抗抑郁药物帕罗西汀,也可以扰乱胆汁酸水平。
次级胆汁酸的损失进一步加剧了胆汁酸的失调,因为胆汁酸的生物合成被上调,导致初级胆汁酸库更大。
几项研究将锻炼与不同的胆汁酸联系起来,但结果存在一些矛盾。
•适度运动可增加胆汁酸的排泄
啮齿类动物研究发现,由于胃肠运动或胆固醇摄取增加,适度运动可增加胆汁酸排泄。然而,令人惊讶的是,这些啮齿动物的胆汁酸合成和信号传导不受运动的影响。
•运动可使循环胆汁酸总体减少
在人类中,持续时间的增加和运动的一致性导致血清和粪便胆汁酸的总体减少。这对结直肠癌和非酒精性脂肪性肝病具有意义,其中循环胆汁酸增加与负面结果相关。
注:除了宿主胆汁代谢外,体育活动还可以逆转非酒精性脂肪性肝患者的微生物群失调,这可能导致不同的次生胆汁酸水平。
此外,对运动训练有生理适应的长跑运动员具有较少的诱变性次生胆汁酸。需要更多的研究来阐明体育活动对肠道和肝脏中胆汁酸的影响。
人们早就知道,饮食,尤其是脂肪和纤维的摄入,可以极大地改变微生物群和胆汁酸代谢。
•胆固醇摄入多会提高胆汁酸总体水平
摄入胆固醇会提高人体内胆汁酸的总体水平,因为胆固醇是胆汁酸的前体,并上调胆汁酸的合成途径。
在高脂肪饮食的人群中,由于表达7α-去羟化酶和表达胆汁酸水解酶的细菌数量的增加,次生胆汁酸和非共轭胆汁酸水平特别高。
•纤维诱导胆汁酸的排泄、抑制其吸收
相比之下,膳食纤维可以在胃肠道消化过程中通过吸附作用促进胆汁酸的排泄以及抑制胆汁酸在肠道的吸收。
高脂肪、低纤维饮食的胆汁酸特性与结肠癌患者有关。最近的一篇文章表明,小鼠的热量限制减少了总胆汁酸,包括石胆酸和去氧胆酸,也减少了总微生物群的含量。
•肝胆系统疾病会导致胆汁酸代谢紊乱
肝胆系统疾病如肝硬化、胆囊疾病等可能会影响胆汁酸的合成和代谢,导致胆汁酸代谢紊乱和相关疾病的发生。
肝细胞受到了损伤,就会出现代谢问题,导致患者的总胆汁酸偏高。
•孕妇的总胆汁酸会偏高
怀孕期间孕妇体内的孕激素水平改变会引起胆汁酸代谢异常,从而使得孕妇总胆汁酸偏高
一些遗传因素也可能会影响胆汁酸代谢,特定的宿主基因变异会影响胆汁酸代谢和肠腔内的胆汁酸含量,导致相关疾病的发生。
▼
人类与微生物群的交流部分依赖于宿主受体对微生物代谢物的反应。微生物群通过几种关键的宿主胆汁酸受体调节胆汁酸的代谢和转运。
肠道菌群调节胆汁酸主要体现在生物转化、生物合成和胆汁酸的运输。
胆汁酸信号传导和网络
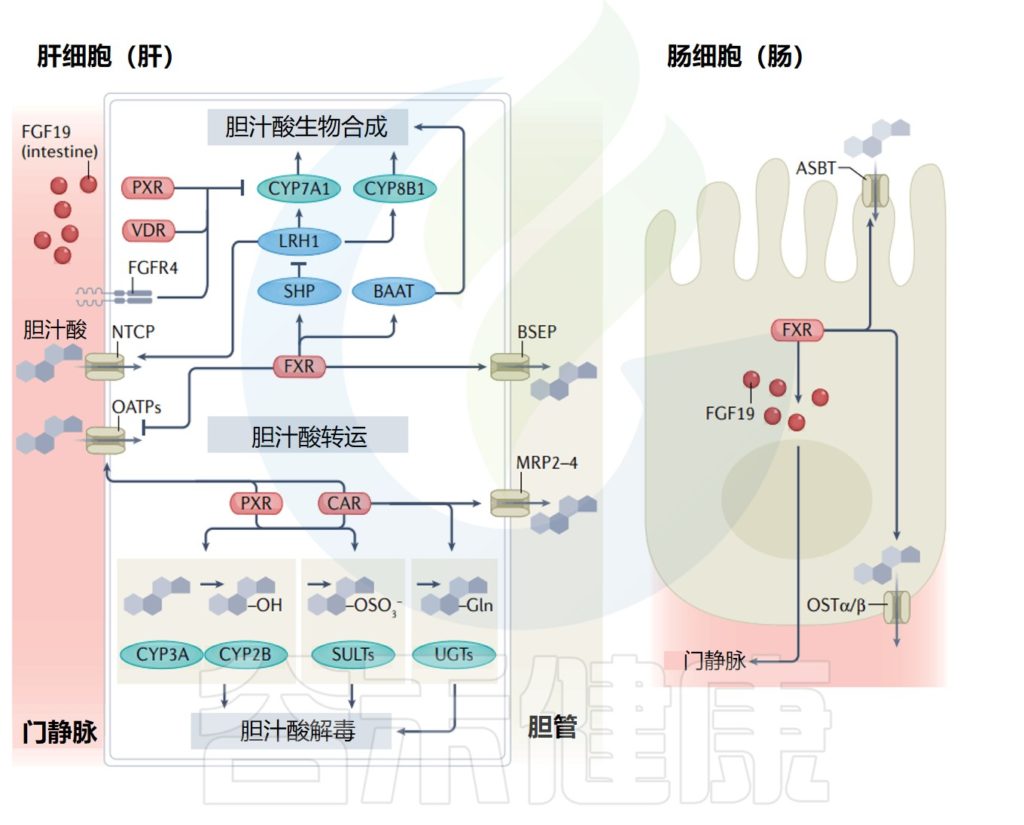
Collins SL,et al.Nat Rev Microbiol.2023
1
肠道菌群调节胆汁酸的生物转化
胆汁酸在肝脏中合成并通过胆道系统释放到肠道,部分释放到远端回肠或结肠,在那里它们被肠道菌群进一步代谢。
✦影响胆汁酸生物毒性和肠道吸收
肠道微生物可以改变胆汁酸,因为它们具有不同的 胆汁酸代谢酶。肠道微生物群通过去结合、7ɑ-脱羟基、异构化、氧化、脱硫和酯化来调节胆汁酸的化学多样性,进而影响其生物毒性和肠道吸收。
去共轭和7ɑ-脱羟基是两个值得注意的反应。
主要通过胆汁酸水解酶进行的微生物去结合是肠道环境中微生物进一步修饰胆汁酸的关键一步。
胆汁酸水解酶编码基因已在各种肠道微生物中检测到并表征,包括双歧杆菌(Bifidobacterium)、乳杆菌(Lactobacillus)、肠球菌(Enterococcus)、梭菌(Clostridium)、拟杆菌(Bacteroides)等。
✦小部分细菌通过脱羟基作用转化胆汁酸
已经表明,怀孕期间分泌胆汁酸水解酶的拟杆菌丰度增加会促进胆汁酸解偶联,从而减少肠细胞胆汁酸的摄取。
胆盐水解酶去结合的胆汁酸随后通过肠道菌群产生的7ɑ-脱羟基作用脱水,并最终转化为次级和三级胆汁酸。
然而,7α-脱羟基作用仅由少数厌氧物种进行,目前的估计表明,只有大约0.0001%的结肠细菌可以进行这种反应,占肠道微生物群总数的不到0.025%。
16S rRNA 序列分析表明,这些细菌主要是梭菌属,包括C.hiranonis、C.scindens、C.hylemonae(簇 XIVa)和C.sordelli(簇 XI)。
在大肠中,梭菌属物种可以通过7α-脱羟基作用将胆酸和鹅去氧胆酸分别转化为去氧胆酸和石胆酸。
✦氧化反应导致差向异构化
氧化和环氧化在胆汁酸的微生物修饰和代谢反应中也受到特别关注,一些肠道微生物合成能够进行可逆氧化还原反应和羟基环氧化的羟基类固醇脱氢酶 (HSDH)。
羟基类固醇脱氢酶(HSDH)活性存在于肠道微生物群的四个主要类别中:
放线菌门(Actinobacteria);
变形菌门(Proteobacteria);
厚壁菌门(Firmicutes);
拟杆菌门(Bacteroidetes)。
这些氧化反应最终会导致差向异构化。然而,由于缺乏合适的分析方法,人们对微生物通过羟基类固醇脱氢酶调节胆汁酸异构化反应的机制知之甚少,但这些反应性胆汁酸的疏水性和毒性是无可辩驳的。
✦肠道微生物的破坏导致胆汁酸代谢紊乱
2019年进行的一项研究支持肠道微生物群的破坏会导致胆汁酸代谢紊乱。这项研究发现,在接受各种抗生素治疗的大鼠的血浆和粪便中,牛磺酸结合的胆汁酸显著增加,而游离胆汁酸减少,胆汁酸的多样性也显著降低。
