 国家高新企业 | ISO9001认证
国家高新企业 | ISO9001认证 二级病原微生物安全实验室
二级病原微生物安全实验室- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
国家高新企业 | ISO9001认证 二级病原微生物安全实验室

谷禾健康

维生素C是一种广泛存在于自然界中的水溶性维生素。维生素C在人体新陈代谢中具有多种重要功能,包括抗氧化、参与胶原蛋白合成、增强铁的吸收等。由于其众多生理益处,维生素C被广泛地应用于修复伤口、治疗感冒、癌症等多种疾病。
人体无法自行合成维生素C,需要从膳食来源(如水果、蔬菜)中获取。维生素C的代谢过程涉及多个酶和转运蛋白。肠道微生物群可能通过影响这些酶和蛋白的活性或表达,来影响维生素C的代谢吸收过程。
维生素C也可以直接调节肠道微生物群,或通过修复肠道屏障、改变氧化还原电位等方式间接对肠道微生物群的平衡起到调节作用。
个体之间的差异、饮食习惯和生活方式等因素各不相同,这些都可能对维生素C与肠道菌群的相互作用产生影响。
本文将从维生素C的结构、功能、吸收和代谢、与肠道菌群的关联等多角度,全面探讨维生素C的作用及其对人体健康的影响,同时介绍了一些维生素C的食物来源、人体需要的剂量、如何补充、注意事项等。
▼
本文主要内容:

编辑
维生素C,也称为抗坏血酸,是一种水溶性维生素,对人体健康非常重要。它在许多身体功能中起着关键作用,包括增强免疫力、抗氧化、胶原蛋白的合成等。
▼
维生素C的化学名称是L-抗坏血酸,它是一种有机化合物。无臭,味酸,易溶于水,微溶于乙醇,不溶于乙醚。
维生素C结构简单,化学式为C6H8O6:
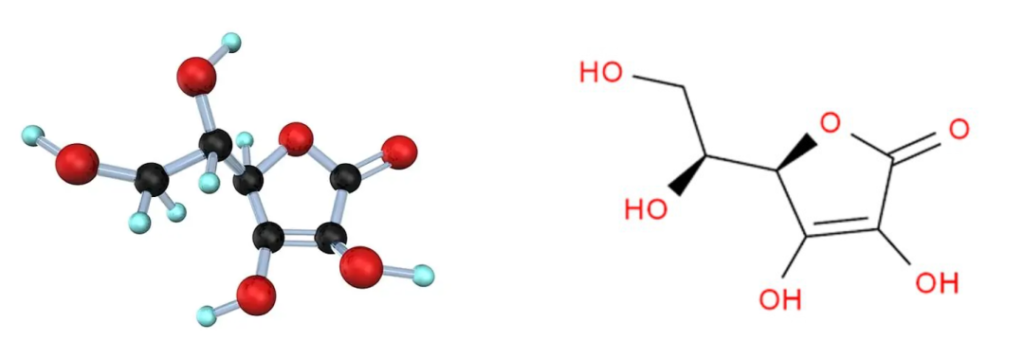
这种结构使维生素C具有抗氧化性质,能够捕捉自由基,并保护细胞免受氧化损伤。
注:体内的分子暴露于环境污染物、吸烟和慢性炎症等情况时,它们会变成自由基。自由基是不稳定的分子,会破坏细胞并导致疾病,维生素C可以通过中和自由基来减缓或预防某些健康问题。
由于其与葡萄糖的结构相似,维生素C可以在许多化学反应中取代葡萄糖,并且可以防止蛋白质的非酶糖基化。
▼
维生素C参与胶原蛋白、激素、肉碱的合成,促进铁离子的吸收,此外,它在免疫系统的功能和调节中发挥着重要作用,对维持内部环境的平衡和中枢神经系统的正常功能极为重要。
维生素C的大部分功能是由于其作为抗氧化剂和辅助因子的能力。由于人类缺乏L-gulono-1, 4-lactone氧化酶,无法自行合成维生素C,因此完全依赖于维生素C的饮食摄入。
▼
从食物中获取营养总是最好的。大约90%的日常需求来自蔬菜和水果,它们是这种维生素的极好来源,例如奇异果、橙子、芒果、草莓、红椒、青椒等。
一些维生素C含量较高的食物
注:单位“杯”是一个常见的非正式计量单位,美规和英规略有区别,大约是237毫升-250毫升左右,涉及到果蔬的份量时,一杯通常是指将果蔬切碎后填满一杯容器的量。
▼
维生素 C 对大脑健康非常重要。大脑在长期缺乏维生素 C 的情况下以牺牲其他组织为代价来保留维生素 C,并且可以维持比其他器官(例如肝脏和肾脏)高很多倍的浓度,如下图。
维生素C的分布在身体各器官之间差异很大
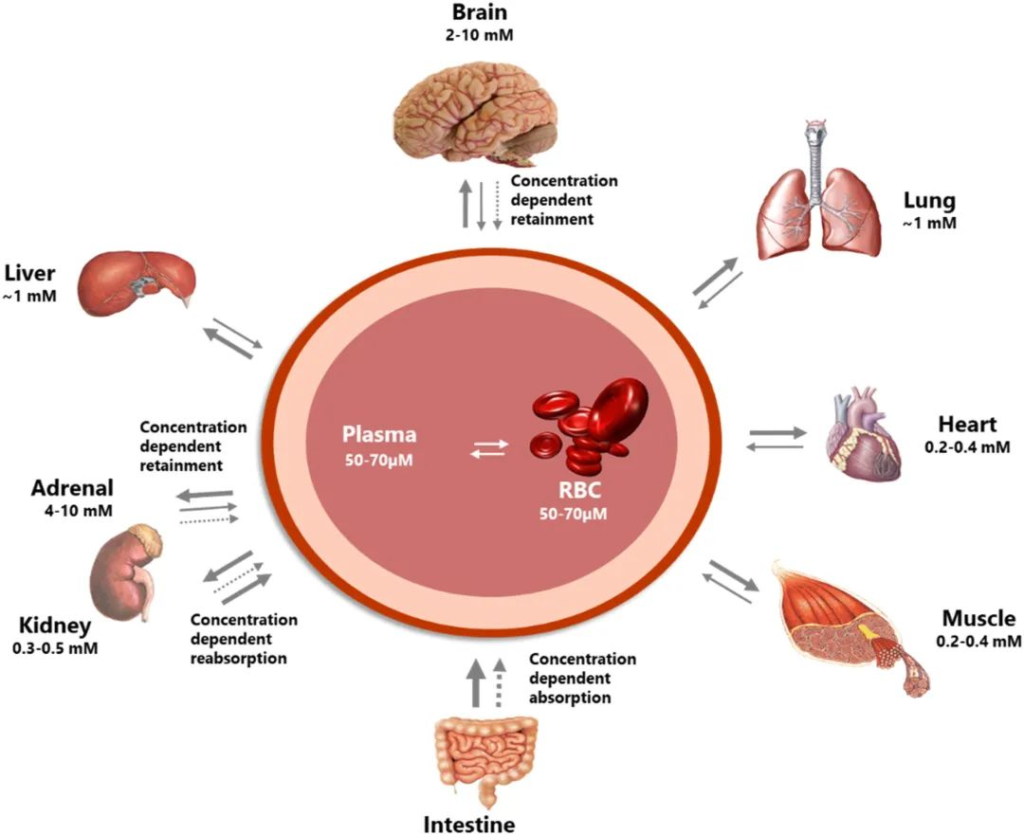
doi.org/10.1016/j.redox.2020.101532
维生素C供应不足时,首先保证大脑里的浓度,那么维生素C对于大脑而言有什么作用?
▼
维生素 C 对胶原蛋白的稳定作用对于形成整个身体的结缔组织框架至关重要;包括皮肤、骨骼、软骨、肌腱、韧带、血管等。
胶原蛋白生产的最后步骤取决于维生素 C,维生素 C 在前胶原脯氨酰和赖氨酰残基的羟基化中充当电子供体。
▼
维生素 C 可增强其他营养素的生物利用度,例如维生素 E 和非血红素铁,这可能会增强含维生素 C 的食物的健康效果。
维生素C经常添加到含铁的口服制剂中,以增加铁的吸收。
▼
维生素 C 是否可以预防或减轻包括普通感冒在内的感染的严重程度是一个有争议的话题。大多数证据都支持其好处。
▼
维生素C对正常骨骼发育至关重要。维生素C水平与骨骼健康之间存在正相关关系,如骨密度、骨折概率、骨转换标志物等。
▼
维生素 C 有助于维持健康的皮肤。
▼
肺部的维生素C水平是血液中的30倍。
维生素C在抵御氧化剂的同时也会被消耗,这表明即使是单剂量的维生素C,也能有效抵御肺部氧化应激的急性增加。
▼
根据现有证据,摄入足够的维生素 C 可能有助于保持健康的情绪。
焦虑
抑郁
▼
人类慢性低维生素 C 状态与神经退行性疾病有关。但是,尚未确定因果关系。
▼
维生素 C 可以通过抑制炎性细胞因子来减轻炎症。
▼
除上述主要的功能之外,在部分小型研究中提到的关于维生素C的功能如下:
助孕育:
助减肥:
降血压、防中风:
降血糖:
助排毒:
助抗癌:
牙周健康:
水溶性维生素在人体内储存较少,从肠道吸收后进入人体的多余的水溶性维生素大多从尿中排出。因此,摄入较多的水溶性维生素一般不会引起中毒现象,但是若摄入量过少,则会很快出现缺乏症状。
▼
刚缺乏的时候症状不明显,饮食中缺乏VC需要大约一个月的时间才会出现症状。
体内维生素C总含量低于300-400mg会出现明显症状。
维生素C的缺乏会出现什么症状?
由于维生素C功能的复杂性及其被不同还原剂的部分替代性,维生素C与坏血病症状的直接联系不容易确定。
如果发展为坏血病,典型症状是:
肌肉无力、牙龈肿胀和出血、牙齿脱落、瘀点出血、自发性瘀斑、贫血、愈合障碍、角化过度、虚弱、肌痛、关节痛和体重减轻(也可能因肿胀而出现矛盾的体重增加),而早期表现包括嗜睡、倦怠、易激惹,甚至呼吸困难等。
在生化上,维生素C血浆水平低于11μM被认为与坏血病的临床症状一致。
在专业医疗人员的指导下,补充维生素C可以轻松有效地逆转坏血病。许多症状可以在几周内轻松解决。富含维生素C的饮食将防止坏血病的发展。
什么人群更容易发生维生素C缺乏?
▼
NIH 认为,成人可耐受的上限是每天 2000 毫克,仅仅靠含有维生素 C 的食物几乎不可能达到这一上限,所以食物可以放心吃。服用补充剂则需注意剂量,可能存在过量的风险。
更高的剂量更有可能导致副作用。
维生素C过量可能会出现什么症状?
单次口服5-10克维生素C会产生短暂的渗透性腹泻和/或腹胀伴疼痛,不建议这样做。
随食物一起摄入可减少这些不良反应。
每天超过 2000 毫克的剂量可能会增加腹泻和肾结石的风险。如果有肾结石病史,每天摄入超过 1000 毫克可能会增加患结石的几率。
那么到底应该怎么补充?每日摄取多少维生素C 合适呢?
▼
科学界对维生素C的最佳剂量方案(摄入量和频率)最健康存在持续争论。
对于大多数健康人来说,通过食物可以获得足量的维生素 C。
维生素C摄入量标准在不同地区有所不同:
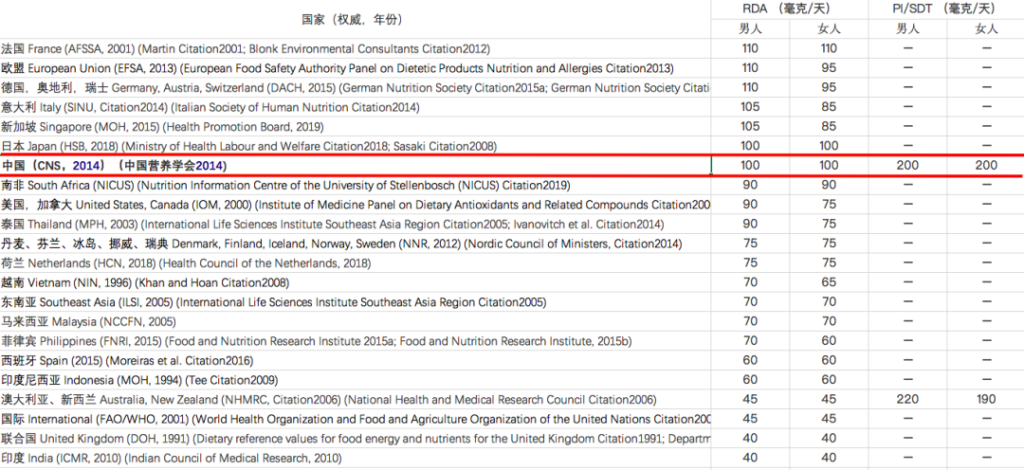
注:RDA – 推荐膳食摄入量,
PI – 建议摄入量,SDT – 建议膳食目标
在中国营养学会编著的《中国居民膳食指南》2022版中对维生素C的推荐摄入量:
中国居民膳食指南2022版
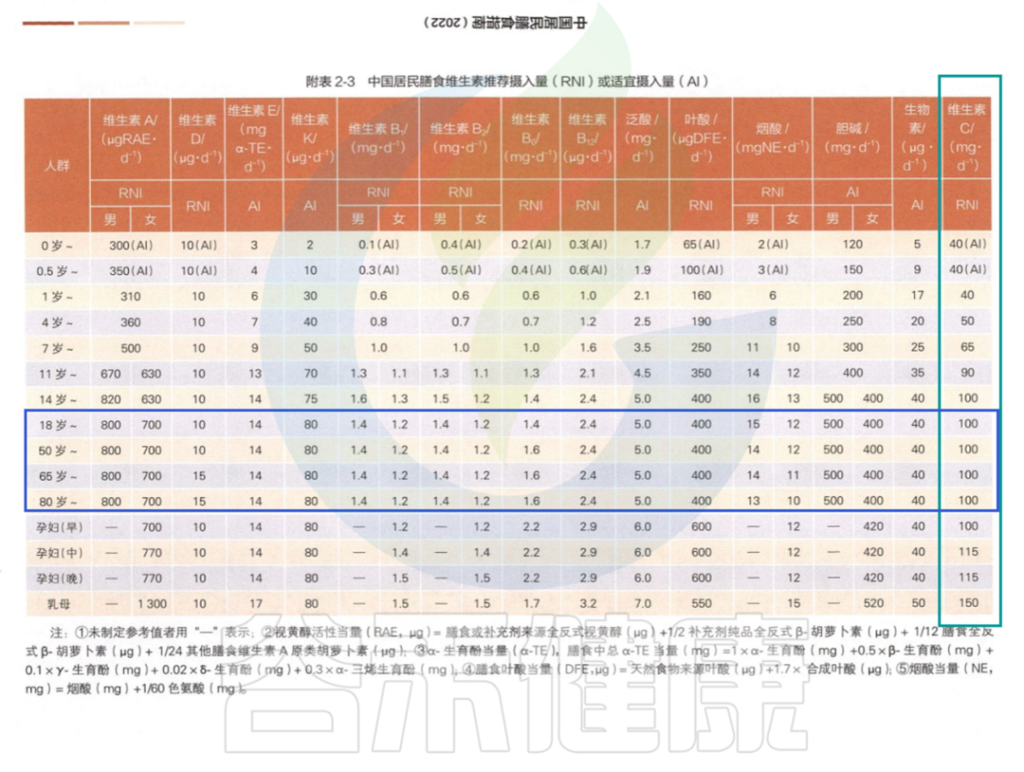
为什么不同地区的标准不一样呢?
这主要是由于RDA标准的基本前提从预防坏血病(~45 mg/d)至健康优化(~200 mg/d)。许多权威机构建议使用最低量的维生素C,但这可能无法满足不同亚群的健康需求。
例如,吸烟者和肥胖者比普通人群有更高的要求。一些国家的吸烟率继续上升,有证据表明,吸烟者每天至少摄入200 mg的维生素C。
随着全球肥胖率的增加,以及与肥胖相关的合并症,如代谢综合征、糖尿病和心血管疾病,需要更多地考虑适当的建议,以优化快速增加的亚健康人群中的维生素C状态。
以上是维生素C的摄入量标准,具体该如何补充,应该注意什么?详见下一章节。
▼
食物:如何才能最大程度地保留其维生素C?
——烹饪方式
长时间烹饪,特别是用大量水煮沸会导致维生素C浸出到水中,显著降低食物中的维生素C含量。
在少量水中蒸或煮,且持续时间较短,是保存维生素C的更温和的方法。
即使将所有外部因素消除到最低限度,也会发生损失,因为由于抗坏血酸氧化酶的存在,材料内部会发生氧化反应。因此,理想的加工方法是用最少量的水快速热灭活酶,然后快速冷却。
——长期保存条件
温度对储存稳定性也有很大影响。随着温度的升高,会出现更显著的损失。然而,在长期储存期间,即使维持短期储存期间仅发生少量损失的条件,维生素C的量也会显著减少。
损失主要是由于酶催化的氧化反应,其程度尤其取决于pH、材料完整性和温度。
总之,为了使水果和蔬菜的维生素C含量保持更长时间,最温和的方法是深度冷冻。
补充剂:一天中分几次服用
食物和许多补充剂中的维生素 C 是一种抗坏血酸的形式。
当肠道面临较低水平的抗坏血酸(即低于约 400 毫克)时,主动运输系统会吸收维生素 C(即,将营养物质通过肠道并进入血液,到身体需要的地方)。
一旦这些主动运输变得不堪重负,被动扩散就会接管吸收其余的维生素 C(这是一个相当低效的过程)。吸收并不像听起来那么容易,事实上抗坏血酸有吸收上限。
身体一次可以处理大约 300 – 400 毫克的纯抗坏血酸形式的维生素 C,更多量一下子难吸收。
所以如果能记得的话,一天中分几次服用比较合适。
▼
维生素C补充剂并非适合所有人。如果遇到以下任何情况,请首先与医生联系:
也不要认为维生素C服用越多越好,每天服用 1000 毫克或更多,实际上会使吸收率降低约 50%。
▼
维生素C可以增加某些药物的吸收,例如:
服用维生素C可以增加含铝药物(如磷酸盐粘合剂)对铝的吸收。这可能对有肾脏问题的人有害。抗酸剂中含铝:不要同时服用维生素C和抗酸剂。服用维生素C后至少等待两个小时,然后再服用抗酸剂。服用抗酸剂后等待四个小时服用维生素C。
维生素C可能会增加左旋甲状腺素的吸收。
补充维生素C会降低一些药物的疗效:
口服维生素C可能会降低这些抗病毒药物的作用。
当与维生素C一起服用时,烟酸和他汀类药物的影响可能会降低,这可能有益于高胆固醇的人。
高剂量的维生素C可能会降低人体对这种抗凝剂的反应。
其他还包括:
维生素C增加或减少药物副作用的风险
如果服用雌激素或基于雌激素的避孕药,维生素C可能会增加激素副作用的风险。这是因为维生素C可能会减缓雌激素离开身体的速度。
一些早期的研究认为, 维生素C可能有助于预防阿司匹林和非甾体抗炎药引起的胃部不适。
▼
维生素 C 和益生菌对肠道健康和免疫力有不同的好处,它们可以很好地互补,这意味着它们可以安全地一起服用。
一项针对学龄前儿童的双盲、随机、安慰剂对照初步研究中,发现益生菌与维生素 C 联合预防呼吸道感染的功效(URTI;33%,P =0.002)。
Lab4 益生菌和维生素 C 组合补充 6 个月的儿童显示,上呼吸道感染症状的发生率和持续时间有所减少,降低感染的严重程度。
注:Lab4 益生菌包含:嗜酸乳杆菌CUL21(NCIMB 30156)和CUL60(NCIMM 30157),双歧杆菌CUL20(NCIMB 30153)和动物双歧杆菌乳亚种CUL34(NCIMM 30172)。
注意:根据说明书剂量服用或遵医嘱。
以上并不是维生素C可能发生的相互作用的完整列表。在开始补充维生素C或调整摄入量之前,请与医生或药剂师沟通,让医生知道你正在服用的所有药物,包括处方药和非处方药、其他维生素或微量营养素、草药补充剂等。
▼
静脉注射和口服给药,这两者可能具有不同的药代动力学特征。
药理学模型显示,口服维生素C,即使是在非常大和频繁的剂量下,也只能适度地增加血浆浓度,从0.07 mM增加到最大0.22 mM。
而静脉注射剂量预计会导致血浆维生素C峰值水平比口服剂量高60倍以上,尿液浓度比口服剂量低140倍。
分子的实际生物利用度由许多因素控制,包括肠道和其他组织的吸收、肾脏的吸收和排泄以及其他患者特异性因素。
除了通过静脉给药和口服给药的浓度差异外,口服给药将维生素C直接输送到肠道微生物组,而不是通过血液;因此,它对肠道微生物的影响可能与动力学和浓度有关,这与影响血浆水平的动力学和浓度完全不同。
静脉注射维生素C常用于临床医疗环境中,用于治疗某些疾病或特殊情况下的高剂量补充,如感染、外伤、手术恢复等。专业医生会给予相应的建议。
口服给药适用于一般的日常维生素C补充,维持正常的维生素C水平。
有几种方法可以评估人体中的维生素C状态。这些包括测量血浆、尿液、组织、粪便中维生素 C 的浓度。
▼
检测血浆维生素C,血浆样品中维生素C的定量测定常见的有两种方法:酶法和色谱法。
酶促维生素C测定
有几种基于维生素C的酶促转化的商业试剂盒,产生可以用光光谱法检测的信号。通常,抗坏血酸氧化酶用于这种类型的测定。这些测定的常见方法是酶联免疫吸附测定(ELISA),它非常适合分批处理样品,但不太方便立即测定少数样品中的值。
根据欧洲外部质量评估计划(Instand EQAS)中报告的方法,基于酶的分析方法在医院中并不常规使用。如果临床对立即测定维生素C的需求增加,由于其直接的技术性质,这些基于酶的测定可用于护理点或集中平台。
色谱法测定维生素C
抗坏血酸和DHA的定量测量目前是通过高效液相色谱(HPLC)方法进行的。如果必须分析具有相似性质的多种化合物,或者如果存在许多可能干扰感兴趣化合物定量的物质,则HPLC方法是优越的。
将酸化样品注射到HPLC仪器中后,通过通过基于化合物的物理性质不同地保留化合物的柱来分离化合物。结果,在分离柱的末端,可以选择性地检测抗坏血酸和DHA,而不受其他化合物的干扰。
目前有两种方法可以检测分离后的抗坏血酸和DHA,一种是电化学检测,另一种是紫外线检测。这两种检测方法给出的结果相同,但由于相对技术简单,紫外线检测更广泛地用于日常检测。
其他检测技术,如荧光检测,需要在柱前对抗坏血酸和DHA进行化学改性,但使用较少。比色/荧光法可能会产生更高的DHA浓度,因为该方法缺乏特异性。
▼
可以查看近期体内维生素状况。
肠道菌群可以影响食物中的营养物质的吸收和利用。肠道菌群的失调可能会影响维生素C吸收,从而出现维生素C缺乏,引发一系列健康问题。因此,检测肠道菌群的状况,可以帮助我们更好地了解维生素C的吸收和利用情况。
与抽血检测不同,肠道菌群的评估更加反映一段时间 ( 一般2周左右 ) 的长期状态,如部分维生素无法在体内留存,需要每日补充,血液检测波动较大。
肠道菌群与维生素C的水平之间存在怎样的关联?
为什么肠道菌群检测报告可以了解维生素情况?
我们来看下一章节。
我们在日常生活会看到,同样吃食物,有些人的维生素吸收状况比较好,有些人就容易缺乏,这是为什么呢?
为什么大剂量补充对一些人的身体有益,而少数人因为过量出现了肾结石呢?
这其实都与肠道微生物群相关。
这里我们分为两个方面来讨论:
一个是肠道微生物群对维生素C的影响,
一个是维生素C的补充对肠道微生物群的影响。
维生素C在人体代谢过程中的吸收和利用,与肠道微生物群相关。了解肠道微生物群对维生素C的吸收和利用的影响,可以帮助我们更好地理解其与人体健康之间的关系。
▼
微量营养素使用各种特定的吸收途径和机制,既可以是被动的,也可以是主动的。
膳食维生素 C 很容易通过钠依赖性维生素 C 转运蛋白(SVCT1 和 SVCT2)在肠道中吸收,其他比如维生素A 、维生素D 的吸收通过小肠中的被动扩散发生。
肠道微生物群是人体肠道中的有效生物反应器,可将各种化合物转化为有益或有害的代谢物,因此对其生物利用度起着至关重要的作用。
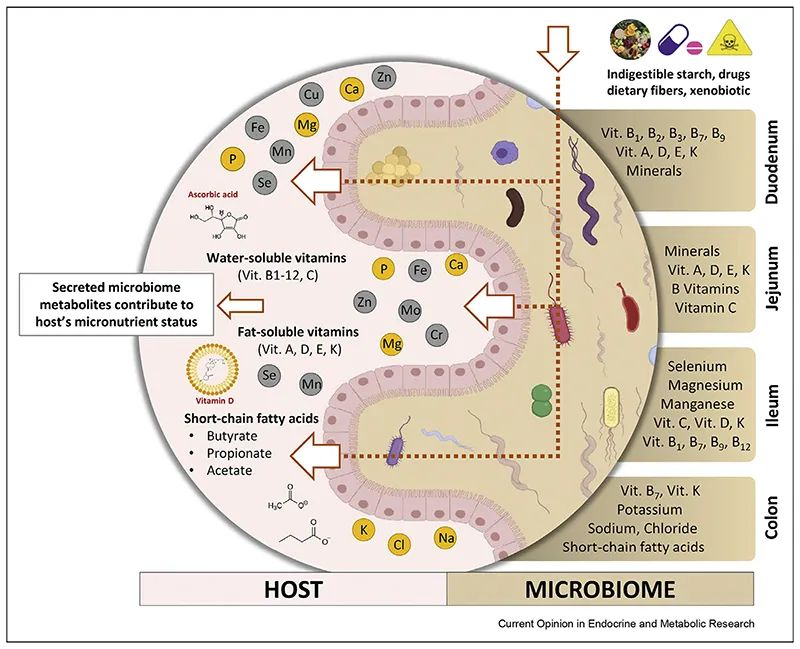
doi: 10.1016/j.coemr.2021.100285
胃肠道各部分理化特性的差异,以及位点特异性受体的存在,使得不同的维生素和矿物质能够沿胃肠道吸收。不同微生物在每个不同部分的定殖会影响当地环境,从而对微量营养素的生物利用度产生积极或消极的影响。
微生物可以干扰维生素C的吸收
微生物可以通过干预生物合成过程和调节吸收,来调节微量营养素的水平,包括维生素C。
来自革兰氏阴性菌的脂多糖降低SVCT-1的表达,进而降低SVCT-1-介导的维生素C的摄取。
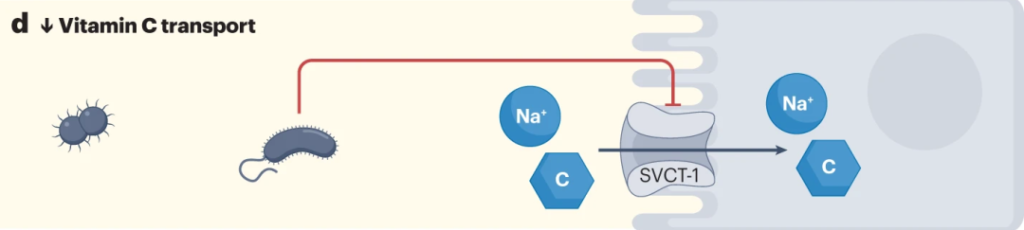
Chmiel JA, et al.,Nat Rev Urol. 2023
大肠杆菌可以通过释放脂多糖来减少宿主对抗坏血酸的摄取,从而增加NF-κB依赖性TNF的产生,其进而通过抑制SLC23A1和SLC23A2启动子(分别编码SVCT1和SVCT2),来降低钠依赖性维生素C转运蛋白SVCT1与SVCT2的表达。从而对维生素C的吸收率产生负面影响。
细菌可以与宿主竞争维生素C
大肠杆菌抗坏血酸转运蛋白对抗坏血酸的亲和力高于哺乳动物SVCT1,这表明细菌可以与宿主竞争维生素C。
因此,肠道菌群中大肠杆菌等革兰氏阴性菌占比较多的情况,可能不利于维生素C的吸收,这在谷禾肠道菌群检测报告中也是可以反映的。
▼
肠道细菌如大肠杆菌和乳酸杆菌代谢维生素C。
利用ula基因簇,大肠杆菌等细菌可以将抗坏血酸代谢为D-木酮糖,宿主细胞可以进一步加工木酮糖以产生草酸盐。
在肺炎链球菌、沙门氏菌、福氏志贺菌、粪肠球菌、肺炎克雷伯菌等病原菌中,也发现了相同的ula基因簇。
带ula的致病菌与草酸盐:携手制造结石
结石形成者(比如肾结石患者)微生物群更常见地富含携带ula基因簇的致病菌,并且这些患者在接受口服维生素C时,草酸盐增加的水平比非结石者增加得更多。高草酸盐水平增加了结石的风险。
因此,如果在肠道菌群检测报告中发现以上提到的致病菌占比较多,则有可能在代谢维生素C的时候产生的草酸盐过多,增加了结石的风险。
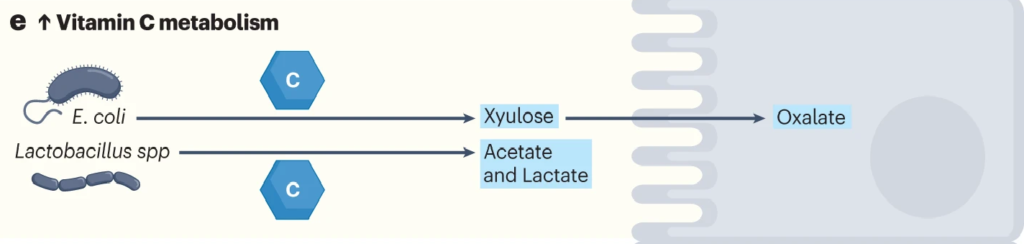
Chmiel JA, et al.,Nat Rev Urol. 2023
“
以上通过微生物群代谢维生素的机制,有助于我们更好地理解结石形成的机制。
既然有促进结石的细菌,自然也有降解的细菌:
一些乳酸杆菌可以将维生素C转化为乙酸盐和乳酸盐,这是一种无毒的代谢产物,通过生物能量途径增加微生物组的功能,并可能促进这些草酸降解细菌的定植。
Oxalobacter formigenes是一种革兰氏阴性厌氧细菌,可降解肠道草酸盐并促进原发性高草酸条件下肠道草酸盐的分泌。该菌在肠道定植可降低尿液或血浆中的草酸盐浓度。
Barone M, et al., Biofactors. 2022
我们看到生活中有人认为不能吃生菠菜,会得肾结石,但有些人每天吃生菠菜也没事…其实可能是因为人家的肠道菌群中致病菌较少,而降解草酸的菌群又在拼命干活…
因此,健康的肠道菌群结构非常重要,菌群在该干活的时候各司其职,井井有条,那么你的身体抵抗疾病的能力也随之提升。
以上是肠道菌群对维生素代谢吸收的影响,反过来,维生素C的补充也可以影响肠道菌群的组成。
二、
补充维生素C可以直接调节肠道微生物群,也可以通过改变氧化还原电位、修复肠道屏障,改善肠道条件,支持部分有益菌生长,防止有害菌泄漏到身体其他部位。
▼
维生素C直接调节肠道菌群
与安慰剂组相比,补充维生素C已被证明可以显着增加微生物生态系统的多样性,以及Collinsella的相对丰度和粪便水平的短链脂肪酸,特别是丁酸盐和丙酸盐。
在健康受试者中,每日高剂量维生素C补充(1000 mg/天):
下列菌群的相对丰度升高
下列菌群的相对丰度下降
一项观察性研究探讨了微量营养素维生素C对肠道微生物组组成和多样性的影响。结果表明,维生素C增加了肠道中双歧杆菌属的丰度,在科水平上,毛螺菌科和双歧杆菌科显著增加。
双歧杆菌属的成员是有益菌,例如增加ATP生成、调节免疫系统、黏膜屏障完整性、短链脂肪酸的产生,对健康有益,维生素C通过增加肠道有益菌促进健康。
研究发现,囊性纤维化患者维生素C摄入量的增加与厚壁菌门的丰度呈正相关,与拟杆菌门的丰度呈负相关。
▼
改变氧化还原电位
调节微生物组的机制类似于维生素B2:通过改变氧化还原电位,改善消化道中的厌氧/耐氧平衡,从而选择性地支持微生物生长,改善肠道条件。
与安慰剂相比,维生素C组粪便样本中的粪便pH值和氧化还原平衡降低。
▼
修复肠道屏障
我们知道,肠道屏障受损,细菌和有害物质有可能会穿过屏障进入血液循环,对人体产生各种负面影响,导致诸如过敏、炎症、自身免疫疾病等多种反应。
关于肠漏可以详见我们之前的文章:
而维生素 C 具有维持肠粘膜屏障完整性和修复粘膜屏障的作用。
这里介绍两种修复肠屏障机制。
——通过激活Notch 信号
Notch 信号影响细胞正常形态发生的多个过程,与许多人类疾病有关,包括IBD,因此被认为是癌症治疗的潜在靶点。Notch 信号通路的激活会改变紧密连接蛋白的表达并影响其分布的连续性,从而降低细胞屏障通透性。
豚鼠结肠组织相关基因检测表明,低剂量的维生素C可强烈激活Notch/Hes-1信号通路,对DSS诱导的结肠炎豚鼠的肠粘膜具有一定的保护作用。肠上皮受损时,Notch-1表达增加可促进上皮细胞增殖,有利于损伤部位的修复和重建。
——通过增加肠道胶原蛋白合成
增加维生素 C 摄入量的另一个潜在好处是肠道胶原蛋白合成增加,从而改善屏障功能。这一提出的机制与抗坏血酸的辅酶功能一致,即羟基化脯氨酸和赖氨酸以交联胶原蛋白。
例如,对具有吲哚美辛诱导的屏障功能障碍的 T84 人隐窝样上皮细胞系的研究表明,细菌通过跨细胞途径穿过上皮细胞,维生素 C 治疗可消除该途径。因此,肠道中抗坏血酸状态不佳可能会加剧屏障功能障碍,从而增加 LPS 衍生的革兰氏阴性菌的易位,从而加剧炎症。
维生素C修复肠道屏障后,肠道屏障可以正常发挥吸收、分解和转换营养物质等功能,同时可以帮助促进健康微生物群栖息和生长,从而促进整体健康。
以上我们了解到,维生素C可以通过多种方式影响肠道菌群,从而促进整体健康。那么当我们看到肠道菌群报告中菌群多样性较低,部分菌群失调,尤其是上面提到的毛螺菌科、肠球菌属、Collinsella、Gemmiger formicilis等,可以考虑通过补充维生素C来调节。
我们首先可以考虑通过食物补充,如卡姆果、针叶樱桃等维生素C含量很高的食物,或者一些常见的蔬菜水果例如:猕猴桃、番石榴(芭乐)、青椒、羽衣甘蓝等。也可以考虑通过维生素C补充剂调节。
人体维生素研究面临着许多挑战,维生素通常是通过食物摄入的,而食物中同时存在多种营养素,这使得难以研究单一维生素的作用;某些高剂量维生素的使用可能存在潜在的风险,进行研究时涉及到人体试验和干预,需要遵循伦理准则,并确保研究的安全性;不同人对维生素的需求和代谢能力存在个体差异,基因、环境和生活方式等因素都可能影响维生素的吸收、利用和代谢。
肠道菌群研究可以揭示不同个体之间菌群组成的差异,这有助于理解个体差异对维生素代谢和利用的影响。可以通过检测肠道菌群的组成和丰度,了解维生素的代谢情况,从而辅助评估维生素水平。
综合运用多种研究方法和技术,结合肠道菌群检测,可以考虑多个因素对维生素代谢和利用的综合影响,进一步理解维生素的作用机制,对人体健康具有重要意义。
注:本账号内容仅作交流参考,不作为诊断及医疗依据。
主要参考文献
Barone M, D’Amico F, Brigidi P, Turroni S. Gut microbiome-micronutrient interaction: The key to controlling the bioavailability of minerals and vitamins? Biofactors. 2022 Mar;48(2):307-314.
Chmiel JA, Stuivenberg GA, Al KF, Akouris PP, Razvi H, Burton JP, Bjazevic J. Vitamins as regulators of calcium-containing kidney stones – new perspectives on the role of the gut microbiome. Nat Rev Urol. 2023 May 9:1–23.
Pham VT, Dold S, Rehman A, Bird JK, Steinert RE. Vitamins, the gut microbiome and gastrointestinal health in humans. Nutr Res. 2021 Nov;95:35-53.
Li XY, Meng L, Shen L, Ji HF. Regulation of gut microbiota by vitamin C, vitamin E and β-carotene. Food Res Int. 2023 Jul;169:112749.
Yang Q, Liang Q, Balakrishnan B, Belobrajdic DP, Feng QJ, Zhang W. Role of Dietary Nutrients in the Modulation of Gut Microbiota: A Narrative Review. Nutrients. 2020 Jan 31;12(2):381.
Steinert RE, Lee YK, Sybesma W. Vitamins for the Gut Microbiome. Trends Mol Med. 2020 Feb;26(2):137-140.
Gomes-Neto JC, Round JL. Gut microbiota: a new way to take your vitamins. Nat Rev Gastroenterol Hepatol. 2018 Sep;15(9):521-522.
Rowland I, Gibson G, Heinken A, Scott K, Swann J, Thiele I, Tuohy K. Gut microbiota functions: metabolism of nutrients and other food components. Eur J Nutr. 2018 Feb;57(1):1-24. doi: 10.1007/s00394-017-1445-8. Epub 2017 Apr 9. PMID: 28393285; PMCID: PMC5847071.
Traber MG, Buettner GR, Bruno RS. The relationship between vitamin C status, the gut-liver axis, and metabolic syndrome. Redox Biol. 2019 Feb;21:101091.
Rozemeijer S, Spoelstra-de Man AME, Coenen S, Smit B, Elbers PWG, de Grooth HJ, Girbes ARJ, Oudemans-van Straaten HM. Estimating Vitamin C Status in Critically Ill Patients with a Novel Point-of-Care Oxidation-Reduction Potential Measurement. Nutrients. 2019 May 8;11(5):1031.
Otten AT, Bourgonje AR, Peters V, Alizadeh BZ, Dijkstra G, Harmsen HJM. Vitamin C Supplementation in Healthy Individuals Leads to Shifts of Bacterial Populations in the Gut-A Pilot Study. Antioxidants (Basel). 2021 Aug 12;10(8):1278.
谷禾健康
锌与肠道微生物
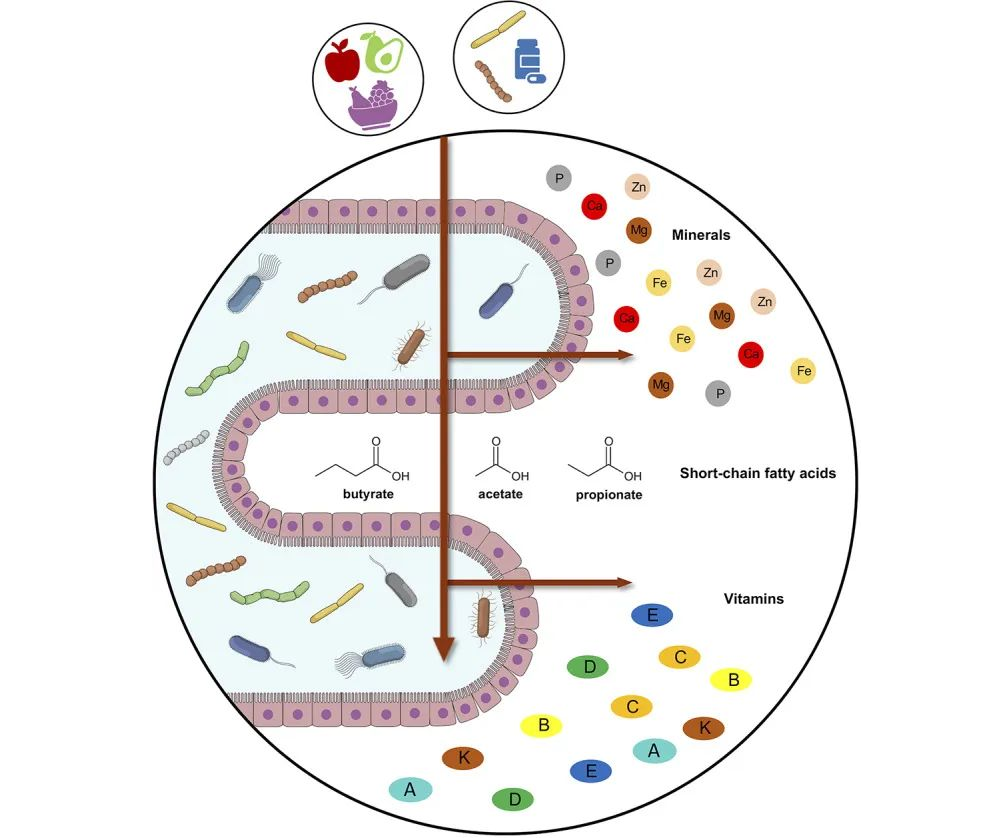
锌(Zn)是人体必需的微量元素,是人体中第二丰富的矿物质。锌在细胞和器官功能中起着关键的催化、调节和结构作用。
★ 膳食锌缺乏或过量均不健康
锌缺乏与发育不良、免疫功能低下、味觉丧失、不良妊娠结局、脱发、皮肤损伤和神经行为异常有关。
世界卫生组织估计,全球三分之一的人口面临锌缺乏症的风险,他们的饮食锌摄入量不足或食物中锌的生物利用度低。
不过需要注意不可在没有医嘱下擅自过量补充,导致锌的摄入量超过既定的营养需求。锌摄入过量也不利于健康,容易导致氧化应激增加、长期记忆受损、肠道吸收和分泌能力改变以及内脏脂肪增加。
近年来的几项体内研究表明,膳食锌摄入缺乏或过量会影响肠道菌群的组成和功能,进而对人体健康产生一定的影响。
本文主要讲述锌缺乏或锌超载情况下细菌类群的一些改变,短链脂肪酸生成变化,细菌金属抗性和抗生素抗性基因增加,以及锌相关转运蛋白的变化。
有助于进一步了解膳食锌对人体肠道微生物组的影响,并阐明利用肠道微生物组作为潜在锌状态生物标志物的适用性。帮助人们更好地控制锌的摄入量来塑造自身健康。
锌(Zinc)是一种化学元素,也是第四”常见”的金属。在现代工业中,锌是电池制造上不可替代、相当重要的金属。
锌还是人体必需的微量元素之一,起着极其重要的作用。
•维持正常食欲,利于生长发育
缺锌会导致味觉下降,锌会影响舌黏膜的正常功能,缺锌时味觉敏感度会比较差,可能还会因此而产生食欲不振或厌食等。长时间厌食会影响营养物质吸收,产生营养不良。
锌有助于生长发育、智力发育,缺乏锌会对我们的身体,特别是生长发育造成严重影响。所以,补充足够的锌是必须的。
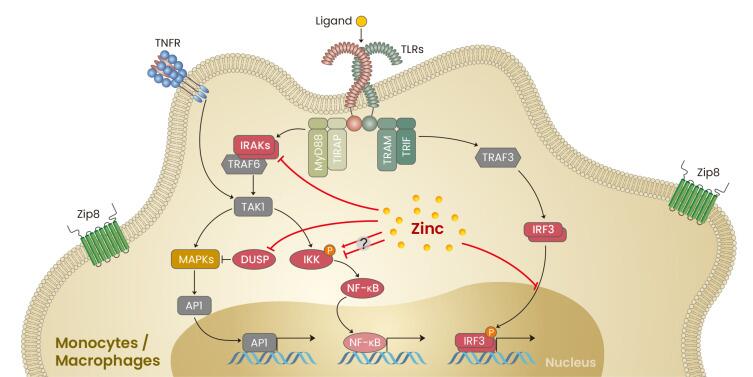
•增强人体免疫力
锌是免疫器官胸腺发育的营养素,只有锌充足才能有效保证胸腺发育,正常分化T淋巴细胞,促进细胞免疫功能。失调的锌稳态会损害整体免疫功能,从而增加对感染的易感性。
锌还以类似于钙的方式作为免疫反应的离子调节剂,通过作为细胞内信使参与信号通路。
锌在单核/巨噬细胞信号传导中的作用

Kim B,et al.Mol Cells.2021
•影响伤口恢复
锌作为许多转录因子和酶系统的辅助因子,包括锌依赖性基质金属蛋白酶,可在伤口修复过程中增强自体清创和角质形成细胞迁移。
缺锌易导致皮肤疾患,患口腔溃疡,受损伤口不易愈合,青春期痤疮等。
★ 锌对肠道微生物有重要影响
肠道微生物在宿主营养状况中发挥着重要作用,在免疫、与宿主细胞相互作用、代谢不可消化的营养素和合成必需维生素等方面发挥着关键作用。
锌结合蛋白占细菌蛋白质组的5%,突出了细菌定植胃肠道的主要锌。锌是一种重要的营养物质和抗菌剂,近年来的研究发现,膳食中锌含量的不同导致肠道微生物的组成和功能产生改变,进而对人体健康产生一系列影响。
膳食锌(锌缺乏或锌过量)对肠道菌群的影响
向上滑动阅览



