-

 CNAS L23010
CNAS L23010

 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业 二级病原微生物安全实验室
二级病原微生物安全实验室- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业 二级病原微生物安全实验室
谷禾健康

消化是人体生命活动的重要组成部分,分解食物提供能量、促进生长发育、参与免疫功能,然而越来越多的人出现了消化不良。
★ 消化不良在人群中很常见
消化不良在全球范围内都是一种常见的疾病,其发病率在不同地区和人群中有所不同。在中国,消化不良的发病率较高,尤其是城市人群中更为常见。据统计,中国城市居民中消化不良的发病率约为20%-30%。
消化不良主要分为器质性消化不良和功能性消化不良(本文主要讲述功能性消化不良)。功能性消化不良其广义上是一种胃肠道多种症状的综合征,主要包括上腹痛或灼热感,餐后饱胀感及早期饱腹感。
随着对功能性消化不良了解的深入,目前研究发现,消化不良病理生理与脑–肠–微生物群轴紊乱、内脏高敏感性、局部低度炎症、胃肠道感染等因素相关。
肠道菌群可以帮助人体消化吸收,同时还可以合成一些对人体有益的物质,在消化过程中扮演着重要角色。肠道菌群作为新的研究切入点,将有助于探索新的改善功能性消化不良的方法。
在了解消化不良前,我们先来看看人体消化的生理过程:
膳食摄入的消化反应

Livovsky DM,et al.Nutrients.2020
在禁食期间,胃肠道会进行周期性的运动,这种运动被称为迁移运动复合体(MMCs)。迁移运动复合体的功能是将残留物从小肠推进到结肠,以便在新的食物进入之前清空肠道。迁移运动复合体包括静止期和强烈的运动和分泌活动期之间的交替,这种刻板模式有助于保持肠道的健康和功能正常。
在进食开始之前,消化系统已经开始做准备工作,例如,预期即将进食的膳食会刺激唾液和胃液的分泌。当食物进入口腔并被吞咽后,消化系统会进入消化间期运动模式。
食物进入胃后,胃壁会收缩并几乎塌陷,这种主动放松被称为胃适应。固体颗粒通过蠕动活动激活胃窦,启动研磨过程,将食物转化为食糜。
餐后阶段胃会逐渐重新收缩,将食糜推入小肠。胃和小肠的活动会适应消化过程的要求,从口腔开始并延伸到末端回肠,食物被消化并随后被吸收。未被吸收的残留物最终到达结肠,成为肠道微生物群的底物,这些微生物会影响宿主的生理和消化功能。
因此,消化系统的正常活动对于人体的健康和营养摄入非常重要。如果消化系统不正常,可能会导致消化问题和其他健康问题。
消化不良是一种常见的临床症状,主要分为器质性消化不良和功能性消化不良。
器质性消化不良是指由某种器官疾病引起的消化不良。一般通过检查,可以明确诊断是由某个器官的病变引起的消化不良症状,例如肝病、胆道疾病、胰腺疾病、糖尿病等。
✦疾病影响消化
这些疾病会影响到消化道内部的消化酶分泌、肠道蠕动、食物吸收等消化过程,导致消化不良症状的出现,如腹胀、腹泻、便秘、恶心、呕吐、胃痛等。

功能性消化不良是最常见的消化系统疾病,大部分消化不良患者都是属于功能性消化不良。功能性消化不良没有明显的器质性病变,但出现了消化不良症状。
由于功能性消化不良患者病情的反复性与迁延不愈,其生活质量明显降低,同时,所带来的社会医疗成本的增加和生产力的下降也不容忽视。
▸ 发病率
在全球范围内,功能性消化不良女性发病率普遍高于男性。不同地区功能性消化不良的患病率也存在明显差异 :西方国家总体较高,约为10%~40%,亚洲国家较低约为5%~30%。
▸高发人群
•饮食不规律的人群:饮食不规律、暴饮暴食、偏食等不良饮食习惯容易导致消化不良。
•长期服用药物的人群:长期服用某些药物,如抗生素、非甾体抗炎药等,可能破坏肠道菌群平衡,导致消化不良。
•精神压力大的人群:精神压力大、情绪波动较大的人群容易出现消化不良症状。
•高龄人群:随着年龄的增长,人体消化功能逐渐下降,高龄人群容易出现消化不良症状。
•婴幼儿:婴幼儿是消化不良的高发人群之一。由于婴幼儿的消化系统尚未完全发育成熟,消化酶的分泌不足,肠道菌群的构成不稳定等因素,使得婴幼儿容易出现消化不良的症状。

▸ 诊断标准与症状
功能性消化不良的诊断标准:
•餐后饱胀感:即在正常量的膳食后感到不舒服;
•早饱:进食后不久即有饱感,以致摄入食物明显减少,无法完成正常量的膳食;
•上腹疼痛或灼痛:上腹胀多发生于餐后,或呈持续性进餐后加重。
注:满足以上一项或多项,并且没有可以解释这些症状的任何结构性疾病(包括上消化道内窥镜检查)的证据;则可以判断为功能性消化不良
根据主要临床症状可大致分为上腹痛综合征和餐后不适综合征两种亚型。
✦持续时间长,并伴有精神症状
在病程中症状也可发生变化,起病多缓慢,经年累月,持续性或反复发作,不少患者有饮食,精神等症状。
早饱和上腹胀常伴有嗳气。恶心、呕吐并不常见,往往发生在胃排空明显延迟的患者,呕吐多为当餐胃内容物。
不少患者同时伴有失眠、焦虑、抑郁、头痛、注意力不集中等精神症状。这些症状在部分患者中与“恐癌”心理有关。
▸ 功能性消化不良和其他胃肠道疾病的关系
伴有肠易激综合征
许多功能性消化不良患者报告伴有肠易激综合征症状,并且这两种症状经常一同出现在更严重的患者中。
注:虽然这两种情况或多或少都与特定的胃肠道症状有关,但没有结构或生化异常可以解释这些症状。
胰腺功能异常
少数但一定人群的功能性消化不良患者存在胰酶异常或胰腺外分泌功能障碍。胰酶异常和胰腺外分泌功能障碍是否直接影响功能性消化不良症状尚不清楚。
肝脏疾病与消化不良直接存在关联
肝脏疾病如肝硬化、肝炎等也可能导致功能性消化不良症状,如腹胀、食欲不振等。这些疾病可能会影响肠道菌群的平衡,导致消化不良。
功能性消化不良的发病机制尚未完全阐明,可能与胃肠运动异常、内脏超敏反应、肠道菌群紊乱、病原微生物感染、遗传因素、社会心理和神经因素、环境因素等多方面调控异常有关。
需要注意的是,肠道菌群数量和紊乱是功能性消化不良的重要病因之一,将在下文中重点讲述。
➤ 1
√功能性消化不良患者胃排空受损
功能性消化不良的发病机制涉及胃调节、胃排空和十二指肠运动的紊乱。
胃排空——食物由胃排入十二指肠的过程称为胃排空。
一项随机、双盲对照研究发现,功能性消化不良患者的症状与胃调节受损之间存在密切关系 。几份报告表明,一些功能性消化不良患者胃排空受损,一项荟萃分析表明,几乎35%的功能性消化不良患者胃排空明显延迟。
√内脏高敏感性影响功能性消化不良
内脏高敏感性是导致功能性消化不良发展的关键病理生理机制。内脏超敏反应可以通过对肠道机械化学刺激的感知增加,这通常表现为疼痛和灼痛感加重。
辣椒素受体(Trpv1)的激活由神经生长因子 (NGF)、热刺激、辣椒素、前列腺素、酸性pH值和炎症介质触发,进一步释放加剧内脏痛觉的神经肽。
那么一般哪些因素会引起或导致人体胃排空受损或内脏高敏感性,常见的原因如下:
1.神经调节失衡:人体内部的神经系统对内脏的感知和调节非常重要。如果神经系统出现失衡,会导致内脏高敏感性,从而引起胃排空受损以及消化不良等症状。
2.饮食不当:食物的种类、质量和摄入量都可能影响胃排空。比如,吃得太快、咀嚼不充分、进食过多或者过少都会导致胃排空受损。
饮食中过多的刺激性食物(如辛辣、油腻的食物)或者过多的咖啡因、酒精等刺激物质,都可能刺激内脏,导致内脏高敏感性。
3.精神压力:长期的精神紧张、焦虑、抑郁等情绪问题,也会影响人体内部神经系统的平衡,进而导致胃排空受损或内脏高敏感性。
4.慢性疾病:一些慢性疾病,如炎症性肠病、胃溃疡等,也会引起胃排空受损以及内脏高敏感性,从而导致消化不良等症状。
5.长期用药:某些药物,如非甾体类抗炎药、抗生素等,长期使用也可能导致内脏高敏感性。
➤ 2
社会心理因素会导致功能性消化不良症状。
√心理对功能性消化不良具有重要影响
一项针对瑞典人群的研究表明,在10年的随访后,焦虑会使患功能性消化不良的风险增加近8倍。在日本,儿童时期的被虐待与功能性消化不良和功能性消化不良症状的严重程度有关。
此外,病理生理学研究表明,社会心理因素和精神障碍可能通过调节大脑中的信号处理在功能性消化不良中发挥作用以及应激激素对痛觉的影响。
社会心理因素和应激激素也会影响胃肠道的其他方面,例如运动、免疫系统激活、渗透性和微生物群。
√功能性消化不良对精神疾病也有反作用
多项研究强调,与健康人相比,功能性消化不良患者的焦虑和抑郁患病率显著增加。这些观察结果表明,精神疾病在功能性消化不良的发病机制中起着重要作用。
另一方面,由于低度肠道炎症中的细胞因子反应,功能性消化不良症状被认为会诱发焦虑或抑郁,这在功能性消化不良患者心理困扰的发展中起着重要作用。
➤ 3
家族史和遗传多态性可能与功能性消化不良相关。许多研究报告了功能性消化不良风险与遗传多态性之间的关联。
遗传多态性是在同一群体中,某个基因座上存在两个或两个以上的等位基因,且等位基因的频率大于0.01的现象。
GNB3 825C>T、SCL6A4 5HTTLPR、CCK-1R 779T>C等基因多态性被认为与功能性消化不良相关。最近的一项荟萃分析发现,GNB3 825C>T中的次要等位基因 (T) 与上腹疼痛综合征亚型的易感性增加有关。
➤ 4
运动不足、睡眠障碍、高脂肪摄入和饮食不规律等生活方式因素与功能性消化不良的病理生理学有关。
√不健康生活方式导致功能性消化不良发病率高
研究发现,睡眠障碍和运动不足与功能性消化不良相关;脂肪摄入过多会加重功能性消化不良的临床症状,不规律的饮食模式也与功能性消化不良相关。
➤ 5
研究表明,沙门氏菌、幽门螺杆菌、空肠弯曲菌、蓝氏贾第鞭毛虫和诺如病毒等病原微生物导致的急性肠胃炎均与功能性消化不良症状有关。
其机制可能与导致胃肠道Cajal间质细胞(ICC)和肌间神经节受损有关,其发生率在功能性消化不良患者约为10%。
√病因微生物感染会加重消化不良症状
最近的一项荟萃分析证实,根除病原体后症状改善比未治疗的对照组效果更好。在持续时间超过4-12周且内镜检查已排除器质性原因的消化不良患者中,成功根除幽门螺杆菌可使症状缓解率提高10%至15%(或至少症状改善)。
➤ 6
来自动物和临床研究的证据表明,肠道菌群在功能性消化不良中起着重要的作用,影响许多致病机制,包括胃肠动力受损、内脏敏感性、免疫激活、粘膜通透性增加和肠脑轴功能改变等。
在人体中,微生物(包括细菌、古细菌、病毒和真菌)的数量远远超过宿主细胞的数量。
微生物组在调节生理功能(包括胃肠运动功能、上皮屏障保护以及肠道和中枢神经系统之间的相互作用)中发挥着重要作用。然而,微生物失调在功能性消化不良患者中非常常见。
•普雷沃氏菌丰度降低
一项研究将功能性消化不良患者的胃液成分与健康对照组进行了比较,报告称与对照组相比,功能性消化不良中普雷沃氏菌属(Prevotella)的频率显著降低。
这种生态失调的原因可能是胃排空延迟,这可能会改变胃的酸度、粘液稠度和部分氧合作用,从而改变胃的细菌定植。
扩展阅读:肠道重要基石菌属——普雷沃氏菌属 Prevotella
•拟杆菌与变形菌比例增加
功能性消化不良患者胃液的微生物群显示拟杆菌与变形菌的比例增加,而未检测到酸杆菌(Acidobacteria)。然而,健康人的胃液中含有酸杆菌,并且拟杆菌与变形菌的比例较低。
•细菌代谢物分泌出现异常
与对照组相比,功能性消化不良患者的胆汁酸阳性胃液样本比例增加更多。由于胆汁酸从十二指肠反流到胃中,生理上发生在胃运动期间,功能性消化不良患者可能会出现胃动力障碍。
物种丰富度的增加表明胃液微生物群的数量和多样性足以使细菌的代谢物和成分影响胃。因此,可能表明肠道的有毒细菌细胞成分,如脂多糖,刺激白细胞产生促炎细胞因子,引发胃部炎症,从而增加粘膜通透性,这可能导致胃肠神经系统功能障碍。
由于脂多糖和胆汁酸会增加粘膜的通透性,因此患者的炎症可能是由含有此类潜在毒性物质的液体回流引起的。
当比较功能性消化不良患者和健康对照组的上消化道微生物组时,功能性消化不良组口腔、食道、胃和十二指肠中的链球菌(Streptococcus)水平较高。
链球菌丰度与上消化道不适呈正相关,表明链球菌与功能性消化不良患者的胃肠道症状之间存在联系。此外,功能性消化不良组表现出更高水平的厚壁菌门。
• 厚壁菌、变形菌丰度增加
根据这些发现,在一项比较功能性消化不良患者和健康受试者十二指肠粘膜微生物群的不同研究中,十二指肠粘膜中最普遍菌属中也有链球菌。
此外,通过分析功能性消化不良和肝郁脾虚综合征大鼠的粪便样本的微生物组成,与对照组相比,该模型中厚壁菌门、变形菌门(Proteobacteria)和蓝藻门(Cyanobacteria)的水平升高,而拟杆菌门的丰度较低。
上述研究表明,功能性消化不良中的微生物改变并不局限于胃肠道中的一个部位,突出了稳态失衡在这些疾病的发病机制中的潜在重要性。
小肠细菌过度生长 (SIFO) 的定义是小肠中存在过量的细菌,并且与胃肠道症状有关。
最近的研究分析了功能性消化不良患者十二指肠的细菌属水平。
链球菌相对丰度增加
澳大利亚的一项研究报道,与对照组相比,功能性消化不良患者的链球菌(Streptococcus)相对丰度增加,尽管不显著(这可能是因为样本量小),并且链球菌的丰度与厌氧属普雷沃氏菌、韦荣球菌(Veillonella)和放线菌(Actinomyces)的丰度之间存在负相关,这些菌群功能性消化不良患者中显著减少。
β多样性发生显著变化
此外,报告的十二指肠的β多样性在患者和对照组之间存在显著差异,而α多样性保持不变,表明该疾病可能涉及更复杂的微生物群结构变化,而不是仅特定属的相对丰度变化。
奈瑟菌和卟啉单胞菌丰度降低
最近的一项研究表明,在使用质子泵抑制剂(PPI)治疗之前,功能性消化不良患者和对照组的十二指肠粘膜奈瑟菌(Neisseria)和卟啉单胞菌(Porphyromonas)丰度降低,但微生物负荷没有差异。
总体而言,研究证实功能性消化不良患者中确实发生了微生物数量和多样性方面的明显变化。
功能性消化不良患者的微生物变化




Brown G,et al.Neurogastroenterol Motil.2022
谷禾还检测了一些功能性消化不良人群的肠道菌群,虽然个体直接存在一定差异,但是共同之处是菌群紊乱,核心菌异常较多,多项病原菌超标。
案例一
基本信息和病症:
1岁3个月,食物过敏,厌食,功能性消化不良,生长发育不良
菌群构成:
重要菌群情况:


注:高亮部分是异常菌属。
总结:菌群失衡,核心菌属异常较多,普雷沃氏菌属(Prevotella)缺乏,多种致病菌超标。
案例二
基本信息和病症:
43岁,每天大便不成形;容易急性肠胃炎;消化不良
菌群构成:
重要菌群情况:
<来源:谷禾健康肠道菌群检测数据库>
总结:菌群失衡,菌群多样性低,核心菌属大量异常,致病菌和病原菌多项超标。
功能性消化不良是一种复杂的多因素导致的功能性胃肠疾病,目前其确切的发病机制尚不明确。
★ 肠道菌群在功能性消化不良中起重要作用
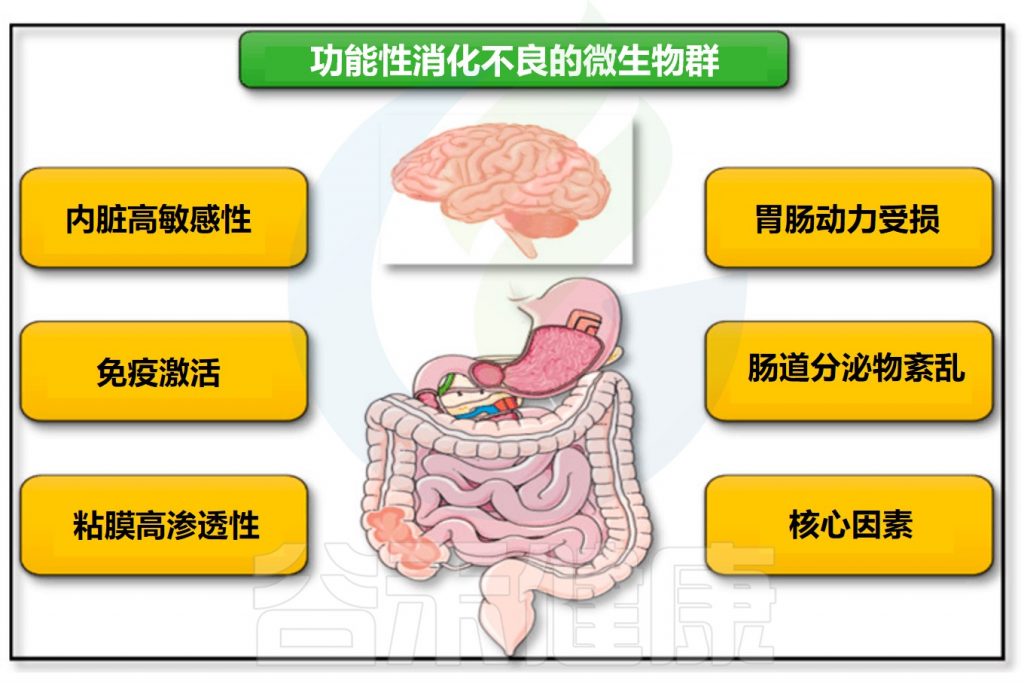
多个研究表明,肠道菌群紊乱在功能性消化不良的发生发展起着重要作用,是功能性消化不良的重要发病机制。影响包括胃肠动力受损、内脏高敏感性、免疫激活、粘膜通透性增加和中枢神经系统疾病。
肠道微生物群参与功能性消化不良发病机制

