-

 国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子
国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子

 二级病原微生物安全实验室
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
谷禾健康
近年来,免疫治疗在癌症治疗领域展现出了巨大的潜力,特别是针对PD1/PDL1通路的免疫检查点抑制剂(ICIs)已在多种实体瘤患者中取得显著疗效。然而,治疗反应的异质性和耐药性的出现依然是当前面临的主要挑战之一。越来越多的研究表明,肠道菌群作为人体内一个庞大的微生态系统,不仅参与机体代谢和免疫调控,而且在调节肿瘤免疫反应方面发挥着关键作用。
让我们快速了解一下肠道菌群如何通过细胞水平、分子信号通路和代谢产物来影响PD1/PDL1肿瘤免疫疗法的疗效。
1 核心菌群与免疫调控机制
多项临床与实验室研究均显示,特定菌群对PD1/PDL1免疫检查点抑制剂治疗具有显著的正向作用。
例如,Bifidobacterium属菌群被发现能显著促进抗PDL1疗法的抗肿瘤作用;而Akkermansia muciniphila在PD1抑制治疗中也表现出与治疗反应正相关的趋势。此外,研究还发现,黑色素瘤患者中较高的微生物多样性和特定菌群如Ruminococcaceae和Faecalibacterium的丰富度均与更长的无进展生存期相关。
肠道菌群对免疫调控的机制主要表现在以下几个方面:
促进树突状细胞成熟:某些菌群(如Bifidobacterium)可通过激活树突状细胞,从而增强抗原呈递能力,提高CD8+ T细胞的活性。
调节T细胞免疫状态:特定菌群通过影响细胞因子分泌和T细胞亚群分布,调节包括CD8+效应T细胞和调节性T细胞(Treg)在内的免疫平衡,从而实现抗肿瘤效应。
通过代谢产物发挥作用:菌群代谢产物,如短链脂肪酸(SCFAs)和次级胆汁酸,对免疫细胞储存、功能激活具有直接调控作用。
这些作用机制不仅在单一免疫治疗模式中发挥效应,同时也对联合治疗(例如PD1/PDL1抑制剂与CTLA-4抑制剂联合使用)产生协同增效作用。
2 关键代谢物及其信号调控路径
肠道菌群通过发酵膳食纤维等底物,产生大量的短链脂肪酸(SCFAs),如丁酸盐和丙酸盐,这些代谢物在调节宿主免疫功能中起到关键作用。研究发现,短链脂肪酸不仅参与维持肠道上皮屏障的稳定,还能调节T细胞分化和促炎/抗炎反应的平衡。此外,菌群代谢的次级胆汁酸也被证明在免疫抑制和调控细胞因子水平中起到重要作用。
值得注意的是,不同菌群通过生成不同的代谢产物,对免疫系统的影响可能存在正负两方面的效应。例如,有研究显示高浓度的丁酸盐和丙酸盐可能会在某些条件下限制CTLA-4抑制剂的疗效,从而提示适度平衡菌群代谢产物对于免疫治疗的成功至关重要。同时,某些菌群如Prevotellaceae和Rikenellaceae则可能通过降低丁酸盐水平来促进促炎性巨噬细胞M1型的极化,从而间接影响免疫治疗的疗效。
3 不同肿瘤类型中的菌群特征与免疫应答
肠道菌群对免疫治疗的影响在不同肿瘤类型中可能存在显著差异。下面介绍一下在黑色素瘤、非小细胞肺癌(NSCLC)、肝细胞癌(HCC)以及结直肠癌(CRC)中的相关发现。
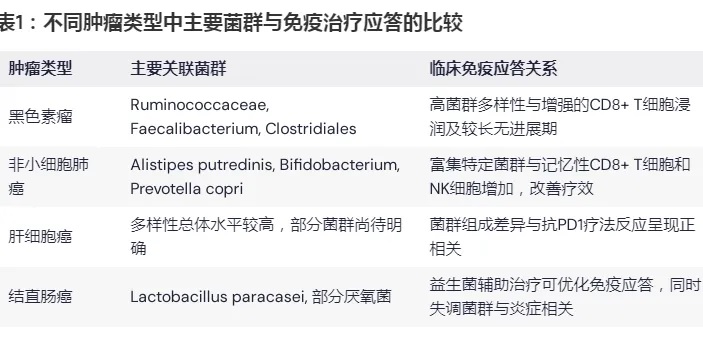
表1说明:各肿瘤类型中显示出菌群多样性和特定菌群丰度与PD1/PDL1免疫治疗效果之间存在明显相关性,该表对比了不同肿瘤中的主要菌群与免疫应答情况。
4 肠道菌群调控机制的分子与细胞通路
肠道菌群通过多条分子信号通路及细胞间相互作用调控宿主的免疫反应,进而影响PD1/PDL1免疫治疗的疗效。
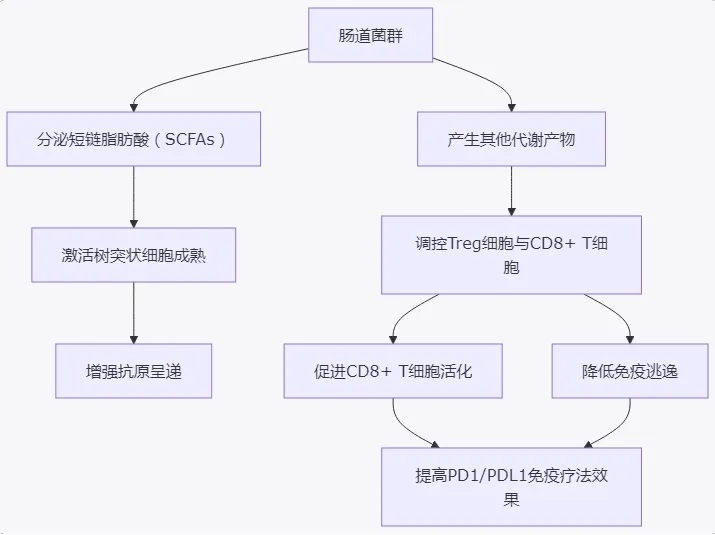
图1:肠道菌群通过分泌代谢产物调控树突状细胞成熟、T细胞活性与调节性T细胞平衡,从而增强PD1/PDL1免疫治疗效果。
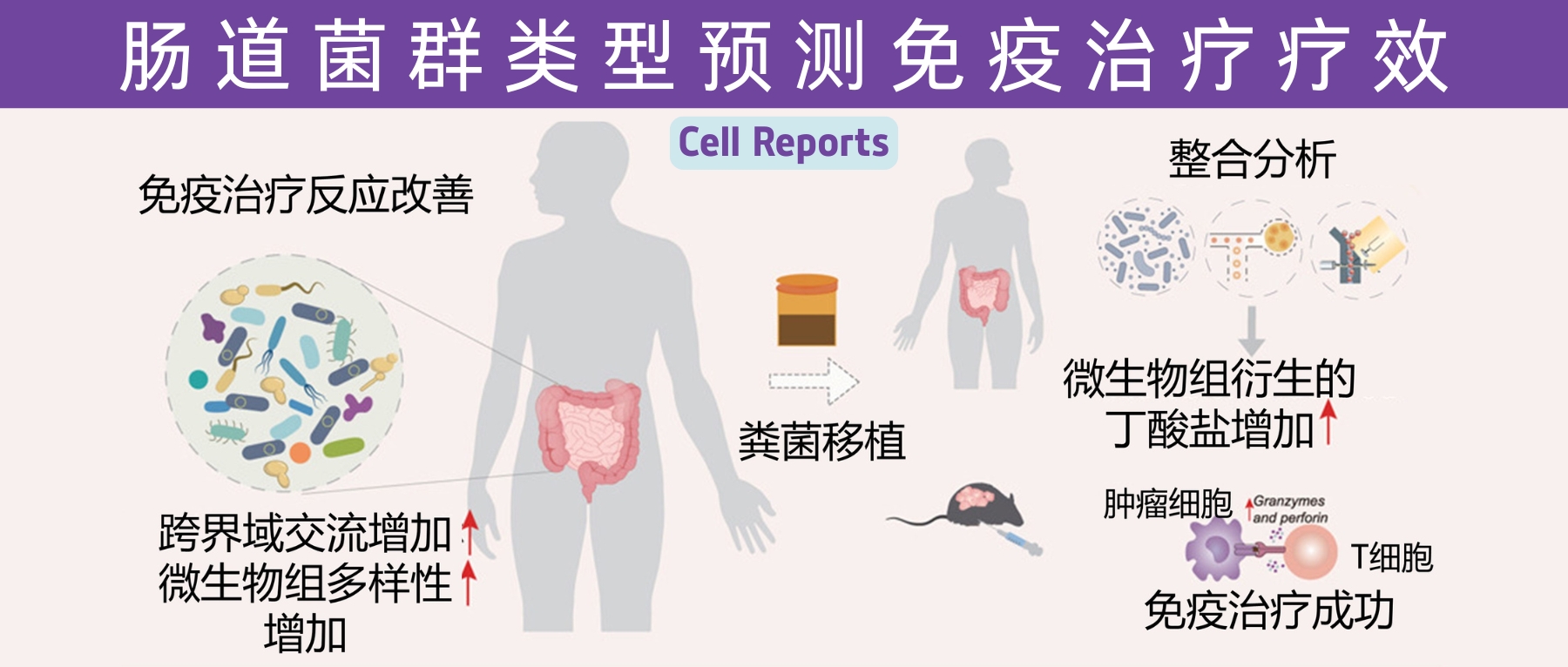
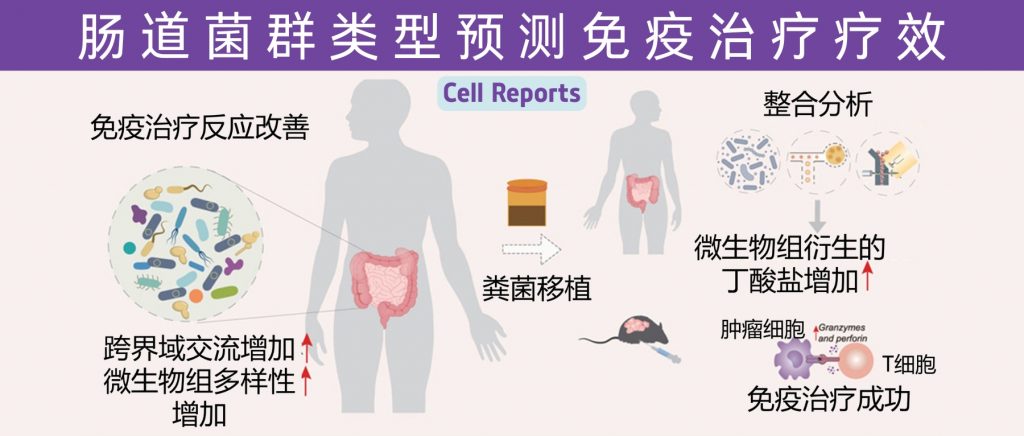
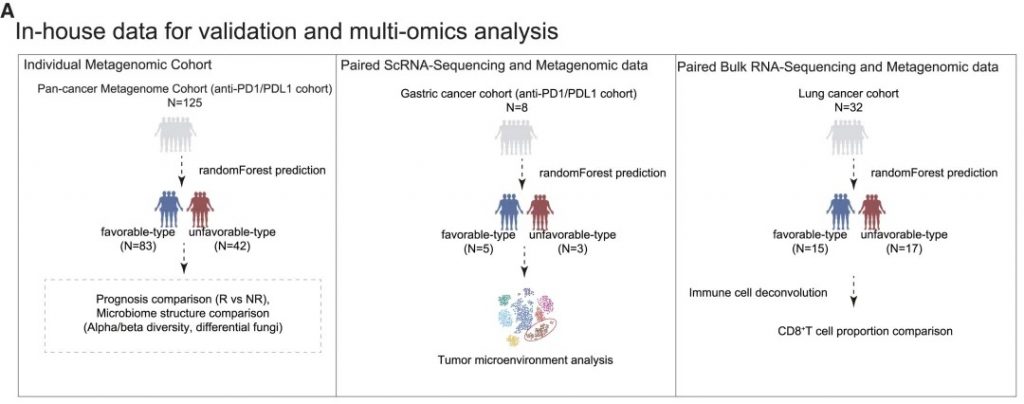
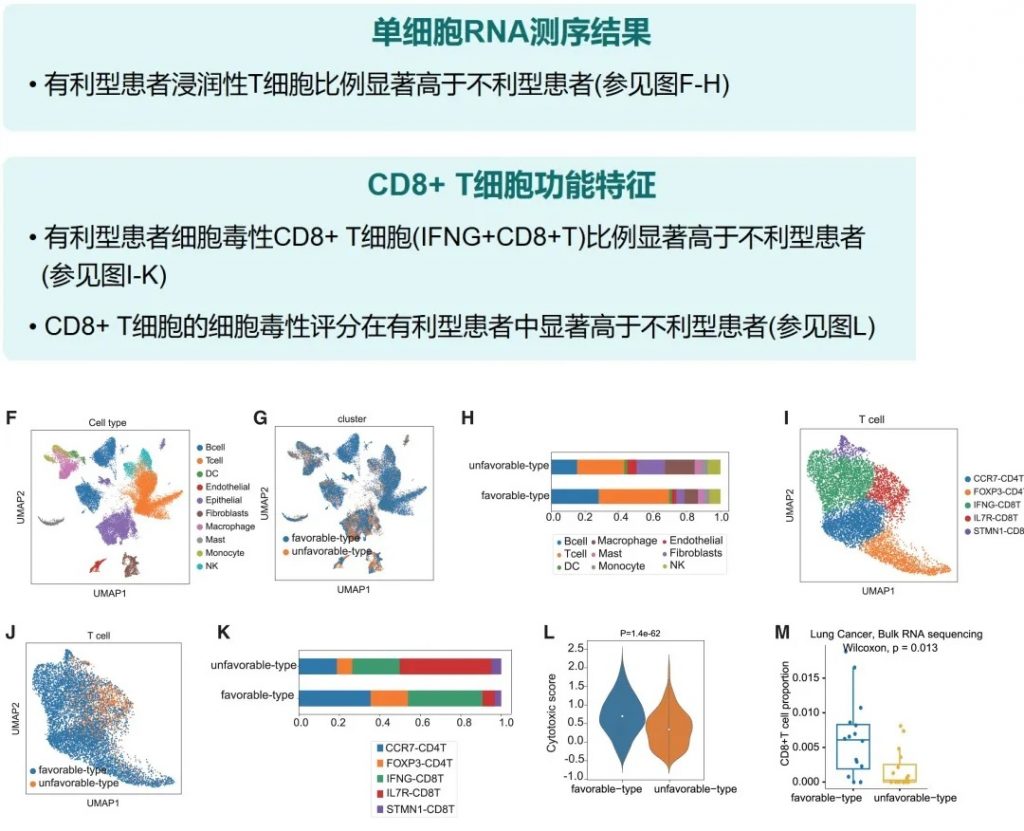
近日,一项来自上海交通大学医学院研究团队的成果发表在《Cell Reports》,通过整合多组学数据与临床队列,揭示了基于肠道菌群分类(有利型与不利型)与抗PD-1/PD-L1免疫治疗疗效的显著关联。

其中,有利型患者表现出更高的真菌与细菌多样性、富集丁酸代谢通路及促免疫菌群(如Faecalibacterium prausnitzii),以及肿瘤微环境中细胞毒性CD8+ T细胞浸润增强的特征,且在多癌种队列中显著关联于抗PD-1/PD-L1治疗的临床缓解与生存获益。
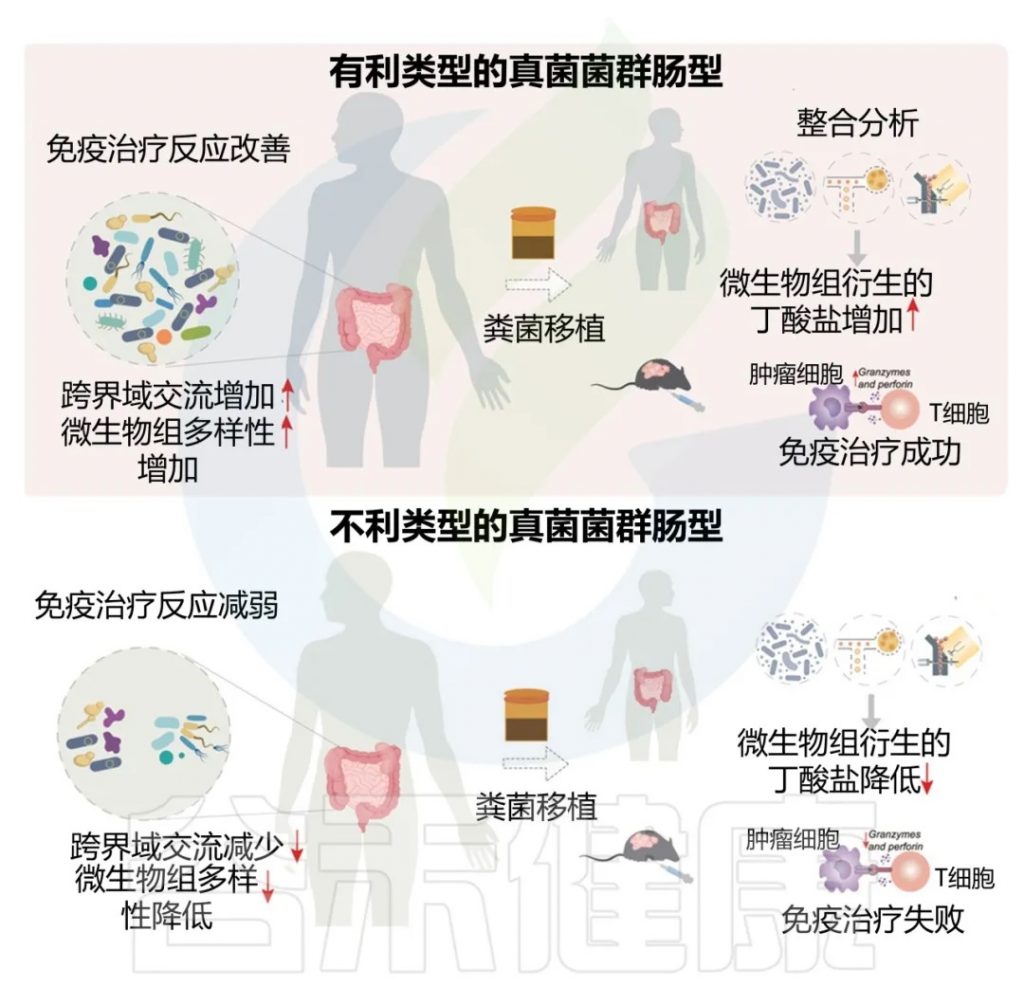
进一步通过FMT实验证实,移植来自有利型供体的粪便可显著提升受体对免疫治疗的敏感性,并重塑肠道菌群代谢功能。
这些发现不仅深化了对肠道真菌组-细菌互作网络的理解,为优化个体化免疫治疗提供了新型生物标志物,更为优化免疫治疗分层策略及FMT供体筛选提供了科学依据,具有重要的临床转化潜力。

多组学关联分析与肠道菌群类型特征
详细对比了两种肠道菌群类型的微生物群落特征差异,包括多样性、产丁酸菌含量、代谢通路等,并展示了它们与临床疗效的关联。
粪便微生物移植实验验证
呈现了临床FMT研究和动物模型验证结果,证明来自有利型供体的FMT可显著提升受体对免疫治疗的敏感性。
总结了研究的创新点及其在预测分层和干预优化方面的临床应用价值。
肠道菌群类型:免疫治疗疗效的新生物标志物
免疫检查点抑制剂(如抗PD-1/PD-L1)显著改变了癌症治疗格局,但患者反应差异大。
该研究团队采用无监督聚类方法分析肠道菌群组成,在多个独立队列中成功识别并验证了两种截然不同的类型——“有利型”与“不利型”,这一发现揭示了肠道菌群对免疫治疗应答的预测价值。

肠道菌群的类型不仅与临床反应相关,还与患者总生存期密切相关,为个体化治疗策略提供了新思路。
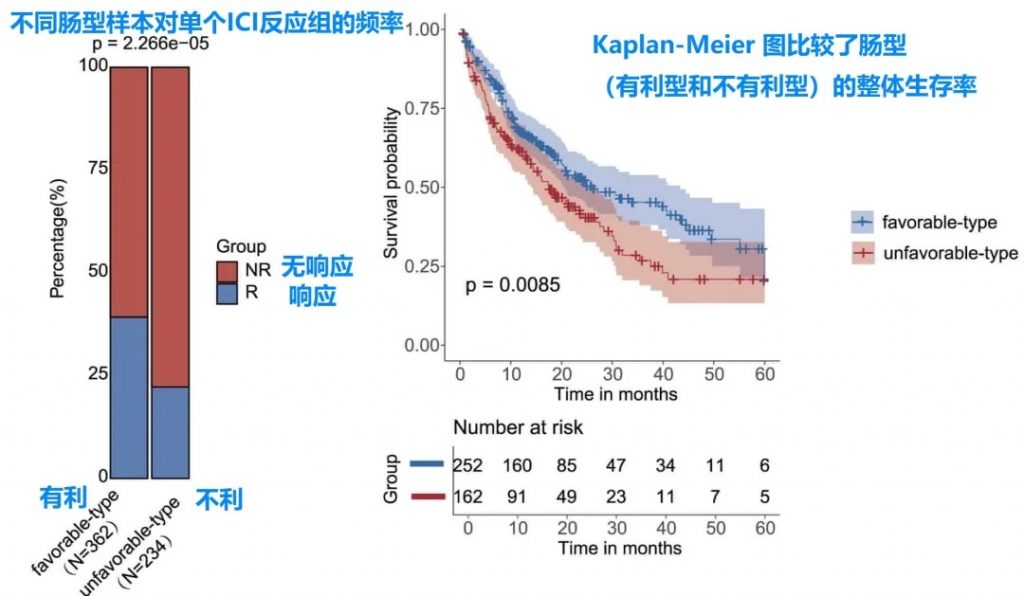
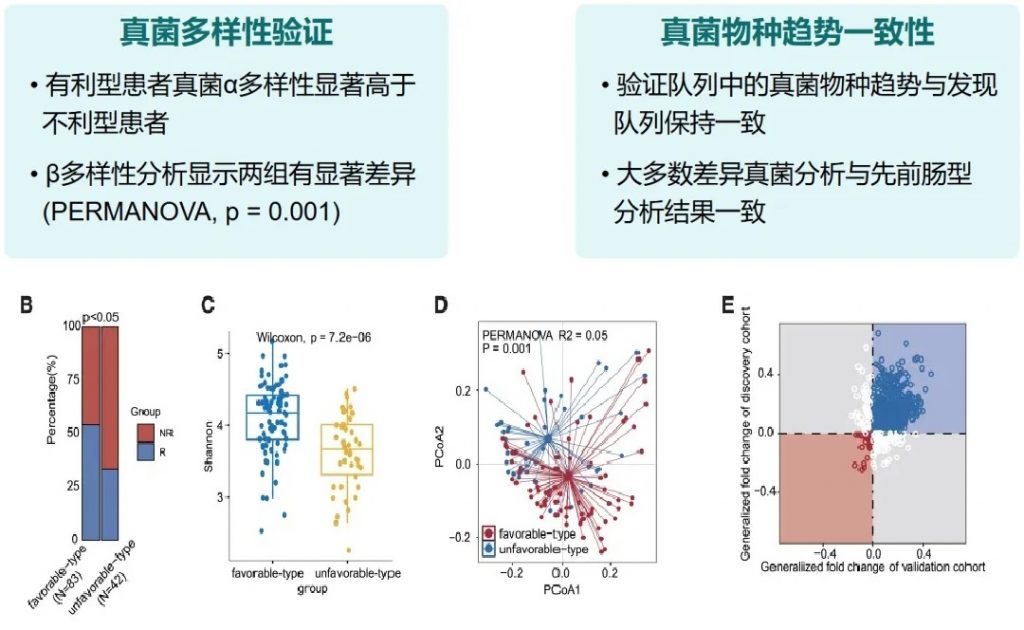
有利型患者对抗PD-1/PD-L1免疫治疗的响应率显著高于不利型 (p=2.266e−5),总生存期也更长 (p=0.0085)。
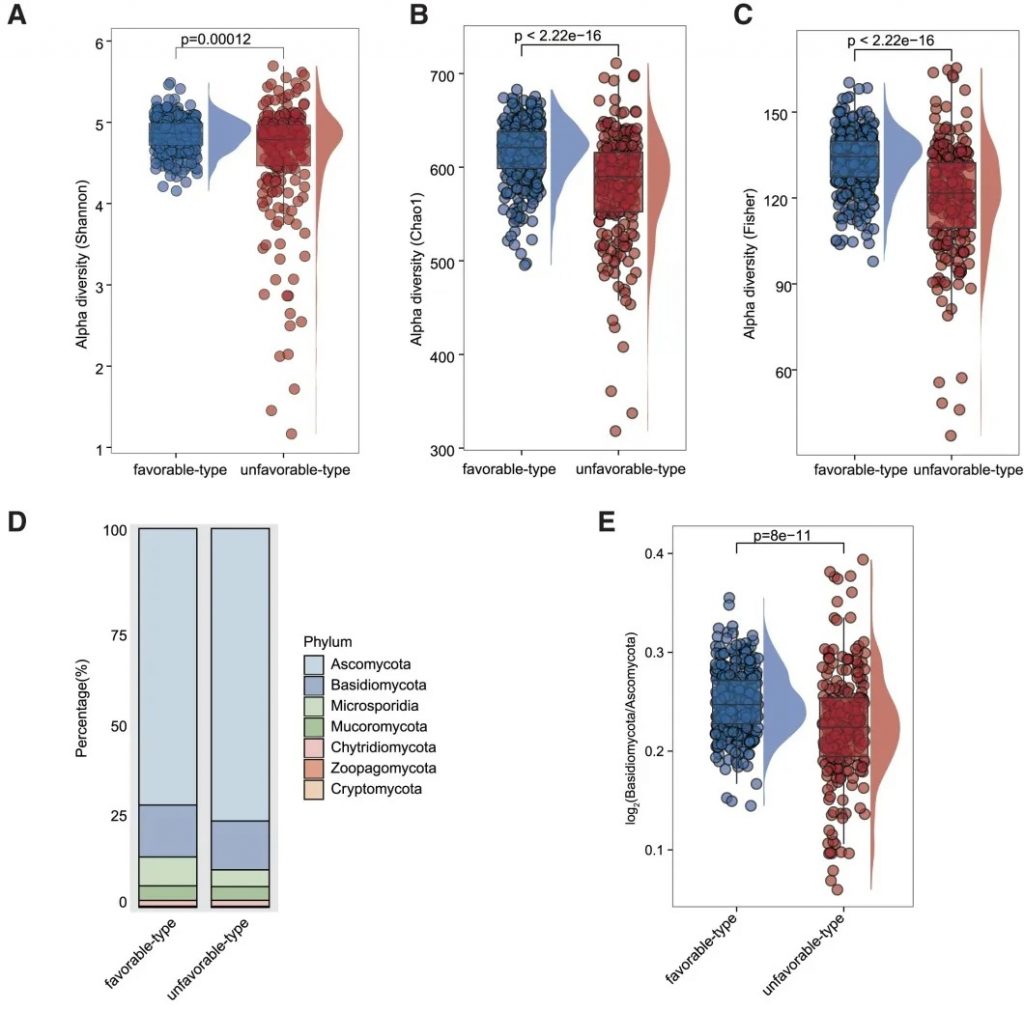
“有利型”和“不利型”肠道菌群差异 细菌、真菌
细菌 多样性
——α多样性差异
有利型患者表现出显著更高的细菌α多样性指数(Shannon、Chao1、Simpson),这与既往研究发现的免疫治疗响应者多样性更高的结论一致。
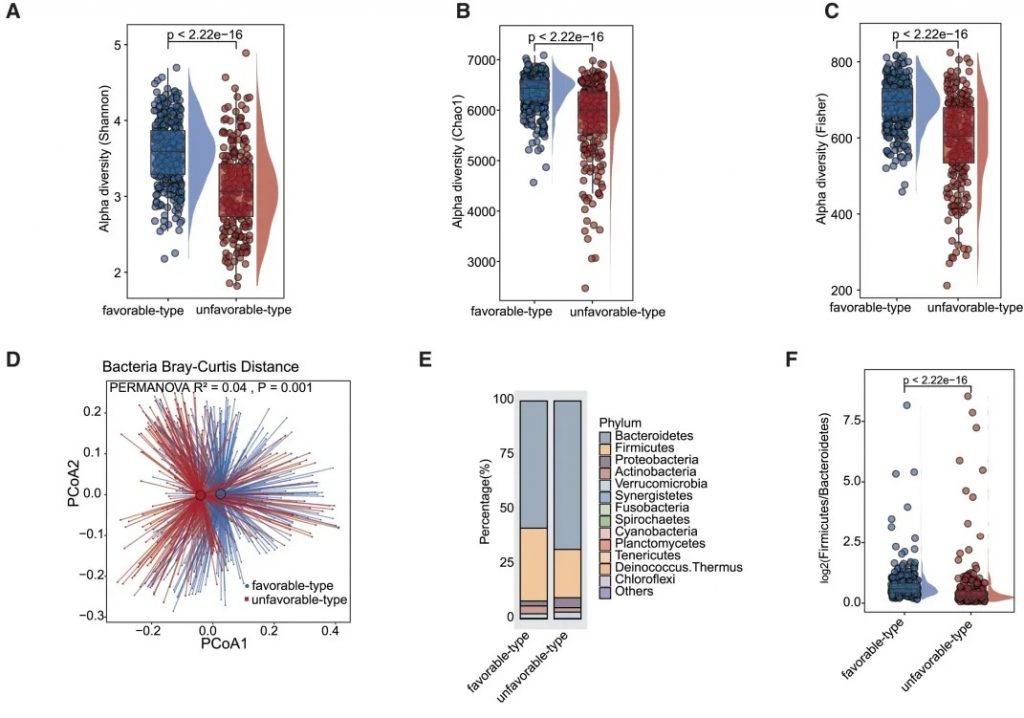
——β多样性差异
细菌 主要构成
有利型患者表现出更高的Firmicutes/Bacteroides比值,这被认为是肠道菌群平衡的重要指标,与免疫治疗的良好反应相关。
有利型的患者肠道中富含产丁酸盐的细菌,特别是Faecalibacterium prausnitzii、Lachnospira eligens,这些细菌通过产生丁酸盐调节肠道微环境,增强抗肿瘤免疫反应。
真菌 多样性
有利型表现出更高的Shannon指数和OTU丰富度,真菌物种多样性更丰富,这与对免疫治疗的更好反应相关联。
真菌 主要构成
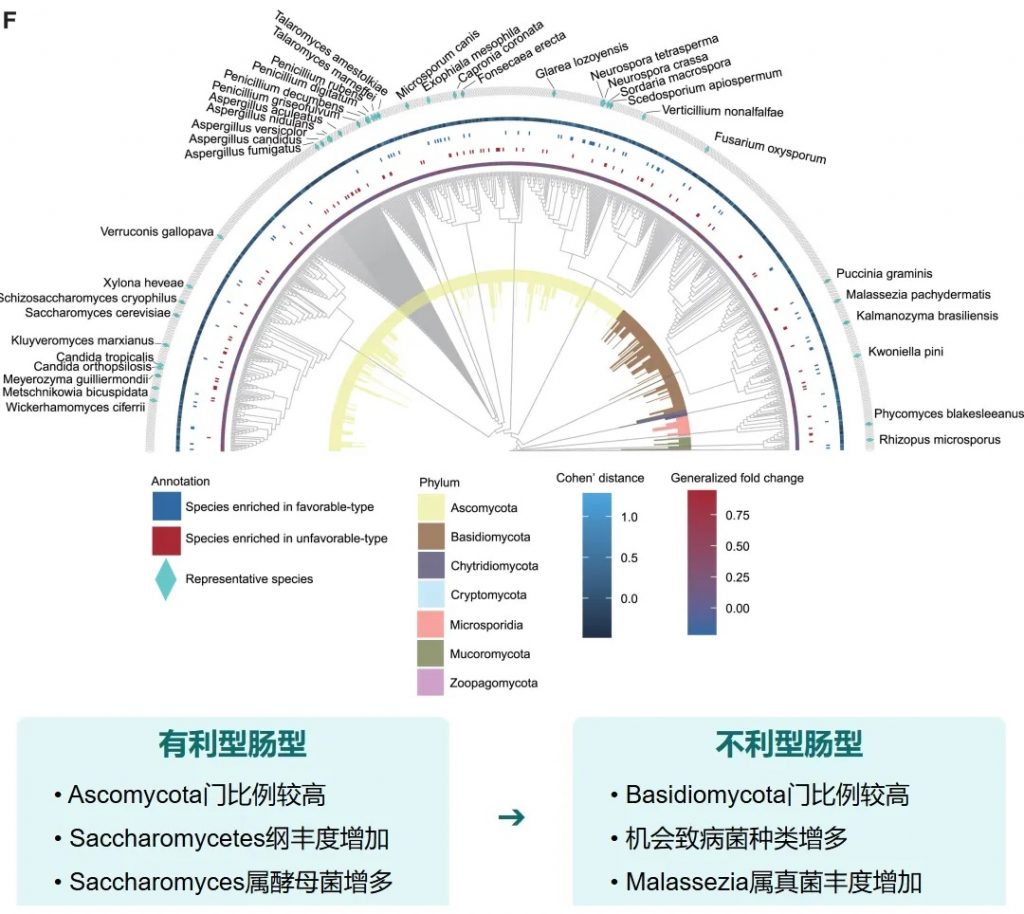
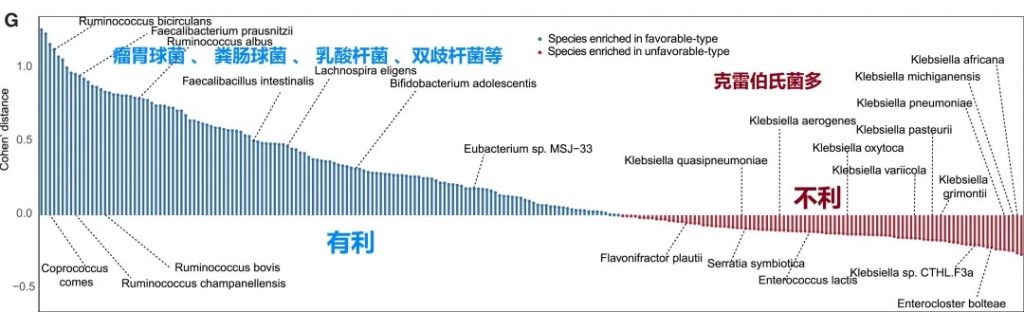
关键真菌与细菌的共现分析
有利型样本在真菌-细菌界中表现出增加的正相互作用,而不利型样本在菌-细菌界表现出更多的负相互作用。
关键真菌-细菌互作对有利型中的关键互作真菌-产丁酸盐细菌正向互作:
有利型的真菌群落结构似乎与某些细菌表现出更积极的共生关系。真菌-细菌间相互作用可能在抗PD-1/PD-L1 ICI的反应中发挥关键作用。
基于特定菌群类型的微生物功能分析
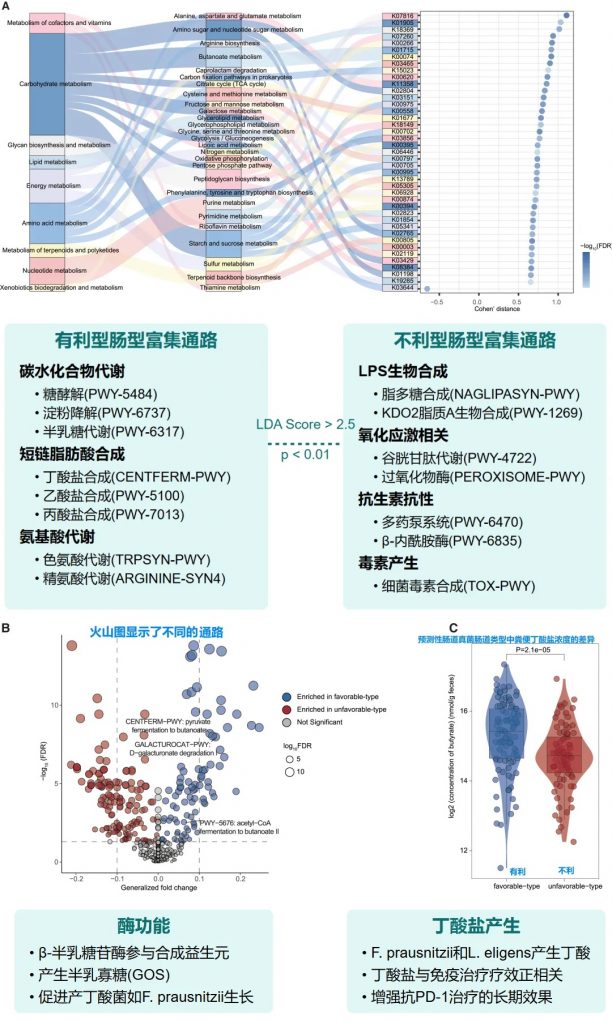
肠道菌群类型的外部验证与多组学分析
验证队列特征
• 125名接受PD-1抗体治疗的泛癌症患者
• 基于随机森林分类器(RandomForestClassifier)进行肠道菌群类型的预测
• 预测结果: 83名有利型患者,42名不利型患者
验证分析方法
• 基于机器学习的肠道菌群类型预测模型
• 随访评估对免疫治疗的临床反应
丁酸可促进CD8+ T细胞浸润肿瘤微环境,直接增强抗肿瘤免疫。
FMT供体选择:优化免疫治疗疗效
整合数据构成
• 3个近期发表的临床试验数据
• 12名FMT供体 (5名有利型, 7名不利型)
• 43名黑色素瘤患者接受者
• 包含宏基因组数据和临床结果
FMT临床疗效分析
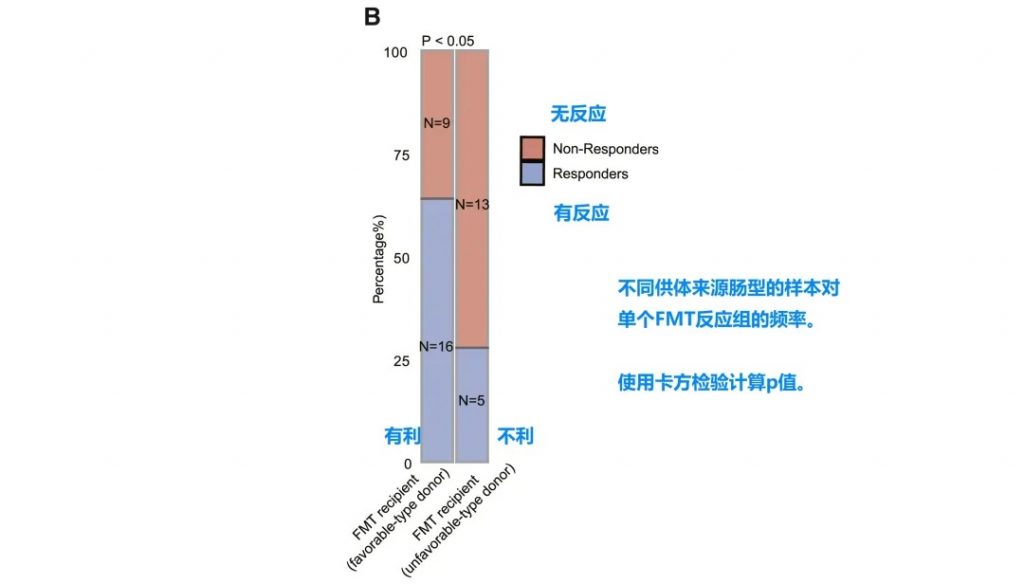
• 接受有利型供体FMT的患者对抗PD-1/PD-L1抗体免疫治疗的反应明显改善
• 卡方检验证实统计学显著差异 (p < 0.05)
接受“有利型”供体FMT的患者显示出对抗PD-1/PD-L1抗体免疫治疗的显著改善反应(p < 0.05)。这一发现具有临床指导意义,为FMT供体筛选提供了新的标准。
FMT前后微生物多样性变化
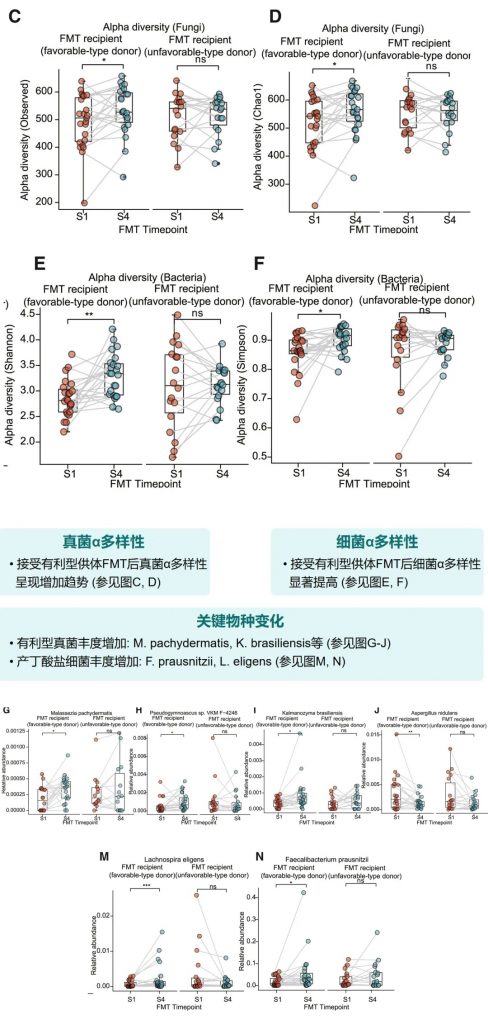
接受有利型供体FMT后,受者的真菌和细菌α多样性均有明显提高。特别是,有利型特征真菌和产丁酸菌的丰度显著增加,这与增强的免疫治疗效果相关。
体内验证实验结果
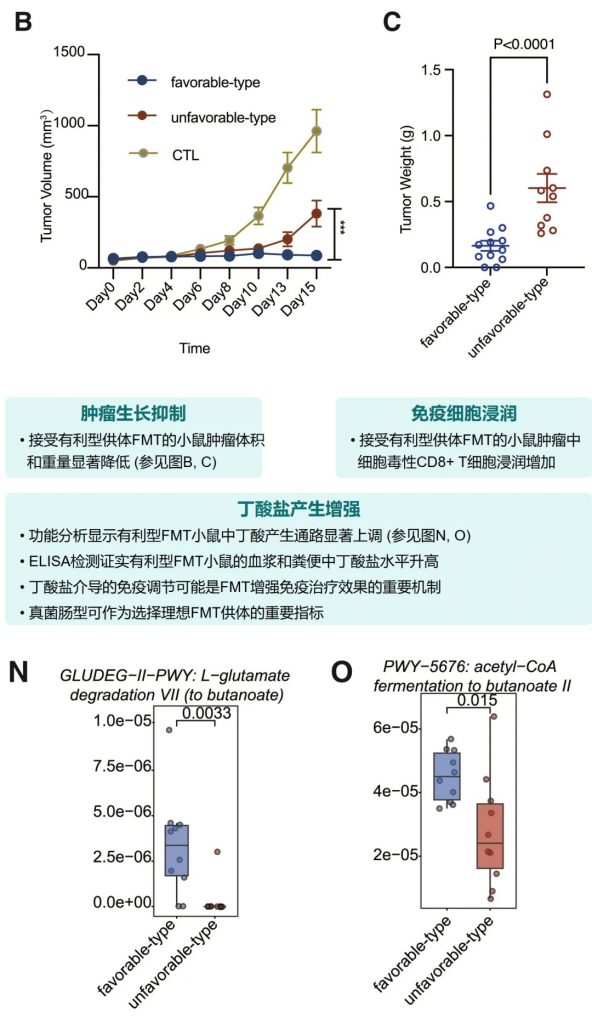
小鼠实验进一步确认了有利型供体FMT可以增强抗PD-1治疗效果,表现为肿瘤生长抑制、CD8+ T细胞浸润增加,以及丁酸盐产生增强。这为FMT增强免疫治疗提供了机制解释。
该研究揭示了肠道微生物群类型与免疫检查点抑制剂(ICI)疗效之间的密切关系,为优化癌症免疫治疗策略提供了新的视角。
肠道菌群与免疫治疗响应之间存在显著关联,且这种关联可通过粪菌移植等干预手段加以调控,为克服免疫治疗耐药提供新策略。
以下是几个具有前景的研究和临床应用方向:
辅助诊断的开发与实施
随着肠道菌群类型与免疫治疗疗效之间关联的确立,开发肠道微生物群分析作为免疫治疗的伴随诊断工具已成为一个明确的临床需求。
粪菌移植(FMT)作为辅助治疗手段
对于被确定为不利型的患者,粪菌移植提供了一种有前景的干预手段,可能显著提高免疫治疗的响应率。
更大规模的临床验证研究
为了将这些发现转化为临床实践,需要进行更大规模的验证研究:
未来,肠道微生物群检测有望成为免疫治疗患者的常规伴随诊断工具,帮助临床医生识别潜在的治疗无应答人群。
也就是说做免疫治疗的人群可以做肠道菌群评估作为辅助诊断,如果免疫治疗没有效果的病人,考虑通过FMT调整菌群后,提高治疗效果。来自有利型供体的粪菌移植可能成为提高治疗效果的有效辅助手段。
这一整合策略不仅能够优化现有免疫治疗的应用,也为开发新型微生物组靶向干预手段提供了方向,最终实现免疫治疗疗效的最大化和个体化。
在癌症精准医疗的框架下,肠道微生物组评估和干预将与基因组学、免疫学等多组学手段一起,共同构成未来肿瘤免疫治疗的个体化决策基础,为患者带来更好的临床获益。
主要参考文献
Hu M, Zhu X, Huang X, Hua L, Lin X, Zhang H, Hu Y, Tong T, Li L, Xuan B, Zhao Y, Zhou Y, Ding J, Ma Y, Jiang Y, Ning L, Zhang Y, Wang Z, Fang JY, Zhang Y, Xiao X, Hong J, Chen H, Li J, Chen H. Optimizing anti-PD-1/PD-L1 therapy efficacy and fecal microbiota transplantation donor selection through gut mycobiome-based enterotype. Cell Rep. 2025 Apr 20;44(5):115589.

谷禾健康

自古以来,中医就有“药食同源”的理念,强调食物与药物之间的内在联系,认为许多食物既可作为日常饮食,又具有调治疾病的作用。随着现代科学的发展,这一古老的智慧得到了新的诠释和应用,逐渐发展成为今天的营养大健康行业的重要组成部分。
药食同源的理念,源于对自然界植物、动物等观察和长期实践。在古代,人们发现某些食物在特定情况下能够缓解或治疗某些症状,这些食物因而被赋予了药用价值。例如,生姜被用来缓解感冒症状,大枣用来滋补身体,这些实践在现代医学中也得到了相应的科学解释和验证。
然而,药食同源的意义远不止于此,它还涉及到食物与人体之间复杂的相互作用,以及食物如何通过影响人体内部环境,特别是肠道菌群,来发挥其健康效益。
近年来,随着研究的深入,人们逐渐认识到肠道菌群对人体健康的重要性。它不仅参与食物的消化和营养吸收,还与免疫系统、神经系统以及许多代谢过程密切相关。因此,肠道菌群的健康状态直接影响到个体的健康状况,而饮食则是影响肠道菌群组成的主要因素之一,这一发现为个性化营养提供了新的视角。
个性化营养的兴起,标志着营养学从“一刀切”的群体指导向“量身定制”的个体化服务转变。在这一过程中,药食同源的概念被赋予了新的生命。通过结合现代营养学、微生物学、分子生物学、数据科学等多学科知识,个性化营养能够更精准地满足个体在不同生理阶段、不同健康状况下的营养需求。这种基于个体差异的精准营养干预,有望提高营养建议的依从性和有效性,从而更好地促进健康和预防疾病。
本文从药食同源的基本概念出发,探讨其在现代营养学中的应用,分析肠道菌群与健康的关系,特别是在肠道菌群检测技术日益成熟的背景下,如何将传统理念与现代科技相结合,为人们提供更科学、更有效的健康解决方案。这里展示药食同源在大健康行业中的潜力和前景,同时指出面临的挑战和未来的发展方向。
在这个营养大健康行业的新时代,我们可以更深入挖掘药食同源的科学意义,利用现代科技手段,为人们提供更加精准、个性化的营养服务。这不仅是对传统智慧的传承和创新,更是对未来健康生活方式的探索和引领。期待能够为营养学界、健康产业等带来相关启示。
本文目录
01 药食同源
药食同源的定义、特点、物质
传统中医中的药食同源理论
现代科学中的药食同源
02 营养基因组学与药食同源:个性化健康新纪元
03 什么原因导致不同个体之间的效果差异?
04 如何实现个性化干预?
肠道菌群参与活性成分的生物转化
药食同源成分调节肠道菌群(多糖、多酚等)
药食同源物质通过肠道菌群影响疾病
消化系统疾病、肝病、内分泌疾病、
神经精神疾病、心血管疾病、其他
05 药食同源干预和开发的注意事项
药食同源-循证依据
如何监测其效用?
药食同源技术创新
展望
药食同源,这一概念最早源于中国古代的医学典籍,其核心思想是某些物质既可以作为日常食物,也可以作为药材使用,即“药食同源”。
在中医理论中,这一理念体现了食物与药物之间的内在联系和相互转化的可能性。
在现代,药食同源的定义得到了进一步的扩展和科学化。它不仅包括了传统意义上的食疗,还涵盖了食物中活性成分的研究、功能性食品的开发以及营养与健康的密切关系。药食同源的物质通常指的是那些在自然界中既可以食用,又具有一定的药理作用,能够在不造成副作用的前提下,通过日常饮食来维护或改善健康的植物、动物或矿物。
▼
药食同源物质的特点
安全性:这些物质应当是长期被人类食用,且对人体没有明显毒性或副作用的。
功能性:含有能够对人体健康产生积极影响的生物活性成分,如多糖、黄酮、皂苷、挥发油等。
营养性:提供必要的营养素,如维生素、矿物质、蛋白质、膳食纤维等。
可食性:这些物质应当能够以食物的形式被人体消化吸收,且在口感、风味上能够被接受。
药食同源的物质在中医中有着广泛的应用,如黄芪、当归、枸杞、山药等,它们既可作为食材加入日常饮食,也可作为药材用于疾病的治疗和调理。
在现代,随着科学研究的深入,越来越多的药食同源物质被发掘和验证,如大蒜、生姜、绿茶、蓝莓等,其健康益处得到了科学界广泛认可。
药食同源的物质 例如:
传统“药食同源”理念的智慧和现代营养学、食品技术的进步,使得药食同源材料,如益智仁、川贝母、苦茶、桂皮、灵芝、生姜、银杏、鱼腥草、甘草、百合、枸杞、黑胡椒、紫苏、茯苓、马齿苋、薏仁、枸杞、大枣等在医药、普通膳食、功能食品和保健品中的应用日益广泛,即从2002年的 86 种增加到2021年的 110 种。

doi: 10.1016/j.jare.2023.05.011
在传统中医中,药食同源理论是一种深植于中国几千年文化和医学实践中的核心理念。中医认为,人体是一个整体,与自然环境息息相关,疾病的产生与人体内外环境的失衡有关。因此,中医治疗的目标是恢复和维持身体的平衡状态,即“阴阳平衡”。在这一理论体系中,食物不仅仅是满足基本营养需求的物质,更是调节身体、预防和治疗疾病的天然药物。
食物的性味归经
中医认为食物具有“四气”(寒、凉、温、热)和“五味”(酸、苦、甘、辛、咸),这些特性决定了食物对人体的作用和影响。
– 寒凉性食物通常具有清热、泻火的作用,适合于热性体质或热性疾病的患者;
– 温热性食物则有温中、补阳的功效,适用于寒性体质或寒性疾病的患者。
此外,食物还根据其对人体脏腑经络的作用被归入不同的“经”,如心经、肝经、脾经等,通过食物的归经特性,可以针对性地调节相应脏腑的功能。
传统中医强调“辨证施食”
根据个体的体质、疾病状态和环境因素来定制饮食方案。这种方法体现了中医个体化治疗的特点,认为没有一种食物或药膳适合所有人。因此,中医会根据患者的具体情况,调整饮食结构,推荐适合的食物和药膳,以达到最佳的治疗效果。例如,某些温热性的食物可能对寒性体质的人有益,但对热性体质的人则可能造成不适。
传统“药食同源”思想的主要特征是:
药食相隐、辨证施食、阴阳平衡、五脏滋养调理、防治结合、顺应自然、适可而止、适度食用。
这些思想体现了一种深度融合医学理论、兼顾个体差异、强调食物与药物功能相辅相成、追求整体平衡与自然和谐的饮食观念。这一理念在当今社会仍然具有重要的指导意义,对提高民众健康意识、倡导健康生活方式有着积极的影响。
现代“药食同源”饮食理念,是在继承和发扬传统“药食同源”思想的基础上,逐步融入了现代遗传学、代谢和营养学理论、生物医学和临床研究成果、先进的工具和技术,以及实际的社会生活经验等。
在中药材理念的基础上,一条集食品、保健品和药品于一体的紧密产业链已经形成。这条产业链不仅包括中药的生产和分销,还包括药食同源食品和营养补充剂的发展。桑葚和枸杞的兴起表明传统和现代健康领域的成功融合。
2022 年,桑葚被创新地用于保健茶,成为药食同源市场中增长最快的成分,价值超过 300 亿元人民币。
枸杞已从中老年人的传统保健选择转变为年轻人中流行的保健元素,预计到 2023 年消费量将达到 138 万吨,市场规模将达到 653 亿元人民币。
由于基因、环境、生活方式等因素的差异,同一种药食同源物质在不同人群中可能产生不同的效果。
例如,大枣这种药食两用的果实,在传统医学中被用于补气养血、安神益智。然而,现代研究发现,大枣的这些效果可能因个体差异而有所不同:
随着人们对健康的追求日益个性化,“药食同源”理念也在不断演变。现代的药食同源不仅包括传统的食疗,还涵盖了功能性食品的开发以及营养与健康的密切关系研究。
然而,这种演变也带来了新的挑战:如何在保持药食同源物质整体效用的同时,实现真正的个性化应用?
融合东西方饮食文化、借助最新技术
融合东西方饮食文化和医疗实践甚至调和/推进两者的现代“药食同源”饮食在当今世界越来越受欢迎。借助人工智能、大数据等新兴技术,基于“药食同源”的个性化营养建议、健康饮食/保健建议、智能饮食生活计划、定制食谱提示、定制膳食建议等远程营养咨询服务应运而生,这将使“药食同源”的体验更具吸引力。

科技赋能传统智慧
全基因组关联研究的进展,包括表观基因组学、代谢组学、转录组学、蛋白质组学和微生物组分析技术(如 DNA 下一代测序)在内的先进“组学”技术等技术突破,使个体营养需求、遗传状况和肠道菌群分布得到全面分析,可以快速识别涉及基因-饮食相互作用的遗传变异及其与各种疾病的潜在联系,从而使“药食同源”饮食更加个性化和精准,从而提高饮食和生活方式建议的有效性。
总的来说,现代“药食同源”的主要特征包括:
科学性、循证性、精准性、标准化、功能性、多样性、生态化、可持续化、智能化、数字化、教育普及化、跨学科协作化、跨文化、国际化。
换言之,现代“药食同源”是一套将传统“药食同源”智慧与现代智能技术、先进的食物、人类和地球知识深度融合的饮食原则,旨在满足当代社会对健康、营养、精神健康、便利性和环境可持续性以及地球健康的多方面饮食需求。
正是在这样的背景下,营养基因组学应运而生。这一领域的出现,为我们理解和应对药食同源效果的个体差异提供了全新的视角和工具。下一章节,我们来探讨营养基因组学如何推动药食同源理论的发展,开启个性化健康的新纪元。
随着人们对饮食及其影响健康的潜力的认识和了解不断增长,导致了营养基因组学出现。
营养基因组学
营养基因组学作为一门新兴的学科,它研究了基因与饮食之间的相互作用,以及如何通过个性化的饮食干预来优化健康。
营养基因组学于 2001 年首次提出,旨在研究食物与遗传基因之间的关系,已应用于开发支持健康和降低饮食相关疾病风险的食品或饮食。
来自世界各地的 1,072 条与营养基因组学相关的研究文献显示,营养基因组学使个性化饮食方法能够恢复体内平衡并消除与饮食相关的疾病,提供了一系列工具来深入了解可能影响基因、蛋白质组或代谢组水平的主要途径以及肠道微生物群和相关代谢物对人类健康的潜在贡献,并提供了重要见解。
通过营养基因组学的研究,可以发现不同人群对药食同源物质的代谢和反应差异,从而为个性化营养提供依据。例如,某些人群可能对某种药食同源物质的代谢能力较弱,需要适当调整摄入量;而另一些人群可能对某种药食同源物质的反应较为敏感,需要谨慎使用。
个性化营养理念与现代人的健康需求不谋而合
现代人更加意识到通过健康的生活方式来预防疾病。人们需要安全、营养充足、身体、精神和情感健康、环境可持续、经济公平和负担得起、文化可接受的食品和饮食。
近年来出现的“个性化食品/饮食”、“个性化营养”、“主动健康”和“天然功能性食品”等术语都反映了人类对基于个人基因、微生物群和生活方式的特定饮食/食品的需求日益增加。
因此,具有特定健康促进特性和定制产品形式的超级成分和功能性食品产品的普及预计将在全球范围内进一步增加。
药食同源物质的机会
自 2023 年以来,人们寻求从方便食品/餐食中获得有意识的享受,这进一步模糊了正餐和零食之间的界限。这为药食两用材料/物质提供了巨大的机会,它们可以作为额外的“补充”加入/强化各种零食类消费食品/餐食,提供维持/促进健康、预防/治疗疾病和延长寿命的效果。
科学知识、现代加工技术和生物技术、组学技术和创新的生物信息学工具的进步,为实现”为人类生命和健康量身定制食品/饮食”提供了可能。这与中医”药食同源”理念的智慧不谋而合,后者强调了”预防”高于”治疗”的首要地位,强调了人与自然的相互影响,以及每个人健康的独特性。
这些理念的现代体现就是”主动护理“或”主动健康”的趋势。
“主动护理”的趋势,重塑可持续健康管理
主动护理(也称为主动健康)是 2019 年疫情以来的主导趋势,在 2020-2021 年成为更为重要的服务模式。在新冠疫情期间和之后,“食物作为药物提供天然的功能”再次得到强调,并得到了显著加强。
人们希望在早期阶段增强对生活压力和不可预测疾病和状况的抵抗力,改善全身健康状况,提高生活质量,延迟和减少进一步医疗和住院治疗的需求,并建立更可持续、更高效的卫生和社会护理体系,从而推动主动护理的转变。
“更安全、温和”的趋势
同时,越来越多的人开始追求更安全、更温和的治疗。预防保健应运而生,以促进生活质量和更健康的生活方式,提倡及早发现健康问题。
当然它也有缺陷,包括不同社会经济群体之间资源可及性的差异、个人的依从性以及不可预见的健康风险和疾病的风险。
根据世界卫生组织(WHO)的数据,草药为全球一半以上的人口提供了初级卫生保健的基础。食物和传统药物的综合使用(例如“人体阴阳平衡和气调”的思想),而不是单独使用药物/药物,已成为古代和现代正常人和亚健康人群预防保健的趋势。

