-

 CNAS L23010
CNAS L23010

 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业 二级病原微生物安全实验室
二级病原微生物安全实验室- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业 二级病原微生物安全实验室

谷禾健康

在我们日常的护肤和美容过程中,我们经常听到关于皮肤的各种话题,从保湿到抗衰老,从痘痘到过敏…
随着科学的不断进步和技术的发展,人们开始逐渐发现,皮肤上隐藏着一个神秘的世界——皮肤微生物群。它在维护我们的皮肤健康方面扮演着举足轻重的角色。
皮肤微生物群由各种细菌、真菌等微生物组成,它们聚集在毛囊、汗腺、皮脂腺等地方,形成一个庞大的生态系统。它们在皮肤表面形成了一道坚固的屏障,阻止了有害菌的入侵。除了提供保护作用外,皮肤微生物群还参与调节角质层的代谢,协助皮肤的水分平衡,并对免疫系统起到了重要的调节作用。
皮肤微生物群的平衡易受到许多因素的干扰。个人的生活方式(过度清洁)、饮食习惯(高糖高脂的饮食)等可能直接影响皮肤微生物的结构和组成,进而引发皮肤问题。外界环境中的污染物、紫外线辐射、气候变化等也会对皮肤微生物群产生影响,从而引发皮肤干燥、过敏、炎症等问题。肠道微生物群的失衡可能导致身体免疫系统的异常反应,进而影响皮肤的健康。
了解皮肤微生物群的特征及其与其他因素的相互关系,对于制定精确的治疗和护肤策略具有重要意义。

图源:Getty Images
本文我们来了解一下整个生命中皮肤微生物组,探讨皮肤微生物群的功能,包括保护屏障、免疫调节等,阐述了皮肤微生物与宿主的相互关系,微生物群在皮肤病中的影响,同时也介绍一些基于微生物群的保持皮肤健康的方法,以及皮肤微生物群在不同领域的应用前景和潜力。
-本文主要内容如下-
编辑
-正文-
皮肤是暴露于外界环境的重要器官,它可以调节体温、防止感染、保护内脏器官等。
皮肤表面是一个酸性、富含盐分、干燥、有氧的环境,而形成毛囊皮脂腺单位的内陷则相对厌氧,甚至富含脂质。
▼
皮肤是身体最大的器官。一个成年人的皮肤平均面积约为1.5-2.0平方米。皮肤除了作为外界与生物体之间的物理和化学屏障的功能外,还作为许多微生物的栖息地。通常,一个人的皮肤上有大约 1000 种细菌。
皮肤微生物组由多种微生物组成,包括细菌、真菌、病毒、螨虫等。
皮肤微生物群通过参与皮肤中发生的基本生理过程,对于维持皮肤屏障、抵御病原体入侵、增强免疫系统、分解天然产物等方面发挥着重要作用。
皮肤和微生物群的结构

编辑
上图可以看到,皮肤由两层组成,即真皮和表皮,具有不同的、专门的生态位或微环境。
▼
皮肤微生物成员和功能可能因皮肤的各种特殊生态位或微环境而异:
具有高密度的毛囊和皮脂腺,例如面部(额头、鼻翼、耳后)、胸部和背部。通常呈高酸性,其特点是细菌可以消耗脂质,需要或可以在厌氧条件下生存,例如:
Corynebacterium minutissimum(微小棒状杆菌)
Cutibacterium
肘部,膝盖,生殖器,肚脐,腹股沟等部位。温和的酸性环境,温度和湿度较高,导致体味的细菌喜欢在这样的环境生活,例如:
Corynebacterium (棒状杆菌)
Staphylococcus (葡萄球菌)
例如手掌等部位。生物量最低,但细菌多样性却最高。
最不稳定的是足部微生物群。足部皮肤上细菌的平均数量从足背表面的103CFU/cm2到第四趾裂处的107CFU/cm2不等。
脚跟底部的真菌居多, 例如:
Malassezia(马拉色菌属)
Aspergillus (曲霉属)
Cryptococcus (隐球菌属)
Rhodotorula (红酵母属)
Epicoccum (附球菌属)
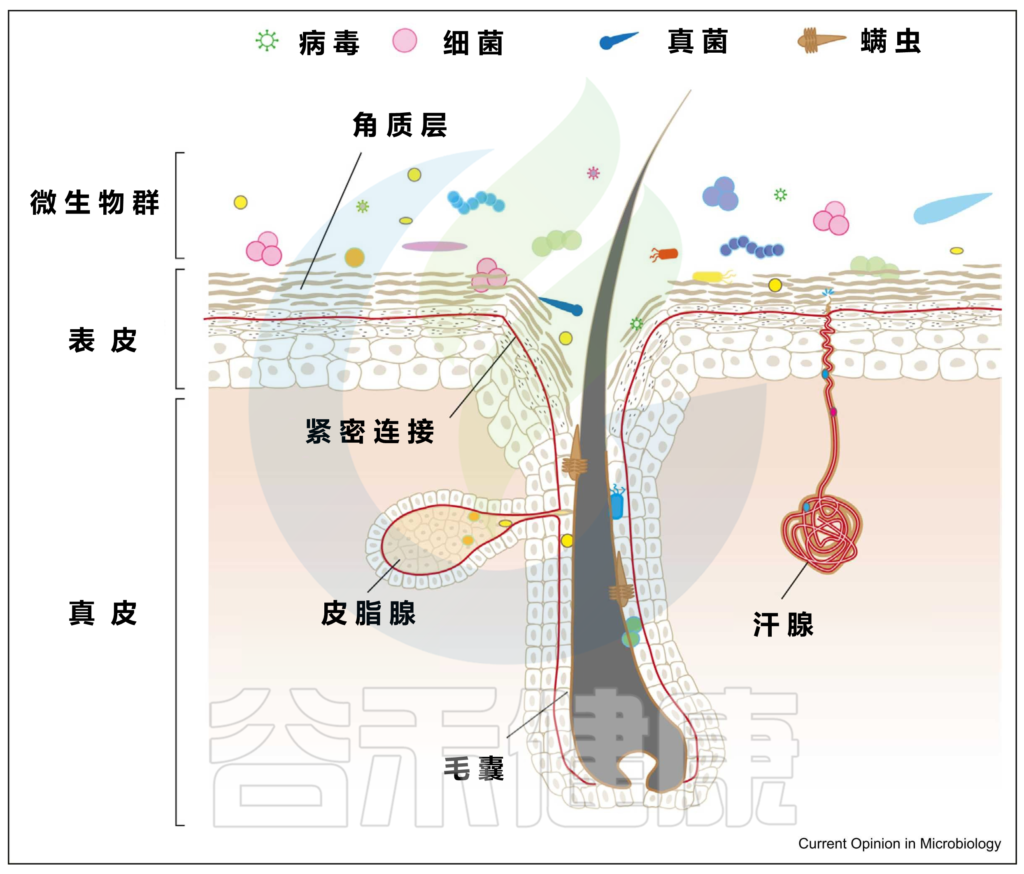
▼
在一生中,随着个人皮肤免疫系统的成熟和激素驱动汗液和皮脂腺的发育,皮肤的生理机能会发生变化。这些变化与突出的皮肤微生物群的相对丰度的变化和整体微生物群落多样性的变化有关。
作为与环境的直接接触面,皮肤也不断地与我们周围的地方和人分享微生物。下图总结了人类一生中皮肤微生物组的变化,并强调了在与年龄相关的关键阶段皮肤微生物组的破坏会影响疾病发展的风险。
皮肤及其微生物组在整个生命周期中的动态平衡
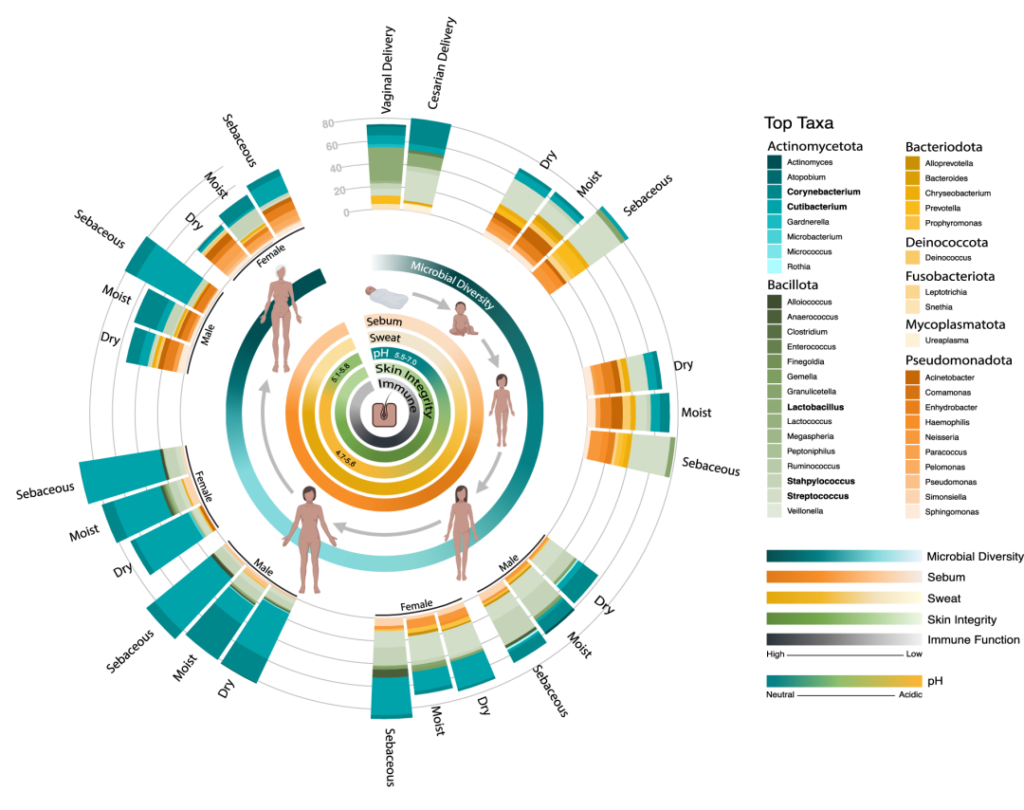
doi.org/10.1042/BST20220216
内圈代表相对微生物多样性、皮脂生成、汗液生成、表面pH值、皮肤完整性和终身免疫功能。微生物组16s测序数据显示了每组前 10 个微生物类群的平均相对丰度。
出生
皮肤微生物组在出生时就已开始定植,并受到多种因素的影响,如:分娩方式、母亲微生物群、抗生素治疗、卫生条件、营养缺乏、住房、动物/宠物接触和环境暴露等。
阴道分娩新生儿的皮肤微生物组以阴道相关菌群为主,主要是乳杆菌,普雷沃氏菌,白色念珠菌。
剖宫产新生儿的微生物群中含有母体皮肤相关微生物,包括葡萄球菌、链球菌、棒状杆菌,Cutibacterium等。
这些初始群落是短暂的,不过物种定殖的顺序和时间会影响菌株后面的相互作用。这些优先效应可以塑造未来的菌群结构,并对皮肤、微生物组和整体健康产生长期影响。
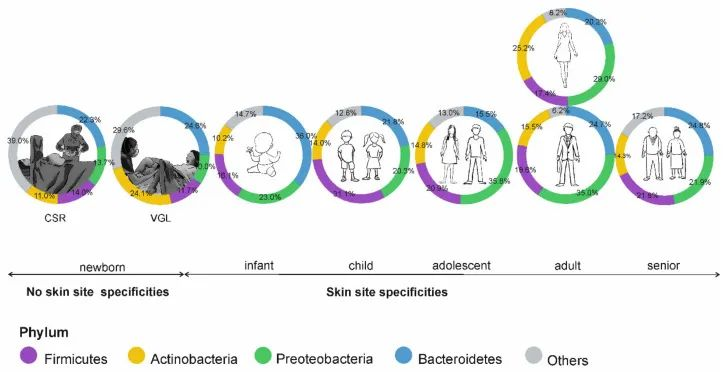
doi: 10.3390/microorganisms9030543
皮肤微生物组的年龄依赖性特异性;CSR剖宫产,VGL阴道分娩。
婴儿期和儿童期
婴儿期,最初接触微生物会促进免疫发育,并通过促进角质形成细胞的适当分化和表皮修复来加强皮肤屏障。
新生儿和婴儿皮肤含水量更高,pH值更高,皮脂生成受到抑制,表皮更新更快,抗菌性能更强。在3-6个月内,微生物分类群与皮肤代谢功能(如脂质生成和pH)之间的联系建立起来。
早期皮脂生成减少与棒状杆菌、Cutibacterium、马拉色菌丰度降低,葡萄球菌、链球菌增加以及以念珠菌为主的真菌生物群落有关。
随着儿童年龄的增长,皮肤进一步酸化并产生更多的皮脂脂质,这促使了酸敏感链球菌(acid-sensitive streptococci)的逐渐减少和整体群落多样性的增加。
在整个儿童时期,皮肤会继续携带来自照顾者的不同微生物群。然而随着年龄的增长,年龄较大的孩子具有更高的皮肤微生物多样性,以及更多来自农村或城市环境的微生物,母婴微生物组之间的相似性逐渐下降。
一旦这种平衡破坏,则可能与更大的炎症有关,并可能增加儿童患特应性皮炎和过敏的风险。
青春发育期
青春期标志着皮肤微生物群的下一个重大转变。驱动身体和性发育的激素也直接促进皮肤的结构和功能变化,如皮脂和顶泌汗液的产生,导致了随后微生物组成的变化。
横断面和纵向研究都表明,Tanner阶段的皮肤微生物组组成发生了明显变化。与V期的年轻人相比,I期的儿童链球菌、拟杆菌和假单胞菌的相对丰度更高,细菌和真菌的多样性也更高。
在年轻成人的皮肤微生物组中主要存在亲脂菌群,如棒状杆菌、痤疮角质杆菌和马拉色菌。与皮脂生成和痤疮相关。
青春期早期和晚期皮肤、微生物组和体味产生的差异
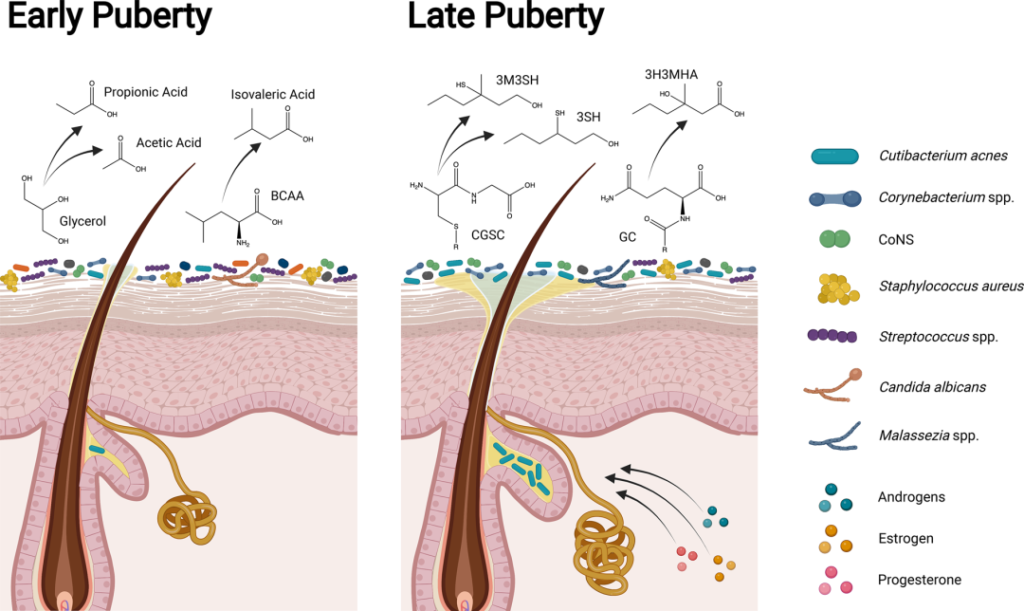
doi.org/10.1042/BST20220216
在儿童期和青春期早期(Tanner阶段I至II),皮肤微生物组高度多样化,体味与凝固酶阴性葡萄球菌属(如表皮葡萄球菌和人型葡萄球菌)产生挥发性脂肪酸(如丙酸、乙酸和异戊酸;酸味)和硫(臭鸡蛋味)有关。随着青春期的发展,类固醇激素促进皮脂腺和顶泌汗腺的发育,改变皮脂中的脂质类型,增强皮肤屏障。
在青春期后期(Tanner IV至V期),脂质生成增加和脂质含量改变与亲脂性类群主导的皮肤微生物组有关。虽然汗液和皮脂成分仍会分解为挥发性脂肪酸,但年轻人的体味与棒状杆菌属更为相关。皮脂和汗液成分代谢为硫烷基烷醇(如3-硫烷基己醇和3-甲基-3-磺基己醇;洋葱味)和挥发性有机化合物(如3-羟基-3-甲基己酸;类孜然味)。
成年期
成年皮肤微生物组在几年内是稳定的。微生物-微生物相互作用网络、持久的成人皮肤生理学和有弹性的皮肤免疫力维持了平衡的成人皮肤微生物群。
成年皮肤微生物群以角质杆菌、棒状杆菌、葡萄球菌、马拉色菌为主。
一旦成年后,成熟和持久的皮肤生理机能,会促进皮脂的产生、汗液成分和表面pH值的一致性,这些共同提供了稳定的身体部位微环境和营养库。免疫系统那时候也成熟了,这些内在特征使皮肤上的大部分微生物群能够在日常环境变化的情况下持续存在。
年龄增长
随着年龄的增长,皮肤会发生明显的变化,包括胶原蛋白合成下降、细胞外基质断裂和皮肤细胞再生减少,皮肤皱纹也就出现了。
随着皮肤屏障的变化,它可能会失去保持水分的能力,导致天然保湿因子(NFM)产生的补偿性增加。NMFs既能吸收水分,又能促进细菌增殖和粘附在皮肤上。随后,NMFs的增加与许多分类群的更丰富有关,如棒状杆菌、微球菌、链球菌、厌氧球菌,同时角质杆菌的减少。皮肤微生物多样性也广泛增加。
女性更年期后皮脂细胞面积和皮脂生成的减少,与角质杆菌的减少以及棒状杆菌、链球菌、不动杆菌和棒状杆菌丰度的增加有关。
在男性中,皮脂分泌下降的速度明显较慢,因此随着年龄的增长,它们保持着更丰富的角质杆菌。
随着年龄增长,免疫系统功能也会慢慢下降。老年人维持低度炎症状态,免疫防御受损和潜在致病菌(如β-溶血性链球菌)增加,皮肤感染的风险大幅增加,难以清除感染。
衰老会改变皮肤结构、功能和微生物定植
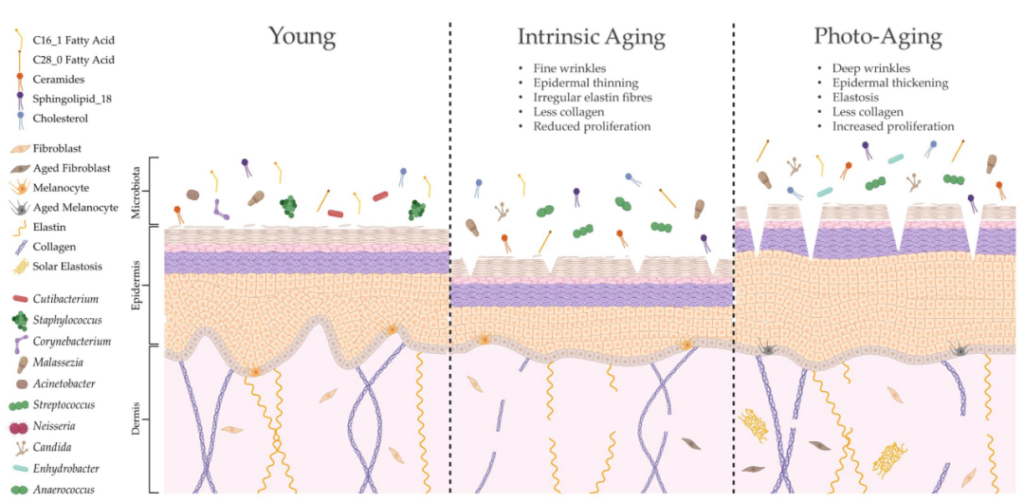
doi.org/10.3390/ijms24043950
内在衰老和光老化会导致皮肤结构和生理的不同变化,导致微生物组成的显著变化。这种改变的皮肤微生物组可能是由脂质成分的特定修饰形成的,这可能进一步导致与年龄相关的皮肤异常。
以上是皮肤微生物组在整个生命周期中的变化情况,那么皮肤微生物组是稳定的吗?它有可能受到哪些因素的影响?我们来看下一章节。
持续暴露于各种外在和内在因素会影响这个皮肤生态系统的平衡。
皮肤结构决定了皮肤微生物组的组成,个体特征取决于宿主的年龄、性别和健康状况等。个人生活方式和所处环境也会影响皮肤上微生物的数量和组成。微生物组的组成可能会随着宿主健康状况的恶化、衰老、甚至居住或职业的改变而改变。皮肤的物理和化学特性影响特定微生物群的优势、它们的比例以及它们之间的相互关系。
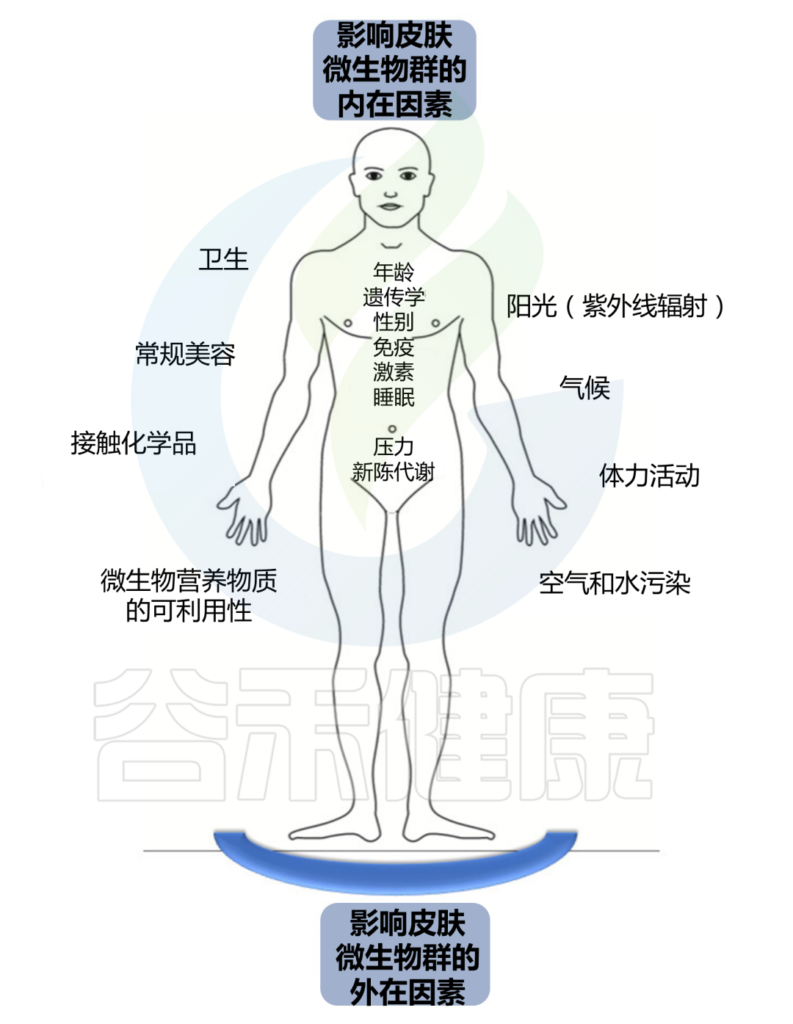
Skowron K, et al., Microorganisms. 2021
我们分为外在因素和内在因素两大块。
外 在 因 素
▼
紫外线辐射,对皮肤细胞有破坏和抗菌作用。大多数与年龄相关的皮肤病是由光老化引起的。皮肤光老化表现为:皱纹、局部色素沉着、毛细血管扩张、干燥和粗糙。这些与表皮和真皮中各种细胞和组织的病理生理变化有关。
皱纹作为光老化最明显的临床特征,主要是由于真皮成纤维细胞减少,以及胶原蛋白和弹性蛋白合成速度减慢但分解速度加快所致。皮肤光老化不仅影响美观,还会损害正常的皮肤屏障功能,增加皮肤炎症性疾病甚至恶性肿瘤的风险。
紫外线:破坏作用
皮肤强烈暴露于紫外线辐射可能会增加其感染的易感性,并加剧相关症状,例如单纯疱疹病毒。
紫外线辐射也可能影响皮肤微生物群的遗传变异,扰乱健康的微生物组结构。
皮肤暴露在紫外线下导致蓝藻菌数量总体增加,乳酸杆菌科和假单胞菌科数量减少。
紫外线:抗菌作用
阳光和紫外线也有效抑制了金黄色葡萄球菌和痤疮丙酸杆菌的生长。痤疮丙酸杆菌数量的减少与卟啉的产生减少有关。
微生物也可以抗紫外线辐射
皮肤微生物组对太阳辐射和紫外线辐射的抵抗力各不相同。一些细菌可以保护皮肤免受紫外线辐射的破坏。皮肤表面的蓝细菌和乳酸杆菌降低了色素沉着的强度和光老化相关损伤的发生。
共生马拉色菌对紫外线辐射表现出高度敏感性,尽管它们有能力合成类似紫外线过滤器的物质——pityriacitrin。
关于紫外线辐射对皮肤微生物群的影响详见谷禾之前的文章:
▼
化学空气污染物,包括 O3、颗粒物(PM 2.5:≤2.5 μm;PM 10:≤10 μm)、挥发性有机化合物和二氧化氮(NO2)等温室气体,是已知的外部暴露组的组成部分,增加过敏性疾病发生和恶化的风险。
空气污染物N2O干扰共生微生物,在对头葡萄球菌和结核棒状杆菌的负面影响大于对金黄色葡萄球菌的负面影响的情况下,有可能发生微生态失调。
烧烤烟雾中较多的成分——多环芳烃,在推动皮肤微生物群分化成不同类型中的作用
多环芳烃来源可以分为自然源和人为源:自然源指火山爆发、森林火灾等自然现象释放到环境介质中的;人为源则是由于人类生产生活活动中化石燃料(煤、油等)不充分燃烧造成的。
我们生活中例如室内外烧烤烟雾中存在较多,在烧烤的过程中,燃料的不完全燃烧或肉类食品脂肪的高温热解均可以产生大量多环芳烃类化合物。
一项研究揭示了多环芳烃暴露与皮肤微生物组分化成不同皮肤类型之间的关联。
皮肤微生物组分化为两种细胞类型(cutotype 1 和 cutotype2)。Cutotype 2与45岁以下受试者的皮肤干燥和色素沉着过度有关。多环芳烃暴露量高与皮肤干燥和cutotype 2有关,cutotype 2富含具有潜在生物降解功能的物种,相关网络结构完整性降低。
cutotype 1中精氨酸生物合成途径中的优势类群、关键功能基因和代谢产物之间的正相关性表明,来自细菌的精氨酸有助于合成聚丝蛋白衍生的天然保湿因子(NMFs),为皮肤提供水合作用,并可解释正常皮肤表型。
这项研究揭示了多环芳烃在推动皮肤微生物群分化成不同类型中的作用,这些类型在分类学和代谢功能上存在广泛的差异,并可能随后导致皮肤与微生物之间的相互作用变化,从而影响人体皮肤的健康。
也就是说:暴露于空气污染后皮肤微生物组组成的变化,可能导致皮肤干燥和炎症的恶化。
▼
全球变暖和极端天气事件等气候变化相关因素,会影响皮肤维持体内平衡的能力,在许多皮肤疾病的发病机制中发挥作用。
全球变暖可能破坏皮肤微生物组
温度和湿度的升高与皮肤上细菌的总体生长有关。
较高的气温和金黄色葡萄球菌的生长之间可能存在关联:
在一项以人群为基础的每月皮肤和软组织感染(SSTI)发病率研究中,SSTI 的时间变化与平均温度和比湿度显着相关。在美国 SSTI 的回顾性分析中(n = 616,375),在气温较高的南部地区,社区获得性耐甲氧西林金黄色葡萄球菌的感染率较高。
温度每升高1˚C,皮脂的产生就会增加10%,这反过来可能会增加微生物的生长,包括角质杆菌和马拉色菌。
极端天气可能引发皮肤病
气候变化导致极端天气事件发生的频率不断增加,包括热浪、干旱、野火、暴雨、洪水和飓风。
例如,洪水的最初影响阶段,经常有创伤与继发性伤口感染的相关风险,包括:嗜水气单胞菌( Aeromonas hydrophila)、创伤弧菌、副溶血性弧菌、Burkholderia pseudomallei等感染。
除了对皮肤病的直接影响外,极端天气事件的额外影响还包括冲突加剧、被迫迁移、心理健康恶化以及传染病的更大传播,所有这些都进一步增加了皮肤病的风险。
▼
农村和城市居民皮肤微生物组的差异,可能与不同程度地接触农业或畜牧业中的土壤、水和生物质中的微生物有关。即使皮肤与土壤和植物材料的短期接触,也会导致手部微生物组的变化以及酸杆菌Acidobacteria和拟杆菌的丰度增加。
在芬兰进行的一项研究结果表明,城市和乡村环境对 1-4 岁儿童的皮肤微生物群有显著影响。这种效应在青少年(14岁)中消失,这直接归因于该年龄段的户外活动时间有限。然而,在其他国家获得的研究结果并未证实这种趋势,表明其他因素(文化差异)也影响皮肤微生物组。
角质杆菌属在农村成年人的背部皮肤上更常见,而Trabulsiella属细菌在城市居民的手和前臂上更丰富。
农村环境的特点是微生物多样性很高
棒状杆菌和角质杆菌属数量的减少,以及假单胞菌和不动杆菌数量的增加,主要发生在与各种农场动物接触的农场工人身上。
封闭空间环境中的微生物有城市和工业区的特点
随着室内城市化的发展,与人类皮肤相关的真菌和细菌的相对丰度也在增加。此外,潜在致病真菌的数量也在增加,包括曲霉菌、马拉色菌、念珠菌等。
由于卫生习惯和西方生活方式,皮肤的细菌多样性降低。许多皮肤共生菌(如表皮葡萄球菌、乳酸杆菌、伯克霍尔德菌Burkholderis、痤疮梭菌)消失,取而代之的是葡萄球菌、棒状杆菌、角质杆菌(Cutibacterium)和微球菌Micrococcus。
▼
不同的动物物种含有独特的微生物群,与动物的持续接触会影响健康人皮肤细菌群落的组成和多样性。例如家养狗和家庭主人共享微生物群。菌群结构受季节的影响,但不受狗的性别、年龄、品种或皮毛类型的影响。
宠物肠道菌群与主人的肠道菌群也会产生关联,详见:
▼
皮肤与衣服的长时间接触也很重要,这会导致微生物的传播,并形成所谓的纺织品和挥发性微生物组。反过来,织物微生物组的组成会受到洗涤和干燥的影响。附着在纤维上的微生物可以利用污垢或皮脂化合物作为基质,并产生挥发性物质作为副产品,从而产生难闻的气味。
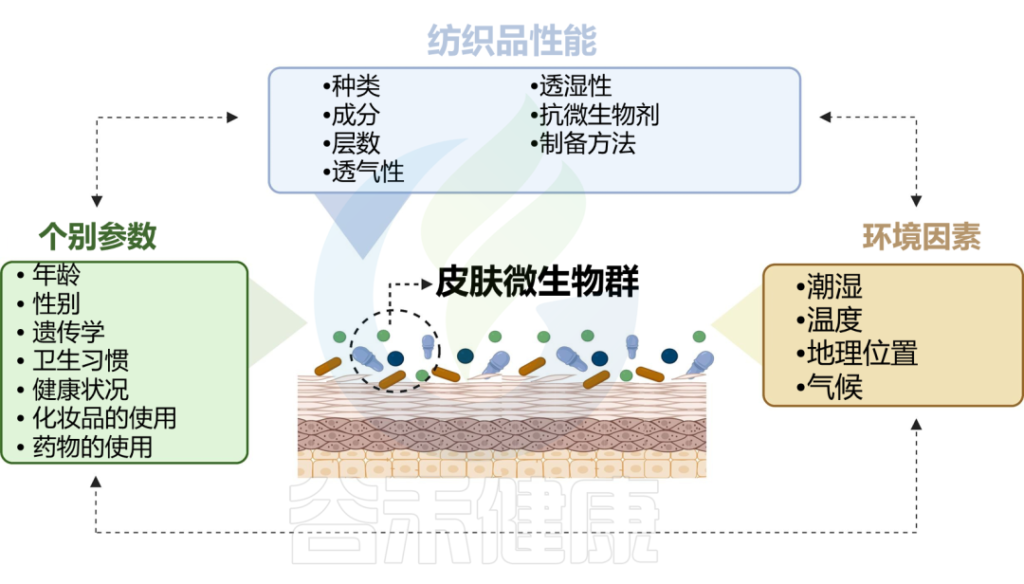
doi.org/10.1016/j.ejpb.2023.05.004
纺织纤维的性质可以直接影响微生物的附着、生长和定植
葡萄球菌属在几乎所有纺织纤维中显示出显着的固定性。Staphylococcus hominis对棉花的亲和力较高,在粘胶纤维和羊毛中不生长。
羊毛促进了许多菌群生长,包括表皮葡萄球菌、Enhydrobacter、角质杆菌、微球菌属。
聚酯为角质杆菌、Enhydrbecter、微球菌属提供了最大的生长环境。
棒状杆菌属无法在棉花、丙烯酸、羊毛、粘胶、尼龙、羊毛和聚酯上进行竞争,这解释了只有少量棒状杆菌属才能从破旧的衣服中分离出来。
合成纤维由于其疏水性和较差的吸附能力,通常抵抗微生物定植。
天然纤维更容易受到微生物定植的影响,因为它们具有高保湿性能,并且它们的聚合物键更容易被微生物酶获取。天然纤维可以以碳水化合物或蛋白质的形式为微生物提供营养和能量来源,支持微生物生长和定植。
与棉花相比,亚麻纺织品对金黄色葡萄球菌和表皮葡萄球菌表现出强烈的抑制作用,同时对角质形成细胞产生细胞毒性。
▼
化妆品旨在改善皮肤,减缓衰老过程。这些产品可能有助于皮肤微生物组的多样化,尤其是当定期或长期使用时。
化妆品中含有的活性成分可能有利于或抑制某些微生物的生长
N-乙酰氨基葡萄糖是刺激皮肤微生物群的化合物之一,它是透明质酸的前体,常见于护肤品中。
保湿产品可以降低皮肤水分流失的强度,并可以增加皮肤微生物群多样性(α多样性高是健康皮肤微生物群的标志),同时减少皮肤细胞剥落。它们的脂质化合物促进亲脂性细菌的生长,如葡萄球菌和角质杆菌。另一方面,皮肤水合水平的提高会降低皮脂含量,并可能减少角质杆菌数量。
化妆品成分的作用持续数周,个体的反应可能差异很大。不合适的化妆品或不合适的应用会减少皮肤微生物组的多样性,从而对其产生负面影响,导致生态失调。洗发水或面霜等化妆品也可能会导致感染,有时会导致严重的健康后果,尤其是在儿童或免疫力下降的人群中使用。
内 在 因 素
▼
皮肤表面呈微酸性(pH值5.6左右)且干燥,但温度比体内低。
表皮细胞自身脱落机制影响菌群组成
表皮外层不断释放角质化皮肤细胞,导致皮肤每四个星期自我更新一次。每小时有 500-3000 个细胞从1cm2的皮肤脱落,这意味着一个成年人每小时释放 600,000-100 万个或更多细胞。由于约 10% 的脱落细胞含有细菌,这种机制可能会显着影响微生物组的组成。
皮肤的厚度、表面褶皱的深度和位置,毛囊和腺体的密度都是影响宿主微生物群的关键因素。腺体释放的分泌物以不同的方式影响微生物,创造刺激或抑制微生物发育的条件。
皮脂腺:确保专性和兼性厌氧菌的最佳环境。这些腺体分泌的皮脂在皮肤上形成保湿、疏水的保护层,并且是微生物使用的脂质的来源。这些脂质水解产生的游离脂肪酸有利于细菌粘附到腺体表面并降低皮肤pH值,抑制金黄色葡萄球菌和化脓性链球菌等病原体的生长。
水分含量:潮湿的区域为许多微生物创造了有利的条件,如棒状杆菌属、葡萄球菌属等。相对干燥且温度波动较大的皮肤部位主要含有变形菌、拟杆菌、放线菌等。微生物的数量随着深层皮肤层中营养物质和水分含量的增加而增加。
▼
男性和女性微生物群之间物种组成的差异是由皮肤的性别特异性特性造成的,即皮肤厚度、毛发、汗液和皮脂腺的数量。女性多样性高于男性。更薄的皮肤、更低的 pH 值和更少的出汗量会导致更多的多样性。
对手部表面的微生物进行的一项研究表明,女性的物种多样性高于男性。在女性手上,肠杆菌和乳杆菌科的数量显著较高(300-400%),而在男性中,观察到更高浓度的角质杆菌和棒状杆菌。
关于不同年龄皮肤菌群构成不同,在前面第一章节已经详细阐述。
▼
在形成皮肤微生物组的遗传因素中,种族是次要的,但也有一定影响。最主要的是不同生活方式的差异。非洲和拉丁美洲男性头皮和腋下的Cutibacterium数量低于其他种族(高加索、非洲裔、东亚和南亚)。中国人皮肤微生物组与其他人群存在差异,比如Enhydrobacter在中国人的皮肤上较为常见。
▼
抑制细菌和减少炎症病变
口服米诺环素(用于治疗痤疮)降低了Cutibacterium、棒状杆菌、普雷沃氏菌、乳酸杆菌和卟啉单胞菌的丰度。
多西环素显著减少痤疮梭菌的数量(治疗6周后为1.96倍)。Snodgrassella alvi的数量也减少了(3.85倍)。另一方面,观察到Cutibacterium granulosum的数量显著增加(4.46倍)。
大环内酯类、四环素类和克林霉素用于治疗痤疮。用利美环素进行的脸颊皮肤治疗减少了角质杆菌的存在,并增加了链球菌、葡萄球菌、微球菌和棒状杆菌的数量。反过来,二甲胺四环素导致微生物组紊乱。
虽然氟喹诺酮类药物(培氟沙星)和大环内酯类药物(红霉素)显著减少了痤疮梭菌的数量,但只有纳氟沙星对凝固酶阴性葡萄球菌表现出抑制活性。
导致出现抗生素耐药性物种
例如痤疮梭菌和表皮葡萄球菌。大环内酯类药物的长期治疗痤疮,增加了痤疮梭菌分离株的数量,但对大环内酯的影响的敏感性降低。
据估计,红霉素和阿奇霉素耐药菌株的比例可能分别达到50%,甚至100%。从感染皮肤分离的G+细菌中,77.5%对青霉素耐药,28%对甲氧西林耐药。在所有测试的菌株中,31.9%对三种以上的抗生素不敏感。
儿童皮肤分析结果显示,36.4%的从皮肤表面分离的金黄色葡萄球菌菌株对甲氧西林有耐药性。此外,耐甲氧西林葡萄球菌(MRSA)是医院感染最常见的原因之一。
▼
肠道内表面和皮肤表面有一些有趣的相似之处:两者都被上皮细胞覆盖,上皮细胞维持着体内与外部环境之间的重要联系,充当第一道防线,在抵御外部病原体、调节免疫反应和抑制分解代谢物方面发挥着重要作用。
肠道和皮肤组织是宿主原核和真核共生微生物的两个主要生态位,因为它们的高细胞周转率决定了定植微生物组的低粘附和感染。
皮肤健康与肠道屏障的完整性有关。一些饮食代谢物可以直接吸收到皮肤中,其他通过肠道微生物代谢来做到这一点,这两者都可能有助于皮肤健康。
由于肠道通透性增加,肠道菌群或其代谢产物可能从肠道迁移到循环系统中并在皮肤中积聚,这可能会损害皮肤屏障并使其容易发炎。
肠道微生物群的变化还可能引发系统性炎症和异常免疫反应,从而破坏皮肤健康。
皮肤或肠道微生物群失调与免疫应答改变密切相关,与多种皮肤病相关,包括特应性皮炎、牛皮癣、寻常痤疮、甚至皮肤癌等,这在下一章节会详细讲述。
饮食强烈影响肠道微生物组的组成,影响代谢和免疫功能,间接影响皮肤健康。关于如何通过饮食调整在最后章节会讲到。
以上是影响皮肤微生物群的外在和内在因素,那么皮肤微生物群会如何影响人体健康呢?我们来看下一章节。
这里我们分为两个部分来阐述:
皮肤微生物群直接影响皮肤健康
我们知道,皮肤是由角质形成细胞的分层角质化上皮组成,这些上皮经历终末分化。这些物理结构通过增强屏障的化学和免疫学特征得到进一步强化。
皮肤微生物群影响皮肤屏障的各个方面,同时也直接与表面遇到的共生微生物和病原微生物相互作用。
皮肤微生物群介导多种屏障功能
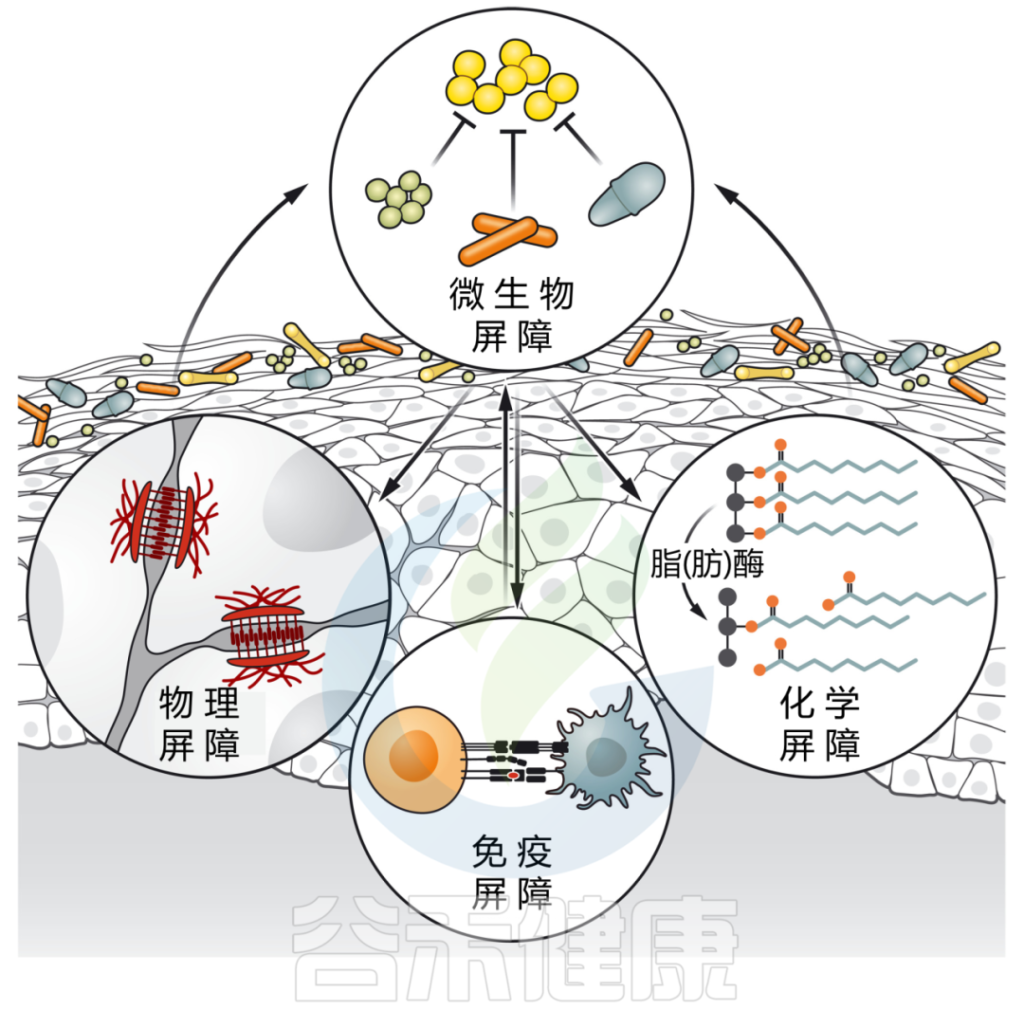
DOI: 10.1126/science.abo0693
微生物群强化皮肤屏障的多个方面:
皮肤微生物通过各种定殖抗性机制,包括资源排斥、直接抑制和/或干扰,形成对抗环境的第一道屏障。
皮肤微生物群也有助于物理皮肤屏障的分化和上皮化。微生物通过产生脂肪酶来增强皮肤的化学屏障,脂肪酶将皮脂甘油三酯消化为游离脂肪酸,从而增强皮肤的酸性,并限制瞬时和致病物种的定植。
最后,微生物刺激先天和适应性免疫防御,如抗菌肽的释放、新生儿耐受性的诱导和保护性免疫的发展。
接下来我们讨论微生物群到底如何与皮肤屏障的微生物、化学以及先天和适应性免疫成分相互作用。
▼
皮肤微生物群本身是抵御外来微生物和病原微生物入侵、定植和感染的屏障。
——直接竞争关系
皮肤微生物争夺资源,并进化出直接对抗对手的机制。
多种CoNS物种(凝固酶阴性葡萄球菌),如人葡萄球菌,产生具有独特化学性质的抗生素,并抑制皮肤病原体金黄色葡萄球菌。
其他物种,如头葡萄球菌,通过干扰金黄色葡萄球菌毒力所需的辅助基因调节因子(agr)群体感应途径来拮抗金黄色葡萄菌。
——拮抗机制与宿主抗菌反应协同作用
人型葡萄球菌和表皮葡萄球菌,可以产生共生衍生的AMPs,其发挥选择性抗菌活性,并与宿主衍生的AMPs协同作用,以抑制皮肤病原体的存活。
痤疮角质杆菌与产生硫肽抗生素角质霉素的特定菌株竞争,以维持其在人类毛皮脂腺单元中的生态位,从而限制金黄色葡萄球菌的定植。
皮肤微生物组内微生物之间的相互作用,可以驱动整体微生物群结构。
主要皮肤菌群产生的抑制其他微生物群,和/或潜在病原体的突出和最近鉴定的抗菌分子汇总在下表,分子作用机制也包括在内。
皮肤上关键的微生物与微生物相互作用