Tziatzios G,et al.Microorganisms.2020
胃肠运动异常是功能性消化不良的基本病理机制,包括胃排空延迟、胃容受性舒张受损及消化期间移行性复合运动(MMC)III期异常。
✦外来菌的增殖抑制胃肠道蠕动
消化期胃和小肠移行性复合运动可将胃肠内容物和致病菌机械性地迁移向远端肠道,而功能性消化不良患者经常出现非传播性和逆传性活动,这可能诱导或加重近端小肠细菌随反流十二指肠液迁移到胃。
而胃排空延迟又导致反流菌能较长时间保留在胃和十二指肠中,引起菌群分布失调的同时,外来菌的增殖可产生内毒素脂多糖刺激免疫应答,进而,抑制胃肠道蠕动,加重功能性消化不良症状。
✦细菌代谢物是胃肠动力的重要来源
短链脂肪酸是肠道共生菌发酵膳食纤维代谢物,除了作为胃肠动力的重要能量来源,也可直接激活肠神经系统以调控肠内分泌细胞合成和分泌某些胃肠激素,如肠激素肽、胆囊收缩素、胰高血糖素样肽等,进而调节胃肠道动力及胃排空。然而在代谢组学研究中发现功能性消化不良大鼠存在短链脂肪酸水平显著降低。
更准确地说,细菌产生的短链脂肪酸不仅调节功能性消化不良中十二指肠碳酸氢盐的分泌,同时它们在十二指肠的快速吸收也可能影响管腔细菌定植抑制。
5-羟色胺影响胃肠道动力
相关研究证实肠道微生物群在调节5-羟色胺(5-HT)合成中起着关键作用,而5-羟色胺水平是影响胃肠道动力的重要因素之一。
大肠杆菌产生的脂多糖延迟胃排空
此外,已发现大肠杆菌(Escherichia coli)产生的细菌脂多糖会导致胃排空显著延迟 ,而双歧杆菌当用作益生菌使用时时可显著增强小肠蠕动。
由上述研究可知,胃肠动力异常可能诱发菌群失调,而菌群失调可反过来进步一步影响胃肠动力,介导功能性消化不良发生发展,但菌群失调与功能性消化不良的因果关系仍需更深入的研究加以验证。
✦心理因素和肠道炎症相互影响
功能性消化不良患者处于十二指肠低度炎症状态。这种慢性低级别的炎症反应会引起疼痛和敏感性。
此外,焦虑或抑郁等心理学因素似乎与慢性炎症反应相互影响,心理压力和由此产生的皮质醇释放激素分泌也会增加十二指肠的局部炎症和全身炎症反应。
✦炎症状态下肠屏障功能发生改变
多项研究报道了功能性消化不良患者肠道中免疫细胞(如肥大细胞、巨噬细胞和嗜酸性粒细胞)数量的增加。
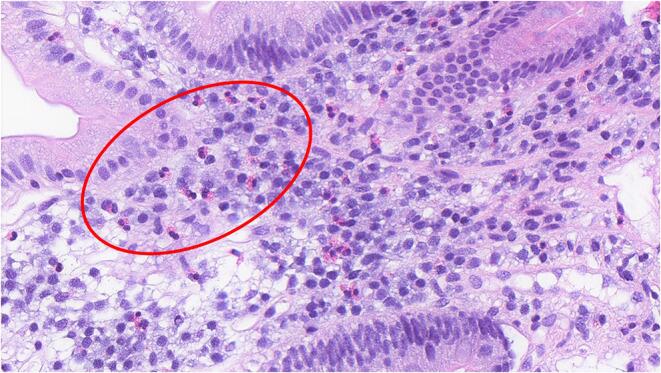
十二指肠活检切片中嗜酸性粒细胞增多

Hari S,et al.Front Allergy.2022
活化的肥大细胞释放细胞因子、组胺、前列腺素和类胰蛋白酶,它们与肠屏障功能障碍和伤害感受通路的改变有关。
嗜酸性粒细胞影响肠道通透性
在生理条件下,十二指肠屏障由粘液层、上皮和下面的固有层组成。在功能性消化不良中,屏障的功能障碍体现为粘膜完整性丧失和通透性增加。
此外,功能性消化不良中嗜酸性粒细胞的募集和激活也发生,这可能使组织损伤和屏障功能障碍持续存在。
正常与功能性消化不良的屏障功能对比
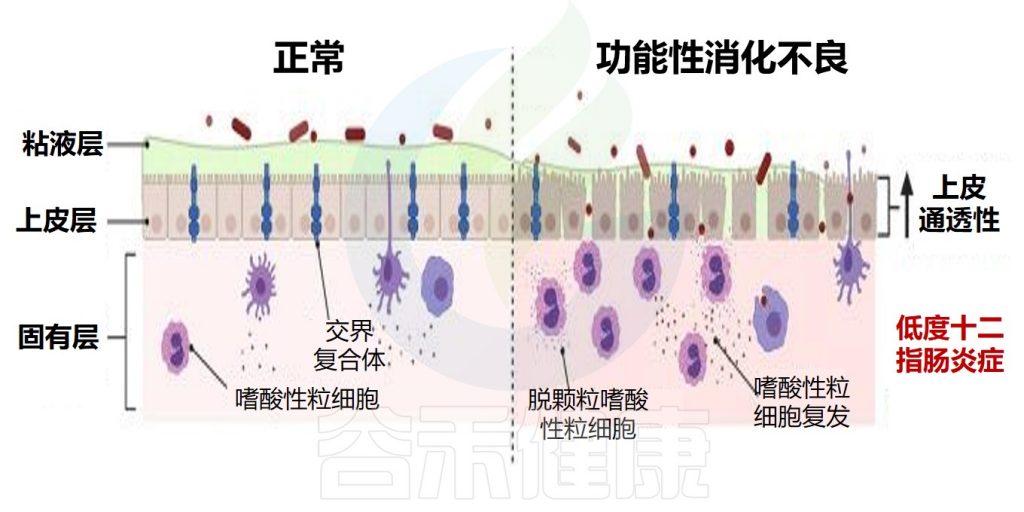

Hari S,et al.Front Allergy.2022
✦肠道菌群也会破坏肠道通透性
肠腔黏膜表面是毒性和免疫原性颗粒面对黏膜相关免疫系统的第一道屏障。肠道微生物菌群失调在功能性消化不良中非常常见。
有害菌破坏紧密连接增加肠道通透性
有研究表明,霍乱弧菌(V.cholera)、艰难梭菌(Clostridium difficile)和产毒素的大肠杆菌的各种菌株已显示可通过直接破坏紧密连接(TJ)、毒素或蛋白酶的产生以及炎症级联反应的活化来增强肠道通透性。
益生菌可以促进屏障完整性
相反,肠道益生菌可以通过增加闭合蛋白、紧密连接蛋白ZO-1和ZO-2的表达来促进屏障完整性。
另外,肠道菌群的某些代谢产物也能够破坏肠道通透性,例如,细菌细胞壁脂多糖易位可以诱导免疫反应和炎症反应,加剧肠道屏障损害并进一步增加肠道的通透性。
扩展阅读:什么是肠漏综合征,它如何影响健康?
除了影响黏膜屏障功能,微生物群也参与调节免疫系统的发育和功能,在先天免疫系统中起关键作用。
✦肠道菌群调节肠上皮免疫功能
研究发现,肠道微生物区系通过激活肠上皮细胞的模式识别受体和内质网应微信号促进免疫系统的发育,从而促进肠上皮细胞的增殖、抗菌肽和黏液的产生,调节与免疫功能相关细胞因子的分泌,如白细胞介素1β、白细胞介素18和白细胞介素25。
✦微生物代谢物也可促进免疫反应
肠道微生物群及其代谢物还通过Toll样受体(TLRs-TLR2、TLR4)干扰信号,并促进促炎细胞因子的产生和免疫反应。
此外,它们的代谢物也可能具有炎症特性或对T细胞分化产生直接影响。
小结
综上所述,肠道菌群失调在影响黏膜生物屏障的同时,可能也导致黏膜机械屏障、化学屏障及免疫屏障等多种屏障功能低下,并可能介导黏膜低度炎症和内脏高敏性的发生。
生物失调的影响
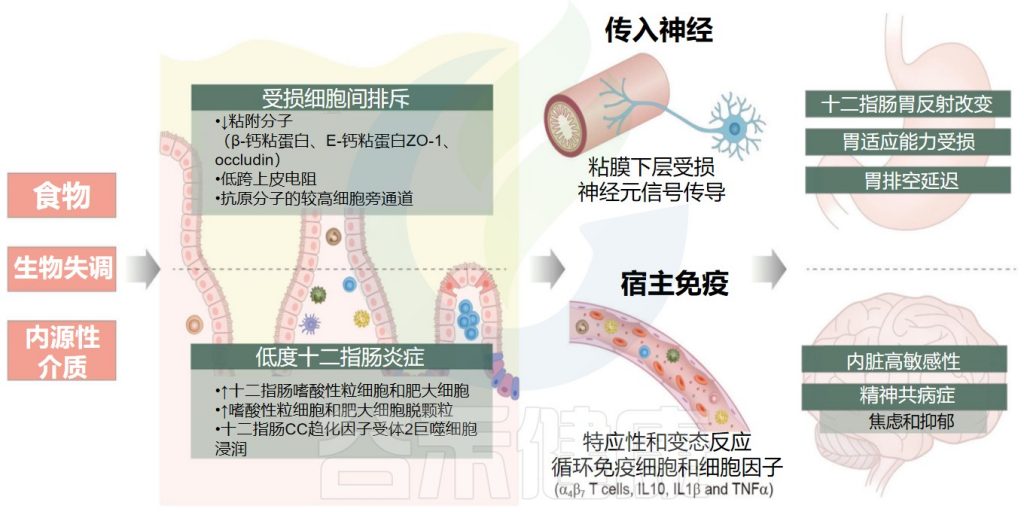

Kim SH.Korean J Gastroenterol.2022
脑-肠轴是连接胃肠道和中枢神经系统的传导通路,包括中枢神经系统、脑和脊髓、自主神经系统、肠神经系统和下丘脑垂体肾上腺轴。
肠道菌群与脑-肠轴存在双向交互作用:
肠道菌群通过神经、内分泌、免疫等至少三种途径与中枢神经系统相通;中枢神经系统调节肠道的运动、分泌及肠道黏膜通透性来影响肠道菌群,或通过肠腔分泌的激素直接调节菌群的基因表达而影响肠道菌群的组成和功能。
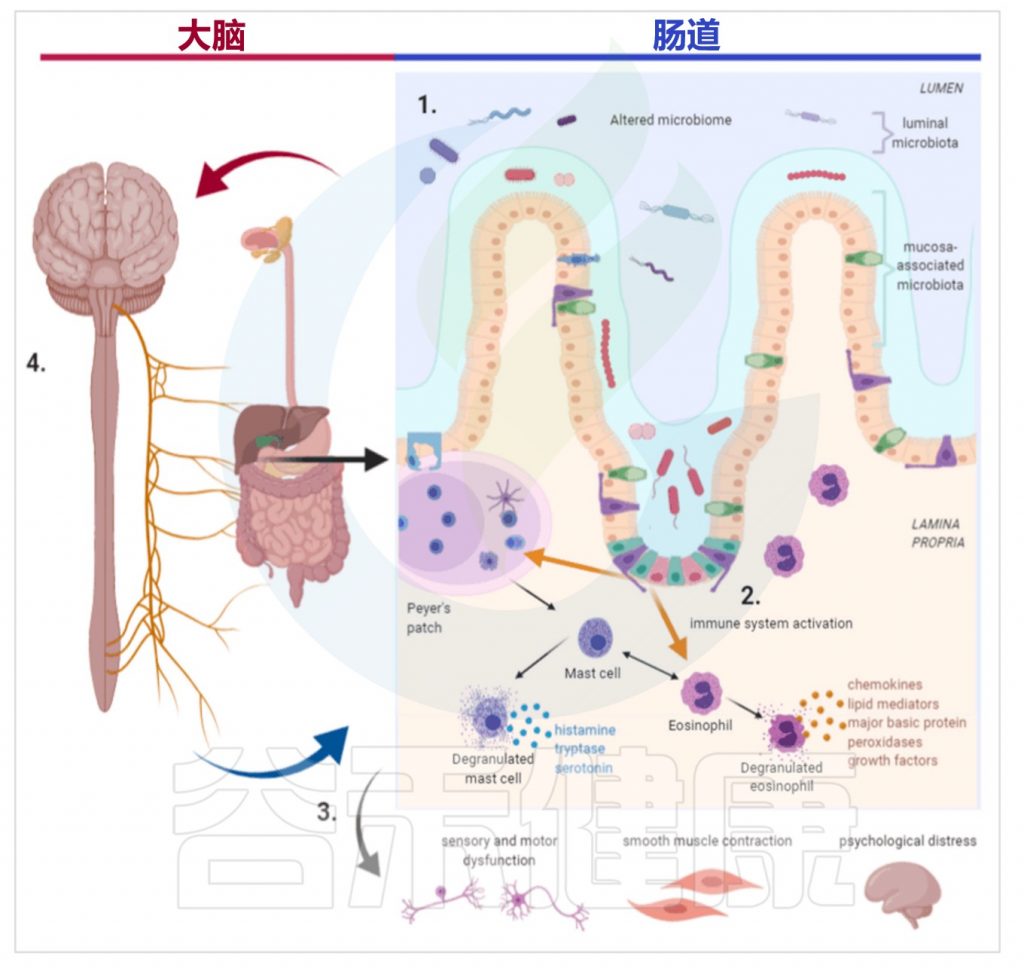

