-

 国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子
国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子

 二级病原微生物安全实验室
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业

谷禾健康
氨基酸(AA)稳态对人体健康至关重要,其紊乱与多种疾病的发生和进展密切相关,如2型糖尿病和炎症性肠病(IBD),并且常常是治疗结果的决定性因素。
肠道微生物群可能通过多种机制调节宿主氨基酸的可用性,例如影响消化酶(如胰蛋白酶)的活性。肠道微生物群定殖还可能改变肠道通透性,从而影响胃肠道中游离氨基酸的运输和吸收。此外,肠道微生物可能直接利用或代谢肠道中的氨基酸,或合成并向宿主提供氨基酸。以往研究表明,肠道微生物组的变化(或其存在)可能影响肠道氨基酸谱。然而,参与调控宿氨基酸稳态的关键菌株及代谢基因仍未完全明确。
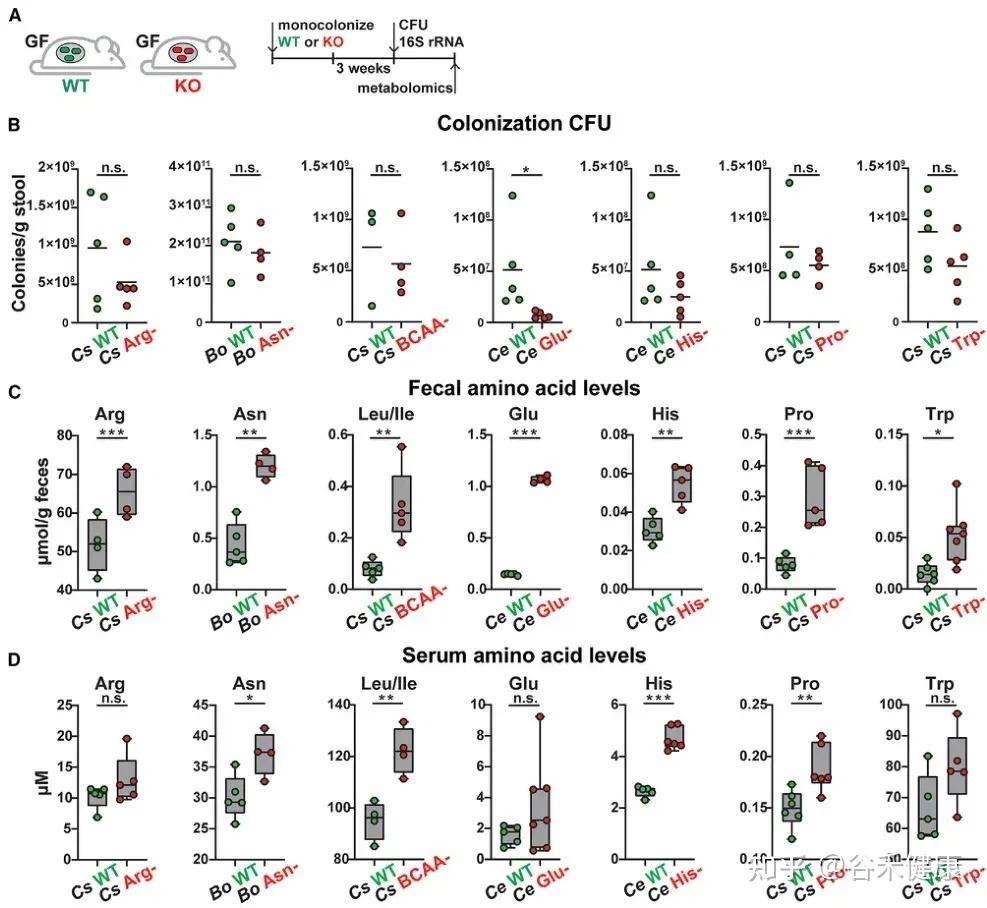
今天分享的这一篇发表在《Cell Host & Microbe》期刊上的研究论文”Microbiota metabolism of intestinal amino acids impacts host nutrient homeostasis and physiology”,通过比较Met无菌(GF)小鼠与无特异病原体(SPF)小鼠的分解物谱已证明,肠道细菌定殖会影响胃肠道(GI)中游离氨基酸谱,还通过高效代谢肠道氨基酸来重塑宿主氨基酸的格局。为确定责任微生物/基因,该研究开发了基于代谢组学的检测法,筛查了104个共生菌,并识别出能高效利用氨基酸的候选基因,并发现这些基因调控了肠道和循环氨基酸的可用性。
结果显示,不同氨基酸具有特定的细菌消费者,消耗效率差异明显。例如,天冬酰胺和谷氨酸被拟杆菌和部分梭菌高效利用,而芳香族氨基酸(色氨酸、酪氨酸)及支链氨基酸(BCAAs)仅为部分厚壁菌门成员所代谢,效率较低。值得注意的是,支链氨基酸与色氨酸代谢相关的微生物基因还能通过调节外周血清素间接影响宿主葡萄糖平衡。
总体而言,该研究首次系统揭示了肠道微生物群编码的氨基酸代谢活动对宿主营养稳态的深远影响,为理解微生物介导的氨基酸利用及其对宿主代谢调控机制提供了关键分子依据。
氨基酸稳态对人类健康至关重要,其紊乱与2型糖尿病和炎症性肠病(IBD)等多种疾病的进展密切相关。虽然传统上认为肠道和肝脏是调控营养代谢的主要器官,但最新研究发现,肠道微生物群通过代谢氨基酸能够深刻重塑宿主的氨基酸谱。
★该研究的核心意义在于:
•揭示了肠道微生物群通过代谢肠道氨基酸影响宿主氨基酸稳态的分子机制;
•鉴定了高效消耗氨基酸的特定肠道微生物及其代谢基因;
•阐明了微生物群氨基酸代谢基因如何通过外周血清素调节宿主葡萄糖稳态;
•为通过调节肠道微生物群代谢活性改善人类健康提供了新的靶点和策略。
▸ 肠道微生物群对宿主氨基酸水平的影响
研究首先通过比较无菌(GF)小鼠、无特定病原体(SPF)小鼠以及接受SPF微生物群移植(FMT)的GF小鼠,证实了肠道微生物群定植显著降低了肠道和循环氨基酸水平。
靶向代谢组学分析显示,SPF小鼠或接受SPF微生物群的GF小鼠肠道和循环氨基酸浓度普遍降低,这与之前的研究结果一致,表明微生物对肠道氨基酸的利用可能是影响宿主氨基酸稳态的主要途径之一。
▸ 高效代谢氨基酸的肠道微生物筛选
为识别高效代谢肠道氨基酸的微生物,研究者建立了一种基于活细胞的高通量代谢组学筛选方法,对104种肠道共生菌进行了系统分析。该方法综合考虑多个因素:
•生长阶段差异:绘制各菌株生长曲线并进行参数拟合,确保在可比生长阶段进行筛选;
•时间动态分析:分别在15、30和60分钟测定氨基酸消耗量,以精确评估不同菌株对特定氨基酸的利用速率;
•营养条件优化:在测定缓冲液中除氨基酸外,补充无机氮源、微量矿物质、维生素及额外碳源(如葡萄糖),以避免因营养限制导致的氨基酸非特异性快速消耗。
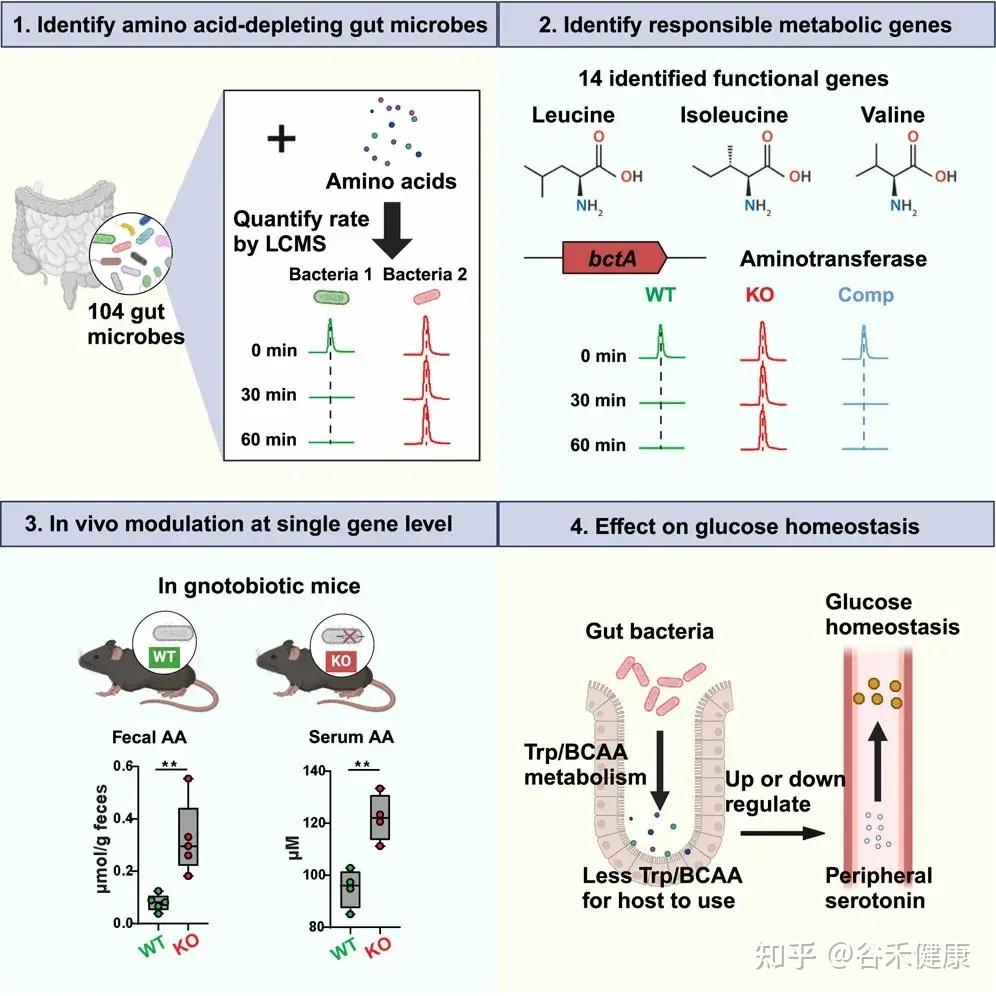
研究流程图展示了从识别消耗氨基酸的肠道微生物、鉴定负责的代谢基因、单基因水平的体内调节到对葡萄糖稳态影响的完整研究路径。
▸ 微生物代谢基因的鉴定与功能验证
研究利用CRISPR-Cas9和ClosTron等基因编辑技术,在系统发育多样的肠道微生物中开展大规模基因缺失分析,以鉴定氨基酸代谢的关键基因。筛选标准包括:
1.代谢基因编码的酶以氨基酸为直接底物;
2.这些基因存在于高效代谢氨基酸的肠道菌株中;
3.携带相关基因的菌株能稳定定植于无菌(GF)小鼠肠道,并降低宿主体内相应氨基酸水平。
通过构建基因敲除和互补菌株并结合代谢组学分析,研究识别出多种氨基酸代谢相关基因,并在单菌定植的GF小鼠模型中验证了其对肠道及血清氨基酸水平的调控作用。
▸ 高效代谢氨基酸的肠道微生物鉴定
通过大规模筛选,研究发现不同肠道微生物对氨基酸的代谢效率和偏好存在显著差异:
鉴定高效利用氨基酸的肠道微生物候选者
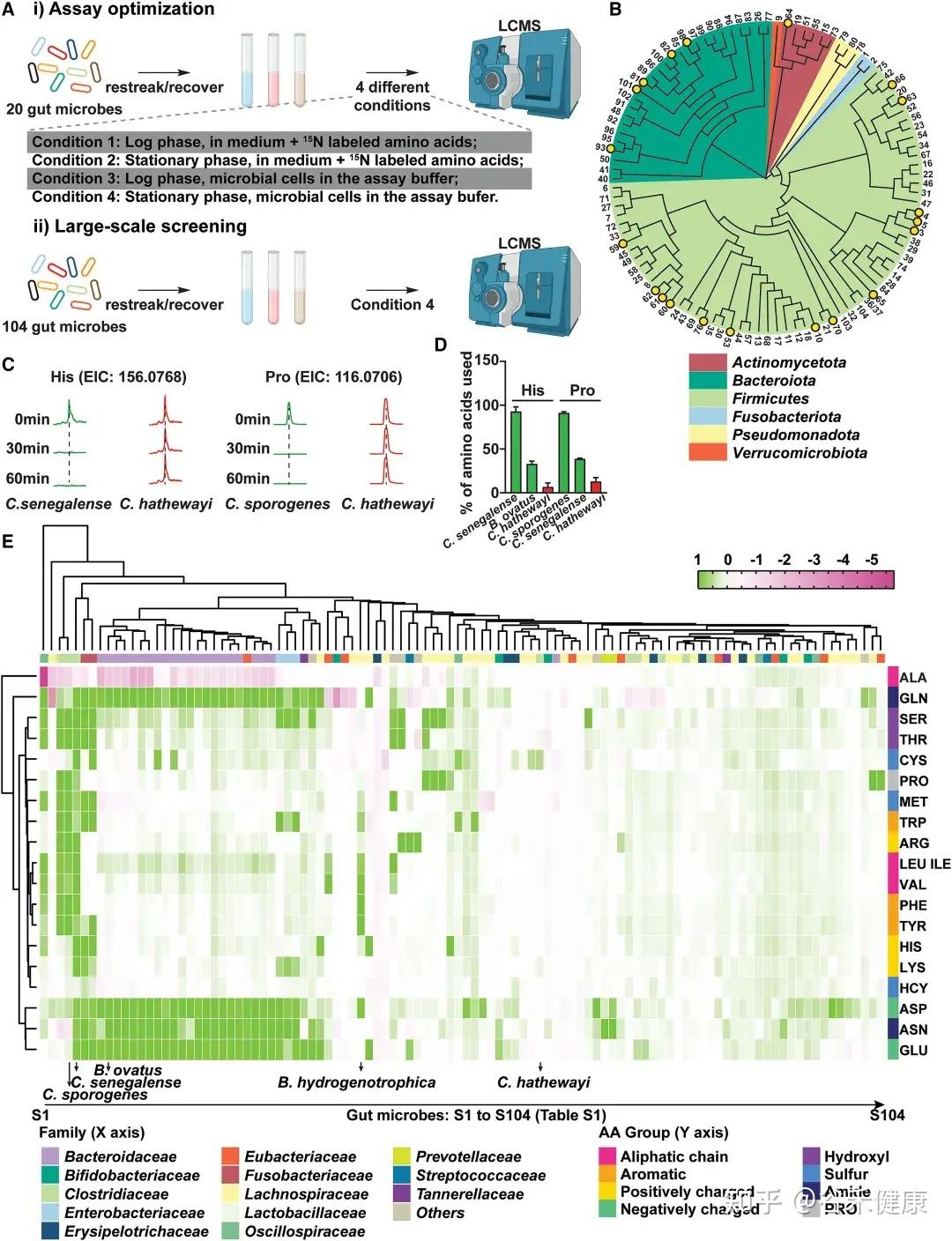
热图展示了104种肠道微生物对不同氨基酸的代谢情况,揭示了微生物系统发育与氨基酸代谢效率之间的复杂关系。
★ 关键发现包括:
•梭菌属微生物:如Clostridium sporogenes ATCC15579和Clostridium senegalense DSM25507,可高效代谢多种氨基酸,包括精氨酸、芳香族氨基酸、支链氨基酸、组氨酸、赖氨酸和脯氨酸等。
•拟杆菌属共生菌:作为肠道多糖的主要利用者,也能有效代谢天冬酰胺(Asn)、天冬氨酸(Asp)和谷氨酰胺(Gln)等氨基酸。
•氨基酸特异性代谢:不同氨基酸具有特定的细菌“消费者”,消耗效率差异明显。天冬酰胺和谷氨酸通常被拟杆菌和部分梭菌快速利用,而芳香族氨基酸(色氨酸、酪氨酸)及BCAA仅被部分厚壁菌门或梭菌较低效地代谢。
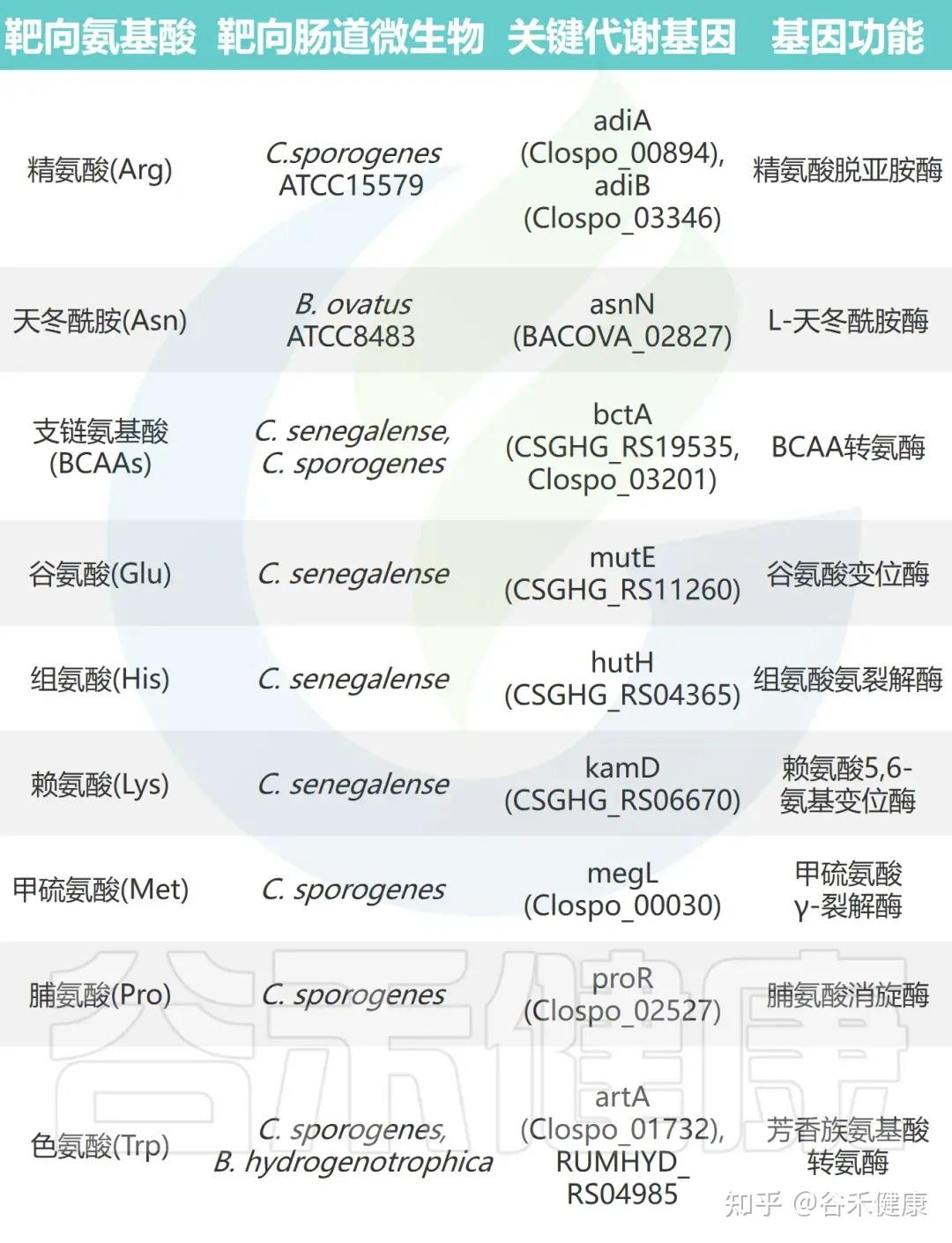
▸ 氨基酸代谢相关微生物基因的鉴定
通过对高效代谢氨基酸的肠道微生物进行大规模基因缺失分析,研究鉴定出多种关键氨基酸代谢基因。这些基因编码的酶包括转氨酶、脱羧酶、消旋酶和氨裂解酶等,具有以下特征:
•功能冗余性:单株细菌常含多个具有相似功能的基因。例如,C. sporogenes 携带两种精氨酸脱亚胺酶基因(adiA 和 adiB),单基因敲除影响较小,而双敲除可完全消除其精氨酸代谢能力。
•底物特异性:代谢酶展现高度底物选择性。例如,C. sporogenes 的转氨酶 ArtA 特异作用于色氨酸和酪氨酸,但对苯丙氨酸代谢影响极小。
•催化多样性:不同类型的酶参与多条代谢途径。支链氨基酸和色氨酸经转氨途径代谢,脯氨酸通过还原反应被 C. sporogenes 利用,而谷氨酸、甲硫氨酸、丝氨酸和苏氨酸则由氨裂解酶快速分解。
•功能可转移性:这些酶的催化功能具有可转移性,在其他肠道共生菌中表达其编码基因可赋予相应氨基酸的消耗能力。
▸ 微生物代谢基因调控宿主氨基酸稳态
通过将野生型菌株及其代谢基因缺失突变体分别单菌定植于GF小鼠,研究发现这些基因显著影响宿主肠道及循环系统中的氨基酸水平:
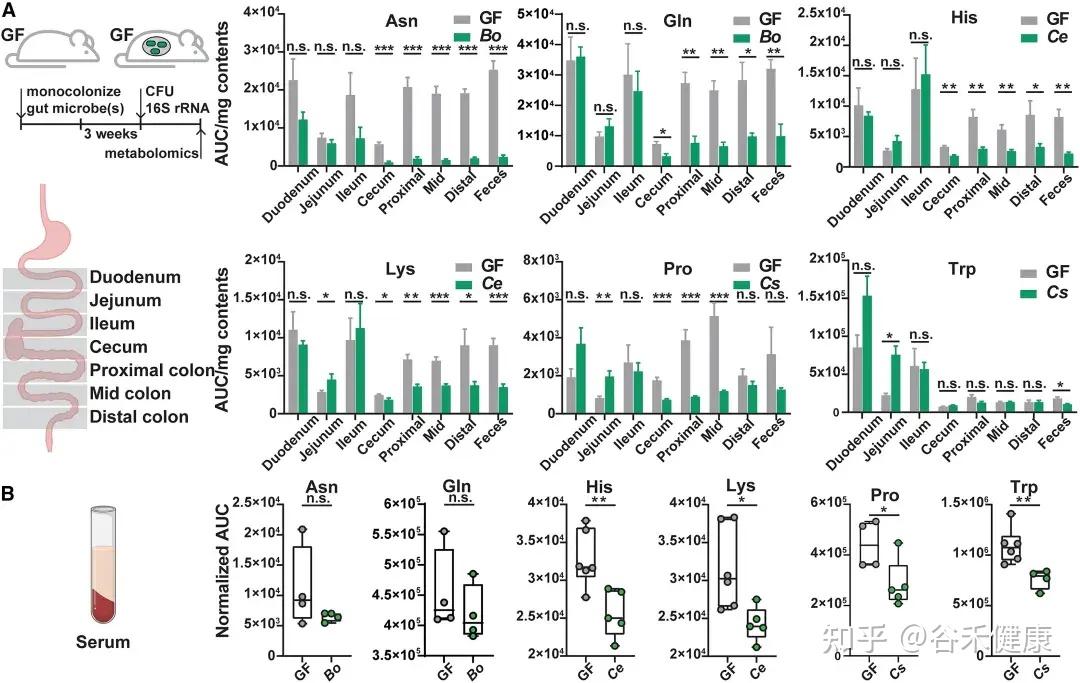
无菌小鼠单一定植B. ovatus、C. senegalense和C.sporogenes野生型菌株后,肠道和血清中特定氨基酸浓度显著降低。
★ 主要发现包括:
•肠道氨基酸稳态调控:多数鉴定出的微生物代谢基因或酶影响肠道氨基酸平衡。由于突变体的相应代谢途径被阻断,肠道内容物和粪便中相关氨基酸显著积累。
•循环氨基酸谱变化:部分代谢基因还影响宿主血清氨基酸水平。定植 Asn、Leu/Ile、Pro 和 His 利用缺陷突变体的无菌小鼠,其血清中对应氨基酸明显升高。
•系统性调节的复杂性:循环氨基酸水平的变化并不总与微生物在肠道中的代谢能力一致。例如,定植 C. sporogenes 精氨酸利用缺陷突变体可显著提高肠道精氨酸水平,但对循环精氨酸影响有限,提示肠上皮或肝脏代谢等额外机制亦参与精氨酸的全身稳态调控。
单基因水平的微生物群氨基酸代谢调控
单基因水平的体内调节实验显示,氨基酸代谢基因缺陷突变体定植的无菌小鼠粪便和血清中相应氨基酸水平显著升高。
▸ 微生物氨基酸代谢通过外周血清素调控宿主葡萄糖稳态
研究发现,微生物中支链氨基酸(BCAAs)和色氨酸代谢相关基因可通过外周血清素间接调节宿主葡萄糖稳态。将 C. sporogenes 野生型、BCAA 缺陷型及色氨酸缺陷型突变体分别单菌定植于 GF 小鼠,并进行口服葡萄糖耐量试验(OGTT),结果显示以下关键发现:
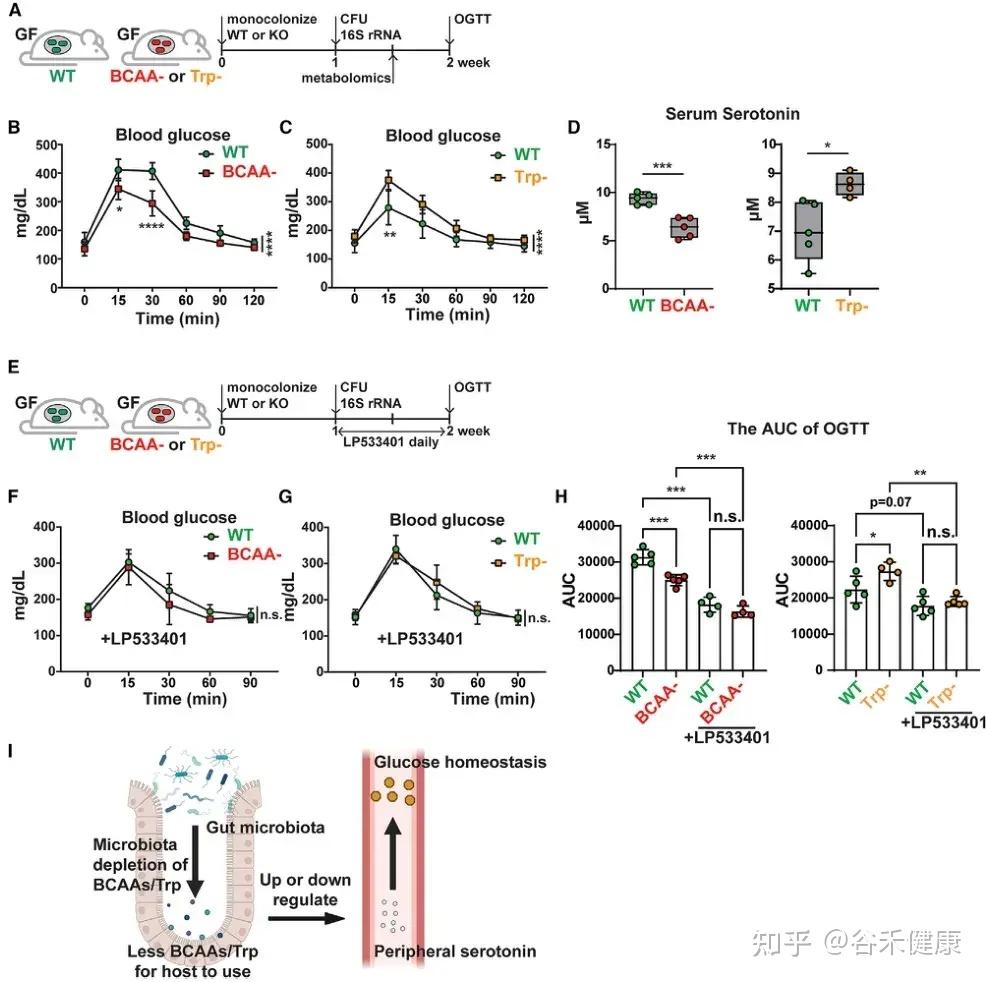
微生物支链氨基酸和色氨酸代谢通过调节外周血清素水平影响宿主葡萄糖稳态的机制示意图及实验验证。
•BCAA代谢与葡萄糖耐受性:定植 BCAA 缺陷突变体的小鼠葡萄糖耐受性显著增强,伴随血清素水平下降。
•色氨酸代谢与葡萄糖耐受性:定植色氨酸代谢缺陷突变体的小鼠表现相反,葡萄糖耐受性下降,血清素水平升高。
•血清素介导机制:应用 Tph1 抑制剂 LP533401 阻断外周血清素合成后,野生型与突变体定植小鼠间的葡萄糖处理差异显著减小,证实微生物氨基酸代谢通过调控外周血清素影响宿主葡萄糖稳态。
这一发现揭示了肠道微生物群、氨基酸代谢、外周血清素及葡萄糖稳态间的复杂关联,为理解代谢性疾病的机制提供了新思路。
!
研究局限性
尽管该研究取得了重要发现,但仍存在一些局限性:
1.微生物氨基酸代谢的多样性:研究主要关注肠道微生物对氨基酸的消耗,而其合成、转运等途径也可能影响宿主氨基酸稳态。体外测定仅反映特定条件下的代谢效率,难以全面代表其在宿主体内的作用。
2.模型简化的局限性:研究采用单菌定植的 GF 小鼠模型,未来应在更接近生理状态的模型(如定植简化微生物群的 GF 小鼠)中验证这些机制。
3.复杂微生物群中基因操作的挑战:虽然本研究成功在宿主定植背景下操纵单一氨基酸代谢途径,但在复杂微生物群中实现精确基因操作仍需进一步技术突破。
4.宿主响应机制研究不足:研究揭示了野生型与突变体共生菌定植小鼠间的生理差异,但尚需深入解析相关宿主细胞类型及信号通路的具体调控机制。
未来的研究在单基因分辨率下揭示了肠道微生物群如何通过代谢肠道氨基酸影响宿主氨基酸稳态。研究鉴定出多种积极代谢肠道氨基酸的细菌,并通过大规模基因缺失分析确定了其关键代谢基因。功能实验显示,这些代谢途径通过直接调控宿主肠道和循环氨基酸的生物利用度,或间接影响外周血清素合成,从而调节宿主的葡萄糖代谢和整体营养稳态。
研究表明,定植微生物群可能以“竞争”模式与宿主共同调控营养平衡,为理解其分子层面的调节机制提供了新视角。更值得关注的是它揭示了一个新的通路:微生物氨基酸利用 → 改变宿主血中氨基酸/代谢产物 → 改变激素/神经递质(如外周 5‑HT)→ 改变葡萄糖代谢等生理功能,所以这些菌在肠道里的存在,更可能是:微调宿主营养和代谢状态,而不是简单的“好/坏”。
参考文献:
Li TT, Chen X, Huo D, Arifuzzaman M, Qiao S, Jin WB, Shi H, Li XV; JRI Live Cell Bank Consortium; Iliev ID, Artis D, Guo CJ. Microbiota metabolism of intestinal amino acids impacts host nutrient homeostasis and physiology. Cell Host Microbe. 2024 May 8;32(5):661-675.e10.
谷禾健康
在快节奏的现代生活中,我们面临着的食品安全挑战也在升级。全球化的食品供应链让美食跨越千山万水来到我们的餐桌,但也为病原菌的传播提供了更多机会。每一餐美食的背后,都可能潜藏着看不见的威胁。从夏日烧烤到家庭聚餐,食源性病原菌如同”隐形杀手”般悄然存在,威胁着我们的健康。
据世界卫生组织统计,全球每年有超过6亿人因食用被污染的食物而患病。在中国,食源性疾病同样影响着数百万人的健康。
有时候我们认为腹泻、呕吐只是”吃坏肚子”的小毛病,忍忍就过去了。然而这可能掩盖了严重的健康风险。食源性病原菌感染不仅会引起急性胃肠炎症状,还可能导致:
脱水和电解质紊乱:特别是对老人和儿童;
肠道菌群失调:破坏肠道微生态平衡;
免疫系统受损:降低机体抵抗力;
慢性健康问题:如肠易激综合征、炎症性肠病等后遗症。
产气荚膜梭菌、蜡样芽孢杆菌、肉毒杆菌、副溶血性弧菌、志贺氏菌等是最常见且危险的食源性病原菌。它们各具特色:有的耐高温,有的耐酸,有的产生致命毒素,有的传播力极强。
通过了解这些常见食源性病原菌的特性和传播规律,有助于我们识别和避免食用易受污染的食物,掌握正确的储存、烹饪和食用方式,同时监测肠道健康状况及时发现菌群异常变化。因为每个人的肠道菌群构成都是独特的,就像指纹一样。这意味着不同的人对同一种病原菌的易感性和抵抗力也不相同。有些人可能对某种病原菌特别敏感,而另一些人则相对耐受。
▶▷▶▷▶▷
▮ 产 气 荚 膜 梭 菌 ▮
产气荚膜梭菌(Clostridium perfringens)是一种革兰氏阳性杆状细菌,会引起食源性腹泻,是工业化国家常见的食源性致病菌之一,全年均有腹泻病例报道。它广泛存在于环境中,对人和动物均具致病性。
作为一种能够形成芽孢的厌氧细菌,产气荚膜梭菌在芽孢形成过程中会产生肠毒素(CPE),其A型到E型菌株与不同疾病相关。其中人类坏死性肠炎是一种致命的危及生命的疾病,会导致小肠溃疡。
该菌能在短期内快速繁殖、在相对较高的温度范围内生长、芽孢能抵抗恶劣环境条件、以及尽管是厌氧菌却具有耐氧能力,这些特性使得产气荚膜梭菌具有很强的环境适应性和致病潜力。
• 梭菌属中具有重要临床意义的致病菌种还有:
破伤风梭菌(Clostridium tetani)
肉毒梭菌(Clostridium botulinum)
巴氏梭菌(Clostridium baratii)
艰难梭菌(Clostridium difficile)
Clostridium argentinese
产气荚膜梭菌的常见栖息地是土壤、水、污泥、污水、食物(主要是生肉和冷冻肉)、受污染的设备等。产气荚膜杆菌也存在于人和动物的肠道中,尤其是老年人的粪便中。
产气荚膜梭菌需要多种氨基酸与维生素,最适生长温度为43–45°C。富含蛋白的肉类尤其适合其繁殖,特别是猪肉、牛肉和家禽产品;用酱料烹制的污染风险很高。
在加热和冷却过程中,如果食物在室温下没有足够的冷却温度,它会增殖并产生毒素。通常,学校食堂,医院,餐馆,监狱和集会是梭菌爆发的高风险场所。
产气荚膜梭菌在应激条件下孢子形成时产生肠毒素,已知可产生14种毒素与5类肠毒素(A–E型)。其还分泌4类细胞外毒素:α、β、ε、iota。
产气荚膜梭菌毒素和降解酶的作用
doi: 10.1080/21505594.2021.1886777.
A型:在人类中引起气性坏疽和腹泻相关的感染性腹泻,在动物中引起肠毒血症、坏死性肠炎和急性胃扩张。
C型:在人类中导致坏死性肠炎(空肠炎),在小猪、羔羊、小牛和鸟类中导致肠毒血症和坏死性肠炎。
B、D、E型:尚未证实致人类疾病;在动物可致肠炎与肠毒血症。
此外,该菌还能产生溶素O(θ毒素)与胶原酶(κ毒素),也需警惕。
与暴发有关的因素是大量准备食物、烹饪温度不够(食物的核心区域没有适当烹饪)、准备好的食物和食用期间受到污染。
集中爆发经常发生在提前准备大量食物的地方,如医院和学校食堂、监狱、聚会和研讨会。
案例调查显示,一道在室温下放置20小时且未再加热的羊肉菜是感染来源。产气荚膜杆菌还从羔羊肉、感染者粪便及尸检样本中被分离出。某校食堂因饮用制备后置于室温的受污染奶昔,导致77例腹泻。
产气荚膜梭菌只有在摄入含有约 10⁷-10⁹个营养细胞/克 的污染食物时才引起感染。
⑴第一阶段:胃肠道定植
-营养细胞抵抗胃酸
-在小肠内生长、增殖和产孢
⑵第二阶段:肠毒素作用机制
靶点结合:大量肠毒素与肠上皮细胞紧密连接处的 claudin蛋白 结合。
复合物形成:结合宿主膜蛋白,经构象变化形成 CPE复合物。
细胞损伤:
-细胞崩解和分离
-肠绒毛尖端损伤
-上皮细胞脱落
-绒毛缩短
膜损伤效应:
-膜上形成孔道
-电解质大量泄漏
-导致细胞溶解
-肠道吸收能力下降
-细胞骨架崩塌
⑶第三阶段:严重并发症
高浓度CPE (>10μg/ml):引起坏死(oncosis)
β-毒素作用:
-引起细胞毒性坏死性肠炎
-影响自主神经系统
-导致肠黏膜出血性坏死
-在真核细胞膜形成孔道
这种多重毒素协同作用使得产气荚膜梭菌食物中毒具有较强的致病性和复杂的临床表现。
主要症状多在食用重度污染食物后8–24小时出现。
常见症状包括腹部绞痛、水样腹泻、呕吐、恶心和发烧,大多数情况下通常在12至24小时内消退。致死率低于0.05%;但婴幼儿、老年人及慢性病患者风险较高;该菌可能侵入并导致严重溃疡,最终因急性脱水而死亡。
婴儿猝死综合征(SIDS)也是由产气荚膜梭菌与睡眠期间大脑的缺陷有关。
检测原理:产气荚膜梭菌食物中毒通过检测粪便样本中的肠毒素和营养细胞/芽孢数量来诊断。
⑴肠毒素检测
培养基法:熟肉培养基 → 巯基乙酸盐流体培养基(10ml) → Duncan-Strong产孢培养基
分子检测方法:通过PCR、多重PCR以及高通量测序手段检测样本中产气荚膜梭菌。例如谷禾健康依托高通量测序的tNGS技术可以特异性的识别粪便样本中的产气荚膜梭菌。
(来源:谷禾健康16S Plus版报告)
生物学方法
小鼠致死试验:评估毒素致死活性
Vero细胞毒性试验:检测细胞毒性效应
⑵免疫学检测
凝胶扩散法
对流免疫电泳
ELISA法
酶联免疫吸附试验进行CPE定量
注意事项:样本蛋白酶活性可降低IgG敏感性,需用1%血清白蛋白保护
⑶分子生物学方法
PCR技术:常规PCR、实时PCR
LAMP:环介导等温扩增
⑴治疗策略
一般治疗
抗生素使用原则
重症治疗
坏死性肠炎需要:
补液治疗
⑵预防控制措施
关键预防原则:烹调后立即食用
时间-温度控制
食品安全措施
▶▷▶▷▶▷
▮ 蜡 样 芽 孢 杆 菌 ▮
蜡样芽孢杆菌(Bacillus cereus)是一种致病性食源性微生物,广泛分布于自然界,如植物、土壤和昆虫及哺乳动物的胃肠道。
也常见于食品加工环境,其耐受性内生孢子可在恶劣条件下存活,污染原料与食品。
该菌可致两型食物中毒:腹泻型(在小肠产多种肠毒素)与呕吐型(摄入含耐热毒素的食物)。亦可引发机会性感染,如菌血症、败血症、肺炎、脑膜炎、胃炎、肝衰竭、肝坏死与脑水肿。
• 其他食源致病性芽孢杆菌:
B. subtilis(枯草芽孢杆菌):可致腹泻、呕吐;
B. licheniformis(地衣芽孢杆菌):常见食物中毒病因;
B. pumilus(短小芽孢杆菌):污染食品致胃肠炎。
• 其他特殊芽孢杆菌:
B. weihenstephanensis:强低温生长能力;
B. anthracis(炭疽芽孢杆菌):炭疽病原,极危;
B. mycoides(蕈状芽孢杆菌):形态特殊;
B. pseudomycoides(假蕈状芽孢杆菌):与蕈状近缘;
B. thuringiensis(苏云金芽孢杆菌):生物杀虫剂,偶可致病。
这些芽孢杆菌都具有形成芽孢的能力,使其在环境中具有很强的抵抗力和传播能力。
⑴基本特征
–革兰阳性菌
–产孢菌
–需氧至兼性厌氧
-有运动性
-杆状细菌
血培养的革兰氏染色蜡样芽孢杆菌
doi: 10.1128/CMR.00073-09.
⑵生长条件
生长温度范围:8-55°C
最适温度:25-37°C
pH范围:4.9-9.3
耐盐浓度达7.5%
内生孢子耐高温、辐射、干燥和消毒剂
⑶污染源
-从原料到包装储存的全程食品加工污染
-生物技术设备和机器污染
-土壤栖息,易传播至蔬菜和农作物
⑷常见污染食品
-大米、小麦、面食、面粉
-乳制品、肉制品、香料
-婴儿食品、鱼类、汤类
-蔬菜和水果
⑸流行病学特点
-全球性分布
-年度持续发生
-与饮食文化相关的地区差异
⑹易感人群
老年人、低胃酸患者
⑺防控挑战
其孢子高耐受性与污染源多样性要求从源头到终端的全程管控,尤其要严格温度控制并预防交叉污染。
蜡样芽孢杆菌产生蛋白质毒素,即肉毒毒素和催吐毒素。
⑴肉毒毒素(腹泻毒素)
当摄入的营养细胞在小肠增殖时产生腹泻毒素。该毒素对蛋白酶敏感,如链霉蛋白酶、胃蛋白酶、胰蛋白酶和胰凝乳蛋白酶。
潜伏期通常为摄入后8–16小时,病程多持续24–48小时。症状以轻度水样腹泻和腹部痉挛为主;少数儿童或免疫低下者可出现血性腹泻与坏死性肠炎,进而致肝功能衰竭和脑水肿。
与肠道疾病相关的三种染色体编码肠毒素包括:
Hbl(溶血素BL):蜡样芽孢杆菌食物中毒的主要毒力因子,在小肠通过渗透溶解形成跨膜孔。
Nhe(非溶血性肠毒素):与 Hbl 同源,亦为三组分成孔毒素。
CytK(细胞毒素K):原型样β-桶成孔毒素,可致出血性腹泻与坏死性肠炎。
⑵呕吐毒素
呕吐型起病更急且更重,由小分子环状耐热肽引起,潜伏期为摄入受污染食物后约2–5小时。
其症状类似金黄色葡萄球菌食物中毒:恶心、呕吐、腹部痉挛,通常持续约24小时。催吐毒素作用机制尚不明,但高度耐热,可经受油炸、烘烤、煮沸和微波等烹调过程。
环境因素(温度、pH、大气成分、营养来源、食物稠度)亦影响毒素的产生。
⑴培养方法
实验室培养基如营养琼脂或血琼脂
⑵ELISA
ELISA技术在商业上被用于测定毒素,但它在评估蜡样芽孢杆菌的产毒活性方面不准确。它只检测一种毒素,溶血素BL或两种无毒蛋白质。
⑶反向被动乳胶凝集(RPLA)肠毒素试验
将样品煮沸以使其在生物学上稳定,从而产生阳性结果。该试验可识别溶血素B组分,但在存在高葡萄糖浓度的情况下,无法检测到毒素。
⑷PCR或高通量测序
通过检测蜡样芽孢杆菌DNA序列,来分析其丰度和毒力基因的存在。
蜡样芽孢杆菌食物中毒多为自限性,通常24–48小时内自行缓解。建议卧床休息并补充液体;重症时可在医生指导下使用抗生素,如克林霉素、万古霉素、庆大霉素或氯霉素。
蜡样芽孢杆菌耐热性强,常规烹调可杀灭营养细胞,但孢子仍可存活,故应在孢子萌发前切断污染。
食品应在储存前迅速冷却、食用前充分加热;芽孢杆菌属难以在高酸性环境存活。保持良好食品处理习惯与卫生。
▶▷▶▷▶▷
▮ 肉 毒 杆 菌 ▮
肉毒杆菌(Clostridium botulinum)是一种生长在缺氧环境下的细菌,在罐头食品及密封腌渍食物中具有极强的生存能力,是毒性最强的细菌之一。
肉毒杆菌中毒是由摄入肉毒毒素(BoNT)导致的致命性食源性麻痹性疾病,虽罕见但病死率高,微量毒素即可致命。
中毒来源包括食物中预先形成的BoNT或肠道内产生的BoNT。已知毒素分为A–G七型,可致人畜发病,其中A、B、E型与人类食源性肉毒中毒最相关。
肉毒杆菌为厌氧、产芽孢的革兰阳性菌,孢子广泛存在于土壤、污水、泥浆及湖海沉积物中,也见于陆生和水生动物肠道。
肉毒杆菌的孢子形成和发芽
doi: 10.1080/21505594.2023.2205251.
其孢子易污染食物并产生毒素。蜂蜜和糖浆曾被报道受其孢子污染,是婴儿肉毒中毒的主要来源。加工不当的罐头亦为重要来源,因孢子可耐受高温。其他高风险食物包括湖鱼、海鱼,以及烟熏、盐腌和发酵肉制品。
A型和B型毒素常见于土壤与生物肥料,故可能污染蔬果,尤其是加热不足的蔬菜制品。
症状通常在摄入后18–38小时出现,先累及胃肠道,常见腹痛、恶心、呕吐、腹泻,偶见便秘。随后出现神经系统表现:弛缓性麻痹、对称性下行无力、构音不清、吞咽困难、眩晕、视物模糊、口干及眼外肌无力。
婴儿肉毒中毒多见于1岁以下,因肠道菌群未成熟,毒素可在肠内产生。表现为哭声微弱嘶哑、上睑下垂、吮吸差、抬头困难与肌无力。
伤口型肉毒中毒为梭菌在伤口、脓肿等处定植,孢子发芽并产毒。其症状包括低血压、呼吸困难、眼肌麻痹、共济失调与瞳孔固定性散大,少见,通常在暴露后3–6天发病。
肉毒杆菌毒素曾在第二次世界大战期间被美国,日本军队用作生化武器。
1991年埃及因食用传统腌鱼 fesaikh 中的E型毒素发生大规模疫情,报告91例、死亡18例。
约90%的肉毒中毒病例见于美国,每年报告约9–10起暴发,常与家庭自制食品相关,尤以自制罐藏、瓶装及油浸食品为主。
口服或肠道内生成的肉毒毒素需先抵御胃酸,经肠道到达十二指肠与空肠。随后经内吞入血,分布至周围神经系统。
毒素进入神经元后作为锌依赖性蛋白酶,切割SNARE蛋白(可溶性N-乙基马来酰亚胺敏感因子附着受体)。其在神经肌肉接头终末结合并抑制乙酰胆碱释放,阻断神经传递,导致弛缓性麻痹。
肉毒毒素中毒的分子步骤
doi: 10.1038/srep23707.
⑴生物测定
小鼠生物测定是检测肉毒毒素最敏感、应用最广的方法:通过注射毒素评估其毒性,小鼠常在4小时内出现腹部颤动、蜂腰样腹部、四肢瘫
一个人表现出类似的肉毒中毒症状应立即置于重症监护与胃肠外营养供应。
如果怀疑有毒素,应在24小时内服用抗毒素,因为它能在到达神经肌肉接头之前中和血液中的所有游离毒素。
在伤口肉毒杆菌中毒的情况下,伤口应该用抗生素治疗,如青霉素。
以上都需要咨询医生确定。
食品应充分加热(≥121°C)并冷藏(≤4°C);加入柠檬酸等酸化剂可降低罐装食品中孢子形成的风险。
避免食用可疑的生或未充分加热食物,并避免其接触伤口。
▶▷▶▷▶▷
▮ 副 溶 血 性 弧 菌 ▮
副溶血性弧菌(Vibrio parahaemolyticus)是导致人类急性肠胃炎的主要病原体之一,通常发生在食用生的、未煮熟的和交叉污染的海鲜产品时。
该菌常见于热带海洋与沿海环境,可在滤食性贝类(牡蛎、蛤蜊、贻贝)肠道黏附并繁殖。除胃肠炎外,还可引起伤口与耳部感染,免疫低下者暴露海水时可发生败血症。
与之相关的人类致病弧菌还包括霍乱弧菌(Vibrio cholerae)与创伤弧菌(Vibrio vulnificus)。在日本及多国亚洲地区,副溶血性弧菌每年占食物中毒病例的约20%–30%。
其在全球范围内是胃肠炎的重要病因之一,亟需关注其流行情况、毒力因子及健康影响,并完善鉴定技术以降低风险。
–革兰氏阴性
-曲线杆状
-非孢子形成者的兼性厌氧菌
-轻度嗜盐,氧化酶阳性
-最佳温度30至35°C
-pH值范围为6.8至10.2。
副溶血性弧菌是海洋嗜盐菌,依靠单极鞭毛在水中游动,并可附着于鱼、蟹、虾、龙虾、浮游动物等水生动物表面。
当食用被副溶血性弧菌污染的生的和未煮熟的海鲜时,它会引起人类急性胃肠炎。当开放性伤口或伤口暴露于受污染的海水时,也会引起感染。
其他来源是副溶血性弧菌强毒株从海产品或设备到其他产品的交叉污染。
副溶血性弧菌按抗原性分为体细胞(O)和荚膜(K)抗原。其主要毒力因子包括粘附素、耐热直接溶血素(TDH)、TDH相关溶血素(TRH)及III型分泌系统(T3SS-1、T3SS-2)。
副溶血性弧菌的结构和毒力因子
doi: 10.3389/fmicb.2015.00437.
此外,它具有两类鞭毛用于运动,并可形成有助于在恶劣环境中生存的荚膜。感染多经粪-口途径,通过细菌表面粘附因子黏附宿主细胞。
TDH通过与红细胞膜结合并在其表面打孔致溶血;同时通过形成通道使细胞外Ca²⁺内流、细胞内Cl⁻分泌增加,导致渗透压升高、细胞形态与功能紊乱,最终细胞膨胀死亡。
TRH与TDH共同介导细胞毒性并诱发肠道感染;不耐热溶血素(TLH)主要用于种属鉴定,对红细胞溶血作用较弱。早期研究还显示,尿素酶是trh⁺菌株的重要辅助毒力因子,有助于在胃肠道定植与致病。
注:尿素酶指可以催化尿素,导致尿素分解成氨和二氧化碳的一种酶,在临床上明确幽门螺杆菌就是人体胃内能够产生大量尿素酶的细菌。
第三型分泌系统(T3SS 1)在组织细胞感染期间诱导自噬和细胞毒性。它连续引起自噬、细胞起泡、溶解最后导致死亡。
在我国东部沿海省份的部分暴发病例中,副溶血性弧菌感染相对较高。并且在亚洲、欧洲、非洲和美洲国家,类似病例的爆发已被频繁报道。
1950年,日本首次爆发副溶血性弧菌病,报告了272例急性胃肠炎,其中20人死亡。
1997年至1998年,美国报告了700多起食用受污染的生牡蛎的病例。
副溶血性弧菌食物中毒多发于夏季(6—10月),因温暖水域有利于其繁殖。
螃蟹、虾、龙虾、贝类、牡蛎、蛤蜊和金枪鱼是高风险的海产品,必须在食用前彻底煮熟。
副溶血性弧菌中毒典型的症状是水样腹泻、恶心、呕吐、腹部痉挛、发烧和发冷。进食受病原体污染的食物后,潜伏期约为12至24小时,并于5至7天内消退。
免疫功能低下的个体可能需要约10至15天的长时间才能恢复,因为疾病是自限性的,不需要药物治疗。
有些严重的患者可能会出现粘液或便血,血压下降,失去知觉,皮肤苍白、发紫,甚至死亡。此时应立即就医。
⑴菌株培养鉴定
使用含十二烷基硫酸钠(SDS)、烷基苯磺酸盐和胆汁盐的选择性增菌培养基可用于副溶血性弧菌的培养与分离。以碱性蛋白胨水为代表的增菌液具最佳pH和较高NaCl浓度,有利于弧菌生长。
⑵分子检测技术
副溶血性弧菌菌株的鉴定也可采用分子技术,如常规表型与生化试验、PCR及DNA检测。高通量宏基因组测序或靶向tNGS可用于分型与菌株区分。
该病多为自限性,但重症时需药物治疗。常用措施包括口服补液及应用四环素等抗生素;充足饮水以补充电解质、卧床休息亦有助于恢复。
副溶血性弧菌食物中毒的预防与控制:
–避免食用生或未熟透的贝类、牡蛎等海产品。
-处理海产品时注重个人卫生,生熟分开,防止交叉污染。
-皮肤有割伤或创口者应避免接触海水或微咸水,并尽量使用防水敷料。
▶▷▶▷▶▷
▮ 志 贺 氏 菌 ▮
志贺氏菌属(Shigella)是引起急性胃肠道感染的原因,这种感染会损害回肠和结肠上皮并引起细菌性痢疾。志贺氏菌感染多见于贫困、欠发达且缺乏卫生与医疗设施的国家和地区。
志贺氏菌病死亡率较高,尤以5岁以下营养不良儿童为甚。患者在腹泻期间因大量丢失电解质和血液而营养受损,可能致死。
• 志贺氏菌属分为四个血清群:
A群:痢疾志贺菌(S.dysenteriae)
B群:福氏志贺菌(S.flexneri)
C群:鲍氏志贺菌(S.boydii)
D群:宋内志贺菌(S.sonnei)
其中,S.dysenteriae 1型(志贺毒素产生菌)可引起严重的流行性痢疾,具有较高的致死率;而其他血清型通常引起相对较轻的痢疾症状。
–革兰阴性杆菌;无芽孢、无鞭毛(不运动)、无荚膜;兼性厌氧。
-最适生长温度为 37°C,生长范围约 10–40°C;可耐受一定盐度与酸性环境。
-低感染剂量:约 10–100 个菌体。
-对巴氏灭菌温度敏感(≥60°C 易被杀灭)。
-对紫外线和化学消毒剂敏感。
-能在不利的物理和化学条件下存活。
⑴病原菌储存宿主
–人类肠道是志贺菌的主要天然储存库;
-动物感染极为罕见,几乎不作为传染源。
⑵传播途径
–粪—口传播为主要途径
–人与人直接接触
–被污染的水和食物
-受污染的灌溉水可污染蔬果
⑶易感人群
–儿童和婴幼儿为高危人群
–免疫力低下者
–密切接触者(如家庭成员、托幼机构)
⑷流行特征
–发展中国家发病率较高
-与卫生设施不完善和个人卫生不良相关
-季节性:夏秋季高发
2003-2013年中国志贺氏菌的流行情况
doi: 10.3389/fcimb.2016.00045.
症状通常在摄入细菌后1–3天出现,视剂量可在12小时至7天内发病。
典型症状包括发热、食欲减退、腹痛、血性或水样腹泻、结肠炎、乏力与不适。大量水样便可致脱水,但较少见。伴随的厌食与营养不良需积极处理,是5岁以下儿童死亡的主要原因。少数患者可出现神经症状,如嗜睡、不自主运动和头痛。
这种疾病一般在5至7天内自行消退,但感染者可能会在很长一段时间内无症状地在粪便中排出细菌,并可能造成传播感染的威胁。
⑴侵入阶段
经口进入消化道
胃酸耐受:可抵抗低 pH
穿越小肠:因上皮更新快、液流迅速,难以定植
最终定植于大肠
⑵细胞侵袭机制
M细胞摄取:进入并被吞噬泡包裹
逃逸入胞:自吞噬泡逃出,进入细胞质
巨噬细胞:被吞噬后诱导其凋亡
⑶细胞内增殖与扩散
在大肠上皮内快速繁殖
经质膜包裹进行细胞间直接传播
继而感染邻近上皮细胞
⑴志贺毒素(主要由S.dysenteriae产生)
作用机制:经胞吞进入宿主细胞
毒性效应:抑制蛋白合成,致细胞死亡
临床表现:出血性腹泻
⑵脂多糖(LPS)
释放时机:细菌死亡破裂时释放
损伤效应:破坏肠道上皮组织
炎症反应:激发强烈的炎症应答
⑶炎症因子
IL-12释放:S. flexneri特别容易诱导IL-12产生
线粒体损伤:导致细胞坏死
组织破坏:引起肠道组织广泛炎症和溃疡
⑷病理损伤特点
组织破坏:主要由宿主免疫反应引起,而非病原菌直接作用
粘膜溃疡:免疫系统过度反应导致的继发损伤
炎症反应:局部组织出现严重炎症和水肿
⑴细菌培养方法
及时采样:出现痢疾症状后立即采集粪便标本
采集时机:感染初期阶段最佳,病原菌体外存活时间有限
样本处理:需要立即检测,避免病原菌死亡
⑵免疫学检测方法
酶免疫测定(EIA):成本低、操作简便。
⑶分子诊断技术
PCR/测序:基因检测快速、可靠,可区分不同志贺菌种。
⑷目标基因
ipaH基因:侵袭相关基因
virA基因:毒力基因
iral基因:调节基因
LPS基因:脂多糖合成基因
质粒DNA:毒力质粒
⑴抗菌治疗
首选:氟喹诺酮类(如环丙沙星)、大环内酯类(如阿奇霉素)
原则:尽早治疗,依据病情调整;监测耐药性
注意:具体用药请遵医嘱
⑵支持治疗
口服补液盐:轻中度脱水
静脉输液:重度脱水或电解质紊乱
营养支持:维持免疫功能
⑶环境与个人卫生
-完善卫生设施与排污系统
-安全处置粪便,防止污染
-保护水源,避免饮用水污染
-手卫生:勤洗手,使用肥皂或消毒剂
⑷食品安全
-生熟分开
-充分加热
-适当冷藏
-果蔬反复清洗
⑸重点人群
儿童与婴幼儿:加强护理与监测
食品从业人员:定期健康检查
集体生活人群:(学校、托幼机构等)
食源性病原菌虽“微小”,却以顽强的生存力、多样的毒力因子与隐匿的传播路径,对公众健康构成持续挑战。产气荚膜梭菌的耐受与产毒、蜡样芽孢杆菌的双型中毒与广泛污染、肉毒杆菌的致命神经毒素、以及志贺氏菌的低感染剂量与高传播力,共同勾勒出从农田到餐桌、从环境到个体的风险图谱。
守护肠道健康,关键在于两端同时用力:一端是系统性防控——完善卫生基础设施、保障饮水安全、规范食品加工与冷链管理、强化时间—温度控制与流程监测;另一端是个人与社区层面的良好习惯——勤洗手、生熟分开、充分加热、规范冷藏、创口防护与及时就医。面向重点人群(婴幼儿、老年人、免疫低下者及集体生活人群),还需更精准的健康教育与常态化监测。
科学认知让风险可见,规范行动让风险可控。以循证为底、以习惯为桥、以制度为盾,我们就能在快节奏与全球化的餐桌上,稳稳地守住每一个人的“肠道安全线”。
主要参考文献
Popoff, M. R.(2014). CLOSTRIDIUM | Detection of Enterotoxin of Clostridium perfringens. Encyclopedia of Food Microbiology, 474–480.
McLauchlin, J., & Grant, K. A. (n.d.). Clostridium botulinum and Clostridium perfringens. Foodborne Diseases, 41–78.
Miyamoto, K., & Nagahama, M. (2016). Clostridium: Food Poisoning by Clostridium perfringens. Encyclopedia of Food and Health, 149–154.
Chukwu, E. E., Nwaokorie, F. O., Coker, A. O., Avila-Campos, M. J., Solis, R. L., Llanco, L. A., & Ogunsola, F. T. (2016). Detection of toxigenic Clostridium perfringens and Clostridium botulinum from food sold in Lagos, Nigeria. Anaerobe, 42, 176–181.
Granum, P. E., & Lund, T. (2006). Bacillus cereus and its food poisoning toxins. FEMS Microbiology Letters, 157(2), 223–228.
Tewari, A., & Abdullah, S. (2014). Bacillus cereus food poisoning: international and Indian perspective. Journal of Food Science and Technology, 52(5), 2500–2511.
Parihar, H. S. (2014). Bacillus cereus. Encyclopedia of Toxicology, 353–354.
Batt, C. A. (2014). BACILLUS | Bacillus cereus. Encyclopedia of Food Microbiology, 124–128.
Carlin, F. (2016). Bacillus cereus and Other Bacillus sp. Causing Foodborne Poisonings, Detection of. Encyclopedia of Food and Health, 301–306.
Drobniewski, F. A. (1993). Bacillus cereus and related species. Clinical Microbiology Reviews, 6(4), 324–338.
Tewari, A., & Abdullah, S. (2014). Bacillus cereus food poisoning: international and Indian perspective. Journal of Food Science and Technology, 52(5), 2500–2511.
Marrollo, R. (2016). Bacillus cereus Food-Borne Disease. The Diverse Faces of Bacillus Cereus, 61–72.
Pavlik, B., Hruska, E., Van Cott, K. et al. Retargeting the Clostridium botulinum C2 toxin to the neuronal cytosol. Sci Rep 6, 23707 (2016).
Notermans, S. H. W., Stam, C. N., & Behar, A. E. (2014). CLOSTRIDIUM | Detection of Neurotoxins of Clostridium botulinum. Encyclopedia of Food Microbiology, 481–484.
Aureli, P. (2017). Botulism. International Encyclopedia of Public Health, 254–262.
McLauchlin, J., & Grant, K. A. (n.d.). Clostridium botulinum and Clostridium perfringens. Foodborne Diseases, 41–78.
Austin, J. W. (2016). Clostridium: Occurrence and Detection of Clostridium botulinum and Botulinum Neurotoxin. Encyclopedia of Food and Health, 155–159.
Lebeda, F. J., Dembek, Z. F., & Adler, M. (2018). Foodborne Botulism From a Systems Biology Perspective. Foodborne Diseases, 275–308.
Horowitz, B. Z. (2005). Botulinum Toxin. Critical Care Clinics, 21(4), 825–839.
Letchumanan, V., Chan, K.-G., & Lee, L.-H. (2014). Vibrio parahaemolyticus: a review on the pathogenesis, prevalence, and advance molecular identification techniques. Frontiers in Microbiology, 5.
Wang R, Zhong Y, Gu X, Yuan J, Saeed AF, Wang S. The pathogenesis, detection, and prevention of Vibrio parahaemolyticus. Front Microbiol. 2015 Mar 5;6:144.
Risk assessment tools for Vibrio parahaemolyticus and Vibrio vulnificus associated with seafood, Food and Agriculture Organization of the Unites Nations, ISSN 1726-5274.
Baker-Austin, C., Jenkins, C., Dadzie, J., Mestanza, O., Delgado, E., Powell, A., … Martinez-Urtaza, J. (2020). Genomic epidemiology of domestic and travel-associated Vibrio parahaemolyticus infections in the UK, 2008–2018. Food Control, 115, 107244.
Yaashikaa, P. R., Saravanan, A., & Kumar, P. S. (2016). Isolation and identification of Vibrio cholerae and Vibrio parahaemolyticus from prawn (Penaeus monodon) seafood: Preservation strategies. Microbial Pathogenesis, 99, 5–13.
Bolen, J. L., Zamiska, S. A., & Greenough, W. B. (1974). Clinical features in enteritis due to vibrio parahemolyticus. The American Journal of Medicine, 57(4), 638–641.
Mumy, K. L. (2014). Shigella. Encyclopedia of Toxicology, 254–255.
Ashkenazi, S. (2018). Shigella Species. Principles and Practice of Pediatric Infectious Diseases, 842–846.e2.

