 国家高新企业 | ISO9001认证
国家高新企业 | ISO9001认证 二级病原微生物安全实验室
二级病原微生物安全实验室- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
国家高新企业 | ISO9001认证 二级病原微生物安全实验室

谷禾健康

每个人的一生都会经历很多,从出生到长大,健康到衰老疾病。你的出生、遗传、家庭环境、很大程度上决定的人生起点,日常的饮食、行为习惯决定你的身体成长,一些不同的选择或意外的事件又会让人生有很多起伏和不同。
每个人的菌群和我们的人生一样也是独一无二的,我们菌群的特点反映着不同人各自生活的烙印。从母亲的腹中开始影响和决定了我们最初的菌群,出生方式、喂养的食物、用药等都决定了我们的菌群基数。当我们开始从喝奶到开始摄入辅食,我们的菌群也同样迎来巨大的演变。当我们生病、感染、运动、饮食、社交、虚弱、衰老这些同样反映在我们菌群的变化和演替上。
相对的,当我们更多的了解我们的菌群,善待和改善它们,同样的变化也会出现在我们的身体和生活中。
越来越多的证据表明,年龄与人类微生物群之间的关联很大,肠道微生物群是许多年龄相关变化的核心,包括免疫系统失调和疾病易感性。几个身体部位的微生物组成可以相对准确地预测人类的年龄。
谷禾健康肠道菌群检测数据库中,也有关于肠道年龄预测:
谷禾健康-肠道年龄预测模型图
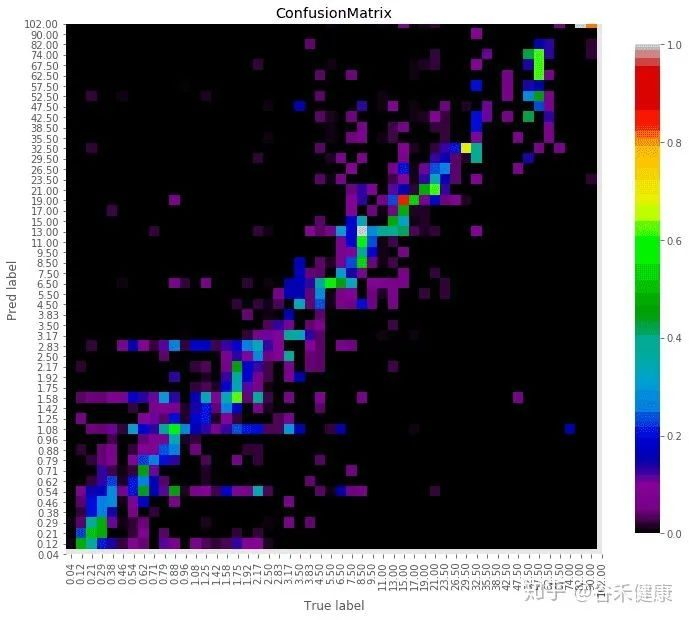
<来源:谷禾健康肠道菌群数据库>
可以看到,肠道年龄和生理年龄基本是符合的。健康人的肠道菌群年龄恰恰是最符合真实年龄的,与真实年龄差异大意味着肠道菌群出现偏离。
健康的人存在更多样化且平衡的肠道菌群。微生物群中与年龄相关的变化归因于生理,生活方式和健康状况。这些因素中的每一个都与某些菌群的相对丰度变化有关。
例如,饮食、卫生、兄弟姐妹、宠物、过敏、儿童疾病和抗生素是影响儿童微生物组的一些突出因素。到了成年期微生物群相对稳定,而到了老年期,一些有益菌开始逐渐下降,菌群又向另一个阶段过渡。
在从出生到死亡和分解的每个生命阶段,微生物群落都是身体的动态组成部分。研究微生物群的自然和诱导变化有可能彻底改变我们对人类生物学的理解。
本文介绍了健康人的微生物群在一生中的变化,讨论了从出生时菌群构成,到疾病或抗生素使用时的变化,再到死亡时的微生物扩展的各个阶段,以及这些阶段在身体部位和组成(细菌、真菌或病毒)上的差异。了解微生物群与年龄关系的未来研究方向,以此对人体微生物群及基于此的干预有更好的了解。
微生物群落存在于人体的每个粘膜表面,人的每个身体部位都有一个独特的生态学。每个人的微生物群像指纹一样,都是独特的。
在个体内,特定的身体部位、地理位置和个体的年龄与健康微生物群具有极强的关系。年龄驱动人类微生物群的α多样性和β多样性。
在了解各个阶段的微生物群变化之前,我们先了解一个概念:微生物演替。
微生物演替是指微生物群落中一种或多种生物的存在、相对丰度或绝对丰度的变化。
在正常或健康衰老期间,微生物演替的三个主要阶段自然发生在人类生活中。
✦初级演替(出生时先锋菌群定植,快速变化直到童年晚期)
第一阶段,初级演替,从先锋物种首次建立群落时开始,随后微生物群落发生快速变化。从出生到童年,变化率降低,许多中间物种存在于出生到童年晚期之间。
初级演替结束于顶级群落的形成,在青春期实现,并在很大程度上持续到成年;该群落的特征是其相对稳定。
虽然成年期的微生物群比儿童期更稳定,但仍然存在变异,这引发了关于人类微生物群中是否存在顶级群落的争论。成年微生物群的自然变异存在于小时(昼夜节律)到年(老化)的时间尺度上,但微生物群相对稳定,除非存在干扰,如饮食或药物的改变。
✦次生演替(菌群的改变,重建)
下一个阶段,即次生演替,发生在一个先前存在的稳定群落一部分被改变或移除之后,然后群落再生到相同的状态或不同的状态。这可以通过抗生素等医疗手段人为实现,也可以通过霍乱弧菌感染等疾病自发实现。
人类的次生演替的特征是至少有一段时间的随机过程占主导地位。在诱导条件下,如单疗程抗生素,群落遵循类似于初级演替的过程,其中现有微生物群落的一部分充当“微生物记忆”,帮助重建一个类似于以前存在的群落。
这一过程被认为是由核心微生物群驱动的,而不是驱动初级演替的先锋微生物。
✦末期演替(自然衰老和死亡阶段)
最终的末期演替是宿主自然衰老和死亡的一部分。在老年期间,微生物群落再次以更高的变化率,成功产生了一个由更少成员组成的群落,通常变形菌门(也称为假单胞菌)的相对丰度增加,有时占总优势。
研究演替的每个阶段使研究人员能够解决与人类相关的微生物群落是如何形成和维持的。通过了解这些过程,我们可以更好地了解微生物群随着年龄的增长的变化及其与人类健康的关系,了解如何管理微生物群。
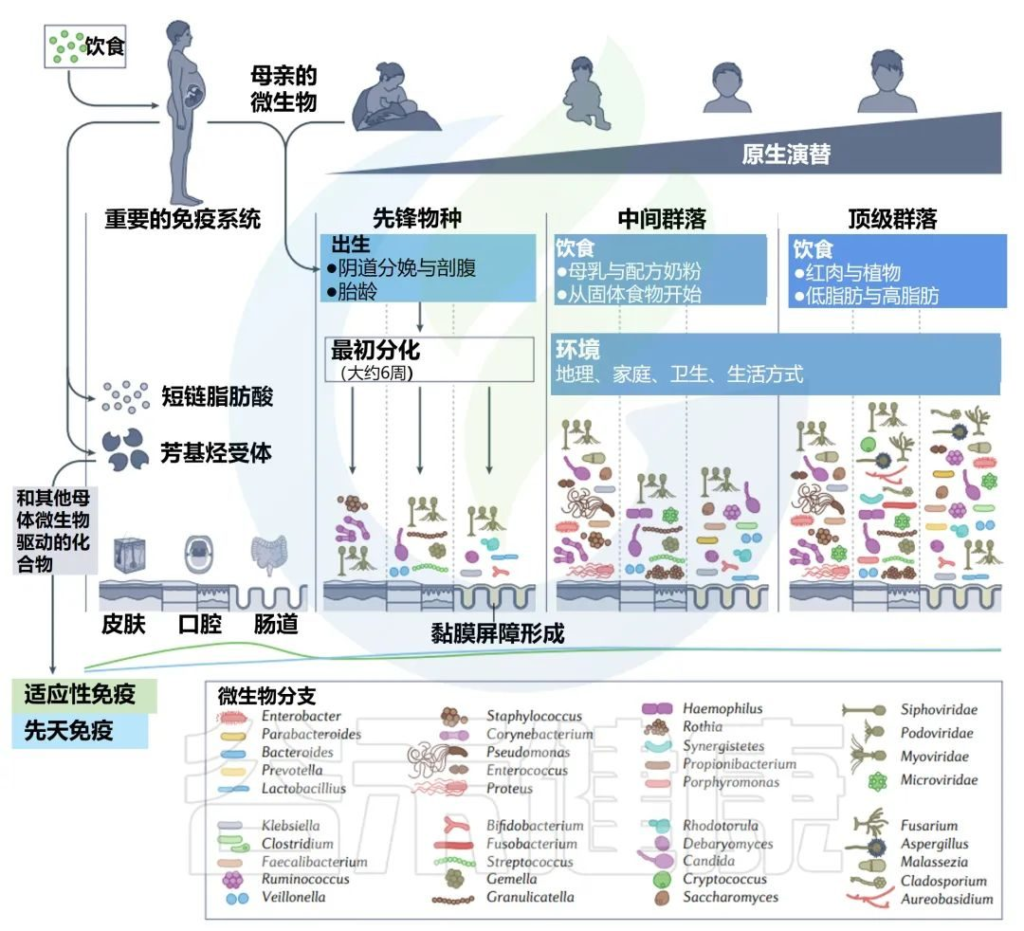
人类相关微生物群从受孕到死亡的变化
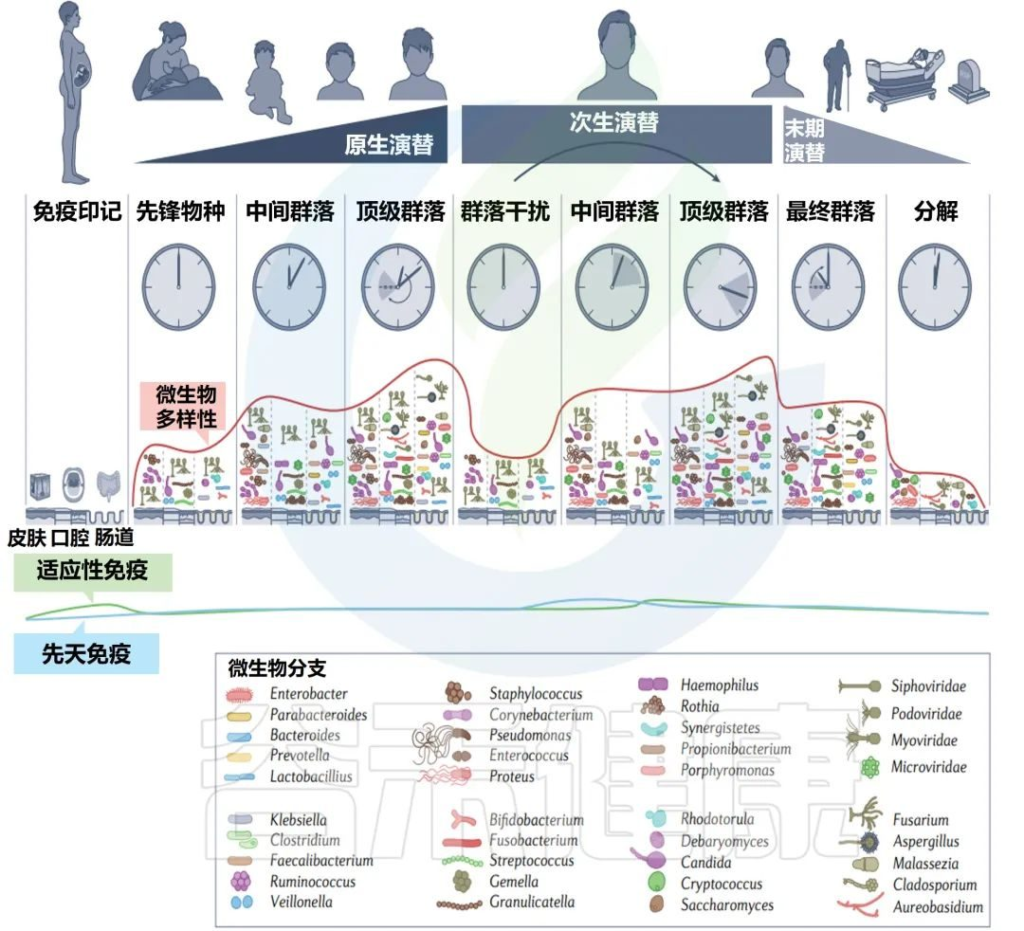
Martino C,et al.Nat Rev Microbiol.2022
常驻细菌、真菌和病毒的多样性在人类生命的各个阶段都会发生变化。模拟时钟代表每个微生物群落阶段发育的宿主年龄的相对时间。
免疫印记在出生前通过母亲的微生物群及其代谢物开始(第一栏)。先锋物种的初始定殖始于出生,身体部位特定的微生物群落出现(第二栏)。这些群落的复杂性不断增加,直到它们达到相对稳定的群落结构(第三列和第四列)。
这些微生物群落的次生演替可能来自内部和外部扰动(第五栏)。中间微生物重新建立初始群落,并再次达到稳定状态(第六列和第七列)。
在晚年,随着寄主接近自然死亡,群落经历了最后的演替和变化(第八栏)。微生物演替的最后阶段发生在腐败和分解阶段。在此阶段,多样性进一步下降,在最初的24-48小时内,许多人类微生物群结构保持不变,但随后很快开始侵蚀分解(第九栏)。
绿线和蓝线分别显示了微生物演替不同阶段的适应性免疫和先天免疫的相对强度。
不同年龄段的细菌多样性测量