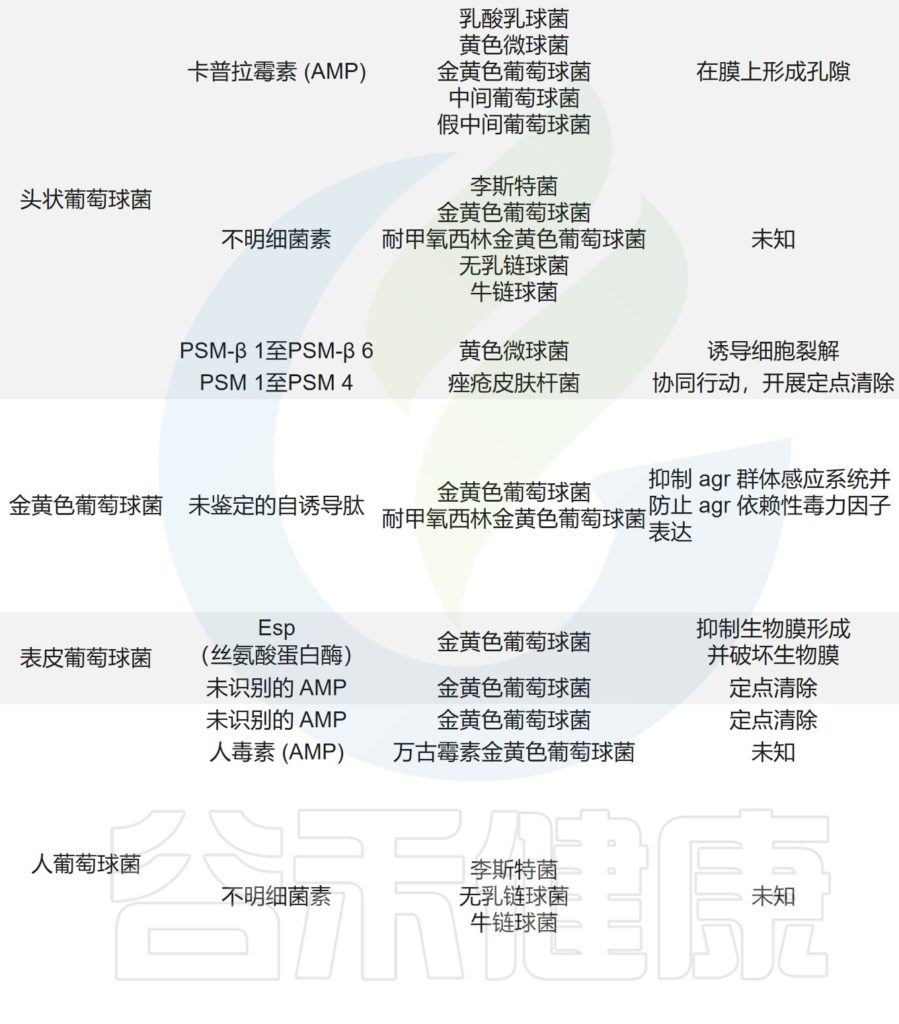
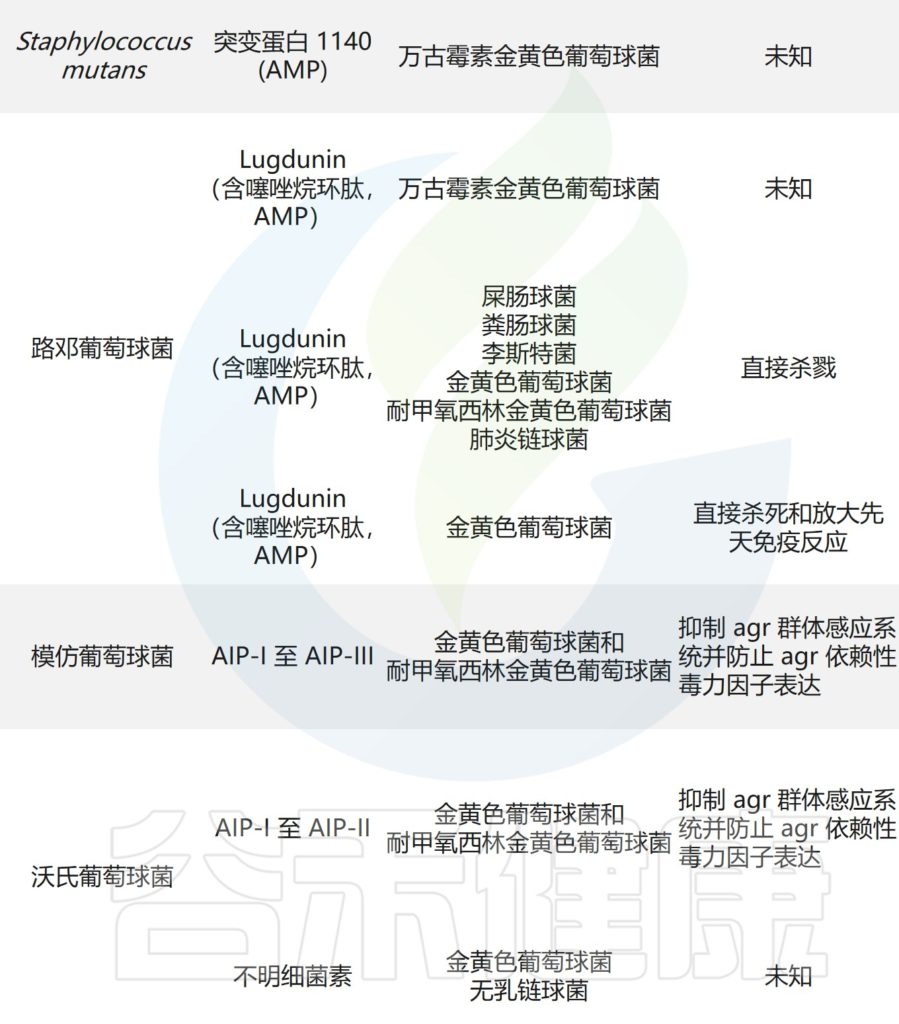
doi.org/10.1042/BST20220216
▼
角质细胞经历严格调控的终末分化程序,形成角质层,该过程由微生物群介导。微生物群通过角质形成细胞芳香烃受体(AHR)的信号传导促进分化和上皮完整性;还分泌鞘磷脂酶,将层状脂质加工成神经酰胺,神经酰胺是角质层的关键成分。
▼
酸性皮肤表面还产生了限制细菌定植的化学环境。痤疮角质杆菌和棒状杆菌都分泌脂肪酶,水解皮脂中甘油三酯中的游离脂肪酸。游离脂肪酸通过直接抑制细菌和刺激人β-防御素2(hBD-2)的表达,进一步增强皮肤免疫力。痤疮角质杆菌也直接与游离脂肪酸结合,这表明游离脂肪酸的存在促进了痤疮角质杆菌的定植。
▼
微生物可以刺激多种与先天免疫反应有关的反应,通常取决于代谢和炎症环境。例如,念珠菌的菌丝和酵母形式在皮肤中刺激不同的免疫反应。S. epidermidis在皮肤中引起的T细胞反应,需要菌体表面特定糖蛋白与宿主先天免疫细胞上的C型凝集素相互作用。
氧气的可用性也会影响皮肤表面宿主与微生物的相互作用。微氧耐性细菌痤疮角质杆菌生成短链脂肪酸,抑制组蛋白去乙酰化酶,后者可作为免疫系统的表观遗传调节因子,从而刺激炎症。
注:在皮肤中,短链脂肪酸具有促炎作用,这点和肠道中不同。SCFAs通过抑制HDAC8和HDAC9以及通过TLR信号通路刺激炎症。
皮肤微生物还通过刺激宿主产生抗菌肽和蛋白来增强皮肤免疫力,这些抗菌肽和蛋白起到天然抗生素的作用。
皮肤微生物群落还在创伤修复过程中协调先天免疫反应。在皮肤中的共生微生物群落会引发I型干扰素(IFN)反应。作为对微生物信号的反应,中性粒细胞会表达CXCL10,吸引活化浆细胞样树突状细胞(pDC)到损伤部位。pDC会产生I型干扰素,通过刺激成纤维细胞和巨噬细胞增长因子反应来加速创伤修复。
实际上,抗原呈递细胞向皮肤的募集是微生物群依赖性的。微生物通过需要IL-1R-MYD88信号传导的过程,在伤口修复和毛囊新生中增强皮肤再生。
▼
皮肤是各种适应性免疫细胞的家园,其中包括大量的常驻记忆T细胞,随时准备对各种环境刺激做出反应,包括致病微生物和共生微生物。
在婴儿早期,暴露于皮肤共生表皮葡萄球菌介导调节性T细胞(Tregs)流入皮肤。这种Treg迁移波与毛囊发育同时发生,需要毛囊角质形成细胞产生趋化因子。Tregs,以及皮肤中的许多其他免疫细胞亚群,最终位于毛囊附近,对在这个发育窗口期间检测到的微生物抗原具有特异性。
在一个平行的过程中,粘膜相关不变T细胞(MAIT)是在婴儿期在类似的时间限制的发育窗口中获得的。MAIT细胞在无菌小鼠中是不存在的,它们的发育需要维生素B2代谢产物,而这些代谢产物仅由细菌和真菌产生,而不是哺乳动物细胞。
在胸腺中,暴露于5-(2-oxopropylideneamino)-6-d-ribitylaminouracil(一种维生素B2的细菌代谢产物,从粘膜部位运输到胸腺),介导MAIT细胞扩增并靶向皮肤和粘膜部位。
微生物细胞表面分子也可以作为宿主的信号。大多数棒状杆菌的细胞膜中含有霉菌酸。棒状杆菌属霉菌酸在稳定状态下可以以IL-23依赖的方式促进γδT细胞的积累。然而,这种相互作用取决于环境,因为高脂肪饮食反而会促进皮肤炎症。因此,微生物暴露时存在的炎症环境影响皮肤内的免疫反应。
这些发现突出了微生物在皮肤免疫细胞的募集和刺激中发挥的关键作用。
以上是皮肤微生物群从物理、化学、免疫等多角度与皮肤之间的关联,如果说上述对局部组织微环境的相互作用,那么接下来我们从更系统的角度来看,皮肤微生物群通过与其他器官的交流,对全身健康产生的影响。
皮肤微生物群通过影响其他器官间接影响
越来越多的证据表明,皮肤损伤和致敏会影响其他屏障部位,如肠道、肺部等。
▼
皮肤和肠道之间存在双向沟通。
为什么浅表皮肤损伤会引起肠道炎症?
浅表皮肤损伤会导致角质形成细胞全身释放 IL-33。IL-33 与 IL-25 协同作用,触发肠道内 ILC2 的激活,产生 IL-4。这反过来又刺激肠道中肥大细胞的扩张,在那里它们准备对食物过敏原做出反应并介导过敏反应。
在模拟炎症性肠病的小鼠模型中,皮肤受伤还会加剧肠道炎症。
皮肤和肠道之间的相互作用取决于损伤期间真皮中产生的透明质酸片段的产生,这些片段刺激肠道成纤维细胞,通过反应性脂肪生成的过程分化为促炎脂肪细胞。这些反应性脂肪细胞通过产生 AMP 和其他炎症介质来传播肠道炎症。
肠道微生物群变化也会影响皮肤炎症
在这两种情况下,肠道免疫网络的激活都会影响皮肤中炎症信号的振幅。
因此,肠道微生物组的改变可能会影响皮肤免疫力。
研究表明,饮食对肠道微生物组的影响,尤其是膳食纤维,对系统免疫有重要影响。皮肤先天免疫反应也与肠道有关,肠道中保护细菌性皮肤感染的AMPs的充分表达,取决于饮食中的维生素A。这些发现加强了我们对饮食在宿主免疫发展中重要性的分子理解。
▼
流行病学证据表明,许多患者经历了“特应性进军”,首先出现特异性皮炎,随后发展为过敏性鼻炎、食物过敏、哮喘。它们的先后出现意味着存在什么样的关联?
皮肤微生物群失调和金黄色葡萄球菌定植增加,与特应性皮炎的发作有关。
表皮暴露于金黄色葡萄球菌刺激角质形成细胞产生IL-36,从而提高血清IgE水平。
而缺乏IL-36受体的小鼠对金黄色葡萄球菌的反应不会产生升高的IgE,并且也可以免受过敏原特异性肺部炎症的影响。这些发现支持了皮肤暴露于微生物病原体作为全身炎症的起始。
微生物组的变化与哮喘、过敏性鼻炎、特应性皮炎和食物过敏的风险有关
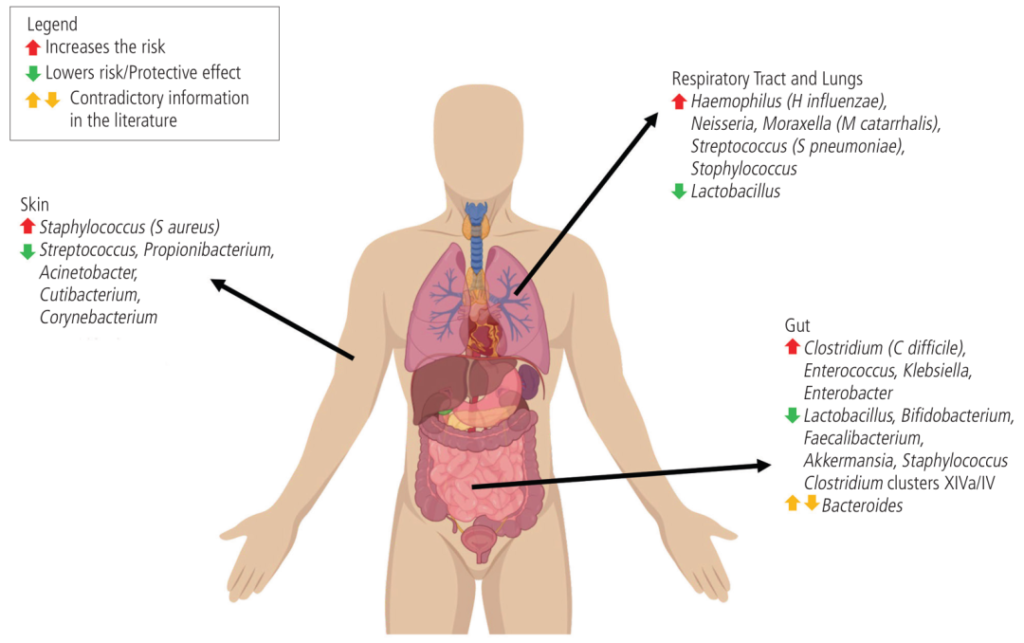
doi: 10.18176/jiaci.0852
在气道中,卡他莫拉克菌(Moraxella catarrhalis)、流感嗜血杆菌(Haemophilus influenzae)和肺炎链球菌水平较高与婴儿哮喘有关。
肠道艰难梭菌的比例高于双歧杆菌,这与更高的食物过敏率有关。
▼
神经免疫相互作用中的皮肤病原体
细菌可以直接激活皮肤中的感觉神经元,并通过产生造孔毒素引起疼痛。菌株水平的变化驱动着可变的反应,这取决于特定毒素和群体感应系统的存在。
关于群体感应,详见:
真菌(白色念珠菌)也可以直接激活皮肤中的感觉神经元。γδT细胞免疫需要刺激才能通过释放神经肽CGRP来控制皮肤念珠菌感染。
相反,引起坏死性筋膜炎的病原体化脓性链球菌,通过分泌链球菌溶血素S直接激活伤害感受器神经元,进而促进神经肽CGRP的释放并抑制化脓性链球菌的杀伤。在这种情况下,CGRP拮抗作用可防止坏死性感染。
皮肤与其他器官系统的交互作用是由微生物群介导的
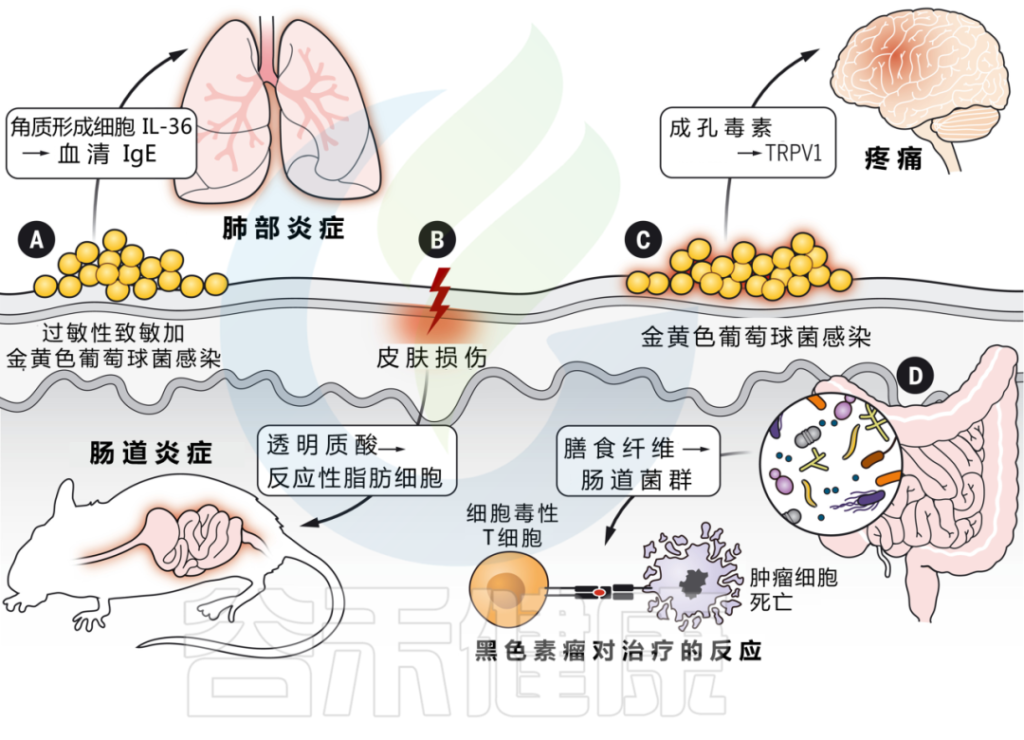
DOI: 10.1126/science.abo0693
微生物与宿主相互作用和皮肤疾病
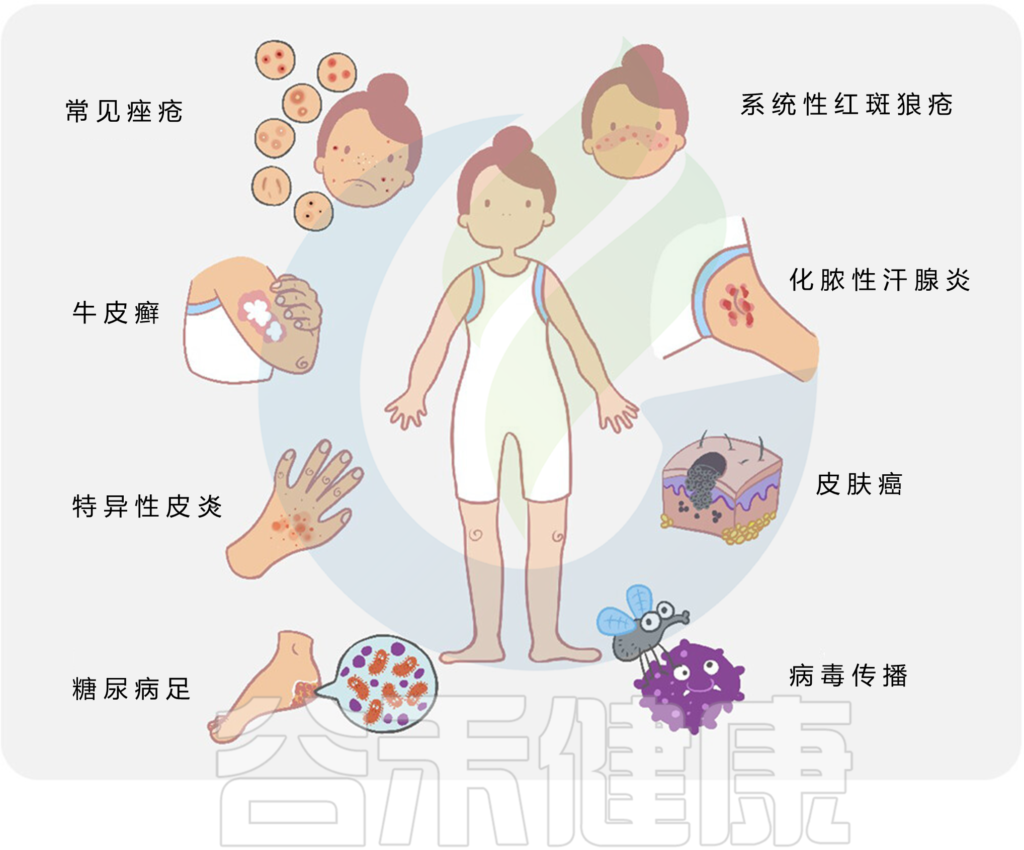
doi.org/10.1002/mlf2.12064
▼
痤疮患者,特别是那些症状严重的患者,表现出α多样性增加,四种革兰氏阴性细菌(即粪杆菌属、克雷伯氏菌属、臭杆菌属和拟杆菌属)的比例更高。
痤疮角质杆菌(C. acnes)的过度生长与痤疮发病机制有着长期的关联。宏基因组分析表明,痤疮患者中痤疮丙酸杆菌的菌株结构与健康个体不同,IV 型和 V 型菌株在受痤疮影响的皮肤中特别普遍。
痤疮丙酸杆菌通过多种不同方式参与痤疮发病机制的调节,它参与:
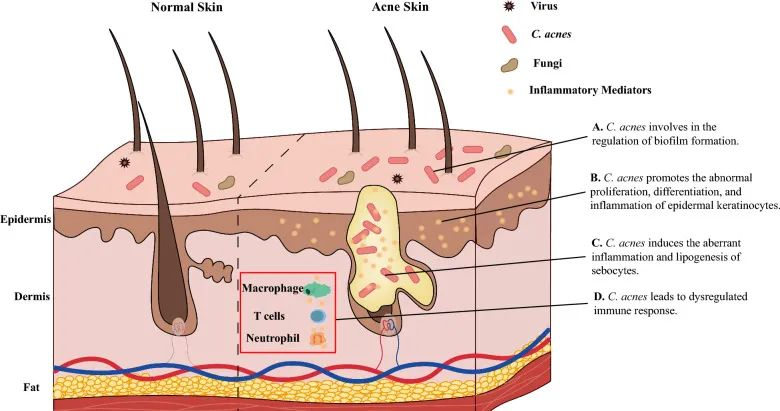
doi: 10.1186/s13578-023-01072-w
痤疮丙酸杆菌和表皮葡萄球菌在痤疮以及炎症后色素沉着过度中具有病理生理作用。
肠道微生物群在皮肤炎症和情绪之间起着中介作用
痤疮和胃肠道功能障碍之间的联系可能起源于大脑。支持这一假设的是压力引起的痤疮加重。实验动物和人类研究表明,压力会损害正常的肠道菌群,尤其是乳酸杆菌和双歧杆菌。心理应激源导致肠道微生物群产生神经递质(即乙酰胆碱、血清素、去甲肾上腺素),这些神经递质穿过肠粘膜进入血流,导致全身炎症。
痤疮中肠-脑-皮肤轴的拟议模型
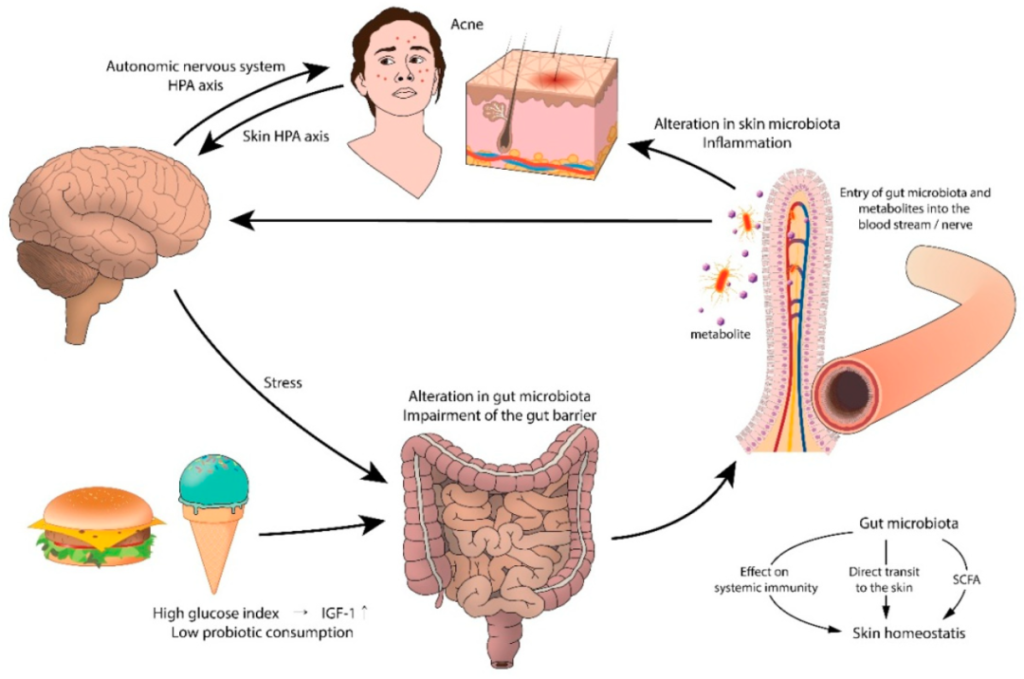
doi.org/10.3390/jcm8070987
西方饮食包括乳制品、精制碳水化合物、巧克力、饱和脂肪等,这些物质可能通过激活营养来源的代谢信号来加重痤疮。高脂肪饮食会降低肠道菌群水平,增加脂多糖的浓度,通过损害结肠上皮完整性和屏障功能、降低粘液层厚度和增加促炎细胞因子的分泌来引起全身炎症。
扩展阅读:
▼
皮肤干燥、斑块发痒和反复出现的湿疹是特异性皮炎的标志。
特异性皮炎引起的皮肤耀斑通常与更多的金黄色葡萄球菌丰度有关,金黄色葡萄球菌定殖的增加与CoNS数量的减少相关,CoNS本来会产生抗菌蛋白,它在特异性皮炎患者中数量少。
金黄色葡萄球菌在病变的真皮中更为普遍,这表明在剥皮过程中更容易接触到更深的皮肤层。
是什么引起金黄色葡萄球菌定植增加呢?
表皮葡萄球菌、痤疮杆菌和棒状杆菌属的丰度降低,它们通常对金黄色葡萄菌的入侵起作用。
特异性皮炎的菌群多样性低。共生细菌数量减少而导致的共生产生的AMPs的缺失,抵御病原体如金黄色葡萄球菌的能力下降,金黄色葡萄球菌定植增加。
与特异性皮炎相关的皮肤屏障缺陷会损害层状膜的完整性,改变皮肤的微生物群,并可能使金黄色葡萄球菌等有害细菌滋生。厌氧微生物的缺乏可能会降低关键的皮肤屏障活性,并促进潜在的感染。
金黄色葡萄球菌分泌毒力因子
金黄色葡萄球菌分泌几种毒力因子,包括纤连蛋白结合蛋白1(FBP1),α-和δ-溶血素,酚溶性调节素(psm)的蛋白家族等,所有这些毒素都会导致更高的炎症反应和更严重的症状。
皮肤稳态取决于复杂的宿主-微生物相互作用,包括金黄色葡萄球菌和特异性皮炎宿主细胞之间的相互作用,微生态失调会导致疾病的发展。
特异性皮炎的其他微生物群变化包括痤疮角质杆菌、棒状杆菌、Dermacoccus、微球菌、CoNS减少,链球菌和一些马拉色菌属增加。这些微生物变化似乎是暂时的,在特异性皮炎发作之前和期间,群落多样性丧失,金黄色葡萄球菌优势更大,在炎症消退后逐渐恢复到基线。
潜在益生菌治疗和预防AD的临床试验

doi.org/10.1016/j.phymed.2023.154824
▼
一般来说,皮肤破裂会导致炎症级联活动;然而,这种免疫反应在糖尿病皮肤中被破坏,也就是无法有效引起免疫反应。微生物组的改变可能会加剧疾病的严重程度。
糖尿病皮肤的菌群特征
糖尿病早期患者的皮肤细菌微生物群与健康人的非常相似。随着疾病的恶化,物种多样性和丰度发生动态变化。总的来说,糖尿病足的皮肤细菌微生物群的多样性低于健康足。因此,不太常见的微生物种类的变化,其中大多数只在健康的足部皮肤中发现,可以用来预测是否患有糖尿病。
糖尿病足皮肤中葡萄球菌的含量通常较低,金黄色葡萄球菌的比例较高。金黄色葡萄球菌的大量存在破坏皮肤微生物群平衡,可能会导致炎症变化,并增加皮肤感染的风险。
慢性溃疡相关菌群
铜绿假单胞菌和厌氧菌通常与深部慢性溃疡有关,但金黄色葡萄球菌通常与急性浅部溃疡有关。比较有慢性感染和没有慢性感染的糖尿病患者的微生物组的研究可以提供有关诊断标志物的信息,这些标志物可以用作发展为慢性损伤的可能性的指标。
▼
牛皮癣患者由于慢性炎症性皮肤病而出现中度至持续性皮肤斑块。许多遗传和环境变量之间的复杂组合导致皮肤过度活跃的炎症反应是病因。
牛皮癣皮肤菌群特征
牛皮癣患者皮肤样本在α、β多样性明显低于正常皮肤。下列菌群相对丰度和分类性能显著下降:
棒状杆菌在牛皮癣的发病机制中发挥重要作用
大量的研究表明,棒状杆菌属丰度上升,棒状杆菌有可能干扰干扰素信号系统,这可能导致皮肤微生物组的微生态失调。
乳制品和糖类的摄入是牛皮癣最常见的诱因之一,而肉类和鸡蛋则被列为次要的常见诱因之一。
关于牛皮癣与肠道菌群之间也存在很多相关性,此处不展开阐述,详见:
▼
皮肤黑色素瘤
黑色素瘤和正常皮肤样本之间的微生物组成和多样性存在显著差异。黑色素瘤样本中的梭杆菌和Trueperella水平较高。
棒状杆菌属与疾病严重程度相关,棒状杆菌水平与IL-17之间存在关系,IL-17可以通过增加IL-6和信号转导器和STAT-3来促进黑色素瘤细胞增殖。
相反,痤疮角质杆菌的细菌上清液增加了黑色素细胞的凋亡。
角质细胞皮肤癌
以皮肤微生物群为代表的生物屏障通过分泌抗微生物肽(AMP)(如组织蛋白酶LL-37和人β-防御素)来抑制病理生物和病原体入侵,从而与角质形成细胞和免疫细胞产生串扰。
研究人员认为金黄色葡萄球菌和鳞状细胞癌之间的联系不是偶然的,皮肤溃疡是有利于外源性搪塞/感染的致病过程的结果。金黄色葡萄球菌也可能参与鳞状细胞癌的发病机制,引起慢性局部炎症,涉及不同的致瘤阶段,包括促进生存、增殖、细胞转化、侵袭、血管生成、转移。
葡萄球菌毒素-α决定了参与炎症过程的局部细胞的分泌,进而导致活化B细胞的NF-Kβ的激活,从而增加不同细胞因子和趋化因子的表达,包括IL-1β、IL-6和IL-12。
其他因素如紫外线辐射(尤其是UVB)也是皮肤癌发生的主要危险因素之一。紫外线照射会改变皮肤微生物群,导致大量形成活性氧、细胞凋亡和炎症,与皮肤癌相关。
总的来说,许多常见的皮肤病,如痤疮、特异性皮炎、牛皮癣、皮肤癌等,都与皮肤微生物群的变化有关。
皮肤病中的关键微生物发现如下:

编辑
doi.org/10.1002/mlf2.12064
●
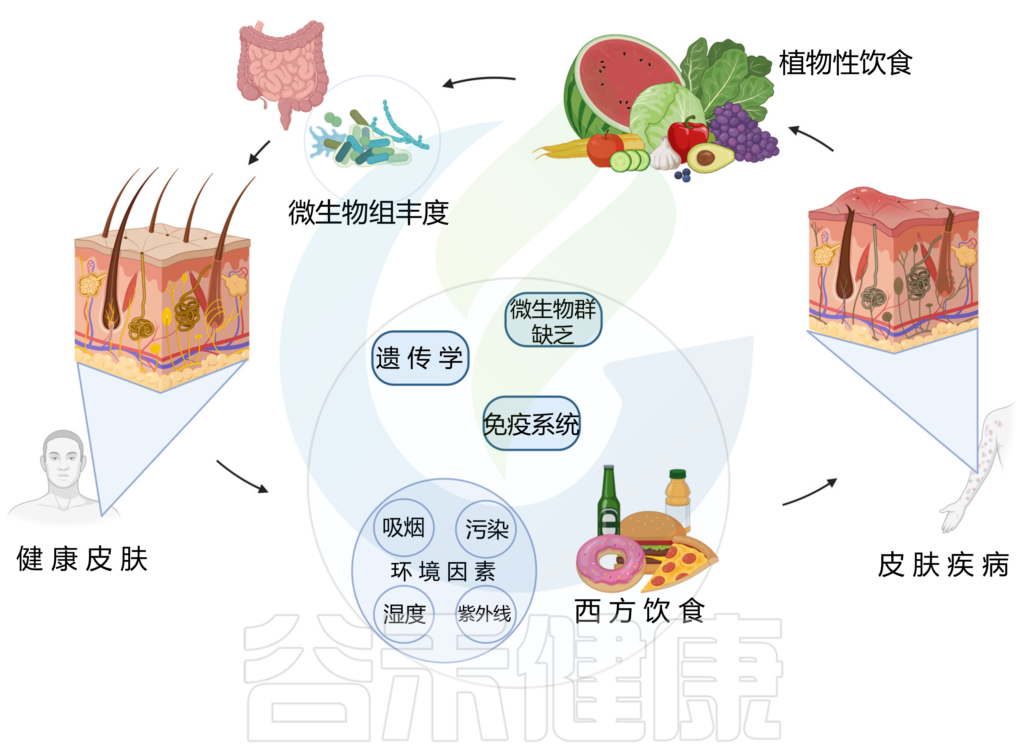
饮食对肠道微生物群的影响较大,皮肤和肠道微生物群是内在相关的,由宿主免疫系统介导。因此,肠道和皮肤可以通过饮食、微生物代谢产物、神经内分泌途径和中枢神经系统等途径相互作用,也就是说,饮食对皮肤也会产生较大影响。
饮食结构
西方饮食已被证明会破坏微生物组并导致皮肤病,从而对皮肤健康产生负面影响。相反,植物性饮食与更健康的皮肤有关。
以植物为基础的饮食是一种由多种蔬菜、水果、豆类、扁豆、豆类、坚果、种子、真菌和全谷物组成的饮食模式,并且限制或不摄入动物产品、加工食品或糖果。
这种饮食的饱和脂肪、反式脂肪和花生四烯酸含量较低,而抗氧化剂和 omega-3 脂肪酸含量较高,再加上其直接治疗作用,可减少炎症和皮肤症状。
植物性饮食与皮肤健康/疾病之间的关联
doi.org/10.3390/nu15132842
多项研究发现,植物性饮食对缓解牛皮癣、特异性皮炎、痤疮等皮肤问题有益。