谷禾健康

如果近期出现鼻子、喉咙、肺部相关症状,如咽喉痛、发烧(体温迅速升高通常超过38.5℃)、咳嗽、流鼻涕、鼻塞、乏力、发冷、浑身疼痛等情况,很有可能是甲流。
根据中国疾病预防控制中心2025年第1周流感监测周报显示(截至2025年1月5日),本周南、北方省份流感病毒检测阳性率继续上升,甲型H1N1流感占比99.4%。
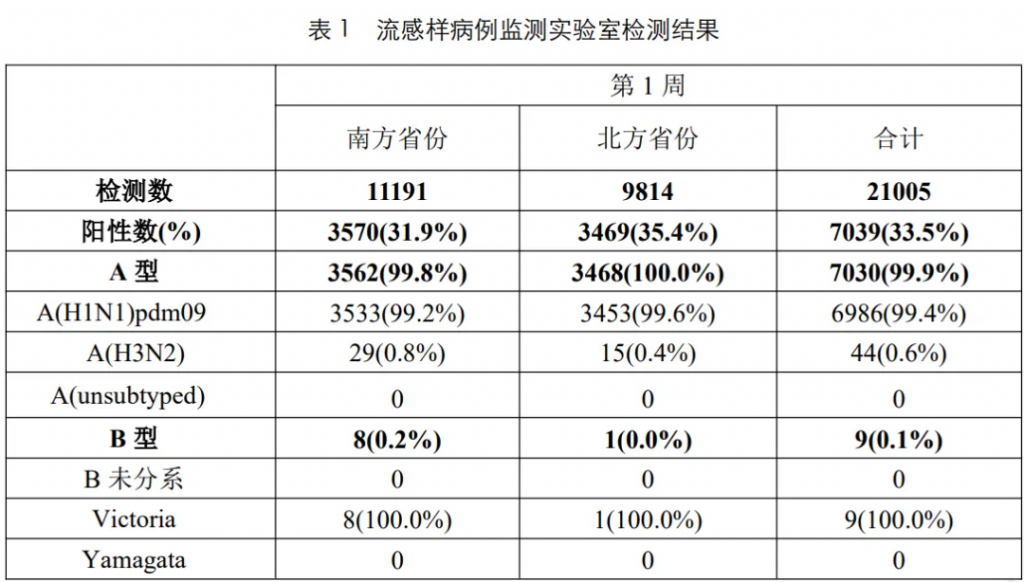
来源:Chinacdc
甲型流感是一种高度传染性的呼吸道感染疾病,其传播速度快、感染范围广,给公共卫生带来了巨大挑战,甲流每年导致300万-500万例严重感染和25万-50万例死亡。甲型流感主要通过咳嗽、打喷嚏或说话时的飞沫传播,症状可持续几天到几周。
近年来的研究发现,甲流感染可能会破坏肠道微生物群的平衡,而肠道生态失调可能会影响甲流患者的疾病进展。
调节肠道菌群可以通过肠-肺轴促进抗甲流病毒的免疫效应。这包括在感染的早期阶段促进甲流病毒清除,并在后期阶段减少炎症损伤。此外,肠道菌群还可以通过维持肠道屏障功能、产生具有保护作用的代谢产物(如短链脂肪酸、氨基酸、胆汁酸)等,促进宿主的免疫反应。
本文我们主要来详细了解一下甲流的症状、原因、潜伏期、危险因素、感染过程,包括肠道菌群及其代谢产物与甲流的关系,以及通过调节肠道菌群及其代谢产物来防治流感的潜在策略,包括益生菌、植物化学物质、中药等干预措施,这为甲流的预防和治疗提供了新的策略。
流感病毒有四种类型:甲(A)、乙(B)、丙(C)、丁(D)。甲型和乙型流感病毒几乎每个冬天都可能会引起季节性疾病流行。
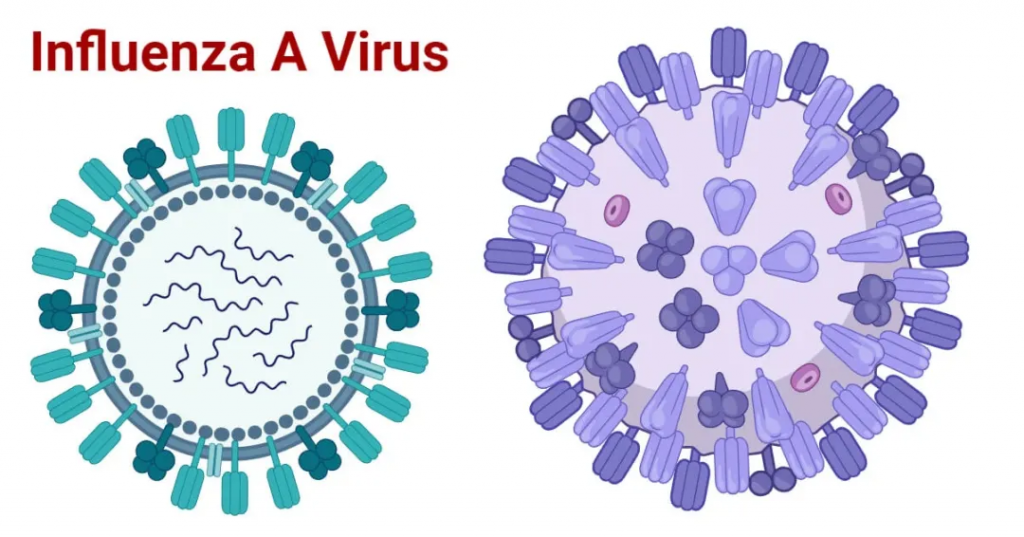
甲流病毒是一种包膜病毒,包膜含有两种糖蛋白,一种叫血细胞凝集素(HA),另一种叫神经氨酸酶(NA)。
注:有18种不同的血凝素亚型和11种不同的神经氨酸酶亚型(分别为H1至H18和N1至N11)。
这两个可以排列组合形成不同的流感亚种,比如说H1N1,就是血细胞凝集素的1型和神经氨酸酶的1型组合。目前经常在人群中传播的甲型流感病毒亚型包括H1N1和H3N2。
甲型流感主要影响呼吸系统,包括鼻子、喉咙、肺。症状可能会突然出现,可能包括:
严重的症状
在严重病例中,甲流引起一系列症状,可能较少见:
图源:mypositiveparenting
并非所有流感都出现呼吸道症状
医生提醒,呼吸道症状是流感的典型症状,但以恶心、呕吐为主的消化道症状患者也大有人在。此外,还有部分患者会出现反应迟钝、嗜睡、躁动等精神系统症状。以上这些症状可能合并出现,也可单独出现,容易给人误导,导致没有及时就诊,延误流感的治疗。
当病毒感染上呼吸道(鼻窦,口腔和喉咙)或下呼吸道(肺部)的组织时,可能会发生甲型流感。病毒可以在这些组织内复制,从而引起症状。
这种流感具有高度传染性。你可以通过咳嗽、打喷嚏或在他人附近说话将病毒传染给他人。
甲流病毒通常在每个流感季节迅速变异。免疫系统可能无法立即抵抗新形式的病毒,即使以前得过流感,你也可能会有几天的症状。
如果被感染,通常会在接触后一到四天(潜伏期)出现流感症状,平均 2 天。
流感可以持续几天到两周。
发烧和身体疼痛等症状可能会突然出现,但通常比其他症状消失得更快。咳嗽或流鼻涕会持续更长时间。
普通感冒由鼻病毒等多种病原体导致,传染性弱,发病没有明显的季节特征,症状表现为鼻塞、流涕、喷嚏、咳嗽等,一般不发热或仅有低热,没有全身性症状,5-7天即可自愈,很少有并发症出现。
甲流传染性强,容易大范围流行,发病季节特征明显。患者表现为高烧、头痛、乏力,并伴有全身肌肉酸痛等。甲流有一定自限性,但容易引起其他并发症,包括肺炎、中耳炎、心肌炎、脑膜炎等严重并发症。
甲流和乙流之间有几个区别:
其他两种流感:
任何人都可能患上甲型流感。某些人出现严重症状和并发症的风险更大,例如:
大多数甲流病例在几周内消退,甲流可能更严重,并导致一些人的并发症,并发症可能包括:
耳部感染:对甲型流感的免疫反应会引起炎症,这会影响并导致咽鼓管中的液体积聚。这根管子把喉咙后部和中耳连接起来。这种液体会吸引可能导致耳部感染的细菌。尤其在儿童中较为常见。
鼻窦感染:甲型流感可能会导致鼻窦充血和炎症,从而吸引可能导致感染的细菌。
脑炎:免疫反应可能导致这种大脑炎症。甲流病毒可以通过血液或神经系统进入脑部,引起脑组织的炎症和损伤。
脑膜炎:表现为头痛、呕吐、意识障碍等症状。
心肌炎: 流感病毒可以直接攻击心肌细胞,引起心肌受损,表现为心悸、胸闷、胸痛、乏力等症状。
肌肉炎症:对流感过度活跃的免疫反应可导致肌肉炎症。症状包括疼痛、触痛和肌肉无力。
肺炎:甲型流感会削弱免疫系统,使其更容易受到细菌的感染,导致肺部感染。表现为高热、咳嗽加剧、呼吸困难等,严重时可导致呼吸衰竭。
继发性细菌性肺炎:它通常发生在病程的后期,在急性疾病观察到一段时间的改善之后,症状和体征是典型的细菌性肺炎。继发性细菌性肺炎比原发性病毒性肺炎更常见。
脓毒症:这是一种严重的、危及生命的感染反应。如果细菌进入血液就会发生。如果不治疗,败血症会导致器官损伤和死亡。
急性呼吸窘迫综合征(ARDS):在重症病例中可能出现,导致严重的呼吸困难。
这些并发症在免疫力低下的人群中更为常见和严重,如老年人、儿童、孕妇以及患有慢性疾病的人。因此,及时就医和治疗对于预防并发症的发生至关重要。
病毒入侵
首先,甲流病毒主要通过呼吸道进入人体。
甲流病毒通过其表面的血凝素(HA)蛋白与呼吸道上皮细胞表面的唾液酸受体结合,从而进入宿主细胞。
病毒复制
病毒进入细胞后,其基因组RNA在细胞核内进行复制,并指导宿主细胞合成病毒蛋白。新合成的病毒颗粒在细胞内组装后,通过细胞膜释放到外界,感染其他细胞。
免疫反应
当甲型流感病毒入侵人体后,免疫系统会迅速识别病毒的病原相关分子模式(PAMPs),激活先天性免疫应答。
巨噬细胞、树突状细胞等免疫细胞被激活,开始分泌多种细胞因子,包括IL-1、IL-6、TNF-α和干扰素等。这些细胞因子一方面协调免疫细胞的募集和活化,增强机体的抗病毒能力;另一方面作用于下丘脑体温调节中枢,导致体温升高,同时还可引起全身性炎症反应,造成肌肉关节疼痛、乏力等症状。
症状形成机制
发热:发热是由于病毒感染触发免疫系统释放的细胞因子(如IL-1、IL-6等)作用于下丘脑温度调节中枢,使体温设定点上调所致,这种发热反应实际上有利于抑制病毒复制并增强免疫功能。
咳嗽:病毒直接感染呼吸道上皮细胞,导致细胞损伤、炎症反应和气道分泌物增多的结果,刺激气道感受器引起咳嗽反射。
全身症状:如肌肉酸痛、乏力、头痛等,主要是由细胞因子介导的系统性炎症反应引起,这些炎症介质可以作用于全身多个器官系统,影响神经-内分泌网络,导致患者出现全身不适。症状的严重程度往往与机体免疫反应的强度直接相关。
病毒变异特点
抗原漂移:点突变导致的小幅变异
抗原转变:基因重配导致的大幅变异
变异使病毒逃避免疫系统识别,是季节性流感和流感大流行的重要原因。
在传统认知中,我们往往将甲流视为一种单纯的呼吸道感染,但随着研究的深入,研究人员发现这种认识可能过于简单化。近年来,越来越多的研究表明,人体的免疫系统是一个高度统一的网络,其中肠道菌群作为人体最大的微生态系统,在调节全身免疫反应中扮演着关键角色。
肠道菌群可以通过调节免疫细胞的分布和功能,影响宿主对流感病毒的免疫反应,显著影响呼吸道感染的疾病进程和预后。此外,肠道菌群的代谢产物,如短链脂肪酸,也被发现能够调节免疫系统,从而可能对甲流的发病机制产生间接影响。接下来,我们进一步探讨肠道菌群是如何参与和调节甲流感染的过程。
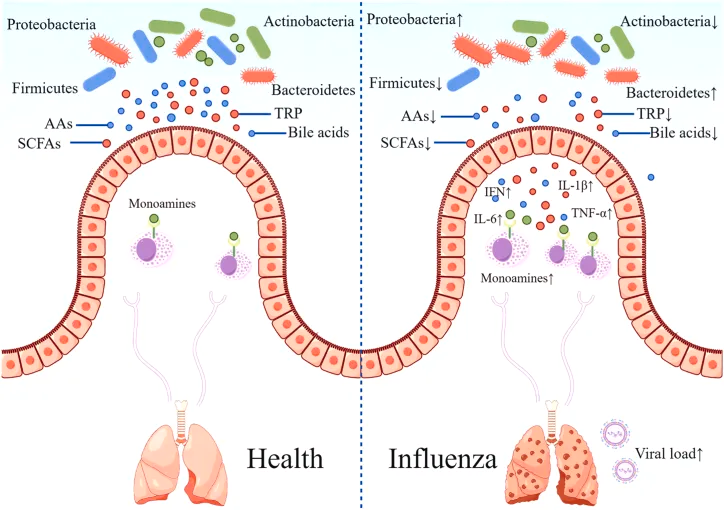
研究表明,肠道微生物群的丰富性和多样性响应于甲流感染发生了显著变化,包括拟杆菌门中细菌数量的增加和厚壁菌门中细菌数量的减少。同样,另一项研究表明,流感感染改变了肠道微生物群,促进肠道特异性厌氧菌的消耗和变形菌门的富集,导致肠道生态失调。
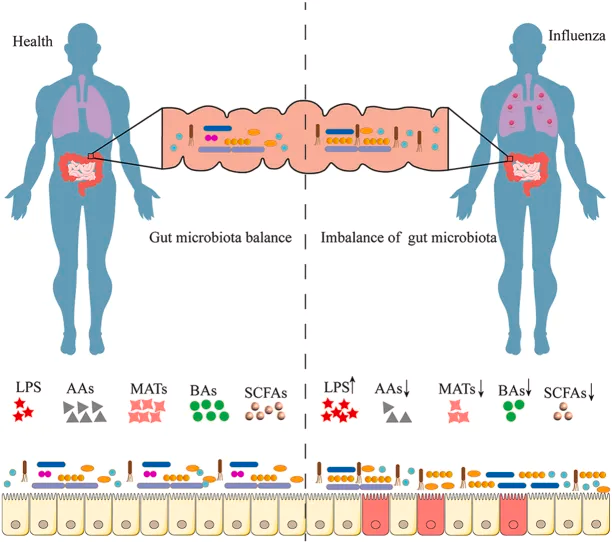
doi: 10.1016/j.heliyon.2024.e37661
▸甲流病毒感染
肠道菌群多样性增加,同时下列菌群增殖较为明显:
而一些有益菌的丰度则降低:
在健康个体中,肠道微生物群以平衡、稳定的方式在肠道中定植,与肠道粘膜屏障和抑菌物质一起维持肠道微生态。 一个平衡良好的肠道菌群能够增强机体的免疫功能,从而降低感染甲型流感的可能性。以下是几个关键机制:
调节免疫系统
在流感病毒感染后,CCR9 + CD4 + T细胞(其是源自肺的效应细胞)被募集到小肠,在小肠中它们分泌干扰素-γ(IFN-γ)。这导致肠道微生物群的不平衡,促进了小肠中的Th17细胞极化。最终导致IL-17 A分泌,其介导免疫损伤。接下来,我们从先天免疫和适应性免疫两方面详细了解肠道菌群对甲流的影响。
先天免疫
肠道微生物主要通过两种方式影响流感病毒感染的先天免疫:
首先,肠道菌群能够影响一种叫做天然淋巴细胞(ILCs)的免疫细胞。这些ILCs细胞在我们的肺部可以对抗病毒,而肠道菌群就像是一个“信号员”,通过释放一些信号物质,告诉肺部的ILCs该怎么做。比如,肠道菌群产生的短链脂肪酸(SCFAs)可以调节肺部ILCs的数量和功能,帮助我们更好地对抗流感病毒。
此外,肠道菌群还能够影响我们的身体识别病毒的能力。它们通过激活一些特殊的受体(比如Toll样受体和RIG-I样受体),帮助我们的身体更快地发现病毒并启动免疫反应。一旦有病毒入侵,就能迅速发出警报,让免疫细胞及时赶到“战场”去消灭病毒。
适应性免疫
肠道微生物群对甲型流感的适应性免疫影响主要体现在三个方面:
1- CD4+T细胞的调节
肠道微生物及其代谢产物可调节CD4+T细胞的分化。
在感染早期,主要促进Th1和Th17细胞分化,有助于病毒清除,乳酸杆菌通过参与色氨酸代谢和产生芳烃受体配体,促进IL-22表达,影响Th17分化;梭菌属通过诱导TGF-β释放,促进结肠Treg细胞分化。
短链脂肪酸(SCFA)通过多种机制促进Treg细胞分化,帮助控制炎症。
抗生素破坏菌群会显著降低CD4+T细胞数量,影响病毒清除。
2- 细胞毒性T淋巴细胞(CTL)的影响
CTL是消灭病毒感染的关键角色,而肠道微生物群能够通过调节树突状细胞(DCs)的活动来增强CTL的细胞毒性作用。
抗生素导致的菌群破坏会降低CTL的免疫应答。
3- 参与调节流感疫苗的免疫效果,促进流感特异性抗体的产生
预防流感的最佳方法是在流行期前接种流感疫苗,以产生特异性抗体。近期的临床试验表明,益生菌制剂可以有效增强流感疫苗的保护效力及提高个体抗体滴度。
然而,动物实验显示,接种前使用抗生素会降低疫苗诱导的抗体滴度及其中和能力,而在无菌小鼠中,恢复肠道植物群能恢复疫苗保护效力。此外,短链脂肪酸(SCFA)的不足会影响B细胞向特异性抗体产生浆细胞的分化。
维护肠道屏障功能
感染甲型流感病毒后,肠黏膜屏障功能障碍与感染的严重程度密切相关。
肠黏膜屏障由肠上皮细胞构成,分泌多种免疫因子并传递细菌抗原,在维持肠道菌群与宿主的共生关系中发挥着重要作用,并作为维持肠道菌群稳定性和生态平衡的控制开关。
流感病毒感染通过破坏肠上皮细胞中紧密连接蛋白和粘附蛋白的功能来影响肠道屏障,这些蛋白调节肠道屏障功能,阻止肠腔内大分子(如细菌和毒素)进入血液。
这些蛋白表达的降低可导致屏障功能受损,从而增加甲型流感病毒的疾病进展和危重症的发生。研究发现,小鼠感染H1N1导致肺和结肠中紧密连接蛋白的表达显著降低。这种减少导致这些器官的屏障结构受损,从而允许肠道细菌易位。随后,通过体液循环发生继发性细菌感染。
产生有益代谢产物
短链脂肪酸
短链脂肪酸包括乙酸盐、丙酸盐和丁酸盐,SCFAs可以通过调整流感病毒感染宿主的肠道菌群结构,如增加双歧杆菌等有益菌的数量,增强免疫适应性等来抵抗流感病毒。
短链脂肪酸能够增强肠道上皮细胞的免疫功能,促进免疫细胞的增殖和分化,同时具有抗炎作用,能够减轻流感病毒感染引起的炎症反应。
在流感期间补充短链脂肪酸,可以减轻肠道病原体如鼠伤寒沙门氏菌的移位,并提高合并感染的存活率。
肠道菌群及其代谢产物对正常人和流感患者的影响
doi: 10.1016/j.heliyon.2024.e37661
氨基酸
支链氨基酸被认为是身体健康的“潜在生物标志物”,在蛋白质合成、分泌和激素(如胰岛素和生长激素)释放中发挥重要作用。 研究表明,庆大霉素治疗后,肠道微生物群紊乱诱导流感小鼠体内支链氨基酸水平升高,从而抑制CD11b + Ly6G +细胞的发育,增加CD8+T细胞水平,导致流感病毒感染程度增加。
色氨酸是必需氨基酸,其缺乏会损害肠道免疫力,通过改善宿主先天免疫应答,降低流感期间的发病率和死亡率,补充色氨酸可能是预防流感的有效策略。
脱氨基酪氨酸(DAT),一种Clostridium orbiscindens菌产生的代谢物,可以通过提高I型干扰素(IFN)的水平,来增强身体对流感病毒的抵抗力,从而减少流感感染小鼠的体重减轻和死亡率。
胆汁酸
初级胆汁酸在肝脏产生,分泌到肠道中,约95%通过肠-肝循环被重吸收。
肠道菌群衍生的代谢产物脱氧胆酸(DCA)通过TGR5受体信号通路,抑制病毒复制,抑制嗜中性粒细胞依赖性组织损伤,增加宿主对流感病毒的抵抗力。
牛磺胆酸是一种胆汁酸。牛磺胆酸钠水合物(STH)抑制流感病毒复制,降低流感病毒RNA、互补RNA和mRNA水平,并对多种流感病毒株(H5N6、H5N1、H1N1、H3N2)表现出广谱抗病毒活性。 此外,STH降低流感病毒感染小鼠中炎症因子(TNF-α、IL-1β、IL-6)的表达,减轻其临床症状,抑制体重减轻,并降低其死亡率,这归因于STH通过抑制NF-B途径的激活而在流感抗性中的作用。
鹅去氧胆酸对甲型流感病毒(H5N1、H9N2和H1N1)具有抑制作用,它能够通过阻断病毒RNA复合物的核输出来抑制甲型流感病毒的复制。
以上我们了解到健康的肠道菌群通过多重免疫调控机制能够增强机体对甲型流感病毒的抵抗力,包括增强免疫系统的警戒能力、提升适应性免疫防护、调节炎症反应等,但在实际临床工作中,准确及时的诊断仍然是控制流感传播和开展针对性治疗的关键。接下来我们来看在临床中,医生可能借助的实验室检测手段。
甲型流感与其他病毒感染(如COVID-19)的症状类似。诊断测试包括:
★ 病毒分离和鉴定
从患者的呼吸道标本中分离出甲型流感病毒,并通过电镜或免疫荧光等方法进行鉴定。
优点:是诊断甲流的金标准。
缺点:操作复杂且耗时较长,一般不作为常规诊断方法
★ 甲型流感病毒核酸检测
通过聚合酶链式反应(PCR)等技术检测病毒的核酸。采集患者的呼吸道标本(如鼻咽拭子、口咽拭子等),进行核酸提取和扩增。
优点:准确率高,是确诊甲流的常用方法。
缺点:需要专业的实验室设备和技术
★ 快速抗原检测
通常使用检测试剂条,从鼻子或喉咙采集组织样本后检测病毒的存在,最快可在15分钟内显示结果,类似于新冠抗原检测。
优点:速度快;
缺点:敏感性和特异性相对较低。
★ 血常规检查
通过检测血液中的白细胞水平等指标,辅助诊断甲流。
优点:可以提供一些间接的诊断信息。
缺点:不能直接确诊甲流
★ 胸部X光
拍摄肺部图像以排除细菌感染或肺炎。
优点:可以排除其他肺部疾病。
缺点:影像学表现无特异性,不能单独用于确诊甲流。
利福昔明通过调节肠道菌群改善甲流病毒感染引起的肺屏障损伤
IAV引起了显着的体重减轻,并破坏了肺和肠的结构。16 S rRNA和代谢组学分析结果表明,感染甲流的小鼠粪便样品中益生菌Lachnoclosdium、Ruminococcocceae_UCG-013和色氨酸代谢产物的水平显著降低。
相比之下,补充50mg/kg利福昔明可以逆转这些变化,包括促进肺屏障的修复,增加粪便中Muribaculum、Papillibacter和色氨酸相关代谢物的含量。此外,利福昔明治疗增加了ILC3细胞数量、IL-22水平以及肺中RORγ和STAT-3蛋白的表达,减轻对肠屏障的损伤,同时增强小肠中AHR、IDO-1和紧密连接蛋白的表达。
在病原体清除方面,用冻干鼠李糖乳杆菌GG和加氏乳杆菌TMC0356进行口服预处理显著降低了PR8感染后6天小鼠肺部的病毒载量。
此外,单独口服煮沸的植物乳杆菌06CC2、植物乳杆菌DK119、热灭活b240和副干酪乳杆菌也降低了甲流病毒感染后BALF中的病毒载量。
口服加氏乳杆菌SBT2055可在感染后5天降低PR8感染小鼠肺部的病毒载量,并上调抗病毒基因Mx1和Oas1a的表达。益生菌治疗降低肺部病毒载量可能与促进对甲流病毒的先天免疫和适应性免疫有关。
煮沸的植物乳杆菌06CC2在感染早期(第二天)增加了抗病毒效应分子IFN-a、IFN-γ、IL-12的水平和NK细胞的活性。
口服植物乳杆菌DK119可增加BALF中IL-12和IFN-γ水平。
副干酪乳杆菌增加了甲流感染后肺组织中树突状细胞的募集。
植物乳杆菌0111口服预处理在H9N2感染后7天上调ISG转录,同时提高脾脏中CD3+CD4+TNF-α+T淋巴细胞百分比和CD3+CD8+TNF-β+T淋巴细胞百分比,增强对病毒的适应性免疫。
在感染的晚期,一些益生菌还可以抑制炎症损伤,促进组织修复。鼠李糖乳杆菌M21可减少甲流感染小鼠肺部的炎症损伤,并增加肺部裂解物中的IFN-γ和IL-2水平。
A. muciniphila可降低H7N9感染小鼠中促炎因子IL-1β和IL-6的水平,并增加炎症调节因子IL-10、IFN-β和IFN-γ的水平。
与益生菌粘膜乳杆菌L. mucosae1025和短双歧杆菌CCFM1026联合治疗可提高甲流感染小鼠盲肠粪便中的丁酸水平,并减轻肺组织中的炎症浸润。
一些口服益生菌可以双向作用,在甲流感染的早期阶段创造一个有利于病毒清除的炎症环境,并在后期抑制过度的炎症激活。
口服Bacteroides dorei在感染早期(第3天)更快地增加了1型干扰素的表达,降低了肺部的病毒载量,而在感染后期(第7天),它降低了1型IFN和其他促炎因子的水平,这有利于组织修复。与此同时,Bacteroides dorei治疗也改变了肠道菌群组成,增加了拟杆菌、普氏菌和乳杆菌,减少了大肠杆菌、志贺氏菌和副拟杆菌。
除此之外,益生菌制剂还可以通过促进甲流特异性抗体的产生来提高流感疫苗的有效性。
植物乳杆菌16和鼠李糖乳杆菌P118共同口服,改善了宿主对甲型流感病毒感染的防御。
研究人员利用C57BL/6和BALB/c小鼠模型的研究结果表明,单独施用植物乳杆菌16(L.plantarum 16)或鼠李糖乳杆菌P118(L.rhamnosus P118)并不能提供足够的流感防护。然而,植物乳杆菌16和鼠李糖乳杆菌P118的联合给药显著降低了呼吸道和肺部的病毒滴度,从而显著缓解了临床症状,改善了预后,降低了死亡率。
这种作用的机制涉及通过共同施用植物乳杆菌16和鼠李糖乳杆菌P118来调节宿主肠道微生物群和代谢,导致厚壁菌门富集和苯丙氨酸相关代谢增强,最终导致抗病毒免疫反应增强。值得注意的是,研究人员发现循环代谢分子2-羟基肉桂酸在抗击流感中起着重要作用。数据表明,植物乳杆菌16和鼠李糖乳杆菌P118这两种细菌或2-羟基肉桂酸在预防流感方面具有潜在的效用。
膳食中添加益生元短链低聚半乳糖和长链低聚果糖会增加小鼠流感疫苗特异性T辅助细胞1反应和mLN中特异性B细胞活化,导致IgG1和IgG2A水平升高。
β-葡聚糖重编程中性粒细胞以促进对甲型流感病毒的疾病耐受
最新研究表明,β-葡聚糖通过重编程造血干细胞来产生具有”调节”功能的中性粒细胞,这些特殊的中性粒细胞具有线粒体氧化代谢特征并能产生IL-10。这种机制通过RoRγt+ T细胞促进中性粒细胞向肺部组织募集,提高了机体对甲流病毒感染的疾病耐受能力,有助于维持肺部组织的完整性,从而降低发病率和死亡率,而这种保护作用并不依赖于抑制病毒载量。
益生元改善肠道菌群,延长流感疫苗抗体效果
接受肠内营养治疗的60岁以上老年人在标准肠内营养配方治疗的基础上,接受了双歧杆菌生长刺激素和半乳糖低聚糖治疗14周,并在治疗后第4周接种了三价季节性流感疫苗(A/H1N1、A/H3N2和B)。在第0、4、6、8和12周测量抗体滴度。结果显示,两组疫苗免疫应答率无差异,但实验组肠道内双歧杆菌数量明显高于对照组,实验组A/H1N1抗体滴度维持时间长于对照组。这表明益生元可能会增加肠道中益生菌的数量,从而维持抗体滴度。
中药在临床应用已有一千多年的历史,最近研究表明,中药在减轻甲流患者肺部炎症、改善临床症状、缩短治疗时间、促进康复等方面具有功效。
感染肺部后,甲流病毒可以改变肠道微生物群的组成和代谢,导致肠粘膜屏障受损、免疫功能受损和炎症因子水平升高。尽管如此,中药干预具有调节肠道微生物群、恢复体内平衡、保持肠粘膜屏障完整性、增强免疫功能和调节炎症反应的潜力。
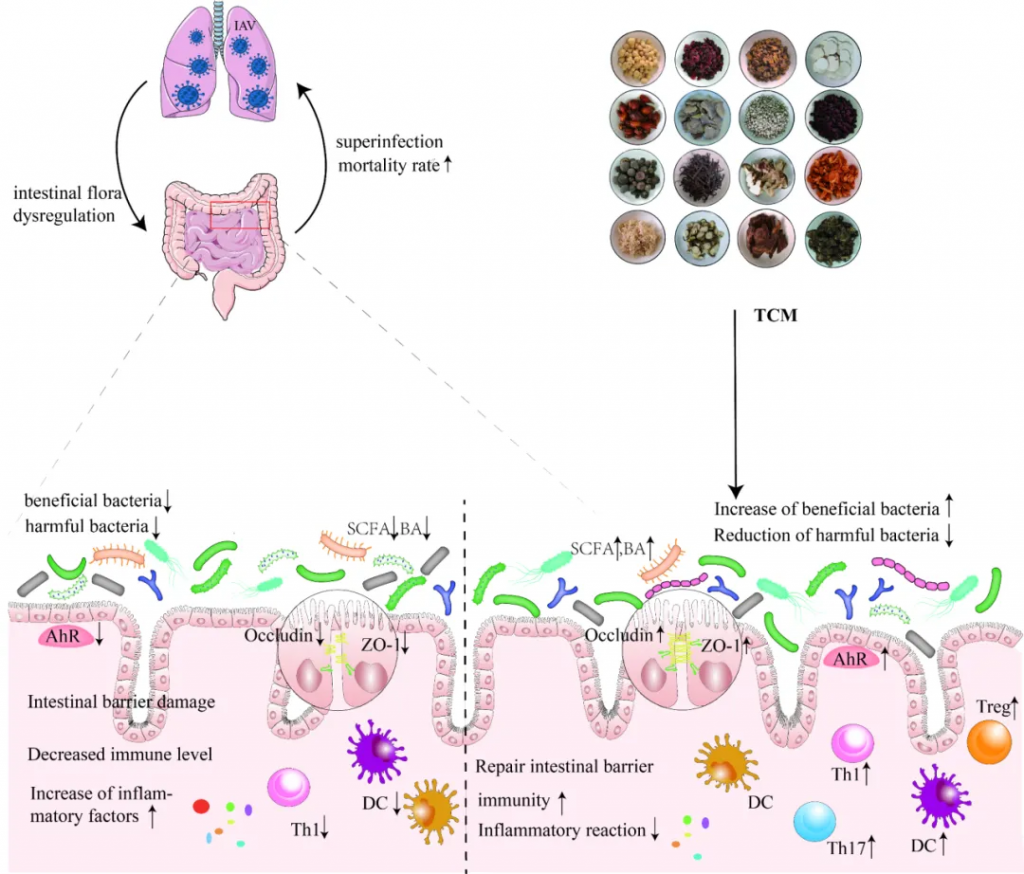
doi: 10.1186/s12985-023-02228-3.
宣肺败毒汤调节肠道微生物群多样性,并与拟杆菌、志贺氏菌、Eubacterium nodatum、Turicibacter、Clostridium sensu stricto 1 的变化呈正相关,而这些变化与 TNF-α 水平相关。
升麻素苷(Prim-O-glucosylcimifugin,POG)是中药防风的提取物,可以通过上调紧密连接蛋白Occludin、Claudin-3和ZO-1的表达水平来调节肠道菌群结构并修复肠道免疫屏障。
999 小儿感冒颗粒可以缓解H1N1感染小鼠的体重减轻,降低IL-6和IL-1β等炎性细胞因子的水平。 降低肺指数和病理损伤,通过维持结肠杯状细胞的数量来保护肠道屏障,降低结肠组织中IL-17 A的表达。
大黄中蒽醌类成分不仅增加了大鼠肠道中一些益生菌和产短链脂肪酸菌的丰度,而且通过上调ZO-1和闭塞素的表达水平来增强肠道屏障功能,从而抑制炎症。
其他中药对菌群及免疫的影响
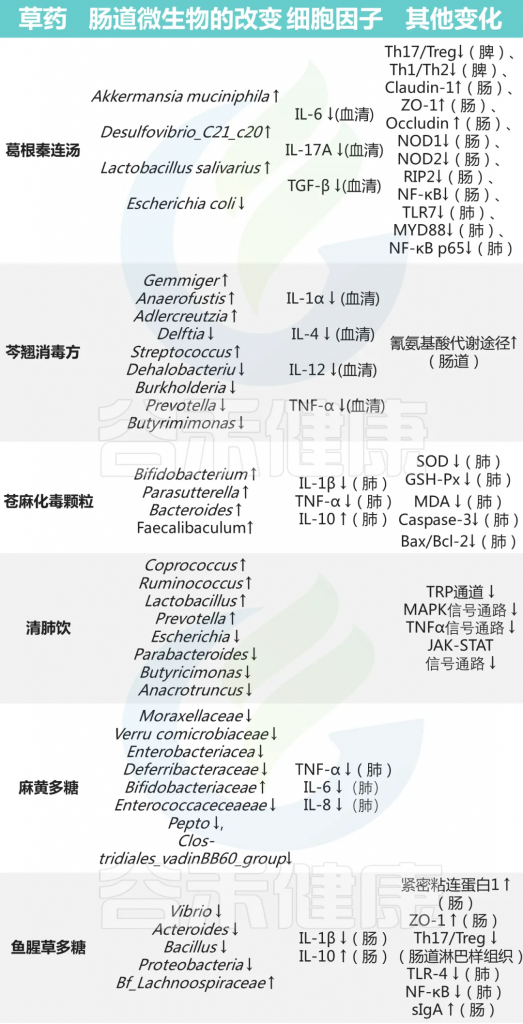
编辑
doi: 10.3389/fimmu.2023.1147724
黄芪多糖具有免疫增强作用,有望成为甲流疫苗的新型佐剂
黄芪多糖组表现出更高的免疫球蛋白G(IgG)、IgG 1和IgG 3水平,以及中和抗体水平。此外,它增加了CD 8+细胞的频率,以增强对致死性感染的抵抗力。
感染后第14天,高剂量黄芪多糖组的存活率(71.40%)高于血凝素组(14.28%),体重恢复更快。
黄芪多糖还能改善肺泡损伤和肠道结构紊乱。上调肠组织紧密连接蛋白Occludin和Claudin-1的表达水平,降低血清TNF-α的表达水平。此外,Colidextribacter、消化球菌科和瘤胃球菌科的群体是病毒感染后黄芪多糖组中的优势肠道微生物群。
doi: 10.1002/ptr.8334