Brown G,et al.Neurogastroenterol Motil.2022
肠道菌群通过肠-脑轴影响5-羟色胺、脑源性神经营养生长因子(BNDF)等神经递质的合成、释放和下丘脑-垂体-肾上腺轴的发育,调节中枢神经系统的发育、功能和行为,影响焦虑、抑郁样情绪障碍和应激反应。
✦功能性消化不良患者伴有精神症状
功能性消化不良属于典型的身心疾病,常有焦虑、抑郁的表现。一项为期12年的前瞻性研究也说明功能性消化不良患者中有1/2~2/3先出现焦虑再有胃肠道症状,提示了功能性消化不良的中枢发病机制。
功能性消化不良患者的中枢神经系统改变
功能性神经影像学,如功能核磁共振,使我们能直接观察到中枢神经系统的改变,证明其与功能性消化不良症状的相关性。
结果显示功能性消化不良患者额叶皮层、感觉皮层、脑岛、前扣带皮层、丘脑、海马体和杏仁核的改变,与功能性消化不良内脏超敏反应、消化不良症状、生活质量及焦虑和抑郁有关。
精神心理应激影响内脏高敏感性
有研究认为功能性消化不良患者在精神心理应激的情况下可通过刺激中枢神经系统的情感活动系统参与患者内脏高敏感性的形成,从而对胃肠道产生影响。
因此,研究人员推测肠道菌群通过脑-肠轴对中枢神经系统的调节,导致消化不良伴焦虑抑郁的发生发展。
✦肠道菌群对肠脑轴具有重要作用
随着研究不断深入,越来越多证据证实肠道菌群与脑-肠-菌群轴功能密切相关。
菌群代谢物影响中枢神经系统信号
一方面,肠道微生物能通过合成和释放短链脂肪酸、次级胆汁酸和色氨酸等代谢物,与肠内分泌细胞,自下而上影响激活内源性中枢神经系统信号机制。
此外,肠道菌群失调会增加脂多糖的产生和分泌,而脂多糖不仅是影响中枢神经系统功能的一个强有力的因素,还会促进其他炎症因子的产生,如肿瘤坏死因子﹣α、白细胞介素﹣1β等,以上炎症因子可影响中枢神经系统的功能, 紊乱神经肽的合成和分泌。
激活肠神经系统信号
另一方面,肠道微生物也可独立完成或促进某些神经活性物质的合成和分泌,如5-羟色胺、γ-氨基丁酸、去甲肾上腺素、多巴胺等,这些小分子物质能通过与肠内分泌细胞相互作用,激活肠神经系统信号机制。
上述研究表明,肠道菌群失调可能诱导脑-肠-微生物群轴功能紊乱,其机制涉及神经、免疫和内分泌信号传导。
✦迷走神经的作用
在生态失调和胃肠道不适的精神状况的背景下,应该注意的一个重要方面是迷走神经的作用。
众所周知,压力会增加肠道通透性并通过各种神经调节剂改变胃肠道微生物组的组成,同时压力可以减少迷走神经活动,这可能会促进胃肠道炎症。
注:虽然目前还没有关于迷走神经刺激对胃肠道微生物组影响的数据,但可以推测迷走神经可能通过其对肠道通透性的影响对肠道微生物组产生影响。
迷走神经的传出可能在肠道中具有抗炎作用,同时降低肠道通透性——这两种作用都可能归因于通过迷走神经活动加强紧密连接。
心理治疗对功能性消化不良患者微生物群和胃肠道症状的影响,也可以用迷走神经的影响来解释。
心理疾病对功能性消化不良的表现有影响
现在人们可能想知道心理疾病的患病率是否在不同的功能性消化不良亚组中有所不同。
已经表明情绪和焦虑障碍在非疼痛主导型功能性消化不良患者中,比在疼痛主导型功能性消化不良患者中更常见。
另一项研究还表明上腹痛与神经质、虐待和躯体化之间存在相关性。
这些发现强调了未来重点关注功能性消化不良患者的心理合并症和特定症状之间的可能联系,研究心理治疗对功能性消化不良患者特别是微生物组的这些有趣影响。
未来随着研究深入,有望更好地理解脑-肠-微生物组轴,功能性消化不良的治疗方法也能更加个性化。
药物经口服途径进入人体,不可避免地与肠道菌群发生相互作用。
多项研究表明,肠道微生物能通过编码多种酶,影响口服药物在体内的吸收、代谢、转化等过程,进而影响药物活性成分的有效性和毒性。
改变微生物群的药物在治疗消化不良中得到了有效证实。
利福昔明是一种广谱抗生素,具有革兰氏阳性、革兰氏阴性、需氧和厌氧覆盖,在胃肠道腔内生物利用度高,全身不良反应极小。
•消化不良症状缓解
在一项随机试验中,利福昔明治疗优于安慰剂治疗,其中79%的患者在服用利福昔明后报告消化不良症状缓解,而安慰剂组在8周时这一比例为47%。
•利于有益细菌生长
在其他胃肠道疾病中,利福昔明治疗已被证明可以保护结肠菌群,增加乳酸杆菌和双歧杆菌(Bifidobacterium)的丰度,从而产生积极效应,利于有益细菌的生长,从而改变整体成分。
扩展阅读:肠道核心菌属——双歧杆菌,你最好拥有它
•益生菌治疗后餐后腹胀改善
益生菌可能在功能性消化不良中发挥治疗作用,三项研究表明,在幽门螺杆菌阳性和阴性消化不良患者中服用益生菌加氏乳杆菌(Lactobacillus gasseri OLL2716)12周后,餐后饱腹感和腹胀症状得到改善。
35.5%的幽门螺杆菌阴性功能性消化不良患者的症状得到了缓解,而安慰剂患者的症状缓解率为17%。
扩展阅读:如果你要补充益生菌 ——益生菌补充、个体化、定植指南
•恢复胃液微生物群
加氏乳杆菌还被证明可以“恢复”功能性消化不良患者异常的胃液微生物群。在服用益生菌12周之前,功能性消化不良患者的胃液显示拟杆菌比变形菌占优势,并且不存在酸杆菌(Acidobacteria),益生菌治疗后,这些比例发生了变化,以反映与健康志愿者相似的微生物群组成。
此外,虽然没有收集小肠细菌数据,但用凝结芽孢杆菌(Bacillus coagulans MY01)和枯草芽孢杆菌(Bacillus subtilis MY02)治疗8周后,功能性消化不良患者的有效率比安慰剂提高了28%。
这些发现提供了初步证据,表明患者症状与肠道微生物群中某些属的丰度有关,这些微生物群可以通过治疗性抗生素和益生菌制剂进行调节。
尽管有这些显著的具体变化,但仍需要对更多不同地理区域的患者群体进行进一步研究,以验证疾病中的微生物组变化。
15项观察性研究和一项随机对照试验的系统回顾发现,高脂肪、小麦、FODMAP(可发酵低聚糖、双糖、单糖和多元醇)和咖啡因等天然食品化学物质含量高的食物与功能性消化不良有关。
扩展阅读:肠道微生物群与健康:探究发酵食品、饮食方式、益生菌和后生元的影响
饮酒与功能性消化不良症状的关联尚不明确,可能取决于酒精的类型。
以下是一些饮食调理建议:
•少食多餐:每天分成5-6餐,每餐食量适中,避免暴饮暴食。
•避免过度饮酒和吸烟:酒精和烟草会刺激胃肠道,加重消化不良症状。
•避免食用刺激性食物:如辛辣、油腻、烧烤等食物,容易引起胃肠道不适。
•增加膳食纤维摄入量:膳食纤维可以促进肠道蠕动,缓解便秘症状。建议多食用蔬菜、水果、全谷类食品等富含膳食纤维的食物。
•注意饮食卫生:避免食用过期食品、生冷食物等容易引起胃肠道感染的食物。
•喝足够的水:保持足够的水分摄入可以促进肠道蠕动,缓解便秘症状。
•食用易消化的食物:如米粥、面条、煮熟的蔬菜等,可以减轻胃肠道负担,缓解消化不良症状。
注意:饮食调理虽然可以缓解功能性消化不良症状,但如果症状持续或加重,建议及时就医进行诊断和治疗。
扩展阅读:20种有效改善肠道健康的科学方法
胃酸抑制的主要药物包括两类药物:质子泵抑制剂(PPI)和组胺H2受体拮抗剂。
•上腹痛和反流症状缓解
一项荟萃分析发现,在两到八周内服用标准剂量的质子泵抑制剂,在减少功能性消化不良症状方面优于安慰剂。
注:治疗益处仅出现在有上腹灼痛和反流样疼痛的患者身上,而不出现在有运动障碍相关问题的患者身上。
•消化不良症状的风险降低
组胺H2受体拮抗剂也是功能性消化不良治疗的一种选择。十二项随机对照试验将H2拮抗剂与安慰剂进行了比较,显示消化不良症状的相对风险降低了23%。
对于以进餐相关症状为特征的功能性消化不良伴餐后窘迫综合征的患者,促胃肠动力药可以减轻症状。
对29项比较促动力药(主要是西沙必利)与安慰剂的试验的回顾发现,功能性消化不良的整体症状显著减少。
对功能性消化不良患者的12项随机对照试验的回顾发现,心理治疗(即认知行为治疗和其他形式的心理治疗)比对照组具有统计学上的显著益处。
认知行为疗法:是一种心理治疗方法,旨在通过调整患者的思维方式和行为习惯,来改善其身体和心理健康问题。在功能性消化不良患者治疗中,认知行为疗法可以帮助患者意识到自己不良的思维和行为习惯,如过度焦虑、负面情绪、饮食习惯不当等,从而通过改变这些不良习惯,减轻和预防消化不良症状的发生。
此外,认知行为疗法还可以帮助患者学会应对压力和情绪问题的技巧,提高其身心健康水平。
•症状严重程度、疼痛强度减弱
认知行为疗法 (CBT)导致症状严重程度、疼痛强度减弱,并进一步降低疾病对患者生活的影响。
•胃动力显著增加
此外,与对照组相比,干预组胃排空率显著增加,胃动力参数发生变化。基于这些结果,认知行为疗法可被视为功能性消化不良患者的有效治疗选择。
注:心理治疗不应作为功能性消化不良的一线治疗,因为数据质量低,缺乏盲法心理干预和主观症状评分导致偏倚风险高。如果药物治疗无效,可以考虑心理治疗。
中药具有多成分、多靶点和多途径的优势,目前中医对功能性消化不良肠道菌群的研究仍处于探索阶段。
中药在治疗功能性消化不良方面具有一定的疗效,可以缓解消化不良症状,改善消化功能,提高生活质量。研究证实,中药能逆转肠道菌群失调,维持肠道微生态平衡。
注:中药治疗功能性消化不良应根据患者的具体情况进行个体化治疗,避免不必要的药物过敏或不良反应。同时,中药治疗应在专业医生的指导下进行。
基于菌群干预后,原先消化不良患者的肠道菌群有了好转:
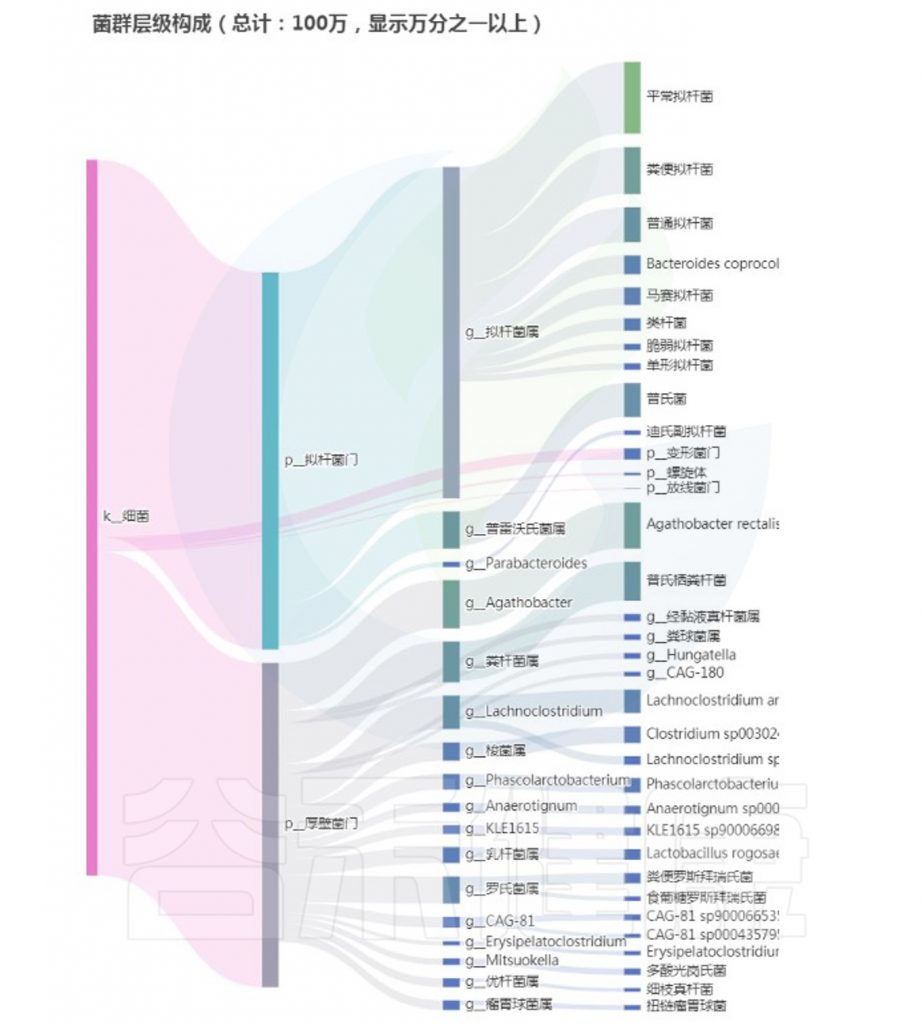

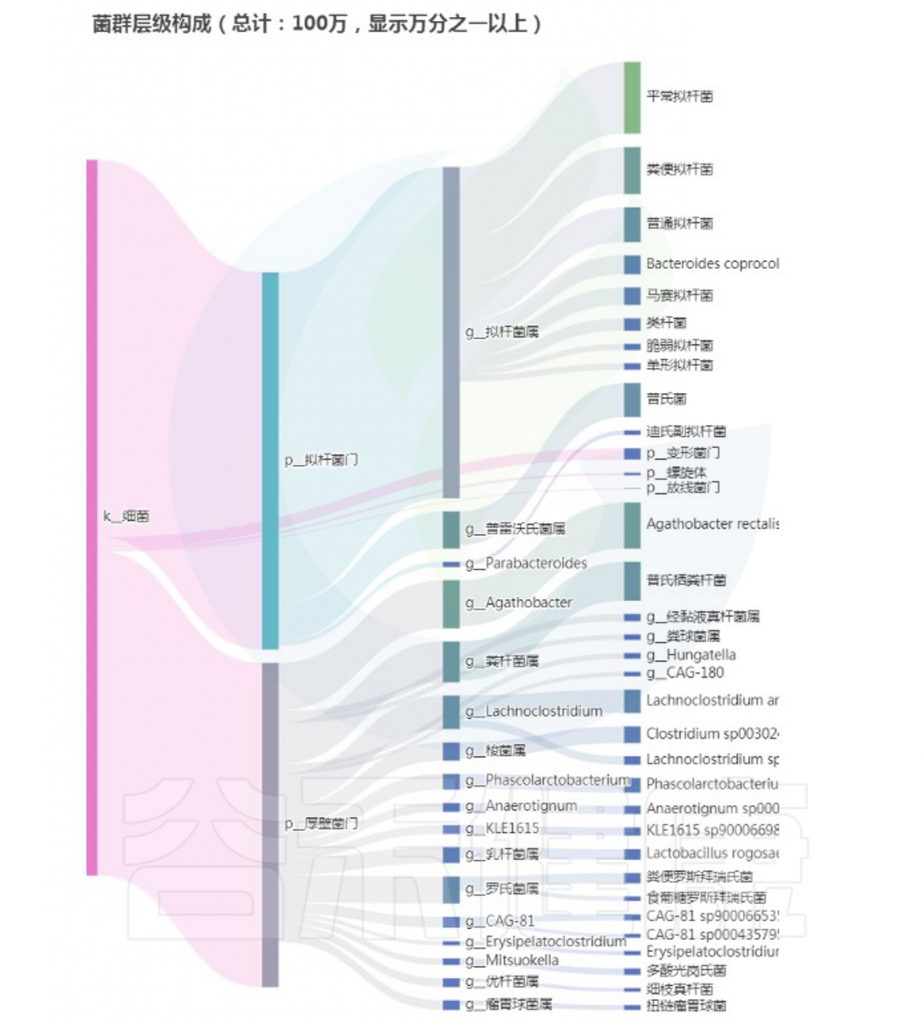






<来源:谷禾健康肠道菌群检测数据库>
从以上菌群评估报告显示,核心菌属和重要菌属没有异常,整体菌群平衡和多样性良好,健康评分显著提升,检测者反馈消化不良症状大幅减轻。
功能性消化不良是一种常见的消化系统疾病,其症状包括腹胀、腹泻、便秘等,给患者带来很大的困扰和痛苦。目前,传统的治疗方法主要是针对症状进行缓解,但效果并不理想。近年来的研究表明,肠道微生物与功能性消化不良密切相关,肠道菌群干预成为了治疗功能性消化不良的新途径。
谷禾的肠道菌群检测经验和研究表明肠道菌群的紊乱和异常特征可以对部分功能性消化不良患者症状背后的病理进行分类和解析,为针对性干预和改善提供了新的可能。对肠道微生物与功能性消化不良之间的关系的进一步深入研究和应用有望带来更有效的治疗方法。
主要参考文献
Singh R, Zogg H, Ghoshal UC, Ro S. Current Treatment Options and Therapeutic Insights for Gastrointestinal Dysmotility and Functional Gastrointestinal Disorders. Front Pharmacol. 2022 Jan 25;13:808195. doi: 10.3389/fphar.2022.808195. PMID: 35145413; PMCID: PMC8822166.
Martinez-Guryn K, Hubert N, Frazier K, Urlass S, Musch MW, Ojeda P, Pierre JF, Miyoshi J, Sontag TJ, Cham CM, Reardon CA, Leone V, Chang EB. Small Intestine Microbiota Regulate Host Digestive and Absorptive Adaptive Responses to Dietary Lipids. Cell Host Microbe. 2018 Apr 11;23(4):458-469.e5. doi: 10.1016/j.chom.2018.03.011. PMID: 29649441; PMCID: PMC5912695.
Shah A, Talley NJ, Holtmann G. Current and Future Approaches for Diagnosing Small Intestinal Dysbiosis in Patients With Symptoms of Functional Dyspepsia. Front Neurosci. 2022 May 6;16:830356. doi: 10.3389/fnins.2022.830356. PMID: 35600619; PMCID: PMC9121133.
Miwa H, Nagahara A, Asakawa A, Arai M, Oshima T, Kasugai K, Kamada K, Suzuki H, Tanaka F, Tominaga K, Futagami S, Hojo M, Mihara H, Higuchi K, Kusano M, Arisawa T, Kato M, Joh T, Mochida S, Enomoto N, Shimosegawa T, Koike K. Evidence-based clinical practice guidelines for functional dyspepsia 2021. J Gastroenterol. 2022 Feb;57(2):47-61. doi: 10.1007/s00535-021-01843-7. Epub 2022 Jan 21. PMID: 35061057; PMCID: PMC8831363.
Mounsey A, Barzin A, Rietz A. Functional Dyspepsia: Evaluation and Management. Am Fam Physician. 2020 Jan 15;101(2):84-88. PMID: 31939638.
Tziatzios G, Gkolfakis P, Papanikolaou IS, Mathur R, Pimentel M, Giamarellos-Bourboulis EJ, Triantafyllou K. Gut Microbiota Dysbiosis in Functional Dyspepsia. Microorganisms. 2020 May 8;8(5):691. doi: 10.3390/microorganisms8050691. PMID: 32397332; PMCID: PMC7285034.
Brown G, Hoedt EC, Keely S, Shah A, Walker MM, Holtmann G, Talley NJ. Role of the duodenal microbiota in functional dyspepsia. Neurogastroenterol Motil. 2022 Nov;34(11):e14372. doi: 10.1111/nmo.14372. Epub 2022 Apr 11. PMID: 35403776; PMCID: PMC9786680.
Hari S, Burns GL, Hoedt EC, Keely S, Talley NJ. Eosinophils, Hypoxia-Inducible Factors, and Barrier Dysfunction in Functional Dyspepsia. Front Allergy. 2022 May 31;3:851482. doi: 10.3389/falgy.2022.851482. PMID: 35769556; PMCID: PMC9234913.
Rupp SK, Stengel A. Bi-Directionality of the Microbiota-Gut-Brain Axis in Patients With Functional Dyspepsia: Relevance of Psychotherapy and Probiotics. Front Neurosci. 2022 Feb 28;16:844564. doi: 10.3389/fnins.2022.844564. PMID: 35295092; PMCID: PMC8919856.

谷禾健康

随着高通量多组学技术的快速创新推动,微生物群,尤其是肠道菌群失调已被明确与许多人类疾病有关,包括 2 型糖尿病和炎症性肠病。
多组学数据的综合分析,包括宏基因组学和代谢组学以及宿主指标的检测和细菌物种的分类,已经确定了许多与疾病相关的细菌和细菌产物。然而,深入了解微生物影响肠道健康的机制需要从关联拓展到因果关系。
目前对肠道微生物群对疾病因果关系的贡献的理解仍然有限,这主要是由于微生物群落结构的异质性、疾病进化的个体差异以及对将微生物群衍生信号整合到宿主信号通路中的机制的不完全理解。
最近,德国慕尼黑工业大学从事肠道菌群和营养研究的Haller 教授团队在《Nature reviews gastroenterology & hepatology》 (自然评论胃肠病学和肝病学)发表评论文章,系统讨论了目前已知的炎症和代谢紊乱相关微生物组的特征和认知,并讨论提高对其作用机制理解的困难所在。
在这里我们将文章整理与大家分享。


关键信息
1、肠道菌群组成的改变和细菌衍生代谢物经宿主加工后的变化与 IBD 和 T2DM 相关,并提供了共同的潜在致病机制。
2、益生菌与 IBD 或 T2DM 之间的因果关系已通过无菌小鼠实验和综合多组学研究明确。
3、对于疾病特异性生物标志物发现的挑战,包括确定观察到的变化的因果关系,了解它们在疾病机制中的功能以及肠道微生物群的地理和种族差异。
4、特定细菌菌株、其编码基因和代谢副产物的大数据细化、测试和验证对于识别疾病生物标志物是必要的。
文章内容
人体消化道含有一系列复杂的微生物,包括细菌、古细菌、病毒和真菌。由于消化道及其微生物组被认为位于免疫和代谢过程的交叉点,本文重点关注炎症性肠病 (IBD) 和 2 型糖尿病 (T2DM) 作为微生物群相关疾病的范例。
IBD 和 T2DM 都被认为是多因素疾病,随着工业化的进展其发病率在全球范围内呈上升趋势。病因涉及遗传易感性、环境诱因和城市生活方式相关因素的复杂相互作用。
在这种共同的背景下,代谢疾病(如 T2DM)的另外特征是肝脏、脂肪组织、肌肉、胰腺和肠道的慢性亚临床炎症,而炎症性胃肠道疾病,如克罗恩病(CD)和溃疡性结肠炎(UC) ,也与炎症驱动的代谢改变有关。
环境触发因素的重要性(肠道菌群)
全基因组关联研究已经确定了大量的遗传变异与 T2DM (143 位点) 或 IBD (>240 位点)的易感性增加相关。然而,这些变异共同解释了这些疾病的一小部分遗传性:T2DM < 10 %,UC < 15 % 和 CD < 50 % 。这种情况表明环境触发因素的重要性,特别是肠道微生物组,作为这些疾病病因的主要贡献者。对大型人群研究和 IBD 或 T2DM 患者队列的多项分析已经确定了与特定疾病表型、复发风险和治疗反应相关的微生物组特征。
IBD 和 T2DM 都与特征性微生物改变有关,特别是随着有益微生物的减少和病原菌的增加而降低群落多样性。尽管它们的病理学不同,IBD 和 T2DM 有几个共同的机制特征。T2DM 的代谢特征伴随着慢性低度炎症和肠道屏障的破坏,IBD 患者的复发性炎症发作与细胞和全身水平的代谢改变共同发生。
这些复杂疾病的治疗仍然具有挑战性,但粪便微生物群移植(FMT) 的对照试验已显示出对T2DM和IBD的临床疗效,包括UC以及较轻的 CD。
FMT对炎症、免疫和代谢疾病有效果但存在差异
FMT 的临床试验还提供了肠道菌群与其他炎症、免疫或代谢疾病之间存在因果关系的证据。例如,FMT 在治疗大约 90% 的艰难梭菌(以前的艰难梭菌)感染患者方面非常有效并已被评估为治疗肥胖和移植物抗宿主病。
在四项随机临床试验中,FMT 在 28% 的 UC 患者中诱导了临床缓解。但很少有临床试验检查过 FMT 在 CD 患者中的疗效,而且结果相当不同。
在一项对 174 名接受 FMT 治疗的 CD 患者进行的研究中,20% 的患者获得了临床缓解,总体而言,43% 的患者获得了临床缓解。
一项单独的随机对照试验发现 FMT 对 CD 患者的临床缓解率没有影响,但供体微生物群的植入增加与维持缓解有关。相反(尽管大量研究表明,特定的菌群失调或特定的微生物群谱与代谢紊乱有关),FMT 对代谢性疾病患者有益的证据尚不明确。具有里程碑意义的研究表明,从较瘦、健康的捐赠者那里接受 FMT 的代谢综合征患者的代谢改善以及肠道微生物组的有益变化。然而,这些影响是不一致和短暂的,这可以通过供体微生物群的有限移植或基线时供体粪便微生物多样性的变化来解释。
有趣的是,口服 FMT 后补充低发酵性纤维可改善肥胖和代谢综合征患者的胰岛素敏感性、增加微生物多样性,并延长供体微生物定植。这些数据强调了微生物调节疗法在逆转代谢功能障碍中的价值。
与这些发现一致,来自代谢受损的肥胖供体的 FMT 会暂时恶化代谢综合征受体的胰岛素敏感性,而胃旁路术后健康供体的 FMT 会导致代谢综合征受体的胰岛素敏感性略有增加。
几项大型队列研究(表 1、表2)通过分析 IBD 患者的肠腔和黏膜微生物群落,研究了肠道微生物群的改变。
总体而言,活跃期IBD 与某些菌群的数量过多有关,如:
肠杆菌科Enterobacteriaceae
梭杆菌属Fusobacterium
咽峡炎链球菌Streptococcus anginosus
肠球菌Enterococcus
巨球菌Megasphaera
弯曲杆菌Campylobacter
Gammaproteobacteria
Deltaproteobacteria
相反,IBD 与有益菌群的缺失有关,例如:
Faecalibacterium prausnitzii
Christensenellaceae
Collinsella
Roseburia
Ruminococcus
其他产丁酸盐的细菌
在我们检测的炎症性肠病的菌群报告中也发现,炎症性肠病风险高的人群中,炎症水平很高,肠杆菌科Enterobacteriaceae,梭杆菌属Ruminococcus gnavus偏高,而Faecalibacterium prausnitzii和Roseburia丰度普遍降低或者缺乏。


