-

 CNAS L23010
CNAS L23010

 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业 二级病原微生物安全实验室
二级病原微生物安全实验室- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业 二级病原微生物安全实验室

谷禾健康
从吃什么到谁在消化
我们常常纠结于吃什么,食物的卡路里、蛋白质或脂肪含量,却可能忽略了一个关键问题:这些食物进入我们体内后,到底发生了什么?
我们吃下的每一口饭,其实都不是我们一个人在享用,在我们肠道里,还住着数万亿的成员——肠道菌群。我们吃进去的营养,需要先经过它们的“再加工”和“巧分配”,才能真正为我们所用。
我们很多人其实已经了解肠道菌群这个系统至关重要,但其内部精密的运作逻辑,对我们而言就像一个黑箱。
关于这些问题,我们用两篇研究来解析这一微观世界的运作机制。
发表在《The ISME Journal》的研究深入微观,揭示了菌群的“社会规则” 。肠道中大部分细菌都存在“氨基酸合成缺陷”,也就是说它们无法独立制造所有生存必需的氨基酸。这种缺陷却不一定是坏事,某种程度上反而驱动微生物群落多样性、稳定性,促成了它们之间广泛的“交叉喂养”(cross-feeding),形成了一个高度依赖、紧密协作的社会网络。
另一篇发表在《Cell》的研究,则是聚焦宏观,首次量化了菌群的“经济贡献” ,也就是说肠道微生物群每天向人体“输送”多少能量以及这一过程受饮食影响的规律。
通过构建一个精密的系统通量模型,该研究精确计算出,肠道微生物群的发酵产物(乙酸、丙酸、丁酸等)可为人体提供每日2%~10%的能量。这个比例受饮食结构(尤其是纤维摄入量)的显著影响,明确了微生物对宿主能量平衡的具体贡献值。
如果说前一篇回答了菌群之间如何协作,那么后一篇是让我们了解了菌群的协作对我们有什么用。这两篇一起能够让我们更系统、更完整地理解肠道菌群不仅是一个随机的微生物集合,而是一个遵循着内部协作规则、并能对外部宿主产生巨大影响的、有组织的生态系统。
本文将立足于这一整体视角,整合两大前沿发现,深入剖析菌群内部的互补协作逻辑与对宿主的代谢贡献,进而揭示“营养-微生物-宿主健康”核心轴线中的多层级调控规律。
氨基酸合成能力缺陷在肠道细菌中非常普遍
人类肠道内有些细菌,自己造不出某些生存必需的营养物质。比如氨基酸或维生素,它们必须依靠外部环境或其他细菌提供这些物质才能活下去,这种情况就叫“营养缺陷型”。这种“专性营养需求”会对单个细菌和整个菌群系统都产生很大影响。
发表在《The ISME Journal》这篇文章,通过对一个包含5414株肠道细菌的基因组目录进行分析后,选取了其中3687个高质量的基因组进行深入研究发现:
氨基酸合成能力缺陷在肠道细菌中非常普遍。例如,近64%的菌株缺乏合成色氨酸的能力。被“丢失”最多的氨基酸(如色氨酸、亮氨酸、异亮氨酸、缬氨酸等),恰好也是人类自身的必需氨基酸。
预测的氨基酸缺陷被分为人类必需和非必需氨基酸
doi.org/10.1038/s41396-023-01537-3
然而,这种看似的缺陷并非生存劣势,而是一种高效的进化策略与生态分工。
更有趣的是,研究还揭示了这种营养缺陷与特定发酵产物生成之间的深刻关联。例如:
这表明,当合成氨基酸的代谢路径被关闭时,其前体物质会被重新导向其他途径,生成对群落乃至宿主健康至关重要的发酵产物。
doi.org/10.1038/s41396-023-01537-3
总而言之,肠道菌群中的营养缺陷并非弱点,而是驱动群落形成社会性分工的关键。
有的细菌不会做饭,只能吃别人做的饭,但因为社区里有人乐意做、多余的饭还能分享,因此大家都能活得更好,整个微生物群落变得更有分工、更稳定。
通过这种交叉喂养,菌群不仅实现了整体的稳定与繁荣,更获得了强大的稳态韧性,使其在面对饮食波动或药物干扰等外界压力时,能更好地维持其核心功能,也增强了微生物与宿主之间的代谢联系。
“营养缺陷型”细菌越多,肠道菌群多样性越高
研究人员发现了一个有意思的现象:肠道菌群中“营养缺陷型”细菌的丰度,与菌群的多样性和长期稳定性呈显著正相关。也就是说,一个人肠道里营养缺陷型细菌越多,他的肠道菌群种类越丰富,也越稳定。
这其实验证了一个早期来自理论生态学的假说,即“代谢交叉喂养能创造新的生态位”。当细菌通过互相提供必需营养物来弥补彼此的合成缺陷时,它们便构筑了共存的基础,避免了直接的生存竞争。
为了验证该假说,研究团队分析了两个独立队列、跨度长达3~4年的纵向人体数据,结果高度一致:
这意味着,一个高度依赖内部“互助网络”的菌群,其整体结构更加稳固,对饮食波动、药物使用等外界干扰具有更强的抵抗力。
研究者引用的另一项关于B族维生素的研究也佐证了此观点:在小鼠模型中,即使科学家改变小鼠饮食中B族维生素的含量,小鼠肠道菌群的组成几乎没变化;B族维生素“缺乏型”的菌群比例也没明显改变。这说明这些菌并不是靠饮食提供的维生素生存,而是靠菌群内部那些“会合成维生素”的菌种提供营养。
因此,这又进一步印证了他们的观点:
肠道中的营养缺陷并非个体的弱点,反而是驱动群落走向更高层次合作与韧性的关键机制。营养缺陷型菌群的越多,越是促进了普遍的交叉喂养,由此形成的复杂互助网络,是维持整个生态系统平衡、健康和具备自我修复能力的核心。
超越肠道:菌群产物的全身性健康效应
研究人员发现肠道里“营养缺陷型”细菌,不仅仅影响菌群结构,也和宿主的代谢状态、体重以及血液代谢物水平之间存在联系。
研究团队注意到以前人们关注焦点是,“肠道菌群的代谢怎么影响人体健康”,但从来没有人专门去了解“这些营养缺陷型细菌的数量会不会和人体健康参数有关”。于是他们第一次统计分析了氨基酸营养缺陷型菌的比例与受试者健康指标的关系。
结果他们发现几个有意思的点:
①
营养缺陷型菌越多,BMI指数越低
他们发现某些氨基酸营养缺陷型菌的总数量跟人体的 BMI 呈负相关,也就是说,一个人的肠道里这些“营养依赖型”细菌比例越高,他的体重(或体脂)往往越低。
虽然不能说明因果,但这暗示了这些细菌群可能和更健康或更瘦的代谢状态有关。
同时,他们没发现这些细菌的丰度和血细胞计数(也就是血象指标)之间有明显关系,所以这种关联主要是代谢方面的,而不是炎症或免疫系统方面。
②
与血液中某些代谢物水平有关
色氨酸营养缺陷型细菌的比例跟血液中 3-吲哚乙酸(3-IAA)和 3-吲哚丙酸(3-IPA)含量呈正相关。
这两种物质都是由色氨酸经过微生物代谢产生的“吲哚类代谢物”,它们已知具有抗氧化、抗炎、神经保护作用,还能调节肠道屏障和免疫系统。
换句话说:这种缺色氨酸合成能力的细菌越多,人体血液里好的代谢产物越多。这说明这些缺陷菌可能刺激或依赖周围菌群代谢色氨酸,从而促进这些有益代谢物的产生。
不止色氨酸,其他几种氨基酸营养缺陷型(比如谷氨酸、天冬氨酸缺陷等)也和这些吲哚类代谢物呈现正相关。这种一致的趋势表明,肠道中代谢互助可能会提高吲哚相关代谢流通量。
③
与其他代谢物也存在相关性(尤其是胆汁酸和含氮代谢物)
他们发现特定氨基酸营养缺陷型细菌的总比例和对甲酚硫酸酯呈正相关。
这种物质是由肠道菌群将酪氨酸分解后经肝脏代谢转化的产物,通常被认为与肠道代谢活性有关。尽管高水平时可能与肾功能或炎症风险相关,但它也代表了一种活跃的芳香族氨基酸代谢通量。
这些胆汁酸变化说明,肠道代谢生态(由菌群活动决定)确实能影响宿主的脂肪吸收、胆盐循环与炎症信号。换句话说,这种“谁缺什么营养、谁提供什么”的群落特征不光影响菌和菌之间的平衡,也会通过代谢物反馈到宿主的肝脏和消化系统。
血清代谢物水平与肠道微生物组中营养缺陷型细菌频率之间的部分斯皮尔曼相关性
④
没有发现明显的氨基酸水平变化
意思是,虽然菌群结构(营养缺陷型比例)跟体重、代谢物水平有关系,但血液中的游离氨基酸浓度本身并没有明显改变。
这意味着这些影响不是因为“血液里多了或少了某种氨基酸”,而是因为菌群间代谢互动带来的代谢网络变化。
这为氨基酸交叉喂养提供了功能性证据:它不仅是“微生物之间共享营养”的现象,更是宿主健康稳态的化学纽带。
以上这些发现为“交叉喂养”提供了强有力的功能性证据。它不仅是微生物之间维持生存的生态策略,更是一条连接菌群内部互作与宿主全身健康稳态的关键化学纽带。
肠道菌群对宿主能量的直接贡献
肠道微生物群与人体的相互作用,主要通过一系列被称为发酵产物的小分子(以乙酸、丙酸和丁酸为主)来介导。
这些分子由大肠中厌氧微生物产生,这些微生物主要以植物性食物中的复合碳水化合物为食,如膳食纤维和抗性淀粉,以及通过小肠和粘蛋白的膳食蛋白质。
产生后,绝大部分发酵产物会被结肠上皮细胞吸收,并整合到宿主的代谢网络中,发挥多重生理效应:
► ▷
我们要讲的这篇发表在《Cell》的研究,其核心突破在于建立了一个“系统级通量模型”。
它是一个强大的计算框架,能够定量分析在真实生活状态下,人体肠道每天究竟产生了多少发酵产物,以及这些产物的代谢去向。
利用这个框架,研究者发现:
超过90%的可被微生物利用的碳水化合物最后会被转化为发酵产物,而这些发酵产物的大部分都被人体吸收。
这意味着发酵产物是肠道微生物与宿主之间最重要的代谢交换物质,对宿主的信号传导、行为和能量平衡都有显著影响。
微生物对能源供应的贡献
在缺氧的大肠中,微生物利用复合碳水化合物(膳食纤维、抗性淀粉)产生短链脂肪酸(乙酸、丙酸、丁酸)。
这些能量丰富的分子随即被结肠上皮细胞高效吸收,直接汇入人体的能量代谢系统。这一过程对宿主总能量的贡献不容小觑,其具体程度与饮食结构紧密相关:
换算成绝对通量,这意味着人体每天从肠道吸收的发酵产物总量约为 450–1500 mmol。这种“微生物供能”机制,因其难以直接测量,在传统营养学评估中常被忽视。而这项研究的定量模型则明确指出,它是一股对人体能量平衡有显著影响的“代谢暗流”。
DOI: 10.1016/j.cell.2025.07.005
饮食与菌群的明确分工:
饮食决定“量”,菌群决定“质”
研究的核心在于厘清了饮食与菌群在肠道发酵过程中的不同角色。结论非常明确:
饮食结构(尤其是微生物可利用碳水化合物 MACs 含量)是决定发酵产物通量的主因。
相比之下,菌群组成的变化对总产量的影响较小(约±10%),但它显著影响发酵产物的具体构成(如丁酸和乳酸的相对比例)。
研究团队首次利用宏基因组与通量模型估算了健康人肠道的发酵产能:
输入
每天约有 50–80克 的可发酵碳水化合物(MACs)进入结肠。
转化
微生物将其高效转化为 450–1500 mmol 的短链脂肪酸。
输出
这为宿主提供了 2%–10% 的总能量需求,具体比例因饮食模式而异。
这些SCFAs并非均质,而是有明确的组成和分工,其中:
这一发现可以用一句话概括:
“吃什么”(饮食)决定了发酵的总能量,而“谁在吃”(菌群)决定了这些能量以何种形式(哪种SCFA)被利用。
例如,高纤维饮食(>35 g/天)能显著增加短链脂肪酸总产量,促进产丁酸菌(如 Roseburia、Faecalibacterium)增殖,从而增强肠道的抗炎屏障。
相反,富集高蛋白与低碳水饮食则会使代谢转向蛋白质分解,增加氨和硫化氢等潜在有害物质的产生,使某些潜在致病菌(如 Bilophila wadsworthia)扩增。
营养-微生物共代谢模型:预测健康的新坐标
核心结论与研究启示
饮食作为核心驱动因素
研究最终明确指出,饮食是决定肠道发酵产物种类与总量的核心驱动力。它通过双重机制发挥作用:一方面,直接决定了可供发酵的底物(主要是MACs)的数量;另一方面,长期饮食模式会选择性地塑造菌群的结构。
研究者强调,理解宿主健康和微生物作用时,必须同时考虑饮食、微生物群组成与发酵产物之间的相互作用轴线。
对研究方法的反思与建议
该研究的一个重要启示是,粪便样本检测存在巨大局限性。由于高达95%的短链脂肪酸在结肠段即被宿主吸收,粪便中的残留量远不能反映肠道内真实的生产通量。因此,准确评估肠道发酵活动,必须超越简单的粪便分析,转而采用结合了饮食记录和微生物代谢通量计算的系统模型。
全球饮食模式与肠道生态的关联
将此模型应用于全球不同人群,研究揭示了饮食模式对肠道发酵功能的深刻影响。
数据显示,工业化国家(如美国的NHANES队列,平均产量286 mmol/天)的发酵产率显著低于仍保持传统高纤维饮食的人群(如哈扎狩猎采集者,产量最大可达~1000 mmol/天)。
发酵产物对宿主的能量贡献率也在 1.7% 至 12.1% 之间大幅波动。这些证据表明,现代饮食的精加工趋势可能正在系统性地削弱人类肠道微生物组的能量代谢功能。
对动物模型外推的审慎态度
研究还发现,实验常用的小鼠模型与人类在肠道发酵上存在显著的物种差异。按体重归一化后,小鼠的SCFA产率(~400 mmol/kg/天)远高于人类(~7 mmol/kg/天),其对微生物发酵的能量依赖也更高(小鼠>21% vs. 人类<12.1%)。
这一差异提醒科研人员,在涉及全身性代谢、能量平衡及行为的研究中,将小鼠数据直接外推至人类需格外谨慎。尽管如此,在局部肠道层面(比如每单位肠道表面积吸收的发酵产物量,人和鼠的数值相近),两者具有可比性,使小鼠在研究局部互作机制时仍具价值。
研究的宏观意义:迈向定量微生物生态学
总结来看,这项工作的核心贡献在于推动微生物组研究从定性的“存在性分析”迈向了定量的“通量分析”。它不再仅仅回答“微生物产生了什么分子”,而是通过整合代谢组学、宏基因组学和饮食数据,精确计算出宿主与微生物之间关键代谢物的交换速率与规模(即“通量”)。
这种对“剂量”的明确计算,为理解发酵产物如何具体影响宿主生理(即“剂量-效应关系”)提供了坚实的数据基础。
作者最后强调,这一强大的定量框架不仅适用于发酵产物,还具有广阔的应用前景,未来可扩展至氮循环、气体代谢、蛋白质降解等其他关键的宿主-微生物相互作用领域,从而开启一个系统化、定量化理解人体共生生态系统的新篇章。
精准氨基酸补充:基于菌群代谢特征
通过外源性氨基酸干预,可有选择地促进益生菌生长:
这种“氨基酸-选择性补养”策略可能成为未来益生营养的新方向。但其具体菌株配对及剂量效应仍需实验验证。
碳源结构优化:调控能量产物比例
通过调控可发酵碳水化合物的含量与结构,可以:
当菌群有充足、优质的碳源(可发酵碳水化合物)作为能量时,它们会优先利用这些碳源,从而减少对蛋白质的分解(即腐败发酵)。这会直接降低氨、硫化氢、部分吲哚和酚类等潜在有害物质的产生。
体化代谢预测:AI + 代谢组学融合
展望未来,我们可以通过整合微生物基因组尺度代谢模型、宿主代谢组学信息以及个体饮食数据,并利用机器学习算法进行深度分析,从而构建一个强大的综合预测模型。
该模型将能够精准评估个体的能量回收效率,预测其罹患肥胖、炎症性肠病(IBD)、2型糖尿病等代谢相关疾病的风险趋势,并模拟不同营养干预方案的个性化反应。这标志着我们正迈向一个能够为精准营养和预防医学提供坚实科学循证依据的新时代。
综合以上两篇《The ISME Journal》与《Cell》的标志性研究成果,一个突破性的生命科学框架逐渐清晰:肠道微生物群落本质上是一个分布式代谢系统 。其核心机制体现为“营养缺陷-代谢互补-发酵能量”的级联功能网络,三者通过动态互作构成人体代谢稳态的关键调控枢纽。
氨基酸营养缺陷型的普遍存在,说明肠道微生物早已从“个体独立的代谢单元”进化为“代谢互助的生态集合体”,代谢功能呈现出模块化和分工化趋势。
这令人联想到人类社会的分工协作:厨师专注烹饪、医生钻研医术、教师传道授业,正是通过放弃全能、追求专精,整个社会系统更能高效运转。菌群的营养缺陷何尝不是如此?
这种“丧失功能以换取依赖”的策略并非退化,而是一种进化经济学的最优解——正如团队成员通过各司其职提升整体效率,微生物通过外部化代谢与共生交换,降低了整个系统的能量成本。
群落层面的发酵产物研究进一步表明,这种微观的营养依赖会在更高层次上塑造整个碳流通量网络。
《Cell》的研究则将这一微观协作的宏观效应进行了精准量化,研究表明,正是通过交叉喂养(cross-feeding)等复杂的内部协作,菌群构建了一个强大的能量生态系统,最终能为宿主贡献每日高达2%-10%的能量。
这是从抽象概念到具体数值的飞跃,证明菌群这个微观社会,集体协作产生的巨大外部价值。这与人类社会的合作共赢理念不谋而合:供应链中上下游企业的互补、科研团队中不同领域专家的协作,都在印证同一个真理——真正的强大是群体互哺所带来的可量化的集体增益。
长期来看,这种“机制解析-模型构建-临床转化”的闭环路径,将重新定义健康管理的范式——从传统的宿主中心视角转向“菌群-宿主生态系统”的动态平衡监测。
未来,随着多组学检测成本的下降与算法迭代,这种“预测-干预-验证”的闭环管理模式有望成为常规临床实践的重要组成部分。结合AI驱动的多组学整合技术,我们有望实现从“疾病治疗”到“代谢稳态预测与维护”的跨越,为个性化营养干预与代谢疾病防治提供全新工具。
参考文献:
Starke S, Harris DMM, Zimmermann J, Schuchardt S, Oumari M, Frank D, Bang C, Rosenstiel P, Schreiber S, Frey N, Franke A, Aden K, Waschina S. Amino acid auxotrophies in human gut bacteria are linked to higher microbiome diversity and long-term stability. ISME J. 2023 Dec;17(12):2370-2380.
Arnoldini M, Sharma R, Moresi C, Chure G, Chabbey J, Slack E, Cremer J. Quantifying the varying harvest of fermentation products from the human gut microbiota. Cell. 2025 Sep 18;188(19):5332-5342.e16.

