-

 CNAS L23010
CNAS L23010

 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业 二级病原微生物安全实验室
二级病原微生物安全实验室- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业 二级病原微生物安全实验室

谷禾健康

群体感应(Quorum Sensing,QS)是一种细胞间的通讯机制,广泛存在于细菌、微生物以及某些多细胞生物中。通过这种机制,细胞能够感知周围环境中同类细胞的密度,并根据细胞数量变化调节基因表达和行为,这种现象在细菌中尤为显著。
在胃肠道等竞争激烈且不断变化的环境中,微生物群已经开发出独特的相互交流方法。肠道菌群产生的群体感应分子调节多种细胞功能,例如毒力基因的表达、生物膜的形成、能力和孢子形成,通常仅在细胞数量达到一定密度时启动这些过程。
肠道细菌产生的群体感应分子在肠道微生物组内建立物种和菌株水平结构方面很重要,但也用于与宿主交流。
微生物之间的对话各不相同,范围从种间交流、自我对话或种内交流到来自一个属的细胞对另一个属产生的信号做出反应。无法产生自身通信信号的细胞正在“监听”其他细胞产生的信号,这种现象类似于为“窃听”。
肠道微生物群使用某些代谢物作为群体感应分子与肠上皮细胞(IEC)进行通讯。例如,金黄色葡萄球菌分泌多种毒力因子,这些因子操纵宿主的免疫系统以保护自身的生存。这些生存策略对宿主的影响通常表现为:
虽然我们目前还不知道哪些群体感应分子与肠道和感觉神经元有最直接的相互作用,但很多证据显示它们会影响我们的生理和心理健康。
例如细菌产生的信号多种多样,包括自诱导剂 (AI)、高丝氨酸内酯(HSL)、喹啉、肽、毒素和蛋白酶。这些信号分子激活细菌细胞壁中的特定受体,并触发细胞质中调节基因表达的传感器。
注:自诱导剂(sAutoinducers)是细菌在群体中相互交流的一种信号分子。它们通过细菌的分泌进入环境,并在细菌密度达到一定水平时被其他细菌重新摄取。它使细菌能够协调其行为,例如生物膜的形成、毒素的产生和其他群体行为。
高丝氨酸内酯(Homoserine Lactones, HSL)是一类特定类型的自诱导剂,主要在革兰氏阴性细菌中发现。当HSL达到一定浓度时,它们会与细菌细胞内的特定受体结合,激活基因表达的调节机制。这种调节可以影响细菌的生长、代谢和致病性等多种生物学过程。
目前,研究很多研究已经建议可以将群体感应分子可用作辅助诊断的生物标志物:
未来,应该有可能使用 QS 生物标志物来诊断神经和精神疾病,例如:焦虑和抑郁、重度抑郁症、精神分裂症、双相情感障碍、自闭症、强迫症等。
微生物群体感应(QS)可以被视为微生物群之间的伙伴关系或协议,本文主要分享和讨论了QS在肠道细菌和肠壁中带来的变化,并探讨和总结这些通信可能对中枢神经系统和心理健康产生的影响。
Interbacterial Communication
革兰氏阴性菌(Gram-Negative Bacteria)
细菌是一类单细胞微生物,它们需要通过各种方式来感知周围环境,并协调自身的行为。革兰氏阴性细菌就利用一类称为自诱导剂 (Autoinducers, AI) 的小分子物质来实现这种细胞间通信。
AHL:细菌间的通用密码
其中最著名的自诱导剂是AHL(N-酰基-高丝氨酸内酯N-Acyl,Homoserine Lactones, AHL)。AHL是一种小的中性脂质分子,由高丝氨酸内酯(Homoserine Lactone, HSL) 部分和4到18个碳原子的酰基侧链组成。
不同细菌产生的AHL可能有不同的酰基侧链长度和取代基,这使得细菌能够区分彼此释放的信号分子。
AHL的合成
AHL的合成需要依赖于S-腺苷甲硫氨酸(S-Adenosyl Methionine,SAM)作为底物,由LuxI或LuxM等合成酶催化产生。有些细菌只有一种AHL合成酶,而另一些细菌则拥有多种AHL合成酶,能够产生多种形式的AHL。
AHL的产生水平受到严格调控,取决于底物的可用性
一些缺乏LuxI型合成酶的细菌会拥有”孤儿”的LuxR型受体,这些受体能够检测到同一环境中其他细菌释放的AHL信号。例如,大肠杆菌(Escherichia coli) 中的SdiA和铜绿假单胞菌(Pseudomonas aeruginosa)中的QscR就是这类孤儿受体的代表。
这些LuxR型受体在不同细菌中高度保守,序列同一性可达67%-84%。它们不仅存在于大肠杆菌,还广泛分布于其他肠道细菌,如肠杆菌(Enterobacter)、柠檬酸杆菌(Citrobacter)、克罗诺杆菌(Cronobacter)、克雷伯菌(Klebsiella)、沙门氏菌 (Salmonella) 、志贺氏菌(Shigella) 中。
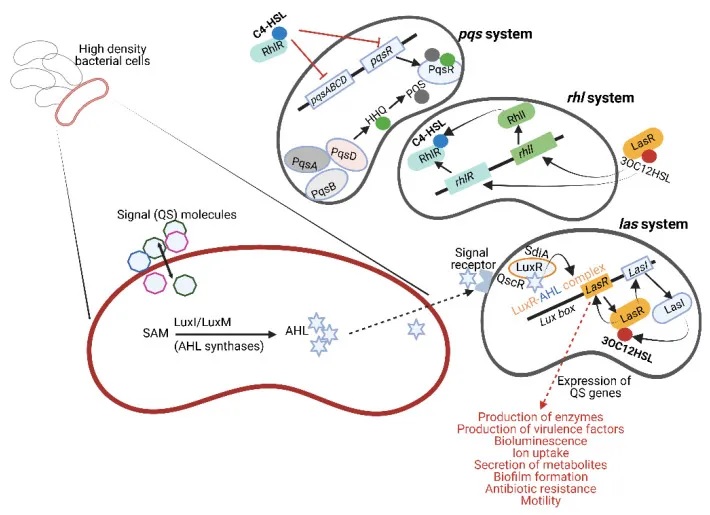
doi.org/10.3390/microorganisms10101969
大肠杆菌的多种群体感应系统及其生理调控
以致病性大肠杆菌(E. coli)为例,它利用多种QS系统来调控自身的生理过程:
其中,大肠杆菌的SdiA蛋白可以被铜绿假单胞菌产生的AHL信号分子激活。这种SdiA/AHL复合物能够增强大肠杆菌中与耐酸性相关基因的表达,对于肠出血性大肠杆菌(EHEC)在胃肠道低pH环境中存活至关重要。
此外,大肠杆菌还利用QS系统调控其毒力基因的表达、生物膜的形成、细菌运动性、III型分泌系统的活性,以及毒素和细菌纤毛的产生。
类似地,沙门氏菌的QS系统也参与调节其致病性,影响侵袭性基因的表达、鞭毛形成,以及与拉链侵袭机制相关的基因。
总之,细菌利用这些精细的群体感应系统,能够在复杂的环境中有效地感知周围情况,并协调自身的生理活动,从而增强其生存和致病能力。这种细菌间的通信机制对于理解细菌的生态和致病机制具有重要意义。
革兰氏阳性菌(Gram-Positive Bacteria)
除了革兰氏阴性菌使用自诱导剂(Autoinducer,AI)进行群体感应(Quorum Sensing, QS)外,革兰氏阳性菌也采用了一种独特的QS机制。
革兰氏阳性菌的独特群体感应机制
革兰氏阳性菌使用由5-17个氨基酸组成的小线性或环状寡肽(Quorum Sensing Peptide, QSP)进行细胞间通信。
其中,研究最多的是芽孢杆菌产生的QS系统,包括能力孢子形成因子(CSF)、五肽和七肽SDLPFEH(PapRIV)等。这些七肽是由48个氨基酸的前体肽经过NprB蛋白酶的裂解而形成的。
QSP在细胞膜上的ATP结合盒转运蛋白的帮助下被分泌到细胞外,并与膜结合的受体或细胞质传感器(如Rap、NprR、PlcR和PrgX蛋白)发生相互作用。
QSP系统在细菌中的实例
以金黄色葡萄球菌为例,其辅助基因调节因子(Agr)就是一个编码肽信号分子Agr的四基因操纵子,起到膜结合传感器的作用。Agr调节多种基因的表达,包括毒力因子、蛋白酶和成孔毒素等。
缺失agr基因(Δagr)的金黄色葡萄球菌菌株会形成生物膜,更容易引起慢性感染和菌血症。
另一种革兰氏阳性菌肠球菌也使用Fsr-QS系统,该系统由四个基因位点fsrABDC控制。一旦肽被裂解激活,就会通过跨膜激酶在细胞内进行磷酸化级联反应,从而诱导靶基因的表达。
总的来说,革兰氏阳性菌利用这种基于寡肽信号分子的QS系统,能够感知周围细菌的数量,并协调自身的生理活动,如毒力因子的表达、生物膜的形成等。这种细菌间的通信机制对于理解它们的生态和致病机制具有重要意义。
革兰氏阳性菌使用的群体感应的一般表示
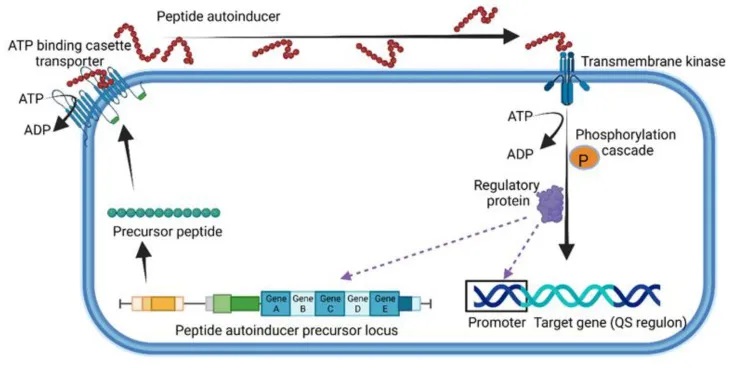
doi.org/10.3390/microorganisms10101969
Interspecies Communication
自身诱导剂-2 (Autoinducer-2, AI-2)是一种广泛存在于细菌中的信号分子,在细菌间的种间通讯和调节特定生物行为中起着关键作用。
AI-2的的合成途径
AI-2是一种呋喃硼酸酯二酯,既可以被革兰氏阴性菌,也可以被革兰氏阳性菌产生。它的合成受到luxS基因的调控。在这个过程中,首先S-腺苷同型半胱氨酸(SAH)被SAH水解酶(SahH)转化为同型半胱氨酸,然后经过Pfs和LuxS两步反应,形成中间体4,5-二羟基-2,3-戊二酮(DPD),最终重排成AI-2分子。
AI-2在细菌中的广泛分布
值得注意的是,在超过三分之一的细菌基因组中都能检测到编码luxS同源物的基因,包括大肠埃希菌(Escherichia coli)、粪肠球菌(Enterococcus faecalis)、空肠弯曲杆菌(Campylobacter jejuni)、金黄色葡萄球菌(S.aureus)、艰难梭菌(Clostridium difficile)、芽孢杆菌属(Bacillus)、链球菌属(Streptococcus)、福氏志贺氏菌(Shigella flexneri)、幽门螺杆菌(Helicobacter pylori)、鼠伤寒沙门氏菌血清型、伤寒沙门氏菌血清型、双歧杆菌属(Bifidobacterium)、乳酸菌属(Lactobacillus)、真杆菌属(Eubacterium)、罗氏菌属(Roseburia) 和瘤胃球菌属(Ruminococcus) 。
这表明:
AI-2系统在细菌的种间通讯中扮演重要角色
例如,大肠杆菌、肺炎链球菌等细菌就利用AI-2系统来调节编码细菌素产生的基因表达。
双歧杆菌也使用AI-2信号来对抗沙门氏菌的感染。此外,AI-2还可能参与调节细菌的运动性、生物膜形成等行为。
有趣的是,在肠道微生物群落中,AI-2信号也被用来帮助细菌应对胃肠道中的各种应激条件。研究发现,通过调控AI-2水平,可以在抗生素治疗后帮助恢复肠道微生物的平衡。但这需要谨慎控制,因为AI-2的细胞内水平受到LsrK激酶的调节。
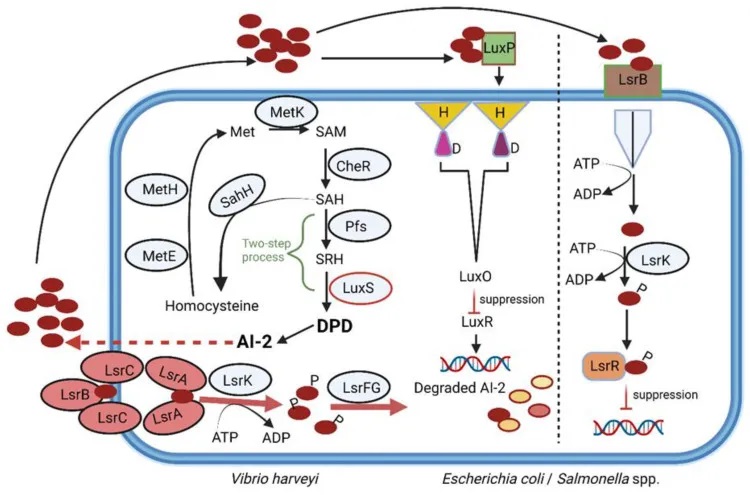
doi.org/10.3390/microorganisms10101969
AI-2在调节肠道微生物群中的应用
一种经改造的大肠杆菌菌株随着 AI-2 的产生增加,导致链霉素抑制的厚壁菌门恢复,并抑制拟杆菌门的生长。从这些发现可以推断,AI-2 可用于在抗生素治疗后恢复肠道微生物群的平衡。如果遵循这种策略,则必须仔细控制,因为 AI-2 的细胞质水平受 LsrK 激酶调节。
厚壁菌门和拟杆菌门种群的变化会改变 SCFA 的水平和组成,进而影响基因表达、细胞因子分泌和调节性 T 细胞诱导。所有这些变化都会影响炎症反应。因此,AI-2 水平的增加可以恢复厚壁菌门和拟杆菌门之间的平衡,并预防或逆转菌群失调、IBD、肥胖、自闭症和压力相关疾病。
利用AI-2来维持肠道微生物平衡,可以但需谨慎
一方面,AI-2确实可以帮助调节肠道细菌的行为,如影响细菌素的产生、生物膜的形成等。但另一方面,高浓度的AI-2也可能上调某些细菌的毒力因子,如增加粪肠球菌噬菌体的释放和基因转移。
在小鼠实验中,AI-2的给药虽然没有影响细胞因子的表达,但却加重了铜绿假单胞菌的肺部感染。这表明,单纯依赖AI-2来维持肠道稳态的做法可能会带来意想不到的后果。
AI-2受体的多样性及其生理影响
目前已经确定了两类AI-2受体,分别是LuxP和LsrB。这两种受体在结构上存在差异,序列相似性仅为11%。它们广泛分布于变形菌门、芽胞杆菌等细菌中。
其他肠道细菌可能也使用类似的受体来感知AI-2信号。有趣的是,有些细菌即使没有这些典型的AI-2受体,也能对AI-2产生反应,这可能涉及其他未知的感知机制。
此外,AI-2信号还可能影响细菌的耐药性等重要生理过程。同时,在结直肠癌相关肿瘤中也检测到AI-2水平的升高,这与肿瘤相关巨噬细胞中TNFSF9基因表达的增加相关。这提示AI-2可能是结直肠癌的一个重要生物标志物,值得进一步研究。
Interkingdom Communication
细菌利用复杂的群体感应系统来感知周围环境,并协调自身的生理活动。除了前面提到的自身诱导剂-2(AI-2)外,细菌还使用其他类型的信号分子进行细胞间通讯。
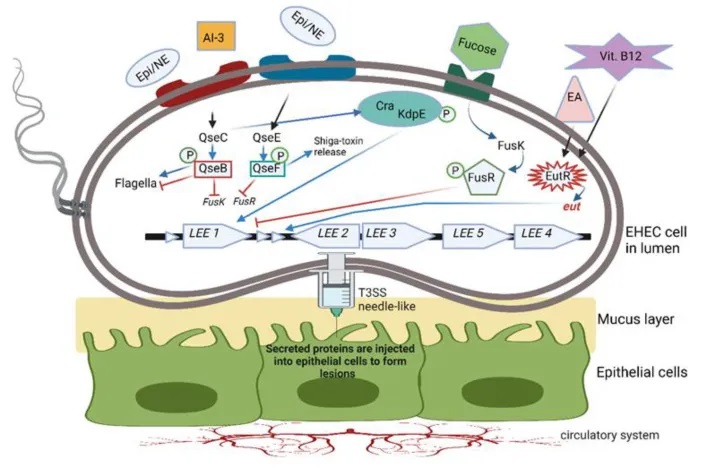
doi.org/10.3390/microorganisms10101969
AI-3信号系统在肠道病原体中的作用
其中,自身诱导剂-3(AI-3)/肾上腺素(Epi)/去甲肾上腺素(NE)界间信号系统在一些重要肠道病原体中发挥关键作用。这种信号系统可以促进鼠伤寒沙门氏菌、啮齿柠檬酸杆菌和肠出血性大肠杆菌(EHEC)等细菌的毒力基因表达。
以EHEC为例,它利用一种双组分QS系统(TCS)来感知和响应这些环境信号分子。该系统由QseBC和QseEF两个TCS组成。当EHEC感知到Epi、NE、AI-3、岩藻糖或乙醇胺等信号分子时,会激活跨膜组氨酸激酶受体,进而激活或抑制相应的反应调节因子。
例如,QseC组氨酸传感器可以激活QseB,QseB则调节鞭毛基因的表达,同时抑制参与岩藻糖代谢和毒力基因表达的fusK/-R基因。
QseC还可以磷酸化KdpE反应调节因子,与Cra蛋白一起刺激LEE操纵子中的基因,从而促进EHEC的黏附和毒力因子的注射。此外,QseC还可以激活QseF,进而诱导志贺毒素的产生。
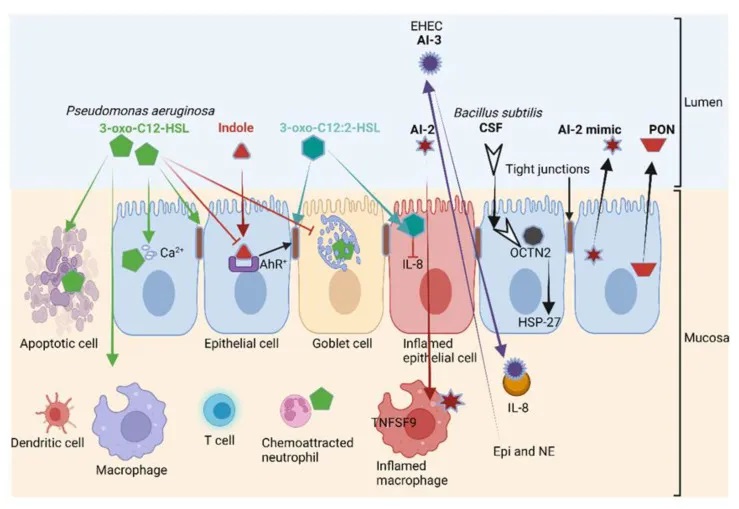
细菌群体感应系统对宿主免疫的影响
另一方面,铜绿假单胞菌(P. aeruginosa)产生的 3-oxo-C12-HSL(下图)在上皮细胞和免疫细胞中积极转运,并通过抑制编码紧密连接蛋白(TJ)的基因表达来破坏肠壁的通透性。这导致occludin、tricellulin、ZO-1、ZO-3、JAM-A、E-cadherin、β-catenin的重新排列(错位),并阻止粘蛋白的产生。
这不仅使上皮细胞受到感染,还会激活粘膜免疫系统,导致白细胞增加和促炎细胞因子的积累。此外,3-oxo-C12-HSL 还抑制肿瘤坏死因子 TNF-α 和 IL-12 的产生,导致 T 辅助细胞-1(Th1)反应功能障碍,并刺激 Th2 产生免疫球蛋白 G1。
抑制 Th1 和 Th2 T 淋巴细胞分化会增加细胞因子的产生 ,加剧氧化应激,刺激细胞凋亡并灭活线粒体。这些细菌间通信机制的研究,有助于我们更好地理解和预防细菌性疾病的发生。
doi.org/10.3390/microorganisms10101969
铜绿假单胞菌产生的 3-oxo-C12-HSL诱导各种细胞类型(包括上皮细胞)的细胞凋亡;破坏紧密连接并减少粘蛋白的产生。
3-oxo-C12-HSL与3-氧代-C12:2-HSL对肠壁影响相反
结构相似的 3-oxo-C12-HSL、3-oxo-C12:2-HSL 对肠壁具有相反的作用。
3-oxo-C12:2-HSL不是破坏上皮细胞的稳定性,而是保护紧密连接蛋白occludin和tricellulin以及细胞质ZO-1免受促炎细胞因子(如干扰素-γ γ)、TNF-α和IL-8的侵害。
3-oxo-C12:2-HSL在肠道免疫细胞中的作用及对IBD的潜在影响
除了一些开创性研究外,3-oxo-C12:2-HSL 对人类肠道免疫细胞的影响在很大程度上仍然未知。
在诊断为IBD的患者中,3-oxo-C12:2-HSL的浓度要低得多。这表明 3-oxo-C12:2 HSL 在保护暴露于免疫冲击的上皮细胞中起重要作用。需要进一步的研究来确定 3-oxo-C12:2-HSL 是否可以用于治疗 IBD。这也需要更好地了解 3-oxo-C12:2-HSL 群体猝灭、AHL 裂解和高丝氨酸内酯(HSL)环水解所涉及的过程。
到目前为止,已在人类和其他哺乳动物的GIT中鉴定出三种参与HSL环水解的对氧磷酶(PON1、PON2和PON3)。其中,PON2 最活跃,主要在空肠中表达 。PON1 和 PON3 在诊断为克罗恩病和溃疡性结肠炎患者中的表达水平较低 。
因此,这些胃肠道疾病有可能通过恢复 PON1 和 PON3 水平来逆转。需要深入研究对氧磷酶在 GIT 不同领域中的作用,以及它们在治疗胃病中的可能应用。
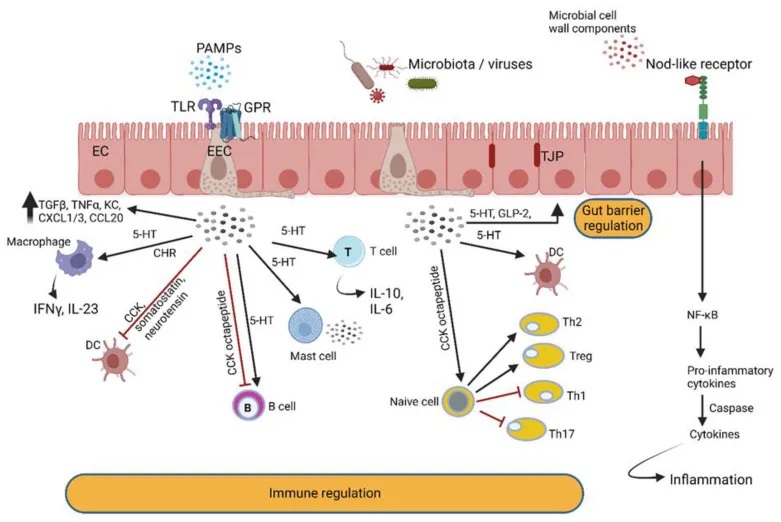
肠道是人体与外界环境直接接触的重要器官,也是细菌与宿主细胞进行密切交流的场所。除了前面提到的自身诱导剂-2(AI-2)和自身诱导剂-3(AI-3)等细菌信号分子外,肠道细胞还能感知和响应其他细菌代谢产物。这些相互作用对维持肠道健康至关重要。
宿主对细菌信号的感知机制
首先,肠道细胞表达一些重要的受体,如:孕烷X受体(PXR)、过氧化物酶体增殖物激活受体(PPAR),这些受体能够感知细菌代谢物,并调节宿主细胞的基因表达,参与解毒、代谢和炎症反应的调控。
例如,PPARγ可以抑制巨噬细胞的活化,减少炎性细胞因子的产生,从而有助于缓解炎性肠病。
细菌-宿主信号相互作用网络
另一方面,细菌也能感知和利用宿主细胞的信号分子。例如,假单胞菌产生的PQS和HHQ信号分子,能够与淋巴细胞、树突状细胞和巨噬细胞相互作用,抑制宿主的先天性和适应性免疫反应。
作为应对,宿主细胞则通过激活芳烃受体(AhR)来感知这些细菌信号,并启动有益的免疫反应,如IL-22和IL-17的表达。
此外,枯草芽孢杆菌产生的CSF信号分子,能够与肠道上皮细胞表面的OCTN2转运蛋白结合,激活HSP-27,从而促进肠道屏障的完整性。一旦进入细胞,CSF还可以作为一种”报告器”,监测肠道微生物群的变化。
总结下,肠道细胞与细菌之间存在复杂的信号交流网络。宿主细胞通过感受细菌代谢物来调节自身的生理活动,而细菌也能利用宿主信号分子来调控自身的行为。这种跨界的“窃听”行为,体现了宿主-微生物之间精细的相互调控机制,对维持肠道健康至关重要。进一步研究这些相互作用过程,有助于我们更好地理解和调控肠道微生态平衡。
PapRIV
由芽孢杆菌产生的 PapRIV 穿过胃肠道,尽管速度很慢,并进入循环系统,大多数肽(87%)从那里以单向方式穿过 BBB(血脑屏障)。从体外研究中可以推断,PapRIV 激活小胶质细胞,因此可能在肠-脑相互作用中发挥作用。
氨基酸
根据2020比利时一项研究,氨基酸天冬氨酸和脯氨酸在小胶质细胞的激活中起关键作用。apRIV还诱导促炎细胞因子IL-6和TNFα的产生,增加细胞内ROS并刺激阿米巴样细胞的增加。丙酮丁醇梭菌(Clostridium acetobutylicum)产生的自体诱导肽(AIP)比肺炎链球菌产生的 AIP 更容易穿过(血脑屏障)。

自诱导肽 AIPs
自诱导肽(Autoinducer peptides, AIPs)是细菌用于细胞间通信的一类信号分子。它们在细菌群体感应中起重要作用。
革兰氏阳性菌穿过肠壁产生的AIPs已在Caco-2细胞的体内研究中得到证实。该研究表明,胃肠道中葡萄球菌(Staphylococcus)、链球菌(Streptococcus)、乳酸杆菌(Lactobacillus)和芽孢杆菌(Bacillus)产生的AIPs对肌肉细胞具有促炎和抗炎作用。
这些屏障的穿越似乎取决于肽的结构和大小。信号肽降解过程中产生的可扩散小分子,称为可扩散信号因子(DSF),也可能起到自诱导剂的作用。
肠道细菌产生的信号由肠壁中的特殊细胞记录(下图),导致身体或心理健康发生暂时或持久的变化。这些细胞通过使用模式识别受体 (PRR)区分本地(地方性)和外来、潜在致病性微生物群产生的信号。
AHL
细菌在生长过程中会产生各种化学信号分子,比如AHL(丙酰基-亮氨酸同系物)。这些信号分子可以被宿主细胞感知到,并引发一些生理反应。
比如,AHL可以增加中性粒细胞的活性,促进成纤维细胞向肌成纤维细胞的转化。这些变化对于组织的修复和再生非常重要。
宿主细胞有一些专门的受体,可以识别这些细菌信号分子。其中包括Toll样受体(TLR)和Nod样受体(NLR),它们可以感知细菌细胞壁的成分。还有一些G蛋白偶联受体(GPR),它们可以感知一些与激素相关的信号分子。
这些受体广泛存在于肠道的免疫细胞、上皮细胞和间质细胞等,在调节炎症反应和维护肠道屏障功能中起重要作用。
总的来说,细菌通过释放各种化学信号分子,可以与宿主细胞进行”对话”(下图),影响宿主的免疫反应和组织修复。这种细菌-宿主之间的相互作用,对于维持肠道健康非常关键。我们需要进一步研究这些复杂的交互机制,以更好地理解和调控肠道微生态平衡。
doi.org/10.3390/microorganisms10101969
肠壁中的肠内分泌细胞 (EEC) 检测肠道细菌和微生物代谢物,并通过分泌与免疫细胞反应的肽激素和细胞因子来做出反应。EEC 产生的激素调节肠道屏障功能并与肠神经反应。后者通过迷走神经与中枢神经系统进行交流。
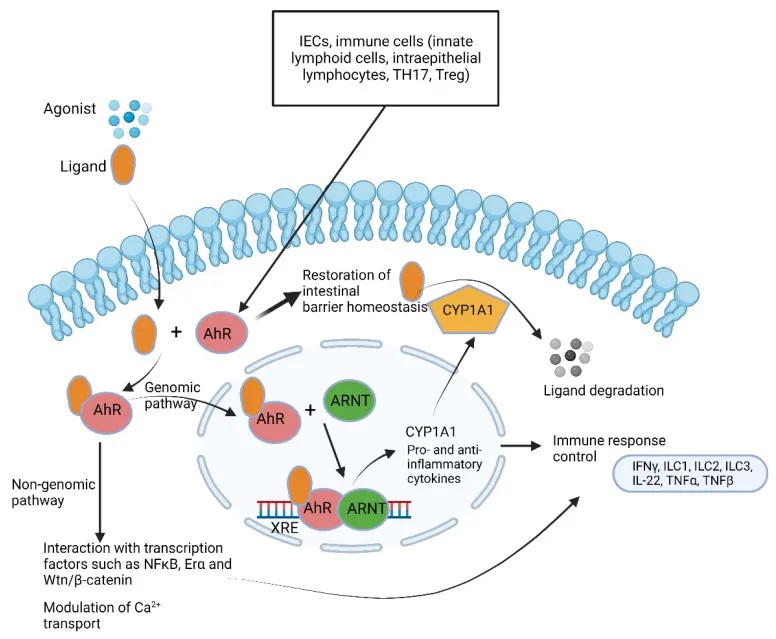
芳香烃受体 AhRs
AhRs 调节免疫反应和发病机制(下图)。大量的AhR由肠上皮细胞和免疫细胞(如先天性淋巴细胞、上皮内淋巴细胞、TH17细胞和Treg细胞)表达,但也存在于肝脏、肺、膀胱和胎盘中。AhR 的关键功能之一是恢复屏障稳态,这是 IBD 中突出的现象。
芳烃受体(AhR) 通路总结
doi.org/10.3390/microorganisms10101969
AhR
芳香烃受体在维护肠道健康中的关键作用
芳香烃受体(Aryl hydrocarbon Receptor, AhR)是一种重要的细胞内受体蛋白,在人体内广泛分布,尤其集中于肝脏、肠道和免疫细胞等组织。
AhR的基本功能
AhR能够感知和结合各种芳香烃类化合物,包括细菌代谢产物、环境污染物等。
当AhR与这些化合物结合后,会进入细胞核内,调控一系列基因的表达,参与解毒代谢、免疫调节、细胞分化等重要生理过程。
AhR与疾病的关联
例如,AhR可以感知细菌产生的PQS信号分子,并激活一些有益的免疫反应,如促进IL-22和IL-17的表达,增强肠道屏障功能,维持肠道微生态平衡。
研究发现,在炎症性肠病(IBD)、代谢综合征或乳糜泻等疾病患者中,粪便中AhR配体的浓度较健康人明显降低,AhR活性也相应降低。而在艰难梭菌感染患者中,则检测到更高的吲哚浓度。
AhR可以被多种细菌代谢产物激活,包括色氨酸降解产物(吲哚、吲哚并[3,2-B]咔唑、吲哚乙酸等)以及假单胞菌产生的2,4-二羟基喹啉、喹诺酮衍生物等。
这些AhR配体的浓度在炎症性肠病、代谢综合征和乳糜泻患者中较健康人有所降低,提示AhR活性的降低可能与这些疾病的发生有关。
AhR在调节肠道屏障和免疫稳态中的作用
AhR参与调节肠道屏障完整性、免疫稳态、上皮内淋巴细胞的产生和存活、炎症反应以及肠道菌群密度等多个关键过程。
通过调节紧密连接蛋白的表达、促进IL-22和IL-10的产生、诱导抗菌肽分泌等,AhR在维持肠道健康中发挥重要作用。
在炎症性肠病患者中,AhR水平降低,尤其是在回肠部位,可能导致ILC1细胞增多,进而促进炎症因子IFN-γ和TNF-α的产生,破坏肠屏障。
AhR配体的来源与饮食影响
色氨酸是AhR配体的重要来源,但人体无法自行合成,需要从饮食中摄取。富含色氨酸的食物包括腰果、向日葵籽、奶酪、鸡胸肉和煮鸡蛋等。
因此,饮食中色氨酸的摄入量可能影响AhR配体的水平,进而影响肠道健康。
耐药菌与群体淬灭技术的兴起
五年前,世界卫生组织公布了一份对目前使用的抗菌剂最具耐药性的病原菌清单。下列物种在名单上名列前茅:
这促使许多科学家研究使用抗QS疗法(称为群体淬灭(quorum quenching,QQ)来预防或控制细菌感染的可能性。近年来,许多已发表的文章报道了有希望的结果,表明与抗生素共同处理时,可以降低微生物的致病性并且更容易根除。

