-

 CNAS L23010
CNAS L23010

 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业 二级病原微生物安全实验室
二级病原微生物安全实验室- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业 二级病原微生物安全实验室

谷禾健康

肠道微生物除了在肿瘤发生过程有重要作用,在抗癌治疗过程中也有着不可忽视的价值。
现有的癌症疗法很多,化疗、放疗等方式虽然有效,但副作用较多;免疫疗法有其局限性,有效率低且针对癌症类型有限。研究表明,肠道微生物群的调节不仅有助于增强机体免疫力,抑制肿瘤细胞的增殖和侵袭,还可以改善治疗的耐受性和减轻不良反应。
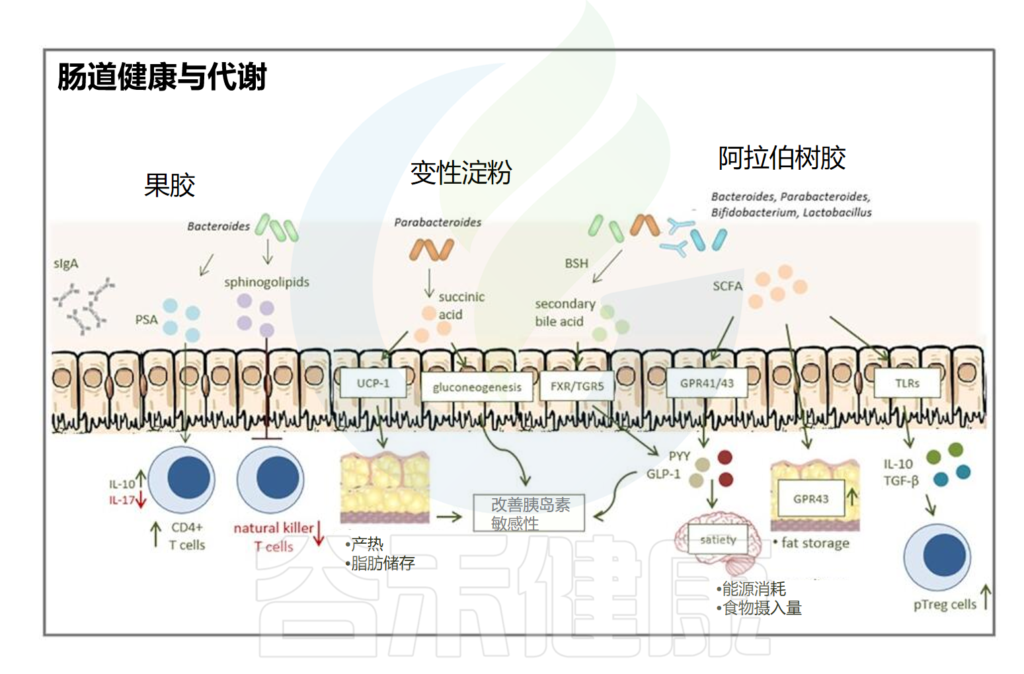
例如,研究发现化疗中伊立替康的毒性取决于肠道微生物群的组成;
毛螺菌科、肠球菌科通过产生短链脂肪酸减轻放疗毒性;
在晚期黑色素瘤患者中,双歧杆菌科、瘤胃球菌科、毛螺菌科等关键细菌与抗PD1治疗后的良好结果有关。
因此,如何通过将肠道微生物群结合现有的癌症治疗方式,更好地促进肿瘤患者的康复和疗效,值得深入挖掘和探讨。
本文我们就来详细了解一下,微生物群与各类癌症疗法,最主要是免疫疗法及其与免疫相关的不良反应。根据患者微生物群组成训练的机器学习方法,可以准确预测患者对免疫疗法的反应能力。
粪菌移植或饮食、益生元等干预可在临床上用于提高癌症患者免疫治疗的成功率,这里讨论了基于微生物群的癌症治疗的新策略。
理解肠道微生物群与肿瘤治疗之间的相互关系,可以为创新的治疗方法提供有力的支持,并为肿瘤治疗领域带来新的突破。
化疗是指使用药物来杀死或控制癌细胞的生长。这些药物可以通过静脉注射、口服或局部应用的方式给予患者。
缺点
由于化疗中使用的药物可能对癌症没有特异性,无法区分细胞好的坏的,会影响身体的其他健康组织,引起副作用,比如脱发、口腔溃疡、恶心、精疲力竭等。
肠道菌群影响化疗药代动力学、疗效和毒性
一个代表性的例子是:伊立替康的毒性取决于肠道微生物群的组成。
注:伊立替康用于治疗结直肠癌的拓扑异构酶 I 抑制剂。
肝脏中:
葡萄糖醛酸转移酶将活性形式的伊立替康 SN-38 解毒成无活性的 SN-38-G。
肠道中:
许多表达 β-葡萄糖醛酸酶的细菌将 SN-38-G 转化回 SN-38,从而引发腹泻。
这种毒性可以通过广谱抗生素或 β-葡萄糖醛酸酶抑制剂来预防。
肠道菌群可优化肿瘤治疗、增强免疫抗癌效果
肠道微生物群可以通过提供有利于药物对癌症细胞毒性作用的肿瘤微环境,并在药物诱导的免疫原性细胞死亡后维持抗癌适应性免疫,从而调节化疗效果。
奥沙利铂、顺铂
无菌或抗生素处理的小鼠中,铂基化合物奥沙利铂和顺铂显示出降低抗肿瘤功效。这些药物仍然可以在微生物群耗尽的小鼠的肿瘤中形成铂-DNA加合物,但没有观察到DNA损伤。肠道微生物群通过NOX2(NADPH氧化酶2)启动肿瘤中的髓细胞产生活性氧(ROS),这是铂治疗诱导的DNA损伤所需的。
肠道微生物发酵的膳食纤维产物——丁酸盐,可以通过 IL-12 信号传导调节 TME 中 CD8+ T 细胞的功能,从而增强奥沙利铂的抗癌作用。
烷化剂环磷酰胺(CTX)
同样,烷化剂环磷酰胺疗效也取决于肠道菌群。
注:烷化剂环磷酰胺既是一种化疗剂,也是一种免疫抑制剂。
CTX减少了小肠中厚壁菌门和螺旋体门,同时增加了其他细菌类群的丰度,其中一些细菌转移到肠系膜淋巴结。CTX诱导免疫原性肿瘤细胞死亡,这依赖于微生物群调节的致病性辅助T细胞17(TH17)和记忆性TH1细胞。
CTX治疗后, Enterococcus hirae移位至淋巴结和Barnesiella intestinihominis在结肠中的积聚促进了癌症免疫。
此外,接受化疗免疫治疗的癌症或卵巢癌患者的存活率与E. hirae和B. intestinihominis特异性TH1反应相关。
吉西他滨
吉西他滨是胰腺导管腺癌常用的化疗药物。肠道微生物参与化疗药物的药代动力学,吉西他滨治疗胰腺导管腺癌的疗效可能受到肠道微生物的影响。
γ-变形菌能够代谢吉西他滨并将其转化为无活性的2′,2′-二氟脱氧尿嘧啶。因此,未来有可能通过将抗γ-变形菌的抗生素与化疗相结合来提高吉西他滨的抗癌效果。
除了负面影响外,肠道微生物代谢产物丁酸盐,还可以通过诱导细胞凋亡来增强吉西他滨对抗癌症细胞的功效。
除了肠道微生物群,肿瘤内细菌促进肿瘤生长并干扰化疗效果,这表明:
肿瘤内微生物群可能是癌症治疗的靶向菌
肿瘤内微生物群已显示在人类乳腺癌、骨癌、胰腺癌、卵巢癌、肺癌、黑色素瘤、胶质母细胞瘤以及原发性和转移性结直肠癌中。肿瘤微生物群可能在肿瘤发生、肿瘤进展和治疗反应中发挥着重要作用。
肿瘤相关γ-变形菌表达细菌胞苷脱氨酶,该脱氨酶使细胞毒性药物吉西他滨失活;使用抗生素环丙沙星可以清除细菌,恢复对吉西他滨的敏感性。然而,吉西他滨治疗也通过增加变形杆菌的丰度来改变肠道微生物群。
↓↓↓
因此,抗生素治疗是否通过对肿瘤相关的γ-变形菌或其他与肠道或肿瘤相关的微生物成分的特异性影响来影响肿瘤生长,目前没有确切的证据。
*************************************************************************
小鼠乳腺癌模型:
通过循环肿瘤细胞携带的肿瘤内细菌,重组了细胞骨架,促进了宿主细胞的存活。通过肿瘤微生物群落中分离的菌株直接注入肿瘤,增强了转移形成。
在原发性人类结肠癌和远处转移中检测到:
——梭杆菌门(Fusobacteria)
使用甲硝唑抗生素治疗可以消除梭杆菌,并减小肿瘤生长速度。
肝癌和肝硬化患者:
肝脏中Stenotrophomonas maltophilia丰度的增加诱导了肝星状细胞的细胞衰老相关的分泌表型(SASP),从而促进了肝癌发生。
↓↓↓
这些数据表明,抗生素有可能靶向肿瘤微生物群,但抗生素治疗的全身效应与肿瘤局部效应可能是拮抗的,特别是考虑到抗生素对免疫检查点阻断的有害影响。
*************************************************************************
细胞内细菌可能逃避RAB11介导的吞噬作用,并寄居于稳定的含有细菌的液泡中,使其对抗生素无效。
↓↓↓
因此,使用细胞穿透肽和核酸可能会最大限度地增强抗生素对肿瘤相关细菌的效力。
肠道微生物群调节癌症治疗,可以有针对性地提高治疗效果并预防不良反应
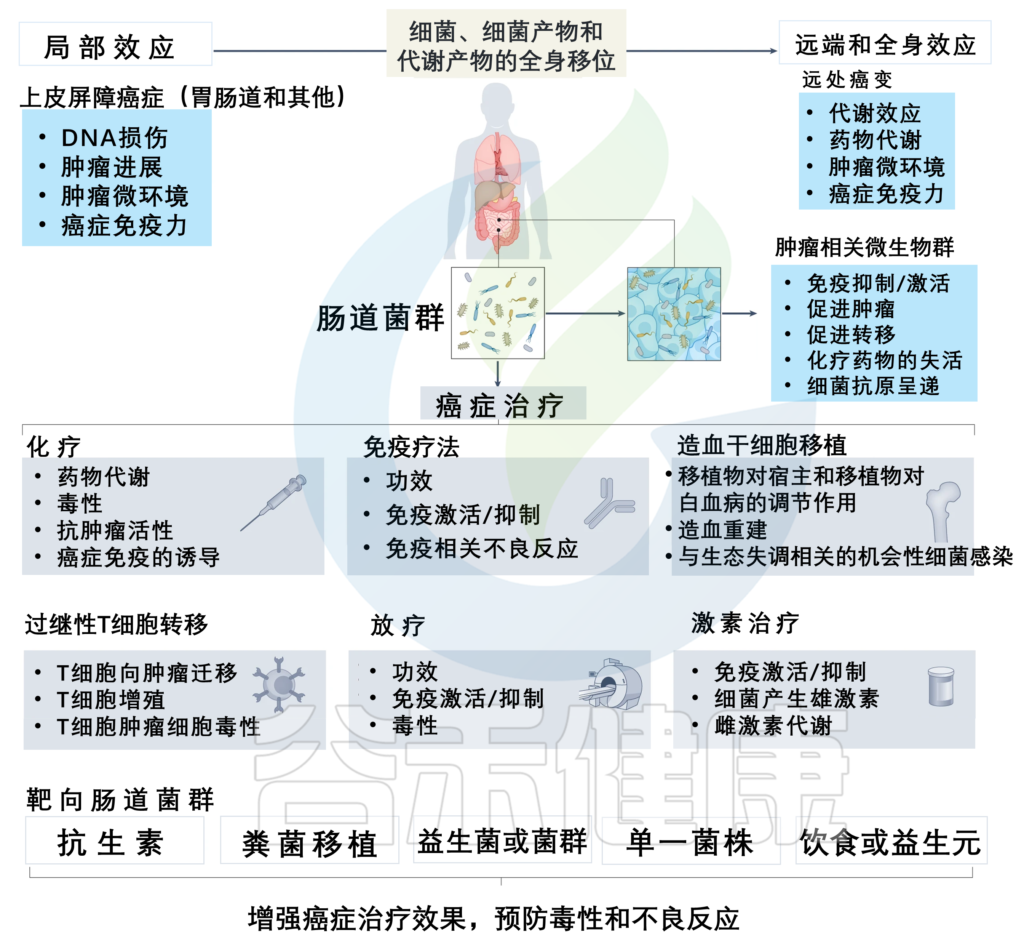
doi: 10.1038/s41568-022-00513-x
放射治疗在于缩小或破坏肿瘤,但过程不同。
辐射是一种高能不可见光波,可以针对癌细胞破坏其遗传物质,最终杀死它们。辐射波会损害癌细胞,癌细胞会随着时间的推移而死亡并被身体清除,从而导致肿瘤缩小。
与化疗不同,放疗通常是一种局部治疗,也就是说一般不会传播到全身。
缺点
如果附近的健康组织在治疗过程中受损,局部放射可能会产生副作用,比如恶心、口腔溃疡、喉咙问题导致进食困难、皮肤干燥、精疲力竭等。
不同癌症类型在毒性和敏感性方面存在差异,这是影响电离放射治疗安全性和有效性的主要障碍。
肠道微生物群的细菌和真菌成分都可能导致患者间的异质性
在实验动物中,对革兰氏阳性细菌有选择性的抗生素万古霉素通过消耗产短链脂肪酸的肠道细菌,提高RTX的有效性,并增强树突状细胞抗原呈递和癌症免疫。
向万古霉素处理的小鼠施用丁酸盐,部分通过抑制树突状细胞功能,包括它们产生IL-12的能力,消除了这种作用。
使用广谱抗生素治疗肠道微生物群会导致酵母菌类真菌肠道扩张,通过β-葡聚糖受体Dectin 1的信号传导抑制肿瘤免疫,从而降低RTX有效性。使用抗真菌药物治疗可提高未经治疗小鼠的RTX有效性,尤其在抗生素治疗的对RTX反应较差的小鼠中。
在乳腺癌患者中,肿瘤内髓细胞中Dectin1的表达与生存率呈负相关,这一观察结果表明真菌免疫抑制作用可能与临床相关。
短链脂肪酸在癌症治疗中的复杂功能:对抗毒性 与 免疫调节
传统饲养的小鼠比无菌小鼠更容易受到全身照射(TBI)毒性的影响,因为微生物群抑制ANGPTL4的表达。
注:ANGPTL4,血管生成素样4,也称为FIAF,是一种支持组织修复的脂蛋白脂肪酶抑制剂。
然而,产短链脂肪酸菌通过诱导ANGPTL4,来保护接受RTX治疗的患者免受结肠炎和粘膜炎的侵袭。
毛螺菌科、肠球菌科→产丙酸→减轻毒性
在全身照射存活的小鼠中,产生色氨酸代谢产物和短链脂肪酸(特别是丙酸盐)的细菌,毛螺菌科(Lachnospiraceae)和肠球菌科(Enterococcaceae)大量存在,并且可以将对TBI毒性的耐药性转移到其他小鼠。
在进行异基因造血干细胞移植(allo-HSCT)预处理过程中,观察到患有白血病的患者的粪便样本中存在着毛螺菌科(Lachnospiraceae)和肠球菌科(Enterococcaceae)。这些患者在接受全身放疗治疗(TBI)时出现的肠道毒性较轻。
这些发现可以解释为什么产生短链脂肪酸的益生菌,如乳酸杆菌和双歧杆菌,可以预防TBI毒性。
总体而言,几项研究表明,不同的短链脂肪酸可能对癌症免疫和治疗毒性产生复杂影响,包括粘膜保护作用,部分由ANGPTL4、IL-18和IL-22的诱导介导,以及通过Treg细胞诱导和抑制树突状细胞功能介导的对立的免疫调节效应。
因此,短链脂肪酸在放疗和其他癌症治疗中的功能似乎很复杂,需要进一步研究。
最新的癌症疗法利用宿主免疫系统来对抗癌症。免疫细胞通过识别和消灭癌细胞来抑制肿瘤的发生。
CTLA-4充当淋巴结中 T 细胞激活的“关闭”开关,而 PD-L1/PD-1降低肿瘤部位的晚期免疫反应。
免疫检查点抑制剂(ICI) 是一种单克隆阻断抗体,可以释放免疫系统的“刹车”,使免疫细胞能够更有效地识别和攻击癌细胞。
这种方法给一些难治癌症患者带来了显著的持久缓解,如晚期和转移性黑色素瘤、肾细胞癌、非小细胞肺癌等。目前用于超过25种不同的肿瘤类型。
缺点
免疫治疗(ICI)每年每个患者的费用超过10万元,成功率仅为20%-40%,同时许多患者会出现与免疫相关的不良事件。
因此,迅速发展的需求突显了将免疫疗法的有效性扩展到所有癌症患者的必要性。
微生物群维持对病原体的抵抗力和疾病耐受性,防止对无害抗原的过度炎症反应。
饮食、生活方式、抗生素和其他药物使用的变化可能会改变这种健康保护生态平衡。与对急性感染的抗原特异性耐药性不同,肿瘤通过劫持疾病耐受机制抑制癌症免疫。因此,肠道微生物群的改变,可能会影响患者对免疫疗法的反应和免疫相关不良事件(irAE)的程度。
过继性T细胞疗法(ACT):万古霉素增强ACT抗肿瘤,其他抗生素降低抗肿瘤
万古霉素的治疗增加了肠道中变形杆菌门的丰度,但耗尽了厚壁菌门和拟杆菌门的大多数物种,增强了ACT在小鼠中的抗肿瘤作用。
而其他抗生素,特别是新霉素和甲硝唑,分别消耗革兰氏阴性需氧菌和厌氧菌,降低了ACT的抗肿瘤作用。
万古霉素修饰的微生物群诱导产生刺激T细胞的IL-12,维持过继转移的细胞毒性T细胞。
Toll样受体激动剂:CpG-ODN、ICB疗法
革兰氏阴性菌与诱导坏死的CpG-ODN的有利先天反应,以及维持适应性CD8+T细胞介导的抗癌免疫的肿瘤微环境的重定向有关。
ICB 疗法增强的适应性免疫(如后面所讨论的)与几种革兰氏阳性细菌的存在相关,包括:
在这种情况下,革兰氏阴性菌与免疫抑制性局部和全身炎症状态有关。
在一项临床试验(NCT03618641)中,证明了 CpG-ODN和ICB组合的功效,抗PD1治疗下,初始CpG-ODN诱导的坏死和维持无复发生存,与革兰氏阴性细菌(变形菌门、拟杆菌门和革兰氏阴性Oscillospiraceae)的粪便丰度相关。
免疫检查点阻断
一般来说,只有20-40%的患者对ICB有反应,因此需要确定响应标志物并开发克服耐药性的治疗方法。
而肠道微生物群已成为预测ICB响应的肿瘤外部生物标志物。
CTLA4
➦ 无菌小鼠对抗CTLA4治疗反应不佳。
➥ 向微生物群耗尽的小鼠注射多形拟杆菌(Bacteroides thetaiotaomicron)或脆弱拟杆菌(Bacteroides fragilis),通过诱导肿瘤中的DC成熟和引流淋巴结中的TH1反应,恢复抗CTLA4反应。
PDL1
相反,小鼠对肿瘤生长的抵抗力和对抗PDL1的反应性与几种双歧杆菌的粪便丰度有关。
在晚期黑色素瘤患者中,放线菌门(双歧杆菌科和Coriobacteriaceae)和厚壁菌门(瘤胃球菌科和毛螺菌科)的关键细菌种类的粪便丰度与抗PD1治疗后的良好结果有关。
在肺癌、肾细胞癌或肝细胞癌患者中,粘蛋白降解的Akkermansia muciniphila在抗PD1治疗中发挥抗肿瘤作用。然而,在一些接受抗生素治疗的患者中,粪便中A. muciniphila的高丰度与抗PD1治疗的耐药性有关,这可能表明AKK菌在抗PD1疗法反应中具有双峰效应。
在抗PD1治疗无反应的患者粪便中,溶血性葡萄球菌和Corynebacterium aurimucosum的更高,在有反应的肾癌和肺癌患者中,E. hirae菌的粪便频率更高。
研究中发现的细菌种类的一致性有限,可能是由于ICB介导的免疫机制的变化。
TIM3
抗生素治疗对抗TIM3癌症治疗的损害,也表明了肠道微生物群在维持这种类型的ICB的疗效中的作用。
肠道菌群、血液相关菌群和肿瘤相关菌群的组成不同,对不同类型的癌症具有特异性,导致不同的细菌种类对免疫治疗的反应产生影响。
对相同类型肿瘤和相同ICB治疗的患者队列的研究,即使仍然受到队列内微生物群异质性的影响,也报告了更一致的结果。
事实上,复杂微生物群落中的未知变异阻碍了稳健的特征识别。未来的研究依赖于细菌亚种和菌株的新特征,并分析单个细菌基因或途径的关联,将更好地识别特定细菌与治疗反应的关联及其作用机制。
肠道菌群对抗PD1免疫检查点阻断反应的调节
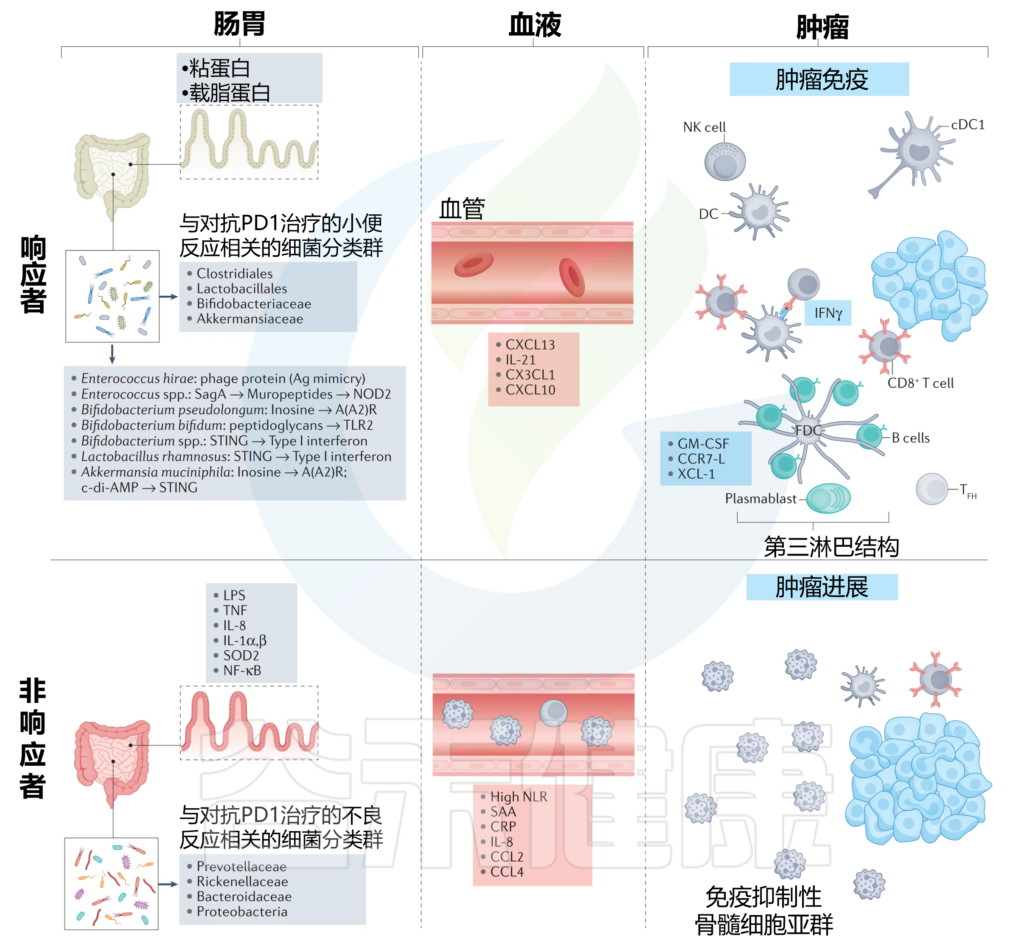
doi: 10.1038/s41568-022-00513-x
在大多数接受抗PD1治疗的癌症患者的研究和荟萃分析中,与有利或不利治疗反应普遍相关的细菌已得到一致定义。
在对抗PD1治疗有反应的患者中观察到一种健康、无炎症的肠粘膜,其特征是编码抗炎载脂蛋白和跨膜粘蛋白的转录物。在这些患者的血液中,参与T细胞和B细胞募集的细胞因子和趋化因子升高,这对抗肿瘤免疫很重要。左上角列出了与抗PD1治疗的有利反应相关的确定细菌种类及其在调节抗肿瘤免疫中的作用机制。
在对抗PD1治疗没有反应的患者中,观察到肠粘膜发炎和炎症细胞脱落,而在他们的血液中,通常存在全身炎症的生物标志物。所列的肠道革兰氏阴性菌群与抗PD1治疗的不良反应以及局部和全身炎症的生物标志物有关,可能反映了免疫抑制肿瘤微环境。
早期临床反应取决于肿瘤内在因素或宿主因素;而肠道菌群的调节作用在治疗开始约1年后更具影响力
一项针对黑色素瘤患者的抗PD1治疗研究报告称,较高的肠道微生物群α多样性与良好反应之间存在关联。但没有更多研究证实,α多样性用于区分健康个体和癌症患者,它不能预测对免疫疗法的反应性。
对已发表的接受抗PD1治疗的黑色素瘤患者队列的数据进行的荟萃分析显示,不仅在单个物种水平上,而且在更大的分类分支中,识别出的细菌之间存在共性。
大多数研究认为有利的微生物群特征,包括放线菌门的多个成员,以及在厚壁菌门中,瘤胃球菌科和Lachnospiraceae科。
而一个不利的特征包括拟杆菌门和变形菌门。
对黑色素瘤患者的免疫治疗反应进行的两项荟萃分析使用了大队列,研究发现,肠道微生物群的基线组成在3个月时进行放射学评估时,区分有反应的患者和没有反应的患者的能力有限,但它可以更好地区分在治疗开始后约1年有进展或没有进展的患者。此外,长期无进展生存期患者在开始抗PD1治疗后数年保持良好的微生物群。
为什么没有在队列中一致地识别出某些菌作为生物标志物?
一项研究的样本来自三个欧洲国家和一些公开的数据集,并包括用抗CTLA4、抗PD1或两者联合治疗的黑色素瘤患者。这里涉及到,地理和治疗类型的异质性。
即使同卵双胞胎,每个个体的肠道微生物组都是独特的,并且这种独特性的很大一部分是在单个菌株水平上编码的。需要分析方法更深入探索这种个体特异性的微生物多样性。
不同地理区域的肠道微生物类型不同,因为人类肠道微生物组受到种族和地理起源的影响。利用美国肠道项目的7000多个样本,对微生物类型进行了地理测绘,揭示了美国各地理区域的分布不均。
接受抗PD1治疗的黑色素瘤患者队列中的微生物类型代表性也不一样,并反映了他们的地理环境。此外,对抗PD1治疗有反应的概率与肠道微生物类型相关。
因此,通过微生物类型视角分析肠道微生物群与免疫疗法反应之间的关系,为先前研究的不一致性提供了可能的解释,并可作为纠正地理和临床中心特异性偏见的工具。
肠道菌群中的肽聚糖酶介导的抗癌
一些肠球菌在小鼠中维持ICB作用的能力是由一簇肽聚糖水解酶介导的,包括分泌抗原a(SagA),它产生NOD2激活的胞壁肽(muropeptides),增强抗PDL1诱导的肿瘤免疫。
当给予小鼠时,SagA转染的乳酸乳球菌抑制肿瘤生长并提高抗PD1反应性。某些与小鼠抗PD1和奥沙利铂癌症治疗协同作用的双歧双歧杆菌菌株富集了肽聚糖合成基因,通过TLR2刺激癌症免疫。
肠道菌群调控干扰素
肠道菌群维持I型干扰素表达,增强肿瘤免疫
在微生物群耗尽的小鼠中,肿瘤内先天免疫细胞中的干扰素信号减弱,改变巨噬细胞极化、树突状细胞分化和DC-自然杀伤(NK)细胞串扰,并使肿瘤微环境从抗肿瘤变为促肿瘤。
肠道微生物通过诱导免疫细胞产生干扰素,通过一系列的受体和信号通路来增强免疫应答。
注:一些列受体如TLR3、TLR4、TLR7、TLR9、Dectin 1、干扰素基因刺激因子(STING)、DDX41、RIG-I等。
微生物群诱导的细胞和基因表达特征激活了小鼠的STING-干扰素途径,并与黑色素瘤患者的良好ICB反应有关。
某些共生菌如AKK菌、双歧杆菌、鼠李糖乳杆菌,通过激活干扰素途径来抑制肿瘤进展,并增强免疫治疗的效果。
干扰素的作用也与肿瘤类型和治疗阶段有关
I型干扰素信号传导并不总是与抑制肿瘤生长有关。肝脏中持续的I型干扰素信号传导,分别通过诱导一氧化氮或改变尿素循环,诱导对抗PD1治疗的耐药性或抑制抗病毒T细胞反应。
不同刺激产生的干扰素还会对肿瘤生长和免疫反应产生不同的影响。在肿瘤治疗中,详细了解干扰素在不同时间点对不同细胞的作用,以及如何影响肿瘤环境和免疫治疗效果,还需深入研究。
肠道促炎:预测ICB疗效不佳
系统性炎症的许多外周血生物标志物,如高中性粒细胞与淋巴细胞比率(NLR)和C反应蛋白(CRP)、IL-8和血清淀粉样蛋白a(SAA)水平升高,是对ICB反应不佳的预测因素。
黑色素瘤患者粪便微生物群中,高NLR与革兰氏阴性拟杆菌门的丰度增加有关。人类粪便转录组分析显示,对抗PD1治疗没有反应的患者肠道中存在促炎特征,主要由脱落的中性粒细胞和其他髓细胞引起,这与LPS和NF-κB信号传导有关,导致产生促炎细胞因子,如TNF、IL-1和IL-8。
革兰氏阴性菌:阻碍ICB反应
总体而言,这些数据表明,肠道微生物群中的革兰氏阴性菌可能通过LPS产生和TLR4信号传导,诱导免疫抑制性局部肠道和全身炎症,从而阻碍ICB反应。
实验诱导的小鼠胆汁淤积诱导革兰氏阴性菌从肠道转移到肝脏,导致TLR4依赖性炎症反应,产生CXCL1,募集免疫抑制性中性粒细胞,抑制抗肿瘤免疫,导致胆管癌生长增强。此外,革兰氏阴性菌Fusobacterium nucleanum与原发性和转移性结直肠癌癌症的关联,通过吸引髓细胞而具有免疫抑制性。
自身抗原和微生物异种抗原之间交叉反应
自身抗原和微生物异种抗原之间的交叉反应在自身免疫性疾病中很常见,可能与癌症免疫有关。身体不仅对病原体产生免疫反应,而且对共生微生物产生免疫反应。尽管这种反应得到了很好的控制,但失调可能会引发免疫病理,并可能在接受ICB治疗的患者中被重新激活,导致免疫相关不良反应,并可能与肿瘤新抗原发生交叉反应。
ICB治疗的患者中鉴定的一些肿瘤新抗原,可能与微生物抗原具有交叉反应
在抗CTLA4处理的小鼠中,通过过继转移B.thetaiotaomin特异性或B.fragilis特异性T细胞导致其活化,进而抑制MCA-205肉瘤肿瘤生长,支持肿瘤和微生物抗原之间分子模拟的观点。
小鼠细菌共生体E.hirae与对环磷酰胺和ICB的有利反应相关的,E.hirae携带表达肽的原噬菌体,针对该肽引发主要组织相容性复合物(MHC)I类反应,并与癌症细胞中过表达的非突变蛋白酶体组分中的表位交叉反应。
在人类中,相同的原噬菌体在肠球菌属中表达。除毛滴虫外,其MHC限制性表位与甘油-3-磷酸脱氢酶1-样蛋白(GPD1L)中的表位发生交叉反应,后者也在肿瘤中过表达。
在人类黑色素瘤转移中,表达人类白细胞抗原(HLA)I类和II类限制性肽的细菌物种存在于造血细胞和肿瘤细胞中。当对表达肿瘤匹配HLA等位基因的EB病毒转化的B细胞进行脉冲时,其中一些肽能够特异性激活肿瘤浸润淋巴细胞。
因此可以推测,在患者中,这些肽通过传统抗原呈递细胞或黑色素瘤细胞的呈递,引发了可能影响抗癌免疫的T细胞免疫反应。
有望成为预测免疫疗法反应的生物标志物
前面第二章节有提到短链脂肪酸与放疗,这里我们主要看短链脂肪酸对接受 ICI 的癌症患者的预测作用。
在日本进行的一项研究中,52名接受抗PD-1抗体治疗的实体瘤患者的粪便短链脂肪酸与良好的临床结果相关。高浓度的粪便乙酸盐、丙酸盐、丁酸盐和戊酸盐以及血清异戊酸盐与PFS的延长有关。
相比之下,另有研究发现,高血清丁酸盐和丙酸盐水平与黑色素瘤患者对抗CTLA4抗体的耐药性和Treg细胞数量的增加有关。
鉴于临床前肿瘤模型也揭示了短链脂肪酸对抗CTLA4抗体或放射治疗的疗效的负面影响,在利用血液或粪便中的任何短链脂肪酸作为ICI疗效的生物标志物之前,需要进一步的研究来验证不同地理位置的大型队列中短链脂肪酸的预测价值。
不同的细菌种类,包括假长双歧杆菌、约氏乳杆菌和粘球菌,产生肌苷及其代谢产物次黄嘌呤。这些分子结合腺苷受体A2a(A2aR),激活抗肿瘤T细胞并提高小鼠的ICB效率,特别是抗CTLA4疗法。
肌苷增强抗PD1治疗效果
除了A2aR介导的T细胞活化外,肌苷是葡萄糖限制下CD8+T细胞的替代碳源,并增强了对不能利用肌苷作为能量来源的肿瘤的抗PD1治疗功效。表明肌苷活性增强了ICB疗效,并且像腺苷一样激活了A2aR,在临床癌症治疗的试验中,A2aR是一个免疫抑制的检查点,增强ICB效果。
肌苷也可能抑制自身免疫
罗伊氏乳杆菌(Lactobacillus reuteri)可以预防ICB诱导的结肠炎,但不会影响ICB在小鼠中的抗肿瘤功效,它通过增强作用于A2aR110的肌苷的产生,来抑制Treg细胞缺陷的Scurfy小鼠的自身免疫。
这些高度矛盾的结果可能可以通过肌苷对T细胞的直接刺激效应与上下文相关的事实来加以解释;在无菌动物中或在没有共生细菌提供的促炎启动的体外,肌苷需要在这些实验条件下分别由CpG-ODN或IFNγ提供的炎症刺激。由于缺乏这种炎症刺激,肌苷确实会抑制T细胞。
因此,在癌症免疫疗法中靶向肌苷依赖性途径需要更好地理解肌苷的这些对比效应,同时考虑到腺苷和肌苷通过A2aR的信号可能不相同,并且肌苷通过A2aR或A3R影响不同的全身炎症模型。
两项研究调查了接受ICIs的晚期癌症患者的犬尿氨酸与色氨酸(Kyn与Trp)比率的临床意义。
Kyn/Trp比率高,与总生存期缩短有关
第一项研究中,早期疾病进展患者(3个月内)的Kyn/Trp比率显著高于有反应者(P = 0.017)。
第二项研究中,在接受nivolumab治疗的黑色素瘤和肾细胞癌患者中,开始治疗4-6周后Kyn与Trp比率的增加(>50%)与中位总生存期的缩短密切相关(P ≤ 0.026)。
有争议的发现
鉴于代谢适应在癌症免疫疗法中的重要性,以及在黑素瘤患者中选择性IDO1抑制剂与ICI联合治疗的疗效不佳(在ECHO-301/KEYNOTE-252第三期试验中),这些发现不仅对未来试验中基于血清肾上腺素/色氨酸比率监测而进行患者分层的必要性提出了质疑,还对ICI治疗期间肠道菌群动态调节的分析提出了质疑。
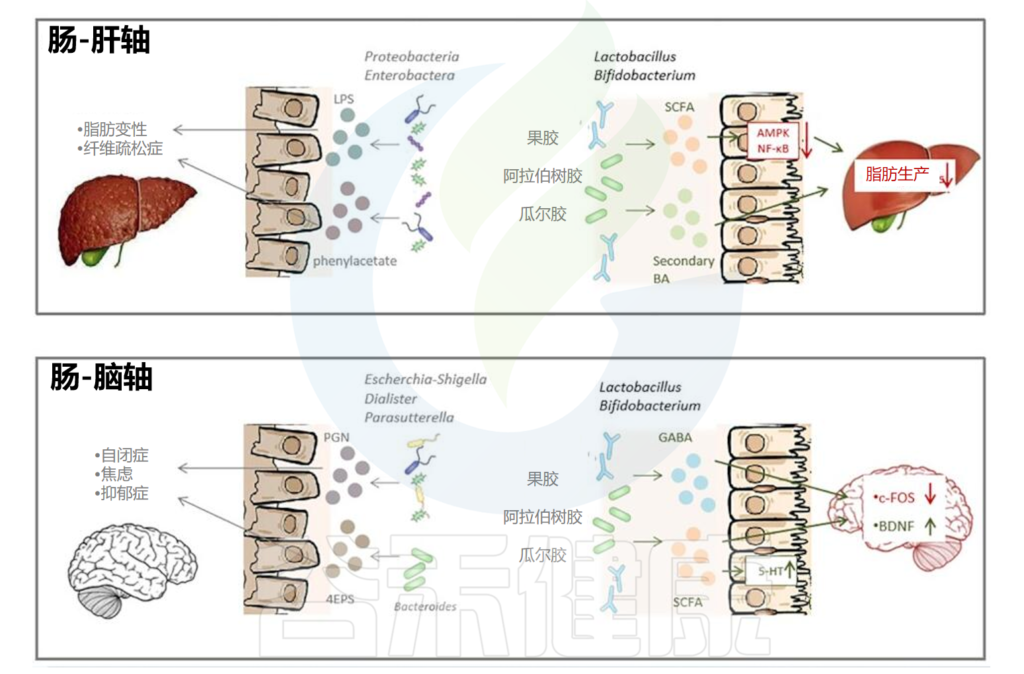
事实上,色氨酸不仅由肿瘤和髓系细胞代谢,而且由肠道菌群中的不同种类代谢成吲哚和吲哚衍生物,包括吲哚乙酸、吲哚-3-醛(I3A)和吲哚丙酸等,这些物质能够与芳香烃受体结合。
可能支持富含色氨酸的饮食,或基于I3A或3-IAA 的益生元与ICI的组合
当罗伊氏乳杆菌与ICI联合给小鼠口服时,它可以从回肠转位到黑素瘤部位,释放I3A,激活CD8阳性T细胞的TCR信号通路,促进IFNγ的释放和抗肿瘤效应。
这些实验发现的临床意义已在一项涉及接受ICI治疗的黑素瘤患者的研究中得到证实,为基于色氨酸的饮食干预在癌症患者中的应用开辟了道路。
细胞内L-精氨酸浓度也直接影响T细胞的代谢适应度和存活率,从而影响它们在小鼠模型中介导有效抗肿瘤免疫反应的能力。
L-精氨酸水平升高诱导了整体代谢变化,包括活化T细胞从糖酵解向氧化磷酸化的转变,并促进了中枢记忆样T细胞的产生。
在发现组和验证组中,低血清L-精氨酸水平(<42 µM)与减少的临床获益率、无进展生存期和总生存期显著独立相关(P = 0.004)。此外,低血清L-精氨酸水平与髓系细胞中PD-L1的表达增加相关。因此,血浆L-精氨酸监测可能成为ICI疗效的适宜预测生物标志物。
doi: 10.1038/s41571-023-00785-8.
尽管广谱抗生素可能会导致真菌过度生长,从而降低放疗效率
万古霉素消耗产短链脂肪酸菌,提高放疗效果
一项正在进行的临床试验(NCT03546829),测试在接受立体定向身体放射治疗的非小细胞癌症(NSCLC)患者中,通过IFNγ的产生评估,万古霉素是否增加抗肿瘤免疫力。
万古霉素还通过增强自然杀伤T细胞(NKT)在肝脏中的迁移和IFNγ的产生,削弱小鼠原发性和转移性癌症的生长。
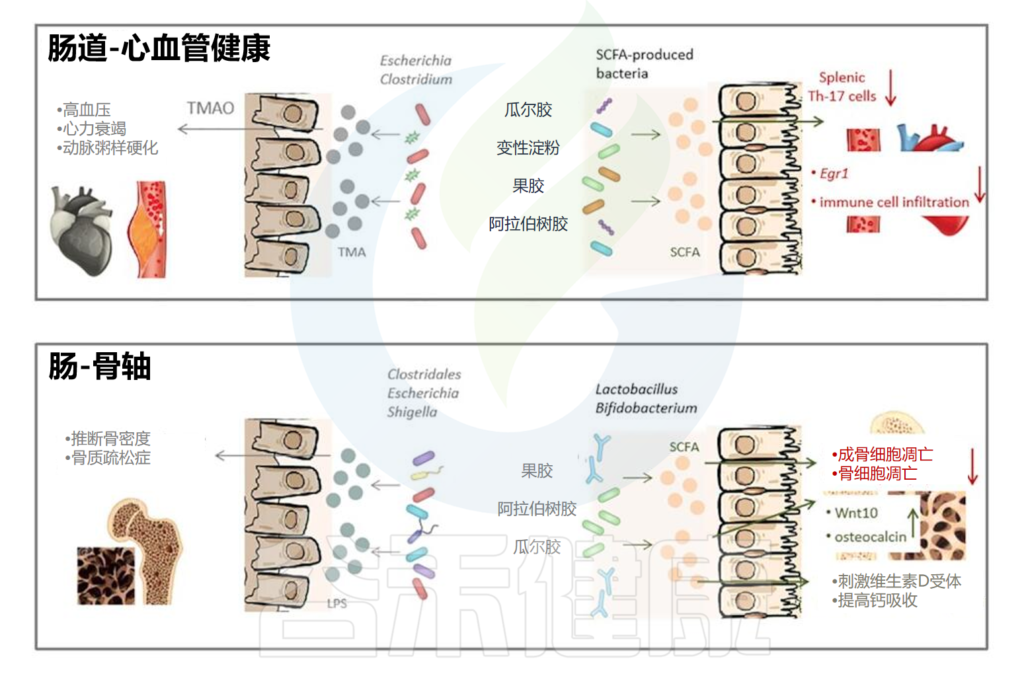
万古霉素耗尽革兰氏阳性菌,调控胆汁酸
万古霉素耗尽革兰氏阳性菌,特别是Clostridium scindens,它将初级胆汁酸转化为次级胆汁酸。微生物群衍生的胆汁酸影响炎症、免疫和致癌物的许多方面。初级胆汁酸,如鹅去氧胆酸,通过肝窦内皮细胞诱导趋化因子CXCL16的产生,该细胞通过CXCR6吸引NKT细胞,而次级胆汁酸,特别是ω-鼠胆酸,抑制CXCL16产生。
还计划进行临床试验(NCT03785210),测试万古霉素是否改善原发性肝癌或转移患者对抗PD1治疗的反应。然而,即使是特定的抗生素也会引起肠道微生物群组成的广泛变化,这可能会对癌症治疗产生相反的影响。
胆汁酸对免疫反应的调节也是复杂的
黑色素瘤患者FMT后,次级胆汁酸水平与抗PD1治疗的良好临床反应有关。
然而在小鼠中,拟杆菌属部分地将抑制TH17细胞的次级胆汁酸3-oxoLCA,转化为激活Treg细胞的isoalloLCA,从而限制自身免疫和癌症免疫。
万古霉素提高ADT疗效
去势抵抗性前列腺癌与促癌的肠道菌群变化有关,使用含有万古霉素的抗生素混合物来消耗小鼠体内产生雄激素的细菌,这种抗生素可以专门减少厚壁菌门的细菌,包括产生雄激素的梭菌属,从而提高ADT(雄激素剥夺疗法)的效果。因此,如果抗生素与ADT一起使用,它们的性质应该是选择性地消耗产生雄激素的细菌,而不是免疫刺激物种。
抗生素治疗仍有挑战,更特异性的方法有待探索
一般来说,在ICB期间或之前使用抗生素对临床反应产生不利影响,很可能是因为它们改变了肠道生态以及有利和不利细菌之间的平衡。尽管通过抗生素来靶向肠道菌群可能具有益处(如上面例子所述),但总体而言,这面临着大部分抗生素的广泛反应性以及可能诱导抗生素耐药性的挑战。
除了抗生素,其他更具特异性的方法,如噬菌体治疗以及更有效、更具特异性的噬菌体介导的CRISPR-Cas3抗微生物药物,可以在我们对直接影响抗肿瘤反应的细菌种类以及由其消除引起的生态变化的认识更加精确之后使用。
FMT可解决80-90%的抗生素耐药性艰难梭菌感染,其他包括各种机会性感染、炎症性肠病等也在研究中。
对免疫疗法后的结肠炎,FMT效果好
抗CTLA4治疗免疫相关不良反应中常见的有结肠炎,这类结肠炎可能比较严重,皮质类固醇或TNF拮抗剂的治疗难以治愈。
两名癌症患者在接受ICB治疗后出现结肠炎,经健康捐赠者FMT治疗后,病情迅速恢复。供体菌群确实发生了定植,厚壁菌门和变形菌门的丰度降低,拟杆菌门和Verrumicrobiae的丰度增加。随后的一份初步报告显示,在FMT治疗的15名癌症患者中,有11人从ICB诱导的结肠炎中恢复。
FMT:提高黑色素瘤患者的抗PD1治疗效果
两项临床试验证明,FMT可以靶向微生物群,以提高黑色素瘤患者的抗PD1治疗效果。由于预测微生物群组成尚不可行,这可能支持抗PD1治疗反应,以选择健康的粪便微生物群供体,两项试验都使用了黑色素瘤患者和对抗PD1疗法有持久反应的患者作为供体。
*****************************************************************************
第一项试验
10名进展中的黑色素瘤患者接受了抗生素治疗(其中6人之前对ICB有阳性反应),并通过结肠镜检查。这10个人做了FMT,供体是两名对抗PD1治疗有反应的捐赠者中的其中一人,随后重复胶囊形式的FMT和抗PD1疗法。
● 在从供体1接受移植的5名患者中,一个完全反应和两个部分反应;
● 在从供体2接受移植的五名患者没有反应。
两个供体的微生物群都包含有利于抗PD1治疗反应的菌群,供体1在梭菌属和放线菌属中更富集。FMT后,供体1受体的微生物组成变化比供体2受体更一致,人类粪便转录组中炎症和抗原呈递特征的表达增加。治疗后,三名有反应的患者表现出更显著的肿瘤排斥反应,其特征是I型干扰素反应和单核细胞、DC、NK细胞和T细胞浸润。
*****************************************
第二项试验
15名对抗PD1治疗原发性无反应的黑色素瘤患者在结肠清洗后(没有抗生素治疗),接受了来自6名抗PD1疗法有反应的供体中的1名,随后接受了持续的抗PD1治疗;1名患者完全缓解,2名患者部分缓解,3名患者病情稳定超过18个月。
这些反应良好的患者的菌群怎么样了?
在反应良好的患者中,FMT后的粪便微生物群变得更像供体,并且针对供体菌群的IgG抗体更显著诱导。有益菌的丰度增加,如毛螺菌科、瘤胃球菌科和双歧杆菌科,有害菌如拟杆菌属减少。
其他指标有什么变化?
在这些有反应的患者的血液中,IL-8水平下降,而CXCR13和IL-21水平上升,表明T卵泡辅助细胞(TFH)有反应。
初级和次级胆汁酸的水平以及苯甲酸盐降解的产物都增加了,苯甲酸盐是微生物群多样性,与抗PD1治疗反应相关的某些菌群的存在的标志。
******************************************************************************
尽管规模小且单臂试验,但这两项初步研究中的整体临床反应率为36%,高于其他联合治疗方案在难治性抗PD-1治疗患者中的反应率,且没有额外严重毒副作用。
FMT后菌群有效定植,干扰时长超1年
总的来说,即使没有抗生素调理,单次结肠镜进行FMT,也会干扰微生物群组成超过1年。宏基因组分析表明,大多数存在于供体而不在受体患者身上的菌属(和微生物基因),可以有效且持久地定植,除非受体接受了抗生素治疗。
供体菌株定植与治疗效果之间可能存在不一致?
事实上,FMT后的微生物群组成不仅反映了供体特异性菌群的定植情况,还反映了FMT引起的肠道生态变化,改变了来自供体或接受者的不同菌群的丰度。
对抗PD1治疗无效的黑色素瘤患者来说,FMT可能只有当患者存在针对肿瘤的免疫反应,但受到免疫抑制或微生物组成不利的影响,导致缺乏免疫促进的微环境时,才会产生对继续抗PD1治疗的反应。FMT可以纠正这种不利的微生物组成。
自体FMT,重建肠道微生物群
供体可以提供更健康、更多样的微生物群,能够重建受体微生物群的多样性。然而,将病原体或抗生素耐药性病原体转移到免疫抑制受体中存在风险,这也说明了患者自体微生物群的储存是合理的。
接受自体FMT的患者每种白细胞谱系的数量都更高,这表明FMT是有利的。Faecaliberium、Ruminococcus、Akkermansia与白细胞数量增加有因果关系的是通过自体FMT重建的。
癌症患者对抗PD1治疗产生抗药性的原因各不相同,可能导致FMT治疗失败
其中原因包括:
1)由于免疫抑制或肿瘤可能缺乏新抗原,无论微生物群组成如何,都无法对肿瘤做出反应
2)FMT供体可能缺乏对抗-PD4治疗做出反应所需的菌群
3)由于供体与受体微生物群的不相容性,FMT不能使微生物群扰动足够大
4)FMT模式的递送方法和频率可能不是最佳的
FMT在癌症免疫治疗中的未来发展
需要重点评估患者预处理和微生物群转移的最佳方案,确定最有可能受益于FMT的患者和合适的供体。
许多计划中以及正在进行的试验将通过评估FMT在不同肿瘤类型中的有效性,以及将FMT与其他方法(如益生菌、饮食和益生元)进行比较或结合,调节肠道微生物群的组成,来确定FMT是否成为治疗对ICB无反应患者的常规方法。
益生菌制剂含有具有抗炎和粘膜保护能力的菌群,如乳酸杆菌属和双歧杆菌属,用于预防化疗和放疗引起的毒性。
临床和小鼠研究,都确定了可能对特定机会性感染产生耐药性的细菌,以及可能纠正微生物群失衡的潜在益生菌。
乳杆菌属:抑制肿瘤生长,改善对ICIs的反应
鼠李糖乳杆菌、嗜酸乳杆菌和发酵乳杆菌对结肠癌发展的缓解作用已在小鼠模型中得到证实。
鼠李糖乳酸杆菌GG可通过cGAS/STING信号传导途径刺激I型干扰素,从而改善对ICIs的反应。
乳酸菌(LAB)可有效减少结直肠癌的发生,这可能归因于炎症因子的减少。此外,LAB还影响肠道微生物群落,其特征是拟杆菌丰度的降低。因此,LAB有利于抑制癌症的发生和发展。
罗伊氏乳杆菌能促进肠上皮的更新和修复,刺激宿主免疫。它能将上皮内CD4+T细胞转化为CD4+CD8αα双阳性上皮内淋巴细胞,从而缓解炎症性肠病,预防部分消化道癌症的发生。
罗伊氏乳杆菌抑制 ICI相关结肠炎的能力,与第 3 组先天淋巴细胞分布的减少有关。
罗伊氏乳杆菌通过产生三种色氨酸降解产物,抑制黑色素瘤、肝癌和结直肠癌的生长:
双歧杆菌属:增强抗PDL1疗效
对无特定病原体或微生物群耗尽的小鼠,给予与ICB反应相关的各种菌群可提高治疗反应。
为了验证双歧杆菌属支持小鼠抗PDL1治疗,施用含有四种双歧杆菌属菌株的非处方商业益生菌制剂可提高对抗PDL1疗法的反应。在对抗PD1治疗有反应的患者中,双歧杆菌也会增加。
只有一些双歧杆菌菌株能够在小鼠中与抗PD1治疗协同作用。这些协同菌株表达参与合成肽聚糖途径的基因,表明它们有助于产生免疫刺激分子。
Faecalibaculum rodentium: 抑制肿瘤生长
F.rodentium和Holdemanella biformis(人类同源物)在肿瘤发生过程中缺失或丢失,这两者都可以产生短链脂肪酸,通过抑制钙调神经磷酸酶和NFATc3激活来控制肿瘤细胞的增殖和蛋白质乙酰化。
当将F.rodentium应用于ApcMin/+小鼠时,80%以上的结直肠癌中会发生结肠腺瘤样息肉(APC)基因突变,或者用偶氮甲烷和右旋糖酐硫酸钠治疗可以减轻小鼠的肿瘤生长。同样,在ApcMin/+模型中,H.biformis在通过丁酸盐抑制肿瘤生长方面似乎与F.rodentium相似。因此,H.biformis可以应用于癌症治疗的设计。
嗜热链球菌:分泌β-半乳糖苷酶,释放叶酸
嗜热链球菌(Streptococcus thermophiles)是一种强大的益生菌,具有消化和免疫益处,通常在结直肠癌患者中被耗尽。更重要的是,嗜热链球菌对肿瘤发生有抑制作用。
具体而言,在结直肠癌小鼠中经口灌胃嗜热链球菌将显著减少肿瘤形成。嗜热链球菌分泌的β-半乳糖苷酶是抑制结直肠癌生长的活性成分;β-半乳糖苷酶可以增加另外两种益生菌乳酸杆菌和双歧杆菌的丰富性,这表明了协同作用。
嗜热链球菌通过释放叶酸来影响肿瘤生长。叶酸是一种主要的膳食元素,在细胞代谢和DNA复制、修复、甲基化和核苷酸合成中发挥重要作用。研究表明,叶酸缺乏在人类中相当普遍,嗜热链球菌释放的叶酸可能与肿瘤抑制有关。此外,嗜热链球菌对淋巴细胞特征、结肠炎的严重程度和调节性T细胞反应有影响。
大肠杆菌Nissle 1917: 减少并发症
大肠杆菌Nissle 1917 菌株可以调节肠道屏障上皮功能,缓解肠道菌群失调,最终减少伊立替康引起的肠道并发症。
丁酸梭菌588菌株:改善抗PD1治疗后患者生存率
相反,在一项回顾性分析中,一种单一的益生菌制剂,在日本和中国广泛用于治疗与胃肠道病理学相关的微生态失调的丁酸梭菌(CBM)588菌株,显著改善了接受抗PD1治疗的小细胞肺癌癌症患者的客观反应、PFS和总生存率。丁酸梭菌通过增加乳酸杆菌和双歧杆菌来改善微生态失调。
然而,一项对接受联合ICB治疗的肾细胞癌症患者的随机试验证实,接受丁酸梭菌治疗的患者临床反应有所改善,但未能检测到双歧杆菌的增加。
多种菌组合,效果 1+1>2
虽然单一益生菌菌株显示出前景,但细菌群落可能更好地维持肠道微生物群内的生态平衡。人类和小鼠种,某些种类的梭状芽孢杆菌与对结肠癌的耐药性有关。
小鼠口服下列四种梭菌菌株:
诱导了激活的CD8+T细胞在肿瘤内的积聚,并成功治疗了化学诱导的结肠癌和可移植结肠癌。
该联合体作为一种独立的治疗方法以及单个物种,在小鼠中比单独的抗PD1治疗更有效。所有四个菌株都产生短链脂肪酸,包括丁酸盐;然而,它们的免疫刺激作用独立于丁酸盐。
鉴定免疫激活细菌群落的另一种方法涉及鉴定罕见的人类共生菌株,该菌株可以激活小鼠肠道中产生IFNγ的CD8+T细胞。一个由11个物种组成的群落在无菌小鼠中对单核细胞增多性李斯特菌感染保持耐药性,并具有与抗PD1治疗相加的抗肿瘤作用。值得注意的是,7种本身没有抗癌活性的拟杆菌目物种确实对该群落的总活性有贡献,这支持了多物种细菌生态学在重建肠道微生物群中的重要性。因此,使用单一菌株和联合体的试验正在进行中。
免疫治疗期间,益生菌的服用也需谨慎
与不服用益生菌的患者相比,服用非处方益生菌的黑色素瘤患者对抗PD1治疗的反应并没有改善,反而有所下降。
这些结果,再加上商业益生菌的配方是异质的,缺乏严格的成分和生存能力质量标准,表明在免疫治疗期间使用这些产品时应谨慎。
饮食的变化会迅速改变肠道微生物群的组成,以及食物发酵衍生的细菌代谢产物的产生,从而产生代谢和免疫后果。
不同的饮食策略,包括热量限制、间歇性禁食、模拟禁食饮食、高纤维饮食、生酮饮食、发酵食物,都已在小鼠或患者中提出或测试,以改善癌症的治疗,至少在某些情况下,显示出通过改变肠道微生物群的组成来影响免疫力。
在T细胞过继转移模型中,小鼠的饮食热量限制促进了记忆性T细胞在骨髓中的积累,并增强了T细胞对细菌感染和肿瘤的免疫力。
发酵食品饮食持续增加了微生物组的多样性,并通过整体抗炎作用改变了其组成。
高纤维饮食对微生物组多样性和组成的影响更为温和,对免疫细胞的频率和细胞因子的表达没有显著影响,但对免疫细胞中的内源性信号传导产生了不同的影响,在一组具有高炎症指数的个体中,内源性信号传导增加,而在另外两组具有低炎症指数特征的个体中则减少炎症指数。
饮食可能通过改变菌群影响抗癌免疫
接受抗PD1治疗的黑色素瘤患者,在纤维充足的饮食超过20g/天时有更好的反应和存活率 ,每5 g纤维的增加相当于进展或死亡风险降低30%。
与另一项研究一致,不同纤维摄入量的人类肠道微生物群在组成或细菌丰富度方面没有显示出重大变化,但从小鼠食物中去除大部分纤维含量导致微生物群迅速变化,双歧杆菌属减少,AKK菌增加。低纤维饮食的小鼠对抗PD1癌症治疗反应较差,与临床结果相似。因此,在无菌小鼠中,高纤维和低纤维饮食不会影响对抗PD1治疗的反应,这表明该饮食可能通过改变微生物群影响抗癌免疫。
跨物种网络分析表明,小鼠的高纤维饮食通过扩大发酵纤维的瘤胃球菌属来维持有效的肿瘤免疫。瘤胃球菌属诱导T细胞的激活和肿瘤浸润,包括表达诱导型T细胞共刺激因子(ICOS)的CD8+和CD4+T细胞。
模拟禁食饮食(FMD)可降低癌症患者的毒性,并改善其对化疗的反应,但大多数关于这种饮食干预的研究尚未解决肠道微生物群改变的作用。
禁食对化疗的积极作用体现在四个方面:
二甲双胍、维生素 C、铂和曲美替尼与禁食联合使用可以增强化疗的效果。
对于三阴性乳腺癌,模拟禁食饮食激活癌细胞中的饥饿逃逸途径,并减少癌症干细胞的表面标志物,从而导致化疗药物的识别增加。
生酮饮食在小鼠中通过诱导由酮体3-羟基丁酸介导的T细胞癌症免疫,来增强抗PD1治疗的抗肿瘤作用。在小鼠和人类中,尽管在低碳水化合物饮食在增加Eisenbergiella massiliensis等细菌种类的丰度方面与抗癌效应相一致,但饮食对肿瘤免疫的影响是由3-羟基丁酸盐的增加介导的,并不依赖于肠道微生物群的改变。
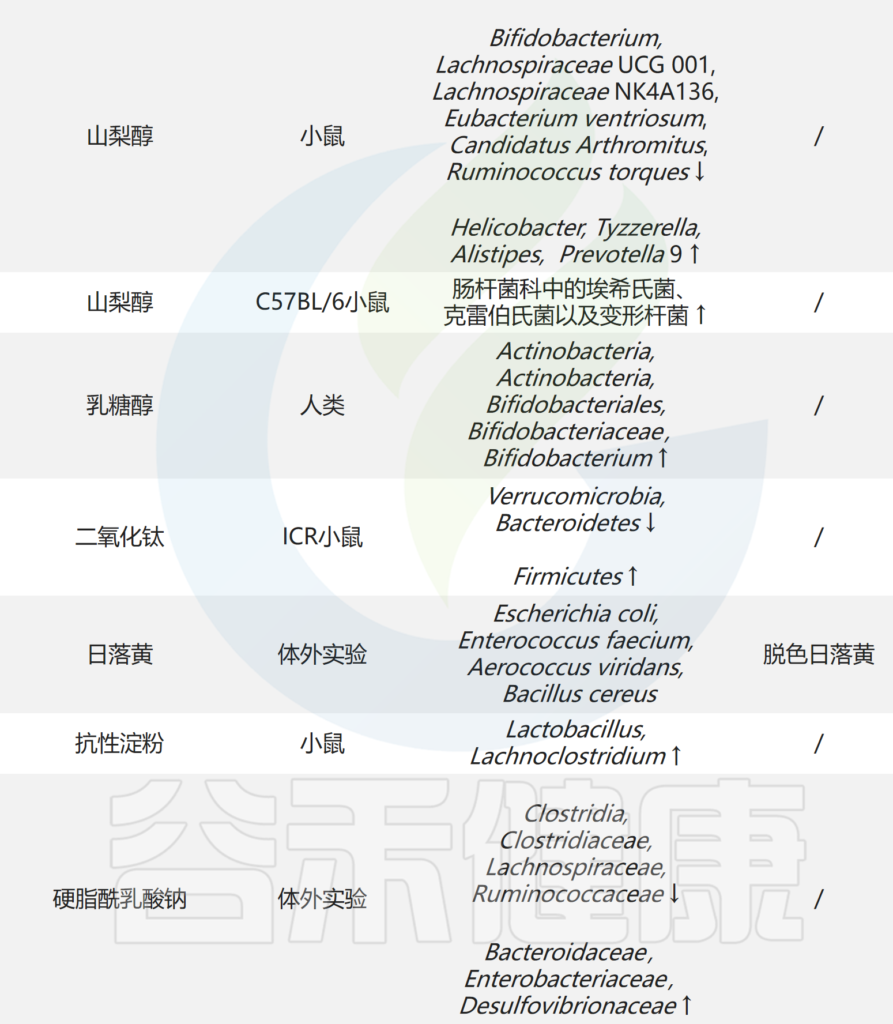
菊粉和果胶是许多蔬菜和水果中天然存在的可溶性纤维,不能被胃肠道酶消化,但可以被细菌发酵。因此,它们通过改变肠道微生物群的组成、增强粘膜屏障、增强上皮完整性以及激活或抑制先天免疫细胞来发挥益生元的作用。
菊粉→产短链脂肪酸菌→增强抗PD1疗效
在小鼠中,菊粉凝胶的口服治疗增加了产短链脂肪酸菌的丰度,并增强了抗PD1疗法的抗肿瘤活性。
↘
果胶增强抗肿瘤免疫,有两种不同的机制。
果胶→产短链脂肪酸菌→增强抗PD1疗效
果胶喂养增强了移植有癌症患者粪便微生物群的小鼠对抗PD1治疗的反应能力,通过扩大纤维喂养和产生短链脂肪酸的Lachnospiraceae和Ruminococceae,通过丁酸盐激活肿瘤免疫。
果胶→AKK菌→激活STING→提高肿瘤免疫
果胶喂养也可以通过导致AKK菌的扩张来提高肿瘤免疫力,从而产生环状二磷酸腺苷(cyclic-di-AMP)并激活STING;由此产生的I型干扰素依赖性免疫肿瘤微环境特征,与小鼠和黑色素瘤患者的免疫介导的肿瘤抵抗相关。
卡姆果→增加多样性,有益菌→增强抗PD1疗效
小鼠研究中,从卡姆果(camu camu)中分离的多酚——栗木鞣花素(castalagin,一种可水解的单宁)的口服给药具有抗肿瘤作用,并通过增加肠道微生物群的多样性,增加有望对抗PD1治疗有益的菌(如瘤胃菌科,颤螺旋菌科Oscillospiraceae和AKK菌)来改变其组成,从而增强小鼠的抗PD1治疗效果。
栗木鞣花素通过影响肠道微生物群组成,重新编程肿瘤免疫微环境,并在一定程度上通过直接结合而不内化于共生细菌(如Ruminococcus bromii)来促进抗癌反应,这可能有助于扩大这种细菌和其他有益细菌的种类,促进抗PD1治疗反应。这表明,栗木鞣花素和其他传统食品添加剂或其成分可能是益生元的来源,通过靶向微生物群来提高免疫疗法的反应性。
人参多糖→增加戊酸→增强抗PD1疗效
人参多糖是一种来源于人参的益生元,可以通过降低犬尿氨酸/色氨酸的比例,增加微生物代谢产物戊酸来增强癌症对PD1抑制剂的反应,从而促进Teff细胞的诱导和调节T细胞的抑制。
肠道微生物群在癌症治疗中,可以通过多种机制影响肿瘤的发展和治疗效果。
研究表明,肠道微生物群可能成为多种癌症类型的预后和预测标记。肠道微生物群有望增强抗癌力,改善放疗化疗效果和增强免疫治疗效果,以降低死亡率和提高生活质量。初步临床试验表明,对抗PD1治疗有反应的患者的FMT可以克服对ICB治疗的原发性和获得性耐药性,这证明可以靶向微生物群增强治疗反应。
然而,在这些临床治疗研究的进展中存在许多障碍和局限性。个体生物学差异可能会阻碍微生物策略的应用。该领域的挑战包括需要在不同的癌症类型、分期、地理区域、年龄、遗传、性别、生活方式、伴随疾病、治疗和合并症中进行前瞻性验证。将小鼠的结果推广到人类也应该谨慎。
虽然存在挑战,但相应的解决方案也在不断研究中。目前应用机器学习方法分析,在考虑了不同群体和地理差异的患者肠道菌群的组成方面取得了进展,这可能很快使肠道菌群成为一个可靠的生物标志物,用于预测患者对治疗的反应,从而实现个性化选择最适合的治疗方法。
将肠道微生物群与独立调节 ICI 功效的其他生物标志物(例如 PD-L1、TMB、IL-8 或某些代谢物等)结合起来,预测免疫疗法效果的能力可能会提高。
未来的临床进展需要更精确地鉴定细菌分类单元对癌症治疗具有积极和负面效应的因果关系,并了解其作用机制,因此需要进行更广泛的试验。
改进的细菌或许可以作为抗癌治疗药物,甚至可以被改造成指导性的“微型机器人”用于药物传递。
总的来说,这一领域的发展为癌症患者提供了新的治疗选择和希望,但同时也需要谨慎评估和监测,确保安全性和有效性。
肠道微生物群对于开发新的抗癌策略的重要性和潜力值得强调,并且有必要探索一种将微生物调节疗法纳入当前癌症管理的整体方法系统。未来我们有望实现更加精准和个性化的肠道菌群调节策略,为癌症治疗带来更大的突破和进展。
主要参考文献
Thomas AM, Fidelle M, Routy B, Kroemer G, Wargo JA, Segata N, Zitvogel L. Gut OncoMicrobiome Signatures (GOMS) as next-generation biomarkers for cancer immunotherapy. Nat Rev Clin Oncol. 2023 Sep;20(9):583-603.
Fernandes MR, Aggarwal P, Costa RGF, Cole AM, Trinchieri G. Targeting the gut microbiota for cancer therapy. Nat Rev Cancer. 2022 Dec;22(12):703-722.
Lee KA, Thomas AM, Bolte LA, Björk JR, de Ruijter LK, Armanini F, Asnicar F, Blanco-Miguez A, Board R, Segata N et al., Cross-cohort gut microbiome associations with immune checkpoint inhibitor response in advanced melanoma. Nat Med. 2022 Mar;28(3):535-544.
Zhao LY, Mei JX, Yu G, Lei L, Zhang WH, Liu K, Chen XL, Kołat D, Yang K, Hu JK. Role of the gut microbiota in anticancer therapy: from molecular mechanisms to clinical applications. Signal Transduct Target Ther. 2023 May 13;8(1):201.
Liu L, Shah K. The Potential of the Gut Microbiome to Reshape the Cancer Therapy Paradigm: A Review. JAMA Oncol. 2022 Jul 1;8(7):1059-1067.
McCulloch JA, Davar D, Rodrigues RR, Badger JH, Fang JR, Cole AM, Balaji AK, Vetizou M, Prescott SM, Fernandes MR, Costa RGF, Yuan W, Salcedo R, Bahadiroglu E, Roy S, DeBlasio RN, Morrison RM, Chauvin JM, Ding Q, Zidi B, Lowin A, Chakka S, Gao W, Pagliano O, Ernst SJ, Rose A, Newman NK, Morgun A, Zarour HM, Trinchieri G, Dzutsev AK. Intestinal microbiota signatures of clinical response and immune-related adverse events in melanoma patients treated with anti-PD-1. Nat Med. 2022 Mar;28(3):545-556.
Guillot N, Roméo B, Manesh SS, Milano G, Brest P, Zitvogel L, Hofman P, Mograbi B. Manipulating the gut and tumor microbiota for immune checkpoint inhibitor therapy: from dream to reality. Trends Mol Med. 2023 Nov;29(11):897-911.