谷禾健康
当我们坐在餐桌前,精心挑选着盘中的食物时,一个常常被忽略的事实是:我们并非独自在享用这顿美餐。在我们的身体内部,一个无比繁华、拥挤而又充满活力的微观世界——肠道菌群,正翘首以盼,等待着我们送去的“补给”。这个由数万亿个细菌、真菌、病毒和古菌组成的复杂生态系统,其细胞总数甚至超过了我们自身的人体细胞,基因数量更是人类基因组的百倍以上。因此,科学家们形象地将其称为人体的“第二基因组”或“隐形器官”。
那么,究竟是什么决定了这个“隐形器官”的构成与功能?虽然遗传、年龄、生活方式及药物等多种因素都有关联,但研究普遍指出,饮食是最关键的驱动力。我们的食物不仅为自身提供能量和营养,也直接影响肠道微生物的“口粮”,不同食物成分会选择性促进不同微生物的生长,正如施用不同肥料会催生出不同的花草组合一样。
在众多营养素中,碳水化合物(尤其是膳食纤维)和脂肪对肠道菌群的影响已有深入研究,而蛋白质这一生命基础营养素,其调节肠道菌群的作用却长期被忽视。
我们都知道蛋白质对构建肌肉、修复组织、维持生命活动至关重要,但我们摄入的蛋白质究竟如何与肠道菌群互动?更重要的是,不同来源的蛋白质——例如,一块牛排(动物蛋白)与一盘扁豆(植物蛋白),会对我们体内的微观生态系统产生截然不同的影响吗?
这正是我们今天要深入探讨的核心问题。本文是基于最近发表在bioRxiv平台、题为《膳食蛋白源塑造肠道微生物结构和预测功能:一项结合机器学习的Meta分析》的重磅研究,为我们揭开提供了迄今为止较为全面、系统化的线索。
该研究巧妙地运用了Meta分析,汇集了全球范围内多项独立研究的数据,并通过前沿的机器学习算法进行深度挖掘,以前所未有的广度和精度,描绘出动物蛋白与植物蛋白在肠道菌群世界里投下的不同“身影”。
接下来,我们将解读这项研究的核心发现,包括动物蛋白和植物蛋白如何影响菌群的多样性与结构,不同饮食下的代表菌种,以及这些变化如何通过代谢功能影响我们的长期健康。
这不仅是对前沿科学的探索,也为我们的饮食选择带来了全新的认知。
在深入探讨研究的具体发现之前,先了解一下此次研究使用的两大“工具”:Meta分析(Meta-Analysis)和机器学习(Machine Learning)。
理解了它们的工作原理,我们才能更深刻地体会到这项研究结论的科学分量与可靠性。
1
Meta分析:站在巨人肩膀上看得更远
关于“不同蛋白质对肠道菌群的影响”,全球许多研究团队曾独立开展实验,
例如:
A研究组可能招募了20名志愿者,让他们分别食用富含牛肉和富含豆类的饮食;
B研究组则可能在小鼠身上测试了乳清蛋白和豌豆蛋白的效果;
C研究组的研究对象则可能是另一群不同文化背景的人。
每一项研究由于其样本量、实验设计、测量方法和地域人群的差异,得出的结论可能不尽相同,甚至有时会相互矛盾。例如,A研究发现动物蛋白显著降低了某种细菌,而B研究可能发现没有显著变化。这会让公众和政策制定者感到困惑:我们到底该相信谁?
Meta分析正是为应对这种问题而设立的统计方法。它并非进行新实验,而是系统收集并整合同一主题下尽可能多的高质量独立研究,利用统计工具合并与再分析这些数据,从而得出更具代表性和可信度的综合结论。
◮ Meta分析有三大优势:
增大样本量:通过合并多个研究,原本几十、几百的样本量可以瞬间扩大到成千上千,极大地提高了统计功效,使得我们能够洞察到在单个小规模研究中难以发现的、更细微但真实存在的效应。
解决不一致性:它可以评估不同研究结果之间的差异程度(即“异质性”),并探究造成这种差异的原因(例如,可能是因为研究对象的年龄不同,或是干预时间长短不一)。
提高结论的普遍性:因为数据来自不同地区、不同人群、不同实验条件,所以Meta分析得出的结论往往比任何单一研究更具普遍适用性。
在本研究中,作者首先进行了大规模文献筛选,在数千篇论文中,依据严格标准最终选取了13项高质量的人类干预研究,构建了总计近600个样本的数据集。
2
机器学习:从海量数据中挖掘模式的智能算法
收集到了海量数据仅仅是第一步。肠道菌群的数据是出了名的高维度、高复杂度。每个人的肠道里都可能有成百上千种不同的细菌,它们的丰度(数量多少)各不相同。面对这样一个由无数变量构成的复杂矩阵,传统的统计方法有时会显得力不从心。
◮ 机器学习对于数据统计和处理更高效
这时,机器学习就是重要的工具了。作为人工智能的一个分支,机器学习算法尤其擅长在看似杂乱无章的海量数据中,自动学习并识别出隐藏的模式和规律。它不像传统统计学那样需要研究者预先设定严格的假设,而是能够以一种更加“开放”和“探索性”的方式,让数据自己“说话”,以上两种方法也是谷禾健康构建人群肠道菌群数据库和识别菌群与健康以及营养的重要工具。
本研究主要采用了“随机森林”模型。可以这样理解:
如果我们想训练医生仅凭肠道菌群组成判断病人偏好动物蛋白还是植物蛋白,可先搭建一棵决策树:医生依次提问,例如“普雷沃氏菌丰度是否超过X?”、“拟杆菌丰度是否低于Y?”,通过一系列“是”或“否”,最终给出饮食类型诊断。
但单棵决策树容易因偶然因素产生偏误,就像初学医生容易过拟合少数病例一样。
为了避免这个问题,“随机森林”模型同时构建了成百上千棵这样的决策树,形成一片茂密的“森林”。它的高明之处在于“随机”二字:
通过这两个“随机”过程,保证了森林里的每一棵树都是独一无二、各具特色的。当需要做出最终判断时,模型会让森林里的每一棵树都进行一次独立的投票,然后采纳“少数服从多数”的原则,得出最终的、最稳健的分类结果。这就像一个由数百位不同背景、不同专长的医生组成的专家委员会进行会诊,其诊断结果自然远比单个医生更加准确和可靠。
◮ 随机森林的特点
样本随机:在构建每一棵树时,都不是使用全部的样本数据,而是从总样本中随机抽取一部分(有放回抽样)。
特征随机:在每个决策节点上选择“提问”的菌种时,也不是从所有菌种里挑选,而是随机抽取一部分菌种作为候选。
更妙的是,随机森林模型在完成分类任务后,还能告诉我们,为了做出准确的判断,它最依赖哪些“线索”。也就是说,它能计算出每一种细菌(即“特征”)对于区分“动物蛋白”和“植物蛋白”饮食模式的重要性得分。得分越高的细菌,就越有可能是区分两种饮食模式的“生物标志物”(Biomarker)。
通过将Meta分析的宏观整合能力与机器学习的微观挖掘能力相结合,这项研究得以在坚实的数据基础上,可以更系统地回答了文章开头提出的核心问题。
现在,让我们正式进入研究的重要发现环节。
在评估一个生态系统的健康状况时,通常会关注两个核心指标:
一是物种的丰富度与均匀度,即生态系统内物种数量多不多,分布均不均匀(对应谷禾健康报告里的多样性分值);
二是物种的组成结构,即这个生态系统主要由哪些类型的物种构成。对于肠道这个微观生态系统而言,这两个指标分别对应着α-多样性(Alpha Diversity)和β-多样性(Beta Diversity)。
1
α-多样性:肠道菌群的“贫富”指数
α-多样性用于衡量单个样本内微生物多样性的“丰富”和“均衡”程度。较高的α-多样性通常代表更健康的肠道生态,意味着功能更稳定、韧性更强,更能抵御外部干扰,如饮食变化或抗生素影响。就像物种繁多的雨林比单一农田更能抵抗病虫害。
那么,动物蛋白和植物蛋白饮食,谁更能促进肠道菌群的“共同富裕”呢?以往,一些小规模的研究在这个问题上存在争议。但这次,通过Meta分析整合了13项研究的庞大数据后,答案变得清晰起来。
◮ 摄入动物蛋白与植物蛋白α多样性的差异不大
研究结果显示,与摄入动物蛋白(如乳清、牛肉、猪肉)相比,摄入植物蛋白(如大豆、豌豆、藜麦)的受试者,其肠道菌群的α-多样性没有发生显著变化,甚至在某些指标上呈现出增加的趋势。
这一发现非常重要。它有力地反驳了一些人担心的“植物蛋白饮食可能会导致肠道菌群变得单一”的观点。恰恰相反,富含植物蛋白的饮食模式,至少在维持甚至提升肠道菌群多样性方面,表现出了与动物蛋白饮食相当甚至更优的潜力。
这背后的原因我们将在后文深入探讨,但一个关键因素在于,植物蛋白往往与一位“黄金搭档”——膳食纤维——相伴相生。这些纤维是许多有益肠道细菌的“顶级美食”,能够有力地支撑起一个物种繁盛的菌群环境。
2
β-多样性:划分肠道菌群的“派系”
如果说α-多样性关注的是菌群的“丰富程度”,那么β-多样性则衡量不同样本间微生物群落结构的差异。如果两组样本(如动物蛋白组和植物蛋白组)β-多样性差异显著,说明各自肠道菌群已经形成了截然不同的“派系”。
◮ 动物蛋白饮食下的菌群与植物蛋白的截然不同
在这项研究中,这正是最显著的发现之一。分析结果明确无误地显示,动物蛋白饮食和植物蛋白饮食塑造出了两种显著不同的肠道菌群结构。这意味着,如果你长期以动物蛋白为主食,你的肠道菌群整体面貌,将会与一个长期以植物蛋白为主食的人,有着清晰可辨的巨大差异。
为直观展示这种差异,研究者使用主坐标分析(PCoA)作图,每个点代表一个人的肠道菌群,点间距离反映差异大小。结果显示,动物蛋白组和植物蛋白组的样本在图上分成两个清晰分离的“星团”。
这一发现具有里程碑式的意义。它证实了膳食蛋白的“来源”,而不仅仅是“数量”,是决定我们肠道菌群生态类型的关键变量。你的餐盘上放的是牛排还是豆腐,正在以一种超乎想象的深度,重塑着你体内那个庞大的微生物帝国。
但问题也随之而来:这两个不同的菌群“派系”,究竟是由哪些具体的细菌成员构成的?又是哪些“关键先生”造成了如此显著的差异?这就要依靠机器学习的强大威力来找出了。
随机森林模型不仅能准确区分两种饮食模式,还给出了区分动物蛋白与植物蛋白饮食最关键的细菌“标志物”名单,这些细菌对于理解不同蛋白来源如何影响肠道菌群至关重要。
研究筛选出多种在两种饮食下丰度明显不同的“核心细菌”,其中值得关注的有:
1
动物蛋白饮食的“拥护者”
在摄入动物蛋白后,一些偏爱在富含蛋白质和脂肪环境中生长的细菌丰度显著增加。这些细菌通常被认为是“蛋白质发酵型”或“腐生菌”,它们擅长分解蛋白质和氨基酸,但其代谢产物有时可能对宿主健康构成潜在风险。
◮ Alistipes和Bilophila——动物蛋白饮食富集
Alistipes(另枝菌属)和Bilophila(嗜胆菌属):这两种菌是动物蛋白饮食下典型的富集菌。Alistipes属的某些菌株与肠道炎症和一些情绪障碍有关联。Bilophila wadsworthia则更为糟糕,它能利用动物性饮食中常见的含硫氨基酸(如牛磺酸,在红肉和海鲜中富含)产生硫化氢(H₂S)。
低浓度的硫化氢对肠道粘膜有保护作用,但过量产生则会破坏肠道屏障,诱发炎症,被认为是炎症性肠病(IBD)的一个潜在触发因素。关于这两种菌详见:
肠道重要菌属——另枝菌属(Alistipes),调节炎症情绪等的潜力菌
肠道重要菌属——嗜胆菌属 (Bilophila)喜欢脂肪、耐胆汁的促炎菌
◮ 动物蛋白饮食还能提升拟杆菌丰度
Bacteroides(拟杆菌属):这是一个庞大且复杂的菌属,主要为革兰式阴性菌,在西方化饮食(高蛋白、高脂肪)人群的肠道中通常占据主导地位。它们是多面手,既能利用碳水化合物,也能分解蛋白质。
动物蛋白饮食能够显著提升它们的丰度。虽然Bacteroides的许多成员是中性甚至有益的共生菌,但它们的过度扩张,特别是当缺乏足够膳食纤维来“喂养”它们时,可能会导致它们转向分解宿主肠道粘液中的糖蛋白,从而削弱肠道的第一道物理防线。
两种饮食结构下,主要菌属的相对丰度
2
植物蛋白饮食的“爱好者”
与动物蛋白饮食形成鲜明对比的是,植物蛋白饮食极大地促进了一批被广泛认可的“有益菌”的生长。这些细菌大多是“糖类发酵型”,它们热衷于分解植物性食物中富含的各种复杂碳水化合物(膳食纤维、抗性淀粉等),并产生对宿主健康至关重要的代谢产物。
◮ 植物蛋白饮食显著提升了双歧杆菌属丰度
Bifidobacterium(双歧杆菌属)和Lactobacillus(乳杆菌属):这两个名字对大众来说可能并不陌生,它们是市面上益生菌酸奶和补充剂中的“常客”。研究发现,植物蛋白饮食显著提升了Bifidobacterium的丰度。双歧杆菌是著名的“健康基石”,尤其在婴幼儿肠道中占据主导地位。它们能够产生短链脂肪酸(SCFAs),抑制病原菌生长,调节免疫系统,增强肠道屏障功能。
◮ 富含纤维的食物还能促进普雷沃氏菌和粪杆菌生长
Prevotella(普雷沃氏菌属)和Roseburia(罗伊氏菌属):Prevotella属偏爱在富含纤维和碳水化合物的饮食环境中生长,在非西方化、以植物为主食的人群中丰度极高。它们是高效的纤维分解者。
而Roseburia则是肠道中产生丁酸盐(一种至关重要的SCFA)的主力军之一。丁酸盐是结肠上皮细胞的首选能源,能直接为肠道细胞“加油”,同时具有强大的抗炎作用。
Faecalibacterium(粪杆菌属):特别是Faecalibacterium prausnitzii(普拉梭菌),被誉为“抗炎明星”。它的丰度降低与多种炎症性疾病(如克罗恩病)直接相关。植物性饮食,通过提供丰富的可发酵底物,为这类有益菌创造了理想的生长环境。
肠道核心菌属——普拉梭菌(Faecalibacterium Prausnitzii),预防炎症的下一代益生菌
编者小结
通过这份“红黑榜”,我们可以清晰地看到,动物蛋白和植物蛋白饮食,在肠道菌群的“物种选择”上,展现出了强烈的偏好性。动物蛋白倾向于富集那些擅长蛋白质发酵、但部分代谢产物可能存在健康隐患的细菌;而植物蛋白则大力扶持了那些以分解纤维、产生有益代谢物而闻名的“健康卫士”。
这不仅仅是细菌名单的改变,更深层次的,是整个肠道微生态系统功能的转变。
肠道菌群结构的变化最终会影响其功能层面,而我们更关注这些细菌在肠道中的具体作用。为此,研究人员采用了PICRUSt2生物信息学工具,通过已知的16S rRNA基因序列,预测菌群的功能基因谱和可能激活或抑制的代谢通路。
分析结果显示,不同饮食模式下肠道内存在截然不同的“代谢模式”。
1
动物蛋白饮食的代谢特征:BCAA代谢亢进
分析发现,动物蛋白饮食组的菌群,其与支链氨基酸(BCAAs)代谢相关的通路被显著上调。BCAAs包括亮氨酸、异亮氨酸和缬氨酸,是人体必需氨基酸,对肌肉生长至关重要。动物蛋白(特别是乳清蛋白)正是BCAAs的极佳来源。
◮ 过量代谢BCAA的细菌可能导致代谢疾病风险
然而,近年来,越来越多的证据表明,循环血液中过高水平的BCAAs及其相关代谢物,与胰岛素抵抗和2型糖尿病的风险增加密切相关。
肠道菌群在BCAA的分解代谢中扮演了重要角色。动物蛋白饮食不仅直接带来了更多的BCAA,还富集了那些能更活跃地代谢BCAA的细菌,这可能导致一系列代谢副产物的产生,这些副产物被认为可能干扰宿主的胰岛素信号通路,从而埋下代谢疾病的隐患。
此外,动物蛋白饮食还与同型半胱氨酸(homocysteine)代谢的上调有关。高水平的同型半胱氨酸是心血管疾病的一个独立风险因子。
2
植物蛋白饮食的代谢特征:SCFA生产菌更丰富
与此相反,植物蛋白饮食组的菌群,展现出了另一幅欣欣向荣的代谢图景。最核心的变化,是与短链脂肪酸(SCFAs)生物合成相关的通路被显著激活。
SCFAs,主要包括乙酸盐、丙酸盐和丁酸盐,是肠道有益菌发酵膳食纤维产生的主要“宝藏”代谢物。它们的功能极其广泛且对健康至关重要:
能源供应:丁酸盐是结肠细胞的主要能量来源,能维持肠道屏障的完整性和健康。
抗炎作用:SCFAs能够抑制炎症反应,对预防和缓解肠道及全身性炎症至关重要。
代谢调节:它们可以通过作用于肠道和远端器官(如肝脏、脂肪组织)的受体,参与调节血糖和血脂代谢,改善胰岛素敏感性。
“肠-脑轴”通讯:SCFAs还能穿过血脑屏障,参与调节食欲、情绪和认知功能。
◮ 植物蛋白饮食促进微生物发酵并产生短链脂肪酸
植物蛋白饮食能够显著促进短链脂肪酸(SCFA)的生成,其主要机制在于植物性食物富含的特定营养素。植物蛋白来源的食物,如豆类、全谷物及坚果,普遍含有高密度的可发酵膳食纤维与抗性淀粉。这些碳水化合物是肠道中特定菌群,例如双歧杆菌属(Bifidobacterium)和罗氏菌属(Roseburia),合成SCFA所偏好的主要底物。
因此,采用植物蛋白为主的饮食模式,实质上是为这些有益微生物提供了一个理想的发酵基质,从而高效促进其代谢活动与增殖。
这一功能差异很可能构成关键的、基于肠道菌群的分子机制,用以解释为何长期坚持植物性饮食与较低的心血管疾病、2 型糖尿病及部分癌症风险相关。
读到这里,一个关键问题值得我们深思:造成上述所有差异的,真的仅仅是蛋白质分子本身的来源不同吗?是牛肉里的氨基酸和豆腐里的氨基酸有什么本质区别吗?
回答并非绝对。
该项荟萃分析的深刻之处,在于其揭示了一个更为宏观的视角:人类摄入的并非孤立的营养素,而是一个完整的“食物基质”(Food Matrix)。不同蛋白质来源之所以产生差异化的生理效应,很大程度上归因于与蛋白质共同摄入的“伴随营养素”所产生的协同作用。
1
动物源性蛋白质的营养基质特征
当摄入动物性食物,如红肉或加工肉制品时,机体获取的是一个复杂的生物活性化合物组合,而非仅仅是动物蛋白。此基质通常包含以下组分:
• 高含量的饱和脂肪与胆固醇:
这类脂质会刺激肝脏合成并分泌更多的初级胆汁酸进入肠道,以辅助脂肪的乳化与吸收。然而,肠道内的特定菌群(如前文提及的嗜胆菌属 Bilophila)能够将这些初级胆汁酸代谢为次级胆汁酸,而后者具有潜在的促炎与致癌特性。
• 丰富的含硫氨基酸(蛋氨酸、半胱氨酸)与牛磺酸:
动物蛋白富含此类化合物,它们为肠道中能够产生硫化氢(H₂S)的细菌提供了充足的反应底物。高浓度的硫化氢可能对肠道细胞产生毒性作用。
• 高浓度的左旋肉碱(L-carnitine)与胆碱:
红肉是这两种物质的重要膳食来源,可被特定肠道菌群代谢为三甲胺(TMA)。TMA被吸收后在肝脏经含黄素单加氧酶氧化为氧化三甲胺(TMAO)。循环中TMAO水平升高已被证实与动脉粥样硬化和心血管疾病风险增加存在因果关系。
• 血红素铁:
尽管红肉中的血红素铁生物利用度高,但过量摄入会作为一种促氧化剂,催化活性氧(ROS)的生成,可能对肠道上皮细胞造成氧化损伤。
• 膳食纤维的缺失:
此为关键区别点。纯动物性食物基质中几乎不含膳食纤维。这使得依赖碳水化合物为主要能量来源的有益菌(即短链脂肪酸生产者)缺乏发酵底物,导致其丰度下降。这种变化为那些进行蛋白质发酵并可能产生有害代谢物的菌群创造了增殖的生态位。
2
植物蛋白的“黄金搭档”
相比之下,植物蛋白的“食物基质”则显得友好得多。当你吃下一碗扁豆、鹰嘴豆或一块豆腐时,你同时摄入的还有:
• 丰富的膳食纤维:
这是植物蛋白最大的优势。各种可溶性和不溶性纤维,是肠道有益菌的“益生元大餐”,直接支撑了SCFA的生产体系,并促进了菌群的多样性。
• 多酚类物质:
植物中含有成千上万种多酚化合物(如黄酮、花青素),它们不仅自身具有强大的抗氧化和抗炎活性,还能被肠道菌群代谢为更具生物活性的小分子物质,同时它们也能反过来调节菌群的组成,抑制有害菌,促进有益菌。
不饱和脂肪酸:
许多植物性食物(如坚果、种子)富含有益心血管健康的单不饱和和多不饱和脂肪酸。
较低的氨基酸密度和不同的氨基酸谱:
植物蛋白的消化速度通常比动物蛋白慢,这使得更多的蛋白质和氨基酸有机会到达结肠,但由于有大量纤维的存在,发酵的模式会更偏向于有益方向。
因此,这场动物蛋白与植物蛋白在肠道中的“较量”,实际上是一场整体饮食模式的较量。
通过对这项结合了Meta分析与机器学习研究的深度解读,我们得以清晰地看到,餐盘中的蛋白质来源,是调控我们肠道微生物的一个强有力杠杆。
★核心结论可以概括为:
结构重塑:膳食蛋白的来源(动物vs.植物)是塑造肠道菌群结构的关键驱动力,能形成两种显著不同的菌群“派系”。
物种筛选:动物蛋白饮食倾向于富集Alistipes、Bilophila等可能与炎症和不良代谢产物相关的细菌;而植物蛋白饮食则大力促进了Bifidobacterium、Roseburia等公认的“有益菌”的繁荣。
功能转向:在功能层面,动物蛋白饮食与支链氨基酸(BCAA)代谢等可能关联胰岛素抵抗的通路被激活相关;而植物蛋白饮食则显著上调了短链脂肪酸(SCFA)的生物合成通路,后者对维持肠道和全身健康至关重要。
这些差异不仅源于蛋白质本身,更重要的是由蛋白质来源食物的整个“食物基质”——包括伴随的脂肪类型、纤维、多酚等共同决定的。
启示和指导
上述结论并非倡导彻底弃用动物蛋白或推行严格纯素,更非妖魔化动物蛋白。科学研究的价值在于提供循证依据,帮助个体做出更明智、均衡的饮食选择。
基于本研究,可提出以下可操作的饮食策略:
1.优化蛋白质来源结构,提升植物蛋白占比:
建议在日常膳食结构中有意识地提升源自植物性食物的蛋白质比例。
优质的植物蛋白来源包括:
豆类: 如小扁豆、鹰嘴豆、黑豆等;
豆制品: 如豆腐、豆豉;
全谷物与准谷物: 如藜麦;
坚果与种子。
可采取循序渐进的策略以实现这一目标,例如,每周设定若干“低动物蛋白日”,或在烹饪时,尝试使用植物蛋白部分或全部替代原有的动物蛋白成分,以逐步调整和优化个人的饮食习惯。
2.优先选择“优质”动物蛋白:
优先选择“优质”动物蛋白:如果选择动物蛋白,可以优先考虑鱼类(富含Omega-3脂肪酸)、禽肉和鸡蛋,适度摄入红肉,并尽量减少加工肉制品(如香肠、培根)的消费。
3.采用基于“食物基质”的整体性评估方法:
在进行膳食选择时,应超越对孤立营养素的关注,建立对食物整体营养构成的认知。评估蛋白质来源时,需系统性地考量其伴随的营养成分。
例如,分析该蛋白质是与有益健康的膳食纤维和植物多酚(如豆类)共同摄入,还是与可能增加代谢负担的饱和脂肪、胆固醇及加工助剂(如加工肉制品)一同进入体内。
4.保证充足膳食纤维摄入:
无论蛋白质来源如何,每日从全谷物、蔬菜、水果、豆类获取足量纤维,是维持肠道微生态的关键。充足的膳食纤维能够为有益菌提供发酵底物,支持其稳态与代谢。
个性化将是未来的大趋势。每个人的初始肠道菌群、遗传背景和生活方式都不同,对同一种蛋白质的反应也可能存在差异。未来的营养学,或许能够根据你的个人菌群特征,为你量身定制最适合你的蛋白质摄入方案。
但在此之前,研究所揭示的宏观规律,已经为我们指明了通往更健康肠道微生态的清晰方向。下一次,当你站在超市的货架前,或在餐厅的菜单上做出选择时,请记住,你的每一次选择,不仅是在喂饱自己,更是在为你体内那个数万亿的微小盟友,投票决定它们的未来,也间接地,决定你自己的健康未来。
参考文献:
Adejumo, Samson, et al. “Dietary Protein Source Shapes Gut Microbial Structure and Predicted Function: A Meta-Analysis with Machine Learning.” bioRxiv (2025): 2025-04.
谷禾健康
儿童呼吸系统疾病,包括呼吸道感染、反复喘息和哮喘,是儿童及其以后年龄发病和死亡的重要原因。
而哮喘是其中比较典型的一种,哮喘是全球最常见的慢性疾病之一,是一种复杂的、异质性的免疫介导的紊乱集合,以气道重塑和慢性气道炎症为特征。
▸ 哮喘的危险因素
哮喘的发病机制仍不清楚,但该疾病与多种遗传、环境、感染和营养因素有关。
哮喘的许多危险因素,包括生命早期的抗菌素暴露、配方奶喂养、以及母体接触抗生素怀孕期间,集中在产前和产后早期,儿童过敏性哮喘的发生可能与微生物和免疫发育关键时期的早期肠道微生物群落有关。
动物模型提供的证据表明,生命早期肠道微生物群的组成可能会影响呼吸道免疫以及对哮喘和呼吸道感染的易感性。
在这里我们总结了婴儿(0-12 个月大)肠道微生物群组成与儿童(0-18 岁)呼吸道疾病(即呼吸道感染、喘息或哮喘)之间的关联。
谷禾健康希望通过研究数据,找到更利于儿童健康的菌群数量与种类,有助于为未来的干预研究提供信息和构建更好的健康。
本文主要内容
●生命早期的肠道微生物
●生命早期肠道微生物群对儿童呼吸道疾病的影响
●早期肠道微生物的调理方法
●青少年哮喘的预防与治疗
每个人都可以被视为一个岛屿,由各种栖息地组成,这些栖息地被微生物群落定殖,并遵循创造和塑造当地组合多样性的规则。
不同婴儿身体部位的微生物群组成
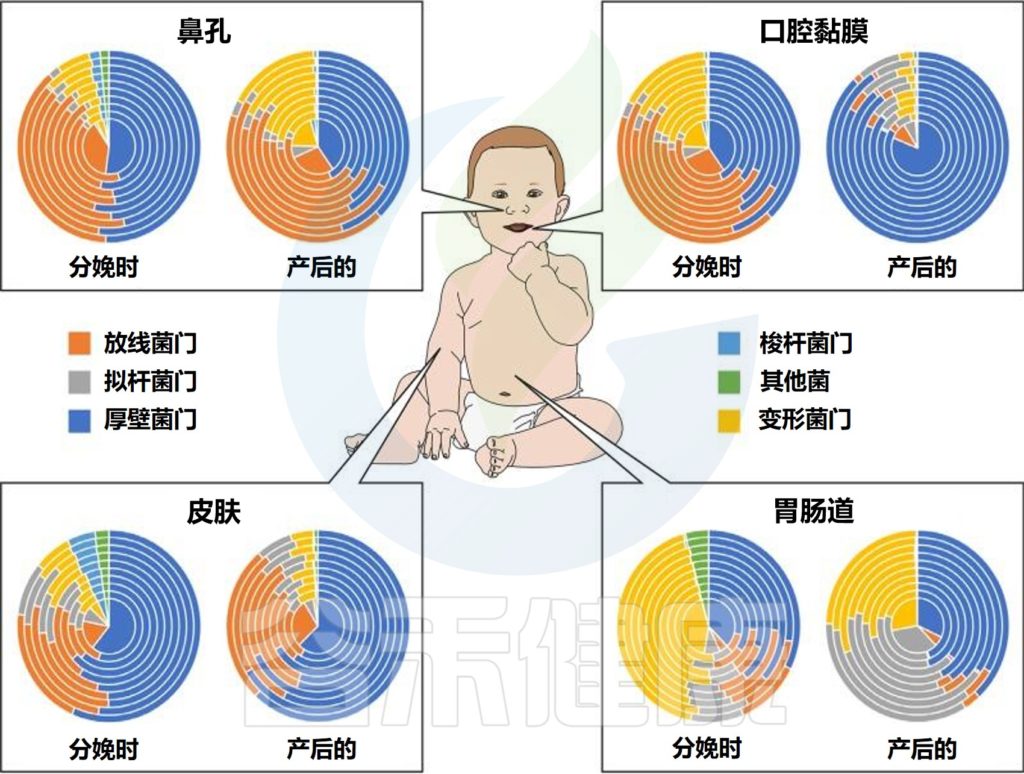
Milani C,et al.Microbiol Mol Biol Rev.2017
该图显示了婴儿微生物群组成的关键门在不同身体部位和生命早期不同阶段的相对丰度的全局概览。同心饼图示意性地表示个体间的可变性。
肠道微生物群是体内最大和最多样化的微生物群,包含数十亿细菌(主要生物)、古细菌、真核生物和病毒。
肠道菌群定植从出生时就开始了,在生命的最初几年是高度动态的,在1-3年后趋于稳定。
婴儿肠道核心微生物群
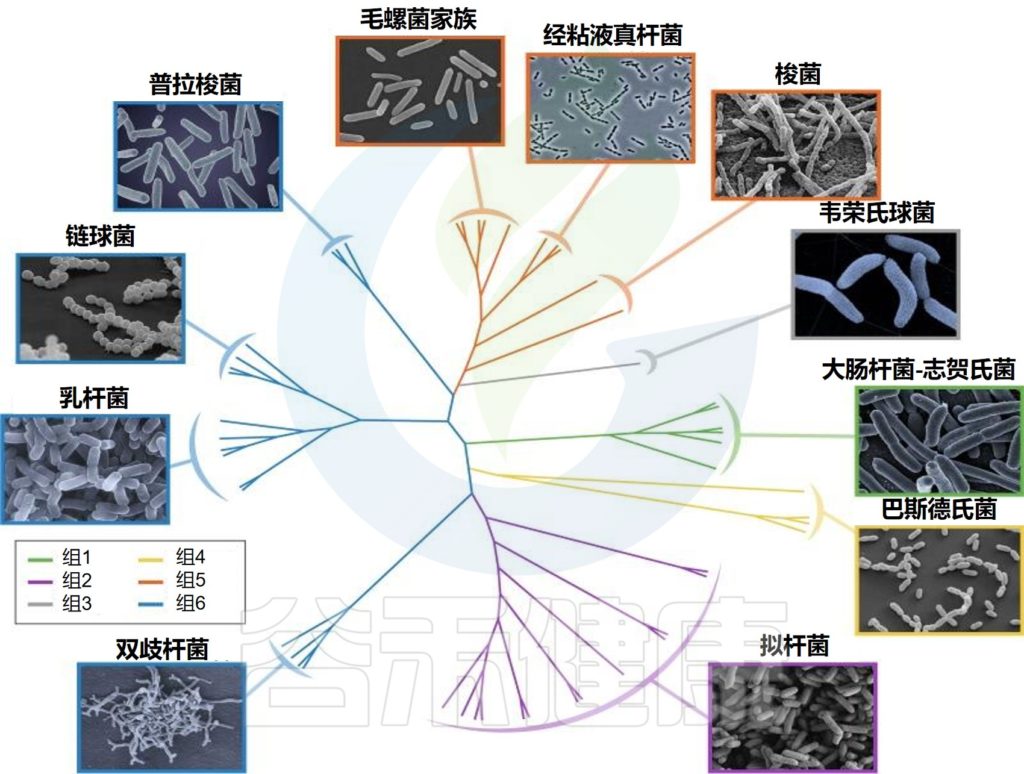
Milani C,et al.Microbiol Mol Biol Rev.2017
该图显示了涉及婴儿细菌核心微生物群的基于 16S rRNA 基因的树。树枝的颜色表示婴儿肠道微生物群的六个主要系统发育组。显示树的每个分支的关键婴儿肠道细菌分类群的电子显微镜图像。
相对于成人或年龄较大的儿童(>1岁)的肠道菌群,婴儿肠道菌群的多样性较低,菌群结构通常不稳定且高度动态。
双歧杆菌通常大量存在于婴儿,特别是母乳喂养的婴儿中,因此被认为是婴儿肠道微生物群的关键成员 。
尽管从婴儿肠道微生物群的初始组合到成人肠道微生物群的建立期间,个体水平的差异很大,但婴儿肠道微生物群可分为六种主要类型。
这种婴儿肠道微生物群的类型是根据肠道微生物群的组成和优势菌群的出现来确定的。详细地说,这些主要群体包括以下:
第1类,由肠杆菌目组成;
第2类,由拟杆菌目和疣微菌目组成;
第3 类,包括Selenomonadales以及梭菌目Pseudoflavonifractor、Subdoligranum和Desulfovibrio的成员;
第4类,包括所有巴斯德氏菌目;
第5类,包括大多数梭菌目;
第6类,包括梭状芽孢杆菌属、厌氧菌属和粪杆菌属、乳酸杆菌属和双歧杆菌属。
Bifidobacterium, Veillonella,
Streptococcus, Citrobacter,
Escherichia, Bacteroides, Clostridium
以上这些菌群在不同个体中主导婴儿肠道微生物群,它们在成人肠道微生物群中也很丰富。
▸ 梭状芽孢杆菌
梭状芽胞杆菌属的成员最近被重新分类为几个属,它们都属于梭状芽胞杆菌纲。这些物种通常存在于婴儿肠道微生物群中的微生物类群中。
▸ 拟杆菌
拟杆菌属。拟杆菌属的成员是成人肠道微生物群的主要成分,尽管它们也可能存在于婴儿肠道微生物群中,它们的存在似乎受到母乳低聚糖(HMO)的调节,其方式类似于双歧杆菌。
母乳低聚糖(HMO)——是母乳中第三丰富的固体成分(仅次于脂肪和乳糖),含量为5~15g/L,具有调节免疫,帮助大脑发育及调节肠道菌群等功能,有助于婴幼儿成长发育。
在小鼠实验中,已显示拟杆菌属的肠道定植。是宿主免疫系统识别和选择的结果,通过Toll样受体 (TLR) 和其他特定微生物-宿主相互作用。该属的成员被归类为能够代谢宿主产生的聚糖(例如HMO和粘蛋白)以及复杂的植物多糖(例如淀粉、纤维素、木聚糖和果胶)的糖破碎细菌。
●拟杆菌的作用
由于细胞外蛋白酶的作用,拟杆菌属物种通常具有蛋白水解活性。拟杆菌属成员利用的其他关键代谢功能包括胆汁酸的去结合。
在拟杆菌属中,脆弱拟杆菌被描述为可以产生多种荚膜多糖的成员,称为多糖A(PSA),是肠道菌群定植、宿主-微生物串扰或免疫调节的重要介质。
在各种拟杆菌属物种中,预计荚膜多糖会改变细胞表面的物理特性,并在宿主细菌共生中发挥关键作用。
▸ 韦荣氏球菌和链球菌
韦荣氏球菌和链球菌是婴儿肠道微生物群的一个次要成分。
这些细菌具有糖分解作用,利用其他婴儿肠道细菌(如链球菌和双歧杆菌)的碳水化合物发酵的最终产物(如乳酸)产生丙酸,形成重要的营养链。
这种短链脂肪酸被认为是肠道菌群的有益产物,因为它表现出抗炎特征,影响葡萄糖和能量稳态,增加胰岛素敏感性。
链球菌属的特定成员也构成婴儿肠道核心微生物群的一部分,并且是婴儿肠道中最早建立的细菌之一,可以在出生后的最初24小时内被识别出来。
▸ 乳酸杆菌
已知乳酸杆菌存在于婴儿肠道微生物群中,尽管它们在大肠中的数量低于上述细菌属,但在分娩后不久就存在。
乳酸杆菌的后续研究表明,与阴道分娩婴儿相比,剖腹产婴儿在生命的前6个月内的不同时间点的乳酸菌属检出率显著降低。
▸ 阿克曼氏菌
阿克曼氏菌自生命早期就存在于人类肠道中,但是水平非常低,阿克曼氏菌的存在与肠道完整性相关,已知其相对丰度和绝对数量会随着年龄的增长而迅速增加,特别是在断奶后。
小鼠实验证实了阿克曼氏菌对肠道屏障功能的影响,并证明其给药可防止饮食引起的肥胖。最近,还提供了涉及通过特定菌毛相关蛋白的TLR信号传导的机制解释。
临床、母体、喂养方式和环境因素共同塑造了生命早期的肠道微生物群。 对身体更方面都产生了一定影响。
考虑了生命早期肠道微生物群与儿童呼吸道疾病(包括呼吸道感染)之间的关联,在这里汇总了一些之前对肠道菌群与呼吸道疾病直接关联的研究。
肠道微生物的测量:通常通过收集粪便样本来测量,并且可以从多样性和丰度方面进行广泛描述。多样性描述了社区内不同分类群的数量。
探索哮喘或特应性喘息的研究
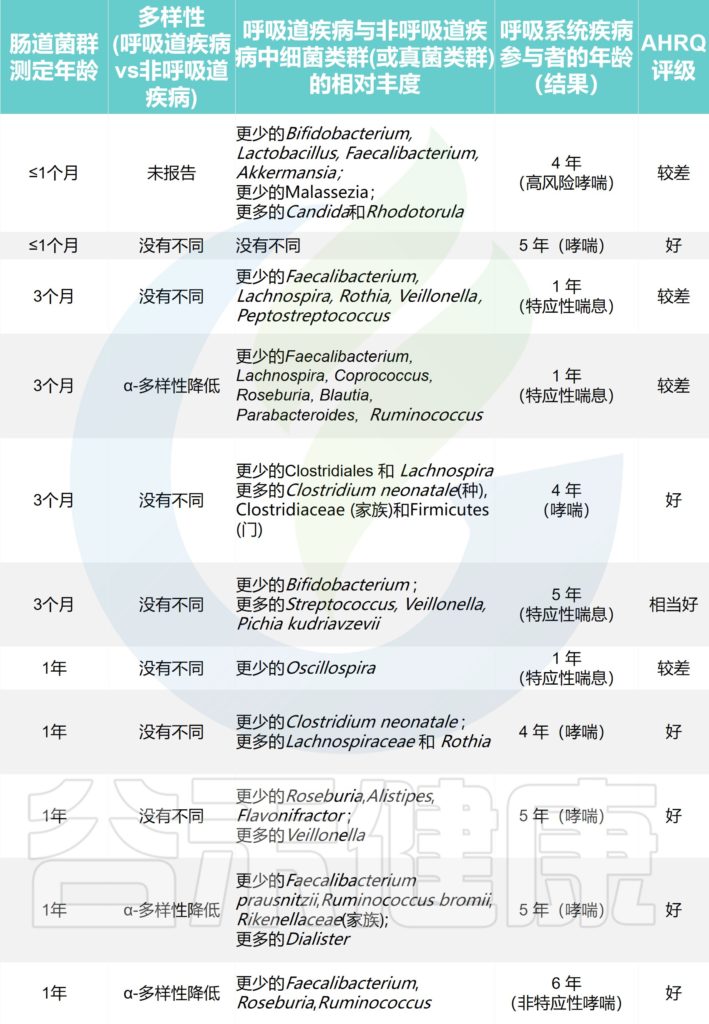
Alcazar CG,et al.Lancet Microbe.2022
探索呼吸道感染的研究