Cheng J,et al.Crit Rev Food Sci Nutr.2023
接下来跟随谷禾了解一下膳食锌缺乏和过量情况下肠道微生物的具体改变
1. 在缺锌条件下,发现能够有效竞争锌的细菌(即肠杆菌科和瘤胃球菌科以及肠球菌属和脱硫弧菌属)增殖。
2. 与碳水化合物和糖代谢相关的微生物代谢途径的表达改变已被发现与锌缺乏相关,其中细菌可以将碳水化合物和糖代谢为短链脂肪酸。
3. 粘蛋白相关细菌丰度的改变(即粘嗜阿克曼氏菌和异源杆菌)也与锌缺乏有关。肠道粘蛋白由粘多糖和糖蛋白组成,细菌可以以这种粘蛋白为食并产生短链脂肪酸。
与锌缺乏相关的潜在结果包括细菌的定植和生长,这些细菌可以有效地竞争锌。
4. 此外,锌缺乏可能导致产生短链脂肪酸的细菌(即Blautia和Ruminococcus)的丰度降低。短链脂肪酸浓度与肠道pH值相关,由于短链脂肪酸含量降低导致的pH值升高会降低锌的溶解度,从而降低锌的生物利用度和吸收。
与锌缺乏相关的潜在宿主和肠道菌群
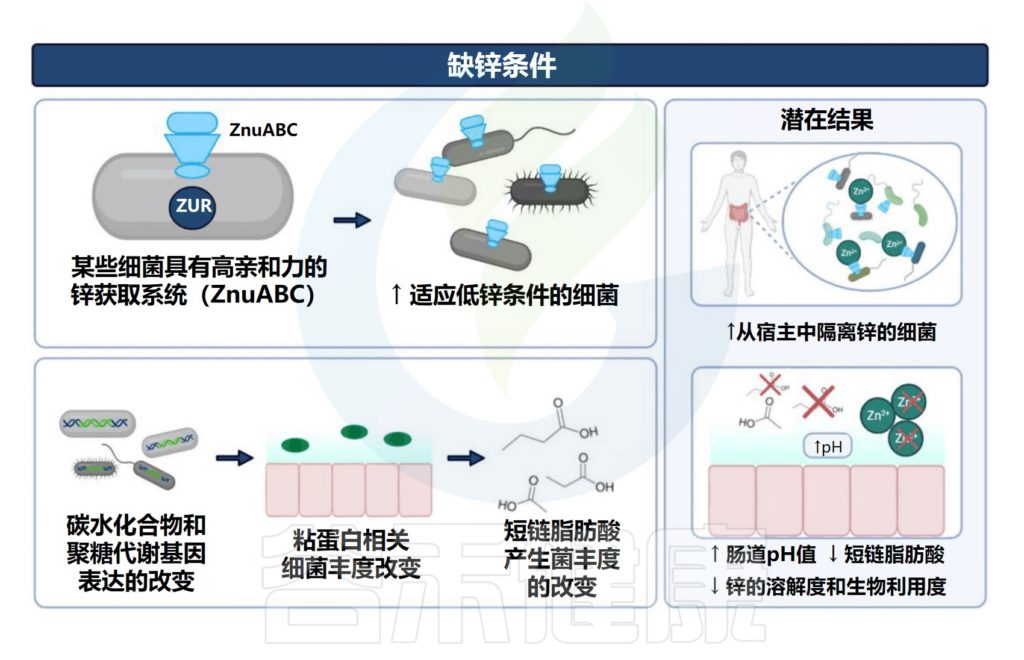
Cheng J,et al.Crit Rev Food Sci Nutr.2023
六项研究比较了锌缺乏相对于锌充足对肠道菌群的影响,其中五项研究通过膳食锌控制诱导锌缺乏。
与锌充足对照组相比,锌缺乏组血浆/血清、指甲、羽毛、肝脏、睾丸和粪便锌含量降低。
锌缺乏组的实验对象大部分生物多样性下降,尤其是粘蛋白相关细菌丰度降低,短链脂肪酸的产量也减少,微生物相关代谢也在一定程度上减弱。
以上实验研究主要是基于小鼠,大鼠,鸡和猪等动物实验,关于人类的临床实验还较少,但是动物的实验也可以为人类提供一定的参考价值。
锌缺乏影响微生物多样性
关于与锌缺乏相关的肠道菌群调节,所有六项研究都评估了α-多样性和β-多样性。
✦α多样性
关于缺锌对肠道菌群α-多样性的影响,六项研究尚无共识。总的来说,三项研究发现α-多样性的改变与缺锌相关,三项研究没有发现变化,其中大多数研究采用了不同的α-多样性测量方法。
缺锌对α多样性的影响具有差异
在锌缺乏的情况下,一项研究发现α-多样性以性别依赖的方式增加或减少(雄性小鼠增加,雌性小鼠减少)。
同样,在人类研究中,缺锌对α-多样性的影响也存在差异。在学龄儿童中,锌缺乏导致α-多样性增加,而在锌缺乏的母乳和配方奶喂养的婴儿中,α-多样性降低。
虽然每项研究中生物多样性变化的差异可能与不同的研究对象有关,但还需要在人类和动物模型中进一步研究锌缺乏对肠道微生物群改变的影响。
✦β多样性变化暂不明确
进行的六项研究中有五项发现了与锌缺乏相关的β多样性变化。
然而,在第六项研究中没有发现β-多样性的变化,该研究使用了θYC距离测量。β多样性的变化并不一定表明细菌类群的有益或消极变化。
●门水平
在门水平上,锌缺乏与变形菌门(Proteobacteria)和黑色素杆菌(Melainabacteria)的增加以及厚壁菌门(Firmicutes)的减少有关。
注:其中变形菌丰度的增加被认为是生态失调和疾病风险的潜在诊断标准,因为在健康的人类肠道中发现了低变形菌丰度。而厚壁菌门的许多成员都是有益菌
这或许可以从一部分角度解释锌缺乏对健康的不利影响
短期缺锌与长期缺锌菌群改变不同
短期缺锌会降低疣微菌门(Verrucomicrobia)的丰度(主要是Akkermansia muciniphila属),而长期缺锌则与疣微菌门丰度的增加有关。
菌群改变出现性别差异
发现与锌充足的野生型小鼠相比,通过ZnT7敲除诱导的锌缺乏导致了敲除(ZnT7-/-)和异质(ZnT7+/-)小鼠的红蝽菌科(Coriobacteriaceae)丰度和Allobaculum属丰度的增加,这种变化仅在雌性小鼠中发现。这表明锌的调节功能可能因性别而异。
注:因此锌对不同性别的人群影响可能不同
●属水平
在属水平上,研究发现双歧杆菌(Bifidobacterium)、希瓦式菌属(Shewanella)、Rheinheimera的减少与血浆锌水平相关。
除此之外还发现粪杆菌属(Faecalibaculum)增加,经黏液真杆菌属(Blautia)、乳杆菌(Lactobacillus)、Negativibacillus减少。
慢性缺锌的情况
在慢性锌缺乏的情况下,以下菌群丰度增加:
肠球菌(Enterococcus) ↑↑↑
肠杆菌(Enterobacteriaceae) ↑↑↑
瘤胃球菌科(Ruminococcaceae) ↑↑↑
而下列菌群丰度下降:
Clostridiales ↓↓↓
消化链球菌科(Peptostreptococcaceae) ↓↓↓
●物种水平
在物种水平上,Lachnospiraceae NK4A136和
真杆菌属(Eubacterium coprostanoligene)的丰度随着锌的缺乏而增加,而Ruminococcus gauvreauii的丰度随着锌的缺乏而降低。
此外还发现了一些菌群的增加和下降
增加的菌群:
吲哚梭菌(Clostridium indolis) ↑↑↑
未分类的某种拟杆菌 ↑↑↑,
减少的菌群:
酸奶瘤胃球菌(Ruminococcus lactaris) ↓↓↓
肠球菌(Enterococcus sp.) ↓↓↓
Clostridium lactatifermentans ↓↓↓
Clostridium clostridioformme ↓↓↓
// 注意
锌缺乏可能使得一些对人体有益的细菌丰度下降,而致病菌的丰度上升,从而对健康产生影响
虽然在上述研究中发现了从门到属水平的显著差异,但在一些学者的研究中,微生物群的相对丰度没有显著差异。
锌缺乏对菌群主要代谢产物——短链脂肪酸的影响
研究了锌缺乏与微生物代谢物(短链脂肪酸)生产和功能的变化。
✦锌缺乏影响短链脂肪酸的产量
锌缺乏时间不同对短链脂肪酸的影响不同
一项小鼠研究发现,短链脂肪酸产量在短期锌缺乏时增加,但在长期锌缺乏时减少。鸡的研究发现,相对于锌充足,锌缺乏会降低短链脂肪酸的产量。
具体而言,短期(4周)饮食锌不足会增加盲肠乙酸,并与各组短链脂肪酸产生细菌(Flavonifractor、Alistipes、Odoribacter、Anaerotruncus、 Alloprevotella、Bifidobacterium adolescentis、Paralactobacillus Leisner)呈正相关,表明在锌匮乏的条件下,肠道微生物群可以增加锌的利用率。
另一方面,长期锌不足(8周)与戊酸盐产量下降有关。发现与碳水化合物、氨基酸、萜类/聚酮、核苷酸代谢、聚糖生物合成和代谢以及异种生物降解和代谢相关的微生物代谢途径随着长期锌缺乏而降低。
✦短链脂肪酸浓度影响锌的利用与吸收
短链脂肪酸浓度与肠腔pH值相关,其中,由于短链脂肪酸含量增加而导致的pH值降低可增加锌的溶解度,从而增加锌的生物利用度和吸收。
说明锌与短链脂肪酸存在相互作用的关系
鉴于短链脂肪酸、宿主营养和肠上皮维持之间的联系,其中短链脂肪酸的产生与黏蛋白的产生有关,细菌发酵碳水化合物和抗性淀粉,短链脂肪酸被用作细菌能量来源。
一组研究人员发现锌缺乏导致乙酸盐产量下降,这可能是由于产生短链脂肪酸的细菌属,如梭状芽胞杆菌和瘤胃球菌的减少。
综上所述,锌缺乏情况下肠道菌群组成的改变及其相关代谢途径影响碳水化合物和糖基代谢、肠道黏蛋白降解,从而影响短链脂肪酸的产生。
锌缺乏与代谢通路变化
✦锌缺乏使微生物相关代谢减弱
在微生物代谢途径中,锌缺乏与碳水化合物代谢相关基因的表达降低有关、以及糖聚糖生物合成和代谢、核苷酸代谢、氨基酸代谢、抗生素耐药性、矿物质吸收、脂肪酸代谢、一级/二级胆汁酸合成和异种生物降解都有联系。
此外,发现与锌吸收、脂肪酸代谢和碳水化合物消化和发酵相关的KEGG途径表达减少,其中最后一种途径也可能导致短链脂肪酸产量减少,这与提高锌生物利用度有关,从而使锌缺乏状态持续存在。
注:基于人类疾病KEGG通路,分别在短期和长期锌缺乏症中发现了更多与物质依赖和心血管疾病相关的基因。
锌缺乏对肠道粘蛋白及相关菌的影响
额外的微生物类群改变与锌缺乏和肠粘蛋白丰富有关。由杯状细胞产生的黏液层由粘多糖和糖蛋白组成,并容纳有助于消化矿物质的肠道微生物群。
粘蛋白——黏蛋白关键特征是其形成凝胶的能力; 因此它们是大多数凝胶状分泌物的关键组成部分,提供润滑,细胞信号通路及化学屏障的功能。
✦锌缺乏影响粘蛋白相关细菌丰度
锌缺乏会影响与粘蛋白相关细菌丰度,例如嗜黏蛋白阿克曼菌的减少不利于肠道屏障的完整性,并在一定程度上影响乙酸和丙酸等短链脂肪酸的产生,从而对健康产生危害。下面是一些具体实验证据:
锌缺乏影响嗜黏蛋白阿克曼菌丰度
共生细菌以黏液蛋白为食并粘附在黏液蛋白上,与宿主相互作用并调节黏液蛋白基因的表达和分泌,与上皮细胞相互作用以维持黏液和上皮内稳态。
在短期锌缺乏的情况下,嗜黏蛋白阿克曼菌(Akkermansia muciniphila)在统计学上显著下降,这与之前的研究一致,这些研究表明嗜黏蛋白阿克曼菌相对丰度较低与锌缺乏相关。
嗜黏蛋白阿克曼菌(A.muciniphila)存在于大肠黏液层,调节肠道屏障的完整性,其丰度与代谢性疾病和炎症性肠病发病率呈负相关。此外,嗜黏蛋白阿克曼菌是一种粘蛋白降解物,可将粘蛋白转化为乙酸和丙酸以供能量使用。
锌缺乏与其他影响粘蛋白丰度的菌群相关
同样,在缺乏锌的情况下,发现经黏液真杆菌属(Blautia)和乳酸杆菌属,以及粪杆菌(Faecalibaculum)属的增加,这些都是可能影响粘蛋白丰度的类群。
乳酸菌的减少可能会减少肠道黏蛋白,因为某些乳酸菌会刺激黏液蛋白2 (MUC2)的产生,黏液蛋白2是杯状细胞表达的主要肠道黏蛋白。
经黏液真杆菌属(Blautia)含量降低,经黏液真杆菌属含有粘蛋白降解基因是一种短链脂肪酸(丁酸和乙酸)生产者,其丰度可能预测锌状态的差异。
异杆菌属丰度增加有利于结肠上皮健康
研究发现,与锌充足野生型(ZnT7+/+)相比,通过敲除ZnT7诱导的锌缺乏导致敲除(ZnT7-/-)小鼠和异质性(ZnT7+/-)小鼠的异杆菌属(Allobaculum)丰度在雌性小鼠中均有所增加。黏蛋白密度(酸性和中性黏蛋白减少,杯状细胞数量减少)与异杆菌属(Allobaculum)相对丰度呈负相关。
异杆菌属(Allobaculum)与发酵碳水化合物(即粘蛋白)有关,以产生短链脂肪酸(乳酸和丁酸),供异源菌群作为碳源利用,这表明异杆菌属(Allobaculum)丰度的增加可能与有利于结肠上皮健康的潜力有关。
小结
总的来说,锌缺乏通常会导致细菌生物多样性、从门到种水平的细菌丰度的降低、短链脂肪酸的产生减少以及与碳水化合物和糖基代谢相关的细菌代谢途径的减弱,锌相关蛋白基因表达也可能在一定程度上减弱。
注:这些变化影响了人体原本正常的代谢,可能导致一些疾病的产生。
注意
所有六项关于锌缺乏和肠道菌群改变影响的研究都使用了膳食锌作为锌源。对于α-多样性和短链脂肪酸产生的变化还缺乏共识,一个可能的解释可能是所使用的动物模型以及模型的生命阶段和性别的差异。
注:只有一项研究使用了大鼠模型,而所有其他研究都使用了小鼠模型。与其他研究相比,所使用的小鼠年龄最大,小鼠肠道菌群随着年龄的增长逐渐稳定。
三项研究仅使用男性受试者,一项研究仅使用女性受试者,两项研究同时使用女性和男性受试者,其中考虑到人类性别之间免疫功能的差异以及锌、免疫功能和肠道微生物组之间的联系,在设计阐明锌缺乏症对肠道微生物组组成和功能的影响的未来研究时,应考虑受试者的性别。
1.与高锌或锌过载相关的潜在结果包括具有高锌条件机制的细菌定殖和生长。
某些细菌具有通过有效的重金属排出系统来抵消高锌浓度的机制。在高锌条件下,发现了对增加金属锌暴露有进化适应的细菌(即梭状芽孢杆菌科、肠杆菌科和大肠杆菌)。
2.增加的细菌进化适应增加的金属暴露(金属抗性),已发现与抗生素耐药性相关的微生物基因表达增加有关。
此外,高锌条件可能会潜在地增加耐药肠道微生物群,其中对高锌的进化适应以及抗生素和金属耐药基因的增选可能会由于水平基因转移而进一步延续这种耐药现象。
与高锌或锌过载相关的潜在宿主和肠道菌
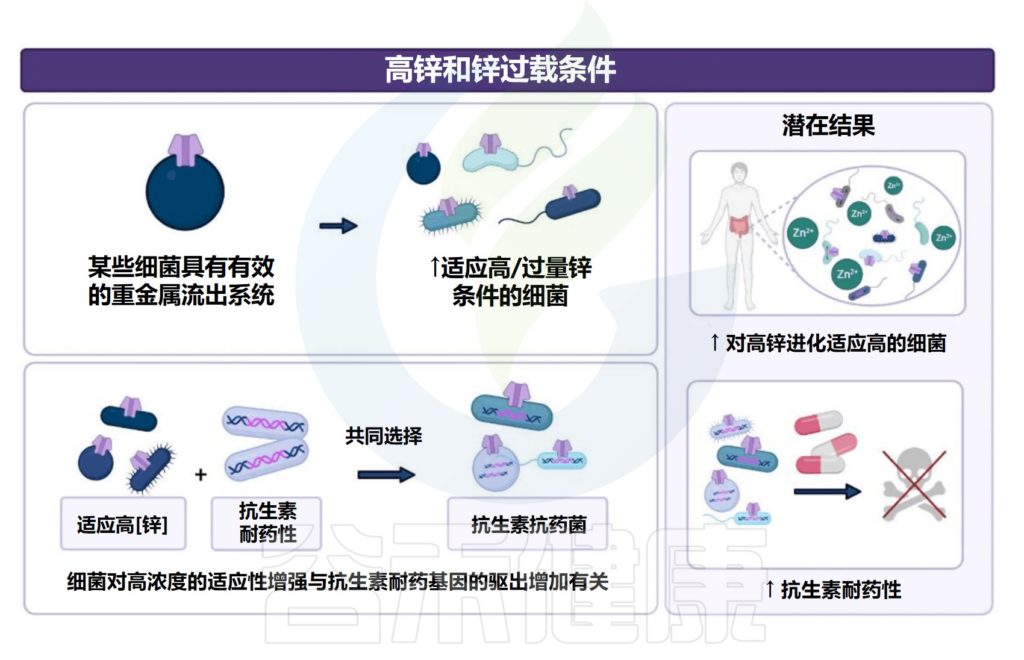
Cheng J,et al.Crit Rev Food Sci Nutr.2023
★锌过量的危害
1.一般情况下,补锌过量可能会导致机体出现锌中毒,大部分人群可能会出现胃肠道症状,如恶心、呕吐等。
2.抵抗力下降:过高的血锌会抑制白细胞的吞噬作用和杀菌作用,使人体的抵抗力下降,容易遭受致病菌侵袭。
3.记忆力下降:对于不缺锌的人,额外补充有可能造成体内锌过量,从而引发代谢紊乱,甚至会对大脑造成损害,可能损伤大脑神经元,导致记忆力下降。
4.贫血:有些家长误把锌制品当作营养品,给婴儿长期大量服用,殊不知补锌过量同样会带来危害,会使体内的维生素C和铁的含量减少,抑制铁的吸收、利用,容易引起、加重缺铁性贫血。
下面具体讲述一下锌过量状态下肠道微生物具体发生的变化
五项研究比较了高膳食锌或锌过载相对于锌充足对肠道菌群的影响。
锌过量与相关代谢基因
与锌充足对照相比,高锌或锌过载导致锌相关蛋白基因表达有所改变。
✦高锌摄入降低锌转运蛋白ZIP4基因表达
ZIP4是锌从肠腔进入肠上皮细胞的主要导入物,由饮食中的锌调节,随着锌摄入的增加,ZIP4的表达下调。
两项研究发现,高锌摄入量会降低ZIP4基因的表达。还发现,高锌摄入的小鼠ZnT2表达增加(仅回肠),MT1和MT2表达增加(回肠和结肠),而其他测试的ZnT(4-7,9)和ZIP(1,3,5-11,13,14)蛋白的基因表达没有变化。
✦脂质代谢基因表达降低
与短链脂肪酸产量降低一致,与脂质代谢相关的微生物基因表达也在高锌和过量锌饲粮中被发现降低。
✦抗性基因表达增强
在微生物代谢途径中,高锌和过量膳食锌与抗生素耐药性相关基因的表达增加有关、传染病和金属离子结合,以及与脂代谢、异种生物降解和酶活性(果胶酯酶、α-葡糖醛酸酶、α-l-阿拉伯糠醛苷酶)相关的基因表达降低。
抗生素耐药性增强
将抗生素抗性基因丰度的增加与肠杆菌的增加联系起来,其中高氧化锌饲粮已普遍用于猪,并导致促进抗生素耐药性。
具体而言,在猪肠道中发现耐多药大肠杆菌的患病率增加,这归因于高氧化锌饮食。
研究发现,饮食中锌过量会增加肠球菌,其中肠球菌具有质粒编码的耐药基因,本质上对许多常见抗菌剂不那么敏感。
尽管没有研究微生物代谢基因,但考虑到细菌类群对锌增加的适应性增加,考虑到金属耐药基因和抗生素耐药基因的共同共存,有可能存在抗生素耐药细菌基因的增加。
总的来说,高锌或锌过载似乎潜在地增加了耐抗生素的肠道微生物群,其中对高锌的进化适应以及抗生素和金属耐药基因的增选可能会由于水平基因转移而进一步延续这种耐抗生素现象,从而导致人体的抗生素和金属耐药性增强。
锌过量影响微生物多样性
评估了与高锌或锌过载相关的肠道菌群调节的多样性测量;4项研究评估了α-多样性,5项研究评估了β-多样性。
✦高锌下α多样性降低
利用Pielou的均匀度α-多样性测量发现,α-多样性随着高锌而降低。通过1/Simpson多样性测量也发现,α-多样性随着Zn过载而降低。
此外,短期(4周)高饲粮锌或锌过载被发现不会显著影响α-多样性,长期高锌被发现不会改变(观察物种)或降低α-多样性,长期(8周)锌过载也不会改变α-多样性。
注:这可能取决于所使用的评估方法
α多样性下降,机会性病原体增多,双歧杆菌和拟杆菌显著减少,兼性厌氧菌、梭杆菌等增多,进而影响人体的的健康。
✦β多样性的变化未达成共识
关于高锌或锌过载对微生物组β-多样性的影响,在五项选定的研究中也没有共识,其中三项研究发现了与高锌或锌过量相关的β-多样性变化,并在其他两项研究中保持不变,尽管不同的研究使用了不同的距离测量。
疣微菌门的丰度与锌过载暴露时间有关
在小鼠中,高饲粮锌和锌过载在短期内增加了疣微菌门(Verrucomicrobia)丰度,但在长期暴露后会降低疣菌菌丰度,此前在仔猪中也有这种情况,其中长期高饲粮锌或锌过载会通过细菌金属间竞争、金属蛋白错金属、或与金属毒性和细菌金属稳态改变相关的氧化还原活性。
高锌显著增加了梭状芽孢杆菌科(Clostridiaceae)的丰度,其中梭状芽孢杆菌科被发现具有通过有效的重金属外排系统来抵消高锌浓度的机制,该系统可能是从携带重金属抗性基因的质粒转移而来的。
梭菌科更适应高锌的浓度
鉴于这些更强的抗锌机制,梭菌科可以适应高锌浓度,因此相对于其他细菌有更多的定殖。最后,在高膳食锌摄入量下,与金属离子结合和金属离子转运相关的微生物基因表达增加,支持了对高膳食锌环境具有特定进化适应的正向选择细菌类群的假设。
注:在所选研究中,在不同的分类水平上研究了肠道微生物群的组成和功能的变化。在这五项研究中都发现了细菌类群的显著变化,这些变化可以归因于微生物群对有效锌浓度增加的适应。
✦门水平
对于细菌类群的变化,在门水平上,两项研究发现高锌没有显著改变细菌丰度。
虽然在两项利用猪模型的研究中拟杆菌门(Bacteroidetes)丰度有所增加,但在门水平上的其他变化没有达成共识。
时间不同菌群丰度变化不同
研究发现短期(4周)高锌和锌过载会增加疣微菌门(Verrucomicrobia)的丰度,而长期暴露(8周)会降低疣微菌门(Verrucomicrobia)的丰度。
另一项研究发现,短期锌过载导致变形菌门丰度增加,长期锌过载导致放线菌门丰度增加。
✦科、属、种水平变化较小
与膳食锌含量过高或过量相关的科、属或种水平变化之间不明显。
在短期(4周)和长期(8周)暴露中,黑色素杆菌(Melainabacteria)与血清锌水平呈负相关,而Desulfovibrio sp. ABHU2SB与血清锌水平呈正相关。
✦锌过载时肠道菌群的具体变化
罗伊氏乳杆菌(L. reuteri)与短期锌过载呈负相关;然而,随着长期锌过载,该属与血清锌无显著相关性。
高锌显著增加了梭菌科(Clostridiaceae)丰度,降低了乳杆菌科(Lactobacillaceae)和肠杆菌科(Enterobacteraceae)丰度。
在利用猪模型的研究中发现, 普雷沃氏菌(Prevotella)和乳酸菌属增加,琥珀酸弧菌属(Succinivibrio) 减少。
另一组研究人员发现锌过载下列菌株丰度增加:
拟杆菌属(Bacteroides) ↑↑↑
副杆菌属(Parabacteroides) ↑↑↑
柯林斯菌属(Collinsella) ↑↑↑
醋弧菌属 (Acetivibrio) ↑↑↑
经黏液真杆菌属(Blautia) ↑↑↑
粪球菌属(Coprococcus) ↑↑↑
普拉梭菌(Faecalibacterium) ↑↑↑
霍尔德曼氏菌(Holdemania) ↑↑↑
Subdoligranulum ↑↑↑
Nestiinimonas ↑↑↑
Lachnoclostridium ↑↑↑
Pseudoflavonifractor ↑↑↑
而以下菌群减少:
巨型球菌属(Megasphaera) ↓↓↓
戴阿利斯特杆菌属 (Dialister) ↓↓↓
氨基酸球菌属(Acidaminococcus) ↓↓↓
瘤胃球菌(Ruminococcus) ↓↓↓
Treponema ↓↓↓
虽然生物多样性变化的差异可能与每项研究中不同的研究对象和不同的研究时间有关,但高膳食锌或锌过量对人类肠道微生物群的影响仍不确定,这突显了进一步研究的必要性。
锌过量对细菌主要代谢产物——短链脂肪酸的影响
✦高锌降低短链脂肪酸含量
长期(8周)高锌与对照组相比,短链脂肪酸总产量更低。具体而言,与锌充足相比,长期高饲粮锌导致盲肠戊酸、丁酸、乙酸、异戊酸、异丁酸和丙酸降低。
短链脂肪酸在管理炎症和促进肠上皮完整性方面发挥着重要作用,高锌除了抑制短链脂肪酸生成外,还发现导致结肠炎症增加。
锌过量与炎症及免疫
虽然锌通常被认为是安全的,因为它不具有氧化还原活性,但大量或过量的锌暴露与肠道功能的改变、免疫功能的改变和炎症有关,因此强调需要确定高剂量和锌过载剂量对肠道微生物组组成和功能的影响。
✦锌过量影响炎症疾病发病率
乳酸菌科的细菌与短链脂肪酸的产生有关,将碳水化合物发酵成乳酸。此外,乳酸菌,产生各种各样的胞外多糖,作为阳离子金属离子(如Zn2+)的结合位点,可以保护这些微生物免受金属毒性。
根据种类的不同,乳酸菌可以是促炎的,也可以是抗炎的。在一项研究中,来自乳酸菌科的细菌丰度增加(Lactobacillus spp.),但在两项研究中(罗伊氏乳杆菌)丰度下降,高膳食锌摄入量。人类罗伊氏乳杆菌丰度的降低与炎症疾病发病率的增加相关。
✦高浓度锌具有抗菌作用
此外,膳食中高浓度的锌在防御致病性大肠杆菌、单核细胞增生李斯特菌和链球菌方面发挥抗菌作用。
小结
总的来说,在大多数选定的研究中,高饲粮锌和锌过载状态与α-多样性降低或不变、短链脂肪酸产量降低以及金属和抗生素抗性基因增加有关。
这些因素都在一定程度上影响人体的健康,对其产生不利的影响。
不足之处
在每一项研究中,并没有量化短链脂肪酸和微生物代谢基因,这本可以更好地了解过量锌摄入相关的微生物群组成和功能。
饮食中高锌或锌过载对人类肠道菌群的影响研究较少。鉴于抗生素耐药性的潜在灾难性影响,未来的研究应了解微生物分类和多样性的变化,以及微生物代谢物产生的变化和驻留菌群的宏基因组潜力。
锌的每日推荐食用量
中国营养学会规定,婴幼儿(0-1岁)5毫克/天,儿童(1-10岁)10毫克/天。青春期男性15毫克/天,女性12毫克/天。
孕妇和哺乳期产妇因为母乳喂养,乳汁中的锌流失比较多,所以锌的摄入量也应多一点,每天20-25毫克。一般锌的摄入量不超过25毫克。
★ 含有锌的食物
海鲜类:如牡蛎、鱼、扇贝、虾仁等,不仅含有较多的锌元素,且含有较多的不饱和脂肪酸,有助于减少动脉硬化的发生风险,从而预防心血管疾病的发生,其中牡蛎的含锌量比较高;
肉禽类:如猪肝、瘦牛肉、猪肉、鸡肉等,通常会含有较多的蛋白质以及微量元素,如锌、铁、磷等,有助于补充锌元素以及预防缺铁性贫血的发生;
蛋类:如鸡蛋、鸭蛋中也含有一定的锌元素,还富含脂溶性维生素、磷脂等,有助于减少口唇干裂、皮炎等疾病的发生;
干果类:如花生、核桃、芝麻、蚕豆、黄豆、绿豆、板栗等,含有钙、锌等多种微量元素,以及胡萝卜素、核黄素等多种维生素和脂肪、碳水化合物,在补充锌元素的同时有助于能量的补充。
膳食锌缺乏与摄入过量都会影响肠道微生物组的组成和功能,进而影响人体正常代谢与健康。
缺锌时,肠道微生物组的改变包括α-多样性的改变、粘蛋白降解细菌丰度的下降、短链脂肪酸产生的减少以及影响碳水化合物和糖基代谢的代谢途径表达减弱。
过量摄入膳食锌的情况下,肠道微生物组变化包括α-多样性没有变化或减少,短链脂肪酸产量减少,细菌金属抗性和抗生素抗性基因的潜在增加。
未来期待更多关于锌等微量营养元素和肠道菌群互作以及影响机理的的人类试验,虽然人类膳食营养的研究中单一变量不太好控制,但是基于大人群的营养调查以及其他协变量控制情况下,去调查这些关键营养素与肠道菌群的扰动可以帮助进一步明确阐明营养在人体健康和疾病扮演的角色和作用机理,对于后续基于菌群或营养素的预防干预治疗有重要的指导价值。
主要参考文献:
Cheng J, Kolba N, Tako E. The effect of dietary zinc and zinc physiological status on the composition of the gut microbiome in vivo. Crit Rev Food Sci Nutr. 2023 Jan 23:1-20. doi: 10.1080/10408398.2023.2169857. Epub ahead of print. PMID: 36688291.
Kim B, Lee WW. Regulatory Role of Zinc in Immune Cell Signaling. Mol Cells. 2021 May 31;44(5):335-341. doi: 10.14348/molcells.2021.0061. PMID: 33986184; PMCID: PMC8175146.
Baholet D, Skalickova S, Batik A, Malyugina S, Skladanka J, Horky P. Importance of Zinc Nanoparticles for the Intestinal Microbiome of Weaned Piglets. Front Vet Sci. 2022 Jun 2;9:852085. doi: 10.3389/fvets.2022.852085. PMID: 35720843; PMCID: PMC9201420.
Chen, X., Y. Jiang, Z. Wang, Y. Chen, S. Tang, S. Wang, L. Su, X. Huang, D. Long, L. Wang, et al. 2022. Alteration in gut microbiotaassociated with zinc deffciency in school-age children. Nutrients 14 (14):2895.
Cheng, J., H. Bar, and E. Tako. 2021. Zinc status index (ZSI) for quantiffcation of zinc physiological status. Nutrients 13 (10):3399.
Chen, L., Z. Wang, P. Wang, X. Yu, H. Ding, Z. Wang, and J. Feng. 2021. Effect of long-term and short-term imbalanced Zn manipulation on gut microbiota and screening for microbial markers sensitive to zinc status. Microbiology spectrum 9 (3):e0048321.
Bombaywala, S., H. J. Purohit, and N. A. Dafale. 2021. Mobility of antibiotic resistance and its co-occurrence with metal resistance in pathogens under oxidative stress. Journal of Environmental Management 297:113315.

谷禾健康

传染病,肠道微生物,营养
传染病和感染目前是许多地区尤其是低收入国家主要死亡原因,也是婴儿和老年人等弱势群体的主要风险。免疫系统在这些感染的易感性、持续性和清除中起着至关重要的作用。由于 70-80% 的免疫细胞存在于肠道中,肠道微生物群、肠上皮层和局部黏膜免疫系统之间存在着错综复杂的相互作用。除了肠道中的局部黏膜免疫反应外,目前越来越多的证据证实:肠道微生物组也会影响全身免疫。
临床医生未来可能会更多地利用关于免疫系统、肠道微生物组和人类病原体之间复杂相互作用的更多知识。现在公认的营养对肠道微生物群组成和免疫系统的影响阐明了营养在改善健康方面可以发挥的作用。
本文将综合介绍维持微生物群、肠道健康、局部免疫反应和全身免疫之间复杂平衡的机制,并将其与一生中的传染病联系起来,并讨论营养在传染病预防和治疗中的影响。

由于卫生条件的改善、疫苗接种和抗生素的使用,传染病的发病率在过去几十年中显着降低。然而,仍有近三分之一的死亡仍与传染病有关,尤其像新冠病毒这样的传播。
此外,感染仍然对婴儿和老年人等弱势群体构成重大风险。上呼吸道感染是医疗保健中的最常见疾病,在老年人中,流感和肺炎仍然是常见的死亡原因 。
世界卫生组织指出,传染性肠道疾病是导致死亡的主要原因之一,根据 2015 年全球疾病负担、伤害和风险因素研究,感染性腹泻是全球主要的死亡原因,尤其是5岁以下儿童。
病原体需要克服三个主要障碍,才能在胃肠道中引起感染:
本小节主要介绍前两个:肠道微生物群和肠上皮层。
肠道微生物群由多物种微生物群组成,由细菌、真菌和病毒组成,它们生活在与宿主协同作用的特定生态位中。肠道微生物和哺乳动物共同进化,因此在微生物获得栖息地繁衍的同时,微生物调节宿主的各种生理功能,包括调节对病原体的保护性免疫。
肠道微生物群的组成受许多因素影响,例如遗传、性别、年龄、社会经济因素、营养、压力、疾病和环境因素(污染物、抗生素等)。
扰乱微生物群落结构和功能的因素,如抗生素的使用,为机会性病原体提供了定殖、生长和持续存在的空间。有几种机制可以确保微生物群防止定植、过度生长、病原体引起的损害以及随后的宿主感染。
定 殖 抗 性
一种机制被称为定殖抗性,其中共生微生物群和入侵微生物在营养或功能空间方面竞争资源可用性或生态位机会。
群 体 感 应
为了允许这种竞争,细菌细胞使用细菌复制过程中积累的信号分子不断地感知环境,从而监测种群密度并相应地调整它们的基因表达,这种机制称为群体感应。
化学信号导致细菌的表型变化,这些变化与粘附、运动和肠道密度或保护性化合物的排泄有关。
群体感应机制被共生体用来确保肠道内稳态,但也被病原体用来最小化宿主免疫反应并增加致病性。可能由饮食、压力、抗生素和药物治疗引起的微生物群落结构或非有益微生物群组成的改变会改变微生物群与宿主之间的整体动态,从而导致低度炎症、降低定植抗性和改变感染易感性。
肠道菌群会影响各种炎症和传染病
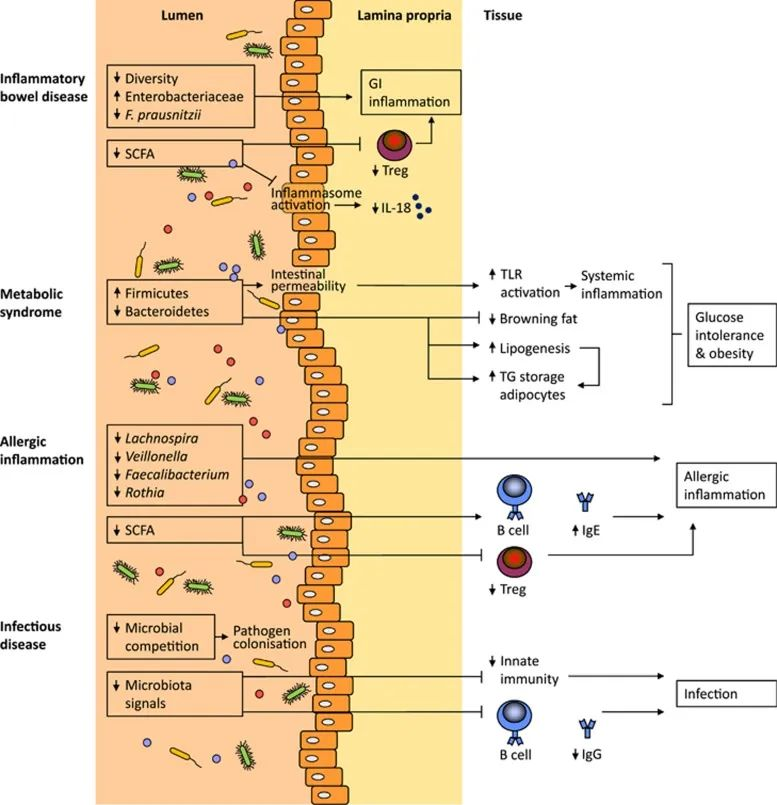
van den Elsen LW et al., Clin Transl Immunology. 2017
除了肠道微生物群,肠道上皮屏障在保护宿主免受病原体感染方面起着至关重要的作用 。
这种将肠道中的共生细菌与下层组织分开的物理屏障是通过紧密连接的蛋白质复合物连接的单层细胞。紧密连接复合物的组装是一个动态过程,某些细菌会通过释放毒素来破坏该过程。
此外,上皮细胞层由一层粘液加强。这种上皮屏障的粘液“衬里”是肠道上皮细胞抵抗细菌入侵的首要防御机制之一,通过防止管腔和粘膜微生物直接与上皮细胞相互作用。
微生物群和抵御感染威胁的肠道屏障
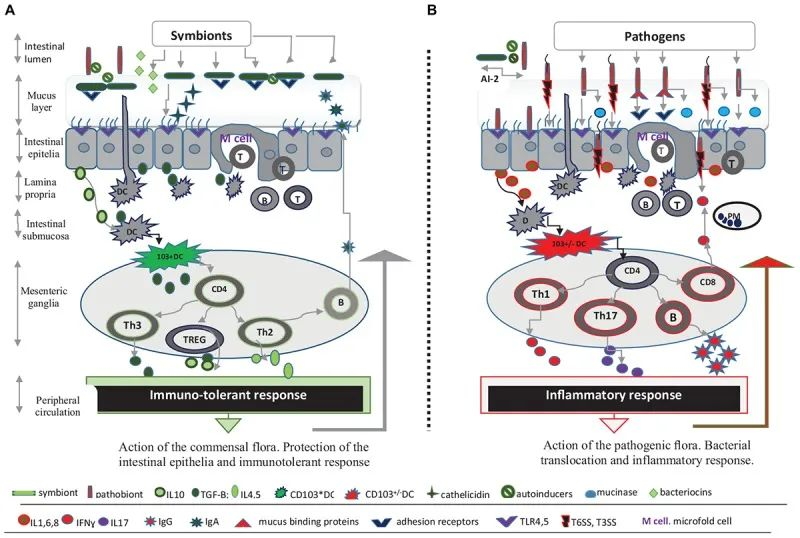
Iacob S, et al., 2019, Front Microbiol.
除了作为生物物理屏障的功能外,粘液还充当宿主产生的抗菌分子(如分泌型 IgA 和防御素)的储存库 。粘液的产生和降解是通过宿主和微生物之间复杂的相互作用来控制的,这种相互作用通过宿主对微生物相关分子模式 (MAMP) 和细菌代谢物的识别来调节,因此容易受到原生微生物群组成变化的影响 。
粘液和微生物之间存在相互关系,宿主炎症状态的变化和微生物群组成的变化都可能导致粘液产生和组成的改变,从而导致感染易感性增加。
短链脂肪酸在抗感染防御中的作用
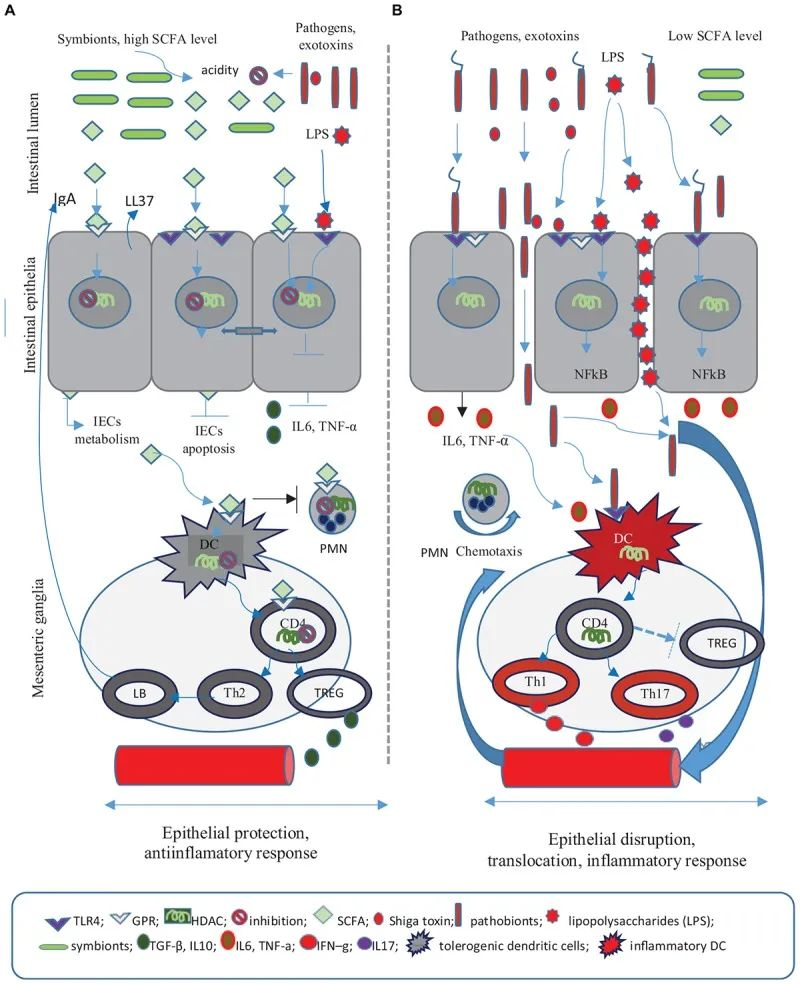
Iacob S, et al., 2019, Front Microbiol.
肠道微生物组成的炎症相关改变导致上皮屏障破坏,使细菌脂多糖能够全身循环。这些代谢变化会导致葡萄糖不耐受和肥胖。
过敏性炎症与特定细菌属的丰度较低有关。耗尽肠道微生物群已被证明会增加 IgE 的产生,这与 Treg 反应减少一起会导致过敏性炎症。
耗尽肠道微生物群会导致肠腔内缺乏微生物竞争已被证明会增加 IgE 的产生,这与 Treg 反应减少一起会导致过敏性炎症。
肠腔内缺乏微生物竞争会导致艰难梭菌定植。
最后,缺乏微生物信号也会降低对微生物的先天免疫和适应性免疫反应,使宿主更容易受到感染。
肠道微生物群和肠道上皮细胞之间的持续相互作用导致持续的免疫信号传导。在共生细菌和入侵病原体存在的情况下,这种免疫反应的调节以及上皮屏障的完整性和通透性对于维持肠道稳态至关重要。如果这个过程受损,可能会导致炎症和感染。
免疫反应在感染的易感性、持续性和清除率中起着至关重要的作用。
免疫系统由两部分组成:
先天免疫系统 通过多种防御机制提供非特异性保护,包括:
适应性免疫系统的细胞,T 和 B 淋巴细胞,识别并响应特定的外来抗原。T 细胞识别已进入宿主细胞的感染因子。这种类型的适应性免疫依赖于细胞的直接参与,因此被称为细胞免疫。
此外,T 细胞在调节 B 细胞功能方面发挥重要作用,B 细胞分泌识别特定抗原的抗体和蛋白质。因为抗体通过体液(即体液)循环,所以由 B 细胞诱导的保护称为体液免疫。
免疫系统的发育和免疫反应的功效与肠道微生物群的发育和组成密切相关
这方面的证据来自比较年龄和性别匹配的没有肠道微生物群的无菌饲养小鼠、相同品系的常规饲养动物和具有特定微生物群的无菌小鼠,即所谓的无菌小鼠。特别是,使用无菌小鼠提高了我们对单一细菌菌株、菌株联合体、特定微生物表达基因和微生物产生的代谢物对肠道稳态以及局部和全身免疫的影响的理解。
这些研究的见解强调,先天免疫在首次识别和响应微生物衍生产物中起着关键作用。
肠道中的先天免疫始于直接暴露于肠腔内容物和微生物产物的单层肠上皮细胞 (IEC)。宿主和微生物之间的关键平衡通过模式识别受体 (PRR) 识别微生物得到安全保护。
PRRs 构成了一个大家族的细胞外和细胞内受体,可识别特定的微生物相关分子模式 (MAMP)。PRR 包括 TLR、C 型凝集素受体 (CLR)、核苷酸结合寡聚结构域 (NOD) 样受体 (NLR) 以及 DNA 和 RNA 的胞质传感器。
MyD88 是 PRR 信号传导下游的重要衔接分子,将 PRR 激活与转录因子 NF-ĸB 的激活联系起来,后者是炎症的主要调节因子。因此,MyD88 的缺乏会导致免疫反应受损和对感染的易感性增加。
然而,PRRs的不当激活可能导致过度的免疫反应,甚至导致炎症性疾病和自身免疫;因此,PRR 反应通过正负反馈回路和交叉调节受到严格调节。
此外,IEC 分泌抗菌肽 (AMP),这是一种具有杀菌、抗炎和抗内毒素特性的先天免疫效应分子。
AMP是先天免疫防御的重要组成部分,可限制病原体与上皮的相互作用。它们的表达可以被某些病原体下调,并因特定微生物的存在而增强,因此,微生物群的组成是形成先天免疫反应的关键。
免疫系统的发育和免疫反应的功效与肠道微生物群代谢产物密切相关
微生物群控制免疫反应的另一种机制是通过肠道微生物群从饮食成分、宿主产物或其他微生物代谢物产生的代谢物的形成。
种类繁多的微生物代谢产物介导了共生细菌的许多保护功能。
具有免疫保护能力的代谢产物包括:
短链脂肪酸、色氨酸代谢物、胆汁酸衍生物等。
➭ 短链脂肪酸
短链脂肪酸通过特殊的肠上皮细胞增强抗菌肽和粘液的产生,并刺激结肠调节性 T 细胞的成熟和扩增,从而抑制对微生物群的局部炎症反应。
短链脂肪酸通过调节上皮屏障来支持结肠内的肠道稳态,并通过诱导这些细胞的增殖和分化来支持肠道细胞的修复 。
短链脂肪酸在先天淋巴细胞 (ILC3) 的增殖中很重要,ILC3 会释放 IL22,这对上皮细胞诱导抗菌分子很重要。
➭ 色氨酸代谢物
色氨酸代谢物,尤其是吲哚类来源于膳食色氨酸的共生发酵,并作为芳烃受体 (AhR) 的配体发挥作用,这是一种对维持肠道稳态很重要的受体,这些代谢物的丢失与炎症性肠病的发生有关疾病。
➭ 胆汁酸衍生物
胆汁酸衍生物通过激活法尼醇 X 受体 (FXR) 和 G 蛋白偶联胆汁酸受体 (TGR5) 来支持肠道稳态并影响大量宿主功能。胆汁酸衍生物是通过特定细菌门表达的细菌胆汁盐水解酶 (BSH) 的作用从胆汁酸代谢而来的,而 BSH 基因丰度的丧失与炎症性肠病的发生有关。
以上表明肠道中的粘膜稳态是肠道微生物群、微生物代谢物和宿主因素之间的微妙平衡。这种持续的相互作用导致严格调节的生理低度炎症状态,维持最佳的宿主防御,从而影响对感染的易感性。
人们越来越认识到,肠道微生物群除了调节局部粘膜免疫系统外,还通过多种机制影响先天性和适应性细胞介导的全身免疫反应。
▸肠道菌群释放微生物可溶性产物,影响外周免疫细胞的激活
一种机制涉及释放微生物可溶性产物,这些产物转移到循环中并影响外周免疫细胞的激活。事实上,肠道远端器官中的常驻免疫细胞可以直接感知循环微生物衍生因子,而微生物群衍生信号分子的缺失会导致免疫功能发生改变,从而导致对全身感染的易感性。
▸肠道菌群对适应性免疫系统(T细胞)的影响
目前,肠道微生物组影响全身免疫反应的最佳表征机制可能是其对适应性免疫系统的 T 细胞室的影响。已经表明,胃肠道微生物群可以影响 T 细胞群分化为 Th1、Th2 和 Th17 细胞或具有调节表型的 T 细胞。
具体而言,丁酸盐作为一种短链脂肪酸可促进外周部位诱导的调节性 T 细胞的这种分化,并且以这种方式能够抑制全身炎症的发展。短链脂肪酸还能够重新编程细胞的代谢活动,从而诱导调节性 B 细胞,并通过戊酸抑制 Th17 细胞的产生,这可能与炎症性肠病和自身免疫性疾病有关。
此外,微生物群来源的 ATP 可以诱导 Th17 细胞的扩增,色氨酸分解产物可以导致上皮内 CD4 + CD8αα + T 细胞的增加,而细菌来源的多糖可以引发调节性 T 细胞。通过其诱导调节群体的能力,微生物组可以支持抑制炎症反应。
▸宿主-病原体相互作用的研究:
结果表明,记忆 T 细胞的共生激活及其向发炎部位的运输,对于防止细菌病原体感染是必要的。
此外,通过共生菌主动控制 IL10 介导的抗炎反应对于防止感染性损伤很重要。这种效应可以使用特定的 toll 样受体 (TLR) 激动剂来重现,这会减少 IL10 的产生,通过增加细菌清除率使小鼠对感染具有更强的抵抗力,并使适当的炎症反应成为可能 。
▸细菌在造血过程中影响调节免疫系统
微生物群释放的信号分子进入循环的能力,也使肠道中的常驻细菌在造血过程中影响免疫细胞发育,调节免疫系统,从而影响对感染的反应。
事实上,短链脂肪酸丁酸盐被证明可以促进骨髓单核细胞从炎症表型分化为更具耐受性的表型。
骨髓细胞也表达多种 PRR(受体),并且对循环中的 MAMP 敏感,其影响由 PRR 表达和 MAMP 可用性决定。例如,在造血干细胞和祖细胞 (HSPCs) 上激活 CLR dectin-1 会导致诱导已经对单核细胞和巨噬细胞进行过训练的免疫。相反,在 HSPCs 上激活 TLR2 会产生具有高抗原呈递共刺激能力的耐受巨噬细胞 。已显示通过 AhR 配体激活 HSPC 可导致产生能够免疫抑制的髓源性抑制细胞。
▸肠道微生物群衍生的信号调节先天免疫防御
除了影响 T 细胞发育和功能外,肠道微生物群衍生的信号被证明还可以通过脾脏中的淋巴刺激、中性粒细胞迁移和功能的调节、巨噬细胞的诱导和活化以及调节自然杀伤 (NK) 细胞的功能来调节先天免疫防御。
▸特定菌群降低皮质酮水平,从而调节炎症反应
最近,研究表明,特定的细菌种类还通过降低血浆皮质酮(肾上腺分泌的皮质醇激素控制炎症“关闭”过程。皮质醇也是身体的减压激素,这个我们下次单独讲)水平来调节炎症反应,这是一种抗炎类固醇,在控制对粘膜损伤的炎症反应中很重要。
综上所述,很明显,肠道微生物群的失调会导致诱导适当的局部和全身免疫反应的能力降低,从而导致局部炎症性疾病,但也会导致远端疾病。
一个远端部位是气道,这两个部位之间的这种特定的直接关系被称为:肠-肺轴。
▸肠道菌群影响肺部:哮喘、过敏性气道疾病、呼吸道感染等,益生菌可改善
事实上,在动物和人类研究中,已经表明抗生素引起的肠道微生物群改变可能与特应性表现、过敏性气道疾病和患哮喘的风险增加有关。
除了影响过敏性气道疾病的发展,已经表明肠道微生物群在防止细菌和病毒呼吸道感染方面发挥着至关重要的作用,因为肠道微生物群直接控制先天性和适应性免疫反应。
事实上,多项人体临床试验表明,使用益生菌可降低呼吸道感染的发生率并改善其健康结果。
▸肠道菌群影响肺部:通过粘膜免疫系统
肠道事件影响肺部疾病的另一种机制是通过常见的粘膜免疫系统,其中在肠道中引发的抗原特异性 B 细胞可以通过胸导管迁移到远端有效部位。
在肠-肺微生物群研究中,确定肠道微生物群变化是疾病的原因还是结果是具有挑战性的。此外,还需要进行纵向研究,以更好地了解肠道微生物群对已确诊肺部疾病的严重程度和病程的影响。
在人群中可以看到,年幼的婴儿和老年人特别容易受到感染。这两个人群的共同点是,在这两个人群中,免疫系统都没有发挥最佳功能。接下来的两个章节,主要介绍婴儿和老年人的免疫系统与肠道菌群的关联。
▸婴儿的免疫系统在出生时受到了极大的抑制,母体抗体提供保护
婴儿的免疫系统在出生时并没有完全发挥作用,这意味着他们的先天和适应性免疫反应都受到了极大的抑制。子宫内环境要求胎儿的免疫系统主动下调并耐受来自母亲的抗原,以避免可能导致终止妊娠的免疫反应。
然而,出生后,暴露于环境抗原,其中许多来自肠道微生物群,需要快速改变免疫反应,以保护婴儿免受病原体入侵。
在生命的最初几个月,母体 IgG 抗体提供了对许多感染的保护,该抗体从母亲转移到婴儿身上;然而,当这些抗体水平下降时,婴儿变得更容易受到感染。
幸运的是,先天免疫细胞提供了抵御入侵病原体的早期第一道防线,在胎儿期就已经发育成熟。但是,这种情况发生在不同的时期,与生命后期相比,新生儿先天免疫所有成分的功能仍然较弱。在抗原暴露的驱动下,适应性免疫系统也在生命的最初几个月迅速发展,从而导致免疫记忆的发展。
▸微生物群和免疫系统密切相关,婴儿的感染率增加也可能与微生物组的变化有关
婴儿最初可能会在子宫内接触细菌,出生后会迅速出现肠道定植。定植模式受以下因素的影响:
有人提出,生命的前 24 个月是建立微生物组的关键发育窗口,甚至可能决定整个生命过程中肠道微生物群的组。由于部分免疫系统发育或成熟需要某些细菌,这两个过程密不可分。
实际上,需要共生微生物来训练免疫系统以区分成为耐受抗原的共生细菌和致病细菌。有缺陷的免疫耐受会加剧自身免疫和炎症性疾病,例如过敏。研究表明,特应性儿童和健康儿童的肠道微生物群组成不同,细菌多样性降低和生态失调与特应性疾病的发展有关。
在年龄范围的另一端,免疫系统也处于次优状态。这种以先天免疫和适应性免疫逐渐下降为特征的免疫系统生物老化是不可逆转的,被称为“免疫衰老”。
▸与年龄相关的免疫系统变化
树突状细胞 (DC) 信号通路中与年龄相关的变化已被证明会影响其功能,从而导致响应病原体的细胞因子分泌模式发生改变。
此外,这些变化会导致吞噬作用降低和呈递抗原的能力受损,并对树突状细胞的迁移能力产生负面影响。同样,研究表明,来自老年人的循环单核细胞、巨噬细胞和迁移性中性粒细胞显示出吞噬功能受损。
在单核细胞、树突状细胞和中性粒细胞中,TLR的表达和功能随着年龄的增长而下降。此外,TLR 的定位受损可诱导细胞因子产生的变化。一个例外是老年人单核细胞上的 TLR5 表达,与年轻人单核细胞中的 TLR5 表达水平相比,它实际上有所增加,并导致老年人中细胞因子的产生增加。
此外,随着年龄的增长,T 细胞会发生复杂的变化,包括表观遗传和代谢变化,这些变化会影响初始 T 细胞、记忆 T 细胞和效应 T 细胞。
此外,T 细胞受体 (TCR) 库减少,功能不活跃的衰老或衰竭 T 细胞的频率增加。与年龄相关的 T 细胞改变的起源可能在于细胞因子产生的调整,因为细胞因子在介导 T 细胞反应中至关重要。确实已经表明,来自老年人的 T 细胞主要表现出 Th2 样表型。
Th17 与调节性 T 细胞的比例似乎也有所增加,这被认为与老年人对感染的反应降低有关。除了 T 细胞区室的变化之外,老年人的 B 细胞库多样性较少,这可能导致老年人更容易受到感染。
▸免疫系统伴随炎症,共同导致慢病
免疫衰老伴随着一种慢性、无菌、低度炎症。有几种先天免疫系统的激活剂会导致炎症。此类刺激包括持续的病毒和细菌感染、细胞分解产物和错误折叠的蛋白质。免疫衰老和炎症共同导致感染、癌症、自身免疫和慢性疾病的患病率增加,以及老年人对疫苗接种的反应不佳。»»»
▸衰老影响肠道菌群稳态
与婴儿肠道微生物群研究以及微生物群对免疫系统的影响相比,较少数量的研究关注衰老过程中肠道微生物群发生的系统发育和功能变化。
尽管老年人的肠道微生物群存在很大差异,但健康的成人肠道微生物群被认为是相当稳定的,直到衰老过程开始影响微生物群的稳态。由此导致的生物多样性减少,尤其是产生抗炎短链脂肪酸的细菌减少,以及肠道微生物群的稳定性受损,通常与感染易感性增加有关。
此外,与肠道菌群变化和免疫衰老相关的慢性低度炎症有利于病菌的生长,病菌是健康肠道菌群的一小部分,在发炎的环境中,可以超过共生菌的生长并导致感染。
▸设计饮食干预实验时,应考虑免疫状态
最近的证据表明,衰老从根本上改变了营养对免疫功能的影响。因此,对调查饮食对免疫功能影响的研究数据的解释在很大程度上取决于受试者的年龄。
在调查膳食成分的功效时,研究设计至关重要,大多数涉及老年人的研究都包括基于病史、实验室测试的严格纳入/排除标准,一般健康状况,通常还有营养状况。
然而,免疫状态很少被考虑在内,但即使在健康的老年人中也可能存在显着差异。有几个明显的例子表明免疫细胞组成、表型和/或功能与年龄相关的变化可以直接改变干预的结果。
随着全球人口迅速老龄化,老年人的健康将越来越受到关注。由于免疫功能下降,与年轻人相比,老年人被认为发生感染的风险增加,严重程度和死亡率增加。尤其是在老年护理机构中,感染容易在居民中传播,预防感染至关重要。
传统上,传染病专家的重点是识别和治疗个体病原体。最有效的治疗策略之一是使用抗生素。然而,抗生素耐药病原体的增加,突出了对替代策略的需求。
▸抗生素破坏肠道菌群结构和功能,给病原体生长的空间
除了刺激耐抗生素病原体的生长外,抗生素还会破坏微生物群的结构和功能,从而使潜在的病原体能够定殖、生长和持续存在。由于对免疫系统、肠道微生物群和病原体之间的复杂平衡和相互作用的认识不断增加,传染病和临床微生物学领域目前正在发生范式转变,临床医生现在开始关注和拓展诊疗和用药信息,这是一个好消息。
▸肠道菌群与传染病之间关系最突出的例子是:抗生素使用后的艰难梭菌感染
在抗生素治疗期间,抗生素敏感细菌被杀死,导致微生物群的信号减少和对艰难梭菌的免疫反应减弱。
此外,由于没有其他细菌,艰难梭菌使用了更多的可用营养物质,这导致在一系列抗生素清除细菌的部位的定植率增加。抗生素的使用与艰难梭菌之间的密切关系使这种感染成为基于微生物组治疗的热门目标。
▸肠道菌群影响病毒感染
同样,多种肠道病毒,包括轮状病毒、诺如病毒(在我们的临床菌群检测报告中也会判别这两种病毒)和脊髓灰质炎病毒,已被证明利用细菌微生物组进行免疫逃避,支持进入肠道并在肠道中复制,从而增加感染率。
使用上述全身免疫机制,肠道微生物组已被证明具有影响全身病毒感染的潜力。例如,微生物群衍生的短链脂肪酸已被描述为通过改变 T 细胞的反应而对流感感染具有保护作用。此外,已证明 HIV 患者肠道内乳酸杆菌目的丰度较高与病毒载量呈负相关,这表明微生物群可以直接或间接地调节 HIV 感染的病理学。
▸ 肠道菌群也可能影响疫苗反应和药物代谢
这是目前有一个非常有意义的领域;然而,这可能也是药物或疫苗特异性的。
在 2017 年一项调查益生元和益生菌对疫苗免疫原性和有效性影响的随机对照试验 (RCT) 的系统评价和荟萃分析中,比较了 13 项使用益生菌的试验和 6 项使用益生元的试验。
然而,应该谨慎解释这项荟萃分析的总体结果,因为它结合了使用不同益生元和益生菌菌株后疫苗反应的数据。查看个别研究中的效果,有时显示没有效果,有时显示干预的积极效果,强调结果高度依赖于干预。
众所周知,营养对肠道微生物群的组成和免疫系统有很大影响,因此可以在健康和疾病的发展中发挥重要作用。
例如,西方饮食通过诱导骨髓祖细胞的表观遗传和转录重编程与炎症反应增强有关,从而直接影响几种非传染性疾病的发展。
增加对肠道微生物群、宿主反应和其他微生物之间关系的理解甚至进一步提供了调节这个三元组的机会。
例如通过营养来帮助维持肠道稳态和抗感染性。应该考虑到不同的膳食成分,如矿物质、碳水化合物、维生素、脂质和蛋白质,都具有特定的特性,它们以不同的方式直接或间接地通过微生物组影响宿主与病原体之间的相互作用。在这些营养素之间建立机制联系,为影响健康提供了多种机会。
饮食干预——
“操纵宿主-微生物轴的宝贵工具”,以帮助维持肠道稳态和感染抵抗力
碳水化合物、脂质、蛋白质、植物化学物质、矿物质和维生素等膳食成分都具有独特的结构和化学(物理化学)特性,可通过微生物组直接或间接影响宿主病原体抵抗力。弥合饮食、宿主和微生物组之间的差距,因为它们与免疫和抗病性有关,是一个多方面的领域,需要了解它们对肠道稳态的综合影响。
饮食促成了微生物群、宿主和病原体之间相互交织的机制的黑匣子
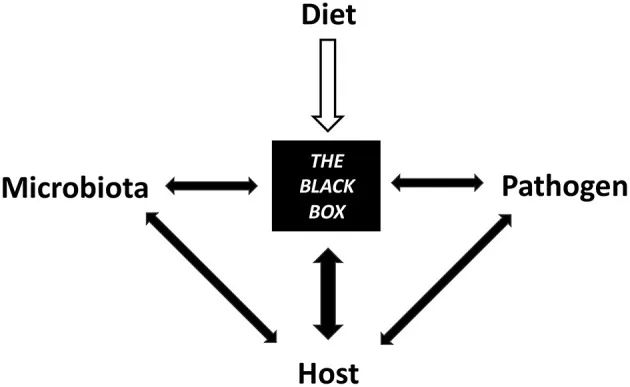
因此,饮食干预应被视为调节传染病风险、防止病原微生物入侵、减轻感染严重程度和支持传染病治疗的宝贵工具;然而,需要在这个迅速兴起的领域进行进一步的研究。
已知有多种营养化合物会对宿主微生物组和免疫系统产生影响,但膳食纤维、益生元和益生菌受到了很多关注。
★ 益生元
益生元是一种不易消化的食物成分,通过选择性地刺激结肠中一种或有限数量的细菌的生长和/或活性来对宿主产生有益的影响。这意味着并非所有的膳食纤维都是益生元,除非有证据表明该纤维被宿主有机体选择性地利用,从而对健康有益。
典型的益生元是人乳低聚糖 (HMO)、菊粉和低聚果糖,以及低聚半乳糖。
★ 膳食纤维
膳食纤维不是典型的益生元,但具有益生元特性,例如,β-葡聚糖、阿拉伯木聚糖 (AX)、果胶和抗性淀粉。益生元和特定的膳食纤维通过充当发酵底物来促进肠道中有益细菌的生长,同时通过排除生态位来抑制病原体的生长。
主要的发酵产物是短链脂肪酸,它们对免疫系统有重大影响,如上所述,因此可以抑制传染病的发展。除了短链脂肪酸,正如 Asadpoor 等人最近审查的那样,益生元和膳食纤维还可以通过排除和抗菌活性直接预防胃肠道感染。
★ 益生元和膳食纤维
益生元和膳食纤维与上皮细胞和免疫细胞的直接相互作用也有助于预防感染。已证明 DFs,如 β-葡聚糖和 AX 可激活 CLR dectin-1,这是一种参与训练免疫诱导的重要受体,可增加对继发感染的免疫反应。
HMO、AX 和果胶还与 TLR 相互作用,从而提高 DC (树突细胞)的功效,通过肠上皮细胞诱导产生耐受性树突系报告,并保护胃肠道免受过度的 TLR 信号传导,但也支持在胃肠道感染后消退炎症。
★ 益生菌
益生菌是活细菌,当以足够的量给药时,可为宿主提供健康益处。使用益生菌的基本原理主要是基于它们改变肠道微生物群的能力,支持共生细菌的生长超过病原菌的生长。
▸ 益生菌通过与病原体竞争营养和功能资源以及产生抗菌物质来塑造微生物群
许多研究调查了益生菌在预防和治疗传染病中的潜在作用;然而,并非所有数据都一致。
✔ 积极作用
在一项关于益生菌在急性腹泻中的疗效的 Cochrane 系统评价中,作者得出结论,益生菌通过缩短急性感染性腹泻的持续时间和减少平均排便频率具有明显的积极作用。
关于呼吸道感染,研究表明,使用益生菌的儿童在出生后第一年的呼吸道感染复发率较低,肺炎和严重急性下呼吸道感染的发生率也有所降低。
○ 单种益生菌未发现积极作用,联合治疗产生效果
一些研究并未发现对下呼吸道感染发病率的影响。例如,一项研究发现接受鼠李糖乳杆菌GG (LGG) 组和对照组之间的中耳炎 (OM) 发生率没有差异。
然而,另一项针对72名婴儿的试验表明,与对照组相比,接受LGG和乳酸双歧杆菌联合治疗的儿童发生OM的几率显著降低。
▸为什么会出现不同的结果?
数据的异质性,主要是由于菌株、剂量、研究环境和测量结果的变化,限制了广泛使用益生菌预防感染的循证建议。
对营养成分(例如益生菌)的影响产生矛盾结果的原因之一,可能是调查营养影响的临床研究通常与调查药物化合物影响的研究类似。这种类似药物的方法可能不适合识别个体对饮食治疗的反应,与药物化合物的靶向作用相比,这可能是多因素的。
如果不报告个体对营养干预的反应,可能会错过对于更好地了解营养、微生物组和宿主之间的相互作用至关重要的信息,而这些信息对于设计个性化的营养方法是必需的。
▸ 有没有什么方法可以深入了解特定营养素对个体特定健康结果的影响?
网络分析、系统生物学和基于机器的学习技术可以整合基于预先存在的大型队列数据集的多个特征(通过多年的积累和持续迭代,谷禾健康已经构建了超过60万例肠道样本数据),可以深入了解特定营养素对个体特定健康结果的影响。
例如,提出了一种机器学习算法来预测对现实生活中膳食摄入反应。然而,由于个人的整体健康结果取决于许多过程和反应,未来还需要调查血液、血糖反应以获得个人健康状况的更多整体情况。
▸ 为什么不同人对营养化合物的反应不同?
个体对营养化合物的反应受个体遗传特征的影响。例如,炎症基因(如 IL1B、IL6 和 TNFA)中的单核苷酸多态性 (SNP) 会导致不同的炎症反应,这可以解释观察到的对营养化合物的反应性差异的部分原因。
还有越来越多的证据表明,个体的遗传特征对于使肠道内的有益细菌定植至关重要,这会影响免疫系统、宿主的整体健康和传染病。
★ 合生元
除了分别使用益生元和益生菌外,还有一些营养概念将益生元和益生菌组合成混合物:合生元。考虑到上述临床研究的局限性,合生元已显示出部分临床上的成功,未来可能有望成为治疗选择之一。
这一点在最近在印度农村进行的一项随机、双盲、安慰剂对照试验中得到印证,该试验显示,在接受为期 7 天的合生元概念干预的新生儿中,败血症和随后的死亡显着减少。
注:败血症是发展中国家新生儿发病率和死亡率的主要原因,这一概念对促进全球儿童健康很有希望。
随着我们更多地了解肠道微生物群影响局部、先天和全身免疫的复杂机制,以及科学家,企业和临床医生开始利用这些信息来开发针对这些过程的方法,以支持改进预防和治疗策略为最终目标的传染性疾病。
目前对抗这些传染原的策略包括疫苗、小分子药物、提高卫生标准和针对特定疾病的干预措施的组合。虽然这些方法有助于大幅降低与感染相关的死亡发生率和人数,但仍需要继续投资于当前战略并开发新的治疗方法,以应对这些全球健康威胁。
这种干预策略应考虑到个体之间微生物组和免疫反应的显着差异,因此需要个性化的方法。饮食干预能够引起微生物组功能和下游免疫反应的快速变化,这一事实可用于开发量身定制的营养概念,从而影响传染病的发展和治疗成功。主要参考文献:
Wiertsema SP, van Bergenhenegouwen J, Garssen J, Knippels LMJ. The Interplay between the Gut Microbiome and the Immune System in the Context of Infectious Diseases throughout Life and the Role of Nutrition in Optimizing Treatment Strategies. Nutrients. 2021 Mar 9;13(3):886.
van den Elsen LW, Poyntz HC, Weyrich LS, Young W, Forbes-Blom EE. Embracing the gut microbiota: the new frontier for inflammatory and infectious diseases. Clin Transl Immunology. 2017 Jan 20;6(1):e125.
Van den Elsen L.W., Poyntz H.C., Weyrich L.S., Young W., Forbes-Blom E.E. Embracing the gut microbiota: The new frontier for inflammatory and infectious diseases. Clin. Transl. Immunol. 2017;6:e125.
Iacob S, Iacob DG, Luminos LM. Intestinal Microbiota as a Host Defense Mechanism to Infectious Threats. Front Microbiol. 2019 Jan 23;9:3328. doi: 10.3389/fmicb.2018.03328. PMID: 30761120; PMCID: PMC6362409.
Kirk M.D., Pires S.M., Black R.E., Caipo M., Crump J.A., Devleesschauwer B., Döpfer D., Fazil A., Fischer-Walker C.L., Hald T., et al. World Health Organization Estimates of the Global and Regional Disease Burden of 22 Foodborne Bacterial, Protozoal, and Viral Diseases, 2010: A Data Synthesis. PLoS Med. 2015;12:e1001921.
Troeger C., Forouzanfar M., Rao P.C., Khalil I., Brown A., Reiner R.C., Jr., Fullman N., Thompson R.L., Abajobir A., Ahmed M., et al. Estimates of global, regional, and national morbidity, mortality, and aetiologies of diarrhoeal diseases: A systematic analysis for the Global Burden of Disease Study 2015. Lancet Infect. Dis. 2017;17:909–948.
Iacob S., Iacob D.G., Luminos L.M. Intestinal Microbiota as a Host Defense Mechanism to Infectious Threats. Front. Microbiol. 2018;9:3328. doi: 10.3389/fmicb.2018.03328.
Lazar V., Ditu L.-M., Pircalabioru G.G., Gheorghe I., Curutiu C., Holban A.M., Picu A., Petcu L., Chifiriuc M.C. Aspects of Gut Microbiota and Immune System Interactions in Infectious Diseases, Immunopathology, and Cancer. Front. Immunol. 2018;9:1830.
Slack E., Hapfelmeier S., Stecher B., Velykoredko Y., Stoel M., Lawson M.A.E., Geuking M.B., Beutler B., Tedder T.F., Hardt W.-D., et al. Innate and adaptive immunity cooperate flexibly to maintain host-microbiota mutualism. Science. 2009;325:617–620

