-

 国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子
国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子

 二级病原微生物安全实验室
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业

谷禾健康
前面的文章中,我们已经了解到,在肝病的发生发展中肠道菌群的变化,详见:
深度解析 | 肠道菌群与慢性肝病,肝癌
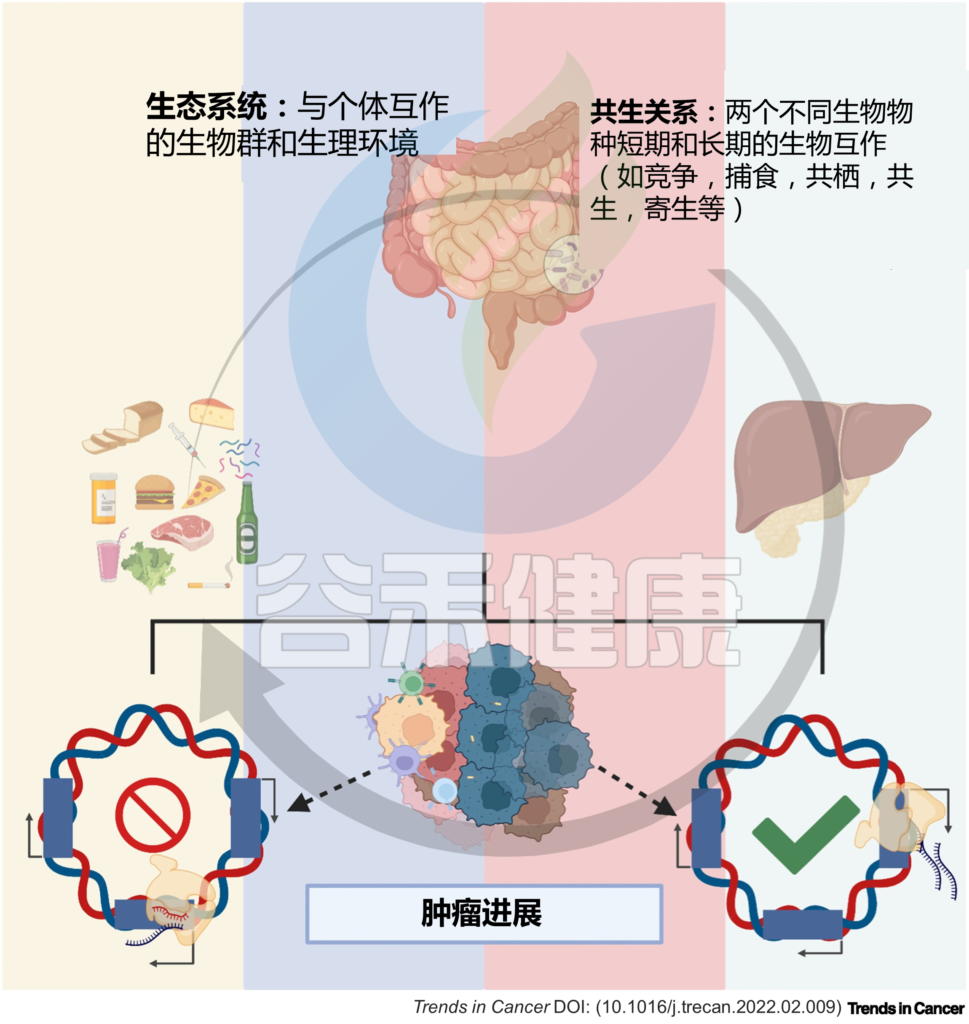
到目前为止,大多数研究都集中在细菌多样性及其代谢物与靶癌细胞表型的关联上,而没有考虑环境的微调。实际上生态系统中,个体在环境中与其他个体相互作用、相互影响和限制。
在肿瘤微环境中,特定的微生物会影响其他细胞(微生物或宿主细胞)并受其影响。
微生物群及其代谢物影响肠-肝轴的组织细胞。肠道微生物群以及肿瘤本身的细菌可以影响肿瘤微环境,包括通过调节癌症、基质和炎症/免疫细胞中的基因转录以及促进或抑制肿瘤进展。
同时,微生物群也受饮食、环境等因素影响,在环境扰动后会适应,从而影响宿主-微生物的相互作用。
本文主要阐述了微生物群在肝脏稳态中的作用,肠道菌群及其代谢物直接和间接地调节肝脏基因表达,导致肠-肝轴失衡,从而促使肝病的发生发展,甚至致癌,并对肿瘤微环境产生影响。
我们的健康和生存能力取决于共生微生物(微生物群)的存在,它们主要存在于上皮细胞界面上,在下消化道中特别丰富。
菌群维持肠道稳态
肠道微生物群中的平衡生态有助于食物加工和吸收,调节宿主新陈代谢,并通过防止病原体和病原菌的扩张或通过调节宿主免疫力和维持肠道上皮的完整性来防止感染。
肠道-肝脏-胰腺轴
在物理上,肝脏和胰腺通过胆管和胰管与肠道相通,而门静脉将肠道菌群产物输送到肝脏。因此,肠道菌群与肝脏和胰腺之间的串扰(肠道-肝脏-胰腺轴)可以将信号整合为一个相互关联的系统。
宿主与微生物群之间复杂而高度协调的相互作用代表了一个自然生态系统。共生相互作用,如互利共生、共栖、捕食、寄生和竞争,是微生物、微生物-宿主和宿主-宿主细胞之间相互作用的基础。
因此,肠-肝轴中的细胞处于稳态平衡,环境扰动会改变这种平衡,从而调节局部和全身的转录反应,并影响健康和疾病,例如癌症。
图1 微生物与宿主的相互作用调节体内平衡和疾病
关于微生物群在致癌中的作用的研究,最初集中在上皮屏障界面的肿瘤上,如胃癌和结肠癌,但胰腺导管腺癌 (PDAC) 为口腔和肠道微生物群以及癌症相关微生物群的作用提供了很多证据。
微生物群对致癌作用的影响,及细菌调节肿瘤微环境的一些机制如下:
胰腺癌研究解决了肠道菌群失调、瘤内细菌和癌症之间的联系
人类胰腺癌前体病变显示被产生 IL-17 的Th17 细胞浸润,加速了癌症的发生和进展。
几项研究表明,胰腺导管腺癌与口腔微生物组的组成、口腔病原体(如牙龈卟啉单胞菌Porphyromonas gingivalis、伴放线聚合杆菌Aggregatibacter actinomycetemcomitans)的丰度增加或口腔微生物抗体的存在有关。胰腺癌中出现了较低的α多样性,产生LPS的细菌增加,产丁酸盐菌减少。
基于与胰腺导管腺癌相关的口腔和肠道微生物群落的概况,已经提出了将胰腺导管腺癌患者与健康个体区分开来的无创诊断模型。
此外,胰腺导管腺癌研究已经确定,胰腺含有与组织相关的细菌和真菌,这些细菌和真菌在胰腺癌中比在正常胰腺组织中的含量更高。胰腺导管腺癌中的细菌主要存在于免疫细胞和癌细胞内。
在分析的每种肿瘤类型中,肿瘤内细菌的组成是不同的,可用于预测肿瘤与正常组织和肿瘤类型。
尽管在大约三分之二的胰腺导管腺癌中观察到细菌,并且数量高于大多数其他肿瘤类型,但与人类细胞相比,它们在肿瘤中的绝对数量仅为约 1/40 – 1/400。
与其他肿瘤相比,胰腺导管腺癌中的肿瘤相关细菌 Gammaproteobacteria较多,尤其是肠杆菌Enterobacterales,与胃癌和结肠癌不同的是,它们的梭杆菌fusobacteria较少。
胰腺导管腺癌相关微生物群的组成与十二指肠微生物群相似,这一事实表明细菌可能是通过胆胰管逆行迁移。
肿瘤相关微生物通过各种机制参与胰腺癌的发生或对治疗产生抵抗
例如,表达胞苷脱氨酶长同工型的细菌(如 Gammaproteobacteria)将吉西他滨代谢成无活性形式,导致胰腺导管腺癌出现耐药性。
瘤内细菌可能通过诱导 MDSCs 和抑制 M1 巨噬细胞分化和 CD4 +和 CD8 + T 细胞活化来重新编程肿瘤微环境;通过抗生素治疗进行的细菌消融可重新编程胰腺肿瘤免疫微环境,防止癌变并使免疫检查点抑制治疗产生反应。
然而,肿瘤内细菌也可能是有益的,并且 胰腺导管腺癌的长期幸存者在肿瘤相关微生物群中表现出更高的微生物 α 多样性和独特的特征(假黄单胞菌属Pseudoxanthomonas、链霉菌属Streptomyces、糖多孢菌属Saccharopolyspora),这可能诱导有效的免疫细胞浸润和抗肿瘤免疫。
最近有多项综述回顾了微生物调节在胰腺导管腺癌中的作用。这些研究支持胰腺导管腺癌患者的肿瘤外和肿瘤内微生物群与发育和临床进展的易感性之间的直接联系。
作为一个相互关联的系统,肠-肝轴中的微生物-宿主串扰有望成为肝癌发生的一个重要因素,就像在胰腺导管腺癌中一样。
细菌通过微生物-宿主和宿主-宿主相互作用参与致癌作用
幽门螺杆菌,已被正式确定为胃癌的明确人类致癌物。然而,新出现的证据表明,细菌,无论是存在于上皮屏障界面上还是存在于肿瘤中,都与局部或远处组织的癌变和肿瘤进展有关。
细菌在肿瘤微环境中产生选择性压力以促进肿瘤发生,部分原因是引发 ROS 的产生,影响对 pH 变化的反应,竞争有限的营养物质,增加 DNA 损伤和诱变,调节癌基因途径,影响化学疗法的代谢药物,或调节免疫。
携带产生大肠杆菌素的聚酮化合物-非核糖体肽合酶操纵子 (pks) 的大肠杆菌菌株在结直肠癌中诱导了明显的突变特征,这为细菌在基因组突变中的作用提供了证据。
微生物群对致癌基因诱导的肿瘤进展的影响得到以下观察结果的支持:
突变的p53仅在远端结肠中致癌,因为存在微生物产生的没食子酸,通过破坏 WNT 通路阻止突变的 p53 作为肿瘤抑制因子。相反,肿瘤会对局部组织细胞施加的竞争压力可能会影响肿瘤微环境、周围组织和肠道中的细菌。
宿主-宿主相邻细胞之间的肿瘤微环境竞争动态是生态系统模型的基础,在结肠和肝脏中均发现有致癌作用,并且可以为细菌调节肿瘤微环境提供底物。
恶性干细胞分泌促进邻近干细胞分化为含有促癌突变的克隆的因子。微生物群的改变可能代表另一个触发因素,结合宿主细胞串扰中涉及的其他多个信号,不仅影响癌症前体靶细胞,而且通过调整局部组织环境影响所有细胞。
此外,肿瘤相关细菌大多存在于癌细胞和免疫细胞的细胞内,可能影响癌细胞的信号传导,并在抗原呈递细胞表面以 MHC 限制性肽的形式呈递,从而刺激宿主免疫。
尽管在改进低生物量正常和肿瘤组织样本中稀有细菌的鉴定和分析技术方面取得了很大进展,但这些结果仍需谨慎解读。不过,这支持肿瘤微环境选择居住的微生物群并反过来受微生物群影响的新概念。
此外,肝脏和胰腺之间的生理联系提出了一个问题,即:最近在胰腺导管腺癌中建立的模型是否可以应用于肝细胞癌 (HCC)?
在此阐述了微生物群在维持肠-肝轴稳态中的作用,并关注环境扰动如何直接(通过诱导微生物相对丰度/多样性的变化)或间接(通过微生物代谢物的作用)触发与肝癌发生相关的基因反应。
微生物群的组成是在婴儿早期建立的,并在成年后保持相对稳定。然而,由于生活方式、饮食、疾病、感染和抗生素的使用,细菌种类的相对丰度可能会迅速改变。
微生物群与宿主之间的串扰对健康和疾病产生关键影响
微生物群通过其扩张/收缩、占据不同的解剖生态位以及遗传物质的突变和交换来适应环境变化并调节宿主反应。
微生物群在宿主从出生开始的先天性和适应性免疫系统发育中发挥着重要作用,免疫系统也塑造了宿主-微生物的相互作用。这个过程依赖于肠道黏膜表面的分隔、微生物群感应和信号传递以及免疫细胞启动,以产生特定的反应并维持体内平衡。
结合起来,上皮屏障、其微生物群落和局部免疫系统不仅可以耐受环境中的共生细菌,还可以使免疫系统对条件致病菌或微生物产物做出反应。
当这些防御机制失败时,例如由于肠道通透性增加(肠道渗漏)或生态失调(与疾病相关或与疾病有因果关系的微生物群组成的变化),就会出现细菌代谢物的涌入或先前被分隔,并且有害微生物可能通过肠道血管屏障(GVB)并通过门静脉循环延伸到肝脏。
微生物群在肝脏稳态中的关键作用
在肝脏中,环境扰动会触发肝脏免疫反应,这种反应依赖于常驻免疫细胞以及来自肠道微生物群的循环抗原和内毒素。由与肠道相关淋巴组织 (GALT) 相关的免疫监视形成,证实了微生物群作为一个相互关联的系统在肝脏稳态中的关键作用。
为什么肠道屏障的紊乱会改变肝脏的微环境?
小肠细菌过度生长(SIBO)也与肠漏和细菌流入肝脏有关。
细菌产物或微生物易位的增加会刺激与慢性肝病相关的促炎反应。相反,肝脏通过胆道在肠道中释放胆汁酸和其他生物活性介质,这些介质可能会被肠道微生物群进行生物转化,然后被吸收并释放到体循环中。
生理组织稳态的改变可能导致癌症等疾病。此外,通过将饮食模式与微生物组对免疫和代谢状态的影响和癌症治疗反应联系起来,这种关系揭示了微生物群和肝脏之间的联系,这是由局部环境扰动引发的一系列相互关联的反应。
总的来说,肠-肝轴生态系统可以触发局部和远处的反应,并勾勒出肠道微生物群与肝脏之间的直接和间接相互作用(图 2)。
图2 肠肝轴的双向关系调节体内平衡
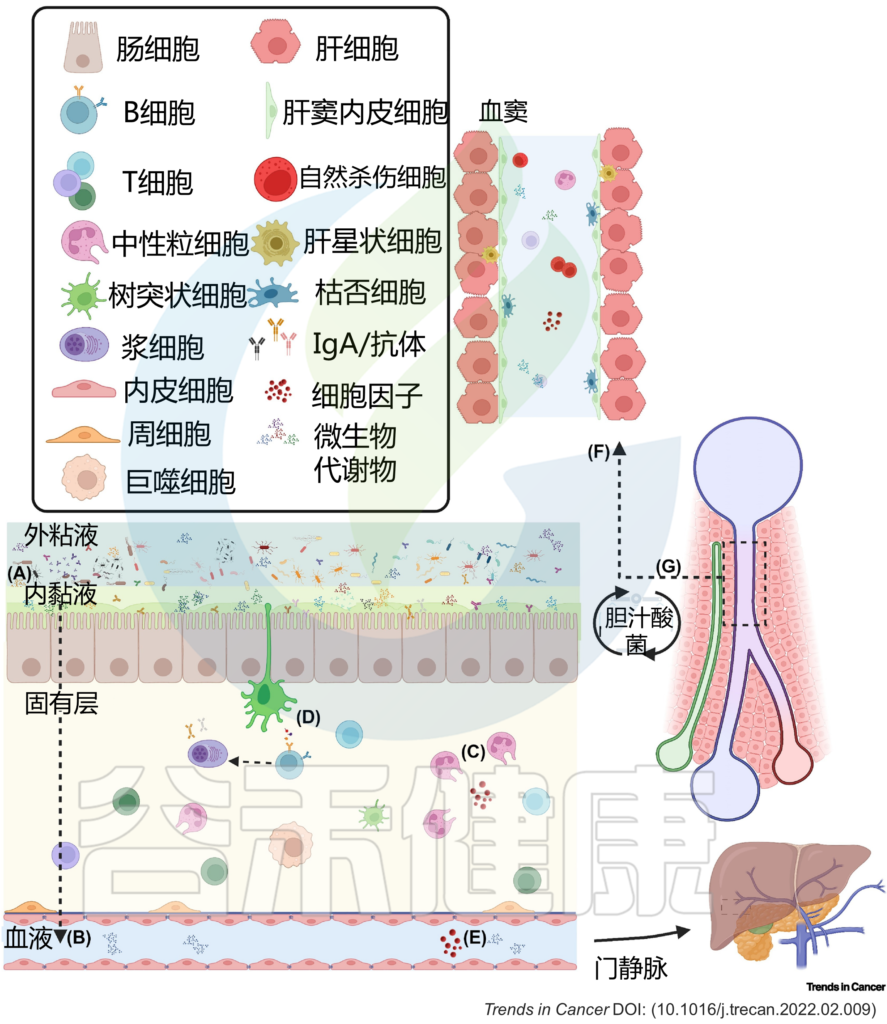
(A) 粘液从物理上将微生物群与上皮衬里分离,而抗菌肽使内部粘液几乎无菌。(B) 因此,微生物群与宿主之间的相互作用主要是间接的,并由代谢产物介导,这些代谢产物可能穿过粘液和上皮屏障,到达固有层中的免疫细胞和基质细胞,或通过淋巴和血管系统到达肝脏和体循环。(C) 免疫细胞通过产生生长因子和细胞因子来巡逻上皮细胞,并加强上皮和粘液屏障。这些产物对微生物产生选择性压力。(D) 树突状细胞感知环境并诱导T细胞和B细胞反应,从而产生IgA抗体,这些抗体转移到管腔,并通过调节微生物组成和多样性来促进粘膜免疫保护。代谢产物通过(E)肠血管屏障通过门静脉流入肝脏。(F) 在肝窦中,免疫细胞扫描异物。(G) 反过来,肝脏通过释放胆汁酸和其他生物活性介质进入胆道与小肠沟通;这些代谢物可被回肠和大肠末端丰富的微生物群生物转化,部分通过门静脉再循环到肝脏,从而可能影响局部和全身功能。
健康的肠道屏障对一些微生物代谢产物是可渗透的,但大多数完整的微生物却不能通过。
然而,饮食和其他环境因素的影响会迅速丰富或消耗特定的营养物质和细菌。这种效应会对微生物代谢物的产生和胆汁酸的转化产生影响,这有可能塑造局部微环境并与包括癌症在内的慢性肝病的发展相关。
例如,酒精会导致肠漏。与 SIBO 一起,它允许内毒素进入循环,导致肝脏疾病。SIBO 是肠道微生物菌群失调的一种表现,其特征是 α 多样性降低,在某些情况下,β 多样性增加,这可能导致全身性炎症。
增加的细菌易位会促进肝硬化进展(肝细胞癌的前体),包括:导致纤维化,通过增加肝细胞与微生物和细菌代谢物的接触,或通过在肝脏中创造转移前的小生境,改变环境以利于转移细胞的募集和增殖,从而促进肝硬化进展。
最后,在某些慢性肝病中,活细菌的易位可能导致 GALT 的免疫麻痹,表明微生物平衡的变化直接影响局部环境的机制。
下文将阐述肠道细菌如何通过扩散到肝脏、影响环境或改变其他细胞中的串扰直接影响肝癌发生。
饮食改变肠道微生物组,对免疫和新陈代谢状况、癌症风险和对癌症治疗的反应产生影响。因此,摄入食物的来源和类型在调节肠道微生物组中起着关键作用,并对宿主-微生物相互作用产生影响。
人类肠道中主要有三种类型(肠型)(拟杆菌属Bacteroides、普氏菌属Prevotella和瘤胃球菌属Ruminococcus)。
在工业化和非工业化人群中观察到的蛋白质和动物脂肪(拟杆菌属Bacteroides)与碳水化合物或植物性食物(普氏菌属Prevotella)的摄入比例不同。工业化与天然富含纤维食品的减少有关。
膳食纤维是饮食中的重要组成部分,分为不溶性(抗发酵)或可溶性(可被肠道微生物群代谢),例如益生元纤维菊粉,在结肠中它被肠道微生物群发酵成短链脂肪酸:乙酸盐、丁酸盐、和丙酸盐。
菊粉 是果糖聚合物的异质混合物。果糖摄入转化为乙酸盐,并通过微生物群衍生的乙酸盐经门静脉到达肝脏触发从头脂肪生成。在肠道中,菊粉对成年人微生物组组成的影响:
下列菌增加:
双歧杆菌Bifidobacterium
厌氧菌Anaerostipes
粪杆菌Faecalibacterium
乳杆菌属Lactobacillus
下列菌减少:
拟杆菌属Bacteroides
普氏菌Prevotella、密螺旋体Treponema、琥珀弧菌属Succinivibrio的丰度增加主要与蔬菜的纤维和碳水化合物发酵相关的饮食有关。
高纤维饮食增加了微生物组编码的聚糖降解碳水化合物活性酶,而不影响群落多样性。相比之下,高发酵食品饮食增加了微生物组的多样性并减少炎症。
从机制上讲,缺乏纤维的饮食可以使结肠粘液降解细菌增多,增强柠檬酸杆菌粘液层翻转和相关的结肠炎。这表明饮食模式可能影响肠-肝轴的多种机制。
健康的植物性食物会影响肠道微生物的多样性和组成,包括产丁酸菌的富集,例如:
Roseburia hominis
Agathobaculum butyriciproducens
普氏粪杆菌Faecalibacterium prausnitzii
厌氧菌Anaerostipes hadrus
丁酸盐是一种短链脂肪酸,通过肠道内膳食纤维的微生物发酵产生。丁酸盐有助于黏膜稳态和肠道内壁的完整性,从而提供肠细胞的大部分能量需求,并通过与几种 G 蛋白偶联受体结合并作为组蛋白脱乙酰酶抑制剂发挥抗炎作用。在小鼠模型中,增加的膳食纤维以微生物群和丁酸盐依赖的方式防止结直肠肿瘤发生。
总的来说,这些研究强调了饮食对微生物群落的深远影响及其对宿主的影响,具体取决于摄入的食物来源。
先前的研究表明,地中海饮食与拟杆菌门Bacteroidetes和某些有益梭菌群Clostridium的富集以及变形杆菌门Proteobacteria和芽孢杆菌门Bacillota的减少有关,可以减少肝脏脂肪,被推荐用于预防非酒精性脂肪性肝病 (NAFLD)。
调节与饮食相关的微生物组是预防肝癌的潜在途径
长期食用可发酵的富含纤维的食物(如可溶性纤维菊粉或富含菊粉的高脂肪饮食)容易导致生态失调的小鼠出现炎症、胆汁淤积和肝细胞癌。总细菌负荷增加,多样性减少,变形杆菌和纤维发酵细菌(如梭菌属)的特定增加。消除这些产生丁酸盐的细菌成功地预防了富含菊粉的肝细胞癌,这表明调节与饮食相关的微生物组是预防肝癌的潜在途径。
由于肠道微生物群失调,高胆固醇/高脂肪饮食依次导致小鼠阶段进展为脂肪变性、脂肪性肝炎、纤维化,最终导致 NAFLD-肝细胞癌。
在每个阶段都富集了不同的微生物群组成,因为在肝细胞癌患者中:
Mucispirillum、Desulfovibrio、Anaerotruncus和 Desulfovibrionaceae依次增加,
而Bifidobacterium、Bacteroides被耗尽。
综合这些发现,饮食模式会通过富集与健康或疾病相关的特定细菌来影响肠道微生物组,从而对肝脏产生潜在影响。
许多微生物群代谢物是肝癌发展的危险因素。考虑到环境塑造和选择特定的微生物群,推测某些物种可能获得哪些竞争优势以及特定微生物群的富集如何影响肝癌的进展。
相似或不同的微生物群组成是否可能参与肝细胞癌和胆管癌 (CCA) 的发展?现在下结论还为时过早,在这部分讨论这两种类型的肝癌。
肝脏中肿瘤和非肿瘤区域之间菌群不同
最近有报道称,病毒和非病毒病因的肝细胞癌中的 16S rRNA 基因测序确定了肝脏中肿瘤和非肿瘤区域之间不同的微生物组成,其中拟杆菌门Bacteroidetes、厚壁菌门Firmicutes、变形杆菌门Proteobacteria的物种占肿瘤相关菌群的主导地位。相关微生物群Ruminococcus gnavus被确定为感染肝炎病毒的 肝细胞癌患者的特征分类群。
肝硬化、肝细胞癌患者的肿瘤微生物群显示出更高丰度的嗜麦芽窄食单胞菌Stenotrophomonas maltophilia,这与肝星状细胞 (HSC) 中的衰老相关分泌表型 (SASP) 相关,证实了菌群失调与肝细胞调节之间的关联。
NAFLD肝硬化中,伴或不伴肝细胞癌的患者菌群不同
肝细胞癌患者的拟杆菌属Bacteroides和瘤胃球菌科Ruminococcaceae丰度增加,双歧杆菌Bifidobacterium丰度降低,这与粪便钙卫蛋白水平升高和全身炎症相关。同样,与肝硬化患者相比,早期肝细胞癌患者的肠道菌群中产生脂多糖 (LPS) 的菌群增加,而产生丁酸盐的菌群减少。
这些数据表明,在患有 NAFLD 和肝硬化的肝细胞癌患者中,肠道菌群组成与全身炎症相关,并可能促进肝癌的发生。
乙肝病毒感染进展的肝细胞癌的菌群特点
从乙型肝炎病毒 (HBV) 感染进展的肝细胞癌患者显示出丰富的抗炎细菌(例如,普氏菌Prevotella、乳酸杆菌Lactobacillus、双歧杆菌Bifidobacterium、粪杆菌Faecalibacterium)和减少的促炎细菌(例如,大肠杆菌-志贺氏菌Escherichia-Shigella、肠球菌Enterococcus),肠道微生物群可能与调节宿主免疫生物学途径的 HBV 感染有关的成分。
这些研究强调了在肝细胞癌中观察到的微生物多样性,这代表了已知风险因素与肝细胞癌发展之间的相关性。
作为饮食和其他环境因素影响微生物组的相互关联的系统,有必要在多个评估部位(肿瘤和非肿瘤)和组织中表征局部微生物群的组成和多样性,与单细胞分析相关并可能相关与环境因素。此外,突出特定细菌物种富集的潜在因果关系的功能分析是超越相关性的关键步骤。
胆管相关的微生物群
一些细菌科,如Dietziaceae、Pseudomonadaceae、Oxalobacteraceae主导了胆管相关微生物群,表明独特的微生物群落存在于这一解剖学定位中。
肝外胆管癌患者有大量肠球菌Enterococcus、链球菌Streptococcus、拟杆菌属Bacteroides、克雷伯氏菌属Klebsiella、锥体杆菌属Pyramidobacter。
此外,与胆总管结石患者相比,胆管癌患者的胆汁样本富含肠杆菌属Enterobacter、假单胞菌属Pseudomonas、窄食单胞菌属Stenotrophomonas。
最后,与肝细胞癌或肝硬化患者和健康个体相比,肝内胆管癌患者肠道菌群中的4个细菌属增加:
乳酸杆菌Lactobacillus、放线菌Actinomyces、消化链球菌Peptostreptococcus、异体卡多菌Alloscardovia。
肠道微生物群特征可以来区分胆管癌和胆石症
在胆管癌中富集的菌群:拟杆菌属Bacteroides、Muribaculaceae_unclassified、Muribaculum、Alistipes属的物种。
而不同的微生物物种在胆石症组中富集,这表明在从良性肝胆疾病到恶性肝胆疾病的演变过程中微生物关联发生了变化。
总的来说,这些过程可能解释了肠道细菌易位直接导致建立有利于肝癌发展和进展的发炎肝脏环境之间的联系。然而,很难根据与微生物相对丰度的相关性来唯一地假设因果关系,微生物相对丰度由于多种环境因素而迅速改变。
因此,全面的跨界网络分析比较肝细胞癌和胆管癌,并将局部肠道和组织微生物群的组成和多样性以及环境因素对代谢、免疫和转录改变的影响联系起来,对于剖析微生物群在肝癌的发生及其作用机制调节中的因果作用至关重要。
“
强调了微生物群变化对肝脏环境的间接影响(可能通过环境中其他细胞或微生物代谢物的串扰)及其与肝癌发生和进展的关系。
为了了解特定微生物群在肝肿瘤中的潜在影响,有必要将潜在机制以及微生物与其他细胞之间的串扰联系起来。
小鼠中肝细胞癌发展的演变
从这个意义上说,最近的一项研究描述了Mdr2 缺陷小鼠中肝细胞癌发展的时间演变,这些小鼠缺乏从肝脏将磷脂分泌到胆汁中的能力,从而经历胆汁淤积和肝细胞癌发展。
在这些代表炎症诱导肝细胞癌的有用模型的小鼠中,肠道菌群失调诱导肠道屏障功能障碍,先于 LPS 介导的肝脏转录改变,从而导致肝细胞癌发展。
此外,肝内炎症基因谱从肝损伤早期的促炎表型转变为肝细胞癌的免疫抑制表型。这种变化与通过微生物组功能从碳水化合物向氨基酸代谢的转变来重新调整能源利用有关。
菌群改变通过代谢和炎症影响肿瘤发生
微生物群的改变通过影响肝脏碳水化合物和脂质代谢调节炎症,从而导致 NAFLD 及其进展为非酒精性脂肪性肝炎 (NASH)。代谢和炎症的调节可能同样影响肝脏肿瘤发生。
肠道微生物组受饮食和其他环境因素的影响,微生物与营养物质的竞争是调节新陈代谢和免疫反应的关键步骤。例如,通过微生物群介导的膳食纤维发酵产生 SCFA 与胆汁淤积型肝细胞癌相关。
有人认为,细菌产物的易位可能会刺激炎症并释放 GALT 中的活性氧 (ROS),从而影响机械和分泌屏障以及局部微生物群。
这些研究强调需要继续进行系统和全球研究,将肠-肝轴中微生物物种的多样性和丰度作为一个生态系统进行表征,同时也需要开始剖析这些表型背后的机制。
最近有人提出,母亲在怀孕期间摄入丁酸盐和谷氨酰胺会影响新生小鼠的粪便微生物群和代谢物,这与拟杆菌和梭状芽胞杆菌的粪便特征有关。
此外,这些新生小鼠对肝脏免疫激活有抵抗力,导致胆管炎症和损伤。
从机制上讲,细菌代谢物在宿主细胞中触发基因反应的影响可能取决于环境中的转录改变。
对急性肝衰竭动物模型中不同的转录特征进行了检查,表明肠道微生物群和 Toll 样受体 (TLR) 信号激活肝星状细胞、枯否细胞和肝窦内皮细胞 (LSEC) 中的 MYC 依赖性转录程序,导致 Ly6C 阳性炎性单核细胞浸润和肝功能衰竭。
图3 微生物代谢产物和多样性是肝癌进展的触发因素