Alcazar CG,et al.Lancet Microbe.2022
喘息是与感染、过敏或后来的哮喘诊断相关的呼吸道症状。我们发现较高的α多样性与的喘息之间存在关联,这主要发生在阴道分娩的婴儿中。
α-多样性是指每个样本检测到的分类群数量,而β-多样性表示样本之间的组成差异。更细微的比较确定了不同分类水平下细菌或真菌的特定相对丰度。
大型研究(>700 名参与者)报告说,高 α 多样性对哮喘和喘息有保护作用。
探索了α-多样性与哮喘或特应性喘息之间的直接关联:与较低的α-多样性相比,生命第一年较高的肠道微生物群α-多样性与1岁时没有特应性喘息显著相关,并且在5岁和6岁时没有哮喘。
一项研究报告称,5周龄时肠道微生物群成熟度增加,肠道微生物群成熟度下降与6-11岁的哮喘高风险相关;还有两项研究报告了12个月时未成熟的肠道微生物群与5-6岁时哮喘风险增加之间存在一定关联。
基于细菌类群组成随时间的变化探索了健康参与者肠道微生物成熟度,并将该微生物群成熟度与儿童呼吸道疾病参与者的微生物群成熟度进行了比较。
总体而言,有证据表明双歧杆菌在3个月前婴儿的粪便中的相对丰度较低,与1岁时的呼吸道感染和4-5岁时的哮喘有关。
在3-12个月时的粪便样本中粪杆菌属、罗氏菌属和瘤胃球菌的丰度较低,与1-6岁时的哮喘和特应性喘息有关。
注意
然而,存在重要的研究限制,包括异质的结果定义和随访时间、残余混杂、小样本量以及异质的生物信息学和统计方法,大多数研究没有报告效果估计。
还有一些研究报告说,在1周龄时,非共生肠道细菌(如克雷伯氏菌和肠球菌)的相对丰度较高与1岁时的呼吸道感染有关; 3 个月时链球菌的相对丰度与 5 岁时的特应性喘息有关;1岁时Rothia或Dialister的高相对丰度与4-5岁时哮喘相关。但是具体作用机制目前还不清楚。
▸ 双歧杆菌增强呼吸道的免疫
双歧杆菌属(Bifidobacterium)是出生后4个月内儿童肠道中最丰富的细菌之一。并且已被证明通过体外和体内的表面相关分子和微生物群衍生代谢物调节个体的全身免疫反应。
在哮喘和呼吸道感染小鼠模型中,特定的双歧杆菌已被证明会影响呼吸道疾病的易感性。
一项研究表明,婴儿双歧杆菌的肠道定植可调节Th1和Th2反应之间的平衡,从而减少诱导小鼠模型中特应性哮喘的症状。
Th1(辅助型T淋巴细胞1)主要是增强吞噬细胞介导的抗感染免疫,特别是抗胞内病原体的感染。
Th2(辅助型T淋巴细胞2)Th2细胞的主要效应是辅助B细胞活化,其分泌的细胞因子可以促进B细胞增殖、分化和抗体的生成。
另一项研究报告称,当受到流感病毒的攻击时,与肠道丰度较低的小鼠相比,肠道丰度较高的双歧杆菌和拟杆菌的小鼠通过增强的CD8 T细胞和调节良好的巨噬细胞反应来提高流感存活率,从而防止过多的气道中性粒细胞流入。
▸ 梭状芽胞杆菌降低呼吸道炎症
Faecalibacterium、Ruminococcus、Lachnospira、Roseburia和Veillonella属于梭状芽孢杆菌类,在4-6个月大的儿童肠道中丰度较高。
已经描述了Roseburia和Faecalibacterium的潜在免疫调节机制,它们产生丁酸盐。
丁酸盐——一种在动物和体外模型中具有抗炎特性的细菌代谢物。
研究发现上呼吸道感染与婴儿粪便样本中丁酸梭菌的丰度降低有关。梭状芽胞杆菌可促进调节性T细胞产生并抑制炎症细胞因子,其中一些与人类全身感染有关。因此,丁酸梭菌对婴儿对感染的免疫反应的潜在抑制作用需要进一步研究。
另一项研究表明,给无菌小鼠接种毛螺菌属、韦荣氏菌属、粪杆菌属和罗氏菌可改善这些小鼠成年后代的气道炎症,但是这些细菌在呼吸系统疾病中的机制作用了解还是较少。
▸ 韦荣氏球菌刺激免疫分化
在我们的研究中,韦荣氏球菌,特别是小韦荣氏球菌(Veillonella parvula),与上呼吸道感染呈正相关,尤其是剖宫产婴儿。Veillonella parvula常见于口腔菌群中,它在口腔和肠道生态系统中都可以观察到。
一项针对120名荷兰婴儿的前瞻性研究发现,在1周大的婴儿中使用 16S V4 rRNA 测序发现了大量的韦荣氏菌操作分类单位,这与出生后第一年的呼吸道感染数量增加有关。
在研究中,Veillonella parvula在人体肠道中产生丙酸盐,这可能会刺激产生IL10的调节性T细胞分化;在小肠中,它会诱导产生IL-8、IL-1β、IL-10和 TNF- α37来影响呼吸道以至于全身的免疫。
▸ 棒状杆菌为呼吸道的致病菌
在剖宫产婴儿中,较高的棒状杆菌属(Corynebacterium)物种相对丰度与较高的上呼吸道感染风险相关。
棒状杆菌属物种通常被列为呼吸道中的致病菌。病例系列表明,痰中的假白喉棒状杆菌是肺部感染的驱动因素,一项来自法国的鼻咽微生物组病例对照研究发现,与健康对照组相比,病毒性呼吸道感染患者的假白喉棒状杆菌富集。
在3个月和1岁收集的粪便中,Faecalibacterium、 Roseburia和Ruminococcus相对丰度较低,与1-6岁的哮喘和特应性喘息相关。
3个月时Lachnospira的相对丰度较低,但1岁时的相对丰度增加也与1-6岁时哮喘和特应性喘息相关。
一项研究显示3个月时Veillonella的相对丰度较低与1岁时的特应性喘息相关,而两项研究报告称3个月和1岁时Veillonella的相对丰度较高与5岁时的哮喘和特应性喘息相关。
✦真菌与哮喘的关系
三项研究探索了真菌和哮喘之间的关系。
在一项研究中,在 1 个月大时测量的念珠菌和红酵母菌的相对丰度较高,而马拉色菌类群的丰度较低。
在另一项研究中,3月龄时Pichia kudriavzevii的相对丰度增加,与4-5岁时的哮喘和特应性喘息相关。
第三项研究却发现真菌成熟度与6岁儿童哮喘之间没有关联。
肠道菌群对哮喘的影响至少部分是由细菌代谢物介导的,这些代谢物可能会影响身体远端的免疫反应。
✦短链脂肪酸降低哮喘致敏性
在人类气道炎症中具有保护作用的最知名代谢物是 短链脂肪酸。1岁时粪便中含有大量丁酸盐和丙酸盐的儿童的特应性致敏性显著降低,并且在3至6岁之间不太可能患哮喘。
✦组胺和氧化脂质影响肺部炎症
与非哮喘志愿者相比,哮喘患者粪便样本中分泌组胺的细菌数量显著高于非哮喘志愿者。
此外,分泌组胺的细菌数量与疾病严重程度相关。然而,在过敏性气道炎症模型中,细菌来源的组胺降低了支气管肺泡液中的总细胞数和肺匀浆中IL-4、IL-5和IL-13的量。
相反,在蟑螂抗原小鼠气道炎症模型中,用12,13-diHOME(一种氧化脂质)对小鼠进行腹腔内治疗会减少肺部调节性T细胞的数量并增加肺部炎症。
建议
越来越多的证据表明细菌在哮喘中的作用,但需要进一步的研究来更清楚地定义所涉及的最重要的物种,并了解哮喘背景下的细菌生态失调是否是疾病的原因或影响。
有必要进行更详细的机制研究,以充分了解生命不同阶段肺和肠道微生物群组成和代谢与特定类型的哮喘炎症之间的复杂关联。
最后,未来的工作应该集中在继续详细描述在哮喘中介导细菌与宿主之间交流的细胞和分子机制。
通常用于预防或治疗不一定由特定病原体引起的感染的抗生素可以有效地消耗肠道微生物群。患有 NEC的新生儿感染肠道微生物的风险很高,抗生素通常用于预防或治疗这些感染。
NEC——新生儿坏死性小肠结肠炎(NEC)为一种获得性疾病,是多种原因引起的肠黏膜损害,使之缺血、缺氧,导致小肠、结肠发生弥漫性或局部坏死的一种疾病。
对于儿童炎症性肠病的治疗,使用单一抗生素对有并发症的患者有益,例如瘘管和脓肿,而广泛的抗生素组合可能会改善临床结果。
注意
在幼儿中使用抗生素存在很大风险。大量证据表明,抗生素会影响我们抵抗感染的能力、免疫系统的功能以及我们加工食物的能力。
肠道微生物群的破坏可能导致长期的健康后果,包括维生素产量减少、营养吸收减少以及糖尿病、哮喘、肥胖和感染风险增加。
口服益生元和益生菌是影响生命早期肠道微生物群发育的最常见方法。
益生元被定义为“选择性刺激肠道微生物群中一种或多种微生物属或物种的生长和活性,从而为宿主带来健康益处”的化合物,而益生菌被定义为“赋予宿主健康益处的活微生物”。摄入足量时会对宿主产生健康影响。
目前可用的益生元包括人乳低聚糖、菊粉、低聚果糖和低聚半乳糖;可用的益生菌包括双歧杆菌和乳酸杆菌属。
✦益生元和益生菌的作用
通过调节肠道微生物群,益生元对宿主产生健康影响。益生菌通过对粘膜和上皮的竞争性粘附、粘膜 IgA反应、抗菌物质的分泌、促炎途径的下调、抗炎细胞因子的产生和免疫系统的调节来增强肠道上皮屏障。
最近的研究表明,益生菌可以预防儿科疾病和障碍的进展,包括过敏、胃肠道感染、肥胖,甚至上呼吸道感染。
干预研究进一步表明,益生菌可以减轻某些疾病的严重程度,但对每种疾病的最佳干预仍然知之甚少。
虽然益生菌在某些情况下可以缓解过敏症状,但它们通常不能有效调节肠道微生物的组成。有证据表明,嗜热链球菌和双歧杆菌的组合可有效预防儿童抗生素相关性腹泻。
此外,合成生物学使益生菌和共生微生物的工程具有新的治疗功能。例如,融合蛋白HSP65-6P277的表达减少了非肥胖糖尿病小鼠中1型糖尿病的发病,而在非肥胖糖尿病鼠中口服重组乳酸乳球菌可改善糖耐量并显著减少胰岛素炎。
建议
尽管益生元和益生菌的应用前景不错,但在未来的研究中,应讨论益生菌和益生素的施用时间、不同菌株和菌株组合的效果、工程、安全性以及这些益生菌与益生素组合是否更有效的确定。
营养素可以通过塑造微生物菌群的组成,对婴儿肠道微生物的定植模式产生短期和长期影响。
越来越多的证据表明,摄入的膳食成分与炎症性肠病、2型糖尿病和动脉粥样硬化的发展有关。肠道菌群的最大变化发生在固体食物的引入,这表明饮食应被视为肠道菌群的核心决定因素。
✦饮食疗法
有趣的是,在中医中,早就有“药食同源”的概念。该概念的一个方面是食物是一种药物,适当的饮食或某些食物可以维持身体的平衡和健康,并预防或减轻某些疾病的发展。
在现代医学中,饮食改变越来越被认为是一种通过改变肠道微生物群来改变全身炎症的相对简单的方法。
早期肠内营养 (EEN) 是一种饮食疗法,已被用作儿童克罗恩病的一线疗法,通过用仅由液体营养素组成的配方代替正常饮食成分,旨在使炎症标志物正常化并诱导临床缓解。
✦高膳食纤维可以减轻呼吸道疾病
此外,另一项研究报告称,喂食高纤维饮食的小鼠可以产生独特的肠道微生物群,从而导致短链脂肪酸乙酸酯水平升高。
高纤维或醋酸盐喂养通过减少与人类哮喘和小鼠哮喘模型相关的某些基因在小鼠胎肺中的表达,显著抑制过敏性气道疾病。
此外,发现肠道微生物不仅与哮喘有关,而且还降低了与哮喘严重程度和炎症表型相关的气道微生物群的多样性和群落组成。
抗炎特性
此外,最近的一些研究证明,膳食纤维具有抗炎特性,这可以部分解释纤维对肠道微生物菌群的影响。 使用临床前模型的几项研究表明,可发酵纤维补充剂通过微生物群诱导的特定抗炎代谢物产生的变化来改变疾病结果。
然而,还需要更多的研究来增加我们对不同饮食如何塑造微生物群和改变健康结果的理解。
粪便微生物群移植(FMT)被定义为将健康供体的粪便悬浮液输注到受体患者的胃肠道中,以恢复肠道微生物群的正常多样性和功能。
粪菌移植的方式:粪便微生物群可通过结肠镜检查、鼻胃管或鼻十二指肠管、灌肠剂或口服胶囊置于患者体内。
由于肠道微生物群宏基因组测序的技术进步以及对其组成和功能的日益了解,粪菌移植近年来引起了越来越多的兴趣和关注。尽管粪菌移植仍然知之甚少,但它不再被认为是一种“替代”和最后的医疗实践,现在作为一种具有生物学合理性的有价值的疗法正在获得主流接受。
此外,这种疗法已被证明能够重建正常运作的微生物群落。通过为患者提供来自合适供体的平衡微生物群,纠正了在艰难梭菌感染 (CDI)发病机制中起重要作用的不平衡肠道微生物菌群。
在一系列关于复发性CDI的研究中,85%接受粪菌移植的患者出现症状缓解。此外,考虑到肠脑轴和肠道微生物之间的相互作用,粪菌移植被认为是治疗某些精神疾病的可能方法,例如自闭症谱系障碍。
建议
然而,粪菌移植的微生物组成尚未完全确定。因此,需要澄清与改善临床结果相关的微生物结构或功能特征,以确定优选的组合。
未来的研究应侧重于确定“健康”微生物菌群的范围以及制定评估最佳组成的标准。
5岁至14岁的哮喘患病率约为10%,使其成为全球儿童时期最普遍的慢性病。尽管下呼吸道感染带来了巨大的健康负担,但目前还没有专门针对它们或儿童哮喘被广泛许可的预防策略,所以暂时只能用一般呼吸道疾病的方法来预防。
✦注意空气卫生
注意室内的清洁和空气流通,因为空气中的灰尘和细菌是哮喘病发的主要致敏原,所以应该勤加打扫,减少空气中的尘埃。尽量减少暴露于空气污染的室内和室外。
✦良好饮食习惯
坚持每天喝水,喝水是排出身体毒素的最佳的方法。在日常生活中注意饮食习惯,一日三餐要按时就餐,少吃油腻。
✦良好生活,避免螨虫
哮喘病人要在日常生活中每天要保持良好的生活态度,放松心情。不要在家里养猫、狗、花、鸟等。经常晾晒被褥、换洗床单,避免螨虫孳生。
✦加强自我管理
对于5岁及以上被诊断为哮喘的儿童或青少年,提供哮喘自我管理计划,包括书面的个性化行动计划和教育。
说明污染会引发或加剧哮喘,并在个性化的行动计划中包含尽量减少暴露于室内和室外空气污染的方法。
注:哮喘好发于青少年和儿童,一旦患病,如防治不当,很容易反复发病,随着发病频度的增加,病情会逐渐加重,必将严重影响生活质量和学习工作能力,给个人家庭和社会造成沉重负担。
我们结合当前的研究与认知,提出了一些适用于儿童和青少年新诊断哮喘或当前治疗无法控制哮喘的治疗建议。
✦药物治疗
•SABA
β2受体激动剂(SABA)是一类能够分布在气道平滑肌上的β2受体产生支气管扩张作用的哮喘治疗药物。这类药物属于支气管扩张药,是哮喘急性发作(气道痉挛)的首选药物,能够迅速改善哮喘急性发作时的呼吸困难、咳嗽等的症状。
对新诊断为哮喘的儿童和年轻人(5至16岁),可以提供SABA作为缓解疗法。
对于患有哮喘的儿童和青少年(5至16岁),他们很少出现短暂的喘息和正常的肺功能,也可以考虑单独使用SABA缓解疗法进行治疗。
•ICS
吸入性糖皮质激素(ICS)是目前控制哮喘病的气道炎症最有效的药物,以定量气雾剂、干粉剂或溶液吸入。
在哮喘炎症表型中,通常接受高剂量吸入性皮质类固醇 (ICS) 的中性粒细胞性哮喘患者表现出较少的细菌负荷,其中嗜血杆菌和莫拉菌属、变形杆菌门的成员相对富集,而链球菌的相对丰度降低。
ICS已经成为目前哮喘治疗的第一线治疗,对病人是最为重要的治疗,任何哮喘患者,只要诊断正确,都应该接受ICS的治疗,这是一个长期维持治疗,可以起到气管局部抗炎的效果,改善病情,预防哮喘急性发作。
为儿童和青少年(5至16岁)提供儿科低剂量ICS作为一线维持治疗。
就诊时出现明显表明需要维持治疗的症状(如导致夜间醒来)或单独使用SABA无法控制的哮喘也应使用ICS治疗。
✦风险分层
同时使用风险分层来识别预后不良风险增加的哮喘患者,并使用此信息优化他们的护理。
根据诸如不依从哮喘药物、心理社会问题和反复发作的哮喘计划外护理等因素进行风险分层。
总体而言,有观察证据表明,在生命的第一年,肠道共生细菌属的低α多样性和相对丰度与随后的呼吸系统疾病,尤其是哮喘有关。因此在婴儿早期关注和了解肠道菌群状况对于后面哮喘发生和预防非常重要,进一步研究哮喘患者的肠道和下呼吸道微生物群可能有助于开发更有效的方法来预防和治疗哮喘。
主要参考文献
Alcazar CG, Paes VM, Shao Y, Oesser C, Miltz A, Lawley TD, Brocklehurst P, Rodger A, Field N. The association between early-life gut microbiota and childhood respiratory diseases: a systematic review. Lancet Microbe. 2022 Aug 18:S2666-5247(22)00184-7. doi: 10.1016/S2666-5247(22)00184-7. Epub ahead of print. PMID: 35988549.
Milani C, Duranti S, Bottacini F, Casey E, Turroni F, Mahony J, Belzer C, Delgado Palacio S, Arboleya Montes S, Mancabelli L, Lugli GA, Rodriguez JM, Bode L, de Vos W, Gueimonde M, Margolles A, van Sinderen D, Ventura M. The First Microbial Colonizers of the Human Gut: Composition, Activities, and Health Implications of the Infant Gut Microbiota. Microbiol Mol Biol Rev. 2017 Nov 8;81(4):e00036-17. doi: 10.1128/MMBR.00036-17. PMID: 29118049; PMCID: PMC5706746.
Zhuang L, Chen H, Zhang S, Zhuang J, Li Q, Feng Z. Intestinal Microbiota in Early Life and Its Implications on Childhood Health. Genomics Proteomics Bioinformatics. 2019 Feb;17(1):13-25. doi: 10.1016/j.gpb.2018.10.002. Epub 2019 Apr 12. PMID: 30986482; PMCID: PMC6522475.
Barcik W, Boutin RCT, Sokolowska M, Finlay BB. The Role of Lung and Gut Microbiota in the Pathology of Asthma. Immunity. 2020 Feb 18;52(2):241-255. doi: 10.1016/j.immuni.2020.01.007. PMID: 32075727; PMCID: PMC7128389.
Ver Heul A, Planer J, Kau AL. The Human Microbiota and Asthma. Clin Rev Allergy Immunol. 2019 Dec;57(3):350-363. doi: 10.1007/s12016-018-8719-7. PMID: 30426401; PMCID: PMC7449604.
Moroishi Y, Gui J, Hoen AG, Morrison HG, Baker ER, Nadeau KC, Li H, Li Z, Madan JC, Karagas MR. The relationship between the gut microbiome and the risk of respiratory infections among newborns. Commun Med (Lond). 2022 Jul 14;2:87. doi: 10.1038/s43856-022-00152-1. PMID: 35847562; PMCID: PMC9283516.