2020年,天津大学的研究团队发现,可以利用QS信号分子丙酰基-亮氨酸同系物(AHL)来控制革兰氏阴性细菌引起的感染。这种方法的关键是要破坏QS对细菌毒力基因表达的调控作用。
群体淬灭的应用效果与风险
当应用群体淬灭时,粘膜相关病原体减少了约60%,主要是梭菌属(Clostridium)。尽管从感染管理的角度来看很有希望,但这种剧烈的变化可能导致促炎性疾病的发展,如囊性纤维化、硬化症和IBD,以及肠球菌(Enterococcus)和艰难梭菌(C. difficile)细胞数量的增加。
也许最令人担忧的是,在luxS QS系统功能失调或缺失的细菌中,细胞聚集和生物膜形成增加的证据,如幽门螺杆菌、霍乱弧菌(Vibrio cholerae)、放线菌集聚杆菌(Aggregatibacter actinomycetemcomitans)、胸膜肺炎放线杆菌(Actinobacillus pleuropneumoniae)、副猪嗜血杆菌(Haemophilus parasuis)、金黄色葡萄球菌(S. aureus)、表皮链球菌(S. epidermidis)、变形链球菌(Streptococcus mutans)、粪肠球菌(Enterococcus faecalis)、蜡样芽孢杆菌(Bacillus cereus)。美罗培南和左氧氟沙星刺激鲍曼不动杆菌(A. baumannii)外排泵的表达,促进AHL的释放,导致QS介导的生物膜形成增加。
群体淬灭技术的局限性与挑战
实际应用中也存在一些挑战。有研究报告,使用群体淬灭方法治疗铜绿假单胞菌和金黄色葡萄球菌感染的效果有限,特别是在生物膜相关感染中。这可能是因为QQ会导致细菌群落结构的剧烈变化,反而促进了一些致病菌的生长,如艰难梭菌等。
此外,一些细菌在缺失LuxS等QS系统时,反而表现出更强的细胞聚集和生物膜形成能力,这可能导致难以根除的持久性感染。一些抗生素如美罗培南和左氧氟沙星,也可能通过刺激细菌外排泵表达,促进AHL释放,增强QS介导的生物膜形成。
因此,单纯使用群体淬灭技术可能并不能完全解决细菌感染问题。研究人员建议,在使用群体淬灭疗法时,还需要结合具体的细菌感染情况,评估其与抗生素联合使用的效果。
此外,细菌也可能通过一些机制,如外排泵基因突变,对群体淬灭疗法产生耐药性。这可能导致难以根治的持久性感染。
群体感应分子对中枢神经系统的影响研究不足。几种QS peptides(群体感应肽分子,QSP)可通过肠粘膜扩散并进入循环系统,从那里它们可以穿透血脑屏障(BBB)。基于这些发现,QSP 可能在肠道微生物组和大脑之间的交流中发挥关键作用。如果是这种情况,QSP 可能会影响神经发育并引发神经退行性疾病。需要进一步的研究来证实这些发现。
金黄色葡萄球菌外毒素对神经系统的影响
金黄色葡萄球菌产生的外毒素激活转录因子辅助基因调节因子(Agr)A,该调节多种基因的表达,包括毒力因子、成孔毒素(PFT) 和细菌蛋白酶。这些毒素会增加细胞内的钙水平,导致感觉神经元的激活。
金黄色葡萄球菌产生的一种毒素叫做PSMS,它能特异性地结合到宿主细胞表面的一种受体叫做FPR(甲酰肽受体样蛋白)。
FPR与另外两种受体b-防御素和MRGPRX2(mas相关G蛋白偶联受体X2)的配体有结构相似性。这表明,MRGPRX2可能也参与了PSMS引起的皮肤过敏等症状。
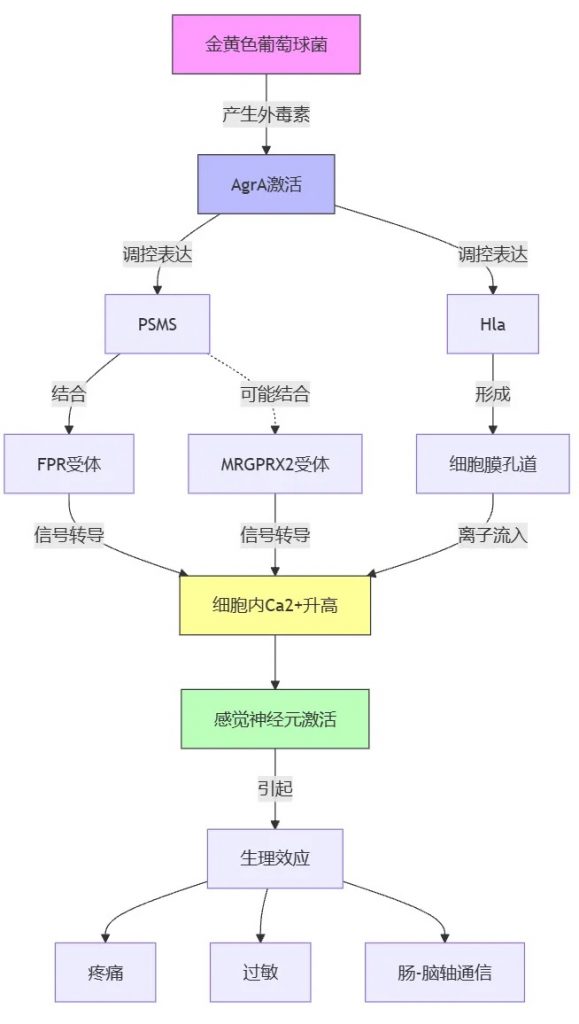
此外,FPR在肠道感觉神经节和背根神经节中有广泛表达,并与肠-脑轴(GBA)中涉及群体感应依赖性通路有关。
此外,金黄色葡萄球菌产生的毒素叫做α-溶血素(Hla),它也能通过增加细胞内钙的转移来激活神经元。与PSMS相比,Hla在细胞膜中形成的孔更小、破坏性也更小。
总之,肠道病原菌类似于金黄色葡萄球菌产生的这些毒素,通过作用于宿主细胞表面的不同受体,如FPR和MRGPRX2,从而引起神经元的激活,导致疼痛、过敏等症状。这些毒素在肠-脑轴通信中也可能发挥重要作用。

根据英国和澳大利亚联合的一项研究结果表明:与PSMS相比,Hla在细胞膜中产生更小、破坏性更小的孔。作者还观察到了 Adam10 的表达,Adam10 是一种在感觉神经元中产生的膜结合金属蛋白酶,Hla 与该酶结合。外毒素在肠脑轴通信中的重要性尚不清楚。
然而,由于金黄色葡萄球菌(S. aureus)与肠易激综合征和食物有关,这些QS分子有可能直接调节肠-脑通讯和肠道反射。

上面这篇研究在6种不同的神经元细胞系上筛选了85个群体感应肽,发现了22个可能对肠脑轴有影响的肽。其中,4 个肽诱导神经突生长,2 个肽抑制神经生长因子(NGF)诱导的神经突生长,8 个肽诱导人 SH-SY5Y 神经母细胞瘤细胞的神经突生长。2 个肽杀死 SH-SY5Y 细胞,6 个肽诱导 IL-6 表达或一氧化氮(NO)产生。
细胞壁成分在肠脑轴中的作用
已经发表了几篇关于细胞壁成分(如脂多糖、多糖和肽聚糖)在神经元激活和肠脑轴中的作用的报道。细胞壁成分还诱导神经肽、ATP 和细胞因子的释放。短链脂肪酸、色氨酸、微量胺和外毒素也具有神经调节剂特性。
5-羟色胺和组胺刺激神经末梢附近的肥大细胞。
阿尔茨海默病(AD)、自闭症谱系障碍(ASD)、多发性硬化症(MS)、帕金森病(PD) 和肌萎缩侧索硬化症(ALS)等神经元疾病与功能失调的小胶质细胞有关。
将患有注意力缺陷多动障碍(ADHD)、阿尔茨海默病和帕金森病的人类粪便移植到小鼠身上,激活了大脑中的小胶质细胞,并加剧了认知和身体损伤。
这些发现以及更多证据表明微生物群失调与神经发育、神经退行性和精神疾病(如自闭症谱系障碍、精神分裂症、阿尔茨海默病、重度抑郁症和帕金森病)之间存在明确联系,促使研究人员更仔细地研究肠脑轴。
有关肠道细菌和神经递质的更多信息,读者可关注谷禾健康以往其他分享文章。
扩展阅读:
肠道菌群检测报告解读——肠道菌群代谢产物包括激素,神经递质等
胃肠道特殊的环境下生存着海量的细菌微生物,因此它们已经开发出与宿主细胞交流的机制也就不足为奇了。
一些群体感应分子是属特异性的,但少数被革兰氏阴性菌和革兰氏阳性菌使用。Epi 和 NE 等激素以及某些碳水化合物(例如岩藻糖和 EA)激活细菌中的特定受体,进而触发细胞质中的传感器以调节基因表达。
在健康的胃肠道中,这些信号分子对于维持稳态状态很重要。一些 QS 分子,如 3-oxo-C12:2-HSL,保护紧密连接蛋白,在治疗肠漏综合征中可能很重要。
一些 QS 分子刺激肿瘤生长并与特定癌症的发展密切相关,而另一些则与神经系统疾病有关。穿透血脑屏障(BBB)的 QSP 构成了一个值得更多研究的领域,特别是因为肠道微生物组越来越被认为是神经精神病学的关键参与者。
通过更深入地了解肠道细菌产生的不同 QS 系统,未来可能能够开发可用于诊断神经和精神疾病的生物标志物,例如焦虑和抑郁、MDD、精神分裂症、双相情感障碍、自闭症和强迫症。
主要参考文献:
Dicks LMT. How does Quorum Sensing of Intestinal Bacteria Affect Our Health and Mental Status? Microorganisms. 2022 Oct 5;10(10):1969.
Janssens Y., Debunne N., De Spiegeleer A., Wynendaele E., Planas M., Feliu L., Quarta A., Claes C., Van Dam D., De Deyn P.P., et al. PapRIV, a BV-2 microglial cell acti-vating quorum sensing peptide. Sci. Rep. 2021;11:10723.
De Spiegeleer A., Elewaut D., Noortgate N.V.D., Janssens Y., Debunne N., Van Langenhove S., Govindarajan S., De Spiegeleer B., Wynendaele E. Quorum sensing molecules as a novel microbial factor impacting muscle cells. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2019;1866:165646.
Miller M.B., Bassler B.L. Quorum Sensing in Bacteria. Annu. Rev. Microbiol. 2001;55:165–199.
Coquant G., Aguanno D., Pham S., Grellier N., Thenet S., Carrière V., Grill J.-P., Seksik P. Gossip in the gut: Quorum sensing, a new player in the host microbiota interactions. World J. Gastroenterol. 2021;27:7247–7270.
Yashiroda Y., Yoshida M. Intraspecies cell–cell communication in yeast. FEMS Yeast Res. 2019;19:foz071.
Prescott R., Decho A.W. Flexibility and Adaptability of Quorum Sensing in Nature. Trends Microbiol. 2020;28:436–444.
Tam K., Torres V.J. Staphylococcus aureus Secreted Toxins and Extracellular Enzymes. Microbiol. Spectr. 2018;7:GPP3-0039-2018.
Fülling C., Dinan T.G., Cryan J.F. Gut Microbe to Brain Signaling: What Happens in Vagus…. Neuron. 2019;101:998–1002.
Krasulova K., Illes P. Intestinal interplay of quorum sensing molecules and human receptors. Biochimie. 2021;189:108–119.
Galloway W.R.J.D., Hodgkinson J.T., Bowden S.D., Welch M., Spring D.R. Quorum Sensing in Gram-Negative Bacteria: Small-Molecule Modulation of AHL and AI-2 Quorum Sensing Pathways. Chem. Rev. 2010;111:28–67.
Wu S, Liu J, Liu C, Yang A, Qiao J. Quorum sensing for population-level control of bacteria and potential therapeutic applications. Cell Mol Life Sci. 2020 Apr;77(7):1319-1343.
Krzyżek P. Challenges and Limitations of Anti-quorum Sensing Therapies. Front. Microbiol. 2019;10:2473.
Wu S., Liu J., Liu C., Yang A., Qiao J. Quorum sensing for population-level control of bacteria and potential therapeutic applications. Cell. Mol. Life Sci. 2019;77:1319–1343.
Janssens Y, Wynendaele E, Verbeke F, Debunne N, Gevaert B, Audenaert K, Van DeWiele C, De Spiegeleer B. Screening of quorum sensing peptides for biological effects in neuronal cells. Peptides. 2018 Mar;101:150-156.


病原微生物和/或有害微生物成功入侵、繁殖并定居于宿主的体内或体内,从而导致健康障碍,称为感染。简单地说,它可以定义为由微生物引起的疾病。感染也被称为传染病或传染病或传染性疾病。
感染每年导致 1300多万 人死亡;2019 年死亡人数为 1370万人(新英格兰医学,2022年统计)。在这 1370 万人死亡中,有 770 万人与细菌感染有关。由于抗菌素耐药性的迅速出现和蔓延,与传染病相关的病例严重程度和死亡率也在增加。
感染和疾病是两个经常互换使用的术语,但它们的含义截然不同。了解两者之间的区别对于有效预防、诊断和治疗至关重要。
感染是指有害微生物(如细菌、病毒或真菌)侵入人体。这些病原体可以通过各种途径进入人体,如呼吸系统、消化系统或皮肤等。感染可以是局部的,也可以是全身性的,程度可轻可重。
疾病是指由于感染或其他原因而出现的症状或异常的表现。感染是原因,而疾病是结果。简而言之,感染是体内存在病原体,而疾病是感染导致的症状的表现。感染可能只是局部的轻微症状,而疾病则是影响身体正常功能的更严重的情况。在某些情况下,如果免疫系统无法控制微生物的传播,感染可能会发展成疾病。
在诊断时,区分感染和疾病非常重要。一个人可能感染了病原微生物,但没有表现出任何疾病症状。另一方面,一个人可能患病但没有活动性感染。在这种情况下,疾病可能是由感染以外的因素引起的,例如遗传易感性或环境诱因。
虽然预防策略主要侧重于避免感染,但防止感染发展为疾病也同样重要。及早发现和适当治疗感染有助于防止发展为严重疾病。比如,长期的呼吸道或肠道感染被认为是相关肿瘤发生的高风险因素之一。
因此,早期识别感染、诊断、治疗和预防是控制感染和疾病的重要组成部分。感染可以用抗生素或抗病毒药物等药物治疗,而疾病可能需要更专业的治疗方案。准确诊断潜在病因对于有效治疗至关重要。
疾病的定义,原因,诊断
// 定义
疾病是一种以身体或精神功能异常为特征的医疗状况。它通常由感染或受伤等外部因素引起,但也可能是由内部失衡或遗传倾向造成的。疾病的严重程度各不相同,会影响身体的不同器官或系统。
// 原因
疾病有多种原因,包括:
// 症状
疾病可表现出各种症状,包括疼痛、疲劳、发烧和身体功能异常,以及癌症。疾病的症状会因具体情况而有很大差异。
常见症状包括发烧、疲劳、疼痛、炎症和身体功能变化。某些疾病还可能导致与受影响器官或系统相关的特定症状,例如呼吸系统疾病的咳嗽或胃肠道疾病的消化问题。
疾病的形成方式多种多样。有些疾病,例如由遗传性疾病引起的疾病,是天生的。其他疾病可能是通过接触有害物质或环境因素而获得的。感染也会导致疾病的发展。当病原体侵入人体并造成伤害时,免疫反应可能会引发炎症、组织损伤和其他导致疾病的变化。
// 诊断和预防
诊断疾病通常需要结合病史、体格检查、实验室检查和影像学检查等。
预防疾病对于保持整体健康至关重要。这可以通过各种方式实现,例如保持良好卫生习惯、接种疫苗、保持健康的生活方式以及避免危险因素。早期发现和治疗感染也有助于防止其发展成更严重的疾病。
疾病的治疗包括药物治疗、手术治疗、其他疗法或这些方法的组合。
总之,疾病涉及身体或精神的异常功能,可能由感染、受伤、遗传因素或失衡引起。识别症状、诊断和了解疾病的原因对于有效预防和治疗疾病至关重要。
感染的定义,原因,诊断
// 原因
感染是一个术语,用于描述有害微生物在体内的入侵和繁殖。这些微生物被称为病原体,包括细菌、病毒、真菌和寄生虫。当这些病原体进入人体时,它们会引起感染。
感染的原因多种多样,包括细菌、病毒、真菌和寄生虫。这些微生物可以通过各种途径进入人体,例如吸入、摄入或通过皮肤破损。
// 症状
虽然感染的严重程度各不相同,但它们通常具有共同的症状。这些症状包括发烧、疲劳、咳嗽、打喷嚏、喉咙痛和炎症。在某些情况下,感染还可能导致更具体的症状,具体取决于所涉及的病原体类型。
感染可能是局部的,即传染源仅感染特定的器官或组织,或者可能是全身性的,即传染源通过血液或淋巴到达身体的不同部位,从而感染不同的器官和组织。
感染可能不会导致疾病等特定症状,因为大多数感染往往是亚临床的。相反,其他感染可能导致严重的症状和并发症。
虽然有些感染可能不被宿主的免疫系统察觉并自行消退,但其他感染可能会引起症状并发展为疾病。
// 预防与治疗
预防是控制感染的一个重要方面。这可以通过保持良好的卫生习惯来实现,例如经常洗手和避免与受感染者密切接触。疫苗接种也可以通过提供针对特定病原体的免疫力来帮助预防某些感染。
预防在降低感染和疾病风险方面发挥着重要作用。保持良好的卫生习惯、接种疫苗和避免与受感染者接触等措施有助于防止感染传播。此外,保持健康的生活方式和增强免疫系统可以降低感染发展为疾病的可能性。
一旦感染,治疗可能涉及使用抗生素、抗病毒药物、抗真菌药物或抗寄生虫药物,具体取决于引起感染的具体微生物。在某些情况下,支持性护理(如休息、补液和缓解症状的药物)可能足以恢复。
// 原因与传播
感染的原因可能因所涉及的微生物类型而异。例如,细菌可通过直接接触受污染的表面、咳嗽或打喷嚏时产生的飞沫,或通过食用受污染的食物或水引起感染。另一方面,病毒通常通过呼吸道飞沫、与受感染者的直接接触或通过受污染的表面传播。
总之,了解感染的原因和传播方式对于预防、诊断和治疗感染至关重要。通过保持良好的卫生习惯、采取适当的预防措施并及时就医,个人可以降低感染风险并最大程度地减少相关疾病的影响。
感染的诊断是基于通过不同的诊断过程对传染源的识别。
某些感染可能会表现出可用于症状诊断的症状,但通常需要进一步确认。感染直接取决于传染源以及宿主对该传染源的免疫反应。
现在我们认为手术是理所当然的,但不久前,即使是最小的手术,如果感染进入体内,也可能是致命的。消毒为我们提供了一种预防手术感染和确保手术安全的方法。
消毒法是使用化学物质(称为防腐剂)来消灭引起感染的细菌的方法。它是由英国外科医生约瑟夫·李斯特发明的。
约瑟夫·李斯特找到了一种预防手术期间和手术后伤口感染的方法。他是第一个将细菌理论的科学应用于外科手术的人。李斯特消毒系统是现代感染控制的基础。
感染和疾病的区别在于症状的严重程度以及对个人整体健康的影响。
感染的定义是细菌、病毒和真菌等微生物在体内的入侵和生长。
感染可能由多种因素引起,包括接触病原体、卫生条件差、免疫系统受损以及食物或水受污染。感染的症状因感染类型和部位而异,但通常包括发烧、疼痛、炎症和疲劳。
疾病是指影响身体或精神功能的特定状况。
疾病可能是由感染引起的,但也可能是由其他因素引起的,例如遗传异常、环境毒素、生活方式选择或自身免疫反应。疾病的症状范围从轻微到严重,并且可能持续很短时间或变成慢性病。
感染的诊断通常涉及检测体液或组织样本以确定病原体的存在。另一方面,疾病的诊断通常需要结合病史、体格检查、实验室检查和影像学检查。
在治疗方面,感染通常使用抗生素、抗病毒药物或抗真菌药物治疗。治疗的目的是消除入侵的微生物并缓解症状。另一方面,疾病可能需要更全面的方法,包括药物治疗、手术、改变生活方式和支持疗法。
大致的区别总结如下:

总之,虽然感染和疾病相关,但它们的定义、原因、症状和治疗方法不同。了解两者的区别可以帮助医护人员和个人采取适当的措施来预防和控制疾病。
感染常见类型
细菌感染是由有害细菌进入人体引起的。它们会影响身体的不同部位,例如呼吸系统、泌尿道或皮肤。常见症状包括发烧、疼痛、肿胀和发红。细菌感染通常用抗生素治疗。
病毒感染是由病毒引起的。它们可导致各种疾病,例如普通感冒、流感或 COVID-19。症状从轻微到严重不等,可能包括发烧、咳嗽、喉咙痛和疲劳。病毒感染通常会自行痊愈,但有些可能需要抗病毒药物。
真菌感染是由真菌引起的,例如酵母菌或霉菌。它们会影响皮肤、指甲或内脏器官。常见的真菌感染包括足癣、酵母菌感染和癣。症状可能包括瘙痒、发红和不适。真菌感染的治疗方法包括抗真菌药物和外用药膏。
寄生虫感染是由寄生在人体内或体表的寄生虫引起的。寄生虫感染的例子包括疟疾、虱子感染和贾第虫病。症状可能因寄生虫类型而异,但可能包括发烧、瘙痒、腹泻和疼痛。寄生虫感染的治疗可能涉及抗寄生虫药物。
性传播感染是通过性接触传播的感染。常见的性传播感染包括衣原体感染、淋病和疱疹。症状范围从轻微到严重,可能包括生殖器分泌物、疼痛和溃疡。性传播感染通常通过检测诊断,可以用抗生素或抗病毒药物治疗。
通过了解不同类型的感染、其症状、原因和治疗方法,个人可以采取预防措施来降低感染风险并在需要时寻求适当的医疗护理。
说到疾病和健康,有各种各样的疾病会影响人体。以下是一些常见的疾病类型:
– 传染性疾病
传染病是由细菌、病毒、真菌和寄生虫等微生物引起的。它们可以通过各种方式在人与人之间传播,包括直接接触、呼吸道飞沫和受污染的食物或水。传染病的例子包括普通感冒、流感、肺结核和艾滋病毒/艾滋病。症状和治疗方法因具体感染而异。
– 慢性疾病
慢性病是一种长期疾病,通常会随着时间的推移而发展,并且没有已知的治疗方法。这些疾病通常有多种原因,并可能受到遗传、生活方式选择和环境因素的影响。慢性病的例子包括心脏病、糖尿病、癌症和慢性阻塞性肺病 (COPD)。治疗通常侧重于控制症状和预防并发症。
其他常见疾病类型
– 自身免疫性疾病
当免疫系统错误地攻击健康细胞和组织时,就会发生此类疾病,从而导致炎症和组织损伤。例如类风湿性关节炎、狼疮和多发性硬化症。
– 心血管疾病
影响心脏和血管。包括冠状动脉疾病、心力衰竭和中风等。
– 呼吸系统疾病
这些疾病会影响肺部和呼吸。一些常见的呼吸系统疾病包括哮喘、慢性支气管炎和肺炎。
– 精神健康障碍
这些障碍会影响一个人的情绪、心理和社会健康。例如抑郁症、焦虑症和精神分裂症。
– 遗传性疾病
这些疾病是由遗传异常基因或突变引起的。这些疾病包括囊性纤维化、镰状细胞病和亨廷顿氏病等。
这些疾病的诊断、治疗和预防策略因具体情况而异。
预防感染
预防是感染和疾病管理的一个重要方面。虽然治疗和诊断很重要,但采取措施预防感染可以大大降低疾病的发病率。
预防感染的一个关键因素是了解病因和传播方式。感染可通过与感染者直接接触、受污染的表面或空气中的颗粒传播。通过了解传播途径,个人可以采取必要的预防措施,将感染风险降至最低。
定期洗手是预防感染最有效的措施之一。用肥皂和水洗手至少 20 秒或使用含酒精的洗手液有助于消除有害微生物。
此外,例如咳嗽或打喷嚏时捂住口鼻,有助于防止呼吸道感染的传播。
接种疫苗是预防感染的另一个重要方面。疫苗刺激免疫系统产生对抗特定病原体的抗体,从而提供免疫力。通过接种疫苗,个人可以保护自己并为社区免疫做出贡献,从而降低传染病的总体流行率。
教育和意识在预防感染方面也发挥着重要作用。了解常见感染的体征和症状可以及早发现并及时治疗,防止进一步的并发症。此外,提倡卫生习惯并提供有关适当预防措施的教育可以使个人能够做出明智的决定并保护自己和他人。
总之,预防是对抗感染和疾病的关键。通过采取积极措施,例如保持良好的卫生习惯、接种疫苗和提高认识,个人可以大大降低感染风险,并为整个社区的健康做出贡献。
预防疾病
预防在减轻疾病负担方面起着至关重要的作用。了解各种感染和疾病的原因、症状和诊断有助于制定有效的预防策略。
预防疾病不仅仅是避免感染。它涉及采取健康的生活方式并做出明智的选择以降低患病风险。这包括保持均衡饮食,定期进行体育锻炼,避免吸烟和过量饮酒等行为,这些行为会增加患某些疾病的风险。
定期筛查和检查对于疾病的早期发现和治疗也很重要。通过早期发现疾病,医护人员可以及时治疗,改善治疗效果并减轻疾病的总体负担。
感染和疾病预防还涉及教育公众预防的重要性,并提供必要的资源和服务。
总之,预防在减少疾病的发生和影响方面起着至关重要的作用。通过了解感染和疾病的原因、症状和诊断,实施预防措施并促进健康行为。
感染治疗
在治疗感染时,了解这些疾病的病因、预防和诊断非常重要。
治疗感染的关键因素之一是防止其传播。
感染的诊断对于有效治疗至关重要。医生可以使用多种方法,包括体检、实验室检查和成像技术来确定感染的原因并确定最合适的治疗方案。
感染的治疗方法取决于感染的类型和严重程度。细菌感染通常用抗生素治疗,而病毒感染则使用抗病毒药物。真菌感染使用抗真菌药物,寄生虫感染则使用抗寄生虫药物。
症状管理
除了针对感染的根本原因外,症状管理也是治疗的一个重要方面。这可能涉及使用非处方药来缓解疼痛、发烧和充血等症状。
治疗持续时间
感染的治疗时间各不相同。有些感染可能需要短期用药,而有些感染可能需要更长时间的治疗。即使症状有所改善,也必须完成处方药物的全部疗程,以确保感染完全根除。
总之,感染的治疗涉及多方面的方法,包括解决根本原因、症状管理和防止感染扩散。通过了解感染的原因、预防和诊断,医疗保健专业人员可以为患者提供最有效的治疗。
疾病治疗
疾病的治疗取决于具体症状和致病因素。
在感染病例中,治疗通常侧重于消除引起感染的生物体。这可能涉及使用抗生素、抗病毒药物或抗真菌药物,具体取决于感染的具体类型。在某些情况下,可以预防性地使用抗病毒或抗真菌药物,以防止感染传播给他人。
另一方面,疾病的治疗不仅仅是治疗感染本身。当疾病被诊断出来后,治疗的目的是缓解症状、控制并发症,并改善整体健康和福祉。
疾病的治疗可能涉及药物治疗、生活方式改变和支持疗法的结合。对于糖尿病或高血压等慢性疾病,主要重点通常是通过药物治疗和生活方式改变(包括定期锻炼和健康饮食)来控制病情。
在某些情况下,可能需要手术干预来治疗疾病。这可能涉及切除受感染或患病的组织、修复受损器官,或在受伤或退化的情况下恢复功能。
预防也是疾病治疗的一个重要方面。通过解决风险因素并实施预防措施,例如接种疫苗、定期健康检查和改变生活方式,可以减少疾病的发生和严重程度。
诊断在确定疾病的最适当治疗方法方面起着至关重要的作用。准确及时的诊断使医疗保健提供者能够根据每位患者的个人需求量身定制治疗计划。
总之,治疗疾病需要综合考虑具体症状、潜在病因和个体因素。通过了解感染和疾病之间的区别,医护人员可以就治疗方案做出明智的决定,并改善患者的治疗效果。
▼
与心血管系统相关的感染,包括血液,即心脏、血管和血液的感染,也称为血流感染或血液循环系统感染。与心血管系统相关的一些常见感染包括:

▼
消化系统是参与食物消化的身体系统。它包括胃肠道 (GI 道) 和相关消化器官。胃肠道两端开放,是外来、可能受污染的物质(包括食物和饮料)的停靠点。这使得消化系统极易受到感染。
幽门螺杆菌感染
消化系统感染包括胃肠道任何部位(从口腔到肛门)和任何消化器官(如肝脏、胰腺和胆囊)的感染。消化系统感染主要包括细菌、病毒、原生动物和寄生虫,真菌病原体感染较少。


▼
呼吸道感染 (RTI) 是指呼吸系统任何器官的感染。它是人类最常见的感染类型。根据感染呼吸道的部位,RTI 可分为上呼吸道感染 (URTI)(即鼻、鼻窦、咽和喉感染)和下呼吸道感染 (LRTI)(即气管、支气管、细支气管和肺感染)。

▼
神经系统感染是指大脑、脊髓和/或人体神经的任何形式的感染。此类神经系统感染可能危及生命。它们是由不同的微生物引起的,主要是病毒和细菌。
▼
泌尿系统包括尿道、膀胱、输尿管和肾脏。这些器官中的任何一个感染都称为尿路感染 (UTI)。UTI 主要由细菌引起,但也有真菌和病毒感染的报道,但这种情况很少见。由于女性尿道较短,因此女性比男性更容易患上 UTI。

▼
生殖系统包括所有参与生殖过程的器官。它在人类中与泌尿系统非常接近。许多引起尿路感染的微生物是造成生殖系统感染的原因。生殖系统感染分为三种类型:

▼
淋巴系统是淋巴管、淋巴结和淋巴器官的网络,淋巴液在其中流动。它是免疫系统的一部分,也是循环系统的一部分。淋巴感染并不常见,但有报道称存在多种细菌和寄生虫感染。

▼
外皮系统包括身体的所有外部覆盖物。它包括皮肤、头发和指甲。外皮系统是我们身体的第一层防御。它是数百万微生物作为正常菌群的家园。病原体在侵入身体之前首先与外皮接触。