(A)一些细菌可能会穿透粘液屏障或参与其降解,在某些情况下导致上皮屏障的破坏或破坏,从而允许(B)微生物和微生物代谢物和免疫细胞之间的直接接触,诱导促炎细胞因子的产生和全身传播。粘液/上皮屏障的破坏也可能促进(C)细菌转移到肝脏(D),这为癌细胞的播散创造了一个有利的生态位。因此,先前划分的细菌和微生物产物的涌入影响了局部肝细胞的基因表达。例如:(E)肝细胞可能表达CXCR1并诱导CRCX2+多形核髓系衍生抑制细胞(PMN-MDSCs)的积累,创造免疫抑制环境促进胆管癌(CCA);(F)激活的肝星状细胞在肝细胞癌(HCC)和癌症转移中发挥多种功能,可能通过CXCL12-CXCR4相互作用破坏肝脏中自然杀伤细胞(NK)的功能,改变NK细胞介导的免疫,促进乳腺向肝脏转移;而(G)其他的肝脏免疫细胞可能通过脂多糖(LPS) – toll样受体4 (TLR4)或脱氧胆酸(DCA) -TLR2调节而被激活,并诱导促进肝细胞癌发生的炎症反应。相比之下,(H)被肠道菌群修饰后的肝脏产生的胆汁酸可能会激活肝脏自然杀伤T细胞(NKTs)在肝脏中的趋化因子依赖性积累,从而控制肿瘤的生长。
LPS调节影响肝细胞癌
循环水平的LPS通过TLR4 激活并诱导肝星状细胞分泌生长因子、调节肝脏慢性炎症状态和抑制细胞凋亡,这些过程与肝细胞癌促进有关。门静脉区域 LPS-TLR4 相互作用下游的转录调节因子 YAP1 的激活调节肝细胞的干性。
因为肝细胞周转的位点定位在激活基础稳态和再生的分子途径中很重要。推测 LPS 也可以通过调节局部微环境重编程来调节这些机制以影响肝细胞癌。
肠道微生物群调节肝细胞的基因表达程序,促进肝细胞癌和胆管癌
在胆管癌中,增加的肠道通透性诱导微生物 LPS 易位进入肝脏,通过 TLR4 依赖性机制诱导肝细胞中 CXCL1 的表达。这种表达反过来导致 CCR2 +多形核髓源性抑制细胞 (MDSCs) 的积累。
肝硬化肝细胞癌患者的瘤内S. maltophilia丰度更高。 通过激活 TLR4/NF-κB/NLRP3 通路诱导衰老肝星状细胞中衰老相关分泌表型SASP因子和促炎因子的表达,从而促进肝纤维化,随之而来的肝纤维化加重并发展为肝细胞癌。
肝星状细胞增殖是肝纤维化发展的关键事件。最后,胆汁酸通过激活表皮生长因子受体诱导肝星状细胞增殖。
饮食会迅速改变人体肠道微生物组。饮食衍生的微生物代谢物对甲酚硫酸盐、4-乙基苯基硫酸盐和 4-甲基儿茶酚会影响肝细胞癌亚型。
人类肠道微生物组编码的代谢途径通过众多生物活性分子不断与宿主基因产物相互作用。例如,营养过剩会增加 IL-17A,进而诱导白色脂肪组织中的中性粒细胞浸润和 NASH 诱导的肝细胞癌。
IL-17A 是一种促肿瘤细胞因子,通过调节Kupffer细胞和骨髓源性单核细胞的炎症反应和脂肪变性肝细胞的胆固醇合成,调节酒精诱导的肝脂肪变性、炎症、纤维化和肝细胞癌的进展。
地高辛,一种类视黄醇孤儿受体 γ t (RORγt) 拮抗剂,降低了 IL-17A 水平并稳定了体重。表明其在代谢紊乱中的关键作用。
此外,TNF和IL-17A 与骨髓来源细胞中NLRP3 炎性体激活诱导的肝脏炎症和纤维化的发展有关。
总的来说,这些研究表明饮食代谢物、细胞因子和肝癌疾病之间存在机制联系。
初级胆汁酸在肝细胞中合成,释放到十二指肠,大部分在小肠中重新吸收。一小部分初级胆汁酸逃逸到结肠,肠道共生细菌将其转化为次级胆汁酸,次级胆汁酸对新陈代谢和宿主先天免疫反应具有多种重要功能。
饮食和微生物胆汁酸代谢物均可调节 RORγt 阳性调节性 T 细胞(Treg) ,有助于维持宿主免疫稳态和改善肠道炎症。此外,胆汁酸代谢物可以通过调节 Th17 和 Treg 细胞的平衡来控制宿主免疫反应。
胆汁酸可以在高脂肪饮食中发挥积极作用
膳食胆固醇诱导肠道细菌代谢物改变,包括增加牛磺胆酸和减少 3-吲哚丙酸,从而在小鼠中驱动 NAFLD-肝细胞癌。因此,胆固醇抑制疗法和肠道菌群操作可能是预防 NAFLD-肝细胞癌的有效策略。
石胆酸 (LCA) 衍生物直接影响 CD4+ T 细胞(3-oxoLCA 和 isoalloLCA)
3-oxoLCA 通过直接结合转录因子 RORγt 抑制 Th17 细胞分化,而 isoalloLCA 增强 Treg 细胞分化,证实肠道微生物群可能控制宿主免疫反应。
对于肝内胆管癌,观察到甘熊去氧胆酸和牛磺脱氧胆酸血浆:粪便比率增加,血浆牛磺胆酸和 IL-4 呈正相关,表明肠道微生物群、代谢物、细胞因子和胆汁酸之间存在相互关系。
肥胖诱导的菌群失调促进肝癌发生
最近的一项综合组学研究揭示了一种胆汁酸代谢物升高、胆固醇代谢失调和与 BMI 增加相关的独特炎症反应的胆管癌亚型,这表明肥胖诱导的肠道微生物群失调促进肝癌发生的模型。
从机制上讲,胆汁酸/致癌轴涉及胆汁酸受体,例如法尼醇 X 受体 (FXR) 和 G 蛋白偶联胆汁酸受体 1,它们可能代表癌症的重要治疗靶点。胆汁酸如脱氧胆酸 (DCA) 被证明可阻断 FXR 的功能及其抑制肠癌干细胞增殖的能力,从而影响肠-肝轴稳态。
此外,胆汁酸传感器 FXR 或 G 蛋白偶联受体 TGR5 的激活通过抑制 NF-κB 依赖性信号通路和 NLRP3 依赖性炎症小体活性来抑制炎症信号传导。因此,饮食-肝脏-胆汁酸-微生物群的串扰在胃肠道炎症以及结直肠癌和肝癌的发生中起重要作用,可用于预防癌症的发生或进展。
菌群利用胆汁酸作为信使影响抗肿瘤免疫
肠道微生物组可以利用胆汁酸作为信使,来控制趋化因子依赖性肝脏自然杀伤 T 细胞 (NKT) 的积累,从而影响肝脏中的抗肿瘤免疫。这一过程是由共生肠道细菌的改变、初级和次级胆汁酸的平衡以及 LSEC 中的 CXCL16 表达介导的,LSEC 是最早暴露于肝脏中肠道衍生代谢物的细胞之一。
相反,NKTs 通过与肝细胞的相互作用与 CD8 T 细胞合作,促进 NASH 和 NASH相关的肝细胞癌。
NK 细胞和肝星状细胞之间的相互作用——癌症休眠和转移的主要开关
基质反应阻碍了 NK 细胞和干扰素 γ 介导的肿瘤细胞休眠的维持,并通过组织损伤和活化的肝星状细胞分泌 CXCL12 的过程诱导肝转移,CXCL12 通过 CXCR4 保留和使 NK 细胞静止,抑制免疫监视和促进转移性生长。
某些与肥胖相关的细菌具有增加次级胆汁酸脱氧胆酸的能力
脱氧胆酸的增加会导致肝星状细胞中的 DNA 损伤,从而诱导衰老相关分泌表型,导致炎症和肿瘤促进因子以及 COX2 诱导的免疫抑制性 PGE2 在肝脏中的产生,从而促进肝细胞癌的发展。
肥胖还与微生物群改变有关,导致 TLR2 激动剂脂磷壁酸 (LTA)这一革兰氏阳性菌的主要细胞壁成分的积累增加。脱氧胆酸增加肝星状细胞上 TLR2 的表达,并与 LTA 协同诱导衰老相关分泌表型因子和 COX2,后者通过 PGE2 诱导免疫抑制,并在化学致癌物暴露后促进肥胖相关的肝细胞癌。
总之,这些研究支持了一个模型,即平衡状态的改变会对环境中的连接细胞产生影响,证实微生物组及其代谢物是肠道和肝脏基因反应的关键影响因素,对肝癌发生具有影响。
本文主要介绍了影响肝细胞癌的微生物群的复杂相互作用,强调了微生物群多样性或肠-肝轴中微生物代谢物的改变触发局部细胞中的基因反应。
与肝细胞癌相比,微生物群和PDAC对患者预后的相关性已得到更彻底的研究,并已明确开始确定特定机制,通过这些机制,微生物群可以局部或系统地影响肿瘤微环境,以及肿瘤进展和对治疗的反应。因此,胰腺导管腺癌研究中使用的一些策略可以应用于肝细胞癌研究。
可以看到,目前的研究分析环境中局部细胞内微生物群介导的直接和间接变化,研究视角已逐渐从靶细胞转向微环境/生态系统。肠道微生物群以及肿瘤本身的细菌通过调节癌症、基质和炎症/免疫细胞中的基因转录程序以及促进或抑制肿瘤进展来影响肿瘤微环境。
癌症干细胞和免疫细胞之间的串扰在癌症进展中起着重要作用,那么癌症干细胞是否容易被微生物/代谢物的改变触发?这也是值得关注的问题。由于物理和生理上的联系,评估微生物群对癌症干细胞分化的影响以及作为一个综合系统对肠-肝轴生态系统的影响也非常重要。
主要参考文献:
Silveira MAD, Bilodeau S, Greten TF, Wang XW, Trinchieri G. The gut-liver axis: host microbiota interactions shape hepatocarcinogenesis. Trends Cancer. 2022 Mar 21:S2405-8033(22)00045-0. doi: 10.1016/j.trecan.2022.02.009. Epub ahead of print. PMID: 35331674.
Komiyama S, Yamada T, Takemura N, Kokudo N, Hase K, Kawamura YI. Profiling of tumour-associated microbiota in human hepatocellular carcinoma. Sci Rep. 2021 May 19;11(1):10589. doi: 10.1038/s41598-021-89963-1. PMID: 34012007; PMCID: PMC8134445.
Zhang T, Zhang S, Jin C, Lin Z, Deng T, Xie X, Deng L, Li X, Ma J, Ding X, Liu Y, Shan Y, Yu Z, Wang Y, Chen G, Li J. A Predictive Model Based on the Gut Microbiota Improves the Diagnostic Effect in Patients With Cholangiocarcinoma. Front Cell Infect Microbiol. 2021 Nov 23;11:751795. doi: 10.3389/fcimb.2021.751795. PMID: 34888258; PMCID: PMC8650695.
Nejman D, Livyatan I, Fuks G, Gavert N, Zwang Y, Geller LT, Rotter-Maskowitz A, et al. The human tumor microbiome is composed of tumor type-specific intracellular bacteria. Science. 2020 May 29;368(6494):973-980. doi: 10.1126/science.aay9189. PMID: 32467386; PMCID: PMC7757858.