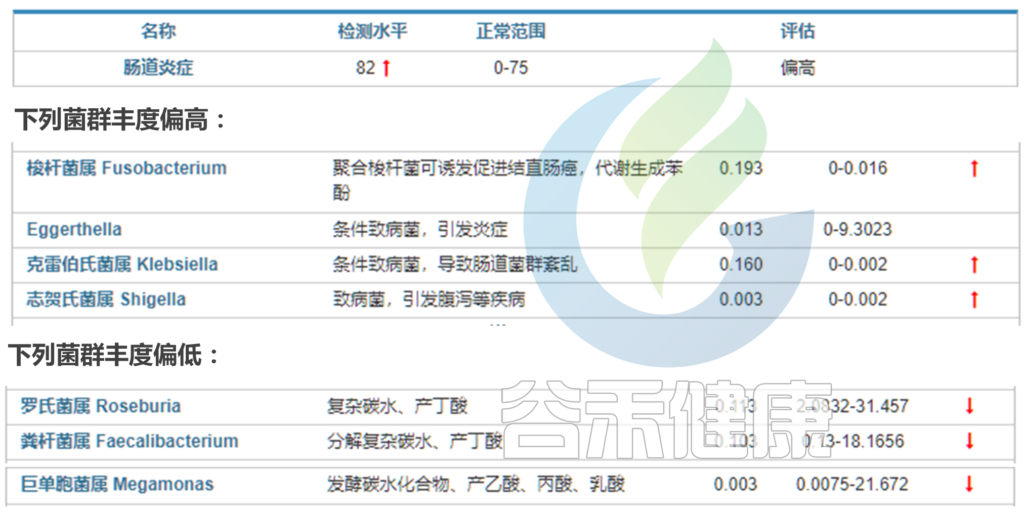
< 选自:谷禾肠道菌群健康检测报告 >
菌群代谢功能
粪便样本的宏基因组学为 IBD 中发生的功能失调和代谢途径的扰动提供了更全面的观点。这些研究表明,参与含硫氨基酸合成、核黄素代谢、谷胱甘肽转运蛋白、氧化应激和营养转运的代谢途径均被上调。
一项能够将微生物群落分解到物种内单个菌株水平的研究显示,与健康对照相比,IBD 或肠易激综合征 (IBS) 患者粪便样本中致病菌的菌株多样性增加,而有益微生物的菌株多样性降低。深入分析表明,219个类群(包括152种)与CD相关,102个类群(包括93种)与UC相关。
CD的主要特征是属于毛螺菌科和瘤胃球菌科的分类群减少和属于肠杆菌科的分类群增加,而对于UC观察到属于拟杆菌科的分类群减少和属于毛螺菌科的分类群增加。与这种异质性一致,在不同的 IBD 研究中仅存在少数物种的共同变化,这表明尽管疾病表型和病程相似,但在 CD 患者群体中仍存在个体间差异。
IBD和肠道菌群的因果关系探究
肠道微生物群在 IBD 中的致病作用的首批临床证据之一源于实验表明,从 CD 患者的小肠发炎段转移粪便流可改善疾病症状。粪便流的恢复和新末端回肠术后暴露于肠腔内容物会诱发炎症,这表明肠道微生物群会引发 CD的术后复发。此外,抗生素治疗对活动性 CD 患者亚群的疗效强调了肠道细菌和 IBD 之间的因果关系。
急性和慢性肠道炎症小鼠模型的机制研究为微生物失调与 IBD 之间因果关系的提供了进一步证据。例如,将IBD 患者的菌群移植到无菌受体小鼠,会将IBD 表型转移到无菌小鼠中。而具有 IBD 遗传易感性的小鼠在无菌条件下不会发生自发性炎症。
免疫响应
IBD 发病的遗传易感小鼠的失调菌群,能够将这种疾病症状转移到无菌受体小鼠。将 IBD 微生物群转移到无菌的野生型小鼠体内会导致肠道 T 细胞反应失衡,肠道 T 辅助 17 (TH17) 细胞和 TH2 细胞数量增加,RORγt +数量减少。同样,肠道微生物群的人类共生细菌脆弱拟杆菌对无菌小鼠的定植诱导CD4 + T 细胞转化为产生IL-10的FOXP3 + T reg细胞,这表明微生物群驱动的存在IBD 的发病机制。
在 T2DM 中也发现了几种细菌类群的丰度变化很大。
例如,据报道,2型糖尿病患者中下列菌相对丰度增加:
大肠杆菌E. coli、
韦荣氏菌属Veillonella、
布劳氏菌属Blautia、
厌氧菌属Anaerostipes、
乳杆菌属Lactobacillus、
粪杆菌属Faecalibacterium、
梭状芽胞杆菌属Clostridiales(等)
相反,下列菌丰度降低:
拟杆菌属Bacteroides
双歧杆菌属Bifidobacterium、
副拟杆菌属Parabacteroides、
颤螺菌属Oscillospira
可降解粘蛋白的阿克曼菌Akkermansia muciniphila
2019 年发表的一项宏基因组和宏蛋白质组学研究分析了来自 254 名中国个体的粪便样本中的肠道微生物群组成和功能,其中包括 77 名未接受治疗的 T2DM 患者、80 名糖尿病前期患者和 97 名葡萄糖耐量正常的对照个体。与代谢健康的对照组相比,T2DM 患者和前驱糖尿病患者的梭菌目Clostridiales细菌丰度较低,而埃氏巨球形菌Megasphaera elsdenii的丰度较高。
菌群代谢功能
在 T2DM 患者和糖尿病前期患者的微生物组中观察到功能差异。与对照个体相比,糖尿病前期个体的肠道微生物群显示出与糖磷酸转移酶系统、细菌分泌系统和氨基酸的 ATP 结合盒 (ABC) 转运蛋白有关的途径富集。这些发现表明,在糖尿病前期患者转变为 T2DM 之前,可以检测到肠道微生物组的疾病特异性变化。
环境因素影响
细菌种类和代谢基因簇谱的差异已被用于确定一组具有正常葡萄糖耐量或 T2DM 的个体的糖尿病状态。然而,包括地理位置、种族、健康状况和用药史在内的混杂因素导致在识别与 T2DM 相关的微生物变化方面不同研究存在不一致。
因果关系研究
几项研究提供了肠道微生物群特定成员与 T2DM 发病机制之间因果关系的证据。例如,A. muciniphila属于在人类和小鼠研究中显示对代谢紊乱具有保护作用的分类群。有趣的是,补充益生元使A. muciniphila的丰度正常化并改善了人类的代谢健康。同样,对喂食高脂肪饮食的小鼠施用A. muciniphila可逆转其增加的脂肪量、代谢性内毒素血症、脂肪组织炎症和胰岛素抵抗。
此外,产生丁酸盐的细菌Anaerobutyricum soehngenii(以前称为Eubacterium hallii菌株 L2-7)显示出丰度增加,这与来自瘦供体的FMT受体的外周胰岛素敏感性改善相关。
对T2DM 患者A. soehngenii菌水平进行管理,在治疗 4 周后改善了外周胰岛素敏感性,这些益处伴随着微生物群组成的改变和胆汁酸代谢的变化。
奇怪的是,特定的细菌分类群在 IBD 和 T2DM 中表现出相似的变化,这表明免疫介导和代谢疾病的共同特征导致微生物群的相似适应。
下列菌丰度下降:
梭状芽孢杆菌属 Clostridium spp.
粪杆菌属 Faecalibacterium
瘤胃球菌属 Ruminococcus
阿克曼氏菌属 Akkermansia
柯林斯氏菌属 Collinsella
罗斯氏菌属 Roseburia
下列菌丰度增加:
肠杆菌科 Enterobacteriaceae
大肠杆菌 E. coli
具核梭杆菌 Fusobacterium nucleatum spp.
这为定义疾病特异性标志物提出了挑战(下图)


Metwaly et al.,Nat Rev Gastroenterol Hepatol. 2022.
例如,一项针对 2,045 名 IBD 患者的研究的作者确定了 8 个分类群的特征,包括克里斯滕森菌科Christensenellaceae和梭杆菌属Fusobacterium的未知成员,它们可以区分 CD 患者和健康个体。
然而,Christensenellaceae 的丰度增加与低 BMI 和体重减轻有关,这是一种在 IBD 患者中经常观察到的分解代谢状况。同样,梭杆菌的富集被认为是转移性结直肠癌患者预后不良的标志。鉴于 IBD 患者患结直肠癌的风险增加,这一提议的微生物组特征可能是一种附带现象,与潜在的疾病机制没有因果关系。
对来自 132 名个体的微生物组、代谢组和转录组数据集的综合网络分析确定了连接关键细菌分类群(即F. prausnitzii 、未分类的Subdoligranulum、Alistipes、大肠杆菌、Roseburia)的某些代谢物(短链脂肪酸、辛酰肉碱和几种脂质)。有趣的是,有和没有 IBD 的研究参与者之间的差异在粪便代谢组中,比在粪便宏基因组、粪便宏转录组或粪便蛋白质组中更明显。
在综合个人组学分析研究 (iPOP) 中,血浆代谢物与来自 106 名个体的纵向样本中的胰岛素抵抗密切相关,这表明宿主代谢组和肠道微生物组之间的相互作用在胰岛素抵抗个体中受到干扰。
许多研究调查了微生物改变作为疾病生物标志物的效用,特别是在 CD 或 UC 患者中。最初试图定义可以作为疾病活动指标的单一细菌分类群。
例如,F. prausnitzii (一种产生丁酸盐的厚壁菌)在 CD患者中被耗尽。CD 患者回肠黏膜活检样本中这种细菌丰度的降低与回肠切除术后内镜下复发风险的增加密切相关。相反,粘附的侵袭性大肠杆菌丰度增加与回肠 CD相关。
然而,由于大多数细菌物种由许多个体菌株组成,这些菌株可以表现出相当大的基因组和蛋白质组变异,因此菌株多样性具有重要的功能,特别是在确定致病性方面。
例如,R. gnavus和大肠杆菌的亚种都与 IBD 的严重程度增加有关。此外, R. gnavus的一个特定亚种表明,来自 IBD 患者的粪便样本中丰度增加含有菌株特异性基因(与改善的细菌定植有关)。这些基因涉及诸如氧化应激反应、细菌粘附、铁获取和宿主粘液利用等功能。同样,不同的脆弱拟杆菌菌株表现出功能差异,导致 IBD 相关小鼠模型中 IgA 诱导水平不同。这些遗传上不同的脆弱拟杆菌菌株在接种到受体小鼠时也表现出不同的致结肠和免疫调节作用。在一项旨在定义用于监测 IBD 患者疾病活动的关键菌群失调的研究中,使用定量 PCR 计算了F. prausnitzii和大肠杆菌的绝对丰度比(也称为 F-E 指数)。F-E 指数的使用提高了 UC 和 IBS 患者与 CD 患者的区分,并有助于区分结直肠癌与其他肠道疾病。然而, F-E 指数无法区分 IBD亚型,这表明单一分类群指标在分类疾病亚型方面的效用有限。
大规模生物标志物分析
基于微生物特征的判别模型
几项研究已经使用机器学习算法来验证横截面和纵向患者队列中复杂的微生物组特征。
例如,2017 年发表的一项研究使用 16S rRNA 微生物群来分析来自大量 IBD 患者和没有 IBD 的对照个体的粪便样本。研究人员使用序列聚类算法根据八种细菌类群的丰度来识别 CD 特异性微生物特征。
此外,另一组研究表明,基线肠道微生物组组成的特征可以预测 IBD 患者在治疗开始后14周对抗整合素治疗的反应。由深度神经网络生成的微生物组特征的受试者工作特征曲线 (AUC) 下面积为 0.87,而基于临床协变量的模型的 AUC 为 0.62。研究小组还评估了微生物组特征作为 IBD 和 T2DM 生物标志物的效用。
在一项研究中,检查了 29 名接受过自体造血干细胞移植的 CD 患者的独特队列中的疾病活动性和对治疗的反应。来自人类供体和人源化小鼠的微生物组和代谢组风险概况的整合将疾病结果的预测模型的性能从 AUC 0.79 提高到 0.96,并确定了与硫代谢相关的疾病相关细菌和代谢物网络。
这些发现听起来很有前景,但重要的是要承认微生物组风险概况是基于来自前瞻性队列研究中的人群或患者组的预测模型,因此比起对于个人的预测结果,对于相似患者组(人群或队列)可能更准确。重要的是记住,预测的风险可能不会直接转化为个体患者,这可能是由于在异质环境中风险概况的普遍性有限。
不忽略混杂变量
另一项研究调查了以德国人群为基础的 1,976 人队列中的代谢健康和肠道微生物群的昼夜节律性。粪便微生物群分析确定了 13 个微生物分类群的风险特征,这些分类群显示 T2DM 患者的昼夜节律性受到破坏。基于这种心律失常风险特征的预测模型成功识别出有患 T2DM 风险的个体,当模型中包含 BMI 时,AUC 为 0.78。
这些例子为微生物组特征在用于诊断和治疗目的的生物标志物发现中的作用提供了证据。然而要注意,生态失调指数不是独立的测量值,需要整合到额外的宿主衍生数据和临床数据中。这些指标的标准化和验证需要大规模究研,包括对潜在生物标志物的纵向评估,并考虑可能的混杂变量,例如饮食、年龄、种族、病史和最后的排便时间,所有这些因素都会影响微生物组的改变。
在寻找疾病生物标志物时,代谢物作为疾病活动的最接近指标,并且与作为疾病机制基础和调节疾病机制的调节信号密切相关。事实上,代谢组和微生物组都随着饮食、环境、衰老和整体健康状况等内源性和外源性因素而波动。
许多研究报告了 IBD或 T2DM患者的肠道代谢物谱的显着变化。
例如,已在 IBD 患者的粪便代谢组中发现中链脂肪酸(如戊酸和己酸水平降低)和 B 族维生素水平降低。相反,据报道,成人和儿童 IBD 患者的粪便和血清中氨基酸、胺和肉碱的含量分别增加。
一项具有里程碑意义的研究结果表明,代谢物分析可以区分 IBD 患者和健康个体。该报告之后有许多其他人一致表明 IBD 患者的代谢物表型与健康个体的不同。有趣的是,代谢物分析还可以区分不同形式的 IBD,例如 CD 和 UC,并且可以进一步将 CD 患者分类为患有回肠或结肠炎症。同样,T2DM 患者的代谢物分析表明代谢途径的活性发生了改变。
在多份报告中,支链和芳香族氨基酸(如亮氨酸、异亮氨酸、缬氨酸、苯丙氨酸、酪氨酸和色氨酸)的血清水平与胰岛素抵抗、肥胖和发生 T2DM的风险相关。
T2DM 患者的代谢物分析还揭示了特定细菌代谢物水平与疾病发作之间的强关联。
例如,色氨酸代谢途径包括几个候选代谢物生物标志物,这些生物标志物由于与人类和小鼠研究中炎症和代谢疾病的发展相关而引起了研究关注。色氨酸是一种从饮食中获得的必需氨基酸,主要在小肠中吸收,尽管一小部分在结肠中分解代谢为吲哚代谢物。
一项纵向队列研究证实,在 213 名中国个体(包括 51 名继续发展为 2 型糖尿病和 162 名保持代谢健康的个体)中,高基线空腹血清色氨酸浓度与患 T2DM 的风险增加有关。此外,色氨酸水平作为生物标志物的预测能力与五种现有氨基酸在区分患有和未患 T2DM 的个体方面的预测能力相当 。
值得注意的是,先前的报告表明,几种氨基酸的血清水平可以以不同的准确性识别来自不同人群的 T2DM 患者。例如,苯丙氨酸和缬氨酸在美国人群中表现最好,而酪氨酸在南亚人群中最准确。这些发现指出了识别区域特异性生物标志物在实现最佳诊断准确性方面的重要性。
有价值的生物标志物必须为从临床信息中获得的分类能力提供额外的分类能力。因此,粪便生物标志物是粘膜疾病诊断标志物的明显来源,因为粪便流与肠粘膜直接接触。
钙卫蛋白
粪便钙卫蛋白是一种可在粪便中检测到的粒细胞衍生的细胞溶质蛋白,由于炎症严重程度与粪便钙卫蛋白水平之间的相关性强,它是最广泛使用的用于炎症性疾病的粪便生物标志物。
两份报告证实了粪便钙卫蛋白水平检测内窥镜炎症的能力,报告的敏感性为70-100% ,特异性为44-100%。这些值的广泛范围可以通过每项研究中应用的截止阈值的变化来解释。
然而,粪便钙卫蛋白水平升高并不是 IBD 特有的,而是反映了肠道炎症状况,这也与其他肠道和代谢疾病(包括 IBS、胃肠道恶性肿瘤、肥胖和 T2DM)有关。
例如,对来自 1,792 个人的粪便样本中肠道微生物群的鸟枪宏基因组分析能够区分 IBD 和 IBS,与单独的粪便钙卫蛋白水平 (AUC) 相比,机器学习算法显着提高了这些预测模型的准确性 (AUC 0.91>0.80)。
重要的是,具有最高预测准确度 (AUC 0.93) 的模型包括粪便钙卫蛋白水平以及前 20 个选定分类群的宏基因组分析。这些结果表明,临床和微生物生物标志物的整合提高了诊断的准确性。这种综合方法已被用于预测 IBD 患者对治疗的反应。
在这项研究中,基线临床数据(包括血清学、内窥镜和临床生物标志物)不足以预测缓解(AUC 0.62),而添加分类学和代谢谱将诊断能力分别提高到 AUC 0.72 和 AUC 0.74。
此外,仅粪便钙卫蛋白水平就能够区分储袋炎患者和无储袋炎患者 (AUC 0.63)。相比之下,微生物组物种模型(有或没有粪便钙卫蛋白水平作为额外的预测因子)实现了 0.78 的 AUC,证实微生物分析在识别储袋炎方面具有优于仅粪便钙卫蛋白水平的诊断性能。
用于诊断 T2DM 的葡萄糖代谢受损的血清学生物标志物包括空腹血糖水平、75g口服葡萄糖激发后 2 小时血糖水平(口服葡萄糖耐量试验)和糖化血红蛋白水平。
一项使用来自两个瑞典队列136数据的研究确定了预测 T2DM 进展的血清学和微生物组生物标志物的组合。
在这项研究中,多变量分析表明胰岛素抵抗程度与微生物组变异之间存在很强的相关性。
有趣的是,使用基于微生物组的机器学习模型来区分验证队列中胰岛素抵抗程度最低和最高的个体的 AUC 为 0.78,这表明肠道微生物群是 T2DM 进展的重要调节因子。
事实上,尽管已经为 T2DM 提出了广泛的诊断生物标志物,但它们中的大多数未能捕捉到这种疾病的复杂性或掌握微生物和代谢的变化。在这方面,已将代谢物生物标志物与已确定的临床风险因素结合使用,以显着改善疾病分类。
微生物特征
了解单个细菌类群(病原菌)和/或复杂微生物群落(生态失调)变化的功能作用和特异性对于解决IBD或T2DM中微生物-宿主相互作用的发病机制至关重要。在这种情况下,肠道微生物组的功能改变可能代表宿主适应的结果。