一项较早的研究还指出,肠道微生物群对胆汁酸有显著影响。无菌小鼠胆汁酸的多样性减少,并且缺乏未结合的和次级胆汁酸。
注:与正常小鼠相比,盲肠、结肠和粪便中的胆汁酸水平降低,但胆囊和小肠中的胆汁酸水平升高。
✦微生物群影响胆汁酸受体信号
实验研究进一步证明了微生物群影响FXR信号。当用tempol处理小鼠时,减少了乳酸杆菌属。及其胆汁酸水解酶改变肠道微生物群的活性,导致厚壁菌门:拟杆菌门比率降低,次级胆汁酸降低,以及牛磺酸-β-鼠胆酸 (T-β-MCA) 积累。
Tempol是一种超氧化物歧化酶(SOD)类似物,可有效中和活性氧。
虽然次级胆汁酸是有效的FXR激动剂,而T-β-MCA是FXR拮抗剂,但抑制胆汁酸受体信号会导致胆汁酸合成增加和胆汁酸库大小增加。
但由于参与胆汁酸脱氢的梭状芽孢杆菌簇XI和XVIa的减少,胆汁酸多样性下降。脱硫弧菌(Desulfovibrionales)也可以逆转这一过程,脱硫弧菌负责从饮食和宿主来源代谢含硫化合物。富含脱硫弧菌的肠道微生物群可以调节胆汁酸的代谢,从而使肠道产生更多的次生胆汁酸。
2
肠道菌群影响胆汁酸的生物合成
✦影响关键酶的表达从而调节胆汁酸的合成
肠道微生物群对胆汁酸形成的调节是复杂的,包括由至少17种不同酶催化的几个反应步骤。然而,更重要的是,肠道微生物群对胆汁酸的代谢会影响许多参与胆汁酸从头合成的关键酶的表达,包括CYP7A1、CYP7B1、CYP8B1和CYP27A1。
在厚壁菌中,可以通过FXR-FGF15/19 反馈机制调节胆汁酸的合成。在肝细胞中,通过来自 SHP/LRH-1/LXRα 的负反馈调节胆汁酸合成。
SHP是LRH-1在肝细胞激活的主要靶基因,胆汁酸通过LRH-1诱导SHP的表达,表达的SHP结合并失活LRH-1,从而抑制胆汁酸合成相关基因及SHP自身的表达。
抑制过程
一旦肝脏FXR被激活,SHP就会立即被触发以抑制LRH转录,从而抑制CYP7A1和CYP8B1转录。在肠道细胞中,FXR通过FXR/FGF19/FGFR4通路抑制胆汁酸合成,其中FXR诱导FGF19/FGF15,后者又与FGFR4和β-klotho复合物结合,触发 MAPK/ERK1/2 通路并最终抑制该基因CYP7A1在肝脏中的表达。
✦没有细菌的情况下无法产生胆汁酸受体激活剂
研究证实肠道在没有细菌的情况下无法产生胆汁酸受体激活剂。肠道微生物群可以通过降低T-MCA水平和促进回肠中FXR依赖性FGF15的表达来抑制CYP7A1和胆汁酸的合成,从而表明肠道微生物群通过FXR-FGF15/19反馈机制调节胆汁酸的合成。
注:正如已经提到的,脱硫弧菌(Desulfovibrionales)衍生的硫化氢除了有利于含7α-脱羟基细菌的生长外,还可以诱导肝脏 FXR 并抑制CYP7A1表达和胆汁酸合成。
3
肠道菌群调节胆汁酸的运输
微生物群还可以通过法尼醇X受体(FXR)反馈机制调节胆汁酸运输。
✦通过影响法尼醇X受体调节胆汁酸的重吸收
肝脏中FXR的激活诱导胆盐输出泵(BSEP)、MRP、OSTα和OSTβ复合物以增强胆汁酸的肝脏消除,同时通过抑制Na+/牛磺胆酸盐对肝细胞中基底外侧NTCP和OATP1B1和OATP1B3的下调来减少胆汁酸重吸收。
肠道中的法尼醇X受体激活上调肠道胆汁酸结合蛋白 ,以促进胆汁盐通过回肠肠细胞,并增强OSTα和 OSTβ表达以帮助胆汁酸从肠道进入门脉循环。
OSTα/β——有机溶质转运蛋白,一种表达在肠上皮细胞基底膜的异二聚体蛋白,主要负责转运胆汁酸入静脉血。
此外,法尼醇X受体可以通过下调ASBT,通过SHP和FGF15/19通路调节肠细胞和胆管细胞对胆汁酸的重吸收。
ASBT——顶端膜钠依赖性胆汁酸转运蛋白
▼
肠道中的胆汁酸被微生物群进一步代谢,进而影响微生物组成。胆汁酸已被证明对肠道微生物群有直接和间接的影响。
胆汁酸是微生物群丰度、多样性和代谢活性的重要决定因素。
1
破坏了胆汁酸不耐受细菌的结构
研究表明,较高的胆汁酸浓度表现出抗菌活性,当胆汁酸不耐受细菌暴露于高浓度胆汁酸时,胆汁酸 会溶解磷脂并分离内在的膜蛋白,导致细胞膜被完全破坏,导致细胞内物质溢出。
除了膜损伤外,胆汁酸还通过干扰RNA二级结构、破坏大分子的稳定性、引起DNA损伤和促进蛋白质错误折叠,从而破坏肠道微生物的组成,从而表现出直接的抗菌活性。
在胆汁酸喂养的大鼠中,厚壁菌门的丰度显著增加,丹毒丝菌(Erysipelotrichi)和梭状芽胞杆菌(Clostridia)中的一些微生物也增加了。
✦胆汁酸的含量对不同细菌作用不同
一般来说,胆汁酸池的减少似乎有利于革兰氏阴性细菌的生长,这些细菌能够产生脂多糖,其中一些具有致病潜力。
相比之下,随着胆汁酸池的增加,观察到革兰氏阳性厚壁菌门的生长,从而促进了次级胆汁酸的产生。
2
影响微生物群的多样性及功能
尽管它们具有毒性作用,但胆汁酸也支持微生物群的多样性。在人体中,牛磺-β-鼠胆酸和牛磺酸胆酸对多种微生物的发展至关重要。
✦胆汁酸的分泌可以提供能量支持微生物多样性
虽然尚未证实其机制,但胆汁酸的分泌可能提供足够的能量来支持大量微生物的多样性。除了成分的改变,胆汁酸还改变了微生物群的功能能力。
例如,去氧胆酸、牛磺胆酸和牛磺酰脱氧胆酸会破坏小鼠中细菌的核苷酸和碳水化合物代谢。
3
抑制细菌的过度生长