doi: 10.2147/IJGM.S361001
维生素C
维生素 C 具有重要的抗炎、免疫调节、抗氧化、抗血栓和抗病毒特性。诺贝尔奖获得者莱纳斯·鲍林从随机对照试验中得出结论,维生素 C 可以预防和缓解感冒。
一项针对儿童上呼吸道感染的荟萃分析发现,维生素C的补充能够减少感染持续时间约1.6天。
口服维生素 C(2-8 克/天)可以减少呼吸道感染的发生率和持续时间。
英国安慰剂对照试验由 168 名志愿者组成,他们在 60 天的冬季期间随机接受安慰剂或维生素 C(每天 2 × 500 毫克)。
维生素 C 组的感冒次数较少(37次 vs.50次,p=0.05)
病毒感染的“感冒”天数更少(85次 vs.178次,p=0.03)
严重症状持续时间较短(1.8天 vs.3.1天,p=0.03)。
试验期间患过两次感冒的参与者人数显着减少(维生素 C 组为 2/84,安慰剂组为 16/84;p=0.04)
维生素D
维生素D缺乏会影响呼吸道感染的发生和哮喘的恶化。
维生素D和肠道微生物组以多种类似的方式影响呼吸道疾病中的免疫系统。它们之间可能存在一些相互作用和/或协同作用。
肠道微生物组可以改变肠道维生素D代谢,1,25(OH)2D (维生素D的活性形式)水平较高的人更有可能拥有有利的肠道微生物群,尤其是更多的产丁酸菌。
维生素D->抗病毒感染->降低哮喘发生和恶化的风险
一项涉及不同国家儿童的队列研究发现,补充维生素D 与 RV感染呈负相关。此外,患有下呼吸道感染的儿童的平均维生素 D 水平明显低于对照组。下呼吸道感染的发生率和严重程度也与维生素 D 水平相关。
维生素D可以双向调节肠道微生物群
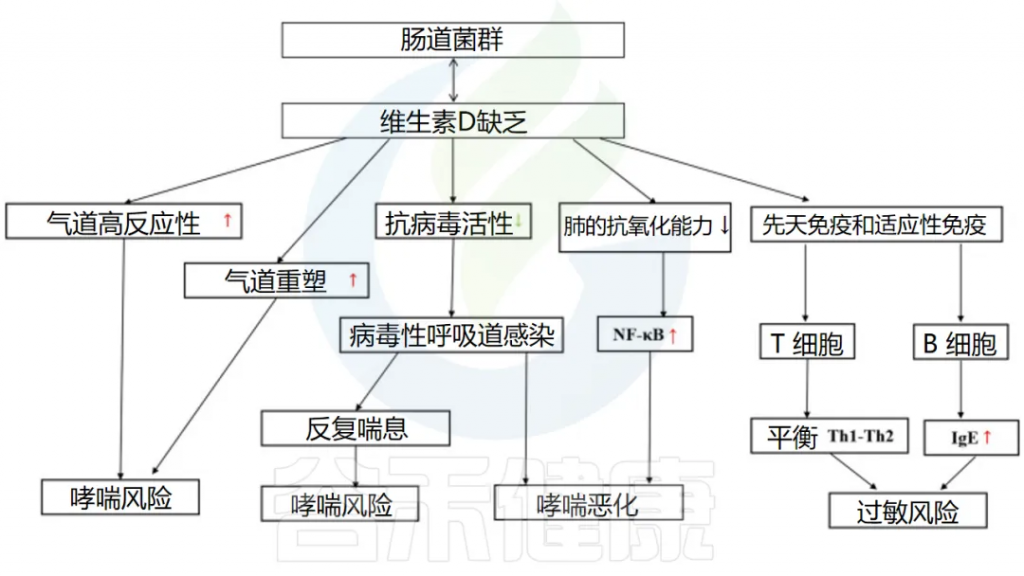
doi: 10.3389/fmicb.2023.1219942
维生素D通过多种途径影响哮喘的发生
因此,保持足够的维生素D水平对于预防和管理哮喘以及呼吸道感染至关重要。
研究表明,甲流病毒主要破坏气道上皮紧密连接,导致急性呼吸窘迫综合征,且胃肠道症状常与甲流病毒感染同时发生,提示肠-肺轴参与了宿主对甲流病毒的反应。在感染甲流病毒的患者和小鼠中发现SCFA(尤其是乙酸盐)显著减少。
在FMT后的小鼠中,这些条件被逆转,减少了肺部炎症损伤,并证实乙酸盐可能是肠-肺轴的重要介质。进一步的实验表明,乙酸盐激活了GPR 43,恢复了一些甲流病毒诱导的气道上皮屏障功能,并降低了TNF-α、IL-6、IL-1β水平。FMT能够有效调节流感感染小鼠肠道菌群结构紊乱。
粪菌移植通过增强I型干扰素通路改善甲流防御
在病毒感染中,I型干扰素(IFNs)非常重要,它们是病原体和宿主之间的重要调节因子。I型干扰素是抵御病毒感染的第一道防线。通过使用感染了甲流病毒的小鼠模型,研究人员发现IFN-κb是最早对H9N2感染做出反应的I型干扰素之一,并且IFN-κb能够有效地抑制多种流感病毒在培养的人类肺细胞中的复制。因此,阻断IFN-κ特异性途径可以作为预防和治疗甲流的依据。
I型干扰素途径对身体很重要。适当调节它既能抵御病毒感染,又能避免免疫或病理对身体造成过度损伤。比如降低IFNAR1水平可抑制流感病毒早期繁殖。
I型干扰素主要会激活一些干扰素刺激基因(ISGs)的表达。ISGs编码的蛋白质具有多种抗病毒功能。抗生素的使用会导致肺间质细胞中ISGs的减少,从而使身体更容易感染流感病毒。
然而,粪菌移植(FMT)可以逆转这一情况,增加肺间质中的I型干扰素信号通路和I型干扰素驱动的抗病毒状态,从而增强对流感病毒感染的防护。
在流感季节很容易感染甲型流感,因为它传染性很强。预防策略可以保护你免受流感或减少并发症的风险。一些常见的预防方法包括:
肠道微生物群是一个复杂而广泛的群落,显著影响各种生物系统,包括免疫系统和代谢功能。最近的研究越来越多地强调肠道微生物群与流感之间的潜在联系,表明肠道微生物群可能影响流感病毒的致病性,从而提供了一种新的治疗靶点。
总之,肠道微生物群表现出对流感的治疗潜力,这可以通过调节肠道微生态来解决。未来的研究应侧重于确定肠道微生物群中的关键菌株、特定代谢物和免疫调节机制,以精确靶向微生物群干预,预防和治疗流感和其他呼吸道病毒感染。
在此基础上,菌群组合的优化研究显得尤为重要。比如说通过研究植物乳杆菌16和鼠李糖乳杆菌P118等益生菌的协同作用,探索不同菌群配比对免疫调节的影响,从而制定个体化的菌群干预方案。
可以进行新型保护性代谢物的筛选与鉴定,深入研究短链脂肪酸、胆汁酸等代谢产物的作用机制,并结合代谢组学分析推进临床应用。
开展一系列临床转化研究,包括建立菌群检测作为辅助诊断的标准化体系,开展益生菌制剂的临床试验,并根据不同患者特点制定个性化预防和治疗方案。同时,预防策略的优化也需要重点关注,特别是探索疫苗免疫效果与肠道菌群的关系,建立预防性菌群调节方案,并为高危人群制定个性化预防策略。
这些研究方向相互关联、相互促进。分子机制的研究为菌群组合优化提供理论指导,代谢产物研究则为临床应用提供新靶点,而临床转化研究的结果又能反馈指导基础研究的方向。通过多维度、多层次的研究,将更全面地理解肠道菌群与甲型流感的关系,为开发新型防治策略提供坚实的科学依据。
主要参考文献
Khan N, Tran KA, Chevre R, Locher V, Richter M, Sun S, Sadeghi M, Pernet E, Herrero-Cervera A, Grant A, Saif A, Downey J, Kaufmann E, Khader SA, Joubert P, Barreiro LB, Yipp BG, Soehnlein O, Divangahi M. β-Glucan reprograms neutrophils to promote disease tolerance against influenza A virus. Nat Immunol. 2025 Jan 8.
Luo C, Yang Y, Jiang C, Lv A, Zuo W, Ye Y, Ke J. Influenza and the gut microbiota: A hidden therapeutic link. Heliyon. 2024 Sep 10;10(18):e37661.
Ma L, Ji L, Wang T, Zhai Z, Su P, Zhang Y, Wang Y, Zhao W, Wu Z, Yu H, Zhao H. Research progress on the mechanism of traditional Chinese medicine regulating intestinal microbiota to combat influenza a virus infection. Virol J. 2023 Nov 13;20(1):260.
Liu X, Wang Q. Effect of natural products on host cell autophagy induced by Influenza A virus infection. Front Cell Infect Microbiol. 2024 Sep 30;14:1460604.
Ho JSS, Ping TL, Paudel KR, El Sherkawi T, De Rubis G, Yeung S, Hansbro PM, Oliver BGG, Chellappan DK, Sin KP, Dua K. Exploring Bioactive Phytomedicines for Advancing Pulmonary Infection Management: Insights and Future Prospects. Phytother Res. 2024 Dec;38(12):5840-5872.
Ou G, Xu H, Wu J, Wang S, Chen Y, Deng L, Chen X. The gut-lung axis in influenza A: the role of gut microbiota in immune balance. Front Immunol. 2023 Oct 20;14:1147724.
Gounder AP, Boon ACM. Influenza Pathogenesis: The Effect of Host Factors on Severity of Disease. J Immunol. 2019 Jan 15;202(2):341-350.
Zhang Y, Chen Y, Xia J, Li L, Chang L, Luo H, Ping J, Qiao W, Su J. Rifaximin ameliorates influenza A virus infection-induced lung barrier damage by regulating gut microbiota. Appl Microbiol Biotechnol. 2024 Sep 19;108(1):469.
Wan C, Yan S, Lu R, Zhu C, Yang Y, Wu X, Yu Z, Jiang M, Peng W, Song W, Wu H, Fang B, He Y. Astragalus Polysaccharide improves immunogenicity of influenza vaccine as well as modulate gut microbiota in BALB/c mice. Microb Pathog. 2024 Oct;195:106893.