▼
肌肉系统包括我们身体的所有肌肉。肌肉感染通常是血源性或传染性传播,但通常很严重,需要立即治疗。

骨骼:
▼
内分泌系统由内分泌腺组成,内分泌腺感染主要是通过血源性播散感染或进行性全身感染而引起。
在当今世界,传染病始终是一个威胁,了解感染的各个阶段对于有效诊断和治疗至关重要。从感染到康复的过程可以粗分为四个不同的阶段,每个阶段都有各自的症状和结果。
了解感染的四个阶段对于确定适当的行动方案至关重要,从预防和早期发现到治疗和康复。识别前驱期及其独特症状有助于早期干预,降低传播率并预防并发症。下面详细了解一下每个时期不同的特征及相关干预。
▼
潜伏期
感染的第一阶段称为潜伏期,在此期间,患者接触到病原体,但可能不会出现任何症状。潜伏期的长短取决于具体感染源,可能为几小时至数周不等。
最初接触病原体的方式多种多样,例如接触受污染的表面、空气中的颗粒物,或直接接触受感染者。在此期间,患者可能并未意识到自己已经接触到病原体。
潜伏期内可能出现前驱期
此阶段的特点是出现轻微症状,这些症状通常不具特异性,容易被忽视或归因于其他因素。前驱期的症状可能包括疲劳、头痛、轻微发烧和全身不适。
值得注意的是,并非所有感染都会经历前驱期。有些感染可能直接从潜伏期进入急性期,这取决于具体病原体和个人的免疫反应。
▼
潜伏期过后,感染进入第二阶段,即入侵和复制。在此阶段,病毒或细菌已成功侵入宿主体内,并开始快速繁殖。这导致体内病原体的浓度较高,从而导致症状的出现。
前驱期是侵袭和复制阶段的一部分,其特点是出现初始症状。这些症状可能是一般性的和非特异性的,例如疲劳、发烧、头痛和肌肉疼痛。前驱期是感染正在发展的警告信号,可让免疫系统为即将到来的战斗做好准备。
随着感染的进展,它会进入急性期,症状会变得更加严重,并且针对特定病原体。人体的免疫系统在各种防御机制的帮助下,试图对抗入侵的病原体。然而,在这个阶段,战斗仍在继续,结果尚不确定。
在慢性期,一些感染可以在体内持续很长一段时间。此阶段的特点是症状较轻,甚至完全没有症状。然而,病原体继续复制并慢慢损害身体组织。如果不及时治疗,慢性感染会导致长期并发症和后遗症。
在某些情况下,随着人体免疫系统成功消灭病原体,感染会自然消退。这会导致康复,症状逐渐消退,患者恢复正常健康状态。然而,在其他情况下,可能会出现并发症,如继发感染或器官损伤,这会延长康复过程,并可能需要额外的医疗干预。
值得注意的是,感染过程的各个阶段可能因具体病原体、个人免疫反应和医疗条件而异。
▼
经过潜伏期和症状出现后,感染者进入前驱期,预示着疾病急性期的开始。
在前驱期,症状逐渐恶化,变得更加明显。感染者可能会出现疲劳、不适和发烧等一般症状,以及特定感染特有的特定症状。
此阶段至关重要,因为它通常表明免疫系统正在积极对抗感染。然而,需要注意的是,并非所有感染都有明确的前驱期。有些人可能会直接从潜伏期进入急性期,而不会出现任何前驱症状。
可能出现并发症
如果不及时治疗或免疫系统无法有效控制感染,前驱期可能会导致进一步的并发症。这些并发症的严重程度可能因感染类型和个人的整体健康状况而异。
前驱期可能出现的一些并发症包括继发感染、器官损伤或感染扩散至身体其他部位。这些并发症会延长病程,增加重症甚至死亡的风险。
前驱期的恢复
对于大多数感染来说,前驱期标志着急性期的开始,在此期间症状达到顶峰。急性期过后,身体逐渐恢复并进入缓解期,症状开始消退,患者开始感觉好些。
从前驱期和随后的急性期恢复可能需要一些时间,因此遵循处方治疗并根据需要休息很重要。在某些情况下,即使感染痊愈后,也可能出现复发或出现长期后遗症,如慢性疲劳或器官损伤。
总体而言,了解前驱期的进展和症状对于有效管理和治疗感染至关重要。认识到这一阶段的开始可以帮助个人寻求适当的医疗护理并采取措施预防进一步的并发症。
▼
潜伏期结束后,感染者进入感染的急性期。在这个阶段,症状会完全显现,感染也会达到高峰。
前驱期是急性期的初始阶段,其特点是出现发烧、疲劳、头痛和肌肉疼痛等症状。在此阶段,感染者还可能感到全身不适。
严重感染的急性期通常持续数天至数周。此阶段的严重程度和持续时间可能因具体病原体和个人的免疫反应而异。
并发症和后遗症
在某些情况下,感染的急性期会导致并发症。这些并发症可能是由于病原体对身体的直接影响或身体对感染的免疫反应而引起的。常见的并发症包括肺炎、器官衰竭和继发感染。
一旦成功控制了急性期,感染者就会进入恢复期。在此阶段,身体会逐渐消除感染,症状开始改善。根据感染的严重程度,恢复可能需要几天到几周的时间。
在某些情况下,感染可能会发展为慢性阶段。当身体无法完全消除病原体时,就会发生这种情况,并且感染会持续很长时间。慢性感染可能导致长期健康问题,可能需要持续治疗。
值得注意的是,并非所有感染都会经历所有阶段,并且进展和症状可能因具体病原体和个体因素而异。
常见症状
在感染的这个阶段,可能会出现常见症状,这些症状可能因具体病原体和个人的免疫反应而异。值得注意的是,并不是所有人都会出现相同的症状,有些人的表现可能比其他人更严重。
在急性期,患者可能会出现发烧、疲劳、身体疼痛和头痛等症状。这些症状通常表明身体对感染产生了免疫反应,可能会持续数天。
急性期过后,患者可能会进入慢性感染期。在此阶段,症状可能会持续很长时间,有时持续数月甚至数年。常见的慢性症状包括持续疲劳、关节疼痛、肌肉无力和认知困难。
有些感染还可能产生后遗症,即感染导致的长期后果或并发症。这些后遗症的严重程度可轻可重,可能影响身体的各个器官或系统。后遗症的例子包括器官损伤、神经系统疾病和免疫功能受损。
复发是另一种可能的结果,尤其是某些病原体引起的感染。经过一段时间的明显缓解后,症状可能会再次出现,表明感染复发。这可能是由于各种因素造成的,例如病原体根除不彻底或休眠感染重新激活。
▼
在感染的呼吸道症状阶段,病毒已到达呼吸系统并开始影响肺部和呼吸道。此阶段通常是在前驱期结束后,发烧和疲劳等一般症状开始消退。
呼吸道症状的严重程度因个人和引起感染的特定病毒而异。常见症状包括咳嗽、呼吸急促、胸痛或不适以及喘息。这些症状通常表明呼吸系统发炎和充血。
在某些情况下,呼吸道症状可能导致肺炎、支气管炎或呼吸衰竭等并发症。这些并发症可能需要额外的医疗干预,并可能延长整体康复过程。如果呼吸道症状恶化或没有随着时间的推移而改善,请务必就医。
一旦呼吸道症状开始改善,患者即被认为处于康复阶段。在此阶段,身体继续抵抗感染,免疫系统努力清除呼吸系统中的病毒。康复阶段的持续时间可能因个人和感染严重程度而异。
在极少数情况下,有些人可能会出现与呼吸道症状相关的后遗症或长期影响。这些可能包括慢性肺损伤、肺功能下降或感染痊愈后仍持续存在的呼吸道疾病。同样,有些人可能会出现呼吸道症状复发,即症状在改善一段时间后再次出现。
为了帮助康复和预防并发症,休息、保持水分充足并遵照处方治疗或药物非常重要。健康的生活方式(包括均衡饮食和定期锻炼)也可以支持免疫系统并促进整体呼吸系统健康。
▼
在感染的第七阶段,即胃肠道阶段,患者可能会出现一系列与消化系统相关的症状。这些症状的严重程度和持续时间可能有所不同,
可能包括:
这些胃肠道症状可能是感染扩散至肠道或人体对感染的免疫反应所致。在某些情况下,这些症状可能会随着时间和休息而自行缓解。但是,可能会出现脱水等并发症,需要医疗干预。
对于处于此阶段的人来说,控制症状并在必要时寻求适当的医疗护理非常重要。根据症状的严重程度,医疗保健专业人员可能会建议休息、补充水分和改变饮食。在某些情况下,可能会开药来缓解症状或治疗并发症。
还要注意的是,虽然有些人可能在此阶段后症状完全消失,但其他人可能会继续经历其他阶段,例如复发或出现后遗症。
▼
神经系统症状可发生在感染的急性期,也可发生在恢复期、复发期和并发症期。这些症状可能是病毒直接入侵的结果,也可能是病毒对神经系统的继发影响。
在感染的急性期,部分患者可能会出现头痛、头晕和精神错乱等神经症状。这些症状通常较轻且短暂,可在数天或数周内缓解。
但在某些情况下,神经系统症状可能会在恢复阶段持续存在。这些症状可能包括持续性头痛、注意力不集中和记忆力问题。
在复发阶段,神经系统症状可能会再次出现或恶化。这可能是病毒重新激活或对神经系统造成进一步损害的结果。
此阶段还可能出现并发症,导致更严重的神经系统症状。这些并发症可能包括脑炎、脑膜炎和中风。
经过适当的医疗护理和治疗,大多数人可以从这些神经症状中恢复过来。然而,有些人可能会出现长期的神经后遗症,如认知障碍或运动功能障碍。
值得注意的是,感染潜伏期也可能出现神经症状。这是接触病毒和出现症状之间的阶段。此阶段的神经症状很少见,但可能包括嗅觉或味觉丧失。
在慢性感染病例中,神经系统症状可能会持续很长一段时间。这些症状可能是间歇性的,严重程度也各不相同。
如果您在感染的任何阶段出现任何神经系统症状,请务必重视。
▼
在感染的急性期,个人通常会出现皮肤症状。皮肤症状是指影响皮肤的任何症状。这些症状的范围从轻微到严重,并可能以各种方式表现出来,具体取决于感染的类型。
在潜伏期,个人可能不会出现任何皮肤症状。然而,随着感染进展到慢性阶段,皮肤症状可能开始出现。这些症状可能包括皮疹、水泡、病变或皮肤变色。这些症状的严重程度因人而异,有些人只会出现轻微症状,而另一些人可能会出现影响日常活动的严重症状。
在某些情况下,皮肤症状可能导致并发症。当感染扩散到身体其他部位或免疫系统对感染反应强烈时,就会出现并发症。这些并发症可能导致更严重的皮肤症状,甚至可能导致其他健康问题。
感染急性期过后,患者可能会出现感染的后遗症或长期影响。这些后遗症可能包括持续的皮肤症状,如疤痕或色素沉着变化。这些长期影响因人而异,取决于感染的严重程度和个人的免疫反应等因素。
在某些情况下,即使感染痊愈,患者也可能会出现皮肤症状复发。如果感染未从体内完全清除,或者免疫系统受损,则可能会出现这种情况。如果患者出现皮肤症状复发,请务必就医,因为这可能表明感染复发或其他潜在健康问题。
总之,皮肤症状可发生在感染的各个阶段,从急性期到慢性期,甚至感染消退后。这些症状可从轻微到严重,并可能对皮肤产生长期影响。个人必须注意这些症状,并在必要时寻求医疗帮助,以确保正确的诊断和治疗。
▼
血液学症状通常出现在感染的恢复期。此阶段发生在急性期之后,急性期的特点是出现症状并出现并发症。
在此阶段,人体的血液系统开始稳定并恢复正常。前驱症状(即感染前出现的一般症状)开始消退。人体开始产生更多的白细胞,负责抵抗感染。
在某些情况下,血液学症状可能会复发。如果感染在急性期没有完全解决,就会出现这种情况。复发可能是由于病毒重新激活或出现新的细菌菌株引起的。
常见的血液学症状:
潜伏期(即从接触感染到症状首次出现之间的时间)也会出现血液学症状。然而,此阶段的症状通常较轻微,可能不明显。
如果血液学症状在恢复期持续存在或恶化,则可能表明出现并发症或发展为慢性感染。在这种情况下,可能需要额外的医疗干预。
在感染的不同阶段密切监测血液学症状对于确保正确的诊断和治疗非常重要。可以定期进行血液检查以跟踪血液系统的变化并指导治疗计划。
▼
在感染的肌肉骨骼症状阶段,患者可能会出现肌肉、关节和骨骼的急性疼痛、僵硬和肿胀。这些症状通常发生在前驱期之后,可能是身体对感染的免疫反应的结果。
肌肉骨骼症状的强度和持续时间会有所不同,具体取决于个人和引起感染的具体病原体。有些人可能只会感到轻微不适,而另一些人则可能会感到剧烈疼痛和活动受限。
在某些情况下,肌肉骨骼症状可能是感染的后遗症,这意味着即使感染已经消退,这些症状仍会持续存在。这在某些病毒感染中更为常见,例如寨卡病毒或基孔肯雅病毒。
注:寨卡病毒属黄病毒科,黄病毒属,单股正链RNA病毒,直径20nm,是一种通过蚊虫进行传播的虫媒病毒,宿主不明确,主要在野生灵长类动物和栖息在树上的蚊子。
基孔肯雅病是由伊蚊传染的一种急性传染病,其临床症状为突然发烧、头疼、呕吐、关节痛及腰下部疼痛等,而最有效的应对途径是采取预防措施,减少蚊虫的滋生。
肌肉骨骼症状的恢复通常包括休息、疼痛管理和物理治疗,以改善力量和活动能力。如果症状严重或持续,可能需要更密集的治疗。
值得注意的是,肌肉骨骼症状也可能是感染的并发症,可能需要额外的医疗干预。并发症可能包括关节炎症、骨骼或软骨损伤或周围组织感染。
在极少数情况下,患者在经过一段时间的缓解或恢复后可能会出现肌肉骨骼症状复发。这可能是由于引起最初感染的病毒或细菌重新激活,或被同一病原体再次感染。
总体而言,感染过程中的肌肉骨骼症状会极大地影响个人的生活质量。如果您出现任何这些症状,建议处理以便获得准确的诊断和适当的治疗。
▼
泌尿生殖系统症状可作为某些传染病发展的一部分出现。这些症状可能表明感染已到达泌尿生殖道,包括生殖系统和泌尿系统的器官。并非所有感染都会发展到这个阶段,并且出现泌尿生殖系统症状并不总是表明感染严重或晚期。
泌尿生殖道阶段通常发生在感染的初期阶段之后,例如急性期和前驱期。在此阶段,病毒或细菌可能已经通过血液或其他身体系统到达泌尿生殖道。
出现泌尿生殖系统症状的患者可能会注意到泌尿系统的变化,例如排尿频率或尿急增加、排尿疼痛、尿液浑浊或带血,或难以完全排空膀胱。影响生殖系统的感染也会导致阴道分泌物、异常出血、盆腔疼痛或男性睾丸疼痛等症状。
在某些情况下,泌尿生殖系统症状可能伴有并发症。这些并发症可能包括肾脏感染、尿路感染、盆腔炎或性传播感染。如果出现这些症状或怀疑有感染,请务必就医。
泌尿生殖系统阶段的持续时间可能因具体感染和个人因素而异。在某些情况下,症状可能会通过适当的休息、补水和对症治疗自行缓解。但是,如果感染未得到适当治疗或存在潜在健康问题,感染可能会发展为慢性阶段或导致复发。
值得注意的是,某些感染会对泌尿生殖系统产生长期影响或后遗症。这些后遗症可能包括不孕症、慢性疼痛、疤痕或其他可能需要持续医疗管理的并发症。
常见的泌尿生殖系统症状:
▼
在感染过程中,个人可能会经历多个阶段。这些阶段包括潜伏期、复发、并发症、缓解,甚至慢性和急性后遗症。然而,一个经常被忽视的阶段是心理症状阶段。
心理症状可发生在感染的任何阶段,程度从轻微到严重不等。这些症状通常表现为情绪、行为和认知的变化。常见的心理症状包括焦虑、抑郁、易怒、困惑和注意力难以集中。
心理症状的存在会对个人的整体幸福感和生活质量产生重大影响。这些症状会影响一个人进行日常活动、维持人际关系甚至工作或上学的能力。为了提供适当的支持和治疗,识别和解决这些症状非常重要。
病原微生物检测在感染管理中的重要性不容忽视。通过病原微生物检测可以有效地控制和预防医院内感染,并加速患者的恢复过程。
随着分子生物学技术的发展,现代医学微生物学检验技术已经取得了显著进步,正在成为指导临床感染辅助诊断和治疗的重要依据。
多种检测手段应用于病原体检测,例如:
在感染的临床管理中,除了传统的病原体检测方法外,肠道菌群检测也是一个重要的发展方向。肠道菌群有助于消化、吸收营养物质,同时还调节人体免疫系统的功能,对人体健康起着至关重要的作用,肠道菌群失衡与多种疾病的发生发展密切相关。
肠道菌群与宿主之间的相互作用对维持内稳态很重要,但这种相互作用一旦受到干扰,就会成为许多慢性疾病的核心驱动因素。
在肠道、相关微生物群和各种器官之间的双向或多向通信连接(轴)
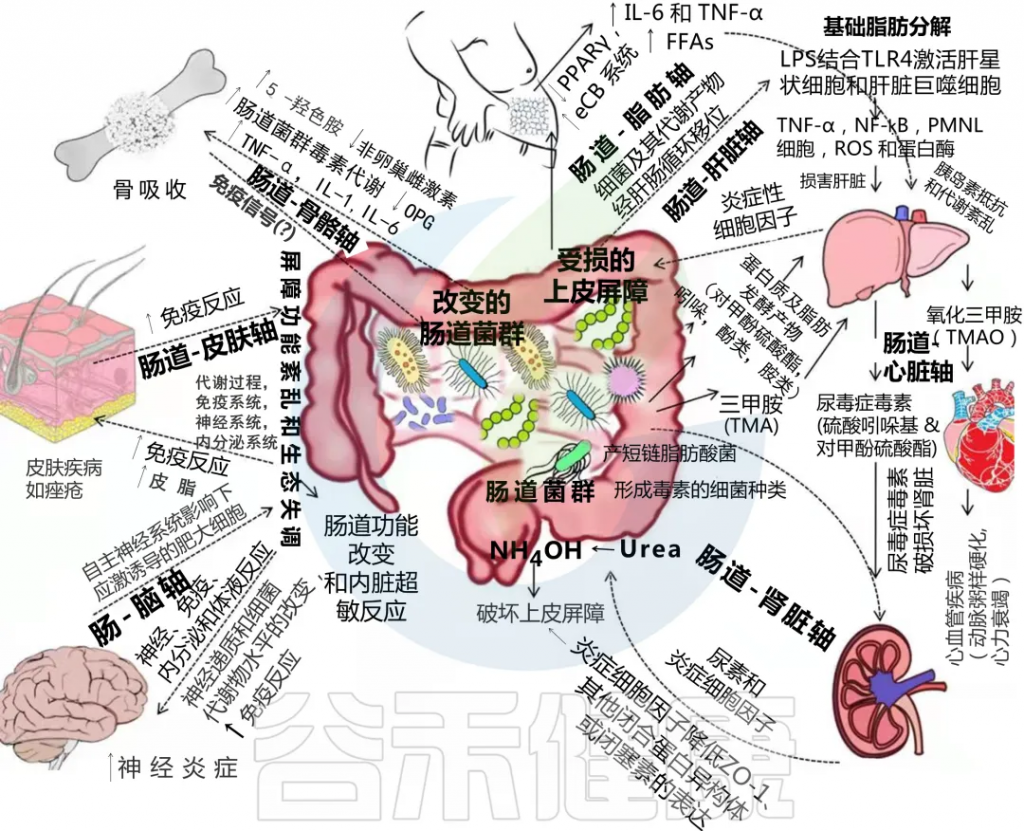
Ahlawat S,et al.,Lett Appl Microbiol. 2021
关于这方面可详见我们之前的文章:
肠道菌群报告中会有致病菌超标等明确指示,此外我们还可以判断整个肠道微生态的健康状况。肠道微生物群的失调可能导致肠道易感性增加,使得易感性疾病如艰难梭菌感染更容易发生。
通过分析肠道微生物组的变化,可以帮助我们更好地理解感染过程,评估治疗效果,并为个性化干预提供依据。
感染,作为全球健康的主要威胁,其影响甚至超过个体的病痛,它触及社会的每一个层面。本文在了解各种感染阶段、症状的同时,也强调了病原体检测在感染管理中的重要性,病原体相关检测手段让我们能够及时识别感染类型,评估其严重程度,并预测可能的并发症。
面对不断演变的病原体和日益严峻的抗生素耐药性问题,我们在推动科研创新的同时,也需提高公众对感染性疾病的认识,强化预防措施,如感染源的识别、个人卫生习惯等,对于控制感染的传播同样至关重要。
在治疗方面,针对病原体的特异性治疗是帮助患者恢复健康的关键。这需要医疗相关专业人员根据相关诊断结果,选择最合适的治疗方案,包括抗生素、抗病毒、抗真菌等。同时,症状管理和支持性护理也是治疗过程中不可或缺的组成部分。
随着科技的进步和医疗知识的积累,整合病原体相关检测,加强跨学科的研究,以及普及健康教育,有望不断提升治疗效果,为患者带来更好的康复和生存机会。
免责声明:本文仅供参考,不构成医疗建议。请务必咨询医疗保健专业人员,以正确诊断和治疗任何感染。
主要参考文献:
Murray, P. R., Rosenthal, K. S., & Pfaller, M. A. (2013). Medical microbiology. Philadelphia: Elsevier/Saunders
Parija S.C. (2012). Textbook of Microbiology & Immunology.(2 ed.). India: Elsevier India.
Sastry A.S. & Bhat S.K. (2016). Essentials of Medical Microbiology. New Delhi : Jaypee Brothers Medical Publishers.
Joseph Lister’s antisepsis system,2018,Science Museum
sciencedirect.com/topics/immunology-and-microbiology/germ-theory-of-disease
biologydictionary.net/germ-theory/
infectioncycle.com/articles/infection-stages-understanding-the-progression-of-infectious-diseases-and-their-impact-on-health
germ theory,Adam Augustyn,2024, 5
National Research Council (US) Committee to Update Science, Medicine, and Animals. Science, Medicine, and Animals. Washington (DC): National Academies Press (US); 2004.
Murray, P.R., Rosenthal, K.S., & Pfaller, M.A. (2015). Medical microbiology (8th ed.). Elsevier.
Fauci, A.S., Braunwald, E., Kasper, D. L., Hauser, S. L., Longo, D. L., & Jameson, J. L. (Eds.). (2008). Harrison’s principles of internal medicine (17th ed.). McGraw-Hill Medical
Mandell, G. L., Bennett, J. E., & Dolin, R. (2010). Mandell, Douglas, and Bennett’s principles and practice of infectious diseases (7th ed.). Churchill Livingstone.
Brook, I. (2013). Microbiology and management of respiratory tract infections. CRC Press.
Control of Communicable Diseases Manual by David Heymann.
G. Authia, S. Fablina, 2022. Global and regional sepsis and infectious syndrome mortality in 2019: a systematic analysis. Published by Elsevier Ltd. Published:March, 2022
Loretta J. Bubenik (2005). Infections of the Skeletal System. , 35(5), 0–1109. doi:10.1016/j.cvsm.2005.05.001
Morrison WB, Kransdorf MJ. Infection. 2021 Apr 13. In: Hodler J, Kubik-Huch RA, von Schulthess GK, editors. Musculoskeletal Diseases 2021-2024: Diagnostic Imaging [Internet]. Cham (CH): Springer; 2021.
Megran DW. Enterococcal endocarditis. Clin Infect Dis. 1992 Jul;15(1):63-71. doi: 10.1093/clinids/15.1.63. PMID: 1617074.
Roberts RB, Krieger AG, Schiller NL, Gross KC. Viridans streptococcal endocarditis: the role of various species, including pyridoxal-dependent streptococci. Rev Infect Dis. 1979 Nov-Dec;1(6):955-66. doi: 10.1093/clinids/1.6.955. PMID: 551516.
Infective endocarditis: A contemporary update. Ronak Rajani, John L Klein. Clinical Medicine Jan 2020, 20 (1) 31-35; DOI: 10.7861/clinmed.cme.20.1.1
Lamas, C. C., & Eykyn, S. J. (2003). Blood culture negative endocarditis: Analysis of 63 cases presenting over 25 years. Heart, 89(3), 258-262.
Smith DA, Nehring SM. Bacteremia. [Updated 2022 Jul 31]. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2023 Jan-.
Britannica, The Editors of Encyclopaedia. “septicemia”. Encyclopedia Britannica, 23 Jun. 2022
Martinez RM, Wolk DM. Bloodstream Infections. Microbiol Spectr. 2016 Aug;4(4).
Akhondi H, Simonsen KA. Bacterial Diarrhea. 2022 Aug 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2023 Jan–. PMID: 31869107.
Bacterial gastroenteritis: Causes, treatment, and prevention (medicalnewstoday.com)
Typhoid (who.int)
Imam Z, Simons-Linares CR, Chahal P. Infectious causes of acute pancreatitis: A systematic review. Pancreatology. 2020 Oct;20(7):1312-1322.

谷禾健康

细菌耐药性
抗生素耐药性细菌感染的发生率正在上升,而新抗生素的开发由于种种原因在制药行业受重视程度下降。
最新在《柳叶刀-微生物》(The Lancet Microbe)上,科学家提出了基于细菌适应性、竞争和传播的生态原则的跨学科研究方法,可能开辟新途径来对抗抗生素耐药性感染。
许多兼性细菌病原体使用人类粘膜表面作为其主要储藏库,并在微生物组和宿主的某些病理状态下诱发传染病,以帮助其横向传播到新的宿主生物体。
有益的细菌共生体可以战胜特定的病原体,从而降低病原体传播和引起严重感染的能力。然而,尽管具有临床相关性,但对其自然栖息地中的共生-病原体相互作用的了解仍然很少。
本文将在人类微生物组和宿主生物学背景下讨论细菌病原体和共生体之间相互作用的研究最新内容,为预防和治疗传染病提供一些新视角。
The Gut Microbiome
对人类微生物组完整性作为健康需求的日益认可,正在极大地改变人们对细菌微生物组成员及其相互作用的评价,无论是微生物之间的互动还是与宿主的互动。这种理解的变化使得微生物生态学的原则成为了预防和治疗主要人类疾病的创新方法的核心。
从海量数据中分析推断出的微生物组特征正被考虑作为不同疾病诊断中的生物标志物,这些疾病包括各种类型的癌症,如结肠癌或乳腺癌,以及自身免疫性疾病,如类风湿性关节炎或银屑病。
粪便微生物移植在治疗艰难梭菌感染方面已被有效使用,而补充微生物组成员(例如AKK菌)或微生物组产品(例如短链脂肪酸),已显示出在改善代谢紊乱和癌症的健康结果方面的潜力。
然而,环境与人类相关的微生物生态系统之间存在根本性差异。许多环境微生物组代表了广阔、不受限制的生态系统,通常具有无限的生命周期,对微生物扩散几乎没有障碍。相比之下,与宿主相关的微生物组,如人类肠道中的微生物组,是受限的生态系统,具有较短的生命周期。
与宿主相关的微生物组需要特定的细菌机制来在个体和世代之间传播,并在其首选的生活条件之外暂时持续存在。
此外,宿主将微生物定殖者暴露于环境栖息地以外的压力源中,特别是通过其粘膜免疫系统。
Microbial Symbiosis and Pathogenicity
▸ 过去,对共生体和病原体的理解不全面
从历史上看,对与宿主相关的微生物生态系统的研究,曾局限于不同研究团队分别对中性或共生的细菌共生体和有害病原体进行的独立调查。因此,对这些不同组的与宿主相关细菌的当前理解高度不对称,且存在对病原体的强烈偏见,这阻碍了对人类微生物组生态的全面理解。
虽然这些细菌组在自然栖息地中有广泛的相互作用,但关于共生菌和病原体相互作用的实验研究却很少。例如,典型的病原体可以被发现几乎是人类微生物组的驯化成员,而共生菌则可以与病原体勾结,这会像多重微生物感染中那样。
▸ 共生菌和致病菌的区分较复杂
尽管在感染医学中,共生菌和致病菌之间的传统区分仍然是一个重要标准,但在生态学上下文中,这两组微生物固有的属性重叠使得这种区分变得不够充分。
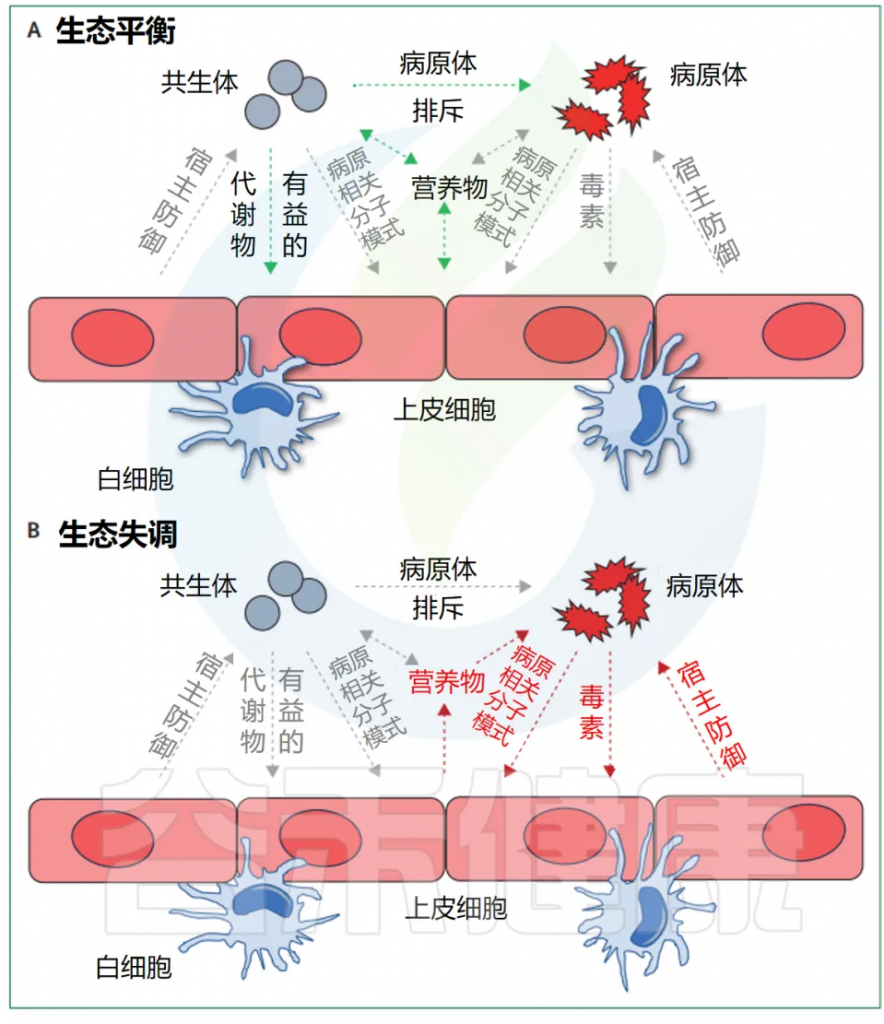
有益的共生现象和对立的致病性代表了细菌行为范围的两个极端(下图),尽管很少有细菌微生物组成员表现出真正的致病行为。多细菌物种或菌株可以根据其环境背景和宿主生理状态动态改变其作为共生体或病原体的能力,这进一步使区别变得复杂。
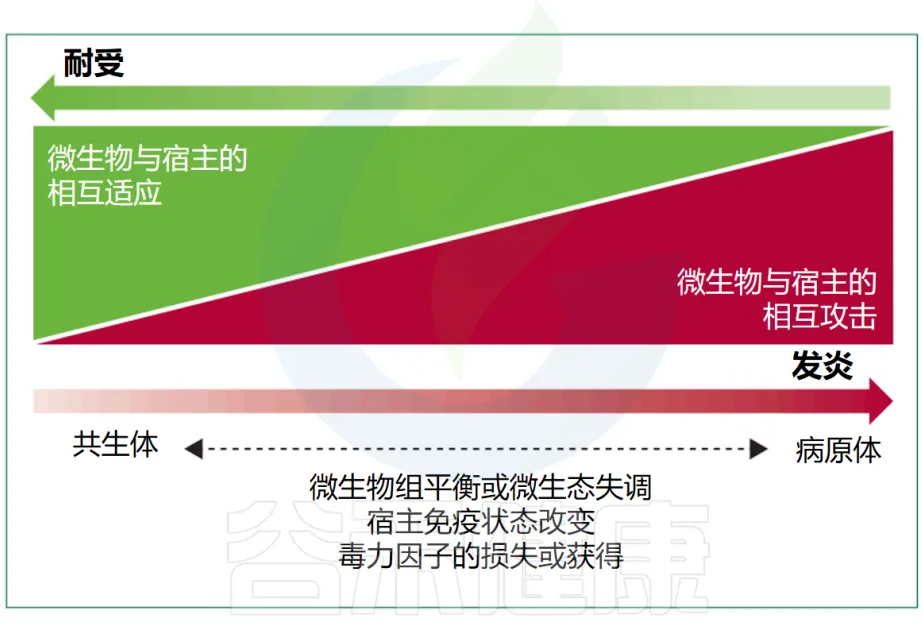
Maier L, et al. Lancet Microbe. 2024
例如,微生物组失调和宿主免疫缺陷,可以将肠球菌(Enterococcus faecium)从一个几乎无害的肠道微生物组成员转变为血流感染的原因。
单个水平基因转移事件可以改变共生生活方式和致病生活方式之间的平衡,例如,当预测的编码毒素成为肠出血性大肠杆菌或皮肤定植和口咽定植的白喉杆菌的主要毒力因子时。这意味着,原本可能是相对无害的微生物,在获得某些特定基因后,其生物学性质和行为可能发生显著变化,从而增加了其致病潜力。这种基因的转移和表达可以使这些微生物在宿主体内表现出更强的侵袭性和病原性,从而对宿主健康构成更大的威胁。
Redefining “Infection”
条件性细菌病原体之间的共生与致病性的动态变化,这挑战了当前对“感染”一词的使用,这种使用往往不一致。
《柳叶刀-微生物》全体作者建议仅将“感染”这个术语,保留用于由特定器官组织中出现的细菌菌株引起的病理状态。例如下列情况:
因此,在这篇个人观点文章中,我们也建议将常规引起感染的细菌称为病原体,而不常引起感染的细菌称为共生菌,尽管这些术语有其局限性,并不能适当描述所有类型的微生物与宿主之间的对抗关系。
目前病原体与共生菌之间的界限模糊,也挑战了科赫的假说,该假说提出了特定微生物与相应疾病之间的单一因果关系。然而,实际上,有些疾病是由多种细菌种类的间接效应引起的,当微生物群落失衡而不仅仅是这些种类的存在,导致了一种特定的病理状态,而这种状态目前并不被视为典型的感染。
Understanding of Bacterial Pathogens and Symbiotic Bacteria
微生物组科学,尤其测序技术的快速进展以及新技术的出现,为微生物学开启了一个新阶段,这一阶段的研究将细菌病原体和共生菌整合在一起,超越了简化的研究方法。
为了探究环境变化如何影响细菌微生物组成员的动态行为变化,来自不同背景的微生物学家应当合作,并结合系统生物学、天然产物化学、粘膜免疫学和临床传染病等互补学科的专业知识。这些新方法可能有助于回答一些最相关和明显的问题:
Future Research on Commensal Bacteria
由于其高度的临床相关性以及培养和操控相对容易,像S. flexneri(志贺氏菌)、S. aureus(金黄色葡萄球菌)和 S. pneumoniae(肺炎链球菌)这样的主要细菌病原体已经被研究了几十年,远超过典型的宿主相关共生菌。相比之下,主要的人类相关共生菌,例如来自属如拟杆菌属、梭菌属(Clostridium)、Cutibacterium,仅被少数实验室研究。
实际上,大多数人类微生物组成员仍然难以培养,并且在遗传上不易处理。在特定情况下可以成为偶发性病原体的一些共生菌,如医院获得性大肠杆菌、粪肠球菌(E.faecium)或表皮葡萄球菌(Staphylococcus epidermidis),已经在一定程度上被研究。但为什么这些特定的细菌比其他更无害的共生菌更频繁地引起侵袭性感染,这一点仍然不清楚(如下表)。