谷禾健康

在研究肠道菌群或复杂微生物样本构成时,“门”(Phylum)是细菌分类的高级分类单位之一。
细菌分类依次为门纲目科属种亚种,最大的分类层面是门,以前写过人群肠道菌群构成主要是以拟杆菌门和厚壁菌门为主,大部分人群占比超90%(个别的变形菌门或放线菌门占比也很高),作为两大“细菌王国”,这两大王国有着不同的生态和代谢属性,又有着相似的营养和生存需求,它们如同天平的两端的砝码,不同的人有着不同倾斜,即使同一个人不同时期不同身体状态下天平的倾斜都不一样。
拟杆菌门被比喻为肠道微生物群落的“多面手”,有超过7000种不同的革兰氏阴性菌组成,主要来自拟杆菌属、副拟杆菌属、普氏菌属和卟啉单胞菌属。它们的脂多糖和鞭毛蛋白与细胞受体相互作用,并通过细胞因子合成增强免疫反应。
拟杆菌门作为多糖降解联盟的成员,尤其是拟杆菌属和普雷沃氏菌属,具有多种可以利用多糖的聚糖和糖苷酶。它们有助于分解膳食纤维和淀粉释放能量,并且它们可能是丙酸盐的主要来源。
拟杆菌门一些细菌的功能还包括胆汁酸的解离。肠道微生物群,特别是肠道拟杆菌,一定程度上也具有使初级胆汁酸解离和脱水并将其转化为人结肠中次级胆汁酸的能力。它们也参与蛋白质分解过程中有毒产物的释放。比如氨,组胺,硫化氢等,这些高浓度的产物会对肠黏膜产生毒性。
该群体的成员具有一些可能有助于抑制炎症的活动,但它们也有可能促进炎症,其中一些已知是机会病原体。
厚壁菌门细菌大多为革兰氏阳性菌,通过短链脂肪酸合成在宿主的营养和代谢中发挥关键作用。
通过其代谢产物,厚壁菌门细菌与其他组织和器官间接连接并调节饥饿和饱腹感。厚壁菌门由大量功能更加多样化的核心细菌组成。从代谢的角度来看,共生梭菌簇 XIVa 和 IV 通过产生短链脂肪酸在宿主和肠道稳态中发挥着重要作用,使肠道通透性正常化,参与脑肠轴调节。许多厚壁菌门的能力与宿主的体重有关。
F/B比率升高或降低被视为生态失调,前者通常与肥胖,代谢障碍相关,这可能与从食物中提取热量增加、脂肪沉积和脂肪生成、胰岛素敏感性受损有关;而后者则与炎症性肠病,抑郁,阿尔茨海默病等相关,这可能与短链脂肪酸尤其丁酸的生成减少,蛋白代谢物组胺,脂多糖累积等相关诱导的免疫炎症反应相关。
目前许多研究工作都集中在识别特定的微生物特征,特别是与肥胖、2型糖尿病和炎症相关的微生物特征。
我们从谷禾肠道菌群检测数据库中随机抽取5万例样本,统计的人群厚壁菌门/拟杆菌门(F/B)比率如下:
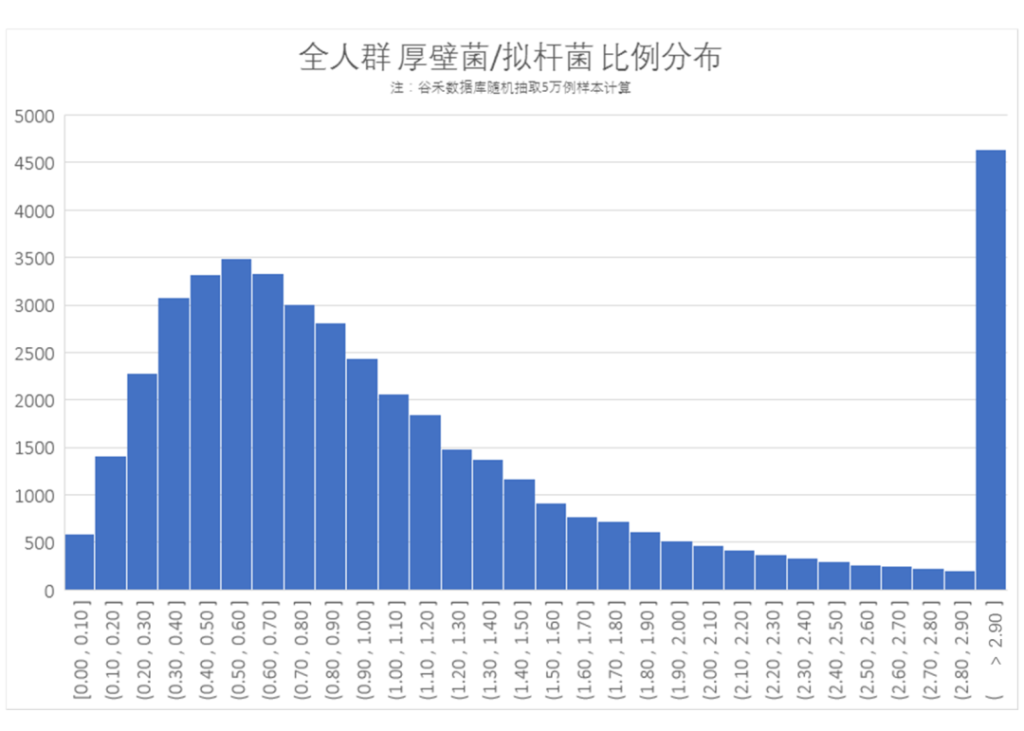
可以看出大部分人群的厚壁菌/拟杆菌比率在0.30-1.0之间(谷禾健康数据库随机抽样5万例样本),表明国人大部分人群肠道菌群的构成中拟杆菌占比多一点;同时抽样人群中1/10人厚壁菌/拟杆菌比率大于2.9,表明也有相当一部分人群厚壁菌门占比较高。
一般而言,从出生到成年,厚壁菌门/拟杆菌门的比例不断增加,并随着年龄的增长而进一步改变。该比率在婴儿、成人和老年人之间存在差异。它可能与生命不同阶段细菌谱的整体变化有关。
厚壁菌门/拟杆菌门比率是一个考虑高水平分类等级(即门)的指数,同一群体的受试者之间厚壁菌门和拟杆菌门的相对丰度差异很大。这可能是由于许多与生活方式相关的因素造成的,包括饮食、疾病状态、食品添加剂和污染物、抗生素消耗、体力活动等影响胃肠道微生物群组成的因素。
由于菌群是一个复杂的生态体系,门层面的简单划分只能提供一个最简单的线索,想要了解肠道菌群需要从最主要的核心菌属,常见的有害菌属以及大量功能菌属的构成来更加准确的分析,此外菌群的代谢通路也提供了不同角度的信息。
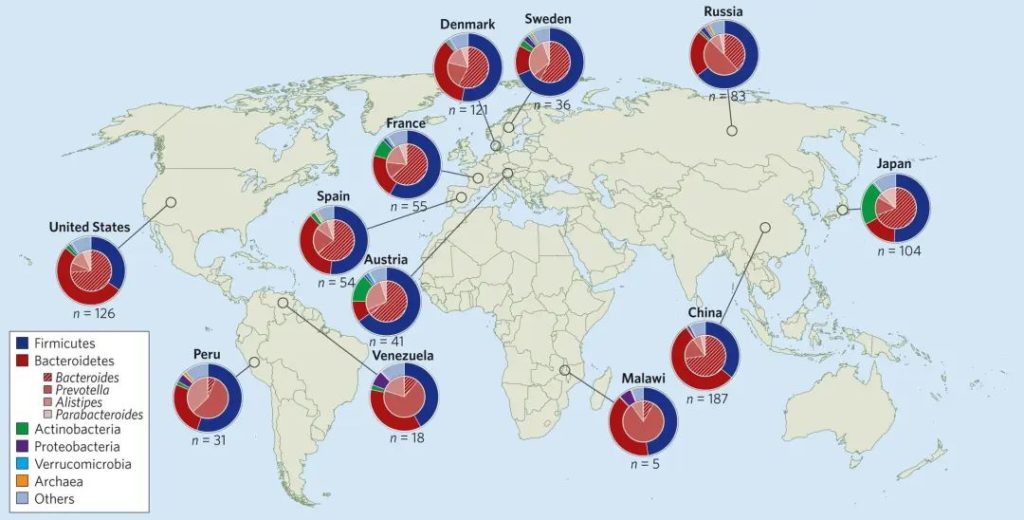
Wexler AG,et al.Nat Microbiol.2017
大多数健康成人微生物群仅由两个细菌门控制——革兰氏阳性厚壁菌门(许多属)和革兰氏阴性拟杆菌门(主要是拟杆菌属、另枝菌属、副拟杆菌属和普氏菌属)——它们共同构成了微生物中的大多数细菌分类群。
此外还有其他分类群包括变形菌门、放线菌门、梭杆菌门、疣微菌门、产甲烷古菌、真核生物(原生生物和真菌)和其他更短暂的定植者。
健康个体中发现的类群在家庭成员(例如父母及其子女)之间往往最相似,而在不同文化和地理空间(例如西方社会与非西方社会)之间则最不相似。
厚壁菌门(Phylum Firmicutes)是细菌的一个门,大多数具有革兰氏阳性细胞壁结构。然而,一些细菌,如巨球菌属、梳状菌属、月单胞菌属和嗜发酵菌属,具有多孔的假外膜,导致它们被染成革兰氏阴性。
科学家们曾经将厚壁菌门分类为包括所有革兰氏阳性细菌,但最近将它们定义为相关形式的核心组,称为低 G+C 组。与放线菌相反。它们有圆形细胞,称为球菌(单球菌),或杆状细胞(芽孢杆菌)。厚壁菌门缺乏脂多糖,而是向宿主呈现厚厚的肽聚糖层;研究人员已经发现了200多种不同类型的属于厚壁菌门的细菌。
★ 厚壁菌门中有许多有益的菌
厚壁菌门的成员包括许多有益的菌。比如乳酸菌是一种常见于酸奶和其他发酵乳制品中的益生菌,属于该门。这些微生物会产生醋酸盐(另一种促进健康的短链脂肪酸)以及乳酸和抗菌物质,以防止病原体定植。
又比如普氏栖粪杆菌(Faecalibacterium prausnitzii),又名:普拉梭菌,是人类肠道菌群中最重要的细菌之一,占健康人粪便样本中检测到的细菌总数的5-15%,是丁酸的重要生产者之一,具有抗炎作用,维持细菌酶的活性,保护消化系统免受肠道病原体的侵害。
★ 厚壁菌门中也存在一些致病菌
但是一些致病菌也属于该门。例如,产气荚膜梭菌是一种引起胃肠道感染的细菌。还有金黄色葡萄球菌,它是一些严重感染的常见原因。
丁酸盐是研究最深入的短链脂肪酸之一,是肠道上皮细胞营养的主要来源。丁酸盐的消耗与肠道屏障完整性受损有关。
注:研究支持,长寿、健康的老年人肠道内的丁酸细菌含量明显较高。
人类肠道包含一个非常密集的代谢活跃微生物群落。然而,只有有限数量的肠道细菌为丁酸生产者。人类肠道中大多数丁酸合成细菌分布于厚壁菌门。在厚壁菌门中,毛螺菌科、瘤胃球菌科、真杆菌科和梭状芽孢杆菌科是四个重要的丁酸合成科。
梭菌目的簇 IV、XIVa、XVI 和 I 包含大多数丁酸盐生产菌种。其中两个重要的物种,直肠真杆菌和普拉梭菌分别属于梭菌簇 XIVa 和梭菌簇 IV,占健康成人粪便样本中肠道菌群总数的12-14% 。此外,还有罗氏菌属,粪球菌属,瘤胃球菌属, Anaerostipes ,丁酸弧菌属和梭菌属。
分布在XIVa 簇中的Butyricicoccus pullicaecorum、Subdolicapsulu variabile、Anaaerotruncus colihominis和分布在 IV 簇中的Papillibacter cinnamivorans是其他产丁酸菌种。
据报道,属于韦荣氏球菌科、热厌氧杆菌科 III 的Megasphaera elsdenii和Caldocellum saccharolyticum也能合成丁酸盐。
最近,对人类样本的高通量宏基因组测序表明,Rawsonibactera saccharolyticus是瘤胃球菌科中的一个新物种,并且肠单胞菌AF211编码丁酰辅酶A:用于合成丁酸的乙酰辅酶A转移酶。
许多厚壁菌门细菌可以产生内生孢子,其耐干燥并且可以在极端条件下生存。孢子在细菌生长中起着重要的作用。细菌的孢子是一种休眠状态,可以在不利环境下存活,并在适宜条件下重新发芽成为活跃的细菌。这种休眠状态使得细菌能够在恶劣条件下存活并传播。
★ 内生孢子可以帮助细菌在恶劣条件下存活
内生孢子由细菌的DNA和部分细胞质组成,周围有一层非常坚韧的外层。它们可以在没有营养的情况下生存,并且能够抵抗紫外线、干燥、高温、极端冷冻和化学消毒剂。
厚壁菌门的代谢偏好
科学家发现,人类肠道微生物群虽然独特,但都可以按照几个分类划分下,称为肠型。第三种肠道型,被称为“谷物爱好者” ,通常以瘤胃球菌(厚壁菌门的成员)为主。
肠型往往可以追溯到长期的饮食习惯。顾名思义,具有“谷物爱好者”微生物组类型的人倾向于消耗更多全谷物中的抗性淀粉和膳食纤维。
✦ 高纤维饮食的人体内厚壁菌更丰富
高纤维饮食可以增加厚壁菌门的丰度并减少拟杆菌的丰度 ,从而增加肠道内短链脂肪酸的浓度。
而葡萄籽原花青素显著降低了肥胖大鼠肠道菌群中厚壁菌门的含量,提高了拟杆菌门的含量,显著降低了厚壁菌门与拟杆菌门比值。
✦厚壁菌偏向从膳食蛋白和循环尿素中获取氮
在厚壁菌门成员中,偏好尿素氮的属往往是菊粉的疯狂使用者,即使用菊粉和尿素合成自己的氨基酸。这包括一些脲酶阴性菌属,它们可能通过交叉喂养获得尿素氮。
此外,在厚壁菌中也看到了一些属更喜欢从膳食蛋白质中获得氮,而其他菌属更喜欢循环尿素。
静脉注射尿素以提高循环尿素浓度后,偏好尿素的厚壁菌以及阿克曼菌的丰度大幅增加。
膳食蛋白质和循环尿素是厚壁菌的主要氮原料,而分泌的宿主蛋白质为拟杆菌提供氮。
!
如果厚壁菌门过高意味着什么?
可能的原因:
– 不良的饮食习惯
– 生态失调
– 消化不良
– 胃酸过少
– 代谢疾病
可能的治疗方法和注意事项:
– 平衡共生细菌
– 当厚壁菌门含量较高时,考虑主要使用双歧杆菌益生菌和布拉氏酵母菌
– 乳杆菌属和芽孢杆菌属(益生菌中发现)可以提高厚壁菌门的数量
– 优化饮食结构:低脂肪饮食可能有助于厚壁菌门水平正常化
滋养肠道内厚壁菌门的食物
纤维和淀粉,是这些细菌的能量来源。富含水果、蔬菜、豆类和全谷物的饮食也是理想的。
研究表明,地中海饮食与更丰富的厚壁菌门有关,包括普拉梭菌。另一方面,富含动物蛋白、脂肪和糖的西方饮食会对有益的这些细菌产生负面影响。
如果想增加肠道中厚壁菌门的丰度,那么应该多吃高纤维食物。以下是一些可以添加到饮食中的食物,以支持您的肠道微生物组并促进厚壁菌门和丁酸盐的产生:
-苹果
-蒜
-大麦
-菊苣根
-燕麦
-亚麻籽
-洋葱
拟杆菌是肠道生态系统中非常成功的竞争者,表现出相当大的营养灵活性以及对宿主和肠道环境施加的压力做出反应的能力。很难权衡肠道拟杆菌对宿主是否产生负面或正面影响。
拟杆菌是复杂碳水化合物的主要降解者,迄今为止调查的所有生态系统中都普遍发现该属,在土壤以及人类和动物肠道中尤其占主导地位。大多数已测序的拟杆菌基因组编码大量多糖降解酶(CAZymes),表明代谢重点是聚糖降解。
CAZymes 在碳水化合物活性酶数据库中分为类别和家族,其中降解酶存在于糖苷水解酶(GH)、多糖裂解酶(PL)、碳水化合物酯酶(CE)和辅助活性酶 (AA) 类别中。
拟杆菌还通过与免疫系统的相互作用,在激活T细胞介导的反应中发挥作用,从而有助于宿主的整体健康。
✦ 防止特定炎症反应
基于多项研究,发现脆弱拟杆菌通过产生称为两性离子多糖的多糖来激活CD4+ T细胞。CD4+ T细胞的产生会刺激白细胞介素10的释放,从而防止特定的炎症反应和脓肿形成。
肠道免疫保护取决于拟杆菌、白细胞介素36信号和巨噬细胞的发育。拟杆菌的这种作用需要其保守的共生定植因子的多糖利用位点。
✦ 抑制胃肠道中的病原体
与胃肠道中发现的其他一些细菌相比,研究表明拟杆菌门的一些成员对各种抗生素和胆汁等物质具有耐药性,这给了他们竞争优势。
除了这种竞争优势之外,这些细菌还被证明在调节胃肠道中其他正常菌群和病原体的数量方面发挥着重要作用。
拟杆菌可以积极改善肠道环境,使其更适合自身和其他微生物。
✦降低肠道内氧水平从而促进厌氧菌生长
例如,许多编码细胞色素bd氧化酶,据推测该酶可以降低细胞内氧水平,进而降低肠道氧水平,从而允许严格厌氧菌生长,否则这些厌氧菌会被氧气的存在杀死。
这种耐受和降低氧气水平的能力可能有助于拟杆菌传播到新宿主,这可能是它们在哺乳动物中如此广泛传播的主要原因。
✦改变肠道的营养状况
它们还可以通过促进宿主的生理变化来诱导某些食物来源(例如岩藻糖基化糖蛋白)的产生,或通过从糖蛋白中释放岩藻糖和唾液酸残基来改变肠道的营养状况,这些残基可以被其他微生物消耗,包括病原体。
通过比较缺乏自身微生物的无菌动物和与单个拟杆菌属菌株和突变体单相关的无菌动物(出生时无菌但后来被微生物定殖),已经鉴定了许多这些拟杆菌属依赖性的肠道环境变化。
✦拟杆菌拥有多糖利用位点有助于代谢
尽管人们可能会想象肠道中的生命充满了微生物的过多食物选择,但在结肠(细菌密度最高的地方),简单、容易获得的糖大部分已经被消耗或吸收。其中的残留物是复杂的长链多糖,它们不能自由吸收,也不能被人类酶促消化。对于许多细菌来说,这些复杂的多糖同样难以处理,并且无法跨膜运输。
然而,早期研究证实拟杆菌是熟练的聚糖降解剂,具有识别和代谢十多种植物和宿主来源的多糖的不同寻常的能力。拟杆菌属物种通过基因簇来实现这一点,这似乎是拟杆菌特有的,称为多糖利用位点(PUL)。
多糖利用位点可以确定拟杆菌可以占据哪些代谢生态位,甚至可以确定它们在肠道内的生物地理位置。多糖利用位点被如此分类是因为它们的susC和susD的同系物,以及其他成分(糖苷水解酶、多糖裂解酶、糖基转移酶、碳水化合物酯酶),它们对于分解多种植物或宿主来源的聚糖很重要。
肠道细菌的其他谱系,包括革兰氏阳性放线菌,降解和输入膳食多糖的途径远不那么复杂,这在很大程度上是由于缺乏外膜屏障。例如,短双歧杆菌使用多结构域、细胞表面锚定酶(而不是一系列单独的酶),可以结合并降解聚合物,例如淀粉、支链淀粉和糖原。
拓展
比较基因组分析已在每个拟杆菌属中鉴定出数十种不同的多糖利用位点。例如,Bacteroides thetaiotaomicron是第一个对其基因组进行测序的拟杆菌属成员,将其 6.26 Mbp 基因组的近五分之一专用于88个不同的多糖利用位点。
拟杆菌不是同时表达所有多糖利用位点以捕获周围环境中可能存在的任何多糖,而是通过大量SusR样调节因子、混合双组分系统和涉及胞质外 σ/抗的跨包膜信号通路来严格调节其表达。
也许是由于在跨外膜运输之前需要部分降解长多糖链,B. thetaiotaomicron和其他肠道拟杆菌已经成功地将适当的多糖利用位点转录与其相应多糖分解中间体的丰度(而不是丰度)耦合起来。
这使得肠道细菌能够快速适应营养可用性的变化。糖分解物质形成甲酸盐、乙酸盐、乳酸盐、丙酸盐和琥珀酸盐作为主要发酵产物。拟杆菌可能是结肠中丙酸盐形成的主要贡献者。
由于多糖利用位点激活是肠道环境的产物,富含植物源性多糖和糖蛋白的饮食将导致多糖利用位点暂时上调,从而降解这些膳食营养素。然而,当这些底物稀缺时,例如在禁食期间或某些低纤维饮食期间(如高糖、高脂肪的西方饮食),亲糖拟杆菌会改变其转录谱以消耗宿主衍生的聚糖,这些暂时的转换可能是导致某些细菌在婴儿期最初在肠道定殖并在我们一生中持续存在的主要因素。
根据最近使用动物模型的研究,拟杆菌属可以在相对较短的宿主婴儿期主要依靠宿主聚糖生存,但当在多个宿主世代的时间内被迫以低纤维饮食为生时,它们最终可能会灭绝。
有的菌会通过破坏中性粒细胞趋化性等机制帮助逃避免疫系统(例如,伤寒沙门氏菌)和逃避宿主补体系统(例如肺炎克雷伯菌)。
相比之下,拟杆菌使用一种称为相变的过程,以开/关方式控制其荚膜多糖生物合成位点的表达。例如,脆弱拟杆菌从八个生物合成位点产生不同的荚膜多糖,其表达受到可逆启动子区域和反式位点抑制剂的严格调控,限制多个位点的同时表达,从而使脆弱拟杆菌的同基因群体共同表现出广泛的多样性。当脆弱拟杆菌漏入通常无菌的腹膜腔时,可能会发生腹膜炎。
注:绝大多数人类腹膜炎病例都是由脆弱拟杆菌引起的。
目前,拟杆菌门中研究的最充分的,包括拟杆菌属和普氏菌属以及卟啉单胞菌属。
✦ 卟啉单胞菌属可导致急性坏死溃疡性牙龈炎
卟啉单胞菌属的成员作为共生体或机会性病原体生活在人类口腔中。牙龈卟啉单胞菌引起临床上侵袭性牙龈炎,可导致急性坏死溃疡性牙龈炎或扩展到口腔和面部组织,这种情况称为坏疽性口炎或口腔癌。
✦ 普雷沃氏菌也是一种条件性致病菌
普氏菌属是一个大属,包括50多个不同的物种。大多数可以从口腔和肠道中分离出来。普雷沃氏菌属存在于人类中,帮助分解蛋白质和碳水化合物食物。也可作为条件致病菌,引起牙周和牙齿、肠道炎症、类风湿性关节炎、细菌性阴道炎等问题。
大型基因组计划,包括美国国立卫生研究院(NIH)资助的人类微生物组项目(http://nihroadmap.nih.gov/hmp/)和欧盟资助的 MetaHIT 项目 (http://www.metahit.eu)对50多种人类来源的拟杆菌门的拟杆菌和普氏菌分离株进行了测序。
基因组草图可从 Genbank 公开获得,并揭示这些物种拥有多种糖苷酶,这些糖苷酶具有预测的对抗植物和宿主来源的多糖的活性,以及维生素和辅因子合成的许多途径。
拟杆菌门的基因组大小相对较大,例如B.thetaiotaomicron 为 6.26 Mb。迄今为止在拟杆菌属中发现的16S rRNA基因的最高拷贝数为7个(在普通拟杆菌中)。
✦ 拟杆菌的组成和代谢受到饮食调节
在全基因组水平上,拟杆菌属与副拟杆菌属共享超过1000个蛋白质家族的核心,但与卟啉单胞菌属和普雷沃菌属共享较少数量的核心家族蛋白质家族。它们的组成和代谢活动在很大程度上受饮食调节。
拟杆菌与高脂肪和蛋白质的摄入量有关,而普雷沃氏菌与富含植物的饮食(高水平的复合碳水化合物和水果和蔬菜的摄入量)有关。因此,拟杆菌属和普雷沃氏菌属物种在人体内具有其特定的栖息地,因为它们大多数是拮抗的。
厚壁菌门与拟杆菌门之间的比率(F/B)与维持体内平衡有关,该比率的变化可能导致各种病理。
例如,特定厚壁菌门或拟杆菌门物种丰度的增加分别导致肥胖和肠道炎症。尽管人们非常关注 F/B 比率,但应该记住,这一比率可能会受到其他门的增加的影响,但是其他门的菌群失调增加或减少并不一定会改变 F/B 比率。
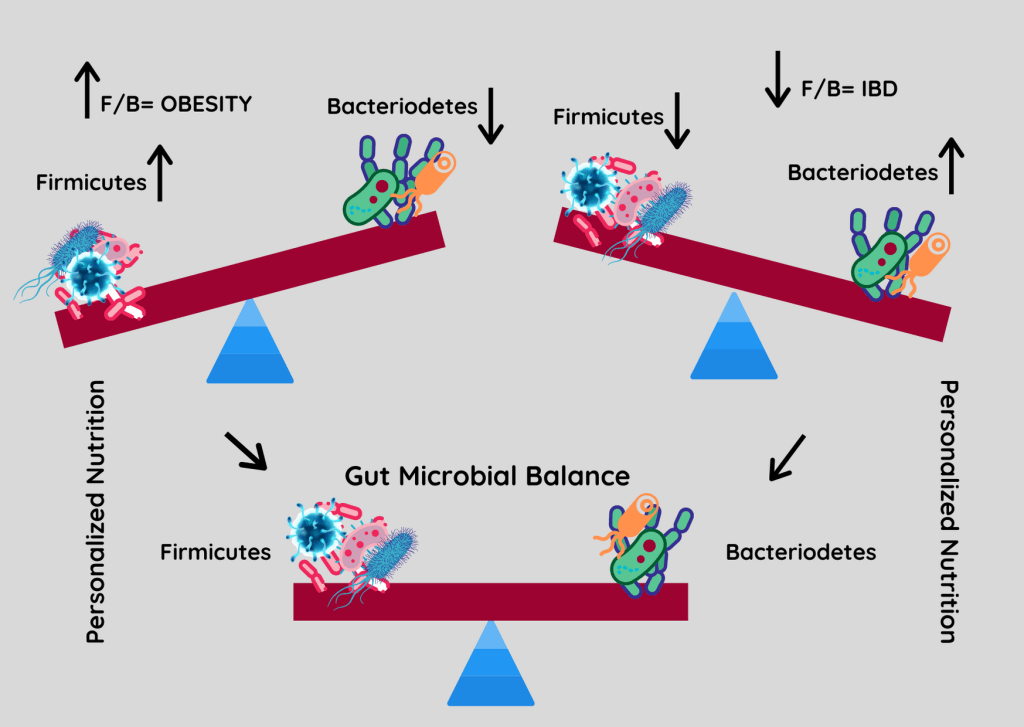
Stojanov S,et al.Microorganisms.2020
肥胖是一种复杂的、多因素的疾病,其原因包括宿主遗传背景、体力活动减少和食物摄入过多等多种因素。在过去的几十年里,肠道微生物群被认为是有利于脂肪储存、体重增加和胰岛素抵抗的额外因素。
世界范围内的研究正在不断寻找其致病因素。厚壁菌门和拟杆菌门的改变首先在肥胖动物和受试者中被描述,这些动物和受试者表现出厚壁菌门丰度增加,而拟杆菌门却减少了。
✦ F/B 比率较高时相同食物会吸收更多能量
当F与B的比例较高或体重增加和肥胖状态时,这两个细菌门影响我们体重的原因是它们调节我们吸收的脂肪量。想象一下两个人都消耗2000卡路里,但F与B的比例不同。F/B 比率较高的一种会吸收更多的卡路里,因此会比另一种增加更多的体重。因此,重点应该是平衡F与B的比例,一方面防止肥胖,另一方面预防炎症性肠病的发生。
当受试者接受热量限制饮食一年后,他们的拟杆菌门丰度增加,厚壁菌门/拟杆菌门比例正常化,同时体重减轻。这些研究得到了对喂食高脂肪或高纤维饮食的动物的研究的支持,这些研究显示厚壁菌门和拟杆菌门的丰度分别较高。
基于这些结果以及从肥胖动物和人类身上获得的其他结果,有人提出厚壁菌门比拟杆菌更能有效地从食物中提取能量,从而促进有效吸收热量和随后的体重增加。
肥胖双胞胎的微生物组富含编码营养转运蛋白的基因,而瘦双胞胎的微生物组富含编码与碳水化合物代谢相关的酶的基因。
最近发表的一项系统综述显示,大多数研究都支持 F/B 比率增加与肥胖之间的关系。
对肥胖和非肥胖日本受试者的粪便样本进行的一项研究显示,他们的 F/B 比率存在显著差异。厚壁菌门的百分比为 37.0 ± 9.1%(非肥胖者)和 40.8 ± 15.0%(肥胖者),而拟杆菌门的百分比为 44.0 ± 9.8%(非肥胖者)和 37.0 ± 14.0%(肥胖者)。
在61名乌克兰成年人中也报告了类似的结果,其中 F/B 比率与体重指数(BMI)显著相关。F/B 比率≥ 1 的个体超重的可能性比F/B 比率<1 的个体高出23%。
同样,卡塔尔受试者(37名肥胖者和36名瘦者)表现出肠道微生物群的改变;肥胖和瘦受试者的 F/B 比率分别为 2.25 ± 1.83 和 1.76 ± 0.58。在哈萨克族和比利时学童中,肥胖组的F/B比值显著高于对照组。
✦ 肥胖人群与正常人群拟杆菌存在差异
然而,两个拟杆菌属的存在差异:与肥胖组相比,正常组中拟杆菌属的高出20%,而肥胖组中普雷沃氏菌属比正常组高出19%。拟杆菌属和普雷沃氏菌属都属于拟杆菌门。
同一群体的受试者之间厚壁菌门和拟杆菌门的相对丰度差异很大。这可能是由于许多与生活方式相关的因素造成的,包括饮食、体力活动、食品添加剂和污染物、抗生素消耗等影响胃肠道微生物群组成的因素。
这种高度变异性可能解释了在比较正常体重和肥胖受试者之间的微生物群时观察到的矛盾结果,使得很难将厚壁菌门/拟杆菌门比率与确定健康状况联系起来。
重要的是要观察肠道微生物种群结构的整体组成,将其作为肥胖和肥胖相关病理(例如GSD)的指标,而不仅仅是厚壁菌门/拟杆菌门的比率。然而,在我们看来,可以修改这个比率,例如引入表征拟杆菌门底下的拟杆菌属或普氏菌属与梭状芽胞杆菌 XIVa 簇菌的比例。
!
F/B 比改变导致肥胖的争议
然而,与这些结果相反,许多研究没有观察到该参数的任何改变,甚至报告肥胖动物和人类中厚壁菌门/拟杆菌门的比例降低。
事实上,在大多数研究中,肥胖患者比瘦受试者表现出更少的细菌多样性,这表明在科、属或种水平上存在其他组成变化,这可能比厚壁菌门/拟杆菌门比率更相关。
✦肥胖增加可能与长期暴露脂多糖有关
关于这一点,代谢性内毒素血症的假说提出,肥胖增加和全身炎症的发展可能是由于长期暴露于脂多糖所致,脂多糖是一种源自革兰氏阴性细菌的促炎分子,它会通过肠腔进入血液。
这一假设与肥胖症中所报道的拟杆菌门丰度下降的情况并不相符,因为该门是肠道微生物群中革兰氏阴性菌的主要群体。
这种差异可以通过这样的事实来解释:来自拟杆菌门细菌的脂多糖的内毒素活性被认为低于来自其他革兰氏阴性细菌(例如属于变形菌门的细菌)的内毒素活性。有趣的是,在肥胖受试者或动物中也观察到变形菌门的增加,并且在无菌小鼠中施用变形菌门成员肠杆菌会导致这些动物出现肥胖和胰岛素抵抗。
✦ F/B比率增加与短链脂肪酸产生无关
另一方面,厚壁菌门/拟杆菌门比例的增加与肥胖个体中观察到的短链脂肪酸产生无关。事实上,据报道,拟杆菌门主要产生乙酸和丙酸,而厚壁菌门则产生更多的丁酸。丁酸盐被认为是一种促进健康的分子,因为它具有增加胰岛素敏感性、发挥抗炎活性、调节能量代谢和增加瘦素基因表达的能力。
结肠中的丙酸盐刺激L-肠内分泌细胞释放GLP-1和 PYY,从而抑制食欲。它还可能到达门静脉循环,主要被肝脏捕获,参与肝糖异生并减少参与脂肪酸和胆固醇从头合成的酶的表达。
乙酸也被吸收并到达全身循环和周围器官,包括脂肪组织、肌肉和大脑。在肝脏中,与丙酸盐相反,它刺激肝脏合成脂质,导致血脂异常。在大脑中,它激活副交感神经系统,分别促进胰腺和胃粘膜分泌胰岛素和生长素释放肽。
这些事件会导致脂肪储存和食欲增加,从而导致肥胖。根据这些结果,醋酸盐通常被认为更容易导致肥胖。肥胖个体中厚壁菌门/拟杆菌门比例的增加意味着这些受试者中丁酸盐的产量较高,丙酸盐和乙酸盐的产量较低,这一发现与这些短链脂肪酸各自的抗肥胖和致肥胖作用部分矛盾。一种解释是,肥胖个体中产生丁酸的细菌减少,并逐渐被属于同一门的其他细菌取代,导致结肠腔中丁酸的产生量降低。例如,葡萄球菌属和罗伊氏乳杆菌的丰度增加(均来自厚壁菌门)
肥胖状态还与A. muciniphila (疣微菌门)丰度较低有关,这是一种粘蛋白降解细菌,有助于稳定肠道屏障功能、分泌抗菌肽和控制炎症。
✦生理及病理状况都会影响肠道微生物群
另一方面,性别、年龄、宿主遗传学的差异。男性和女性的肠道微生物群存在差异,例如男性中拟杆菌-普雷沃氏菌群的水平较高,而女性中厚壁菌门的比例较高。然而,个别研究也发现女性中拟杆菌门的丰度最高。
此外,存在病理状况(糖尿病、心血管疾病、炎症性肠病等)。例如,Oddi括约肌松弛与胆管结石有关,可能是由于肠内容物回流增强改变了微环境所致。
治疗(抗生素、二甲双胍等)通过富集Akkermansia muciniphila以及几种产生短链脂肪酸的微生物群(丁酸弧菌、两歧双歧杆菌等)来改变肠道微生物群组成。
总之,这些导致厚壁菌门/拟杆菌门比率并不是与肥胖相关的微生物群失调的有力标志。
我们建议与肥胖相关的肠道微生物组研究不应该研究肥胖本身的分类标志物,而应侧重于确定用于将患者分层为亚组的分类标志物。引入微生物组患者分层将通过直接操纵患者微生物组来制定个性化治疗决策,从而改善肥胖的管理。
炎症性肠病代表小肠和大肠炎症反应引起的肠道疾病,由两种疾病组成:克罗恩病(CD)和溃疡性结肠炎(UC)。
溃疡性结肠炎是由结肠和直肠(从大肠末端开始到肛门结束的腔室)的长期炎症引起的,而克罗恩病是由结肠或回肠(小肠的最后部分)或两者的长期炎症引起的。
✦ 炎症性肠病患者厚壁菌门丰度下降
肠道微生物群会导致这些疾病。具体来说,当厚壁菌门减少或拟杆菌门相对于厚壁菌门增加时(两种情况下肠道菌群失调),就会导致慢性炎症和消化系统疾病,如克罗恩病和溃疡性结肠炎。
一项研究检查了克罗恩病和溃疡性结肠炎患者以及健康对照活检组织中的微生物多样性。观察到厚壁菌门丰度下降;更准确地说,克罗恩病患者中梭状芽胞杆菌类细菌减少,但溃疡性结肠炎患者或健康个体中则没有减少。
相反,与溃疡性结肠炎患者和健康个体相比,克罗恩病患者的拟杆菌丰度显著增加(分别为74.97%、64.31%和67.41%)。
✦肠道微生物变化与炎症性肠病严重程度相关
肠道微生物群的变化也与克罗恩病和溃疡性结肠炎的疾病活动度和严重程度相关。例如,与患有非活动性疾病的患者相比,患有活动性疾病的溃疡性结肠炎患者中厚壁菌门的含量较低。同样,与患有非侵袭性疾病的克罗恩病患者相比,患有侵袭性疾病的克罗恩病患者的厚壁菌门丰度显著降低。
具有抗炎作用的普氏粪杆菌减少
普氏粪杆菌(F. prausnitzii)是一种厚壁菌门物种,在炎症性肠病患者中减少,其减少与克罗恩病术后六个月复发相关。
普氏粪杆菌在体内和体外均表现出抗炎作用。在患有诱导性结肠炎的小鼠中,活普氏粪杆菌(或其上清液)降低了结肠炎的严重程度并纠正了生态失调。此外,普氏粪杆菌对血液单核细胞的体外刺激导致IL-10分泌增加以及IL-12和干扰素-γ(IFN-γ)分泌减少。
拟杆菌在炎症性肠病中增加
相比之下,拟杆菌在炎症性肠病中通常会增加,并与其进展和发展相关。对炎症性肠病患者和健康个体肠道炎症和非炎症区域的粘膜活检显示,炎症性肠病样本中厚壁菌门减少,拟杆菌门丰度增加,而肠杆菌科仅在克罗恩病患者中增加。活检还显示,与非发炎区域相比,发炎区域的细菌失调程度更高。
另一项对克罗恩病和溃疡性结肠炎患者进行粘膜活检和粪便菌群构成的研究证明了肠道微生物群和 F/B 比率的改变。克罗恩病和溃疡性结肠炎患者的F/B比值均较对照组显著降低。
此外,种、属和科的组成也发生了变化。与克罗恩病患者和对照组相比,溃疡性结肠炎患者中拟杆菌和乳酸菌的丰度较高;与对照组相比,溃疡性结肠炎患者中大肠杆菌的丰度也有所增加。相反,克罗恩病患者中球状梭菌减少。
✦ 不同拟杆菌种类与不同炎症性肠病程度相关
不同的拟杆菌与不同程度的炎症性肠病相关。在经过抗生素预处理且存在转化生长因子(TGF)βRII和IL-10R2信号传导缺陷的小鼠系中,在用普通拟杆菌和多形拟杆菌灌胃后观察到严重的溃疡性疾病。
与显示 F/B 比率对肥胖影响不大的研究类似,并不是在所有炎症性肠病病例中都观察到 F/B 比率降低。炎症性肠病患者粪便微生物分析显示,拟杆菌门、厚壁菌门和疣微菌门的丰度下降,而变形菌门、放线菌门和梭杆菌门的丰度增加。在48名克罗恩病患者中,粪便样本中的拟杆菌细菌丰度不到健康个体的10%,梭菌亚群 XIVab(厚壁菌门)也显著降低。
然而,大多数研究表明,拟杆菌因内毒素而表现出促炎特性,并影响细胞因子的产生,从而导致炎症性肠病。此外,厚壁菌门细菌具有抗炎作用,可以缓解炎症性肠病的进展。
2型糖尿病(T2DM)是一种代谢性疾病,其主要原因是与肥胖相关的胰岛素抵抗。然而,其他一些因素,如精神压力、感染和遗传倾向也可能导致糖尿病。
肥胖和糖尿病的特征都是慢性低度炎症状态,并伴有多种炎症介质(例如肿瘤坏死因子和白细胞介素)的异常表达和产生。
最近的研究基于大规模16S rRNA 基因测序、定量实时PCR (qPCR) 和荧光原位杂交 (FISH),显示了肠道微生物群组成与糖尿病等代谢疾病之间的关系。
有人提出,肠道微生物群引导增加肠道对单糖的摄取,并指示宿主增加肝脏产生与胰岛素抵抗相关的甘油三酯。
✦ 2型糖尿病患者中产丁酸厚壁菌门丰度降低
权威研究显示主要的丁酸盐合成菌厚壁菌门在2型糖尿病患者中显著减少。多项人体研究支持这样一个事实:与健康受试者相比,2型糖尿病受试者中合成丁酸的厚壁菌门丰度显著降低。
对这些患者的粪便微生物群分析显示,与正常耐糖受试者相比,新诊断或长期患有糖尿病的受试者中瘤胃球菌科和毛螺菌科等科的丰度有所下降。
还报告了罗氏菌属(Roseburia)和普拉梭菌(Faecalibacter prausnitzii)的丰度在健康对照中比2型或1型糖尿病患者更高。
对粪便样本进行深入的宏基因组学和宏蛋白质组学分析证实,在患有糖尿病前期 (Pre-DM) 或初治2型糖尿病 (TN-T2D) 的个体中,产生丁酸盐的菌含量显著降低。与正常耐糖个体相比, 患有糖尿病前期个体中的普拉梭菌的水平低于正常耐糖个体和初治2型糖尿病个体。
肠道微生物组可以反映整个身体的健康状况。厚壁菌门和拟杆菌门是结肠的主要门,可以影响与肥胖相关的疾病,这些疾病也是乳腺癌的危险因素。
✦ 乳腺癌患者F/B比值比健康人群低很多
因此,研究对乳腺癌患者和健康对照者的血清中提取细菌细胞外囊泡,使用微生物组测序分析厚壁菌门和拟杆菌门。分析乳腺癌的预后因素和血清学检测结果与 F/B 比值的相关性。发现乳腺癌患者的 F/B 比值比健康对照者低三倍。
此外,乳腺癌的危险因素,如空腹血糖,被发现与F/B比值有关。F/B比值可以作为乳腺癌的危险因素,并可能作为解释影响乳腺癌发生的潜在机制的线索之一。
健康对照者和乳腺癌患者中 F/B 的比率
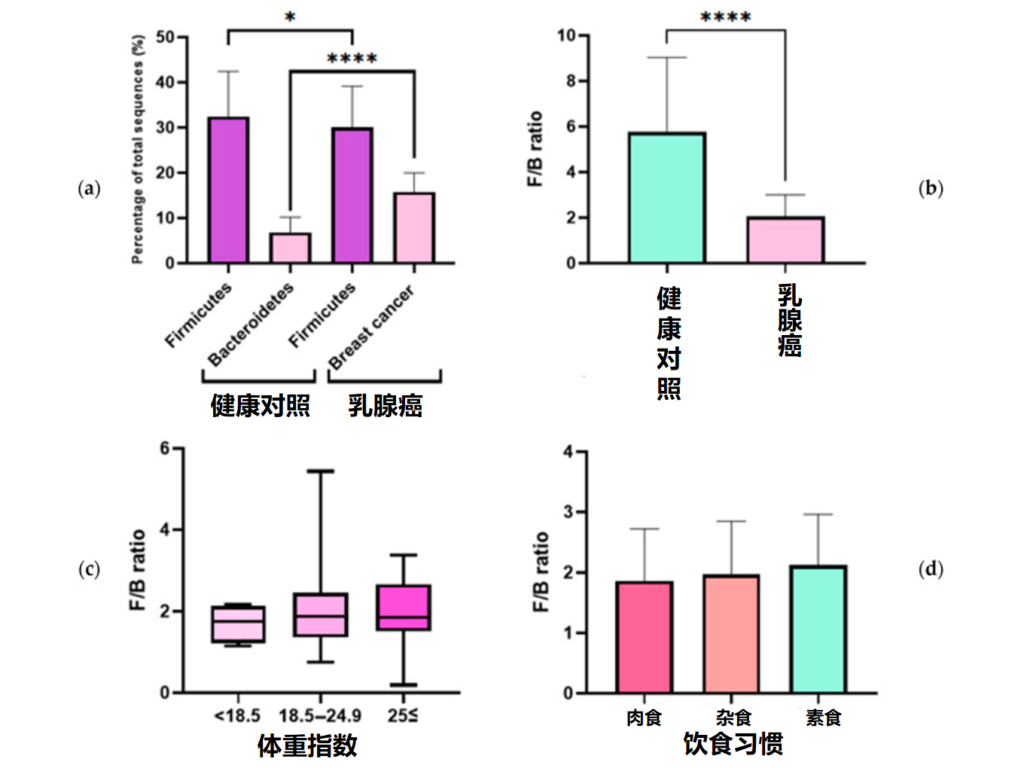
An J,et al.J Clin Med.2023
(a)健康对照组和乳腺癌患者中厚壁菌门和拟杆菌门占总微生物群的比例;(b)健康对照者和乳腺癌患者的F/B比率;(c)F/B比与体重指数之间的关系;(d)F/B比与饮食习惯的关系(*p≤0.05,**** p≤0.0001)。
调查研究发现中国饮食中的动物脂肪和蛋白质含量高于印度饮食中的全谷物和植物性素食。而印度参与者微生物组中拟杆菌的比例几乎是中国参与者的四倍,分别为16.39%和4.27%(p=0.001)。
✦ 动物产品食用较少与拟杆菌数量较多有关
据推测,印度人体内拟杆菌门的丰度较高是由于他们对动物产品的消耗较少;表明饮食中动物产品含量较低与拟杆菌数量较多有关。
对欧洲儿童和布基纳法索儿童的微生物群的比较也显示了由于饮食习惯不同而导致的微生物群组成的差异,欧洲儿童的饮食富含动物蛋白、糖、淀粉和脂肪,纤维含量低。以素食为主的饮食主要由碳水化合物、纤维和非动物蛋白组成。欧洲儿童微生物组富含厚壁菌门和变形菌门,而放线菌门和拟杆菌门在非洲儿童中较多。
✦ 营养不良儿童的肠道微生物多样性较低
研究发现,与同一地区中高收入家庭的健康儿童相比,孟加拉国贫困社会经济地位家庭的营养不良儿童的肠道微生物群多样性较低,其特点是拟杆菌门的相对丰度较低,变形菌门占主导地位。
注:拟杆菌门的存在率较低众所周知,它们可以消化复杂的膳食材料,从而提高从各种食物中提取能量,而潜在致病性变形菌的较高含量可能有助于解释营养不良儿童的健康状况不佳。
✦ 增加纤维摄入量可以增加厚壁菌门的数量
一项研究将西方饮食的美国儿童与食用大米、面包和扁豆等植物性饮食的孟加拉国儿童进行了比较。在食用西方饮食的美国儿童中,厚壁菌门/拟杆菌门的比例减少了三倍。
另一项研究要求参与者增加纤维摄入量并避免西方减肥食品。研究表明厚壁菌门:拟杆菌门比率增加了。
通过分析三名健康受试者6-12个月的肠道宏蛋白质组,进一步证实了这些结果。在这项研究中,参与碳水化合物运输和代谢的蛋白质占检测到的蛋白质的10%以上,构成了所有测试对象中发现的核心宏蛋白质组的一部分。
糖酵解途径尤其引人注目,并鉴定出几种相关酶。在为每个门的每个COG功能类别分配光谱命中后,很明显,厚壁菌门和放线菌门负责活跃的碳水化合物代谢,而拟杆菌门则显示出更多的混合功能。
这些例子反映了将某些门与特定饮食广泛联系起来主要挑战是需要考虑涵盖整个微生物组的微生物的状态和相互作用动态。
✦ 当F/B比率增加时
即更多厚壁菌门,拟杆菌较少。或两者兼而有之,厚壁菌门增长更快。可能导致以下疾病:
-肥胖(BMI较高)
-高血压中的肠道菌群失调
-自闭症谱系障碍
-慢性疲劳综合症
-肠易激综合症
-干燥综合症
-类风湿性关节炎
✦当F/B比率下降时
即厚壁菌门较少,更多拟杆菌。或两者兼而有之,厚壁菌门减少得更快。可能导致以下疾病:
-系统性红斑狼疮
-乳糜泻
-克罗恩病
-老龄化
我们审查评估了饮食干预如何影响微生物组成的相关文章。总体而言得出以下建议供参考:
✦ 增加植物蛋白摄入,避免摄入过量动物蛋白
总体而言,蛋白质摄入量,尤其植物蛋白的摄入与肠道拟杆菌门,细菌多样性呈正相关。乳清和豌豆蛋白可以增加双歧杆菌和乳酸菌的数量,而乳清还可以减少脆弱拟杆菌和产气荚膜梭菌的数量,这两种细菌都是致病性的。
此外,豌豆蛋白被发现可以增加短链脂肪酸水平,短链脂肪酸被认为具有抗炎作用,并在粘膜完整性中发挥作用。然而,一项研究发现,高动物蛋白/低碳水化合物饮食会导致罗氏菌属和直肠真杆菌(这两种有益细菌)减少。动物蛋白可能含有大量脂肪,而脂肪也会影响肠道微生物群。
✦ 尽量摄入不饱和脂肪
当研究人员观察脂肪时,发现并不是所有的脂肪都是平等产生的。在一项研究中,小鼠被喂食猪油或鱼的脂肪。喂猪油的小鼠中拟杆菌属和嗜胆菌属(Bilophila)增加,而双歧杆菌和Adlercreutzia、乳酸菌(乳杆菌和链球菌)和疣微菌属(Akkermansia muciniphila)在鱼油喂养的小鼠中增加。
猪油喂养的小鼠还出现白色脂肪组织炎症和胰岛素敏感性受损。喂食鱼油的小鼠总体表现更好,研究人员得出结论,肠道微生物群促进了高饱和脂肪饮食的整体炎症反应。
✦ 高纤维的饮食可以减少炎症
碳水化合物,包括可消化的(淀粉和糖)和不可消化的(纤维),可能是研究最多的常量营养素,因为它们具有改变肠道微生物组的能力。
人类喂食大量葡萄糖、果糖和蔗糖,双歧杆菌水平增加,拟杆菌减少。相反,当喂食糖精(一种人工甜味剂)时,拟杆菌丰度增加,罗伊氏乳杆菌减少,微生物发生与天然糖完全相反的变化。
纤维被认为是一种不易消化的膳食成分,有时被称为益生元,因为它通过刺激有益肠道细菌的生长而对宿主有益。
来源包括大豆、菊粉、未精制的小麦和大麦、生燕麦和不可消化的低聚糖,例如果聚糖、聚葡萄糖、低聚果糖 (FOS)、低聚半乳糖 (GOS)、低聚木糖 (XOS) 和阿拉伯低聚糖 (AOS)。
低纤维饮食与较低的细菌丰度有关,而富含这些不易消化的碳水化合物的饮食会增加双歧杆菌和乳酸杆菌。较高的纤维摄入量导致促炎细胞因子IL-6和胰岛素抵抗减少。
✦ 发酵食品可以增加双歧杆菌和乳杆菌水平
除了常量营养素和纤维对肠道细菌的影响外,含有乳酸菌的发酵食品(例如发酵乳制品)也被发现可以增加双歧杆菌和乳酸杆菌的水平。
✦ 多酚具有抗氧化活性能够减少一些致病菌
最后但并非最不重要的是多酚,包括儿茶素、黄酮醇、黄酮、花青素、原花青素和酚酸(存在于水果、种子、蔬菜、茶、可可制品和葡萄酒等食品中),被发现可以增加拟杆菌,减少金黄色葡萄球菌、鼠伤寒沙门氏菌和梭菌等致病菌。
拟杆菌门的成员被假设可以降低细胞内的氧水平,从而有利于厌氧菌的生长,从而促进肠道平衡的维持,并且它们被认为是关键的聚糖降解细菌比厚壁菌门等其他类群更能代谢多酚。
多酚可以改变肠道环境,它们可以被肠道细菌分解代谢,将它们转化为多种化合物比原始化合物具有更强的抗氧化活性。
厚壁菌门/拟杆菌门(F/B比率)作为一个指标在很多关于肠道菌群的研究中被提及和表征,从疾病发生进展,到药物饮食干预,涉及肠道菌群的研究,首先会描述细菌门水平的变化,而人类或动物肠道细菌约90%都属于厚壁菌门与拟杆菌门,而它们的比率也被认为是肠道菌群动态平衡的标志。
拟杆菌门主要为革兰氏阴性菌,能激活T细胞介导的免疫反应,降解多糖保护肠道和其他病原菌,厚壁菌门主要为革兰氏阳性菌,尤其产丁酸的菌,但是更容易吸收能量。
但是肠道菌群受到抗生素,药物,年龄以及分析注释数据库等的影响,此外随着肠道菌群研究队列的扩大,以及对菌群构成与不同疾病状态和药物饮食等因素的相关性研究的深入,F/B比率作为一个相对粗放的指标已经无法准确反映复杂的肠道菌群状况。我们需要更加深入和全面的通过菌属甚至菌种的构成变化来解释和分析肠道菌群特征,并进而更准确的解释肠道菌群和健康及生活方式的关联。
主要参考文献
Stojanov S, Berlec A, Štrukelj B. The Influence of Probiotics on the Firmicutes/Bacteroidetes Ratio in the Treatment of Obesity and Inflammatory Bowel disease. Microorganisms. 2020 Nov 1;8(11):1715.
Mariat D, Firmesse O, Levenez F, Guimarăes V, Sokol H, Doré J, Corthier G, Furet JP. The Firmicutes/Bacteroidetes ratio of the human microbiota changes with age. BMC Microbiol. 2009 Jun 9;9:123.
Gui X, Yang Z, Li MD. Effect of Cigarette Smoke on Gut Microbiota: State of Knowledge. Front Physiol. 2021 Jun 17;12:673341.
Grigor’eva IN. Gallstone Disease, Obesity and the Firmicutes/Bacteroidetes Ratio as a Possible Biomarker of Gut Dysbiosis. J Pers Med. 2020 Dec 25;11(1):13.
Vaiserman A, Romanenko M, Piven L, Moseiko V, Lushchak O, Kryzhanovska N, Guryanov V, Koliada A. Differences in the gut Firmicutes to Bacteroidetes ratio across age groups in healthy Ukrainian population. BMC Microbiol. 2020 Jul 22;20(1):221.
Indiani CMDSP, Rizzardi KF, Castelo PM, Ferraz LFC, Darrieux M, Parisotto TM. Childhood Obesity and Firmicutes/Bacteroidetes Ratio in the Gut Microbiota: A Systematic Review. Child Obes. 2018 Nov/Dec;14(8):501-509.
Hidalgo-Cantabrana C, Delgado S, Ruiz L, Ruas-Madiedo P, Sánchez B, Margolles A. Bifidobacteria and Their Health-Promoting Effects. Microbiol Spectr. 2017 Jun;5(3).
An J, Kwon H, Kim YJ. The Firmicutes/Bacteroidetes Ratio as a Risk Factor of Breast Cancer. J Clin Med. 2023 Mar 13;12(6):2216.

谷禾健康

就在前不久,世界卫生组织将阿斯巴甜列为可能致癌物,引发了人们不小的热议。阿斯巴甜作为人工甜味剂的一种,不提供任何卡路里,就可以提供蔗糖几百倍的甜度。
人工甜味剂几乎不提供热量,或只提供极少的热量,所以深受健身人士及减肥人群的喜爱。人工甜味剂相关产业发展迅速,许多无糖饮料、无糖冰淇淋、巧克力等食品中都有其身影。
除了人工甜味剂外,乳化剂、着色剂、防腐剂等食品添加剂也是现代食品产业的重要组成部分,用于维持或提高食品的口味、安全性、新鲜度和外观。
但随着食品中的添加剂越来越多,其对健康的影响正引起人们的担忧。在此背景下,食品添加剂对微生物群的影响受到广泛研究。
事实上许多国家允许的食品添加剂并不会直接对人产生有害影响,但是会改变肠道微生物及其代谢。近年来出现的证据表明食品添加剂和微生物群之间存在相互作用,这可能会间接影响宿主健康。
例如盐是用于食品保鲜的最常见天然添加剂之一。高盐摄入会改变肠道微生物群组成和粪便短链脂肪酸的产生,通过调节辅助T细胞17影响肠道免疫轴并促进局部和全身组织炎症,可能导致高血压和肥胖。
此外,人工甜味剂有可能引起糖代谢障碍。随着人工甜味剂摄入量的增加,患心血管疾病的风险上升,尤其是阿斯巴甜与脑血管疾病,安赛蜜和三氯蔗糖与冠动脉性心脏病的风险有关。
本文主要总结了食品添加剂中的人工甜味剂、多元醇、乳化剂、色素、防腐剂对人体肠道微生物的影响,并间接导致了一些宿主代谢的变化,已有研究发现过量摄入食品添加剂与肠道炎症之间存在关联。在最后,我们提出了一些有助于减轻食品添加剂对身体的损害的小建议。
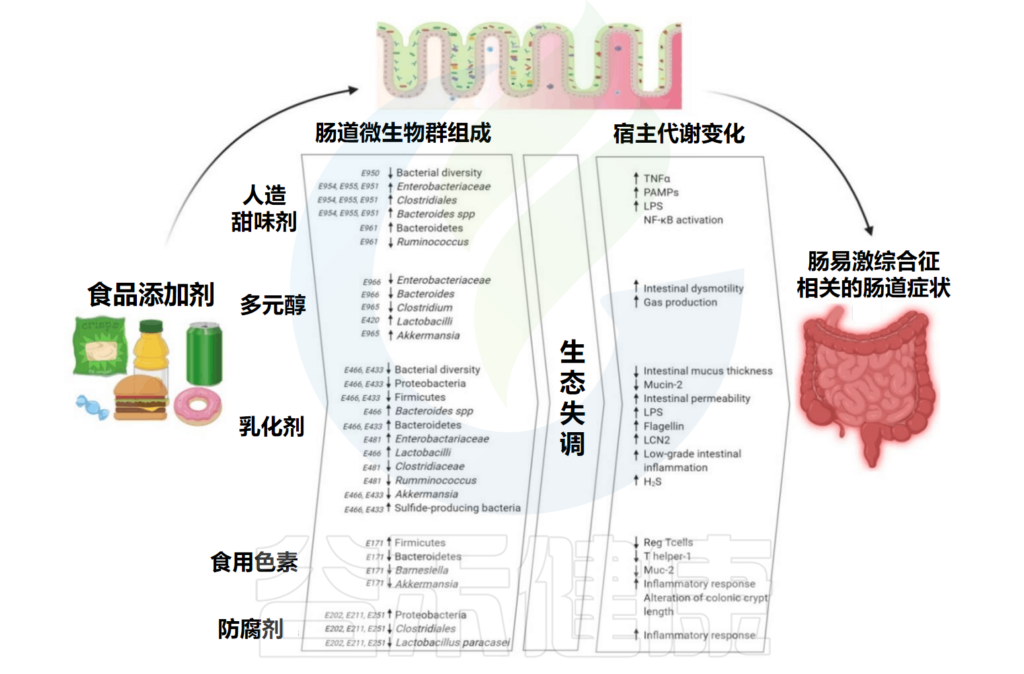
Rinninella E,et al.Int J Environ Res Public Health.2020
甜味剂与食品风味和人体健康密切相关,由于现在许多人们重视对身材的管理,无热量人工甜味剂逐渐兴起,其是蔗糖的替代品,特点是能够在不增加热量摄入的情况下具有更高的甜味强度,不会增加热量或引发血糖反应。
其中包括阿斯巴甜、安赛蜜、三氯蔗糖、糖精、纽甜和相应的化合物,主要存在于无糖饮料、无糖糖果和乳制品中。目前的研究已观察到人工甜味剂引起的肠道微生物群和代谢物的改变。
阿斯巴甜是一种低热量、强效的人造甜味剂。
•阿斯巴甜会改变人体肠道微生物的多样性
在人类中,阿斯巴甜可能不会改变粪便微生物群的丰度,但会改变其多样性。产生短链脂肪酸的细菌(例如双歧杆菌和另一项研究中检测到Blautia coccoides和拟杆菌/普雷沃氏菌比例下降。
•阿斯巴甜会影响短链脂肪酸浓度
在动物实验中,经阿斯巴甜处理后,厚壁菌门和柔嫩梭菌属(Clostridiumleptum)的丰度较高,肠球菌和副萨特氏菌 (Parasutterella)的丰度较低。
注:厚壁菌门和柔嫩梭菌都是重要的产丁酸菌属。
而在另一项人体研究中,随着阿斯巴甜的摄入,血清、粪便和盲肠内容物中的短链脂肪酸(包括丙酸和丁酸)浓度有所增加。
鉴于阿斯巴甜在不同动物和体外研究中存在一定的差异,阿斯巴甜对人体短链脂肪酸的影响还需要更多的实验研究,但目前可以确定的是,阿斯巴甜的摄入对人体短链脂肪酸的含量有影响。
阿斯巴甜存在一定的安全隐患!
需要注意的是,苯丙酮尿症患者苯丙氨酸代谢困难,应控制阿斯巴甜的摄入量。并且在此之前,美国一项研究发现,阿斯巴甜还会导致小鼠产生焦虑行为,这种影响还会跨代遗传。
阿斯巴甜对革兰氏阴性菌N-酰基高丝氨酸内酯(AHL)具有显著的抑制作用,通过群体感应抑制影响肠道微生物群落的平衡,从而促进消化系统疾病的进展。
此外,法国一项大规模前瞻性队列研究的结果表明,较高的人工甜味剂消耗量(尤其是阿斯巴甜)与心血管疾病风险增加之间存在潜在的直接关联。

安赛蜜,又称AK糖,是一种常用的人造甜味剂,也适合1-3岁儿童用于特殊医疗项目。
安赛蜜的可接受每日摄入量为9毫克/公斤体重
•安赛蜜在改变肠道微生物时具有性别特异性
在一项研究中,安赛蜜的摄入量在改变肠道微生物及其代谢物方面表现出高度的性别特异性。
在女性中,观察到乳杆菌属(Lactobacillus)和梭菌属(Clostridium)的丰度较低,Mucispirillum的丰度较高;而在男性中,拟杆菌属、萨特氏菌(Sutterella)、Anaerostipes有所增加。
•过量的安赛蜜会引起肠道炎症
在怀孕和哺乳期间将小鼠暴露于三氯蔗糖和安赛蜜会改变其后代肠道微生物群的α和β多样性,厚壁菌门增加和潜在抗炎细菌Akkermansia muciniphila的极度减少。
抗生素治疗后施用安赛蜜还会诱导硫酸盐还原菌脱硫弧菌的扩增以及结肠中促炎细胞因子的较高表达。
在炎症性肠病患者的粪便样本中,安赛蜜治疗后抗炎细菌减少,并伴有一些细菌功能改变,如脂多糖和胆汁酸合成。这表明安赛蜜诱导的肠道微生物组和代谢物扰动可能是破坏肠道稳态的因素,这可能会增加炎症性肠病的风险。
此外,安赛蜜可能会通过菌群失调引起吲哚美辛诱导的肠道损伤。
三氯蔗糖又名三氯半乳蔗糖,其甜度是蔗糖的600-650倍。
•长期摄入三氯蔗糖会改变肠道菌群并影响胰岛素水平
在人类研究中,摄入三氯蔗糖并不会在短期中影响肠道微生物,而年轻人摄入十周可能会导致Blautia coccoides增加和嗜酸乳杆菌减少,并改变肠道菌群和血清中的胰岛素和葡萄糖水平。
注:B. coccoides是一种参与促炎途径的细菌。
•三氯蔗糖的摄入导致体内多种代谢物改变
三氯蔗糖的摄入导致了多种代谢物的改变。酪氨酸水平升高,而对羟基苯乙酸和肉桂酸水平降低。这些化合物可以抑制参与色氨酸代谢的活性氧的产生。
此外,胆汁酸含量受损,盲肠内容物中胆酸浓度较高,胆酸/鹅去氧胆酸比率较高。研究人员认为这些代谢物的变化将导致引发和维持肝脏炎症。
▷大胆推测:三氯蔗糖会增加结肠炎的易感性
根据现有证据,我们认为三氯蔗糖对结肠炎有害,会增加促炎细菌的丰度。
三氯蔗糖会增加2,4,6,三硝基苯磺酸诱发的结肠炎易感性,并导致拟杆菌门和变形菌门的水平升高,厚壁菌门和放线菌门的数量减少。
最近的一项调查还发现,三氯蔗糖可能会增加患结肠炎相关结直肠癌的风险。
邻苯甲酰磺酰亚胺,俗称糖精。是一种热量为0的甜味剂,其甜度为蔗糖的300~500倍,但吃起来会有轻微的苦味和金属味。大部分糖精会被吸收并最终通过尿液排出,而未被吸收的糖精则通过粪便排出体外。
•糖精的消耗导致黏膜炎症细胞富集并改变肠道通透性
最近的一项研究发现,糖精的消耗会导致粘膜炎症细胞的富集并改变小鼠的肠道通透性。
此外,用抗生素预处理的糖精暴露小鼠可诱导较低水平的粘膜炎症和肠道屏障功能障碍。这些发现表明肠道菌群失调被认为是介导这些异常的原因。
•大量食用糖精会导致菌群失调
盲肠内容物中糖精的高含量与需氧菌群的增加有关。糖精暴露会导致严重的菌群失调,从而导致葡萄糖不耐受。许多增加的类群属于拟杆菌门和梭菌目,但厚壁菌门和蓝藻门减少了。
在无菌小鼠中无法诱导代谢变化,但在移植肠道微生物群后却发生了代谢变化。糖精还会导致肝脏炎症,导致阿克曼氏菌、棒状杆菌(Corynebacterium)和Turicibacter增加,并减少Anaerostipes、瘤胃球菌(Ruminococcus)和Dorea。
纽甜的味道比蔗糖甜7000-13000倍。纽甜的摄入降低了粪便微生物组的α多样性并改变了β多样性。
观察到厚壁菌门数量急剧下降。拟杆菌门,尤其是拟杆菌属的丰度增加。值得注意的是,毛螺菌科和瘤胃球菌科的多种成分显著减少,包括Blautia、Dorea、Oscillospira和瘤胃球菌属。
•纽甜的摄入改变了丁酸等物质的代谢
纽甜的摄入还改变了肠道微生物组的两种丁酸发酵途径。其中一条途径包括编码t4-羟基丁酰辅酶A脱水酶、丁酰辅酶A脱氢酶和乙酸辅酶A转移酶的三个基因的减少。这些基因参与了琥珀酸发酵为丁酸的过程。
另一方面,有关丙酮酸发酵丁酸的上游基因也被下调。此外,氨基酸代谢、脂多糖生物合成和抗生素生物合成增强,而脂肪酸和碳水化合物代谢途径减少。
为了方便大家观看,谷禾将一些食品添加剂与肠道微生物群之间的相互作用整理在了下表:




Liu C,et al.Nutrients.2022
人工甜味剂对肠道功能和代谢的影响
多项研究探讨了人工甜味剂对肠道微生物群和宿主生理功能的影响。他们证明人工甜味剂消耗与胰岛素抵抗和葡萄糖耐受不良增加之间的关联。
•富含人工甜味剂的饮食可能导致病态肥胖
研究表明,与对照小鼠相比,食用糖精的小鼠的聚糖降解显著增加。鉴于肠道细菌将聚糖发酵成短链脂肪酸,接触糖精可能会减少短链脂肪酸的产生。
丁酸盐通过肠-脑神经回路降低食欲和激活棕色脂肪组织而具有关键的抗肥胖作用。因此富含人工甜味剂的饮食可能与成年人的病态肥胖有关。
这些结果强调了一些人工甜味剂对聚糖发酵和短链脂肪酸产生的潜在不利影响,从而导致代谢紊乱。
•长期食用人工甜味剂会诱导炎症反应
此外,分析了给予糖精6个月后的小鼠肝脏,报告显示,与对照组相比,糖精处理的小鼠肝脏出现明显的肝脏炎症,且TNF-α基因表达升高。
注:TNF-α是炎症中的关键细胞因子,TNF-α的表达可以激活NFκB通路并诱导细胞损伤和炎症反应。
如前所述,在动物模型中,人工甜味剂的摄入可能会增加拟杆菌和肠杆菌科。肠杆菌科,如大肠杆菌、克雷伯菌属和变形杆菌属,位于粘膜上皮附近。
病原体感染、化学诱导的结肠炎或宿主免疫力不足可能导致肠道炎症,并刺激肠杆菌科的生长。因此,人工甜味剂似乎与肠道微生态失调、肠道炎症以及肠易激综合征相关的症状有关。
谷禾在下图中汇总了各种食品添加剂对肠道代谢产物的影响:



Liu C,et al.Nutrients.2022
多元醇,也叫糖醇,是一种有机物。主要种类有赤藓糖醇、异麦芽酮糖醇、山梨糖醇和木糖醇。它们天然存在于一些水果、蔬菜和蘑菇中。
其热量约为白糖的一半,并且只会导致血糖轻微变化。它们中的大多数在宿主中不能被很好地吸收或代谢,并且一部分会被结肠中的微生物群发酵。
糖醇对肠道微生物和代谢物的影响
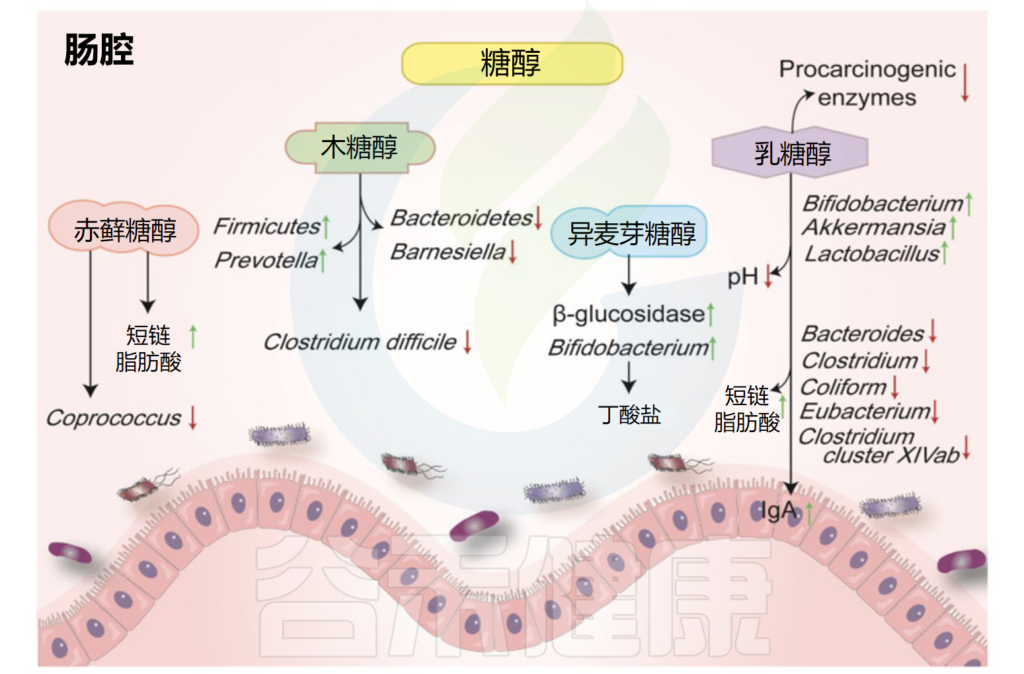
Liu C,et al.Nutrients.2022
赤藓糖醇在自然界中广泛存在,葡萄、梨、蘑菇、地衣中都有赤藓糖醇。摄入赤藓糖醇后,约90%可被小肠吸收,但代谢率很低,并主要以原形通过尿液排出体外。
•摄入赤藓糖醇增加了体内短链脂肪酸含量
在结肠中,肠道菌群可以代谢未吸收的部分。研究证明,摄入赤藓糖醇会导致丁酸和戊酸增加。考虑到到达大肠的赤藓糖醇数量有限,该化合物仅对肠道微生物群的组成产生轻微影响。
然而,赤藓糖醇会增强肠道微生物群以产生短链脂肪酸,从而减轻肠道炎症。赤藓糖醇被发现可以改善高脂肪饮食中的小肠炎症,并诱导粪球菌属丰度降低。
注:研究发现摄入赤藓糖醇后血清、粪便和白色脂肪组织中短链脂肪酸的浓度明显升高。
▷目前认为赤藓糖醇是一种对肠道稳定有利的物质
尽管缺乏赤藓糖醇对炎症性肠病的直接证据,但它被认为是一种对细菌友好的多元醇,可以稳定肠道微环境,并且可以降解为对炎症性肠病友好的代谢物。
异麦芽酮糖醇长期以来一直在食品和制药工业中用作甜味剂。微生物可以很容易地降解结肠中未吸收的异麦芽酮糖醇。
•异麦芽酮糖醇提高双歧杆菌的丰度
最近的研究认为它是一种益生元。在人类肠道中发酵的异麦芽酮糖醇可提高双歧杆菌的丰度并抑制致病菌,此外还降低了细菌β-葡萄糖苷酶的活性。
粪便中的短链脂肪酸、乳酸、胆汁酸、中性甾醇、苯酚和对甲酚也发生了变化。一些双歧杆菌菌株可以在体外发酵异麦芽,产生更高含量的丁酸盐。然而,暴露于异麦芽后没有发现不同的基因表达。
尽管很少有临床试验探讨异麦芽酮糖醇对肠道炎症的影响,但异麦芽酮糖醇可能被认为是一种双歧多元醇,是肠道稳态和微环境的“亲密朋友”。
木糖醇是是从白桦树、橡树、玉米芯、甘蔗渣等植物原料中提取出来的一种天然甜味剂。木糖醇被认为是一种非致龋性甜味剂,常被应用于口香糖中。
•木糖醇有一定的抗炎作用并能抑制变型链球菌
据报道,木糖醇会影响肠道菌群,并对脂多糖诱导的炎症细胞因子表达具有抑制作用。在一项人体研究中,摄入包括木糖醇在内的低消化性碳水化合物会导致Anaerostipes spp和粪便中的丁酸盐显著升高。
木糖醇降低了粪便拟杆菌(Bacteroidetes)和Barnesiella属的水平,而厚壁菌门和普雷沃氏菌(Prevotella)的丰度增加。较低的木糖醇浓度还可以抑制有害的变形链球菌。
注:变型链球菌是口腔天然菌群中占比例最大的链球菌属中的一种,是龋病的主要致病菌。
因此,我们认为木糖醇可能有利于肠道菌群的生长和代谢,而不会对肠道产生低有害刺激。
其他食品添加剂对肠道菌群及其功能的影响