谷禾健康

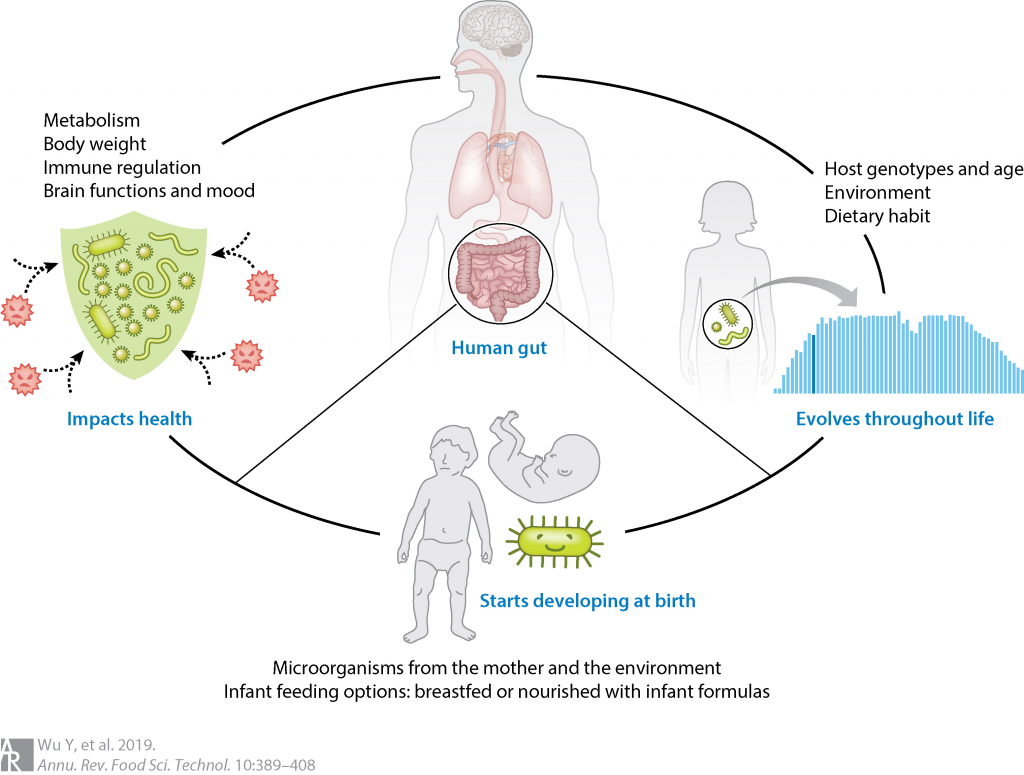
每个人的一生都会经历很多,从出生到长大,健康到衰老疾病。你的出生、遗传、家庭环境、很大程度上决定的人生起点,日常的饮食、行为习惯决定你的身体成长,一些不同的选择或意外的事件又会让人生有很多起伏和不同。
每个人的菌群和我们的人生一样也是独一无二的,我们菌群的特点反映着不同人各自生活的烙印。从母亲的腹中开始影响和决定了我们最初的菌群,出生方式、喂养的食物、用药等都决定了我们的菌群基数。当我们开始从喝奶到开始摄入辅食,我们的菌群也同样迎来巨大的演变。当我们生病、感染、运动、饮食、社交、虚弱、衰老这些同样反映在我们菌群的变化和演替上。
相对的,当我们更多的了解我们的菌群,善待和改善它们,同样的变化也会出现在我们的身体和生活中。
越来越多的证据表明,年龄与人类微生物群之间的关联很大,肠道微生物群是许多年龄相关变化的核心,包括免疫系统失调和疾病易感性。几个身体部位的微生物组成可以相对准确地预测人类的年龄。
谷禾健康肠道菌群检测数据库中,也有关于肠道年龄预测:
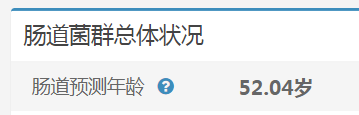
谷禾健康-肠道年龄预测模型图
<来源:谷禾健康肠道菌群数据库>
可以看到,肠道年龄和生理年龄基本是符合的。健康人的肠道菌群年龄恰恰是最符合真实年龄的,与真实年龄差异大意味着肠道菌群出现偏离。
健康的人存在更多样化且平衡的肠道菌群。微生物群中与年龄相关的变化归因于生理,生活方式和健康状况。这些因素中的每一个都与某些菌群的相对丰度变化有关。
例如,饮食、卫生、兄弟姐妹、宠物、过敏、儿童疾病和抗生素是影响儿童微生物组的一些突出因素。到了成年期微生物群相对稳定,而到了老年期,一些有益菌开始逐渐下降,菌群又向另一个阶段过渡。
在从出生到死亡和分解的每个生命阶段,微生物群落都是身体的动态组成部分。研究微生物群的自然和诱导变化有可能彻底改变我们对人类生物学的理解。
本文介绍了健康人的微生物群在一生中的变化,讨论了从出生时菌群构成,到疾病或抗生素使用时的变化,再到死亡时的微生物扩展的各个阶段,以及这些阶段在身体部位和组成(细菌、真菌或病毒)上的差异。了解微生物群与年龄关系的未来研究方向,以此对人体微生物群及基于此的干预有更好的了解。
微生物群落存在于人体的每个粘膜表面,人的每个身体部位都有一个独特的生态学。每个人的微生物群像指纹一样,都是独特的。
在个体内,特定的身体部位、地理位置和个体的年龄与健康微生物群具有极强的关系。年龄驱动人类微生物群的α多样性和β多样性。
在了解各个阶段的微生物群变化之前,我们先了解一个概念:微生物演替。
微生物演替是指微生物群落中一种或多种生物的存在、相对丰度或绝对丰度的变化。
在正常或健康衰老期间,微生物演替的三个主要阶段自然发生在人类生活中。
✦初级演替(出生时先锋菌群定植,快速变化直到童年晚期)
第一阶段,初级演替,从先锋物种首次建立群落时开始,随后微生物群落发生快速变化。从出生到童年,变化率降低,许多中间物种存在于出生到童年晚期之间。
初级演替结束于顶级群落的形成,在青春期实现,并在很大程度上持续到成年;该群落的特征是其相对稳定。
虽然成年期的微生物群比儿童期更稳定,但仍然存在变异,这引发了关于人类微生物群中是否存在顶级群落的争论。成年微生物群的自然变异存在于小时(昼夜节律)到年(老化)的时间尺度上,但微生物群相对稳定,除非存在干扰,如饮食或药物的改变。
✦次生演替(菌群的改变,重建)
下一个阶段,即次生演替,发生在一个先前存在的稳定群落一部分被改变或移除之后,然后群落再生到相同的状态或不同的状态。这可以通过抗生素等医疗手段人为实现,也可以通过霍乱弧菌感染等疾病自发实现。
人类的次生演替的特征是至少有一段时间的随机过程占主导地位。在诱导条件下,如单疗程抗生素,群落遵循类似于初级演替的过程,其中现有微生物群落的一部分充当“微生物记忆”,帮助重建一个类似于以前存在的群落。
这一过程被认为是由核心微生物群驱动的,而不是驱动初级演替的先锋微生物。
✦末期演替(自然衰老和死亡阶段)
最终的末期演替是宿主自然衰老和死亡的一部分。在老年期间,微生物群落再次以更高的变化率,成功产生了一个由更少成员组成的群落,通常变形菌门(也称为假单胞菌)的相对丰度增加,有时占总优势。
研究演替的每个阶段使研究人员能够解决与人类相关的微生物群落是如何形成和维持的。通过了解这些过程,我们可以更好地了解微生物群随着年龄的增长的变化及其与人类健康的关系,了解如何管理微生物群。
人类相关微生物群从受孕到死亡的变化
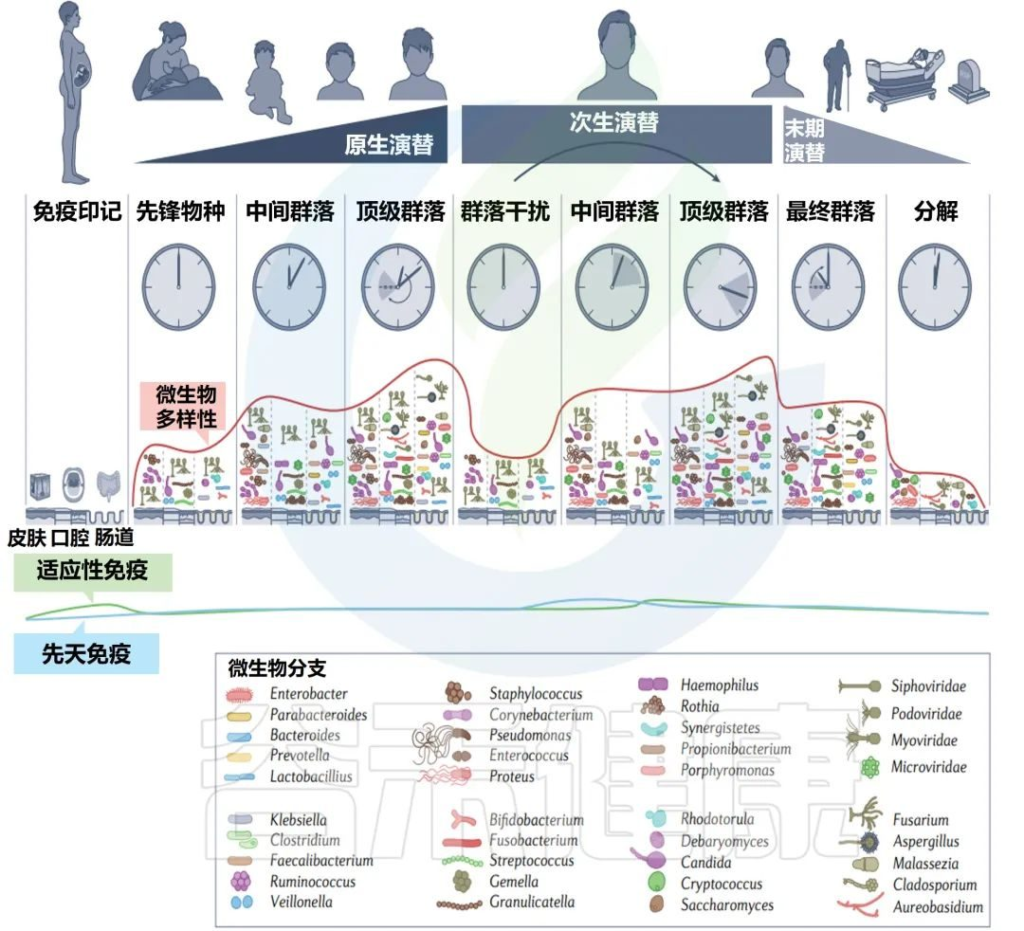
Martino C,et al.Nat Rev Microbiol.2022
常驻细菌、真菌和病毒的多样性在人类生命的各个阶段都会发生变化。模拟时钟代表每个微生物群落阶段发育的宿主年龄的相对时间。
免疫印记在出生前通过母亲的微生物群及其代谢物开始(第一栏)。先锋物种的初始定殖始于出生,身体部位特定的微生物群落出现(第二栏)。这些群落的复杂性不断增加,直到它们达到相对稳定的群落结构(第三列和第四列)。
这些微生物群落的次生演替可能来自内部和外部扰动(第五栏)。中间微生物重新建立初始群落,并再次达到稳定状态(第六列和第七列)。
在晚年,随着寄主接近自然死亡,群落经历了最后的演替和变化(第八栏)。微生物演替的最后阶段发生在腐败和分解阶段。在此阶段,多样性进一步下降,在最初的24-48小时内,许多人类微生物群结构保持不变,但随后很快开始侵蚀分解(第九栏)。
绿线和蓝线分别显示了微生物演替不同阶段的适应性免疫和先天免疫的相对强度。
不同年龄段的细菌多样性测量
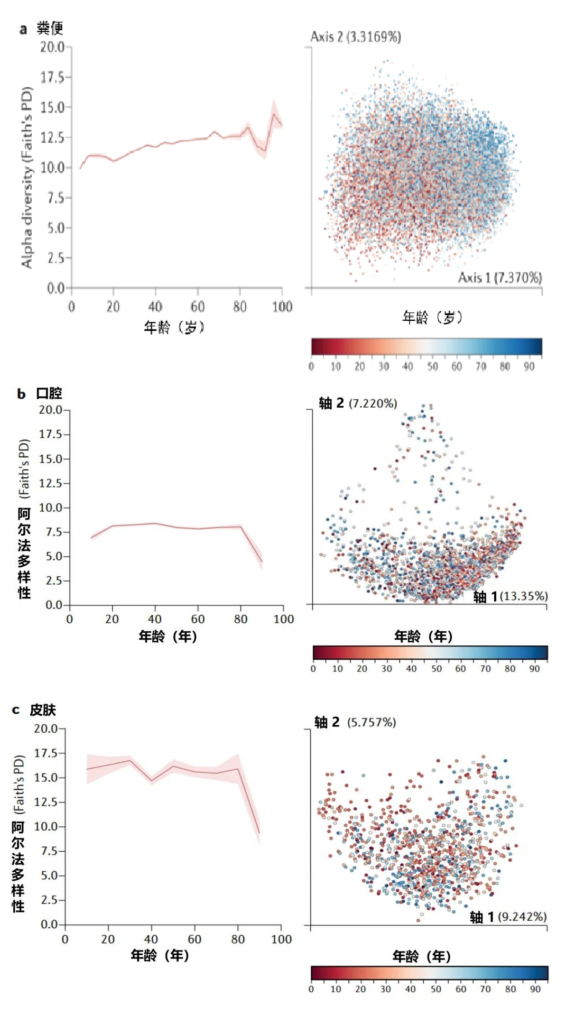
Martino C,et al.Nat Rev Microbiol.2022
美国一个肠道项目集中测量了从儿童到老年的人类粪便(a部分)、口腔(b部分)和皮肤(c部分)微生物群的细菌多样性和系统发育史,该项目包含21919个粪便、1920个口腔和998个皮肤微生物群样本,带有16S核糖体RNA基因扩增子序列。
α多样性,一种对样本中不同类型微生物数量的定量测量,通过Faith的系统发育多样性(PD)α多样性度量跨年龄测量。
UniFrac β多样性主坐标分析,一种用于比较微生物群落相似性的方法,其中空间上接近的点表示相似的样本,空间上远离的点表示不同的样本,按年龄着色。
✦胎儿时期——菌群及代谢物影响免疫发育
塑造人类微生物群的第一个因素来自胎儿发育过程中的母亲。
胎儿通过胎盘接触到母亲微生物群落产生的代谢物,这些代谢物会影响其免疫系统,并会影响正常微生物群和后期病理学的各个方面。代谢物,如短链脂肪酸(乙酸盐)和其他微生物化合物,可以通过胎盘转移到胎儿体内,并影响免疫发育。母亲的饮食和健康也会影响这些代谢物。
胎儿组织中的乙酸盐影响与成人调节性T细胞生成相关的表观遗传印记,其与防止生命后期哮喘的发展相关。
✦出生后——菌群受出生模式,饮食,环境等影响
出生后,微生物群落根据身体部位迅速分化。
在最初的时候,先锋物种和未来4年的群落发展可能会受到出生模式和妊娠时间的影响。中间群落由饮食影响,如母乳或配方奶粉的消费,以及环境。
最后,饮食和环境再次塑造了稳定的顶级群落。主要由真菌、细菌和病毒组成。
子宫内和生命早期的主要演替
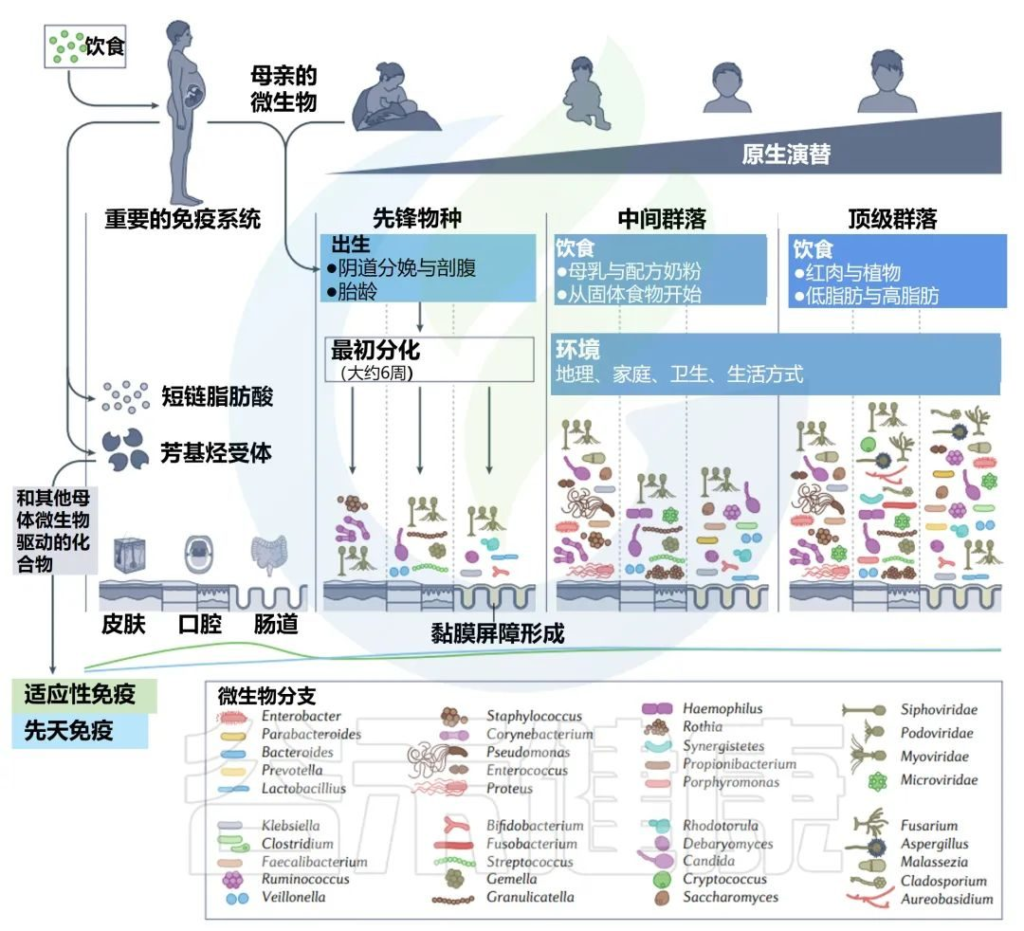
微生物代谢物和配体调节宿主芳基烃受体,这有助于塑造新生儿微生物和免疫发育。母亲使用抗生素和胃肠道相关疾病,如炎症性肠病,也被认为会通过胎儿免疫系统的印记增加后代的病理风险。
然而,这些联系仅在非人类实验中研究过。在一个案例中,由患有炎症性肠病的孕妇或其新生儿的微生物群所定殖的无菌小鼠继续发展出异常微生物群和指示炎症性肠病的免疫发育。
✦怀孕期间母体的微生物群与免疫系统的变化
在怀孕期间,母亲的微生物群和免疫系统也发生了改变。母亲的阴道微生物群变得更加多样化,通常由在其他身体部位发现的许多微生物群组成。
孕期母体免疫系统与胎儿形成协同作用,包括通过胎盘转移IgG抗体。
关于出生时获得的微生物群是否通过混合来源于阴道和粪便,或者阴道微生物群本身在出生时是否具有多能性,是否是微生物先驱的主要来源,存在一些争议。
无论确切的母体来源如何,这一阶段的特征是先锋细菌种类。包括下列菌群:
然后这些细菌定居在常规身体部位:肠道、口腔和皮肤。
许多先锋细菌是兼性厌氧菌,它们会消耗氧气,从而使专性厌氧菌能够在以后的每个环境中定居。起初,新生儿的每个身体部位都相对未分化,但先锋微生物很快开始启动身体部位依赖性微生物多样性的级联,至少在生命的第4到第6周,每个部位的细菌都可以很容易地区分。
先锋细菌进驻后,生命早期的微生物群逐渐开始形成。接下来的章节我们来了解生命早期的肠道,口腔,皮肤等各部位的微生物群(包括细菌、真菌、病毒等)。
✦肠道细菌群——双歧杆菌主导
人类肠道细菌群落的发展已经得到了很好的研究。
双歧杆菌属一直占主导地位,直到在生命的第一年结束时,它们被双歧杆菌、梭状芽孢杆菌和拟杆菌属的组合所取代。拟杆菌属的丰度增加,而双歧杆菌属等物种的丰度相对减少。
双歧杆菌分解母乳低聚糖,开始终生影响免疫系统
最近,一项研究发现,双歧杆菌等细菌含有母乳低聚糖分解代谢所需的基因,与婴儿免疫发育之间存在功能联系。特别是,接受Bifidobacterium infantis EVC001极化初始T细胞的婴儿的粪便水与来自对照组的粪便水平不同,其方式与减少肠道炎症有关。
其他菌属也可降解母乳低聚糖(如拟杆菌、阿克曼菌)
到3-6岁时,肠道细菌群落汇聚到整个成年期持续的顶级群落。这一微生物群是已知的密度最大、多样性最强的生态群落之一。通常,在这段时间内,普通健康人中只有两个细菌门占优势:厚壁菌门和拟杆菌门。
✦肠道其他微生物群——真菌、古细菌、病毒
在人类肠道发育过程中,对病毒组、真菌组和古菌组的研究远远少于细菌组。在整个生命周期中,真菌群落所占的总数远远少于细菌组或病毒组。
//真菌群落
真菌群落在生命的最初几天含有大量的Rhodotorula和Debaryomyces,接下来的一个月则是CandidaCryptococcus和Saccharomyces spp.。
到成年时,主要的真菌属是Aspergillus, Candida和Saccharomyces。
//古细菌群落
发育期间肠道的古细菌群落尚不清楚,但古细菌是一些最早的移生菌落,但丰度较低。
早期定植的古细菌包括Methanosphaera和Methanobrevibacter。
// 病毒群落:噬菌体家族在出生后就开始流行
主要由噬菌体组成的病毒群落在出生后的第一周数量众多。噬菌体家族Siphoviridae、Podoviridae和Myoviridae在出生后立即流行,主要以溶原形式整合到细菌基因组中。
到生命的第四个月,有尾噬菌体目大量生长,成员更常为裂解型(传染性噬菌体颗粒或主动复制的噬菌体)。
在成人中,Caudovirales和Microviridae在肠道噬菌体群落中占主导地位,但噬菌体肠道病毒组对个体具有高度特异性,其演替仍有许多未知之处。
与噬菌体不同,感染真核病毒的肠道病毒组主要与儿童和成人的病理相关。最近,在健康儿童和健康成人中也观察到一些感染真核细胞的病毒丰度较低,但其发生时间和流行率尚不清楚。
✦口腔细菌群:出生后几个月逐渐趋于稳定,牙齿形成后再次转变
在出生时,口腔细菌群在以下菌属中的流行率很高:
在接下来的几个月里,Lactobacillus和Fusobacterium也开始流行。Staphylococcus的丰度在出生后3个月左右达到峰值,然后稳步下降,让位与更高丰度的GemellaGranulicatella, Haemophilus和Rothia spp.
牙齿形成后,口腔微生物群再次转变,在成年期具有更高丰度的梭杆菌门, Synergistetes, Tenericutes, Saccharibacteria (TM7), SR1 。
✦口腔其他微生物:成年口腔含产甲烷菌,最常见的噬菌体群是尾状病毒
口腔真菌群落被认为比皮肤和内脏的真菌多样性少。Candida spp.是口腔的第一批真菌定植菌。对中级口腔真菌群落知之甚少,但成年人CandidaCladosporiumAureobasidium
AspergillusFusarium和Cryptococcus spp.的丰度较高。
发育过程中的口腔古菌体尚不清楚,但成年口腔中含有许多古菌产甲烷菌,包括甲烷杆菌属。
目前对人类婴儿口腔中病毒的知之甚少。在成年人中,与肠道类似,最常见的噬菌体群是尾状病毒。
口腔病毒群在本质上通常被视为病理性的(例如柯萨奇A病毒、麻疹病毒、红疹病毒和人乳头瘤病毒),并且没有对病毒群落组成进行纵向研究。然而,在无症状和健康成人中也观察到许多真核病毒分类群。
✦皮肤细菌群落:出生时母亲阴道乳杆菌属占据较多,4-5周与成人相似
皮肤细菌群落在出生时含有大量的母亲阴道乳杆菌属。到第4-5周,婴儿皮肤微生物群与成人皮肤微生物群相似,但在青春期继续变得更具位点特异性。
Staphylococcus和Corynebacterium在不同位点PseudomonasEnterobacterEnterococcus,
Proteus和Klebsiella在特定位点(如腋窝与前臂)。
✦皮肤其他微生物:马拉色菌占比较高,古细菌占4%左右
在皮肤真菌群落中,MalasseziaCandida和 Saccharomyces在生命的前30天最为普遍。对于中间群落的确切组成知之甚少,但成年真菌群落中Malassezia的丰度通常很高,估计约占真菌群落总组成的75%至90%。
关于皮肤古细菌群落的发育情况了解较少,但古细菌约占成年人菌群的4%。大体上,成年人皮肤古细菌群由Thaumarchaeota门和Euryarchaeota门代表。在成人皮肤上也发现了Halobacteriaceae和 Methanobrevibacter。
与肠道和口腔不同,健康的皮肤微生物群拥有相对较少的已知病毒多样性,很少有对其进行研究,可能是由于与低生物量样本相关的技术限制。不过,皮肤上有一些自然存在的病毒群。
以上了解关于生命早期肠道、口腔、皮肤的微生物群,那么哪些因素会给生命早期的微生物群发展带来影响?
在生命的最初几年中,有几个因素塑造并区分了微生物群落的发展。
✦出生方式和母体抗生素的使用
出生方式和母体抗生素的使用是影响人类微生物群落的研究最好、最清楚的因素之一。然而,微生物的发育可能会导致独特的结果,即使是在同居的同卵双胞胎中,这可能是由于许多未知或随机的过程。
通过剖腹产和围产期和新生儿抗生素暴露,自然微生物群落的建立过程可能会在所有身体部位受到干扰。这一发现突出了阴道微生物群落的重要性,阴道微生物群落自然含有大量Lactobacillus spp.,但在青春期发生改变,对女性健康至关重要。
一些最佳样本的婴儿发育研究,通常缩写为DIABIMMUNE ECAM和TEDDY,在婴儿出生后的前2年和3年进行了随访,重点关注抗生素使用或出生方式的影响。
在上述所有研究中,阴道分娩的婴儿的拟杆菌属相对丰度高于剖腹产婴儿。
由于缺乏建立微生物群落的天然先锋微生物群,导致可变的群落组成被认为是由随机过程而不是确定性过程驱动的,出生模式对微生物群落组成的影响直到生命的第四年仍然可见。
出生模式影响的一个例外是早产,可能是由于在出生后的头几天大量使用抗生素,其特点是无论出生模式如何,微生物发育都不稳定。婴儿微生物群自然发育的这种改变与感染、免疫疾病、肥胖和神经内分泌异常的风险增加相关。
✦母乳喂养:母乳低聚糖给菌群带来稳定性
其次,与其他因素相比,母乳喂养对微生物群的发育有很大影响。与母乳喂养相比,配方奶粉的使用导致了更高的多样性和更不确定的微生物群落。
例如,考虑到出生时肠道中双歧杆菌科的自然优势,缺乏某些母乳低聚糖作为主要营养源可能会导致初始定植的不稳定性。然而,微生物群、牛奶代谢组和免疫系统发育的多组学整合是一个活跃且快速发展的研究领域。
除了母乳低聚糖,母乳还含有其他免疫调节化合物,例如革兰氏阴性细菌的脂多糖、分泌性IgA、先天免疫因子、抗菌肽和益生元因子。
最后,所有这些因素都会影响人类免疫发育。微生物相关分子模式识别受体与微生物群衍生分子相互作用,代谢物如短链脂肪酸(与GPR43、GPR41和GPR109相互作用)和次级胆汁酸(与FXR相互作用)直接影响免疫发育。
//
这些因素加在一起,有助于形成一个独特的、相对稳定的细菌、真菌和病毒微生物群落,这种微生物群落在人类生命的大部分时间都持续存在。
前面章节了解了婴儿期初级演替期间发生的巨大变化,与之相比,成年期微生物群基本上是稳定的(15-65岁),但该群落可能会受到干扰,因此本章节从以下三方面展开讨论:
健康成年人中某些细菌的基因组随着时间的推移而进化,表明在次生演替中,功能和组成进化以稳定状态发生。
• 昼夜节律影响菌群变化
成人微生物群也会发生自然的短期变化,时间尺度为一天到数月或数年。
短期变化的一个典型例子是微生物群落组成的昼夜节律。与昼夜节律相关的人类基因表达和免疫激活,以及肠道微生物群中细菌的丰度和组成也遵循这种模式。
在小鼠中表现出昼夜循环的细菌家族包括瘤胃球菌科、毛螺菌科、Muribaculaceae和疣微菌科,但对人体的等效周期知之甚少。
青春期和成年生活中的二次演替
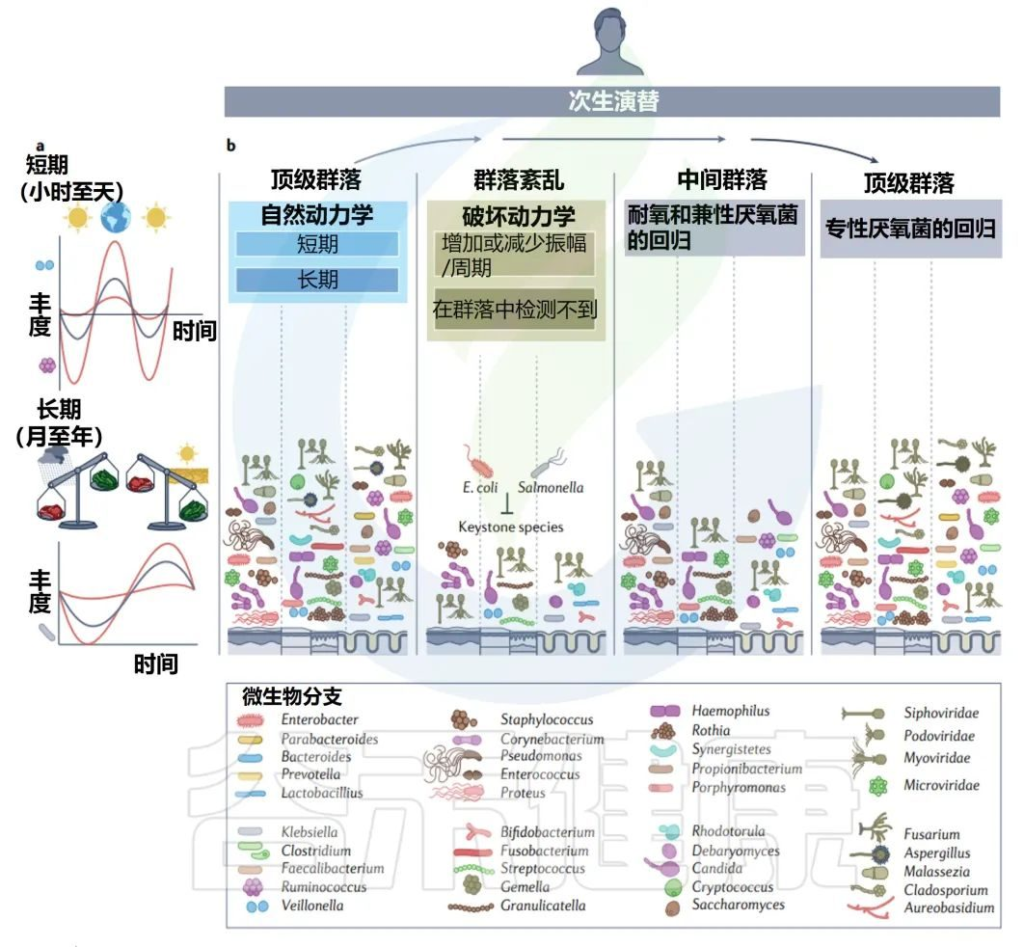
Martino C,et al.Nat Rev Microbiol.2022
• 口腔和皮肤的微生物群随清洗而变化
在口腔中,整组真菌和细菌的每日振幅与刷牙频率一致。在皮肤上,真菌和细菌每天的变化也与洗涤频率一致,并依赖于个人护理产品。
• 饮食会影响肠道微生物群
一个经过充分研究的发生在几周到几年范围内的变化的例子是饮食驱动的肠道微生物群的改变。饮食对微生物群落有很大影响,可以包括群落中的自然和可逆变化。
例如,坦桑尼亚哈扎部落在旱季食用富含肉类和块茎的饮食,但在雨季食用富含蜂蜜和浆果的饮食,在拟杆菌等属中表现出较大的季节波动。
饮食对微生物群形成的巨大影响也可能在人类健康中发挥作用,许多工作致力于了解特定的饮食成分和总体饮食模式如何影响微生物群及其对健康的影响。
肠道细菌喜欢大量的水果、蔬菜、全谷物、橄榄油等健康食物。研究表明,饮食主要由富含纤维的食物(如地中海饮食)组成的人具有更大的微生物组多样性,并且通常更健康。
此外例如,西方饮食中红肉含量高,这与全因死亡率有关。肠道微生物群可能以有害的方式将红肉中富含的左旋肉碱转化为三甲胺,而肝脏则将三甲胺转化为三甲胺氮氧化物,据推测这会促进动脉粥样硬化。
肠道微生物群也可以起到保护作用,例如,在红肉被肠道吸收之前将其分解,以防止炎症。除了饮食,还有许多其他因素有助于形成成年微生物群,包括遗传学、地理、宿主因素,如代谢病和药物。
扩展阅读:深度解析 | 炎症,肠道菌群以及抗炎饮食
• 抗生素对微生物群的影响巨大
由于微生物群的破坏而发生的次生演替已被广泛研究和审查。在破坏微生物群的众多因素中,抗生素是最强的,治疗后的恢复率往往各不相同。
抗生素治疗后肠道微生物群反弹的能力被认为取决于特定的群落成员,如拟杆菌和青春双歧杆菌。
扩展阅读:抗生素对微生物组及对人体健康的影响
细菌的天敌抗生素,如何用好这把救命的双刃剑?
疾病本身也会破坏微生物群,无论这种变化是由微生物群落内部、宿主还是多种因素共同引起的。
• 疾病破坏菌群
——肠道:炎症破坏菌群
肠道中的许多其他疾病,如炎症性肠病,破坏了微生物群落,但没有达到新的稳定群落组成,而是在没有干预的情况下继续长期不稳定。
——皮肤:炎症引起金黄色葡萄球菌大量增殖
在皮肤上,特应性皮炎的特征是免疫介导的炎症引起的金黄色葡萄球菌大量繁殖和细菌多样性减少。在金黄色葡萄球菌大量繁殖期间观察到马拉色菌属的数量减少,反之亦然,真菌数量增加导致金黄色葡萄菌数量减少,这部分可能是由于真菌产生蛋白酶的能力,蛋白酶消化金黄色葡萄球菌生物膜并降低细菌逃避免疫系统的能力。
——口腔:细菌和真菌间的竞争和协同
口腔中也存在类似的跨界相互作用;例如,真菌白色念珠菌的定殖依赖于细菌生物膜,但同时,Pseudomonas和Staphylococcus等细菌属分别形成竞争和协同关系。
这些例子强调了微生物群落的相互作用和演替是如何跨域和与宿主作用的,但由于其高阶相互作用的复杂性质,仍然没有完全理解。
干扰后微生物群落恢复的障碍导致许多研究人员探索有针对性地恢复微生物群落的干预措施的可能性。微生物群落恢复包括定向重新播种或某些物种的富集或耗竭,旨在促使微生物群落恢复到接近扰动前的水平。
这可以通过益生菌、益生元、抗生素或其他药物、从健康个体移植完整的微生物联合体或这些的组合来尝试。
尽管这些疗法在某些特征明确的环境中可以非常有效地恢复健康的微生物群落,但它们往往因缺乏与现有群落相互作用的机理知识,或因其仅短暂移植的能力而受到限制。
为了解决这些,研究集中在两个领域:
第一个领域涉及更好地了解群落是如何组合的。例如,对人类发育的研究有助于确定微生物群落在发育过程中如何聚集,以及这种聚集在生命后期的影响。
其次,正在开发新方法,通过探索微生物群落相互作用来确定机制,包括计算和实验,包括高通量共培养和微生物群落的基因组编辑。
为了解决瞬时性问题,采用了两种主要方法:
首先,微生物群疗法的短暂和个性化影响是由每个人的微生物群的个体性质决定的。因此,精准医学将群落改变的目标定位于每个人独特的微生物群,前景广阔。例如,基于微生物群落组成的个性化营养在盲法随机对照干预中有效地改善了餐后血糖。
另外,超越细菌组,探索病毒组和真菌群落及其之间的相互作用,具有巨大的前景。例如,噬菌体疗法已经用于严重的耐药细菌感染,并且对目标细菌菌株具有高度特异性。但大多数此类干预措施仍处于初步研究阶段,且规模成本高昂。
前面章节我们了解了成年微生物群的变化,以及变化后的恢复情况等,成年稳定微生物群在老年时转变为最终群落,本章节来详细了解老年微生物群。
“老年”的确切时间尺度取决于其他几个与宿主相关的因素,如疾病,但迄今为止大多数文献将“老年人”定义为65岁及以上的人。
接近寿命终点的晚期演替
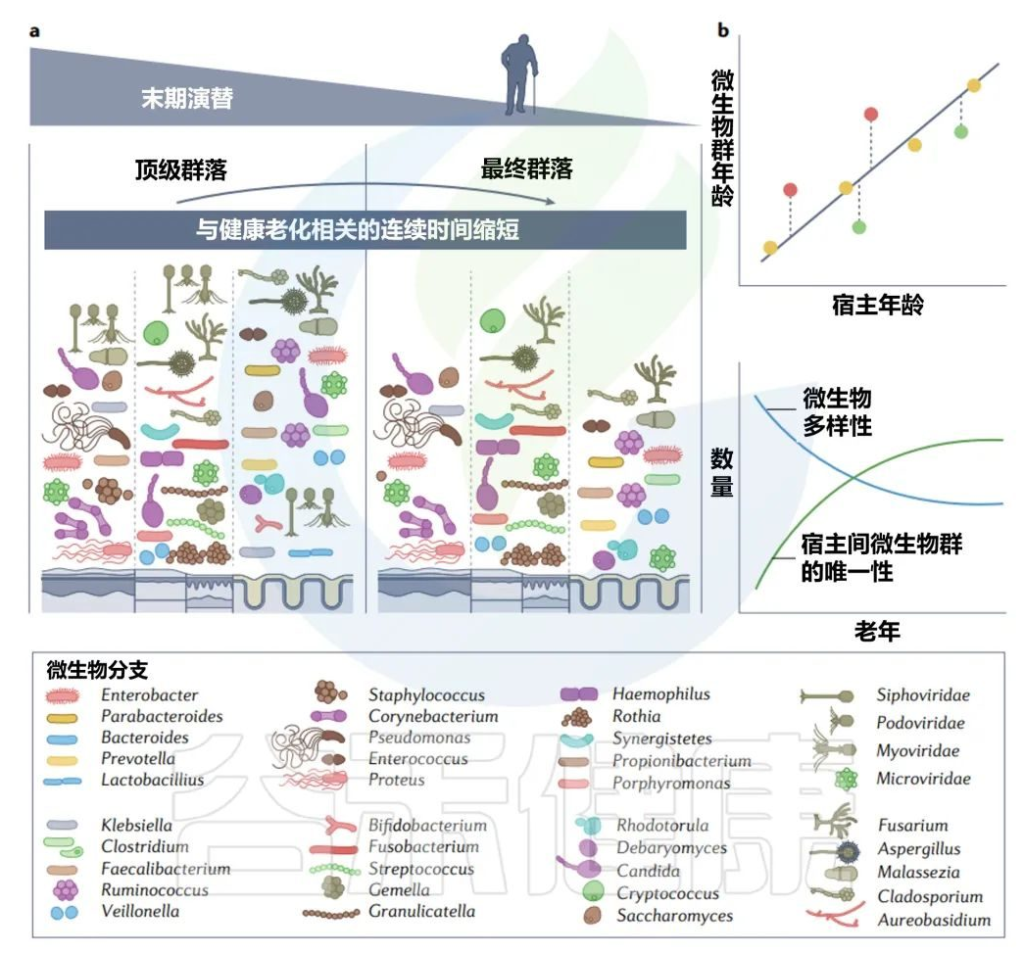
由于生物编程和生命中损伤的累积而导致的衰老影响细胞功能的各个方面,微生物群也不例外。随着年龄的增长,肠道微生物群α多样性减少,β多样性增加。
关于老年微生物群,仍有许多未知之处,而文献也有些矛盾(一项报告称65岁及以上成年人拟杆菌数量增加,与其他研究相矛盾),大多数研究都集中在肠道细菌上。
老年微生物群:年轻优势菌丰度减少
一般而言,肠道中观察到的群落演替是年轻成年人中占优势和普遍的细菌属丰度减少,如Bifidobacteria, Bacteroides, Lactobacillus, 抵御机会细菌爆发的能力降低。
• 皮肤
在65岁及以上的人群中,genera Cutibacterium和Staphylococcus的皮肤细菌数量减少,同时观察到的Corynebacterium。
• 口腔
在口腔部位,Rothia和Streptococcus spp.是核心口腔细菌群落,PorphyromonasTreponema和Faecalibacterium spp.的数量持续减少。
• 肠道
老年期肠道真菌群落的特征是Penicillium, CandidaAspergillus和Saccharomyces spp.的优势度增加。
在皮肤和口腔部位的研究很少,但老年期皮肤上的Malasseziaspp.和口腔内的Candidaspp.丰度减少。
在肠道噬菌体中,成年期的Siphoviridae占主导地位,而老年期的Microviridae和Podoviridae则占主导地位。与肠道细菌、真菌和噬菌体群体相比,真核病毒的多样性在童年后和整个余生中保持不变。
研究重点
由于个体之间的高度变异性,老年微生物演替的研究重点主要是比较健康和不健康的衰老。
目前尚不清楚微生物群是否在健康衰老中起着机械作用,还是仅仅是其他变量的一个有力指标,如饮食、运动和药物。然而,在那些长寿健康的人中,可以观察到在健康成年人中高度流行的菌群的持续保留方面的共同点。
然而,百岁老人表现出更独特的微生物群,α多样性增加,群落组成的个体间差异更大,使“健康”和“不健康”年龄之间的比较复杂化。次生胆汁酸在百岁老人中含量丰富,也可能在健康老龄化中发挥作用。尽管前景看好,但这一研究领域仍处于起步阶段。
扩展阅读:肠道菌群与健康长寿
• 微生物的演替不会随着个体的死亡而结束
宿主的死亡可以视为微生物群的生态干扰。心脏停止后,组织立即因缺氧而开始分解。细胞功能持续,直到所有剩余的氧气耗尽,二氧化碳不再能够从组织中运输为止。细胞内二氧化碳的积累创造了一个缺氧的酸性环境,导致细胞破裂。
细胞成分,例如酶会泄漏到周围环境中,在被称为“自溶”的过程中进一步促进组织分解。自溶通过消除免疫系统、松开细胞连接并为微生物群提供营养,触发了一系列负责组织分解的微生物过程。
死亡后的微生物群
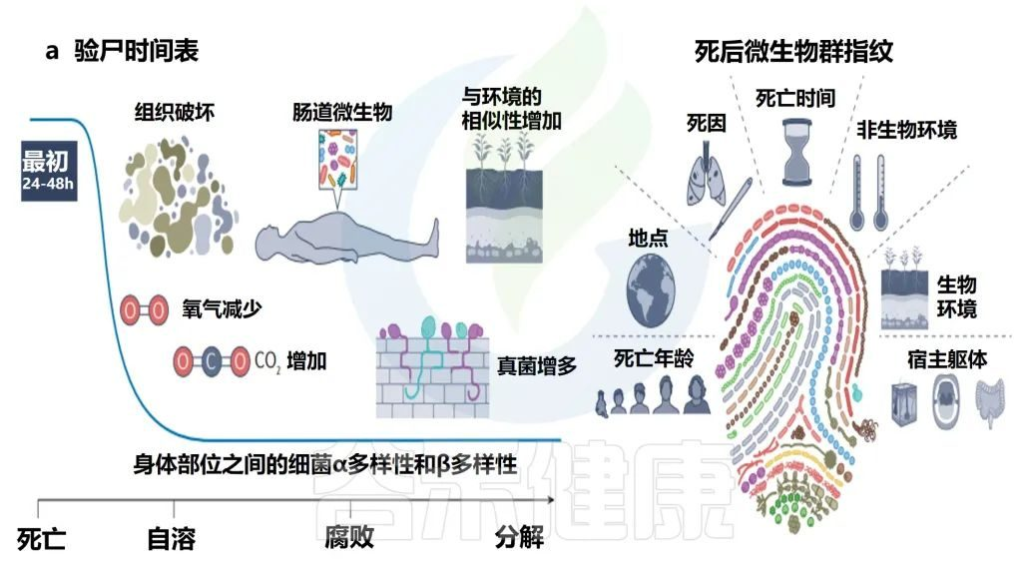
Martino C,et al.Nat Rev Microbiol.2022
• 死亡后微生物群分解
人类微生物群在死亡后的前24-48小时内相对稳定,具有不同的身体部位微生物生态、年龄的α多样性模式和可识别的个性化皮肤微生物群特征。
在分解的最初几天到几周内,腐败主要由细菌进行,但随着分解的进行,真菌的作用增加。然而,在这个过程中,对病毒组的演替和功能作用了解甚少。
随后,环境变化促进了微生物的演替,改变了人体和微生物群,不再像活着的个体(除非身体被冷冻)。
由于缺乏宿主生活中先前遇到的环境限制,使得微生物的相对丰度发生了快速变化以及在身体各部位的移动。迁移的细菌群成为从肠道转移到肠外部位的先锋物种,根据身体部位参与初级演替或次级演替。
• 死亡微生物群——生物指示器
死亡微生物群因其对法医调查的影响而引起了越来越多的关注。与多个个体和身体部位相关的一致的时间序列模式证明,死后微生物群可以作为死后间隔的生物指示器。
每个尸体的死后微生物群都是独一无二的,并且根据死亡时间、死因、环境、死亡地点和年龄以及开始时身体部位之间的差异,尸体之间的微生物群是不同的。
当微生物演替包括群落成员的快速更替时,在分解的早期阶段(即死亡后的前2-3周),死后时间间隔估计更为准确,但在分解的后期阶段(例如骨骼)仍然有用,因为几乎没有证据可以估计死后时间间期。
• 死亡原因与微生物群存在联系
还证明了与死亡原因和微生物群存在的联系。例如,在死于心脏病的个人的口腔微生物群中发现了的Rothia spp.。
此外,皮肤微生物群脱落可能通过将个人与他们接触过的物品联系起来,从而有助于追踪证据;然而,这一独特特征能够准确匹配到个体的时间取决于对象的材料和用途。
人类微生物群是动态的。考虑到这一点,设计一种能够捕捉微生物群的时间和空间变异性的采样策略非常重要,特别是当这些波动与所提出的科学问题相关时。
✦测量时间不同:多个时间点的样本采集
横断面研究从每个个体收集一个样本,而重复测量研究在多个时间点或身体部位收集样本。随着时间的推移,采样频率应该调整到研究人员试图观察的现象。
例如,小鼠昼夜节律研究通常每2-4小时收集一次粪便样本;而在炎症性肠病中,在一周内对患者进行三到五次采样可以改善疾病分类。
在其他应用中,例如研究特定治疗对个体微生物群的影响,这可能与进行“一对一”研究有关,在该研究中,同一参与者被反复检测其微生物群的结果变化;治疗前采集的样本被视为个体水平的对照。
✦测量空间不同:城市化/农村环境不同
同样重要的是要考虑到人口的微生物群高度依赖于地理和种族。
例如,在一个大型中国群体中,一种与年龄高度相关的微生物在一个美国大型群体中根本没有检测到。
另一个具体的例子涉及城市化社会的“建筑环境”;城市化人群通常较少接触环境微生物,更多地使用家用抗菌剂,与来自农村社会的人类微生物群相比,这导致了重大变化。
这些考虑因素与微生物群领域尤其相关,因为大多数公共微生物群数据来自城市化的北美和欧洲人。因此,现有数据集的结论可能无法很好地推广到全球人口。
从人类微生物群和微生物群研究中生成的测序数据的主要类别是扩增子测序数据和鸟枪测序数据。
✦扩增子测序
在扩增子测序中,对已建立的高变区的PCR产物(扩增子)进行深度测序,从而能够通过与个体“条形码”匹配来识别和测量群体成员。
这里有两种选择:要扩增的基因和该基因的哪一部分要扩增。微生物基因组的常见扩增区域包括:细菌的16S核糖体RNA基因、真核微生物的18S核糖体DNA基因和真菌的内部转录间隔区。
每个特定基因中高变区的选择取决于要捕获的特定微生物,但广泛使用的高变区包括来自地球微生物组项目的V4区。
✦肠道微生物群参与人体的调节
在鸟枪测序中,所有微生物DNA都被测序,而不仅仅是PCR产物,从而能够对微生物进行更具体的分类。由于鸟枪测序不依赖于任何标记基因,因此与扩增子测序相比,它对某些微生物的偏向性较小。
然而,鸟枪测序的成本要高得多,并且需要更大的计算能力,这使得在不需要提高鸟枪序列分辨率的情况下,扩增子测序具有吸引力。
结合其他技术进行扩增子或宏基因组测序可以丰富对微生物群和宿主的理解。定量PCR和荧光激活细胞分选等技术通过将相对丰度锚定到可靠的绝对丰度测量值,为相对丰度提供了更多的背景。
酶联免疫吸附试验和单细胞测序可以通过提供宿主细胞类型或宿主免疫信息与宏基因组测序很好地配对。
培养组学使研究人员能够通过实验验证功能或活性的基因组预测,并将微生物转化为益生菌。微生物产生的代谢物或蛋白质,即微生物群的下游效应物,可以分别通过代谢组学和蛋白质组学进行探测。
最后,宿主基因组学和转录组学越来越多地与扩增子或宏基因组学数据配对,以深入了解宿主基因表达和微生物群之间的联系。
最后,从被调查的参与者那里收集数据至关重要。一般微生物群研究的一些重要元数据类别包括人口统计、临床信息和饮食信息;然而,使用的确切元数据因研究而异。应采用产生标准化元数据的实践,以便结果可重复使用和再现。
本文描述了目前对不同年龄和不同身体部位的人类常驻微生物群落组成的研究现状。
人类健康与微生物群组成之间存在许多联系,对肠道菌群的干预可能改善健康。侧重于整个微生物群而不是单一物种的富集或消除的干预措施,需要了解这些群落是如何形成和维持的。
不同人群年龄,不同部位的微生物群需要依托于大样本数据库的构建,这为微生物群研究的准确性提供了保障。
通过研究人类整个生命周期中的微生物群,我们可以更好地了解这些微生物群复杂的相互作用,以及如何有效地将微生物群推向宿主所需的组成。此外也正应用于除人类健康外的其他领域,如法医学。随着微生物群的相关研究不断突破,将给人类生命健康和生产生活带来巨大的影响。
主要参考文献:
Martino C, Dilmore AH, Burcham ZM, Metcalf JL, Jeste D, Knight R. Microbiota succession throughout life from the cradle to the grave. Nat Rev Microbiol. 2022 Jul 29. doi: 10.1038/s41579-022-00768-z. Epub ahead of print. PMID: 35906422.
Lim, A. I. et al. Prenatal maternal infection promotes tissue-specific immunity and inflammation in offspring. Science 373, eabf3002.
Al Nabhani, Z. & Eberl, G. Imprinting of the immune system by the microbiota early in life. Mucosal Immunol. 13, 183–189.
Helve, O. et al. 2843. Maternal fecal transplantation to infants born by cesarean section: safety and feasibility. Open. Forum Infect. Dis. 6, S68.
Seppo, A. E. et al. Infant gut microbiome is enriched with Bifidobacterium longum ssp. infantis in old order mennonites with traditional farming lifestyle. Allergy 76, 3489–3503.

谷禾健康
菌群多样性
✦
关于菌群多样性,小伙伴们常会问到:
菌群多样性是如何形成的?
菌群多样性越高越好吗?
它与健康的关系如何?
该怎么去改善呢?
……
✦
✦
本期推文就为大家统一解答这类问题。
本文要点
在了解前面问题之前。我们首先要明确,菌群多样性是什么?
首先,我们知道肠道微生物群代表了一个不断变化的生态系统。
为了便于理解,我们可以把肠道菌群比作一块草地,健康的肠道菌群包含多种菌群,它们之间相对平衡,就好比一块生长完好的草地,生长各种类型的草,整体和谐 ↓↓

一旦遇到疾病,药物等干扰之后,一些菌群被杀死,也就是说少了很多种类型的草,这块“草地”会逐渐变秃,甚至部分沦为荒漠 ↓↓


之后如果不加任何看管,没有很好得到恢复的话,部分菌群可能会野蛮生长,破坏了原有的平衡 ↓↓


以上,我们可以看到关于多样性的几个特点。
一个是种类:
成年人群中肠道菌群的种类参考范围在100~2000种,种类数量越多多样性越高。类比图1那样。
一个是均匀性:
即各个菌种的含量丰度较为均一,如果出现单一菌种占据绝大部分的情况,类比以上图3,那么多样性也不理想。
多样性的评估一般通过一个叫做香农-维纳多样性指数的指标来进行评估,计算公式为:
H=-∑(Pi)(log2Pi)
其中Pi为每个菌的占比例,值越大代表物种种类越多,均匀性也更好相应的多样性也越高。正常人群中香浓指数在2~9之间,一般大于3以上表明具有一定多样性。
肠道菌群多样性表现在:
微生态系统的稳定性,以及面对外界致病菌等入侵的抵御能力。
这要追溯到生命早期。
在胎儿发育和婴儿期,菌群多样性通常很低。
由于在生命的早期阶段,肠道含有氧气,此时肠道内的微生物大多是耐氧的。
随着不断发育,这些菌群开始逐渐被厌氧菌取代。肠道菌群的这种进化是迅速的。
研究表明,婴儿在出生后第一周,肠道内发现的病毒序列有56%,在第二周后就不存在了。
这种多样性在生命的前3个月继续迅速扩大,其中约95%的微生物随着时间的推移而保存下来。
婴儿的微生物群组成具有特征丰富的Akkermansia muciniphila、拟杆菌属、韦荣氏菌属、球状梭菌属和肉毒梭菌属等。肠道微生物多样性延迟与过敏、哮喘和营养不良有关。
肠道菌群变得稳定,微生物群落多样性随着年龄的增长而增加,直到它成为一个稳定的成年微生物群,主要由三个细菌门组成:
厚壁菌门
(Lachnospiraceae和ruminococaceae)
拟杆菌门
(拟杆菌科、Prevotellaceae和Rikenellaceae)
放线菌门
(双歧杆菌科和Coriobacteriaceae)
从出生到2.5岁的微生物群发育期可能非常关键,婴儿早期肠道微生物群的破坏可能会严重影响人类健康。
大约三岁时,儿童肠道微生物群的组成和多样性与成人最为相似。
在个体的整个生命过程中,它继续以更稳定、更慢的速度进化。

研究还发现,青春期之前儿童,肠道微生物群富含可能参与持续发育功能的菌群,如叶酸和维生素B12的合成方面也很丰富。
青少年微生物群,梭状芽胞杆菌属、双歧杆菌属的丰度显著高于成人。
健康成人肠道菌群以厚壁菌门和拟杆菌门为主,也包括较小比例的Verrucomicrobia、放线菌门(Actinobacteria)、变形菌门(Proteobacteria)。
一般来说,老年微生物群的特点是微生物多样性下降,机会性病原体增多,双歧杆菌和拟杆菌显著减少,兼性厌氧菌、梭杆菌门、梭状芽胞杆菌和真细菌Eubacteria增加,产生短链脂肪酸菌群减少。衰老显著影响老年人微生物群的组成和功能。
是不是所有老人的肠道菌群多样性都会降低?
不是,长寿老人的肠道菌群多样性水平高。
研究人员检测了一群健康的长寿老人的肠道微生物组,来自中国四川都江堰市的长寿老人,包括“90-99岁”和“≥100岁”两个年龄段。
他们发现长寿人群的肠道菌群比年轻成年人的肠道菌群更多样化,这与传统观点相矛盾。
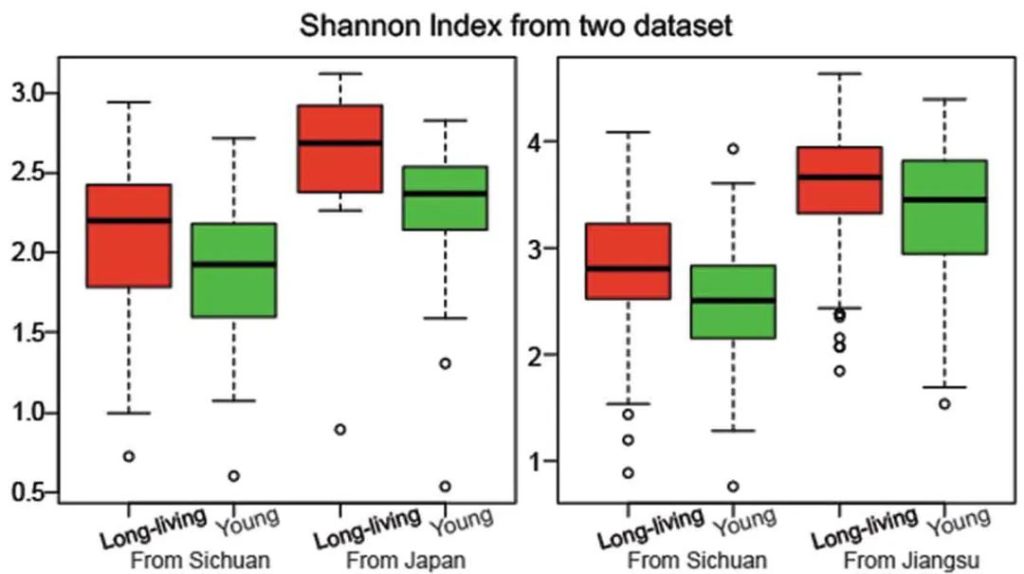
Kong F, et al., Gut Microbes, 2018
他们还发现了产短链脂肪酸菌在长寿老人中开始增加,例如梭状芽胞杆菌XIVa。
国内外研究结果一致
为了验证他们的发现,他们分析了来自一个意大利小组的独立数据集。
Kong F, et al., Gut Microbes, 2018
出现一致的结果:长寿的意大利人也比年轻的人群的肠道菌群多样化水平更高 。
庆幸的是,谷禾肠道菌群数据库中也有比较长寿的老人肠道菌群数据。
我们抽取其中一例相对较为健康的长寿老人的数据:
编号:083*****97 ,98岁(谷禾肠道菌群数据库)
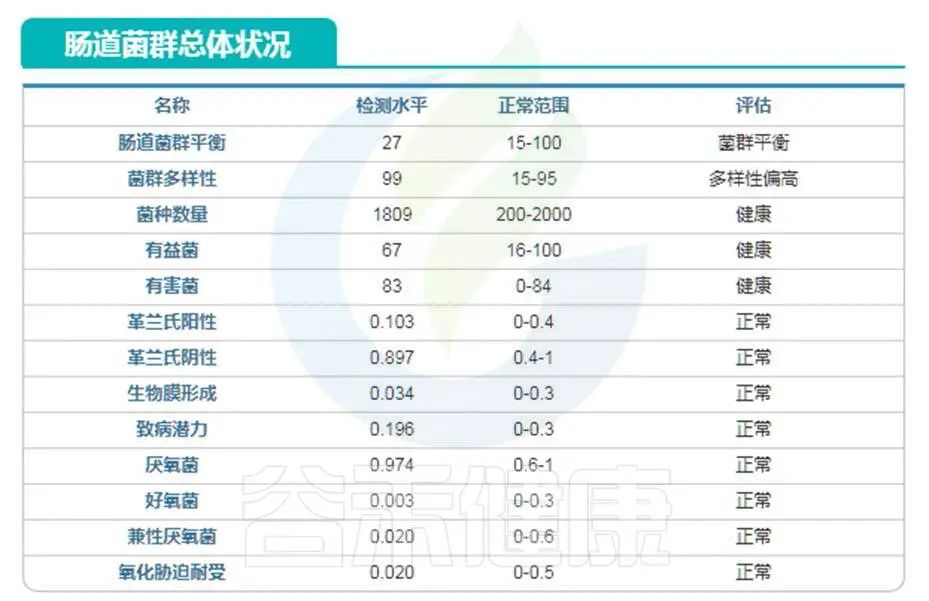
可以看到肠道菌群多样性水平也是明显增高,与文献报道相符。大部分指标都处于正常水平。
那么,菌群多样性是不是仅随年龄的变化而变化呢?
不是的,我们会发现同一年龄层人群的多样性还是有差别的。也就是说除了年龄这个因素之外,肠道菌群变化与某些其他因素也存在关联。
下一章节我们逐一展开了解一下其他相关因素。
前一章节,我们发现年龄会影响菌群多样性变化,其他也可能是由遗传、饮食习惯、生活方式、生理变化、地理位置、药物等因素共同决定的。
▼ 遗传(双胞胎菌群特征相似)
研究发现,分开生活多年的同卵双胞胎在微生物特征上表现出高度的相似性,而生活在相同环境和饮食习惯相似的配偶则没有。


厚壁菌门、放线菌门、Tenericutes、Euryarchaeota的遗传性更强,而拟杆菌门的遗传性很低。
以上这些发现有力地证明,遗传因素在塑造肠道微生物群方面提供了强大的力量。
其他也有研究证实了猪、人和小鼠之间的遗传分类群和候选基因的功能类别的高度相似性,这表明宿主基因对哺乳动物肠道微生物群的影响机制相似。
▼ 地理位置(一方水土养一方人)
肠道菌群中每一个门的比例随着地理位置的变化而变化。这些可能是受到各种不同生活方式有关的大气、基因、饮食或其他因素影响。
在非洲农村布吉纳法索村的儿童(高纤维饮食,脂肪和动物蛋白含量低),马拉维和委内瑞拉的儿童和成人中富含普雷沃特菌,他们吃的主要是玉米、木薯和其他植物源性多糖。


研究表明,普氏杆菌与碳水化合物和单糖有关(在农业社会普遍存在)。
在西方国家,拟杆菌门与厚壁菌门的比例更高。
拟杆菌与长期饮食中,富含动物蛋白、几种氨基酸和饱和脂肪有关(在美国和欧洲普遍存在);


西方国家,肠道菌群多样性的普遍减少,引发了人们的思考,更广泛人群中重要共生菌该如何维持?全球饮食趋势是否会导致有益菌群永久消失?