Maier L, et al. Lancet Microbe. 2024
The Important Role of Symbiotic Bacteria in Human Health
越来越多的证据表明,一些共生细菌对人类健康至关重要。
▸ Blautia producta 抑制粪肠球菌
例如,Blautia producta 的共生菌株通过产生特定的抗菌化合物,可以抑制并排除粪肠球菌(E.faecium),而Staphylococcus lugdunensis的共生菌则可以抑制并排除金黄色葡萄球菌。
▸ 枯草芽孢杆菌阻止金黄色葡萄球菌
肠道中的枯草芽孢杆菌(Bacillus subtilis)释放一种抑制性化合物,可以阻止金黄色葡萄球菌的定植能力,或抑制肠球菌(Enterococcus faecalis)的毒力因子表达。
一些共生菌还能产生对宿主有直接益处的化合物,例如促进肿瘤疗法的成功。病原体的研究可以帮助理解这些有益共生菌的生物学特性,并利用它们来对抗细菌感染。
共生菌的许多重要特性在不同菌株间可能会有所不同,这取决于例如获得或丢失移动遗传元素,如编码抗性或适应性特征的基因岛。用于病原体菌株特异性分类的现有方法,如序列分型方案,也可以应用于共生菌。这些方法可以帮助提升当前的诊断策略,达到个性化感染医学的新水平,不仅监测特定有害病原体的存在,还监测如那些保护免受潜在病原体定植的特定有益共生菌的缺失。
bacterial pathogens and symbiotes
▸ 共生菌的宿主特异性适应
与许多主要的细菌病原体一样,很多共生菌也特定于特定的宿主物种。这种适应过程导致宿主特异性,可能作为一种策略来增加细菌在与其他适应性较差的微生物竞争中的生存能力。这些基础机制对于病原体来说理解得很少,对于非致病性共生菌来说则基本上是不清楚的。
▸ 共生菌的粘附与持续定殖机制
特定宿主的持续定殖往往依赖于有效地粘附到上皮细胞的结合基序,如表面蛋白、蛋白多糖或糖脂。尽管在许多主要病原体中已经在一定程度上研究了相应的细菌粘附素,但现在才开始探索共生微生物中的这些机制。
▸ 共生菌与宿主免疫系统的相互作用
细菌在上皮表面的生存受到粘膜宿主防御机制的限制,这些机制包括IgA、抗菌肽和脂质以及活性氧和氮化合物的产生。先天和适应性免疫机制,通过感知与微生物相关的分子模式分子或通过粘膜白细胞检测微生物抗原来启动,有助于粘膜免疫过程,导致促炎或抗炎信号和效应分子的释放。
宿主的持续定殖依赖于细菌耐受抗菌免疫效应分子的能力或通过诱导免疫耐受来减少它们的表达。具有对抗菌宿主效应分子增强耐受性的细菌甚至可以诱导和利用抗菌宿主反应来消除更易感的竞争者。
根据宿主免疫反应,与微生物群其他成员的拮抗干扰已在表皮葡萄球菌和鼠伤寒沙门氏菌中得到记录。广泛的研究揭示了主要病原体的免疫逃避机制,但共生菌是否使用类似或不同的策略仍然未知。
▸ 共生菌免疫调节的复杂性及其潜在影响
例如,一些肠道共生菌通过产生非炎症性鞭毛蛋白来钝化先天免疫反应,这些蛋白抑制人类的Toll样受体5,或产生可以在粘膜表面诱导调节性T细胞的短链脂肪酸,以促进免疫耐受。然而,这些潜在的控制机制可能会被干扰,并可能在宿主无法启动耐受并以炎症反应的情况下导致疾病,例如当口腔共生菌异位定殖在肠道时。
doi.org/10.1016/S2666-5247(24)00049-1
总的来说,宿主相关生态系统中的细菌间的相互作用很大程度上依赖于分泌的因子,这些因子可以通过专门的分泌系统以单独的可溶性分子的形式释放出来,或者作为膜囊泡的组成部分释放出来(上图)。
因此,细菌病原体的分泌毒力因子已经被广泛研究。相比之下,共生菌分泌的初级或次级代谢产物或蛋白质介质如何调节与病原体和宿主的相互作用的研究较少。这些研究显示,例如一些共生菌可以释放杀菌素等分子,消灭金黄色葡萄球菌等病原体,或产生营养物促进艰难梭菌等病原体的扩展,由此凸显了宿主相关微生物生态系统中的一个新的复杂层面。
Fitness mechanisms of facultative human pathogens
许多主要的人类细菌病原体不是专性病原体,而是作为常见微生物组成员定植于人类或动物体表面,但不会引起疾病(下表)。

doi.org/10.1016/S2666-5247(24)00049-1
事实上,在大多数兼性病原体的共生生活方式中,急性感染是罕见的。然而,研究主要集中在S.flexneri、金黄色葡萄球菌、肺炎链球菌等病原体的毒力机制上,而在共生行为期间引导这些生物体与其他微生物组成员竞争的适应性的机制仍然被忽视。
从微生物的角度来看,感染是否以及哪些类型确实对细菌有利,或者应该被视为偶然事件,而不利于它们在几代宿主中的长期进化成功,目前尚不清楚。
Importance of a Deep Understanding of Bacterial Infections
现在比以往任何时候都更需要深入理解细菌感染,因为全球抗菌药物抗性和细菌感染的负担日益增加。目前对细菌感染的了解主要限于少数模型病原体,如金黄色葡萄球菌和肺炎链球菌。相比之下,一些臭名昭著的抗生素抗性细菌种类,根据以下病原体种属的首字母被命名为ESKAPE病原体,包括粪肠球菌、金黄色葡萄球菌、肺炎克雷伯菌、鲍曼不动杆菌、假单胞菌和肠杆菌,这些病原体的研究细节远不够充分。
▸ ESKAPE病原体的持久抗性与适应性进化
抗生素抗性机制在某些情况下可能会给细菌带来适应性的负担,即抗性可能会影响细菌的其他生存能力,但在没有抗生素的环境中,这种抗性通常会消失。然而,一些被称为ESKAPE病原体的细菌已经进化出了一种能力,使它们能够在医疗环境之外保持这种抗性特征。补偿性突变可以帮助这些细菌克服由于抗性而带来的适应性负担,从而促进抗性菌的成功和抗性的传播。
▸ 社区相关的抗性细菌扩散
这就是为什么社区相关的甲氧西林抗性金黄色葡萄球菌(MRSA)和万古霉素抗性屎肠球菌(VRE)等细菌,能够如此有效地在社区中扩散和扩张,甚至以牺牲它们对抗生素敏感的同类为代价,这一现象仍然是科学界未解之谜。这表明这些细菌可能已经发展出了一些未知的机制来维持和增强其抗性,即使在抗生素使用减少的环境中也是如此。这种现象需要进一步的研究来解明其背后的具体机制和因素。
The Relationship Between Bacterial and Ecological Success
生态学概念在普通微生物学和环境微生物学领域很常见,但尚未广泛应用于对病原菌的理解。
细菌在与其他微生物组成员的竞争中的成功依赖于多种机制,包括利用生长限制性营养物质的能力,从其他细菌那里获取促进生长的共同物质如聚合物水解酶或微量金属捕获剂,抵抗由微生物群体成员释放的抗菌分子,或者附着在少数上皮细胞附着点上。
一个细菌种类或菌株的长期生态成功来自于该种类在特定宿主体内的增殖和向新宿主体传播的综合效应。为了研究一个种类在特定宿主体内的增殖,应采用一系列研究策略,从共生菌的研究到对主要专业和偶发病原体的调查。
细菌有两种主要的传播方式:垂直传播和横向传播。
每个人体内都有大量细菌,它们组成了独特的微生物组。这些细菌有的倾向于垂直传播,有的倾向于横向传播,不同细菌的传播方式和速度是不一样的。
2022年的一项研究就发现,微生物组中的细菌在这两种传播方式的偏好上有所不同。我们已经对一些常见的致病细菌的横向传播有了较多了解,比如幽门螺杆菌、肺炎链球菌、金黄色葡萄球菌等。它们能在人群中快速传播,传播过程也被很好地监测和记录。但目前对这些细菌是如何在人体外存活,又是如何进入新的人体的,还不太清楚。
The association between the epidemic spread of pathogens
主要病原体的流行扩散很可能与它们引起的感染类型和严重程度有关(下图)。
▸ 感染类型
疾病特性对细菌传播的贡献在引起腹泻的病原体(如志贺氏菌)中最为明显,这类病原体通过污染的污水传播而获益。
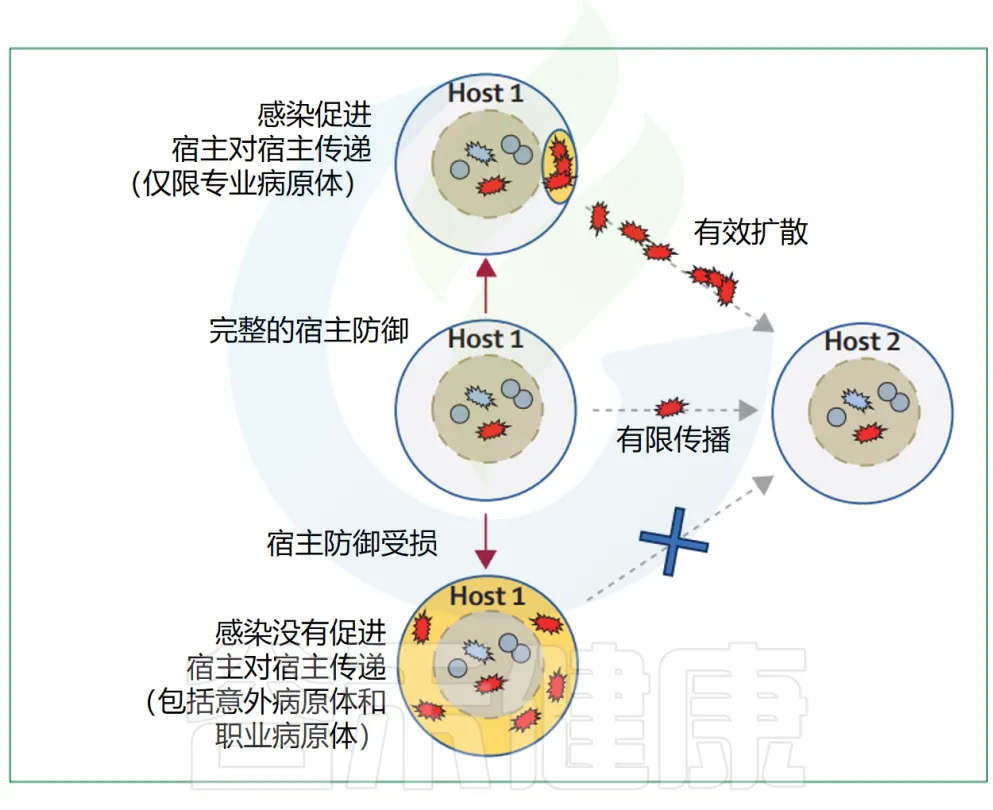
其他常见的感染表现可能也具有类似的作用。例如,金黄色葡萄球菌引起的典型感染——化脓性皮肤和伤口感染,会导致大量的S.aureus细胞在体表出现,从而支持通过皮肤接触的宿主间传播。
类似的机制也可以使尿生殖道病原体,如淋病奈瑟菌Neisseria gonorrhoeae在宿主间迅速传播。引起呼吸道感染的病原体,如百日咳博德特氏菌Bordetella pertussis、化脓性链球菌Streptococcus pyogenes或肺炎链球菌S pneumoniae则通过咳嗽或打喷嚏的个体释放的气溶胶传播。
▸ 严重程度
关于疾病的严重程度如何通过支持传播来增强病原体的适应性已有讨论,例如在SARS-CoV-2的背景下,但尚未对细菌病原体进行系统性评估。值得注意的是,大多数专业化的细菌病原体并非人类微生物组的核心成员,它们只是暂时性地定植在人类体内(见表1),这就需要它们具备有效的横向传播机制。
专业人类病原体(如志贺氏菌和淋病奈瑟菌N.gonorrhoeae)的持续定植在人群中较为罕见,或仅在人群的一小部分(如金黄色葡萄球菌)或特定年龄组(如化脓性链球菌或肺炎链球菌)中发现,这表明维持广泛的致病力涉及与其他微生物组成员竞争时的重大适应性负担。
诸如粪肠球菌(E.faecium)和表皮葡萄球菌( S.epidermidis )等偶然病原体表达的因子有助于其免疫逃避能力,但几乎不表达任何侵袭性毒素,这可能导致这些病原体在人类微生物组中比大多数专业病原体具有更高的患病率和持久性。
由于偶发性病原体主要在免疫力低下的个体中引起感染,它们也常被称为机会性病原体。然而,这一术语也经常用于诸如金黄色葡萄球菌和肺炎链球菌等专业病原体,这些病原体在免疫力低下的个体中引起的感染类型不同且更为严重(通常是血流感染),而在免疫力正常的个体中则不然。因此,专业病原体有时也可能引起偶发性感染,这种感染并不促进病原体的传播。偶发性人类感染也可以由适应于非人类宿主的病原体如军团菌(Legionella pneumophila)或霍乱弧菌引起,这些病原体只在特定的环境条件下感染人类。
The association between the epidemic spread of pathogens
在自然环境中同时研究共生菌和病原体,可以帮助我们更好地控制微生物,对抗感染以及其他与微生物组相关的疾病。
共生细菌对传染病的影响远比先前假设的要复杂得多。一些共生菌种采用主动防御策略,如释放抗菌肽或通过依赖接触的V型、VI型或VII型分泌系统消除其他细菌,这些系统在针对特定目标物种的特异性上可能有很大差异。此外,共生菌还可以使用更微妙的抑制策略来增加其在抗击病原体定植方面的生态成功,这基于代谢干扰。
共生细菌群落可以以依赖于群落多样性及其与病原体的代谢重叠的协作方式,阻止诸如克雷伯肺炎菌和鼠伤寒沙门氏菌等病原体获取营养物。
病原体的排除可能是由于共生菌释放的金属载体螯合必需的微量金属,或产生抑制性代谢产物而产生的。例如,一些肠道共生菌可以将原生胆酸转化为抑制肠道病原体艰难梭菌C difficile孢子生长的次生胆酸代谢产物。
Challenges in Fecal Microbiome Transplantation Treatment
粪便微生物组移植(FMT)是治疗艰难梭菌感染的一种有效策略。粪便微生物组移植成功至少部分是由于恢复了有益细菌群及其代谢产物,这些代谢产物能抑制艰难梭菌的生长。然而,粪便微生物组移植难以标准化,其对抗其他病原体的效果仍不明确。
目前,临床前和临床试验正在评估使用基于共生菌的生命治疗产品来预防处于风险中的个体被抗生素耐药病原体定植。然而,那些防止病原体定植的共生菌往往比目标病原体更易受抗生素影响,这可能会消除共生菌的有益效果,并在抗生素治疗期间加剧与菌群失调相关的疾病。
因此,研究人员需要平衡使用共生菌治疗产品的风险和好处。深入了解不同共生菌菌株对抗生素的敏感性,以及这些菌株如何与目标病原体相互作用。只有通过深入的研究和严格的监测,才能确保共生菌的治疗潜力得到充分发挥,同时避免不良后果的发生。
系统评估关键共生菌群的抗生素敏感性,并将这些知识纳入个性化的、基于微生物组的抗生素管理方案中,对于最小化广谱抗生素对微生物组的附带伤害至关重要。
除了我们通常使用的抗生素之外,许多针对人类的其他药物也可能意外地影响到人体肠道中的有益细菌。这种药物的副作用在未来的医疗实践中需要被特别注意,特别是对于那些需要同时使用多种药物治疗多种疾病的患者。此外,人体内的微生物群体本身可能是一种新型抗菌剂的重要来源。通过系统地研究和描述这些微生物产生的创新化合物,我们可能会发现新的药物,这些药物有望用于预防或消除病原体在人体内的定植。
Antibiotic Resistance Issues in Treatment
在现代医疗中,我们常用的广谱抗生素不仅攻击病原体(引起病症的微生物),还会影响到人体内的有益共生细菌。这种广泛的攻击会导致细菌之间的抗药性基因传递,特别是通过一些可以在不同细菌间转移的元素,如质粒和噬菌体。这种基因的传递可能导致抗生素更难以消灭病原体。
由于这种抗药性基因的传递,需要更好地监控不仅是病原体的抗药性发展,也要监控那些可能是抗药性传播者的共生菌。这样可以更有效地控制病原体的传播和抗药性的发展。
此外,通过理解这些微生物之间的生态关系,可以更好地发现病原体的弱点,从而开发出新的治疗方法。
这些方法不仅限于传统的抗生素,还包括能够利用有益共生菌或宿主防御机制的新型药物。这些新药物的开发需要在模拟真实生态环境的实验模型中进行,比如使用人类挑战模型,这种模型允许我们在受控的环境中测试病原体和治疗方法。
总之,通过更好地理解微生物之间的相互作用和生态平衡,可以开发出更有效的策略来对抗病原体,同时减少对有益共生菌的损害。这种方法有助于限制病原体的传播和疾病的爆发。
主要参考文献:
Maier L, Stein-Thoeringer C, Ley RE, Brötz-Oesterhelt H, Link H, Ziemert N, Wagner S, Peschel A. Integrating research on bacterial pathogens and commensals to fight infections-an ecological perspective. Lancet Microbe. 2024 Apr 9:S2666-5247(24)00049-1. doi: 10.1016/S2666-5247(24)00049-1. Epub ahead of print. PMID: 38608681.

谷禾健康

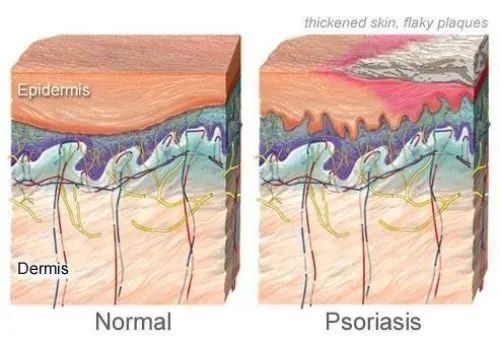
银屑病,又叫牛皮癣,会导致出现皮疹伴发痒的鳞状斑块,最常见于膝盖、肘部、躯干和头皮。通常呈周期性发展,发作数周或数月,然后消退一段时间,长期的发作和复发会给患者带来很大的痛苦和困扰,严重影响到生活质量和日常工作。银屑病影响着全球超过 6000 万成人和儿童。
银屑病是一种免疫介导的炎症性皮肤病,涉及遗传和环境因素之间复杂的相互作用。免疫学和遗传学研究确认IL-17和IL-23是银屑病发病机制中的关键驱动因子。然而,由于银屑病的反复发作和难治性质,目前还没有简单治愈方法。
在有遗传倾向的个体中,非遗传因素也是触发牛皮癣发病和复发所必需的,其中包括感染、皮肤和肠道微生物群失调、脂质代谢失调、性激素失调、精神疾病等,其他环境因素也可能诱发牛皮癣,例如皮肤创伤、不健康的生活方式和药物。
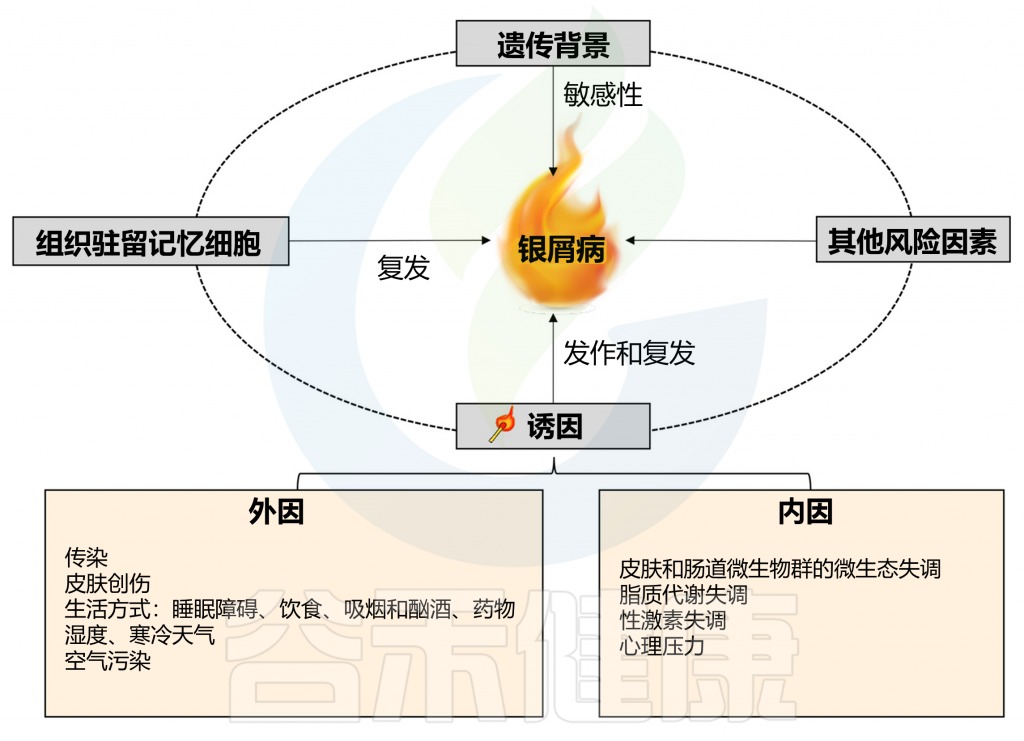
Liu S, et al., Cell Communication and Signaling, 2024
大量研究发现,银屑病不再被认为是仅影响皮肤的疾病,而是被视为一种全身性炎症性疾病,这与多种共病相关,包括结肠直肠癌、代谢综合征、肥胖、克罗恩病和溃疡性结肠炎等。微生物感染被认为是银屑病的重要病因,尤其是链球菌上呼吸道感染。
肠道和皮肤之间存在很强的双向联系,在肠道-皮肤轴中,皮肤炎症会影响肠道健康,而肠道微生物群也会影响皮肤健康,与健康对照相比,牛皮癣患者的肠道菌群失调,多样性降低,Firmicutes/Bacteroidetes(F/B)异常,大肠杆菌水平往往更高,Lachnospira、Akkermansia muciniphila、Blautia、Faecalibacerium减少,这种变化与人类结肠微生物群中丁酸盐代谢和产丁酸菌相关。丁酸与调节多种炎症因子有关,包括脂多糖、TNF-α、IL-10、IL-1β等。
扩展阅读:
其他菌群变化也与银屑病患者中与炎症相关标志物的程度相关联,特别是IL-2受体,其与Phascolarctobacterium呈正相关,与Dialister呈负相关。Phascolarctobacterium和Dialister的相对丰度可作为银屑病活动的预测指标。详见:
肠道核心菌属——考拉杆菌属(Phascolarctobacterium)
近日,来自华中科技大学同济医学院附属协和医院皮肤科的研究人员,总结了牛皮癣发病和复发的触发因素,发表在《Cell Communication and Signaling》上。

本文结合牛皮癣与肠道菌群的关联,更新了有关这些因素如何引发该疾病的潜在机制的当前证据。了解这些触发因素如何在银屑病的发病和复发中发挥作用,可以深入了解银屑病的发病机制,并更好地进行临床治疗。
银屑病是一种常见的红斑鳞屑性皮肤病,具有多种皮肤表现和全身参与,可影响任何皮肤部位,发生于任何年龄和任何地理区域。
在牛皮癣患者中,免疫系统错误地攻击了健康的皮肤细胞,导致皮肤细胞生长过快,形成皮肤表面的鳞屑。
银屑病的常见体征和症状包括:
图源:WebMD
根据银屑病临床表现的不同,通常分为以下类型:
其他还包括:
牛皮癣和湿疹一样吗?
牛皮癣和湿疹是两种不同的皮肤病。这两种情况都会引起类似的症状,如皮肤变色、皮疹和瘙痒。
牛皮癣会传染吗?
牛皮癣不会传染。接触他人的牛皮癣皮疹不会感染牛皮癣。
牛皮癣是一种慢性炎症性皮肤病,世界卫生组织最近的报告指出,牛皮癣的患病率正在上升,在发达国家从 1.5% 到 5% 不等。
据统计,中国的牛皮癣发病率在1.3%-2.4%之间,约有1000万-2000万的患者。男性的发病年龄在30-39岁和60-69岁,而女性的发病时间大约要早十年,年轻人群体中发病率也有上升的趋势。
许多银屑病患者可能持续数年毫无症状,直至某种环境因素诱发这种疾病。常见的银屑病诱发因素包括:
银屑病关节炎
一般来说,银屑病越严重,患银屑病关节炎的风险就越高。2015 年欧洲一项研究发现,十分之一的银屑病患者患有未确诊的银屑病关节炎。未确诊的银屑病关节炎患者的生活质量受到很大影响。
心血管疾病
研究表明牛皮癣与心血管疾病之间存在联系,慢性炎症可能是心血管疾病和中风风险增加的原因,因为随着时间的推移,炎症会对动脉造成损害。
代谢综合征
根据一项研究,患有牛皮癣的人患代谢综合征的风险,是其他没有牛皮癣的人的六倍。
肥胖
研究表明,与一般人群中的其他人相比,牛皮癣患者更有可能肥胖。
炎症性肠病
牛皮癣和炎症性肠病 之间存在联系。《JAMA Dermatology》杂志 2018 年的系统综述和荟萃分析结果表明,牛皮癣与炎症性肠病相关。
抑郁症
抑郁症是银屑病的首要合并症。患有皮肤问题会导致抑郁。对于某些人来说,治疗牛皮癣可以帮助减轻抑郁症状并提高生活质量。
以上我们了解了关于银屑病的基本情况,接下来,我们从感染,微生物群失调,脂质代谢失调,心理压力和精神障碍,性激素失调,其他环境因素等方面来了解银屑病的相关触发机制。
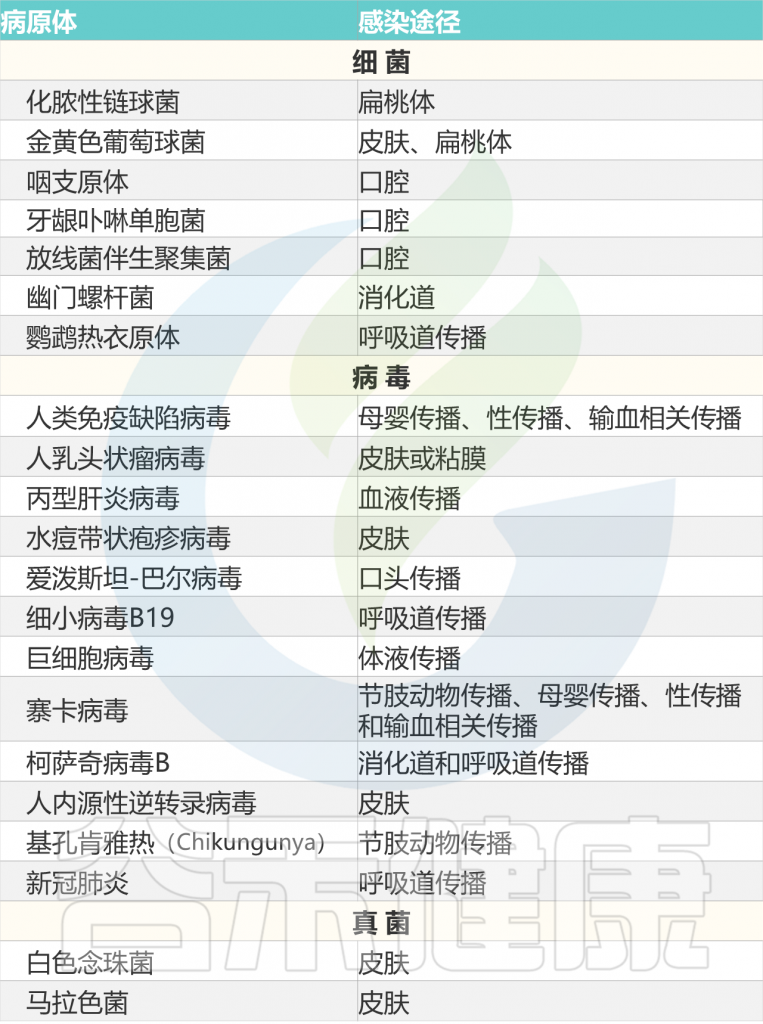
多种微生物是银屑病的触发因素,见下表。
Liu S, et al., Cell Communication and Signaling, 2024
化脓性链球菌引起的扁桃体感染,可引发或加剧斑块状和滴状银屑病中的银屑病皮肤病变。
研究人员通过遗传关联研究将链球菌咽喉感染与银屑病联系起来,表明公认的银屑病风险等位基因HLA-C06:02是链球菌扁桃体炎的风险因素,而推断的银屑病风险单倍型HLA-C06:02/HLA-B*57:01是扁桃体炎最强的风险因素。
儿童银屑病与复发性扁桃体炎密切相关
一项临床队列研究还报道,年龄从10岁-11岁的儿童银屑病与复发性扁桃体炎密切相关。在银屑病患者的皮肤和扁桃体组织中发现了相同的T细胞克隆,提示在链球菌感染后的疾病中扁桃体内产生致病性T细胞。因此,建议将扁桃体切除术作为解决银屑病的干预措施,这可以减少循环T细胞的数量。
但仍应进行长期随访,验证扁桃体切除术的适应症和长期益处。同时,没有确凿的证据表明抗链球菌干预措施的有效性。有趣的是,肛周链球菌感染也会引发喉结型银屑病,但它不如喉咙感染常见。
化脓性链球菌喉咙感染和银屑病之间发病联系
链球菌扁桃体炎可能通过以下多种机制来引发牛皮癣:
a) 化脓性链球菌(SP)的M蛋白模仿人类 K17
CD8 + T细胞识别银屑病患者中链球菌M蛋白和人角蛋白17(K17)共享的表位,K17可以成为CD8的自身抗原和靶点 + 以HLA-C*06:02限制性模式浸润银屑病皮肤病变的T细胞。
b) SP超抗原激活IL-12的释放,然后促进CD4+T 细胞中皮肤归巢CLA的表达
皮肤寻求性皮肤淋巴细胞相关T细胞(CLA + T细胞)与化脓性链球菌的相互作用,为理解银屑病的免疫发病机制提供了新的概念。通过刺激IL-12的产生途径,化脓性链球菌超抗原诱导T细胞上皮肤特异性归巢受体(CLA抗原)的表达,并促进CLA+ T细胞向皮肤的迁移。
此外,在携带HLA-C*06:02的银屑病患者中,如果他们患有化脓性链球菌引起的扁桃体炎,其CLA+ T细胞和表皮细胞的培养中观察到高Th17反应。化脓性链球菌能够在斑块型和滴状银屑病中诱导循环CLA+ T细胞产生IL-17,这进一步在CLA+ T细胞迁移到皮肤后诱导银屑病自身抗原(如ADAMTS样蛋白5和LL-37)的产生。在体外银屑病模型中,化脓性链球菌提取物诱导CLA+ T细胞产生IL-9,从而上调IL-17A的产生。
c) 化脓性链球菌肽聚糖 (PG) 的辅助作用
含有PG的巨噬细胞与PG特异性CD4+密切接触银屑病病变中的T细胞,然后是PG特异性CD4 + T细胞以HLA-DR等位基因限制的方式增殖并产生IFN-γ。此外,PG固有识别的改变增强了T细胞对化脓性链球菌的反应,并诱导银屑病。
图2 感染和皮肤微生物失调可能诱发银屑病
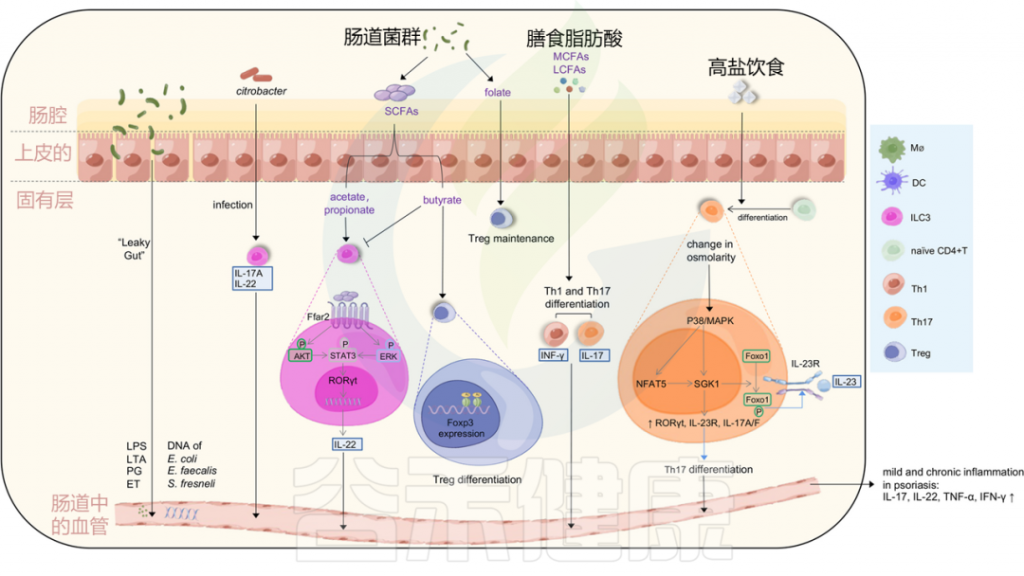
Liu S, et al., Cell Communication and Signaling, 2024
金黄色葡萄球菌在大约60%的银屑病患者的皮损和鼻孔中,而在5%至30%的健康个体中定植。
从22名银屑病患者中的11名患者的喉咙中分离出金黄色葡萄球菌。
一项研究显示,与健康对照组相比,银屑病受试者对超级抗原毒素的炎症皮肤反应增加,银屑病表皮中TNF-αmRNA水平增加。
然而,在银屑病病变中没有发现表达特异性T细胞受体Vβ的T细胞的选择性扩增,这是超抗原刺激的标志。这种T细胞依赖性反应可能是由于角质细胞中HLA-DR的高表达,增强了皮肤对超级抗原的炎症反应。
此外,银屑病的严重程度与葡萄球菌肠毒素的产生显著相关。
牙周炎患者患银屑病的风险较高
牙周炎期间,口腔微生物群可能影响银屑病的发展和恶化。一项涉及13项研究的荟萃分析显示,牙周炎患者患银屑病的风险高于对照组。
口腔微生物(尤其是咽支原体)在银屑病发展中的作用
一名患者最初患有滴状银屑病,后来发展为斑块型银屑病,其皮肤感染了咽支原体(Mycoplasma faucium),属于口腔软壁菌门,这种细菌出现在银屑病棘层角质形成细胞中,以及银屑病损害上层真皮的细胞外。
与对照组(骨关节炎)相比,在银屑病关节炎(PsA)患者的血清和滑液中也发现了更多种类和更高浓度的口腔细菌(牙龈卟啉单胞菌和Prevotella nigrescens)DNA。
牙龈病原体(尤其是牙龈卟啉单胞菌)在牙周炎与银屑病发病机制中的桥梁作用
牙龈卟啉单胞菌和放线菌聚集杆菌(Aggregatibacter actinomycetemcomitans)是与牙龈炎相关的病原体,可激活人CD14 + 单核细胞在体外增强Th17分化和IL-17产生。牙龈卟啉单胞菌蛋白酶可以通过降解其他关键细胞因子如IL-12,增强Th17系列反应,而髓样抗原呈递细胞(APCs)被触发产生与Th17相关的细胞因子IL-1β、IL-6、IL-23。然而,与健康受试者相比,牙周炎患者的牙龈组织中IL-17+细胞的频率增加,而在外周血中没有增加。
牛皮癣的皮肤炎症可以通过宿主的抗病毒免疫反应失调由病毒感染引发。
视黄酸诱导基因I (RIG-I)是病毒的主要细胞质传感器。通过激活RIG-I抗病毒信号传导,病毒感染可以在遗传易感个体中触发CD11c+树突细胞产生IL-23的表达,从而导致银屑病的发展。
HIV
与普通人群相比,人类免疫缺陷病毒(HIV)患者的银屑病标准化发病率更高。HIV可以直接作为超抗原或抗原呈递的共刺激因子触发银屑病,在HIV感染期间,活化的CD8 +T细胞产生更多的IFN-γ。神经肽物质P可以从HIV感染的免疫细胞中释放,然后调节炎症和免疫反应,刺激角质细胞的增殖。
HPV
人乳头瘤病毒(HPV)也与银屑病相关。一项针对66274名HPV感染患者的全国性人群队列研究显示,HPV感染后银屑病的患病率更高。
SARS-CoV2
严重急性呼吸综合征冠状病毒2型(SARS-CoV2)也被认为是银屑病恶化的原因。在严重急性呼吸系统综合征冠状病毒感染患者的血浆中炎性细胞因子水平升高,粒细胞集落刺激因子和TNF-α的浓度与疾病严重程度相关。
此外,一些接种新冠肺炎疫苗的患者患有慢性免疫介导的皮肤病,如牛皮癣,但皮肤反应通常轻微且自限性的。
NLRP1炎症小体
NLRP1是已鉴定的炎症小体形成模式识别受体(PRRs)之一,先天免疫系统可以通过该受体检测病原体。在正链RNA病毒感染过程中产生的长双链RNA(dsRNA),例如Semliki Forest病毒,可以结合并激活人类角质形成细胞中的NLRP1炎症小体。NLRP1炎症体在促进银屑病发病的过程中发挥作用,无论是通过增加对银屑病的易感性,还是通过促炎细胞因子(包括IL-1β和IL-18)的失调释放。
同样,NLRP1具有感知细菌病原体外毒素的能力,如铜绿假单胞菌分泌的外毒素A和白喉棒状杆菌分泌的白喉毒素,并诱导细胞死亡和IL-1β / IL-18分泌。
注:NLRP1,核苷酸结合域和富含亮氨酸重复pyrin结构域的蛋白质1(NLRP1),一种在人体免疫系统中发挥作用的蛋白质。
银屑病皮肤中的多种真菌已被鉴定为通过遗传易感个体的先天免疫系统激活银屑病。
白色念珠菌
来自白色念珠菌等微生物的超级抗原可能导致感染患者银屑病的恶化。接触白色念珠菌也可在银屑病皮肤中引发对IL-17的临床相关反应。银屑病CLA + T细胞/表皮细胞共培养物通过增加IL-9、IL-17A和IFN-γ的产生,对白色念珠菌提取物产生反应。
此外,皮肤白色念珠菌感染通过产生IL-17的CD4 + TRM诱导复发性银屑病。在小鼠模型中,CD4 + TRM在感染30天后成为IL-17的主要来源。
除白色念珠菌外,马拉色菌可能与头皮银屑病的恶化有关。
幽门螺杆菌
与对照组相比,幽门螺杆菌感染在中度和重度银屑病患者中显著增加,但在轻度银屑病患者中没有增加。在幽门螺杆菌感染的银屑病患者中,银屑病面积和严重程度指数(PASI)评分较高,银屑病相关细胞因子IL-1β、IL-6、IL-8和TNF-α的粘膜水平也较高。然而,2015年的一项发现表明,银屑病患者的幽门螺杆菌患病率没有比健康对照组增加。还需进一步研究。
目前,许多研究都致力于人类微生物组在银屑病发病机制中的作用,特别是皮肤和肠道微生物组之间的关系,即“肠道-皮肤轴”。
银屑病患者厚壁菌门增加
几位研究人员推测,银屑病可能与宿主皮肤微生物群的失调密切相关(图2)。与非病变皮肤和对照组相比,银屑病病变皮肤的细菌载量更高,但细菌多样性更低。厚壁菌门和放线菌门分别是银屑病患者和健康对照组中最常见的细菌门,病变皮肤中厚壁菌的增加和放线菌的相应减少是显著的。然而,另一项研究报道了银屑病病变中放线菌门和厚壁菌门都增加。这种差异可能是由于采样方法、皮肤部位、药物和分析方法的多样性。
棒状杆菌丰度与银屑病严重程度相关
根据新的证据,与未受影响和健康的皮肤相比,银屑病病变的棒状杆菌(Corynebacterium)浓度更高,Cutibacterium更低。棒状杆菌丰度与疾病严重程度相关,大多数种类的棒状杆菌在小鼠皮肤中诱导强烈的IL-23依赖性反应。在小鼠皮肤上涂抹假白喉棒状杆菌后,皮肤IL-1β蛋白水平升高,真皮中γδT17细胞增多。
银屑病耳朵皮肤显示葡萄球菌过度表达
较低丰度的表皮葡萄球菌和痤疮丙酸杆菌可能促进金黄色葡萄球菌在银屑病中的定植,这可以刺激小鼠模型中的Th17极化并引发IL-17介导的皮肤炎症。
为了解释银屑病的发病机制,提出了从屏障破坏到微生物群紊乱,再到病变加重的循环来解释银屑病的发病机制。
具有表皮屏障缺陷的小鼠具有增加的细菌载量和抗微生物肽(AMP)表达。在施用局部抗生素后,通过减少皮肤上的细菌负荷,以及减少IL-17和IL-22的产生,可以缓解小鼠的银屑病样表型。
除了皮肤微生物群的微生态失调外,紊乱的肠道微生物群也会影响银屑病的病理生理学(图3)。与健康对照组相比,银屑病患者的肠道微生物群在组成和功能潜力方面都发生了变化。银屑病患者的肠道微生物群特征显著紊乱,细菌多样性低,厚壁菌门/拟杆菌门(F/B)比率升高,并与PASI评分呈正相关。除了厚壁菌门和拟杆菌门外,银屑病患者和健康对照组16种属水平上的类群也存在显著差异。
银屑病患者肠道微生物的相关研究