编辑
Zhou X,et al.Molecules.2023
多元醇引起的菌群变化对肠道健康有益
多元醇属于所谓的“FODMAP”(可发酵低聚糖、二糖和单糖以及多元醇)饮食类别。近年来,FODMAP饮食已成为肠易激综合征患者的宝贵治疗选择,对其限制将改善肠易激综合征治疗的结果。
“FODMAP”一词于2004年创建,用于定义高度可发酵的碳水化合物和多元醇。
乳化剂在日常生活中被广泛应用,因为它们能够稳定乳液并通过防止储存期间分离来延长保质期。它们还可以通过食品中的脂肪分子充当胶凝剂和表面活性剂。
常见的乳化剂包括羧甲基纤维素、聚山梨酯、卡拉胶等。它们存在于各种加工食品中,如酱汁、布丁、人造黄油和冰淇淋,在西方饮食中非常普遍。研究发现乳化剂会通过肠道微生物影响人体健康。
乳化剂通过肠道微生物诱发结肠炎的机制
编辑
Liu C,et al.Nutrients.2022
羧甲基纤维素(CMC)和聚山梨酯80(P80)常见于食用油、冰淇淋、蛋糕粉、糖霜和巧克力糖浆中。然而,接触乳化剂CMC和P80会对肠道微生物群产生负面影响。
•可能通过微生物群驱动肠道炎症
有益菌如短链脂肪酸的生产菌减少
聚山梨酯80的施用引起了与炎症性肠病类似的人类肠道微生物群的改变,导致有益的双歧杆菌属、重要的短链脂肪酸生产者如粪杆菌属和罕见小球菌属 (Subdoligranulum)以及Clostridium leptum的减少。
炎症性肠病相关病原菌增长显著
在小鼠中,摄入P80会加剧吲哚美辛诱发的回肠炎,从而降低肠道微生物群的α多样性。包括肠杆菌科在内的硫化物生产者的生长和炎症性肠病相关物种奇异变形杆菌的群集行为得到显著促进。
群集是一种鞭毛介导的运动,需要营养细胞分化成一种称为群集细胞的特殊细胞类型。奇异变形杆菌的群集能力与其在炎症性肠病中的发病机制高度相关。
增加病原菌的入侵
研究发现长期摄入羧甲基纤维素或聚山梨酯80会诱发低度肠道炎症,并促进易患这种疾病的小鼠出现严重结肠炎,最终导致细菌入侵增加。
卡拉胶,又名角叉菜胶,是从麒麟菜、石花菜、鹿角菜等红藻类海草中提炼出来的亲水性胶体。它们常见于调味牛奶、冰咖啡、乳制品冰淇淋和冷冻甜点中。
卡拉胶的代谢主要由宿主肠道微生物群进行。因此,肠道微生物直接与卡拉胶相互作用,影响肠道稳态。有趣的是,不同种类和分子量的卡拉胶对宿主有不同的影响。
卡拉胶可分为低分子量或高分子量、降解或未降解卡拉胶。
•低分子量的卡拉胶会增加肠道通透性并与结肠炎相关
低分子量卡拉胶已被证明可以增加肠道通透性并与结肠炎的发生有关。据报道,角叉菜胶会诱发和加重肠道炎症,改变肠道微生物群组成。
卡拉胶的摄入导致变形菌门和脱铁杆菌门(Deferribacteres)的增加,以及厚壁菌门、放线菌门和拟杆菌门的减少。
卡拉胶的不同异构体均被认为会对肠道生态产生有害影响。对于人类肠道微生物群,每种异构体都会诱导α-多样性的变化并增加微生物群的促炎潜力。
κ-卡拉胶和λ-卡拉胶显著富集了拟杆菌,而ι-卡拉胶减少了Faecaliberium,处理后鞭毛蛋白含量更高。κ-卡拉胶在高脂饮食模型中诱导了结肠炎,显著增加了志贺菌并减少了双歧杆菌。两种炎症相关细菌Alistipes finegoldii和Bacteroides acidifaciens的丰度也显著增加。
然而,高分子量卡拉胶可能具有良好的抗肿瘤和抗氧化活性。此外,之前的研究认为,肠道微生物组的改变仅在饮用水中添加卡拉胶时观察到,而在补充卡拉胶的饮食中则未观察到,这表明与蛋白质等其他食物成分的结合可能改变其构象并消除其对某些细菌的生物利用度。
▷推测:食用卡拉胶不利于肠道炎症患者恢复
在一项研究中,两种协同菌株:Bacteroides xylanisolvens 38F6A4和E. coli 38F6C1是从健康人的粪便中获得的,并给予无菌小鼠,与卡拉胶一起食用时会加剧肠道炎症。
对于缓解期的溃疡性结肠炎患者,在含卡拉胶的饮食治疗后复发率较高。鉴于卡拉胶对炎症性肠病患者和肠道菌群的直接有害作用,我们推测卡拉胶可能由于肠道菌群和肠道稳态紊乱而对炎症性肠病患者产生有害影响。
!
乳化剂可能通过肠道微生物影响宿主心理
研究结果表明,膳食乳化剂也可能对肠脑轴产生影响,并通过微生物群依赖机制诱导暴露者的心理/行为障碍。
肠道微生物群中聚山梨酯80和羧甲基纤维素依赖性的变化也是小鼠性别特异性行为和神经改变的原因。特别是,雌性获得了反社会行为,而雄性表现出更高的焦虑水平。
这两种变化都与微生物群特征和α-黑素细胞刺激素水平的明显变化相关,这两个神经肽参与调节焦虑相关行为、食欲和能量。
小结
乳化剂中羧甲基纤维素和聚山梨酯80过量食用会导致生态失调,粘液降解细菌过度生长,以及白细胞介素10或Toll样受体5进一步缺乏。
乳化剂可能还会增加肠上皮屏障中病原微生物的易位,引起肠道炎症,从而导致炎症性肠病发病率增加。
除此之外,乳化剂等胶体还会通过肠脑轴、肠肝轴、肠道与骨骼、心血管之间的联系进一步影响人体代谢和健康。
其他乳化剂在体内的生理影响



Tan H,Nie S.FEMS Microbiol Rev.2021
乳化剂和相关微生物代谢对人体的影响
编辑
Tan H,Nie S.FEMS Microbiol Rev.2021
食品着色剂又称食品色素,是以食品着色为主要目的,赋予食品色泽和改善食品色泽的物质。
食品着色剂主要添加到奶酪、酱汁、冰淇淋、糕点、糖果、巧克力和口香糖中。
二氧化钛(TiO2)是我国常用的食品着色剂,通常用作食品中的增白剂或增亮剂。
•二氧化钛影响肠道保护细菌的丰度
近年来,各种研究试图确定口服二氧化钛对小鼠和人类肠道微生物群组成的影响。在用二氧化钛处理的小鼠中,与对照组相比,观察到厚壁菌门显著增加,拟杆菌门减少。特别是,巴恩斯氏菌(Barnesiella),一种关键的保护性肠道细菌,其丰度受到二氧化钛暴露(160mg/kg/天,持续28天)的显著影响。
巴恩斯氏菌(Barnesiella)可以清除肠道中的有害细菌。消除对万古霉素耐药的肠球菌的定植,并抑制抗生素耐药细菌的传播。它还可以改善环磷酰胺等抗癌化合物的性能。
巴恩斯氏菌(Barnesiella)的减少可能与炎症性肠病的发病机制有关。此外,它们导致结肠中的炎症浸润和线粒体异常,粪便中Turicibacter属和格氏乳杆菌(Lactobacillus gasseri)和Lactobacillus NK4A136_group的增加。
•高剂量的二氧化钛会诱导肠道炎症加剧
在人类中,添加二氧化钛会导致肠道微生物群多样性适度下降,并导致卵形拟杆菌(Bacteroides ovatus)和Clostridium cocleatum减少。
卵形拟杆菌(Bacteroides ovatus)被誉为新一代益生菌,在肠道免疫、肿瘤治疗、膳食纤维代谢方面发挥重要作用。
剂量反应实验表明,单独使用微量二氧化钛对两组巨噬细胞的吞噬作用没有影响。然而,高剂量的二氧化钛与细菌抗原协同作用,可促进巨噬细胞产生 IL-8、肿瘤坏死因子-α和IL-10,并损害其转化生长因子-β分泌/吞噬细胞活性。
据报道,喂食富含二氧化钛饮食的小鼠肠道炎症加剧。特别是,微生物群组成发生了显著变化,活性氧释放增强,NLRP3炎症小体激活。
!
食品着色剂会影响肠道稳态
二氧化钛等食品着色剂可能会影响肠道稳态。事实上,各种研究已经证明了二氧化钛暴露与对肠道微生物群功能性不利影响之间的潜在关联。
炎症反应加剧
在口服二氧化钛一周(10mg/kg体重/天)的大鼠中,在派尔贴片的免疫细胞和参与炎症反应的调节性T细胞中检测到二氧化钛。具体而言,在二氧化钛暴露后,从Peyer贴片中分离的免疫细胞的刺激显示辅助性T细胞减少,IFN-γ分泌增加,Th1/Th17炎症反应增加。
注:更进一步研究表明二氧化钛暴露与啮齿动物肠道疾病和结直肠癌的发展之间可能存在关联。
短链脂肪酸水平降低
研究了小鼠体内不同剂量的二氧化钛(2、10、50 mg/kg体重/天),并报告了高剂量暴露后短链脂肪酸水平的降低、粘液相关基因表达的降低、炎症反应的增加和结肠隐窝长度的改变。
不同食品着色剂对粘膜屏障和肠道微生态的影响
Liu C,et al.Nutrients.2022
所有这些发现表明,由于暴露于二氧化钛等食品着色剂而导致的菌群失调可能导致微生物群失调,其中特定科和属细菌的变异可能参与炎症性肠病的发病机制。
食品防腐剂可以延缓食品的降解,延长食品的保质期,限制微生物的生长,抑制食品的氧化。
防腐剂的益处和安全性存在争议,许多常见的防腐剂,如苯甲酸、山梨酸钾、亚硝酸钠和亚硫酸钠以及银纳米颗粒被报道可以诱导肠道微生物群的改变。
美国儿科学会警告说,防腐剂可能与恶化的多动行为或致癌风险有关。
苯甲酸和苯甲酸钠均可作为食品防腐剂,因为它们能够限制病原微生物的生长。所有吸收的苯甲酸均可完全降解为马尿酸。
•适量的苯甲酸会提高有益微生物的水平
最近的研究发现苯甲酸和苯甲酸钠通过调节氧化状态和免疫状态对肠道屏障功能和肠道微生物群的有益影响。
苯甲酸处理增强了回肠微生物群的生物多样性,以剂量依赖性方式降低了总需氧细菌的丰度。它还会减少十二指肠中革兰氏阴性菌的丰富度。在小猪中,苯甲酸处理增加了肠绒毛的高度,提高了有益微生物(例如双歧杆菌和乳杆菌)的水平,并减少了有害微生物的数量。
•过量摄入可能会破坏肠道屏障
然而,过量摄入苯甲酸可能会通过氧化还原状态破坏肠道屏障。
最近的研究报告称,苯甲酸增加了小鼠中诱导变形菌失调的易感性。值得注意的是,变形菌的增殖被认为是生态失调的潜在诊断标志物,并且与炎症性肠病等疾病的风险相关。
虽然现有的研究对苯甲酸表现出相互矛盾的观点,仍需要更多的研究来阐明其对肠道菌群和肠道炎症的影响。
山梨酸钾是一种低毒防腐剂,能强烈抑制腐败细菌和霉菌,其对感官特性影响轻微。
•山梨酸钾会降低肠道微生物多样性
先前的研究报道,山梨酸钾通过改变宿主免疫来抑制肠道微生物群的生存能力。暴露于山梨酸钾显著降低了斑马鱼肠道中IgG、IL-1β和TNF-α的含量,并激活了免疫系统。
在属水平上,有益菌粪杆菌(Faecalibacterium)以及病原菌气单胞菌(Aeromonas)和甲基杆菌(Methylobacterium)的含量呈现显著下降趋势。在体外,施用山梨酸钾后,普氏粪杆菌的生长也显著下降。
•革兰氏阳性菌可能阻碍山梨酸钾的传递
值得注意的是,山梨酸钾的亲脂性在调节不同类型的肠道微生物群中发挥着重要作用。革兰氏阴性细菌细胞壁中含有大量脂质,从而抑制转录和碳水化合物代谢途径。然而,革兰氏阳性菌含有较高的肽聚糖含量,可能会阻碍山梨酸钾的传递。大肠杆菌已被证明通过其外排泵机制对山梨酸钾具有抗性。
亚硫酸钠是常见的食品防腐剂之一。
•亚硫酸钠对人体有一定的危害
它强烈抑制普拉梭菌(Faecalibacterium prausnitzii)的生长。干酪乳杆菌、鼠李糖乳杆菌和植物乳杆菌因具有产生乳酸的能力而被认为是有益的肠道细菌,但它们也减少了。
在另一项研究中,亚硫酸钠持续降低奇异变形杆菌、Escherichia fergusonii、摩氏摩根菌(Morganellamorganii)的活力。亚硫酸钠对肠道有益菌较强的抗菌能力,可能诱发肠道疾病。
•具有抗炎特性的细菌可能对防腐剂更敏感
在一项体外研究中,发现人类肠道微生物对亚硝酸钠、苯甲酸钠和山梨酸钾高度敏感,尤其是酪丁酸梭菌(Clostridium tyrobutyricum)或副干酪乳杆菌(Lactobacillus paracasei),具有已知的抗炎特性,与具有促炎或致大肠杆菌特性的粪肠球菌或多形拟杆菌相比,对这三种防腐剂更敏感。
减轻食品添加剂对身体损害的一些建议
•选择天然食物:尽量选择天然食物,例如新鲜水果、蔬菜、全谷物和未经加工的肉类。这些食物通常不含添加剂。
•阅读食品标签:在购买加工食品时,仔细阅读食品标签。避免购买添加剂含量较高的食品,特别是那些含有大量人工色素和防腐剂的食品。
•自制食物:尽量选择自制食物,这样可以控制所使用的成分和添加剂的量。自制食物可以使用新鲜的食材,并尝试使用天然的调味料来增添风味。
•限制加工食品摄入量:减少加工食品的摄入量,特别是那些高度加工和包装的食品。这些食品通常含有更多的添加剂。
•多样化饮食:保持饮食的多样性,摄入各种不同类型的食物。这样可以减少对某一种食品及其潜在添加剂的过度依赖。
需要注意的是,并非所有的食品添加剂都对每个人都会产生负面影响。食品添加剂经过严格的监管和安全评估,以确保在合理摄入量下的安全性。
然而,对于敏感人群来说,或在过量摄入的情况下,某些食品添加剂可能会引起问题。因此,适度和平衡地摄入食品添加剂是很重要的。
越来越多的研究调查肠道微生物群与食品添加剂之间的相互作用,在总结当前研究中收集的信息后,我们得出结论:食品添加剂对肠道微生物群和肠道稳态产生多种影响,这可能与肠道炎症的发病和进展有关。
食品添加剂显著介导细菌功能的改变。总而言之,安赛蜜、三氯蔗糖和糖精等甜味剂可能通过提高细菌炎症潜力而诱发或加剧结肠炎。
羧甲基纤维素和聚山梨酯80等乳化剂被认为会改变肠道微生物群的多样性并增加细菌的侵入,从而对肠道健康有害。
食品着色剂的安全性应重新评估,因为其会引发肠道紊乱和菌群失调;然而,多元醇似乎通过改善肠道微生物的结构和功能而对肠道微生物群有益。
根据目前的证据和法规,必须谨慎考虑对炎症性肠病患者不利的食品添加剂,例如食品着色剂和乳化剂,包括羧甲基纤维素和聚山梨酯80。
尽管研究显示食品添加剂对肠道微生物组和肠道炎症有不同的影响,但将体外或动物模型中获得的相同结果归因于人类是不可行的。因此,未来的研究应该基于生物相关模型来复制人类的生理条件。
总之,需要更多的研究来阐明食品添加剂、肠道微生物群和人体健康之间的关系,并了解后代由于当代人的消费习惯而可能面临的炎症性肠病风险。
相关阅读:
你的焦虑可能与食品添加剂有关,警惕食品添加剂引起的微生物群变化
肠道微生物群与健康:探究发酵食品、饮食方式、益生菌和后生元的影响
主要参考文献
Zhou X, Qiao K, Wu H, Zhang Y. The Impact of Food Additives on the Abundance and Composition of Gut Microbiota. Molecules. 2023 Jan 7;28(2):631.
Rinninella E, Cintoni M, Raoul P, Gasbarrini A, Mele MC. Food Additives, Gut Microbiota, and Irritable Bowel Syndrome: A Hidden Track. Int J Environ Res Public Health. 2020 Nov 27;17(23):8816.
Tan H, Nie S. Functional hydrocolloids, gut microbiota and health: picking food additives for personalized nutrition. FEMS Microbiol Rev. 2021 Aug 17;45(4):fuaa065.
Liu C, Zhan S, Tian Z, Li N, Li T, Wu D, Zeng Z, Zhuang X. Food Additives Associated with Gut Microbiota Alterations in Inflammatory Bowel Disease: Friends or Enemies? Nutrients. 2022 Jul 25;14(15):3049.
Del Pozo S, Gómez-Martínez S, Díaz LE, Nova E, Urrialde R, Marcos A. Potential Effects of Sucralose and Saccharin on Gut Microbiota: A Review. Nutrients. 2022 Apr 18;14(8):1682.
Ravash N, Peighambardoust SH, Soltanzadeh M, Pateiro M, Lorenzo JM. Impact of high-pressure treatment on casein micelles, whey proteins, fat globules and enzymes activity in dairy products: a review. Crit Rev Food Sci Nutr. 2022;62(11):2888-2908.
Laudisi F, Stolfi C, Monteleone G. Impact of Food Additives on Gut Homeostasis. Nutrients. 2019 Oct 1;11(10):2334.
Abiega-Franyutti P, Freyre-Fonseca V. Chronic consumption of food-additives lead to changes via microbiota gut-brain axis. Toxicology. 2021 Dec;464:153001.

谷禾健康

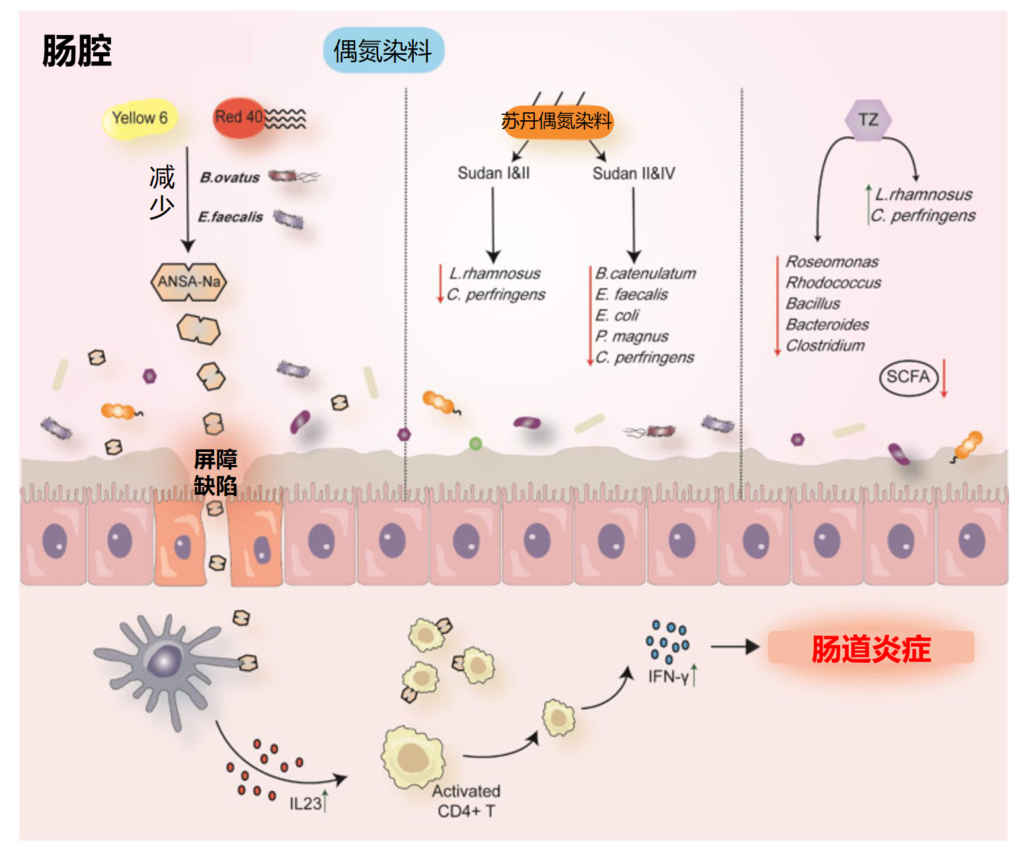
营养不良的微生物 Malnourished Microbes
儿童营养不良是全球主要的健康负担,营养干预措施只能部分解决这一问题。儿童营养不良的慢性和急性形式均以多种生物系统紊乱为特征,包括新陈代谢、免疫和内分泌系统。
越来越多的证据支持肠道微生物组在调节这些影响早期生命生长的途径中的作用。观察性研究报告了营养不良儿童肠道微生物组的变化,而临床前研究表明,这可能引发肠病,改变宿主代谢,并破坏免疫介导的针对肠道病原体的抵抗力,这些都会导致早期生命生长不良。
肠道微生物组在健康和疾病中发挥着不可或缺的作用,而饮食是其组成、多样性和功能的主要驱动因素。鉴于婴儿和儿童肠道微生物组的动态发育,解决两个主要问题至关重要:
a) 饮食能否改变肠道微生物组的组成、多样性或功能
b) 这种修改是否会影响功能/临床结果,包括免疫功能、认知发展和整体健康?
本文汇编了临床前和临床研究的证据,包括生命早期肠道微生物组影响宿主代谢、免疫、肠道功能、内分泌调节和其他导致儿童营养不良的途径的新兴病理生理学途径。
我们综合了儿童青少年的营养、营养干预措施和肠道微生物群之间联系的最新研究,该年龄段儿童是一个生长阶段,对疾病的易感性各不相同。还讨论了营养素如何单独或组合如何影响肠道微生物组的机制。
同时综合并讨论了新兴的微生物组导向疗法,并考虑未来的研究方向,以确定和针对儿童营养不良的微生物组治疗目标。
2013年发表在《柳叶刀》上文章统计,儿童营养不良导致全球儿童死亡人数的 45%,并对健康产生终生影响。全世界超过五分之一的5岁以下儿童至少表现出一种形式的慢性或急性营养不良。
首先我们来了解一下发育迟缓、消瘦的定义。
发育迟缓[定义为年龄别身长 Z 得分 (LAZ) < −2],影响着全球 22% 的儿童,与认知发展较差、终身综合能力下降以及成年后慢性病风险增加有关。
消瘦[定义为身高体重 Z 得分 (WHZ) < –2] 影响着全世界近 7% 的儿童,除了长期认知和健康缺陷不佳之外,还与高死亡率相关。
▸ 目前的治疗方法无法完全解决这一问题
营养疗法不足以完全恢复生长缺陷和营养不良的相关病理生理后果。标准生命早期营养补充剂仅可将发育迟缓降低 12-14%。
而严重消瘦后会出现高死亡率、再入院和长期生长缺陷。因此,存在与儿童营养不良相关的隐藏的病理生理学负担,目前的治疗方法无法完全解决这一问题。
▸ 儿童营养不良与多个生命早期生物系统紊乱有关
发育迟缓和消瘦儿童的肠道功能受到环境肠道功能障碍(EED)的影响。这种肠道病理学损害营养吸收,并可能导致全身炎症,从而损害早期生命生长。
在营养不良的儿童中宿主免疫力广泛缺陷,导致抵抗感染能力受损,特别是病原体密集的环境。
营养良好和营养不良的儿童之间存在激素差异,尤其是与生长 [生长激素 (GH) 和胰岛素样生长因子1 (IGF-1)] 和食欲调节(瘦素和生长素释放肽)相关的荷尔蒙差异。
营养不良时宿主代谢发生改变,氨基酸、脂质和能量代谢受到干扰。这些受到干扰的生理系统表明,儿童营养不良背后存在复杂的病理生理学,涉及相互关联的生物系统。
▸ 微生物群影响生命早期的生长发育
生命早期接触微生物也可能导致儿童营养不良中这些扰动途径。肠道病原体携带量高与线性生长和体重生长下降以及 EED 相关,而腹泻和呼吸道感染也与儿童营养不良密切相关。
除了病原体之外,共生肠道微生物群也会影响早期生命的生长。生命早期复杂微生物组的组装对于免疫训练、对病原体的定植抵抗、母乳寡糖(HMO) 和其他营养物质的代谢、肠道结构和内分泌信号传导至关重要。
因此,儿童时期肠道微生物组的破坏可能会损害这些有助于儿童健康成长的途径。
▸ 儿童在出生时就获得了一些的微生物组
除了环境获得的微生物外,这些微生物组主要源自母体肠道、阴道、口腔和皮肤微生物组。
在纯母乳喂养的儿童中,最初的婴儿肠道微生物群的多样性仍然很低,主要由大肠杆菌、双歧杆菌和拟杆菌组成物种。
在接下来2-3 年里,肠道微生物群会经历一种模式化的组装,这种组装主要是由纯母乳喂养和随后的补充食品的引入所形成的,这导致肠道微生物群的快速扩张和多样化。
▸ 许多环境因素会影响微生物,从而影响生长
包括地理位置、分娩方式、抗生素暴露、胎龄以及其他母亲和家庭因素。这些微生物群在出生后在胃肠道内的积累,除了肠道结构和对病原体的定植抵抗力之外,还推动了免疫、代谢和内分泌途径的成熟,这些都有助于儿童的正常生长。
如果微生物演替因营养不足、卫生或抗生素暴露而受到干扰,这些生长决定途径可能会受到损害。很少有研究纵向研究在营养匮乏的环境中保持良好营养的儿童与消瘦或发育迟缓的儿童的肠道微生物群的演替和组装情况。
因此,这种依赖高收入环境的微生物组数据库的现象,限制了低收入和中等收入国家中个体所产生的营养不良微生物组的解释性(这也是构建本地数据库和谷禾长期追踪重点人群纵向数据的主要原因),这些国家的肠道微生物组特征不太明确。
来自中低收入国家的横断面和短期纵向研究的越来越多的证据揭示了微生物组的差异,这些差异将营养不良与健康生长区分开来。
▸ 发育迟缓——多因素驱动
“发育迟缓”是一种慢性营养不良的形式,由多种遗传和环境因素共同驱动。高达30%的发育迟缓发生在子宫内,这可以通过出生体重对发育迟缓风险的强烈影响来证明。
之后一系列环境因素有助于产后线性生长。很少有研究纵向研究健康婴儿与发育不良婴儿肠道微生物组的获取和组装情况。
▸ 横断面研究:肠道菌群特征破坏导致发育迟缓
对马拉维和孟加拉国队列的二次分析发现,微生物组多样性减少和Acidaminococcus(氨基酸球菌)丰度增加分别与发育迟缓严重程度和未来线性生长缺陷相关。
其他一些小型横断面研究已经发现,在一系列中低收入国家队列中,发育不良儿童与健康儿童的粪便微生物组组成存在不同差异。来自中非共和国和马达加斯加的一项更大规模的多点横断面分析发现,发育不良儿童的粪便微生物群中大肠杆菌/志贺氏菌和弯曲杆菌种类丰富,而产生丁酸的种类较少。
▸ 发育迟缓的儿童:小肠细菌过度生长
在发育迟缓的儿童中观察到小肠细菌过度生长的比率较高,其特征是十二指肠中口咽微生物的富集。发现一群来自孟加拉国的发育迟缓儿童的小肠细菌载量与 LAZ 之间呈负相关。韦荣球菌属、链球菌属和Rothia mucilaginosa的十二指肠丰度也与 LAZ 呈负相关。
▸ 肠道菌群的功能潜力——预测儿童生长的指标
对 335 名津巴布韦农村的 1 至 18 个月大儿童的肠道微生物组的组成和功能发育的跟踪发现,肠道微生物组的功能宏基因组组成(而非分类组成),可以预测孩子已达到的线性生长和未来的生长速度,其中 B 族维生素和核苷酸生物合成途径是最具预测性的特征之一。这些数据表明,肠道微生物组的功能潜力是儿童生长的更强大的指标。
来自马拉维的一个纵向队列发现了类似的结果,即 16S 测序无法识别成分多样性、成熟度或物种丰度与 LAZ 之间的关联。秘鲁对 6-24 个月大的儿童(作为营养不良和肠道疾病队列的一部分)进行的一项纵向研究表明,成分多样性与线性生长之间存在一定关联,仅针对出生时发育迟缓的儿童。
该研究组另外一项研究究表明,肠道微生物组可能介导腹泻和弯曲杆菌感染对线性生长的影响。除了肠道微生物组细菌成分的影响之外,发育迟缓婴儿与非发育迟缓婴儿中噬菌体(可以调节微生物组组成的病毒)的丰度也有所不同。
总的来说,发育不良儿童的肠道微生物群与营养良好的儿童不同。然而,这因地理群体而异。因此,目前不存在一致的发育迟缓的组成或功能微生物组特征。
▸ 儿童口腔微生物组与发育迟缓有关
发育迟缓的儿童在十二指肠和粪便中表现出更丰富的口咽微生物,包括唾液乳杆菌,这表明胃肠道的隔室化可能导致营养不良儿童中肠道菌群变化,并通过减少营养吸收而导致生长不良或口腔微生物在其生态位之外的炎症作用。
▸ 怀孕期间母亲的口腔微生物组预测儿童营养不良
牙周炎是一种口腔生态失调疾病,会导致局部免疫反应功能失调,并与低出生体重呈负相关。在怀孕期间的母亲口腔中,Actinomyces naeslundii(内氏放线菌)的相对丰度与出生体重和妊娠持续时间呈负相关,而Lactobacillus casei(干酪乳杆菌)则呈正相关,两者都强烈预测儿童营养不良。
▸ 早产的阴道微生物组,与新生儿低出生体重和随后的发育迟缓密切相关
高度多样化、缺乏乳杆菌的阴道微生物群与西方地区的早产以及撒哈拉以南非洲地区新生儿 LAZ 的减少密切相关。阴道微生物组的改变很可能与早产的经典感染触发因素同时发生,包括绒毛膜羊膜炎、细菌性阴道病和泌尿生殖感染,从而导致泌尿生殖环境炎症,这些共同可能限制胎儿生长或引发早产。
▸ 消瘦及其分类:营养不良形式的反映
消瘦是营养不良的一种形式,可以反映慢性营养缺乏和急性疾病,通常与发育迟缓同时发生。
消瘦可分为中度[中度急性营养不良(MAM)]或重度[严重急性营养不良(SAM)],取决于 WHZ 阈值、中上臂围 (MUAC) 和/或水肿的存在。
▸ 水肿性SAM、非水肿性SAM、复杂SAM
SAM 还以两种主要形式存在:
如果出现急性感染、食欲不振、休克或水肿(也称为复杂 SAM)。
患有水肿性和非水肿性 SAM 的儿童具有不同的临床结果,其中非水肿性 SAM 儿童在某些情况下接受 SAM 住院治疗后表现出更高的死亡率和再入院率。
尽管治疗 SAM 的营养疗法有所改进,但需要住院治疗的复杂 SAM 的死亡率在 10% 至 40% 之间,而生长缺陷可能会持续长达7年,此外,日后认知缺陷和慢性病风险也会增加。
▸ 在SAM 儿童肠道中肠道微生物组具有影响力
患有 SAM 的儿童肠道微生物多样性降低,肠杆菌科细菌增多,而与健康生长一致的双胞胎相比,SAM不一致的双胞胎肠道病毒组受到干扰。
▸ 年龄微生物群Z评分——微生物群成熟度指标
2014 年,孟加拉国的一项具有里程碑意义的研究创建了微生物群成熟度指标,称为年龄微生物群Z评分(MAZ),该评分在 SAM 儿童中显着降低,并且与术后营养恢复的人体测量指标高度相关。这项研究和最近的后续研究使得所谓的微生物组生态群得以细化,该生态群由 15 种细菌类群组成,这些细菌类群在不同地理环境中在生命的头 2 年中表现出一致的共变,可用作微生物群落的指标。
不成熟的微生物组发育并衡量MAM 或 SAM 后微生物组的恢复程度。根据 MAZ 的评估,标准治疗性往往只能暂时恢复微生物组的成熟度,表明 MAZ 可能有潜力作为未来 MAM/SAM 复发的指标,或者针对肠道微生物组的营养疗法可能有助于营养恢复。
事实上,这些微生物群成熟度指标为 MAM 中新的微生物群导向营养干预措施提供了信息,在试点研究中,与标准治疗食品相比,这些干预措施在更大程度上改善了营养恢复。
▸ 在水肿性SAM 和非水肿性 SAM 儿童差异
这可能部分解释了他们营养恢复的差异。与水肿性SAM儿童相比,非水肿性 SAM 儿童的α多样性显著降低,并且与普雷沃氏菌科(Prevotellaceae)、毛螺菌科 (Lachnospiraceae) 、瘤胃菌科(Ruminoccoaceae)相对丰度的减少有关。
总的来说,与健康成长的儿童相比,SAM 儿童的微生物组始终不成熟且多样性降低。这种失调的微生物组可能是急性营养状态以及与治疗相关的合并症和抗生素治疗的结果。
如上所述,越来越多的人类观察研究证据表明,营养良好和营养不良的儿童之间的微生物组组成存在差异。
临床前和临床研究已开始揭示,生命早期肠道微生物群紊乱可能导致儿童生长不良的病理生理机制。其中包括肠道微生物组对免疫、新陈代谢、肠道功能和内分泌信号传导等途径的影响(如下图)。
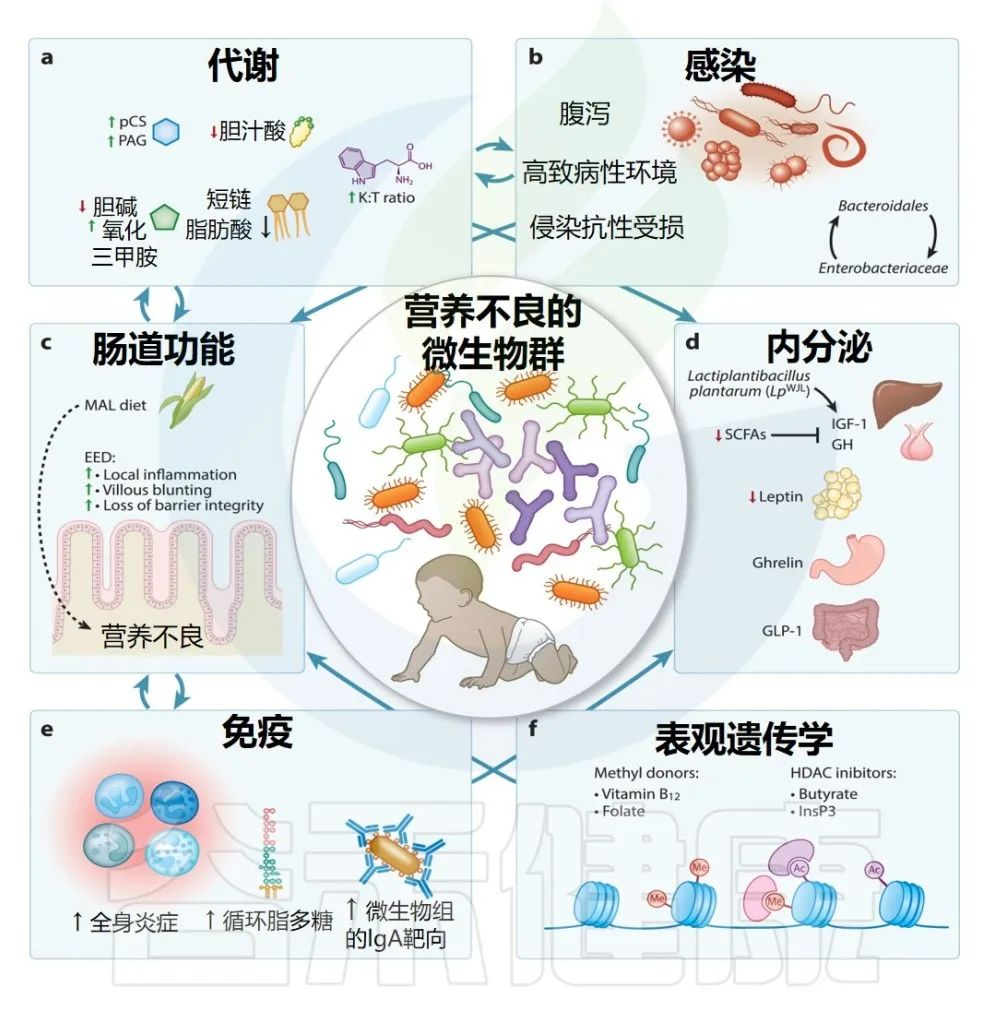
doi.org/10.1146/annurev-nutr-061121-091234
下面我们逐一来了解营养不良的微生物群带来的多方面影响。
共生肠道微生物群的存在,对于生命早期肠道屏障的正常结构、功能和转录发育至关重要。临床前研究的大量证据表明,无菌小鼠幼崽表现出肠道屏障成熟延迟,这与自发性结肠炎、更容易肠道感染和营养吸收不良有关。
生命早期长期接触肠道病原体会破坏肠道结构和功能。环境肠道功能障碍(EED)是一种胃肠道现象,在营养匮乏的环境中很常见,并且经常在营养不良的儿童中观察到(我们以前也专门写过儿童EED的文章)。
▸ 什么是环境肠道功能障碍(EED)?
EED 通常是亚临床的,局限于小肠,其特征是肠道通透性增加、绒毛萎缩和变钝、粘膜屏障变化、吸收不良、局部炎症和隐窝伸长。屏障功能的丧失还导致微生物及其产物(最常研究的是脂多糖)从肠道转移到体循环中,从而刺激全身炎症。据推测,营养吸收不良、肠道炎症和慢性全身炎症的结合会阻碍儿童的健康成长。
▸ 环境肠道功能障碍(EED)与什么相关?
EED 通常归因于在水、环境卫生和个人卫生 (WASH) 较差的环境中长期接触肠道病原体。然而,在此类环境中改善怀孕期间和早期生活中的 WASH 的大型随机试验未能减少幼儿 EED 的常见生物标志物,这可能是由于同时未能减少其肠道病原体携带。
除了病原体携带之外,共生微生物组的破坏也可能导致 EED。对611 名儿童( 6、18 和 30 个月大)大型分析发现,微生物群多样性/成熟度与 EED 的三种生物标志物之间存在负相关:
表明肠道微生物群的不成熟可能会导致肠道炎症和屏障功能障碍。特定菌群的丰度升高,包括Megasphaera、Mitsuokella、Sutterella也与 EED 有关。
注:REG1B,Regenerating family member 1 beta 是一种蛋白质,它是一种由胰腺细胞产生的蛋白质,在胰岛细胞再生和胰岛功能调节中起着重要作用。
REG1B在胰腺细胞再生过程中被表达,且参与了胰岛细胞增殖和分化,对胰岛细胞的生长和修复有促进作用,在胰岛素分泌和血糖调节中发挥一定的调节作用。
EED 动物模型有助于深入了解可能驱动该疾病表型的机制。一些模型能够通过应用低蛋白饮食和 LPS 或吲哚美辛等肠道损伤来复制 EED 的某些方面。
注:吲哚美辛:用于治疗关节炎,癌性疼痛,痛风,滑囊炎、肌腱炎及肩周炎等非关节软组织炎症;可用于高热的对症解热,恶性肿瘤引起的发热或其他难以控制的发热。
▸ 肠道微生物对 EED 的潜在贡献
利用异常微生物状态和改变的营养环境之间相互作用的新模型已经成功地概括了几种 EED 表型。例如,肠道生理学和微生物组通过血管紧张素 I 转换 2 酶 (ACE2) 的表达联系起来。
注:ACE2是一种在分化上皮细胞的管腔表面表达的蛋白质。
与野生型小鼠相比,缺乏 ACE2 的小鼠在受到DSS攻击时,表现出类似 EED 的病理学、氨基酸代谢紊乱和肠道微生物组紊乱。DSS 攻击期间的 EED 表型可以在从 ACE2 缺陷小鼠进行粪便移植后转移到野生型动物中,这表明微生物群被破坏。
注:DSS,右旋糖酐硫酸钠,一种破坏肠上皮屏障并导致结肠炎的刺激物
膳食色氨酸及其代谢物烟酰胺可以通过哺乳动物雷帕霉素靶点 (mTOR) 活性刺激抗菌肽的产生,从而挽救 ACE2 缺陷小鼠的表型,从而维持微生物组稳态。
▸ 微生物结合营养物质影响EED
低蛋白、低脂肪饮食会导致小鼠小肠微生物群遭到破坏,其特点是小肠内物种丰富度升高,此外拟杆菌门和变形菌门也会扩张。
将营养不良的饮食与拟杆菌和大肠杆菌结合起来作者产生了 EED 的表型,包括肠道通透性、绒毛和隐窝萎缩增加、空肠细胞因子 (IL-6 和 MCP-1)释放到腔内增加,以及细菌对小肠上皮细胞的粘附增加。这种表型依赖于微生物组的改变和营养不良的饮食,这表明 EED 表型是复杂的微生物与营养物质相互作用的结果。
总体表明,肠道微生物群破坏在营养不良相关肠道功能障碍的病理学中具有潜在的因果作用。
▸ 腹泻和有症状的呼吸道感染与发育迟缓和消瘦有关
关于腹泻和儿童营养不良的大型多中心研究的有力证据表明,肠道病原体的高负担与生命早期生长不良有关。
健康且多样化的肠道微生物群通过生态位排除提供针对肠道病原体的定植抵抗力。因此,受损的肠道微生物组可能为病原体定植提供了环境。
肠道微生物组也可能介导远处部位的感染易感性,包括肺部。因此,生命早期健康肠道微生物组可能有助于在卫生条件差的环境中抵抗病原体的定植,从而防止感染负担和相关的生长不良。
▸ 发育迟缓:肠道菌群多样性减少——预示腹泻发病率的增加
来自秘鲁的发育不良儿童在生命的前两年表现出肠道微生物组多样化轨迹受损。在此期间,发育迟缓的儿童的微生物多样性显着减少,腹泻后多样性的恢复也较慢。因此,肠道微生物组的组装受损可能会加剧感染-发育迟缓的循环。
来自同一队列的一项研究发现,弯曲杆菌的携带与肠道微生物群的改变有关。此外,肠道微生物组内与弯曲杆菌携带相关的物种与 LAZ 的减少独立相关,这表明肠道微生物组组成受损会促进病原体定植和并发生长缺陷。
▸ 营养不良:微生物组对病原体定植产生影响
与营养良好的小鼠相比,由营养不良饮食和肠道微生物群紊乱引起的 EED 小鼠模型对感染的敏感性更高,这表明与营养不良相关的微生物群破坏可能会导致病原体定植和感染负担。
蓝氏贾第鞭毛虫感染后生长缺陷的易感性也取决于动物模型中的肠道微生物组。人类研究报告称,贾第鞭毛虫和相关的肠侵袭性病原体,除了那些涉及粘膜破坏的病原体外,对全身炎症、肠道炎症和生长受损具有最深远的影响。
在小鼠中,贾第鞭毛虫与小肠中的肠杆菌科细菌相互作用,以限制蛋白质营养不良期间的生长,这种作用在用没有抗贾第鞭毛虫活性的抗菌药物治疗后消失,表明失调的微生物群在此过程中发挥着关键作用。
这种相互作用表明特定的病原体或病原体依赖于共生肠道微生物群的成员来定殖和增殖。这在人类发育迟缓研究中很明显,即肠杆菌科细菌和拟杆菌目细菌之间的交叉喂养与儿童发育迟缓有关。
▸ 在体外,肠杆菌科和拟杆菌目之间协同生长
特别是在蛋白质和铁含量低的营养不良条件下。在这种情况下拟杆菌属利用饮食和粘蛋白衍生的糖和肠杆菌科细菌提高铁的生物利用度,铁是特定病原体增殖所必需的微量营养素。
拟杆菌科和肠杆菌科在营养不良的儿童中具有很强的相关性,但在营养良好的儿童中则没有。另一种拟杆菌属的脆弱拟杆菌,还依赖周围的微生物群来诱导小鼠的生长效应。含有bft毒素的脆弱拟杆菌的产肠毒素菌株会导致无菌小鼠体重减轻和能量代谢受损,这些小鼠被来自发育迟缓儿童的微生物群定植,但对于那些被来自营养良好儿童的微生物群定植的小鼠则不会。
这些数据表明,病原体和感染对儿童营养不良的影响可能依赖于共生肠道微生物群的组成和功能,在儿童营养不良患病率高的环境中减少病原体负担的策略除了考虑病原体负担外,还必须考虑共生微生物群。
营养不良儿童的宿主蛋白质、脂肪和能量代谢紊乱早已有报道。然而,肠道微生物组的代谢能力大大扩展了哺乳动物宿主的生物转化能力,扩大了可加工底物的多样性,并增加了宿主接触的分子范围。
许多微生物衍生的代谢物可以在肠道和肠壁中发挥局部作用,并且在吸收后还可以影响周围组织中的宿主过程。因此,儿童早期肠道微生物组的变化可能会通过改变提供给儿童的代谢副产物和宿主代谢途径本身来影响代谢表型,从而影响生长。
▸ 成熟度与微生物代谢
与用于绘制儿童微生物组成熟度的MAZ评分类似,代谢成熟度模型,称为年龄表组 Z 评分 (PAZ),是根据秘鲁、坦桑尼亚和孟加拉国儿童的尿液样本构建的,并用于追踪生化年龄及其与生长的关系。用于计算 PAZ 的八种代谢物中的三种——对甲酚硫酸盐 (pCS)、苯乙酰谷氨酰胺 (PAG) 和马尿酸,与肠道微生物代谢相关。
有趣的是,与未生长受限的婴儿相比,生长受限的婴儿早在出生后 3 个月就出现生化不成熟(即,其生化年龄低于实际年龄)。这凸显了微生物群在宿主整体代谢能力发展及其对生长的后续影响中的重要性。
健康肠道微生物群的一个主要功能是发酵宿主无法消化的膳食底物。在母乳喂养期间,这主要涉及HMO的微生物消化。患有发育不良婴儿的母亲的母乳中 HMO 含量明显较低,结构相似的牛乳低聚糖可以恢复营养不良模型中小鼠的生长。
然而,这种效果取决于微生物群,因此补充乳寡糖后,无菌小鼠的生长不会恢复。断奶后,肠道微生物群转向不可消化的碳水化合物、抗性淀粉、粘蛋白和蛋白质的代谢。
▸ 糖分解产生短链脂肪酸,满足能量需求
糖分解活性(即碳水化合物的分解)的主要最终产物是短链脂肪酸。这些都有助于宿主日常的能量需求。丁酸盐是结肠细胞的主要能量来源,提供其所需能量的 80%,而乙酸盐在肌肉等全身区域代谢,丙酸盐在肝脏中用于产生 ATP。
这种微生物活动提供了一种从饮食中释放能量的机制,可用于支持生长。此外,短链脂肪酸还具有其他有益作用,例如降低结肠 pH 值以增强对潜在病原体的定植抵抗力并提高矿物质吸收。此外,丁酸盐已被证明可以通过增加 AMP 激活蛋白激酶 (AMPK) 活性和紧密连接蛋白的组装来促进肠道屏障的发育。
▸ 营养不良:从碳水化合物发酵和短链脂肪酸生产转向蛋白水解代谢
例如,与营养良好的儿童相比,印度尼西亚中度营养不良儿童的粪便丙酸盐和丁酸盐含量较低,粪便 pH 值较高。
在营养不良的猪模型中,肠道微生物组产生的丁酸盐较少,并且与肝脏脂肪酸代谢(β-氧化)减少有因果关系。同样,在对巴西东北部儿童进行的代谢组学研究中,发育不良的婴儿排出了更多由微生物降解氨基酸而产生的微生物-宿主共代谢物。
其中包括 pCS 和 PAG,它们分别源自酪氨酸和苯丙氨酸的微生物分解。营养不良病原体相关的EED导致肠道吸收不良可能会推动这一观察。这增加了小肠中氨基酸的可用性,从而导致发育不良儿童中的小肠细菌过度生长,并增加到达结肠进行细菌代谢的氨基酸。
▸ 营养不良:微生物群与发育中的宿主竞争氨基酸
有趣的是,虽然 SCFA 促进上皮屏障完整性,但 pCS 被发现会损害它,可能导致营养不良个体中观察到的肠漏表型。
此外,pCS由艰难梭菌和其他病原体产生,限制肠道微生物群的生物多样性。可能导致营养不良婴儿中观察到的微生物群失调。另外两种氨基酸代谢物,吲哚乳酸和N-乙酰谷氨酸,分别源自色氨酸和谷氨酸代谢,在来自孟加拉国的一组儿童中,它们与 2-24 个月龄之间的线性和体重生长指标呈正相关。长双歧杆菌也与生长呈正相关,它编码了参与这些化合物生产的大部分微生物代谢途径,这些化合物可能有助于支持早期生命的生长。
▸ 膳食胆碱的微生物分解增加还导致蛋白质营养不良和发育迟缓
在蛋白质缺乏的小鼠模型中,尿胆碱减少,而胆碱代谢的微生物产物三甲胺和二甲胺增加。
同样,在马拉维儿童中,血清中三甲胺-N-氧化物(TMAO)与胆碱的比率与线性生长障碍呈正相关。
胆碱和甜菜碱排泄量降低与巴西婴儿发育迟缓有关。胆碱的可用性不仅对于肌肉获得很重要,对于S-腺苷甲硫氨酸 (SAMe)的生成也是必要的,而 S-腺苷甲硫氨酸 (SAMe) 是 DNA 甲基化和发育的关键。
重要的是,胆碱是骨骼肌的必需营养素以及神经发育和大脑功能。胆汁酸是宿主肝脏和肠道微生物群组合代谢产生的代谢物的一个例子,在消化中发挥重要作用。
初级胆汁酸,例如胆酸和鹅去氧胆酸,在肝脏中合成,然后通常与甘氨酸或牛磺酸结合,并分泌到胆汁中。释放到肠道后,一小部分胆汁酸可以到达结肠,在那里微生物群将它们分解并转化为次级胆汁酸,例如脱氧胆酸。这些修饰的胆汁酸可以通过粪便排出或再循环回肝脏。
与没有 EED 的儿童相比,患有 EED 的儿童的血清总胆汁酸含量较低,其中牛磺鹅去氧胆酸、牛磺鼠胆酸和甘氨酰去氧胆酸存在特定差异。除了消化之外,胆汁酸在代谢调节中也发挥着重要作用。通过充当全身表达的受体(例如 FXR、PXR、VDR 和 TGR5)的配体,胆汁酸可以调节多种宿主途径和功能的表达,例如能量稳态、葡萄糖和脂质代谢。
▸ 这些细菌相关代谢物还具有抗菌特性,既能抑制又能促进特定病原体的生长
例如,胆汁酸代谢的变化,特别是细菌胆汁盐水解酶活性,被认为是隐孢子虫卵囊排泄的重要触发因素。此外,观察到未患阿米巴病的儿童粪便中次级胆汁酸、脱氧胆酸的含量较低。研究表明,给小鼠注射这种胆汁酸足以增加粒细胞-单核细胞祖细胞的数量,并提供针对阿米巴病的保护。这表明胆汁酸可能在预防营养不良环境中常见的不同肠道感染和对这些感染的易感性方面发挥潜在作用。
▸ 从微生物代谢,到宿主代谢的改变
除了微生物代谢的改变外,在营养不良的儿童中还观察到宿主代谢的改变,这可能部分是由菌群失调引起的。例如,从发育迟缓儿童的小肠中分离出的口腔微生物在体外和体内模型中会损害脂质吸收,从而提供小肠微生物组和小肠细菌过度生长在营养不良中的潜在因果作用。
肠道屏障功能的丧失会导致微生物群及其产物(包括脂多糖)从肠道进入体循环的易位增加。LPS 易位可导致慢性全身炎症,从而通过吲哚胺 2,3-双加氧酶诱导激活色氨酸-犬尿氨酸途径。事实上,秘鲁和坦桑尼亚儿童的血浆犬尿氨酸与色氨酸 (K:T) 比率与血浆 LPS 和 LPS 结合蛋白 (LBP) 相关,并且还与线性增长缺陷相关。
扩展阅读:
犬尿氨酸途径的激活可以抑制炎症并促进耐受性,但可能会失调色氨酸途径,而色氨酸途径对于血清素的产生以及 NAD +和烟酰胺的生成非常重要,而后者是生长的关键。免疫耐受性的增加也可能对增强对病原体的有效反应产生影响,这也可能受到色氨酸剥夺的影响,并可能导致此类感染在营养不良的肠道中持续存在。
儿童营养不良与内分泌信号中断有关,包括介导食欲和能量代谢的瘦素和生长素释放肽,以及共同构成生长轴的GH 和IGF-1 。
发育迟缓儿童与非发育迟缓儿童相比,在童年后期血浆 IGF-1 较低以及生命的前 18 个月,一种与慢性炎症相关的现象。
▸ 瘦素水平低可预测SAM儿童的死亡率
新数据表明,肠道微生物组直接和间接影响肠道局部激素的产生,例如胰高血糖素样肽-1 (GLP-1),并通过介导瘦素、生长素释放肽和 IGF-1 的产生来系统地进行,每一个都调节新陈代谢和营养状况。
▸ 微生物组→产生SCFA→GLP-1 的产生→抑制胰高血糖素原的表达
GLP-1 是一种肠道源性激素,负责刺激胰岛素分泌、减少胃排空和增加饱腹感,从而影响新陈代谢和营养状况。
在没有微生物组的情况下,在无菌小鼠中,GLP-1 的产生在一种假设的适应性机制中升高,当微生物组被破坏或不存在时,GLP-1 的产生会增加营养物质的吸收,而 SCFA 给药可以逆转这种效应。
▸ 微生物衍生的SCFA也会影响 IGF-1 的产生
IGF-1 在肝脏和脂肪组织中产生,影响骨形成、骨量和骨骼生长,尤其是在生命早期。当无菌小鼠被微生物定植时,血浆中血清 IGF-1 的浓度会增加。
同样,抗生素的使用会降低 IGF-1 浓度,而补充 SCFA 可恢复抗生素治疗期间 IGF-1 的浓度。不产生短链脂肪酸的细菌种类也可以通过刺激生长轴来刺激实验动物的生长。植物乳杆菌的特定菌株可恢复果蝇和小鼠的线性生长、股骨长度、IGF-1 产生和活性以及外周组织对 GH 的敏感性。
植物乳杆菌WJL ( Lp WJL ) 菌株对 IGF-1 产生和生长的活性取决于肠上皮细胞中含有核苷酸结合寡聚化结构域的蛋白 2 (NOD2) 的刺激。
▸ 肠道共生细菌还可以防止感染时的肌肉萎缩
共生大肠杆菌在骨骼肌中维持 IGF-1 信号传导,独立于宿主代谢、热量吸收或炎症,从而防止肠道或肺部感染后肌肉萎缩。
▸ 与饱腹感和新陈代谢有关的激素与特定共生菌群丰度相关
在动物模型中,双歧杆菌和乳酸菌丰度与瘦素浓度呈正相关,而生长素释放肽则与拟杆菌和普雷沃氏菌丰度呈正相关。
在营养不良模型中,生长素释放肽水平还与双歧杆菌、乳酸菌和球状球菌-直肠真杆菌呈负相关。一项针对 6 至 24 个月大的冈比亚儿童( n = 60)的横断面观察队列研究发现,严重营养不良儿童的肠道微生物群与代谢激素浓度之间存在许多密切相关性,使用网络分析模拟肠道微生物组和激素相互作用。埃希氏菌/志贺氏菌通过其与生长素释放肽和生长素释放肽受体的相互作用,在区分健康对照和营养不良患者方面具有高度预测性,而粘膜乳杆菌与瘦素/瘦素受体之间的相互作用也对营养不良和营养良好的儿童具有高度区分性。肠杆菌科细菌和 IGF-1 的相互作用也区分了 MAM 和 SAM。
这些相互作用可能是双向的,瘦素可以刺激粘蛋白的产生并改变肠道微生物组的组成,而特定的产生脂多糖的肠杆菌科细菌的炎症活性可能会损害 IGF-1 和 GH 信号传导。
微生物群在整个生命过程中与宿主免疫系统保持着相互的、动态的沟通,而这种沟通会因营养不良而被破坏。
越来越多的证据表明,营养不良的儿童在免疫学上与营养充足的同龄人不同,胸腺较小,免疫细胞组成和分布在质量和数量上不同,全身和肠道环境中的促炎介质长期升高,以及由不利因素驱动的免疫基因上的表观遗传标记。
产前和产后暴露。迄今为止,大多数营养不良儿童的免疫表型都是在感染和病原体携带的背景下进行研究的。然而,新出现的数据表明,肠道微生物组内共生和条件致病微生物的多样性可能会驱动营养不良儿童的独特免疫表型,而与病原体无关。
▸ 营养不良引起的肠道炎症的特征——EED生物标志物
例如钙卫蛋白和髓过氧化物酶,这些生物标志物是由肠道组织中微生物激活的先天免疫细胞产生的,这些细胞可能会被破坏的肠道微生物组激活。
EED 动物模型表明,营养缺乏的小鼠与营养充足的小鼠相比,肠道菌群失调会导致小肠上皮内淋巴细胞数量增加和促炎细胞因子分泌增加。尽管空肠免疫激活增加,但营养缺乏的动物在肠道内抑制鼠伤寒沙门氏菌感染的能力较差,导致细菌传播到脾脏和肝脏,并导致肝脏分泌促炎介质。
▸ 儿童营养不良还与慢性全身炎症有关
尽管反复出现症状的感染可以引发全身炎症表型,但小鼠模型表明,在没有明显感染的情况下,肠道微生物群和/或其成分系统性扩散到功能失调的肠道屏障进入循环也可能发生这种情况,如 EED 中观察到的那样。
与营养充足的幼鼠相比,断奶后采用缺乏蛋白质、铁、锌的饮食的健康小鼠出现了微生物群失调和体重减轻,这与向结肠给药时未能含有脂多糖有关;全身、盲肠和结肠(但不包括回肠)促炎细胞因子升高;在没有感染的情况下,LPS 攻击后促炎细胞因子的分泌增多。
在该模型中,通过针对革兰氏阴性(LPS 阳性)而非革兰氏阳性(LPS 阴性)共生体的抗生素治疗,使体重增加和促炎性免疫反应的差异正常化。
该模型中抗生素治疗效果的选择性支持了对革兰氏阴性微生物群成分的播散性促炎性免疫反应的作用,这些微生物群成分从肠道转移到循环系统中,从而导致营养不良的体重和身高缺陷。
由微生物易位驱动的慢性全身炎症可能影响人体测量的一种方式是通过抑制生长因子信号传导和骨/骨骼肌生长。来自动物研究的证据表明,依赖于微生物群代谢的唾液酸化HMO会增加成骨细胞驱动的骨形成,并且该过程是由 Th2 极化免疫反应介导的,即增加大肠中的嗜酸性粒细胞募集和嗜酸细胞趋化因子浓度。
中低收入国家儿科队列研究一致的证据表明,炎症介质也与IGF-1呈负相关。
注:前面讲过,营养良好和营养不良的儿童之间存在激素差异,尤其是与生长[生长激素 (GH) 和胰岛素样生长因子 1 (IGF-1)] 和食欲调节(瘦素和生长素释放肽)相关的激素差异。
包括 sCD14、补体蛋白 2、巨噬细胞炎症蛋白 1B 和 LBP 在内的炎症介质与SAM入院后康复儿童的体重和/或 MUAC 增加呈负相关。
▸ 炎症介质对生长的直接影响
无菌小鼠持续过度表达促炎细胞因子 IL-6 和循环 IGF-1 减少,生长速度较慢,并且比非无菌同窝小鼠小 30-50%,这种表型可以通过给予 IL-6 来部分挽救。
研究发现,IL-1β 和TNFα 与 IL-6 一起下调人肝细胞中生长激素受体 (GHR) 的表达、通过细胞因子信号传导抑制因子 3 (SOCS3) 产生的 GH 信号传导以及 IGF-1 的产生体外细胞系和小鼠肝脏体内细胞系。
▸ 微生物群:或将使炎症驱动的生长缺陷正常化
在一项以豆类为基础的治疗性营养的临床试验中,已经探索了将微生物群作为使炎症驱动的生长缺陷正常化的一种手段的潜力,该营养旨在增强营养不良的孟加拉国儿童肠道微生物组的多样性和成熟度。
与之前的观察结果一致,30 名 SAM 儿童在基线时可以通过血浆蛋白质组与 21 名健康儿童区分开来,该血浆蛋白质组与较低的骨化和成骨细胞分化以及较高的急性期炎症反应有关,包括 C 反应蛋白 (CRP)、IL- 6、促炎核因子 kappa B (NF-κB) 信号通路中的中间蛋白。干预后,尽管证据表明总体肠道病原体携带或绝对粪便微生物群多样性存在差异,但 WHZ 和微生物群成熟度有所增加;这些变化与血浆蛋白质组向健康生长和减少炎症特征的转变有关,表明肠道微生物组针对性的干预措施可以减少营养不良相关的炎症。
菌群失调引起肠道和全身炎症的能力也可能取决于抗体介导的肠道微生物组成的控制。肠道中的分泌性免疫球蛋白,特别是免疫球蛋白 A (IgA),选择性地结合微生物组成分,从而调节微生物组组成。
扩展阅读:
▸ 发育不良:IgA靶向的细菌比例更高
研究发现,与来自马达加斯加和中非共和国的非发育迟缓对照儿童相比,发育不良儿童的粪便中 IgA 所靶向的细菌比例更高。除了 IgA 靶向肠道微生物群的数量差异外,IgA 还靶向健康儿童与 SAM 儿童不同的肠道微生物子集。其中 IgA 倾向于靶向SAM 儿童中的肠杆菌科细菌,同时针对健康儿童中更广泛的共生生物。
从患有 SAM 的儿童身上提取的IgA +微生物群在移植到小鼠体内时会诱发类似 EED 的病理学和全身炎症,这种效应依赖于营养不良的饮食。
在另一个模型中,给小鼠喂食低蛋白、低脂肪饮食,并进行或不进行细菌灌胃来模拟 EED,这些小鼠的 IgA 对乳杆菌属的靶向作用受损。尽管空肠中乳酸杆菌的相对丰度、粪便 IgA 总量以及 IgA 目标细菌总数的百分比与营养良好的对照组相似。
这些差异是由于乳杆菌碳水化合物代谢对饮食变化的适应,而不是 IgA 丰度或亲合力的差异。总的来说,这些观察结果强调了肠道屏障处免疫-微生物群相互作用的可塑性,有证据表明,微生物群和组织驻留免疫细胞的适应可能会损害 EED 中微生物及其抗原的免疫介导遏制,从而导致生长缓慢。
▸ 营养不良儿童的口服疫苗反应受损并反映营养不良中存在的一些免疫缺陷
小鼠模型为营养不良时微生物组失调如何损害口服疫苗的免疫原性提供了一些线索。然而,这些范式需要在人类队列研究中进一步探索。
在针对产肠毒素大肠杆菌(ETEC)进行初免加强口服疫苗接种后,由于饮食缺乏和大肠杆菌粘附定植而诱导出现 EED 样表型的小鼠,其小肠中的疫苗特异性 CD4 + T 细胞较低,且疫苗水平较低-相对于饲喂营养充足饮食的小鼠和饲喂营养不足或充足饮食的未定植小鼠的特异性 IgA。这些差异取决于微生物群,因为 ETEC 定植小鼠的小肠疫苗特异性 T 细胞数量在广谱抗生素治疗 3 周后恢复正常。
EED 表型中疫苗反应受损是由于微生物群依赖性 RORγT + FOXP3 +Treg 的扩张,这些细胞能够抑制小肠中的疫苗反应;小肠中疫苗特异性 CD4 + T 细胞增殖和 IgA 丰度得到恢复,并且在有条件地消耗 Tregs 后体重增加百分比增加。
总的来说,这些研究为微生物群靶向疗法如何恢复营养不良儿童受损的免疫表型(包括受损的疫苗反应)提供了概念证明,但也强调了固有的免疫和微生物异质性,在设计转化研究和人群时应考虑到这一点。
许多研究开始探索食物或食物成分对整个童年时期肠道微生物组的影响。
纳入的试验采用了涉及天然食品的干预措施;微量营养素补充或强化;食品成分,例如牛乳脂肪球膜(MFGM);以及特殊配方食品,例如早期限量配方食品(ELF)、即用治疗食品、即用补充食品(RUSF)和脂类营养补充剂(LNS)。
母乳
纯母乳喂养的婴儿往往含有较高比例的双歧杆菌和乳酸菌属。与配方奶喂养的婴儿相比,他们的肠道微生物组往往含有更高丰度的潜在致病菌,例如变形菌门成员。
双歧杆菌属丰度更高对婴儿健康有益,而肠球菌属的比例较高与不良的健康结果相关。
从瓶子中挤出的母乳,与潜在病原体的富集和双歧杆菌的消耗相关。
▸ 母乳是一种复杂的基质,有助于双歧杆菌定植
母乳含有许多成分,包括乳糖、400 多种不同的脂肪酸、蛋白质(乳清)、核苷酸、维生素、矿物质、乳脂肪球 (MFG) 、HMO等。
HMO 可以与牛奶中的其他化合物(例如糖复合物)结合,形成母乳聚糖 (HMG),其完整地到达婴儿结肠并驱动细菌定植,特别是双歧杆菌的定植。
HMG 逃脱肠道消化并转运到结肠,进入那里常驻双歧杆菌的发酵循环,使这些物种能够增殖并在 HMO 的双歧杆菌效应的过程中占据主导地位。特别是,婴儿双歧杆菌(ATCC 15697) 、短双歧杆菌、长双歧杆菌和双歧双歧杆菌(PRL2010) 在婴儿期占主导地位,是更健康肠道微生物组的标志。
▸ 母乳本身也含有细菌
在几组哺乳期妇女中,发现母乳样本最常见有葡萄球菌、链球菌和假单胞菌,其他分类单元因女性和环境而异;这些细菌可能有助于在婴儿肠道中播种。
配方奶
▸ 配方奶与对肠道微生物群的潜在负面影响有关
a ) pH 值的变化,导致大肠杆菌和沙门氏菌等细菌过度生长,从而破坏肠道完整性,可能是由于蛋白质类型和含量的差异
b ) 牛乳中缺乏母乳中存在的游离氨基酸和某些生物活性化合物。
食用牛奶配方奶粉的婴儿往往具有较高的肠道微生物多样性和丰富的厚壁菌门、梭状芽胞杆菌、肠球菌属、肠杆菌科和拟杆菌属和双歧杆菌属的丰度较低。
在配方奶喂养的婴儿中存在的双歧杆菌属中,有更多的青春双歧杆菌和假链状双歧杆菌,这两种细菌都与成人肠道微生物群有关。
▸ MFGM 对肠道菌群的影响:嗜血杆菌水平较低
MFGM (牛乳脂肪球膜)因其在婴儿肠道成熟、免疫以及大脑结构和功能中的潜在作用而受到关注。MFGM 源自乳腺上皮,包含 60% 蛋白质和 40% 脂质,使 MFG 稳定为乳液。为了复制母乳,婴儿配方奶粉中添加了牛 MFGM。
在 2 至 6 个月龄期间,MFGM 配方奶粉组的几种氨基酸及其分解产物的含量较低,乳酸和琥珀酸的含量也较低。12 个月时,MFGM 组的婴儿嗜血杆菌水平较低与接受标准配方奶粉的婴儿相比,该属含有多种病原体。
然而,与母乳喂养(非随机)参考组相比,MFGM 配方奶粉组的微生物群与标准配方奶粉组的微生物群更相似,因此,将 MFGM 和标准组的结果汇总进行分析,并与来自 MFGM 配方奶粉组的结果进行比较。
▸ ELF:改善健康的同时,尽量不破坏肠道菌群
母乳和婴儿配方奶粉中乳清蛋白和酪蛋白的含量各不相同。婴儿配方奶粉中牛奶的比例较高,较难消化。水解牛奶可以提高其消化率,水解牛奶用于早期限量配方食品(ELF),用于纯母乳喂养的新生儿作为干预措施,以增加肠内摄入量并避免并发症,例如胆红素血症或脱水。
与纯母乳喂养相比,ELF 不会导致α 多样性或群落结构出现显着差异,也不会导致乳杆菌丰度降低或更高丰度的梭状芽胞杆菌。这些发现表明,ELF 可能有助于改善体重大幅减轻的新生儿的健康,同时不会对肠道微生物群产生不利影响。
碳水化合物
碳水化合物包括简单的单糖和二糖以及复杂的寡糖和多糖,例如淀粉和纤维素。简单的糖在小肠 (SI) 中被消化和吸收,而复杂的或非血糖碳水化合物在到达结肠之前仍未被消化。
后者已被证明可以通过聚糖的降解来调节肠道微生物群,特别是以纤维和寡糖的形式。
关于单糖对肠道微生物群影响的研究有限,因为它们通常不会到达大肠;更常见的是针对人工甜味剂影响的研究或高糖、高脂肪的西方饮食的组合对肠道微生物群的影响。
注:单糖,如葡萄糖、半乳糖和果糖;二糖,例如乳糖、蔗糖、麦芽糖和海藻糖;和糖醇(多元醇)
▸ 果糖
一项小鼠研究中,果糖可以逃避肠道吸收并到达结肠,从而降低多形拟杆菌的丰度。
在一项针对 12-19 岁青少年的队列研究中,高果糖摄入量与真杆菌和链球菌含量降低有关,而这些细菌被认为有利于碳水化合物代谢。这些研究的结果提供了一些证据,证明在随机试验中检查单糖对人类肠道微生物组的影响是有益的,这些试验尚未在儿童中进行。
▸ 复杂碳水化合物
包括寡糖、多糖和膳食纤维,可以起到益生元的作用,被定义为“被宿主微生物选择性利用并赋予健康益处的底物”。这些分子不会被人类消化,而是被完整地运送到结肠,在那里微生物可能会代谢和发酵它们。
碳水化合物益生元包括HMO、低聚果糖、低聚半乳糖、甘露低聚糖、低聚木糖和膳食纤维,例如不溶性纤维,包括纤维素和半纤维素,例如β-葡聚糖(来自燕麦)。碳水化合物益生元还包括可溶性纤维,例如果胶和菊粉,两者都是微生物发酵罐的底物,包括长双歧杆菌亚种、婴儿杆菌、拟杆菌、拟杆菌、乳杆菌和普氏粪杆菌。
▸ 益生元通过直接机制发挥作用
简而言之,结构与宿主聚糖相似的碳水化合物分子可以直接阻断与宿主细胞的粘附;β-葡聚糖可以与吞噬细胞和自然杀伤细胞上的受体结合,引发中性粒细胞的吞噬作用;
菊粉型果聚糖可以通过 Toll 样受体、C 型凝集素受体和半乳糖凝集素作为肠道树突状细胞的受体,诱导抗炎细胞因子。
▸ 益生元可以间接地选择有益共生细菌的增殖
其代谢副产物可以直接影响肠道环境或宿主基因表达。例如,与潜在致病性大肠杆菌和梭菌相比,双歧杆菌属、拟杆菌和厚壁菌门中的物种将到达结肠的未消化碳水化合物发酵成微生物副产物,宿主或其他共生细菌可以将其用作底物。
▸ 麸皮添加:微生物群的变化因基线饮食差异而异
麸皮(小麦、燕麦和大米等谷物的外皮)是益生元可溶性纤维以及其他营养素(如植物化学物质、脂肪酸和酚类物质)的主要来源。
在一项试验中,生活在尼加拉瓜 ( n = 47) 和马里 ( n = 48)的 6 个月大婴儿每天补充米糠(每天 1-5 克),添加到断奶食品中,或者不进行干预 6 个月。
不同国家的基线肠道微生物群落结构(通过非度量多维尺度(NMDS)绘制的布雷-柯蒂斯差异来测量)存在差异,这种差异导致 8 个月和 12 个月龄时微生物对米糠补充剂的反应越来越明显;例如,马里样本在两个时间点的群落结构差异(NMDS 图上的分离)更为明显。对米糠摄入量敏感的属也因国家而异:在尼加拉瓜,8 个月和 12 个月龄时,双歧杆菌属、乳杆菌和韦荣球菌的相对丰度增加,而在马里乳杆菌在 8 个月和 12 个月大时,对米糠的反应丰度有所增加。
这些反应性类群被公认为碳水化合物消化者。这项研究在比较这些人群时的一个局限性是,整个马里队列都是纯母乳喂养和阴道分娩,而尼加拉瓜样本包括配方奶喂养和大多数剖腹产婴儿;这些基线饮食差异可能导致了微生物对米糠的不同反应。
引入补充食品,包括低聚糖的植物性食物来源,例如大豆和其他豆类、水果和蔬菜,帮助婴儿肠道微生物群多样化并增加其丰富度,使其达到类似成人的组成。
▸ 蜂蜜:双歧双歧杆菌有变化
蜂蜜是单糖(即葡萄糖、果糖、蔗糖和麦芽糖)以及益生元低聚果糖的来源。
在早产儿肠道微生物群研究中,埃及早产儿 ( n = 40) 连续 14 天食用添加到牛奶配方奶粉中的 5、10、15 或 0 克医用级三叶草蜂蜜。从定量实时聚合酶链反应 (qRT-PCR) 结果来看,双歧双歧杆菌似乎以剂量依赖性方式对蜂蜜做出反应,而乳酸杆菌种不会。10g 组的计数最高,5g、15g 和 0g 组变化不大;这可能与这些细菌的底物代谢差异有关。
一些局限性包括样本量小(每组仅 10 名婴儿)、通过 qRT-PCR 定量特定细菌(相对于整个细菌群落的下一代测序)、干预持续时间短以及研究结果的适用性(考虑到不推荐蜂蜜)对于 12 个月以下的儿童,因为担心它可能含有导致婴儿肉毒杆菌中毒的细菌。
然而,作为一项试点研究,这些发现为进一步研究蜂蜜和单糖对老年人群肠道微生物组的潜在影响提供了支持。除了豆类外,没有其他试验研究过碳水化合物在天然食品中的作用,包括天然存在的益生元,对足月婴儿或年龄较大的儿童或青少年人群的作用。
脂 肪
脂肪酸,包括单不饱和脂肪酸 (MUFA)、多不饱和脂肪酸 (PUFA)(例如亚麻酸 ω-3 和亚油酸 ω-6 脂肪酸)和饱和脂肪酸 (SFA),构成脂质分子的一部分,例如甘油三酯、磷脂、胆固醇和植物甾醇。
膳食脂肪在整个胃肠道中被分解。大多数甘油三酯的消化和吸收发生在小肠(特别是十二指肠)中,并且依赖于胰脂肪酶、肝脏中的胆汁盐和胆囊中的胆汁。
初级胆汁盐,包括胆酸盐和鹅脱氧胆酸盐,是由胆固醇合成的;共生微生物群从初级胆汁盐合成次级胆汁盐、脱氧胆酸盐和石胆酸盐,这些盐进一步被细菌用作底物或被肝细胞修饰。大约 7% 的游离脂肪酸完好无损地到达结肠,它们可能会影响体内的微生物群。食物通常含有短链、中链和长链脂肪酸的混合物。
脂肪酸影响肠道微生物群的机制仍然是一个活跃的研究领域,并且关于 SFA、MUFA 和 PUFA 对人类肠道微生物组的影响存在相互矛盾的证据。
▸ 促炎:饱和脂肪酸、ω-6 多不饱和脂肪酸; 抗炎:ω-6多不饱和脂肪酸、单不饱和脂肪酸
一些研究表明,SFAs 和 ω-6 PUFA 会促进炎症和氧化应激,而 ω-3 PUFA 和 MUFA 则具有积极作用,例如增加乳酸杆菌和双歧杆菌属。由于膳食脂肪的代谢需要氧气,因此脂肪不太可能成为厌氧肠道细菌的能量来源。然而,较高的脂肪摄入量可能会取代饮食中的膳食纤维和碳水化合物;因此,对肠道微生物群的影响可能是由于饮食替代或较低的碳水化合物底物可用性,而不是较高的脂肪摄入量。膳食脂肪可能对细胞膜有杀菌作用,导致细菌丰度降低。此外,膳食脂肪可以诱导细菌增加胆汁酸代谢,以应对增加的脂肪。
一项研究旨在分析鱼和红花油的混合物对接受小肠肠造口术的早产儿(n = 16)肠道微生物群的影响(即在腹壁上开一个开口,以便在出生后进行肠内喂养)因炎症性疾病而需要进行腹部手术,例如坏死性小肠结肠炎 (NEC),与标准护理相比这是早产儿的常见病症。
与标准营养疗法相比,干预措施持续 9 周,结果增加了微生物α 多样性,降低了潜在致病菌(肠杆菌科、梭状芽胞杆菌)的丰度,并丰富了碳水化合物代谢的预测基因功能。
总之,这些研究表明,ω-3 LCPUFA 干预可能会改善婴儿肠道微生物群多样性、组成和预测功能,特别是患 NEC 的早产儿,但母亲在怀孕期间摄入 LCPUFA 不会影响婴儿肠道微生物组。
蛋白质
膳食蛋白质由形成肽的氨基酸 (AA) 组成,肽是组织的基本组成部分。氨基酸对于骨骼肌功能、生长、健康、发育和生存至关重要,可以为宿主和肠道微生物群提供能量,而肠道微生物群可以改变宿主对氨基酸的生物利用度。
膳食蛋白质的消化从胃开始,蛋白酶将蛋白质切割成大肽。小肠中的肽酶分解这些肽,以便随后由肠上皮细胞和腔细菌进行转运和分解代谢。
增加蛋白质摄入量通常会导致结肠消化率降低和可发酵底物增加。细菌将游离氨基酸转化为多肽有助于肠道中氨基酸的代谢和生物利用度,而 L-谷氨酰胺等氨基酸的可用性可以调节必需和非必需氨基酸的小肠细菌代谢。
结肠细菌,特别是拟杆菌、梭状芽胞杆菌和大肠菌,将摄入的蛋白质和来自宿主酶、粘蛋白和脱落的肠细胞的内源蛋白质转化,产生副产物,例如较短的肽、AA、脂肪酸(例如 SCFA)和气体例如氨和硫化氢。
整个肠道的蛋白水解活性各不相同:例如,体外实验发现,与回肠微生物群相比,结肠微生物群可以更有效地降解结肠中的牛血清白蛋白,具体取决于 pH 水平、碳水化合物可用性和肠道模型保留时间。
▸ 较高的蛋白质摄入量:促进潜在致病菌
较高的蛋白质摄入量(例如西方饮食中的蛋白质)被认为会增加 pH 值,从而减少双歧杆菌等严格厌氧菌的生长。并促进蛋白质发酵,潜在致病性兼性厌氧细菌。
这些细菌,例如大肠杆菌和沙门氏菌,可以通过形成降解产物(例如三甲胺氧化物以及芳香族和支链氨基酸)来破坏肠道屏障和免疫系统;这些副产品与胰岛素抵抗和 2 型糖尿病有关。除了摄入水平之外,蛋白质来源、浓度和 AA 组成也会影响肠道微生物群的组成和功能。
▸ 蛋白质来源会导致微生物组成和功能的变化
由于植物细胞壁的原因,豆类等植物蛋白不能被宿主酶完全消化,导致它们作为微生物发酵的益生元运输到结肠。
难解析出植物蛋白来源中的蛋白质、纤维或其他化学物质的个体效应,但总的来说,这些蛋白质来源往往与细菌组成的有益变化有关,包括乳酸菌的生长和更大的微生物多样性。
进行了两项试验,利用 V4 区的 16S 测序来检查植物性蛋白质来源(包括豇豆、黄豌豆和普通豆)对肠道微生物群的影响。
一项试验比较了烤豇豆粉(25% 蛋白质,21% 纤维)、烤普通豆粉(25% 蛋白质,28% 纤维)和挤压煮熟的玉米大豆混合 (CSB) 面粉(13% 蛋白质,8% 纤维) ,6 个月大的儿童每天服用 6 个月,豇豆导致双歧杆菌属的比例更高。
与普通豆和 CSB 相比, 9 至 12 个月期间普雷沃氏菌丰度较低,6 至 9 个月期间埃希氏菌/志贺氏菌丰度较低;这是出乎意料的,因为与普通豆相比,豇豆的纤维含量较低。
另一项针对儿童(6-10 岁)的研究检查了四种不同剂量的补充微量营养素的豆类蛋白质(黄豌豆)(即 6、8、10 或 12 克),连续 1 个月每天食用两次,在厚壁菌门和拟杆菌门中发现了 9 个有区别的类群,其中许多与植物多糖发酵和 SCFA 生产有关——对应于肠道微生物组的变化。
三个类群(普雷沃菌属、颤旋菌属、普氏镰刀菌)与儿童线性生长的增加显着相关,并且所有类群的相对丰度发生变化,特别是在最低蛋白质剂量组(6 g)和最高蛋白质剂量组(12 g)。这些结果强调了豆类对肠道微生物群的潜在剂量反应效应,并确定了与功能结果(例如儿童线性生长)相关的分类群以进一步研究。
与植物蛋白相比,动物来源的蛋白质在近端肠道更容易消化,因此运输到结肠的蛋白质较少,导致结肠微生物群调节减少,并可能抑制病原体。
维生素A
维生素A是一类具有全反式视黄醇生物活性的类视黄醇化合物,包括视黄醛(视黄醛)、视黄酸、视黄酯(主要是棕榈酸视黄酯)和类胡萝卜素化合物,如β-胡萝卜素、α-胡萝卜素和β-胡萝卜素。
▸ 维生素 A 的功能
维生素 A 对于维持肠道屏障的完整性和受损粘膜上皮的再生至关重要,特别是肠上皮细胞的正常增殖和产生粘液的杯状细胞分化,这会影响肠道微生物群的定植和粘附。
膳食维生素 A 可以作为植物来源的维生素 A 原(类胡萝卜素)或动物来源的预制维生素 A(视黄酯)形式食用。
▸ 维生素 A在体内的加工
消化从咀嚼开始,包括通过胆汁盐和胰脂肪酶的胃作用,胆汁盐和胰脂肪酶在肠腔中从膳食脂质形成混合胶束以供吸收。虽然预先形成的维生素 A 来源更容易被宿主肠细胞吸收以运输至储存或循环,但维生素 A 原类胡萝卜素必须进一步加工才能转化为类视黄醇并被宿主使用。
▸ β-胡萝卜素的吸收
根据食物基质的不同,大约 5-50% 的 β-胡萝卜素被吸收。例如,膳食脂质的存在会增加小肠中类胡萝卜素的吸收;如果β-胡萝卜素被纤维吸附,它可以绕过上肠的吸收并被转运到结肠。结肠微生物群可以消化纤维,释放β-胡萝卜素供结肠细胞吸收。在结肠粘膜中发现了类胡萝卜素,主要是在升结肠中。
一项研究发现,大肠杆菌表达一种与 β-胡萝卜素单加氧酶 1 同源的耐盐酶,使大肠杆菌能够积累 β-胡萝卜素;然而,没有证据表明这种β-胡萝卜素被裂解形成视网膜以供进一步使用。
▸ 维生素A与普通拟杆菌:此消彼长
在一项针对小鼠的研究中,普通拟杆菌(Bacteroides vulgatus)先前被证明是一种生长歧视类群,随维生素 A 补充而丰度降低,并因维生素 A 缺乏而丰度增加,可能与胆汁酸代谢的改变有关。作者报告说,这些变化是由于普通双歧杆菌视黄醇流出系统的破坏以及随后对视黄醇和胆汁辅助敏感性的影响造成的。
▸ 肠道共生菌与视黄酸代谢
另一项针对小鼠的研究发现,粘膜树突状细胞 (DC) 对肠道共生婴儿双歧杆菌进行摄取,导致其从维生素 A 向视黄酸的转化增加,并且具有更高数量的具有耐受性的粘膜 DC,例如抑制 1 型 T 辅助细胞(Th1) 和 Th17 细胞。这种效果也随着宿主视黄酸状态的不同而变化。然而,还需要对人类进行进一步的研究,因为啮齿类动物的类胡萝卜素代谢效率比人类高得多,并且没有其他动物模型能够完全代表人类类胡萝卜素代谢。
最近的一项研究检查了补充维生素 A 对新生儿肠道微生物群的影响,结果发现对细菌组成(包括大双歧杆菌属)有积极的以及性别特异性的影响。
与安慰剂相比,入组时单剂量 50,000 国际单位剂量不会改变α-多样性,但在 6-15 周龄时对雄性产生双歧效应,但对雌性没有影响;作者推测,这可能有助于解释本研究中男性死亡率低于女性的原因。
维生素E
维生素 E 主要通过 α-生育酚活性发挥抗氧化剂的作用。
α-生育酚可来源于坚果、植物油、乳制品、奶酪和鸡蛋,也可以作为酯化 α-生育酚的补充剂形式食用。
作为自由基清除剂,α-生育酚可破坏细胞膜和血浆脂蛋白中的自由基链,以维持 LCPUFA 的完整性。维生素E与膳食脂质一起被吸收到肠道细胞中,融入乳糜微粒中,分泌到淋巴系统中,并转运到肝脏,然后被极低密度脂蛋白吸收并分泌到与α-生育酚转移蛋白结合的血液中; 然而,其吸收和贩运的许多细节基本上不为人所知。
经过额外的代谢步骤后,未结合的维生素 E 异构体通过粪便和尿液排出体外。维生素 E 与铁一起,与氧化电位降低有关;因此,维生素 E 被认为可以改善铁相关炎症的影响,改善肠道微生物群的结果。
一项针对美国缺铁 6 个月大婴儿的研究发现,每天两次服用 18 毫克维生素 E 和 15 毫克元素铁,持续 2 个月,并不会导致细菌多样性的变化,但确实会导致相对细菌多样性的变化出现差异。随着时间的推移,维生素 E 与铁的结合导致拟杆菌门(尤其是拟杆菌科)数量减少,而厚壁菌门(尤其是毛螺菌科)和罗氏菌属(Roseburia)数量增加。Roseburia产生丁酸盐,可以刺激结肠血流。
铁
铁是人体内最丰富的微量营养素,对于人体的氧运输、氧化还原反应、新陈代谢和电子传递链机制以及许多细菌的新陈代谢和毒力功能至关重要。
膳食血红素铁 (10%) 和非血红素铁 (90%) 的摄取和吸收存在不同的机制。牛肉是血红素铁的来源;美国的一项对测序研究使用牛肉作为干预措施。从 16S测序数据来看,以牛肉作为补充食品的群体中双歧杆菌属没有下降。与基线相比,4 周后细菌种类丰富度更高;4 个月后,放线菌和 XIVa 族梭菌(丁酸盐生产者)的丰度增加,并且与铁强化婴儿谷物相比,拟杆菌门较低。
差异可能是由于肉类中铁的生物利用度较高,以及未吸收的铁进入结肠,以及铁强化谷物类中存在植酸盐。
▸ 铁的生物利用度
铁的生物利用度随着植酸盐和多酚的存在而降低,而抗坏血酸(维生素 C)则提高铁的生物利用度。胃酸和蛋白水解可能会从食物基质中释放非血红素铁,需要更多的消化才能释放血红素和铁蛋白。一些细菌可以通过特殊机制获取食物中的铁源,例如解没食子链球菌或路邓葡萄球菌降解多酚单宁酸。
营养干预措施对肠道菌群影响的潜在机制
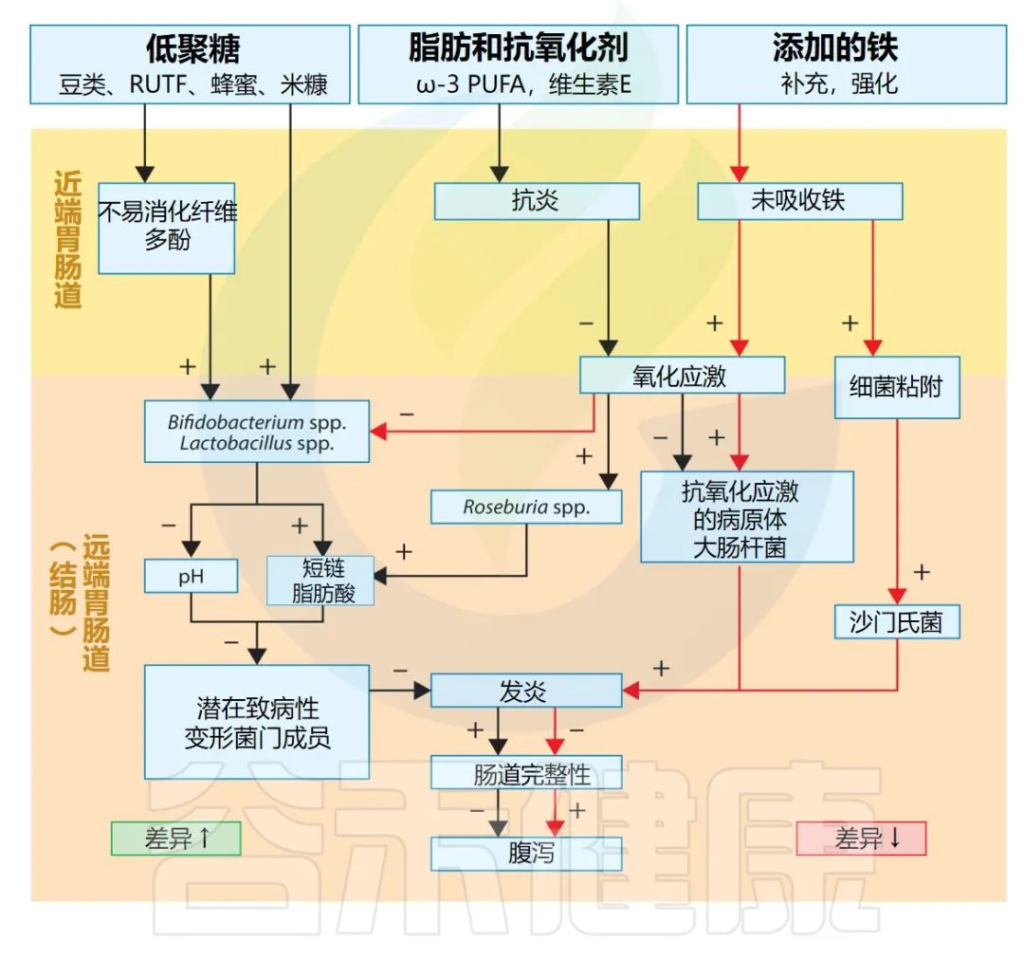
doi.org/10.1146/annurev-nutr-021020-025755
▸ 未吸收的铁被认为会刺激肠道中细菌病原体的生长和毒力
例如大肠杆菌,而宿主铁的状态会影响细菌免疫防御和宿主炎症反应,包括乳杆菌科和伯氏疏螺旋体在内的一些细菌不依赖铁生长,而是利用锰。
链球菌属可以使用铁或锰,具体取决于可用性。大肠杆菌使用 Feo 摄取系统来摄取二价铁,而三价铁在摄取之前首先被细胞外还原酶还原为二价铁,或者被摄取为柠檬酸铁或与细菌铁载体结合。
▸ 感染期间可用铁量的变化向毒力基因发送信号
低铁会导致铁载体抑制和毒素上调,而较高的铁可用性会诱导细菌粘附到肠上皮细胞,如肠沙门氏菌的情况鼠伤寒血清型。缺铁还会减少肠道细菌(包括Roseburia 、直肠真杆菌和梭状芽胞杆菌IV 族成员)等产生的短链脂肪酸。
在易缺铁的人群中,婴儿和幼儿最常需要口服铁剂治疗,例如补充剂,尽管他们的肠道微生物群仍在成熟。
▸ 铁干预措施对腹泻:负面影响
多项研究表明铁干预措施对腹泻等胃肠道症状有负面影响以及肠道微生物群的变化,例如较高的肠杆菌科和较低的乳杆菌科和双歧杆菌属。
▸ 铁补充剂,取决于基线铁或贫血状态
为了帮助抵消铁的这些影响,儿童随机试验检查了铁与维生素 E 等抗氧化剂结合使用的效果或冻干覆盆子作为抗氧化剂来源。例如,在加拿大一项研究中,与仅接受铁强化谷物的婴儿相比,接受含冻干覆盆子的电解铁强化米粉的婴儿在2 -4周后肠道微生物组的丰富度和多样性有所增加。
然而,接受冻干覆盆子的组活性氧并没有显着减少,与胃肠道中过量铁相关的炎症过程的引发剂;这可能是由于冻干覆盆子的消耗量很小,或者与新鲜覆盆子中的抗氧化剂含量相比可能较低,而新鲜覆盆子没有被任何群体食用。
在未来的研究中,考虑其他抗氧化剂来源(例如新鲜浆果或提取物)和不同剂量的这些抗氧化剂来源,以及对大量贫血婴儿的研究或铁充足,将有助于建立铁和抗氧化剂之间潜在的相互作用。
▸ 使用铁剂联合干预的随机试验
包括叶酸, 锌和多种微量营养素(MMN),无论是片剂的形式还是作为微量营养素粉(MNP)的一部分。铁的形式包括硫酸亚铁、富马酸亚铁、NaFeEDTA 和电解铁,它们可能对儿童肠道微生物群产生不同的影响;其他变量包括给药剂量、基线铁和贫血状况、抗氧化剂和其他微量营养素等辅助干预措施以及研究环境。
大多数检查 MNP 的研究都是在肯尼亚针对开始添加辅食的 6 个月大婴儿进行的;MNP 与玉米粥一起服用 3 至 4 个月。MNP 的微量营养素成分各不相同,但都含有维生素 A、叶酸和维生素 C,并且除了一种还含有 B 族维生素、维生素 D、铜、碘、硒和锌。
根据靶向 qRT-PCR 结果,MNP ( MixMe ) 加上 2.5 mg NaFeEDTA 4 个月,导致大肠杆菌/志贺氏菌丰度增加,以及肠杆菌科:双歧杆菌属比例增加。与单独使用相同MNP产生的丰度相比,10个月大时的比例增加。
在一项比较 MNP(Sprinkles)加 12.5 毫克富马酸亚铁与单独使用相同 MNP 的试验中,以及当含铁组组合并与仅使用 MNP 的组进行比较时,也观察到了类似的结果。
在另一项试验中,与单独使用相同的 MNP 相比,MNP 中的铁含量(2.5 mg NaFeEDTA 和 2.5 mg富马酸亚铁)对微生物组组成有显着影响,包括较低丰度的有益菌(如乳杆菌属和双歧杆菌属)以及较高比例的梭菌目,而拟杆菌门在各组之间保持相似。在一项辅助研究中,抗生素治疗并没有抵消铁的作用,铁仍然导致肠杆菌科细菌的增加。
最后,在另一项试验中,与单独的 MNP 相比,含铁的 MNP(12.5 毫克富马酸亚铁)在多样性或相对丰度方面没有组间差异。然而,接受含铁 MNP 的组梭状芽胞杆菌增加,双歧杆菌减少,而仅接受 MNP 的组大肠杆菌减少。
尽管 MNP 配方和 16S 测序区域存在差异,但这些研究一致发现铁会导致有益细菌减少和可能有害细菌增加,这为铁对肠道微生物组的影响提供了多种证据。需要进一步的研究来检查铁在其他环境中对肠道微生物组的影响。
▸ 铁强化和补充剂:对肠道菌群的影响
在科特迪瓦 6-14 岁儿童中(n = 60),铁强化饼干(20 毫克电解铁)不会导致微生物多样性发生变化,肠杆菌科细菌增加,乳杆菌属细菌减少,并且双歧杆菌属没有差异。6个月后这些发现与之前对婴儿进行的研究一致。
在南非进行的另一项针对 6-11 岁儿童的研究中,与安慰剂组(两组均服用维生素 C)相比,饮用含 50 毫克硫酸亚铁的饮料会导致组内相对含量存在差异;然而,尽管铁剂量较高,但没有发现组间差异。这些发现可能是由于肠道病原体负担和卫生设施的差异造成的,包括优质水和卫生设施、饮食质量和感染负担。
▸ 存在于全食物基质中的铁可能有利于铁的吸收
如LNS,不会产生与铁相关的负面影响,例如增加潜在致病菌、炎症和腹泻等胃肠道疾病症状。
LNS由植物油、花生、奶粉、糖、维生素和矿物质组成,食用量约为54g小袋,或在少量的情况下,食用量为20g(~6g脂肪),以帮助解决大量营养素和能量不足问题,并满足微量营养素需求。
已经进行了三项随机试验来检验直接给予婴儿 LNS 的影响或在怀孕期间给予母亲,然后给予婴儿。在同一队列的两项研究中,6 个月大的儿童接受基于牛奶蛋白的 LNS、基于大豆蛋白的 LNS、CSB 或 12 个月内不接受干预。通过 16S rRNA V4 区域测序评估肠道微生物群,双歧杆菌属的定植率或计数没有差异。与未接受营养补充剂的对照组相比,接受 LNS 和 CSB 补充剂的组中发现了金黄色葡萄球菌或其他肠道细菌。
在另一项试验中,将 SQ-LNS(20 克铁)给予孕妇(妊娠 <29 周),然后给予 6 至 18 个月大的婴儿;对照组包括给怀孕期间和哺乳期前 6 个月的母亲服用含有 20 毫克铁的 MMN,或者给怀孕期间的母亲服用标准护理铁和叶酸(含有 60 毫克铁)加上安慰剂哺乳期的前 6 个月。
12 个月时,与任一对照组相比,SQ-LNS 组的婴儿肠道微生物α 多样性和均匀度更高,而β 多样性没有差异或观察年龄微生物群 z 评分 (MAZ)。
然而,由于干预组之间干预的类型、数量、接受者和给药方案存在差异(只有 SQ-LNS 组中的婴儿接受直接补充),因此很难确定任何一种特定的营养素或干预类型可能会造成这些影响。总之,这些发现表明,怀孕和哺乳期间补充铁可能不会影响新生儿和婴儿的微生物组。
锌
微量矿物质锌在体内发挥催化、结构和调节作用,包括肠道健康和免疫健康,并已广泛用于治疗严重腹泻。
锌的吸收主要发生在人体的小肠中,受到食品中含锌物质是否存在以及食品加工方法的影响。例如,植物性锌来源(如种子、根和块茎)中的植酸盐会抑制锌的吸收,而铁等其他微量营养素会竞争性地抑制锌的吸收,而谷物的发酵可以提高锌的吸收。
此外,由于存在组氨酸等氨基酸增强锌的溶解度,动物源产品中的锌更容易被利用。过量的锌,包括未吸收的膳食锌和内源性锌,以受控机制通过粪便排出体外,以维持体内平衡。由于体内没有锌的储存,低锌饮食后可能会迅速出现锌缺乏症。
▸ 共生菌和病原菌在肠道中的定植和功能可能受到锌的调节
锌对于许多细菌(例如毒力因子)至关重要,并因其抗菌作用而用于动物生产。
在大肠杆菌中,锌受到严格调控,并在 300 多种蛋白质中发挥结构和催化作用,并且许多细菌拥有重金属外排系统或进行特定重金属质粒的基因转移。乳酸杆菌等细菌对膳食锌的抗性也不同。
关于锌和肠道微生物群的研究主要在动物模型中进行;这些研究的作者发现了细菌组成的差异,特别是乳杆菌属、梭菌属和肠杆菌科。
一项针对患有继发于肺炎的抗生素相关性腹泻的 2-36 个月大儿童的研究,检查了锌补充剂与益生菌的作用,为期 14 天,与单独益生菌相比,双歧杆菌属的计数没有显着差异。与基线相比,每个干预组中大肠杆菌减少。
另一项针对 6 个月大儿童食用铁和锌强化或仅铁强化谷物的研究发现,锌可以抵消铁强化对肠道微生物群的潜在不利影响。
上面的证据表明肠道微生物组在生命早期生长中的作用,为儿童营养不良的营养和药物治疗提供了新的目标。
目前预防发育迟缓和消瘦的营养策略并不能完全解决短期和长期生长缺陷或相关的临床结果。6-24 个月龄小剂量脂质营养补充剂可将发育迟缓发生率降低12%,将严重发育迟缓发生率降低 17%。
使用即用型治疗食品 (RUTF) 对 SAM 进行社区管理可显着改善营养恢复;然而,复杂的 SAM 后仍然存在高死亡率和长期生长缺陷。
总的来说,目前的治疗方法通过抗生素解决感染负担,并通过单糖、脂质和微量营养素恢复宿主营养;然而,这些治疗方法并不专门针对肠道微生物组,而肠道微生物组可能有助于改善生长。
事实上,目前对复杂 SAM 的建议(包括在所有情况下进行抗生素治疗)可能会损害肠道微生物组的恢复。而越来越多的试验报告了针对微生物群的干预措施对儿童营养不良的临床有益效果。
益生菌
少数益生菌试验报告了在营养不良负担较高的环境中益生菌对儿童生长的不同影响。对 795 名 SAM 儿童进行的一项大型随机临床试验发现,尽管观察到门诊死亡率有降低的趋势,但多物种益生菌和益生元联合治疗对营养恢复、死亡率或相关临床症状没有影响。较小规模的试验表明,其他益生菌种类对 SAM 和其他形式的营养不良具有潜在益处。
长双歧杆菌亚种由于婴儿益生菌与资源匮乏环境中的生长呈正相关,因此它具有作为促进生长的潜在益生菌的巨大潜力。在孟加拉国开展的一项 SAM 儿童试验,在临床稳定和急性期管理后,将 62 名参与者随机分为婴儿双歧杆菌益生菌(婴儿双歧杆菌 EVC001)、益生菌和纯化 HMO(乳-N-新四糖)或安慰剂,为期4周在医院。
在开始治疗 8 周后的研究终点,益生菌组的年龄别体重 Z 评分 (WAZ) 和 MUAC 显着更高。鼠李糖乳杆菌GG (LGG) 和动物双歧杆菌亚种组合的试验。SAM 治疗期间的乳酸菌和 LGG 也证明了感染发生率和门诊腹泻减少的证据。
由于患有复杂 SAM 的儿童的临床不稳定和高死亡率,特定益生菌用作 SAM 标准治疗的潜力将在很大程度上取决于成本、可持续性和安全性。
此外,需要合理选择益生菌,以确定那些能够适当定植于营养不良儿童受损肠道的益生菌,并针对可以改善生长恢复的特定途径。
来自动物研究的进一步数据可能有助于为未来合适益生菌的临床试验提供信息,例如针对内分泌生长途径的益生菌。
即食治疗性食品(RUTF)
针对肠道微生物群的改良营养疗法有潜力成为儿童营养不良的可持续且经济有效的疗法。SAM 的证据表明,RUTF 喂养后微生物组成熟度的恢复是短暂的,并且往往在治疗后 3-4 个月恢复到不成熟状态。
因此,针对 SAM 恢复中肠道微生物组和预防发育迟缓的改良补充食品可能有助于促进生长。
一项试点试验评估了在 SAM 治疗期间向 F75 和 F100 治疗奶中添加可微生物发酵的碳水化合物菊粉或豇豆粉,发现与对照组相比,对营养恢复没有改善作用;然而,补充营养可以防止住院治疗期间物种多样性的暂时丧失,这部分是由抗生素治疗引起的。
豇豆辅助喂养可以显着减少有发育迟缓风险的儿童的一些 LAZ 缺陷;然而,它仅引起肠道微生物组组成的适度变化,包括双歧杆菌增加和大肠杆菌/志贺氏菌减少,表明对生长的有益影响可能与肠道微生物组无关。
来自孟加拉国的一个队列的研究,与标准 RUTF 疗法相比,微生物群导向的补充食品 (MDCF) 对 MAM 恢复儿童生长的影响显着增强。通过合理设计由当地可用的、文化上可接受的食物组成的补充食品组合 (MDCF-2),该组合可促进人源化动物模型中早期生命肠道微生物组的成熟。
这些有希望的数据表明,与目前的疗法相比,针对微生物群的补充食品可以在更大程度上促进儿童营养不良后的生长恢复。从长远来看,这些小但显着更大的生长改善是否与持续的生长改善、认知益处或感染和慢性病风险的降低相对应,目前尚不清楚。
来自高收入环境的临床试验越来越多的证据支持针对微生物群的治疗在各种感染、胃肠道和代谢疾病中的功效。
由于世界上超过五分之一的儿童发育迟缓或消瘦,旨在进一步表征导致儿童营养不良的微生物介导的病理生理途径的研究对于为更好的治疗提供信息至关重要。
儿童营养不良涉及许多生理系统的紊乱,包括代谢、免疫和内分泌系统,其中许多系统与肠道微生物组密切相关,因此可能适合针对微生物组的干预措施。
未来潜在的微生物组靶向疗法将补充针对宿主营养需求和感染负担的现有疗法。此外,必须根据国家或地区的具体要求调整这些干预措施,以制定具有成本效益且本国或本地区上可接受的可持续干预措施。
Tips
在这篇文章中综合了营养干预试验的现有证据,这些试验测试了多种饮食干预措施对婴儿和儿童微生物组相关结果的影响,并讨论了来自实验室和动物研究的支持证据,以及来自人类研究的观察数据,旨在指导未来的研究并增强可解释性,为饮食相关政策提供信息。
可比性较差,因为纳入的研究在干预类型和持续时间上有所不同,是在有或没有既往疾病的不同儿科人群中进行的,并且是在可能影响基线微生物组成的各种环境中进行的。
虽然调查婴儿微生物群的研究发现,母亲在怀孕期间的饮食变化几乎没有影响儿童肠道微生物群。但是通过单独检查研究而不是包括整个全球背景,很难将研究偏差与真实的生物现象分开。可以使用 Qiita 等工具尝试进行此类分析,前提是研究数据与符合标准的样本元数据是公开的,以促进未来分析中的跨研究可比性。
在进行 16S rRNA 测序的 19 项研究中,只有两项对 16S V4 区域进行测序的试验可用于 Qiita 的进一步分析。作者将这两个试验人群的肠道微生物群与 2012 年具有里程碑意义的微生物组队列研究的亚组进行了比较。
在 2012 年的研究中发现,马拉维出生的婴儿和美国出生的婴儿之间存在显着的微生物分离,但在纳入最近的两项试验时,缺乏数据来深入研究营养干预措施的效果与2012年的研究对比。
尽管微生物组生物信息学取得了很大进展,并且研究的人群类型和健康状况的广度不断扩大,但在营养和肠道微生物组领域,特别是儿童,仍需要解决许多研究空白和问题。
为了解决基于发现的差距,未来的研究人员将需要:
a)识别饮食细分成分响应微生物群并确定它们是否影响宿主的生理结果;
b)与最适合现实世界应用的特定膳食成分相比,了解全食干预措施的作用;
c)提高复制性和再现性,例如在不同人群中使用相同的饮食干预措施,以及在同一人群中测试不同的饮食干预措施;
d)确定食品加工和准备的作用,了解熟食或生食如何影响肠道微生物群(烹饪的影响最近已被研究,但仍是一个研究不足的领域);
e)考虑到依从性的差异,设计和使用饮食干预措施,这需要大量的队列和长时间的随访;
f)将研究结果和序列数据整合到一个数据库中,用于对类似处理的序列进行汇总分析并了解全局现象,同时最大限度地减少研究偏差。
介入/治疗差距
未来的研究应解决以下五个介入和治疗差距。
首先,需要解决稳定性和可塑性问题。虽然短期饮食变化可以改变肠道微生物群,但长期饮食模式与更稳定的微生物群构象相关,并且很难改变。仍有待确定的是哪种饮食模式具有最大或最小的可塑性,以及这种可塑性是否也取决于基线微生物种群和过去的饮食模式。
第二个差距是干预研究中针对不同人群的营养素和食物研究不足。尽管本次综述确定了几项微量营养素粉 (MNP) 试验,而关于补充个别维生素、矿物质和天然食品的试验相对较少,但仍然需要在中国以及其他国家重复研究中对 MNP 进行调查。
第三,应进行纵向、长期的人体研究。需要对 2-18 岁婴儿、青春期、成年早期和晚年等人群进行长期喂养研究,以了解衰老微生物群的动态。
第四,解决精准营养健康问题。
第五,未来健康建议应考虑大局。建议食物和饮食(生活方式)改变需要全面的方法和来自营养师和营养科学家、心理学家、医生和生物统计学家的多方面团队的支持。
小编寄语
中国在医疗和队列研究方面的发展仍有待提升。正如首都医科大学附属北京天坛医院院长王拥军教授所指出的:“中国目前还缺乏具有全球影响力的标志性队列研究,尤其是能够改变临床指南的里程碑试验”。
举例来说,美国的弗明汉心脏研究和英国的UK Biobank人群队列都对临床研究产生了重大影响。弗明汉心脏研究自1948年开始,最初只有5200名参与者,但经过70年的发展,参与者数量增加到15000人,产生了3698篇文章。这项研究改变了几乎所有心血管疾病的临床危险因素的认知。
另外,英国的UK Biobank人群队列从2006年开始策划,至今已发表超过3000篇文章。该队列成为世界上其他国家难以逾越的临床队列研究之一。
因此,谷禾也愿意和期待与各大医疗机构合作构建大型研究队列,尤其是关于青少年儿童的。通过对这些队列进行全面多组学以及生活营养指标的调查分析,将为我国医疗研究带来新的机遇和突破,为改善人们的健康水平做出重要贡献。
主要参考文献
Jones HJ, Bourke CD, Swann JR, Robertson RC. Malnourished Microbes: Host-Microbiome Interactions in Child Undernutrition. Annu Rev Nutr. 2023 Aug 21;43:327-353.
Acosta A, De Burga R, Chavez C, Flores J, Olortegui M, et al. 2017. Relationship between growth and illness, enteropathogens and dietary intakes in the first 2 years of life: findings from the MAL-ED birth cohort study. BMJ Glob. Health 2(4):e000370
Alves da Silva AV, de Castro Oliveira SB, Di Rienzi SC, Brown-Steinke K, Dehan LM, et al. 2019. Murine methyl donor deficiency impairs early growth in association with dysmorphic small intestinal crypts and reduced gut microbial community diversity. Curr. Dev. Nutr. 3(1):nzy070
Mehta S, Huey SL, McDonald D, Knight R, Finkelstein JL. Nutritional Interventions and the Gut Microbiome in Children. Annu Rev Nutr. 2021 Oct 11;41:479-510.
Amadi B, Fagbemi AO, Kelly P, Mwiya M, Torrente F, et al. 2009. Reduced production of sulfated glycosaminoglycans occurs in Zambian children with kwashiorkor but not marasmus. Am. J. Clin. Nutr. 89(2):592–600
Amadi B, Zyambo K, Chandwe K, Besa E, Mulenga C, et al. 2021. Adaptation of the small intestine to microbial enteropathogens in Zambian children with stunting. Nat. Microbiol. 6(4):445–54
Ansaldo E, Farley TK, Belkaid Y. 2021. Control of immunity by the microbiota. Annu. Rev. Immunol. 39:449–79
Attia S, Versloot CJ, Voskuijl W, van Vliet SJ, Di Giovanni V, et al. 2016. Mortality in children with complicated severe acute malnutrition is related to intestinal and systemic inflammation: an observational cohort study. Am. J. Clin. Nutr. 104:1441–49
Barratt MJ, Nuzhat S, Ahsan K, Frese SA, Arzamasov AA, et al. 2022. Bifidobacterium infantis treatment promotes weight gain in Bangladeshi infants with severe acute malnutrition. Sci. Transl. Med. 14(640):eabk1107
Bartelt LA, Bolick DT, Mayneris-Perxachs J, Kolling GL, Medlock GL, et al. 2017. Cross-modulation of pathogen-specific pathways enhances malnutrition during enteric co-infection with Giardia lamblia and enteroaggregative Escherichia coli. PLOS Pathog. 13(7):e1006471
Bartz S, Mody A, Hornik C, Bain J, Muehlbauer M, et al. 2014. Severe acute malnutrition in childhood: hormonal and metabolic status at presentation, response to treatment, and predictors of mortality. J. Clin. Endocrinol. Metab. 99(6):2128–37
Bhattacharjee A, Burr AHP, Overacre-Delgoffe AE, Tometich JT, Yang D, et al. 2021. Environmental enteric dysfunction induces regulatory T cells that inhibit local CD4+ T cell responses and impair oral vaccine efficacy. Immunity 54(8):1745–57.e7
Black RE, Victora CG, Walker SP, Bhutta ZA, Christian P, et al. 2013. Maternal and child undernutrition and overweight in low-income and middle-income countries. Lancet 382(9890):427–51
Bourdon C, Lelijveld N, Thompson D, Dalvi PS, Gonzales GB, et al. 2019. Metabolomics in plasma of Malawian children 7 years after surviving severe acute malnutrition: “ChroSAM” a cohort study. EBioMedicine 45:464–72
Aakko J, Grzeskowiak L, Asukas T, Paivansade E, Lehto KM, et al. 2017. Lipid-based nutrient supplements do not affect gut Bifidobacterium microbiota in Malawian infants: a randomized trial. J. Pediatr. Gastroenterol. Nutr. 64:610–15
Aly H, Said RN, Wali IE, Elwakkad A, Soliman Y, et al. 2017. Medically graded honey supplementation formula to preterm infants as a prebiotic: a randomized controlled trial. J. Pediatr. Gastroenterol. Nutr. 64:966–70
Amarri S, Benatti F, Callegari ML, Shahkhalili Y, Chauffard F, et al. 2006. Changes of gut microbiota and immune markers during the complementary feeding period in healthy breast-fed infants. J. Pediatr. Gastroenterol. Nutr. 42:488–95
Andrews SC, Robinson AK, Rodriguez-Quinones F. 2003. Bacterial iron homeostasis. FEMS Microbiol. Rev. 27:215–37
Arimond M, Zeilani M, Jungjohann S, Brown KH, Ashorn P, et al. 2015. Considerations in developing lipid-based nutrient supplements for prevention of undernutrition: experience from the International Lipid-Based Nutrient Supplements (iLiNS) Project. Matern. Child Nutr. 11:31–61
Arrieta MC, Stiemsma LT, Amenyogbe N, Brown EM, Finlay B. 2014. The intestinal microbiome in early life: health and disease. Front. Immunol. 5:427
Black RE, Victora CG, Walker SP, Bhutta ZA, Christian P, de Onis M, Ezzati M, Grantham-McGregor S, Katz J, Martorell R, Uauy R; Maternal and Child Nutrition Study Group. Maternal and child undernutrition and overweight in low-income and middle-income countries. Lancet. 2013 Aug 3;382(9890):427-451.
谷禾健康
偏头痛
偏头痛是一种常见且使人衰弱的神经系统疾病,也是世界经济的重大负担。大约14%的成年人患有偏头痛。其特点是反复出现一侧或两侧严重头痛(抽痛),并伴有独特的相关症状,包括畏光、恐声、恶心、呕吐等。
偏头痛在儿童晚期/青春期早期发病,在 40 多岁的人群中发病率最高,此后发病率下降。在所有年龄段中,女性偏头痛的发病率始终高于男性。
潜在危险因素包括压力、饮食、急性药物过量、激素波动、睡眠、环境、天气等。
对偏头痛根本原因的研究已经跨越了几个世纪,涉及外周和中枢机制等,但其根本机制仍未完全了解。已经确定三叉神经系统的激活和敏化在偏头痛发作期间至关重要。
胃肠道系统和中枢神经系统之间存在双向关系。多项研究表明偏头痛与某些胃肠道疾病有关,例如幽门螺杆菌感染、肠易激综合征(IBS)、胃轻瘫、乳糜泻等。
越来越多的证据表明微生物介导的肠-脑串扰可能导致偏头痛的发病机制。在偏头痛患者中,肠道微生物群的组成发生了改变,包括肠道菌群多样性下降,产丁酸菌减少,梭状芽孢杆菌增加等。
偏头痛患者肠道和大脑之间的相互作用,可能受到多种因素的影响,如炎症介质(IL-1β、IL-6、IL-8 和 TNF-α)、肠道微生物群、肠道菌群代谢产物、血清素途径、谷氨酸途径、神经肽(包括 CGRP、P物质、PACAP、NPY)等。
本文主要介绍了偏头痛相关症状、发病人群、四个阶段,基于偏头痛的发病机制,阐述肠道微生物群如何通过各个途径参与偏头痛的病理生理学,并探讨了潜在的干预策略。