在发展中国家保持个体菌群(特别是在农业文化中),可能有助于保存人类微生物群中潜在的重要组成部分。
此外,特殊环境,如医院的ICU病房、更衣室等消毒严格的地方,也可能导致环境菌群多样性下降。
▼ 饮食 (调节菌群怎么离得开吃?)
都知道饮食对肠道微生物群的调节起着至关重要的作用,这其中可能起到有益的作用,也可能是有害的。
为什么要把婴儿单独拎出来讲呢?因为对他们来说,人生重要事儿就是吃吃睡睡,那么吃就占了很重要的一块。饮食可以说是塑造肠道菌群的关键驱动力。
婴儿早期,肠道微生物群富含与母乳中低聚糖消化有关的基因,后来,随着其他辅食的引入,富含与多糖和维生素代谢相关的基因。
母乳喂养的婴儿表现出放线菌的过度生长以及厚壁菌和变形菌的抑制。母乳中含有可被这些细菌有效代谢的低聚糖,导致短链脂肪酸增加,从而使免疫系统增加免疫球蛋白G的表达。
配方奶粉喂养的婴儿表现出梭菌、链球菌、拟杆菌和肠杆菌的增加。婴儿期后,肠道微生物群继续发展,饮食成为组织肠道微生物群结构、形状和多样性的主要关键。
日常强调的水果和蔬菜,在决定α多样性方面发挥了最重要的作用。研究发现α多样性或微生物丰富度与长期水果和蔬菜摄入呈正相关(p<0.05)。

膳食纤维的摄入对于保持肠道粘膜屏障功能的完整性至关重要。高纤维饮食可以改善2型糖尿病患者的血糖控制和促进更健康的代谢状况。
素食与健康、肠道菌群多样性的变化以及厚壁菌和拟杆菌的优势有关。
高蛋白质和高脂肪的饮食与大量耐胆汁的菌群有关,如拟杆菌、嗜胆汁菌和厚壁菌,以及对厚壁菌的抑制。不过需要注意的是,这种饮食习惯,可能会导致免疫力降低、感染易感性增加和代谢性疾病的发展。因此,减肥人士也需谨慎。
具体如何选择可见后面章节。
▼ 季节 (果蔬也有属于它们的生长季节)
食物供应的季节性变化的影响菌群多样性。
首先,初步研究将坦桑尼亚的哈扎狩猎采集人与城市化人进行了比较,结果与意大利城市居民相比,哈扎人的饮食富含复杂的多糖,并表现出更高水平的微生物多样性,表现为拟杆菌的比例较高。
后续宏基因组分析显示碳水化合物活性酶的富集,这与觅食、富含多糖的饮食一致。肠道微生物组保留的代谢复杂碳水化合物和蛋白质的功能潜力,可能是由于食物供应的季节性变化。
哈扎人肠道菌群在组成上,经历了与湿季和旱季同时发生的季节性循环,其中 Prevotellaceae 、 Spirochaetaceae 是 2 个最具季节性变化的菌群。Prevotellaceae 的比例在雨季下降,这与宏基因组中存在的碳水化合物活性酶显着减少有关,特别是植物碳水化合物。
另一项研究针对北达科他州的社区哈特派人群,他们的夏季饮食富含高纤维的新鲜水果和蔬菜。与冬季相比,北达科他州的夏季肠道微生物群与拟杆菌门的丰度显着增加相关,而放线菌和厚壁菌门的丰度相应减少。
当然,在城市化人群中并没有这种现象。
总的来说,研究城市化程度、地理位置和特定食物供应的全球饮食模式,有助于在宏观层面上形成全球肠道微生物群的巨大异质性。
▼ 生活方式 (除了吃还有什么能改变菌群?动!)
——运动锻炼
在人类研究中,运动增加了肠道菌群的多样性,与蛋白质摄入量和肌酸激酶水平呈正相关。
与非运动员相比,运动员的拟杆菌水平较低,厚壁菌数量较多。
对运动产生反应的细菌种类包括乳酸杆菌、双歧杆菌和阿克曼菌,而其他种类则有所减少,如变形杆菌、Turicibacter、Rikenellaceae.
此外,在运动的反应中,产短链脂肪酸菌(尤其产丁酸菌),如梭菌Clostridiales、罗斯氏菌Roseburia、毛螺菌科Lachnospiraceae、Erysipelotrichaceae增加,并在调节运动对宿主肠道菌群的影响中发挥重要作用。

运动增强丰富菌群的机制是什么?
涉及内部和外部因素。
【内】对耐力训练的内在适应,如血流量减少、组织缺氧和转运增加,会导致胃肠道的变化。
【外】运动型个体暴露于他们的环境生物圈中,遵循整体健康的生活方式,促进更丰富的微生物群。
与对照组相比,运动员表现出较低的炎症和改善的代谢标志物,并且运动与较低的慢性炎症导致的发病率降低有关。
有研究人员提出,运动可以作为一种治疗支持,可用于治疗与失调相关的疾病,如肥胖和其他一些胃肠道疾病。
关于运动详见:《运动如何影响肠道微生物群,如何正确运动》
《肠道微生物组如何影响运动能力,所谓的“精英肠道微生物组”真的存在吗?》
——吸烟
在戒烟的健康个体的粪便微生物群中也观察到显著的变化,包括厚壁菌门和放线菌门相对丰度的增加,拟杆菌门和变形菌门的减少。
研究证实,吸烟者和非吸烟者的口腔菌群存在显著差异。特别是在吸烟者的口腔中,卟啉单胞菌和奈瑟菌的繁殖增加,Gemella减少。


▼ 药物
抗生素的使用是一把双刃剑:它摧毁致病菌的同时,可能也伤害了有益菌,导致肠道微生物群失调,多样性大幅下降,并且需要一段时间才能恢复。
广谱抗生素导致厚壁菌门和拟杆菌门之间的不平衡。在治疗过程中,菌群整体多样性降低,这些细菌的丰度也降低。
微生物组组成的改变取决于抗生素类别、剂量、暴露时间、药理作用和目标细菌等。例如,连续两年服用克林霉素会导致肠道微生物群发生变化,而拟杆菌种类不会恢复。
在幽门螺杆菌治疗中使用克拉霉素会导致放线菌数量减少,而环丙沙星会导致治疗半年后仍未恢复的瘤胃球菌数量减少。
研究表明,氨苄青霉素、万古霉素、甲硝唑和新霉素具有长期的负面影响,可引起肠道菌群多样性的不可逆变化,从而增加宿主疾病的风险。
关于抗生素对菌群的影响详见:
《抗生素对微生物组及对人体健康的影响》
《细菌感染和抗生素使用》
《细菌的天敌抗生素,如何用好这把救命的双刃剑?》
其他药物也会降低菌群多样性,如治疗胃溃疡和反酸的质子泵类药物也会导致菌群多样性降低。
以上是常见的菌群多样性的决定因素,那么菌群多样性是如何对身体产生影响的?下一章节我们探讨这部分内容。
微生物组的多样性和平衡在人类健康中发挥着重要作用。
▼ 消化系统疾病
以IBD为例。
在 IBD 患者中,肠道菌群的多样性减少,这主要是由于厚壁菌门的多样性减少。在厚壁菌中,梭菌减少,尤其是普拉梭菌,普拉梭菌是产丁酸盐的菌群之一。这些菌群少了,产出来的丁酸盐就少,丁酸有什么用呢?
它能对肠道防御机制具有多阶段调节作用,包括通过促进肠上皮中的紧密连接蛋白保护肠粘膜屏障,支持先天性和适应性免疫反应,刺激成熟的结肠细胞,抑制未分化的恶性细胞和干细胞,通过降低环氧合酶-2(COX-2)水平抑制氧化应激,并通过诱导过氧化氢酶,改善过氧化氢(H2O2)的解毒作用。
丁酸也能刺激芳香烃受体(AhR)、GPR41和GPR109A受体,并抑制不同细胞类型的HDAC,从而稳定肠道屏障功能并减少炎症过程。
丁酸少了,肠道屏障可能受损,炎症反应增加,就会出现IBD等肠道疾病。
在克罗恩病(CD)、肠易激综合征(IBS)等消化系统疾病中,无论是否伴有腹泻和结直肠癌,都会持续观察到多样性降低。
乳糜泻患者的十二指肠相关微生物群多样性也不太理想。
艰难梭菌结肠炎复发的危险因素之一就是多样性降低。
扩展阅读:
《深度解析 | 炎症,肠道菌群以及抗炎饮食》
《炎症性肠病中宿主与微生物群的相互作用》
▼ 神经系统疾病
目前越来越多的研究表明,在抑郁症、帕金森病 、阿尔茨海默病等神经系统疾病中,肠道菌群多样性减少。
抑郁症、帕金森病和AD患者肠道菌群的变化
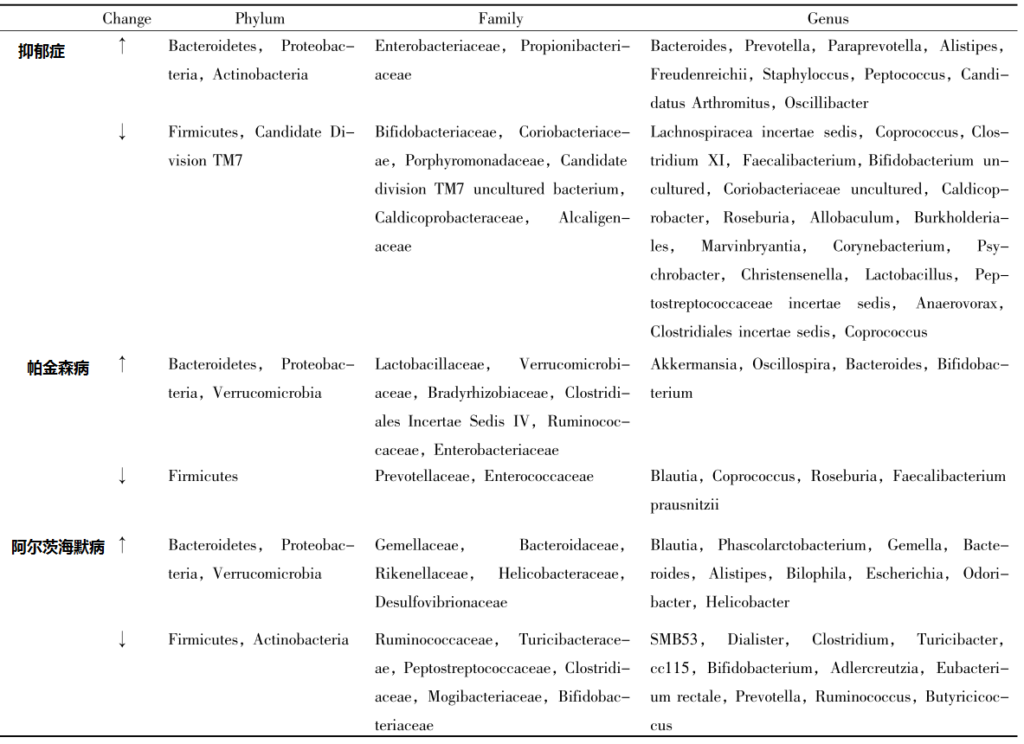
MU Xin et al., Chinese Pharmacological Bulletin 2019
扩展阅读:《肠道微生物组在人类神经系统疾病中的作用》
《最新研究速递 | 柳叶刀:肠道微生物群在神经系统疾病中的作用》
《深度解读 | 肠道菌群和中枢神经系统的关系》
▼ 癌症
以乳腺癌为例。
乳腺癌患者多样性减少。一项基于人群的病例对照试验研究中,调查了绝经后妇女的粪便微生物群与乳腺癌之间的关系。发现与对照组患者相比,病例患者的微生物群组成显着改变(β-多样性,P = 0.006)和较低的α-多样性(P = 0.004)。
肠道菌群失调可诱发慢性炎症,因此有可能影响乳腺癌的发展。
扩展阅读:《肠道微生物群与五种癌症的相互作用:致癌 -> 治疗 -> 预后》
《结直肠癌防治新策略——微生物群》
《“隐藏高手” 胰腺癌的新出路——微生物》
《肺癌最新研究进展,与肠肺微生物息息相关》
《深度解析 | 肠道菌群与慢性肝病,肝癌》
▼ 代谢类疾病
肥胖
一般来说,肥胖个体的微生物多样性低于瘦的个体。饮食干预可以改善微生物的丰富度和相关的临床表型,例如改善代谢和炎症状态。
扩展阅读:《体重增长:目前为止我们所知道的一切(更新你的减肥工具箱)》
糖尿病
1 型糖尿病患者的肠道微生物群,高水平的拟杆菌,大量产生乳酸和丁酸的细菌以及细菌功能多样性降低。
2型糖尿病前期,菌群多样性就已经存在下降趋势。
扩展阅读:《2型糖尿病如何做到可防可控?肠道菌群发挥重要作用》
▼ 儿童发育
对来自孟加拉国的营养不良儿童和营养良好的健康儿童的肠道微生物群进行了横向比较研究,结果发现,营养不良儿童微生物群的多样性显著降低。
扩展阅读:《生命早期营养不良和微生物群影响大脑发育和行为》
《真实案例 | 儿童发育迟缓肠道菌群检测的应用》
《发育迟缓/营养不良不容忽视,问题很有可能在肠道》
▼ 心脑血管疾病
高血压个体的肠道微生物多样性减少,厚壁菌门/拟杆菌门比率增加。
扩展阅读:《认识肠道微生物及其与高血压的关系》
《饮食-肠道微生物群对心血管疾病的相互作用》
▼ 免疫系统疾病
以过敏为例。
在出生第一年甚至更早的时候,饮食多样性可能对预防过敏产生积极影响。早期接触多种食物抗原可能会促进免疫耐受的发展。
扩展阅读:《婴幼儿过敏有望改善,与肠道菌群密切相关》
《生命早期微生物接触和过敏风险:如何预防》
《微生物群对三大过敏性疾病发展的影响》
▼ 睡眠
人类肠道微生物群可以通过脑-肠道微生物群轴影响健康。
一项研究使用活动描记术量化睡眠测量,并结合肠道菌群采样,以确定肠道菌群与各种睡眠生理测量之间的相关性。测量了免疫系统生物标记物,并进行了神经行为评估,因为这些变量可能会改变睡眠和肠道菌群组成之间的关系。
总微生物组多样性与睡眠效率、总睡眠时间的增加呈正相关,睡眠越好,肠道菌群多样性越丰富,与睡眠开始后的觉醒呈负相关。
总微生物组多样性与IL-6之间存在正相关。微生物组分分析显示,拟杆菌和厚壁菌门内的丰富性与睡眠效率、IL-6浓度和抽象思维呈正相关。研究人员也发现了几个分类群(Lachnospiraceae, Corynebacterium, Blautia)与睡眠指标呈负相关。
肠道菌群多样性高有助于减轻压力,改善睡眠。
扩展阅读:
《肠道菌群与睡眠:双向调节》
《深度解析|睡眠健康与肠道健康之间的双向联系》
以上是菌群多样性与疾病的关联,了解这些我们可以有针对性地去改善菌群多样性。
我们在实际检测中会发现有这样一种情况:
多样性指标虽然很高,但是整体看起来健康总分并不理想。甚至还有很多慢性疾病风险,这是为什么呢?
这种情况可能是核心菌群丰度不够,核心菌群在代谢、免疫等方面都发挥重要作用,一旦核心菌群丰度下降,则可能造成外源物质侵入。感染、旅行等可能会出现这种情况。
如果发现肠道菌群检测报告中多样性很低,该如何判别具体情况,这要看以下几种情况:
✦
总的来说,在一定范围内,更高的多样性通常代表饮食更加丰富多样,同时也意味着更健康的身体状况。
✦
一个健康、有弹性的肠道微生物群依赖于高丰富性和生物多样性,也就是说,当你的肠道菌群多样性相对较高时,会更强大、更稳定。
就好比你拥有一支队伍,有很多不同类型的、不同专长的人可供你使用时,整体的能力就会很强大。


前面我们已经知道,菌群多样性的各种决定因素,例如,饮食、年龄、遗传、环境等。
多样性低不代表一定有疾病,但是更容易受到饮食,环境或疾病的影响,包括更易发生水土不服或更容易因饮食不洁导致腹泻等。菌群多样性低可能是肠道菌群不健康不稳定的一种表现形式。
当然,也不能过度追求菌群多样性高。
随着对饮食-微生物组-健康轴的理解逐渐加深,最近创造了个词叫“食品药学”,即“食品作为药物”的概念逐渐得到推广,基于个体微生物组特征的“食物疗法”的个性化得到越来越多的关注。
饮食干预引起的改变可能在几周甚至几天就会发生。对调节肠道菌群而言,饮食调节是一个非常重要且纯天然的方式。那到底该吃什么?如何吃?
我们可以从食物种类,饮食结构,饮食质量等多个角度去优化日常饮食,从而调节菌群。
✔ 食物种类:
膳食纤维是饮食中的重要组成部分,分为不溶性(抗发酵)或可溶性(可被肠道微生物群代谢),例如益生元纤维菊粉,在结肠中它被肠道微生物群发酵成短链脂肪酸:乙酸盐、丁酸盐、和丙酸盐。
食用膳食纤维时,各种菌群可以利用这些膳食纤维作为基质,扩大其种群,从而增加微生物组的总体多样性。
许多住在结肠和盲肠内的厌氧细菌,依赖于复杂碳水化合物的代谢来产生短链脂肪酸,短链脂肪酸是肠粘膜的重要能量来源,在宿主代谢、免疫功能和细胞增殖中发挥关键作用。
丁酸盐是肠道中一种丰富的短链脂肪酸,在结肠癌中起着复杂的作用,如最近的两项临床前研究所示,丁酸盐似乎是浓度和环境依赖性的。丁酸盐可以抑制肿瘤的发生,因为丁酸盐受体Grp109a缺乏的小鼠增加了炎症刺激或APC突变促进的肿瘤发生,并且通过Grp109a的信号传导抑制了这些刺激诱导的肿瘤发生。
▸ 富含膳食纤维的食物在哪里?
膳食纤维可以在各种水果、蔬菜、坚果、种子、豆类和谷类中找到。

吃洗过的生水果和蔬菜,这些水果和蔬菜含有环境微生物(如果难以消化生蔬菜,可以榨汁),包括益生元,以促进多样化的微生物群,不要只吃某一种蔬菜或水果。
▸ 足量的膳食纤维该吃多少?
推荐的纤维摄入量(充足摄入 克/天)
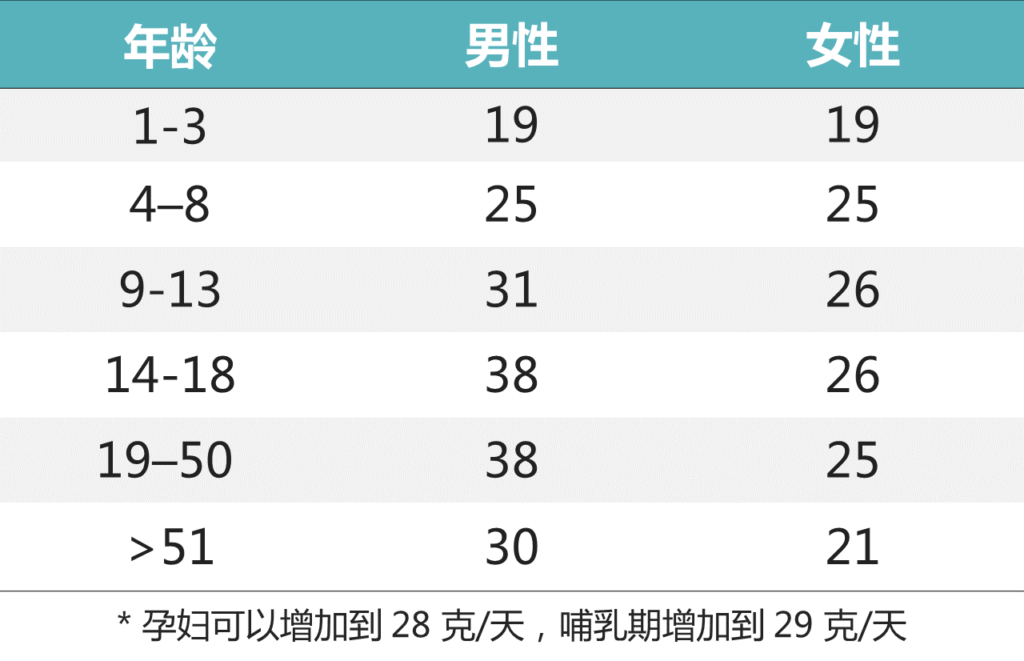
▸ 具体哪些水果蔬菜中的富含膳食纤维?
各类水果蔬菜中每标准份量膳食纤维的含量及热量
数据来源:U.S. Department of Agriculture, Agricultural Research Service. FoodData Central
同时,水果蔬菜里面一般富含多酚,多酚可以塑造肠道细菌的组成。
Vemana Gowd, et al.,Trends in Food Science & Technology, 2019
白藜芦醇
食用白藜芦醇可减轻结肠炎小鼠模型中的炎症状态并恢复微生物群多样性。
新的证据表明,红酒中的白藜芦醇能通过支持短链脂肪酸的产生来预防阿尔茨海默病 (AD),短链脂肪酸会干扰有毒的 β-淀粉样蛋白聚集体形成。在AD背景下,白藜芦醇的消耗与较高比例的有益菌有关,例如 F. prausnitzii、Akkermansia 物种和具有脑抗炎特性的产丁酸盐细菌。
姜黄素
姜黄素能够部分逆转多样性的变化 肠道微生物群,增加 F/B 比并降低 Anaerotruncus 和 Helicobacter 属的丰度。
已证明,姜黄素治疗降低了在癌症患者中发现的与癌症相关的菌群丰度(例如普氏菌属、冠状杆菌属和瘤胃球菌属)。
富含多酚的食物包括:


此外,饮食中添加全麦大麦、糙米或这两种的混合物会增加菌群多样性。
发酵食品是益生菌的另一个重要来源。
2021 年 7 月的《细胞》杂志上,健康成年人被分为两组,21人食用水果、全谷物、豆类、蔬菜、坚果和种子等高纤维食品,18人食用发酵食品。
在试验前的三周、指定饮食后的10周内,以及研究结束后的四周内,当参与者想吃什么就吃什么时,从受试者身上收集粪便样本。还采集了血液样本以测量炎症和免疫功能。
结果发现食用发酵食品的人,除了增加肠道微生物群的多样性外,同时对四种免疫细胞的激活程度较低,包括IL-6在内的几种炎症蛋白水平也降低。
然而,在大多数情况下,发酵食品对我们肠道微生物群的改变是暂时的,因此,持续摄入发酵食品更有利于对于维持高水平的有益菌。
▸ 发酵食物有哪些?
坚果(尤其是核桃)对健康有益,食用坚果可以降低血脂水平。核桃如何改变菌群?
在一项随机对照试验(交叉设计)中,96名健康参与者接受了为期8周的富含核桃的饮食,然后改用无坚果饮食。第二组98名参与者按相反顺序遵循饮食模式,收集粪便样本做16S rRNA测序分析。
在α多样性方面没有发现差异,但在β多样性方面,观察到核桃和对照组的明显聚集,也就是说,他们的菌群发生明显变化。
与对照饮食相比,核桃饮食仅解释了观察到的差异的5%左右。在核桃组,与对照组相比,瘤胃球菌Ruminococcaceae和双歧杆菌的丰度显著增加,同时梭状芽孢杆菌属簇XIVa物种的数量减少。
因此,摄入核桃可能会促进肠道微生物群向潜在的益生菌和产短链脂肪酸菌群的组成转变。核桃含有丰富的多酚和n-3 脂肪酸,这两种脂肪酸都具有益生元特性。
以上是各类食物对菌群多样性的影响。那么这些食物如何搭配在一起可以对菌群产生更好的影响?
这就涉及到日常饮食结构。
以现代地中海饮食金字塔为代表的高品质饮食,包括大量食用水果和蔬菜、豆类、坚果和全麦,适量橄榄油、鱼、家禽和酒,尽可能少吃红肉、糖果和加工肉类等食品。
坚持地中海饮食与降低疾病风险和改变微生物多样性有关。
地中海饮食评分与拟杆菌、普氏杆菌丰度较高呈正相关,与双歧杆菌/大肠杆菌的比例、粪便丙酸盐和丁酸盐浓度较高呈正相关。
地中海饮食中存在的典型食物成分与肠道微生物群中特定菌株的存在相关。
例如,全谷物与双歧杆菌、粪杆菌、 Tenericutes、Dorea的存在有关。橄榄油和红酒的食用量与Faecalibacterium的存在有关,蔬菜的食用量与Rikenellaceae, Dorea, Alistipes Ruminococcus存在有关,豆类与Coprococcus的存在有关。同一作者还观察到饮食中的多酚含量(通常在地中海饮食中较高)与特定梭菌(XIVa)和粪杆菌簇的存在之间存在相关性,这些梭菌簇能够合成丁酸,可能具有抗炎作用(如Akkermansia,也更多地与地中海饮食相关)。