doi.org/10.3390/nu15132842
食 物
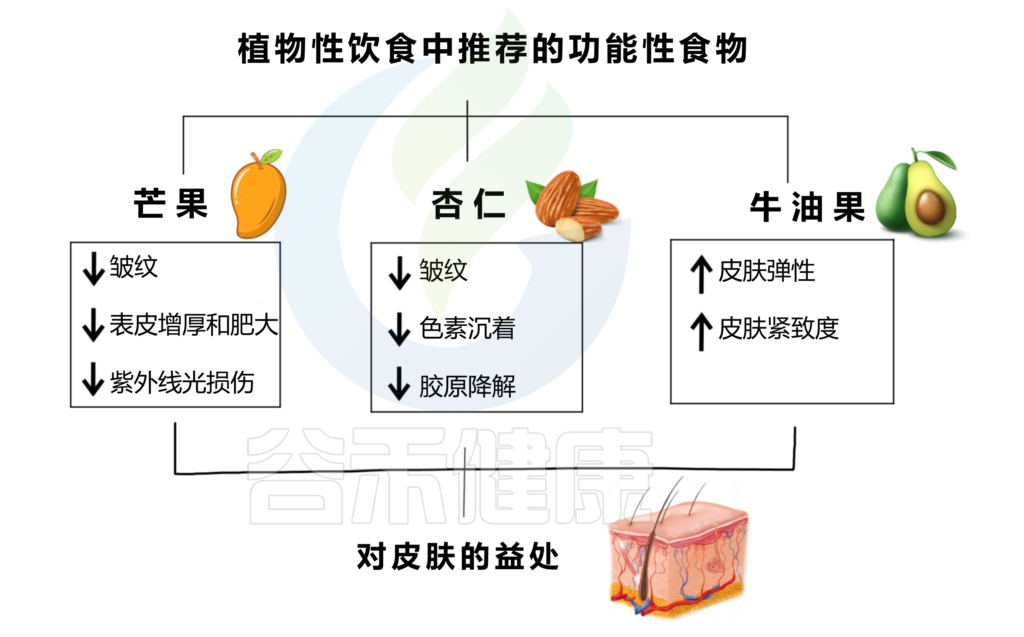
植物性功能性食品增强皮肤健康,减少皮肤老化迹象,并改善整体外观。下图是芒果、杏仁、牛油果及其对皮肤健康的积极影响。
编辑
doi.org/10.3390/nu15132842
芒果能够减少皱纹、表皮变薄和肥厚,防止 UVB 损伤。无论是果肉还是果汁,芒果酚酸的抗氧化特性和生物利用度都会得到保留,而果汁的呈现可能会增强其特性。从芒果干中提取的芒果提取物也可以减少UVB辐射引起的皱纹的形成。
杏仁富含α-生育酚(或维生素E)、脂肪酸、多酚,因此是一种具有抗氧化特性的食物,可以减少皱纹、色素沉着和胶原蛋白降解。
牛油果含有类胡萝卜素、单不饱和脂肪酸、酚类化合物,某些基因的表达,如胶原蛋白和弹性蛋白基因,在进行饮食调整后被诱导,因此可以促进皮肤弹性和紧致度的增加。
限制饮酒和甜食
酒精会使你的身体和皮肤脱水,这可能会使皮肤看起来更加干燥或有皱纹,许多含有酒精的混合饮料也富含糖,这都不利于皮肤健康。糖可能使胶原蛋白变硬,从而使皮肤老化,也可能带来炎症。
喝 水
对于每日饮水量较低的人(即那些本来就脱水的人)来说,增加饮水量对皮肤外观有积极影响,有助于维持皮肤水合水平。同时,尝试在食物中多加入黄瓜、芹菜、西葫芦、西瓜、草莓和花椰菜等,也可以适当补水。
●
适当的清洁和保湿可用于维持皮肤的生理pH值。据报道,早在1995年,与使用 pH8 的普通肥皂相比,使用酸性合成皂(pH 5.5-5.6)可显著减少非炎症和炎症病变。
从那时起,pH 值的变化通过皮肤屏障的完整性与痤疮的发病机制联系起来,建议使用 pH 值约为 5.5 的皮肤清洁剂。
为什么洗脸很重要?
脸每天面对风吹、紫外线、化妆/护肤品、屏幕等刺激,会积聚污垢、油脂和其他碎屑,如果不及时清除,可能会导致刺激和其他皮肤问题。
合适的洗脸方式
注意:
应该多久洗一次脸?
没有既定的指导方针,一般来说,最好每天洗两次脸。
如果皮肤干燥或敏感,可以在晚上用清洁去除污垢,然后在早上用温水冲洗脸。
即使当天不化妆或者不出门,污垢、油脂和其他不需要的碎屑仍然会在一天中积聚在皮肤上,因此最好在睡觉之前洗脸。
如果刚在健身房、参加高温瑜伽课或在户外徒步旅行,并且出汗较多,最好马上洗脸。
如果存在敏感问题或其他特殊的皮肤状况,请与医生沟通。
●
痤疮是一种慢性炎症性皮肤病,对于痤疮,护肤品有多种作用机制,包括:
1) 保护和改善皮肤屏障
2) 保护皮肤微生物组
3) 维持健康的皮肤 pH 值
4) 抵御紫外线伤害
保护皮肤屏障是皮肤化妆品改善痤疮管理的重要机制,临床上,屏障功能障碍表现为皮肤干燥、刺痛/烧灼/刺痛、紧绷、疼痛或刺激性皮炎等形式。这些被认为与经表皮失水 (TEWL) 有关,并且可以通过使用保湿剂至少部分缓解。
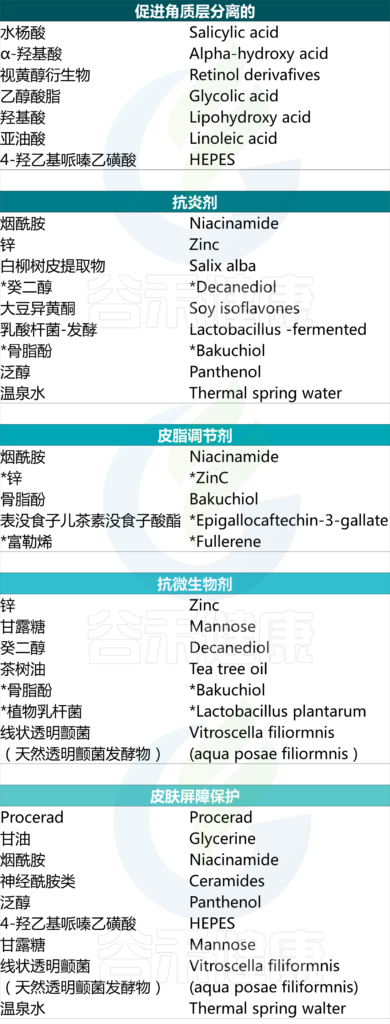
特别适用于痤疮的成分包括烟酰胺、视黄醇衍生物、水杨酸、神经酰胺、甘油、温泉水、泛醇等。
护肤品中可能存在的活性成分及其针对性作用
Kurokawa I, et al., Dermatol Ther (Heidelb). 2023
而皮肤微生物组的核心作用与表皮屏障功能一起,为优化护肤提供了强有力的支持。可以通过尝试恢复微生物组的多样性并通过下调先天免疫来抑制炎症。
总的来说,现有文献结果的总体趋势表明,护肤品可以改善整体皮肤健康,减少痤疮皮损,在处方治疗后维持痤疮清除,并且可能对减少表面皮肤油腻具有有益作用。
然而需要进一步研究才能更好地理解这一作用。在此情况下可能有益的成分包括但不限于:乙醇酸、LHA、亚油酸、烟酰胺、锌、吡罗克酮乙醇胺、procerad、Vitroscella filiformis.
注意:
痤疮的治疗管理需谨慎,一些基于类维生素A的治疗方案,可能会加剧皮肤干燥和刺激,这不仅可能导致屏障功能改变,而且还会增加深色皮肤患者继发性妊娠高血压综合征的可能性。
●
温泉水已被证明对膜流动性、皮肤屏障修复、抗自由基、抗氧化、抗炎和免疫调节特性以及增殖活性和衰老和保湿过程的调节有影响。
温泉水的水微生物群中的生物活性化合物可以改善特应性皮炎或红斑痤疮等皮肤病,并改善瘙痒和干燥症;还能增加对紫外线的防护,强化屏障功能,维持皮肤防御良好的稳态,修复受损皮肤,促进伤口愈合,改善皮肤状况,减少皮肤色素不均匀,防止皮肤老化。
未来,护肤品研发人员、水文学家、温泉中心之间的合作,将推动该行业更好地了解温泉水的水生生物群落对皮肤病的作用,并考虑将这种水生生物群落的衍生物纳入皮肤病配方(以发酵罐、裂解物、提取物等形式)。
●
某些益生菌菌株及其代谢物可能带来许多益处,如:改善皮肤屏障功能、减少炎症、改善易长粉刺或易湿疹的皮肤、抗皮肤光老化等。因此,近年来益生菌已成为护肤品中的流行成分。
益生菌分为口服和外涂。
口服益生菌
肠道微生物群的变化可能会引发全身炎症和异常免疫反应,从而破坏皮肤健康。口服益生菌直接作用于肠道微生物群,帮助恢复肠道微生物群的稳态,这在皮肤稳态中发挥着至关重要的作用。
益生菌在皮肤光老化中的作用
含有约氏乳杆菌和营养类胡萝卜素的膳食补充剂的摄入对紫外线暴露的长期和反复影响有益处,并且对光老化更有针对性。
含有长双歧杆菌和低聚半乳糖的膳食补充剂由于其抗炎和抗氧化特性,保护皮肤免受UVB诱导的光老化。它们也提高了血清中短链脂肪酸和乙酸盐的水平,可以增加和激活依赖于组蛋白乙酰化的皮肤固有Treg。
口服植物乳杆菌HY7714通过抑制JNK/AP-1信号通路的激活,降低了UVB损伤细胞中过量的MMP-13转录水平和MMP-2和MMP-9的活性。
口服清酒乳杆菌可以通过阻断MAPK信号通路来抑制AP-1的表达,以增加真皮成纤维细胞中的胶原蛋白并延缓皮肤光老化。
益生菌通过多种途径对抗皮肤光老化的作用
编辑
doi: 10.2147/CCID.S388954
局部益生菌(外涂)
局部益生菌于1912年首次被提出作为皮肤疾病的治疗方法。
在特定条件下,益生菌可以持续存在并成功定殖皮肤:
虽然益生菌对皮肤健康有一定益处,但益生菌使用的安全性可能存在一些限制,主要针对免疫系统较弱的人群,如婴儿、孕妇、老年人等。需要进一步的研究来证明益生菌作为皮肤病治疗和护肤品的功效,作用机制以及主要是局部使用益生菌的安全性。
●
过氧化苯甲酰 (BPO) 治疗可以调节痤疮患者的皮肤微生物群,治疗后细菌种类的数量和多样性均减少,接近健康组。也有研究人员认为,BPO治疗虽然降低了GAGS评分并降低了微生物多样性,但它也损害了痤疮的表皮屏障,这可以被认为是一种副作用。
抗生素
针对痤疮丙酸杆菌的抗生素一直是痤疮治疗的支柱。其中,大环内酯类、克林霉素和四环素类药物的处方最为广泛。
红霉素、罗红霉素、克拉霉素和阿奇霉素是大环内酯类药物。常用于痤疮的四环素类药物是多西环素、四环素和米诺环素。
异维甲酸是一种全反式维甲酸原药,是严重顽固性痤疮患者的最终选择,它抑制皮脂生成,它可以使痤疮患者的C.acnes/TLR-2介导的先天免疫反应正常化,也就是说,异维甲酸会间接影响皮肤微生物。
其他,比如抗生素治疗会降低皮肤伤口中的细菌密度并改变细菌组成,其次是RegIIIγ表达的降低,这可能有助于延迟伤口修复。
●
几项研究表明,互惠共生对维持微生物物种之间的新陈代谢很重要。我们不仅需要关注微生物组的转移,还需要关注潜在的交叉喂养和共同居住。
有研究表明,整个原始皮肤微生物组从一个皮肤部位移植到另一个部位。研究人员将能够在腋下产生气味的细菌转移到受试者的前臂,前臂上培养的双菌群样本产生强烈的气味,这表明引起气味的细菌可以从腋下传播到前臂。
这项研究表明,通过皮肤微生物组移植重塑人类气味,降低对传染病媒介的吸引力,从而阻断病毒传播,为传染病预防和控制提供了一条新的途径。
●
尽可能选择天然纤维材料,如棉、亚麻或丝绸等,这些材料通常具有良好的透气性和吸湿性,可以帮助皮肤保持干爽。同时,天然纤维材料也更加温和,减少与皮肤的摩擦和刺激。
避免合成纤维材料:尽量减少使用合成纤维材料,如尼龙或涤纶等。
保持衣物的清洁和卫生非常重要。定期清洗和消毒衣物,避免细菌、真菌和其他有害微生物的滋生。
●
吸烟会使皮肤最外层的微小血管变窄,从而减少血液流动并使皮肤更苍白。这也耗尽了皮肤对皮肤健康很重要的氧气和营养物质。吸烟还会损害胶原蛋白,也可能会增加患鳞状细胞皮肤癌的风险。
压力会通过肠-脑-皮肤轴影响皮肤健康,皮肤细胞的代谢和分泌作用可能发生变化,导致皮肤微生物群失去原本的平衡状态,皮肤更加敏感,并引发痤疮爆发和其他皮肤问题。
长期的压力会导致免疫系统的紊乱,使皮肤更容易受到各种外界因素的侵害,从而引发炎症、过敏等症状。
睡眠可以促进皮肤细胞的修复和再生,有助于提升肤色的均匀度。同时,在深度睡眠状态下,身体会产生更多的生长激素,促进皮肤中胶原蛋白和弹力纤维的生成,有助于减少皱纹的产生。因此,保证充足的睡眠对皮肤健康也相当重要。
适度的身体运动可以促进血液循环和新陈代谢,有助于清除毒素和废物,使皮肤更加健康。选择适合自己的运动方式,如散步、跑步、瑜伽等,坚持每周进行几次。
随着年龄的增长,我们的皮肤微生物群组成发生了很大变化且可预测。未来的研究将继续阐明动态皮肤微生物组在整个生命周期中的积极作用。
皮肤微生物群扮演着重要角色,它不仅是阻止致病菌进入皮肤的屏障,还可以通过调节免疫系统减少炎症等方式对皮肤疾病起到积极作用。肠道微生物组的研究为我们探索皮肤微生物群在其他生理系统中的作用提供了途径。确定哪些微生物及其代谢产物对维持人类健康和疾病至关重要。
了解皮肤微生物群和肠道微生物群之间的相互关系可以帮助我们更好地理解肠-皮肤轴的作用机制。
未来的研究需要更深入地了解皮肤微生物组的分子基础,包括微生物-微生物相互作用、微生物-宿主-微生物相互作用、环境因素-微生物相互作用以及不同细菌菌株之间的互动对宿主健康的影响,为调控皮肤微生物群的紊乱提供更专业的治疗方法。
微生物群研究的相关应用领域
护肤品可以改变皮肤上的分子和菌群多样性及微生物在皮肤上的动力学和结构。微生物群在护肤中的应用是一种先进、前沿的方法。
服装和皮肤之间的关系已成为探索纺织品如何通过调节皮肤微生物群,来治疗或缓解皮肤疾病的新领域。
考虑到生物活性纺织品的广泛应用,需要寻找创新技术和产品。对于抗菌纺织品,根据其对纺织材料和人体皮肤微生物群的主要影响,可以在下述生物医学研究方向中找到有价值的用途:
这些应用使得生物活性纺织品更加绿色环保、安全、高性能,能够提供更多的功能性和舒适性,满足人们对健康和舒适的需求。
通过利用皮肤微生物穿透皮肤屏障的能力,或许可以开发微生物活化免疫细胞来传递细胞因子、小分子化学物质或疫苗。
微生物在皮肤上引发免疫反应,并与皮肤免疫细胞进行相互作用,调节免疫应答和免疫耐受。这为开发新的免疫调节治疗策略提供了新的思路。
通过调节肠道微生物组来靶向皮肤健康是一种很有前景的替代疗法。对于一些慢性皮肤病患者来说,肠道微生物群的紊乱可能是病情加重或难以治愈的原因之一,而通过调整饮食、生活习惯、环境等因素,同时加入益生菌或其他补充剂等,调节肠道微生物群的平衡,增强皮肤对各种病原体的抵抗力;同时通过肠道菌群调节影响自身免疫反应,改善皮肤疾病的症状。
通过皮肤、肠道微生物群联合检测可以探索和评估微生物之间的相互关系,并在皮肤问题的中提供更全面有效、个性化的治疗方案。
主要参考文献:
Townsend EC, Kalan LR. The dynamic balance of the skin microbiome across the lifespan. Biochem Soc Trans. 2023 Feb 27;51(1):71-86.
Kurokawa I, Kobayashi M, Nomura Y, Abe M, Kerob D, Dreno B. The Role and Benefits of Dermocosmetics in Acne Management in Japan. Dermatol Ther (Heidelb). 2023 Jul;13(7):1423-1433.
Harris-Tryon TA, Grice EA. Microbiota and maintenance of skin barrier function. Science. 2022 May 27;376(6596):940-945.
Belzer A, Parker ER. Climate Change, Skin Health, and Dermatologic Disease: A Guide for the Dermatologist. Am J Clin Dermatol. 2023 Jul;24(4):577-593.
Patra V, Bordag N, Clement Y, Köfeler H, Nicolas JF, Vocanson M, Ayciriex S, Wolf P. Ultraviolet exposure regulates skin metabolome based on the microbiome. Sci Rep. 2023 May 3;13(1):7207.
Skowron K, Bauza-Kaszewska J, Kraszewska Z, Wiktorczyk-Kapischke N, Grudlewska-Buda K, Kwiecińska-Piróg J, Wałecka-Zacharska E, Radtke L, Gospodarek-Komkowska E. Human Skin Microbiome: Impact of Intrinsic and Extrinsic Factors on Skin Microbiota. Microorganisms. 2021 Mar 5;9(3):543.
Trompette A, Ubags ND. Skin barrier immunology from early life to adulthood. Mucosal Immunol. 2023 Apr;16(2):194-207.
Alashkar Alhamwe B, López JF, Zhernov Y, von Strandmann EP, Karaulov A, Kolahian S, Geßner R, Renz H. Impact of local human microbiota on the allergic diseases: Organ-organ interaction. Pediatr Allergy Immunol. 2023 Jun;34(6):e13976.
Xu H, Li H. Acne, the Skin Microbiome, and Antibiotic Treatment. Am J Clin Dermatol. 2019 Jun;20(3):335-344.
Zubeldia-Varela E, Barker-Tejeda TC, Obeso D, Villaseñor A, Barber D, Pérez-Gordo M. Microbiome and Allergy: New Insights and Perspectives. J Investig Allergol Clin Immunol. 2022 Oct;32(5):327-344.
De Almeida, C.V.; Antiga, E.; Lulli, M. Oral and Topical Probiotics and Postbiotics in Skincare and Dermatological Therapy: A Concise Review. Microorganisms 2023, 11, 1420.
Flores-Balderas, X.; Peña-Peña, M.; Rada, K.M.; Alvarez-Alvarez, Y.Q.; Guzmán-Martín, C.A.; Sánchez-Gloria, J.L.; Huang, F.; Ruiz-Ojeda, D.; Morán-Ramos, S.; Springall, R.; et al. Beneficial Effects of Plant-Based Diets on Skin Health and Inflammatory Skin Diseases. Nutrients 2023, 15, 2842.
Suellen Ferro de Oliveira C, Kekhasharú Tavaria F. The impact of bioactive textiles on human skin microbiota. Eur J Pharm Biopharm. 2023 Jul;188:66-77.
Santiago-Rodriguez, T.M.; Le François, B.; Macklaim, J.M.; Doukhanine, E.; Hollister, E.B. The Skin Microbiome: Current Techniques, Challenges, and Future Directions. Microorganisms 2023, 11, 1222.
Fernandes A, Rodrigues PM, Pintado M, Tavaria FK. A systematic review of natural products for skin applications: Targeting inflammation, wound healing, and photo-aging. Phytomedicine. 2023 Jul;115:154824.
Lee, H.-J.; Kim, M. Skin Barrier Function and the Microbiome. Int. J. Mol. Sci. 2022, 23, 13071.
Ito Y, Amagai M. Dissecting skin microbiota and microenvironment for the development of therapeutic strategies. Curr Opin Microbiol. 2023 Apr 3;74:102311.
Mourelle, M.L.; Gómez, C.P.; Legido, J.L. Hydrobiome of Thermal Waters: Potential Use in Dermocosmetics. Cosmetics 2023, 10, 94
Azzimonti, B.; Ballacchino, C.; Zanetta, P.; Cucci, M.A.; Monge, C.; Grattarola, M.; Dianzani, C.; Barrera, G.; Pizzimenti, S. Microbiota, Oxidative Stress, and Skin Cancer: An Unexpected Triangle. Antioxidants 2023, 12, 546
Leung MHY, Tong X, Shen Z, Du S, Bastien P, Appenzeller BMR, Betts RJ, Mezzache S, Bourokba N, Cavusoglu N, Aguilar L, Misra N, Clavaud C, Lee PKH. Skin microbiome differentiates into distinct cutotypes with unique metabolic functions upon exposure to polycyclic aromatic hydrocarbons. Microbiome. 2023 Jun 1;11(1):124.
Kengmo Tchoupa A, Kretschmer D, Schittek B, Peschel A. The epidermal lipid barrier in microbiome-skin interaction. Trends Microbiol. 2023 Jul;31(7):723-734.

谷禾健康

与人类密切相关的微生物
我们的世界大到浩瀚宇宙,小到微观下的生物分子。我们总说漫天繁星,其实身边微生物数量可能更多。动物、植物、真菌、细菌、病毒等,共同构成了丰富多彩的生命世界。
细菌、真菌、病毒是其中的三个大类,虽然它们都体型微小,但是相互之间可以说是天差地别。并且它们与我们的生活以及健康息息相关,有对人体有益的益生菌,也有对人体危害极大的病毒。

谷禾在本文中介绍了细菌、真菌、病毒的一些特征,它们的分类及繁殖方式,以及有致病性的微生物和感染后的一些症状与免疫过程。
在最后,针对一些病原体的感染。我们提出了一些预防的方法,以及目前技术条件下的治疗手段。微生物检测技术能够帮助人们更好地了解就在我们体内或身边的微生物,有助于塑造更健康的身体及生活。
本文主要从以下几个方面讲述
●什么是细菌、真菌、病毒?
●细菌、真菌、病毒的分类与繁殖方式
●细菌、真菌、病毒的致病性与感染症状
●细菌、真菌、病毒在体内的免疫反应
●病原体感染的预防及治疗方法
细菌
细菌(Bacteria)是生物的主要类群之一,属于细菌域。广义的细菌即为原核生物, 是指一大类细胞核无核膜包裹,只存在拟核区(或拟核)的裸露DNA的原始单细胞生物,包括真细菌(eubacteria)和古细菌(archaea)两大类群。
注:其中除少数属古细菌外,多数的原核生物都是真细菌。
✦结构简单、个体小
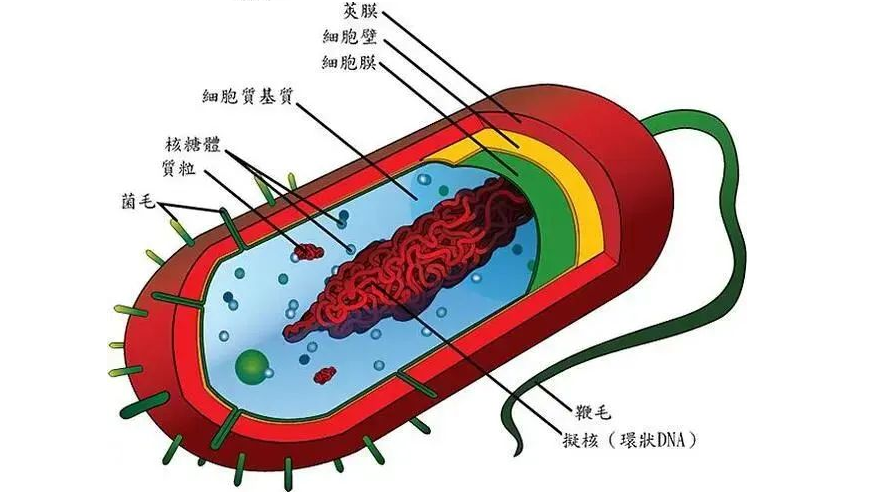
图片来源:百度
细菌为原核微生物的一类,是一类形状细短,结构简单,多以二分裂方式进行繁殖的原核生物。细菌一般是单细胞,主要由细胞壁、细胞膜、细胞质、核质体等部分构成,有的细菌还有荚膜、鞭毛、菌毛等特殊结构, 缺乏细胞核、细胞骨架以及膜状胞器,例如粒线体和叶绿体。
细菌的个体非常小,绝大多数细菌的直径大小在0.5~5μm之间。目前已知最小的细菌只有0.2微米长,因此大多情况只能在显微镜下看到它们。但处于有利环境中时,细菌可以形成肉眼可见的集合体,例如菌簇。
✦数量众多、分布广泛
细菌是所有生物中数量最多的一类,据估计,其总数约有5×10的三十次方个。
细菌广泛分布于土壤和水中,或者与其他生物共生。人体是大量细菌的栖息地;可以在皮肤表面、肠道、口腔、鼻子和其他身体部位找到。据估计,人体内及表皮上的细菌细胞总数约是人体细胞总数的十倍。
此外,也有部分种类分布在极端的环境中,例如温泉,甚至是放射性废弃物中,它们被归类为嗜极生物,其中最著名的种类之一是海栖热袍菌(Thermotoga maritima),科学家是在意大利的一座海底火山中发现这种细菌的。
★ 常见的细菌
大肠埃希氏菌(Escherichia coli)
大肠杆菌是短杆菌,两端呈钝圆形,属革兰氏阴性菌,于1885年首次被发现。
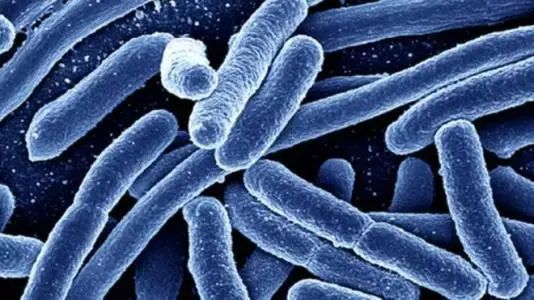
图片来源:百度百科
大肠杆菌是条件致病菌,在一定条件下可以引起多种疾病,如腹泻,肠炎,尿路感染,呼吸道感染、菌血症和其他临床感染(如新生儿脑膜炎)。
金黄色葡萄球菌(Staphylococcus aureus)
金黄色葡萄球菌也称“金葡菌”,隶属于葡萄球菌属,是革兰氏阳性菌代表,为一种常见的食源性致病微生物。
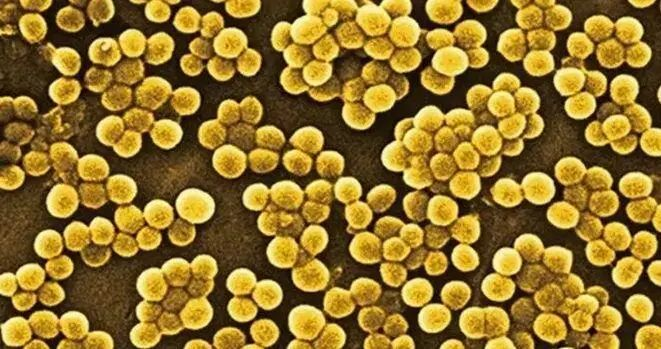
图片来源:百度
该菌最适宜生长温度为37℃,pH为7.4,耐高盐,可在盐浓度接近10%的环境中生长。金黄色葡萄球菌常寄生于人和动物的皮肤、鼻腔、咽喉、肠胃、痈、化脓疮口中,空气、污水等环境中也无处不在。
金黄色葡萄球菌是最臭名昭著、分布最广的细菌病原体之一。这种病原体可引起多种疾病,从中度严重的皮肤感染到致命的肺炎和败血症。
每年在全球范围内造成难以估计数量的无并发症皮肤感染,并可能导致数十万至数百万更严重的侵入性感染。
双歧杆菌属(Bifidobacterium)
双歧杆菌是一种革兰氏阳性、不运动、细胞呈杆状、一端有时呈分叉状、严格厌氧的细菌属,广泛存在于人和动物的消化道、阴道和口腔等生境中。双歧杆菌属的细菌是人和动物肠道菌群的重要组成成员之一。
双歧杆菌是一种重要的肠道有益微生物。双歧杆菌作为一种生理性有益菌,对人体健康具有生物屏障、营养作用、抗肿瘤作用、免疫增强作用、改善胃肠道功能、抗衰老等多种重要的生理功能。
★ 细菌对人类有利有弊
细菌也对人类活动有很大的影响。一方面,细菌是许多疾病的病原体,可以通过各种方式,如接触、消化道、呼吸道、昆虫叮咬等在正常人体间传播疾病,具有较强的传染性,对社会危害极大。
另一方面,人类也时常利用细菌,例如乳酪及酸奶和酒酿的制作、部分抗生素的制造、废水的处理等,都与细菌有关。在生物科技领域中,细菌也有着广泛的运用。
真菌
真菌(Fungus)是一种真核生物。在生物学分类上属于藻菌植物中真菌超纲。微生物中只有真菌具有真正的细胞核和完整的细胞器,故又称真核细胞型微生物。
最常见的真菌是各类蕈类,另外真菌也包括霉菌和酵母。现在已经发现了七万多种真菌,估计只是所有存在的一小半。大多真菌原先被分入动物或植物,现在成为自己的界,分为四门。
✦结构
菌体由菌丝组成,无根、茎、叶的分化,无叶绿素,不能自己制造养料,以寄生或腐生方式生活的低等生物。
真菌菌丝呈管状,多数菌丝有隔膜,此类菌丝为多细胞,隔膜中央有小孔,使细胞质、细胞核得以通过。有些真菌的菌丝无隔膜,为多核细胞。
✦分布
真菌广泛分布于全球各带的土壤、水体、动植物及其残骸和空气中,营腐生、寄生和共生生活。
★ 常见的真菌
蘑菇(Agaricus campestris)