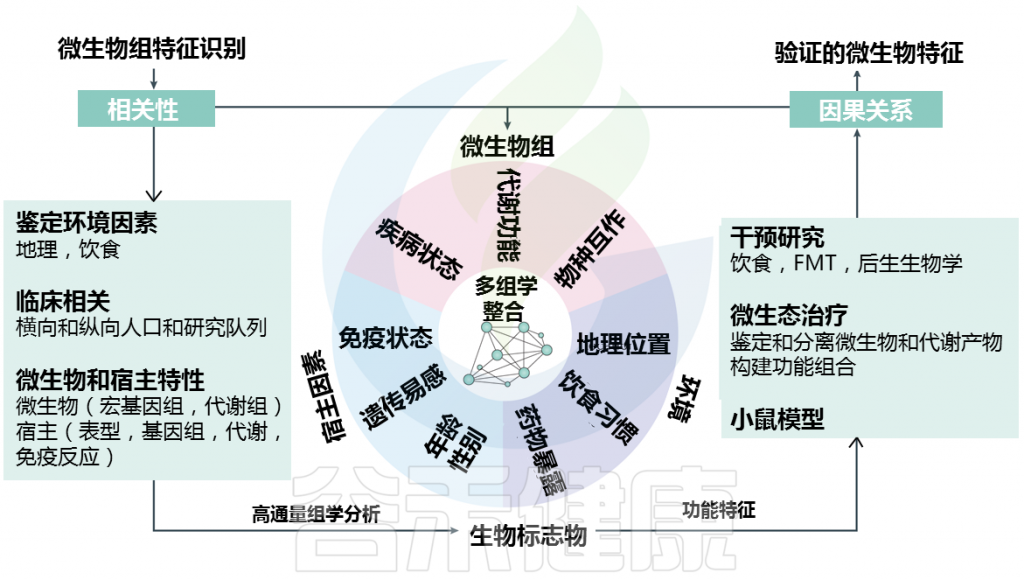
Metwaly et al.,Nat Rev Gastroenterol Hepatol. 2022.
肠道微生物群与多种疾病之间存在因果关系,已在小鼠实验中得到证实。无菌小鼠模型可以选择性地用单一细菌菌株、最小细菌聚生体或来自人类粪便或其他供体材料的定义复杂的肠道微生物生态系统进行定植,以研究它们对宿主表型的影响。在 IBD 中,无菌小鼠模型的单菌株定植有多种共生细菌,包括大肠杆菌、粪肠球菌、普通拟杆菌和Bilophila wadsworthia使我们能够了解疾病引发或保护的一些潜在机制。已有的研究工作表明,肠道菌群是驱动结肠炎小鼠模型炎症所必需的,而这些细菌与 CD患者的复发风险相关。肠道微生物群移植到无菌小鼠体内导致了几种疾病状态,从而揭示了与炎症有关的共享功能代谢途径。同样,以前的工作表明葡萄糖耐量和胰岛素抵抗受肠道微生物组组成的影响,已通过一系列 FMT 研究得到证实。
在过去的二十年里,人类和小鼠研究的证据揭示了肠道微生物组在炎症和代谢疾病(如 IBD 和 T2DM)的发病机制中的基本作用。肠道微生物生态系统结构和功能的变化(失调),与这些疾病患者的疾病活动、复发风险或对治疗的反应有关。然而,大多数这些疾病的复杂性和多因素发病机制,以及人类研究中存在多种混杂因素,依然对微生物组特征在诊断、预测预后和治疗决策中的临床应用提出重大挑战。
当前的微生物组研究不仅仅局限于描述微生物群落结构和疾病关联,还在了解肠道菌群在复杂慢性病发病机制中的致病作用方面取得进展。预计这些努力将增强微生物组建模,并推进可用于临床环境的基于菌群特征和/或疾病风险的模型开发。
参考文献:
Metwaly A, Reitmeier S, Haller D. Microbiome risk profiles as biomarkers for inflammatory and metabolic disorders. Nat Rev Gastroenterol Hepatol. 2022 Feb 21. doi: 10.1038/s41575-022-00581-2. Epub ahead of print. PMID: 35190727.

谷禾健康

随着工业革命的发展,城市化进程的加快和经济的发展导致能源消耗和废物排放激增,在过去的几十年里,环境污染物已成为一种日益普遍的健康危害。
最近的研究还表明,人类微生物组可以代谢环境化学物质,进而可能会受到化学物质暴露的影响。
这种直接相互作用表明,在风险评估中可能需要考虑微生物组。
目前越来越多的证据表明,接触这些环境化学物质是导致多种健康障碍发展的重要因素之一。例如,体外、体内和流行病学研究已将人类暴露于内分泌干扰的化学物质与肥胖、代谢综合征和 2 型糖尿病联系起来。
此外,接触环境污染物与产前缺陷、呼吸系统疾病、心脏病和精神障碍以及患癌可能性增加和预期寿命缩短有关,而胃肠道微生物群对各种宿主代谢和免疫功能起着至关重要的作用。
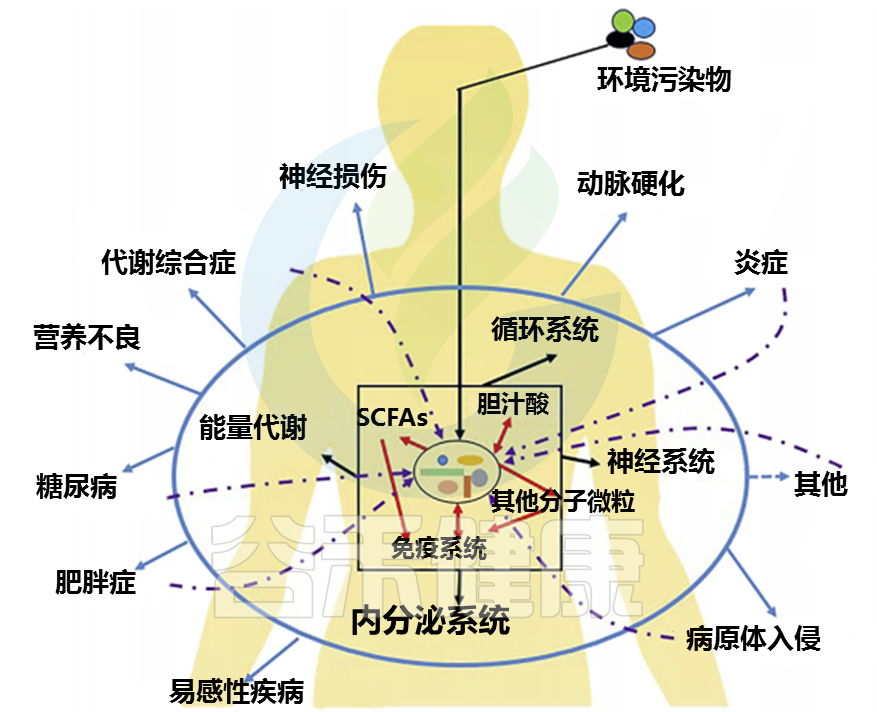
Jin Y et al. Environ Pollut. 2017.
尽管大多数环境污染物并不直接针对肠道微生物群,但一些污染物可以通过不同途径进入人体并与肠道微生物群相互作用。
先前的多项研究表明,暴露于环境污染物会改变肠道微生物群的组成,导致能量代谢、营养吸收和免疫系统功能障碍或产生其他毒性症状。有人提出胃肠道微生物可能通过改变环境化学物质的吸收、处置、代谢和排泄来影响肥胖和糖尿病等疾病。
肠道微生物群对药物、饮食以及环境污染物都非常敏感,包括抗生素、重金属、持久性有机污染物、杀虫剂、纳米材料和食品添加剂等。
环境污染物可能对肠道微生物群的影响及其对健康的产生后续影响。
我们每个人的肠道中都携带着数千种细菌以及一些其他类型的微生物。虽然说所有人都具有非常相似的微生物群,但没有两个人的肠道中细菌种类的组成完全相同,换句话说,每个人的微生物群可能会像指纹一样独特。
不过,不同的细菌物种在人与人之间可能存在很大差异,但这些物种往往编码相同的代谢途径。
通过查阅文献得出肠道微生物具有广泛的代谢环境化学物质的能力,污染物的细菌依赖性代谢会调节宿主的毒性。相反,来自各种化学家族的环境污染物已被证明会改变胃肠道细菌的组成和/或代谢活动,这可能是影响个体微生物群的重要因素。
直接代谢排出
几十年来,人们已经知道胃肠道微生物参与异生物质的生物转化。
注:异生物质,是指由人工合成的化学物质。如农药、洗涤剂及化工原料等,其中各种合成农药占绝大多数。异生物质是对身体来说是异物的物质。
早在 1973 年,Scheline 就提出,胃肠道微生物群代谢外来化合物的潜力与肝脏不差上下。此后已为胃肠道微生物群鉴定了 40 多种药物底物,并强调肠道微生物对药物进行多种化学转化的能力,包括:
还原、水解、琥珀酸基团的去除、脱羟基、乙酰化、脱乙酰、N-氧化物键的裂解、蛋白水解、脱硝、解偶联、噻唑开环、去糖基化、去甲基化
胃肠道是异生物质进入人体的主要途径。细菌代谢的速率和程度受到达远端肠道的异生物质数量的影响,在那里细菌丰度最高。环境化学物质在摄入后可能吸收不良,随后被蠕动扫到远端小肠和盲肠,或者,它们及其代谢物可能会穿过肠壁从血液中分离出来。因此,许多化学物质可以被胃肠道微生物群直接代谢。
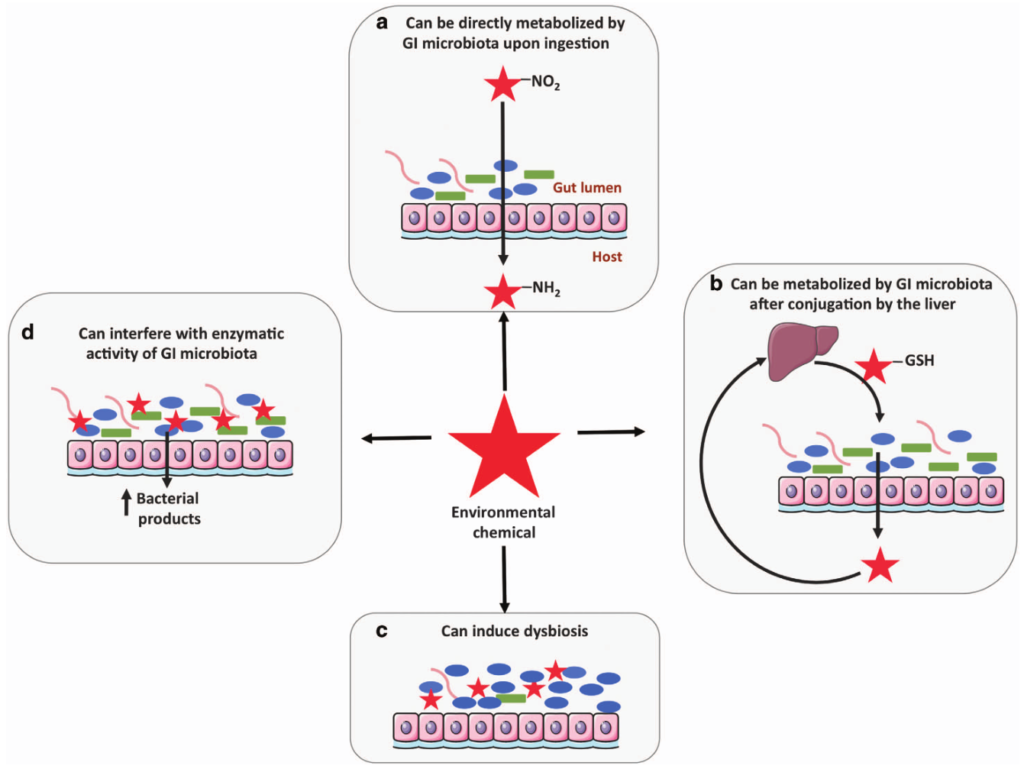
环境化学物质(或其代谢物)也可能从胆汁中排出。大多数外源性物质是非极性的,因此在胃肠道中被吸收并通过门静脉血输送到肝脏进行解毒。肝脏通常氧化异生素并产生葡萄糖醛酸、硫酸盐或谷胱甘肽结合物。在大多数情况下,结合反应促进排泄,结合物从尿液中消除。
然而,结合物也可以在胆汁中排泄。决定一种化学物质是否排泄到胆汁中的因素尚不完全清楚,一般规则是低分子量化合物 (<325 kDa) 很少排泄到胆汁中,而较高分子量 (>325) 的化合物可以显着排出。分泌到胆汁中的结合物进入小肠,那些未被吸收的向下移动到大肠,在那里它们可能被微生物群代谢。
Claus SP et al. NPJ Biofilms Microbiomes, 2016
解偶联,形成肝脏循环
胃肠道微生物群可以通过解偶联并减少肝脏异生物质代谢物,导致形成低分子量的非极性分子,这些分子很容易被重吸收。这些非极性分子的重吸收和它们返回肝脏被称为“肠肝循环”。肝肠循环控制着体内内源性底物(如胆汁酸和类固醇)的储存和再利用。然而,它也延迟了体内环境化学物质的消除。
胃肠道细菌介导的环境异生物质转化的例子, 如下:
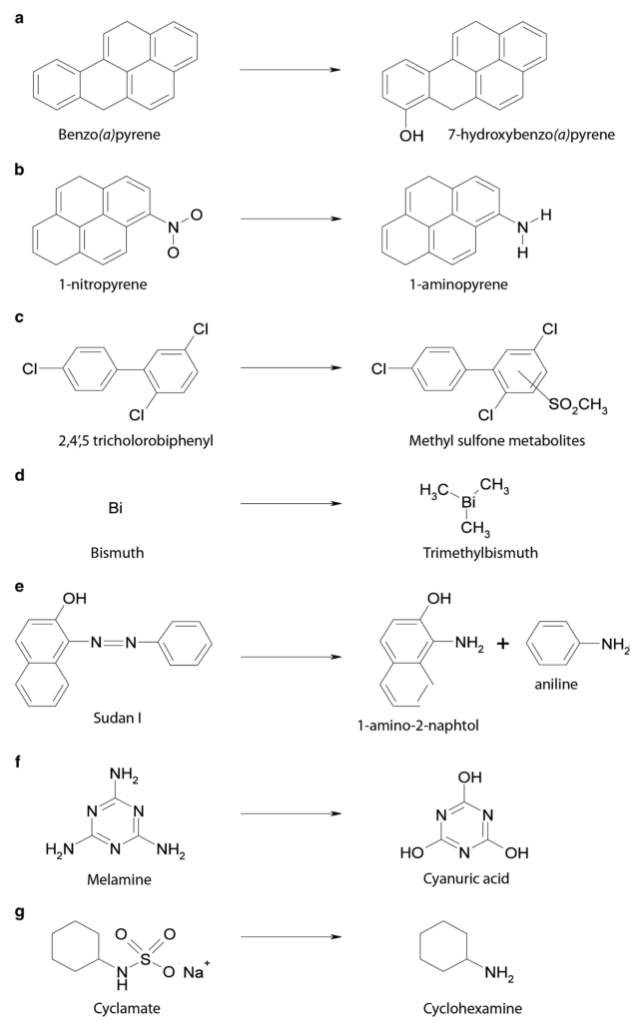
Claus SP et al., NPJ Biofilms Microbiomes, 2016
胃肠道微生物群的异生物质代谢酶
尽管已经将各种化学反应归因于胃肠道微生物,但只有少数酶家族被确定为胃肠道微生物异生物质代谢。
主要偶氮还原酶、nitroreductases、β-葡糖醛酸酶、硫酸酯酶和β-裂解酶。
胃肠道微生物群的异生物质代谢酶,如下:
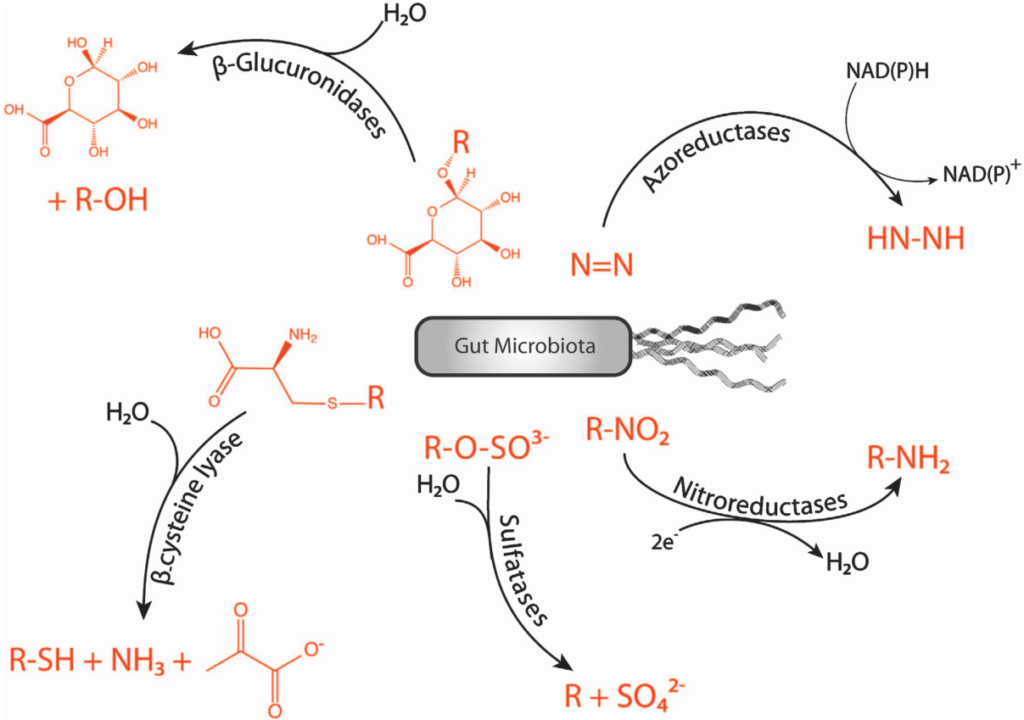
(i) 偶氮 (N=N) 键的还原裂解由细菌偶氮还原酶进行。
(ii) 细菌硝基还原酶还原硝基 (–NO2) 官能团到相应的胺。
(iii) 内源性硫酸酯在胃肠道中被细菌来源的硫酸酯酶水解。
(iv) 异生素的谷胱甘肽结合物也大量排泄在胆汁中。它们被各种哺乳动物酶(γ-谷氨酰转肽酶和羧肽酶)降解,形成半胱氨酸结合物。这些半胱氨酸结合物可能会到达含有 ß-裂解酶活性的胃肠道的一部分,并转化为相应的硫醇。
(v) β-葡糖醛酸酶存在于整个胃肠道中,并在异生物质葡糖醛酸苷的水解中发挥作用,异生物质葡糖苷酸是胆汁中排泄的最大一类异生物质结合物。肠道微生物被认为是 ß-葡萄糖醛酸酶的主要来源。
肠道细菌和人体细胞合作,制造生命所需的酶
据 NIEHS 科学家领导的一个团队称,人类肠道中的细菌与人体细胞合作,制造出一种生命必不可少的分子。烟酰胺腺嘌呤二核苷酸 (NAD) 分子是细胞生存必须完成的数百个生化反应所必需的。
研究人员还发现,一些肿瘤内的细菌保护肿瘤免受靶向 NAD 的抗癌药物的侵害。研究结果于2020年3 月 3 日在线发表在 Cell Metabolism 上。
该研究的主要作者指出,NAD 水平会随着哺乳动物的年龄增长而降低。他说有些人服用补充剂来促进 NAD 的产生。Shats 解释说,他的小组的最新研究表明,这些化合物的功效取决于肠道微生物群。
“如果你的肠道细菌没有发现负责促进 NAD 合成的酶,那么补充剂的效率就会低得多”。
内分泌干扰化学物质 (EDC) 由一大类化学物质组成,这些化学物质会改变内分泌系统并影响人类的发育和繁殖。EDC 的来源包括工业化学品、药品和个人护理产品、除草剂、有机氯杀虫剂和添加剂等。
EDC 被美国环境保护署描述为外源性因素,因为它可以干扰内源性激素,从而解除对发育过程的管制。由于内分泌系统对器官的影响,EDCs对身体具有深远的全身影响。EDC 已通过消化道对动物和人类健康产生不利影响,例如炎症、代谢紊乱、先天性异常,菌群失调。
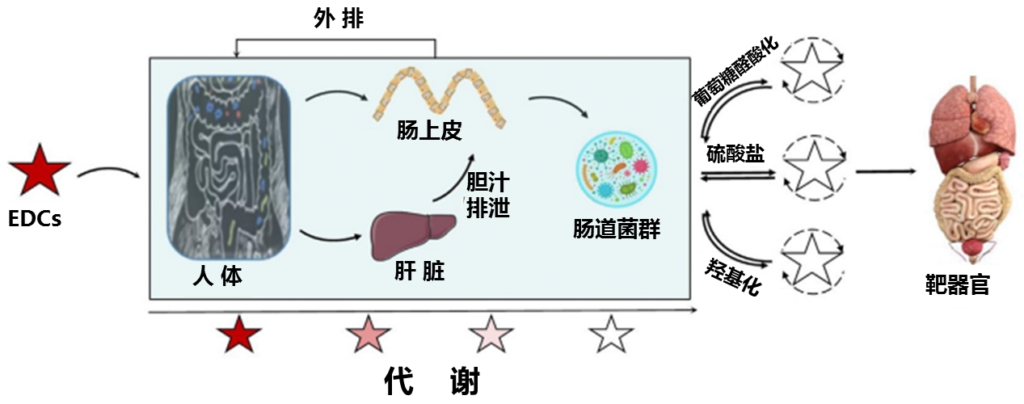
Li N, et al. Environ Pollut. 2021
抗生素:直接改变或者杀死微生物
我们现在知道人类和动物除了故意给药外,还可以通过多种途径被动摄入抗生素。通常,抗生素可以对微生物造成直接的影响。因此,即使某些抗生素的浓度非常低,动物肠道微生物群的组成也可能发生变化。人们越来越注意到,环境抗生素引起的肠道菌群失调与人类健康密切相关。
在过去的 50 年中,全世界开发了大量抗生素用于人类和兽医学。由于它们在人和动物体内的吸收不完全,部分摄入的抗生素通过粪便或尿液排出到环境中。
越来越多的研究发现,在中国和其他国家的河流和湖泊沉积物、地表水、农业土壤和废水等自然环境中发现了高浓度的各种抗生素。