谷禾健康

随着大家陆续“阳康”,大家逐渐恢复以往的生活,城市的烟火气回来了。
然而阳康后真的万事大吉了吗?
还是有很多朋友处于这样的状态:感觉恢复了,又好像没有完全恢复,身体多少有点不适,开始关心:
这种新冠肺炎后遗症,也就是临床上常说的“长新冠”,已经成为部分人不得不面临的事实。
“长新冠”是一种多系统疾病,据估计,全球至少有 6500 万人患有长新冠,病例每天都在增加。由于许多未记录在案,这个数字实际可能要高得多。
“长新冠”与所有年龄段和急性期疾病严重程度相关,许多患者在多个器官系统中经历了数十种疾病,包括心血管、血栓、脑血管疾病、2型糖尿病、自主神经功能障碍等。基于对其超过 2 年的研究,如果不采取行动任其发展,很大一部分患有长新冠的人可能会有严重后果。
新出现的临床前和临床研究表明,肠道微生物群可能有助于理解 COVID-19 的发病机制和疾病结果;SARS-CoV-2 感染与肠道微生物群的改变有关,并与炎症和免疫反应相关。
本文基于多篇文献报道,主要介绍关于“二次感染”以及“长新冠”问题的最新研究进展,同时也着重介绍了多种与新冠恢复期调理相关的干预措施。
本文由于篇幅较长,主要分为上、中、下三大篇章,主要包括以下内容:
上篇: 关于“二次感染”
中篇: 关于“长新冠”
下篇: 关于“干预措施”
上 篇
关于 “二次感染”
已经阳康的朋友,可能关心会不会马上又来一波,出现“二次感染”?
我们先了解一下,二次感染指的是什么?
“二次感染” 也叫“再感染”或者“重复感染”,是指一个人感染了新冠之后,经过足够的时间,然后再次感染。一般而言,如果一个人在第一次阳性检测后 90 天或更长时间再次检测呈阳性,则被视为二次感染。
注:也有部分研究定义间隔>30天。
再感染后可能出现临床症状,患者因核酸载量高而具有传染性。
与没感染过的人群相比,已经感染过的人群再感染风险低87%
先看一组数据:
➪
一项Meta分析综合了先前感染SARS-CoV-2人群再感染风险的全球证据:先前感染SARS-CoV-2的人可能会再次感染,他们的感染风险比之前没有感染的人低。
SARS-CoV-2再感染发病率为:
0.7 / 10,000人日(标准差0.33).
以前感染过的人比从未感染过的人再次感染的可能性低87%(HR = 0.12)。
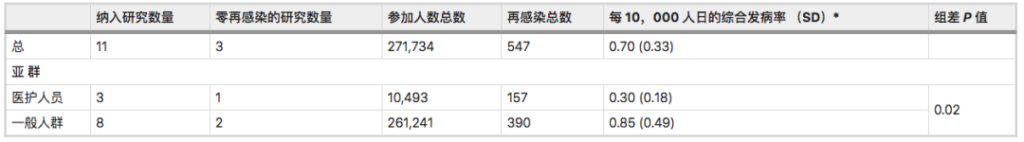
注:该研究荟萃分析和系统审查分别包括11项研究和11份病例报告(样本量都不小于100)。
看到这里是不是觉得可以放心了,再次感染的概率很低?
并不一定。需要注意的是,该研究虽在近期发表,但研究的数据是基于出现在2021年4月及之前的毒株,当时奥密克戎变种尚未出现。
研究表明,奥密克戎变异株的免疫逃逸,导致比其他变异株具有更高的传播。那么,有没有关于奥密克戎毒株的数据?
有。
与感染过德尔塔毒株的人群相比,在奥密克戎阶段,再感染风险显著高于德尔塔时期
➪
一项回顾性观察性研究,分析来自意大利北部利古里亚地区的感染者,在研究期间(2021年9月至2022年5月),利古里亚记录了335117例SARS-CoV-2感染病例,其中15715例再次感染。在奥密克戎阶段(从2022年1月3日开始占主导地位),再感染的风险是德尔塔阶段的4.89倍(p<0.001)。
其他也有来自意大利的研究(2021年8月至2022年3月)认为,奥密克戎时期的再感染风险比德尔塔时期高18倍。
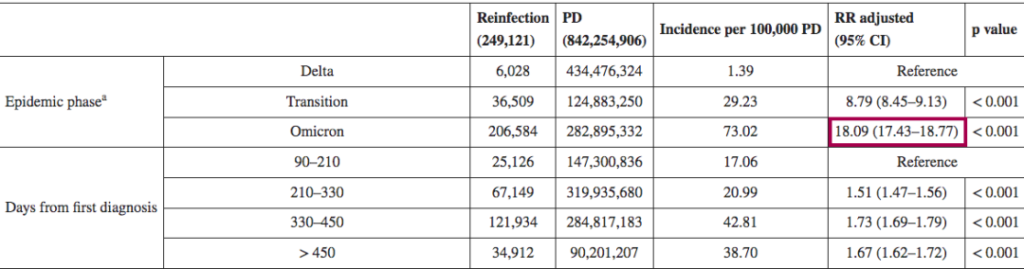
DOI: 10.2807/1560-7917.ES.2022.27.20.2200372
➪
英国爱丁堡大学的研究人员发现,奥密克戎变异株的可能再感染率约为德尔塔变异株的10倍。
➪
来自圣保罗大学医院281名医护人员的再感染率(2020年3月10日至2022年3月10日):
奥密克戎时期再感染率显著增加(0.8%到4.3%;相对风险5.45 [95%IC 3.80–7.81];p < 0.001)
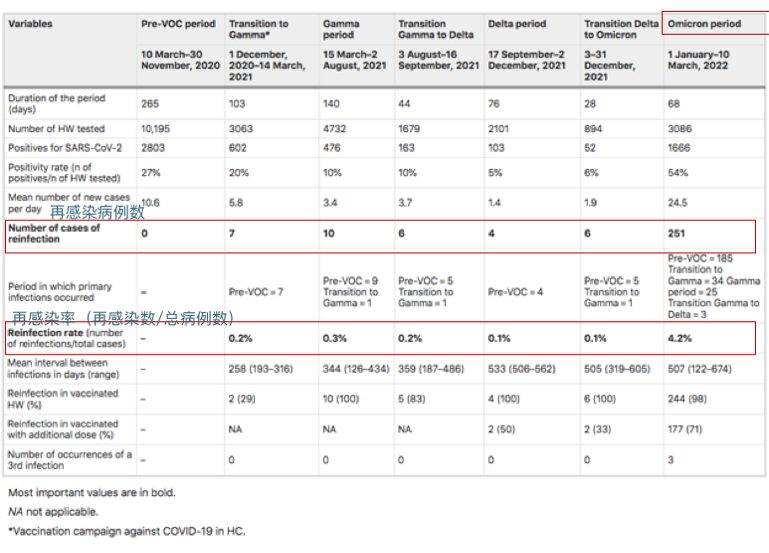
Guedes AR, et al., Sci Rep. 2023
关于奥密克戎再感染率上升的证据相当多,此处就不一一列举了。既往感染虽然可以在一定程度上防止新冠的再次感染,但对于奥密克戎来说,之前的抗体保护作用较弱,并且在一定期限后迅速减弱,接下来我们来看一下,既往感染的保护期限大概能维持多久。
队列研究证实,既往感染对奥密克戎变异株(BA.1、BA.2、BA.4 和 BA.5)再感染的保护作用低于对德尔塔和较旧变异株再感染的保护作用。
➪
50项研究共纳入118名再感染患者,从首次感染到再感染的最短时间为19日,最长为293日。
● 针对奥密克戎BA.1和BA.2毒株:
先前感染德尔塔变异株可将症状性感染的风险降低50%至67%.
➪
在丹麦的一项队列研究中,如果先前感染发生在3至6个月前,则对奥密克戎BA.1或BA.2的保护率为43.1%,如果先前感染至少发生在6个月前,则为22.2%.
● 针对奥密克戎BA.4和BA.5毒株:
BA.4 和 BA.5 这两个变体与 BA.2 的相似性高于 BA.1 菌株,携带着它们自己独特的突变,包括病毒刺突蛋白中 L452R 和 F486V 的变化,这些变化可能会调整其锁定宿主细胞并避开某些免疫反应的能力。
对卡塔尔人群的分析提供了关于奥密克戎BA.4和BA.5防护的详细信息:
在未接种疫苗的人群中,既往感染奥密克戎BA.1或BA.2可使任何感染奥密克戎BA.4或BA.5的风险降低至少68.7%(CI,64.0%至72.9%),而如果先前感染发生在奥密克戎变异株出现之前,则仅为27.7%(CI,19.3%至35.2%).
针对BA.4或BA.5免受BA.1或BA.2感染的保护作用在4个月内是强大的,但这种保护可能会迅速减弱。
关于再感染的风险和保护期限的研究
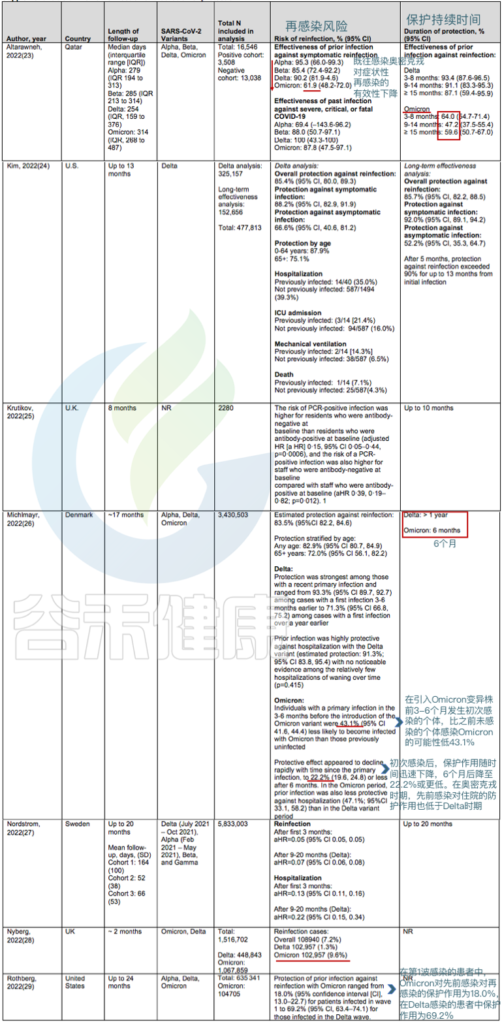
doi:10.7326/M22-1745
高龄和抑郁状态是奥密克戎再感染的危险因素
一项前瞻性队列研究,纳入了 933 名被诊断为 Omicron BA.2.2 感染且治疗后检测呈阴性的成年患者。
最终,683例符合标准,进行研究分析。
注:数据来源以及筛选标准:
患者来自四川省临水县,数据由四川省疾病预防控制中心提供。如果患者符合以下任何标准,则被排除在外:
感染前诊断为精神疾病,伴随需要住院治疗的严重疾病,以及交流障碍或拒绝参与。
在683名奥密克戎感染患者中,出院后30天内有116例再阳性,总体再阳性率为16.4%.
预测再阳性风险变量的单变量逻辑回归
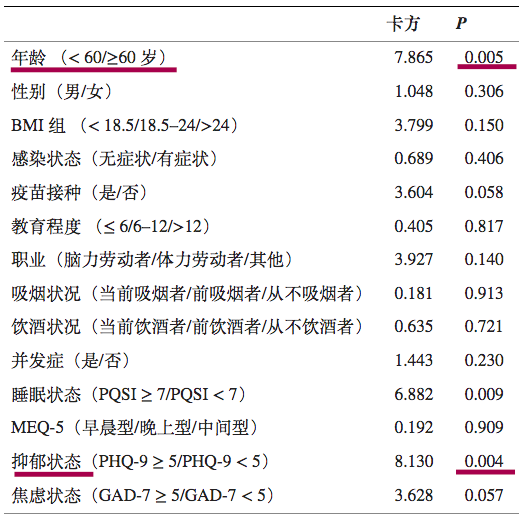
DOI:10.3389/fpubh.2022.1014470
注:PHQ-9,患者健康问卷九。PHQ-9量表的总分范围为0至27分,其中0至5分表示没有抑郁,得分>5表示抑郁状态;评分越高,抑郁症状越严重。
比较分析显示:
慢性肾功能衰竭、心血管疾病、支气管肺病、神经病变和自身免疫性等疾病患者再感染的风险相对增加
来自意大利北部地区的数据显示:
女性个体的再感染风险比男性高17%(OR为1.17,95%CI为1.13-1.21,p <0.0001)
在再感染者中,60岁及以上患至少一种潜在慢性病人群的风险比其他年龄组高7倍。
在慢性心血管疾病中,心力衰竭和脑血管病变与再感染风险的相关性最高,风险为1.24(95%CI为1.04-1.47,p = 0.0184)和1.49(95%CI为1.29-1.71,p <0.0001)是 未再感染个体的1.24倍(95%CI为1.29-1.71,p 0.0001).
在慢性肾衰竭患者中,接受透析的患者再感染风险几乎高出 3 倍(OR 为 2.77,95% CI 为 1.76–4.38,p < 0.0001).
与未再感染者相比,哮喘和呼吸衰竭/氧疗患者的再感染风险分别增加 1.17 倍(95% CI 为 1.05 –1.33,p = 0.0070)和 1.67 倍(95% CI 为 1.23–2.25,p = 0.0009).
在神经病变患者中,癫痫患者、帕金森病和阿尔茨海默病的再感染风险几乎是未再感染个体的两倍。
痴呆患者的风险大约高出四倍(OR为3.71,95%CI为3.04-4.52,p <0.0001).
在患有自身免疫性疾病的个体中,桥本甲状腺炎是再感染个体中最相关的疾病(OR为1.19,95%CI为1.00-1.44,p = 0.0479)
2021年9月至2022年5月期间与SARS-CoV-2再次感染患者有关的主要合并症的详细信息:
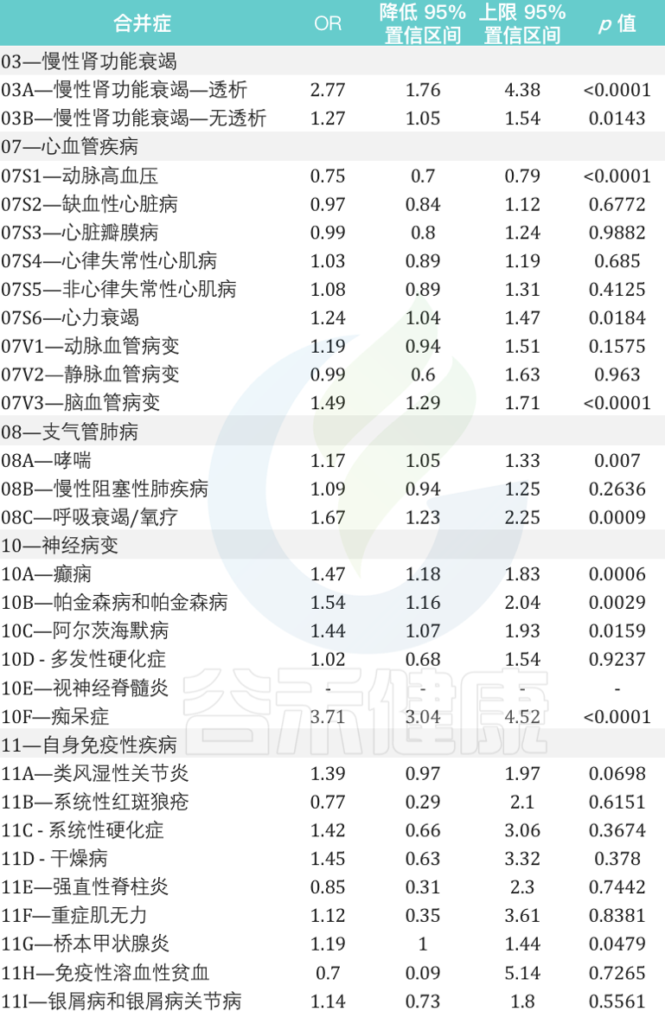
doi: 10.3390/vaccines10111885
此外,2022年发表的一篇系统评价报告了2019年12月1日至2021年9月1日的数据,发现高血压和肥胖是再感染患者中最常见的,其次是终末期肾衰竭、哮喘、慢性阻塞性肺病、痴呆、血脂异常和2型糖尿病。
其他研究也报道,终末期肾衰竭、高血压、糖尿病、慢性呼吸系统疾病、肝病和心血管疾病病史患者的再感染风险更高。
二次感染依然有症状
大多数再感染患者表现出临床症状,只有少数研究报告患者在第一次和继发感染时均无症状。
其他包括50项研究的数据显示再感染患者特征:


注:以上数据是2019年12月1日至2021年9月1日期间的研究
➪
二次感染相对首次感染严重程度有所减轻
韩国疾病控制和预防局于2020年1月20日至2022年5月7日在流行病学调查中通过综合系统报告的新冠肺炎病例数据库与健康保险审查和评估服务系统合并。使用具有二项分布的广义线性模型估计二次感染发作时与一次感染时的严重性比值比(SOR)。
结果发现,在所有患者中,再次感染的SOR为0.89(95%置信区间[CI]:0.82–0.95),与首次感染发作相比,严重程度有所减轻。
➪
一项研究显示,截至 2022 年 11 月 9 日,估计 94%(95% CrI,79%-99%)的美国人口至少感染过一次 SARS-CoV-2。
2022年11月针对SARS-CoV-2感染和重症的有效保护大大高于2021年12月
结果发现,2022年11月9日,在美国,对奥密克戎变异株SARS-CoV-2感染的保护估计为63%(51%-75%),对重症的保护率为89%(83%-92%).
11 月的人口免疫力高于最初奥密克戎激增后立即出现的情况。
对二次感染后的重症情况,不同研究结果不一致。
➪
发表在《Nature Medicine》的一项研究建立了一个包括5819264人的队列,其中SARS-CoV-2感染者(n = 443588)、再感染(两次或多次感染,n = 40947)和非感染对照(n = 5334729). 使用逆概率加权生存模型来估计死亡、住院和事件后遗症的风险和6个月的负担。
结果发现,与无再感染相比,再感染增加了死亡风险(危险比(HR) = 2.17,95%置信区间(CI)1.93–2.45),住院风险(HR = 3.32,95%可信区间3.13-3.51)和后遗症风险,包括肺部、心血管、血液学、糖尿病、胃肠道、肾脏、精神健康、肌肉骨骼和神经系统疾病。无论疫苗接种状况如何,风险都很明显。
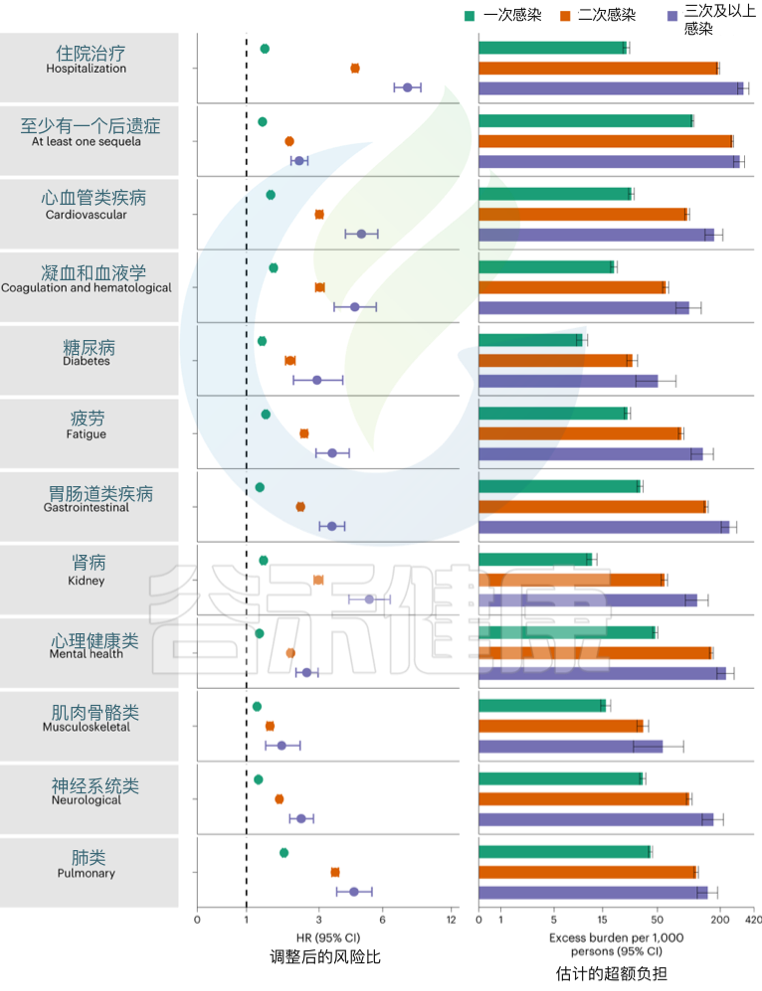
Bowe B, et al., Nat Med. 2022
风险在急性期最为明显,但在6个月后持续存在。与未感染的对照组相比,反复感染的累积风险和负担随着感染次数的增加而增加。
注:该研究数据主要来自美国退伍军人事务部国家医疗保健数据库,选取时间在2020年3月1日至2022年4月6日期间。退伍军人事务部的人口大多由老年人和男性组成,该队列也包括 10.3% 的女性,12% 在 38.8 岁以下(2021 年美国人口的中位年龄)。
关于未来:
英国牛津大学研究病毒进化的科学家认为,奥密克戎及其分支继续占据主导地位的时间越长,慢性感染产生全新变种的可能性就越小。
流行病学研究表明,连续的 COVID-19 浪潮正在变得温和。但这种趋势不应该被视为理所当然。
随着全球对反复接种疫苗和感染后免疫力的增强,研究人员预计 SARS-CoV-2 浪潮的频率将会放缓。
也有研究人员认为,SARS-CoV-2 的一个可能未来是,随季节起伏,通常在冬季达到顶峰,通常每三年左右重新感染一次。
关于其带来的症状是否会越来越轻等问题,研究人员还将持续关注。
总的来说,COVID-19疫情是一个高度动态的全球事件,各种流行病学驱动因素随着时间的推移而变化(包括新变异株的出现、疫苗吸收的增加和免疫力的减弱等各种因素),再感染的流行病学及其健康后果也可能随着时间的推移而改变。
对于已经感染过一次的人来说,继续保持警惕,降低再感染的风险,对于整体健康来说可能很重要。
以上是针对阳康后担心自己是否会二次感染的朋友的问题,然而也有一些朋友到现在,仍然觉得身体没有完全恢复,那就要考虑新冠可能带来的后遗症,也就是临床上说的“长新冠”,接下来章节,我们来了解一下关于“长新冠”及其持续时间,什么人群容易发生,会影响哪些器官功能,具体形成机制,与肠道菌群的关联等。
中 篇
关于“长新冠”
▾◆▽◆▽◆▾
►
“长新冠”是如何定义的?
2021年10月,世界卫生组织将其定义为:
有疑似或确诊新冠感染史,通常发生于起病三个月后,症状通常至少持续两个月,且不能被其他诊断所解释的症状。
►
“长新冠”会有哪些表现?
常见症状包括:
也包括其他一些症状,通常或多或少会影响日常生活。
症状可能是:
从新冠急性发作初步恢复后新出现的症状;
也可能从最初的疾病中持续存在;
也可能随着时间的推移而波动或复发。
►
“长新冠”什么时候发生发展?持续多长时间?
症状的发作和时间过程因个体和症状类型而异。
神经系统症状通常延迟发作数周至数月:在有认知症状的参与者中,43%的人报告在COVID-19至少1个月后才出现认知症状,延迟发作与年龄较轻有关。一些神经认知症状会随着时间的推移而恶化,并且往往会持续更长时间。而胃肠道和呼吸道症状更容易解决。
关节、骨骼、耳朵、脖子和背部的疼痛在1年时比2个月时更常见,感觉异常、脱发、视力模糊以及腿、手和脚肿胀也是如此。
麻痹症在初次感染后平均3个月发病;与其他神经认知症状不同,它通常会随着时间的推移而减少。
很少有长新冠患者能够完全康复,一项研究发现,在最初感染2个月后出现症状的患者中,85%在症状出现1年后出现症状。尽管ME/CFS和自主神经障碍的诊断通常是终身的,但未来的预后仍不确定。
►
有多少人会患“长新冠”?
据保守估计,全世界至少有 6500万 人患有长新冠,病例每天都在增加。
注:有许多未登记病例,这个数字实际上可能要高的多。
据估计,非住院病例的发病率为10-30%,住院病例为50-70%,接种疫苗病例为10-12%。
长冠肺炎与所有年龄段和急性期疾病严重程度相关,36岁至50岁之间的确诊率最高,大多数长冠肺炎病例发生在患有轻度急性疾病的非住院患者中,因为该人群占新冠肺炎总病例的大多数。
►
哪些人群更有可能患“长新冠”?
风险因素可能包括:
注:三分之一的长期 COVID 患者没有确定的原有疾病。
COVID-19 可能对任何患者产生长期影响,包括无症状或轻症患者。相比儿童和青少年,“长新冠”看起来在成人中更为常见。
►
“长新冠”的发病机制?
简单来说,潜在机制包括:
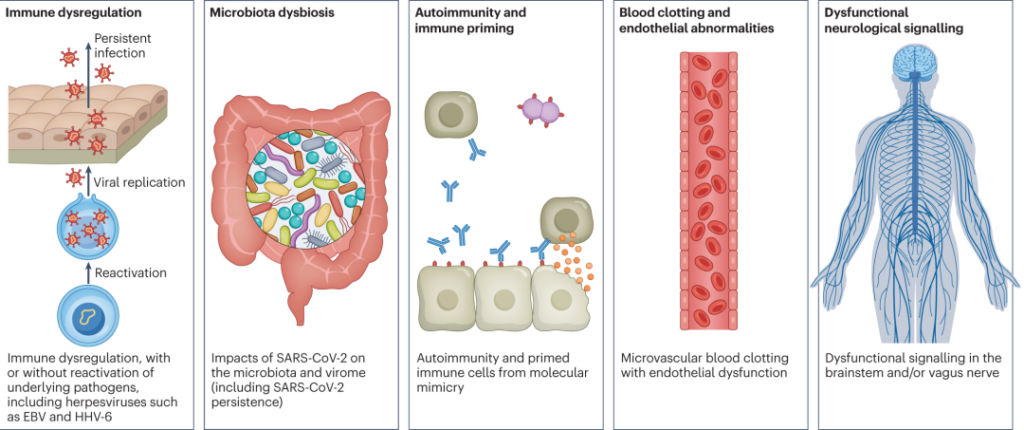
DOI: 10.1038/s41579-022-00846-2
以上是关于“长新冠”的一些基本科普,具体关于“长新冠”带来的多种不良后果,需要关注的疾病,形成机制,与菌群的关联等,我们将在接下来的小节详细介绍。
▾◆▽◆▽◆▾
长新冠包括多种不良后果,常见的新发疾病包括:心血管、血栓和脑血管疾病, 2型糖尿病、肌痛性脑脊髓炎/慢性疲劳综合征(ME/CFS)和自主神经功能障碍,体位性心动过速综合征 (POTS).
长新冠症状及其对多种不同病理器官的影响
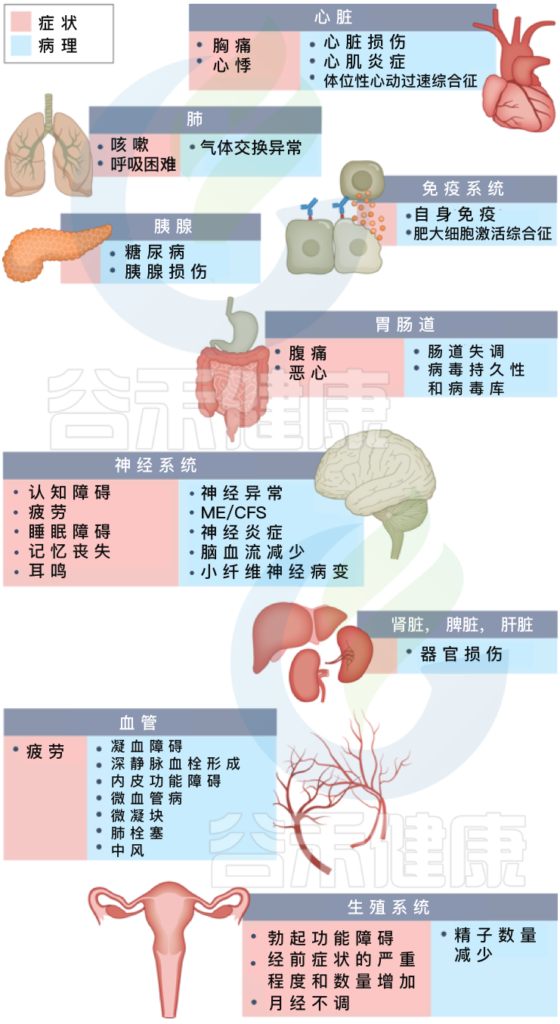
Davis HE, et al., Nat Rev Microbiol. 2023
★
血管问题和器官损伤
SARS-CoV-2会损害许多器官系统。在不同组织中已经证明的损伤主要归因于免疫介导的反应和炎症,而不是病毒对细胞的直接感染。
循环系统破坏包括内皮功能障碍和随后的下游影响,以及深静脉血栓形成、肺栓塞和出血事件的风险增加。
微凝块-> 促进血栓形成
在急性新冠肺炎和长冠肺炎中检测到的微凝块有助于血栓形成,在长新冠中也发现了血细胞大小和硬度的长期变化,有可能影响氧气输送。感染18个月后,长新冠患者的血管密度长期降低,特别影响小毛细血管。
心血管疾病风险增加
对美国退伍军人事务部数据库的分析显示,SARS-CoV-2感染1年后超过150000人患多种心血管疾病的风险显著增加,包括心力衰竭、心律失常和中风,与最初出现新冠肺炎的严重程度无关。
持久的心脏异常
心脏MRI研究显示,在100名既往有新冠肺炎发作史的患者中,78%的患者(感染后平均71天进行调查)和58%的长期冠状病毒感染者(感染后12个月进行研究)存在心脏损害,这增强了心脏异常的持久性。
新冠与多器官损害有关
除了心脏之外,新冠还与其他多种器官受损相关。
一项针对低风险个体的前瞻性研究,观察了心脏、肺、肝脏、肾脏、胰腺和脾脏,发现201名患者中70%至少有一个器官受损,29%有多个器官受损。
在同一研究小组对536名参与者进行的一项为期一年的随访研究中,研究作者发现,59%的人有单器官损伤,27%的人有多器官损伤。
一项针对VA数据的专门肾脏研究,包括89000多名患有新冠肺炎的患者,发现许多肾脏不良结果的风险增加。
另一项VA数据分析,包括181000多名患有新冠肺炎的患者,发现感染也会增加2型糖尿病的风险。
长新冠患者的心血管疾病风险增加,此外,器官损伤似乎是持久的,长期影响仍然未知。
★
神经和认知系统
神经和认知系统的症状是长新冠的主要特征,包括感觉运动症状,记忆丧失、认知障碍、感觉异常、头晕和平衡问题、对光和噪音的敏感性、嗅觉或味觉丧失(或幻觉)以及自主神经功能障碍,影响日常生活。长新冠的前庭听觉表现包括耳鸣、听力损失和眩晕。
认知障碍程度,类似酒驾上限
一项荟萃分析中,在感染后12周,32%的新冠肺炎患者出现疲劳,22%的患者出现认知障碍。长新冠患者的认知障碍会使人衰弱,与英国酒后驾驶限制的醉酒程度或认知衰老10年的程度相同,并且可能会随着时间的推移而增加。
犬尿氨酸途径的激活可能与认知障碍相关
一项研究发现,16%的患者在感染后2个月出现,26%的患者在感染后12个月出现。犬尿氨酸途径的激活,特别是代谢产物喹啉酸、3-羟基邻氨基苯甲酸和犬尿氨素的存在,已在长新冠中确认,并与认知障碍相关。
部分患者可能没有意识到认知障碍
在新冠康复的个体中也发现了认知障碍,当使用客观与主观测量时,认知障碍的比率更高,这表明有一部分认知障碍患者可能没有意识到和/或报告他们的障碍。
认知障碍是一种独立于焦虑和抑郁等心理健康状况的特征
认知障碍在住院和非住院患者中发生率相似。一份有130多万新冠肺炎患者参与的报告显示,随着时间的推移,焦虑和抑郁等心理健康状况恢复正常,但认知障碍(脑雾)、癫痫、痴呆、精神病和其他神经认知疾病的风险增加至少持续了2年。
神经病理学的可能机制包括:神经炎症、凝血病和内皮功能障碍对血管的损伤以及神经损伤
长新冠患者脑脊液异常,年龄较小可能延迟发作
研究发现,患有长新冠的患者存在阿尔茨海默病样信号,脑和脑干代谢低下与特异性症状和长新冠患者中的异常脑脊液发现相关,且年龄较小与神经症状延迟发作相关。
长新冠患者——“化疗脑”
在最近的一份预印本中,轻度感染的长新冠患者的多线细胞失调和髓磷脂丢失,其小胶质细胞反应性类似于化疗,即“化疗脑”。
即使非住院患者,认知能力也可能下降
英国生物银行(UK Biobank)的一项研究,包括新冠流行前后相同患者以及对照组的大脑成像,显示眶额皮层和海马旁回(初级嗅觉皮层相关区域的组织损伤标志物)的灰质厚度减少,与对照组相比,即使是非住院患者,新冠后患者的大脑整体缩小,认知能力下降更大。
尽管该研究将新冠感染者与对照组进行了比较,但并不是特别长的新冠患者,这可能对长新冠的认知成分有影响。在中枢神经系统中发现了线粒体蛋白以及SARS-CoV-2刺突蛋白和核衣壳蛋白的异常水平。在长新冠中也发现了四氢生物蝶呤缺乏症和氧化应激。
眼睛不适?可能与病毒在视网膜感染和复制有关
在眼睛中,在长新冠患者中发现角膜小神经纤维丢失和树突细胞密度增加,瞳孔光反应显著改变,视网膜微循环受损。SARS-CoV-2可以在视网膜和大脑类器官中感染和复制。长新冠的其他表现包括视网膜出血、棉絮斑和视网膜静脉闭塞。
焦虑和抑郁人群比例上升
在爱尔兰,研究报告了20-28%的普通人群出现焦虑和抑郁症状,多达4%的人有自残或自杀的想法。而在2018年爱尔兰健康调查(一项全国性的代表性调查),结果显示,在新冠大流行之前,自我报告的抑郁和焦虑发生率为6%.
仓鼠模型:持续炎症与焦虑和抑郁行为相关
轻度感染的小鼠模型显示小胶质细胞反应性和CCL11水平升高,这与认知功能障碍和受损神经发生有关。
仓鼠模型表现出持续的炎症状态,包括T细胞和髓细胞活化、产生促炎细胞因子和干扰素反应,与仓鼠的焦虑和抑郁行为相关,在新冠肺炎患者的组织中发现了类似的转录特征。轻度疾病感染的非人灵长类动物表现出神经炎症、神经元损伤和凋亡、脑微出血、慢性低氧血症和脑缺氧。
血液皮质醇水平较低,持续症状超过1年
最近的报告表明,与对照组相比,长新冠患者的血液皮质醇水平较低,持续症状超过1年。肾上腺产生的低皮质醇应通过垂体产生的促肾上腺皮质激素(ACTH)来补偿,但事实并非如此,这支持下丘脑-垂体-肾上腺轴功能失调。这也可能反映了潜在的神经炎症过程。此前,ME/CFS患者的皮质醇水平较低。
注:ME/CFS——肌痛性脑脊髓炎/慢性疲劳综合征,是一种多系统神经免疫性疾病。将在下一小节详细阐述。
★
ME/CFS、自主神经功能障碍和相关疾病
ME/CFS通常在病毒或细菌感染后发病。标准包括至少6个月的“疾病前从事职业、教育、社会或个人活动的能力大幅降低或受损”,伴随着无法通过休息缓解的严重疲劳,以及运动后不适、睡眠不足和认知障碍或直立不耐受(或两者兼有)。
高达75%的ME/CFS患者不能全职工作,25%的人患有重度ME/CFS,这通常意味着他们卧床,对感官输入极度敏感,并依赖他人照顾。
大约一半的长新冠患者符合ME/CFS标准
许多研究人员评论了ME/CFS与长新冠之间的相似性;据估计,大约一半的长新冠患者符合ME/CFS标准,在测量运动后不适的主要ME/CFS症状的研究中,大多数长新冠患者报告经历了运动后不舒服。
一项对长新冠患者和ME/CFS患者的直立压力的研究发现,与健康个体相比,两组患者的血流动力学、症状和认知异常相似。
• ME/CFS中一致的异常发现
包括自然杀伤细胞功能减弱、T细胞衰竭和其他T细胞异常、线粒体功能障碍以及血管和内皮异常,包括红细胞变形和血容量减少。
其他异常包括运动不耐受、耗氧量受损和无氧阈值降低,以及代谢异常,包括脂肪酸和氨基酸的使用改变。还观察到神经功能改变,包括神经炎症、脑血流减少、脑干异常和心室乳酸水平升高,以及眼睛和视力异常。反应性疱疹病毒(包括EBV、HHV-6、HHV-7和人巨细胞病毒)也与ME/CFS相关。
• 长新冠患者中观察到上述这些发现
长新冠研究发现,线粒体功能障碍包括线粒体膜电位丧失和可能的线粒体代谢失调、脂肪酸代谢改变和线粒体依赖性脂质分解代谢失调,与运动不耐受、氧化还原失衡、运动不耐受和氧提取受损的线粒体功能障碍一致。
研究还发现了内皮功能障碍、脑血流异常和代谢变化(即使是POTS症状减轻的长新冠患者)、广泛的神经炎症、疱疹病毒重新激活、红细胞变形以及其他地方讨论的许多发现。不仅在长新冠患者中,而且在ME/CFS患者中也发现了微裂纹和过度活化的血小板。
自主神经障碍,特别是POTS,通常与ME/CFS共病,也常伴有病毒性发作
POTS与G蛋白偶联的肾上腺素能受体和毒蕈碱乙酰胆碱受体自身抗体、血小板储存库缺陷、小纤维神经病变和其他神经病变有关。POTS和小纤维神经病变通常在长新冠中发现,一项研究发现67%的长新冠队列中存在POTS。
注:POTS——体位性心动过速综合征,一种随着姿势的改变而心率增加的情况,例如躺着坐起来或站着。这会导致头晕或昏厥。
肥大细胞激活综合征也通常与ME/CFS共病
与新冠前患者和对照组相比,长新冠患者肥大细胞激活综合征症状的数量和严重程度显著增加,组胺受体拮抗剂可改善大多数患者的症状。
注:肥大细胞活化综合征(MCAS)是一种由肥大细胞异常活化导致的慢性多系统性疾病.肥大细胞广泛分布于胃肠道,因此MCAS易累及胃肠道并出现相应症状。
其他可能共病的疾病
其他通常与ME/CFS共病的疾病包括结缔组织疾病,包括Ehlers–Danlos综合征和高移动性、神经矫形脊柱和颅骨疾病以及子宫内膜异位症。
长新冠中观察到与ME/CFS,自主神经障碍,肥大细胞激活综合征等疾病类似的发现,其他疾病也可能与长新冠合并。应进一步探讨病毒后条件与这些条件的重叠。
★
生 殖 系 统
长新冠中经常报告对生殖系统的影响。与无冠状病毒病史的女性以及患有新冠肺炎但不长时间冠状病毒的女性患者相比↓↓↓
患长新冠的女性更有可能发生月经改变
月经和月经前一周已被患者确定为长新冠症状复发的诱因。
在新冠肺炎患者中观察到卵巢储备下降和生殖内分泌紊乱,初步理论表明SARS-CoV-2感染会影响卵巢激素的产生和/或子宫内膜反应,因为卵巢和子宫内膜组织中ACE2受体丰富。
与那些没有月经变化的人相比,同时患有新冠和月经改变的人更容易出现疲劳、头痛、身体疼痛和气短,最常见的月经变化是月经不规律、经前症状增加和月经不频繁。
ME/CFS与多种妇科疾病存在关联
对ME/CFS的研究表明,ME/CFS与经前焦虑障碍、多囊卵巢综合征、月经周期异常、卵巢囊肿、绝经早期和子宫内膜异位症之间存在关联。妊娠、产后变化、围绝经期和月经周期波动会影响ME/CFS,并影响代谢和免疫系统变化。长新冠的研究应该关注这些关系,以更好地理解病理生理学。
病毒在阴茎组织中的持续存在,勃起功能障碍的风险也增加
这可能是由内皮功能障碍引起的。在一项研究中,与对照组相比,长新冠患者的精子计数、精液体积、活力、精子形态和精子浓度受损,并与细胞因子水平升高以及精液中胱天蛋白酶8、胱天蛋白酶9和胱天蛋白酶3的存在相关。
长新冠患者女性月经变化(月经不规律等),更容易出现疲劳,头痛等症状,男性精子质量变化,长期影响仍需进一步研究。
★
呼吸系统
呼吸系统疾病是长新冠的常见表现型,在一项研究中,新冠肺炎幸存者的发病率是普通人群的两倍。
呼吸短促和咳嗽是最常见的呼吸道症状,分别在40%和20%的长新冠患者中持续至少7个月。
几项包括长新冠患者的非住院患者的影像学研究显示了肺部异常,包括空气潴留和肺部灌注。
对感染后3-6个月的患者进行的免疫学和蛋白质组学研究表明,气道中的细胞凋亡和上皮损伤,但血液样本中没有。
进一步的免疫学特征比较了长新冠肺炎患者和新冠肺康复者,发现肺功能下降、全身炎症和SARS-CoV-2特异性T细胞之间存在相关性。