药食两用的水果举例:龙眼
龙眼就是一个很好的例子,它是一种药食两用的水果。龙眼在中国和日本传统医学中用于补心健脾、养血安神,以及治疗健忘、失眠、气血不足等症状。
现代研究表明,龙眼通过抑制NF-κB 和 AP-1 信号通路来抑制脂多糖刺激的巨噬细胞一氧化氮的产生,其多糖能提供免疫调节作用。
在小鼠实验中龙眼能够增强学习和记忆能力(部分由脑源性神经营养因子表达和未成熟神经元存活介导),并改善认知能力、减轻病理损伤 [部分通过抑制RAS/丝裂原活化蛋白激酶(MEK)/细胞外信号调节激酶(ERK)信号通路介导]。
药食同源理论和个性化营养的发展为我们提供了全新的健康管理视角。然而,在实际应用中,有些人可能对龙眼的免疫调节作用特别敏感,而另一些人则可能在认知功能改善方面受益更多。
这不仅关系到药食同源理论的实际应用效果,也是个性化营养和精准医疗领域的核心挑战。要回答这个问题,我们需要深入探讨导致个体差异的多种因素,包括遗传背景、生活环境、饮食习惯、肠道菌群等。
要回答这个问题之前,我们先了解一下有哪些食物或者说药材及其成分。
2020年,国家卫健委、国家市场监督管理总局联合发布的《关于开展党参等9种按传统既是食品又是中药材的物质管理试点工作的通知》指出:
众多药食同源的中药材中含有当归、八角、豆蔻、丁香、小茴香、白芷、地茯苓、火麻仁、人参、人参果、人参叶、山楂、马苋菜、刀豆、甘草、人参、木瓜、梅干、白果、山药、木薯等84种成分。
这些药食同源物只要使用得当,几乎没有任何危险的副作用,且含有生物碱、长链脂肪酸、多酚、多糖、蛋白质、三萜类化合物、有机酸和维生素等生物活性物质,具有抗氧化、抗炎、抗菌、抗癌、抗高血压、增强免疫力和保护肾脏等多种生理功能。
例如,枸杞含有丰富的维生素C、维生素E、矿物质和多糖,具有增强免疫力和抗氧化能力,促进患者康复的功效。
现代科学研究:药食同源物质中的活性成分
现代科学研究揭示了许多传统药食同源物质中的活性成分,如多酚类、黄酮类、皂苷类、生物碱等,这些成分具有抗氧化、抗炎、抗菌、抗病毒等多种生物活性。例如:
正如我们所了解的,药食同源物质含有多种生物活性成分,这些成分对健康有着广泛的影响。然而,这些成分在不同个体中的作用效果会有差异。这种差异性主要是由以下因素造成的:
遗传差异
药食同源物质含有多种成分,个体基因的不同会影响这些成分的代谢途径。例如,某些人可能有更高效的酶来代谢这些成分,从而影响其效果。有些人可能对某些药食同源物质更敏感,即使是很小的剂量也可能产生显著效果。例如,某些人对大蒜中的蒜素特别敏感,可能会有更强烈的生理反应。
健康状况
个体的健康状况,特别是肝脏和肾脏功能,对药食同源物质的代谢和排泄至关重要。例如,肝脏功能不佳可能会影响甘草中甘草酸的代谢,进而影响其抗炎效果。
年龄和性别
不同年龄和性别的人可能对药食同源物质有不同的反应。随着年龄的增长,新陈代谢减慢可能会影响某些成分的效果。
生活习惯
饮食、运动、睡眠和压力管理等生活习惯也会影响药食同源物质的效果。例如,高脂肪饮食可能会影响绿茶中儿茶素的吸收,而规律运动可能增强黄芪的抗疲劳效果。
剂量
药食同源物质的剂量也会影响其效果,不同个体可能需要不同的剂量以达到最佳效果。例如,不同个体对人参的敏感性不同,所需的最佳剂量也会有所不同。
药食同源成分——相互协同作用
中医认为这些同源植物具有“四气”(寒、热、温、凉)和“五味”(酸、苦、甘、辣、咸),有助于调节人体各方面的机能和阴阳平衡,相互促进和制约,促进脏腑功能和气血协调,从而预防或治疗疾病。例如:
张锡纯在《中西医结合》一书中指出:“食疗既治病又充饥,既充饥又可口,对症下药,病自然会慢慢好,即使不对症下药,也不会有其他问题。”
这些思想强调食疗与药疗的协同作用,在现代依然适用,比如说:
药食同源物质/成分中的营养成分和生物活性物质是可靠的,在人类治疗某些健康状况或疾病方面有着悠久的安全历史,副作用较小,尤其适用于慢性病的长期治疗。这些药食同源物质/成分的组合应经过量身定制,以最大限度地发挥其对人体阴阳平衡和免疫力的调节作用,以达到治疗某些疾病或在药物治疗过程中起到辅助作用。
基于这些药食同源物种的特点,对食物和药物进行个性化选择,可以使两者共同发挥作用,提高药物的疗效和生物利用度,同时抵消部分药物引起的不良反应。
肠道菌群不仅参与消化过程,帮助我们分解食物中的营养素,还参与了药物和食物成分的代谢,影响其生物利用度和效果。
因此,当我们考虑药食同源物质对每个人效果的差异时,必须将肠道菌群作为一个关键因素纳入考量。
个性化的营养和健康管理策略应当考虑到个体的肠道菌群组成,以及它们如何与药食同源物质相互作用,从而优化这些天然资源的健康效益。
不同的厨师用同样的食材,可以做出不同口味的菜肴,那么同样,不同人的身体体质,基础条件,代谢能力等都不一样,药食同源物质活性成分发挥的效果就不一样。这些差异不仅体现在生理和遗传层面,还涉及到很重要的——肠道微生物群。不同的肠道菌群可能通过不同的代谢途径处理相同的活性成分,产生不同的代谢产物。
肠道菌群会分泌出一些酶,这些酶能够改变这些天然材料的结构。一旦结构被改变了,这些天然材料的物理化学性质和生物活性也会跟着变化,这样的变化能让它们在身体里发挥更好的治疗效果。
肠道微生物群转化,比化学合成有什么优势?
与化学合成对这些天然产物的修饰相比,肠道菌群的生物转化具有独特的优势:
1) 能干
肠道菌群分泌的催化酶驱动的大多数反应,难以通过化学合成完成;
2) 精准
酶促反应选择性强,也就是特别挑,只对特定部位下手,不用担心误伤到不用改变的部分;
3) 温和
生物转化反应条件相对温和,不会像高温、高压那样粗暴,这样能更好地保护那些珍贵的活性成分不受损。
下面具体来看,一些药食同源的成分经过肠道菌群代谢后,如何产生更容易吸收的或者更有益于人体的物质。
呋喃香豆素—经黏液真杆菌属—更容易吸收代谢
白芷里就含有多种呋喃香豆素,比如花椒毒素、佛手柑内酯、欧前胡素和异欧前胡素等等。
呋喃香豆素是一种在食物和草药里常见的活性成分,抗炎、抗癌、保护神经。但是,如果我们吃了含有呋喃香豆素的东西再去晒太阳,可能会有光毒性,就是皮肤对光特别敏感,容易发炎。
呋喃香豆素还会影响肝脏里的一种叫做CYP3A的酶,这种酶负责代谢药物,一旦被抑制了,药物在身体里待的时间就会变长。
肠道菌群是怎么发挥作用的?
肠道菌群中的经黏液真杆菌属(Blautia),能产生一种叫做 Co O-甲基转移酶MRG-PMF1,裂解甲基芳基醚,这种酶能把花椒毒素和香柠檬烯这些成分变成花椒毒醇和香柠檬醇,重要的是,异戊二烯基芳醚基团以相同的方式裂解。
去甲基化和去异戊烯化
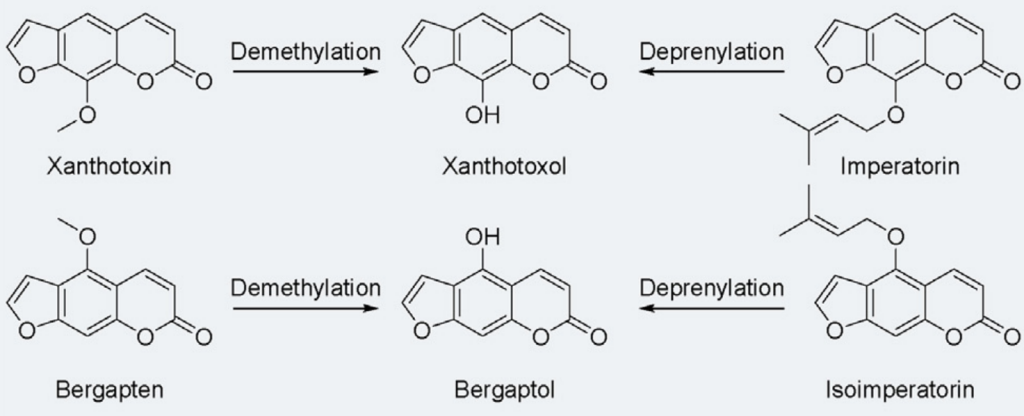
doi: 10.1016/j.jare.2023.05.011
欧前胡素和异欧前胡素经肠道微生物转化脱异戊烯基化也产生了上述产物。
这些溶解性更好的脱甲基和脱异戊烯基酚代谢物可以迅速随尿液排出。
也就是说,肠道内经黏液真杆菌高的人,可能更有效地代谢和排出体内的某些毒素,减少因体内毒素积累引起的炎症反应,从而降低慢性炎症性疾病的风险。
人参皂苷—双歧杆菌——抗癌、抗疲劳效果↑
人参皂苷的生物利用度通常约为给药量的 0.1% ~ 0.5%。某些肠道菌群如拟杆菌、双歧杆菌、真杆菌、普氏菌、梭杆菌,可以代谢人参皂苷并产生具有促进健康作用和显著提高生物利用度的新化合物。
比如,人参皂苷Rb1在双歧杆菌的作用下,先变成Rd,继续去糖基化,再变成抗癌和抗糖尿病效果更强的Compound K。而且,Rg1去糖基化变成的20(S)-protopanaxatriol(PPT),抗疲劳效果就更强。
那么,有些肠道内双歧杆菌含量较高的人,对他们来说,吃人参的效果可能更显著。
黄芪中毛蕊异黄酮—拟杆菌属13—抗病毒效果↑
黄芪中的主要异黄酮类化合物有毛蕊异黄酮-7-O-β-D-葡萄糖苷 (CG) 。
来自人类肠道细菌的拟杆菌属 13对 CG 进行去糖基化,获得了糖苷配基毛蕊异黄酮,它比相应的糖苷具有更强的抗病毒能力,可以保护 PC12 细胞免受损伤。
去糖基化
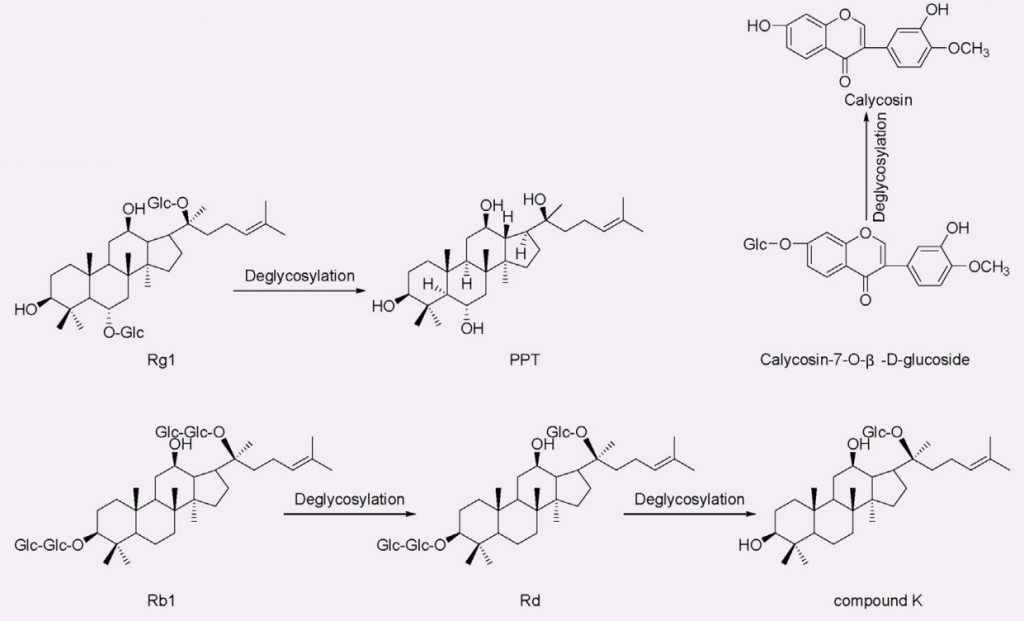
doi: 10.1016/j.jare.2023.05.011
毛蕊异黄酮—拟杆菌58—芒柄花素预防癌症
芒柄花素是一种植物雌激素,用于预防和治疗乳腺癌、前列腺癌、结肠癌和骨质疏松症。
来自人类肠道细菌的拟杆菌属58能够使毛蕊异黄酮脱水并将其转化为芒柄花素。
此外,毛蕊异黄酮的乙酰化是由菌株梭菌属21-2完成的。
脱水和乙酰化
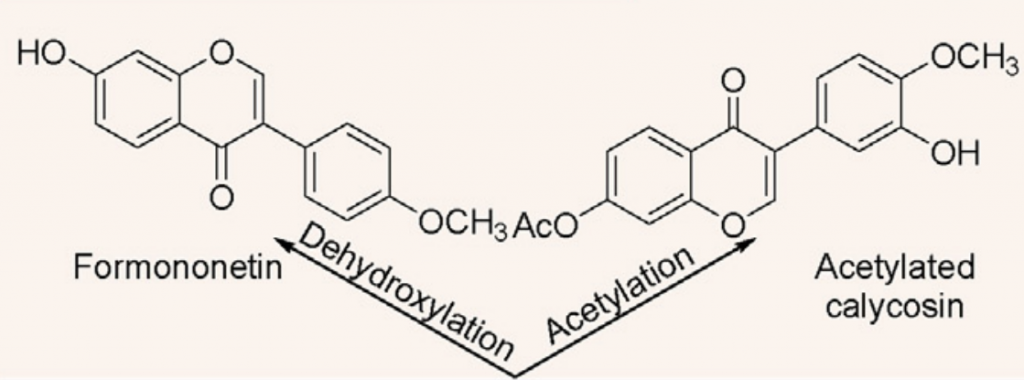
doi: 10.1016/j.jare.2023.05.011
药食同源物质对肠道菌群的调节作用是现代营养学和传统医学交汇点上的一个研究热点。
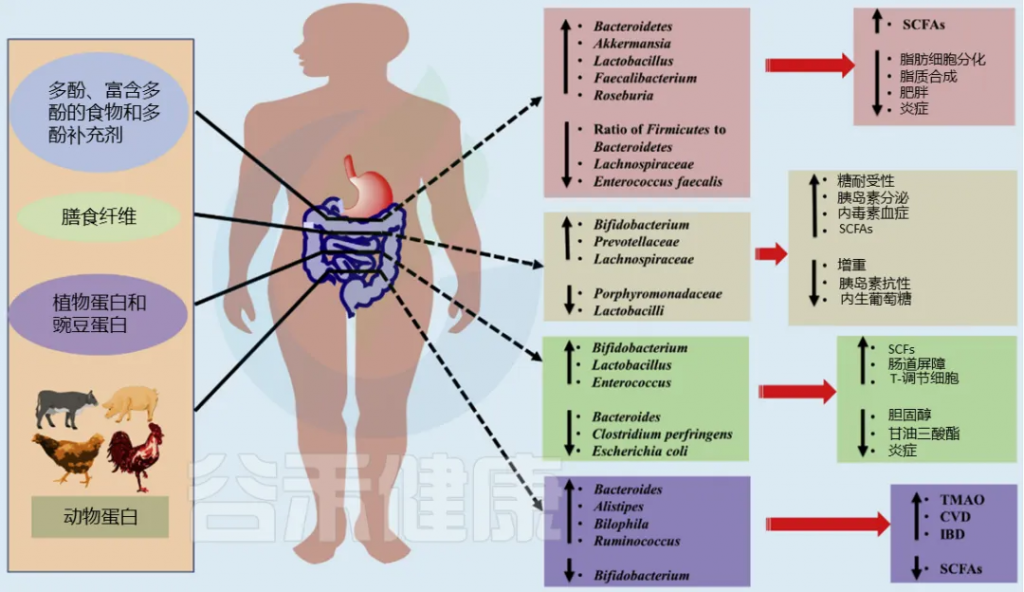
药食同源物质,如中药材、功能性食品和富含生物活性成分的天然食品,已被证明能够对肠道菌群产生积极影响。这些物质中含有的多糖、多酚、挥发油、皂苷等生物活性成分,都可以调节肠道菌群。
这些生物活性成分不仅为肠道菌群提供营养,还能选择性地促进某些有益菌的生长,同时抑制有害菌的繁殖,从而帮助维持肠道微生态的平衡。
多糖
促进有益菌,减少有害菌,比如说,麦冬多糖可以通过调节肠-肝轴显著保护非酒精性脂肪性肝病。
麦冬多糖显著降低了一些有害细菌的相对丰度,包括乳球菌、肠杆菌、Turicibacter、Tyzzerella、Oscillibacter等,并增加一些有益菌,如Alistipes、Ruminiclostridium、Rikenella。这种多糖还显著增加了两种产短链脂肪酸菌( Butyricimonas、Roseburia )丰度以及乙酸和戊酸的水平,从而改善了炎症反应和肝脏脂质代谢。
促进短链脂肪酸(SCFAs)的产生
枸杞多糖补充剂可降低厚壁菌门与拟杆菌门的比例,增加产短链脂肪酸菌,如Lacticigenium、Butyricicoccus、Lachnospiraceae等,从而改善肥胖小鼠的肠道菌群失调。
改善代谢,缓解疾病
银耳多糖通过多途径调节肠道菌群及其代谢物,改善了DSS诱导的溃疡性结肠炎。银耳多糖可以增加Lactobacillus的丰度,从而改善色氨酸的分解代谢。这导致黄嘌呤酸、KA和吲哚衍生物(如5-羟吲哚、5-羟吲哚-3-乙酸、5-羟吲哚乙酰酸)的增加。
天然多糖因其在缓解运动性疲劳中的潜在效果和较少的副作用而受到关注。研究表明,肌肉功能与肠道菌群的多样性和组成密切相关,而天然多糖如决明子、灵芝、枸杞、冬虫夏草等可通过不同机制发挥抗疲劳作用。
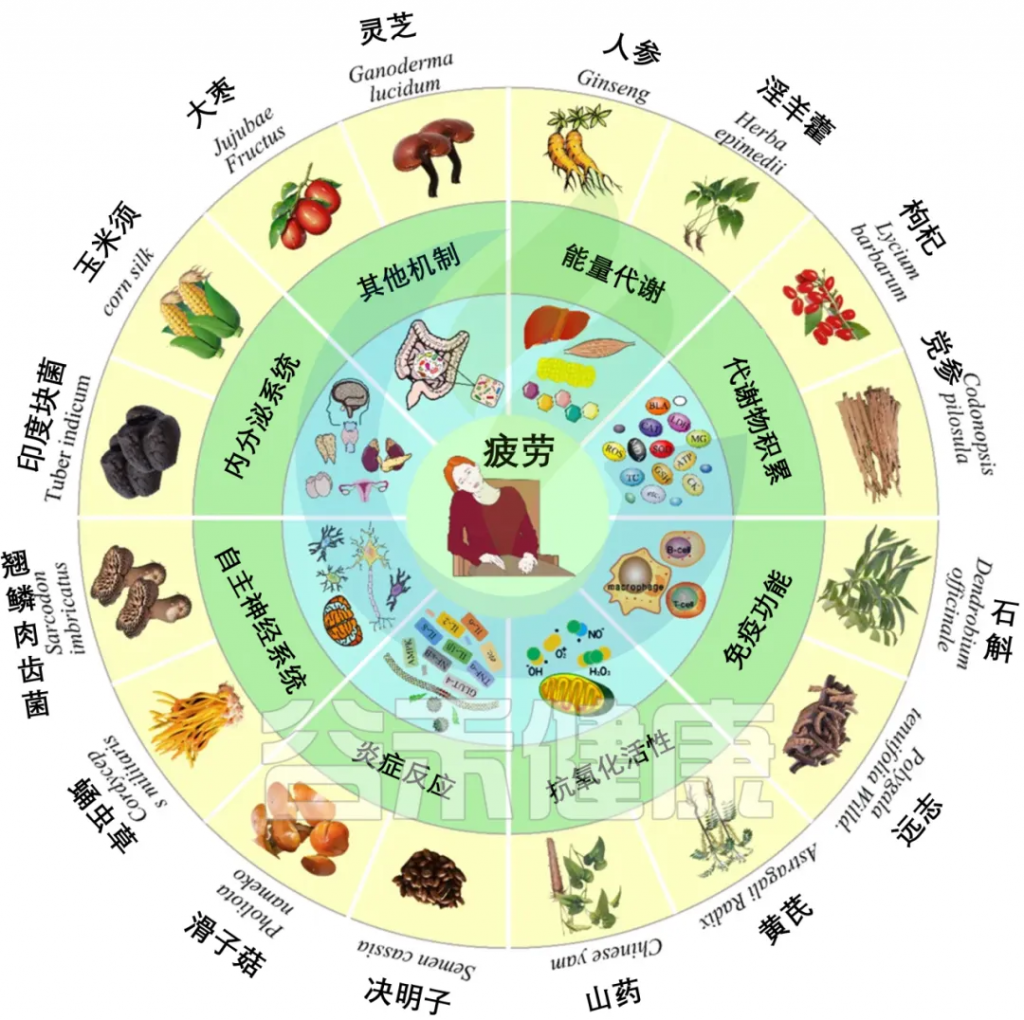
doi.org/10.3390/foods12163083
多酚
多酚是药食同源食物中含量丰富的活性化合物,膳食多酚类化合物具有预防慢性疾病的抗氧化剂,而临床试验中证据很弱,主要因为个体间的差异很大。多酚的生物利用度低(5%–10% 被小肠吸收)。这些化合物的生物利用度严格取决于肠道微生物的酶促机制。
肠道微生物群通过改变糖苷配基、糖苷和结合物的结构来影响多酚的生物利用度。
一般来说,口服多酚在上消化道的吸收相对较低;很大一部分多酚积聚在结肠中,影响和改变肠道微生物群的组成。摄入多酚可能通过促进有益细菌的生长,抑制有害细菌的生长,从而使宿主受益。
随机,双盲,安慰剂对照的人类试验表明,经口摄入没食子儿茶素-3-没食子酸酯和白藜芦醇,持续12周超重男性显著降低拟杆菌的粪便丰度和趋向于降低Faecalibacterium prausnitzii 的丰度。
在高脂饮食中补充膳食葡萄多酚导致肠道微生物群落结构发生显著变化,包括厚壁菌门与拟杆菌门的比例降低以及AKK菌的大量繁殖。膳食多酚促进肠道细菌Akkermansia muciniphila 的生长并减轻高脂饮食诱导的代谢综合征。
Vemana Gowd, et al.,Trends in Food Science & Technology, 2019
下面按照疾病类型分类分为消化系统疾病、肝病、内分泌疾病、神经精神疾病、心血管疾病等,逐一了解药食同源如何通过肠道菌群影响健康。
■ 消化系统疾病
炎症性肠病(IBD)
炎症性肠病(IBD)是一种肠道的慢性病,它会反复发生,造成肠道发炎和损伤。肠道屏障是保护肠道不受病菌侵害的关键。肠道菌群失衡可能会引起IBD。特别是,如果肠道里的革兰氏阴性细菌多了,它们细胞壁上的脂多糖(LPS)也会增加,这可能会引发过度的免疫反应,导致IBD。
人参→拟杆菌、疣微菌、变形菌↓→LPS↓→抗炎
枸杞→副拟杆菌属,Parasutterella,梭菌属↓ 螺杆菌科↑
玉竹提取物→脱硫弧菌科↓
除了上面提到的,还有很多药食同源的植物也能调节肠道菌群,帮助治疗IBD,比如山楂、蜂蜜、山药、菊花、酸枣仁、姜黄、黄芪和灵芝。这些植物可能成为治疗IBD的新药物。
多糖治疗IBD的机制
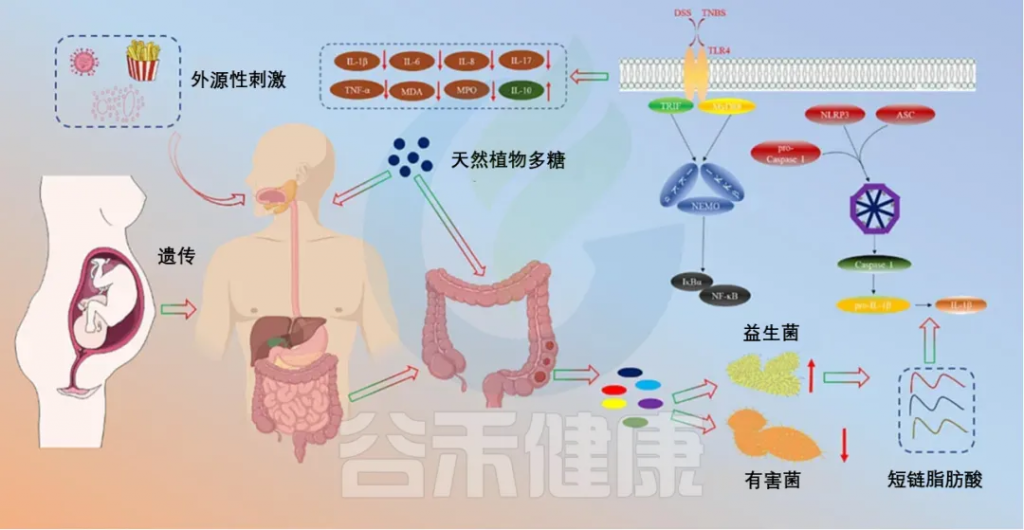
doi.org/10.1016/j.ijbiomac.2023.126799
结直肠癌(CRC)
结直肠癌(CRC)是全球常见的癌症之一,占全球每年所有癌症诊断和所有癌症相关死亡的 10%。长期患有 IBD 和不健康的生活方式导致肠道菌群紊乱的患者患结直肠癌的风险更高。因此,发基于药食同源物种的新型、有效但低毒的产品用于预防和治疗结直肠癌变得越来越重要。
灵芝→厚壁菌门、双歧杆菌、乳酸杆菌↑ 拟杆菌门、脱硫弧菌、颤杆菌↓ →预防结直肠癌
西洋参→疣微菌↓→抗炎、抗肿瘤
大枣→双歧杆菌、拟杆菌、乳酸杆菌↑→预防肿瘤
■ 肝 病
酒精性肝病(ALD)
酒精性肝病(ALD)与长期喝酒太多有关,它包括脂肪肝、酒精性肝炎和酒精性肝硬化等不同的肝脏问题。喝酒太多不仅会伤害肝脏,还会让肠道菌群不平衡,破坏肠道屏障,这会影响肠道和肝脏之间的正常联系,导致肝病。
枸杞→阿克曼氏菌、瘤胃球菌↑→保护肝脏
灵芝→瘤胃球菌、普雷沃氏菌、颤杆菌克属↑→改善脂肪代谢、抑制炎症
对沙棘、葛根、黄芪及其相关加工产品的药食同源物种的研究表明,它们通过肠道菌群在预防和减轻ALD方面发挥越来越多作用。
非酒精性脂肪性肝病
非酒精性脂肪性肝病(NAFLD)肝脏中脂肪积累过多,这与胰岛素抵抗和氧化应激有关,可能导致肝细胞损伤和炎症。虽然NAFLD的肝脏变化与酒精性肝病(ALD)相似,但NAFLD患者并不饮酒过量。目前,除了运动和饮食调整外,还没有特定的药物被批准用于治疗NAFLD。
黄芪多糖→厚壁菌门/拟杆菌门比例↓→抗炎
决明子中的蒽醌+益生菌 DM9054 和 86066→预防 NAFLD
其他药食同源的物质,如砂仁、茯苓、山楂、葛根,也显示出通过调节肠道菌群来预防和治疗NAFLD的潜力,可能通过激活相关的信号通路来发挥作用。
其他肝病
其他肝病还包括其他亚型,比如肝纤维化和肝硬化,它们可能由炎症或外界因素引起。
覆盆子→双歧杆菌、Turicibacter↑→肝功能改善
马齿苋→阿克曼氏菌和粪杆菌↑→减轻肝肾损伤
这些草药通过调节肠道菌群,显示出在肝病治疗中的潜力。
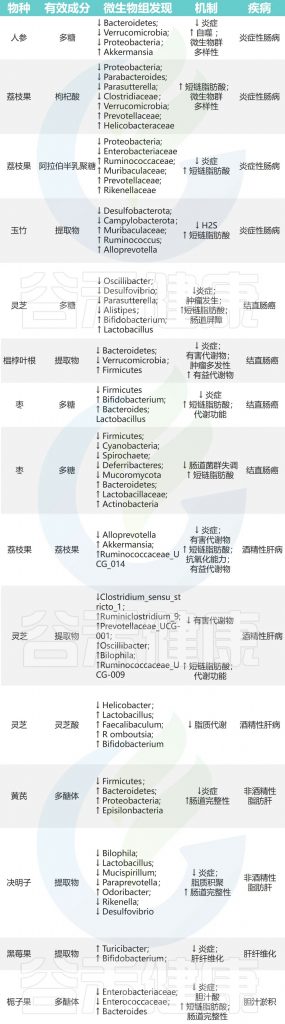
doi: 10.1016/j.jare.2023.05.011
■ 神经和精神疾病
阿尔茨海默病(AD)
阿尔茨海默病(AD)的大脑中淀粉样斑块和神经纤维缠结的形成,会导致大脑损伤和认知能力下降,目前还没有治愈的方法。近年来,研究表明肠道菌群,也就是我们体内的“第二大脑”,可能通过肠-脑轴影响阿尔茨海默病的发展。
茯苓→乳酸杆菌↑产内毒素菌↓→产GABA
黄芪→Roseburia、Lactobacillus↑→延缓淀粉样蛋白的沉积
天麻→约翰逊乳杆菌、鼠乳杆菌、罗伊氏乳杆菌↑→改善认知障碍
西洋参和黄芪→异杆菌属、嗜粘蛋白阿克曼氏菌、乳酸杆菌↑→有助于治疗AD
茯苓在阿尔茨海默治疗中作用
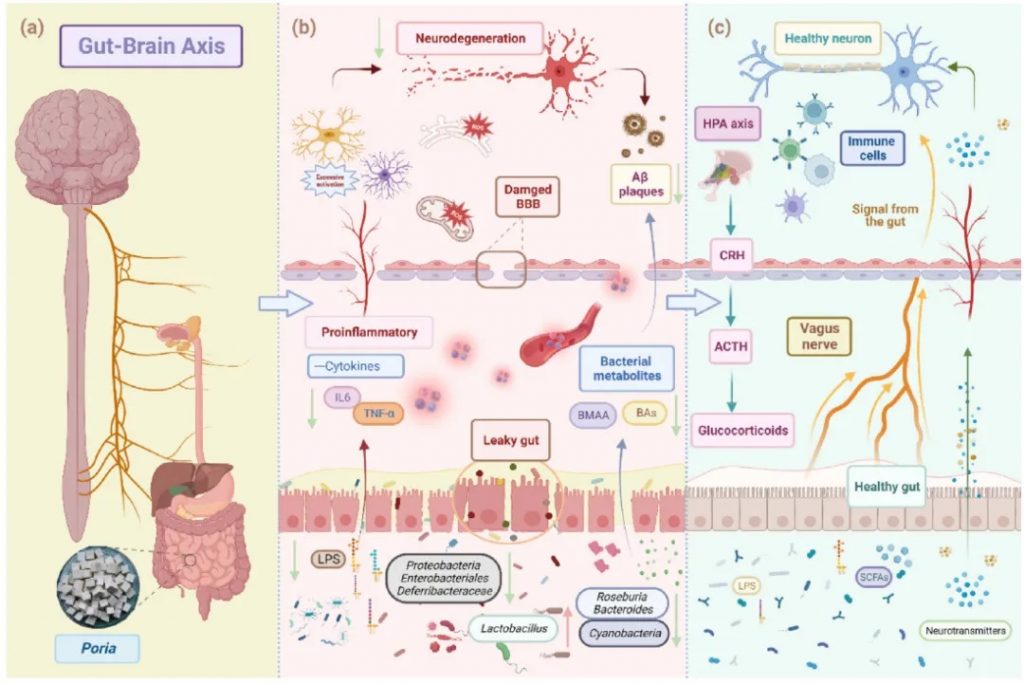
doi: 10.1016/j.jare.2023.05.011
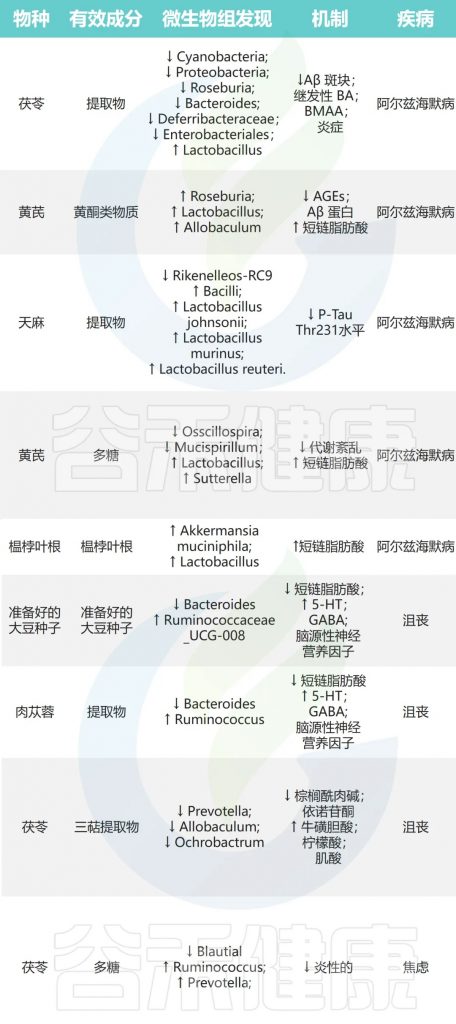
抑郁症
抑郁症是一种常见的精神障碍,影响很多人的生活。科学家认为,大脑中某些化学物质,比如GABA、去甲肾上腺素(NE)和血清素(5-HT)水平降低,可能会导致抑郁症。还有一种叫做脑源性神经营养因子(BDNF)的物质,它在调节抑郁症状中也很重要。
豆豉→瘤胃球菌↑ 拟杆菌↓→抗抑郁
肉苁蓉→瘤胃球菌↓ 拟杆菌↑→缓解抑郁症
茯苓→普雷沃氏菌、异杆菌、苍白杆菌↓→抗抑郁
焦虑症
焦虑症是一种常见的精神障碍,主要症状是过度紧张、担忧、失眠等,严重影响患者生活质量。
茯苓→Blautia↓瘤胃球菌、普雷沃氏菌↑→抗焦虑
doi: 10.1016/j.jare.2023.05.011
■ 心血管疾病
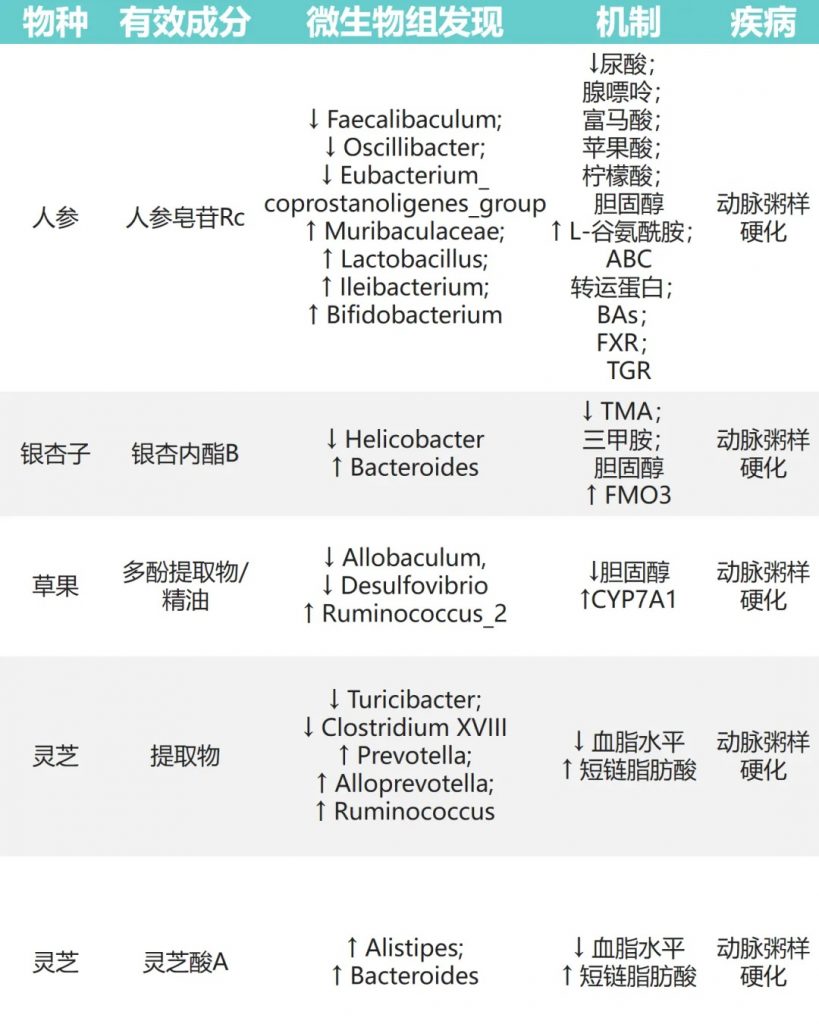
动脉粥样硬化(AS)
动脉粥样硬化(AS)是一种心血管疾病,可能因为血脂异常等问题引起,是全球主要的死亡原因之一。研究发现,一些天然物质可以通过调节肠道菌群来帮助缓解动脉粥样硬化。
人参皂苷→Muribaculaceae、乳杆菌、双歧杆菌↑
氧化三甲胺 (TMAO) 可以预测动脉粥样硬化早期阶段,它与胆固醇代谢和氧化应激有关。
银杏内酯B→拟杆菌↑、幽门螺杆菌↓→TMAO↓
草果→异杆菌、脱硫弧菌↓ Ruminococcus_2↑→降低胆固醇
灵芝→普氏菌、产短链脂肪酸菌↑→改善血脂异常
这些研究表明,通过调节肠道菌群,天然物质可能在预防和治疗动脉粥样硬化方面发挥作用。
doi: 10.1016/j.jare.2023.05.011
■ 内分泌疾病
糖 尿 病
糖尿病是一种代谢紊乱,高血糖是由于胰岛素分泌不足和/或作用不足造成的。根据国际糖尿病联合会2021年的数据,全球有 5.37 亿人患有糖尿病。研究表明,肠道菌群在控制宿主的生理和代谢方面至关重要,其失调与糖尿病的进展有关。
枸杞→瘤胃球菌科、Intestinimonas↑→GLP-1、PYY↑→促进胰岛素的分泌
罗汉果→Elasimicrobium↑、脱硫弧菌、大肠埃氏菌属-志贺氏菌属↓
桑叶→ Romboutsia、Oscillatoriales cyanobacterium↓ Alloprevotella、副拟杆菌、Muribaculaceae↑→缓解糖尿病
马齿苋→ 支链氨基酸↓→ 减缓糖尿病
肥 胖
肥胖已成为全球严重的健康问题,药食同源物种通过调节肠道菌群和相关信号发挥其抗肥胖活性。
这些研究表明,通过调节肠道菌群,药食同源物种可能成为管理肥胖的有效策略。

doi: 10.1016/j.jare.2023.05.011
■ 其他疾病
慢性非细菌性前列腺炎 <茯苓多糖>
茯苓多糖(PPs)可以通过调节肠道菌群来缓解慢性非细菌性前列腺炎。茯苓多糖显著增加副拟杆菌属、梭杆菌属、Parasutterella等有益菌,同时改变结肠上皮中关键基因表达,调节激素水平,抑制前列腺炎症。
恶性黑色素瘤 <黄芪多糖>
黄芪多糖(AP)能够通过调节肠道菌群来降低髓系抑制细胞(MDSC)的数量和相关分子的表达,增加CD8 T细胞的数量,削弱MDSC抑制CD8 T细胞杀死肿瘤细胞的能力。黄芪多糖干预下,与假长双歧杆菌呈负相关、与约氏乳杆菌呈正相关的L-谷氨酸和肌酸显著上调,揭示了黄芪多糖抑制肿瘤生长的治疗能力。
乳腺癌 <茯苓>
茯苓能够通过增加有益细菌如乳酸杆菌和双歧杆菌,减少有害细菌如硫酸盐还原菌脱硫弧菌和Mucispirillum等,来改善乳腺癌小鼠的菌群失调。
骨质疏松症 <杜仲叶提取物>
杜仲叶提取物补充剂能够促进保加利亚乳杆菌的生长,增加粪便和血清中的短链脂肪酸浓度,表现出抗骨质疏松症活性。
慢性肾脏疾病 <茯苓补充剂>
茯苓补充剂可以通过调节肠道菌群及其代谢产物来延缓慢性肾脏疾病的发展。
药食同源领域的研究正处于快速发展阶段,它融合了传统智慧与现代科学,为我们提供了一个全面的健康新视角。随着研究的深入,我们对药食同源物质的安全性、独立性效益以及它们如何影响肠道菌群有了更深入的理解。
药食同源物质的安全性是其研究的首要关注点。历史上,许多药食同源物质已被人类长期安全食用。这块还可以探索这些物质在不同剂量、不同人群中的使用效果,以及可能的长期影响。
如何通过科学的方法来证明药食同源物质的疗效和安全性?
它涉及到的是研究设计、实验方法、数据分析等,目的是建立药食同源物质功效的科学证据基础。这通常包括以下几个方面:
文献回顾:查找和分析现有的研究,了解药食同源物质的历史和当前的知识状态。
实验室研究:在细胞和动物模型中测试药食同源物质的生物活性。
临床试验:通过随机对照试验等方法,在人体中测试药食同源物质的效果。
横断面研究:在某一时间点对大量人群进行调查,了解药食同源物质的使用情况及其与健康状况的关联。
系统评价和荟萃分析:综合多项研究的结果,以评估药食同源物质的整体效果。
长期跟踪和安全性评估:对药食同源物质进行长期跟踪,评估其长期使用的效果和潜在的副作用。通过毒理学研究和临床监测,确保药食同源物质的安全性。
法规和标准化:遵循相关法规和指南,同时应确保药食同源物质的质量,包括纯度、稳定性和成分标准化。
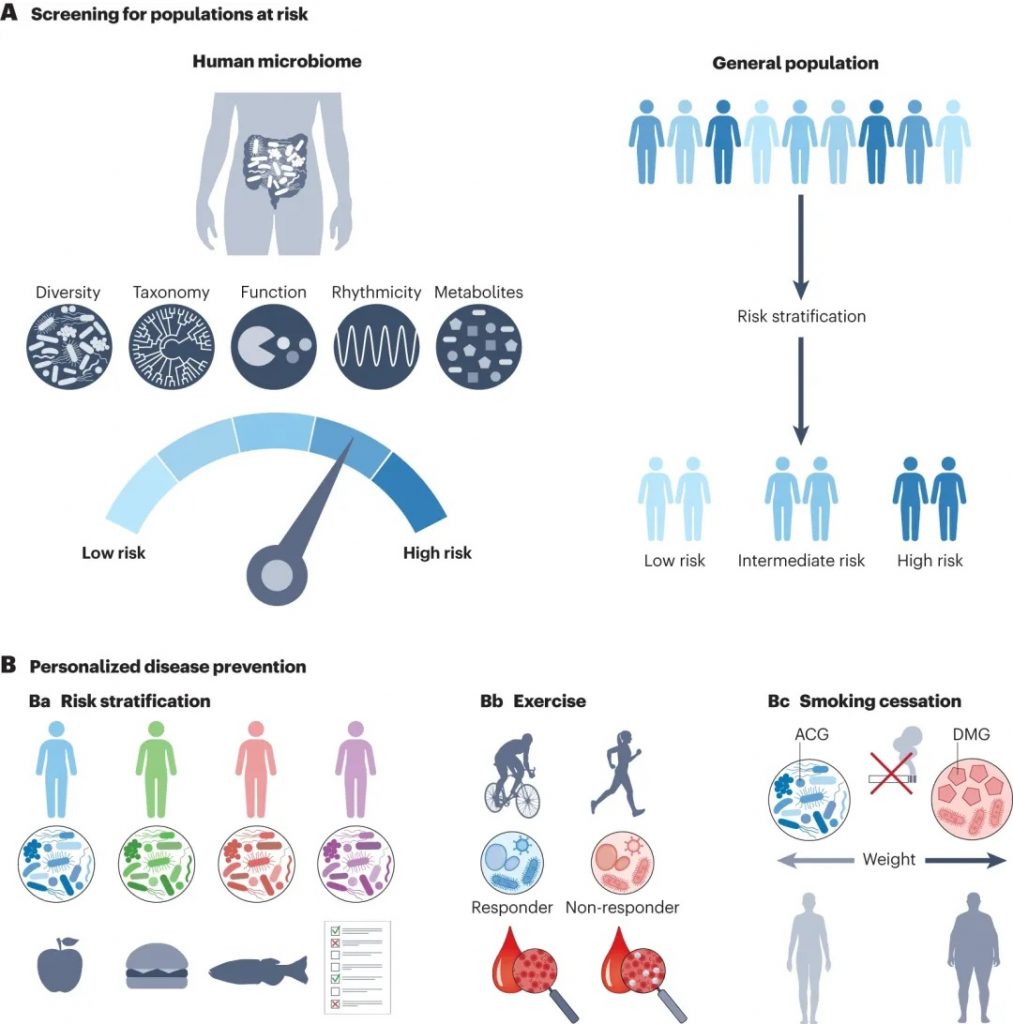
利用微生物组数据进行疾病筛查和个性化预防
doi.org/10.1038/s41579-023-00998-9
目前常用的监测手段包括生理指标监测(心率、血压、血糖等)、生物标志物检测(血液、尿液等)、肠道菌群检测、自我报告和问卷调查等。
症状记录:记录使用药食同源物质前后的症状变化,如疲劳感、消化问题、睡眠模式等。
免疫功能检测:例如,灵芝多糖可以通过增加NK细胞活性和T细胞增殖来增强宿主的抗肿瘤免疫反应。可以检测外周血中NK细胞活性和T细胞亚群比例来评估灵芝多糖的效用。
肠道通透性检测:例如,枸杞多糖可以通过上调紧密连接蛋白(如Occludin和ZO-1)的表达来增强肠道屏障功能。可以通过检测血清中细菌内毒素(LPS)的水平来间接评估肠道通透性,从而评估枸杞多糖的效用。谷禾肠道菌群检测报告中也有相关详细的菌群代谢产物水平提示。
肠道菌群检测:可以用来评估特定药食同源物质(如中药材、功能性食品)对肠道菌群组成的影响。通过比较干预前后的菌群变化,可以了解这些物质如何改变肠道微生物的多样性、丰度等,这有助于监测治疗效果和疾病进展。同时,结合大数据分析与生物信息学技术,能够识别出一些特定菌群的变化,疾病风险评估,根据肠道菌群评估营养状况,找到一些个性化的健康管理方案。
doi.org/10.1002/imt2.230
提高生物利用度
例如,基于纳米技术的人参皂苷Rb1递送系统,可以显著提高其在肠道中的稳定性和吸收率。这种新型递送系统不仅可以提高药食同源物质的效果,还可能降低所需剂量,减少潜在的副作用。
新的药食同源物开发
利用大数据分析技术,可以从大量的药食同源物质中筛选出具有潜在药用价值的候选物质。通过分析这些物质的化学成分、生物活性和用途,结合肠道菌群检测技术和药食同源的研究,为药食同源物质的深入研究提供科学依据,推动理论创新和技术进步,发现新的生物活性物质和药物靶点,同时可以推动新药食同源物质和/或功能性食品的开发。
药食同源物质与现代药物的协作
药食同源物质与现代药物协同作用的研究,例如,灵芝多糖可以增强某些化疗药物的抗肿瘤效果,同时减轻其副作用。这种协同作用的研究不仅可以提高现有治疗方案的效果,还可能为开发新的联合治疗策略提供依据。
污染问题监测的新型解决方案
药食同源产品的生产和加工过程中可能会遇到食品安全和污染的问题,如农药残留、重金属污染等。确保产品的安全性是药食同源产品开发的重要挑战。
肠道菌群检测可以作为评估和解决这一问题的有效工具。
每个人的基因、代谢类型、生活方式、健康状况都不同,对营养的需求也各异。随着科技的发展,药食同源可以与肠道菌群检测、人工智能、大数据分析、基因编辑等前沿科技相结合,为实现精准、高效、可持续的个性化营养提供更多创新的可能。比如,根据个人的肠道菌群特点选择合适的药食同源物质,满足个性化的营养需求,从而提高营养干预的有效性和依从性。
药食同源的研究涉及营养学、医学、微生物学、遗传学等多个学科,推动了跨学科研究的发展。这种跨学科合作有助于深入理解食物、营养和健康之间的复杂关系,有助于制定促进全民健康的膳食指南。
主要参考文献
Xia X, Xiao J. Natural Ingredients from Medicine Food Homology as Chemopreventive Reagents against Type 2 Diabetes Mellitus by Modulating Gut Microbiota Homoeostasis. Molecules. 2021 Nov 17;26(22):6934.
Yang, Minmin, et al. “Advances in understanding of health‐promoting benefits of medicine and food homology using analysis of gut microbiota and metabolomics.” Food frontiers 1.4 (2020): 398-419.
Zhong, Huan, et al. “The evolution and significance of medicine and food homology.” Acupuncture and Herbal Medicine 4.1 (2024): 19-35.
Zuo WF, Pang Q, Yao LP, Zhang Y, Peng C, Huang W, Han B. Gut microbiota: A magical multifunctional target regulated by medicine food homology species. J Adv Res. 2023 Oct;52:151-170.
Neu, J. The evolution of personalized nutrition. Nat Med 30, 1826–1827 (2024).
Sun-Waterhouse, et al. “Transformation from traditional medicine-food homology to modern food-medicine homology.” Food & Medicine Homology 1.1 (2024).
Doris Vandeputte, Personalized Nutrition Through The Gut Microbiota: Current Insights And Future Perspectives, Nutrition Reviews, Volume 78, Issue Supplement_3, December 2020, Pages 66–74
Chen, Jiali, et al. “Dietary therapies interlinking with gut microbes toward human health: Past, present, and future.” iMeta: e230.
Singar S, Nagpal R, Arjmandi BH, Akhavan NS. Personalized Nutrition: Tailoring Dietary Recommendations through Genetic Insights. Nutrients. 2024 Aug 13;16(16):2673.
Guess, N. Big data and personalized nutrition: the key evidence gaps. Nat Metab 6, 1420–1422 (2024).
Simon, Marie‐Christine, et al. “Gut microbiome analysis for personalized nutrition: the state of science.” Molecular Nutrition & Food Research 67.1 (2023): 2200476.
谷禾健康
2024年6月19日,广东省人民医院儿科杨敏团队取得了一项备受瞩目的成果。该团队关于“肠道菌群失调通过糖原贮积病中的 CCL4L2-VSIR 轴引发炎症性肠病”,在国际著名期刊《Advanced Science》(影响因子IF=15.1)发表,针对这一通路不仅可以为 GSD 患者提供有益的治疗,还可以为结肠炎和其他形式的 IBD 患者提供有益的治疗。
谷禾健康技术团队参与并支持本研究的菌群测序和分析部分。
这一突破性研究为糖原贮积病及其相关并发症的治疗提供了新的视角,这也是我国儿科在罕见病领域的研究中取得的重大突破。