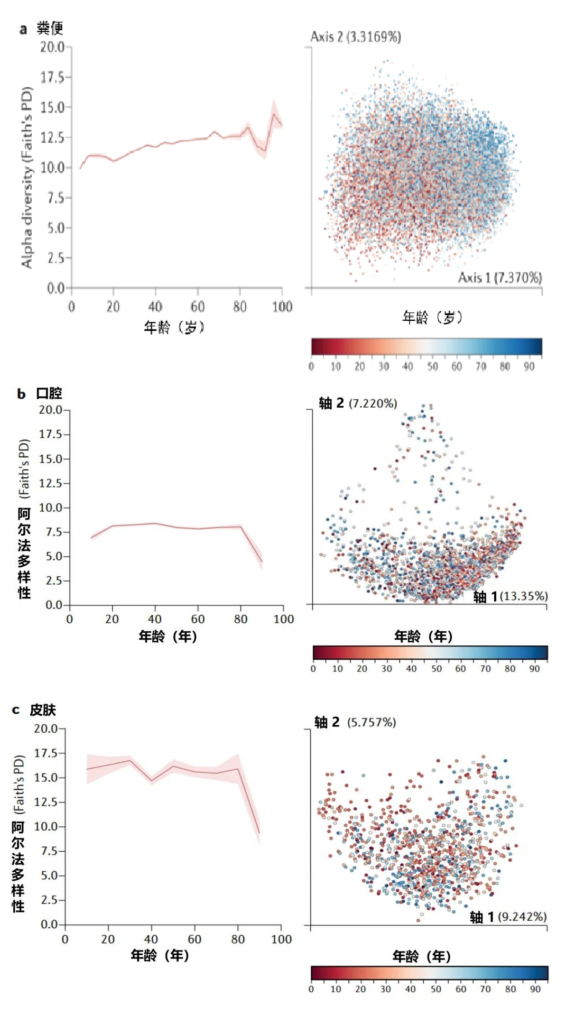
Martino C,et al.Nat Rev Microbiol.2022
美国一个肠道项目集中测量了从儿童到老年的人类粪便(a部分)、口腔(b部分)和皮肤(c部分)微生物群的细菌多样性和系统发育史,该项目包含21919个粪便、1920个口腔和998个皮肤微生物群样本,带有16S核糖体RNA基因扩增子序列。
α多样性,一种对样本中不同类型微生物数量的定量测量,通过Faith的系统发育多样性(PD)α多样性度量跨年龄测量。
UniFrac β多样性主坐标分析,一种用于比较微生物群落相似性的方法,其中空间上接近的点表示相似的样本,空间上远离的点表示不同的样本,按年龄着色。
✦胎儿时期——菌群及代谢物影响免疫发育
塑造人类微生物群的第一个因素来自胎儿发育过程中的母亲。
胎儿通过胎盘接触到母亲微生物群落产生的代谢物,这些代谢物会影响其免疫系统,并会影响正常微生物群和后期病理学的各个方面。代谢物,如短链脂肪酸(乙酸盐)和其他微生物化合物,可以通过胎盘转移到胎儿体内,并影响免疫发育。母亲的饮食和健康也会影响这些代谢物。
胎儿组织中的乙酸盐影响与成人调节性T细胞生成相关的表观遗传印记,其与防止生命后期哮喘的发展相关。
✦出生后——菌群受出生模式,饮食,环境等影响
出生后,微生物群落根据身体部位迅速分化。
在最初的时候,先锋物种和未来4年的群落发展可能会受到出生模式和妊娠时间的影响。中间群落由饮食影响,如母乳或配方奶粉的消费,以及环境。
最后,饮食和环境再次塑造了稳定的顶级群落。主要由真菌、细菌和病毒组成。
子宫内和生命早期的主要演替
微生物代谢物和配体调节宿主芳基烃受体,这有助于塑造新生儿微生物和免疫发育。母亲使用抗生素和胃肠道相关疾病,如炎症性肠病,也被认为会通过胎儿免疫系统的印记增加后代的病理风险。
然而,这些联系仅在非人类实验中研究过。在一个案例中,由患有炎症性肠病的孕妇或其新生儿的微生物群所定殖的无菌小鼠继续发展出异常微生物群和指示炎症性肠病的免疫发育。
✦怀孕期间母体的微生物群与免疫系统的变化
在怀孕期间,母亲的微生物群和免疫系统也发生了改变。母亲的阴道微生物群变得更加多样化,通常由在其他身体部位发现的许多微生物群组成。
孕期母体免疫系统与胎儿形成协同作用,包括通过胎盘转移IgG抗体。
关于出生时获得的微生物群是否通过混合来源于阴道和粪便,或者阴道微生物群本身在出生时是否具有多能性,是否是微生物先驱的主要来源,存在一些争议。
无论确切的母体来源如何,这一阶段的特征是先锋细菌种类。包括下列菌群:
然后这些细菌定居在常规身体部位:肠道、口腔和皮肤。
许多先锋细菌是兼性厌氧菌,它们会消耗氧气,从而使专性厌氧菌能够在以后的每个环境中定居。起初,新生儿的每个身体部位都相对未分化,但先锋微生物很快开始启动身体部位依赖性微生物多样性的级联,至少在生命的第4到第6周,每个部位的细菌都可以很容易地区分。
先锋细菌进驻后,生命早期的微生物群逐渐开始形成。接下来的章节我们来了解生命早期的肠道,口腔,皮肤等各部位的微生物群(包括细菌、真菌、病毒等)。
✦肠道细菌群——双歧杆菌主导
人类肠道细菌群落的发展已经得到了很好的研究。
双歧杆菌属一直占主导地位,直到在生命的第一年结束时,它们被双歧杆菌、梭状芽孢杆菌和拟杆菌属的组合所取代。拟杆菌属的丰度增加,而双歧杆菌属等物种的丰度相对减少。
双歧杆菌分解母乳低聚糖,开始终生影响免疫系统
最近,一项研究发现,双歧杆菌等细菌含有母乳低聚糖分解代谢所需的基因,与婴儿免疫发育之间存在功能联系。特别是,接受Bifidobacterium infantis EVC001极化初始T细胞的婴儿的粪便水与来自对照组的粪便水平不同,其方式与减少肠道炎症有关。
其他菌属也可降解母乳低聚糖(如拟杆菌、阿克曼菌)
到3-6岁时,肠道细菌群落汇聚到整个成年期持续的顶级群落。这一微生物群是已知的密度最大、多样性最强的生态群落之一。通常,在这段时间内,普通健康人中只有两个细菌门占优势:厚壁菌门和拟杆菌门。
✦肠道其他微生物群——真菌、古细菌、病毒
在人类肠道发育过程中,对病毒组、真菌组和古菌组的研究远远少于细菌组。在整个生命周期中,真菌群落所占的总数远远少于细菌组或病毒组。
//真菌群落
真菌群落在生命的最初几天含有大量的Rhodotorula和Debaryomyces,接下来的一个月则是CandidaCryptococcus和Saccharomyces spp.。
到成年时,主要的真菌属是Aspergillus, Candida和Saccharomyces。
//古细菌群落
发育期间肠道的古细菌群落尚不清楚,但古细菌是一些最早的移生菌落,但丰度较低。
早期定植的古细菌包括Methanosphaera和Methanobrevibacter。
// 病毒群落:噬菌体家族在出生后就开始流行
主要由噬菌体组成的病毒群落在出生后的第一周数量众多。噬菌体家族Siphoviridae、Podoviridae和Myoviridae在出生后立即流行,主要以溶原形式整合到细菌基因组中。
到生命的第四个月,有尾噬菌体目大量生长,成员更常为裂解型(传染性噬菌体颗粒或主动复制的噬菌体)。
在成人中,Caudovirales和Microviridae在肠道噬菌体群落中占主导地位,但噬菌体肠道病毒组对个体具有高度特异性,其演替仍有许多未知之处。
与噬菌体不同,感染真核病毒的肠道病毒组主要与儿童和成人的病理相关。最近,在健康儿童和健康成人中也观察到一些感染真核细胞的病毒丰度较低,但其发生时间和流行率尚不清楚。
✦口腔细菌群:出生后几个月逐渐趋于稳定,牙齿形成后再次转变
在出生时,口腔细菌群在以下菌属中的流行率很高:
在接下来的几个月里,Lactobacillus和Fusobacterium也开始流行。Staphylococcus的丰度在出生后3个月左右达到峰值,然后稳步下降,让位与更高丰度的GemellaGranulicatella, Haemophilus和Rothia spp.