图片来源:百度
蘑菇属于腐生真菌中的一种,其体内并没有叶绿素的存在,因此不能直接在光照下进行光合作用。蘑菇生长过程中,主要是将培养料中的各类营养物质作为营养来源,从而实现生长发育。
酵母(Saccharomyces)
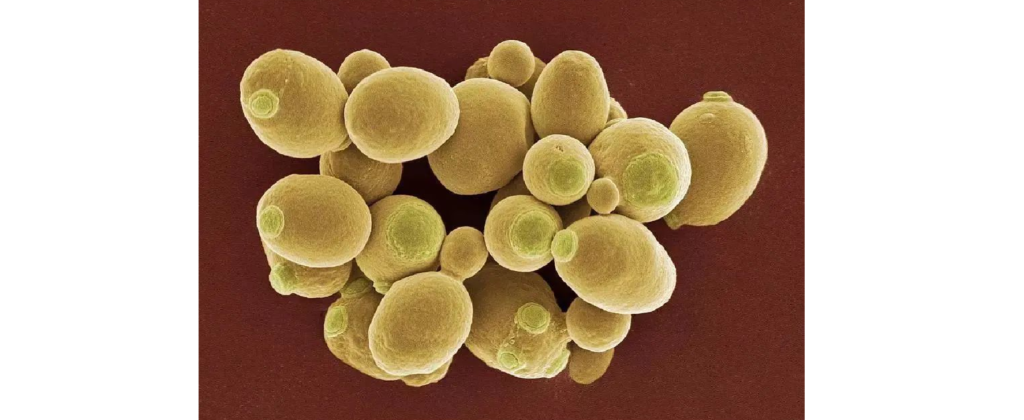
图片来源:百度
酵母菌是单细胞微生物。它属于高等微生物的真菌类。有细胞核、细胞膜、细胞壁、线粒体、相同的酶和代谢途径。酵母无害,容易生长,空气中、土壤中、水中、动物体内都存在酵母。有氧气或者无氧气都能生存。
酵母菌的作用
1、在面包、蛋糕、饼干和烤饼等这种一般的粮食制品掺入酵母菌可以提高食品的营养价值,酵母菌还在婴儿食品以及健康食品中作为食品营养的强化剂。
2、利用酵母菌可以将糖类发酵成酒精,在白酒、啤酒、果酒、黄酒等的酿造过程中都会加入酵母菌。
3、此外,大量的酵母菌还能令酒免受外界杂菌的侵害,在酵母菌发酵完成之后有澄清酒体的作用。
4、酵母菌还有入药价值,入药后的酵母菌不仅在治疗克山病和大骨节病起到辅助作用,还可以具有一定的防衰老作用。
酵母菌的危害
1、个别的酵母菌会危害生物或家庭用具,例如红酵母菌会生长在浴帘或者一些潮湿的家具上,会慢慢地腐蚀这些家具。
2、酵母菌种类中的白色假丝酵母菌,也就是俗称的白色念珠菌,主要出现在口腔、肠道、尿道和阴道等部位的粘膜上,会引起鹅口疮或尿道炎等感染疾病。
病毒
病毒(virus)是一种可以利用宿主细胞系统进行复制的微小, 无完整细胞结构的亚显微粒子。病毒不具细胞结构,无法独立生长和复制, 但病毒可以感染所有的具有细胞的生命体, 具有遗传、复制等生命特征。
✦无细胞结构
病毒主要由核酸和蛋白质外壳组成。有些病毒有囊膜和刺突,如流感病毒。病毒基因同其他生物的基因一样,也可以发生突变和重组,因此也是可以演化的。
对于病毒到底是一种生命形式,还是仅仅是一种能够与生物体作用的有机结构,人们的观点各不相同。
病毒有高度的寄生性,完全依赖宿主细胞的能量和代谢系统,获取生命活动所需的物质和能量,离开宿主细胞,它只是一个大化学分子,停止活动,可制成蛋白质结晶,为一个非生命体,遇到宿主细胞它会通过吸附,进入、复制、装配、释放子代病毒而显示典型的生命体特征,所以病毒是介于生物与非生物之间的, 一种处于“生命边缘的生物体”。
★ 常见的病毒
狂犬病毒(Rabies virus)
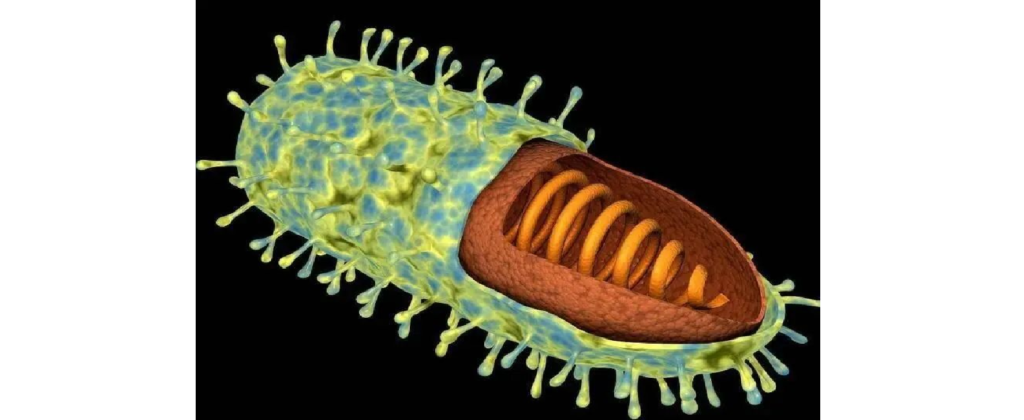
图片来源:百度
狂犬病病毒为弹状病毒,其头部为半球形,末端常为平端,形态呈典型的子弹状,长约130-240nm,直径65~80nm,内含有单链RNA。
狂犬病病毒是引起狂犬病的病原体。狂犬病毒具有两种主要抗原:一种是病毒外膜上的糖蛋白抗原,能与乙酰胆碱受体结合使病毒具有神经毒性,并使体内产生中和抗体及血凝抑制抗体,中和抗体具有保护作用;另一种为内层的核蛋白抗原,可使体内产生补体结合抗体和沉淀素,无保护作用。
冠状病毒
图片来源:百度
冠状病毒属的病毒是具囊膜、基因组为线性单股正链的RNA病毒,是自然界广泛存在的一大类病毒。
冠状病毒直径约80~120nm,基因组全长约27-32kb,是已知RNA病毒中基因组最大的病毒。
冠状病毒是成人普通感冒的主要病原之一,在儿童可以引起上呼吸道感染,一般很少波及下呼吸道。
冠状病毒还可以引起婴儿、新生儿急性肠胃炎,主要症状是水样大便、发热、呕吐,每天10余次,严重者可以出现血水样便。
2019新型冠状病毒(2019-nCoV ,引发新型冠状病毒肺炎COVID-19)是已知的第7种可以感染人的冠状病毒。
注:第一个已知的病毒是烟草花叶病毒,由马丁乌斯·贝杰林克于1899年发现并命名,如今已有超过5000种类型的病毒得到鉴定。研究病毒的科学被称为病毒学,是微生物学的一个分支。
细菌和真菌的名称中均有一个“菌”字,同属微生物,但两者在生物类型、结构、大小、增殖方式上却有着诸多不同。比较如下:
✦生物类型不同
一是就有无成形的细胞核来看:细菌没有核膜包围形成的细胞核,属于原核生物;真菌有核膜包围形成的细胞核,属于真核生物。
二是就组成生物的细胞数目来看:细菌全部是由单个细胞构成,为单细胞型生物;真菌既有由单个细胞构成的单细胞型生物(如酵母菌),也有由多个细胞构成的多细胞型生物(如食用菌、霉菌等)。
✦细胞结构不同
细菌和真菌都具有细胞结构,属于细胞型生物,在它们的细胞结构中都具有细胞壁、细胞膜、细胞质,但却存在诸多不同,具体表现在:一是细胞壁的成分不同:细菌细胞壁的主要成分是肽聚糖,而真菌细胞壁的主要成分是几丁质。
二是细胞质中的细胞器组成不同:细菌只有核糖体一种细胞器;而真菌除具有核糖体外,还有内质网、高尔基体、线粒体、中心体等多种细胞器。
三是细菌没有成形的细胞核,只有拟核;真菌具有。细菌没有染色体,其DNA分子单独存在;真菌细胞核中的DNA与蛋白质结合在一起形成染色体。
✦细胞大小差异
细胞大小:原核细胞一般较小,直径一般为1μm-10μm;真核细胞较大,直径一般为10μm-100μm。
✦增殖方式不同
细菌是原核生物,为单细胞型生物,通过细胞分裂而增殖,具有原核生物增殖的特有方式(二分裂);真菌为真核生物,细胞的增殖主要通过有丝分裂进行,因真菌种类的不同其个体增殖方式主要有出芽生殖(如酵母菌)和孢子生殖(食用菌)等方式。
注:尽管在细菌和真菌的名称中都有一个菌字,但细菌的名称中一般含有:球、杆、弧、螺旋等描述细菌形态的字眼,只有乳酸菌例外(实为乳酸杆菌);而真菌名称中则不含有。
✦体积差异大
细菌和病毒同属于微生物,只有在显微镜下才能看到。但两者是截然不同的东西。
细菌和病毒均属于微生物。在一定的环境条件下,细菌和病毒都可以在人体中增殖,并可能导致疾病发生。细菌较大,用普通光学显微镜就可看到,它们的生长条件也不高。病毒则较小,一般要用放大倍数超过万倍的电子显微镜才能看到。
注意:有一点值得指出的是,在人们身体的许多部位都有细菌的增殖。医学上称之为正常菌群,它们与我们和平相处,互惠互利。而在任何情况下从机体中发现病毒都非正常状况。因为只有侵入我们的活组织细胞中这些病毒才能存活。
✦结构不同
细菌是和植物一样,有细胞壁,而人的细胞是没有细胞壁的,这就是很多抗生素杀菌的原理。比如破坏它的细胞壁或者阻止合成细胞壁,细菌就死掉了,而人没有这个结构,所以对人无影响。
病毒与细菌不同之处是,病毒没有细胞结构,可以说是最低等的生物,但是它的能耐可不小,人类的疾病从小的感冒到大的癌症都和它有关系。
病毒构造很简单,外面是一层蛋白质,称为病毒外壳。蛋白质外壳内部包裹着病毒的遗传物质,可以是DNA,也可以是RNA。病毒自己不能完成新陈代谢,也不能完成繁殖,需要寄生在其它细胞内完成。
病毒没有自己的生长代谢系统,它的生存靠寄生在宿主(如人)和细胞中依赖他人的代谢系统。也是因为如此,目前抗病毒的特殊药物不多。
✦按形状分类
细菌具有不同的形状,并可根据形状分为三类,即:球菌、杆菌和螺旋菌(包括弧菌、螺菌、螺杆菌)。
✦不同生存条件
按细菌的生活方式来分类,分为两大类:自养菌和异养菌,其中异养菌包括腐生菌和寄生菌。
按细菌对氧气的需求来分类,可分为需氧(完全需氧和微需氧)和厌氧(不完全厌氧、有氧耐受和完全厌氧)细菌。
按细菌生存温度分类,可分为喜冷、常温和喜高温三类。
✦按细胞壁组成分类
细菌的结构十分简单,原核生物,没有成形的细胞核,没有膜结构的细胞器例如线粒体和叶绿体,但是有细胞壁,有的细菌还有鞭毛和荚膜,根据细胞壁的组成成分,细菌分为革兰氏阳性菌和革兰氏阴性菌。
“革兰氏”来源于丹麦细菌学家革兰,他发明了革兰氏染色。
革兰氏阳性菌具有较厚的肽聚糖细胞壁结构,在革兰氏染色试验中呈紫色/蓝色;
革兰氏阴性菌的细胞壁较薄,在革兰氏染色试验中呈红色至粉红色。
革兰氏阳性菌特征
革兰氏阳性细菌的主要特征是它们的结构。一般有以下特点:
●没有外膜。革兰氏阳性细菌没有外膜,但革兰氏阴性细菌有。
●复杂的细胞壁。包围细胞质膜的细胞壁由肽聚糖、多糖、磷壁和蛋白质组成。它很容易吸收外来物质。
●厚的肽聚糖层。在革兰氏阳性细菌中,肽聚糖有40到80层厚。
●某些表面附属物。革兰氏阳性细菌可能有鞭毛,可以帮助它们移动。它们很少有被称为菌毛的毛发状结构。
革兰氏阴性菌特征
革兰氏阳性菌和革兰氏阴性菌具有不同的结构。通常,革兰氏阴性菌具有以下特征:
●外脂质膜
●肽聚糖薄层(2-3纳米)
●通常不含磷壁酸
●可以有鞭毛或毛
小结
主要的区别是外脂膜。它很难渗透,这给了革兰氏阴性细菌额外的保护。革兰氏阳性细菌没有这种特征。
由于这种差异,革兰氏阴性细菌更难杀死。这意味着革兰氏阳性菌和革兰氏阴性菌需要不同的处理方法。
真菌较高层级的分类系统仍有很大争议,新理论不断被提出,各个分类阶层的名称均常有变动。且同一种真菌还可能在生活史的不同阶段,例如无性与有性世代拥有数种不同的学名,使真菌分类更加复杂。目前将真菌界分为4门和1类。
✦壶菌门
壶菌门(Chytridiomycota)壶菌门是游动细胞具有“9+2”结构的鞭毛,并能在水中游动的一类真菌,游动孢子具有一根后生尾鞭式鞭毛。
壶菌多水生,大多腐生在动植物残体上或寄生于水生植物、藻类、小动物和其他真菌上,少数寄生于高等种子植物上。大多数种类能分解纤维素和几丁质。
✦接合菌门
接合菌门(Zygomycota)是由低等的水生真菌发展到陆生种类,由游动的带鞭毛的孢囊孢子发展为不游动的孢囊孢子——静孢子或单孢孢子囊的分生孢子。
接合菌门菌物共同特征是有性生殖产生接合孢子。接合菌营养体为单倍体,大多是很发达的无隔菌丝体,少数菌丝体不发达,较高等的种类菌丝体有隔膜。有的种类菌丝体可以分化形成假根和匍匐丝。细胞壁的主要成分为几丁质。
✦子囊菌门
子囊菌门(Ascomycota)是真菌中最大的类群,与担子菌被称为高等真菌,生殖菌丝细胞出现较短双核阶段,其区别于其他真菌的一个特征是产生子囊。
子囊菌大都陆生,营养方式有腐生、寄生和共生。腐生的子囊菌可以引起木材、食品、布匹和皮革的霉烂以及动植物残体的分解。
✦担子菌门
担子菌门(Basidiomycota)是一类高等真菌,构成双核亚界,包含2万多种,包括蘑菇、木耳等主要食用菌。
担子菌门包括以下组:蘑菇,马勃,鬼笔科,和人体致病酵母隐球菌属等等。
担子菌门的真菌基本全为陆生品种,主要特征是由多细胞,有横隔膜的菌丝体组成,菌丝分为两种,初生菌丝体的细胞只有一个细胞核,次生菌丝体的细胞有两个核,两个核的次生菌丝体可以形成一种子实体,称为担子果,经过有性繁殖过程,在担子上生成担孢子;也可以经过无性繁殖过程生成无性孢子或出芽繁殖。
✦半知菌类
半知菌类(Deuteromycota)是一种已废止的生物分类,指在子囊菌、担子菌的同伴之中,还未发现有性繁殖阶段而在分类学上位置不明的一种临时分类。
只进行无性繁殖的菌类被称作不完全型,这一阶段被称为无性阶段。进行有性繁殖的被称为完全型,该阶段被称作有性阶段,通常有性阶段的菌类也是同时进行无性生殖的。
从遗传物质分类:DNA病毒、RNA病毒、蛋白质病毒(如:朊病毒)
RNA病毒和DNA病毒在结构、成分、复制能力和致病力等方面都不同。RNA病毒就是遗传物质是RNA的一种病毒。DNA病毒也称为脱氧核苷酸病毒,是一种生物病毒,属于原发病毒。
✦DNA病毒和RNA病毒的区别
1、结构:RNA病毒是单链病毒,比较容易发生变异。DNA病毒是双链结构的病毒,不容易发生变异,更稳定。
2、组成成分:RNA病毒是由核糖和磷酸组成,一般只有数百个或者数千个核苷酸。DNA病毒是由去氧核糖和磷酸组成,通常有上百万个核苷酸单位。
3、复制能力:RNA病毒感染人体后,在人体细胞中复制非常活跃,而复制过程中发生变异后也不修复,而是继续复制下去。这样就使RNA病毒变异非常快,不容易被攻破。比如甲型流感H1N1病毒和SARS病毒以及新冠病毒等,都是RNA病毒。
4、致病力:DNA病毒相对不容易变异,致病比较单一。由于RNA病毒相对比较容易变异,因此较容易致病。
从病毒结构分类:真病毒(Euvirus,简称病毒)和亚病毒(Subvirus,包括类病毒、拟病毒、朊病毒)
从寄主类型分类:噬菌体(细菌病毒)、植物病毒(如烟草花叶病毒)、动物病毒(如禽流感病毒、天花病毒、HIV等)
从性质来分:温和病毒(例如HIV)、烈性病毒(例如狂犬病毒)。
✦无性二分裂方式
细菌主要以无性二分裂方式繁殖,即细菌生长到一定时期,在细胞中间逐渐形成横隔,由一个母细胞分裂为两个大小相等的子细胞。
细胞分裂是连续的过程,分裂中的两个子细胞形成的同时,在子细胞的中间又形成横隔,开始细菌的第二次分裂。有些细菌分裂后的子细胞分开,形成单个的菌体,有的则不分开,形成一定的排列方式,如链球菌、链杆菌等。
分裂过程
采用电子显微镜研究细菌的分裂过程表明:细菌细胞分裂大致可经过核物质与细胞质分裂、横隔壁形成和子细胞分离等过程。
细菌细胞分裂时,核质DNA与中介体或细胞膜相连,首先DNA复制并向细胞两端移动,与此同时,细菌细胞膜向内凹陷并形成一垂直于细胞长轴的细胞质隔膜,使细胞质和核质均匀分配到两个子细胞中。
其次细胞形成横隔壁,在细胞膜不断内陷,形成子细胞各自的细胞质膜同时,母细胞的细胞壁也从四周向中心逐渐延伸。最后,逐渐形成子细胞各自完整的细胞壁。接着,子细胞分裂,形成两个大小基本相等的子细胞。
✦繁殖速度快
细菌繁殖速度快,一般细菌约20-30min便分裂一次,即为一代。接种子肉汤培养中的细菌在适宜的温度下迅速生长繁殖,肉汤很快即可变浑浊,表明有细菌的大量生长。不过也有些细菌,如结核分枝杆菌(M.tuberculosis)的繁殖速度较慢,需要15-18小时才能繁殖一代。
真菌的繁殖方式分为无性繁殖和有性繁殖两种。
✦无性繁殖
无性繁殖是指营养体不经过核配和减数分裂产生后代个体的繁殖。它的基本特征是营养繁殖通常直接由菌丝分化产生无性孢子。
常见的无性孢子有三种类型:
(1)游动孢子:形成于游动孢子囊内。游动孢子囊由菌丝或孢囊梗顶端膨大而成。游动孢子无细胞壁,具1-2根鞭毛,释放后能在水中游动。
(2)孢囊孢子:形成于孢囊孢子囊内。孢子囊由孢囊梗的顶端膨大而成。孢囊孢子有细胞壁,水生型有鞭毛,释放后可随风飞散。
(3)分生孢子(conidium)产生于由菌丝分化而形成的分生孢子梗(conidiophore)上,顶生、侧生或串生,形状、大小多种多样,单胞或多胞,无色或有色,成熟后从孢子梗上脱落。有些真菌的分生孢子和分生孢子梗还着生在分生孢子果内。孢子果主要有两种类型,即近球形的具孔口的分生孢子器(pycnidium)和杯状或盘状的分生孢子盘(acervulus)。
✦有性繁殖
真菌生长发育到一定时期(一般到后期)就进行有性生殖。有性生殖是经过两个性细胞结合后细胞核产生减数分裂产生孢子的繁殖方式。
多数真菌由菌丝分化产生性器官即配子囊,通过雌、雄配子囊结合形成有性孢子。其整个过程可分为质配、核配和减数分裂三个阶段。
第一阶段:质配阶段
即经过两个性细胞的融合,两者的细胞质和细胞核(N)合并在同一细胞中,形成双核期(N+N)。
第二阶段:核配阶段
核配阶段,就是在融合的细胞内两个单倍体的细胞核结合成一个双倍体的核(2N)。
第三阶段:减数分裂阶段
双倍体细胞核经过两次连续的分裂,形成四个单倍体的核(N),从而回到原来的单倍体阶段。
经过有性生殖,真菌可产生四种类型的有性孢子。
(1)卵孢子(oospore):卵菌的有性孢子。是由两个异型配子囊——雄器和藏卵器接触后,雄器的细胞质和细胞核经授精管进入藏卵器,与卵球核配,最后受精的卵球发育成厚壁的、双倍体的卵孢子。
(2)接合孢子(zygospore):接合菌的有性孢子。是由两个配子囊以配子囊结合的方式融合成1个细胞,并在这个细胞中进行质配和核配后形成的厚壁孢子。
(3)子囊孢子(ascospore):子囊菌的有性孢子。通常是由两个异型配子囊——雄器和产囊体相结合,经质配、核配和减数分裂而形成的单倍体孢子。子囊孢子着生在无色透明、棒状或卵圆形的囊状结构即子囊内。
每个子囊中一般形成8个子囊孢子。子囊通常产生在具包被的子囊果内。子囊果一般有四种类型,即球状而无孔口的闭囊壳,瓶状或球状且有真正壳壁和固定孔口的子囊壳,由于座溶解而成的、无真正壳壁和固定孔口的子囊腔,以及盘状或杯状的子囊盘。
(4)担孢子(basidiospore):担子菌的有性孢子。通常是直接由“+”、“-”菌丝结合形成双核菌丝,以后双核菌丝的顶端细胞膨大成棒状的担子。在担子内的双核经过核配和减数分裂,最后在担子上产生4个外生的单倍体的担孢子。
此外,有些低等真菌如根肿菌和壶菌产生的有性孢子是一种由游动配子结合成合子,再由合子发育而成的厚壁的休眠孢子(restingspore)。
✦自我复制
病毒繁殖借助宿主细胞为其提供的原料、能量和酶等必要条件,以自我复制的方式进行增殖,利用宿主细胞的核苷酸和氨基酸来自主地合成自身的一些组件,装配下一代个体。
流感病毒自我复制过程
1.病毒体附着到宿主细胞表面并通过胞吞进入细胞;2.衣壳分解后,病毒核糖核蛋白转运入核;3a.病毒基因组转录;3b.病毒基因组复制;4.新合成的病毒mRNA出核并完成翻译;5a.合成的核蛋白入核与新复制的核酸结合;5b.合成的病毒表面蛋白进入高尔基体完成翻译后修饰并转运上膜;6.新形成的核衣壳进入细胞质并与插有病毒表面蛋白的细胞膜结合;7.新生成的病毒体通过出泡方式离开宿主细胞。
✦细菌因素
主要与病原菌的毒力和数量有关。毒力强或数量多的致病菌进入机体,引起败血症的可能性较大。
注:败血症是指各种致病菌侵入血液循环,并在血中生长繁殖,产生毒素而发生的急性全身性感染。
✦人体因素
细菌侵入人体后是否引起感染,与人的防御、免疫功能有关。
•皮肤和黏膜是抵御细菌的有效武器
完整的皮肤和粘膜是防止细菌侵入人体的天然屏障,破损后细菌易于从此处侵入体内,挤压皮肤炎症部位或脓肿时细菌侵入的可能性更大。
严重烧伤时,创面为细菌敞开门户,皮肤坏死、血浆渗出又为细菌繁殖提供了良好环境,故极易发生感染。尿路、胆道、胃肠道、呼吸道粘膜受破坏后,若同时有内容物积滞、压力增高,细菌更易进入血中。保留导尿管、静脉等血管内留置导管、人工辅助呼吸时插管等,也使细菌易于侵入。
•免疫细胞能清除细菌
人体免疫功能正常时,进入血中的细菌迅速被血中免疫细胞如单核细胞、嗜中性粒细胞等所清除,而患肝硬变、糖尿病、血液病、结缔组织病等慢性病者,可因代谢紊乱、体液免疫及细胞免疫功能减低,易导致细菌感染发生;各种免疫抑制药物的使用、放射治疗亦是导致细菌感染发病率高的原因。
注:广谱抗菌药物使用后,对药物敏感的细菌虽被抑制或杀灭,而一些耐药菌乘机繁殖,亦可酿成细菌感染。
★细菌病
由细菌引起的疾病有许多,如:伤寒和副伤寒、细菌性食物中毒、 细菌感染性腹泻、霍乱、弯曲菌感染(弯曲菌肠炎、幽门螺杆菌感染) 细菌性痢疾、鼠疫、炭疽、白喉、百日咳、猩红热、流行性脑脊髓膜炎、结核病、人感染猪链球菌病、破伤风、败血症等。
✦症状
原发炎症:各种病原菌所引起的原发炎症与其在人体的分布部位有关。原发炎症的特点是局部的红、肿、热、痛和功能障碍。
皮疹:见于部分患者,以瘀点最为多见,多分布于躯干、四肢、眼结膜、口腔粘膜等处,为数不多。
关节症状:可出现大关节红、肿、热、痛和活动受限,甚至并发关节腔积液、积脓,多见于革兰阳性球菌、脑膜炎球菌、产碱杆菌等败血症的病程中。
感染性休克:约见于1/5~1/3败血症患者,表现为烦燥不安,脉搏细速,四肢厥冷,皮肤花斑,尿量减少及血压下降等,是严重败血症所致。
除外伤性、手术后、挤压疮疖后发生的败血症有较明显的潜伏期外,大多发病急骤。
注意
由于新生儿及老年患者具有不同的生理特点,其败血症亦各有特征。
•新生儿免疫力弱,发病率高
新生儿的皮肤粘膜屏障功能、淋巴及单核吞噬细胞系统功能尚不健全,补体尚缺乏,体液免疫水平低,细胞免疫也未完善;脐带残端为细菌入侵创造了有利条件,孕母泌尿生殖道感染或全身感染等均可使新生儿败血症发病率高、表现复杂、并发症多。
•老年人发病往往比较严重
老年人败血症的发病率有增高的趋势,由于机体反应性差,早期临床表现较隐蔽,热型往往不规则;又因免疫功能低下,病情常较严重,进展迅速且不易控制;老年人脏器功能多有减退或原有慢性病,败血症极易诱发脏器功能衰竭。
故新生儿及老年败血症预后差、死亡率亦高。及时发现新生儿和老年败血症,主要依据年龄特点和提高警惕。
真菌感染性疾病根据真菌侵犯人体的部位分为4类:浅表真菌病、皮肤真菌病、皮下组织真菌病和系统性真菌病;前二者合称为浅部真菌病,后二者又称为深部真菌病。
真菌感染多为继发性感染,由机会致病性真菌引起,特别是深部真菌感染多是由于各种诱因使机体免疫功能显著下降所致。
某些真菌如白假丝酵母菌、烟曲霉中可产生高分子强毒素或低分子毒素,这些毒素也会在治病中起到一定作用。另外,真菌的黏附能力,对免疫系统功能的抑制及胞壁中的酶类也与致病性有关。
诱发因素:发烧、创伤、肿瘤、严重其他微生物感染等。
✦浅部真菌感染
主要是由于人体接触所致,如皮肤廯菌、角层癣菌等皮肤感染真菌,多具有嗜角质性,可分解细胞的角蛋白和脂质,还可通过机械刺激和代谢产物作用,引起局部病变。
✦深部真菌感染
多发生于人体抵抗力下降或菌群失调时,为继发性感染,常见菌属有白色念珠菌、新生隐球菌、肺孢子菌、曲霉及毛菌等,因患者抵抗力较差,治疗效果往往不佳。
✦系统性真菌感染
在机体抵抗力低下时致病,如患有白血病、淋巴瘤、糖尿病等疾病或有长期大量广谱抗生素、激素使用史时,许多条件致病菌、如念珠菌、曲霉、毛霉等感染后在体内发展繁殖,从而引起系统性真菌感染。
✦症状
•浅表真菌病
感染仅仅局限于皮肤角质层的最外层,极少甚至完全没有组织反应,感染毛发时也只累及毛发表面,很少损伤毛发。
主要包括:花斑癣、掌黑癣和毛结节菌病。
•皮肤真菌病
感染累及皮肤角质层和皮肤附属器,如毛发、甲板等,能广泛破坏这些组织的结构并伴有不同程度的宿主免疫反应;这类真菌感染中最常见的是皮肤癣菌病,其他真菌引起的感染还包括皮肤念珠菌病等。
皮肤癣菌病根据不同的发病部位可以分为足癣(俗称”脚气”)、手癣、体癣、股癣、甲癣以及头癣等各类癣病;在世界范围内广泛发生,是最常见的真菌性疾病,发病率高。
•皮下真菌病
感染皮肤、皮下组织,包括肌肉和结缔组织,一般不会经血液流向重要脏器播散;但有些感染可以由病灶向周围组织缓慢扩散蔓延,如足菌肿等;也有些则沿淋巴管扩散,如孢子丝菌病、着色芽生菌病。免疫受损患者的皮下真菌具有潜在的播散全身的危险。
•系统性真菌病
除侵犯皮肤和皮下组织外,还累及组织和器官,甚至引起播散性感染,又称为侵袭性真菌感染。
近年来,随着高效广谱抗生素、免疫抑制剂、抗恶性肿瘤药物的广泛应用,器官移植、导管技术以及外科其他介入性治疗的深入开展,条件致病性真菌引起的系统性真菌病日益增多,新的致病菌不断出现,病情也日趋严重。
主要包括念珠菌病、曲霉病、隐球菌病、接合菌病和马内菲青霉病等。
病毒通过多种途径侵入机体,并在易感的宿主细胞中增殖。
✦水平传播
水平传播是指病毒在人群中不同个体之间的传播,包括病毒从动物到人的传播。常见的水平传播方式有以下几种。
(1)经呼吸道传播:病毒经空气、飞沫等吸入感染,如流感病毒、风疹病毒等。
(2)经消化道传播:病毒污染了食物和水源,经口食入而感染。如甲型肝炎病毒、脊髓灰质炎病毒等。
(3)经泌尿生殖道传播:由直接性接触而感染,如人类免疫缺陷病毒、单纯疱疹病毒等。
(4)经皮肤伤口传播:经昆虫媒介的叮咬、动物咬伤或皮肤伤口直接接触病毒而感染。如流行性乙型脑炎病毒、狂犬病病毒等。
(5)经血液传播:经输血或血液制品,包括经注射、器官移植等途径引起的感染,如乙型肝炎病毒、人类免疫缺陷病毒等。
✦垂直传播
病毒经胎盘、产道、哺乳由母亲传给胎儿或新生儿的方式,称为垂直传播。可经垂直传播的病毒有风疹病毒、人类免疫缺陷病毒、乙型肝炎病毒等。
✦症状
机体感染病毒后,可表现出不同的临床类型。依据有无症状,可分为显性感染和隐性感染;依据病毒滞留时间及症状持续时间长短,又可分为急性感染和持续性感染。
•隐性感染
由于侵入机体的病毒数量较少、毒力较弱或机体的抵抗力较强,病毒在宿主细胞内增殖,但机体不出现明显的临床症状,称为隐性感染。
隐性感染可使机体获得对该病毒的特异性免疫,保护机体免受该病毒的再次感染。隐性感染虽不出现临床症状,但病毒仍在体内增殖并向外界传播病毒,成为重要的传染源。
•显性感染
由于侵入机体的病毒数量较多、毒力较强或是机体的抵抗力较弱,病毒在宿主细胞内大量增殖,出现明显的临床症状,称为显性感染。显性感染根据感染持续时间长短。分为急性感染和持续性感染。
急性感染:病毒侵入机体后,其潜伏期短、发病急、病程数日至数周,病后常可获得特异性免疫力,机体可通过自身的免疫机制把病毒完全清除出体外,如甲型肝炎病毒。
持续性感染:病毒侵入机体后,在体内持续存在数月、数年,甚至数十年,机体可出现临床症状,也可不出现临床症状而长期带有病毒,成为重要的传染源。持续感染按病程、致病机制的不同,可分为三种。
①慢性感染:病毒侵入机体后,长期存在于血液或组织中,机体可出现症状,也可不出现症状。在整个病程病毒均可被查出,如乙型肝炎病毒引起的慢性肝炎。
②潜伏感染:原发感染后,病毒基因潜伏在机体一定的组织或细胞中,但不复制增殖出具有感染性的病毒,此时机体既没有临床症状,也不会向体外排出病毒。在某些条件下病毒可被激活而急性发作,并可检测出病毒,如单纯疱疹病毒。
③慢发病毒感染:经显性或隐性感染后,病毒长时间潜伏在机体内,潜伏期可长达数月至数年,此时机体一般无症状,一般也检测不出病毒。一旦发病,则呈亚急性进行性加重直至死亡,如人类免疫缺陷病毒的感染。
抗细菌感染的免疫是指机体抵御细菌感染的能力,是由机体的非特异性免疫和特异性免疫共同协调来完成的。
先天具有的非特异性免疫包括机体的屏障结构,吞噬细胞的吞噬功能和正常组织及体液中的抗菌物质;后天获得的特异性免疫包括以抗体作用为中心的体液免疫和致敏淋巴细胞及其产生的淋巴因子为中心的细胞免疫。
病原菌侵入机体后,由于其生物学特性的不同,致病物质的不同。机体对它们的免疫反应也各有差别。
✦宿主体表的防御功能
(一)机械的阻挡和排除作用
健康和完整的皮肤与粘膜能有效地阻挡细菌的侵入。
呼吸道粘膜上皮细胞的纤毛向上颤动,可将细菌咳出或咽下;随粪便每日约排菌1012个;小便可清除尿道上皮的细菌。
(二)分泌液中化学物质的局部抗菌作用
汗腺分泌的乳酸,皮脂腺分泌的脂肪酸均有一定的抗菌作用。
胃酸能杀死寒杆菌、痢疾杆菌和霍乱弧菌。阴道分泌物中的酸类亦有抗菌作用。前列腺分泌的精素是正常精液中存在的对革兰氏阳性细菌有效的抑制物。泪液、唾液、乳汗和呼吸道分泌物中广泛分布的溶菌酶能溶解革兰氏阳性细菌。
(三)正常菌群的拮抗作用
人体表以及与外界相通腔道中的正常菌群,可以通过它们的代谢产物对抗病原菌入侵。
例如皮肤上的痤疮丙酸菌(Propionibacterium acnes)能产生抗菌性脂类、抑制金黄色葡萄球菌和化脓性链球菌在皮肤上生长;肠道中的某些厌氧菌能产生脂肪酸阻止沙门氏菌在局部生存;肠道中大肠杆菌产生的大肠菌毒和酸性产物能抑制痢疾杆菌、金黄色葡萄球菌;咽部的草绿色链球菌(Viridans Streptococci)似能阻止肺炎球菌在局部生长;鼻腔的表皮葡萄球菌和类白喉杆菌能妨碍金黄色葡萄球菌定居等。当这种拮抗作用受影响时,则可发生菌群失调症。
✦机体抗毒性免疫
抗毒性免疫是一种以体液抗体为主的免疫应答。许多以外毒素致病的病原菌造成的感染,如白喉、破伤风、气性坏疽及内毒中毒等,机体的免疫应答,主要表现为抗毒素(lgG)中和毒素的作用。
由抗毒素与外毒素特异结合形成的复合物,可被吞噬细胞吞噬,并将其降解消除。抗毒素与毒素结合,可以通过空间阻碍使毒素不能吸附到敏感的宿主细胞(受体)上,或者使毒素生物学活性部位(酶)被封闭,从而使毒素不能发生毒性作用。
注意:抗毒素不能对已与组织结合的毒素起中和作用。
// 建议
根据外毒素的免疫特点,可应用类毒素进行预防接种,应用抗毒素血清进行早期治疗与紧急预防,使用时要保证“早期足量”。
✦机体的抗菌性免疫
病原侵入机体后,由于其生物学特征的不同,可分为胞外菌感染和胞内菌感染两类,机体对这两类感染的免疫反应是有差别的。
(一)胞外寄生菌的抗感染免疫
1.抗体对细菌繁殖的抑制作用:抗体与细菌结合,可以出现凝集和鞭毛制动现象,但一般而言,对细菌的活力只有微弱的影响,甚至没有影响。如果抗体的结合能抑制细菌的重要酶系统或代谢途径,则可能抑制细菌的生长。
例如,某些细菌(例如败血巴氏杆菌)从血清转铁蛋白摄取铁的能力可被特异性抗体封闭,从而导致细菌生长受抑制。
2.抗体对细菌吸附作用的抑制:病原菌吸附到粘膜上皮细胞是造成感染的先决条件。粘膜表面的抗体,在防止病原菌对粘膜的侵犯中具有更重要的作用。
在粘膜表面起这种作用的抗体主要是SlgA它是局部免疫的主要因素。SlgA抗细菌感染可有以下几种方式:在补体和溶菌酶的参与下溶解某些细菌;在肠道局部增强吞噬作用;防止细菌对粘膜上皮细胞的吸附。
例如SlgA能阻止链球菌、致病性大肠杆菌、霍乱弧菌、淋球菌、百日咳杆菌等对粘膜表面的吸附。
3.抗体和补体对细菌的溶解作用:在许多感染中,机体能产生相应抗体(lgG、lgM、lgA),当细菌表面抗原和lgG、lgM结合的免疫复合物一旦通过经典途径使补体活化或由分泌型 lgA或聚合的血清lgA通过替代途径活化补体,即可引起细胞膜的损伤,最终发生溶菌。
实验证明补体的溶菌作用仅对革兰氏阴性菌,其中包括霍乱弧菌、大肠杆菌、痢疾杆菌、伤寒杆菌等发挥作用。但这种作用往往并不彻底,仅使杆菌菌体膨大或变为球形,不引起溶解。但若于试验中系统中加入适量的溶菌酶,则可出现溶菌现象。
4.抗体和补体对吞噬作用的调理:抗体和补体单独能适当的靶细胞起调理吞噬作用,若两者联合作用效应更加强大。中性粒细胞和单核吞噬细胞表面具有lgG的Fc受体。当lgG通过其特异性抗原结合部位(Fab)与细菌表面相应抗原结合后,其Fc段可与吞噬细胞表面相应Fc受体结合,即可在细菌与吞噬细胞间形成抗体“桥梁”,这不仅能促进吞噬细胞对细菌的吞噬,而且有助于强化细胞内的杀菌作用。
注:中性粒细胞和单核细胞表面还有C3b 受体。因此,细菌与所有能结合补体的抗体(lgg 、lgM )形成的复合物,均可激活补体形成活化产物C3B,从而发挥调理吞噬作用。尤以lgM 的作用更强,此作用在感染的早期特别重要,因为此时lgM抗体占优势。
(二)胞内寄生菌的细胞免疫
凡侵入人体后大部分时间停留在宿主细胞内并繁殖的病原菌称胞内寄生菌。例如结核杆菌、麻风杆菌、布氏杆菌等均属此类。
由于抗体不能进入细胞内,所以体液免疫对这类细菌感染的作用受到限制,对胞内感染的防御功能主要靠细胞免疫。例如机体初次感染结核杆菌,由于细胞免疫尚未建立,吞噬细胞虽可将它们吞噬,但不能有效地消化杀灭,因此病原菌容易随吞噬细胞在体内扩散,蔓延,而造成全身感染。
但在传染过程中,机体在病原菌的刺激下逐渐形成细胞免疫,通过致敏淋巴细胞释放的各种淋巴因子,激活吞噬细胞,可增强其吞噬消化能力,抑制病原菌在吞噬细胞内生存,从而获得防御同种病种原菌再感染的免疫力。
✦非特异性免疫
人类对真菌感染有天然免疫力。包括皮肤分泌短链脂肪酸和乳酸的抗真菌作用,血液中转铁蛋白扩散至皮肤角质层的抑真菌作用;中性粒细胞和单核巨噬细胞的吞噬作用,以及正常菌群的拮抗作用。
注意:许多真菌病受生理状态影响,如婴儿对念珠菌病易感,学龄前儿童易患头癣。
✦特异性免疫
•细胞免疫排菌杀菌
真菌感染中细胞免疫是机体排菌杀菌及复原的关键,T细胞分泌的淋巴因子对加速表皮角化和皮屑形成,随皮屑脱落,将真菌排除;以T细胞为主导的迟发型变态反应引起免疫病理损伤能局限和消灭真菌,以终止感染。
一般反应强度与体内菌量呈反比,如阴性则菌量增加,病情严重,而经治疗又转阳性,说明治疗见效,预后良好。
•体液免疫具有保护作用
体液免疫对部分真菌感染有一定保护作用,如特异性抗体可阻止真菌转为菌丝相以提高吞噬细胞的吞噬率;抗白色念珠菌抗体与菌表面甘露醇蛋白质复合物结合,阻止本菌粘附宿主细胞;全身性白色念珠菌感染,尽管其迟发型变态反应阳性,或通过被动转移致敏淋巴细胞,还必须同时输入特异抗体才起保护作用。
注意:而DTH反应阴性者即使有抗体,不能引起保护作用,表明抗体须在具有良好的细胞免疫基础的机体内才发生保护作用。
DTH指的是迟发型超敏反应,主要是T细胞与相应抗原作用后,引起单个核细胞浸润以及组织细胞损伤为主的炎症反应。
✦先天性免疫
人体抵御病毒的第一道防线是先天性免疫系统。这一免疫系统由能够抵御非特异性病毒感染的细胞和其他机制组成,即以一种通用方式来对入侵的病原体做出识别和反应,但不同于获得性免疫系统,这一免疫系统并不产生持久的或保护性的免疫。
RNA干扰是对抗病毒的一种重要的先天性防御机制。
✦体液免疫
当人体的获得性免疫系统探测到病毒时,会产生特异性的抗体来与病毒结合并使其失去感染性,这种作用被称为体液免疫。
其中,有两类抗体非常重要。第一类被称为IgM(免疫球蛋白M),它能高效地使病毒去活,但免疫系统的细胞产生IgM的时间只有几个星期。第二类被称为IgG(免疫球蛋白G),它能够被免疫系统不停地制造出来。
IgM存在于宿主的血液中是用于急性感染的情况,而IgG的存在则表明过去曾经受到某种感染(用于防御以后的同类感染)。进行免疫性测试时,通常是对体内的IgG型抗体进行测量。
✦细胞免疫
人体对抗病毒的另一道防线是细胞免疫,包括了被称为T细胞的免疫细胞。人体中的细胞不断地将其内部蛋白质的片断展示在细胞表面(抗原呈递)供T细胞来进行检查,一旦T细胞识别出可能的病毒片断,那么对应的细胞就会被病毒特异性T细胞扩增所消灭。
诸如巨噬细胞在内的一些细胞专门负责抗原呈递。制造干扰素是一种重要的宿主防御机制。干扰素是病毒感染之后由机体所产生的一种激素,它在免疫中的作用较为复杂,可以确定的是它能够通过杀死受感染细胞及其邻近细胞来逐步阻止病毒的复制。
注:并非所有的病毒感染都会引起保护性免疫反应。例如,艾滋病毒可以通过不断地变换其病毒体表面蛋白的氨基酸序列来逃避免疫系统的打击。这些顽固的病毒采用多种方式来逃脱免疫系统的控制,如隔离、阻断抗原呈递、产生细胞因子抗性、逃避自然杀伤细胞的作用、逃脱细胞凋亡以及抗原转移。其他一些病毒,如向神经病毒,可以通过神经来传播,而在神经系统中免疫系统可能无法接触到它们。
✦预防
细菌感染的预防需要重点关注防止感染,对于具有传播性质的疾病,应从传染源、传播途径和易感人群三个基本环节中进行防控。
1、注意卫生:日常生活中注意饮食卫生,家庭居所最好日常进行消毒、灭菌,防止带入外界环境的致病菌。
2、接种疫苗:接种疫苗后可使机体产生免疫应答,产生特异性抗体,使机体获得针对病原疫苗的免疫力,应及时接种相关疫苗提高自身抗细菌感染能力。
3、体育锻炼:适当从事体育锻炼可以提高免疫力,可根据自身情况选择打太极拳、健身、跑步、郊游等,劳逸结合。
4、远离传染源:有传染性疾病的人群应做好隔离措施,其他人群也应做好防护措施,避免与其接触,避免到细菌感染流行病区。
小结
经常保持皮肤和粘膜的清洁和完整,避免创伤,控制慢性病,合理使用免疫抑制剂和抗生素类药物,烧伤病房应严格消毒等措施,均可预防细菌感染。
一切明显的或隐匿的化脓性病灶如能及早予以清除,感染的发生就可以减少。小儿时常见的传染病如麻疹、流行性感冒、百日咳等每易继发较重的呼吸道细菌感染,从而发生细菌感染。对这类病儿,必须加强保护。对不论多么细小的皮肤创伤必须予以重视,早作适当处理。
✦治疗
一般治疗:卧床休息,加强营养,补充适量维生素。维持水、电解质及酸碱平衡。必要时给予输血、血浆、白蛋白和丙种球蛋白。高热时可给予物理降温,烦躁者给予镇静剂等。
病原治疗:及时选用适当的抗菌药物是治疗的关键。应注意早期、足量并以杀菌剂为主;一般两种抗菌药物联合应用,多自静脉给药;首次剂量宜偏大,注意药物的半衰期,分次给药;疗程不宜过短,一般三周以上,或热退后7~10天方可酌情停药。
局部病灶的处理:化脓性病灶不论原发性或迁徙性,均应在使用适当、足量抗生素的基础上及时行穿刺或切开引流。化脓性胸膜炎、关节脓肿等可在穿刺引流后局部注入抗菌药物。胆道及泌尿道感染有梗阻时应考虑手术治疗。
建议
关键在于及时选用适当的抗菌药物,并予以休息及适量的营养。诊断基本肯定后应尽早治疗,在培养未获阳性结果前可根据细菌入侵途径及临床表现推测致病菌的种类给药,若获阳性培养而治疗效果欠佳时,则可按药物敏感试验选用适宜抗菌药物。
革兰氏阳性球菌感染者可选用青霉素、红霉素、头孢菌素等;革兰氏阴性杆菌感染则选用庆大霉素、丁胺卡那霉素、头孢菌素及半合成广谱青霉素;厌氧菌感染则首选甲硝唑,也可选用青霉素、氯霉素、氯洁霉素等;败血症确为真菌所致则应选用二性霉素。
此外,正确处理局部病灶及各阶段的突出矛盾(如感染性休克、弥漫性血管内凝血、心肾功能不全)亦很重要。
✦预防
1.保持皮肤干燥和清洁。
2.穿着宽松的衣服。
3.避免共用发刷,梳子和毛巾,因为它们可能含有与皮肤真菌菌落片段。
4.为避免脚气,应该使用备用鞋,每2、3天换洗。
5.尽量选择天然纤维制品的衣服,如棉花、蚕丝,使皮肤呼吸。
6.糖尿病患者应控制血糖水平。疾病有可能会导致免疫功能低下,增加了真菌感染的概率。
✦治疗
•大多真菌感染症状较轻微
除头癣和甲癣外,大多数真菌感染症状都较轻微,常用抗真菌霜剂治疗。一般不用抗真菌粉剂。抗真菌药物的活性成分有咪康唑、克霉唑、益康唑和酮康唑等。
一般霜剂每天涂敷两次,治疗持续到皮损消退后7~10天。如果霜剂停用太快,感染并未消除,皮疹又会复发。
抗真菌霜剂要在使用几天后才显效,其间可用皮质类固醇霜剂缓解瘙痒和疼痛。严重的或顽固性感染,可用灰黄霉素治疗几个月,有时同时用抗真菌霜剂。
•部分口服药物会引起副作用
口服灰黄霉素很有效,但可引起副作用,如头痛、胃肠道功能紊乱、光敏、水肿和白细胞减少等。停用灰黄霉素后,感染可能复发。皮肤真菌感染也可用酮康唑治疗。与灰黄霉素一样,口服酮康唑也有严重的副作用,包括肝脏损害。
保持感染部位清洁、干燥有助于抑制真菌繁殖,促进皮肤愈合。感染处应经常用肥皂和水清洗,擦干后扑撒滑石粉。避免使用含玉米粉的粉剂,因为它容易促进真菌生长。
注意
如果真菌感染有渗液,可能并发了细菌感染。需要用抗生素治疗。涂敷抗生素霜剂或口服抗生素。稀释醋酸铝溶液或怀特菲尔德软膏也可用来使渗液的皮肤干燥。
由于病毒使用了宿主细胞来进行复制并且寄居其内,因此很难用不破坏细胞的方法来杀灭病毒。现在最积极的对付病毒疾病的方法是疫苗接种来预防病毒感染或者使用抗病毒药物来降低病毒的活性以达到治疗的目的。
部分病毒感染以对症支持治疗为主,如普通感冒,甲型肝炎、病毒性胃肠炎等,可通过补液维持体内水分、电解质和酸碱平衡。
✦疫苗接种预防感染
疫苗接种是一种廉价而又有效的防止病毒感染的方法。早在病毒被发现之前,疫苗就已经为人们用于预防病毒感染。随着疫苗接种的普及,病毒感染相关的一些疾病(如小儿麻痹、痳疹、腮腺炎和风疹)的发病率和死亡率都大幅度下降,而曾经是致命疾病的天花已经绝迹。
目前各类疫苗可以预防超过30种对人体的病毒感染,而有更多的疫苗被用于防止动物受到的病毒感染。
疫苗的成分可以是活性降低或死亡的病毒,也可以是病毒蛋白质(抗原)。活疫苗包含了活性减弱的可致病的病毒,这样的病毒被称为“减毒”病毒。
注意
虽然活性减弱,但活疫苗对于那些免疫力较弱或免疫缺陷的人可能是危险的,对他们注射活疫苗可能反而会导致疾病。
生物技术和基因工程被用于改造病毒疫苗,改造后的疫苗(即亚单位疫苗)只含有病毒的衣壳蛋白,如乙肝疫苗。由于不含有病毒核酸,因此亚单位疫苗对于免疫缺陷的病人是安全的。
对于活疫苗的安全性也有一些例外,如黄热病毒疫苗,虽然是一种减毒病毒株(被称为17D),却可能是目前所有疫苗中最安全和最有效的。
✦治疗
•抗病毒药物
在过去的二十年间,抗病毒药物的发展非常迅速。艾滋病的不断蔓延推动了对抗病毒药物的需求。抗病毒药物常是核苷类似物,当病毒复制时如果将这些类似物当作核苷用于合成其基因组就会产生没有活性的病毒基因组(因为这些类似物缺少与磷相连能够相互连接形成DNA“骨架”的羟基,会造成DNA的链终止),从而抑制病毒的增殖。
核苷类似物作为抗病毒药物的例子包括阿昔洛韦,可用于抑制单纯疱疹病毒感染,和拉米夫定,可用于治疗艾滋病和乙型肝炎。阿昔洛韦是最早出现也是最经常被指定使用的抗病毒药物。其他使用中的抗病毒药物是针对病毒生活周期的不同阶段。艾滋病毒需要依赖一种被称为HIV-1蛋白酶的作用来获得完整的感染能力;而通过使用大量的蛋白酶抑制剂类的药物可以使这种酶失活。
•“吃病毒”生物
近日,美国的研究人员称,他所在的研究团队于近日首次发现了有生物会把“病毒”作为“食物”。他们研究后发现,两种浮游生物——“Halteria”和“Paramecium”可以主动食用病毒并茁壮成长。
研究发现,“Halteria”在两天内就有明显成长的迹象,纤毛的种群在两天内增长了约15倍,而氯病毒含量则下降了100倍以上。而在没有氯病毒的对照样本中,“Halteria”与初始状态相差无几。“Paramecium”也有类似表现,同样把氯病毒作为营养来源。并且,标记在氯病毒DNA的荧光绿移动痕迹证实病毒被“吃掉”了。
从科学上讲,这是人类第一次改变看待病毒的方式:病毒不仅是导致机体发生病变的“病原体”,还可以是自然界食物链中的一环。这可能对未来治疗病毒相关疾病具有重要作用。
细菌、真菌、病毒等生物共同构成了我们生活的世界。它们基本上无处不在,也时刻影响着我们的生命活动。而人体内的肠道菌群是其中数量最庞大的一类,其对人们的影响巨大。
微生物检测技术可以较为清晰地发现生活中的一些细菌和其他微生物,有助于我们判断健康状况,并根据此来做出一些调整。
随着测序技术和其他体外诊断技术的快速发展,新标志物的发现等新技术的发展将能更快更精准的区分和诊断感染病原,甚至大大提高用药的针对性和减少无效用药和耐药性,将有利于人类和微生物更好的共存。
主要参考文献:
Kwiecinski JM, Horswill AR. Staphylococcus aureus bloodstream infections: pathogenesis and regulatory mechanisms. Curr Opin Microbiol. 2020 Feb;53:51-60. doi: 10.1016/j.mib.2020.02.005. Epub 2020 Mar 12. PMID: 32172183; PMCID: PMC7244392.
Fisher CR, Streicker DG, Schnell MJ. The spread and evolution of rabies virus: conquering new frontiers. Nat Rev Microbiol. 2018 Apr;16(4):241-255. doi: 10.1038/nrmicro.2018.11. Epub 2018 Feb 26. PMID: 29479072; PMCID: PMC6899062.
Riley LW. Distinguishing Pathovars from Nonpathovars: Escherichia coli. Microbiol Spectr. 2020 Dec;8(4). doi: 10.1128/microbiolspec.AME-0014-2020. PMID: 33385193.
Cheung GYC, Bae JS, Otto M. Pathogenicity and virulence of Staphylococcus aureus. Virulence. 2021 Dec;12(1):547-569. doi:
10.1080/21505594.2021.1878688. PMID: 33522395; PMCID: PMC7872022.
Liu N, Pang X, Zhang H, Ji P. The cGAS-STING Pathway in Bacterial Infection and Bacterial Immunity. Front Immunol. 2022 Jan 13;12:814709. doi: 10.3389/fimmu.2021.814709. PMID: 35095914; PMCID: PMC8793285.
Klein EY, Monteforte B, Gupta A, Jiang W, May L, Hsieh YH, Dugas A. The frequency of influenza and bacterial coinfection: a systematic review and meta-analysis. Influenza Other Respir Viruses. 2016 Sep;10(5):394-403. doi: 10.1111/irv.12398. Epub 2016 Jun 24. PMID: 27232677; PMCID: PMC4947938.
Piepenbring M, Maciá-Vicente JG, Codjia JEI, Glatthorn C, Kirk P, Meswaet Y, Minter D, Olou BA, Reschke K, Schmidt M, Yorou NS. Mapping mycological ignorance – checklists and diversity patterns of fungi known for West Africa. IMA Fungus. 2020 Jul 7;11:13. doi: 10.1186/s43008-020-00034-y. PMID: 32699745; PMCID: PMC7341642.
Wu HY, Chang PH, Huang YS, Tsai CS, Chen KY, Lin IF, Hsih WH, Tsai WL, Chen JA, Yang TL, Lee CY, Ho TS, Wang HW, Huang SF, Wu AY, Chen HJ, Chen YC, Chen WC, Tseng CH, Lin PC, Yang CH, Hong PL, Lee SS, Chen YS, Liu YC, Wang FD; Infectious Disease Society of Taiwan; Medical Foundation in Memory of Dr. Deh-Lin Cheng; Foundation of Professor Wei-Chuan Hsieh for Infectious Diseases Research and Education; CY Lee’s Research Foundation for Pediatric Infectious Diseases and Vaccines,; 7th Guidelines Recommendations for Evidence-based Antimicrobial agents use in Taiwan (GREAT) working group; Members of the expert panel and board members of the IDST are listed in alphabetical order. Recommendations and guidelines for the diagnosis and management of Coronavirus Disease-19 (COVID-19) associated bacterial and fungal infections in Taiwan. J Microbiol Immunol Infect. 2022 Dec 21:S1684-1182(22)00284-5. doi: 10.1016/j.jmii.2022.12.003. Epub ahead of print. PMID: 36586743; PMCID: PMC9767873.
Dickson RP, Singer BH, Newstead MW, Falkowski NR, Erb-Downward JR, Standiford TJ, Huffnagle GB. Enrichment of the lung microbiome with gut bacteria in sepsis and the acute respiratory distress syndrome. Nat Microbiol. 2016 Jul 18;1(10):16113. doi: 10.1038/nmicrobiol.2016.113. PMID: 27670109; PMCID: PMC5076472.
de Wilde AH, Snijder EJ, Kikkert M, van Hemert MJ. Host Factors in Coronavirus Replication. Curr Top Microbiol Immunol. 2018;419:1-42. doi: 10.1007/82_2017_25. PMID: 28643204; PMCID: PMC7119980.

谷禾健康

对微生物组的深入研究,有望为困扰我们的健康问题提供新的解决之道。
然而目前有大多数微生物基因组尚未被培养,就算在已发现的基因组序列中,也有很多无法进行功能注释。因此还不能充分捕捉微生物系统发育树的功能多样性,这限制了我们对生物序列高级特征进行建模的能力。而模型的构建又是微生物组研究中重要的一块。
这里我们介绍一个新发布的深度学习模型,为大家的微生物研究提供一些新的思路。

研究人员搭建了LookingGlass程序,应用RNN循环神经网络和LSTM长短期记忆神经网络学习方法,学习序列中的每个核苷酸字符,以达到能预测分类不同功能、同源性和环境起源的reads的目的。
LookingGlass模型还具有迁移学习的能力,经过微调后可以执行一系列不同的任务:例如识别新的氧化还原酶,预测酶的最适温度,以及识别氨基酸序列。
LookingGlass模型能够对其他未知和未注释的序列进行功能相关表征,从而挖掘微生物暗物质。
代表性细菌和古细菌基因组序列的分类由GTDB51(89.0 版)确定。
完整的基因组序列通过 NCBI Genbank ftp下载。这产生了 24,706 个基因组,包括 23,458 个细菌基因组和 1248 个古细菌基因组。
为了确定它们实际的序列长度,使用MetaSeek API下载了它们的测序元数据。去除长度<60bp或>300bp的样本,最终获得了平均序列长度为136bp的共计7909个样本。
LookingGlass模型的训练集、验证集和测试集都是在纲水平上划分的,在该分类水平下三者之间没有重叠的部分。
主要应用RNN循环神经网络和LSTM长短期记忆神经网络。
LookingGlass使用三层LSTM编码器模型,每个隐藏层有1152个单元,根据超参数调整的结果,embedding大小为104。
LookingGlass以自我监督的方式进行训练,根据序列中前面的核苷酸的上下文,预测一个被掩盖的核苷酸。
对于训练集序列中的每个reads,考虑多个训练输入,将被掩盖的核苷酸沿序列长度从第二位置移动到最后位置。因为它是一个字符级模型,线性解码器从可能的词汇“A”、“C”、“G”和“T”中预测序列中的下一个核苷酸,并带有“开始阅读”的特殊标记、“未知核苷酸”(对于不明确序列的情况)、“读取结束”(在LookingGlass训练期间仅对“读取开始”进行标记)和“填充”标记(仅用于分类)。
LSTM 的正则化和优化利用 dropout 和梯度下降方法以获得最佳性能,使用fastai 库进行训练。
硬件方面,LookingGlass在 Microsoft Azure 上,内存为 16GB的 Pascal P100 GPU 进行训练。总共训练了 12 天,共 75 个 epoch,根据超参数优化的结果逐渐降低学习率:15 个 epoch,学习率为 1e-2,15 个 epoch,学习率为2e-3,并以 1e-3 的学习速率进行 45 个 epoch。
通过随机搜索调整超参数,主要调整:
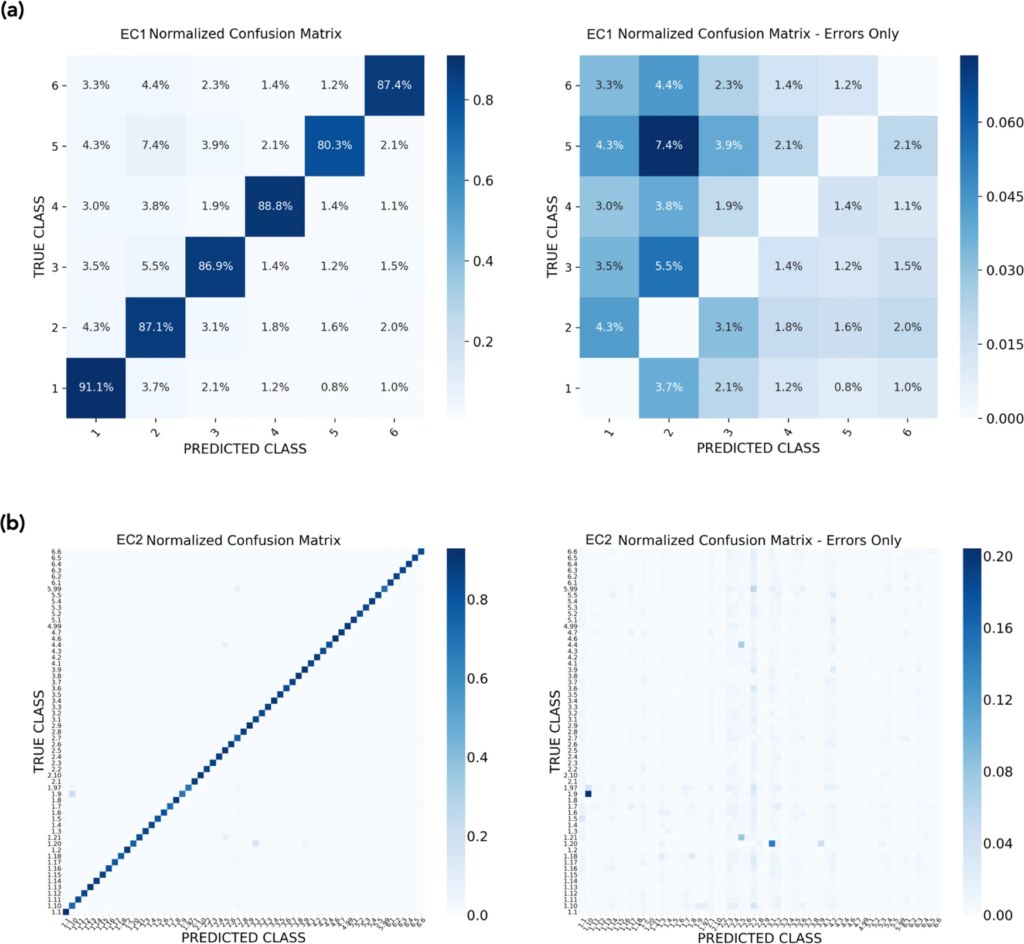
功能注释预测的多分类混淆矩阵。横轴表示真实值,纵轴表示预测值。方块内的数值为归一化后的预测百分比,左边为预测正确的,右边为预测错误的。
图a表示对验证集中EC功能编号的第一个位置的预测,图b表示对第二个位置的预测,显示准确率都在80%以上。

LookingGlass在门水平上识别同源序列对。蓝色为同源(Homologous),红色为非同源(Nonhomologous)。
图a为embedding相似性度的组间比较,同源组显著高于非同源组,embedding相似度计算为embedding向量之间的余弦相似度。
图b为准确度、精确度、召回率和 F1分值的变化,可见在embedding相似性阈值为0.62时其准确度(accuracy)最高,达到了66.4%,这是指门水平的。文中表示在纲水平上达到了68.3%,在目水平上达到了73.2%,在科水平上达到了76.6%,在属水平上达到了78.9%。LookingGlass使用embedding方法区分同源和非同源序列,而不依赖它们的序列相似性(Smith-Waterman比对)。
图c比较了这两种方法在搜索同源物时的结果,图中的黑框表示被LookingGlass 正确识别的同源序列,但使用比对时遗漏了。可见许多同源物具有非常低的序列相似性(bit score<50),不能被基于比对的方法捕获到,但LookingGlass可以。LookingGlass识别同源基因的高精度,与它们的序列相似性无关,表明它捕捉到了高水平的特征,可能反映了序列之间的在系统发育上的关系。

来自100个不同环境样本的宏基因组功能注释集作为验证集。从中对每个环境组别中随机抽取20000个序列计算embedding相似度。发现组间的embedding相似性通常低于组内的,即来自相同环境背景的序列通常聚集在一起。
以LookingGlass的底层架构作为起点,微调预训练模型,以执行不同任务。
1. 预测分类氧化还原酶

对LookingGlass功能注释分类模型进行微调后,执行氧化还原酶的预测分类任务。
上图为对序列相似性(bit score<50)的序列,预测分类是否为氧化还原酶的编码基因的准确度、精确度、召回率和 F1分值的变化,结果表示默认阈值为0.5时,其准确度(accuracy)最高,为82.3%。
基于LookingGlass模型能够区分不同环境背景下的序列这一优势,研究人员使用来自16个海洋宏基因组作为测试集,样品覆盖范围从纬度(从-62 度到 76 度),海洋深度(从表层~5米到中层~ 200-1000米)以及氧浓度(包括来自氧最低区的4个中上层样品),并从中对每个宏基因组随机抽取 2000 万reads。
挖掘其中的氧化还原酶序列,并证明LookingGlass对氧化还原酶的分类优于传统的基于同源性的方法。

图a为LookingGlass在海洋表层区(surface)、中层区(mesopelagic)以及氧浓度最低区(OMZ)预测分类为氧化还原酶序列的比例。
图b为在海洋表层区组中,纬度与氧化还原酶的相关性,结果表示存在显著相关(R2 = 0.79,P = 0.04)。
图c为分别使用LookingGlass、MG-RAST和 mi-faser工具搜索氧化还原酶序列,并统计其预测为氧化还原酶(oxidoreductases)、非氧化还原酶(not oxidoreductases)和未注释(unannotated)的序列比例。结果为MG-RAST 注释了 26.7-50.3% 的reads,其中 0.01-4.0% 被鉴定为氧化还原酶。Mi-faser 注释了 0.17-2.9% 的reads,其中 0.04-0.59% 被鉴定为氧化还原酶。可见,LookingGlass更具优势。
2. 使用LookingGlass识别氨基酸序列
LookingGlass直接从CDS预测翻译帧起始位置(1、2、3、-1、-2 或 -3),准确率达到了97.8%,但目前仅用于非编码DNA比例较低的原核序列。
3. 从 DNA 序列片段预测酶的最佳温度
酶的最佳温度部分取决于 DNA 序列特征,但难以预测,尤其是短读长。将温度划分为嗜冷(<15°C)、嗜温(20-40°C)或嗜热(>50°C),微调LookingGlass程序后,输入序列预测最佳温度类别,准确率达70.1%。
LookingGlass程序能够不通过比对参考数据库来预测表征DNA序列,从而获得功能注释和系统发育相关信息。同时,LookingGlass的迁移学习框架能够快速学习、训练和收敛以适用不同分类任务,这对于未来复杂生物系统建模提供了一些贡献。
预测分类氧化还原酶可以挖掘位置序列的潜在功能,未来还会扩大可预测的酶类。预测酶的最佳温度可以用于指导蛋白质设计所需的功能和最佳温度。
总而言之,这是一个不错的探索。作者已经将本文所用到的模型功能封装为python库—fastBio,能够直接使用本文使用到的数据集进行模型训练,可在以下地址获得:github.com/ahoarfrost/fastBio/
主要参考文献:Hoarfrost A, Aptekmann A, Farfañuk G, Bromberg Y. Deep learning of a bacterial and archaeal universal language of life enables transfer learning and illuminates microbial dark matter. Nat Commun. 2022 May 11;13(1):2606. doi: 10.1038/s41467-022-30070-8. PMID: 35545619; PMCID: PMC9095714.