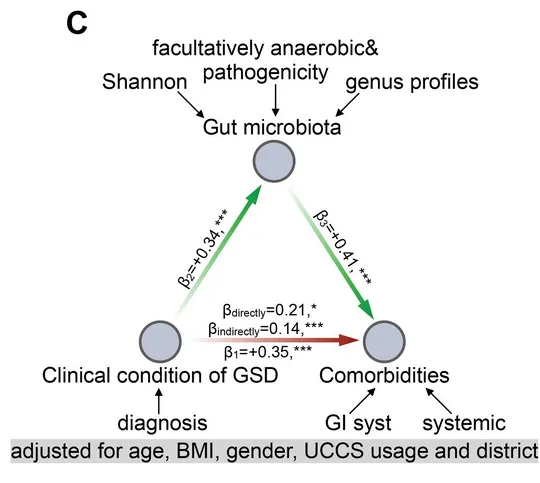
糖原贮积病(GSD),是一类由于先天性酶缺陷所造成的糖原代谢障碍疾病,多数属常染色体隐性遗传,发病因种族而异。该病类型多样,其中GSD-Ib型患者常伴有炎症性肠病。
在GSD-Ib型患者中,肠道菌群失调通过CCL4L2-VSIR轴影响炎症性肠病(IBD)。研究发现,与典型IBD不同,GSD-Ib患者展现出独特的消化道症状和肠道菌群特征。
肠道菌群失调主要由特定病原菌引起,导致肠道巨噬细胞过度激活,CCL4L2-VSIR轴的过度激活促进上皮细胞特定基因的表达,从而推动IBD的进展。
研究结果表明,靶向肠道菌群失调或CCL4L2-VSIR轴可能成为治疗GSD相关IBD的潜在策略。
糖原是肝脏和骨骼肌中储存的葡萄糖形式,其分解提供稳定的葡萄糖供应。当特定酶的活性缺失时,会导致糖原积累,进而影响组织的功能,导致一系列健康问题,包括低血糖、肌肉痉挛、疲劳、肝脏肿大等。
糖原贮积病的类型多样,根据受影响的酶和组织不同,可以分为几种主要类型。例如:
糖原贮积病的治疗主要是通过调节血糖水平和预防低血糖来管理。对于GSD-Ia和GSD-Ib,过去十多年的治疗方法包括间歇性未煮玉米淀粉或通过胃管给予夜间葡萄糖输注。给GSD-III患者高蛋白饮食,以增加儿童的生长速率。
GSD不仅影响儿童,成人也可能受到影响。未经最佳终身饮食葡萄糖治疗的成人GSD患者的长期预后不佳,可能会出现多种并发症,成人GSD患者常见的并发症包括骨质疏松症和骨折、肾结石、肾盂肾炎等。
▼
正常情况下,人体摄入的碳水化合物会被转化为葡萄糖,一部分葡萄糖会被立即利用以提供能量,而多余的葡萄糖则会在肝脏和肌肉等组织中合成糖原储存起来。当身体需要能量时,糖原又会分解为葡萄糖以供使用。
然而,在糖原贮积病患者中,由于参与糖原合成或分解的某些酶存在缺陷,导致糖原的合成、分解或储存出现异常,从而引起一系列的健康问题,包括低血糖、肌肉痉挛、疲劳、肝脏肿大等。
这类疾病有一个共同的生化特征,即是糖原贮存异常,绝大多数是糖原在肝脏、肌肉、肾脏等组织中贮积量增加。
▼
糖原贮积病是一种罕见病,总体发病率相对较低,但具体的发病率因不同类型的糖原贮积病而有所差异。
《世界胃肠病学杂志》资料显示,发病率约为20000-43000名新生儿中的1人。
由于其症状的多样性和隐匿性,一些病例可能未被准确诊断,实际的发病率可能被低估。
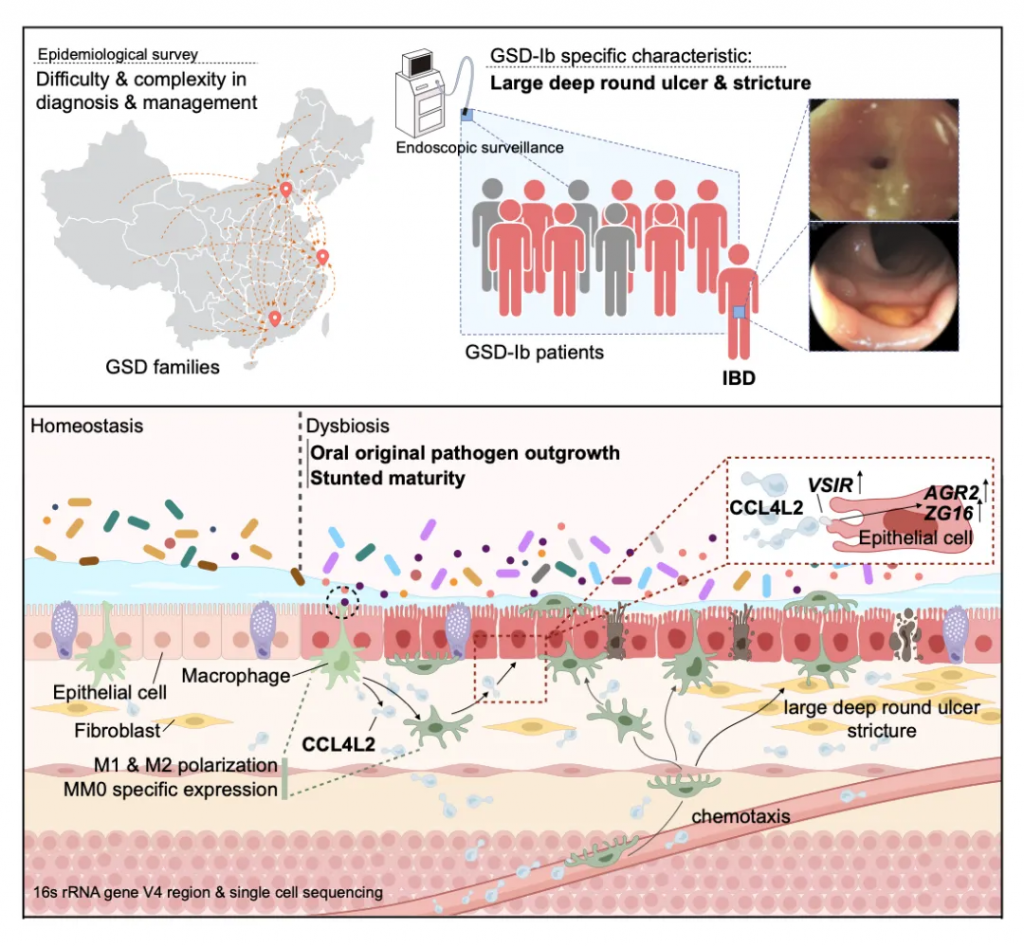
为了调查中国大陆GSD的流行病学和临床特征,杨敏及其团队进行了两项大型多中心研究,并通过问卷调查报告了2020年10月至2021年6月的209例GSD病例(如图)。
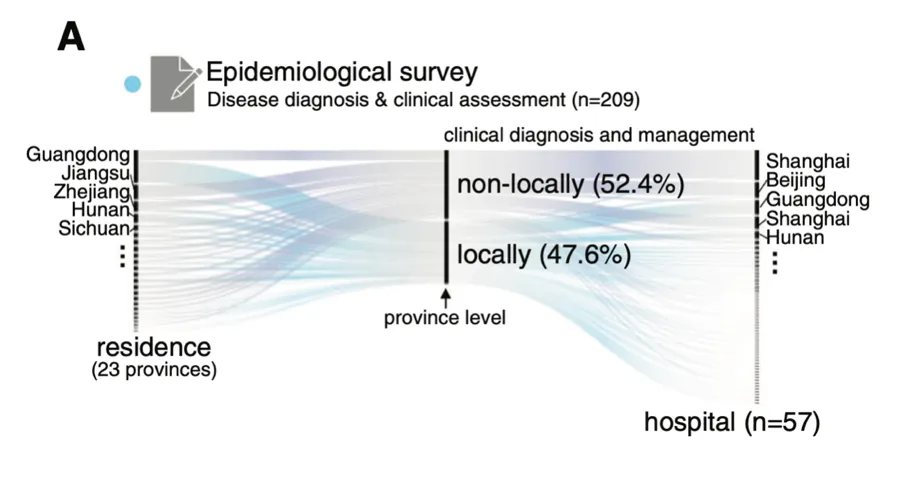
关于患者的就医情况,其中最显著的一个特点是超半数家庭(52.4%)为“省级”外来病例,就医的医院主要来自相对发达地区。且就诊地主要集中在上海、北京、广东等医疗资源相对发达的城市。
▼
糖原合成和分解代谢中所必需的各种酶至少有8种,由于这些酶缺陷所造成的临床疾病有15型,其中:
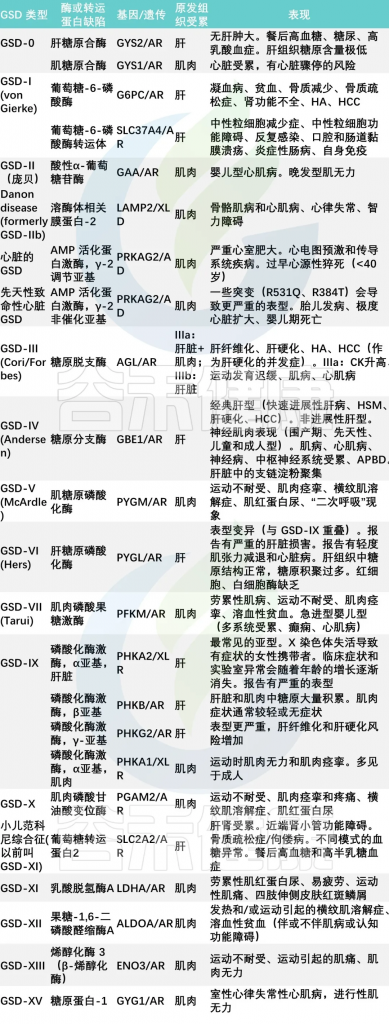
doi: 10.3748/wjg.v29.i25.3932
杨敏团队的调查研究中,主要包括6种类型的GSD,即:
其中最常见的是I型(66.9%)。
▼
不同类型的糖原贮积病临床表现各异,但常见的症状包括:
低血糖、肝肿大、酸中毒(乳酸性酸中毒)、高脂血症、高尿酸血症、疲劳、肌肉无力等。
由于糖原无法正常分解为葡萄糖,患者在空腹或长时间运动后容易出现低血糖症状,如头晕、乏力、出汗、心慌等。
糖原在肝脏中过度积累,导致肝脏体积增大。
长期的低血糖和代谢紊乱可能影响儿童的生长发育,导致身高、体重增长缓慢。
某些类型的糖原贮积病会影响肌肉功能,表现为肌肉无力、疼痛、痉挛等。
如Ⅱ型糖原贮积病可能导致心脏肥大、呼吸肌无力等,严重影响心肺功能。
其中:
口疮性口炎、肛周病变、胃肠炎、呼吸道感染和皮肤感染等症状在I型GSD中更常见;
厌食、呕吐、腹泻、粘液/血便、腹痛和腹胀是 GSD 患者常见的胃肠道并发症,尤其是在 GSD-Ib 组(77.8%)。
造成患者家庭问诊困难的一个主要原因是该疾病表型的复杂,例如下图:
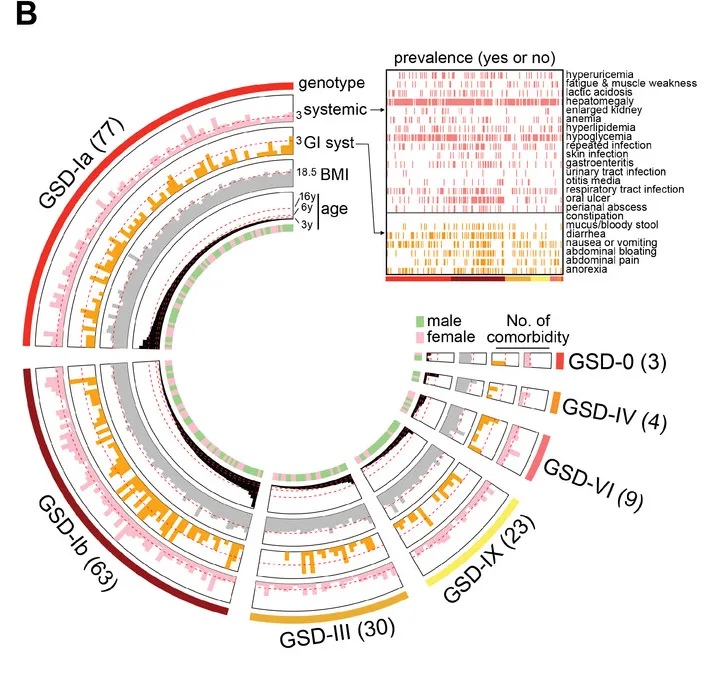
一种奇怪的糖原贮积病亚型:许多GSD-lb患者也患有慢性 IBD
杨敏教授表示,他们关注 GSD相关IBD 已有近10年了。多中心内镜监测研究中,他们招募了 32 名 GSD 患者。这些患者在内镜检查期间表现出活动性胃肠道症状。在这 32 名患者中,27 名患有 GSD-Ib,25 名最初诊断为 IBD。
其中GSD-Ib型患儿消化道症状比例最高(77.8%)且最为特殊,表现为单个或多发散在深圆形溃疡、炎性假息肉、梗阻和狭窄(如下图)。
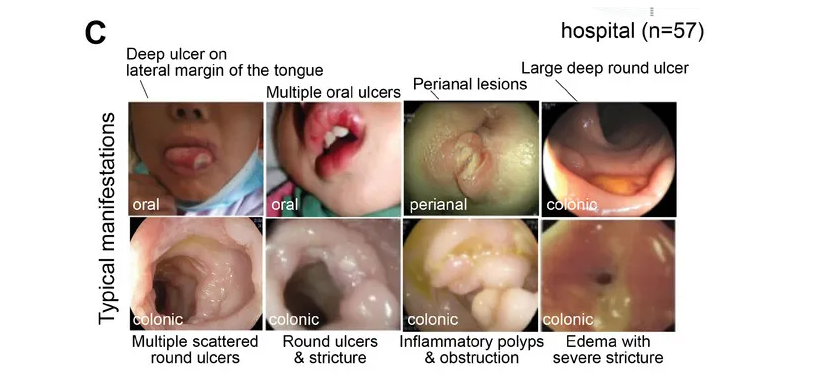
目前人们对 GSD-lb 中 IBD 的发病机制和发病原因知之甚少,超过 70% 的病例会出现严重的消化道症状,而这种症状在其他疾病亚型中偶尔才会出现。
如果不具体了解这些机制以及免疫系统与肠道微生物群之间的关系,科学家将无法开发出针对与 GSD 相关的 IBD 症状的有效治疗方法或干预措施。杨敏及其团队希望改变这一现状,开始了他们的实验探索过程。
▼
为了探究GSD群体消化系统异常,特别是IBD高发的原因,杨敏及其团队在全国23个省份招募了150名GSD患者(共涵盖Ia、Ib、III、IX、VI、IV及0型共7种类型)采集粪便样本, 327个健康对照者的粪便样本。
注:这327个对照样本包括:137 名家庭对照者和 190 名在山东、浙江、湖南等地收集的无关健康对照者。
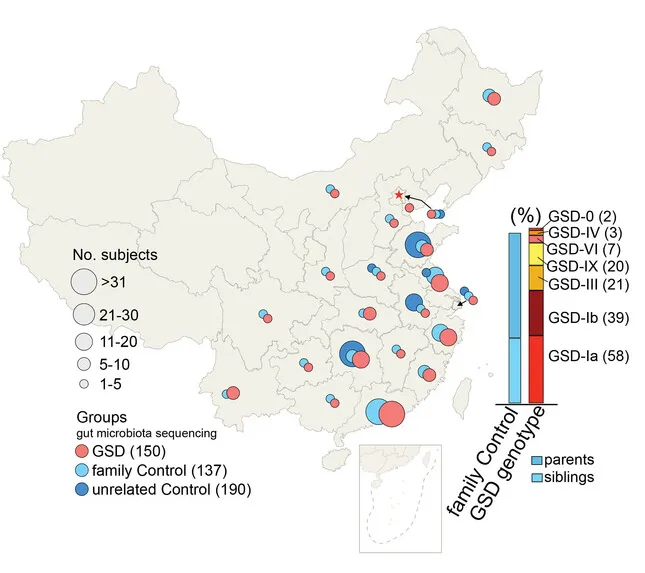
GSD基因型是肠道菌群的主要影响因素
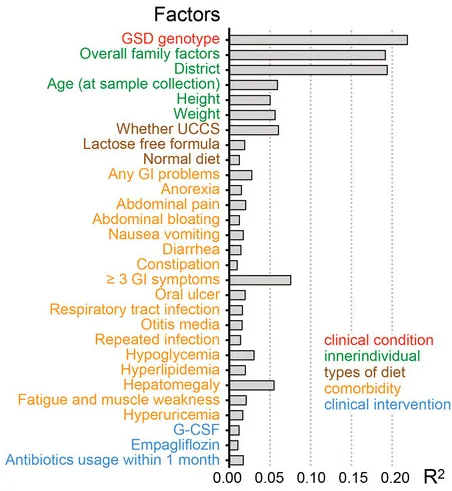
肠道菌群受多种因素影响,包括饮食、地域和医疗条件等。
通过结合个人临床信息与肠道菌群数据的EnvFit分析发现(如下图),共有45个个人因素与肠道菌群的变异显著相关,如疾病情况(基因型)、家庭因素、年龄、身高体重、共患病等等。
值得注意的是,在这个队列中,GSD基因型主要影响肠道菌群组成。总体而言,家庭因素(家庭组配对比较)和居住地区是影响肠道菌群组成的第二和第三重要因素。
这在肠道菌群研究中颇为少见,因为多数研究认为地域因素极大且显著地影响个体菌群构成。这说明,特殊的基因型对于塑造个体肠道菌群构成起到决定性作用。
具体而言,研究人员将患者分为三组:
GSD-Ia、GSD-Ib、GSD-no IaIb
▼
GSD引起了个体肠道菌群多样性显著降低(下图H)及潜在致病菌显著升高(下图G),特别是GSD-Ia和GSD-Ib。这些肠道菌群的变化在校正性别、年龄、BMI和地域等因素后,仍然存在。
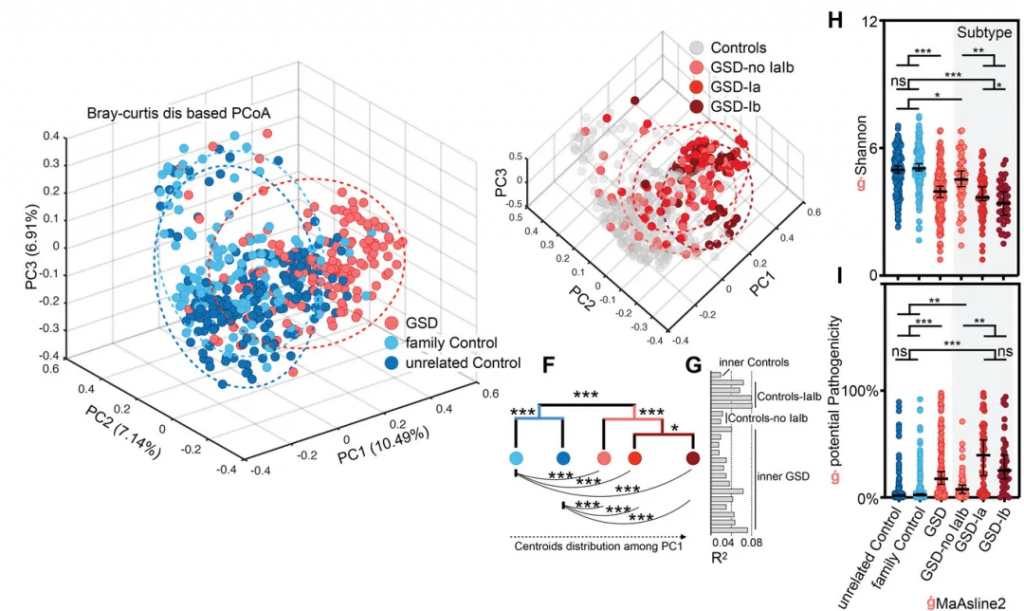
厚壁菌门和拟杆菌门仍是主要菌群;但变形菌门和放线菌门的比例显著增加。
GSD 患者共有31个属发生显著改变。其中18个属减少,而其余13个增加。这31个属中28个属在校正掉性别、年龄、BMI和地域等因素影响后,仍与GSD显著相关(下图D)。
有22物种的丰度和GSD显著关联(下图E),其中10 个增加和12个降低。
这31个显著改变的属的总丰度在不同分组人群中的总占比达到60-70%,种水平则为~25%,表明这些GSD相关的属种变化,是肠道菌群构成主体的紊乱。
▼
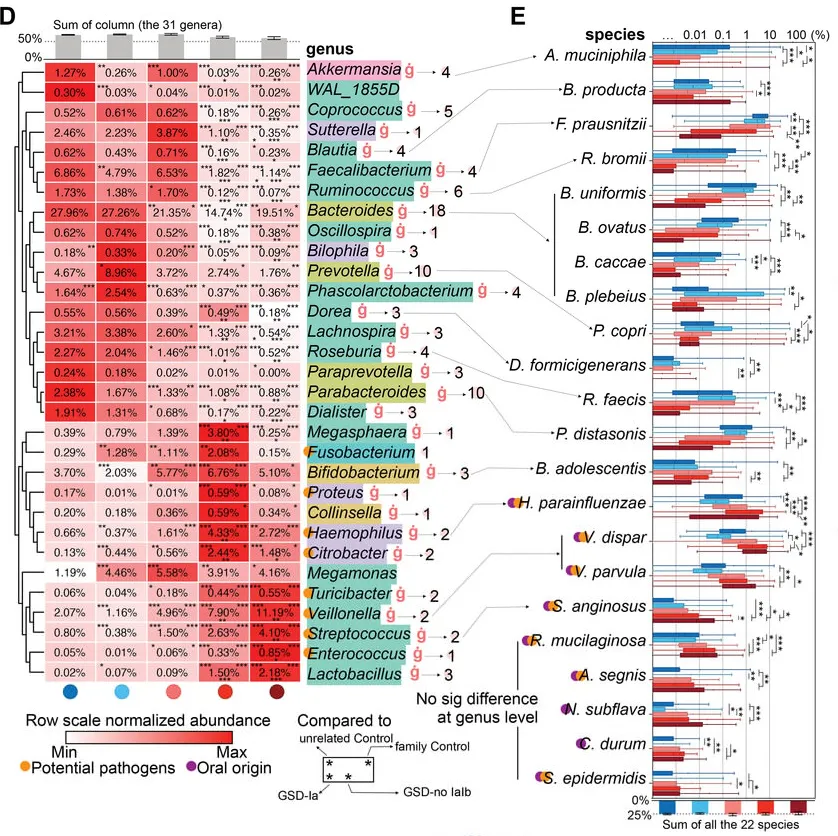
GSD-Ia和GSD-Ib患者表现出更严重的菌群紊乱,特别是那些最常见、功能性的微生物类群在其肠道内丢失,例如下列菌群在GSD-Ia和-Ib几乎完全耗尽:
丁酸可增强上皮氧合,有利于结肠中的低氧微环境并抑制病原体定植。然而,主要丁酸生产菌,如拟杆菌属、粪杆菌属和瘤胃球菌,以及相关代谢途径在GSD中显著降低,这可能导致兼性厌氧病原菌的延伸。
具体到每个菌来说:
AKK菌与肠道屏障的维持和抗炎作用有关,其减少可能会削弱肠道屏障,增加肠道炎症的风险,从而可能与IBD症状的出现有关。
粪杆菌参与肠道内的发酵过程,对维持肠道健康和免疫功能有重要作用。在糖原贮积病患者中,由于糖原代谢异常可能影响了肠道环境,进而影响了粪杆菌的定殖和功能,导致其数量减少。这可能会减少短链脂肪酸的产生,影响肠道健康。
拟杆菌在肠道内参与碳水化合物的分解,产生短链脂肪酸,糖原贮积病患者拟杆菌的减少可能导致肠道炎症反应加剧,与IBD症状的出现有关。
瘤胃球菌是产生短链脂肪酸的重要菌群,对肠道健康和宿主能量代谢有重要作用。瘤胃球菌的减少可能会导致短链脂肪酸的产生下降,影响肠道屏障功能和抗炎作用,从而可能与IBD症状的出现有关。以动物为基础的饮食会增加有害菌Ruminococcus gnavus。
GSD患者中存在口腔源的潜在病原体激增,例如:
口腔中的致病菌可以在肠道中定植和繁殖,说明肠道为这些菌群的生长繁殖提供了条件和环境。
链球菌,已被证明可通过GasderminA依赖性细胞焦亡诱导肠道损伤;可能通过产生超抗原等物质激活宿主T细胞免疫反应进而引发肠道炎症。
肠球菌的成员已显示出促炎作用;肠球菌可以通过分泌金属蛋白酶分解上皮钙黏蛋白破坏肠屏障。
韦荣氏球菌属在IBD中延伸,细小病毒通过产生硝酸盐诱导炎症。韦荣氏球菌属和链球菌的组合抑制了IL-12p70的生物合成,随后增强了IL-8、IL-6和TNFα的炎症反应。
菌群该定植时未定植,该退出时却长期占据
值得注意的是,双歧杆菌、韦荣球菌和乳杆菌这些本应该在健康儿童生命早期保持较高比例的细菌,在GSD患者的肠道中扩张,并且这些细菌在 GSD 患者的整个童年和成年期都会扩增,这可能抑制了其他正常菌群的定植。
▼
GSD患者肠道菌群发育落后于实际年龄
使用微生物组年龄Z分数(MAZ)评估菌群发育状态,随机森林回归和调整的深度神经网络分析均显示GSD患者的MAZ显著降低,特别是在GSD-Ia和Ib患者中。
这表明GSD患者的肠道菌群发育落后于实际年龄,呈现发育延迟状态。
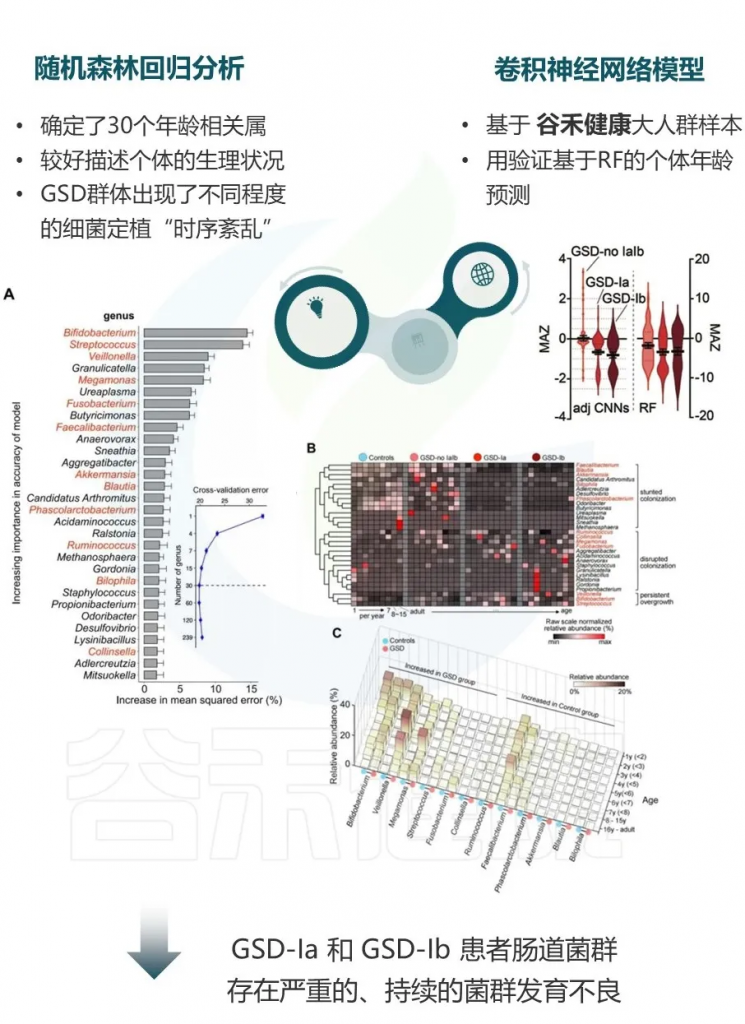
▼
从数据分析的角度,研究团队进一步分析了GSD患病情况、共患病与肠道菌群紊乱的中介关系。
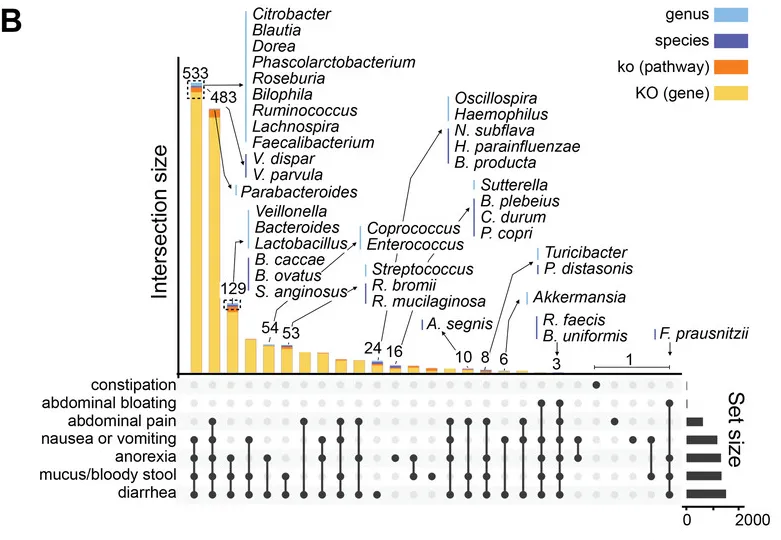
他们用最具代表性的肠道微生物特征和临床元数据构建了一个中介模型,在校正年龄、性别、BMI、地域及特殊治疗性饮食(生玉米淀粉)的影响后,肠道菌群仍然是GSD背景下,患者消化道异常的显著中介因子(下图)。说明GSD会通过影响肠道菌群构成最终造成消化道异常。
GSD 中这些细菌可能相互依赖资源和生存,形成一个复杂的相互支持网络,肠道菌群失调可能通过细菌的“群体犯罪”引起 GSD 患者的肠道上皮功能障碍。
下面我们来看一下该研究的另一个重要内容——肠道微生物群和免疫细胞之间的“串扰”。
▼
为了明确GSD相关的肠道菌群紊乱,如何引起个体IBD表性的产生。研究者们采集3名GSD-Ib患者的结肠粘膜组织(下图G),并使用单细胞测序技术分析患者结肠粘膜单细胞层面的转录水平变化。通过转录水平注释发现,相较于健康人、典型IBD和结肠炎的肠粘膜组织,GSD-Ib患者肠粘膜上有非常高的巨噬细胞聚集(下图H)。
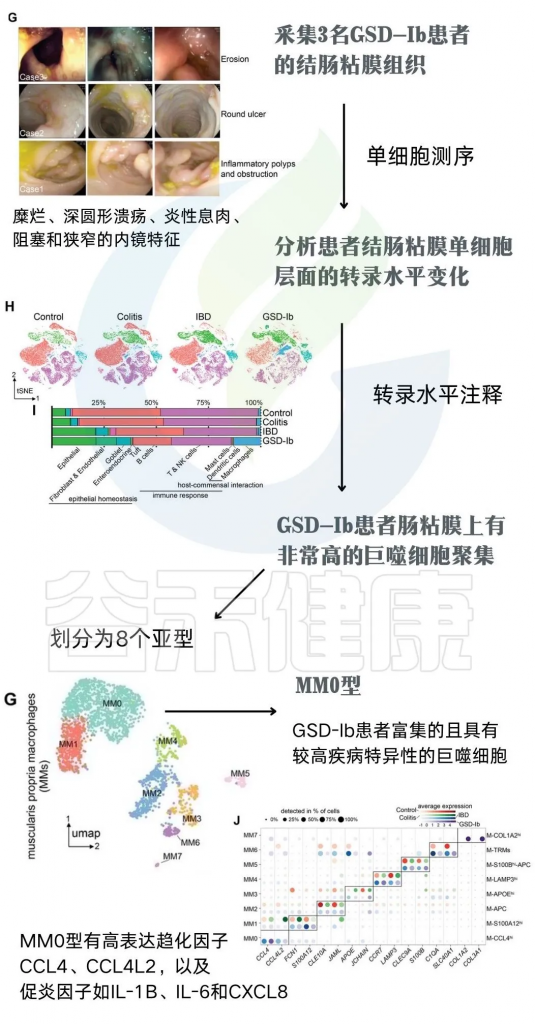
注:巨噬细胞中差异表达基因的KEGG通路注释显示,GSD-Ib中调节抗菌体液免疫反应和趋化因子介导的信号通路的基因集显著富集。GSD-Ib组中上调的基因主要参与环境信息处理和人类疾病。
“
更深入的分析确定了一种巨噬细胞亚型(MM0)的重要性,这种亚型产生一种名为CCL4L2的趋化因子,负责激活其他免疫细胞对抗有害细菌的生长。
虽然CCL4L2的自然功能通常是保护性的,但杨敏及团队怀疑,由于GSD-Ib患者相关肠道细菌的存在,CCL4L2的过度或失调刺激可能是他们出现IBD症状的可能原因。
这一点最初在实验室中得到了支持,其中用有害细菌感染的细胞获得的细胞培养基处理的巨噬细胞,比用对照培养基处理的巨噬细胞更加“激活”。
下面我们来看一下他们选用了哪些菌群进行相关实验,来刺激巨噬细胞:
▼
研究人员选择了三种GSD相关菌群来刺激巨噬细胞,即:
暴露于粪肠球菌、小弧菌和咽峡炎链球菌以及混合感染 (EVS) 的巨噬细胞中 CCL4L2 的表达在 mRNA 和蛋白质水平上显著增加,但不影响 IL-1 β和 IL-6 的表达。
来自EVS感染的条件培养基(CM)与对照组相比,诱导了显著的巨噬细胞迁移趋化性,而这种趋化性部分被CCL4L2中和所减弱。
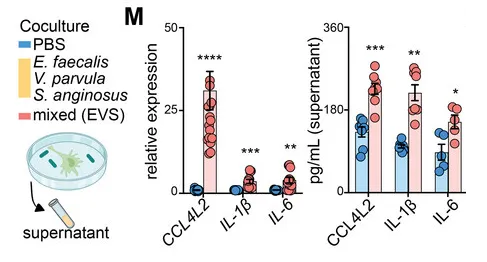
因此,是特定的肠道病原菌(如E. faecalis、V. parvula、S. anginosus)引发了CCL4L2的高表达,那么具体是如何影响炎症性肠病的进展?我们接着看。
“
然后,团队在从GSD-Ib患者取得的活检样本中寻找CCL4L2,并发现CCL4L2蛋白与一种叫做VSIR的受体结合,这种受体存在于肠道细胞壁中。
在IBD的小鼠模型中也发现了类似的结果,科学家们发现与健康对照组相比,巨噬细胞的水平更高,并且在用一种旨在抑制VSIR的抗体处理后,他们发现小鼠的症状加重了。
巨噬细胞通过CCL4L2-VSIR配体-受体信号与肠道上皮细胞相互作用,以促进损伤修复。
具体研究如下:
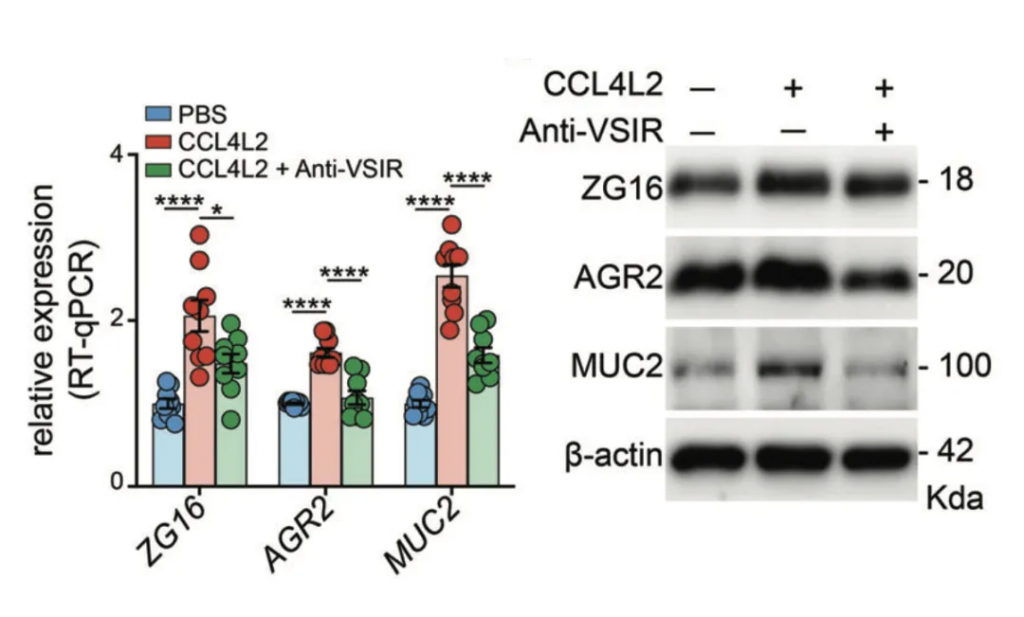
调节核糖体、氧化磷酸化途径和抗病原体反应的基因集在GSD-Ib上皮细胞中特异性富集。
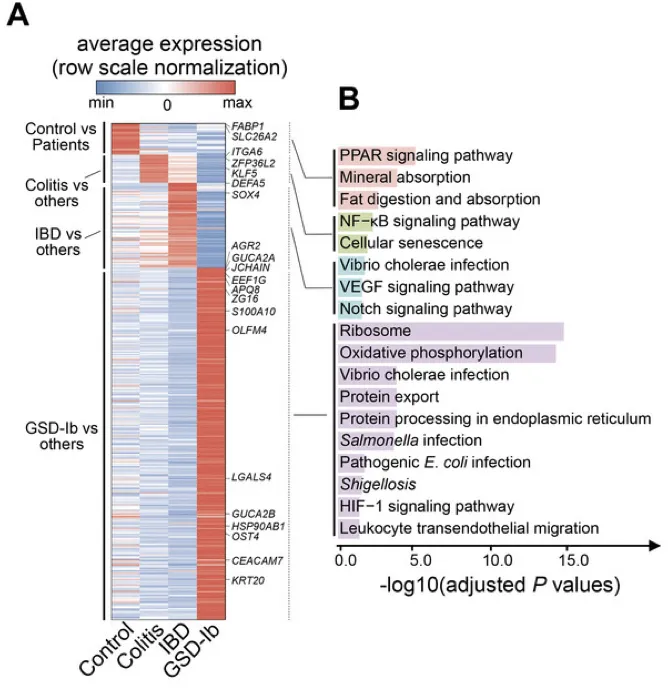
上皮细胞中典型的差异表达基因是AQP8、ZG16、GUCA2C、SLC26A3、AGR2,它们负责吸收、分泌、代谢、上皮屏障完整性、pH稳态、粘液屏障功能。
▼
免疫荧光染色显示GSD-Ib患者肠道组织中CCL4L2和VSIR的显著共定位,进一步验证了CCL4L2在巨噬细胞中的高表达及其与VSIR的相互作用。
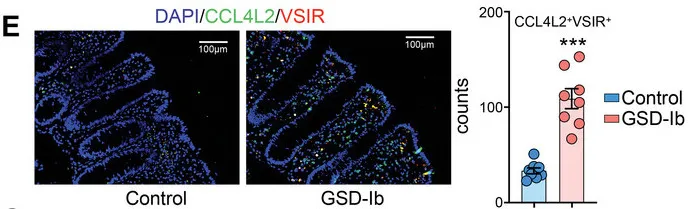
CCL4L2与其受体VSIR形成的CCL4L2-VSIR轴主导参与了MM0型巨噬细胞与其他类型细胞交互,特别是OLFM4+上皮细胞(这类细胞是GSD特异性增加的,同时也被报道在克罗恩病中较高)。
预测MM0巨噬细胞与其他细胞类型之间的相互作用
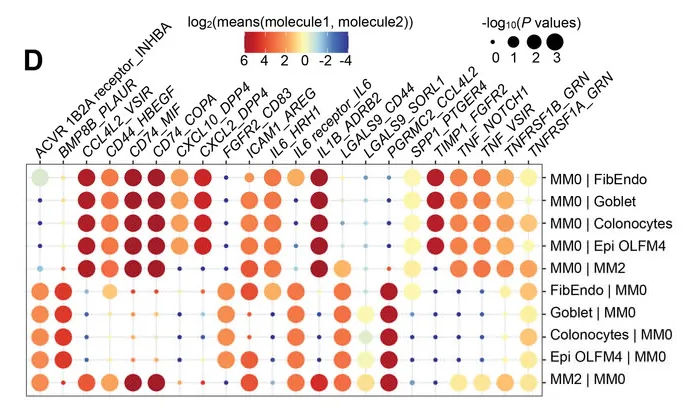
而这些受体细胞的基因表达情况,决定了患者肠道组织的疾病发展方向,如AGR2、ZG16、MUC2等。
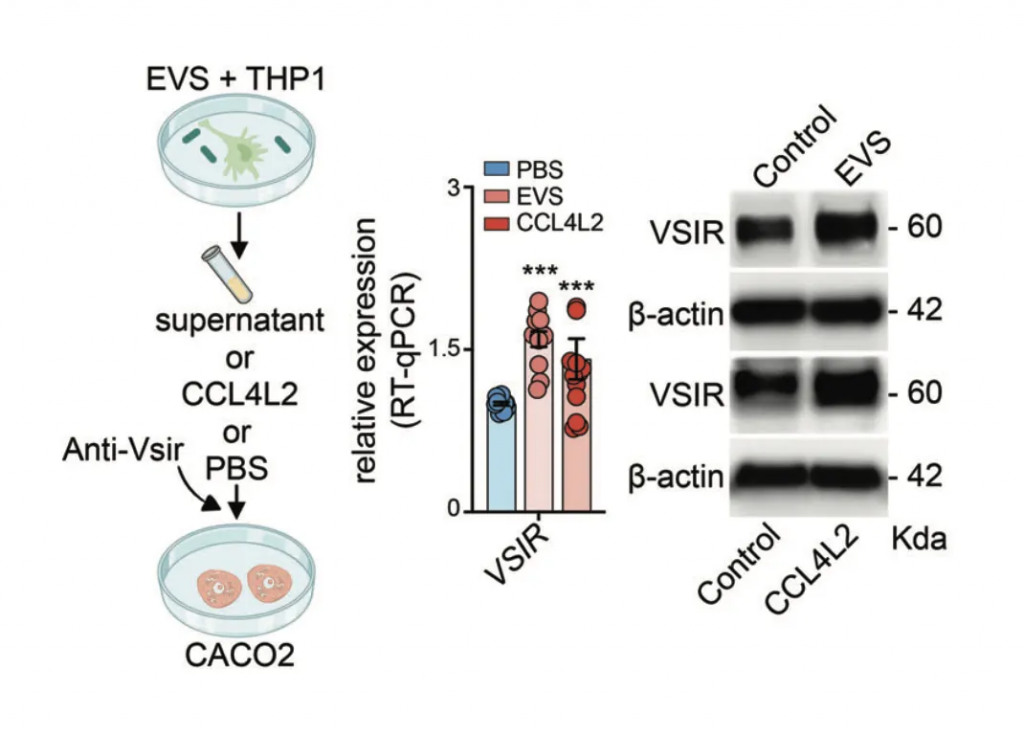
表明GSD相关的肠道菌群紊乱,通过引起肠上皮巨噬细胞高表达趋化因子CCL4L2,激活其他下游细胞膜蛋白VSIR表达,从而引起了GSD独有的消化系统IBD表型。这个现象进一步在C57BL/6小鼠中得到验证。
▼
研究人员在C57BL / 6小鼠中进行了选择性细菌移植,通过口服灌胃的方式将EVS移植到小鼠体内,并用DSS诱导结肠炎。研究发现,抗生素治疗显著降低了EVS灌胃诱导的小鼠血清CCL4L2水平,而抗VSIR则显示出完全相反的趋势。
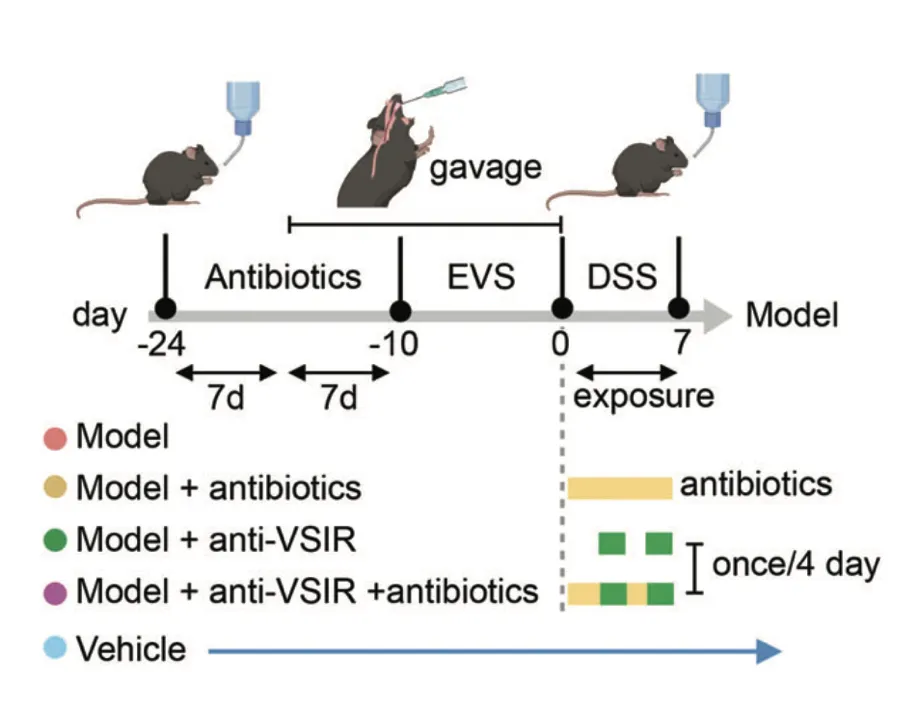
免疫荧光染色显示,模型组中MM0巨噬细胞增强,但在给予抗生素或抗VSIR注射后减少,这进一步导致模型组中肠上皮细胞的MUC2、AGR2和ZG16表达显著下调,引发粘膜损伤。
研究结果表明,EVS 增强的 MM0 巨噬细胞可以通过调节 CCL4L2-VSIR 轴来介导结肠肠上皮稳态。因此,靶向CCL4L2-VSIR通路可用于结肠炎和IBD的广泛应用。
肠道驻留免疫细胞和上皮之间的串扰,对于胃肠道稳态、抗原致敏调节、预防感染和 IBD 发展至关重要。在该研究中 GSD相关IBD 与典型的 IBD 相比具有不同的特征,特别是复杂的内镜表现及其特殊的解构:大量巨噬细胞积聚和上皮细胞增殖。
从病因上讲,肠道致病菌激活粘膜巨噬细胞导致胃肠道内形成炎症环境。在 GSD-Ib 中,一群独特的巨噬细胞由致病菌诱导,并表达高水平的趋化因子CCL4L2。
关于CCL4L2的有益方面:
潜在有害方面:
值得注意的是,CCL4L2-VIR通路的激活可能促进严重细胞因子风暴中的T细胞耗竭,这与新冠肺炎的不良预后有关。
靶向CCL4L2-VSIR可以抑制各种炎症性疾病、自身免疫性疾病和肿瘤转移。在这里,研究人员揭示了胃肠道中CCL4L2-VSIR轴的异常激活,最终驱动GSD-Ib中的非典型IBD。该发现为诊断和治疗GSD-Ib以及潜在的GSD相关IBD引入了一个新的靶点,并为VSIR相关研究提供了证据。
总的来说,这项研究证实了在糖原贮积病群体中独有的肠道菌群紊乱,即:
而这样的肠道菌群紊乱会引起个体结肠巨噬细胞高表达CCL4L2,并通过其受体蛋白VSIR介导上皮细胞的异常基因表达,最终造成GSD相关的IBD表型。
该研究的意义及未来方向:
该研究通过识别GSD-Ib与IBD之间的特定生物学联系,推动了精准医疗在代谢性疾病和消化系统疾病中的应用。
目前杨敏团队正在研究 CCL4L2-VSIR 通路的作用机制,并将探索其在患者治疗中的潜力。该团队预计,针对这一通路不仅可以为 GSD 患者提供有益的治疗,还可以为结肠炎和其他形式的 IBD 患者提供有益的治疗。
CCL4L2-VSIR轴的发现也可能促进新药开发,特别是针对GSD-Ib相关IBD的治疗。
这些研究结果也可应用于个性化治疗方案(基于肠道菌群特征),为患者提供更有效的治疗选择,有助于改善GSD-Ib患者及更广泛消化系统疾病患者的生活质量。
主要参考文献
Lan J, Zhang Y, Jin C, Yang M et al., Gut Dysbiosis Drives Inflammatory Bowel Disease Through the CCL4L2-VSIR Axis in Glycogen Storage Disease. Adv Sci (Weinh). 2024 Jun 18:e2309471.
Gümüş E, Özen H. Glycogen storage diseases: An update. World J Gastroenterol. 2023 Jul 7;29(25):3932-3963.
Hannah WB, Derks TGJ, Drumm ML, Grünert SC, Kishnani PS, Vissing J. Glycogen storage diseases. Nat Rev Dis Primers. 2023 Sep 7;9(1):46.
Zhong J, Gou Y, Zhao P, Dong X, Guo M, Li A, Hao A, Luu HH, He TC, Reid RR, Fan J. Glycogen storage disease type I: Genetic etiology, clinical manifestations, and conventional and gene therapies. Pediatr Discov. 2023;1(2):e3.
Wang Y, Liu H, Dong F, Xiao Y, Xiao F, Ge T, Li D, Yu G, Zhang T. Altered gut microbiota and microbial metabolism in children with hepatic glycogen storage disease: a case-control study. Transl Pediatr. 2023 Apr 29;12(4):572-586.

谷禾健康

“胖!可怎么办?”《柳叶刀》发布的一项报告称,截止2022年,全球超过10亿人患有肥胖症,超过20亿人存在超重。从1990年到2022年间,全球患肥胖症的成年人增加了一倍多,患肥胖症的儿童和青少年(5至19岁)更是增加了约3倍。超重和肥胖已成为全球日益严重的流行病。
超重和肥胖的不良影响可能不会马上体现,有时会延迟十年或更长时间。流行病学研究证实,超重和肥胖程度的增加是寿命缩短的重要预测因素。在一项心脏研究中,在30岁到42岁之间,体重每增加一磅(0.45公斤),26年内死亡的风险增加1%,在50岁到62岁之间,死亡风险增加2%。
肥胖还会引起或加剧许多健康问题,这些问题既有独立的,也有与其他疾病相关的。例如超重和肥胖增加了患高血压、冠心病和中风等心血管疾病的风险。肥胖还是2型糖尿病主要风险因素;此外,肥胖和超重人群更容易患上睡眠呼吸暂停、哮喘和其他呼吸系统问题。过重的身体还会给骨骼和关节造成巨大压力,增加骨质疏松症和关节炎的风险。
“那么我们应该如何改善或是预防肥胖呢?”除了常规的“管住嘴,迈开腿”,谷禾在最近与减肥前后相关的检测中发现,在不改变其他生活方式和饮食的情况下,额外补充一定量的膳食纤维对减肥的帮助效果显著。
肠道菌群已越来越多地被认为是宿主生理和病理的重要调节器。肠道菌群调节炎症、脂肪储存和葡萄糖代谢进而影响体重和代谢健康。
而对肠道微生物群影响最大的是饮食,人类从食物中提取和储存卡路里的能力至少部分受到肠道微生物的影响。这使得饮食成为驱动人类肠道微生物群组成的重要因素。
膳食纤维是一种来自植物的复合多糖,在消化过程中不被小肠吸收,并通过为微生物生长提供底物来改变肠道微生态环境。膳食纤维可能会改变微生物群的丰度、多样性和代谢,包括短链脂肪酸的产生。最近的研究表明,富含膳食纤维的饮食具有预防肥胖的潜力。
首先,膳食纤维的物理化学性质(粘性、可发酵性等)具有预防肥胖的作用,因为粘稠的纤维可以延长胃排空和小肠运输时间,从而增加饱腹感,减少进食频率,降低热量的摄入。
其次,膳食纤维可通过增加代谢相关有益肠道菌群的丰度和多样性来改善能量稳态并预防肥胖,降低门水平上厚壁菌门/拟杆菌门的比例(F/B比)并增加罗氏菌属(Roseburia)的相对丰度。
第三,膳食纤维可被肠道菌群发酵产生短链脂肪酸 (SCFA),通过降低肠道腔内pH值、抑制致病或有害肠道细菌、减少脂多糖(LPS)和代谢有害化合物,在能量代谢方面发挥重要作用。同时,研究还表明,在高脂膳食中添加纤维可降低外周血炎症水平,在本文一起了解下肥胖的原因,哪些肠道菌群参与肥胖及其作用机理,此外,我们还分享了几个根据检测报告针对性调整和补充膳食纤维合生元等干预措施减重改善健康的案例。
肥胖不仅仅是因为吃得多这一个原因造成的,而是由多种因素共同导致的慢性代谢性疾病。
肥胖受到遗传、环境、生活方式、社会心理因素、内分泌和健康状况、个体的微生物和营养差异等因素的影响,这些因素通过能量摄入和消耗的生理机制相互作用,最终导致了肥胖。
变胖最直接的原因就是能量摄入与能量消耗的不平衡。我们知道,要维持健康的体重,需要保持食物摄入和能量消耗的平衡。

★ 摄入大于消耗时,变成脂肪慢慢堆积
当一天中人体的能量摄入大于能量消耗时,多余的能量就会被储存为脂肪、糖原或蛋白质,而体重就会上涨;相反,当摄入量小于消耗量时,机体就会通过转化储存能量(大部分来源于脂肪)来弥补热量差,体重就会下降。
简单来说就是:当你吃的比消耗的多时,能量储存=能量摄入-能量消耗
尽管在过去的20年里,环境变化导致了肥胖率的上升,但家庭和双胞胎研究表明,遗传因素在肥胖的发展中也起着关键作用。
★ 瘦素等基因突变易导致肥胖
迄今为止,已确定十几个基因的变异是肥胖的单基因原因;包括瘦素、瘦素受体、黑皮质素3受体和黑皮质素4受体(MC4R)基因。
最著名的肥胖相关基因是瘦素(ob)及其受体(db)基因。洛克菲勒大学的研究首次发现ob基因突变和缺乏ob基因的小鼠导致严重肥胖,能量摄入增加(暴饮暴食),但能量消耗减少(代谢率、产热和身体活动减少)。
注:该基因产物后来被描述为循环因子,命名为瘦素。瘦素主要由脂肪细胞分泌,循环至全身并穿过血脑屏障屏障来传递饱腹信号,从而减少食物摄入。
★ 肥胖的遗传原因大致可分为:
1)单基因原因:由单个基因突变引起,主要位于瘦素-黑皮质素通路。许多基因,如PYY(食欲促进基因)或MC4R(黑皮质素4受体),被发现与单基因肥胖有关,这些基因会破坏食欲和体重的调节系统以及位于下丘脑弓状核的受体感知激素信号(胃促生长素、瘦素、胰岛素)。
罕见的单基因缺陷与高饥饿水平有关,并可能导致幼儿严重肥胖。
2)综合征性肥胖是由神经发育异常和其他器官/系统畸形引起的严重肥胖。这可能是由单个基因或包含多个基因的较大染色体区域的改变引起的。
3)多基因肥胖是由许多基因的累积作用引起的。这些类型的基因存在会导致热量摄入增加、饥饿感增加、饱腹感减少、储存身体脂肪的倾向增加以及久坐不动的倾向增加。
★ 有肥胖家族史的人患肥胖风险较高