doi: 10.3390/nu13010007
地中海饮食它没有指定份量大小或具体数量。由每个人根据自身情况设定每餐吃多少,大致比例参考以上金字塔。
目前认为的健康的饮食结构越来越偏向于,强调植物性食物和少吃红肉的饮食。
在地中海饮食结构中,我们可以看到,这类饮食结构的特点是:水果、蔬菜、高纤维食物类占比较多。
大多数研究表明,以植物为基础的饮食可以促进微生物的多样性。与杂食动物相比,纯素食者和素食者菌群具有显着更高的 α 多样性,也就是丰富度。
健康的植物性食物会影响肠道微生物的多样性和组成,包括产丁酸菌的富集,例如:
Roseburia hominis
Agathobaculum butyriciproducens
普氏粪杆菌Faecalibacterium prausnitzii
厌氧菌Anaerostipes hadrus
一些研究表明,高纤维饮食可以降低体重增加的风险。高纤维饮食随后有助于形成高度多样的微生物群。
高纤维饮食的摄入会降低饮食的能量密度,从而导致体内的短链脂肪酸促进体内更高水平的糖异生。这会导致肠促胰岛素的形成,从而使人更快、更长时间地拥有饱腹感。
地中海饮食与肠道菌群生长之间的相关性
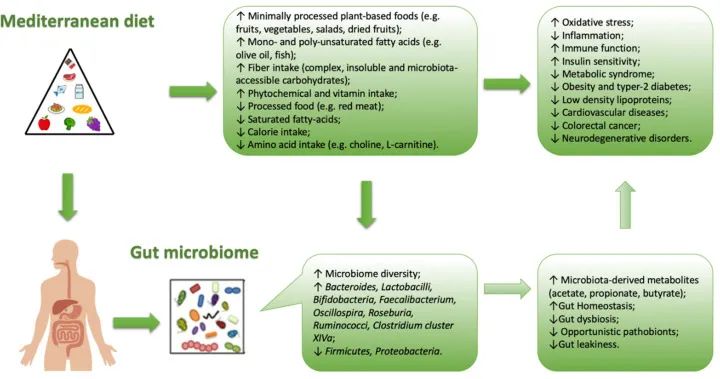
doi: 10.3390/nu13010007
此外,我们看到,地中海饮食结构中红肉类,加工肉类等是需要少吃,那么脂肪是否足够?能不能满足人体需求?
不必担心,地中海饮食中含有大量植物脂肪,这些脂肪来自橄榄油(主要是特级初榨橄榄油)和坚果,它们属于高脂肪能量密集型。
★ 地中海饮食中含有优质脂肪
榛子、 杏仁、夏威夷果、开心果、腰果等,都是单不饱和脂肪酸的重要来源。此外,核桃、碧根果和夏威夷果还富含植物性ω-3脂肪酸。核桃中的ω-3脂肪酸与ω-6脂肪酸的比例极好。
研究表明这些脂肪源与健康代谢状况直接相关,对改善大脑健康、消减炎症、降低各种原因的死亡风险和预防心脏病等有明显的作用,主要归因于它们的特定脂肪成分和生物活性分子含量。
ω-3脂肪酸,在肠道微生物群中的作用,可以调节炎症反应,改善结肠炎时的上皮屏障。
注:炎症反应是几种慢性非传染性退行性疾病的基础,如动脉粥样硬化、癌症、神经退行性疾病、慢性肾功能衰竭、糖尿病、男性肥胖、继发性性腺功能减退等。
地中海饮食中,除了坚果之外,鱼类、海鲜等食物都富含ω-3脂肪酸。显然优质的脂肪更有利于健康。
除了以上脂肪之外,地中海饮食中强调,“鱼和海鲜、鸡蛋、家禽类和高脂奶制品的摄入量每周几次;红肉每月只吃几次”,那么,蛋白质的量是否足够满足人体需求?
我们来看看地中海饮食中,蛋白质对菌群的影响。需要考虑多方面,比如蛋白质的数量、质量、加工方式和来源。
★ 高蛋白饮食可能不利于健康,地中海饮食中蛋白比例更合适
研究表明,与正常蛋白质饮食(20% 蛋白质,56% 碳水化合物)相比,喂给 Wistar 大鼠的高蛋白饮食(45% 蛋白质,30% 碳水化合物)对结肠微生物群有不利影响。
在高蛋白饮食中,链球菌、大肠杆菌/志贺菌和肠球菌分别增加了5.36倍、54.9倍和31.3倍,其丰度与与疾病发病相关的基因和代谢物(包括代谢物尸体)呈正相关,它来源于赖氨酸的脱羧作用,大量使用已被证明会引起氧化应激和DNA损伤。
有益菌在高蛋白饮食中大量减少,包括丁酸生产菌F.prausnitzii(减少3.5倍)、瘤胃球菌(减少8.04倍)和Akkermansia(在高蛋白饮食组中未检测到)。
慢性肾病下,长期低蛋白饮食更有利于改善健康。
研究表明,六个月的低蛋白饮食(0.6 g/kg/天)可以降低非透析慢性肾病患者的血清尿毒症毒素水平,包括对甲酚硫酸盐。低蛋白饮食也与肾功能改善、总胆固醇和低密度脂蛋白胆固醇降低有关。
★ 植物来源的蛋白比动物来源蛋白可能更有利于菌群和健康
在喂食大豆、猪肉、牛肉、鸡肉、鱼类和酪蛋白(后者作为对照)的大鼠中进行了为期14天的喂食试验,结果显示,到第2天,尤其是红肉(猪肉和牛肉)和白肉(鱼和鸡肉)之间的变化。主成分分析显示,在第7天和第14天有不同的微生物群,因此大豆蛋白组与肉类和对照组分开,也就说菌群特征显著变化。
在另一项类似的研究中,与喂食白肉、红肉或酪蛋白的大鼠相比,大豆蛋白与大鼠粪便短链脂肪酸增加有关。大豆组中拟杆菌和普氏杆菌的相对丰度也较高,它们是丙酸盐和其他短链脂肪酸的主要生产者。
另外研究发现,与接受牛奶分离蛋白的仓鼠相比,大豆喂养的仓鼠的血脂显著降低,至少部分原因是大豆蛋白引起的肠道微生物群变化。在高脂饮食制度下,大豆分离蛋白减少了高脂饮食诱导的小鼠体重增加和脂肪组织质量积累,并减轻了肝脏脂肪变性,而乳蛋白没有观察到这种情况。
荞麦蛋白抑制大肠杆菌的生长,促进乳酸杆菌、肠球菌和双歧杆菌的生长,后者与血脂密切相关。荞麦喂养的小鼠粪便中总胆汁酸和短链脂肪酸的排泄量显著增加。
绿豆蛋白在降低高脂饮食诱导的小鼠体重增加方面也优于酪蛋白。绿豆蛋白导致GLP-1分泌增加,盲肠和粪便胆汁酸池增加,次级和初级胆汁酸比率显著升高;在无菌小鼠中被消除的效应。就肠道微生物群而言,食用绿豆作为高脂肪饮食的一部分,导致了瘤胃菌科的扩张,并导致拟杆菌门分类群的增加和厚壁菌门丰度的减少。
乳清蛋白提取物和豌豆蛋白已被证明可以增加多样性、双歧杆菌和乳酸杆菌,其中,乳清蛋白提取物减少拟杆菌和梭菌。
以上这些研究清楚地表明,植物源性蛋白质在促进有益微生物群方面优于动物源性蛋白质,对宿主代谢具有积极影响。
蛋白质的加工(包括热加工)及其对蛋白质功能的影响(包括微生物组的调节)尚不完全清楚,需要进一步研究。
因此,在我们的饮食干预中,不能认为高蛋白饮食一定是健康的,还取决于其来源、加工方式等方面,对待“高蛋白饮食”需谨慎,尤其是在长期或反复饮食实践中。
我们可以看到,地中海饮食结构中,来源于动物蛋白只占金字塔上部(比例较小),无论是对慢病患者或者普通人群,都是一种更为安全、稳妥、健康的选择。
以上是地中海饮食结构对菌群多样性及健康的影响。那么知道吃什么,如何搭配之后,还需要注重什么?
✔ 饮食质量
同样是植物性饮食,有些高温下高油高盐等烹饪方式也会影响其营养。
为了防止破坏大量维生素、矿物质和纤维,加工烹饪的时候应尽量简化。天然食物的本味,对味蕾和身体是一种滋养。
在有条件的情况下,可以尽可能选用本地的、应季的新鲜蔬果作为食材。
高度加工的食物,富含脂肪、精制糖、盐等,与肠道微生物多样性降低有关,对宿主健康产生负面影响。
此外,各类添加剂比如,羧甲基纤维素 (CMC) 和聚山梨醇酯 80 (P80) 这两种乳化剂会降低肠道的微生物多样性并显着改变微生物群组成。
关于食品添加剂对菌群影响详见:《你的焦虑可能与食品添加剂有关,警惕食品添加剂引起的微生物群变化》
服用含有益生元的食物,乳糖不耐症的症状减轻。
低聚半乳糖是益生元,通过刺激肠道中的有益细菌对人体有益。对于乳糖不耐症患者,当低聚半乳糖诱导的微生物群变化增加了双歧杆菌、粪杆菌、乳酸杆菌、Roseburia等乳糖发酵物种的丰度时,它们会起到有益作用。
一项研究观察到,通过向参与者喂食高纯度低聚半乳糖(>95%),乳糖不耐受相关症状得到改善。在有反应的受试者中,观察到微生物组的组成发生了显著变化,但是,双歧杆菌、乳酸杆菌和粪杆菌,所有乳糖发酵菌均出现增加,50%的原本腹痛的受试者报告在使用低聚半乳糖治疗结束时没有腹痛。在将乳制品重新引入饮食后,喂低聚半乳糖的受试者在治疗后,乳糖耐受性的可能性也增加了六倍。
总的来说,以上这些研究强调了饮食对肠道菌群及其对人体的深远影响,具体取决于摄入的食物种类、饮食结构、饮食质量等。除了饮食之外,其他方式也能对菌群产生影响,具体如下:
益生菌
益生菌通过多种机制发挥作用,对肠道菌群的多样性产生积极影响,并改变人体的免疫反应,与改善多种慢性疾病密切相关。
研究发现,益生菌会抑制肠杆菌科(志贺氏菌和埃希氏菌)的生长,同时促进厚壁菌门的大量增殖,尤其是厌氧菌属。益生菌具有帮助从抗生素治疗中恢复个体菌群的显著能力。
益生菌可以直接作用于肠黏膜层和下层上皮,影响肠屏障功能和黏膜免疫;也可以通过调节常驻微生物种群结构或酶活性对宿主产生间接影响。
益生元
益生元本质上是一种肠道菌群的肥料,可以滋养有益菌群。例如,菊粉等益生元支持双歧杆菌的平衡。
益生元通过多种方式与潜在病原体、免疫系统的直接相互作用,调节肠道环境,从而有利于人体健康。
关于益生菌、益生元这部分内容由于在前面的文章中多次提到,此处就不详述了,详见:
《如何调节肠道菌群?常见天然物质、益生菌、益生元的介绍》
《益生菌的靶向递送:研究和商业化前景》
《益生菌的简单入门指南》
补充维生素D可显着增加肠道微生物多样性。具体而言,拟杆菌/厚壁菌的比例增加,益生菌类Akkermansia和双歧杆菌的丰度增加。拟杆菌属和普氏杆菌属显著变化,表明补充后肠型发生了变化。
详见:《维生素D与肠道菌群的互作》
缺铁可能导致微生物群组成的显著重组,微生物多样性降低。
详见:《人与菌对铁的竞争吸收 | 塑造并控制肠道潜在病原菌的生长》
如果从婴儿时期开始,住在农村或者养宠物,可能对儿童菌群多样性的建立比较有益。
另外,多去户外活动,亲近大自然。
一项研究对城市环境生物多样性进行人为干预试验,以检测其对儿童共生菌群和免疫调节的影响。在为期28天的生物多样性干预期间,对标准城市托儿所和自然托儿所的儿童进行对比分析,研究儿童皮肤和肠道微生物群以及血液免疫标记物的变化。
干预使环境和皮肤Gammaproteobacteria群落多样化,这反过来又与血浆TGF-β1水平和调节性T细胞比例的增加有关。
该研究发现,生物多样性干预增强了免疫调节途径,降低城市社会中免疫介导疾病的风险。
这可能为未来的预防方法提供了很大的参考价值。
写在最后
跟任何生态系统一样,我们的肠道菌群处于一种微妙的平衡。在不断变化的环境中,我们永远不知道自己会遇到什么新的威胁。
肠道菌群多样性高,能使整体菌群更有能力去抵抗外界的干扰,例如压力,抗生素药物等。
当然,对于“菌群多样性”这个指标,我们也应该理性看待,多样性出现异常高,如果是各类致病菌大量定植,核心菌群难以生存,那么也不利于健康…
肠道菌群检测报告可以帮助我们从更多角度去看待肠道菌群的整体状况,利用机器学习等最新技术,发现潜在的疾病风险,反映不同营养物质摄入比例,应该多吃什么、尽量少吃什么…
每个人对于要达到健康这个目的,可能有不同的路径,饮食需求不能一概而论,“个性化饮食建议”可以成为我们的健康指向标。我们可以利用好它,结合自身健康状态,有针对性地做出饮食或其他干预措施,从而达到更接近理想的健康状态。
主要参考文献:
Fatima Enam, Thomas J Mansell, Prebiotics: tools to manipulate the gut microbiome and metabolome, Journal of Industrial Microbiology and Biotechnology, Volume 46, Issue 9-10, 1 October 2019, Pages 1445–1459
Rinninella, Emanuele et al. “What is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases.” Microorganisms vol. 7,1 14. 10 Jan. 2019, doi:10.3390/microorganisms7010014
Wu Y, Wan J, Choe U, Pham Q, Schoene NW, He Q, Li B, Yu L, Wang TTY. Interactions Between Food and Gut Microbiota: Impact on Human Health. Annu Rev Food Sci Technol. 2019 Mar 25;10:389-408. doi: 10.1146/annurev-food-032818-121303. PMID: 30908952.
Muralidharan J, Galiè S, Hernández-Alonso P, Bulló M, Salas-Salvadó J. Plant-Based Fat, Dietary Patterns Rich in Vegetable Fat and Gut Microbiota Modulation. Front Nutr. 2019;6:157. Published 2019 Oct 11. doi:10.3389/fnut.2019.00157
Fitzgerald E, Lambert K, Stanford J, Neale EP. The effect of nut consumption (tree nuts and peanuts) on the gut microbiota of humans: a systematic review. Br J Nutr. 2021 Mar 14;125(5):508-520. doi: 10.1017/S0007114520002925. Epub 2020 Jul 27. PMID: 32713355.
Azcarate-Peril MA, Ritter AJ, Savaiano D, Monteagudo-Mera A, Anderson C, Magness ST, Klaenhammer TR. Impact of short-chain galactooligosaccharides on the gut microbiome of lactose-intolerant individuals. Proc Natl Acad Sci U S A. 2017 Jan 17;114(3):E367-E375. doi: 10.1073/pnas.1606722113. Epub 2017 Jan 3. PMID: 28049818; PMCID: PMC5255593.
Smith RP, Easson C, Lyle SM, et al. Gut microbiome diversity is associated with sleep physiology in humans. PLoS One. 2019;14(10):e0222394. Published 2019 Oct 7. doi:10.1371/journal.pone.0222394
Spolidoro GCI, Azzolino D, Cesari M, Agostoni C. Diet Diversity Through the Life-Course as an Opportunity Toward Food Allergy Prevention. Front Allergy. 2021;2:711945. Published 2021 Sep 24. doi:10.3389/falgy.2021.711945
Ecklu-Mensah G, Gilbert J, Devkota S. Dietary Selection Pressures and Their Impact on the Gut Microbiome. Cell Mol Gastroenterol Hepatol. 2022;13(1):7-18. doi:10.1016/j.jcmgh.2021.07.009
Merra G, Noce A, Marrone G, et al. Influence of Mediterranean Diet on Human Gut Microbiota. Nutrients. 2020;13(1):7. Published 2020 Dec 22. doi:10.3390/nu13010007
Elena Mengheri, Diet Quality Is Associated with Microbial Diversity and Host Health, The Journal of Nutrition, Volume 149, Issue 9, September 2019, Pages 1489–1490
Wilson, Annette S et al. “Diet and the Human Gut Microbiome: An International Review.” Digestive diseases and sciences vol. 65,3 (2020): 723-740. doi:10.1007/s10620-020-06112-w
Huang C, Feng S, Huo F, Liu H. Effects of Four Antibiotics on the Diversity of the Intestinal Microbiota. Microbiol Spectr. 2022 Mar 21:e0190421. doi: 10.1128/spectrum.01904-21. Epub ahead of print. PMID: 35311555.
Shreiner AB, Kao JY, Young VB. The gut microbiome in health and in disease. Curr Opin Gastroenterol. 2015;31(1):69-75. doi:10.1097/MOG.0000000000000139
Sakkas H, Bozidis P, Touzios C, Kolios D, Athanasiou G, Athanasopoulou E, Gerou I, Gartzonika C. Nutritional Status and the Influence of the Vegan Diet on the Gut Microbiota and Human Health. Medicina (Kaunas). 2020 Feb 22;56(2):88. doi: 10.3390/medicina56020088. PMID: 32098430; PMCID: PMC7073751.

谷禾健康
幽门螺杆菌(helicobacterpylori,H.pylori)是一种独特的,能持续定植于人类胃粘膜并能引起胃感染的细菌。
幽门螺杆菌是革兰氏阴性,螺旋形,微需氧细菌,定居于人类胃粘膜中。世界上有超过一半的人感染了幽门螺杆菌,但很多没有临床症状。幽门螺杆菌及其患病率在某些人群中高达80%。
据推测,幽门螺杆菌可能是人类土著微生物组的一部分,它与人类宿主之间有着很复杂的关系。
本文主要讨论幽门螺杆菌是如何与人类共同进化的,可能的机制可以解释基于人群的研究中幽门螺杆菌感染与几种疾病的发展之间的正相关性和负相关性,以及炎症和/或微生物组的变化是如何联系的各自的结果。
大多数幽门螺杆菌感染者并没有明显症状,但当症状出现时,一般是以下几种:
恶心,胃痛或胃灼热,空腹时腹部疼痛加剧。
怎样的情况有可能感染此菌?
幽门螺杆菌可能通过接触感染者的唾液、呕吐物或粪便传播,食用受污染的食物或水也会感染。
大多数感染幽门螺杆菌的人都是在儿童时期感染的。
有些因素可能会增加感染幽门螺杆菌的风险:
比如在拥挤的空间生活,没有干净的水,和患有幽门螺杆菌的人一起生活等。
什么情况下需要看医生?
当感觉有不寻常的胃痛或不适时,尤其是持续或反复发作的胃痛;
吞咽困难;
血腥,黑色或柏油样的粪便;
呕吐物带血或看起来像咖啡渣;
…
以上情况,最好去看医生。
( 如自行随意服用抗生素,效果不好的同时很可能带来抗生素耐药 )
如何诊断幽门螺杆菌?
目前最常见也实惠的方法是呼气检测,从分析到出结果只需数分钟,简单准确。
胃镜检查:做胃镜检查也包括幽门螺杆菌检测,因为可以取到胃里活性组织做病理切片检查。
抗体检查:通过抽血检查,以确定是否感染过幽门螺杆菌以及目前处于感染状态。
幽门螺杆菌会导致并发症?
包括溃疡,胃炎和胃癌。
溃疡约有10%的幽门螺杆菌患者会发展为胃溃疡(疼痛)。当幽门螺杆菌破坏保护胃和小肠内壁的粘膜时,就会发生这种情况,胃酸会渗透到衬里并造成伤害。
幽门螺杆菌可导致90%以上的肠道溃疡和80%的胃溃疡。
溃疡本身也会导致严重的并发症:
• 内部出血胃酸或溃疡渗入血管可能导致出血。
•阻塞溃疡会阻止食物离开您的胃。
•穿孔溃疡可能会深入并穿透胃或肠壁。
•腹膜炎 当溃疡引起感染或发炎时,可能发生腹膜炎症(腹膜)。
胃炎胃炎是胃黏膜炎症。幽门螺杆菌是处于患有这种病症的风险增加。
如果不治疗胃炎,可能会导致严重失血,并可能增加患胃癌的风险。胃癌是世界上与癌症相关的死亡的第二大最常见原因。
此外,幽门螺杆菌感染还与其他疾病有关,例如缺血性心脏病,2型糖尿病,贫血,肥胖受试者的不良代谢特征和胰岛素抵抗等。
幽门螺杆菌(Hp)感染会在胃微环境中产生剧烈变化,进而影响胃微生物群组成,并可能与肠道微生物群变化有关。可能会触发肠道共生稳态的重大改变,从而出现新的胃肠道平衡。
同时,用于根除幽门螺杆菌的治疗策略可以调节这种生理共生,但也可能相反,受到其特性的影响。感染,饮食,抗生素和/或生活方式会干扰这种共生关系。
幽门螺杆菌感染对胃微生物群的干扰
下一代测序对胃液和活检标本中的人体胃微生物群进行了研究,结果表明胃中存在多种细菌类群,主要由五个门组成,包括放线菌门、拟杆菌门、厚壁菌门、梭菌门和变形菌门。幽门螺杆菌感染极大地改变了胃微生物群的特征。
一项初步研究报道,幽门螺杆菌定植改变了胃微生物群,降低了微生物多样性,根除幽门螺杆菌可以恢复微生物多样性。
幽门螺杆菌可以利用几种机制来调节胃微环境。通过干扰质子泵的表达,幽门螺杆菌可以调节胃腔的酸度,从而使微环境与通常不能在胃中生长的微生物相容。
评估幽门螺杆菌感染对胃微生物群影响的研究
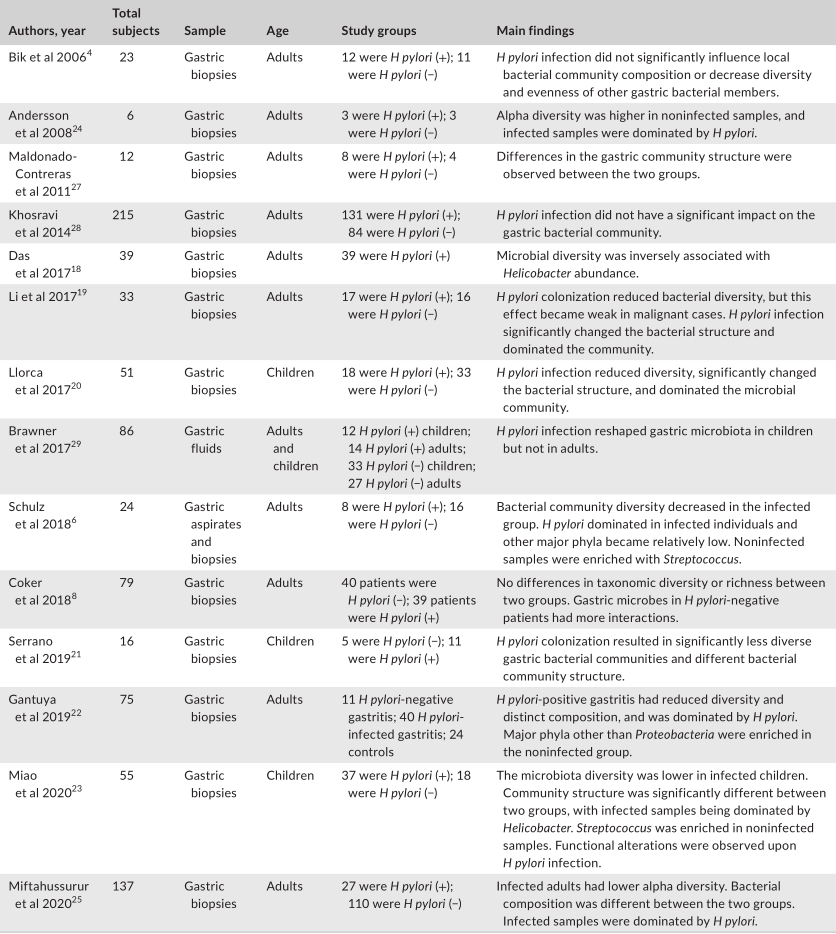
Tao et al., Helicobacter, 2020
此外,由于幽门螺杆菌的存在而改变的免疫反应可能会影响其他微生物群。幽门螺杆菌还可能通过动员抗菌肽或通过营养竞争来改变其他微生物群的生长。一般来说,研究人员表明,幽门螺杆菌的定植与α多样性的显著降低有关。微生物多样性和幽门螺杆菌丰度之间存在反比关系。
幽门螺杆菌感染对肠道微生物群的影响
与幽门螺杆菌阴性对照组相比,胃内有大量幽门螺杆菌的个体具有不同的肠道菌群,这表明胃肠段之间存在相互作用。
大多数研究表明,幽门螺杆菌感染与肠道微生物多样性之间的呈正相关。由于较高的微生物多样性通常与总体较好的健康状况有关,因此,探讨幽门螺杆菌感染后肠道微生物多样性的增加是否对宿主有一些有益的影响将是一个有趣的问题。
幽门螺杆菌感染对肠道微生物群影响的研究分析
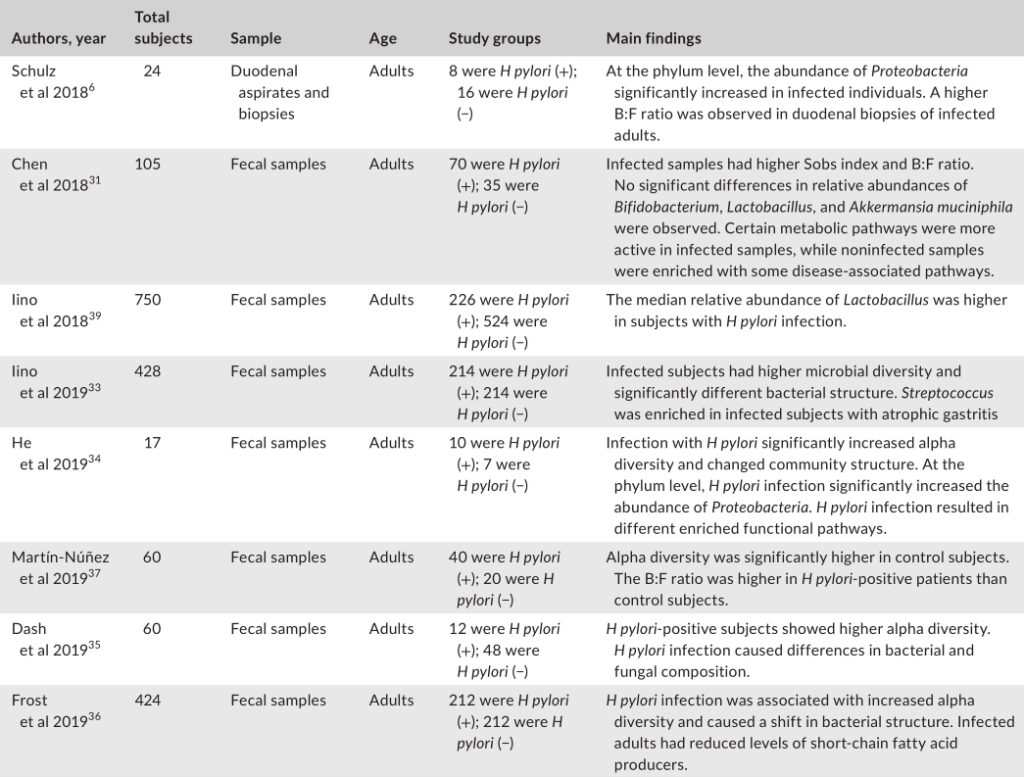
Tao et al., Helicobacter, 2020
幽门螺杆菌感染组和阴性对照组的微生物群落结构存在显著差异。
在门水平上的成分分析发现,受感染个体中变形菌的丰度显著增加,这可能是由于幽门螺杆菌从胃转移到肠腔所致。值得注意的是,拟杆菌与厚壁菌(B:F)的比率在幽门螺杆菌阳性组中更高。改变的B:F比值与临床相关,因为发现厚壁菌和拟杆菌与宿主的脂质代谢和能量平衡有关。
受感染与没感染组相比,双歧杆菌、乳酸杆菌和嗜粘液阿克曼菌的丰度没有显著差异。其他研究人员发现,幽门螺杆菌感染者体内乳酸杆菌的丰度更高。
一项研究发现,在幽门螺杆菌感染的患者中,萎缩性胃炎患者的肠道微生物群中链球菌更为丰富。
另一项研究发现,幽门螺杆菌感染的个体表现出短链脂肪酸(SCFA)产生者(如丁酸盐)水平的下降,SCFA对宿主产生有益的代谢作用。
此外,鉴于SCFAs受体在免疫细胞中的普遍表达,这些代谢物被认为在调节肠道内稳态中发挥重要作用。因此,幽门螺杆菌感染时抑制SCFAs的输出可能对人类健康有害。
有功能分析指出,在幽门螺杆菌阴性人群中,疾病相关途径更活跃。相比之下,许多代谢途径在感染的患者中的比例过高。因此,微生物群中的功能变化可能是特定于部位的,因为在肠道中观察到的变化与在胃中观察到的变化呈现不同的趋势。
根除疗法对胃微生物群的影响
Tao et al., Helicobacter, 2020
充分的证据表明,成功清除幽门螺杆菌后,胃微生物多样性显著增加,但显示治疗失败后没有改善。研究人员认为恢复可能需要一定的时间,因为他们认为多样性从第0周到第6周和第26周逐渐增加。
此外,研究表明,成功根除幽门螺杆菌后,α多样性可以完全恢复到未感染对照组的水平。相反,尽管在清除幽门螺杆菌后,群落结构也可以部分恢复,但对于是否仍存在显著差异存在争议在根除后组和阴性对照组之间。
一些研究人员发现,接受抗幽门螺杆菌治疗的患者在治疗2个月后,胃微生物群落的组成恢复到未感染儿童的水平,然而,其他研究确定,成功治疗6个月后的成人样本仍显示出不同的菌群结构,阴性对照组则不同。
纳入分析的不同年龄组可能解释了相反的结论,因为另一项招募儿童的研究也发现根除组的菌群结构在治疗4周后与幽门螺杆菌阴性组的菌群结构接近。
根除治疗后评估胃微生物群变化的研究

Tao et al., Helicobacter, 2020
成分分析揭示了幽门螺杆菌的相对丰度治疗后幽门螺杆菌显著下降,而其他主要菌门,包括放线杆菌、拟杆菌、厚壁菌和梭杆菌,则增加。
在属水平上,治疗后乳酸杆菌和双歧杆菌(两种公认的益生菌)的数量显著增加。
两项研究的功能分析表明,成功治疗后,蛋白质和碳水化合物代谢途径上调。因此幽门螺杆菌感染期间,胃微生物群营养代谢功能受损,根除治疗可部分恢复。然而,预测的功能改变还需要进一步的验证。
根除治疗后肠道微生物群的变化
通过降低酸度,质子泵抑制剂(PPIs)已被证明对肠道微生物群产生显著影响。因此,抗幽门螺杆菌治疗对肠道微生物群的影响是PPIs、抗生素、铋联合应用的共同结果。总的趋势是,α多样性在根除后立即下降,随后又恢复。
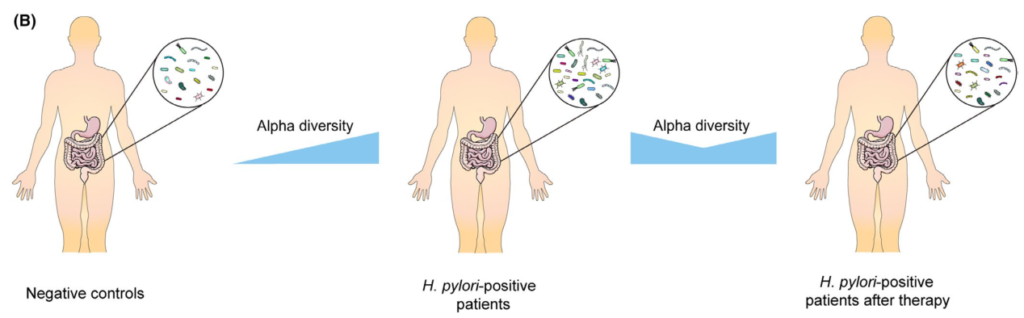

Tao et al., Helicobacter, 2020
具体来说,恢复需要多长时间仍有待阐明。大多数研究收集了随访2个月内的信息显示α多样性降低,而此后报告与基线无显著差异。
然而,一项研究报告,从第0周到第6周和第26周,Sobs指数和Chao指数都在下降没有达到统计学意义。值得注意的是,另一项研究发现,不同的治疗方案对肠道微生物群的影响可能不同。
在临床试验中,三种不同的治疗方案,包括三联疗法、联合疗法和铋四联疗法。结果表明,所有组的α多样性在第2周时都有所下降,但只有接受三联疗法的患者在第8周时才具有基本α多样性,而其他两组的α多样性在治疗后1年也无法恢复。
希望将来的研究验证根除疗法是否会导致肠道菌群的持续紊乱,以及不同的治疗方案如何影响可逆性。
分析根除疗法对肠道微生物群影响的研究
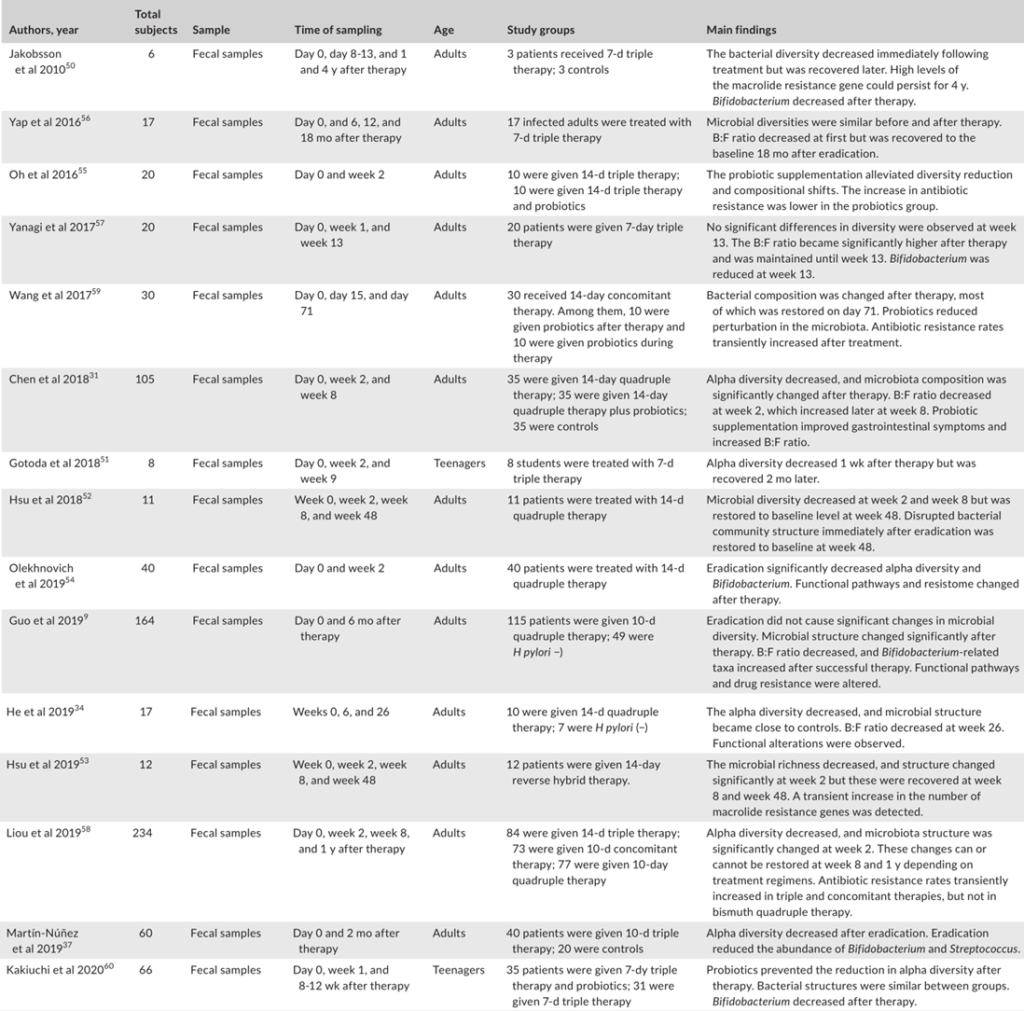

Tao et al., Helicobacter, 2020
益生菌的补充可以缓解抗生素引起的多样性和结构紊乱
益生菌,已被证明可稳定微生物群并改善胃肠道症状。在门水平上,治疗6个月后观察到厚壁菌群增加和拟杆菌减少,表明在幽门螺杆菌感染的个体中观察到的高B:F比率逆转。在另一项研究中报告了9个类似的结果。
临床证据支持肥胖与B:F比率降低相关。因此,这些研究中检测到的B:F比率降低可能为根除治疗后体重增加提供了一种可能的解释。然而,一些研究人员说明,在根除18个月后,降低的B:F比率可以逆转。有趣的是,益生菌补充被证明与根除治疗后较高的B:F比率相关。值得注意的是,报告的结果不一致。
一项研究表明,治疗后B:F比率立即增加,并在第13周保持高于基线水平。宏基因组研究的分类图谱显示,治疗后双歧杆菌的相对丰度下降。然而,另一项研究表明,双歧杆菌在成功根除6个月后增加。
在幽门螺杆菌阳性萎缩性胃炎的肠道微生物群中富集的链球菌在根除治疗后显著减少。
尽管不同研究中肠道微生物群的长期变化不同,但根除引起肠道内稳态的短期紊乱似乎是一致的。
根除疗法对肠道菌群抗生素耐药性的影响
随着根除疗法的广泛应用,抗生素耐药性的出现已成为临床实践中的一个重要问题。
根除幽门螺杆菌感染后,抗生素耐药性的上升以及根除治疗后的长期安全性是重要的问题。
通过药敏试验指导的个性化治疗可以在一线治疗中提供可靠的优异根除率,但贵且无法广泛获得。根据局部地区的抗生素耐药性,针对特定人群的经验疗法可能是一种替代策略。
已证明会改变幽门螺杆菌相关疾病风险的宿主因素包括宿主的年龄、在幽门螺杆菌的获得和幽门螺杆菌感染之间的持续时间,但更重要的是宿主免疫介质(多态性)和酸分泌状态。
除宿主因素外,吸烟和饮食等环境因素还会影响幽门螺杆菌定植的生态位,因此可能会影响疾病风险。

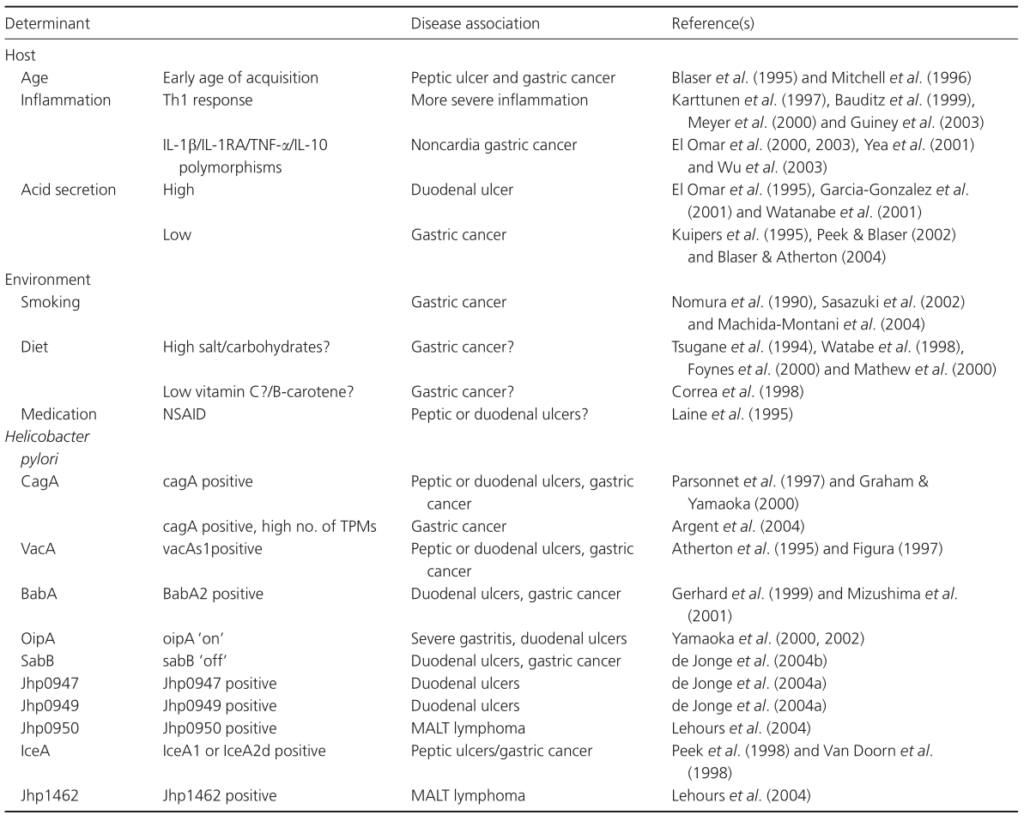
Karin et al., FEMS Microbiol Rev. 2006
在年幼时期感染幽门螺杆菌会增加患胃溃疡和胃癌的风险。 胃溃疡和胃癌的发展需要幽门螺杆菌的长期感染,因此,推测这些疾病是在慢性感染幽门螺杆菌的患者中更为常见。
此外,这种关联被认为是由不同年龄组的宿主免疫反应的差异所决定的。
感染幽门螺杆菌后,宿主胃上皮细胞释放细胞因子IL-8。这种细胞因子参与了巨噬细胞、中性粒细胞、肥大细胞、B细胞和T细胞募集到炎症部位。这些细胞通过分泌其他炎症介质,如干扰素(IFN-g)、肿瘤坏死因子(TNF-a)和白细胞介素(IL-1b),进一步增强免疫反应。
幽门螺杆菌相关发病机制的示意图