谷禾健康

口服酶可以对肠道微生物群的组成产生深远的影响,并且可以作为一种有吸引力的替代调节剂。
本文总结了酶影响肠道微生物群的三种方式,并讨论了选择合适的酶来调节肠道微生物群的挑战。
酶:一种被忽视的调节肠道微生物群的因子
人类肠道容纳多达100万亿微生物,包括细菌、古细菌、真菌等。
最近的一项研究发现,有178种肠道微生物可产生400多种高度丰富的代谢物,其中许多可通过肠道屏障迁移,进入血液循环,并对宿主进行免疫、代谢和神经元调节。
目前用于调节肠道微生物群的大多数策略都集中在化学品和整个微生物细胞上。而酶,作为具有催化功能的特殊蛋白质,却没有被广泛考虑。
几乎在所有生化反应中都需要酶。在畜牧业中,各种酶被广泛添加到饲料中,以促进动物的生长。它们的有益作用与肠道菌群的变化有关,并可能部分归因于肠道菌群的变化,这强烈表明可以有意选择酶来调节肠道菌群。
事实上,在小鼠和其他实验动物中,酶无疑已被证明对肠道微生物群有潜在影响。
酶可以通过三种方式影响肠道微生物群。
1
// 一些酶可以杀死肠道微生物
溶菌酶、类溶菌酶糖苷水解酶和细菌噬菌体溶菌酶直接降解细菌细胞壁的主要成分肽聚糖。

Su X, Yao B. Trends Microbiol. 2022
在小鼠中,口服赖氨酸-防御素嵌合蛋白可显著减少粪便艰难梭菌孢子,从而降低死亡率。酶可以产生对肠道微生物有害的反应产物。
在肠道中,管腔葡萄糖(浓度为几十毫摩尔)、L-氨基酸(回肠中的几毫摩尔),甚至细菌衍生的游离D-氨基酸(盲肠内容物中的约200-500 nmol/g)都可以被酶氧化,释放过氧化氢杀菌。在小鼠体内,D-氨基酸氧化酶导致霍乱弧菌(Vibrio cholera)在小肠中的定植显著减少。
2
// 酶可以刺激肠道微生物生长
在人类中,食物中约一半的木聚糖被肠道微生物木聚糖酶降解为低聚木糖,低聚木糖支持某些肠道微生物(如拟杆菌、双歧杆菌和乳酸杆菌属)的生长。

Su X, Yao B. Trends Microbiol. 2022
木聚糖酶的刺激谱可以通过肠道微生物的生态网络进一步扩展。
酶还可以通过催化去除有害化学物质来刺激肠道微生物生长。
肠道碱性磷酸酶(IAP)是一种由肠上皮细胞分泌的内源性酶,通过降低抑制细菌的管腔核苷酸三磷酸的浓度来促进特定肠道微生物的生长。这与减轻肠道屏障损伤和减轻酒精诱导的肝脂肪变性等疾病密切相关。
3
// 酶能通过干扰微生物网络影响肠道菌群
群体感应(QS)是微生物群形成网络的一种方式。通过感应信号分子,微生物同步生成生物膜并排出有毒分子。

Su X, Yao B. Trends Microbiol. 2022
通过N-酰基高丝氨酸乳糖等酶去除QS分子(群体猝灭)可以调节金鱼的肠道微生物群,这可以通过增加变形菌的丰度和减少肠道中致病性嗜水气单胞菌(Aeromonas hydrophila)来证明。
分散素B也证明了微生物网络的干扰,它水解生物膜稳定剂1,6-N-乙酰-D-葡萄糖胺,从而显著减少铜绿假单胞菌(Pseudomonas aeruginosa)在肠道中的定植。
研究表明,有充分的机会利用酶来调节肠道微生物群,进而促进宿主的健康。
这其中最大的挑战可能是该使用什么酶。
以治疗慢性疾病为例。由于整合多组学分析的能力不断发展,慢性病中的致病菌群正逐渐被识别出来。
因此,酶可以作为有选择地富集或减少有害致病微生物的宝贵工具。
因为酶能够通过将致病微生物与微生物结合模块(如抗体)连接起来,从而瞄准致病微生物。
为了制造这种具有精确调节功能的酶,可以从拟杆菌的“自私”策略中获取经验。这种细菌编码细胞表面附着的甘露聚糖酶,以帮助它在竞争激烈的肠道生态位中靠甘露聚糖繁衍生息。
此外,成功地将赖氨酸与细菌素融合,使酶能够杀死革兰氏阴性细菌,可以减少不必要的致病菌。
酶可以作为已知化学物质或微生物的增强剂,影响致病微生物。
例如,2型糖尿病患者(50%)对两种精心设计的高纤维饮食没有积极的反应。在那些从饮食中受益的人中,利用低聚糖的细菌,如双歧杆菌属,属于少数“公会”致病微生物。在这些饮食中添加纤维特异性酶可以释放更多低聚糖,从而改善细菌的生长,从而有助于缓解更多患者的疾病。
然而,许多其他疾病与肠道微生物群变化之间的机制联系目前仅限于关联而非因果关系。因为肠道中几乎所有的成分都是酶的潜在底物,除了上述几种酶之外,还有大量其他候选酶。
从理论上讲,直接受酶影响的肠道微生物可以从酶的作用模式中推断出来,根据已知的机械关联,可以使用该模式选择酶作为进一步测试的候选酶。
酶可以用来帮助释放真正的效应剂
这在使用化学药品和微生物来调节肠道微生物群的组成时是不容易做到的。例如,溶菌酶通过杀死乳酸乳球菌(Lactococcus lactis)和释放细胞内超氧化物歧化酶来缓解结肠炎。
总的来说,这些结果表明,通过询问与上述三种酶以及其他类似的候选酶,并对其进行系统地测试,可以发现在有效调节肠道微生物群方面的隐藏酶。或者,将有关酶、饮食和宿主基因型的累积信息集成到现有数据库中,如Amadis (http://gift2disease.net/GIFTED/)将肠道微生物群与疾病联系起来,可能会有更合理的选择,并减少需要筛选的酶的数量。
尽管生产、储存和口服都很容易,但在选择合适的酶时仍有重要的考虑因素。
1、考虑酶的耐受条件,相应增加剂量
口服给药时,酶必须能够耐受肠道内的恶劣条件,包括酸性pH值和蛋白酶消化,这表明应选择候选酶来满足抵抗力,最初计算的剂量应在实验验证的基础上相应增加,以补偿储存和使用过程中任何可能的活性损失。
2、考虑酶的多效作用甚至危害影响
虽然酶不具有水平基因转移和引入多药耐药生物体甚至与基因工程益生菌和粪便微生物群移植相关的病原体的安全风险,但酶可以发挥不必要的多效作用,甚至产生有害影响。例如,磷脂酶将磷脂酰胆碱水解为胆碱,可被肠道细菌进一步转化为代谢产物三甲胺,三甲胺与不良疾病相关。
3、同一家族中不能调节菌群的酶需要排除
同一家族中的一些酶略有不同。那些不能调节目标微生物的也应该被排除在外。例如,木聚糖酶释放大量不同的低聚木糖,肠道细菌如Roseburia intestinalis和Bacteroides ovatus对具有不同糖链长度的低聚木糖的反应非常不同。
酶影响肠道微生物群,进而影响宿主健康,再加上大量候选者的可用性和可感知的优势,使其成为调节肠道微生物群的一种不错的方式。了解酶的功能和催化机制可以更好地调节肠道菌群,指导药物使用,治疗人类疾病。
然而,由于酶与其底物、肠道微生物群和宿主之间的复杂相互作用,开发酶等新试剂并非易事。需要针对不同情况选择合适的酶。此外,酶并非排他性的,它们可以与化学物质和微生物合作,改变肠道微生物群的组成,进一步促进宿主的健康。
主要参考文献:
Su X, Yao B. Exploiting enzymes as a powerful tool to modulate the gut microbiota. Trends Microbiol. 2022 Feb 1:S0966-842X(22)00003-8. doi: 10.1016/j.tim.2022.01.003. Epub ahead of print. PMID: 35120774.
Jia B, Han X, Kim KH, Jeon CO. Discovery and mining of enzymes from the human gut microbiome. Trends Biotechnol. 2022 Feb;40(2):240-254. doi: 10.1016/j.tibtech.2021.06.008. Epub 2021 Jul 22. PMID: 34304905.
Neves ALA, Yu J, Suzuki Y, Baez-Magana M, Arutyunova E, O’Hara E, McAllister T, Ominski KH, Lemieux MJ, Guan LL. Accelerated discovery of novel glycoside hydrolases using targeted functional profiling and selective pressure on the rumen microbiome. Microbiome. 2021 Nov 23;9(1):229. doi: 10.1186/s40168-021-01147-1. PMID: 34814938; PMCID: PMC8609826.
谷禾健康
慢性肾病 (CKD) 影响着全球约 13.4% 的人口,是一个日益严重的全球健康负担。成人中,高血压和糖尿病是慢性肾病的主要原因,而先天性肾脏和泌尿生殖道异常占儿童慢性肾病病因的大部分。慢性肾病与心血管疾病、神经系统并发症、不良妊娠结局和高钾血症等严重健康状况的发展有关。在儿童中,慢性肾病会影响神经认知能力、学校表现、成长、生活质量等。
而许多研究已证明,慢性疾病过程与人类肠道微生物群及其代谢物之间存在关联。
那么慢性肾病与肠道微生物群之间存在什么关系?
肠道菌群代谢产物在慢性肾病中起着什么样的作用?
肠道失调是如何启动炎症过程并导致菌群代谢产物泄漏到血液中的?
哪些饮食方式可以对其进行干预?
本文一起来了解一下。
概 要
· 慢性肾病与菌群关系是双向的;肠源性代谢物和毒素影响慢性肾病的进展,尿毒症环境影响微生物群。
· 微生物代谢物和毒素的积累与肾功能丧失和死亡风险增加有关,但短链脂肪酸和胆汁酸等肾脏保护代谢物有助于恢复肾功能和提高慢性肾病患者的存活率。
· 改变肠道微生物组的特定饮食干预可改善慢性肾病患者的临床结果。
·低蛋白和高纤维饮食增加了产生短链脂肪酸和抗炎菌的丰度。
· 尿液微生物组的波动与感染易感性和抗生素耐药性的增加有关。
01
慢性肾病是什么
肾脏的存在好处多多。肾可以帮助调控血液稳态,维持电解质平衡,调控全身水平衡,甚至可以产生激素。
慢性肾病是一个泛指,包含多种肾功能轻微下降的症状,且肾功能下降和微结构改变持续时间超过3个月,而小于3个月的肾功能恶化则是急性肾损伤。
慢性肾病的病理生理学
慢性肾病包括一系列与肾功能异常和肾小球滤过率(GFR)逐渐下降相关的病理生理过程。
注:单位时间内两肾生成滤液的量称为肾小球滤过率,正常成人为100-125ml/min/1.73㎡。
但是呢,由于GFR的测量麻烦、经济与时间成本较高,临床实践中相对较少使用。于是就出现了eGFR(Estimated Glomerular filtering Rate,即估算的肾小球滤过率),临床上一般用这个指标来衡量肾脏的工作情况。可以对慢性肾病患者的疾病严重程度进行分级,分级越高,滤过率越低,病情越严重。