一项研究发现,如果父母一方肥胖,孩子成年后患肥胖的风险会提高3倍;而如果父母双方都肥胖,那么孩子成年后患肥胖的风险会提高10倍。(当然这在一定程度上也可能与相同的生活方式有关)
一项针对260名儿童(139名女性、121名男性,年龄分别为2.4岁至17.2岁)的横断面观察研究表明,心脏代谢疾病家族史和肥胖是儿童期肥胖严重程度的关键危险因素。
生活方式等环境因素在肥胖发展中也发挥作用。肥胖患病率的显著增加与饮食和生活方式的改变密切相关。
在工业化国家,教育程度较低和收入较低的人超重和肥胖的发生率较高,尽管发展中国家的情况可能正好相反。
例如,生活在美国的皮马印第安人平均比生活在墨西哥的皮马印第安人重25公斤。生活在美国的非洲人也出现了类似的趋势。
在男性和女性中,超重和肥胖的患病率随着年龄的增长而增加,直到50至60岁;这在20岁到40岁之间尤为明显。结婚后,超重的趋势也会增加。
★ 高糖的零食和饮料加剧了肥胖
在众多导致肥胖的因素中,过量食用高热量食物是罪魁祸首之一。目前,无论是发达国家还是发展中国家,高热量食物在食品行业的大规模生产和营销方面都相当成功。这类食物在商店、餐馆、超市和家庭中随处可见。
两餐之间(尤其是晚餐后)吃零食,以及每天饮用果汁、碳酸饮料、糖果和高糖食物。这些不健康的饮食与肥胖风险增加密切相关。
★ 运动量的减少也导致肥胖率增加
体育锻炼应该是人们日常生活的一部分,但是随着青少年的学业负担增加、中年人的工作负担沉重,运动量大幅减少。如果平时不注意运动,长时间久坐,会导致脂肪堆积,从而出现肥胖的现象。
其他可能导致身体活动减少的因素包括花在电子游戏和移动设备上的时间增加,而花在户外活动上的时间减少。这些不良习惯都会损害人们的健康。
★ 睡眠不足导致皮质醇升高也不利于减肥
缺乏充足睡眠会引发皮质醇水平的升高,这对身体的减肥过程是不利的,因为高水平的皮质醇可能会干扰新陈代谢和影响体重管理的效果。
你也许见过,本来身材挺匀称的一个人,因为生病或是吃了一些药物而迅速变胖,这就是内分泌紊乱进而导致肥胖的结果。
许多外周激素参与中枢神经系统(CNS)对食欲和食物摄入、食物奖励或成瘾的控制。美味的食物和一些药物都能激活中脑边缘多巴胺(DA)奖励系统,而该系统对于调节人类和动物的成瘾至关重要。
★ 瘦素、胰岛素等激素会影响进食行为
来自脂肪组织的瘦素、胰腺的胰岛素和胃肠道的胆囊收缩素、胰高血糖素样肽-1(GLP-1)、肽YY3-36(PYY3-36)和生长素释放肽参与通过主要针对下丘脑和脑干的神经激素肠脑轴传递有关饥饿和饱腹信号,这些信号可能会直接或间接地调节食欲,影响个体的进食行为。
瘦素和瘦素受体的基因缺陷可导致儿童早发性严重肥胖。同时当内分泌系统出现紊乱时,可能会导致个体无法有效地控制食欲,进而对食物上瘾或是出现严重的暴饮暴食症。
肠道能量吸收、饱腹感调节和全身炎症
DOI:10.1016/B978-0-12-407825-3.00011-3.
★ 内分泌紊乱影响代谢更易导致肥胖
一些疾病如甲状腺功能减退会影响基础代谢率,导致能量代谢异常,进而影响体重控制,甚至引发肥胖。
胰岛素抵抗是指身体细胞对胰岛素的反应下降,导致血糖不能有效地被细胞吸收利用。这可能导致胰岛素分泌增加,促使脂肪细胞更多地吸收葡萄糖并转化为脂肪,从而引起肥胖。
多囊卵巢综合征(PCOS)一种影响女性生殖系统的疾病,患者常伴有胰岛素抵抗和雄激素水平异常。这些因素会导致体重增加和脂肪堆积,使得患者更容易发展为肥胖。
有证据表明,胎儿在宫内发育期间的营养不良可能决定了肥胖、高血压和2型糖尿病的后期发病,而这与基因遗传无关。这种现象表明,由于宫内生长的改变,可能存在基因表达的长期编程。
★ 胎儿时期的营养不良可能导致中老年时的肥胖
研究人员假设,子宫内不良的营养环境会导致身体器官发育缺陷,从而导致“程序化”的易感性,这种易感性与后来的饮食和环境压力相互作用,在几十年后导致明显的疾病。
这表明胎儿的生长和新陈代谢为了适应出生后营养不足的预期。这可能在子宫内具有生存优势,因为它将可用的营养物质定向到重要器官,并在以后的生活中,通过增加以脂肪形式储存能量的能力,为食物短缺时提供能量储备。
有报告显示腹部脂肪与出生体重呈负相关,一项研究提供了一些有说服力的证据,证明妊娠早期和晚期是随后肥胖发展的关键时期。与没有在怀孕期间接触过饥荒的对照组相比,那些胎儿在怀孕的前两个月接触过饥荒等到成年后的肥胖患病率明显更高。
肠道微生物群在肥胖中的作用是多方面的,并且与肥胖的发生和发展密切相关。肠道微生物群参与能量稳态的调节,并通过影响营养吸收、食欲和脂肪组织功能来影响肥胖的发生。
接下来我们详细了解下肠道微生物群在肥胖中的作用和角色。
越来越多的研究和谷禾实践检测数据表明肠道微生物群与肥胖之间存在着许多相互作用,虽然大人群水平来看,肠道菌群只能解释其中部分的肥胖原因,但是在个体水平上,肠道菌群的状况和构成对于肥胖以及肥胖相关的能量摄入,炎症反应,食物消化等都息息相关。
肠道菌群在肥胖中的作用简单分为两个层面,直接参与与间接影响。
一,菌群是直接影响致肥胖途径还是通过其成分/代谢产物影响致肥胖途径?
二,是否存在导致肥胖的特定宿主-微生物信号传导机制?
队列人群和小鼠研究试验均表明,肥胖个体的肠道微生物群种类和丰富度与正常体重个体存在明显差异。
★ 肥胖人群与健康个体之间占主导的肠道微生物不同
大量研究集中于拟杆菌门和厚壁菌门水平变化与肥胖和减肥之间的动态关系。通过比较瘦人和肥胖个体的肠道菌群,发现大人群水平上肥胖个体的拟杆菌门比例降低,而厚壁菌门水平升高。更有趣的是,经过饮食治疗后,拟杆菌门的相对丰度增加,而厚壁菌门的相对丰度降低。
在谷禾的检测案例里也发现,同一个人减肥前后的肠道菌群构成(如下桑基图展示)发生了较大的变化,有的肠型也会发生改变。
减肥前后的主要肠道菌群构成变化
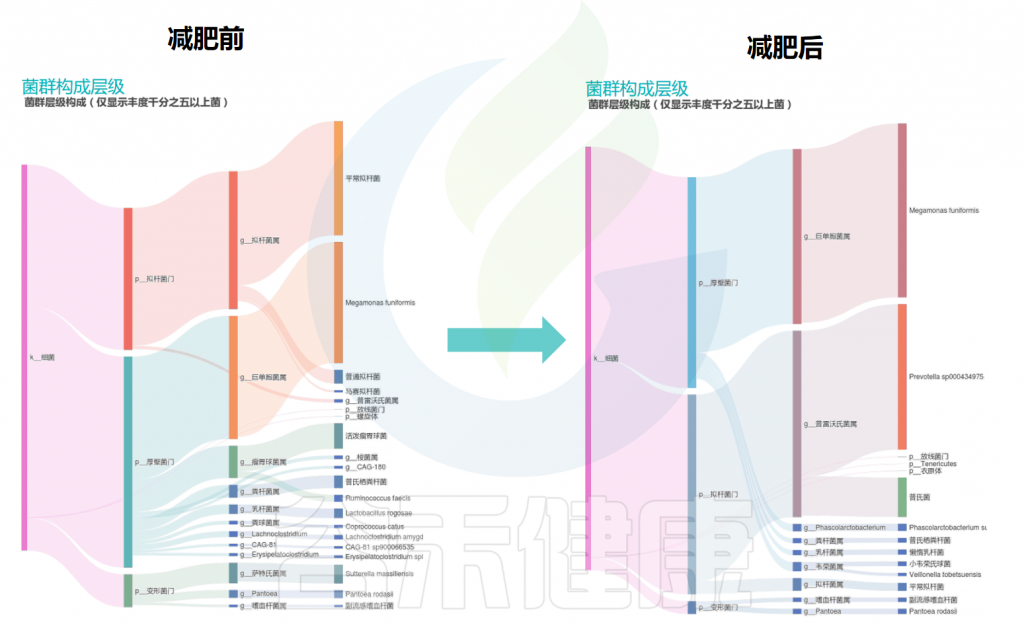
通过上图的变化可以看到:减肥之后拟杆菌门的比例增加,与此同时,普雷沃氏菌属的丰度也大大增加,其他菌属的构成也有一定程度的变化。
该案例通过饮食管理和膳食纤维补充,减肥后普雷沃氏菌占比为主可能是水果蔬菜、高纤维豆类的饮食摄入相关。
★ 减肥的效果与基线肠道菌群相关
2021年,发表在《Gut Microbes》和《Gastroenterology》两篇研究分别以饮食控制和维生素给予为变量,同时都研究了基线时和干预后肠道菌群的变化。研究结论表示作为节食前个人体重减轻轨迹的预测指标,基线肠道微生物的作用超过了其他因素。
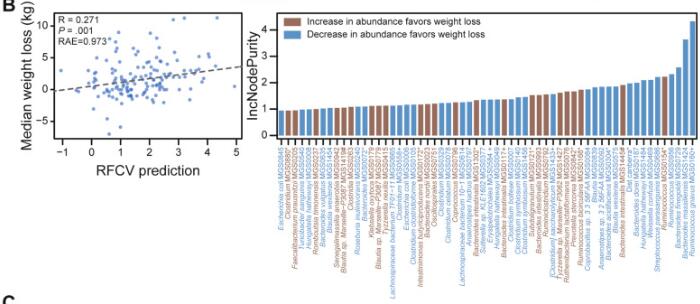
doi: 10.1053/j.gastro.2021.01.029.
同时使用随机森林算法,基于相对于基线的种水平物种变化来预测体重下降,预测精度R=0.271,发现R. gnavus (MGS0160), Bacteroides massiliensis (MGS1424)和Bacteroides finegoldii (MGS0729)这三个物种在模型中贡献度最大。
此外,基线菌属如普雷沃氏菌,罗氏菌属(Roseburia)的丰度也会影响饮食营养干预的减肥效果。
这些研究同时确定有哪些饮食因素与个体的体重下降相关。使用GLMMLASSO模型,结果如下图,当coef为非零时被认为是显著的。发现,在所有受试者中,体重下降与卡路里摄入量(系数=-0.153)、膳食中大量营养素组成(脂肪,系数=-0.161;碳水化合物,系数=-0.055;蛋白质,系数=0.084;纤维,系数=0.1)、膳食微量营养素含量和体力活动之间的具有弱相关。
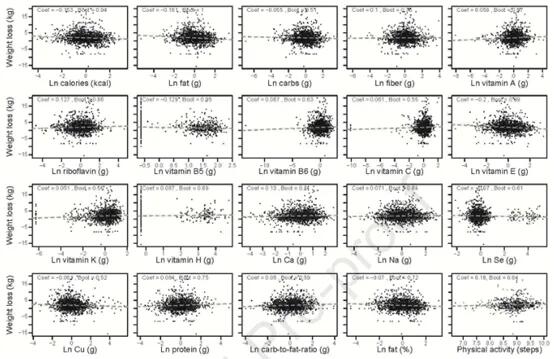
Jie Zhuye, et al., .Gastroenterology, 2021
但是在个体间有很大差异。例如在受试者F00161中,纤维摄入量的增加与减肥之间存在正相关关系,而在F00147中,受试者的脂肪摄入量增加与体重减轻之间存在负相关关系。相反,尽管纤维或脂肪摄入量发生变化,F00203人仍然对体重变化不敏感。所以,即使是相似的膳食大量营养素,体重反应也是高度个性化的。
饮食控制期间肠道菌群的组成变化是否会影响减肥轨迹?
研究发现许多参与者的肠道菌群组成发生了实质性的变化,0-3个月的微生物群的变化幅度与体重下降的程度呈正相关,有15个个体肠型被改变了,体重变化范围从20公斤到+7公斤。这两个时间段都是一致的。这一发现表明,在饮食控制计划中,饮食和肠道菌群之间存在着持续的相互作用。如下图:
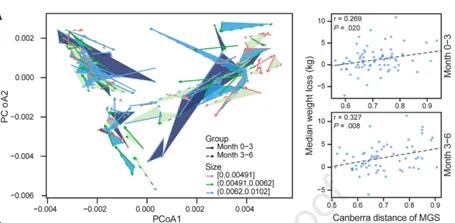
Jie Zhuye, et al., .Gastroenterology, 2021
这些结果表明可以基于基线肠道菌群组成和饮食数据建立个性化的体重预测模型。我们可以利用肠道菌群的分布做个性化的饮食推荐,以此调节体重,进而促进宿主健康。
此外,谷禾根据权威研究和检测大数据库,在菌群检测报告中给出了肥胖,便秘,失眠,过敏等症状相关菌(包括正相关,负相关菌,证据强度,菌的说明和异常菌的个性化干预调整措施)。
如下是谷禾菌群报告里肥胖相关菌的截图展示:
与肥胖症状相关的菌属
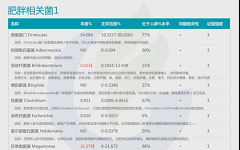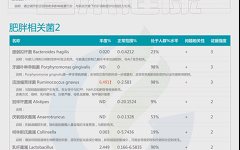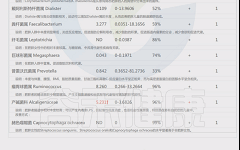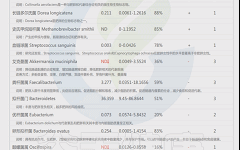
这些菌里,包括有益菌,核心菌,以及有害菌和致病菌等。部分菌在以往文章中详细介绍过:
★ 普雷沃氏菌——在摄入膳食纤维后对减脂更有利
普雷沃氏菌丰度高的健康超重成人在食用富含全谷类和纤维的随意饮食6周后,比普雷沃氏菌丰度低的受试者减脂更多。
普雷沃氏菌的高水平不仅与肥胖有关,且与非糖尿病患者的BMI指数、胰岛素抵抗、高血压和非酒精性脂肪性肝显著相关。
★ Blautia——治疗炎症肥胖相关的潜力菌
Blautia是肠道中常见的乙酸生产者,可通过激活G蛋白偶联受体 GPR41 和 GPR43 来抑制脂肪细胞中的胰岛素信号传导和脂肪积累,进而促进其他组织中未结合的脂质和葡萄糖的代谢,从而减轻肥胖相关疾病。
Blautia是有效减肥组女性肠道菌群中的优势菌属,但在减肥无效组中则不然。Blautia,特别是B. luti和B. wexlerae,可能有助于减少与肥胖相关的炎症。
肠道核心菌属——经黏液真杆菌属(Blautia),炎症肥胖相关的潜力菌
★ Bifidobacterium——减轻体重和减少体脂
双歧杆菌对糖尿病、肥胖症和高脂血症的有益作用也得到了研究,证据显示其对普通人群的血糖水平和胰岛素抵抗具有有益作用,同时还能降低孕妇妊娠糖尿病的发病率。
★ Phascolarctobacterium——帮助减肥
比较容易减肥的人体肠道内考拉杆菌属(Phascolarctobacterium)水平较高,因此该菌也被认为可以用来预测肥胖指标。在代谢综合征女性中观察到的Phascolarctobacterium属的丰度高于代谢综合征男性。
肠道核心菌属——考拉杆菌属(Phascolarctobacterium),与减肥相关?
★ Ruminococcus——含量过多与炎症和肥胖有关
瘤胃球菌(Ruminococcus)在新陈代谢中起着至关重要的作用。
一项研究包括肥胖组(BMI≥40kg/m2)和对照组(BMI18.5~ 24.9kg/m2之间)的粪便菌群,其中Ruminococcus bromii, Ruminococcus obeum 在肥胖患者中丰度较高。
瘤胃球菌喜欢植物中的多糖。如果肠道中有过多的瘤胃球菌,细胞可能会吸收更多的糖,导致体重增加。
★ Desulfovibrio——含量过高与肥胖相关
脱硫弧菌属(Desulfovibrio)里的一种能够引起炎症的细菌,当其含量过多时与肥胖相关。并在便秘型肠易激综合征患者,帕金森,系统性硬化症患者患者富集,可产生硫化氢对肠道上皮具有毒性,会导致人体腹泻。
★ 嗜胆菌属——喜欢脂肪、耐胆汁的促炎菌
嗜胆菌属(Bilophila)是变形菌门,脱硫弧菌科的一种厌氧、革兰氏阴性、耐胆汁,该菌是“喜欢动物脂肪喜欢胆汁”的微生物——在以动物为基础的饮食,尤其富含肉类和乳制品脂肪时,其肠道中Bilophila丰度会增加。
《Nature》杂志的一项研究发现,当人们从素食转变为以肉类和奶酪为主的饮食结构上时,他们肠道里的细菌Bilophila几乎立即增加,但植物性为主的饮食结构可以降低该菌群的数量。
Bilophila是机会致病菌,其丰度的增加与肠道炎症相关。其代表菌种Bilophila wadsworthia增加了高脂饮食诱导的代谢综合征,这是一种与低程度全身炎症相关的疾病,伴随着较高的体重指数。
肠道重要菌属——嗜胆菌属 (Bilophila)喜欢脂肪、耐胆汁的促炎菌
★ 脆弱拟杆菌——肥胖儿童中含量较高
宿主的生活方式和生理状态也会影响肠道脆弱拟杆菌的丰度。例如,缺乏运动可能会导致脆弱拟杆菌和其他拟杆菌属物种显著富集。
以往的研究表明,脆弱拟杆菌过多与肥胖呈正相关,肥胖儿童中脆弱拟杆菌的丰度高于瘦儿童。
扩展阅读:
★ 短链脂肪酸影响饱腹感、促进能量消耗
菌群关键代谢物短链脂肪酸(SCFA)可以说是研究最广泛的微生物代谢物,对人体代谢有许多影响。
短链脂肪酸(SCFA)在维持宿主健康和影响代谢方面发挥着广泛的作用。SCFA调控体内各种生理过程,包括维持结肠上皮和粘液水平的先天性肠道屏障的完整性、调节肠道蠕动以及控制重要肠道激素的分泌,如肽YY(PYY)、血清素、胃抑制肽和胰高血糖素样肽1(GLP-1) 。
短链脂肪酸参与L细胞产生的肽YY和胰高血糖素样肽1(GLP1)激素的调节。这两种激素都调节神经系统的饱腹感,GLP1在葡萄糖刺激的胰岛素敏感性和分泌中也起作用。
-乙酸盐
乙酸盐的健康影响还存在争议。一些研究将其与通过GPR41/43相互作用减少食欲、减轻体重以及增强胰岛素敏感性联系起来,而另一些研究则表明其作为肝脏和脂肪组织脂肪产生的底物,在促进肥胖方面发挥着作用。
-丙酸盐
丙酸可由拟杆菌属、考拉杆菌属(Phascolarctobacterium succinatutens)、戴阿利斯特杆菌属(Dialister)和韦荣氏球菌属通过琥珀酸途径产生;或是由埃氏巨球形菌属(Megasphaera elsdenii)、粪球菌属(Coprococcus catus)、沙门氏菌属(Salmonella spp.)、Roseburia inulinivorans和Ruminococcus obeum通过丙烯酸途径产生。
人体研究表明,丙酸具有整体抗肥胖作用,因为它可以增加餐后GLP-1和PYY水平,减少体重增加、腹部脂肪和肝细胞内脂质含量,并预防胰岛素敏感性问题。丙酸还通过减少中性粒细胞释放白细胞介素8(IL-8)和肿瘤坏死因子α(TNF-α)而表现出抗炎特性。
-丁酸盐
在胃肠道发酵产生的所有短链脂肪酸中,丁酸尤其值得注意。重要的产丁酸属和种有Coproccocus 属、Anaerostipes属、真杆菌属(Eubacterium)、普拉梭菌(Faecalibacterium prausnitzii)和罗氏菌属。
丁酸盐是成熟结肠细胞的主要能量来源,支持结肠健康,并且是一种具有强效抗炎特性的微生物代谢物,局部和系统性作用均有。此外,丁酸盐在调节局部和全身免疫、维持粘膜完整性和抑制细胞水平的肿瘤改变方面起着至关重要的作用。丁酸盐与丙酸盐类似,具有通过刺激食欲抑制激素的释放和促进瘦素的合成来减少肥胖的作用。
★ 胆汁酸影响脂肪代谢
许多研究报告了肠道微生物组、胆汁酸和肥胖或肥胖相关疾病之间存在联系。胆汁酸在肠道中与脂肪分子结合,形成复合物,促进脂肪的分解和吸收。适当的胆汁酸可以帮助提高脂肪代谢效率,减少脂肪在体内的堆积。
此外,胆汁酸在消化系统中可以影响胃液分泌,从而影响饱腹感和食欲调节。适当的胆汁酸水平可以帮助控制饥饿感,减少摄入的热量,有助于减轻体重。
★ 吲哚——抗肥胖特性
一项研究发现较高的血浆吲哚丙酸水平与降低患2型糖尿病的风险之间存在关联。
另一项研究发现,与瘦对照相比,患有2型糖尿病的肥胖受试者的吲哚丙酸水平降低。吲哚丙酸显示通过与孕烷X受体结合并随后下调肿瘤坏死因子α来调节炎症。
吲哚丙酸也被证明在小鼠中具有抗肥胖活性,微生物衍生的吲哚乙酸进一步限制了巨噬细胞中脂肪酸的积累和炎症标志物的产生。
★ 谷氨酸过量与肥胖潜在危害有关
谷氨酸是一种多功能氨基酸,谷氨酸在生物体内的蛋白质代谢过程中占重要地位。根据对肥胖和瘦受试者的队列进行的全基因组关联分析显示,谷氨酸盐具有潜在危害。
通过进行途径分析,谷氨酰胺/谷氨酸转运系统在肥胖个体中高度富集。这与拟杆菌属(包括B.thetaiotaomicron)的物种呈负相关。事实上,与瘦受试者相比,肥胖者体内这种细菌的数量减少。因此谷氨酸与人体肥胖之间也存在一定联系。
★ 肥胖人群的肠道微生物能够更多地获取能量
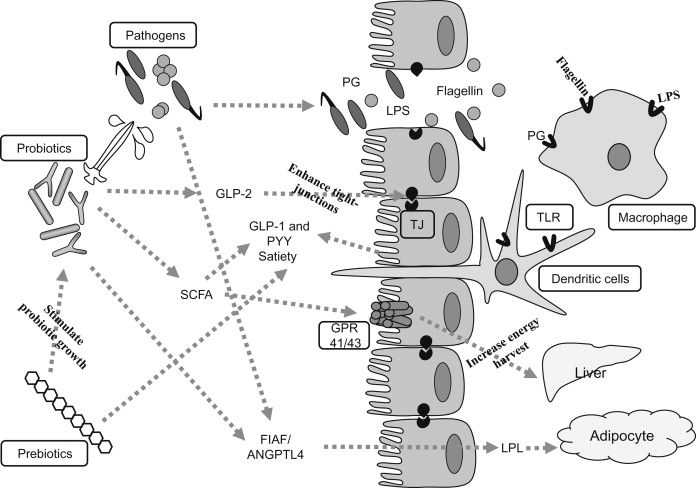
对肠道微生物群影响膳食能量收集和储存过程的探索揭示了两种关键机制:肥胖人群具有分解难以消化的膳食细菌多糖水解酶从而对多糖降解;以及抑制一种名为禁食诱导脂肪因子(FIAF)或血管生成素样4(ANGPTL4)的脂蛋白脂肪酶(LPL)抑制剂的肠道基因表达。
一项比较肥胖小鼠及瘦小同窝小鼠的功能性宏基因组的研究发现,肥胖微生物群富含八个能够水解膳食多糖的糖苷水解酶家族。第二种机制涉及微生物对FIAF的抑制,通过影响LPL活性导致脂肪堆积增加。
细菌多糖水解产生脂肪形成底物,即单糖和短链脂肪酸。肠道微生物群还通过增强钠/葡萄糖转运蛋白-1(SGLT1)的表达以及使小肠毛细血管密度加倍来促进有效的单糖吸收。这些脂肪形成底物到达肝脏后促进了肝脏甘油三酯的合成。
★ 肠道微生物群的组成影响从食物中获取的能量
“能量获取理论”最近也在人类受试者身上进行了测试。招募了12名瘦弱和9名肥胖的成年男性,在最初3天的体重维持饮食之后,以随机交叉的方式分配到2400或3400千卡/天的饮食,持续3天。
在初始体重维持饮食中,瘦人和肥胖个体的三大细菌门(厚壁菌门、拟杆菌门和放线菌门)存在显著差异。作为对改变的热量负荷的反应,肠道厚壁菌门增加、拟杆菌门减少与宿主能量吸收增加约150kcal有关。
瘦人似乎对增加热量摄入更为敏感,在2400vs. 3400 kcal/天饮食中,粪便能量损失和肠道微生物组成的变化均显著减少。而肥胖个体没有出现类似的变化。我们认为,与体重维持饮食相比,能量摄入的差异程度可能会通过肠道微生物群影响饮食中能量的吸收效率。
暴饮暴食是肥胖的主要诱因,是由调节食物摄入的过程失衡造成的,包括“饥饿”、“食欲”和“饱腹感”,以及环境因素。
★ 饱腹和饥饿信号影响人们的进食行为
正常的饮食行为在我们感到饱腹(饱腹感)时就会停止,并在感到饥饿一段时间后再次开始。两餐之间的时间是饱腹感的指标。有许多肠道激素会向大脑传递“饱腹”或“饥饿”的信号。肠内分泌L细胞分泌的胆囊收缩素(CCK)和胰高血糖素样肽1(GLP-1)是与饱腹感相关的肠道激素,可传递“饱腹”信号。
肠内分泌L细胞分泌的肽YY(PYY)和肠内分泌K细胞分泌的葡萄糖依赖性胰岛素促泌多肽(GIP)也传达“饱腹感”,而胃细胞分泌的生长素释放肽则传达决定开始进餐的“饥饿”信号。
★ 短链脂肪酸可以影响激素水平进而调节饱腹感
研究表明,肠道微生物群和其发酵产物短链脂肪酸可以影响肠道激素水平,进而调节饱腹感。肠内分泌L细胞分泌的GLP-1和PYY含有短链脂肪酸受体GPR41和GPR43,暗示结肠短链脂肪酸的生成与食欲或摄食量存在关联。
膳食纤维可选择性地支持肠道中有益细菌的生长和短链脂肪酸的产生。与饲喂对照饮食或高蛋白饮食的大鼠相比,饲喂高纤维饮食的大鼠血浆GLP-1和PYY水平较高,血浆GIP水平较低,结肠PYY mRNA水平增加5倍,胰高血糖素原mRNA水平增加11倍。
菊粉型果聚糖已被证明可以提高血浆GLP-1水平和结肠胰高血糖素原基因表达,保护免受高脂肪饮食引起的肥胖。一项人体研究表明,每天摄入21克低聚果糖(FOS)可降低超重成人的生长素释放肽并增加PYY,同时减少卡路里摄入量,有助于减肥。
★ 肠道微生物通过影响免疫也会导致贪食、肥胖
另一个影响食物摄入和肥胖的因素是肠道微生物群与先天免疫系统之间的相互作用。Toll样受体5(TLR5)是被广泛研究的受体之一。TLR5在小鼠肠粘膜中高表达,可以识别细菌鞭毛蛋白作为病原相关分子模式(PAMP),与肠道炎症和肠道微生物群的变化有关。
最新研究表明,TLR5基因敲除小鼠(T5KO)表现出贪食、肥胖、代谢综合征以及相关的高脂血症、高血压和胰岛素抵抗。T5KO小鼠的贪食/肥胖表型与肠道微生物群组成的细菌物种水平变化相关,T5KO小鼠有116种来自不同门类的细菌属丰富或减少。
将T5KO小鼠肠道微生物移植到健康小鼠中,导致健康小鼠发展代谢综合征,表明T5KO小鼠的肠道微生物影响代谢健康。
总的来说,肠道微生物群通过短链脂肪酸信号传导或与先天免疫系统相互作用,在调节食欲、肥胖和糖尿病方面发挥着重要作用。
因此,可发酵纤维和益生元等膳食成分以及肠道内生成的微生物代谢产物可以作为对抗这些疾病的简单而有效的手段。
肥胖与慢性、轻度全身性炎症相关。脂肪细胞和肥胖下的脂肪组织内的巨噬细胞都释放炎症细胞因子。这种轻度全身性炎症部分来源于细菌脂多糖(LPS)从肠道进入血液循环。
血浆中脂多糖浓度增加两到三倍,被称为“代谢性内毒血症”,与肥胖、胰岛素抵抗、糖尿病和动脉粥样硬化等多种慢性疾病相关。
★ 高脂饮食会加剧炎症状态
肠道脂多糖倾向于通过乳糜微粒携带,这些是脂蛋白颗粒,用于输送膳食脂质。因此,高脂饮食可能诱发或加剧代谢性内毒血症。
肠道通透性可能导致脂多糖位移,肠道微生态失调或引发肠道壁炎症可能增加通透性。双歧杆菌和乳酸杆菌的几种菌株及其代谢产物(如短链脂肪酸)已被证实能促进紧密连接蛋白的表达,这些蛋白维持有效的肠道屏障。而肠道中双歧杆菌相对缺乏与肠道通透性增加相关。
★ 肥胖状态下促炎巨噬细胞比例增加
引起脂肪组织炎症加剧的一个关键因素是巨噬细胞的渗入。产生促炎细胞因子的脂肪组织主要来自渗入的巨噬细胞。动物研究表明,巨噬细胞在肥胖中逐渐积累,瘦小鼠脂肪组织中不到10%的细胞是巨噬细胞,而肥胖小鼠中这一比例超过50%。
巨噬细胞本身表现出不同表型,可极化为促炎的“M1”或抗炎的“M2”表型。肥胖状态改变了M1和M2巨噬细胞的比例,导致M1巨噬细胞增加。
注:有趣的是,TLR4缺乏可减轻脂肪组织炎症,促进脂肪组织和腹膜巨噬细胞极化为M2型,但不会影响全身胰岛素敏感性。
另一项研究比较了无菌小鼠和大肠杆菌单个定植小鼠,结果显示单个定植小鼠脂肪组织中存在脂多糖依赖性巨噬细胞积累。大肠杆菌的定植还增加了巨噬细胞的极化,使其转变为促炎的M1型,并导致葡萄糖和胰岛素耐受性下降。
小结
肠道微生物组影响肥胖和相关代谢状态的三种机制,即通过能量获取、食欲调节和炎症状态,综合起来看,每条途径都是协同作用而非独立作用。这些调节过程的核心是宿主、微生物组和饮食之间复杂而动态的三向相互作用。
人体的消化过程和细菌作用在肠道中协同处理膳食成分。产生的代谢产物的特征取决于宿主遗传、宿主生理和肠道微生物的组成。微生物与营养消化产物结合,可以通过与宿主细胞受体的特定相互作用或以非特定方式进入一般血液循环,对宿主代谢产生不同的影响。
以下是来自君好美健康科技公司的3个案例,采取“膳食纤维+后生元”君好美膳食片作为主食,三餐吃饱——肉鱼蛋奶豆制品蔬菜充分吃,水果干果控制吃,在基本不改变在原有饮食情况下,一段时间后,个案的体重体脂明显下降、肠菌菌群得到有效改善。
案例一
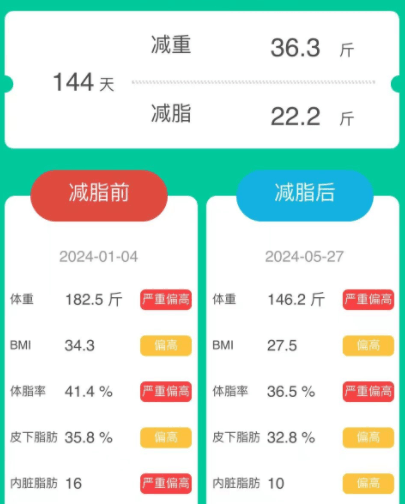
女,44岁,原本体重为182.5斤,属于严重偏胖。
经历144天的改善后,体重减轻了36.3斤,减脂了22.2斤,减肥效果非常明显。体脂率、皮下脂肪和内脏脂肪都有所减少。
身体数值的各项变化
编辑
改善前后的报告对比
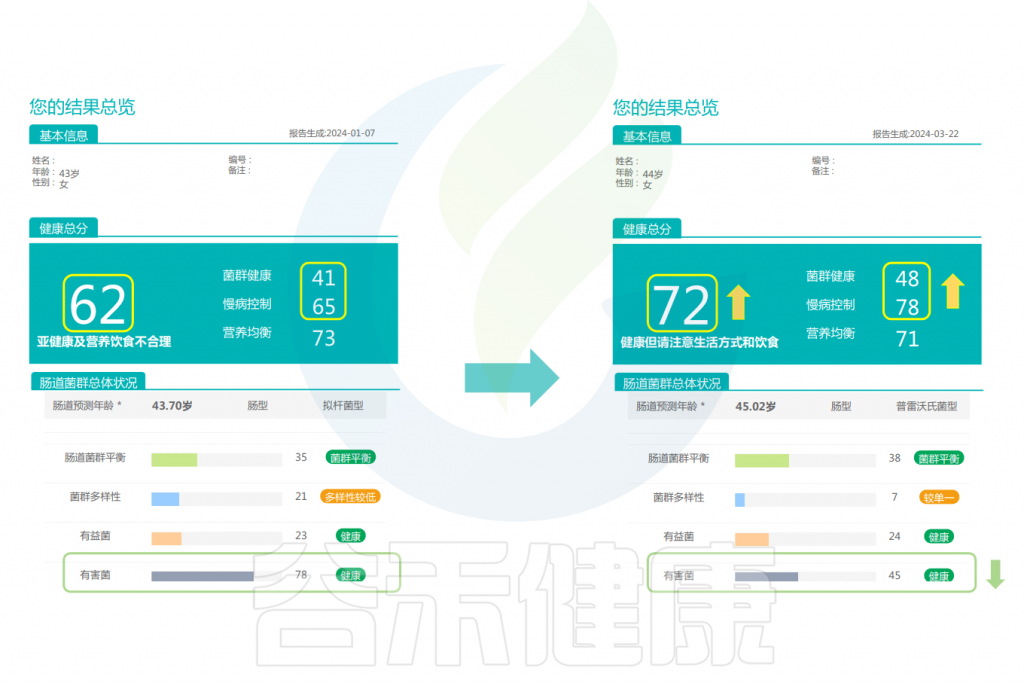
可以看到:在额外补充膳食纤维后,健康总分升高,肠道微生物的结构更健康,通过增加膳食纤维来增加微生物的数量和多样性是非常有效的。有益菌的分值提高,有害菌明显减少,肠型由原来拟杆菌型变成了普雷沃氏菌型,慢病风险总分下降。
▸ 具体到菌种

活泼瘤胃球菌与短链脂肪酸的产生有关。与非酒精性脂肪肝病和肥胖等疾病存在正相关。在改善后由原本的过高丰度,降低到了正常水平。
干预前:

干预后:

颤螺菌属也是与肥胖相关的菌群,过多或过少都可能影响肥胖。在改善后由原本的过低,丰度有所增加,恢复到了正常水平。
干预前:
干预后:

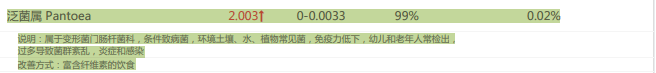
泛菌属是一种条件致病菌,过多导致菌群紊乱,炎症和感染。而在补充膳食纤维后丰度有所降低(尽管还有一点偏高)。
干预前:
干预后:
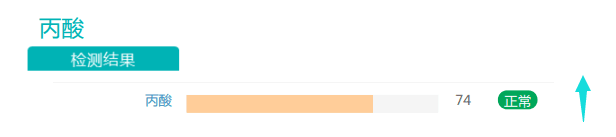
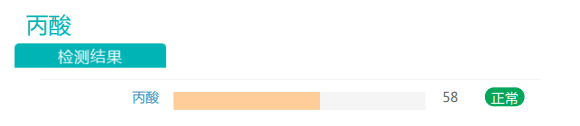
此外,丙酸盐的丰度也有一定程度的提高。有充分证据表明,随着膳食纤维摄入量的增加,产生的短链脂肪酸(SCFA)也增加。SCFA能够激活游离脂肪酸受体,从而促使厌食激素(如瘦素和肽YY)的分泌。在减肥中发挥益处。
根据以上这些变化,可以看出在额外补充膳食纤维后,可以在一定程度上改善人体的肥胖和代谢健康。
案例二
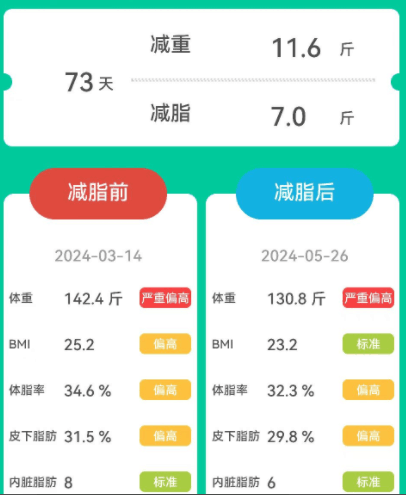
女,52岁,原本体重为142.4斤,属于严重偏胖。
经历73天的改善后,体重减轻了11.6斤,减脂了7斤,虽然没有上一个人减重多,但是BMI恢复到了正常水平。体脂率、皮下脂肪和内脏脂肪也均有所减少。
身体数值的各项变化
改善前后的报告对比
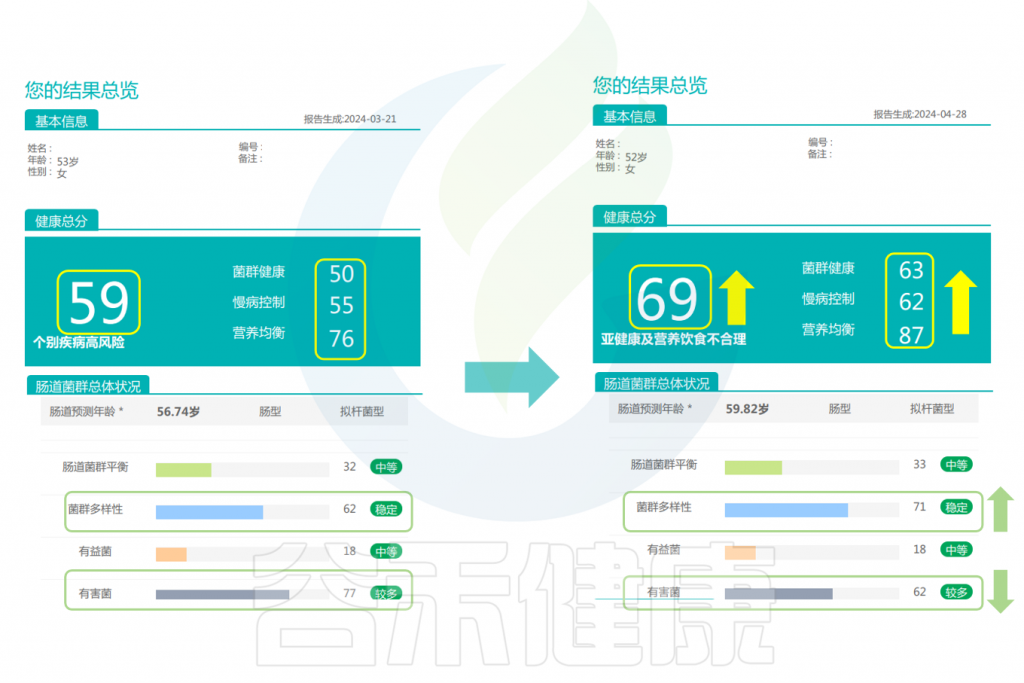
可以看到,该女性在经过干预后,健康总分也升高了。并且菌群变得更健康、对于慢病的控制评分更高,营养也更均衡了。肠道菌群多样性提高,有害菌明显减少。
与此同时,核心菌属也发生了一定程度的变化:
干预前:

干预后:
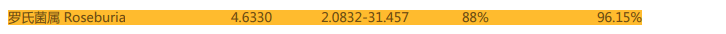
重要的肠道基石菌,产短链脂肪酸菌属罗氏菌在改善后由原来的丰度过低变成了正常丰度。罗氏菌具抗炎特性,有助于分解不可消化的碳水化合物如膳食纤维,对健康有利。
干预前:

干预后:

过多会导致菌群紊乱的副拟杆菌属由原本的过高丰度降低到了正常值。
肠道核心菌——副拟杆菌属(Parabacteroides),是否是改善代谢减轻炎症的黑马?
干预前:

干预后:

能够产生脂多糖,代谢生成苯酚,引发感染和并发症的梭杆菌属丰度由原来的过高水平恢复到了正常水平。
梭杆菌属Fusobacterium——共生菌、机会致病菌、致癌菌
干预前:

干预后:

过高时会导致肥胖和2型糖尿病的副萨特氏菌属在额外补充膳食纤维后丰度也有所下降(尽管仍然高于正常值)。
干预前:
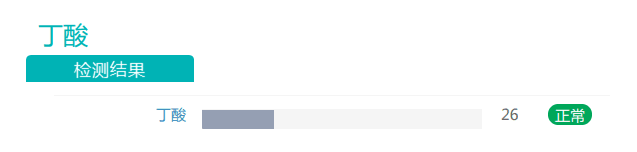
干预后:
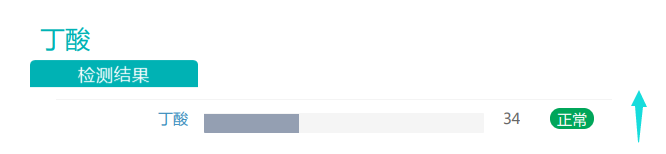
随着膳食纤维摄入量的增加,丁酸盐的丰度有所上升。丁酸对于维持肠道内环境温度预防结直肠癌发生具有重要作用,是肠上皮细胞最重要能量来源,对肠粘膜有营养作用,利于代谢健康的重要物质。
案例三
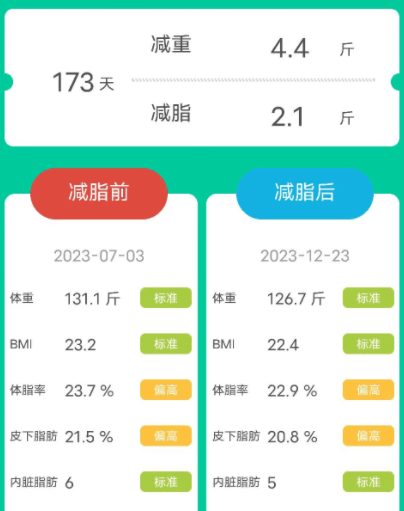
男,54岁,原本体重为131.1斤,属于标准体重,本来健康状态就比较好。因此在经历173天后,体重只下降了4.4斤,减脂2.1斤。体脂率、皮下脂肪、内脏脂肪也稍微有所下降,但都变得更健康了。
此外,三个案例可以看出膳食纤维对不同基础体重人群的作用效果不太一样,大体重人群在额外补充膳食纤维后减重更多。
身体数值的各项变化
改善前后的报告对比
<来源:谷禾肠道菌群健康检测数据库,下同>
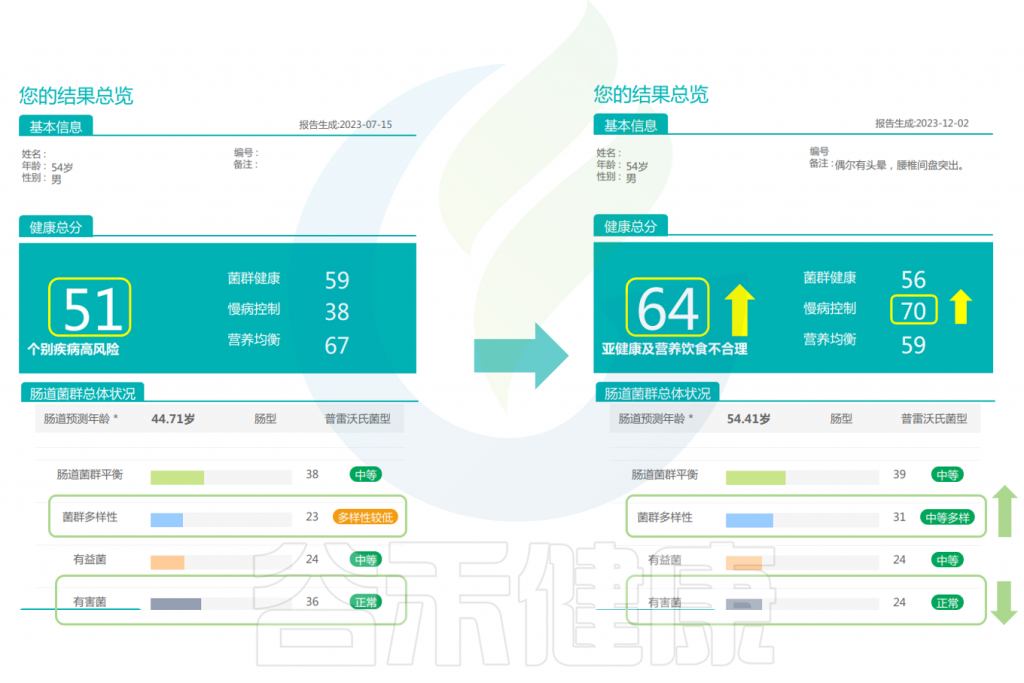
根据谷禾的健康报告前后对比可以看到,该男性在经过干预后,健康总分升高,对于慢病控制的评分显著升高。肠道微生物的多样性变得更丰富,有害菌减少明显。
干预前:

干预后:

过高时与肥胖相关,可能通过抑制乙酸水平来加速肥胖的脆弱拟杆菌在额外补充膳食纤维后丰度降低到了正常水平。
干预前:

干预后:
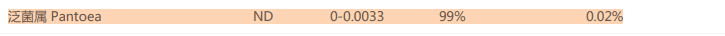
肠道的条件性致病菌泛菌属,过多导致菌群紊乱,炎症和感染。在补充额外的膳食纤维后水平也下降到正常值。
肠道微生物群多样性和高纤维摄入量与长期体重增加较低有关
此外一项对1632个人的研究也发现,微生物群多样性会影响膳食纤维与体重增加之间的关系。在微生物群多样性较高的人群中,纤维摄入量与体重增加风险降低有显著相关性。

膳食纤维对体重的有益作用可能在微生物组多样性较高的个体中更为明显。实验表明,纤维摄入会降低饮食的能量密度,由此产生的短链脂肪酸促进肠道糖异生、肠促胰岛素形成并随后产生饱腹感,同时短链脂肪酸也会向宿主输送能量并影响脂肪异生。
TIPs: 膳食纤维与益生元的区别
膳食纤维和益生元虽然都是植物性食物中的复杂碳水化合物,但它们之间存在一些区别:
1.膳食纤维的定义: 膳食纤维是指植物性食物中不被人体消化酶分解的碳水化合物,包括不溶性纤维和可溶性纤维。不溶性纤维,如纤维素,主要作用是增加大便体积,促进肠道蠕动,帮助消化系统健康。可溶性纤维,如果胶和树胶,可以溶解在水中,形成凝胶状物质,有助于降低血糖和胆固醇。
2.益生元的定义: 益生元是一种可溶于水的可溶性纤维,它作为益生菌的食物,可以被肠道中的有益细菌发酵,从而促进有益细菌的生长和活动。益生元主要包括低聚果糖(FOS)菊粉、低聚半乳糖(GOS)等。
3.作用机制: 膳食纤维的作用更广泛,包括促进肠道蠕动、帮助排便、降低血糖和胆固醇等。而益生元的主要作用是喂养和促进益生菌的生长,尤其是那些能够产生短链脂肪酸(如丁酸盐)的细菌,这些短链脂肪酸对肠道健康至关重要。
4.种类: 膳食纤维的种类很多,包括纤维素、半纤维素、果胶、树胶、抗性淀粉等。而益生元的种类相对较少,主要是一些特定的低聚糖和某些类型的多糖。
5.健康益处: 膳食纤维对整体消化系统健康有益,而益生元则更专注于通过促进益生菌的生长来改善肠道微生物群的平衡。
总的来说,膳食纤维是一个更广泛的概念,包括了益生元但它们的作用和重点略有不同。膳食纤维对消化系统的整体健康有益,而益生元则专门针对促进一些细菌的生长。
拓展:不同纤维对肠道菌群的影响

一项研究了不同谷物纤维(包括麦麸)对健康成人肠道菌群组成的影响。
-食用小麦纤维对肠道微生物的影响:
食用小麦纤维或麸皮纤维对肠道菌群有显著影响,以下门类的菌群均显著增加:
双歧杆菌(Bifidobacteria) ↑↑↑
乳酸杆菌(Lactobacillus) ↑↑↑
奇异菌属(Atopobium) ↑↑↑
肠球菌(Enterococci) ↑↑↑
梭状芽孢杆菌(Clostridia) ↑↑↑
毛螺菌科(Lachnospiraceae) ↑↑↑
埃格氏菌(Eggerthella) ↑↑↑
柯林斯菌(Collinsella) ↑↑↑
棒状杆菌(Corynebacterium) ↑↑↑
拟杆菌(Bacteroides) ↑↑↑
普氏菌属(Prevotella) ↑↑↑
-食用大麦纤维对肠道微生物的影响:
所有研究都表明大麦纤维对肠道菌群标志物有显著影响:要么改变微生物群落 ,要么改变发酵代谢物。发现厚壁菌门和放线菌(特别是罗氏菌属、Dialister、真杆菌和双歧杆菌)显著增加,拟杆菌减少。
其余3项研究测量了发酵标志物,结果显示,在食用大麦纤维后,总短链脂肪酸、丁酸盐和乙酸盐显著增加,呼气氢显著增加。由于发酵代谢物的积极作用,血糖反应同时改善。
-食用燕麦纤维对肠道微生物的影响:
燕麦粒的外层含有不溶性和可溶性(β-葡聚糖)纤维的混合物,这两种纤维都为肠道微生物群提供了食物来源。已证实可溶性燕麦β-葡聚糖有助于降低血液胆固醇水平。
对患有轻度高血糖症或高胆固醇血症的参与者研究了每天早餐食用全麦燕麦片与精制谷物片的影响。据报道,食用全麦燕麦片后,粪便总细菌、乳酸杆菌和双歧杆菌数量显著增加,而食用精制谷物片后,总细菌数量和双歧杆菌数量均下降。
-食用玉米纤维对肠道微生物的影响:
玉米的淀粉含量相对于其他谷物来说较高,麸皮含量较低。
一项急性干预研究比较了单次食用48克全麦玉米早餐麦片(14.2克纤维)对肠道菌群的影响,并与48克低纤维玉米早餐麦片(0.8克纤维)进行了比较。
3周后,高纤维组和低纤维组均报告粪便双歧杆菌增加,高纤维组的增幅更大,但未达到显著性,乳酸杆菌、肠球菌和奇异菌属物种的增加不显著。
-混合全谷物对肠道微生物的影响:
全麦谷物包括谷物的胚乳、胚芽和麸皮成分,因此其营养成分与谷物的麸皮纤维部分不同,这可能会影响肠道微生物群。
共计357人的研究发现,增加完整谷物纤维的摄入具有显著的益生元作用,细菌多样性、放线菌、双歧杆菌、梭菌、毛螺菌显著增加;但阿克曼氏菌、罗氏菌、乳酸杆菌和肠球菌的增加趋势不显著。促炎性肠杆菌科细菌的水平也显著下降。
作者认为,对高纤维干预的反应取决于基线肠道微生物丰富度——由于膳食纤维的增加,基线微生物丰富度有限的人会随着时间的推移表现出更大的微生物群变化。
如今全球肥胖症的流行,寻求新的有效解决方案已成为研究的重中之重。这主要是因为大多数人无法长期坚持既定的饮食和身体活动方案,从而无法达到并保持健康的体重。
肠道微生物群由于位于宿主营养/能量代谢的关键位置并能够影响它,因此已成为一种有希望的新治疗靶点。
饮食、肠道微生物群和肥胖之间的相互关系是高度互动和动态的。虽然饮食热量摄入是能量平衡方程的主要因素,研究估计现在知道某些肠道微生物可以从饮食中获取“额外”的能量,大约150千卡/天;相当于一年内可能增加6.8公斤的体重。
除了热量含量外,饮食成分也是与肥胖相关的饮食-微生物相互作用的重要因素,例如肠道微生物失调以及高脂饮食引起的全身炎症。
肠道微生物还通过调节食欲、食物摄入和饱腹感来影响肥胖:这是能量平衡的一部分。膳食纤维、合生元是调节肠道微生物组成的有效方法,这些干预措施不仅降低了体重,体脂,而且同时改善了与肥胖和相关代谢状况相关的生物标志物,例如血糖水平、胰岛素敏感性、血浆脂联素等。
总体而言,这些结果表明,通过膳食纤维,合生元以及饮食调节肠道微生物群可以作为人类减肥的独立方法,同时可以有助于减轻风险并控制肥胖相关疾病(如心血管疾病、胰岛素抵抗和2型糖尿病)的症状。
在当前肥胖流行和健康挑战的背景下,个体化肠道菌群干预显得尤为重要。每个人的肠道微生物群独一无二,受基因、生活方式和环境等多方面影响。因此,针对个体的肠道菌群进行精准干预,可以更有效地调节体重、改善健康状况。
主要参考文献
Li H, Zhang L, Li J, Wu Q, Qian L, He J, Ni Y, Kovatcheva-Datchary P, Yuan R, Liu S, Shen L, Zhang M, Sheng B, Li P, Kang K, Wu L, Fang Q, Long X, Wang X, Li Y, Ye Y, Ye J, Bao Y, Zhao Y, Xu G, Liu X, Panagiotou G, Xu A, Jia W. Resistant starch intake facilitates weight loss in humans by reshaping the gut microbiota. Nat Metab. 2024 Mar;6(3):578-597.
Jie Zhuye,Yu Xinlei,Liu Yinghua et al. The Baseline Gut Microbiota Directs Dieting-Induced Weight Loss Trajectories.[J] .Gastroenterology, 2021.
Pham Van T,Fehlbaum Sophie,Seifert Nicole et al. Effects of colon-targeted vitamins on the composition and metabolic activity of the human gut microbiome- a pilot study.[J] .Gut Microbes, 2021, 13: 1-20.
Corica D, Aversa T, Valenzise M, Messina MF, Alibrandi A, De Luca F, Wasniewska M. Does Family History of Obesity, Cardiovascular, and Metabolic Diseases Influence Onset and Severity of Childhood Obesity? Front Endocrinol (Lausanne). 2018 May 2;9:187.
Bartsch M, Hahn A, Berkemeyer S. Bridging the Gap from Enterotypes to Personalized Dietary Recommendations: A Metabolomics Perspective on Microbiome Research. Metabolites. 2023 Dec 2;13(12):1182.
Zhang M, Liu J, Li C, Gao J, Xu C, Wu X, Xu T, Cui C, Wei H, Peng J, Zheng R. Functional Fiber Reduces Mice Obesity by Regulating Intestinal Microbiota. Nutrients. 2022 Jun 28;14(13):2676.
Menni C, Jackson MA, Pallister T, Steves CJ, Spector TD, Valdes AM. Gut microbiome diversity and high-fibre intake are related to lower long-term weight gain. Int J Obes (Lond). 2017 Jul;41(7):1099-1105.
Mayengbam S, Lambert JE, Parnell JA, Tunnicliffe JM, Nicolucci AC, Han J, Sturzenegger T, Shearer J, Mickiewicz B, Vogel HJ, Madsen KL, Reimer RA. Impact of dietary fiber supplementation on modulating microbiota-host-metabolic axes in obesity. J Nutr Biochem. 2019 Feb;64:228-236.
Jefferson A, Adolphus K. The Effects of Intact Cereal Grain Fibers, Including Wheat Bran on the Gut Microbiota Composition of Healthy Adults: A Systematic Review. Front Nutr. 2019 Mar 29;6:33.
Zhang, Y.; Liu, J.; Yao, J.; Ji, G.; Qian, L.; Wang, J.; Zhang, G.; Tian, J.; Nie, Y.; Zhang, Y.E.; et al. Obesity: Pathophysiology and Intervention. Nutrients 2014, 6, 5153-5183.
Lin X, Li H. Obesity: Epidemiology, Pathophysiology, and Therapeutics. Front Endocrinol (Lausanne). 2021 Sep 6;12:706978.Lin X, Li H. Obesity: Epidemiology, Pathophysiology, and Therapeutics. Front Endocrinol (Lausanne). 2021 Sep 6;12:706978. Pathophysiology, and Therapeutics. Front Endocrinol (Lausanne). 2021 Sep 6;12:706978