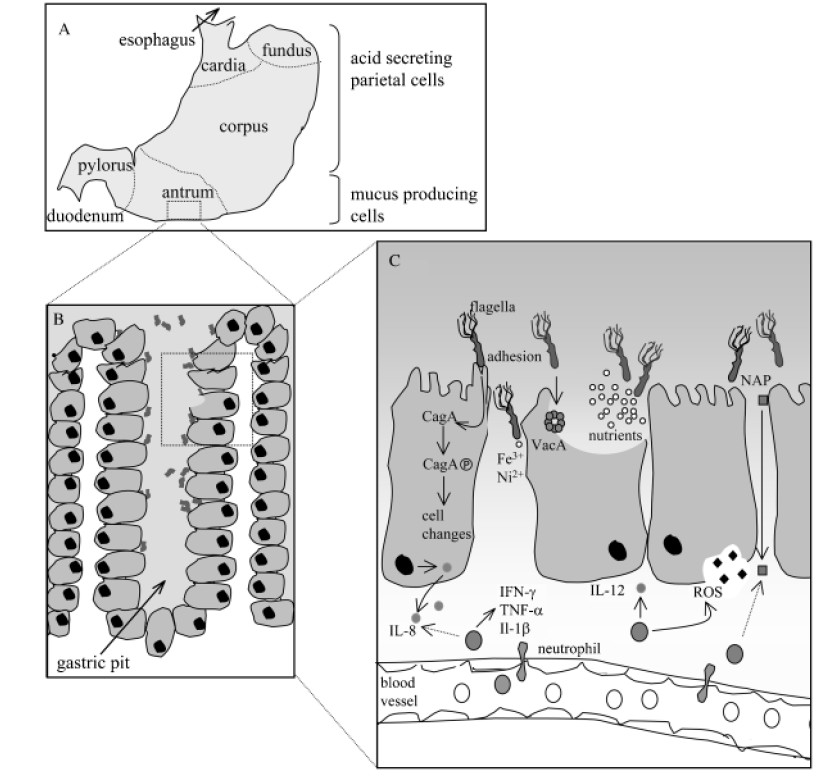
Karin et al., FEMS Microbiol Rev. 2006
A)在人类胃中,幽门螺杆菌主要定植于缺乏胃酸分泌壁细胞的胃窦中;
B)在胃小凹中,幽门螺杆菌尤其在上皮细胞紧密连接处附近增殖;
C)幽门螺杆菌对定植、持续和疾病发展具有重要作用的因子。
针对幽门螺杆菌的免疫反应进一步表现为活化的人类中性粒细胞中的氧化爆发。这种爆发会导致活性氧(ROS)的释放,活性氧是一种低分子量的代谢物,可破坏包括核酸在内的重要生物分。ROS可能损伤控制细胞生长的基因;并刺激癌症的发展。因此,免疫介质的存在会导致胃上皮的完整性受损。这种损伤是引起大多数与幽门螺杆菌感染相关的病理学,而不是直接的幽门螺杆菌活动。
幽门螺杆菌感染期间存在的免疫细胞主要是促炎细胞,而不是抗炎细胞。促炎介质由辅助性T细胞1(Th1)分泌。原始T细胞受多种因素驱动分化为Th1细胞,如TNF-a和IL-12。
在幽门螺杆菌刺激下,单核细胞大量产生细胞因子IL-12。因此,通过刺激IL-12的释放,幽门螺杆菌增强了促炎反应。这导致更多的损伤胃上皮,这容易发展胃萎缩。
一些编码免疫调节因子的宿主基因含有多态区域。这些基因的多态性将改变它们的转录,从而影响炎症过程。因此,幽门螺杆菌相关疾病的风险可能会改变。到目前为止,IL-1b、IL-1受体拮抗剂、IL-10和TNF-a基因的多态性已被证明与远端胃癌显著相关。
人类白细胞抗原(HLA)通过将病原体衍生的肽片段呈递给适当的T细胞,有助于消除病原体。HLA基因具有高度多态性,某些HLA等位基因与胃腺癌的发生有关。幽门螺杆菌感染也与胃腺癌的发生有关,但其存在与这些HLA等位基因的存在无关。因此,HLA等位基因在幽门螺杆菌介导的腺癌中的重要性仍然存在争议。
因此,遗传上预定的显性促炎性Th1反应与幽门螺旋杆菌疾病相关。
另一方面,Th1反应的强度可能受到抗炎Th2细胞因子IL10的产生的限制,从而延长感染时间。Th2型反应是由寄生虫引起的。因此,感染幽门螺杆菌的人的炎症反应和病理改变逐渐减弱。这可能导致这些个体慢性胃炎进展为胃癌的时间延迟。另外,不同的促炎反应可能直接导致幽门螺杆菌相关病理学的不同途径。
前面提到Th1反应导致胃壁细胞抑制其酸分泌。酸性较低的环境可能促进幽门螺杆菌的传播和持续。此外,当胃酸水平较低时,幽门螺杆菌以外的微生物可能在人体胃中生长,从而增强局部炎症反应。促炎细胞因子和胃酸分泌减少的联合作用增加了患消化性溃疡病和胃癌的风险。
另一方面,当酸的分泌减少时,十二指肠溃疡就不太可能发生。当酸的分泌增强时,十二指肠溃疡发生的风险增加。然而,同样在低酸性条件下,幽门螺杆菌感染后十二指肠溃疡的风险可能会增加。
胃壁细胞控制其产酸的能力在幽门螺杆菌相关疾病的发展中是重要的。幽门螺杆菌感染主要发生在人类胃中酸性较弱的部位,如胃窦。因此,胃酸分泌量高的宿主胃窦内幽门螺杆菌密度高,且主要发展为胃窦性胃炎。随后,胃体中完整的壁细胞被刺激分泌酸,从而诱导十二指肠的胃化生。幽门螺杆菌定植于这种胃化生,这可能导致十二指肠溃疡。
另一方面,胃酸分泌量低的宿主不仅胃窦易受幽门螺杆菌感染,而且胃体也易受幽门螺杆菌感染。感染的幽门螺杆菌可进一步抑制胃壁细胞的酸分泌。这可刺激胃上皮细胞持续增殖,导致胃腺进行性丧失,最终导致胃萎缩和癌症。
吸烟
一些环境因素与幽门螺杆菌相关的发病机制有关。吸烟是胃癌发展的主要因素。烟草烟雾含有致癌的亚硝胺,引发癌症的发展。
高盐饮食
一些饮食因素与幽门螺杆菌疾病有关。高盐饮食与幽门菌定植率、胃炎和胃癌风险增加。高盐浓度导致壁细胞萎缩,粘膜屏障破坏。然而,只有长期高盐摄入才会增加幽门螺杆菌相关疾病的风险。
高米饭摄入量
高米饭摄入量与胃癌之间存在正相关关系。米饭含有碳水化合物,可能会刺激胃粘膜。
水果和蔬菜摄入量低
水果和蔬菜摄入量低可能会增加胃癌的风险。水果和蔬菜含有抗氧化剂维生素C和β-胡萝卜素,通过中和活性氧来防止致癌。
因此,尽管有几个环境因素被认为与幽门螺杆菌相关疾病的发生有关,但它们的作用往往是有争议的。据推测,不同环境因素以及宿主和细菌因素之间的复杂相互作用妨碍了对研究结果的解释。
幽门螺杆菌因素在疾病发展中的总体影响是复杂的。到目前为止,已经描述了几种与幽门螺杆菌相关疾病的发生有关的幽门螺杆菌特异性蛋白。这些蛋白质被认为是导致幽门螺杆菌持续存在的原因。作为副作用,这些幽门螺杆菌蛋白诱导和改变炎症过程,损伤胃上皮,从而决定幽门螺杆菌感染后的结局。
CagA
与感染cagA阴性幽门螺杆菌分离株的患者相比,携带细胞毒素相关基因A(cagA)的幽门螺杆菌分离株的定植与严重胃炎、消化性溃疡病和远端胃腺癌的风险增加有关。
幽门螺杆菌cagA基因的存在或不存在经常被用作幽门螺杆菌中致病岛(PAI)存在或不存在的标记。 cag PAI由大约30个基因组成,并且在所有幽门螺杆菌分离株中占50%–70%。 位于cag PAI侧翼的两个31 bp重复序列之间的重组可能导致整个cag区域的缺失或获得。
cag PAI上的18个基因对于产生IV型分泌系统至关重要。 IV型分泌系统将细菌细胞中的蛋白(与毒力相关)转运到宿主细胞的胞质溶胶中。 这通常导致上皮细胞反应的级联反应的启动,例如细胞骨架的变化和细胞因子IL-8的分泌。
在幽门螺杆菌中,IV型分泌系统的成分将CagA蛋白转运到胃上皮细胞中。 随后,CagA蛋白被磷酸化。 然后,这种磷酸化的CagA与宿主磷酸酶SHP-2相互作用,引起细胞骨架的重排。 这导致宿主细胞形态发生变化,也称为“蜂鸟”表型。该表型的特征在于细胞扩散,上皮细胞的延长生长以及片状脂蛋白和丝状伪足的存在。
作为抵消事件,磷酸化的CagA与宿主Src激酶相互作用,从而减弱宿主SHP-2磷酸酶的信号传导。 这个过程减少了细胞骨架的重排,并阻止了CagA的进一步磷酸化。
最近,已经证明幽门螺杆菌菌株在体内诱导较高水平的CagA磷酸化。上皮细胞诱导更多的细胞骨架变化,并且更可能与胃癌有关。CagA磷酸化水平的差异是由cagA基因的30个区域内酪氨酸磷酸化基序(TPM)数量的差异引起的。
幽门螺杆菌人群,需要最低限度的免疫识别。因此,在萎缩的生态位中,这些TPM数量减少的亚克隆诱导了较弱的宿主免疫反应。
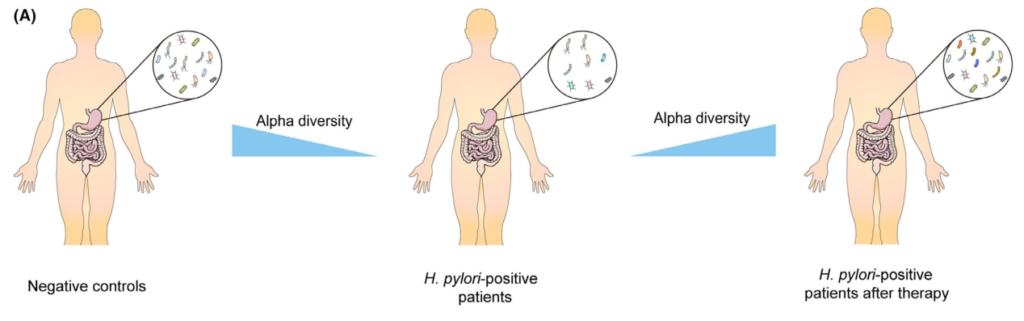
另一方面,较高水平的CagA磷酸化可引起强烈的炎症反应。 这种炎症会导致萎缩,并可能在限制高酸输出的生态位酸应激中发挥作用。
通过观察发现,缺乏cagA基因的幽门螺杆菌菌株是从消化性溃疡或胃癌患者中分离出来的,尽管其频率比cagA阳性幽门螺杆菌菌株的频率更低。
cagA和幽门螺杆菌相关疾病之间的关联在不同的地理区域中有所不同。 这种变异可能与CagA表达的差异有关。
然而,CagA并不是唯一的。幽门螺杆菌蛋白负责屏障功能障碍,而这一过程并没有随着CagA的清除而丢失。下一节讨论的VacA蛋白也很重要。
VacA
最初显示真核细胞在带有幽门螺杆菌的体外测试系统中经历空泡化和连续变性。 后来证明这种现象是由幽门螺杆菌毒素诱导的,现在被称为空泡细胞毒素A(VacA)。
VacA或CagA不能单独用作临床结果的决定因素。
VacA如何发挥作用,导致细胞死亡?
幽门螺杆菌VacA是一种高度免疫原性的95-kDa蛋白。VacA结合上皮细胞的顶端部分,形成阴离子选择性孔。
通过这些孔,碳酸氢盐,氯化物和尿素从细胞质中释放出来,然后,VacA被内吞进入晚期的内体区室,并改变这些区室的通透性。
在弱碱(例如氨)的存在下,这会导致水涌入,从而导致囊泡肿胀和液泡形成。 细胞内内吞途径的损伤最终导致细胞死亡。
这导致了上皮细胞抗性的降低,因此低分子量分子如Fe3+和Ni2+可以很容易地穿过上皮细胞层。Fe3+和Ni2+分子分别是幽门螺杆菌生长和脲酶活性的关键因子。
因此,推测通过降解上皮细胞屏障,幽门螺杆菌可以更容易地获得这些关键因子。
幽门螺杆菌VacA也与壁细胞的降解有关,这导致酸分泌减少,使宿主容易患上胃癌。 持续感染过程中VacA表达水平的差异导致毒性改变。 因此,溃疡的消长可以通过随时间变化的VacA表达变化来解释。
在体外在胃上皮细胞附近诱导vacA基因的转录。 这一发现证实了VacA与宿主细胞相互作用的重要性。 此外,VacA可以逃避适应性免疫反应,从而增强幽门螺杆菌在胃粘液层中的持久性。
幽门螺杆菌外膜蛋白(HOPs)
在革兰氏阴性细菌中,外膜介导与其周围环境的相互作用。 在感染期间,假定存在于幽门螺杆菌外膜上的蛋白质被改变,使得宿主免疫系统的识别作用降至最低。
幽门螺杆菌分离物含有约30种不同的外膜蛋白(HOP)。 其中几种是粘附素。 在革兰氏阴性细菌中,粘附素最常形成聚合菌毛结构。 但是,在幽门螺杆菌中,这些粘附素已经适应了胃环境,其中酸性条件可能会使这种聚合物菌毛结构解聚。
岩藻糖基化的糖蛋白和唾液酸化的糖脂都已被证明是胃上皮中幽门螺杆菌的结合位点。
与宿主细胞的粘附可保护幽门螺杆菌免受蠕动和粘膜脱落。
此外,推测粘附力可以使幽门螺杆菌更好地获取从胃上皮释放的营养物质,并更有效地将细菌毒素传递至宿主细胞。
另一方面,在剧烈炎症的部位,粘附特性的丧失可能使幽门螺杆菌逃脱宿主免疫细胞的杀伤。目前为止,一些HOP与疾病的发展有关。
幽门螺杆菌BabA (HopS)介导幽门螺杆菌粘附于人Leb血型抗原,这些抗原存在于胃上皮细胞上。BabA黏附促进幽门螺杆菌定植,增加上皮细胞分泌IL-8,导致粘膜炎症增强。
BabA调节的两种机制都有助于促进慢性感染的动态反应。
在慢性炎症期间,选择增加或减少粘附的周期可在所有血型的人群中获得所有类型的Leb结合。
宿主粘膜糖基化模式导致BabA进化,使幽门螺杆菌菌株适应其个体宿主,这有助于避免宿主反应,并在全世界范围内造成幽门螺杆菌感染的异常长期性。
幽门螺杆菌可塑性区域毒力因子
两个单独菌株基因组序列的比较表明,一个菌株中存在的幽门螺杆菌基因中约有6%-7%不存在于另一个菌株中,反之亦然。
大约一半的菌株特异性基因存在于高变区;可塑性区。 位于这种可塑性区域的基因通常与毒力增加相关。
在幽门螺杆菌中,可塑性区确实编码了与IV型分泌有关的Vir型ATP酶。 在幽门螺杆菌中,该分泌系统在例如分泌CagA细胞毒素中很重要。目前为止,一些位于可塑性区的幽门螺杆菌基因与幽门螺杆菌相关的疾病有关。
幽门螺杆菌Jhp0947和Jhp0949与十二指肠溃疡疾病相关。这两个基因均位于幽门螺杆菌菌株J99可塑性区,在幽门螺杆菌介导的IL-12释放中很重要。
在体外测试系统中从单核细胞中提取。 细胞因子IL-12对于使免疫反应偏向促炎性Th1应答是必不可少的,并且与十二指肠溃疡的形成密切相关。
由jhp0947和jhp0949编码的蛋白质的功能未知。 然而,已经假设这些蛋白通过与单核细胞的相互作用诱导促炎细胞因子IL-12的产生。
IL-12水平升高会导致幼稚T细胞分化为活化的Th1细胞,从而导致炎症反应增强和组织损伤增加。
幽门螺杆菌Jhp0950,编码一种未知功能的蛋白质,也是J99可塑性区的一部分。该基因的存在与cagA、cagE、vacA s1m1、babA2、hopQ T1、oipA有关。
在幽门螺杆菌J99基因组中,Jhp0950与Jhp0949相邻。与Jhp0949不同,Jhp0950与十二指肠溃疡风险增加无关。然而,它与疾病的关联支持了位于可塑性区域的幽门螺杆菌基因可能与毒性相关的观点。
其他幽门螺杆菌毒力因子
在美国和荷兰,幽门螺杆菌iceA1基因(iceA基因的两个等位基因变体之一)在幽门螺杆菌感染的消化性溃疡患者中比仅在幽门螺杆菌感染的胃炎患者中更为普遍。
与许多其他与毒力相关的基因相似,IceA表达与幽门螺杆菌相关疾病之间的联系是群体依赖性的。
在东南亚,未发现iceA1与幽门螺杆菌相关疾病之间的关联。 有趣的是,对iceA1呈阳性的南非幽门螺杆菌分离株在幽门螺杆菌感染的胃癌患者中比仅在幽门螺杆菌感染的胃炎患者中更普遍。
注:幽门螺杆菌iceA1最初是在与胃上皮细胞接触后转录上调后被鉴定的,iceA1基因编码的核酸内切酶与乳球菌中的限制性核酸内切酶NlaIIIR非常相似,伴随的高度保守的脱氧核糖核酸腺嘌呤甲基转移酶的活性,由幽门螺杆菌编码,似乎与控制幽门螺杆菌的基因表达有关。
幽门螺杆菌Jhp1462与MAL T淋巴瘤、十二指肠溃疡和胃腺癌的风险增加相关。由Jhp1462编码的蛋白质的功能是未知的,其在严重胃十二指肠疾病发展中的重要性仍有待阐明。
以上,我们看到幽门螺杆菌通过各种方式给人体带来种种不利影响,它似乎不应该在人体中长期生存,然而,幽门螺杆菌在人体内存在有着的悠久的历史,那么幽门螺杆菌是如何保护自己在人体中安稳生存下来?
幽门螺杆菌在疾病发生之前会长期感染人胃粘膜。 因此,导致幽门螺杆菌定植和持续存在的因素与幽门螺杆菌相关的发病机理具有内在联系。 在胃腔进入后,幽门螺杆菌必须应付胃酸。
幽门螺杆菌通过其耐酸性在这种酸性条件下得以生存:然后穿过粘液层到达其接近胃上皮细胞的位置。趋化性,运动性和粘附性是胃上皮细胞定殖的重要过程。
为了在人胃粘膜中长期持久存在,幽门螺杆菌还不断需要宿主提供营养。 这是通过降解粘液层和下面的胃上皮细胞的完整性来实现的。 此外,通过抑制宿主免疫系统,抗原变异和抗原拟态避免了幽门螺杆菌的清除。
耐酸性
幽门螺杆菌生长的下限pH值是5.0-5.5,具体取决于测试的分离物。在胃粘膜层中,pH也是酸性的,大约在4-6.5之间变化。而且,当粘液层被破坏时,pH值可能会偶尔下降。
酸冲击(pH<3)后幽门螺杆菌的存活取决于幽门螺杆菌蛋白脲酶的活性,该酶将尿素转化为氨和碳酸氢盐。这导致幽门螺杆菌的周质或细胞质被中和。幽门螺杆菌尿素酶活性对于在酸性pH下体外存活以及在动物模型中胃粘膜定植至关重要,这证明了尿素酶在幽门螺杆菌感染中的重要性。
在中性pH下高度活跃的脲酶和酸调节的尿素通道的结合解释了为什么幽门螺旋菌在人类胃部的生存能力是独特的。有效抑制UreI蛋白,将提供一种从正常的,分泌酸的胃中根除幽门螺杆菌的方法。
除耐酸性外,脲酶还具有与胃定植有关的其他基本功能
尽管尿素酶对于酸休克的生存是必不可少的,但是幽门螺杆菌尿素酶在处理胃上皮细胞附近的慢性酸性环境时还是不够的。 用酸抑制剂治疗不能恢复尿素酶阴性突变体定殖在生侏儒仔猪胃中的能力。
脲酶参与幽门螺杆菌的氮代谢,脲酶活性产生的氨和碳酸氢盐被认为会影响宿主的许多细胞过程,包括细胞裂解。
除脲酶外,非脲酶基因也有助于耐酸,并且在酸性pH下的存活和生长与脲酶无关。 在这些非脲酶基因中,已显示三种调节蛋白(Fur,NikR和HP0166)对幽门螺杆菌的适应性有贡献。 这表明严格调节耐酸机理的重要性。
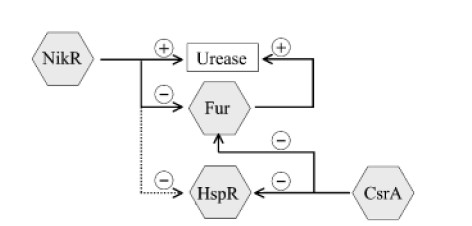
Karin et al., FEMS Microbiol Rev. 2006
NikR通过Fur调节系统直接和间接调节脲酶的表达。CsrA调节Fur和HspR调节系统的表达。通过不同基因调节蛋白的相互作用,幽门螺杆菌能够感知和响应多种信号。
细胞的趋化作用与运动
胃上皮细胞层覆盖着一层厚厚的粘液。假定该层甚至对于小分子也相对不可渗透。幽门螺杆菌的能动性和趋化性是有效穿透这一粘液层的关键。
在小鼠感染模型中,几乎任何运动和趋化性系统基因的诱变都消除了幽门螺杆菌感染胃粘膜的能力,强调了这些因素对定殖的重要性。
运动性
幽门螺杆菌的运动性是通过存在两到六个极性的带鞘鞭毛而实现的。鞭毛由三个结构元素组成:基体,钩子和细丝。迄今为止,超过50种蛋白质与鞭毛的结构或调控组织有关。
鞭毛丝由两个亚基FlaA和FlaB组成。幽门螺杆菌flaA和flaB突变体无法在感染仔猪的模型中定植,表明幽门螺杆菌鞭毛对定殖是必需的,幽门螺杆菌的毒力也因此而增加。
幽门螺杆菌motB基因编码鞭毛运动旋转蛋白。 幽门螺杆菌motB基因敲除突变体仍包含鞭毛,但不能运动。该突变体在小鼠中定植的能力降低,证实了运动性是完全传染性所必需的。
鞭毛基因转录的调控不同于许多其他细菌。与许多其他带鞭毛的革兰氏阴性细菌相反,幽门螺杆菌没有转录调控因子来调控鞭毛基因的转录。
据推测,幽门螺杆菌的运动不需要严格的调节,因为它在任何时候都是不可或缺的。然而,在幽门螺杆菌中仍然可以看到一些转录调控。例如,蛋白质FlhA和FlhF,基体的组成部分,需要在中后期鞭毛基因转录之前出现。
趋化性
除了运动性,趋化性对于幽门螺杆菌的定植也是必不可少的。趋化性允许幽门螺杆菌将其运动导向胃上皮细胞层。
由HP0099编码的幽门螺杆菌趋化性受体传感器识别碳酸氢盐和精氨酸作为引诱剂。幽门螺杆菌外膜上这种传感器蛋白与反应调节剂CheY偶联。
反应调节剂CheY调节鞭毛运动蛋白的表达,使幽门螺杆菌游向其生态位。
应答调节因子CheY的一个敲除突变体未能在非生物小猪体内定殖,强调了趋化性在最初定殖中的重要性。
反应调节因子CheY及其组氨酸激酶的敲除突变体显示出对粘蛋白的趋化性反应减弱。
免疫逃避
在长期的定殖过程中,宿主对多种幽门螺杆菌抗原产生强大的免疫反应。然而,这些抗原通常没有表面暴露。
另一方面,表面暴露的抗原通常高度多样化,使宿主清除幽门螺杆菌变得更加困难。幽门螺杆菌感染极少产生足够的免疫反应,能够清除细菌。 实际上,幽门螺杆菌能够有效规避先天性和适应性免疫反应。
先天的免疫反应不能实质上识别幽门螺杆菌。大肠杆菌脂多糖(LPS)通过TLR4激活先天免疫应答。然而,幽门螺杆菌LPS远不是TLR4介导的基因表达的有效诱导剂。
在幽门螺杆菌中,TLR5介导的免疫应答激活效率也较低。 与肠沙门氏菌血清型鼠伤寒鞭毛蛋白相比,幽门螺杆菌鞭毛蛋白没有被释放,并且在通过TLR5激活IL-8分泌中也没有那么强。
先天性免疫反应未能识别幽门螺杆菌可能导致适应性免疫反应失败以随后清除感染。
自适应免疫系统的逃避至少由两个重要机制介导:抗原模拟和免疫抑制。抗原模拟导致宿主免疫系统无法正确识别幽门螺杆菌。例如,幽门螺杆菌LPS上的Le血型抗原也在人类上皮细胞上表达。结果,幽门螺杆菌LPS免疫原性差,从而支持幽门螺杆菌的持久性。在其LPS上缺少O抗原的幽门螺杆菌菌株在小鼠中定殖的能力显着降低。因此,幽门螺杆菌LPS的典型结构在定植中很重要。
除了通过抗原模拟来逃避免疫外,免疫抑制也有助于幽门螺杆菌感染的慢性化。
免疫应答可能偏向更抗炎的应答,或者下调(VacA)。幽门螺杆菌VacA可以阻断巨噬细胞中的吞噬体-溶酶体融合,导致巨噬细胞无法杀死幽门螺杆菌。
此外,VacA表达阻断T细胞活化并因此增殖,活化T细胞凋亡,并抑制T细胞中的抗原呈递。
B淋巴细胞的抗原加工可能被阻止,可能是通过将内体的成熟限制在发生抗原加载的MHC II类区室中。因此,VacA限制了针对幽门螺杆菌的T细胞和B细胞应答。
幽门螺杆菌的存在可以预防某些疾病?
通过以上各种方式,幽门螺杆菌得以在人体中生存下来,从某种程度上来说,幽门螺杆菌并不总是对人类宿主有害。
研究表明,幽门螺杆菌的存在可能抑制引起结核的细菌(结核分枝杆菌),预防哮喘,克罗恩病,食管反流,腹泻病以及食道癌。 幽门螺杆菌的发病率下降与儿童过敏性疾病和自身免疫性疾病,多发性硬化,腹腔疾病的发病率上升相关。
因此,从某种程度上来说,幽门螺杆菌有其存在的价值。这引起了关于是否需要根除幽门螺杆菌的讨论。
需要明确的是,幽门螺杆菌感染后不会立即对人体产生致死性危害和严重后果,所导致的慢性胃炎、消化道溃疡和胃癌等疾病是长期存在并慢性与其他因素共同作用的结果。
第五次全国幽门螺杆菌感染处理共识报告中有如下陈述:
根除幽门螺杆菌的获益在不同个体之间存在差异
根除幽门螺杆菌促进消化性溃疡愈合和降低溃疡并发症发生率,根除幽门螺杆菌可使约80%早期胃 MALT 淋巴瘤获得缓解。与无症状和并发症的 幽门螺杆菌感染者相比,上述患者根除幽门螺杆菌的获益显然更大。胃癌发生高风险个体有胃癌家族史、早期胃癌内镜下切除术后和胃黏膜萎缩和(或)肠化生等。根除幽门螺杆菌预防胃癌的获益高于低风险个体。

在做出可靠的功能性消化不良诊断前,必须排除幽门螺杆菌相关消化不良。
幽门螺杆菌胃炎伴消化不良症状的患者,根除幽门螺杆菌后可使部分患者的症状获得长期缓解,是优选选择。
幽门螺杆菌感染是消化性溃疡主要病因,不管溃疡是否活动和是否有并发症史,均应该检测和根除幽门螺杆菌。
…
总的来说,对于无抗衡因素 ( 高龄、伴存疾病、社区再感染率、卫生经济因素等 ) 的个体,根除幽门螺杆菌治疗利大于弊。
儿童:
不推荐对 14 岁以下儿童行常规检测幽门螺杆菌。推荐对消化性溃疡儿童进行幽门螺杆菌检测和治疗,因消化不良行内镜检查的儿童建议行幽门螺杆菌检测与治疗。
与成人相比,儿童根除治疗不利因素较多,包括抗菌素选择余地小,对药物不良反应耐受性低。此外,儿童幽门螺杆菌感染有一定自发清除率,根除后再感染率也可能高于成人。
老年人:
老年人根除幽门螺杆菌治疗药物不良反应风险增加,因此对老年人根除幽门螺杆菌治疗应该进行获益-风险综合评估,个体化处理。
问卷调查显示,多数临床医生对老年人根除幽门螺杆菌治疗的态度趋向保守。一般而言,老年人(年龄>70 岁)对根除幽门螺杆菌治疗药物的耐受性和依从性降低,发生抗生素不良反应的风险增加;另一方面,非萎缩性胃炎或轻度萎缩性胃炎患者根除幽门螺杆菌预防胃癌的潜在获益下降。
合理看待幽门螺杆菌感染后果至关重要,对自然人群中幽门螺杆菌感染是否需要进行干预,需要综合权衡利弊。对于有根除治疗指征的患者,需要给予规范治疗,提高首次根除率,减少耐药性发生。
在慢性感染过程中,多株菌株同时定植于一个宿主,并且幽门螺杆菌分离株的遗传变异使幽门螺杆菌相关的发病机理难以从单个因素中预测。 幽门螺杆菌与其宿主的共同进化使它得以终生定居。
随着现代卫生环境改善及抗生素的大量使用,幽门螺杆菌的生长和传播越来越难,年轻一代中感染率较低,在未来几十年中幽门螺杆菌感染率将进一步下降。
更好地理解幽门螺杆菌和其他胃肠道微生物群之间的对话可能为预防或治疗其感染提供新的方向。如在肠道菌群健康检测报告中发现幽门螺杆菌感染,则应引起重视。当需要治疗时,应评估新的治疗策略和药物,或者应该制定替代策略来控制细菌的毒力,从而避免出现溃疡和胃癌而不消除其积极影响。
附录: 幽门螺杆菌的发现史
在1980年代初,幽门螺杆菌首次被发现是引起胃部疾病的原因。该发现帮助研究人员将幽门螺杆菌感染与胃炎,溃疡和胃癌等联系起来。
1982年,两名澳大利亚研究人员巴里·马歇尔和罗宾·沃伦描述了幽门螺杆菌感染的作用。
然而,沃伦和马歇尔关于幽门螺杆菌功能的理论并未立即被医学界接受。当时,大多数科学家都不相信细菌可以在人的胃中生活。
马歇尔无法在实验室小鼠中进一步测试他的工作,也不允许在人身上进行实验。马歇尔这位勇敢的科研工作者想到了一个办法。
他决定亲自吃下幽门螺杆菌。他从病人的肠道中提取该菌,将其放入肉汤中,然后饮用。
几天后,马歇尔出现了肠道症状,例如恶心,呕吐,口臭和疲劳。他做了胃活检,且能够培养幽门螺杆菌,证明是细菌引起了他的症状。
医学界最终接受了幽门螺杆菌引起溃疡和其他肠道问题的观念,但是接受的道阻且长。
1994年,美国国立卫生研究院共识发展会议正式承认幽门螺杆菌与溃疡之间的紧密联系。该小组建议对溃疡患者进行抗生素治疗。
1996年,FDA批准了首个针对幽门螺杆菌引起的溃疡的抗生素疗法。
一年后,疾控中心(CDC)与其他政府机构和学术机构合作发起了一项全国运动,为医患相关人员提供有关幽门螺杆菌与溃疡之间的联系。
2005年,马歇尔和沃伦获得了诺贝尔生理学或医学奖。溃疡不再被视为无法治愈的慢病,而是一种可以通过短期的抗生素和酸分泌抑制剂治疗的疾病。
【参考文献】
Burucoa C, Axon A. Epidemiology of Helicobacter pylori infection. Helicobacter. 2017 Sep;22 Suppl 1. doi: 10.1111/hel.12403. PMID: 28891138.
Camilo V, Sugiyama T, Touati E. Pathogenesis of Helicobacter pylori infection. Helicobacter. 2017 Sep;22 Suppl 1. doi: 10.1111/hel.12405. PMID: 28891130.
Eusebi LH, Zagari RM, Bazzoli F. Epidemiology of Helicobacter pylori infection. Helicobacter. 2014 Sep;19 Suppl 1:1-5. doi: 10.1111/hel.12165. PMID: 25167938.
Mentis A, Lehours P, Mégraud F. Epidemiology and Diagnosis of Helicobacter pylori infection. Helicobacter. 2015 Sep;20 Suppl 1:1-7. doi: 10.1111/hel.12250. PMID: 26372818.
Thorell K, Lehours P, Vale FF. Genomics of Helicobacter pylori. Helicobacter. 2017 Sep;22 Suppl 1. doi: 10.1111/hel.12409. PMID: 28891132.
Julie Marks.Robert Jasmer, MD What Is H. Pylori? Symptoms, Causes, Diagnosis, Treatment, and Prevention. 2020.9
History of H. Pylori: What We Do and Don’t Know Before the discovery of the bacterium, doctors thought stomach ulcers were due to stress and spicy foods.
Burucoa C, Axon A. Epidemiology of Helicobacter pylori infection. Helicobacter. 2017 Sep;22 Suppl 1. doi: 10.1111/hel.12403. PMID: 28891138.
Kamboj AK, Cotter TG, Oxentenko AS. Helicobacter pylori: The Past, Present, and Future in Management. Mayo Clin Proc. 2017 Apr;92(4):599-604. doi: 10.1016/j.mayocp.2016.11.017.
Eusebi LH, Zagari RM, Bazzoli F. Epidemiology of Helicobacter pylori infection. Helicobacter. 2014 Sep;19 Suppl 1:1-5. doi: 10.1111/hel.12165. PMID: 25167938.
Yang JC, Lu CW, Lin CJ. Treatment of Helicobacter pylori infection: current status and future concepts. World J Gastroenterol. 2014 May 14;20(18):5283-93. doi: 10.3748/wjg.v20.i18.5283.
Sun Y, Zhang J. Helicobacter pylori recrudescence and its influencing factors. J Cell Mol Med. 2019 Dec;23(12):7919-7925. doi: 10.1111/jcmm.14682. Epub 2019 Sep 19.
Tao Zhi-Hang,Han Ji-Xuan,Fang Jing-Yuan,Helicobacter pylori infection and eradication: Exploring their impacts on the gastrointestinal microbiota.[J] .Helicobacter, 2020, 25: e12754.
Algood HM, Cover TL. Helicobacter pylori persistence: an overview of interactions between H. pylori and host immune defenses. Clin Microbiol Rev. 2006 Oct;19(4):597-613. doi: 10.1128/CMR.00006-06.
Kori M, Daugule I, Urbonas V. Helicobacter pylori and some aspects of gut microbiota in children. Helicobacter. 2018 Sep;23 Suppl 1:e12524. doi: 10.1111/hel.12524. PMID: 30203591.
van Amsterdam K, van Vliet AH, Kusters JG, van der Ende A. Of microbe and man: determinants of Helicobacter pylori-related diseases. FEMS Microbiol Rev. 2006 Jan;30(1):131-56.
Waskito LA, Yamaoka Y. The Story of Helicobacter pylori: Depicting Human Migrations from the Phylogeography. Adv Exp Med Biol. 2019;1149:1-16. doi: 10.1007/5584_2019_356. PMID: 31016625.
Pereira-Marques J, Ferreira RM, Pinto-Ribeiro I, Figueiredo C. Helicobacter pylori Infection, the Gastric Microbiome and Gastric Cancer. Adv Exp Med Biol. 2019;1149:195-210.
RU Nan, DU Yi-qi, LI Zhao-shen. 幽门螺杆菌根除指征的演变[J]. 中国实用内科杂志, 2019, 39(006):515-519.
中华医学会消化病学分会幽门螺杆菌和消化性溃疡学组, 全国幽门螺杆菌研究协作组, 刘文忠,等. 第五次全国幽门螺杆菌感染处理共识报告[J]. 胃肠病学, 2017(6).