谷禾健康
人的身体拥有数千种细菌,这些细菌在维持健康方面发挥着重要作用。当这些细菌失控繁殖并侵入身体的其他部位或将有害细菌引入身体的系统时,可能会发生细菌感染。
细菌感染的严重程度取决于所涉及的细菌类型和所感染的部位等。细菌最常感染肠道、皮肤和呼吸系统,包括肺、泌尿道和阴道。
目前应对细菌感染的主要治疗方法还是使用抗生素,但是抗生素管理,或改进抗生素的处方和使用方式,对于优化感染患者的治疗、保护患者免受伤害和对抗抗生素耐药性至关重要。
今天我们重点了解下什么是细菌感染,感染的症状、原因和类型,包括诊断测试和治疗方案,抗生素什么时候使用,什么时候不用,抗生素的耐药性如何产生以及使用抗生素的注意事项等。
了解什么是细菌感染以及如何治疗细菌感染,我们应该先了解细菌的概念。
来源:MedicineNet
地球上所有的生物都是由两种基本类型的细胞中的一种组成:
真核细胞,其遗传物质被包裹在核膜内;
原核细胞,其遗传物质与细胞的其他部分不分离。
传统上,所有的原核细胞都被称为细菌,被归为原核生物界。
1970年代后期,美国微生物学家卡尔·沃斯(Carl Woese)率先在分类上进行了重大变革,将所有生物分为真核生物、细菌(原名真细菌)和古细菌(原名古细菌)三个领域,以反映三条古老的进化路线。原核生物以前被称为细菌,然后被分为两个领域,细菌和古细菌。

published by Woese et al.
细菌和古细菌在表面上是相似的,例如,它们没有细胞内的细胞器,它们有环状DNA。但是,它们在本质上是截然不同的,它们的分离是基于其古老而又独立的进化谱系的遗传证据,以及其化学和生理学的根本差异。这两个原核域的成员彼此之间的区别与它们和真核细胞中的区别一样。
细菌细胞在几个方面不同于动物细胞和植物细胞。一个根本的区别是细菌细胞缺乏动物细胞和植物细胞中都存在的细胞内细胞器,例如线粒体,叶绿体和细胞核。细菌体积小,设计简单,代谢能力强,使它们能够迅速生长和分裂,并在几乎任何环境中生存和繁衍。
细菌,动物和植物细胞的比较

除此之外,原核和真核细胞在许多其他方面有所不同,包括脂质组成,关键代谢酶的结构,对抗生素和毒素的反应以及遗传信息的表达机制。
真核生物包含多个线性染色体,这些染色体的基因比编码蛋白质合成所需的基因大得多。遗传信息的核糖核酸(RNA)副本(脱氧核糖核酸或DNA)的大部分被丢弃,剩余的信使RNA(mRNA)在被翻译成蛋白质之前已被充分修饰。相反,细菌具有一个包含所有遗传信息的环形染色体,它们的mRNA是其基因的精确副本,不会被修饰。
细菌细胞在结构上比真核细胞小得多,也简单得多,但细菌是一个在大小、形状、生境和代谢上都有差异的极其多样化的有机体群体。
许多关于细菌的知识来自对致病细菌的研究,这些细菌比许多自由生活的细菌更容易在纯培养中分离出来,也更容易被研究。必须注意的是,许多自由生活的细菌与适应作为动物寄生虫或共生体生活的细菌有很大的不同。因此,关于细菌的组成或结构没有绝对的规则,任何说法都会有许多例外。
大多数细菌对地球上的生命都是有益的,甚至是生命所必需的,但少数细菌却对人类有害。目前,没有一种古细菌被认为是病原体,但是包括人类在内的动物不断遭到大量细菌的“轰击和居住”。
口腔,肠道和皮肤被大量特定类型的细菌定殖,这些细菌适应这些栖息地的生活。这些微生物在正常情况下是无害的,并且仅在它们以某种方式穿过身体的屏障并引起感染时才变得危险。
一些细菌擅长入侵宿主,被称为病原体或疾病产生者。一些病原体作用于人体的特定部位,例如:
在食物中传播的许多致病菌的毒素在摄入时会引起食物中毒。
由金黄色葡萄球菌(Staphylococcus aureus)产生的毒素,能引起迅速、严重但有限的胃肠道不适。
肉毒梭菌(Clostridium botulinum)的毒素,通常是致命的。在密封前未完全煮熟的罐装非酸性食品中可能产生肉毒杆菌毒素。肉毒梭菌形成耐热孢子,可以发芽为营养细菌细胞,在厌氧环境中茁壮成长,这有利于产生其极强的毒素。
其他食源性感染实际上是由受感染的食物处理者传播的,包括伤寒、沙门氏菌病(沙门氏菌属)和志贺氏菌病(痢疾志贺氏菌属)。
关于食物中毒相关细菌详见:正值夏季,警惕食源性疾病,常见的食物中毒的病原菌介绍
细菌感染是指病原菌侵入宿主组织。当有害细菌进入人体或伤口并繁殖,导致疾病、器官损伤、组织损伤或疾病时,就会发生细菌感染。细菌可以感染身体的任何部位。
细菌感染很常见,但它们并不完全相同。细菌有很多种,每种对身体都有不同的影响。广义上来讲,细菌感染也可以是由任何细菌引起的任何内部或外部疾病的广义术语。
细菌存在于与人体的各种关系中。它们在体表内定殖并提供益处,正常情况下细菌可以与人类身体共存不会产生有害的影响(在共生关系中)。但是有时候,通常无害地存在于我们体内或皮肤上,而不会造成任何异常问题的细菌会失控并导致感染。这种情况通常发生在我们的免疫系统不够强大,无法保持它们的平衡,或者我们的微生物区系的组成发生了一些变化,对某些细菌生存繁殖更有利。
细菌感染是由体内的坏细菌引起的。一些细菌感染可能危及生命。标准的医学治疗通常涉及一剂抗生素。细菌感染很容易与病毒感染混淆。但是他们并不一样,后面我们会单独解释。
当皮肤粘膜有破损或发生化脓性炎症时,细菌则容易侵入体内。
人体的免疫反应可分为非特异性免疫反应及特异性免疫反应两种,后者又可分为细胞免疫与体液免疫两方面。当机体免疫功能下降时,不能充分发挥其吞噬杀灭细菌的作用时,即使入侵的细菌量较少,致病力不强也能引起感染;条件致病菌所引起的医源性感染也逐渐增多。
人体免疫功能正常时,进入血中的细菌迅速被血中防御细胞如单核细胞、嗜中性粒细胞等所清除,而患肝硬变、糖尿病、血液病、结缔组织病等慢性病者,可因代谢紊乱、体液免疫及细胞免疫功能减低,易招致败血症发生;各种免疫抑制药物的使用、放射治疗亦是导致败血症发病率高的原因。广谱抗菌药物使用后,对药物敏感的细菌虽被抑制或杀灭,而一些耐药菌乘机繁殖,亦可酿成败血症。
细菌侵入人体后是否引起感染,除了与人的防御、免疫功能,还与细菌的毒力及数量有关。毒力强或数量多的致病菌进入机体,引起感染的可能性较大。
严重烧伤时,创面为细菌敞开门户,皮肤坏死、血浆渗出又为细菌繁殖提供了良好环境,故极易发生感染。
尿路、胆道、胃肠道、呼吸道粘膜受破坏后,若同时有内容物积滞、压力增高,细菌更易进入血中,保留导尿管、静脉等血管内留置导管、人工辅助呼吸时插管等,也使细菌易于侵入。
细菌是微观的,通常是单细胞生物,到处都可以找到(例如,在我们体内、皮肤上、空气中、水里、土壤中或我们吃的食物里)。细菌具有传染性,因此,它可以通过与患者的密切接触在人与人之间传播。接触受污染的表面、食物、水以及打喷嚏和咳嗽就是例子。
发生感染的几个条件:
此外,细菌还可以导致急性感染(快递治疗)和慢性感染(持续很长时间甚至终生),以及潜伏感染(最初可能没有任何明显迹象或症状但可以过段时间自然发展)。症状可以从轻度、中度到重度不等。在严重的情况下,导致数百万人死亡;例如,许多人死于黑死病或鼠疫。
细菌感染的常见特征:
常见的各部位细菌感染的症状

细菌性感冒
细菌性感冒(Bacterial cold)是指继发细菌感染引起的感冒。感冒在习惯上分为病毒性感冒和细菌性感冒。
病毒性感冒有:普通感冒、流行性感冒和病毒性咽炎等。其主要不同是致病因素不同,病毒性感冒是由于病毒所致,而细菌性感冒是由于细菌所致。
注:流行性感冒(流感通过病毒传播,不是细菌性感冒,注意区分),是由流感病毒引起的急性呼吸道传染病。
细菌的感染还取决于受感染细菌的类型,常见的细菌感染例如:
细菌感染具有高度传染性,因此需要特别注意避免通过洗手、打喷嚏和咳嗽时遮盖以及不共用杯子和饮料瓶来传播感染。
细菌有多种传播方式,包括:

当你忍不住咳嗽或打喷嚏时,用纸巾捂住口鼻或用手肘内侧。
在下述情况下需要洗手,至少洗20秒:
如果没有肥皂和水,可以使用酒精含量至少为 60% 的酒精类洗手液。
除此之外,其他需要注意的:
接种疫苗。如轮状病毒是一种导致婴儿和幼儿严重肠胃炎的病毒。所有婴儿都应在 6 周和 4 个月大时接种轮状病毒疫苗。
流感疫苗。当患上流感时,免疫系统会被削弱。这使细菌有机会侵入肺部并引起感染。每年注射一次流感疫苗可以防止感染肺部细菌。
肺炎球菌多糖疫苗。美国疾病控制与预防中心 (CDC) 推荐 65 岁及以上的人接种肺炎球菌多糖疫苗 (PPSV)。它也适用于免疫系统较弱或患肺炎风险较高的成年人。
治疗取决于感染的严重程度、患者的年龄、免疫系统有多强、是否存在任何并存症状以及现有的医疗条件。
治疗可能包括:
扩展阅读:
细菌素——对抗感染、保存食品、重塑肠道菌群
还在滥用抗生素?15种天然抗生素助你调节肠道菌群
这里我们主要来了解一下,用于细菌感染的药物——抗生素。
目前抗生素是对抗细菌感染的常用药物。它们通过破坏细菌细胞生长和增殖所需的过程来发挥作用。
抗生素根据抗菌谱可以分为:
广谱抗生素,中谱抗生素和窄谱抗生素。
✓ 广谱抗生素
广谱抗生素可对抗多种细菌。广谱抗生素可治疗革兰氏阳性菌和阴性菌,因此如果医生不确定有哪些细菌,他或她可能会开出其中一种抗生素。
阿莫西林、奥格门汀、头孢菌素(第 4 代和第 5 代)、四环素氨基糖苷类和氟喹诺酮类(环丙沙星)是广谱抗生素的例子。
✓ 中谱抗生素
中谱抗生素针对一组细菌。青霉素和杆菌肽是流行的中谱抗生素。
✓ 窄谱抗生素
窄谱抗生素用于治疗一种特定的细菌。多粘菌素属于这一小类抗生素。当确定患有哪种细菌感染时,治疗会更容易和更有效。
抗菌活性通常分为五种机制:
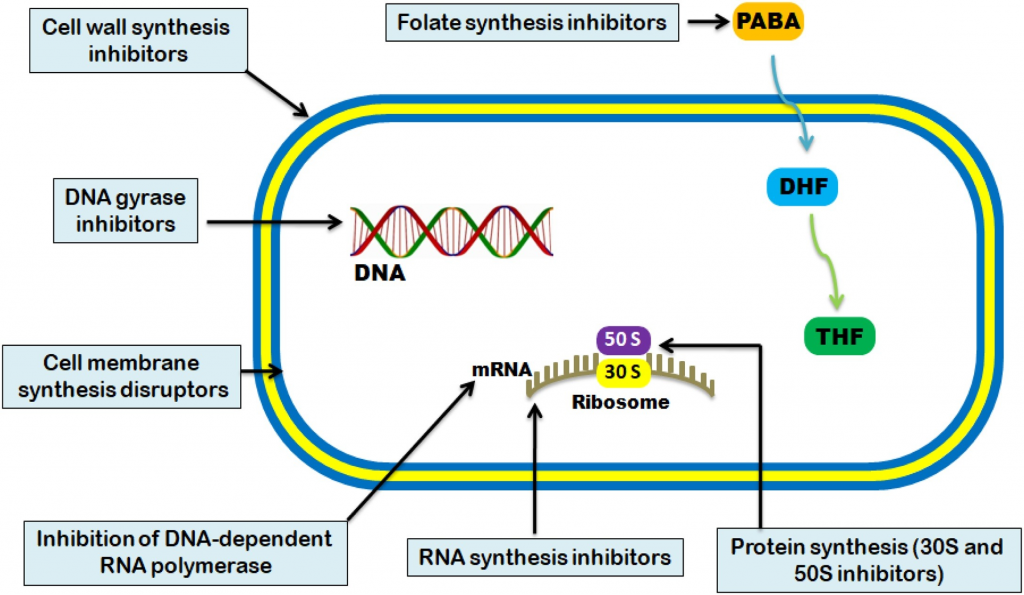

Uddin TM, et al., J Infect Public Health. 2021
不同种类抗生素作用的方式


Uddin TM, et al., J Infect Public Health. 2021
青霉素
最初的青霉素仅用于革兰阳性菌感染的治疗,由于金黄色葡萄球菌很快产生一种青霉素酶对青霉素产生了耐药,人们研制出了以甲氧西林、苯唑西林为代表的耐酶青霉素,用于产青霉素酶的葡萄球菌( 甲氧西林耐药者除外) 感染。
之后,以氨苄西林、阿莫西林为代表的广谱青霉素由仅对革兰阳性(G+) 菌有效扩展到对革兰阴性(G-) 菌( 主要为肠杆菌科细菌) 也具有抗菌活性。在广谱青霉素的基础上又发展为对铜绿假单胞菌等非发酵菌也具抗菌活性者,当前应用主要为脲基类青霉素,包括: 哌拉西林、阿洛西林、美洛西林。
代表药物: 阿莫西林(amoxicillin)
阿莫西林是一种抗细菌的青霉素抗生素。
阿莫西林用于治疗由细菌引起的多种不同类型的感染,例如扁桃体炎、支气管炎、肺炎以及耳、鼻、喉、皮肤或泌尿道感染。
阿莫西林有时还与另一种称为克拉霉素( Biaxin ) 的抗生素一起用于治疗由幽门螺杆菌感染引起的胃溃疡。这种组合有时与一种称为兰索拉唑(Prevacid)的胃酸减少剂一起使用。
四环素
四环素是一类抗生素,可用于治疗由易感微生物引起的感染,例如革兰氏阳性菌和革兰氏阴性菌、衣原体、支原体、原生动物或立克次体。
它们是在 1940 年代发现的,第一个四环素是从链霉菌中获得或衍生的。
四环素抑制微生物 RNA 中的蛋白质合成(一种重要的分子,作为 DNA 的信使)。它们主要是抑菌剂,这意味着它们可以防止细菌繁殖,但不一定会杀死它们。
四环素类药物现今应用的品种除四环素外,更常用有多西环素( 强力霉素) 和米诺环素( 二甲胺四环素) 。由于常见病原菌对本类药物耐药性普遍升高及其不良反应多见,当前本类药物临床应用已受到很大限制。
代表药物:强力霉素(doxycycline)
强力霉素是一种四环素抗生素,可以对抗体内的细菌。
强力霉素用于治疗许多不同的细菌感染,例如痤疮、尿路感染、肠道感染、呼吸道感染、眼部感染、淋病、衣原体、梅毒、牙周炎(牙龈疾病)等。
强力霉素也用于治疗由红斑痤疮引起的瑕疵、肿块和痤疮样病变。
某些形式的强力霉素用于预防疟疾、治疗炭疽或治疗由螨虫、蜱虫或虱子引起的感染。
什么是林可霉素衍生物?
林可霉素衍生物是一小类抗生素,可抑制细菌蛋白质的合成,而细菌蛋白质对细菌的生存至关重要。林可霉素衍生物保留用于治疗由肺炎球菌、葡萄球菌和链球菌的敏感菌株引起的感染。一种衍生物也可用于治疗恶性疟原虫(疟疾)。
林可霉素类包括林可霉素及克林霉素。克林霉素的体外抗菌活性优于林可霉素,主要可应用于敏感肺炎链球菌、其他链球菌属( 肠球菌属除外) 及甲氧西林敏感金葡菌所致的各种感染。该类药物对厌氧菌有良好的抗菌活性常与其他抗菌药物联合用于腹腔感染及盆腔感染,也由于其骨组织浓度较高适用于骨和关节的G + 菌感染。使用本类药物时,应注意假膜性肠炎的发生,如有可疑应及时停药。本类药物有神经肌肉阻滞作用,应避免与其他神经肌肉阻滞剂合用,应注意静脉滴注速度宜缓慢滴注,不可静脉推注。
代表药物:克林霉素(Clindamycin)
克林霉素是一种抗生素,可以对抗体内的细菌。
克林霉素用于治疗由细菌引起的严重感染。
服用此药前 如果对克林霉素或林可霉素过敏,则不应使用该药。
为确保克林霉素的安全,请告诉医生是否曾经有过:结肠炎、克罗恩病或其他肠道疾病; 湿疹或皮肤过敏反应; 肝病; 哮喘或对阿司匹林的严重过敏反应; 对黄色食用色素过敏。
什么是喹诺酮类药物?
喹诺酮类是一种抗生素。抗生素杀死或抑制细菌的生长。
有五种不同的喹诺酮类。此外,另一类抗生素,称为氟喹诺酮类,是从喹诺酮类通过氟修饰其结构衍生而来的。氟喹诺酮类抗生素可导致严重或致残的副作用,这些副作用可能不可逆。
喹诺酮类药物和氟喹诺酮类药物有许多共同点,但也有一些区别,例如它们对哪些生物有效。有些人互换使用喹诺酮和氟喹诺酮这两个词。
喹诺酮类和氟喹诺酮类对细菌产生的两种酶(拓扑异构酶 IV 和 DNA 促旋酶)的功能产生不利影响,因此它们不能再修复 DNA 或帮助其制造。
现今临床主要应用为氟喹诺酮类,常用有诺氟沙星、环丙沙星等主要用于单纯性下尿路感染或肠道感染。但应注意,现今国内尿路感染的主要病原菌大肠埃希菌中,耐药株已达半数以上。环丙沙星尚与β-内酰胺类抗生素联合用于治疗中、重度革兰阴性杆菌感染包括铜绿假单胞菌的感染。
近年来研制的新品种左氧氟沙星、莫西沙星等对肺炎链球菌、化脓性链球菌等G + 球菌的抗菌作用增强,对衣原体属、支原体属、军团菌等细胞内病原或厌氧菌的作用亦有增强,但对G-菌活性增强不明显,对铜绿假单胞菌的抗菌活性仍以环丙沙星为最强。左氧氟沙星、莫西沙星除可用于尿路感染或肠道感染外也可用于呼吸道感染,特别适合于肺炎链球菌( 包括耐青霉素肺炎链球菌) 、支原体、衣原体、军团菌等所致社区获得性肺炎的治疗,此外亦可用于皮肤软组织感染。
在治疗腹腔、胆道感染及盆腔感染时除莫西沙星外需与甲硝唑等抗厌氧菌药物合用。部分品种可与其他药物联合应用作为治疗耐药结核分枝杆菌和其他分枝杆菌感染的二线用药。
代表药物:环丙沙星(Ciprofloxacin)和左氧氟沙星(Levofloxacin)
环丙沙星和左氧氟沙星是一种氟喹诺酮抗生素,用于治疗不同类型的细菌感染。也用于治疗接触过炭疽或某些类型鼠疫的人。环丙沙星和左氧氟沙星应仅用于无法用更安全的抗生素治疗的感染。
左氧氟沙星、莫西沙星:对肺炎链球菌、A 组溶血性链球菌等革兰阳性球菌、衣原体属、支原体属、军团菌等细胞内病原或厌氧菌的作用强。
环丙沙星、左氧氟沙星:主要适用于肺炎克雷伯菌、肠杆菌属、假单胞菌属等革兰阴性杆菌所致的下呼吸道感染。
头孢霉素
头孢菌素是一大类从霉菌顶孢菌(以前称为头孢菌素)中提取的抗生素。头孢菌素具有杀菌作用(杀死细菌),其作用方式与青霉素相似。它们结合并阻断负责制造肽聚糖的酶的活性,肽聚糖是细菌细胞壁的重要组成部分。它们被称为广谱抗生素,因为它们对多种细菌有效。
自 1945 年发现第一个头孢菌素以来,科学家们一直在改进头孢菌素的结构,以使其对更广泛的细菌更有效。每次结构发生变化,都会产生新的“一代”头孢菌素。迄今为止,头孢菌素已有五代。所有头孢菌素均以 cef、ceph 或 kef 开头。请注意,该分类系统在不同国家/地区的使用并不一致。
第一代头孢菌素是指发现的第一组头孢菌素。它们的最佳活性是对抗革兰氏阳性细菌,如葡萄球菌和链球菌。它们对革兰氏阴性菌几乎没有活性。
第二代头孢菌素尚可应用于由流感嗜血杆菌、大肠埃希菌、奇异变形杆菌等中的敏感株所致的尿路感染、皮肤软组织感染、败血症、骨及关节感染和腹腔、盆腔等感染。但如用于腹腔感染和盆腔感染时需与抗厌氧菌药合用。头孢呋辛尚可用于脑膜炎球菌、流感嗜血杆菌所致脑膜炎的治疗,也可作为围术期预防用药。
第三代头孢菌素:对肠杆菌科细菌等革兰阴性杆菌具有强大抗菌作用,头孢他啶和头孢哌酮对铜绿假单胞菌亦具较强抗菌活性;注射品种有头孢噻肟、头孢曲松、头孢他啶、头孢哌酮等,口服品种有头孢克肟和头孢泊肟酯等,口服品种对铜绿假单胞菌均无作用。应注意的是,不是所有的三代头孢菌素都可用于非发酵菌( 如铜绿假单胞菌、不动杆菌等) 感染的治疗,除了头孢哌酮和头孢他啶;外,如头孢曲松和头孢噻肟不可以用于非发酵菌感染的治疗。
第四代头孢菌素:常用者为头孢吡肟,对肠杆菌科细菌作用与第三代头孢菌素大致相仿,对铜绿假单胞菌的作用与头孢他啶相仿,对革兰阳性球菌的作用较第三代头孢菌素略强。其不同于三代头孢菌素之处为对产头孢菌素酶( AmpC) 的肠杆菌属有效,其次是增强了对作用靶位细菌青霉素结合蛋白( PBP) 的亲和力以及其抗革兰阳性菌的抗菌活性亦较三代头孢菌素增强。
第五代(或下一代)头孢菌素头孢洛林( Teflaro ) 对耐甲氧西林金黄色葡萄球菌(MRSA)具有活性。Avycaz 含有β-内酰胺酶抑制剂avibactam。
代表药物:头孢氨苄(Cephalexin)
头孢氨苄是一种头孢菌素(SEF 一种低孢子)抗生素。它通过对抗体内的细菌起作用。
头孢氨苄用于治疗由细菌引起的感染,包括上呼吸道感染、耳部感染、皮肤感染、尿路感染和骨骼感染。
头孢氨苄用于治疗成人和至少 1 岁儿童的感染。
注意事项:
如果对头孢氨苄或类似抗生素(如Ceftin、Cefzil、Omnicef等)过敏,则不应使用该药。如果对任何药物(尤其是青霉素或其他抗生素)过敏,请告诉医生。
对任何药物(尤其是青霉素)过敏;
肝脏或肾脏疾病;要么肠道问题,例如结肠炎,请告诉医生。
头孢氨苄可以进入母乳。如果正在哺乳婴儿,请咨询医生。
β-内酰胺酶抑制剂
β-内酰胺酶抑制剂是一类阻断 β-内酰胺酶(也称为 β-内酰胺酶)活性,防止 β-内酰胺类抗生素降解的药物。本类药物适用于因产β-内酰胺酶的细菌感染,其抗菌谱主要依据原有抗生素的活性而酶抑制剂仅有对抗细菌产酶的作用,一般不增加抗菌活性。
β-内酰胺酶由以下细菌的某些菌株产生:拟杆菌属、肠球菌属、流感嗜血杆菌、卡他莫拉菌、淋病奈瑟菌和葡萄球菌属,无论是组成型还是暴露于抗微生物剂。
β-内酰胺酶裂解易感青霉素和头孢菌素的 β-内酰胺环,使抗生素失活。一些抗微生物药物(如头孢唑啉和氯唑西林)对某些 β-内酰胺酶具有天然耐药性。β-内酰胺类:阿莫西林、氨苄青霉素、哌拉西林和替卡西林,可以通过与β-内酰胺酶抑制剂组合来恢复和扩大其活性。
克拉维酸、舒巴坦和他唑巴坦都是β-内酰胺酶抑制剂。
大环内酯类
大环内酯类药物主要作为青霉素过敏患者的替代药物,用于治疗β-溶血性链球菌、肺炎链球菌中的敏感菌株所致的上、下呼吸道感染、敏感β-溶血性链球菌引起的猩红热及蜂窝织炎等感染。
大环内酯类是从红糖多孢菌(原名红链霉菌)中提取的一类抗生素,一种土壤传播的细菌。
红霉素是第一个发现的大环内酯类;其他大环内酯类包括阿奇霉素、克拉霉素和罗红霉素。
它们的作用主要是抑菌,但在高浓度时可能具有杀菌作用,或取决于微生物的类型。
代表药物:红霉素
红霉素为大环内酯类原型代表药物,由于其胃肠道反应大、口服吸收差使其疗效受到影响。当下在临床应用的主要为大环内酯类新品种罗红霉素、阿奇霉素和克拉霉素等,其他大环内酯类疗效多不如该三种药物。
罗红霉素主要是改善了其药代动力学,口服生物利用度明显提高、给药剂量减小、不良反应明显减少,但其抗菌谱没有明显的改善,是红霉素的替代药物。
氨基糖苷类
临床常用的氨基糖苷类药物主要有: 链霉素、卡那霉素、丁胺卡那霉素、庆大霉素、妥布霉素、奈替米星、依替米星。
其中链霉素当前主要用抗结核治疗,其次可用于治疗鼠疫及布鲁菌病; 庆大霉素、妥布霉素、奈替米星、依替米星肠杆菌科细菌和铜绿假单胞菌等G - 杆菌具强大抗菌活性,对葡萄球菌属亦有良好作用者。所有氨基糖苷类药物对肺炎链球菌、溶血性链球菌的抗菌作用均差。
链霉素、卡那霉素:对肠杆菌科和葡萄球菌属细菌有良好抗菌作用,但对铜绿假单胞菌无作用者。
庆大霉素、妥布霉素、奈替米星、阿米卡星、异帕米星、小诺米星、依替米星:对肠杆菌科细菌和铜绿假单胞菌等革兰阴性杆菌具强大抗菌活性,对葡萄球菌属亦有良好作用。
碳青霉烯类
碳青霉烯类药物是当下抗菌药物中抗菌谱最广的药物,对各种革兰氏阳性球菌、革兰氏阴性杆菌( 包括铜绿假单胞菌的非发酵菌) 和多数厌氧菌具强大抗菌活性,对大多数β-内酰胺酶高度稳定,但对甲氧西林耐药葡萄球菌和嗜麦芽窄食单胞菌等抗菌作用差。
当下在国内应用的碳青霉烯类抗生素有亚胺培南-西司他丁、美罗培南、帕尼培南-倍他米隆、比阿培南。厄他培南与上述品种不同对非发酵菌缺乏抗菌活性,而其他抗菌作用与上述药品基本相同,适合于治疗社区与医院早期特别是G-杆菌感染的重症患者。
碳青霉烯类对各种革兰阳性球菌、革兰阴性杆菌(包括铜绿假单胞菌、不动杆菌属)和多数厌氧菌具强大抗菌活性,对多数β-内酰胺酶高度稳定,但对甲氧西林耐药葡萄球菌和嗜麦芽窄食单胞菌等抗菌作用差。
现用于细菌感染的抗生素被认为是医学史上最重要的突破之一。不幸的是,细菌的适应性很强,抗生素的过度使用让许多人对抗生素产生了耐药性,造成了严重的问题,尤其是在医院的环境里。
抗生素耐药性是细菌抵抗抗生素杀伤力的能力。换句话说,以前治疗感染的抗生素没有效果了,或者说根本无法杀死细菌。
由于过度使用抗生素,抗生素耐药率持续上升,新的抗菌药物开发缓慢。感染耐药细菌可能导致住院时间更长、费用更高,并增加死于感染的风险。
抗生素耐药性是对全球健康的紧迫威胁,包括后来逐渐出现危险耐药细菌——“超级细菌”。
疾病控制和预防中心 (CDC)发布了对美国的前 18 种耐药性威胁的清单,至少包括:
注:以上这些并未全面列出所有耐药菌。抗生素耐药性模式不断演变,细菌可能并不总是对每位患者的选择抗生素表现出耐药性。在正常情况下,抗生素的选择应基于感染部位和由医疗保健专业人员评估的临床表现、培养/敏感性和其他所需的实验室结果、局部耐药性/敏感性模式以及患者的特定特征。
细菌通过以某种方式调整其结构或功能作为防御机制,从而对抗生素产生抗药性。
当细菌以某种方式发生变化时,细菌会抵抗药物。这种变化可以保护细菌免受药物的影响或限制药物接触细菌。或者这种变化可能导致细菌改变药物或破坏药物。
细菌可以用以下方式与抗生素抗争:
如果说以上列举的是抗生素产生耐药性的内因,那么除细菌本身外的因素也会导致抗生素耐药性的出现。
早前,医生有时会依赖不可靠或不准确的知识,开具抗生素“以防万一”,或在特定窄谱抗生素可能更合适时却开出广谱抗生素。这些情况加剧了选择压力并加速了抗菌素耐药性。
当医生不清楚是细菌或病毒加剧了感染时,他们可能会开抗生素。抗生素对病毒感染不起作用,可能会产生耐药性。
抗生素自我治疗(SMA)与药物使用不当的可能性有关,这会使患者面临药物不良反应的风险,掩盖潜在疾病的迹象,并在微生物中产生耐药性。
过度使用抗生素,尤其是在不正确治疗的情况下服用抗生素,会促进抗生素耐药性。根据疾病控制和预防中心的数据,人类使用大约三分之一的抗生素既不需要也不合适。
抗生素可以治疗由细菌引起的感染,但不能治疗由病毒引起的感染(病毒感染)。例如,抗生素是可以治疗由细菌引起的链球菌性咽喉炎。但对于大多数由病毒引起的喉咙痛,并不是正确的治疗方法。
每天都有成千上万的患者、工作人员和访客到达医院,每个人的衣服上和身体上/体内都有自己的一套微生物组。如果医院没有适当的程序和规程来帮助保持空间清洁,细菌就会传播。
抗生素被用作动物的生长补充剂和生长促进剂。在家畜中发现的耐抗生素细菌可能对人类致病,很容易通过食物链传播给人类,并通过动物粪便在生态系统中广泛传播。在人类中,这可能会导致复杂的、无法治疗的和长期的感染。
由于技术挑战、缺乏知识、对抗细菌生理学的重大困难,制药业对新抗生素的发明在很大程度上放缓了,当新抗生素普及时,耐药性的发展(在相对较短的时间内)几乎是不可避免的。
世界范围内抗生素的过度使用和滥用正在导致抗生素耐药性的全球医疗问题。可能会发生抗生素耐药性感染,在最坏的情况下,可能会没有有效的抗生素。这种情况在严重感染时可能会危及生命。
大多数病毒性疾病不需要特殊药物并且是“自限性”的,也就是说患者自身的免疫系统可以抵抗疾病。病毒性疾病的患者可以休息、多喝水并使用对症治疗。
由病毒引起的疾病包括:
有时,在复杂或长期的病毒感染中,细菌也可能侵入,并导致所谓的“继发感染”。在这些情况下,如果需要,可以使用抗生素。
在感染由病毒引起的疾病中,例如咳嗽、感冒或流感,患者不应要求医生开抗生素。抗生素不能治愈病毒感染,患者可能会因不必要的药物而产生副作用。医生可以用其他方法来帮助患有病毒性疾病的患者进行治疗。
如果感染没有得到正确治疗,抗生素耐药细菌也可能传播给其他人。
——有以下几个原因:
它可能不是治疗感染的正确抗生素
它可能已过时且无效
整个疗程可能没有足够的药物
如果新疾病是病毒感染,则不需要抗生素
一些疫苗可以预防细菌性疾病。抗病毒疫苗,例如流感疫苗或COVID-19 疫苗,可以帮助预防可能与继发性细菌感染(如严重的肺部肺炎)相关的原发性疾病。
在治疗的最初几天感觉好些了的时候,仍应完成整个抗生素疗程。
抗生素使用注意事项
✓ 按照指示使用抗生素
按照医生的指示治疗感染。医生会选择最能对抗导致感染的特定细菌的抗生素类型。目前市面上有许多不同种类的抗生素,大部分需要医生开出的非处方抗生素。
✓ 了解抗生素
有的抗生素需要进行皮试,比如青霉素,以防过敏,不同的抗生素杀菌的作用原理不尽相同,比如青霉素是通过能破坏细菌的细胞壁并在细菌细胞的繁殖期起杀菌作用的一类窄普抗生素,杀菌作用强,可用于敏感菌所致的严重感染,由于该类药物作用的靶位为细菌细胞壁,而人体的细胞没有细胞壁,因此毒性低,用于老年人,新生儿和孕妇时安全性相对较高。
✓ 确保清楚抗生素的使用方式
确保明确知道应该服用多少抗生素,以及何时服用。有些抗生素需要与食物一起服用,有些需要在晚上服用等。特别注意要用完整的病征疗程抗生素,有些人担心抗生素对身体有副作用,病情稍微好转就会停止服用或注射抗生素,但是这可能导致体内的致病细菌未完全清楚,再次发生感染,或引起形成抗生素抗药性,使得以后的感染治疗变得困难。
✓ 清洁伤口以防止细菌感染
通过立即正确清洁和包扎伤口来预防皮肤感染。适当的急救治疗对于帮助预防细菌感染至关重要,但不应尝试自己治疗严重的皮肉伤口。如果伤口很深、很宽或出血很多,应该立即寻求医疗帮助。
治疗伤口前先洗手。如果你用脏手治疗伤口,会增加细菌感染的机会。用温水和抗菌肥皂洗手 20 秒,然后擦干。如果有条件可以戴上手套。
细菌感染性疾病起病急、进展快,重症疑难患者病原细菌复杂,诊断及治疗困难,可危及生命。早期、准确地明确病原微生物对感染判定至关重要,有助于改善预后。
准确诊断细菌感染对于避免不必要的抗生素使用和集中适当的治疗至关重要。
细菌感染是细菌的存在与炎症或全身功能障碍的结合;因此,通常需要不止一种诊断方式进行确认,确定患者是否符合临床病例定义的病史和检查。
不同的病原微生物检测技术对感染的判定价值不同,临床上也会根据患者的病情,可能感染的部位,可供采集的标本等选择适宜的检测技术。
当前,病原微生物检测技术层出不穷,能识别的微生物种类越来越多,但如何正确利用和分析微生物检测结果仍是临床关注的难题。值得注意的是,感染首先是个临床诊断,所有的判定必须基于临床。
定植容易对感染判定产生混淆。正常生理状态下人体口腔、胃肠道、呼吸道及体表等部位均有细菌、真菌存在,患者无感染症状时称为定植菌。当定植菌的致病力改变或机体防御能力下降时,可大量繁殖或被带入机体深部引起感染成为致病菌。
临床医生应如何判断所获取的病原体为定植还是感染,是长久以来的难题,且未形成成熟统一的解决方案。
呼吸道定植菌的综合分析:
就呼吸道定植菌而言,既往认为可根据菌落数量或菌种拷贝数来判定定植与感染,但结果存在偏差。越来越多的学者认为,临床上判定是定植还是感染,需结合患者症状、体征及其他检查结果综合分析。
若患者存在与检测阳性结果相匹配的临床症状和体征,如发热、咳嗽、咳痰等,感染指标升高,肺部出现新发病灶,应考虑感染并给予相应的抗感染治疗。如仅培养阳性,患者无任何感染相关临床表现,则倾向于定植菌。
念珠菌定植的综合分析:
念珠菌可广泛定植于呼吸道、胃肠道及泌尿生殖道,正常屏障破坏、免疫功能受损或局部菌群失调时,定植的念珠菌可生长繁殖引起感染。痰和尿标本中分离出的念珠菌亦应根据临床症状和体征鉴别是定植还是感染,念珠菌血培养阳性时要高度警惕念珠菌血症。
此外,病原微生物检测结果阳性,并不意味着患者一定存在感染或感染一定由检出的病原体所致,需结合标本质量、采集部位、病史及其他检测结果综合分析。
不同部位标本检测出同一种病原体,其代表的临床意义可能并不相同。
以肺炎克雷伯氏菌为例,从肠道粪便样本中检出肺炎克雷伯氏菌,其中丰度占比超过1%的人群有3765例,占比28.2%【谷禾健康数据库】。但是当肺炎克雷伯菌进入血液并感染时,通常会发生细菌血症。症状可能包括发冷,发烧,发抖,肌肉酸痛,疲劳和嗜睡。当肺炎克雷伯菌感染肝脏时,会引起脓性病变,也被称为化脓性肝脓肿。这种类型的感染更常见于患有糖尿病一段时间或已经服用抗生素多年的人。
因病原微生物种类不同,微生物检测方法的选择也不相同;即便检测同一种微生物,因采集部位不同所选择的检测方法也有所差别。
如大肠埃希菌,血培养阳性时需考虑血流感染,怀疑致病性大肠埃希菌感染肠道时需特殊培养或者血清学、PCR,16s测序检测等。因普通大肠埃希菌可寄居在肠道中,粪便普通培养有大肠埃希菌生长并不能直接考虑大肠埃希菌肠道感染。
病原微生物检测技术快速发展,能识别的微生物种类越来越多,但感染首先是个临床诊断,所有的判定必须基于临床。随着检测技术革新带来的是临床辅助手段的进步和多样化,更快、更准确地判定感染一直都是病原学检测的挑战,不同方法各有优劣。当前,并没有一项技术可通过从标本中识别微生物来直接判定机体是否感染该病原体,甚至依靠检测的阴性结果来排除感染都很难实现。
在面对细菌感染诊疗过程中始终需要思考3个问题:患者是否存在感染?如果有感染,致病病原体是什么?如何治疗干预?
充分利用现有的病原微生物检测方法和其他实验室检查手段,综合判定,给出合适的抗菌治疗或其他综合治疗方法,是科学可持续的手段。
主要参考文献:
Uddin TM, Chakraborty AJ, Khusro A, Zidan BRM, Mitra S, Emran TB, Dhama K, Ripon MKH, Gajdács M, Sahibzada MUK, Hossain MJ, Koirala N. Antibiotic resistance in microbes: History, mechanisms, therapeutic strategies and future prospects. J Infect Public Health. 2021 Dec;14(12):1750-1766. doi: 10.1016/j.jiph.2021.10.020. Epub 2021 Oct 23. PMID: 34756812.
Cheung GYC, Bae JS, Otto M. Pathogenicity and virulence of Staphylococcus aureus. Virulence. 2021 Dec;12(1):547-569. doi: 10.1080/21505594.2021.1878688. PMID: 33522395; PMCID: PMC7872022.
Tshibangu-Kabamba E, Yamaoka Y. Helicobacter pylori infection and antibiotic resistance – from biology to clinical implications. Nat Rev Gastroenterol Hepatol. 2021 Sep;18(9):613-629. doi: 10.1038/s41575-021-00449-x. Epub 2021 May 17. PMID: 34002081.
BRM, Mitra S, Emran TB, Dhama K, Ripon MKH, Gajdács M, Sahibzada MUK, Hossain MJ, Koirala N. Antibiotic resistance in microbes: History, mechanisms, therapeutic strategies and future prospects. J Infect Public Health. 2021 Dec;14(12):1750-1766. doi: 10.1016/j.jiph.2021.10.020. Epub 2021 Oct 23. PMID: 34756812.
Carmen Fookes, BPharm, Bacterial Infection. January 13, 2020 Medicine
Spagnolo F, Trujillo M, Dennehy JJ. Why Do Antibiotics Exist? mBio. 2021 Dec 21;12(6):e0196621. doi: 10.1128/mBio.01966-21. Epub 2021 Dec 7. PMID: 34872345; PMCID: PMC8649755
Baquero F, Coque TM, Cantón R. Counteracting antibiotic resistance: breaking barriers among antibacterial strategies. Expert Opin Ther Targets. 2014 Aug;18(8):851-61. doi: 10.1517/14728222.2014.925881. Epub 2014 May 31. PMID: 24881465.
武洁, 王荃. 病原微生物检测在感染判定的意义[J]. 中国小儿急救医学, 2020, 27(3):6.
李丹鹤, 荣爱国, 马瑞芝,等. 病原微生物检验在抗感染经验治疗中的临床意义[J]. 医学理论与实践, 2019, 32(09):127-129.
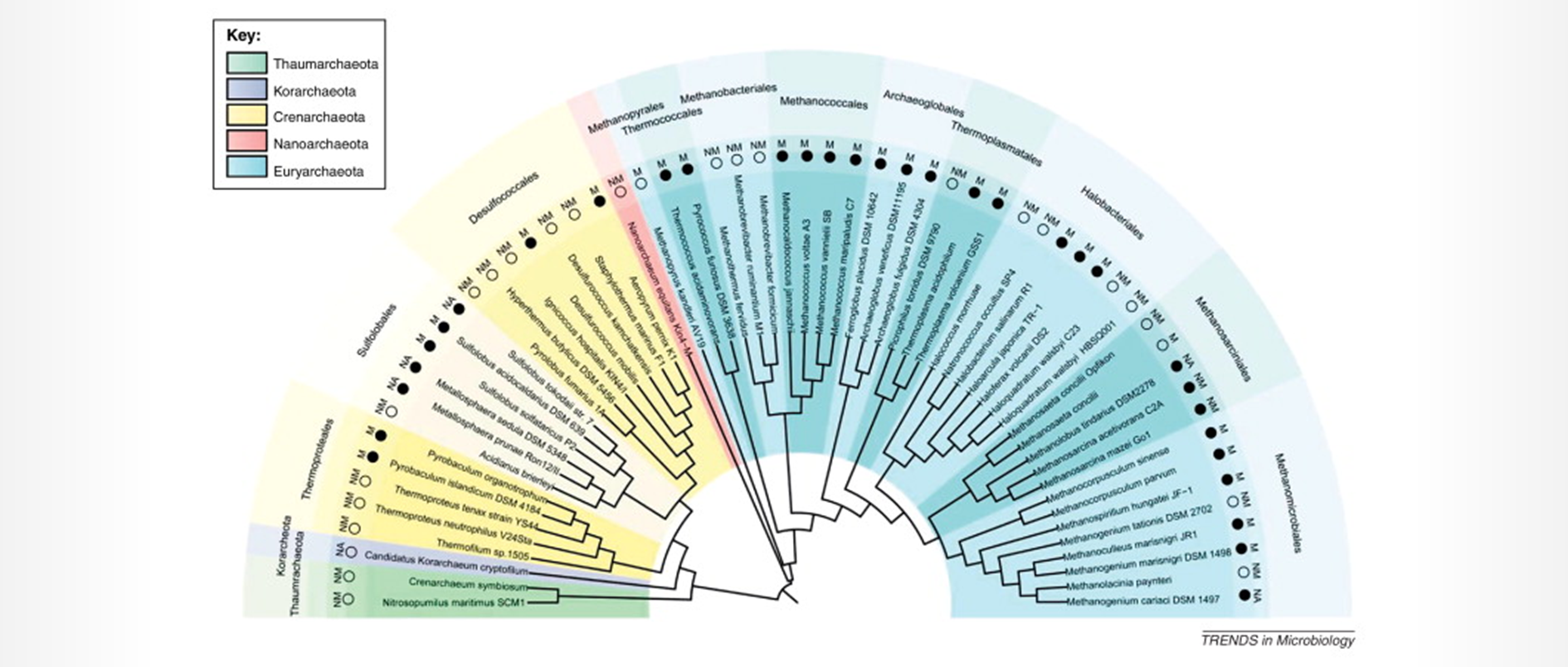
谷禾健康
随着基因组测序的发展,大量的数据为分类学提供了更全面的框架。基因组序列的空前可用性意味着分类学可以建立在一个全面的进化框架上,这是分类学家的长期目标。
然而,采用单一的框架来保持分类自由存在阻碍,而且从未培养的原核生物衍生的基因组数量不断增加,威胁着目前基于特征分离物的命名实践。
接下来的挑战是就分类学框架达成共识,并对现有的命名密码进行调整和扩展,或者创建一个新的密码,以系统地将未培养的分类单元纳入所选择的框架。