谷禾健康

膀胱癌主要是老年人的疾病,约75%的新诊断发生在65岁以上的患者,约45%发生在75岁以上的患者。膀胱癌对男性和女性的影响比例为3:1,但随着年龄的增长,男性和女性的发病率都显著增加。
年龄增长与膀胱癌的发病率和死亡率升高有关。高龄还与慢性炎症标志物升高以及肠道和尿液微生物群紊乱有关。
老年人膀胱癌发病率和死亡率增加的一个原因可能是,微生物组中与年龄相关各种变化,引起全身代谢变化,导致免疫失调,具有潜在的致癌作用。
膀胱癌患者的肠道和尿道微生物群可能失调,尽管这些变化的影响尚不清楚。
提高对免疫系统、肠道和尿液微生物群的年龄相关改变的理解,可能有助于深入了解老年人膀胱癌发生和发展的风险。在膀胱癌患者中,提高对微生物群的认识也可能为治疗干预提供潜在的靶点。
目前的数据支持炎症、免疫和微生物失调可能是已知的与衰老相关的膀胱癌发病率增加的原因,但需要在这一领域开展工作,以直接检验这一假设,以及更好地了解肠道中微生物成分的组成,膀胱癌期间膀胱和膀胱肿瘤微生物组及其对免疫系统的影响。
从这个角度出发,本文讨论了炎症、免疫、衰老和癌症之间的复杂联系。研究了表明老龄化和炎症可促进膀胱癌发展的数据,老龄化如何导致或促进多个区域的菌群失调,以及菌群失调如何影响对膀胱癌治疗的反应。
本文主要从以下几个方面讲述
本文出现的专业名词缩写
非肌肉浸润性膀胱癌——NMIBC
肌肉浸润性膀胱癌——MIBC
肿瘤坏死因子——TNF
损伤相关的分子模式——DAMPs
病原相关的分子模式——MAMPs
血清中性粒细胞:淋巴细胞比值——NLR
免疫组织化学——IHC
肠血管屏障——GVB
高龄不仅与患膀胱癌的风险增加有关,还与发生侵袭性肿瘤有关,这些肿瘤复发和发展为侵袭性疾病的可能性增加。
这些数据很复杂,因为“年轻”和“老年”的定义在研究中高度可变,可以作为连续或分类报告变量。此外,癌症特异性生存率增加背后的原因可能是多因素的,但仍然包括这样的事实:年轻患者通常比老年患者的肿瘤侵袭性更小、级别更低。
✦ 非肌肉浸润性膀胱癌:年轻患者肿瘤侵袭性更小,级别低,不易复发
年轻患者,如年龄小于30岁的患者,其乳头状、低级别Ta、非肌肉浸润性膀胱癌(NMIBC)的发病率高于年龄大于30岁的患者,其肿瘤通常较小、单发且不太可能复发。
尽管数据表明年轻患者和老年患者的肿瘤分期相似,即使按年龄<30岁或>30岁分组。膀胱癌基因表达数据的荟萃分析结果表明,在多个肿瘤亚型中,年轻化与激活成纤维细胞生长因子受体(FGFR)突变率增加相关,这一发现与Ta肿瘤相关,Ta肿瘤比高级别肿瘤具有更有利的预后。
在晚期膀胱癌中,与FGFR激活突变相关的有利结果丢失,因为FGFR3在化疗背景下可能与较差的结果相关。
在对肿瘤亚型的同一荟萃分析中,与老年患者相比,年轻患者的肿瘤中claudin的表达水平比老年患者高,claudin是一种肿瘤转移的反向标记物。
✦ 肌肉浸润性膀胱癌:发病率随年龄增长而增加
相比之下,在普通人群中,肌肉浸润性膀胱癌(MIBC)的发病率随着年龄的增长而增加。85岁以上的患者中有30%以上发生MIBC,而10岁以下的患者中有23%发生MIBC。
随着年龄的增长,肿瘤抑制因子TP53、APC和PTEN12突变的可能性增加,这在MIBC中很常见,并与不良预后相关。
最后,随着人类年龄的增长,他们通常会累积共病和虚弱指数增加,这导致了与年轻患者的总体预后差异,特别是与使用根治性膀胱切除术的手术干预有关。
有证据表明,当仔细选择时,许多老年患者是手术治疗的合适人选,但老年患者较不积极的治疗也会影响预后。
所有这些数据都广泛支持这样一种观点,即年轻患者的膀胱癌出现在疾病的较低阶段,临床上侵袭性较低,最终,年轻患者比他们的老年对手更有可能存活。
膀胱癌中与年龄和微生物组相关的变化
在衰老过程中,炎症系统和肠道和尿道中的微生物都发生了大量的变化。
✦ 膀胱癌
•老年患者的膀胱癌预后比年轻患者差,年轻患者通常表现为低级肿瘤。
•在膀胱癌亚型中,年轻患者比老年患者表达FGFR3突变的比例更高,连接蛋白水平也更高
•在膀胱癌亚型中,老年患者更有可能发生肿瘤,肿瘤抑制蛋白p53, APC和PTEN突变
•在24个月的随访点,老年患者(80岁以上)的非肌肉浸润性膀胱癌患者对卡介苗的反应低于年轻患者(61-70岁) (39%对61%的绝对反应率)
✦ 肠道微生物组
• 老年个体(90岁以上)β-多样性较年轻个体减少,α-多样性没有变化
• 老年个体(90岁以上)与年轻个体相比肠道中以下菌群丰度增加:
Akkermansia ↑↑↑
Escherichia ↑↑↑
Bifidobacterium ↑↑↑
Christensenellaceae ↑↑↑
• 老年个体(90岁以上)与年轻个体相比肠道中以下菌群丰度降低:
Faecalibacterium ↓↓↓
Bacteroides ↓↓↓
Lachnospiracae ↓↓↓
• 百岁老人(100岁)比非百岁老人群体有的Akkermansia,通常与改善健康状况有关。
关于AKK菌详见:AKK菌——下一代有益菌
✦ 尿液微生物群
• 70岁及以上的个体与年轻个体相比尿道中:
saccharofermentans ↑↑↑
Proteiniphilum ↑↑↑
Jonquetella ↑↑↑
Parvimonas ↑↑↑
• 70岁及以上的男性比年轻男性有的细菌属
• 70岁及以上的男性比年轻男性体内的细菌总数要少
• 与年轻女性相比,70岁以上的女性在细菌种类或数量上没有差异
✦ 肠道上皮屏障
• 与4或12月龄小鼠相比,21月龄小鼠血清脂多糖增加与肠道通透性增加相关
• 在老年小鼠模型中,IL-6和肿瘤坏死因子随肠道通透性线性增加
• 与10岁及以下的狒狒相比,18岁及以上的狒狒IL-6、IL-1β和IFNγ水平显著升高
• 18岁及以上的狒狒与10岁及以下的狒狒相比,occludin、claudin和紧密连接分子减少
• 与年轻人相比,老年人的短链脂肪酸(如丁酸、丙酸、乙酸)水平较低,这些脂肪酸被认为可以保持粪便中的肠道-上皮完整性
先天和适应性免疫系统在衰老过程中都出现失调。
衰老主要通过两种途径导致免疫系统失调事件:
在膀胱癌患者中,先天免疫活性的血清和组织标志物升高预后较差,可能与肿瘤生长有关。
适应性免疫系统在膀胱癌中的作用尚不清楚,基于所研究免疫细胞的差异、其相对丰度和抗原肿瘤标记物的存在,结果相互矛盾。
先天免疫系统和适应性免疫系统都可能影响膀胱癌的进展,与衰老相关的免疫系统变化会对肿瘤清除产生有害影响。
衰老:免疫系统逐渐破坏、炎症增加
随着年龄的增长,细胞分解的无菌产物的积累,包括代谢废物,如细胞游离DNA或错误折叠或氧化的蛋白质,以及异常分布的病原相关的分子模式(MAMPs),如脂多糖(LPS),长期激活先天免疫系统。
当衰老伴随着其他对人类健康有害的问题,如营养过剩、慢性惰性感染、癌症或肠道失调时损伤相关的分子模式(DAMPs)和错误折叠蛋白的积累会加速,并导致免疫系统中与衰老相关的破坏。
这种破坏至少部分是由巨噬细胞和中性粒细胞中的细胞信号传导介导的:巨噬细胞上进化保守的toll样受体(TLRs)识别升高的内源性MAMPs和DAMPs。
在衰老过程中,MAMPs和DAMPs在组织中积累,检测它们诱导IL-6、IL-8和tnf等细胞因子的表达,导致先天免疫活性升高,并从常驻免疫细胞向组织释放活性氧(ROS)。
临床前癌症模型支持这一观点,即局部炎症的增加会导致细胞因子的产生和导致DNA损伤的局部活性氧水平的增加,并可能会在某些肿瘤中刺激肿瘤生长和进展。
异常的DNA修复和增强的炎症也与老年小鼠中与肿瘤形成增加相关的DNA损伤有关。
先天免疫信号升高——膀胱癌不良预后
临床上,先天免疫信号升高似乎与NMIBC和MIBC患者的不良预后相关。这种作用不仅限于膀胱癌,因为肿瘤相关的巨噬细胞(TAMs)和肿瘤浸润性中性粒细胞(TINs)可能促进结直肠癌和肺癌的发生。
在膀胱癌中,使用免疫组织化学(IHC)对296个肿瘤的泛巨噬细胞标志物CD68和泛T细胞标志物CD3的组织芯片分析表明,巨噬细胞与T细胞的比例是不良生存的最大预测因子。
CD163+TAMs的存在:复发率增加
一项对13项来自人类膀胱组织的meta分析结果显示,CD163+TAMs的存在与复发率增加相关,而CD68+TAMs的存在与复发率无关。
值得注意的是,这一发现可能与这些巨噬细胞的炎症状态有关,因为CD163是抗炎巨噬细胞的标志,与肿瘤免疫逃逸相关。相比之下,CD68并不能描述巨噬细胞的炎症状态,并由大多数巨噬细胞亚型表达。
TINs水平升高:预后不良,生存率较差
对102个人膀胱癌肿瘤的免疫组化分析表明,TINs水平的升高与NMIBC复发率的升高和较差的总生存率呈正相关。
此外,在MIBC患者中,TIN水平升高与总生存期降低和化疗反应不良相关。然而,所有这些研究都使用了免疫组化相关分析,限制了推断因果关系的程度。
由于肿瘤浸润免疫细胞的测量需要在组织中进行评估,因此确定系统性炎症标志物是否可以作为组织相关炎症蛋白或细胞的替代物是有用的。
NLR高分与无复发和无进展生存期相关
系统性炎症的一个指标,血清中性粒细胞:淋巴细胞比率(NLR),可以预测NMIBC和MIBC的疾病复发和进展。
在MIBC中,两项回顾性研究结果显示,NLR评分大于3.0和2.5的患者5年无复发较差(HR 1.49;95% CI 1.12-2.0),总体(危险比1.67;95% CI 1.17-2.39)和无病生存期(危险比1.946;95% CI 1.03-3.66)高于NLR评分较低的患者。
在NMIBC中,对6项回顾性研究的荟萃分析结果显示,NLR高分与无复发和无进展生存期相关。
在一项对899例NLR分界点为2.7的MIBC患者的大型回顾性研究中,年龄的增加与NLR评分的增加相关,年龄的增加与免疫功能障碍的增加有关。
免疫变化在 NMIBC 中任何年龄都可能很重要
NMIBC的研究结果大部分显示膀胱癌患者的年龄与NLR评分之间没有显著的相关性,但仍显示NLR可以预测结果,这表明在NMIBC中,无论年龄如何,免疫改变可能都是重要的,年龄、NLR和预后之间的联系在MIBC中比在NMIBC中更强。
一项针对NMIBC患者的单一前瞻性研究发现,校正年龄后,NLR与无复发和无进展生存期之间的差异消失,这表明年龄是主要决定因素,应与NLR状态一起测量和校正,这进一步复杂化了高龄、NLR和膀胱癌预后之间的潜在关系。
当考虑到先天免疫系统在肿瘤微环境中的潜在致瘤作用时,显然,需要进一步的研究来确定膀胱局部先天免疫信号是否与系统性免疫失调相关,这可以很容易地测量和潜在地用于预后。
年龄、NLR和预后在MIBC中的相关性似乎比NMIBC更强,这并不奇怪,因为MIBC由于侵袭组织而被认为是更具有炎症性的疾病。
先天免疫调节膀胱癌:活性氧产生,增强致癌性
来自小鼠模型的数据支持先天免疫调节膀胱癌的观点。在膀胱癌模型中,与仅用亚硝胺(BBN)治疗的小鼠相比,致癌物n-丁基-n(4-羟基丁基)亚硝胺(BBN)和膀胱内脂多糖治疗的小鼠体内活性氧浓度与肿瘤发生增加呈正相关。
脂多糖注射促进了大鼠模型膀胱癌的形成,支持了先天免疫激活增加有助于肿瘤发展的观点。
脂多糖激活的中性粒细胞增强大鼠尿路上皮细胞系的恶性转化,这与脂多糖在其他肿瘤中的致瘤作用的数据一致,表明先天免疫系统产生的活性氧可以通过氧化应激和基因突变促进肿瘤的形成。
在缺乏铜/锌超氧化物歧化酶的老年小鼠中进行的研究结果表明,与年龄匹配的对照组相比,这些小鼠在肝癌模型中活性氧水平和致癌性增加,这支持了先天免疫系统不受控制的活性氧产生可增强致癌性的观点。
先天免疫系统与免疫疗法的复杂作用
在治疗上,炎症抑制剂,如塞来昔布,可以阻断促炎前列腺素的形成,减少大鼠膀胱癌模型中的肿瘤数量和体积,并减少体外细胞系中的细胞增殖。
在膀胱癌小鼠模型中,先天免疫系统似乎也调节了卡介苗的反应;在膀胱癌小鼠原位模型中,中性粒细胞减少消除了卡介苗介导的免疫治疗的反应。
这些数据表明,先天免疫系统的作用可能是复杂的,因为它可能需要通过启动免疫反应对卡介苗免疫疗法作出适当反应,但如果不加以检查,也会促进癌症的进展,这取决于肿瘤微环境和肿瘤的状态。
值得注意的是,这种效应在本研究中归因于中性粒细胞,但这一结论是在发现用于消耗中性粒细胞的抗体也会消耗单核细胞之前得出的,因为该抗体识别单核细胞上的LY6C和中性粒细胞上的LY6C和LY6G。
因此,单核细胞也可能影响模型中免疫治疗的疗效,突出了改善治疗结果的另一个潜在调控靶点。事实上,区分中性粒细胞和单核细胞在膀胱癌中的作用可能是增强患者免疫治疗和了解疾病进展的一种手段。
总之,这些数据支持这样一种观点:
在多种实体肿瘤类型中,与衰老相关的异常先天免疫导致了致瘤前表型。
然而,目前还缺乏利用解决潜在机制的转化模型来理解浸润肿瘤的先天免疫细胞在膀胱癌中的作用。这些模型是有根据的,因为阻断中性粒细胞募集也可以减少肿瘤的生长和其他肿瘤的转移进展,如乳腺、结肠和肝脏。
通过使先天免疫系统的慢性激活正常化,减少老年先天免疫过度刺激的新方法可能是减缓膀胱肿瘤发病率和复发的潜在手段,特别是在炎症指数自然增加的老年患者中。
先天免疫反应可以在老年时被长期激活,但适应性免疫的组成部分,如初始CD4和CD8 T细胞,由于免疫衰老而数量下降,这是由细胞衰老引起的免疫细胞随时间的变化,整体多样性下降。
随着年龄增长,老年人适应性免疫反应受损,肿瘤的感知和清除能力下降
随着个体年龄的增长,他们的初始T细胞逐渐减少,T细胞受体多样性减少,T细胞上的共刺激效应分子CD28的水平也比年轻的T细胞上的CD28低。这些变化损害了老年人的适应性免疫反应。
随着适应性免疫反应的下降,肿瘤的感知和清除能力也在下降。这些改变在很大程度上导致了观察到的免疫差异。
将适应性免疫细胞蛋白(如CD3和CD8)和肿瘤浸润淋巴细胞(TILs)浓度与临床结果相关联的尝试产生了相互矛盾的结果。
免疫组化分析显示,肿瘤突变负担增加的膀胱肿瘤CD8效应因子评分相应增加,T细胞浸润增强。对膀胱切除术后MIBC肿瘤的研究结果,支持CD3+或CD8+ TILs的增加与生存率的提高相关的观点。
MIBC中TILs的存在是否受年龄的影响尚不清楚。在一项对>50岁患者的MIBC膀胱切除术样本的研究中,没有发现患者的年龄、性别或T细胞标记CD4、CD8或CD56存在相关性。
然而,从更广泛的免疫衰老和衰老意义上来说,所使用队列中的患者的年龄范围相当有限,并没有在特定年龄组之间进行比较。
在更有针对性的比较队列(<40岁和>80岁)中,需要了解免疫衰老的变化如何影响免疫治疗,以提高对膀胱癌中衰老对适应性免疫的影响的理解。
衰老与基础先天免疫活动的增加有关,数据表明,在人类组织和小鼠模型中,先天免疫细胞(如中性粒细胞)的增加是肿瘤形成的促进剂。
然而,尽管T细胞在肿瘤免疫中的非常重要,但是适应性免疫反应在衰老和肿瘤发生中的作用仍不清楚。还需要确定炎症如何促进膀胱癌的决定性实验。
肠道微生物群
人体的肠道微生物组估计有超过1000种不同的细菌和100万亿个单独的细菌细胞。
这些不同菌群的代谢产物通过一层肠细胞、肠道粘液和微折叠免疫细胞与全身循环分离。肠道微生物的产物被腔细胞摄取后,运输到全身,对全身免疫系统产生广泛影响。
由肠道菌群组成的变化引起的宿主和肠道菌群间稳态的破坏,是菌群失调,会导致炎症、肿瘤进展和肿瘤对乳腺癌、肺癌和结直肠癌治疗的反应。
菌群失调如何影响免疫
菌群失调破坏肠道屏障,导致释放促炎症的MAMPs,如脂多糖和未甲基化的CpG DNA,加速年龄相关的免疫系统失调。
肠道物质的泄漏也发生在肠道血管屏障(GVB)。对肝脏疾病的研究表明,酒精性和非酒精性脂肪性肝炎等情况可破坏肠血管屏障,导致微生物物种的全身传播。
我们知道,衰老会导致上皮完整性屏障的破坏,因此,除了肠道-上皮完整性的丧失外,肠血管屏障的完整性的丧失可能是肠道功能障碍中微生物或微生物组分系统性传播的一个尚未被充分研究的方面。据推测,这些物质的泄漏会以对人体健康有害的方式广泛影响免疫力。
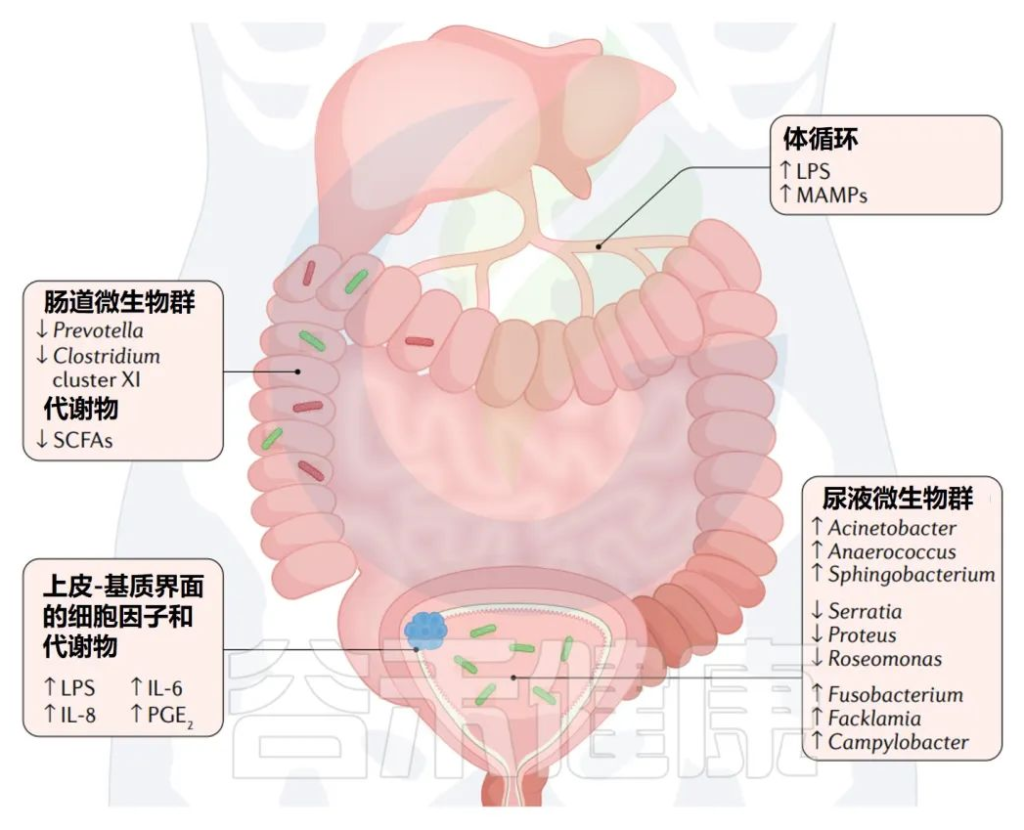
膀胱癌中的微生物群及其的潜在联系
Martin A,et al.Nat Rev Urol.2022
多种微生物群可能会影响或被膀胱癌的形成所改变,包括肠道微生物群的改变、尿液微生物群的改变和上皮基质界面的改变。
与健康人相比,膀胱癌肠道微生物群的变化包括Prevotella和Clostridium的丰度降低。
膀胱癌患者尿微生物组中以下菌群的丰度高于健康人群:
同时以下三个菌群的丰度较健康人群偏低:
肠道失衡与结直肠癌之间的正相关已经得到了很好的证实,但是肠道失衡在多大程度上影响其他肿瘤的进展,包括膀胱癌,还不清楚。
此外,肠上皮完整性的破坏在多大程度上是由肠道菌群的非共生变化驱动的,反之亦然,这一点也存在争论。
膀胱癌患者:普雷沃氏菌、梭菌属减少,丁酸水平降低
一项研究包括26名膀胱癌患者和16名年龄匹配的健康参与者通过定量PCR分析证明,普雷沃氏菌和梭状菌群显著减少 。
在同一研究中,膀胱癌患者粪便中的丁酸盐(一种短链脂肪酸)水平降低,而血清中的脂多糖水平升高。鉴于丁盐酸的抗肿瘤作用以及脂多糖和其他与肠道功能障碍相关的MAMPs的促肿瘤作用,这些数据表明微生物群的变化可能是疾病发生的相关因素。
迄今为止只进行了这种单一的、小规模的试验。这些发现是否代表肠道失衡、炎症和膀胱癌发生之间的潜在因果联系,或仅是相关的,还需要进一步的研究来确定。
需要在临床前模型中采用直接介入方法来了解短链脂肪酸在肠道失衡和膀胱癌中的作用,更重要的是,需要进行更大规模的试验,在膀胱癌患者的分子水平上调查肠道功能障碍,以验证任何发现。
需要进一步调查来确定,这些发现是否代表肠道菌群失调、炎症和膀胱癌肿瘤发生之间的潜在因果关系。
年龄相关的失调机制和对系统性炎症标志物的影响
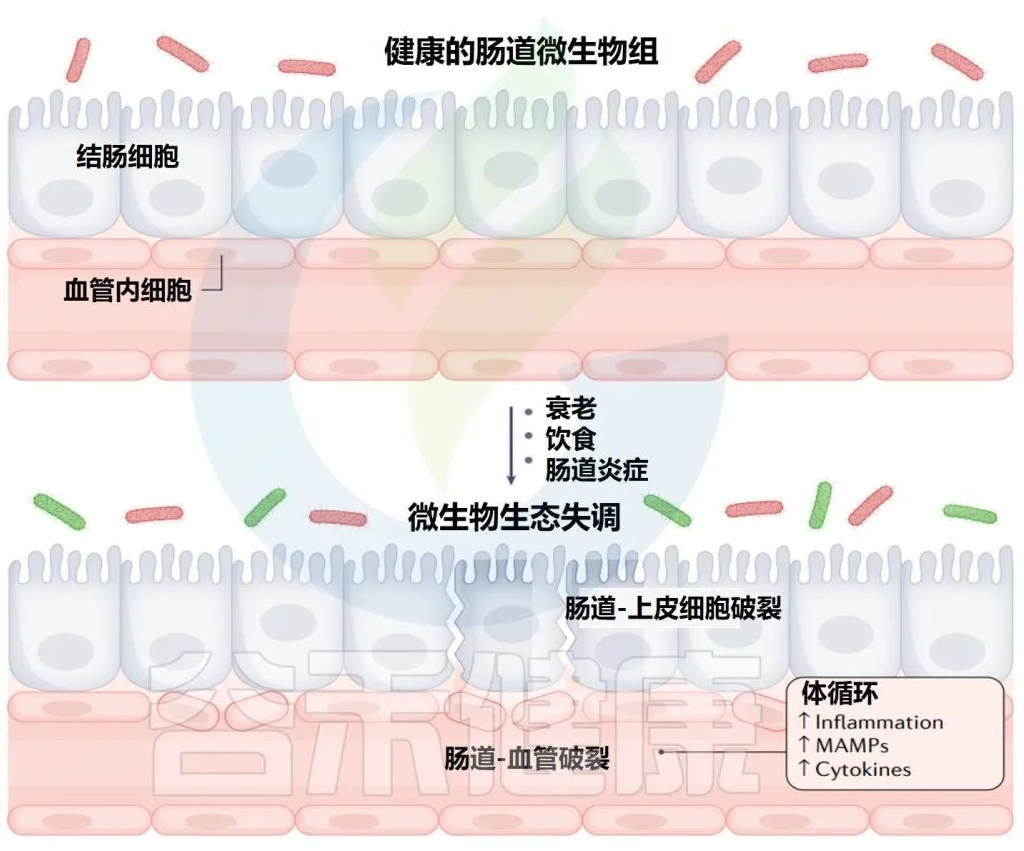
Martin A,et al.Nat Rev Urol.2022
衰老和饮食都可以影响肠道微生物群,导致微生物失调和肠道上皮细胞的破坏。
这种分解导致微生物相关分子模式(MAMPs)的释放增加和短链脂肪酸(SCFAs)的产生减少,并可导致体循环中MAMP和细胞因子的炎症增加。
大多数研究的结果都认为,当肠道菌群失调发生时,肠上皮细胞的分解通常与之相关,尽管特定细菌亚群的变化及其与肠上皮细胞的相互作用并不完全清楚。了解这些变化对于确定哪些菌群可能对免疫系统产生负面的全身影响非常重要。
首先出现的问题是菌群失调或上皮破裂,这一点很复杂,因为它们之间可能有着不可分割的联系,因此,一个或另一个的刺激可能会促成这两个事件。
对健康百岁老人肠道菌群的研究,为通过恢复肠道健康进行治疗干预提供了潜在的途径。这一观点得到了一项研究的支持,该研究显示,健康状况良好的30-100岁人群的微生物含量具有很强的相似性,这表明存在健康的微生物组,并可能对不符合这一类别的患者进行调节。
到目前为止,结果还没有表明任何特定细菌种类的丰度与长寿是绝对相关的。
相反,尽管老年人有效对抗感染的能力明显下降,但长寿的人会形成促炎和抗炎细菌的平衡。
对27项研究的系统综述结果表明,长寿个体在β-多样性(微生物环境间的多样性程度)方面与年轻个体存在差异;然而,通常不会观察到 α 多样性的差异,即每个微生物系统的多样性程度。
总的来说,8项研究涉及百岁老人,4项研究比较了高龄人群和59-85岁人群,4项研究中有2项报告了高龄人群和低龄人群微生物环境多样性程度的差异,而没有研究发现α-多样性的差异。
▸ 老年人的菌群丰度
与年轻个体相比,老年人至少有两个菌群的丰度更高:
与此同时下列菌群的丰度更低:
然而,研究设计存在相当大的差异,关于哪些菌群构成良好的健康,以及老年人存在哪些变化仍未确定的一致意见。
▸ 长寿老人:AKK菌丰度高
值得注意的是,在系统综述中,Akkermansia在高龄人群(99岁以上)中最常见,这可能与此相关,因为该菌在适当水平时与肠道通透性降低、内毒素血症减少和对癌症治疗的免疫反应改善有关。
阿克曼菌(Akkermansia,简称AKK菌)还促进Treg分化,限制肠道功能障碍高脂饮食模型中细胞因子的产生,并通过类似机制预防衰老相关炎症。没有保留阿克曼菌的患者可能会增加疾病发展的风险,包括膀胱癌,或者可能对化疗反应不好。
▸可能存在:AKK菌促进健康的“正确”水平或必要的平衡
抗生素的使用与免疫治疗的不良结果相关,矛盾的是,与不使用抗生素的患者相比,许多服用抗生素的患者AKK菌增加,而且,AKK菌高水平的存在与总生存率呈负相关(使用抗生素的患者P = 0.011,不使用抗生素的患者P = 0.018)。
然而,AKK菌只是可能对微生物功能至关重要的许多不同实体之一,单一实体治疗不太可能像基于平衡微生物组的更强大的改变(如粪菌移植)那样有效。
肠道菌群随年龄或并发疾病的变化可能是膀胱癌的危险因素,可通过直接给予益生元和益生菌或通过粪菌移植减轻。
▸ 短链脂肪酸
短链脂肪酸(SFCAs)由肠道细菌产生,是肠细胞的主要能量来源,是G蛋白偶联的游离脂肪酸受体配体。作为组蛋白去乙酰化酶抑制剂,可通过预防致瘤前表观遗传变化直接限制肿瘤生长。
短链脂肪酸主要存在于肠道中,但它们也可能对治疗膀胱癌有用。在体外,短链脂肪酸在很大程度上抑制先天免疫系统。
由于它们的多种作用和潜在的抗肿瘤活性,产生有益的短链脂肪酸可能是肠道维持健康环境的一种手段,也可能有助于预防结肠癌。
丁酸盐减少癌细胞生长,帮助维持健康的上皮屏障
由于癌细胞与非恶性细胞之间代谢的Warburg效应型变化(有氧糖酵解),丁酸盐通过氧化代谢和能量产生促进非恶性细胞的生长,但减少了癌细胞的生长,因为癌细胞不能有效代谢丁酸盐,丁酸盐在细胞核中积累,在细胞核中发挥组蛋白去乙酰化酶抑制剂的作用。因此,丁酸盐可以帮助维持健康的上皮屏障。
百岁老人肠道菌群中的短链脂肪酸水平上调
在一项研究中,80-99岁的老年人群中健康的老人总短链脂肪酸浓度升高;另一项研究结果显示,与老年患者相比,百岁老人的肠道菌群中产生短链脂肪酸的代谢途径(如乙酸和丙酸)表达水平上调。
除了存在一些与短链脂肪酸产生相关的细菌外,与小于80岁的人群相比,老年人粪便中的细菌数量通常减少。短链脂肪酸水平随着年龄的增长逐渐下降,而百岁老人开始恢复Akkermansia水平。
粪菌移植期间补充Akk菌可能有助于短链脂肪酸的保留
另一项研究的结果表明,在粪菌移植期间用巴氏消毒Akkermansia治疗小鼠,会导致肠道中短链脂肪酸的形成短暂增加,因此,如果这项技术能够完善,在粪菌移植期间补充Akkermansia或其他产生短链脂肪酸的细菌可能有助于改善短链脂肪酸的保留,并预防或治疗癌症。
在脂多糖处理的骨髓源性巨噬细胞中,丁酸、丙酸和乙酸均以剂量依赖的方式降低IL-6和IL-12的产生。丁酸和丙酸也会降低脂多糖处理单核细胞中单核细胞趋化蛋白-1的表达。
通过这些作用,短链脂肪酸可能缓和过度的免疫激活,促进免疫调节。
短链脂肪酸体外作用似乎具有抗炎作用,但其对膀胱癌适应性免疫系统的作用尚不清楚。
特别是丁酸盐,通过诱导调节性T细胞产生IL-10和激活 FAS 介导的T细胞凋亡,下调适应性免疫反应。
短链脂肪酸高可能影响免疫治疗效果
丁酸盐和丙酸盐均通过抑制抗原呈递细胞产生IL-12来抑制CD8+T细胞活化,全身水平的短链脂肪酸高与转移性黑色素瘤细胞毒性T淋巴细胞抗原4(CTLA4)免疫治疗的不良结果有关。
直接通过短链脂肪酸或间接通过前列腺素E2下调T细胞对免疫治疗和顺铂化疗均有不利影响,因为传统的肌肉浸润性膀胱癌新辅助顺铂化疗会刺激效应T细胞。因此,降低T细胞的疗效也可能改变化疗。
这些观察结果可以部分解释在肿瘤床上升高的前列腺素E2表达与不适当抑制的免疫反应和化学耐药性有关。
总之,这些发现表明短链脂肪酸在宿主免疫功能中参与发挥抗炎作用,尽管这一假设可能过于简单化,短链脂肪酸还可以刺激中性粒细胞趋化,因此可能有其他重要的调节作用。
然而,文献强烈支持短链脂肪酸的抗肿瘤作用,尤其在结直肠癌中,可能是因为短链脂肪酸在结肠中的浓度增加,以及介导丁酸差异效应的代谢变化,对癌细胞具有直接作用。
在衰老过程中,维持体内稳态需要抗炎和促炎信号,而短链脂肪酸除了自身具有抗肿瘤作用外,还可能是调节这一系统的一种手段。这些发现已经在体外和体内观察到,但迄今为止没有研究将人类短链脂肪酸水平与膀胱癌结果直接相关。
鉴于与肠道和全身短链脂肪酸水平相关的多种影响,短链脂肪酸可能充当免疫调节剂,因此,塑造使短链脂肪酸产生正常化的微生物含量,是增强或促进免疫治疗的潜在手段。
肠道微生物群的变化可能成为膀胱癌和其他癌症的治疗靶点。应该在这些群体和其他群体之间进行更广泛的比较,以了解哪些类型的微生物生态系统对人类的健康和寿命最有益。
当前的数据支持这样一种观点,即与年龄相关的微生物群变化可能最终影响疾病进展、结果或治疗反应等因素。
衰老在肠道微生物变化中的作用尚不清楚;此外,一些证据表明,环境因素对肠道微生物群有实质性的影响。
一项包括来自中国同一小区域的1000多名极健康个体的研究结果表明,当研究人员试图控制总体健康时,30至100岁的个体粪便微生物群组成几乎没有变化,虽然在30岁以下的个体和30岁以上的个体中发现了相当大的差异。
通过主成分分析评估,30-100岁非常健康的个体的粪便微生物群高度一致,支持了其他研究的结果,这些研究表明环境,而不是年龄或遗传,是微生物组的主要影响者。
值得注意的是,该研究的入选标准具有极高的选择性,通过严格的生活方式分析,几乎99%的患者被拒绝入选;选择标准包括吸烟状况、饮酒、饮食和其他与肠道功能障碍相关的因素。
因此,尽管这些数据表明年龄是肠道微生物群变化的促成因素,但这一观察结果可能仅与这些高选择性标准所代表的一小部分人有关。然而,本文提出了一个假设,即衰老本身可能不如与衰老相关的饮食变化或生活方式变化重要。
在小鼠、狒狒和果蝇模型中,肠道菌群的变化与年龄有关,并与肠道屏障完整性的破坏有关。
在小鼠中,用荧光素异硫氰酸酯(FITC)标记的葡聚糖测量肠道通透性,其血清水平与IL-6和肿瘤坏死因子(TNF)水平一起随年龄线性增加。
肠道菌群组成——肠道屏障功能障碍
在果蝇中,观察到老年果蝇肠道中细菌组成的改变和细菌总负荷的增加,与年轻果蝇相比,肠道中不可吸收蓝色染料(肠道屏障功能障碍的替代品)浓度也相应增加。
结肠完整性的破坏——年龄
在狒狒中,结肠完整性的破坏与年龄有关,与10岁以下的狒狒相比,年长的狒狒(年龄≥18岁)结肠组织的免疫组化分析显示封闭蛋白(occludin)、连接蛋白(claudin)和紧密连接粘附分子减少,同时大分子辣根过氧化物酶(肠通透性的另一标志)在结肠上皮的通量增加。
此外,18岁狒狒的结肠活检中IL-6、IL-1β和IFNγ水平高于10岁灵长类动物。
肠道功能障碍——细菌成分的全身释放
另一项比较21月龄小鼠与4或12月龄小鼠肠道通透性的研究结果显示,老年小鼠血清FITC-右旋糖酐水平的增加与体循环中脂多糖水平的增加相关,将肠道功能障碍和细菌成分的全身释放联系起来。
LPS 和其他 MAMP 与 IL-1β、IL-6 和 TNF 等细胞因子的增加密切相关,提供了肠道菌群失调、慢性炎症和肿瘤发生之间可能的机制联系。
所有这些数据都支持这样的观点,即肠道功能障碍随着年龄的增长而增加,与其他哺乳动物的饮食或生活方式的变化无关,但由于适当控制人类饮食的复杂性,尚未在人类中进行评估。
免疫功能障碍——衰老——微生物群
衰老过程中,循环细胞因子的增加在具有14个月或以上老年动物的微生物群的小鼠模型中加剧,将免疫功能障碍与衰老和微生物群联系起来。
相比之下,无菌小鼠在衰老过程中通常不会表现出,老年特异性无病原体小鼠所表现出的循环细胞因子的增加,并且在很大程度上可以防止衰老相关炎症和巨噬细胞功能障碍,除非它们再次暴露于微生物群。
衰老——肠道通透性——炎症——菌群
将无菌小鼠暴露于14月龄动物的菌群中,比无菌小鼠暴露于幼龄小鼠的菌群中更能促进肠道通透性和细胞因子释放。
这一机制依赖于肿瘤坏死因子,使用抗肿瘤坏死因子拮抗剂治疗可以改善衰老引起的微生物群的变化。因此,由肠道功能障碍介导的MAMP或DAMP释放与老年炎症表型之间发生了实质性的相互作用,从而导致炎症系统的慢性全身激活。
为了将这些发现转化为膀胱癌,除了了解局部微生物群(包括尿液微生物群和肿瘤本身)之间的联系外,还需要进一步了解膀胱癌背景下的肠道微生物群。
到目前为止,对尿液微生物群的研究还缺乏一个关于什么是正常尿液微生物群的明确共识:由于收集方法的差异和相对较小的样本量,目前关于健康尿液微生物群的定义存在很大差异。
目前对尿液和膀胱微生物群及其改变的了解有限。局部微生物环境的失衡与多种上皮性癌症的肿瘤进展有关,包括乳腺癌、宫颈癌和子宫内膜癌。
健康的尿液微生物群
目前的初步研究一致认为,健康成年志愿者的尿液微生物中至少包含四种细菌种群:乳酸菌和链球菌是尿液微生物中最稳定存在的属,与年龄和采样方法有关,而厚壁菌门和不动杆菌门是两性尿液微生物中最丰富的门。表明这些是该人群中最有可能构成健康尿液微生物组的成分。
女性尿液中乳酸杆菌种类的含量高于男性
根据对健康志愿者进行的两项独立研究的结果,尿液微生物群的性别差异很明显。研究表明,女性尿液中乳酸杆菌种类的含量高于男性尿液中的含量。
我们还不清楚这一观察结果与癌症的关系,但这一发现的一个潜在的重要方面是,男性与女性泌尿系统微生物群的差异或女性泌尿系统微生物群的个体间差异可能与女性泌尿系统感染的流行有关,或她们膀胱癌发病率的降低,因此可能会影响膀胱的整体微环境。
菌群在不同年龄的差异
在同一项包括23名健康成年人的研究中,下列四种菌群仅在>70岁的个体中发现:
此外,在男性中,属的数量会随着年龄的增长而增加,而细菌总数减少。性别或年龄差异会影响尿液微生物群。
尿液微生物:个体间差异极高
研究结果还表明,个体间差异极高,限制了更深入的分析。由于这种高变异性,对泌尿系统微生物的调查缺乏类似的大型多种族和多种族肠道微生物群研究的力量,因此饮食、基因、男性和女性短期或长期激素变化的重要性,以及泌尿系统微生物的卫生还没有确定。
需要大型的、多中心的、多种族的、具有共识方法论和清晰统一的分析管道的研究来完善该领域的假设并产生共识。
多项研究报道尿失禁、间质性膀胱炎和泌尿生殖系统癌患者尿液微生物群落的物种多样性和相对丰度存在差异。
健康人群与膀胱癌人群的微生物组差异
在膀胱癌方面,一项包括12例膀胱癌患者和11名年龄匹配的志愿者的研究结果显示,与健康成年人相比,膀胱癌患者富集了以下四种菌群:
与间质性膀胱炎和慢性前列腺炎等疾病相比,使用Simpson指数测量的健康成人和膀胱癌患者之间的物种多样性(α-多样性)在种或科水平上没有总体差异,这表明膀胱癌的尿液微生物群可能不像其他炎症性疾病状态那样受影响,尽管这一观察结果可能是因为该研究力量不足。
然而,在特异性操作分类单元(OTUs)水平上进行差异分析,可以减少测量参数的数量,从而识别差异,表明相互关联的物种可能在肿瘤发生中发挥作用。
一项针对31名膀胱癌男性的研究结果显示,与18岁匹配的健康个体相比,下列三种菌群的丰度增加:
同时下列几种菌群降低:
由于个体数量少,这些研究不足以完全确定特定细菌种群的变化。对于尿液微生物菌群研究在多大程度上反映了膀胱内的微生物群也知之甚少。
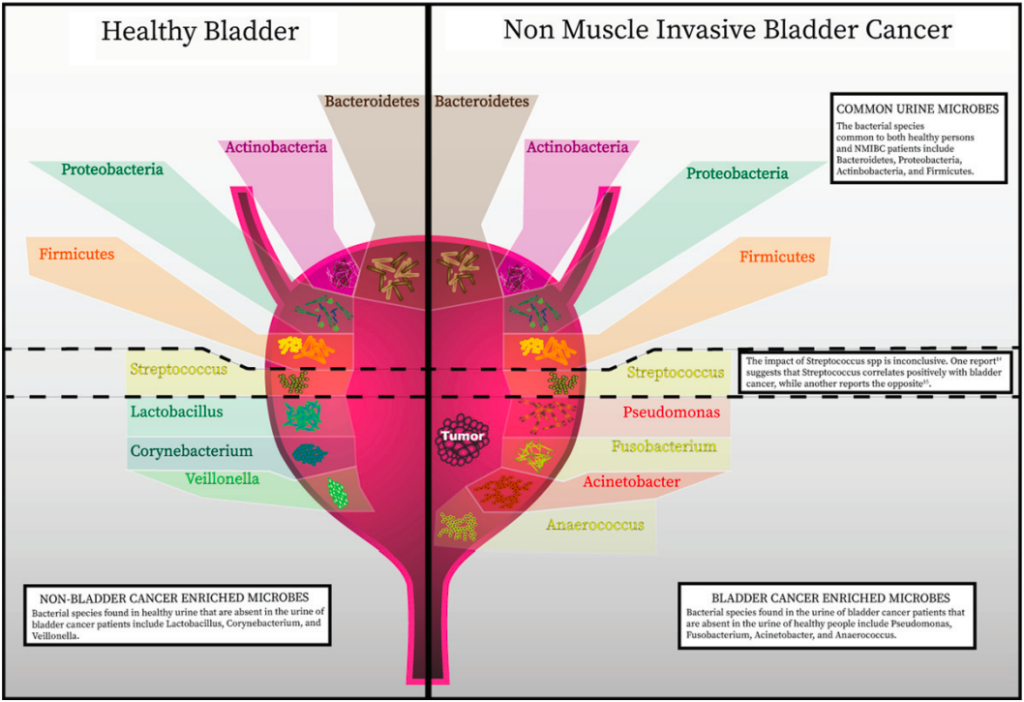
Andolfi C, et al., Bladder Cancer. 2020
关于膀胱肿瘤微生物组的研究仅限于两项研究。两者的结果都表明,肿瘤与非恶性组织中的 α 多样性导致物种丰富度相对降低。一项研究发现,膀胱微生物组的门(如厚壁菌门和放线菌门)的相对丰度与之前在其他研究中报道的尿液微生物组值相似。
联合测量来自同一患者尿液和膀胱肿瘤样本的微生物群目前在文献中还没有。这些因素支持了对肿瘤分级或分期与泌尿系统微生物组之间差异的调查,以及使用更大的患者队列进行明确的、多机构的、控制良好的研究的必要性。
尿液微生物群的影响尚不完全清楚,衰老和肠道生态失调导致的免疫失调的直接影响可能会增加膀胱癌的发病风险。
然而,没有研究直接比较同一患者的尿液微生物群和肠道微生物群的变化。需要进行研究来评估与衰老和炎症相关的肠道生态失调是否也发生在泌尿系统的微生物群水平上,或任何其他含有微生物的解剖部位。
如果存在潜在的机制,尿液微生物的失调可能是衰老、炎症、失调和膀胱癌风险之间的联系。
系统性炎症、尿路上皮增生与膀胱癌之间的假设联系

Martin A,et al.Nat Rev Urol.2022
衰老引起的炎症变化以及肠道上皮完整性的降低可导致循环促炎介质水平的增加,例如细胞因子和微生物激活分子模式 (MAMP).
LPS介导的toll样受体4 (TLR4)激活可导致骨形态发生蛋白4 (BMP4)、音猬因子 (SHH)、WNT -β-catenin和STAT3通路的激活,促进尿路上皮的增殖。BMP4、SHH、WNT -β-catenin和STAT3通路在膀胱癌中调控异常,但还没有研究直接将LPS或MAMP介导的BMP4、SHH、WNT -β-catenin和STAT-3的激活与膀胱癌的发生联系起来。当转化发生时,带有肿瘤相关巨噬细胞(TAMs)和肿瘤浸润性中性粒细胞(TINs)的尿路上皮的慢性炎症阻止了适当的免疫反应和肿瘤清除,并可能通过释放细胞因子、生成活性氧(ROS)和促进免疫抑制环境进一步加剧肿瘤的形成。PGE2,前列腺素E2.
由尿路上皮细胞和局部感染性细菌引起病原相关分子模式(MAMPs)驱动和损伤相关分子模式驱动炎症可能是年龄、肠道上皮破坏和在许多膀胱肿瘤中发现的先天免疫信号之间的一种机制联系。
脂多糖增加炎症因子的释放
在体外人类和小鼠模型中,来自尿路致病性大肠杆菌的脂多糖通过TLR4信号通路增加尿路上皮细胞中IL-6、IL-8和PGE2细胞因子的释放。
此外,脂多糖激活尿路上皮细胞上的电压门控钾通道,介导IL-6释放。脂多糖通过磷酸化途径激活巨噬细胞和泌尿黏膜中的TLR4,这与膀胱癌的发展和化疗耐药有关。
STAT3促肿瘤
免疫组织化学(IHC)分析发现,与非侵袭性发光型肿瘤相比,局部侵袭性肿瘤中STAT3磷酸化水平升高,抑制STAT3可降低膀胱癌细胞系的增殖。在小鼠中,阻断尿路上皮细胞中的 STAT3 可降低膀胱癌的发生和侵袭性。
IL-8和PGE2促肿瘤
脂多糖介导的尿路上皮IL-8和PGE2的增加在膀胱癌中也具有促肿瘤作用。在经尿道膀胱肿瘤标本切除的新鲜冷冻组织中,与非侵入性肿瘤相比,浸润性肿瘤中IL-8和基质金属蛋白酶9水平升高。
前列腺素E2促肿瘤
此外,前列腺素E2增加高级别膀胱癌细胞系的增殖,在接受吉西他滨和顺铂标准化疗方案治疗的小鼠中使用塞来昔布抑制前列腺素E2,产生了持续的治疗反应,并通过消除患者来源的肿瘤异种移植模型中CK14+细胞的形成克服了化疗耐药性。
这些研究表明尿路感染期间脂多糖相关炎症与膀胱癌进展之间存在多种趋同途径。
尿路感染后信号通路的改变
重要的是,对尿路感染的生理尿路上皮反应受到音猬因子重组蛋白(SHH)、骨形态发生蛋白4 (BMP4)和WNT -β-catenin信号通路的高度调控,这些信号通路在恶性转化过程中经常发生改变。
尿路感染后的尿路上皮再生是由尿路上皮基底层分泌SHH的干细胞增殖驱动的。感染后,下游SHH信号通路通过BMP4和WNT-β-catenin信号通路诱导尿路上皮和基质细胞增殖和终末分化。
这种作用在前列腺素合成抑制剂治疗后被逆转。
这些再生途径中的每一个的丧失或改变都与膀胱癌发生有关。在 BBN 小鼠模型中,SHH 阳性基底细胞是膀胱癌的主要祖细胞。SHH 的丧失和进展为膀胱癌可能是通过 BMP 基质信号传导的丧失而发生的。
免疫组化分析显示,与非恶性尿路上皮黏膜相比,在人膀胱癌肿瘤中观察到WNT糖蛋白活性升高。这一发现在小鼠模型中得到了进一步的支持,在功能获得突变体中β-连环蛋白的构成性激活导致了12周后30%的小鼠发生低级别膀胱肿瘤。
有趣的是,在具有构成型活性β-连环蛋白的小鼠中,肿瘤的发生是性别依赖的,45%的雄性小鼠发生肿瘤,而雌性小鼠只有3%,这最终与雄激素受体(AR)信号有关。
在人类和普通小鼠模型中都发现了膀胱癌的性别二态,但其原因是多因素的,也与表观遗传机制中的染色体差异有关。
这些发现呈现了尿路上皮在响应急性脂多糖介导的炎症反应和尿路上皮癌的发展之间呈现出许多重叠特征。
年龄相关的肠屏障破坏,诱发炎症;免疫系统变化,增加肿瘤风险
然而,尿路感染与膀胱癌风险的增加并没有特别的关联。当与衰老和肠上皮完整性丧失相结合时,由于与年龄相关的肠屏障破坏,长期暴露于基底尿路上皮层的脂多糖或其他损伤相关分子可能会诱发慢性膀胱炎症,并增加细胞周转,以及与年龄相关的免疫系统变化,增加肿瘤转化的可能性。
慢性膀胱炎症诱导的肿瘤转化可能是女性特有的一种机制,因为尿路感染在女性中更为突出,而膀胱癌在男性中更为常见。
关于衰老相关炎症、微生物群和膀胱癌发生的研究仍然不完整,需要更多的假设检验来确定这些不同实体(衰老、微生物功能障碍、炎症改变)在这一过程中导致的具体变化和肿瘤相关结果。
研究肠道和尿液微生物群对浅表性癌症治疗反应的影响,可能为微生物群和膀胱癌进展提供额外的见解。
新的证据表明,浅表膀胱癌治疗受到肠道和尿道微生物群的影响。
目前,高级别非肌肉浸润性膀胱癌的一线治疗是细菌疫苗株卡介苗。卡介苗的疗效随着年龄的增长而降低,早期数据表明,疗效还受尿微生物中变形菌群丰度的影响。
一项包括1106例患者的卡介苗干扰素膀胱内治疗的II期多中心试验的数据表明,年龄在61-70岁的患者在卡介苗干扰素治疗后的无癌生存期优于80岁或以上的患者(3 年无癌生存率为 65%对 47% ,P = 0.036)或卡介苗单药治疗(3 年无癌生存率 55% 对 32%,P < 0.003)。
令人惊讶的是,在这项研究中,小于50岁的患者的预后较差;然而,在多变量分析中,年龄仍然是反应的独立危险因素。
在一项包括805名Ta、T1或CIS高分级患者的独立研究中,年龄为>70岁的患者的无癌生存期低于年龄<70岁患者(37% 对 27%,P = 0.005)。
在接受益生元或益生菌治疗的个体中,非肌肉浸润性膀胱癌复发减少。最后,肠道菌群介导了实体肿瘤(如黑色素瘤)中对PD1治疗的反应,这可能与晚期膀胱癌中免疫治疗具有共同机制。
关于卡介苗抑制肿瘤复发的机制尚不清楚,一种假设是卡介苗诱导膀胱内的局部促炎性先天免疫反应,进而刺激对卡介苗和肿瘤抗原的适应性免疫反应。
在治疗后的前3年,由于未知的原因,卡介苗未能阻止肿瘤复发的患者占30%,年龄可能是无反应的原因之一。
年龄越大,卡介苗疗效越差
接受卡介苗治疗后,61-70岁的患者中有61%在24个月时无复发,而>80岁的患者中有39%无复发。
这些结果得到了另一项研究的支持,该研究表明70岁以下(27%)和70岁以上(37%)患者5年后无病生存率的绝对差异为10%;然而,这两个年龄组中与年龄相关的死亡风险没有报道。
因此,年龄可能是非肌肉浸润性膀胱癌复发的独立危险因素,但与年龄相关的免疫系统衰退是否相关尚不清楚。
卡介苗应答取决于多种因素,不止年龄
在一项小型研究中,31名卡介苗应答者与32名卡介苗无应答者进行了比较,结果表明,肿瘤PDL1状态而不是T细胞PD1状态是卡介苗应答的决定因素,这表明卡介苗的总体应答可能取决于许多不同的因素,而不仅仅是年龄。
尿液微生物群也可能在卡介苗反应中起作用
在一项包括31例接受卡介苗治疗的高危非肌肉浸润性膀胱癌患者的研究中,观察到卡介苗应答者和无应答者在OTU水平上的尿液微生物群差异。
卡介苗治疗6个月后无病的患者比复发患者尿厚壁菌门(如乳杆菌和变形杆菌)丰度更高。
鉴于非肌肉浸润性膀胱癌中益生菌和饮食的潜在保护作用,对卡介苗有反应的人群中乳酸杆菌丰度的增加尤其引人注目。
年龄、免疫状态和尿液微生物组与卡介苗反应之间的确切联系尚未确定。
考虑到有相当多的非肌肉浸润性膀胱癌患者最终会进行膀胱切除术,对年龄相关卡介苗反应降低的机制进行更好的了解,将对评估复发风险和改善对现有治疗的反应非常有益。
在日本,肠道微生物组长期以来一直是浸润性膀胱癌治疗的目标,部分原因是发酵乳制品中广泛使用的乳酸菌,以及报道的小鼠口服乳酸菌制剂的抗肿瘤特性。
★ 乳酸杆菌
乳酸杆菌与膀胱癌复发减少或低发病率相关
一项包括138名患者的双盲随机对照试验结果显示,口服干酪乳杆菌制剂与安慰剂相比,男性和女性浸润性膀胱癌复发减少。
这些结果得到了一项病例对照研究的支持,该研究包括180名膀胱癌患者和445名年龄和性别匹配的健康志愿者,他们有10-15年的发酵牛奶的习惯摄入史(发酵奶是高脂肪的天然来源),每周1-2次乳酸杆菌(OR 0.46,95 % CI 0.27–0.79)或3-4次乳酸杆菌(OR 0.61,95% CI 0.38–0.99)与膀胱癌发病风险的降低有关。
这些发现得到了一项前瞻性试验结果的进一步支持,该试验包括202例经尿道膀胱肿瘤切除术后的浸润性膀胱癌患者,他们被随机分为两组,一组接受六次膀胱内表阿霉素治疗,另一组接受六次膀胱内表阿霉素治疗,外加每日3克干酪乳杆菌口服制剂,为期1年。
接受表阿霉素加每日制剂乳酸杆菌的患者3年无复发生存率为74.6%,而单独表阿霉素组仅为59.9% 。
这些研究没有调查粪便微生物组是否发生了变化,因此可以推断因果关系的程度有限。需要通过研究确定益生菌相关的变化,以及这些变化如何影响膀胱癌相关结果,来验证这些发现。
乳酸杆菌与肠道黏膜炎症减少有关
然而,在葡聚糖硫酸钠诱导的小鼠结肠炎症模型中,乳酸杆菌种类与肠道黏膜炎症减少有关。
在体外实验中,唾液乳杆菌降低肠上皮细胞系中TLR受体的激活,导致上皮细胞之间紧密连接的组装增加,可能减少系统性脂多糖和慢性炎症。
还需要在膀胱癌动物模型中进行更多的研究,以直接确定全身的病原相关分子模式水平是否也与全身炎症相关。
扩展阅读:如何调节肠道菌群?常见天然物质、益生菌、益生元的介绍
★ 萝卜硫素
除了乳酸菌,萝卜硫素也可能在恢复肠道屏障健康和降低膀胱癌发展风险方面发挥保护作用。
萝卜硫素饮食的小鼠丁酸水平增加
用亚硝基胺(BBN)治疗的小鼠和喂食高萝卜硫素饮食的小鼠丁酸水平增加,但与单独用亚硝基胺治疗的小鼠相比没有表现出增加的存活率。
肿瘤发生时间延长,发病率降低
然而,有趣的是,饲喂西兰花提取物(萝卜硫素的主要来源)的大鼠在暴露于亚硝基胺8周后,与未接受西兰花提取物的大鼠相比,肿瘤的发生时间延长了24周,其膀胱肿瘤发病率降低了58%.
重要的是,口服西兰花提取物导致小鼠在喂食12小时后尿液中生物活性异硫氰酸盐(包括萝卜硫素)的浓度比血清中高70% 。
这些化合物也会干扰亚硝基胺的代谢,由于它们在尿液中积累,其功能主要是通过改善肠道健康还是直接作用于尿路上皮细胞的其他机制尚不清楚。需要新的研究来调查它们对肠道健康、随后的炎症和微生物差异的影响,以改进对确切机制的定义。
总之,这些研究的结果表明,改变饮食,无论是口服乳酸杆菌或萝卜硫素,都可以降低膀胱癌的风险。这两种方法都与血清中细菌脂多糖浓度的降低相关,脂多糖是小鼠模型中肠道炎症的系统性标记物。
然而,关于乳酸杆菌或萝卜硫素在多大程度上诱导与治疗反应相关的肠道或尿液微生物组的积极改变,目前还缺乏数据。
菌群失调对PDL1-PD1轴的影响是一个有趣的话题。2016年首次批准PD1抑制剂用于顺铂难治性晚期膀胱癌。
总的来说,阿克曼菌Akkermansia、肠球菌、梭状芽孢杆菌、瘤胃球菌、粪杆菌和双歧杆菌的丰度增加与肺癌、肾癌和黑色素瘤等癌症对PDL1治疗的反应改善有关。
在转移性黑色素瘤患者中,研究中定义的患者粪便中“有益”与“无益”OTU数量的增加与抗PD1治疗反应的改善呈正相关。
PD1响应者粪菌移植的小鼠有更强的抗肿瘤反应
在对肠道微生物组和免疫治疗反应的三项独立研究中,从PD1治疗反应者到无菌小鼠的粪便移植产生了比无菌小鼠更强的抗肿瘤反应。
在一项研究中,研究了转移性黑色素瘤患者肠道微生物组和肿瘤微环境之间的相关性,以评估肿瘤微环境的发现是否可以预测对PD1免疫疗法的治疗反应。
对肿瘤活检的免疫组化分析显示,对PD1免疫治疗反应良好的患者,其肿瘤床上的CD8+ T细胞密度更大,调节性T细胞更少。
此外,响应者的物种多样性和瘤胃球菌科细菌的相对丰度高于无响应者。考虑到准确定义和匹配供体材料的重要性,必须了解粪菌移植如何改善抗免疫抑制分子或抗PDL1治疗的反应。
鉴于这些变化,作者得出结论,为 FMT 和其他微生物疗法准确匹配和定义微生物群可能需要额外的匹配,如年龄和性别,并准确定义理想的供体-宿主关系。
总之,这些发现强烈表明,在PD1免疫治疗的情况下,肠道微生物群可能会影响对癌症的全身免疫反应。
所有关于肠道微生物组和对PD1抑制剂的反应的初步研究都比较了相对物种丰度和处理反应。这种方法不能解释细菌总负荷的变化以及体循环中代谢物浓度的潜在差异。
微生物组是否通过直接的抗原相互作用对免疫系统产生影响,或者微生物组的代谢产物,包括潜在的有益成分和潜在的有害成分,是否间接地影响免疫系统,目前尚不清楚。
这种区别很重要,因为如果微生物组的主导作用是通过体循环中的代谢物间接发生的,那么微生物组可以被广泛地认为是一个更大的代谢产物网络的代表。
在这个模型中,许多不同的细菌组合有可能创造一个治疗性代谢网络,这与结果一致,表明促炎和抗炎成分对健康的老龄化都是必要的。
如果微生物组主要通过直接的免疫原性相互作用发挥作用,使免疫细胞对抗特定抗原,那么某些细菌种群的丰度是至关重要的。
膀胱癌的发病率与衰老密切相关。在老年患者中常见的炎症导致先天免疫系统失调,持续激活与活性氧表达和细胞因子释放相关的细胞。
与年龄相关的肠道微生物组变化可能有助于防止老年患者肿瘤的发生和肿瘤的治疗,但这些变化是否在功能上与老年患者的致瘤性炎症特征有关还有待确定。
许多膀胱癌患者以及其他尿道疾病患者与健康人群相比,尿液微生物群出现紊乱。需要更多大样本的基础研究,来确定膀胱癌或尿道疾病相关的菌群变化机制,从而更好地帮助治疗疾病。
没有确定的方法可以预防膀胱癌。但可以做一些有助于降低风险的措施。
虽然说没有一种食物本身可以绝对防止患癌症。但研究表明 ,富含各种水果、蔬菜、豆类、全谷物和其他植物性食物的饮食可以降低患多种癌症的风险,包括膀胱癌。
绿叶蔬菜
根据BCAN的数据,研究人员将每份蔬菜与膀胱癌风险减少10%联系起来。绿叶蔬菜含有抗氧化剂,抗氧化剂修复细胞损伤,同时还具有抗炎特性,甚至可以帮助预防癌症。例如:西兰花、卷心菜、南瓜、小麦胚芽、菠菜、羽衣甘蓝等
水果
研究人员还认为水果可以降低10%的膀胱癌风险。柠檬、橙子、葡萄柚和酸橙等柑橘类水果具有抗氧化特性,可以起到保护作用。其他还包括浆果,梨,苹果等。
茶
BCAN 表示,每天喝一杯茶,患膀胱癌的风险降低 6%,对动物的研究表明,多酚化合物会阻碍膀胱肿瘤的生长。绿茶中含有表没食子儿茶素-3-没食子酸酯,具有抗癌特性。
注意:红肉等加工食品中的化学物质会加剧膀胱癌的病情,此外还应避免酒精,零食等。尽可能均衡、健康地饮食。
西兰花芽丸、绿茶或姜黄素补充剂已经成为接受膀胱癌治疗的流行天然补充剂。然而,研究未能提供它们对人类有益的结论性证据。
在膀胱癌治疗期间服用姜黄素补充剂似乎可以使小鼠的肿瘤减小。
西兰花芽提取物对膀胱癌细胞的体外生长有抑制作用。
维生素E
据MayoClinic称,大量摄入富含维生素E的食物可以降低膀胱癌恶化的风险。
硒
硒是指一种具有抗氧化特性以防止细胞损伤的矿物质。因此,专家认为通过食用富含硒的食物可能帮助控制膀胱癌。如巴西坚果、核桃和燕麦片等。
限制场所
化工场所包括橡胶、皮革、印刷材料、纺织,油漆,铝,芳香胺、砷,染料,地毯,美发,打印等行业,接触时间过长可能患膀胱癌的风险更高,如在这些行业工作,尽可能遵守安全规则,做好防护措施。
尽可能减少吸烟
吸烟的人患膀胱癌的可能性至少是不吸烟的人的 3 倍。
多喝水
多喝水可能降低患膀胱癌的风险。如果喝白开水有困难,可以在水中加入新鲜柠檬片等来保持水分。
注:本账号内容仅作交流参考,不作为诊断及医疗依据。
主要参考文献:
Martin A, Woolbright BL, Umar S, Ingersoll MA, Taylor JA 3rd. Bladder cancer, inflammageing and microbiomes. Nat Rev Urol. 2022 Jul 7. doi: 10.1038/s41585-022-00611-3. Epub ahead of print. PMID: 35798831.
Babjuk M. Bladder Cancer in the Elderly. Eur Urol. 2018 Jan;73(1):51-52. doi: 10.1016/j.eururo.2017.04.018. Epub 2017 May 4. PMID: 28478044.
Andolfi C, Bloodworth JC, Papachristos A, Sweis RF. The Urinary Microbiome and Bladder Cancer: Susceptibility and Immune Responsiveness. Bladder Cancer. 2020 Sep 21;6(3):225-235. doi: 10.3233/BLC-200277. PMID: 33195783; PMCID: PMC7605348.
Kacew A, Sweis RF. FGFR3 Alterations in the Era of Immunotherapy for Urothelial Bladder Cancer. Front Immunol. 2020 Nov 5;11:575258. doi: 10.3389/fimmu.2020.575258. PMID: 33224141; PMCID: PMC7674585.
Palumbo C, Knipper S, Pecoraro A, Rosiello G, Luzzago S, Deuker M, Tian Z, Shariat SF, Simeone C, Briganti A, Saad F, Berruti A, Antonelli A, Karakiewicz PI. Patient frailty predicts worse perioperative outcomes and higher cost after radical cystectomy. Surg Oncol. 2020 Mar;32:8-13. doi: 10.1016/j.suronc.2019.10.014. Epub 2019 Oct 25. PMID: 31683158.
Zhang Y, Hou Q, Ma C, Zhao J, Xu H, Li W, Wang Y, Ma H, Zhang H, Sun Z. Lactobacillus casei protects dextran sodium sulfate- or rapamycin-induced colonic inflammation in the mouse. Eur J Nutr. 2020 Jun;59(4):1443-1451. doi: 10.1007/s00394-019-02001-9. Epub 2019 May 23. PMID: 31123864.