谷禾健康
过完今天,这个魔幻的2020就要过去了。面临工作、学习、出行等方方面面一而再地按下暂停键,很多人都希望这一年能够重启。
这一年发生了太多故事
每一个都足以让人痛惜
“活着”、“健康”这样的字眼在这一年显得尤为珍贵。简单的道理往往在经历重大的事件后才会尤为深刻。
古有求仙药,今有各种医疗技术的不断革新,大家对于长寿有着一致的追求。当基本的生活有了一定的保障后,人们不仅想要活得长久,还要健健康康地活着。
我们可以看到,人类的预期寿命已开始延长,并且仍在继续提高。“健康长寿”不再停留于一句祝福语,而是可行的人生目标。
长寿是多种变量复杂组合的结果。由于不同地区自然环境、社会制度、社会经济发展状况和人口构成等因素千差万别。
根据相关研究,影响人类寿命的因素至少包括:
遗传因素如线粒体状态、染色体稳定性、端粒长短、疾病、干细胞活性;
环境因素如肠道微生物、饮食、运动、空气质量以及生活环境;
其他因素如情绪压力、社交爱情、目标成就、投入预防等等。
人们在迈向健康长寿的过程中处于不同的阶段和水平。若干年后,也许有相当多的人进入百岁人生。
在2020年的最后一天,我们就来聊聊肠道菌群和长寿的故事。

伊卡里亚岛
伊卡里亚岛
该岛位于萨摩斯岛西南约10海里,这是爱琴海的一个小岛,是世界上寿命最长的地区之一。他们是90岁以上人群中世界上人口最多的国家,百岁老人在岛上已有400多年的历史了。科学家将那里长寿的机会与空气,水,社区精神,饮食习惯和遗传倾向等因素相关联。

意大利
提起意大利,你想到的是足球还是意大利面,其实这个国家还盛产长寿老人,根据欧盟统计,意大利为欧洲第一长寿国,女性平均寿命为84岁,男性平均寿命为78.3岁。
研究发现100岁意大利老人的肠道菌群种类分布与30岁意大利人相比,出现了较明显的变化,厚壁菌门中的拟杆菌XIVa明显减少,而芽孢杆菌上升,身体的炎症反应状况高,因此科学家得出长寿的关键因素:菌群种类的改变,更好的应对和调节炎症反应。

中国 新疆和田,广西巴马,四川都江堰青城山等
中国新疆和田,广西巴马以及四川都江堰市青城山等地区都很大比例的长寿健康老人。动物遗传育种研究所李英团队在《Current Biology》发表的一项关于寿老人和年轻人群肠道菌群研究发现长寿老人肠道菌群多样性和菌群丰度显著高于年轻组,这一结论在意大利相应人群中也得到了证实,提示更多有益菌群以及更丰富的菌群多样性可能是人类健康长寿的重要原因之一。
对长寿的研究可能有助于我们理解人类是如何延缓衰老,如何战胜与年龄相关的疾病。
肠道微生物群被认为是监测和可能支持健康衰老的变量之一。事实上,宿主-肠道微生物体内平衡的破坏与炎症和肠道通透性以及骨骼和认知健康的普遍下降有关。肠道微生物群作为健康衰老可能的介质,通过对抗炎症、肠道通透性以及认知和骨骼健康的恶化来保持宿主环境的稳态。
健康老年人的肠道菌群如何定义?
考虑到大多数老年人都患有肠道相关合并症,因此在该人群中定义健康的肠道微生物组极具挑战性。肠道环境的变化,例如炎症,肠道渗漏,活性氧的产生以及药物的使用,都可能影响肠道微生物组。在这方面,健康百岁老人一直被用作健康老龄化的典范,因为他们有能力推迟或避免慢性疾病。因此,该队列中的肠道微生物组可用于定义健康的肠道微生物组。
长寿者肠道菌群多样性水平高
一般认为,随着年龄增长时,肠道微生物多样性通常会降低。可能是由于生理,饮食,药物和生活方式的变化所致。
是不是所有老人的肠道菌群多样性都会降低?
研究人员检测了一群健康的长寿老人的肠道微生物组,来自中国四川都江堰市的长寿老人,包括“90-99岁”和“≥100岁”两个年龄段。
他们发现长寿人群的肠道菌群比年轻成年人的肠道菌群更多样化,这与传统观点相矛盾。
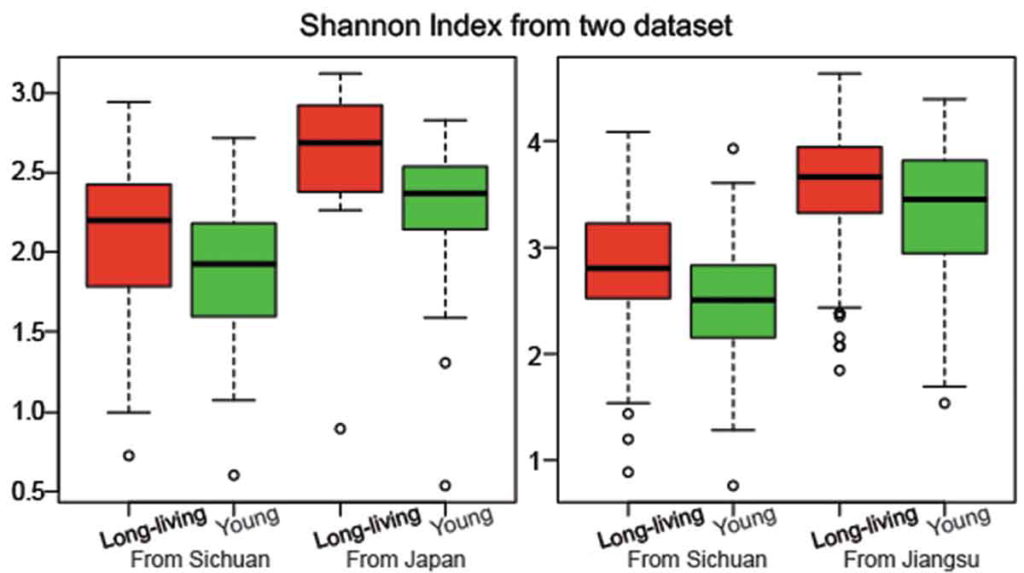
他们还发现了产短链脂肪酸菌在长寿老人中开始增加,例如梭状芽胞杆菌XIVa。
国内外研究结果一致
为了验证他们的发现,他们分析了来自一个意大利小组的独立数据集。
出现一致的结果:长寿的意大利人也比年轻的人群的肠道菌群多样化水平更高 。
庆幸的是,谷禾肠道菌群数据库中也有比较长寿的老人肠道菌群数据。
我们抽取其中一例相对较为健康的长寿老人的数据:
编号:083*****97 ,98岁(谷禾肠道菌群数据库)
谷禾健康数据库
可以看到肠道菌群多样性水平也是明显增高,与文献报道相符。大部分指标都处于正常水平。
长寿者产短链脂肪酸菌增多
结合意大利和中国的数据集,发现尽管肠道微生物群结构存在显著差异(可能是由于饮食、基因和环境的差异),但区分长寿个体和年轻群体的前50种细菌特征中,有11种特征是相同的。同样,这些特征包括肠道菌群多样性水平更高和几个产短链脂肪酸菌丰度更高。
一项后续研究中,另外两个独立的队列中也观察到了长寿人群中更大的肠道微生物组多样性:一个来自中国江苏省,另一个来自日本。
以上这些研究都清楚地表明,健康长寿的人存在更多样化且平衡的肠道菌群,而在患有不同合并症的老年人中观察到肠道菌群紊乱。
因此,研究人员假设调节肠道微生物组(如通过饮食、益生菌)来维持健康的肠道微生物组将有利于健康地衰老。
进一步假设,在患有慢性疾病的老年人中,将紊乱的肠道菌群调节为健康的肠道菌群将减轻他们的症状,提高他们的生活质量。
肠道微生物组和健康衰老的有效假设
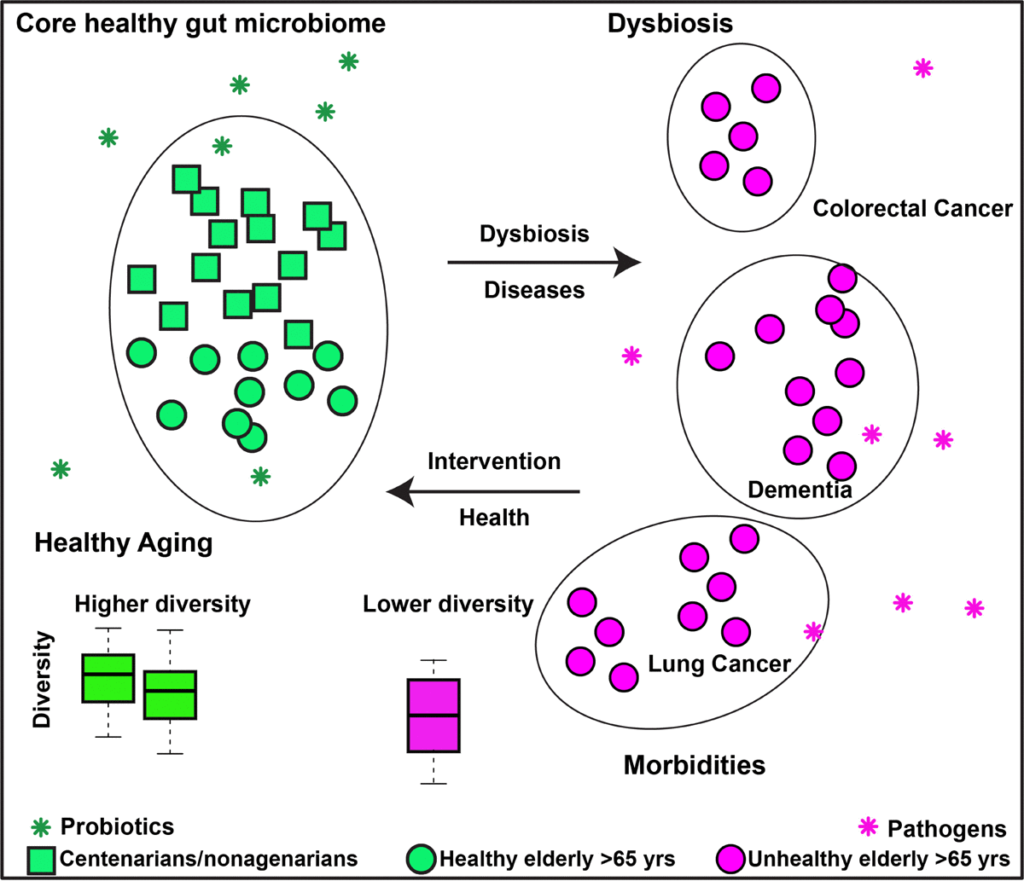
该假设背后的一个基本原理是慢性炎症,即老年人中慢性低度炎症的增加,这与不同的慢性疾病有关。
短链脂肪酸对维持肠道止血很重要。短链脂肪酸为结肠上皮细胞提供主要能量,并具有抗炎特性。这些产短链脂肪酸菌在长寿老人中的富集表明,这些细菌可能会减轻炎症及由此造成的损害,这可能是他们能够健康衰老的原因。
以上我们知道长寿老人的产短链脂肪酸菌增多,那么其他菌群会有什么样的变化?
在门类水平上,大多数研究都证明了变形菌丰度的增加。
长寿者菌群变化,潜在有益菌较多
· 不同地区比较:
一项研究分析并比较了长寿村庄中百岁老人与同一地区和城市化城镇中的老年人和成年人的肠道菌群。采集长寿村的百岁老人、老年人和年轻人的粪便样本,以及来自韩国城市城镇的老年人和年轻人的公共数据库获得肠道菌群数据。
与城镇化老年人相比,长寿村老人:
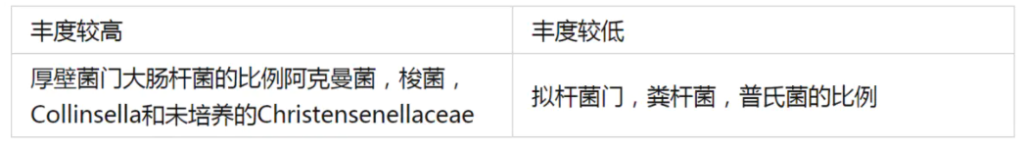
康复医院百岁老人的肠道菌群也不同于居家。这些差异可能是由于饮食方式和生活环境的差异。
· 不同年龄比较:
我们来看一项研究,对62个人的粪便微生物组进行宏基因组测序,年龄从22岁至109岁不等。
下图可以看到,随着年龄的增长,肠道微生物群发生了变化。
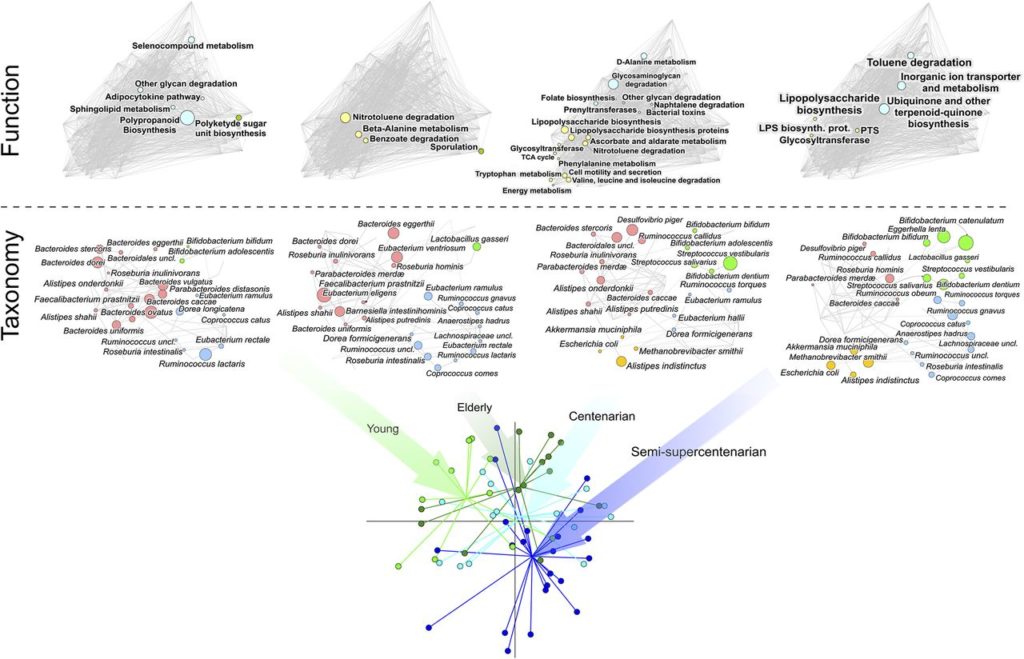
注:4个年龄组的肠道微生物组:
11个年轻人 (22 – 48岁,young);
中年13人 (65 – 75岁,elderly);
15名百岁老人 (99至104岁, centenarian);
23名半超百岁(105至109岁,semisupercentenarian)
研究人员发现与年轻人相比,长寿者菌群变化如下:

编号:083*****97 ,98岁(谷禾肠道菌群数据库)
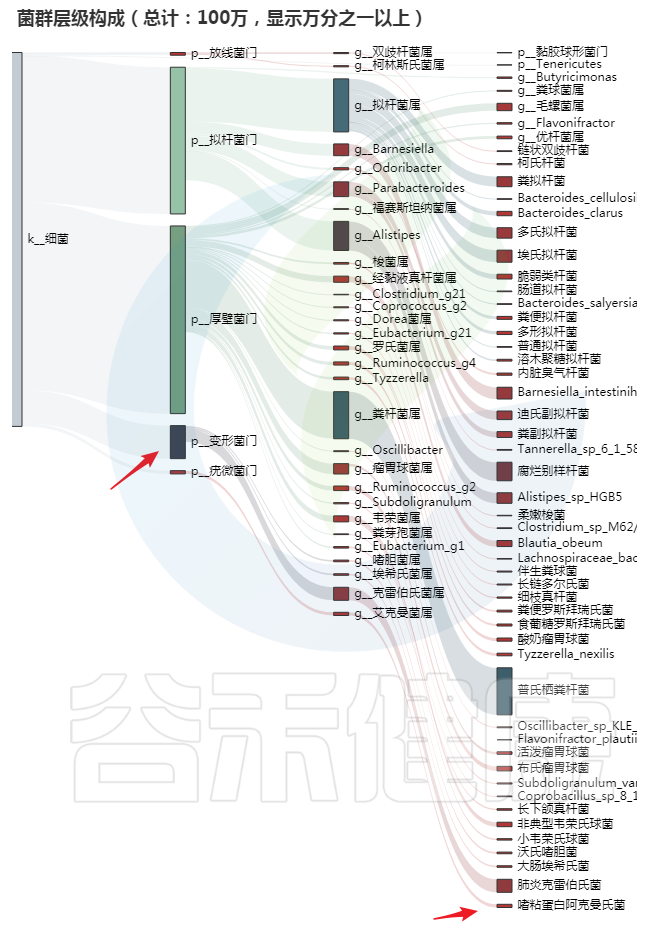
我们发现同样,变形菌门增加,另外有益菌如阿克曼菌增多。
长寿者碳水化合物代谢相关基因减少
有趣的是,当研究人员将分析集中在功能规模上时,发现与碳水化合物代谢有关的基因减少。
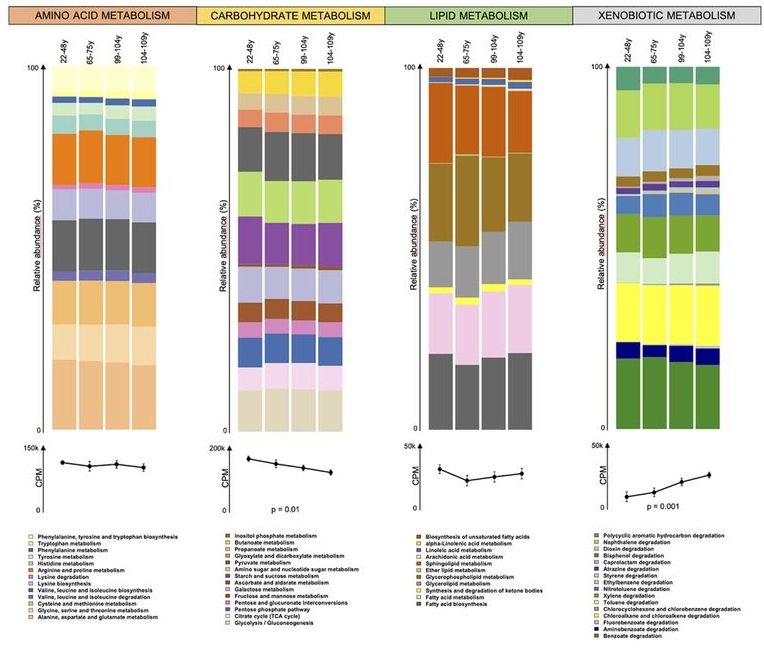
这种功能重塑在百岁老人和半超百岁老人的肠道微生物组中更为明显,研究人员观察到淀粉和蔗糖(KEGG途径编号ko00500),磷酸戊糖(ko00030)以及氨基糖和核苷酸糖(ko00520)途径的贡献减少。
异种生物降解有关的基因数量增加
同时,研究人员发现了和甲苯(ko00623),乙苯(ko00642),己内酰胺(ko00930)以及氯环己烷和氯苯(ko00361)降解途径的随之增加。
乙苯,氯苯,氯环己烷,甲苯是主要来源于工业生产和城市排放的普遍化学物质,由于其毒性作用,是世界各地监测的主要环境污染物之一。这些分子的主要人造来源实际上是汽车和废气汽车的排放,以及香烟烟雾。
此外,众所周知,它们是在加工精制石油产品(如塑料)的过程中产生的,并包含在普通消费产品(如油漆和漆、稀释剂和橡胶产品)中。
己内酰胺是尼龙的原料,用于生产合成纤维、树脂、合成皮革、增塑剂等多种室内产品。先前的研究表明,这些分子在室内的负担比在室外环境中更高,并强调了室内暴露对人类健康的特殊重要性。
生活在强人为下的环境中,例如意大利的艾米利亚-罗马涅区(工业发达),导致持续不断地暴露于这些普遍的异生物质中,促进它们在身体组织(包括肠道)中的维持和累积。
研究人员认为,这可以为人类宿主创造合适的条件,以选择能够解毒此类化合物的肠道微生物组成分,就微生物组和宿主在人类环境中的适应性而言互惠互利。
百岁和半百岁的人都是长寿的人,他们接触异生生物刺激的时间更长,他们的微生物群更适合降解这些异生生物。
脂质代谢基因变化
除了异物降解基因和糖代谢相关基因外,我们还发现了其他代谢途径中与年龄相关的差异,包括与脂质代谢有关的差异。
百岁老人和半超百岁老人显示出更多的α-亚油酸(KEGG途径编号ko00592)和甘油脂(ko00561)代谢的信息。另一方面,年轻人显示鞘脂(ko00600)和甘油磷脂(ko00564)代谢相关基因的贡献更大。
鉴于已知甘油磷脂和鞘脂在动物源性食品中更为丰富,而α-亚油酸主要来自植物源食物,这些特征可能与饮食习惯有关,特别是长寿者的植物源性脂肪摄入量高于年轻人的动物脂肪摄入量。
氨基酸代谢基因变化
此外,涉及氨基酸代谢的功能途径:
色氨酸(ko00380),酪氨酸(ko00350),甘氨酸,丝氨酸和苏氨酸(ko00260)的代谢基因随着年龄的增长而逐渐增加。
另一方面,发现年轻人中丙氨酸,天冬氨酸和谷氨酸代谢的基因(ko00250)更为丰富。色氨酸和酪氨酸的代谢被认为是蛋白水解代谢增强的指标。
此外,血清中色氨酸的生物利用度降低,以及尿液中酪氨酸代谢引起的酚类代谢产物水平升高。
慢性炎症水平低
研究人员发现随着衰老,脂多糖生物合成基因(ko00540)逐渐增加,这可能与病原菌(即肠杆菌科的成员)的存在和慢性炎症的水平低有关。
然而,更长的寿命并不一定等于健康的衰老。随着年龄的增长,人们更有可能患上各种疾病,如心脏病、中风、高血压、认知障碍、癌症等。
前面章节有一项研究(长寿村老人与城镇化老人肠道菌群)提到,来自不同地区的老人,虽然都是长寿,但菌群情况不尽相同。
因此,我们想更具体地了解,同样是长寿老人,健康长寿和不健康长寿具体到个人,在哪些方面会有区别。我们抽取谷禾肠道菌群数据库中两例报告来进行直接比较分析。
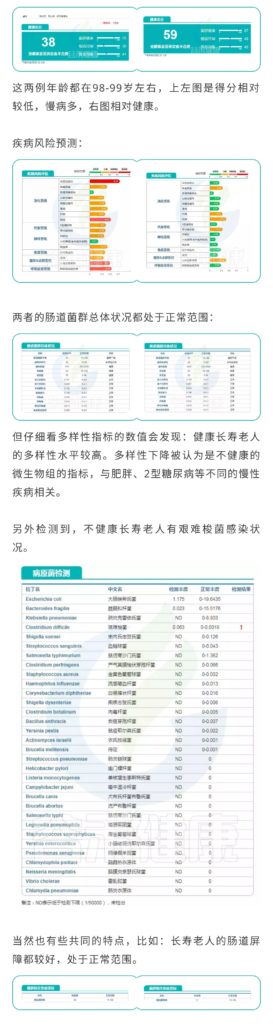
以上只是数据库中的两个案例,在经过谷禾肠道菌群数据库筛查后,我们总结了一些关于长寿老人(90岁以上)的肠道菌群的趋势,分享关于报告中的一些指标判断,供大家参考。
健康总分
健康总分能很好的反映一个人的总体健康水平,有慢病或其他问题的老人一般低于55分。
菌群多样性
菌群多样性健康长寿老人的菌群多样性水平最好能高于50,菌种数量在1000~1800左右较好,超过2000则可能会伴有病原菌感染的情况。
慢病情况
主要是心脑血管及糖尿病和部分消化道疾病,涉及慢性炎症和代谢疾病。
病原菌
病原菌感染是老人中最常出现的问题,包括呼吸道和肠道病原菌,随着衰老,肠杆菌科的部分机会致病菌比例会上升,需要注意饮食健康,以及呼吸道健康和口腔健康。
肠道屏障及炎症水平
长寿老年人中Akk菌水平普遍较中年人群较高,Akk菌有助于降低肥胖等代谢疾病,但是Akk菌丰度过高会导致肠粘膜黏蛋白降解,破坏肠道屏障,也是需要注意的指标。
短链脂肪酸水平
短链脂肪酸生成的菌的水平与短链脂肪酸和炎症水平密切相关,短链脂肪酸缺乏通常是慢性炎症的推手。
益生菌水平
在谷禾检测的90岁以上人群中,益生菌水平普遍较高,基本超过人群平均水平。
06 长寿者避雷专区——谣言粉碎机
信息爆炸的时代,我们可以轻易获得大量关于营养保健的信息,然而其中大多数可能是不正确或者过时的观念。
1. 减肥仅靠控制热量?
我们都知道,减肥需要燃烧比摄入更多的能量,但这不是唯一。那些遵循“卡路里摄入,卡路里消耗”方法的人通常只专注于食物的卡路里值,而不是其营养价值。这对于整体健康而言,并非最佳选择。
如果出现体内激素失调,甲状腺功能低下,代谢状况,药物使用等健康问题,可能即使在严格饮食下也难以减轻体重。
2. 高脂食物不健康?
许多人仍然担心高脂肪的食物并遵循低脂肪的饮食习惯,认为减少脂肪的摄入有益于整体健康。
膳食脂肪对于保持最佳健康至关重要。另外,低脂饮食与包括代谢综合征在内的健康风险更高有关,并且可能导致胰岛素抵抗和甘油三酸酯水平升高,这是已知的心脏病危险因素。
而且,在鼓励减肥方面,高脂肪饮食已被证明比低脂肪饮食有效(甚至更高)。
当然,无论是低脂还是高脂饮食,任何一种极端情况都可能危害健康。尽可能遵循“中庸之道”。
3.非营养性甜味剂是健康的?
市场上出现越来越多的非营养性甜味剂(NNS)的产品有所增加。显然,高糖饮食会大大增加疾病的风险,但摄入NNS也会导致不良的健康后果。
例如,摄入NNS可能会引起肠道菌群产生负面变化并促进血糖失调,从而增加2型糖尿病的发病率。
该领域的研究仍正在进行中,未来需要高质量研究来确认这些潜在的联系。
4. 你必须很瘦才能健康?
我们知道,肥胖与许多健康状况相关,包括2型糖尿病,心脏病,抑郁症,某些癌症等。
尽管如此,降低疾病风险并不是说要你必须要达到模特身材。最重要的是营养饮食并保持积极的生活方式,因为这些行为通常会改善体重和体内脂肪百分比。
5. 所有食物都用低脂和减肥食品来代替?
去超市你会发现各种标有“清淡”,“低脂”,“无脂”的产品。虽然这些产品对那些想要减少体内多余脂肪的人来说很诱人,但它们通常是不健康的选择。
研究表明,许多低脂和减肥食品所含的糖和盐要比普通脂肪食品多得多。最好不要经常食用这些产品,有时候也可以享受一下正常食物,例如全脂酸奶,奶酪和坚果黄油。
6. 钙补充剂对骨骼健康必不可少?
很多人听说添加钙补充剂以保持骨骼系统健康。但是,目前的研究表明,补充钙可能弊大于利。
例如,一些研究已将钙补充剂与心脏病风险增加联系起来。此外,研究表明,它们不会降低骨折或骨质疏松症的风险。
当然,如果你担心缺钙,最好注意下钙的饮食来源,例如全脂酸奶,沙丁鱼,豆类和种子食物等。
7.所有果汁和果汁都是健康的 ?
某些果汁营养丰富。例如,主要由非淀粉类蔬菜制成的新鲜果汁可以是增加维生素,矿物质和抗氧化剂摄入量的好方法。
然而,要知道外面买到的大多数果汁中都含有糖和卡路里。如果过量食用,会促进体重增加和其他健康问题,例如蛀牙和血糖失调。

8. 每个人都需要补充益生菌?
益生菌的概念现在越来越火。但是,研究表明,不是所有人补充益生菌都有用,搞不好有副作用。
某些人的消化系统对益生菌的定殖有抵抗力,而且通过补充剂引入益生菌可能会导致肠道细菌产生负面变化。另外,与益生菌使用相关的小肠细菌过度生长会导致腹胀,气体和其他不良副作用。
益生菌不应作为千篇一律的补充剂,而应更加个性化,最好在做完肠道菌群检测之后再确定是否需要补充益生菌,补充哪一类益生菌,这样才能真正让身体恢复健康。
9. 减肥很容易?
你可能看过很多生动的减肥前后的图片,甚至还有传奇的故事,几乎不费吹灰之力就可以迅速减肥的故事,不要随意相信。
减肥其实并不容易。它需要坚持不懈,自律,努力和耐心。另外,由于遗传或其他药物因素使某些人的减肥困难很大,我们需要正视这一切,面对它,慢慢来,给自己多一点耐心,找到一种对你有效的可持续的模式最重要。

10. 纤维补充剂是高纤维食品的良好替代品?
许多人缺乏足够的膳食纤维,这就是为什么纤维补充剂如此受欢迎的原因。尽管纤维补充剂可以改善肠蠕动和血糖控制,从而有益于健康,但它们不应代替真正的食物。
高纤维全食(例如蔬菜,豆类和水果)包含营养物质和植物化合物,它们可以协同工作以促进您的健康,并且不能完全由纤维补充剂替代。
LONGEVITY
随着时代的不断发展,旧的观念也在不断更新。曾经的认知也许是“七十古来稀”,而现如今更多的是百岁人生。
我们的追求也会越来越高,不仅是长寿,更是健康的长寿。可以预见,长寿时代将促使健康产业结构升级。
是的,微生物产业作为健康领域的其中一块,发展迅速。值得庆幸的是,在应对突如其来的疫情下,肠道微生态也在被应用于治疗,技术的革新为提高健康水平提供有力支撑,各个制度完善也在为健康领域的可持续发展构建强大保障,人类命运共同体正彰显其感召力。

而谷禾健康作为微生物产业的一员,自2012年创立起,对于科研事业一直贡献着自己的力量,与此同时,这么多年来,谷禾一直希望将科研真正服务于大众,将科研成果带给每一个人,这是我们的使命。
现如今,我们也已经看到阶段性硕果。曾经,“肠道菌群”还只是一个概念,谷禾健康从肠道菌群的研究构思,到取样专利的落地,肠道菌群检测报告逐步完善,再到样品运输的细节管理,我们都在经历从0到1的过程,勇于创新,不断摸索,在微生物产业的道路上,迈着坚定的步伐。
令我们感到欣慰的是,“肠道菌群”现已逐渐从研究过渡到临床甚至普通人群,并且从模糊的健康概念走向精准检测甚至个性化辅助治疗。
2021寄语
愿你所有努力都有回报
所有的美好都如期而至
参考文献:
Kong F, Deng F, Li Y, Zhao J. Identification of gut microbiome signatures associated with longevity provides a promising modulation target for healthy aging. Gut Microbes.
2019;10(2):210-215. doi: 10.1080/19490976.2018.1494102. Epub 2018 Aug 24. PMID: 30142010; PMCID: PMC6546316.
Rampelli S, Soverini M, D’Amico F, Barone M, Tavella T, Monti D, Capri M, Astolfi A, Brigidi P, Biagi E, Franceschi C, Turroni S, Candela M. Shotgun Metagenomics of Gut Microbiota in Humans with up to Extreme Longevity and the Increasing Role of Xenobiotic Degradation. mSystems. 2020 Mar 24;5(2):e00124-20. doi: 10.1128/mSystems.00124-20. PMID: 32209716; PMCID: PMC7093822.
Deng F, Li Y, Zhao J. The gut microbiome of healthy long-living people. Aging (Albany NY). 2019 Jan 15;11(2):289-290. doi: 10.18632/aging.101771. PMID: 30648974; PMCID: PMC6366966.
Kim BS, Choi CW, Shin H, Jin SP, Bae JS, Han M, Seo EY, Chun J, Chung JH. Comparison of the Gut Microbiota of Centenarians in Longevity Villages of South Korea with Those of Other Age Groups. J Microbiol Biotechnol. 2019 Mar 28;29(3):429-440. doi: 10.4014/jmb.1811.11023. PMID: 30661321.