doi.org/10.3390/ijms25041984
肠道屏障完整性与银屑病严重程度呈正相关
肠道脂肪酸结合蛋白(FABP)是肠道屏障完整性的生物标志物,其水平与银屑病的严重程度呈正相关。肠道微生物群失调可能通过减少粘液层的厚度、干扰肠道上皮细胞的增殖和代谢以及影响AMPs的产生来增加肠道通透性,也称为“肠漏”。
肠道微生物失调导致肠漏,促进细菌和抗原转移
肠道细菌可以通过DC通过上皮细胞之间的过程逃逸到血液中,而不影响紧密连接功能,或者通过覆盖在Peyer贴片上的微折叠细胞,将微生物产物呈递给APC。渗漏的肠道促进细菌移位,并允许外部抗原从肠腔进入血液和淋巴循环,从而驱动局部和全身免疫反应。
银屑病患者中细菌DNA易位与全身炎症的增加
与其他患者和健康对照组相比,斑块型银屑病患者血液样本中细菌DNA易位增加,主要是由肠道细菌引起的,包括大肠杆菌、粪肠球菌和弗氏志贺菌。细菌DNA易位的患者也表现出更高水平的全身炎症反应。
另一项研究还报道,在25%的斑块型银屑病患者的血液中观察到细菌DNA,与其他肠道类型的患者相比,肠型2(普雷沃氏菌占优势)的患者更容易发生细菌移位。这些微生物在被重新激活后可能会释放出高效的炎症因子,如脂多糖(LPS)和脂磷壁酸(LTA),这可能会导致宿主机体中的轻度和慢性炎症,银屑病患者会受此困扰。从肠道吸收的细菌内毒素(ET)和PGs也会加剧银屑病,银屑病的治疗已通过阻止其吸收或破坏内毒素来证明这一点。
短链脂肪酸促进Treg分化,调节免疫活性
微生物群可以通过肠道中的微生物代谢产物来改变免疫活性。短链脂肪酸(SCFAs)是肠道微生物组不可消化碳水化合物的主要发酵产物,主要包括乙酸盐、丁酸盐和丙酸盐。其中,丁酸盐可增强Foxp3基因座启动子区的组蛋白H3乙酰化,表明其有可能影响Treg细胞的分化。
微生物群对T细胞分化的影响:叶酸与色氨酸代谢途径
叶酸来自肠道微生物群和饮食,而饮食中的叶酸对维持Foxp3+Tregs有选择性作用。作为宿主色氨酸代谢途径之一,犬尿氨酸途径可以通过吲哚胺2,3双加氧酶将成熟的DC转化为耐受性DC,从而增强Tregs并抑制效应T细胞。这些结果可能表明,微生物群代谢产物通过触发肠道T细胞的分化,成为银屑病的非感染性风险因素。
固有淋巴细胞ILC3的调控:微生物代谢产物的双重作用
近年来,第3组固有淋巴细胞(ILC3)的功能受到了广泛关注。ILC3主要在肠道和皮肤中发现,并通过产生IL-17A和IL-22在银屑病中发挥致病作用。
当身体感染某些细胞外病原体(如柠檬酸杆菌)时,ILC3产生IL-22和/或IL-17,用于对抗病原体的粘膜免疫。ILC3的功能也可以受到微生物代谢产物的调节,如短链脂肪酸。丁酸盐可由厚壁菌门产生,而乙酸盐和丙酸盐主要由拟杆菌门产生。
图3 肠道微生物失调和饮食可能会在银屑病中诱导轻度和慢性炎症
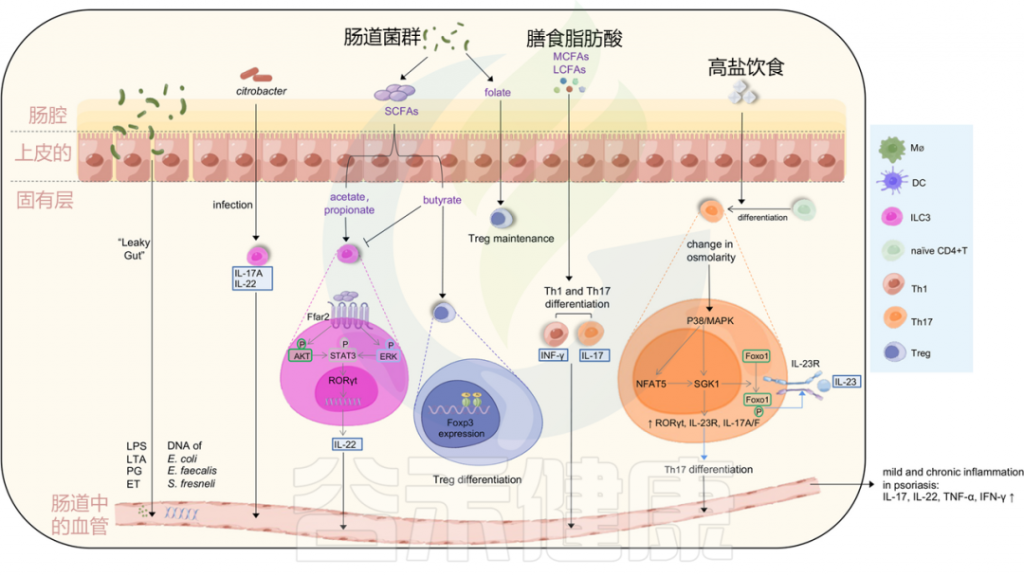
Liu S, et al., Cell Communication and Signaling, 2024
肠道通透性增加导致的“渗漏性肠病”可以诱导脂多糖(LPS)、脂蛋白(LTA)、外毒素(ET)和肽聚糖(PG)等强效炎症介质的释放,以及肠道细菌DNA进入血液。
柠檬酸杆菌感染可以刺激ILC3s产生IL-22和IL-17A,从而有助于黏膜免疫。ILC3s的功能也受到微生物代谢产物短链脂肪酸(醋酸、丁酸和丙酸)的影响。
乙酸和丙酸与结肠ILC3s上的FFAR2受体相互作用,激活AKT或ERK信号通路,并通过STAT3轴释放IL-22;
而丁酸则降低ILC3s的数量,同时,丁酸增强Treg细胞的分化,叶酸有助于维持Treg细胞,中长链脂肪酸支持Th1和Th17细胞的分化。
高盐饮食导致渗透压改变激活P38/MAPK通路,进而上调下游靶点NFAT5和SGK1,这反过来驱动转录因子RORγt、IL-23R、IL-17A和IL-17F的表达,导致Th17分化。上调的SGK1通过磷酸化Foxo1促进IL-23R的表达和稳定Th17分化。
益生菌的治疗已证明在改善银屑病方面具有潜在的益处,尽管尚未制定标准化的治疗方法。粪菌移植提供了另一种可能的治疗策略,因为它们通过允许健康粪便微生物组的“真生物”在受影响患者的肠道中重新定居来缓解自身免疫性疾病。未来更好地了解微生物群失调无疑将有助于缓解银屑病的治疗方法,我们在后面第八章也有具体的介绍。
许多研究已经表明肥胖和血脂异常与银屑病的关系,肥胖是银屑病的一个重要风险因素。而且,肥胖会加重已有的银屑病,这种自身免疫性疾病与肥胖之间的可能联系依赖于脂肪组织。
一般来说,脂肪组织可能通过肥大脂肪细胞分泌大量激素和细胞因子(也称为脂肪因子),包括IL-6、TNF-α和瘦素,在脂质代谢失调的患者中充当免疫器官,这些因子调节炎症途径和免疫细胞的活动。各种银屑病RNA-seq数据集表明,脂质代谢途径与银屑病的发病机制密切相关。
图4 将血脂异常与银屑病联系起来的潜在机制
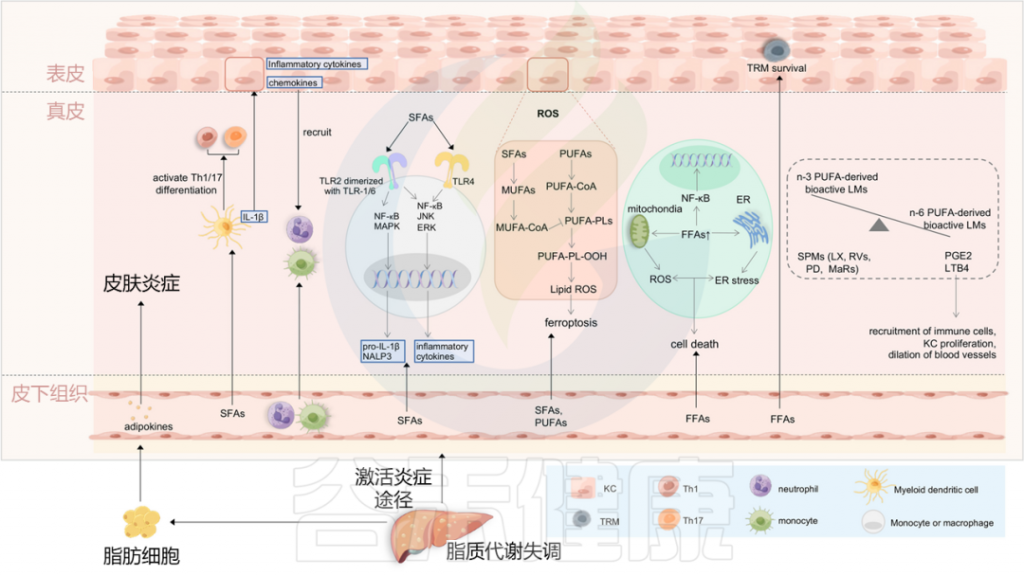
Liu S, et al., Cell Communication and Signaling, 2024
最近的研究表明,与肥胖相关参数无关的饮食成分,可能在银屑病的恶化中发挥关键作用。一项使用小鼠银屑病模型的研究提供了证据,证明饮食中的游离脂肪酸(FFAs),特别是饱和脂肪酸(SFAs),是银屑病皮炎的关键放大器。
饱和脂肪酸:加重银屑病皮炎的可能潜在机制
循环中的饱和脂肪酸被转运到皮肤中,并诱导骨髓源性树突细胞产生各种促炎细胞因子,如IL-1β。这些促炎细胞因子随后促进角质形成细胞分泌趋化因子和炎性细胞因子,导致中性粒细胞和单核细胞募集到皮肤,并导致银屑病皮炎放大。
饱和脂肪酸还可以通过激活TLR2和TLR4,来调节单核细胞或巨噬细胞中的NALP3炎症小体和炎症小体介导的IL-1β分泌。饱和脂肪酸在银屑病皮炎恶化中的另一种机制是通过激活DC来促进Th1/Th17分化。
体内游离脂肪酸的增加可能通过支持表皮中TRM细胞的存活而促使银屑病的复发。此外,膳食游离脂肪酸、中链和长链脂肪酸(MCFA和LCFA)的成分指导肠形Th细胞分化,这与上述丁酸盐诱导的Treg分化相反(图3)。
多不饱和脂肪酸(PUFA)及其衍生的生物活性脂质介质
除饱和脂肪酸外,多不饱和脂肪酸(PUFA)和多不饱和脂肪酸衍生的生物活性脂质介质(LMs)也可介导银屑病的炎症反应。
其中,生物活性LMs来源于两种多不饱和脂肪酸,亚油酸(LA,n-6 PUFA)和α-亚麻酸(ALA,n-3 PUFA),它们分别因其在银屑病中的促银屑病和抗炎特性而闻名。
n-6
作为代表性的n-6 多不饱和脂肪酸衍生的LMs,LTB4有助于中性粒细胞趋化,PGE2有助于角质细胞增殖。
n-3
衍生自 n-3 多不饱和脂肪酸的专门的促分解脂质介质(SPMs),包括脂氧素(Lxs)、resolvins (Rvs)、保护蛋白(PD) 和Maresins (MaRs),在银屑病中具有抗炎和免疫调节功能。
一个研究小组专注于基于液相色谱-串联质谱(LC–MS/MS)分析鉴定人类银屑病中的生物活性LMs和SPMs。根据他们的研究,衍生自n-6 多不饱和脂肪酸的生物活性LMs在银屑病皮肤中丰富,同时resolving D1(RvD1)、resolving D5(RvD5)、保护蛋白D1(PD1)及其双加氧异构体10S,17S diHDHA(又名PDx),阿司匹林触发形式的脂氧素A4和脂氧素B4(AT-LXA4和AT-LXB4)可能是解决银屑病病理生理学中炎症反应的专门的促分解脂质介质。
脂质代谢与铁死亡(Ferroptosis)密切相关,铁死亡促进银屑病角质形成细胞引发炎症,这涉及一系列连续事件,即活性氧(ROS)的积累导致脂质过氧化并进一步诱导铁死亡。
PUFA促进银屑病角质形成细胞铁死亡和炎症的机制
在银屑病期间,脂质过氧化与Th22/Th17通路在单细胞水平上呈正相关,证明了脂质过氧化的增强。
角质形成细胞对铁死亡也具有时间和浓度依赖性。在角质形成细胞的脂质代谢中,循环中积累的PUFA被催化为关键底物PUFA-CoA,最终酯化为PUFA-PLs,当存在生物活性铁时,PUFA-PL-OH可以过氧化为PUFA-PL-OH。随后,PLOOH可以通过产生脂质羟基自由基和脂质过氧自由基使细胞对铁死亡敏感。
MUFA-CoA和Fer-1通过抑制脂质过氧化抑制银屑病铁死亡
相反,来自饱和脂肪酸的单不饱和脂肪酸(MUFA)的产物MUFA-CoA可以通过抑制PUFA-PLs的过氧化来减少脂质过氧化的可用底物,从而抑制铁死亡。
此外,各种研究表明,Ferrostatin-1(Fer-1)是一种有效的脂质过氧化抑制剂,可以抑制铁死亡并阻断银屑病中的炎症反应。
除了氧化应激外,脂质还可以启动内质网(ER)应激,这具有双向作用:
饱和游离脂肪酸通过诱导ROS和ER应激参与银屑病发病机制
在非脂肪细胞中,过量的饱和游离脂肪酸通过脂质代谢和信号通路诱导ROS和ER应激。随后的线粒体和ER功能障碍是导致终末细胞死亡的关键步骤。此外,长期的内质网应激可导致氧化应激,脂质诱导的ROS也可能间接触发内质网应激,尽管确切机制尚不清楚。
脂质抗原激活限制性T细胞促进银屑病炎症反应
脂质自身抗原在驱动血脂异常相关自身免疫性疾病中的作用也引起了关注。银屑病病变含有高水平的磷脂酶A2(PLA2),其参与新脂皮肤抗原的产生。在IFN-α的诱导下,细胞质PLA2组IVD(PLA2G4D)可以以外泌体的形式从银屑病肥大细胞中释放,并转移到邻近的表达CD1a的Langerhans细胞。然后,新脂质抗原被脂质特异性CD1a反应性T细胞识别,释放IL-22和IL-17A。
自身脂质反应性T细胞在高脂血症与银屑病关联中的角色
除了CD1a限制性T细胞外,其他CD1分子(如CD1b和CD1c)限制性T淋巴细胞也对自身脂质产生反应并诱导细胞因子的产生。
在一项研究中,通过皮肤病变中积累的磷脂和胆固醇的一些自身抗原直接激活了CD1b自身反应性HJ1 T细胞。在患有高脂血症血清的小鼠中,CD1b+树突细胞增加了IL-6的产生,以及HJ1 T细胞分泌IL-17A,表明高脂血症和银屑病之间的潜在联系可能在于自身脂质反应性T细胞。
心理压力与银屑病的发病、严重程度和复发之间可能存在相关性
在31-88%的患者报告压力是银屑病的诱因,在过去12个月内经历压力事件的受试者中,银屑病的发病率更高。
另一项病例对照研究利用Holmes和Rahe的社会适应评定量表来评估压力生活事件,得出结论,压力在银屑病的发展中起着重要作用,特别是在复发和延长方面。
然而,一项荟萃分析报告称,没有令人信服的证据表明应激事件与银屑病之间存在这种关联。因此,未来应在基于人群的研究中前瞻性地审查这种关系,利用标准化的压力测量工具,并结合额外的生理和生化压力标志物。
心理压力激活HPA轴促进银屑病发病
压力可能通过下丘脑-垂体-肾上腺(HPA)轴、免疫途径和外周神经系统在银屑病的病理生理学中发挥作用。(图5)
作为对心理压力的反应,下丘脑产生促肾上腺皮质素释放激素(CRH),进一步激活垂体促肾上腺皮质激素(ACTH)和肾上腺皮质醇的分泌。CRH能够抑制角质细胞的凋亡,这是银屑病的典型现象。
另一方面,CRH通过刺激血管内皮生长因子(VEGF)增强血管生成,并增加血管通透性,促进银屑病斑块中炎症细胞的渗透。肥大细胞(MC)也可以被CRH激活,然后释放细胞因子和趋化因子,包括IL-1、IL-6、IL-31、TNF和CXCL-8。
此外,压力刺激皮肤外周神经末梢神经肽的释放,导致MC激活后神经源性炎症的发展。这些神经肽包括神经降压素(NT)、物质P(SP)、神经生长因子(NGF)和垂体腺苷酸环化酶激活多肽(PACAP)。
重度抑郁、PTSD、焦虑
除压力外,重度抑郁障碍或创伤后应激障碍患者患银屑病的风险显著高于对照组。
甚至父母常见的精神障碍(焦虑和抑郁)与后代患银屑病风险之间的关系也已确定。
双相情感障碍
另一项研究报告称,一名患有双相情感障碍的女性随后发展为银屑病,并在每次躁狂发作期间银屑病病变加重。
自闭症
根据自闭症谱系障碍患者银屑病炎症的临床升高,自闭症小鼠模型中高水平全身炎症,表明自闭症与银屑病活动之间存在联系。此外,先前的研究已经证实,严重精神障碍和银屑病之间存在遗传重叠。
越来越多的研究阐明了性激素对皮肤的多种生物和免疫调节作用。银屑病的自然病程似乎受到妊娠、月经和更年期的调节,从而暗示女性激素诱导的机制可能参与调节皮肤炎症。此外,研究表明,与女性相比,男性银屑病的患病率和严重程度更高,尤其是在雌激素充足的年龄,这表明不同性激素对银屑病的调节作用不同。
雌激素对银屑病具有保护作用
研究发现,雌激素通过下调中性粒细胞和巨噬细胞产生的IL-1β,这一过程通过雌激素受体α和β(ERα和ERβ)介导,具有潜在的抗银屑病作用。
同样,一项体内研究表明,雌二醇通过调节中性粒细胞和巨噬细胞的功能,在咪喹莫特(IMQ)诱导的小鼠银屑病炎症中发挥保护作用。体外,17β-雌二醇阻断了10 kDa的IFN-γ/干扰素诱导蛋白(IP-10)的正反馈回路,支持银屑病中Th1介导的炎症。
雌激素在银屑病中也可能具有促炎特性
一些孕妇的银屑病症状有所改善,而另一些孕妇的症状则有所恶化。
一份病例报告表明,一名患者在接受三苯氧胺治疗(一种抗中风药物)后,银屑病症状得到缓解,但在围绝经期症状恶化。值得注意的是,与对照组相比,男性银屑病患者的血清雌二醇水平显著升高,这表明雌激素可能参与银屑病的发展。
此外,一项使用咪喹莫特诱导的银屑病模型的体内研究也表明,雌激素通过Erα诱导IL-23,在银屑病中发挥促炎作用。
总之,这些证据支持雌激素在特定情境下对银屑病有双重影响,这导致了偶尔出现的相互矛盾的观察结果。
孕激素在银屑病中具有保护作用
临床观察证明银屑病通常在妊娠期间改善或消退,但在分娩后再次出现。
一项病例对照研究确定了孕妇受影响体表面积的改善与雌二醇、雌三醇、雌激素与孕酮比例升高之间的相关性。一些研究人员已经证明,角质细胞通过在银屑病皮肤中表达孕酮受体(PR)而成为孕酮的靶标。此外,孕酮在妊娠期间诱导转录改变,这些改变富含与银屑病相关的基因。STAT1和STAT3显著下调,其下游靶点,包括IL-12β、OSM和CXCL10,受到影响。
睾酮
一些研究强调雄性激素在牛皮癣中的作用。他们表明,严重的牛皮癣与低血清睾酮水平有关。无论患者年龄(40 岁以上或以下)如何,总睾酮和游离睾酮与 PASI 之间均呈负相关。
睾酮可以作为一种免疫抑制因子,下调树突状细胞的活性,干扰B细胞的分化,减少促炎细胞因子的释放。此外,在动物模型中,睾酮可以通过降低 IL-17 的水平来充当肝脏炎症的抑制剂,IL-17 是一种已知在银屑病发展中发挥关键作用的白细胞介素。
雄激素剥夺疗法(ADT)在晚期前列腺癌中作为加剧或缓解银屑病因素
一份病例报告显示一名前列腺癌患者ADT后银屑病加重。相反,一项调查发现ADT与银屑病风险降低之间存在相关性。另一项研究显示,无论年龄组如何,总睾酮或游离睾酮与PASI之间存在显著的负相关。
催乳素:刺激角质形成细胞的增殖
催乳素是垂体前叶分泌的一种多肽激素,刺激角质形成细胞的增殖,充当血管生成的诱导剂,并促进 Th1 细胞浸润银屑病病变。
此外,催乳素在调节免疫反应方面具有复杂的作用。它可以作为体液和细胞免疫反应以及 Th1 和 Th17 反应的刺激剂。
催乳素可以刺激角质形成细胞的增殖。最近的一项荟萃分析支持了激素在银屑病发病机制中发挥重要作用,该分析评估了循环催乳素水平与银屑病严重程度之间的关系。该荟萃分析包括 12 项研究,总共涉及 446 名银屑病患者和 401 名健康受试者。银屑病患者的循环催乳素水平显著较高,并且与疾病严重程度呈正相关。
总之,目前探索性激素对银屑病影响的研究主要依赖于观察性研究,缺乏深入的机制探索。这些关于雌激素和雄激素的矛盾发现表明,需要更多高质量的证据来更好地理解性激素与银屑病发病机制之间的复杂联系。
一些触发因素的潜在机制无法归类为感染因素、皮肤微生物群失调、肠道微生物群失调或脂质代谢失调,如下图。
图5 与银屑病发生相关的其他诱因
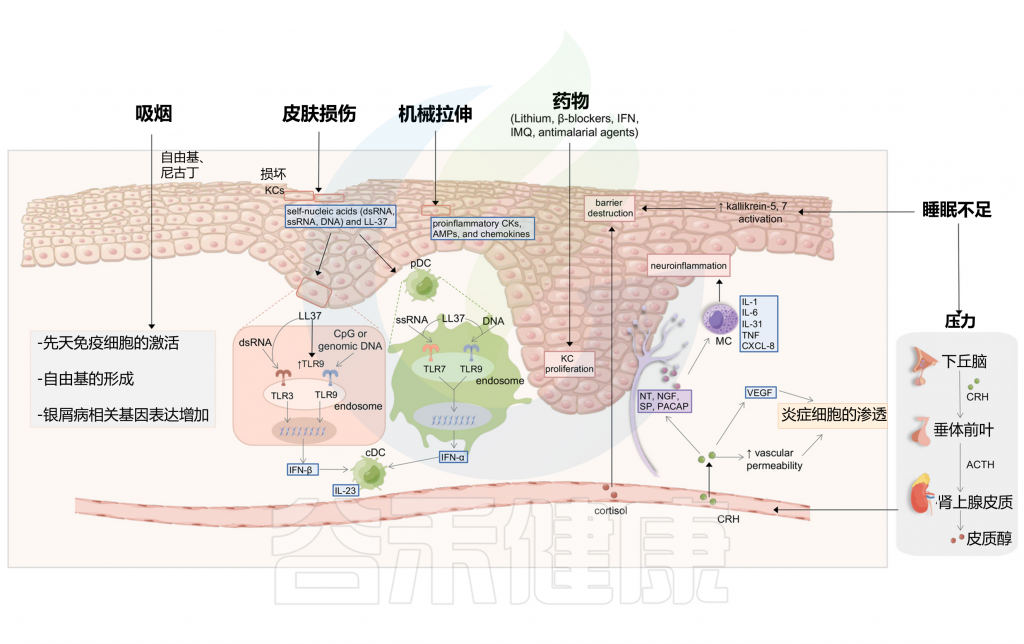
Liu S, et al., Cell Communication and Signaling, 2024
科布内氏现象与银屑病:皮肤损伤诱发疾病
皮肤创伤或压力可引发银屑病,称为科布内氏现象(Koebner phenomenon,KP)。
注:科布内氏现象,也称同形反应,是指在某些皮肤病患者健康皮肤区域受到损伤后出现的与原发皮肤病相同的皮损。
拔罐疗法与银屑病:传统治疗方法的争议
拔罐疗法作为一种传统的中医药,曾被用于治疗银屑病,但现在却存在争议,因为一些银屑病患者通过同形反应产生局部皮肤病变,而没有达到预期的治疗效果。
在银屑病患者中,拔罐治疗会导致拔罐部位出现同形反应,而Hijama(中东国家进行的一种湿拔罐)只会导致切口区域出现同形反应。
皮肤损伤激活角质细胞和免疫细胞参与银屑病发病
在皮肤损伤过程中,受损的角质细胞释放自身核酸,包括dsRNA、单链RNA(ssRNA)和DNA,并诱导LL-37的表达。
LL-37能够通过TLR7或TLR9在浆细胞样树突状细胞(pDC)中识别ssRNA或DNA,这最终导致IFN-α的分泌。LL-37暴露也可以通过DNA-LL-37复合物非依赖性机制或通过TLR3对dsRNA的识别诱导IFN-β的产生。对于前一种机制,LL-37增加TLR9的表达,从而促进TLR9配体的识别,如CpG或基因组DNA。浆细胞样树突状细胞的IFN-α和角质细胞的IFN-β促进常规树突状细胞的成熟。银屑病在创伤部位的复发归因于创伤部位TRM细胞的积聚和再激活。
机械拉伸与皮肤损伤:银屑病病变潜在触发因素
一份病例报告描述了一名患寻常型银屑病的妇女在插入组织扩张器后发展出新的银屑病病变。机械拉伸被怀疑会触发角质细胞释放ATP(5’-三磷酸腺苷),并随后产生Th17极化细胞因子,如前IL-1β和IL-6。被释放的ATP也可以激活表皮兰氏细胞。
在小鼠皮肤扩张模型中,观察到表皮过度增殖、皮肤屏障功能受损以及表皮角质细胞中银屑病相关细胞因子的上调。在人类角质细胞中,连续拉伸可以导致银屑病相关促炎细胞因子、抗微生物肽和趋化因子的产生。
除了拉伸,角质细胞的抓伤损伤可以通过细胞因子或趋化因子CCL20触发科布内氏现象,在较小程度上,这种触发作用与抓痕线数有关,也与CXCL8有关。
吸烟强度和持续时间与银屑病有关
与普通人群相比,银屑病患者中吸烟患病率更高,并揭示了吸烟开始和戒烟对银屑病的因果影响。吸烟强度和持续时间可能对银屑病的发病率具有剂量依赖性影响。作为银屑病发展的一个独立风险因素,吸烟对银屑病患者有许多负面影响,包括较高的PASI评分、指甲受累增加和心血管疾病的发展。
吸烟可能通过炎症、氧化和遗传机制引发银屑病
尼古丁通过释放炎性细胞因子刺激先天免疫细胞,如DC、巨噬细胞和角质细胞。此外,吸烟会引发自由基的形成,从而激活银屑病相关的蛋白质信号通路。
吸烟上调银屑病相关基因表达
在遗传学方面,吸烟上调银屑病相关基因的表达,包括HLA-C*06:02、HLA-DQA1*0201和CYP1A1。
烟草与银屑病:保护作用or风险因素?
最近的一项研究阐明了烟碱受体基因CHRNA5在银屑病的发展和发病机制中的作用。沉默的CHRNA5可以抑制人类角质细胞的增殖和迁移。有趣的是,吸烟也会增加普通人群患银屑病的风险,但吸烟在银屑病患者中表现出保护作用,这被称为“吸烟悖论”。然而,最近的一项孟德尔随机化研究涵盖了105912名具有生活方式因素、生物化学和基因型数据完整信息的个体,该研究表明吸烟是银屑病的一个独立风险因素,但不是银屑病的因果风险因素。
睡眠障碍:促炎因子显著增加,影响表皮屏障,加剧银屑病
睡眠障碍通常是银屑病的危险因素之一。一个具有全国代表性的基于人群的数据集表明,当发生阻塞性睡眠呼吸暂停时,银屑病和精神分裂症的风险增加。睡眠不足可能通过失眠的心理压力改变屏障稳态和角质层完整性。研究人员发现,银屑病小鼠在睡眠不足后,促炎细胞因子(IL-1β、IL-6和IL-12)显著增加,抗炎细胞因子(如IL-10)减少。
睡眠不足也促进了银屑病皮肤中kallikrein-5和kallikrein-7的活性,从而影响表皮屏障,导致银屑病的发展。此外,皮质醇在失眠等一些睡眠障碍中会增加。皮质醇刺激皮肤MC,破坏皮肤屏障功能,并上调促炎细胞因子,从而进一步加剧银屑病。
饮食因素在银屑病发病机制中的作用
73%的患者在坚持无麸质饮食三个月后出现临床改善,银屑病真皮中的Ki67淋巴细胞也减少了。
除麸质外,氯化钠(NaCl)摄入量的增加被认为对银屑病的发病机制有潜在影响(图3)。在高盐条件下,活化的p38/MAPK通路可以上调下游靶向活化T细胞核因子5(NFAT5)和血清/糖皮质激素调节激酶1(SGK1)。靶基因的上调可以驱动转录因子RORγt、IL-23R、IL-17A和IL-17F的表达,从而导致银屑病Th17细胞从初始CD4 + T细胞分化。SGK1对于通过Foxo1的磷酸化促进IL-23R表达和稳定Th17细胞分化至关重要。
银屑病与饮酒之间存在着复杂的多因素关系
银屑病患者的过度饮酒率高于普通人,酗酒会增加银屑病的严重程度并降低治疗效果。中重度银屑病患者的死亡风险也会增加,酒精是一个主要因素。然而,一项调查报告称,饮酒与银屑病的发展没有显著联系,孟德尔随机化研究也没有发现饮酒与银屑病之间的因果关系。目前还没有足够的证据来确定饮酒是否与银屑病的发病和复发有关。
许多药物都能引发银屑病,如锂、β受体阻滞剂、抗疟药、非甾体抗炎药、血管紧张素转换酶抑制剂、IFN、IMQ、特比萘芬、他汀类药物、贝特类药物和抗程序性细胞死亡蛋白1(PD-1)或抗程序性死亡配体1(PD-L1)抗体。在罕见病例中,TNF抑制剂也可能自相矛盾地诱发银屑病。
银屑病是锂的最常见皮肤不良反应
锂诱发和加重银屑病的发病率范围从3.4%至45%不等。
β受体阻滞剂治疗引发的角质细胞过度增殖及T细胞活性变化
当用β受体阻滞剂治疗时,负责促进细胞分化和抑制增殖的细胞内信使CAMP在表皮显示减少,最终导致角质细胞过度增殖。
此外,银屑病T细胞与对照组在蛋白酪氨酸磷酸化活性方面存在重要差异,蛋白酪氨酸激酶的诱导对包括淋巴细胞和角质细胞在内的细胞的激活和增殖至关重要。
抗PD-1/PD-L1诱导银屑病及潜在疗法
随着抗PD-1或PD-L1抗体在晚期恶性肿瘤治疗中的应用迅速增长,抗PD-1/PD-L1诱导的银屑病病例逐渐被报道。有研究者认为,PD-1的抑制通过加速表皮CD8 + 的浸润而促进皮肤炎症参与与角质细胞致病性串扰的T细胞。他们进一步证明了IL-6靶向疗法对抗PD-1/PD-L1诱导的银屑病的潜在疗效。
其他药物也通过不同的机制与银屑病有关
银屑病也可以由生物制剂引发
这被认为是一种自相矛盾的反应。一项研究显示,216例疑似TNF抑制剂诱导或加重的银屑病报告病例,英夫利昔单抗治疗银屑病的发生率更高,在克罗恩病和类风湿关节炎治疗的第一年最为普遍。对TNF抑制剂诱导的银屑病患者的回顾性分析也得出了一致的结果,表明英夫利昔单抗是主要的触发因子,而克罗恩病和类风湿关节炎是最常见的原发性疾病。这种自相矛盾的反应可能与通过抑制易感个体TNF活性而诱导的免疫改变有关。其发病机制也与IL-23/Th17轴参与TNF抑制的设定有关。
其中一个悬而未决的重大谜团是,在靶向TNF-α、IL-23和IL-17A/IL-17RA的生物制剂停用后,银屑病病变往往在相同的区域复发。目前最流行的观点是,现有生物制剂主要用于抑制致病性免疫细胞的活性,而不是完全消除它们。
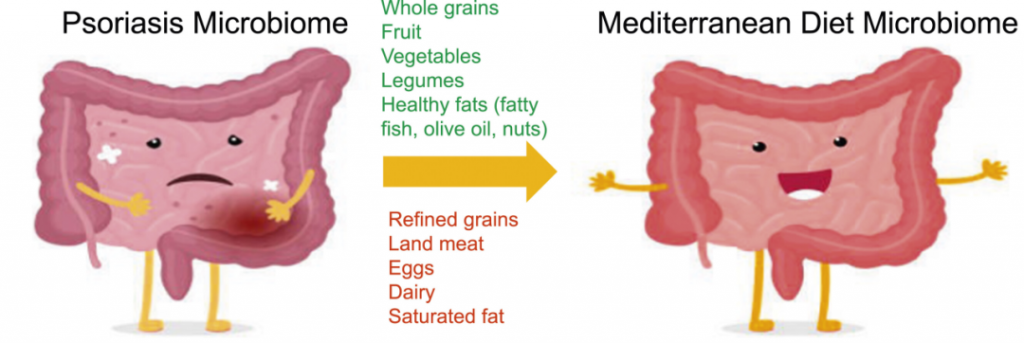
地中海饮食(MeD),因为它具有抗炎作用。重点是增加蔬菜、橄榄油和豆类等富含营养和抗氧化剂的食物的消费,同时限制肉类和奶制品的摄入。
一项针对 3557 名银屑病患者的队列研究发现,与地中海饮食更一致的饮食模式与较低的银屑病严重程度相关。
一种方式是通过增加拟杆菌门细菌来降低F/B比例,从而帮助恢复短链脂肪酸的平衡,并促进丁酸盐的产生,这反过来又可以减轻肠道和全身炎症,对牛皮癣具有重要影响。
肠道中其他产丁酸菌包括Akkermansia和普拉梭菌,地中海饮食通过增加这些细菌也促进了短链脂肪酸的产生。
doi.org/10.1177/24755303241226626
食用特级初榨橄榄油含有高含量的多酚,已在健康患者中显示出能减少IFNy和IL7R的基因表达,表明减少银屑病患者炎症的潜在途径。
间歇性禁食可能为难以遵循低热量饮食的患者提供另一种策略。尽管关于这种饮食的研究仅限于斋月禁食,这增加了几个混杂因素,但总的来说有益处。以减重为重点的饮食改善可能的原因可能是由于腹部脂肪和脂肪率的减少,随后降低脂肪因子水平并减少银屑病进展所依赖的炎症。
血清脂肪因子之一的脂联素与PASI评分呈正相关,IL-22也是如此。肥胖加剧银屑病的机制因此可能是由于脂肪因子诱导Th-17相关细胞因子的增加。对于这些建议,应避免严重的饮食改变,以防止营养不良。低热量饮食和间歇性禁食可能推荐给有适当动机的患者。
无麸质饮食意味着避免摄入小麦、大麦和黑麦中的这种蛋白质,这对患者有益。银屑病患者食用无麸质饮食 3 个月后发表了积极结果,显著改善了PASI 评分。其他研究小组证实,一年的无麸质饮食可以改善抗麦胶蛋白肽 IgA 水平较高的群体的 PASI 评分。
高脂饮食破坏IL-23和胆汁酸信号通路的稳态,促进皮肤的γδT细胞浸润并增强其产生IL-17A的能力来诱发银屑病样皮炎。也会通过改变粘液屏障和肠道微生物群来加剧银屑病皮肤炎症,从而导致全身 IL-17 反应增强,从而加剧银屑病。
极低热量生酮饮食(VLCKD)可以显著减少炎症,是缓解肥胖银屑病患者症状的有效手段,可能与微生物群-肠道-皮肤轴有关,但无论短期还是长期都有一定的副作用。
鱼油已被加入到许多银屑病患者用来缓解症状的补充剂中。2014 年,对几项有关在银屑病患者中补充鱼油的研究进行荟萃分析发现,虽然一些研究显示了中等结果,但其他研究尚未发现任何相关性。
最近,在一大群个体(超过 25,000 名男女登记受试者)中,在他们的饮食中补充了维生素 D(2000 IU/天)和/或 omega-3 脂肪酸(1000 毫克/天),并且该组随访 5 年,记录了银屑病 与其他自身免疫性疾病的发病率。研究结果表明,在补充组中,包括银屑病在内的自身免疫性疾病的发病率降低了 22% 以上。
姜黄素是一种抗炎的天然化合物,口服后会在胃肠道中积聚,并可能通过调节肠道微生物群的微生物多样性和组成来发挥其调节作用。一项人类随机安慰剂对照试验,补充组每天接受6000 mg姜黄提取物,并在治疗开始时和8周后进行微生物群分析。大多数梭菌属、拟杆菌属、柠檬酸杆菌属、Cronobacter、肠杆菌属、肠球菌属、克雷伯菌属、拟副杆菌属和假单胞菌属在反应参与者中均匀增加。
产生丁酸的细菌数量和粪便丁酸水平增加,对银屑病患者口服补充姜黄素12周进行评估,结果PASI评分显著降低,IL-22血清水平降低。
在斑块型银屑病患者的局部光疗的同时口服姜黄素似乎可以诱导对治疗的更快、更渐进的治疗反应。考虑到这些炎症发病机制与银屑病受试者的发病机制相似,补充姜黄素可能代表了该病理管理的未来前景。
白藜芦醇是一种抗炎的非黄酮类多酚化合物。在小鼠实验中也发现了白藜芦醇对肠道菌群的有利效果。口服白藜芦醇能够增强肠道屏障功能,同时减少通透性和炎症。白藜芦醇治疗后肠道菌群拟杆菌属、Alistipes、Rikenella、Odoribacter、Parabacteroides和Alloprevotella分类的丰度水平增加,白藜芦醇治疗恢复了小鼠的菌群失调。
此外,白藜芦醇的给药,8周内每公斤400毫克白藜芦醇,增加了高脂饮食喂养的大鼠中产丁酸菌Blautia和Dorea。因此,白藜芦醇补充可能改善患者肠道微生物多样性。
槲皮素是一种植物黄酮醇,它可能存在于各种水果、蔬菜、叶子、种子和谷物中。以其抗炎、细胞保护和免疫抑制特性闻名。
最近的一项研究针对亚甲基丙烯酰亚胺诱导的小鼠,探讨了槲皮素补充(30、60和120 mg/kg)的效果,显示PASI评分显著降低,银屑病斑块改善。此外,槲皮素成功降低了小鼠血清中的TNF-α、IL-6和IL-17水平,加强了抗炎效果。
此外,研究表明口服补充槲皮素,减少了亚甲基丙烯酰亚胺诱导的小鼠类银屑病样皮炎,显著降低了角质形成细胞的增殖和异常分化,以及炎症细胞的浸润。口服槲皮素后,观察到IL-23/Th17轴上的细胞因子表达降低和Th17细胞反应减少。
其他中草药如雷公藤等,也对银屑病患者有用,改善了他们的临床参数,可作为辅助治疗。
回顾性分析显示,83.7% 的银屑病患者在接受唾液链球菌K-12治疗 24 周后,PASI 评分改善了 100%,并且随着治疗持续时间的延长,疗效持续改善。
一项随机对照试验发现,持续口服婴儿双歧杆菌35624(B.fantis)可显著改善银屑病的进展,并降低C反应蛋白和肿瘤坏死因子TNF-α的表达,显示出免疫调节作用。人体微生物群的作用不仅限于粘膜免疫系统,还延伸到全身免疫系统。
在明确单一益生菌的作用机制后,益生菌的组合应用以及相应的标准和方法仍需探索。不同的饮食和环境,可能会导致肠道微生物群的差异。
一项为期12周的开放标签、单中心临床试验评估了益生菌[印度芽孢杆菌(HU36)、枯草杆菌(HU58)、凝结芽孢杆菌(SC208)、草绿芽孢杆菌(SL307)和克劳氏芽孢杆菌(SC109)]和精确益生元(低聚果糖、低聚木糖和低聚半乳糖)对接受局部治疗的银屑病患者的有效性。接受抗银屑病局部治疗和益生菌及益生元补充的患者,在疾病活动性的测量中表现更好,包括银屑病面积和严重性指数、皮肤病生活质量指数、炎症标志物和皮肤厚度。肠道菌群发生了有利的变化,转变为抗炎的特征。
喂食富含菊粉的高纤维饮食(HFD)降低了咪喹莫特诱导的皮炎的增厚和总严重程度评分,并降低了表皮厚度、炎症浸润,包括Ly6G+中性粒细胞和表皮Ki67+增殖细胞。
富含菊粉的高纤维饮食增加了拟杆菌门和拟杆菌属的相对丰度,增加了血清和粪便中丙酸盐的浓度。膳食菊粉补充剂部分通过丙酸盐改善银屑病样皮炎,可能是一种很有前途的银屑病辅助治疗方法。
当然,何时使用益生菌、益生元以及针对不同个体的精准应用,还需结合肠道菌群检测进一步研究。
FMT是纠正肠道菌群失调、重建肠道微生态平衡的方法。近年来,这种疗法已用于治疗牛皮癣、系统性红斑狼疮、肠易激综合征、帕金森病疾病等。
FMT治疗的效果取决于不同的疾病、移植的形式和数量、给药途径以及所使用的供体。
在一项为期五周的干预性临床试验中,一名患有斑块型银屑病和IBS的受试者通过内窥镜和结肠镜接受了两次FMT。身体表面积、PASI评分、皮肤病学生活质量指数、肠道症状和血清TNF-α水平在干预后都有所改善,没有观察到不良反应。
然而,在周围性银屑病关节炎患者中,FMT在治疗活动性疾病方面并不那么有效。FMT可能在减轻银屑病患者的疾病严重程度方面带来一些益处,但需要更多的临床试验来证明这一点。
其他相关预防干预措施,皮肤护理等详见我们之前的文章:
以上我们了解了银屑病发病和复发的危险因素和潜在病理机制。银屑病的发展是复杂的,可能是由多种触发因素引起的。这些触发事件可能在不同条件下独立发生,或者表现出累积或协同效应。因此,很难确切地将这种疾病归因于特定的诱因。
鉴于银屑病是由遗传基础上的环境风险因素引发的,疾病的预防和管理值得应有的重视。根据这些明确的触发因素,可以制定银屑病风险评估和疾病管理指南,有助于轻症或非典型病例的早期诊断和银屑病的精准管理。例如,感染史(不仅是化脓性链球菌感染,还包括这里列出的其他感染)、肥胖和高血脂水平、过度的心理压力、吸烟、睡眠障碍、高盐饮食、服用特定药物史都应被视为银屑病的风险因素。
从患者的角度来看,消除这些风险因素对于他们个人对疾病的管理至关重要。结合肠道菌群检测报告查看相关菌群超标或缺乏情况例如链球菌,AKK菌,产丁酸菌等,以及整体菌群平衡情况,对于疾病预警有着重要作用。通过及时调整生活方式、饮食习惯和适当药物治疗,可以有效地降低银屑病的发病风险,提高患者的生活质量。
从治疗角度来看,患者可能受益于早期治疗,包括但不限于结合菌群的抗生素治疗、标准化益生菌补充、抗高脂血症治疗等方式。了解触发因素在银屑病发病机制中的作用,与肠道菌群的关联,也将为开发针对银屑病发病和复发期间触发机制的新疗法提供线索。
注:本账号内容仅作交流参考,不作为诊断及医疗依据。
主要参考文献
Zhu Q, Wu K, Yang Q, et al. Advances in psoriasis and gut microorganisms with co-metabolites[J]. Frontiers in Microbiology, 2023, 14: 1192543.
Liu S, He M, Jiang J, et al. Triggers for the onset and recurrence of psoriasis: a review and update[J]. Cell Communication and Signaling, 2024, 22(1): 108.
Kranyak A, Haran K, Smith P, et al. The Mediterranean Diet as a Potential Solution to the Gut Microbiome Dysbiosis in Psoriasis Patients[J]. Journal of Psoriasis and Psoriatic Arthritis®, 2024: 24755303241226626.
Ryguła I, Pikiewicz W, Grabarek B O, et al. The Role of the Gut Microbiome and Microbial Dysbiosis in Common Skin Diseases[J]. International Journal of Molecular Sciences, 2024, 25(4): 1984.
Buhaș, M.C.; Candrea, R.; Gavrilaș, L.I.; Miere, D.; Tătaru, A.; Boca, A.; Cătinean, A. Transforming Psoriasis Care: Probiotics and Prebiotics as Novel Therapeutic Approaches. Int. J. Mol. Sci. 2023, 24, 11225.
Constantin, C.; Surcel, M.; Munteanu, A.; Neagu, M. Insights into Nutritional Strategies in Psoriasis. Nutrients 2023, 15, 3528.
Tampa, M.; Mitran, M.I.; Mitran, C.I.; Matei, C.; Georgescu, S.R. Psoriasis: What Is New in Markers of Disease Severity? Medicina 2024, 60, 337.
Zang C, Liu J, Mao M, Zhu W, Chen W, Wei B. Causal Associations Between Gut Microbiota and Psoriasis: A Mendelian Randomization Study. Dermatol Ther (Heidelb). 2023 Oct;13(10):2331-2343.