先前的研究发现,抗生素治疗并没有减少肠道微生物群的总数,而是改变了人类和动物中某些物种的相对数量。此外,抗生素暴露通常会通过增加或减少多样性来改变微生物组的多样性。
更重要的是,抗生素对人类肠道微生物群的影响可持续数年。如克拉霉素和甲硝唑治疗持续改变肠道微生物群组成长达 4 年。婴儿完成抗生素治疗后,虽然微生物群组成的某些方面恢复到治疗前的水平,但某些细菌种类的丰度发生了永久性改变。这种变化对婴儿有害,扰乱他们的早期发育。值得注意的是,不同的环丙沙星治疗对个体微生物群影响也略有不同。
在生命早期向小鼠施用低剂量的青霉素,可通过降低乳酸菌Lactobacillus, Candidatus, Arthromitus, Allobaculum的水平来增强代谢表型并促进脂质积累。另一份报告显示,给小鼠服用青霉素 G、红霉素或两者结合可以增加脂质积累并诱导炎症反应。所有这些处理都降低了拟杆菌属/厚壁菌门的比率。
接触某些抗生素可能会加重某些疾病的严重程度。例如,艰难梭菌是抗生素引起的腹泻的主要原因,它会大大增加住院患者的发病率和死亡率。
用克林霉素和氨苄青霉素这两种最常用的抗生素进行治疗,通过减少 Clostridium scindens菌(胆汁酸代谢的二级调节剂)来增加患者对艰难梭菌感染的易感性。
链霉素治疗可以增加Porphyromonadaceae 和 Bacteroidaceae 两个科的水平,它们的存在与小鼠对肺炎和结肠炎的易感性或超敏性呈正相关。
甲硝唑治疗减少了小鼠结肠的内部粘液层,从而通过减少厌氧拟杆菌的组成,以及增加包括乳酸杆菌在内的耐氧细菌的组成来增加对柠檬酸杆菌引起的结肠炎的易感性。
抗生素治疗还增加了大肠杆菌等病原菌的入侵,抗生素治疗后肠道微生物群中富集了抗生素抗性基因,造成抗生素抗性。
由于使用抗生素治疗,一些非微生物驱动的疾病会更频繁地发生。

在生命早期给小鼠施用万古霉素会通过增加乳杆菌科和疣微菌科的丰度来增强过敏性哮喘,并且这些微生物群的变化与 Treg 细胞减少有关。
最近,一项研究报告称,口服链霉素、粘菌素和氨苄青霉素的组合或单独使用万古霉素治疗会增加小鼠胰腺 b 细胞死亡和 I 型糖尿病的发病率。
万古霉素处理小鼠在属水平上增加了埃希氏菌、乳杆菌和 Sutterella 的水平,并减少了属于梭菌目和毛螺菌科、普氏菌科和 Rikenellaceae 科的细菌。联合抗生素治疗导致肠道微生物群几乎完全消失。这些微生物组结构变化与回肠和结肠固有层中产生白细胞介素 (IL)17 的细胞的出现密切相关,表明肠道微生物群在免疫稳态中发挥着非常重要的作用。
然而,抗生素引起的肠道微生物群的改变有时对疾病有益。囊性纤维化是一种常染色体隐性疾病,可导致肠道细菌过度生长。链霉素治疗小鼠 9 周通过减少肠道细菌过度生长、通过降低乳酸杆菌丰度来调节 T 细胞谱以及增加肺和肠系膜淋巴结中的免疫细胞水平来改善这种疾病。
对肝硬化患者施用利福昔明可降低韦荣球菌科的丰度并增加真杆菌科的丰度。这些微生物组成的变化改变了患者的代谢特征并降低了疾病的严重程度。诺氟沙星可以减少肝硬化患者的细菌移位,改善肠道屏障功能,从而降低自发性细菌性腹膜炎的风险。
总之,抗生素很容易引起肠道微生物群失调,并对健康产生不利影响。人类和动物可能会长期被动地暴露于环境中低浓度的抗生素中。因此,环境衍生的抗生素对人和动物的健康风险不容忽视。
重金属:导致肠道微生物群失调
近年来,环境中的重金属已成为严重的健康风险。作为一种常见的环境污染形式,重金属与多种毒性作用有关,包括致癌作用、氧化应激和 DNA 损伤,以及对免疫系统的影响。最近,几项研究表明,重金属暴露也可能导致肠道微生物群失调。
砷 (As) 是一种已知的致癌物质,是一种常见的有毒污染物,广泛分布在我们的环境中。砷通常以砷酸盐 [As(V)] 的形式附着在固体表面上。摄入 As(V) 后,肠道微生物群暴露于砷会诱导 As 硫醇化和甲基化。

据此前报道,砷暴露与多种疾病有关,如皮肤癌、膀胱癌、肺癌和肝癌;糖尿病; 和心血管疾病。此外,暴露于 10 mg/L 砷 4 周会显着扰乱小鼠的微生物组组成和代谢组学特征。一般而言,As 暴露后厚壁菌门的丰度显着下降,而拟杆菌门的丰度显着增加。
还观察到这些变化与脂质代谢物、含吲哚的代谢物、异黄酮代谢物和胆汁酸代谢物密切相关。长期接触砷会导致结肠微生物组的结构和组成发生变化,并促进雄性小鼠宿主的氮和氨基酸代谢。
镉 (Cd) 经常且广泛地用于制造许多产品,例如电池、金属电镀、颜料和塑料。在一些国家,尤其是中国等发展中国家,在水生系统、沉积物和土壤中观察到高浓度的 Cd。
据报道,镉毒性与致癌作用、肝毒性、氧化应激和免疫毒性有关。最近,研究报告雄性小鼠亚慢性暴露于低剂量 Cd(饮用水中 10 毫克/升)10 周会降低厚壁菌门和变形菌门的相对丰度,并增加盲肠和粪便中拟杆菌门的相对丰度。肠道微生物组组成的这些变化与雄性小鼠血清中脂多糖 (LPS) 水平升高、肝脏炎症甚至能量代谢失调有关。
铅 (Pb) 也是一种剧毒金属,疾病控制和预防中心非常重视。事实上,环境铅是一种普遍存在的全球健康危害,因为它广泛用于各种消费品,例如汽油中的四乙基铅。由于它在空气、土壤、水、旧油漆和食物中的持久性,人类和动物可以通过摄入、吸入和皮肤吸收吸收铅。铅中毒还通过其能量产生和其他代谢过程的中断而与肥胖的发展有关。
最近的一份报告显示,围产期铅暴露(饮用水中 32 毫克/升)持续 40 周会导致成年雄性小鼠体重增加,但不会导致雌性小鼠体重增加。接触铅后微生物组中没有观察到拟杆菌/厚壁菌的比例下降; Desulfovibrionaceae、Barnesiella 和 Clostridium XIVb的丰度在科水平上有所增加;接触铅后,乳球菌、肠杆菌和柄杆菌Caulobacterales的丰度下降。此外,Akkermansia nuciniphila在暴露于铅的小鼠中。
Li N, et al. Environ Pollut. 2021
持久性有机污染物:改变微生物组成,导致炎症
持久性有机污染物 (POPs) 包括有机氯农药、多氯联苯 (PCBs)、多溴联苯醚和多环芳烃 (PAHs) 是合成化合物,具有持久性、半挥发性和高毒性,可在生物组织中积累。
持久性有机污染物受到了特别关注,因为它们是导致近年来全球肥胖、糖尿病、自身免疫疾病和一些发育障碍发病率上升的众多因素之一。摄入食物和水被认为是主要的 POP 接触途径。
PCB 是一种致癌物质,因其介电特性和化学稳定性而被用于制造电容器、变压器、冷却液、液压油和润滑剂。PCBs 可以在小鼠暴露后 2 天内显着改变肠道微生物群的组成。
肠道中的产气荚膜梭菌(Clostridium perfringens)和拜氏梭菌(Clostridium beijerinckii )可以通过去除氯原子和打开苯环来降解多氯联苯。
暴露于一种 PCB,2, 3, 7, 8 四氯二苯并呋喃 (TCDF),以 24 毫克/公斤体重的剂量连续 5 天会降低厚壁菌门/拟杆菌门的比率,增加黄杆菌(Flavobacteria)的水平,并降低盲肠中的梭菌。
这些微生物群组成的变化与胆汁酸代谢的改变有关。此外,在分子水平上,TCDF 还可以抑制法尼醇 X 受体 (FXR) 信号通路,由于细菌发酵而引发宿主体内显着的炎症和代谢紊乱。此外,TCDF 诱导的变化以芳烃受体 (AHR) 依赖性方式发生。
PAH 被认为是高度优先的环境污染物,因为它们对人类具有毒性、致癌性和假定的雌激素或抗雌激素特性。人类接触高分子量多环芳烃的途径主要是通过口服炭烤、烤和熏肉或摄入未清洁干净的蔬菜。
摄入的多环芳烃到达小肠肠细胞和肝脏肝细胞并作为 AHR 配体。此外,结肠中的微生物群可以催化多环芳烃转化为雌激素。这种生物活化可能是 PAH 毒性的潜在机制。另一种 AHR 调节剂二恶英也有可能影响小鼠肠道微生物组的组成。
最近, 据报道,2, 3, 7, 8-四氯二苯-对-二恶英 (TCDD) 给药(6 毫克/千克体重)26 周显着增加厚壁菌门的丰度并降低拟杆菌门的丰度。在科水平上,TCDD暴露后粪便中乳酸杆菌科和脱硫弧菌科的含量显着增加,普氏菌科的含量下降。这些肠道微生物组的变化与链脲佐菌素诱导的高血糖小鼠的肝脏和免疫毒性相关。
农药/杀虫剂:改变微生物组成
农药残留是一个长期存在且严重的环境问题,因为在食品材料、水和土壤中检测到了大量农药。近年来,关于农药对动物影响的健康问题也有所增加。由于某些杀虫剂的抗菌活性,杀虫剂有可能改变肠道微生物组并诱发动物的其他症状。
氯菊酯 (PEM) 是最具代表性的拟除虫菊酯化合物之一,在低剂量和长期治疗的情况下(Nasuti 等人,2016)可以降低拟杆菌、普氏菌和卟啉单胞菌的丰度,并增加肠杆菌科和乳杆菌的丰度。肠道微生物群的这些变化可能导致 PEM 的神经毒性。
五氯苯酚 (PCP) 是世界范围内广泛使用的农药。即使在非常低的浓度下,五氯苯酚暴露也会导致水生动物的遗传和生殖毒性。PCP可在鱼类肝脏和肠道内蓄积,抑制鱼类生长,引起肝脏氧化和组织病理学损伤。此外,在金鱼中,接触五氯苯酚 (100 毫克/升) 28 天也通过降低厚壁菌门/拟杆菌门的比率来改变微生物组的组成。拟杆菌的相对丰度增加,金黄色杆菌、微杆菌、节杆菌和军团菌的相对丰度在属水平上下降。
环氧环唑 (EPO) 是一种广谱唑类杀菌剂,可有效防治小麦叶斑病 (Septoria tritici) 和锈病 (Puccinia triticina),被广泛用作农业抗真菌剂。据报道,EPO 对大鼠具有胎儿毒性作用,这与母体血浆中睾酮增加和雌二醇减少密切相关。
当大鼠以高剂量 EPO饮食喂养 90 天时,粪便中厚壁菌门的相对丰度下降,拟杆菌门和变形菌门的相对丰度增加。此外,毛螺菌科和肠杆菌科的相对数量被选择性地富集。作者进一步指出这些由 EPO 引起的微生物群组成的改变与肝功能受损有关。
毒死蜱 (CPF) 是一种有机磷杀虫剂,常用于处理水果、蔬菜作物和葡萄园。
CPF 可以被肝脏和肠道中的细胞色素 P450 酶代谢。围产期 CPF 暴露降低了幼鼠的体重和身长,并抑制了它们的肠道发育。
此外,CPF 诱导较高水平的拟杆菌、肠球菌和梭状芽孢杆菌,同时导致较低水平的乳杆菌属和双歧杆菌属。CPF 诱导的微生物群失调会破坏粘膜屏障,增加细菌易位,并刺激先天免疫系统。
多菌灵(CBZ;2-苯并咪唑氨基甲酸甲酯)是一种内吸性广谱苯并咪唑杀菌剂,被广泛用于防治真菌病害。 CBZ 对健康有多种不利影响,例如肝脏氧化应激和水肿、生殖毒性和内分泌紊乱。
在最近的一项研究中,我们观察到,以高剂量向小鼠口服 CBZ 28 天会增加肝脏脂质积累和体重,此外还会增加血清中促炎细胞因子 IL-1b 和 IL-6 的水平。
这些变化与 CBZ 诱导的肠道微生物群失调有关,包括在门水平上拟杆菌的相对水平降低和厚壁菌、变形菌和放线菌的相对水平增加。在科水平上,脱硫弧菌科、瘤胃球菌科、毛螺菌科和去铁杆菌科的相对丰度增加,而S24-7、副普雷沃氏菌科、瑞肯菌科、普雷沃氏菌科、拟杆菌科、卟啉单胞菌科、金银藻科(Christensencealae)的相对丰度减少。
最近,口服暴露(100 毫克/公斤体重)至 IMZ 28 天可导致小鼠肠道微生物群失调和结肠炎症。在盲肠中,49.3% 的细菌在 IMZ 暴露后发生了变化。在门水平上,拟杆菌、变形菌和蓝藻的丰度下降,而绿弯菌Chloroflexi、厚壁菌、放线菌和酸杆菌的丰度增加。在属水平上,梭菌、毛螺菌科、螺杆菌科和螺杆菌的丰度显着增加,而S24-7、理肯菌科、普氏菌、厌氧菌和柠檬酸杆菌的丰度显着降低。
微塑料:诱发肠道菌群失调和炎症
全球塑料污染问题太普遍了,因为塑料的大规模生产在过去 70 年左右的时间里呈爆炸式增长,从每年约 200 万吨增加到约 3.8 亿吨。每年约有 800 万吨塑料到海洋中,塑料碎片为水生环境中的微生物提供了稳定的基质和新的生态位,被称为“塑料圈”。