慢性肾病的潜在病因因年龄、合并症、急性肾损伤反复发生和蛋白尿水平而异。
无论潜在的病因如何,剩余肾单位的过度过滤和肥大、肾小管间质纤维化、肾素-血管紧张素-醛固酮系统的激活以及内皮屏障的破坏都很常见,并导致肾排泄功效和eGFR下降。
从一个等级到下一个等级的转变通常伴随着肾脏内分泌功能的丧失。特别是,患有心血管病的慢性肾病患者表现出肾功能恶化和严重炎症。
肾小管间质间隙中免疫细胞的浸润和免疫衍生成分的积累促进慢性肾病的进展。
慢性肾病治疗的一个关键目标是防止患者进展到疾病的下一阶段。
02
慢性肾病中的肠道菌群失调
最近的研究表明,肠道微生物群失调在慢性肾病的病理生理学中起着关键作用,并导致严重的慢性肾病。
慢性肾病中的菌群变化
双歧杆菌和乳酸杆菌与慢性肾病进展和长期生存率呈负相关。
一项对223名终末期肾病患者的研究表明,与对照组相比,慢性肾病患者的次级胆汁酸和尿毒症毒素水平升高与Eggerthella lenta、Fusobacterium nucleatum、Alistipes shahii呈正相关。在这项研究中,作者表明,Faecalibacterium prausnitzii(普拉梭菌)、Roseburia、Prevotella(产短链脂肪酸菌)的存在与疾病进展和尿毒症毒素积累呈负相关。
另一项对92例慢性肾病患者的研究报告称:
慢性肾病队列中的Paraprevotella,Pseudobutyrivibrio(假丁酸弧菌属),Collinsella数量增加;这一发现使作者提出,这个特征可以用来区分慢性肾病患者(甚至是处于疾病早期的患者)和健康人。
肠道菌群失调引发慢性肾病的两种机制
其一:影响肠道屏障
微生物群组成的变化增强了肠道氨的产生,从而提高了肠腔的生理 pH 值,导致粘膜刺激并破坏了结肠上皮屏障。这导致肠道通透性增加,通常称为“肠漏”。
因此,内毒素和细菌产物易位进入循环并诱导局部炎症,由免疫细胞激活和促炎细胞因子和趋化因子的释放引起,以及慢性全身炎症,加剧肾功能的恶化。
其二:影响血压变化
肠道菌群失调可能促进慢性肾病进展的另一个机制是通过肠道生态失调在内皮功能障碍、血管收缩反应和随后的高血压发展中的作用。
肠道中乳酸杆菌的较低丰度与高血压和肾脏疾病的发生有关。与正常饮食的小鼠相比,高盐饮食的小鼠具有异常的微生物群;这些变化与T淋巴细胞活化和血压升高有关。
肠道微生物群的变化可能是慢性肾病通过一系列免疫反应改变、血压改变、代谢变化和长期炎症进展的起点。
上述是菌群失调影响慢性肾病,反过来,慢性肾病也影响菌群失调。
肠道菌群 ⇋ 慢性肾病
菌群失调与慢性肾病的发病机制之间存在双向关系。
吃进去的营养物质被分解代谢最终产物中的氨,通过肝脏代谢转化为尿素,并释放到循环中。尿素主要通过肾脏排出,部分通过结肠排出。
肾功能的恶化将主要排泄部位从肾脏转移到结肠。结肠中尿素的持续存在会触发产脲酶菌的增殖,导致肠道生态失调。
肠道微生物组与慢性肾脏病之间的关系是双向的
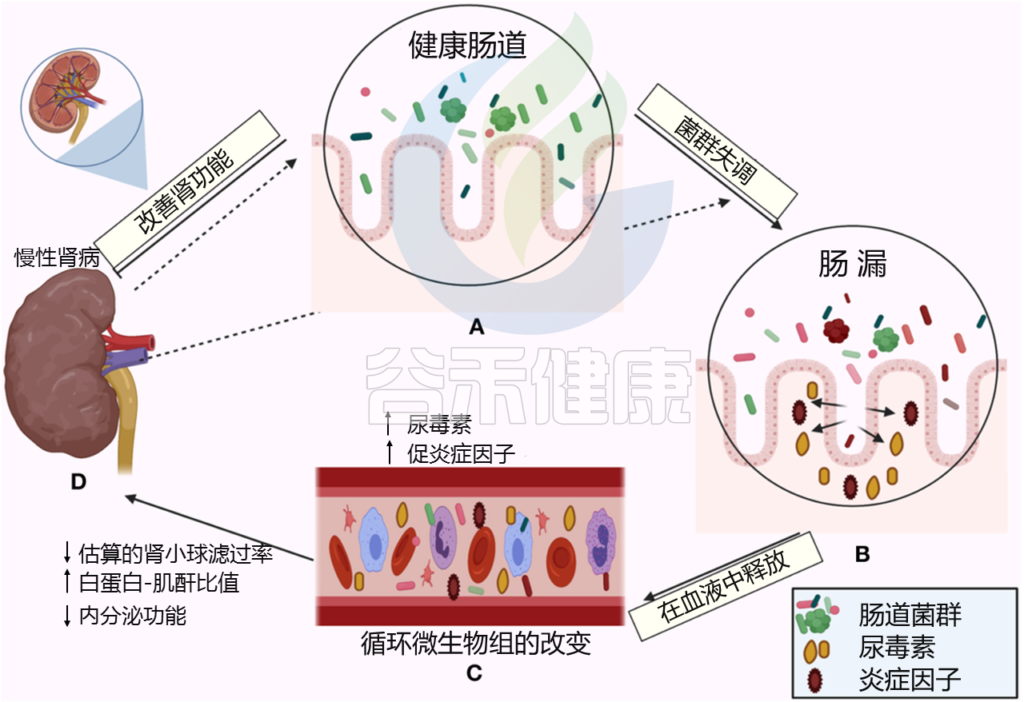
Al Khodor D, et al., Frontiers in Medicine,2022
← 在一个方向上,肠道菌群影响肾脏:
(A)健康的肠道
(B)肠道微生物失调和破坏粘膜层
(C)释放血液中炎性因子和炎症级联的开始,尿毒症毒素积累
(D)估计的肾小球滤过率下降(eGFR),白蛋白肌酐比值(ACR)升高,肾脏内分泌功能丧失
→ 在另一个方向,慢性肾病驱动肠道内的生态失调(虚线箭头所示),并引发炎症级联
03
慢性肾病中的微生物代谢物
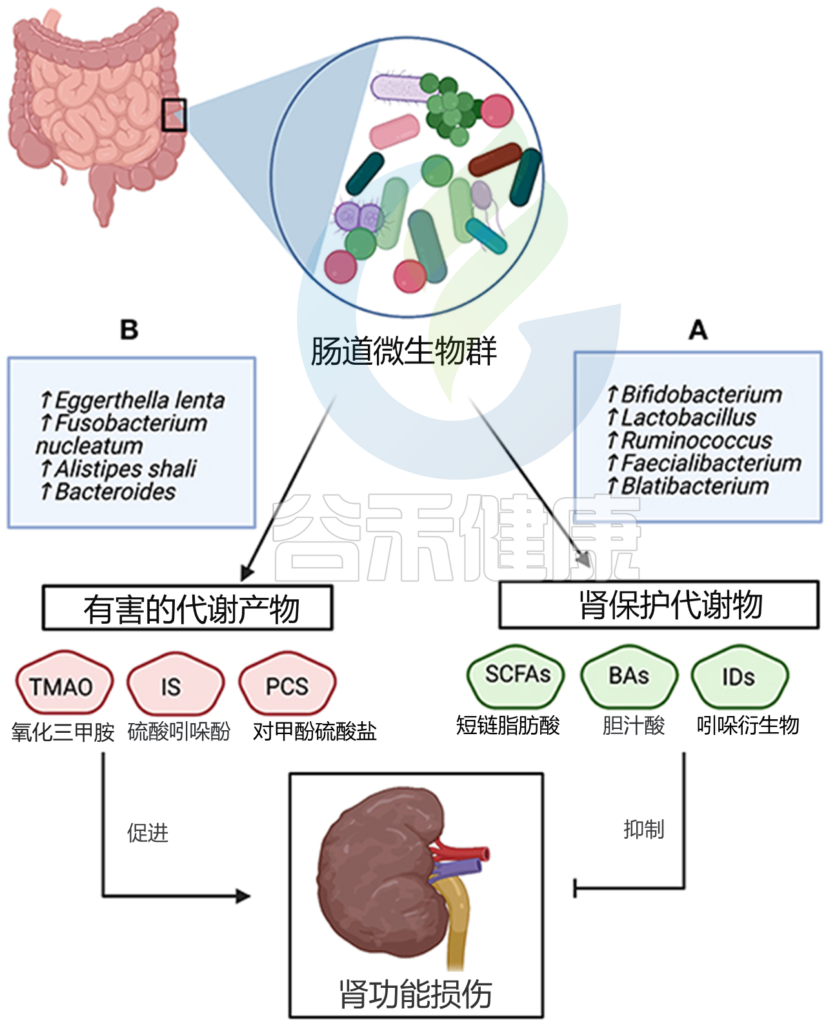
一般来说,与慢性肾病相关的微生物代谢物分为两类;有害和肾脏保护代谢产物。
一方面,有害代谢物的水平增加,包括三甲胺 N-氧化物 (TAMO)、硫酸吲哚酚和对甲酚硫酸盐与肾纤维化、内皮功能障碍、估计肾小球滤过率 (eGFR) 下降、心血管并发症以及慢性肾病死亡率和发病率增加有关。此外,5-甲氧基色氨酸和硫酸吲哚酚的血清水平与慢性肾病进展呈正相关。
另一方面,包括短链脂肪酸在内的肾脏保护代谢物通过抑制上皮屏障的破坏和调节抗炎反应来预防慢性肾病的进展。来自肠道菌群的吲哚丙酸水平与慢性肾病患者的对甲酚硫酸酯和吲哚硫酸酯浓度呈负相关。
菌群代谢产物和慢性肾病的关系
Al Khodor D, et al., Frontiers in Medicine,2022
一些人类和动物研究已经证明了TAMO对肾脏的有害影响,表现为肾间质纤维化、eGFR下降、内皮功能障碍和心血管疾病风险增加。
我们知道,肾脏的功能就是代谢身体废物,它的功能类似一个“清洁工”,肾脏生病也就是清洁工罢工,那么代谢废物就清除不出去了,于是在血浆中累积。
慢性肾病患者死亡率和发病率的增加归因于毒素的积累:硫酸吲哚酚和硫酸对甲酚。这些毒素与血浆蛋白具有很高的亲和力,从而减轻了它们通过透析膜的清除。
TAMO、硫酸吲哚氧基和硫酸对甲酚分别参与SMAD信号传导、色氨酸代谢和酪氨酸途径。
广泛的尿毒症毒素和其他微生物代谢物积聚在慢性肾病患者的生物样本中,包括血浆、粪便和尿液等常见生物样本中的毒素和其他微生物代谢物,也包括呼出气中的挥发性代谢物和粪便培养物中收集的气体。例如,慢性肾病患者体内会积聚气体代谢物,包括异戊二烯、醛、二甲基二硫、二甲基三硫和硫酯。
04
慢性肾病中的饮食干预
慢性肾病患者存在微生物失调和肠道代谢物积累。
益生菌
对慢性肾病患者进行的随机对照临床试验表明,益生元和益生菌治疗后肠道微生物群组成的变化改善了疾病结果,并降低了尿毒症毒素水平。
双歧杆菌和乳酸杆菌含量高的患者血清中尿毒症毒素水平较低,炎症环境减轻,肾功能改善。
益生元
益生元是不易消化的膳食成分,如膳食纤维和耐消化淀粉。它们存在于谷物、水果、牛奶、蜂蜜和蔬菜中,或者可以作为膳食补充剂。益生元发酵通过增加双歧杆菌和乳酸杆菌的丰度,降低类杆菌、梭状芽孢杆菌和肠杆菌的水平,有益地改善肠道细菌。
不利肾脏的食物:
摄入富含胆碱和 L-claritin的食物(TMAO的前体),如蛋黄、内脏、肉类和牛奶,与尿毒症毒素的大量积累和肾小球滤过率的下降相关。
低蛋白饮食减少炎症菌
一项前瞻性交叉临床试验将60例慢性肾病患者随机分为不同的饮食干预组;与常规饮食组相比,极低蛋白饮食组的肠道放线菌丰度增加,炎性变形菌减少。
膳食纤维降低慢性肾病风险
在慢性肾病患者中,膳食纤维摄入可降低循环中促炎细胞因子的水平,减缓eGFR的下降,降低尿毒症毒素的血浆水平,并将慢性肾病相关的心血管风险降至最低。
抗性淀粉降低血浆毒素
研究人员研究了补充抗性淀粉(16克/天)对慢性肾病患者的影响;他们观察到尿毒症毒素(硫酸吲哚氧基和硫酸对甲酚)、IL-6和硫代巴比妥酸反应物质的血浆水平降低。
乳果糖糖浆降低血清肌酐
这些结果与另一项将32例慢性肾病患者随机分为两组的研究一致;接受乳果糖糖浆治疗8周的组,肠道微生物群中双歧杆菌和乳酸杆菌的含量更高,血清肌酐水平降低。
虽然这些研究表明益生菌和益生元对慢性肾病有有益的作用,但也有其他研究表明循环肠道菌群代谢物或慢性肾病结果没有显著变化。
重要的是要指出,现有的研究是异质的;他们使用不同的膳食补充剂,有不同的干预持续时间,并对其他共病患者、肾病严重程度和潜在病因不同的患者进行管理。这种异质性使得从这些研究中得出结论非常困难。这就是说,在儿童饮食干预研究中可能会获得更好的结果,因为其他共同因素是最小的。
总之,这些研究表明,饮食干预疗法有可能调节微生物组组成及其代谢产物,从而改善慢性肾病并发症和慢性肾病进展率。然而,需要进一步设计良好的前瞻性研究来明确证明营养疗法对慢性肾病的益处。
05
尿和血液微生物群在慢性肾病中的作用
微生物组学领域的大部分注意力集中在肠道微生物组及其代谢产物上;然而,尿液微生物组正受到更多的关注。
新一代测序技术的发展使研究表明,健康个体的尿路由不同种类的微生物控制,这些微生物的分布模式影响尿路健康。
泌尿系微生物组的波动发生在泌尿系感染中,并与抗生素耐药性有关。肾移植后,尿液微生物组发生变化,这些改变被认为是导致同种异体移植功能障碍和增加感染易感性的原因。此外,慢性肾病患者尿液微生物多样性与eGFR值相关。
健康个体的循环微生物群包含多种细菌类群,其中以变形菌门为主。血液中循环的肠源性内毒素可改变血液微生物组。
一项研究调查了399名参与者的血液代谢组与肠道微生物群α多样性之间的相关性,结果表明,对甲酚和TAMO等肠道菌群代谢物反映了肠道细菌的香农多样性,可能是反映肠道健康的生物标记物。
使用16S rRNA靶序列对血液样本进行的病例对照研究表明,与对照组相比,慢性肾病患者的肠杆菌科和假单胞菌科的多样性较高,这也与较低的eGFR相关。
因此,肠道微生物群通过不同途径对慢性肾病的结局产生最终影响。
06
结 语
双歧杆菌、乳酸杆菌和胆汁酸成分水平较低与慢性肾病患者的不良后果有关。TAMO、硫酸吲哚氧基、硫酸对甲酚和其他有害微生物代谢产物在慢性肾病患者体内积累,这些代谢产物的水平与疾病进展相关。
肠道、泌尿道和血液微生物群以及相关代谢物之间的复杂相互作用可能协调慢性肾病发病机制中的亚临床变化,并促进疾病的发生。
通过饮食干预调节肠道微生物群可以改善慢性肾病患者的临床结果。
随着肠道微生物群的深入研究,可为慢性肾病的病因、代谢途径和潜在治疗提供线索。
未来可在以下方面深入开展研究:
主要参考文献:
Al Khodor D, Wehedy E, Shatat I F. The human microbiome in chronic kidney disease: a double-edged sword[J]. Frontiers in Medicine, 2986.
Mertowska P, Mertowski S, Wojnicka J, et al. A Link between Chronic Kidney Disease and Gut Microbiota in Immunological and Nutritional Aspects. Nutrients. 2021;13(10):3637. Published 2021 Oct 17. doi:10.3390/nu13103637
Feng Z, Wang T, Dong S, et al. Association between gut dysbiosis and chronic kidney disease: a narrative review of the literature. J Int Med Res. 2021;49(10):3000605211053276.
Giordano L, Mihaila SM, Eslami Amirabadi H, Masereeuw R. Microphysiological Systems to Recapitulate the Gut-Kidney Axis. Trends Biotechnol. 2021 Aug;39(8):811-823. doi: 10.1016/j.tibtech.2020.12.001. Epub 2021 Jan 6. PMID: 33419585.
谷禾健康
10.29-11.1谷禾管理后台数据库升级,涉及到数据迁移和测试,期间可能会在后台显示信息缺失,数据并无问题仅因数据库迁移可能导致显示不正常,预计11.2日会完成升级工作。
特此通知
杭州谷禾信息技术有限公司