谷禾健康

最近新型冠状病毒肺炎(COVID-19)感染人数再次开始上升,Covid-19危机和近期频发的流感(甲流、乙流、支原体感染等),可能让免疫学进入一个新的时代。
现在人们因 Covid-19 感染住院或死亡的可能性降低了,那么危险已经过去了吗?如果您第二次、第三次甚至第四次感染,还有理由担心吗?
专家表示,感染 Covid-19 的风险比以前要小,但仍有充分的理由不要随意对待它。
在大流行的破坏中,医学研究和科学家收集了关于免疫系统的新见解,这些见解正在重塑一些长期以来关于人类免疫的观念。
这场全球性的大流行不仅提升了公众对免疫系统工作原理的认识,也推动了科学界对免疫学研究的投资和兴趣。在这个时期,肠道菌群和微生态管理成为了其中串联的角色,因为它们在人体免疫系统中扮演着至关重要的角色。随着对肠道微生态和宿主健康关系的深入理解,免疫学的疆界被进一步拓展,带来了研究和治疗上的新机遇。在这个新时代,我们可以预见到几个关键的发展趋势。
首先,疫苗开发将更加快速和高效。COVID-19疫苗的研发打破了速度记录,mRNA技术的成功应用预示着未来可以对抗更多疾病的疫苗将以更快的速度被开发出来。同时,针对肠道菌群的疫苗也将成为可能,直接通过调节菌群来提高免疫力。
其次,免疫疗法的研究和应用将得到扩展。COVID-19疫情期间,对于如何调节免疫系统以治疗严重病例的研究取得了进展。这些知识的积累将有助于治疗其他免疫炎症相关的疾病,如自身免疫疾病、炎症性疾病、过敏性疾病以及某些类型的癌症。免疫疗法也将受益于对肠道微生态的深入研究。
第三,全球免疫监测和疾病预防策略将得到加强。大流行暴露了许多国家在疾病监测和应对方面的不足。因此,未来可能会看到更多的资源被投入到预防性策略和监测系统的建立上,以便更快地识别和遏制新的疫情。
此外,精准医疗在免疫学领域的应用也将增加。通过对个体免疫系统的深入了解,医生可以为患者量身定制更加精确的治疗方案。基因组学、蛋白质组学和细胞组学等技术的结合,将使得医生能够更好地预测疗法的效果,减少副作用,并提高治疗成功率。在个性化医疗时代,肠道菌群检测也将成为重要的诊断工具,用以预测疗效、指导用药。这可以大大提高治疗的精准性,减少不必要的试错。
最后,公共卫生政策和教育将更加注重免疫健康。从学校教育到成人继续教育,对免疫系统的知识将被纳入课程,以提高人们对于预防疾病的认识。这需要从基础教育做起,向公众宣传合理膳食、生活习惯的重要性。如果社会环境本身就是一个促进健康的生态系统,有望尽早打造出一个更加健康的世界。
总之,COVID-19疫情虽然给全球带来了巨大的挑战,但也催生了免疫学领域的重大进展。在这个新的黄金时代,我们有理由相信,通过科学的力量,人类将更加有能力应对未来可能出现的健康危机。
✦ 疫苗带来的免疫保护可能会逐渐减弱
从这次大流行中得出的一个关键见解是认识到 mRNA 疫苗引起的免疫反应逐渐减弱可能并不是这些疫苗所独有的。对于以前被认为能诱导终身免疫的其他疫苗来说,情况也可能如此。
mRNA疫苗是一种全新的免疫方式。它能对有症状的新冠病毒产生快速、有力但短暂的反应。虽然 mRNA 疫苗针对死亡和严重疾病的保护作用随着时间的推移仍然很强,但针对症状感染的保护作用不太持久。这主要是因为病毒不断变异,因此它可以逃避免疫系统的防御——宿主和病原体之间的进化竞赛。
不过,这也是因为人体免疫系统需要更多时间来形成针对病原体的长期免疫记忆。每次免疫系统遇到病毒时,无论是通过疫苗还是自然暴露,这种记忆都会增强。
✦ 接种了疫苗并不意味着终生免疫
水痘和麻疹也可能发生类似的情况。你得了麻疹或水痘,或者你接种了疫苗,你以为就终身免疫了么?有新的证据表明情况并非如此。
例如,最近的一项研究发现,带状疱疹的发病率在过去60年里有所上升,但其原因仍知之甚少。引起带状疱疹的病原体水痘带状疱疹病毒也是儿童感染水痘的罪魁祸首。
一种可能的解释可能是近年来传播的病毒水平相对较低,而在过去,这些病毒会导致亚临床感染,从而自然地“增强”先前感染或接种疫苗的个体的免疫系统。
✦ 人体可能在多次感染中增强了免疫,而不仅仅是因为接种了疫苗
在过去,我们会接种疫苗或从感染中恢复,但感染仍然是地方性的,因此我们都可能被再次感染并多次加强,而往往是在不知情的情况下。
我们大多数人可能多次被许多病毒再次感染,并产生了非常好的免疫力。因此,一次接触病原体就会导致终生免疫的想法可能需要重新考虑。疫苗也是如此——一些疫苗和病毒暴露所产生的保护作用必然会随着时间的推移而减弱。
疫苗、mRNA 等可以挽救生命。然而可能需要放弃这样一种观念,即它们提供一次性的保护。疫苗仍然是增强我们免疫防御的最重要且最安全的工具。
然而免疫力是单独来自疫苗还是由于疫苗加上暴露后的自然增强?对于一种非常容易传播的病原体变异后,我们能否获得完全的免疫力?这些问题还值得深入研究,但确定的是我们可以通过疫苗接种来预防重大疾病。
新型疫苗有望成为癌症等疾病治疗的新平台
COVID mRNA 疫苗的工作原理是诱导人体产生对自己安全的SARS-CoV-2刺突蛋白,并训练免疫系统识别和禁用它,然后在遇到实际病毒时执行相同的操作。
这种方法首次成功用于对抗 SARS-CoV-2,已成为利用人体免疫反应产生针对各种传染病以及多种癌症、心脏病、罕见疾病和某些自身免疫性疾病的“疗法”蓝图。
这次大流行使人们对免疫印记(免疫的核心特征)有了更细致的了解。
当我们的身体第一次遇到病毒时,它们会对其形成长期记忆。这种最初的遭遇不仅影响了对同一种病毒的后续反应,也影响了对看起来相似的病原体的后续反应。这种形式的免疫印记通常被描述为“原始抗原原罪”,指的是第一次遇到特定抗原,随后使免疫系统偏向于该病原体。
✦ 免疫系统对新病毒和曾感染过的相似病毒具有不同反应
当免疫系统遇到一种新病毒时,它不会快速产生针对这种新病毒的高度特异性抗体,而是默认对最初的相似病毒的记忆——就像一个视力不佳的人在远处看到模糊的熟悉的面孔,当他们只是看起来像熟人的陌生人时,他们会像朋友一样打招呼。
免疫印记可以对类似于原始抗原的病毒产生保护性免疫反应,因为这些新病毒的某些部分看起来像创建记忆的原始抗原。然而,免疫印记也会干扰针对新病毒或先前见过的病毒突变部分的快速免疫反应的形成。它在一定程度上削弱了初始防御的准确性。
✦ 曾感染过的病毒突变可能会更容易受到感染
疫情是一件可怕的事情,但抛开这一点,看看我们的免疫系统如何处理信息并看到生物学在起作用,这对我们今后的健康至关重要。
初次接触记忆会更强烈,并可能导致终生对病毒原始毒株的偏见。事实上,即使随后的暴露扩大了免疫反应以涵盖更新的毒株,对原始病毒的反应性仍然较高。这可能意味着,当以前遇到的病毒发生足够的突变而无法被产生抗体的免疫细胞识别时,个体会变得更容易受到感染。
但是,生命早期接触病毒往往也能提供持久的保护,防止类似病毒株引起的严重疾病,即使晚年的反应可能不足以预防有症状的感染。
再次感染(COVID-19)会怎么样?
有证据表明,当感染过SARS-CoV-2病毒原始毒株或接种疫苗的个体遇到后来的变种时,与原始毒株相比,他们产生了更高水平的针对原始毒株的抗体,而针对新毒株的抗体水平则更少。
然而,研究表明,任何先前通过感染或接种疫苗接触 SARS-CoV-2 的情况都会降低随后感染的疾病严重程度。
此外,先前的疫苗接种并不排除对病毒突变部分形成新的反应。免疫印记的局限性可以通过在疫苗中使用更高的抗原剂量、通过对一种以上变体产生反应的多价疫苗或通过用佐剂(增强免疫反应的物质)增强疫苗来克服。
最重要的是,当快速反应比高度精确的反应更重要时,基于过去暴露的免疫回忆的抗体生产是一种快速、经济的方法,可以在不确定的情况下建立快速保护。
免疫系统具有广泛的功能和对过去接触的长期记忆,具有进化优势。这是一种对抗未来病原体遭遇风险的方法。
之前认为免疫系统可以释放记忆并删除带有先前病毒遭遇记忆的细胞,但事实并非如此。保留这些旧的“文件”增加了至少一些长寿的抗体产生细胞识别前所未见的病毒的某些部分的可能性。制造高度适应新病毒的新抗体需要时间和精力,这在反应速度至关重要的高风险情况下是宝贵的资源。
因此,免疫系统部署了两种反应路线:一种是快速但不精确的反应,依赖于先前存在的防御;另一种是缓慢但更精确的反应,逐渐磨练其武器库来攻击新病毒。
免疫系统足够聪明,知道它需要速度和精度,但为了做到这两点,它最初会放弃一点精度,转而追求速度。
研究人员早就知道病毒和宿主免疫系统处于永久的竞争中,但为什么这场战斗在大多数人身上达到停滞状态,而在其他人身上却造成严重感染呢?
更重要的是,这场战斗在细胞和分子层面上到底是如何展开的?这个问题一直困扰着科学家,并阻碍了对预防或治愈感染的治疗的探索。
哈佛医学院的研究人员最近使用实验室工程细胞进行了一项研究,并发表在《美国国家科学院院刊》上,揭示了宿主和病原体在争夺细胞主导地位时所使用的精确策略。
此外,该研究还展示了免疫系统如何在细胞控制中心(细胞核)发生的战斗中阻止病毒。
✦ 干扰素信号蛋白阻止病毒感染
该研究揭示了一组称为干扰素的信号蛋白的关键作用,它们招募其他保护性分子并阻止病毒建立感染。
一旦进入宿主体内,单纯疱疹病毒就会利用宿主的遗传机制在细胞核内复制自身,从而进行繁殖。为此,病毒必须战胜宿主的免疫系统。
干扰素因其干扰病原体感染细胞的能力而得名,是当免疫系统检测到微生物(例如病毒)存在时释放的信号分子。干扰素发出的求救信号会激活该细胞和其他产生蛋白质的细胞中的基因,从而首先阻止病毒建立感染。
干扰素用于阻止细胞质(充满细胞的凝胶状液体)内的病毒的几种不同机制是众所周知的。但干扰素如何对抗DNA病毒——那些在细胞核内发起攻击的病毒——仍然是个谜。
✦ 干扰素阻止病毒复制
在这项新研究中发现,干扰素会招募一种名为 IFI16 的宿主蛋白,以多种方式帮助阻止病毒复制。
IFI16 用来抵御病毒的策略之一是在病毒 DNA 基因组周围构建和维持分子外壳。这种分子“气泡膜”可以防止病毒展开。当病毒被包裹起来时,它就无法激活其 DNA 来表达其基因并复制自身。
注:然而,为了对抗这些保护性策略,病毒会产生称为VP16和ICP0的分子,这些分子可以去除包裹物,使宿主细胞的保护性分子失活,并使病毒能够繁殖。
IFI16 对抗病毒感染的另一种机制是中和VP16和 ICP0。正常情况下,当细胞未准备好击退病毒入侵者时,细胞核内会存在一些IFI16。但IFI16的水平不足以对抗病毒辅助蛋白并保持病毒的包裹和抑制。
如果没有干扰素向细胞发出更多的IFI16信号,病毒就会赢得军备竞赛并感染细胞。然而,实验表明,当干扰素信号招募更高水平的IFI16时,免疫系统就会获胜。
目前的这项研究与类似的发现相呼应,即在组织的临床样本中发现 IFI16 水平升高,在这些组织中,免疫系统似乎成功地控制了密切相关的病毒症状,这提供了有关避免病毒爆发的分子机制的重要见解。
最新研究显示,五分之一的人在服用抗病毒药物后出现新冠病毒反弹。病毒反弹是指在对病毒性疾病进行治疗后,病毒水平暂时下降但之后再次上升的现象。
Paxlovid 是一种用于治疗 COVID-19 的口服抗病毒药物。此前的研究表明,该药物可有效减少重症 COVID-19 病例的住院率和死亡人数。自从 Paxlovid 纳入 COVID-19 治疗以来,一些患者报告了病毒学反弹。此前一项名为 EPIC-HR 的 3 期临床试验表明,服用 Paxlovid 的患者中只有1%至2%出现病毒学反弹。
✦ 特定药物可能增加了新冠病毒反弹
哈佛医学院研究人员进行的一项新研究发现,在接受尼尔马瑞韦-利托那韦疗法(俗称 Paxlovid)治疗 COVID-19 严重症状的人中,有五分之一的检测结果呈阳性,并且已痊愈初次康复且检测呈阴性后,存在潜在传染性的活病毒。相比之下,不服用 Paxlovid 的人只有大约2%出现反弹。
研究人员指出,这些发现不应阻止临床医生开出药物,但应促使他们向服用药物的患者提供关于病毒反弹和将病毒传播给他人的风险的咨询。
新研究表明,Paxlovid 引起的病毒反弹比之前认为的更为常见。结果表明,病毒反弹的发生频率可能比之前认为的要高,这引发了人们对那些确实经历病毒复发的人传播病毒风险的质疑。
✦ 病毒反弹使一些患者在康复后可能具有传染性
超过20%服用 Paxlovid 的人出现这种现象——而且个体在出现反弹时会释放活病毒,这意味着他们在最初康复后可能具有传染性。
研究人员警告说,该研究依赖于观察,而不是随机对照试验,因此科学家们不能确定服用 Paxlovid 的人的反弹率增加仅仅是由于使用该药物所致。
该团队使用阳性病毒培养物作为传播病毒风险的标志,但没有正式测量经历病毒学反弹的人的传染性有多大。尽管如此,他们指出,活病毒脱落是其他呼吸道病毒传播的一个公认因素,因此临床医生应建议患者在反弹期间保持警惕并进行隔离。
✦ 出现反弹的个体传染性持续时间也较长
出现反弹的个体病毒排出时间也较长,平均为14天,而没有出现反弹的个体则少于5天,这表明他们的传染性可能会持续更长时间。令人欣慰的是,没有证据表明病毒在反弹的患者中会对药物产生耐药性。
最初的EPIC-HR 研究仅在两个时间点评估患者的结果,而新研究则更频繁地跟踪患者。当研究人员将他们的数据与第一个 EPIC-HR 研究的选定时间点进行比对时,他们也发现只有2.4%的参与者出现了病毒学反弹,这一发现表明之前的研究并未捕捉到病毒学反弹的全部范围。
这项研究在未来还需要知道为什么有些人会经历反弹,而另一些人却没有。他们计划研究 Paxlovid 反弹现象背后的生物学机制,并确定改变治疗持续时间是否有助于对抗反弹效应。
一些研究探讨肠道细菌在抵抗病毒感染中的作用,肠道微生物组在疾病和健康中的作用已得到充分证实。然而,我们肠道中的细菌如何保护我们免受病毒感染尚不清楚。
✦ 肠道微生物调节免疫以抵御病毒入侵
哈佛医学院的研究人员首次描述了这种情况在小鼠身上发生的情况,并确定了调节局部和全身免疫反应以抵御病毒入侵的特定肠道微生物群。
这项研究于11月18日发表在《细胞》杂志上,精确定位了一组肠道微生物以及其中的一个特定物种,这些微生物会导致免疫细胞释放被称为1型干扰素的病毒排斥化学物质。
研究人员进一步确定了该组中许多肠道细菌所共有的精确分子,该分子可以解锁免疫保护级联反应。研究人员指出,这种分子可以成为增强人类抗病毒免疫力药物的基础。
“鉴于干扰素在疾病和健康中发挥的关键作用,我们鉴定出一种可以诱导干扰素保护信号的细菌分子,这为开发治疗化合物提供了一种有前景的新方法,这种化合物可以增强抗病毒免疫力,从而降低病毒感染的风险。”该研究的资深作者、哈佛医学院免疫学教授丹尼斯·卡斯珀说。
✦ 脆弱拟杆菌通过产生干扰素β进行抗病毒保护
在对细胞和动物进行的一系列实验中,研究人员发现,大多数人类肠道中存在的一种微生物——脆弱拟杆菌,会启动信号级联反应,诱导结肠中的免疫细胞释放一种名为干扰素-β的蛋白质——一种重要的免疫化学物质,通过两种方式提供抗病毒保护:它诱导病毒感染的细胞自我毁灭,并刺激其他类别的免疫细胞攻击病毒。
具体来说,实验证明细菌表面的一种分子通过激活所谓的TLR4-TRIF信号通路来触发干扰素β的释放。这种细菌分子会刺激由九种Toll样受体(TLR)之一启动的免疫信号通路,TLR是先天免疫系统的一部分。
当免疫细胞表面的蛋白质识别出各种感染性生物体表面的某些明显的分子模式时,该途径就会被激活,并针对这些入侵者组织免疫防御。
✦ 其他肠道细菌可能也具有免疫保护信号传导作用
由于解锁这一级联的特定表面分子并非脆弱拟杆菌所独有,并且也存在于同一家族的多种其他肠道细菌中,因此研究人员测试了携带该分子的其他细菌物种是否可以触发类似的免疫信号传导。
在一组小鼠中进行的一组实验表明,在拟杆菌属细菌家族的多个其他物种中发现的含有这种分子的膜可以成功地启动类似的信号传导——这一发现表明,广泛的肠道细菌共有更广泛的免疫保护信号传导。
为了确定脆弱拟杆菌是否可以保护动物免受感染,研究人员测试了两组小鼠,一组使用抗生素治疗以耗尽肠道微生物群,另一组则保持完整的肠道微生物群。
接下来,研究人员将接受治疗和未接受治疗的动物暴露于水疱性口炎病毒(VSV)中,这种生物体几乎感染所有哺乳动物,但在人类中导致大部分无症状感染。与未接受抗生素且肠道微生物群完整的小鼠相比,经过抗生素治疗且肠道微生物群耗尽的小鼠在接触病毒后更有可能出现活动性感染,并且在感染后病情更严重。
结果证明了肠道微生物在诱导保护性干扰素-β信号传导和增强对病毒感染的自然抵抗力方面的作用。观察结果证实,共生微生物群正是通过干扰素-β信号传导发挥其保护作用。
大多数时候,病毒的变化是渐进的。从本质上讲,它只改变了其表面的一部分,但这还不足以在它试图闯入时完全掩盖我们的免疫系统或疫苗。
• 病毒发生巨大变化时会导致疫苗和免疫系统无法识别
有时,它会进行改造。可以说,它剪了头发、染了头发、做了整容手术,还减掉了很多体重。这些巨大的变化使得我们的免疫系统无法识别它,有时我们用来抵御它的疫苗和药物也无法识别它。
这发生在奥密克戎(Omicron)毒株的第一波浪潮中,它迅速在世界范围内传播,感染接种疫苗的人和以前感染过的人,仍然引起了Covid-19。2021年冬季,Omicron在全球范围内造成大量感染,令人瞠目结舌。
另一种新冠变异病毒也已经出现在世界舞台上,它被称为BA.2.86,它的刺突蛋白有30多个氨基酸变化,这使得它在基因上与其下一个最近的祖先 BA.2 相距甚远,就像最初的 Omicron 变体来自 SARS 的祖先菌株一样。
• 疫苗和人体的免疫也需要不断加强
目前尚不清楚 BA.2.86 的具体来源。科学家认为,它所携带的突变模式是一种在慢性感染者体内发生变化的病毒特征。通常,这些患者的免疫功能减弱,无法完全清除体内的病毒,但他们有足够的免疫力,这会给病毒带来压力,使其不断变化以求生存。
新型冠状病毒可以以这种方式变异,这意味着我们可能需要不断更新我们的疫苗和免疫力才能跟上步伐。
新冠病毒感染者担心三件事:“‘我会死吗?我最终会住院吗?但对于大多数人来说,问题是‘我会生病并病上好几个月吗?我会长期感染新冠病毒吗?”
• 长期并发症的风险正逐渐下降
“实际上,对于大多数人来说,这是最重大的风险。”
专家指出,根据我们现有的科学知识,长期并发症的风险似乎正在下降。
澳大利亚最近的另一项研究对2022年7月至8月期间近 23000名确诊感染Covid-19的人进行了研究,结果发现,在超过11000名接受调查的人中,有18%符合长期 Covid 的病例定义。研究人员将其定义为感染 Covid-19 后 90 天以上出现的任何新症状或持续症状。
• 中老年群体更有可能长期感染新冠病毒,即使接种过疫苗也不例外
这也是一个高度接种疫苗的群体。超过94%的受访者至少接种了三剂Covid-19疫苗。在这项研究中,女性、年龄在50岁至69岁之间的人、居住在农村地区的人以及疫苗剂量较少的人更有可能报告长期感染新冠病毒。
• 但接种疫苗可以降低感染的风险和病情程度
根据治疗长期新冠患者的经验表示,人们接种疫苗后,感染新冠病毒的风险会从大约10%到20%下降到个位数。他说,如果他们使用Paxlovid等抗病毒药物,感染率会进一步下降。
“一般原则是,你的病情越重,你的免疫越弱,出现急性和慢性并发症的机会就越高,多项研究表明,通常在大流行的早期,随着最初的变异,人们有更多的急性和慢性并发症。”加州大学旧金山分校传染病专家 Peter Chin-Hong 博士说。
新冠病毒以这种方式引起长期疾病的能力似乎仍然是独一无二的,感染流感的人有可能会患上长期流感的问题,但最终患上这种疾病的人比例约为1%。
传染病专家表示,这种风险足够大,不应忽视。她说:“即使是轻微的新冠病例也可能导致症状持续数周、四个月或更长时间,所以不应该掉以轻心。”
• 再次感染可能会出现更严重的危害
在整个人群中,感染导致慢性并发症的风险可能会下降,但每一次感染都会增加一个人可能面临持久伤害的机会。最近发表在《自然医学》杂志上的一项研究发现,再次感染不是良性的。每一次额外的感染都会增加一个人的死亡、住院和其他长期问题的风险。
美国国立卫生研究院最近资助的一项研究通过梳理数百万份患者记录来查找首次和第二次感染Covid-19的人,该研究支持了这些发现。
研究人员发现,在超过30万名再次感染的人中,第二次感染更严重疾病的风险略高。
此外,新冠对儿童的危害也不容忽视,可能还会出现一些特定的症状。
• 儿童感染新冠病毒后可能出现罕见并发症
在大流行早期,儿科传染病专家一直在寻找儿童感染 Covid-19 的一种罕见并发症,称为儿童多系统炎症综合征(MIS-C)。
MIS-C在Covid-19感染后两到六周开始。据美国疾病控制与预防中心称,它可能会导致持续发烧以及以下一种任意症状:胃痛、眼睛充血、腹泻、头晕、皮疹或呕吐。
注:MIS-C可能很严重,但大多数孩子通过医疗护理会有所好转。
最近的研究表明,随着时间的推移,全球MIS-C病例的数量和严重程度均有所下降。而接种疫苗可进一步降低风险,达90%以上。
过去,研制传统疫苗需要四到二十年的时间,而针对新冠病毒的新型信使RNA (mRNA)疫苗来说,这是创纪录的11个月。这个过程可能永远改变了药物开发的方式。
支持COVID-19疫苗开发的是对 mRNA 疫苗十多年的研究,该疫苗教会人类细胞如何制造触发特定免疫反应的蛋白质。
• 未来mRNA疫苗还可能治疗心脏病等复杂疾病
有一天,疫苗可能会治疗心脏病等疾病。在不久的将来,mRNA 技术可能会带来更好的流感疫苗,随着流感病毒随季节的变化而快速更新,或者开发出可能有效数年的“通用”流感疫苗。
它也可能针对我们最大的杀手。未来的mRNA疗法可以帮助衰竭心脏的肌肉再生,并通过个性化癌症疫苗针对个体癌症的独特遗传学。医生将能够对你的肿瘤进行测序,并用它来制造疫苗,唤醒你的免疫系统来对抗它。mRNA 疫苗也将使我们为未来的流行病做好准备。
与此同时,请使用我们现有的疫苗,不要跳过目前为老年人推荐的流感、肺炎、带状疱疹等常规疫苗。仅流感疫苗一项每年就可以挽救数万人的生命,并将因流感住院的风险降低28%。
自我保健是指个人为了维持和提升健康而采取的一系列主动行为和习惯。这包括合理膳食、适量运动、良好睡眠、避免有害物质、管理压力和保持积极心态等方面。
为什么要关心自我保健?呵护对于您自己和周围人的幸福至关重要。让人感到放松的活动对我们的健康和平衡至关重要,而且这种自我保健的心态可能会持续下去。
• 自我保健有助于提高免疫,更好地抵御病毒
无论是长时间的泡澡,在后院修建花草,享受下午茶,还是晒晒太阳。对自己好一点,可以让我们免受来自外面威胁的任何恐惧。舒适将帮助我们轻松恢复生活,更好地抵御病毒的入侵。
通过自我保健,个人可以在很大程度上控制和改善自己的健康状况,减少对医疗系统的依赖。因此,推广自我保健意识和实践对于提高公众健康水平至关重要。
你的健康与生活方式直接相关——营养、身体活动、健康的体重和恢复性的睡眠。”
• 高龄会导致致命感染的风险增加
然而年龄带来的风险也较高,合并症(本质上是同时存在两种或多种健康问题)更为重要。具体来说,75 岁及以上的健康人发生致命感染的风险是所有65岁以下参与者的四倍。但如果你比较所有75岁及以上的人——包括那些患有高血压、肥胖或肺病等慢性疾病的人问题,可能性增加了十三倍。
• 而生活方式的改善可以降低老年人患新冠风险
研究发现:健康的75岁老人死于冠状病毒的可能性是患有多种慢性健康问题的65岁老人的三分之一。年龄会影响患上新冠肺炎重症的风险,但应该更加注重避免慢性健康问题。生活方式的改变可以改善您的整体健康状况,这可能会直接降低您患上严重新冠病毒或死于新冠病毒的风险。
锻炼仍然至关重要,英国对387109名40岁至60岁成年人进行的一项研究发现,避免体力活动的人患重症新冠病毒的风险要高出38%。活动能力应该被视为健康的重要标志之一。
从新冠疫情中学到的另一点,孤独是每个人的问题。这不仅发生在老年人身上,也发生在我们所有人身上。
孤独有多致命?一项令人震惊的调查,在大流行的前5个月里,旨在保护患有痴呆症的老年人和弱势成年人的疗养院封锁导致比往年增加了13200人死亡。
• 新冠疫情期间隔离造成孤独问题需要正确对待
隔离可能成为新常态。密歇根大学的一项民意调查发现,56%的50岁以上成年人表示,他们在2020年感到孤独,是2018年(新冠疫情爆发前)感到孤独的人数的两倍。根据约翰·霍普金斯大学在《美国医学会杂志》上发表的一项调查,随着疫情的加剧,所有成年人的心理困扰率都在上升,年轻人的心理困扰率增加了六倍,30至54岁的人则增加了四倍。
新冠疫情期间实施的隔离措施导致了广泛的孤独感,这对人们的健康产生了负面影响。孤独不仅影响心理健康,如增加抑郁和焦虑的风险,还可能对身体健康造成损害,比如增加心脏病和免疫系统功能下降的风险。
因此,新冠时期隔离措施虽然是控制疫情传播的必要手段,但其带来的孤独感需要被认真对待,并采取措施减轻其对健康的不良影响。
如果说老年人的生活围绕着定期亲自看医生,那么由于大流行给我们的医疗保健系统带来的压力,从某种程度上来说,医疗保健系统已经开始转变:远程医疗逐渐兴起并应用在医疗中,特别是对于那些患有慢性疾病的人来说。
• 远程医疗能够更早发现问题并及时调整
告别例行就诊,注册远程血糖监测的患者使用支持蓝牙的仪表,通过智能手机应用程序将结果直接传输到他们的健康记录中。远程监控使我们能够及早发现何时应该调整治疗。
但这需要意愿和大量装备,例如支持蓝牙的血压监测仪,以及医生方面存储和分析数据的系统。“人们需要使用设备,医疗保健系统必须准备好处理所有这些数据。”
• 集体就诊有助于提高医疗效率
集体就诊是指一组人在同一时间和地点接受医疗服务,通常是由一名医生或医疗团队为他们提供服务。
集体就诊可能是一种先进的方式。这种方式不仅有助于提升医疗服务的效率,还能促进患者之间的信息交流和经验分享,有助于患者更好地理解自身的病情和治疗方法。
参考链接:
medicalxpress.com/news/2024-01-pandemic-immune.html
www.aarp.org/health/conditions-treatments/info-2021/lessons-from-covid.html
www.cnn.com/2023/08/28/health/risk-from-catching-covid-wellness/index.html