研究发现小鼠肠道微生物群落过度增殖和细菌易位,通过使用口服胆汁酸,以诱导法尼醇X受体激活,从而抑制细菌过度生长。
✦胆汁酸受体诱导抗菌肽和宿主免疫反应
这是因为法尼醇X受体可以诱导参与肠道保护的基因,法尼醇X受体诱导抗菌肽产生和宿主免疫反应调节来塑造微生物组。
在回肠中缺乏法尼醇X受体受体表达细胞的小鼠也表现出肠道微生物数量的变化,拟杆菌和厚壁菌的数量分别增加和减少。其主要原因是法尼醇X受体敲低后胆汁酸的合成增加,这表明胆汁酸可以抵消法尼醇X受体信号通路,从而影响肠道微生物的组成。
小结
一般来说,肠道微生物群和胆汁酸之间存在双向相互作用。
肠道微生物群可以调节胆汁酸的合成和代谢,相反,胆汁酸可以改变肠道菌群的组成。
因此,维持胆汁稳态对肠道微生态非常重要。然而,还需要更多的研究来了解胆汁酸对肠道微生物群的功能操作如何影响宿主-微生物群的交流。
微生物群和胆汁酸之间的相互作用会影响肠道屏障功能的维持,调节先天免疫和适应性免疫,并调节定植抵抗力。
胆汁酸对宿主细胞的影响主要由膜相关和胆汁酸受体介导,包括法尼醇X受体(FXR)、G蛋白偶联受体5(TGR5)、孕烷X受体(PXR)和维生素D受体(VDR)。
FXR和TGR5在肝脏、回肠远端和结肠、上皮细胞、内皮细胞和免疫细胞中高度表达。除了在调节胆汁酸合成中的作用外,这两种受体对于维持肠道屏障完整性和限制炎症都是必不可少的。
注:胆汁酸激活TGR5的能力不同,顺序为石胆酸>去氧胆酸>鹅去氧胆酸>熊去氧胆酸>胆酸。
肠上皮细胞形成紧密连接的能力对于肠屏障的形成和维持至关重要。几项研究支持胆汁酸在调节紧密连接功能中的作用。
胆汁酸介导的肠道屏障功能调节
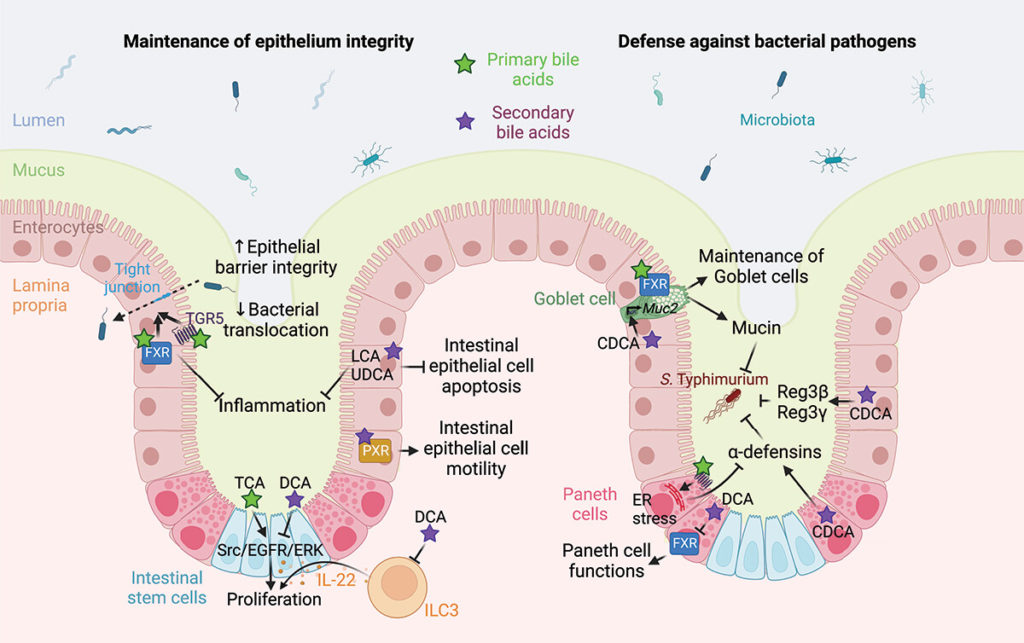
Larabi AB,et al.Gut Microbes.2023
▷肠道通透性与胆汁酸浓度有关
在喂食高脂肪饮食的小鼠和大鼠中,肠道通透性增加和紧密连接蛋白表达减少与盲肠和血浆胆汁酸浓度的改变有关,总胆汁酸库和次级胆汁酸增加。
法尼醇X受体缺失会增加肠道通透性
胆汁酸对肠上皮完整性的调节是由其激活受体的能力介导的。在小鼠和大鼠胆管结扎胆汁流阻塞模型中,法尼醇X受体的缺失增加了肠道通透性和细菌移位,并降低了紧密连接蛋白的表达。
同样,在化学诱导的结肠炎小鼠模型中,法尼醇X受体的激活限制了上皮屏障的通透性并防止了肠道炎症。
法尼醇X受体在肠上皮稳态中的作用是通过FGF蛋白介导的。补充喂食去氧胆酸的小鼠会出现生态失调,这会减少胆汁酸去结合,从而限制FXR-FGF15轴的激活并损害粘膜屏障功能。
G蛋白偶联受体5缺陷小鼠肠道通透性增加
此外,与野生型小鼠相比,G蛋白偶联受体5缺陷小鼠表现出紧密连接的表达改变、肠道通透性增加并且更易患化学诱导的结肠炎,表明这种胆汁酸受体在维持肠道屏障中的作用。
▷诱导上皮细胞增殖
胆汁酸还诱导肠上皮细胞增殖并限制细胞凋亡。在小鼠中,次级胆汁酸石胆酸和熊去氧胆酸可防止肠道炎症并限制上皮细胞凋亡。胆汁酸通过作用于肠道干细胞中的G蛋白偶联受体5受体来促进上皮再生。
牛磺胆酸诱导肠上皮细胞增殖
在体外,化学激动剂对法尼醇X受体的刺激增加了肠上皮细胞的运动和伤口闭合。牛磺胆酸通过表皮生长因子受体(EGFR)和胞外调节蛋白激酶(ERK)激活在体外诱导肠上皮细胞增殖,而去氧胆酸通过法尼醇X受体依赖性机制抑制细胞增殖。
最后,高脂肪饮食诱导的去氧胆酸增加通过减少3型先天淋巴细胞的数量来减少肠道干细胞的增殖和分化,从而减少潘氏细胞和杯状细胞。
▷调节粘液层的形成和组成
胆汁酸还调节粘液层的形成和组成,粘液层由浸泡在抗菌药物(如防御素)中的粘蛋白组成。
补充鹅去氧胆酸粘蛋白转录增加
在化学诱导的结肠炎小鼠模型中,法尼醇X受体的激活可防止产生粘蛋白的杯状细胞的损失。补充鹅去氧胆酸饮食的小鼠表现出潘氏细胞α-防御素的表达增加,杯状细胞Muc2(粘蛋白2-编码基因)的转录增加,回肠上皮C型凝集素Reg3β和Reg3γ的合成增强。
次级胆汁酸去氧胆酸具有刺激作用,而熊去氧胆酸在体外抑制人β-防御素-1和β-防御素-2的表达和分泌,这可能对维持肠道稳态也有影响。
微生物群产生的胆汁酸调节免疫的不同方面,包括诱导炎症基因募集先天性和适应性免疫细胞。
微生物衍生的胆汁酸调节肠道先天性和适应性免疫
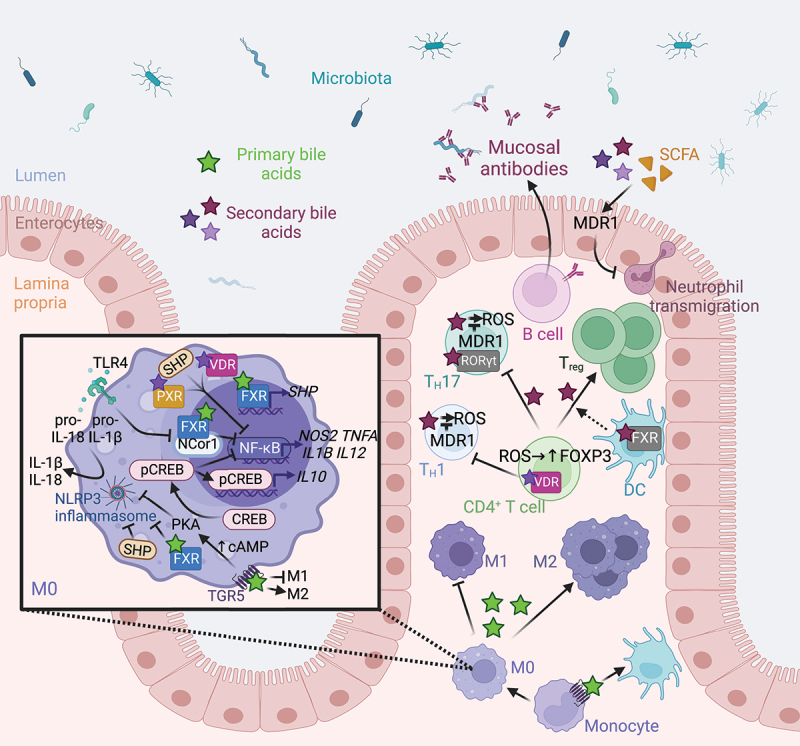
Larabi AB,et al.Gut Microbes.2023
▷调节促炎基因表达
胆汁酸受体FXR、TGR5和PXR调节促炎基因表达。在化学诱导的结肠炎小鼠模型中,FXR缺陷恶化,而FXR激动剂奥贝胆酸治疗可防止粘膜炎症并促进抗菌基因的表达。
胆汁酸受体影响促炎基因的表达
此外,法尼醇X受体可以通过直接结合促炎基因的启动子,以不依赖于SHP的方式调节促炎基因的表达。
病原体相关分子模式激活Toll样受体4会导致促炎基因启动子释放NCor1,从而激活其转录。最后,孕烷X受体和维生素D受体直接抑制NF-κB信号,从而减少促炎反应。
▷限制炎性体的产生
胆汁酸也限制炎症小体的激活。FXR和SHP通过与NLRP3炎症小体和胱天蛋白酶-1的物理相互作用来抑制炎症小体的组装,而TGR5环磷酸腺苷(cAMP)途径的激活通过诱导其泛素化来阻断NLRP3炎性小体的激活,这最终限制了白细胞介素-1β和白细胞介素-18的产生。