谷禾健康
人一生中平均有三分之一的时间在睡觉。睡眠对于我们是必不可少的存在,它能够促进我们的幸福感。

然而目前,睡眠障碍已经成为了一种流行病。尤其是在新冠肺炎流行之后,进一步加剧了睡眠问题。
随着睡眠障碍的患病率越来越高,人们开始深入研究——是什么因素决定了睡眠的质量?
众所周知,各种环境因素和其他因素(如心理压力和饮食)的共同作用会影响睡眠质量和睡眠持续时间(下图)。
图1 各种因素的相互作用影响睡眠质量和持续时间
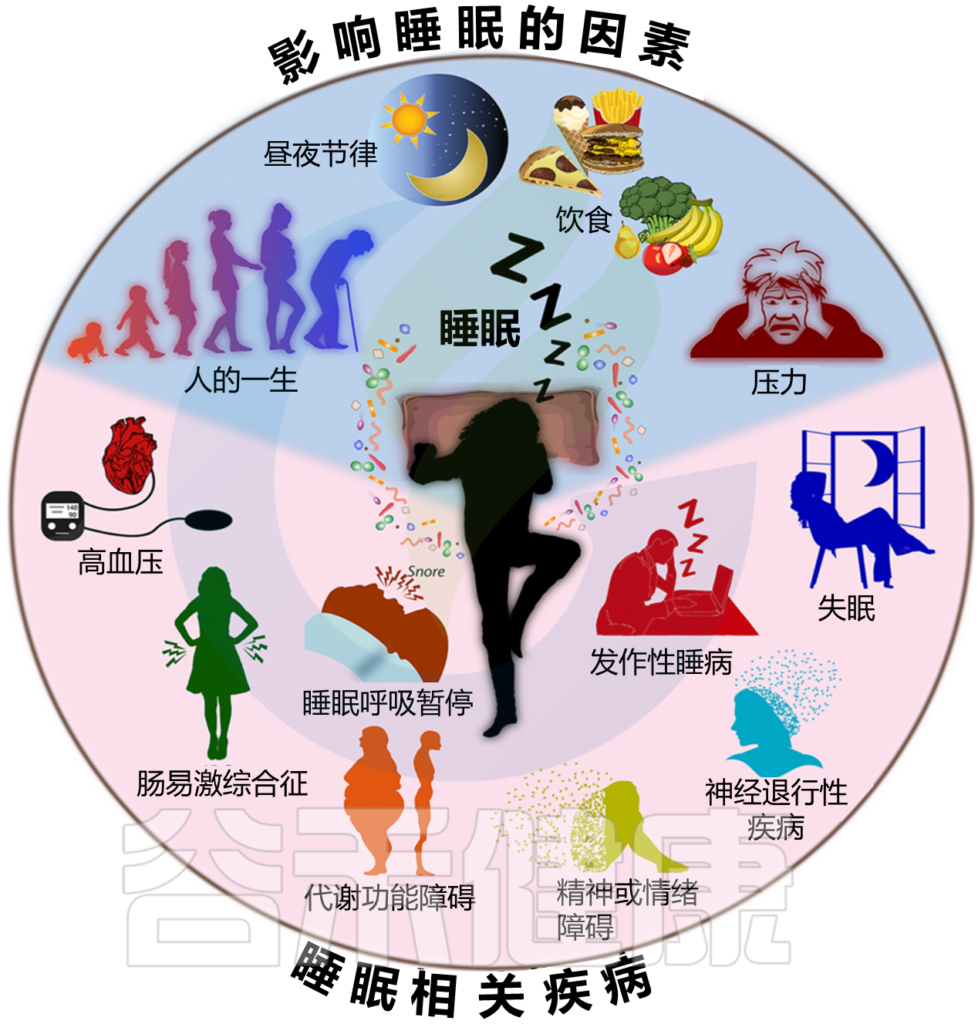
Sen P, et al.,Trends Mol Med. 2021
各种因素对睡眠质量和持续时间的相互作用影响。已知各种不同的因素会影响睡眠。其中包括饮食、压力、昼夜节律调整以及整个生命周期的各个阶段(上图,蓝色背景)。最近的研究表明,这些因素与肠道微生物群相互作用,影响睡眠。
反过来,睡眠质量和持续时间也影响各种因素。睡眠质量差或睡眠不足可导致免疫、代谢功能障碍、血压升高,并可发展为肠易激综合征、各种神经退行性疾病和精神障碍、睡眠呼吸暂停和发作性睡病(下窗格,粉红色背景)。睡眠不足可能通过影响肠道菌群组成而导致这些疾病的发展。
本文从微生物群的角度探讨各种因素如何与肠道微生物群相互作用影响睡眠。包括睡眠与微生物群具有相互作用的可能机制。
此外,还提出了常见的伴随睡眠障碍的疾病以及针对各种睡眠障碍时微生物群对肠道和大脑进行调节改变的现有证据。
睡眠质量与肠道菌群组成之间存在双向关系。使用抗生素干扰肠道微生物群会导致更大的非快速眼动(NREM)睡眠碎片化,从而降低睡眠质量,睡眠中断也会导致肠道微生物群组成的变化。不过,关于睡眠障碍和微生物组成之间关系的结果缺乏一致性。
此外,大多数睡眠研究只关注单一时间点的肠道微生物群。由于肠道菌群每天动态变化,睡眠和肠道菌群之间的关系可能在一天的不同时间有所不同。
尽管如此,越来越多的研究表明,微生物-肠道-大脑轴与睡眠生理和行为有关。
表1 探索肠道微生物如何影响睡眠的各种研究

Sen P, et al.,Trends Mol Med. 2021
睡眠和微生物群组成:交流机制
肠道微生物群通过微生物群-肠道-大脑轴与大脑沟通的各种可能机制。
图2 影响睡眠的微生物群-肠道-大脑轴交流模式

谷禾健康
2型糖尿病是一种血糖水平升高(高血糖)的慢性代谢疾病,主要由胰岛素抵抗和胰岛素分泌不足引起。2型糖尿病全球患病率在不断上升。
截止到2020年,根据美国糖尿病学会评估标准,糖尿病患病率已达到12.8%。其实在我国成年人中,已经有将近一半的人存在血糖异常,未来极有可能发展为2型糖尿病。如果不加干预治疗,会引发许多不良后果。
很多研究表明,肠道菌群在宿主的代谢和疾病状态中起着重要的作用。特别是2型糖尿病,其病因复杂,包括肥胖、慢性低度炎症,受肠道微生物群和微生物代谢产物的调节。
本文将从肠道菌群的角度了解其与2型糖尿病的关系;肠道菌群及其代谢物对葡萄糖代谢、免疫系统的影响;
从影响肠道菌群变化的因素来了解各种干预措施,包括药物,饮食,益生菌/益生元,粪菌移植,间接性禁食等方式。
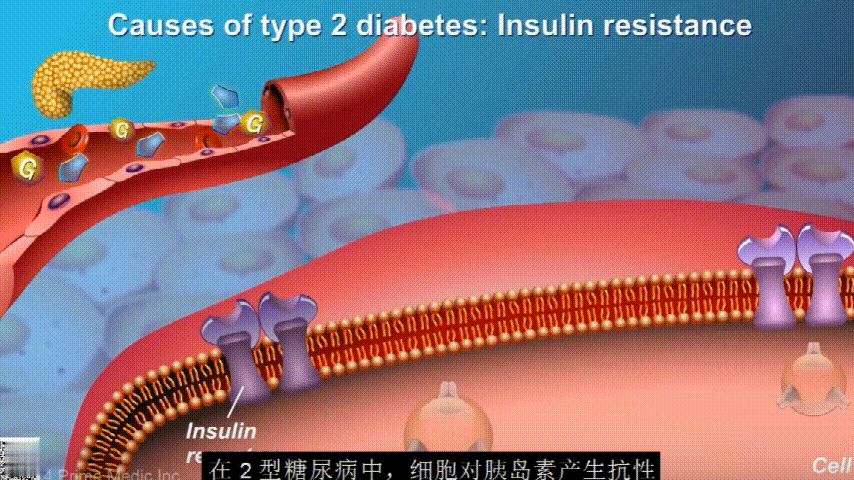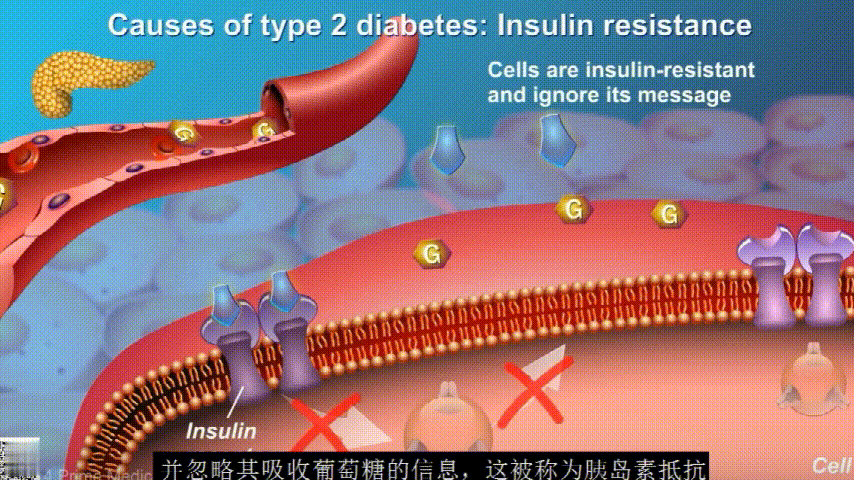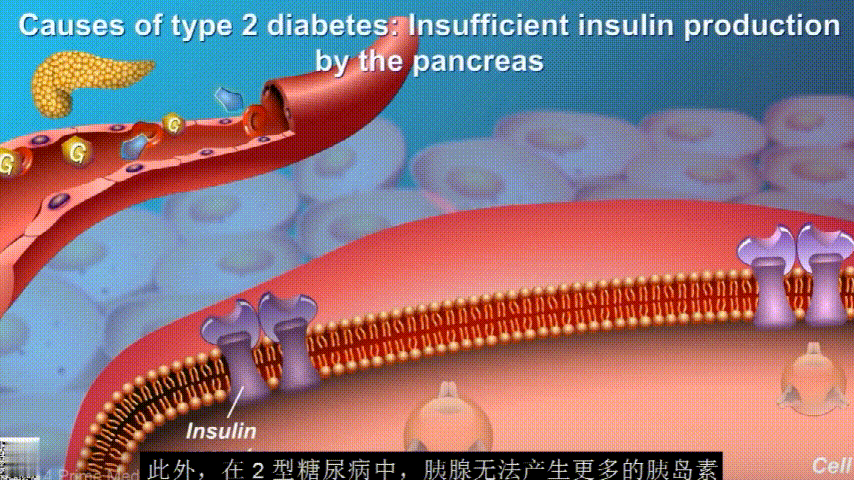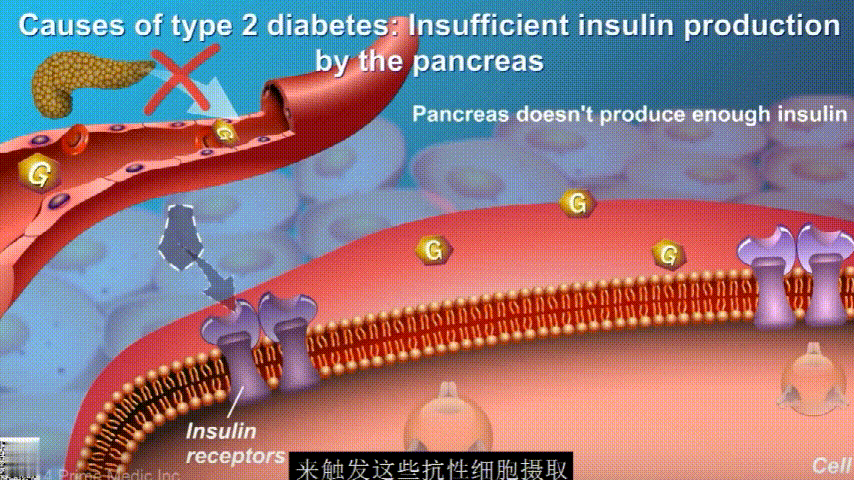
葡萄糖主要通过食用的食物和饮料进入人体。胰腺会产生一种叫做胰岛素的激素,以帮助血液中的葡萄糖进入肌肉、脂肪和肝脏,用作能量。
胰岛素通过血液循环进入不同组织。组织大多数细胞表面有一种结构,叫胰岛素受体。当胰岛素流过时,会附着在胰岛素受体上,就像钥匙打开锁一样,糖可以进入细胞内,细胞获得能量维持正常运作,血液中糖分就会回落到正常水平。
在2型糖尿病患者身上,细胞对胰岛素没有反应,这种情况就叫胰岛素抵抗。胰岛素没有了“钥匙”的作用,不能有效开启细胞上的“锁”,或者“锁”的数量也减少了,由此,糖分就不能进入细胞有效利用,被挡在了细胞外面,只好在血液中积累,血糖就升高了。
刚开始的时候,胰腺会觉得自己使不上力,于是不甘心开始拼命工作,产生更多的胰岛素来克服这种抵抗。
随着时间的推移,胰腺开始累了,力不从心,不能产生更多的胰岛素,这时候就是2型糖尿病开始了。
肠道微生物群可以直接或间接地影响人类健康,肠道微生物群失调可能会增加促炎疾病的患病率,如肥胖、炎症性肠病、2 型糖尿病、关节炎和癌症。
2型糖尿病人群中个体微生物群的差异
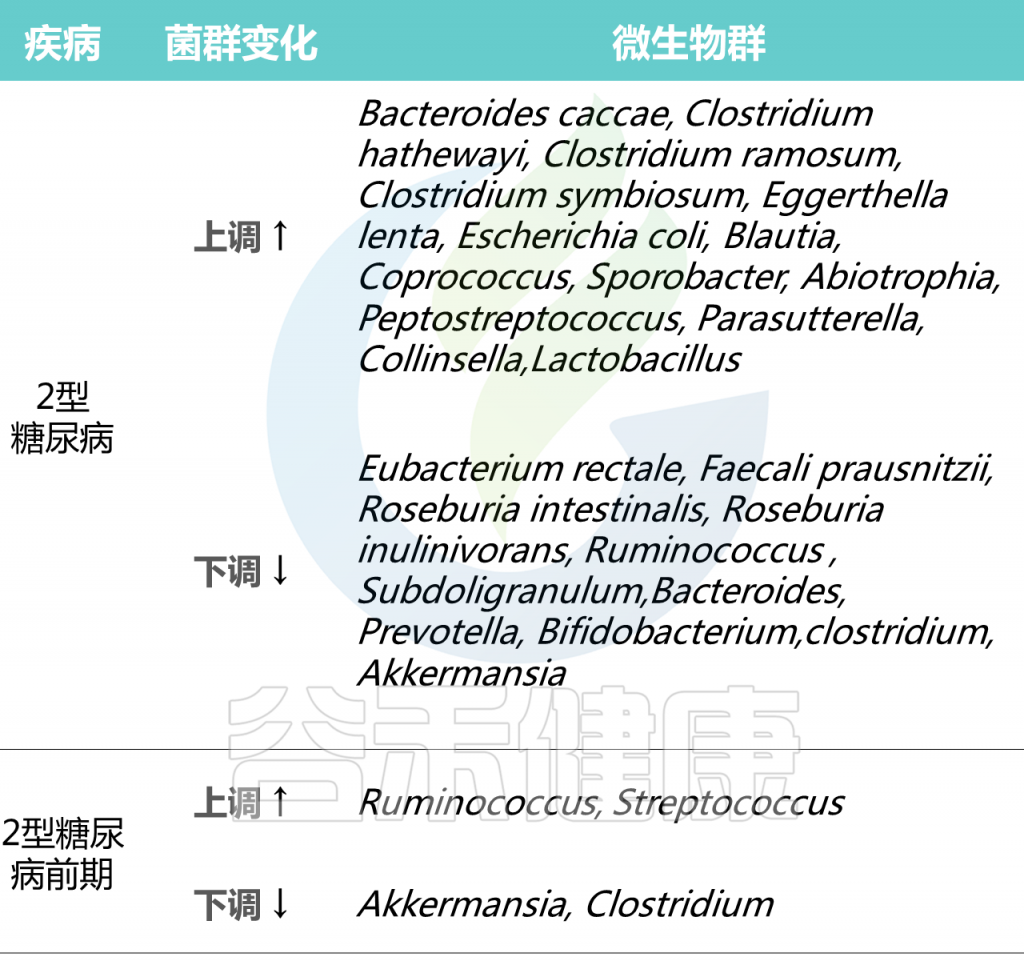

Cunningham A L et al., Gut Pathog, 2021
在2型糖尿病患者普遍具有相对高丰度的特定属:Blautia、Coprococcus、Sporobacter、Abiotrophia、Peptostreptococcus、Parasutterella、Collinsella。
2型糖尿病患者中,产生丁酸菌特别缺乏,特别是梭菌目,包括:
Ruminococcus、Subdoligranulum,Eubacterium rectale、Faecali prausnitzii、Roseburia intestinalis 、
Roseburia inulinivorans
2型糖尿病患者中,其他明显较少的菌属有:
拟杆菌属、普雷沃氏菌属、双歧杆菌属

谷禾健康


曾经,央视网也有过类似的报道 ↓↓↓
军团菌到底是什么?
军团病为什么会这么严重?
夏季吹空调怎样才能避免?
今天我们就详细了解一下
——“军团病”
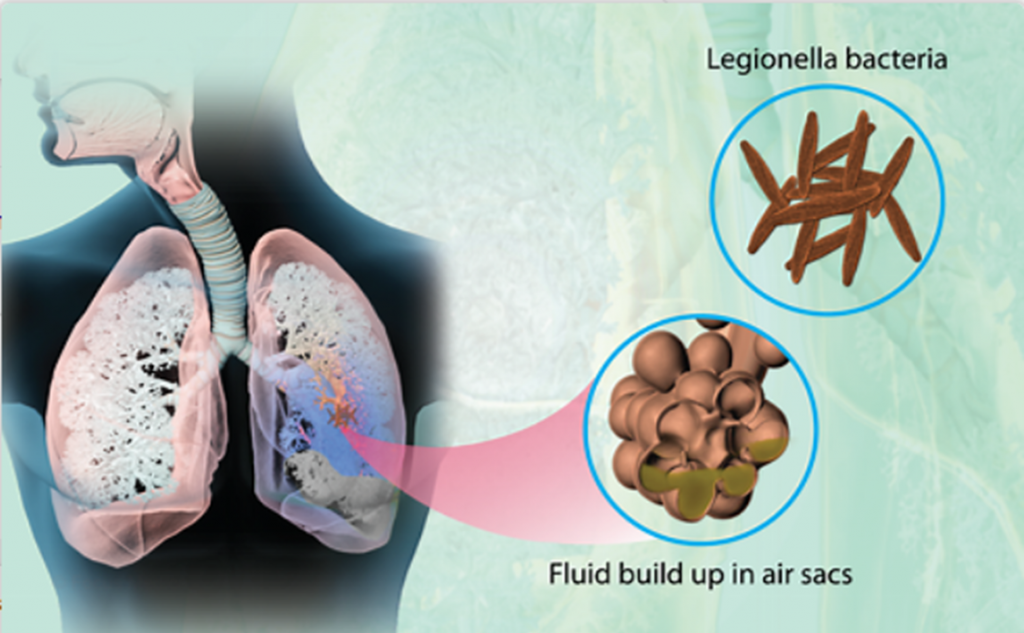
是一种由军团菌引起的严重肺炎。大多数病例都是由嗜肺军团菌引起的,但其他种类的军团菌也能引起这种疾病。它之所以被命名为军团病,是因为1976年,在宾夕法尼亚州费城参加美国退伍军人大会的人群中爆发肺炎后,首次发现了这种疾病。
军团菌是一种小型,水性的,未被包裹的芽孢杆菌,革兰氏阴性菌,过氧化氢酶呈阳性,专性需氧菌。该菌通过雾化水颗粒传播,不会在干燥的表面上存活。
军团菌科有超过42种,其中嗜肺军团菌L. pneumophila 是最常见的,已鉴定出嗜肺军团菌的15个血清群,其中血清组1、4和6被确定为人类疾病的病因。血清组1被认为是造成80%报告病例的原因。
军团菌广泛存在于自然水系统中,例如河流和池塘。然而,人们从这些来源感染疾病的情况很少。
最有可能发生感染的情况,包括淋浴器、水龙头、浴缸、装饰喷泉、游泳池或通过大型建筑(如酒店、办公楼、医院、游轮)通风系统的水滴。