近日,一篇发表在The ISME Journal(10.1038/s41396-021-00941-x)题为“Prokaryotic taxonomy and nomenclature in the age of big sequence data” 的文章,针对微生物分类学和命名法进行完整概述,主要侧重于细菌和古菌,从过去到现代研究其发展脉络,并探讨了如何利用基因组测序技术来让微生物分类学更加全面和完善。
将生命形式分类为一个等级系统(taxonomy 分类学),并将名称应用于这个等级系统(nomenclature 命名法)是微生物学领域的一个重要转折点。
最早在1700年代,Linnaeus在工作中就引入了现代生物分类学 taxonomy(将动植物分类为等级类别)和命名法 nomenclature(命名动植物分类组的规则)的原理,这些原则如今已成为生物分类的基础。
最初的分类法是基于共同的特性,发育过程和行为,后来统称为表型,这些特征用于区分遗传信息(基因型)。
Taxonomy 分类学在生物学中最常被定义为科学的一个分支,它根据共同的特性对生物体进行命名和分类。
生物学家认同分类学应该建立在进化关系的基础上,因为进化关系是排列生物的最自然的方式。由于微生物的表型特征大部分都没有揭示其共同祖先,所以微生物仍是系统发育框架中最有争议的分类单元 。
第一次尝试根据表型特性对细菌进行分类,始于1923年Bergey 写的第一版确定性细菌学手册,细菌分类为嵌套的层次分类,以指示不同的相关度。最初,排名从最高(最远亲)到最低(最紧密相关)排名。关键主要依赖于形态、培养条件和致病特征,其主要目标是在物种水平上进行分离菌株的实际鉴定,而不是构建一个进化框架。
数值分类法,由Sokal和Sneath于1962年提出,用于定量比较包含几十种特征的细菌的表型特性。虽然在原则上,数字分类法可以纳入系统发育信息,但在实践中,它主要用于鉴定,缺乏一个严格的进化框架。
Zuckerkandl 和 Pauling 通过使用信息大分子来推断进化关系,打破了表型预测的局限性。Woese 发现核糖体可以用来构成所有生命进化框架的基础。最著名的是小亚基核糖体RNA(16S / 18S rRNA),基于其高度的序列保守性,并且和可变区域穿插在一起。而对小亚基rRNA的比较分析能够为整个生命树提供客观的进化框架。 Woese和他的同事的分析的亮点是发现古细菌。

图源:scienceterms.net
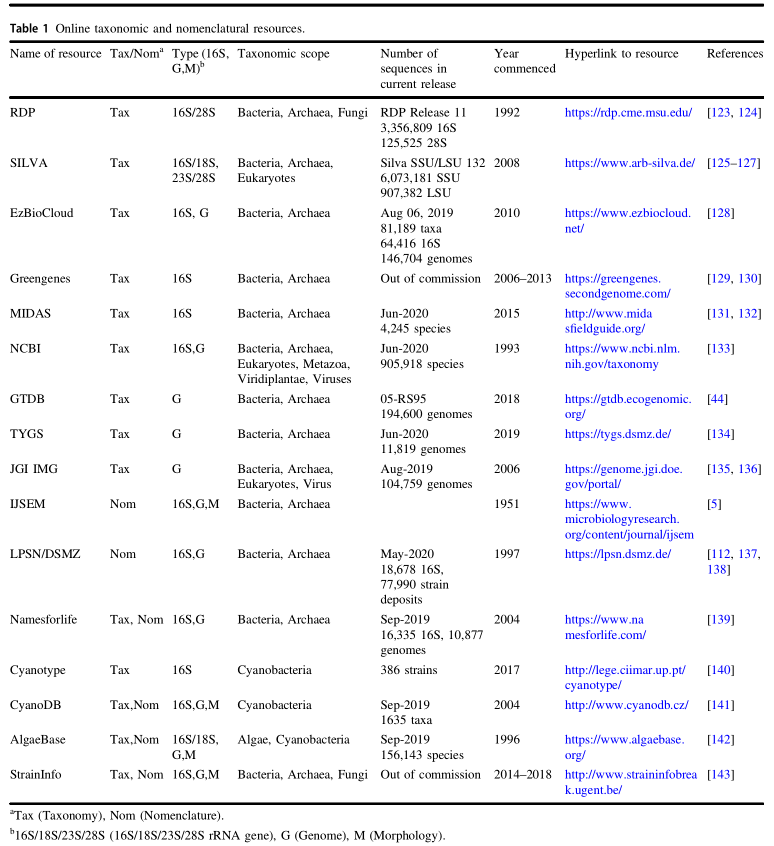
传统培养方法遗漏的大量微生物可以通过16S rRNA基因获取。Pace和同事首次通过使用高度保守的“通用”引物,直接从环境中获得的16S rRNA序列表征微生物。这些引物随后用于从提取的基因组环境DNA中PCR扩增16S rRNA基因。 然后将混合的扩增子克隆并测序,来获得微生物群落。随着测序技术的改进,大量的样本从各种环境被提取出来,从而开发出各种数据库和工具,用于分析和分类16S rRNA基因序列(表1)。
Hugenholtz P ,et al. ISME J,2021
多相分类法整合了表型和基因型的一种共识分类法,该分类法最能反映生物体的多种属性。
随着16S rRNA分析的出现,系统发育分类得到发展,由于16S rRNA基因的高序列保守性,对多相分类进行了分层,以使16S rRNA树可为属及以上分类水平属提供分类信息。
而化学分类学方法(如多基因座酶电泳和全细胞蛋白质分析)可以更好地划分物种和亚种水平。
全基因组测序的出现以及近年来技术的进步,加速了细菌和古细菌分类学上的发展,从基于16S rRNA的分类向基于基因组分类过渡。
与16S rRNA基因一样,基因组序列也可用于构建系统发育框架,并以此为基础进行系统分类。
基于基因组的分类比16S rRNA基因(仅占平均3-Mbp原核基因组的0.05%)对最古老和最近的关系提供了更高的分辨率,这是因为在比较中使用了更大比例的基因组,从而提供了更好的系统发育信号。
基于基因组序列构建进化树的方法有两种:超级树 supertrees 和 超级矩阵 supermatrices。
超级树:创建独立的基因树,然后将其组合以产生生物之间的系统发育关系。
超级矩阵:将基因连接到比对序列的系统发育矩阵中,然后从该矩阵中推断出树。
两种方法尽管基于不同的标记基因集,它们在分类学上的一致性为98.2%。
基于基因组序列的其他方法包括:成对的基因组之间进行相似性测量,或在编码蛋白质水平,或在核酸水平和DNA-DNA杂交,这些方法主要用于定义和识别物种。


基因组测序也可以对未培养的物种进行分析。随着测序技术和计算方法的改进,如今可以从环境DNA中获得天然微生物种群近乎完整的基因组序列,即所谓的宏基因组组装的基因组(MAG)。由于相对容易从单个基因组中获得多个MAG,因此可用MAG的数量迅速超过了分离基因组的数量。
未培养的分类单元的基因组数据的快速积累给分类学带来巨大挑战,根据系统发育树的指标,估计未培养的分类单元占微生物多样性的85%以上,意味着过去几十年建立的分类框架在其中存在重大差距。这个问题在病毒世界中更加明显。
众所众知,原核微生物分类学在系统发育上存在不一致的情况(多系统分类学)由以下原因导致:沿用历史上的表型数据、基于16S rRNA基因序列嵌合来自环境的PCR产物、基于早期的系统发育结论缺乏合适群体。来自未培养分类群的基因组使得问题更加复杂。
因此,已经开发了一些数据库和工具,试图通过建立可靠的微生物分类系统发育框架来解决这些缺点,首先使用16srRNA基因序列,最近使用宏基因组序列。但这必须建立在数十万个序列提供的微生物多样性全局视图上,对于单个基因来说已经很困难了,对于宏基因组来说更为困难。随着信息的不断增长,这些数据库能否继续扩张还有待观察。
从历史上看,基于表型数据的等级定义非常主观,特别是对于物种以上等级。基于基因组的分类能实现量化操作更加客观,并且可实现定义属和更高等级的机会。
2014年,Yarza及其同事提出了基于16S rRNA基因序列同一性的标准化阈值,用于定义从属到门的原核世系。尽管可以肯定地消除了分类学分类中的许多不一致之处,并且可以容纳未培养的分类单元,但这种方法并未考虑谱系之间的系统发育关系和可变的进化速率。
命名法的发展
图1:过去100年中原核生物分类学和命名法中的关键事件
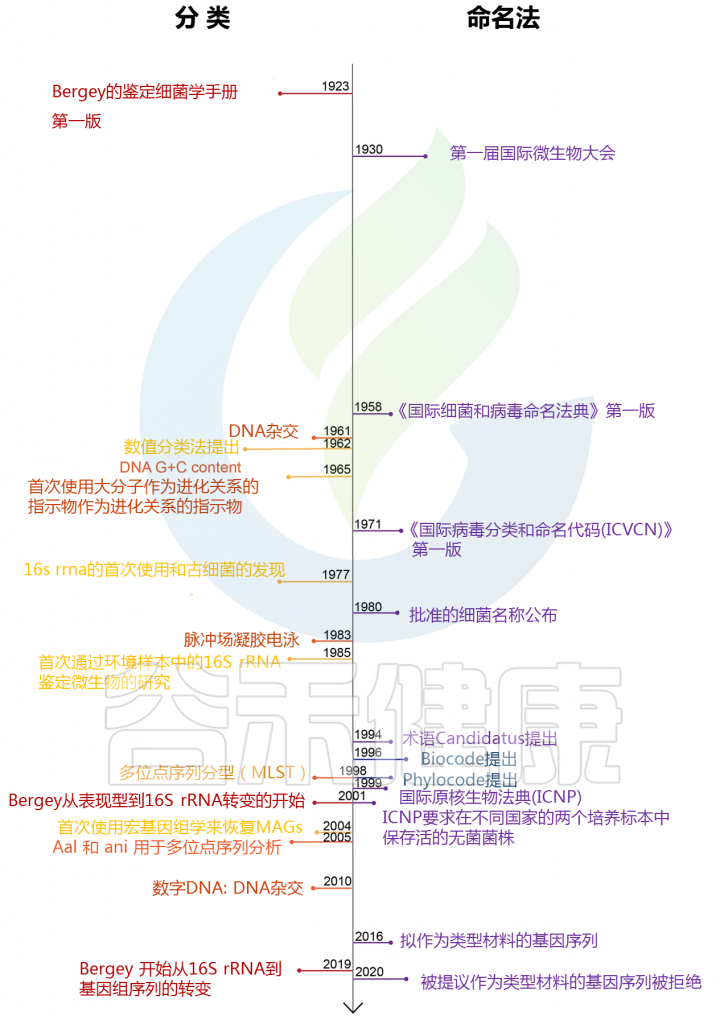

Hugenholtz P ,et al. ISME J,2021
缺乏未培养的多样性的命名法
由于技术的进步,未培养微生物的详细分子表征是一项相对较新的创新(请参见16S rRNA和基于基因组的分类)。这类生物对原核生物的编码构成了挑战,因为它们的名称不能被有效命名,这是由于物种描述必须建立在纯培养型菌株的基础上。
1994年,通过引入Candidatus的临时状态介绍,提出了一种未培养类群的早期命名规则。Candidatus一词作为任何等级的共同名称的前缀,以表示该分类单元的临时性质。
Candidatus最初是在16S rRNA环境调研时提出的。与分离株相比,它们的描述范围是受限的。然而随着MAGs和SAGs的出现,微生物不需要分离就能描述其功能。在2016年,有人提出,基因序列作为类型材料,因为它们能够用于命名法提供明确的参考点,特别是全基因组序列。鉴于掺入未培养的微生物进原核代码的命名法的困难,建议创建一个独立的编码命名这些类群。
命名定标问题
最近的一项估计显示,全球原核生物的数量在220万—430万之间,低于之前估计的数万亿。即使是这个向下修正的估计,在数百万种物种和目前有效公布的物种数量(~21K)和基因组描述的物种数量(~25K)之间仍存在巨大差距。
在未来的几十年里,作者很可能在基因组表达方面弥补这一差距,但通过原核密码验证如此大量的新物种的名称目前还不可能用于未培养的类群。并不是所有被识别的物种都需要用拉丁名称,前提是建立和维持一个系统的分类框架,该框架具有基因组限定物种的唯一和永久对象标识符。只有对科学界有足够兴趣的物种才会被更深入地描述和命名。
原核命名和微生物生态学间争论
微生物生态学家一直认为有必要为他们研究的微生物命名,然而,大多数生态学家并不太熟悉命名规则。这导致了两个学科之间的一些争论点,一旦在原核代码或新代码下更正式地考虑到未培养的分类群,这些争论点可能会扩大。
首先,代码要求严格遵守正确的拉丁语语法,在作为原始文章或验证列表发表在《国际系统与进化微生物学杂志》(IJSEM)上之前,由一小群专家定期检查名称的词源正确性。
第二,自1975年《原核生物密码法》修订以来,有一项要求,即从属名的词干和标准化后缀组成更高级别的属名。
此外,要求在从属属茎上形成更高等级的名称,导致有人提议完全改变一些更高分类单元的名称,尽管法典中有保留这一要求之前的较早名称的余地。
原核生物的分类和命名处在一个有趣的十字路口。从积极的方面来看,从来没有像现在这样,利用新兴的微生物基因组测序资源,以客观的进化关系为基础来发展一种分类方法。随着时间的推移,微生物分类法随着方法的改进而不断演变,有人认为,出于这个原因,应该避免正式的分类法,以防止它在方法上过时。然而,基因组是生命最基本的蓝图,因此不太可能出现一种被广泛接受的替代方法,从而产生一种完全不同的、改进的分类方法。尽管开发一个全面的基于基因组的分类法在生物信息学上面临挑战,但使用不同标记基因组合的独立项目之间的高度一致性预示着一个稳健的进化框架,可以形成一个稳定的分类法的基础。
虽然分类学的多相方法的想法是可以理解的,特别是使用多种特征来定义生态一致单位的目标,但作者认为,单独的基因组序列,特别是保守的垂直遗传核心操作基因的子集,应该构成分类学框架的基础。所有其他表型、基因型和生态数据都可以有效地叠加到这个框架上,以便了解它们相对于树种的个体分布和进化轨迹。
科学界普遍接受的单一一致分类法的好处是多方面的,包括改进互操作性和通信。这是发展GTDB的动力,GTDB非常强调包含性(即,使用尽可能多的来自培养和未培养分类群的高质量序列数据)和系统化(例如,定义物种代表性和等级的统一和可复制的方法,提供从领域到物种的完整分类学分配)。
一个标准化的分类学框架需要一个相似的、可复制的、客观的、能随手头的任务而扩展的命名法。官方的原核命名法是在大规模基因组测序和未培养分类群特征化出现之前制定的,因此不包括未培养微生物的大多数。
要克服这一点,要么开发一种以基因组序列为类型材料的单独命名法,要么对原核代码中分类群的管理规则进行重大修改。如果有必要开发一个单独的命名法,它可以提供一个机会,利用原核生物代码的最佳元素,简化陷入历史遗留问题的其他部分,这些部分对用户不友好,不能很好地适应大序列数据的挑战。一个例子是简化或自动形成拉丁语或希腊语的名称,并具有正确的词源,否则全世界只有少数实践者能够确保这一点。

消极的一面是,采用统一的标准化分类法将不可避免地伴随着越来越多的漏洞。一些行业已经开始制定特定的分类法和相关的命名法,它们不一定遵循进化框架。开发一个新的命名法,它确实带有与现有原核代码互操作性挑战的潜在负面影响。然而,这并不是史无前例的,例如蓝细菌的案例,因此应该通过命名委员会之间的公开对话来管理。
通过精心管理和充足的资源,对于新一代使用现代方法研究微生物世界的研究人员而言,基于基因组的分类和简化的命名将会受到欢迎。
谷禾健康