谷禾健康

营养不良的微生物 Malnourished Microbes
儿童营养不良是全球主要的健康负担,营养干预措施只能部分解决这一问题。儿童营养不良的慢性和急性形式均以多种生物系统紊乱为特征,包括新陈代谢、免疫和内分泌系统。
越来越多的证据支持肠道微生物组在调节这些影响早期生命生长的途径中的作用。观察性研究报告了营养不良儿童肠道微生物组的变化,而临床前研究表明,这可能引发肠病,改变宿主代谢,并破坏免疫介导的针对肠道病原体的抵抗力,这些都会导致早期生命生长不良。
肠道微生物组在健康和疾病中发挥着不可或缺的作用,而饮食是其组成、多样性和功能的主要驱动因素。鉴于婴儿和儿童肠道微生物组的动态发育,解决两个主要问题至关重要:
a) 饮食能否改变肠道微生物组的组成、多样性或功能
b) 这种修改是否会影响功能/临床结果,包括免疫功能、认知发展和整体健康?
本文汇编了临床前和临床研究的证据,包括生命早期肠道微生物组影响宿主代谢、免疫、肠道功能、内分泌调节和其他导致儿童营养不良的途径的新兴病理生理学途径。
我们综合了儿童青少年的营养、营养干预措施和肠道微生物群之间联系的最新研究,该年龄段儿童是一个生长阶段,对疾病的易感性各不相同。还讨论了营养素如何单独或组合如何影响肠道微生物组的机制。
同时综合并讨论了新兴的微生物组导向疗法,并考虑未来的研究方向,以确定和针对儿童营养不良的微生物组治疗目标。
2013年发表在《柳叶刀》上文章统计,儿童营养不良导致全球儿童死亡人数的 45%,并对健康产生终生影响。全世界超过五分之一的5岁以下儿童至少表现出一种形式的慢性或急性营养不良。
首先我们来了解一下发育迟缓、消瘦的定义。
发育迟缓[定义为年龄别身长 Z 得分 (LAZ) < −2],影响着全球 22% 的儿童,与认知发展较差、终身综合能力下降以及成年后慢性病风险增加有关。
消瘦[定义为身高体重 Z 得分 (WHZ) < –2] 影响着全世界近 7% 的儿童,除了长期认知和健康缺陷不佳之外,还与高死亡率相关。
▸ 目前的治疗方法无法完全解决这一问题
营养疗法不足以完全恢复生长缺陷和营养不良的相关病理生理后果。标准生命早期营养补充剂仅可将发育迟缓降低 12-14%。
而严重消瘦后会出现高死亡率、再入院和长期生长缺陷。因此,存在与儿童营养不良相关的隐藏的病理生理学负担,目前的治疗方法无法完全解决这一问题。
▸ 儿童营养不良与多个生命早期生物系统紊乱有关
发育迟缓和消瘦儿童的肠道功能受到环境肠道功能障碍(EED)的影响。这种肠道病理学损害营养吸收,并可能导致全身炎症,从而损害早期生命生长。
在营养不良的儿童中宿主免疫力广泛缺陷,导致抵抗感染能力受损,特别是病原体密集的环境。
营养良好和营养不良的儿童之间存在激素差异,尤其是与生长 [生长激素 (GH) 和胰岛素样生长因子1 (IGF-1)] 和食欲调节(瘦素和生长素释放肽)相关的荷尔蒙差异。
营养不良时宿主代谢发生改变,氨基酸、脂质和能量代谢受到干扰。这些受到干扰的生理系统表明,儿童营养不良背后存在复杂的病理生理学,涉及相互关联的生物系统。
▸ 微生物群影响生命早期的生长发育
生命早期接触微生物也可能导致儿童营养不良中这些扰动途径。肠道病原体携带量高与线性生长和体重生长下降以及 EED 相关,而腹泻和呼吸道感染也与儿童营养不良密切相关。
除了病原体之外,共生肠道微生物群也会影响早期生命的生长。生命早期复杂微生物组的组装对于免疫训练、对病原体的定植抵抗、母乳寡糖(HMO) 和其他营养物质的代谢、肠道结构和内分泌信号传导至关重要。
因此,儿童时期肠道微生物组的破坏可能会损害这些有助于儿童健康成长的途径。
▸ 儿童在出生时就获得了一些的微生物组
除了环境获得的微生物外,这些微生物组主要源自母体肠道、阴道、口腔和皮肤微生物组。
在纯母乳喂养的儿童中,最初的婴儿肠道微生物群的多样性仍然很低,主要由大肠杆菌、双歧杆菌和拟杆菌组成物种。
在接下来2-3 年里,肠道微生物群会经历一种模式化的组装,这种组装主要是由纯母乳喂养和随后的补充食品的引入所形成的,这导致肠道微生物群的快速扩张和多样化。
▸ 许多环境因素会影响微生物,从而影响生长
包括地理位置、分娩方式、抗生素暴露、胎龄以及其他母亲和家庭因素。这些微生物群在出生后在胃肠道内的积累,除了肠道结构和对病原体的定植抵抗力之外,还推动了免疫、代谢和内分泌途径的成熟,这些都有助于儿童的正常生长。
如果微生物演替因营养不足、卫生或抗生素暴露而受到干扰,这些生长决定途径可能会受到损害。很少有研究纵向研究在营养匮乏的环境中保持良好营养的儿童与消瘦或发育迟缓的儿童的肠道微生物群的演替和组装情况。
因此,这种依赖高收入环境的微生物组数据库的现象,限制了低收入和中等收入国家中个体所产生的营养不良微生物组的解释性(这也是构建本地数据库和谷禾长期追踪重点人群纵向数据的主要原因),这些国家的肠道微生物组特征不太明确。
来自中低收入国家的横断面和短期纵向研究的越来越多的证据揭示了微生物组的差异,这些差异将营养不良与健康生长区分开来。
▸ 发育迟缓——多因素驱动
“发育迟缓”是一种慢性营养不良的形式,由多种遗传和环境因素共同驱动。高达30%的发育迟缓发生在子宫内,这可以通过出生体重对发育迟缓风险的强烈影响来证明。
之后一系列环境因素有助于产后线性生长。很少有研究纵向研究健康婴儿与发育不良婴儿肠道微生物组的获取和组装情况。
▸ 横断面研究:肠道菌群特征破坏导致发育迟缓
对马拉维和孟加拉国队列的二次分析发现,微生物组多样性减少和Acidaminococcus(氨基酸球菌)丰度增加分别与发育迟缓严重程度和未来线性生长缺陷相关。
其他一些小型横断面研究已经发现,在一系列中低收入国家队列中,发育不良儿童与健康儿童的粪便微生物组组成存在不同差异。来自中非共和国和马达加斯加的一项更大规模的多点横断面分析发现,发育不良儿童的粪便微生物群中大肠杆菌/志贺氏菌和弯曲杆菌种类丰富,而产生丁酸的种类较少。
▸ 发育迟缓的儿童:小肠细菌过度生长
在发育迟缓的儿童中观察到小肠细菌过度生长的比率较高,其特征是十二指肠中口咽微生物的富集。发现一群来自孟加拉国的发育迟缓儿童的小肠细菌载量与 LAZ 之间呈负相关。韦荣球菌属、链球菌属和Rothia mucilaginosa的十二指肠丰度也与 LAZ 呈负相关。
▸ 肠道菌群的功能潜力——预测儿童生长的指标
对 335 名津巴布韦农村的 1 至 18 个月大儿童的肠道微生物组的组成和功能发育的跟踪发现,肠道微生物组的功能宏基因组组成(而非分类组成),可以预测孩子已达到的线性生长和未来的生长速度,其中 B 族维生素和核苷酸生物合成途径是最具预测性的特征之一。这些数据表明,肠道微生物组的功能潜力是儿童生长的更强大的指标。
来自马拉维的一个纵向队列发现了类似的结果,即 16S 测序无法识别成分多样性、成熟度或物种丰度与 LAZ 之间的关联。秘鲁对 6-24 个月大的儿童(作为营养不良和肠道疾病队列的一部分)进行的一项纵向研究表明,成分多样性与线性生长之间存在一定关联,仅针对出生时发育迟缓的儿童。
该研究组另外一项研究究表明,肠道微生物组可能介导腹泻和弯曲杆菌感染对线性生长的影响。除了肠道微生物组细菌成分的影响之外,发育迟缓婴儿与非发育迟缓婴儿中噬菌体(可以调节微生物组组成的病毒)的丰度也有所不同。
总的来说,发育不良儿童的肠道微生物群与营养良好的儿童不同。然而,这因地理群体而异。因此,目前不存在一致的发育迟缓的组成或功能微生物组特征。
▸ 儿童口腔微生物组与发育迟缓有关
发育迟缓的儿童在十二指肠和粪便中表现出更丰富的口咽微生物,包括唾液乳杆菌,这表明胃肠道的隔室化可能导致营养不良儿童中肠道菌群变化,并通过减少营养吸收而导致生长不良或口腔微生物在其生态位之外的炎症作用。
▸ 怀孕期间母亲的口腔微生物组预测儿童营养不良
牙周炎是一种口腔生态失调疾病,会导致局部免疫反应功能失调,并与低出生体重呈负相关。在怀孕期间的母亲口腔中,Actinomyces naeslundii(内氏放线菌)的相对丰度与出生体重和妊娠持续时间呈负相关,而Lactobacillus casei(干酪乳杆菌)则呈正相关,两者都强烈预测儿童营养不良。
▸ 早产的阴道微生物组,与新生儿低出生体重和随后的发育迟缓密切相关
高度多样化、缺乏乳杆菌的阴道微生物群与西方地区的早产以及撒哈拉以南非洲地区新生儿 LAZ 的减少密切相关。阴道微生物组的改变很可能与早产的经典感染触发因素同时发生,包括绒毛膜羊膜炎、细菌性阴道病和泌尿生殖感染,从而导致泌尿生殖环境炎症,这些共同可能限制胎儿生长或引发早产。
▸ 消瘦及其分类:营养不良形式的反映
消瘦是营养不良的一种形式,可以反映慢性营养缺乏和急性疾病,通常与发育迟缓同时发生。
消瘦可分为中度[中度急性营养不良(MAM)]或重度[严重急性营养不良(SAM)],取决于 WHZ 阈值、中上臂围 (MUAC) 和/或水肿的存在。
▸ 水肿性SAM、非水肿性SAM、复杂SAM
SAM 还以两种主要形式存在:
如果出现急性感染、食欲不振、休克或水肿(也称为复杂 SAM)。
患有水肿性和非水肿性 SAM 的儿童具有不同的临床结果,其中非水肿性 SAM 儿童在某些情况下接受 SAM 住院治疗后表现出更高的死亡率和再入院率。
尽管治疗 SAM 的营养疗法有所改进,但需要住院治疗的复杂 SAM 的死亡率在 10% 至 40% 之间,而生长缺陷可能会持续长达7年,此外,日后认知缺陷和慢性病风险也会增加。
▸ 在SAM 儿童肠道中肠道微生物组具有影响力
患有 SAM 的儿童肠道微生物多样性降低,肠杆菌科细菌增多,而与健康生长一致的双胞胎相比,SAM不一致的双胞胎肠道病毒组受到干扰。
▸ 年龄微生物群Z评分——微生物群成熟度指标
2014 年,孟加拉国的一项具有里程碑意义的研究创建了微生物群成熟度指标,称为年龄微生物群Z评分(MAZ),该评分在 SAM 儿童中显着降低,并且与术后营养恢复的人体测量指标高度相关。这项研究和最近的后续研究使得所谓的微生物组生态群得以细化,该生态群由 15 种细菌类群组成,这些细菌类群在不同地理环境中在生命的头 2 年中表现出一致的共变,可用作微生物群落的指标。
不成熟的微生物组发育并衡量MAM 或 SAM 后微生物组的恢复程度。根据 MAZ 的评估,标准治疗性往往只能暂时恢复微生物组的成熟度,表明 MAZ 可能有潜力作为未来 MAM/SAM 复发的指标,或者针对肠道微生物组的营养疗法可能有助于营养恢复。
事实上,这些微生物群成熟度指标为 MAM 中新的微生物群导向营养干预措施提供了信息,在试点研究中,与标准治疗食品相比,这些干预措施在更大程度上改善了营养恢复。
▸ 在水肿性SAM 和非水肿性 SAM 儿童差异
这可能部分解释了他们营养恢复的差异。与水肿性SAM儿童相比,非水肿性 SAM 儿童的α多样性显著降低,并且与普雷沃氏菌科(Prevotellaceae)、毛螺菌科 (Lachnospiraceae) 、瘤胃菌科(Ruminoccoaceae)相对丰度的减少有关。
总的来说,与健康成长的儿童相比,SAM 儿童的微生物组始终不成熟且多样性降低。这种失调的微生物组可能是急性营养状态以及与治疗相关的合并症和抗生素治疗的结果。
如上所述,越来越多的人类观察研究证据表明,营养良好和营养不良的儿童之间的微生物组组成存在差异。
临床前和临床研究已开始揭示,生命早期肠道微生物群紊乱可能导致儿童生长不良的病理生理机制。其中包括肠道微生物组对免疫、新陈代谢、肠道功能和内分泌信号传导等途径的影响(如下图)。
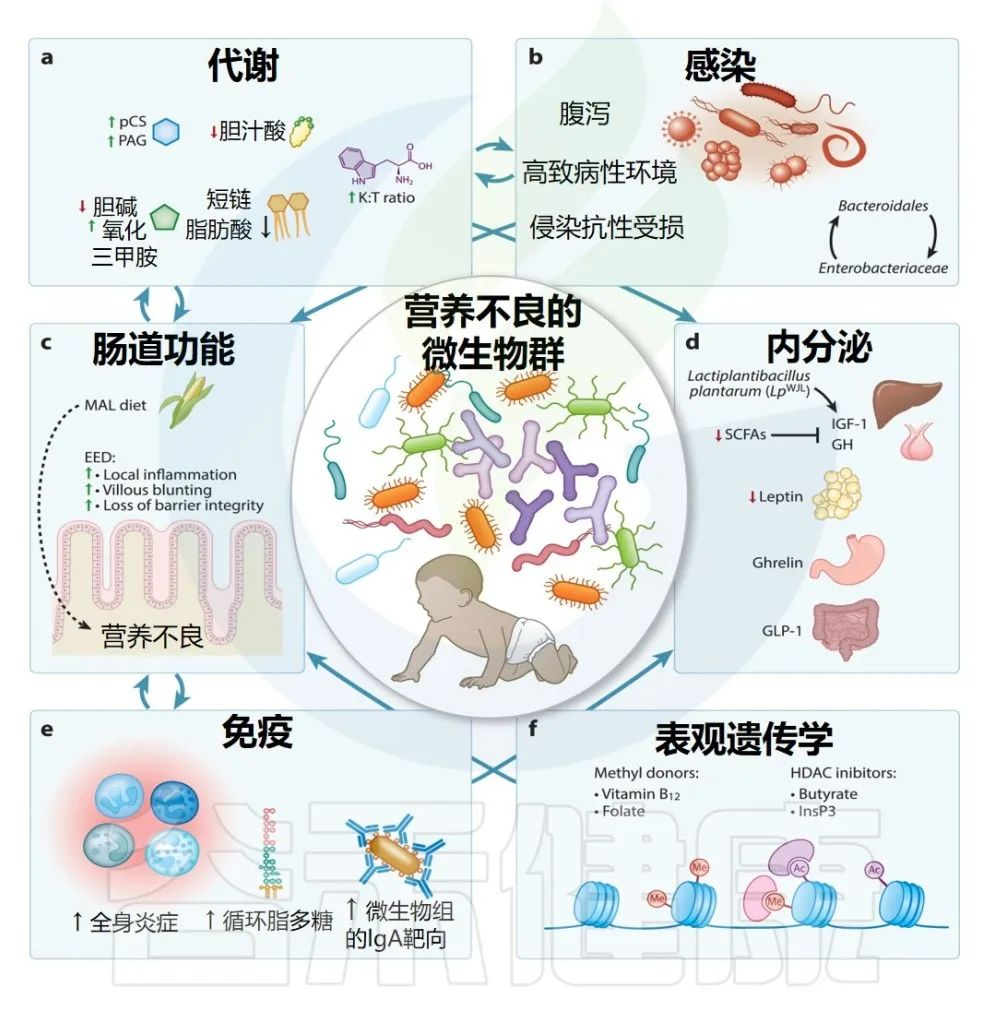
doi.org/10.1146/annurev-nutr-061121-091234
下面我们逐一来了解营养不良的微生物群带来的多方面影响。
共生肠道微生物群的存在,对于生命早期肠道屏障的正常结构、功能和转录发育至关重要。临床前研究的大量证据表明,无菌小鼠幼崽表现出肠道屏障成熟延迟,这与自发性结肠炎、更容易肠道感染和营养吸收不良有关。
生命早期长期接触肠道病原体会破坏肠道结构和功能。环境肠道功能障碍(EED)是一种胃肠道现象,在营养匮乏的环境中很常见,并且经常在营养不良的儿童中观察到(我们以前也专门写过儿童EED的文章)。
▸ 什么是环境肠道功能障碍(EED)?
EED 通常是亚临床的,局限于小肠,其特征是肠道通透性增加、绒毛萎缩和变钝、粘膜屏障变化、吸收不良、局部炎症和隐窝伸长。屏障功能的丧失还导致微生物及其产物(最常研究的是脂多糖)从肠道转移到体循环中,从而刺激全身炎症。据推测,营养吸收不良、肠道炎症和慢性全身炎症的结合会阻碍儿童的健康成长。
▸ 环境肠道功能障碍(EED)与什么相关?
EED 通常归因于在水、环境卫生和个人卫生 (WASH) 较差的环境中长期接触肠道病原体。然而,在此类环境中改善怀孕期间和早期生活中的 WASH 的大型随机试验未能减少幼儿 EED 的常见生物标志物,这可能是由于同时未能减少其肠道病原体携带。
除了病原体携带之外,共生微生物组的破坏也可能导致 EED。对611 名儿童( 6、18 和 30 个月大)大型分析发现,微生物群多样性/成熟度与 EED 的三种生物标志物之间存在负相关:
表明肠道微生物群的不成熟可能会导致肠道炎症和屏障功能障碍。特定菌群的丰度升高,包括Megasphaera、Mitsuokella、Sutterella也与 EED 有关。
注:REG1B,Regenerating family member 1 beta 是一种蛋白质,它是一种由胰腺细胞产生的蛋白质,在胰岛细胞再生和胰岛功能调节中起着重要作用。
REG1B在胰腺细胞再生过程中被表达,且参与了胰岛细胞增殖和分化,对胰岛细胞的生长和修复有促进作用,在胰岛素分泌和血糖调节中发挥一定的调节作用。
EED 动物模型有助于深入了解可能驱动该疾病表型的机制。一些模型能够通过应用低蛋白饮食和 LPS 或吲哚美辛等肠道损伤来复制 EED 的某些方面。
注:吲哚美辛:用于治疗关节炎,癌性疼痛,痛风,滑囊炎、肌腱炎及肩周炎等非关节软组织炎症;可用于高热的对症解热,恶性肿瘤引起的发热或其他难以控制的发热。
▸ 肠道微生物对 EED 的潜在贡献
利用异常微生物状态和改变的营养环境之间相互作用的新模型已经成功地概括了几种 EED 表型。例如,肠道生理学和微生物组通过血管紧张素 I 转换 2 酶 (ACE2) 的表达联系起来。
注:ACE2是一种在分化上皮细胞的管腔表面表达的蛋白质。
与野生型小鼠相比,缺乏 ACE2 的小鼠在受到DSS攻击时,表现出类似 EED 的病理学、氨基酸代谢紊乱和肠道微生物组紊乱。DSS 攻击期间的 EED 表型可以在从 ACE2 缺陷小鼠进行粪便移植后转移到野生型动物中,这表明微生物群被破坏。
注:DSS,右旋糖酐硫酸钠,一种破坏肠上皮屏障并导致结肠炎的刺激物
膳食色氨酸及其代谢物烟酰胺可以通过哺乳动物雷帕霉素靶点 (mTOR) 活性刺激抗菌肽的产生,从而挽救 ACE2 缺陷小鼠的表型,从而维持微生物组稳态。
▸ 微生物结合营养物质影响EED
低蛋白、低脂肪饮食会导致小鼠小肠微生物群遭到破坏,其特点是小肠内物种丰富度升高,此外拟杆菌门和变形菌门也会扩张。
将营养不良的饮食与拟杆菌和大肠杆菌结合起来作者产生了 EED 的表型,包括肠道通透性、绒毛和隐窝萎缩增加、空肠细胞因子 (IL-6 和 MCP-1)释放到腔内增加,以及细菌对小肠上皮细胞的粘附增加。这种表型依赖于微生物组的改变和营养不良的饮食,这表明 EED 表型是复杂的微生物与营养物质相互作用的结果。
总体表明,肠道微生物群破坏在营养不良相关肠道功能障碍的病理学中具有潜在的因果作用。
▸ 腹泻和有症状的呼吸道感染与发育迟缓和消瘦有关
关于腹泻和儿童营养不良的大型多中心研究的有力证据表明,肠道病原体的高负担与生命早期生长不良有关。
健康且多样化的肠道微生物群通过生态位排除提供针对肠道病原体的定植抵抗力。因此,受损的肠道微生物组可能为病原体定植提供了环境。
肠道微生物组也可能介导远处部位的感染易感性,包括肺部。因此,生命早期健康肠道微生物组可能有助于在卫生条件差的环境中抵抗病原体的定植,从而防止感染负担和相关的生长不良。
▸ 发育迟缓:肠道菌群多样性减少——预示腹泻发病率的增加
来自秘鲁的发育不良儿童在生命的前两年表现出肠道微生物组多样化轨迹受损。在此期间,发育迟缓的儿童的微生物多样性显着减少,腹泻后多样性的恢复也较慢。因此,肠道微生物组的组装受损可能会加剧感染-发育迟缓的循环。
来自同一队列的一项研究发现,弯曲杆菌的携带与肠道微生物群的改变有关。此外,肠道微生物组内与弯曲杆菌携带相关的物种与 LAZ 的减少独立相关,这表明肠道微生物组组成受损会促进病原体定植和并发生长缺陷。
▸ 营养不良:微生物组对病原体定植产生影响
与营养良好的小鼠相比,由营养不良饮食和肠道微生物群紊乱引起的 EED 小鼠模型对感染的敏感性更高,这表明与营养不良相关的微生物群破坏可能会导致病原体定植和感染负担。
蓝氏贾第鞭毛虫感染后生长缺陷的易感性也取决于动物模型中的肠道微生物组。人类研究报告称,贾第鞭毛虫和相关的肠侵袭性病原体,除了那些涉及粘膜破坏的病原体外,对全身炎症、肠道炎症和生长受损具有最深远的影响。
在小鼠中,贾第鞭毛虫与小肠中的肠杆菌科细菌相互作用,以限制蛋白质营养不良期间的生长,这种作用在用没有抗贾第鞭毛虫活性的抗菌药物治疗后消失,表明失调的微生物群在此过程中发挥着关键作用。
这种相互作用表明特定的病原体或病原体依赖于共生肠道微生物群的成员来定殖和增殖。这在人类发育迟缓研究中很明显,即肠杆菌科细菌和拟杆菌目细菌之间的交叉喂养与儿童发育迟缓有关。
▸ 在体外,肠杆菌科和拟杆菌目之间协同生长
特别是在蛋白质和铁含量低的营养不良条件下。在这种情况下拟杆菌属利用饮食和粘蛋白衍生的糖和肠杆菌科细菌提高铁的生物利用度,铁是特定病原体增殖所必需的微量营养素。
拟杆菌科和肠杆菌科在营养不良的儿童中具有很强的相关性,但在营养良好的儿童中则没有。另一种拟杆菌属的脆弱拟杆菌,还依赖周围的微生物群来诱导小鼠的生长效应。含有bft毒素的脆弱拟杆菌的产肠毒素菌株会导致无菌小鼠体重减轻和能量代谢受损,这些小鼠被来自发育迟缓儿童的微生物群定植,但对于那些被来自营养良好儿童的微生物群定植的小鼠则不会。
这些数据表明,病原体和感染对儿童营养不良的影响可能依赖于共生肠道微生物群的组成和功能,在儿童营养不良患病率高的环境中减少病原体负担的策略除了考虑病原体负担外,还必须考虑共生微生物群。
营养不良儿童的宿主蛋白质、脂肪和能量代谢紊乱早已有报道。然而,肠道微生物组的代谢能力大大扩展了哺乳动物宿主的生物转化能力,扩大了可加工底物的多样性,并增加了宿主接触的分子范围。
许多微生物衍生的代谢物可以在肠道和肠壁中发挥局部作用,并且在吸收后还可以影响周围组织中的宿主过程。因此,儿童早期肠道微生物组的变化可能会通过改变提供给儿童的代谢副产物和宿主代谢途径本身来影响代谢表型,从而影响生长。
▸ 成熟度与微生物代谢
与用于绘制儿童微生物组成熟度的MAZ评分类似,代谢成熟度模型,称为年龄表组 Z 评分 (PAZ),是根据秘鲁、坦桑尼亚和孟加拉国儿童的尿液样本构建的,并用于追踪生化年龄及其与生长的关系。用于计算 PAZ 的八种代谢物中的三种——对甲酚硫酸盐 (pCS)、苯乙酰谷氨酰胺 (PAG) 和马尿酸,与肠道微生物代谢相关。
有趣的是,与未生长受限的婴儿相比,生长受限的婴儿早在出生后 3 个月就出现生化不成熟(即,其生化年龄低于实际年龄)。这凸显了微生物群在宿主整体代谢能力发展及其对生长的后续影响中的重要性。
健康肠道微生物群的一个主要功能是发酵宿主无法消化的膳食底物。在母乳喂养期间,这主要涉及HMO的微生物消化。患有发育不良婴儿的母亲的母乳中 HMO 含量明显较低,结构相似的牛乳低聚糖可以恢复营养不良模型中小鼠的生长。
然而,这种效果取决于微生物群,因此补充乳寡糖后,无菌小鼠的生长不会恢复。断奶后,肠道微生物群转向不可消化的碳水化合物、抗性淀粉、粘蛋白和蛋白质的代谢。
▸ 糖分解产生短链脂肪酸,满足能量需求
糖分解活性(即碳水化合物的分解)的主要最终产物是短链脂肪酸。这些都有助于宿主日常的能量需求。丁酸盐是结肠细胞的主要能量来源,提供其所需能量的 80%,而乙酸盐在肌肉等全身区域代谢,丙酸盐在肝脏中用于产生 ATP。
这种微生物活动提供了一种从饮食中释放能量的机制,可用于支持生长。此外,短链脂肪酸还具有其他有益作用,例如降低结肠 pH 值以增强对潜在病原体的定植抵抗力并提高矿物质吸收。此外,丁酸盐已被证明可以通过增加 AMP 激活蛋白激酶 (AMPK) 活性和紧密连接蛋白的组装来促进肠道屏障的发育。
▸ 营养不良:从碳水化合物发酵和短链脂肪酸生产转向蛋白水解代谢
例如,与营养良好的儿童相比,印度尼西亚中度营养不良儿童的粪便丙酸盐和丁酸盐含量较低,粪便 pH 值较高。
在营养不良的猪模型中,肠道微生物组产生的丁酸盐较少,并且与肝脏脂肪酸代谢(β-氧化)减少有因果关系。同样,在对巴西东北部儿童进行的代谢组学研究中,发育不良的婴儿排出了更多由微生物降解氨基酸而产生的微生物-宿主共代谢物。
其中包括 pCS 和 PAG,它们分别源自酪氨酸和苯丙氨酸的微生物分解。营养不良病原体相关的EED导致肠道吸收不良可能会推动这一观察。这增加了小肠中氨基酸的可用性,从而导致发育不良儿童中的小肠细菌过度生长,并增加到达结肠进行细菌代谢的氨基酸。
▸ 营养不良:微生物群与发育中的宿主竞争氨基酸
有趣的是,虽然 SCFA 促进上皮屏障完整性,但 pCS 被发现会损害它,可能导致营养不良个体中观察到的肠漏表型。
此外,pCS由艰难梭菌和其他病原体产生,限制肠道微生物群的生物多样性。可能导致营养不良婴儿中观察到的微生物群失调。另外两种氨基酸代谢物,吲哚乳酸和N-乙酰谷氨酸,分别源自色氨酸和谷氨酸代谢,在来自孟加拉国的一组儿童中,它们与 2-24 个月龄之间的线性和体重生长指标呈正相关。长双歧杆菌也与生长呈正相关,它编码了参与这些化合物生产的大部分微生物代谢途径,这些化合物可能有助于支持早期生命的生长。
▸ 膳食胆碱的微生物分解增加还导致蛋白质营养不良和发育迟缓