谷禾健康

你可知道,为什么明明摄入了很多的蛋白质,仍然可能出现氨基酸缺乏的情况吗?谷禾今天就带你深入了解这一现象背后的各种奥秘和原因,帮助你更好地理解蛋白质的摄入与氨基酸利用关系。
造成这种情况的原因可以主要分为几个方面,我们将逐一详细讲述:
一、内源性氨基酸的产生不足。这些氨基酸是在体内合成的,主要通过蛋白质的分解与转化而来。然而,内源性氨基酸的生成受到多种因素的影响,比如身体的整体代谢状态、营养状况、肝脏的功能等。如果这些影响因素出现问题,就会导致体内内源性氨基酸的产生不足,从而影响整体的氨基酸平衡。特别值得注意的是,肠道菌群在这个过程中扮演着关键角色,它们不仅参与氨基酸的合成,还能影响宿主的代谢状态。如果肠道菌群失衡,可能会影响内源性氨基酸的产生效率。
二、外源性蛋白质的消化和吸收率不高。外源性蛋白质来自我们摄入的食物,经过消化过程转化为氨基酸。但这个过程的效率不总是很高,因素如食物的种类、加工方式、个人的消化功能等都会影响外源性蛋白质的消化和氨基酸的吸收。健康的肠道菌群能够提高蛋白质的消化效率,产生有益的代谢物,并维持肠道屏障功能,从而促进氨基酸的吸收,如果肠道菌群失衡,消化吸收不良,即使摄入了足够的蛋白质,身体也无法有效利用其中的氨基酸。
三、氨基酸的消耗过大,导致消耗量超过了摄入量。这就像一个游泳池,放水速度比注水速度快,那么即使持续地注入水,也很难增加池内的水量。而肠道菌群的代谢活动也会影响氨基酸的消耗速率,某些菌群可能会竞争利用宿主的氨基酸资源。
每个人对氨基酸的需求和消耗情况是不同的,这和个体的活动水平、身体状况以及生理需求都有很大关系,同时,每个人的肠道菌群组成都是独特的,这种个体差异会影响到氨基酸的代谢和利用效率。有些人可能会在日常生活中消耗掉大量氨基酸,导致最终的总量不足。因此,在摄入氨基酸时,需要综合考虑个体需求的差异,关注肠道健康状况,以确保氨基酸的充分吸收和利用,满足身体所需。
本文我们一起来了解一下,蛋白质在胃肠道内的消化过程,详细讨论影响氨基酸消化和吸收的关键因素,肠道菌群如何影响氨基酸的吸收,探讨根据不同人群的特点(如素食者、运动人群、孕妇等)来优化蛋白质的摄入策略,以及如何通过调节肠道菌群来提高氨基酸的利用效率,从而更好地把握蛋白质摄入与氨基酸利用的关系,制定有效的营养策略。
▸ 氨基酸的定义
氨基酸(amino acid)是含有氨基和羧基的一类有机化合物,是促进人体生长,维持身体正常代谢,影响生命活动的重要物质,也是合成机体所需抗体、激素、酶类的原料。
如果氨基酸代谢异常,可能导致生长发育、全身稳态受损,甚至死亡。总的来说,氨基酸作为生命体的最基本组成单位,在人体扮演着重要角色。
▸ 氨基酸与蛋白质的关系
蛋白质我们都知道,人体必需的营养素之一。它是有机大分子,是构成细胞的基本有机物,是生命活动的主要承担者。可以这么说,没有蛋白质就没有生命。
而氨基酸是蛋白质的基本组成单位。蛋白质和氨基酸之间的关系是:氨基酸通过脱水缩合形成肽键,进而连接形成肽链,肽链折叠盘旋形成具有空间结构的蛋白质。
图源:National Human Genome Research Institute
氨基酸的种类、数目、排列方式、多肽链的盘曲、折叠方式及其空间结构的不同导致了蛋白质结构的多样性,其结构的多样性导致了功能的多样性。
通俗地讲:氨基酸是构成蛋白质的小分子“积木”。想象一下,蛋白质就像一串珠子项链,而每颗珠子就是一个氨基酸。它们通过不同的组合和排列形成各种蛋白质。
通常,我们体内的氨基酸以游离氨基酸总量计算,包括两部分,一是内源氨基酸,即体内组织蛋白降解产生或自身合成的非必需氨基酸;二是外源氨基酸,即食物蛋白质经消化吸收而来。两者共同分布于身体各处,参与代谢,总称为氨基酸代谢库。
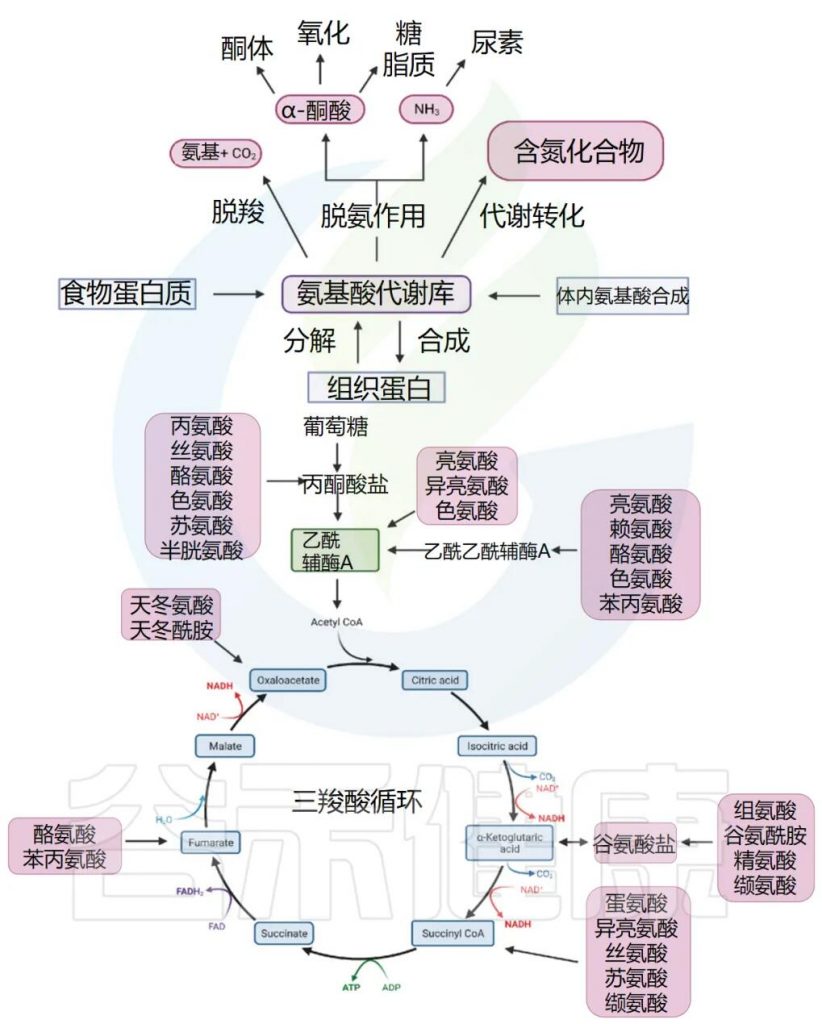
1
内源性氨基酸的合成
◆ 大多数非必需氨基酸可通过转氨基作用合成
转氨基作用指的是一种α-氨基酸的α-氨基转移到一种α-酮酸上的过程,可以看成是氨基酸的氨基与α-酮酸的酮基进行了交换。生成相应的α-氨基酸;而原来的α-氨基酸则转变为相应的α-酮酸。
例如,天冬氨酸是三羧酸循环中间产物草酰乙酸经转氨作用得到,谷氨酸是三羧酸循环中间产物α-酮戊二酸经转氨作用得到,丙氨酸是糖酵解生成的丙酮酸经转氨作用得到。
一般有以下几类:
1) 谷氨酸族:以a-酮戊二酸为起始物,可合成Glu,GIn,Pro,Arg(称为谷氨酸族);
2) 天冬氨酸族:以草酰乙酸为起始物,可合成Asp,Asn,Met,Thr,Lys,le(称为天冬氨酸族);
3)丙氨酸族:以丙酮酸为起始物,可合成Ala,Val,Leu(称为丙氨酸族);
4)丝氨酸族:以甘油酸-3-磷酸为起始物,可合成Ser,Gly,Cys(丝氨酸族);
5)芳香族氨基酸:以赤藓糖-4-磷酸、磷酸烯醇式丙酮酸为起始物,可合成芳香族氨基酸(Phe,Tyr,Trp);
6)组氨酸(His):由核糖-5-磷酸衍生,核糖-5-磷酸通过磷酸戊糖途径产生。
◆ 通过必需氨基酸转化
从上述合成的过程可以发现,氨基酸的种类发生了改变,数量并没有发生改变。那么,有没有不是通过转氨基途径合成氨基酸呢?答案肯定是有的,如下所示是半胱氨酸的合成示意图:
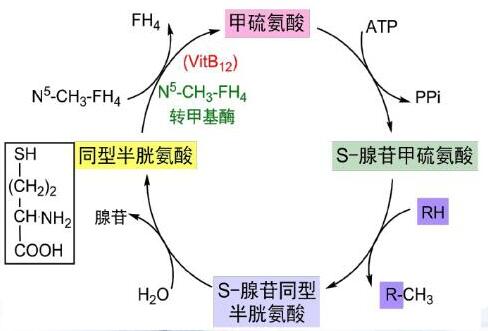
半胱氨酸在体内可以由甲硫氨酸转化而成,而这一过程必须有维生素B6参与。
必需氨基酸还可以通过苯丙氨酸羟化酶催化合成非必需氨基酸酪氨酸,如果缺少这种酶,转化过程很难进行,苯丙氨酸就会转化为苯丙酮酸,这就是苯丙酮尿症的原因。
2
影响内源性氨基酸合成的因素
▸ 营养因素
•必需氨基酸的充足供应
虽然非必需氨基酸可以在体内合成,但合成过程中往往需要以必需氨基酸为前体或底物。如果必需氨基酸缺乏,可能会影响非必需氨基酸的合成途径,导致非必需氨基酸合成减少。
例如,当饮食中缺乏蛋氨酸时,可能会影响半胱氨酸的合成,因为半胱氨酸可以由蛋氨酸转化而来。
•能量供应
非必需氨基酸的合成需要消耗能量,因此充足的能量供应对于非必需氨基酸的合成至关重要。如果能量摄入不足,身体会优先利用能量来维持生命活动的基本需求,从而可能减少非必需氨基酸的合成。
在饥饿或能量摄入不足的情况下,身体会分解蛋白质来提供能量,这会导致非必需氨基酸的合成减少。
•维生素和矿物质的供应
某些维生素和矿物质是参与非必需氨基酸合成的酶的辅助因子或激活剂。例如,维生素B6是参与氨基酸代谢的多种酶的辅酶,缺乏维生素B6可能会导致谷氨酸合成谷氨酰胺的过程受阻。锌、镁等矿物质也可能参与某些氨基酸合成酶的活性调节。
▸ 生理因素
•年龄和生长发育阶段
不同年龄和生长发育阶段的人对非必需氨基酸的合成能力可能不同。婴幼儿和青少年时期,身体生长发育迅速,对蛋白质和氨基酸的需求较大,非必需氨基酸的合成能力也相对较强。
随着年龄的增长,身体的代谢功能逐渐下降,非必需氨基酸的合成能力可能会减弱。因此老年人可能需要更多的优质蛋白质补充来满足身体对氨基酸的需求。
•健康状况和疾病状态
某些疾病可能会影响非必需氨基酸的合成。例如,肝脏疾病可能会影响氨基酸的代谢和转化,导致非必需氨基酸的合成减少。肾脏疾病可能会影响氨基酸的排泄和重吸收,从而影响体内氨基酸的平衡。
此外,一些慢性疾病如糖尿病、癌症等也可能影响非必需氨基酸的合成,因为这些疾病会导致身体的代谢紊乱和营养消耗增加。
•激素水平
激素可以调节身体的代谢过程,包括氨基酸的合成和分解。例如,生长激素可以促进蛋白质的合成和细胞的生长,从而可能增加非必需氨基酸的合成。胰岛素可以促进葡萄糖的摄取和利用,同时也可以促进氨基酸的摄取和蛋白质的合成,可能对非必需氨基酸的合成有一定的促进作用。
相反,一些应激激素如皮质醇可能会促进蛋白质的分解,从而可能减少非必需氨基酸的合成。
▸ 环境因素
•饮食结构和饮食习惯
长期的不良饮食结构和饮食习惯可能会影响非必需氨基酸的合成。例如,高糖、高脂肪、低蛋白的饮食可能会导致蛋白质摄入不足,从而影响非必需氨基酸的合成。过度饮酒、吸烟等不良习惯也可能对身体的代谢功能产生不良影响,从而影响非必需氨基酸的合成。
此外,一些特殊的饮食限制,如素食主义、低蛋白饮食等,需要更加关注非必需氨基酸的合成问题,以确保身体的正常代谢需求。
•运动和体力活动
适度的运动和体力活动可以促进身体的代谢功能,运动可以增加肌肉的蛋白质合成,从而可能增加对非必需氨基酸的需求和合成。
然而,过度的运动和体力活动可能会导致蛋白质分解增加,从而影响非必需氨基酸的合成。此外,运动过程中如果没有及时补充足够的营养物质,也会影响非必需氨基酸的合成。
•环境压力和应激
长期处于高压力、高应激的环境中,会影响身体的代谢功能,从而影响非必需氨基酸的合成。在应对环境压力和应激时,身体会优先利用能量和营养物质来维持生命活动的基本需求,从而可能减少非必需氨基酸的合成。
当这些因素受到影响时,会导致内源性氨基酸的产生不足。这种不足会进一步影响人体内氨基酸的整体水平,使得总氨基酸的水平较低。
还有一些氨基酸人体不能合成或合成速度远不能适应人体需要,必须通过食物中的蛋白质补充,也称为必需氨基酸。
1
蛋白质和多肽在肠道内的消化
以前,蛋白质消化和氨基酸吸收被认为是截然不同的独立过程;现在人们认识到,蛋白质分解成更小的单位,随后被肠细胞吸收,并将氨基酸释放到门静脉血液中,这是一个高度组织和紧密结合的过程。
消化涉及食物基质、自身物理过程和哺乳动物及细菌酶的化学过程之间的多种相互作用,所有这些都发生在胃肠道的复杂环境中。
•大分子蛋白质先被胃蛋白酶水解成多肽
摄入的食物在口腔中经过咀嚼发生物理分解,食物与唾液等液体混合,产生可被吞咽进入胃部的食物丸。胃蛋白酶在不同的裂解点水解蛋白质,形成更小的多肽,胃酸使蛋白质变性并部分展开,帮助胃蛋白酶更好地进入它们的键。
随后,剩余的蛋白胨和多肽被转移到小肠中,在小肠中胰腺产生和分泌多种蛋白酶和肽酶,如肠黏膜的胰蛋白酶、羧肽酶。上皮细胞的刷状缘膜结合酶包括许多氨基肽酶和一种羧肽酶,并作用于胰腺消化后留下的任何寡肽。
•多肽再被切割成各个氨基酸
胰腺蛋白酶将多肽切割成寡肽和氨基酸。胃蛋白酶、胰蛋白酶、凝乳胰蛋白酶和弹性酶都属于内肽酶,它们攻击某些内部键,产生大的多肽。
注:胃蛋白酶在芳香氨基酸如苯丙氨酸、色氨酸和酪氨酸旁切割消化键,而胰蛋白酶在碱性氨基酸精氨酸和赖氨酸旁切割键。
在这些内肽酶作用后剩余的寡肽受到外肽酶的攻击,如羧基肽酶A和B,它们每次从多肽或蛋白质的羧基或氨基端切割一个氨基酸。
羧基肽酶从肽链的羧基端切除氨基酸,羧基肽酶A优先释放缬氨酸、亮氨酸、异亮氨酸和丙氨酸,羧基肽酶B优先释放碱性氨基酸如精氨酸和赖氨酸。
2
氨基酸的吸收
蛋白质消化的产物,主要是游离氨基酸、二肽和三肽,通过几种运输机制从小肠腔吸收。
肽和氨基酸通过不同的转运体被小肠肠细胞吸收。空肠近端是肽和氨基酸吸收的主要部位,但小肠的其他部位也具有显著的运输能力。
短肽(二肽和三肽)通过肠细胞顶膜中的H+依赖性肽转运体(如PepT1)转运进入肠细胞,并在上皮细胞中进一步水解为氨基酸。
•不同的氨基酸有特定的转运系统
游离氨基酸(FAAs)被碱性、阳离子、中性氨基酸以及亮氨酸和甘氨酸转运体以梯度依赖的方式通过肠上皮细胞的刷状边界膜吸收。有许多刷状边缘(顶端膜)和基底侧膜转运系统,特定于特定的氨基酸或氨基酸群。
例如,浓缩氨基酸转运蛋白在顶膜上表达,用于吸收中性氨基酸(B0AT1,广义中性氨基酸转运蛋白 1,SLC6A19)、阳离子氨基酸(囊胚中性和阳离子氨基酸转运蛋白,SLC7A9)、甘氨酸和脯氨酸 (PAT1,质子氨基酸转运蛋白1,SLC36A1;SIT,系统亚氨基转运蛋白,SLC6A20) ,阴离子氨基酸(兴奋性氨基酸转运蛋白3(EAAT3),SLC1A1)和β-氨基酸(PAT1,SLC36A1,TauT,牛磺酸转运蛋白,SLC6A6)。
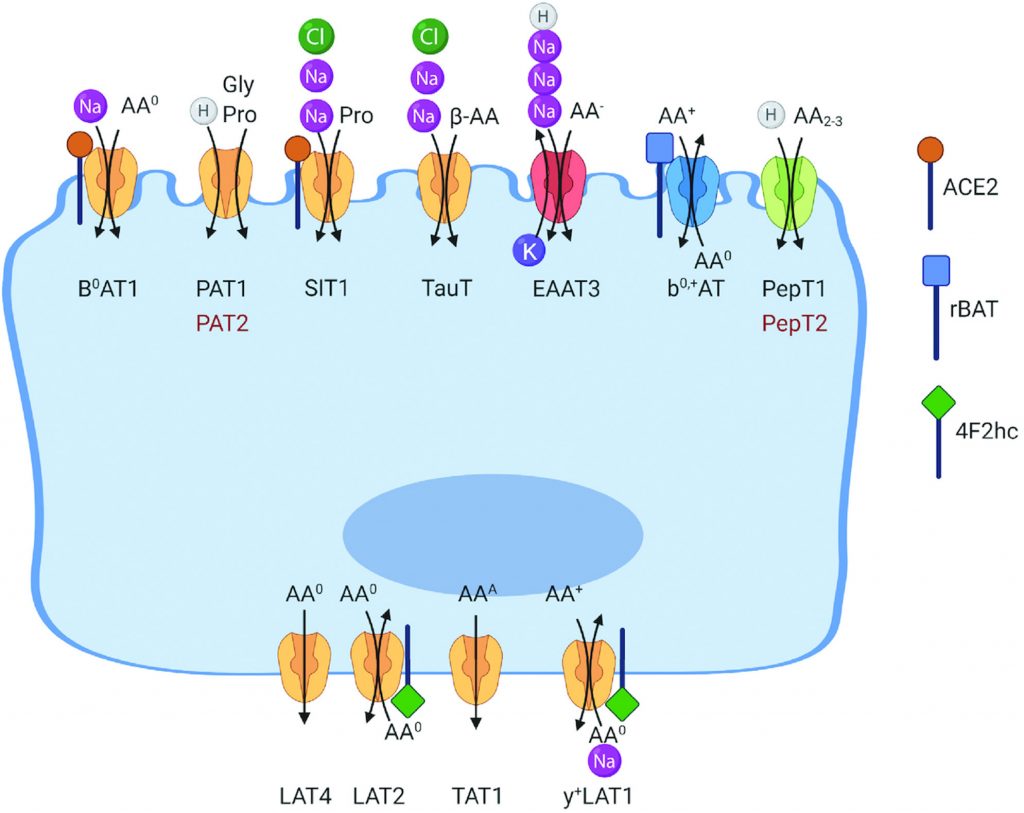
doi: 10.1093/jn/nxab342.
•小肠可以适应性地上调氨基酸吸收能力
人体从膳食蛋白质中获得氨基酸的净正流量是至关重要的,因此小肠具有适应性地上调其氨基酸吸收的能力。个体的膳食蛋白质摄入量和生理状态(影响氨基酸需求)都可能随着时间的推移而改变,氨基酸吸收机制可以在组织和细胞水平上进行相应的调整。例如转运蛋白上调和粘膜增生的共同作用可导致氨基酸吸收能力比禁食状态增加数倍。
继基底外侧膜转运到间质液后,氨基酸通过绒毛毛细血管进入门静脉。门静脉直接供应肝脏,在那里氨基酸可以进一步代谢或运输到其他器官和组织。
3
影响膳食氨基酸消化和吸收的因素
虽然膳食蛋白质通常在胃肠道内被消化,但它们并不一定被完全消化和吸收。此外,不同的氨基酸从蛋白质中释放出来,并在不同程度上被肠道吸收。因此,有必要了解膳食蛋白质和氨基酸的消化和吸收率。影响膳食氨基酸消化和吸收的因素主要有以下几个方面:
▸ 食物因素
•蛋白质的来源和性质
不同来源的蛋白质其氨基酸组成和结构不同,消化和吸收的难易程度也有所差异。例如,动物蛋白(如肉类、蛋类、奶类)通常含有较高比例的优质蛋白质,其氨基酸组成与人体需求较为接近,且消化率较高。
蛋白质的消化是一个有效的过程,从鸡蛋中粗蛋白的97%消化率到谷物中的≥70%不等。
蛋白质的结构也会影响消化。例如,胶原蛋白等结构紧密的蛋白质较难被消化酶分解,而球状蛋白等结构较为松散的蛋白质则相对容易消化。研究表明,与酪蛋白相比,乳清蛋白的消化和吸收速度更快。
•食物的加工和烹饪方式
适当的加工和烹饪可以提高蛋白质的消化率。例如,加热可以使蛋白质变性,使其结构变得松散,更容易被消化酶作用。此外,研磨、破碎等加工方式可以增加蛋白质与消化酶的接触面积,促进消化。
然而,过度加工和烹饪可能会破坏蛋白质的结构和营养成分,降低其消化率。例如,高温油炸、长时间炖煮等可能会导致蛋白质发生美拉德反应,产生难以消化的化合物,同时也会破坏一些必需氨基酸。当被吸收时,通常不被利用,而是被分解并随尿液排出。
•抗营养因子
抗营养因子是一类能够干扰人体对营养素的吸收和利用的物质。食品中的胰蛋白酶抑制剂、单宁和植酸盐等化合物会显著降低蛋白质和氨基酸的消化率。此外,还可能会与蛋白质结合,形成难以消化的复合物,影响蛋白质的消化和吸收。
▸ 消化系统因素
•消化酶的分泌和活性
胃液中的胃蛋白酶、胰液中的胰蛋白酶、糜蛋白酶等消化酶在蛋白质的消化过程中起着关键作用。消化酶的分泌量和活性受到多种因素的影响,如年龄、健康状况、饮食等。老年人的消化酶分泌量可能会减少,导致蛋白质消化能力下降。某些疾病(如胰腺疾病、胃肠道疾病)也会影响消化酶的分泌和活性。
此外,消化酶的活性还受到pH值、温度等环境因素的影响。例如,胃蛋白酶在酸性环境下活性较高,而胰蛋白酶等则在碱性环境下活性较高。
•胃肠道的生理状态
胃肠道的蠕动、分泌、吸收等生理功能对氨基酸的消化和吸收至关重要。胃肠道蠕动过慢会导致食物在胃肠道内停留时间过长,影响消化和吸收;而蠕动过快则可能使食物来不及充分消化就被排出体外。
胃肠道的分泌功能也会影响消化。例如,胃液、胰液、胆汁等消化液的分泌不足会影响蛋白质的消化。此外,胃肠道的吸收面积和功能也会影响氨基酸的吸收。例如,小肠黏膜的损伤会导致吸收面积减少或吸收功能障碍。
▸ 个体因素
•年龄
婴幼儿的消化系统尚未发育完全,消化酶的分泌和活性较低,胃肠道的吸收能力也较弱。随着年龄的增长,消化系统的功能逐渐成熟,但老年人的消化和吸收功能又会逐渐下降。
不同年龄段的人对蛋白质的需求和消化吸收能力不同,因此在饮食中应根据年龄特点选择合适的蛋白质来源和摄入量。
•健康状况
一些疾病会影响蛋白质的消化和吸收。例如,胃肠道疾病(如胃炎、胃溃疡、肠炎等)、肝脏疾病、肾脏疾病等都可能会影响消化酶的分泌、胃肠道的蠕动和吸收功能。此外,一些慢性疾病(如糖尿病、心血管疾病等)也可能通过影响代谢和营养状态间接影响蛋白质的消化和吸收。
营养不良、消瘦、肥胖等也可能影响蛋白质的消化和吸收。营养不良可能导致消化酶的合成减少、胃肠道黏膜萎缩等,从而影响消化和吸收功能;而肥胖可能会引起胃肠道激素分泌异常、肠道菌群失调,也会影响蛋白质的消化和吸收。
•遗传因素
个体的遗传差异可能会影响蛋白质的消化和吸收。例如,某些人可能存在特定的基因突变,导致消化酶的活性或结构异常,从而影响蛋白质的消化。此外,遗传因素还可能影响胃肠道的生理功能、肠道菌群的组成等,进而影响蛋白质的消化和吸收。
▸ 其他因素
•饮食中的其他成分
饮食中过多的膳食纤维、脂肪等成分可能会影响蛋白质的消化和吸收。膳食纤维可以促进肠道蠕动,但过多的膳食纤维可能会吸附蛋白质和消化酶,影响蛋白质的消化。脂肪可以延缓胃排空,使蛋白质在胃中停留时间延长,有利于胃蛋白酶的作用,但过多的脂肪可能会引起消化不良。
•药物和补充剂
某些药物会影响蛋白质的消化和吸收。例如,抗生素可能会破坏肠道菌群,影响消化酶的合成和活性;抗酸药会改变胃肠道的pH值,影响消化酶的活性。一些补充剂,如蛋白粉、氨基酸补充剂等,如果使用不当也可能会影响蛋白质的消化和吸收。
在使用药物和补充剂时,应注意其对蛋白质消化和吸收的影响,并在医生或营养师的指导下合理使用。
•肠道菌群
肠道微生物群在氨基酸的调节、消化和吸收过程中发挥着至关重要的作用,它们通过多种方式来影响氨基酸的利用效率。在下一章节中,我们将详细探讨肠道微生物群如何在氨基酸的消化、吸收及利用中发挥作用,并分析其对整体健康的影响。
氨基酸对各种生物活性化合物的合成至关重要,这些化合物在信号通路和代谢中发挥关键的调节作用。肠道微生物群在促进氨基酸调节以及氨基酸消化和吸收过程中起着不可缺少的作用。
doi: 10.2174/1389203719666180514145437.
1
参与氨基酸代谢和吸收
过去对肠道微生物群和氨基酸相互作用的研究表明,许多肠道细菌参与了胃肠道中蛋白质的消化和氨基酸的吸收。
•一些肠道细菌能够产生肽酶,用于降解蛋白质
蛋白质的转换主要发生在肠道中,结肠细菌降解内源性或外源性蛋白质的效率较高。拟杆菌属(Bacteroides)、梭状芽胞杆菌属(Clostridium)、丙酸杆菌属(Propionibacterium)、梭杆菌属(Fusobacterium)、乳酸杆菌属(Lactobacillus)和链球菌属(Streptococcus)在蛋白质水解过程中发挥着重要作用。
其中一些细菌可以直接代谢氨基酸,并具有分泌各种蛋白酶和肽酶的能力。发酵过程中,复杂蛋白质首先被各种细菌肽酶、蛋白酶和内肽酶切割,释放出游离氨基酸和短肽。然后对氨基酸和短肽进行发酵。产生支链脂肪酸(2-甲基丁酸酯、异丁酸酯、异戊酸酯)、有机酸、气体(H2和CO2)以及微量酚、胺、吲哚和氨。
肠道内的梭菌属细菌(赖氨酸或脯氨酸利用的基础细菌)是氨基酸发酵的关键驱动因素,而消化链球菌属细菌是谷氨酸或色氨酸利用的关键驱动因素。

Yadav M,et al.Arch Microbiol.2018
其他例如瘤胃细菌,Selenomonas ruminantium, Megasphaera elsdenii, Prevotella ruminicola, Misuokella multiacidas, Butyrivibrio fibrisolvens, Streptococcus bovis等含有极为活跃的二肽基肽酶和二肽酶。
•代谢氨基酸的基因广泛分布于人类肠道细菌

研究了来自不同系统发育群体的肠道细菌中相关代谢酶及其同源物的分布情况。对代表10个门的380种肠道细菌的基因组做了blast比对。
结果表明,大多数这些酶广泛分布在肠道细菌中,但有些酶表现出不同的分布模式。例如,L-天冬酰胺酶在拟杆菌门(Bacteroidetes)和假单胞菌门(Proteobacteria)中普遍存在,而芳香族氨基酸转氨酶和精氨酸脱亚胺酶主要存在于厚壁菌门(Firmicutes)和放线菌门(Actinomycetota)中。
支链氨基酸转氨酶仅在拟杆菌门和厚壁菌门中观察到,而前消旋酶主要在厚壁菌门中普遍存在。此外,大多数拟杆菌表现出快速消耗天冬酰胺(Asn)的能力,而L-脯氨酸降解菌仅在属于厚壁菌门的毛螺菌科和梭菌科中发现。
这些结果表明,氨基酸代谢基因广泛分布在人类肠道微生物中,并编码消化代谢蛋白质氨基酸的关键酶。
2
从头合成氨基酸
肠道菌群除了参与蛋白质的降解外,还可以合成氨基酸,包括从头合成。
体外研究表明,瘤胃中的细菌种类可以在不同的肽浓度下合成氨基酸。包括Streptococcus bovis,Selenomonas ruminantium, Prevotella bryantii。
doi: 10.2174/1389203721666200212095503.
此外,体内研究还表明,微生物衍生的赖氨酸(一种必需氨基酸)被吸收并并入宿主蛋白质。在无菌和常规化大鼠体内,将15NH4CL中的15N并入赖氨酸的比较表明,检测到的所有15N赖氨酸均来自微生物来源。在后续研究中,这些研究人员确定约75%的微生物15N标记的赖氨酸被小肠吸收。
对18岁以上的人样本进行了检查,这些人被提供了氮充足的饮食,并发现微生物衍生的赖氨酸和苏氨酸对游离血浆赖氨酸和苏氨酸库有显著贡献。此外,有报告说,大肠中微生物群的富集是通过与必需氨基酸(EAA)生物合成相关的基因进行的,该生物合成基于人血浆池产生的前体。
3
对氨基酸吸收的影响
肠道微生物群还可以通过以下几种方式影响氨基酸的吸收。
▸ 调节肠道环境影响氨基酸吸收
•改变肠道pH值
肠道菌群的代谢活动可以改变肠道的pH值。不同的氨基酸在不同的pH值下具有不同的离子化状态,从而影响其吸收。例如,在酸性环境下,一些氨基酸更容易以阳离子形式存在,而在碱性环境下则更容易以阴离子形式存在。
某些肠道菌群如乳酸菌可以产生乳酸,降低肠道pH值,使一些氨基酸更容易被吸收。而另一些细菌如大肠杆菌等则可以产生碱性物质,升高肠道pH值,可能会影响某些氨基酸的吸收。
•影响肠道通透性
肠道菌群可以通过调节肠道上皮细胞之间的紧密连接来影响肠道通透性。正常情况下,肠道上皮细胞之间的紧密连接可以阻止大分子物质和有害物质进入体内,但也会限制一些氨基酸的吸收。
一些有益的肠道菌群如双歧杆菌可以增强肠道上皮细胞之间的紧密连接,维持肠道屏障功能,防止有害物质进入体内,同时也可以促进一些氨基酸的选择性吸收。而一些有害的致病菌则可能破坏肠道上皮细胞之间的紧密连接,增加肠道通透性,导致氨基酸吸收紊乱和肠道炎症。
▸ 与宿主相互作用影响氨基酸吸收
•竞争吸收位点
肠道菌群和宿主细胞可能会竞争氨基酸的吸收位点。一些肠道细菌可以表达与宿主细胞相似的氨基酸转运蛋白,从而与宿主竞争氨基酸的吸收。
例如,某些大肠杆菌可以表达与人体小肠上皮细胞相似的赖氨酸转运蛋白,从而竞争赖氨酸的吸收。这种竞争会导致宿主明明摄入了较多的蛋白质,但实际对氨基酸的吸收量却不多。
•调节宿主基因表达
肠道菌群可以通过产生代谢产物、分泌信号分子等方式调节宿主基因的表达,从而影响氨基酸的吸收。
例如,一些短链脂肪酸如丁酸可以作为信号分子,调节肠道上皮细胞中氨基酸转运蛋白的表达。此外,肠道菌群还可以通过调节肠道免疫系统、内分泌系统等间接影响氨基酸的吸收。
4
补充益生菌增加了植物蛋白的吸收

该实验研究了益生菌补充剂对植物蛋白(如豌豆蛋白)中氨基酸吸收的影响。
研究发现,副干酪乳杆菌LP-DG®(每天50亿 CFU)给药显著增加了摄入豌豆蛋白后蛋氨酸(+20.0%)、组氨酸(+40.4%)、缬氨酸(+21.5%)、亮氨酸(+23.3%)、异亮氨酸(+26.0%)、酪氨酸(+16.0%)、总支链氨基酸(+22.8%)和总必需氨基酸(+16.0%)的含量。
而副干酪乳杆菌 LPC-S01(每天50亿 CFU)给药显著增加了蛋氨酸(+16.3%)、组氨酸(+49.2%)、缬氨酸(+24.7%)、亮氨酸(+25.2%)、异亮氨酸(+26.1%)、酪氨酸(+11.6%)、总支链氨基酸(+26.8%)和总必需氨基酸(+15.6%)最大浓度。
这项研究表明,通过补充益生菌可能是一种重要的营养策略,可以改善餐后血液氨基酸的变化,并克服植物蛋白的组成缺陷。益生菌可以通过优化肠道菌群结构,提高植物蛋白的消化吸收率。
氨基酸在人体营养和生理活动中至关重要,人们对蛋白质的需求实际上是对氨基酸的需求。我们每天从食物中摄取蛋白质,正是为了满足这些生理活动所必需的氨基酸。那么氨基酸被人体吸收后主要有哪些去处呢?让我们继续来看。
1
氨基酸吸收后的利用
人体的游离氨基酸主要有三条去路:
一是合成人体所需的多肽和蛋白质,多肽和蛋白质是机体生命活动的执行者。蛋白质合成过程较复杂,需根据特定基因上携带的遗传信息,经复制、转录、翻译等一系列过程,以各种氨基酸为原料装配成人体蛋白质。
二是经脱氨、脱羧等分解代谢途经生成氨、α-酮酸和胺类、CO2。其中,生成的α-酮酸可进一步代谢,经氨基化作用生成非必需氨基酸供机体利用。或氧化供能(当非蛋白质能量来源不足时,氨基酸可能被用于能量代谢,以满足能量需求)。氨基酸脱氨作用产生的游离氨,在正常情况下,主要在肝脏中合成尿素,随尿排出体外,少部分氨在肾脏中以铵盐形式由尿排出。
三是转变成其它含氮化合物,比如经一系列酶催化转变成嘌呤、嘧啶、肌酸等(赖氨酸合成肉碱,精氨酸合成肌酐,蛋氨酸作为甲基供体)。
!
注意
一些氨基酸代谢产生出具有特殊重要生理功能的小分子化合物,比如:谷氨酸脱氨分解生成γ-氨基丁酸(GABA),GABA是抑制性神经递质,可抑制中枢神经;组氨酸脱羧分解产生组胺,组胺是一种强烈的血管扩张剂,引起平滑肌收缩,并增加毛细血管通透性;色氨酸脱羧分解产生5-羟色胺(又叫血清素),也是一种抑制性神经递质,直接影响神经传导。
•不是所有氨基酸都能被吸收利用
人体内的氨基酸除了被利用外,也会有部分无意流失。每天摄入大量的蛋白质进入胃肠道;虽然其中一些物质被消化和吸收,但相当数量的氨基酸在胃肠道中分解代谢或被纳入微生物蛋白质,这些蛋白质与一些未消化的蛋白质一起在粪便中从体内流失。
2
氨基酸的重吸收
生物体氨基酸稳态的一个重要因素是发生在肾皮层的肾小球滤过/肾小管重吸收机制。
每天都会不可避免地流失一些氨基酸,如尿素、肌酐和氨。当蛋白质成分增加超过必需替代或氨基酸用于糖异生时,尿素的量会增加。
•肾小管重吸收受损会导致氨基酸的流失增加
肾小管重吸收是氨基酸循环的一部分。它由于肠道中发现的几乎相同的一组转运蛋白介导,当肾小管重吸收受损时,可能导致氨基酸流失增加,即使摄入了较多的蛋白质,氨基酸的总含量仍缺乏。
3
不同人群氨基酸需求不同
多年来,人们进行了大量的工作来确定人类对膳食氨基酸的需求。粮农组织、世卫组织和联合国建议,每公斤体重0.83克优质蛋白质是每日膳食蛋白质的“安全”摄入量,即体重70公斤的参考男性每天摄入58克蛋白质,体重57公斤的参考女性每天摄入47克蛋白质。
但越来越多的研究和谷禾数据表明,由于个体消化吸收差异以及不同食物和饮食习惯的代谢影响,每个人对蛋白质的需求量各不相同。
•不同人群对氨基酸的需求和利用都不同
每个人的不同状态对蛋白质的需求量也都不同:在成长中的儿童中,需要氨基酸来维持体内蛋白质和生成新的组织,而在成人中,需要氨基酸来补充体内丢失的氨基酸。对于孕妇和哺乳期妇女来说,还需要额外的氨基酸来促进胎儿的生长和产奶量。健身人群则需要大量的蛋白质来合成肌肉。
因此,成人所需的膳食氨基酸不是固定值,每日最佳摄入量取决于你的身高体重、健康状况、目标、身体活动水平。
•消化率较低的人群需要摄入更多的蛋白质
素食主义者对蛋白质的需求较高,因为植物蛋白在生物利用度上通常不如动物蛋白。植物蛋白消化率为60-80%,而动物蛋白消化率超过90%。
此外一些植物含有抑制蛋白质消化和吸收的抗营养因子,如胰蛋白酶抑制剂、凝集素、植酸盐和单宁、硫苷,会导致素食主义者的蛋白质消化和吸收率偏低。而消化率较低的人群需要摄入更多的蛋白质,以确保满足身体的营养需求。
•注意摄入蛋白的氨基酸构成
另外,一些蛋白质也含有限制性氨基酸,限制性氨基酸是指食物所含必需氨基酸的量与人体所需的蛋白质必需氨基酸的量相比,比值偏低的氨基酸。
一种食物里特别缺少某一种氨基酸,即使其它的氨基酸含量很高,因为这个氨基酸导致它的各个氨基酸比例不平衡。人体对这种食物的吸收依然不理想。多样性饮食结合不同的膳食蛋白可以帮助弥补它们各自的不足。
比如说,赖氨酸是最常见的限制性氨基酸,特别是在谷类中,如小麦和水稻。坚果往往也含有赖氨酸作为限制性氨基酸。另一方面,豆类含有足够的赖氨酸,可以弥补,但缺乏硫氨基酸,如蛋氨酸和半胱氨酸。
通过整合微生物组学、宏基因组学、代谢组学和蛋白质组学数据,我们可以构建更完整的个体营养代谢图谱,有望建立更精确的营养-微生物-代谢互作网络模型,为精准营养干预提供依据。
益生菌在提高氨基酸吸收率方面的应用潜力。新型益生菌的筛选和开发将更加注重其对氨基酸代谢的调节作用,有望开发出更有针对性地提高氨基酸的生物利用度的微生态制剂。
随着精准医疗的发展,个性化的氨基酸需求评估和补充策略将成为研究热点,这需要我们更深入地了解个体差异对氨基酸代谢的影响。基于机器学习的预测模型将帮助我们更准确地评估个体的氨基酸需求,这需要综合考虑肠道菌群特征、代谢指标和临床表型等多维度数据。通过这些数据的综合分析,我们可以构建出更加个性化的营养干预方案,优化个体健康和疾病预防。
主要参考文献
Li TT, Chen X, Huo D, Arifuzzaman M, Qiao S, Jin WB, Shi H, Li XV; JRI Live Cell Bank Consortium; Iliev ID, Artis D, Guo CJ. Microbiota metabolism of intestinal amino acids impacts host nutrient homeostasis and physiology. Cell Host Microbe. 2024 May 8;32(5):661-675.e10.
Jäger R, Zaragoza J, Purpura M, Iametti S, Marengo M, Tinsley GM, Anzalone AJ, Oliver JM, Fiore W, Biffi A, Urbina S, Taylor L. Probiotic Administration Increases Amino Acid Absorption from Plant Protein: a Placebo-Controlled, Randomized, Double-Blind, Multicenter, Crossover Study. Probiotics Antimicrob Proteins. 2020 Dec;12(4):1330-1339.
Neis EP, Dejong CH, Rensen SS. The role of microbial amino acid metabolism in host metabolism. Nutrients. 2015 Apr 16;7(4):2930-46.
Hou Y, Yao K, Yin Y, Wu G. Endogenous Synthesis of Amino Acids Limits Growth, Lactation, and Reproduction in Animals. Adv Nutr. 2016 Mar 15;7(2):331-42.
Gorissen SHM, Trommelen J, Kouw IWK, Holwerda AM, Pennings B, Groen BBL, Wall BT, Churchward-Venne TA, Horstman AMH, Koopman R, Burd NA, Fuchs CJ, Dirks ML, Res PT, Senden JMG, Steijns JMJM, de Groot LCPGM, Verdijk LB, van Loon LJC. Protein Type, Protein Dose, and Age Modulate Dietary Protein Digestion and Phenylalanine Absorption Kinetics and Plasma Phenylalanine Availability in Humans. J Nutr. 2020 Aug 1;150(8):2041-2050.
Bröer S, Gauthier-Coles G. Amino Acid Homeostasis in Mammalian Cells with a Focus on Amino Acid Transport. J Nutr. 2022 Jan 11;152(1):16-28.
Mariotti F, Gardner CD. Dietary Protein and Amino Acids in Vegetarian Diets-A Review. Nutrients. 2019 Nov 4;11(11):2661.
Abdallah A, Elemba E, Zhong Q, Sun Z. Gastrointestinal Interaction between Dietary Amino Acids and Gut Microbiota: With Special Emphasis on Host Nutrition. Curr Protein Pept Sci. 2020;21(8):785-798.
Fan P, Li L, Rezaei A, Eslamfam S, Che D, Ma X. Metabolites of Dietary Protein and Peptides by Intestinal Microbes and their Impacts on Gut. Curr Protein Pept Sci. 2015;16(7):646-54.


短链脂肪酸这一词经常出现在谷禾的文章和报告中,那你真的了解短链脂肪酸吗?短链脂肪酸(SCFA)主要是肠道微生物群在结肠内通过发酵碳水化合物(包括膳食和内源性碳水化合物,主要是抗性淀粉和膳食纤维)和一些微生物可利用的蛋白质而产生的。
短链脂肪酸主要是乙酸、丙酸和丁酸,在结肠中的浓度比大致为60-70%:20-30%:10-20%。这些代谢产物能够被宿主利用,尤其是丙酸和丁酸,它们发挥一系列促进健康的功能。它们能被肠粘膜有效吸收,作为能量来源,还能作为基因表达调节剂以及特定受体识别的信号分子,对宿主生理产生重要影响。近几年的研究还发现其能够调节免疫细胞发育并抑制炎症。
然而,三种主要短链脂肪酸——乙酸盐、丙酸盐和丁酸盐——它们在体内的功能和组织分布不同,对宿主生理的影响存在显著差异。丁酸盐优先被肠粘膜用作能量来源,还具有抗炎特性,可增强肠道屏障功能和粘膜免疫。丙酸盐有助于抵抗肝脏中的脂肪形成、降低胆固醇、也有一定的抗炎和抗致癌作用。而乙酸在血液中浓度最高,可用作肝脏胆固醇和脂肪酸合成的底物,增加结肠血流量和氧气吸收,并通过影响回肠收缩来增强回肠运动。
其次,它们与宿主蛋白(如丁酸和丙酸抑制组蛋白去乙酰化酶)和受体的相互作用也不同。因此,特别需要考虑这些短链脂肪酸的微生物来源。与此同时,饮食结构和肠道生理变化如何影响这些脂肪酸的相对产量和在结肠中的浓度也是至关重要的。
本文将重点介绍丙酸盐和丁酸盐,因为这两种短链脂肪酸被认为对健康有重要影响,包括丁酸预防结直肠癌,丙酸促进饱腹感并降低胆固醇。而乙酸是大多数肠道厌氧菌的主要发酵产物,也可由还原性乙酸生成,而丙酸和丁酸则由不同的肠道细菌群产生。
我们将探讨在人类结肠微生物群中已知的形成这两种短链脂肪酸的途径,以及各种饮食和环境因素对其产生的调节可能性。详细了解肠道微生物群的短链脂肪酸代谢及其生理功能对于制定个性化的健康营养方案是必不可少的。
丙酸盐和丁酸盐在结构、来源、生理功能以及对健康的影响方面存在一些差异。
▸ 结构上的差异:
丙酸盐(Propionate)含有三个碳原子,羟基(-OH)位于第二个碳原子上。
丁酸盐(Butyrate)含有四个碳原子,羟基(-OH)位于第四个碳原子上。
▸ 来源上的差异:
丙酸盐通常由肠道细菌通过发酵L-鼠李糖、聚葡萄糖、阿拉伯木聚糖、D-塔格糖、甘露寡糖、昆布多糖等糖类物质产生。
丁酸盐可以通过肠道微生物群发酵富含抗性淀粉和果聚糖的食物来增加,如菊粉、马铃薯、洋葱等。
▸ 功能上的差异:
丙酸盐在体内的主要功能是作为肝脏中糖原合成的前体物质,有助于调节血糖水平,影响食欲。
丁酸盐是结肠上皮细胞的首选能源,有助于维持肠道屏障,发挥免疫调节和抗炎作用。丁酸盐还可以通过减少NF-kB信号传导和诱导凋亡来促进神经保护。
▸ 对健康的影响:
丙酸盐的健康益处主要是调节能量代谢和改善代谢综合征,可能对调节肠道菌群平衡也有积极作用,但其具体影响取决于个体的肠道菌群组成。
丁酸盐对健康的影响更为广泛,包括维持肠道稳态、促进肠道屏障的完整性、刺激绒毛的生长、促进粘蛋白的产生。以及改善认知功能、促进睡眠、调节社交行为和在糖尿病中的潜在益处。
丙酸盐是许多生物(从细菌到人类)的代谢副产物,产生丙酸的代谢途径可分为三类。主要发酵途径将不同的碳源分解代谢为丙酸盐(图A);分解代谢途径则能将多种氨基酸降解为丙酸盐(图B);此外,通过与从丙酮酸或二氧化碳生成生物质前体相关的合成代谢途径也可以生产丙酸盐(图C)。
产生丙酸盐的代谢途径
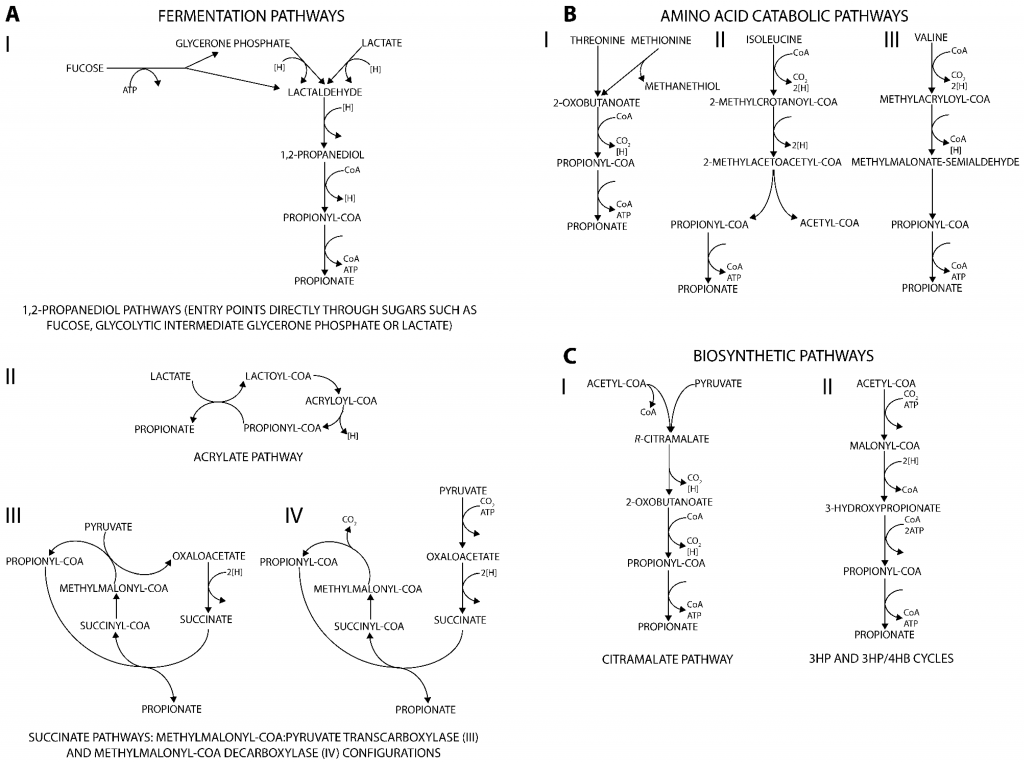
doi.org/10.3390/fermentation3020021
肠道微生物通过发酵不可消化的碳水化合物产生的丙酸盐是体内丙酸盐的主要来源。下面将讲述肠道中丙酸盐通过发酵碳水化合物的三种主要产生途径以及参与这些途径的微生物。
与氨基酸降解和生物合成途径相比,发酵途径不仅提供能量,还帮助消耗由糖分解代谢产生的还原辅因子。它们在能量产生和维持氧化还原平衡中的作用,使这些途径能够与细胞生长相耦合。
能够产生丙酸盐的微生物种类