很多人在听到偏头痛这个词的时候,会想到严重的头痛,但头痛并不直接等于偏头痛,头痛只是偏头痛的一种症状。
偏头痛是一种常见的神经系统疾病,会引起多种症状,最明显的是头部一侧的搏动性头痛,伴随着头部不适感和其他一些症状。
偏头痛是原发性头痛,这意味着它不是由其他疾病引起的。偏头痛的发作通常会对患者的日常生活和工作造成较大的干扰,那么偏头痛发作时,具体会有什么表现?
偏头痛可能会有以下症状:
日常体力活动、运动,甚至咳嗽或打喷嚏都会加剧头痛。
偏头痛最常发生在早晨,尤其是刚醒来时。有些人会在可预测的时间出现偏头痛,例如月经前或一周紧张工作后的周末。许多人在偏头痛后感到疲惫或虚弱,但在两次发作之间通常没有症状。
偏头痛的频率可以是每年一次、每周一次、或介于两者之间的任意时间。
每月发生两到四次偏头痛是最常见的。
未经治疗的发作持续 4~72 小时。
大多数偏头痛持续约 4 小时,但严重的可能会持续 3 天以上。
偏头痛的两种主要类型是:
先兆偏头痛
以前称为经典偏头痛,包括视觉障碍和其他神经系统症状,这些症状在实际头痛前约10-60分钟出现,通常持续不超过一小时。个人可能会暂时失去部分或全部视力。这种先兆可能会在没有头痛的情况下出现,并且随时可能发作。
其他典型症状包括:
说话困难;身体一侧感觉异常、麻木或肌肉无力;手或脸有刺痛感,意识混乱。头痛之前可能会出现恶心、食欲不振以及对光、声音或噪音的敏感性增加。
无先兆偏头痛
或普通偏头痛是更常见的偏头痛形式。症状包括毫无征兆地出现头痛,通常在头部的一侧感觉到,并伴有恶心、意识模糊、视力模糊、情绪变化、疲劳以及对光、声音或噪音的敏感性增加。
其他类型包括:
经期偏头痛
这些通常发生在经期开始前 2 天,并持续到经期后 3 天。可能还会在每月其他时间出现其他类型的偏头痛,但月经前后的偏头痛通常没有先兆。
沉默偏头痛
会出现前兆症状,但没有头痛。事实上,前兆通常是这种类型偏头痛的主要警告信号。但可能还会出现恶心和其他偏头痛症状。它通常只持续约20-30分钟。
前庭偏头痛
有平衡问题、眩晕、恶心、呕吐,伴或不伴头痛。这种情况通常发生在有晕车病史的人身上。
腹部偏头痛
可能会导致胃痛、恶心和呕吐。它经常发生在儿童身上,随着时间的推移可能会变成典型的偏头痛。
偏瘫偏头痛
身体一侧出现短暂瘫痪 (偏瘫)或无力。可能还会感到麻木、头晕或视力变化。这些症状也可能是中风的征兆,因此请立即寻求医疗帮助。
眼部偏头痛
也称为眼部或视网膜偏头痛。它会导致一只眼睛短暂、部分或完全丧失视力,并伴有眼睛后面的钝痛,这种疼痛可能会扩散到头部的其他部位。请立即就医检查这些症状。
带脑干先兆的偏头痛
头痛之前可能会出现头晕、意识混乱或失去平衡。疼痛可能会影响头后部。这些症状通常突然出现,并可能伴有说话困难、耳鸣和呕吐。这种类型的偏头痛与激素变化密切相关,主要影响年轻的成年女性。请立即就医检查这些症状。
持续偏头痛
这种严重类型的偏头痛可持续超过 72 小时。疼痛和恶心非常严重需要去医院。有时,药物或停药可能会导致它们出现。
眼肌麻痹性偏头痛
这会导致眼睛周围疼痛,包括周围肌肉麻痹。这是医疗紧急情况,因为这些症状也可能是由眼睛后面的神经受压或动脉瘤引起的。其他症状包括眼睑下垂、复视或其他视力变化。
活动性偏头痛的高峰发病年龄为 40 岁左右,其中 35%左右的女性和15%左右的男性受到影响。
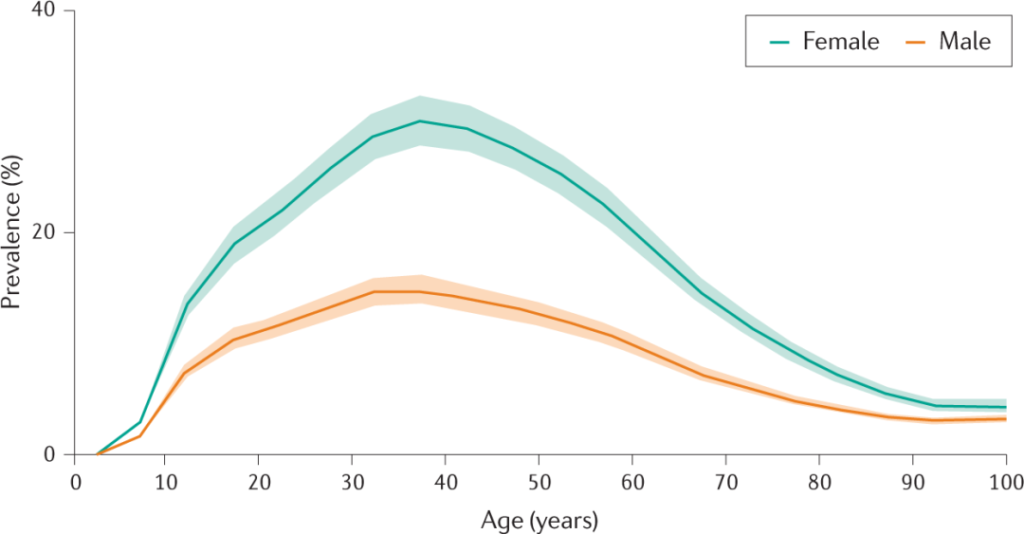
doi: 10.1038/s41572-021-00328-4.
一些常见的偏头痛诱因包括:
许多女性注意到她们在月经期间、怀孕期间或排卵期间会感到头痛。症状也可能与更年期、使用激素避孕或激素替代疗法有关。
当感到压力时,大脑会释放化学物质,这些化学物质会导致血管变化,从而可能导致偏头痛。
某些食物和饮料,例如陈年奶酪、酒精、硝酸盐(意大利辣香肠、热狗和午餐肉中的硝酸盐)、味精等食品添加剂,高盐和加工食物,可能对部分人造成影响。
摄入过多或未达到习惯的摄入量都会导致头痛。咖啡因本身可以治疗急性偏头痛。
风暴锋、气压变化、强风或海拔变化都可能引发偏头痛。
大声的噪音、明亮的灯光和强烈的气味都会引发偏头痛。
口服避孕药、血管扩张剂可以扩张血管,从而引发这些症状。
这包括体育锻炼和性生活等。
当睡眠过多或不足时,可能会感到头痛。
不良习惯包括不吃饭(或推迟进餐),吸烟,酗酒等,还有晕车也都可能引发偏头痛。
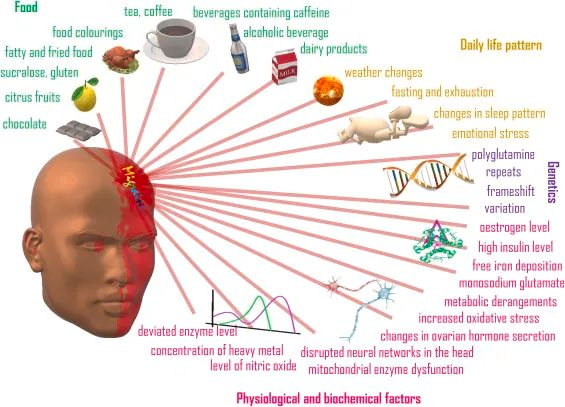
doi.org/10.1016/j.biopha.2021.111557
前驱症状阶段
有时称为头痛前阶段,此阶段的特点是在偏头痛出现前数小时或数天出现,特点是没有疼痛的症状。这些包括情绪波动、对食物的渴望、颈部僵硬、严重口渴、腹胀、便秘或腹泻、注意力不集中、排尿增多等。
先兆阶段
先兆是指偏头痛之前或期间发生的感觉障碍。这些障碍会影响一个人的视力、触觉或言语,但并不是每个偏头痛患者都会出现视觉异常期。例子包括:
视力模糊、看到黑点、波浪线、闪光或不存在的东西、渐渐扩大的盲点、手臂麻木、手臂和腿部有沉重感、口齿不清或语句混乱。
头痛阶段
这是通常出现疼痛的阶段,疼痛程度可能从轻微到无法忍受。
偏头痛通常以钝痛开始,然后发展为抽痛,有时候是钻孔感。也可能出现颈部疼痛、僵硬、鼻塞、恶心、呕吐、失眠等。
体力活动以及接触光、声音和气味可能会加剧疼痛。然而,有些人在发作偏头痛时并不会出现头痛。
恢复阶段
最后阶段是疼痛消退时。在此阶段,人们可能会感到疲惫、困惑或总体不适。
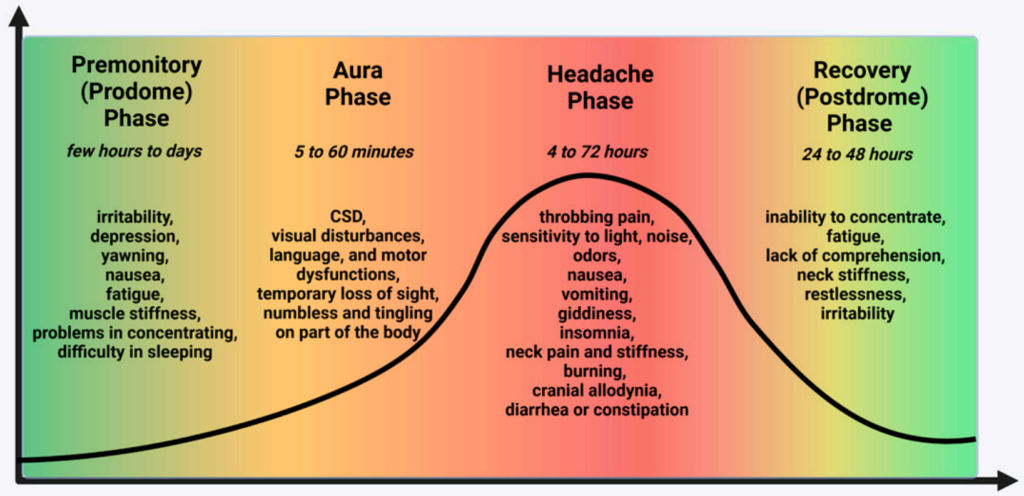
doi.org/10.3390/neurolint15030073
简单来说,在偏头痛发作之前几天,可能会感到疲劳,情绪变化,打哈欠,出现睡眠问题,恶心等情况,对光和声音敏感,这些预警信号与大脑中某个特殊的区域——下丘脑有关。下丘脑通常控制体内激素平衡,昼夜节律等,偏头痛发作之前下丘脑很活跃。
偏头痛还有常见的症状,视觉变化,刺痛感,言语困难等,这些感觉与细胞膜电位变化相关,导致脑活动和血流变化在大脑中扩散,电位变化快速在大脑表面传播,它传向哪里,哪里就出问题(*>﹏<*) …当它侵入视觉皮层,可能出现视觉盲点。
在头痛阶段,三叉神经起到关键作用,三叉神经会传递来自脸部皮肤,头皮,覆盖大脑皮层的血管和膜层的触觉,温度等,一旦被激活,三叉神经会传递痛觉信号,当偏头痛发作时,这个痛觉通路会变得敏感化,也就是说能触发痛觉的阈值下降了,那么平时不太会引起疼痛的也变得容易痛了,比如说,咳嗽,弯腰,一些强光,噪声等都会引起疼痛。
具体来说,关于偏头痛的发病机制,目前主要有以下几种理论。
I) 血管学说
该学说认为由血管舒缩功能障碍引起。基于先兆期间大脑血管收缩引起的三叉神经过度兴奋,随后产生炎症神经肽释放引起血管舒张,导致偏头痛。
II)神经学说 (皮层扩散性抑制)
该学说认为偏头痛是原发性神经功能紊乱疾病。其中通过皮层-三叉神经通路,大脑皮层的过度神经兴奋(异常的神经放电)会增加细胞外谷氨酸、钙离子、钠离子和钾离子的释放,从而引发去极化作用,造成皮质扩散性抑制(CSD)。
皮质扩散抑制是什么意思呢?
它其实是一种缓慢传播的大脑活动改变波,涉及神经、血管功能的巨大变化,皮质扩散抑制可能是偏头痛先兆的原因。这些慢电波可以激活负责感知大脑周围覆盖物(“脑膜”)疼痛的神经,并改变血管功能,这两者都可能导致偏头痛。
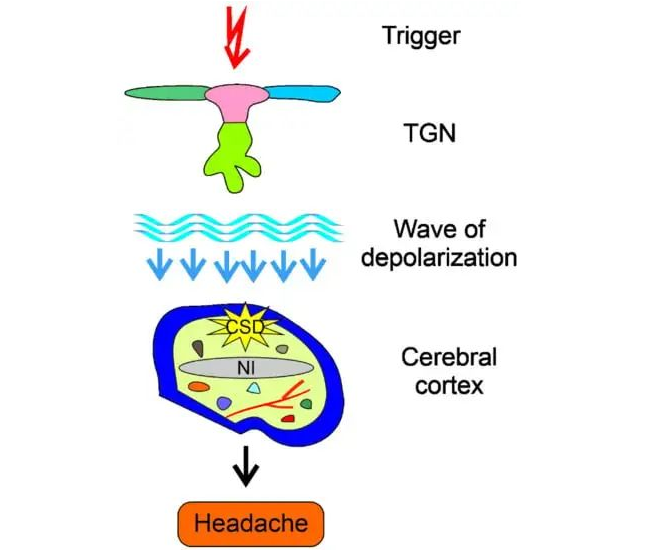
doi.org/10.3390/ijms221810134
III)三叉神经血管学说 (近年流传较广)
三叉神经感觉纤维的激活(传递疼痛信号),导致脑膜血管(硬脑膜)发生痛性神经源性炎症,此过程通过炎症神经肽(CGRP、SP、NK-A)的释放介导,这表明CGRP释放与神经源性硬脑膜血管舒张有关,这可能在偏头痛的发生中很重要。
注:关于CGRP在第四章节会详细介绍。
简单来说,激活三叉神经血管通路:
激活三叉神经元 → 释放P物质、CGRP等血管活性神经肽 → 血管扩张和促炎分子(组胺、缓激肽)的释放 → 神经源性炎症 → 激活脑膜痛觉受体
偏头痛中的 CGRP 和三叉血管系统
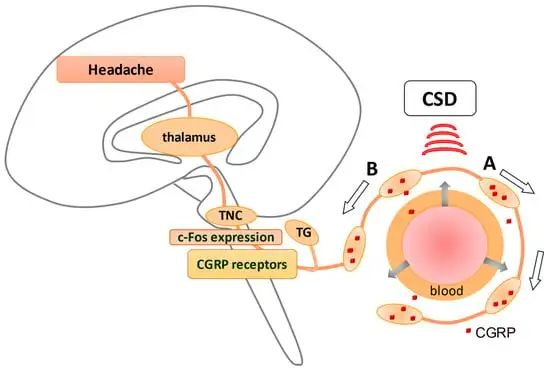
doi.org/10.3390/ijms22168929
三叉血管系统是一组感知面部和大脑覆盖层疼痛的神经细胞。三叉血管系统的反复激活会导致神经系统过敏和持续疼痛。
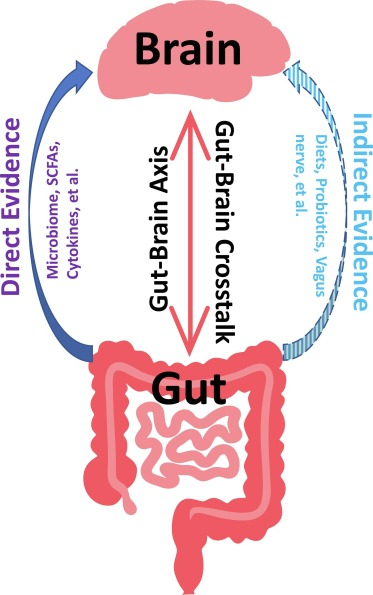
IV)肠脑轴(GBA)理论
肠-脑轴作为肠道功能和维护的协调系统,将大脑的情绪中心与肠外周机制连接起来,包括肠反射、肠道通透性、免疫反应和肠内分泌信号传导等。
肠道微生物群组成的改变可能会影响肠脑轴并引起炎症,从而可能导致偏头痛。
通过肠-脑轴影响偏头痛
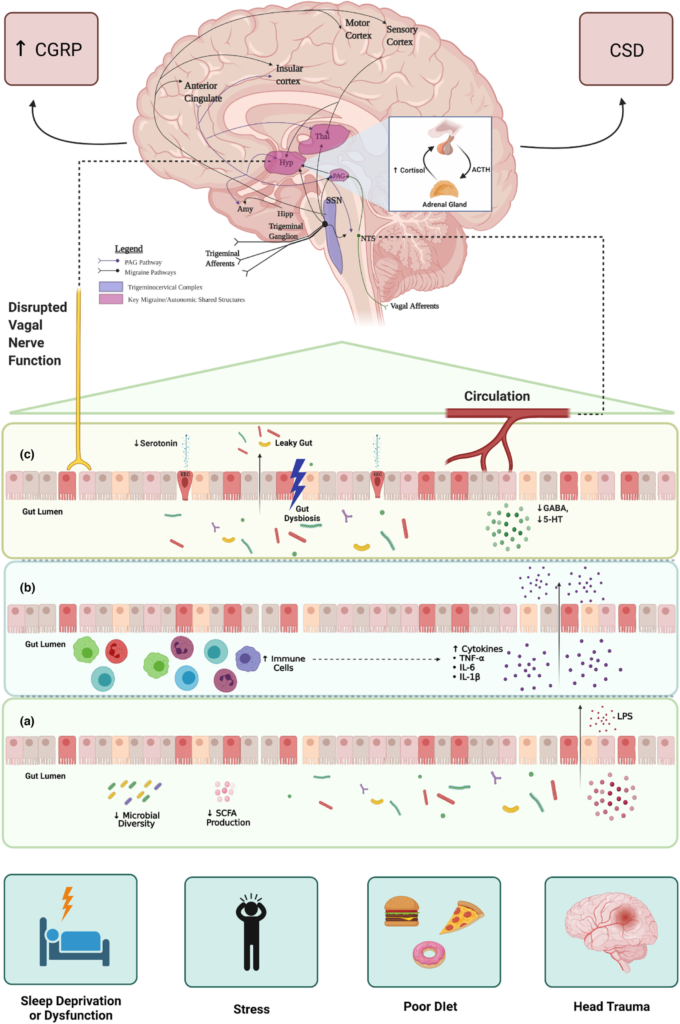
doi.org/10.1111/ene.15934
众所周知,睡眠不足和压力是原发性头痛障碍的常见诱因,不良饮食会增加偏头痛的严重程度和频率。
a) 上述因素导致肠道微生物组多样性和组成的改变,导致肠道细菌产生的短链脂肪酸减少,LPS转移到系统循环中。
b) 此外,这些因子激活免疫细胞,产生的促炎细胞因子也进入系统循环。
c) 这反过来又促进了肠道微生态失调,增加了肠道上皮的通透性,并将分子转移到系统循环中。
肠道微生物组的功能障碍,会导致与头痛病理生理学有关的必需神经递质GABA和血清素的耗竭。这反过来又改变了迷走神经功能。
而系统性促炎细胞因子和LPS,改变了HPA轴的功能,并激活了孤束核,孤束核介导了大脑中的疼痛处理途径,包括前扣带、岛叶皮层、运动皮层和感觉皮层,导致疼痛感知的改变。
神经炎症刺激三叉神经纤维释放CGRP,并驱动皮层扩散性抑郁(CSD),改变兴奋性和抑制性传递,激活三叉神经-颈复合体,加剧头痛严重程度并介导头痛频率。
接下来,我们主要就以上肠脑轴理论展开讨论一下,这其中涉及到的肠道菌群及其代谢产物与偏头痛有什么样的关联。
偏头痛常伴随一些胃肠道症状,包括恶心、呕吐、腹泻、便秘和消化不良。一些研究表明,肠道微生物群失调,可能会下调免疫系统功能,改变宿主代谢,破坏肠-脑轴,导致代谢和神经疾病的发生。胃肠道中的炎症和神经免疫调节可能在偏头痛的发病机制中发挥重要作用。
偏头痛患者的肠道微生物群的物种多样性(α多样性)和代谢功能降低,产丁酸菌减少。
而梭状芽孢杆菌增加,如:
硝酸盐已被确定为一种普遍的偏头痛诱因。
与没有偏头痛的人相比,在偏头痛患者的口腔和粪便样本中观察到能够减少硝酸盐、亚硝酸盐和一氧化氮的细菌丰度更高,如嗜血杆菌属和Rothia属。
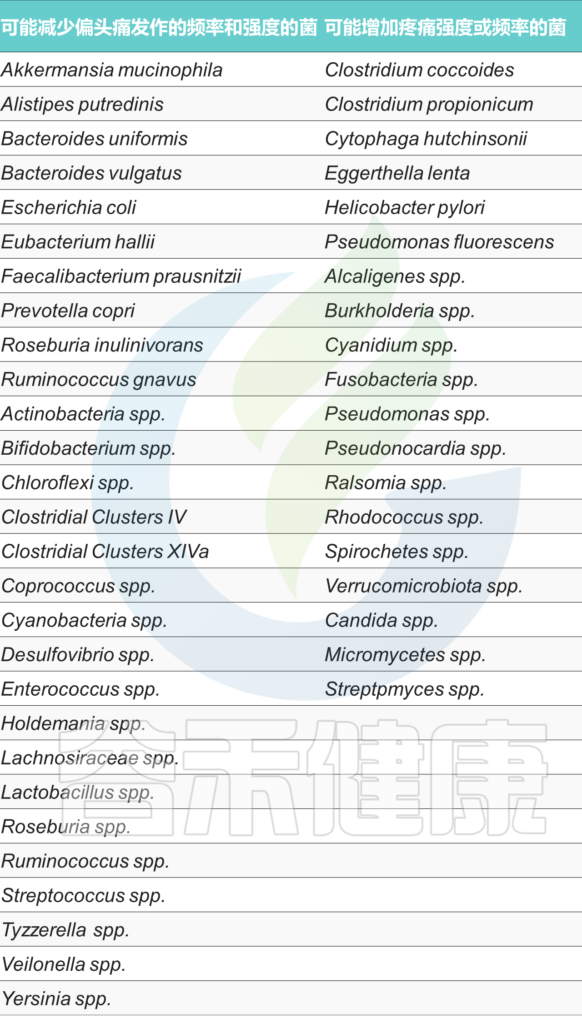
编辑
doi.org/10.3390/microorganisms11082083
特殊年龄段的偏头痛研究队列:
儿童和青少年
偏头痛在儿童和青少年中表现出多种形式,估计全世界儿童和青少年的总患病率为 10%。儿童双侧偏头痛更为常见,且持续时间较短。偏头痛儿童会出现多种胃肠道症状,包括腹痛、恶心、呕吐、腹泻、便秘。
在381名 7-18 岁儿童的队列中(40 名患有偏头痛,341 名没有偏头痛,平均年龄 11.5 岁),在偏头痛儿童和健康对照儿童有明显的菌群差异。
与健康对照相比,偏头痛儿童下列菌群升高:
老年患者

一项针对108位老年女性(54 名偏头痛患者、 54 名健康受试者)的宏基因组关联研究发现偏头痛患者的细菌特征如下:
厚壁菌门,尤其是梭菌属显著增加。
在健康对照中有更多有益菌,例如:
普氏粪杆菌、青春双歧杆菌、史密斯甲烷杆菌。
同时,该研究发现犬尿氨酸降解、谷氨酸降解和γ-氨基丁酸(GABA)合成途径较多,提示神经传递和代谢可能发生转变。
接下来我们看这些肠道菌群代谢产物会怎样参与到偏头痛的发病机制中。
谷氨酸和γ-氨基丁酸(GABA)等都参与疼痛感知。肠道中的这些神经递质参与了几种信号通路,这些信号通路除了调节疼痛外,还调节促炎细胞因子的释放。
★ 谷 氨 酸
谷氨酸作为一种兴奋性神经递质,通过不同的作用,包括皮质扩散抑制、中枢敏化和刺激三叉血管系统,在偏头痛病理生理学中发挥作用。
研究证实,各种细菌菌株,包括在环境中发现的或用于食品发酵的菌株,都能够产生谷氨酸。
偏头痛患者谷氨酸水平升高
由于谷氨酸可以沿着三叉神经血管通路对伤害性神经元产生刺激作用,因此它可能在偏头痛和偏头痛相关的中枢敏化的病理生理学中至关重要。这一理论得到了进一步的证明,偏头痛患者的血液谷氨酸水平在间歇期和发作期都有升高,在血浆和脑脊液中均升高。
有研究表明,血浆谷氨酸水平可能是慢性偏头痛和阵发性偏头痛的潜在指标。