牙齿形成后,口腔微生物群再次转变,在成年期具有更高丰度的梭杆菌门, Synergistetes, Tenericutes, Saccharibacteria (TM7), SR1 。
✦口腔其他微生物:成年口腔含产甲烷菌,最常见的噬菌体群是尾状病毒
口腔真菌群落被认为比皮肤和内脏的真菌多样性少。Candida spp.是口腔的第一批真菌定植菌。对中级口腔真菌群落知之甚少,但成年人CandidaCladosporiumAureobasidium
AspergillusFusarium和Cryptococcus spp.的丰度较高。
发育过程中的口腔古菌体尚不清楚,但成年口腔中含有许多古菌产甲烷菌,包括甲烷杆菌属。
目前对人类婴儿口腔中病毒的知之甚少。在成年人中,与肠道类似,最常见的噬菌体群是尾状病毒。
口腔病毒群在本质上通常被视为病理性的(例如柯萨奇A病毒、麻疹病毒、红疹病毒和人乳头瘤病毒),并且没有对病毒群落组成进行纵向研究。然而,在无症状和健康成人中也观察到许多真核病毒分类群。
✦皮肤细菌群落:出生时母亲阴道乳杆菌属占据较多,4-5周与成人相似
皮肤细菌群落在出生时含有大量的母亲阴道乳杆菌属。到第4-5周,婴儿皮肤微生物群与成人皮肤微生物群相似,但在青春期继续变得更具位点特异性。
Staphylococcus和Corynebacterium在不同位点PseudomonasEnterobacterEnterococcus,
Proteus和Klebsiella在特定位点(如腋窝与前臂)。
✦皮肤其他微生物:马拉色菌占比较高,古细菌占4%左右
在皮肤真菌群落中,MalasseziaCandida和 Saccharomyces在生命的前30天最为普遍。对于中间群落的确切组成知之甚少,但成年真菌群落中Malassezia的丰度通常很高,估计约占真菌群落总组成的75%至90%。
关于皮肤古细菌群落的发育情况了解较少,但古细菌约占成年人菌群的4%。大体上,成年人皮肤古细菌群由Thaumarchaeota门和Euryarchaeota门代表。在成人皮肤上也发现了Halobacteriaceae和 Methanobrevibacter。
与肠道和口腔不同,健康的皮肤微生物群拥有相对较少的已知病毒多样性,很少有对其进行研究,可能是由于与低生物量样本相关的技术限制。不过,皮肤上有一些自然存在的病毒群。
以上了解关于生命早期肠道、口腔、皮肤的微生物群,那么哪些因素会给生命早期的微生物群发展带来影响?
在生命的最初几年中,有几个因素塑造并区分了微生物群落的发展。
✦出生方式和母体抗生素的使用
出生方式和母体抗生素的使用是影响人类微生物群落的研究最好、最清楚的因素之一。然而,微生物的发育可能会导致独特的结果,即使是在同居的同卵双胞胎中,这可能是由于许多未知或随机的过程。
通过剖腹产和围产期和新生儿抗生素暴露,自然微生物群落的建立过程可能会在所有身体部位受到干扰。这一发现突出了阴道微生物群落的重要性,阴道微生物群落自然含有大量Lactobacillus spp.,但在青春期发生改变,对女性健康至关重要。
一些最佳样本的婴儿发育研究,通常缩写为DIABIMMUNE ECAM和TEDDY,在婴儿出生后的前2年和3年进行了随访,重点关注抗生素使用或出生方式的影响。
在上述所有研究中,阴道分娩的婴儿的拟杆菌属相对丰度高于剖腹产婴儿。
由于缺乏建立微生物群落的天然先锋微生物群,导致可变的群落组成被认为是由随机过程而不是确定性过程驱动的,出生模式对微生物群落组成的影响直到生命的第四年仍然可见。
出生模式影响的一个例外是早产,可能是由于在出生后的头几天大量使用抗生素,其特点是无论出生模式如何,微生物发育都不稳定。婴儿微生物群自然发育的这种改变与感染、免疫疾病、肥胖和神经内分泌异常的风险增加相关。
✦母乳喂养:母乳低聚糖给菌群带来稳定性
其次,与其他因素相比,母乳喂养对微生物群的发育有很大影响。与母乳喂养相比,配方奶粉的使用导致了更高的多样性和更不确定的微生物群落。
例如,考虑到出生时肠道中双歧杆菌科的自然优势,缺乏某些母乳低聚糖作为主要营养源可能会导致初始定植的不稳定性。然而,微生物群、牛奶代谢组和免疫系统发育的多组学整合是一个活跃且快速发展的研究领域。
除了母乳低聚糖,母乳还含有其他免疫调节化合物,例如革兰氏阴性细菌的脂多糖、分泌性IgA、先天免疫因子、抗菌肽和益生元因子。
最后,所有这些因素都会影响人类免疫发育。微生物相关分子模式识别受体与微生物群衍生分子相互作用,代谢物如短链脂肪酸(与GPR43、GPR41和GPR109相互作用)和次级胆汁酸(与FXR相互作用)直接影响免疫发育。
//
这些因素加在一起,有助于形成一个独特的、相对稳定的细菌、真菌和病毒微生物群落,这种微生物群落在人类生命的大部分时间都持续存在。
前面章节了解了婴儿期初级演替期间发生的巨大变化,与之相比,成年期微生物群基本上是稳定的(15-65岁),但该群落可能会受到干扰,因此本章节从以下三方面展开讨论:
健康成年人中某些细菌的基因组随着时间的推移而进化,表明在次生演替中,功能和组成进化以稳定状态发生。
• 昼夜节律影响菌群变化
成人微生物群也会发生自然的短期变化,时间尺度为一天到数月或数年。
短期变化的一个典型例子是微生物群落组成的昼夜节律。与昼夜节律相关的人类基因表达和免疫激活,以及肠道微生物群中细菌的丰度和组成也遵循这种模式。
在小鼠中表现出昼夜循环的细菌家族包括瘤胃球菌科、毛螺菌科、Muribaculaceae和疣微菌科,但对人体的等效周期知之甚少。
青春期和成年生活中的二次演替
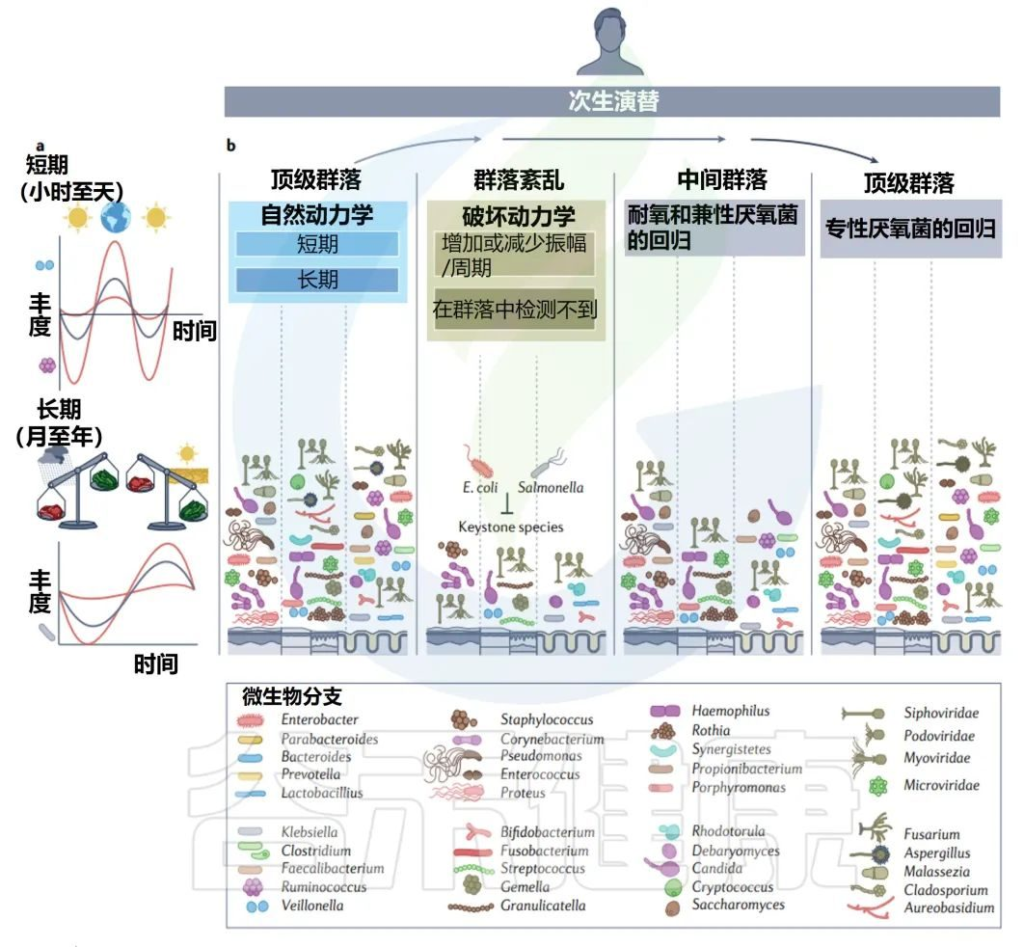
Martino C,et al.Nat Rev Microbiol.2022
• 口腔和皮肤的微生物群随清洗而变化
在口腔中,整组真菌和细菌的每日振幅与刷牙频率一致。在皮肤上,真菌和细菌每天的变化也与洗涤频率一致,并依赖于个人护理产品。
• 饮食会影响肠道微生物群
一个经过充分研究的发生在几周到几年范围内的变化的例子是饮食驱动的肠道微生物群的改变。饮食对微生物群落有很大影响,可以包括群落中的自然和可逆变化。
例如,坦桑尼亚哈扎部落在旱季食用富含肉类和块茎的饮食,但在雨季食用富含蜂蜜和浆果的饮食,在拟杆菌等属中表现出较大的季节波动。
饮食对微生物群形成的巨大影响也可能在人类健康中发挥作用,许多工作致力于了解特定的饮食成分和总体饮食模式如何影响微生物群及其对健康的影响。
肠道细菌喜欢大量的水果、蔬菜、全谷物、橄榄油等健康食物。研究表明,饮食主要由富含纤维的食物(如地中海饮食)组成的人具有更大的微生物组多样性,并且通常更健康。
此外例如,西方饮食中红肉含量高,这与全因死亡率有关。肠道微生物群可能以有害的方式将红肉中富含的左旋肉碱转化为三甲胺,而肝脏则将三甲胺转化为三甲胺氮氧化物,据推测这会促进动脉粥样硬化。
肠道微生物群也可以起到保护作用,例如,在红肉被肠道吸收之前将其分解,以防止炎症。除了饮食,还有许多其他因素有助于形成成年微生物群,包括遗传学、地理、宿主因素,如代谢病和药物。
扩展阅读:深度解析 | 炎症,肠道菌群以及抗炎饮食
• 抗生素对微生物群的影响巨大
由于微生物群的破坏而发生的次生演替已被广泛研究和审查。在破坏微生物群的众多因素中,抗生素是最强的,治疗后的恢复率往往各不相同。
抗生素治疗后肠道微生物群反弹的能力被认为取决于特定的群落成员,如拟杆菌和青春双歧杆菌。
扩展阅读:抗生素对微生物组及对人体健康的影响
细菌的天敌抗生素,如何用好这把救命的双刃剑?