施用胆汁酸可减轻部分炎症
将去氧胆酸和石胆酸直肠给药于各种小鼠结肠炎模型可减轻炎症,部分是通过作用于TGR5受体。因此,溃疡性结肠炎患者体内菌群失调引起的次级胆汁酸缺乏可能会促进炎症,而炎症可以通过恢复次级胆汁酸水平得到缓解。
相反,另一项研究报告称,结肠中的去氧胆酸给药可部分通过刺激组织蛋白酶B释放来激活 NLRP3 炎性体,这会增加巨噬细胞分泌白细胞介素-1β并加剧结肠炎。
鉴于结肠炎模型中次级胆汁酸的相反作用,需要进一步的研究来更好地了解其在肠道炎症中的作用。
▷影响免疫细胞的募集和分化
胆汁酸还指导各种免疫细胞的募集和分化。与野生型小鼠相比,法尼醇X受体缺陷小鼠在结肠炎期间表现出炎症细胞募集减少。
接受鹅去氧胆酸补充剂的小鼠表现出向肠粘膜募集的单核细胞、巨噬细胞和嗜中性粒细胞减少,其中鼠伤寒沙门菌(S.Typhimurium)和柠檬酸杆菌(C.rodentium)感染期间B细胞的相对数量增加。
最近的研究表明,次级胆汁酸的氧代、异代和同种异体调节T细胞分化。
在体内,微生物群衍生的IsoDCA通过限制法尼醇X受体活性增加树突状细胞的免疫刺激特性,从而间接促进结肠调节性T细胞的分化。
▷通过抑制中性粒细胞迁移限制肠道炎症
暴露于固有层中的胆汁酸会驱动效应细胞TH1和TH17中的氧化应激。效应T细胞在迁移到回肠时通过上调外源性转运蛋白多药耐药蛋白1(MDR1,也称为P-糖蛋白)的表达来适应,以限制胆汁酸驱动的氧化应激。
胆汁酸与短链脂肪酸协同作用限制了肠道炎症
胆汁酸和微生物群衍生的短链脂肪酸协同作用,调节MDR1的表达,抑制中性粒细胞迁移,从而限制肠道炎症。
总之,肠道微生物组产生次级胆汁酸的能力是调节炎症以及先天和适应性免疫细胞募集、分化和激活的重要因素。
相反,适应性免疫调节微生物群和次级胆汁酸的产生。因此,维持这些因素之间的平衡对于维持肠道稳态是必要的。
微生物群通过竞争资源和产生限制细菌生长的代谢产物,如短链脂肪酸,来保护细菌免受机会性感染。
微生物群衍生的限制细菌生长的代谢产物可以被视为选择最适合环境的代谢特征的栖息地过滤器。微生物群通过与宿主合作来防止机会病原体在肠道定植。
微生物群介导的胆汁酸代谢和对病原体的防御
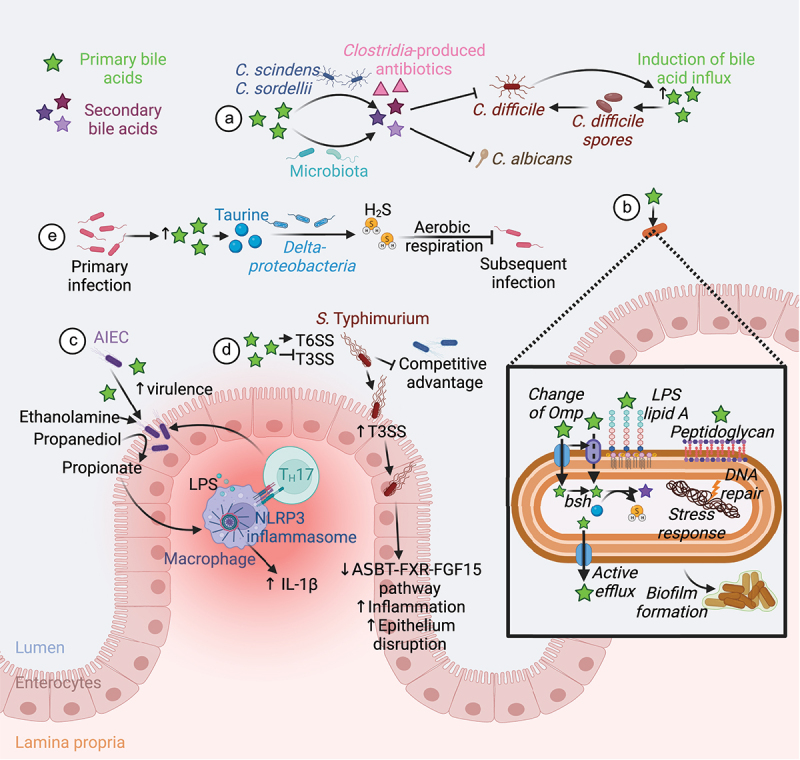
Larabi AB,et al.Gut Microbes.2023
▷增强了对病原体的防御
微生物群对胆汁酸的调节在保护宿主免受致病性感染方面发挥作用。
在人类中,肠道微生物群中胆汁酸水解酶的丰度和活性越高,通过降解激活病原体毒力基因表达的牛磺胆酸,对霍乱弧菌(Vibrio cholerae)感染的抵抗力就越强。
次级胆汁酸对艰难梭菌具有定值抗性
将初级胆汁酸转化为次级胆汁酸的共生细菌提供了对艰难梭菌(C. difficile)的定植抗性。初级胆汁酸可诱导艰难梭菌孢子萌发,而次级胆汁酸对营养细胞有毒。
编码操纵子的共生梭菌通过产生抑制艰难梭菌萌发、生长和毒素产生的次级胆汁酸来保护艰难梭菌不受感染。
石胆酸和去氧胆酸对白色念珠菌具有抵抗性
在体外,次级胆汁酸石胆酸和去氧胆酸对白色念珠菌具有直接的抗真菌活性。相反,通过改变微生物群组成和减少肠道单核吞噬细胞和TH17细胞的数量,向抗生素治疗的小鼠施用牛磺胆酸会加剧白色念珠菌的定植和传播。
因此,微生物群对胆汁酸的作用调节了定植抗性,直接通过抑制病原体生长或间接通过调节粘膜固有和适应性反应。
▷作为致病菌和病原体的环境信号
由于胆汁酸及其代谢产物是肠道环境的关键特征,许多机会性病原体利用这些线索来调节肠道定植所需的毒力因子的表达。
粘附侵袭性大肠杆菌(AIEC) 是一种独特的常驻粘膜相关病理细菌,克罗恩病患者中富含这种细菌。利用特定的肠道环境来增加它们的复制并诱发炎症。
在管腔中,胆汁酸促进AIEC毒力基因的表达,例如有利于细菌在肠道中持久存在的鞭毛蛋白FliC,以及促进细菌与潘氏斑相互作用和生长的长极性菌毛LpF。
此外,胆汁盐的存在激活了次级代谢途径,使AIEC能够使用乙醇胺作为氮源,使用丙二醇作为碳源,从而赋予这些菌株相对于其他共生细菌的竞争优势。但也与脂多糖协同作用,触发白细胞介素-1β的产生和TH17细胞的激活,从而促进T细胞依赖性肠道炎症。
影响病原菌的毒力和运动性
鼠伤寒杆菌(S.Typhimurium)暴露于胆汁会增加其VI型蛋白分泌系统(T6SS)的活性,将具有抗菌活性的效应蛋白输送到邻近细胞,从而杀死共生细菌并成功地在肠道定植。
在霍乱弧菌(V. cholerae)中,初级胆汁酸增加了毒力和运动性。艰难梭菌(C. difficile)在宿主定植过程中诱导胆汁酸快速流入肠道,这有助于孢子萌发和生长。
▷会记住过去的感染,以便在未来更好地抵抗
肠道病原体感染会破坏胆汁酸的回肠吸收和胆汁酸产生的内分泌调节。
表现出更强的定值抗性
实验小鼠在感染假结核耶尔森菌(Y.pseudotuberculosis)数周后,表现出对肺炎克雷伯菌(Klebsiella pneumoniae)更强的定植抗性。
潜在的机制是感染假结核耶尔森菌(Y.pseudotuberculosis),一种侵入回肠派尔氏斑块的肠道病原体,增加了肠道微生物群中Deltaproteobacteria的丰度。