此外,有证据表明谷氨酸在皮质扩散性抑制中起主要作用,皮质扩散性抑制被假设为偏头痛先兆的生理底物。基于这些,谷氨酸能神经传递可能是偏头痛和微生物组之间的联系。
扩展阅读:
★ 血 清 素
血清素存在于大脑、肠道和血液的组织中,其中肠道含有~95%的人体总血清素。血清素能神经元存在于中缝背核和整个疼痛通路中。
它在体内有很多作用,例如在神经细胞之间发送信号并帮助身体控制血管收缩的方式。它还在控制情绪、睡眠、饮食和消化方面发挥作用。
血清素从血小板释放到血浆中可能与偏头痛先兆期的病理生理学有关。偏头痛发作期间储存在血小板中的血清素被耗尽;血清素血浆水平降低,而相应代谢物羟基吲哚乙酸 (5-HIAA) 水平升高。
血清素既是血管扩张剂又是血管收缩剂,可作为伤害性疼痛的调节剂
偏头痛患者经常报告说,呕吐后头痛就会停止,呕吐会刺激肠道蠕动并提高血液血清素水平。而血清素的合成与肠道菌群有关,例如肠球菌、链球菌、大肠杆菌等。
血清素可能具有促伤害感受和抗伤害感受特性,具体取决于亚型
现在估计至少有七种不同的受体家族(5-HT1-7),其中许多具有亚型,偏头痛的发生涉及不止一种亚型。由于色氨酸是血清素的前体并且可以穿过血脑屏障,因此大脑中血清素的量取决于色氨酸的水平。
接下来我们看看关于色氨酸的代谢。
★ 色 氨 酸
色氨酸在人体内以三种主要途径代谢:血清素(5-HT),犬尿氨酸(l-kyn)和微生物群相关的吲哚途径。
一些报告表明,增加膳食色氨酸的摄入量可预防偏头痛的发生或减轻其胃肠道相关症状,如恶心、呕吐以及畏光。
色氨酸的代谢途径在五个肠道门中最有效:放线菌门、厚壁菌门、拟杆菌门、变形菌门、梭杆菌门。梭菌属、Burkholderia、链霉菌属、假单胞菌属、芽孢杆菌属5个属也具有较大的肠道内色氨酸代谢潜力。
色氨酸代谢的犬尿氨酸途径,可能对偏头痛具有治疗潜力
色氨酸代谢产物可能作用于谷氨酸能系统,该系统参与疼痛传递、中枢敏化和皮质扩散性抑制,谷氨酸能离子型和代谢型受体参与偏头痛的发病机制,KYNA(犬尿喹啉酸)是NMDA受体的竞争性拮抗剂,其抑制作用可防止谷氨酸诱导的兴奋性毒性,因此有理由推测色氨酸代谢的犬尿氨酸途径可能对偏头痛具有治疗潜力。
注:NMDA受体是离子型谷氨酸受体的一个亚型
犬尿氨酸途径是色氨酸转化的主要途径,因为它在正常情况下约占色氨酸代谢的95%,并且与神经系统疾病的行为和认知症状有关。
犬尿氨酸(l-kyn)途径(KP)将l-色氨酸转化为几种神经活性化合物。犬尿氨酸和喹啉酸的产生,耗尽了作为血清素前体的色氨酸和色胺的供应。

doi.org/10.3390/ijms221810134
犬尿氨酸衍生物具有有效的镇痛作用,其机制包括抑制CGRP。犬尿氨酸途径产物是芳烃受体(AhR)的重要配体,其激活与胃肠道炎症和偏头痛的发病机制有关。
AhR、TLR 和 MyD888 激活可能强调犬尿氨酸参与偏头痛和功能性胃肠道疾病。
扩展阅读:
★ 短 链 脂 肪 酸
短链脂肪酸 (SCFA) 是胃肠道细菌的代谢产物,对葡萄糖稳态、饱腹感、抗炎作用和大脑信号传导有许多局部和全身影响。
通过调节免疫细胞和细胞因子→影响神经炎症
如短链脂肪酸的施用可以减少硝酸甘油诱导引起的痛觉过敏,并导致肠道促炎细胞因子如 TNF-α 和 IL-1β 的释放减少。
丙酸可以保护血脑屏障免受氧化应激
一些菌群如:普通拟杆菌、B.uniformis、Alistipes putredinis、Prevotella copri、Roseburia inulinivorans、Veilonella、Akkermansia mucinophila等,具有很高的丙酸盐生产能力。
丁酸和丙酸等短链脂肪酸通过迷走神经,穿过血脑屏障,然后激活可能改变多巴胺能和血清素能信号传导的受体。
神经保护特性:如丁酸盐
丁酸盐影响肠嗜铬细胞释放血清素,前面我们已经了解到血清素在偏头痛发病中的重要性。
与产丁酸相关的菌群,例如:
快速和极端的饮食变化直接影响肠道微生物群,影响肠道微生物群的β多样性。在高脂肪饮食中添加益生元(可发酵纤维)可以恢复产丁酸菌和双歧杆菌水平,这也突出了饮食对肠道微生物群组成的关键影响。
丁酸盐刺激齿状回的细胞增殖和分化,并增强脑源性神经营养因子(BDNF)和胶质源性神经营养因子(GDNF)的表达。丁酸盐还通过抑制 TNF-α 的合成而在大脑中显示出抗炎作用,TNF-α 是由内毒素脂多糖 (LPS) 通过抑制NF-κB 诱导的。
下面我们来看看炎症因子在偏头痛发病中扮演怎样的角色。
炎症介质
在炎症性疾病和偏头痛中都观察到炎症免疫反应增强。促炎性免疫反应的主要触发因素是由于肠道通透性增加而导致脂多糖进入循环,那么,炎症反应可能出现在身体的各个部位,当三叉神经上伤害感受器的激活时,偏头痛就出现了。
研究表明偏头痛发作期间,下列炎症因子升高:
这些炎症因子是如何在偏头痛发病中发挥作用的呢?
IL-1β 是一种主要细胞因子,可驱动大脑和全身炎症,激活普遍存在的细胞表面受体 IL-1R1。
IL-1β/IL-1R1 轴激活一系列炎症分子,包括其他细胞因子和趋化因子。IL-1β 激活脑膜伤害感受器并使之敏感,是皮质扩散性抑制后三叉神经激活的关键介质。也就是说,IL-1β/IL-1R1 轴在外周TG和中枢神经系统的三叉血管通路中尤其发挥着至关重要的作用。
IL-1和IL-6 是由调节性B细胞响应微生物群的紊乱而释放的。
研究表明,肠道微生物群失调通过三叉神经伤害感受系统中的TNF-α水平上调,促进了类似偏头痛疼痛的持续性。
肠道菌群通过间接信号传导维持肠脑轴正常平衡的作用机制
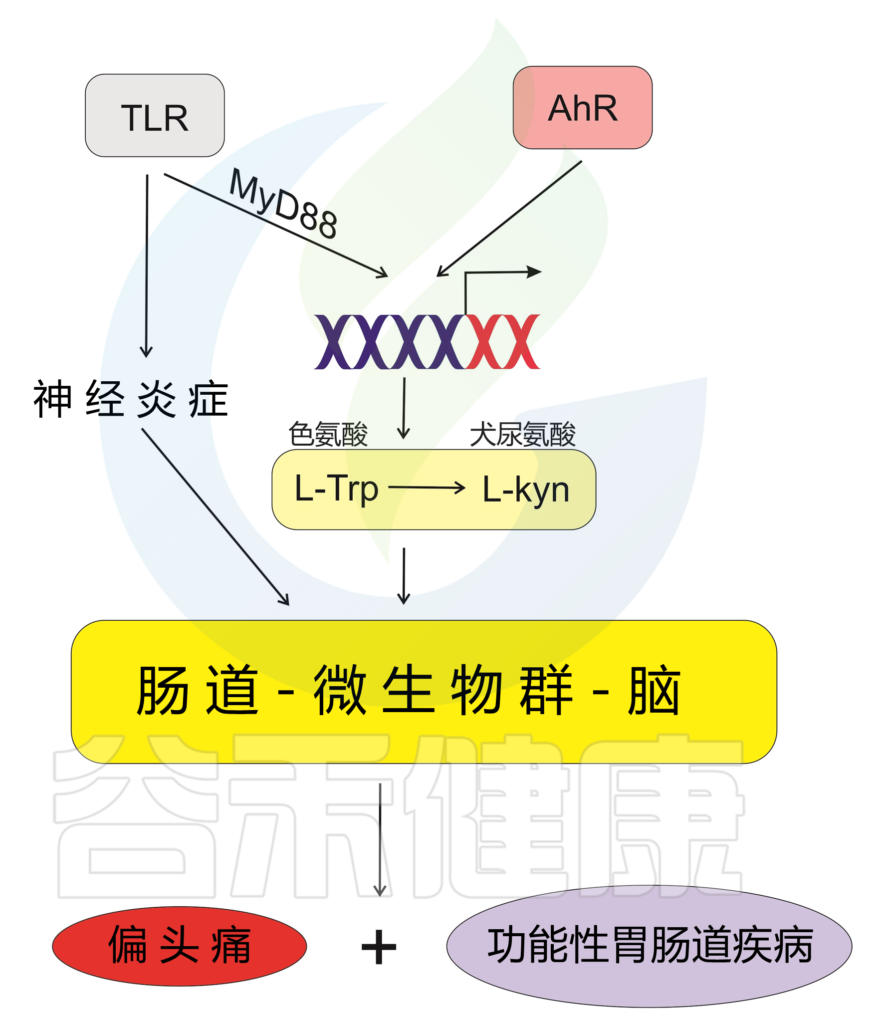
doi: 10.1186/s10194-020-1078-9
IL-8的表达,涉及到神经肽,与P物质有关,P物质可以帮助调节免疫细胞及其产生的各种细胞因子。
接下来,我们来看偏头痛中重要的——神经肽。
★ CGRP(降钙素基因相关肽)
对于患有慢性偏头痛的患者,CGRP 水平在偏头痛发作期间持续升高,并且在两次发作之间继续升高。CGRP是我们反复提到的词,我们来看看它在偏头痛疾病中发挥什么样的作用。
CGRP,降钙素基因相关肽 ,一种由37个氨基酸组成的肽,可能通过胃肠道免疫细胞和肠道运动与肠道微生物有关。在偏头痛的情况下,它在三叉神经节、三叉神经尾核、杏仁核、丘脑和皮质中浓度很高。
CGRP有两种形式:
CGRP升高如何与偏头痛关联起来:
星形胶质细胞和神经元参与了CGRP合成释放过程的正反馈循环,并维持了增加的炎症和致敏状态。
CGRP 可以作为一种有价值的治疗标志物
目前,许多用于治疗和预防偏头痛药物的原理,就是针对这个CGRP或其受体的,抑制 CGRP 转录并减少其释放,从而缓解偏头痛。
★ P 物 质
P物质(substance P),是一种广泛分布于细神经纤维内的神经肽,主要作用是传递痛觉信息。
呕吐是最常见的胃肠道反应之一,受P物质的控制
在中枢神经系统中,延髓有面突区和孤束核的区域,这两个区域控制呕吐反射并含有高水平的P物质。P物质释放结合到NK1R上会触发呕吐反应。
P物质的一个功能是通过改变细胞信号通路作为神经递质和疼痛感知调节剂
有一个假说是P物质有助于突触后神经元对谷氨酸的敏感性,有助于将疼痛信号传递到大脑的体感区域。在许多其他中枢神经系统病理中通过神经源性炎症引起早期血脑屏障的破坏。
P物质的关键作用是双向调节免疫细胞及其产生的各种细胞因子
IL-1、IL-4和IFN-γ可以诱导巨噬细胞表达NK1R。诱导细胞因子释放会导致巨噬细胞和树突状细胞的招募,这有助于刺激IL-8的表达。P物质间接参与了免疫细胞的招募和迁移。P物质能刺激和诱导肥大细胞脱颗粒化。
★ PACAP
在与偏头痛病因机制相关的几个结构中发现了PACAP(垂体腺苷酸环化酶激活肽)的存在,包括硬脑膜、脑血管、三叉神经节、三叉神经原球和颈椎脊髓。
PACAP在神经调节、神经源性炎症和痛觉中起着重要作用。PACAP在感觉神经元炎症后上调。PACAP与CGRP在一些硬膜神经纤维中共同表达,也与疼痛的中枢敏化和情绪负荷有关。
PACAP对革兰氏阳性菌、革兰氏阴性菌和真菌具有抗菌活性,因此它可以直接影响膜的杀菌功能。有效的防御能力和较低的细菌耐药性使抗微生物神经肽成为一类有吸引力的新型抗生素。
★ NPY(神经肽Y)
NPY是去甲肾上腺素能系统功能的可能指标,通过脑循环调节影响脑血流量。值得一提的是,在偏头痛发作期检测到较高水平的NPY。
NPY已经在肠-脑轴的各个层面上被检测到。NPY通路也有助于改变胃肠道功能及其血流、免疫系统和炎症状态、疼痛、能量稳态、情绪、情绪以及行为和大脑认知功能等。
★ 胆囊收缩素 (CCK)
胆囊收缩素(CCK)由小肠粘膜内层(I 细胞)、哺乳动物大脑(例如皮质、丘脑、中脑边缘、导水管周围灰质和中脑)和脊髓的肠内分泌细胞合成。这种肽抑制胃排空和胃酸分泌,刺激胆囊收缩和胰腺分泌,并通过 CCK1 受体在大脑中激发饱腹感。
CCK在导水管周围灰质中产生,可能是偏头痛内源性疼痛信号系统及其水平升高的原因。CCK也存在于三叉神经节中,因此刺激三叉神经节会导致局部CCK增加。
CCK——肥胖和偏头痛之间的桥梁之一
肥胖患者偏头痛发作的频率和严重程度明显更高。偏头痛和肥胖之间联系的一种可能解释是,是高脂饮食引起的CCK分泌,因为十二指肠内游离脂肪酸会刺激CCK的分泌。
关于CCK可以详见我们之前肠道激素的文章:
当考虑到性激素时,我们知道偏头痛在女性中的发病率是男性的2-3倍,女性的发作时间更长、更严重,恢复时间也更长。
无先兆偏头痛的风险在月经前 2 天和月经期间最高,这也是生殖道微生物群多样性最低的时候。周期中期雌激素和孕酮水平高与无先兆偏头痛的风险较低以及生殖道内微生物群落的稳定性较高有关。
众所周知,雌激素会改变肠道内的紧密连接和粘蛋白保护,改变肠道通透性。
雌激素受体广泛表达于中枢神经系统,有证据表明雌激素调节痛觉传导,雌激素敲除模型在脊髓背角水平导致CGRP升高,从而改变了发起偏头痛攻击所需的阈值。临床上支持这一点的是黄体晚期偏头痛发作的可能性增加。虽然孕酮撤退似乎不会触发偏头痛发作,但它可能在疼痛敏感性和病程演化中起作用。
关于雌激素与肠道菌群详见我们之前的文章:
睾酮降低了对CSD的易感性,增加了血清素能调,并具有抗炎特性。较高水平的睾酮与肠道微生物群多样性增加有关,研究表明,患有偏头痛的男性存在雄激素缺乏和雌二醇水平增加。
因此,肠-脑轴可能介导性激素和头痛疾病之间的关系。
以上我们阐述了肠道菌群及其代谢产物等参与偏头痛发病机制,包括炎症介质、肠道微生物群、神经肽、血清素途径等。有证据表明偏头痛和胃肠道疾病之间存在共同的病理生理学,下面我们来了解一下它们之间的关联。
偏头痛和胃肠道合并症的病理生理学都很复杂,尚未完全阐明;然而,科学证据确实表明这些复杂疾病之间的病理生理学重叠。研究这种重叠有助于阐明常见的病理生理异常或生物学机制。
特别是,自主神经系统(ANS)被认为在偏头痛和胃肠道功能障碍之间的关联中起重要作用,因为它们的症状特征相似,包括恶心,呕吐,消化不良和胃轻瘫。
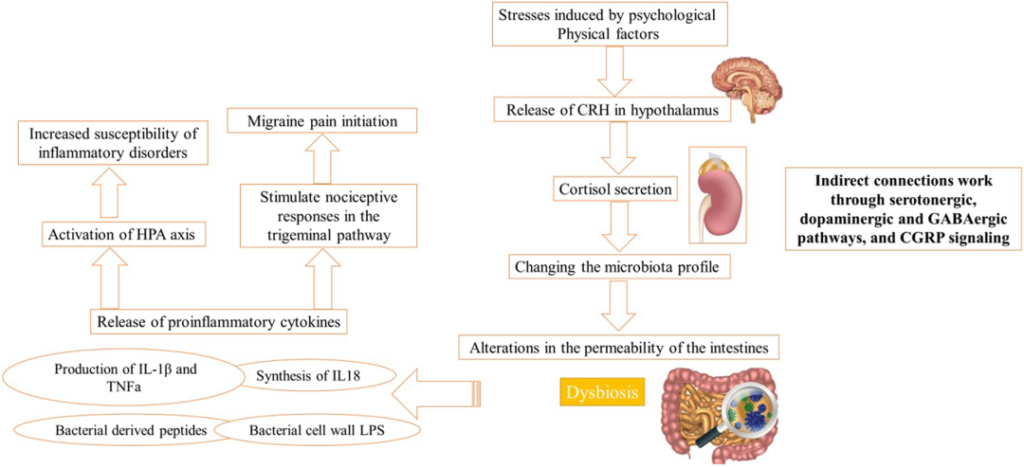
doi: 10.1111/head.14099
肠易激综合征(IBS)和炎症性肠病(IBD)的患者中,偏头痛的发生率增加。两者都是严重的肠道疾病,与肠道通透性增加和微生物引起的炎症有关。此外,中枢性、内脏性和热性皮肤过敏在IBS和偏头痛中很常见。患有长期和更频繁头痛的偏头痛患者更有可能被诊断为IBS。
偏头痛与IBS有一些共同的重要特征,这两种疾病都是慢性疼痛相关疾病,在女性中比男性更普遍,并且在纤维肌痛、慢性疲劳综合征、间质性膀胱炎、失眠、抑郁等方面与心理合并症有关。据估计,约60%的偏头痛患者患有异常性疼痛,除内脏超敏反应外,大多数IBS患者还报告异常性疼痛。
在IBS患者的粪便微生物群中评估出偏头痛的严重程度与肠道菌群失调之间存在强正相关。约一半没有先兆的年轻女性偏头痛患者患有肠道微生物群失调。
美国肠道计划队列研究发现,偏头痛与口腔微生物中具有还原硝酸盐、亚硝酸盐和一氧化氮功能活性的菌群水平呈正相关关系。
IBD是一种慢性复发-缓解炎症性疾病,由克罗恩病和溃疡性结肠炎组成。关于偏头痛和IBD之间可能关系的数据很少。
虽然机制不明确,IBD中存在的自身免疫性炎症反应、吸收不良、内皮功能障碍以及为该疾病开具的免疫抑制治疗可能参与了炎症性肠病和偏头痛之间的病理生理关联。
扩展阅读:
根据罗马IV标准,功能性消化不良分为餐后不适综合征(PDS)和上腹疼痛综合征(EPS)。
偏头痛是功能性消化不良和餐后症状患者的常见合并症。
扩展阅读:
基于人群的研究,个体主要是女性,最常见的症状(无论是持续性还是偶发性)是恶心和呕吐,但也可能包括腹痛、腹胀、体重减轻、餐后饱腹感和早饱感。
在患有特发性胃轻瘫和腹痛的个体中,腹痛严重的人比症状较轻的人更有可能患有重叠偏头痛。
在胃轻瘫中,观察到胃排空时间与偏头痛发作患者头痛、恶心和对光敏感的严重程度显著相关。多潘立酮是一种多巴胺受体拮抗剂,可用于治疗胃轻瘫,并已被证明在早期高剂量给药时可预防大多数偏头痛发作。另一种多巴胺受体拮抗剂甲氧氯普胺可用于治疗胃轻瘫和恶心,作为偏头痛的急性静脉注射治疗是有效的。
根据 5 项病例对照研究的荟萃分析结果,约 45% 的偏头痛患者患有幽门螺杆菌感染,而健康对照组的患病率估计约为 33% 。有证据表明根除幽门螺杆菌可能与缓解偏头痛症状有关。
幽门螺杆菌感染与持续的慢性炎症状态有关,这反过来可能导致炎症介质和血管活性化合物的产生增加。因此,偏头痛和幽门螺杆菌感染之间的关联机制可能包括诱导免疫、炎症和血管反应,以及随后免疫细胞、炎症和血管活性物质释放到胃粘膜中,最终可能导致大脑疼痛敏感结构的超敏反应。
扩展阅读:
正确认识幽门螺杆菌
乳糜泻具有多种神经系统表现,如癫痫、共济失调、小脑性共济失调、情绪障碍、脑炎、周围神经病变、神经肌肉疾病、痴呆、学习障碍、发育迟缓和偏头痛。
与没有这种疾病的人相比,被诊断患有乳糜泻(一种由谷蛋白肽引发的自身免疫性疾病)的人表现出更高的偏头痛发生率,据估计,大约 21-28% 的乳糜泻患者患有偏头痛。
相反,患有偏头痛的人乳糜泻的患病率更高。临床医生可能会考虑在存在此类改变的偏头痛受试者中寻找乳糜泻,特别是如果存在其他提示乳糜泻的症状。
偏头痛和乳糜泻之间的联系可归因于几种同时存在的机制,包括面筋引起的促炎细胞因子的激活、吸收不良导致的重要维生素和必需元素的缺乏、血管张力的紊乱、神经系统敏感性的增加、脑灌注不足、血管周围炎症等。
✔ 消除饮食
消除饮食需要先确定首先引发头痛的饮食成分,然后从常规饮食中排除这些成分。这是一种个人方法,个人可以识别引发或加剧头痛的成分。
如果某种食物在接触后 1 天内引起头痛,则可将其视为触发因素。大多数患者存在多种触发因素;因此,很难识别单一成分。况且有些食物含有不止一种成分,因此要完全识别出来并不简单。
✔ 综合饮食
富含ω-3和ω-6脂肪酸的低脂、高叶酸和生酮饮食对偏头痛患者有益。
改良的阿特金斯饮食提供神经保护,改善血清素能功能障碍,抑制神经炎症,改善线粒体功能。
生酮饮食会增加产生的酮体浓度,这对头痛患者有帮助。
低血糖饮食也能改善炎症状态。保持ω-6脂肪酸和ω-3摄入之间的微妙平衡可以减少炎症,维持血管张力,增强血小板功能。
偏头痛发作频率与坚持“健康”饮食模式(大量食用水果、蔬菜、鱼类、豆类、发酵食物等)成反比(P=0.04)。与坚持“西方”饮食方式(大量食用可乐、咸坚果、加工肉和快餐)成正比(P=0.02).
生姜、薄荷和辣椒都具有天然的止痛功效。生姜和薄荷有助于减少与偏头痛相关的恶心。
薰衣草可以减少血管炎症,通常是治疗焦虑、压力和压力引发的偏头痛有效的草药。
✔ 低钠饮食 / 高钠饮食
对脑脊液的分析显示,与对照组相比,偏头痛患者的钠含量较高,尤其是在头痛发作时。然而,低钠饮食的效果取决于许多因素,因此在考虑低钠饮食之前应详细评估其效果。
低钠饮食可以有效减少老年高血压患者头痛的发生。除了控制血压之外,低钠饮食还可以降低头痛的发生率。
相比之下,在没有高血压的低至正常体重指数的年轻女性人群中,高钠饮食有助于降低偏头痛发作的频率。
因此,低/高钠饮食应该根据患者群体进行调整。
低热量饮食 or 生酮饮食?
有研究认为,生酮饮食对偏头痛患者更有益,低热量饮食对偏头痛患者不利 (n = 108) 。在另一项研究中,生酮饮食显着改善了这些患者的头痛特征,并减少了发作频率和药物使用。停止生酮饮食后 2 个月观察到持续改善。
虽然看起来生酮饮食患者的头痛频率有所减少,但过渡期的头痛频率更严重。然而,经过3个月和6个月的随访后,低热量饮食组的头痛频率显著改善。
✔ 低血糖饮食(LGD)
通常从碳水化合物中获取总热量的 50-55% 是安全饮食。饮食转向低血糖指数碳水化合物已被证明可以增加短链脂肪酸水平。在某些情况下,LGD 是一种重要的替代方案,例如高脂血症、糖尿病、癫痫和体重管理。
罗马进行的一项队列研究中报告称,增加全麦面包(P = 0.04)和全麦面食(P = 0.004)的消费,以及减少白面包(P = .004)的消费,与偏头痛发作频率和每月使用药物救治的统计显著减少有关。
✔ 低脂饮食
一项研究调查了54名成年人减少脂肪饮食对偏头痛治疗的影响。患者被告知他们的脂肪消耗量将降至<20 g/天(12周)。发现头痛药物的发生率、严重程度和需求显著降低。
在一项为期 12 周的试验中,患有慢性偏头痛的成年人被随机分配接受高omega-3 / 低omega-6 饮食或低 omega-6 饮食。与采用低omega-6 饮食的偏头痛患者相比,采用高omega-3/低 omega-6 饮食的人的头痛改善程度更高。
可能与此相关:omega-6 和 omega-3 之间的平衡有助于炎症控制。omega-6 脂肪酸促进血管舒张。
谨慎选择下列食物
▸
味精在高浓度和溶解在液体中可能会引发偏头痛。并不是说引发每个患者的偏头痛发作,不能一概而论。
▸
咖啡因可能缓解偏头痛或者引发偏头痛。与阿司匹林和对乙酰氨基酚结合,咖啡因是一种高效的止痛药。咖啡因戒断会引发咖啡因使用者的偏头痛发作。需要注意自己的剂量。
咖啡因与偏头痛频率之间的联系没有定论。一些研究数据表明,不同的偏头痛和慢性日常头痛在咖啡因使用者中比不摄入这种物质的人更常见。也有研究认为偏头痛频率和咖啡因摄入之间没有关联。
▸
饮酒是偏头痛发作最常见的原因之一。酒精可以刺激三叉神经节中的脑膜伤害感受器,触发疼痛信号,然后在三叉神经脊核传递到丘脑核,最后传递到体感皮层。也可能涉及其他机制,例如血管舒张作用、脱水、毒性等。除了酒精,酒精饮料还含有某些化合物(酒精发酵的副产品),这些化合物都可能会引发偏头痛。
▸
许多研究提出了巧克力摄入与头痛之间的联系,但其确切的生理机制尚不清楚。可能是巧克力中的黄烷醇刺激内皮一氧化氮合酶(eNOS)活性,这可以通过增加一氧化氮(NO)的产生导致血管舒张。
也有研究认为巧克力含有许多维生素和矿物质(例如镁和核黄素),用于预防偏头痛。富含可可的饮食通过抑制CGRP的表达来预防三叉神经节神经元的炎症反应。
▸
酪胺(Tyramine),是一种衍生自酪氨酸的胺化合物,存在于各种食品中,包括陈奶酪、腌肉、熏鱼、啤酒、发酵食品和酵母提取物等。酪胺有可能通过促进去甲肾上腺素的释放并对α-肾上腺素受体产生激动作用而引发头痛。
▸
阿斯巴甜是一种人造甜味剂。几项研究表明,它会引起各种神经或行为症状;也会引起头痛,尤其是对于长期使用中等或高剂量(900–3000 mg/天)的人。
基于以上所述,识别和避免饮食中的偏头痛诱因至关重要,因为这有助于降低偏头痛的频率,让偏头痛患者能够控制让他们感到疲惫和无助的情况。
B族维生素
维生素B2对氧化代谢至关重要,可以降低偏头痛的风险。根据随机对照试验,服用维生素B2可以显著减少每月偏头痛天数1-3天。通过增加产短链脂肪酸菌丰度,维生素B2和其他B族维生素有助于对肠道微生物组产生正面影响。
扩展阅读:
维生素C
水溶性维生素C是最重要的非酶抗氧化剂之一,参与神经代谢。维生素C的膳食来源是水果和蔬菜。
维生素C可能调节偏头痛过程中的神经炎症和氧化应激活性的影响。
一项开放标签研究中,患者接受了含有120 mg粉红树皮提取物、60 mg维生素C和30 IU维生素E的胶囊(每粒胶囊)。在完成治疗期的患者中,得到了改善,包括头痛频率和头痛严重程度的降低。该研究人员还进行了一项开放标签研究,以检查维生素C(150 mg)和辐射松树皮提取物对偏头痛症状的影响。患者服用上述剂量的维生素C和辐射松树皮提取物3个月。治疗后,患者表现出显著改善:头痛频率和头痛严重程度减轻。
扩展阅读:
维生素D3
补充维生素D3会影响肠道微生物群的组成。为健康人补充8周维生素D3也显著减少了幽门螺杆菌的数量。几项研究报告称,血清维生素D水平可能与偏头痛/头痛风险增加有关。
偏头痛患者中这种维生素缺乏/不足的患病率可能更高。补充维生素D对偏头痛发作的强度和频率也有良好的影响。
镁
镁参与多种酶促反应,还维持细胞膜的平衡,影响其通透性并减少自发去极化的可能性。它影响外周和中枢神经系统的兴奋性和神经传导,因此在偏头痛病程中起着重要作用。
镁缺乏导致皮质扩散抑制或谷氨酸能神经传递异常是镁与偏头痛关系的一个可能机制。镁不足可能会影响 NMDA 受体阻断、钙通道、谷氨酸和 NO 活性,以及血清素受体的亲和力。
许多临床医生认为,以正确的剂量和形式补充镁可以成为预防偏头痛发作的治疗方法。补充镁也可以配合其他药物。
益生菌是偏头痛的另一种潜在疗法,它与饮食干预一样能够改变肠道微生物群。
在 NTG 诱导的偏头痛小鼠模型中,发现口服益生菌混合物显著抑制抗生素治疗引起的偏头痛样疼痛延长。
在一项随机双盲对照试验中观察到,与安慰剂对照相比,在 40 名发作性偏头痛患者和 39 名慢性偏头痛患者中,为期 8 周的益生菌试验显著降低了偏头痛的严重程度、频率和药物使用量。
其中含有14种益生菌菌株:
益生菌组偏头痛发作的平均频率显著降低(平均变化:-9.67 与 -0.22;p ≤ 0.001)。补充益生菌显著降低了严重程度(平均变化为-2.69;p≤0.001.),降低慢性偏头痛患者的发作持续时间(平均变化:-0.59;p≤0.034),减少每天服用药物数量(平均变化为-1.02;p<0.001)。
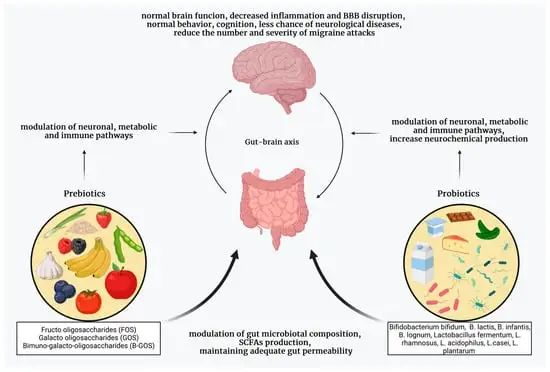
doi.org/10.3390/neurolint15030073
将芽孢杆菌、双歧杆菌、乳酸杆菌和链球菌菌株的混合物应用8-10周,可以降低发作性偏头痛患者的发作严重程度和频率,并降低慢性偏头痛患者的频率、严重程度、持续时间和每天服用的药物数量。
姜黄素
姜黄素是从姜黄根茎中提取的主要姜黄素,也是最著名的植物多酚之一。姜黄素具有许多有益的特性,如抗氧化、抗动脉粥样硬化、抗菌、免疫调节、抗衰等特性。
姜黄素可以显著抑制 IL-1β 和 TNF-α 表达、氧化应激和蛋白质积累。
对偏头痛治疗进行了一项研究,比较了单独使用舒米曲普坦(ST)和与姜黄素一起使用的效果,采用了硝酸甘油诱导的偏头痛大鼠模型。
姜黄素以下面两种形式静脉注射,因为姜黄素的肠道吸收较低:
a)酒精溶液(用生理盐水稀释)
b)脂质体的形式
当姜黄素以脂质体的形式进行注射时,其抗氧化作用更为显著。作者将所得结果归因于姜黄素能够清除羟基或过氧自由基、直接与过氧化物自由基相互作用以及抑制参与炎症性因子NF-κBs的活性。
偏头痛发病机制的一个考虑的机制涉及肿瘤坏死因子α(TNF-α)的参与,通过引发神经元超兴奋性、刺激伤害感受器和前列腺素的产生,从而引发神经炎症。
一项研究,旨在确定姜黄素和ω-3脂肪酸对TNF-α基因表达的协同作用。接受两种物质治疗的患者在血浆中显示出TNF-α mRNA水平的降低(反映了TNF-α表达的减少)。在只接受其中一种分析物的患者中,没有发现这种情况。
辅酶Q10
除了抗氧化特性外,辅酶Q10还表现出抗炎特性,参与嘧啶合成(作为辅因子),进而参与DNA复制和RNA修复过程。由于其潜在的积极作用,是许多神经系统疾病的膳食补充剂。
酶Q10被列为预防偏头痛最常用的治疗方法之一。给予偏头痛患者辅酶Q10会导致CGRP的降低。
儿童和青少年中研究了补充辅酶Q10与偏头痛发生之间的关系。患者每天服用100毫克辅酶Q10,为期4个月。结果显示,在主要结果方面,两组之间没有统计学上的显著差异。然而,与对照组相比,偏头痛患者的研究组头痛发作频率呈下降趋势。此外,在补充辅酶Q10的前四周,观察到发作性偏头痛患者头痛的严重程度降低。
在辅酶Q10的抗氧化特性中有以下方面,即超氧化物歧化酶活性的调节(导致细胞抗氧化防御的改善)、氧化应激OS的减少或增加谷胱甘肽还原形式(用作抗氧化剂)浓度的能力。上述机制可缓解偏头痛,降低偏头痛发作的频率,缩短偏头痛发作的持续时间。
银杏内酯B
银杏内酯B是从银杏叶中提取的一种草药成分。银杏叶提取物具有神经保护特性,对记忆有有益作用,也有抗氧化剂的某些特性。
一项为期六个月的开放标签多中心研究,证明了银杏内酯B对每月至少发作一次的偏头痛患者治疗先兆偏头痛的有效性。
槲皮素和银杏内酯B对减轻偏头痛症状有特殊作用。银杏叶提取物成分可降低氧化应激,抑制血小板活化因子,并影响谷氨酸能传递。
所有生物学机制都会增加神经保护作用,从而降低偏头痛的频率,减轻偏头痛,缓解先兆症状。
小白菊
小白菊(Tanacetum parthenium)属菊科植物,广泛分布于南美洲。其特征是具有抑制醛糖还原酶活性的高潜力,并显示出必要的抗氧化特性。
小白菊用于治疗多种疾病,不仅可以缓解偏头痛的症状,还可以缓解另一种来源的疼痛,如炎症、恶心和呕吐。
一项双盲、多中心、安慰剂对照研究,以确定小白菊稳定提取物(MIG-99)在降低偏头痛发作频率方面的有效性。最显著的效果记录为6.25mg剂量的MIG-99。三年后,同一作者发表了剂量为6.25 mg的MIG-99制剂的有效性和安全性结果。在服用该制剂的组中,偏头痛发作频率每月减少1.9次(安慰剂组每月减少1.3次)。
粪菌移植可以通过改变肠道菌群,帮助恢复肠道微生物平衡,使肠道微生物恢复更加有效和持久,理论上可减轻偏头痛发作和偏头痛严重程度,超过340项临床试验正在进行中。肠道菌群的改变可导致炎症介质和血清素途径的正常化,从而缓解偏头痛的频率和强度。
扩展阅读:
吴茱萸汤是一种临床用于治疗和预防偏头痛的配方,可anaerostipes和acidifaciens丰度,以恢复细菌的比例。由于5-HT合成与微生物群、功能和代谢紊乱相关,吴茱萸汤可以通过影响中枢和外周5-HT来缓解痛觉过敏。研究表明,在基因敲除小鼠和豚鼠中,5-HT1D受体具有抑制神经肽释放的作用,从而改变硬脑膜神经源性炎症反应。
良好的生活习惯
建立规律的睡眠时间。每天同一时间起床和上床睡觉,甚至周末也是如此。一天结束时放松一下。
减少吸烟。偏头痛患病率与每日吸烟的相关性为正相关(Spearman系数,rs=0.49)。
定期运动
身体活动期间,身体会释放某些化学物质来阻隔脑部疼痛信号。这些化学物质还有助于缓解焦虑和抑郁症,这两种状况都可能加剧偏头痛。
肥胖也会增加慢性头痛的风险。通过运动和饮食维持健康体重可能也有助于管理偏头痛。
值得尝试的有氧运动可能包括快走或慢跑、骑自行车、瑜伽、游泳等。
缓解压力
简化生活,明确管理时间,适当休息,可以选择接触大自然,做深呼吸(专注且缓慢),冥想,瑜伽等放松的方式,保持积极心态,享受乐趣,每天抽出至少 15 分钟做自己喜欢的事。
写偏头痛日记
记下偏头痛发作时周围的环境条件。写下感官刺激(明亮的灯光、响亮的音乐、奇怪的气味等)、压力的原因、饮食习惯和睡眠习惯。几次发作后,回顾一下日记,找出偏头痛发作前通常存在的常见情况。这些情况可能是“触发因素”,下次尽量避免。
当偏头痛发作时的缓解小方法:
关灯、静音
光线和声音会使偏头痛加剧,尽量在昏暗且安静的房间里放松,闭目休息,能睡就睡吧,让眼睛和大脑得到放松,帮助缓解头痛的不适感。
找一个安静的环境,避免嘈杂的声音,减轻对偏头痛的刺激。
如果不能马上找到这样的地方,可以考虑戴上太阳镜和耳塞。或者尝试舒缓的音乐。
试试温度疗法
热敷或冷敷头部或颈部。
按摩、放松
用双手的食指和中指,以小幅度打圈的方式轻轻按摩太阳穴、颈侧、颈后。
通过深呼吸放松身体和心理,减轻紧张和疼痛感。适量饮水,保持身体水分平衡,有助于缓解头痛。
偏头痛是由多种因素共同引起的,由遗传、内分泌、代谢和/或环境因素相互交织促成的。基于各种初始因素,广泛接受的观点是皮质扩散抑制假说和三叉神经血管假说。前者指出了大脑与神经炎症之间的相互关系,后者指出神经和血管是发生的关键因素。偏头痛复杂的机制和病理生理学仍在广泛研究中。
本文我们了解到,肠道微生物群通过参与肠-脑轴各个途径,肠道微生物群和大脑通过各种途径相互作用,包括免疫系统、色氨酸代谢、迷走神经和肠神经系统等,可能发挥着关键作用,或将作为偏头痛的有前途的生物标志物和治疗靶点。了解这种疾病的多系统联系,极大地扩展了对偏头痛机制的理解,并提供了更广泛的治疗蓝图。
偏头痛本身会产生一些诊断问题,这些问题可能会因合并症而加剧,包括功能性胃肠道疾病等。同时,复杂的疼痛通路,食物和药物代谢、免疫反应等造成的个体差异,会给在偏头痛的临床管理治疗过程中带来难度,目前药物并不能完全缓解症状或满足要求。
考虑以互补的方式将临床前和临床研究结合起来,同时包括科研人员、临床医生、健康管理以及其他相关机构的成员之间的相互交流、协作,在此过程中纳入肠道菌群健康检测,集结众多力量的合作,从预防偏头痛辅助判别其潜在风险,到偏头痛发作的疾病管理,再到预后减少发病次数等多个阶段,帮助降低偏头痛和偏头痛合并的各种胃肠道疾病的风险,改善偏头痛患者的生活质量,为他们带来更多希望。
注:本账号内容仅作交流参考,不作为诊断及医疗依据。
主要参考文献
Ferrari MD, Goadsby PJ, Burstein R, Kurth T, Ayata C, Charles A, Ashina M, van den Maagdenberg AMJM, Dodick DW. Migraine. Nat Rev Dis Primers. 2022 Jan 13;8(1):2.
Khan J, Asoom LIA, Sunni AA, Rafique N, Latif R, Saif SA, Almandil NB, Almohazey D, AbdulAzeez S, Borgio JF. Genetics, pathophysiology, diagnosis, treatment, management, and prevention of migraine. Biomed Pharmacother. 2021 Jul;139:111557.
Arzani M, Jahromi SR, Ghorbani Z, Vahabizad F, Martelletti P, Ghaemi A, Sacco S, Togha M; School of Advanced Studies of the European Headache Federation (EHF-SAS). Gut-brain Axis and migraine headache: a comprehensive review. J Headache Pain. 2020 Feb 13;21(1):15.
Aurora SK, Shrewsbury SB, Ray S, Hindiyeh N, Nguyen L. A link between gastrointestinal disorders and migraine: Insights into the gut-brain connection. Headache. 2021 Apr;61(4):576-589.
Zhou Y, Pang M, Ma Y, Lu L, Zhang J, Wang P, Li Q, Yang F. Cellular and Molecular Roles of Immune Cells in the Gut-Brain Axis in Migraine. Mol Neurobiol. 2023 Sep 11.
He Q, Wang W, Xiong Y, Tao C, Ma L, Ma J, You C; International Headache Genetics Consortium. A causal effects of gut microbiota in the development of migraine. J Headache Pain. 2023 Jul 17;24(1):90.
Kim JH, Lee Y, Kwon YS, Sohn JH. Clinical Implications of the Association between Respiratory and Gastrointestinal Disorders in Migraine and Non-Migraine Headache Patients. J Clin Med. 2023 May 12;12(10):3434.
Cámara-Lemarroy CR, Rodriguez-Gutierrez R, Monreal-Robles R, Marfil-Rivera A. Gastrointestinal disorders associated with migraine: A comprehensive review. World J Gastroenterol. 2016 Sep 28;22(36):8149-60.
Slavin M, Li HA, Frankenfeld C, Cheskin LJ. What is Needed for Evidence-Based Dietary Recommendations for Migraine: A Call to Action for Nutrition and Microbiome Research. Headache. 2019 Oct;59(9):1566-1581.
Park CG, Chu MK. Interictal plasma glutamate levels are elevated in individuals with episodic and chronic migraine. Sci Rep. 2022 Apr 28;12(1):6921.
Sgro M, Ray J, Foster E, Mychasiuk R. Making migraine easier to stomach: the role of the gut-brain-immune axis in headache disorders. Eur J Neurol. 2023 Jun 17.
Do TP, Hougaard A, Dussor G, Brennan KC, Amin FM. Migraine attacks are of peripheral origin: the debate goes on. J Headache Pain. 2023 Jan 10;24(1):3.
Pleș, H.; Florian, I.-A.; Timis, T.-L.; Covache-Busuioc, R.-A.; Glavan, L.-A.; Dumitrascu, D.-I.; Popa, A.A.; Bordeianu, A.; Ciurea, A.V. Migraine: Advances in the Pathogenesis and Treatment. Neurol. Int. 2023, 15, 1052-1105.
Kappéter, Á.; Sipos, D.; Varga, A.; Vigvári, S.; Halda-Kiss, B.; Péterfi, Z. Migraine as a Disease Associated with Dysbiosis and Possible Therapy with Fecal Microbiota Transplantation. Microorganisms 2023, 11, 2083.
Yamanaka, G.; Suzuki, S.; Morishita, N.; Takeshita, M.; Kanou, K.; Takamatsu, T.; Suzuki, S.; Morichi, S.; Watanabe, Y.; Ishida, Y.; et al. Role of Neuroinflammation and Blood-Brain Barrier Permutability on Migraine. Int. J. Mol. Sci. 2021, 22, 8929.
Martami F, Togha M, Seifishahpar M, Ghorbani Z, Ansari H, Karimi T, Jahromi SR. The effects of a multispecies probiotic supplement on inflammatory markers and episodic and chronic migraine characteristics: A randomized double-blind controlled trial. Cephalalgia. 2019 Jun;39(7):841-853.
Fila, M.; Chojnacki, J.; Pawlowska, E.; Szczepanska, J.; Chojnacki, C.; Blasiak, J. Kynurenine Pathway of Tryptophan Metabolism in Migraine and Functional Gastrointestinal Disorders. Int. J. Mol. Sci. 2021, 22, 10134.
Goschorska, M.; Gutowska, I.; Baranowska-Bosiacka, I.; Barczak, K.; Chlubek, D. The Use of Antioxidants in the Treatment of Migraine. Antioxidants 2020, 9, 116.
Spekker, E.; Nagy-Grócz, G. All Roads Lead to the Gut: The Importance of the Microbiota and Diet in Migraine. Neurol. Int. 2023, 15, 1174-1190

谷禾健康
不知道大家有没有这样的体验:紧张时会想上厕所,伤心时会食欲不振。任何情绪变化,好像都会反映在消化系统。
与此同时,你相不相信一个人的肠道微生物可以影响其认知、性格、气质、社交情况、甚至是责任心。
有大量证据表明神经系统与肠道微生物的组成和功能有关。目前,已经描述了各种肠-脑串扰途径,包括通过迷走神经途径的免疫、内分泌和神经回路。此外,中枢神经系统控制肠道菌群的失衡,构成双向通讯系统。
肠道菌群失衡,也许是导致精神问题的潜在原因。一些研究发现,肠道菌群改变(生态失调)与阿尔茨海默病、自闭症和帕金森病等神经精神疾病以及焦虑、抑郁和认知功能障碍等其他心理疾病有关。
肠道菌群可以分泌神经递质,例如乙酰胆碱、γ-氨基丁酸(GABA)和色氨酸;此外肠道中有数百万个神经元,可以通过神经递质向大脑发送信号。
肠脑轴功能障碍被认为是精神障碍的病理生理学;因此,恢复受干扰的微生物生态系统为管理或预防这些神经系统疾病提供了一种新颖且有前途的方法。
本文对人类肠道微生物群失调与几种精神和心理疾病之间联系进行了概述。此外,还展示了一些使用精神药物与调节肠道微生物相结合的方式以改善精神疾病或心理障碍的干预措施。肠道微生物群在未来可能成为缓解压力与治疗精神疾病的新工具。
人类肠道含有多样化的微生物群落,据估计,肠道中的微生物数量可以达到数万亿个。微生物群的密度在肠道不同部位也有所不同,结肠和直肠的微生物密度较高,而小肠的微生物密度较低。
肠道微生物由多种微生物类群组成,包括细菌、病毒、真菌、古细菌和原生动物,其中最主要的是细菌。
•古细菌
在健康的人类肠道中,古细菌的数量很少,其中史密斯产甲烷短杆菌(Methanobrevibactersmithii)是最常见的物种。
•真菌
肠道中最常检测到的真核微生物是真菌,例如念珠菌(Candida)和酵母菌(Saccharomyces)。这些真菌维持生态和免疫肠道微生物组的平衡。
•病毒
人体病毒组主要由噬菌体组成,它们在肠道中的作用是作为细菌组的调节剂。
•原生生物
一些原生动物,例如芽囊原虫属(Blastocystis),已在人类肠道微生物群中检测到,它们的存在通常与胃肠道疾病有关。
✦细菌是人类肠道中最常见的微生物
细菌是人体肠道中最常见的微生物,一般属于以下8个门:放线菌门 (Actinobacteria)、芽孢杆菌门 (Bacillota)(又名厚壁菌门)、拟杆菌门 (Bacteroidetes)、弯曲菌门 (Campylobacterota)、梭杆菌门(Fusobacteria)、变形菌门(Proteobacteria)、热脱硫杆菌门(Thermodesulfobacteriota)和疣微菌门(Verrucomicrobia)。
下图显示了健康人肠道微生物群中主要的门、科和属。
Borrego-Ruiz A,et al.Prog Neuropsychopharmacol Biol Psychiatry.2023
✦肠道不同部位的微生物群组成具有差异
肠道区室中的微生物有特定的空间分布。上消化道中最主要的是孪生球菌属(Gemella)、韦荣氏球菌属(Veillonella)、奈瑟菌属(Neisseria)、梭杆菌属(Fusobacterium)、链球菌属(Streptococcus)、普雷沃氏菌属(Prevotella)、假单胞菌属(Pseudomonas)和放线菌属(Actinomyces)。
而在下消化道中则有粪杆菌属(Faecalibacterium)、拟杆菌属(Bacteroides)和瘤胃球菌属(Ruminococcus)。
十二指肠
研究发现人类十二指肠粘膜富含芽孢杆菌(Bacillus)、Solibacillus、赖氨酸芽孢杆菌(Lysinibacillus)、微小杆菌属(Exiguobacterium)、海洋芽胞杆菌属(Oceanobacillus)和类芽孢杆菌(Paenibacillus)。
空肠
在空肠中,变形菌门和厚壁菌门是最常见的门,而埃希氏菌属、乳杆菌属和肠球菌属是空肠中最常见的属。
回肠
链球菌属(Streptococcus spp.)、大肠杆菌(Escherichia coli)和梭菌属(Clostridium spp.),是回肠中最常见的物种。
结肠
结肠是一个厌氧环境,以拟杆菌门、放线菌门和厚壁菌门以及毛螺菌科(Lachnospiraceae)、瘤胃球菌科(Ruminococcaceae)、普雷沃氏菌科(Prevotellaceae)、拟杆菌科(Bacteroidaceae)和理研菌科(Rikenellaceae)为主,以及已鉴定的主要细菌属乳杆菌属、双歧杆菌属和粪杆菌属。
研究了不同结肠区域的细菌属优势,他们发现乙状结肠中存在拟杆菌属、肠球菌属。近端结肠和远端结肠为粪杆菌和大肠杆菌。
阑尾中微生物多样性较高,以放线菌门、梭杆菌门、变形菌门、拟杆菌门和厚壁菌门为主;以及毛螺菌科、肠杆菌科、拟杆菌科、梭杆菌科和双歧杆菌科。
尽管人类肠道微生物群的组成因年龄、营养和抗生素使用等因素而在分类学和功能上存在差异,但人类肠道微生物群主要是由乳杆菌属、芽孢杆菌属、梭菌属、肠球菌属、瘤胃球菌属、粪杆菌属、罗氏菌属(Roseburia)、经黏液真杆菌属(Blautia)、Dorea属、拟杆菌属、普雷沃氏菌属、双歧杆菌属和埃希氏菌属。
✦肠道微生物对于消化非常重要
肠道微生物群为人类宿主提供了多种积极影响,包括免疫和代谢功能。肠道微生物是消化的关键调节者,通过营养物质的吸收以及多种代谢物的合成,包括脂质、氨基酸、维生素和短链脂肪酸。
✦肠道微生物有助于预防病原体感染
此外,肠道微生物群通过多种竞争过程在预防病原菌感染方面发挥着关键作用,并且还有助于维持肠上皮的完整性。
✦肠道微生物产生的化合物可作为信号分子
肠道微生物群从未消化的食物中产生多种分泌化合物。其中一些可以作为细菌间通讯的信号分子,影响细菌调节、稳态、生长、毒力和生物膜形成等。
色氨酸代谢物
细菌还可能分泌其他生物活性化合物,包括色氨酸分解代谢物、短链脂肪酸、多胺和组胺。色氨酸的代谢产物[5-羟色胺(血清素)、犬尿氨酸和吲哚]与生理和神经元活动有关。
吲哚
吲哚调节肠促胰岛素的分泌,调节肠道屏障通透性,并且可以作为微生物和宿主细胞之间细胞间通讯的信号分子。此外,吲哚及其衍生物被描述为神经炎症的抑制化合物。
短链脂肪酸
最重要的定量代谢物是由微生物降解不可消化的膳食纤维、蛋白质和糖蛋白产生的短链脂肪酸。如丁酸盐、乙酸盐和丙酸盐,可以作为信号分子,局部调节从十二指肠到结肠的肠道功能,并且通过肠内分泌细胞,它们还可以控制肝脏、肌肉和大脑的代谢,影响肠道功能及宿主能量稳态。
此外,短链脂肪酸通过诱导神经炎症反应呈现神经活性特性。短链脂肪酸是强大的表观遗传调节剂,控制DNA甲基化遗传物质的可及性并抑制组蛋白脱乙酰化。
该机制依赖于10-11易位 (TET) 蛋白,该蛋白催化胞嘧啶残基羟基化为5-羟甲基胞嘧啶,介导主动DNA去甲基化。确定丁酸盐诱导前额皮质中许多行为相关基因的变化,特别是影响涉及神经元兴奋或抑制的基因。几种兴奋性神经递质和神经元激活标记基因的减少,以及抑制性神经递质基因的增加表明丁酸盐促进抑制途径转录物的转录。
✦细菌代谢产物会影响神经发育
最近的研究表明,细菌代谢产物会影响神经发育。一些研究人员还报道了短链脂肪酸影响神经发生基因的表达。
使用无微生物的斑马鱼模型,观察到无微生物培养的胚胎中神经基因表达显著下降,并通过添加斑马鱼代谢物重新建立了该模型,识别出超过300个下调的基因。
基因分析表明,这些基因参与重要的神经发育途径,包括转录调控和Wnt信号传导。事实上,Wnt信号通路在发育和疾病中发挥着重要作用,可以推测其调控与多细胞真核生物的细菌定植共同进化。
下图展示了肠道微生物群合成的一些关键神经递质及其与心理和精神疾病的关联。这些神经活性化合物通过内在或外在传入神经通路将信息从肠腔传输到肠神经系统、肠神经胶质细胞和中枢神经系统。
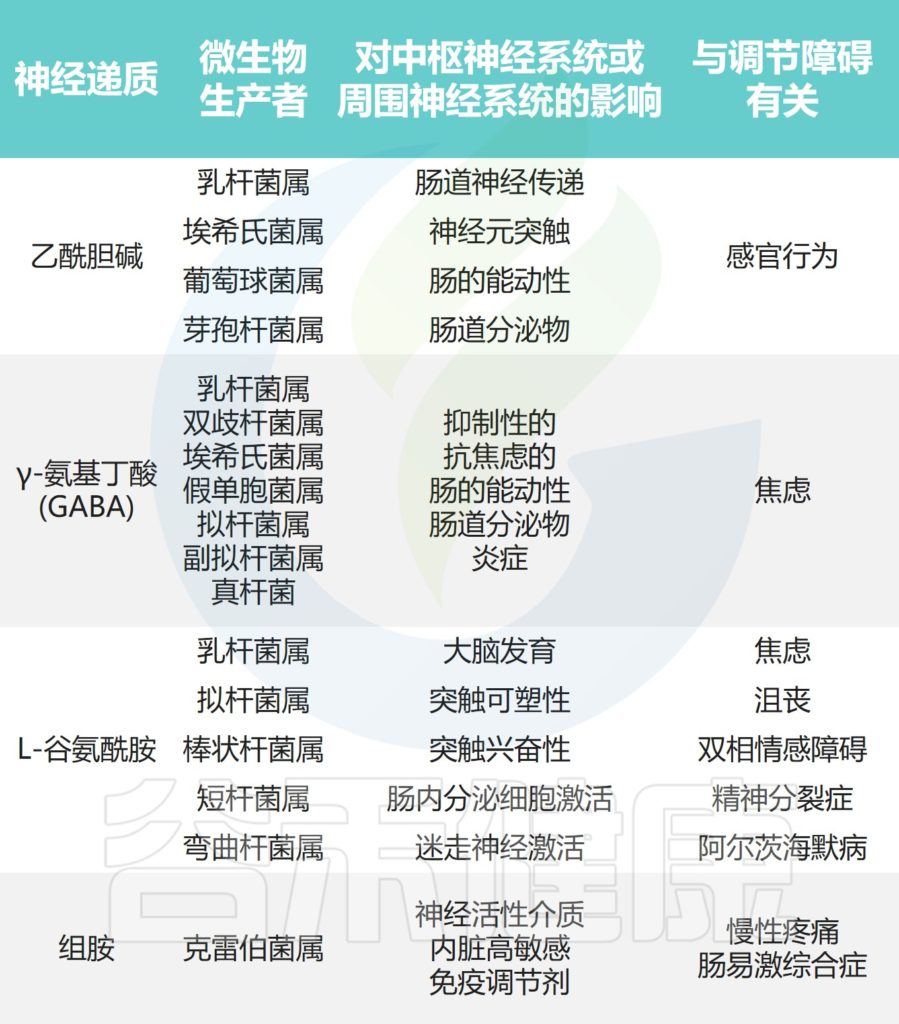

Borrego-Ruiz A,et al.Prog Neuropsychopharmacol Biol Psychiatry.2023
阿尔茨海默病(AD)是一种慢性神经退行性疾病,其中记忆和执行功能进行性丧失,与突触损伤和神经损失有关,主要发生在海马体和大脑皮层。
多项研究已明确β-淀粉样肽(Aβ)在阿尔茨海默病中的作用。最近的研究表明,细菌来源的淀粉样蛋白,例如curli、Csg A、tau、TasA、FapC和酚溶性调节蛋白,在Aβ寡聚体的聚集和促进错误折叠中发挥作用。
细菌淀粉样蛋白通过与Toll样受体2相互作用激活宿主免疫系统,诱导促炎介质刺激大脑中神经元淀粉样蛋白的产生。
•肠道微生物失调与阿尔兹海默病存在关联
其他证据表明肠道微生物群失调与阿尔茨海默病的早期阶段有关,导致细胞因子分泌增强、免疫衰老、神经炎症和氧化应激。
此外,肠道微生物失调会诱导脂多糖的分泌,从而破坏胃肠道通透性和血脑屏障,调节炎症途径,促进神经炎症和认知能力下降,并导致阿尔茨海默病中的神经元死亡。
在大鼠中,腹膜脂多糖给药导致炎症因子(例如IL-1、IL-6和TNF-α)水平升高,表明微生物组在阿尔茨海默病先天免疫反应的启动中发挥作用。
•阿尔兹海默病中短链脂肪酸水平降低
研究表明,阿尔茨海默病小鼠的微生物群组成和多样性受到干扰,短链脂肪酸水平降低,这预示着30多种代谢途径的改变,这可能与淀粉样蛋白沉积有关。
肠道菌群失调可能会增加氧化三甲胺(TMAO)浓度,这与不同的功能有关,例如β-淀粉样蛋白的形成、外周免疫反应的激活和氧化应激。此外,还与肠粘膜屏障功能障碍、血脑屏障通透性以及胆汁酸和胆固醇进入大脑的通道增加有关。
•阿尔茨海默病患者体内促炎菌群比例增加
研究发现阿尔茨海默病患者中大肠杆菌/志贺氏菌等促炎菌群的比例增加,而直肠真杆菌(eubacterium rectale)、霍氏大肠杆菌(E.hallii)、普拉梭菌(F. prausnitzii)和脆弱拟杆菌等产生丁酸的细菌则减少。
这些细菌与外周炎症状态有关,这是轻度认知障碍(MCI)和脑淀粉样变性患者的典型症状。
注:淀粉样脑血管病主要是由淀粉样物质在软脑膜以及大脑皮质小动脉中层沉积所导致的疾病,多见于80岁以上的老年人,可能会导致血管壁受损。
•阿尔茨海默病患者肠道微生物多样性较低
据报道阿尔茨海默病患者肠道菌群的微生物多样性较低,厚壁菌门和双歧杆菌种类减少。
此外,发现门水平上拟杆菌门细菌成员增加,放线菌门减少,以及科水平上瘤胃球菌科、肠球菌科和乳杆菌科增加,毛螺菌科、拟杆菌科和韦荣氏菌减少。
促炎细菌类群增加,同时产生丁酸盐的细菌减少,例如丁酸弧菌(B.proteoclasticus和B.Hungatei)、梭菌属、真杆菌属(E.Hallii、E.rectal和E.eligens)、普拉梭菌和罗斯拜瑞氏菌 (R.hominis)。
几种精神障碍疾病中的肠道菌群失调
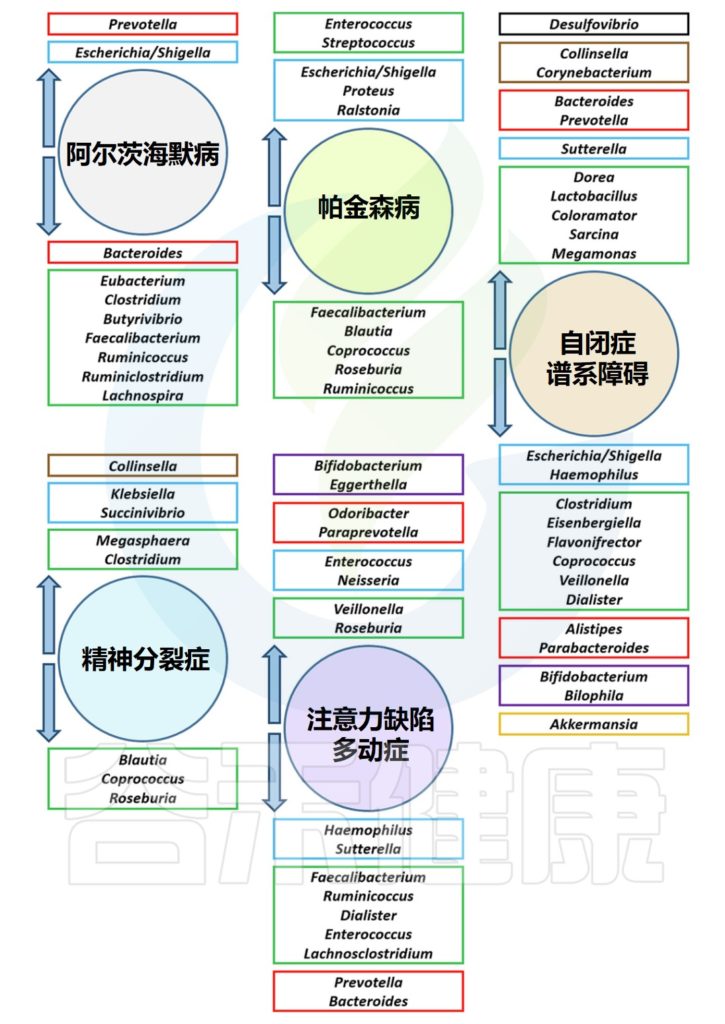
Borrego-Ruiz A,et al.Prog Neuropsychopharmacol Biol Psychiatry.2023
自闭症谱系障碍(ASD)是一种大脑发育障碍,其特征是刻板行为以及沟通和社交互动缺陷。
注:儿童自闭症是儿童精神类疾病当中最为严重的一种。
自闭症谱系障碍的确切病理学和病因很难确定,研究重点是调查遗传原因、免疫系统失调、环境毒物暴露以及肠道微生物群的改变。
•自闭症儿童的肠道微生物多样性较高
据报道自闭症儿童的肠道微生物多样性较高。自闭症谱系障碍样本中最常见的属如下:
拟杆菌属(Bacteroides),
脱硫弧菌属(Desulfovibrio),
柯林斯氏菌属(collinsella),
棒状杆菌属 (Corynebacterium),
乳杆菌属(Lactobacillus),
普雷沃氏菌属(Prevotella),
萨特氏菌属(Sutterella),
八叠球菌(Sarcina),
罗氏菌属(Roseburia),
粪杆菌属(Faecalibacterium),
巨单胞菌(Megamonas),
Caloramator,
Dorea;
此外以下属的存在减少:
双歧杆菌(Bifidobacterium) ↓↓↓
另枝菌属(Alistipes) ↓↓↓
嗜胆菌属(Bilophila) ↓↓↓
韦荣氏球菌属(Veillonella) ↓↓↓
阿克曼菌(Akkermansia) ↓↓↓
粪球菌属(Coprococcus) ↓↓↓
埃希氏杆菌属-志贺氏杆菌 ↓↓↓
梭菌属(Clostridium) ↓↓↓
艾森伯格氏菌(Eisenbergiella) ↓↓↓
嗜血杆菌属(Haemophilus)↓↓↓
Dialister ↓↓↓
Parabacteroides ↓↓↓
Flavonifractor ↓↓↓
这些发现强烈表明肠道微生物群失调与自闭症行为表现之间存在联系。
•有害菌会产生神经毒素影响自闭症
在自闭症行为表现患者中发现双歧杆菌丰度较低,而双歧杆菌是产生GABA神经递质的关键,双歧杆菌丰度过低会导致焦虑、认知缺陷和行为障碍。
产生神经毒素的艰难梭菌和溶组织梭菌的积累与自闭症谱系障碍症状相关。这些释放的有毒分子会影响血清素信号传导,可能导致自闭症谱系障碍行为模式,例如社交能力下降、对疼痛的反应减弱、语言异常以及自虐或重复行为。
受自闭症行为表现影响的个体表现出其他潜在有毒化合物水平的改变,例如抑制多巴胺-β-羟化酶的对甲酚和对甲酚硫酸盐。此外,在受自闭症谱系障碍影响的个体中检测到梭菌衍生的代谢物3-(3-羟基苯基)-3-羟基丙酸增加,这可能反映了儿茶酚胺代谢的改变。
•短链脂肪酸对自闭症的作用存在矛盾结果
短链脂肪酸在自闭症谱系障碍中的作用存在矛盾的结果。丙酸盐会诱导小胶质细胞激活、神经毒性细胞因子产生、基因表达改变、海马组织学异常和神经行为异常,例如重复动作和社交互动受损。
然而,丁酸盐对一种自闭症谱系障碍样小鼠模型的社交和重复行为具有有益影响。丁酸盐改善血脑屏障的不渗透性可能是丁酸盐可以恢复丙酸盐诱发的自闭症样疾病异常的另一种机制。
帕金森病(PD)是老年人中常见的一种痴呆症,其主要疾病症状包括神经炎症、中脑多巴胺能神经元丧失以及伴有非运动症状的异常运动。
•微生物失调引起的炎症可能导致帕金森病发展
一些研究人员认为,微生物群失调引起的炎症反应可能会导致帕金森病病理学的发展或恶化。这些反应可以促进脂多糖和α-突触核蛋白在肠道和大脑中的积累,并且由于促炎细菌活性导致氧化应激增加,从而促进α-突触核蛋白病理学通过迷走神经从头侧向尾侧区域扩散。
•帕金森病患者体内短链脂肪酸显著减少
另一方面,微生物短链脂肪酸可以防止肠粘膜通透性增加,从而减少细菌移位。帕金森病中短链脂肪酸显著减少。
短链脂肪酸在帕金森病病理学中的作用尚未完全阐明,报道短链脂肪酸的消耗导致肠神经系统中路易体的形成;相反,有研究表明短链脂肪酸能够降低血脑屏障的通透性。
注:路易体主要是以帕金森病为代表的脑门特征性标志物,患者常常会出现功能障碍。
考虑到短链脂肪酸缺乏与神经炎症和小胶质细胞激活以及便秘、肠漏和结肠炎症等胃肠道特征有关,这些研究结果支持了这样的假设:短链脂肪酸缺乏可能是帕金森病的病因。
•帕金森病患者有抗炎作用的菌群减少
帕金森病患者经常出现属于乳杆菌科、巴氏杆菌科和肠球菌科的某些细菌种类水平升高。
此外发现,帕金森病患者粪便和粘膜中产生抗炎丁酸的细菌水平显著降低(普雷沃氏菌、布氏菌、粪球菌属和罗氏菌属),以及粪杆菌属的水平显著降低。
此外,像罗尔斯通氏菌(Ralstonia)这样的促炎细菌在帕金森病患者的肠粘膜中更为丰富。
拓展
基于肠杆菌科细菌滴度与帕金森病症状严重程度之间的正相关性,这些作者认为肠道微生物群可能与帕金森病表型有关。
相关分析显示疾病严重程度和帕金森病持续时间与纤维素降解剂呈负相关,但与致病生物呈正相关,可能导致短链脂肪酸产生减少以及神经毒素和内毒素增加,可能与帕金森病病理学的发展有关。
此外,普雷沃菌科丰度的降低和乳杆菌科丰度的增加与胃饥饿素浓度的降低有关,胃饥饿素是一种与维持正常多巴胺功能有关的肠道激素,帕金森病患者的胃饥饿素分泌发生了改变。
精神分裂症是一种复杂的精神障碍,患者对现实的理解异常,经常出现焦虑和严重抑郁症状。
这种疾病的起源是有争议的,表明是遗传起源。然而,值得注意的是,通过表观遗传学机制,如神经传递基因甲基化、核苷酸修饰或非编码RNA的作用,一些环境因素可能在其易感性和疾病的发展中发挥作用。
•肠道菌群代谢物影响精神分裂症风险
短链脂肪酸、色氨酸分解代谢物和神经递质(GABA、谷氨酸)的差异被称为精神分裂症相关的肠脑模块。
微生物产生的多巴胺与精神分裂症有关,而与精神分裂症相关的胃肠道炎症增加强烈表明肠道微生物群可能在患精神分裂症或其表现的风险中发挥作用。
•精神分裂症患者肠道微生物群显著改变
研究表明抗生素治疗引起的肠道微生物群失调与精神分裂症的发病率之间存在直接关系。精神分裂症患者中巨球型菌属、琥珀酸弧菌属、梭菌属、柯林斯氏菌属、甲烷短杆菌属和克雷伯菌属的丰度显著增加,而与健康人相比,精神分裂症患者中经黏液真杆菌属、粪球菌属和罗氏菌属的滴度下降。
•抗精神病药物治疗后肠道微生物群发生改变
几项研究已经确定抗精神病药物治疗与肠道微生物群之间的联系。精神分裂症患者在接受抗精神病药物治疗后发现微生物群丰度发生了显著变化。
在其他研究中,接受抗精神病药物治疗的患者的厚壁菌门与拟杆菌门的比例逐渐增加,与体重指数的上升相关。此外,接受治疗的个体的肠道微生物群富含短链脂肪酸和血清素代谢的基因。
注意缺陷与多动障碍(ADHD)是一种神经发育障碍,其特征是高度多动冲动和注意力问题。尽管可能涉及环境和遗传因素,一些证据表明注意缺陷与多动障碍的发展或症状可能与饮食成分对肠道微生物组的调节有关。
•ADHD儿童体内有害菌丰度较高,有益菌较少
最近发现,食用加工饮食的注意缺陷与多动障碍儿童体内有害细菌(如肠杆菌、大肠杆菌和梭菌菌株)的丰度明显较高,而有益细菌(如双歧杆菌和瘤胃球菌菌株)的丰度明显较低。
食用加工食品的注意缺陷与多动障碍患者的肠道微生物群α多样性和短链脂肪酸水平显著低于对照组,研究人员认为不平衡的饮食会扰乱结肠微生物平衡,并可能成为多动症患病的潜在风险因素。
•患者肠道菌群失衡影响激素和神经递质水平
宿主-微生物组的相互作用对激素和神经递质水平产生影响,被认为与注意缺陷与多动障碍的病理生理学有关。肠道菌群失调加上持续接触微生物病原体引起的免疫功能障碍可能会导致受影响的多动症患者出现过度活跃的行为。
肠道菌群提供多巴胺和去甲肾上腺素的前体
注意缺陷与多动障碍与预测的多巴胺和去甲肾上腺素合成异常有关,其前体由肠道细菌(主要是双歧杆菌)提供。
压力被定义为由困难情况或环境压力源引起的非特定情绪或身体反应。这种反应与心理和生物因素有关,例如性激素、高情绪反应性、被动应对技巧、糖皮质激素抵抗以及中枢和外周免疫激活。
✦压力会影响肠道微生物组成与活动
不同类型的心理和社会压力源已被证明可以通过神经元、免疫细胞和结肠嗜铬细胞释放信号分子、激素或神经递质来调节肠道微生物群的组成和活动。
压力对肠道微生物群的影响可能直接通过宿主肠道微生物群信号传导介导,也可能间接通过肠道生态系统的变化介导,并涉及炎症反应、微生物栖息地的改变、肠道运动和粘蛋白分泌。
✦产生短链脂肪酸的细菌减少可能是导致焦虑的关键
暴露于压力会导致肠道微生物群组成的实质性变化,肠杆菌科细菌的增加,而乳杆菌属的细菌减少。拟杆菌属减少,梭菌属增加。以及大肠杆菌和假单胞菌属的增加。
某些微生物群可能参与应激性焦虑和一般性焦虑症,其特征是社交和职业功能下降。研究人员报道,在一般性焦虑症患者中,粪杆菌(Faecalibacterium)、直肠真杆菌(E.rectale)、毛螺菌属(Lachnospira)、丁酸球菌(Butyricicoccus)和萨特氏菌(Sutterella)较低。这些都是短链脂肪酸的重要生产者。
应激性焦虑是指在强烈的精神或躯体应激事件的刺激下,产生的过度焦虑、恐惧等情绪。患者可出现紧张、坐立不安、心慌、呼吸急促等症状,还可伴有睡眠障碍、食欲改变等表现。
大胆猜想
短链脂肪酸产生菌的减少是焦虑和抑郁单胺类假说的关键。
单胺假说——指科学家在抗抑郁药领域提出的假说,即抑郁患者神经突触间隙可有效利用的单胺类神经递质浓度明显下降,而升高突触间隙单胺递质浓度(主要是血清素)能发挥抗抑郁作用。
抑郁症是一种与压力相关的情绪障碍,涉及神经免疫-神经内分泌失调,与促炎细胞因子水平升高相关。
促炎细胞因子与高水平的皮质醇有关,通过抑制四氢生物蝶呤酶发挥重要作用,四氢生物蝶呤对于合成多巴胺、血清素和去甲肾上腺素至关重要。
多巴胺、血清素和去甲肾上腺素是人体内重要的神经递质,它们在神经系统中发挥着重要的作用。
✦肠道微生物的多样性与抑郁严重程度负相关
几项研究报告称,不同抑郁症患者的肠道微生物群组成差异较大,粪便微生物多样性的增加与抑郁症症状的严重程度呈负相关。
研究人员发现,Gelria、Turicibacter、Anaerofilum、Paraprevotella、Holdemania和Eggerthella属在抑郁症患者中普遍存在,而普雷沃氏菌(Prevotella)和戴阿利斯特杆菌(Dialiste)的存在减少。
抑郁症患者体内有益菌大量减少
后来的研究还发现,与对照组相比,抑郁症患者肠道微生物总体减少。肠道微生物群中的双歧杆菌和乳酸菌减少。Dorea、瘤胃球菌属和阿克曼氏菌属的显著减少,以及副拟杆菌属(Parabacteroides)、普雷沃氏菌属(Prevotella)和放线菌属(Actinobacteria)的增加与抑郁个体相关。
✦短链脂肪酸有助于释放神经递质和调节血脑屏障
几项研究表明,重度抑郁症患者的丁酸盐、乙酸盐和丙酸盐减少,表明短链脂肪酸可能通过直接刺激神经通路或通过神经内分泌和免疫激活的间接中枢效应改变行为。
短链脂肪酸也有助于结肠嗜铬细胞合成和释放外周神经递质(血清素和乙酰胆碱),并有助于交感神经元合成和释放去甲肾上腺素。
除了局部作用外,短链脂肪酸还可以直接作用于血脑屏障的中心受体。此外,体外研究表明,丙酸盐和丁酸盐,而不是乙酸盐,都可以调节血脑屏障的通透性,防止脂多糖引起的通透性增加。
强迫症是一种慢性且持久的疾病,会被迫出现侵入性、自我张力障碍的想法或冲动或重复行为。
尽管只有少数研究涉及肠道微生物群与强迫症的关系,但压力和抗生素治疗都会影响微生物群组成,已被认为是与强迫症症状同时发生的因素。
✦强迫症患者肠道微生物丰富度较低
据报道,患有强迫症的个体肠道微生物群的物种丰富度较低,产生丁酸盐的属(颤旋菌属、Odoribacter和Anaerostipes)的相对丰度较低。
最近还报道强迫症患者粪便样本显示文肯菌科(Rikenellaceae)(另枝菌属)丰度增加,而普雷沃氏菌和毛螺菌科(Lachnospiraceae)丰度降低。
几种心理障碍中的肠道菌群失调
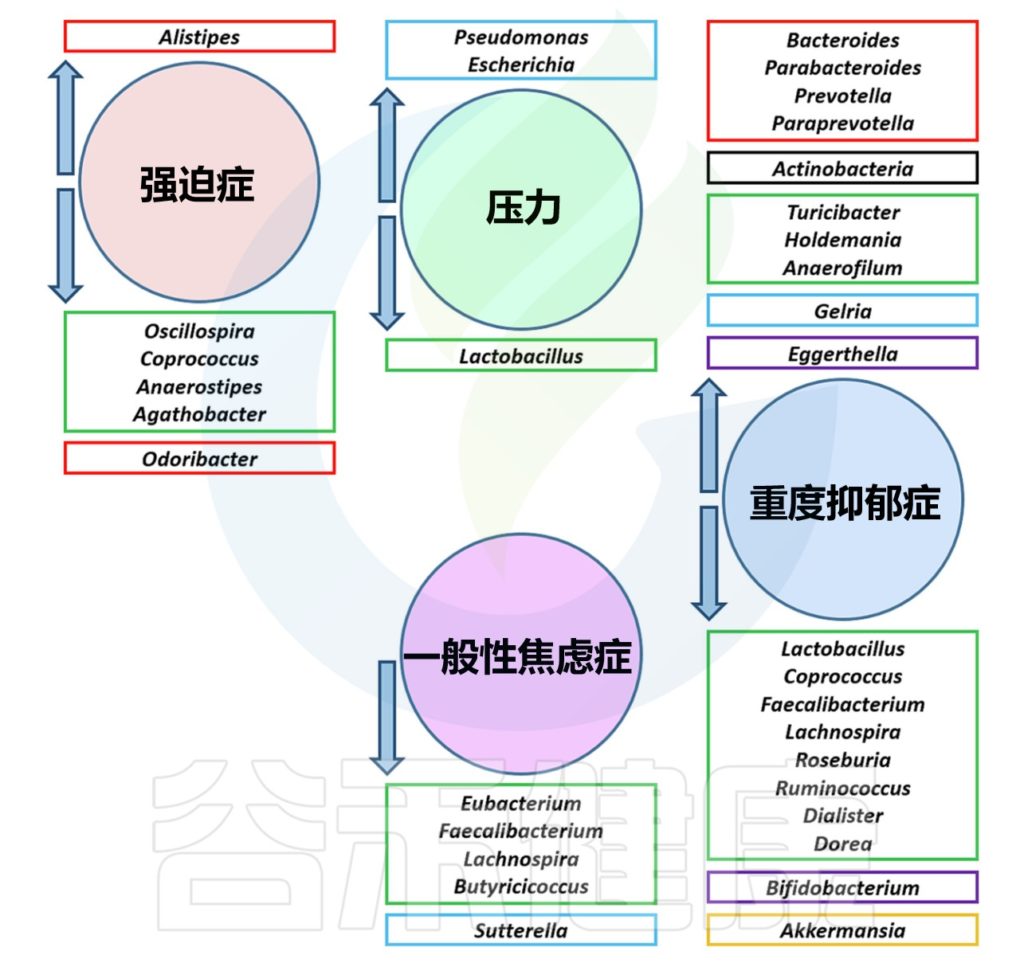
Borrego-Ruiz A,et al.Prog Neuropsychopharmacol Biol Psychiatry.2023
杏仁核在与压力相关的情绪、行为反应和情绪调节中发挥着关键作用,它显著受到肠道微生物群的影响。
杏仁核,又名杏仁体,呈杏仁状,是边缘系统的一部分。
√肠道微生物的变化可以调节认知功能
在动物模型中,有一些证据支持肠道微生物群组成的变化可以影响调节认知功能,尽管只有少数研究关注人类认知。
拟杆菌水平较高的人群拥有较好的认知表现
最近,根据粪杆菌(Faecalibacterium)、拟杆菌(Bacteroides)和瘤胃球菌科(Ruminococcaceae)的丰度建立了三组受试者,并得出结论,较好的认知表现与较高水平的拟杆菌相对应。
使用抗生素会影响认知功能,故不可滥用
用抗生素治疗剥夺或破坏肠道微生物群也会影响认知功能,包括语言学习能力以及工作和空间记忆。
√肠道微生物会影响一个人的气质和性格
由于人格特征和行为模式以及生理和心理健康结果之间存在很强的关联,一些研究已经证明肠道微生物群与气质和性格之间存在关联,这些特征甚至可以通过粪菌移植在受试者之间传播。
研究了肠道微生物的组成与儿童气质之间的关系,发现更大的暴躁/外向和恐惧与系统发育多样性呈正相关,包括戴阿利斯特杆菌属 (Dialister)、文肯菌科(Rikenellaceae)、瘤胃球菌科(Ruminococcaceae)和约氏副拟杆菌(Parabacteroides)的丰度变化。
产丁酸细菌更丰富的人可能有更高的责任心
发现高度神经质和低外向性与γ-变形菌纲丰度增加有关。低责任性与变形菌丰度增加和毛螺菌科丰度减少有关,而开放程度高的人与更大的系统发育多样性和丰富度相关。
另一方面,高度的责任心与产生丁酸的细菌滴度的增加有关,这些细菌主要属于毛螺菌科。
√多样化的肠道微生物有助于社交和好心情
最近有研究表明,特定细菌属的丰度与人格特征显著相关。对肠道微生物组多样性的分析表明,具有较高社交网络的个体呈现出多样化的肠道微生物组,而焦虑和压力与肠道微生物组的组成改变和多样性降低有关。
研究还发现某些细菌属,包括阿克曼氏菌、脱硫弧菌、乳球菌、颤螺菌和萨特氏菌可能与行为密切相关。
已经使用了几种方法来确定肠道微生物群对中枢神经系统功能的作用,包括抗生素治疗、无菌动物模型和粪菌移植。
此外,精神抗生素和益生元已被用作精神和心理疾病的辅助治疗。精神抗生素的类别可以定义为“一种活的有机体,通过与共生肠道细菌相互作用,为患有精神疾病的患者带来心理健康益处”。这些药物通过调节和改善肠道微生物群发挥作用。
为了让大家有更清晰的认识,谷禾在下表整理了使用益生菌或益生元治疗对心理或精神疾病的影响。





Borrego-Ruiz A,et al.Prog Neuropsychopharmacol Biol Psychiatry.2023
•基因集富集分析展示下肠道菌群与精神疾病的关联
使用基因集富集分析 (GSEA) 能够检测肠道微生物群与精神疾病之间的以下显著关联:注意力缺陷与多动障碍常伴有脱硫弧菌和梭状芽孢杆菌富集;自闭症谱系障碍与拟杆菌属和脱硫弧菌有关;对于重度抑郁障碍,观察到脱硫弧菌、梭状芽孢杆菌目、钩端螺旋菌科和拟杆菌类的关联。
最近报道了普雷沃氏菌与自闭症谱系障碍以及放线菌属与精神分裂症之间的特定关联。还有一组研究人员提出了放线菌和假单胞菌与重度抑郁障碍之间的关系。
肠道微生物群和中枢神经系统通过两条途径进行交流:
(1)通过迷走神经途径的神经免疫和代谢回路;
(2)通过微生物群合成的代谢产物、肠道激素和内分泌肽。
先前的研究表明,肠道微生物群的失调是几种精神疾病的病理生理学。因此,恢复受干扰的微生物生态平衡可能为管理或预防神经系统疾病提供一种新的、有前景的方法。
另一方面,心理干预已证明在治疗功能性胃肠道疾病如肠易激综合征方面是有效的;然而还需要进一步的研究来确定心理干预对肠道微生物群变化的具体影响。
通过粪菌移植、健康的饮食和生活方式或使用心理生物药物等干预措施重建失调的肠道微生物群,可能在未来调节微生物群肠脑轴功能和促进心理精神健康方面具有重大潜力。
特别是,心理干预可能对某些神经系统疾病具有潜在的治疗作用。然而,这些干预措施的有效性并不相同,取决于给药方法、干预时间、使用的微生物菌株的数量和具体类型以及宿主的生理状况等因素。
在未来,单独或与抗精神病药物联合的个性化精神微生物干预可能成为临床患者的一种新的治疗策略。
主要参考文献
Borrego-Ruiz A, Borrego JJ. An updated overview on the relationship between human gut microbiome dysbiosis and psychiatric and psychological disorders. Prog Neuropsychopharmacol Biol Psychiatry. 2023 Sep 8:110861.
Abuaish S, Al-Otaibi NM, Abujamel TS, Alzahrani SA, Alotaibi SM, AlShawakir YA, Aabed K, El-Ansary A. Fecal Transplant and Bifidobacterium Treatments Modulate Gut Clostridium Bacteria and Rescue Social Impairment and Hippocampal BDNF Expression in a Rodent Model of Autism. Brain Sci. 2021 Aug 5;11(8):1038.
Bear T, Dalziel J, Coad J, Roy N, Butts C, Gopal P. The Microbiome-Gut-Brain Axis and Resilience to Developing Anxiety or Depression under Stress. Microorganisms. 2021 Mar 31;9(4):723.
Ansari F, Pourjafar H, Tabrizi A, Homayouni A. The Effects of Probiotics and Prebiotics on Mental Disorders: A Review on Depression, Anxiety, Alzheimer, and Autism Spectrum Disorders. Curr Pharm Biotechnol. 2020;21(7):555-565.
Bharwani A, Mian MF, Foster JA, Surette MG, Bienenstock J, Forsythe P. Structural & functional consequences of chronic psychosocial stress on the microbiome & host. Psychoneuroendocrinology. 2016 Jan;63:217-27.
Cerovic M, Forloni G, Balducci C. Neuroinflammation and the Gut Microbiota: Possible Alternative Therapeutic Targets to Counteract Alzheimer’s Disease? Front Aging Neurosci. 2019 Oct 18;11:284.
Cryan JF, O’Riordan KJ, Sandhu K, Peterson V, Dinan TG. The gut microbiome in neurological disorders. Lancet Neurol. 2020 Feb;19(2):179-194.