疾病本身也会破坏微生物群,无论这种变化是由微生物群落内部、宿主还是多种因素共同引起的。
• 疾病破坏菌群
——肠道:炎症破坏菌群
肠道中的许多其他疾病,如炎症性肠病,破坏了微生物群落,但没有达到新的稳定群落组成,而是在没有干预的情况下继续长期不稳定。
——皮肤:炎症引起金黄色葡萄球菌大量增殖
在皮肤上,特应性皮炎的特征是免疫介导的炎症引起的金黄色葡萄球菌大量繁殖和细菌多样性减少。在金黄色葡萄球菌大量繁殖期间观察到马拉色菌属的数量减少,反之亦然,真菌数量增加导致金黄色葡萄菌数量减少,这部分可能是由于真菌产生蛋白酶的能力,蛋白酶消化金黄色葡萄球菌生物膜并降低细菌逃避免疫系统的能力。
——口腔:细菌和真菌间的竞争和协同
口腔中也存在类似的跨界相互作用;例如,真菌白色念珠菌的定殖依赖于细菌生物膜,但同时,Pseudomonas和Staphylococcus等细菌属分别形成竞争和协同关系。
这些例子强调了微生物群落的相互作用和演替是如何跨域和与宿主作用的,但由于其高阶相互作用的复杂性质,仍然没有完全理解。
干扰后微生物群落恢复的障碍导致许多研究人员探索有针对性地恢复微生物群落的干预措施的可能性。微生物群落恢复包括定向重新播种或某些物种的富集或耗竭,旨在促使微生物群落恢复到接近扰动前的水平。
这可以通过益生菌、益生元、抗生素或其他药物、从健康个体移植完整的微生物联合体或这些的组合来尝试。
尽管这些疗法在某些特征明确的环境中可以非常有效地恢复健康的微生物群落,但它们往往因缺乏与现有群落相互作用的机理知识,或因其仅短暂移植的能力而受到限制。
为了解决这些,研究集中在两个领域:
第一个领域涉及更好地了解群落是如何组合的。例如,对人类发育的研究有助于确定微生物群落在发育过程中如何聚集,以及这种聚集在生命后期的影响。
其次,正在开发新方法,通过探索微生物群落相互作用来确定机制,包括计算和实验,包括高通量共培养和微生物群落的基因组编辑。
为了解决瞬时性问题,采用了两种主要方法:
首先,微生物群疗法的短暂和个性化影响是由每个人的微生物群的个体性质决定的。因此,精准医学将群落改变的目标定位于每个人独特的微生物群,前景广阔。例如,基于微生物群落组成的个性化营养在盲法随机对照干预中有效地改善了餐后血糖。
另外,超越细菌组,探索病毒组和真菌群落及其之间的相互作用,具有巨大的前景。例如,噬菌体疗法已经用于严重的耐药细菌感染,并且对目标细菌菌株具有高度特异性。但大多数此类干预措施仍处于初步研究阶段,且规模成本高昂。
前面章节我们了解了成年微生物群的变化,以及变化后的恢复情况等,成年稳定微生物群在老年时转变为最终群落,本章节来详细了解老年微生物群。
“老年”的确切时间尺度取决于其他几个与宿主相关的因素,如疾病,但迄今为止大多数文献将“老年人”定义为65岁及以上的人。
接近寿命终点的晚期演替
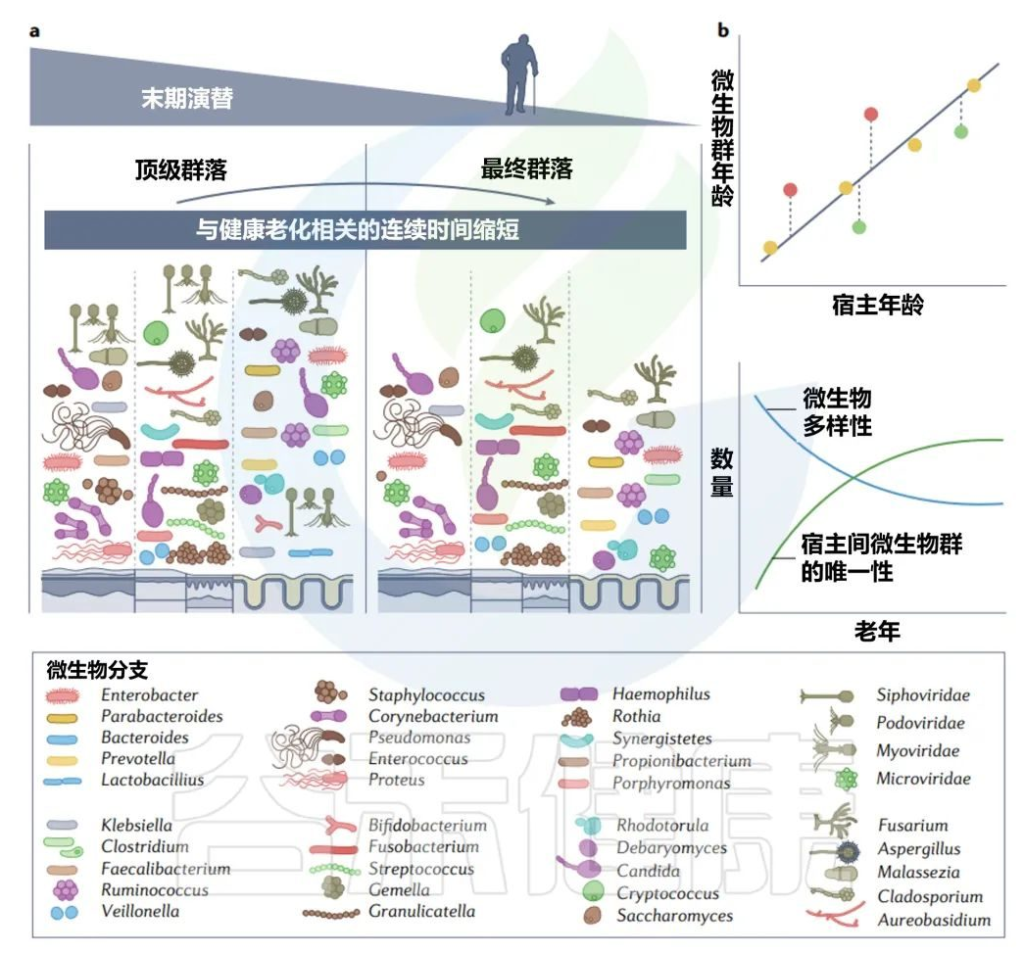
由于生物编程和生命中损伤的累积而导致的衰老影响细胞功能的各个方面,微生物群也不例外。随着年龄的增长,肠道微生物群α多样性减少,β多样性增加。
关于老年微生物群,仍有许多未知之处,而文献也有些矛盾(一项报告称65岁及以上成年人拟杆菌数量增加,与其他研究相矛盾),大多数研究都集中在肠道细菌上。
老年微生物群:年轻优势菌丰度减少
一般而言,肠道中观察到的群落演替是年轻成年人中占优势和普遍的细菌属丰度减少,如Bifidobacteria, Bacteroides, Lactobacillus, 抵御机会细菌爆发的能力降低。
• 皮肤
在65岁及以上的人群中,genera Cutibacterium和Staphylococcus的皮肤细菌数量减少,同时观察到的Corynebacterium。
• 口腔
在口腔部位,Rothia和Streptococcus spp.是核心口腔细菌群落,PorphyromonasTreponema和Faecalibacterium spp.的数量持续减少。
• 肠道
老年期肠道真菌群落的特征是Penicillium, CandidaAspergillus和Saccharomyces spp.的优势度增加。
在皮肤和口腔部位的研究很少,但老年期皮肤上的Malasseziaspp.和口腔内的Candidaspp.丰度减少。
在肠道噬菌体中,成年期的Siphoviridae占主导地位,而老年期的Microviridae和Podoviridae则占主导地位。与肠道细菌、真菌和噬菌体群体相比,真核病毒的多样性在童年后和整个余生中保持不变。
研究重点
由于个体之间的高度变异性,老年微生物演替的研究重点主要是比较健康和不健康的衰老。
目前尚不清楚微生物群是否在健康衰老中起着机械作用,还是仅仅是其他变量的一个有力指标,如饮食、运动和药物。然而,在那些长寿健康的人中,可以观察到在健康成年人中高度流行的菌群的持续保留方面的共同点。
然而,百岁老人表现出更独特的微生物群,α多样性增加,群落组成的个体间差异更大,使“健康”和“不健康”年龄之间的比较复杂化。次生胆汁酸在百岁老人中含量丰富,也可能在健康老龄化中发挥作用。尽管前景看好,但这一研究领域仍处于起步阶段。
扩展阅读:肠道菌群与健康长寿
• 微生物的演替不会随着个体的死亡而结束
宿主的死亡可以视为微生物群的生态干扰。心脏停止后,组织立即因缺氧而开始分解。细胞功能持续,直到所有剩余的氧气耗尽,二氧化碳不再能够从组织中运输为止。细胞内二氧化碳的积累创造了一个缺氧的酸性环境,导致细胞破裂。
细胞成分,例如酶会泄漏到周围环境中,在被称为“自溶”的过程中进一步促进组织分解。自溶通过消除免疫系统、松开细胞连接并为微生物群提供营养,触发了一系列负责组织分解的微生物过程。
死亡后的微生物群
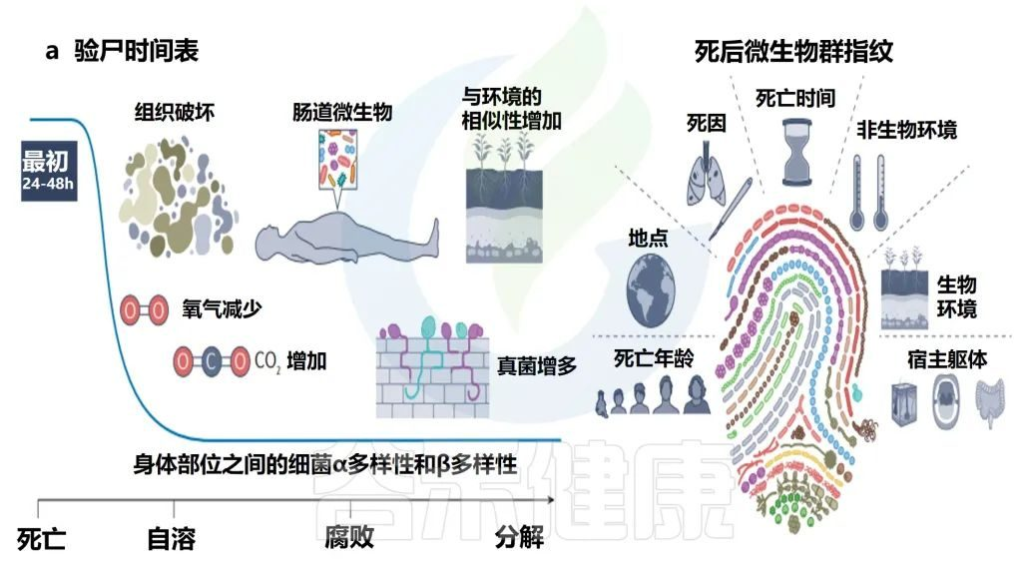
Martino C,et al.Nat Rev Microbiol.2022
• 死亡后微生物群分解
人类微生物群在死亡后的前24-48小时内相对稳定,具有不同的身体部位微生物生态、年龄的α多样性模式和可识别的个性化皮肤微生物群特征。
在分解的最初几天到几周内,腐败主要由细菌进行,但随着分解的进行,真菌的作用增加。然而,在这个过程中,对病毒组的演替和功能作用了解甚少。
随后,环境变化促进了微生物的演替,改变了人体和微生物群,不再像活着的个体(除非身体被冷冻)。
由于缺乏宿主生活中先前遇到的环境限制,使得微生物的相对丰度发生了快速变化以及在身体各部位的移动。迁移的细菌群成为从肠道转移到肠外部位的先锋物种,根据身体部位参与初级演替或次级演替。
• 死亡微生物群——生物指示器
死亡微生物群因其对法医调查的影响而引起了越来越多的关注。与多个个体和身体部位相关的一致的时间序列模式证明,死后微生物群可以作为死后间隔的生物指示器。
每个尸体的死后微生物群都是独一无二的,并且根据死亡时间、死因、环境、死亡地点和年龄以及开始时身体部位之间的差异,尸体之间的微生物群是不同的。
当微生物演替包括群落成员的快速更替时,在分解的早期阶段(即死亡后的前2-3周),死后时间间隔估计更为准确,但在分解的后期阶段(例如骨骼)仍然有用,因为几乎没有证据可以估计死后时间间期。
• 死亡原因与微生物群存在联系
还证明了与死亡原因和微生物群存在的联系。例如,在死于心脏病的个人的口腔微生物群中发现了的Rothia spp.。
此外,皮肤微生物群脱落可能通过将个人与他们接触过的物品联系起来,从而有助于追踪证据;然而,这一独特特征能够准确匹配到个体的时间取决于对象的材料和用途。
人类微生物群是动态的。考虑到这一点,设计一种能够捕捉微生物群的时间和空间变异性的采样策略非常重要,特别是当这些波动与所提出的科学问题相关时。
✦测量时间不同:多个时间点的样本采集
横断面研究从每个个体收集一个样本,而重复测量研究在多个时间点或身体部位收集样本。随着时间的推移,采样频率应该调整到研究人员试图观察的现象。
例如,小鼠昼夜节律研究通常每2-4小时收集一次粪便样本;而在炎症性肠病中,在一周内对患者进行三到五次采样可以改善疾病分类。