doi: 10.3389/fimmu.2021.686029
★
胃肠道系统
长新冠的胃肠道症状包括恶心、腹痛、食欲不振、胃灼热、便秘等。
持续的呼吸道和神经系统症状都与特定的肠道病原体有关。
SARS-CoV-2 RNA存在于新冠肺炎患者的粪便样本中,一项研究表明,12.7%的参与者在新冠肺炎确诊4个月后粪便中持续存在,3.8%的参与者在确诊7个月后持续存在。大多数感染7个月后出现长新冠症状和炎症性肠病的患者在肠粘膜中存在抗原持久性。
与没有长新冠或SARS-CoV-2阴性对照的患者相比,长新冠肺炎患者的血浆中发现了来自肠道和/或肺上皮的较高水平的真菌易位,可能会诱导细胞因子的产生。
长新冠患者的胃肠道症状可能与肠道菌群变化相关
与非新型冠状病毒肺炎对照组(疫情爆发前)相比,长新冠感染者中发现了较高水平的Ruminococcus gnavus和普通拟杆菌(Bacteroides vulgatus)和较低水平的普氏粪杆菌(Faecalibacterium prausnitzii),肠道失调持续至少14个月;低水平的产丁酸盐细菌与6个月时的长新冠密切相关。
将长新冠患者的肠道细菌转移到健康小鼠体内,导致小鼠认知功能丧失,肺部防御受损。
为什么长新冠患者的肠道菌群会发生变化?
胃肠道和呼吸道症状怎样将微生物群与SARS-CoV-2感染联系起来?
病毒是如何感染肠道的?
感染后的肠道菌群及宿主免疫会受到什么样的影响?
……
我们将在下一小节详细介绍。
▾◆▽◆▽◆▾
要了解病毒是如何感染,为什么会与肠道菌群相关联,我们来先从病毒的结构说起:
SARS-CoV-2是新冠肺炎的病原体。它是一种正向单链RNA病毒。它编码膜蛋白(M蛋白)、核衣壳蛋白(N蛋白)、刺突蛋白(S蛋白)和包膜结构蛋白(E蛋白)和多种非结构蛋白。
呼吸道胃肠道是人类微生物群的主要栖息地,也是SARS-CoV-2感染的目标
⇓⇓⇓
SARS-CoV-2病毒如何实现感染?
病毒利用表面的刺突蛋白和人体细胞上的ACE2(血管紧张素转化酶2)进行结合,从而实现感染。
在病毒体上,刺突蛋白(S蛋白)是包含S1和S2亚基的同源三聚体:
病毒劫持宿主细胞表面蛋白酶,如跨膜丝氨酸蛋白酶2(TMPRSS2),TMPRSS2反过来激活病毒S蛋白,切割ACE2受体,并促进病毒与宿主细胞膜的结合。
除了ACE2和TMPRSS2介导的进入外,SARS-Cov-2还可以利用宿主细胞的吞噬作用或内吞作用侵入某些免疫细胞类型,如巨噬细胞。
为什么SARS-CoV-2可感染肠道?
ACE2和TMPRSS2在呼吸道和胃肠道中强烈表达。由于后者与外部环境沟通,它们是SARS-CoV-2入侵的主要目标。
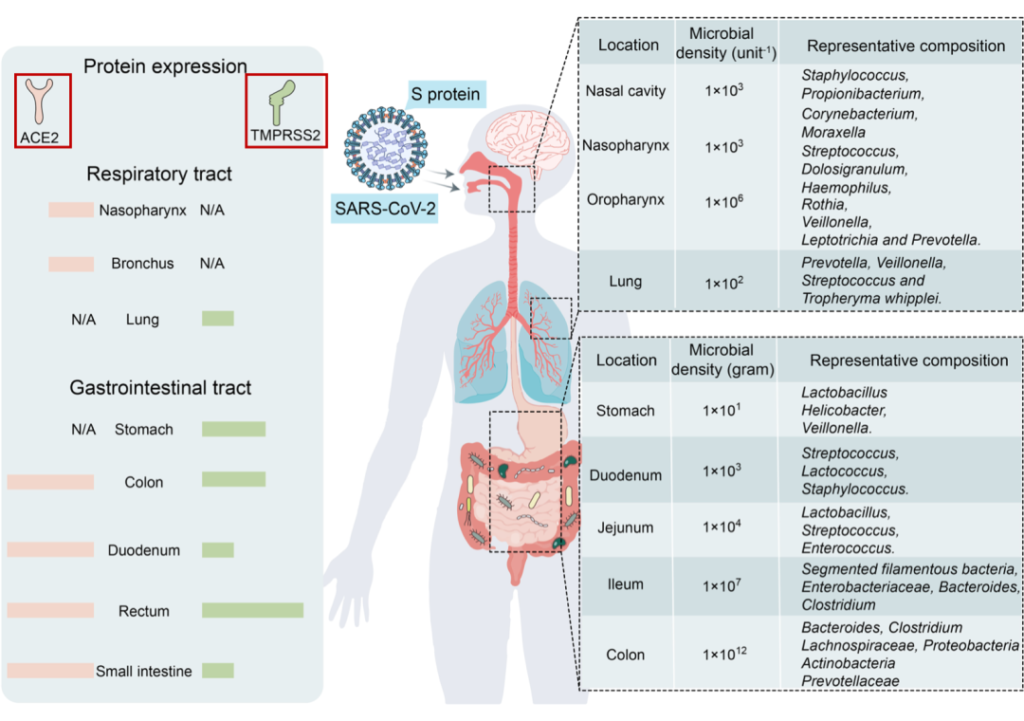
Wang B, et al., Signal Transduct Target Ther. 2022
SARS-CoV-2受体ACE2和TMPRSS2主要在呼吸道和胃肠道中表达,为微生物提供了许多合适的栖息地。
由于ACE2和TMPRSS2在胃肠道中高度表达,SARS-CoV-2也以肠道为靶点。一些研究报告称,新冠肺炎患者粪便样本中SARS-CoV-2病毒RNA呈阳性。
对一名新冠肺炎患者进行的活组织检查显示,SARS-CoV-2蛋白涂层存在于胃、十二指肠和直肠中。因此,SARS-CoV-2可感染肠道。
COVID-19的胃肠道和呼吸道症状将微生物群与SARS-CoV-2感染联系起来
⇓⇓⇓
几项临床研究报告,11-39% 的 COVID-19 患者有胃肠道症状,包括恶心、呕吐、腹泻和腹痛。
智利进行的一项研究报告称,在 7016 名 COVID-19 患者中,有 11% 表现出胃肠道症状。
在浙江的651例COVID-19患者中,8.6%表现出腹泻,4.15%表现出恶心或呕吐。
胃肠道症状与相对较高的住院风险和/或更高的疾病严重程度相关。在重症和/或危重患者中,疾病进展并引起并发症,例如急性呼吸窘迫综合征(ARDS),败血症,继发性病原体肺炎和终末期器官衰竭。
由于微生物群维持呼吸道和胃肠道稳态和健康,上述COVID-19相关症状可能将微生物群与SARS-CoV-2感染联系起来。
一些研究证实,肠道菌群有助于调节肠道免疫稳态和病原体感染。因此,肠道细菌可能对SARS-CoV-2感染的宿主免疫反应至关重要。
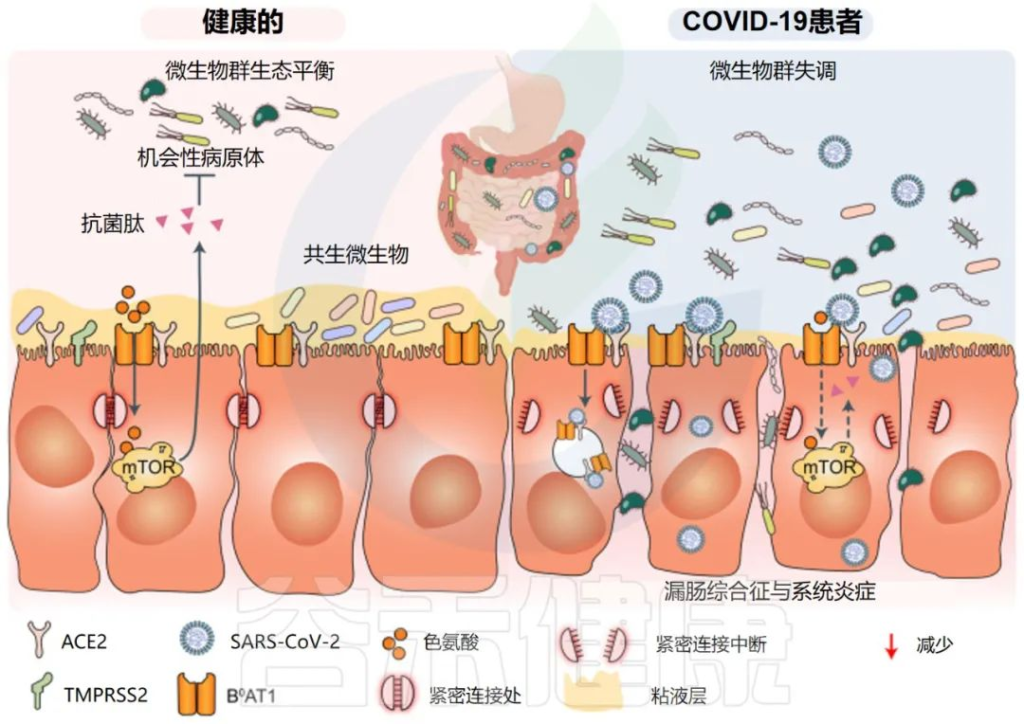
一些研究小组报告称,与新冠完全康复的患者相比,长新冠患者的肠道微生物群存在差异。在诊断时检测到微生物差异,但在6个月后被夸大。特别是,长新冠患者出院后,微生物群的丰富度没有恢复到正常状态。
持续的症状可能与免疫隔离组织中存在少量残余病毒有关,特别是身体中不受抗体直接保护的区域,如肠道。
一个重要的观察结果显示,病毒清除后,肠道失调持续了数月。与作为对照的健康个体相比,从新冠肺炎中恢复的患者在3个月时细菌多样性和丰富度降低,同时有益共生菌丰富度较低,机会性病原体丰富度较高。在随访6个月时,与对照组相比,COVID-19患者中双歧杆菌和瘤胃球菌的相对丰度显著降低(P < 0.001).
一项前瞻性研究追踪了香港106名新冠肺炎住院患者肠道微生物群的纵向动态,大约四分之三的患者在感染后6个月出现”长新冠”症状(通常是疲劳、记忆力差和焦虑)。
粪便样本的Shotgun宏基因组分析显示,与没有长新冠的个体和作为对照的健康个体相比,长新冠患者微生物多样性显著降低,细菌类型减少。
长新冠患者的Bacteroides vulgatus和Ruminococcus gnavus 的丰度增加,而P.prausnitzii的丰度减少。
有趣的是,与健康个体(n = 11)相比,PI-IBS患者(n = 11)的粪便样本中Bacteroides vulgatus也显示出6倍的升高;这一发现表明,Bacteroides vulgatus可能与长新冠和PI-IBS的发病机制有关。
注:因感染而导致的IBS被称为感染后肠易激综合征(PI-IBS)
doi: 10.1038/s41392-022-00986-0
此外,SARS-CoV-2感染后6个月出现呼吸道症状与Streptococcus vestibularis和Streptococcus anginosus等机会性致病物种的水平升高有关。
而疲劳和神经精神症状与医院内病原体有关,如:Clostridium innocuum 和 Actinomyces naeslundii。
在脱发患者中,产丁酸盐细菌显著减少,某些细菌,如Bifidobacterium pseudocatenulatum和F. prausnitzii,与长新冠的发展具有最大的负相关。入院时的细菌种类,包括长双歧杆菌(B.longum)和Blautia wexlerae,与6个月时长新冠的发展呈负相关,这意味着这些细菌在恢复期具有潜在的保护作用。
相比之下,在长新冠患者中富集的菌有:
Atopobium parvulum
Actinoomyces johnsonii
Actinomyces sp. S6 Spd3
这些发现表明,一个人在感染时的肠道微生物组组成能会影响其对新冠长期并发症的敏感性。尽管如此,这些变化可能代表长新冠的反应性变化,未来的研究需要包括从感染到症状发展的非住院患者的前瞻性纵向研究,以描述肠道菌群失调对长新冠症状的确切影响。
持久战——免疫细胞数量变化
针对患有轻度急性新冠肺炎的长冠肺炎患者的免疫失调进行的研究发现,T细胞改变,包括耗尽的T细胞、CD4+和CD8+效应记忆细胞数量减少以及中央记忆细胞PD1表达升高,持续至少13个月。
研究还报道了高度活化的先天免疫细胞,缺乏初始T细胞和B细胞,I型和III型干扰素(IFN-β和IFN-λ1)的表达升高,持续至少8个月。
一项综合研究将长新冠患者与未感染者和无长新冠的感染者进行了比较发现,在感染后中位数14个月,长新冠患者的非经典单核细胞、活化B细胞、双阴性B细胞以及分泌IL-4和IL -6的CD4+ T细胞数量增加,常规树突状细胞和T细胞数量减少,皮质醇水平降低。
疲劳和神经认知功能障碍可能与细胞因子变化和EBV再激活相关
细胞毒性T细胞的扩增已被发现与长冠的胃肠道表现有关。更多的研究发现细胞因子水平升高,特别是IL-1β、IL-6、TNF和IP10,最近的预印本报道了CCL11水平的持续升高,这与认知功能障碍有关。
在长新冠患者中发现了包括EBV和HHV-6在内的反应性病毒(已在ME/CFS45中发现),并导致线粒体断裂,严重影响能量代谢。最近的一份预印本报告,在长新冠患者中,EBV再激活与疲劳和神经认知功能障碍相关。
抗体生成量低,可预测长新冠
几项研究表明,在新冠肺炎急性期,无论是住院患者还是非住院患者,SARS-CoV-2抗体生成量低或无,以及其他免疫反应不足,都可以预测6至7个月后长新冠。
这些不足的免疫反应包括IgG的低基线水平、受体结合域和刺突特异性记忆B细胞的低水平、核衣壳IgG的低水平和刺突特异性IgG的低峰值。
在最近的一份预印本中,发现严重长新冠患者的CD4+T细胞和CD8+T细胞应答较低或缺失,另一项研究发现,与无长新冠的感染对照组相比,长新冠患者的CD8+T淋巴细胞表达CD107a的水平较低,核衣壳特异性干扰素-γ-产生的CD8+T细胞下降。
二次感染可能与抗体水平低相关
长新冠患者体内高水平的自身抗体与保护性新冠肺炎抗体呈负相关,这表明高水平自身抗体的患者更有可能发生突破性感染。
SARS-CoV-2病毒在肠道内反弹,可能是由于病毒持续存在,也与受体结合域IgA和IgG抗体的水平较低和产生较慢有关。
长新冠驱动因素:病毒持续存在
一些报告指出,病毒持续存在可能是长新冠的驱动因素;在生殖系统、心血管系统、大脑、肌肉、眼睛、淋巴结、阑尾、乳腺组织、肝组织、肺组织、血浆、粪便和尿液中发现了病毒蛋白和/或RNA。胃肠道活检后的多份报告表明存在病毒,提示某些患者体内存在持久性病毒库。
ACE2——病毒感染人体细胞的“钥匙”
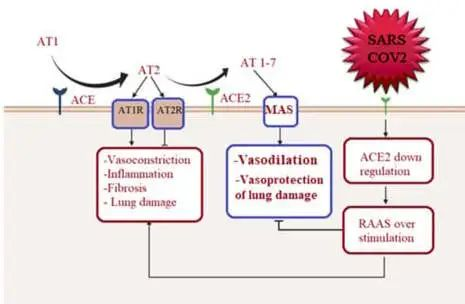
前面我们已经知道,病毒利用表面的刺突蛋白和人体细胞上的ACE2(血管紧张素转化酶2)进行结合,从而实现感染,可见,ACE2作为病毒进入的一个入口点,扮演者重要的角色。
ACE2能做什么?
ACE2在肾素-血管紧张素系统(RAS)中起主要作用,除此之外,其活性在肺部也有作用,因为它通过抑制des-Arg9-缓激肽调节缓激肽1受体信号传导,从而减少血管舒张和血管通透性。
ACE也在肠道肠细胞中表达,它可以调节微生物生态、先天免疫和饮食氨基酸稳态。
ACE2亦敌亦友
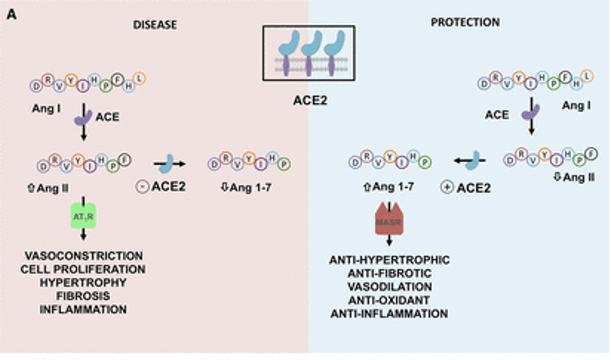
ACE2在肺部的保护作用可能是COVID-19中的一把双刃剑:
ACE2在肾素-血管紧张素系统(RAS)中的作用以及严重急性呼吸综合征SARS-CoV-2诱导的细胞表面ACE2表达下调的机制如下:
doi: 10.1161/CIRCULATIONAHA.120.047049
ACE2平衡肾素-血管紧张素系统的2个轴:
➭ 增加的ACE2促进保护性ACE2/Ang 1-7/MASR
➭ ACE2的丢失导致向以ACE/血管紧张素II(Ang II)/AngII受体1型(AT1)受体轴(AT1R)过度活性为特征的疾病状态转变。
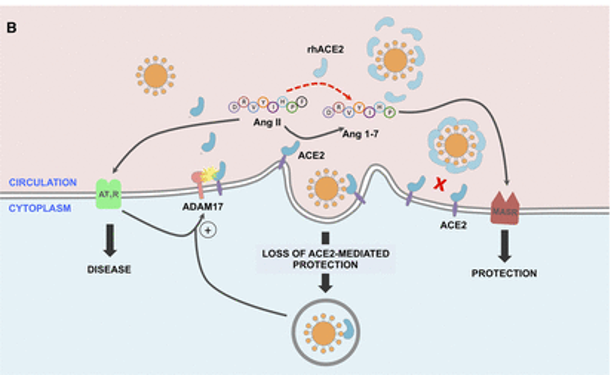
doi: 10.1161/CIRCULATIONAHA.120.047049
SARS-CoV-2的病毒刺突糖蛋白与细胞表面ACE2相互作用,并通过内吞作用一起内化,导致表面ACE2表达降低。ACE2的丢失导致Ang II的积累,Ang II通过AT1受体也上调ADAM17,导致细胞表面ACE2的进一步裂解。
肠道微生物群对肠道ACE2表达的影响?
动物研究中,无菌小鼠的肠道和肺部ACE2表达水平明显高于常规小鼠。以不同微生物群定殖的促性腺激素小鼠显示出肠道ACE2表达的可变性,这可能部分归因于微生物组编码的蛋白酶和肽酶类型的差异。
2021发表的一项研究确定了调节肠道ACE2表达的转录因子,包括已知受肠道微生物群调节的GATA4。这些数据表明,肠道微生物群可能在调节ACE2表达中发挥作用。
特定肠道菌群抑制ACE2表达
在小鼠模型中,特定的细菌种类,如多氏拟杆菌(Bacteroides dorei)和长双歧杆菌,可以抑制结肠ACE2的表达。
与小鼠结肠ACE2表达下调相关的四种拟杆菌显示出显著性差异(P < 0.05),与粪便SARS-CoV-2载量呈负相关。
“阳”之前肠道菌群已经异常,促进易感性
死于新冠肺炎的高危人群通常是那些患有糖尿病、心血管疾病和肥胖症等共病的人群,这些疾病也与微生物异常有关,其特征是细菌多样性减少。
这一发现表明,感染前的肠道微生物群可能促进宿主对SARS-CoV-2和ACE表达的易感性。
拟议的感染前、中、后肠道菌群变化模型及新冠措施如何影响人一生中的菌群多样性
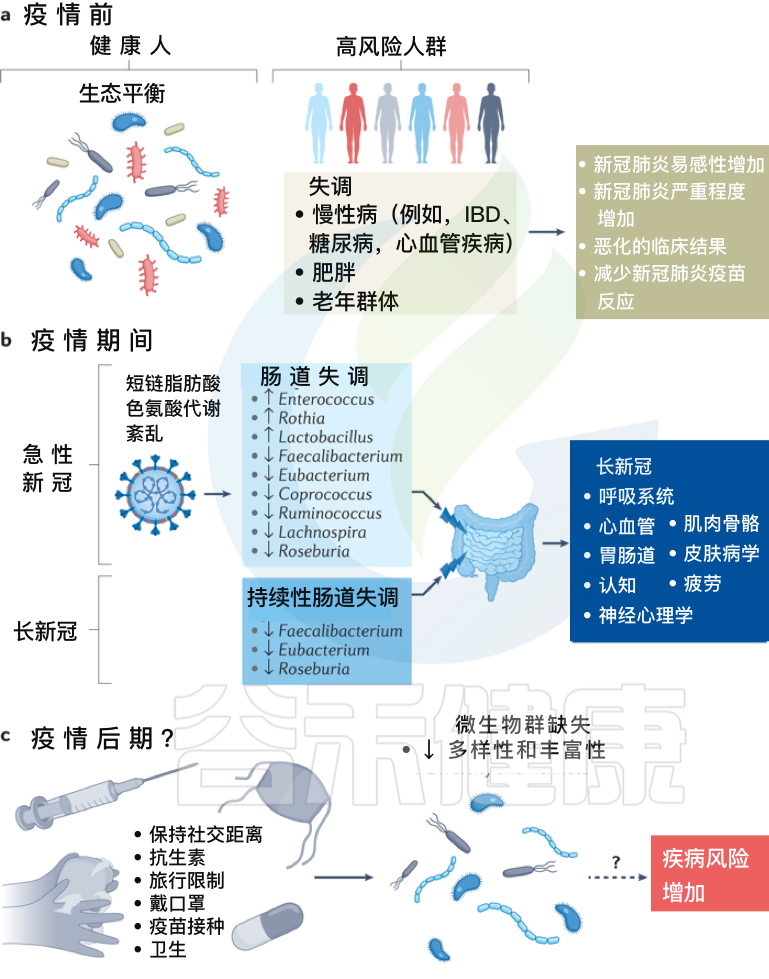
Zhang F, et al., Nat Rev Gastroenterol Hepatol. 2022 Oct
a、在新冠疫情之前,健康个体的肠道微生物群以“生态平衡”为特征,这是一个具有丰富微生物多样性的平衡肠道生态系统。
而某些个体,包括老年群体和患有炎症性肠病、糖尿病、心血管疾病和肥胖等慢性病的个体,肠道生态系统发生了变化,微生物多样性降低,肠道微生物组成失调可能导致严重急性呼吸综合征SARS-CoV-2感染的易感性增加,新冠肺炎严重程度增加,临床结局恶化和/或新冠肺炎疫苗反应降低。
b、在疫情期间,急性新冠感染与持续的肠道微生物群组成变化、短链脂肪酸生物合成受损和色氨酸代谢紊乱有关。
最初感染中出现的功能障碍也与急性后新冠肺炎综合征有关,包括慢性呼吸道症状(例如咳嗽或气短)、心血管症状(例如胸痛或心悸)、胃肠道症状(例如食欲不振或腹泻)、神经精神症状(例如焦虑或失眠)、肌肉骨骼症状(例如关节疼痛或肌肉无力)和皮肤病症状(例如皮疹或脱发)。
在急性新冠肺炎疫情后阶段,肠道微生物群持续受到破坏,其特征是产短链脂肪酸的细菌粪杆菌、真杆菌和Roseburia持续耗竭。急性期肠道菌群组成的改变也与多器官急性后新冠肺炎综合征有关。
c、除了大流行之外,现有的疫情控制做法,包括严格执行社交距离、广泛的卫生消毒措施、定期接种疫苗和限制旅行,可能会对婴儿的微生物组多样性产生负面影响,并对肠道中的早期细菌定植产生重大影响,对疾病风险产生未知后果。
需要做更多的工作来调查和确认新冠、微生物群丧失和未来疾病风险之间的潜在联系。未来导致微生物损失的感染控制措施需要与促进微生物多样性的战略相平衡,以确保子孙后代的健康益处。
迄今为止,大多数临床研究表明 SARS-CoV-2 感染与肠道菌群改变之间存在关联,但尚不清楚肠道菌群改变是感染的原因还是影响。
为此研究人员提出了两种假设,并推断了相应的机制。
SARS-CoV-2感染 ▬► 肠道菌群失调
那么研究人员假设了 SARS-CoV-2 可导致肠道菌群失调,推断了几种可能的机制:
1) SARS-CoV-2侵入肺部可导致组织损伤,可以激活以NF-κB和TNF途径上调为特征的强促炎途径,激活模式识别受体(TLR、RLR、NLR),这些受体被先天免疫细胞识别,导致各种促炎细胞因子的释放。
或者肠道感染可导致肠道结构的直接损伤和肠上皮屏障的破坏,可能损害肠道通透性,并促进肠道炎症。
激活的全身炎症和肠道炎症可能导致破坏肠道微生物群平衡,导致:
• 机会病原体(例如肠杆菌科和肠球菌)的丰度增加
• 共生菌(例如Faecaliberium、Eubacterium和Roseburia)的丰度降低
关于Faecaliberium详见:
肠道核心菌属——普拉梭菌(F. Prausnitzii),预防炎症的下一代益生菌
关于Eubacterium 详见:
肠道核心菌属——优/真杆菌属(Eubacterium),你为什么要关心它?
关于Roseburia 详见:
2) SARS-CoV-2感染可下调肠上皮细胞管腔表面ACE2和B0AT1(分子ACE2伴侣)的表达,这可能促进病原菌的生长。
3) 一项体外研究发现SARS-CoV-2可能直接感染细菌。揭示了SARS-CoV 2 影响肠道微生物群的另一种可能机制。
肠道菌群特征 ▬► 严重程度和免疫反应
动物研究提供了证据,SARS-CoV-2感染可能在驱动肠道微生物群生态变化中发挥作用。
➪
当用SARS-CoV-2攻击恒河猴和食蟹猴的非人灵长类动物模型时:
➪
仓鼠模型能够重现人类严重新冠的一些特征:
研究人员提出SARS-CoV-2感染点的特定内在“微生物组特征”,可以通过几种假设机制影响感染的严重程度和宿主免疫反应:
i) 增加的机会性病原体可能会被先天性淋巴细胞进一步识别,并强化肠道促炎反应。
ii)机会性病原体和毒素可能转移到循环系统中,导致菌血症,加剧系统炎症和疾病严重程度。
iii)有益菌逐渐减少耗尽,可能会对免疫细胞的募集产生负面影响,如激活的粘膜相关不变性T细胞(MAIT),从而影响呼吸道感染的易感性和严重性。
肠道菌群可能导致免疫反应功能失调和 COVID-19严重程度的潜在机制
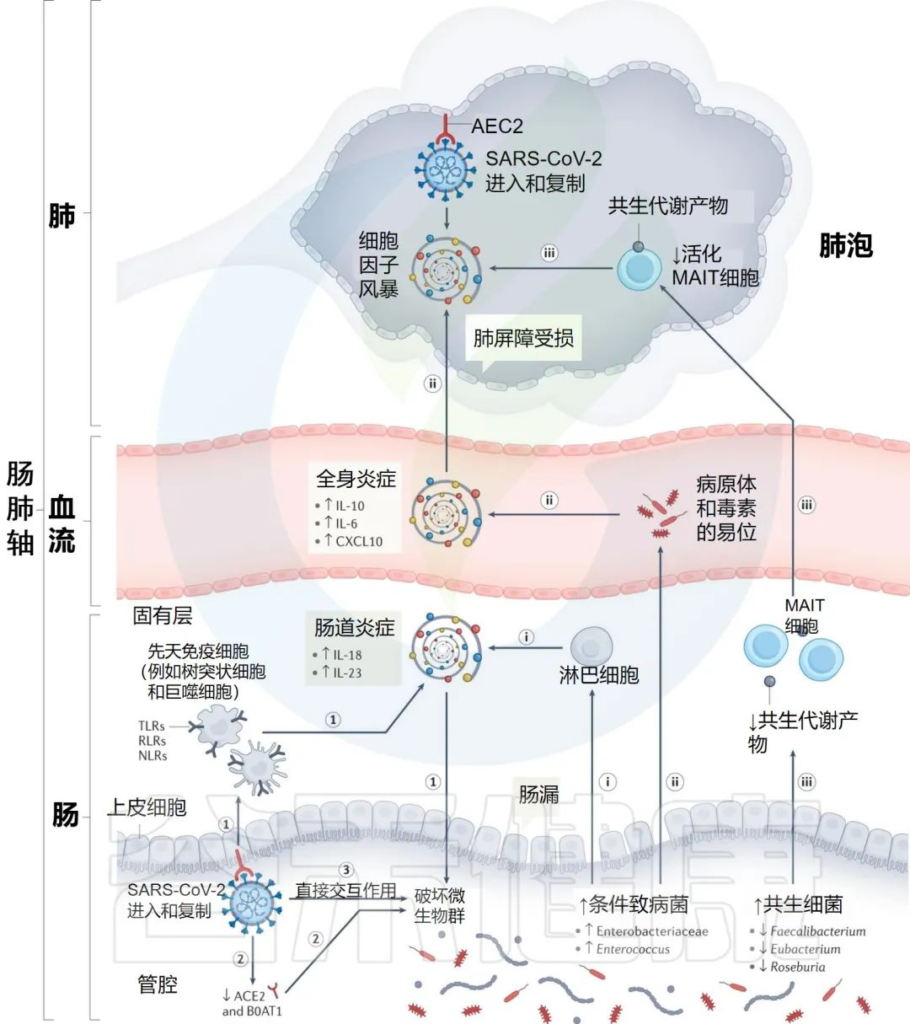
Zhang F, et al., Nat Rev Gastroenterol Hepatol. 2022 Oct
肠道菌群代谢产物
SARS-CoV-2感染与肠道微生物群的碳水化合物、脂质和氨基酸代谢改变有关。
——短链脂肪酸:合成受损
多项研究表明,COVID-19 患者的粪便样本中的短链脂肪酸生物合成受损。
在对66名新冠肺炎抗生素无效患者和70名未感染新冠的患者进行的宏基因组分析中,SARS-CoV-2感染患者的肠道微生物群合成短链脂肪酸的能力降低,这与疾病严重程度和血浆中促炎细胞因子IL-10和趋化因子CXCL10浓度升高呈负相关。
通过粪便代谢物的测量,19名与新冠相关的严重和/或危重疾病患者的粪便中短链脂肪酸(包括乙酸、丙酸、丁酸、戊酸和己酸)浓度持续下降。
短链脂肪酸可以激活免疫细胞的抗炎反应,抑制炎症信号通路,并保持肠道屏障的完整性,以防止肠道内毒素和细菌进入循环,从而减轻局部和全身炎症反应。
鉴于短链脂肪酸在调节宿主免疫反应中的重要性,新冠肺炎中短链脂肪酸生物合成不足可能与疾病发病机制和严重程度有关。然而,短链脂肪酸缺失是否是新冠感染的原因或后果尚待阐明。
——色氨酸代谢:受到干扰
一些测量新冠患者血浆代谢物的研究表明,与健康人作为对照组相比,色氨酸代谢受到干扰,与色氨酸新陈代谢有关的犬尿氨酸途径的激活增强。
色氨酸代谢通过调节调节性T细胞与TH17细胞的比率和B细胞活性与自身免疫、病毒感染和肠道健康相关。
在人类和动物研究中,犬尿氨酸途径代谢产物进入大脑的增加可能会引发疲劳、记忆力差和抑郁等症状,这是“长新冠”的常见症状。重要的是,色氨酸代谢产物是宿主-微生物群界面的关键介质。
根据人类和动物研究的证据,肠道微生物群可以直接使用色氨酸作为底物,并影响宿主色氨酸的吸收和代谢,以调节宿主的生理和免疫反应。内源性宿主色氨酸代谢产物可以深刻影响肠道微生物群的组成和功能,如阿克曼氏菌和乳杆菌。
综上所述,这些数据表明色氨酸代谢是肠道微生物群参与新冠肺炎的一种可能机制。
——胆汁酸代谢:菌群失调影响胆汁酸代谢,胆汁酸浓度升高破坏肠道屏障,引起炎症
初级胆汁酸由胆固醇、胆酸和鹅去氧胆酸通过与甘氨酸或牛磺酸结合在肝脏中合成。然后,它们被分泌到小肠,在那里它们被肠道菌群转化为次级胆汁酸,主要是脱氧胆酸(DCA)和石胆酸(LCA)。
次级胆汁酸在上皮细胞和内皮细胞以及肝细胞中充当核受体FXR、VDR和PXR的配体。它们还与TGR5相互作用。
注:TGR5是一种在肠、胰腺、淋巴组织和大脑中表达的膜结合受体。DCA和LCA都能够通过上述受体调节免疫系统。
研究人员根据疾病严重程度将新冠肺炎患者从无症状患者到处于关键阶段的患者进行分组。随着疾病严重程度的增加,来自患者的肠道菌群中厚壁菌/拟杆菌的比例逐渐受到更大的影响。
来自无症状患者的肠道菌群保留了有益菌种类,这些细菌与新冠肺炎的不良后果呈负相关:
此外,与危重症患者的微生物群不同,他们的微生物群的特征在于参与次级胆汁酸生物合成代谢途径的基因的高表达。
Harry Sokol等人观察了SARS-CoV-2感染对灵长类微生物群的影响,并发现总胆汁酸的数量随疾病严重程度而增加。值得注意的是,初级/次级胆汁酸的比率也明显较高。
这些数据表明,SARS-CoV-2感染对肠道菌群的破坏随着疾病的严重程度而加剧。随着菌群失调程度的增加,回肠的内在功能进一步改变,导致肠内转运增加,从而阻止胆汁酸的完全重吸收,从而增加其在结肠中的浓度。此外,重症新冠肺炎患者的肠道菌群功能有限,因此胆汁酸集中在这些患者的粪便中。
血清胆汁酸谱显示,急性呼吸窘迫综合征(ARDS)患者符合这些观察结果,因此可以推断,在严重的新冠肺炎中,胆汁酸浓度升高可能会破坏肠道屏障,并通过血流到达包括肺、心脏、肾和内皮在内的外周组织。它们的细胞毒性活性可能会损伤外周组织的细胞膜,导致局部和全身炎症反应,并在临床上表现出来。
——鞘脂:肠道微生物鞘脂代谢改变
据报道,新冠肺炎患者血清和粪便中鞘脂浓度降低,肠道微生物鞘脂代谢改变。鞘磷脂是生物膜的组成部分,介导信号转导和免疫激活。
拟杆菌产生的鞘脂可以增加外源鞘脂,从而增强体外或体内研究中观察到的调节性T细胞的分化,这可能抑制冠状病毒的复制。
这一观察支持肠道微生物群衍生的鞘脂可能调节宿主对SARS-CoV-2感染的防御的假设。
——蔗糖、葡萄糖:异常
与47名健康人作为对照组相比,56名新冠肺炎患者的粪便蔗糖水平升高,粪便葡萄糖水平降低。蔗糖和葡萄糖的异常水平可能与蔗糖酶-异麦芽糖酶活性受损有关。
这种变化可能与新冠肺炎常见的肠道症状有关,如腹泻、呕吐、肠胃气胀和腹痛。胀气通常是由细菌在肠道中发酵未吸收的碳水化合物引起的。
蔗糖水平的增加与放线菌和Streptococcus parasanguinis水平的增加有关,这意味着COVID-19 中的生态失调,可能会破坏肠道发酵并导致胃肠道症状。
▾◆▽◆▽◆▾
累积的证据表明,肠道菌群失调与 COVID-19 感染的严重程度和疾病恢复后的长期多系统并发症有关。
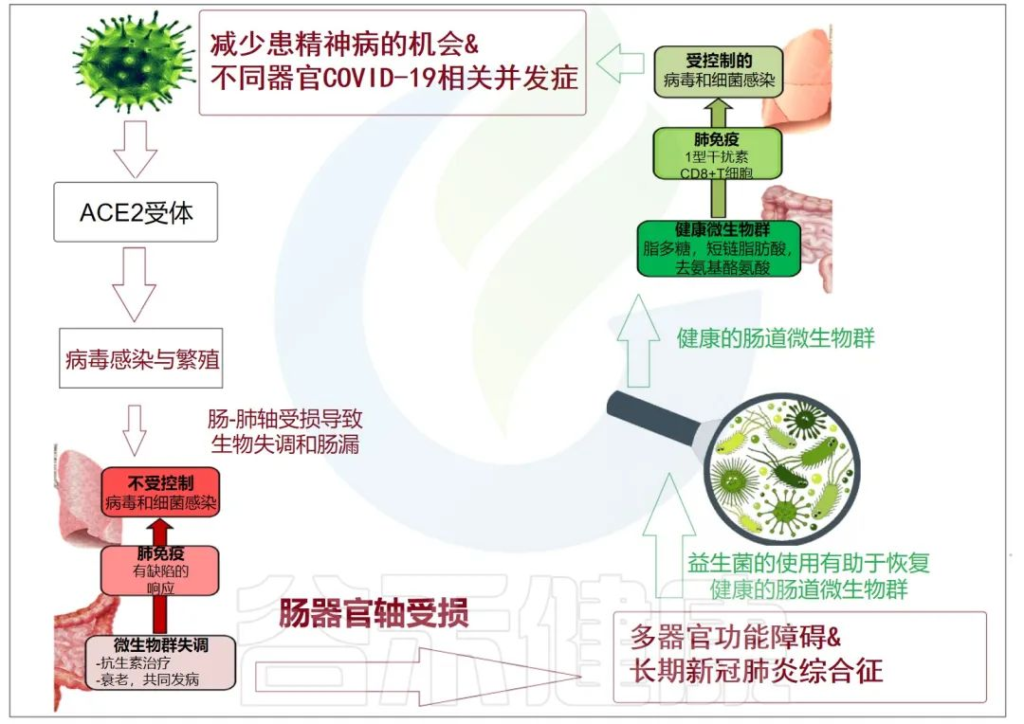
doi: 10.3390/metabo12100912
肠-肝轴是指肠道及其微生物群和肝脏之间的双向通路。这种相互作用是由门静脉建立的,通过门静脉,肠道菌群产物直接运输到肝脏,肝脏将胆汁和抗体反馈到肠道。
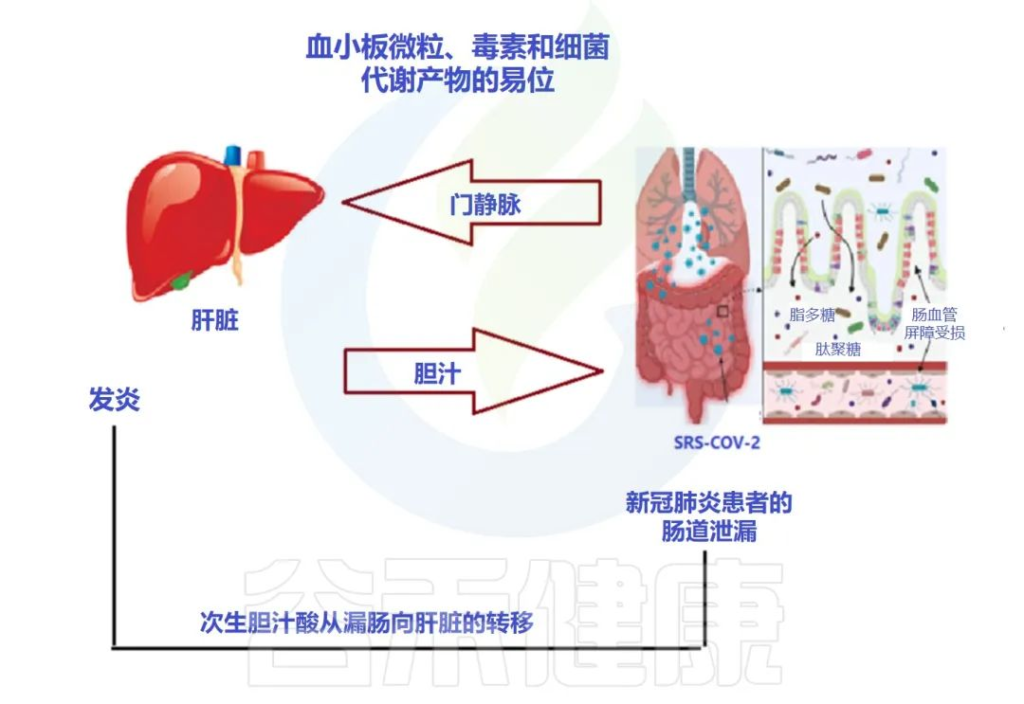
doi: 10.3390/metabo12100912
最近的两项研究表明:
代谢功能障碍相关性脂肪肝(MAFLD)是进展为严重和长期COVID-19的主要危险因素。
MAFLD征象的患者风险高
两项研究均证明,有MAFLD征象的患者发生呼吸系统疾病进展的风险高于无MAFLD的患者,年轻COVID-19患者的风险远高于老年COVID-19患者。
风险增加与病毒影响肠道通透性有关
研究人员认为,在MAFLD患者中观察到的风险增加可能与SARS CoV-2感染对肠道的影响有关,SARS CoV-2感染使肠道通透性和粘膜炎症恶化,从而加剧全身免疫功能障碍,这是严重COVID-19的特征。
当然,这个过程也可以阐明肥胖、2型糖尿病甚至炎症性肠病中 COVID-19 进展的较高风险,这与肠道微生物群改变、粘膜炎症和肠道通透性增加有关。
大量研究表明,腹泻、呕吐和腹痛等胃肠道症状在COVID-19患者中很常见,胃肠道症状的严重程度与呼吸系统疾病和肝功能障碍同时增加。
胃肠道中病毒进入受体表达的增加
已发现ACE-2 SARS CoV-2受体在肠细胞细胞上表达,因为粪便中高水平的SARS CoV-2病毒表明肠道是病毒感染和炎症的合理部位。
用于SARS-CoV-2进入的跨膜丝氨酸蛋白酶2在肠道细胞中也广泛表达。基于此,胃肠道中病毒进入受体表达的增加以及胃肠道症状的早期发作,意味着胃肠道异常可能是由病毒的直接恶化肠漏引起的,而不是对上呼吸道感染的继发性免疫致病反应的结果。
胃肠道症状的临床表现与肝功能不全的生物标志物呈正相关,支持了PAMPs向肝脏传播增加的观点。
SARS-CoV-2 感染会破坏肠道屏障,导致全身细菌脂多糖和肽聚糖升高,并有助于增强全身炎症。因此,肠漏和肠道菌群失调可能导致COVID-19重症患者发生细胞因子风暴。
基于此,已经开发的用于治疗肠漏的治疗方法,例如用于肠道粘膜保护/再生的益生菌和益生元,可以最大限度地减少进展为严重和长新冠的 MAFLD/肥胖/T2D 患者的数量。此外,在SARS CoV-2病毒感染期间,应避免使用干扰肠道微生物群的药物,例如抗生素。
充分证据表明,肠道微生物群改变和肠道细菌多样性减少,在心力衰竭合并冠状动脉疾病患者中很常见。
肠漏导致炎症,和心血管疾病相关
功能失调的肠道屏障会延缓菌群产物的被动泄漏,其中包括促炎脂多糖(LPS)进入血液,这可以通过炎症小体激活导致全身炎症。LPS结合蛋白(LBP)作为老年男性心血管风险高预测生物标志物的血浆水平显著升高,证明了这一点。
有趣的是已经发现,肠漏和炎症小体激活,与肌钙蛋白作为心肌损伤的标志物呈正相关。
肠道菌群-心轴在长新冠综合征中的作用
相当大比例的COVID-19住院患者有心脏问题。早期心血管疾病和心血管疾病危险因素(如肥胖)似乎是发生严重且长新冠并发症的关键危险因素。然而,高比例的COVID-19患者既往无心血管疾病的心脏受累。
在COVID-19患者中,心脏问题也被视为一个危及生命的实质性问题,从MI和心肌炎到伴有心脏应激的肺动脉高压。这种心脏受累的机制尚不清楚。
心肌感染可能与ACE2受体相关
ACE2在多个器官中表达,除肺、心脏和肾脏组织外,ACE2也在肠道中表达,肠细胞中的ACE2表达区可作为SARS-CoV-2进入和提示肠道感染的位点。继发于下调ACE2(SARSCoV-2受体)的抗炎和心脏保护性血管紧张素(AT)-1-7通路下调,通过表达ACE2的心脏细胞引导心肌感染,导致心脏炎症。
doi: 10.3390/metabo12100912
心脏成像的长期随访,结合肠道菌群分析,是进一步测试长冠状病毒肺炎患者肠道-心脏轴潜在影响的必要后续步骤。
肠道微生物群与肾脏疾病之间的致病性相互关联
肠道菌群参与广泛的临床表现,如慢性肾脏病(CKD),急性肾损伤(AKI)和高血压。
在肠漏的情况下,活细菌经常从肠道转移到其他肠外位置,例如肾脏。这种细菌易位可能伴有菌群失调、病原菌过度生长和宿主免疫系统低下。
在慢性肾病的情况下,肠道微生物群会产生许多毒素和尿毒症溶质,例如对甲酚硫酸盐 (PCS)、硫酸吲哚酯和三甲胺 (TMA) N-氧化物。另一方面,尿素水平升高可能导致肠道微生物群的改变(图3)。
尿毒症毒素可能导致慢性肾病患者出现疲乏、矿物质骨疾病、神经系统疾病和心血管损害。
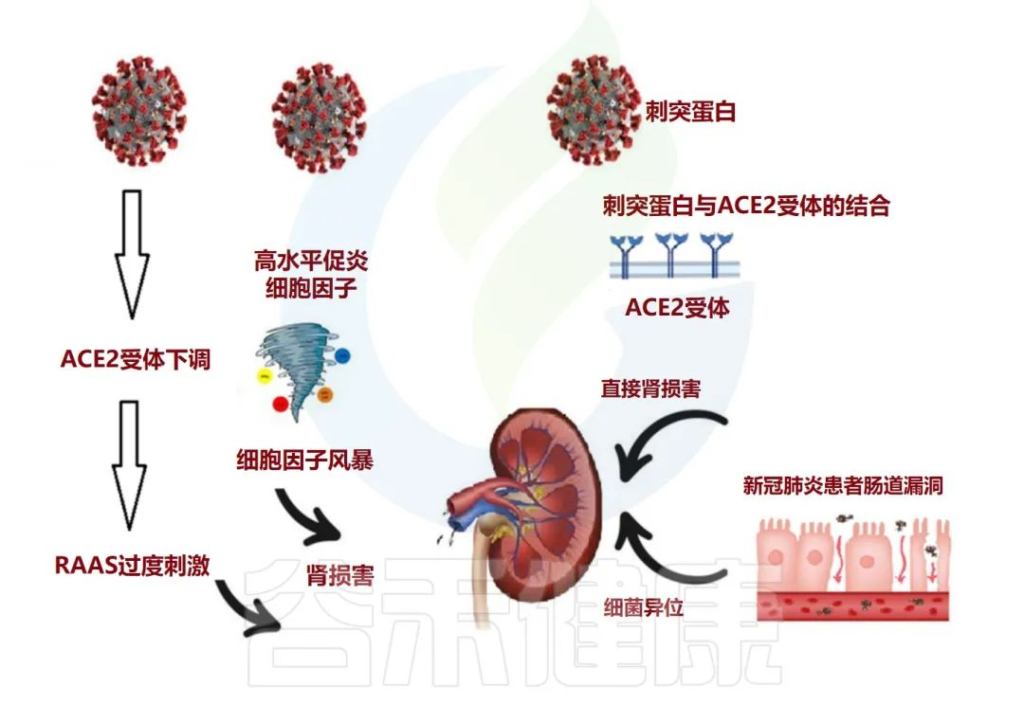
doi: 10.3390/metabo12100912
肠道生态失调在与 COVID-19 相关的长期肾脏问题中的作用
急性肾损伤 (AKI) 通常被视为 COVID-19 患者的并发症。除了先前存在的慢性肾病与COVID-19中的重症或死亡有关外,值得注意的是,解决SARS CoV-2通过AEC2受体进入肾脏,并诱导临床表现的不同途径。
人们普遍认为,该病毒可直接进入肾脏并复制,导致功能障碍,并且通过肾素-血管紧张素-醛固酮系统(RAAS)稳态的局部紊乱影响肾脏。
研究显示,既存慢性肾病患病率较高的群体可能更容易发生急性肾损伤。新出现的证据还表明,COVID-19的肾脏表现与长期严重COVID-19相关肾脏并发症的风险增加有关。
在菌群失调型COVID-19患者中,有益菌(主要是双歧杆菌和乳酸杆菌)逐渐消失,并且由于微生物群改变和病原体优势,观察到短链脂肪酸和胆汁酸水平下降。
短链脂肪酸,特别是丁酸盐是结肠细胞的重要能量来源,在上皮完整性中也起着重要作用。此外,短链脂肪酸受体GPR109A的激活与几种促炎介质的抑制有关。这可能解释了在 COVID-19 患者中出现的显著长期并发症。
作者报告说,慢性肾病患者的厌氧菌群减少,而有氧菌群增加,以肠杆菌科为主。所有这些机制都可以解释一些COVID-19患者的长期肾脏并发症。
在COVID-19住院期间监测肾功能,有助于识别后果更严重风险的患者,有助于早期和更有效的干预。
尽管 COVID-19 的主要临床表现与呼吸系统有关,但也会带来脑相关的问题,引发急性脑血管问题和颅内感染。
约35%的患者和高达85%的重症患者报告神经系统症状,包括头痛、头晕、肌痛或味觉和嗅觉丧失。
COVID-19感染可能导致神经系统疾病以及大脑结构和功能改变的机制有很多。
部分感染后的患者存在一致的记忆缺陷模式
认知问题是最常报告的症状之一,影响10%-25%的COVID-19患者,表现为SARS-CoV感染后的慢性疾病。作者发现,经历过COVID-19感染的人存在一致的记忆缺陷模式,随着自我报告的持续症状的严重程度,记忆缺陷也在增加。
此外,他们报告说,最初疾病期间的疲劳/混合症状和持续的神经系统症状可以预测认知能力。
doi: 10.3390/metabo12100912
COVID-19与神经损伤有关,主要见于神经系统症状
对COVID-19死亡患者的尸检显示,有缺血性损伤的指征和神经炎症的证据是病因机制。许多研究记录了不同脑区的功能和结构畸形,如出血性损伤和癫痫样放电。
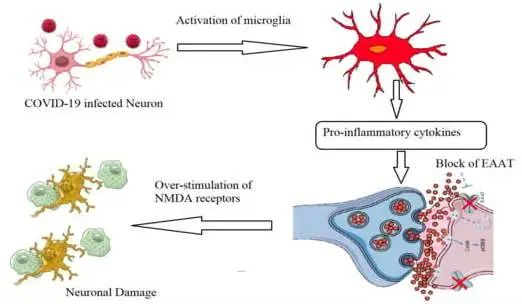
谷氨酸兴奋性毒性的作用:诱导促炎细胞因子产生
关于神经炎症,谷氨酸兴奋性毒性的作用应考虑为导致长新冠相关神经系统症状的原因。Ahmed等人(2020)报道,由于谷氨酸兴奋性毒性,SARS-CoV感染可诱导促炎细胞因子的产生和神经元变性显著增加。
简单地说,谷氨酸作为神经系统中的主要兴奋性神经递质,主要由神经元产生并在突触间隙中排出,之后它与配体依赖性AMPA受体(α-氨基-3-羟基-5-甲基-4-异恶唑丙酸受体)结合。这有助于钠离子的进入和神经冲动通过突触后神经元,导致N-甲基-D-天冬氨酸受体(NMDA)的激活,从而诱导钙离子进入。
新冠引起的神经元感染,扰乱谷氨酸稳态的控制
在新冠病毒引起的神经元感染过程中,小胶质细胞产生促炎细胞因子(TNF-α、IL-1β和IL-6),下调星形胶质细胞和突触前神经元上的谷氨酸转运蛋白1(GLT-1)。这将降低谷氨酸有效再摄取的速率,并导致谷氨酸 / GABA神经递质的不平衡和NMDA受体的过度刺激。这些事件扰乱了谷氨酸稳态的控制,突触间隙中谷氨酸的过量产生诱导神经元兴奋性毒性,钙显著进入,最终导致神经细胞变性和损失。
关于谷氨酸与大脑健康以及肠道菌群之间的关联,我们在之前的文章也有写过,详见:
新冠引起的肠道菌群失衡➔影响肠道屏障和血脑屏障通透性 ➔ 菌群代谢产物进入大脑➔脑功能障碍
肠道菌群维持肠上皮屏障、免疫稳态和防止病原体入侵。此外,肠道菌群可以通过增加紧密连接蛋白的表达来影响血脑屏障 (BBB) 的完整性和通透性。ACE2与肠道微生物稳态密切相关。
SARS-CoV-2 感染引起的肠道感染和 ACE2 表达下调可导致肠道菌群组成异常,包括乳酸杆菌和双歧杆菌等菌群水平降低。微生物失衡和肠道炎症势必会影响肠道屏障功能和血脑屏障的完整性和通透性,从而导致肠道细菌、毒素等肠道微生物代谢产物易位,通过血液循环进入大脑,最终导致脑功能障碍。
SARS-CoV-2侵入肠神经系统,出现异常,通过迷走神经影响大脑
迷走神经与肠神经系统中的神经元形成突触连接,并将肠道信息传输到NTS,在那里信息被整合并传递到大脑。
肠神经系统的异常不仅会导致胃肠功能障碍,还会通过肠-脑轴影响大脑功能。先前的研究表明,嗜神经病毒持续感染肠神经系统,并导致肠功能障碍。
SARS-CoV-2进入肠道后,通过与ACE2结合侵入肠神经系统,然后通过迷走神经进入大脑,影响中枢神经系统功能。
更重要的是,肠神经系统损伤导致肠道运动障碍、肠道血流异常和上皮屏障功能障碍,从而进一步促进肠道微生物和细菌代谢产物的毒素进入血液,加重大脑损伤。
越来越多的证据表明,肠道菌群失调也被发现是增加骨质流失的关键因素,而骨质流失又反过来促进了几种骨相关疾病的发展,例如类风湿性关节炎和骨质疏松症。
一系列报告显示,肠道菌群通过改变骨组织的质量来影响骨强度。肠道菌群调节骨骼健康的机制是通过促进调节代谢产物的生成,如吲哚衍生物、三甲基胺N-氧化物(TMAO:氧化胺)、短链脂肪酸和气体递质,如硫化氢(H2S)。
在卵巢切除术(ovx)诱导的绝经后骨质疏松小鼠模型中,H2S供体化合物GYY4137通过激活Wnt10b生成,从而增加骨形成,减少小梁骨损失,从而增强骨健康。
短链脂肪酸如乙酸、丙酸和丁酸诱导骨吸收细胞的代谢重编程,导致糖酵解增强,从而降低破骨细胞的特异性基因,如NFATc1和TRAF6,这是骨骼稳态的有效调节因子。
肠道菌群失调会加剧COVID-19疾病的严重程度和骨质流失
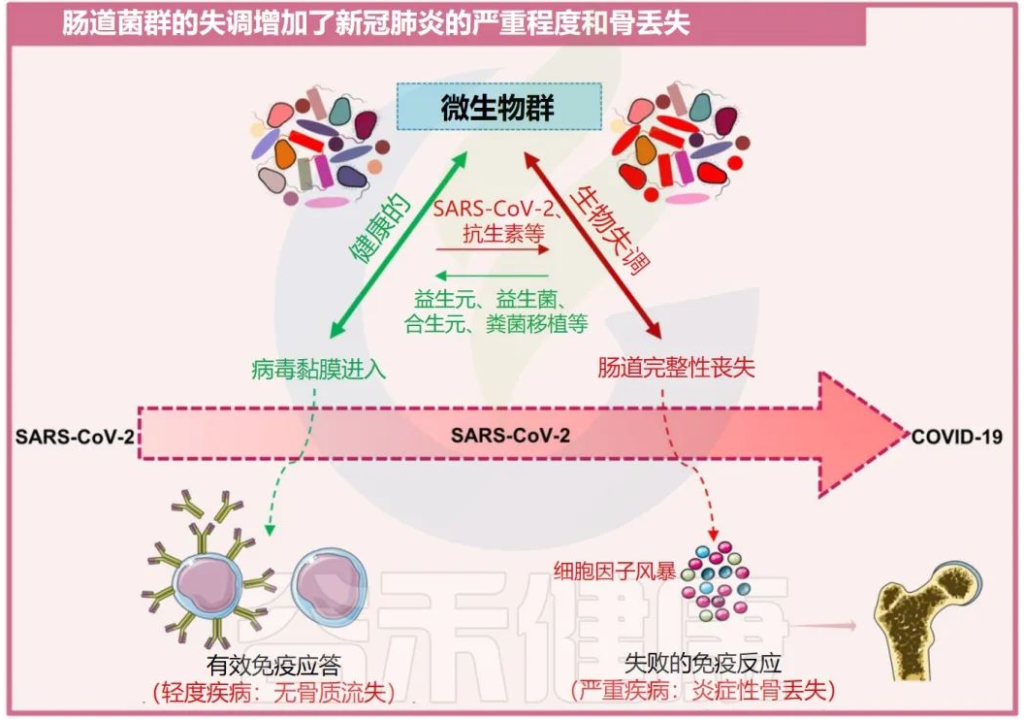
Sapra L, et al., Inflamm Res. 2022
还需要在该领域进行更多研究,并对康复的 COVID-19 感染患者进行长期随访,以确定 COVID-19 引起的骨病变的机制。
★ 肠-脾轴
SARS-CoV-2 被证明可能通过 ACE-2 受体诱导脾脏的特定嗜性。人们认为脾功能障碍与其他机制一起导致B细胞和T细胞淋巴细胞减少,这是感染后COVID-19的典型特征。
与健康对照组相比,脾切除术或脾功能障碍患者的革兰氏阴性菌产物LPSs丰度较高,因此肠道微生物群组成改变是血浆LPS升高的主要原因,可能与长期COVID-19并发症有关。
下 篇
干 预 措 施
饮 食 模 式
坚持高质量的饮食模式与较低的COVID-19感染和住院风险相关
一项Meta分析纳入5项研究,包括4023663名受试者(3149784名高质量饮食者和873881名对照组)。
高质量饮食模式对SARS-CoV-2感染和住院的有效性分别为28%(95% CI 19% ~ 36%)和62% (95% CI 25% ~ 80%)。
基于不同类型的高质量饮食和COVID-19感染风险的亚组分析显示:
一项病例对照研究调查了来自六个国家(法国、德国、意大利、西班牙、英国、美国)的2884名一线医护人员的饮食模式与新冠肺炎之间的关系。
注:这些医护人员是根据与新冠肺炎患者的大量接触情况进行筛选的。这项研究依赖于主要由男性医生组成的自我报告人群,不包括受最严重COVID-19病例影响的个体。
报告食用植物性饮食或鱼素饮食(包括益生元食品在内的)的人,中度至重度 COVID-19 的几率分别降低了 73% 和 59%。
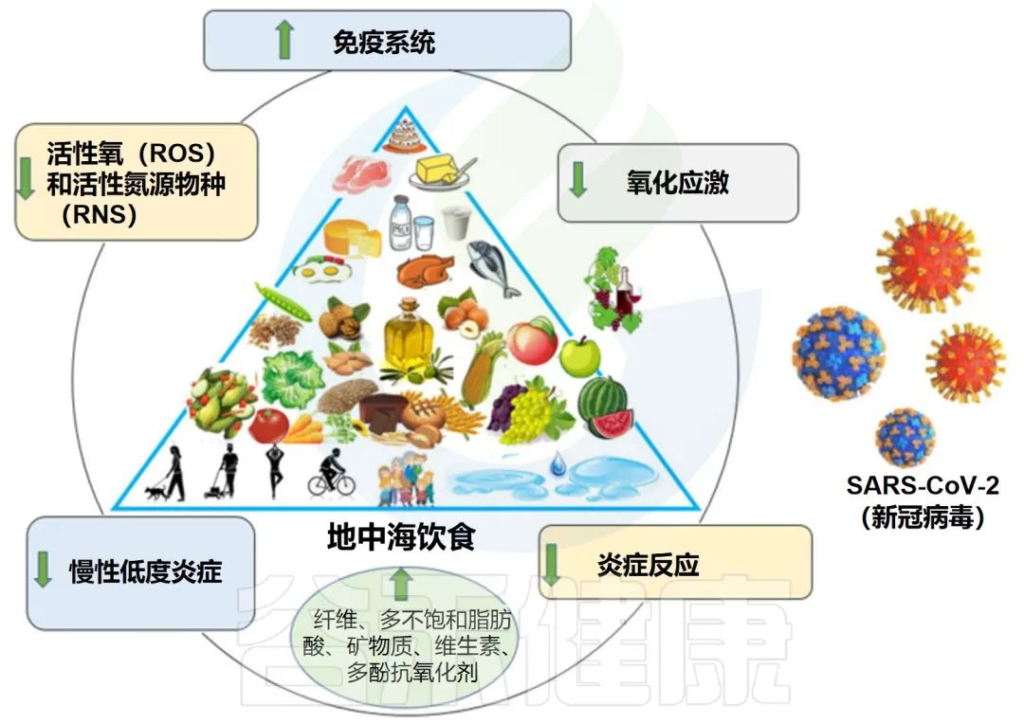
地中海饮食特征:
地中海饮食的依从性高,COVID-19严重程度和症状的可能性降低
一项横断面研究中,对 250 名年龄在 18 至 65 岁之间的 COVID-19 患者进行了检查。结果发现,对地中海饮食的依从性越高,COVID-19严重程度和症状的可能性降低,住院和康复时间缩短,炎症生物标志物也越短。
地中海饮食和COVID-19风险及相关死亡呈负相关
一项仅针对欧洲国家的生态学研究表明,地中海饮食与COVID-19相关死亡之间存在显着的负相关(r2= 0.771, p = 0.030).
一项观察性病例对照研究探讨了不同饮食模式与 COVID-19 事件和结局之间的可能关联。结果显示,病例的地中海饮食评分平均值(p=0.009)低于对照组,表明地中海饮食与COVID-19风险呈负相关。
地中海饮食有益地调节肠道微生物群和免疫系统
地中海饮食富含益生元物质,如半乳聚糖、果聚糖、纤维和菊粉。大量报道表明,这些化合物被宿主微生物使用,支持有利细菌的生长并促进有益代谢物的产生。
膳食纤维是影响复合碳水化合物对炎症影响的重要因素。研究表明,纤维消耗量的增加(约30g/d)与hs-CRP浓度的显着降低有关。膳食纤维摄入的另一个优点是对肠道微生物组组成更有利,可降低肠道和全身炎症。
多酚是地中海饮食中最丰富的次生植物化合物或植物化学物质之一,可能通过抑制NF-κB和AP-1以及激活Nrf2发挥许多抗氧化和抗炎作用。对肠道微生物群具有益生元作用。
地中海饮食:对抗冠状病毒感染的潜在策略
DOI:10.3390/medicina57121389
植物性饮食并不完全等于素食。在少量摄入动物源性食物的基础上,健康植物性饮食更倾向于新鲜蔬菜、坚果等健康的植物源性食物;而不健康的植物性饮食则更倾向于腌制蔬菜、糖等不健康的植物源性食物。
研究报告了植物性饮食是营养丰富的,包括高浓度的多酚,类胡萝卜素,纤维,维生素A,C和E,叶酸,铁,钾和镁。植物性饮食在预防高血压和心血管等疾病方面有益处。素食植物性饮食还可以增强免疫系统,减少炎症和氧化应激,并可能有助于预防慢性肾脏疾病和保持肾功能。
一项流行病学前瞻性队列研究表明,植物性食物与较低的COVID-19严重程度有关。
对来自美国和英国的五十多万参与者进行了一项研究,其中记录了 31815 例 COVID-19 病例。那些食用健康植物性饮食的人患 COVID-19 的风险降低了 10%,COVID-19 的严重程度降低了 40%.
对于年龄较大的COVID-19患者,研究人员发现,非素食饮食与COVID-19严重程度的风险较高有关。
对 2021 年 5 月至 2021 年 8 月期间在一家医疗中心被诊断为 COVID-19 的 509 名患者进行了回顾性评估。患者根据疾病严重程度分为三组。对于 ≥65 岁的患者,COVID-19 症状严重程度与坚持素食有统计学意义且呈负相关(p = 0.013).
水果、蔬菜摄入的重要性
一项针对COVID-19住院患者的横断面研究表明,水果、蔬菜和纤维的摄入量增加与 COVID-19 的严重程度、临床症状、住院和康复持续时间以及炎症标志物的浓度呈负相关。
水果和蔬菜富含纤维,是抗炎和增强免疫力的维生素,矿物质和抗氧化剂的良好来源。
为了确定饮食习惯对全球大流行期间COVID-19风险和严重程度的影响,需要进行进一步的研究。
间歇性禁食是一种潜在的补充疗法,不仅影响慢性病风险,而且有充分证据表明对传染病有影响。
SARS-CoV-2可能诱发肠道微生物群失调,导致致病菌的增殖增强,并导致有害的继发性病原体从肠道进入血流。
没有研究直接评估禁食对SARS-CoV-2感染者微生物组的影响,但由于其他观察结果,禁食塑造微生物组以支持与最佳代谢健康和低肥胖风险相关的物种,因此可以预期在预防不良变化方面产生影响。禁食期间,肠道微生物生长及其副产物(例如三甲胺N-氧化物)的产生受到抑制。
间接性禁食改变了微生物群丰度
间歇性禁食改变了各种微生物的丰度,例如脱硫弧菌科、Hydrogenoanaerobacterium、阿克曼氏菌、瘤胃球菌科等。
一项针对两种间歇性禁食(即限时进食和隔日禁食)的系统评价报道,禁食对微生物组有明显影响,例如改变厚壁菌/拟杆菌的比例,两种方案均增加了代谢保护微生物(如乳杆菌属和 Akkermansia municiphila)的丰度。
周期性禁食患者的死亡、住院和新诊断心力衰竭的风险也较低。特别是,禁食会在禁食期间急剧大幅降低葡萄糖水平,并在长期内降低基础葡萄糖水平,这在两种情况下都会使葡萄糖在感染期间减少使用,因此应该抵消SARS-CoV-2对糖酵解的刺激。
注意:禁食的安全问题包括轻微的潜在副作用,如饥饿、疲劳、头晕、便秘、头痛等,对健康的人是安全的。禁食会降低血糖,并可能导致低血糖。也可能导致脱水。2型糖尿病患者应谨慎禁食。其他安全问题可能包括身体过度紧张和营养缺乏等。
然而不可忽视的是间歇性禁食的一系列机制,使受损的人体免疫系统自我修复。在感染之前开始间歇性禁食方案可能对预防 COVID-19 等疾病的严重后果最有利。
总的来说,整体健康的饮食可能在预防SARS-CoV-2感染和降低感染严重程度方面发挥作用。需要更多的研究来证实这些发现,未来的研究应确定饮食质量与COVID-19感染风险之间关系的生物学机制。
生 活 方 式
注意,这里的运动并不是剧烈运动,特别是爬山、徒步等运动量大的活动更要注意量力而行。仍要加强健康监测。
刚阳康后需要进行一个休息阶段,尤其是老年“阳康”者,建议可循序渐进,先进行低等强度的温和运动,如适当散步、拉伸等。可以配合一些温和的养生方法,例如广播操,太极拳,八段锦等。
⇘⇘
太极拳作为中国传统运动之一,其特点是动作连贯、柔和,既能增强体质,调和身心,又能改善肺功能,避免运动时的呼吸短促。太极拳作为一种慢性病辅助治疗方法,已被临床证实能改善和缓解慢性阻塞性肺病。慢性阻塞性肺病,是一种常见的导致呼吸困难的慢性肺部疾病,被认为是COVID-19患者病情恶化的一个风险因素。
一项Meta分析共纳入11项随机对照实验,共纳入708名慢性阻塞性肺疾病患者。中期(3-6个月)或长期(12个月)太极拳运动对改善肺通气功能有效,且对改善慢性阻塞性肺疾病患者肺通气功能效果而言,干预时间在12个月以上的太极拳运动相较于中期运动更明显。
太极拳的简单易操作更容易让疫情期间老年人群体做出选择,同时也因为负荷量小更能够保证老年人的锻炼需求,从而达到免疫力提升的效果。
⇘⇘
八段锦具有运动强度小、动作幅度舒缓等特征,符合新冠肺炎患者年龄偏大、身体乏力、行动不便的需求,对于预防期新冠肺炎的发生以及预后呼吸功能的回复具有很好的预防和恢复作用。
90名亚健康状态学生为研究对象,随机分为对照组、散步组、八段锦组,30例/组。结果:八段锦组亚健康恢复率为50.00%(15/30),高于对照组的13.33%(4/30)和散步组的30.00%(9/30),差异有统计学意义(P<0.01). 八段锦练习能够显著改善亚健康状态,提高亚健康恢复率,且效果优于日常散步活动。
吸烟被认为与不良疾病预后有关,因为大量证据强调了吸烟对肺部健康的负面影响。吸烟会增加肺部感染的风险和严重程度,因为它会损害上呼吸道并降低肺部免疫功能。
吸烟已被证实是 COVID-19 负面进展的一个危险因素,尤其是在疾病严重程度和死亡方面。
Meta分析纳入 40 项研究,目前吸烟和以前吸烟都会显著增加疾病严重程度的风险(分别为:OR=1.58;95%CI:1.16–2.15,p=0.004;OR=2.48;95%CI:1.64–3.77,p<0.001),表现出中度异质性。
前吸烟者患疾病严重程度的几率是从不吸烟者的 1.58 倍。前吸烟者患疾病严重程度的几率是从不吸烟者的 2.48 倍。
同样,目前吸烟和以前吸烟也会显著增加死亡风险(分别为OR=1.35;95%CI:1.12–1.62,p=0.002;OR=2.58;95%CI:2.15–3.09,p<0.001;),并出现中度异质性。
对于死亡结果,当前和以前吸烟也分别使死亡风险显著增加 1.35 倍和 2.58 倍。
冥想练习调节注意力和情绪,向外关注特定的身体和感官刺激,向内转向精神体验和身体体验的躯体感觉。
由于冥想对包括抑郁症、焦虑症、慢性疼痛和药物滥用在内的精神病理学的特定领域产生积极影响,以及它与注意力障碍、创伤性压力、饮食失调和严重精神疾病的关联,冥想越来越多地被纳入心理健康干预措施。
与当地招募的对照组相比,长期冥想的佛教僧侣的肠道微生物群组成发生了显著变化。在属水平上,普氏菌属和拟杆菌属在冥想组中显着丰富。根据 LEfSe 分析,两个有益细菌属(巨型单胞菌属和粪杆菌属)在冥想组中显着增加。功能预测分析进一步表明,包括聚糖生物合成、新陈代谢和脂多糖生物合成在内的几种途径在冥想组中显着丰富。
这种改变的肠道微生物群组成可以降低焦虑和抑郁的风险,并改善身体的免疫功能。生化标志物概况表明冥想可以降低心身医学中心血管疾病的风险。这些结果表明,长期深度冥想可能对肠道微生物群产生有益影响,使身体保持最佳健康状态。
来自麻省理工学院、加州大学圣地亚哥分校、乔普拉综合研究图书馆和哈佛大学的研究人员探索后认为,某些冥想、瑜伽体式(姿势)和调息(呼吸)练习可能是治疗和/或预防SARS-CoV-2感染的有效辅助手段。
来自 13 个国家的 44 项研究,共 4023 人,时间范围从 20 到 4800 分钟不等。主要研究结果表明,瑜伽、冥想和调息,无论是单独使用还是组合使用,都可以通过调节抗炎和促炎生物标志物来有效提高健康和临床人群的免疫力,可能有效降低 IL-6、皮质醇和 TNF-α 患者的水平。
⇘⇘
晒太阳
时间可以选择在11点-15点,晒5-30分钟,尽量不隔着玻璃,夏季应注意避开紫外线最强的时候。
2022 年 1 月发表在《韩国家庭医学杂志》上的一项研究着眼于气候如何影响 COVID-19。研究人员发现,当涉及较高的湿度、气温和暴露在阳光下时,COVID-19 病例较少。
⇘⇘
卫生措施
阳康后依然有必要进行日常的防护措施,包括出门戴口罩,回家后勤洗手。
衣服上的病毒会传染吗?
2020 年 11 月发表的一项研究发现,虽然活病毒在皮肤上存在长达四天,但在衣服上,病毒存活不到八小时。
美国疾病控制与预防中心指出,COVID-19 主要通过三种方式传播:
受污染的衣服(或其他材料)不被视为主要传播方式。
马里兰州约翰霍普金斯健康安全中心资深学者、传染病专家认为,病毒的生长和存活在很大程度上取决于环境条件——温度和湿度。总的来说,衣服不是 SARS-CoV-2 的“主要传播媒介”。
衣服上的病毒可以被洗掉吗?
莱斯特德蒙福特大学的研究表明,使用洗涤剂在洗衣机中用热水洗衣服可以完全消除病毒。
⇘⇘
保暖
此外,天气寒冷时去户外应注意防寒保暖,天气寒冷时可适当减少外出。
以上是针对饮食、生活方式的调整,是所有调理方式的基石。当然可能还有许多没有列举出来的其他健康的生活方式,可以慢慢探索,找到一种适合自己的健康的生活方式,“病毒”就难以伤害你。
肠道菌群对免疫调节至关重要。针对肠道菌群的干预措施可能对COVID-19患者产生全身性有益作用。
益生菌和益生元是我们饮食中会影响微生物组的两个组成部分。营养状况和饮食在COVID-19中起着至关重要的作用,主要是由于肺和肠道微生物群之间的双向相互作用。
下图描述了人体肠道和肺部之间的相互作用以及益生菌和益生元引发的潜在阳性免疫反应。
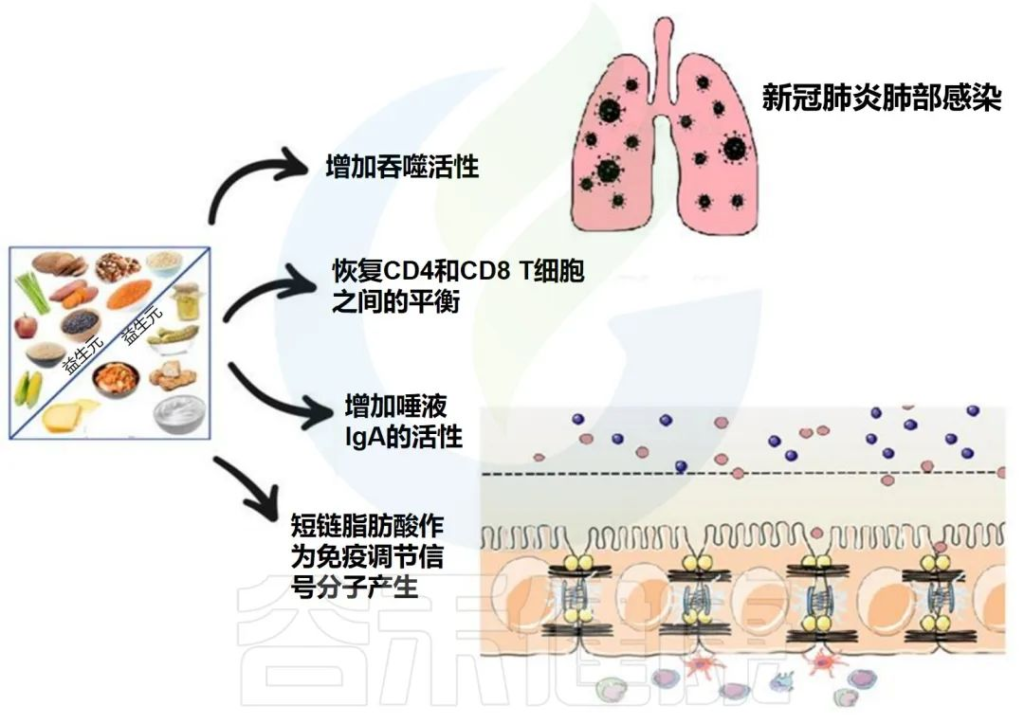
doi.org/10.3390/metabo12100912
益生菌和益生元都可以增强巨噬细胞的吞噬活性,平衡T细胞免疫以有利于更多的调节状态,增加唾液IgA的活性,并通过产生短链脂肪酸作为重要的信号分子,来发挥免疫调节细胞外和细胞内功能。
在临床研究中,使用益生元和益生菌操纵肠道微生物群是治疗肺部疾病的一种有前途的方法。
国家卫生健康委员会和国家中医药管理局指南建议重症COVID-19感染患者在常规治疗的同时食用益生菌,以改善肠道菌群平衡并预防继发细菌感染。
益生菌可能会改变肠道微生物群的组成,并在维持肠道微生物群的生态系统中发挥至关重要的作用。
益生菌通过调节肠道菌群有助于COVID-19治疗
益生菌有利于增强上皮屏障功能和改善肠道微生物多样性。此外,益生菌可对抗和阻断肠道中的有害细菌菌株或增强有益的信号通路。
尽管细菌引起的免疫反应与病毒引起的免疫反应相对不同,但许多临床研究得出结论,益生菌有助于治疗COVID-19。
注:已有超过 25 项注册临床试验旨在调查益生菌给药对 COVID-19 管理的生物学和治疗作用。
在迄今为止发表的有限试验中,关于益生菌给药的主要发现是:症状的更快改善,疲劳减少,并可能解决胃肠道问题。
已发表的研究:服用益生菌对COVID-19及其相关后遗症的影响