塑料降解的自然过程非常缓慢。例如,一个 PET 瓶的寿命可能长达数百年。这为环境中存在的各种微生物对这些化合物做出反应提供了足够的进化时间,并且在之前的研究中发现了许多不同的酶,它们具有降解不同塑料的能力。
在塑料污染严重的地方,更多的微生物可以降解塑料。具有降解塑料能力的微生物酶的数量正在增加,这与当地塑料污染水平有关。
最近发表在科学杂志mBIO 上的这项新研究分析了来自世界各地数百个地点的环境 DNA 样本。研究人员使用计算机建模来搜索具有塑料降解潜力的微生物酶,在污染最严重的地区增加酶的数量增加,换句话说,塑料降解酶的数量和多样性正在增加,直接响应当地的塑料污染水平。
总共发现了超过 30,000 种酶“同源物”,它们有可能降解 10 种不同类型的常用塑料。同源物是具有相似特性的蛋白质序列的成员。一些含量最高的地点是众所周知的高污染地区,例如来自地中海和南太平洋的样本。
这么多不同的微生物和环境栖息地中发现如此多的酶。这是一个令人惊讶的发现,真正说明了问题的严重性,这也说明对抗全球塑料危机的潜在途径。
塑料造成的环境污染已成为公共卫生问题。然而,微塑料对肠道微生物群、炎症发展及其潜在机制的影响尚未得到很好的表征。
用高浓度的微塑料处理增加了肠道微生物种类的数量、细菌丰度和菌群多样性。喂养组显示葡萄球菌丰度显着增加,同时副杆菌属显着减少丰度,与空白(未处理)组相比。此外,所有喂养组血清白细胞介素1α水平均显着高于空白组。
值得注意的是,微塑料处理降低了 CD4 +细胞中 Th17 和 Treg 细胞的百分比,而在 Th17/Treg 细胞比率方面,空白组和处理组之间没有观察到显着差异。喂食高浓度微塑料的小鼠的肠道(结肠和十二指肠)表现出明显的炎症和较高的 TLR4、AP-1 和 IRF5 表达。因此,聚乙烯微塑料可诱发肠道菌群失调和炎症,为微塑料相关疾病的防治提供了理论依据。
大多数塑料碎片上定植的细菌多样性高于周围水域。塑料碎片也可能比周围的水积累更多的潜在病原体。源头追踪结果表明,下游塑料圈从上游继承的细菌类群比例远高于浮游细菌。下游水中约 92.3-99.7% 的细菌群落不是来自上游,而是来自下游人类活动的输入。相反,下游塑料圈中高比例的细菌分类群与上游密切相关。塑料圈比浮游细菌具有更高的生态功能多样性。
城市和下游农业和水产养殖业的密集人类活动对乌鲁木齐河微生物群结构和功能群有很大影响。Pastisphere 群落比浮游细菌更能抵抗人类干扰。与周围的水相比,塑料圈增加了上游微生物结构和功能的遗传,也增加了下游水中病原体的存活和繁殖,其中含有高浓度的抗生素。
Li N, et al. Environ Pollut. 2021
空气污染物:增加肠道通透性
空气污染是指空气中可能由自然和人类活动导致的有害物质的存在。空气污染是一种复杂的气体混合物(包括臭氧、二氧化碳、二氧化硫、一氧化碳和二氧化氮)、颗粒物,包括化石燃料/汽车尾气的燃烧、多环芳烃/PAHS、花粉、孢子、微生物颗粒、矿物粉尘、有机碳、硝酸盐和硫酸盐。
大气颗粒物,尤其是空气污染是世界性的环境问题,会引发多种健康问题。颗粒物质其直径范围为2.5 μM-10 μM。
事实上,世界不同地区的几项基于队列的研究已经解决了长期暴露于空气污染与死亡率之间的相关性。他们已经证明长期暴露于细颗粒物与心血管和肺部疾病风险增加以及肺癌风险增加之间存在关联。空气污染可能与炎症性肠病的病理生理有关,从而与胃肠道疾病相关。

作为空气污染成分的颗粒物和臭氧现在被认为具有严重的健康问题,因为臭氧和颗粒物会增加肠道通透性,而且它们可能会破坏肠道细胞壁中存在的紧密连接。由于空气污染对肠道微生物群的影响而发生的变化知之甚少。当颗粒物被摄入时,它会被肠道微生物群代谢成其他一些对整个肠道有害的有毒代谢物,如果这种代谢物进入循环,那么它可能会引起其他一些影响。
生物学研究的最新进展表明,空气污染正在导致肠道微生物群的组成和生理发生改变。人类中,吸入的 PM 会迅速从肺部清除并运输到肠道,在那里它可能会导致菌群发生改变。
在炎症性肠病 (IBD) 的小鼠模型中,口服环境 PM10的浓度代表了在高水平空气污染期间可能发生的剂量,已被证明会显着影响肠道微生物群。厚壁菌门的比例增加,而拟杆菌门减少,炎症反应和肠道通透性得到提高。
肠道微生物群相对浓度的这种显着变化导致支链脂肪酸(异丁酸和异戊酸)的形成,从而导致丁酸浓度降低。丁酸是结肠细胞和肠黏膜细胞必需的脂肪酸,丁酸的减少会导致肠道屏障受损,也会导致黏膜炎症。
另一项研究表明,当小鼠暴露于另一种污染物,即多氯联苯 (PCB) 时,与肠道微生物群相关的组成和代谢过程也会发生改变。
流行病学证据表明,空气污染物也与人类 IBD 风险增加有关。有人提出,一般而言,空气污染和 PM 可能会促进人类的胃肠道疾病。
除了 IBD 外,暴露于空气污染与 2 型糖尿病和肥胖症有关,这可能是由于对肠道微生物群的影响。与 NOX暴露相关的特定肠道细菌家族,Bacteroidetes增加,而 Coriobacteriaceae减少。这些变化与发展为 2 型糖尿病的空腹血糖水平升高有关。
纳米材料和食品添加剂:直接改变或者杀死微生物
纳米粒子 (NP)被广泛用于制造许多产品,例如电子元件、无刮痕涂料、药品、运动器材、化妆品、食品容器和膳食补充剂。
由于它们的体积非常小,NPs 能够通过吸入、摄入、皮肤渗透或注射进入人体。作为一种新的环境污染物,NPs 会诱导氧化应激,这被认为是最具毒性的细胞内和体内机制。一些纳米颗粒在体外显示出抗菌特性。例如,银纳米颗粒 (Ag NPs) 对细菌、病毒和真菌生物体具有明显的细胞毒性。摄入后,Ag NPs 可以被小肠吸收并被回肠组织内的派尔氏斑吸收。碳纳米管与细胞壁结合,破坏细胞膜,并对球状或杆状形态的革兰氏阳性菌和革兰氏阴性菌具有抗菌活性。
考虑到 NPs 的生物分布和抗菌活性的潜力,NPs 有可能通过口服和非口服途径影响肠道微生物群。然而,关于纳米颗粒对肠道微生物群的影响的研究很少。
银纳米颗粒(Ag NPs): 最近的一项研究报告称,给予 Ag NPs(1 mg/mL 在 2 mM 柠檬酸盐缓冲液中或在 100 mg/mL 聚乙烯吡咯烷酮水溶液中持续 28 天)不会改变小鼠肠道微生物组的组成、结构或多样性。然而,另一项研究表明,暴露于 Ag NPs13 周会降低门水平厚壁菌的丰度,并扰乱大鼠回肠中免疫调节基因的表达。
此外,Ag NPs 对果蝇幼虫有毒,因为它们会降低幼虫肠道微生物群的多样性。需要更多证据确定 NPs 是否具有抗菌活性或对肠道微生物群的其他影响。
无热量人造甜味剂 (NAS)可为不含高热量糖分的食物提供甜味,是无糖汽水、谷物和无糖甜点中的常见成分,推荐用于减肥和患有葡萄糖耐受不良的人 或 2 型糖尿病。
最近的一份研究表明,长期给小鼠喂食 NAS(包括糖精、三氯蔗糖和阿斯巴甜)会导致更高的葡萄糖耐受性,并且与肠道中拟杆菌属和梭菌属细菌的丰度增加有关。这种葡萄糖耐受不良的增加依赖于肠道微生物群;NAS 在无菌小鼠或抗生素治疗小鼠中均未引起葡萄糖耐受不良;然而,将肠道微生物群从 NAS 处理的小鼠或体外暴露于 NAS 的细菌转移到无菌小鼠或抗生素处理的小鼠,会损害葡萄糖稳态。
乳化剂是食品添加剂,通常存在于加工食品(如面包)中,以延长保质期和新鲜度。

最近的一项研究表明,乳化剂的类似洗涤剂的特性可能会对肠道微生物群和肠道组织的完整性产生负面影响。给小鼠喂食羧甲基纤维素或聚山梨醇酯-80(乳化剂)12 周,增加了粘液溶解细菌(如瘤胃球菌)的丰度,并降低了粪便中拟杆菌的丰度。微生物群的这些变化伴随着肠道粘液厚度的减少、轻度炎症和代谢综合征的症状,包括肥胖增加和葡萄糖失调。有趣的是,乳化剂对无菌小鼠没有影响。
此外,这些变化可以通过从乳化剂处理的小鼠身上转移微生物群来转移到无菌小鼠身上。此外,作者观察到乳化剂在 IL-10 缺陷和 Toll 样受体 5 (TLR5) 缺陷小鼠中促进了强烈的结肠炎,这表明乳化剂可能会促进易感宿主的强烈结肠炎,并诱发低度炎症。
大规模的环境污染和由此产生的抗健康问题是当前主要的公共卫生问题。
当环境污染物改变肠道微生物群组成时,微生物群代谢物,包括短链脂肪酸和胆汁酸,将首先受到影响。这些特性支持短链脂肪酸影响能量代谢、肠道免疫、癌症甚至神经系统功能。
环境污染物会直接影响肠道微生物群并导致代谢物和免疫系统发生变化。生物多样性的丧失和污染物的暴露最终会导致免疫系统失衡,并增加威胁公众健康的炎症性疾病。
环境污染对肠道微生物群的影响及其后续影响
Li N, et al. Environ Pollut. 2021
需要注意的是,环境毒理性研究大多数来自动物模型,主要是大小鼠。确定动物结果对推测人类的重要性并不容易,因为它们没有相同的环境暴露以及物种差异。没有人能排除环境污染物可能对人类肠道菌群和表型产生长期影响的可能性,而这些影响迄今为止在动物身上不存在,短期使用是规则。
总之,动物实验和流行病学研究并不完全平行。因此,即使有足够的证据支持基于动物模型结果的“三向相互作用”理论,仍需要更多的流行病学证据来验证。
几十年来,评估环境化学品健康风险的核心要素保持不变(参见相关报道)。迄今为止,还没有研究方法明确考虑人类微生物组的潜在影响。因此,微生物组在改变人类对环境相关暴露的毒性易感性方面的作用在很大程度上仍不确定。
值得庆幸的是,许多已知的微生物转化必须与酶和基因相关联,为临床研究中的诊断标志物提供特定目标。对肠道菌群异物代谢的分子理解将为毒理学风险评估提供信息,并为环境污染中毒的治疗提供新的靶点。
建议今后应该确定以下几方面:
许多微生物在人类存在之前就已经存活在大自然中,它们也在不断进化从而适应环境。微生物群与人类、环境和它们自身相互作用或许远超出我们想象。
微生物群检测是我们人类得以了解它们的一个通道,对于环境微生物群的深入了解,或许能成为我们了解微生物群与人体关系的一块重要拼图。
主要参考文献
Claus SP, Guillou H, Ellero-Simatos S. The gut microbiota: a major player in the toxicity of environmental pollutants? [published correction appears in NPJ Biofilms Microbiomes. 2017 Jun 22;3:17001]. NPJ Biofilms Microbiomes. 2016;2:16003. Published 2016 May 4. doi:10.1038/npjbiofilms.2016.3
Li N, Li J, Zhang Q, Gao S, Quan X, Liu P, Xu C. Effects of endocrine disrupting chemicals in host health: Three-way interactions between environmental exposure, host phenotypic responses, and gut microbiota. Environ Pollut. 2021 Feb 15;271:116387. doi: 10.1016/j.envpol.2020.116387. Epub 2020 Dec 24. PMID: 33401209.
Jin Y, Wu S, Zeng Z, Fu Z. Effects of environmental pollutants on gut microbiota. Environ Pollut. 2017 Mar;222:1-9. doi: 10.1016/j.envpol.2016.11.045. Epub 2017 Jan 11. PMID: 28086130.
Anwar H, Iftikhar A, Muzaffar H, Almatroudi A, Allemailem KS, Navaid S, Saleem S, Khurshid M. Biodiversity of Gut Microbiota: Impact of Various Host and Environmental Factors. Biomed Res Int. 2021 May 12;2021:5575245. doi: 10.1155/2021/5575245. PMID: 34055983; PMCID: PMC8133857.

谷禾健康

肠道菌群是居住在肠道中各种微生物。微生物群的建立甚至在个体出生之前就开始了,随着时间的推移而改变,并在体内持续存在,直到个体死亡。这些微生物群的组成是宿主特定的,在个体的一生中不断进化,并且容易受到各种因素的影响。
胎龄、分娩方式、饮食(母乳与配方奶)、卫生、抗生素,激素、疾病、衰老等都会影响并塑造肠道菌群。肠道菌群的定植、发育、成熟、稳定,老化与我们人类发育以及免疫成熟等高度吻合,不同阶段不同部位的菌群的构成以及丰度有不同特征。
肠道细菌是免疫系统发育和功能的重要组成部分。肠道菌群的变化可能是许多炎症性疾病发生的重要因素。而生活方式的改变可能改变了肠道菌群的初始发育或稳定维持。
本文从肠道菌群的功能,初始构建,到菌群健康/失衡的状态,以及相应的改善措施等进行全面阐述。
肠道菌群,居住在肠道(宿主)内的所有本土细菌的总和,被视为一个器官,执行着一系列重要的、对健康至关重要的功能,而这些功能无法通过任何其他方式复制。
研究人员通过比较无菌实验动物(没有任何肠道菌群)和正常菌群对照动物来确定这些功能。这些发现也在人群中得到了证实。
以下是肠道菌群最重要功能的简要概述:
◥ 粪便中的水分滞留
粪便主要是由水组成(平均水含量75%;各项研究的平均范围为63-86%)。单细胞生物,如细菌,主要含有水,被无法穿透的膜包围。
◥ 形成正常粪便
由于细菌是正常粪便中最主要的成分,它们的缺失可能会导致持续性慢性腹泻。
◥ 生产必需维生素
细菌会合成各种物质,包括某些复合维生素 B、维生素 B12 和维生素 K,这些物质对血液正常凝固至关重要。
◥ 保护肠道上皮(粘膜)免受病原体侵害
正常的肠道菌群控制着不良细菌的数量,例如白色念珠菌(酵母)或大肠杆菌的感染性菌株。保护机制有很多种,食物供应的竞争、对肠粘膜的粘附、维持所需的 pH 值平衡以及产生过氧化物和酶,从而杀死外来细菌。
◥ 组织发育和再生
与健康动物相比,无菌实验动物的肠粘膜(上皮)和淋巴组织(派尔氏斑)发育不良,肠道健康黏膜薄弱、免疫淋巴组织不发达等存在许多缺点。
◥ 免疫
正常肠道细菌负责实现吞噬作用:吞噬细胞在全身范围内破坏致病细菌、病毒、过敏原和其他异物,吞噬细胞是负责非特异性(抗体前)免疫系统防御的专门血细胞。
生命的前三年是可塑性增强的时期,肠道微生物群的发育很容易受到环境因素的影响。在婴儿期人与人之间的肠道微生物组差异变化最大,在成年期变得更加相似。
★ 婴幼儿
当婴儿出生后,在几口初乳后,大肠得到“培养”,初乳是一种淡黄色的液体,包含母亲的细菌,富含必需的营养。初乳先于富含脂肪和蛋白质的母乳流出。这个过程在母乳中继续,新生儿的肠道菌群在第6个月开始逐渐成熟,直到“成人”状态。
肠道微生物群的发育及主要影响环境因素
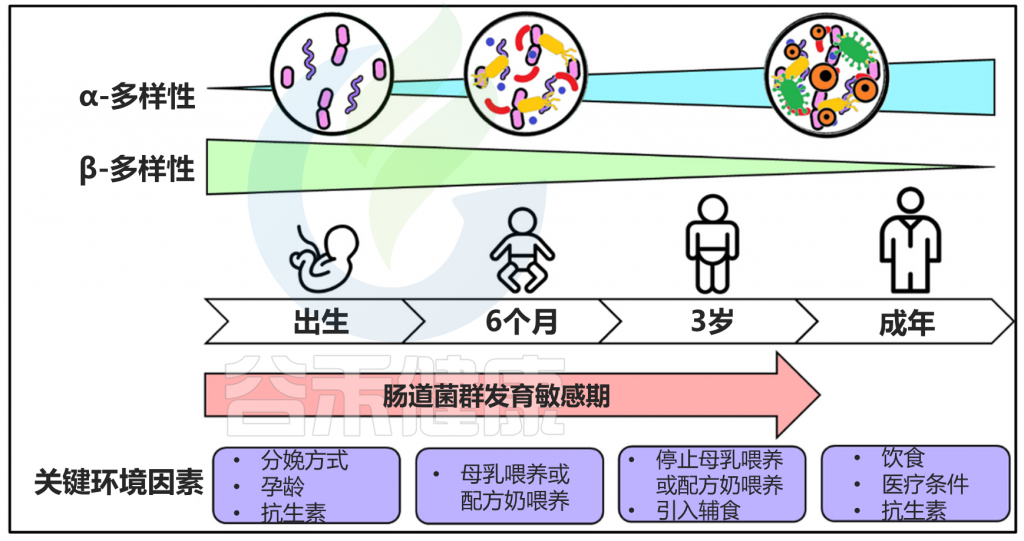

Parkin K et al., Microorganisms,2021
谷禾健康
在与细菌性疾病的斗争中,抗生素是不可避免的。抗生素是人类使用了几十年的治疗细菌感染的救命药物。

虽然抗生素挽救了数百万人的生命,但它们也会消灭一些常驻细菌。现有文献已经确定了与抗生素相关的肠道微生物群失调(多样性降低)。也就是说过度使用抗生素会影响肠道健康。
抗生素对肠道菌群的不同影响
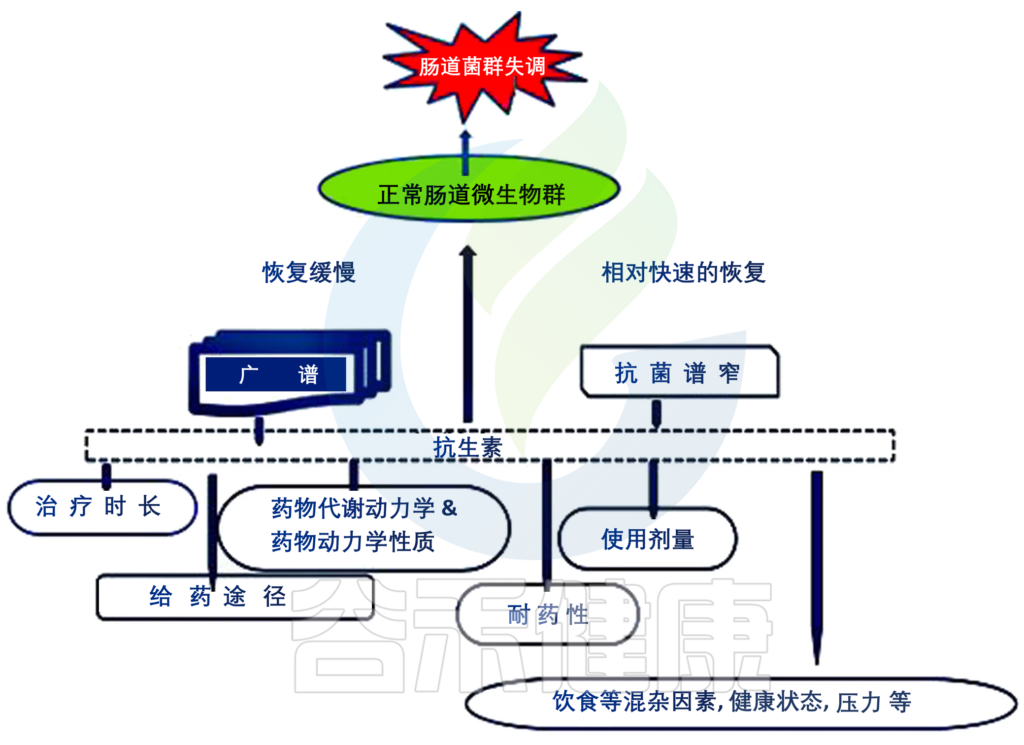
Yang L, et al., AMB Express. 2021
药物使用时间越短,肠道菌群恢复越快,反之亦然。然而,虽然一些物种将在几个月后恢复,但有些灭绝的物种在试验期间没有恢复。每个疗程中抗生素浓度的比例恢复是一种现象,它将从体积上说明所使用的抗生素的疗效。