在其他应用中,例如研究特定治疗对个体微生物群的影响,这可能与进行“一对一”研究有关,在该研究中,同一参与者被反复检测其微生物群的结果变化;治疗前采集的样本被视为个体水平的对照。
✦测量空间不同:城市化/农村环境不同
同样重要的是要考虑到人口的微生物群高度依赖于地理和种族。
例如,在一个大型中国群体中,一种与年龄高度相关的微生物在一个美国大型群体中根本没有检测到。
另一个具体的例子涉及城市化社会的“建筑环境”;城市化人群通常较少接触环境微生物,更多地使用家用抗菌剂,与来自农村社会的人类微生物群相比,这导致了重大变化。
这些考虑因素与微生物群领域尤其相关,因为大多数公共微生物群数据来自城市化的北美和欧洲人。因此,现有数据集的结论可能无法很好地推广到全球人口。
从人类微生物群和微生物群研究中生成的测序数据的主要类别是扩增子测序数据和鸟枪测序数据。
✦扩增子测序
在扩增子测序中,对已建立的高变区的PCR产物(扩增子)进行深度测序,从而能够通过与个体“条形码”匹配来识别和测量群体成员。
这里有两种选择:要扩增的基因和该基因的哪一部分要扩增。微生物基因组的常见扩增区域包括:细菌的16S核糖体RNA基因、真核微生物的18S核糖体DNA基因和真菌的内部转录间隔区。
每个特定基因中高变区的选择取决于要捕获的特定微生物,但广泛使用的高变区包括来自地球微生物组项目的V4区。
✦肠道微生物群参与人体的调节
在鸟枪测序中,所有微生物DNA都被测序,而不仅仅是PCR产物,从而能够对微生物进行更具体的分类。由于鸟枪测序不依赖于任何标记基因,因此与扩增子测序相比,它对某些微生物的偏向性较小。
然而,鸟枪测序的成本要高得多,并且需要更大的计算能力,这使得在不需要提高鸟枪序列分辨率的情况下,扩增子测序具有吸引力。
结合其他技术进行扩增子或宏基因组测序可以丰富对微生物群和宿主的理解。定量PCR和荧光激活细胞分选等技术通过将相对丰度锚定到可靠的绝对丰度测量值,为相对丰度提供了更多的背景。
酶联免疫吸附试验和单细胞测序可以通过提供宿主细胞类型或宿主免疫信息与宏基因组测序很好地配对。
培养组学使研究人员能够通过实验验证功能或活性的基因组预测,并将微生物转化为益生菌。微生物产生的代谢物或蛋白质,即微生物群的下游效应物,可以分别通过代谢组学和蛋白质组学进行探测。
最后,宿主基因组学和转录组学越来越多地与扩增子或宏基因组学数据配对,以深入了解宿主基因表达和微生物群之间的联系。
最后,从被调查的参与者那里收集数据至关重要。一般微生物群研究的一些重要元数据类别包括人口统计、临床信息和饮食信息;然而,使用的确切元数据因研究而异。应采用产生标准化元数据的实践,以便结果可重复使用和再现。
本文描述了目前对不同年龄和不同身体部位的人类常驻微生物群落组成的研究现状。
人类健康与微生物群组成之间存在许多联系,对肠道菌群的干预可能改善健康。侧重于整个微生物群而不是单一物种的富集或消除的干预措施,需要了解这些群落是如何形成和维持的。
不同人群年龄,不同部位的微生物群需要依托于大样本数据库的构建,这为微生物群研究的准确性提供了保障。
通过研究人类整个生命周期中的微生物群,我们可以更好地了解这些微生物群复杂的相互作用,以及如何有效地将微生物群推向宿主所需的组成。此外也正应用于除人类健康外的其他领域,如法医学。随着微生物群的相关研究不断突破,将给人类生命健康和生产生活带来巨大的影响。
主要参考文献:
Martino C, Dilmore AH, Burcham ZM, Metcalf JL, Jeste D, Knight R. Microbiota succession throughout life from the cradle to the grave. Nat Rev Microbiol. 2022 Jul 29. doi: 10.1038/s41579-022-00768-z. Epub ahead of print. PMID: 35906422.
Lim, A. I. et al. Prenatal maternal infection promotes tissue-specific immunity and inflammation in offspring. Science 373, eabf3002.
Al Nabhani, Z. & Eberl, G. Imprinting of the immune system by the microbiota early in life. Mucosal Immunol. 13, 183–189.
Helve, O. et al. 2843. Maternal fecal transplantation to infants born by cesarean section: safety and feasibility. Open. Forum Infect. Dis. 6, S68.
Seppo, A. E. et al. Infant gut microbiome is enriched with Bifidobacterium longum ssp. infantis in old order mennonites with traditional farming lifestyle. Allergy 76, 3489–3503.

谷禾健康
虽然每个人都会老去,但是衰老的速度却不一定相同。生理年龄是每个人的遗传,环境和生活方式的影响。
人的一生,从出生到成年到老年,微生物组在免疫系统成熟,功能和调节中起着基本作用。免疫系统和微生物组形成一种互惠关系。
随着年龄的增长,免疫系统和肠道微生物组的组成和功能都会发生重大变化,这与对传染病的易感性增加有关。
导读
01)免疫衰老的一般标志
02)衰老和肠道微生物组
03)微生物老化的衡量指标
04)微生物老化加快免疫衰老和虚弱
05)肠道微生物组对疫苗反应的影响
06)对抗与年龄相关宿主免疫力下降的营养策略
07)恢复肠道微生物稳态、减少炎症和免疫衰老
08)健康长寿的普适方式
免疫衰老
“免疫衰老”是指在老年人中观察到的功能障碍,免疫反应有缺陷或异常。
与年龄相关的免疫反应的质量和数量的变化导致触发有效抗体和细胞反应抵御感染和疫苗的能力逐渐下降。
衰老的T淋巴细胞生物学研究十分突出,但所有造血源性细胞都显示出衰老的特征,包括功能失调的抗体产生B细胞、抗原呈递细胞、自然杀伤细胞和中性粒细胞。最早的造血祖细胞中也报告了表观遗传学改变,这可能解释了观察到的年龄相关的髓样细胞偏斜。
主要淋巴器官的老化(骨髓和胸腺磨损)、慢性抗原过载(如CMV)、肠道失调或炎症是免疫衰老的驱动因素,这些因素与遗传缺陷、细胞应激和/或细胞衰竭的累积一起,会导致免疫适应度随着年龄的增长而下降。
炎症
炎症是一种高度控制的生理过程,对对抗病原体、清除碎屑和愈合损伤至关重要。
随着年龄的增长,由于基因、环境和生活方式因素的复杂和不断变化的相互作用,促炎和抗炎之间的动态平衡下降。
慢性炎症状态是发病和死亡的一个重要危险因素。包括慢性感染、缺乏运动、内脏肥胖、饮食、心理压力、睡眠不足或肠道失调等多种因素都会引发和维持炎症。
长期暴露于应激源会加速细胞衰老和先天免疫失调,这是炎症的一个主要特征,反映在局部和全身炎症介质与白细胞介素-6(IL-6)、肿瘤坏死因子TNF-α 水平持续升高,IL-1β,C-反应蛋白(CRP)在老年人中普遍存在。
尽管先天性单核-巨噬细胞网络的失调可能是炎症的中心,但新的证据表明衰老细胞(包括T细胞和B细胞)通过其衰老相关的分泌表型参与慢性低度炎症的关键作用。
而慢性表达或暴露于炎症刺激可能使预先激活的免疫细胞对进一步的刺激难以耐受,从而导致观察到的老年人感染性疾病的频率和严重程度增加。
因此,先前存在的炎症已被证明是疫苗反应性的一个重要决定因素。
此外,炎症可能通过助长年龄相关疾病(包括代谢综合征、心血管疾病、肌细胞减少症、癌症和神经退行性疾病)而产生多种健康后果,因为大多数(如果不是所有)年龄相关疾病都具有炎症特征。
尽管衰老、炎症和慢性病之间存在着共同性,但生物医学研究继续巨额花费来单独解决这些疾病状态。这就提出了一个具有挑战性的问题,即:
针对慢性低度炎症或引起炎症的机制是否可能减缓衰老及其相关疾病?