编辑
doi: 10.1016/j.clnesp.2022.08.023
益生菌通过调节宿主免疫系统带来益处
除了改善肠道微生物平衡外,最近的证据表明,益生菌还可以通过调节宿主免疫功能对宿主产生有益作用。一些研究报告了益生菌与ACE2相互作用的潜力,ACE2是SARS-COV2 进入宿主的受体。
例如,据报道,几种益生菌(主要是益生菌乳酸菌)在牛奶发酵过程中释放出对ACE2具有高亲和力的肽。同样,益生菌也可能通过ACE2途径改善呼吸道感染。
益生菌还可以提高肺免疫系统中自然杀伤细胞(NK细胞)、I.型干扰素、T和B淋巴细胞以及APC的水平。
注:NK细胞在针对病毒感染的早期免疫反应中起重要作用,主要是通过清除病毒感染。
先前的一项研究表明,益生菌会改变IL-10的表达并降低炎性细胞因子的表达。
益生菌还抑制其他促炎细胞因子,如TNF-a、CRP、IL-1b、IL2、IL-6、IL7、MCP1和LDH等。
益生菌通过细菌素抑制病毒与其受体的结合
除了抗炎作用、免疫调节和调节微生物组的机制外,益生菌还可以通过细菌素抑制 SARS-CoV-2 与其受体的结合。
益生菌植物乳杆菌(LPG)的独特菌株可以通过增强干扰素信号传导和抑制凋亡和炎症途径,在有效阶段和记忆阶段促进SARS-CoV-2特异性免疫反应。
由植物乳杆菌等益生菌分泌的植物素等细菌素参与抑制SARS-CoV-2的进入和复制
doi: 10.1016/j.clnesp.2022.08.023
分子对接研究预测,植物素结构可能通过靶向S蛋白或结合RNA依赖RNA聚合酶(RdRP)来阻碍病毒的进入,从而阻碍基因组的转录。
植物乳杆菌的PlnE和PlnF可以通过在解旋酶的ssRNA或ATP结合位点结合来抑制SARS CoV-2复制。
益生菌并不是治疗COVID-19的灵丹妙药,益生菌的功效和安全性在文献中存在争议,例如在接受Bacillus clausii 治疗的免疫功能低下患者中出现了菌血症等现象。因此,对一个病人有益的配方可能对另一个病人有害。还需要进行更多的研究。
此外,在推断临床试验结果时,重要的是要意识到混淆因素,如不同年龄组,不同免疫系统状态,不同季节,益生菌的成分,剂量,营养状况,正在服用的其他补充剂等。因此,也可能需要根据症状进行调整。
更有针对性的方法,包括肠道菌群检测等措施,以充分了解微生物群的作用及其在饮食和外部应激源之间的相互作用,以对抗SARS-CoV-2感染,可能会带来个性化治疗,以便益生菌能给每个需要的人带来真正益处。
益生元包括多不饱和脂肪酸、抗性淀粉、阿拉伯寡糖、低聚糖、果聚糖、低聚糖、半乳甘露聚糖、车前子、蔗糖乳糖、乳糖酸、多酚等。
大多数益生元是从植物多糖中合成或分离的,是低聚糖,例如:
含有益生元的食物,如纤维、低聚糖和多酚,可以改善细菌的生长。例如,富含菊粉的饮食刺激双歧杆菌和拟杆菌的生长;全麦谷物可以改变细菌谱,增加双歧杆菌和乳酸杆菌的相对数量。
益生元以与益生菌类似的方式调节肠道微生物群,从而抑制病原体并刺激免疫系统。同样,益生元通过直接和间接机制,对免疫系统和宿主的健康产生有益的改变。
益生元为益生菌的生长提供能量
此外,益生元选择性地刺激益生菌的有利生长并增强益生菌的活性。益生元通过增强益生菌的生长和生存能力,对 COVID-19 感染具有潜在作用。益生元也可能通过阻断ACE2对COVID-19引起的胃肠道症状产生潜在影响。
益生元明显降低了促炎IL-6的水平,这似乎是迄今为止描述的COVID-19严重预后的主要原因,并改善了抗炎IL-10的水平。
母乳低聚糖:在COVID-19中的潜在应用
母乳低聚糖在母乳中固体成分的比例排名第三。母乳低聚糖可以发挥多种功能,即抗感染(针对细菌和病毒),信号传导,抗炎/免疫调节和益生元作用。
母乳低聚糖对抗SARS-CoV-2的潜在作用模式
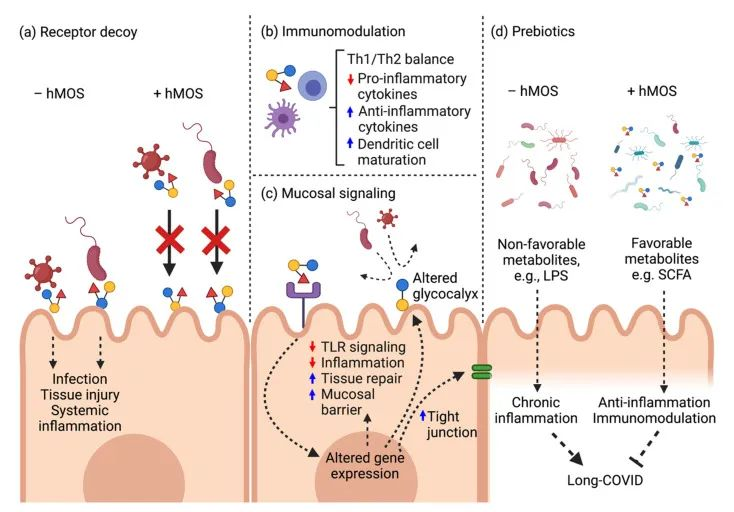
doi: 10.3390/biomedicines10020346
a) 母乳低聚糖分子结构类似于HBGA,并充当受体诱饵以阻止病毒进入。
b) 母乳低聚糖诱导局部防御和免疫调节。
c) 母乳低聚糖减弱TLR4介导的信号通路以维持粘膜稳态。
d) 母乳低聚糖在长新冠中缓解肠道菌群失调并恢复健康的肠道微生物群。
地中海饮食多酚:对 COVID-19 引起的炎症的潜在用途
研究人员发现,地中海饮食中存在的主要酚类化合物作为COVID-19预防/治疗剂的潜在用途,基于其抗氧化和抗炎作用。
目前的证据支持羟基酪醇、白藜芦醇、黄酮醇(如槲皮素)、黄烷醇(如儿茶素)和黄烷酮可能对 COVID-19 产生的潜在益处。
茶多酚:具有抗病毒固体和抗氧化特性,可能有助于降低出现严重COVID-19症状的风险
肠肺轴在SARS-CoV-2感染中起着重要作用,因此靶向肠肺轴治疗COVI-19尤为重要。茶多酚被认为是多功能生物活性分子,除了抗菌和调节肠道菌群以增强免疫功能外,还具有抗病毒作用。因此,茶多酚对COVID-19具有潜在的预防和治疗作用。
茶多酚降低 COVID-19 合并症风险