谷禾健康

氨基酸(Amino Acid, 简称AA ) 是蛋白质合成的基石。它们是正常细胞生长、分化和功能所必需的细胞结构元素和能量来源。
蛋白质是长链氨基酸。人的身体有数千种不同的蛋白质,每种蛋白质都有重要的作用。每种蛋白质都有自己的氨基酸序列。该序列使蛋白质呈现不同的形状并在体内具有不同的功能。
可以将氨基酸视为字母表中的字母。当你以不同的方式组合字母时,会组成不同的单词。氨基酸也是如此——当以不同的方式组合它们时,就会产生不同的蛋白质。
氨基酸代谢紊乱与许多病理状况有关,包括代谢疾病、心血管疾病、免疫疾病和癌症。就肿瘤而言,氨基酸代谢的改变不仅可以用作癌症进展的临床指标,还可以用作治疗策略。
由于肿瘤的生长发育依赖于外来氨基酸的摄入,因此越来越多的研究针对肿瘤相关氨基酸的代谢来选择性杀伤肿瘤细胞。此外,免疫相关研究证实,氨基酸代谢调节效应T细胞和调节性T细胞的功能,影响免疫细胞的功能。因此,确定氨基酸代谢途径中的靶点可能有助于疾病的治疗。
近日,浙江大学医学院附属第一医院,国家卫健委多器官联合移植重点实验室,中国医学科学院器官移植诊治重点实验室,以及器官移植重点实验室联合发表在《Signal Transduct Target Ther》综述成果详细阐述和讨论氨基酸代谢相关的代谢性疾病、心血管疾病和免疫相关疾病的研究及临床研究进展。
本文就该篇文章为主要参考,分享围绕肿瘤相关疾病中的氨基酸代谢研究,结合讨论肠道微生物群在蛋白质和氨基酸代谢中的作用,以期为靶向氨基酸代谢管理治疗提供理论依据。
官方氨基酸定义包括任何同时含有羧基和氨基的有机化合物(如下图)。它们都具有共同的结构特征氨基 (-NH3+)、羧酸根 (-COO–) 基团和与同一碳原子键合的氢键。
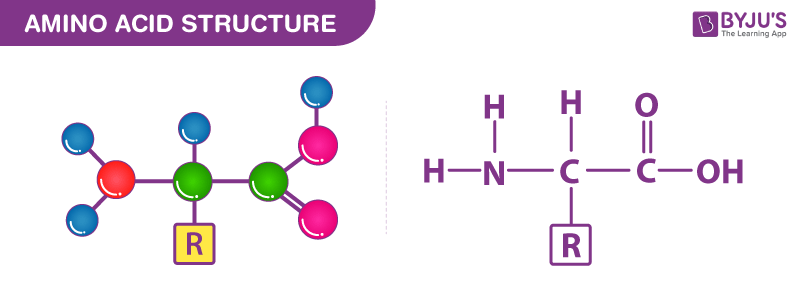
氨基酸有多少种?
氨基酸一共有20种,每种氨基酸在体内都发挥着非常特殊的作用,并通过其各自的氨基酸侧链来区分。从细菌到人类,都是由同一组 20 种氨基酸构成。
每个氨基酸都有 4 个不同的基团连接到 α-碳。它们的不同之处在于称为 R 基团的侧链。如下氨基酸的R基团不同:
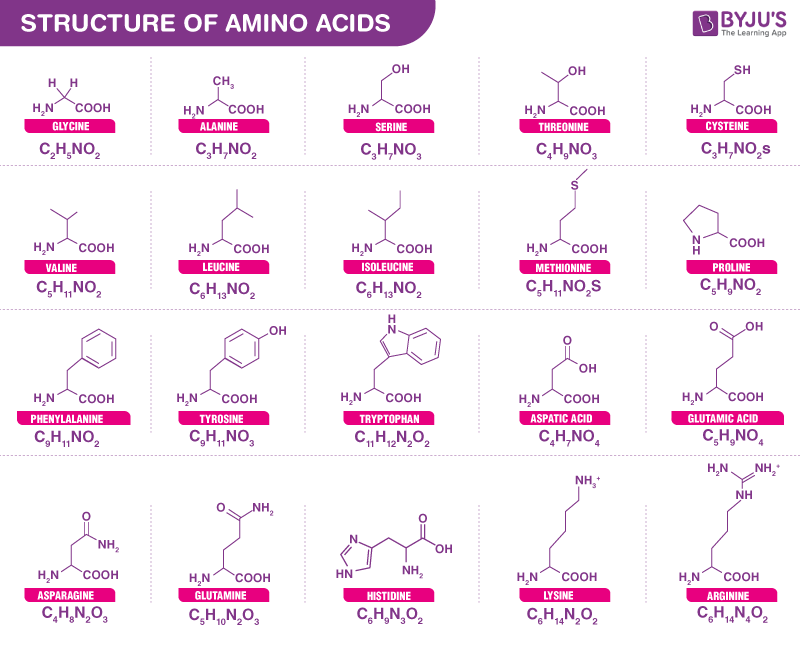
来源:byjus
氨基酸还可以根据其结构和侧链分为其他组,
包括:
根据每种氨基酸中“R”基团的性质,氨基酸可以分为四个大类。
氨基酸可以是极性的、非极性的、带正电的或带负电的。
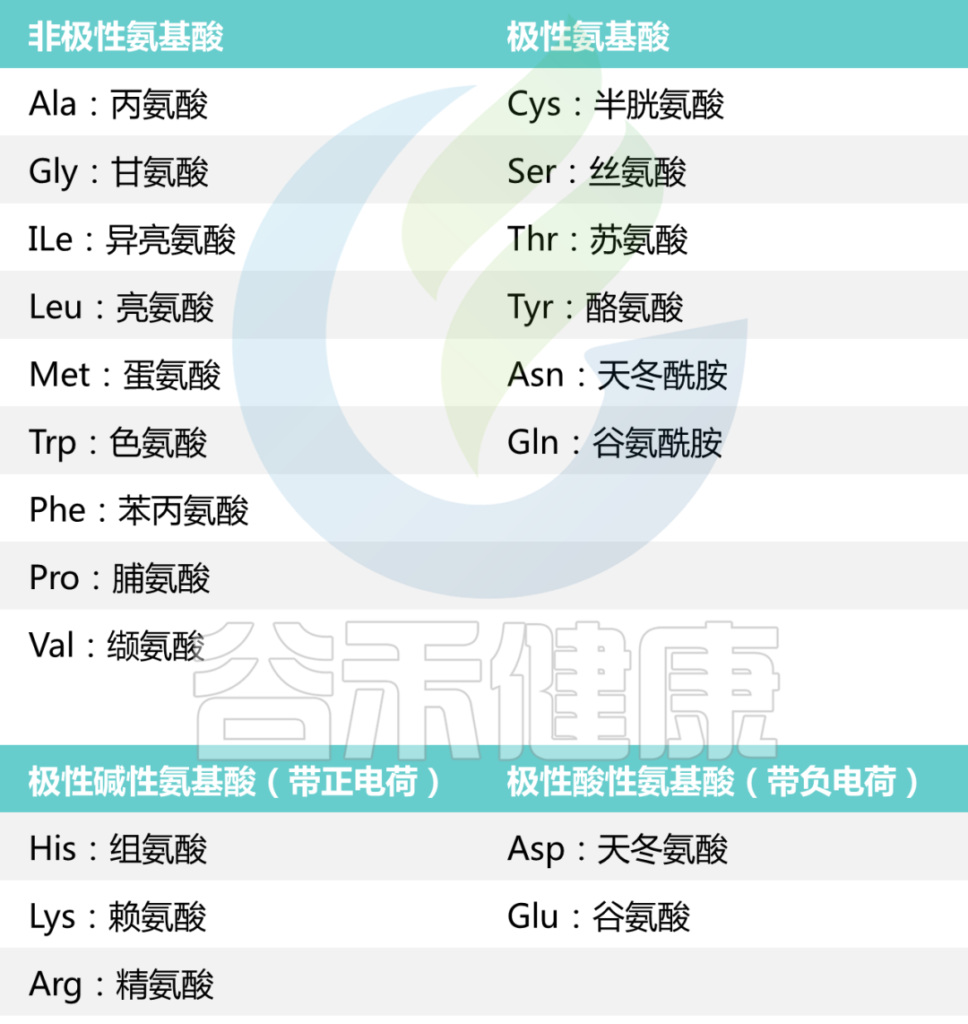
氨基酸是作为蛋白质构建块的基本分子。蛋白质由一条或多条氨基酸链(称为多肽)组成,其序列由基因编码。
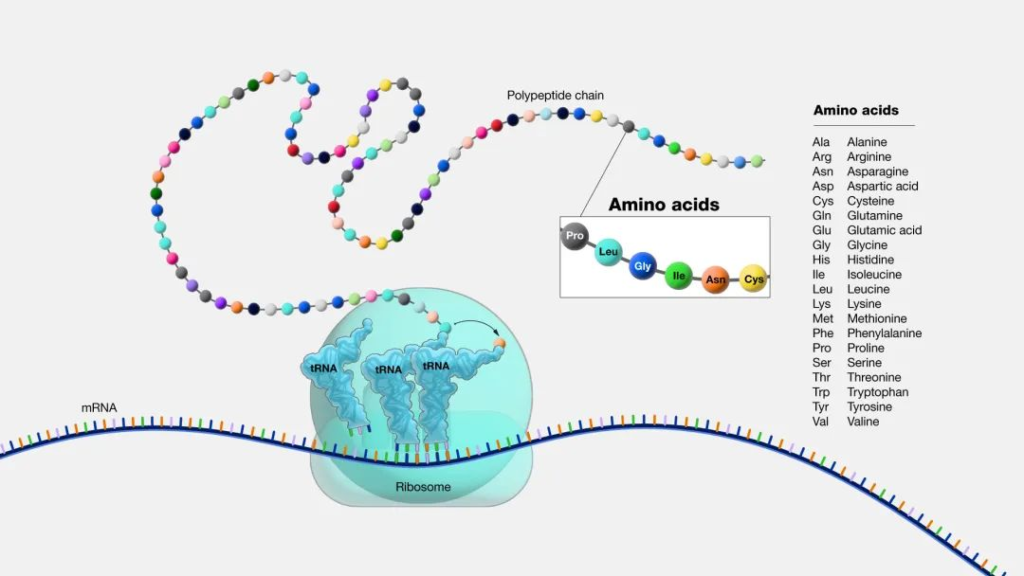
图源:National Human Genome Research Institute
这些氨基酸参与几乎所有生物过程并有助于:
伤口愈合、激素产生、免疫功能、肌肉生长、能源生产等。
身体需要所有氨基酸才能发挥作用并茁壮成长。有些可以在体内产生,而另一些则需要从食物中获取。
氨基酸有着有趣而悠久的历史。天冬酰胺是第一个被发现的氨基酸,由法国化学家 Louis Nicolas Vauquelin 和 Pierre Jean Robiquet 于 1806 年从芦笋营养中分离出来。
身体所需的 20 种氨基酸可进一步分为两个不同的类别:必需氨基酸和非必需氨基酸。
人体无法合成必需氨基酸,这意味着必须从食物中获取它们才能满足需求。
必需氨基酸有多少种?
必须通过饮食获得九种必需氨基酸,包括:
赖 氨 酸
赖氨酸是生长和组织修复以及多种激素、蛋白质和酶的产生所必需的。赖氨酸对于促进抗体、激素和酶的形成以及骨骼中钙的形成和固定是必需的。
亮 氨 酸
这种必需氨基酸参与蛋白质合成、伤口愈合、血糖控制、生长激素和新陈代谢。
异 亮 氨 酸
研究表明异亮氨酸有助于解毒、免疫功能和激素排泄。此外,在血红蛋白的形成、刺激胰腺合成胰岛素以及将氧气从肺部输送到各个部位方面起着至关重要的作用。
色 氨 酸
色氨酸是最著名的氨基酸之一,也是功能性氨基酸,参与维生素 B3 和血清素激素的产生。血清素是调节情绪、疼痛、食欲和睡眠的神经递质。
苯 丙 氨 酸
这种氨基酸有助于产生其他氨基酸以及多巴胺和去甲肾上腺素等神经递质,有助于维持健康的神经系统和增强记忆力。
苏 氨 酸
研究表明苏氨酸是胶原蛋白和弹性蛋白等结缔组织的基础,这些蛋白质为皮肤和结缔组织提供结构。它们还有助于形成血凝块,从而有助于防止出血。苏氨酸在脂肪代谢和免疫功能中也发挥着重要作用。
缬 氨 酸
缬氨酸支持大脑功能、肌肉协调和平静。缬氨酸参与肌肉生长、组织再生和产生能量。
组 氨 酸
这种氨基酸维持人体内髓鞘的健康,保护神经细胞免受损伤。参与许多酶促过程以及红细胞和白细胞的合成。组氨酸有助于制造一种称为组胺的大脑化学物质(神经递质)。组胺在身体的免疫功能、消化、睡眠和性功能中发挥着重要作用。
蛋 氨 酸
研究表明蛋氨酸可以保持皮肤弹性,并有助于强化头发和指甲。还用于治疗肾结石、保持皮肤健康,还用于控制病原菌的入侵。蛋氨酸有助于身体的组织生长、新陈代谢和解毒。蛋氨酸还有助于吸收必需矿物质,包括锌和硒。
在饮食中摄入多种氨基酸对于维持整体健康至关重要。任何这些关键必需氨基酸的缺乏都会导致严重的副作用,影响健康的几乎各个方面,包括免疫功能、肌肉质量、食欲等。
相比之下,非必需氨基酸可以由身体产生,这意味着通过吃的食物获取它们并不那么重要。
非必需氨基酸列表中共有 11 种氨基酸,包括:
精 氨 酸
精氨酸可刺激免疫功能、抗疲劳并优化心脏健康。一水肌酸在肝脏、胰腺和肾脏中形成,主要是在精氨酸、甘氨酸和蛋氨酸的帮助下形成的。有助于促进蛋白质和激素的合成、肾脏解毒、愈合伤口和维持健康的免疫系统。
丙 氨 酸
研究表明丙氨酸有助于新陈代谢,为肌肉、大脑和中枢神经系统提供能量。此外可以帮助清除体内毒素并参与葡萄糖和其他氨基酸的生产。
半 胱 氨 酸
半胱氨酸是头发、皮肤和指甲中发现的主要蛋白质类型,作为一种抗氧化剂,对于胶原蛋白的产生和皮肤健康至关重要。
谷 氨 酸
这种非必需氨基酸在中枢神经系统中充当神经递质,主要参与人脑的发育和功能。
天 冬 氨 酸
天冬氨酸有助于产生其他几种氨基酸,包括天冬酰胺、精氨酸和赖氨酸。主要参与将氮输送到身体细胞中、形成嘌呤和嘧啶以合成DNA、神经系统的发育和提高我们的身体耐力。
甘 氨 酸
甘氨酸有助于维持适当的细胞生长及其功能,并且在愈合伤口方面也起着至关重要的作用。此外,甘氨酸作为神经递质支持大脑健康。
脯 氨 酸
脯氨酸存在于胶原蛋白中,有助于促进关节健康、新陈代谢和皮肤弹性。主要参与胶原蛋白形成过程中的组织修复,防止动脉壁增厚和硬化(动脉硬化)以及新皮肤的再生。
丝 氨 酸
丝氨酸是脂肪代谢、免疫功能和肌肉生长所必需的。
酪 氨 酸
甲状腺激素-T3 和 T4 的产生以及合成一类神经递质和黑色素(我们的眼睛、头发和皮肤中发现的天然色素)方面发挥着至关重要的作用。
谷 氨 酰 胺
研究表明,谷氨酰胺支持许多代谢过程并为体内细胞提供能量。促进健康的大脑功能,是合成核酸(DNA 和 RNA)所必需的。
天 冬 酰 胺
天冬酰胺起到利尿剂的作用,可以优化大脑和神经细胞的功能。
氨基酸列表中的一些化合物也被认为是“有条件必需的”。这意味着它们通常不是身体所需要的,但在某些情况下可能变得必不可少,例如极端疾病或压力。
氨基酸缺乏的一些主要症状可能包括:
蛋白质缺乏症会影响任何饮食中氨基酸摄入不足的人。老年人和患有癌症等慢性病的人缺乏蛋白质的风险特别高,因为他们通常对蛋白质的需求增加而食物摄入量减少。
那些遵循纯素或素食饮食的人还应该仔细计划他们的饮食,以确保通过食用各种植物性蛋白质食物来满足他们的蛋白质需求。
富含氨基酸的食物包括植物性产品,如西兰花、豆类、甜菜根、南瓜、卷心菜、坚果、干果、奇亚籽、燕麦、豌豆、胡萝卜、黄瓜、绿叶蔬菜、洋葱、大豆、全谷物、花生豆类、扁豆等。
富含氨基酸的水果有苹果、香蕉、浆果、无花果、葡萄、甜瓜、橙子、木瓜、菠萝、石榴等。
其他动物产品包括乳制品、鸡蛋、海鲜、鸡肉、肉类、猪肉等。
从富含蛋白质的食物中摄入大量蛋白质不太可能引起任何负面影响。然而,有可能过量摄入蛋白质,尤其是蛋白质补充剂。
摄入过多蛋白质可能产生的一些副作用包括体重增加、肾脏问题、便秘和口臭,炎症以及肠道菌群的失调。
氨基酸是含有氨基和羧基的有机化合物,根据核心结构官能团的位置可分为α-、β-、γ-、δ-氨基酸,其中大部分参与蛋白质合成的是 α-氨基酸。
氨基酸参与生物合成、神经传递和其他生命过程。肽键连接氨基酸形成多肽链,多肽链经过翻译后修饰,有时与其他多肽链结合形成蛋白质。
九种氨基酸不能由其他化合物合成,必须从食物中获取,这些氨基酸从食物中被人体摄入后,除了用于蛋白质等生物分子的合成外,还可以通过氧化途径氧化为尿素和二氧化碳作为能量来源。
氧化途径以转氨酶介导的脱氨作用开始,将氨基转移至α-酮戊二酸,形成谷氨酸进入尿素循环。另一种产物酮酸进入柠檬酸循环,为生命活动提供能量(下图)。该循环提供某些氨基酸的前体,以及还原剂NADH,用于许多其他反应。
doi: 10.1038/s41392-023-01569-3
人体可以通过食物消化吸收、组织分解、内部合成三种途径获得氨基酸。
氨基酸代谢库中的氨基酸可脱酸,产生氨基酸和二氧化碳,或在代谢产物的转化中参与嘌呤、嘧啶等含氮化合物的合成;或者脱氨基产生α-酮酸和NH3。
根据不同的酶和途径,α-酮酸可以产生酮体,或参与氧化供能或糖和脂质的合成;NH3进入尿素循环。
细胞或细胞器对氨基酸的摄取需要氨基酸转运蛋白(AAT)的参与。不同的氨基酸依赖于特定的AAT,但氨基酸和转运蛋白并不是一一匹配的。多个AAT可以转运一个氨基酸,同一个转运蛋白也可以转运多个底物。AAT 除了充当氨基酸进出细胞的通道外,还充当感测氨基酸水平的探针和营养信号的引发剂。
根据结构和功能的多样性,AAT可分为不同的家族,其中溶质载体(SLC)超家族约占人类基因组编码的所有膜蛋白的20%,是最大的膜转运蛋白超家族。
根据底物特异性,AAT可分为中性、碱性和酸性类别,以及进一步的子类别,包括钠依赖性和钠非依赖性类型。从机制上讲,由于哺乳动物细胞(包括人类)细胞内液中的氨基酸浓度通常高于细胞外液中的氨基酸浓度,因此 AAT 通过离子共轭或氨基酸交换转运氨基酸以产生钠离子。
氨基酸除了是肽和蛋白质的组成部分外,还参与维持细胞生长、代谢和免疫的关键途径。
例如,哺乳动物雷帕霉素靶蛋白 (mTOR) 信号通路是调节蛋白质合成的主要机制。mTOR 系统包含雷帕霉素敏感复合物 1 (mTORC1) 和雷帕霉素不敏感复合物 2 (mTORC2)。mTORC1 被谷氨酰胺 (Gln)、精氨酸 (Arg) 和亮氨酸 (Leu) 激活,并通过 eIF4E 结合蛋白 1 (4E-BP1) 和核糖体蛋白 S6 激酶 1 (S6K1) 的磷酸化激活蛋白质合成。
此外,丙氨酸(Ala)可以通过抑制丙氨酸激酶来调节糖异生和糖酵解,从而维持饥饿肝脏产生的葡萄糖量。
TIPs:
糖异生是指非糖物质(如某些氨基酸、乳酸、丙酮酸和甘油等)在人体的肝脏和肾脏等器官中某些酶的催化下转变成糖原或葡萄糖的过程。简单说来,就是非糖物质转化为糖的过程。
糖分解是生物体取得能量的主要方式。生物体中糖的氧化分解主要有3条途径:糖的无氧氧化、糖的有氧氧化和磷酸戊糖途径。其中,糖的无氧氧化又称糖酵解(glycolysis)。生物在无氧条件下,从糖的降解代谢中获得能量的途径,也是大多数生物进行葡萄糖有氧氧化的一个准备途径。
糖酵解/糖原异生的差异比较
在某种程度上,糖异生可能被认为是糖酵解的逆过程,因为糖酵解将葡萄糖分解成两个丙酮酸分子以获得能量并供给三羧酸循环,而糖异生将两个丙酮酸分子构建成一个葡萄糖分子。然而,仅仅把糖异生看作糖酵解是不太正确的,因为这两者之间有一些重要的区别。
糖酵解是一种古老的代谢途径,为所有形式的细胞生命所共有,并且可以由所有细胞进行以获得能量(每个葡萄糖净2个三磷酸腺苷)。
相比之下,糖异生更具特异性:在人类中,它主要在肝细胞中进行,在某种程度上也在肾上腺皮质中进行。它还有一个更具体的目标:当血糖水平较低时,产生葡萄糖分泌到血液中。
在免疫方面,氨基酸参与免疫细胞增殖、分化和功能激活。例如,T细胞激活会上调多种氨基酸转运蛋白,包括SLC7A5,而SLC7A5的缺失会导致mTOR信号通路的激活和转录因子MYC的上调,从而抑制T细胞增殖。
当T细胞缺乏色氨酸(Trp)和 精氨酸 (Arg)时,活化的T细胞无法进入S期,这证明Trp和Arg是T细胞进入细胞周期的关键物质。此外,亮氨酸(Leu) 和异亮氨酸 (ILe) 的消耗会诱导 T 细胞进入 S-G1 期,然后停止分裂并死亡。
综上所述,氨基酸是维持生命所必需的有机化合物,是生物合成的原料,也是生命活动的能量来源。
接下来,我们针对支链氨基酸、天冬氨酸、谷氨酰胺、精氨酸、蛋氨酸五种氨基酸的代谢展开详细讨论,包括它们在癌症或其他疾病中的作用,希望给大家呈现一个更全面的视角,为疾病的发生发展和临床治疗方向带来更多的启示。
★
支链氨基酸(BCAA)是一类具有一个分支的脂肪侧链氨基酸,包括亮氨酸、异亮氨酸和缬氨酸。三种支链氨基酸占人体必需氨基酸肌肉中必需氨基酸的35%。
所有物种中支链氨基酸的分解过程都是相似的,最初通过支链氨基酸转移酶(BCAT)形成支链α-酮酸(BCKA),并将氮转移到氮受体(最常见的氮受体是α-酮戊二酸) (α-KG) 形成谷氨酸)。
第二步是支链α-酮酸脱氢酶(BCKDH)催化的不可逆限速反应,该反应被特定激酶BCKDH激酶(BCKDK)磷酸化并失活,并被蛋白磷酸酶1K(PPM1K)去磷酸化和激活。然后产物通过进一步氧化参与不同的生理活动。
促进肝脏和其他组织中的蛋白质合成
支链氨基酸参与多种生理过程。在代谢和信号通路研究方面,支链氨基酸尤其是亮氨酸,是mTOR信号通路的有效激活剂。亮氨酸可以与 Sestrin2(mTORC1 活性的负调节因子)结合,促进 mTORC1 激活,从而促进肝脏和其他组织中的蛋白质合成。
促进肝脏和骨骼肌对糖原的吸收,淋巴细胞增殖
支链氨基酸还可以促进肝脏和骨骼肌对糖原的吸收,并增强糖原的合成。
支链氨基酸对于免疫系统中免疫细胞的正常功能至关重要,通过氧化分解免疫细胞表达的脱氢酶和脱羧酶,促进淋巴细胞增殖和细胞毒性 T 细胞活化。
癌症患者的支链氨基酸循环水平发生了变化。
胰腺导管腺癌
最近的代谢组学回顾性研究表明,血浆支链氨基酸水平升高与胰腺癌风险增加相关,这一点在胰腺导管腺癌 (PDAC) 基因工程小鼠模型中得到了验证。可能是由于系统性蛋白质分解以满足致瘤期生长所需的支链氨基酸所致。
非小细胞肺癌 v.s. 胰腺导管腺癌
一项研究表明KRAS突变可以促进支链氨基酸代谢。尽管非小细胞肺癌 (NSCLC) 和 胰腺导管腺癌 (PDAC) 中存在 KRAS 激活和 P53 缺失,这两种肿瘤尽管初始事件相同,但对支链氨基酸的利用却不同。胰腺导管腺癌细胞倾向于分解并利用细胞外蛋白质获取氨基酸,而非小细胞肺癌细胞则通过分解循环支链氨基酸来提取氮。
为什么胰腺导管腺癌的支链氨基酸代谢低于周围正常组织?
胰腺导管腺癌细胞中的CBP和SIRT4结合BCAT2的K44位点,使该位点乙酰化,从而进一步促进BCAT2通过泛素蛋白途径降解,降低代谢胰腺导管腺癌中支链氨基酸的比例,进而抑制肿瘤细胞的生长。
注:CBP——cAMP反应元件结合(CREB)结合蛋白
KRAS和USP1也可以通过泛素-蛋白酶途径调控PDAC中BCAT2的表达:KRAS通过抑制脾酪氨酸激酶(SYK)和E3泛素化连接酶TRIM21对BCAT2的泛素化来稳定PDAC中BCAT2的表达;而USP1则通过去泛素化BCAT2的K229位点,并通过GCN2-eIF2a途径促进USP1蛋白的翻译水平,促进支链氨基酸表达。
另一项研究发现,USP1 和 BCAT2 的表达水平在基因编辑小鼠和临床样本中一致呈正相关,阐明了为什么胰腺导管腺癌的支链氨基酸代谢低于周围正常组织,进而转向其他途径获取氮。
癌症中的支链氨基酸代谢
doi: 10.1038/s41392-023-01569-3
血浆支链氨基酸升高是胰腺癌的危险因素
在胰腺癌进展的早期阶段,在人类和小鼠胰腺癌模型中观察到循环支链氨基酸升高,并且由于胰腺癌周围组织中蛋白质过度分解,血液中支链氨基酸水平升高。
肿瘤微环境中的支链氨基酸代谢
doi: 10.1038/s41392-023-01569-3
肺部肿瘤支链氨基酸摄取量高于胰腺导管腺癌
对标记的支链氨基酸代谢物的分析显示,非小细胞肺癌细胞中标记的 α-酮异己酸 (α-KIC) 和亮氨酸衍生的支链α-酮酸(BCKA)较多。同时BCKDK在非小细胞肺癌中高表达并调节细胞内ROS的产生,影响细胞存活。
代谢性疾病
支链氨基酸及其代谢物是肥胖、胰岛素抵抗和 2 型糖尿病等代谢性疾病的最强生物标志物。支链氨基酸及其代谢物升高是2型糖尿病等代谢性疾病早期进展的关键。
每种支链氨基酸都有独特的代谢作用
低异亮氨酸饮食→快速恢复肥胖小鼠代谢健康
异亮氨酸可以作为代谢健康的调节剂,低异亮氨酸饮食可以改善肥胖对代谢的不利影响。
肥胖可通过增加肝细胞中BCKDK(BCKDH激酶)/ PPM1K(BCKDH去磷酸化酶)的比例,抑制肝脏对支链氨基酸的利用,导致BCKDH失活。在肥胖和胰岛素抵抗的小鼠模型中,通过限制支链氨基酸饮食或调节 BCKDK/PPM1K 比率可以逆转这种现象。
肝脏和肾脏疾病
肝硬化:循环支链氨基酸降低 ↓↓
非酒精性脂肪肝:循环支链氨基酸升高↑↑
肝硬化患者中,高氨血症环境中支链氨基酸分解代谢增强、谷氨酸合成增加以及循环支链氨基酸水平降低被认为是该疾病的标志,并与肝性脑病风险增加相关。
在非酒精性脂肪肝 (NAFLD) 中检测到循环支链氨基酸升高。
此外,这种支链氨基酸代谢紊乱与 2 型糖尿病的发展具有协同作用。其他研究表明,补充支链氨基酸有助于恢复慢性肝病患者的葡萄糖稳态并增强免疫系统功能。
慢性肾病:循环支链氨基酸降低 ↓↓
在肾脏疾病中,慢性肾功能衰竭患者的循环支链氨基酸水平显著降低。这种现象在慢性肾脏病 (CKD) 患者中已观察到,一项 II 期慢性肾病队列研究发现,与正常对照相比,慢性肾病患者血浆亮氨酸 和缬氨酸显著降低。
这可能是由于慢性肾病患者长期营养不良和血液透析导致支链氨基酸水平下降。代谢性酸中毒还会增强支链氨基酸脱氢酶 (BCKD) 活性并加速蛋白质分解。然而,给慢性肾功能衰竭患者补充支链氨基酸和其他必需氨基酸,有助于维持蛋白质平衡,减少尿毒症毒性。
★
天冬氨酸(Asp)是一种用于蛋白质合成的 α-氨基酸,具有 α-氨基、α-羧酸基和侧链甲酰胺。它是一种非必需氨基酸,因为人体可以合成它。
草酰乙酸——天冬氨酸的前体
转氨酶将氨基从谷氨酸转移到草酰乙酸,产生 α-酮戊二酸和天冬氨酸。在天冬酰胺合成酶介导的酶反应中,谷氨酰胺提供氨基,与 β-天冬氨酸-AMP 结合形成天冬酰胺 (Asn) 和 AMP。
天冬酰胺——大脑发育必需的氨基酸
由于血液中的天冬氨酸不能直接通过血脑屏障,神经细胞的发育依赖于其在大脑中的合成。
当脑内天冬氨酸合成酶水平不足时,脑细胞的增殖就会受到限制,甚至导致细胞死亡。
反过来,在分解代谢过程中,天冬酰胺被天冬氨酸酶水解为天冬氨酸,然后用 α-酮戊二酸胺化,形成谷氨酸和草酰乙酸。然后草酰乙酸进入柠檬酸循环(下图)。
天冬氨酸、精氨酸和蛋氨酸代谢
doi: 10.1038/s41392-023-01569-3
天冬氨酸也是尿素循环的代谢物
在苹果酸-天冬氨酸穿梭中携带还原当量,在肌苷合成中提供氮原子,并在 ATP 合成中充当氢受体。
天冬氨酸也是四种必需氨基酸(蛋氨酸、苏氨酸、赖氨酸和异亮氨酸)的前体。天冬氨酸还可以作为氨基酸交换因子,成为氨基酸进出细胞的介质,特别是组氨酸、精氨酸和丝氨酸。天冬氨酸通过氨基酸交换因子功能调节丝氨酸代谢、核苷酸合成和 mTORC1 活性。
TP53是人类癌症中突变频率最高的基因。该基因编码的蛋白p53通过调节细胞周期、细胞凋亡、基因组稳定性等途径抑制肿瘤的发展。
结肠癌
结肠癌细胞系中的天冬氨酸(Asp)和 天冬酰胺 (Asn) 可通过与 LKB1(编码丝、苏氨酸激酶,并直接磷酸化蛋白质产物以激活 AMPK)来抑制其活性,从而抑制 AMPK 介导的 p53 激活。
淋巴瘤和结直肠肿瘤
在淋巴瘤和结直肠肿瘤模型中,p53 的激活会破坏 天冬氨酸-天冬酰胺 稳态,并促使细胞衰老和周期停滞。缺氧条件下,天冬氨酸是肿瘤生长的限制因素。
缺氧会抑制电子传递链 (ETC),影响能量和天冬氨酸的合成。研究了肿瘤细胞对线粒体 ETC 抑制剂的敏感性,发现对 ETC 抑制不敏感的肿瘤细胞通过 Asp/谷氨酸转运蛋白 SLC1A3 维持细胞内的Asp 浓度,这使肿瘤细胞具有生存优势。
膀胱癌
在另一项关于肿瘤代谢的研究中,发现当环境中缺乏氧气时,天冬氨酸合成是膀胱癌生长的限制因素。在膀胱癌细胞中,天冬氨酸细胞的渗透性差,阻碍了肿瘤细胞从环境中摄取天冬氨酸。虽然细胞对天冬酰胺的通透性高于天冬氨酸,但膀胱癌细胞中天冬酰胺酶的活性不足,无法将天冬酰胺转化为天冬氨酸。
使用豚鼠天冬酰胺酶 1 (gpASNase1) 促进肿瘤细胞中天冬酰胺转化为 Asp 后,肿瘤细胞的生长速度显著增加,表明 Asp 获取是 Asp 获取困难的肿瘤的内源性代谢限制。有人提出,Asp是体内某些肿瘤生长的内在限制,突破这一限制将促进肿瘤生长。
天冬氨酸-谷氨酸转运蛋白 SLC1A3 与 ETC 抑制剂的作用密切相关,并且 SLC1A3 位点在非神经胶质上皮肿瘤亚簇中扩增,从而对抗天冬氨酸限制。
乳腺癌
发现SLC1A3促进乳腺癌细胞对L-天冬酰胺酶(ASNase)的抵抗。而且,SLC1A3可以补充ASNase对天冬氨酸和谷氨酸的消耗,从而消除ASNase的抑制作用,促进肿瘤的发展。
此外,另一种氨基酸转运蛋白SLC25A22可以促进KRAS突变结直肠癌(CRC)细胞中天冬氨酸的合成,激活AMPK通路并减少氧化应激。
这些研究表明,AAT 是肿瘤代谢重编程的潜在靶标。目前正在测试的针对 AAT 的药物见(见后面的表格)。
实体瘤中的天冬氨酸代谢
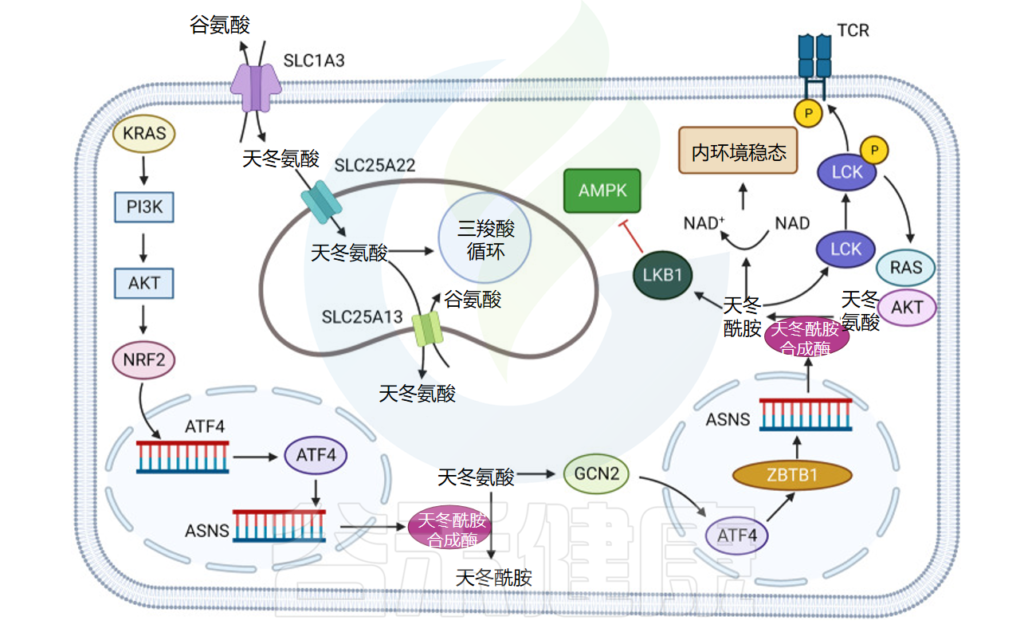
doi: 10.1038/s41392-023-01569-3
天冬酰胺作为癌症治疗的新靶点受到广泛关注
报道指出乳腺癌中天冬酰胺合成酶(ASNS)的表达水平与转移复发密切相关,抑制ASNS或限制饮食天冬酰胺可以减少肿瘤转移。
非小细胞肺癌
在非小细胞肺癌中,激活转录因子 4 (ATF4) 可以通过 KRAS 下游的 AKT 和 NRF2 改变氨基酸摄取并增加天冬酰胺合成。此外,将AKT抑制剂与细胞外天冬酰胺(ASN)消耗结合使用可以显著抑制肿瘤生长。
天冬酰胺在免疫细胞的生长和功能中发挥关键作用
发现CD8 + T细胞在CD8 + T细胞激活的早期阶段几乎不表达天冬酰胺合酶(ASNS),并且CD8 + T细胞的生长、激活和代谢重编程在天冬酰胺剥夺的情况下被破坏。
还证明,活化的 CD8 + T 细胞中的天冬酰胺水平增加,并与 SRC 家族酪氨酸激酶 LCK 结合,协助 LCK 在 Tyr394 和 505 处磷酸化,增强 LCK 活性和 T 细胞受体信号传导。
血液恶性肿瘤
天冬酰胺在血液恶性肿瘤中也发挥着关键作用。目前,细菌源性 L-天冬酰胺酶 (ASNase) 在儿童急性淋巴细胞白血病 (ALL) 中的应用已显著提高了治愈率。然而,在实体瘤中,多项临床试验表明会出现药物相关的毒副作用,如胰腺炎、中性粒细胞减少和低蛋白血症。
这些毒副作用至少部分是由L-天冬酰胺酶中谷氨酰胺酶的协同活性引起的。基于提高L-天冬酰胺酶在血液恶性肿瘤中的疗效、扩大L-天冬酰胺酶的用途、减少副作用的目的,新一代L-天冬酰胺酶正在开发中。
临床试验中靶向氨基酸代谢的药物
doi: 10.1038/s41392-023-01569-3
免疫疾病
自身免疫性疾病中免疫细胞代谢异常可促进炎症细胞的趋化性和炎症因子的产生。
类风湿性关节炎:T细胞天冬氨酸的丰度降低↓↓
在类风湿性关节炎 (RA) 中,细胞因子肿瘤坏死因子 (TNF) 的过量产生是发病机制中的一个核心事件,而富含内质网 (ER) 的 T 细胞是发炎关节中 TNF 的主要释放者。
发现类风湿性关节炎(RA)T细胞中线粒体天冬氨酸(Asp)的丰度降低,从而抑制NAD +周转,导致NAD +/NADH 比值降低以及NAD+蛋白质的ADP核糖基化减少依赖。
类风湿性关节炎和其他自身免疫性疾病的治疗策略是使用抗体来阻断细胞因子或其受体。最新的小分子抑制剂是靶向 Janus 激酶 (JAK) 抑制剂。这些治疗策略旨在阻断炎症途径的下游实践。然而,这些下游信号通路广泛分布在免疫细胞以外的细胞类型中,这会导致血栓形成等不良事件。因此,对自身免疫性疾病上游炎症的研究有助于从源头上预防疾病的发展。
神经系统疾病
天冬酰胺合成障碍是一种新发现的神经系统疾病,与染色体 7q2 上的 ASNS 基因突变有关。
自闭症:血清和脑脊液中天冬酰胺降低↓↓
自闭症谱系障碍(ASD)严重影响早期神经发育,导致智力障碍、发育迟缓、顽固性癫痫发作、进行性脑萎缩和呼吸系统缺陷。目前,该疾病中有一小部分人的血清和脑脊液中天冬酰胺水平可检测到降低,这阻碍了该测试用于初步筛查。由于血脑屏障的存在,天冬酰胺不会在大脑中积极积聚,因此大脑中 ASNS 活性的降低被认为是导致这种疾病的原因。
迄今为止,已报道了15种与ASD相关的突变,其中一些突变破坏了蛋白质结构,降低了ASNS的底物结合能力和催化效率。
例如,R49Q是位于N端结构域的谷氨酰胺 (Gln)结合袋中的突变,该突变不仅导致第二个β-折叠的氢键丢失,而且导致Gln的氢键丢失。此外,G289A和T337I突变位于C端结构域的ATP结合袋附近,G289A会引起与Ser293的空间冲突,而T337I会在蛋白质表面引起疏水斑并降低蛋白质溶解度。
补充天冬氨酸部分有效,需进一步研究
在治疗方面,膳食补充天冬氨酸并没有想象中那么有效,人为升高血液天冬氨酸可能会因竞争共转运蛋白而影响其他氨基酸的吸收。目前的治疗方法仅部分有效,需要进一步了解该疾病的机制来开发有效的药物。
★
谷氨酰胺(Gln)是一种用于蛋白质合成的 α-氨基酸。它的结构与谷氨酸类似,但侧链的羧酸基团被酰胺取代。谷氨酰胺是从食物中获取的非必需氨基酸,也是消耗最多的氨基酸,参与合成所有非必需氨基酸 (NEAA) 和蛋白质。
哪里产生谷氨酰胺?
肌肉组织在人体内产生最多的谷氨酰胺,约占全部合成谷氨酰胺的90%。
大脑和肺部也能释放少量的谷氨酰胺。肝脏虽然也能合成谷氨酰胺,但其主要功能是调节从肠道吸收的大量谷氨酰胺。
哪里消耗谷氨酰胺?
肠道细胞、肾细胞、激活的免疫细胞和各种肿瘤细胞是谷氨酰胺最迫切的消耗者。
具体通路
谷氨酰胺通过氨基酸转运蛋白 ASCT2/SLC1A5 进入细胞,并通过涉及谷氨酰胺酶 (GLS) 的脱氨反应在线粒体中转化为谷氨酸。然后谷氨酸在谷氨酸脱氢酶(GDH)或谷氨酸转氨酶或天冬氨酸转氨酶(TAs)的催化下产生α-酮戊二酸(α-KG)。α-KG是TCA循环的中间产物。
在缺氧或线粒体功能障碍下,α-KG可通过异柠檬酸脱氢酶(IDH2)催化的羧化反应转化为柠檬酸,用于氨基酸和脂肪酸的合成以及还原剂NADPH的生产。
肿瘤细胞是谷氨酰胺的紧急消耗者。信号分子Akt、Ras和AMPK可以通过激活糖酵解引起Warburg效应来诱导乳酸产生,促使肿瘤细胞通过谷氨酰胺代谢来满足能量需求。
在各种肿瘤中,谷氨酰胺代谢受癌基因/抑癌基因(例如 c-Myc 和 p53)的调节。癌基因 c-Myc 通过 GLS 和 SLC1A5 基因的转录激活上调谷氨酰胺代谢。
GLS 驱动的谷氨酰胺代谢是前列腺癌 (PCa) 放疗耐受性的调节因子,并且 GLS 1 和 c-MYC(谷氨酰胺的关键调节因子)的高表达与接受治疗的前列腺癌患者的无进展生存期缩短显著相关与放射治疗。
谷氨酰胺代谢可以通过α-KG依赖性染色质双加氧酶维持前列腺癌干细胞(CSC)。抑制谷氨酰胺代谢可降低体内 CSC 群体的频率以及小鼠模型中肿瘤生长的速率。
结肠癌
在结肠癌中,谷氨酰胺剥夺刺激p53激活,促进天冬氨酸/谷氨酸转运蛋白SLC1A3的表达,从而促进谷氨酸、谷氨酰胺和核苷酸合成,维持电子传递链和三羧酸循环活性。
SLC1A3 的缺失会降低肿瘤细胞对谷氨酰胺饥饿的抵抗力并抑制肿瘤细胞的生长。此外,研究表明,高表达胱氨酸/谷氨酸抗转运蛋白SLC7A11/xCT的肿瘤细胞高度依赖谷氨酰胺代谢。
在缺乏胱氨酸等氨基酸的情况下,细胞通过通用控制非阻遏物 2 (GCN2) -真核起始因子 (eIF2a) 信号通路促进 ATF4 的翻译,从而促进参与氨基酸代谢和应激反应的基因转录,包括 SLC7A11,使细胞能够应对氨基酸饥饿。
肿瘤中的谷氨酰胺代谢
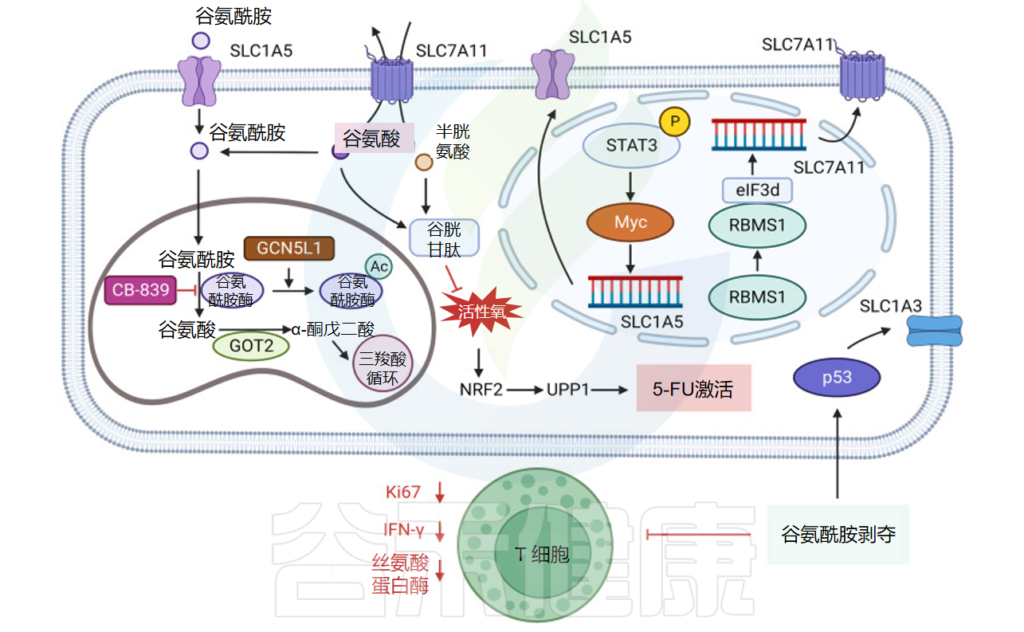
doi: 10.1038/s41392-023-01569-3
由于肿瘤细胞通过 SLC7A11 将细胞内的谷氨酸交换为细胞外的胱氨酸,细胞内的谷氨酸被消耗,导致细胞吸收更多的谷氨酰胺,并激活谷氨酰胺酶来补充细胞内的谷氨酸,使得 SLC7A11 高表达的细胞成为谷氨酰胺依赖型。
SLC7A11在三阴性乳腺癌、肺癌、胰腺导管腺癌、肾癌和肝癌中高表达
在三阴性乳腺癌 (TNBC) 中,与其他乳腺癌细胞相比,SLC7A11 高表达的细胞消耗更多的谷氨酰胺,并且对谷氨酰胺饥饿更敏感。
研究发现,SLC7A11 的缺失对小鼠正常胰腺组织的发育没有影响,但会严重损害 KRAS 驱动的胰腺导管腺癌生长。
生理条件下 SLC7A11 的非必需性以及 SLC7A11 在肿瘤中的高表达使 SLC7A11 成为癌症治疗的有希望的靶标。
谷氨酰胺酶可将谷氨酰胺水解为谷氨酸,是谷氨酰胺代谢中的关键酶。
谷氨酰胺酶的表达具有组织特异性。谷氨酰胺酶在汇管周围肝细胞、肾上皮细胞和中枢神经系统中活跃表达,用于合成尿素和神经递质。
人类谷氨酰胺酶的四种异构体分为两种由 GLS1 编码的高活性肾谷氨酰胺酶类型和两种由 GLS2 编码的低活性肝谷氨酰胺酶类型。
不同肿瘤中GLS1和GLS2表达的异质性表明恶性肿瘤细胞对谷氨酰胺代谢的需求不同。发现肝癌细胞线粒体中氨基酸合成5样1(GCN5L1)的通用控制可以促进GLS1和GLS2异构体的乙酰化和失活,从而抑制mTORC1激活和细胞增殖。
卵巢透明细胞癌
在卵巢透明细胞癌 (OCCC) 中,谷氨酰胺酶抑制剂 CB-839 抑制 ARID1A(富含 AT 的相互作用结构域蛋白 1A)突变的 PDX 肿瘤生长。
肾细胞癌
一项针对晚期肾细胞癌 (RCC) 的随机、双盲、对照 II 期试验证明,谷氨酰胺酶抑制剂 telaglenastat (CB-839) 和 mTOR 抑制剂依维莫司 (TelaE) 联合使用具有协同抗癌作用,且患者耐受性良好既往接受过 TKI 治疗的患者。此外,与安慰剂加依维莫司 (PboE) 相比,TelaE 可以改善无进展生存期 (PFS)。
另一项 Ib 期临床试验也显示 TelaE 或 telaglenastat 联合卡博替尼 (TelaC) 治疗 RCC 具有良好的耐受性和临床活性。
胰腺炎
补充谷氨酰胺降低重症死亡率,改善IBS,利于伤口愈合
谷氨酰胺可作为多种疾病的营养补充剂。多项荟萃分析发现,补充谷氨酰胺可以降低重症胰腺炎患者的死亡率、并发症发生率和总住院时间。
一项随机、双盲、安慰剂对照临床研究表明,在低发酵低聚单糖和多元醇 (FODMAP) 饮食中补充谷氨酰胺可改善肠易激综合征 (IBS) 症状。
在促进伤口愈合方面,研究发现,补充精氨酸和谷氨酰胺对伤口愈合有积极影响,补充谷氨酰胺显著影响患者的氮平衡,减少住院时间和死亡率。
补充谷氨酰胺并没有显著影响烧伤患者的预后
在一项招募 1200 名患者的双盲、随机、安慰剂对照试验中,谷氨酰胺补充剂组的出院生存期为 40 天,而安慰剂组为 38 天。谷氨酰胺组的死亡率为 17.2%,与安慰剂组的 16.2% 没有显著差异,并且补充谷氨酰胺并没有减少住院时间。
在他们的研究中,表明了补充谷氨酰胺的益处和风险,而烧伤和其他疾病的临床试验却显示出相互矛盾的结果。
补充谷氨酰胺对各种疾病的益处和风险仍需要更多临床试验数据。
心血管疾病
在心血管疾病中,Myc 和 Myc 相关因子 X (Max) 上调肺动脉高压中的谷氨酰胺转运蛋白 SLC1A5 和 SLC7A5 以及线粒体苹果酸,从而促进谷氨酰胺分解诱导的右心室肥厚。
氧化应激下,谷氨酰胺显著下降
在氧化应激下,心肌细胞中的谷胱甘肽(GSH)水平下降 60-70%,谷氨酰胺、谷氨酸和 α-酮戊二酸(α-KG)水平也显著下降,同时将谷胱甘肽转化为谷胱甘肽的 GLS 酶活性显著降低。
调节葡萄糖稳定性的关键氨基酸
2 型糖尿病是心血管疾病发生的主要危险因素。糖尿病患者骨骼肌代谢失调会影响胰岛素敏感性和葡萄糖稳态。研究发现,谷氨酰胺是调节葡萄糖稳定性和胰岛素敏感性的关键氨基酸,谷氨酰胺水平影响骨骼肌的炎症反应并调节胰岛素信号抑制剂适应性蛋白GRB10的表达。此外,谷氨酰胺的全身升高可改善肥胖小鼠模型中的胰岛素敏感性并恢复葡萄糖稳态。
补充谷氨酰胺——心脏保护的新希望
蒽环类抗生素阿霉素(DOX)是一种广泛应用于实体恶性肿瘤的抗肿瘤药物;然而,这种疗法可能因自由基和氧化应激而导致严重的心脏毒性。补充谷氨酰胺显著降低心脏脂质过氧化物水平,增加过氧化物酶和谷胱甘肽水平,保护 DOX 治疗的大鼠模型的心脏功能。
针对心脏谷氨酰胺代谢的药物正在开发中。冬凌农 (Ori) 是一种从植物冬凌草 (Hemsl.) 中提取的天然萜类化合物,在大鼠心肌损伤模型中,可提高心脏谷氨酰胺水平并抑制 ATP/ADP 比值下降,保护心肌细胞并减少梗塞面积。
严重急性呼吸道综合症
严重急性呼吸综合征冠状病毒 2 (SARS-CoV-2) 是 2019 年冠状病毒病 (COVID-19) 的病因。该疾病通过人与人之间的密切接触或感染者的呼吸道分泌物传播。
COVID-19:谷氨酰胺降低↓↓
COVID-19的危险因素包括心血管疾病和糖尿病,此类高危人群表现出谷氨酰胺、NAD +水平低和透明质酸(HA)过量产生的共同代谢特征。
谷氨酰胺和 NAD +水平会导致 SIRT1 失调
SIRT1 是透明质酸合酶 2 (HAS2) 基因的关键负调节因子。这些代谢改变最终导致 HA 和纤溶酶原激活剂抑制剂 1 (PAI-1) 的过量产生以及 Tregs 和骨髓源性抑制细胞 (MDSC) 群体的扩张。因此,谷氨酰胺缺乏导致了 COVID-19 高危人群的免疫功能障碍和 HA 过量产生。
HA可以通过PAI-1激活STAT3。由于 SIRT1、STAT3 和 O-GlcNacylation 失调,通过激活 HAS2 诱发透明质酸风暴。
谷氨酰胺消耗加剧
此外,尽管SARS-CoV-2疫苗显著减少了COVID-19病例,但SARS-CoV-2感染后细胞处于强烈的氧化应激条件下,从而促进谷氨酰胺的消耗以合成谷胱甘肽。这一过程加剧了高危人群的谷氨酰胺缺乏,并可能诱发代谢功能障碍。同时,它还能引起STAT3通路失活和PAI-1激活,导致部分人出现严重的COVID-19并发症。
补充谷氨酰胺降低患者严重程度
小型临床试验表明,补充谷氨酰胺可以降低 COVID-19 患者感染后的严重程度。然而,这部分研究需要扩大,以更准确地评估谷氨酰胺在治疗 COVID-19 中的价值。
★
精氨酸,又称L-精氨酸,是蛋白质合成的原料,也是尿素和一氧化氮循环的中间产物。精氨酸被归类为条件必需氨基酸,其需求量取决于发育阶段和健康状况。
在人类中,小肠上皮细胞将谷氨酰胺和谷氨酸转化为瓜氨酸,然后通过循环系统转运至肾近端肾小管细胞,其中精氨酸由尿素循环中的精氨酸-琥珀酸合成酶和精氨酸-琥珀酸裂合酶合成。
当小肠和肾功能受损时,精氨酸合成就会受损,从而产生对精氨酸的饮食需求。在其他细胞类型中,瓜氨酸合成的精氨酸非常低,但当诱导型一氧化氮合酶 (NOS) 增加时,精氨酸合成会急剧增加。
在这些条件下,一氧化氮合成的副产物瓜氨酸可以通过精氨酸-瓜氨酸途径回收精氨酸。精氨酸对于细胞分裂、伤口愈合和免疫功能很重要。
蛋白质中的精氨酸可被 PAD 酶催化为瓜氨酸,这一过程称为瓜氨酸化,是正常免疫过程的一部分。另一种类型的翻译后修饰是精氨酸甲基转移酶 (PRMT) 的甲基化,其中精氨酸可以甲基化为单甲基化精氨酸或二甲基化精氨酸。
精氨酸甲基转移酶可分为以下三类:
精氨酸甲基化通常发生在富含甘氨酸和精氨酸的“GAR基序”中。许多精氨酸甲基化蛋白质已被证明可以与 DNA 或 RNA 相互作用,并且精氨酸残基充当磷酸主链的重要氢供体。
此外,精氨酸甲基化还会影响各种细胞过程中涉及的蛋白质-蛋白质相互作用,例如蛋白质运输、信号转导和转录调节。
正常细胞中的瓜氨酸和天冬氨酸可通过尿素循环中的精氨酸-琥珀酸合成酶 1 (ASS1) 和精氨酸-琥珀酸裂解酶 (ASL) 转化为精氨酸。
精氨酸琥珀酸合成酶 1 (ASS1) 转录抑制发生在各种肿瘤中,产生对外部精氨酸的依赖,并实现精氨酸剥夺疗法。
在GBM中使用精氨酸消耗剂聚乙二醇化精氨酸脱亚胺酶ADI-PEG20可以增加一氧化氮(NO)合成并产生细胞毒性过亚硝酸盐,增加肿瘤细胞对电离辐射的敏感性,显著增强放疗对GBM的效果。
精氨酸剥夺促进HCC细胞中GCN2依赖性周期停滞,而精氨酸剥夺HCC细胞中GCN2的抑制促进细胞衰老并增加衰老化合物的功效。
Ass 缺陷的前列腺癌和胰腺癌也被证明对 ADI-PEG20 敏感,而 ADI-PEG20 通过诱导自噬和细胞凋亡来促进细胞死亡。
肿瘤细胞中的精氨酸代谢
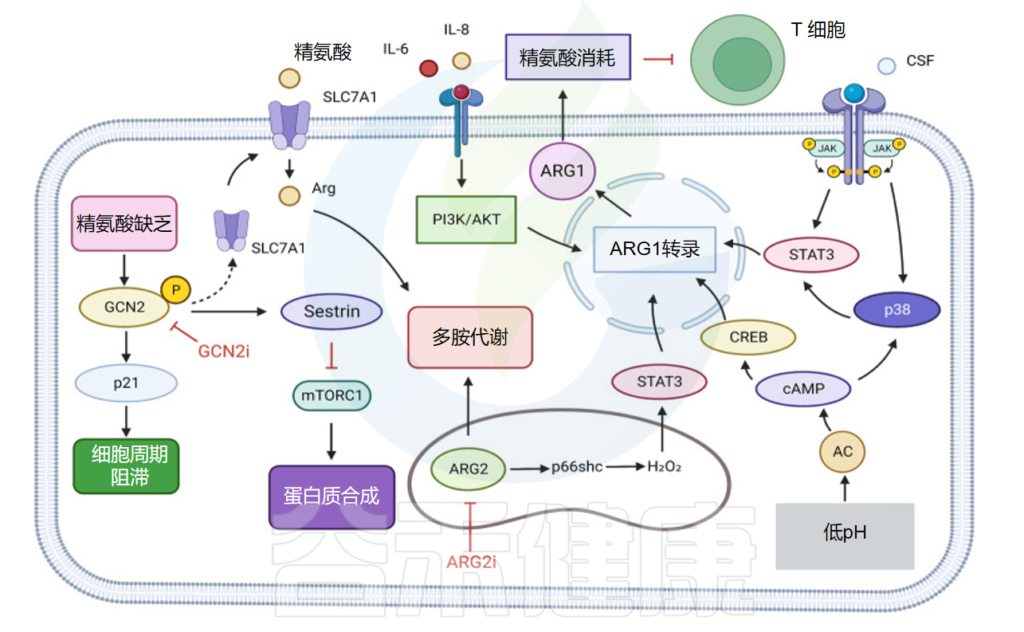
doi: 10.1038/s41392-023-01569-3
精氨酸耗竭可以增加肝细胞癌细胞中GCN2的磷酸化水平,激活GCN,增加SLC7A11的表达水平,增加精氨酸的摄取。
伤口愈合
在伤口愈合过程中,精氨酸通过精氨酸-NO途径参与炎症因子的反应。此外,精氨酸酶降解精氨酸产生的鸟氨酸和尿素在此过程中是必需的,并且在胶原蛋白和多胺的合成中具有关键作用。
精氨酸可以通过 GPRC6A-ERK1/2 和 PI3K/Akt 信号通路促进成纤维细胞增殖。
精氨酸调节免疫
精氨酸可以在炎症早期阶段增加单核细胞迁移和外周血中促炎因子的产生;在炎症后期,精氨酸还可以抑制免疫细胞的活性,调节免疫状态。总之,精氨酸及其代谢物对于伤口愈合至关重要,并参与伤口愈合的多个阶段,包括胶原蛋白形成、细胞增殖和免疫调节。
补充精氨酸有助于各种伤口愈合
膳食补充精氨酸是最方便的方法,对伤口愈合有多种好处。补充精氨酸可以增强人体 DNA 的合成。在结肠炎模型中,精氨酸补充剂可抑制炎症因子和趋化因子的表达,抑制炎症反应,促进受损组织的修复。
遭受创伤/失血性休克的患者由于胶原蛋白合成减少而难以实现伤口愈合。相反,补充精氨酸可以显著缓解上述问题并增加伤口强度。
糖尿病伤口愈合过程中,补充精氨酸可以逆转NO合成不足的情况,恢复受损组织中NO的浓度,促进伤口愈合。精氨酸也被用来降低压疮的风险,在压疮(褥疮)高风险患者中补充精氨酸可以显著加速压疮愈合。
阿尔茨海默病(AD)
阿尔茨海默病 (AD) 的特征是由淀粉样蛋白-β 和磷酸化 tau 沉积引起的老年斑和神经原纤维缠结 (NFT)。
精氨酸治疗改善阿尔茨海默病
晚期糖基化终末产物 (AGE) 会修饰蛋白质,导致其功能障碍。AMPK-γ 亚基的糖基化会抑制 AMPK 功能,精氨酸治疗可保护 AMPK-γ 免受糖基化并增加阿尔茨海默病小鼠模型中的 AMPK 磷酸化,从而改善阿尔茨海默病。
与其他药物联合使用,改善认知
在轻度阿尔茨海默病 /认知障碍 (MCI) 患者中,联合使用 L-精氨酸、HMG-CoA 抑制剂辛伐他汀和四氢生物蝶呤可增强内皮一氧化氮合酶 (eNOS) 通路,从而适度增加脑血流量并改善认知。
此外,据报道PRMT4催化的不对称二甲基精氨酸(ADMA)作为配体与NOS结合,导致NOS功能障碍,导致脑血流量减少,加重阿尔茨海默病,通过抑制PRMT4可以逆转这种情况。
肺部疾病
哮喘是一种多变的、反复发作的、长期的呼吸道炎症性疾病。精氨酸和一氧化氮代谢失衡与哮喘的病理生理学有关。
哮喘:精氨酸水平显著降低↓↓
对哮喘儿童血浆代谢质谱分析显示,与非哮喘易感组相比,哮喘易感组的精氨酸(Arg)、赖氨酸(Lys)和蛋氨酸(Met)水平显著降低。
还显示,哮喘和阻塞性睡眠患者的血清不对称二甲基精氨酸 (ADMA) 浓度显著增加,一氧化氮合酶 (NOS) 抑制增强,精氨酸分解代谢增强,鸟氨酸 (Orn) 和脯氨酸 (Pro) 水平增加,以及精氨酸/鸟氨酸比值降低呼吸暂停(OSA)。
在标记的哮喘药物中添加 L-精氨酸并不能显著减少哮喘发作
这可能是由于 IL-4 和 IL-13 诱导的精氨酸酶活性显著增加,以及过敏原刺激的肺部下游产物腐胺显著增加。
在肥胖哮喘患者的药物中添加 L-瓜氨酸(L-精氨酸循环和 NO 合成的前体),可能有助于控制哮喘并改善 NO 排泄分数 (FeNO) 水平。
心血管疾病
不对称二甲基精氨酸 (ADMA) 升高↑↑
患有高胆固醇血症和血管疾病的患者通常会出现不对称二甲基精氨酸 (ADMA) 升高,这与 NO 合成受损和内皮功能障碍的早期标志物有关。
ADMA 是一种内源性一氧化氮合酶 (NOS) 抑制剂,可显著减少血管舒张剂 NO 的合成,导致心血管疾病的发生。
PRMT1是催化ADMA的主要酶。它通过调节心肌肌动蛋白启动子区域的组蛋白甲基化修饰来调节基因激活。PRMT1的消融可以下调心肌素等收缩基因的表达,并显著降低主动脉的收缩力和血管平滑肌细胞(VSMC)的牵引力。
PRMTs的异常活性导致ADMA和MMA增加,从而增加心血管疾病的风险。
针对 PRMT 的抑制剂正在开发和实验测试中。精氨酸(Arg)甲基化酶抑制剂 (AMI) 是对称磺化尿素,可特异性抑制 PRMT 活性,并在大鼠模型中抑制环氧合酶 2 (COX-2) 表达并抑制炎症。
★
蛋氨酸(Met)是一种必需氨基酸,也是其他氨基酸的前体,例如半胱氨酸 (Cys) 和牛磺酸,以及 S-腺苷-L-甲硫氨酸 (SAM) 和谷胱甘肽 (GSH)。蛋氨酸生物合成的骨架主要来自天冬氨酸。
在分解代谢中,蛋氨酸被蛋氨酸腺苷转移酶 (MAT) 催化为 SAM。作为甲基供体,SAM 参与各种甲基转移反应,并在反应中转化为 S-腺苷高半胱氨酸 (SAH)。蛋氨酸可以增加细胞内谷胱甘肽的浓度,促进细胞氧化还原调节,并通过与氧化代谢物结合来保护细胞。
蛋氨酸作为一种必需氨基酸,在肿瘤的生长和代谢中具有重要作用。除外源供应外,蛋氨酸补救途径是唯一的蛋氨酸来源。该途径需要甲基腺苷磷酸化酶 (MTAP) 和 蛋氨酸合酶 (MS) 的活性。
MTAP 位于肿瘤抑制因子细胞周期蛋白依赖性激酶抑制剂 2 A (CDNK2A) 的外围,这两个基因的共同缺失发生在约 15% 的癌症中,导致高度侵袭性肿瘤且预后不良。这些酶在恶性肿瘤中常常下调,导致细胞对外部环境中蛋氨酸摄入的强烈依赖。
肿瘤细胞中的蛋氨酸代谢
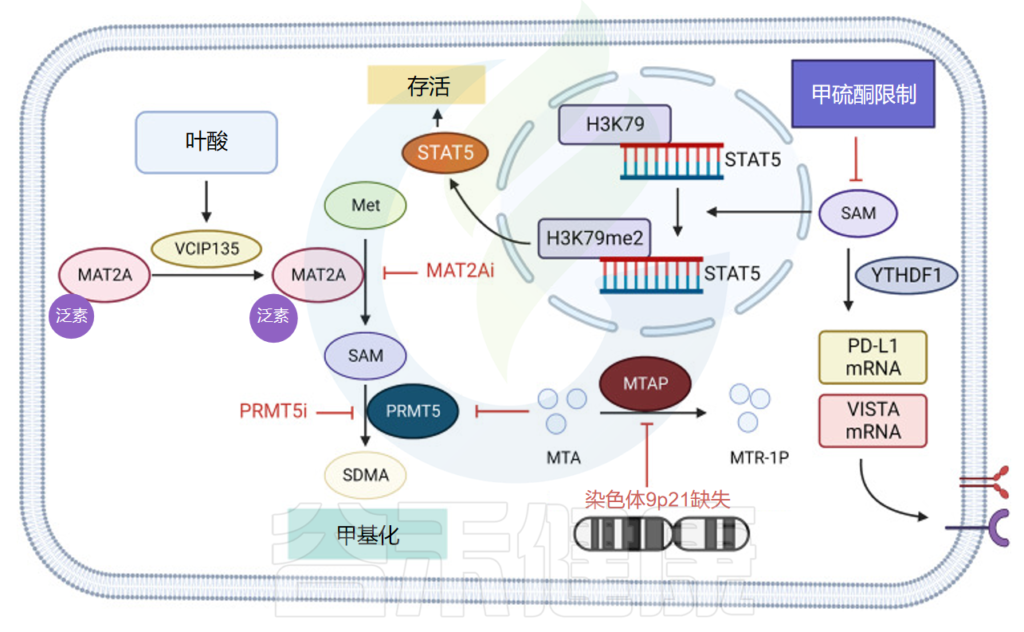
doi: 10.1038/s41392-023-01569-3
肿瘤细胞竞争消耗蛋氨酸→T细胞蛋氨酸缺乏
肿瘤细胞可以通过SLC43A2的高表达来增加蛋氨酸的摄入,竞争性消耗环境中的蛋氨酸,导致T细胞蛋氨酸缺乏。T细胞蛋氨酸限制可抑制细胞内正常甲基化,导致STAT5基因转录受阻,影响T细胞存活和功能。另一方面,蛋氨酸代谢抑制 PD-L1 和 T 细胞激活的 V 结构域 Ig 抑制因子 (VISTA) 免疫检查点翻译。
由于其在甲基化中的核心作用,蛋氨酸被认为是由 10-11 易位 (TET)、异柠檬酸脱氢酶 (IDH) 蛋白、甲基转移酶和其他表型修饰剂驱动的肿瘤治疗的候选靶点。
无蛋氨酸饮食的抗肿瘤作用,抑制肿瘤免疫逃逸
无蛋氨酸饮食的抗肿瘤作用首次在患有 Walker-256 肉瘤的 Sprague-Dawley 大鼠中被报道。
Met限制饮食可以减少肿瘤细胞中的N6-甲基腺苷(m6A)甲基化和免疫检查点翻译,例如PD-L1和T细胞激活的V域Ig抑制因子(VISTA)。
此外,它还能增加肿瘤浸润CD8 + T细胞的数量,从而抑制肿瘤免疫逃逸。补充蛋氨酸或抑制肿瘤细胞中SLC43A2的表达可以逆转上述T细胞功能抑制,激活肿瘤免疫。
这些研究表明,蛋氨酸限制不仅抑制肿瘤细胞代谢,还涉及免疫细胞,并且肿瘤细胞本身具有复杂的优先途径来调节蛋氨酸代谢。因此,应针对肿瘤细胞蛋氨酸代谢开发更精准、更有针对性的治疗方法。
脂肪肝疾病
非酒精性脂肪性肝病(NAFLD)是一种由于代谢途径异常导致甘油三酯(TG)在肝脏积聚而引起的疾病。肥胖和2 型糖尿病是 NAFLD 的重要危险因素。
高脂饮食、蛋氨酸和胆碱缺乏饮食(MCD)可以模拟人类疾病的组织学和代谢异常,通常用于建立NAFLD小鼠模型。
非酒精性脂肪性肝病早期,蛋氨酸水平降低↓↓
在评估两种方法构建NALFD/NASH模型的差异时发现,MCD饮食可在2-4周内自发导致肝纤维化,并显著影响肝纤维化途径相关基因的表达。高脂饮食的这种作用直到胰岛素抵抗后 24 周才观察到,从而减少了肝纤维化。
临床数据显示,NAFLD 早期阶段蛋氨酸水平降低,且较高的蛋氨酸摄入量与纤维化风险呈负相关。甲基供体补充通过激活 AMPK 信号通路增加脂肪酸消耗来减少肝脏脂肪积累。
肾脏疾病
常染色体显性多囊肾病:蛋氨酸水平升高↑↑
常染色体显性多囊肾病(ADPKD)是一种常见的单基因疾病,其特征是肾囊肿增大。在ADPKD模型中,蛋氨酸(Met)和SAM的水平升高,从而诱导Mettl3的表达。
Mettl3 可以增加 c-Myc 和 Avpr2 mRNA 修饰,激活 c-Myc 和 cAMP 通路,并加速囊肿生长。限制蛋氨酸饮食可能会减缓常染色体显性多囊肾病的进展。
在蛋氨酸限制 (MR) 小鼠损伤模型中,肾损伤标志物簇蛋白和胱抑素 c 显著下降。与正常喂养模型相比,MR模型中Emr1、Nos2、Tnfa等肾脏炎症基因下调,嗜碱性粒细胞聚集程度较低。
糖尿病
2 型糖尿病:循环蛋氨酸水平升高 ↑↑
糖尿病肾病:循环蛋氨酸水平升高 ↑↑
在2 型糖尿病和糖尿病肾病 (DKD) 中可以检测到循环蛋氨酸、乙酰天冬氨酸和天冬酰胺水平升高。此外,循环蛋氨酸(Met)水平升高可用于预测患糖尿病的风险。
蛋氨酸代谢调节 Cys 和内源性硫化氢 (H2S) 水平。H2S 抑制胰腺 β 细胞中葡萄糖诱导的胰岛素释放和脂肪组织中胰岛素刺激的葡萄糖摄取。
胱硫醚γ-裂解酶(CSE)是H2S合成中的关键酶,CSE抑制剂的使用会增加脂肪细胞对葡萄糖的摄取。
总之,蛋氨酸限制 (MR) 可以通过干扰葡萄糖稳态、增加胰岛素敏感性和炎症反应来缓解糖尿病。
氨基酸水平和代谢影响多个层面的细胞代谢和许多细胞过程,从蛋白质合成到表观遗传调控。这些生理过程与维持细胞稳态和正常功能密切相关。因此,氨基酸代谢异常可能导致疾病的发展。
综上,一些肿瘤病人比如,肝硬化、肺癌、乳腺癌、食管癌、肾癌、胃癌、甲状腺癌、头颈部肿瘤等病人血浆中氨基酸浓度与健康人血浆中氨基酸浓度相比表现出异常,对氨基酸代谢变化的分析可以用来辅助诊断疾病。氨基酸代谢的改变对肿瘤细胞及肿瘤免疫微环境有着显著影响。肿瘤细胞通常依赖于外源氨基酸的供应,相对于肿瘤细胞,正常细胞对氨基酸的需求较低,干扰氨基酸的可获得性,通过氨基酸消耗疗法来诱导癌细胞凋亡也是近年来备受瞩目的研究热点之一。
支链氨基酸在肿瘤和代谢疾病中起重要作用,但不同肿瘤之间对支链氨基酸的需求存在差异。支链氨基酸代谢对促炎CD4+和CD8+T细胞以及免疫抑制调节性Treg细胞的功能同样重要,Treg细胞在代谢性疾病、肝脏和肾脏疾病中发挥着关键作用。
天冬氨酸在某些肿瘤生长中是一个内在的限制因素,其转化依赖于天冬酰胺酶的活性。转氨产物天冬酰胺比天冬氨酸更具渗透性。然而,膀胱癌细胞缺乏天冬酰胺酶,导致转化为天冬氨酸的功能障碍。SLC1A3在某些实体瘤中高表达,可能为解决天冬氨酸酶耐药提供解决方案。
谷氨酰胺在肠、肾、免疫和肿瘤细胞中广泛消耗,与癌基因c-Myc和STAT3有关。SLC7A11在多种癌症中显著高表达,可作为有前景的靶标。GLS是肾小管氨基酰胺代谢的关键酶,CB-839是其抑制剂,显示出良好的抗肿瘤活性。
精氨酸调节免疫,缺乏ASS1的细胞对精氨酸具有依赖性。精氨酸甲基转移酶(PRMT)在肿瘤发生发展中的调节作用也受到广泛关注,多种 PRMT 抑制剂表现出良好的抗肿瘤活性。
蛋氨酸作为必需氨基酸的地位及其在转甲基化过程中的作用注定了细胞对蛋氨酸代谢的依赖。甲硫腺苷磷酸化酶 (MTAP) 基因缺失在一些实体瘤中发生,针对缺乏MTAP的肿瘤细胞的治疗策略是抑制蛋氨酸代谢途径中的关键酶MAT2A。蛋氨酸限制饮食可以抑制肿瘤免疫逃逸。
但是,氨基酸代谢靶向治疗面临挑战,血液氨基酸代谢是一个复杂的生物过程,涉及多个代谢途径和相互作用。因此,准确评估血清氨基酸水平对于疾病筛查和治疗有主要的意义。同时,目前血清各类氨基酸测定方法,一般是空腹抽取这增加了检测的复杂性。而且检查前一周熬夜、饮酒、劳累、服药、情绪抑郁以及进食油腻食物等都会影响氨基酸的评估。
肠道菌群在蛋白质和氨基酸代谢中扮演着重要的角色。通过评估肠道菌群的组成和功能,可以更从另外一个维度了解氨基酸代谢的状态。
一些研究发现,通过调节肠道菌群可以影响氨基酸代谢和相关疾病的进展。通过了解个体的肠道菌群状态,可以针对性地选择合适的综合治疗方法,如益生菌、益生元和饮食干预,以优化氨基酸代谢和减少治疗的不良副作用,比如抗体来阻断细胞因子或其受体导致的血栓形成等不良事件。
除本文常见的20种蛋白质氨基酸外,还包括在各个代谢途径中较为重要的氨基酸及短肽,比如牛磺酸、γ-氨基丁酸(GABA)、肌酸、肌肽等。它们在某些方面对人体的重要作用一样不比蛋白质氨基酸逊色,后续我们再和大家分享。
主要参考文献:
Ling ZN, Jiang YF, Ru JN, Lu JH, Ding B, Wu J. Amino acid metabolism in health and disease. Signal Transduct Target Ther. 2023 Sep 13;8(1):345.