虽然临床前研究表明炎症是导致年龄相关疾病和免疫反应性降低的一个因素,但证实这一假设的人类数据在很大程度上是缺失的。
而最近一项CANTOS的研究(Canakinumab Anti-inflammatory Thrombosis Outcomes Study)为老年人带来了巨大的希望。
对10000多名既往有心肌梗死史的稳定患者进行卡那单抗(Canakinumab,一种针对白细胞介素-1β的人单克隆抗体)治疗,可显著降低参与者的全身低度炎症。
重要的是,治疗方案可以预防复发性血管事件和肺癌的发生。虽然这些研究唤起了人们对有效抗衰老疗法的希望,但通过靶向阻断关键炎症介质来减少全身炎症需要在对传染病的易感性方面谨慎平衡。
这些免疫指标包括基于炎症年龄评分的iAGE、依赖于免疫球蛋白糖基化的聚糖生长或南丁格尔健康指数,其中糖蛋白乙酰化的全身炎症预测呼吸道感染、心血管疾病和全因死亡率的长期风险。
随着老龄化社会的健康管理成为一个日益增长的经济负担,这种免疫指标代表了有希望的工具,以确定风险个人的早期药物或营养干预。
为了应对不断变化的环境,肠道微生物通过菌种组成和代谢功能的变化动态响应。这个过程受到宿主免疫系统的严格调控,想象宿主免疫系统是一位建筑师,通过允许共生细菌生长和占据粘膜生态位,同时选择性地消除或中和有害微生物,从而塑造肠道微生物群。
随着年龄的增长,免疫适应度逐渐下降,宿主与微生物动态信号交换的监测受到损害,从而对宿主健康和免疫造成广泛的功能后果(图1)。
图1 模型:微生物老化及其对宿主免疫的相关影响
Nabil Bosco & Mario Noti,Genes & Immunity, 2021
(1) 年龄相关的肠道微生物群落变化和相关的肠道组织功能下降可能加剧炎症
(2) 由全身低度炎症引起的慢性免疫刺激、代谢组和微生物刺激的变化导致免疫衰老
(3) 免疫功能受损(如胸腺退化、造血功能改变),导致老年人感染风险增加,老年人接种疫苗反应差
在人类中,年龄相关的菌群失调(微生物老化)特征是梭状芽孢杆菌和双歧杆菌数量减少,变形菌门以及肠杆菌科等病原菌数量过多。
肠道微生物群落结构的组装也可能取决于宿主器官的功能。考虑到肠道的组织功能和完整性受到与年龄相关的显著变化,如再生能力、上皮屏障形成、黏液层组成和蠕动的改变,随着我们年龄的增长,粘膜生态位的变化很可能导致了失调状态。
这种肠道完整性的亚临床改变可能会促进微生物向全身部位渗出,全身性低度炎症,慢性炎症的发生和过早死亡。然而,确定人类的因果关系仍然具有挑战性,下文将进行讨论。
虽然,我们已经开始了解细菌的分类组成和多样性是如何随着年龄的变化而变化的,但我们对细菌进化和与宿主适应性相关的功能后果的了解仍然有限。有两种可能的情况:
1 肠道微生物群落结构中与年龄相关的变化是宿主衰老过程中发生的生理组织适应的简单结果
2 与年龄相关的失调是细菌进化的产物,通过使特定菌群逃避免疫监视,直接触发宿主衰老
因此,更好地了解老化的宿主微生物群,对于推进以微生物组为基础的治疗方法来对抗老化和与年龄相关的疾病至关重要。
我们需要先了解这样的概念。
生理年龄:指人达到某一时序年龄时生理和其功能所反映出来的水平,是从医学、生物学角度来衡量的。
肠道年龄:是基于健康人群每个年龄段样本的菌群构成使用深度神经网络模型提取特征菌属并构建的预测模型。
谷禾健康-肠道年龄预测模型图
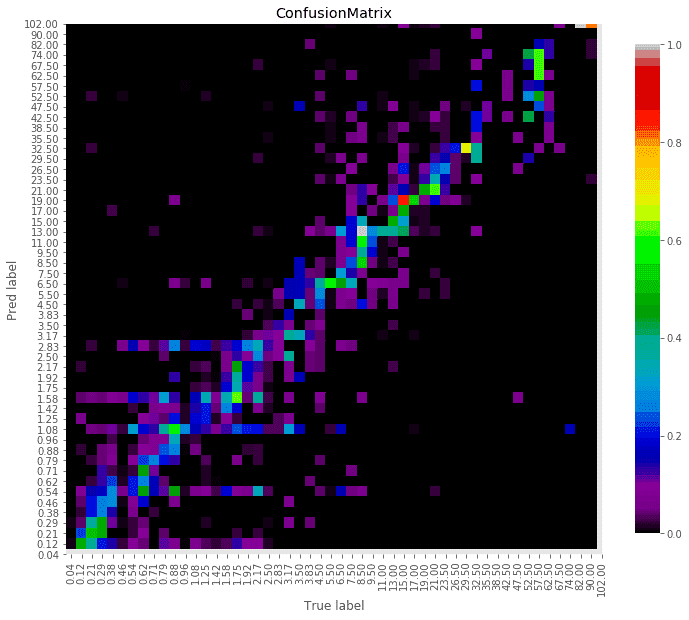

注: 谷禾报告中的肠道年龄是基于超过6万人群队列的深度学习模型构建的,伴随年龄的变化肠道菌群也会相应的改变。对于0~2岁的儿童,肠道年龄通常偏差小于3个月,3~5岁偏差在6个月以内,6~15岁偏差在1岁左右,16~50岁人群正常肠道年龄的偏差在3岁以内,50岁以上正常偏差在5岁以内。在正常偏差内年龄差异可以理解为年轻或衰老,超出正常偏差的年龄无论是超过或低于都可能是菌群异常或健康状况不佳。
真实年龄与肠道预测年龄在范围内的差异可以反映其肠道菌群的发育和衰老状况。
以下情况可能会导致肠道预测年龄完全偏离真实年龄,包括:肠道菌群紊乱,菌群结构过于单一,近期服用包括抗生素等可能严重干扰肠道菌群的药物,病原菌感染或者处于疾病状态。
肠道菌群失调可以触发先天性免疫反应和慢性低度炎症,导致许多与年龄有关的退化性疾病和不健康的衰老。肠道菌群通过各种生物分子,营养素信号独立途径和表观遗传机制与宿主进行交流。与年龄有关的肠道菌群失调会干扰这些交流,从而影响宿主的健康和寿命。
与年龄相关的肠道微生物组成的改变不仅发生在人类身上,也发生在实验室模型生物身上。这些控制遗传、年龄、饮食和微生物组本身的模型系统提供了强有力的科学证据,表明宿主生理或微生物进化的内在改变足以促进一种失调状态。
这些模式生物已经证明肠道微生物群有潜力有益地调节衰老过程,以促进宿主的健康和寿命。
黑腹果蝇
对黑腹果蝇的研究(一个经常被用来研究微生物动力学、肠道生理中与年龄相关的变化和生物体健康之间相互作用的衰老模型)——已经完美地证明了肠道生态失调不仅是年龄相关肠道屏障功能障碍、系统免疫激活和机体死亡的前兆,而且还预测了这些疾病的发生。
结果表明,在整个生命过程中保持在无菌条件下的果蝇显示出较低的衰老率,这表明在这些模型设置中防止与年龄相关的菌群失调可以限制炎症,改善免疫稳态,促进健康。
非洲绿松石鳉鱼
利用自然短命的脊椎动物非洲绿松石鳉鱼(Nothobranchius furzeri)进一步证实了肠道微生物群在调节衰老过程中的影响。年轻的微生物群的慢性定植在中年鱼,诱导了长期有益的系统效应,导致脊椎动物的寿命延长。将年轻的供体微生物群定植在年老的鱼,与已知产生代谢产物的关键细菌属的存在有关,这些代谢产物既能维持免疫系统健康,又有抗炎作用。
小鼠
年轻小鼠菌群 → 早衰小鼠:特定菌发挥作用
将年轻供体的粪便微生物群移植到早衰小鼠中,可改善后者的健康状况和寿命,并将疣微菌门verrucomicrobia,Akkermansia muciniphila移植到早衰小鼠中,从而充分发挥有益作用。
此外,与年龄相关的Akkermansia muciniphila缺失与肠道完整性受损和胰岛素抵抗相关——这是一个通过微生物组-单核细胞-B细胞轴介导的过程。
除了对宿主代谢的影响,Akkermansia muciniphila还参与了调节抗原特异性T细胞反应和抗体产生来调节宿主免疫功能。
小鼠菌群 → 无菌小鼠:T细胞活化增加
用年轻或年老小鼠的肠道微生物群定植无菌(GF)小鼠。将老年供体微生物组移植给年轻小鼠足以促进肠道炎症、微生物产物向循环的渗漏和慢性低级别炎症的发生。作为全身低度炎症的结果,全身免疫区的T细胞活化增加。
年老菌群→年轻小鼠:巨噬细胞功能失调
另一项研究报告了类似的发现,在年轻的GF接受者中,移植一种老年微生物组可促进全身低度炎症。这些情况下的炎症进一步与巨噬细胞功能失调相关,巨噬细胞是炎症细胞因子的有力来源,其细菌杀灭活性较差。此外,基因或抗体介导的肿瘤坏死因子α(低度炎症的标志性细胞因子)的减少,可以预防年龄相关的肠道失调和相关的全身低度炎症。
年龄相关肠道免疫力下降:增加M细胞可恢复
最近的一项研究强调,年龄相关肠道免疫力下降可以通过操纵肠道微生物组增加M细胞数量来恢复。年老小鼠暴露于年轻的微生物群或鞭毛蛋白刺激都足以观察到这种效应:Peyer’s补丁中的M细胞成熟恢复,增强抗原摄取,年老小鼠肠道IgA反应增加。基于微生物组干预的M细胞修复确实依赖于肠道干细胞功能的改善,这表明修复老年肠道的再生能力可能对提高肠道免疫力有额外的好处。这些发现可能与改善口服疫苗应答或预防老年人胃肠道感染有关。
年龄相关肠道菌群改变影响造血
除了局部免疫调节外,年龄相关的肠道群落结构变化也可能对造血产生直接影响。对小鼠的研究表明,肠道微生物组的改变与造血系统的多谱系改变和多能祖细胞的抑制有关。
鉴于肠道微生物群通过促进造血功能密切参与了细菌感染的控制,与年龄相关的肠道微生物群平衡和多样性的变化可能导致老年人造血功能受损、更易感染和减少疫苗接种应答。然而,为了更好地理解年龄相关的生态失调对造血系统调节的影响,还需要更多的研究来支持这一假设。
总的来说,这些临床前模型系统的发现表明,校正与年龄相关的肠道菌群失调是有益的,为基于微生物组的治疗方法改善免疫系统功能、对抗衰老及相关疾病提供了理论依据。
虽然粪便微生物群移植(FMT)在动物模型中具有抗衰老特性(见上文),并且FMT已成功用于治疗男性复发性艰难梭菌感染,但在临床环境中提出FMT作为抗衰老策略仍存在一些障碍。更好地了解健康微生物组的特征(包括病毒组和真菌组)对于确保接受者的长期安全至关重要。
尽管我们对整个生命中肠道微生物群的理解取得了很大进步,但在疫苗学上的潜力尚未实现。鉴于微生物组严格调控免疫细胞的发育和功能,它可能最终影响疫苗的疗效。因此,由环境、社会经济、营养或卫生条件引起的肠道微生物群落结构的变化可以解释疫苗应答中的地理异质性。
对微生物群如何促进疫苗应答的机制更好地理解,可能有助于制定新的策略,以减少老年人感染性死亡。
直到最近,表明肠道微生物组影响疫苗接种反应的最佳证据来自使用无菌或缺乏微生物的小鼠的临床前模型系统。
研究人员探讨不同抗生素方案对抗原特异性体液免疫应答的影响,研究了克拉霉素、强力霉素和氨苄西林对小鼠破伤风类毒素(TT)、肺炎球菌多糖疫苗(PPV)、乙型肝炎病毒表面抗原(HBsAg)疫苗和减毒沙门氏菌活疫苗(Ty21a)的一级和二级抗体应答的影响。
有趣的是,抗原和抗生素的特异性反应都受到了影响。
克拉霉素和强力霉素抑制典型的T细胞依赖性和T细胞非依赖性抗体反应,而氨苄西林的效果较差或无效果。
此外,所有三种抗生素,特别是氨苄西林增强了对Ty21a-a减毒细菌粘膜疫苗模型的体液反应。在GF小鼠中进行疫苗接种实验的进一步工作证实了这些初步发现。
到目前为止,现有的研究主要集中在细菌,以及包括病毒、真菌、原生动物和古生菌在内的微生物群的其他组成部分。然而,仍然存在两个重要问题:
(一) 微生物群参与宿主免疫反应(特别是疫苗反应)的机制是什么?