注:Deltaprotoebacteria是一类代谢胆汁酸衍生的牛磺酸的细菌。
Deltaproterobacteria摄入牛磺酸会导致硫化氢的释放,可以通过有氧呼吸抑制肺炎克雷伯菌生长。
Deltaproteobacteria丰度的增加也增强了对柠檬酸杆菌(C.rodentium)的定殖抗性,是一种需要氧气才能在肠道环境中生长的病原体。
这些研究表明微生物群衍生的硫化氢限制了兼性厌氧机会性病原体的生长。
注意
尽管结肠中胆汁酸浓度的增加可能有利于增强定植抵抗力,但胆汁酸浓度过高与炎症性疾病和结直肠癌有关。
因此,需要对胆汁酸代谢进行精细调节,以增强定植抵抗力,同时限制对宿主的有害影响。
肠道菌群和胆汁酸在人体健康和疾病中发挥着重要的作用。肠道菌群可以代谢胆汁酸,影响其在体内的水平和作用,从而影响脂质代谢、肝胆功能和肠道健康等方面。
同时,胆汁酸也会影响艰难梭菌感染、癌变与细胞周期失调。下面谷禾列举了胆汁酸在影响人类疾病中的一些例子。
宿主和微生物群依赖性胆汁酸对人类疾病的影响
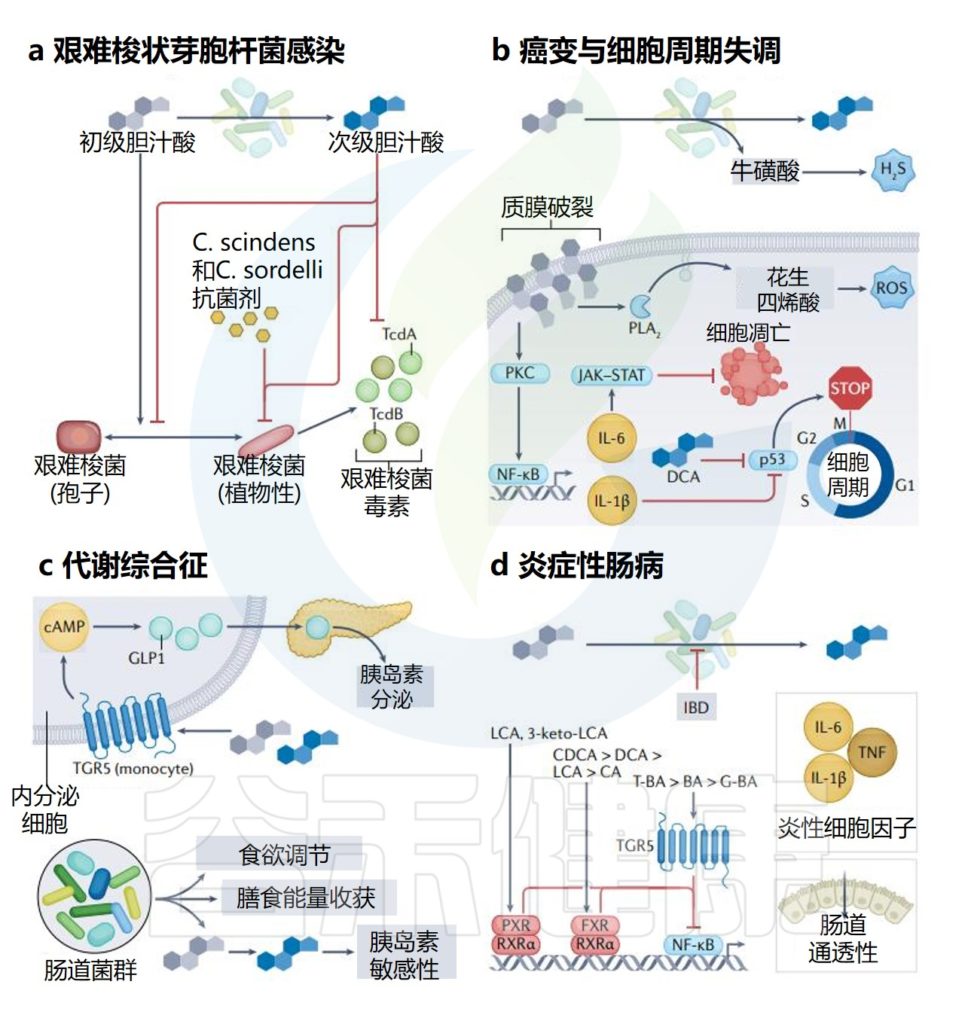
编辑
Collins SL,et al.Nat Rev Microbiol.2023
代谢综合征是指至少有以下三种体征的个体:高血压、肥胖、血液甘油三酯水平升高或血糖水平升高。
患有代谢综合征的人有多种疾病的风险,包括糖尿病、中风、心脏病和结直肠癌。
√肠道菌群改变胆汁酸对代谢稳态非常重要
遗传、饮食和环境都在代谢性疾病中起作用,其中包括微生物群和胆汁酸组成。微生物群通过增加膳食能量收获、调节食欲和改变胆汁酸池对代谢稳态至关重要。
√次级胆汁酸与非酒精性脂肪肝相关
抗生素相关的厚壁菌门细菌丰度减少和次级胆汁酸的消耗与代谢综合征患者胰岛素敏感性降低相一致,而微生物群依赖的次级胆汁酸水平增加与非酒精性脂肪肝相关,强调了微生物群相关胆汁酸对代谢健康的重要性。
√次级胆汁酸影响脂质和葡萄糖稳态
虽然次级胆汁酸可能有助于缓解代谢综合征,但其机制尚不清楚。胆汁酸受体如FXR和TGR5维持脂质(胆固醇和甘油三酯)和葡萄糖稳态。
被胆汁酸激活的受体增加胰岛素敏感性
被胆汁酸激活的TGR5通过诱导胰高血糖素样肽-1(GLP1)的分泌而增加胰岛素敏感性。虽然FXR的激活也会增加GLP1的分泌,但它在代谢性疾病中的作用是复杂的,似乎是特定于环境的。
在一些研究中,激活FXR对胆固醇、甘油三酯和葡萄糖水平有有益的影响。例如,肝脏FXR激活通过抑制SREBP1c脂质生成途径来抑制甘油三酯的产生。
√胆汁酸通过法尼醇X受体活性影响代谢
包括奥贝胆酸在内的强效合成FXR激动剂的临床试验显示,非酒精性脂肪肝患者体重减轻,可能是通过刺激棕色脂肪分化和代谢。
然而,关于奥贝胆酸对非酒精性脂肪肝患者胰岛素抵抗的影响,有相互矛盾的报道。此外,在一些研究中,敲除小鼠体内的法尼醇X受体与激活法尼醇X受体有相似的效果(防止体重增加和增加胰岛素敏感性)。
FXR基因敲除小鼠的益处依赖于微生物群,因为微生物群从这些小鼠转移到无菌小鼠可以减轻体重,降低体脂率和胰岛素敏感性。
因此,在检查胆汁酸对肥胖和代谢性疾病的影响时,应仔细考虑肠道和肝脏FXR活性。
艰难梭菌感染(CDI)是全世界范围内最常见的感染之一,一部分感染者为无症状携带者,严重的患者可能出现腹泻、发烧,甚至会死亡。
当正常的微生物群被抗生素耗尽时,就会出现艰难梭菌感染。抗生素消耗的常驻微生物群减少了分泌抗菌肽的梭状芽胞杆菌(Clostridium spp.)的数量,而梭状芽胞杆菌通常控制艰难梭菌的数量。
√微生物胆汁酸代谢对预防艰难梭菌感染有益
此外,原生微生物群将原发性胆汁酸转化为继发性胆汁酸对于预防艰难梭菌感染至关重要。事实上,抗生素相关的原发性胆汁酸升高和继发性胆汁酸水平降低与复发性艰难梭状芽胞杆菌感染有关。
一些初级胆汁酸会诱导艰难梭菌到产毒状态
微生物胆汁酸代谢对预防艰难梭菌感染的益处是多方面的。某些初级胆汁酸,包括胆酸、牛磺胆酸和甘氨胆酸,可诱导艰难梭菌从孢子萌发到活跃的产毒状态。
次级胆汁酸能直接抑制艰难梭菌的生长
相反,去氧胆酸和石胆酸已被证明能直接抑制艰难梭菌的萌发和生长。这部分解释了为什么编码胆汁酸的梭状芽胞杆菌(如C. scindens)产生次生胆汁酸可以保护艰难梭菌感染。
注:C.scindens和Clostridium也分泌抗艰难梭菌的抗菌剂,与去氧胆酸和石胆酸结合使用更有效。
次级胆汁酸可以降低艰难梭菌毒性
次级胆汁酸也直接结合并隔离艰难梭菌毒素B(TcdB)以降低其毒性。细菌胆汁酸代谢对预防艰难梭菌感染很重要,这一认识可能会提供新的治疗选择。
粪菌移植作为抗生素的替代方法已被证明可以通过恢复次级胆汁酸来成功减轻艰难梭菌感染。
肠道菌群相关的胆汁酸会影响胃肠道炎症和肿瘤的发展。
早在20世纪30年代,去氧胆酸就被证明会引起小鼠注射部位的肿瘤。特别是与高脂肪饮食相结合,两种主要的次级胆汁酸(去氧胆酸和石胆酸)长期以来都与胃肠道癌症有关,特别是结直肠癌和肝细胞癌。
√胆汁酸影响致癌作用
微生物群的组成和胆汁酸合成潜力对炎症和癌症的进展至关重要。
高脂肪饮食更容易发生肝细胞癌