谷禾健康

胃肠道除了在食物消化和吸收中发挥作用外,还是一个主要的内分泌器官,产生多种生物活性肽和胺类物质,作为肠道激素。
大多数肠道激素来源于肠内分泌细胞,但来源于其他肠上皮细胞类型的生物活性肽也与代谢调节有关。
为什么说肠道激素重要呢?我们在日常生活中胃口好不好都可能与它相关,它能影响食欲和食物摄入,通过影响大脑中的食欲中枢,减轻食欲或刺激饱腹感。
其实,肠道激素在多种器官的多种代谢活跃组织中协调关键的生理过程,包括胰腺、肝脏、脂肪组织、肠道和中枢神经系统等。从局部控制营养吸收和调节肠道生长、肠道运动和液体分泌,到远距离调节胰岛素释放、脂肪细胞功能。
近日,来自英国剑桥大学威康Trust-MRC代谢科学研究实验室团队发表了一篇文章在《Nature Reviews Gastroenterology & Hepatology》期刊,主要讲述了参与代谢调节的肠道激素,讨论了它们在胃肠系统内外的作用。

肠道激素由多种肠内分泌细胞和非肠内分泌细胞产生,它们对不同刺激的反应不同,并分泌特定的激素组合。
肠内分泌细胞通过一系列感觉转运蛋白、离子通道和受体感知各种营养和非营养刺激,使其成为营养吸收和肠道激素反应之间的有效联系。
这些肠道激素信号传导过程可以作为治疗靶点;例如,GLP2受体激动剂用于治疗短肠综合征,并且GLP1受体激动器用于治疗2型糖尿病和肥胖。此外,激素类药物的组合可用于最大限度地提高疗效,同时最大限度地减少不良反应。
因此,深入了解肠道内分泌系统介导的复杂器官间串扰,有助于设计基于或靶向肠道激素及其受体的更有效药物,并将其治疗潜力扩展到肥胖和糖尿病之外的疾病。
肠内分泌细胞沿着胃肠上皮的长度广泛分布。肠道内的一些细胞会分泌一些激素来应对不同的刺激。
■ 肠内分泌细胞可分为两大类:
以前我们认为这些肠内分泌细胞只会分泌单一类型的激素,但现在我们知道,实际上它们通常会同时分泌多种不同的激素,所以它们之间的分泌有一些重叠的部分。
■ 肠道中不同部位的激素表达也会有所不同,如:
肠道特定区域的激素产生与激素的功能作用及其反应的感觉输入有关。
决定肠道不同区域肠内分泌细胞激素特征的因素在很大程度上仍然未知。来自不同肠段的肠道类器官似乎保留了与起源相关的激素特征,这表明肠道干细胞的效力有限。
■ 肠道细胞功能受限的机制与激素分泌的发育演化
然而,随着细胞的发育成熟,肠内分泌细胞的激素特征会发生变化,并沿着隐窝-绒毛轴分化。
举个例子,回肠L细胞在肠隐窝中共表达GLP1和肽YY(PYY),但当这些细胞到达绒毛时,前胰高血糖素转录下调,分泌素转录上调。一旦这些激素分泌出来,它们有两条路:
■ 激素在肠道中的分泌调节及其影响
肠道激素控制多种代谢活性组织的广泛生理过程,包括胰腺、肝脏、脂肪组织、肠道和中枢神经系统。这些激素大多具有不止一种生理作用,许多生理作用由不止一种激素发挥。如下表:
调节新陈代谢的主要肠道激素的特征

Bany Bakar R, et al., Nat Rev Gastroenterol Hepatol. 2023
■ 肠内分泌细胞如何与营养物质密切接触
这就需要我们先来了解一下,肠内分泌细胞的结构特点,大多数肠内分泌细胞是开放型细胞,也就是说它们有一个顶端表面,带有向肠腔延伸的微绒毛,以及一个基底外侧表面,负责在刺激时通过胞吐释放分泌颗粒。
因此,这些细胞与消化和吸收的营养物质直接接触,把它们推向产生肠道激素反应的理想位置,将营养吸收的变化传达给身体其他部位。
它怎么传达呢?
肠内分泌细胞的化学感受能力归因于它们表达一系列感觉转运蛋白、离子通道和受体,这些受体解释了它们对各种营养刺激的反应能力,包括葡萄糖、消化脂质、碳水化合物和蛋白质,以及非营养刺激,如胆汁酸、肠道激素、肠道神经递质,微生物产物和炎性胞质分裂。
■ GPCR——肠内分泌细胞通过其检测营养物质吸收
G蛋白偶联受体(GPCR)检测到广泛的营养和非营养刺激,肠内分泌细胞差异表达一系列GPCR,这些GPCR主要位于其基底外侧表面,能够检测出胆汁酸、游离脂肪酸(FFAs)和氨基酸在上皮层内的消化和吸收过程中的变化。
与肠道激素释放密切相关的GPCR包括:
各种脂质感应受体,包括:
相反,葡萄糖在通过顶端膜上的钠偶联葡萄糖转运蛋白SGLT1吸收时激活肠内分泌细胞。SGLT1对钠离子与葡萄糖的共转运直接触发质膜去极化,进而激活电压门控钙通道。由此产生的钙流入进一步使肠内分泌细胞质膜去极化,并启动分泌颗粒的释放(下图)。
调节肠内分泌细胞活性的营养诱导机制
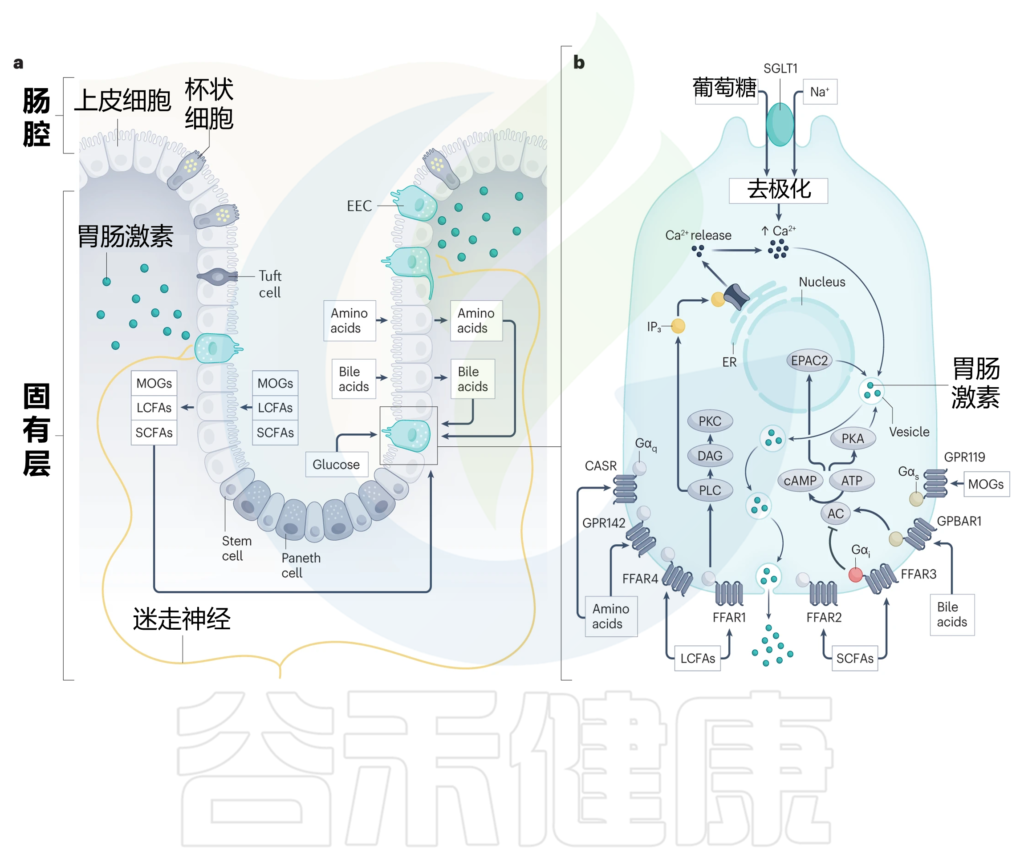
Bany Bakar R, et al., Nat Rev Gastroenterol Hepatol. 2023
肠道激素GDF15、FGF19(小鼠中的FGF15)、鸟苷素和尿鸟苷素不是由肠内分泌细胞产生的,而是由肠细胞和其他上皮谱系产生的。对这些激素生物合成和释放的途径了解相对较少。
肠内分泌细胞衍生的肽激素是由激素原转化酶的活性产生的,这些酶共同储存在囊泡中,囊泡与细胞膜融合以响应分泌刺激;而其他细胞类型中激素的产生则不同,它们并不经过传统的囊泡激素原转化酶和调节的囊泡分泌机制。
■ 肠道激素与二甲双胍疗效的关联
这些激素的生物合成与它们的mRNA水平有关,mRNA对特定刺激有反应。举例来说,FGF19(小鼠中的FGF15)的表达仅限于回肠末端,并通过核法尼糖样受体FXR被胆汁酸上调。在人体中,饭后胆汁释放后90-120分钟内FGF19的循环水平上升。在小鼠中,GDF15在多个器官中表达,但其在肠道中的表达受到细胞应激途径的增加的影响,就像在二甲双胍治疗过程中观察到的那样。实际上,治疗2型糖尿病患者的二甲双胍可能导致循环GDF15水平升高,从而产生该药物减肥效果。
■ Proguanylin:胃切除后水平升高?
Proguanylin在成熟肠细胞、一些杯状细胞和Paneth细胞中表达,在接受胃切除术或肠移植的患者中检测到的循环水平高于健康人。
Proguanylin释放到肠腔中,在那里被切割形成成熟的生物活性鸟苷肽。鸟苷由两个分子内二硫键稳定,这两个键对消化酶提供了一些保护,并从顶端方向靶向上皮膜鸟苷环化酶受体。Proguanylin和 prouroguanylin 也进入血液,但这些形式是如何分泌和激活的尚不清楚。
肠道激素控制许多胃肠道功能,这些功能涉及协调对食物摄入的反应,并确保摄入的食物被有效消化和吸收。
肠内分泌细胞检测管腔内容物的化学成分和局部营养吸收率,以及它们释放的激素控制过程,如胃酸分泌的调节、胃排空、胆囊收缩、胰腺酶和电解质的分泌、肠道运动和上皮屏障功能。
▸ 胃酸分泌
胃中特殊的壁细胞分泌酸有助于食物的消化和矿物质的吸收,并降低胃肠道感染的风险。胃酸的产生受到严格调节,以确保有效的食物消化,而不会损害胃和十二指肠粘膜。
胃窦G细胞分泌的胃泌素作用于肠嗜铬细胞上的胃泌素受体(称为CCKB或CCK2受体),刺激组胺的释放,组胺是一种强效的盐酸促分泌剂。胃窦和胃底D细胞的生长抑素对该系统进行微调,除了直接抑制壁细胞的酸释放外,生长抑素还能抑制胃泌素和组胺的释放。
其他几种肠道激素,包括胆囊收缩素(CCK)、氧调素、PYY和分泌素,通过不完全表征的机制抑制胃酸分泌,并募集涉及生长抑素的旁分泌回路。
▸ 胃排空
胃排空的速度决定了胃内容物输送到十二指肠的速率,并受到各种因素的调节,包括食物特性、肠道神经信号、肠道激素和血糖水平。胃排空率与食物消化率和营养吸收率密切相关,并受到调节,使得胃排空率仅反映小肠消化和吸收所接收营养的能力。
如果胃排空速率超过十二指肠吸收能力,营养物质会进入小肠并触发回肠制动,从而导致胃排空的反馈抑制,从而恢复平衡。主要位于回肠和结肠的肠道激素GLP1和PYY在回肠制动中起主要作用。回肠制动导致食物滞留在胃中,导致胃胀,从而激活发出饱腹信号的机械敏感迷走神经传入神经元。
▸ 胰腺和胆囊功能
一旦胃内容物进入小肠,其他激素就会被吸收以促进消化。由十二指肠 I 细胞产生的CCK刺激胰腺酶分泌,并通过刺激胆囊神经节中传出迷走神经神经递质的释放来促进胆囊排空。
分泌素在各种成熟的肠内分泌细胞中产生,刺激胰腺碱性电解质分泌,也与餐后热遗传学有关。鸟苷激活肠细胞中的鸟苷酸环化酶-cGMP信号,从而触发上皮氯化物和液体分泌。
这种途径被热稳定肠毒素劫持,是它们引发分泌性腹泻的能力的基础。鸟苷素-cGMP信号通路也与结直肠癌的抑制有关。
▸ 肠道运动
幽门以外胃肠道的运动由协调纵向蠕动波的肠神经系统和几种肠道激素之间的相互作用调节。
其中一种激素是胃动素,它存在于人类十二指肠中(尽管啮齿类动物不存在),并在胃中启动迁移运动复合体,特别是在餐间状态。胃动素受体激动剂,如低剂量红霉素,对胃轻瘫有治疗作用,一项研究表明,胃动素接收器的功能缺失变体在胃轻瘫患者中富集。
与调节小肠和大肠运动有关的其他激素包括:
粘膜血清素对肠道运动和液体分泌的重要性受到质疑,因为缺乏产生粘膜血清素所需的色氨酸羟化酶1的小鼠没有强烈的表型。然而,在一个模型中,产生血清素的肠嗜铬细胞被有条件地切除,这一发现强调了这些细胞确实在肠道转运控制中发挥作用。
▸ 肠道功能
除了控制肠道通道外,肠道激素还调节其他重要的肠道过程。例如:
GLP2刺激上皮生长,从而增加给定肠段的吸收能力。这种作用可能有助于GLP2治疗短肠综合征的疗效,
GLP1–GLP2双激动剂更有效。与血清素和神经降压素等其他肠道激素一样,GLP1和GLP2也会影响屏障功能并调节免疫系统。分泌血清素的肠嗜铬细胞与内脏疼痛有关,对肠道炎症、肝脏病理生理学和焦虑具有重要意义。
内 分 泌 胰 腺
我们知道饭后一段时间,人体内胰岛素的分泌量会增加,这其实也跟肠道激素有关。GLP1和GIP是两种主要的肠促胰岛素,占餐后胰岛素分泌的70%。
肠促胰岛素对葡萄糖刺激的胰岛素释放的影响可以通过以下事实来证明:口服葡萄糖比静脉注射葡萄糖(不刺激肠促胰岛素释放)能引起更大的胰岛素分泌——这种效应被称为肠促胰岛素效应。从生理上讲,肠促胰岛素调节对碳水化合物摄入的反应,并确保餐后健康的葡萄糖耐受性,无论摄入的葡萄糖负荷如何。
GLP1和GIP如何增加胰岛素分泌?
GLP1和GIP分别通过直接作用于GLP1受体和GIP受体来增加胰岛素分泌,这两种受体在胰腺β细胞中高度表达。
在β细胞中,GLP1和GIP受体主要募集Gαs并触发cAMP的升高。cAMP影响通过激活蛋白激酶A(PKA)和由cAMP-2激活的交换蛋白(EPAC2)调节胰岛素释放的几种下游途径。这两种效应物都调节离子通道活性以及参与葡萄糖依赖性胰岛素分泌的囊泡运输和融合。
GLP1和GIP具有额外的促胰岛素作用,但两者对肠促胰岛素作用的贡献有争议
GLP1和GIP对β细胞的作用不仅限于调控胰岛素分泌,还包括上调PDX1,促进胰岛素基因转录,增加β细胞质量并通过细胞周期和MAPK的激活促进新生。
基于这些作用,GLP1受体激动剂的长期治疗被认为是2型糖尿病的一种疾病改良治疗方法,可以通过增加或保留功能性β细胞质量来减轻β细胞功能障碍的进展。
注:迄今为止还没有证据表明GLP1受体促动剂调节人类的β细胞质量。
虽然说GLP1和GIP都刺激胰岛素分泌,但它们对胰腺α细胞分泌胰高血糖素的作用相反。
GIP刺激胰高血糖素分泌,而GLP1抑制分泌。
有证据表明,GLP1对胰岛素和胰高血糖素分泌的相反作用都有助于降低血浆葡萄糖。
GLP1通过激活GLP1受体介导的内在(非旁分泌)途径抑制α细胞,从而减少了胰高血糖素的分泌。
然而,其他研究结果表明,GLP1通过涉及胰岛内生长抑素的旁分泌机制间接抑制胰高血糖素的释放,并且GLP1和GIP的不同作用可以通过GIP对α细胞具有强大的直接刺激作用来解释,该作用掩盖了局部生长抑素的抑制作用。
肠促胰岛素的作用通常在2型糖尿病中受损,这种作用是糖尿病的后果而非原因。
在这种情况下,即使GIP的水平是超生理的,其作用也会减弱或缺失,而GLP1的促胰岛素作用相对保留。然而,GIP在2型糖尿病中保留刺激胰高血糖素分泌的能力,这会导致餐后高血糖。在各种研究中,GIP对胰腺β细胞的作用减弱归因于GIP受体表达减少、GIP受体下调或细胞内信号机制的受体后缺陷。
值得注意的是,2型糖尿病患者的血糖正常化提高了对GIP和GLP1的敏感性,这也许解释了为什么在双激动剂药物中在GL1中添加GIP对胰岛素分泌有额外的有益作用。
2型糖尿病是否与GLP1或GIP分泌缺陷有关尚不清楚。对2型糖尿病患者循环GIP水平的研究产生了矛盾的结果——高、低和不变的水平都有报道。相比之下,几项研究表明,2型糖尿病患者在混合餐后GLP1的分泌减少。
总的来说,2型糖尿病对肠道激素分泌的影响显然相对较小,这可能反映了这样一个事实,即:
肠内分泌细胞从隐窝干细胞中不断更新,在不利的代谢条件下,细胞损伤几乎没有时间积累。
然而,2型糖尿病对肠道运动和营养吸收的影响可能通过改变营养和胆汁酸刺激的局部递送而间接影响肠内分泌细胞的释放。
GLP1对胰岛素释放的影响比GIP大,GLP1已被开发为2型糖尿病的主要治疗靶点,并改变了2型糖尿病和肥胖患者血糖控制和体重减轻的临床治疗。
肝 脏
来自动物研究和临床试验的数据强调了GLP1和GLP1受体激动剂对肝脏代谢的多种有益作用。
GLP1受体是否在肝脏中表达有争议:
因此,在将GLP1对肝脏病理和功能的任何观察到的影响归因于肽对肝脏本身的直接影响之前,必须谨慎。
GLP1和GLP1受体激动剂对肝功能的间接作用
GLP1和GLP1受体激动剂对肝功能的间接作用是预期的,因为它们能够刺激胰岛素释放、降低胰高血糖素水平、改善葡萄糖稳态、减少胃肠道中乳糜微粒的产生以及减少热量摄入和体重。GLP1对肝脏影响的复杂性体现在它可以抑制肝脏葡萄糖产生并刺激肝脏葡萄糖摄取的发现上。20世纪90年代发表的研究将这些观察结果归因于胰岛素水平改变的间接影响,但后来的研究表明,胰岛素不是关键因素。
GLP1对肝脏脂质代谢有几种有益作用:
肝脏脂质代谢的这些改善可能至少部分归因于食物摄入的减少和随之而来的体重减轻。然而,一些研究结果表明,GLP1在调节肝脏脂质代谢方面具有额外的直接作用。
GLP1是否具有与体重减轻、胰岛素和胰高血糖素相关的肝脏作用之外的肝脏作用是一个有趣的问题。一项研究令人惊讶地表明,GLP19-36是GLP1的主要循环代谢产物,它独立于胰岛素的变化而抑制肝葡萄糖的产生,当GLP1受体被阻断时,这种作用仍然存在。这种作用的机制尚不清楚。一些证据表明,它涉及一种未鉴定的膜受体的激活,而其他研究结果表明,GLP19–36的降解产物(GLP128–36和GLP132–36)靶向PKA–β-连环蛋白–Wnt信号通路;这些降解产物可以在不涉及特定受体的情况下穿透质膜。需要进一步调查,以澄清和验证所涉及的机制。
无论GLP1对肝脏的有益作用是直接的还是间接的,GLP1受体激动剂正在被研究用于治疗非酒精性脂肪肝(NAFLD),这是肝脂代谢紊乱的最常见的慢性肝病之一。
GLP1受体激动剂:治疗非酒精性脂肪肝的希望
GLP1受体激动剂可以通过多种途径的组合来改善与非酒精性脂肪肝的发作和进展相关的基本代谢变化。
对非酒精性脂肪肝和非酒精性脂肪性肝炎(NASH)小鼠模型的研究表明,GLP1受体激动剂可降低肝脏脂肪含量,逆转脂肪变性,发挥抗炎作用,并改善纤维化的组织学严重程度。
此外,GLP1受体激动剂治疗非酒精性脂肪肝的临床试验(主要包括T2DM患者)产生了有希望的结果。几项研究表明,用GLP1受体激动剂治疗可降低肝脏脂肪含量,导致非酒精性脂肪性肝炎的组织学消退,并减缓纤维化的进展。然而,在这些临床研究中观察到体重和血红蛋白A1c水平显著降低,因此GLP1受体激动剂的作用是否独立于体重减轻和血糖控制的改善仍不清楚。
双激动剂疗法:研究非酒精性脂肪肝的新选择
双激动剂疗法,其中GLP1受体激动剂与直接抑制脂肪酸合成或靶向纤维化的第二分子结合,正在作为非酒精性脂肪肝的治疗选择进行研究;一个例子是GLP1–胰高血糖素组合肽。
其他肠内分泌激素(包括胃饥饿素和胃肠肽)对肝脏的影响尚不清楚。几项研究表明,GIP调节肝脏脂质含量,并可能参与非酒精性脂肪肝的发病机制。然而,GIP是否独立于其对胰岛素和脂肪组织的影响而直接影响肝脏中的脂质积聚(见“脂肪组织”)仍存在争议,因为GIP受体在肝脏中的表达仍未得到证实。几项动物研究表明,GIP信号传导的遗传或药理学破坏可以减少肝脏脂质积聚,但这些干预措施也可以防止饮食诱导的体重增加,因此研究结果不能证明GIP对肝脏有直接作用。
FGF19的多重代谢作用
与肠内分泌细胞衍生的肽不同,FGF19(小鼠中的FGF15)在回肠末端上皮中对胆汁酸吸收产生反应,是调节肝胆汁酸生物合成的重要抑制性反馈回路的基础。它还通过激活肝糖原合成和抑制糖异生等途径具有更广泛的代谢作用。
在不同器官中由肠道激素调节的过程
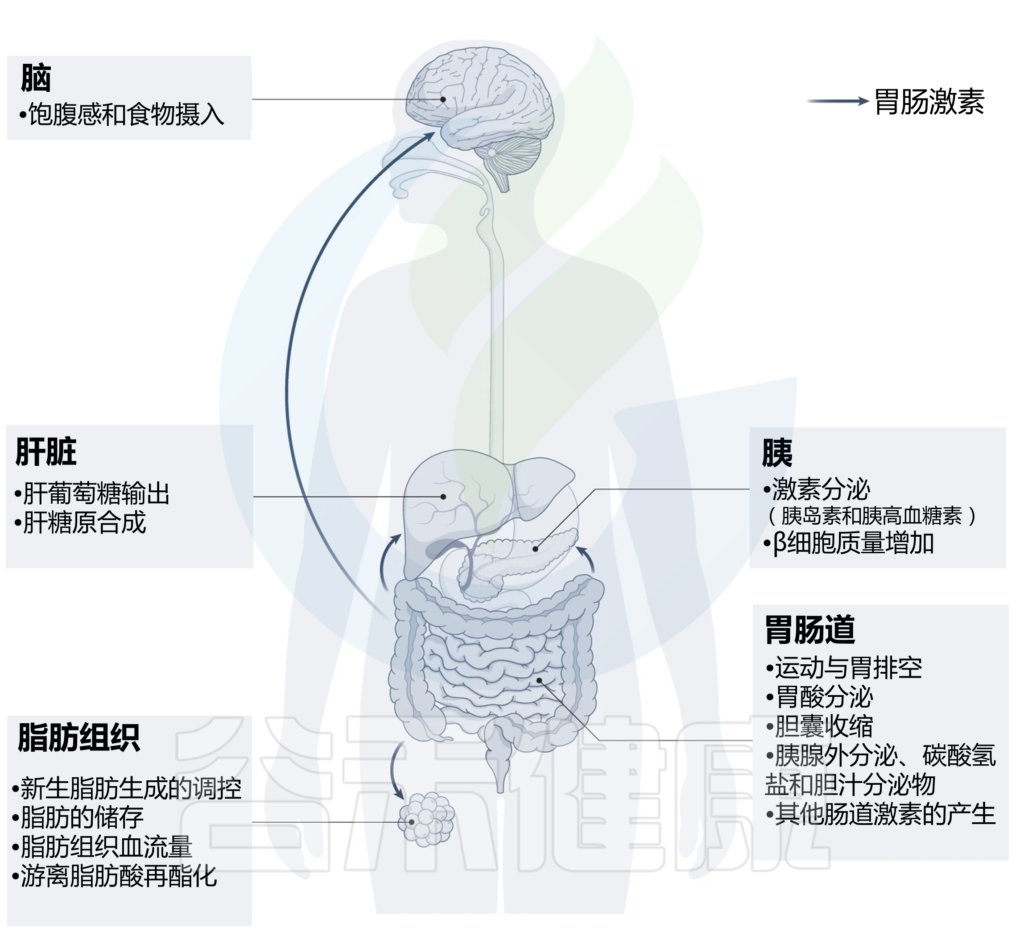
Bany Bakar R, et al., Nat Rev Gastroenterol Hepatol. 2023
脂 肪 组 织
脂肪细胞是哺乳动物的主要燃料储存室,它们的调节可以产生重要的代谢后果。在肠内分泌激素中,GIP与脂肪细胞功能的关系最为密切。
GIP与脂肪细胞功能:代谢调节的关键
几项研究表明,GIP促进白色脂肪组织(WAT)中的脂质储存,包括在大鼠中观察到,GIP通过增加脂肪组织葡萄糖摄取、从葡萄糖重新生成脂肪和将FFAs重新酯化为甘油三酯来支持脂肪组织中的脂质存储,以及在小鼠中敲除GIP受体可以预防饮食诱导的肥胖。
几项研究表明,脂肪组织中的GIP受体激活与胰岛素协同作用,增加脂质积累、葡萄糖摄取、新生脂肪生成和FFA再酯化。
脂肪细胞中的GIP受体类型:研究争议与需解决的问题
然而,这些发现需要在脂肪组织中哪些细胞类型表达GIP受体的争议中进行解释。单核转录组学表明,GIP受体不在成熟脂肪细胞中表达,而是在周细胞和间充质细胞中表达。考虑到许多脂肪细胞模型是由可以表达GIP受体的未成熟细胞在体外产生的,这些模型中的GIP受体表达是否能概括天然成熟脂肪细胞中的表达值得怀疑。
另一个必须考虑的因素是,GIP受体与Gαs偶联,Gαs下游cAMP水平升高通常与脂解而非脂肪生成有关,例如β3-肾上腺素能受体激活的脂解作用。
GIP的脂肪生成作用有益,改善脂质储存,减少异位脂肪积聚
有趣的是,GIP受体在小鼠和人类脂肪组织周细胞中表达的发现与GIP在体内显著增加脂肪组织血流量的证据一致,这一过程可以间接促进胰岛素依赖性作用,比如:清除甘油三酯、摄取葡萄糖。
无论潜在机制如何,GIP的脂肪生成作用都是有益的,因为它改善了白色脂肪组织中健康的长期脂质储存,减少了肝脏、心脏和胰腺等外周组织中的脂质溢出和异位脂肪积聚。
在2型糖尿病和肥胖的GLP1受体激动剂上添加GIP受体激动剂的代谢益处
早期的研究表明,GIP纯粹是致肥胖的,因此有利于GIP拮抗作用的翻译策略来实现减肥,而自从2018年以来发表的研究表明双重GIP受体-GLP1受体激动剂,替西帕肽在2型糖尿病患者中实现血糖控制和减轻体重方面比单独使用任一受体的激动剂更有效,该治疗改善了白色脂肪组织健康。替西帕肽治疗显著降低了肝脏脂肪含量以及内脏和皮下脂肪组织的体积,因此可能成为非酒精性脂肪肝或非酒精性脂肪性肝炎的重要治疗选择,尤其是在同时患有2型糖尿病或肥胖的患者中。
然而,对于是激动还是拮抗GIP受体,仍存在不清晰性;强有力的证据表明,任何一种方式都可以减轻肥胖和体重。
注:已经提出了几个假设来调和这些自相矛盾的观察结果,但还需要更多的研究来澄清这一领域。
尽管GIP在肠道白色脂肪组织轴中的确切作用机制仍有争议,但GIP在脂肪组织中的生理作用是明确的。在啮齿类动物中,棕色脂肪细胞组织(BAT)中GIP受体的表达已得到充分证实,给予GIP或替西帕肽可促进产热;替西帕肽也与支链氨基酸分解代谢增加有关。
然而,小鼠棕色脂肪细胞组织中GIP受体的条件性敲除并不能像全局敲除GIP受体那样防止饮食诱导的肥胖,这表明GIP对棕色脂肪细胞组织的影响在生理上并不是主要的。那主要的会是什么呢?
其他激素在肠道棕色脂肪细胞组织轴中发挥重要作用
例如,血清素抑制棕色脂肪细胞组织活性,而缺乏血清素产生所需的色氨酸羟化酶1的小鼠可以免受饮食诱导的肥胖。然而,鉴于血浆5-羟色胺是由血栓细胞缓冲的,肠道来源的5-羟色胺的内分泌作用值得怀疑,而且关于血浆5-羟色胺水平相对于用餐时间的日变化的报道一直不一致。相反,分泌素激活棕色脂肪细胞组织,这种激活与食欲的改变有关。
脑
肠-脑轴是胃肠道和大脑之间传递信息的渠道,包括神经、激素和营养信号;下丘脑核团和后脑是检测和整合这些外周信号的主要中心。肠-脑轴的紊乱已在多种疾病中被发现,包括炎症和功能性胃肠道疾病以及饮食障碍。
肠道激素在大脑的作用可以解释一些饮食失调和进食障碍的发生?
肠道向大脑发送关于饮食营养摄入状态的实时信号,肠道激素是肠道和大脑之间复杂相互作用的主要介质,这些相互作用是饱腹感(两餐之间不饿的感觉)和饱足感(一餐结束时吃得足够的感觉)的基础。许多肠道肽及其受体也在中枢神经系统中表达,并调节中枢回路,包括与食欲有关的回路。这种在中枢神经系统中的表达可能会使涉及肠道激素受体的药理学或全局遗传操作的研究的解释复杂化,因为其结果可能归因于对中枢神经系统回路的影响,而与肠道分泌的激素无关。
传入迷走神经神经元的细胞体位于结节神经节,并投射到孤束核(NTS),是介导肠道激素对食物摄入影响的主要途径。肠内分泌细胞通过局部升高的肠道激素水平以及可能的突触连接与传入迷走神经和内脏神经末梢进行交流。到达心室周围器官(例如,脊后和正中隆起区)和邻近细胞核(分别为下丘脑的NTS和弓状核)的循环肠道激素与这些信号汇合,以调节进食行为。几种肠道激素对大脑有着重要而独特的影响,下文将对此进行讨论。
CCK是在食欲调节中发挥作用的肠道激素。餐后释放CCK通过与CCK1受体结合激活传入迷走神经,向大脑提供反馈信号,从而减少进食。然而,根据小鼠研究,反馈信号的性质仍不清楚,因为动物可能因各种原因停止进食。在人类中,输注生理浓度的CCK可以抑制能量摄入并增加幽门收缩,而不会引起恶心;食物摄入减少被认为是幽门收缩的结果。在小鼠中,CCK通过传入迷走神经发出信号,激活中枢奖赏通路,并增加脂肪和糖的消耗。
CCK似乎会触发相对短暂的饱腹信号,PYY与餐后食物摄入的长期抑制有关。已知PYY的两种不同形式:PYY1-36和PYY3-36。PYY1–36通过二肽基肽酶IV转化为PYY3–36。两种形式的PYY都参与调节食物摄入,但它们表现出不同的特性和受体偏好。
GLP1和GLP1受体激动剂可以抑制食欲
GLP1受体激动剂主要通过作用于中枢神经系统内的GLP1受体来抑制食物摄入,但肠道来源的GLP1在大脑中是否达到足够高的水平以在生理条件下激活GLP1受体仍不清楚。
几项研究表明,迷走神经传入神经元的末端暴露于胃肠道中高水平的局部生物活性GLP1,介导内源性肠道衍生的GLP1的饱腹作用。
肠道来源GLP1控制食物摄入——不是主要的
在小鼠中,迷走神经神经元表达GLP1受体,而且响应机械刺激,外源性GLP1也可以增强由胃和肠道拉伸引起的饱腹感。然而,敲除肠道来源的GLP1不会影响小鼠的食物摄入或体重,这表明肠道来源的GLP1对控制食物摄入并不重要。
脑源性GLP1控制食物摄入——明显作用
虽然去除后脑GLP1表达也不会影响自由进食行为,但当化学遗传学激活时,它确实削弱了表达GLP1的神经元减少进食的能力,证明了脑源性GLP1在控制食物摄入方面的明显作用。
GLP1受体与氧调素的互动:探索胰岛素类似物的潜力
GLP1受体也被氧调素激活,氧调素是一种由肠道L细胞分泌的37个氨基酸的略微延伸的胰高血糖素类似物,在胰岛素受体处也具有活性。氧调节蛋白在体重管理和葡萄糖稳态方面具有有益作用,这一作用正开始通过使用GLP1受体和胰岛素受体双激动剂而得到利用;鉴于目前还没有发现特异性的尿囊素受体,这些作用被认为是由其对这两种受体的双重作用介导的。
减肥手术后,餐后GLP1、氧调节蛋白和PYY水平显著升高;循环水平足以对食物摄入和葡萄糖耐受性产生有益影响。然而,在小鼠中,Roux-en-Y胃旁路术的减肥作用在缺乏神经肽Y受体2型和GLP1受体的动物中仍然存在,这表明这些激素作用之外的机制也有助于减肥手术的益处。
神经降压素也与减肥手术的效果有关
神经降压素(Neurotensin)是一种13个氨基酸的肽,主要存在于大脑和胃肠道中。在啮齿类动物中,神经降压素直接进入大脑时会减少食物摄入,这种作用主要由NTSR1(神经降压素受体1)介导。
在一些研究中,Ntsr1敲除小鼠食欲增加,体重增加,对神经降压素的食欲抑制作用没有反应。
然而,在其他研究中,Ntsr1敲除降低了进食量并增加了活性,但对体重或组成没有影响。
脑源性神经降压素抑制食欲,外周的神经降压素对代谢的影响尚不清楚
一些研究表明,神经降压素的外周给药会导致啮齿动物的食物摄入量暂时减少。一些证据表明,这种影响可能归因于肠道中脂肪吸收的减少,但减少进食的确切机制尚不清楚。
另一种与减肥手术结果有关的激素是分泌素,因为它在手术后的餐后水平也会升高,尽管它在这些患者中的生理重要性尚不清楚。
来自非肠道内分泌细胞的肠道激素(GDF15、FGF19、uroguanylin)似乎也通过中枢途径调节食欲的作用。
最有力的证据是GDF15,它靶向后脑中有限的神经元群体,并导致由一系列毒性刺激(如化疗剂)诱导的恶心,这种恶心发生在癌症恶病质和妊娠剧吐中。
GDF15反应神经元表达GDF15受体,即GFRAL,并与脑干GLP1受体和CCK回路密切相关。有趣的是,GIP可以通过表达GIP受体的海马后区GABA能神经元抑制GDF15(和其他肠道激素,包括GLP1和PYY)引起的恶心。
Ghrelin:非凡食欲刺激者
胃饥饿素(Ghrelin)是唯一被广泛接受的刺激食物摄入的肠道激素。根据这种生物活性,血液中胃饥饿素的水平在用餐前升高,在进食后下降。
其生物学效应是通过生长激素促分泌受体(GHSR)介导的,GHSR存在于全身,包括胰腺、迷走神经传入神经元和大脑。几项研究表明,Ghrelin通过在下丘脑弓状核的中央和迷走神经传入神经的外周发挥其食欲产生活性。
然而,后一种机制是有争议的,一些研究表明,胃饥饿素的食欲产生作用不需要迷走神经传入信号。一些研究表明,胃动素也能促进食物摄入,但胃动素的生理学特征很差,因为它不存在于啮齿类动物中。
注:胃动素是一种与胃饥饿素有关的肽,由十二指肠中重叠的肠内分泌细胞群体产生。
INSL5:微弱但可能的食欲促进者
INSL5由远端结肠和直肠L细胞的一个子集分泌,对小鼠的研究表明它它具有食欲促进作用。它通过RXFP4(松弛素-胰岛素样家族肽受体4)发挥作用,该受体在大脑的关键进食中心表达。
INSL5的增食欲作用远低于胃饥饿素,这在一些研究中没有观察到,并且可能在体内被PYY所淹没,PYY是从相同的肠内分泌细胞平行分泌的。这些观察结果让人怀疑外周释放的INSL5是否在控制食物摄入方面具有重要的生理作用。然而,一项研究已经确定了腹内侧下丘脑中表达RXFP4的神经元在调节食物摄入和偏好方面的潜在作用。
胃肠道和身体其他活跃的组织之间有着双向的信息传递,而这个信息传递的改变与多种代谢性疾病有关。目前对于胃肠激素如何与这个复杂的调节网络相互作用的理解正在逐渐加深,但仍需要进一步的研究来完全揭开这个谜团。
肠道菌群的异常变化可能影响肠道激素的正常分泌,从而导致代谢相关疾病发生。因此,肠道菌群的稳定和平衡对于保持肠道激素的正常分泌和代谢健康非常重要。这些发现为我们深入了解肠道菌群和肠道激素的相互作用、开发相关治疗方法提供了新的方向。通过调节肠道菌群的结构和功能,我们可能能够干预肠道激素的分泌,从而实现对代谢相关疾病的治疗和预防。
通过开发创新、微创且耐受性良好的治疗方法,我们可以利用胃肠激素受体的转化潜能。GLP-1双重和三重激动剂疗法的临床试验已经证明了这种方法的价值。这个领域研究进展迅速,很可能会进一步细化现有的认识,解决关键的机制问题。期待揭示出胃肠激素控制的多个信号通路,可以成功地用于治疗2型糖尿病和肥胖,并且将这些基于胃肠激素的药物在其他代谢性疾病中推广应用。
主要参考文献:
Bany Bakar R, Reimann F, Gribble FM. The intestine as an endocrine organ and the role of gut hormones in metabolic regulation. Nat Rev Gastroenterol Hepatol. 2023 Aug 25.