谷禾健康

帕金森综合征
帕金森病(Parkinson’s disease,PD)是全球增长最快的神经系统疾病之一,原因有很多,包括人口老龄化和环境影响。老年人多见,平均发病年龄为60岁左右,40岁以下起病的青年帕金森病较少见。
我国流行病学调查显示,65岁以上人群帕金森病的患病率大约是1.7%。估计全国每年新发患者数达10万以上,我国现有帕金森病患者人数近300万。大部分帕金森病患者为散发病例,仅有不到10%的患者有家族史。
帕金森病最主要的病理改变是中脑黑质多巴胺(dopamine, DA)能神经元的变性死亡,由此而引起纹状体多巴胺含量显著性减少而致病。
虽然这种神经退行性疾病目前主要由临床运动特征(运动迟缓、僵硬和静止性震颤)来定义,但非运动特征,如自主神经功能障碍、认知行为问题和睡眠障碍,大大增加了总体疾病负担。
200多年前,詹姆斯·帕金森首次正式描述帕金森病时,胃肠功能障碍被认为是帕金森病临床表现的一部分。值得注意的是,他还推测帕金森病可能起源于胃肠系统,并评论道“胃肠紊乱状态可能会导致脊髓的一部分发生病态行为”。
肠道微生物群对神经系统过程(如神经发生、髓鞘形成和小胶质细胞激活)至关重要,能够调节行为和认知,并越来越多地参与调节神经退行性疾病和神经精神疾病的易感性和进展。
帕金森发病过程中肠道和大脑之间的串扰被认为受到肠道微生物失衡(失调)以及由此引起的细菌代谢活性改变的重大影响。
本文概述了帕金森病的肠-脑轴,包括迷走神经、肠神经系统中的α-突触核蛋白、改变的肠道通透性和炎症以及肠道微生物及其代谢活动的潜在作用和相互关系。阐述了肠道相关因素(包括肠道菌群)在帕金森病治疗进展中的作用和作为治疗靶点的作用,并基于研究文献讨论了常见的一些帕金森疗法。
关键词解释
α-突触核蛋白
是一种在中枢神经系统突触前及核周表达的可溶性蛋白质,它与帕金森病的发病机制和相关功能障碍密切相关,是路易小体的主要成分。
路易小体
是以帕金森病为代表的路易体病患者脑内的特征性标志物。显微镜下为圆形粉红色均质状结构。
▸ 震颤
震颤通常始于手或手指,其他包括手臂、腿、下巴或头部都可能出现震颤。可能会来回摩擦拇指和食指,类似弹丸滚动的震颤。手在休息时可能会颤抖。
▸ 肌肉僵硬
肌肉长时间保持收缩。尤其是在疾病的早期阶段,可能被误认为关节炎或骨科问题。
▸ 运动缓慢
随着时间的推移,帕金森病可能会让人减慢运动速度,使简单的任务变得困难且耗时。走路时步数可能会变短,可能拖着脚。可能很难从椅子上站起来。
▸ 平衡和协调受损
可能会变得弯腰,或者可能因帕金森病而出现平衡问题,有时会导致跌倒。
▸ 认知变化
一项研究发现,认知障碍是帕金森病常见且进展迅速的症状。在平均患有帕金森病五年且在研究开始时认知正常的参与者中,约有一半在确诊帕金森病后的六年内(约11年)出现轻度认知障碍。
少数出现轻度认知障碍的患者在五年内发展为痴呆。帕金森痴呆症患者可能有严重的记忆和思维问题,影响日常生活。
▸ 抑郁、焦虑等
可能会经历抑郁症,有时是在非常早期的阶段。
大约 40%-50% 的帕金森患者会出现抑郁症。可能还会经历其他情绪变化,例如恐惧、焦虑或失去动力等。也包括可能出现的精神病,例如幻觉或妄想。
▸ 吞咽、咀嚼和说话困难
由于吞咽缓慢,唾液可能会积聚在口腔中,导致流口水。晚期帕金森病会影响口腔中的肌肉,使咀嚼变得困难。这会导致窒息和营养不良。
▸ 排尿问题或便秘
帕金森病可能会导致膀胱问题,包括无法控制尿液或排尿困难。许多帕金森病患者出现便秘,主要是由于消化道较慢。
▸ 睡眠问题
帕金森病患者经常有睡眠问题,包括整夜频繁醒来、早起或白天想睡觉。
▸ 其他影响
其他还包括嗅觉障碍、疲劳、皮肤问题(黑色素瘤)、性功能障碍、疼痛、头晕目眩、眼睛和视力问题、体重减轻等问题。
帕金森氏症的症状和进展速度因人而异。这种疾病的早期症状很微妙,并且逐渐发生。例如:
有时候朋友或家人可能是第一个注意到早期帕金森氏症患者的变化的人。他们可能发现患者:
当疾病发展到晚期帕金森病时,症状变得更加严重且相当复杂,行动变得局限。其中包括:
从1980年代帕金森病患者肠道神经系统(ENS)中发现α-突触核蛋白沉积开始,进行了尸检研究,提出突触核蛋白病起源于迷走神经背运动核(DMNV)和前嗅核,帕金森病病理学肠-脑传播的“上升解剖学理论”获得了广泛的关注。
根据这一理论,作用于肠道的损伤会触发α-突触核蛋白的错误折叠和聚集,α-突触核蛋白开始于肠上皮细胞,然后通过朊病毒样细胞间转移传播到大脑。
这些结果表明,需要进一步的研究才能得出α-突触核蛋白的肠-脑支持是否是帕金森病的关键致病步骤。
鉴于帕金森病的复杂性和异质性以及肠-脑相互作用的多方面性质,目前的观点是,这种相互作用可能在帕金森病中起作用,但在个别患者中的作用程度不同。
▸▸
在过去十年中,多项临床和临床前研究提供了帕金森病患者肠道通透性改变(“肠道渗漏”)和炎症的证据。
Toll样受体4(TLR4)信号通路可感知受损或坏死组织释放的革兰氏阴性细菌脂多糖和内源性分子,被认为在帕金森病的肠道和脑部炎症中起着至关重要的作用。
虽然研究普遍显示帕金森病患者存在肠道炎症的证据,但这些变化仅在一部分患者中明显,胃肠道通透性研究的结果也具有高度异质性。
肠道相关因素在某些但并非所有患者的神经退行性变中起着重要作用。已知炎症可导致肠屏障功能障碍,在炎症性肠病(IBD)患者中,抗肿瘤坏死因子(anti-TNF)治疗可抑制肠道炎症,使胃肠道通透性恢复正常。
然而,屏障缺陷有时可以作为疾病的早期事件发生,与炎症无关。
一些支持和反对肠道优先参与PD的证据
▸流行病学研究
支持证据:
• 便秘、胃食管反流和胃轻瘫等胃肠道症状可先于帕金森病运动症状几十年。
• 大量研究表明,完全切断迷走神经干与显著降低帕金森病的发病风险相关。
反对证据:
• 胃肠功能障碍等非运动特征可能比运动特征更早出现,因为副交感神经系统(PNS)和肠神经系统(ENS)对神经退行性变的补偿能力不如中枢神经系统(CNS)。
• 一项大型纵向研究发现迷走神经切断术后帕金森病风险没有显著降低。
▸神经病理学研究
支持证据:
• 尸检研究表明,几乎所有帕金森病患者的肠神经系统中都存在突触核细胞病变。
• 在帕金森病诊断前20年,在食道、胃、小肠和结肠的活检中检测到α-突触核蛋白沉积。
反对证据:
• 对老年人的尸检研究发现,在没有脑α-突触核蛋白参与的情况下,没有“仅肠道”路易病理学,尽管不能排除在胃肠道迅速传播或高度局限性病理学的可能性。
• 其他活检研究发现,神经健康个体的胃肠道中存在α-突触核蛋白积聚。
▸动物研究
支持证据:
• 在啮齿动物模型中已经证明,肠-脑α-突触核蛋白传递,伴有黑质多巴胺能神经变性和帕金森病样运动和非运动症状,可能与年龄有关。
• 在鱼藤酮帕金森病小鼠模型中,半迷走神经切断术阻止了α-突触核蛋白在迷走神经背侧运动核(DMNV)中的积聚以及与该方法同侧的黑质多巴胺能细胞死亡。
•暴露于胃肠道中的微生物淀粉样蛋白(CULLI)会增加老年大鼠肠道和大脑中的α-突触核蛋白病理学和炎症,以及转基因α-突触核蛋白过度表达的小鼠,并伴有相应的运动缺陷。
反对证据:
• 涉及将大量α-突触核蛋白注射到胃肠道的动物模型可能无法准确再现人类帕金森病。
• 中枢神经系统紊乱也会改变肠道功能;例如,大鼠的黑质病变导致肠道运动障碍、炎症和微生物组改变。
半透性肠道屏障调节营养吸收,允许免疫感应,同时限制有害物质的运输。
在帕金森病中,“渗漏”的肠道允许微生物和分子(包括细菌和毒素)通过肠道上皮细胞的流量增加。表达脂多糖(LPS)的失调的肠道微生物群可以激活上皮细胞、免疫细胞和肠胶质细胞上表达的Toll样受体,并促进肠道渗漏以及肠道和脑炎症。
正如在患者结肠活检和粪便样本中观察到的那样,肠道炎症部分通过系统机制驱动疾病发病,包括细胞因子产生增加、血脑屏障破坏、炎性细胞迁移到大脑和小胶质细胞激活,最终导致神经功能障碍或丢失。
恶性循环可能发生,生物失调、通透性增高、炎症和α-突触核蛋白聚集相互延续。
这些因素的重要性和时间顺序可能因患者而异,这取决于环境影响(如影响微生物组的饮食)、衰老和宿主遗传学,导致不同的途径更容易出现功能障碍。
α-突触核蛋白参与先天性和适应性免疫
炎症环境增强了α-突触核蛋白的表达、错误折叠和聚集,进而诱导局部促炎免疫反应,从而形成促进α-突触核蛋白扩散的反馈环。
帕金森患者肠道屏障、炎症和α-突触核蛋白聚集
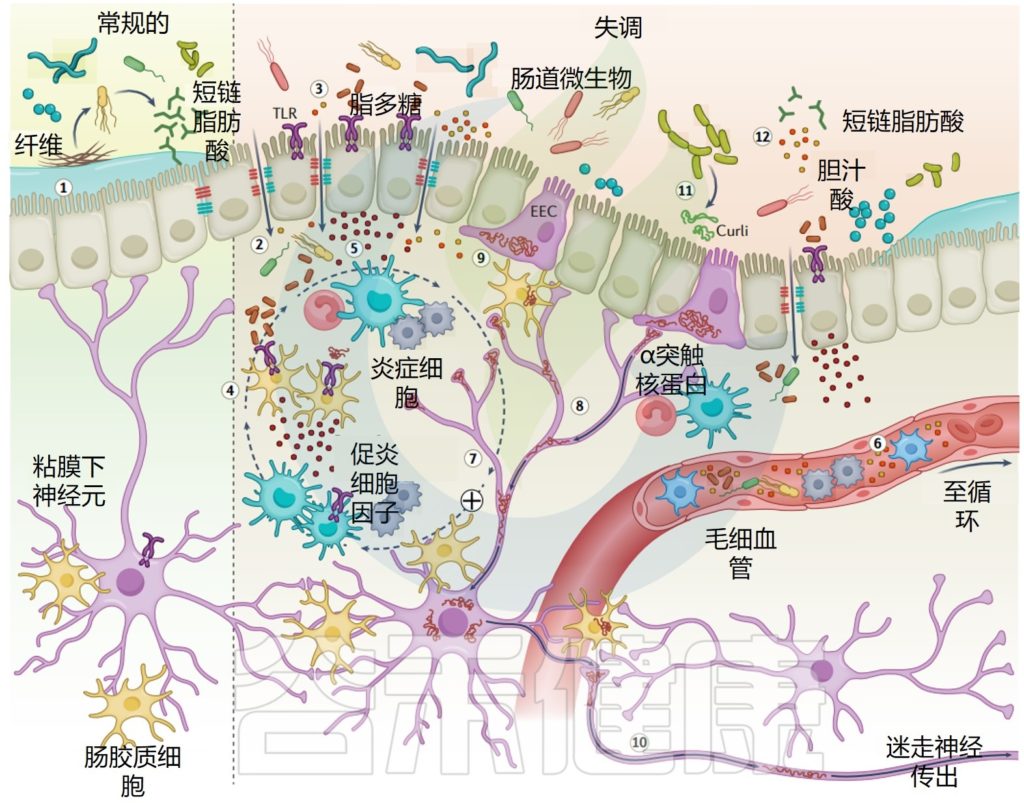
Tan AH, et al.,Nat Rev Neurol. 2022
值得注意的是,轴突终止于肠腔的粘膜下神经元和具有直接管腔和神经连接的肠内分泌细胞可以表达α-突触核蛋白,可能是α-突触核蛋白聚集的来源,并通过迷走神经扩散到大脑。微生物卷曲被建议在肠道中模板α-突触核蛋白聚集。
虽然肠道炎症和α-突触核蛋白表达可能是常见事件,但人们认为只有在存在其他促发因素(如宿主遗传脆弱性或衰老)时,帕金森才会发生。潜在有益的肠道代谢物的改变,如短链脂肪酸也可能在帕金森病发病中发挥作用。
帕金森病中微生物-肠-脑轴的示意图
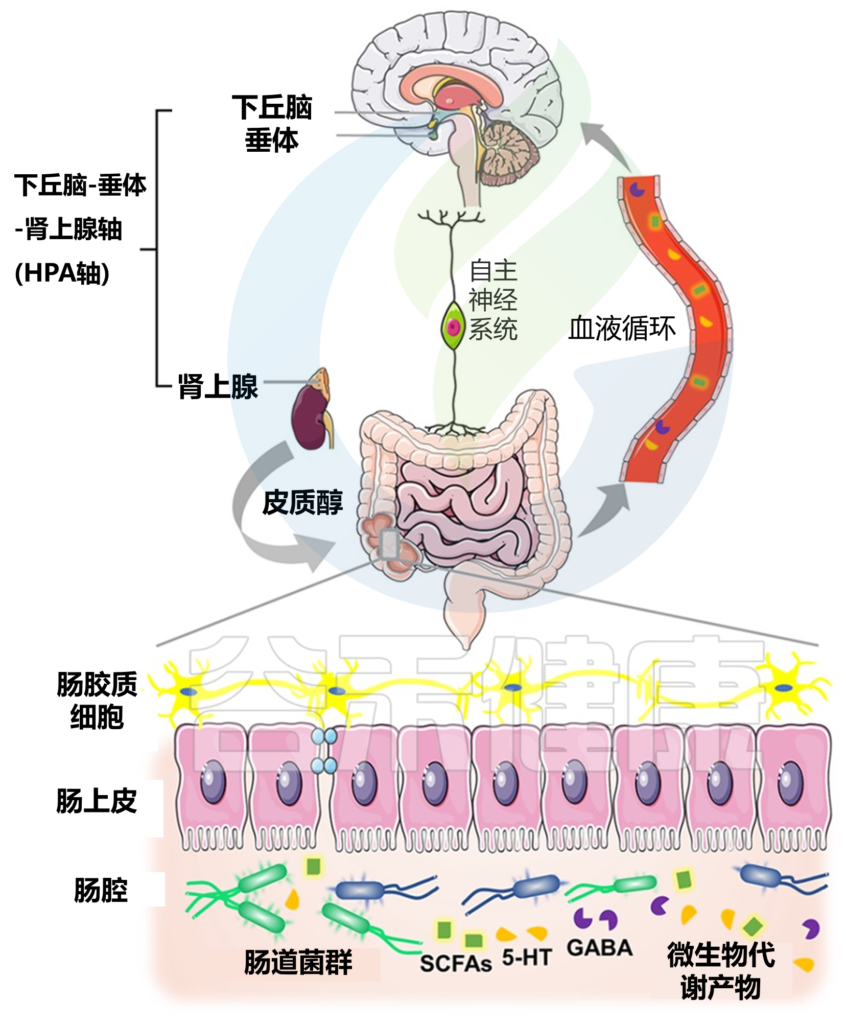
Wang Q, et al., Brain. 2021
在肠神经系统中,肠道菌群通过释放各种信号分子来调节大脑功能,这些信号分子可能进入体循环并到达目标部位。
中枢神经系统通过自主神经系统 (ANS) 下丘脑-垂体-肾上腺 (HPA) 轴在生理上支配胃肠道;反过来,肠道向大脑提供反馈以建立双向交流。
肠道微生物与宿主共同进化,形成复杂的互惠关系:胃肠道为其微生物群落提供滋养环境,而微生物群发挥着广泛的重要功能,影响宿主的生理机能。
在解释与疾病相关的肠道微生物组组成和功能改变时,宿主-微生物组相互作用的多维度和微生物组的个体变异是需要考虑的重要因素。
各种内在和外在因素改变了肠道微生物组,导致微生物组成的显著个体间差异。
影响肠道菌群的因素
Tan AH, et al.,Nat Rev Neurol. 2022
微生物组适应环境和宿主因素的变化并不总是产生有害后果,宿主-微生物和微生物-微生物相互作用的动力学是整个肠道微生物组成和功能的重要决定因素。
事实上,健康个体之间的微生物组成可能差异很大,但仍保持着非常相似的功能能力,这为肠道健康提供了更好的衡量标准。因此,对肠道失调的研究不仅应关注与疾病相关的微生物组成变化,还应考虑影响疾病病因、发病机制、诊断、预后和治疗的功能变化。
衰老是帕金森病病理生理学的一个重要因素,并对肠道微生物生态系统有重要影响。在讨论PD相关变化之前,我们先概述与年龄相关的肠道微生物组变化。
与年龄相关的肠道菌群变化可归因于肠道生理衰老(例如,黏膜变薄、屏障破坏、肠道运动能力下降和免疫衰老)、生活方式的改变(例如,饮食、生活条件和体育活动)和健康状况(例如,共病、药物和虚弱)。
关于肠道微生物群在衰老过程中发生变化详见我们之前的文章:
肠道微生物群的老化及其对宿主免疫力的影响
健康长寿的步伐永不停歇
值得注意的是,肠道菌群具有调节炎症信号和与其他器官相互作用的能力,越来越多地认为它在“炎症”中发挥着核心作用。
来自ELDERMET联盟的开拓性研究表明,老年人肠道微生物组个体间具有高度的差异性,居住在社区的老年人与居住在长期护理机构的老年人之间存在显著差异,后者的微生物组多样性和时间稳定性较差。
长期护理患者的微生物组变化也与饮食因素、营养状况受损以及炎症和虚弱标志物的增加有关。
▸ 百岁老人的肠道菌群多样性较高,有益菌富集
在寻找健康老龄化和长寿的微生物标志的过程中,对百岁老人和超级百岁老人的研究几乎无一例外地揭示出,与其他老年人和年轻人相比,他们的微生物组多样性水平很高,不同地域不同人群研究表明长寿个体会富集例如瘤胃球菌(Ruminococcaceae)和阿克曼菌(Akkermansia)在内的几个细菌类群,同时,两组shotgun宏基因组的功能分析表明,长寿人群的微生物群具有较高的碳水水化合物和脂类代谢能力,以及外源生物降解能力。
▸ 炎症变化导致菌群变化
随着年龄的增长,可能出现相关炎症,引起肠道生理和营养环境变化,这时候,能够适应炎症的肠道环境的菌群,具有竞争优势,而其他微生物可能在炎症状态下丢失。
整合了来自 11 个国家的 30 项独立病例对照 NGS 研究,几乎所有的研究都发现,帕金森病患者和对照组之间的总体肠道微生物组组成存在显著差异。
重要的是,一些研究发现了可能影响肠道微生物组的潜在混杂因素,包括年龄、饮食因素(可能受到吞咽困难或 PD 嗅觉和味觉改变的影响)、便秘、虚弱和帕金森病药物(COMT 抑制剂和左旋多巴),虽然在控制这些因素后,帕金森病患者和对照组之间的差异仍然很大,不过这些差异的影响大小是在适度范围内的。
事实上,研究方法和地理位置可以解释微生物组组成中比PD状态更大的方差比例(在18-54%之间)。与之前关于微生物多样性减少与不健康衰老之间关系的观点相反,大多数研究发现帕金森病患者与对照组相比要么没有差异,要么总体多样性增加。
帕金森病:AKK菌、双歧杆菌、乳酸杆菌增加
据报道,帕金森病患者与对照个体之间至少有42科、102属和44种细菌存在差异。然而,只有大约四分之一的研究结果在一项或多项研究中得到了重复。
帕金森病最一致的发现是阿克曼菌(Akkermansia)的丰度增加(30项研究中的14项),该属属于疣状菌科(12项研究中也发现帕金森病增加)。
其他组成特征包括双歧杆菌(Bifidobacterium)属和乳酸杆菌(lactobacillus)属的丰度增加,丁酸生产菌(Roseburia)、粪杆菌属和布劳特氏菌(Blautia)的丰度减少。
帕金森病患者在物种或亚种水平上的研究仍然很少,在两项鸟枪式宏基因组研究和一些具有较高测序分辨率的16S rRNA研究中进行了初步探索。
Akk菌增加可能的原因
阿克曼菌(Akkermansia)、双歧杆菌(Bifidobacterium)和乳酸杆菌(lactobacillus) 被广泛视为“有益”细菌,后两个属通常包含在益生菌制剂中。
它们在帕金森病含量增加的原因尚不清楚,但可能与它们在改变的肠道环境中生存的适应性更强有关。值得注意的是,Akkermansia的增多与结肠运输缓慢和体重或脂肪量低有关,这两个特征在帕金森病患者中普遍存在。
其他神经系统疾病中也有Akkermansia丰度增加的报道,包括多系统萎缩(MSA)、进行性核上性麻痹、阿尔茨海默病和多发性硬化症,表明这一特征并非帕金森病特有。
扩展阅读:Nature | AKK菌——下一代有益菌
引人注目的是,对跨越十种疾病的28项病例对照研究进行的荟萃分析发现,与疾病相关的细菌属之间存在明显的重叠,表明某些变化代表了对健康和疾病的共同反应。
菌株特异性
细菌的功能可能具有菌株特异性,同一物种的不同菌株可能产生相反的作用。因此,根据目前在属和种水平上的知识推断细菌在帕金森病中的潜在作用可能信息不足。
此外,人类肠道微生物群中的大多数基因尚不能进行功能分配,其动态转录和翻译活动尚待阐明。
在统一的人类胃肠道基因组目录中,从超过200000个人类微生物基因中鉴定出的物种中,有70%以上仍有待培养或实验鉴定,对这些物种内的菌株和功能变异性的了解仍极为有限。
至关重要的是,单一微生物分类群的作用不能在隔离中看待,但应在社区环境中考虑,考虑到具有类似代谢功能的微生物与具有对抗作用的微生物的净效应,以及微生物-微生物和微生物-宿主相互作用。
因此,使用粪便转录组学、代谢蛋白质组学和代谢组学对代表肠道菌群集体功能输出的活性细菌代谢途径和产物进行直接测量,可能比细菌分类学描述更具相关性,以便更深入地了解肠道菌群在健康和疾病中的作用。
帕金森病患者:短链脂肪酸下降
帕金森病患者粪便短链脂肪酸的水平(乙酸盐,丙酸盐,丁酸盐)一直在下降。短链脂肪酸在微生物群-肠-脑串扰中发挥关键作用,部分通过调节肠道和血-脑屏障的完整性、炎症级联、内分泌信号和神经元存活。
除了作为结肠上皮的主要基质外,实验性帕金森模型中还发现丁酸通过组蛋白脱乙酰酶抑制发挥神经保护作用,包括减轻运动损伤和多巴胺能细胞死亡。
在MSA中也发现粪便短链脂肪酸和胆碱代谢产物水平降低,表明这种情况与帕金森病之间存在共同的微生物反应。
帕金森病患者肠道微生物组和代谢物的临床相关性在后面章节关于帕金森病风险和进展中的肠道相关因素中进行了讨论。
在几种帕金森病啮齿动物模型中已证实肠道微生物群的改变,包括:通过腹腔注射或皮下注射的神经毒素模型等,大多数研究还发现,与野生同窝鼠相比,患病啮齿动物的胃肠道(肠道运动障碍、炎症、屏障破坏和神经功能丧失)或大脑(运动功能障碍、炎症和多巴胺能细胞丧失)发生病理生理学变化。
在其中一些研究中,观察到肠道变化先于帕金森病样运动特征的出现;然而,肠道微生物对观察到的变化的贡献尚不清楚。
与帕金森病相关的菌群改变的因果关系推断
几项使用无菌小鼠(出生和饲养时未接触活微生物)、粪便微生物移植(FMT)或使用广谱抗生素治疗的研究,可以推断出与人类帕金森病相关的微生物组改变的因果关系。
与无菌ASO小鼠相比,含有复杂微生物群的ASO小鼠运动功能和便秘更差,伴有明显的神经炎症反应和脑内α-突触核蛋白沉积。
值得注意的是,帕金森病患者肠道微生物群的移植加剧了无菌ASO小鼠的运动功能障碍。有趣的是,补充短链脂肪酸(SCFA)通常被认为是有益的,与无菌ASO小鼠中的小胶质细胞激活和α-突触核蛋白聚集有关。
然而,无菌小鼠小胶质细胞缺陷的观察结果使这些发现与人类帕金森病的解释复杂化。短链脂肪酸的作用也可能取决于剂量和类型;肠道中不同短链脂肪酸浓度和比率的差异可能导致不同的健康结果。
在另一项研究中,长期口服鱼藤酮会导致常规饲养小鼠的肠道通透性增高和运动功能障碍,但在无菌小鼠中则没有。
在另外两项针对啮齿动物的研究中,抗生素治疗耗尽了肠道微生物负荷和多样性,似乎可以减轻MPTP和六羟基多巴(6-OHDA)诱导的多巴胺能神经毒性。
综上所述,这些实验模型提供了证据,证明肠道微生物的存在、组成和代谢产物在帕金森病样发病机制中具有致病或促进作用。
肠道微生物及其代谢产物的高度个人化和复杂性质使微生物组因果成分的鉴定变得复杂,这些微生物及其代谢产物可能单独发挥作用,或在微生物群中发挥增强或抵消作用。
此外,大多数研究中使用的供体数量较少,无法捕获人类肠道微生物群的广泛个体间差异。至关重要的是,未能解释宿主-微生物相互作用和内环境平衡及其在个人整个生命周期中的演变,以及饮食、生活方式、衰老和其他因素的变化,是这些实验的主要限制。
几项大型流行病学和临床研究将肠道疾病、肠道干预和饮食因素与随后的帕金森病发展和恶化风险联系起来。
便秘是帕金森病的典型前驱症状,20多年前发表了第一项基于社区的长期研究,证明患有这种疾病的个体患帕金森病的风险升高。
帕金森病的便秘是由一系列因素引起的,包括结肠传输缓慢和肛门直肠功能障碍。2021发表的一项研究表明,便秘的不同亚型与帕金森病的不同发病风险相关,慢传输比排便协同障碍的发病风险略高。
便秘也预示着帕金森病的恶化趋势
一项针对465名社区帕金森病患者的研究发现,患者在发病时便秘的存在和严重程度与痴呆更快进展之间存在关系。
这些患者从诊断开始就接受了长达8年的监测。严重便秘患者平均得痴呆症的时间为6.0年,而无便秘的患者则为7.5年。排便协同障碍是便秘患者排便过程中出现的一种反常肛门括约肌收缩障碍。
同样,使用来自帕金森病进展标志物倡议423名新诊断帕金森病患者的数据,这些患者接受了长达5年的监测发现,胃肠道症状的综合得分越高(肠蠕动、大便硬和失禁的频率越低),认知能力越差,轻度认知障碍和痴呆的发生率越高。
在这两项研究中,认知功能与非胃肠道自主症状无关,这表明这种关联是胃肠道系统特有的,而不是自主功能障碍本身。
帕金森病患者便秘的相关因素
在一个单独队列的3年随访研究中,基线便秘严重程度与体脂质量损失独立相关。反过来,帕金森病患者的体重减轻与营养不良、骨折、压疮和死亡的风险增加有关。
2021发表的一项粪便代谢组学研究发现,帕金森病患者的便秘严重程度与丁酸盐(一种潜在有益的SCFA)水平降低相关。
另一项初步研究发现,帕金森病和便秘患者外周血中辅助性T细胞的数量增加。这种具有强大炎症潜能的CD4+T辅助细胞亚群可由特定的肠道细菌诱导,并与帕金森病(以及炎症性肠病)的发病机制有关。
长期以来,人们推测结肠淤滞会导致细菌产物和其他毒素更多地转移到循环中;尽管这一点尚未得到令人信服的证明,但粪便稠度被认为对肠道微生物群组成有重大影响。
如上所述,炎症与帕金森病的发病机制有关,帕金森病患者的一些结肠活检和粪便结果与炎症性肠病患者相似。
迄今为止最大的一项研究调查了美国1.7亿多人的索赔数据,发现炎症性肠病患者的帕金森病发病率增加了28%。
荟萃分析表明,当分别分析克罗恩病患者和溃疡性结肠炎患者时,帕金森病风险的增加仍然显著。研究人员还发现,抗TNF疗法可以显著降低这种风险,这表明炎症性肠病的有效治疗可能会降低帕金森病的发病风险。
一项利用瑞典全国组织病理学数据库进行的研究还发现,显微镜下结肠炎(另一种慢性肠道炎症疾病)与帕金森病之间存在强烈的相关性。
遗传学研究表明,帕金森病和炎症性肠病患者(尤其是克罗恩病)的发生有共同的生物学机制。LRRK2基因中的突变是单基因(常染色体显性)帕金森病最常见的原因之一。
随后的几项研究确定了LRRK2变异体与散发性帕金森病和克罗恩病之间的关联,还报告了帕金森病和炎症性肠病的其他重叠风险位点。这些位点进一步暗示了免疫反应和微生物感应(例如,NOD2基因和HLA位点),以及溶酶体功能障碍,这是两种情况下的常见机制。
约有50个和10个基因分别与单基因炎症性肠病和帕金森病相关,但导致这两种疾病的单基因形式的罕见变异迄今尚未报道。
最后,在2022年发表的一项研究中,与无结肠炎的小鼠相比,患有结肠炎的小鼠的运动功能降低,多巴胺能神经元丢失加重,进一步支持LRRK2、结肠炎和帕金森病之间的联系。
肠易激综合征(IBS)症状,如腹痛和肠道习惯改变,在帕金森病患者中很常见,并与粪便微生物群的变化相关。
在肠易激综合征患者身上观察到的病理生理变化,包括失调、肠道通透性增加和轻度粘膜炎症,可能会使胃肠道更容易受到帕金森病病理的影响。
肠易激综合征与帕金森病之间的关联
在一项来自台湾的全国性注册研究中,涉及约120000人,肠易激综合征与帕金森病风险增加相关。
然而,在芬兰进行的一项类似研究中,在前2年的随访中,风险显著升高,导致作者得出结论,肠易激综合征与帕金森病之间的关联可归因于反向因果关系(即帕金森病的前驱胃肠道症状被误解为肠易激综合征症状);或检测偏差(即,对肠易激综合征症状患者进行更密切的临床监测可提高诊断帕金森病的可能性)。
韩国的一项研究包括约285000名受试者,他们接受了长达11年的监测,发现患有肠易激综合征的老年人(65岁及以上)患帕金森病的风险增加。
迄今为止最大的一项涉及瑞典170万参与者的研究发现,专家诊断肠易激综合征与帕金森病风险增加相关。
然而,在同一研究者进行的补充队列研究中,肠易激综合征的诊断基于自我报告的症状,结果没有统计学意义。因此,这种常见的肠-脑相互作用障碍(影响全球十分之一的人)与帕金森病之间的可能联系仍然没有定论。
迷走神经
迷走神经起源于延髓,在腹腔内为胃、小肠和大肠提供副交感神经支配,直至脾曲。
在两项大型研究中,完全躯干(但非选择性)迷走神经切断术与患帕金森病的风险显著降低相关。
另一项研究分析了相同的丹麦数据库,但包含了较长时间内的大量病例,并使用了不同的编码分类和统计方法,发现躯干或其他类型的迷走神经切断术没有显著降低风险。
相互矛盾的结果可能部分归因于迷走神经切断术的混合效应:虽然这种干预可能限制α-突触核蛋白病理学从肠道向大脑的传播,但由于胆碱能抗炎途径的破坏,它也可能导致免疫稳态向促炎反应的转变。
阑尾切除术
虽然阑尾以前被认为是一个退化器官,但现在人们认识到它富含多种免疫细胞,可以影响胃肠道的炎症和自身免疫,它还充当肠道微生物组的蓄水池和调节器。
研究发现,除了迷走神经背侧运动核的密集神经支配外,阑尾在帕金森病发病机制中的可能作用还表现在该器官中大量聚集的α-突触核蛋白。
在一项研究中,发现年轻人和老年人(从婴儿到84岁)的阑尾中都有大量截短形式的α-突触核蛋白,这些截短形式的α-突触核蛋白是路易小体的主要成分,在实验模型中被证明是α-突触核蛋白聚集的有效种子,包括那些没有神经系统疾病的人,表明阑尾是α-突触核蛋白致病形式的储库。
在2021发表的一项研究中,53%的帕金森病患者和8%的对照组患者的CT扫描显示有慢性阑尾炎的病变。
一小群7名患者接受了阑尾切除术,对切除组织的组织病理学分析显示,阑尾粘膜和粘膜下层存在α-突触核蛋白聚集。
在一些研究中,阑尾切除术与帕金森病风险降低相关,但在大多数研究中,未发现任何影响,甚至风险轻度升高。
有趣的是,一项针对12名帕金森病患者的小型研究采用多组学方法研究了阑尾微生物群,发现了脂质和胆汁酸代谢紊乱的证据,包括阑尾中微生物源性(次生)胆汁酸升高5.6–18.7倍。这些胆汁酸具有毒性作用,并与肠道炎症有关。
与上文所述的对帕金森病下消化道失调的理解的扩展相反,关于帕金森病上消化道失调的研究主要集中在幽门螺杆菌感染和小肠细菌过度生长(SIBO)。
有幽门螺杆菌感染史的人帕金森风险高
幽门螺杆菌感染与帕金森病之间的联系始于20世纪60年代,一项观察性研究显示,患有这种疾病的患者中消化性溃疡的发生率很高,通常在8-10年前出现帕金森症状。
一项来自台湾的研究发现,有感染史的人患帕金森病的风险增加了一倍。
来自丹麦的另一项研究发现,患者根除幽门螺杆菌的治疗史与5年或更长时间后患帕金森病的风险增加45%相关。
帕金森病患者的幽门螺杆菌感染率在不同地理区域的研究中有所不同,从呼吸和粪便抗原检测的25-33%到血清学检测的48-50%不等。这些比率与当地人口的比率相当。
帕金森患者的小肠细菌过度生长患病率范围广
小肠细菌过度生长可作为幽门螺杆菌诱导的胃酸过少的后遗症;其他原因包括使用质子泵抑制剂、小肠运动障碍和免疫反应受损。
研究发现,帕金森病患者的小肠细菌过度生长患病率范围广泛(25–67%),这可能是由于研究参与者的异质性和呼吸测试协议,这些协议提供了小肠细菌过度生长的间接评估,准确性有限。
幽门螺杆菌感染和小肠细菌过度生长均与更严重的运动严重性和更严重的运动反应并发症(延迟和缩短时间)相关。
致病原理
幽门螺杆菌感染和小肠细菌过度生长可能通过降低左旋多巴的生物利用度,或通过诱导慢性外周炎症状态来加剧神经退行性变过程,从而导致帕金森病患者的运动功能障碍。
例如,幽门螺杆菌诱导的细胞毒素可以改变宿主的免疫反应并触发大量促炎物质的释放,这可能导致血脑屏障的破坏、小胶质细胞的激活,最终导致神经元损伤和死亡。
在一项小型研究中,幽门螺杆菌阳性的帕金森病患者被发现对神经功能至关重要的蛋白质的自身抗体水平升高,包括核因子I亚型A、血小板衍生生长因子B和真核翻译起始因子4A3。
另一方面,肠道运动障碍(以及更普遍的自主神经功能障碍)可能是严重帕金森病的表现,并可能导致小肠细菌过度生长,导致观察到这些疾病之间的关联。
几项研究调查了肠道微生态系统与帕金森病风险或前驱症状之间的关系,包括睡眠行为障碍(RBD),它可能在 PD 运动诊断之前 20 年或更长时间。
肠道菌群
对来自德国和日本的队列(包括46名特发性睡眠行为障碍(RBD)患者、223名帕金森病患者和137名对照个体)进行的荟萃分析表明,睡眠行为障碍患者与帕金森病患者具有不同的微生物成分,尽管观察到一些重叠(例如,两种情况下Akkermansia的丰度增加)。
有趣的是,与对照组相比,在一小群睡眠行为障碍患者中,产生短链脂肪酸的细菌数量并没有减少,
作者认为短链脂肪酸可能在孤立性快速眼动睡眠行为障碍向帕金森病的转化中发挥作用。同时,在包括666名个体的大趋势队列研究中,一些帕金森病的风险或前驱标记物与微生物组组成的改变相关,尽管其影响很小。
如上所述,病例对照研究已经确定了帕金森患者中存在大量差异丰富的菌群。然而,在单独的研究中,只有少数与运动特征(MDS-UPDRS评分、运动反应并发症或姿势不稳定和步态困难(PIGD)表型)或非运动特征(认知、抑郁、焦虑或便秘)相关。
在一项研究中,肠杆菌科细菌的丰度与更严重的运动表型相关,这与大肠杆菌通过产生脂多糖和卷曲菌毛在帕金森病发病机制中的可能作用有关。
纵向研究的结果也不一致。在25名新发PD患者的3年随访研究中,更快的PD运动进展(使用Hoehn和Yahr评分量表或UPDRS测量)与较低的基线Roseburia丰度相关;在36名PD189患者的2年随访研究中,使用19种细菌分类群的定量PCR,与双歧杆菌和Atopobium 的基线丰度相关。
短链脂肪酸
帕金森病患者肠道微生物群衍生代谢的临床研究仍然很少。低水平的粪便丁酸盐与较差的认知功能和步态困难评分以及更早出现运动和非运动症状相关,而低水平的粪便乙酸盐与低BMI和排便频率降低相关。
帕金森病患者的血浆乙酸盐和丙酸盐水平高于对照组。一些研究人员推测,由于肠道屏障破坏和结肠上皮细胞对肠道短链脂肪酸的利用或清除减少,肠道短链脂肪酸会泄漏到体循环中。
血浆丙酸水平与运动严重程度呈负相关,这与短链脂肪酸对帕金森病有益的总体临床趋势一致。
氧化三甲胺(TMAO)
另一种对帕金森病越来越感兴趣的肠道代谢物是氧化三甲胺(TMAO),根据实验研究,氧化三甲胺可以通过促进正确的蛋白质折叠和防止病理性不溶性纤维的形成来防止神经变性。
如上所述,一些研究发现帕金森病患者粪便和血浆样本中的氧化三甲胺水平均降低,尽管其他研究发现帕金森病患者的血浆氧化三甲胺水平高于对照组。血浆氧化三甲胺水平与帕金森病进展之间的相关性也得到了相互矛盾的结果。
饮食是决定微生物组成和功能的关键因素。肠道微生物将摄入的营养物质用于基本的生物过程,反过来,它们的代谢产物被认为对人类健康有重要影响。
尽管每种主要的宏量营养素和微量营养素都能改变肠道微生物群,但为大肠微生物提供主要能量来源的不可消化碳水化合物(例如纤维和抗性淀粉)在饮食-微生物群相互作用中发挥着特别突出的作用。
这些聚糖的低消耗与微生物多样性降低、短链脂肪酸产量减少以及肠道微生物对粘液的觅食增加有关,导致肠道屏障完整性中断。
许多研究调查了帕金森病风险与饮食成分(包括大量营养素、微量营养素、特定食物和饮食模式)之间的关系。
饮食与帕金森病风险和进展
Tan AH, et al.,Nat Rev Neurol. 2022
研究结果往往相互矛盾,前瞻性研究和病例对照研究之间存在差异。虽然前瞻性研究比病例对照研究更好地确定饮食和疾病之间的因果关系,但它们不能完全消除反向因果关系,即患者因疾病早期迹象改变饮食习惯。
此外,病例对照研究受到选择和饮食回忆偏差的限制。然而,令人惊讶的是,咖啡因研究的结果具有显著的可重复性,这使得咖啡因成为最能降低帕金森病风险的环境暴露之一。
地中海饮食
近年来,地中海饮食已成为一种潜在的“健康”饮食模式,与降低慢性病(包括帕金森病)的风险相关,以及几项前瞻性研究中的总体死亡率减低。
这种饮食通常包括相对较高的水果、蔬菜、全谷物和不饱和脂肪的摄入量,相对较低的肉类(包括家禽)和乳制品的摄入量,以及适度的酒精摄入。
据推测,地中海饮食的有益健康效应部分是通过保护机体免受氧化应激和炎症的影响,以及通过肠道微生物代谢产物改善宿主代谢的调节来实现的。
与其他慢性病一样,大量研究与缺乏减少帕金森病发展或进展的决定性饮食建议之间仍存在明显差距。对饮食化合物反应的个体间差异,可能源于肠道微生物群和宿主基因组成的差异,进一步使饮食与疾病之间的关联复杂化,基于人群的研究结果可能对个别病例的普遍性有限。帕金森的饮食干预研究仍处于起步阶段,将在后面的治疗策略部分进行讨论。
前面介绍了LRRK2基因、帕金森病和炎症性肠病之间相互关系的概念。LRRK2蛋白在免疫细胞中高度表达,并在接触微生物病原体后上调。
与克罗恩病患者的非炎症结肠组织相比,炎症结肠组织中检测到更高水平的LRRK2,与匹配对照组相比,散发性帕金森病患者的外周免疫(血单个核)细胞中检测到更高水平的LRRK2。
LRRK2过度表达也显示出增加结肠炎的严重程度,因此,LRRK2被认为在一定程度上通过介导肠道中的异常炎症反应参与帕金森病的发病机制。然而,其他作者推测,通过增强肠道细菌清除和减轻肠道炎症,增加LRRK2激酶活性会产生有益的效果。
免疫细胞中高水平的LRRK2是否对帕金森病产生保护作用或有害作用,目前尚不清楚。
就动物模型而言,缺乏PTEN基因诱导的假定激酶1(PINK1)的小鼠的肠道革兰氏阴性细菌感染,该激酶参与线粒体内环境平衡,并由帕金森病的某些单基因形式突变的基因编码,触发外周和大脑中自反应性线粒体特异性细胞毒性CD8+T细胞的产生,导致纹状体多巴胺能丧失和运动障碍。
左旋多巴是帕金森病的主要治疗药物,必须到达原代小肠才能被吸收。
胃轻瘫是一种在没有机械阻塞的情况下胃排空延迟的情况,导致左旋多巴留在胃中并转化为多巴胺,而多巴胺不能穿过血脑屏障进入大脑。
反过来,胃中的多巴胺可能进一步损害胃运动。通过使用无线运动胶囊(“智能药丸”),这是一项相对较新的技术,35%患有各种胃肠道症状的帕金森病患者发现胃排空时间延长,20%有小肠传输延迟,8%有合并传输异常。
胃轻瘫通过降低左旋多巴的生物利用度,导致反应波动,如延迟开启状态,甚至剂量失效。因此,胃轻瘫的治疗,以及“绕过”胃的新疗法,可能会缓解帕金森病的运动波动和症状。
饮食蛋白质限制可以减少摄入的氨基酸和左旋多巴之间的竞争,以实现通过肠道上皮和血脑屏障的可饱和主动运输;然而,由于人们认识到体重减轻、营养不良、肌肉减少和骨质疏松症在帕金森病中已经很常见,并且可能被这种方法放大,因此现在人们往往不鼓励这种做法。
胃肠功能和左旋多巴药代动力学
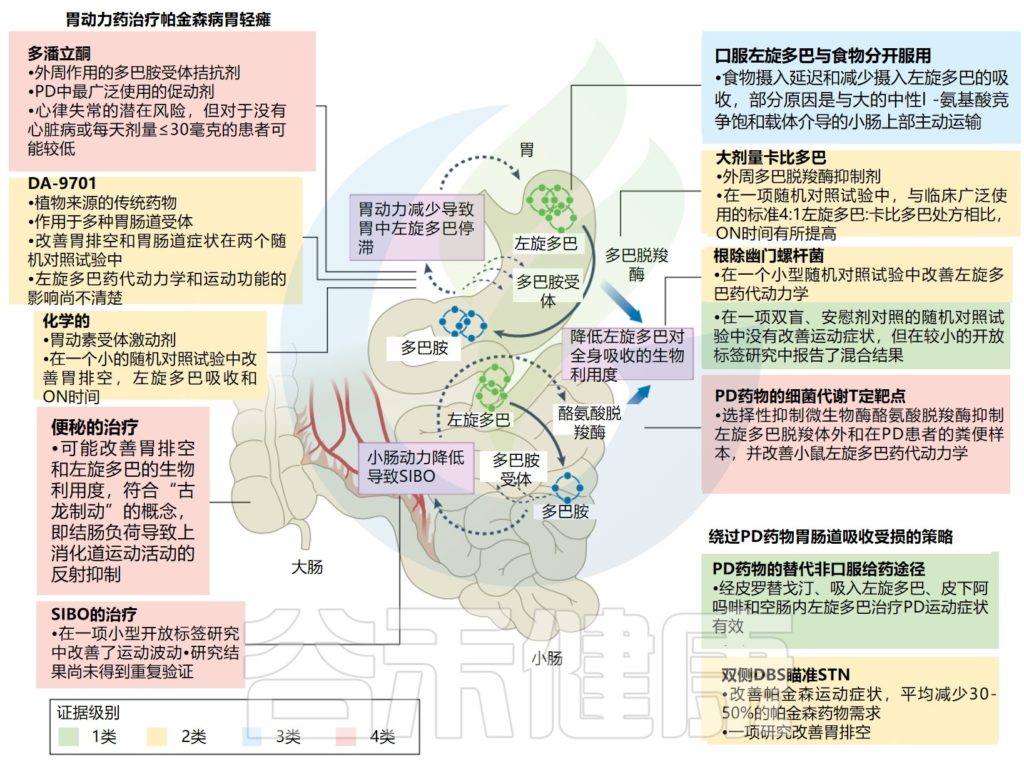
Tan AH, et al.,Nat Rev Neurol. 2022
该图说明了胃轻瘫和小肠微生物活性对帕金森病患者左旋多巴药代动力学的影响,并概述了改善胃肠功能和提高左旋多巴生物利用度的治疗策略。
促动力药物,尤其是多潘立酮,是治疗帕金森病相关胃轻瘫的主要药物,临床试验证明了DA-9701和Camicinal (胃动素受体激动剂)等新型促动力药物的潜在益处,而其他药物,如5-羟色胺5-HT4受体激动剂莫沙必利和普鲁卡因丙利、组胺激动剂尼扎替丁和胃饥饿素(ghrelin)激动剂对帕金森病的益处的证据仍然有限。
从食物中单独口服左旋多巴或与大剂量卡比多联合使用可以帮助提高左旋多巴的生物利用度。一些临床研究探讨了幽门螺杆菌感染、小肠细菌过度生长(SIBO)和便秘治疗对左旋多巴生物利用度和帕金森病运动反应并发症的影响。
肠道微生物在左旋多巴代谢中的作用也得到强调,并可能开辟新的治疗途径来改善帕金森病药物的吸收。绕过胃肠道的策略,包括非口服形式的多巴胺替代品和DBS,是治疗帕金森病运动症状的可能选择。
高度多样化的肠道微生物群及其丰富的酶库可以将许多药物转化为活性更高或活性更低的代谢物,现已被认为是药物药理、疗效和毒性个体间差异的主要因素。
在一项开创性的研究中,在271种不同的口服药物中,三分之二的药物被76种人类肠道细菌分离物中的至少一种显著耗尽,每种菌株代谢11-95种不同的药物。
细菌酪氨酸脱羧酶的发现可以在小肠中将左旋多巴转化为多巴胺,从而限制其生物利用度并可能导致外周多巴胺能不良反应,这为帕金森病患者对左旋多巴观察到的显著个体间反应提供了一种解释。
粪肠球菌(Enterococcus faecalis)是负责左旋多巴脱羧的主要微生物。考虑到小肠细菌过度生长与运动严重程度和波动之间的关联,推测小肠中的细菌过度生长会导致酪氨酸脱羧酶或其他代谢左旋多巴的细菌酶的产生增加。
值得注意的是,左旋多巴肠道细菌代谢产生的多巴胺也会损害肠道运动,可能导致恶性循环,增加小肠细菌过度生长。
然而,目前几乎没有直接证据支持这些观点,部分原因是进入小肠的技术困难。帕金森病的所有研究都使用呼吸试验评估小肠细菌过度生长,如前所述,缺乏标准化。
一项针对18名帕金森病患者的小型研究发现,使用利福昔明进行西博治疗不会影响左旋多巴的药物动力学,但会改善治疗的及时性(但6个月后复发率为43%)。
肠道微生物也可以通过间接机制与左旋多巴相互作用,包括肠道环境的变化(例如,粘膜损伤和炎症),导致细菌降解产物与左旋多巴之间的无效吸收和竞争,以在小肠中进行主动转运。
例如,幽门螺杆菌相关性胃十二指肠炎伴胃低氯酸盐被认为可减少左旋多巴的吸收,左旋多巴在酸性降低的环境中溶解性较低。然而,幽门螺杆菌感染对左旋多巴药代动力学的影响尚不清楚。
一项小型研究(n=34)发现根除幽门螺杆菌后左旋多巴药代动力学得到改善;然而,另一项较大的研究(n=73)没有发现帕金森病幽门螺杆菌阳性和阴性患者之间左旋多巴的药代动力学有任何差异。
此外,尽管几项小型开放标签研究发现根除幽门螺杆菌后运动结果显著改善,但2020年发布的一项双盲、安慰剂对照、随机临床试验(DBPC-RCT)在短(12周)或长(52周)随访后,未发现运动功能或运动波动(使用MDS-UPDRS和可穿戴传感器评估)或生活质量的改善。小肠细菌过度生长状态不影响本研究的结果。
虽然在本研究中根除幽门螺杆菌与任何临床意义的改善无关,但从长远来看或在疾病早期阶段,仍有可能改变根除幽门螺杆菌对帕金森病的影响。
各种微生物导向疗法有望在各种情况下进行症状管理或疾病治疗,但临床试验患者人数相对较少。
最好的证据来自粪菌移植(FMT)对严重梭状芽胞杆菌感染的治疗,证明了约90%的疗效,并为这些方法的临床应用提供了概念证明。
前面我们已经知道,饮食可以通过微生物群-肠-脑轴影响肠道微生物群的组成和中枢神经系统的神经活动。
★ 地中海饮食
前面已经提到,地中海饮食与帕金森的关联。
也有研究表明,坚持地中海饮食与前驱帕金森病的低发病率显着相关。
最近的一项随机临床试验调查了地中海饮食对帕金森病患者认知功能的影响,发现坚持地中海饮食可显着提高执行功能、语言、注意力、记忆力并改善认知功能。因此,地中海饮食可能是减少帕金森病发病和发展的有前景的干预策略。

地中海饮食
地中海饮食是以大量植物性食物为特征的健康饮食模式,如水果、蔬菜、谷类、豆类、坚果等,中低量的乳制品,以橄榄油为主,少量和中等量的鱼和肉,以及少量的红肉,用香料代替盐。
高膳食纤维是地中海饮食的特点之一。膳食纤维结构的细微变化可能强烈影响肠道微生物群的组成和功能。
扩展阅读:你吃的膳食纤维对你有帮助吗?
将富含抗氧化剂的食物(对整体大脑健康很重要)加入饮食中。包括颜色鲜艳的和深色的水果和蔬菜。前瞻性研究发现膳食维生素 E 和胡萝卜素的摄入可以降低患帕金森病的风险。这些抗氧化剂可以中和氧自由基的作用,减少氧化损伤。
橄榄油是地中海饮食的主要脂肪来源,主要含有单不饱和脂肪酸,具有抗氧化活性并抑制α突触核蛋白聚集。橄榄油中的酚类化合物具有抗炎、抗氧化、神经保护和免疫调节活性。
补充水分,每天至少应饮用 8 杯水。
需要限制或避免摄入的:
避免食用含有大量胆固醇或饱和脂肪的食物,少吃肉类(尤其是红肉类)也很重要。
糖、盐、饮料的摄入量也应该严格限制。
尽量限制加工食品的摄入。
★ 禁食疗法
除了我们熟悉的地中海饮食,与预防或治疗帕金森病相关的饮食疗法还包括禁食疗法。
模拟禁食的饮食可以减少 MPTP 诱导的帕金森病小鼠黑质中多巴胺能神经元的损失。
间歇性禁食可以保护疾病动物模型中的神经元,减少多巴胺能神经元的数量并改善帕金森病的运动功能。
关于间歇性禁食详见我们上一篇文章:
间歇性禁食 & 肠道菌群 & 心血管代谢疾病
药物治疗时需要注意避免混合在一起的:
在服用药物治疗帕金森氏症时最好避免饮酒,因为酒精可能会干扰药物治疗。
对于一些帕金森病患者来说,如果服药时间与蛋白质或铁(以铁补充剂的形式)的摄入时间接近,那么服药时间的安排尤其成问题。这些人对药物的吸收很敏感。
总之,饮食可以调节肠道菌群的改变,并通过抗炎、减少氧化应激和神经保护作用对大脑产生影响。饮食介导的治疗可以作为治疗和预防帕金森病的新策略。
益生元主要是不易消化的碳水化合物,短链脂肪酸是由其发酵产生的。益生元是宿主微生物(或施用的益生菌菌株)利用的基质,具有健康益处。
在一项小型开放标签研究(n=19)中,富含不溶性纤维(一种常见益生元食物来源)的饮食改善了便秘、血浆左旋多巴生物利用度和运动功能。
此外,一项有87名参与者参与的开放性研究报告称,在接受益生元干预的帕金森病患者中,使用抗药性进行益生元干预后,非运动症状评分得到改善,粪便丁酸显著增加,粪便钙卫蛋白水平降低。
在仅接受饮食指导的患者中未观察到此类变化,并且两种干预措施均未导致微生物成分或便秘严重程度的显著变化。
帕金森病肠道微生物组的治疗调节
Tan AH, et al.,Nat Rev Neurol. 2022
益生菌是活的微生物,当剂量足够时,可带来健康益处。
两项研究为多系列益生菌的价值提供了I级证据(作为添加益生元纤维的发酵乳服用(n=120)或作为每日一次的胶囊配方(n=72))用于治疗帕金森病便秘,据报道,肠道频率和粪便组成有所改善。
益生菌配方中包括各种乳酸杆菌(Lactobacillus)、双歧杆菌(Bifidobacterium)和肠球菌(enterococcus)菌株。由于这两项研究的干预持续时间仅为4周,因此需要进一步研究,以检查益生菌治疗帕金森病的长期疗效和安全性。
事实上,大多数益生菌制剂中都含有双歧杆菌和乳酸杆菌,但它们是帕金森病患者中已经普遍升高的两个属。肠球菌有时也包括在益生菌中,但已显示出强大的左旋多巴降解能力,强调在PD患者中使用这些益生菌时需要谨慎。
粪菌移植(FMT)对肠道微生物群的调节作用可能通过免疫、内分泌、代谢和神经通路对神经系统疾病的症状或进展产生影响。
粪菌移植在帕金森病患者便秘治疗中的潜在价值已在6名患者的病例系列和11名患者的开放标签研究中显示。
从供体向患者传播传染病或其他微生物介导的特征是与这种方法相关的潜在风险。
在两项研究中,供体粪便菌群均采用内窥镜管理,无重大不良事件发生。目前正在研究粪菌移植(FMT)对帕金森病运动和非运动症状的影响。
抗菌剂减少了许多肠道微生物的数量,可能无法区分有害微生物和有益微生物。此外,反复治疗(例如,反复出现的小肠细菌过度生长)也有产生抗生素耐药性的风险。
上文讨论了根除幽门螺杆菌或治疗帕金森病SIBO的抗生素。尽管在研究后1年的随访中,根除幽门螺杆菌并没有改善临床特征,但潜在的疾病改善效应仍然存在。
在几项研究中发现的安慰剂干预后小肠细菌过度生长状态的自发变化和治疗后相对较高的复发率突出了与研究和试图根除帕金森病患者小肠细菌过度生长相关的挑战。
后生元是指赋予健康益处的无生命(杀死的)微生物和/或其成分的制剂。作用于肠道相关靶点的小分子、生物制剂或代谢物可能包括抑制肿瘤坏死因子、Toll样受体或富含亮氨酸的重复激酶2信号、短链脂肪酸和肠道屏障恢复治疗,如紧密连接调节剂或粘液稳定剂。
其他有可能使帕金森病患者受益的微生物导向药物,包括小分子药物和生物制剂,尚未在临床环境中进行广泛探索。
给予短链脂肪酸和靶向抑制左旋多巴代谢细菌酶似乎具有特别的前景,但仍需在人类干预研究中进行测试。
免疫调节和肠道屏障恢复方法也值得进一步关注。抗TNF治疗可降低炎症性肠病患者的帕金森病发病率,已证明可使炎症性肠病患者的粪便微生物群组成向健康人的粪便微生物群组成转变,丁酸盐和相关底物的粪便水平与临床缓解有关。
针对肠道神经系统中帕金森病相关分子发病机制(例如,涉及α-突触核蛋白或葡萄糖脑苷酶)的策略可能代表另一个新的前沿;在一项开放性试验中,口服角鲨胺(建议以静电方式取代肠道神经元中的α-突触核蛋白聚集体)是安全的,并能显著改善帕金森病患者的便秘症状。
发表在《神经病学》杂志上的一项研究发现,患有早期帕金森病的人每周两次定期通过散步或园艺等活动进行一到两个小时的适度体育锻炼,他们的平衡、行走能力、力量和耐力可能会有所改善。
五年以上经常锻炼的早期帕金森患者,在认知测试中表现更好,症状进展较慢。
每周至少进行4小时中等强度到剧烈运动(比如走路或跳舞)的人,与没有进行那么多运动的人相比,平衡能力下降的速度要晚5年。
有规律的活动,即使是小量的活动,也会有产生影响。
不同形式的体力活动似乎有不同的效果:
研究表明,89% 的帕金森病患者会出现言语和声音障碍,包括声音轻柔、单调、呼吸沙哑和发音不确定。因此,患有帕金森的人不太能参与正常对话,或者对社交环境没有信心。
言语障碍会逐渐降低帕金森患者的生活质量。一个人越早接受基线言语评估和言语治疗,就越有可能随着疾病的进展保持沟通技巧。沟通是帕金森患者生活质量高、充满信心的关键因素。
音乐疗法作为一种发展相对成熟、安全、方便实施而又成本低廉的非药物干预方法,被广泛应用于帕金森病人的康复中。
★ 措施
根据音乐疗法的性质及其应用方式可将音乐疗法分为 3 种类型:
① 主动音乐疗法:
大多采用治疗师与病人合作的方式,通过让病人唱歌、跳舞等形式调节病人情绪,逐步提高病人适应外界的能力,是国外康复机构主要治疗方法之一。
② 被动音乐疗法:
多注重治疗师的引导作用,常在病人休息时进行,让病人在欣赏和感受音乐的过程中,通过音乐的节奏、音色等因素影响人的神经系统,从而达到治疗的作用。
③ 综合疗法:
综合疗法往往是主动、被动双管齐下,并不限于哪种方法的使用,如在绘画时听音乐以陶冶情绪,或用音乐导引练习气功等。通过综合考虑音乐风格、人格类型、文化背景 、兴趣爱好 、职业等因素开出音乐处方实行辨证施乐。
★ 效果
改善运动障碍,增加运动协调性
对11例男性帕金森病人实施音乐疗法方案,用维也纳测试系统(VTS)测量病人在听音乐前后的身体运动协调性,研究发现实施音乐疗法后病人肢体瞄准和直线追踪方面有显著改善。
从华盛顿大学医学院招募了 23 例帕金森病人进行音乐疗法实验,研究者让病人边走边唱自己熟悉且韵律感强的歌曲,结果发现当病人以与舒适的步态步调相匹配的节奏唱歌时,可以改善步态的协调性和稳定性,同时还增强了病人的语言清晰度和声音强度。
18 例帕金森病人随机分为干预组和对照组,干预组采用音乐疗法,病人根据音乐节拍做一些特定的舞蹈动作或跟着音乐唱歌,对照组则采用传统理疗方式,结果显示干预组病人在身体灵活性、步态等方面较对照组有显著改善,病人的跌倒次数也显著减少。
研究平板训练联合音乐疗法对帕金森病人冻结步态的影响,结果显示平板训练联合音乐干预能够显著缓解帕金森病人冻结步态,减少摔跤次数。
提高认知能力,改善吞咽功能和言语障碍
有研究招募了 25 例帕金森病高龄病人为研究对象,实验组病人接受音乐疗法 24 周,对照组不进行任何干预,结果发现实验组病人认知能力、处理速度、注意力、记忆能力较对照组有明显改善。
美国一项研究对 27 例帕金森病人进行音乐疗法干预后,病人吞咽功能明显改善,同时说话吐字也更加清晰,但在实验干预的过程中使用多个研究人员进行干预和评估,无法保证被测病人的 2 次评估是由相同的研究人员完成,因此不同的评估者的主观因素可能会影响评估结果。
对 4 例帕金森病人进行了为期 1 个月的音乐干预,结果显示音乐疗法提高了帕金森病人的声音强度和语言清晰度。和谐而又有节奏感的音乐能够有效改善帕金森病人的认知能力,同时也有利于病人的言语康复。
改善负面情绪,提高病人生活质量
将80 例帕金森病人随机分为对照组和实验组各 40 例,对照组接受常规治疗,实验组病人在常规治疗的基础上接受音乐干预,通过汉密尔顿抑郁量表评估干预后的效果,结果显示实验组抑郁评分明显下降,并激发了病人积极的情绪状态。
对 32 例帕金森病人进行为期 3 个月的随机对照实验发现,接受音乐干预的帕金森病人调节情绪的能力明显提高,同时病人也对音乐疗法提供的社交和创造性的交流方式表达了欣赏和喜爱,幸福指数也明显增高。
镁
矿物镁的摄入量低会使过量的重金属和毒素在大脑中沉积,从而导致帕金森和阿尔茨海默病。
当有足够的镁存在时,重金属不能在小肠中被吸收。镁在保护神经元免受铝和汞中毒的致命影响方面起着至关重要的作用。
镁也是治疗震颤最重要的自然疗法之一。研究人员发现,每天服用镁补充剂可以缓解手部震颤的症状。
富含镁的食物:
B族维生素
缺乏维生素 B9 和 B12可能导致最初表现为抑郁、焦虑甚至精神病的大脑问题。其他 B 族维生素可用于防止与年龄有关的脑萎缩,并可能防止记忆丧失。
谷胱甘肽
谷胱甘肽是抗氧化剂,能有效中和自由基损伤,并大大减少破坏神经元的氧化应激。
葡萄籽提取物
具有超强抗氧化作用,可减少大脑中的 DNA 碎片。它能够进入大脑,保护脑细胞免受自由基损伤。
帕金森病微生物组研究的当前差距和未来方向
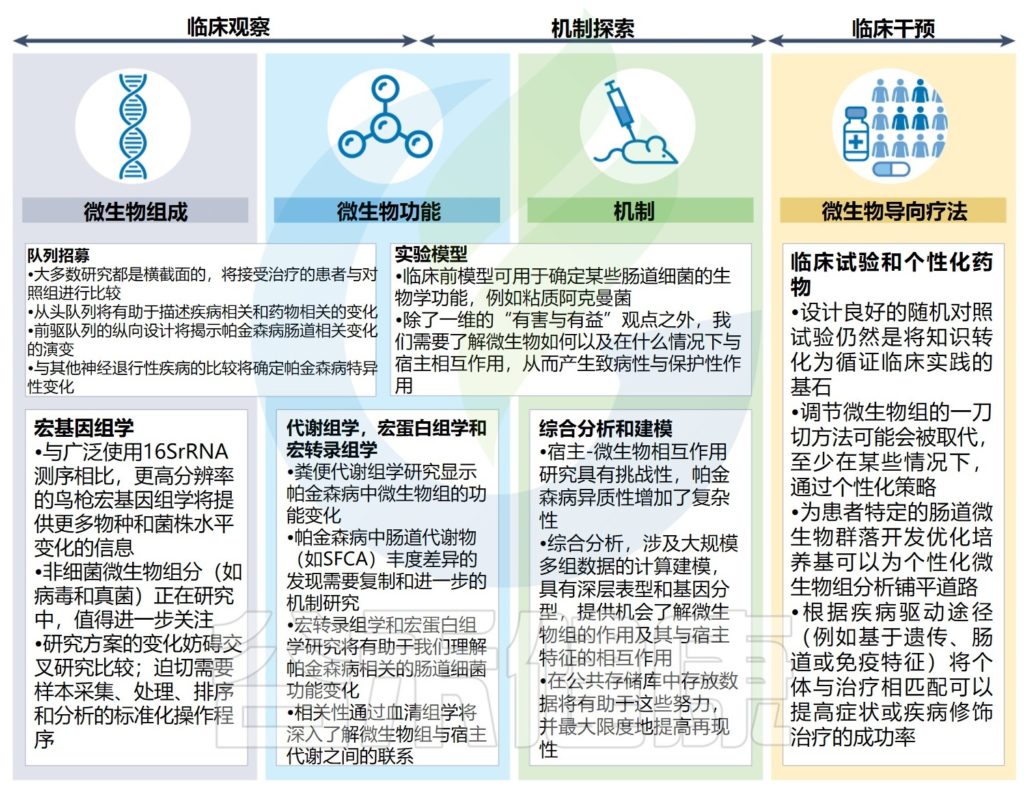
Tan AH, et al.,Nat Rev Neurol. 2022
帕金森病微生物组研究重点正在从研究微生物组结构和潜力(即肠道中存在哪些微生物及其相对丰度和代谢潜力)转向通过使用多组学平台和机理研究来了解微生物功能。
根据观察性研究和基本机制研究的结果,以回答有关肠道微生物和代谢物在帕金森病发病、维持和进展中的作用的基本问题,该领域有望发展到微生物导向疗法的临床试验,从而将微生物组学转化为这种疾病患者的临床现实。
目前,有关肠道相关因素与人类帕金森病之间关系仍然存在很大的相关性,现在需要从观察性研究设计过渡到干预性研究设计。
然而,一些问题仍然存在,确定改变是否是疾病过程的因果关系或促成因素将是至关重要的。例如,与肠道微生物组分的相关性——可能反映了对疾病因素(如转运时间改变或炎症)的次要或共同的非特异性反应。
总之,关于胃肠道参与神经退行性路径,许多疑难杂症仍有待解开。我们希望这一领域的科学进步可以给人们带来新的诊断、预后和治疗方法,最终将改善帕金森病患者及其家人的生活质量。主要参考文献:
Tan AH, Lim SY, Lang AE. The microbiome-gut-brain axis in Parkinson disease – from basic research to the clinic. Nat Rev Neurol. 2022 Jun 24. doi: 10.1038/s41582-022-00681-2. Epub ahead of print. PMID: 35750883.
Wang Q, Luo Y, Ray Chaudhuri K, Reynolds R, Tan EK, Pettersson S. The role of gut dysbiosis in Parkinson’s disease: mechanistic insights and therapeutic options. Brain. 2021 Oct 22;144(9):2571-2593. doi: 10.1093/brain/awab156. PMID: 33856024.
Menozzi, E., Macnaughtan, J. & Schapira, A. H. V. LRRK2 parkinsonism: does the response to gut bacteria mitigate the neurological picture? Mov. Disord. 36, 71–75 (2021).
Nishiwaki, H. et al. Short- chain fatty acid- producing gut microbiota is decreased in Parkinson’s disease but not in rapid- eye-movement sleep behavior disorder. mSystems 5, e00797-20 (2020).
Vascellari, S. et al. Clinical phenotypes of Parkinson’s disease associate with distinct gut microbiota and metabolome enterotypes. Biomolecules 11, 144 (2021).
马春潮.帕金森的防治[J].开卷有益-求医问药,2021(09):35.
Koutzoumis,D.N. et al. Alterations of the gut microbiota with antibiotics protects dopamine neuron loss and improve motor deficits in a pharmacological rodent model of Parkinson’s disease. Exp. Neurol. 325, 1 13159 (2020).