高脂肪饮食喂养的小鼠更容易发生肝细胞癌,部分原因是革兰氏阳性群体增加,它们通过7α-去羟基化酶产生去氧胆酸。
由于其疏水性,次级胆汁酸如去氧胆酸和石胆酸比初级胆汁酸更具有致癌性。在肝细胞癌和结直肠癌中,疏水性胆汁酸的癌症加速是基于它们损伤细胞和诱导炎症的能力。
未结合的疏水性去氧胆酸和石胆酸更容易作为清洁剂破坏细胞膜并诱导细胞损伤反应。
抑制法尼醇X受体活性进一步加剧了次级胆汁酸的致癌潜力。许多研究已经观察到人类结直肠癌中法尼醇X受体激活降低,法尼醇X受体敲除小鼠易患结肠和肝脏肿瘤。
作为胆汁酸的受体,法尼醇X受体是癌症进展过程中脂质代谢和细胞信号中断事件之间的重要联系。
炎症会降低法尼醇X受体的激活从而影响癌变
炎症,特别是通过NF- κB信号通路分泌肿瘤坏死因子和白细胞介素-1β,降低了法尼醇X受体的激活。因此,上述细胞对去氧胆酸和石胆酸的促炎反应会促进癌变。
法尼醇X受体失活还会增加胆汁酸的合成并减少肝细胞的胆汁酸输出,进一步加剧胆汁酸积累介导的损伤和炎症。
细菌水解的牛磺酸偶联胆汁酸的促癌作用的另一种机制已被提出。去偶联释放的牛磺酸最终通过磺酸代谢为硫化氢,这是一种强效致癌物。
√炎症性肠病中胆汁酸代谢失调
炎症性肠病是胃肠道慢性炎症性疾病的集合,最常见的是克罗恩病和溃疡性结肠炎。溃疡性结肠炎只影响结肠,而克罗恩病可以同时影响小肠和结肠。
在炎症性肠病发作期间,肠上皮发炎并失去屏障的完整性。虽然炎症性肠病的病因尚不完全清楚,但与肠道菌群的大规模变化有关。
炎症性肠病中胆汁酸代谢明显失调,特别是当微生物群被破坏时。活动性炎症性肠病患者粪便中共轭胆汁酸水平升高,次级胆汁酸水平降低。
注:一项利用计算预测炎症性肠病患者和健康人微生物群的解偶联和修饰潜能的研究发现,炎症性肠病患者微生物群的胆汁酸代谢潜能受到干扰。
次级胆汁酸受体促进肠上皮再生并预防肠道炎症
次级胆汁酸激活G蛋白偶联受体5(TGR5)可促进肠上皮细胞再生并预防肠道炎症。胆汁酸受体FXR和 TGR5被胆汁酸衍生物或其他激动剂激活,刺激靶向通路以调节肠道稳态,预防炎症和肿瘤发生。
炎症性肠病和结直肠癌中的胆汁酸受体
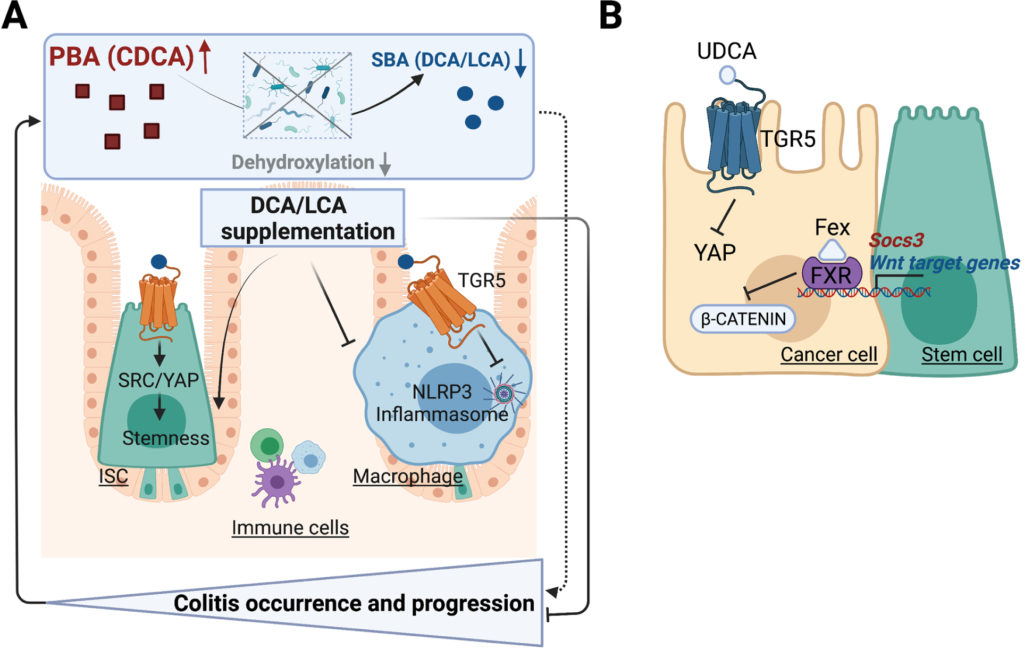
Cai J,et al.Cell Host Microbe.2022
然而,一些因素使胆汁酸、微生物群和炎症性肠病之间的关系复杂化。特别是,活动性炎症性肠病患者肠道转运率的增加减少了总微生物群数量,限制了代谢胆汁酸的足够反应时间。
胆汁酸能预测炎症性肠病的早期缓解
胆汁酸不仅与疾病发生高度相关,而且能够预测炎症性肠病的早期缓解。在最近发表的一项研究中,进行了粪便宏基因组学、血清代谢组学和蛋白质组学,以揭示预测炎症性肠病中抗细胞因子和抗整合素治疗的不同反应的微生物决定因素。
血清胆汁酸,如甘氨石胆酸钠、甘氨脱氧胆酸和熊去氧胆酸的富集可预测接受抗细胞因子治疗的炎症性肠病患者的早期病情缓解。
此外,微生物种类的多样性与与抗细胞因子反应相关的缓解可能性呈正相关,这表明微生物特征可能有利于炎症性肠病的生物治疗反应。
√胆汁酸与结直肠癌发病相关
根据2020年美国癌症协会的统计数据,在美国,结直肠癌是男性和女性癌症死亡的第三大原因。炎症性肠病和结直肠癌密切相关,炎症性肠病被认为是结直肠癌的主要风险因素。
与炎症性肠病类似,结直肠癌也是一种与肠道菌群密切相关的异质性疾病。鉴于肠道微生物群与结肠肿瘤发生之间的联系以及结直肠癌的异质性,多组学技术已被应用于了解疾病的病因学、发现预后或预测性生物标志物以及开发治疗或预防药物。
人体炎症性肠病和结直肠癌中的胆汁酸
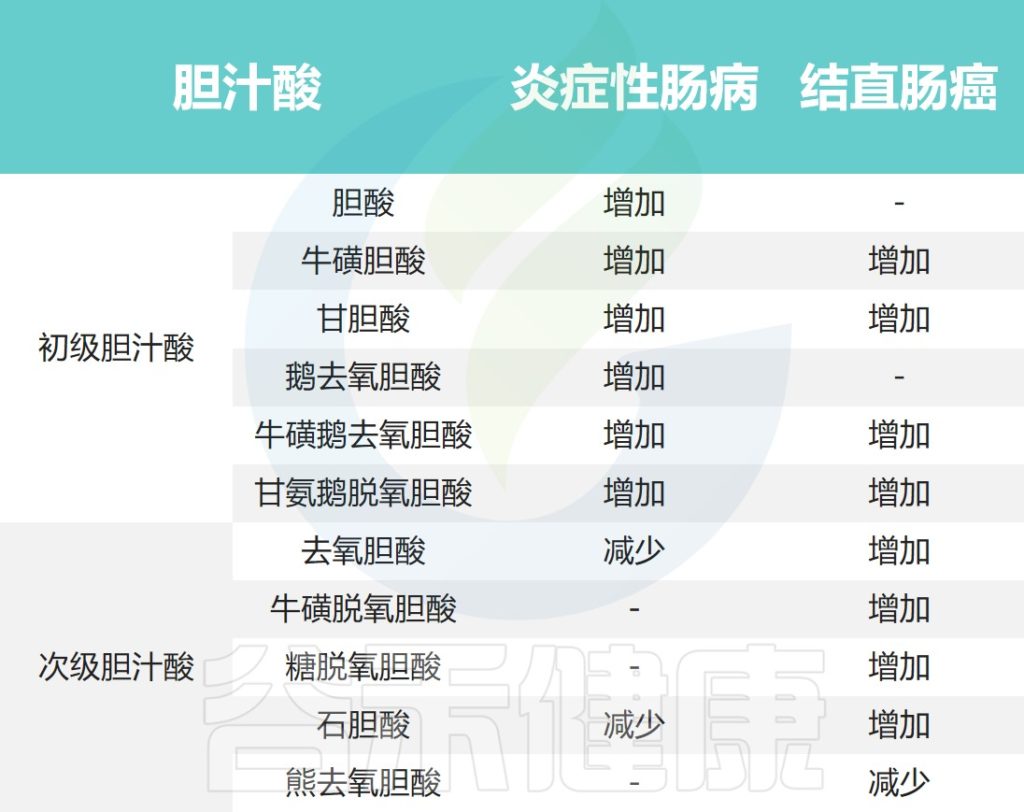
Cai J,et al.Cell Host Microbe.2022
胆汁酸合成基因与结直肠癌显著相关
研究发现胆汁酸合成相关基因的丰度与人类结直肠癌显著相关。
在对来自七个不同国家的八项结直肠癌研究进行的无偏倚荟萃分析中,胆汁酸基因在基因组和转录组水平上都在结直肠癌患者的粪便中高度富集,这可能被用作结直肠癌诊断的替代微生物组标志物。
胆盐水解酶与结直肠癌存在联系
在另一项关于人类微生物组的全球宏基因组研究中,主要来自厚壁菌门的胆汁酸水解酶基因的丰度与结直肠癌呈负相关,这与之前显示结直肠癌患者中厚壁菌门减少的研究一致, 表明胆盐水解酶丰度与人类结直肠癌之间的关系。
胆汁酸含量与肿瘤风险相关的研究:
在一项全基因组鸟枪法宏基因组学和代谢组学研究中,粪便样本取自结直肠肿瘤不同阶段的患者,研究了多步肿瘤发生过程中的微生物组和代谢组学变化。多发性息肉样腺瘤患者的去氧胆酸显著增加。
在欧洲癌症与营养前瞻性调查队列中的一项病例对照研究中,七种血浆胆汁酸的水平,包括甘氨胆酸、牛磺胆酸、牛磺去氧胆酸和甘胆酸等与结肠癌风险呈正相关,而未结合的胆汁酸和三级胆汁酸与癌症风险无关。
其他研究发现,在粪便代谢组学研究中,血清胆汁酸在腺瘤组中升高,并与拟杆菌(Bacteroides)呈正相关而与健康对照相比,结直肠癌患者粪便样本中的熊去氧胆酸有所减少。
一些研究表明去氧胆酸可能有助于结直肠癌的发病机制,不过还需要更多的研究来确定其他胆汁酸,尤其是原发性胆汁酸在人类结直肠癌中的作用。
胆固醇结石病 (GS) 是一种世界范围内流行的疾病,尤其是在西方国家。最近的调查发现,它在中国上海的发病率超过12%。