还有当喝水时咳嗽或意外呛到,这时候如果吸入的水含有军团菌,那么也有可能发展为军团病或庞蒂亚克热。
注:庞蒂亚克热是一种比军团病更温和的感染,类似流感。症状主要是发烧和肌肉酸痛。症状可在接触细菌后数小时至 3 天内开始出现,通常持续不到一周。庞蒂亚克热通常会自行痊愈,但军团病需要治疗。
大多数接触军团菌的人不会患上军团病。生病的人通常在接触后2~10天内出现症状,但也可能需要更长时间(20天)。最初的症状可能包括头痛、寒战、肌肉疼痛和温度可达40℃或更高的发烧。
其他症状通常在最初症状后1~2天出现,可能包括咳嗽、气短、胸痛、关节疼痛、食欲不振、腹泻、恶心和呕吐,甚至意识混乱、协调性丧失。军团病主要影响肺部,但有时也会导致身体其他部位的感染,如心脏或身体伤口。

谷禾健康
围产期营养是健康生长和长期发育轨迹的早期决定因素。关键发育时期的营养不良可以显著、持续地改变许多身体系统的发育过程,包括神经系统。
营养不良 是指热量、大量营养素或微量营养素的缺乏、过量或差异比例的改变。
研究表明,肠道微生物组可以调节饮食对宿主生理的影响,且调节神经系统的发育和功能。
本文围绕营养不良、肠道微生物群和神经发育三者之间的关系,讨论了围产期营养不良改变孩子大脑发育的证据,并研究了母体和新生儿微生物群作为一个潜在的促成因素,介导营养不良和大脑发育之间的关系。
常量营养素
生命早期蛋白摄入:与运动,智商,抑郁等都有关
营养不良的幼儿在社会行为上有缺陷。尤其是出生后早期蛋白质摄入与早产儿的运动和认知评分以及总脑容量呈正相关,与髓鞘形成相关缺陷、脑萎缩和心室扩张、皮质树突发育不良以及枕大池扩大和室周白质异常呈负相关。
出生后第一年低蛋白摄入的青少年表现出认知功能下降,包括儿童时期的智商和注意力下降,青春期抑郁症状增加。
生命早期脂肪摄入:影响大脑发育,甚至持续到成年
其他常量营养素也同样至关重要:在早产儿中,出生后早期脂肪摄入与更大的小脑、基底神经节、丘脑和全脑有关,此外还有内囊、辐射状晕、丘脑放射、后纵束和皮质脊髓束的分数各向异性增加。重要的是,大脑发育与后来的精神运动功能有关。
一些神经缺陷可能会持续到成年:在荷兰饥荒出生队列的118名成员中,暴露于产前营养不良与成年后期脑白质灌注减少,男性扣带皮层血流量减少,与男性衰老相关的大脑特征相关。
来自巴巴多斯的成年人在出生后第一年受到蛋白质限制,有持续的注意力缺陷,执行功能下降,包括认知灵活性、工作记忆和视觉空间整合。
微量营养素
出生后早期缺乏微量营养素,如维生素B12、叶酸和维生素K,也与脑萎缩和胼胝体变薄、精神疾病和颅内出血风险增加有关。
营养过剩
母亲营养过剩(怀孕前和/或怀孕期间的高BMI)与婴儿在认知和语言发展领域得分下降、视觉运动技能受损、社交和学习能力下降(尤其是男孩)、胎儿丘脑和皮质连接性改变有关。
实际上,营养需求以及这些需求得不到满足的后果可能比以前的模型所显示的更加复杂和相互依存,可能构成一个连续体,而不是营养不足和营养过剩的二元对立现象。
为了进一步了解营养不良作为一个连续体,研究产前营养不良的特定亚型破坏大脑和行为发育的基本机制至关重要。
肠道微生物群是饮食对宿主生理影响的关键决定因素,具有调节大脑发育和行为的能力。
饮食和营养的变化,改变了肠道微生物群的结构和功能,尤其是在发育的关键时期。
营养不良——α多样性下降:
孟加拉国2岁以下儿童出生后严重急性营养不良与微生物α多样性(物种丰富度和均匀度)下降相关,表明菌群不成熟的情况即使在饮食干预后也持续存在。在一个类似的队列中,α多样性降低的原因是发育迟缓和非发育迟缓儿童之间噬菌体(细菌病毒)的差异。
营养不良不同亚型,α多样性下降程度不同:
在塞内加尔和尼日尔的幼儿中,出生后营养不良亚型之间的微生物多样性差异显著:
不同亚型之间的菌群构成差异:
出生后严重急性营养不良儿童体内的微生物群减少,代谢和营养吸收相关的功能途径减少,毒力相关基因增加。还显示肠道氧化还原电位增加,这可能是微生物群落组成改变的功能结果。
营养不良——α多样性升高:
谷禾健康
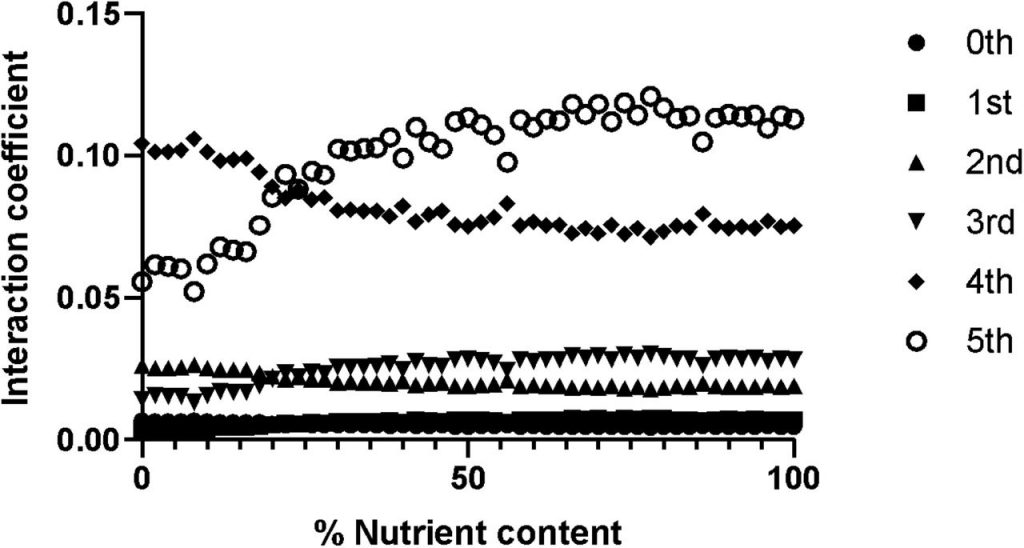
生态学的一个长期目标是捕获多种物种相互作用的多样性,生态系统中存在多种物种时产生的不可预测的影响。例如,动物具有多样的微生物群落,它们的组成是可变的,受随机过程控制,这会影响系统的整体行为。许多涉及复杂网络结构的生态研究通常集中于成对相互作用,忽略了涉及三个或更多的分类单元之间的高阶交互作用(higher-order interactions)带来的潜在影响。
高阶交互作用
高阶交互作用(higher-order interactions):高阶相互作用一直是遗传学领域中相对严格的研究对象,在遗传学领域中,它们被讨论为上位性,或基因和突变之间的非线性相互作用。尤其是高阶上位性,因为这些相互作用包含了理解和研究其他系统甚至微生物中的高阶相互作用的所有复杂性和挑战。
上位性(epistasis)是指某一基因受不同位点上别的基因抑制而不能表达的现象。现也将在群体遗传学和数量遗传学中非等位基因的遗传效应为非相加性时统称之为上位性。也就是位于不同座位上的基因间的非相加性相互作用。
与基因组不同,肠道微生物群是由单个实体或信息群(在这种情况下,是微生物群中的组成类群)之间的相互作用所定义的复杂系统。因此,科学家会预测微生物群中的生物群之间的更高阶相互作用可能是微生物群相关生物表型的基础。
例如一项研究了多物种相互作用对确定群落功能(即淀粉酶表达)的贡献。在存在更高阶的交互作用时,加性无效模型(没有交互作用)在预测社区功能方面的预测能力会降低。但是,通过将行为和人口动力学效应都计入其无效模型,高阶交互确实为社区功能提供了良好的预测。
最近发表的一篇关于此研究的文章,作者利用一种简单的数学方法——Hadamard-Walsh变换,研究了昆虫肠道菌群中的高阶相互作用。以此说明不同环境下肠道菌群之间的高阶相互作用是如何影响宿主表型的。希望能打破仅关注群落水平或两两互作关系的局限性,更多关注三阶、四阶以至多阶的分类群之间的互作关系。


一个由五个核心类群组成的肠道群落中。发现根据宿主特征,三向,四向和五向相互作用占所有可能病例的13-44%。然而,低阶相互作用(2对)仍然占系统中所有观察到的表型的至少一半。Sanchez-Gorostiage等人的工作研究了多物种相互作用对确定群落功能(即淀粉酶表达)的贡献。在存在更高阶的交互作用时,加性无效模型(没有交互作用)在预测社区功能方面的预测能力会降低。但是,通过将行为和人口动力学效应都计入其无效模型,高阶交互确实为社区功能提供了良好的预测。前述研究提供了可如何测量高阶相互作用的示例,并表明它们与理解微生物分类群如何影响某些表型有关。尽管多样性和主体互动的重要性很明显,但据我们所知,还没有研究试图明确区分环境环境中高阶互动的影响。
Hadamard-Walsh沃尔什-阿达玛变换使人们能够量化潜在相互作用的物体或地块之间相互作用的显著程度。它的主要输出是沃尔什系数(Walsh coefficient),该系数传达给定交互的大小(交互有多大)和符号(正交互或负方向)。
该方法以向量的形式实现表型(本研究中指宿主感染风险)值,然后将其重新格式化为Hadamard矩阵(然后通过对角线矩阵进行缩放)。输出的是与分类群之间相互作用的强度相对应的系数的集合。
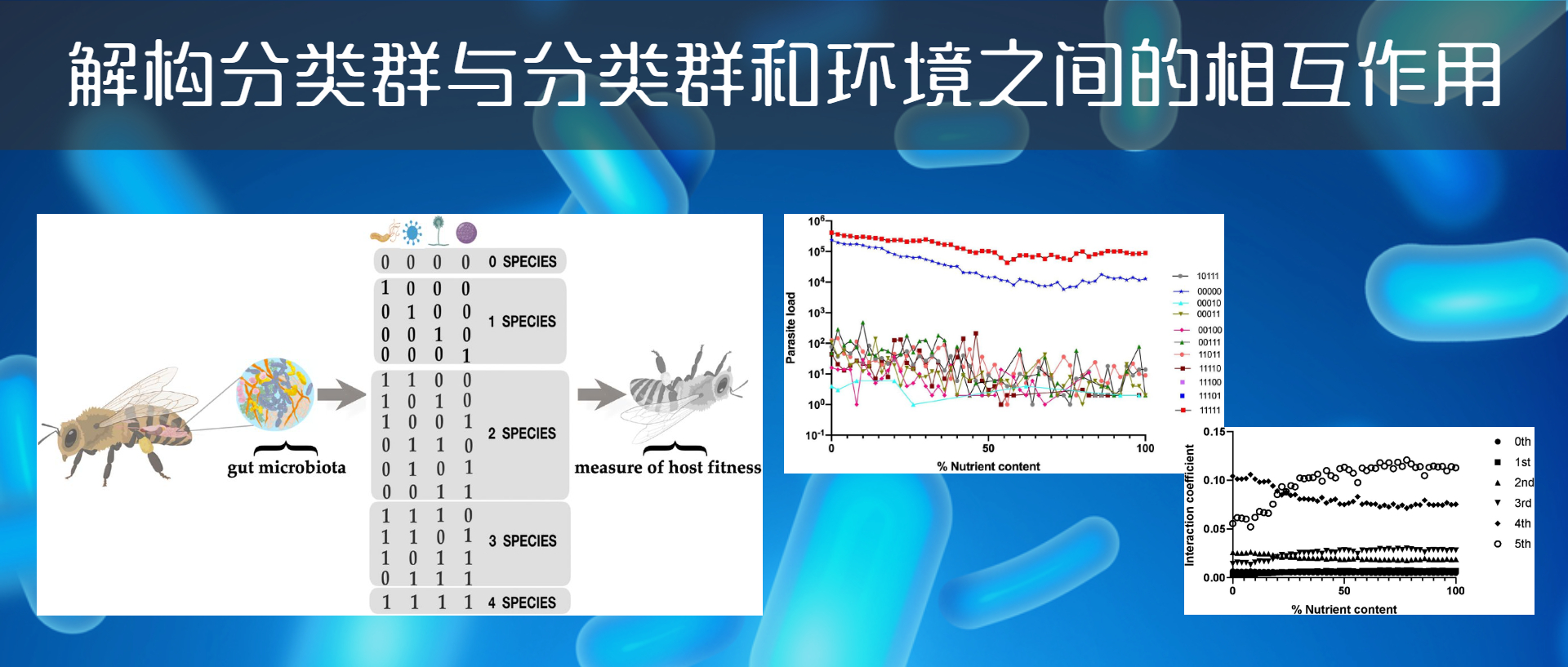
作者将沃尔什系数以二进制形式重新编码,例如: *B*DE 被编码为 01011
“*”表示不存在的分类群,在这个示例中存在BDE三个分类群,并将这三者之间的相互作用贴上“三阶相互作用”的标签,以此类推零阶即为*****或00000,五阶即为ABCD或11111。对于一个庞大的微生物群落,可以计算三阶相互作用的强度,并将它们与四阶相互作用的强度进行比较。
作者构建了一个昆虫肠道菌群的理论环境。用于研究饮食中不同品质的营养对宿主肠道菌群中寄生虫的易感性的影响。
利用从优质饲料(食物培养基中不含甲基纤维素)到低品质饲料(用甲基纤维素取代食物培养基的10%、20%、30%、40%、50%、60%、70%、80%、90%)的不同资源水平,对不同营养环境对鳞翅目害虫抗寄生性的影响进行了实证研究。
0%的饮食相当于极低质量的营养饮食,而100%的饮食相当于由昆虫的标准食物量组成的高质量饮食。
数据集来自用于生成理论上的适应度地形(fitness landscapes,遗传学算法中的名词,指由一系列基因型组成的三维的地形图)的原始数据。
这些原始数据由5位字符串组成。这些数据集可用于研究群体遗传学的高级课题,比如高阶上位性。数据被定义为字符串(例如,01011或11001),每个字符串具有对应的“表现型”( phenotype)值。由此组成了含161个微生物群的昆虫肠道环境。如下图:
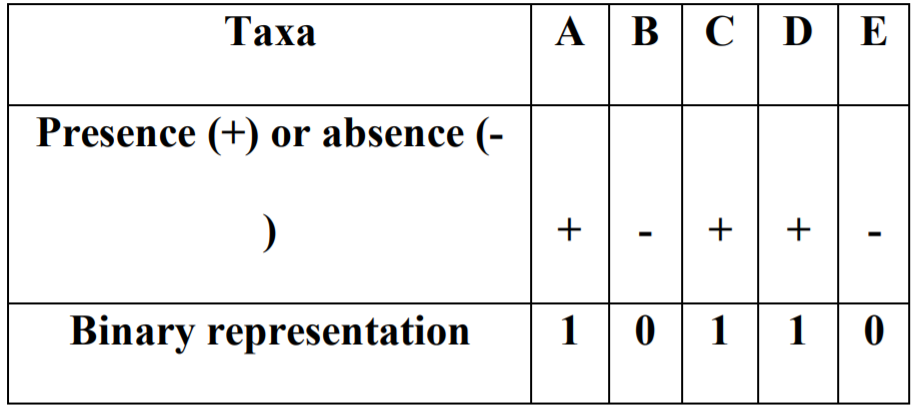