谷禾健康

嗜黏蛋白阿克曼菌(Akkermansia muciniphila, 简称A. muciniphila, Akk菌)的缺乏或减少与多种疾病(如肥胖、糖尿病、肝脂肪变性、炎症和对癌症免疫治疗的反应)有关。
关于AKK菌,我们之前的一篇文章也有详细介绍过,点击详见:
肠道重要菌属——Akkermansia Muciniphila,它如何保护肠道健康
谷禾肠道样本大数据库显示A. muciniphila缺乏或未检出情况在人群中很常见,尤其是那些有肠道问题的人。在健康个体中其约占肠道微生物群总数的0.5%–3%。
现如今,关于AKK菌的研究正在从动物模型转向人类验证试验,AKK菌与疾病之间的研究不仅仅停留在相关性,更是开始向因果性及具体产生作用的机制方面深入探讨。
– A. muciniphila –
本文我们将依次介绍A. muciniphila的起源,主要特性,以及它与不同疾病间的联系,并解释产生有益作用的主要机制。
一所在瓦格宁根(Wageningen)的微生物实验室的研究人员使用一种专门用于分离优势细菌的策略,即稀释到消亡(即基于连续稀释的分离),从一名健康成人身上分离出一株高度丰富的粘液降解菌株。
该菌株似乎是疣微菌门(Verrucomicrobiota)中一个新属的新种,被命名为嗜黏蛋白-阿克曼菌(Akkermansia muciniphila, A. muciniphila, Akk菌),并以典型菌株MucT为代表。
粘液主要由粘蛋白组成,粘蛋白是一种由粘多糖组成的保护肠道细胞的糖蛋白。
长期以来,粘液的降解被认为是一种可能导致宿主紊乱的不良特性。然而,对于结肠微生物群来说,粘蛋白在肠道中大量分泌,因此提供了持续的宿主产生的碳、能量和氮源。粘蛋白的代谢转换需要一组酶,如唾液酸酶和硫酸酯酶,这些酶参与顺序降解。
粘蛋白降解“专家”
A. muciniphila 是人类早期生命中存在的独特的粘蛋白降解“专家”。此外,对无菌小鼠的单体型关联研究表明,A. muciniphila MucT不会损害宿主,并在结肠中显示出特异性的代谢和免疫信号。其2.7Mb的基因组预测了粘蛋白降解的酶机制,比较生长分析表明,A. muciniphila是体外利用粘蛋白最有活性的菌株。
A. muciniphila MucT也可以利用人乳寡糖作为能量来源。值得注意的是,在纯培养中,只有少数其他底物被发现能刺激其生长,包括二甲双胍、甜菜碱和色氨酸。
由于A. muciniphila是人类肠道中疣微菌门的唯一代表,许多报道这一门的16S rRNA基因序列调查往往代表A. muciniphila 。最近的一项研究分析了2,000多个Akkermansia基因组,表明A. muciniphila是迄今为止具有高度相似(超过98%同一性)16S rRNA 序列的优势物种。
非工业化人群A. muciniphila下降
在这个大型宏基因组数据库中对四个已确定的A. muciniphila亚种的详细分析表明,它们的基因组通常存在于西方和中国人群中。对哈扎部落和其他非工业化人群的肠道微生物群与工业化世界个体的肠道微生物群进行分析比较,发现拟杆菌纲和疣微菌门微生物群的丰度减少。这些结果表明,非工业化种群的A. muciniphila丰度水平有所下降,但这一观察结果是否与粪便样本保存、DNA提取或测序深度的技术问题有关仍有待调查。
值得注意的是,A. muciniphila宏基因组已在非人类灵长类动物中发现,包括野生和圈养。类似地,与模式菌株MucT具有高度基因组同源性的A. muciniphila已从同样生活在野外的各种动物中分离出来。
部分A. muciniphila菌株产维生素B12
长期以来,A. muciniphila型菌株MucT是唯一可用的人类分离株然而,在过去的 5 年中,从不同的肠道环境中分离到了其他A. muciniphila菌株,包括野生哺乳动物和圈养哺乳动物,特别是人类。在临床前试验中报告了人类相关 A. muciniphila菌株之间的一些变异性,但尚不清楚观察到的差异的稳定性和可重复性如何。
迄今为止,观察到的菌株之间最显著的代谢差异是产生维生素B12的能力,而维生素B12是产生丙酸盐所必需的。大约三分之一的A. muciniphila分离株能产生类似于A. glycaniphila的维生素B12。多种结肠微生物可以产生可供A. muciniphila利用的维生素B12,正如霍氏真杆菌(Eubacterium hallii)(重命名为Anaerobutyricum soehn genii)所显示的那样,在共培养实验中,发现该细菌与菌株MucT形成微生物网络,导致从粘液中产生丙酸盐和丁酸盐。
代谢紊乱
由于观察到益生元补充剂(即低聚果糖)对肥胖发育的保护作用与啮齿动物疣微菌门的主要繁殖相吻合,因此引起了人们对A. muciniphila和代谢紊乱的兴趣。随后在人类和啮齿动物上进行的观察研究都表明,与瘦小的对应物相比,患有肥胖症的个体肠道中的A. muciniphila反复出现表达不足。代谢紊乱,包括肥胖、T2DM、非酒精性脂肪性肝病(NAFLD)和心血管疾病,都与Akkermansia spp.丰度减少有关。
研究发现,11名超重者和38名肥胖者体内A. muciniphila的丰度与更健康的代谢状态以及热量限制后较好的临床结果呈正相关。通过在小鼠和人类身上注射该模式菌株,研究了A. muciniphila在肥胖过程中的因果作用及其潜在的有益影响。首次证明,小鼠每天服用活的A. muciniphila MucT可以逆转高脂饮食引起的代谢紊乱,包括脂肪增加、代谢内毒素、脂肪组织炎症和胰岛素抵抗。
在合成培养基中生长的A. muciniphila MucT的巴氏杀菌增强了它减少小鼠脂肪团发育、胰岛素抵抗和血脂异常的能力。事实上,A. muciniphila MucT可以降低小鼠的胆固醇水平和血清甘油三酯水平。
代谢健康背景下黏质阿克曼菌的代谢效应及主要影响因素
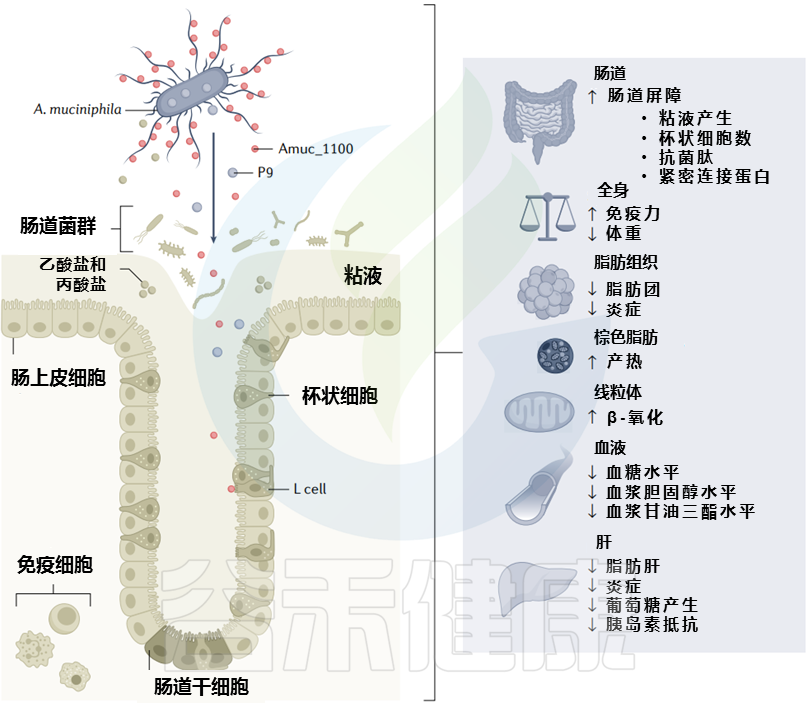
Cani PD, et al. Nat Rev Gastro Hepat. 2022
影响肝脏脂肪代谢
研究发现A. muciniphila MucT补充剂通过调节与脂肪合成有关的基因的表达(例如,降低肝脏中固醇调节元件结合蛋白的表达)和不同的炎症标志物(例如,降低IL-1β和IL-6的表达,ALT和髓过氧化物酶活性),可以积极地影响小鼠的肝脏脂肪代谢,预防非酒精性脂肪性肝病。
减轻动脉粥样硬化
在动脉粥样硬化发展的小鼠模型(载脂蛋白E缺陷(ApoE−/−)小鼠)中,A. muciniphila MucT的注射似乎减轻了动脉粥样硬化的损害。
降低糖尿病发病率
有趣的是,除了T2DM,在啮齿类动物和人类中,A. muciniphila通过降低肠道通透性、减少炎症和胰岛炎症的机制减少了T1DM的发病,从而有助于胰岛的保存。早期应用万古霉素可增加非肥胖型糖尿病小鼠(T1DM小鼠模型)的A. muciniphila丰度,降低糖尿病发病率。
神经退行性疾病
2016年至2020年间发表的几份报告表明,A. muciniphila在帕金森病和多发性硬化症中的作用,因为其相对丰度与受影响患者的疾病严重程度呈正相关。然而,尽管将患者的粪便微生物移植到无菌小鼠体内会导致疾病某些方面的发展,但这些研究都没有检测到受体小鼠粪便中的A. muciniphila,从而表明其他细菌可能参与了该病的发生表型。
多发性硬化症
对于多发性硬化症,在一项纳入62名复发缓解性疾病患者的研究中,作者发现脑脊液中的抗-Akkermansia IgG水平高于健康同行,并且局部脑脊髓特征与残疾评分呈正相关,而在同一个人的血液中检测到IgG水平没有明显的改变。
虽然在这些研究中观察到的抗-Akkermansia IgG水平的增加与肠道中嗜粘蛋白A. muciniphila丰度的改变无关,但另一项研究提供了支持 A. muciniphila在多发性硬化症中的积极和有益作用的机制解释。
这项研究表明,在多发性硬化症实验性自身免疫性脑脊髓炎小鼠模型的粪便和未治疗的多发性硬化症高峰期患者的粪便中发现的A. muciniphila的大量繁殖与miR-30d的富集有关。
有人发现,从这些小鼠或人类身上收集的粪便的转移导致A. muciniphila的丰度增加,并有利于调节性T (Treg)细胞的扩增,进而控制效应T细胞以抑制疾病症状。研究人员表明在患有多发性硬化症的小鼠中接种A. muciniphila可降低疾病评分、减少轴突损失的脱髓鞘和增加Treg细胞群。
相反,A. muciniphila衍生肽被确定为环境因素,与多发性硬化症(即HLA-DR15单倍体型)最强的遗传风险易感因素相结合,可以介导免疫调节介导的患者自身反应性T细胞的激活。
更具体地说,从患者体内分离出的 HLA-DR-SP反应性 CD4+ T细胞可以被某些外来因子(例如Epstein-Barr病毒和从A. muciniphila中分离出的一些肽)激活,然后对血液中的潜在致病多肽或自身抗原(如髓鞘碱性蛋白)产生反应,可能还会在脑脊液和/或脑中发生交叉反应,从而针对脑组织。
帕金森病
文献中关于阿尔茨海默病和A. muciniphila的发病机制存在差异,相关研究显示阿尔茨海默病患者的A. muciniphila含量较高,而干预研究清楚地报告了这种细菌在病理学中的有益作用。阿尔茨海默病是一种疾病,其最强的病理标志之一是淀粉样蛋白β肽42 (Aβ42)在老年斑中的积累。
由于肥胖和T2DM是痴呆的重要危险因素,一项机制研究探讨了在喂食高脂肪饮食的阿尔茨海默病小鼠模型中施用A. muciniphila的影响。除了抗肥胖作用外,无论饮食如何,接种都能显着减少大脑中的 Aβ并改善认知测试的表现。A. muciniphila给药还恢复了生命早期(生命3周时)暴露于高脂肪饮食的小鼠的认知能力下降和海马发育障碍。
癌症免疫治疗反应
免疫疗法是一种利用免疫系统对抗肿瘤形成细胞的癌症疗法,并已发展成为治疗各种癌症的成功方法。
肠道微生物组的相关性是增强对检查点抑制剂治疗的临床反应的标志物和辅助剂。
该疗法通过阻断程序性死亡受体1(PD1)等免疫检查点来恢复对抗癌细胞的活性。对100名对抗-PD1抗体反应良好的非小细胞肺癌和肾细胞癌患者的微生物组进行了详细的表征,结果显示A. muciniphila富集。此外,在疾病进展迅速的个体和后来死亡的个体中,A. muciniphila的患病率最低(34%)。
A. muciniphila菌株在用抗-PD1 药物治疗后,在移植无应答者粪便的无菌小鼠和预先暴露于抗生素的小鼠中改善了抗肿瘤活性。一项具有统计学意义的研究旨在专门评估338例非小细胞肺癌患者的A. muciniphila 基线检测与临床反应之间的关联。证实了A. muciniphila的存在与临床反应之间的密切关联。
在另一项研究中,42名主要使用抗-PD1药物治疗的转移性黑色素瘤患者的肠道微生物群也被证实富含多种分类群,包括Akkermansia。2020 年发表的一项研究确定肌苷是抗癌细胞活性的潜在因素,尽管这种核苷是由许多其他细菌产生的。
总之,这些研究表明,A. muciniphila是一种有望提高对检查点抑制剂免疫疗法的临床反应的候选菌。
肠道屏障和肠道炎症
最初发现A. muciniphila MucT 可以通过恢复小鼠的粘液层厚度以及抗微生物肽Reg3g的肠道表达来改善肠道屏障功能,这在肥胖和代谢紊乱期间都会发生改变。后来,在A. muciniphila MucT补充剂中观察到的粘液层厚度增加与小鼠粘液产生细胞数量的增加有关。
此外,来自A. muciniphila MucT (AmEVs)的细胞外囊泡,也显示通过调节小鼠的紧密连接来降低肠道通透性。
肠道炎症的情况下保护的肠道屏障
已有研究表明,与健康人相比,克罗恩病 (n = 26)和溃疡性结肠炎(n = 20和n = 15 ) 患者相比,A. muciniphila显著减少。这种相关性已在临床前模型中进行了因果研究。
改善结肠炎
已发现A. muciniphila MucT在结肠炎中的有益作用,AmEVs可保护葡聚糖硫酸钠 (DSS) 诱导小鼠结肠炎的进展。
随后,几项研究观察到A. muciniphila MucT细胞恢复肠道屏障功能并改善了:
改善衰老引起的变化
衰老是另一种肠道屏障减少和炎症增加的情况。各种人类研究报告称,与年轻成人(<50岁)和百岁老人相比,老年人(>65岁)的A. muciniphila丰度较低。
这一发现导致了旨在评估A. muciniphila MucT给药对不同衰老小鼠模型中几个年龄相关参数的影响的研究。
总之,这些研究表明,施用A. muciniphila MucT改善了一些与年龄相关的变化,包括炎症、屏障完整性和行为。
代谢综合征
在啮齿动物中,A. muciniphila被认为是健康的生物标记物。在啮齿类动物中,许多对A. muciniphila种群具有生长促进作用的膳食补充剂因其促进健康的作用而受到广泛关注。
在患有代谢综合征的人类中,给药被证实是可行的、安全的和有良好的耐受性。事实表明,与炎症、血液学、肾脏、肝脏和肌肉功能相关的任何标志物的变化,都可以用任何A. muciniphila MucT制剂来观察。这一结果是在不考虑所使用的形式以及短期(即2周)和长期(即3个月暴露)的情况下观察到的。
巴氏灭菌Akk菌比活菌效果好
以前的结果表明,高压灭菌灭活A. muciniphila可消除其对小鼠代谢综合征的保护作用。对植物乳杆菌或干酪乳杆菌菌株进行的研究表明,不剧烈的热诱导灭活,如巴氏杀菌,可以使细菌在稳定它们的同时保留部分有益的特性。
因此,有人在相同的饮食诱导肥胖小鼠模型上,通过比较活菌和巴氏灭菌菌的给药效果,检验A. muciniphila巴氏灭菌的效果。结果是,接受巴氏灭菌菌株的小鼠在体重、脂肪质量增加、血脂和胰岛素抵抗标志物方面的降幅甚至比接受活细菌的小鼠更大。除了提高A. muciniphila粘液的有效性,巴氏杀菌还具有提高菌稳定性和延长其保质期的好处,从而便于给人服用。
在未经治疗的患者中,给予巴氏灭菌A. muciniphila MucT成功地防止了与代谢综合征相关的参数的自然恶化。与补充安慰剂的志愿者相比,补充已过活的A. muciniphila MucT显著改善了胰岛素敏感性,降低了胰岛素血症和血浆总胆固醇水平。
为了进一步评估巴氏灭菌A. muciniphila MucT的安全性,对大鼠进行了一项强有力的毒理学长期评估。结果表明,口服90天,即使在测试的最高剂量(每公斤体重9.6×1010A. muciniphila MucT细胞,没有观察到不良反应的水平)下,也没有转化为亚慢性毒性,而体外遗传毒性试验显示阴性结果。
如前面所述,A. muciniphila可能有多种作用模式,所有这些都已用 MucT菌株进行了研究。当使用巴氏杀菌细菌时,可以观察到A. muciniphila MucT的几种有益效果和令人惊讶的功效增加。
产短链脂肪酸,作用于宿主代谢
最明显的答案是短链脂肪酸(如丙酸盐)的一般作用,因为A. muciniphila MucT在维生素B12存在下被称为丙酸盐生产者。虽然A. muciniphila对肠道中丙酸盐总水平的贡献可能相对于其他主要的丙酸盐生产者(例如拟杆菌)相对较低,尤其是在禁食条件下。但 Akkermansia似乎在动物和人类中都上调,并且可能对生产这种短链脂肪酸做出重大贡献。
在这种情况下,某些影响可能与肠道上皮细胞和肠道黏膜中存在的不同类型免疫细胞上表达的FFAR3和FFAR2受体(也分别称为GPR41和GPR43)有关。然而,值得注意的是,巴氏杀菌的 A. muciniphila MucT也可以作用于宿主代谢,不会改变丙酸盐的产生。
通过外膜上的蛋白质Amuc_1100发挥作用
2017年,有人确定了A. muciniphila MucT可以在宿主健康中发挥作用的特定和独特的分子机制。他们发现存在于A. muciniphila外膜中的一种特定蛋白质,称为Amuc_1100,概括了这种细菌的有益作用。A. muciniphila的所有有益作用是否都可归因于Amuc_1100目前尚不清楚,但其每个细胞的生产水平足以解释临床前模型中的作用。
重要的是,他们发现这种蛋白质在用于巴氏杀菌的温度下仍保持其活性构象,从而解释了为什么巴氏杀菌的A. muciniphila MucT在小鼠和人体实验中保持活性。
此外,发现Amuc_1100可激活Toll样受体2 (TLR2),单独给药可复制A. muciniphila MucT细胞的大部分有益作用,包括在肠道炎症和结肠癌的特定疾病模型中的作用。
这一发现表明,即使死亡(即巴氏杀菌后),A. muciniphila MucT仍然可以改善宿主健康,并反对需要潜在的分泌代谢物来观察细菌的有益作用。
外膜上的蛋白质 P9,刺激GLP1水平增加
2021年的一项研究确定了A. muciniphila MucT产生的另一种蛋白质。发现A. muciniphila处理的饮食诱导的肥胖小鼠在口服葡萄糖挑战期间表现出循环GLP1水平略有增加。
通过体外实验,作者鉴定了一种84kDa的蛋白质,命名为蛋白 9(P9;由早期鉴定的 Amuc_1831 基因编码),它负责体外刺激GLP1。
在体内,他们测试了 8周以非常高的剂量口服P9的效果,发现体重增加和脂肪量增加都有所减少,胰高血糖素前体(即GLP1的前体)的肠道表达增加。
这些影响怎么与较高的循环 GLP1 水平联系起来?
研究发现P9的药理作用激活 ICAM2,这些作用与可能的GLP1分泌有关,因为抗-ICAM2抗体在体外部分消除了P9对GLP1分泌的影响。
此外,A. muciniphila MucT增加了小鼠回肠中IL-6的表达。除了证实IL-6是一种促炎细胞因子外,这种细胞因子还在体外剂量依赖性地增加GLP1的分泌。
因此,有人假设A. muciniphila MucT也可以通过 IL-6-GLP1信号传导发挥作用。为了进一步支持他们的发现,研究人员使用了IL-6敲除小鼠,发现这些小鼠对P9诱导的GLP1分泌没有反应,而阻断GLP1受体也消除了P9对产热的影响。尽管这些发现是相关的,但应评估其他因素,例如P9的生理剂量和位置(可能被分泌)。
与Akkermansia或相关分子在疾病中的作用相关的主要机制
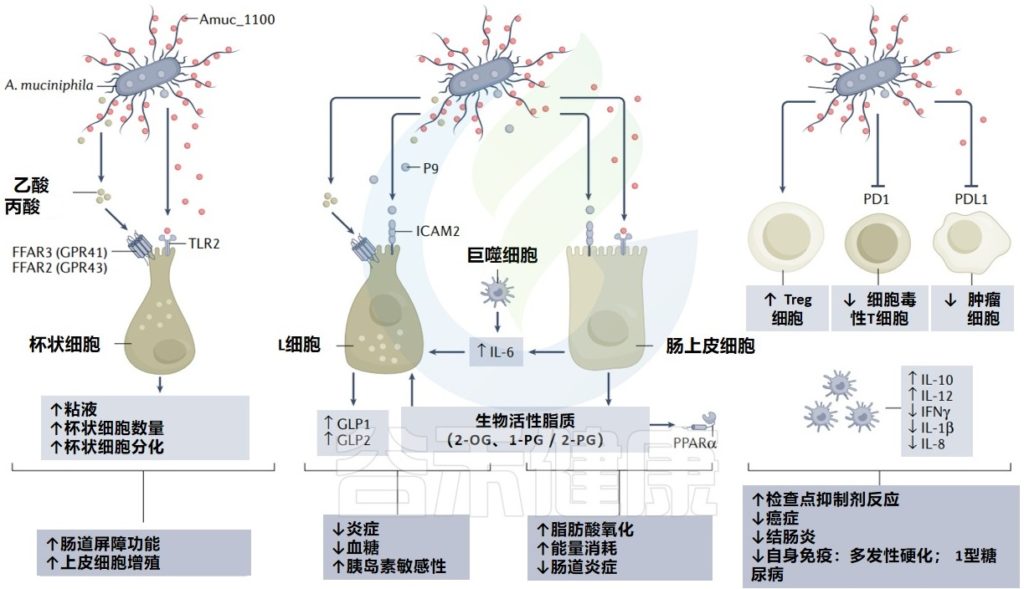
Cani PD, et al. Nat Rev Gastro Hepat. 2022
A. muciniphila MucT表达的酶潜在益处
一项研究证明天冬氨酸蛋白酶Amuc_1434*通过TRAIL介导的细胞凋亡途径抑制LS174T细胞活力。然而,没有进一步的位置、热稳定性或体内数据报告,因此尚无法评估这一发现的潜在和生理相关性。
第二项研究表明,A. muciniphila MucT表达的β-乙酰氨基己糖苷酶Amuc_2109*保护小鼠免受DSS诱导的结肠炎;然而,尚未报告热稳定性数据。
此外,已在人类身上表明,活的或巴氏杀菌的A. muciniphila MucT 改善了几种心脏代谢风险因素,包括胰岛素抵抗,而不增加 GLP1 的循环水平,也不影响 DPP4 活性,DPP4 是一种参与 GLP1 降解的酶。
有趣的是,无论使用何种形式的A. muciniphila MucT——活的或巴氏杀菌的,甚至是蛋白Amuc\U 1100,文献中的所有数据都通过作用于不同的关键标记物来强化肠道屏障。
事实上,所有的治疗都指向更高的粘液产量,紧密连接蛋白的恢复,抗菌因子的恢复,以及最终加强肠道屏障。
特异性调节
此外,一部分人通过对用活的或巴氏杀菌的 Akkermansia治疗的人体进行脂质组学和代谢组学分析。已经发现A. muciniphila MucT治疗诱导了不同生物活性脂质的特异性调节,这些被鉴定为 PPARα 激动剂(2-PG和1-PG)。
重要的是,在人类中使用非靶向代谢组学分析,他们已经能够重建一条指向通过β-氧化激活脂肪酸氧化的代谢途径,并且所有鉴定的代谢物都在PPARα的控制下趋向于增加线粒体活性,这些数据也在啮齿动物身上得到了证实。
除了对肠道屏障的影响,这可能涉及对几种疾病(即糖尿病、肥胖症、NAFLD和/或非酒精性脂肪性肝炎、炎症性肠道疾病、多发性硬化症)的保护作用,A. muciniphila对癌症的作用也是与依赖免疫系统的其他机制有关。
例如,使用检查点抑制剂(抗-PD1)的免疫疗法与动物数据中涉及 IL-12 依赖性效应的特定机制方面有关。此外,数据表明A. muciniphila菌株对抗-PD1反应的辅助作用增加了特定T细胞向肿瘤床的募集。
在炎症性肠病、T1DM或肠癌的小鼠模型中,A. muciniphila MucT或特定蛋白增加Treg细胞群的分化或降低结肠中浸润性巨噬细胞和CD8+细胞毒性T淋巴细胞的水平。
A. muciniphila MucT的代谢和抗炎作用是强大的,因为来自不同研究团队的许多结果已经证实了A. muciniphila菌株的各种健康特性。
A. muciniphila的生理和有益作用是多效性的(例如,能量、脂质、葡萄糖代谢、炎症、免疫、脑功能),重要的是要强调有许多汇聚的作用模式,可能是由于其特异性特性与其粘蛋白代谢的特化有关。
事实上,已经确定了几种常见的途径,都指向调节肠道屏障功能(即粘液产生和免疫系统)。肠道屏障的恢复,也有助于恢复几种改变的途径的正常功能,包括例如线粒体活性、肝脏代谢、脂肪组织和大脑活动。
最后,A. muciniphila MucT的有益作用已从临床前观察转化为代谢综合征背景下的人类干预。这种情况是独一无二的,是其他下一代微生物无法比拟的。
需要注意的是,虽然A. muciniphila具备各种有益特性,但是该菌丰度并不是越多越好,要保持在合适范围。谷禾肠道菌群检测结果中曾经有案例,发现该菌丰度过高,占比超50%。
该菌过量将过度消耗粘液蛋白而存活下来,这是大多数其他细菌所缺乏的生存优势,该菌增殖异常,从而可能导致肠道屏障损伤,诱发肠道炎症、LPS 进入血液的增加、自身免疫性疾病有关。
该样本检测报告也显示菌群多样性低,肠炎和几项慢病注意风险:
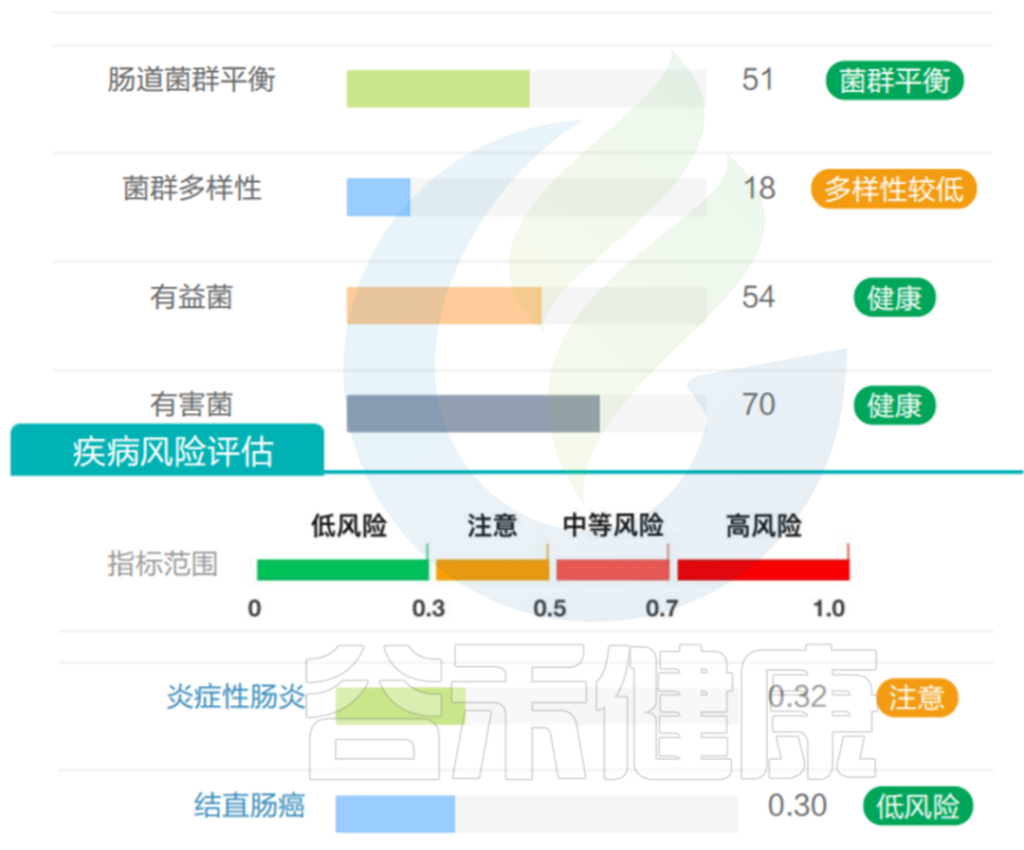
<来源:谷禾健康数据库>
总的来说,在合理范围内,A. muciniphila 带来有益影响。
当然也需要更多的研究来支持A. muciniphila的有益特性及临床治疗应用:
主要参考文献
Cani PD, Depommier C, Derrien M, Everard A, de Vos WM. Akkermansia muciniphila: paradigm for next-generation beneficial microorganisms. Nat Rev Gastroenterol Hepatol. 2022 May 31. doi: 10.1038/s41575-022-00631-9. Epub ahead of print. PMID: 35641786.
Kostopoulos I, Elzinga J, Ottman N, Klievink JT, Blijenberg B, Aalvink S, Boeren S, Mank M, Knol J, de Vos WM, Belzer C. Akkermansia muciniphila uses human milk oligosaccharides to thrive in the early life conditions in vitro. Sci Rep. 2020 Aug 31;10(1):14330. doi: 10.1038/s41598-020-71113-8. PMID: 32868839; PMCID: PMC7459334.
Yin J, Song Y, Hu Y, Wang Y, Zhang B, Wang J, Ji X, Wang S. Dose-Dependent Beneficial Effects of Tryptophan and Its Derived Metabolites on Akkermansia In Vitro: A Preliminary Prospective Study. Microorganisms. 2021 Jul 14;9(7):1511. doi: 10.3390/microorganisms9071511. PMID: 34361945; PMCID: PMC8305782.
Paone P, Cani PD. Mucus barrier, mucins and gut microbiota: the expected slimy partners? Gut. 2020 Dec;69(12):2232-2243. doi: 10.1136/gutjnl-2020-322260. Epub 2020 Sep 11. PMID: 32917747; PMCID: PMC7677487.
谷禾健康
现状
全球肥胖患病率的上升是一个主要的社会经济负担,肥胖与许多疾病的风险增加有关,包括糖尿病、心血管疾病和癌症。
尽管人们努力改善生活方式选择,提高对潜在病因的认识,但在预防和治疗肥胖方面的长期成功似乎有限,因为饮食诱导的体重减轻在5年随访后仅维持约25%。
近年来,在了解肠道微生物群作为宿主能量和底物代谢调节器参与肥胖和相关心脏代谢并发症方面取得了进展。因此,通过肠道微生物群靶向宿主代谢可能是饮食干预减轻体重的一项重要策略。
过去十年中,关于肠道微生物组对宿主代谢影响的研究数量呈指数增长,研究的数量和质量都在迅速发展,这些研究表明,基线微生物组成可以预测包括肥胖在内的代谢综合征。然而,研究同时表明微生物群组成的调节不可能会在所有条件下对人体代谢产生重要积极的影响,而这种影响取决于个体的特征,例如年龄、习惯性饮食、代谢表型和基线肠道微生物谱。
肠道微生物群的组成由可遗传、人口统计和环境因素决定,包括出生时的分娩方式、年龄、性别、胃肠道转运时间和药物使用。但是诸多因素中,饮食已成为塑造和定义肠道微生物组的关键因素。
饮食尤其是膳食纤维等引起的肠道微生物群组成和功能变化与肥胖和相关疾病的发展有关。这些研究结果发现肠道微生物群的个体间差异可以作为对抗肥胖代谢疾病的更精确饮食方法的基础。
本文将介绍有关饮食成分、肠道微生物组和宿主代谢之间相互作用的知识和研究成果,以及如何整合这些知识来制定基于精确的营养策略,以改善人类的体重控制和代谢健康。
厚壁菌/拟杆菌门
肠道微生物群影响免疫功能和上皮完整性、能量和底物代谢以及葡萄糖稳态。初步研究表明,与瘦个体相比,肥胖的人类和啮齿动物的厚壁菌门与拟杆菌门的比例增加,但也有个别研究未能观察到这种差异,甚至报告了比例下降。
多样性和微生物基因丰富度
在代谢健康与不健康个体的比较中,代谢不健康组的α多样性较低。而且重度肥胖症患者的低微生物基因丰富度比例高达75%,而瘦或超重/中度肥胖症患者的低微生物基因丰富度比例为23%-40%。
(小编推测可能是由于中重度肥胖人群其饮食比较丰富且量大,微生物不需要太多多余的基因就可以代谢获得生存繁殖的食物,而较瘦的个体食物不太丰富,那么菌需要更多的基因才获取生存的食物和繁殖生存)
具体菌属
具体而言,颤螺菌属(Oscillospira)和 红蝽菌科(Coriobacteriaceae)的细菌与良好的代谢健康相关。 在一项包含正常体重和超重/肥胖人群的研究人群中,特定菌属的丰度与代谢特征相关。 例如,产气柯林氏菌、Dorea formicigenans 和 Dorea longicatena 在超重/肥胖人群中的丰度更高。
Akkermansia属的细菌是最有说服力的证据,它与患肥胖症和代谢综合征的风险呈负相关。在超重/肥胖患者中,为期 3 个月的 Akkermansia muciniphila 补充剂可改善胰岛素敏感性并降低肝功能障碍和炎症的血液标志物。
基线菌属
另一项研究表明,在瘦肉型个体中,嗜粘菌A.muciniphila和Alistipes obesi显著富集,而在肥胖型个体中,Ruminococcus gnavus显著富集。该研究还确定,当在基线检查时高丰度存在的菌,如Blautia wexlerae 和 Bacteroides dorei 减肥前以高丰度存在时将有助于减肥。此外,基线普雷沃菌属 (Prevotella)普氏菌丰度可以预测肥胖人群在膳食纤维干预减肥中是否可以成功。
此外,与健康个体相比,II型糖尿病患者和代谢受损个体表现出微生物功能改变和发酵能力降低,尤其是产丁酸盐细菌丰度较低的个体。此外,胰岛素抵抗个体的肠道微生物组可能具有增加的生物合成潜力,并减少了支链氨基酸(BCAA,主要由Prevotella copri,B. vulgatus驱动)的吸收和分解代谢,这与有害代谢效果有关。
总之,代谢受损个体的微生物基因丰富度和多样性降低。肠道微生物群组成和功能的个体差异与饮食干预的反应变化有关。
在当前的西方世界,习惯性饮食结构已转向高能量密集型食物,包括相对较高的饱和脂肪和简单碳水化合物含量,以及较低的膳食纤维含量。尤其是膳食纤维的消耗,以及大量营养素的质量和消耗量都会强烈影响肠道微生物群的组成和功能。基于人群的宏基因组分析揭示了微生物组成和多样性与60多种饮食因素的习惯饮食之间的关联。这些因素包括能量和大量营养素的摄入,以及面包和软饮料等特定食品的消耗。这些数据证实了饮食对塑造肠道微生物群的重要性。
饮食塑造肠型
在一项纵向单卵双生子研究中,粪便微生物群分析表明,能量的习惯性摄入、不饱和脂肪酸(FA)的类型和可溶性纤维会影响微生物群的组成,尤其是拟杆菌属和双歧杆菌的丰度。微生物肠道类型与长期习惯性饮食密切相关,尤其是蛋白质和动物脂肪(拟杆菌属)与碳水化合物摄入(普雷沃氏菌属)相比。
与此一致,长期坚持地中海饮食与特定分类群以及肠道微生物谱的功能有关。肠道微生物组的组成是地中海饮食与心脏代谢疾病风险之间保护性关联的调节因素。当比较习惯性高脂肪饮食和高碳水化合物饮食时,高脂肪饮食的微生物多样性似乎较低。此外,与高(饱和)脂肪饮食和高碳水化合物/纤维饮食相比,微生物多样性似乎更低。这种饮食诱导的失调被认为是肥胖症代谢障碍的诱因。
饮食干预菌群变化较快,但是整齐菌群结构稳定
虽然主要在动物模型中得到证实,但数量有限的人体研究表明,饮食干预引起的微生物组成和功能改变可能已经在饮食摄入改变后的几周甚至几天内发生。在人类中,在严格转向完全以植物或动物为基础的饮食后,发现了适度的微生物变化。这些相当极端的饮食干预形式提供了对饮食-肠道微生物组相互作用的潜在机制的见解,并表明饮食干预引起的微生物变化可能会非常迅速地发生。
与此一致,一项小型控制喂养研究显示,在开始高脂肪/低纤维或低脂肪/高纤维饮食后 24 小时内微生物组组成发生了变化,尽管在整个为期 10 天的研究中肠型特征保持稳定。这些研究结果表明,成年人存在微生物复原力的趋势,这可能与长期习惯性饮食摄入有关。然而,由于缺乏对肠型动力学和复原力的理解,细菌肠型的概念受到了其他几项研究的质疑。
一项为期 1 年的干预研究比较了限制能量的地中海饮食和增加体力活动与等热量地中海饮食对超重/肥胖成年人的影响,结果显示两组之间肠道菌群组成的变化存在显著差异。尽管如此,两种饮食的微生物转移趋势是相同的。这表明饮食模式对于肠道微生物的整齐迁移起关键作用。
饮食与肠道和宿主代谢中的糖酵解和蛋白水解发酵之间的相互作用
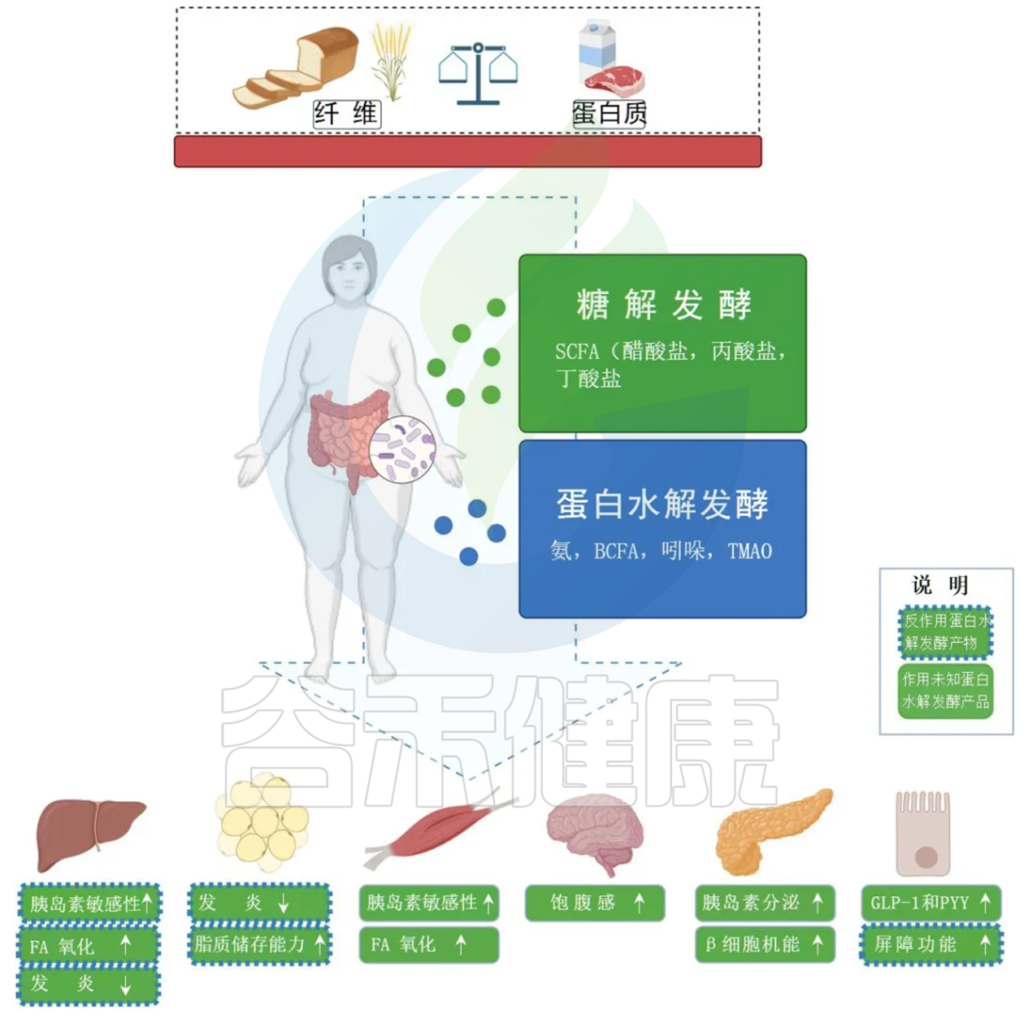
Jardon KM et al., Gut. 2022
膳食纤维的发酵主要发生在近端结肠并产生 SCFA,既可以用作肠细胞的燃料,也可以充当外周信号分子。SCFA 通过影响 GLP-1 和 PYY 的分泌,参与集中调节食物摄入和能量消耗。
蛋白质发酵主要发生在远端结肠并产生更多样化的代谢物,包括与肠道和代谢健康有害影响的 BCFA。
绿框表示 SCFAs 对周围器官代谢过程的影响。
蓝色边框表示蛋白水解发酵产物的相反方向位点方向(虚线)或未知方向(无线)的影响。
BCFA,支链脂肪酸;FA,脂肪酸;GLP-1,胰高血糖素样肽 1;PYY,肽YY;SCFA,短链脂肪酸;TMAO,三甲胺 N-氧化物。
成人肠道微生物组的塑造在生命早期就已经开始,这取决于诸如暴露于母体微生物组、分娩方式和早期暴露于膳食成分等因素。在所有生命阶段影响肠道微生物群组成和功能的众多因素中,饮食是调节特定细菌种类及其功能的丰度的关键。反之亦然,个人对某种饮食或饮食成分的反应可能在很大程度上受肠道微生物群特征的影响。
肠道微生物群能够发酵宿主无法获得的食物成分。小肠中不能被酶分解的膳食纤维和其他复杂碳水化合物可以(部分)被大肠中的细菌发酵,这一般是细菌作为首选能源,发酵后产生微生物产品,如短链脂肪酸(主要是乙酸盐、丙酸盐和丁酸盐)。
主要的产丁酸菌属于厚壁菌门,尤其是:
Faecalibacterium prausnitzii、Clostridium leptum、Eubacterium rectale 、Roseburia.
其他短链脂肪酸的产生由双歧杆菌等细菌介导,双歧杆菌在碳水化合物发酵过程中产生乙酸盐和乳酸。此外,A. muciniphila 物种同时产生丙酸盐和乙酸盐。
稳定同位素技术与13 C标记的短链脂肪酸可根据呼吸、尿液和血液分析对体内结肠产生的短链脂肪酸进行量化。短链脂肪酸主要在结肠中形成,其中约95%随后被吸收。
短链脂肪酸的作用
丁酸盐主要用作结肠细胞的主要能量来源,而丙酸盐和乙酸盐则通过门静脉进入肝脏。特别是,乙酸盐在进入体循环后也能到达外周组织,引起多种代谢和饱腹感相关效应。
短链脂肪酸可与G蛋白偶联受体(GPRs)结合。研究最好的受体包括GPR41、GPR43、GPR109a和GPR164,它们在大量细胞中表达,包括结肠上皮、胰腺β细胞、免疫细胞和周围组织,如脂肪组织。
短链脂肪酸对外周组织的影响包括脂肪生成、抑制脂肪组织脂肪分解(尤其是通过乙酸盐)和减轻脂肪细胞炎症、骨骼肌脂质氧化能力增加、胰腺胰岛素分泌和β细胞功能增加,肝脏的胰岛素敏感性和脂质氧化增加并改变肠-脑相互作用。但是注意这些数据主要来自体外和啮齿动物研究。
短链脂肪酸减脂(人类研究)
在人类研究中发现,长期结肠丙酸盐输送可防止体重增加,减少腹部肥胖和肝细胞内脂质含量,并防止超重成年人胰岛素敏感性的恶化。与这些发现一致,人体内数据表明,在超重或肥胖的成年人中,饮食诱导微生物短链脂肪酸产生变化或直接结肠短链脂肪酸输注后,空腹脂质氧化和静息能量消耗增加。
碳水化合物的消化是一个复杂的过程,涉及摄入的碳水化合物类型的特定酶。大多数可消化的膳食碳水化合物在小肠中被消化和吸收,而某些不可消化的碳水化合物,包括抗性淀粉和膳食纤维,很容易被结肠中含量最高的肠道微生物发酵。
膳食纤维对肠道菌群的有益影响
膳食纤维已被证明对与健康益处相关的肠道微生物群的组成和功能具有显著影响。这些因膳食纤维的结构、物理和化学特性可能会有所不同,例如水溶性、粘度、粘合和膨胀能力以及发酵性。高度可发酵的纤维,如 β-葡聚糖、菊粉和低聚半乳糖,在对微生物群组成和肠道代谢物产生的影响方面得到了很好的定义,而不溶性纤维虽然部分发酵,但大多数人都知道它们对粪便的有益作用一致性和结肠传输时间。

摄入高纤维饮食有益地影响宿主的健康,其中包括影响葡萄糖和脂质代谢。重要的机制包括调节营养吸收或产生短链脂肪酸,但有关膳食纤维对健康影响的数据存在争议。
对于膳食纤维研究中不一致发现的解释:
首先,在大多数人体研究中,只补充了一种特定的可发酵纤维,因此只刺激了一种或几种个体(潜在有益的)细菌属。后者的后果可能是其他必需细菌或核心菌属的丰度减少,这可能导致微生物生态系统的不平衡。因此,结合刺激多种不同细菌属的不同纤维可能对维持微生物丰富度以及对免疫状态和代谢健康产生更显著的(相加或协同)影响很重要,所以多样化膳食纤维和饮食摄入对于健康益处的微生物调节更有用。
有趣的是,一项研究表明,结肠中产生短链脂肪酸的部位可能是代谢健康的决定因素。急性远端结肠乙酸盐给药增加了超重男性的循环乙酸盐浓度,增加了脂肪氧化和刺激饱腹感激素 PYY,并降低了血浆肿瘤坏死因子-α。与远端输注相比,近端结肠中的乙酸盐给药不影响代谢特征。因此,通过结合不同的膳食纤维和/或更复杂的膳食纤维,针对远端结肠中微生物物种的膳食纤维可用性和短链脂肪酸形成,可能是改善免疫和代谢健康的有前景的策略。
TIPs
短链脂肪酸在一定范围内是越高越好,但是超过一定范围,也会产生害处。例如,高纤维饮食增加丁酸盐,诱导Stx受体球形三酰神经酰胺表达从而促进致病大肠杆菌定植。
此外,有益的短链脂肪酸一般需要通过结肠部位的菌群发酵产生,如果外源性的补充摄入,例如,丙酸盐有助于防止食物上霉菌,被广泛使用于烘焙食物、动物饲料和人造调味品中。如果长期摄入过量含有丙酸盐的食物,可能会增加人类患糖尿病和肥胖症的风险。
其次,到目前为止,大多数膳食纤维干预研究都没有考虑基线微生物组或代谢表型。基线肠道微生物组的特征可能与饮食干预结果密切相关。例如,已经表明肠道微生物群对膳食纤维(抗性淀粉与非淀粉多糖)的反应可以根据肥胖男性的基线微生物多样性来预测。高微生物多样性与微生物群的较低膳食反应性相关,这可能支持肠道微生物的更高多样性与微生物生态系统的稳定性有关的假设。
与此一致,与基因计数低的个体相比,基线时的高微生物基因计数与对减肥饮食的不太明显的反应有关。在低基因计数组中,基因丰富度和临床参数有所改善,尽管在基因丰富度低的个体中炎症标志物的变化不太明显。
一项针对肥胖个体的研究表明,不是基线微生物多样性而是厚壁菌门的基线丰度预测了个体微生物群的饮食反应。总之,这些发现表明微生物多样性并不总是饮食反应性的预测指标,这意味着需要进一步研究以更好地了解复杂的饮食-微生物组-宿主代谢相互作用。
作为对菊粉型果聚糖益生元的反应,具有高习惯性膳食纤维摄入量的健康个体的肠道菌群组成发生了更大的变化,而习惯性纤维摄入量低的人肠道菌群似乎更能适应变化。在II型糖尿病患者中进行的一项研究表明,膳食纤维促进了一组精选的产生短链脂肪酸的菌株,而许多其他微生物,包括蛋白水解发酵中的微生物,要么减少要么不变,表明微生物基因丰富度总体下降。粪便短链脂肪酸增加,尤其是丁酸盐,伴随着葡萄糖稳态的改善。因此,如几项人类纤维膳食干预研究所示,更高的微生物基因丰富度本身可能无益,但生理结果可能更依赖于微生物网络的功能。
在一项调查 6 周全麦饮食对体重变化影响的研究中,普雷沃氏菌属的高基线丰度与超重、健康成年人的体重减轻程度较高相关。这些发现表明,作为对特定饮食干预的反应,肠道微生物群的影响调节剂具有预测能力。
此外,发现超重、前驱糖尿病个体与瘦个体相比,对短期施用长链菊粉和抗性淀粉的微生物多样性和餐后胰岛素敏感性的变化的反应降低。与此一致,最近的研究表明,基线肠道微生物特征可以预测补充 3 个月长链菊粉后 BMI 的变化,这种效应在不同个体的粪便微生物群定植的小鼠中得到了复制。
有趣的是,可溶性菊粉纤维已被证明可以降低空腹血糖受损人群的胰岛素抵抗,但不能降低葡萄糖耐量受损的人群。鉴于空腹血糖受损与肝脏胰岛素抗性密切相关的发现,后一发现可能表明纤维 – 肠道微生物群 – 宿主代谢串扰中的组织特异性。
总体而言,益生元膳食纤维对代谢健康结果的有效性可能取决于几个参数,包括基线微生物组成以及微生物发酵的部位。
在低膳食纤维的西方饮食人群中,结肠远端的微生物群更擅长于利用剩余肽和蛋白质的发酵,因为首选的燃料,可发酵碳水化合物,已经在近端结肠中被人体大量使用。这种蛋白水解发酵过程的产物包括气体产物,如氢、甲烷、二氧化碳和硫化氢;BCFAs异丁酸酯、2-甲基丁酸酯和异戊酸酯(源自BCAAs发酵)、酚类和吲哚类化合物(源自芳香族氨基酸微生物发酵)以及较小的、未知的短链脂肪酸。
与糖解发酵产物相比,大多数蛋白水解发酵产物被认为对宿主肠道和代谢健康有害,尽管一些动物数据表明吲哚和硫化氢对肠道和外周组织功能有益。
例如,一些只能由肠道细菌(吲哚)或哺乳动物宿主(酪胺、色胺和短链脂肪酸)产生的氨基酸衍生化合物通过影响GLP-1和肠内分泌细胞血清素的分泌,直接影响哺乳动物的饱腹感和肠道运动。
然而,大多数这些化合物对宿主肠道和周围组织的生理作用仍不清楚。许多此类化合物的人类来源和细菌来源之间的区别尚未完全确定,需要进一步的体内研究来验证此类效应。
结肠中糖酵解和蛋白水解发酵之间的平衡,以及对宿主生理的假定有益和有害调节之间的平衡,可能对制定饮食干预策略很有意义。
一些研究表明,增加膳食纤维的摄入量,特别是缓慢发酵纤维的摄入量,会减少肠道微生物群仅产生有害的蛋白水解代谢物,使得整体发酵平衡向更有益的糖酵解发酵转变。
摄入的膳食蛋白质首先在小肠中被胰酶和来自肠细胞的肽酶消化。然后,大量的寡肽和氨基酸通过肠细胞转运蛋白被转运到门静脉血流中,在那里它们被用作蛋白质合成的氨基酸前体或被代谢为燃料或肠粘膜代谢物必需的前体。
由于远端小肠和近端结肠中的大多数细菌优先使用可发酵碳水化合物而不是蛋白质,因此大多数氨基酸作为能量来源的发酵发生在碳水化合物被耗尽的远端结肠。
摄入的蛋白质到达大肠的百分比也可能取决于蛋白质质量,估计约为 10%。由于植物的细胞壁不易消化,源自植物的蛋白质的消化率较低,而源自动物的蛋白质更容易在大肠中消化,这表明功能结果存在潜在差异。
酪蛋白是一种从动物产品中提取的相对缓慢消化的蛋白质,是防止高脂肪/高蛋白饮食小鼠体重增加和脂肪量增加的最有效蛋白质来源。
蛋白水解和糖酵解发酵之间的平衡可能决定对生活方式干预的反应情况,因此应在未来的研究中加以考虑。


流行病学研究还表明,摄入乳制品和素食蛋白质来源与预防肥胖有关,而大量摄入肉类(尤其是红肉)则预示着体重增加会更高。
尽管研究较少,但蛋白质摄入已被证明会影响微生物群的组成和功能。效果取决于蛋白质的氨基酸组成和消化率,而蛋白质的来源和摄入量会影响它们。
蛋白质摄入影响微生物组成
在大鼠研究中,高蛋白饮食与C. coccoides, C. leptum, F. prausnitzii 减少有关,而超重或肥胖雄性中Roseburia, E. rectale, C. aerofaciens, Bacteroides, Oscillibacter 减少。
值得注意的是,以等热量的方式比较高脂肪/高蛋白饮食与中等蛋白质或低蛋白质饮食会导致饮食之间碳水化合物或脂肪含量的差异。因此,对于所有的等热量膳食宏量营养素交换研究,很难确定导致肠道微生物群组成变化的主要膳食因素,这可能归因于一种(宏量)营养素的增加或另一种营养素的减少。
膳食脂肪已被广泛研究与饮食相关的代谢疾病(如肥胖)相关,但其对人类肠道微生物群的影响尚不明确,而且研究通常会得出相反的结果。
不同类型的脂肪酸(饱和、单不饱和、多不饱和脂肪酸)、碳链长度和饱和度可能对肠道微生物群组成有明显影响。
横断面研究表明,食用富含动物蛋白和脂肪的饮食与拟杆菌属肠型有关,而高纤维、水果和蔬菜的摄入与健康成年人的普氏菌肠型有关。

此外,主要饱和脂肪酸(SFA)的高摄入量与成人和婴儿肠道微生物丰富度和多样性的降低有关。在超重和肥胖人群中, 主要饱和脂肪酸与肠单胞菌属呈负相关,而主要饱和脂肪酸与Roseburia呈正相关,后者在体重正常的个体中也非常丰富。在这项研究中,根据 BMI,习惯性 主要饱和脂肪酸摄入量与产丁酸菌表现出相反的关联特征。
总体而言,应该注意的是,与膳食纤维相比,膳食脂肪-微生物组-宿主生理学相互作用的研究较少,而且其机理知识主要基于动物研究。根据人类生理学比较难解释这些发现,应进一步研究。
多酚主要作为酚类化合物存在于水果和蔬菜中,以其作为抗氧化、抗炎、心脏保护、癌症预防和神经保护剂的有益作用而闻名。
补充天然存在于茶中的表没食子儿茶素-3-没食子酸酯(epigallocatechin-3-gallate) 2个月,对肥胖小鼠胆汁酸代谢和疣微菌科Verrucomicrobiaceae丰度均有影响,促进了A. muciniphila丰度的增加。在其他研究中,后者与有益的代谢作用有关。
此外,虽然也在动物模型中,但 8 周的多酚补充剂可防止饮食引起的肥胖和肠道炎症,这与Akkermansia的丰度增加有关。在健康、超重或肥胖的个体中,12 周的白藜芦醇和表没食子儿茶素-3-没食子酸酯联合补充剂改善了男性的代谢参数并减少了拟杆菌门,但女性没有。
以上两项研究都表明存在性别特异性微生物反应,在评估干预反应时应考虑这一点。

总体而言,在饮食中添加膳食多酚似乎可以促进肠道和代谢健康,尽管仍然需要对人体研究的机制见解。
基于微生物组的精准营养预测代谢健康参数,如血糖反应和变异性,或用于抵消代谢紊乱,目前已受到很大关注。
该领域的一项具有里程碑意义的研究表明,尽管餐后血糖反应的人际差异很大,但在机器学习算法的帮助下创建的个性化饮食(基于习惯性饮食、身体活动和肠道微生物群)可能会成功降低血糖反应和不良代谢健康,还有助于减肥。
研究测试在对不同类型面包的血糖反应中发现了显著的人际差异,并且这种血糖反应可以通过基线微生物组特征来预测。值得注意的是,这些研究主要基于他们对急性膳食挑战和短期干预的反应,而不是长期干预反应。
肠道微生物组的预测能力正变得越来越明显,特别是在检查纤维和粪便微生物群移植效果的研究中。在长期的肠道菌群检测经验实践中也证实,基线微生物特征是对饮食干预(例如,膳食纤维或复合蛋白质)的反应性的有趣生物标志物,也是个性化健康管理的应该纳入的指标基础。
微生物组-宿主代谢轴可能对胰岛素抵抗患者的饮食干预存在抗性,这表明干预可能需要更长的时间,或者需要摄入的功能性膳食成分(如膳食纤维)来诱导有益的效果。特定功能微生物群的特点是对膳食成分的不同消化能力,导致微生物代谢物(如 短链脂肪酸)的不同产生,随后影响宿主代谢的调节。
总的来说,在评估饮食模式和常量营养素组成不同的饮食时,重要的是要同时考虑饮食成分的数量和质量,由于与宿主的微生物和代谢表型的不同相互作用,在整体饮食方法中要考虑到微量营养素和生物活性成分,如多酚。
对饮食干预的反应不仅取决于肠道微生物群的特征,还取决于饮食、生活方式和环境因素以及代谢表型等临床特征之间复杂的多因素相互作用。
Jardon KM, et al., Gut. 2022
为了将基于精确的策略转化为医疗保健实践或指南,我们需要彻底了解为什么人们对饮食的反应不同,差异反应和相关表型是否长期保持,以及开发的算法在多大程度上是可重复的。
在饮食干预研究中通过最先进的方法进行详细的微生物和代谢表型分析至关重要。显然,鉴于复杂性,除了生活方式和环境因素的详细信息外,还需要详细的信息,包括出生方式、病史、药物使用情况(尤其是抗生素)、身体活动、心理压力和睡眠质量等。这也意味着需要先进的统计和建模方法来梳理不同因素的重要性。

主要参考文献:
Jardon KM, Canfora EE, Goossens GH, Blaak EE. Dietary macronutrients and the gut microbiome: a precision nutrition approach to improve cardiometabolic health. Gut. 2022 Feb 8:gutjnl-2020-323715. doi: 10.1136/gutjnl-2020-323715. Epub ahead of print. PMID: 35135841.
Agus A, Clément K, Sokol H. Gut microbiota-derived metabolites as central regulators in metabolic disorders. Gut. 2021 Jun;70(6):1174-1182. doi: 10.1136/gutjnl-2020-323071. Epub 2020 Dec 3. PMID: 33272977; PMCID: PMC8108286.
Jie Zhuye,Yu Xinlei,Liu Yinghua et al. The Baseline Gut Microbiota Directs Dieting-Induced Weight Loss Trajectories.[J] .Gastroenterology, 2021
Jie Z, Yu X, Liu Y, Sun L, Chen P, Ding Q, Gao Y, Zhang X, Yu M, Liu Y, Zhang Y, Kristiansen K, Jia H, Brix S, Cai K. The Baseline Gut Microbiota Directs Dieting-Induced Weight Loss Trajectories. Gastroenterology. 2021 May;160(6):2029-2042.e16. doi: 10.1053/j.gastro.2021.01.029. Epub 2021 Jan 20. PMID: 33482223.