DOI:10.3389/fnut.2022.899842
茶多酚可以促进肠道中有益细菌的生长,并抑制肠道中病原微生物的生长,从而调节肠道菌群的组成。
研究人员研究了茶多酚对回肠损伤和肠道菌群紊乱的治疗和预防作用。结果表明,茶多酚可以减少炎症和氧化应激标志物,提高抗氧化酶和紧密连接蛋白的水平,有效改善肠道菌群失衡,减少对肠粘膜的损害,增强机体免疫力。
使用茶多酚预防和治疗COVID-19并发症
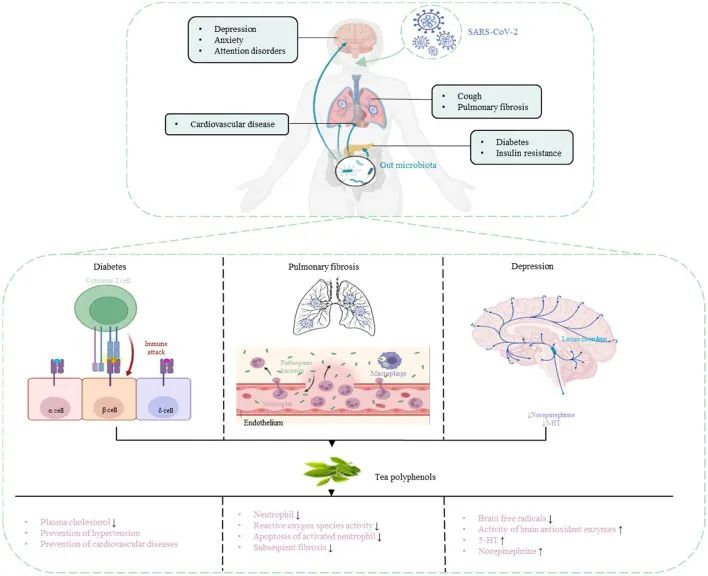
DOI:10.3389/fnut.2022.899842
在一项针对200名医护人员的随机双盲试验中,每天六粒胶囊(包括378毫克儿茶素和270毫克EGCG)持续5个月,在预防流感病毒方面优于安慰剂。
注意:在确定使用茶多酚治疗COVID-19之前,仍然需要大量实验来确认具体的药物给药(绿茶饮料、粉状绿茶提取物、儿茶素混合物、单独儿茶素)、剂量方案(不同剂量、不同治疗持续时间)和给药途径管理(饮食口服、饮料口服)。
针对COVID-19住院患者研究中,接受合生元患者临床症状缓解的比例更高
在一项针对55名COVID-19住院患者的开放标签研究中,与标准治疗组相比,接受双歧杆菌菌株和益生元的合生元配方(SIM01)4周的患者的临床症状得到缓解的比例更高(88%对63.3%),抗SARS-CoV-2的IgG抗体增加,IL-6、CCL2、M-CSF、TNF和IL-1RA等血液促炎标志物减少。
在接受SIM01的个体肠道菌群中,共生菌(如双歧杆菌、真杆菌和粪杆菌)的丰度也有所增加,而机会致病菌(如大肠杆菌和拟杆菌)的丰度则有所下降。
合生元配方 SIM01 可加速针对 SARS-CoV-2 的抗体形成,降低鼻咽病毒载量,减少促炎免疫标志物,并恢复住院 COVID-19 患者的肠道生态失调。
合生元辅助治疗两周可以有效调节针对COVID-19感染的炎症反应
一项随机安慰剂对照试验招募了 78 名确诊 COVID-19 感染的住院患者。干预组和对照组分别每天两次接受合生元或安慰剂胶囊,持续两周。
注:合生元胶囊含有多种菌株益生菌,如鼠李糖乳杆菌 、瑞士乳杆菌、干酪乳杆菌、乳双歧杆菌、嗜酸乳杆菌、短双歧杆菌、保加利亚乳杆菌、长双歧杆菌、植物乳杆菌、双歧双歧杆菌、格氏乳杆菌和嗜热链球菌,以及低聚果糖益生元剂。
结果发现:
后生元:在宿主中具有生物活性的微生物的非活细菌产物或代谢产物。也有研究人员称之为“幽灵益生菌”、“灭活益生菌”、“非活性益生菌”等。
短链脂肪酸、微生物细胞组分、功能蛋白、细胞外多糖(EPS)、细胞裂解物、替胆酸、肽聚糖衍生的多肽和毛状结构等多种代谢产物都算后生元。
后生元对抗 COVID-19 的抗病毒机制可以与以下作用相关联:
(a) 抗病毒抑制代谢物的产生
(b) 改善肠上皮衬里屏障功能
(c) 调节先天性和适应性免疫系统
(d) 对肠脑轴的影响
(e) 缓解继发性真菌感染
各种后生元对肠道屏障完整性的调节
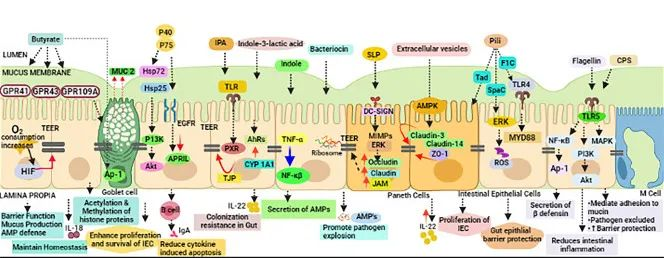
doi: 10.1007/s12602-021-09875-4
人们越来越热衷于利用益生菌,尽管由于宿主微生物组和侵入性病毒之间的固有复杂性和串扰,其详细的作用机制仍在研究中。后生元在降低致命的SARS-CoV-2感染严重程度方面可能比益生菌具有显着优势。
然而从治疗和调节的角度来看,还有一些关键问题需要回答,例如:最适合提取它们的方法,它们在宿主内的有效传递方法,它们的稳定性和保质期,后生元商业化的生物处理策略等。
未来,根据病原体变异的流行情况,将后生元用作个性化疗法或许是一种可行的选择。基于后生元在增强或调节个体免疫力方面的作用,也可以探索降低对其他病毒感染的易感性或严重程度。
在粪菌移植中,来自健康供体的体外培养或粪便物质纯化的粪便或复杂微生物群落被接种到患者的肠道中。粪菌移植已证明对结肠炎、糖尿病和复发性艰难梭菌感染有效。
11名COVID-19患者中有5名报告胃肠道症状有所改善,血液免疫标志物和肠道微生物群组成有良好的改善,双歧杆菌和粪杆菌的丰度增加。
在两名合并复发性艰难梭菌感染的 COVID-19 患者中,FMT 治疗似乎是安全的,并且 COVID-19 相关呼吸道症状在 FMT 后 1 个月内迅速消退。
一项注册临床试验(ClinicalTrials.gov 标识符号 NCT04824222),试图验证粪菌移植作为免疫调节风险降低剂,在与细胞因子风暴和炎症升级相关的COVID-19疾病进展中的功效。
对照组接受标准的药物治疗,而实验组也接受口服FMT,剂量为30–50,双层,耐胃酸,肠溶性冷冻60-g胶囊。
➳ 主要的结果指标是:
给药后第30天安全试验组的不良事件发生率。
➳ 另一个结果指标是:
研究组和对照组中需要升级无创氧疗方式的患者百分比
如增加FiO2、给予高流量鼻插管氧治疗(HFNOT)、持续气道正压(CPAP)或有创通气、呼吸机和/或ICU住院治疗,对应于新冠肺炎表现状态量表中5-7级疾病恶化。
这个试验仍在进行中。然而,考虑到肠道微生物群在免疫调节中的重要作用,研究人员认为FMT是抑制新冠肺炎诱导的细胞因子风暴和炎症的一种可能的治疗选择。
益生菌、益生元和FMT旨在增加有益细菌的丰度,而抗生素则用于抑制有害微生物群的丰度。世卫组织建议,患有可能患有严重急性呼吸道感染(SARI)和败血症的COVID-19患者可能需要广谱抗生素,这可以覆盖尽可能多的致病菌。
密歇根州38家医院新冠肺炎患者的随机数据显示,至少50%的患者接受了早期经验性抗生素治疗。抗生素在危重症患者中的应用也占很大比例,并在抑制患者继发感染方面发挥关键作用。
然而,抗生素治疗可以不加区别地消除正常的共生微生物群,同时消除病原体,导致肠道菌群失调。
与健康对照组相比,新冠肺炎患者的抗生素治疗对肠道微生物群有显著影响,有益于宿主免疫的共生菌较少,包括粪杆菌、粪球菌和直肠真杆菌,而铜绿假单胞菌和分枝杆菌,这已被证实与新冠肺炎的严重程度增加呈正相关。
特别是在疫情期间,抗生素的使用面临巨大挑战。
在早期大流行中,抗生素被普遍使用。一项荟萃分析估计,全球四分之三的新冠肺炎患者曾服用过抗生素,这一比例明显高于新冠肺炎中细菌合并感染的估计发生率,后者仅为8.6%。
45名中度新冠肺炎患者的临床结果显示,服用和未服用抗生素的患者之间没有差异,这表明抗生素对改善新冠肺炎的临床结局并无益处。
据报道,在住院期间接受抗生素治疗的新冠肺炎患者出现明显的肠道失调,抗生素诱导的肠道失调损害了人类对季节性流感疫苗的免疫反应。
一项对200名新冠肺炎患者的纵向研究表明,新冠肺炎爆发前一年抗生素摄入量的减少与疾病严重程度的减轻和SARS-CoV-2的快速清除有关。
因此,抗菌药物管理对于预防抗生素诱导的失调、严重的新冠肺炎和新冠肺炎患者的抗菌药物耐药性风险至关重要。抗菌治疗必须有明确严格的适应症,应谨慎选择病原学检测后敏感的抗生素,并根据患者的具体情况适当调整用药时间。
➤ 多 酚
多酚是源自植物的酚类化合物,富含抗氧化和抗炎特性。膳食多酚大致分为四类,包括酚酸、木脂素、二苯乙烯和类黄酮。
有大量证据强调了多酚对肠道的益生元作用。这可能有助于纠正据报道由SARS-CoV-2感染引发的肠道微生物群的生态失调。
——槲皮素
槲皮素被认为有助于预防严重的COVID-19症状,因为它具有已知的抗炎、抗氧化和免疫调节特性。分子对接和体外研究通过多种机制揭示了强大的抗病毒潜力,包括阻止与ACE2受体的附着和阻断病毒复制。
槲皮素的临床试验结果很有希望。在一项随机、开放标签试验中,研究人员测试了用向日葵磷脂配制的槲皮素提高吸收的有效性。
与标准护理相比,槲皮素组的住院风险降低了68.2%,住院时间缩短了76.8%,对氧气治疗的需求减少了93.3%。此外,槲皮素组没有患者入住ICU或死亡,对照组分别有10.5%和3.9%的患者入住ICU或死亡。同一组的进一步随访研究证实,槲皮素显著改善了病毒清除率,缩短了症状时间,改善了炎症标志物,与对照组的19%相比,槲皮素组的57%在7天后完全康复。
最近的一项小型研究证实了这些结果。槲皮素组的炎症标志物和住院时间显著减少。此外,槲皮素组入住ICU的患者数量减少,ICU天数减少,无死亡,而对照组有3例死亡。在所有情况下,这些结果都接近显著性,但很可能由于样本量小而没有统计学意义。
虽然缺乏大型临床试验,存在局限性,上述研究仍显示了症状减轻、进展至严重疾病和死亡率方面的益处。鉴于槲皮素在短期服用时具有良好的安全性,以及其广泛的可用性和众多其他健康益处,可以与患者讨论使用。
槲皮素可以从几种水果和蔬菜中获得,如浆果、芦笋,红叶生菜、洋葱、苹果、莳萝、萝卜、刺山柑、香菜、银杏叶、葡萄、葱、西红柿、西兰花、青椒、豌豆等。也可以从含有槲皮素或其一些合成衍生物的补充片剂中补充。
——姜黄素
姜黄素是姜黄中存在的生物活性化合物,具有多机制作用模式。
它可以抑制病毒进入细胞,包裹病毒和病毒蛋白酶。它调节各种信号通路。
姜黄素可能在治疗 COVID-19 感染中发挥有益作用,因为它能够调节负责 SARS-CoV-2 在许多器官(如肾脏、肝脏和心血管系统)中的附着和内化的各种靶点。它还可以抑制COVID-19感染时触发的纤维化相关通路和肺水肿。
研究表明,口服姜黄素可降低死亡率、恢复时间、对氧气的需求、机械通气、住院时间以及存在几种炎症标志物。
在三项安慰剂对照试验中,每天服用160 mg纳米姜黄素或1050 mg姜黄素和胡椒碱的患者的死亡率分别降低了80%、50%和82%。最近的一项荟萃分析表明,姜黄素治疗患者的死亡率总体降低了77%.
六项临床试验中,有五项也发现轻度、中度和重度新冠肺炎患者的症状持续时间显著缩短。
虽然还需要进行更大规模的试验,但现有数据表明姜黄素对降低新冠肺炎的严重程度非常有效。
鉴于几乎所有先前的临床试验都表明姜黄素补充剂是安全且耐受性良好的,即使剂量高达8000 mg/d,建议在症状出现时或首次阳性试验时每天服用1000 mg 较为合适。
虽然补充剂是达到试验中同等剂量的最佳方法,但用于烹饪的姜黄粉含有约3%的姜黄素。因此,一茶匙(5克)的姜黄粉含有大约150毫克。虽然姜黄素的低生物利用度一直存在问题,但胡椒碱(一种存在于黑胡椒中的化合物)已被证明可将生物利用度提高20倍,因此想要提高饮食中姜黄素摄入量的做法,可以在加姜黄粉的同时加入黑胡椒。
——白藜芦醇
白藜芦醇是一种多酚,白藜芦醇可能与SARS-CoV-2相互作用,至少部分是通过触发Nrf2,而Nrf2作为宿主防御机制对某些呼吸道病毒疾病(如呼吸道合胞病毒病)具有重要的调节作用。
白藜芦醇可以降低氧化应激,通过谷胱甘肽过氧化物酶的调节。这增强了谷胱甘肽的产生,并抵消了氧化应激介导的组织损伤。
白藜芦醇通过抗氧化和抗炎机制对SARS-CoV-2诱导的损害的主要潜在保护作用
doi: 10.1007/s13105-022-00926-0.
白藜芦醇通过eIF2α和NADPH氧化酶途径降低氧化应激水平。
高血压和动脉粥样硬化是新冠病毒感染的两个危险因素。在这方面,白藜芦醇调节SIRT1和Nrf2通路的能力,以及ROS的产生,导致更大的一氧化氮(NO)生物利用度。因此,白藜芦醇介导的NO增加很可能是多酚的血管扩张剂和抗血小板作用的基础,这反过来又可以减轻许多患者的COVID-19严重程度。
白藜芦醇在内皮细胞中积累,由于其潜在的抗血栓作用,能够保护内皮屏障。
由于其抗氧化活性,白藜芦醇可以减轻与ROS介导的氧化应激相关的炎症反应。较低的ROS水平导致NF-κB和细胞外信号调节激酶/丝裂原活化蛋白激酶(ERK/MAPK)的抑制。
对随机临床试验的各种荟萃分析强调了白藜芦醇的抗炎作用,这可能有助于缓解新冠肺炎特有的所谓“炎症形式”。
项研究利用网络药理学方法和生物信息学基因分析探讨了白藜芦醇对新冠肺炎患者作用的潜在机制。该研究表明,白藜芦醇可以通过抑制IL-17、TNF和NF-κB信号通路来减轻SARS-CoV-2产生的过度炎症。
白藜芦醇,自然存在于不同的食物来源中,浓度较低,典型的地中海饮食,包括葡萄、红酒、浆果和坚果。
关于白藜芦醇与肠道菌群的关联详见:
——羟基酪醇
羟基酪醇,存在于橄榄中,在橄榄成熟过程中由于橄榄苦苷水解而增加。它是从橄榄叶和果实中提取出来的,在特级初榨橄榄油中含量尤其丰富。
注:橄榄油是地中海饮食中最具特色的食物之一。
羟基酪醇的抗病毒能力是众所周知的,部分原因是其抗炎作用。不同的研究表明,这种酚类化合物抑制MMP-9和COX-2酶的活性。
MMP-9循环水平升高被认为是COVID-19患者呼吸衰竭的早期指标。在急性肺损伤(例如COVID-19中发生的肺损伤)中,MMP-9从中性粒细胞中释放出来,从而产生炎症和肺泡毛细血管屏障的退化,进而促进炎症细胞的迁移,导致肺组织的进一步破坏。
羟基酪醇诱导的MMP-9抑制可以通过该途径减轻和/或部分预防COVID-19产生的肺损伤。
羟基酪醇通过抗炎机制对 SARS-CoV-2 诱导的肺泡组织损伤的潜在保护作用
doi: 10.1007/s13105-022-00926-0.
➤ 类胡萝卜素
类胡萝卜素是四萜类。它们包括由植物、藻类和细菌产生的橙色、红色和黄色有机色素。一些常见的类胡萝卜素是α-和β-胡萝卜素、叶黄素、玉米黄质和番茄红素。类胡萝卜素以其抗氧化特性和抑制ROS而闻名。
因此,低水平的类胡萝卜素与氧化应激有关。叶黄素、胡萝卜素和玉米黄质的抗病毒作用已被报道。
一些类胡萝卜素可作为维生素A的前体,与免疫调节功能直接相关。据报道,类胡萝卜素可缓解COVID-19感染期间导致肺损伤的炎症反应。
当通过饮食提供必要的微量营养素(如维生素 A、B 和 D)以及硒、锌和铜等必要质量和数量时,可以达到最佳健康状况。
随着新冠的持续,很明显,最容易感染的人是那些失去生理营养状况和免疫系统平衡的人。这种不平衡使SARS-CoV-2病毒得以发展并导致疾病的不同临床形式(无症状,轻度,中度和重度)。
SARS-CoV-2感染中个体反应的微量营养素调节
DOI:10.1007/s12011-022-03290-8
维生素A缺乏导致第一道防御屏障缺乏先天免疫的重要成分,从而增强了病毒的反应。
维生素B和硒在感染期间抵抗氧化应激的免疫和抗氧化反应中发挥作用。维生素B、D、硒、锌和铜以非常重要的方式参与,促进抑制促炎细胞因子的合成,促炎细胞素引发细胞因子风暴,从而通过抑制Th1反应和促进Th2细胞产生细胞因子来调节适应性免疫应答。
锌本身参与细胞信号转导,因此参与细胞和病毒基因表达模式,从而避免病毒突变。
➤ 维 生 素 A
维生素A,一种脂溶性维生素。是身体许多部位正常生长和功能所必需的,包括眼睛、皮肤和免疫系统。
维生素A或视黄酸在全身水平上作为一种激素,并通过核视黄酸受体(RAR和RXR)信号调节免疫系统中的I型干扰素(IFN)合成。此外,它还通过维甲酸诱导基因I(RIG-I)信号通路负责对病毒感染的永久性免疫系统反应。另外调节NF-kB的活化。
德国进行的一项多中心、观察性、横断面、前瞻性分析调查中,研究人员发现维生素A缺乏与ARDS的发展和死亡率之间存在很强的关联(OR 5.21 [1.06–25.5],p = 0.042)。
注:ARDS,呼吸窘迫综合征
该研究纳入了40名SARS-CoV-2感染住院患者,并被诊断为中度、重度和危重度ARDS。对照组由47例症状较轻且不需要住院治疗的康复患者组成,结果显示,随着ARDS严重程度的增加,维生素A缺乏症更高,康复组的维生素A缺乏症明显降低(p <0.01至p <0.001)。
最近一项针对155名老年患者(18-95岁)的研究显示,36.5%的患者缺乏维生素A (< 0.343 mg/L); 作者认为,COVID-19疾病患者耗尽了血清维生素A储存。
免疫反应机制从先天转变为适应性,阻止了维甲酸的使用,这表明炎症平衡了COVID-19严重程度和维生素A水平之间的关系。
维生素A缺乏会降低影响上皮机械屏障功能的先天免疫应答,并增强呼吸道和肠道感染。这些上皮中的粘蛋白产生受视黄酸调节;因此,中等剂量维生素A补充剂通过调节上皮生长因子和相关细胞因子的基因表达来改善屏障完整性。
食物来源:肝脏,鱼类,鸡蛋,乳制品等,还包括橙黄色蔬菜、绿叶蔬菜、西红柿、水果等。
注意:维生素 A 补充剂可能与某些类型的避孕、抗癌药物、痤疮治疗和血液稀释剂相互作用,服用任何这些类型药物的人在服用维生素A补充剂之前应咨询医生。
➤ B 族 维 生 素
B族维生素是多种细胞反应的辅助因子,介导氨基酸的合成。包括维生素 B1、B2、B3、B5、B6、B7、B9、B12等,对免疫系统对抗感染反应至关重要。
B族维生素在激活先天性和适应性免疫应答中起关键作用,下调促炎细胞因子和炎症的产生,并显著改善呼吸功能。B族维生素还可以减少胃肠道问题,预防高凝状态,并缩短COVID-19患者的住院时间。
一项研究的311名法国受试者中,与7455名健康受试者相比,SARS-CoV-2阳性患者的维生素B9摄入量较低(OR = 0.84 (0.72,0.98),p = 0.02)。
在以色列,162例诊断为重症COVID-19的患者叶酸水平低于中轻度病例(分别为9.6ng/mL vs 12.9ng/mL vs 18.2ng/mL,p=0.005),其中12%为免疫抑制,9%需要无创氧合,15%为插管。
一组来自新加坡的患者纳入了43例50岁以上的COVID-19患者,发现联合补充维生素B12、维生素D和镁入院后与疾病严重程度的降低有关。在补充的患者中,与未接受这些微量营养素补充剂的患者相比,对氧疗的需求减少(17.6 vs 61.5%,p = 0.006)和重症监护需求减少有关。
——维生素B12
维生素B12对DNA合成和调节至关重要。转钴胺素发挥其抗氧化机制,促进还原性谷胱甘肽在细胞质中的生物利用度,从而促进氧化性谷胱甘肽的合成。钴胺素由肠道微生物群产生,有助于调节肠-脑轴,防止肠道生态失调,并有利于产生适当比例的微生物代谢物。这些过程对DNA合成、细胞稳态、造血和免疫至关重要。
在生理条件下,VB12调节抗炎细胞因子和生长因子的表达,减轻全身炎症。此外,通过促进NK细胞和CD8 + T细胞的增加,提高免疫反应,调节抗病毒反应。
当维生素B12缺乏时,感染的风险更大,严重程度也会增加。
一些因素如年龄和某些药物的使用与维生素B12缺乏的风险较高有关。在老年人中,内因子(intrinsic factor)的产生减少导致B12吸收不良、营养不良或尿和肠损失增加。
二甲双胍作为2型糖尿病的治疗也与VB12吸收不良引发的缺乏有关。因此,这可能会使这些患者更容易受到感染。
➤ 维 生 素 C
维生素C在免疫功能和伤口愈合中很重要,并具有抗氧化,抗病毒和抗炎的特性。维生素C已被证明可以增加中性粒细胞向感染部位的迁移,引发活性氧(ROS)和吞噬作用的产生。
据报道,饮食中的抗坏血酸可以降低c反应蛋白的浓度。维生素C被认为通过增加干扰素蛋白的产生而表现出抗病毒活性。
维生素C被认为是严重疾病的潜在治疗方法,也是降低感染风险的预防措施,因为它能够调节免疫细胞活性并减少炎症。
小规模研究显示,在对ICU入院后24小时内循环维生素C水平的分析中发现,高达82%的COVID-19患者缺乏维生素C,许多患者的维生素C水平仅为0.1mg/dL(正常值为0.4-2mg/dL),18%的维生素C水平检测不到(低于0.1mg/dL)。
在 50 例 COVID 病例中进行了一项临床试验,其中给予了高剂量维生素C 静脉干预,结果显示 COVID 患者的氧合指数发生了积极变化。
有研究表明,高剂量(8000mg/d)口服维生素补充剂可使COVID-19恢复率提高70%.
仍需要进一步的临床试验来充分描述维生素 C 对 COVID-19 感染的影响。
维生素 C 静脉注射被 FDA 归类为药物,只有口服剂量才能被评估并用作膳食补充剂,儿童(取决于年龄)每天 400 至 1800 毫克,成人每天 2000 毫克被认为是安全的。
富含维生素C的食物包括:猕猴桃,橙子,辣椒,草莓,樱桃,苹果,西兰花,菠菜,青椒,甜菜,花椰菜等。
➤ 维 生 素 D
除了对钙的吸收和骨骼强度的强大影响外,维生素D对于维持免疫力至关重要。它还减慢病毒复制,降低炎症活动,并增加体内 T 调节细胞的数量。
维生素 D 还可以缓解传染病、感染性休克和 ARDS 疾病(由于维生素 D 水平低)等并发症,这可能有助于预防新型冠状病毒作为辅助补充剂。
维生素D通过中性粒细胞和巨噬细胞合成组织蛋白酶和防御素来提高先天免疫力。这些作用控制病毒复制,并下调肿瘤坏死因子TNFα和IFN-γ的表达。
维生素D在增强免疫力中的作用
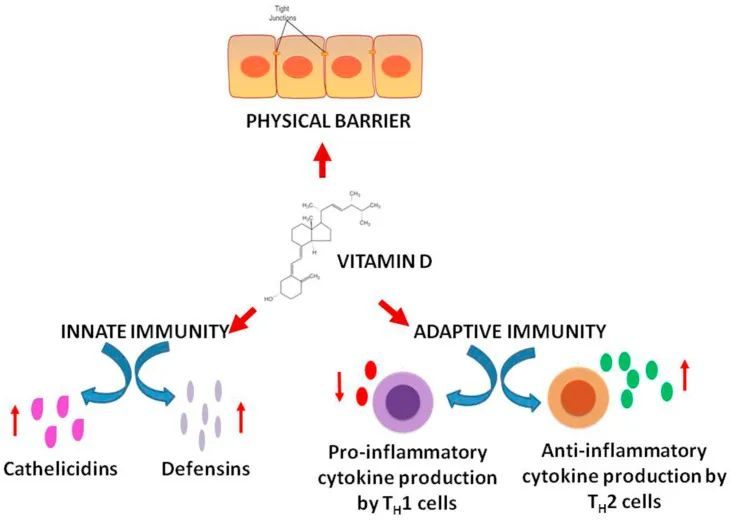
DOI:10.1016/j.clnesp.2022.04.007
低水平的维生素D(<20 ng/mL)与慢性疾病的发展和进展有关,如心血管疾病、2型糖尿病、癌症和抑郁症,也与骨骼健康不良有关。一些研究认为,维生素 D 水平低的人更容易患上 COVID-19 并患有更严重的疾病。维生素 D 缺乏会导致宿主体内的 SARS-CoV-2 病毒存活和复制。
与维生素D充足(感染率8.1%)或最高(感染率5.9%)的患者相比,缺乏维生素D的患者更有可能感染COVID-19(感染率12.5%)。
一项研究比较了中国335名COVID-19患者和560名健康志愿者的血液25(OH)D水平,发现COVID-19患者的25(OH)D浓度(中位数为26.5 nmol·L−1 [10.6 ng·mL−1])明显低于健康志愿者(中位数为32.5 nmol·L−1 [13 ng·mL−1])。
维生素D不足[具体定义为血清25(OH) D低于30 nmol·L−1 (12 ng·mL−1)]在COVID-19患者中比健康志愿者更常见。
与轻度COVID-19相比,重度COVID-19症状与最显著的维生素D缺乏相关。
土耳其的95名住院COVID-19患者,向他们补充了25-羟基维生素D,显示纤维蛋白原浓度降低,补充维生素D减少了住院时间,即使存在合并症。
在对13项研究的荟萃分析中,Pal等人计算出补充维生素D的人患严重疾病的风险降低了73%。在COVID-19发病后进行补充的研究中,风险降低最为显著(降低88%,OR 0.12)。
对SARS-CoV-2感染确诊病例的回顾性分析表明,维生素D水平低与较高的死亡风险显着相关,特别是在既往患病的老年人和男性个体中。
补充维生素D可以帮助25(OH) D水平低于25 nmol·L−1 (10 ng·mL−1) 的患者避免呼吸道感染。
补充维生素D,可以通过多晒太阳;饮食补充富含脂肪的鱼类如鲑鱼,鳟鱼,鲫鱼,金枪鱼,鳗鱼等,其他包括蘑菇,肝脏,蛋黄,新鲜水果蔬菜等。
注:冬季日照不够的情况可以通过维生素D补充剂,推荐最低剂量是每日400IU.
➤ 维 生 素 E
维生素E是一种脂溶性维生素,包括生育酚和生育三烯酚。是一种有效的抗氧化剂。可以影响免疫系统细胞,因为它具有抗氧化活性、蛋白激酶C(PKC)抑制和通过酶调节的信号转导。
在巨噬细胞中,维生素E修饰环氧化酶活性,从而控制过氧亚硝酸盐的合成。这导致前列腺素E2的产生降低,T细胞介导的T淋巴细胞反应上调。此外,它通过一氧化氮调节提高自然杀伤(NK)细胞活性。
与健康对照组相比,COVID-19患者的维生素E水平较低。
补充维生素E既往已被证明可增加免疫活性,特别是T细胞增殖,改善老年患者对疫苗接种的反应,并将老年肺炎患者的再住院风险降低63%.
维生素E来源:如小麦胚芽、葵花籽、杏仁、榛子、松子、鳄梨和甜红辣椒等。
扩展阅读:
➤ 铁
先天免疫反应精心控制铁代谢。铁稳态的破坏与感染,癌症,心血管疾病,肾脏疾病和血液疾病密切相关。
铁在病毒感染中也起着重要作用。在病毒复制过程中,需要ATP,而ATP合成需要铁。此外,铁调节素铁调素的表达增加几种细胞因子(例如,IL-6、IL-1),已知铁调素水平升高与血浆铁水平低有关(一种基于铁剥夺入侵病原体的非特异性宿主防御机制)。
注:铁调素通过结合和介导铁转运蛋白的降解来发挥其作用,铁转运蛋白是唯一已知的细胞铁输出剂,存在于肠细胞和巨噬细胞的细胞膜中,从而防止铁从这些细胞外排到血浆中。
铁缺乏在预测从轻度到重度疾病的转变方面很有价值,低血清铁水平是COVID-19患者死亡的独立危险因素。
汇总研究结果显示,COVID-19患者、严重程度状态和非幸存者的铁水平分别明显低于对照组、非严重程度状态和幸存者。
最近的研究中,与对照组(1.87±0.66mg/L)相比,COVID-19患者的血清铁(重症患者为1.33±0.7mg/L)显著降低。
此外,铁稳态的紊乱可在疾病发作后持续数月,并与肺部病变未消退和体能受损密切相关。
细胞内铁水平的升高和降低都是危险信号,分别通过NF-κB和HIF-1激活炎症和抗菌途径。
与中度病例相比,重症病例的血红蛋白、红细胞计数较低,铁蛋白水平较高,而非幸存者的铁蛋白水平高于幸存者。
铁过量也被认为是COVID-19发病机制的一个因素,因为它在活性氧的产生中发挥了作用,但更重要的可能是由于诱导非凋亡细胞死亡的铁死亡。
富含铁的食物包括:肝脏、动物血、红肉、扇贝、干、木耳、紫菜、菠菜、海带、黑芝麻、李子、桃、杏、苹果等。
扩展阅读
➤ 锌
锌是最普遍和最重要的微量元素,它参与细胞代谢的许多方面和近 100 种酶的催化活性。
它刺激适应性和固有免疫力,以及有助于维持呼吸上皮等组织屏障的抗病毒和抗炎特性。初步证据表明,ACE-2表达受Sirtuin 1(SIRT1)调节。由于锌能够下调SIRT1活性,这可能导致ACE-2表达降低,并减少SARS-CoV 2 进入细胞。
锌缺乏会阻碍淋巴细胞的产生、刺激和成熟,导致免疫功能下降。它还影响免疫介质,抑制 T 细胞、白细胞介素-2 合成以及抑制自然杀伤和细胞毒性 T 细胞活性。此外,它的缺失与更多的促炎介质有关,这可能会增加对疾病和炎症性疾病的易感性,尤其是那些影响肺部的疾病和炎症性疾病。
补充锌可能减轻COVID-19症状,因为这种金属抑制细胞内SARS-CoV 2复制的pH依赖性步骤,增加细胞内小泡的pH.
COVID-19典型的细胞因子过度产生(即“细胞因子风暴”),能够影响多个器官,似乎与短暂缺锌显著相关。
在2020年报告的一项研究显示,COVID-19患者的高剂量锌盐(每天<200毫克)改善了治疗后24小时的氧合并减少了发烧。尽管评估的病例数量很少,但这项工作确定了锌在SARS-CoV-2病毒控制和COVID-19并发症中可能产生的有益作用。
一项横断面对照研究报道,血清锌水平相对于疾病严重程度下降(中位数56.61μg/dL,n=200),重度COVID-19患者的降幅最大。
锌的典型每日剂量为15-30毫克锭剂,直接保护上呼吸道。补锌前检测确定锌水平至关重要。
➤ 硒
硒(Se)具有广泛的作用,如抗氧化和抗炎。硒在病毒感染中的氧化还原信号传导、氧化还原稳态和抗氧化防御中具有重要作用。
硒在保护呼吸系统,特别是抵御病毒感染方面发挥着重要作用。大量证据表明,硒缺乏与RNA病毒感染的易感性和不良结局有关。
硒与辅酶Q10联合使用可降低非特异性炎症反应和心血管疾病的死亡率。当与乙酰半胱氨酸一起服用时,硒有助于实现细胞内GSH(还原型谷胱甘肽)的正常水平,这是GPX(谷胱甘肽过氧化物酶是最强的抗氧化硒酶之一)的最佳水平的原因。GPX模拟依布硒(ebselen,一种合成硒化合物)是主要SARS-CoV-2蛋白酶的强抑制剂。
硒在正常免疫系统功能中的作用与其抗氧化特性和增加白细胞介素IL-2产生的能力有关,白细胞介介素IL-2具有免疫调节特性;例如,它根据当前需求刺激或抑制免疫反应。
适当剂量的硒对SARS-Cov-2感染患者的免疫系统的有益作用很重要,因为它调节IL-6的分泌,IL-6在疾病的病理生理学中起关键作用。硒的剂量建议为每天100至200µg。
➤ 铜
铜(Cu)是一种必需的微量元素,具有两种主要的生物学功能,第一种是酶的结构/催化辅因子,第二种是关键转录因子的辅激活因子。
铜转运基因参与巨噬细胞介导的宿主防御,其缺乏会降低IL-2和T细胞的增殖,并降低循环中性粒细胞的数量及其产生超氧阴离子的能力。在病毒感染和ROS启动氧化反应时,铜下调NF-κB表达,导致炎症细胞因子、趋化因子和粘附分子受到抑制。
一项对铜消费的荟萃分析显示,中国儿童反复呼吸道感染的发生与免疫功能、遗传因素和营养状况有关。在这项研究的结果中,锌和铜缺乏可能是儿童易患呼吸道感染的因素。

编辑
DOI:10.1007/s12011-022-03290-8
微量元素锌和铜之间存在拮抗作用和竞争吸收关系。当锌摄入量长期超标时,会导致铜缺乏,反之亦然。
自行服用锌补充剂的COVID-19患者,极有可能缺铜。作为一种必需的微量元素,铜是维持生物机制和细胞稳态所必需的;因此,有必要注意保持平衡。
关于微量元素的补充,应考虑锌与其他微量金属和维生素的相互作用。锌、铜、铁的吸收和生物利用度取决于元素之间的竞争,当以等比例给予时,铁和铜的吸收约减少40%。因此,过量补充锌可能导致铜或铁不足。
从目前的角度来看,除临床试验外,COVID-19中的维生素和微量元素补充剂不应超过一般人群和年龄组的推荐剂量。
药用植物,作为抗病毒、抗感染、抗炎、抗氧化、退热和肺肠道免疫增强剂,已被用于治疗与新冠肺炎相关的症状。
药用植物,在免疫刺激和维持平衡肠道微生物组中具有支持作用,可能是管理COVID-19的有效策略。
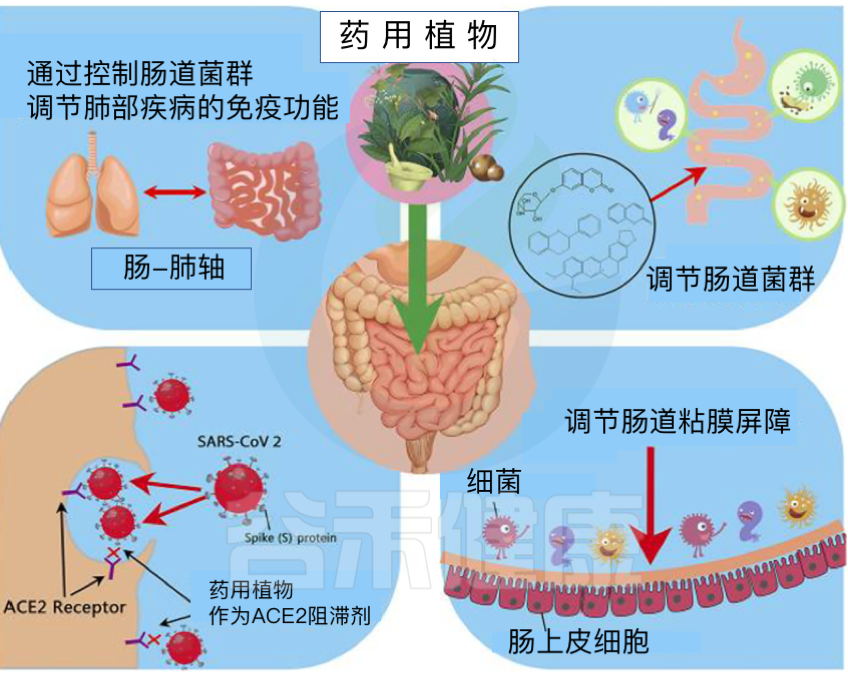
doi: 10.1186/s43088-022-00277-1.
➦ 吉 洛 伊 (Giloy,Guduchi)
由于其免疫调节特性,粉末状吉洛伊植物在前病毒期和后病毒期被彻底使用。此外,吉洛伊植物也可用作抗氧化剂,因此可以降低体内的整体氧化应激。这进一步防止了体内重要生物分子的氧化分解。
在慢性新冠肺炎感染期间,吉洛伊表现出抗炎作用,这在冠状病毒诱导的肺炎和肺部炎症中有很大的作用。血液中的氧饱和度低于正常范围,导致缺氧、疲劳,甚至死亡。
临床试验评估其对新冠肺炎确诊的无症状至轻度症状患者的疗效。91名年龄在18岁至75岁之间的患者,其中11.7%的对照组在平均1.8天后出现轻微症状,试验组没有任何其他症状。研究人员从研究中建议,Guduchi Ghan Vati可以作为无症状新冠肺炎患者的预防和治疗药物(NCT04480398)。
➦ 甘 草 (Mulethi)
由豆科甘草的根和细根组成,其活性化学成分是甘草酸。甘草是一种阿育吠陀药,以其外围和中枢作用的抗咳特性而闻名。
研究表明,甘草酸苷具有止咳作用,可以抑制咳嗽和相关的呼吸短促;也就是新冠肺炎感染的主要症状。它已被证明可以阻止被SARS感染的细胞释放HMGB1核蛋白(高运动性基盒蛋白1)。这使巨噬细胞失效,巨噬细胞在激活免疫中发挥积极作用,并负责产生促炎细胞因子。因此,避免了“细胞因子风暴”的开始及其对健康的不利影响。
在伊朗进行了一项单中心、开放标签、随机、平行组临床试验,以评估甘草对60名年龄≥18岁(体重≥35 kg)的新冠肺炎患者的抗炎作用(IRCT20200506047323N2)。
➦ 茯 苓 (Poria cocos)
其活性成分为茯苓酸。蛋白酶负责生物在宿主细胞中的复制。在冠状病毒中发现的一种蛋白酶是Mpro酶。Mpro是SARS-CoV-2的关键酶,参与介导病毒复制和转录。茯苓的主要活性成分茯苓酸与Mpro酶结合并抑制,从而阻止生物体在宿主细胞中的进一步复制和生长。
在COVID-19重症病例中,很少患者可能出现肺炎并发症,65岁或以上人群的并发症甚至更高。在这种情况下,茯苓酸的抗炎特性可以被解释为提供缓解。在一项研究中,用茯苓酸治疗肺炎大鼠肺部,结果显示通过NF-κB通路抑制炎症因子的表达。
➦ 蒙古紫云英(Mongolian milkvetch)
它由豆科黄芪的根、茎和叶组成。活性植物化学物质为黄芪甲苷、熊竹素和毛蕊异黄酮。黄芪甲苷是黄芪根、茎和叶中发现的多糖,可调节促炎因子如细胞因子、TNFα、IL-1β和NFATc4的表达,从而发挥免疫调节作用。它已被发现有助于避免细胞因子风暴的状况。
熊竹素在一定程度上也能成功抑制Mpro酶。作为一种免疫调节剂,在COVID-19的情况下,作为一种预防措施,可以使用蒙古紫云英来增强免疫力。
研究人员对小鼠血浆进行了代谢组学研究。结果表明,蒙古紫云英与人参同时食用可提高大鼠的脾、胸腺指数、脾淋巴细胞增殖和NK细胞毒活性。
➦ 其 他
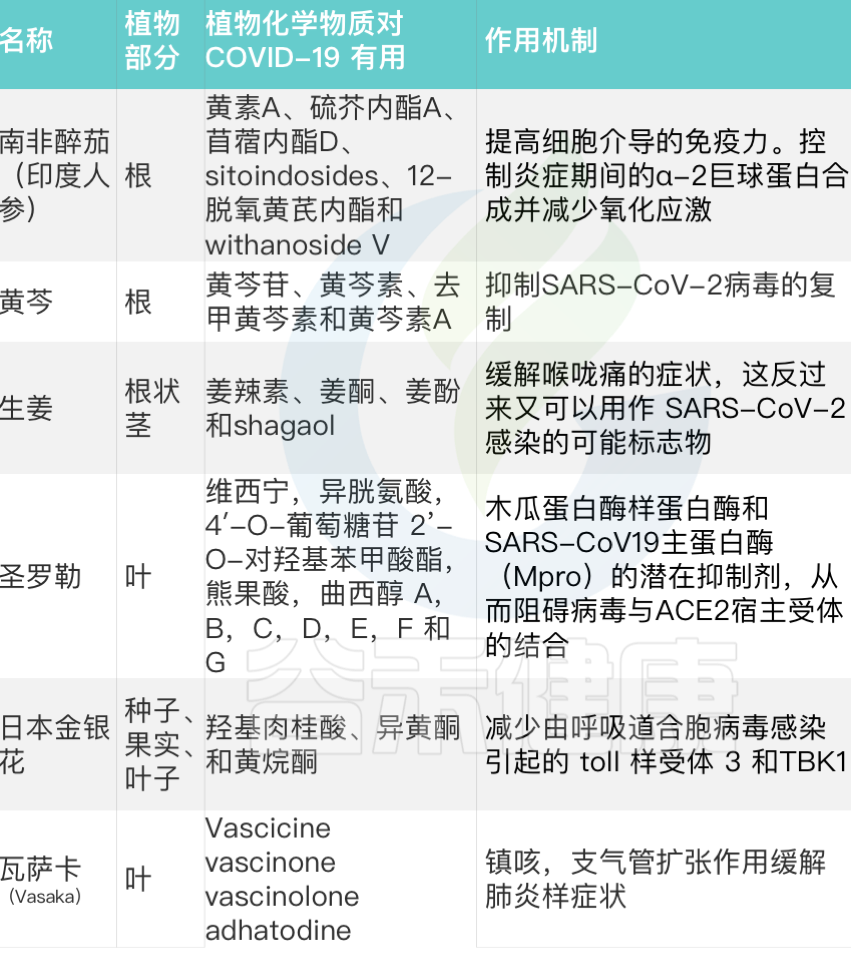
doi: 10.1016/j.ccmp.2022.100021.
改变的肠道微生物群或菌群失调可以作为全身炎症活动的调节剂,并可以通过多个肠道器官轴影响不同的器官。肠道通透性增加或者说肠漏,使细菌代谢物和毒素进入循环系统,并进一步恶化全身炎症反应,导致不同的新冠肺炎并发症。
肠道微生物群不仅显着影响COVID-19的发展和疾病的严重程度,而且还反映了COVID-19患者对长期并发症的易感性。
初步临床研究揭示了益生菌对 SARS-CoV-2 感染及恢复期的潜在调节作用。鉴于针对COVID-19的特定药物仍然是个谜,疫苗是有效的预防和控制策略;然而,病毒的持续突变加剧了这一难题。因此,通过益生菌、益生元、膳食补充剂、药用植物、FMT等多种方式,进行肠道菌群干预,是未来治疗COVID-19的有前途的补充策略。
尽管大规模临床试验有限,但相信这个领域正在发展,并蕴藏着巨大的机遇,一个个新诞生的研究可以在疫情的不同浪潮中推动微生物群的发现走向临床应用。
在此,也向一直奋斗在前线的医务工作者、在该领域辛勤耕耘的科研人员、其他相关工作者致敬。
对于个人而言,通过多种方式增强体质提高自身免疫力,是抵抗病毒侵袭的最好选择。
在新开启的2023里,希望大家都能找到属于自己的健康生活。
注:本账号内容仅作交流参考,不作为诊断及医疗依据。
主要参考文献:
Li M, Peng H, Duan G, Wang J, Yu Z, Zhang Z, Wu L, Du M, Zhou S. Older age and depressive state are risk factors for re-positivity with SARS-CoV-2 Omicron variant. Front Public Health. 2022 Oct 4;10:1014470. doi: 10.3389/fpubh.2022.1014470. PMID: 36268004; PMCID: PMC9576942.
Sacco C, Petrone D, Del Manso M, Mateo-Urdiales A, Fabiani M, Bressi M, Bella A, Pezzotti P, Rota MC, Riccardo F; Italian Integrated Surveillance of COVID-19 study group. Risk and protective factors for SARS-CoV-2 reinfections, surveillance data, Italy, August 2021 to March 2022. Euro Surveill. 2022 May;27(20):2200372. doi: 10.2807/1560-7917.ES.2022.27.20.2200372. PMID: 35593164; PMCID: PMC9121659.
Deng L, Li P, Zhang X, Jiang Q, Turner D, Zhou C, Gao Y, Qian F, Zhang C, Lu H, Zou H, Vermund SH, Qian HZ. Risk of SARS-CoV-2 reinfection: a systematic review and meta-analysis. Sci Rep. 2022 Dec 1;12(1):20763. doi: 10.1038/s41598-022-24220-7. PMID: 36456577; PMCID: PMC9714387.
Hansen CH, Friis NU, Bager P, Stegger M, Fonager J, Fomsgaard A, Gram MA, Christiansen LE, Ethelberg S, Legarth R, Krause TG, Ullum H, Valentiner-Branth P. Risk of reinfection, vaccine protection, and severity of infection with the BA.5 omicron subvariant: a nation-wide population-based study in Denmark. Lancet Infect Dis. 2022 Oct 18:S1473-3099(22)00595-3. doi: 10.1016/S1473-3099(22)00595-3. Epub ahead of print. PMID: 36270311; PMCID: PMC9578720.
Guedes AR, Oliveira MS, Tavares BM, Luna-Muschi A, Lazari CDS, Montal AC, de Faria E, Maia FL, Barboza ADS, Leme MD, Tomazini FM, Costa SF, Levin AS. Reinfection rate in a cohort of healthcare workers over 2 years of the COVID-19 pandemic. Sci Rep. 2023 Jan 13;13(1):712. doi: 10.1038/s41598-022-25908-6. PMID: 36639411; PMCID: PMC9837751.
Holmer HK, Mackey K, Fiordalisi CV, Helfand M. Major Update 2: Antibody Response and Risk for Reinfection After SARS-CoV-2 Infection-Final Update of a Living, Rapid Review. Ann Intern Med. 2023 Jan;176(1):85-91. doi: 10.7326/M22-1745. Epub 2022 Nov 29. PMID: 36442059; PMCID: PMC9707440.
Bowe B, Xie Y, Al-Aly Z. Acute and postacute sequelae associated with SARS-CoV-2 reinfection. Nat Med. 2022 Nov;28(11):2398-2405. doi: 10.1038/s41591-022-02051-3. Epub 2022 Nov 10. PMID: 36357676; PMCID: PMC9671810.
Davis HE, McCorkell L, Vogel JM, Topol EJ. Long COVID: major findings, mechanisms and recommendations. Nat Rev Microbiol. 2023 Jan 13:1–14. doi: 10.1038/s41579-022-00846-2. Epub ahead of print. PMID: 36639608; PMCID: PMC9839201.
O’ Mahony L, Buwalda T, Blair M, Forde B, Lunjani N, Ambikan A, Neogi U, Barrett P, Geary E, O’Connor N, Dineen J, Clarke G, Kelleher E, Horgan M, Jackson A, Sadlier C. Impact of Long COVID on health and quality of life. HRB Open Res. 2022 Apr 22;5:31. doi: 10.12688/hrbopenres.13516.1. PMID: 36101871; PMCID: PMC9440374.
Zhang F, Lau RI, Liu Q, Su Q, Chan FKL, Ng SC. Gut microbiota in COVID-19: key microbial changes, potential mechanisms and clinical applications. Nat Rev Gastroenterol Hepatol. 2022 Oct 21:1–15. doi: 10.1038/s41575-022-00698-4. Epub ahead of print. Erratum in: Nat Rev Gastroenterol Hepatol. 2023 Jan 12;: PMID: 36271144; PMCID: PMC9589856.
Sun K, Tempia S, Kleynhans J, von Gottberg A, McMorrow ML, Wolter N, Bhiman JN, Moyes J, Carrim M, Martinson NA, Kahn K, Lebina L, du Toit JD, Mkhencele T, Viboud C, Cohen C; PHIRST-C group. Rapidly shifting immunologic landscape and severity of SARS-CoV-2 in the Omicron era in South Africa. Nat Commun. 2023 Jan 16;14(1):246. doi: 10.1038/s41467-022-35652-0. PMID: 36646700; PMCID: PMC9842214.
Wang B, Zhang L, Wang Y, Dai T, Qin Z, Zhou F, Zhang L. Alterations in microbiota of patients with COVID-19: potential mechanisms and therapeutic interventions. Signal Transduct Target Ther. 2022 Apr 29;7(1):143. doi: 10.1038/s41392-022-00986-0. PMID: 35487886; PMCID: PMC9052735.
Vaezi M, Ravanshad S, Rad MA, Zarrinfar H, Kabiri M. The effect of synbiotic adjunct therapy on clinical and paraclinical outcomes in hospitalized COVID-19 patients: A randomized placebo-controlled trial. J Med Virol. 2023 Jan 5. doi: 10.1002/jmv.28463. Epub ahead of print. PMID: 36602047.
Chutipongtanate S, Morrow AL, Newburg DS. Human Milk Oligosaccharides: Potential Applications in COVID-19. Biomedicines. 2022 Feb 1;10(2):346. doi: 10.3390/biomedicines10020346. PMID: 35203555; PMCID: PMC8961778.
Nasir Ahmed M, Hughes K. Role of ethno-phytomedicine knowledge in healthcare of COVID-19: advances in traditional phytomedicine perspective. Beni Suef Univ J Basic Appl Sci. 2022;11(1):96. doi: 10.1186/s43088-022-00277-1. Epub 2022 Aug 4. PMID: 35966214; PMCID: PMC9362587.
Gang J, Wang H, Xue X, Zhang S. Microbiota and COVID-19: Long-term and complex influencing factors. Front Microbiol. 2022 Aug 12;13:963488. doi: 10.3389/fmicb.2022.963488. PMID: 36033885; PMCID: PMC9417543.
Xu L, Ho CT, Liu Y, Wu Z, Zhang X. Potential Application of Tea Polyphenols to the Prevention of COVID-19 Infection: Based on the Gut-Lung Axis. Front Nutr. 2022 Apr 14;9:899842. doi: 10.3389/fnut.2022.899842. PMID: 35495940; PMCID: PMC9046984.
Xu L, Yang CS, Liu Y, Zhang X. Effective Regulation of Gut Microbiota With Probiotics and Prebiotics May Prevent or Alleviate COVID-19 Through the Gut-Lung Axis. Front Pharmacol. 2022 Apr 25;13:895193. doi: 10.3389/fphar.2022.895193. PMID: 35548347; PMCID: PMC9081431.
Rahmati M, Fatemi R, Yon DK, Lee SW, Koyanagi A, Il Shin J, Smith L. The effect of adherence to high-quality dietary pattern on COVID-19 outcomes: A systematic review and meta-analysis. J Med Virol. 2023 Jan;95(1):e28298. doi: 10.1002/jmv.28298. Epub 2022 Nov 18. PMID: 36367218.
Pandey M, Bhati A, Priya K, Sharma KK, Singhal B. Precision Postbiotics and Mental Health: the Management of Post-COVID-19 Complications. Probiotics Antimicrob Proteins. 2022 Jun;14(3):426-448. doi: 10.1007/s12602-021-09875-4. Epub 2021 Nov 22. PMID: 34806151; PMCID: PMC8606251.
Horne BD, Bunker T. Pathogenic Mechanisms of the Severe Acute Respiratory Syndrome Coronavirus 2 and Potential Direct and Indirect Counteractions by Intermittent Fasting. Nutrients. 2022 Dec 21;15(1):20. doi: 10.3390/nu15010020. PMID: 36615679; PMCID: PMC9823718.
Hou YC, Su WL, Chao YC. COVID-19 Illness Severity in the Elderly in Relation to Vegetarian and Non-vegetarian Diets: A Single-Center Experience. Front Nutr. 2022 Apr 29;9:837458. doi: 10.3389/fnut.2022.837458. PMID: 35571931; PMCID: PMC9101048.
Karupaiah T, Lu KC. Editorial: Nutraceuticals for the recovery of COVID-19 patients. Front Nutr. 2022 Nov 14;9:1054632. doi: 10.3389/fnut.2022.1054632. PMID: 36451742; PMCID: PMC9703639.
Itsiopoulos C, Mayr HL, Thomas CJ. The anti-inflammatory effects of a Mediterranean diet: a review. Curr Opin Clin Nutr Metab Care. 2022 Nov 1;25(6):415-422. doi: 10.1097/MCO.0000000000000872. Epub 2022 Aug 30. PMID: 36039924.
Yue Y, Ma W, Accorsi EK, Ding M, Hu F, Willett WC, Chan AT, Sun Q, Rich-Edwards J, Smith-Warner SA, Bhupathiraju SN. Long-term diet and risk of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) infection and Coronavirus Disease 2019 (COVID-19) severity. Am J Clin Nutr. 2022 Dec 19;116(6):1672-1681. doi: 10.1093/ajcn/nqac219. PMID: 35945354; PMCID: PMC9384672.
Zargarzadeh N, Tadbir Vajargah K, Ebrahimzadeh A, Mousavi SM, Khodaveisi H, Akhgarjand C, Toyos FMP, Cerqueira HS, Santos HO, Taghizadeh M, Milajerdi A. Higher Adherence to the Mediterranean Dietary Pattern Is Inversely Associated With Severity of COVID-19 and Related Symptoms: A Cross-Sectional Study. Front Med (Lausanne). 2022 Jul 19;9:911273. doi: 10.3389/fmed.2022.911273. PMID: 35928288; PMCID: PMC9343686.
Xu J, Ren Z, Cao K, Li X, Yang J, Luo X, Zhu L, Wang X, Ding L, Liang J, Jin D, Yuan T, Li L, Xu J. Boosting Vaccine-Elicited Respiratory Mucosal and Systemic COVID-19 Immunity in Mice With the Oral Lactobacillus plantarum. Front Nutr. 2021 Dec 22;8:789242. doi: 10.3389/fnut.2021.789242. PMID: 35004816; PMCID: PMC8733898.
Gualtieri P, Marchetti M, Frank G, Cianci R, Bigioni G, Colica C, Soldati L, Moia A, De Lorenzo A, Di Renzo L. Exploring the Sustainable Benefits of Adherence to the Mediterranean Diet during the COVID-19 Pandemic in Italy. Nutrients. 2022 Dec 26;15(1):110. doi: 10.3390/nu15010110. PMID: 36615768; PMCID: PMC9824251.
Ferro Y, Pujia R, Maurotti S, Boragina G, Mirarchi A, Gnagnarella P, Mazza E. Mediterranean Diet a Potential Strategy against SARS-CoV-2 Infection: A Narrative Review. Medicina (Kaunas). 2021 Dec 20;57(12):1389. doi: 10.3390/medicina57121389. PMID: 34946334; PMCID: PMC8704657.
Chavda VP, Patel AB, Vihol D, Vaghasiya DD, Ahmed KMSB, Trivedi KU, Dave DJ. Herbal Remedies, Nutraceuticals, and Dietary Supplements for COVID-19 Management: An Update. Clin Complement Med Pharmacol. 2022 Mar;2(1):100021. doi: 10.1016/j.ccmp.2022.100021. Epub 2022 Feb 5. PMID: 36620357; PMCID: PMC8816850.
Nasir Ahmed M, Hughes K. Role of ethno-phytomedicine knowledge in healthcare of COVID-19: advances in traditional phytomedicine perspective. Beni Suef Univ J Basic Appl Sci. 2022;11(1):96. doi: 10.1186/s43088-022-00277-1. Epub 2022 Aug 4. PMID: 35966214; PMCID: PMC9362587.
Renata RN, Arely GA, Gabriela LA, Esther MM. Immunomodulatory Role of Microelements in COVID-19 Outcome: a Relationship with Nutritional Status. Biol Trace Elem Res. 2022 Jun 6:1–19. doi: 10.1007/s12011-022-03290-8. Epub ahead of print. PMID: 35668151; PMCID: PMC9170122.
Li Y, Luo W, Liang B. Circulating trace elements status in COVID-19 disease: A meta-analysis. Front Nutr. 2022 Aug 12;9:982032. doi: 10.3389/fnut.2022.982032. PMID: 36034929; PMCID: PMC9411985.
Bego T, Meseldžić N, Prnjavorac B, Prnjavorac L, Marjanović D, Azevedo R, Pinto E, Duro M, Couto C, Almeida A. Association of trace element status in COVID-19 patients with disease severity. J Trace Elem Med Biol. 2022 Dec;74:127055. doi: 10.1016/j.jtemb.2022.127055. Epub 2022 Aug 4. PMID: 35985069; PMCID: PMC9349050.
Milton-Laskibar I, Trepiana J, Macarulla MT, Gómez-Zorita S, Arellano-García L, Fernández-Quintela A, Portillo MP. Potential usefulness of Mediterranean diet polyphenols against COVID-19-induced inflammation: a review of the current knowledge. J Physiol Biochem. 2022 Nov 8:1–12. doi: 10.1007/s13105-022-00926-0. Epub ahead of print. PMID: 36346507; PMCID: PMC9641689.
Sun Y, Ju P, Xue T, et al., Alteration of faecal microbiota balance related to long-term deep meditation. General Psychiatry 2023;36:e100893. doi: 10.1136/gpsych-2022-100893
Callaway E. What Omicron’s BA.4 and BA.5 variants mean for the pandemic. Nature. 2022 Jun;606(7916):848-849. doi: 10.1038/d41586-022-01730-y. PMID: 35750920.
Wang K, Gheblawi M, Oudit GY. Angiotensin Converting Enzyme 2: A Double-Edged Sword. Circulation. 2020 Aug 4;142(5):426-428. doi: 10.1161/CIRCULATIONAHA.120.047049. Epub 2020 Mar 26. PMID: 32213097.
Sapra L, Saini C, Garg B, Gupta R, Verma B, Mishra PK, Srivastava RK. Long-term implications of COVID-19 on bone health: pathophysiology and therapeutics. Inflamm Res. 2022 Sep;71(9):1025-1040. doi: 10.1007/s00011-022-01616-9. Epub 2022 Jul 28. PMID: 35900380; PMCID: PMC9330992.
Alenazy MF, Aljohar HI, Alruwaili AR, Daghestani MH, Alonazi MA, Labban RS, El-Ansary AK, Balto HA. Gut Microbiota Dynamics in Relation to Long-COVID-19 Syndrome: Role of Probiotics to Combat Psychiatric Complications. Metabolites. 2022 Sep 27;12(10):912. doi: 10.3390/metabo12100912. PMID: 36295814; PMCID: PMC9611210.
Shi Y, Li Z, Yang C, Liu C. The role of gut-brain axis in SARA-CoV-2 neuroinvasion: Culprit or innocent bystander? Brain Behav Immun. 2021 May;94:476-477. doi: 10.1016/j.bbi.2021.01.024. Epub 2021 Feb 15. PMID: 33600935; PMCID: PMC7883713.
De R, Dutta S. Role of the Microbiome in the Pathogenesis of COVID-19. Front Cell Infect Microbiol. 2022 Mar 31;12:736397. doi: 10.3389/fcimb.2022.736397. PMID: 35433495; PMCID: PMC9009446.