(二) 这些发现对人类的影响是什么?
迄今为止,最令人信服的数据表明,肠道微生物群提供了佐剂的天然来源,能够调节宿主的系统和黏膜疫苗反应。
这一观点最初是在Nakaya等人的系统生物学报告中提出的,他们描述了在非佐剂流感疫苗接种三价灭活疫苗(TIV)后,人类toll样受体-5 (TLR5)的早期(第3天)和瞬时基因上调。TLR5的表达与TIV诱导的血凝抑制(HAI)滴度(滴度是稀释度的倒数)呈正相关。
随后,该研究小组证实TLR5对小鼠肠道微生物分泌的鞭毛蛋白的感应与TIV介导的应答有关。虽然直接给予鞭毛蛋白或有鞭毛的细菌移植可以提供天然佐剂来改善非佐剂流感疫苗的应答,但它不能与其他佐剂或活疫苗如破伤风-白喉-百日咳(Tdap)、黄热病(YF-17D)或重组乙型肝炎抗原(Recombivax HB)配合使用。
除TLR5外,另一种模式识别受体核苷酸结合寡聚结构域2 (NOD2)的特异性贡献,人血清白蛋白(HSA)抗原和霍乱毒素(CTX)佐剂鼻内刺激诱导的粘膜疫苗应答小鼠模型。在GF小鼠、抗生素处理小鼠或基因修饰缺乏NOD2信号的突变小鼠中,HSA特异性IgG反应持续下降。当这些动物接受一种被NOD2或表达MDP的细菌识别的肽聚糖MDP(muramyl dipeptide, MDP)时,HSA特异性IgG反应恢复。
虽然尚未使用人体相关疫苗,但这第二项工作表明,常驻微生物也可以增强用于粘膜疫苗接种方案的CTX的佐剂效果。需要进一步研究揭示肠道微生物群、其细胞壁成分或代谢物作为内源性疫苗佐剂的免疫能力,以放大对特定病原体的适应性免疫应答。
临床前和临床报告均观察到膳食纤维对功能免疫参数的积极影响,包括疫苗应答(表1),进一步支持基于其组成(细菌的性质)和或活性(分泌的代谢物的性质)的肠道微生物群的重要免疫调节潜力。
表1 益生元和/或益生菌临床干预试验探讨老年人免疫相关结果


Nabil Bosco & Mario Noti,Genes & Immunity, 2021
关于纤维消化产生的短链脂肪酸(SCFAs)免疫调节活性的大量文献证明了这一点。这些代谢物在局部或全身释放,参与宿主的一般生理过程。
肠道微生物组组成和功能的动态变化也可以解释疫苗应答中观察到的地理异质性。研究人员还报道了成人和老年人接种流感疫苗后体液免疫反应的显著差异(超过100倍)。
宿主遗传、营养状况、母乳喂养习惯以及卫生条件和/或之前接触过病原体,被提出用来解释疫苗免疫原性的差异。然而,近年来研究宿主微生物群的技术进步为这一领域提供了新的思路。
粪便微生物群特征研究表明,肠道菌群组成可能影响口服脊髓灰质炎、卡介苗、破伤风类毒素和乙肝病毒的疫苗接种效果。
此外,在某些国家的儿童中,小肠细菌过度生长(SIBO)的流行率高。SIBO还可能限制疫苗的性能,因为相关的吸收不良、肠道微生物群和宿主免疫细胞之间争夺关键营养物质以及或通过渗漏的肠道系统释放微生物分子。从南非收集的样本中观察到反应迟钝的全血细胞(标准TLR刺激试验),并将来自北美和欧洲儿童的年龄匹配样本进行了比较。
上述人体研究仅存在相关性。
最近进行了三项研究,以确定婴儿和成人肠道微生物群失调和疫苗应答之间的因果关系。
在第一个大型(n = 754)和良好对照的研究中,广谱抗生素治疗(阿奇霉素)减少了致病性肠道细菌的流行,但没有改善印度婴儿的口服脊髓灰质炎疫苗接种。
接下来,Harris等人在开放标签试验中使用广谱抗生素(万古霉素)或广谱抗生素(万古霉素、环丙沙星和甲硝唑)治疗66名荷兰健康成人,并研究TT、口服轮状病毒(RV)和多糖肺炎球菌(Pneumo23)疫苗的应答。虽然RV疫苗观察到一些积极的效果,抗生素治疗没有改善TT或肺炎23价反应。
最后,Hagan和他的同事进行了一项研究,22名年轻的成年人接受广谱抗生素(万古霉素、新霉素和甲硝唑)治疗,然后接受TIV疫苗。虽然这种治疗对以前接种过流感疫苗的成年人影响有限,但在11名以前没有接触过流感(接种疫苗或自然感染)的健康个体中进行的第二次试验提供了突破性的发现。那些首次接受抗生素治疗的人的体液免疫反应,特别是对H1N1流感毒株的免疫反应大大减弱。
这些数据证实了早期的小鼠研究,并证明抗生素驱动的生态失调导致了非佐剂TIV引起的疫苗应答的显著改变。用系统生物学方法进行的更深层次的分子图谱也揭示了一种特定的炎症基因特征,即与抗生素治疗相关的更活化的髓系树突状细胞。
合理设计以微生物为内源性佐剂的疫苗在疫苗学领域具有广阔前景。这些方法可能需要个性化和工程可逆性来管理脆弱个体的疗效和潜在并发症。鉴于饮食是塑造肠道微生物群的最有力因素之一,营养干预与益生菌和益生元促进多样化的微生物群来维持健康受到了相当大的关注。
食品的发展是为了促进健康或减少疾病风险,1980年代中期,出现了“功能食品”一词:希望支持以营养为基础的预防性方法,以提高生活质量,减少与老龄化人口相关的医疗成本。
由于肠道微生物与宿主免疫系统的密切相互作用,临床试验研究的提高老年人免疫力的功能食品主要包括益生元(如纤维)、益生菌、两者的结合(即共生)或分泌可溶性代谢物(也称为后生元,如短链脂肪酸)。
随着年龄的增长,免疫适应能力下降,老年人无力抵抗感染,无力对抗原挑战作出反应,大多数这些试验探讨了感染流行程度、严重程度和持续时间或特定的疫苗接种结果。
在已确定的400项临床研究中,研究人员根据其相关性(免疫读数和目标人群为60岁的老年人)保留了31项研究,并将其总结在表1中。
这些研究是在社区居民、疗养院居民或住院患者中进行,这些患者可以服用口服补充剂或需要管饲。
大多数研究(20项研究)是随机双盲安慰剂对照组,考虑经典的混杂因素,如年龄、性别,有时还有医疗条件、营养状况、感染史或疫苗接种史。
预防感染性并发症,特别是危重症或选择性手术患者中艰难梭菌相关腹泻是第二个有意思的领域。
在对老年人进行的13项研究中,6项呈阳性。它们积极的性质也可能表明,疾病预防可以通过人体免疫系统的不同方面来实现。
疫苗挑战研究通常测试适应性免疫系统的功能,但没有解决某些细菌感染中与年龄相关的先天免疫下降的后果。然而,最近在英国23家养老院进行的规模最大的研究,即“益生菌减少养老院居民感染”试验,LGG和BB12胶囊每天服用长达1年,对预防感染没有效果。在感染症状、抗生素使用、住院或死亡率方面没有发现差异。
总的来说,可能有多种原因,研究之间缺乏一致性,如菌株特定差异或剂量,以及益生元性质和数量,或受试者的年龄和医疗条件。
这些研究中没有一项对干预前和干预后的患者微生物群进行了广泛的分析。虽然这些数据可能作为独立报告存在,以描述成分的特性,在未来,对干预前后的人体微生物群和免疫参数进行系统的平行评估是至关重要的,以揭示相关的相互作用或因果关系。
虽然实际年龄是不可逆转的,但随着年龄的增长,与肠道微生物群、炎症和免疫衰老相关的衰老变化是免疫介导的慢性疾病的共同帮凶,这可能是维持免疫和健康的目标。
当消炎药和衰老疗法选择性地消除衰老细胞(衰老抑制剂)或抑制衰老相关分泌表型(衰老表型)时,作为抗衰老疗法的临床试验正在迅速发展(图2),它们的长期使用需要谨慎地平衡对传染病的易感性和潜在的其他副作用。
为什么减少全身低度炎症可以促进疫苗应答,而疫苗学的主流观点是佐剂通过促进局部炎症来改善疫苗应答,这一悖论还需要进一步的研究来解释。
图2 恢复肠道微生物稳态、减少炎症和免疫衰老以支持老年人免疫功能的方法
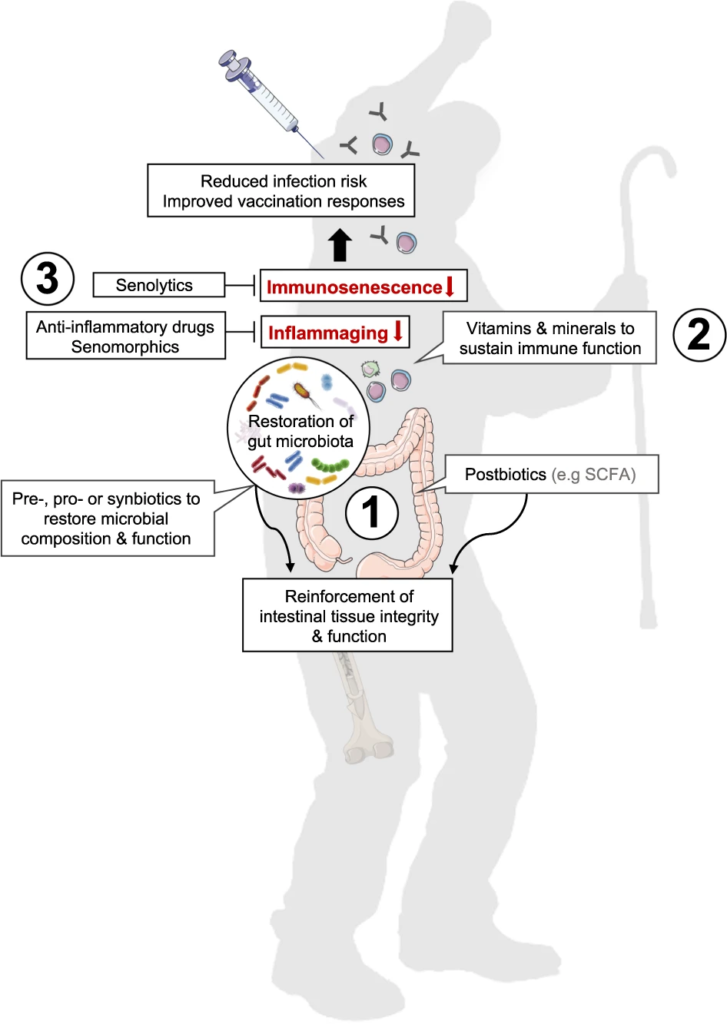
Nabil Bosco & Mario Noti,Genes & Immunity, 2021
(1)益生菌或益生元或合生元、后生元(如SCFAs)等进行干预可能有助于恢复与年龄相关的肠道微生物组成和功能下降
(2) 补充维生素和矿物质有助于正常的免疫细胞功能。
(3) 消炎药或老年药可能有助于减少炎症,而老年药直接消除老化细胞,燃料炎症。
单独或联合使用可能有助于增强宿主免疫,更好地控制感染,并随着我们年龄的增长产生适当的疫苗接种反应。
鉴于肠道微生物对宿主免疫系统的影响越来越大,可以合理推测,通过个性化营养或补充,恢复与年龄相关的肠道微生物丰富度和功能的下降,可能是一种对抗免疫适应性功能下降的预防措施。