图一

图二
数据集是5个分类群(A-D)的存在和缺失(+或-)的组合。这种组合的可能性相当于四种不同昆虫环境中2的5次方=32个分类群的理论组合。
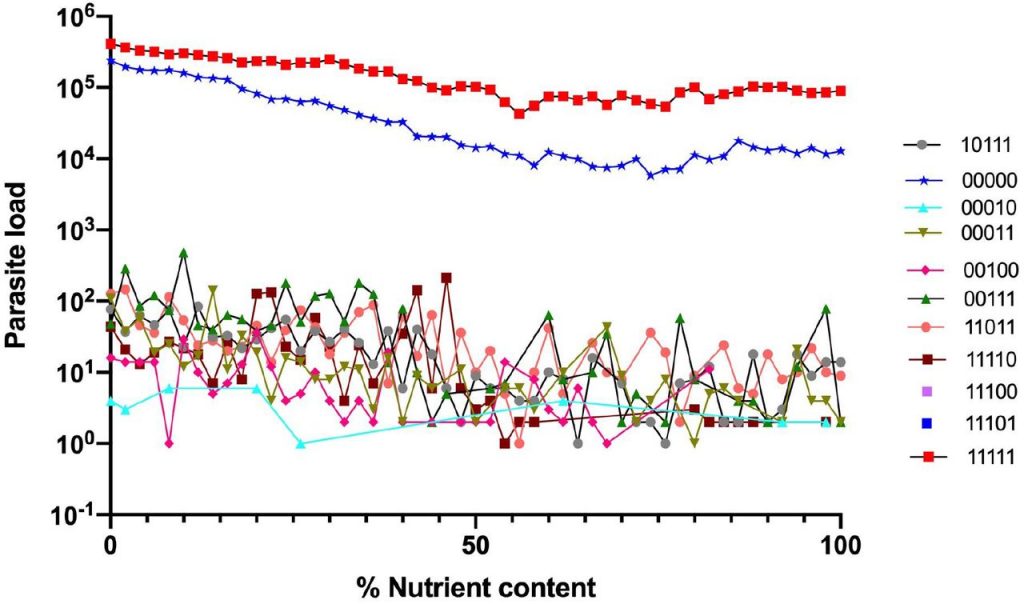
图一表示营养含量的消耗(x轴)与寄生虫含量(y轴)之间的反应关系。
结果显示与其他昆虫微生物区系组合相比,对应于00000(不含分类群) 和11111(存在各种分类群)的两个昆虫肠道最易感染寄生虫。相比之下所有其他昆虫微生物群组合(包括组合00100;11011;11101),感染的寄生虫含量显著较低。
图二说明了阶级变化与相互作用强度是相关的,比如低阶的如零一二三,它们的相互作用强度也较低,相反,高阶的较高。以及营养含量对相互作用模式的影响,比如营养含量为20%时,可以明显看见五阶分类群的相互作用强度开始上升,四阶的下降。

图三

图四
上图结果表明,高阶微生物群的相互作用限制了寄生虫在营养环境中的入侵。肠道菌群的营养状况对宿主的健康起着重要作用。
使用高阶建模方法能够捕捉到来自复杂生态相互作用的丰富信息,将这些方法应用于实验数据的分析,将会推动微生物群动力学研究。
相关阅读:
Science|180种动物肠道菌群测序结果有哪些信息值得我们关注
参考文献
Deconstructing taxa x taxa x environment interactions in the microbiota: A theoretical examination
Senay Yitbarek, John Guittar, Sarah A. Knutie, C. Brandon Ogbunugafor
bioRxiv 647156; doi: https://doi.org/10.1101/647156

谷禾健康
每个人身上都有“独特的味道”。有些人身上的味道几乎感受不到,而有些人身上却会出现令人尴尬的“体臭”。
慢性体臭可能带来许多不便,焦虑,自尊降低和生活质量下降,社交困难,甚至导致严重的心理问题。

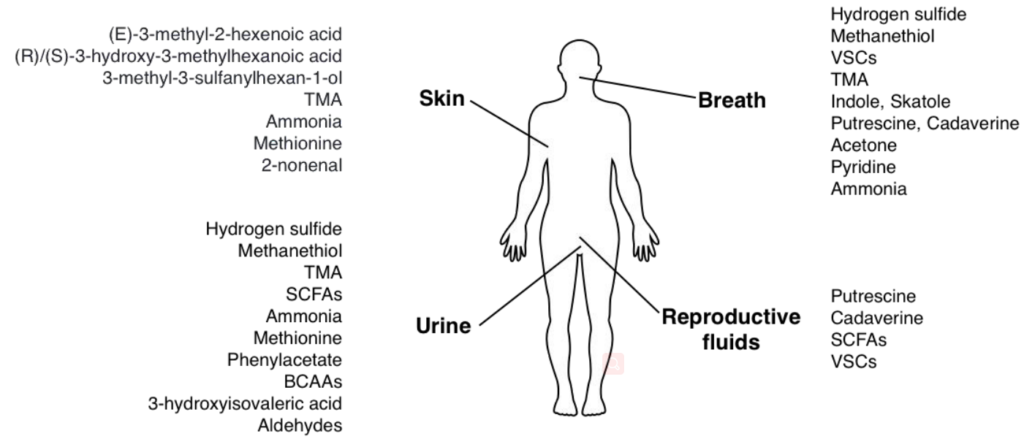
人体会通过呼吸,唾液,汗水(皮肤),尿液或生殖器官液体释放出有气味的物质,主要的气味物质是小的挥发性化合物。
越来越多的证据表明,微生物群在人类的生理过程中起着至关重要的作用。它可能通过产生大量有气味的分子(例如氨,挥发性硫化合物或三甲胺)来加剧人体臭味。
本文我们来讨论下关于体臭最常见的原因,以及潜在的治疗选择。
化合物的过度积累会产生难闻的气味。与体液中细菌代谢物积聚相关的体臭可能由以下一种或多种原因引起:

主要的细菌气味物质及其散发部位
Mogilnickaet al., Int J Mol Sci, 2020
接下来,我们针对身体各个部位气味及其原因逐一解读。
口臭(口腔之味或呼吸之味)是一种以口腔内或口腔外的口臭为特征的疾病。全世界约有20-50%的成人或青少年患有口臭。[更精确的数字,为31.8%(95%可信区间24.6–39.0%)]