√胆汁酸分泌异常与胆结石形成相关
导致胆结石的先决生化障碍是胆囊中胆固醇过饱和胆汁的形成。研究中,我们发现胆固醇结石患者和胆结石易感小鼠的粪便中富含脱硫弧菌(Desulfovibrio),诱导胆结石形成。
肝脏胆固醇积聚和胆汁胆固醇分泌增强
我们进一步证明,携带脱硫弧菌与肠道微生物群活化胆汁酸脱羟基作用导致的次级胆汁酸产生增加有关,导致胆汁酸疏水性增加,从而促进肠道胆固醇吸收,导致肝脏胆固醇积聚和胆汁胆固醇分泌增强。
微生物群促进胆固醇结石形成的潜在机制
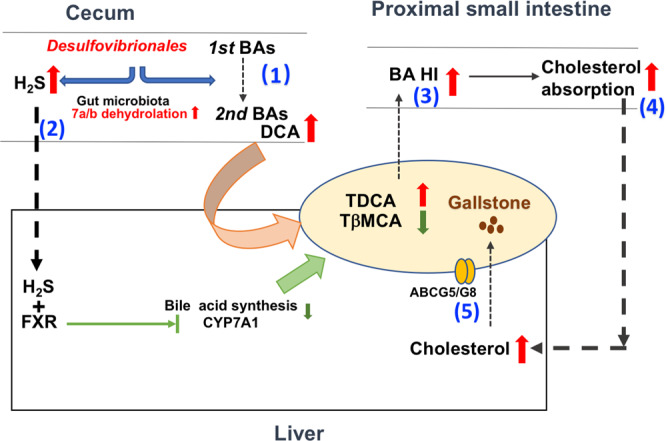
Hu H,et al.Nat Commun.2022
√易患胆结石的微生物群调节胆汁酸的方式:
易患胆结石的微生物群(富含脱硫弧菌)通过以下方式调节肝胆汁酸代谢:
(1) 增加7α-脱羟基细菌在盲肠中产生更多的次级胆汁酸;
(2) 产生硫化氢并调节肝FXR-CYP7A表达,这些物质影响胆汁酸的合成以及在肝脏中的重吸收;
(3) 导致去氧胆酸增加,进而提高胆汁酸的疏水性指数,抑制肝胆汁酸合成;
(4) 促进肠道胆固醇吸收,导致肝脏胆固醇超载;(5) 促进小管胆固醇分泌到胆汁中并诱导胆固醇胆结石形成。
通过改变微生物群的组成或微生物群代谢某些胆汁酸的能力,或通过施用次生胆汁酸,来控制微生物胆汁酸代谢,具有提供健康益处的潜力。
随着研究阐明了特定胆汁酸对健康的确切影响,治疗可以从更普遍的改变微生物群的方法(例如,粪便移植、益生菌和益生元)转向专门针对单个胆汁酸。
粪便微生物群移植是将健康供体的粪便微生物转移到受体患者,最初是为了恢复完整的微生物群而开发的。
•粪菌移植可以治疗艰难梭菌感染
粪菌移植治疗艰难梭菌感染的成功可能部分是由于恢复了依赖胆汁酸水解酶的次级胆汁酸生产。
尽管一些初级胆汁酸,如牛磺胆酸,促进艰难梭菌孢子萌发,但次级胆汁酸去氧胆酸和石胆酸抑制艰难梭菌生长和艰难梭菌孢子发芽,并且它们的水平在艰难梭菌感染抗性个体中升高。
•减轻肠道炎症等相关疾病
虽然粪菌移植在治疗溃疡性结肠炎方面的效果不一,但石胆酸的衍生物3-oxo LCA的恢复与粪菌移植相关,并表明次级胆汁酸在减轻疾病方面的作用。
减少产生胆汁酸水解酶的细菌缓解肠易激综合征
粪菌移植可以通过减少胆汁酸水解酶活性细菌的数量来缓解与腹泻型肠易激综合征相关的次级胆汁酸过量产生。
•粪菌移植也可用于治疗生态失调
粪菌移植也在小鼠身上进行了研究,以治疗观察到微生态失调的各种其他情况(例如衰老)。
粪菌移植缺乏特异性
粪菌移植广泛应用的主要限制之一是其在调节微生物群方面缺乏特异性,这可能导致意想不到的不良反应。
在完整的细菌群落及其对人类健康的影响尚不清楚的情况下,设计一种成功的粪菌移植治疗方法仍然具有挑战性。
具有明确胆汁酸代谢特性的活微生物可用于改变生态失调和疾病中的胆汁酸库。
•使用益生菌更具针对性
与粪菌移植相比,益生菌的优势在于它们更具针对性,具有明确的作用机制。具有7α-去羟基化活性的梭状芽胞杆菌(Clostridium spp.),特别是C.scindens,被证明可以通过产生抑制性的次级胆汁酸来消除艰难梭菌。
注:通过基因工程,来自具有7α-脱羟基活性的细菌的操纵子已被移植,以将这些益处赋予共生产孢梭菌菌株。这种方法可以将胆汁酸合成归因于各种共生细菌,并精确地生成需要的胆汁酸。
生孢梭菌——能够成为具有发展前景的癌症治疗手段
•益生菌可以调节胆汁酸受体
益生菌的使用可以调节胆汁酸受体如法尼醇X受体、维生素D受体的激活。
•益生菌通过调节胆汁酸降低胆固醇水平
益生菌混合物VSL#3可以治疗肠易激综合征和溃疡性结肠炎,但也含有表达胆汁酸水解酶的细菌,可以增加胆汁酸的解结和排泄。
VSL#3通过抑制FXR-FGF15途径上调肝脏胆汁酸生物合成。这种作用可能有利于降低循环中的胆固醇水平。
•益生菌还可以减少肠道炎症和肿瘤形成
然而,同样的益生菌混合物在结直肠癌患者中可能具有相反的恢复法尼醇X受体激活的效果。事实上,施用VSL#3可防止结直肠癌相关的胆汁酸下调,并减少炎症和肿瘤形成。
另一种活性益生菌,罗伊氏乳杆菌NCIMB 30242,在人类中具有类似的抗炎和降胆固醇作用,这表明这些细菌的有益作用依赖于将共轭胆汁酸转化为次级胆汁酸。
与其给药产生胆汁酸的细菌,不如直接给药具有生物活性的次级胆汁酸,因为它们具有相似的预期效果。
•施用熊去氧胆酸可以抑制胃肠道癌症
熊去氧胆酸主要用作抗胆汁淤积剂,是原发性胆道自身免疫性疾病(如原发性胆道胆管炎)患者的主要治疗选择,但也可能抑制胃肠道癌症(如结直肠癌和肝细胞癌)的进展。
注:熊去氧胆酸减轻结直肠癌的发生是由胆汁酸膜受体TGR5介导的。
•施用次级胆汁酸降低了结肠炎的严重程度
胆汁酸也可能有助于结肠炎患者的症状缓解。石胆酸代谢物通过调节转录因子RORγt和产生线粒体活性氧,支持T细胞分化为调节性T细胞而不是促炎性T辅助17细胞。
这种次级胆汁酸介导的对调节性T细胞分化的促进降低了结肠炎的严重程度,并且部分地解释了细菌联合治疗小鼠结肠炎的益处。
•改善肥胖患者胆固醇及甘油三酯含量
熊去氧胆酸和石胆酸均为法尼醇X受体激动剂,可通过FXR-FGF15途径减轻代谢综合征。由于FXR-FGF15的激活,肥胖小鼠在接受熊去氧胆酸和石胆酸治疗后,胆固醇、血浆甘油三酯和血浆脂肪酸水平降低了。