doi.org/10.3390/fermentation3020021
▸
丙二醇相关途径
• 鼠伤寒沙门氏菌和Roseburia inulinivorans在这一过程可以产生丙酸
丙酸盐在肠胃中的生成是由1,2-丙二醇(PDO)发酵菌和PDO消耗菌组成的微生物联合作用的结果。已知一些生物体(如鼠伤寒沙门氏菌和Roseburia inulinivorans)能同时进行这两种过程。
在二醇脱水酶和两种常与乙酸代谢相关的混杂酶(辅酶A依赖性醛脱氢酶磷酸转酰基酶和乙酸激酶)的共同作用下,PDO分解代谢为丙酸,同时生成一个ATP和一个还原辅因子。
然而,通过该途径产生丙酸取决于可用于其他细菌生长的碳水化合物,据报道岩藻糖和鼠李糖是丙酸的前体。
通过1,2-丙二醇形成丙酸的微生物途径
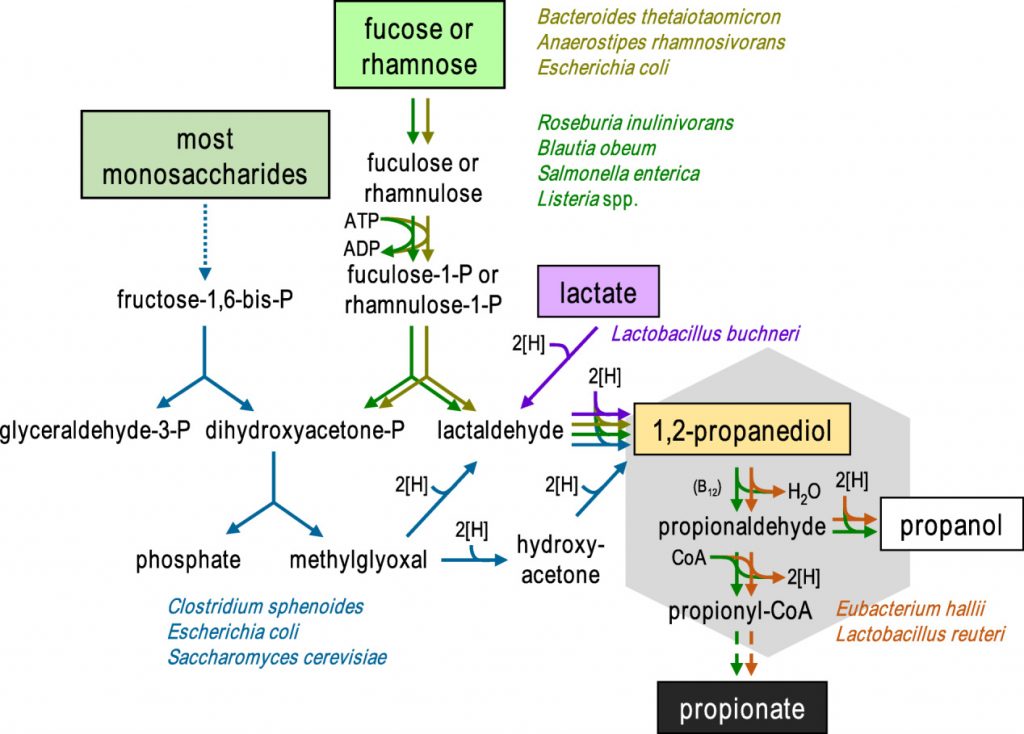
doi: 10.1111/1462-2920.13589.
霍氏大肠杆菌(E.hallii)和罗伊氏乳杆菌(Lactobacillus reuteri)虽然无法在岩藻糖或鼠李糖上生长,但仍然能够利用1,2-丙二醇产生丙酸和丙醇。此外,宏基因组研究表明,其他肠道厌氧菌,包括Flavonifractor plautii、Intestinimonas butyriproducens和Veillonella spp. 也可能能够从这种底物中产生丙酸。因此,不同细菌之间中间体 1,2-丙二醇可能在脱氧糖生产丙酸中发挥重要作用。
▸ 丙烯酸酯途径
丙烯酸酯途径在消耗NADH的情况下,使乳酸在ATP中性条件下转化为丙酸。该途径存在于几种细菌中,包括丙酸梭菌(Clostridium propionicum)、埃氏巨球菌(Megasphaera elsdenii)和瘤胃普氏菌(Prevotella ruminicola)。
虽然多种底物可以分解为丙酸和乙酸,包括乳酸、丝氨酸、丙氨酸和乙醇,但葡萄糖发酵在这过程中似乎不会导致任何天然生产者产生丙酸,这可能是因为葡萄糖发酵不会触发启动循环所需的乳酸消旋酶的表达。
▸ 琥珀酸途径
琥珀酸途径主要存在于拟杆菌门和厚壁菌门中,拟杆菌门的一些细菌从膳食碳水化合物生成丙酸,并且拟杆菌门的相对丰度与人类粪便中丙酸盐的相对水平相关。
• 琥珀酸转化为丙酸盐还需要维生素B12
琥珀酸是丙酸的前体,但在高pCO2(二氧化碳分压)和高稀释率等条件下,它可在磷酸烯醇式丙酮酸羧激酶受到抑制的拟杆菌属培养物中积累。琥珀酸转化为丙酸还需要维生素B12,如果缺乏B12,琥珀酸可能无法转化成丙酸盐。
人类结肠中的一些厚壁菌门细菌(例如Phascolarctobacterium succinatutens)能将琥珀酸转化为丙酸;其他革兰氏阴性菌通过琥珀酸途径(如韦荣氏球菌属)或丙烯酸酯途径(Megasphaera elsdenii)将乳酸转化为丙酸盐。
▸ 氨基酸降解产生丙酸
缬氨酸、苏氨酸、异亮氨酸和蛋氨酸的降解可导致通过丙酰辅酶A产生丙酸和ATP。
由于氨基酸的合成和随后的分解代谢途径存在于多种微生物中,因此可以使用氨基酸合成代谢和分解代谢途径的组合从葡萄糖生产丙酸。
几种拟杆菌在蛋白水解和肽形成丙酸方面发挥重要作用。
苏氨酸的产生和分解代谢已在大肠杆菌中得到广泛研究,作为丙酰辅酶A的可能来源,可用于生产各种化学物质,包括丙醇、红霉素和3-羟基戊酸。此外,考虑到氧化还原和能量平衡,缬氨酸/异亮氨酸和苏氨酸途径的组合具有协同作用,可以略微提高丙酸产量。
除此之外,可以利用丙酮酸经过柠檬酸途径产生丙酸,由于和菌群关联较小,在这就不展开描述了。
▸ 增加丙酸盐生成的底物
一些益生元等化合物可以影响丙酸盐的生产,不过由于浓度以及肠道微生物群落构成的不同,这些化合物的调节丙酸盐产生时具有一定差异。
• L-鼠李糖能够明显增加丙酸盐产量
L-鼠李糖或6-脱氧-L-甘露糖是一种天然脱氧糖。它存在于多种动物、植物和细菌多糖中。在短期体外实验中,L-鼠李糖已被证明能使丙酸产量增加四倍于乳果糖。
在一项人体体内研究中也获得了类似的结果,其中受试者在三个不同的时间被给予25克L-鼠李糖、乳果糖或D-葡萄糖。摄入后24小时测量血清丙酸,L-鼠李糖的血清丙酸明显高于乳果糖或D-葡萄糖的血清丙酸。
一项长期研究也证实了L-鼠李糖诱导丙酸的作用,该研究结果表明,与摄入D-葡萄糖作为对照相比,摄入25克L-鼠李糖可显著提高人体28天内的血清丙酸水平。
• 抗性淀粉有助于增加丙酸
抗性淀粉对淀粉酶降解具有抗性,但它会发酵成丁酸或丙酸。特别是,来自大米的抗性淀粉与丙酸产量增加有关。
研究了抗性淀粉在大鼠体内的发酵情况,饲喂抗性淀粉(630g/kg饲料)的大鼠的肝脏甘油三酯和总胆固醇浓度显著低于对照组。与此同时,血清丙酸浓度也显著增加。
• 菊粉对于增加丙酸和丁酸非常有效
菊粉属于果聚糖家族,主要由β-(2,1)连接的果糖基组成。它天然存在于菊苣和菊芋等开花植物中。作为益生元,菊粉已被证明对增加丁酸和丙酸的产量非常有效。使用人体肠道微生物体外模拟研究了菊粉增加丙酸的效果。
补充菊粉1周(5g/d)后观察到短链脂肪酸产生的代谢变化。较高浓度的短链脂肪酸源于丙酸和丁酸产量的增加。
此外一项针对喂食菊粉(10%)的大鼠的体内研究也导致丙酸产量大幅增加,高达 58.4mmol/。
• 聚葡萄糖
聚葡萄糖是一种支链、随机聚合的多糖,主要由葡萄糖合成,在胃肠道上部不会被消化。使用结肠模拟器研究了这种底物对结肠微生物组成和代谢活性的调节作用。与对照糖木糖醇(8.3mmol/L)相比,短链脂肪酸产量显著增加,尤其是丙酸盐(22.9mmol/L)。
• 阿拉伯木聚糖
阿拉伯木聚糖是许多谷物中发现的主要非淀粉多糖,是膳食纤维的一部分。
在体内研究中,比较了54只大鼠,这些大鼠分别喂食对照饮食(含710g/kg小麦)、阿拉伯木聚糖补充饮食(610g/kg小麦淀粉加100g/kg玉米阿拉伯木聚糖)和胆固醇补充饮食(不含或含2g/kg 胆固醇)。由于短链脂肪酸的积累,尤其是丙酸(摩尔百分比>45%),盲肠pH值从7降至6。然而,丁酸的产生不受影响。
车前草是一种可溶性纤维来源,可提供与麦麸阿拉伯木聚糖相当的多糖。在一项大鼠体内研究中,比较了车前草(5%)对盲肠和结肠发酵的影响与麦麸 (10%) 的影响。研究发现,车前草发酵可产生更高的短链脂肪酸,尤其是盲肠和所有结肠中的丙酸更多。
除此之外,D-塔格糖、甘露寡糖、昆布多糖等物质也可以增加人体丙酸盐的产生。
丙酸盐已被证明具有抗脂肪形成和降低胆固醇的作用。它还对体重控制和进食行为有很强的影响。此外,有研究表明,丙酸和丁酸一样,对结肠癌细胞具有抗增殖作用。
丙酸盐对健康的影响
doi: 10.1111/j.1753-4887.
然而,与结肠细胞用作能量来源的丁酸不同,丙酸在血液循环中的浓度较高。因此,丙酸的生物活性可能不仅限于结肠本身,还扩展到人体的其他部位。需要强调的是,了解体内结肠丙酸浓度或短链脂肪酸浓度不足以推断健康状况。
▸ 影响肝细胞的脂质合成
肝脏的脂质合成包括将饮食来源的脂肪酸和甘油转化为具有不同脂肪酸组成的胆固醇和甘油三酯。然后,这些肝脏脂质分子被结合到脂蛋白中,从而通过循环分布到各种组织中。
• 脂质合成受到短链脂肪酸的强烈影响
有趣的是,肝细胞中的脂质合成受到肠道纤维发酵产生的短链脂肪酸的数量和类型的强烈影响。丙酸盐已被确定在其中一些过程中发挥重要作用。
对肝脏脂质合成的饮食调节早期观察显示,膳食纤维摄入与肝脏脂质合成密切相关。这种影响的部分原因如下:1)粪便中胆固醇和胆汁酸从肠道排出的增加;2)胆固醇向胆汁酸的肝脏转化率较高;3)通过减少乳糜微粒的大小和降低胆固醇在乳糜微粒中的掺入来优化脂蛋白的外周代谢。
• 降低血清胆固醇水平、抑制脂肪酸合成
短链脂肪酸作为微生物碳水化合物发酵的产物,在肝脏脂质合成中发挥着特定作用,已被证明在喂食大鼠时可降低血清胆固醇水平。对分离的大鼠肝细胞的体外研究表明,丙酸对脂肪酸合成有抑制作用,但对胆固醇合成没有抑制作用。
其他大鼠实验表明,加入纤维的饮食可降低肝和血浆胆固醇水平及血浆甘油三酯中的胆固醇,而肝甘油三酯没有受到影响。
尽管这些研究的结果令人信服,但其他研究并不总是能够证实丙酸盐对脂质代谢的抑制作用。例如,每天在面包中补充9.9克丙酸盐不会改变6名健康志愿者的脂质代谢,甚至会导致5名受试者的甘油三酯浓度升高。
在另一项研究中,比较了丙酸盐对人和大鼠肝细胞脂质代谢的影响。发现浓度为0.1mmol/L的丙酸盐对大鼠乙酸盐合成脂质有抑制作用。然而,在人类肝细胞中,需要更高浓度的丙酸盐(约10-20 mmol/L)才能获得同样的抑制作用。该值比门静脉血中丙酸盐的浓度高100-200倍,表明大鼠模型不能完全外推到人类的情况。
▸ 作为影响饱腹感的分子
丙酸盐不仅具有降低胆固醇和抗脂肪生成的作用,还可能通过刺激饱腹感来控制体重。已有研究表明,短链脂肪酸(如乙酸、丙酸、丁酸)具有诱发饱腹感的作用。
• 丙酸盐影响肠道激素的形成进而影响饱腹感
现有证据表明,细菌调节肠道激素(如胰高血糖素样肽1(GLP-1)和肽YY(PYY))的形成,是通过短链脂肪酸介导的。乙酸、丙酸和丁酸的生理浓度,以及pH值从7.5降至6.0,会显著增加肠内分泌结肠细胞系STC-1中的胰高血糖素原和PYY。
GLP-1和PYY是刺激饱腹感的激素,由L细胞(主要位于回肠和结肠)响应营养摄入而释放。GLP-1促进胰岛素分泌和胰腺β细胞增殖,同时控制肌肉细胞中的糖原合成;而PYY则减缓胃排空。相反,生长素释放肽刺激食欲,主要由胃中的P/D1细胞产生。
不易消化的碳水化合物,如低聚果糖、乳糖醇和抗性淀粉,通过调节肠道肽GLP-1、PYY和生长素释放肽的产生,有效地诱导饱腹感,这一机制还涉及肠道微生物群落。
• 丙酸盐对摄食行为有显著影响,诱导瘦素产生
在短链脂肪酸中,丙酸盐被重点研究作为一种饱腹感诱导剂,对能量摄入和摄食行为有显著影响。人体和动物试验表明,丙酸盐给药(体内范围为130-930mmol/L,体外范围为0.01-10mmol/L)显著增强饱腹感并降低进食欲望。
丙酸盐触发的饱腹感信号之一是瘦素,这是一种强效的厌食激素,通过中枢神经系统中表达的受体抑制食物摄入。研究显示,每天服用500µmol的丙酸盐几乎使小鼠血浆中的瘦素浓度翻了一倍。
在另一项研究中,浓度为3mmol/L的丙酸在mRNA和蛋白质水平上诱导了人内脏脂肪组织中瘦素的产生。这些数据表明,肠道微生物群对肥胖的调节作用可能部分由短链脂肪酸(特别是来自微生物碳水化合物发酵的丙酸)介导。
▸ 影响心脑血管健康
丙酸盐通过与肠道受体GPR 41和GPR 43(也称为脂肪酸受体FFAR2和FFAR3)相互作用,对心脑血管健康具有一些潜在的影响。
• 脑血管患者体内的丙酸盐含量较低
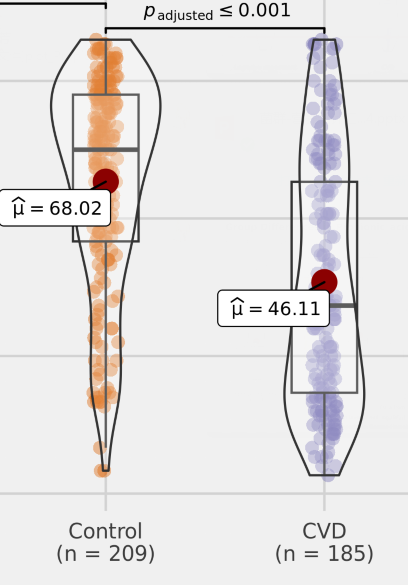
编辑
谷禾的数据中发现,与健康对照相比,脑血管疾病患者中的丙酸盐含量较低,并且具有明显的统计学差异。因此丙酸盐的含量可能是影响心脑血管健康的一个重要指标。
• 较高的丙酸盐浓度患冠状动脉硬化风险较低
最近一项大型横断面研究表明血浆丙酸浓度与冠状动脉疾病存在关联,较高浓度的丙酸盐与较低的冠状动脉粥样硬(CAD)风险相关,且与已知的心血管风险因素无关。
如上所述,越来越多的实验数据表明,丙酸盐可能对高血压、内皮功能障碍和高胆固醇血症等心血管风险因素产生有益影响。
▸ 丙酸盐在癌症中的潜在作用
短链脂肪酸对癌症(尤其是结肠癌)的影响已被广泛研究。丁酸能够调节基因表达,并对细胞凋亡和细胞周期的关键调节因子产生影响。
几种机制促成了丁酸对基因表达的调节作用。这些机制包括组蛋白和非组蛋白的过度乙酰化以及DNA甲基化的改变,从而增强了转录因子对核小体DNA的可及性。
• 丙酸盐诱导结直肠癌细胞凋亡
在一项研究中,丙酸盐和乙酸盐(浓度分别为26-40和9-16mmol/L)在人类结直肠癌细胞系中诱导了典型的细胞凋亡迹象。这些迹象包括线粒体跨膜电位的丧失、活性氧的产生、胱天蛋白酶3加工和核染色质凝聚。
• 抑制结肠癌细胞系的生长
短链脂肪酸对结肠上皮细胞增殖具有矛盾作用。虽然这些阴离子刺激正常隐窝细胞,但它们抑制结肠癌细胞系的生长。
丁酸盐和丙酸盐也是诱导分化和细胞凋亡的最有效脂肪酸。因此,它们通常可以预防癌症的发展,尤其是预防结直肠癌。虽然丁酸比丙酸更有效,但它主要被结肠细胞吸收作为能量来源。相比之下,丙酸和乙酸盐进入血液循环的浓度比丁酸高得多,并且它们被肝脏大量吸收(约60%)。由于这些阴离子在肝脏中的浓度很高,它们很可能会影响肝癌细胞以及已知会导致肝脏转移的其他典型癌细胞,例如乳腺癌和结肠癌。
一项针对猝死患者的研究表明,外周血中的短链脂肪酸数量可以量化。因此,这种循环中的丙酸盐、乙酸盐和丁酸盐的抗癌作用非常值得研究;例如,这种影响会在多大程度上延伸到小肠、大肠和肝脏之外,从而影响不同的组织?
!
• 丙酸盐过量可能与阿尔茨海默病相关
最近的几项研究提供了丙酸盐与阿尔茨海默病(AD)之间联系的证据。例如,分析了阿尔茨海默病患者和健康对照者的唾液样本,发现阿尔茨海默病患者的丙酸水平比健康对照者高出1.35倍。分析了轻度认知障碍者、阿尔茨海默病患者和健康对照者的唾液样本,也发现AD患者的丙酸水平显著升高。
几项啮齿动物研究也将粪便和循环中丙酸盐水平与阿尔茨海默病联系起来。接受AD患者粪便微生物群移植的小鼠丙酸水平高于对照组。
此外,发现阿尔茨海默病小鼠海马体中的丙酸增加了1.23倍。也发现阿尔茨海默病小鼠前额叶皮层中的丙酸浓度显著高于野生型小鼠,6个月大的阿尔茨海默病小鼠粪便中的丙酸浓度也显著更高。
• 丙酸血症的丙酸盐代谢异常
丙酸血症是人类最常见的有机酸代谢紊乱,是一种由丙酰辅酶A羧化酶基因缺陷引起的先天性代谢错误,丙酸过量且无法通过丙酰辅酶A转化。
丙酸血症患者的体内丙酸及其代谢物水平显著升高,会引起代谢性酸中毒、血氨升高,可能还会诱导一些严重的并发症如脑损伤、心肌病发生。
•有研究认为过多的丙酸盐可能导致肥胖和糖尿病
2019年哈佛大学公共卫生学院发表的一篇文献称:小鼠长期接触丙酸将导致肝细胞产生更多葡萄糖,形成高血糖症,这是糖尿病的一个典型特征。此外,研究人员还发现,长期服用丙酸盐剂量的小鼠,其体重会显著增加,胰岛素抗性也会增大。
还对14名健康人开展对照试验,受试者被随机分为两组,其中一组的餐食中添加1克丙酸盐,另一组添加安慰剂。结果显示,进餐后不久,丙酸盐组受试者血液中的去甲肾上腺素水平显著升高,胰高血糖素等激素水平也有所增长。这表明丙酸盐可能会作为“代谢破坏者”,增加人类患糖尿病和肥胖症的风险。
而这与前文丙酸盐会影响脂质合成与摄食行为相矛盾,因此,丙酸盐对人体的影响可能取决于其含量以及人体独特的肠道菌群结构与代谢能力。
内源性丙酸盐与外源性丙酸盐存在区别
内源性丙酸:指的是人体内部产生的丙酸,通常情况下,人体中的微生物能够在结肠中通过发酵未完全消化的碳水化合物来产生丙酸,这些丙酸对人体是有益的。
外源性丙酸:指的是从外部来源摄入的丙酸,比如通过食物摄入或作为补充剂。以及在一些药物中也可能含有丙酸盐。
★ 外源性丙酸盐摄入过多对代谢健康有害
但已经有各种研究表明,外源性地摄入和自体产生的丙酸作用并不一样,外来的丙酸对个体的许多代谢过程有不利影响。
哈佛大学公共卫生学院的研究人员完成了一项实验,发现从食品中摄入的丙酸在短期内会导致高血糖、胰岛素偏高,而且长期摄入将会造成肥胖和胰岛素抵抗等更严重的症状。
我们通过口腔摄入含丙酸添加剂的食物后,会激活我们的交感神经系统,并促使体内胰高血糖素和脂肪酸结合蛋白4等激素的升高。受到激素影响,肝糖原持续分解,血糖升高。身体开始分泌更多胰岛素进行代偿降低血糖浓度,最终导致胰岛素抵抗和肥胖产生。
这项研究表明,自身产生的丙酸和外界摄入的丙酸具有不同的功能,我们推测,这是因为从食物中摄入会极大地增加细胞和丙酸的接触,而细菌产生的仅仅只会局限在结肠部分。
在人体肠道中,丁酸由膳食纤维经细菌发酵产生,通过两种代谢途径。在第一条途径中,丁酰辅酶A被磷酸化形成丁酰磷酸,并通过丁酸激酶转化为丁酸。在第二条途径中,丁酰辅酶A的辅酶A部分通过丁酰辅酶A:乙酸辅酶A转移酶转移到乙酸盐,从而形成丁酸和乙酰辅酶A。其中丁酰辅酶A:乙酸辅酶A转移酶途径占主导地位。
对人类粪便微生物群的放射性同位素分析也表明,肠道中的大多数丁酸盐是由碳水化合物通过Embden-Meyerhof-Parnas(糖酵解途径)由乙酰辅酶A产生的。
• 少量的丁酸盐也可由蛋白质合成
除了碳水化合物,少量的丁酸也可以通过谷氨酸、赖氨酸、戊二酸和γ-氨基丁酸途径由蛋白质合成。厚壁菌种也对氨基酸表现出高活性,特别是肠单胞菌(Intestinimonas AF211),它通过不同的途径将葡萄糖和赖氨酸发酵成丁酸。
形成丁酸盐的微生物途径
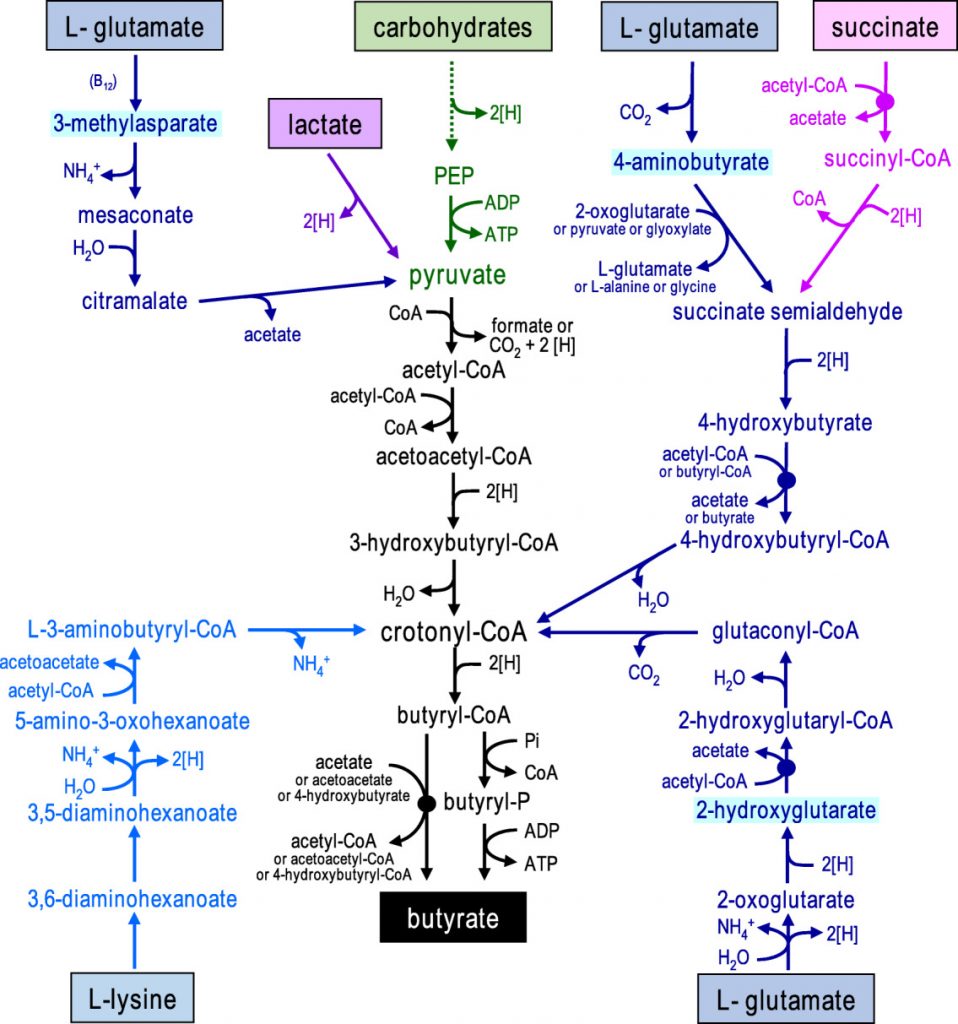
doi: 10.1111/1462-2920.13589.
碳水化合物通过糖酵解发酵为丙酮酸显示为绿色,从乙酰辅酶A形成丁酸盐显示为黑色,氨基酸发酵途径显示为蓝色(以中间体命名不同的谷氨酸途径),乳酸和琥珀酸发酵分别显示为紫色和粉色。
• 人结肠中的瘤胃球菌和毛螺菌是产丁酸的主力
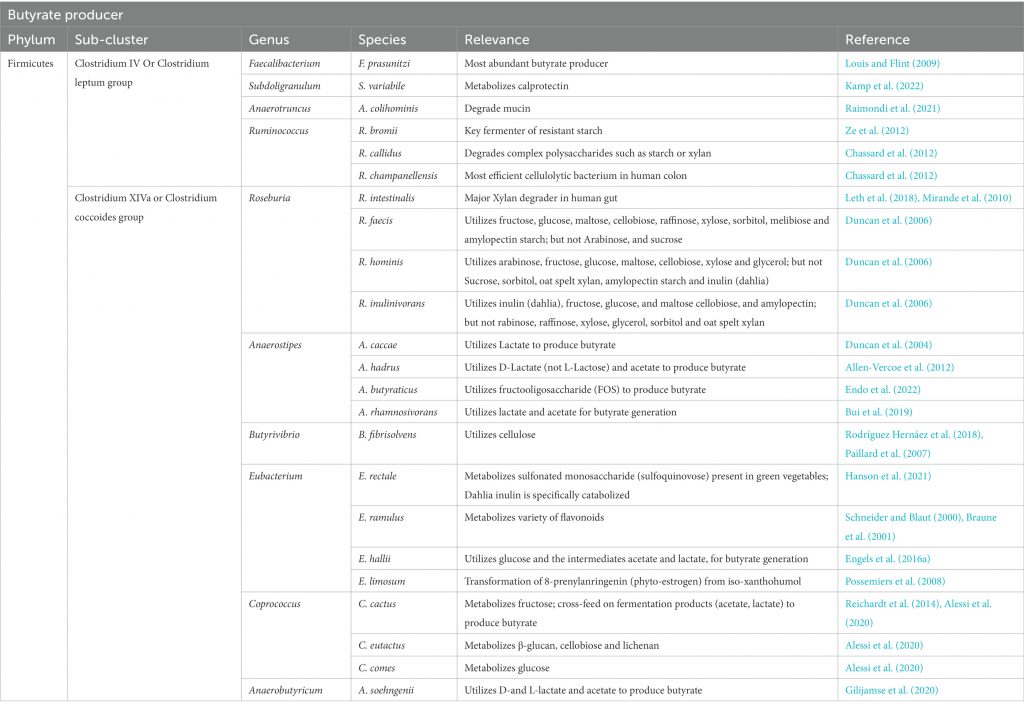
除食源性丁酸,人体内丁酸主要是由盲肠和结肠的厌氧型细菌发酵产生,而由胃和小肠产生的丁酸含量极低。大肠产丁酸的菌种主要是梭菌属XIVa和IV族,以及真杆菌属和梭杆菌属
盲肠和结肠中丁酸的产生速度和数量主要取决于肠道微生物组成、日常膳食中可利用发酵成分组成等。在人的结肠中,厚壁菌门中的瘤胃球菌科(Ruminococcaceae)和毛螺菌科(Lachnospiraceae)这两个主要科,以及包括丹毒丝菌科(Erysipelotrichaceae)和梭菌科(Clostridiaceae)在内的其他厚壁菌门物种,都发现了丁酸生产菌种与非生产菌种的交替存在。
但应注意,许多人类结肠的主要厚壁菌门(例如Blautia spp.、Eubacterium eligens、Ruminococcus spp.)缺乏从碳水化合物中生成丁酸的能力。
产生丁酸盐的细菌被认为在生命的第一年内定植于宿主,并且在成年时占总细菌群落的20%以上。在消化道中发现的已知丁酸盐生产者中,大多数似乎属于毛螺菌科和瘤胃球菌科。我们将简要介绍两个丁酸生产菌。
▸ 普拉梭菌
普拉梭菌(Faecalibacterium prausnitzii)是健康人类微生物群中最丰富的菌种之一,也是主要的丁酸盐生产者之一。它通过丁酰辅酶A:乙酸辅酶A转移酶产生丁酸,并消耗乙酸。
虽然F.prausnitzii菌株是专性厌氧菌,但在核黄素(维生素B2)和还原性化合物(如半胱氨酸或谷胱甘肽)存在的情况下,低浓度氧气也能促进其生长。
氧气消耗会伴随丁酸形成的减少。F. prausnitzii分离株在利用食物多糖(如淀粉和半纤维素)生长方面能力有限,但一些菌株可以利用菊粉和果胶衍生物,并且普遍利用糖醛酸。
• 肠道炎症患者产生丁酸会减少
F. prausnitzii在炎症性肠病患者中很少见,尤其是克罗恩病,有证据表明它具有抗炎作用,因此引起了人们对其作为潜在治疗药物的兴趣。同样,据报道,Butyricicoccus pullicaecorum在炎症性肠病患者中较少见,并且也可能具有治疗潜力。
注:其他瘤胃球菌科细菌也能产生丁酸,但人们对大多数此类生物知之甚少。关于Faecalibacterium prausnitzii,详见:肠道核心菌属——普拉梭菌(F. Prausnitzii),预防炎症的下一代益生菌
▸ 毛螺菌科
直肠真杆菌(Eubacterium rectale)和密切相关的罗氏菌属(Roseburia)构成了产丁酸毛螺菌科的主要群体,它们具有相同的丁酰辅酶 A:乙酸辅酶 A 转移酶途径来生产丁酸,并且其丁酸合成基因的基因组组织也相同,从乙酰辅酶A到丁酰辅酶A。
• 罗氏菌属通过代谢饮食多糖产生丁酸
在某些Roseburia菌株中,特别是在弱酸性pH值下,丁酸几乎是唯一产生的发酵酸,乙酸的净消耗通常伴随着丁酸的形成。一些其他菌株和物种除了产生丁酸外,还产生甲酸和乳酸。基因组分析表明,该群体具有相当大的利用饮食来源的多糖的能力,包括淀粉、阿拉伯木聚糖和菊粉,不同菌株和物种之间的差异很大。
其他拥有丁酰辅酶A:乙酸辅酶A转移酶基因的毛螺菌科包括Eubacterium hallii、Anaerostipes hadrus、Coprococcus catus、与分离株SS3/4和M62/1有关的未鉴定物种。
• 一些毛螺菌科细菌利用乳酸和乙酸来产生丁酸
某些毛螺菌科能够在乳酸和乙酸盐存在下生长并产生丁酸,其总净化学计量为4mol乳酸和2mol乙酸盐产生3mol丁酸 。
这包括仅使用D-乳酸的物种Anaerostipes hadrus和能够利用两种乳酸异构体的E. hallii。
此外,双歧杆菌等常见益生菌配方中的微生物与丁酸盐生产者之间的交叉喂养相互作用已被证明是可以产丁酸盐的。
人体肠道中的主要丁酸生产者
doi: 10.3389/fmicb.2022.1103836.
▸ 增加丁酸盐生成的食物
饮食对人体微生物群有显著影响,通过饮食干预可显著改变细菌数量并增加微生物多样性。富含纤维的饮食对丁酸盐的产生特别有益,因为它可以滋养产生丁酸盐的细菌。
• 高纤维饮食能够有效增加丁酸盐的生成
蔬菜、水果、豆类和全谷物,它们含有膳食纤维,可被肠道细菌发酵成丁酸盐等有机化合物。通过食物增强微生物群是促进消化系统健康和丁酸盐生成的有效且安全的方法。因此,增加丁酸盐产量的最佳方法是通过高纤维饮食。
• 高脂、低纤维饮食不利于丁酸盐的生成
高蛋白、高脂肪、低碳水化合物的饮食已被证明会破坏微生物组中丁酸盐的产生。在一项研究中,研究人员分析了短期饮食限制碳水化合物摄入量的肥胖参与者的微生物组,从而限制了他们对植物性膳食纤维的消耗。
在低碳水化合物饮食(每天24克)和中等碳水化合物饮食(每天164克)4 周后,短链脂肪酸的浓度低于高碳水化合物饮食(每天399克)。具体来说,当碳水化合物摄入量减少时,丁酸盐浓度会降低。
同时还发现,厚壁菌门细菌Roseburia和E.Rectale的密度与丁酸盐浓度之间存在联系,两者都随着碳水化合物摄入量的减少而降低。
一些食源性物质产生短链脂肪酸的量
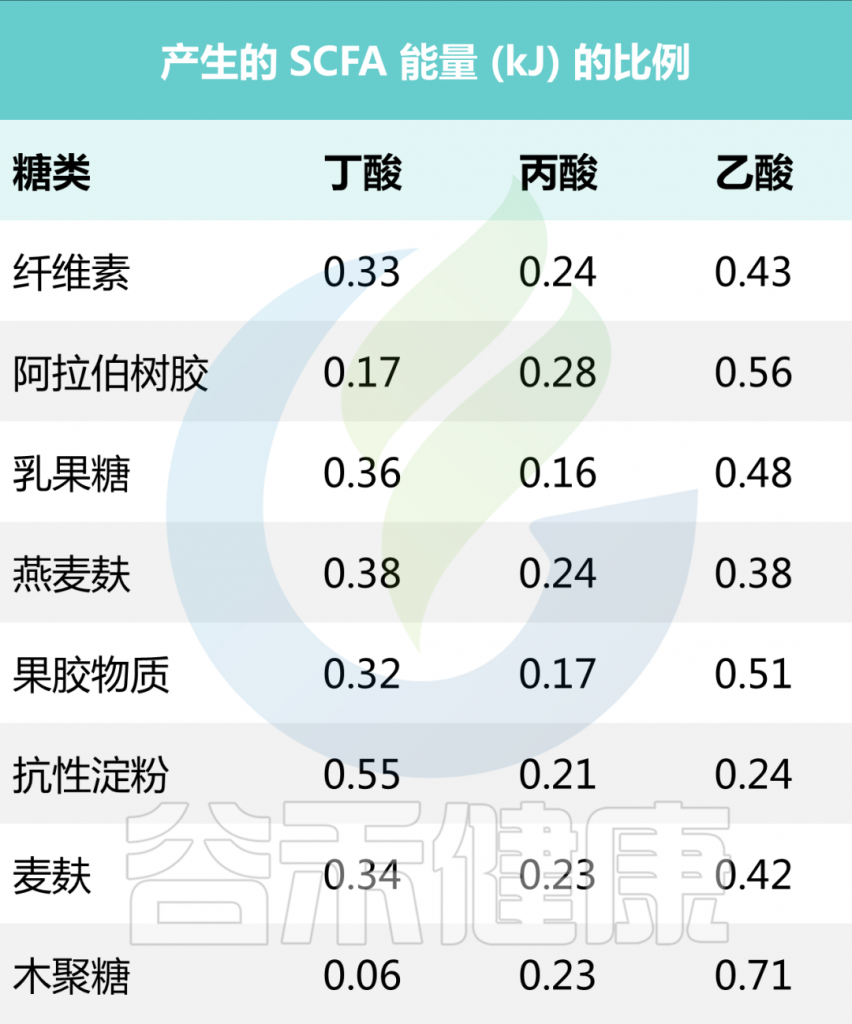
• 抗性淀粉
研究表明,从生马铃薯、高直链淀粉玉米和全谷物中提取的阿拉伯木聚糖中,含有高含量2型抗性淀粉的食物显著增加了丁酸盐的产量。
在174名健康年轻人的饮食中添加马铃薯抗性淀粉后,丁酸盐产量增加。玉米、菊苣和玉米中的抗性淀粉也进行了测试,但只有当食用土豆中的抗性淀粉时,粪便中的丁酸总量才会显著增加。
• 果聚糖(菊粉)
许多研究表明,菊粉可以增加短链脂肪酸的产量,包括丁酸盐。这可能解释了香蕉在溃疡性结肠炎和克罗恩病患者的饮食中如此有效的原因之一。然而,链长较短的果聚糖通常比链长的果聚糖(如菊粉)更容易喂养产丁酸菌。
也就说,抗性淀粉和果聚糖(短链低聚果糖和长链菊粉)在肠道中发酵时会产生丁酸盐。
富含抗性淀粉的食物:全麦面包、燕麦、大麦和糙米等全谷物;黑豆、红豆、绿豆等豆类;马铃薯、玉米、菊苣根、牛蒡根、魔芋根、亚麻籽。
富含果聚糖的食物:洋葱、菊苣、香蕉、朝鲜蓟、芦笋、大蒜、韭菜、西兰花、开心果。
需要注意的
由于人与人之间的常驻微生物存在显著差异,并且它们消化某些底物的能力不同,因此比较明智的选择是食用包含多种抗性淀粉和果聚糖的饮食。例如以马铃薯抗性淀粉、燕麦麸皮纤维或车前草种子或短链低聚半乳糖、长链低聚果糖和谷氨酰胺的混合物的形式短期补充,增加丁酸盐水平。
丁酸盐作为一种重要的的调节因子,是宿主-微生物串扰的关键介体。丁酸盐在体内可以通过脂肪酸氧化为机体供应能量,是肠道上皮细胞的主要供能物质。
丁酸盐还与机体健康密切相关,对调节肠道健康、修复肠道屏障、抑制炎症及癌症等病症意义重大。在养殖业中常添加丁酸盐保护动物健康生长,如预防断奶仔猪腹泻、调节鸡肠道菌群并增强其免疫力等。
▸ 为肠道细胞提供能量
丁酸盐是结肠细胞的主要能量来源,结肠细胞是构成肠道内壁的细胞。
与身体中使用糖(葡萄糖)作为主要能量来源的大多数其他细胞不同,肠道内壁细胞(结肠细胞)主要使用丁酸盐。如果没有丁酸盐,这些细胞就无法正确执行其功能。
• 丁酸盐为结肠细胞提供能量,细胞利于产丁酸盐细菌的生长
厚壁菌属的成员以产生丁酸盐而闻名,像Roseburia,Faecalibacterium prausnitzii,直肠真杆菌(E.rectale)等。
这种关系是相互的。丁酸盐为结肠细胞提供燃料,作为回报,这些细胞有助于提供一个无氧环境,有益的肠道微生物在其中茁壮成长。这可以控制炎症,保持肠道细胞健康,并使肠道细菌保持健康。
▸ 促进肠道运动
实验研究表明,丁酸盐通过作为短链脂肪酸受体的配体和激活剂,诱导肠道激素肽YY或介导肠嗜铬细胞释放5-羟色胺来促进肠道运动。
丁酸盐还可以通过上调Na+—H+交换器和诱导ATPase离子交换器基因来增强水和电解质的吸收。并且可能有益于预防某些类型的腹泻。
▸ 抗炎、抗癌特性
丁酸盐对肠道具有抗炎和抗癌功能。
肠道内壁会保持低水平的炎症,以防与微生物群接触的粘膜表面发生任何变化。低水平的炎症受到严格控制,但如果它被破坏,会导致氧化损伤,并可能在很长一段时间内导致癌症。
• 丁酸盐减少促炎细胞因子的产生
丁酸盐会阻止体内的一些促炎物质发挥作用。丁酸盐的抗炎作用可减少氧化应激并控制自由基造成的损害。
丁酸盐的抗炎特性,部分原因是其抑制核转录因子(NF-κB)的激活,通过下调NF-κB信号通路,丁酸盐可以调节促炎细胞因子的产生。
• 丁酸盐阻止结肠癌细胞的生长
丁酸盐也是一种组蛋白脱乙酰酶(HDAC)抑制剂。组蛋白脱乙酰酶是大多数癌症中产生的酶。因为丁酸盐是一种抑制剂,它实际上会改变基因表达,抑制细胞增殖,诱导细胞分化或凋亡。因此,它可以阻止癌细胞的发展。
有证据表明,结直肠癌(CRC)患者的微生物组图显示主要产丁酸菌属减少,包括罗氏菌属(Roseburia)、Clostridiales、Faecalibacterium和Lachnospiraceae科成员,而使用产丁酸的丁酸梭菌可有效减少癌细胞增殖并增强癌细胞凋亡。
▸ 修复肠道屏障,防止肠漏
肠上皮细胞的屏障功能是重要的第一道防线,确保上皮层具有适当的通透性。研究发现丁酸盐能够修复和增强肠上皮细胞的屏障功能。
• 丁酸盐上调粘蛋白的表达,增强粘液屏障
最新研究表明,丁酸对肠屏障功能具有保护作用。例如,丁酸能够上调粘蛋白2(MUC2)的表达。粘蛋白2是肠粘膜表面主要的粘蛋白,可增强粘膜层,从而限制有害物质通过肠道。
• 丁酸盐对肠道屏障的影响可能具有浓度依赖性
此外,丁酸还能增加三叶因子(TFF)的表达,这是一种粘蛋白相关肽,有助于维持和修复肠粘膜。丁酸还调节紧密连接蛋白的表达,以降低细胞旁通透性。其中一种机制是通过激活单分子层中的AMP活化蛋白激酶。
体外模型研究表明,丁酸对肠道屏障功能的影响可能具有浓度依赖性。丁酸在低浓度(≤2 mM)时可促进肠道屏障功能,但在高浓度(5或8 mM)时可能通过诱导细胞凋亡来破坏肠道屏障功能。
根据哺乳动物胃肠道中的生理浓度,目前体外模型中推荐使用的丁酸浓度为0-8mM。然而,考虑到大多数丁酸在结肠上皮中被代谢为能量底物,体内和体外模型中治疗剂量可能存在差异。
▸ 抗氧化能力
说起抗氧化,我们先了解一下自由基。它基本上是体内化学反应产生的废物。另一方面,抗氧化剂是身体抵御它们的防御措施。大量自由基会造成损害并压倒身体的修复系统。我们称之为氧化应激。氧化应激被认为是导致衰老和疾病的一个重要因素。
• 丁酸盐增加谷胱甘肽,可以中和自由基
结肠或大肠是身体产生的废物的储存容器。较高的丁酸盐水平已被证明会增加谷胱甘肽的水平,谷胱甘肽是一种在人体细胞中产生的抗氧化剂,可以中和肠道中的自由基(自由基与炎症和许多疾病有关),从而起到抗氧化作用。
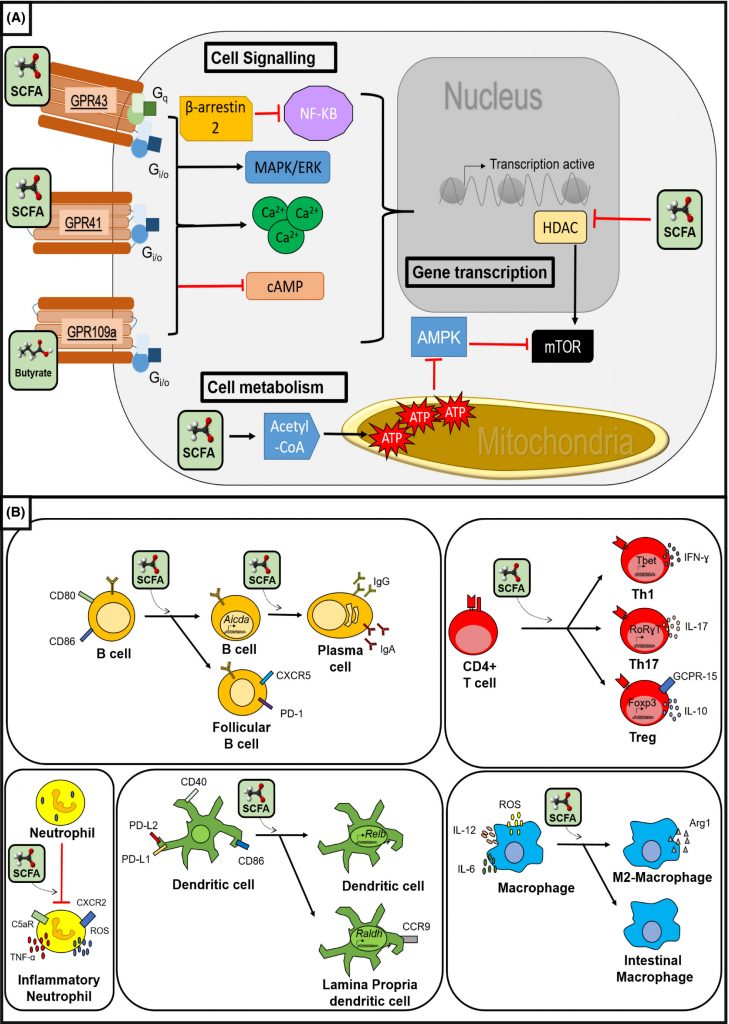
▸ 调节肠道免疫
• 维持厌氧环境,阻止有害菌生长
丁酸通过增强结肠细胞的氧消耗和稳定缺氧诱导因子(HIF)来维持结肠内的厌氧环境,而丁酸的缺失则会促进潜在有害细菌和分子的积聚,例如沙门氏菌、大肠杆菌和一氧化氮(NO)。
肠道中丁酸水平降低,这促进了肠道上皮氧合和鼠伤寒沙门氏菌(S.Typhimurium)的生长,鼠伤寒沙门氏菌是食源性肠道炎症和腹泻的已知病因。
• 影响免疫细胞迁移、粘附
除了营造无氧环境外,短链脂肪酸,尤其是丁酸盐,还可以作为免疫细胞趋化和粘附的调节剂。丁酸可以调节肠上皮细胞介导的中性粒细胞向炎症部位的迁移,并且这种作用是浓度依赖性的。
此外,丁酸在细胞增殖和凋亡中发挥作用。丁酸刺激细胞生长和DNA合成,并诱导细胞周期G1期的生长停滞。虽然低浓度的丁酸会增强细胞增殖,但高浓度的丁酸会诱导细胞凋亡。
丁酸盐等短链脂肪酸可以在先天反应过程中通过影响巨噬细胞和粒细胞以及树突状细胞的抗原呈递发生,也可能在适应性免疫反应过程中通过影响T细胞和B细胞功能发生。
短链脂肪酸的免疫调节作用
doi: 10.1111/apt.14689.
▸ 影响代谢健康
• 对肥胖存在一定影响,但还不能完全确定
包括丁酸盐在内的短链脂肪酸可通过激活肠细胞内的FFAR来降低食欲和体重。这促进胰高血糖素样肽1(GLP-1)和肽YY的释放,前者促进胰岛素分泌并抑制胰高血糖素分泌,后者降低食欲并减缓胃排空。
短链脂肪酸还能减少所谓的“饥饿激素”——胃促生长素(ghrelin)的分泌;FFAR2存在于ghrelin分泌细胞上,包括丁酸盐和丙酸盐在内的FFAR2激动剂可减少ghrelin分泌。
丁酸盐影响脂质代谢的机制
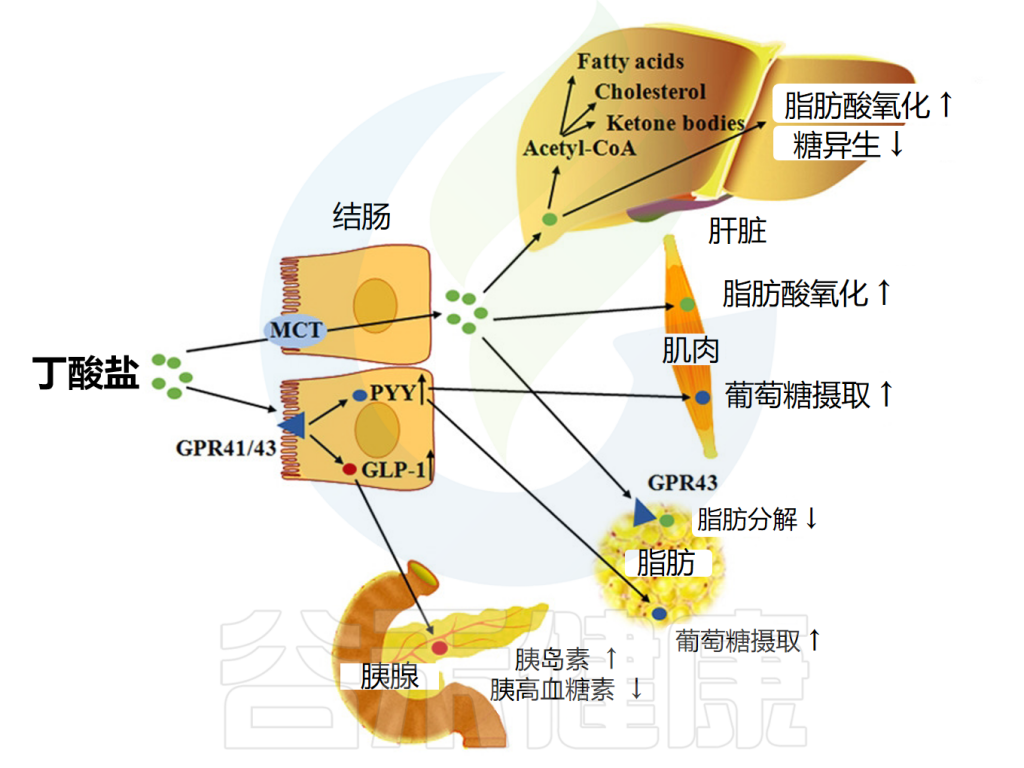
doi: 10.1093/advances/nmx009.
也有证据表明短链脂肪酸作用于交感神经系统,交感神经节中FFAR3的激活导致能量消耗增加。
然而,与丙酸盐一样,关于丁酸盐对食物摄入的影响,有相互矛盾的结果报道。
• 丁酸盐对控制血糖水平可能有潜在好处
研究发现,糖尿病患者和糖尿病前期受试者中丁酸盐水平降低。
丁酸在肠内分泌细胞(EEC)中与游离脂肪酸受体(FFAR)FFAR2和FFAR3结合,调节肠道激素释放,如胰高血糖素样肽1(GLP-1)和肽YY(PYY)。
丁酸盐通过多种途径影响糖代谢的调节
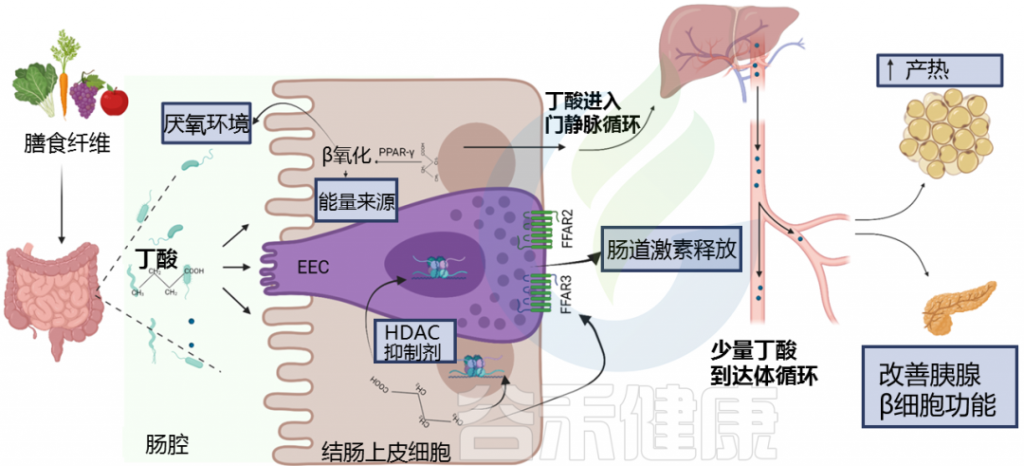
doi: 10.3389/fendo.2021.761834.
这些激素共同作用以保持血糖水平稳定。当血糖过高时,胰岛素会告诉身体的肌肉和脂肪细胞吸收多余的葡萄糖,所以说这些激素对肥胖和糖尿病很重要。丁酸盐增加这些肠道激素的释放,表明对控制血糖水平和防止体重增加有潜在的好处。
!
丁酸盐过低可能会导致以下危害:
•肠道屏障功能受损:丁酸盐有助于维持结肠的“生理性缺氧”,并提供能量给结肠细胞,有助于维持肠道稳态。丁酸盐的减少可能导致肠道屏障功能受损。
•增加慢性疾病风险:变形菌门的兼性厌氧细菌的扩张几乎总是伴随着产丁酸盐细菌丰度的减少,并与许多慢性疾病有关,包括炎症性肠病、肠易激综合症、结直肠癌、2型糖尿病、肥胖等。
•神经炎症:丁酸盐具有抗炎特性,对大脑健康具有重要意义。丁酸盐过低可能导致炎症增加,影响大脑健康。
•睡眠问题:丁酸盐可能作为细菌源性促睡眠信号,丁酸盐过低可能影响睡眠,导致睡眠问题。
•肠道菌群失衡:丁酸盐过低可能导致肠道菌群失衡,因为变形菌门的扩张伴随着产丁酸盐细菌的减少。
•不利于肠道损伤修复:在轻微炎症或轻度溃疡部位添加丁酸盐可以促进肠道损伤的修复,而丁酸盐过少可能不利于溃疡的恢复。
!
丁酸盐并不总是越多越好,低浓度丁酸促进细胞增殖和生长,高浓度丁酸反而抑制细胞增殖和生长,增加肠道的通透性。
• 不同部位对丁酸盐的耐受阈值存在差异
胃肠道不同部位对丁酸盐的耐受阈值也存在一定差异,胃和小肠对丁酸的耐受阈值低,结肠和盲肠耐受阈值高。
添加普通丁酸钠制剂(主要在肠道前端被吸收利用),反而造成肠道炎症、菌群失调。
• 肠道严重溃疡时丁酸盐可能加剧症状
更有意思的是,轻微炎症或者轻度溃疡部位添加丁酸盐可以促进肠道损伤的修复,在严重溃疡肠道部位添加,不利于溃疡的恢复,甚至加剧整个溃疡。
有学者发现溃疡部位的粘膜组织对丁酸的代谢降低,甚至只有正常粘膜组织的一半,主要是由于其转运载体和氧化相关的酶活降低。
主要参考文献
Louis P, Flint HJ. Formation of propionate and butyrate by the human colonic microbiota. Environ Microbiol. 2017 Jan;19(1):29-41.
Gill PA, van Zelm MC, Muir JG, Gibson PR. Review article: short chain fatty acids as potential therapeutic agents in human gastrointestinal and inflammatory disorders. Aliment Pharmacol Ther. 2018 Jul;48(1):15-34.
Hosseini E, Grootaert C, Verstraete W, Van de Wiele T. Propionate as a health-promoting microbial metabolite in the human gut. Nutr Rev. 2011 May;69(5):245-58.
Arora T, Tremaroli V. Therapeutic Potential of Butyrate for Treatment of Type 2 Diabetes. Front Endocrinol (Lausanne). 2021 Oct 19;12:761834.
Singh V, Lee G, Son H, Koh H, Kim ES, Unno T, Shin JH. Butyrate producers, “The Sentinel of Gut”: Their intestinal significance with and beyond butyrate, and prospective use as microbial therapeutics. Front Microbiol. 2023 Jan 12;13:1103836.
Gonzalez-Garcia, R.A.; McCubbin, T.; Navone, L.; Stowers, C.; Nielsen, L.K.; Marcellin, E. Microbial Propionic Acid Production. Fermentation 2017, 3, 21.
Reichardt N, Duncan SH, Young P, Belenguer A, McWilliam Leitch C, Scott KP, Flint HJ, Louis P. Phylogenetic distribution of three pathways for propionate production within the human gut microbiota. ISME J. 2014 Jun;8(6):1323-35.
Liu H, Wang J, He T, Becker S, Zhang G, Li D, Ma X. Butyrate: A Double-Edged Sword for Health? Adv Nutr. 2018 Jan 1;9(1):21-29.