谷禾健康
今年是新冠病毒影响的第三年了,在病毒的影响下,我们的生活方式发生了很大的变化。近日,多地政府颁布了疫情防控的新政策,除了一些特殊场所,出入其他场所不用扫场所码了。
从一方面说,这样的情况方便了我们日常的生活,但是从另一方面来讲,对于病毒的防范没有那么严格了。最终如果全面放开的话,我们可能需要通过自身的免疫来抵抗病毒,最终实现与病毒的共存。

★ 面对新冠病毒,健康仍是第一位
新冠疫情给全球带来了不可估量的损失。经济发展缓慢,生产制造受阻,人们精神压力增大。但是无论如何,健康仍然是第一位的,健康乃生死大事,是众多其他所愿所求之根本。
在疫情防范严格时尚且有那么多人感染,那么在逐渐放宽的今天,我们应该如何避免新冠病毒感染,并在感染后将影响降到最低呢?免疫在这其中发挥了关键的作用。
本文从人体内的免疫作用概述开始,讲述了免疫系统如何抵抗细菌及病毒的感染,影响免疫的诸多因素、新冠病毒下身体的免疫,以及微生物群对免疫的影响。
在文章的最后,提出了一些有助于提高免疫力的方法。希望可以帮助人们更好地了解免疫,以便在当下和未来拥有健康的身体,不受病毒所侵害。这对未来以微生物群为中心的预防和治疗方法也具有启示意义。
本文主要从以下几个方面讲述
●免疫与免疫系统
●免疫力的高低与评估
●新冠病毒在人体中的免疫反应
●影响免疫的因素
●肠道菌群对免疫的影响
●提高免疫力的方法
我们常说“一个人的免疫力好,就不容易生病”。那么,什么是免疫力呢?
免疫力
免疫力是人体自身的防御机制,是人体识别和消灭外来侵入的任何异物(病毒、细菌等);处理衰老、损伤、死亡、变性的自身细胞以及识别和处理体内突变细胞和病毒感染细胞的能力。
现代免疫学认为,免疫力是人体识别和排除“异己”的生理反应。数百万年来,人类生活在一个既适合生存又充满危险的环境,人类得以存续,也获得了非凡的免疫力。所以说免疫力是生物进化过程的产物。

▸ 非特异性免疫
非特异性免疫,也叫固有免疫。它和特异性免疫都是人类在漫长进化过程中获得的一种遗传特性,但是非特异性免疫是人一生下来就具有,而特异性免疫需要经历一个过程才能获得。
当细菌侵入到机体的不同部位,机体的免疫系统通过皮肤黏膜屏障作用对病原体的侵入开始攻击。当病原体突破体表和黏膜层进入机体内部遇到固有免疫细胞及分子所介导的固有免疫应答。
固有免疫应答是指固有免疫分子和细胞在遇到细菌后,被即刻激活且发挥生物学效应,将病原体和异物清除的过程。
固有免疫应答出现在宿主抗感染应答的早期阶段,以抗原非特异性方式识别和清除细菌。
✦特点
•作用范围广:机体对入侵抗原物质的清除没有特异的选择性。
•反应快: 抗原物质一旦接触机体,立即遭到机体的排斥和清除。
•有相对的稳定性: 既不受入侵抗原物质的影响,也不因入侵抗原物质的强弱或次数而有所增减。
•有遗传性:生物体出生后即具有非特异性免疫能力,并能遗传给后代。
▸ 特异性免疫
特异性免疫又称获得性免疫或适应性免疫,这种免疫只针对一种病原。是获得免疫经后天感染(病愈或无症状的感染)或人工预防接种(菌苗、疫苗、类毒素、免疫球蛋白等)而使机体获得抵抗感染能力。
一般是在微生物等抗原物质刺激后才形成的(免疫球蛋白、免疫淋巴细胞),并能与该抗原起特异性反应。
✦特点
•具有特异性(或称专一性):机体的二次应答是针对再次进入机体的抗原,而不是针对其他初次进入机体的抗原。
•有免疫记忆:免疫系统对初次抗原刺激的信息可留下记忆。
•有多种细胞参与:针对抗原刺激的应答主要是T细胞和B细胞,但在完成特异性免疫的过程中,还需要其他一些细胞(巨噬细胞、粒细胞等)的参与。
•有个体的特征:特异性免疫是机体出生后,经抗原的反复刺激而在非特异性免疫的基础上建立的一种保护个体的功能,这种功能个体上具有差别,不同于非特异性免疫。
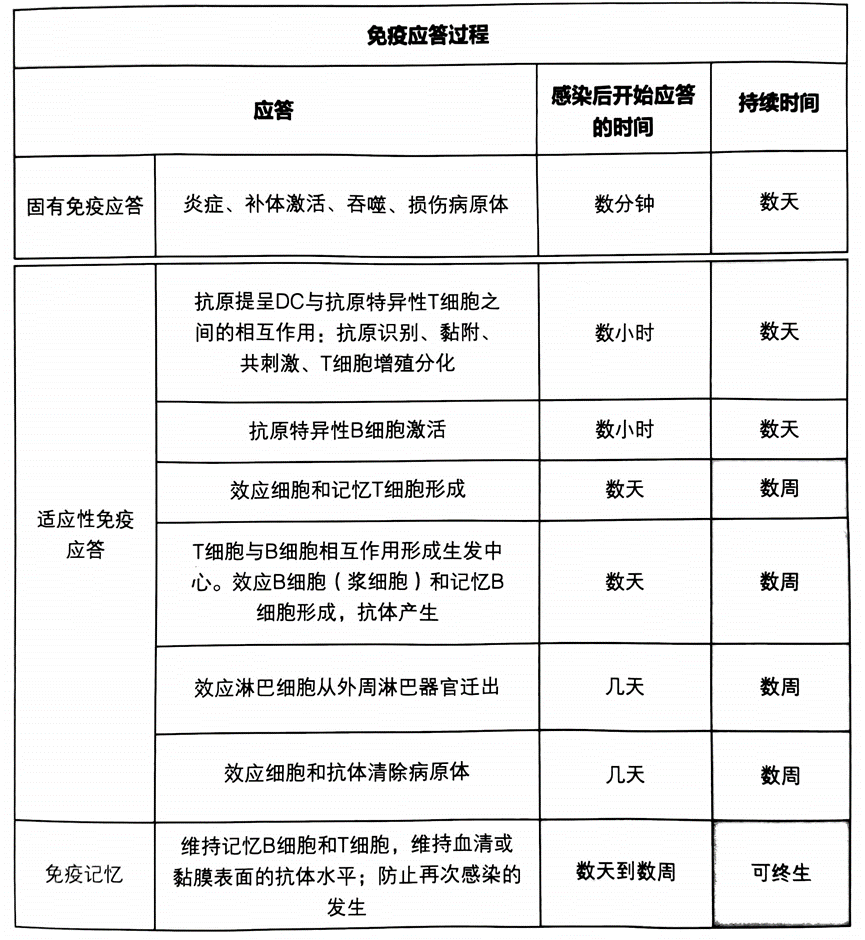
来源:詹韦免疫生物学(原书第九版)
提到免疫力,不得不提到的就是人类的免疫系统,免疫系统包括了免疫器官、免疫细胞和免疫活性物质三大类。
•免疫器官
常见的免疫器官有:骨髓、胸腺、扁桃体、脾、淋巴结、阑尾等。
免疫器官遍布全身
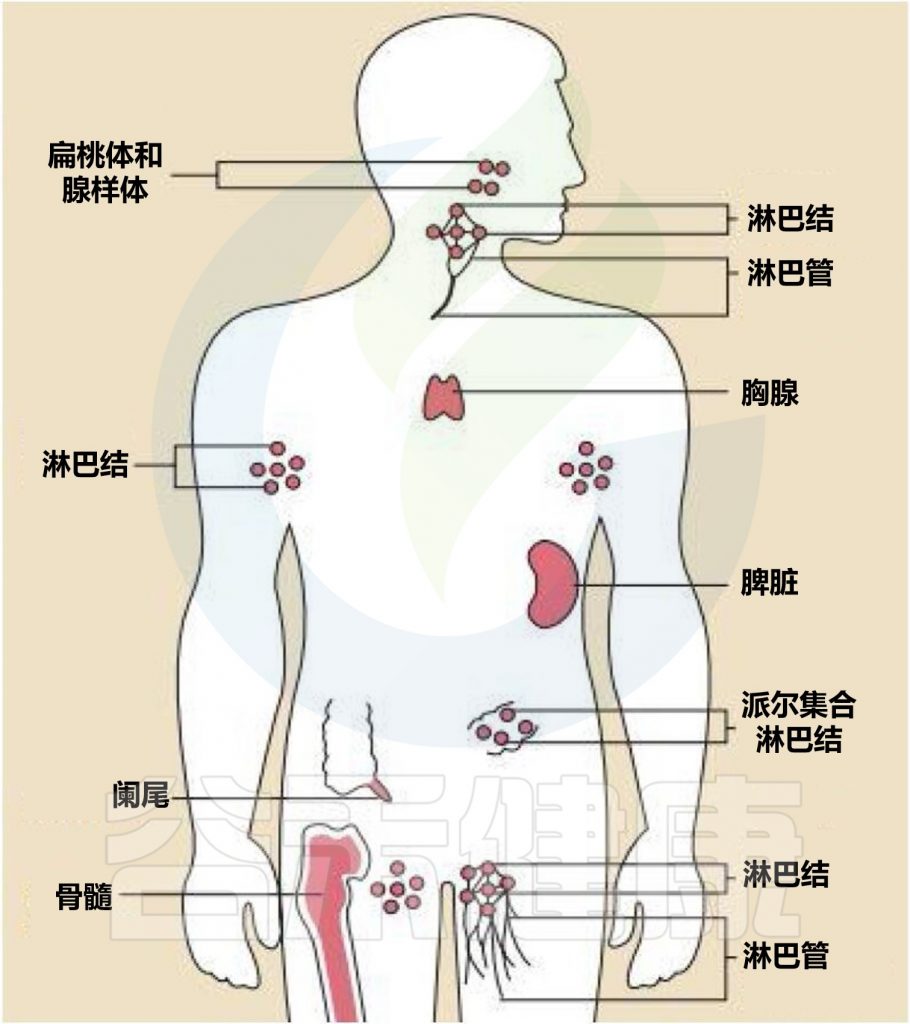

Chowdhury MA,et al.J Infect Public Health.2020
作用
免疫器官主要的作用就是产生、分化、成熟和储存免疫细胞,也是免疫应答发生的场所,还包括合成某些活性物质、建立和维持自身免疫耐受、对免疫细胞进行调节等作用。
•免疫细胞
常见的免疫细胞有:淋巴细胞、T细胞、B细胞、自然杀伤细胞、白细胞、吞噬细胞等。
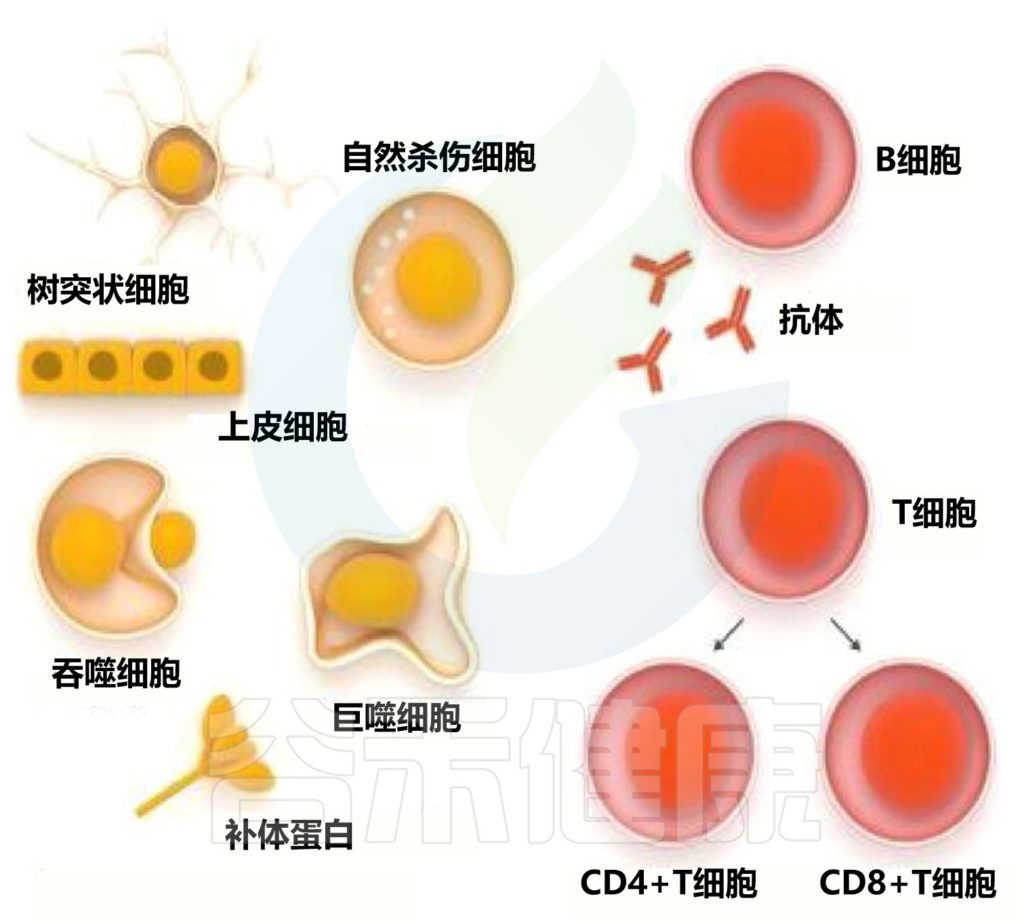
Chowdhury MA,et al.J Infect Public Health.2020
作用
免疫细胞就是战场上的“士兵”,负责巡视人体,一旦发现入侵的病原体,就会立刻展开吞噬和清除工作。
作用过程
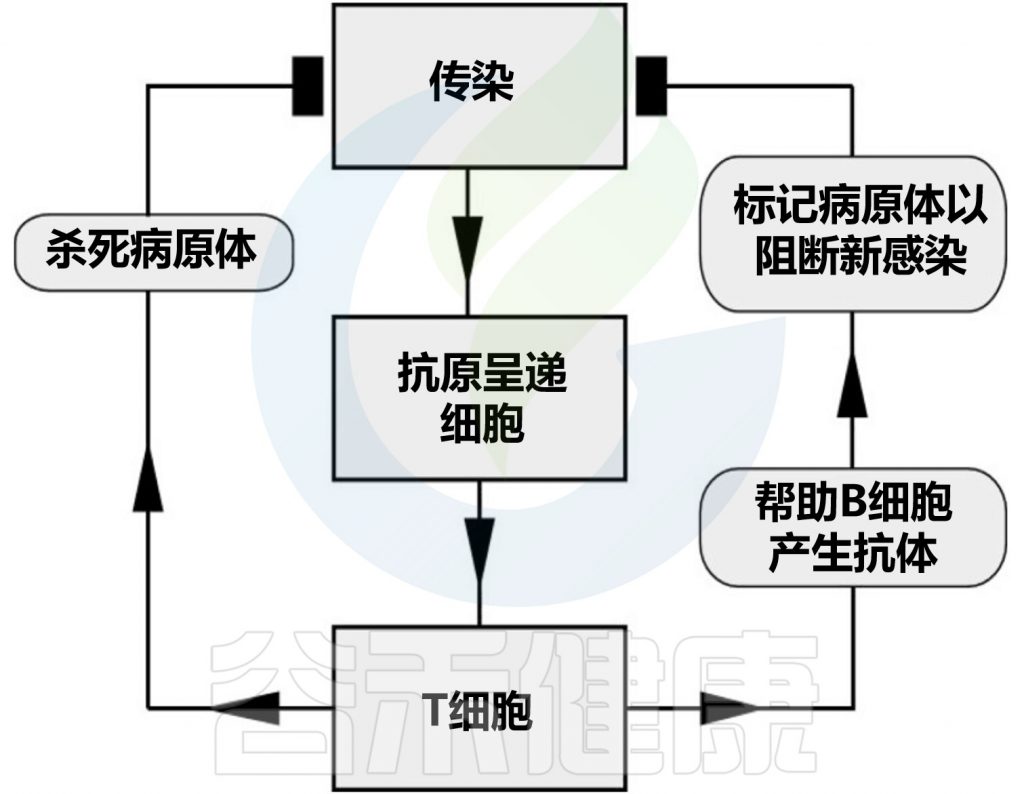

由巨噬细胞首先发起进攻,将病原体吞噬、分解,将分解的片段显示在细胞表面,并且提示T细胞,T细胞与巨噬细胞交流过后,就会向整个免疫系统发出“敌人入侵警报”,免疫系统收到后,会派出杀伤性T细胞,杀伤性T细胞找到并清除已经被感染的人体细胞,防止被感染的细胞继续繁殖,同时杀伤性T细胞会派出B淋巴细胞,在感染病原体的人体细胞被摧毁的同时,B淋巴细胞产生抗体,与细胞内的致病微生物结合,使其失去致病作用。
•免疫活性物质
常见的免疫活性物质有:抗体、免疫球蛋白、干扰素、细胞因子等。
作用
免疫活性物质可以辅助免疫细胞,使免疫性细胞的作用发挥的更强,放大其作用效果。
免疫系统的功能主要表现为三方面,即防御功能、稳定功能及免疫监视作用,这些功能一旦失调,即产生免疫病理反应。
•免疫防御
防御病原微生物侵害机体。就是人体抵御病原体及其毒性产物侵犯,使人免患感染性疾病。当该功能过于亢进,发生超敏反应;当该功能过于低下,发生免疫缺陷病。
•免疫自稳
人体组织细胞时刻不停地新陈代谢,随时有大量新生细胞代替衰老和受损伤的细胞。免疫系统能及时地把衰老和死亡的细胞识别出来,并把它从体内清除出去,从而保持人体的稳定。该功能异常时,发生自身免疫病。
•免疫监视
免疫系统具有识别、杀死并及时清除体内突变细胞,防止肿瘤发生的功能,称为免疫监视。免疫监视是免疫系统最基本的功能之一。
宿主通常采取三种策略应对病毒等微生物带来的威胁:
“规避”机制包括解剖学屏障和行为矫正,可防止机体暴露于微生物环境中。
机体被感染后,“抵抗”可减少或消除病原体。为了抵御不同种类的微生物,免疫系统包含众多调控分子和功能性细胞,称为免疫介质或效应机制。
“耐受”是指增强组织抵抗微生物诱导损伤的能力。“免疫耐受”是指阻止针对宿主自身组织的免疫应答。
注:“耐受性”被广泛用于植物的易感性而非动物免疫。例如,植物应对损伤的常见“耐受”机制是通过激活休眠的分生组织来促进更新,即未分化细胞分化为植物的新生部分。这应与术语“免疫耐受”进行区分。
解剖学和化学屏障是抵抗感染的最初屏障。
皮肤和黏膜属于一种“规避”策略,可防止机体内部组织暴露于病毒等微生物环境中。
在大多数解剖学屏障中,“抵抗”策略进一步增强了宿主的防御能力。例如,黏膜表面会产生多种抗菌蛋白,可作为天然抗生素防止病毒等微生物进入人体。
如果这些屏障被破坏,固有免疫系统的其他组分将立即发挥作用,比如前面提到的补体。补体系统由30余种协同作用的蛋白质分子组成,是血清和间质组织中最重要的免疫分子之一。
补体不仅能与抗体协同作用,还可以在没有特异性抗体存在时直接结合外来抗原,因此,它在固有免疫应答和适应性免疫应答中都发挥重要作用。
抵抗病原体的防御等级
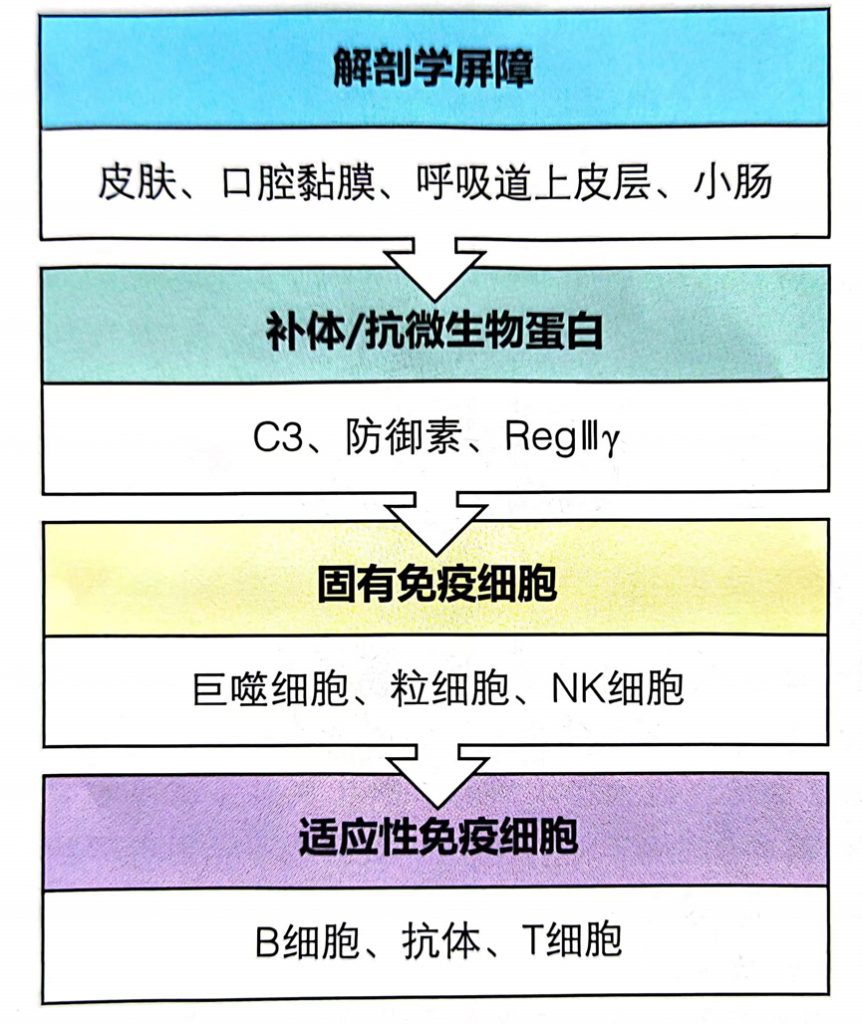

来源:詹韦免疫生物学(原书第九版)
既然免疫对我们如此重要,那是不是越高越好呢?免疫是否会影响一些疾病?以及我们如何了解自身的免疫能力呢?
各种原因使免疫系统不能正常发挥保护作用,在此情况下,极易招致细菌、病毒、真菌等感染,因此免疫力低下最直接的表现就是容易生病。
✦免疫力低下与生病容易恶性循环
免疫力低下或免疫力不健全,容易经常患病,加重了机体的消耗,所以一般有体质虚弱、营养不良、精神萎靡、疲乏无力、食欲降低、睡眠障碍等表现,生病、打针吃药便成了家常便饭。而营养不良这些又会使免疫力下降,形成恶性循环。
每次生病都要很长时间才能恢复,而且常常反复发作。长此以往会导致身体和智力发育不良,还易诱发重大疾病。
免疫力是越高越好吗?
答案是否定的。
随着医学专家对新冠肺炎的研究不断深入,又有一个专业名词“细胞因子风暴”,也就是“炎症风暴”,走进大众视野。它是指在患者体内结束潜伏期后爆发的病毒激发人体内免疫系统的强烈抵抗,会引起急性呼吸窘迫综合征和多器官衰竭,夺走患者的生命。
✦免疫力过高引起过敏
而且,免疫力过高容易使人体处于高度敏感状态,所有物质都可成为变应原,引发变应性鼻炎、过敏性哮喘、食物过敏等情况,严重的可致对身体内部自己的组织细胞产生反应,患自身免疫病。
如类风湿关节炎、系统性红斑狼疮、恶性贫血等疾病。
因此,健康适度的免疫力需要在清除外源物质和降低自身损伤之间做一个平衡。
免疫力的过高或过低以及在体内的不平衡都会引起一系列疾病
✦过敏性反应
无害的环境或饮食抗原触发2型免疫反应(涉及嗜酸性粒细胞、肥大细胞、固有淋巴样细胞、辅助T细胞和IgE),导致暴露组织的炎症和损伤。例如过敏性接触性皮炎、过敏性鼻炎、过敏性哮喘和食物过敏。
✦同种异体免疫
来自同一物种的非自抗原触发免疫反应,即体液(抗体介导)或细胞介导的免疫反应,导致外来组织或器官的细胞毒性和破坏。
反之亦然,移植到宿主体内的外来免疫细胞会攻击和损伤宿主组织。例如,移植物抗宿主病、输血反应和器官移植排斥反应
✦自身免疫
自我抗原被自身反应性的T和B细胞不适当地靶向,从而逃避耐受机制,导致抗体或细胞介导的组织损伤。自身免疫可分为器官特异性或系统性,根据主要影响一个或多个器官的临床表现。
例如,类风湿关节炎、干燥综合征和系统性红斑狼疮(系统性例子),I 型糖尿病、多发性硬化症和自身免疫性肝炎(器官特异性例子)。
✦自体炎症
在没有传染性药物的情况下,由于天然免疫细胞激活(中性粒细胞和巨噬细胞)而导致组织损伤后的自身炎症性疾病。
与自身免疫相反,损伤发生在没有明显的T和B细胞参与或自身抗体的情况下。自身炎症主要发生在单基因综合征和晶体驱动的疾病,如痛风,但也发生在一些自身免疫性疾病,具有突出的自身炎症成分(特别是炎症性肠病和脊椎关节病)。
例如,家族性地中海热和肿瘤坏死因子受体相关周期综合征。
✦食物不耐受引起的自身免疫性疾病
不适当的饮食成分在遗传易感性的个人中,导致自身免疫反应。这些疾病将与食物不耐受区分开来,而不依赖于免疫反应(例如,酶缺乏引起的乳糖不耐受)或复杂的非免疫超敏反应综合征(例如肠易激综合征)的功能障碍。
食物过敏的主要区别是饮食抗原引起的适应性免疫反应类型,例如腹腔疾病。
为什么要做免疫评估?
人的免疫系统就像一支精锐的军队24小时昼夜不停地守护着人类健康。免疫功能衰退会引发身体各种亚健康症状,各种疾病也随之到来。近年来随着人均寿命的延长、城市化的加快、不良生活习惯、职业压力、环境污染、遗传易感基因等因素的影响,身体的免疫力也会随之发生变化。
每个人的免疫系统都不一样,免疫能力自然也不一样,先天免疫能力较差的个体,应该在后天生活中选择更加健康的生活方式,以促进机体免疫系统的强大,每个人对营养的需求也都不相同,免疫力评估可以指导你如何为了更加健康而选择怎样的生活方式。
免疫力评估是一项全面评估免疫系统的检查,其中不仅包括了血常规中最基础的检查项目,还包括了免疫细胞的比例数目和T细胞亚群的检测,通过以上检测,可以评估人体三大防线的情况,还可以评估免疫的平衡作用,适合所有人群。判断免疫系统的强弱,指导更加健康的生活方式。

✦抽血查免疫球蛋白、补体
免疫球蛋白是检查机体体液免疫功能的一项重要指标,免疫球蛋白、补体的检测可以反映机体的免疫力。
✦淋巴细胞功能检查
淋巴细胞转化实验是为了检测免疫细胞的活性,根据淋巴细胞转化程度测定机体免疫应答功能,淋巴细胞转化率的高低可以反映机体细胞免疫水平。
形态学方法参考值:淋巴细胞转化率(LTT)为60.1%±7.6%。
✦细胞因子检测
细胞因子是由免疫细胞和一些非免疫细胞经刺激而合成分泌的一类具有广泛生物学活性的小分子蛋白质,调控免疫应答。
临床上有细胞因子6项检测,或细胞因子12项检测,包括白细胞介素-6、白细胞介素-2、干扰素、炎症细胞因子(TNF)等等。
小结
免疫系统评估通常包含了基因差异带来的先天免疫能力的不同,可以评估免疫的基础水平、对病毒的抵抗能力等,充分了解自身的免疫系统风险;还可以评估目前受检机体的营养吸收、代谢水平,根据基因找到最适宜的睡眠节律,结合以上检测,可以制定最适宜个体的健康方案。
COVID-19是指2019新型冠状病毒感染导致的肺炎。该病毒是一种具有冠状外观的RNA病毒。其直径约为60–140nm。我们一般称其为新冠病毒。
✦病毒传播
它通过咳嗽和打喷嚏产生的呼吸道飞沫传播,并通过吸入进入鼻腔系统并开始复制。研究表明,佩戴口罩等外在防护可以有效降低病毒的传播率。
ACE2是COVID-19病毒的主要受体。COVID-19表面存在的刺突蛋白(S蛋白),与ACE2受体结合。
接下来,病毒开始在体内传播,可以通过鼻咽拭子检测到。然后病毒传播并到达呼吸道,在那里它面临更强大的先天免疫反应。在此阶段,疾病具有临床表现,先天反应细胞因子可能预示着随后的临床过程。
✦病毒的影响

•大部分患者症状轻微
对于90%的受感染患者,这种疾病将是轻微的,并且主要局限于上呼吸道。通过保守的对症治疗即可治愈,这些人可能会在家中接受监测。
最常见的症状是发烧和呼吸道不适。也有胃肠道感染的报道,症状包括腹泻、恶心、呕吐、腹痛和食欲不振等。
•小部分患者症状比较严重
一小部分感染患者会出现肺部浸润,其中一些会出现非常严重的疾病。根据中国疾病预防控制中心的流行病学研究,新冠重症患者的死亡率可高达49%。
•患病率与年龄有关
在武汉,对292名COVID-19患者进行了研究。年龄是重症患者的危险因素。重症患者年龄每增加5岁,风险增加15.15%。COVD-19患者多为重症组老年患者,有基础疾病。慢性阻塞性肺疾病、高血压、恶性肿瘤、冠心病、慢性肾脏病在重症组中的发生率高于轻症组。
145例重症患者中,51例(34.69%)死亡,90.2%的死亡患者年龄在60岁以上。51例死亡患者中有40例患有基础疾病 (78.43%),大部分还是高血压的患者。
✦死亡原因
COVID-19患者死亡的主要原因是急性呼吸窘迫综合征引起的呼吸衰竭。继发性噬血细胞性淋巴组织细胞增生症 (sHLH) 的特征是伴有多器官衰竭的暴发性和致命性高细胞因子血症,并且未被充分认识。
病毒感染引发sHLH,发生在3.7%–4.3%的成人败血症病例中。sHLH,类似于细胞因子谱,与 COVID-19疾病严重程度相关,其特征是白细胞介素2、白细胞介素7、干扰素诱导蛋白10、粒细胞集落刺激因子、巨噬细胞炎症蛋白1、单核细胞增加趋化蛋白1和肿瘤坏死因子。
COVID-19感染的进展和潜在的辅助干预措施
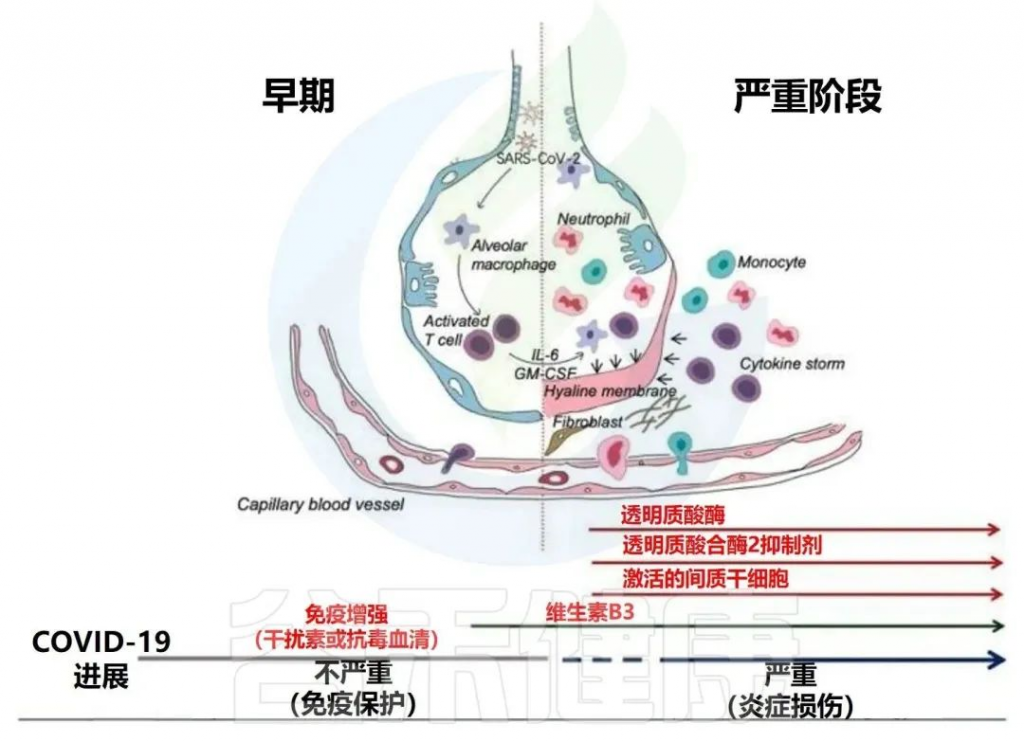

Chowdhury MA,et al.J Infect Public Health.2020
最近一项针对中国武汉150例COVID-19确诊病例的回顾性死亡预测多中心研究包括升高的铁蛋白和白细胞介素6,这表明死亡可能是由于病毒驱动的过度炎症所致。
✦第一次感染时无及时、有效的免疫
免疫的三种类型是先天免疫(快速反应)、适应性免疫(慢反应)和被动免疫。当身体第一次遇到病毒时,免疫系统无法正常运作,就会生病。这种情况就是COVID-19的情况。
注:按照获得方式的不同,可分为天然被动免疫和人工被动免疫。前者是人或动物在天然情况下被动获得的免疫力。例如,母体内的抗体可经胎盘或乳汁传给胎儿,使胎儿获得一定的免疫力。后者是用人工方法给人或动物直接输入免疫物质(如抗毒素、丙种球蛋白、抗菌血清、抗病毒血清)而获得免疫力。
当免疫系统的细胞收到信号时,它们通过在中央和外周淋巴器官之间再循环并通过血液从损伤部位迁移来完成它们的工作。
血液将幼稚和受过训练的免疫细胞从一个部位带到另一个部位,因为它流经全身,并充当免疫系统的管道。细胞再次进入血流,在通过传出淋巴管离开这些节点后被运送到全身组织。
✦病毒刺激免疫细胞进行分化工作
受病毒影响后免疫反应介导抗体。B细胞在T细胞的协助下分化为浆细胞,然后浆细胞产生针对病毒抗原的特异性抗体。中和性抗体能有效完全阻断病毒进入宿主细胞,限制感染,在感染后期起到很强的保护作用,防止感染复发。
相比之下,可以在受感染的细胞内观察到由T淋巴细胞介导的细胞免疫反应。整体适应性免疫反应由辅助性T细胞指导,细胞毒性T细胞在病毒感染细胞的清除和清洁中起着至关重要的作用。

✦免疫过程
•巨噬细胞先进行非特异性吞噬
当病菌再次进入到人体后,免疫系统中的巨噬细胞首先发起进攻,将它们吞噬到“肚子“里,然后通过酶的作用,把他们分解成一个个片断,并将这些微生物的片断显现在巨噬细胞的表面,成为抗原,表示自己已经吞噬过入侵的病菌,并让免疫系统中的T细胞知道。
•激活T细胞免疫
T细胞与巨噬细胞表面的微生物片断,或者说微生物的抗原,连着相遇后如同原配的锁和钥匙一样,马上发生反应。这时,巨噬细胞便会产生出一种淋巴因子的物质,他最大的作用就是激活T细胞。T细胞一旦“醒来”便立即向整个免疫系统发出“警报”,报告有“敌人”入侵的消息。这时,免疫系统会出动一种杀伤性T淋巴细胞,并由它发出专门的B淋巴细胞,最后通过B淋巴细胞产生专一的抗体。
•杀伤性T细胞直接作用,B淋巴细胞产生抗体
杀伤性T淋巴细胞能够找到那些已经被感染的人体细胞,一旦找到之后便像杀手那样将这些受感染的细胞摧毁掉,防止致病微生物的进一步繁殖。
在摧毁受感染的细胞的同时B淋巴细胞产生抗体,与细胞内的致病微生物结合使其失去治病作用。通过以上一系列复杂的过程,免疫系统终于保卫住我们的身体。
注意:当第一次的感染被抑制住以后,免疫系统会把这种致病微生物的所有过程用具的记录下来。如果人体再次受到同样的致病微生物入侵,免疫系统已经清楚地知道该怎样对付他们,并能够很容易、很准确、很迅速的作出反应,将入侵之敌消灭掉。
新冠病毒是RNA病毒,病毒是简单,也是多样的病原体,借助宿主细胞实现自身复制和传播。
病毒的遗传物质可以激活细胞内的PRR,进而引发固有和适应性免疫应答裂解被感染的细胞,也可以诱导 I 型干扰素反应,激活细胞内固有的级联反应限制病毒的复制。
尽管多种细胞可以产生 I 型干扰素,但pDC 是一种在病毒感染早期专门产生大量 I 型干扰素的细胞,它与NK 细胞一起,在适应性反应的的早期病毒宿主防御中发挥核心作用。后者涉及适应性免疫的各个方面:
病毒拮抗宿主免疫防御的策略复杂多样,这与病毒基因的类型有关,RNA病毒缺乏利用校对功能的RNA聚合酶来复制,所以与DNA病毒相比,这类病毒具有较高的突变率且基因组较小,有利于RNA病毒快速改变抗原表位,而抗原表位是适应性免疫靶向位点,是RNA病毒免疫逃逸的机制之一。
另外,一些RNA病毒基因组是分节段的,病毒复制有利于病毒基因组重组。流感病毒是一种常见的季节性病原体,引起急性感染并造成全球爆发,其利用以上2中策略实现免疫逃逸。
建议
佩戴口罩和一些外在防护,以及提前接种新冠疫苗,可以有效地降低病毒的感染率。并且接种疫苗的人群,在感染了病毒后,由于存在特异性免疫,身体会快速有效地做出反应,及时地清除病原体,将对身体的影响降到最低。
为了自己和他人的健康,建议大家外出佩戴口罩,尽量接种疫苗。
人体的免疫功能会受到多种因素影响:包括遗传因素以及很多环境因素,如年龄、温度、运动、压力等。
人体的免疫功能不论是体液免疫还是细胞免疫都与遗传有着密切的关系,由于遗传因素,子代体内抗体T淋巴细胞、吞噬细胞等数量减少或功能降低时,都可以使抗感染能力下降,而引起严重的反复感染,称之为原发性的免疫缺陷病。
主要有以下几种:
一,体液性免疫缺陷病即丙种球蛋白缺乏症。
二,细胞性免疫缺陷病,是由于胸腺发育不良,或缺乏胸腺而引起的体内T淋巴细胞缺乏。
三,联合性免疫缺陷病,也就是T淋巴细胞和B淋巴细胞都缺乏。
四,吞噬细胞缺陷病,是患儿吞噬细胞内缺乏溶菌酶,而不能将吞噬细胞杀死,临床表现为婴幼儿反复发生严重的化脓性感染。
同一个体不同发育阶段(年龄)、营养、健康状况都对免疫有影响。
✦年龄
•刚出生时抵抗力弱
刚出生或出生不久的动物对许多抗原的刺激通常不能激起有效地免疫反应。其原因:一是机体免疫应答的能力差;二是从母体获得的母源抗体。
所谓母源抗体,是指动物通过胎盘,初乳、卵黄等途径从母体获得的抗体。母源抗体方面可保护幼龄儿童免于感染但同时也能抑制或中和相应抗原,使其对机体的刺激强度大为减弱,从而削弱了机体对抗原的反应能力。
•衰老抵抗力变弱
另一方面,随着年龄的增长,身体的各种组织也以不同的方式衰老。其中也包括我们的免疫器官。
抗体由B细胞产生,并且产生途径非常复杂,涉及了许多前体细胞类型。该过程的所有早期步骤都发生在骨髓中,也正是在骨髓中从造血(产生血液)的干细胞产生了前体B细胞。
年龄增长的过程强烈影响了这些早期步骤,减少了前体B细胞的数量,也减少了这些细胞向分泌抗体的成熟B细胞的发育。重要的是,这降低了抗体库的多样性。
由于每个B细胞产生不同的抗体,这本身就像一个数字游戏:拥有的B细胞前体越少,产生能够应对任何感染产生抗体的成熟B细胞的可能性就越小。
✦病理因素
先天性的免疫球蛋白缺陷、自身免疫性疾病、艾滋病(获得性免疫缺陷综合征)、血液系统疾病、感染及慢性病等疾病都会导致患者的免疫功能受到影响。
甲状腺功能亢进症、白血病、肺炎、糖尿病,这些疾病会导致T淋巴细胞等细胞数量减少,使其功能降低,导致免疫功能失调,就会使患者免疫力受到影响。
注:艾滋病病毒是一种能攻击人体免疫系统的病毒。它把人体免疫系统中最重要的CD4T淋巴细胞作为主要攻击目标,大量破坏该细胞,使人体丧失免疫功能。
✦心理及生理
学习或者工作压力大,身体过渡劳累、睡眠不足、过度紧张,会加重植物性神经的负担,内分泌系统紊乱,植物性神经和内分泌系统及免疫系统有着紧密的联系,会带来不良影响,从而造成免疫力下降。
✦饮食对免疫非常重要
饮食具有很重要的影响力,因为有些食物的成分能够协助刺激免疫系统,增强免疫能力。如果缺乏这些重要营养素成分,会严重影响身体的免疫系统机能。
饮食失衡,饮食混乱、进食时间不规律、挑食等,都会使提供人体免疫系统所需的营养不足。
(在后面我们会介绍一些有助于增强免疫力的食物,帮助大家在平时增加免疫力)
✦适量运动增强免疫力
运动能够提高身体的免疫功能,一方面人体通过不断的长期运动,能够促进全身的血液循环,从而促进新陈代谢,包括免疫细胞的更新迅速,比如白细胞,淋巴细胞等,能够使这些免疫细胞的活性比较强。一旦身体有被细菌、病毒等病原体入侵,这些免疫细胞能够快速聚集起来,杀灭这些病原体。
另外在运动过程中,长时间的运动能够提高人体脏器的机能,比如肺功能会提高,心脏功能也会提高,这样会使人体的免疫反应增强,能够使心脏和肺脏的耐受力增强,从而最后提高人体整体的免疫力。
运动不足导致体力下降,体力跟不上就难以抵抗劳累,进而造成免疫力下降。
注:过量的运动使机体疲惫,在一定程度上也会降低免疫力,所以在剧烈运动后要注意自身防护。
免疫细胞监测并响应环境以及各种内源性触发因素,导致其功能改变。人类和动物研究表明,不同的环境温度可以改变细胞和体液方面的免疫反应。
环境温度对小鼠和人类的代谢和免疫影响


Wang H,et al.FEBS J.2022
✦寒冷使活性降低,免疫反应受限
寒冷降低了单核细胞上的主要组织相容性复合物II类(MHCII)并使其活性降低,这反过来又抑制了自身免疫过程中致病性T细胞的启动。
这导致T细胞细胞因子表达减少,从而减轻神经炎症。数据表明,由于小鼠免疫系统的能量可用性降低,资源优先用于产热,导致免疫反应受限。
✦热中性环境下增强了免疫细胞
此外,热中性环境增强了免疫细胞在肿瘤微环境中的渗透。这与不断积累的证据一致,即反复寒冷暴露会抑制小鼠的免疫活动,而温暖会激发更大的抗病毒免疫反应。
有趣的是,处于热中性环境的小鼠在骨髓中积累LyG6+单核细胞,但在循环血液中减少,从而对动脉粥样硬化产生保护作用。
寒冷期间对某些病毒感染的易感性也会增加,免疫的变化可以从一定程度上解释夏天新冠疫情的感染率较低,而到了冬天就比较严重。
抗原的性质对免疫应答的影响是多方面的。例如注射类毒素主要引起体液免疫,而细胞内慢性病毒感染则主要激发细胞免疫;可溶性抗原如类毒素的免疫潜伏期较长,而颗粒性抗原如细菌免疫潜伏期则简短。
✦通过接种疫苗来增强免疫
我们可以通过接种疫苗来增强对某一特殊疾病的预防免疫作用。疫苗的主要目的就是通过疫苗接种所产生的免疫应答,诱导机体产生对特异性抗原的特异性抗体,以便对这种特殊性的疾病,有着积极的预防作用。
注:活苗产生的保护力及免疫应答的持续时间都较死苗优越。
•抗原的次数和剂量都会影响免疫
在一定的限度内,抗体的产量随抗原用量的增加而相应增加,但超过了一定限度,抗体的行程反而受到抑制,这种现象称为“免疫麻痹”。剂量过小,不足以刺激机体产生抗体。
所以在进行疫苗接种时,疫苗的剂量必须严格按照规定,不能随意增减。
仅注射一次疫苗,抗体出现较慢,而且效价低,持续时间段。如果要建立强大的免疫,最好间隔一段时间连续注射2-3次。
注:一般菌苗需要间隔7-10天,类毒素至少间隔6周;注射弱活毒苗,由于活微生物可以在局部适当繁殖,能较长久地在机体内存在,对机体刺激较强,一次注射便可达到目的。
除了上述的因素,越来越多的研究发现肠道菌群对免疫有很大的影响。宿主与微生物相互作用是免疫系统发展的基础。
微生物群在免疫系统中起着基础性作用
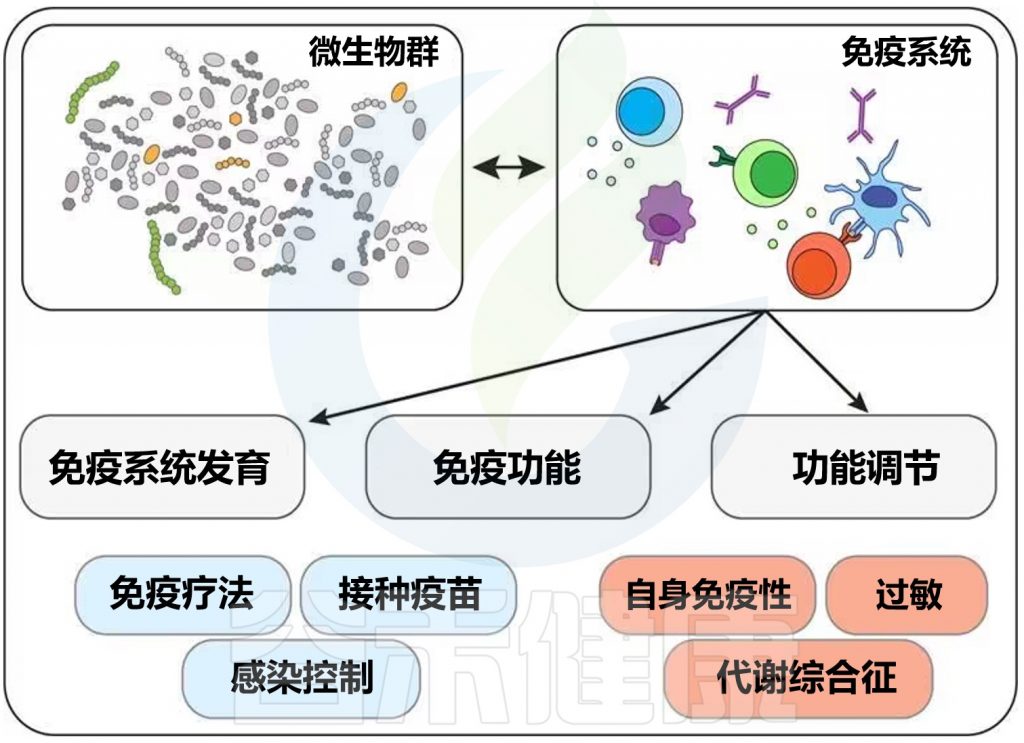

Belkaid Y,et al.Immunity.2017
哺乳动物免疫系统的进化与复杂微生物群的获得同时发生,证明了宿主免疫系统与其共生微生物群之间的共生关系。
这种共生状态的崩溃可能导致慢性炎症性疾病,包括自身免疫、过敏和代谢综合征。相反,微生物群的选择性调节在增强肿瘤免疫治疗、疫苗接种和对抗生素耐药微生物的耐药性方面具有巨大的治疗潜力。
什么是肠道菌群?
人体由大约30万亿个细胞组成,同时还有39万亿个细菌,真菌,病毒共同构成的巨大的微生物群。
这些菌群存在于我们人体的各个部位,有多达2000种不同的种类。其中最多的微生物是聚集在肠道里的细菌,我们称它们为肠道菌群。
肠道菌群并非人体与生俱来的。在母体子宫内,胎儿所处环境几乎是无菌的。在婴儿出生时,其肠道暂时处于无菌状态。
出生三个月后,细菌通过哺乳时口腔摄入、空气吸入等途径进入婴儿体内,并逐渐开始在肠道内定值,兼性厌氧菌首先定值,其后是厌氧菌。随着生长发育和集体功能的不断完善,肠道菌群种类与结构逐渐稳定,进而形成成熟的肠道菌群。在人体中起着至关重要的作用。
✦肠道菌群是肠道屏障的重要组成
肠道固有菌和肠黏膜在肠道内共同构成一道免疫屏障, 阻止细菌、病毒和抗原入侵。
防止产生过度免疫反应的最明显的方法是保持上皮屏障。然而,屏障需要有选择性地渗透,使营养物质进入宿主,有益的微生物与免疫细胞相互作用,以促进成熟和分化。
屏障功能是由复杂的细胞生物学过程维持的,这些过程受到严格的调控。多种微生物信号促进肠道稳态、屏障完整性和免疫成熟,部分是通过先天和适应性免疫信号来实现的。
例如,通过TLR途径激活强直性微生物是维持肠道屏障稳态的必要条件。
✦细菌代谢物影响屏障稳态
然而,影响适应性免疫成熟的共生物质的另一个例子是微生物衍生的短链脂肪酸促进抗炎环境,以诱导辅助性T细胞并促进屏障稳态。适应性调节机制的丧失、屏障功能障碍或先天免疫缺陷都是免疫介导的疾病的发病机制。
肠道屏障宿主-微生物群稳态的破坏增加了对微生物的过度免疫反应的可能性,从而促进炎症性肠病。同样,在这种情况下,对饮食成分的异常反应可能会发生,引发食物过敏和腹腔疾病。
✦影响免疫应答和免疫交流
免疫应答和免疫交流受到肠道菌群组成与数量的影响,在细菌入侵或抗原激活时,机体主要通过上皮内淋巴细胞维持肠道内环境稳定。
研究表明,白细胞介素-22诱导产生的固有淋巴细胞对一直肠道菌群的大面积弥散起重要作用。当健康的小鼠缺乏固有淋巴细胞时,可观察到炎性细胞扩散,同时远端多器官出现炎症,而补充白细胞介素-22后,炎症细胞扩展手限制且炎症消失。
•肠道微生物与免疫之间相互作用
在一项关于肠道菌群免疫应答关系的研究中,研究人员使每个参与者的免疫细胞受三种细菌刺激物刺激(共生细菌艰难梭菌、常见病原体金黄色葡萄球菌、大肠杆菌),进而分析了500名参与者的血液和粪便样本,通过分析病原体免疫应答的个体差异,肠道差异及两因素的相互影响,发现了肠道微生物群体及其功能与免疫应答之间的相互作用。
✦肠道菌群调节宿主免疫稳态
肠道菌群免疫保护作用的证据来自最近的研究,表明抗真菌药物对肠道真菌的靶向扰动对宿主免疫和健康产生持久影响。
巨噬细胞和单核细胞对真菌细胞壁成分(如β-葡聚糖和几丁质)具有记忆特性,可以保护小鼠免受真菌的二次攻击。
✦影响细菌和真菌病原体的感染
通过用白色念珠菌(Candida albicans)的人血液分离物在肠道定植,可以在小鼠体内诱导训练有素的免疫力通过抗生素治疗小鼠的胃肠道 。
该免疫保护机制效果持续时间短,依赖于白细胞介素6,并且可以在T细胞和B细胞缺陷小鼠中重现。然而,在类似的模型中,人类血液白色念珠菌分离株也可以防止随后的全身性白色念珠菌感染,这主要是由于诱导了Th17适应性免疫反应。
除了防止系统性真菌感染外,白色念珠菌在肠道中的定植还可以防止系统性金黄色葡萄球菌(Staphylococcus aureus)感染;这种保护是否取决于经过训练的免疫力或适应性念珠菌特异性Th17细胞,或两者的结合,仍然未知。
这些结果表明,经典的适应性免疫反应和先天免疫记忆都可以由肠道中的白色念珠菌(Candida albicans)引发,并表明菌株特异性特征可能会告知宿主免疫反应。
先天性和适应性免疫机制可能会协同维持体内平衡。用白色念珠菌或酵母菌对无菌小鼠进行单一定植,促进肠道稳态的建立。
✦益生菌促进免疫反应
共生细菌促进效应和调节性T细胞反应
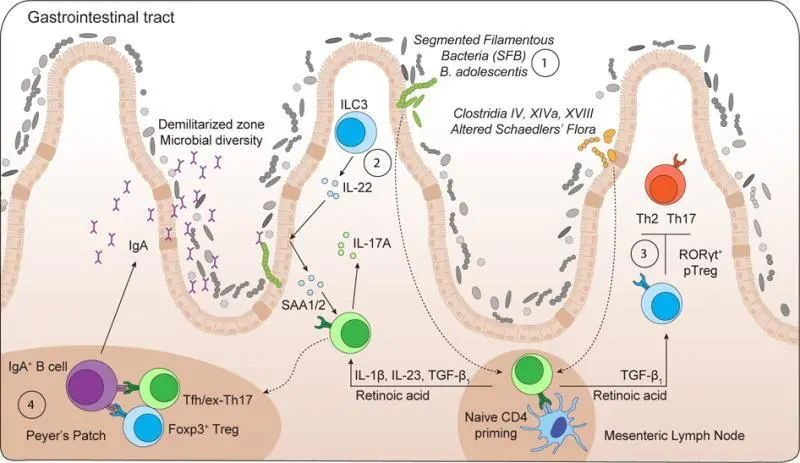
Belkaid Y,et al.Immunity.2017
除了微生物配体或代谢物在免疫系统功能中的多效性之外,现在越来越清楚的是,特定的微生物或细菌群可以在稳定状态下对免疫系统产生显著影响。
在生态系统中,那些具有最重要影响的生物被称为“关键物种”。分段丝状细菌 (SFB) 代表了胃肠道中关键物种的原型。这种形成孢子的革兰氏阳性厌氧菌定殖在小鼠回肠末端,通过促进小肠中Th17和Th1细胞的积累并驱动IgA的产生,对粘膜免疫系统产生显著影响。
•抵抗胃肠道病原体
宿主产生强大的分段丝状细菌特异性反应,这种现象与其促进抵抗胃肠道病原体的能力有关。
分段丝状细菌通过与淋巴结和上皮细胞的紧密粘附与粘膜组织密切相互作用,在接触部位诱导这些细胞的细胞骨架重组。这种与上皮细胞的密切接触是少数共生生物共有的特性,据信可以解释特定微生物提高组织免疫力的能力。
•青春双歧杆菌促进Th17细胞积累
最近的一项研究发现青春双歧杆菌(Bifidobacterium adolescentis)是一种人类共生体,也是一种可以促进Th17细胞在单克隆化小鼠肠道中积累的微生物。
值得注意的是,青春期双歧杆菌引发了一个不同于分段丝状细菌的转录程序,支持Th17细胞(淋巴细胞的一个子集,对屏障位点的稳态至关重要)的诱导可以通过不同或重叠的途径发生。
•皮肤微生物影响T细胞积累
皮肤微生物群还控制产生白细胞介素17的T细胞的积累。特定的皮肤微生物在调节适应性免疫系统不同分支的能力方面表现出高度专业化。
用确定的表皮葡萄球菌(Staphylococcus epidermidis)分离株定植小鼠,表皮葡萄球菌特异性T细胞通过其产生白细胞介素17的能力促进角质形成细胞产生AMP,从而促进针对真菌感染的异源保护。
肠杆菌(Enterobacteriaceae)定植可能有助于酿酒酵母对硫酸葡聚糖(DSS)诱导的结肠炎的保护作用,表明预先存在的肠道微生物群(真菌和细菌)可能进一步调节肠道炎症和保护性免疫的结果。
注:这些发现如何适用于不断暴露于真菌和细菌抗原的人类,仍不清楚。
小结
大量研究表明益生菌能够刺激人类分泌型免疫球蛋白A细胞增殖, 阻止肠道微生物及其毒素分子对肠黏膜的攻击。
益生菌对机体细胞免疫的影响, 主要包括激活巨噬细胞、B淋巴细胞和自然杀伤细胞, 促进白细胞介素和干扰素等细胞因子的产生。增强我们体内的免疫应答。
微生物在免疫疾病发病机制中的作用