谷禾健康

健康的身体,匀称的身材,充沛的精力,想必是很多人所向往的,有一个物质在其中起到了重要的作用,它就是亮氨酸。
亮氨酸——我们有时会在一些报告和补剂中看见这个名字,那么它究竟是什么?对我们人体又有什么作用?
亮氨酸,又称白氨酸,是支链氨基酸之一(支链氨基酸是人体合成蛋白质的重要物质)。亮氨酸因其促进肌肉生长和提高运动表现的能力而受到运动员的喜爱。不仅如此,它还可以减缓老年人的肌肉退化,并有助于控制血糖。
近年来,亮氨酸引起了相当大的关注,部分原因是它对肌肉合成的促进作用以及对葡萄糖耐量、胰岛素敏感性的有益影响。此外亮氨酸还可以影响体内和体外的脂质和能量代谢,从而有利于减少肥胖。
亮氨酸可以调节线粒体功能障碍,有人认为,通过增加亮氨酸的存在率和活性来增加能量消耗和去除有毒脂质可能是治疗肥胖及其后续疾病(如胰岛素抵抗、糖尿病和心血管疾病)的一种有前景的治疗策略。
亮氨酸与肠道微生物之间也存在着密切的关系。首先,亮氨酸可以作为一种营养物质提供给肠道菌群,促进有益菌的生长和繁殖。其次,亮氨酸会在肠道中被一些细菌代谢为有益的代谢产物,如短链脂肪酸,影响肠道菌群的活性及人体健康。此外,肠道菌群的失衡也会影响亮氨酸的代谢和利用。
本文总结了亮氨酸的生物学功能及其在哺乳动物能量代谢中的作用,特别是促进肌肉生长和脂质分解,并为代谢疾病、肌肉减少症、神经系统疾病的治疗和食品行业及畜牧业的科学应用提供一定参考。
亮氨酸,又称白氨酸,化学名称为α-氨基异己酸,于1819年首次从奶酪中分离出来。后来从肌肉和羊毛的酸水解产物中将其结晶出来,并将其命名为Leu(亮氨酸)。
✦亮氨酸一般分为L-亮氨酸和D-亮氨酸
L-亮氨酸是氨基酸的天然形式,存在于体内的蛋白质中,是用作补充剂的主要形式。
注:人体中的亮氨酸基本都是L-亮氨酸。
D-亮氨酸是L-亮氨酸的镜像,它是在实验室中产生的,也可用作补充剂。一般用于工业。
★ 亮氨酸在代谢中起重要作用
亮氨酸具有很强的氧化能力,其主要生理功能包括调节蛋白质代谢和氧化能量的供应。这种能量供应可用于特殊生理时期,如饥饿、哺乳和运动,以及调节免疫功能和脂质代谢。
亮氨酸也可以直接分解成乙酰辅酶A,使其成为体内最重要的生酮氨基酸之一。(在葡萄糖不能供能的情况下动用脂肪产生酮体),可以通过提高血液胰岛素水平直接或间接促进蛋白质的合成,并且可以抑制骨骼肌蛋白质的分解。
注意:亮氨酸通常被认为比其他支链氨基酸更可取,因为它分解和吸收更快,使其比其他类型的氨基酸更容易使用。
★ 亮氨酸是一种必需氨基酸,但广泛存在于食物中
作为一种必需氨基酸和支链氨基酸,亮氨酸广泛存在于动物蛋白(牛肉、马肉、虾米)和乳制品(全脂奶粉)以及豆类中。一些坚果如杏仁和腰果中的含量也较高。
必需氨基酸——人体所必需的氨基酸,同时人体自身不能合成,或合成速度远不足以满足人体的需要,从而必需由食物提供的氨基酸。
支链氨基酸——指具有支链侧链结构的氨基酸,支链氨基酸是蛋白质中三种常见的氨基酸,另两种支链氨基酸是缬氨酸和异亮氨酸。
亮氨酸是一种氨基酸,它在人体中具有多种重要功能。当亮氨酸偏低或偏高时,可能都会对身体产生一些不良影响。
↓↓ 亮氨酸偏低的后果:
蛋白质合成受阻
亮氨酸是蛋白质合成的重要组成部分,如果亮氨酸偏低,可能会影响身体内蛋白质的合成,导致肌肉组织的损失和功能受损。
免疫功能下降
亮氨酸对免疫系统的正常功能至关重要。亮氨酸偏低可能会导致免疫功能下降,增加感染的风险。
能量供应不足
亮氨酸在体内可以被转化为能量,供给身体使用。亮氨酸偏低可能导致能量供应不足,影响身体的正常代谢和功能。
引起疲劳、头痛
膳食摄入亮氨酸不足导致身体缺乏时,还会引起一系列症状,如疲劳和头痛等。有些情况下,还会导致晕眩和易怒。
▸ 容易亮氨酸偏低的人群
营养不良:饮食中缺乏蛋白质或亮氨酸含量不足,可能导致亮氨酸水平下降。
肝脏疾病:肝脏是合成蛋白质的重要器官,如果肝脏功能受损,可能会影响蛋白质合成和亮氨酸的产生。
消化系统问题:某些肠道疾病或吸收问题可能会影响亮氨酸的吸收和利用。
慢性疾病:某些慢性疾病,如肾脏疾病或肠道疾病,可能会导致亮氨酸水平下降。
其他因素:某些药物、代谢异常或遗传性疾病也可能导致亮氨酸水平降低。
↑↑ 亮氨酸偏高的后果:
氨基酸失衡
亮氨酸偏高可能导致其他氨基酸的相对不足,破坏氨基酸的平衡,影响蛋白质合成和身体的正常功能。
氮代谢紊乱
亮氨酸偏高会导致氮代谢紊乱,增加肾脏的负担,对肾功能产生不良影响。
毒性效应
亮氨酸在高浓度下可能对神经系统产生毒性效应,引起神经系统的损伤。引起谵妄和神经损害,并可危及生命。
引发一些疾病
亮氨酸浓度过高会引起亮氨酸血瘀、痛风、枫糖尿病等疾病。
▸ 容易亮氨酸偏高的人群
•遗传性疾病:某些遗传性疾病会导致亮氨酸的代谢异常,这些疾病通常是由于亮氨酸代谢酶的缺陷引起的。例如枫糖尿病或遗传性亮氨酸血症。
•肝脏疾病:肝脏是身体中重要的代谢器官,如果肝脏功能受损,可能会导致亮氨酸等物质在体内积累过多。
•营养补充过量:过量摄入亮氨酸的营养补充剂或蛋白质粉末可能会导致亮氨酸水平升高。
因此,在使用任何营养补充剂之前,最好咨询医生或专业营养师的建议。
亮氨酸的偏高或偏低通常是由于饮食不平衡、基因突变、代谢紊乱等原因引起的。值得注意的是,亮氨酸的需求量会因个体差异和特定状况而有所不同。例如,运动员和肌肉量较大的人可能需要更多的亮氨酸来支持肌肉生长和修复。谷禾的健康检测报告中也能评估体内亮氨酸的含量,并给出个性化的建议。
前面讲述了许多亮氨酸的重要性,那么在日常生活中我们应该如何补充它呢?亮氨酸作为必需氨基酸和支链氨基酸,通常被认为比其他支链氨基酸更可取,因为它的分解和吸收速度更快,比其他类型(如异亮氨酸和缬氨酸)更容易使用。
亮氨酸也可以直接分解成乙酰辅酶A,使其成为体内最重要的生酮氨基酸之一。虽然大多数其他氨基酸被转化为葡萄糖,但由亮氨酸形成的乙酰辅酶A可用于制造酮体。
运动与亮氨酸的摄入
✦ 运动后亮氨酸摄入的持续时间和效果
运动后的合成窗口是运动后身体特别容易吸收营养的时期,持续30分钟到两小时。在这个时间内摄取亮氨酸可以增加肌肉蛋白质合成率20-30%,具体取决于个体差异和运动的具体情况。
✦ 运动前摄入亮氨酸的潜在副作用
运动前摄入亮氨酸有其缺点。由此产生的胰岛素激增可能会导致运动中的血糖水平下降,导致头晕甚至昏厥。
此外,运动前的高亮氨酸水平可能会重新引导血流到消化系统,减少流向正在使用的肌肉的血流。
不同来源的亮氨酸含量
✦ 不同蛋白质来源,亮氨酸含量不同
你应该吃什么来获得足够的蛋白质和亮氨酸?这取决于你选择的蛋白质来源。亮氨酸作为必需氨基酸,只能从食物或补充剂中获得。
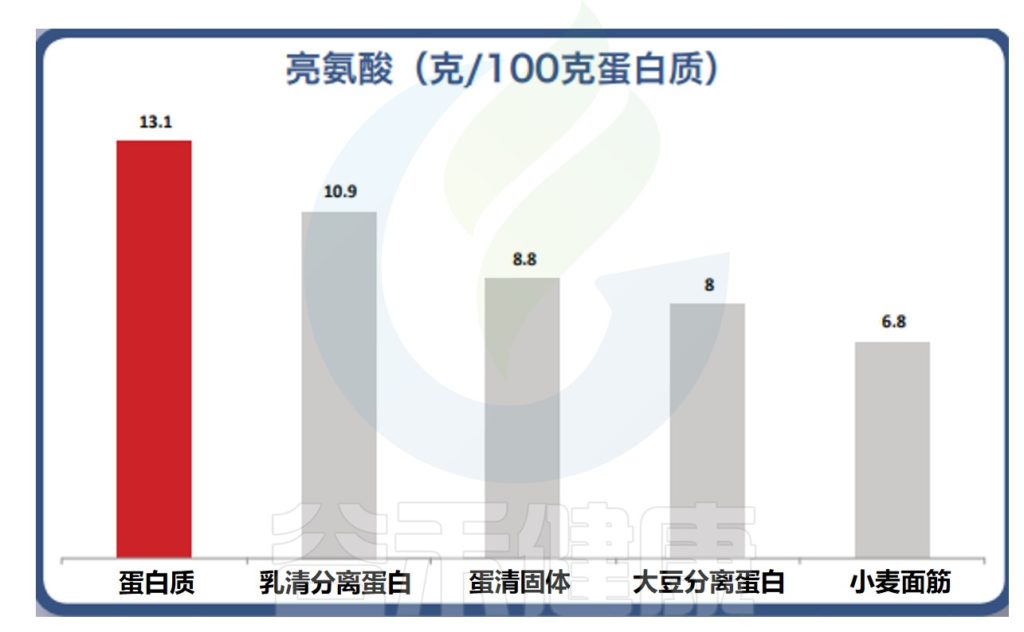
✦动物性蛋白的亮氨酸含量更高
植物性蛋白缺乏一种或多种必需氨基酸,对肌肉蛋白合成的刺激效果较差,而乳制品和动物源蛋白具有较高的亮氨酸百分比。
最近的一项研究,受试者每天摄入19克分离乳清蛋白或26克分离大豆蛋白——两者都提供大约1.8克亮氨酸。
谷禾在此整理了一些常见食物中的亮氨酸含量表,以供大家参考(mg/100g可食用部分)
★ 摄入建议
世界卫生组织建议成年人每天至少需要补充39mg/kg亮氨酸,每餐建议摄入1-3g亮氨酸。婴儿、发育期的青少年以及运动员可以根据需求适量多补充一些亮氨酸。一般来说正常饮食的人亮氨酸不会缺乏。
需要注意的是,最近有研究指出,老年人群亮氨酸推荐量应该更高,因为与年轻人相比,老年人的亮氨酸代谢存在差异:老年人内脏中亮氨酸的滞留量是年轻人的两倍,并且亮氨酸对肌肉蛋白质合成能力的刺激作用随着年龄的增长而降低。
✦ 禁忌
补充亮氨酸会降低血糖水平,从而导致患有以下疾病的人血糖水平不健康:
•低血糖症(低血糖人群)
•糖尿病,服用降血糖药物的患者
关于亮氨酸的拓展知识和认知
亮氨酸的分解代谢是生理衰老的保守调节因子,参与多种生理和病理过程。
•亮氨酸的分解代谢过程
哺乳动物中亮氨酸的分解很复杂,涉及两个过程。最初,摄入的亮氨酸被支链氨基酸转移酶(BCAT)催化生成α-酮异己酸(KIC)和β-羟基-β-甲基丁酸(HMB)的前体;这种转氨作用是快速且双向的。
随后,KIC进入两种代谢途径之一,产生异戊酰辅酶A(占亮氨酸代谢的90-95%)或HMB(占亮氨酸代谢的5-10%)。
后者中,α-酮异己酸通过α-酮异己酸双加氧酶不可逆地代谢为HMB;前者中,α-酮异己酸通过支链α-酮酸脱氢酶(BCKD)催化的一系列反应,发生不可逆且限速的氧化脱羧。
最终,亮氨酸转化为乙酰乙酸和乙酰辅酶A,这是三羧酸循环的中间体。过量的α-酮异己酸可以被释放到循环中,并被肝脏和脂肪组织等其他器官吸收,然后在那里被重新合成为支链氨基酸或氧化生成腺苷三磷酸(ATP)。
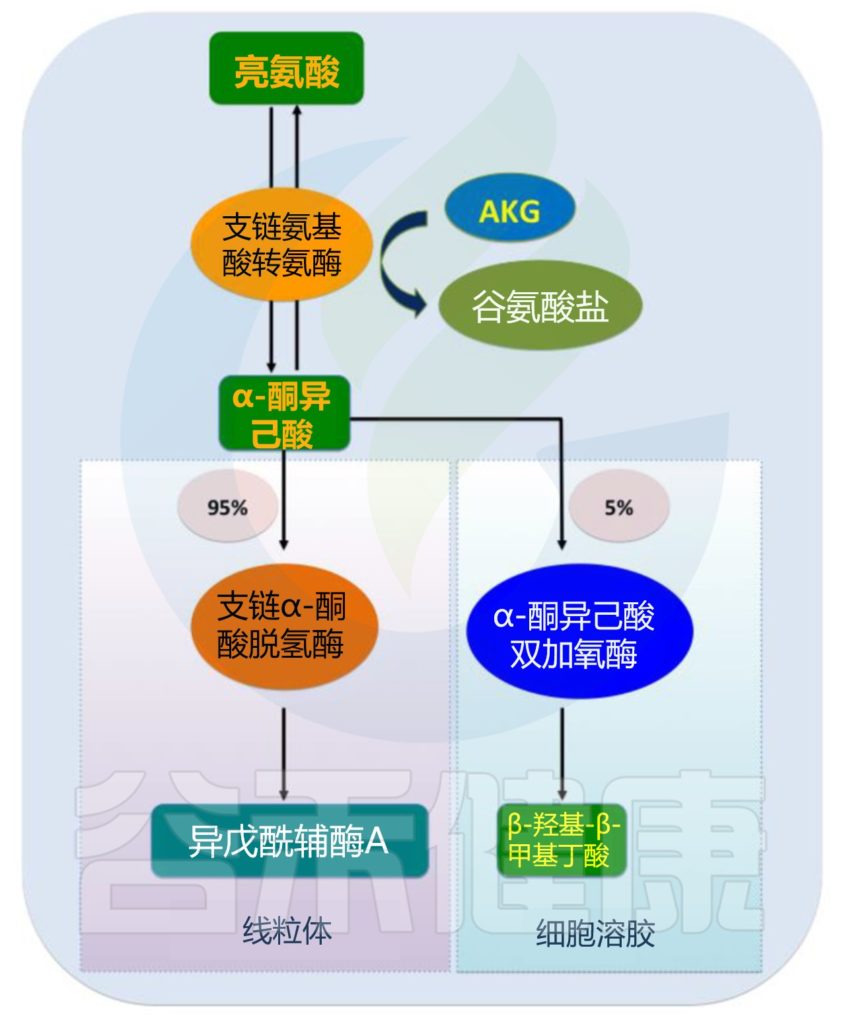
Zhang L,et al.Nutrients.2020
★ 亮氨酸的代谢物可以影响能量稳态调节
亮氨酸及其代谢产物被假设为能量稳态的调节信号。研究表明,亮氨酸代谢产物而不是亮氨酸本身可能是mTOR激活的信号。
mTOR——哺乳动物雷帕霉素靶蛋白,是细胞生长和增殖的重要调节因子。
除了亮氨酸,β-羟基-β-甲基丁酸和α-酮异己酸是沉默信息转录调节因子1(SIRT1)酶的直接激活剂。很明显,亮氨酸在将能量从脂肪组织分配到骨骼肌方面发挥着关键作用,导致脂肪细胞中的能量储存减少,肌肉中脂肪酸的利用增加。
亮氨酸的代谢物对能量调节的作用
这里介绍两种:
α-酮异己酸(KIC)、β-羟基-β-甲基丁酸(HMB)
α-酮异己酸(KIC)
充分考虑现有文献表明,KIC在激活mTOR信号传导和SIRT1方面比亮氨酸更有效。KIC和亮氨酸均抑制脂肪细胞中的脂质合成代谢,同时促进脂肪酸氧化 (FAO)。
此外,KIC处理增加了培养的C2C12肌管中支链氨基酸的氧化。KIC通过抑制BCKD激酶增加骨骼肌中的完整脂肪酸氧化,导致BCKD复合物的强烈激活和通过支链氨基酸氧化途径的流量增加。
游离脂肪酸氧化的增加会降低葡萄糖的利用率,而增加肌肉质量可以有效增强脂肪的氧化。然而,值得注意的是,KIC可能是一把双刃剑,在促进生长的同时,通过下调腺苷5′-单磷酸激活蛋白激酶 (AMPK) 的磷酸化来增加脂肪酸合成,从而导致负面影响脂肪组织中脂质代谢。
β-羟基-β-甲基丁酸(HMB)
体外和体内证据表明,亮氨酸向HMB(β-羟基-β-甲基丁酸) 的内源转化效率约为5-10%。尽管如此,HMB作为膳食补充剂的影响一直是近期脂质代谢研究的焦点。
HMB是一种有趣的运动补充剂。在人体试验中,耐力训练中摄入HMB对减少脂肪量具有有利作用。由于运动员试图维持一定的体重(主要是通过降低脂肪组织的量),因此HMB供应可能是他们的合适选择,可以对他们的身体表现产生积极影响。
在一项特别有趣的研究中,饮食诱导的肥胖小鼠接受低剂量(2g/kg饮食)或高剂量(10g/kg饮食)HMB治疗6周,导致脂肪SIRT1活性增加,肌肉葡萄糖摄取增加和棕榈酸酯的摄取、胰岛素敏感性以及炎症应激生物标志物的改善和肥胖的减少。
膳食中补充HMB可以调节脂肪组织功能,包括脂肪酸和脂肪分解,同时增加血清脂联素浓度。这些效应可能部分由AMPKα–mTOR通路介导,并与线粒体生物合成、AMPK–SIRT1–增殖物激活受体γ共激活因子-1α (PGC-1α) 轴和肌因子相关。
值得注意的是,HMB在调节线粒体功能方面也发挥着关键作用,线粒体功能与许多疾病有关,如衰老、神经退行性疾病、肥胖、糖尿病和心血管疾病。
用一定剂量的HMB(50mM) 处理肌管24小时显著增加线粒体质量、呼吸能力和生物发生,并且优于用亮氨酸处理观察到的效果。
综上所述,这些结果表明β-羟基-β-甲基丁酸(HMB)可能通过脂肪组织中的AMPKα-SIRT1-PGC-1α轴调节线粒体生物合成和脂肪酸氧化。
亮氨酸可降低脂肪酸转运与合成相关蛋白的活性,抑制脂肪酸的合成。可调控相关信号因子的表达和刺激胰高血糖素样肽的分泌,从而促进脂肪分解。
•亮氨酸利于脂肪分解,并减少肥胖
据报道,亮氨酸可以抑制脂肪生成,促进脂肪分解和脂肪酸合成,并通过mTOR信号通路显著增加脂肪细胞中的瘦素分泌,有利于减少肥胖。
在脂肪细胞分化过程中增加亮氨酸会降低脂滴周围的脂滴涂层蛋白水平,提高激素敏感脂肪酶的磷酸化水平,并促进脂肪分解。
•亮氨酸可以降低糖尿病风险
膳食亮氨酸可降低高脂肪饮食引起的高血糖和高胆固醇,降低体内脂肪和脂肪产生率,并增加胰岛素敏感性。
研究发现补充亮氨酸可明显改善糖耐量,并呈良好的剂量-效应关系。同时,亮氨酸可以抑制摄入淀粉后的血糖上升,能有效抑制餐后血糖的升高,其机制与促进胃肠道激素胰高血糖素样肽-1(GLP-1)密切相关。
GLP-1具有调节胰岛素释放和糖代谢的作用。当补充亮氨酸后,大量葡萄糖以糖原形式贮存在肝脏和肌肉中,有效降低血中葡萄糖浓度。对治疗头晕有作用。1型和2型糖尿病患者都可以从富含亮氨酸的饮食获得好处,这可以尽量减少碳水化合物摄入。
•亮氨酸可以降低心血管疾病风险
此外,研究人员还总结说,人类饮食中的亮氨酸代谢物会导致甘油三酯和低密度脂蛋白胆固醇降低,从而改善心血管功能。
在运动员中,补充β-羟基-β-甲基丁酸(HMB)进行4周的阻力训练可显著降低心血管危险因素,如低密度脂蛋白或总胆固醇和甘油三酯。
最近,一些研究人员强调,亮氨酸氧化可能是mTOR激活所必需的,mTOR是一种胞质丝氨酸/苏氨酸蛋白激酶,似乎介导脂肪酸氧化,因此亮氨酸可能通过KIC或HMB调节脂肪组织的代谢,充当营养物质传感器。
脂肪组织的脂肪酸氧化能量供应主要用于肌肉组织中蛋白质的周转,这也是亮氨酸减少脂肪沉积、减轻体重的原因之一,但这些现象背后的确切机制还需要进一步研究。
膳食亮氨酸对脂肪组织脂质代谢的影响
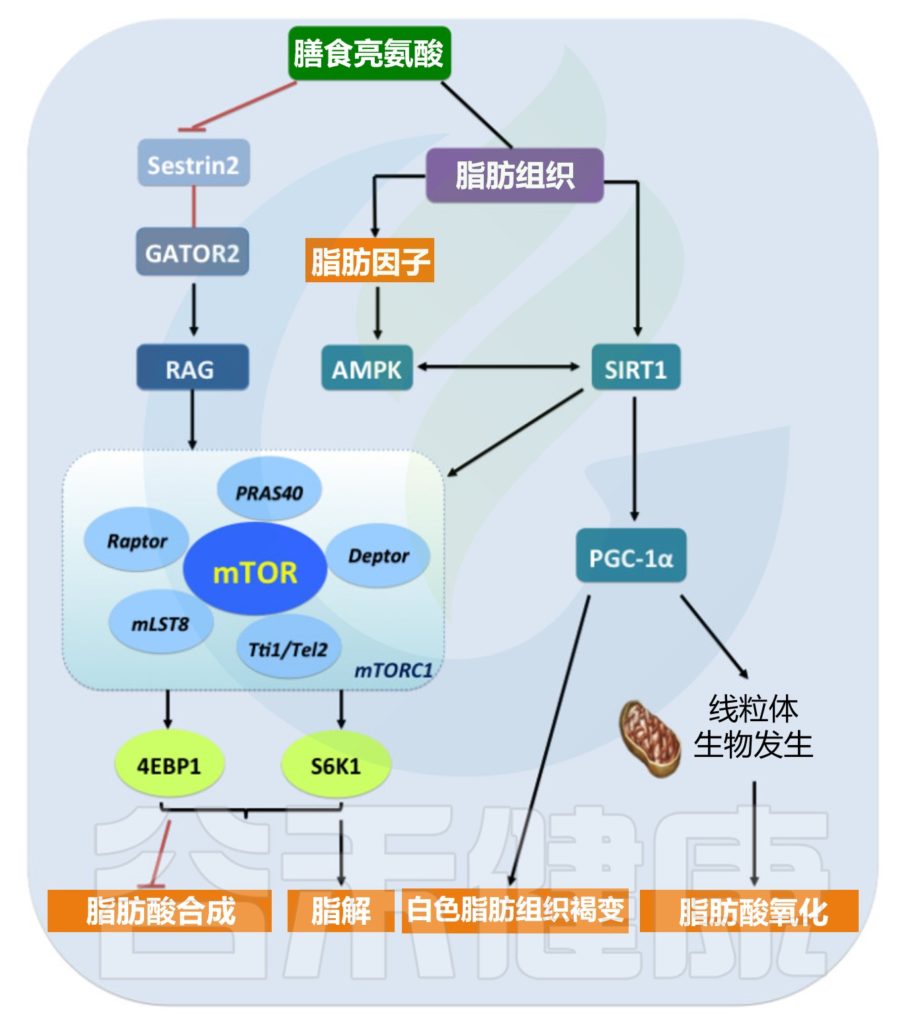
Zhang L,et al.Nutrients.2020
亮氨酸可能通过SIRT1-AMPK-PGC-1α轴促进白色脂肪组织 (WAT) 中的褐变和线粒体生物合成。
•亮氨酸通过促进脂肪组织褐变改善代谢健康
脂肪组织通过白色脂肪细胞的能量储存和褐色脂肪细胞的能量消耗在调节全身能量代谢中发挥重要作用。事实上,过量白色脂肪组织的积累会对代谢健康产生有害影响,而褐色脂肪组织的激活有助于平衡血糖水平并增加能量消耗,从而对肥胖、胰岛素抵抗和高脂血症产生有益影响。
已知亮氨酸对于褐色脂肪细胞分化是必不可少的。因此,在刺激白色脂肪组织中白色脂肪细胞的发育时,“褐变”可能会减少白色脂肪组织的不利影响,并可能有助于改善代谢健康。
•亮氨酸可以通过肠道微生物影响脂肪组织
最近,肠道微生物群已被证明可以调节白色脂肪组织的褐变和褐色脂肪组织的活性;该活性可以由亮氨酸调节。亮氨酸补充已被证明可诱导白色脂肪组织中解偶联蛋白1(UCP-1)(一种棕色脂肪特异性基因)的mRNA表达增加近四倍。
越来越多的证据表明线粒体可能在调节脂肪细胞脂质代谢中发挥关键作用。具体而言,线粒体是底物氧化和ATP生成所必需的,ATP为细胞功能提供能量。此外,脂肪组织中核因子‑红细胞2相关因子2过表达诱导的线粒体丰度增加,增加了脂联素的合成,并已被证明能刺激脂肪酸氧化。
注:核因子‑红细胞2相关因子2是细胞内调节抗炎、抗凋亡和抗氧化基因表达的重要分子。
•亮氨酸调节脂肪细胞的信号通路比其他氨基酸更有效
在脂肪组织中,mTOR通路似乎在前脂肪细胞的分化、脂肪组织形态发生、肥大生长和瘦素分泌中发挥重要作用。新鲜分离的脂肪细胞含有一个亮氨酸刺激的识别位点,该位点与mTOR信号传导耦合,并调节哺乳动物生理学其他方面的脂质代谢,包括饱腹感、胰岛素分泌和线粒体生物发生。
这种活性在很大程度上是由于mTORC1蛋白激酶(一种主要生长控制器)的激活,它由亮氨酸传感器Sestrin2调节,一种抑制mTORC1信号传导的相互作用蛋白。事实上,亮氨酸在体外和体内均可调节脂肪细胞中的mTOR信号通路,并且比其他氨基酸更有效。
•亮氨酸促进线粒体生物合成从而调节脂质代谢
此外,mTOR通过以不依赖于蛋白激酶B的方式控制线粒体的氧化功能来平衡能量代谢。BCAT激活mTOR 信号通路,促进线粒体生物发生和ATP产生,并通过调节相关基因的表达来防御氧化应激。也就是说,亮氨酸促进线粒体生物合成,从而调节脂质代谢。
线粒体参与亮氨酸调节的能量代谢
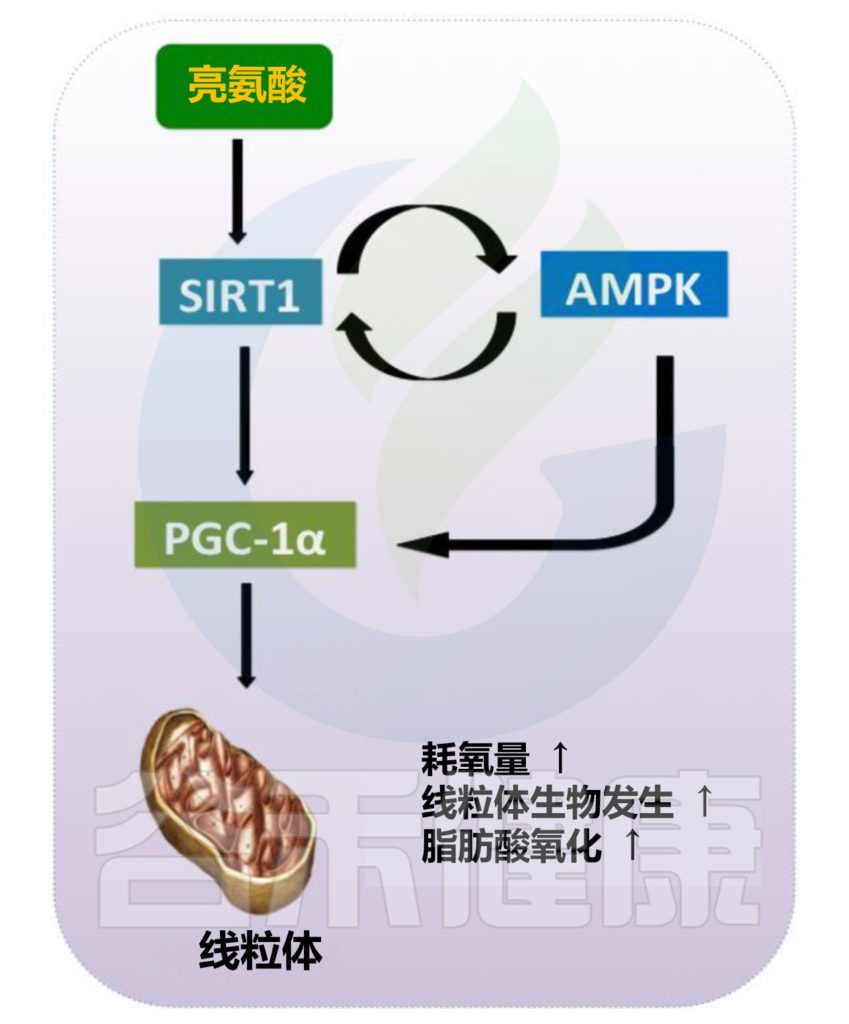
Zhang L,et al.Nutrients.2020
亮氨酸能够刺激脂肪细胞中雷帕霉素敏感的4EBP1磷酸化。简而言之,亮氨酸通过调节mTOR信号传导,调节脂肪细胞组织成组织样结构,并从脂肪组织合成/分泌瘦素。
综上所述,亮氨酸通过调节SIRT1和mTOR信号通路在稳定能量和代谢方面发挥重要作用,从而促进线粒体生物发生并调节脂质代谢。
骨骼肌是利用葡萄糖和脂肪酸的主要部位,也是导致肥胖和2型糖尿病中胰岛素抵抗的主要组织之一。骨骼肌通过清除血清游离脂肪酸、全身脂肪酸和脂质利用在能量稳态中发挥着至关重要的作用。
人体用氨基酸来制造肌肉。这个过程被称为肌肉蛋白合成(MPS),对于修复由运动、受伤和衰老引起的身体压力造成的肌肉纤维至关重要。
亮氨酸是蛋白质合成过程中的必需氨基酸之一,但在特殊生理时期(饥饿、泌乳、应激和运动等)能作为能量来源。
由于蛋白质合成和蛋白质降解不断发生,肌肉蛋白质处于持续更新状态。“合成代谢状态”是指肌肉蛋白质的净增加。相反,“分解代谢状态”是指肌肉蛋白的净损失。
肌肉蛋白由二十种氨基酸组成,所有这些氨基酸都是合成新肌肉蛋白所必需的。其中九种氨基酸必须通过膳食蛋白质来源获得。支链氨基酸是九种必需氨基酸中的三种,对于肌肉蛋白质代谢非常重要。特别是亮氨酸被认为是蛋白质代谢的重要介质。
骨骼肌中支链氨基酸的分解代谢
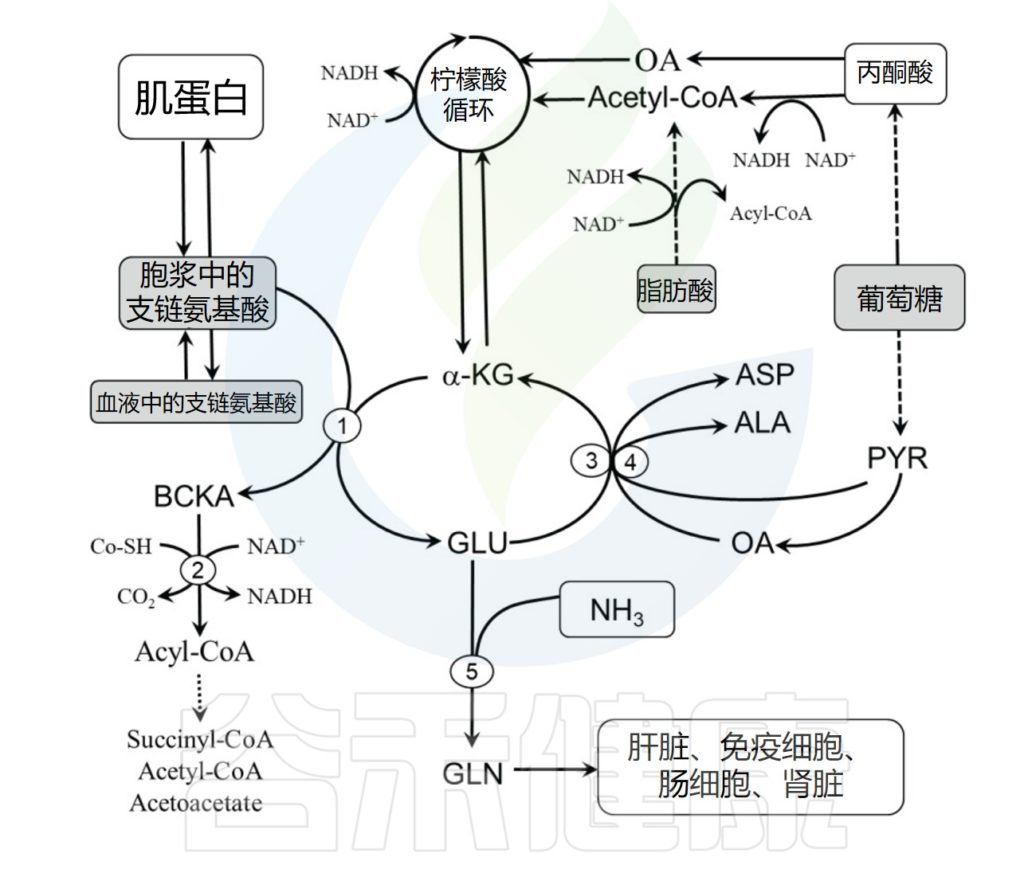
Holeček M.Physiol Res.2021
•亮氨酸通过刺激肌肉蛋白合成来提高运动表现
亮氨酸是唯一能够通过刺激肌肉蛋白合成来提高运动表现的。研究发现,亮氨酸向胰岛素发出信号,最终导致促进蛋白质合成和防止肌肉分解的途径得到更大的激活。
研究发现,年轻人和老年人群的急性亮氨酸补充可以增加肌肉蛋白质的合成。
•运动员应该补充足够的亮氨酸
研究还认为,更多的亮氨酸摄入可以降低体脂。这意味着运动员应该补充足够的蛋白,其中含有高亮氨酸含量,以优化肌肉蛋白合成。
亮氦酸是骨骼肌与心肌唯一可调节蛋白质周转的氨基酸,可以调节机体内氮的利用,进而促进机体蛋白质的合成,促进胰岛素的分泌,抑制胰高血糖素分泌,从而抑制糖原异生,减缓肌肉蛋白的分解。
亮氨酸增加蛋白质合成高达50%,抑制分解为25%。亮氨酸抑制分解主要通过α-KIC促进胰岛素的分泌,从而抑制糖原异生,减缓肌肉蛋白的分解。
亮氨酸的代谢产物α-酮戊己酸(α-KIC)和β-羟基-β-甲基丁酸(HMB)也具有调节蛋白质代谢的作用。此外,β-羟基-β-甲基丁酸(HMB)作为亮氨酸的代谢产物,还具有缓解疲劳的作用。
众所周知,哺乳动物的雷帕霉素复合物1(mTORC1) 靶点是细胞生长和代谢的重要调节因子。当mTORC1被激活时,它会促进合成代谢并抑制分解代谢。
mTORC1的活性受到多种信号分子的影响,包括氨基酸。特别是,已知亮氨酸会影响mTORC1活性。在亮氨酸丰度状态下,亮氨酸传感蛋白通过多种机制被激活,然后引起mTORC1复合蛋白的募集和亚细胞定位,从而导致其激活和蛋白翻译的上调。
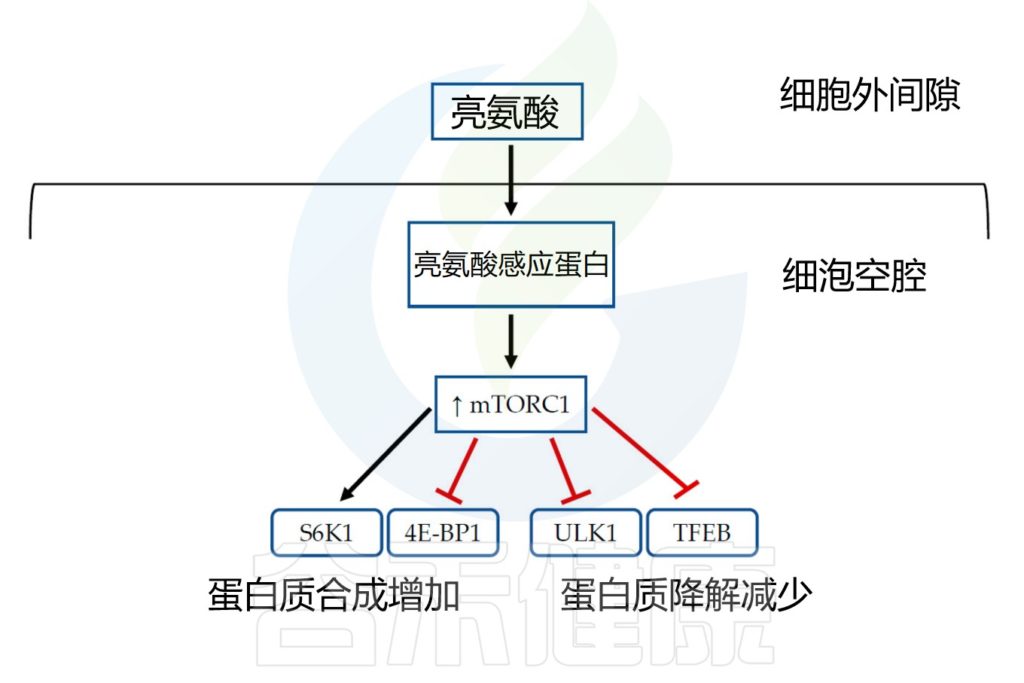
Beaudry AG,et al.Nutrients.2022
•亮氨酸促进能量分配到肌肉细胞
其主要代谢场所在肌肉中,在转氨酶作用下,亮氨酸将氨基转移给酮戊二酸生成谷氨酸,谷氨酸将氨基转移给丙酮酸生成丙氨酸。
亮氨酸通过调节线粒体生物发生来调节能量代谢,并促进脂肪酸氧化和线粒体生物发生。亮氨酸促进能量从脂肪细胞分配到肌肉细胞,导致脂肪细胞中的脂质储存减少并增加肌肉中的脂肪利用率。
此外,饮食中的亮氨酸通过促进骨骼肌中的脂肪酸合成来增加胰岛素敏感性。表明亮氨酸能够通过促进酶与其底物和烟酰胺腺嘌呤二核苷酸(NAD)+的亲和力来直接激活SIRT1,从而导致脂肪细胞和肌肉中的线粒体生物发生和脂肪酸氧化升高。
亮氨酸还促进相关激素的释放,如生长激素、IGF-1(胰岛素样生长因子-1),对于儿童和青少年的生长发育至关重要。
•饮食补充亮氨酸增强肌肉能量代谢,减少肥胖
研究证明亮氨酸可以防止高脂饮食诱导的线粒体损伤,并通过SIRT1介导的机制刺激线粒体生物发生和从脂肪细胞到肌肉细胞的能量分配。
与这些观察结果一致,已发现亮氨酸显著增强骨骼肌中的氧化能力并增加线粒体密度,部分是通过SIRT1依赖性途径AMPK介导的PGC-1α增加。亮氨酸增加线粒体生物发生和PGC-1α表达,表明亮氨酸可能部分通过调节PGC-1α表达来调节骨骼肌能量代谢。
此外,用低剂量白藜芦醇与亮氨酸或β-羟基-β-甲基丁酸联合喂养饮食诱导的肥胖小鼠,可以增加脂肪SIRT1活性、肌肉葡萄糖、棕榈酸摄取和胰岛素敏感性。改善炎症应激生物标志物,并减少肥胖。
在另一项研究中,发现补充亮氨酸可以减少体内脂肪,同时瘦素浓度也成比例下降。通过饮用水摄入亮氨酸导致高脂饮食喂养的小鼠肥胖减少25%,减少了饮食引起的肥胖、高血糖和高胆固醇血症。
肌肉减少症,是指因持续骨骼肌量流失、强度和功能下降而引起的综合症。
•补充亮氨酸可以增加肌少症患者的肌肉质量
目前的荟萃分析中,在不考虑体育锻炼的情况下,补充乳清蛋白、亮氨酸和维生素D可以有效增加肌少症患者的四肢肌肉质量。
•补充亮氨酸伴随锻炼可以更好改善身体表现
根据是否存在伴随体育锻炼对结果进行分析表明,补充乳清蛋白、亮氨酸和维生素D并伴有体育锻炼可以显著改善肌少症患者的握力和身体表现。相反,当体育锻炼不与乳清蛋白、亮氨酸或维生素D补充剂相结合时,食用它并没有显著改善肌少症患者的握力和身体机能。但无论是否伴随的体育锻炼,食用亮氨酸后四肢肌肉质量都会显著增加。
因此,我们认为,肌少症患者除了食用乳清蛋白、亮氨酸、补充维生素D外,还应结合体育锻炼。单独补充乳清蛋白、亮氨酸和维生素D,而不同时进行体育锻炼,对肌肉减少症患者产生积极治疗效果的能力有限。
•摄入亮氨酸对于老年患者保持肌肉质量很重要
充足的蛋白质摄入对于老年患者保持肌肉质量和功能至关重要。
据报道,摄入乳清蛋白和亮氨酸可提供足够量的必需氨基酸,尤其是亮氨酸,它对于适当的肌肉合成至关重要。
此外,维持血清25-羟基维生素D浓度的最佳水平(50-75nmol/L)对于维持四肢肌肉的足够力量和增强蛋白质合成代谢是必要的。因此,我们相信补充乳清蛋白、亮氨酸和维生素D有助于保持肌肉减少症患者的肌肉质量并改善肌肉功能。
25羟维生素D是维生素D在体内的主要存在形式。
建议
总之,食用乳清蛋白、亮氨酸和维生素D补充剂可以增加肌肉减少症患者的四肢肌肉质量。此外,将体育锻炼与补充乳清蛋白和亮氨酸相结合可显著提高肌肉力量和表现。
因此,临床医生可以考虑建议肌少症患者补充乳清蛋白、亮氨酸和维生素D,同时进行体育锻炼。
肠道微生物在亮氨酸的代谢和利用中扮演着重要的角色。接下来将讲述肠道微生物群与亮氨酸直接可能对存在的相互作用。
人们普遍认为代谢健康和代谢稳态的许多方面都受到肠道的影响。肠道屏障在预防感染方面发挥着重要作用,是抵御细菌和病毒抗原的第一道防线。
•补充亮氨酸可以改善肠上皮细胞的能量代谢
亮氨酸对肠道脂质和能量代谢的影响已开始引起人们的关注。在肠上皮细胞中,亮氨酸已被证明可以激活多种参与生物体代谢调节的信号传导途径。
特别是,发现补充亮氨酸可以通过降低活性氧水平来显著调节断奶小猪模型肠上皮细胞的能量代谢,活性氧的稳态对猪的健康至关重要。
活性氧的减少是通过mTOR缺氧诱导因子-1α途径从氧化磷酸化转变为糖酵解来诱导的。
•亮氨酸调节肠道脂肪酸氧化
肠道脂肪酸氧化是一条关键的代谢途径,不仅有助于肠道细胞的能量稳态,而且有助于整个生物体的能量稳态。
研究发现,抗肥胖小鼠的小肠脂质分解代谢能力较高,摄入亮氨酸后对肠道脂肪酸氧化的刺激与体重减轻有关。
事实上,肠道脂肪酸氧化受亮氨酸调节。具体来说,研究表明,补充亮氨酸可能会减缓人十二指肠粘膜中的脂肪酸氧化,涉及四种与脂质代谢相关的蛋白质:HADHA(三功能酶亚基α)、ACADVL(酰基辅酶A脱氢酶超长链)、CPT2(肉碱O-棕榈酰转移酶)和FABP1(肝脏脂肪酸结合蛋白)。
哺乳动物肠道微生物群由数万亿个微生物组成,其中一些微生物有利于宿主健康,包括赋予对胃肠道疾病的定植抵抗力。
人们关注到饮食(如膳食亮氨酸)调节肠道微生物群的组成和功能,从而调节脂质代谢并影响健康的机制。
•亮氨酸通过影响肠道微生物的组成来减少肥胖
临床和临床前研究表明,饮食中的支链氨基酸,尤其是亮氨酸,对人类发挥有益作用,包括增加线粒体呼吸作用、减少氧自由基的产生、影响脂质代谢,并最终延缓肠道微生物群的年龄依赖性变化。
膳食亮氨酸、宿主和微生物群之间的作用是相互的,膳食亮氨酸对肠道微生物群落的组成具有关键影响;反过来,微生物会影响从摄入的食物中获取亮氨酸的效率。
亮氨酸通过塑造肠道微生物组的组成来发挥抗肥胖作用,这往往会逆转短链脂肪酸的改变并产生丙酸,同时改善微生物多样性并介导脂质代谢。
•亮氨酸通过短链脂肪酸调节微生物群
三种支链短链脂肪酸(异丁酸、2-甲基丁酸和异戊酸)主要来源于支链氨基酸的分解代谢。这些短链脂肪酸已被广泛认为既可以作为宿主能量来源,又可以作为信号分子,将肠道微生物群的代谢活动与宿主能量稳态联系起来,特别是通过调节脂质代谢。
亮氨酸可能通过支链短链脂肪酸调节微生物群和肠道脂质代谢,这种现象必须进一步研究。短链脂肪酸刺激一些细胞因子的表达,例如瘦素、IL-10和 IL-18。
此外,短链脂肪酸通过G蛋白偶联受体调节脂肪细胞中的脂肪分解,随后对脂质代谢产生进一步影响。高脂饮食引起的肥胖可以通过调节肠道微生物群来缓解,因为高脂饮食喂养的小鼠中短链脂肪酸减少,同时肠道微生物失调和脂质积累加重。
•肠道微生物群协助肠上皮细胞干预亮氨酸代谢
肠道微生物群在一定程度上改变氨基酸代谢,这被认为会影响许多生理功能。研究证明肥胖受试者的肠道微生物群诱导循环支链氨基酸显著增加,这是肥胖状态的特征。
亮氨酸诱导的脂质代谢与肠道微生物群的脂质代谢密切相关,因为肠道微生物可能协助肠上皮细胞吸收并干预亮氨酸代谢。
最近,粪便微生物群移植已成为肥胖症的一个重要研究领域和治疗方法。这种研究方法也可用于研究脂质代谢——移植产生亮氨酸代谢物的微生物已被观察到以依赖饮食的方式发挥抗肥胖作用。
然而,目前尚不清楚亮氨酸如何靶向肠道微生物群来调节脂质代谢和预防肥胖,这表明需要进一步研究来确定其潜在机制。
亮氨酸缺乏会导致动物胸腺和脾萎缩,淋巴组织受损,并使免疫球蛋白、补体C3和铁转运蛋白水平降低,断奶猪仔缺乏亮氨酸会导致合成特异性抗体的能力下降。
补体C3是血清中含量最高的补体成分,主要由巨噬细胞和肝脏合成,在补体经典激活途径和旁路激活途径中均发挥重要作用。补体C3的升高或降低,对于患者疾病的诊断、预后恢复情况以及病情的监测具有重要意义。
研究发现,添加瘤胃保护性α-KIC(0.05%)可增强羔羊抗猪红细胞的抗体反应与淋巴细胞增殖反应;绵羊饲养中添加亮氨酸与α-KIC,可调节其T-淋巴细胞亚群的免疫功能。
★
哺乳动物雷帕霉素靶蛋白(mTOR)是免疫功能的重要调节剂。mTOR被认为是一个信号中枢,它感知和整合来自免疫微环境的信息,以组织与细胞生长、增殖和死亡相关的反应。
近年来,mTOR被确定为适应性免疫的主要调节因子,如记忆CD8+和CD4+ T细胞分化和人类树突细胞的文献中所述。
mTOR信号通路也在先天免疫中发挥作用,被认为在先天免疫细胞(如先天样自然杀伤细胞、单核细胞、中性粒细胞、肥大细胞、巨噬细胞和树突细胞)激活后触发效应反应。
•亮氨酸可能是一种潜在的抗炎剂
有证据表明mTOR通路信号在促炎和抗炎细胞因子的调节中发挥作用。雷帕霉素在Toll样受体刺激过程中抑制mTORC1已被证明可以通过增强NF-κB信号传导增加促炎细胞因子IL-12的表达,并通过抑制STAT3信号传导减少抗炎性IL-10的表达。
基于mTOR通路信号传导的潜在抗炎作用,支链氨基酸喂养可能会减少疾病或肌肉损伤期间的炎症。因此,亮氨酸可能通过其对mTOR通路的影响而成为潜在的抗炎剂。
然而,亮氨酸如何通过mTOR激活特异性介导炎症信号传导还需要进一步探索。
除了促进肌肉的合成和脂肪的分解以及一些代谢类疾病外,有研究发现亮氨酸在其他疾病中也有影响作用,下面两章将为大家讲述亮氨酸在其他一些疾病中的作用。
亮氨酸在神经疾病中的作用越来越受到重视。在了解抑郁症中的作用之前,我们先来了解一下亮氨酸是如何影响神经系统的。
亮氨酸是合成重要神经递质的前体物质之一,例如多巴胺、去甲肾上腺素和肾上腺素。这些神经递质在神经系统中起着调节心情、注意力、情绪和认知功能的作用。
亮氨酸还可以影响神经传递的速度。研究表明,亮氨酸可以增加神经传递物质的释放和神经元之间的通讯速度,从而促进神经信号的传递效率。
有研究发现亮氨酸在改善抑郁症社交回避行为中起到一定作用。

营养对情绪状态的保持至关重要。临床研究发现,抑郁症患者血清中许多必需氨基酸发生改变,如色氨酸、苏氨酸、亮氨酸、异亮氨酸和缬氨酸。目前,已有一些必需氨基酸的补充被认为有助于改善抑郁行为,但某种必需氨基酸的缺乏是否会对抑郁行为造成影响呢?
为了探讨抑郁和社交回避行为的潜在机制,研究人员使用MetaboAnalyst 5.0网站对差异代谢物进行分析。选择KEGG数据库中的Mus musculus pathway library来支持结果(图A)。y轴的-log(P)表示显著性,-log(P)值越大表示差异越显著。Impact越大表示节点命中,节点命中表示代谢属于该通路的比例有多大。
当使用KEGG数据库支持分析时,排名前十的途径包括:(1)氨基酰基trna生物合成;(2)缬氨酸、亮氨酸和异亮氨酸生物合成;(3)甘氨酸、丝氨酸和苏氨酸代谢;(4)精氨酸和脯氨酸代谢;(5)苯丙氨酸、酪氨酸和色氨酸生物合成;(6)嘌呤代谢;(7)缬氨酸、亮氨酸和异亮氨酸降解;(8)苯丙氨酸代谢;(9)精氨酸生物合成;(10)泛酸和辅酶a生物合成。
可以看到,同样的途径集中在氨基酸代谢和脂肪酸代谢。富集分析用于支持上述结果(图B)。
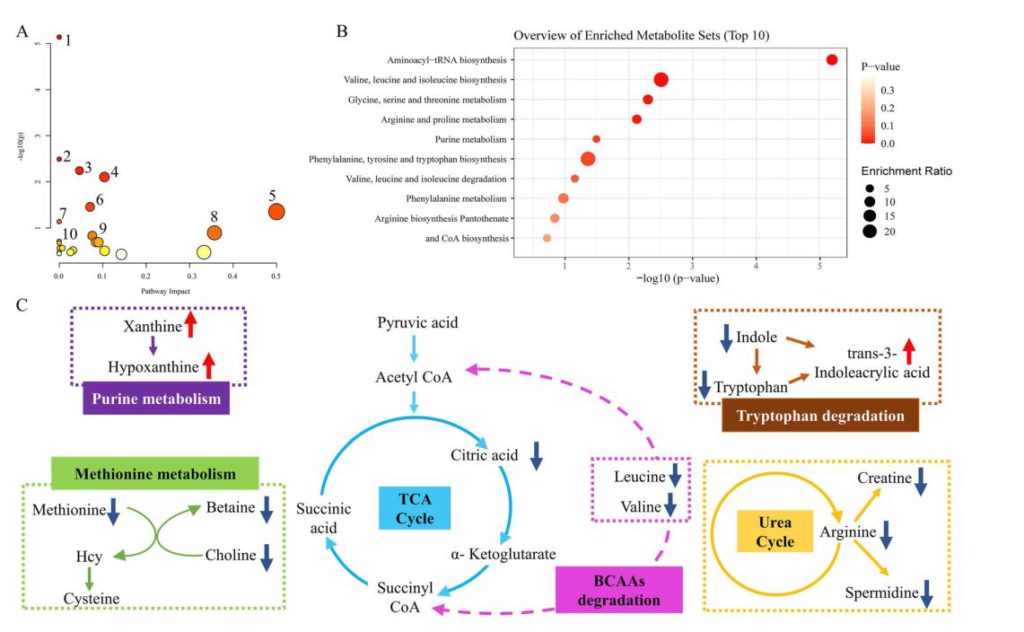
Wang Q,et al.Food Funct.2023
结果还表明,氨基酸代谢和脂肪酸代谢在其潜在机制中起重要作用。如图C所示,氨基酸代谢占很大比例,如色氨酸降解、支链氨基酸降解、蛋氨酸降解。还有其他几种代谢途径,包括嘌呤代谢,TCA循环和尿素循环。因此,进一步研究了支链氨基酸和色氨酸含量的变化。
亮氨酸是社交回避行为相关的关键代谢物
进行Spearman相关分析以确定与社交回避和抑郁样表现相关的关键代谢物。在六个行为指标中,SIR被认为是评估社交回避行为的最佳指标。
结果表明,亚精胺、肌酸、缬氨酸、柠檬酸、黄嘌呤、亮氨酸、甜菜碱、尿酸、苯丙氨酸、PEG-4、棕榈酰肉碱、血小板活化因子、亚油酰胺、油酰胺、醋酸视黄酯、花生四烯酸等16种代谢物与SIR呈显著正相关,其中亮氨酸与SIR呈特异且极显著正相关。
其他15种代谢物虽然与SIR相关,但也与其他行为指标相关。
补充外源性亮氨酸可改善社交回避和抑郁行为
采用CSDS模型(慢性社会挫败应激模型)验证外源性亮氨酸补充,进一步评估其在社交回避和抑郁行为中的作用。
选择盐酸帕罗西汀作为阳性对照药物。适应期结束时,四组间体重无显著差异。在CSDS应激后,亮氨酸可以逆转CSDS引起的体重缓慢增加,其效果略低于阳性药物,但无显著差异。
此外,亮氨酸改善了小鼠的社交回避行为,在社交互动测试中,在社交区停留的时间比例显著高于MOD组。低亮氨酸组的社交回避行为发生率略高于PCM组。同时,亮氨酸也能在一定程度上改善抑郁引起的兴趣和流动性下降。
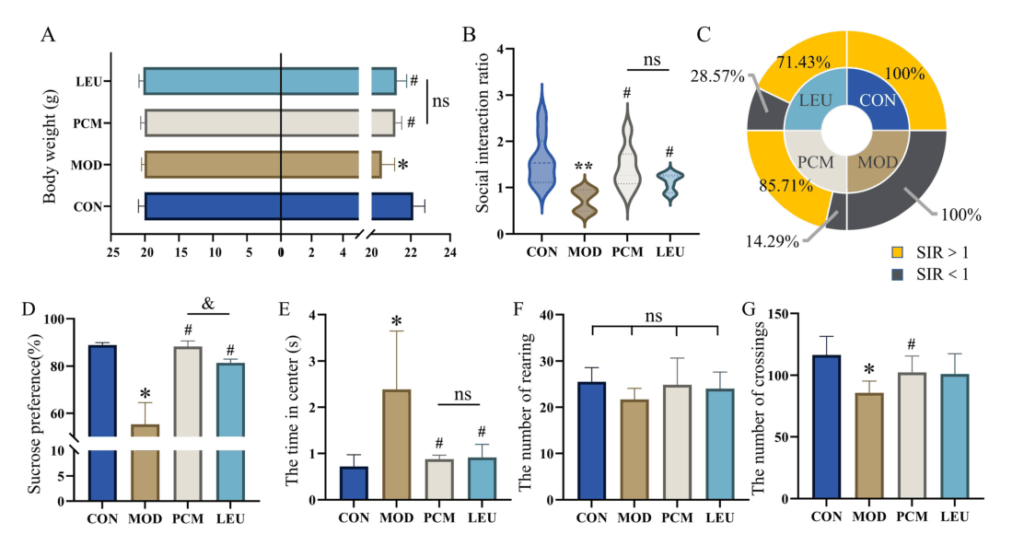
Wang Q,et al.Food Funct.2023
综上所述,外源性补充亮氨酸可在一定程度上调节小鼠抑郁样行为,改善小鼠社交回避行为。在先前的基础上,亮氨酸可能是抑郁和社交回避行为过程中重要的小分子化合物。
亮氨酸与抑郁症相关的横向证据
支链氨基酸和色氨酸是竞争性抑制关系,它们会穿过血脑屏障并影响相应的大脑功能。亮氨酸的活性在支链氨基酸中最高,我们发现亮氨酸在外周和中枢神经系统中的含量降低。
在更高浓度下,亮氨酸触发下丘脑神经元中的mTOR信号级联。几项研究表明,mTOR在抑郁症患者和模型动物中的表达降低。上述信息是亮氨酸与抑郁症之间潜在关系的横向证据。
因此,当体内亮氨酸减少时,更多的色氨酸会通过LAT-1转化到大脑中。CDSD小鼠的研究也表明,当色氨酸进入大脑时,海马中的色氨酸更多地代谢为犬尿氨酸。
最近的一项研究表明,亮氨酸与LAT-1联合给药阻碍了犬尿氨酸向大脑的转运,从而阻止了对脂多糖反应时抑郁样行为的发展。犬尿氨酸途径由炎症因子触发,激活并破坏其神经保护和神经毒性分支之间的平衡。色氨酸向犬尿氨酸的代谢增加也会降低其向5-羟色胺的代谢,进一步促进抑郁症的发作。
思考
总之,这项研究验证了亮氨酸对抑郁状态和社交回避行为的积极影响。同时,它为亮氨酸在神经疾病中的重要作用提供了新的证据。
▸ 什么是癌症恶病质?
癌症恶病质是一种身体消瘦和进行性功能衰退的复杂综合征,其中骨骼肌和脂肪量持续减少。癌症恶病质会降低癌症治疗的耐受性和有效性,同时还会导致严重的疲劳和虚弱;此外,它还会降低预期寿命和生活质量。
★ 补充亮氨酸改善癌症恶病质对机体的影响
与饥饿不同,恶病质不能通过单独增加能量摄入来逆转。尽管如此,有针对性的营养支持是多模式综合征管理的必要组成部分。有趣的是,亮氨酸被发现可以通过mTORC1通路激活来增加蛋白质合成并减少蛋白质降解。多项临床前研究探讨了补充亮氨酸对恶病质肿瘤宿主的影响。
癌症恶病质动物模型的亮氨酸补充研究



Beaudry AG,et al.Nutrients.2022
在14项临床前癌症恶病质研究中,13项表明富含亮氨酸的喂养可能是癌症恶病质的有益附加治疗。
关于亮氨酸对癌症恶病质的影响存在局限性
目前,所有专门研究癌症恶病质期间富含亮氨酸喂养应用的文献仅限于临床前啮齿动物研究。尽管由于各种原因它非常有益,但临床前恶病质研究具有固有的局限性。模型之间存在差异,例如恶病质发生机制(癌细胞注射、致癌物质暴露、肿瘤移植等)、宿主的年龄和性别、啮齿动物的类型和品系、恶病质发生率、肿瘤位置(异位与原位)和负担、肿瘤是否转移以及抗癌药物的使用。这种变异性使得临床前癌症恶病质的研究变得困难,同时也限制了对人群的可转化性。
此外,目前所有与癌症恶病质背景下亮氨酸补充相关的临床前文献都是使用雄性或雌性啮齿动物进行的。没有研究同时使用两种性别。因此,无法探究性别差异。
检查动物研究时的另一个重要考虑因素是蛋白质代谢的种间差异。与人类相比,啮齿动物的蛋白质周转率更高,估计快大约10倍。总之,这些因素使当前工作的综合变得复杂,并使结果的整体解释变得困难。需要进行更标准化的工作,以进一步探索癌症恶病质背景下补充亮氨酸的安全性和有效性。
亮氨酸作为必需氨基酸之一,意味着你的身体无法合成它,所以必须通过食物来源获得。
亮氨酸可以通过服用补充剂的形式获得,也可以通过食用肉类、海鲜、家禽、鸡蛋、奶制品、坚果等食物获得。
搭配营养丰富且全面的饮食一般不会亮氨酸缺乏。大多数人可以安全地食用食物中的这种氨基酸,反而高剂量的补充剂会引起副作用和毒性。
迄今为止,亮氨酸及其代谢物已被证明通过直接或间接增加脂肪酸的氧化和改善代谢健康,它还被证明可以促进和保持肌肉质量、改善运动表现、促进脂肪减少、稳定血糖水平并支持肌肉恢复。
此外,亮氨酸可能具有减轻线粒体功能障碍的附加作用,代表了一种针对衰老、神经退行性疾病、肥胖、糖尿病和心血管疾病的新治疗方法。
亮氨酸有潜力用作人类健康和工业生产的功能性添加剂。然而,有必要进行更多的实验来深入了解其调控机制。
关于亮氨酸的应用
1
临床应用
亮氨酸是临床上选用的复合氨基酸静脉注射制剂中不可缺少的原料,对维持危重病人的营养需求,挽救病人的生命等方面起到了积极的作用。
亮氨酸还应用于肝病及肝性脑病的治疗;此外,亮氨酸在一些神经系统疾病的治疗中可能起到一定的作用。例如,亮氨酸被用于治疗帕金森病,因为它可以作为多巴胺的前体物质,增加多巴胺的合成。
2
食品应用
亮氨酸可以应用于运动饮料补充剂,以减轻运动所引起的肌肉损伤,增强运动能力及减少疲劳。还可作为食品添加剂改善食品风味。
亮氨酸还用于增加食品的营养价值,例如蛋白质补充剂的配方中,以增加蛋白质的含量和完整性。
3
畜牧养殖
亮氨酸在动物生产性能、机体免疫功能和肉品质方面有较多的研究和应用。例如,饲料中添加适量的亮氨酸能够促进生长中期草鱼的生长,改善肌肉品质。
亮氨酸对畜禽动物的繁殖效果也有积极影响。它可以提高动物的生殖能力,增加繁殖成功率。
4
化妆品应用
亮氨酸在化妆品和护肤品中也有应用、提亮肤色、改善皮肤质地、减少色斑。
亮氨酸具有抗氧化作用,可以帮助减少自由基对皮肤的损害,从而延缓皮肤衰老的过程。它可以用于抗衰老产品,如抗皱霜、紧致精华等。
亮氨酸还可以促进皮肤细胞的修复和再生,加速伤口愈合和皮肤恢复。它常被用于修复霜、面膜等产品中。
总的来说,亮氨酸作为一种必需氨基酸,在人体中具有多种重要功能,例如促进生长发育,调节能量代谢,影响机体免疫等,广泛应用于食品、化妆品、医药和生物技术等领域。
肠道菌群检测报告中亮氨酸指标过高或过低的结果能够提供有关肠道菌群代谢功能以及与健康风险相关的信息,当发现亮氨酸水平异常时,可能意味着肠道菌群的代谢功能出现阶段性波动或者紊乱,也可能与本文中提到的一些代谢过程或者疾病的发生和发展相关。
主要参考文献
Chang MC, Choo YJ. Effects of Whey Protein, Leucine, and Vitamin D Supplementation in Patients with Sarcopenia: A Systematic Review and Meta-Analysis. Nutrients. 2023 Jan 19;15(3):521.
Zhang L, Li F, Guo Q, Duan Y, Wang W, Zhong Y, Yang Y, Yin Y. Leucine Supplementation: A Novel Strategy for Modulating Lipid Metabolism and Energy Homeostasis. Nutrients. 2020 May 2;12(5):1299.
Beaudry AG, Law ML. Leucine Supplementation in Cancer Cachexia: Mechanisms and a Review of the Pre-Clinical Literature. Nutrients. 2022 Jul 9;14(14):2824.
Wang Q, Wu Z, Xiang H, Zhou Y, Qin X, Tian J. Revealing the role of leucine in improving the social avoidance behavior of depression through a combination of untargeted and targeted metabolomics. Food Funct. 2023 Jul 17;14(14):6397-6409.
Millward DJ. Leucine requirements for the elderly. Am J Clin Nutr. 2021 Apr 6;113(4):1056-1057.
Holeček M. The role of skeletal muscle in the pathogenesis of altered concentrations of branched-chain amino acids (valine, leucine, and isoleucine) in liver cirrhosis, diabetes, and other diseases. Physiol Res. 2021 Jul 12;70(3):293-305.
Szwiega S, Pencharz PB, Rafii M, Lebarron M, Chang J, Ball RO, Kong D, Xu L, Elango R, Courtney-Martin G. Dietary leucine requirement of older men and women is higher than current recommendations. Am J Clin Nutr. 2021 Feb 2;113(2):410-419.
Wilkinson K, Koscien CP, Monteyne AJ, Wall BT, Stephens FB. Association of postprandial postexercise muscle protein synthesis rates with dietary leucine: A systematic review. Physiol Rep. 2023 Aug;11(15):e15775.

谷禾健康

拟杆菌是革兰氏阴性、专性厌氧菌,在人类肠道微生物群中含量丰富,构成了肠道菌群中大部分比例。
Bacteroides uniformis (一般简称为 B.uniformis),翻译为单形拟杆菌或普通拟杆菌,是拟杆菌属的主要成员,在肠道中起着重要的作用。也被鉴定为阴道微生物组的组成部分。
Bacteroides uniformis是与人类粪便中异黄酮染料木黄酮降解相关的特定细菌物种,可以产生纤维素酶和淀粉酶,降解复杂多糖。
Bacteroides uniformis与肠炎、厌食、肥胖、糖尿病、腹胀、认知发育与情绪运动、多动症、慢性肾病、骨质疏松等相关,是可引起人类或动物疾病的物种,不太可能在人群中传播,并且存在足够的预防或治疗方法。
对单形拟杆菌进行深入研究,有助于我们更好地理解其在人体肠道中的功能,并为相关疾病的预防和治疗提供新的思路。
Bacteroides uniformis是一种革兰阴性细菌,呈杆状。它们通常具有弯曲或弯钩形态,并且没有芽胞。

图源:Microbe Canvas
生理特性
Bacteroides uniformis是一种厌氧菌,不能在氧气存在的环境中生长。它们通常能够利用多糖类物质作为碳源,如淀粉、纤维素和其他复杂碳水化合物。还可以产生一些酶,如纤维素酶和淀粉酶,用于降解消化这些复杂碳水化合物。
分布和生存环境
Bacteroides uniformis是人体肠道,阴道中常见的细菌之一,也可以在其他动物的肠道中找到。它们在肠道中扮演重要的生态角色,参与食物消化、营养吸收和免疫调节等过程。Bacteroides uniformis能够适应肠道的微生态环境,与其他肠道细菌相互作用。
功能和代谢
Bacteroides uniformis在肠道中具有多种功能和代谢活性。它们参与葡萄糖和其他碳水化合物的代谢,并产生短链脂肪酸(如丙酸和乙酸),这些酸有益于肠道健康。此外,Bacteroides uniformis还参与胆固醇代谢、氨基酸代谢和维生素合成等生物化学过程。
肥胖和肥胖相关疾病构成了世界范围内的重大公共卫生问题。除了传统的减肥药物外,下一代益生菌(NGP)似乎非常有希望作为潜在的肥胖预防和治疗药物。
主要的候选NGP已在肥胖和肥胖相关疾病的临床前模型中显示出前景,例如:
B.uniformis已被证明具有对抗肥胖和代谢紊乱的有益特性。在动物模型中, B. uniformis与减肥、降低血清胆固醇和甘油三酯水平以及改善肝脂肪变性有关。此外,它还与血清瘦素水平降低、空腹血糖浓度降低和糖耐量改善有关。
推测的机制包括调节肠道菌群和改善肠道菌群失调、改善肠道屏障功能、减少慢性低度炎症和调节肠道肽分泌。
一项对41名体重指数(BMI)正常的个体肠道菌群和内脏脂肪面积 (VFA)的研究显示,内脏脂肪面积高/低(VFA)两组的肠道菌群存在显著差异,在 L-VFA 组中,B. plebeius 和B.uniformis含量很高。此外,B.uniformis与血清 LDL-胆固醇水平呈负相关。
另一项对32名肥胖成年人(14 名接受了腹腔镜袖状胃切除术 (LSG))和30名基线健康对照者的肠道菌群以及身体指标的研究,并在术后 6 个月进行了随访。通过磁共振成像测量腹部内脏脂肪面积(VFA)和皮下脂肪面积(SFA)。腰围、臀围、腰臀比(WHR)和体重指数(BMI)被列为简单的肥胖参数。通过宏基因组测序分析肠道微生物组。在与腹部内脏脂肪面积(VFA)负相关的物种中,Eubacteria eligens相关性最强,其次是Clostridium citroniae、C. symbiosum、Bacteroides Uniformis、E. ventriosum、C. hathewayi等。
一项涉及 45 名儿童(6 至 12 岁)的横断面研究调查了肠道微生物群与儿童肥胖之间的关系。发现两个BMI组之间的多样性存在差异。这种差异包括真杆菌属(Eubacterium)、普拉梭菌(Faecalibacterium prausnitzii)、Dialister、Monoglobus pectinilyticus、Bifidobacterium pseudocatenulatum、Intestinibacter bartlettii、 Bacteroides intestinalis、Bacteroides uniformis (单形拟杆菌)和 史密斯甲烷杆菌(Methanobrevibacter smithii)。
一项针对减肥手术是否显著改善体重的研究显示单形拟杆菌的丰度可能影响手术后的效果。
术前属于拟杆菌门和厚壁菌门的成员丰度存在差异无应答组:
富含拟杆菌科、拟杆菌属、单形拟杆菌、Alistipes Finegoldii、Alistipes alistipes、Dorea formicigenerans 、瘤胃球菌。
应答组:
富含消化链球菌科、 Gemmiger、Gemiger formicilis、Barnesiella、Prevotellaceae 、 Prevotella。
线性判别分析 >2;p < 0.05。与有反应组相比,无反应组中普氏菌与拟杆菌的比率显着较低 (p = 0.048)。
神经性厌食症(Anorexia nervosa, AN)的特点是一种神秘的食物回避模式,导致严重的营养不良,这种疾病在青春期早期发展。近期,在《Nature Microbiology》上发表的文章研究了77名患有神经性厌食症的成年女性和70名被认为是健康体重的对照组。
利用宏基因组学,作者对微生物组的细菌和病毒成分进行了深入分析。细菌基因组中的结构变异,特别是Bacteroides uniformis(单形拟杆菌)中的结构变异,可能造成了硫胺素生产的破坏。
硫胺素(或称维生素B1)缺乏会影响人体健康,包括记忆丧失、焦虑、抑郁、易怒、失眠、食欲减退和胃肠道不适。这也可以解释在神经性厌食症患者中普遍观察到的硫胺素缺乏的现象。
认知发展
一项针对健康以色列阿拉伯儿童的横断面研究显示一些菌群分类学差异与儿童智商测试FSIQ评分显着正相关,包括Prevotella(普雷沃氏菌属)、Dialister属、Sutterella(萨特氏菌属)、Ruminococcus callidus和Bacteroides uniformis(单形拟杆菌)。
情绪相关
一项针对长期封闭环境中心理变化和肠道菌群的研究,鉴定出四种潜在的“精神抗生素”(单形拟杆菌Bacteroides Uniformis、Roseburia inulinivorans、Faecalibacter prausnitzii、Eubacterectale )。
根据宏基因组、宏蛋白质组和代谢组学分析,四种潜在的精神抗生素主要通过与神经系统功能相关的三种途径改善情绪:
首先,通过发酵膳食纤维,它们可能产生短链脂肪酸,例如丁酸和丙酸;
其次,它们可能调节天冬氨酸、谷氨酸、色氨酸等氨基酸代谢途径(例如,将谷氨酸转化为γ-氨基丁酸;将色氨酸转化为血清素、犬尿酸或色胺);
第三,它们可能调节其他途径,例如牛磺酸和皮质醇代谢。此外,动物实验的结果证实了这些潜在的精神抗生素对情绪的积极调节作用和机制。
运动相关
肠道微生物群可能有助于改善运动表现。日本庆应义塾大学先进生命科学研究所通过分析25名男性长跑运动员的比赛成绩后发现,运动员粪便中单形拟杆菌(Bacteroides uniformis)的丰度与其3000米比赛成绩间存在相关性。
在随后的一项随机、双盲、安慰剂对照研究中,研究人员让健康男性志愿者服用α-环糊精,以增加肠道中的单形拟杆菌总量。结果表明,补充α-环糊精可以提高人体的耐力运动能力。
此外,在小鼠实验中,食用单形拟杆菌显著延长了动物游泳的持续时间;动物盲肠短链脂肪酸浓度以及肝脏中与糖异生相关酶的基因表达均上升,肝糖原含量降低。该研究表明,单形拟杆菌可能通过促进肝脏内源性葡萄糖生成,增强个体的耐力运动能力。
妊娠糖尿病
妊娠合并糖尿病包括孕前糖尿病(PGDM)和妊娠期糖尿病(GDM)。 一项针对234份孕妇粪便样本的16S rRNA测序数据显示,随着妊娠的进展,妊娠糖尿病PGDM 组的肠道细菌种类数量有所减少。巨单胞菌、拟杆菌和 Roseburia Enteris 在 PGDM 组中富集,而 Bacteroides vulgatus、Faecalibacter prausnitzii、Eubacterectale、Bacteroides uniformis(单形拟杆菌)、Eubacteria eligens、Subdolilegum在对照组中比例更高,这些菌血糖与血糖特征呈负相关。
二型糖尿病
一项针对亚洲人的包括36个不同二型糖尿病T2DM研究(健康,n = 3378;T2DM,n = 551)的meta分析显示,T2DM 组中普通杆菌菌、单形拟杆菌和普氏粪杆菌较低。
另一项包括29名T2DM患者和28名健康对照者研究显示,Megamonas funiformis 和 Escherichia 的丰度在T2DM患者的肠道中显着增加。相反,Bacteroides stercoris、单形拟杆菌和Phascolarctobacterium faecium的丰度降低。
单形拟杆菌 (Bacteroides uniformis) 还可能调节动物肠道稳态,减轻小鼠结肠炎。一项研究使用B.uninis JCM5828 及其代谢物对患有葡聚糖硫酸钠盐 (DSS) 诱导的结肠炎的雌性 C57BL/6J小鼠进行治疗。
Bacteroides uniformis治疗显著减轻了结肠炎的进展并恢复了机械和免疫屏障蛋白的表达。此外,Bacteroides uniformis还增加了共生菌双歧杆菌和阴道乳杆菌的丰度,同时减少了致病性埃希氏菌、志贺氏菌的丰度,并调节了肠道胆汁酸代谢。
Bacteroides uniformis在很大程度上调节结肠组织中NF-κB和丝裂原激活蛋白激酶(MAPK)信号通路关键调节蛋白的表达以及TH17细胞的分化。然而,Bacteroides uniformis在体外不能直接抑制TH17细胞分化;它应该是通过参与胆汁酸代谢和调节关键代谢物(α-鼠胆酸、猪去氧胆酸和异石胆酸)来调节固有层的过程,从而调节肠道免疫反应。
此外在一项溃疡性结肠炎(UC)患者的菌群研究中,六种拟杆菌属表现出显着较低的相对丰度,十二种拟杆菌属被发现与至少一项疾病活动指标显着相关。五个物种( Alistipes putredinis、Bacteroides stercoris、Bacteroides Uniformis、Bacteroides rodentium、Parabacteroides merdae )的相对丰度与溃疡性结肠炎活性的四个指标中的三个相关。
12 个关键物种表达了与 UC 发病机制相关的5 个基因(TARP、C10ORF54、ITGAE、TNFSF9和LCN2 )。作为潜在生物标志物的关键物种的丧失可能会加剧 UC 活动。
比较功能性腹胀和膨胀(FABD)患者和12健康个体粪便微生物群的多样性和组成,并评估小肠细菌过度生长 (SIBO) 。结果显示FABD 组的微生物多样性显着低于对照组。与健康对照相比,FABD 患者中变形菌的比例显着升高,放线菌的比例显着降低。与健康对照相比,FABD患者中粪杆菌的比例显着升高,而SIBO(+)FABD患者中普氏菌和粪杆菌的比例也显着升高。与健康对照相比,FABD 患者的普氏粪杆菌 (Faecalibacter prausnitzii) 的丰度显着增加,但单形拟杆菌 (Bacteroides uniformis)和青春双歧杆菌数量明显减少。
乳糜泻患者肠道中的 Bacteroides distasonis、Bacteroides fragilis / Bacteroides thetaiotaomicron、Bacteroides Uniformis、Bacteroides ovatus比对照组更少(p<0.05)。
骨质疏松
一项研究中,与对照组相比,几个物种的丰度发生了变化,在骨质疏松组中发现了特定的功能途径,骨质疏松组中下列菌群的水平下降:
类风湿性关节炎
一项研究包括 76 名类风湿关节炎患者的粪便宏基因组和血浆代谢组的分期概况。
骨关节炎
骨关节炎是一种病因不明的慢性退行性疾病。一项研究针对骨关节炎患者(n = 32)和正常对照组(n = 57)肠道菌群检测,宏基因组测序显示,骨关节炎患者中Bacteroides stercoris、Bacteroides vulgatus、Bacteroides uniformis在物种水平上的丰度显著降低,大肠杆菌、肺炎克雷伯菌、Shigella flexneri、唾液链球菌在骨关节炎中显著增加。
特应性皮炎AD
使用 16S rRNA 测序分析了 93 名志愿者的粪便样本,其中包括 44 名 AD 患者和 49 名健康对照受试者,年龄为 6-22 岁。
卟啉单胞菌科、Blautia、Parabacteroides、Bacteroides ovatus、Bacteroides uniformis 和 Prevotella stercorea 的相对丰度显著较高(P < 0. 05) AD 患者高于健康对照者。与 AD 患者相比,健康对照受试者中梭状芽胞杆菌和粪疟原虫含量较高 (P < 0.05)。
儿童多动症
对11项儿童多动症研究的meta分析显示,拟杆菌属与多动和冲动水平相关。在物种水平上,多动症儿童Faecalibacterium丰度较低,但Bacteroides uniformis、肠球菌、双歧杆菌和Odoribacter增加,可能导致中枢神经系统多巴胺相关功能受损。
此外,多动症儿童肠道中Faecalibacterium丰度的降低可能导致炎症细胞因子的渗透性和交叉性更高。
一项调查多动症儿童及其饮食模式和肠道菌群的研究显示,在注意缺多动障碍(ADHD)组中,粪肠拟杆菌(B. coprocola)的相对丰度降低,而单形拟杆菌(B.uniformis)、卵形拟杆菌(B.ovatus)和粪类杆菌(S. stercoricanis)的相对丰度升高。
结直肠癌转移组
转移性结直肠癌组和非转移性结直肠癌组患者的肠道菌群存在差异。在属水平上,拟杆菌属菌种脆弱拟杆菌和单形拟杆菌组成在转移组中显着高于非转移组。
PD-1免疫疗法反应
肠道微生物群被认为是调节免疫检查点抑制剂治疗功效的关键因素。一项波兰黑色素瘤患者队列中PD-1疗法反应与基线肠道微生物组的研究显示,普氏菌和单形拟杆菌的丰度与反应有关,而无反应者的肠道微生物群富含普氏粪杆菌和脱硫弧菌以及一些未分类的厚壁菌门。
慢性肾病
一项台湾地区人群肠道菌群研究显示,在慢性肾病 (CKD)患者中发现的最丰富的属是拟杆菌属,在物种水平上,血液透析患者中的B. ovatus、B. caccae 、 B.uniformis表现出显著丰度,而腹膜透析患者中的Blautia表现出较高的丰度。
白塞病
在活动性白塞病BD患者其单形拟杆菌丰度显著高于其匹配的健康对照和处于非活动状态的疾病患者 ( p = 0.038)。
单形拟杆菌可以消化多种膳食纤维,作为一种代谢膳食纤维产生乙酸的肠道菌已有大量研究表明日常膳食纤维摄入会影响单形拟杆菌的丰度。
一项横断面研究中检查了全谷物和精制谷物、纤维和麸质的习惯摄入是否与肠道微生物群相关。对779 名参与者137 项食物频率的调查问卷和肠道菌群的分析显示。最高全谷物摄入量四分位与较高丰度的普通拟杆菌、普氏粪杆菌、Blautiaproducta和Rikenellaceae以及较低丰度的单形拟杆菌有关。
一项对不同特异性(低-低聚果糖、低至中-2 型抗性淀粉、中-果胶和高-不溶性 β-1,3-葡聚糖特异性)纤维的微生物发酵研究显示,高特异性不溶性β-葡聚糖促进目标细菌大量增加,Anaerostipes sp. 平均从0.3%增加到16.5%,单形拟杆菌平均为 2.5% 至 17.9%,这与这些细菌所在的每个微生物群落中相关代谢物(分别为丁酸盐和丙酸盐)比例的增加有关。
水溶性醋酸纤维素 (WSCA) 是一种合成纤维来源,可以导致拟杆菌属成员的丰度增加,尤其是单形拟杆菌。在四种具有代表性的人类肠道拟杆菌属中,只有单形拟杆菌在WSCA上的纯培养物中生长并活跃地产生乙酸。
一项对健康志愿者进行了为期 4周的营养干预分析天然单宁提取物混合物对肠道微生物变化的研究显示。从前两周起就可以观察到微生物群的组成和功能的变化,测序显示以下菌相对丰度增加,例如:
利用同位素标记发,发现食用菊粉富集的前三种细菌为单形拟杆菌(Bacteroides uniformis)、产酸拟杆菌(Bacteroides acidifaciens)和狄氏副拟杆菌(Parabacteroides distasonis)。
一项研究评估富含黄酮类化合物的蔓越莓多酚(CP)和龙舌兰(AG)(一种高支链龙舌兰衍生的新果聚糖)对C57BL6雄性小鼠的心脏代谢反应、肠道微生物群组成、代谢内毒素血症和粘膜免疫调节的影响,该雄性小鼠喂食致肥胖高脂肪高糖(HFHS)饮食9周。AG,单独或与CP(CP+AG)联合,主要刺激聚糖降解细菌:
与未经治疗的小鼠相比,接受AG的肥胖小鼠中聚糖降解细菌的增加与丁酸水平的显著增加一致。
总的来说,研究发现高特异性不溶性β-葡聚糖、天然单宁提取物混合物、菊粉、蔓越莓多酚和龙舌兰能增加Bacteroides uniformis的丰度。
主要参考文献:
Vallianou NG, Kounatidis D, Tsilingiris D, Panagopoulos F, Christodoulatos GS, Evangelopoulos A, Karampela I, Dalamaga M. The Role of Next-Generation Probiotics in Obesity and Obesity-Associated Disorders: Current Knowledge and Future Perspectives. Int J Mol Sci. 2023 Apr 4;24(7):6755.
Nie X, Chen J, Ma X, Ni Y, Shen Y, Yu H, Panagiotou G, Bao Y. A metagenome-wide association study of gut microbiome and visceral fat accumulation. Comput Struct Biotechnol J. 2020 Sep 20;18:2596-2609.
Murga-Garrido SM, Ulloa-Pérez EJ, Díaz-Benítez CE, Orbe-Orihuela YC, Cornejo-Granados F, Ochoa-Leyva A, Sanchez-Flores A, Cruz M, Castañeda-Márquez AC, Plett-Torres T, Burguete García AI, Lagunas-Martínez A. Virulence Factors of the Gut Microbiome Are Associated with BMI and Metabolic Blood Parameters in Children with Obesity. Microbiol Spectr. 2023 Feb 14;11(2):e0338222.
Lapidot Y, Maya M, Reshef L, Cohen D, Ornoy A, Gophna U, Muhsen K. Relationships of the gut microbiome with cognitive development among healthy school-age children. Front Pediatr. 2023 May 19;11:1198792.
Cheng M, Zhao Y, Cui Y, Zhong C, Zha Y, Li S, Cao G, Li M, Zhang L, Ning K, Han J. Stage-specific roles of microbial dysbiosis and metabolic disorders in rheumatoid arthritis. Ann Rheum Dis. 2022 Aug 19;81(12):1669–77.
Shirvani-Rad S, Ejtahed HS, Ettehad Marvasti F, Taghavi M, Sharifi F, Arzaghi SM, Larijani B. The Role of Gut Microbiota-Brain Axis in Pathophysiology of ADHD: A Systematic Review. J Atten Disord. 2022 Nov;26(13):1698-1710.
Wang LJ, Yang CY, Chou WJ, Lee MJ, Chou MC, Kuo HC, Yeh YM, Lee SY, Huang LH, Li SC. Gut microbiota and dietary patterns in children with attention-deficit/hyperactivity disorder. Eur Child Adolesc Psychiatry. 2020 Mar;29(3):287-297.
Hao Z, Meng C, Li L, Feng S, Zhu Y, Yang J, Han L, Sun L, Lv W, Figeys D, Liu H. Positive mood-related gut microbiota in a long-term closed environment: a multiomics study based on the “Lunar Palace 365” experiment. Microbiome. 2023 Apr 24;11(1):88.
Ren Y, Hao L, Liu J, Wang P, Ding Q, Chen C, Song Y. Alterations in the Gut Microbiota in Pregnant Women with Pregestational Type 2 Diabetes Mellitus. mSystems. 2023 Apr 27;8(2):e0114622.
Wu X, Park S. Fecal Bacterial Community and Metagenome Function in Asians with Type 2 Diabetes, According to Enterotypes. Biomedicines. 2022 Nov 21;10(11):2998.
Lv M, Li L, Li W, Yang F, Hu Q, Xiong D. Mechanism research on the interaction regulation of Escherichia and IFN-γ for the occurrence of T2DM. Ann Palliat Med. 2021 Oct;10(10):10391-10400.
Yan Y, Lei Y, Qu Y, Fan Z, Zhang T, Xu Y, Du Q, Brugger D, Chen Y, Zhang K, Zhang E. Bacteroides uniformis-induced perturbations in colonic microbiota and bile acid levels inhibit TH17 differentiation and ameliorate colitis developments. NPJ Biofilms Microbiomes. 2023 Aug 14;9(1):56.
Nomura K, Ishikawa D, Okahara K, Ito S, Haga K, Takahashi M, Arakawa A, Shibuya T, Osada T, Kuwahara-Arai K, Kirikae T, Nagahara A. Bacteroidetes Species Are Correlated with Disease Activity in Ulcerative Colitis. J Clin Med. 2021 Apr 17;10(8):1749.
Noh CK, Lee KJ. Fecal Microbiota Alterations and Small Intestinal Bacterial Overgrowth in Functional Abdominal Bloating/Distention. J Neurogastroenterol Motil. 2020 Sep 30;26(4):539-549.
Wang H, Zhang K, Wu L, Qin Q, He Y. Prediction of Pathogenic Factors in Dysbiotic Gut Microbiomes of Colorectal Cancer Patients Using Reverse Microbiomics. Front Oncol. 2022 Apr 27;12:882874.
Ye S, Yan F, Wang H, Mo X, Liu J, Zhang Y, Li H, Chen D. Diversity analysis of gut microbiota between healthy controls and those with atopic dermatitis in a Chinese population. J Dermatol. 2021 Feb;48(2):158-167.
Zhou P, Dai Z, Xie Y, Li T, Xu Z, Huang Y, Sun D, Zhou Y. Differences in tissue-associated bacteria between metastatic and non-metastatic colorectal cancer. Front Microbiol. 2023 Jun 9;14:1133607.
Shivani S, Kao CY, Chattopadhyay A, Chen JW, Lai LC, Lin WH, Lu TP, Huang IH, Tsai MH, Teng CH, Wu JJ, Hsieh YH, Wang MC, Chuang EY. Uremic Toxin-Producing Bacteroides Species Prevail in the Gut Microbiota of Taiwanese CKD Patients: An Analysis Using the New Taiwan Microbiome Baseline. Front Cell Infect Microbiol. 2022 Apr 26;12:726256.
Kim JC, Park MJ, Park S, Lee ES. Alteration of the Fecal but Not Salivary Microbiome in Patients with Behçet’s Disease According to Disease Activity Shift. Microorganisms. 2021 Jul 6;9(7):1449.
Um CY, Peters BA, Choi HS, Oberstein P, Beggs DB, Usyk M, Wu F, Hayes RB, Gapstur SM, McCullough ML, Ahn J. Grain, Gluten, and Dietary Fiber Intake Influence Gut Microbial Diversity: Data from the Food and Microbiome Longitudinal Investigation. Cancer Res Commun. 2023 Jan 11;3(1):43-53.
Cantu-Jungles TM, Bulut N, Chambry E, Ruthes A, Iacomini M, Keshavarzian A, Johnson TA, Hamaker BR. Dietary Fiber Hierarchical Specificity: the Missing Link for Predictable and Strong Shifts in Gut Bacterial Communities. mBio. 2021 Jun 29;12(3):e0102821.
Molino S, Lerma-Aguilera A, Jiménez-Hernández N, Rufián Henares JÁ, Francino MP. Evaluation of the Effects of a Short Supplementation With Tannins on the Gut Microbiota of Healthy Subjects. Front Microbiol. 2022 Apr 27;13:848611.