细 菌
单一细菌,是一种微观的单细胞微生物,生活在地球上的几乎每个角落,从深海喷口到地表以下再到人类的消化道,都存在于其中。
结核分枝杆菌
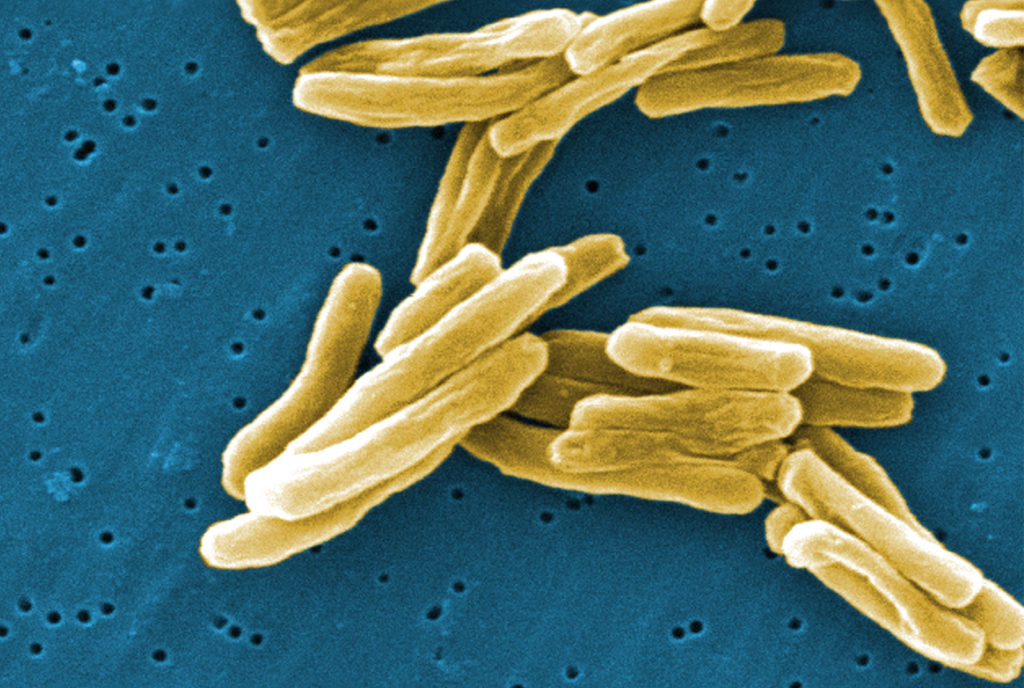
图源:Centers for Disease Control and Prevention
革兰氏阳性结核分枝杆菌细菌(结核病的原因)的扫描电子显微照片
细菌细胞
细菌缺乏与膜结合的细胞核和其他内部结构,因此被称为原核生物。原核生物是地球上占主导地位的生物,在地球历史上大约有四分之三的时间都存在,并且已经适应了几乎所有可用的生态栖息地。
作为一个群体,它们表现出极其多样化的代谢能力,几乎可以使用任何有机化合物和一些无机化合物作为食物来源。一些细菌可以引起人类、动物或植物的疾病,但大多数是无害的,是有益的生态因子,其代谢活动维持更高的生命形式。
其他细菌是植物和无脊椎动物的共生体,它们对宿主起着重要的作用,如固氮和纤维素降解。没有原核生物,土壤就不会肥沃,死去的有机物腐烂的速度也会慢得多。一些细菌被广泛用于食品、化学品和抗生素的制备。对不同细菌群之间关系的研究,不断为地球生命起源和进化机制提供新的见解。
细菌——原核生物
地球上所有的生物都是由两种基本类型的细胞中的一种组成:一种是真核细胞,其遗传物质被包裹在核膜内;另一种是原核细胞,其遗传物质不与细胞的其他部分分离。传统上,所有的原核细胞都被称为细菌,被归为原核生物界。但是,它们的分类为Monera,在分类学上与其他王国(植物界,动物界,真菌和原生生物)相当,这低估了原核细胞相对于真核细胞表现出的显着遗传和代谢多样性。
1970年代后期,美国微生物学家卡尔·沃斯(Carl Woese)率先在分类上进行了重大变革,将所有生物分为真核生物、细菌(原名真细菌)和古细菌(原名古细菌)三个领域,以反映三条古老的进化路线。原核生物以前被称为细菌,然后被分为两个领域,细菌和古细菌。细菌和古细菌在表面上是相似的;例如,它们没有细胞内的细胞器,它们有环状DNA。但是,它们在本质上是截然不同的,它们的分离是基于其古老而又独立的进化谱系的遗传证据,以及其化学和生理学的根本差异。 这两个原核域的成员彼此之间的区别与它们与真核细胞中的区别一样。
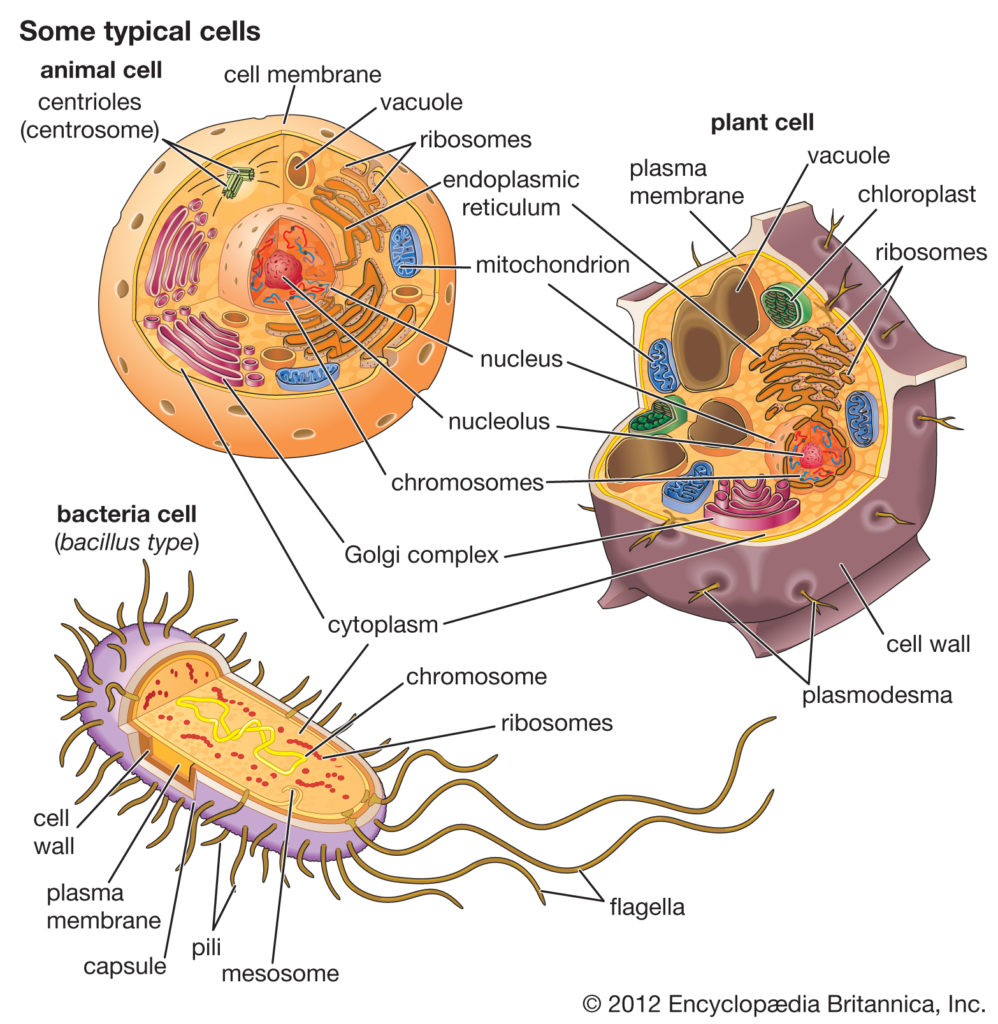
细菌,动物和植物细胞的比较
细菌细胞在几个方面不同于动物细胞和植物细胞。一个根本的区别是细菌细胞缺乏动物细胞和植物细胞中都存在的细胞内细胞器,例如线粒体,叶绿体和细胞核。
原核细胞(即细菌和古细菌)与构成其他生命形式的真核细胞有根本的不同。原核细胞的定义比真核细胞要简单得多。最明显的简化是缺乏细胞内的细胞器,这是真核细胞的特征。所有由细胞器执行的活动也发生在细菌中,但它们不是由专门的结构执行的。此外,原核细胞通常比真核细胞小得多。细菌体积小,设计简单,代谢能力强,使它们能够迅速生长和分裂,并在几乎任何环境中生存和繁衍。
杆菌型细菌细胞
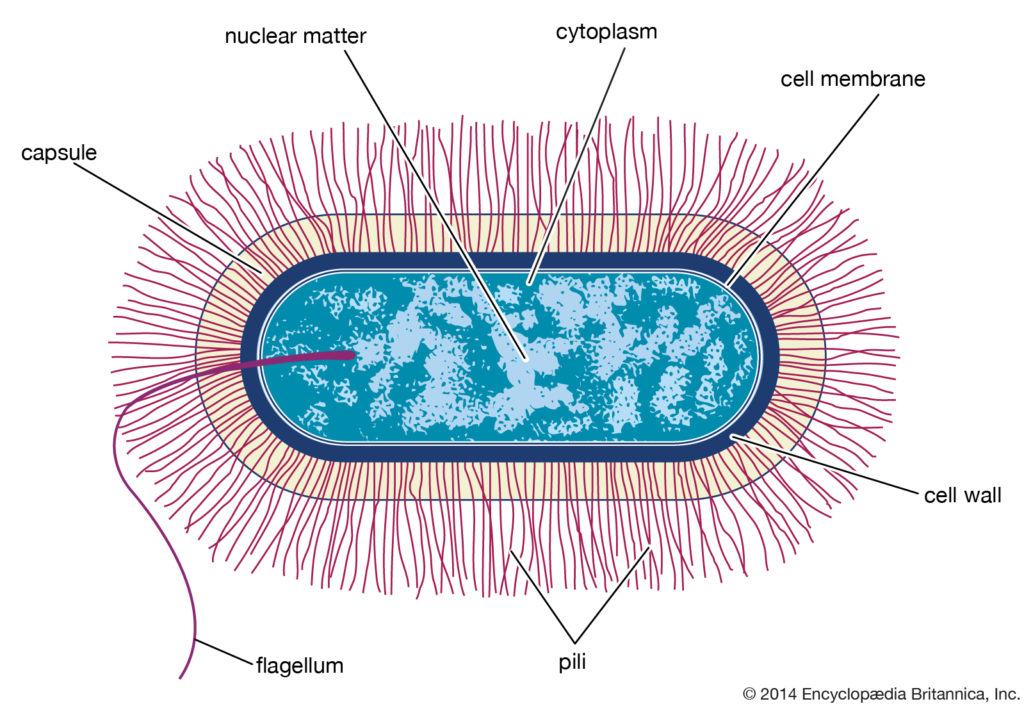
典型的芽孢杆菌属细菌细胞的结构示意图
原核和真核细胞在许多其他方面有所不同,包括脂质组成,关键代谢酶的结构,对抗生素和毒素的反应以及遗传信息的表达机制。 真核生物包含多个线性染色体,这些染色体的基因比编码蛋白质合成所需的基因大得多。 遗传信息的核糖核酸(RNA)副本(脱氧核糖核酸或DNA)的大部分被丢弃,剩余的信使RNA(mRNA)在被翻译成蛋白质之前已被充分修饰。相反,细菌具有一个包含所有遗传信息的环形染色体,它们的mRNA是其基因的精确副本,不会被修饰。
细菌结构的多样性
尽管细菌细胞在结构上比真核细胞小得多,也简单得多,但细菌是一个在大小、形状、生境和代谢上都有差异的极其多样化的有机体群体。许多关于细菌的知识来自对致病细菌的研究,这些细菌比许多自由生活的细菌更容易在纯培养中分离出来,也更容易被研究。必须注意的是,许多自由生活的细菌与适应作为动物寄生虫或共生体生活的细菌有很大的不同。因此,关于细菌的组成或结构没有绝对的规则,任何一般的说法都会许多例外。
单个细菌可以呈现三种基本形状之一:球形(球菌)、棒状(芽孢杆菌)或弧形(弧菌、螺旋体或螺旋体) 。在细菌的实际形状中可以看到相当大的变化,细胞可以在一维中拉伸或压缩。细胞分裂后不分离的细菌形成有助于鉴定的特征性簇。
例如,有些球菌主要成对出现,包括肺炎链球菌Streptococcus pneumoniae(一种引起细菌性大叶性肺炎的肺炎球菌)和淋病奈瑟菌Neisseria gonorrhoeae(一种引起性传播疾病淋病的淋病球菌)。大多数链球菌类似于一长串珠子,而葡萄球菌则形成随机的团块(“葡萄球菌”这个名字来源于希腊语staphyle,意思是“葡萄簇”)。
此外,有些球菌呈方形或立方形。杆状杆菌通常单独出现,但有些菌株形成长链,如棒状杆菌的杆状杆菌,通常以任意角度彼此相连。有些杆菌的末端是尖的,而有些杆菌的末端是方的,有些杆菌的杆弯曲成逗号形状。这些弯曲的杆状病毒通常被称为弧菌,包括霍乱弧菌,它能引起霍乱。
其他形状的细菌包括弯曲和弯曲的螺旋形螺线管和螺旋形螺线管,螺旋形螺线管类似于开瓶器,其中细胞体包裹在称为轴向细丝的中央纤维周围。
变形链球菌
图源:David M. Phillips/Visuals Unlimited
变形链球菌细菌是球形(球菌)细菌的一个例子。这种细菌通常会聚集成对和短链。
细菌是最小的生命体。一种细菌的平均大小,如杆状大肠杆菌,是人类和动物肠道的“正常居民”,长约2微米(μm;百万分之一米),直径0.5μm,金黄色葡萄球菌的球形细胞直径可达1μm。
少数细菌种类甚至更小,如肺炎支原体是最小的细菌之一,其宽度约为0.1至0.25μm,长度约为1至1.5μm;百日咳的病原体为棒状百日咳杆菌,其直径为0.2至0.5μm,长度为0.5至1μm;梅毒病原体螺旋形梅毒螺旋体直径仅0.1~0.2μm,长度6~15μm。蓝藻聚球藻平均直径约0.5~1.6μm。有些细菌相对较大,例如固氮菌,其直径为2至5μm或更大;无色菌,其最小宽度为5μm,最大长度为100μm,视物种而定。用肉眼可以看到巨大的细菌,例如平均直径为750μm的Thiomargarita namibiensis和长度在30到600μm之间的杆状Epulopiscium fishelsoni。
细菌是单细胞微生物,因此通常不会组织起来。 每个细菌的生长和分裂都独立于其他任何细菌,尽管常会发现细菌的聚集体,有时包含不同物种的成员。许多细菌可以形成称为生物膜的聚集结构。
生物膜中的生物通常表现出与处于个体状态或浮游状态的同一生物基本不同的特性。 聚集到生物膜中的细菌可以传达有关种群大小和代谢状态的信息。 这种类型的通信称为群体感应,通过产生称为自动诱导剂或信息素的小分子来进行操作。
群体感应分子(最常见的是肽或酰化高丝氨酸内酯(AHL;特殊的信号化学物质))的浓度与生物膜中相同或不同物种的细菌数量有关,有助于协调生物膜的行为。
革兰氏染色
细菌如此之小,直到1677年才被首次发现,当时荷兰科学家安东尼·列文虎克借助原始显微镜(在设计上与现代放大镜比现代显微镜更相似)在各种物质中看到了微生物,其中一些能放大200倍以上。现在细菌通常在能放大1000倍以上的光学显微镜下进行检查;然而,只有借助更强大的透射电子显微镜才能观察到其内部结构的细节。除非使用特殊的合适的显微镜,否则细菌必须用有色染料染色,以使它们从背景中脱颖而出。
肺炎克雷伯菌
图源:AW Rakosy /Encyclopædia
从肺炎患者的肺脓肿中分离出的革兰氏阴性杆菌肺炎克雷伯菌。
对细菌最有用的染色反应之一叫革兰氏染色,由医生汉斯·克里斯蒂安·格兰姆(Hans Christian Gram)于1884年开发。
悬浮液中的细菌通过短暂加热固定在玻片上,然后暴露在两种染料中,这些染料会在每个细胞内结合形成大的蓝色染料复合物。 当用酒精溶液冲洗载玻片时,革兰氏阳性菌会保留蓝色,而革兰氏阴性菌会失去蓝色。然后用一种较弱的粉红色染料将玻片染色,这种粉红色的染料会使革兰氏阴性菌变成粉红色,而革兰氏阳性菌则保持蓝色。革兰氏染色剂对细菌细胞表面结构的差异起反应,当在电子显微镜下观察细胞时,这种差异是显而易见的。
金黄色葡萄球菌
图源:AW Rakosy /Encyclopædia
实验室培养中的革兰氏阳性球菌金黄色葡萄球菌
细胞包膜
细菌细胞表面(或包膜)的结构变化很大,它在细胞的性质和能力中起着核心作用。所有细胞的一个特征是细胞质膜,它将细胞内部与外部环境分开,调节营养物质的流动,维持适当的细胞内环境,并防止细胞内容物的丢失。细胞质膜执行许多必要的细胞功能,包括能量产生、蛋白质分泌、染色体分离和有效的营养物质主动运输。它是一种典型的由蛋白质和脂类组成的单位膜,与包围所有真核细胞的膜基本相似。在电子显微照片中,它是由脂质和蛋白质组成的三层结构,完全包围细胞质。
水螺螺旋藻的肽聚糖层
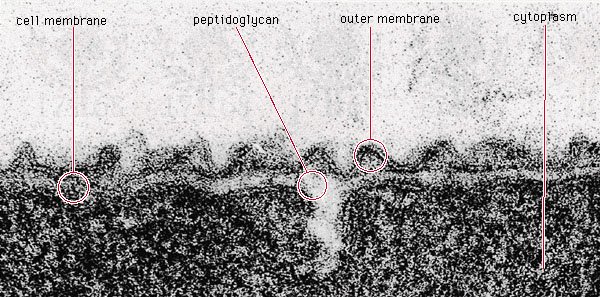
图源:AW Rakosy /Encyclopædia
革兰氏阴性细菌蛇形藻(Aquaspirillum serpens)具有薄的肽聚糖层,位于细胞膜和外膜之间。
肽聚糖只存在于细菌中(没有细胞壁的细菌除外,如支原体)。肽聚糖是两个重复糖(N-乙酰氨基葡萄糖和N-乙酰壁酸)的长链聚合物,其中相邻的糖链通过肽桥相互连接,从而提供刚性稳定性。
肽桥的性质在不同种类的细菌之间有很大差异,但一般由四种氨基酸组成:L-丙氨酸与D-谷氨酸相连,在革兰氏阴性细菌中与二氨基丙酸相连,或在革兰氏阳性细菌中与赖氨酸、L-鸟氨酸或二氨基丙酸相连,最后与D-丙氨酸相连。
在革兰氏阴性菌中,肽桥将一条链上的D-丙氨酸连接到另一条链上的二氨基丙酸。在革兰氏阳性细菌中,可以有一个额外的肽链延伸交叉连接的范围;例如,在金黄色葡萄球菌中有一个额外的五个甘氨酸桥。
肽聚糖的合成是许多有用的抗菌剂的目标,包括阻断肽桥交联的β-内酰胺类抗生素(如青霉素)。动物合成的一些蛋白质作为天然抗菌防御因子攻击细菌的细胞壁。例如,一种名为溶菌酶的酶分解肽聚糖分子的主链糖链。这些药物中的任何一种都会削弱细胞壁并破坏细菌。
在革兰氏阳性细菌中,细胞壁主要由一层厚的肽聚糖网状结构组成,该网状结构与被称为磷壁酸(来自希腊单词teichos,意思是“壁”)的其他聚合物以及一些蛋白质或脂质交织在一起。相反,革兰氏阴性菌有一个复杂的细胞壁,由多层组成,其中一个外膜层位于一个薄肽聚糖层的顶部。这种外膜由磷脂和脂多糖组成,磷脂是一种含有磷酸盐分子的复合脂质,脂多糖是一种复合脂质,通过脂质末端锚定在细胞外膜上,并有一长链糖从细胞延伸到培养基中。
脂多糖,通常被称为内毒素,对动物和人类是有毒的;它们在血液中的存在会导致发烧、休克,甚至死亡。
对于大多数革兰氏阴性细菌来说,外膜形成了一道屏障,阻碍了许多对细菌有害的化学物质的通过,例如通常溶解细胞膜的染料和洗涤剂。对油溶性化合物的不渗透性在其他生物膜中是不存在的,这是由于膜中存在脂多糖和外膜蛋白的不寻常特性造成的。作为外膜抵抗恶劣环境的能力的证据,一些革兰氏阴性细菌在浮油、喷气燃料箱、酸性矿井排水甚至是瓶装消毒剂中生长良好。
古细菌的表面结构与细菌明显不同。它们没有肽聚糖;相反,它们的膜脂是由支链类异戊二烯通过乙醚键与甘油相连组成的。一些古菌的壁材料与肽聚糖相似,只是与氨基酸桥相连的特定糖不是壁酸而是塔罗糖胺酸。许多其他古细菌使用蛋白质作为其细胞壁的基本组成部分,有些则缺乏坚硬的细胞壁。
胶囊和粘液层
许多细菌细胞以荚膜或粘液层的形式分泌一些细胞外物质。黏液层松散地与细菌结合在一起,很容易被洗掉,而胶囊则紧紧地附着在细菌上,并有明确的边界。通过将细胞置于印度墨水的悬浮液中,在光学显微镜下可以看到胶囊。胶囊排除了墨汁,在细菌细胞周围出现了清晰的光晕。
胶囊通常是单糖(多糖)的聚合物,尽管炭疽杆菌的胶囊是由聚谷氨酸制成的。大多数胶囊都是亲水的,可以通过防止水分流失帮助细菌避免干燥(脱水)。胶囊可以保护细菌细胞免受白细胞的吞噬和破坏。虽然逃避吞噬作用的确切机制尚不清楚,但可能是因为胶囊使细菌表面成分更光滑,帮助细菌逃避吞噬细胞的吞噬。肺炎链球菌中存在的胶囊是导致肺炎的最重要因素。肺炎链球菌的突变株失去了形成荚膜的能力,很容易被白细胞吸收,不会引起疾病。在许多其他种类的细菌中也发现了毒力和荚膜形成的联系。
醋酸钙不动杆菌
图源:T.J. Beveridge
这些细菌周围的荚膜材料(钙不动杆菌)显示在印度墨水的悬浮液中,并通过光学显微镜(放大约2500倍)观察。
胞外多糖材料的包膜层可以将许多细菌包裹成一个生物膜,并具有多种功能。引起龋齿的变形链球菌Streptococcus mutans,会分解食物中的蔗糖,并利用其中一种糖类来构建它的荚膜,使之紧紧地附着在牙齿上。被困在胶囊中的细菌利用其他糖来促进新陈代谢,并产生一种强酸(乳酸),攻击牙釉质。当铜绿假单胞菌定植于囊性纤维化患者的肺部时,它会产生一种厚的海藻酸囊膜聚合物,从而导致根除细菌的困难。Zoogloea属的细菌分泌纤维素纤维,将细菌缠绕成漂浮在液体表面的絮状物,使细菌暴露在空气中,这是该属新陈代谢的需要。一些杆状细菌,如Sphaerotilus,分泌长而复杂的管状鞘,这些鞘包围了大量的细菌。这些细菌和许多其他环境细菌的外壳会被铁或锰氧化物包裹。
变形链球菌
图源:Kateryna Kon / Shutterstock
变形链球菌是一种在口腔中发现的细菌,可导致蛀牙。
鞭毛、菌毛
许多细菌是运动的,能够在液体介质中游动,或在固体表面滑动或聚集。游动和成群的细菌拥有鞭毛,鞭毛是运动所需的细胞外附属物。
鞭毛是由单一类型的蛋白质组成的长而螺旋的细丝,位于杆状细胞的末端,如霍乱弧菌或铜绿假单胞菌,或遍布细胞表面,如大肠杆菌。鞭毛可以在革兰氏阳性和革兰氏阴性杆菌上找到,但在球菌上很少见,并且被困在螺旋体的轴向丝中。
鞭毛在其基部附着在细胞膜的基体上。在膜上产生的原动力被用来转动鞭毛丝,就像涡轮一样,由氢离子流通过基体进入细胞。当鞭毛以逆时针方向旋转时,细菌细胞以直线游动;顺时针方向旋转导致以相反方向游动,或者,如果每个细胞有多个鞭毛,则随机翻滚。趋化性使细菌能够调整自己的游泳行为,使其能够感觉到并向不断增加的引诱性化学物质或远离排斥性化学物质的方向迁移。
细菌不仅能够游向或滑向更有利的环境,而且它们还有附属物,使它们能够粘附在表面,防止被流动的液体冲走。有些细菌,如大肠杆菌和淋病奈瑟菌,会产生笔直、坚硬的尖刺状突起,称为菌毛,它们从细菌表面延伸出来,附着在这些菌株的其他细胞上的特定糖上,肠上皮细胞或泌尿道上皮细胞。菌毛只存在于革兰氏阴性菌中。某些菌毛是用来让一个细菌识别和坚持另一个在有性交配过程中称为接合。许多水生细菌产生一种酸性粘多糖固着物,使它们能够紧紧地附着在岩石或其他表面。
细胞质
虽然细菌在表面结构上有很大的不同,但它们的内部内容物非常相似,显示出相对较少的结构特征。
所有细胞的遗传信息都存在于DNA超长分子的含氮碱基序列中。与真核细胞中的DNA不同的是,细菌细胞中的DNA并没有被隔离在膜结合的细胞器中,而是呈长螺旋状分布在细胞质中。
在许多细菌中,DNA是以单个环状染色体的形式存在的,尽管有些细菌可能含有两条染色体,而且在某些情况下DNA是线性的而不是环状的。可变数量的较小的,通常是圆形的(虽然有时是线性的)DNA分子,称为质粒,可以携带辅助信息。
DNA中的碱基序列已经为数百种细菌所确定。细菌染色体的DNA含量从生殖支原体的580000碱基对到大肠杆菌的4700000碱基对,再到黄色粘球菌的9450000碱基对。粘液杆菌纤维素(Sorangium cellulosum),拥有最大的细菌基因组之一,包含超过1300万个碱基对。大肠杆菌染色体的长度,如果从细胞中取出并拉伸到最大限度,约为1.2毫米,考虑到细胞的长度约为0.001毫米,这蛮惊人的。
与所有生物体一样,细菌DNA含有四种含氮碱基腺嘌呤(A)、胞嘧啶(C)、鸟嘌呤(G)和胸腺嘧啶(T)。 双链DNA分子碱基配对的规则要求腺嘌呤和胸腺嘧啶碱基的数目相等,胞嘧啶和鸟嘌呤碱基的数目也相等。
G和C碱基对的数量与A和T碱基对的数量之间的关系是生物体内进化和适应性遗传变化的重要指标。
G + C的比例或摩尔比可以用G + C除以所有碱的总和(A + T + G + C)乘以100%来测量。生物体之间G + C比率的变化程度可能是相当大的。在动植物中,G + C的比例约为50%。 在原核生物中,G + C的比例范围更广,从大多数支原体的约25%到大肠杆菌中的约50%,到微球菌,放线菌和结实的粘菌中的近75%。但是,单个属中某个物种内的G + C含量非常相似。
细菌的细胞质包含高浓度的酶,代谢产物和盐。 另外,细胞的蛋白质是在分散在整个细胞质中的核糖体上制造的。 细菌核糖体与真核细胞中的核糖体不同之处在于它们更小,具有更少的成分(由三种类型的核糖体RNA和55种蛋白质组成,而真核生物中则由四种类型的rRNA和78种蛋白质组成)并且与作用于真核核糖体的抗生素相比,它们受到不同抗生素的抑制。
细菌细胞质中有许多包涵体或颗粒。这些物体从不被膜封闭,而是作为储存容器。糖原是葡萄糖的聚合物,是碳水化合物和能量的储备。 Volutin或变色颗粒含有聚合磷酸盐,是无机磷酸盐和能量的储存形式。许多细菌具有含有聚β-羟基丁酸聚合物酯或相关化合物的脂滴。这与真核生物不同,真核生物利用脂滴储存甘油三酯。在细菌中,储存颗粒是在有利的生长条件下产生的,并在营养物质从培养基中耗尽后被消耗掉。 许多水生细菌产生气体空泡(或叫气囊),这是一种蛋白质结合的结构,含有空气,其中包含空气,并使细菌能够调节其浮力。 细菌也可以有内部的膜结构,作为细胞质膜的外生物形成。
细菌的生物类型
致病细菌不断地与宿主的免疫系统搏斗,这一事实可能解释了属于同一物种但可通过血清学试验加以区分的不同菌株或类型的细菌数量之多,令人困惑。微生物学家通常通过细胞表面的特定分子来识别细菌,这些分子是用特定的抗体检测出来的。抗体是一种血清蛋白,在免疫反应中与外来分子(抗原)紧密结合,以清除或破坏抗原。抗体具有显着的特异性,蛋白质中甚至一个氨基酸的取代都可能阻止该蛋白质被抗体识别。
对于许多细菌种类,有成千上万种不同的菌株(称为血清变异体,用于血清学变异体),它们主要或仅在其脂多糖、鞭毛或荚膜的抗原特性上彼此不同。例如,肠内细菌的不同血清型,如大肠杆菌和沙门氏菌,常常被发现与栖息在不同宿主动物或引起不同疾病的能力有关。这些众多血清型的形成反映了细菌对免疫系统强烈的防御行为作出有效反应的能力。
生殖过程
二分裂法
大多数原核生物通过二分裂过程进行繁殖,细胞体积不断增大,直至分裂成两半,产生两个完全相同的子细胞。每个子细胞可以继续以与父细胞相同的速度生长。为了实现这一过程,细胞必须在其整个表面生长,直到细胞分裂时,在细胞中部的分裂隔膜处形成一个新的半球形极。
在革兰氏阳性细菌中,隔膜沿着细胞的中点从质膜向内生长;在革兰氏阴性细菌中,细胞壁更具弹性,当侧壁向内挤压时形成分裂隔膜,将细胞一分为二。为了使细胞分裂成两半,肽聚糖的结构必须在半球形帽中不同于在细胞壁的笔直部分,并且不同的壁交联酶必须在隔膜处比在其他地方更活跃。
出芽生殖
一群环境细菌通过出芽繁殖。在这个过程中,在母细胞的一端或丝上形成一个小的芽,称为前列腺癌。随着生长的进行,母细胞的大小保持不变,但芽会扩大。当芽的大小和母细胞差不多时,它就会分离。这种繁殖方式类似于萌芽真菌,如啤酒酵母(Saccharomyces cerevisiae)。分裂和出芽的一个区别是,在后者中,母细胞通常具有与后代不同的特性。在某些巴氏杆菌菌株(Pasteuria strains)中,子芽有鞭毛,并能运动,而母细胞没有鞭毛,但有长的菌毛,但在芽对面的末端具有长的菌毛和固定的附肢。在浮游生物中发现的相关浮萍菌在芽对面的末端具有长的纤维状茎。 在Hyphomicrobium中,菌丝(prostheca)从细胞的一端长出,芽从prostheca的尖端长出,与母细胞相距相对较长的距离。
多形生丝单胞菌
图源:Biological Photo Service
在萌芽的多形性hyphomonas细菌中,芽长在称为prostheca的细丝末端。
孢子繁殖
许多环境细菌能够产生稳定的休眠或休眠形式,作为其生命周期的一个分支,以增强其在不利条件下的生存能力。这些过程不是细胞生命周期的一个强制性阶段,而是一种中断。这种休眠形式被称为内生孢子、胞囊或异胞囊(主要见于蓝藻),这取决于孢子形成的方法,而孢子形成的方法因细菌群而异。
在许多属的细菌中都发现了形成内生孢子的能力,这些细菌主要是革兰氏阳性菌,包括需氧芽孢杆菌(aerobic rod Bacillus),微需氧菌芽孢杆菌(microaerophilic rod Sporolactobacillus),厌氧菌梭状芽孢杆菌(anaerobic rods Clostridium)和脱硫芽孢杆菌(Desulfotomaculum),球菌孢子菌(coccus Sporosarcina)和丝状嗜热放线菌(filamentous Thermoactinomyces)。
孢子的形成是对营养缺乏的反应。 因此,在营养物质可用之前,内生孢子不具有代谢活性,这时它们能够从孢子分化为营养细胞。 在孢子形成过程中,每个细菌细胞内部仅形成一个孢子。孢子的形成始于细菌染色体拷贝周围细胞质膜的内陷,从而使较小细胞的内容物与母细胞分离。
母细胞的膜吞噬了其细胞质内的较小细胞,有效地提供了两个同心的单元膜来保护正在发育的孢子。 薄的孢子膜和厚的肽聚糖皮质位于两个单元膜之间。 皮质外部形成坚硬的孢子皮,包围整个孢子结构。孢子皮具有类似于角蛋白的特性,能够抵抗热,干燥(脱水),冷冻,化学药品和辐射的致死作用。 内生孢子抵抗这些有害物质的能力可能来自于孢子内部极低的水含量。 甲基肌球菌属中的甲烷氧化细菌(Methylosinus)也会产生耐干燥性的孢子,称为外孢子。
包囊是由固氮菌(Azotobacter), 蛭弧菌(Bdellovibrio,蛭包囊)和粘球菌(Myxococcus,粘孢子)的休眠成员产生的厚壁结构。它们对干燥和其他有害条件有抵抗力,但程度低于内生孢子。在固氮固氮菌的包囊中,细胞分裂后形成一层厚厚的多层细胞壁和外壳,包围着静息细胞。
丝状放线菌产生两类生殖孢子:分生孢子,是在地上或基质菌丝体上形成的多个孢子链;孢子囊孢子,是在称为孢子囊的特殊囊中形成的。
遗传信息交流
细菌在其生命周期中没有一个专有性生殖阶段,但它们可以非常活跃地交换遗传信息。DNA中携带的遗传信息可以从一个细胞转移到另一个细胞;然而,这不是真正的交换,因为只有一个’同伴’接收到新的信息。
此外,转移的DNA量通常只是染色体的一小部分。发生这种情况有几种机制。在转化过程中,细菌吸收漂浮在培养基中的游离DNA片段。为了有效地吸收DNA,细菌细胞必须处于一种有能力的状态,这是由细菌结合DNA自由片段的能力来定义的,并且仅在有限数量的细菌中自然形成,例如嗜血杆菌(Haemophilus)、奈瑟菌(Neisseria)、链球菌(Streptococcus)和芽孢杆菌(Bacillus)。在实验室条件下,例如通过暴露于氯化钙(CaCl2)溶液,可以使许多其他细菌,包括大肠杆菌,变得人为地具有竞争力。转化是重组DNA技术中的一个主要工具,因为来自一个生物体的DNA片段可以被另一个生物体摄取,从而使第二个生物体获得新的特性。
细菌之间交换遗传物质
图源:Encyclopædia
细菌DNA可以通过结合和转导过程从一个细胞传递到另一个细胞。
转导是指通过一种称为噬菌体的细菌感染病毒将DNA从一种细菌转移到另一种细菌。转导是在细菌之间转移DNA的有效手段,因为封闭在噬菌体中的DNA受到保护,免受物理降解和环境中酶的攻击,并通过噬菌体直接注入细胞中。 然而,通过转导的广泛基因转移具有有限的意义,因为将细菌DNA包装到病毒中效率低下,并且噬菌体通常在它们可感染的细菌种类范围内受到高度限制。 因此,通过转导进行种间转移的DNA很少。
图源:Encyclopædia
接合是通过质粒(非染色体DNA分子)介导的细胞间直接接触的DNA转移。接合质粒编码一种极其有效的机制,它介导自己从供体细胞到受体细胞的转移。因为仅供体细胞包含结合质粒,所以该过程在一个方向上发生。在革兰氏阴性细菌中,供体细胞产生特定的质粒编码菌毛,称为性菌毛,该菌毛将供体细胞附着到受体细胞上。 连接后,两个细胞将直接接触,并形成一个偶联桥,DNA通过该桥从供体转移到受体。 许多接合质粒可以在大量不同的革兰氏阴性细菌之间转移,并在其中繁殖。 质粒的大小不等,从几千个到超过100,000个碱基对。 后者有时被称为巨质粒。
巴氏杀菌如何保护食物
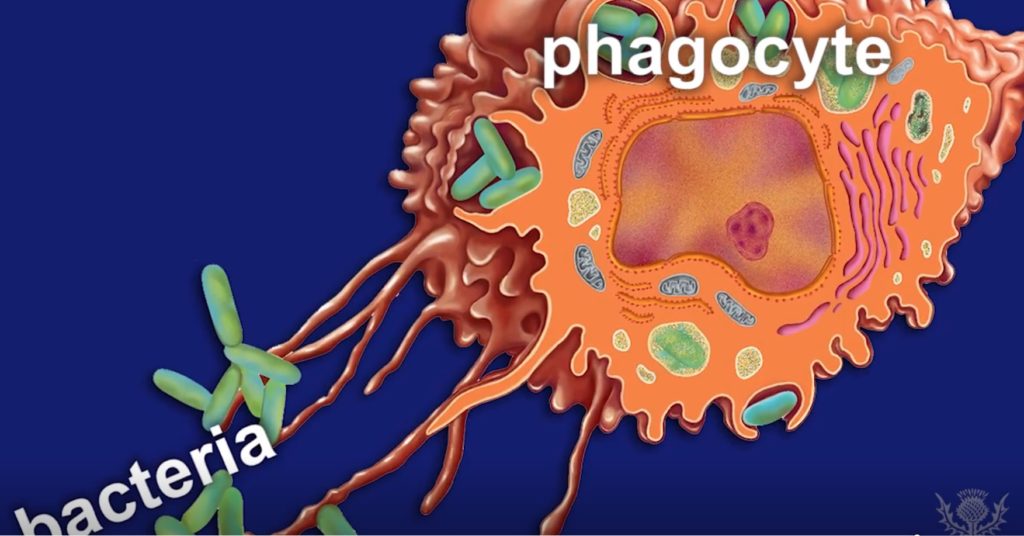
图源:Encyclopædia
细菌可能是致病的(引起疾病),并可能产生毒素,使食物变质并引起食物中毒。细菌引起的某些疾病包括肉毒杆菌中毒,伤寒和肺炎。
细菌的种群增长
细菌培养物的生长被定义为一个群体中细菌数量的增加,而不是单个细胞大小的增加。细菌种群的生长是以几何或指数方式发生的:每一个分裂周期(世代),一个细胞产生2个细胞,然后是4个细胞,然后是8个细胞,然后是16个细胞,然后是32个细胞,依此类推。生成一代所需的时间,即生成时间(G),可通过以下公式计算:
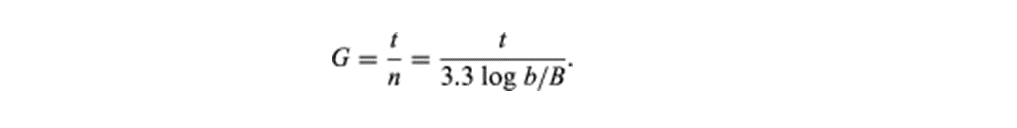
在公式中,B是观察开始时出现的细菌数量,B是时间段t后出现的细菌数量,n是世代数。这种关系表明,平均产生时间是恒定的,细菌数量增加的速度与任何给定时间的细菌数量成正比。这种关系只在种群以指数方式增长的时期有效,称为对数增长期。因此,显示细菌培养物生长的图形被绘制为细胞数的对数。
枯草芽孢杆菌生长周期
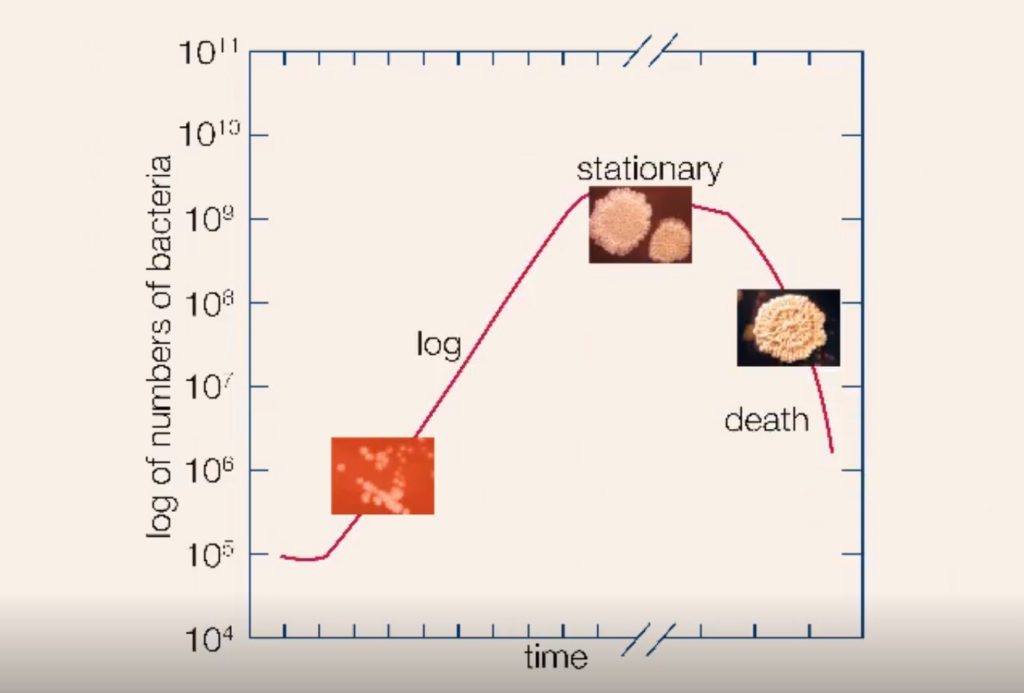
图源:AW Rakosy /Encyclopædia
细菌菌落通过生长的四个阶段进行:滞后阶段,对数阶段,静止阶段和死亡阶段。
细菌的产生时间因细菌而异,受许多环境条件和细菌种类的性质所控制。例如,生长最快的细菌之一产气荚膜梭状芽胞杆菌的最佳生成时间约为10分钟。 大肠杆菌每20分钟可以翻一倍; 生长缓慢的结核分枝杆菌的产生时间为12至16小时。 一些研究人员建议,某些生活在地球表面深处的细菌种群可能以极慢的速度生长,每隔几千年才繁殖一次。 “生长培养基” 例如摄入食物的组成是控制生长速率的主要因素。 当培养基提供更好的能源和细胞原本必须自己制造的更多生物合成中间体时,生长速率会增加到最大。
细菌生长曲线
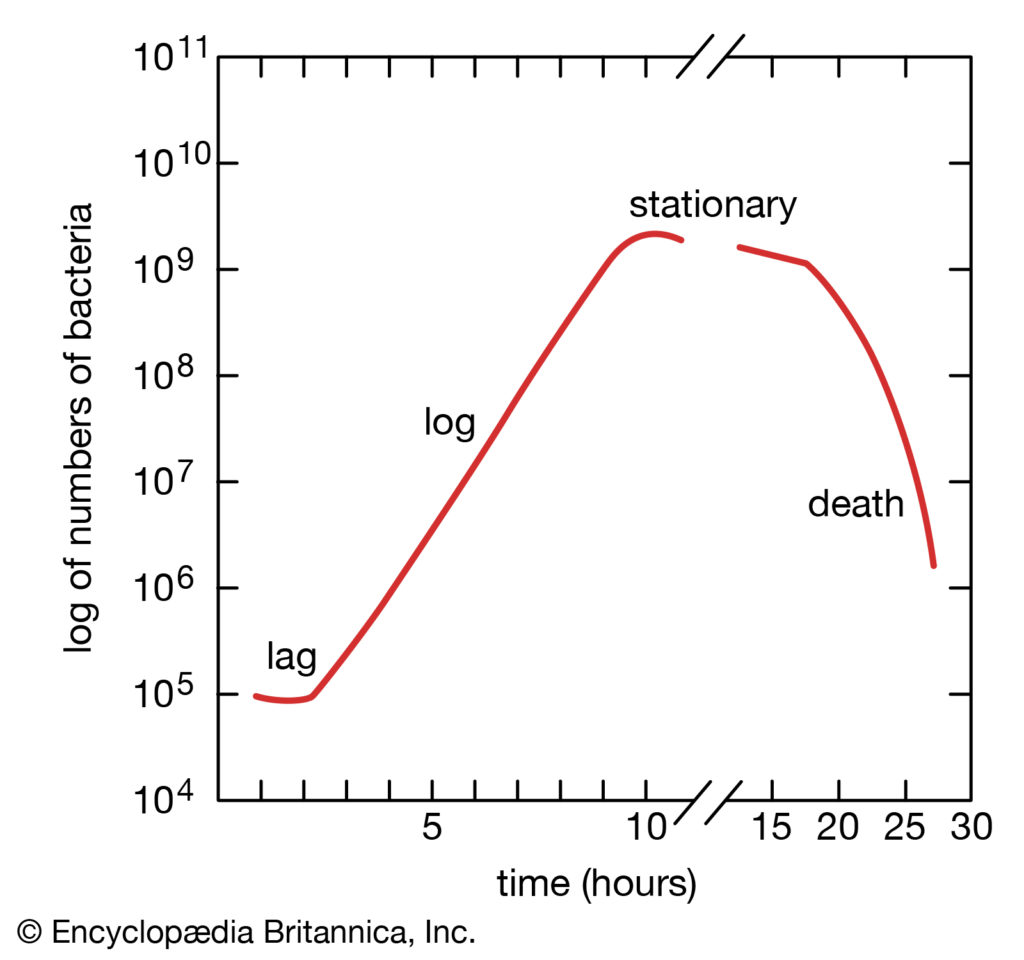
广义细菌生长曲线显示了细菌菌落生长的各个阶段。
当细菌被放置在一个能提供其生长所需的所有营养物质的培养基中时,菌群表现出四个生长阶段,这四个阶段代表了典型的细菌生长曲线。一旦接种到新的培养基中,细菌不会立即繁殖,种群大小保持不变。在这一时期,称为滞后期,细胞代谢活跃,只增加细胞大小。它们还在新的环境条件下合成细胞分裂和种群增长所需的酶和因子。
然后,种群进入对数阶段,细胞数量以对数方式增加,每一代细胞与前一代细胞在相同的时间间隔内发生,从而使每一代细胞的成分均衡增加。对数阶段一直持续到营养物质耗尽或有毒物质积累,此时细胞生长速度减慢,一些细胞可能开始死亡。在最佳条件下,对数末期某些细菌的最大种群密度可达每毫升100亿至300亿个细胞。
枯草芽孢杆菌
图源:AW Rakosy /Encyclopædia
枯草芽孢杆菌的细菌菌落在37℃下进入生长的对数期培养18-24小时后,(98.6°F;放大约6倍)
细菌生长的对数阶段之后是稳定阶段,在这个阶段,细菌种群的大小保持不变,即使一些细胞继续分裂,而另一些细胞开始死亡。稳定期之后是死亡期,在这一阶段,群体中细胞的死亡超过了新细胞的形成。死亡阶段开始前的时间长短取决于物种和培养介质。即使在缺乏营养的情况下,细菌也不一定会死亡,并且它们可以长期存活。
大自然中的分布
原核生物在地球表面无处不在。 从极地冰到冒泡的温泉,从山顶到海底,从动植物到森林土壤,都可以找到它们。 有些细菌可以在接近冰点的温度下在土壤或水中生长,而另一些细菌则可以在接近沸腾(100°C 的温度下繁殖。 每种细菌都适合生活在特定的环境环境中,无论是海洋表面,泥浆沉积物,土壤还是其他生物的表面。 空气中的细菌含量很低,但是当灰尘被悬浮时含量会升高。 在未受污染的自然水体中,细菌数可以是每毫升数千。 在肥沃的土壤中,细菌数可以达到每克数百万。 在粪便中,细菌数量可能超过每克数十亿。

细菌在有机分解中的作用是从垃圾填埋场和水中去除不需要的生物材料的过程的一部分。
细菌是其栖息生态系统中的重要成员。尽管它们的体积很小,但它们的数量之多意味着它们的新陈代谢在外部环境中的元素转化中起着巨大的作用,有时有益,有时有害。可能每一种自然产生的物质,以及许多人工合成的物质,都能被某些种类的细菌降解(代谢)。奶牛最大的胃——瘤胃是一个发酵室,细菌在其中消化草料和饲料中的纤维素,将其转化为脂肪酸和氨基酸,这是奶牛使用的基本营养素,也是奶牛产奶的基础。污水或堆肥堆中的有机废物被细菌转化为适合植物新陈代谢的营养物质,或转化为气态甲烷(CH4)和二氧化碳。包括动植物在内的所有有机物质的残余物最终通过细菌和其他微生物的活动转化为土壤和气体,从而可供进一步生长。
许多细菌生活在溪流和其他水源中,它们在水样中的低种群密度并不一定表明水不适合饮用。然而,含有大肠杆菌等细菌的水,是人类和动物肠道的正常居民,表明污水或粪便物质最近污染了该水源。这类大肠菌群本身可能是病原体(致病生物),它们的存在表明其他不太容易检测到的细菌和病毒病原体也可能存在。净水厂中使用的沉淀、过滤和氯化程序旨在去除这些以及任何其他可能存在于人类饮用水中的微生物和传染源。水中微生物分解有机物质消耗氧气(生化需氧量),导致氧气水平下降,这对接收废水的溪流和湖泊中的水生生物非常有害。污水处理的一个目标是在有机物排放到水系统之前尽可能地氧化它们,从而降低废水的生化需氧量。为此,污水消化池和曝气装置专门利用细菌的代谢能力。
土壤细菌通过转化土壤的各种物质、腐殖质和矿物质,在影响生物化学变化方面极为活跃。对生命至关重要的元素,如碳、氮和硫,被细菌从无机气态化合物转化为可供植物和动物使用的形式。
细菌还将动植物代谢的最终产物转化为细菌和其他微生物可以利用的形式。氮循环可以说明细菌在影响各种化学变化中的作用。氮在自然界中以几种氧化状态存在,如硝酸盐、亚硝酸盐、二氮气体、几种氮氧化物、氨和有机胺(含有一种或多种取代烃的氨化合物)。固氮是将大气中的二氮气体转化为可供生物利用的形式。一些固氮细菌,如固氮菌、巴氏梭菌和肺炎克雷伯菌,是自由生活的,而根瘤菌则与豆科植物密切相关。
土壤中的根瘤菌生物识别并侵入其特定植物宿主的根毛,进入植物组织,并形成根瘤。 这个过程导致细菌失去许多自由活动特性。 它们变得依赖于植物所提供的碳,并且作为碳的交换,它们将氮气转化为氨,氨被植物用于其蛋白质的合成和生长。 另外,当硝酸盐用作电子受体时,许多细菌可以将硝酸盐转化为胺,以合成细胞材料,或将其转化为氨。反硝化细菌将硝酸盐转化为氮气。 氨或有机胺向硝酸盐的转化是通过需氧生物硝化单胞菌和硝化细菌的联合活动来完成的,它们利用氨作为电子供体。
固氮细菌 (右)豌豆植物(Pisum sativum)的根部,其根瘤内藏有固氮细菌(Rhizobium)。(左)根瘤是根瘤菌与植物根毛之间共生关系的结果。细菌识别根毛并开始分裂(A),通过感染线(B)进入根部,使细菌进入根细胞,然后分裂成结节(C)
在碳循环中,二氧化碳被植物和自养原核生物转化为细胞物质,有机碳通过异养生物返回大气。 微生物分解的主要分解产物是二氧化碳,它是由呼吸需氧生物而形成的。
甲烷是碳代谢的另一种气体最终产物,是全球碳循环中相对次要的组成部分,但在当地情况下具有重要意义,是供人类使用的可再生能源。甲烷的产生是由高度专业化和专性厌氧产甲烷原核生物进行的,它们都是古细菌。产甲烷菌利用二氧化碳作为末端电子受体,从氢气(H2)中接收电子。其他一些物质可以被这些生物转化为甲烷,包括甲醇、甲酸、乙酸和甲胺。
尽管产甲烷菌可利用的物质范围极窄,但在许多有机物质(包括纤维素、淀粉、蛋白质、氨基酸、脂肪、醇和大多数其他底物)的厌氧分解过程中,甲烷的生成非常普遍。从这些物质中生成甲烷需要其他厌氧菌将这些物质降解为醋酸盐或二氧化碳和氢气,然后由产甲烷菌(methanogens)使用。产甲烷菌通过去除甲烷代谢活动中产生的氢气来支持混合物中其他厌氧菌的生长。消耗氢气会刺激其他细菌的新陈代谢。
尽管产甲烷菌的代谢能力非常有限,而且对氧非常敏感,但它们在地球上很普遍。大量的甲烷是在厌氧环境中产生的,比如沼泽和沼泽,但是土壤和反刍动物也会产生大量的甲烷。大气中至少80%的甲烷是由产甲烷菌作用产生的,其余的则是从煤层或天然气井中释放出来的。
食物中的细菌
健康奶牛的牛奶最初含有很少的细菌,这些细菌主要来自奶牛的皮肤和处理牛奶的程序。牛奶是许多细菌的极好生长介质,除非牛奶经过适当加工,否则细菌的数量会迅速增加。如果存在致病菌,细菌生长会破坏牛奶,甚至对健康造成严重危害。可以从被感染的牛传播的疾病包括结核病(结核分枝杆菌,Mycobacterium tuberculosis),不发烧的发热(流产的布鲁氏菌Brucella abortus)和发热的病原体(柯氏杆菌Coxiella burnetii)。
此外,伤寒可通过受感染的牛奶处理者的牛奶传播。 巴氏杀菌程序将牛奶的温度升高30分钟至63°C(145°F)或15秒升高至71°C(160°F),这可以杀死可能存在的任何病原菌,尽管这些程序不能杀死所有微生物。
某些细菌将牛奶转化成有用的乳制品,如酪乳、酸奶和奶酪。商业培养的酪乳是从接种了乳酸菌(Lactococcus)发酵剂(通常是乳酸乳杆菌或乳酪乳杆菌)的牛奶中制备的。酸奶和其他发酵乳制品是用不同的细菌培养物以类似的方式生产的。许多奶酪也是通过细菌的作用制成的。产酸细菌如乳乳酸杆菌在牛奶中的生长会使酪蛋白沉淀成凝乳。除去水分和加盐后,凝乳在其他微生物的作用下成熟。不同的细菌赋予食物不同的风味和特性;例如,干酪乳杆菌、嗜热链球菌和舍曼丙酸杆菌的混合物负责瑞士奶酪的成熟,并产生其特有的味道和大气泡。其他类型的细菌长期以来被用于制备和保存通过细菌发酵生产的各种食品,包括腌制产品、泡菜和橄榄。
在食物中传播的许多致病菌的毒素在摄入时会引起食物中毒。其中包括一种由金黄色葡萄球菌(Staphylococcus aureus)产生的毒素,这种毒素能引起迅速、严重但有限的胃肠道不适,或者肉毒梭菌(Clostridium botulinum)的毒素,这种毒素通常是致命的。在密封前未完全煮熟的罐装非酸性食品中可能产生肉毒杆菌毒素。肉毒梭菌形成耐热孢子,可以发芽为营养细菌细胞,在厌氧环境中茁壮成长,这有利于产生其极强的毒素。其他食源性感染实际上是由受感染的食物处理者传播的,包括伤寒、沙门氏菌病(沙门氏菌属)和志贺氏菌病(痢疾志贺氏菌属)。