谷禾健康

氨基酸(Amino Acid, 简称AA ) 是蛋白质合成的基石。它们是正常细胞生长、分化和功能所必需的细胞结构元素和能量来源。
蛋白质是长链氨基酸。人的身体有数千种不同的蛋白质,每种蛋白质都有重要的作用。每种蛋白质都有自己的氨基酸序列。该序列使蛋白质呈现不同的形状并在体内具有不同的功能。
可以将氨基酸视为字母表中的字母。当你以不同的方式组合字母时,会组成不同的单词。氨基酸也是如此——当以不同的方式组合它们时,就会产生不同的蛋白质。
氨基酸代谢紊乱与许多病理状况有关,包括代谢疾病、心血管疾病、免疫疾病和癌症。就肿瘤而言,氨基酸代谢的改变不仅可以用作癌症进展的临床指标,还可以用作治疗策略。
由于肿瘤的生长发育依赖于外来氨基酸的摄入,因此越来越多的研究针对肿瘤相关氨基酸的代谢来选择性杀伤肿瘤细胞。此外,免疫相关研究证实,氨基酸代谢调节效应T细胞和调节性T细胞的功能,影响免疫细胞的功能。因此,确定氨基酸代谢途径中的靶点可能有助于疾病的治疗。
近日,浙江大学医学院附属第一医院,国家卫健委多器官联合移植重点实验室,中国医学科学院器官移植诊治重点实验室,以及器官移植重点实验室联合发表在《Signal Transduct Target Ther》综述成果详细阐述和讨论氨基酸代谢相关的代谢性疾病、心血管疾病和免疫相关疾病的研究及临床研究进展。
本文就该篇文章为主要参考,分享围绕肿瘤相关疾病中的氨基酸代谢研究,结合讨论肠道微生物群在蛋白质和氨基酸代谢中的作用,以期为靶向氨基酸代谢管理治疗提供理论依据。
官方氨基酸定义包括任何同时含有羧基和氨基的有机化合物(如下图)。它们都具有共同的结构特征氨基 (-NH3+)、羧酸根 (-COO–) 基团和与同一碳原子键合的氢键。
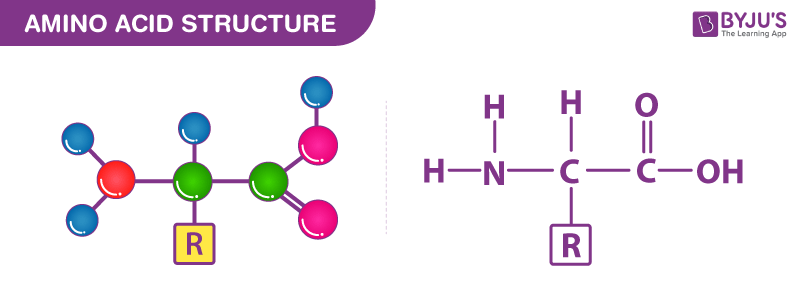
氨基酸有多少种?
氨基酸一共有20种,每种氨基酸在体内都发挥着非常特殊的作用,并通过其各自的氨基酸侧链来区分。从细菌到人类,都是由同一组 20 种氨基酸构成。
每个氨基酸都有 4 个不同的基团连接到 α-碳。它们的不同之处在于称为 R 基团的侧链。如下氨基酸的R基团不同:
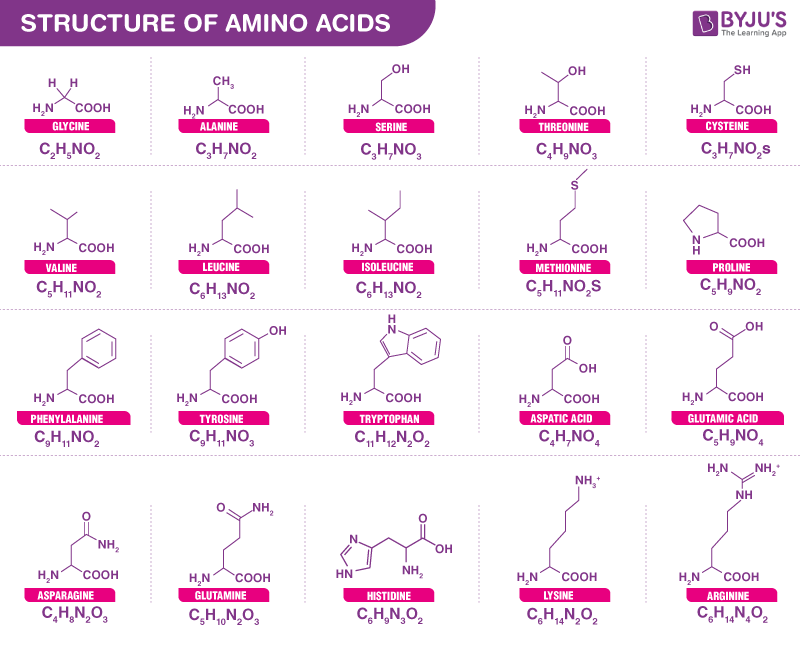
来源:byjus
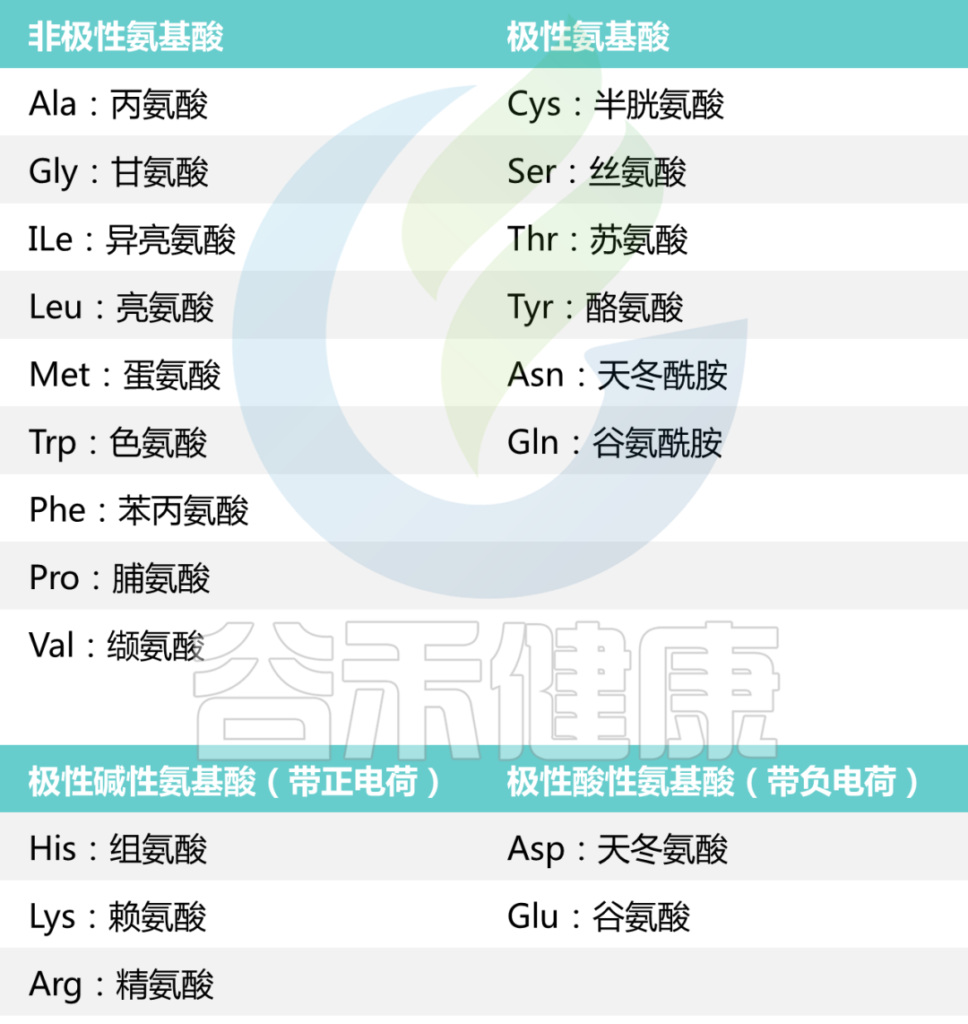
氨基酸还可以根据其结构和侧链分为其他组,
包括:
根据每种氨基酸中“R”基团的性质,氨基酸可以分为四个大类。
氨基酸可以是极性的、非极性的、带正电的或带负电的。
氨基酸是作为蛋白质构建块的基本分子。蛋白质由一条或多条氨基酸链(称为多肽)组成,其序列由基因编码。
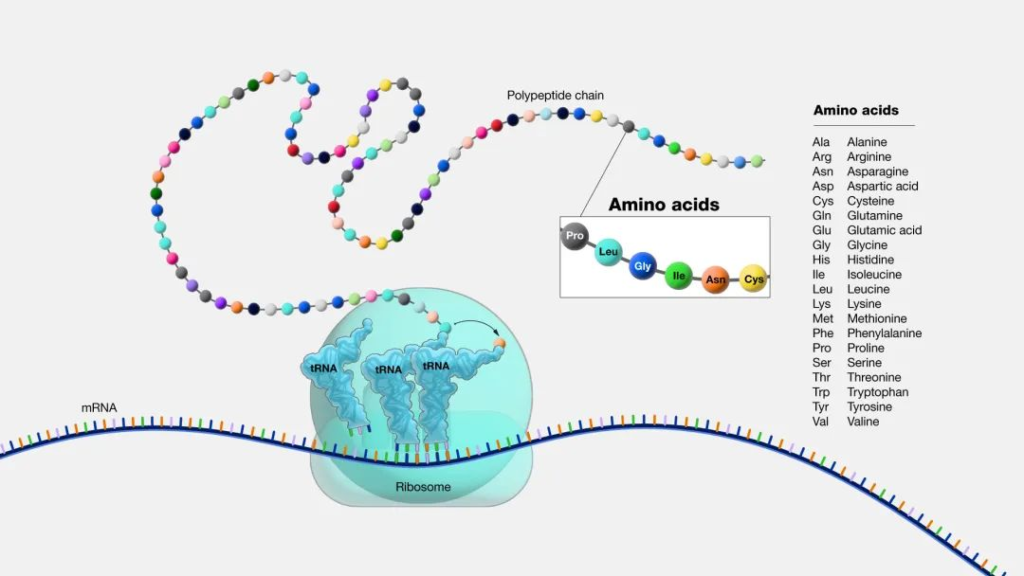
图源:National Human Genome Research Institute
这些氨基酸参与几乎所有生物过程并有助于:
伤口愈合、激素产生、免疫功能、肌肉生长、能源生产等。
身体需要所有氨基酸才能发挥作用并茁壮成长。有些可以在体内产生,而另一些则需要从食物中获取。
氨基酸有着有趣而悠久的历史。天冬酰胺是第一个被发现的氨基酸,由法国化学家 Louis Nicolas Vauquelin 和 Pierre Jean Robiquet 于 1806 年从芦笋营养中分离出来。
身体所需的 20 种氨基酸可进一步分为两个不同的类别:必需氨基酸和非必需氨基酸。
人体无法合成必需氨基酸,这意味着必须从食物中获取它们才能满足需求。
必需氨基酸有多少种?
必须通过饮食获得九种必需氨基酸,包括:
赖 氨 酸
赖氨酸是生长和组织修复以及多种激素、蛋白质和酶的产生所必需的。赖氨酸对于促进抗体、激素和酶的形成以及骨骼中钙的形成和固定是必需的。
亮 氨 酸
这种必需氨基酸参与蛋白质合成、伤口愈合、血糖控制、生长激素和新陈代谢。
异 亮 氨 酸
研究表明异亮氨酸有助于解毒、免疫功能和激素排泄。此外,在血红蛋白的形成、刺激胰腺合成胰岛素以及将氧气从肺部输送到各个部位方面起着至关重要的作用。
色 氨 酸
色氨酸是最著名的氨基酸之一,也是功能性氨基酸,参与维生素 B3 和血清素激素的产生。血清素是调节情绪、疼痛、食欲和睡眠的神经递质。
苯 丙 氨 酸
这种氨基酸有助于产生其他氨基酸以及多巴胺和去甲肾上腺素等神经递质,有助于维持健康的神经系统和增强记忆力。
苏 氨 酸
研究表明苏氨酸是胶原蛋白和弹性蛋白等结缔组织的基础,这些蛋白质为皮肤和结缔组织提供结构。它们还有助于形成血凝块,从而有助于防止出血。苏氨酸在脂肪代谢和免疫功能中也发挥着重要作用。
缬 氨 酸
缬氨酸支持大脑功能、肌肉协调和平静。缬氨酸参与肌肉生长、组织再生和产生能量。
组 氨 酸
这种氨基酸维持人体内髓鞘的健康,保护神经细胞免受损伤。参与许多酶促过程以及红细胞和白细胞的合成。组氨酸有助于制造一种称为组胺的大脑化学物质(神经递质)。组胺在身体的免疫功能、消化、睡眠和性功能中发挥着重要作用。
蛋 氨 酸
研究表明蛋氨酸可以保持皮肤弹性,并有助于强化头发和指甲。还用于治疗肾结石、保持皮肤健康,还用于控制病原菌的入侵。蛋氨酸有助于身体的组织生长、新陈代谢和解毒。蛋氨酸还有助于吸收必需矿物质,包括锌和硒。
在饮食中摄入多种氨基酸对于维持整体健康至关重要。任何这些关键必需氨基酸的缺乏都会导致严重的副作用,影响健康的几乎各个方面,包括免疫功能、肌肉质量、食欲等。
相比之下,非必需氨基酸可以由身体产生,这意味着通过吃的食物获取它们并不那么重要。
非必需氨基酸列表中共有 11 种氨基酸,包括:
精 氨 酸
精氨酸可刺激免疫功能、抗疲劳并优化心脏健康。一水肌酸在肝脏、胰腺和肾脏中形成,主要是在精氨酸、甘氨酸和蛋氨酸的帮助下形成的。有助于促进蛋白质和激素的合成、肾脏解毒、愈合伤口和维持健康的免疫系统。
丙 氨 酸
研究表明丙氨酸有助于新陈代谢,为肌肉、大脑和中枢神经系统提供能量。此外可以帮助清除体内毒素并参与葡萄糖和其他氨基酸的生产。
半 胱 氨 酸
半胱氨酸是头发、皮肤和指甲中发现的主要蛋白质类型,作为一种抗氧化剂,对于胶原蛋白的产生和皮肤健康至关重要。
谷 氨 酸
这种非必需氨基酸在中枢神经系统中充当神经递质,主要参与人脑的发育和功能。
天 冬 氨 酸
天冬氨酸有助于产生其他几种氨基酸,包括天冬酰胺、精氨酸和赖氨酸。主要参与将氮输送到身体细胞中、形成嘌呤和嘧啶以合成DNA、神经系统的发育和提高我们的身体耐力。
甘 氨 酸
甘氨酸有助于维持适当的细胞生长及其功能,并且在愈合伤口方面也起着至关重要的作用。此外,甘氨酸作为神经递质支持大脑健康。
脯 氨 酸
脯氨酸存在于胶原蛋白中,有助于促进关节健康、新陈代谢和皮肤弹性。主要参与胶原蛋白形成过程中的组织修复,防止动脉壁增厚和硬化(动脉硬化)以及新皮肤的再生。
丝 氨 酸
丝氨酸是脂肪代谢、免疫功能和肌肉生长所必需的。
酪 氨 酸
甲状腺激素-T3 和 T4 的产生以及合成一类神经递质和黑色素(我们的眼睛、头发和皮肤中发现的天然色素)方面发挥着至关重要的作用。
谷 氨 酰 胺
研究表明,谷氨酰胺支持许多代谢过程并为体内细胞提供能量。促进健康的大脑功能,是合成核酸(DNA 和 RNA)所必需的。
天 冬 酰 胺
天冬酰胺起到利尿剂的作用,可以优化大脑和神经细胞的功能。
氨基酸列表中的一些化合物也被认为是“有条件必需的”。这意味着它们通常不是身体所需要的,但在某些情况下可能变得必不可少,例如极端疾病或压力。
氨基酸缺乏的一些主要症状可能包括:
蛋白质缺乏症会影响任何饮食中氨基酸摄入不足的人。老年人和患有癌症等慢性病的人缺乏蛋白质的风险特别高,因为他们通常对蛋白质的需求增加而食物摄入量减少。
那些遵循纯素或素食饮食的人还应该仔细计划他们的饮食,以确保通过食用各种植物性蛋白质食物来满足他们的蛋白质需求。
富含氨基酸的食物包括植物性产品,如西兰花、豆类、甜菜根、南瓜、卷心菜、坚果、干果、奇亚籽、燕麦、豌豆、胡萝卜、黄瓜、绿叶蔬菜、洋葱、大豆、全谷物、花生豆类、扁豆等。
富含氨基酸的水果有苹果、香蕉、浆果、无花果、葡萄、甜瓜、橙子、木瓜、菠萝、石榴等。
其他动物产品包括乳制品、鸡蛋、海鲜、鸡肉、肉类、猪肉等。
从富含蛋白质的食物中摄入大量蛋白质不太可能引起任何负面影响。然而,有可能过量摄入蛋白质,尤其是蛋白质补充剂。
摄入过多蛋白质可能产生的一些副作用包括体重增加、肾脏问题、便秘和口臭,炎症以及肠道菌群的失调。
氨基酸是含有氨基和羧基的有机化合物,根据核心结构官能团的位置可分为α-、β-、γ-、δ-氨基酸,其中大部分参与蛋白质合成的是 α-氨基酸。
氨基酸参与生物合成、神经传递和其他生命过程。肽键连接氨基酸形成多肽链,多肽链经过翻译后修饰,有时与其他多肽链结合形成蛋白质。
九种氨基酸不能由其他化合物合成,必须从食物中获取,这些氨基酸从食物中被人体摄入后,除了用于蛋白质等生物分子的合成外,还可以通过氧化途径氧化为尿素和二氧化碳作为能量来源。
氧化途径以转氨酶介导的脱氨作用开始,将氨基转移至α-酮戊二酸,形成谷氨酸进入尿素循环。另一种产物酮酸进入柠檬酸循环,为生命活动提供能量(下图)。该循环提供某些氨基酸的前体,以及还原剂NADH,用于许多其他反应。
doi: 10.1038/s41392-023-01569-3
人体可以通过食物消化吸收、组织分解、内部合成三种途径获得氨基酸。
氨基酸代谢库中的氨基酸可脱酸,产生氨基酸和二氧化碳,或在代谢产物的转化中参与嘌呤、嘧啶等含氮化合物的合成;或者脱氨基产生α-酮酸和NH3。
根据不同的酶和途径,α-酮酸可以产生酮体,或参与氧化供能或糖和脂质的合成;NH3进入尿素循环。
细胞或细胞器对氨基酸的摄取需要氨基酸转运蛋白(AAT)的参与。不同的氨基酸依赖于特定的AAT,但氨基酸和转运蛋白并不是一一匹配的。多个AAT可以转运一个氨基酸,同一个转运蛋白也可以转运多个底物。AAT 除了充当氨基酸进出细胞的通道外,还充当感测氨基酸水平的探针和营养信号的引发剂。
根据结构和功能的多样性,AAT可分为不同的家族,其中溶质载体(SLC)超家族约占人类基因组编码的所有膜蛋白的20%,是最大的膜转运蛋白超家族。
根据底物特异性,AAT可分为中性、碱性和酸性类别,以及进一步的子类别,包括钠依赖性和钠非依赖性类型。从机制上讲,由于哺乳动物细胞(包括人类)细胞内液中的氨基酸浓度通常高于细胞外液中的氨基酸浓度,因此 AAT 通过离子共轭或氨基酸交换转运氨基酸以产生钠离子。
氨基酸除了是肽和蛋白质的组成部分外,还参与维持细胞生长、代谢和免疫的关键途径。
例如,哺乳动物雷帕霉素靶蛋白 (mTOR) 信号通路是调节蛋白质合成的主要机制。mTOR 系统包含雷帕霉素敏感复合物 1 (mTORC1) 和雷帕霉素不敏感复合物 2 (mTORC2)。mTORC1 被谷氨酰胺 (Gln)、精氨酸 (Arg) 和亮氨酸 (Leu) 激活,并通过 eIF4E 结合蛋白 1 (4E-BP1) 和核糖体蛋白 S6 激酶 1 (S6K1) 的磷酸化激活蛋白质合成。
此外,丙氨酸(Ala)可以通过抑制丙氨酸激酶来调节糖异生和糖酵解,从而维持饥饿肝脏产生的葡萄糖量。
TIPs:
糖异生是指非糖物质(如某些氨基酸、乳酸、丙酮酸和甘油等)在人体的肝脏和肾脏等器官中某些酶的催化下转变成糖原或葡萄糖的过程。简单说来,就是非糖物质转化为糖的过程。
糖分解是生物体取得能量的主要方式。生物体中糖的氧化分解主要有3条途径:糖的无氧氧化、糖的有氧氧化和磷酸戊糖途径。其中,糖的无氧氧化又称糖酵解(glycolysis)。生物在无氧条件下,从糖的降解代谢中获得能量的途径,也是大多数生物进行葡萄糖有氧氧化的一个准备途径。
糖酵解/糖原异生的差异比较
在某种程度上,糖异生可能被认为是糖酵解的逆过程,因为糖酵解将葡萄糖分解成两个丙酮酸分子以获得能量并供给三羧酸循环,而糖异生将两个丙酮酸分子构建成一个葡萄糖分子。然而,仅仅把糖异生看作糖酵解是不太正确的,因为这两者之间有一些重要的区别。
糖酵解是一种古老的代谢途径,为所有形式的细胞生命所共有,并且可以由所有细胞进行以获得能量(每个葡萄糖净2个三磷酸腺苷)。
相比之下,糖异生更具特异性:在人类中,它主要在肝细胞中进行,在某种程度上也在肾上腺皮质中进行。它还有一个更具体的目标:当血糖水平较低时,产生葡萄糖分泌到血液中。
在免疫方面,氨基酸参与免疫细胞增殖、分化和功能激活。例如,T细胞激活会上调多种氨基酸转运蛋白,包括SLC7A5,而SLC7A5的缺失会导致mTOR信号通路的激活和转录因子MYC的上调,从而抑制T细胞增殖。
当T细胞缺乏色氨酸(Trp)和 精氨酸 (Arg)时,活化的T细胞无法进入S期,这证明Trp和Arg是T细胞进入细胞周期的关键物质。此外,亮氨酸(Leu) 和异亮氨酸 (ILe) 的消耗会诱导 T 细胞进入 S-G1 期,然后停止分裂并死亡。
综上所述,氨基酸是维持生命所必需的有机化合物,是生物合成的原料,也是生命活动的能量来源。
接下来,我们针对支链氨基酸、天冬氨酸、谷氨酰胺、精氨酸、蛋氨酸五种氨基酸的代谢展开详细讨论,包括它们在癌症或其他疾病中的作用,希望给大家呈现一个更全面的视角,为疾病的发生发展和临床治疗方向带来更多的启示。
★
支链氨基酸(BCAA)是一类具有一个分支的脂肪侧链氨基酸,包括亮氨酸、异亮氨酸和缬氨酸。三种支链氨基酸占人体必需氨基酸肌肉中必需氨基酸的35%。
所有物种中支链氨基酸的分解过程都是相似的,最初通过支链氨基酸转移酶(BCAT)形成支链α-酮酸(BCKA),并将氮转移到氮受体(最常见的氮受体是α-酮戊二酸) (α-KG) 形成谷氨酸)。
第二步是支链α-酮酸脱氢酶(BCKDH)催化的不可逆限速反应,该反应被特定激酶BCKDH激酶(BCKDK)磷酸化并失活,并被蛋白磷酸酶1K(PPM1K)去磷酸化和激活。然后产物通过进一步氧化参与不同的生理活动。
促进肝脏和其他组织中的蛋白质合成
支链氨基酸参与多种生理过程。在代谢和信号通路研究方面,支链氨基酸尤其是亮氨酸,是mTOR信号通路的有效激活剂。亮氨酸可以与 Sestrin2(mTORC1 活性的负调节因子)结合,促进 mTORC1 激活,从而促进肝脏和其他组织中的蛋白质合成。
促进肝脏和骨骼肌对糖原的吸收,淋巴细胞增殖
支链氨基酸还可以促进肝脏和骨骼肌对糖原的吸收,并增强糖原的合成。
支链氨基酸对于免疫系统中免疫细胞的正常功能至关重要,通过氧化分解免疫细胞表达的脱氢酶和脱羧酶,促进淋巴细胞增殖和细胞毒性 T 细胞活化。
癌症患者的支链氨基酸循环水平发生了变化。
胰腺导管腺癌
最近的代谢组学回顾性研究表明,血浆支链氨基酸水平升高与胰腺癌风险增加相关,这一点在胰腺导管腺癌 (PDAC) 基因工程小鼠模型中得到了验证。可能是由于系统性蛋白质分解以满足致瘤期生长所需的支链氨基酸所致。
非小细胞肺癌 v.s. 胰腺导管腺癌
一项研究表明KRAS突变可以促进支链氨基酸代谢。尽管非小细胞肺癌 (NSCLC) 和 胰腺导管腺癌 (PDAC) 中存在 KRAS 激活和 P53 缺失,这两种肿瘤尽管初始事件相同,但对支链氨基酸的利用却不同。胰腺导管腺癌细胞倾向于分解并利用细胞外蛋白质获取氨基酸,而非小细胞肺癌细胞则通过分解循环支链氨基酸来提取氮。
为什么胰腺导管腺癌的支链氨基酸代谢低于周围正常组织?
胰腺导管腺癌细胞中的CBP和SIRT4结合BCAT2的K44位点,使该位点乙酰化,从而进一步促进BCAT2通过泛素蛋白途径降解,降低代谢胰腺导管腺癌中支链氨基酸的比例,进而抑制肿瘤细胞的生长。
注:CBP——cAMP反应元件结合(CREB)结合蛋白
KRAS和USP1也可以通过泛素-蛋白酶途径调控PDAC中BCAT2的表达:KRAS通过抑制脾酪氨酸激酶(SYK)和E3泛素化连接酶TRIM21对BCAT2的泛素化来稳定PDAC中BCAT2的表达;而USP1则通过去泛素化BCAT2的K229位点,并通过GCN2-eIF2a途径促进USP1蛋白的翻译水平,促进支链氨基酸表达。
另一项研究发现,USP1 和 BCAT2 的表达水平在基因编辑小鼠和临床样本中一致呈正相关,阐明了为什么胰腺导管腺癌的支链氨基酸代谢低于周围正常组织,进而转向其他途径获取氮。
癌症中的支链氨基酸代谢
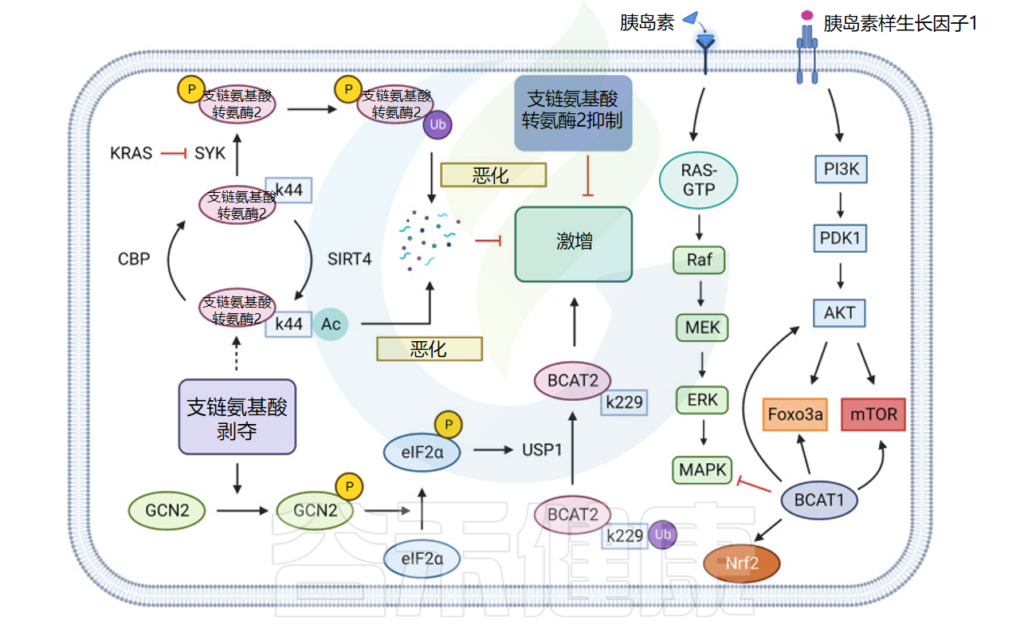
doi: 10.1038/s41392-023-01569-3
血浆支链氨基酸升高是胰腺癌的危险因素
在胰腺癌进展的早期阶段,在人类和小鼠胰腺癌模型中观察到循环支链氨基酸升高,并且由于胰腺癌周围组织中蛋白质过度分解,血液中支链氨基酸水平升高。
肿瘤微环境中的支链氨基酸代谢
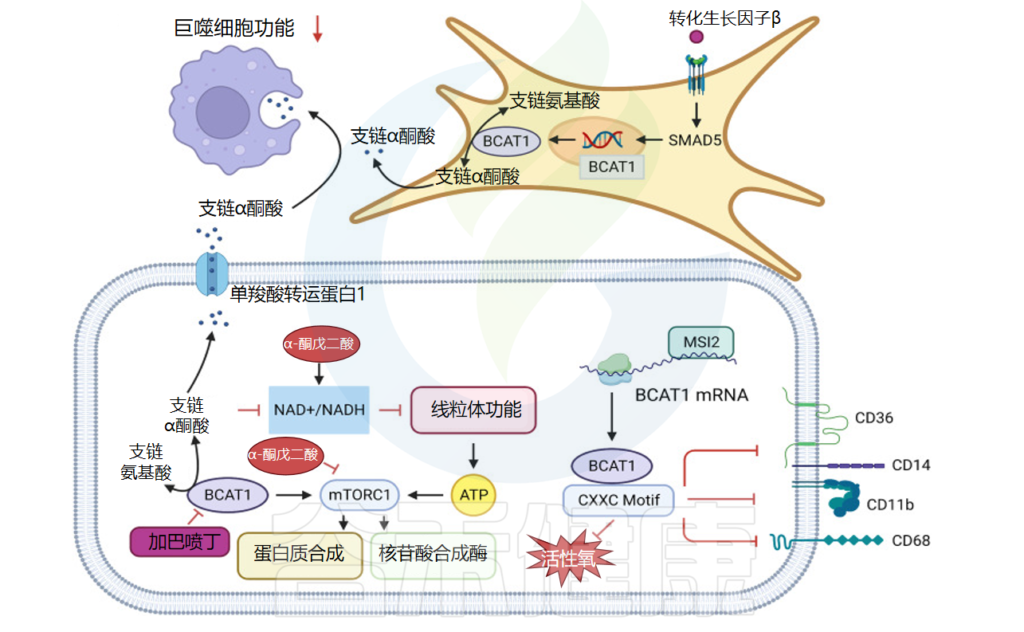
doi: 10.1038/s41392-023-01569-3
肺部肿瘤支链氨基酸摄取量高于胰腺导管腺癌
对标记的支链氨基酸代谢物的分析显示,非小细胞肺癌细胞中标记的 α-酮异己酸 (α-KIC) 和亮氨酸衍生的支链α-酮酸(BCKA)较多。同时BCKDK在非小细胞肺癌中高表达并调节细胞内ROS的产生,影响细胞存活。
代谢性疾病
支链氨基酸及其代谢物是肥胖、胰岛素抵抗和 2 型糖尿病等代谢性疾病的最强生物标志物。支链氨基酸及其代谢物升高是2型糖尿病等代谢性疾病早期进展的关键。
每种支链氨基酸都有独特的代谢作用
低异亮氨酸饮食→快速恢复肥胖小鼠代谢健康
异亮氨酸可以作为代谢健康的调节剂,低异亮氨酸饮食可以改善肥胖对代谢的不利影响。
肥胖可通过增加肝细胞中BCKDK(BCKDH激酶)/ PPM1K(BCKDH去磷酸化酶)的比例,抑制肝脏对支链氨基酸的利用,导致BCKDH失活。在肥胖和胰岛素抵抗的小鼠模型中,通过限制支链氨基酸饮食或调节 BCKDK/PPM1K 比率可以逆转这种现象。
肝脏和肾脏疾病
肝硬化:循环支链氨基酸降低 ↓↓
非酒精性脂肪肝:循环支链氨基酸升高↑↑
肝硬化患者中,高氨血症环境中支链氨基酸分解代谢增强、谷氨酸合成增加以及循环支链氨基酸水平降低被认为是该疾病的标志,并与肝性脑病风险增加相关。
在非酒精性脂肪肝 (NAFLD) 中检测到循环支链氨基酸升高。
此外,这种支链氨基酸代谢紊乱与 2 型糖尿病的发展具有协同作用。其他研究表明,补充支链氨基酸有助于恢复慢性肝病患者的葡萄糖稳态并增强免疫系统功能。
慢性肾病:循环支链氨基酸降低 ↓↓
在肾脏疾病中,慢性肾功能衰竭患者的循环支链氨基酸水平显著降低。这种现象在慢性肾脏病 (CKD) 患者中已观察到,一项 II 期慢性肾病队列研究发现,与正常对照相比,慢性肾病患者血浆亮氨酸 和缬氨酸显著降低。
这可能是由于慢性肾病患者长期营养不良和血液透析导致支链氨基酸水平下降。代谢性酸中毒还会增强支链氨基酸脱氢酶 (BCKD) 活性并加速蛋白质分解。然而,给慢性肾功能衰竭患者补充支链氨基酸和其他必需氨基酸,有助于维持蛋白质平衡,减少尿毒症毒性。
★
天冬氨酸(Asp)是一种用于蛋白质合成的 α-氨基酸,具有 α-氨基、α-羧酸基和侧链甲酰胺。它是一种非必需氨基酸,因为人体可以合成它。
草酰乙酸——天冬氨酸的前体
转氨酶将氨基从谷氨酸转移到草酰乙酸,产生 α-酮戊二酸和天冬氨酸。在天冬酰胺合成酶介导的酶反应中,谷氨酰胺提供氨基,与 β-天冬氨酸-AMP 结合形成天冬酰胺 (Asn) 和 AMP。
天冬酰胺——大脑发育必需的氨基酸
由于血液中的天冬氨酸不能直接通过血脑屏障,神经细胞的发育依赖于其在大脑中的合成。
当脑内天冬氨酸合成酶水平不足时,脑细胞的增殖就会受到限制,甚至导致细胞死亡。
反过来,在分解代谢过程中,天冬酰胺被天冬氨酸酶水解为天冬氨酸,然后用 α-酮戊二酸胺化,形成谷氨酸和草酰乙酸。然后草酰乙酸进入柠檬酸循环(下图)。
天冬氨酸、精氨酸和蛋氨酸代谢
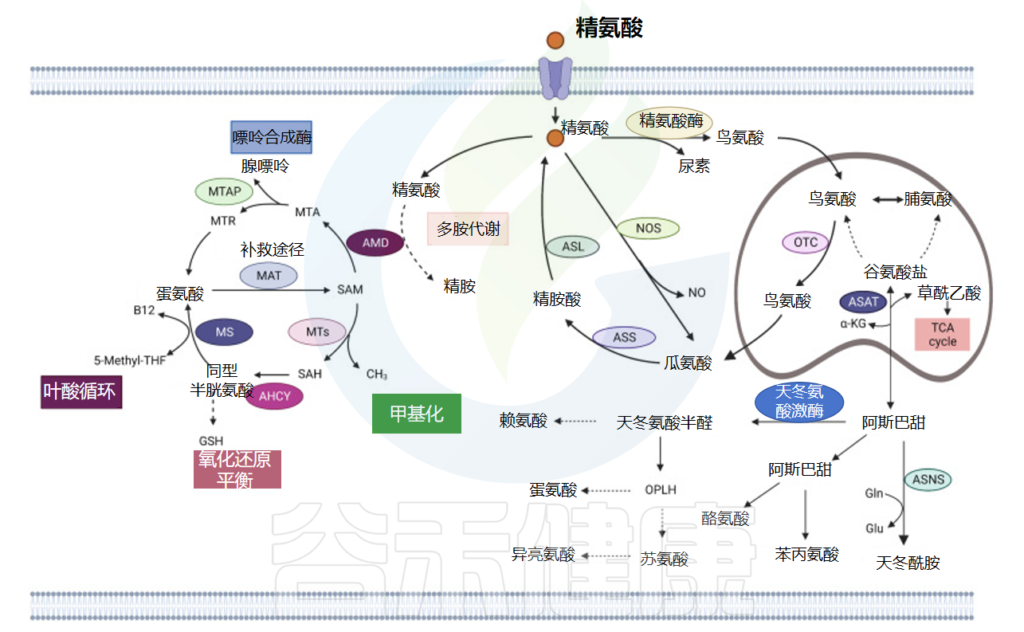
doi: 10.1038/s41392-023-01569-3
天冬氨酸也是尿素循环的代谢物
在苹果酸-天冬氨酸穿梭中携带还原当量,在肌苷合成中提供氮原子,并在 ATP 合成中充当氢受体。
天冬氨酸也是四种必需氨基酸(蛋氨酸、苏氨酸、赖氨酸和异亮氨酸)的前体。天冬氨酸还可以作为氨基酸交换因子,成为氨基酸进出细胞的介质,特别是组氨酸、精氨酸和丝氨酸。天冬氨酸通过氨基酸交换因子功能调节丝氨酸代谢、核苷酸合成和 mTORC1 活性。
TP53是人类癌症中突变频率最高的基因。该基因编码的蛋白p53通过调节细胞周期、细胞凋亡、基因组稳定性等途径抑制肿瘤的发展。
结肠癌
结肠癌细胞系中的天冬氨酸(Asp)和 天冬酰胺 (Asn) 可通过与 LKB1(编码丝、苏氨酸激酶,并直接磷酸化蛋白质产物以激活 AMPK)来抑制其活性,从而抑制 AMPK 介导的 p53 激活。
淋巴瘤和结直肠肿瘤
在淋巴瘤和结直肠肿瘤模型中,p53 的激活会破坏 天冬氨酸-天冬酰胺 稳态,并促使细胞衰老和周期停滞。缺氧条件下,天冬氨酸是肿瘤生长的限制因素。
缺氧会抑制电子传递链 (ETC),影响能量和天冬氨酸的合成。研究了肿瘤细胞对线粒体 ETC 抑制剂的敏感性,发现对 ETC 抑制不敏感的肿瘤细胞通过 Asp/谷氨酸转运蛋白 SLC1A3 维持细胞内的Asp 浓度,这使肿瘤细胞具有生存优势。
膀胱癌
在另一项关于肿瘤代谢的研究中,发现当环境中缺乏氧气时,天冬氨酸合成是膀胱癌生长的限制因素。在膀胱癌细胞中,天冬氨酸细胞的渗透性差,阻碍了肿瘤细胞从环境中摄取天冬氨酸。虽然细胞对天冬酰胺的通透性高于天冬氨酸,但膀胱癌细胞中天冬酰胺酶的活性不足,无法将天冬酰胺转化为天冬氨酸。
使用豚鼠天冬酰胺酶 1 (gpASNase1) 促进肿瘤细胞中天冬酰胺转化为 Asp 后,肿瘤细胞的生长速度显著增加,表明 Asp 获取是 Asp 获取困难的肿瘤的内源性代谢限制。有人提出,Asp是体内某些肿瘤生长的内在限制,突破这一限制将促进肿瘤生长。
天冬氨酸-谷氨酸转运蛋白 SLC1A3 与 ETC 抑制剂的作用密切相关,并且 SLC1A3 位点在非神经胶质上皮肿瘤亚簇中扩增,从而对抗天冬氨酸限制。
乳腺癌
发现SLC1A3促进乳腺癌细胞对L-天冬酰胺酶(ASNase)的抵抗。而且,SLC1A3可以补充ASNase对天冬氨酸和谷氨酸的消耗,从而消除ASNase的抑制作用,促进肿瘤的发展。
此外,另一种氨基酸转运蛋白SLC25A22可以促进KRAS突变结直肠癌(CRC)细胞中天冬氨酸的合成,激活AMPK通路并减少氧化应激。
这些研究表明,AAT 是肿瘤代谢重编程的潜在靶标。目前正在测试的针对 AAT 的药物见(见后面的表格)。
实体瘤中的天冬氨酸代谢
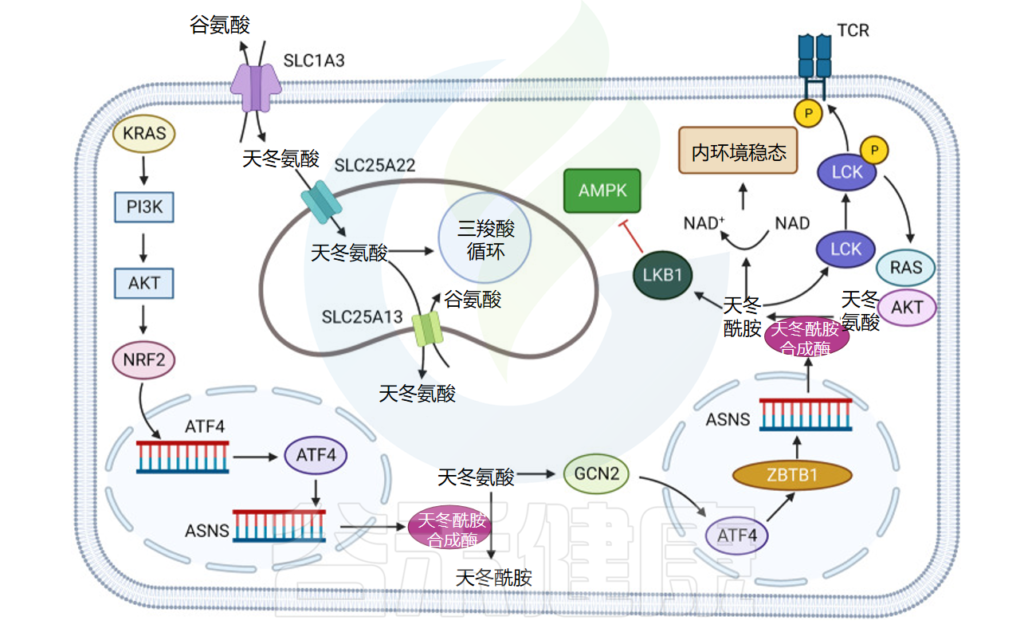
doi: 10.1038/s41392-023-01569-3
天冬酰胺作为癌症治疗的新靶点受到广泛关注
报道指出乳腺癌中天冬酰胺合成酶(ASNS)的表达水平与转移复发密切相关,抑制ASNS或限制饮食天冬酰胺可以减少肿瘤转移。
非小细胞肺癌
在非小细胞肺癌中,激活转录因子 4 (ATF4) 可以通过 KRAS 下游的 AKT 和 NRF2 改变氨基酸摄取并增加天冬酰胺合成。此外,将AKT抑制剂与细胞外天冬酰胺(ASN)消耗结合使用可以显著抑制肿瘤生长。
天冬酰胺在免疫细胞的生长和功能中发挥关键作用
发现CD8 + T细胞在CD8 + T细胞激活的早期阶段几乎不表达天冬酰胺合酶(ASNS),并且CD8 + T细胞的生长、激活和代谢重编程在天冬酰胺剥夺的情况下被破坏。
还证明,活化的 CD8 + T 细胞中的天冬酰胺水平增加,并与 SRC 家族酪氨酸激酶 LCK 结合,协助 LCK 在 Tyr394 和 505 处磷酸化,增强 LCK 活性和 T 细胞受体信号传导。
血液恶性肿瘤
天冬酰胺在血液恶性肿瘤中也发挥着关键作用。目前,细菌源性 L-天冬酰胺酶 (ASNase) 在儿童急性淋巴细胞白血病 (ALL) 中的应用已显著提高了治愈率。然而,在实体瘤中,多项临床试验表明会出现药物相关的毒副作用,如胰腺炎、中性粒细胞减少和低蛋白血症。
这些毒副作用至少部分是由L-天冬酰胺酶中谷氨酰胺酶的协同活性引起的。基于提高L-天冬酰胺酶在血液恶性肿瘤中的疗效、扩大L-天冬酰胺酶的用途、减少副作用的目的,新一代L-天冬酰胺酶正在开发中。
临床试验中靶向氨基酸代谢的药物
doi: 10.1038/s41392-023-01569-3
免疫疾病
自身免疫性疾病中免疫细胞代谢异常可促进炎症细胞的趋化性和炎症因子的产生。
类风湿性关节炎:T细胞天冬氨酸的丰度降低↓↓
在类风湿性关节炎 (RA) 中,细胞因子肿瘤坏死因子 (TNF) 的过量产生是发病机制中的一个核心事件,而富含内质网 (ER) 的 T 细胞是发炎关节中 TNF 的主要释放者。
发现类风湿性关节炎(RA)T细胞中线粒体天冬氨酸(Asp)的丰度降低,从而抑制NAD +周转,导致NAD +/NADH 比值降低以及NAD+蛋白质的ADP核糖基化减少依赖。
类风湿性关节炎和其他自身免疫性疾病的治疗策略是使用抗体来阻断细胞因子或其受体。最新的小分子抑制剂是靶向 Janus 激酶 (JAK) 抑制剂。这些治疗策略旨在阻断炎症途径的下游实践。然而,这些下游信号通路广泛分布在免疫细胞以外的细胞类型中,这会导致血栓形成等不良事件。因此,对自身免疫性疾病上游炎症的研究有助于从源头上预防疾病的发展。