在绝大多数(80-90%)持续口臭的患者中,口臭的原因是口腔原因,其中舌苔和牙周病最为普遍。
在口臭的口外(非口)原因中,还包括糖尿病酮症酸中毒,先天性代谢性疾病,胃肠道或呼吸疾病等。
当然,尽管口臭可能是一些严重疾病的征兆,但食用洋葱或大蒜等几种食物以及吸烟也可能导致口臭。
口臭患者呼吸中存在的主要物质是挥发性硫化合物,如硫化氢(H2S)、乙硫醇、硫代乙酸乙酯、二乙基二硫化物、二甲基硫化物((CH3)2S) 和甲硫醇(CH3SH或甲硫醇)。
一些革兰氏阴性菌与食物或唾液中含硫底物产生挥发性硫化合物有关,如:
福赛拟杆菌Bacteroides forsythus、牙龈卟啉单胞菌Porphyromonas gingivalis、放线共生放线杆菌Actinobacillus actinomycetemcomitans 、中间普雷沃氏菌Prevotella intermedia.
最常见的细菌气味
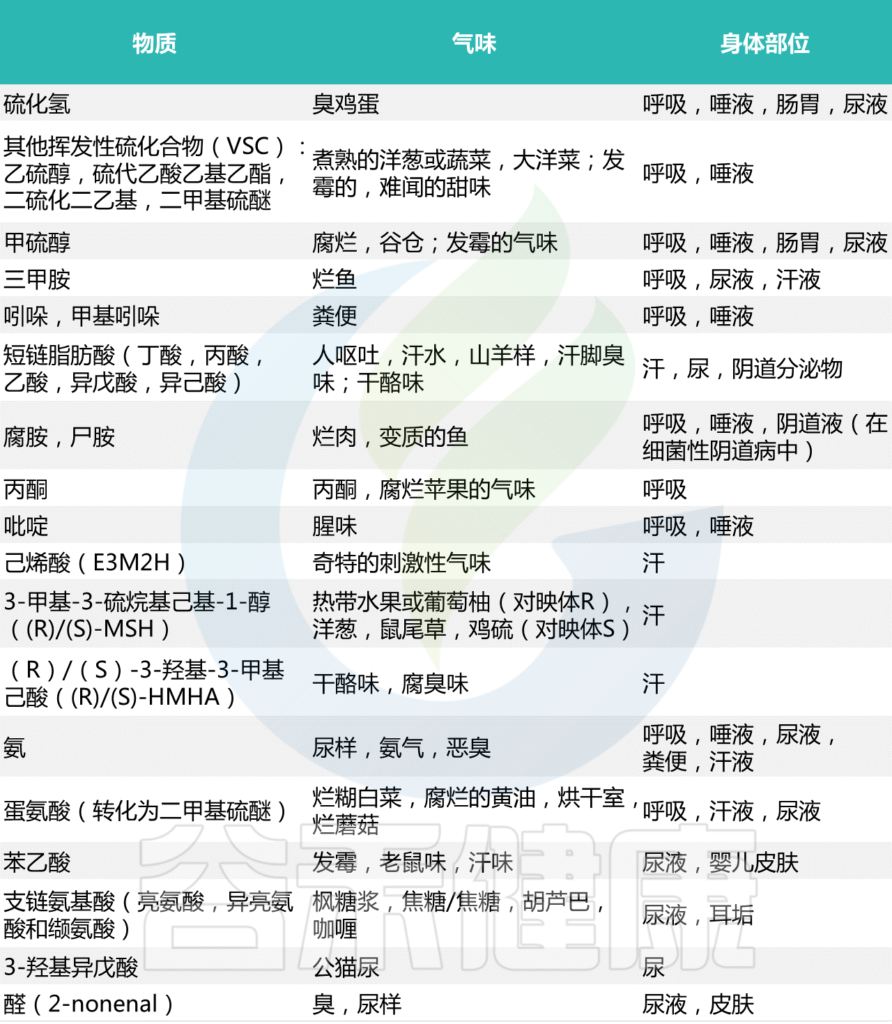
Mogilnickaet al., Int J Mol Sci, 2020
与恶臭相关的疾病
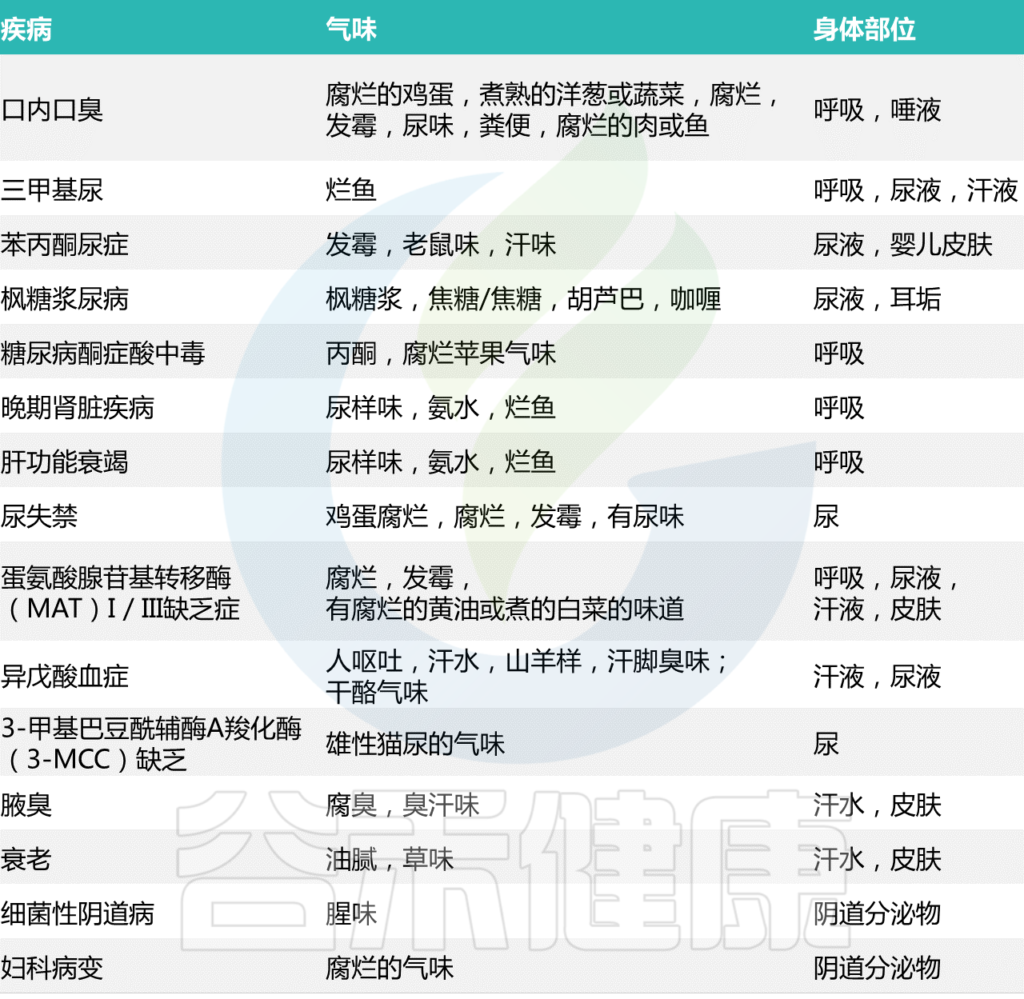
Mogilnickaet al., Int J Mol Sci, 2020
H2S——臭鸡蛋的气味
在低浓度下,硫化氢(H2S)是一种重要的生物介质,类似于其他气体递质,如一氧化氮或一氧化碳。然而,在较高的浓度下,H2S最出名的是它的毒性作用和特殊的臭味,经常被比作臭鸡蛋的气味。H2S即使浓度很低,也会产生明显的气味。
众所周知,口腔中存在的厌氧菌(龈下微生物群)可能会将含有氨基磺酸半胱氨酸的脱硫物质降解为有气味的挥发性硫化合物,而H2S是难闻呼吸气味的重要来源。
几十年前,已经报道了产生硫化氢的菌属,如:
多齿拟杆菌Treponema denticola 和中间拟杆菌Bacteroides intermedius,以及其他属如消化链球菌Peptostreptococcus、真细菌Eubacterium和梭杆菌Fusobacterium。
多年后确定,口腔异味患者以韦荣氏球菌属Veillonella、放线菌属Actinomyces、普雷沃氏菌属 Prevotella 为主。
另一方面有研究认为,在呼出高浓度H2S的患者口腔中占优势的菌:奈瑟氏球菌属 Neisseria、梭杆菌属Fusobacterium、卟啉单胞菌属 Porphyromonas.
而以CH3SH为主的患者口腔中以韦荣氏球菌属Veilonella和普雷沃氏菌属Prevotella为主。
在某些食品中,可能存在由细菌还原酶生产H2S的底物。例如,洋葱,大蒜,葡萄酒,卷心菜,花椰菜,蘑菇,坚果,土豆和干果含有硫化物和亚硫酸盐(用作抗菌和抗氧化剂)。
甲硫醇(CH3SH/MT/MeSH) ——腐败、霉味
在口臭患者的呼吸中检测到的另一种分子是甲硫醇。研究表明CH3SH是口腔内口臭的主要致病因素。MT的气味常被形容为腐臭、霉味,可比作谷仓气味。
事实上,口臭患者的恶臭通常更类似于臭鸡蛋的气味(H2S的特征)。客观存在的阈值为0.5 nM (12 ppb) 。与H2S类似,口外(血源性)口臭患者的口腔空气中也检测不到甲基叔丁基醚,而其他口腔鳞状细胞癌在其中起着至关重要的作用。
研究人员报告说,在口臭患者唾液中,普雷沃氏菌 Prevotella、韦荣氏球菌Veillonella、阿托波毕姆菌Atopobium、巨形球菌Megasphaera和硒单胞菌Selenomonas的丰度很高,并怀疑这些菌群参与CH3SH的产生。
对口腔恶臭患者口腔气体中的金属硫蛋白含量测量,发现在患有牙周病的个体中显著增加的浓度。
其他挥发性硫化合物——熟洋葱的甜味和霉味
口臭患者的呼吸和唾液中检测到的其他挥发性含硫化合物包括乙硫醇、硫代乙酸乙酯、二乙基二硫化物和二甲基硫化物。这些化合物与蔬菜独特的、甜的、发霉的气味有关,通常类似于熟洋葱的气味。
二甲基硫(DMS,CH3SCH3)是血液传播(口外)臭味的主要来源。相比之下,口腔内口臭患者口腔和鼻子呼吸中的二甲基硫浓度在正常范围内,未达到气味阈值。
血液中存在二甲基硫的可能原因是代谢紊乱。文献中报道的其他原因包括肝衰竭、高硫氨酸血症(一种主要与甲硫氨酸水平升高相关的遗传性甲基化障碍)和治疗性摄入二甲亚砜或半胱胺。
三甲胺——鱼腥味
三甲胺(TMA)是一种挥发性的脂肪族叔胺,因其腐烂鱼的特有气味和高浓度的毒性作用而闻名。它是由肠道细菌产生的过量胆碱和其他含TMA的膳食营养素形成的。
已经表明,许多肠道中的共生细菌通过代谢膳食中含有TMA的物质来产生TMA,
例如,厌氧球菌属Anaerococcus、普罗维登夏氏菌属Providencia、爱德华氏菌属Edwardsiella、梭菌属Clostridium、柯林塞拉氏菌属Collinsella、脱硫弧菌属Desulfovibrio、乳杆菌属Lactobacillus 和 变形杆菌属Proteus.
从肠道吸收后,TMA被肝脏氧化成几乎无味的三甲胺氧化物(TMAO)。在三甲基胺尿症(也叫TMAu或“鱼味综合征”)中,由于缺乏含鸟苷的单加氧酶3 (FMO3),一种氧化TMA的肝酶,TMA积累并排泄到体液中。
继发性(获得性)TMau在患有严重肝脏或肾脏疾病患者中。文献中还描述了儿童(与含胆碱的食物补充剂摄入有关)和妇女(与月经有关)的短暂形式。这种代谢障碍导致底物超载和独特的体臭,可以在包括呼吸在内的多个身体部位检测到。
吲哚和甲基吲哚——粪便的气味
吲哚代表一组微生物来源的化合物,由色氨酸产生,色氨酸是一种必需氨基酸,是色胺、血清素和褪黑激素内源性合成的前体。
被怀疑导致口臭的吲哚包括吲哚和甲基吲哚,它们是由口腔内厌氧革兰氏阴性菌产生的,如:
中间卟啉单胞菌Porphyromonas intermedia、具核梭杆菌Fusobacterium nucleatum和牙龈卟啉单胞菌Porphyromonas gingivalis.
它们的气味可以等同于粪便的气味,而这两种化合物的气味更浓。与挥发性含硫化合物相比,吲哚和甲基吲哚的挥发性要低得多,因此,它们对口臭的影响相当小。
然而,有些口臭患者呼吸中可能存在无法检测到的VSCs浓度以及高水平的吲哚,在这样一组患者中,常用的用于测定口气气味的工具(如检测VSCs的口臭计)可能不够充分并延误诊断。
腐胺和尸胺——腐肉或鱼的气味
腐胺和尸胺这两种二胺一直被怀疑是造成口臭的原因,都与牙菌斑中的细菌引起的食物腐败有关。
它们在唾液中通过氨基酸脱羧(分别是赖氨酸和鸟氨酸)或转氨作用产生。
腐胺的气味常被比作变质的鱼或腐肉的气味,而尸胺的气味除了上述气味之外,还可能会让人联想到尿液或精液的气味。口臭和唾液中存在尸胺有强相关性。
丙酮——水果味
丙酮是一种三碳挥发性酮,来源于乙酰乙酸酯(通过脱羧或酶促转化)。长期以来,它的水果味(常被比作烂苹果的味道)一直与糖尿病(DM)有关。
高浓度的呼吸丙酮与糖尿病酮症酸中毒有关。它也随着禁食、高脂肪或生酮饮食而增加。
吡啶——鱼腥味、汗味
吡啶是一种芳香的含氮挥发性化合物,有一种鱼腥味和汗味,可能会导致口臭。在中度和重度牙周病患者的培养唾液中发现了吡啶及其类似物(2-,3-和4-甲基吡啶)。相比之下,这些分子在口腔健康患者的样本中是不存在的。
氨——类似尿液的气味
氨(NH3)以铵离子(NH4+)的形式存在于所有体液中,但其高浓度是有毒的,因此它由尿素循环精确调节。氨在肝脏和肾脏疾病以及口臭中作为生物标志物发挥着重要作用。
去除舌苔和牙菌斑后呼吸中氨水平的降低,这可能表明口腔中存在的一些微生物是口腔内口臭中氨产生的原因。
在患有终末期肾病(ESRD)的患者中,尿毒症呼吸气味是由唾液中的高浓度尿素引起的,尿素被分解成氨。多达34.1%的ESRD患者主诉尿毒症恶臭。
肝衰竭历来与尿样胎儿肝性脑病(肝性呼吸)有关。比较肝硬化患者和非肝硬化患者的血氨和呼吸氨水平,患有肝性脑病的患者的呼吸氨水平明显高于对照组。此外,呼吸和血氨随着高氨血症的治疗而降低。
肠道细菌是哺乳动物体内氨形成的重要因素。胃肠道细菌(主要是革兰氏阴性肠杆菌科)产生脲酶,将尿素水解成二氧化碳和氨。其他细菌菌株,例如大肠杆菌和肠道沙门氏菌,能够通过半胱氨酸脱硫酶从半胱氨酸形成氨。
此外,大肠杆菌还可以将硝酸盐还原成氨。源自肠道的氨被肠道细菌用于蛋白质再合成,通过GBB(肠道-血液屏障)被吸收到循环中,或者随粪便排出体外。在正常情况下,肠道中产生的氨在肝脏中代谢。然而,在肝衰竭状态下,它不能转化为无毒衍生物,如尿素或谷氨酰胺。
因为氨也是在蛋白质分解代谢过程中形成的,所以可以假设富含蛋白质的饮食可能会增加其血液水平,并导致更多的口呼出氨。研究表明,蛋白质的摄入导致血清和唾液尿素增加,从而导致呼吸氨浓度增加。这一点需要认识到,尤其是因为许多流行的、不健康的时尚饮食是基于高蛋白的摄入。
尿液成分受身体新陈代谢的影响,但也受消耗的食物和饮料的影响。因此,需要强调的是,并不是尿液中发现的每一种气味都是不好的。
例如,在摄入芦笋后不久,有些人的尿液可能会有明显的硫磺味(想象煮熟的卷心菜)。虽然导致这种气味的确切分子还没有被明确地鉴定出来,但是一些类似甲硫醇或二甲基硫醚的挥发性物质是可疑的。
尿液中特殊气味化合物的鉴定可能有助于诊断苯丙酮尿症,高硫氨酸血症或枫糖尿症。值得一提的是,多种有气味的物质是由肠道菌群产生的,它们可以在从肠道吸收到循环系统后出现在尿液中。
下面将介绍几种影响尿液气味的物质。
H2S ——臭鸡蛋的味道
在尿失禁(UI,不自觉的尿液泄漏,显著影响生活质量)患者中,出现难闻的气味,也是造成社会尴尬的重要原因。
研究尿液和吸收性失禁垫主要挥发性气味物质,与甲硫醇和醛类等其他分子一起,硫化氢被检测到高于气味阈值。此外,在尿路感染中,大肠杆菌是恶臭硫化氢的常见产生者。
甲硫醇 ——腐败、霉味
健康人尿液中的气态甲硫醇低于检测阈值。在用半胱胺治疗的患者,肝甲硫氨酸腺苷转移酶缺乏症和UI 中发现水平升高。甲硫醇产生的主要因素是肠道细菌群,如大肠杆菌、柠檬酸杆菌和变形杆菌。甲硫醇从肠道吸收,进入循环,然后可以随尿液排出。
三甲胺——鱼腥味
在三甲胺尿中,过量的三甲胺不能被氧化成无味的TMAO,分泌到包括尿液在内的多种体液中。需要强调的是,一些患者只有间歇性TMAu,这使得建立正确的诊断更加困难,因为尿液测试可能在气味不明显的期间呈阴性,必须重复进行。
例如,来月经的女性应该在月经期间或之前进行测试,以最大限度地增加检测TMAu的机会,因为在此期间TMA的排泄会增加。
尿中TMA的另一个来源可能是肠道菌群,主要是
厌氧球菌Anaerococcus,普罗维登夏氏菌 Providencia, 爱德华氏菌Edwardsiella,梭状芽孢杆菌Clostridium, 柯林塞拉氏菌Collinsella, 脱硫弧菌Desulfovibrio,乳杆菌Lactobacillus,变形杆菌Proteus.
短链脂肪酸——干酪味
异戊酸血症(IA)通常与“汗脚”或人体呕吐物的特殊干酪样体臭有关,这在代谢危机期间可以注意到。这种疾病是异戊酰辅酶a脱氢酶(IVD)缺乏导致亮氨酸代谢异常的结果。在IA患者的尿液中可以检测到的病理物质是异戊酰甘氨酸。
氨——类似尿液的气味
氨是第一批被认为会在尿失禁患者周围引起恶臭的分子之一。根据这一概念,细菌尿素酶(主要是大肠杆菌、奇异变形杆菌Proteus mirabilis和粪肠球菌 Enterococcus faecalis)会将尿素分解成难闻的氨。然而,也有研究人员对这一假设提出质疑。
蛋氨酸及其代谢物——腐臭黄油或煮卷心菜的气味
甲硫氨酸腺苷转移酶(MAT) I/III缺乏是甲硫氨酸代谢的遗传性错误,主要在新生儿筛查中发现。它是由导致蛋氨酸及其代谢物积累的MAT1A基因突变引起的。是持续性孤立性高硫血症最常见的原因。
苯乙酸盐——霉味、鼠味
另一个导致尿液有独特的类似老鼠气味的原因是苯丙酮尿症(PKU)。
支链氨基酸(亮氨酸、异亮氨酸和缬氨酸)及其酮酸——焦糖或枫糖浆的气味
枫糖浆尿病(MSUD,亮氨酸病)是另一种代谢紊乱,其症状之一是有明显的尿气味。
3-羟基异戊酸——雄性猫尿的气味
在3-甲基巴豆酰辅酶a羧化酶(3-MCC)缺乏的其他症状中,一些研究人员报告了尿液的特殊气味。
醛类(乙醛、丁醛、异戊醛)——类似尿液的气味
在尿失禁患者的尿液样本和尿失禁卫生垫中检测到少量有气味的醛类物质。据报道,乙醛、丁醛、异戊醛(具有恶臭、类似尿液的气味)的浓度高于检测阈值,可以假设它们有效地增加了UI患者的气味强度。尿醛的来源至少部分是肠道及其微生物群。
从皮肤表面散发出的气味分子大多来自汗液,汗液是汗腺分泌的产物。人体汗腺主要分三种类型:大汗腺、小汗腺和大小汗腺(混合型)。
小汗腺产生大量的汗液,主要含有水和电解质,它们几乎分布在整个身体表面。
大汗腺大多位于腋窝、会阴、生殖器区域和乳头周围。它们在青春期后变得活跃,分泌的汗液比外分泌腺少。
大汗腺分泌的汗液类似于小汗腺产生的汗液,但其分泌速率较高,尤其出现在腋窝部位。这些特征使得大汗腺成为腋窝出汗的重要原因。
顶泌汗液是无味的,它只有被皮肤表面的微生物分解后才会变得有气味,如微球菌科Micrococcaceae、好氧类白喉菌aerobic diphtheroids 和丙酸杆菌Propionibacteria。
激素平衡的变化、消耗的食物和新陈代谢的变化可能会对汗水的数量和质量构成产生影响。
皮肤微生物群的任何变化以及细菌感染都可能改变汗液的成分,通常会产生独特的气味,因为人类有机体与几种能够转化汗液化合物的微生物物种保持共生关系。
据报道,在链球菌性皮炎中,患者皮肤有一种独特的臭味。
此外,许多代谢疾病的特征是皮肤散发出各种气味。其中一些疾病包括苯丙酮尿症,甲硫氨酸吸收不良综合征,高硫氨酸血症,或TMAu。
下面将讨论已知会导致皮肤散发气味的物质。
E3M2H,HMHA,MSH——发臭、发臭的汗味
体臭,是一种令人尴尬痛苦的疾病,特别是在腋窝、生殖器或脚等部位。
这三种类型的汗腺都在这种疾病的发病机制中起作用。过度出汗后,细菌分解汗液成分,会产生难闻的汗味。与小汗腺腋臭相反,大汗腺腋臭在青春期后出现,是这种疾病最常见的形式。
细菌将顶泌汗液分解成许多挥发性分子,如氨和短链脂肪酸,例如(E)-3-甲基-2-己烯酸(E3M2H) ,这是一种C7支链和不饱和酸。它有一种非常强烈的刺鼻气味。
Natsch等人报道了有气味的E3M2H及其水合类似物,(R)/(S)-3-羟基-3-甲基己酸((R)/(S)-HMHA)是由位于腋窝皮肤上的共生棒状杆菌物种的特定锌依赖性N-α-酰基-谷氨酰胺氨酰酶(N-AGA)从谷氨酰胺缀合物(存在于腋窝分泌物中)中释放出来的。
HMHA(以腐臭、奶酪味为特征)是最丰富的。3-甲基-3-磺酰己-1-醇的(S)-异构体((R)/(S)-MSH)及其洋葱味和类似鼠尾草的气味是引起腋臭的另一种颗粒。
此外,多种因素与小汗腺腋臭有关。这些包括摄入某些食物产品,如大蒜或洋葱,角蛋白的细菌降解,代谢紊乱和多汗症。
三甲胺——鱼腥味
在TMAu患者中,过量的未代谢的三甲胺也会从皮肤表面渗出(伴有汗液),引起特有的鱼腥味体臭,无论患者的个人卫生状况如何,都会有这种体臭。
在对患有特发性恶臭的患者进行的一项研究中,约三分之一的患者三甲基胺尿检测呈阳性。在这些个体中,自我识别的体臭是主要症状(29.9%),其次是体臭和口臭(21.4%)。
然而,只有5%的TMAu阳性患者确实在手掌上检测到明显的恶臭,并且他们中没有人在社交距离处发出明显的体臭。在摄入胆碱(合成TMA的一种底物)后,多达10%受试者在社交距离内有明显的体臭。
氨——类似尿液的气味
研究表明,汗液中含有氨。然而,汗液中的氨来源尚未得到证实。一些研究人员认为它是从血浆中运输的,而其他人则认为它直接来自汗腺。
比较呼吸和汗液中氨浓度的研究结果也没有定论。一项研究表明,有气味的氨的皮肤释放量高于呼气。另一项研究则认为,皮肤气体中的氨含量比呼吸中的低。
蛋氨酸及其代谢物——腐臭黄油或煮卷心菜的气味
在新生儿筛查中未确诊的蛋氨酸腺苷转移酶I/III缺乏症(导致高蛋氨酸血症)患者,不仅在呼吸或尿液中,而且在汗液中可能会发现类似水煮卷心菜或腐臭黄油的特殊气味。这种独特的气味很可能是由蛋氨酸形成的有气味的二甲基硫引起的。
2-壬烯醛–油腻、青草味
自然身体气味的化学成分、强度和宜人性在一生中都会发生变化。众所周知,老年人有一种特殊的体臭,有时称“老人气味”。
研究人员对与衰老相关的体臭变化进行研究,发现了一种特殊的粒子,2-壬烯醛,它是老年人和中年人身体气味的特征。
这种不饱和醛具有独特的油腻和青草气味,是由皮肤表面脂质中的ω-7单不饱和脂肪酸降解产生的。

正常阴道分泌物:几乎无气味或有类似酸奶的气味。
与生殖液体一起释放的干酪味或鱼腥味可能是位于生殖器区域的感染(例如,细菌性阴道病、滴虫病或念珠菌病)以及非感染疾病(例如尿失禁、恶性溃疡、三甲基铵尿症或慢性便秘)的症状。
细菌性阴道病是阴道气味最常见的原因。其他症状包括阴道分泌物、瘙痒和刺激。这种情况下阴道液的异常腥味是由阴道加德纳氏菌Gardnerella vaginalis 等细菌产生的挥发性胺(腐胺、尸胺、TMA)引起的。在大多数情况下,细菌性阴道病可以用抗生素迅速治愈。然而,在大约三分之一的患者中,常规治疗并不能改善症状。
阴道气味的非感染性原因比感染性原因更不常见,因此它们构成了更大的临床挑战。医生也应该认识到,阴道的气味可能有不同的来源,如肛管或尿道。生殖器汗味的难闻气味也可能被误认为是阴道气味。
胃肠疾病,如慢性便秘和大便失禁,也应被视为肛门生殖器区域散发恶臭的可能原因。在这些状态下,挥发性有气味的化合物包括硫化氢、甲硫醇和二甲基硫化物,它们是造成屁和粪便气味的原因。这些患者的阴道检查不会显示任何异常。
最后,一些妇科肿瘤和病变患者抱怨阴道分泌物带有恶臭。例如,伴有坏死的外阴恶性溃疡可能导致腐烂气味,这可能是由于细菌形成腐胺、尸胺、短链脂肪酸(异戊酸和丁酸)和含硫化合物。
遗憾的是,缺乏管理体臭的循证指南,也没有普遍的治疗方法。医学文献中已经讨论了几种临时解决方案,如刷牙、漱口、嚼口香糖或经常用抗菌肥皂洗澡和使用除臭剂。然而,这些方法不能解决潜在的问题,而是掩盖或减少不愉快的气味到可接受的水平。只有把原因考虑进去,才能取得令人满意的结果。
一般来说,与体液中细菌代谢物积累相关的恶臭是由气味物质的合成和排泄之间的不平衡引起的。
可能是由以下一个或多个因素引起的:
(1) 含有气味物质或其产生底物的饮食
(2) “前气味”细菌组成和细菌的代谢活性
(3) 增加从肠道对气味物质或其前体的吸收(增加肠道转运时间,增加肠道-血液屏障通透性)
(4) 减少气味物质的肝脏代谢
(5) 减少尿排泄
基于以上所述,除了标准的卫生程序之外,还可以建议一些一般的但不是基于证据的预防措施。
1. 减少产生气味的食物基质。
2. 经常排便以减少通过时间(缩短肠道细菌代谢时间和细菌代谢物吸收时间),治疗便秘。
3. 益生菌和益生元治疗(尝试改变肠道细菌组成)。
4. 多喝水,以增加代谢物随尿液的排泄。
下表总结了与细菌气味剂有关体味的治疗选择。根据气味类型进行管理:

Mogilnickaet al., Int J Mol Sci, 2020
目前为止,已经确定了几种导致体臭的化合物,其中大多数是细菌来源的。虽然没有基于证据的管理身体恶臭的指南,一些针对饮食、肠道菌群组成的治疗措施可能会减轻症状。需要提高临床认识以及进一步的研究来解决体臭问题。
相关阅读:
肠道微生物组如何影响运动能力,所谓的“精英肠道微生物组”真的存在吗?
参考文献:
Li, M.; Al-Sarraf, A.; Sinclair, G.; Frohlich, J. Fish odour syndrome. CMAJ 2011, 183, 929–931.
Rath, S.; Heidrich, B.; Pieper, D.H.; Vital, M. Uncovering the trimethylamine-producing bacteria of the human gut microbiota. Microbiome 2017, 5, 54
Piotr, K.; Marcin, U. Indoles – Gut Bacteria Metabolites of Tryptophan with Pharmacotherapeutic Potential.Curr. Drug Metab. 2018, 19, 883–890
Mogilnicka Izabella,Bogucki Pawel,Ufnal Marcin,Microbiota and Malodor-Etiology and Management.[J] .Int J Mol Sci, 2020, 21.
Bielinska, K.; Radkowski, M.; Grochowska, M.; Perlejewski, K.; Huc, T.; Jaworska, K.; Motooka, D.; Nakamura, S.; Ufnal, M. High salt intake increases plasma trimethylamine N-oxide (TMAO) concentration and produces gut dysbiosis in rats. Nutrition 2018, 54, 33–39
Suzuki, N.; Nakano, Y .; Watanabe, T.; Yoneda, M.; Hirofuji, T.; Hanioka, T. Two mechanisms of oral malodorinhibition by zinc ions. J. Appl. Oral Sci. 2018, 26, e20170161