Ruff W E,et al.Nature Reviews Microbiology.2020
✦肠相关疾病
•鸡肠球菌会导致多种免疫炎症
鸡肠球菌向肝脏的转移促进了自身免疫性肝炎,并与肺炎克雷伯菌和奇异变形杆菌一起,通过包括TH17细胞分化在内的多种机制导致原发性硬化性胆管炎。鸡肠球菌诱导的肝芳香烃受体(AhR)和内源性逆转录病毒(ERV)也参与了小鼠狼疮性肾炎的系统性抗ERV反应。
Leptotrichia goodfellowii能与小鼠Ⅰ型糖尿病自身抗原胰岛特异性葡萄糖-6-磷酸酶催化亚单位相关蛋白(IGRP)发生交叉反应。在小鼠Ⅰ型糖尿病中,未定义的微生物群易位到胰腺淋巴结并激活NOD2,而来自微生物群的NOD1配体的易位驱动自身免疫性胰腺炎。
NOD2作为一种胞内的模式识别受体在机体免疫系统抵抗胞内菌感染过程中起重要作用。
✦非肠相关疾病
微生物群对非肠道自身免疫性疾病的影响

Ruff W E,et al.Nature Reviews Microbiology.2020
鸡肠球菌影响多种免疫疾病
鸡肠球菌和乳杆菌(Lactobacillus)在系统性红斑狼疮模型中向肠系膜淋巴结、肝脏和脾脏转移,从而激活先天性(浆细胞样树突状细胞)途径。鸡肠球菌还刺激狼疮患者滤泡辅助因子和自身抗体的产生。
抗磷脂综合征是一种凝血性自身免疫性疾病,鸡肠球菌传染性支气管炎也加重了这种疾病,它诱导肝脏中的自身抗原β2GPI。
瘤胃球菌加重狼疮性肾炎
瘤胃球菌在狼疮性肾炎时扩张,可与狼疮双链DNA发生交叉反应。
粘附性侵袭性大肠杆菌可导致克罗恩病相关性脊柱炎。在关节炎模型中,分段丝状细菌还使CD4+T细胞向TFH细胞倾斜,并通过携带双TCRs的TH17细胞在同一模型中引发间质性肺病。
其他细菌
Ro60直系表达细菌在狼疮患者中引起Ro60自身免疫反应;肠道中的类泰奥托米克龙杆菌、口腔或肠道中的大量放线菌(Actinomycetes)和阿米巴棒状杆菌,除了狼疮外,还可能导致干燥综合征,丙酸杆菌与亚急性皮肤狼疮皮损通过交叉反应。
✦免疫特异性疾病
嗜黏蛋白阿克曼菌(Akkermansia muciniphila)在多发性硬化症中增强TH1反应,并且未定义的微生物群与GDP-l-岩藻糖合酶(一种神经炎症疾病的自身抗原)交叉反应。
未定义的小鼠肠道微生物群与TH17细胞交叉反应,识别自身免疫性葡萄膜炎的自身抗原——视黄酸结合蛋白(IRBP)。
重要的是,微生物失调本身通过各种机制引起失调的免疫反应,包括辅助性T细胞倾斜、旁观激活、表位扩散、交叉反应和双T细胞受体(TCR)刺激,它们分别作用于疾病的易感性、引发和传播。
✦新冠患者肠道微生物改变
病毒感染可以改变胃肠道微生物群。一份报告确认了COVID-19患者粪便样本中乳酸杆菌和双歧杆菌(Bifidobacterium)的减少。同样,使用抗生素会干扰COVID-19患者的微生物组。
在新冠病毒清除后的30天内,肠道微生物组的改变依然存在,主要体现在以下菌属的减少:
普拉梭菌(Faecalibacterium Prausnitzii) ↓↓↓
直肠真杆菌(Eubacterium rectale) ↓↓↓
双歧杆菌(Bifidobacterium) ↓↓↓
✦微生物的变化导致免疫变化
某些微生物的变化可以增加对先天性免疫受体的刺激,例如核苷酸结合寡聚化域样受体和Toll样受体。该受体的刺激触发几个促炎症信号和细胞因子和趋化因子的产生,它们调节适应性免疫系统,影响局部和全身免疫反应。

✦肠外接种卡介苗增强肺部免疫力
最新的一项研究发现:肠外接种卡介苗,通过分枝杆菌(Mycobacterium)传播,引起肠道微生物组、屏障功能和微生物代谢物的时间依赖性改变,以及随后循环和肺代谢物的变化。
这些变化导致记忆巨噬细胞的诱导和肺部免疫力的增强。
注:这些数据确定了远端粘膜组织先天免疫记忆发育的肠道微生物群介导途径,并对开发针对呼吸道病原体的下一代疫苗策略具有重要意义。
小结
总而言之,这些研究表明,平衡的肠道菌群有助于维持宿主免疫稳态,而微生物失调本身通过各种机制引起失调的免疫反应。
可以说肠道菌群在促进免疫系统发育,维持正常免疫功能,协同拮抗病原菌入侵方面,发挥着重要作用。
既然免疫功能对我们如此重要,我们应该如何拥有一个健康的免疫系统来保卫我们的身体呢?谷禾在这里列举了一些可以提高免疫力的方法。
疫苗是将病原微生物,比如细菌、病毒等、以及细菌病毒的代谢产物经过人工减毒、灭活或利用转基因等方法,制成的用于预防传染病的自动免疫制剂。
预防传染病的疫苗已经彻底改变了医疗保健,延长了数百万人的生命。疫苗方法是安全、持久和有效的。鉴于致病菌与传染源具有相似的免疫靶点,这种方法可能对免疫介导的微生物感染性疾病也有用。
✦接种疫苗后免疫反应迅速高效
疫苗的作用是使身体产生专门对外部病毒、细菌或其他微生物的进行特异性免疫。通常,疫苗通过激活患者的免疫系统产生相应的抗体,在人体免疫系统进行特异性记忆,如果有同一种病毒或细菌攻击身体,免疫系统可以迅速反应,控制病毒、细菌或其他致病微生物。能有效的起到预防疾病、增强免疫力等作用。
疫苗在保护人类健康方面的影响是巨大的,疫苗使人类在面对传染病的威胁时,首次化被动为主动,在降低死亡率和提高人均预期寿命方面发挥了不可替代的作用。
在这里建议还未接种新冠疫苗的人们尽量都去接种,可以有效地保护我们的身体。
饮食和营养可以直接影响免疫系统或通过肠道菌群介导宿主免疫。饮食还可以加强肠道屏障,从而改善微生物群和宿主之间的界面。
✦营养不均衡导致免疫力下降
要吃出健康,进而增强身体的免疫力,最重要的就是营养充足及均衡,任何一种营养的缺乏都会导致经常性或长期性的疾病。
摄入不足也是不可取的,当人感觉到饥饿时,身体会分泌肾上腺素;如果体重每周减轻850g以上,抵御疾病和外来病毒的T细胞就会受到抑制。
专家建议,在搭配膳食时,要食用一些含不饱和脂肪酸的食物,摄入足够的蛋白质,多吃水果和蔬菜,多喝果汁和开水。而号称能提高免疫力的保健品作用并不明显。


✦维生素有助于增强免疫力
每天适当补充维生素和矿物质。专家指出,身体抵抗外来侵害的武器,包括干扰素及各类免疫细胞的数量与活力都和维生素与矿物质有关。例如维生素B、C、E等微量营养素,可以保护免疫细胞免受自由基的氧化损伤,维护上皮细胞组织,增强免疫力。
蔬菜、水果类
身体健康的人最好通过吃水果、蔬菜来提高免疫力,现在市场上销量较大的西红柿、草莓、胡萝卜、橘橙等都含有大量的维生素。
柑橘类水果:柑橘类水果富含维生素 C,能够促进免疫系统的发育和激活,例如:橘子、橙子、西柚、葡萄柚等。
浆果莓果:包括蓝莓、草莓、树莓、黑莓、蔓越莓、巴西莓、樱桃等,都是具备抗氧化,抗炎等功效的食物。
其他富含维生素或抗炎类的水果,例如:苹果、猕猴桃、番石榴(红心芭乐)、牛油果、葡萄、西瓜、哈密瓜、香蕉、杏、梨、芒果、山楂、桃子等。
蔬菜类食物:蔬菜类食物富含多种微量元素和维生素,例如:羽衣甘蓝、甜菜、菠菜、花菜、卷心菜、西兰花、芝麻菜、芥菜、芦笋、甜椒、抱子甘蓝、萝卜、香菜、甜叶菊、生菜、萝卜、番茄等都是对提高免疫力非常不错的食物。
豆类食物:豆类食物富含蛋白质和多种维生素,能够促进免疫系统的发育和激活,例如:鹰嘴豆、黑豆、扁豆等。
全谷类食物:全谷类食物富含纤维素和多种微量元素,能够提高免疫系统的活性。
鱼虾类食物:鱼虾类食物富含蛋白质和不饱和脂肪酸,能够促进免疫系统的发育和激活。例如:三文鱼、沙丁鱼、金枪鱼、鳟鱼、鲭鱼、鲱鱼等。
注意:有些鱼含有汞和其他污染物,孕妇和婴儿遵医嘱。
大蒜、洋葱
大蒜和洋葱都是热性食物,对改善体质有良好的作用。大蒜具有杀菌杀毒功能,吃大蒜最好生食,因为生蒜具有抗病毒、提高机体免疫力的作用。大蒜中所含的具有增强免疫力功能的有效成分大蒜素,在加热的过程中会失去功效。洋葱也是一种天然的杀菌杀毒食物,可以有效地抵抗病毒和细菌。
含锌食物
锌是人体不可缺少的微量元素,人体中许多种酶必须有锌参与才能发挥作用,锌对调节免疫功能十分重要。此外,它还有另一个功能,就是抗感染。每天摄入50~100毫克的锌,就可以预防流感。海产品、瘦肉、粗粮和豆类食品都富含锌。
食用菌类
蘑菇、猴头菇、草菇、黑木耳、银耳、百合等:都有明显增强免疫力的作用;
香菇所含的香菇多糖能增强人体免疫力。
鸡汤
喝鸡汤能够预防感冒和流感等上呼吸道感染性疾病。鸡肉中含有人体所必需的多种氨基酸,营养丰富,特别是其中所含的的半胱氨酸,可以增强机体的免疫力。此外,喝鸡汤对感染后加速痊愈也有积极作用。
姜黄
姜黄素是姜黄中的活性成分,研究表明它可以帮助对抗炎症、稳定血糖水平,甚至可以防止癌细胞生长。
生姜
生姜具有强大的药用价值,含有维生素C、镁和钾,与姜黄有着密切的关系。它还具有强大的抗炎和抗氧化特性。
蜂产品
蜂王浆:能提高机体免疫力及内分泌的调节能力,并含具有防癌作用的蜂乳酸(10-HDA);
蜂胶:蜂胶是蜜蜂采集胶源植物新生腋芽分泌物和蜜蜂自身分泌物(如蜂蜡)混合而成的天然产物,含有最为丰富的黄酮类化合物及其它生物活性成分。药理及临床证明,蜂胶能有效提高机体免疫功能,克服癌症的神奇功效。另外,长期服用蜂胶可减少成人慢性病的发生,对糖尿病、心脑血管病有改善的功效。
总之,多吃上述食物可以有效增强免疫力,帮助人体更好地抵抗外来物质的侵害。建议在饮食中搭配多种食物,以获得更多的营养,提高免疫力。
烹饪方式
相对健康的方式一般认为是:少糖、少盐、少人工调味料,运用各式香草、坚果、特级初榨橄榄油、适量海盐来调味。
总之,把握食材新鲜、多样的原则,简单清爽保留食物原味,避免过度烹调。
更多详见:
✦饮食对肠道菌群的影响也很大
在健康个体中,肠道微生物群的组成很容易因饮食而改变。有趣的是,饮食不仅会迅速影响人类肠道菌群,还会影响肠道病毒,这也可能有助于研究饮食如何影响免疫介导的疾病。
总体而言,特定宿主中饮食与微生物群落相互作用的复杂性和个体间差异使得为免疫介导的疾病制定特定饮食方案具有挑战性。然而,饮食调整仍然是一种提高免疫力的方法。
✦免疫需要充足的睡眠
睡眠与人体免疫力密切相关。良好的睡眠可使体内的两种淋巴细胞数量明显上升。睡眠时人体会产生一种称为胞壁酸的睡眠因子,此因子促使白血球增多,巨噬细胞活跃,肝脏解毒功能增强,从而将侵入的细菌和病毒消灭。
在黑暗中人体才会产生抵御疾病褪黑激素。睡眠不足或晚上长时间处于灯光下,都会减少褪黑激素的释放量,同时使雌激素的分泌增加,这样就很容易患乳腺癌。

✦适量的运动增强免疫
有规律、长期的适度锻炼能够增强身体的抵抗力。
研究显示,每天运动30到45分钟,每周5天,持续12周后,免疫细胞数目会增加,抵抗力也相对增加。
体育锻炼不仅有益于心血管,对改善情绪、减轻压力也很有好处。然而,活动强度要适当,如果感觉到吃力,超出了平常的承受能力,就要及时调整。
✦压力大抑制免疫作用
压力会使人体分泌类固醇激素和肾上腺素,这些荷尔蒙对人体免疫系统有抑制作用。这就很好地解释了为什么当人们处于压力状态下,如面临期末考试、情感出现危机时特别容易感冒。
噪音不仅伤害人们的听力,还会造成肌肉紧张、心跳加速、血管收缩和消化系统的不适———这和受到惊吓或感到压力时身体的反应是一样的。人如果长期处于噪音之中会导致血压、胆固醇水平和免疫功能的不良变化。
与朋友在一起时,由于感受到友情和社会认同,人的免疫系统会处于良好的状态。
为了证明此观点,巴黎身心健康中心的研究人员让参加实验的人与感冒病毒接触,然后由研究人员跟踪监测他们的社会交往情况。结果发现,和朋友、家人、同事相处时间越长的人感冒的几率越小。同时,拥抱、抚摸等身体接触也能使人体的免疫细胞更加活跃。
建议
免疫系统会受到思想和感觉的暗示,所以,我们应该保持积极向上的心态。最近的研究中也发现,消极、情绪低落、容易感觉到压力的人对肝炎病毒的抵抗力比乐观的同龄人弱得多。
注:美国的研究人员还发现,年轻时乐观的人比悲观的人多活12年。
考虑到宿主与微生物群相互作用对免疫的影响,针对微生物群的疗法在一定程度上也可以增强免疫能力。
✦益生菌、益生元疗法
益生菌或益生元或合生元、后生元(如短链脂肪酸)等单独或联合使用可能有助于增强宿主免疫,更好地控制感染,并随着我们年龄的增长产生适当的免疫反应。
益生菌产生代谢物可影响共生微生物群,肠屏障,免疫系统。益生菌还可以帮助预防或治疗细菌和病毒感染。益生菌的使用可以提高感染流感病毒的小鼠的存活率。除流感病毒外,研究还显示出对呼吸道合胞病毒感染的有益保护作用。
扩展阅读:
✦噬菌体
病毒以噬菌体为主导,而噬菌体与微生物群的细菌成员共同进化。天然噬菌体和合成噬菌体都可以代表一种针对性强的方法,与抗药性病原体类似,可以消灭免疫介导疾病中的致病菌。
例如,针对参与炎症性肠病发病机制的肺炎克雷伯菌菌株的噬菌体目前正被生物技术公司评估用于干预。
最近在动物模型中证明了在酒精性肝病中成功地将噬菌体靶向肠球菌。该方法对于其他与免疫介导的疾病有关的肠球菌属也是可行的,特别适合那些易患免疫介导疾病并被“关键病理生物”定植的个体。
还可以构想组合噬菌体疗法,以解决涉及免疫介导疾病的几种不同菌株,例如三个物种协同作用:肺炎克雷伯氏菌,变形杆菌和鸡肠球菌。
注:与疫苗接种方法类似,噬菌体疗法可能仍然存在理论上的风险,即在不同的情况下去除菌株可能是有益的。然而,这两种方法都比传统的抗生素更有针对性,会导致多种脱靶效应。
小结
鉴于肠道微生物对宿主免疫系统的影响越来越大,可以合理推测,通过个性化营养或补充,恢复相关的肠道微生物丰富度和功能,可能是一种对抗免疫适应性功能下降的预防措施。
免疫在身体中发挥着重要的作用。身体免疫力提高后能够阻断病毒的入侵,减少疾病发生,即使在生病以后恢复时间也比较快。
肠道微生物群是一个有待挖掘的宝藏,尤其在抵抗病原体包括病毒等方面。正如许多临床前研究所证明的那样,保持强壮丰富健康的微生物群可以通过维持免疫和营养需求等来提高宿主的活力和生存力。
注:本账号内容仅作交流参考,不作为诊断及医疗依据。
主要参考文献:
Yeoh YK, Zuo T, Lui GC, Zhang F, Liu Q, Li AY, Chung AC, Cheung CP, Tso EY, Fung KS, Chan V, Ling L, Joynt G, Hui DS, Chow KM, Ng SSS, Li TC, Ng RW, Yip TC, Wong GL, Chan FK, Wong CK, Chan PK, Ng SC. Gut microbiota composition reflects disease severity and dysfunctional immune responses in patients with COVID-19. Gut. 2021 Apr;70(4):698-706. doi: 10.1136/gutjnl-2020-323020. Epub 2021 Jan 11. PMID: 33431578; PMCID: PMC7804842.
Ruff W E, Greiling T M, Kriegel M A. Host–microbiota interactions in immune-mediated diseases[J]. Nature Reviews Microbiology, 2020: 1-18.
Chowdhury MA, Hossain N, Kashem MA, Shahid MA, Alam A. Immune response in COVID-19: A review. J Infect Public Health. 2020 Nov;13(11):1619-1629. doi: 10.1016/j.jiph.2020.07.001. Epub 2020 Jul 14. PMID: 32718895; PMCID: PMC7359800.
Li XV, Leonardi I, Iliev ID. Gut Mycobiota in Immunity and Inflammatory Disease. Immunity. 2019 Jun 18;50(6):1365-1379.doi:10.1016/j.immuni.2019.05.023. PMID: 31216461; PMCID: PMC6585451.
Belkaid Y, Harrison OJ. Homeostatic Immunity and the Microbiota. Immunity. 2017 Apr 18;46(4):562-576. doi: 10.1016/j.immuni.2017.04.008. PMID: 28423337; PMCID: PMC5604871.
Kenneth Murphy,Casey Weaver 著,周洪 译。詹韦免疫生物学(原书第九版),科学出版社,2022.
Allert S, Forster TM, Svensson CM, Richardson JP, Pawlik T, Hebecker B, Rudolphi S, Juraschitz M, Schaller M, Blagojevic M, et al.(2018). Candida albicans-Induced Epithelial Damage Mediates Translocation through Intestinal Barriers. mBio 9.

谷禾健康
虽然每个人都会老去,但是衰老的速度却不一定相同。生理年龄是每个人的遗传,环境和生活方式的影响。
人的一生,从出生到成年到老年,微生物组在免疫系统成熟,功能和调节中起着基本作用。免疫系统和微生物组形成一种互惠关系。
随着年龄的增长,免疫系统和肠道微生物组的组成和功能都会发生重大变化,这与对传染病的易感性增加有关。
导读
01)免疫衰老的一般标志
02)衰老和肠道微生物组
03)微生物老化的衡量指标
04)微生物老化加快免疫衰老和虚弱
05)肠道微生物组对疫苗反应的影响
06)对抗与年龄相关宿主免疫力下降的营养策略
07)恢复肠道微生物稳态、减少炎症和免疫衰老
08)健康长寿的普适方式
免疫衰老
“免疫衰老”是指在老年人中观察到的功能障碍,免疫反应有缺陷或异常。
与年龄相关的免疫反应的质量和数量的变化导致触发有效抗体和细胞反应抵御感染和疫苗的能力逐渐下降。
衰老的T淋巴细胞生物学研究十分突出,但所有造血源性细胞都显示出衰老的特征,包括功能失调的抗体产生B细胞、抗原呈递细胞、自然杀伤细胞和中性粒细胞。最早的造血祖细胞中也报告了表观遗传学改变,这可能解释了观察到的年龄相关的髓样细胞偏斜。
主要淋巴器官的老化(骨髓和胸腺磨损)、慢性抗原过载(如CMV)、肠道失调或炎症是免疫衰老的驱动因素,这些因素与遗传缺陷、细胞应激和/或细胞衰竭的累积一起,会导致免疫适应度随着年龄的增长而下降。
炎症
炎症是一种高度控制的生理过程,对对抗病原体、清除碎屑和愈合损伤至关重要。
随着年龄的增长,由于基因、环境和生活方式因素的复杂和不断变化的相互作用,促炎和抗炎之间的动态平衡下降。
慢性炎症状态是发病和死亡的一个重要危险因素。包括慢性感染、缺乏运动、内脏肥胖、饮食、心理压力、睡眠不足或肠道失调等多种因素都会引发和维持炎症。
长期暴露于应激源会加速细胞衰老和先天免疫失调,这是炎症的一个主要特征,反映在局部和全身炎症介质与白细胞介素-6(IL-6)、肿瘤坏死因子TNF-α 水平持续升高,IL-1β,C-反应蛋白(CRP)在老年人中普遍存在。
尽管先天性单核-巨噬细胞网络的失调可能是炎症的中心,但新的证据表明衰老细胞(包括T细胞和B细胞)通过其衰老相关的分泌表型参与慢性低度炎症的关键作用。
而慢性表达或暴露于炎症刺激可能使预先激活的免疫细胞对进一步的刺激难以耐受,从而导致观察到的老年人感染性疾病的频率和严重程度增加。
因此,先前存在的炎症已被证明是疫苗反应性的一个重要决定因素。
此外,炎症可能通过助长年龄相关疾病(包括代谢综合征、心血管疾病、肌细胞减少症、癌症和神经退行性疾病)而产生多种健康后果,因为大多数(如果不是所有)年龄相关疾病都具有炎症特征。
尽管衰老、炎症和慢性病之间存在着共同性,但生物医学研究继续巨额花费来单独解决这些疾病状态。这就提出了一个具有挑战性的问题,即:
针对慢性低度炎症或引起炎症的机制是否可能减缓衰老及其相关疾病?
虽然临床前研究表明炎症是导致年龄相关疾病和免疫反应性降低的一个因素,但证实这一假设的人类数据在很大程度上是缺失的。
而最近一项CANTOS的研究(Canakinumab Anti-inflammatory Thrombosis Outcomes Study)为老年人带来了巨大的希望。
对10000多名既往有心肌梗死史的稳定患者进行卡那单抗(Canakinumab,一种针对白细胞介素-1β的人单克隆抗体)治疗,可显著降低参与者的全身低度炎症。
重要的是,治疗方案可以预防复发性血管事件和肺癌的发生。虽然这些研究唤起了人们对有效抗衰老疗法的希望,但通过靶向阻断关键炎症介质来减少全身炎症需要在对传染病的易感性方面谨慎平衡。
这些免疫指标包括基于炎症年龄评分的iAGE、依赖于免疫球蛋白糖基化的聚糖生长或南丁格尔健康指数,其中糖蛋白乙酰化的全身炎症预测呼吸道感染、心血管疾病和全因死亡率的长期风险。
随着老龄化社会的健康管理成为一个日益增长的经济负担,这种免疫指标代表了有希望的工具,以确定风险个人的早期药物或营养干预。
为了应对不断变化的环境,肠道微生物通过菌种组成和代谢功能的变化动态响应。这个过程受到宿主免疫系统的严格调控,想象宿主免疫系统是一位建筑师,通过允许共生细菌生长和占据粘膜生态位,同时选择性地消除或中和有害微生物,从而塑造肠道微生物群。
随着年龄的增长,免疫适应度逐渐下降,宿主与微生物动态信号交换的监测受到损害,从而对宿主健康和免疫造成广泛的功能后果(图1)。
图1 模型:微生物老化及其对宿主免疫的相关影响
Nabil Bosco & Mario Noti,Genes & Immunity, 2021
(1) 年龄相关的肠道微生物群落变化和相关的肠道组织功能下降可能加剧炎症
(2) 由全身低度炎症引起的慢性免疫刺激、代谢组和微生物刺激的变化导致免疫衰老
(3) 免疫功能受损(如胸腺退化、造血功能改变),导致老年人感染风险增加,老年人接种疫苗反应差
在人类中,年龄相关的菌群失调(微生物老化)特征是梭状芽孢杆菌和双歧杆菌数量减少,变形菌门以及肠杆菌科等病原菌数量过多。
肠道微生物群落结构的组装也可能取决于宿主器官的功能。考虑到肠道的组织功能和完整性受到与年龄相关的显著变化,如再生能力、上皮屏障形成、黏液层组成和蠕动的改变,随着我们年龄的增长,粘膜生态位的变化很可能导致了失调状态。
这种肠道完整性的亚临床改变可能会促进微生物向全身部位渗出,全身性低度炎症,慢性炎症的发生和过早死亡。然而,确定人类的因果关系仍然具有挑战性,下文将进行讨论。
虽然,我们已经开始了解细菌的分类组成和多样性是如何随着年龄的变化而变化的,但我们对细菌进化和与宿主适应性相关的功能后果的了解仍然有限。有两种可能的情况:
1 肠道微生物群落结构中与年龄相关的变化是宿主衰老过程中发生的生理组织适应的简单结果
2 与年龄相关的失调是细菌进化的产物,通过使特定菌群逃避免疫监视,直接触发宿主衰老
因此,更好地了解老化的宿主微生物群,对于推进以微生物组为基础的治疗方法来对抗老化和与年龄相关的疾病至关重要。
我们需要先了解这样的概念。
生理年龄:指人达到某一时序年龄时生理和其功能所反映出来的水平,是从医学、生物学角度来衡量的。
肠道年龄:是基于健康人群每个年龄段样本的菌群构成使用深度神经网络模型提取特征菌属并构建的预测模型。
谷禾健康-肠道年龄预测模型图

注: 谷禾报告中的肠道年龄是基于超过6万人群队列的深度学习模型构建的,伴随年龄的变化肠道菌群也会相应的改变。对于0~2岁的儿童,肠道年龄通常偏差小于3个月,3~5岁偏差在6个月以内,6~15岁偏差在1岁左右,16~50岁人群正常肠道年龄的偏差在3岁以内,50岁以上正常偏差在5岁以内。在正常偏差内年龄差异可以理解为年轻或衰老,超出正常偏差的年龄无论是超过或低于都可能是菌群异常或健康状况不佳。
真实年龄与肠道预测年龄在范围内的差异可以反映其肠道菌群的发育和衰老状况。
以下情况可能会导致肠道预测年龄完全偏离真实年龄,包括:肠道菌群紊乱,菌群结构过于单一,近期服用包括抗生素等可能严重干扰肠道菌群的药物,病原菌感染或者处于疾病状态。
肠道菌群失调可以触发先天性免疫反应和慢性低度炎症,导致许多与年龄有关的退化性疾病和不健康的衰老。肠道菌群通过各种生物分子,营养素信号独立途径和表观遗传机制与宿主进行交流。与年龄有关的肠道菌群失调会干扰这些交流,从而影响宿主的健康和寿命。
与年龄相关的肠道微生物组成的改变不仅发生在人类身上,也发生在实验室模型生物身上。这些控制遗传、年龄、饮食和微生物组本身的模型系统提供了强有力的科学证据,表明宿主生理或微生物进化的内在改变足以促进一种失调状态。
这些模式生物已经证明肠道微生物群有潜力有益地调节衰老过程,以促进宿主的健康和寿命。
黑腹果蝇
对黑腹果蝇的研究(一个经常被用来研究微生物动力学、肠道生理中与年龄相关的变化和生物体健康之间相互作用的衰老模型)——已经完美地证明了肠道生态失调不仅是年龄相关肠道屏障功能障碍、系统免疫激活和机体死亡的前兆,而且还预测了这些疾病的发生。
结果表明,在整个生命过程中保持在无菌条件下的果蝇显示出较低的衰老率,这表明在这些模型设置中防止与年龄相关的菌群失调可以限制炎症,改善免疫稳态,促进健康。
非洲绿松石鳉鱼
利用自然短命的脊椎动物非洲绿松石鳉鱼(Nothobranchius furzeri)进一步证实了肠道微生物群在调节衰老过程中的影响。年轻的微生物群的慢性定植在中年鱼,诱导了长期有益的系统效应,导致脊椎动物的寿命延长。将年轻的供体微生物群定植在年老的鱼,与已知产生代谢产物的关键细菌属的存在有关,这些代谢产物既能维持免疫系统健康,又有抗炎作用。
小鼠
年轻小鼠菌群 → 早衰小鼠:特定菌发挥作用
将年轻供体的粪便微生物群移植到早衰小鼠中,可改善后者的健康状况和寿命,并将疣微菌门verrucomicrobia,Akkermansia muciniphila移植到早衰小鼠中,从而充分发挥有益作用。
此外,与年龄相关的Akkermansia muciniphila缺失与肠道完整性受损和胰岛素抵抗相关——这是一个通过微生物组-单核细胞-B细胞轴介导的过程。
除了对宿主代谢的影响,Akkermansia muciniphila还参与了调节抗原特异性T细胞反应和抗体产生来调节宿主免疫功能。
小鼠菌群 → 无菌小鼠:T细胞活化增加
用年轻或年老小鼠的肠道微生物群定植无菌(GF)小鼠。将老年供体微生物组移植给年轻小鼠足以促进肠道炎症、微生物产物向循环的渗漏和慢性低级别炎症的发生。作为全身低度炎症的结果,全身免疫区的T细胞活化增加。
年老菌群→年轻小鼠:巨噬细胞功能失调
另一项研究报告了类似的发现,在年轻的GF接受者中,移植一种老年微生物组可促进全身低度炎症。这些情况下的炎症进一步与巨噬细胞功能失调相关,巨噬细胞是炎症细胞因子的有力来源,其细菌杀灭活性较差。此外,基因或抗体介导的肿瘤坏死因子α(低度炎症的标志性细胞因子)的减少,可以预防年龄相关的肠道失调和相关的全身低度炎症。
年龄相关肠道免疫力下降:增加M细胞可恢复
最近的一项研究强调,年龄相关肠道免疫力下降可以通过操纵肠道微生物组增加M细胞数量来恢复。年老小鼠暴露于年轻的微生物群或鞭毛蛋白刺激都足以观察到这种效应:Peyer’s补丁中的M细胞成熟恢复,增强抗原摄取,年老小鼠肠道IgA反应增加。基于微生物组干预的M细胞修复确实依赖于肠道干细胞功能的改善,这表明修复老年肠道的再生能力可能对提高肠道免疫力有额外的好处。这些发现可能与改善口服疫苗应答或预防老年人胃肠道感染有关。
年龄相关肠道菌群改变影响造血
除了局部免疫调节外,年龄相关的肠道群落结构变化也可能对造血产生直接影响。对小鼠的研究表明,肠道微生物组的改变与造血系统的多谱系改变和多能祖细胞的抑制有关。
鉴于肠道微生物群通过促进造血功能密切参与了细菌感染的控制,与年龄相关的肠道微生物群平衡和多样性的变化可能导致老年人造血功能受损、更易感染和减少疫苗接种应答。然而,为了更好地理解年龄相关的生态失调对造血系统调节的影响,还需要更多的研究来支持这一假设。
总的来说,这些临床前模型系统的发现表明,校正与年龄相关的肠道菌群失调是有益的,为基于微生物组的治疗方法改善免疫系统功能、对抗衰老及相关疾病提供了理论依据。
虽然粪便微生物群移植(FMT)在动物模型中具有抗衰老特性(见上文),并且FMT已成功用于治疗男性复发性艰难梭菌感染,但在临床环境中提出FMT作为抗衰老策略仍存在一些障碍。更好地了解健康微生物组的特征(包括病毒组和真菌组)对于确保接受者的长期安全至关重要。
尽管我们对整个生命中肠道微生物群的理解取得了很大进步,但在疫苗学上的潜力尚未实现。鉴于微生物组严格调控免疫细胞的发育和功能,它可能最终影响疫苗的疗效。因此,由环境、社会经济、营养或卫生条件引起的肠道微生物群落结构的变化可以解释疫苗应答中的地理异质性。
对微生物群如何促进疫苗应答的机制更好地理解,可能有助于制定新的策略,以减少老年人感染性死亡。
直到最近,表明肠道微生物组影响疫苗接种反应的最佳证据来自使用无菌或缺乏微生物的小鼠的临床前模型系统。
研究人员探讨不同抗生素方案对抗原特异性体液免疫应答的影响,研究了克拉霉素、强力霉素和氨苄西林对小鼠破伤风类毒素(TT)、肺炎球菌多糖疫苗(PPV)、乙型肝炎病毒表面抗原(HBsAg)疫苗和减毒沙门氏菌活疫苗(Ty21a)的一级和二级抗体应答的影响。
有趣的是,抗原和抗生素的特异性反应都受到了影响。
克拉霉素和强力霉素抑制典型的T细胞依赖性和T细胞非依赖性抗体反应,而氨苄西林的效果较差或无效果。
此外,所有三种抗生素,特别是氨苄西林增强了对Ty21a-a减毒细菌粘膜疫苗模型的体液反应。在GF小鼠中进行疫苗接种实验的进一步工作证实了这些初步发现。
到目前为止,现有的研究主要集中在细菌,以及包括病毒、真菌、原生动物和古生菌在内的微生物群的其他组成部分。然而,仍然存在两个重要问题:
(一) 微生物群参与宿主免疫反应(特别是疫苗反应)的机制是什么?
(二) 这些发现对人类的影响是什么?
迄今为止,最令人信服的数据表明,肠道微生物群提供了佐剂的天然来源,能够调节宿主的系统和黏膜疫苗反应。
这一观点最初是在Nakaya等人的系统生物学报告中提出的,他们描述了在非佐剂流感疫苗接种三价灭活疫苗(TIV)后,人类toll样受体-5 (TLR5)的早期(第3天)和瞬时基因上调。TLR5的表达与TIV诱导的血凝抑制(HAI)滴度(滴度是稀释度的倒数)呈正相关。
随后,该研究小组证实TLR5对小鼠肠道微生物分泌的鞭毛蛋白的感应与TIV介导的应答有关。虽然直接给予鞭毛蛋白或有鞭毛的细菌移植可以提供天然佐剂来改善非佐剂流感疫苗的应答,但它不能与其他佐剂或活疫苗如破伤风-白喉-百日咳(Tdap)、黄热病(YF-17D)或重组乙型肝炎抗原(Recombivax HB)配合使用。
除TLR5外,另一种模式识别受体核苷酸结合寡聚结构域2 (NOD2)的特异性贡献,人血清白蛋白(HSA)抗原和霍乱毒素(CTX)佐剂鼻内刺激诱导的粘膜疫苗应答小鼠模型。在GF小鼠、抗生素处理小鼠或基因修饰缺乏NOD2信号的突变小鼠中,HSA特异性IgG反应持续下降。当这些动物接受一种被NOD2或表达MDP的细菌识别的肽聚糖MDP(muramyl dipeptide, MDP)时,HSA特异性IgG反应恢复。
虽然尚未使用人体相关疫苗,但这第二项工作表明,常驻微生物也可以增强用于粘膜疫苗接种方案的CTX的佐剂效果。需要进一步研究揭示肠道微生物群、其细胞壁成分或代谢物作为内源性疫苗佐剂的免疫能力,以放大对特定病原体的适应性免疫应答。
临床前和临床报告均观察到膳食纤维对功能免疫参数的积极影响,包括疫苗应答(表1),进一步支持基于其组成(细菌的性质)和或活性(分泌的代谢物的性质)的肠道微生物群的重要免疫调节潜力。
表1 益生元和/或益生菌临床干预试验探讨老年人免疫相关结果


Nabil Bosco & Mario Noti,Genes & Immunity, 2021
关于纤维消化产生的短链脂肪酸(SCFAs)免疫调节活性的大量文献证明了这一点。这些代谢物在局部或全身释放,参与宿主的一般生理过程。
肠道微生物组组成和功能的动态变化也可以解释疫苗应答中观察到的地理异质性。研究人员还报道了成人和老年人接种流感疫苗后体液免疫反应的显著差异(超过100倍)。
宿主遗传、营养状况、母乳喂养习惯以及卫生条件和/或之前接触过病原体,被提出用来解释疫苗免疫原性的差异。然而,近年来研究宿主微生物群的技术进步为这一领域提供了新的思路。
粪便微生物群特征研究表明,肠道菌群组成可能影响口服脊髓灰质炎、卡介苗、破伤风类毒素和乙肝病毒的疫苗接种效果。
此外,在某些国家的儿童中,小肠细菌过度生长(SIBO)的流行率高。SIBO还可能限制疫苗的性能,因为相关的吸收不良、肠道微生物群和宿主免疫细胞之间争夺关键营养物质以及或通过渗漏的肠道系统释放微生物分子。从南非收集的样本中观察到反应迟钝的全血细胞(标准TLR刺激试验),并将来自北美和欧洲儿童的年龄匹配样本进行了比较。
上述人体研究仅存在相关性。
最近进行了三项研究,以确定婴儿和成人肠道微生物群失调和疫苗应答之间的因果关系。
在第一个大型(n = 754)和良好对照的研究中,广谱抗生素治疗(阿奇霉素)减少了致病性肠道细菌的流行,但没有改善印度婴儿的口服脊髓灰质炎疫苗接种。
接下来,Harris等人在开放标签试验中使用广谱抗生素(万古霉素)或广谱抗生素(万古霉素、环丙沙星和甲硝唑)治疗66名荷兰健康成人,并研究TT、口服轮状病毒(RV)和多糖肺炎球菌(Pneumo23)疫苗的应答。虽然RV疫苗观察到一些积极的效果,抗生素治疗没有改善TT或肺炎23价反应。
最后,Hagan和他的同事进行了一项研究,22名年轻的成年人接受广谱抗生素(万古霉素、新霉素和甲硝唑)治疗,然后接受TIV疫苗。虽然这种治疗对以前接种过流感疫苗的成年人影响有限,但在11名以前没有接触过流感(接种疫苗或自然感染)的健康个体中进行的第二次试验提供了突破性的发现。那些首次接受抗生素治疗的人的体液免疫反应,特别是对H1N1流感毒株的免疫反应大大减弱。
这些数据证实了早期的小鼠研究,并证明抗生素驱动的生态失调导致了非佐剂TIV引起的疫苗应答的显著改变。用系统生物学方法进行的更深层次的分子图谱也揭示了一种特定的炎症基因特征,即与抗生素治疗相关的更活化的髓系树突状细胞。
合理设计以微生物为内源性佐剂的疫苗在疫苗学领域具有广阔前景。这些方法可能需要个性化和工程可逆性来管理脆弱个体的疗效和潜在并发症。鉴于饮食是塑造肠道微生物群的最有力因素之一,营养干预与益生菌和益生元促进多样化的微生物群来维持健康受到了相当大的关注。
食品的发展是为了促进健康或减少疾病风险,1980年代中期,出现了“功能食品”一词:希望支持以营养为基础的预防性方法,以提高生活质量,减少与老龄化人口相关的医疗成本。
由于肠道微生物与宿主免疫系统的密切相互作用,临床试验研究的提高老年人免疫力的功能食品主要包括益生元(如纤维)、益生菌、两者的结合(即共生)或分泌可溶性代谢物(也称为后生元,如短链脂肪酸)。
随着年龄的增长,免疫适应能力下降,老年人无力抵抗感染,无力对抗原挑战作出反应,大多数这些试验探讨了感染流行程度、严重程度和持续时间或特定的疫苗接种结果。
在已确定的400项临床研究中,研究人员根据其相关性(免疫读数和目标人群为60岁的老年人)保留了31项研究,并将其总结在表1中。
这些研究是在社区居民、疗养院居民或住院患者中进行,这些患者可以服用口服补充剂或需要管饲。
大多数研究(20项研究)是随机双盲安慰剂对照组,考虑经典的混杂因素,如年龄、性别,有时还有医疗条件、营养状况、感染史或疫苗接种史。
预防感染性并发症,特别是危重症或选择性手术患者中艰难梭菌相关腹泻是第二个有意思的领域。
在对老年人进行的13项研究中,6项呈阳性。它们积极的性质也可能表明,疾病预防可以通过人体免疫系统的不同方面来实现。
疫苗挑战研究通常测试适应性免疫系统的功能,但没有解决某些细菌感染中与年龄相关的先天免疫下降的后果。然而,最近在英国23家养老院进行的规模最大的研究,即“益生菌减少养老院居民感染”试验,LGG和BB12胶囊每天服用长达1年,对预防感染没有效果。在感染症状、抗生素使用、住院或死亡率方面没有发现差异。
总的来说,可能有多种原因,研究之间缺乏一致性,如菌株特定差异或剂量,以及益生元性质和数量,或受试者的年龄和医疗条件。
这些研究中没有一项对干预前和干预后的患者微生物群进行了广泛的分析。虽然这些数据可能作为独立报告存在,以描述成分的特性,在未来,对干预前后的人体微生物群和免疫参数进行系统的平行评估是至关重要的,以揭示相关的相互作用或因果关系。
虽然实际年龄是不可逆转的,但随着年龄的增长,与肠道微生物群、炎症和免疫衰老相关的衰老变化是免疫介导的慢性疾病的共同帮凶,这可能是维持免疫和健康的目标。
当消炎药和衰老疗法选择性地消除衰老细胞(衰老抑制剂)或抑制衰老相关分泌表型(衰老表型)时,作为抗衰老疗法的临床试验正在迅速发展(图2),它们的长期使用需要谨慎地平衡对传染病的易感性和潜在的其他副作用。
为什么减少全身低度炎症可以促进疫苗应答,而疫苗学的主流观点是佐剂通过促进局部炎症来改善疫苗应答,这一悖论还需要进一步的研究来解释。
图2 恢复肠道微生物稳态、减少炎症和免疫衰老以支持老年人免疫功能的方法
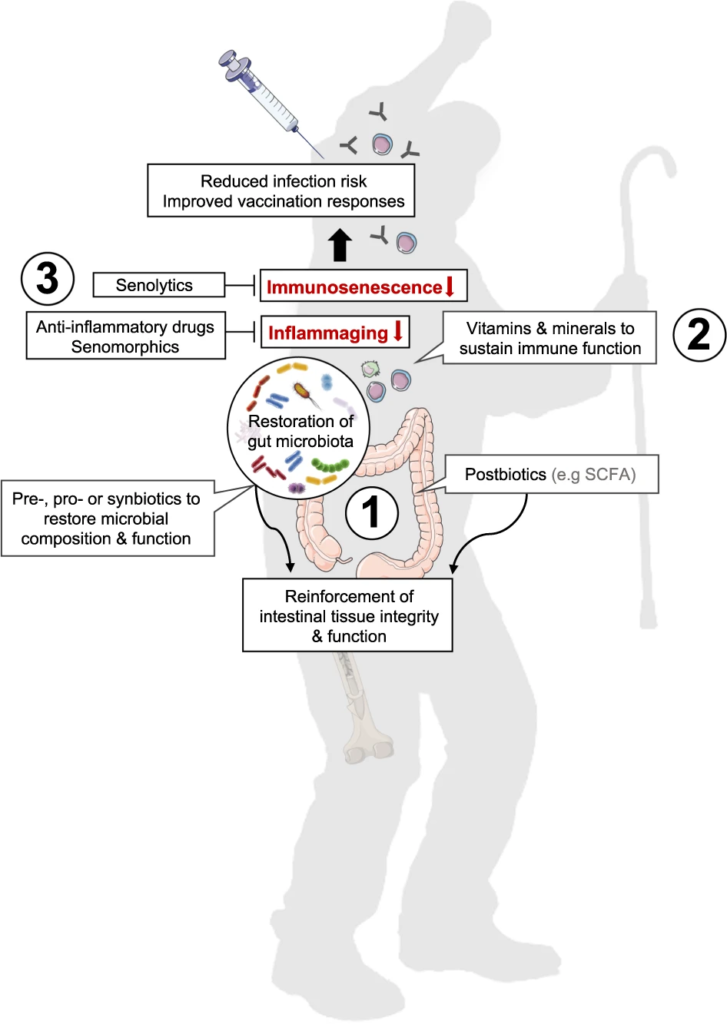
Nabil Bosco & Mario Noti,Genes & Immunity, 2021
(1)益生菌或益生元或合生元、后生元(如SCFAs)等进行干预可能有助于恢复与年龄相关的肠道微生物组成和功能下降
(2) 补充维生素和矿物质有助于正常的免疫细胞功能。
(3) 消炎药或老年药可能有助于减少炎症,而老年药直接消除老化细胞,燃料炎症。
单独或联合使用可能有助于增强宿主免疫,更好地控制感染,并随着我们年龄的增长产生适当的疫苗接种反应。
鉴于肠道微生物对宿主免疫系统的影响越来越大,可以合理推测,通过个性化营养或补充,恢复与年龄相关的肠道微生物丰富度和功能的下降,可能是一种对抗免疫适应性功能下降的预防措施。
在这种情况下,已在临床环境中测试了能够通过支持肠道屏障完整性或调节炎症过程来增强免疫力的益生菌、益生元、后生元或合生元(表1)。
然而,由于研究、菌株特异性差异或剂量、益生元的性质和数量或受试者的年龄和医疗条件之间缺乏一致性,因此很难验证这种方法在增强与年龄相关的宿主免疫适应性下降方面的有效性。
肠道微生物群是一个有待挖掘的宝藏,衰老学也不例外。正如许多临床前研究所证明的那样,恢复年轻的微生物群可以通过维持免疫和健康跨度来恢复老年宿主的活力。
因此,更好地了解肠道微生物群落结构和相关代谢组的动态变化,这些改变如何影响细胞免疫网络,以及这些通路如何被治疗靶向,将对未来加强甚至恢复老化免疫系统的策略产生广泛的影响。随着人口老龄化的加剧,迫切需要这样的解决方案来支持健康老龄化,减缓不断增长的医疗费用。
健康饮食
Omega-3脂肪酸
随着年龄的增长,神经细胞萎缩,向大脑提供的营养丰富的血液供应减少,Omega-3脂肪酸,尤其是二十二碳六烯酸(DHA)可以促进神经细胞之间的有效电信号传递,减少炎症,甚至可以改善精神集中度并与记忆力丧失作斗争。
较小的脑体积与阿尔茨海默氏症以及正常衰老有关。研究人员发现,血液中omega-3脂肪酸EPA和DHA水平较高的绝经后妇女的脑体积也更大。
富含Omega-3脂肪酸的食物:
鱼类:鲱鱼,沙丁鱼,鲭鱼,鲑鱼,大比目鱼,鳟鱼
绿叶蔬菜: 抱子甘蓝,菠菜,芝麻菜,薄荷,羽衣甘蓝和豆瓣菜
油: 亚麻籽油,正大籽油,鳕鱼肝油和磷虾油
其他:蛋、核桃等
黄酮类化合物
几乎所有的水果,蔬菜都含有类黄酮,具有许多健康益处,包括减少炎症,降低心脏病风险和湿疹症状,对衰老的大脑有益。有研究发现,食用大量浆果(类黄酮含量较高)的老年妇女将记忆力下降的时间推迟了两年以上。
类黄酮含量高的食物:
浆果: 蓝莓,草莓和黑莓
绿叶蔬菜: 菠菜,羽衣甘蓝和豆瓣菜
彩色农产品:胡桃南瓜,牛油果,李子,红葡萄
其他:咖啡,黑巧克力,红酒
维生素E
防止细胞损伤。已有几项研究发现,维生素E可以延缓轻度到中度阿尔茨海默病的进展
富含维生素E的食物:
坚果和种子:杏仁,山核桃,花生酱,花生,榛子,松子和葵花籽
油:小麦胚芽油,葵花籽油,红花油,玉米油,大豆油
绿叶蔬菜:菠菜,蒲公英嫩叶,唐莴苣,萝卜叶
运动
走楼梯:
瑞士一项研究发现,久坐不动的人把乘电梯换成走楼梯,过早死亡的风险可以降低15%。哈佛早期的研究显示,与每周爬楼梯少于10层的人相比,每周能爬35层或更多的楼梯能显著提高寿命。
骑车:
骑自行车上班是一种环保又健康的方式,在一天中挤出锻炼时间在户外,还能节省点油钱。
游泳:
一项数据发现,经常游泳的男性比久坐不动的男性死亡率低50%,游泳者的死亡率也比运动时走路或跑步的男性低。
每天锻炼15分钟:
2011年的一项研究发现,与久坐不动的人相比,每天15分钟锻炼可以平均延长3年的寿命。
保护牙齿
如果牙齿一直很敏感,那么随着年龄的增长,牙齿会越来越容易脱落,这会影响老年生活品质。请保持日常牙科护理的最佳状态,防止蛀牙。
保持良好的生活习惯
保证充足的睡眠:
NIH的数据显示,每晚持续睡眠少于7个小时的成年人患高血压,心脏病,肥胖,糖尿病和抑郁症的风险更高。另外,晚上睡不好会抑制重要激素的释放,这些激素可以修复细胞和组织,抵抗疾病和感染,睡不好会使身体无法自然康复。
尽可能避免吸烟:
戒烟可以减少许多疾病风险。
适当缓解压力:
压力容易让人变老。尝试放弃明显的压力源,适当安排休息时间,
经常锻炼记忆力
记忆就像肌肉一样可以训练;如果充分利用自己的记忆并定期使用它,记忆力可以磨练到老。比如每天尝试做一个填字游戏,以帮助建立和维护词汇量和记忆力。
《阿尔茨海默氏病杂志》上的一项研究显示,吃蓝莓可以增强记忆力和学习能力,喝苹果汁可以通过防止重要的神经递质的衰退来改善记忆力。
选择适宜的养老居住地
可以选择气候宜人的地方居住,良好的空气质量有助于保持健康,环境条件佳也有利于保持好心情。此外尽量选择拥有良好的经济和医疗体系的地方,帮助维持积极长寿的生活。
做好健康管理
定期进行肠道菌群健康检测或其他检查,及时了解自身健康状况,包括哪些风险,需要注意事项等,做好健康管理,享受健康幸福的晚年生活。
相关阅读:
主要参考文献:
Hägg S, Jylhävä J, Wang Y, Xu H, Metzner C, Annetorp M, et al. Age, frailty, and comorbidity as prognostic factors for short-term outcomes in patients with coronavirus disease 2019 in geriatric care. J Am Med Dir Assoc. 2020;21:1555–9.e2.
Khan SS, Singer BD, Vaughan DE. Molecular and physiological manifestations and measurement of aging in humans. Aging Cell. 2017;16:624–33.
Crooke SN, Ovsyannikova IG, Poland GA, Kennedy RB. Immunosenescence and human vaccine immune responses. Immun Ageing. 2019;16:25–16.
Butler CC, Lau M, Gillespie D, Owen-Jones E, Lown M, Wootton M, et al. Effect of probiotic use on antibiotic administration among care home residents: a randomized clinical trial. Jama. 2020;324:47–56.
Bosco Nabil,Noti Mario,The aging gut microbiome and its impact on host immunity.[J] .Genes Immun, 2021
Book: Mayo Clinic Family Health Book, 5th Edition
Wang J, Varghese M, Ono K, Yamada M, Levine S, Tzavaras N, Gong B, Hurst WJ, Blitzer RD, Pasinetti GM. Cocoa extracts reduce oligomerization of amyloid-β: implications for cognitive improvement in Alzheimer’s disease. J Alzheimers Dis. 2014;41(2):643-50.
James V. Pottala, Kristine Yaffe, Jennifer G. Robinson, Mark A. Espeland, Robert Wallace, William S. Harris Neurology Feb 2014, 82 (5) 435-442
Dysken MW, Sano M, Asthana S, et al. Effect of Vitamin E and Memantine on Functional Decline in Alzheimer Disease: The TEAM-AD VA Cooperative Randomized Trial. JAMA. 2014;311(1):33–44.
Berryman CE, West SG, Fleming JA, et al. Effects of Daily Almond Consumption on Cardiometabolic Risk and Abdominal Adiposity in Healthy Adults With Elevated LDL-Cholesterol: A Randomized Controlled Trial. Journal of the American Heart Association. January 2015.