神经系统疾病
天冬酰胺合成障碍是一种新发现的神经系统疾病,与染色体 7q2 上的 ASNS 基因突变有关。
自闭症:血清和脑脊液中天冬酰胺降低↓↓
自闭症谱系障碍(ASD)严重影响早期神经发育,导致智力障碍、发育迟缓、顽固性癫痫发作、进行性脑萎缩和呼吸系统缺陷。目前,该疾病中有一小部分人的血清和脑脊液中天冬酰胺水平可检测到降低,这阻碍了该测试用于初步筛查。由于血脑屏障的存在,天冬酰胺不会在大脑中积极积聚,因此大脑中 ASNS 活性的降低被认为是导致这种疾病的原因。
迄今为止,已报道了15种与ASD相关的突变,其中一些突变破坏了蛋白质结构,降低了ASNS的底物结合能力和催化效率。
例如,R49Q是位于N端结构域的谷氨酰胺 (Gln)结合袋中的突变,该突变不仅导致第二个β-折叠的氢键丢失,而且导致Gln的氢键丢失。此外,G289A和T337I突变位于C端结构域的ATP结合袋附近,G289A会引起与Ser293的空间冲突,而T337I会在蛋白质表面引起疏水斑并降低蛋白质溶解度。
补充天冬氨酸部分有效,需进一步研究
在治疗方面,膳食补充天冬氨酸并没有想象中那么有效,人为升高血液天冬氨酸可能会因竞争共转运蛋白而影响其他氨基酸的吸收。目前的治疗方法仅部分有效,需要进一步了解该疾病的机制来开发有效的药物。
★
谷氨酰胺(Gln)是一种用于蛋白质合成的 α-氨基酸。它的结构与谷氨酸类似,但侧链的羧酸基团被酰胺取代。谷氨酰胺是从食物中获取的非必需氨基酸,也是消耗最多的氨基酸,参与合成所有非必需氨基酸 (NEAA) 和蛋白质。
哪里产生谷氨酰胺?
肌肉组织在人体内产生最多的谷氨酰胺,约占全部合成谷氨酰胺的90%。
大脑和肺部也能释放少量的谷氨酰胺。肝脏虽然也能合成谷氨酰胺,但其主要功能是调节从肠道吸收的大量谷氨酰胺。
哪里消耗谷氨酰胺?
肠道细胞、肾细胞、激活的免疫细胞和各种肿瘤细胞是谷氨酰胺最迫切的消耗者。
具体通路
谷氨酰胺通过氨基酸转运蛋白 ASCT2/SLC1A5 进入细胞,并通过涉及谷氨酰胺酶 (GLS) 的脱氨反应在线粒体中转化为谷氨酸。然后谷氨酸在谷氨酸脱氢酶(GDH)或谷氨酸转氨酶或天冬氨酸转氨酶(TAs)的催化下产生α-酮戊二酸(α-KG)。α-KG是TCA循环的中间产物。
在缺氧或线粒体功能障碍下,α-KG可通过异柠檬酸脱氢酶(IDH2)催化的羧化反应转化为柠檬酸,用于氨基酸和脂肪酸的合成以及还原剂NADPH的生产。
肿瘤细胞是谷氨酰胺的紧急消耗者。信号分子Akt、Ras和AMPK可以通过激活糖酵解引起Warburg效应来诱导乳酸产生,促使肿瘤细胞通过谷氨酰胺代谢来满足能量需求。
在各种肿瘤中,谷氨酰胺代谢受癌基因/抑癌基因(例如 c-Myc 和 p53)的调节。癌基因 c-Myc 通过 GLS 和 SLC1A5 基因的转录激活上调谷氨酰胺代谢。
GLS 驱动的谷氨酰胺代谢是前列腺癌 (PCa) 放疗耐受性的调节因子,并且 GLS 1 和 c-MYC(谷氨酰胺的关键调节因子)的高表达与接受治疗的前列腺癌患者的无进展生存期缩短显著相关与放射治疗。
谷氨酰胺代谢可以通过α-KG依赖性染色质双加氧酶维持前列腺癌干细胞(CSC)。抑制谷氨酰胺代谢可降低体内 CSC 群体的频率以及小鼠模型中肿瘤生长的速率。
结肠癌
在结肠癌中,谷氨酰胺剥夺刺激p53激活,促进天冬氨酸/谷氨酸转运蛋白SLC1A3的表达,从而促进谷氨酸、谷氨酰胺和核苷酸合成,维持电子传递链和三羧酸循环活性。
SLC1A3 的缺失会降低肿瘤细胞对谷氨酰胺饥饿的抵抗力并抑制肿瘤细胞的生长。此外,研究表明,高表达胱氨酸/谷氨酸抗转运蛋白SLC7A11/xCT的肿瘤细胞高度依赖谷氨酰胺代谢。
在缺乏胱氨酸等氨基酸的情况下,细胞通过通用控制非阻遏物 2 (GCN2) -真核起始因子 (eIF2a) 信号通路促进 ATF4 的翻译,从而促进参与氨基酸代谢和应激反应的基因转录,包括 SLC7A11,使细胞能够应对氨基酸饥饿。
肿瘤中的谷氨酰胺代谢
doi: 10.1038/s41392-023-01569-3
由于肿瘤细胞通过 SLC7A11 将细胞内的谷氨酸交换为细胞外的胱氨酸,细胞内的谷氨酸被消耗,导致细胞吸收更多的谷氨酰胺,并激活谷氨酰胺酶来补充细胞内的谷氨酸,使得 SLC7A11 高表达的细胞成为谷氨酰胺依赖型。
SLC7A11在三阴性乳腺癌、肺癌、胰腺导管腺癌、肾癌和肝癌中高表达
在三阴性乳腺癌 (TNBC) 中,与其他乳腺癌细胞相比,SLC7A11 高表达的细胞消耗更多的谷氨酰胺,并且对谷氨酰胺饥饿更敏感。
研究发现,SLC7A11 的缺失对小鼠正常胰腺组织的发育没有影响,但会严重损害 KRAS 驱动的胰腺导管腺癌生长。
生理条件下 SLC7A11 的非必需性以及 SLC7A11 在肿瘤中的高表达使 SLC7A11 成为癌症治疗的有希望的靶标。
谷氨酰胺酶可将谷氨酰胺水解为谷氨酸,是谷氨酰胺代谢中的关键酶。
谷氨酰胺酶的表达具有组织特异性。谷氨酰胺酶在汇管周围肝细胞、肾上皮细胞和中枢神经系统中活跃表达,用于合成尿素和神经递质。
人类谷氨酰胺酶的四种异构体分为两种由 GLS1 编码的高活性肾谷氨酰胺酶类型和两种由 GLS2 编码的低活性肝谷氨酰胺酶类型。
不同肿瘤中GLS1和GLS2表达的异质性表明恶性肿瘤细胞对谷氨酰胺代谢的需求不同。发现肝癌细胞线粒体中氨基酸合成5样1(GCN5L1)的通用控制可以促进GLS1和GLS2异构体的乙酰化和失活,从而抑制mTORC1激活和细胞增殖。
卵巢透明细胞癌
在卵巢透明细胞癌 (OCCC) 中,谷氨酰胺酶抑制剂 CB-839 抑制 ARID1A(富含 AT 的相互作用结构域蛋白 1A)突变的 PDX 肿瘤生长。
肾细胞癌
一项针对晚期肾细胞癌 (RCC) 的随机、双盲、对照 II 期试验证明,谷氨酰胺酶抑制剂 telaglenastat (CB-839) 和 mTOR 抑制剂依维莫司 (TelaE) 联合使用具有协同抗癌作用,且患者耐受性良好既往接受过 TKI 治疗的患者。此外,与安慰剂加依维莫司 (PboE) 相比,TelaE 可以改善无进展生存期 (PFS)。
另一项 Ib 期临床试验也显示 TelaE 或 telaglenastat 联合卡博替尼 (TelaC) 治疗 RCC 具有良好的耐受性和临床活性。
胰腺炎
补充谷氨酰胺降低重症死亡率,改善IBS,利于伤口愈合
谷氨酰胺可作为多种疾病的营养补充剂。多项荟萃分析发现,补充谷氨酰胺可以降低重症胰腺炎患者的死亡率、并发症发生率和总住院时间。
一项随机、双盲、安慰剂对照临床研究表明,在低发酵低聚单糖和多元醇 (FODMAP) 饮食中补充谷氨酰胺可改善肠易激综合征 (IBS) 症状。
在促进伤口愈合方面,研究发现,补充精氨酸和谷氨酰胺对伤口愈合有积极影响,补充谷氨酰胺显著影响患者的氮平衡,减少住院时间和死亡率。
补充谷氨酰胺并没有显著影响烧伤患者的预后
在一项招募 1200 名患者的双盲、随机、安慰剂对照试验中,谷氨酰胺补充剂组的出院生存期为 40 天,而安慰剂组为 38 天。谷氨酰胺组的死亡率为 17.2%,与安慰剂组的 16.2% 没有显著差异,并且补充谷氨酰胺并没有减少住院时间。
在他们的研究中,表明了补充谷氨酰胺的益处和风险,而烧伤和其他疾病的临床试验却显示出相互矛盾的结果。
补充谷氨酰胺对各种疾病的益处和风险仍需要更多临床试验数据。
心血管疾病
在心血管疾病中,Myc 和 Myc 相关因子 X (Max) 上调肺动脉高压中的谷氨酰胺转运蛋白 SLC1A5 和 SLC7A5 以及线粒体苹果酸,从而促进谷氨酰胺分解诱导的右心室肥厚。
氧化应激下,谷氨酰胺显著下降
在氧化应激下,心肌细胞中的谷胱甘肽(GSH)水平下降 60-70%,谷氨酰胺、谷氨酸和 α-酮戊二酸(α-KG)水平也显著下降,同时将谷胱甘肽转化为谷胱甘肽的 GLS 酶活性显著降低。
调节葡萄糖稳定性的关键氨基酸
2 型糖尿病是心血管疾病发生的主要危险因素。糖尿病患者骨骼肌代谢失调会影响胰岛素敏感性和葡萄糖稳态。研究发现,谷氨酰胺是调节葡萄糖稳定性和胰岛素敏感性的关键氨基酸,谷氨酰胺水平影响骨骼肌的炎症反应并调节胰岛素信号抑制剂适应性蛋白GRB10的表达。此外,谷氨酰胺的全身升高可改善肥胖小鼠模型中的胰岛素敏感性并恢复葡萄糖稳态。
补充谷氨酰胺——心脏保护的新希望
蒽环类抗生素阿霉素(DOX)是一种广泛应用于实体恶性肿瘤的抗肿瘤药物;然而,这种疗法可能因自由基和氧化应激而导致严重的心脏毒性。补充谷氨酰胺显著降低心脏脂质过氧化物水平,增加过氧化物酶和谷胱甘肽水平,保护 DOX 治疗的大鼠模型的心脏功能。
针对心脏谷氨酰胺代谢的药物正在开发中。冬凌农 (Ori) 是一种从植物冬凌草 (Hemsl.) 中提取的天然萜类化合物,在大鼠心肌损伤模型中,可提高心脏谷氨酰胺水平并抑制 ATP/ADP 比值下降,保护心肌细胞并减少梗塞面积。
严重急性呼吸道综合症
严重急性呼吸综合征冠状病毒 2 (SARS-CoV-2) 是 2019 年冠状病毒病 (COVID-19) 的病因。该疾病通过人与人之间的密切接触或感染者的呼吸道分泌物传播。
COVID-19:谷氨酰胺降低↓↓
COVID-19的危险因素包括心血管疾病和糖尿病,此类高危人群表现出谷氨酰胺、NAD +水平低和透明质酸(HA)过量产生的共同代谢特征。
谷氨酰胺和 NAD +水平会导致 SIRT1 失调
SIRT1 是透明质酸合酶 2 (HAS2) 基因的关键负调节因子。这些代谢改变最终导致 HA 和纤溶酶原激活剂抑制剂 1 (PAI-1) 的过量产生以及 Tregs 和骨髓源性抑制细胞 (MDSC) 群体的扩张。因此,谷氨酰胺缺乏导致了 COVID-19 高危人群的免疫功能障碍和 HA 过量产生。
HA可以通过PAI-1激活STAT3。由于 SIRT1、STAT3 和 O-GlcNacylation 失调,通过激活 HAS2 诱发透明质酸风暴。
谷氨酰胺消耗加剧
此外,尽管SARS-CoV-2疫苗显著减少了COVID-19病例,但SARS-CoV-2感染后细胞处于强烈的氧化应激条件下,从而促进谷氨酰胺的消耗以合成谷胱甘肽。这一过程加剧了高危人群的谷氨酰胺缺乏,并可能诱发代谢功能障碍。同时,它还能引起STAT3通路失活和PAI-1激活,导致部分人出现严重的COVID-19并发症。
补充谷氨酰胺降低患者严重程度
小型临床试验表明,补充谷氨酰胺可以降低 COVID-19 患者感染后的严重程度。然而,这部分研究需要扩大,以更准确地评估谷氨酰胺在治疗 COVID-19 中的价值。
★
精氨酸,又称L-精氨酸,是蛋白质合成的原料,也是尿素和一氧化氮循环的中间产物。精氨酸被归类为条件必需氨基酸,其需求量取决于发育阶段和健康状况。
在人类中,小肠上皮细胞将谷氨酰胺和谷氨酸转化为瓜氨酸,然后通过循环系统转运至肾近端肾小管细胞,其中精氨酸由尿素循环中的精氨酸-琥珀酸合成酶和精氨酸-琥珀酸裂合酶合成。
当小肠和肾功能受损时,精氨酸合成就会受损,从而产生对精氨酸的饮食需求。在其他细胞类型中,瓜氨酸合成的精氨酸非常低,但当诱导型一氧化氮合酶 (NOS) 增加时,精氨酸合成会急剧增加。
在这些条件下,一氧化氮合成的副产物瓜氨酸可以通过精氨酸-瓜氨酸途径回收精氨酸。精氨酸对于细胞分裂、伤口愈合和免疫功能很重要。
蛋白质中的精氨酸可被 PAD 酶催化为瓜氨酸,这一过程称为瓜氨酸化,是正常免疫过程的一部分。另一种类型的翻译后修饰是精氨酸甲基转移酶 (PRMT) 的甲基化,其中精氨酸可以甲基化为单甲基化精氨酸或二甲基化精氨酸。
精氨酸甲基转移酶可分为以下三类:
精氨酸甲基化通常发生在富含甘氨酸和精氨酸的“GAR基序”中。许多精氨酸甲基化蛋白质已被证明可以与 DNA 或 RNA 相互作用,并且精氨酸残基充当磷酸主链的重要氢供体。
此外,精氨酸甲基化还会影响各种细胞过程中涉及的蛋白质-蛋白质相互作用,例如蛋白质运输、信号转导和转录调节。
正常细胞中的瓜氨酸和天冬氨酸可通过尿素循环中的精氨酸-琥珀酸合成酶 1 (ASS1) 和精氨酸-琥珀酸裂解酶 (ASL) 转化为精氨酸。
精氨酸琥珀酸合成酶 1 (ASS1) 转录抑制发生在各种肿瘤中,产生对外部精氨酸的依赖,并实现精氨酸剥夺疗法。
在GBM中使用精氨酸消耗剂聚乙二醇化精氨酸脱亚胺酶ADI-PEG20可以增加一氧化氮(NO)合成并产生细胞毒性过亚硝酸盐,增加肿瘤细胞对电离辐射的敏感性,显著增强放疗对GBM的效果。
精氨酸剥夺促进HCC细胞中GCN2依赖性周期停滞,而精氨酸剥夺HCC细胞中GCN2的抑制促进细胞衰老并增加衰老化合物的功效。
Ass 缺陷的前列腺癌和胰腺癌也被证明对 ADI-PEG20 敏感,而 ADI-PEG20 通过诱导自噬和细胞凋亡来促进细胞死亡。
肿瘤细胞中的精氨酸代谢
doi: 10.1038/s41392-023-01569-3
精氨酸耗竭可以增加肝细胞癌细胞中GCN2的磷酸化水平,激活GCN,增加SLC7A11的表达水平,增加精氨酸的摄取。
伤口愈合
在伤口愈合过程中,精氨酸通过精氨酸-NO途径参与炎症因子的反应。此外,精氨酸酶降解精氨酸产生的鸟氨酸和尿素在此过程中是必需的,并且在胶原蛋白和多胺的合成中具有关键作用。
精氨酸可以通过 GPRC6A-ERK1/2 和 PI3K/Akt 信号通路促进成纤维细胞增殖。
精氨酸调节免疫
精氨酸可以在炎症早期阶段增加单核细胞迁移和外周血中促炎因子的产生;在炎症后期,精氨酸还可以抑制免疫细胞的活性,调节免疫状态。总之,精氨酸及其代谢物对于伤口愈合至关重要,并参与伤口愈合的多个阶段,包括胶原蛋白形成、细胞增殖和免疫调节。
补充精氨酸有助于各种伤口愈合
膳食补充精氨酸是最方便的方法,对伤口愈合有多种好处。补充精氨酸可以增强人体 DNA 的合成。在结肠炎模型中,精氨酸补充剂可抑制炎症因子和趋化因子的表达,抑制炎症反应,促进受损组织的修复。
遭受创伤/失血性休克的患者由于胶原蛋白合成减少而难以实现伤口愈合。相反,补充精氨酸可以显著缓解上述问题并增加伤口强度。
糖尿病伤口愈合过程中,补充精氨酸可以逆转NO合成不足的情况,恢复受损组织中NO的浓度,促进伤口愈合。精氨酸也被用来降低压疮的风险,在压疮(褥疮)高风险患者中补充精氨酸可以显著加速压疮愈合。
阿尔茨海默病(AD)
阿尔茨海默病 (AD) 的特征是由淀粉样蛋白-β 和磷酸化 tau 沉积引起的老年斑和神经原纤维缠结 (NFT)。
精氨酸治疗改善阿尔茨海默病
晚期糖基化终末产物 (AGE) 会修饰蛋白质,导致其功能障碍。AMPK-γ 亚基的糖基化会抑制 AMPK 功能,精氨酸治疗可保护 AMPK-γ 免受糖基化并增加阿尔茨海默病小鼠模型中的 AMPK 磷酸化,从而改善阿尔茨海默病。
与其他药物联合使用,改善认知
在轻度阿尔茨海默病 /认知障碍 (MCI) 患者中,联合使用 L-精氨酸、HMG-CoA 抑制剂辛伐他汀和四氢生物蝶呤可增强内皮一氧化氮合酶 (eNOS) 通路,从而适度增加脑血流量并改善认知。
此外,据报道PRMT4催化的不对称二甲基精氨酸(ADMA)作为配体与NOS结合,导致NOS功能障碍,导致脑血流量减少,加重阿尔茨海默病,通过抑制PRMT4可以逆转这种情况。
肺部疾病
哮喘是一种多变的、反复发作的、长期的呼吸道炎症性疾病。精氨酸和一氧化氮代谢失衡与哮喘的病理生理学有关。
哮喘:精氨酸水平显著降低↓↓
对哮喘儿童血浆代谢质谱分析显示,与非哮喘易感组相比,哮喘易感组的精氨酸(Arg)、赖氨酸(Lys)和蛋氨酸(Met)水平显著降低。
还显示,哮喘和阻塞性睡眠患者的血清不对称二甲基精氨酸 (ADMA) 浓度显著增加,一氧化氮合酶 (NOS) 抑制增强,精氨酸分解代谢增强,鸟氨酸 (Orn) 和脯氨酸 (Pro) 水平增加,以及精氨酸/鸟氨酸比值降低呼吸暂停(OSA)。
在标记的哮喘药物中添加 L-精氨酸并不能显著减少哮喘发作
这可能是由于 IL-4 和 IL-13 诱导的精氨酸酶活性显著增加,以及过敏原刺激的肺部下游产物腐胺显著增加。
在肥胖哮喘患者的药物中添加 L-瓜氨酸(L-精氨酸循环和 NO 合成的前体),可能有助于控制哮喘并改善 NO 排泄分数 (FeNO) 水平。
心血管疾病
不对称二甲基精氨酸 (ADMA) 升高↑↑
患有高胆固醇血症和血管疾病的患者通常会出现不对称二甲基精氨酸 (ADMA) 升高,这与 NO 合成受损和内皮功能障碍的早期标志物有关。
ADMA 是一种内源性一氧化氮合酶 (NOS) 抑制剂,可显著减少血管舒张剂 NO 的合成,导致心血管疾病的发生。
PRMT1是催化ADMA的主要酶。它通过调节心肌肌动蛋白启动子区域的组蛋白甲基化修饰来调节基因激活。PRMT1的消融可以下调心肌素等收缩基因的表达,并显著降低主动脉的收缩力和血管平滑肌细胞(VSMC)的牵引力。
PRMTs的异常活性导致ADMA和MMA增加,从而增加心血管疾病的风险。
针对 PRMT 的抑制剂正在开发和实验测试中。精氨酸(Arg)甲基化酶抑制剂 (AMI) 是对称磺化尿素,可特异性抑制 PRMT 活性,并在大鼠模型中抑制环氧合酶 2 (COX-2) 表达并抑制炎症。
★
蛋氨酸(Met)是一种必需氨基酸,也是其他氨基酸的前体,例如半胱氨酸 (Cys) 和牛磺酸,以及 S-腺苷-L-甲硫氨酸 (SAM) 和谷胱甘肽 (GSH)。蛋氨酸生物合成的骨架主要来自天冬氨酸。
在分解代谢中,蛋氨酸被蛋氨酸腺苷转移酶 (MAT) 催化为 SAM。作为甲基供体,SAM 参与各种甲基转移反应,并在反应中转化为 S-腺苷高半胱氨酸 (SAH)。蛋氨酸可以增加细胞内谷胱甘肽的浓度,促进细胞氧化还原调节,并通过与氧化代谢物结合来保护细胞。
蛋氨酸作为一种必需氨基酸,在肿瘤的生长和代谢中具有重要作用。除外源供应外,蛋氨酸补救途径是唯一的蛋氨酸来源。该途径需要甲基腺苷磷酸化酶 (MTAP) 和 蛋氨酸合酶 (MS) 的活性。
MTAP 位于肿瘤抑制因子细胞周期蛋白依赖性激酶抑制剂 2 A (CDNK2A) 的外围,这两个基因的共同缺失发生在约 15% 的癌症中,导致高度侵袭性肿瘤且预后不良。这些酶在恶性肿瘤中常常下调,导致细胞对外部环境中蛋氨酸摄入的强烈依赖。
肿瘤细胞中的蛋氨酸代谢
doi: 10.1038/s41392-023-01569-3
肿瘤细胞竞争消耗蛋氨酸→T细胞蛋氨酸缺乏
肿瘤细胞可以通过SLC43A2的高表达来增加蛋氨酸的摄入,竞争性消耗环境中的蛋氨酸,导致T细胞蛋氨酸缺乏。T细胞蛋氨酸限制可抑制细胞内正常甲基化,导致STAT5基因转录受阻,影响T细胞存活和功能。另一方面,蛋氨酸代谢抑制 PD-L1 和 T 细胞激活的 V 结构域 Ig 抑制因子 (VISTA) 免疫检查点翻译。
由于其在甲基化中的核心作用,蛋氨酸被认为是由 10-11 易位 (TET)、异柠檬酸脱氢酶 (IDH) 蛋白、甲基转移酶和其他表型修饰剂驱动的肿瘤治疗的候选靶点。
无蛋氨酸饮食的抗肿瘤作用,抑制肿瘤免疫逃逸
无蛋氨酸饮食的抗肿瘤作用首次在患有 Walker-256 肉瘤的 Sprague-Dawley 大鼠中被报道。
Met限制饮食可以减少肿瘤细胞中的N6-甲基腺苷(m6A)甲基化和免疫检查点翻译,例如PD-L1和T细胞激活的V域Ig抑制因子(VISTA)。
此外,它还能增加肿瘤浸润CD8 + T细胞的数量,从而抑制肿瘤免疫逃逸。补充蛋氨酸或抑制肿瘤细胞中SLC43A2的表达可以逆转上述T细胞功能抑制,激活肿瘤免疫。
这些研究表明,蛋氨酸限制不仅抑制肿瘤细胞代谢,还涉及免疫细胞,并且肿瘤细胞本身具有复杂的优先途径来调节蛋氨酸代谢。因此,应针对肿瘤细胞蛋氨酸代谢开发更精准、更有针对性的治疗方法。
脂肪肝疾病
非酒精性脂肪性肝病(NAFLD)是一种由于代谢途径异常导致甘油三酯(TG)在肝脏积聚而引起的疾病。肥胖和2 型糖尿病是 NAFLD 的重要危险因素。
高脂饮食、蛋氨酸和胆碱缺乏饮食(MCD)可以模拟人类疾病的组织学和代谢异常,通常用于建立NAFLD小鼠模型。
非酒精性脂肪性肝病早期,蛋氨酸水平降低↓↓
在评估两种方法构建NALFD/NASH模型的差异时发现,MCD饮食可在2-4周内自发导致肝纤维化,并显著影响肝纤维化途径相关基因的表达。高脂饮食的这种作用直到胰岛素抵抗后 24 周才观察到,从而减少了肝纤维化。
临床数据显示,NAFLD 早期阶段蛋氨酸水平降低,且较高的蛋氨酸摄入量与纤维化风险呈负相关。甲基供体补充通过激活 AMPK 信号通路增加脂肪酸消耗来减少肝脏脂肪积累。
肾脏疾病
常染色体显性多囊肾病:蛋氨酸水平升高↑↑
常染色体显性多囊肾病(ADPKD)是一种常见的单基因疾病,其特征是肾囊肿增大。在ADPKD模型中,蛋氨酸(Met)和SAM的水平升高,从而诱导Mettl3的表达。
Mettl3 可以增加 c-Myc 和 Avpr2 mRNA 修饰,激活 c-Myc 和 cAMP 通路,并加速囊肿生长。限制蛋氨酸饮食可能会减缓常染色体显性多囊肾病的进展。
在蛋氨酸限制 (MR) 小鼠损伤模型中,肾损伤标志物簇蛋白和胱抑素 c 显著下降。与正常喂养模型相比,MR模型中Emr1、Nos2、Tnfa等肾脏炎症基因下调,嗜碱性粒细胞聚集程度较低。
糖尿病
2 型糖尿病:循环蛋氨酸水平升高 ↑↑
糖尿病肾病:循环蛋氨酸水平升高 ↑↑
在2 型糖尿病和糖尿病肾病 (DKD) 中可以检测到循环蛋氨酸、乙酰天冬氨酸和天冬酰胺水平升高。此外,循环蛋氨酸(Met)水平升高可用于预测患糖尿病的风险。
蛋氨酸代谢调节 Cys 和内源性硫化氢 (H2S) 水平。H2S 抑制胰腺 β 细胞中葡萄糖诱导的胰岛素释放和脂肪组织中胰岛素刺激的葡萄糖摄取。
胱硫醚γ-裂解酶(CSE)是H2S合成中的关键酶,CSE抑制剂的使用会增加脂肪细胞对葡萄糖的摄取。
总之,蛋氨酸限制 (MR) 可以通过干扰葡萄糖稳态、增加胰岛素敏感性和炎症反应来缓解糖尿病。
氨基酸水平和代谢影响多个层面的细胞代谢和许多细胞过程,从蛋白质合成到表观遗传调控。这些生理过程与维持细胞稳态和正常功能密切相关。因此,氨基酸代谢异常可能导致疾病的发展。
综上,一些肿瘤病人比如,肝硬化、肺癌、乳腺癌、食管癌、肾癌、胃癌、甲状腺癌、头颈部肿瘤等病人血浆中氨基酸浓度与健康人血浆中氨基酸浓度相比表现出异常,对氨基酸代谢变化的分析可以用来辅助诊断疾病。氨基酸代谢的改变对肿瘤细胞及肿瘤免疫微环境有着显著影响。肿瘤细胞通常依赖于外源氨基酸的供应,相对于肿瘤细胞,正常细胞对氨基酸的需求较低,干扰氨基酸的可获得性,通过氨基酸消耗疗法来诱导癌细胞凋亡也是近年来备受瞩目的研究热点之一。
支链氨基酸在肿瘤和代谢疾病中起重要作用,但不同肿瘤之间对支链氨基酸的需求存在差异。支链氨基酸代谢对促炎CD4+和CD8+T细胞以及免疫抑制调节性Treg细胞的功能同样重要,Treg细胞在代谢性疾病、肝脏和肾脏疾病中发挥着关键作用。
天冬氨酸在某些肿瘤生长中是一个内在的限制因素,其转化依赖于天冬酰胺酶的活性。转氨产物天冬酰胺比天冬氨酸更具渗透性。然而,膀胱癌细胞缺乏天冬酰胺酶,导致转化为天冬氨酸的功能障碍。SLC1A3在某些实体瘤中高表达,可能为解决天冬氨酸酶耐药提供解决方案。
谷氨酰胺在肠、肾、免疫和肿瘤细胞中广泛消耗,与癌基因c-Myc和STAT3有关。SLC7A11在多种癌症中显著高表达,可作为有前景的靶标。GLS是肾小管氨基酰胺代谢的关键酶,CB-839是其抑制剂,显示出良好的抗肿瘤活性。
精氨酸调节免疫,缺乏ASS1的细胞对精氨酸具有依赖性。精氨酸甲基转移酶(PRMT)在肿瘤发生发展中的调节作用也受到广泛关注,多种 PRMT 抑制剂表现出良好的抗肿瘤活性。
蛋氨酸作为必需氨基酸的地位及其在转甲基化过程中的作用注定了细胞对蛋氨酸代谢的依赖。甲硫腺苷磷酸化酶 (MTAP) 基因缺失在一些实体瘤中发生,针对缺乏MTAP的肿瘤细胞的治疗策略是抑制蛋氨酸代谢途径中的关键酶MAT2A。蛋氨酸限制饮食可以抑制肿瘤免疫逃逸。
但是,氨基酸代谢靶向治疗面临挑战,血液氨基酸代谢是一个复杂的生物过程,涉及多个代谢途径和相互作用。因此,准确评估血清氨基酸水平对于疾病筛查和治疗有主要的意义。同时,目前血清各类氨基酸测定方法,一般是空腹抽取这增加了检测的复杂性。而且检查前一周熬夜、饮酒、劳累、服药、情绪抑郁以及进食油腻食物等都会影响氨基酸的评估。
肠道菌群在蛋白质和氨基酸代谢中扮演着重要的角色。通过评估肠道菌群的组成和功能,可以更从另外一个维度了解氨基酸代谢的状态。
一些研究发现,通过调节肠道菌群可以影响氨基酸代谢和相关疾病的进展。通过了解个体的肠道菌群状态,可以针对性地选择合适的综合治疗方法,如益生菌、益生元和饮食干预,以优化氨基酸代谢和减少治疗的不良副作用,比如抗体来阻断细胞因子或其受体导致的血栓形成等不良事件。
除本文常见的20种蛋白质氨基酸外,还包括在各个代谢途径中较为重要的氨基酸及短肽,比如牛磺酸、γ-氨基丁酸(GABA)、肌酸、肌肽等。它们在某些方面对人体的重要作用一样不比蛋白质氨基酸逊色,后续我们再和大家分享。
主要参考文献:
Ling ZN, Jiang YF, Ru JN, Lu JH, Ding B, Wu J. Amino acid metabolism in health and disease. Signal Transduct Target Ther. 2023 Sep 13;8(1):345.