在这种情况下,已在临床环境中测试了能够通过支持肠道屏障完整性或调节炎症过程来增强免疫力的益生菌、益生元、后生元或合生元(表1)。
然而,由于研究、菌株特异性差异或剂量、益生元的性质和数量或受试者的年龄和医疗条件之间缺乏一致性,因此很难验证这种方法在增强与年龄相关的宿主免疫适应性下降方面的有效性。
肠道微生物群是一个有待挖掘的宝藏,衰老学也不例外。正如许多临床前研究所证明的那样,恢复年轻的微生物群可以通过维持免疫和健康跨度来恢复老年宿主的活力。
因此,更好地了解肠道微生物群落结构和相关代谢组的动态变化,这些改变如何影响细胞免疫网络,以及这些通路如何被治疗靶向,将对未来加强甚至恢复老化免疫系统的策略产生广泛的影响。随着人口老龄化的加剧,迫切需要这样的解决方案来支持健康老龄化,减缓不断增长的医疗费用。
健康饮食
Omega-3脂肪酸
随着年龄的增长,神经细胞萎缩,向大脑提供的营养丰富的血液供应减少,Omega-3脂肪酸,尤其是二十二碳六烯酸(DHA)可以促进神经细胞之间的有效电信号传递,减少炎症,甚至可以改善精神集中度并与记忆力丧失作斗争。
较小的脑体积与阿尔茨海默氏症以及正常衰老有关。研究人员发现,血液中omega-3脂肪酸EPA和DHA水平较高的绝经后妇女的脑体积也更大。
富含Omega-3脂肪酸的食物:
鱼类:鲱鱼,沙丁鱼,鲭鱼,鲑鱼,大比目鱼,鳟鱼
绿叶蔬菜: 抱子甘蓝,菠菜,芝麻菜,薄荷,羽衣甘蓝和豆瓣菜
油: 亚麻籽油,正大籽油,鳕鱼肝油和磷虾油
其他:蛋、核桃等
黄酮类化合物
几乎所有的水果,蔬菜都含有类黄酮,具有许多健康益处,包括减少炎症,降低心脏病风险和湿疹症状,对衰老的大脑有益。有研究发现,食用大量浆果(类黄酮含量较高)的老年妇女将记忆力下降的时间推迟了两年以上。
类黄酮含量高的食物:
浆果: 蓝莓,草莓和黑莓
绿叶蔬菜: 菠菜,羽衣甘蓝和豆瓣菜
彩色农产品:胡桃南瓜,牛油果,李子,红葡萄
其他:咖啡,黑巧克力,红酒
维生素E
防止细胞损伤。已有几项研究发现,维生素E可以延缓轻度到中度阿尔茨海默病的进展
富含维生素E的食物:
坚果和种子:杏仁,山核桃,花生酱,花生,榛子,松子和葵花籽
油:小麦胚芽油,葵花籽油,红花油,玉米油,大豆油
绿叶蔬菜:菠菜,蒲公英嫩叶,唐莴苣,萝卜叶
运动
走楼梯:
瑞士一项研究发现,久坐不动的人把乘电梯换成走楼梯,过早死亡的风险可以降低15%。哈佛早期的研究显示,与每周爬楼梯少于10层的人相比,每周能爬35层或更多的楼梯能显著提高寿命。
骑车:
骑自行车上班是一种环保又健康的方式,在一天中挤出锻炼时间在户外,还能节省点油钱。
游泳:
一项数据发现,经常游泳的男性比久坐不动的男性死亡率低50%,游泳者的死亡率也比运动时走路或跑步的男性低。
每天锻炼15分钟:
2011年的一项研究发现,与久坐不动的人相比,每天15分钟锻炼可以平均延长3年的寿命。
保护牙齿
如果牙齿一直很敏感,那么随着年龄的增长,牙齿会越来越容易脱落,这会影响老年生活品质。请保持日常牙科护理的最佳状态,防止蛀牙。
保持良好的生活习惯
保证充足的睡眠:
NIH的数据显示,每晚持续睡眠少于7个小时的成年人患高血压,心脏病,肥胖,糖尿病和抑郁症的风险更高。另外,晚上睡不好会抑制重要激素的释放,这些激素可以修复细胞和组织,抵抗疾病和感染,睡不好会使身体无法自然康复。
尽可能避免吸烟:
戒烟可以减少许多疾病风险。
适当缓解压力:
压力容易让人变老。尝试放弃明显的压力源,适当安排休息时间,
经常锻炼记忆力
记忆就像肌肉一样可以训练;如果充分利用自己的记忆并定期使用它,记忆力可以磨练到老。比如每天尝试做一个填字游戏,以帮助建立和维护词汇量和记忆力。
《阿尔茨海默氏病杂志》上的一项研究显示,吃蓝莓可以增强记忆力和学习能力,喝苹果汁可以通过防止重要的神经递质的衰退来改善记忆力。
选择适宜的养老居住地
可以选择气候宜人的地方居住,良好的空气质量有助于保持健康,环境条件佳也有利于保持好心情。此外尽量选择拥有良好的经济和医疗体系的地方,帮助维持积极长寿的生活。
做好健康管理
定期进行肠道菌群健康检测或其他检查,及时了解自身健康状况,包括哪些风险,需要注意事项等,做好健康管理,享受健康幸福的晚年生活。
相关阅读:
主要参考文献:
Hägg S, Jylhävä J, Wang Y, Xu H, Metzner C, Annetorp M, et al. Age, frailty, and comorbidity as prognostic factors for short-term outcomes in patients with coronavirus disease 2019 in geriatric care. J Am Med Dir Assoc. 2020;21:1555–9.e2.
Khan SS, Singer BD, Vaughan DE. Molecular and physiological manifestations and measurement of aging in humans. Aging Cell. 2017;16:624–33.
Crooke SN, Ovsyannikova IG, Poland GA, Kennedy RB. Immunosenescence and human vaccine immune responses. Immun Ageing. 2019;16:25–16.
Butler CC, Lau M, Gillespie D, Owen-Jones E, Lown M, Wootton M, et al. Effect of probiotic use on antibiotic administration among care home residents: a randomized clinical trial. Jama. 2020;324:47–56.
Bosco Nabil,Noti Mario,The aging gut microbiome and its impact on host immunity.[J] .Genes Immun, 2021
Book: Mayo Clinic Family Health Book, 5th Edition
Wang J, Varghese M, Ono K, Yamada M, Levine S, Tzavaras N, Gong B, Hurst WJ, Blitzer RD, Pasinetti GM. Cocoa extracts reduce oligomerization of amyloid-β: implications for cognitive improvement in Alzheimer’s disease. J Alzheimers Dis. 2014;41(2):643-50.
James V. Pottala, Kristine Yaffe, Jennifer G. Robinson, Mark A. Espeland, Robert Wallace, William S. Harris Neurology Feb 2014, 82 (5) 435-442
Dysken MW, Sano M, Asthana S, et al. Effect of Vitamin E and Memantine on Functional Decline in Alzheimer Disease: The TEAM-AD VA Cooperative Randomized Trial. JAMA. 2014;311(1):33–44.
Berryman CE, West SG, Fleming JA, et al. Effects of Daily Almond Consumption on Cardiometabolic Risk and Abdominal Adiposity in Healthy Adults With Elevated LDL-Cholesterol: A Randomized Controlled Trial. Journal of the American Heart Association. January 2015.