图源:Encyclopædia
工业中的细菌
各种细菌的厌氧糖发酵反应会产生不同的最终产物。用酵母生产乙醇已经被酿酒工业开发了几千年,并被用于燃料生产。在醋的生产过程中,特定的细菌将酒精氧化成醋酸。其他的发酵过程可以制造出更有价值的产品。有机化合物,如丙酮、异丙醇和丁酸,是由各种梭状芽孢杆菌(Clostridium)在发酵过程中产生的,可以在工业规模上制备。在极端环境的生物中也发现了其他细菌产物和反应。 从嗜热细菌中分离出的酶引起了极大的兴趣,在这些酶中,由于可以在较高的温度下进行反应,因此可以以较高的速率进行反应。
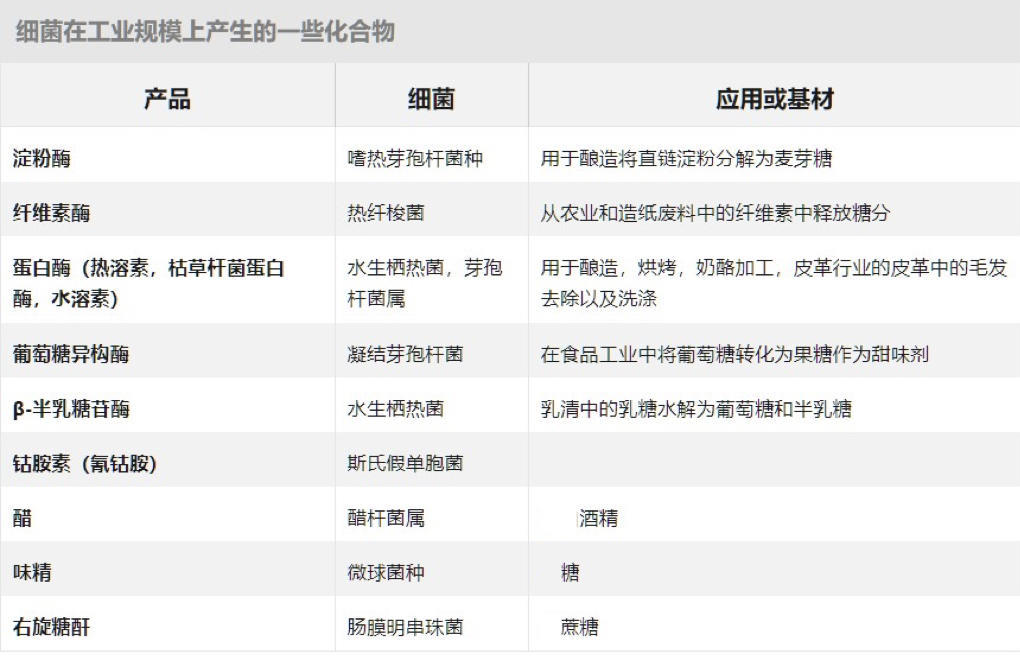
来源:Encyclopædia
烃类氧化细菌和真菌对石油产品的微生物分解具有重要的生态学意义。石油的微生物分解是一个好氧过程,如果石油沉淀到底部的厌氧沉积物层(厌氧环境中的天然石油沉积物有数百万年的历史),那么这一过程就会被阻止。碳氢化合物氧化细菌附着在水面上漂浮的油滴上,它们的作用最终将油分解为二氧化碳。 将此类细菌及其生长因子喷洒到溢油上以提高非挥发性脂族和芳族烃的降解速度已成为一种常见的做法。
医学中的细菌
菌性疾病在人类历史上起着主导作用。 霍乱和鼠疫的广泛流行使世界上某些地区的人口减少了三分之一以上。细菌性肺炎可能是老年人死亡的主要原因。 也许有更多的军队被伤寒,痢疾和其他细菌感染击败,而不是被武力击败。 随着管道和卫生设备的现代发展,细菌疫苗的发展以及抗菌素的发现,细菌性疾病的发生率已降低。 然而,细菌没有继续作为传染因子消失,因为它们继续进化,产生了越来越强的毒株并获得了对许多抗生素的抗性。
尽管大多数细菌对地球上的生命都是有益的,甚至是生命所必需的,但少数细菌却对人类有害。 目前,没有一种古细菌被认为是病原体,但是包括人类在内的动物不断遭到大量细菌和各种细菌的轰击和居住。 与动物接触的大多数细菌都可以通过宿主的防御系统迅速消除。 口腔,肠道和皮肤被大量特定类型的细菌定殖,这些细菌适应这些栖息地的生活。 这些生物在正常情况下是无害的,并且仅在它们以某种方式穿过身体的屏障并引起感染时才变得危险。
一些细菌擅长入侵宿主,被称为病原体或疾病产生者。 一些病原体作用于人体的特定部位,例如脑膜炎球菌(Neisseria meningitidis),它会侵袭并刺激脑膜,大脑和脊髓周围的膜,并刺激脑膜。白喉细菌(Corynebacterium diphtheriae),最初感染喉咙; 以及在肠道中繁殖的霍乱细菌(霍乱弧菌),其产生的毒素导致该霍乱的大量腹泻。 其他能感染人类的细菌包括葡萄球菌(主要是金黄色葡萄球菌),它能感染皮肤引起疖子(疖子),血液引起败血症(血液中毒),心脏瓣膜引起心内膜炎,或骨头引起骨髓炎。
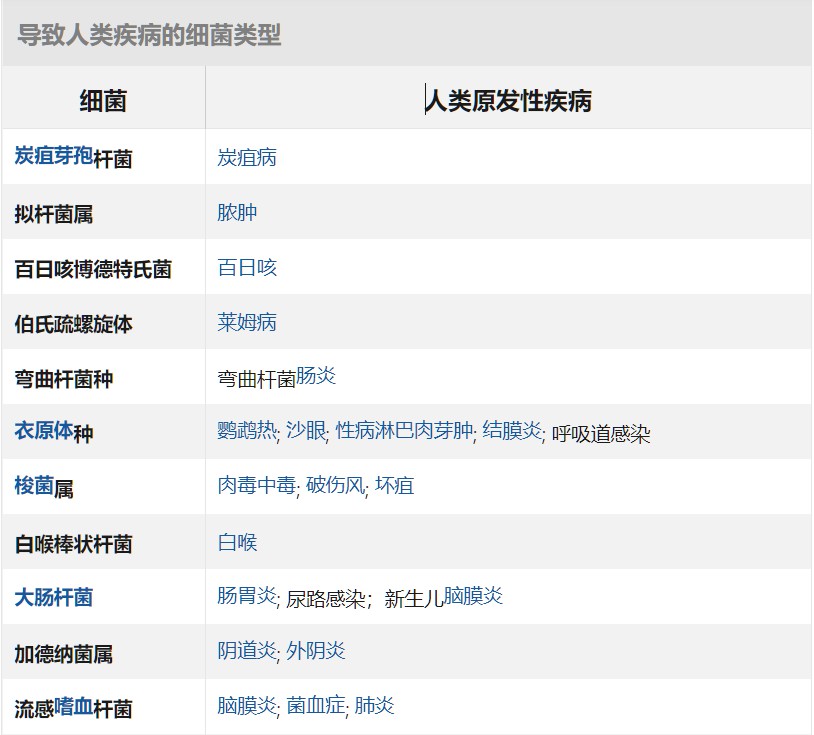
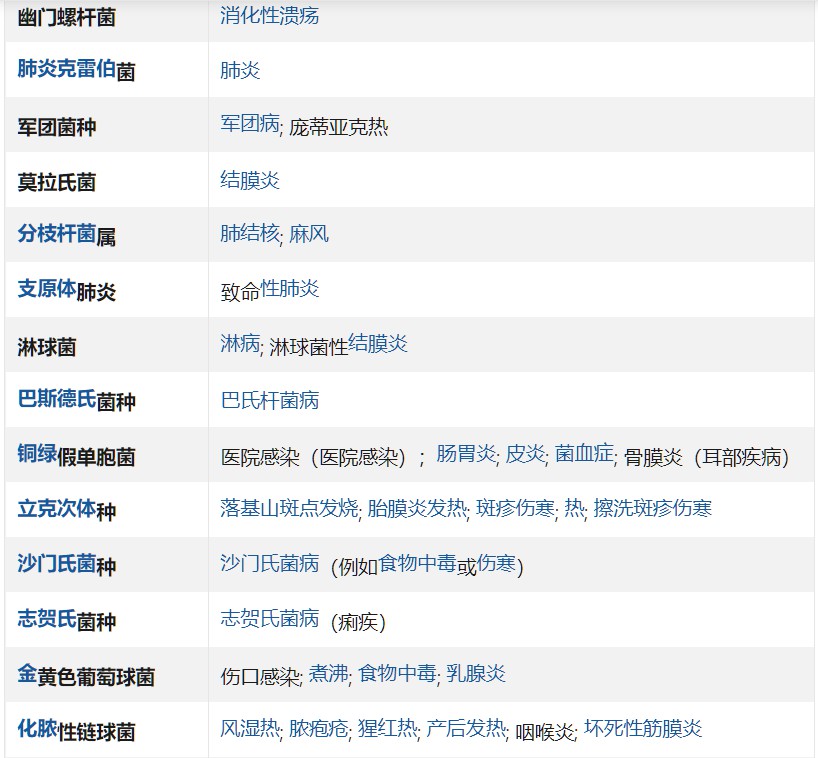
来源:Encyclopædia
人体中的细菌
图源:Encyclopædia
人体屏障(例如皮肤和肠道内壁)中的天然细菌在正常的人类生理过程中起着重要的作用。
侵入动物血液的病原菌可以利用多种机制中的任何一种来逃避宿主的免疫系统,包括形成长的脂多糖链,从而对一组血清免疫蛋白(称为补体)产生抵抗力,补体通常会抑制细菌的生长。细菌表面蛋白的致病性重组阻止动物产生的抗体识别病原体,在某些情况下使病原体能够在吞噬性白细胞中存活和生长。许多病原菌产生毒素,协助它们入侵宿主。这些毒素包括蛋白酶,分解组织蛋白质的酶,以及脂肪酶,分解脂质(脂肪)并通过破坏细胞膜来损伤细胞的酶。其他毒素通过在细胞膜上形成孔或通道来破坏细胞膜。一些毒素是修饰参与蛋白质合成或控制宿主细胞代谢的特定蛋白质的酶;例如白喉、霍乱和百日咳毒素。
补体激活途径
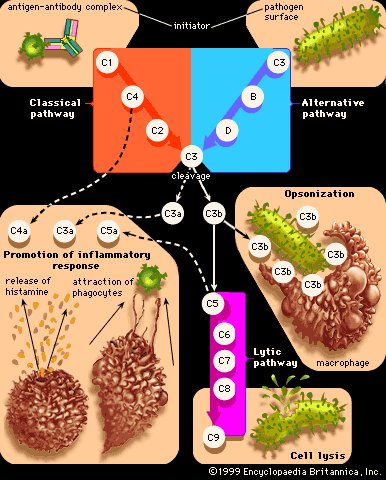
补体蛋白的主要功能是通过刺穿病原体的外膜(细胞裂解)或使它们对吞噬细胞(例如巨噬细胞)更具吸引力来帮助破坏病原体(这一过程称为调理作用)。一些补体成分还通过刺激细胞释放组胺和将吞噬细胞吸引到感染部位来促进炎症。
一些病原细菌在宿主体内形成区域,在这些区域中它们被封闭并不受免疫系统的保护,例如葡萄球菌形成的皮肤疖子和结核分枝杆菌形成的肺部空洞。脆弱拟杆菌(Bacteroides fragilis)大量存在于人体肠道中,但是一般不会对宿主造成任何困难。如果这种细菌通过损伤进入体内,细菌囊会刺激身体将细菌隔离成脓肿,从而减少细菌的传播。在许多情况下,细菌感染的症状实际上是免疫系统过度反应的结果,而不是细菌产生毒性因子的结果。
对抗致病性细菌感染的其他方法包括使用生物治疗剂或益生菌。这些无害的细菌会干扰病原菌的定殖。另一种方法是利用噬菌体,即杀死细菌的病毒,来治疗特定细菌病原体的感染。此外,20世纪80年代发展起来的重组DNA技术使细菌合成几乎任何蛋白质成为可能,大肠杆菌是这一过程中常见的宿主。重组DNA技术用于廉价、大规模生产极为稀缺和有价值的动物或人类蛋白质,如激素、凝血因子,甚至抗体。
细菌自地球生命史以来就已经存在。 在岩石中发现的细菌化石至少可以追溯到泥盆纪时期(4.192亿至3.589亿年前),并且令人信服的论点是自前寒武纪大约35亿年前就已经存在细菌。细菌至少在大约18亿年前的古元古代末期就已在地球上广泛传播,当时由于蓝细菌的作用,大气中出现了氧气。 因此,细菌有足够的时间来适应其环境,并产生了许多后代。
关于生命起源所涉及的原始祖先的性质,人们有相当多的猜测。有人认为原始细胞可能使用RNA作为其遗传物质,因为研究表明RNA分子可以具有多种催化功能。在这个时期,细菌和古菌很早就从它们共同的前体中分化出来。这两类原核生物倾向于栖息在不同类型的环境中,并以不同的速度产生新的物种。许多古菌喜欢高温生态位。古细菌树的一个主要分支仅由嗜热物种组成,而另一个主要分支中的许多产甲烷菌可以在高温下生长。相反,没有一个主要的真细菌分支仅仅由嗜热菌组成。细菌和古细菌都含有能够在高温下生长的成员,以及其他能够在低温下生长的物种。另一个显著的区别是细菌广泛地适应好氧条件,而许多古细菌是专性厌氧菌。没有古细菌是专性光合作用的。也许古细菌是一种较原始的生物体,对变化的环境条件的遗传反应受损。 适应新情况的能力有限可能会将古细菌限制在恶劣的环境中,与其他生命形式的竞争较少。
根据三域系统的生命之树
有机体必须不断进化或适应不断变化的环境,很明显,突变,即有机体DNA中核苷酸序列的变化,在所有有机体中不断发生。DNA序列的变化可能会导致该DNA片段编码的蛋白质的氨基酸序列发生变化。因此,改变后的蛋白质可能更适合或不太适合在当前条件下发挥功能。尽管DNA中可能发生的许多核苷酸变化对细胞的适应性没有影响,但如果核苷酸变化能促进细胞的生长,哪怕是很小的程度,那么突变型细胞就能够增加其在群体中的相对数量。然而,如果核苷酸的改变阻碍了细胞的生长,那么突变形式就会被其他细胞所取代而丧失。
在生物体之间传递遗传信息的能力是适应环境变化的一个主要因素。DNA交换是高等真核生物生命周期的重要组成部分,在所有真核生物中都有发生。基因交换也发生在整个细菌世界,虽然转移的DNA量很小,但这种转移可以发生在远缘生物之间。携带在质粒上的基因可以进入细菌染色体,成为细菌遗传的稳定部分。生物体通常拥有称为转座子的可移动遗传元件,可以重新排列染色体上任何基因的顺序和存在。转座子可能在帮助加快进化速度方面发挥作用。
细菌快速进化的例子很多。在20世纪40年代以前,抗生素还没有用于医疗实践。当抗生素最终投入使用时,大多数致病菌对它们很敏感。然而,从那时起,细菌对一种或多种抗生素的耐药性增加,以至于以前有效的抗生素对某些类型的细菌不再有用。大多数致病细菌对抗生素耐药的例子并不是由于突变改变了抗生素攻击的蛋白质,尽管这种机制可能发生。相反,抗生素抗性通常涉及细菌产生的酶,这些酶会改变抗生素并使之失活。抗生素耐药性传播的一个主要因素是可传播的质粒,它将药物灭活酶的基因从一个细菌携带到另一个细菌。尽管这些酶的基因的原始来源尚不清楚,但可移动的基因元件(转座子)可能在它们的出现中起了作用,也可能允许它们转移到其他细菌类型。
影响细菌生长的因素
细菌在最佳生长所需的条件上有很大的不同。就营养需求而言,所有细胞都需要碳、氮、硫、磷、多种无机盐(如钾、镁、钠、钙和铁)以及大量其他微量元素(如锌、铜、锰、硒、钨和钼)。碳是细菌最需要的元素,因为氢和氧可以从水中获得,这是细菌生长的先决条件。同时还需要一种能量来源来促进细菌的新陈代谢。组织细菌的一种方法是基于这些基本的营养需求:碳源和能源。
细胞可用于碳的来源有两种:无机化合物和有机化合物。使用无机化合物二氧化碳(CO2)作为碳源的生物称为自养生物。 需要碳的有机源(例如糖,蛋白质,脂肪或氨基酸)的细菌称为异养生物(或有机营养菌)。 许多异养生物,例如大肠杆菌或铜绿假单胞菌,都由简单的糖(例如葡萄糖)合成所有细胞成分,因为它们具有必要的生物合成途径。 其他异养生物失去了其中一些生物合成途径。 为了生长,它们要求其环境包含化学上完整的特定氨基酸,含氮碱或维生素。
铜绿假单胞菌
除了碳,细菌还需要能量,能量几乎总是通过电子从电子供体转移到电子受体来获得的。有三种基本的能源:光、无机化合物和有机化合物。光营养细菌利用光合作用从光能中产生三磷酸腺苷(ATP)形式的细胞能量。化学营养体从化学物质(有机和无机化合物)中获取能量;化学石营养体从与无机盐的反应中获取能量;化学异养体从有机化合物中获取碳和能量(能量源也可作为这些生物体的碳源)。
在大多数情况下,细胞能量是通过电子转移反应产生的,其中电子通过一种途径从电子或有机供体分子移动到受体分子,从而保留了电子在转移过程中释放出的能量,其形式为电子俘获 细胞可用于其化学或物理工作的物质。 从电子传输中捕获的主要能量形式是ATP。 分解有机分子以产生能量的代谢过程称为分解代谢反应。 相反,合成分子的代谢过程称为合成代谢反应。
许多细菌可以使用大量的化合物作为碳和能源,而其他细菌的代谢能力受到很大限制。尽管碳水化合物是真核生物的常见能源,但是由于大多数细菌不具有代谢这些通常复杂的分子所必需的酶,因此这些分子仅通过有限数量的细菌进行代谢。 相反,许多细菌都依赖于其他能源,例如氨基酸,脂肪或其他化合物。 对细菌重要的其他化合物包括磷酸盐,硫酸盐和氮。 由于许多细菌无法合成磷酸盐,因此在许多环境中,尤其是在水中,磷酸盐含量低可能是细菌生长的限制因素。 另一方面,大多数细菌可以将硫酸盐或硫化物转化为蛋白质合成所需的有机形式。 生命有机体从氨中吸收氮的能力是广泛存在的,细菌将其他形式的氮(例如土壤中的硝酸盐或大气中的二氧化氮转化为细胞物质的能力也有所不同。
有些细菌是专性寄生虫,只在活的宿主细胞内生长。例如,立克次体(Rickettsia)和衣原体(Chlamydia), 在真核细胞中生长,蛭弧菌(Bdellovibrio)在细菌细胞中生长。梅毒螺旋体(Treponema pallidum)很难(即使不是不可能)在培养中生长,可能是因为它需要低的氧张力和低的氧化还原水平,这是由于存在动物细胞而不是任何特定的营养素所致。因为有些细菌可能只以动物或植物寄生虫的形式生长,或者只在牛奶等营养丰富的来源中生长,所以它们在自然界中不可能以游离细菌的形式生长。许多来自自然环境的细菌与其他细菌联合存在,很难从该伙伴关系的其他成员中分离和培养。
对于不同的细菌类型来说,最适合细菌生长的物理条件有很大的不同。作为一个群体,细菌在不同环境中的生存能力在所有生物中表现出最大的差异。
氧 气
细菌之间最显著的区别之一是它们对大气氧(O2)的需求和反应。实际上,所有的真核生物都需要氧气才能茁壮成长,而许多种类的细菌可以在厌氧条件下生长。需要氧气才能生长的细菌称为专性需氧细菌。在大多数情况下,这些细菌需要氧气才能生长,因为它们产生能量和呼吸的方法依赖于电子向氧气的转移,而氧气是电子传递反应中的最终电子受体。
专性需氧菌包括枯草芽孢杆菌(Bacillus subtilis)、铜绿假单胞菌(Pseudomonas aeruginosa)、结核分枝杆菌(Mycobacterium tuberculosis)和氧化亚铁硫杆菌(cidithiobacillus ferrooxidans)。
只有在缺氧条件下才能生长的细菌,如梭状芽孢杆菌、类杆菌和产甲烷的古细菌(产甲烷菌)被称为专性厌氧菌,因为它们产生能量的代谢过程与氧气的消耗相无关。事实上,氧气的存在实际上会毒害它们的一些关键酶。有些细菌(肺炎链球菌)是微需氧或耐氧厌氧菌,因为它们在低浓度的氧气中生长得更好。在这些细菌中,氧气通常会刺激微小的代谢过程,从而增强能量产生的主要途径。兼性厌氧菌可以根据氧气的存在改变其代谢过程,在氧气存在时使用更有效的呼吸过程,在缺氧时使用效率较低的发酵过程。兼性厌氧菌包括大肠杆菌和金黄色葡萄球菌。
破伤风梭状芽胞杆菌
图源:Centers for Disease Control and Prevention
细菌对氧的反应不仅仅取决于它们的代谢需要。氧是一种非常活泼的分子,会形成一些有毒的副产物,如超氧物(O2-)、过氧化氢(H2O2)和羟基自由基(OH·)。有氧生物产生的酶可以使这些氧气产物解毒。
最常见的排毒酶是分解过氧化氢的过氧化氢酶和分解超氧化物的超氧化物歧化酶。 厌氧细菌通常不产生过氧化氢酶,其超氧化物歧化酶的水平随细胞对氧气的敏感性而成比例地变化。 许多厌氧菌对氧气过敏,短时间接触就会被杀死,而其他厌氧菌,包括大多数梭状芽孢杆菌,对氧的存在更具耐受性。这些酶的联合作用,以消除过氧化氢和超氧物是重要的,因为这些副产品与铁一起形成极为活跃的羟基自由基,这是能够杀死细胞。
温度
细菌已适应多种温度。 在低于约15°C(59°F)的温度下生长的细菌是嗜冷菌。细菌在低温下生长的能力并不意外,因为温带土壤的平均地下温度约为12°C(54°F),90%的海洋温度为5°C(41°F)或更低。专性嗜冷菌是从北极和南极海水和沉积物中分离出来的,其最适生长温度约为10℃(50℉),如果暴露在20℃(68℉)下,则无法存活。
大多数嗜冷细菌属于革兰氏阴性菌属假单胞菌属、黄杆菌属、无色杆菌属和产碱菌属。中温细菌是指在20至45°C(68至113°F)之间发生最佳生长的细菌,尽管它们通常可以在10至50°C(50至122°F)的温度下存活和生长。动物病原体一般是嗜中性的。
嗜热原核生物可以在高于60°C(140°F)的温度下生长。这些温度在腐烂的堆肥堆、温泉和海洋地热喷口中都会遇到。在温泉的径流中,在温度下降到70℃左右的源头附近发现了嗜热菌,如水热菌(生长的最适温度为70℃[158℉];最高温度为79℃[174℉])。
蓝藻合成球藻和光营养滑动细菌氯曲菌的厚席在径流较冷的部分发育。古生酸硫菌对酸性条件有很高的耐受性,允许在pH值约为1.0至6.0的范围内生长,最适温度为80°C(176°F)。许多细菌和古细菌适应于50至70°C(122至158°F)的温度范围,包括芽孢杆菌属、热放线菌属、甲烷杆菌属、甲基球菌属和硫菌属的一些成员。
最引人注目的是上世纪80年代中期在深海海底营养丰富、温度极高的热液喷口中发现了细菌和古细菌。焦菌属的古细菌在80到110°C(176到230°F)的温度范围内繁衍生息,在这种温度下,水只有在极高的压力下才能保持液态。
pH值
大多数细菌在中性pH值范围内(5-8之间)生长,尽管有些物种已经适应了酸性或碱性极端条件下的生活。 嗜酸细菌的一个例子是氧化铁曲霉。 当煤层通过采矿作业暴露于空气中时,黄铁矿硫化物沉积物会受到A.ferrooxidans的攻击而生成硫酸,从而将pH值降低至2.0甚至0.7。 但是,铁氧化农杆菌的耐酸性仅适用于硫酸,因为这些细菌在暴露于等浓度的其他酸(例如盐酸)时会死亡。许多细菌不能耐受酸性环境,特别是在厌氧条件下,结果,植物聚合物在酸性沼泽(pH值在3.7和5.5之间),松树林和湖泊中会缓慢降解。 与嗜酸菌相反,嗜碱菌能够在高达10至11的pH值的碱性浓度下生长。嗜碱菌已从土壤中分离出来,大多数是革兰氏阳性芽孢杆菌属。
盐和水
水是生命的基本需求。有些细菌喜欢咸的环境,因此被称为嗜盐菌。极端嗜盐菌,如嗜盐杆菌,在含盐量为20%至30%的条件下表现出最佳生长,如果盐含量降低,它们会溶解(破裂)。这种细菌存在于死海、盐水池中,有时也存在于咸鱼和兽皮上。中度嗜盐细菌在含盐量为5%至20%的条件下生长,在盐卤和泥浆中也有发现。
死海中的盐柱

图片来源:Peter Carmichael/ASPECT
细菌代谢
异养代谢
如上所述,异养(或有机营养)细菌需要有机分子来提供它们的碳和能量。产生能量的分解代谢反应可以有许多不同的类型,尽管它们都涉及电子转移反应,其中电子从一个分子到另一个分子的运动与产生ATP的能量捕获反应相耦合。一些异养细菌可以代谢糖或复杂的碳水化合物来产生能量。这些细菌必须产生许多特定的蛋白质,包括将多糖降解为其组成糖单位的酶,在细胞内积累糖的运输系统,以及将糖转化为代谢中间产物之一的酶,如葡萄糖-6-磷酸。糖酵解的Embden-Meyerhof途径和磷酸戊糖途径都存在于真核细胞中。有些细菌拥有Entner-Doudoroff途径,主要将葡萄糖转化为丙酮酸,还有一些途径可以用较少的酶催化步骤将葡萄糖转化为较小的化合物。
糖代谢通过发酵和呼吸两个不同的过程为细胞产生能量。发酵是在没有任何外部电子受体的情况下进行的厌氧过程。有机化合物,如糖或氨基酸,被分解成更小的有机分子,这些分子接受在能量源分解过程中释放的电子。
这些分解代谢反应包括几个步骤,直接形成ATP。当葡萄糖被分解成乳酸时,就像某些乳球菌和乳酸杆菌以及高等真核生物的肌肉细胞中发生的那样,每个葡萄糖分子只产生两个ATP分子,相当数量的葡萄糖必须被降解以提供足够的能量供细菌生长。由于有机分子在发酵过程中仅被部分氧化,发酵细菌的生长会产生大量的有机终产物,而消耗的每个葡萄糖分子的能量输出相对较小。
很少有细菌只产生乳酸,乳酸对细菌毒性很大,限制了菌落的生长。特定细菌利用多种额外的发酵途径分解葡萄糖;这些途径的特征性终产物有助于细菌的鉴定。这些最终产物通常比乳酸毒性小,或者是利用额外的代谢能量形成的。例如,大肠杆菌混合酸发酵的产物包括乳酸、琥珀酸、乙酸、甲酸、乙醇、二氧化碳和氢气。产气肠杆菌能产生大部分相同的发酵产物,以及大量的2,3-丁二醇,它是非酸性的,允许更多的细菌生长。
细胞可以从呼吸中获得相当多的能量,在这个过程中,糖分子的电子不是转移到另一个有机分子,而是转移到一个无机分子上。最常见的呼吸过程(有氧呼吸)使用氧气作为最终的电子受体。糖被完全分解成二氧化碳和水,每分子葡萄糖最多产生38个ATP分子。
电子通过电子传递链转移到氧中,电子传递链是位于细胞膜上的酶和辅助因子系统,其排列方式使电子沿着链的传递与质子(氢离子)穿过膜并离开细胞的运动相耦合。电子传输引起带正电荷的氢离子向细胞外移动,带负电荷的离子向细胞内移动。这种离子梯度导致外部介质的酸化和带电的质膜,其电荷为150至200毫伏。离子梯度的产生,包括质子动力(质子梯度),是所有生物能量产生和储存的一个共同方面。
质子的梯度被细胞直接用于许多过程,包括营养物质的主动运输和鞭毛的旋转。质子还可以通过一种被称为F1F0质子转位ATP酶的膜酶从细胞外部进入细胞质,该酶将质子运动与ATP合成结合,其过程与真核细胞线粒体中发生的过程相同。
能够利用呼吸作用的细菌每一个糖分子产生的能量比发酵细胞多得多,因为能量源的完全氧化(分解)可以完全提取所有可用的能量,如呼吸生物的ATP产量比发酵细菌的ATP产量大得多所示。呼吸有机体利用一定量的营养物质获得更高的细胞物质产量;它们也产生更少的有毒最终产品。然而,氧在水中的溶解度是有限的,需氧菌群的生长和存活与可用的氧供应量成正比。只有与空气接触的细菌才能获得连续的氧气供应,当细菌能够漂浮在暴露在空气中的表面上时,或者当细菌所在的介质被剧烈搅拌时,就会发生这种情况。
在厌氧条件下,呼吸也可以通过称为厌氧呼吸的过程发生,其中最终的电子受体是无机分子,如硝酸盐(NO3-)、亚硝酸盐(NO2-)、硫酸盐(SO42-)或二氧化碳(CO2)。在硫酸盐和二氧化碳的情况下,使用这些受体的细胞可获得的能量产量比氧气的呼吸低得多,但它们仍然比发酵可获得的能量产量高得多。一些细菌利用无机分子进行无氧呼吸的能力可能具有环境意义。大肠杆菌可以使用氧气,硝酸盐或亚硝酸盐作为电子受体,斯图氏假单胞菌在反硝化,硝酸盐转化为亚硝酸盐和氮气(N2)方面具有重要的全球意义。脱硫弧菌和脱硫单胞菌分别还原硫酸盐和元素硫,产生硫化物(S2-),而木醋杆菌和产甲烷古细菌,如嗜热甲烷杆菌,将二氧化碳还原为乙酸盐和甲烷。古细菌通常使用氢作为电子供体,使用二氧化碳作为电子受体以产生甲烷,或者使用硫酸盐作为电子受体以产生硫化物。
自养代谢
自养细菌以二氧化碳为碳源合成所有的细胞成分。从二氧化碳合成有机化合物的最常见途径是还原性戊糖磷酸(Calvin)循环,还原性三羧酸循环和乙酰辅酶A途径。 由美国生物化学家梅尔文·卡尔文(Melvin Calvin)阐明的卡尔文循环是这些途径中分布最广泛的途径,在植物,藻类,光合细菌和大多数需氧自养细菌中起作用。
卡尔文循环的关键步骤是核酮糖1,5-二磷酸与二氧化碳的反应,产生两个分子的3-磷酸甘油酯,葡萄糖的前体。这种循环在能量方面对细胞来说是极其昂贵的,因此一个甘油醛-3-磷酸分子的合成需要消耗九个ATP分子和六个电子供体分子的氧化,即烟酰胺腺嘌呤二核苷酸磷酸(NADPH)的还原形式。自养行为取决于细胞进行光合作用或有氧呼吸代谢的能力,这是唯一能够提供足够能量来维持碳固定的过程。
卡尔文循环
细菌使用加尔文循环来合成有机化合物。核糖1,5-二磷酸(RuBP)与二氧化碳的反应导致生成3-磷酸甘油酸酯(PGA)分子,该分子通过多次中间反应转化为一分子3-磷酸甘油醛(Gal3P)。然后可以将3-磷酸甘油醛转化为其他分子,包括糖或淀粉。
需氧非光合自养菌是指不仅以二氧化碳为唯一碳源,而且以氧为电子受体的无机化合物(电子供体)为能源的细菌。这些细菌在分类上是多样的,通常由它们使用的电子供体来定义。例如,欧洲亚硝基单胞菌将氨(NH4+)氧化为亚硝酸盐,而Nitrobacter winogradskyi将亚硝酸盐氧化为硝酸盐。 硫杆菌将硫代硫酸盐和元素硫氧化为硫酸盐,而氧化铁曲霉将亚铁离子氧化为三价铁。
这种不同的氧化能力使氧化亚铁杆菌能够耐受高浓度的许多不同离子,包括铁、铜、钴、镍和锌。所有这些类型的细菌似乎都是专性的岩性滋养菌,不能在很大程度上利用有机化合物。一氧化碳(CO)被寡糖carboxidovorans氧化成二氧化碳,而氢气(H2)被真核产碱杆菌(Alcaligenes eutrophus)氧化,在较小程度上也能被其他许多细菌氧化。
代谢能是由这些电子供体的氧化提供的,其方式基本上与呼吸异养生物相同,异养生物将电子从有机分子转移到氧气中。当电子沿着电子传递链传递到氧时,质子在细胞膜上产生梯度。这个梯度被用来产生ATP分子。岩石自养生物中存在的其他反应是用于从无机供体中去除电子和固定二氧化碳的反应。
光营养代谢
地球上的生命依靠光合作用将太阳能转化为细胞能量。光合作用的一般过程是利用叶绿素来吸收来自太阳的光能,并释放出一个能量更高的电子。这种电子通过电子传递链,通过形成质子梯度和伴随的ATP合成产生能量。电子最终返回叶绿素。这种循环反应路径可以满足电池的能量需求。
然而,为了使细胞生长,二氧化碳固定的卡尔文循环必须被激活,电子必须转移到辅助因子NADP以形成NADPH,这是循环运行所需要的大量的NADPH。因此,光营养细胞的生长需要一个电子源来取代生物合成反应中消耗的电子。
光合生物根据这些电子来源的性质分为两大类。其中一类包括高等植物、真核藻类和蓝藻(蓝绿藻);这些生物含有色素叶绿素a,在产生氧气的反应中以水为电子源。
据认为,到18亿年前,蓝藻的前身已经在全球产生了足够的氧气,开始允许更高形式的生命的发展。析氧光合作用需要两个独立的光吸收系统的作用,将水中电子的能量提高到足以转移到NADP的水平。因此,在这些生物体中存在两个不同的光反应中心,一个用于产氧反应,另一个用于能量产生的循环过程。
在蓝藻中,两个光反应中心都含有叶绿素a。它们的光合器官还含有其他光吸收色素,这些色素作为触角来捕获光能并将其转移到反应中心。蓝藻触角包括额外的叶绿素a分子和藻胆体,叶绿素a分子将能量转移到循环反应中心,藻胆体是一种蛋白质色素,它们吸收短、高能波长的光,并将能量传递到析氧反应中心。在几乎所有的蓝藻中,光合器都包含在一个广泛的细胞内扁平膜囊系统中,称为类囊体,其外表面布满规则排列的藻胆体颗粒。这种色素聚集体存在于类囊体表面的排列称为光系统。
其他光合细菌只含有一种不同色素的单一反应中心,称为细菌叶绿素,它能吸收长而低能波长的光。这些生物需要除水以外的电子供体,并且不释放氧气。绿色细菌(绿藻科)和紫色硫细菌(藻科)使用元素硫、硫化物、硫代硫酸盐或氢气作为电子供体,而紫色非硫细菌使用氢或有机基质中的电子。这些细菌需要厌氧条件才能进行光合作用。绿色细菌的光系统与高等植物的光系统Ⅰ有关,而紫色细菌的光系统Ⅱ则与光系统Ⅱ有关,后者提供了从细菌到植物的进化轨迹。
细菌的生物合成途径
许多原核生物能够将任何给定的碳源转化为生物合成的组成部分,例如氨基酸、嘌呤、嘧啶、脂类、糖和酶辅因子。在这些生物合成途径中,每种酶的数量和活性都受到严格的调节,因此细胞在任何时候只产生所需的任何化合物。
在进化过程中,一些细菌失去了编码某些生物合成反应的基因,因此可能需要营养补充。例如,支原体的DNA含量约为大肠杆菌的四分之一,它有许多营养需求,甚至丧失了形成细胞壁的能力。
支原体
图源:Centers for Disease Control and Prevention
革兰氏阴性人型支原体和在琼脂培养基上生长的常见机会性T株支原体的分离株。
细菌是肉眼看不到的单细胞微生物,是地球上最古老的生物之一,它们无处不在且数量惊人,难以逃避。由于我们看不到它们并且对它们的了解相对较少,因此细菌的世界对我们来说是神秘的,并且通常不为大众所重视。
人类每天使用细菌。细菌可用于保存食物,为农作物增添养分,制作奶酪和酸奶以及消除固体废物。这些用途有益于人类的日常生活。一些细菌在生物研究以及药物和疫苗的开发中非常有用。有的细菌帮助管理化粪池系统和清理漏油,或制作酸奶,奶酪和葡萄酒。人体中的细菌可以使身体受益,这取决于它们是细菌的类型。一些细菌有助于训练免疫系统,并有助于预防过敏。其他的则可以保护人体免受有害的致病细菌的侵害。细菌负责帮助我们消化食物,有些细菌会产生对人体非常有用的酶,激素和维生素。
细菌还会导致人患病或死亡。当人打喷嚏或咳嗽时,传染病(例如普通感冒和流感)可能会从细菌中传播。粪便中的细菌也会使人生病。如果一个人上完洗手间后没有正确洗手,他们可能会感染大肠杆菌和自己。这些特性使人类需要花更大精力来研究和重视细菌。
客观全面认识细菌,是我们正确管理和利用它们的第一步。
附录:
常见革兰氏阳性菌属:
Corynebacterium 棒杆菌属 , Propionibacterium 丙酸杆菌属, Alloiococcus 差异球菌属, Oscillospira 颤螺菌属, Enterococcus 肠球菌属, Dialister 戴阿利斯特杆菌属, Anaerostipes 丁酸弧菌属, Butyrivibrio 丁酸弧菌属, Mobiluncus 动弯杆菌属, Planomicrobium 动性杆菌属, Brevibacillus 短芽孢杆菌属, Actinomycetales 放线菌目, Actinomyces 放线菌属, Mycobacterium 分枝杆菌属, Finegoldia 芬戈尔德菌属, Faecalibacterium 粪杆菌属, Coprococcus 粪球菌属, Coprobacillus 粪芽孢菌属, Gordonia 戈登氏菌, Holdemania 霍尔德曼氏菌属, Rhodococcus 红球菌属, Arthrobacter 节杆菌属, Blautia 经黏液真杆菌属, Rubrobacter 红色杆菌属, Cetobacterium 鲸杆菌属, Catonella 卡托氏菌属, Lachnobacterium 毛螺旋菌属, Collinsella 柯林斯氏菌属, Nocardioides 类诺卡氏菌属, Paenibacillus 类芽孢杆菌属, Streptococcus 链球菌属, Ruminococcus 瘤胃球菌属, Roseburia 罗氏菌属, Rothia 罗氏菌属, Vagococcus 漫游球菌属, Lachnospira 毛螺菌属, Moryella 毛螺旋菌属, Oribacterium 毛螺旋菌属, Leuconostoc 明串珠菌属, Ureaplasma 尿枝原体属, Dermabacter 皮杆菌属, Staphylococcus 葡萄球菌属, Aerococcus 气球菌属, Carnobacterium 肉杆菌属, Lactobacillus 乳杆菌属, Lactococcus 乳球菌属, Slackia 史雷克氏菌属, Bifidobacterium 双歧杆菌属, Fusobacterium 梭杆菌属, Parvimonas 微单胞菌属, Weissella 魏斯氏菌属, Peptostreptococcus 消化链球菌属, Peptococcus 消化球菌属, Sporosarcina 芽孢八叠球菌属, Bacillus 芽孢杆菌属, Anaerococcus 厌氧球菌属, Alicyclobacillus 脂环酸杆菌属, Dactylosporangium 指孢囊菌属
常见革兰氏阴性菌属:
Escherichia 埃希氏菌属, Eikenella 艾肯氏菌属, Acidaminococcus 氨基酸球菌属, Hymenobacter 薄层菌属, Phenylobacterium 苯基杆菌属, Burkholderia 伯克霍尔德氏菌属, Acinetobacter 不动杆菌属, Filifactor 产丝菌属, Asticcacaulis 不粘柄菌属, Ochrobactrum 苍白杆菌属, Oxalobacter 草酸杆菌属, Flavobacterium 产黄菌属, Enterobacter 肠杆菌属, Comamonas 丛毛单胞菌属, Acetobacter 醋杆菌属, Brevundimonas 短波单胞菌属, Capnocytophaga 二氧化碳嗜纤维菌属, Delftia 代尔夫特菌属, Paracoccus 副球菌属, Arcobacter 弓形菌属, Devosia 德沃斯氏菌属, Mesorhizobium 根瘤菌属, Citrobacter 枸橼酸杆菌属, Azospirillum 固氮螺菌属, Stenotrophomonas 寡养单胞菌属, Marinobacter 海杆菌属, Rhodobacter 红细菌属, Rhodoplanes 红游动菌属, Porphyromonas 红棕色单胞菌属, Vibrio 弧菌属, Methylobacterium 甲基杆菌属, Pseudomonas 假单胞菌属, Succiniclasticum 解琥珀酸菌属, Chryseobacterium 金黄杆菌属, Kingella 金氏菌属, Megasphaera 巨球形菌属, Megamonas 巨单胞菌属, Klebsiella 克雷伯氏菌属, Lautropia 口动菌属, Aggregatibacter 凝聚杆菌属, Bradyrhizobium 慢生根瘤菌属, Agrobacterium 农杆菌属, Sphingomonas 鞘氨醇单胞菌属, Rikenella 理研菌属, Myroides 类香味菌属, Helicobacter 螺杆菌属, Chloronema 绿丝菌属, Erwinia 欧文氏菌属, Bilophila 嗜胆菌属, Treponema 密螺旋体属, Moraxella 莫拉氏菌属, Neisseria 奈瑟氏菌属, Sphingobacterium 鞘氨醇杆菌属, Bacteroides 拟杆菌属, Prevotella 普雷沃氏菌属, Sporocytophaga 生孢噬纤维菌属, Psychrobacter 嗜冷杆菌属, Hyphomicrobium 生丝微菌属, Prosthecobacter 突柄杆菌属, Cytophagales 嗜细胞菌目, Pectinatus 梳状菌属, Lysobacter 溶杆菌属, Sutterella 萨特氏菌属, Serratia 赛氏杆菌属, Cupriavidus 贪铜菌属, Leptotrichia 纤毛菌属, Haemophilus 嗜血杆菌属, Cytophaga 噬纤维菌属, Leptolyngbya 瘦鞘丝藻属, Campylobacter 弯曲杆菌属, Desulfobulbus 脱硫叶菌属, Enhydrobacter 水栖菌属, Pedomicrobium 土微菌属, Desulfovibrio 脱硫弧菌属, Veillonella 韦荣菌属, Shewanella 希瓦氏菌属, Gemmatimonas 牙单胞菌属, Selenomonas 月形单胞菌属, Phormidium 席蓝细菌属, Nitrospira 硝化螺菌属, Cardiobacterium 心杆菌属, Anaerovibrio 厌氧弧菌属, Halomonas 盐单胞菌属, Anaerobiospirillum 厌氧螺菌, Bdellovibrio 蛭弧菌属
以上信息整理来自谷禾健康,若需转载,请备注出处。
相关阅读:
主要参考资料:
Robert J. Kadner, Kara Rogers, Yamini Chauhan Swati, Yamini Chauhan, Swati Chopra, Aakanksha Gaur, Parul Jain, Robert Lewis, Gloria Lotha, Deepti Mahajan, Richard Pallardy, Chelsey Parrott-Sheffer, Marco Sampaolo, Veenu Setia, Shiveta Singh, Grace Young. Science Biology Archaea & Bacteria Bacteria. (1999-2020)
GEORGE M. GARRITY et al. (eds.), Bergey’s Manual of Systematic Bacteriology, 5 vol., 2nd ed. (2001–12)
MICHAEL T. MADIGAN et al., Brock Biology of Microorganisms, 14th ed. (2015)
JOANNE M. WILLEY, LINDA M. SHERWOOD, and CHRISTOPHER J. WOOLVERTON, Prescott’s Microbiology, 10th ed. (2015)
LUCY SHAPIRO and RICHARD LOSICK (eds.), Cell Biology of Bacteria (2011)
THOMAS D. BROCK, The Emergence of Bacterial Genetics (1990)
LARRY SNYDER et al., Molecular Genetics of Bacteria, 4th ed. (2013)
CLIVE EDWARDS (ed.), Microbiology of Extreme Environments (1990)
CHARLES GERDAY and NICOLAS GLANSDORFF (eds.), Physiology and Biochemistry of Extremophiles (2007)

谷禾健康
大部分人都遭受过头皮屑的困扰。头皮屑虽然看起来不是严重的疾病,但有时候足以令人头疼和尴尬,影响整体生活质量。

头皮屑是一种非常常见的皮肤病,无论年龄,性别等,任何人都可能会有。
头皮屑通常看起来像油腻的皮肤片,如果飘落在深色衣服上更加明显。在头皮上,它也可能表现为皮肤的鳞片状斑点,有时伴有发红、发痒或发炎。
头皮表面为微生物提供了独特的微环境,主要来自宿主的生理条件,包括皮脂含量,水分,pH值等。
头皮屑与多种可能的原因有关,包括卫生差,压力大和饮食不当,当然头皮屑的产生也离不开微生物群。
微生物包括真菌和细菌。
真菌
少量脱落实际上是正常的,因为皮肤细胞会死亡,以便形成新的皮肤。然而,在某些情况下,由于一种或多种潜在的问题可能会使头皮真菌的过量生长,导致剥落异常大量。
头皮表面发现的最多的真菌是马拉色菌(malassezia)。这种真菌生活在头皮上,以皮肤油脂为食。
马拉色菌里有限制性马拉色菌、球形马拉色菌等,不同种类的马拉色菌的致病性不同,并适应人体皮肤的不同部位,这主要归因于宿主的生理条件(皮脂,免疫反应,汗液等),环境(温度,湿度,紫外线暴露)以及其他微生物的存在。
其中限制性马拉色菌与球形马拉色菌的比例较低与健康的头皮有关。
当球形马拉色菌利用脂肪酶代谢其消耗的油脂时,会产生一种副产物——油酸,该副产物渗透到头皮并引起头皮屑。
功能分析表明,真菌菌群主要通过与头皮屑头皮细胞-宿主粘附有关的途径富集。马拉色菌的粘附与头皮角质层屏障功能受损有关,可能进一步有助于头皮屑区马拉色菌的发育。
与健康头皮相比,头皮屑头皮中还发现了大量与一般功能和遗传信息处理相关的通路(硫中继系统、蛋白酶体通路、酵母细胞周期和减数分裂等),且与限制性马拉色菌呈显著正相关。
“酵母细胞周期”和“减数分裂”是参与真菌细胞周期和增殖的重要通路,这两通路在头皮屑头皮上的较高数量表明,它们在头皮屑头皮上的马拉色菌的增殖中起作用。
“蛋白酶体通路”与包膜的形成有关,包膜是真菌的主要致病决定因素。

细菌
头皮表面发现的最丰富的两个细菌属是Cutibacterium菌(健康头皮)和葡萄球菌(头皮屑头皮)。
研究表明,与健康人群相比,头皮屑患者的限制性马拉色菌和葡萄球菌的丰度较高(p <0.05)。
不同的葡萄球菌丰度变化也不同。
与健康人群相比,头皮屑患者头皮上表皮葡萄球菌的丰度减少,而同时头状葡萄球菌的丰度显著增加。头状葡萄球菌丰度的增加足以驱动头皮屑损伤部位葡萄球菌绝对物种丰度的总体增加。
细菌菌群的功能分析显示出,健康头皮与以下代谢相关通路明显高于头皮屑头皮:
氨基酸:丙氨酸、天冬氨酸、谷氨酸、精氨酸、脯氨酸、赖氨酸、组氨酸、半胱氨酸、蛋氨酸代谢;
维生素:维生素B6,B7
其他辅助因子:卟啉,叶绿素,烟酸和烟酰胺代谢,泛醌和其他萜醌的生物合成。
这些头皮微生物群合成的维生素、生物素、氨基酸等被头皮角质形成细胞吸收,为头皮提供营养,在保持头发和头皮健康以及控制头皮屑方面起着有益的作用。
可能引起头皮屑的一些潜在潜在问题包括:
•皮肤状况-例如脂溢性皮炎,湿疹和牛皮癣,会增加头皮屑的风险。
•神经系统疾病-发现患有帕金森氏症等神经系统疾病的人更容易患头皮屑。
•洗护用品的刺激-购买的洗护品含有许多化学物质,可能会刺激头皮并导致发炎,发痒和脱落。
•不良的饮食习惯-摄入含锌和B族维生素的食物不足,可能会导致头皮屑。
•年龄-虽然说任何年龄的任何人都可能患有头皮屑,但往往在年轻人和中年人中更为普遍。
•气候-冬季的寒冷天气可能促进头皮中酵母样真菌的生长。
•激素-体内激素的变化可能会影响头皮的天然皮脂分泌。激素失调的人更容易有头皮屑。

为了想要尽快清除头皮屑,有人会选择含药的洗发水。但是,药用洗发水真的可以治愈头皮问题吗?
首先,含药的洗发香波只有在您持续使用时才有效-它们不能彻底治愈头皮屑。这就意味着,必须要每周使用几次这种洗发水才能看到效果,一旦停止使用,头皮屑很可能卷土重来。

这些洗发剂大多数都含有抗真菌成分,例如酮康唑。虽然可以有效控制头皮屑,但连续使用会刺激头皮,并使头发变得蓬松干燥。
其他药用去屑洗发水也可能含有煤焦油,煤焦油是通过延迟皮肤细胞再生的过程来控制头皮屑。但是,煤焦油是已知的致癌物质,因此最好避免含有这种成分的洗发水。
皮肤会吸收施加的任何物质,自然疗法是最佳选择。
盐
快速去除头皮屑且不使用任何有害化学物质的最安全方法之一是:用盐来去除片状头皮屑。
在洗澡之前,将盐轻轻按摩到干头皮上,使头发在漂洗时更容易去除。
椰子油
椰子油可以帮助改善水合作用,减少刺激感,并防止真菌在头皮上生长。恢复头皮的水分有助于止痒。

根据一项临床试验的结果,在117名1-13岁的儿童中,将纯净的椰子油涂在皮肤上可使特应性皮炎症状减少68.23%。当然也需要进行更多研究,以评估椰子油在治疗头皮屑中的作用。
椰子油的保湿和抗菌特性,使其成为抵抗干燥的绝佳自然选择。
茶树油
使用茶树油治疗头皮屑也是比较快速简便的。有研究人员认为茶树油中的化合物可以有效控制表皮葡萄球菌。

只需在头皮上按摩几滴茶树油,可减少瘙痒和多余油脂的产生。也可以在洗发水中混合几滴茶树油。
芦荟凝胶
芦荟是一种多汁植物,以其愈合特性而闻名。叶片中的凝胶含有几种生物活性化合物,例如氨基酸和抗氧化剂,可减少头皮屑。
2019年一项研究检查了23项涉及芦荟的临床试验。这些研究的结果表明,芦荟凝胶可以改善皮肤中的水分保留并促进伤口愈合。芦荟还可以减轻炎症,这可以帮助有头皮屑症状的人减轻症状,例如发痒。
2015年的研究表明,芦荟的抗菌性能可预防头皮屑。

饮 食
头皮屑通常与不良饮食有关,首先请开始调整饮食习惯。
Omega-3脂肪酸
Omega-3脂肪酸可以降低血压,增加“良好的” HDL胆固醇水平,并支持心脏和大脑健康。这种脂肪酸的缺乏会导致不良症状,例如头皮屑,指甲变脆和皮肤干燥。
锌
锌是支持人体免疫系统并促进细胞生长的矿物质。可以从动物蛋白,坚果等获取锌。

根据NIH研究表明,严重的锌缺乏与脱发、腹泻、皮肤损伤等有关。
2016年的一研究将缺锌列为脂溢性皮炎和头皮屑的潜在诱因。
减少糖和谷物
应该减少糖和谷物的摄入,因为已知这些会促进酵母或真菌的过度生长。
水果
水果和蔬菜含有许多必需的维生素,矿物质和抗氧化剂,可以帮助减轻炎症。

一项涉及4379人的观察性研究称,多吃水果的人患脂溢性皮炎的可能性较小。
研究结果还表明,典型的西方饮食可能会增加女性脂溢性皮炎的风险。
生物素
2018年的一项审查显示,生物素缺乏症可能导致几种皮肤疾病,包括脂溢性皮炎。生物素,也称为维生素B7,在支持健康的头发,指甲和皮肤方面发挥作用。富含生物素的食物包括:肝、蛋黄、坚果、三文鱼等。

晒太阳
还有一种非常简单的方法是:经常晒晒太阳。阳光可以防止真菌在头皮上繁衍。同时可以增加维生素D的含量,从而有助于改善皮肤状况。

解 压
此外,压力是头皮屑的最常见诱因之一。学习如何正确处理压力也很重要。适当培养一些兴趣爱好,定期放松解压,尽可能远离压力源。

其 他
最后,要保持良好的个人卫生习惯,避免过多抓挠,发痒最初是由头皮屑引起的刺激引起的,抓挠会增加刺激性并导致恶性循环。
避免使用过多的护发产品,防止头皮发炎。
摆脱头皮屑需要时间,精力和耐心,才能获得持久的效果。
Q & A
如何区分头皮屑和虱子?
如果白色颗粒在吹动或梳理头发时掉落,痒的程度相对较轻,则很可能是头皮屑。头虱在头皮部叮咬吸血,主要症状头皮部剧烈瘙痒,斑点粘在发干上,则可能是虱子或虱子空卵。
头皮屑可以治愈吗?
没有永久治愈的方法,但合理的方式可以控制住头皮屑。一些自然疗法包括茶树油、椰子油等。
头皮屑会导致脱发吗?
头皮屑本身不会直接导致脱发。然而,因为头皮屑抓挠发痒的头皮,久而久之可能会导致发丝脱落。
相关阅读:
主要参考文献:
Grimshaw SG, Smith AM, Arnold DS, Xu E, Hoptroff M, Murphy B. The diversity and abundance of fungi and bacteria on the healthy and dandruff affected human scalp. PLoS One. 2019;14(12):e0225796. Published 2019 Dec 18.
Saxena R, Mittal P, Clavaud C, et al. Comparison of Healthy and Dandruff Scalp Microbiome Reveals the Role of Commensals in Scalp Health. Front Cell Infect Microbiol. 2018;8:346. Published 2018 Oct 4.
National Health Service, Dandruff
Lorch JMP J.M.; Vanderwolf K.J.; Schmidt K.Z.; Verant M.L.; Weller T.J.; Blehert D.S. Malassezia vespertilionis sp. nov.: a new cold-tolerant species of yeast isolated from bats. Persoonia. 2018; 41:56–70. 10.3767/persoonia.2018.41.04
Symptom Find, Dandruff
Lin Qingbin,Panchamukhi Ananth,Li Pan et al. Malassezia and Staphylococcus dominate scalp microbiome for seborrheic dermatitis.[J] .Bioprocess Biosyst Eng, 2020
Zinc, Fact Sheet for Health Professionals
Borda, L. J., & Wikramanayake, T. C. (2016). Seborrheic dermatitis and dandruff: A comprehensive review.
Hekmatpou, D., et al. (2019). The effect of aloe vera clinical trials on prevention and healing of skin wound: A systematic review.
Kim, S., et al. (2017). Enhanced barrier functions and anti-inflammatory effect of cultured coconut extract on human skin.
Omega-3 fatty acids fact sheet for consumers. (2018).
Sanders, M. G. H., et al. (2019). Association between diet and seborrheic dermatitis: A cross-sectional study.
Varma, S. R., et al. (2019). In vitro anti-inflammatory and skin protective properties of virgin coconut oil.