谷禾健康

随着一日三餐米面肉蛋菜等一些列食物的食用,数百种化学成分会进入我们的消化道。在那里,它们被肠道微生物组进一步代谢,这是数千种微生物物种的独特集合。
因此,肠道微生物组在决定营养如何影响健康方面发挥着重要作用。然而到目前为止,微生物组中的许多微生物的代谢能力仍然是未知的。这意味着我们不知道它们以什么物质为食,以及它们是如何处理这些物质的。
近期,来自普林斯顿大学的研究人员在《CELL》期刊上发表了最新的文章:
“Gut bacterial nutrient preferences quantified in vivo”,研究人员使用同位素追踪定量研究了小鼠肠道微生物群的输入和输出。

微生物碳水化合物发酵的主要输入是膳食纤维,支链脂肪酸和芳香代谢物的主要输入为膳食蛋白质。此外,循环宿主乳酸、3-羟基丁酸和尿素(但不是葡萄糖或氨基酸)为肠道微生物群提供食物。
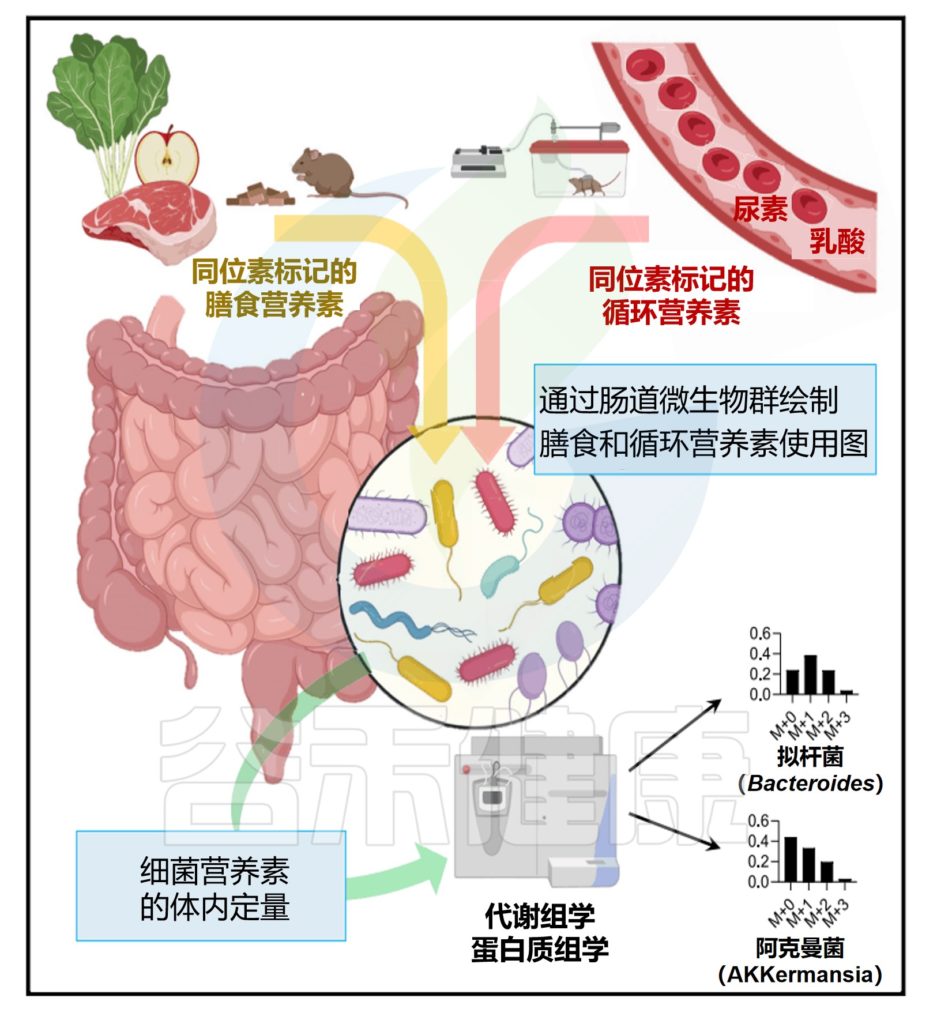
肠道菌群拥有巨大的酶多样性,超过哺乳动物基因组的数量100多倍。这些酶的能力能使摄入的膳食营养物质加工成一些列微生物代谢物。
为了复制自身和释放代谢产物,肠道细菌需要营养输入。这些形式包括摄入的食物、宿主合成的肠道粘液和宿主循环代谢物。
//
在本文中,研究人员通过对肠道菌群及其进入宿主循环系统的代谢物进行了大规模的定量评估。
研究了膳食淀粉、纤维和蛋白质的贡献以及宿主粘液的贡献,也研究了大多数主要的循环宿主营养素,发现乳酸、3-羟基丁酸和尿素在从宿主传递到肠道微生物群中表现突出。基于对细菌特异性肽序列的测量,评估了不同细菌属的营养偏好,并表明这些偏好与响应改变饮食的微生物组分变化一致。
同位素追踪能够定量测量代谢物和生物量的输入。与质谱检测相结合的稳定同位素示踪剂,使得能够测量特定下游产物的标记。通过注入氮标记的苏氨酸来标记宿主粘液,研究人员能够比较饮食和粘液蛋白对肠道微生物群的贡献,并观察到喂食低蛋白饮食的小鼠中粘液贡献的变化。
从小鼠尾部静脉抽取血样;
使用注射器从小鼠膀胱采集尿液;
所有血清样品在没有抗凝剂的情况下置于冰上 15 分钟,并在 4°C 下以 16,000 x g 离心 15 分钟。
用预冷的Wollenberger钳在液氮中快速分离并快速冷冻(< 5秒)获得组织;夹紧前取出肠内容物;盲肠内容物取样时,先将小鼠盲肠取出并在表面切开,然后用镊子将盲肠内容物挤出。
取新鲜粪便,轻揉小鼠腹部诱导排便。将血清、组织和粪便样本保存在 -80 ºC 直至进一步分析。
为了测定血清和组织样本中的代谢物浓度,进行了同位素标配(isotope spike-in)或标准标配(standard spike-in )。
对于前者将已知浓度的同位素标记标准品加入血清或组织提取液中,通过标记与未标记代谢物的比值计算浓度。
当没有同位素标准品时,加入连续稀释的非标记标准品,测量的总离子计数与加入的标准品浓度之间产生线性拟合。然后通过拟合线的x截距确定内源代谢物的浓度;蛋白质氨基酸组成采用酸水解法测定。
首先,使用13C同位素标记的不同营养物质,通过口服管饲法对小鼠进行灌胃采集小鼠的血清、组织和粪便样本。对粪便和肠内容物进行16S rRNA测序获得细菌分类。
首先使用代谢组学方法测定盲肠内容物中游离氨基酸13C-或15N标记。
然后,对于每个肽,模拟了未标记(Iunlabeled)和由游离盲肠氨基酸(Ifree)合成的肽的同位素包膜模式。标量γ可以通过将测量的肽同位素分布(Imeasured)与Iunlabeled和Ifree的线性组合拟合来确定。
注意,当一个菌属使用的特定营养素超过该营养素对盲肠游离氨基酸的贡献时,γ将大于1。
具体来说,测量的每个肽的γ如下:
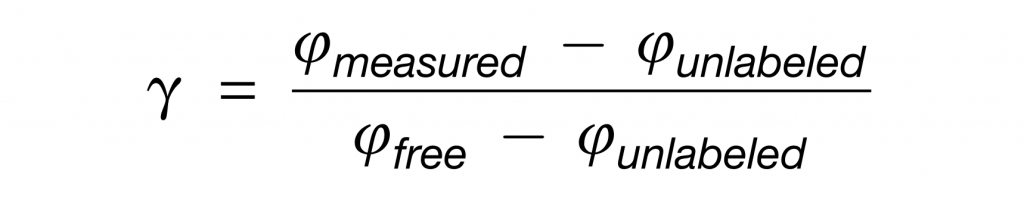

对于细菌属水平的原料贡献程度的测量,分析中只保留测量超过3个肽的属,多肽的中位数为γ-genus。
对于细菌科水平,仅分析在蛋白质组学中始终检测到的属,以及在 16S rRNA 基因扩增子测序中检测到 (> 0.5%) 的属的上一级科。
每种营养物质对菌属的贡献程度的定量公式如下:
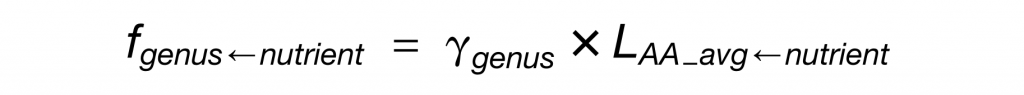

LAA_avg-nutrient为各营养物质对细菌蛋白质的贡献程度,其计算公式如下:
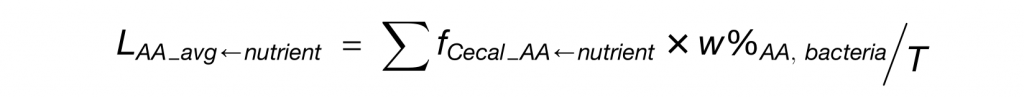

1 微生物组消耗较少的可消化膳食成分
微生物群影响宿主生理学的主要机制是通过分泌代谢产物。研究人员在门静脉和体循环以及盲肠内容物中测量了微生物衍生的50多种代谢产物的绝对浓度。
微生物群相关代谢物的绝对浓度和来源
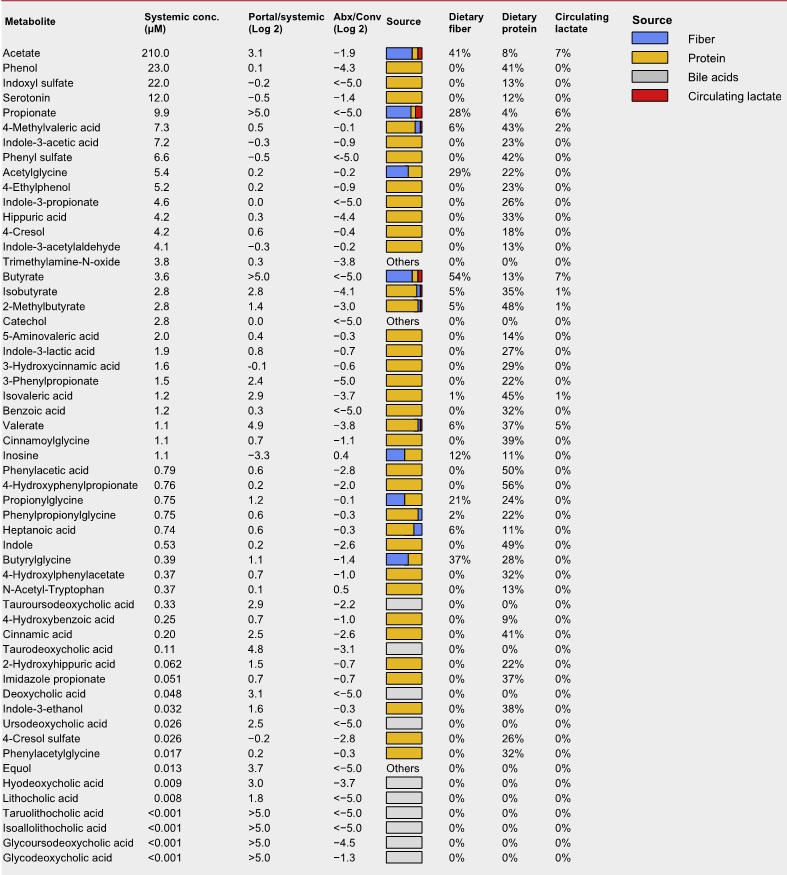
上表可以看到,与全身血液相比,大多数在门静脉循环中升高,除两种(肌苷和N-乙酰色氨酸主要来源于宿主)外,其余均被抗生素治疗耗尽。
门静脉血中主要排泄产物是短链脂肪酸。
其他相对丰富的微生物群产物是芳香族氨基酸发酵产物(苯酚、吲哚硫酸盐和3-苯丙酸盐)和支链脂肪酸(戊酸盐、异戊酸盐,4-甲基戊酸、异丁酸盐和2-甲基丁酸盐)。
探索肠道微生物产物的膳食输入:淀粉、菊粉
研究人员通过口服管饲法、淀粉(易消化葡萄糖聚合物)和菊粉(易消化果糖聚合物,即可溶性纤维)喂养小鼠:
13C淀粉灌胃后,标记的葡萄糖、乳酸和丙氨酸迅速出现在门脉循环中,并占大多数淀粉碳(约75%)。
13C菊粉和13C淀粉有什么不同?
13C菊粉灌胃后,没有观察到大量标记的果糖、葡萄糖、乳酸和丙氨酸,取而代之的是标记的门静脉代谢产物以短链脂肪酸的形式缓慢出现,约40%的菊粉碳成为短链脂肪酸,其余未消化并随粪便排出。
膳食菊粉,而不是淀粉,在盲肠内容物中广泛标记糖酵解和TCA中间体和氨基酸。
藻类蛋白大量标记了微生物群衍生的门静脉代谢物:短链脂肪酸、支链脂肪酸和芳烃(吲哚、吲哚-3-丙酸盐和3-苯丙酸盐)。
“难以消化的碳水化合物和蛋白质直接为微生物组提供营养,并通过微生物产物间接为宿主提供营养。”
研究中发现宿主循环系统中的乳酸,3-羟基丁酸以及尿素能为肠道细菌提供营养。
如图A,将同位素标记的营养物质通过静脉输注到小鼠的全身血液循环中。 2.5 小时后收集血清和粪便以量化每种营养物质对相应菌群代谢物的碳贡献。
图BCD表示了13C标记的各种营养物质在小鼠的血液和粪便中的含量,可见乳酸和 3-羟基丁酸有进入肠道菌群中,而其余大部分营养物质如柠檬酸盐、葡萄糖、氨基酸等都没有进入到肠道菌群中。
图F为15N标记的营养物质,可见尿素也同样被菌群大量利用。
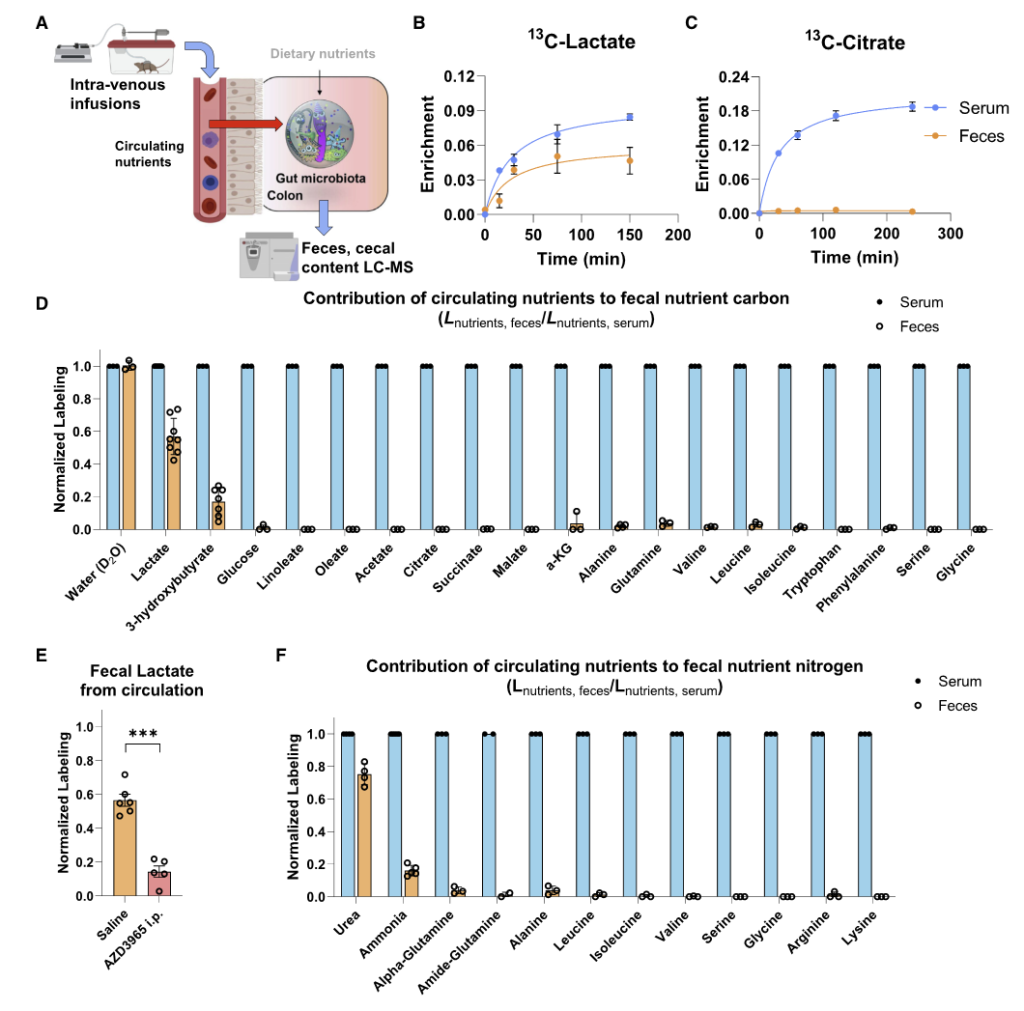
为了定量确定微生物代谢物的来源,研究人员给小鼠喂食部分纤维、脂肪或蛋白质13C标记的标准食物,盲肠标记在12小时内达到稳定状态。
为了说明循环营养输入,研究人员还注入了13C乳酸或3-羟基丁酸。
这些研究确定了大多数微生物群中心代谢物中的碳供给:
接下来,研究人员检查了微生物组游离氨基酸的输入,并用15N标记的膳食蛋白和注入的尿素进行追踪。
与哺乳动物不同,大多数肠道细菌具有合成所有20种蛋白质氨基酸的生物合成能力。
然而,研究人员观察到“必需氨基酸”主要来源于膳食蛋白质,哺乳动物无法制造,需要在细菌中表达广泛的生物合成途径。
“非必需氨基酸”主要在肠道微生物群中合成,使用膳食菊粉和循环乳酸作为碳源。
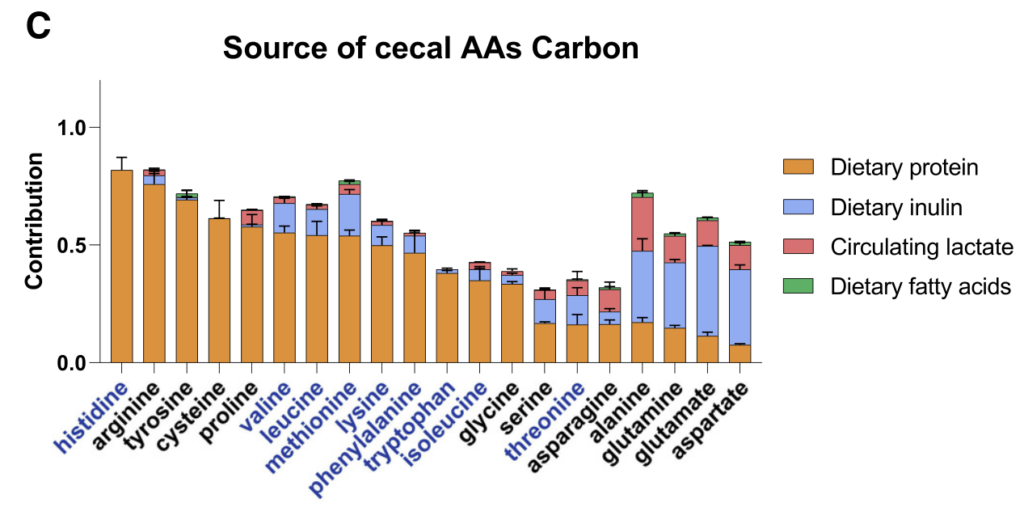
抗生素或无菌小鼠中的微生物群消耗有利于盲肠中氨基酸的积累(基于同位素追踪研究),这些氨基酸主要来自膳食蛋白质和微生物合成的氨基酸的消耗。
膳食蛋白质是必需氨基酸和非必需氨基酸的主要氮源,宿主尿素对非必需氨基酸也有很大贡献。
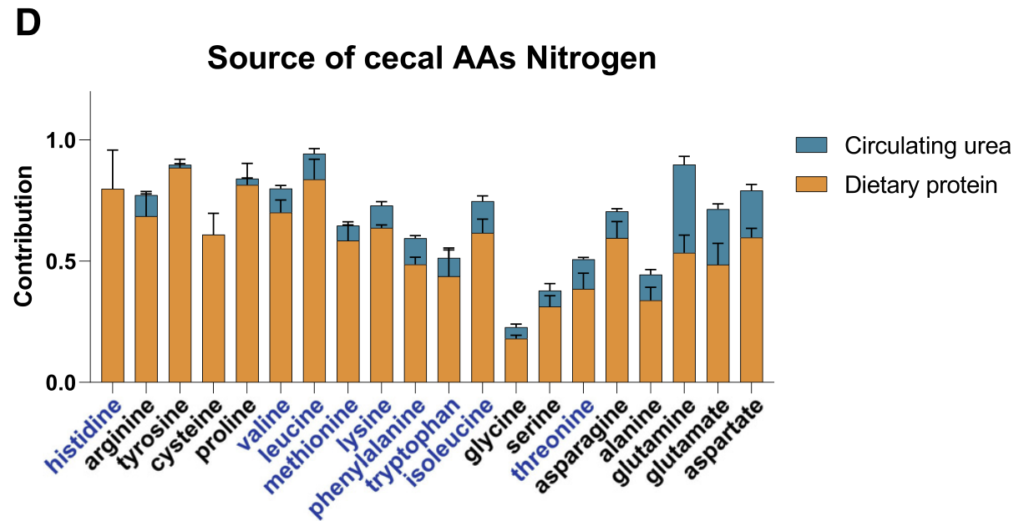
研究人员的发现如下:
【1】必需氨基酸,尽管能够由微生物群合成,但主要来自饮食,不经历任何碳重排;
【2】与TCA连接最紧密的非必需氨基酸基本上由微生物群合成,使用来自纤维的碳,通过中心代谢反应与其他碳争夺;
【3】转氨反应部分地将来自饮食衍生氨基酸的氮与来自宿主尿素的氮混合。
研究人员发现,许多微生物来源的代谢物来源于到达结肠的未吸收膳食蛋白。假设这些代谢物的循环水平将取决于膳食蛋白质到达结肠微生物群的程度。
为了控制这一点,研究人员给小鼠喂食的食物中,一部分蛋白质(酪蛋白,部分到达结肠微生物群)被游离氨基酸(基本上在小肠中完全吸收)取代。
2周后对全身血液进行代谢组学研究。含有较少完整蛋白质和更多游离氨基酸的饮食往往会增加循环氨基酸水平。
重要的是,蛋白质衍生的循环微生物代谢物(酚类、吲哚类和酰基甘氨酸)串联下降。
“微生物代谢物营养来源的知识可用于操纵其系统水平。”
研究人员通过结合13C营养标记和蛋白质组学来定量不同微生物的碳原料。
每种13C标记的营养素(膳食菊粉、膳食藻蛋白或循环乳酸)提供24小时,这足以在肠道细菌中实现稳态标记。
如同B-D,分别计算了在膳食中使用的菊粉和蛋白质以及乳酸在各细菌内的喜好程度,这个喜好程度也就是将在细菌特异性肽上被同位素标记的程度进行了量化。
结果可见:
拟杆菌属和梭状芽胞杆菌利用菊粉的程度是 Akkermansia、Muribaculum 或 Alistipes 的 4 倍多。
总体而言,厚壁菌门下的菌属比拟杆菌门的使用膳食中的蛋白质(厚壁菌0.237±0.052;拟杆菌0.175±0.031,p=0.02)。
Akkermansia通常被认为是一种促进健康的肠道微生物,使用的菊粉和蛋白质最少。相比之下,它使用了来自宿主的循环乳酸最多。
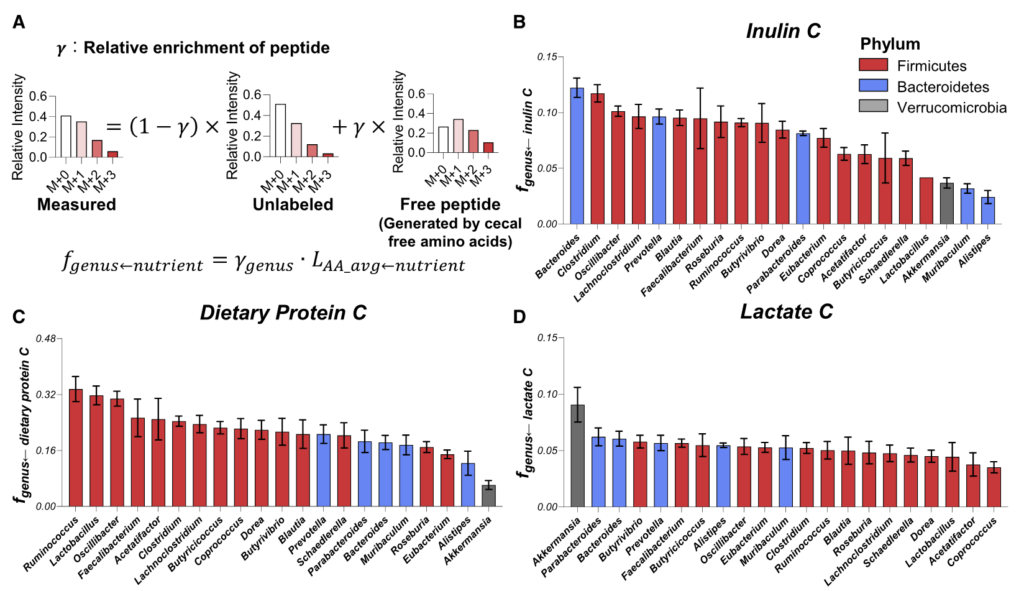
为了知晓这些细菌的营养偏好是否能预测饮食变化后的肠道菌群的组成变化。研究人员给小鼠喂食富含菊粉或藻类蛋白的饮食 2 天,并通过 16S rRNA 测序测量微生物组的组成。
结果如图F和I:
利用最多菊粉的拟杆菌属在高菊粉饮食后增加了4倍;
另一种利用较多菊粉的梭状芽胞杆菌也增加了2倍;
利用较少菊粉的菌属要么没有变化,要么略有下降;
富含藻类蛋白饮食的实验结果同理。
图G和J计算了这两种营养物与对其利用程度最高的前两名菌属相对丰度的相关性,p<0.05呈显著相关。
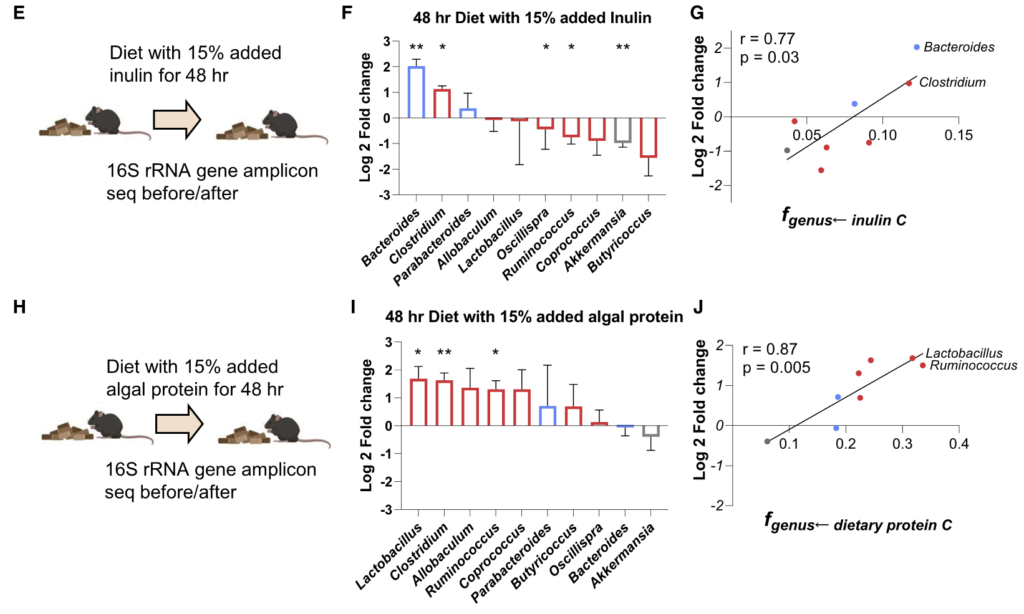
“不同肠道细菌的营养偏好有助于解释饮食操作后微生物组分的变化。”
最后,研究人员转向不同肠道细菌的氮源偏好,比较15N标记的膳食蛋白喂养和15N尿素输注。
高度利用膳食蛋白质中碳的细菌属也高度利用膳食蛋白中的氮,这与细菌蛋白质组中完整吸收的膳食蛋白质中的氨基酸一致。
厚壁菌喜欢从膳食蛋白质获取氮
在厚壁菌门成员中,偏好尿素氮的属往往是菊粉的疯狂使用者,即使用菊粉和尿素合成自己的氨基酸。这包括一些脲酶阴性菌属,它们可能通过交叉喂养获得尿素氮。
此外,在厚壁菌中也看到了一些属更喜欢从膳食蛋白质中获得氮,而其他属更喜欢循环尿素。
静脉注射尿素以提高循环尿素浓度后,偏好尿素的厚壁菌以及阿克曼菌的丰度大幅增加。
拟杆菌喜欢从宿主分泌的蛋白质中获取氮
与厚壁菌相比,拟杆菌对膳食蛋白质和循环尿素氮的利用率较低,这提出了一个关键问题:
拟杆菌如何获得氮?
肠道微生物群的一些成员(如拟杆菌和阿克曼菌)能够消化宿主分泌的蛋白质,如粘蛋白。
假设宿主分泌的蛋白质是拟杆菌氮的关键来源。为了探索这种可能性,研究人员进行了长期15N标记的赖氨酸和精氨酸输注(12、18和36小时),以标记结肠中的宿主蛋白。
尽管没有直接给微生物组喂食,但在36小时输注后,赖氨酸和精氨酸确实起作用,这与通过宿主蛋白进行的标记一致。这种标记优先发生在拟杆菌和阿克曼菌中。
膳食和分泌宿主蛋白的氮贡献呈负相关,与某些肠道细菌优先消耗膳食蛋白和其他宿主蛋白一致。
“膳食蛋白质和循环尿素是厚壁菌的主要氮原料,而分泌的宿主蛋白质为拟杆菌提供氮。”
研究人员开发了定量同位素追踪方法来测量肠道细菌的营养偏好。除了膳食纤维和分泌的宿主蛋白外,还将膳食蛋白和循环宿主乳酸、3-羟基丁酸和尿素确定为喂养肠道细菌的重要营养素。排除了其他循环宿主营养素(如葡萄糖和氨基酸)对结肠微生物群的直接贡献。
一项关键技术成就是能够从不同碳源和氮源追踪到细菌特异性肽,从而揭示复杂和竞争性肠腔环境中不同细菌的营养偏好。
厚壁菌门倾向于从膳食蛋白质获得氨基酸,而拟杆菌门更多地依赖宿主分泌蛋白。同样,一些厚壁菌门(如梭菌属)大量利用纤维(菊粉),而其他厚壁菌门则不利用纤维。
动物饮食干预实验发现,拟杆菌属和梭菌属是转化纤维最活跃的菌属。宿主循环代谢物水平也可能影响微生物组的营养获取和最终组成。
本文提供了关于哪些营养素喂养肠道微生物群以及哪些细菌更喜欢哪些营养素的基础知识。
文中所开发的方法具有广泛的应用前景,最终将有助于全面和定量地了解饮食-微生物-健康的关系。
参考文献:Zeng X, Xing X, Gupta M, Keber FC, Lopez JG, Lee YJ, Roichman A, Wang L, Neinast MD, Donia MS, Wühr M, Jang C, Rabinowitz JD. Gut bacterial nutrient preferences quantified in vivo. Cell. 2022 Sep 1;185(18):3441-3456.e19. doi: 10.1016/j.cell.2022.07.020. PMID: 36055202; PMCID: PMC9450212.