谷禾健康


嗜粘蛋白-阿克曼氏菌(Akkermansia muciniphila,A.muciniphila),简称为 AKK,于 2004 年发现,是一种从人类粪便中分离出来的椭圆形革兰氏阴性细菌。
该细菌以荷兰微生物生态学家 Anton DL Akkermans 的名字命名,以表彰他对该领域的贡献。A. muciniphila 是人类肠道共生菌,并可以依靠肠粘液层的黏蛋白生存。
在过去十年中,越来越多的研究证明,在糖尿病、心血管疾病、疾病性肠病、神经疾病发现A. muciniphila丰度降低。最近的干预研究还证实如茶或富含多酚水果的饮食可增加其丰度,有助于改善糖尿病和肥胖个体的代谢功能。
总的来说,来自动物和人类研究的越来越多的证据表明A. muciniphila将成为下一代有临床应用前景的益生菌。尤其是它在预防和治疗糖尿病、肥胖症及癌症,这对未来的研究具有重要意义和发展。
A. muciniphila是一种厌氧、革兰氏阴性、无运动、没有内生孢子的卵圆形肠道细菌,其最适生长温度是37℃,最适生长pH为6.5,该菌的倍增时间大约是1.5 h。A. muciniphila属于疣微菌门,事实上,它是人类粪便样本中发现的属于这个特殊门的唯一物种。
值得注意的是,A. muciniphila具有一定的耐氧性,其暴露在空气中24 h,仍然有1%以上的存活率。此外,研究还发现低氧气浓度(纳摩尔)下可以显著促进A. muciniphila的生长。
A. muciniphila单独或者成对存在,很少成链生长,其代谢产物为乙酸盐、丙酸盐、1,2-丙二醇等。
在提供蛋白源的基础培养基中,加入N-乙酰氨基葡萄糖、N-乙酰半乳糖胺和葡萄糖作为能源,A. muciniphila可以生长,但是在果糖和纤维二糖等其他糖类作为能源的培养基中A. muciniphila不能生长,说明该菌的生长繁殖需要蛋白质。
A. muciniphila的特别有趣之处在于,与大多数其他有益细菌不同,它不完全依赖于我们的饮食来喂养。即使消化道中没有营养物质,它也会以粘膜层中的粘蛋白为食。
Akkermansia Muciniphila一般通过母体获得, 通常在儿童中大量存在,并随着年龄和疾病而减少。将其恢复到最佳水平可以带来许多好处。
肠道中低浓度的Akkermansia muciniphila可能表明黏液层较薄,从而导致肠道屏障功能减弱,此外还会增加细菌毒素的易位,炎症性肠病、哮喘、肥胖症和 II 型糖尿病患者的Akkermansia muciniphila浓度往往较低。
我们知道整个胃肠道 (GI) 的粘膜完整性对于维持人体健康至关重要。这其中,粘液屏障的完整性是胃肠道的第一道防线。肠道黏液层的周转包括黏液的合成、分泌和降解,这是一个需要调节和平衡的微妙过程,以确保黏液保持最佳的保护功能。
粘液由多种成分组成:水 (90%–95%)、电解质、脂质 (1%–2%)、蛋白质等。这种粘液是一种稀的、水性和粘弹性的分泌物,这要归功于被称为粘蛋白的特定粘液蛋白,它是粘液中存在的主要结构和功能成分,浓度为 1%–5%。
粘蛋白是一个大的、复杂的、糖基化的蛋白质家族,其特征是一个重要的元素,即“粘蛋白结构域”。它由一个蛋白质核心组成,该核心由含有氨基酸残基脯氨酸 (Pro)、苏氨酸 (Thr) 和丝氨酸 (Ser) 的序列组成,称为富含 PTS 的序列,通常串联重复,其中 Ser 和 Thr 广泛为 O-糖基化并赋予“瓶刷”状构象。
MUC2(黏糖蛋白2)的化学结构和肠道粘液的合成
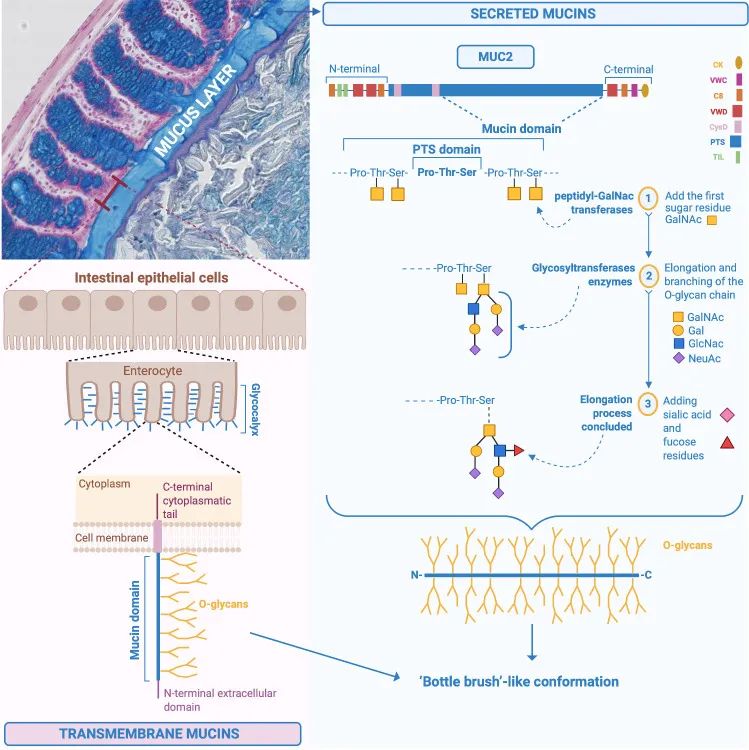

Paone P & Cani PD. Gut. 2020
肠道菌群在肠道中的分布梯度随其位置不同而变化;微生物密度从近端到远端肠道增加,每克肠道内容物的微生物细胞数量大约如下:
十二指肠中103 个
空肠中 104 个
回肠中 107个
结肠中1012 个
此外,微生物密度从上皮细胞向管腔增加,在后者中发现的细菌数量最多,与管腔相比,实际上很少有细菌物种能够很好地粘附和驻留在粘液层中。除了粘附这一重要功能外,肠道微生物群对调节肠道粘液层有很大贡献。
需要普及一个认知就是,当我们谈到构成微生物组的各种细菌物种时,有些菌似乎比其他细菌具有更显着的影响,比如肠道基石菌,核心菌,益生菌等,确保这些个体物种的最佳水平——不要太低,也不要太高——可以产生对健康深远的影响。这其中包括有益细菌的关键物种之一 Akkermansia Muciniphila。
谷禾肠道样本大数据库显示A. muciniphila缺乏或未检出情况在人群中很常见,尤其是那些有肠道问题的人。在健康个体中其约占肠道微生物群总数的0.5%–3%。
但是也有例外,在我们在最近的检测的一个56岁女士肠道内该菌的丰度占比超50%,菌群构成如下:


然而,好事太多也可能是坏事。它的名字——“muciniphila”的意思是粘液爱好者,指的是它独特的能力,降解黏蛋白和粘液层。过量的Akkermansia 将过度消耗粘液蛋白而存活下来,这是大多数其他细菌所缺乏的生存优势。在这种情况下,非粘液消耗物种的数量显着减少,导致物种多样性减少, Akkermansia 增殖异常,从而可能导致肠道屏障损伤,诱发肠道炎症、LPS 进入血液的增加、自身免疫性疾病有关,神经退行性疾病等。
该样本检测报告也显示菌群多样性低,肠炎和几项慢病注意风险:
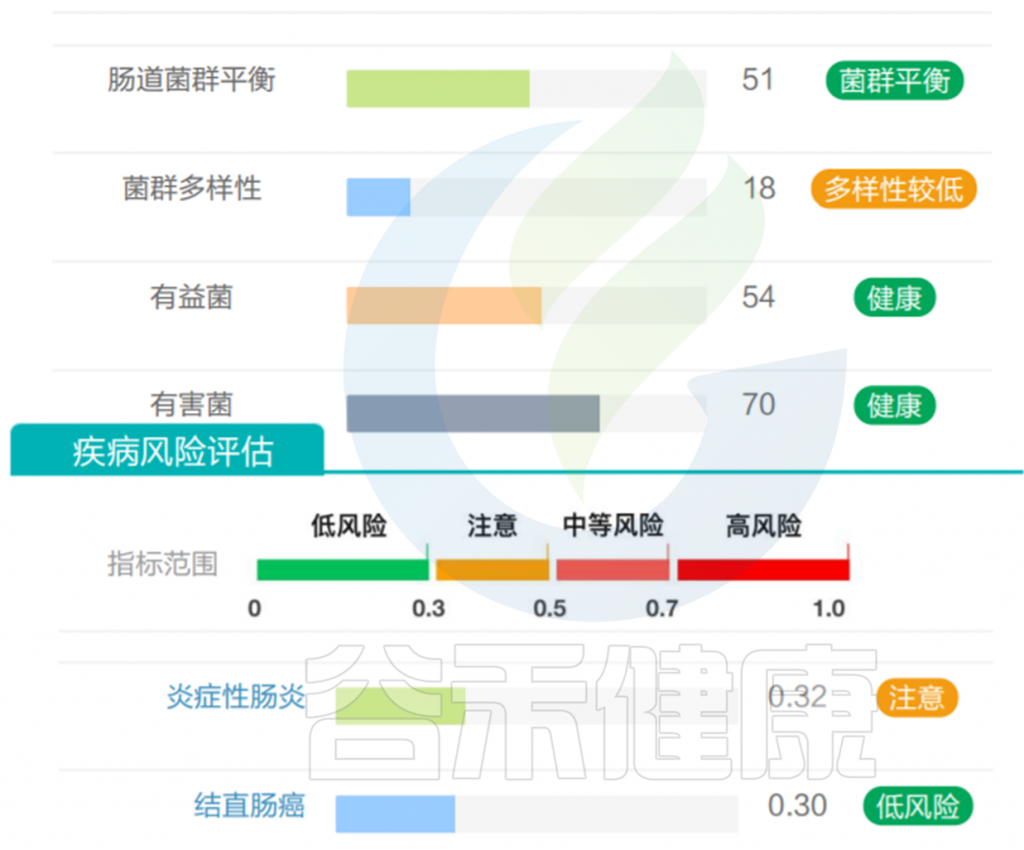

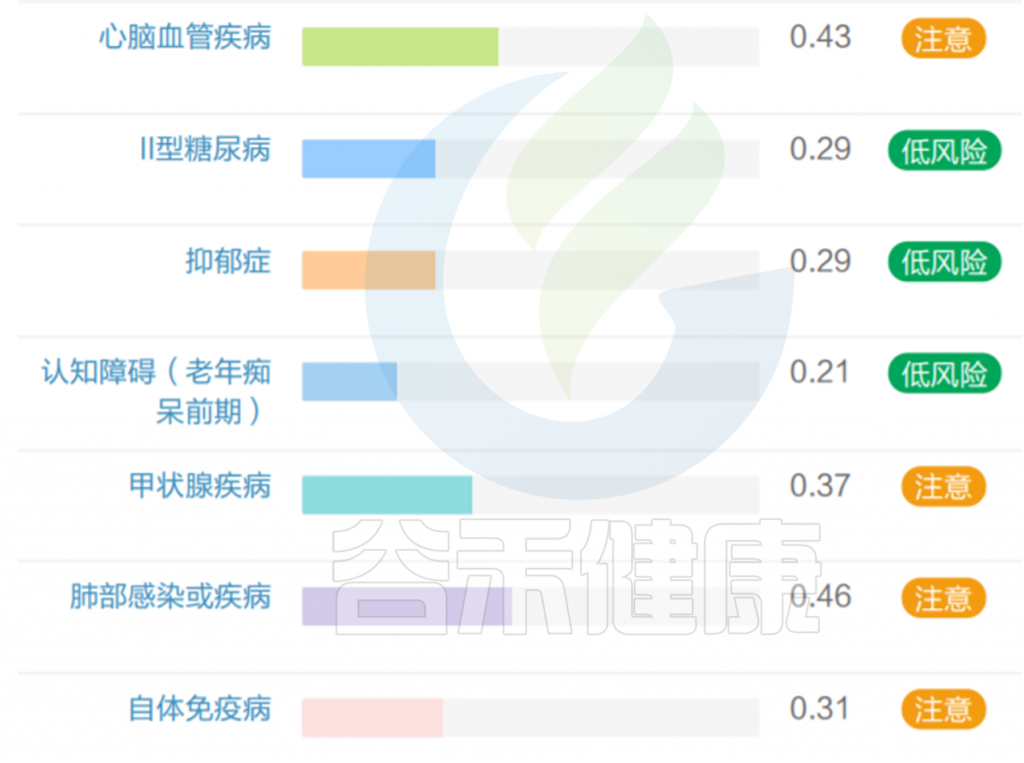

此外,在其它研究文献中也得出Akkermansia muciniphila丰度高的负面研究,如:
在中国人中,A. muciniphila菌的丰度与2型糖尿病呈正相关。
血红素在肠道中的过度增生,与A. muciniphila的粘膜溶解活性有关,在特定情况中,A. muciniphila参与促进代谢综合征。
北京协和团队发现Akkermansia muciniphila可能参与了系统性红斑狼疮的发生发展,致自身免疫亢进特征。
通过细菌16S rRNA测序表明其属于疣微菌门(Verrucomicrobia),与Verrucomicrobium spinosum有92%序列相似性,是现在唯一已知的人类肠道疣微菌门成员。其中文译名尚无统一,在文献中以阿克曼黏细菌、艾克曼黏细菌等名字出现。
为了揭示A. muciniphila的功能潜力,对其基因组进行了测序和注释。发现Akkermansia muciniphila在其相对较小的基因组中编码了大量黏蛋白降解酶,黏蛋白被其降解后其下游聚糖副产物可以交叉饲养其他肠道细菌。
根据DNA-DNA杂交(Digital DNA-DNA hybridiztion,dDDH)将23个Akkermansia属的菌株分为4种,分别为:
Akkermansia sp.CAG
Akkermansia sp.KLE
Akkermansia. muciniphila
Akkermansia. glycaniphila
但是发现它包含许多候选粘蛋白酶编码基因,但缺乏编码典型粘液结合域的基因。
种内存在多样性。在整个基因组中发现的大量噬菌体相关序列,表明病毒在该物种的进化中发挥了重要作用。此外,发掘了 37 个胃肠道宏基因组中,确定Akkermansia序列的存在和遗传多样性。在 37 个中,11 个包含 16S 核糖体 RNA 基因序列,与A. muciniphila 的相同 > 95% , 称为Akkermansia-like 微生物。
瓦赫宁根大学微生物实验室从蟒蛇体内分离培养出于A. muciniphila 基因序列具有94.4%相似度的新菌株,并且发现它们也具有相似的特性,将该微生物命名为Akkermansia glycaniphila。
Akkermansia muciniphila是一种肠道中的黏蛋白降解细菌。粘蛋白是肠上皮黏液层的重要组成部分。粘蛋白由杯状细胞产生肠上皮,是高度糖基化的分子,还包括丝氨酸、苏氨酸和半胱氨酸,它们在肠组织表面形成透明的粘液层,是大量肠道微生物栖息的主要部位。
A. muciniphila主要定植在胃肠道的外黏液层,以胃肠道的黏蛋白作为自身生长的碳和氮来源,其消耗黏蛋白与杯状细胞再生黏蛋白能够达到动态平衡,从而维持黏液层稳定。A.muciniphila以及其分泌物,如囊泡(Extracellular vesicles, EV)通过与结肠上皮细胞Toll受体(Toll-like receptor,TLR)结合及调节紧密连接蛋白的表达等来维持肠道稳态,从而改善高脂饮食诱导的肥胖和炎症性肠病等疾病。
此外,粘蛋白还覆盖在呼吸道、消化道和泌尿生殖道的细胞表面。许多癌症(胰腺癌、肺癌、乳腺癌、卵巢癌、结肠癌和其他组织)都会增加粘蛋白的产生。粘蛋白也在肺部疾病中过度表达,例如哮喘、支气管炎、慢性阻塞性肺病 (COPD) 或囊性纤维化。
Akkermansia muciniphila 高度适应从粘蛋白降解中生存——例如,它需要苏氨酸才能生长,并且具有多种适合利用粘蛋白寡糖中的特殊糖类的酶,如唾液酸酶和岩藻糖苷酶,甚至硫酸酯酶。表明宿主动物和AKK菌之间的共同进化进入了高级阶段,并暗示了这些疣状微生物对宿主的潜在功能意义。由于其向其他细菌物种提供粘蛋白降解产物,该物种也是肠道重要的共生菌属。
Akkermansia muciniphila 能够使用粘蛋白作为碳、氮和能量的唯一来源。尽管宿主肠道层中的许多黏液降解微生物可能具有致病性,但A. muciniphila不一样。Akkermansia muciniphila从粘蛋白发酵中产生乙酸盐、丙酸盐和乙醇,以调节宿主的生物学功能,包括宿主免疫反应和脂质代谢。由于A. muciniphila保留在外黏膜层并且不会渗透到内层,因此它的定植是有益的,因为细菌会促进外黏蛋白层的自然更新。
除了基因序列多样性外,这种细菌还表现出多种生物学功能,包括:
促进肠道屏障完整性;
调节免疫反应;
抑制炎症;
交叉喂养他微生物群;
降低某些心脏病风险,如胰岛素抵抗、总血胆固醇和脂肪组织储存;
防止体重增加。
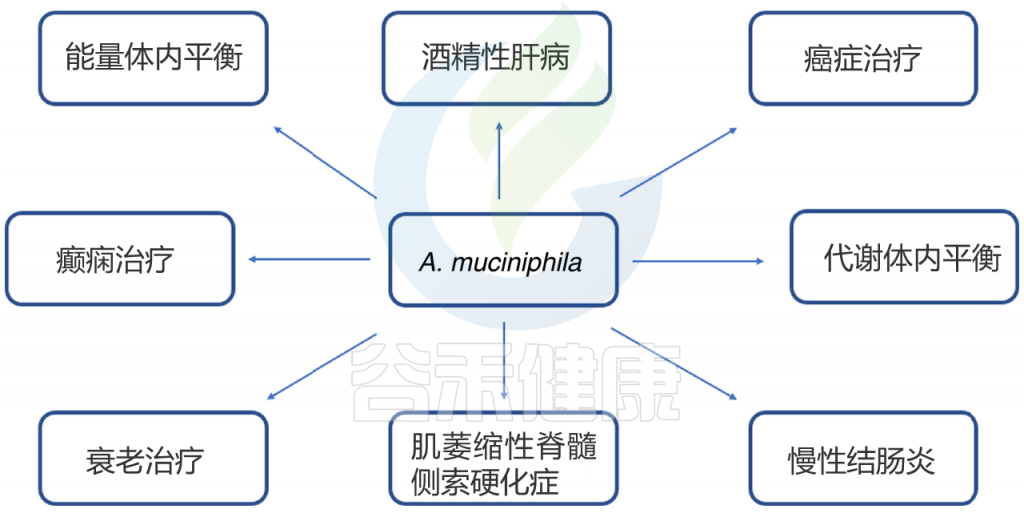

DOI: (10.1111/jam.14911)
促进肠道屏障
肠道屏障是一个多层复杂的系统,它允许营养吸收,同时防止微生物及其产物的易位。肠道屏障的破坏导致管腔内容物进入血流,激活免疫反应并诱发炎症。
粘液覆盖外肠上皮细胞层,起到物理保护作用,防止微生物和有害化合物的渗透。除了降解粘蛋白外,A. muciniphila还被发现可以刺激粘蛋白的产生 。
A. muciniphila在防止人类肠道致病菌增殖中的作用
Kalia VC, et al., Indian J Microbiol. 2021
今年发表在Gut Microbes研究得出,虽然A. muciniphila 是一种粘蛋白降解细菌,但是通过促进分泌型 IEC 的分化,反而增加了粘液的产生。A. muciniphila处理以 Gpr41/43 依赖性方式增强了ISC 增殖,加速肠上皮再生,促进 IEC 发育并维持肠道稳态。
注释:哺乳动物肠上皮细胞(IEC)更新较快,每 3-5 天补充一次。所有类型的 IEC 都源自肠干细胞(ISC)
粘蛋白的周转率为 6-12 小时,内层的周转率约为 1 小时,据信其分泌受神经、激素和旁分泌作用的调节。
在动物模型中,A. muciniphila补充剂使结肠粘液层的厚度增加了大约 3 倍,明显高于由有益细菌植物乳杆菌引起的厚度增加。此外,在体外,发现A. muciniphila通过直接与肠细胞结合来改善肠细胞单层完整性 。
A. muciniphila如何促进粘液厚度尚不清楚。原因之一可能是A. muciniphila通过从降解的粘蛋白中制造短链脂肪酸来刺激粘液周转率,这是合成和分泌粘蛋白的宿主上皮的优选能源。事实上,A. muciniphila补充剂增加了小鼠产生粘蛋白的杯状细胞的数量。
抑制炎症
有证据表明A. muciniphila可以调节炎症。在加速衰老的小鼠模型中补充这种细菌可以减轻炎症 。其他研究也显示了A. muciniphila在不同小鼠模型中的抗炎特性,包括无菌模型、肝损伤模型和肥胖模型。
A. muciniphila可以减少由牙龈卟啉单胞菌引起的瘦或肥胖小鼠的炎症。
此外,A. muciniphila分泌的囊泡能够降低TLR4(Toll-like receptor 4, TLR4)的表达,从而调节NF-κB通路,减少促炎因子IL-6、IL-8的分泌。
调节免疫
A. muciniphila在小鼠体内平衡期间,在诱导肠道驻留 T 细胞方面发挥着宿主相关的作用。
在一项针对围产期 HIV 感染儿童和青少年的横断面研究中报告说,A. muciniphila丰度与升高的IL-6 和可溶性 CD14 相关 。
A. muciniphila降解黏蛋白,释放可以被周围非黏液降解细菌利用的短链脂肪酸(SCFAs)、寡糖等代谢物。SCFAs能抑制组蛋白脱乙酰基酶,从而调节NF-κB通路,减少炎症因子的表达。丁酸可以促进Treg细胞的外周活化,增加结肠中Treg细胞亚群的数量,抑制促炎免疫细胞CD4+T淋巴细胞、CD8+T淋巴细胞活性。
免疫相关的皮肤病——牛皮癣
银屑病(俗称牛皮癣)是一种免疫介导的慢性炎症性皮肤病。研究表明银屑病和肠道炎症之间存在关联。一项研究评估了来自巴西的21名银屑病患者与24名健康对照者的肠道微生物群组成和多样性。与对照组相比,银屑病患者的 Akkermansia muciniphila 数量有所减少。
扩展阅读:牛皮癣看似皮肤病,实则关系到肠道
交叉喂养
除了A. muciniphila外,粘液层中还常见其他微生物。产生丁酸盐的细菌不具有降解粘液的能力,而是利用被粘蛋白降解的物种(如A. muciniphila)降解的碳和氮,这些微生物包括细菌如Faecalibacterium prausnitzii,罗斯氏菌等其产生抗炎短链脂肪酸(SCFA)。
产生的乙酸能够促进粪厌氧棒状菌(Anaerostipes caccae),霍氏真杆菌(Eubacterium hallii)。
报道共培养A. muciniphila与非粘液降解丁酸盐产生细菌F. prausnitzii、A. caccae和Eubacteriumhallii导致同养生长和丁酸盐的产生。因此,A. muciniphila不仅自身在保护肠道上皮方面发挥重要作用,而且还支持抗炎肠道微生物群。
A. muciniphila和其他细菌之间的相互作用
Hagi T, et al., Appl Microbiol Biotechnol. 2021
有些细菌不能降解粘蛋白,因此非粘蛋白降解细菌受益于粘蛋白降解细菌。
A. muciniphila产生来自粘液和短链脂肪酸的糖,如乙酸盐和丙酸盐。非粘液降解细菌,如Anaerostipes caccae, Eubacterium hallii and Faecalibacterium prausnitzii ,利用A. muciniphila从粘液中降解的糖进行生长。
A. muciniphila的粘蛋白降解支持了Anaerostipes caccae的生长和丁酸生产。Anaerostipes caccae诱导AKK中粘蛋白降解基因的表达增加,核糖体基因的表达减少。
防止体重增加
普遍发现A. muciniphila在瘦个体中更为丰富。在超重个体中,细菌的丰度较低。
在小鼠中,与肥胖小鼠的丰度相比,瘦动物粪便中A. muciniphila丰度增加之间的相关性表明了这种意义,并且观察到在数周内每天向高脂肪饮食诱导的肥胖小鼠喂食Akkermansia显然可以逆转这种诱导的肥胖以及改善上皮完整性和相关效应。
也有一些数据表明,A. muciniphila干预可能成为人类受试者肥胖改善方案的基础。但是强调只有活的Akkermansia菌才能在小鼠中产生这种效果,但是发现高温预处理杀菌的细胞甚至纯化的Akkermansia蛋白,与活细胞治疗的效果相比,Akkermansia的热杀死细胞似乎对高脂肪饮食肥胖小鼠具有相同甚至增强的有益效果。
此外,A. muciniphila通过分解粘蛋白产生醋酸盐这样的 SCFAs,对我们的体重产生积极影响。因为乙酸盐可以阻止我们感到饥饿,从而防止体重增加。
A.muciniphila治疗不会显著改变饮食诱导的肥胖小鼠的肠道微生物组成,但它可以逆转HFD诱导的代谢紊乱,包括脂肪质量增加、代谢内毒素血症、脂肪组织炎症和胰岛素抵抗,这表明它可能用于预防或治疗肥胖和相关的代谢紊乱。
人类肠道微生物群中丰富的A.muciniphila可预防疾病
Kalia VC, et al., Indian J Microbiol. 2021
改善代谢
A. muciniphila补充剂能够恢复肥胖和 2 型糖尿病小鼠的粘液厚度,其中高脂肪饮食治疗破坏了肠道粘液;该治疗还导致血清脂多糖 (LPS) 显着减少,这是一种代谢性内毒素血症,并改善了代谢状况。
对肥胖小鼠施用Akkermansia还通过减少循环中的炎性脂多糖和减轻胰岛素抵抗而导致观察到的“代谢性内毒素血症”减少作为小鼠肥胖的心脏代谢并发症,如动脉硬化。至少来自小鼠模型的结论Akkermansia效应得到了许多不同研究的支持。
A. muciniphila及其衍生产物对宿主代谢的影响
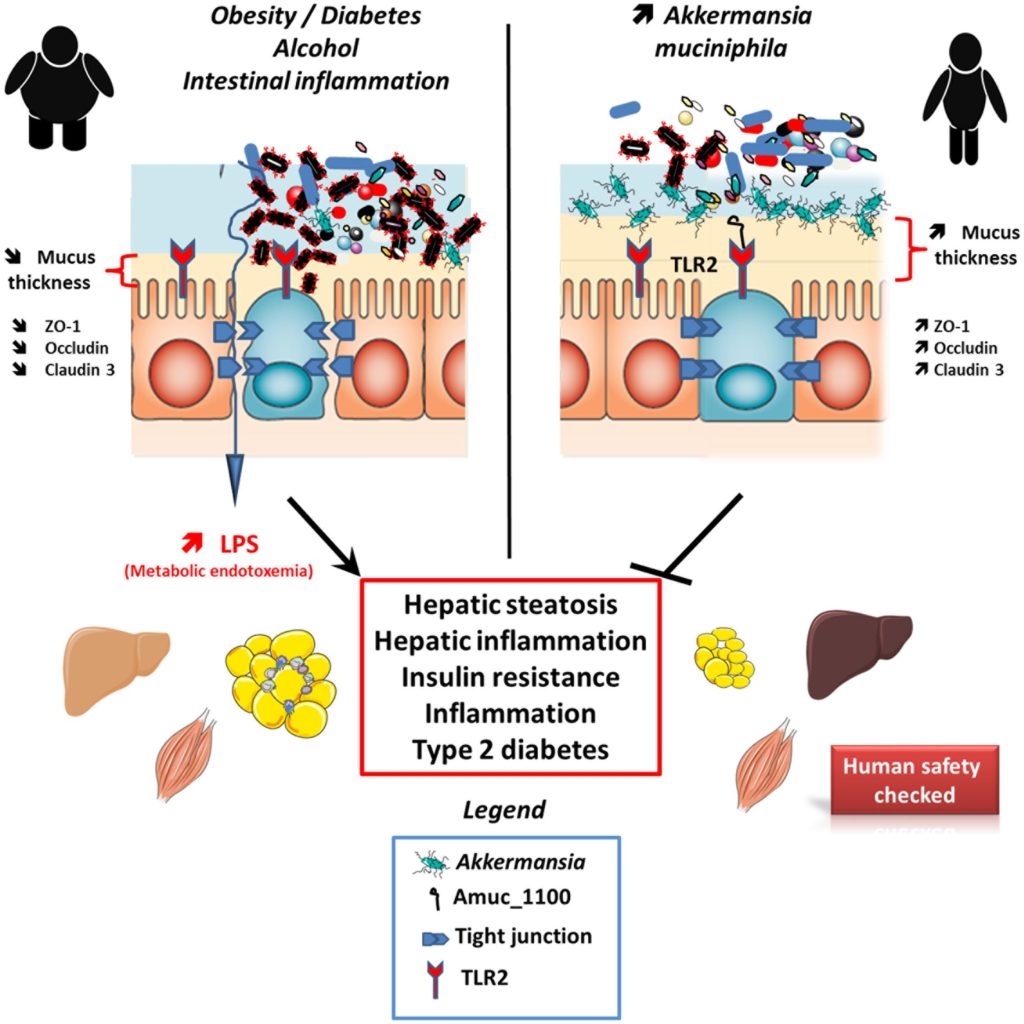
Cani PD,et al.,Front Microbiol. 2017
非酒精性脂肪性肝
与野生型小鼠相比,在NAFLD小鼠中观察到A. muciniphila丰度显著降低,而导致A.muciniphila丰度增加的干预措施改善了代谢参数。使用益生元(如低聚果糖)可以恢复A.muciniphila的丰度,并改善相关疾病。
酒精性肝病
酒精性肝病(ALD)由脂肪变性、纤维化和肝硬化发展为急性酒精性脂肪性肝炎,死亡率高,但治疗方法非常有限。结果发现,内毒素血症水平升高的酒精性肝病患者肝损伤程度增加。乙醇消耗引起的肠道紧密连接的破坏导致炎症信号通过病原体相关的分子模式,如脂多糖。
研究ALD患者队列时,发现与健康受试者相比,ALD患者粪便A. muciniphila数量减少。给乙醇喂养的小鼠口服粘液粘菌A. muciniphila可恢复该细菌的消耗,减少肠漏,增加Claudin-3和Occludin的紧密连接表达。因此,炎症和肝损伤促进细胞因子白介素1β (IL-1β)和肿瘤坏死因子α (TNF-α)明显减少,导致MPO +中性粒细胞浸润,改善肝损伤。
扩展阅读:深度解析 | 肠道菌群与慢性肝病,肝癌
改善血糖和胰岛素抵抗
A. muciniphila在糖耐量正常的人群中比糖尿病前期或 II 型糖尿病人群中的数量更多。
事实上,这些细菌已被证明可以增强称为二甲双胍的抗糖尿病药物的作用。在喂食高脂肪饮食的小鼠中,二甲双胍治疗增加了A. muciniphila的数量 并改善了它们的血糖水平。
同样,喂食高脂肪饮食的小鼠口服A. muciniphila但不服用二甲双胍,对葡萄糖的耐受性也有所提高。因此,这种细菌通过改变肠道微生物群,可能有助于在未来找到控制 2 型糖尿病的新方法。
扩展阅读:2型糖尿病如何做到可防可控?肠道菌群发挥重要作用
癌症免疫治疗
Akkermansia muciniphila也可以积极影响宿主对癌症免疫疗法的反应。研究发现,抗生素的消耗与对免疫治疗 PD-1 阻断剂的反应较差有关。进一步研究确定较低水平的Akkermansia muciniphila确实对反应率有直接影响,给抗生素治疗的小鼠口服细菌补充剂,发现它恢复了它们对免疫疗法的反应。根据这项研究,有人建议维持健康的肠道菌群,主要包括Akkermansia muciniphila,对于提高癌症免疫治疗有效性很重要。
此外,Akkermansia muciniphila通过刺激 DCs 产生 IL-12 并促进 CCR9 + CXCR3 + CD4 +的积累来增强 ICB 功效 上皮肿瘤和淋巴结中的 T 细胞。
根据美国癌症研究中心新研究显示Akkermansia muciniphila提升免疫检查点抑制剂的效果,还能产生大量调节T细胞反应的肌苷。食用高纤维食物可以有效地调节肿瘤微环境中的单核巨噬细胞的组成,而食用得越多,Akkermansia muciniphila的肠道菌数量就会更多。
该菌还能产生大量环二腺苷酸,即使是无菌小鼠,仅仅植入Akkermansia muciniphila,它们也能有更强的抗癌反应。未来,或许一种通过食物和环二腺苷酸联合的促进抗癌疗法就在眼前。
神经退行性疾病
肌萎缩性侧索硬化症(amyotrophic lateral sclerosis, ALS)是一种复杂的神经退行性疾病,患者通常会出现运动神经元的过早死亡,导致诊断后的平均生存时间不足5年。然而,针对肌萎缩性侧索硬化症的药物所做的巨大努力却收效甚微。
急性ALS小鼠模型经抗生素治疗后显示,微生物群失调在渐冻症的进展和恶化中起着重要作用。
通过挑选11株与ALS相关的微生物菌株,包括A.muciniphila和单接种到经抗生素治疗的ALS小鼠中,只有A.muciniphila与运动神经元改善相关。
其他疾病,例如,帕金森病与对照组的肠道微生物组组成显著不同, Akkermansia 丰度增加。
年龄
年龄和地理环境等因素影响人体A. muciniphila的数量。从婴幼儿时期开始A. muciniphila在人类胃肠道定植,并在一年后迅速接近成年人的水平。随着年龄增长A. muciniphila在人体内的数量逐渐降低。
不同的是,另一研究发现大于60岁人群A. muciniphila的数量升高。此外,另外研究发现在百岁长寿老人中A. muciniphila数量显著增加,提示A. muciniphila水平的升高可能有助于延缓机体的衰老。
对两种类型的早衰综合征小鼠模型,外用该菌A. muciniphila可显著提高小鼠的健康寿命。
地理位置
另外,不同地理位置A. muciniphila在人群中定植也存在差别。如通过细菌16S rRNA分析发现,智利健康受试者粪便中的微生物菌群和巴布亚新几内亚受试者有明显不同;但与美国和阿根廷受试者接近;A. muciniphila在智利受试者的丰富度最高。
中国南方的男性和女性定植率没有差异,但是中国南方人群的A. muciniphila定植率低于欧洲人群。暨南大学理工学院在广州地区人群的A. muciniphila定植率为89.31%,平均丰度为5.825 lg(CFU/mL)。肠道消化模拟不同时间点的RT PCR结果显示,A. muciniphila在模拟人类肠道微生物菌群环境中具备生存能力;对Bifidobacterium属和Bacteroides属具有抑制效果,对Lactobacillus属具有促进作用。
2021年在Nature发表的文章中,研究人员将古肠道样本数据和一组来自于已测序的工业化及非工业化生活方式下现代人群的粪便样本数据进行对比。比起非工业化样本和古粪便,包括Akkermansia muciniphila(可降解人体粘液)在内的BloSSUM类群在工业化样本中更丰富。
工业化生活下微生物群落中的粘液降解菌和基因更普遍,这可能是由西方饮食导致的。
药物
药物是影响肠道A.muciniphila丰度的重要因素之一。首先,抗菌药物作用后可以直接影响肠道A.muciniphila的数量。
在小鼠出生后早期用万古霉素进行干预可以有效预防1型糖尿病,显著降低小鼠血糖值以及减轻胰岛炎症;而对8周龄小鼠未见明显预防效果。万古霉素作用于幼儿时期的小鼠使A. muciniphila成为胃肠道优势菌群,该研究提示A. muciniphila可能是推迟1型糖尿病的关键菌群。
对60例6~11个月的婴儿使用阿奇霉素治疗3 d后,婴儿A. muciniphila数量显著减少。此外Dubourg等的研究中,2例成人患者经过抗生素(强力霉素、哌拉西林、羟氯喹、亚胺培南等)治疗后,A. muciniphila数量显著增加,在肠道菌群的比例均大于40%。
有趣的是,在另外一项小鼠模型中也发现类似现象,给小鼠灌胃四联抗生素(氨苄青霉素,万古霉素,新霉素和甲硝唑)可显著逆转免疫抑制剂导致的A. muciniphila数量减少。这些研究表明抗菌药物等药物使用会影响肠道内A. muciniphila数量的变化,其对人体代谢产生的影响需进一步研究。
最新的研究发现,代谢综合征相关治疗药物对A.muciniphila丰度的影响可能参与其作用机制。如二甲双胍通常用作治疗代谢综合征(如肥胖和2型糖尿病)的一线用药。
对14例服用二甲双胍的糖尿病患者进行肠道菌群分析,与未服用二甲双胍的糖尿病患者以及健康受试者相比,A. muciniphila显著增加。在体外实验中,收集二甲双胍治疗组和对照组小鼠的粪便,发现二甲双胍培养组A. muciniphila的比例显著增加。
在另外一项研究使用另外一种代谢病治疗药物利拉鲁肽(Liraglutide)。对高脂饮食诱导的肥胖小鼠进行皮下注射给药,连续15 d给药后处死小鼠,收集肠道内容物进行菌群分析,发现治疗小鼠肠道中A. muciniphila显著增加。此外,抗肿瘤药物也可以影响肠道A.muciniphila的丰度。如在胰腺导管腺癌(Pancreatic ductal adenocarcinoma, PDAC)异种移植的小鼠模型中发现,吉西他滨(Gemcitabine)治疗后使A. muciniphila在肠道中的数量显著增加(5%~33%)。
饮食方式
饮食对肠道A.muciniphila影响最显著。
与正常饮食组相比,高蛋白饮食显著降低大鼠肠道A. muciniphila。
Akkermansia 的数量随着短期(3 周)低FODMAPs饮食而减少。这不足为奇,因为 FODMAP 基本上与益生元纤维相同。
Akkermansia在生酮饮食中显着增加,并且与 Parabacteroides一起,可能是减少酮症癫痫发作的原因。
小鼠实验表明,A.muciniphila数量受到高脂饮食的强烈影响,高脂饮食三周后A.muciniphila丰度开始下降,并且A.muciniphila丰度与体脂呈负相关。
与高脂高糖饮食组相比,小鼠食用热带水果8周后,可以显著增加A. muciniphila在胃肠道中的丰度;同时干预后可以降低高脂高蔗糖饮食喂养小鼠的内毒素血症,改善其胰岛素敏感性。
饱和脂肪含量高的饮食与较高的炎症发生率有关。而富含多不饱和脂肪酸的饮食,如亚麻籽和鱼油中的那些,已被证明可以抵抗炎症,并且与更瘦的人有关。
这些膳食脂肪也会影响肠道中A. muciniphila的水平。

在一项研究中,老鼠被喂食由不同脂肪组成的不同饮食。一组喂猪油,另一组喂鱼油十一周,结果很有趣。
在鱼油喂养组中,A. muciniphila的丰度增加,乳酸杆菌的丰度也增加。
然后将两组的粪便材料移植到新的小鼠组中,在接受鱼油喂养小鼠粪便移植的小鼠中,它们的A. muciniphila水平增加,炎症水平降低。然而,接受猪油喂养小鼠移植的小鼠的炎症水平很高。
因此,摄入的膳食脂肪类型也会极大地影响肠道中A. muciniphila的数量。
我们已经知道多酚可以减少炎症并有助于预防许多疾病,因此高多酚食物可以增加Akkermansia.
多酚是强大的抗氧化剂,可以对抗体内的自由基(与多种疾病有关的化合物,包括糖尿病、心脏病和癌症)。
有许多不同类型的多酚。其中一些不能很好地被身体吸收留在肠道中,这对 Akkermansia来说是个好消息,它们吞噬了多酚并因此增强其力量,从而达到双赢。
此外,丁酸钠和菊粉、北极浆果提取物、绿茶提取物和异麦芽寡糖、膳食多酚等这些益生元也能增加Akkermansia丰度。
含有高水平多酚和鱼油的食物是提高A. muciniphila菌最好的食物。
高多酚食品
多酚使食物一般颜色丰富多彩,尝起来略带苦味或酸味。泡了很长时间的茶尝起来很苦:浸泡时间越长,茶多酚含量越高。浓橄榄油的青草味或红酒的味道也是很好的例子。

在自然界中,苦味可以抵御想要攻击植物的昆虫。有超过8000种多酚。一些例子是红酒中的白藜芦醇、辣椒中的辣椒素、百里香中的百里酚、肉桂中的肉桂酸、迷迭香中的迷迭香酸。
食物中多酚的含量在很大程度上取决于其生长的土壤、成熟度以及种植、运输和制备方式,因此以下列表仅供参考。
高多酚食品:
蔓越莓、石榴、山莓、黑莓、蓝莓、草莓、樱桃、李子、亚麻种子、黑巧克力、栗子、红茶、绿茶、苹果汁、苹果、黑麦面包、核桃、榛子、红酒。

其他常见的高多酚食物包括葡萄、橄榄、菠菜、李子和桃子。一般来说,颜色越深越好,所以选择绿色的黑橄榄和葡萄。
油性鱼
Akkermansia 喜欢鱼油,但目前还不清楚它们喜欢鱼油的特定成分——例如脂肪酸——或者它们喜欢所有的部分。所以目前,最好吃鱼。
尽管鱼油现在是有争议的,一些科学家认为它的好处被夸大了。研究人员发现,食用整条鲑鱼的一组人的 DHA(一种脂肪酸)水平是食用鱼油的一组人的 9 倍。食用鱼类的同时既增加鱼油,同时增加优质蛋白质摄入,均有助于A. muciniphila。
试着每周吃两到三份鱼:
沙丁鱼、鲭鱼、鲱鱼、三文鱼、鳟鱼、鳀鱼、旗鱼、金枪鱼

补充膳食纤维,选择有机产品。
含纤维的食物:
生菊苣根、绿色蒲公英 、麦麸 、芦笋 、大蒜、香蕉 、洋葱 、韭菜,也存在于燕麦片、红酒、蜂蜜、枫糖浆、豆类和其他食物中。
避免高脂肪饮食和酒精
微生物多样性是我们肠道健康的主要目标。实现这一目标的最佳方法是均衡饮食,吃能让肠道菌群快乐的食物,从而你的感觉也会良好。
难点
A. muciniphila与机体健康联系紧密,有潜力成为新型益生菌。但目前A. muciniphila是专性厌氧菌且难以培养,成为新型益生菌,还有以下几点重要问题亟待解决:
首先,需研发高效快速培养A. muciniphila的新技术。目前实验室或小规模已经成功在实验室中获得活性A. muciniphila,并且能够稳定,但是这只是实验室规模生产,大规模的工业生产仍是需要解决的问题。比如培养基中存在蛋白质来源问题,是否会对人体可能有过敏反应,工业化生产时需研发合理的工业化生产培养基。
其次,需确定安全有效的治疗人群。目前A. Muciniphila 与改善肝功能、降低葡萄糖毒性、减轻氧化应激和抑制炎症有强关联,但是,在炎症性肠病、动脉粥样硬化等其他疾病中的干预,不同研究中存在不同的结果,对于其作为药物面对的安全问题,其应用仍待商榷。
最后,需要优化A. muciniphila的补充或给药方式。在体外模拟胃肠道的释放实验中,发现包封的 A. muciniphila活性比直接分散在模拟胃肠道的系统中活性更高,并且能够定向在肠道释放细菌,显著增加A. muciniphila对胃的抵抗力,对于实现在肠道的定向给药还需要更多的实验进行优化。
最后,尽管已经有研究证明有活性的A. muciniphila或者是巴氏消毒的A. muciniphila对人体是安全的,但目前A. muciniphila的临床研究较少,在临床上应用还需要更多的实验验证。
主要参考文献
Hasani A, Ebrahimzadeh S, Hemmati F, Khabbaz A, Hasani A, Gholizadeh P. The role of Akkermansia muciniphila in obesity, diabetes and atherosclerosis. J Med Microbiol. 2021 Oct;70(10). doi: 10.1099/jmm.0.001435. PMID: 34623232.
Zhang T, Ji X, Lu G, Zhang F. The potential of Akkermansia muciniphila in inflammatory bowel disease. Appl Microbiol Biotechnol. 2021 Aug;105(14-15):5785-5794. doi: 10.1007/s00253-021-11453-1. Epub 2021 Jul 27. PMID: 34312713.
Druart C, Plovier H, Van Hul M, Brient A, Phipps KR, de Vos WM, Cani PD. Toxicological safety evaluation of pasteurized Akkermansia muciniphila. J Appl Toxicol. 2021 Feb;41(2):276-290. doi: 10.1002/jat.4044. Epub 2020 Jul 28. PMID: 32725676; PMCID: PMC7818173.
Kalia VC, Gong C, Shanmugam R, Lin H, Zhang L, Lee JK. The Emerging Biotherapeutic Agent: Akkermansia. Indian J Microbiol. 2021 Dec 16:1-10. doi: 10.1007/s12088-021-00993-9. Epub ahead of print. PMID: 34931096; PMCID: PMC8674859.
Yu Y, Lu J, Sun L, Lyu X, Chang XY, Mi X, Hu MG, Wu C, Chen X. Akkermansia muciniphila: A potential novel mechanism of nuciferine to improve hyperlipidemia. Biomed Pharmacother. 2021 Jan;133:111014. doi: 10.1016/j.biopha.2020.111014. Epub 2020 Nov 26. PMID: 33246225.
Cheng D, Xie MZ. A review of a potential and promising probiotic candidate-Akkermansia muciniphila. J Appl Microbiol. 2021 Jun;130(6):1813-1822. doi: 10.1111/jam.14911. Epub 2020 Nov 15. PMID: 33113228.
Hagi T, Belzer C. The interaction of Akkermansia muciniphila with host-derived substances, bacteria and diets. Appl Microbiol Biotechnol. 2021;105(12):4833-4841. doi:10.1007/s00253-021-11362-3
Anhê, F. F et al. (2016). Triggering Akkermansia with dietary polyphenols: A new weapon to combat the metabolic syndrome?. Gut microbes, 7(2), 146–153.
Schade L, Mesa D, Faria AR, Santamaria JR, Xavier CA, Ribeiro D, Hajar FN, Azevedo VF. The gut microbiota profile in psoriasis: a Brazilian case-control study. Lett Appl Microbiol. 2021 Dec 12. doi: 10.1111/lam.13630. Epub ahead of print. PMID: 34897759.
Belzer, C et al. (2017). Microbial Metabolic Networks at the Mucus Layer Lead to Diet-Independent Butyrate and Vitamin B12 Production By Intestinal Symbionts. American Society for Microbiology: 8(5).
Caesar, R et al. (2015). Crosstalk Between Gut Microbiota and Dietary Lipids Aggravates WAT Inflammation Through TLR Signalling. Cell Metab: 22(4), pp 658-668.
Dao, M, C et al. (2015). Akkermansia muciniphila and Improved Metabolic Health During a Dietary Intervention in Obesity: Relationship with Gut Microbiome Richness and Ecology. Gut.
de Vos, W, M. (2017). Microbe Profile: Akkermansia muciniphila: A Conserved Intestinal Symbiont that Acts as The Gatekeeper of our Mucosa. Microbiology: 163(5).
Derrien, M et al. (2017). Akkermansia muciniphila and its Role in Regulating Host Functions. Microbial Pathogenesis: 106, pp 171-181.
Everard, A et al. (2013). Cross-Talk Between Akkermansia muciniphila and intestinal Epithelium Controls Diet-Induced Obesity. PNAS: 110(22), pp 9066-9071.
Geerlings, S, Y et al. (2018). Akkermansia muciniphila in the Human Gastrointestinal Tract: When, Where, and How? Microorganisms: 6(3).
Krishnan, A. (2016). Akkermansia muciniphila: What You Need to Know About this Bacterium. uBiome.
Naito, Y et al. (2018). A Next-Generation Beneficial Microbe: Akkermansia muciniphila. J. Clin. Biochem. Nutr: 63(1), pp 33-35.
Ottman, N et al. (2017). Pili-Like Proteins of Akkermansia muciniphila Modulate Host Immune Responses and Gut Barrier Function. PLOS One.
Pierre, J, F et al (2015). Cranberry Proanthocynaidins Improve the Gut Mucous Layer Morpohology and Function in Mice Receiving Elemental Enteral Nutrition. JPEN J Parenter Enteral Nutr: 37(3), pp 401-409.
Shin, N, R et al. (2013). An Increase in the Akkermansia spp. Population Induced by Metformin Treatment Improves Glucose Homeostasis in Diet-Induced Obese Mice. Gut: 63, pp 706-707.
Zhang, T et al. (2019). Akkermansia muciniphila is a promising probiotic. Microbial Biotechnology.
Ouyang J, Lin J, Isnard S, et al. The Bacterium Akkermansia muciniphila: A Sentinel for Gut Permeability and Its Relevance to HIV-Related Inflammation. Front Immunol. 2020;11:645. Published 2020 Apr 9. doi:10.3389/fimmu.2020.00645
Zhou K. Strategies to promote abundance of Akkermansia muciniphila, an emerging probiotics in the gut, evidence from dietary intervention studies. J Funct Foods. 2017;33:194-201. doi:10.1016/j.jff.2017.03.045
Naito, Y., Uchiyama, K. & Takagi, T. A next-generation beneficial microbe: Akkermansia muciniphila. J Clin Biochem Nutr 63, 33–35 (2018).
Derrien, M., Vaughan, E. E., Plugge, C. M. & de Vos, W. M. Akkermansia muciniphila gen. nov., sp. nov., a human intestinal mucin-degrading bacterium. Int. J. Syst. Evol. Microbiol. 54, 1469–1476 (2004).
Hansson, G. C. Role of mucus layers in gut infection and inflammation. Curr. Opin. Microbiol. 15, 57–62 (2012).
van der Lugt, B. et al. Akkermansia muciniphila ameliorates the age-related decline in colonic mucus thickness and attenuates immune activation in accelerated aging Ercc1-/Δ7 mice. Immun. Ageing 16, 6 (2019).
Lukovac, S. et al. Differential modulation by Akkermansia muciniphila and Faecalibacterium prausnitzii of host peripheral lipid metabolism and histone acetylation in mouse gut organoids. MBio 5, (2014).
Dao, M. C. et al. Akkermansia muciniphila and improved metabolic health during a dietary intervention in obesity: relationship with gut microbiome richness and ecology. Gut 65, 426–436 (2016).
Depommier, C. et al. Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: a proof-of-concept exploratory study. Nat. Med. 25, 1096–1103 (2019).
Png, C. W. et al. Mucolytic bacteria with increased prevalence in IBD mucosa augment in vitro utilization of mucin by other bacteria. Am. J. Gastroenterol. 105, 2420–2428 (2010).
Anhê, F. F. et al. A polyphenol-rich cranberry extract protects from diet-induced obesity, insulin resistance and intestinal inflammation in association with increased Akkermansia spp. population in the gut microbiota of mice. Gut 64, 872–883 (2015).
Pierre, J. F. et al. Cranberry proanthocyanidins improve the gut mucous layer morphology and function in mice receiving elemental enteral nutrition. JPEN J Parenter Enteral Nutr 37, 401–409 (2013).
Vinson, J. A. & Cai, Y. Nuts, especially walnuts, have both antioxidant quantity and efficacy and exhibit significant potential health benefits. Food Funct 3, 134–140 (2012).
Mao, B. et al. Effects of different doses of fructooligosaccharides (FOS) on the composition of mice fecal microbiota, especially the bifidobacterium composition. Nutrients 10, (2018).