因此,产生这些次生胆汁酸的细菌,如狄氏副拟杆菌(Parabacteroides distasonis),有助于降低高脂血症。
注:合成的FXR激动剂奥贝胆酸是一种被批准用于治疗原发性胆道胆管炎的药物,并且在治疗非酒精性脂肪性肝炎和肥胖症方面也取得了成功。
提示
虽然直接补充胆汁酸可以产生对致癌、结肠炎和代谢综合征有益的结果,但这些效果可能只在治疗期间持续。相反,改变微生物群以增加胆汁代谢可能有助于延长效果。因此调节肠道微生物群的平衡十分重要。
细菌转化胆汁酸的失衡是代谢性、炎症性、感染性和肿瘤性疾病的重要因素,主要通过胆汁酸受体的失调来实现。
由于许多胆汁酸受体在胃肠道以外的组织中表达(例如,脑,T细胞和平滑肌),因此值得进一步研究胆汁酸在其他部位的影响。
随着对肠道菌群和胆汁酸的深入研究,我们将更好地了解它们在人类健康中的作用机制,以及它们与人类疾病的关系。
同时,我们也将探索更多的方法来维护肠道菌群和胆汁酸的平衡,如饮食、运动、药物等。这些研究将为预防和治疗相关疾病提供新的思路和方法,为人类健康事业做出更大的贡献。
主要参考文献
Collins SL, Stine JG, Bisanz JE, Okafor CD, Patterson AD. Bile acids and the gut microbiota: metabolic interactions and impacts on disease. Nat Rev Microbiol. 2023 Apr;21(4):236-247. doi: 10.1038/s41579-022-00805-x. Epub 2022 Oct 17. PMID: 36253479.
Yu L, Liu Y, Wang S, Zhang Q, Zhao J, Zhang H, Narbad A, Tian F, Zhai Q, Chen W. Cholestasis: exploring the triangular relationship of gut microbiota-bile acid-cholestasis and the potential probiotic strategies. Gut Microbes. 2023 Jan-Dec;15(1):2181930. doi: 10.1080/19490976.2023.2181930. PMID: 36864554; PMCID: PMC9988349.
Cai J, Sun L, Gonzalez FJ. Gut microbiota-derived bile acids in intestinal immunity, inflammation, and tumorigenesis. Cell Host Microbe. 2022 Mar 9;30(3):289-300. doi: 10.1016/j.chom.2022.02.004. PMID: 35271802; PMCID: PMC8923532.
Hu H, Shao W, Liu Q, Liu N, Wang Q, Xu J, Zhang X, Weng Z, Lu Q, Jiao L, Chen C, Sun H, Jiang Z, Zhang X, Gu A. Gut microbiota promotes cholesterol gallstone formation by modulating bile acid composition and biliary cholesterol secretion. Nat Commun. 2022 Jan 11;13(1):252. doi: 10.1038/s41467-021-27758-8. PMID: 35017486; PMCID: PMC8752841.
Poland JC, Flynn CR. Bile Acids, Their Receptors, and the Gut Microbiota. Physiology (Bethesda). 2021 Jul 1;36(4):235-245. doi: 10.1152/physiol.00028.2020. PMID: 34159805; PMCID: PMC8526331.
Larabi AB, Masson HLP, Bäumler AJ. Bile acids as modulators of gut microbiota composition and function. Gut Microbes. 2023 Jan-Dec;15(1):2172671. doi: 10.1080/19490976.2023.2172671. PMID: 36740850; PMCID: PMC9904317.
McCarville JL, Chen GY, Cuevas VD, Troha K, Ayres JS. Microbiota metabolites in health and disease. Annu Rev Immunol [Internet] 2020; 38:147–170.