♞ 抗生素滥用正导致耐药性危机日益增长
构成抗生素滥用的一种形式与食用动物有关。由于抗生素的过度使用,微生物的遗传能力随着耐药基因的增加而增加。显然,细菌对几乎所有可用的抗生素都产生了多种机制的耐药性。
抗生素滥用所造成的神经毒性效应,既有局部作用,也有更显著的全身作用。
滥用抗生素的不良反应除了增加微生物耐药性之外,还有微生物群异常,所谓微生物群异常也就是菌群失调。
☟ 菌群失调
可导致自身免疫性疾病、胃肠道疾病、过敏、感染、关节炎、哮喘、癌症、肥胖等疾病。
过量服用抗生素会改变肠道微生物群,据研究,这与神经和精神疾病的发展有关。在控制抗生素滥用和耐药性的同时,为了减少抗生素的使用,抗菌肽(AMPs)与一些传统抗生素联合使用是有前景的。
从最近的研究来看,水平基因转移(HGT)和抗生素耐药基因在胃肠道中大量发生。这带来了许多临床意义,因为益生菌、共生菌和条件致病菌可以通过转移耐药基因在抗菌药物中生存。
◢ 在耐药性癫痫患者中的菌群失调
由于肠道生态失调被发现与黏液消耗、炎症、血脑屏障破坏导致神经调节剂改变有关,与药物敏感患者和健康对照相比,耐药性癫痫患者的肠道微生物组组成发生了改变。
低碳水化合物饮食疗法通过改变肠道微生物组减少一些耐药性癫痫患者的癫痫发作。益生菌补充剂通过改变微生物群的组成,可以减少某些患者的癫痫发作。
◢ 健康人肠道肠杆菌科出现替加环素耐药性
在新加坡健康人群肠道微生物群的队列研究中,通过替加环素灭活酶Tet (X4)对宿主的作用检测到高水平的替加环素耐药性。大肠杆菌2EC1-1和94EC分别携带Tet(X4)、IncI1型质粒p2EC1-1和p94EC-2。产 Tet(X4) 肠杆菌科细菌的粪便携带率为 10.1%(95% CI = 5.1%-17.3%),表明 Tet(X4) 广泛分布于新加坡健康个体的肠道微生物群。
接下来我们详细了解下,目前常用的抗生素对肠道菌群会带来哪些影响。
人类肠道微生物群在平均稳定的群落中是的,而对所有物种来说都是没有弹性的。从某种意义上说,就像突变的效果一样,一旦基因发生突变,就不会有恢复。因此,在使用后果不明的药物削弱肠道微生物群时,必须高度谨慎。
表1 抗生素使用对肠道菌群变化的不同影响
克林霉素
克林霉素是常用的广谱抗生素之一。它主要针对对维持肠道健康至关重要的厌氧菌。

谷禾健康
中风是全球范围内最具破坏性的最严重的脑部疾病之一,治疗选择非常有限。中风和其他神经系统疾病与微生物群破坏有关。反过来,中风诱发的菌群失调会通过调节免疫反应而对疾病结果产生重大影响。
了解肠细菌-脑串扰的性质可能会导致基于微生物组的治疗方法,从而可以改善患者的康复。
本文主要介绍关于肠道微生物组在中风中的作用的最新知识,以及肠-免疫-脑串扰的潜在机制。
肠道微生物群和周围器官之间的信号传导是由微生物组相关分子模式(MAMP)和微生物组分泌的代谢产物介导的,这些代谢产物可以与粘膜上皮和肠道免疫细胞相互作用,刺激迷走神经或到达全身循环以向大脑发出信号和可能调节神经元和免疫应答。
反过来,支配肠壁的副交感神经和交感神经纤维会从大脑传递输入信号,从而影响肠蠕动,免疫细胞活性,并能诱导肠成分改变。
01
中风改变了微生物群的组成
我们知道,肠道微生物群稳态的破坏,就会容易患病。
实验和临床研究中越来越多的证据表明中风会改变肠道菌群的组成。对中风患者肠道菌群的分析显示,与健康个体相比,微生物组成发生了变化,而菌群失调程度与病变的严重程度相关。
同样,观察小鼠的粪便微生物丰度的变化,发现严重中风要比轻微中风更为明显。但是,在分析菌群多样性和特定菌群时,在临床和实验研究中有些不同结果。
为什么会出现不同结果?
Yin 等人发现与无症状对照组相比,中风患者粪便样本的多样性增加,Stanley等人发现假手术组和中风小鼠之间的微生物群多样性没有变化。相反,其他研究人员则在实验和临床环境中观察到某些菌减少和其他细菌的过度繁殖导致多样性降低,表明多样性程度不一定能预测中风的严重程度。
不同的研究发现,沙门氏菌总体上减少了,同时伴有拟杆菌的过度生长。然而,当在较低的生物分类等级上分析微生物群变化时,在临床和实验性中风中特定细菌变化方面缺乏一致性。
可能的混淆因素
粪便采样位置的差异(在小鼠的胃肠道和患者的粪便中),用于基因组DNA提取和16S rRNA基因分析的方法,中风模型的严重程度以及基线的差异微生物群组成(人类的小鼠起源或个体间和个体内变异)可能是造成矛盾结果的混淆因素。
影响肠道菌群组成的另一个混杂因素是饮食类型。
严重的中风病例总是住院治疗,他们将接受药物治疗并接受医院控制的饮食,而实验性中风后对小鼠的干预很少,这将以啮齿动物和人类的不同方式影响微生物群的组成。
总体而言,临床和实验数据表明中风后微生物群组成发生了变化。然而,迄今为止,尚不清楚哪种类型的肠道细菌参与中风的病理生物学机制。
年龄对微生物群状态的重要性
衰老会改变肠道菌群的组成,并与炎症和肠道通透性增加有关。中风后,与年轻小鼠相比,有高龄微生物群的小鼠具有更高的死亡率,运动功能受损和促炎性细胞因子反应。
粪便微生物群移植(FMT)可以在年轻和老年小鼠中恢复中风的结果。这些实验强调了年龄对微生物群状态的重要性及其对疾病发展的影响,以及微生物群的操纵如何能够导致非侵入性疗法以预防和改善中风预后。
虽然实验结果表明与年龄、肠道生物失调和中风结果相关,但其他风险因素(糖尿病、高血胆固醇水平、饮酒、高脂肪饮食、缺乏锻炼)和性别差异对肠道微生物群和中风结果的影响仍有待实验和临床研究解决。
脑损伤改变微生物组组成的机制
由于中风后自主神经系统(ANS)的失调而降低肠道运动性和粘膜屏障的完整性。脑损伤后,肠道内胆碱能活性下降,肾上腺素能信号传导增加,这与肠屏障的破坏有关。
因此,使用β受体阻滞剂抑制β肾上腺素能活性可显着恢复中风诱导的肠道通透性,并减少细菌向周围器官的扩散。
在这方面,Singh等人证明了回肠瘫痪本身就可以驱动微生物群变化,这是一种中枢神经系统介导的生态失调。
肠道细菌可以感觉到来自胃肠道的激素和神经递质,并且在体外培养中,在应激激素去甲肾上腺素(NE)存在下,某些条件致病菌能够增殖更多。
有趣的是,在实验性中风后,盲肠中的去甲肾上腺素释放增加,这可能是某些病原体或共生菌过度生长的原因。
另外,ANS调节肠粘液分泌,大脑对肠信号的扰动可能对肠粘液层的大小和质量产生重要影响。
Houlden等人观察到,中风后盲肠内,粘液蛋白产生的失调和产粘液的杯状细胞降低。中风后粘液层的形成受损可能使共生膜与上皮直接接触,转移并引发免疫反应,以及中风后感染。此外,还需要更多关于细菌移位导致实验性中风感染的直接影响的数据。
脑损伤触发肠道中胆碱能和肾上腺素能信号的失衡,增加释放的NE的浓度,降低肠道运动性,并改变与微生物群失调有关的肠脑屏障。所有这些事件都将影响肠道微生物的功能输出,并可能进一步扰乱肠道的免疫稳态。
02
中风的肠-免疫-脑轴
迄今为止,使用动物模型进行的研究提供了最有力的证据,并有助于阐明中风后肠-免疫-脑相互作用的潜在机制。
T淋巴细胞的关键作用
已知T淋巴细胞在脑损伤后继发的组织损伤中起关键作用。促炎性T辅助1细胞(Th1),T辅助17细胞(Th17)和γδT-IL-17+细胞与中风后神经毒性有关,而调节性T细胞(Treg)发挥作用抗炎和神经保护特性。
越来越多的证据,支持微生物群通过调节胃肠道中促炎Th17细胞与抗炎Treg细胞之间的平衡来维持免疫稳态。
通过诱导促炎Th1和Th17介导的反应,具有中风后遗症粪便含量的无菌(GF)小鼠的定殖导致中风结果的恶化,这表明中风诱导的微生物失调触发促炎性免疫反应,从而加剧脑损伤。
这组作者证明,来自派尔氏淋巴集结(Peyer’s patches)的T细胞会侵袭梗塞周围组织,并在中风的急性期产生免疫反应。
肠道菌群的免疫调节作用对于中风恢复至关重要
此外,通过健康捐献者的FMT恢复肠道微生物群的稳态,减少了损伤大小并增加了缺血性脑中Treg细胞的数量。
然而,单独存在的微生物群具有神经保护作用,因为与定殖小鼠相比,完全根除GF小鼠的肠道细菌导致更大的梗塞体积。
同样,这种微生物群介导的对中风的有益作用取决于淋巴细胞反应,表明肠道细菌的免疫调节作用对于中风恢复至关重要。通过显示在缺血性损伤之前使用抗生素对微生物群组成进行控制可减少梗塞体积并改善感觉运动功能,从而证明了肠-脑双向沟通在中风中的关键参与。
在这项研究中,他们表明神经保护作用是由微生物依赖性的肠道树突状细胞(DC)引发的,介导小肠Treg的扩增,通过抑制促炎性γδT-IL-17+细胞发挥作用。
此外,他们还认为IL-10是Treg细胞介导的γδT细胞增殖抑制的主要协调器,因为IL-10-/- 小鼠不受保护作用的影响。由于较少的促炎IL-17 +γδT细胞积聚在脑膜中,因此肠道中的这种抗炎环境会传导到大脑,这与较小的梗死面积有关。
有趣的是,肠道T细胞从肠道迁移到脑膜和脑实质中,表明中风有直接的肠脑沟通途径。
另外,肠道微生物群组成的修饰可通过致耐受性DC使肠道免疫应答偏向抗炎环境。
这些研究表明中风后微生物群受到影响,中风后细菌数量的变化会促进促炎性T细胞反应,肠道免疫细胞迁移至脑膜,这可能会参与继发性脑损伤并恶化中风结果。
也突显了通过肠道淋巴细胞从肠道运输到大脑的过程,沿着肠道-脑轴的直接联系,在那里它们调节了对中风的神经炎症反应。
当然,对于肠免疫细胞是否直接导致脑损伤或修复机制还不是很清楚。重要的是,这种肠道细菌-DC之间相互作用的分子途径仍然难以捉摸。
图 肠道代谢物——中风中的免疫系统干扰
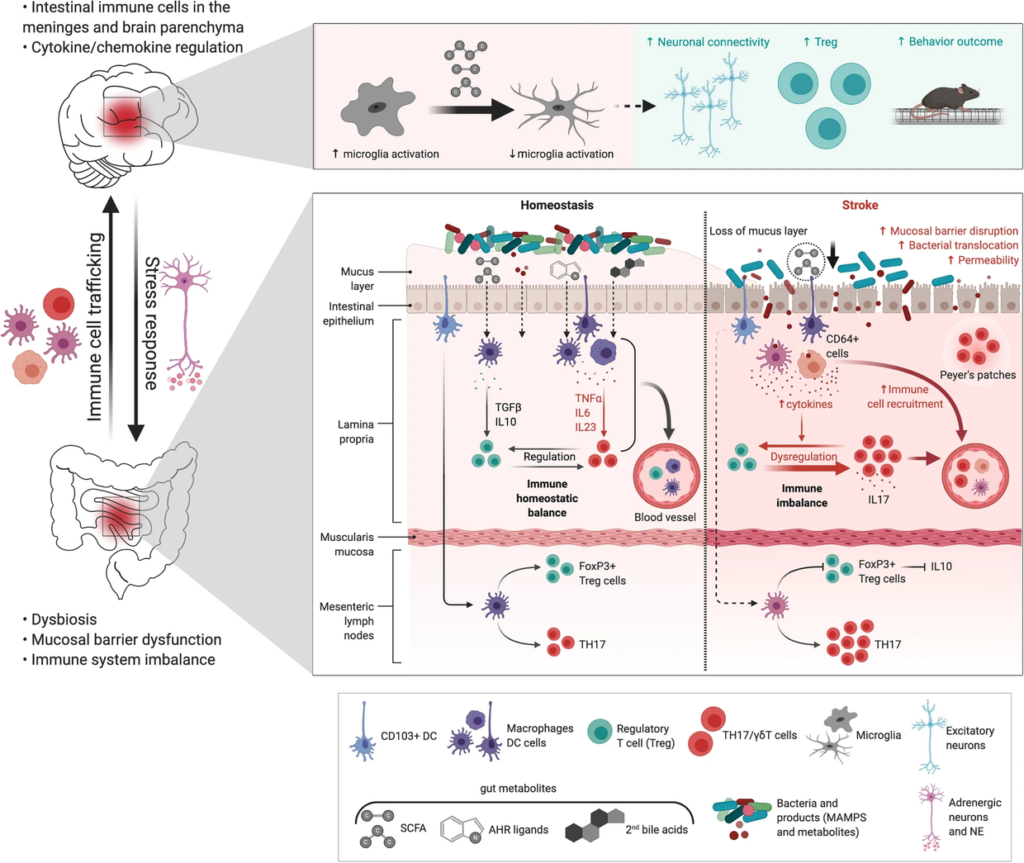
Delgado.et al., Neuromolecular Med, 2020.
中风后肠道微生物对免疫系统的调节。
下图:在肠道中,中风引起生物失调、粘膜屏障功能障碍、肠道通透性增加、细菌移位、中风后感染和通过树突细胞的促炎性T细胞反应(DC)。中风后,免疫细胞,特别是T细胞、CD64 +巨噬细胞和树突状细胞从肠道迁移到脑膜和大脑。肠道代谢物(AHR,次生胆汁酸,MAMPs)作为肠道免疫细胞免疫调节剂的作用尚未明确。中风后微生物来源的SCFA减少,可能引发γδT-IL-17 +细胞和调节性T细胞(Treg)的失衡。
上图:补充SCFA调节神经元活动和突触密度,并且与小胶质细胞激活减少和Tregs增加以及中风后更好的恢复相关。
03
中风肠道菌群的操纵
Benakis等人证明了在诱导缺血性损伤之前用氨苄西林或万古霉素治疗小鼠具有神经保护作用,而使用新霉素则不能保护小鼠免于中风。这种神经保护作用与微生物组组成中的抗生素特异性变化有关,变形菌和厚壁菌门增多,而拟杆菌属的减少。
此外,当他们分析与有益菌群变化相关的预测酶途径时,他们发现异种/芳香族化合物代谢可预测缺血性病变的大小。
微生物代谢产物是细菌-宿主串扰的主要沟通渠道之一
与中风的肠道免疫脑轴有关的一个可能的缺失环节可能是仅由细菌产生的代谢产物。
已知必需芳香族氨基酸色氨酸的代谢产物可通过在DC和T细胞中表达的芳烃受体(AHR)调节免疫细胞功能。一些菌群能够将肠道中的色氨酸分解为AHR的配体。
在小鼠中,微生物群衍生的吲哚激活AHR通过诱导免疫细胞中IL-22促进肠道稳态。在星形胶质细胞的AHR通过激活在多发性硬化的小鼠模型中色氨酸代谢物的神经保护作用的驱动器。
然而,保护作用是配体依赖,其他色氨酸代谢产物已报告加重病程。有趣的是,负责色氨酸分解代谢的内源性犬尿氨酸途径的活性被上调下列缺血性中风和已与炎性反应和恶化的结果。因此,AHR受体的药理和遗传阻断改善了中风恢复。
这些发现暗示了色氨酸衍生化合物在中风中的关键作用。尚不清楚微生物源性色氨酸代谢产物在缺血性损伤中的免疫调节作用。
短链脂肪酸(SCFA)是细菌发酵的生物产物,具有免疫调节作用:它们能够根据免疫情况直接诱导T细胞分化为效应细胞和调节细胞。
小鼠中风后血浆SCFA的水平显着降低,这与恶化的结果有关。
中风之前在小鼠中补充SCFA改善了行为恢复,改善了皮质网络的连通性,并改变了突触可塑性的组织学标志,这与改善中风的长期中风有关。这些作用与小胶质细胞形态向稳态状态的改变和侵袭性淋巴细胞的减少有关。
菊粉改善中风后神经功能缺损
在另一项研究中,他们观察到,与年幼小鼠相比,口服菊粉(产生SCFA的细菌和制造SCFA的细菌基质),可减少老年小鼠中风后的神经功能缺损和改善抑郁样行为。
此外,这些变化伴随着缺血性脑中IL-17 +γδT细胞百分比的降低,但SCFA是否直接影响T细胞极化和迁移尚未解决。出人意料的是,在两项研究中,梗死面积均不受SCFA干预的影响。
所有这些证据表明,微生物代谢产物SCFA在中风后恢复中起重要作用,并且可能参与中风后肠道菌群发挥的免疫调节作用。
04
结 语
本文总结了微生物群组成如何通过调节免疫反应影响中风预后的最新发现。肠道菌群的变化引起主要的促炎性T细胞反应,这与更大的梗塞体积和恶化的预后相关。
由于肠道菌群是一个非常复杂的生态系统,受环境和宿主的影响,具有冗余功能和协同关系,因此,研究微生物组代谢组学特征以将其转化为临床实验研究是一种更好的策略。
然而,尚无实验数据将特定微生物群合成的化合物与脑损伤后的免疫反应之间的因果关系联系起来。为了在中风患者中成功应用这些发现,有必要进一步研究中风后共生细菌和免疫系统之间相互作用的分子机制。
相关阅读:
参考文献:
Singh, V., Roth, S., Llovera, G., Sadler, R., Garzetti, D.,Stecher, B.,et al. (2016). Microbiota dysbiosis controls the neuroinflammatory response after stroke. Journal of Neuroscience, 36(28), 7428–7440.
Houlden, A., Goldrick, M., Brough, D., Vizi, E. S., Lénárt, N., Martinecz, B., et al. (2016). Brain injury induces specific changes in the caecal microbiota of mice via altered autonomic activity and mucoprotein production. Brain, Behavior, and Immunity, 57, 10–20.
Lee, J., d’ Aigle, J., Atadja, L., Quaicoe, V., Honarpisheh, P., Ganesh, B. P., et al. (2020). Gut microbiota-derived short-chain fatty acids promote poststroke recovery in aged mice. Circulation Research, 127(4), 453–465.
Delgado Jiménez Rosa,Benakis Corinne,The Gut Ecosystem: A Critical Player in Stroke.[J] .Neuromolecular Med, 2020.
Yin, J., Liao, S. X., He, Y., Wang, S., Xia, G. H., Liu, F. T., et al. (2015). Dysbiosis of gut microbiota with reduced trimethylamine-N-oxide level in patients with large-artery atherosclerotic stroke or transient ischemic attack. Journal of the American Heart Association.
Stanley, D., Mason, L. J., Mackin, K. E., Srikhanta, Y. N., Lyras, D., Prakash, M. D., et al. (2016). Translocation and dissemination of commensal bacteria in post-stroke infection. Nature Medicine.
Benakis, C., Brea, D., Caballero, S., Faraco, G., Moore, J., Murphy, M., et al. (2016). Commensal microbiota affects ischemic stroke outcome by regulating intestinal γδ T cells. Nature Medicine, 22(5), 516–523.
Lamas, B., Natividad, J. M., & Sokol, H. (2018). Aryl hydrocarbon receptor and intestinal immunity. Mucosal Immunology, 11(4), 1024–1038.