在蛋白质缺乏的小鼠模型中,尿胆碱减少,而胆碱代谢的微生物产物三甲胺和二甲胺增加。
同样,在马拉维儿童中,血清中三甲胺-N-氧化物(TMAO)与胆碱的比率与线性生长障碍呈正相关。
胆碱和甜菜碱排泄量降低与巴西婴儿发育迟缓有关。胆碱的可用性不仅对于肌肉获得很重要,对于S-腺苷甲硫氨酸 (SAMe)的生成也是必要的,而 S-腺苷甲硫氨酸 (SAMe) 是 DNA 甲基化和发育的关键。
重要的是,胆碱是骨骼肌的必需营养素以及神经发育和大脑功能。胆汁酸是宿主肝脏和肠道微生物群组合代谢产生的代谢物的一个例子,在消化中发挥重要作用。
初级胆汁酸,例如胆酸和鹅去氧胆酸,在肝脏中合成,然后通常与甘氨酸或牛磺酸结合,并分泌到胆汁中。释放到肠道后,一小部分胆汁酸可以到达结肠,在那里微生物群将它们分解并转化为次级胆汁酸,例如脱氧胆酸。这些修饰的胆汁酸可以通过粪便排出或再循环回肝脏。
与没有 EED 的儿童相比,患有 EED 的儿童的血清总胆汁酸含量较低,其中牛磺鹅去氧胆酸、牛磺鼠胆酸和甘氨酰去氧胆酸存在特定差异。除了消化之外,胆汁酸在代谢调节中也发挥着重要作用。通过充当全身表达的受体(例如 FXR、PXR、VDR 和 TGR5)的配体,胆汁酸可以调节多种宿主途径和功能的表达,例如能量稳态、葡萄糖和脂质代谢。
▸ 这些细菌相关代谢物还具有抗菌特性,既能抑制又能促进特定病原体的生长
例如,胆汁酸代谢的变化,特别是细菌胆汁盐水解酶活性,被认为是隐孢子虫卵囊排泄的重要触发因素。此外,观察到未患阿米巴病的儿童粪便中次级胆汁酸、脱氧胆酸的含量较低。研究表明,给小鼠注射这种胆汁酸足以增加粒细胞-单核细胞祖细胞的数量,并提供针对阿米巴病的保护。这表明胆汁酸可能在预防营养不良环境中常见的不同肠道感染和对这些感染的易感性方面发挥潜在作用。
▸ 从微生物代谢,到宿主代谢的改变
除了微生物代谢的改变外,在营养不良的儿童中还观察到宿主代谢的改变,这可能部分是由菌群失调引起的。例如,从发育迟缓儿童的小肠中分离出的口腔微生物在体外和体内模型中会损害脂质吸收,从而提供小肠微生物组和小肠细菌过度生长在营养不良中的潜在因果作用。
肠道屏障功能的丧失会导致微生物群及其产物(包括脂多糖)从肠道进入体循环的易位增加。LPS 易位可导致慢性全身炎症,从而通过吲哚胺 2,3-双加氧酶诱导激活色氨酸-犬尿氨酸途径。事实上,秘鲁和坦桑尼亚儿童的血浆犬尿氨酸与色氨酸 (K:T) 比率与血浆 LPS 和 LPS 结合蛋白 (LBP) 相关,并且还与线性增长缺陷相关。
扩展阅读:
犬尿氨酸途径的激活可以抑制炎症并促进耐受性,但可能会失调色氨酸途径,而色氨酸途径对于血清素的产生以及 NAD +和烟酰胺的生成非常重要,而后者是生长的关键。免疫耐受性的增加也可能对增强对病原体的有效反应产生影响,这也可能受到色氨酸剥夺的影响,并可能导致此类感染在营养不良的肠道中持续存在。
儿童营养不良与内分泌信号中断有关,包括介导食欲和能量代谢的瘦素和生长素释放肽,以及共同构成生长轴的GH 和IGF-1 。
发育迟缓儿童与非发育迟缓儿童相比,在童年后期血浆 IGF-1 较低以及生命的前 18 个月,一种与慢性炎症相关的现象。
▸ 瘦素水平低可预测SAM儿童的死亡率
新数据表明,肠道微生物组直接和间接影响肠道局部激素的产生,例如胰高血糖素样肽-1 (GLP-1),并通过介导瘦素、生长素释放肽和 IGF-1 的产生来系统地进行,每一个都调节新陈代谢和营养状况。
▸ 微生物组→产生SCFA→GLP-1 的产生→抑制胰高血糖素原的表达
GLP-1 是一种肠道源性激素,负责刺激胰岛素分泌、减少胃排空和增加饱腹感,从而影响新陈代谢和营养状况。
在没有微生物组的情况下,在无菌小鼠中,GLP-1 的产生在一种假设的适应性机制中升高,当微生物组被破坏或不存在时,GLP-1 的产生会增加营养物质的吸收,而 SCFA 给药可以逆转这种效应。
▸ 微生物衍生的SCFA也会影响 IGF-1 的产生
IGF-1 在肝脏和脂肪组织中产生,影响骨形成、骨量和骨骼生长,尤其是在生命早期。当无菌小鼠被微生物定植时,血浆中血清 IGF-1 的浓度会增加。
同样,抗生素的使用会降低 IGF-1 浓度,而补充 SCFA 可恢复抗生素治疗期间 IGF-1 的浓度。不产生短链脂肪酸的细菌种类也可以通过刺激生长轴来刺激实验动物的生长。植物乳杆菌的特定菌株可恢复果蝇和小鼠的线性生长、股骨长度、IGF-1 产生和活性以及外周组织对 GH 的敏感性。
植物乳杆菌WJL ( Lp WJL ) 菌株对 IGF-1 产生和生长的活性取决于肠上皮细胞中含有核苷酸结合寡聚化结构域的蛋白 2 (NOD2) 的刺激。
▸ 肠道共生细菌还可以防止感染时的肌肉萎缩
共生大肠杆菌在骨骼肌中维持 IGF-1 信号传导,独立于宿主代谢、热量吸收或炎症,从而防止肠道或肺部感染后肌肉萎缩。
▸ 与饱腹感和新陈代谢有关的激素与特定共生菌群丰度相关
在动物模型中,双歧杆菌和乳酸菌丰度与瘦素浓度呈正相关,而生长素释放肽则与拟杆菌和普雷沃氏菌丰度呈正相关。
在营养不良模型中,生长素释放肽水平还与双歧杆菌、乳酸菌和球状球菌-直肠真杆菌呈负相关。一项针对 6 至 24 个月大的冈比亚儿童( n = 60)的横断面观察队列研究发现,严重营养不良儿童的肠道微生物群与代谢激素浓度之间存在许多密切相关性,使用网络分析模拟肠道微生物组和激素相互作用。埃希氏菌/志贺氏菌通过其与生长素释放肽和生长素释放肽受体的相互作用,在区分健康对照和营养不良患者方面具有高度预测性,而粘膜乳杆菌与瘦素/瘦素受体之间的相互作用也对营养不良和营养良好的儿童具有高度区分性。肠杆菌科细菌和 IGF-1 的相互作用也区分了 MAM 和 SAM。
这些相互作用可能是双向的,瘦素可以刺激粘蛋白的产生并改变肠道微生物组的组成,而特定的产生脂多糖的肠杆菌科细菌的炎症活性可能会损害 IGF-1 和 GH 信号传导。
微生物群在整个生命过程中与宿主免疫系统保持着相互的、动态的沟通,而这种沟通会因营养不良而被破坏。
越来越多的证据表明,营养不良的儿童在免疫学上与营养充足的同龄人不同,胸腺较小,免疫细胞组成和分布在质量和数量上不同,全身和肠道环境中的促炎介质长期升高,以及由不利因素驱动的免疫基因上的表观遗传标记。
产前和产后暴露。迄今为止,大多数营养不良儿童的免疫表型都是在感染和病原体携带的背景下进行研究的。然而,新出现的数据表明,肠道微生物组内共生和条件致病微生物的多样性可能会驱动营养不良儿童的独特免疫表型,而与病原体无关。
▸ 营养不良引起的肠道炎症的特征——EED生物标志物
例如钙卫蛋白和髓过氧化物酶,这些生物标志物是由肠道组织中微生物激活的先天免疫细胞产生的,这些细胞可能会被破坏的肠道微生物组激活。
EED 动物模型表明,营养缺乏的小鼠与营养充足的小鼠相比,肠道菌群失调会导致小肠上皮内淋巴细胞数量增加和促炎细胞因子分泌增加。尽管空肠免疫激活增加,但营养缺乏的动物在肠道内抑制鼠伤寒沙门氏菌感染的能力较差,导致细菌传播到脾脏和肝脏,并导致肝脏分泌促炎介质。
▸ 儿童营养不良还与慢性全身炎症有关
尽管反复出现症状的感染可以引发全身炎症表型,但小鼠模型表明,在没有明显感染的情况下,肠道微生物群和/或其成分系统性扩散到功能失调的肠道屏障进入循环也可能发生这种情况,如 EED 中观察到的那样。
与营养充足的幼鼠相比,断奶后采用缺乏蛋白质、铁、锌的饮食的健康小鼠出现了微生物群失调和体重减轻,这与向结肠给药时未能含有脂多糖有关;全身、盲肠和结肠(但不包括回肠)促炎细胞因子升高;在没有感染的情况下,LPS 攻击后促炎细胞因子的分泌增多。
在该模型中,通过针对革兰氏阴性(LPS 阳性)而非革兰氏阳性(LPS 阴性)共生体的抗生素治疗,使体重增加和促炎性免疫反应的差异正常化。
该模型中抗生素治疗效果的选择性支持了对革兰氏阴性微生物群成分的播散性促炎性免疫反应的作用,这些微生物群成分从肠道转移到循环系统中,从而导致营养不良的体重和身高缺陷。
由微生物易位驱动的慢性全身炎症可能影响人体测量的一种方式是通过抑制生长因子信号传导和骨/骨骼肌生长。来自动物研究的证据表明,依赖于微生物群代谢的唾液酸化HMO会增加成骨细胞驱动的骨形成,并且该过程是由 Th2 极化免疫反应介导的,即增加大肠中的嗜酸性粒细胞募集和嗜酸细胞趋化因子浓度。
中低收入国家儿科队列研究一致的证据表明,炎症介质也与IGF-1呈负相关。
注:前面讲过,营养良好和营养不良的儿童之间存在激素差异,尤其是与生长[生长激素 (GH) 和胰岛素样生长因子 1 (IGF-1)] 和食欲调节(瘦素和生长素释放肽)相关的激素差异。
包括 sCD14、补体蛋白 2、巨噬细胞炎症蛋白 1B 和 LBP 在内的炎症介质与SAM入院后康复儿童的体重和/或 MUAC 增加呈负相关。
▸ 炎症介质对生长的直接影响
无菌小鼠持续过度表达促炎细胞因子 IL-6 和循环 IGF-1 减少,生长速度较慢,并且比非无菌同窝小鼠小 30-50%,这种表型可以通过给予 IL-6 来部分挽救。
研究发现,IL-1β 和TNFα 与 IL-6 一起下调人肝细胞中生长激素受体 (GHR) 的表达、通过细胞因子信号传导抑制因子 3 (SOCS3) 产生的 GH 信号传导以及 IGF-1 的产生体外细胞系和小鼠肝脏体内细胞系。
▸ 微生物群:或将使炎症驱动的生长缺陷正常化
在一项以豆类为基础的治疗性营养的临床试验中,已经探索了将微生物群作为使炎症驱动的生长缺陷正常化的一种手段的潜力,该营养旨在增强营养不良的孟加拉国儿童肠道微生物组的多样性和成熟度。
与之前的观察结果一致,30 名 SAM 儿童在基线时可以通过血浆蛋白质组与 21 名健康儿童区分开来,该血浆蛋白质组与较低的骨化和成骨细胞分化以及较高的急性期炎症反应有关,包括 C 反应蛋白 (CRP)、IL- 6、促炎核因子 kappa B (NF-κB) 信号通路中的中间蛋白。干预后,尽管证据表明总体肠道病原体携带或绝对粪便微生物群多样性存在差异,但 WHZ 和微生物群成熟度有所增加;这些变化与血浆蛋白质组向健康生长和减少炎症特征的转变有关,表明肠道微生物组针对性的干预措施可以减少营养不良相关的炎症。
菌群失调引起肠道和全身炎症的能力也可能取决于抗体介导的肠道微生物组成的控制。肠道中的分泌性免疫球蛋白,特别是免疫球蛋白 A (IgA),选择性地结合微生物组成分,从而调节微生物组组成。
扩展阅读:
▸ 发育不良:IgA靶向的细菌比例更高
研究发现,与来自马达加斯加和中非共和国的非发育迟缓对照儿童相比,发育不良儿童的粪便中 IgA 所靶向的细菌比例更高。除了 IgA 靶向肠道微生物群的数量差异外,IgA 还靶向健康儿童与 SAM 儿童不同的肠道微生物子集。其中 IgA 倾向于靶向SAM 儿童中的肠杆菌科细菌,同时针对健康儿童中更广泛的共生生物。
从患有 SAM 的儿童身上提取的IgA +微生物群在移植到小鼠体内时会诱发类似 EED 的病理学和全身炎症,这种效应依赖于营养不良的饮食。
在另一个模型中,给小鼠喂食低蛋白、低脂肪饮食,并进行或不进行细菌灌胃来模拟 EED,这些小鼠的 IgA 对乳杆菌属的靶向作用受损。尽管空肠中乳酸杆菌的相对丰度、粪便 IgA 总量以及 IgA 目标细菌总数的百分比与营养良好的对照组相似。
这些差异是由于乳杆菌碳水化合物代谢对饮食变化的适应,而不是 IgA 丰度或亲合力的差异。总的来说,这些观察结果强调了肠道屏障处免疫-微生物群相互作用的可塑性,有证据表明,微生物群和组织驻留免疫细胞的适应可能会损害 EED 中微生物及其抗原的免疫介导遏制,从而导致生长缓慢。
▸ 营养不良儿童的口服疫苗反应受损并反映营养不良中存在的一些免疫缺陷
小鼠模型为营养不良时微生物组失调如何损害口服疫苗的免疫原性提供了一些线索。然而,这些范式需要在人类队列研究中进一步探索。
在针对产肠毒素大肠杆菌(ETEC)进行初免加强口服疫苗接种后,由于饮食缺乏和大肠杆菌粘附定植而诱导出现 EED 样表型的小鼠,其小肠中的疫苗特异性 CD4 + T 细胞较低,且疫苗水平较低-相对于饲喂营养充足饮食的小鼠和饲喂营养不足或充足饮食的未定植小鼠的特异性 IgA。这些差异取决于微生物群,因为 ETEC 定植小鼠的小肠疫苗特异性 T 细胞数量在广谱抗生素治疗 3 周后恢复正常。
EED 表型中疫苗反应受损是由于微生物群依赖性 RORγT + FOXP3 +Treg 的扩张,这些细胞能够抑制小肠中的疫苗反应;小肠中疫苗特异性 CD4 + T 细胞增殖和 IgA 丰度得到恢复,并且在有条件地消耗 Tregs 后体重增加百分比增加。
总的来说,这些研究为微生物群靶向疗法如何恢复营养不良儿童受损的免疫表型(包括受损的疫苗反应)提供了概念证明,但也强调了固有的免疫和微生物异质性,在设计转化研究和人群时应考虑到这一点。
许多研究开始探索食物或食物成分对整个童年时期肠道微生物组的影响。
纳入的试验采用了涉及天然食品的干预措施;微量营养素补充或强化;食品成分,例如牛乳脂肪球膜(MFGM);以及特殊配方食品,例如早期限量配方食品(ELF)、即用治疗食品、即用补充食品(RUSF)和脂类营养补充剂(LNS)。
母乳
纯母乳喂养的婴儿往往含有较高比例的双歧杆菌和乳酸菌属。与配方奶喂养的婴儿相比,他们的肠道微生物组往往含有更高丰度的潜在致病菌,例如变形菌门成员。
双歧杆菌属丰度更高对婴儿健康有益,而肠球菌属的比例较高与不良的健康结果相关。
从瓶子中挤出的母乳,与潜在病原体的富集和双歧杆菌的消耗相关。
▸ 母乳是一种复杂的基质,有助于双歧杆菌定植
母乳含有许多成分,包括乳糖、400 多种不同的脂肪酸、蛋白质(乳清)、核苷酸、维生素、矿物质、乳脂肪球 (MFG) 、HMO等。
HMO 可以与牛奶中的其他化合物(例如糖复合物)结合,形成母乳聚糖 (HMG),其完整地到达婴儿结肠并驱动细菌定植,特别是双歧杆菌的定植。
HMG 逃脱肠道消化并转运到结肠,进入那里常驻双歧杆菌的发酵循环,使这些物种能够增殖并在 HMO 的双歧杆菌效应的过程中占据主导地位。特别是,婴儿双歧杆菌(ATCC 15697) 、短双歧杆菌、长双歧杆菌和双歧双歧杆菌(PRL2010) 在婴儿期占主导地位,是更健康肠道微生物组的标志。
▸ 母乳本身也含有细菌
在几组哺乳期妇女中,发现母乳样本最常见有葡萄球菌、链球菌和假单胞菌,其他分类单元因女性和环境而异;这些细菌可能有助于在婴儿肠道中播种。
配方奶
▸ 配方奶与对肠道微生物群的潜在负面影响有关
a ) pH 值的变化,导致大肠杆菌和沙门氏菌等细菌过度生长,从而破坏肠道完整性,可能是由于蛋白质类型和含量的差异
b ) 牛乳中缺乏母乳中存在的游离氨基酸和某些生物活性化合物。
食用牛奶配方奶粉的婴儿往往具有较高的肠道微生物多样性和丰富的厚壁菌门、梭状芽胞杆菌、肠球菌属、肠杆菌科和拟杆菌属和双歧杆菌属的丰度较低。
在配方奶喂养的婴儿中存在的双歧杆菌属中,有更多的青春双歧杆菌和假链状双歧杆菌,这两种细菌都与成人肠道微生物群有关。
▸ MFGM 对肠道菌群的影响:嗜血杆菌水平较低
MFGM (牛乳脂肪球膜)因其在婴儿肠道成熟、免疫以及大脑结构和功能中的潜在作用而受到关注。MFGM 源自乳腺上皮,包含 60% 蛋白质和 40% 脂质,使 MFG 稳定为乳液。为了复制母乳,婴儿配方奶粉中添加了牛 MFGM。
在 2 至 6 个月龄期间,MFGM 配方奶粉组的几种氨基酸及其分解产物的含量较低,乳酸和琥珀酸的含量也较低。12 个月时,MFGM 组的婴儿嗜血杆菌水平较低与接受标准配方奶粉的婴儿相比,该属含有多种病原体。
然而,与母乳喂养(非随机)参考组相比,MFGM 配方奶粉组的微生物群与标准配方奶粉组的微生物群更相似,因此,将 MFGM 和标准组的结果汇总进行分析,并与来自 MFGM 配方奶粉组的结果进行比较。
▸ ELF:改善健康的同时,尽量不破坏肠道菌群
母乳和婴儿配方奶粉中乳清蛋白和酪蛋白的含量各不相同。婴儿配方奶粉中牛奶的比例较高,较难消化。水解牛奶可以提高其消化率,水解牛奶用于早期限量配方食品(ELF),用于纯母乳喂养的新生儿作为干预措施,以增加肠内摄入量并避免并发症,例如胆红素血症或脱水。
与纯母乳喂养相比,ELF 不会导致α 多样性或群落结构出现显着差异,也不会导致乳杆菌丰度降低或更高丰度的梭状芽胞杆菌。这些发现表明,ELF 可能有助于改善体重大幅减轻的新生儿的健康,同时不会对肠道微生物群产生不利影响。
碳水化合物
碳水化合物包括简单的单糖和二糖以及复杂的寡糖和多糖,例如淀粉和纤维素。简单的糖在小肠 (SI) 中被消化和吸收,而复杂的或非血糖碳水化合物在到达结肠之前仍未被消化。
后者已被证明可以通过聚糖的降解来调节肠道微生物群,特别是以纤维和寡糖的形式。
关于单糖对肠道微生物群影响的研究有限,因为它们通常不会到达大肠;更常见的是针对人工甜味剂影响的研究或高糖、高脂肪的西方饮食的组合对肠道微生物群的影响。
注:单糖,如葡萄糖、半乳糖和果糖;二糖,例如乳糖、蔗糖、麦芽糖和海藻糖;和糖醇(多元醇)
▸ 果糖
一项小鼠研究中,果糖可以逃避肠道吸收并到达结肠,从而降低多形拟杆菌的丰度。
在一项针对 12-19 岁青少年的队列研究中,高果糖摄入量与真杆菌和链球菌含量降低有关,而这些细菌被认为有利于碳水化合物代谢。这些研究的结果提供了一些证据,证明在随机试验中检查单糖对人类肠道微生物组的影响是有益的,这些试验尚未在儿童中进行。
▸ 复杂碳水化合物
包括寡糖、多糖和膳食纤维,可以起到益生元的作用,被定义为“被宿主微生物选择性利用并赋予健康益处的底物”。这些分子不会被人类消化,而是被完整地运送到结肠,在那里微生物可能会代谢和发酵它们。
碳水化合物益生元包括HMO、低聚果糖、低聚半乳糖、甘露低聚糖、低聚木糖和膳食纤维,例如不溶性纤维,包括纤维素和半纤维素,例如β-葡聚糖(来自燕麦)。碳水化合物益生元还包括可溶性纤维,例如果胶和菊粉,两者都是微生物发酵罐的底物,包括长双歧杆菌亚种、婴儿杆菌、拟杆菌、拟杆菌、乳杆菌和普氏粪杆菌。
▸ 益生元通过直接机制发挥作用
简而言之,结构与宿主聚糖相似的碳水化合物分子可以直接阻断与宿主细胞的粘附;β-葡聚糖可以与吞噬细胞和自然杀伤细胞上的受体结合,引发中性粒细胞的吞噬作用;
菊粉型果聚糖可以通过 Toll 样受体、C 型凝集素受体和半乳糖凝集素作为肠道树突状细胞的受体,诱导抗炎细胞因子。
▸ 益生元可以间接地选择有益共生细菌的增殖
其代谢副产物可以直接影响肠道环境或宿主基因表达。例如,与潜在致病性大肠杆菌和梭菌相比,双歧杆菌属、拟杆菌和厚壁菌门中的物种将到达结肠的未消化碳水化合物发酵成微生物副产物,宿主或其他共生细菌可以将其用作底物。
▸ 麸皮添加:微生物群的变化因基线饮食差异而异
麸皮(小麦、燕麦和大米等谷物的外皮)是益生元可溶性纤维以及其他营养素(如植物化学物质、脂肪酸和酚类物质)的主要来源。
在一项试验中,生活在尼加拉瓜 ( n = 47) 和马里 ( n = 48)的 6 个月大婴儿每天补充米糠(每天 1-5 克),添加到断奶食品中,或者不进行干预 6 个月。
不同国家的基线肠道微生物群落结构(通过非度量多维尺度(NMDS)绘制的布雷-柯蒂斯差异来测量)存在差异,这种差异导致 8 个月和 12 个月龄时微生物对米糠补充剂的反应越来越明显;例如,马里样本在两个时间点的群落结构差异(NMDS 图上的分离)更为明显。对米糠摄入量敏感的属也因国家而异:在尼加拉瓜,8 个月和 12 个月龄时,双歧杆菌属、乳杆菌和韦荣球菌的相对丰度增加,而在马里乳杆菌在 8 个月和 12 个月大时,对米糠的反应丰度有所增加。
这些反应性类群被公认为碳水化合物消化者。这项研究在比较这些人群时的一个局限性是,整个马里队列都是纯母乳喂养和阴道分娩,而尼加拉瓜样本包括配方奶喂养和大多数剖腹产婴儿;这些基线饮食差异可能导致了微生物对米糠的不同反应。
引入补充食品,包括低聚糖的植物性食物来源,例如大豆和其他豆类、水果和蔬菜,帮助婴儿肠道微生物群多样化并增加其丰富度,使其达到类似成人的组成。
▸ 蜂蜜:双歧双歧杆菌有变化
蜂蜜是单糖(即葡萄糖、果糖、蔗糖和麦芽糖)以及益生元低聚果糖的来源。
在早产儿肠道微生物群研究中,埃及早产儿 ( n = 40) 连续 14 天食用添加到牛奶配方奶粉中的 5、10、15 或 0 克医用级三叶草蜂蜜。从定量实时聚合酶链反应 (qRT-PCR) 结果来看,双歧双歧杆菌似乎以剂量依赖性方式对蜂蜜做出反应,而乳酸杆菌种不会。10g 组的计数最高,5g、15g 和 0g 组变化不大;这可能与这些细菌的底物代谢差异有关。
一些局限性包括样本量小(每组仅 10 名婴儿)、通过 qRT-PCR 定量特定细菌(相对于整个细菌群落的下一代测序)、干预持续时间短以及研究结果的适用性(考虑到不推荐蜂蜜)对于 12 个月以下的儿童,因为担心它可能含有导致婴儿肉毒杆菌中毒的细菌。
然而,作为一项试点研究,这些发现为进一步研究蜂蜜和单糖对老年人群肠道微生物组的潜在影响提供了支持。除了豆类外,没有其他试验研究过碳水化合物在天然食品中的作用,包括天然存在的益生元,对足月婴儿或年龄较大的儿童或青少年人群的作用。
脂 肪
脂肪酸,包括单不饱和脂肪酸 (MUFA)、多不饱和脂肪酸 (PUFA)(例如亚麻酸 ω-3 和亚油酸 ω-6 脂肪酸)和饱和脂肪酸 (SFA),构成脂质分子的一部分,例如甘油三酯、磷脂、胆固醇和植物甾醇。
膳食脂肪在整个胃肠道中被分解。大多数甘油三酯的消化和吸收发生在小肠(特别是十二指肠)中,并且依赖于胰脂肪酶、肝脏中的胆汁盐和胆囊中的胆汁。
初级胆汁盐,包括胆酸盐和鹅脱氧胆酸盐,是由胆固醇合成的;共生微生物群从初级胆汁盐合成次级胆汁盐、脱氧胆酸盐和石胆酸盐,这些盐进一步被细菌用作底物或被肝细胞修饰。大约 7% 的游离脂肪酸完好无损地到达结肠,它们可能会影响体内的微生物群。食物通常含有短链、中链和长链脂肪酸的混合物。
脂肪酸影响肠道微生物群的机制仍然是一个活跃的研究领域,并且关于 SFA、MUFA 和 PUFA 对人类肠道微生物组的影响存在相互矛盾的证据。
▸ 促炎:饱和脂肪酸、ω-6 多不饱和脂肪酸; 抗炎:ω-6多不饱和脂肪酸、单不饱和脂肪酸
一些研究表明,SFAs 和 ω-6 PUFA 会促进炎症和氧化应激,而 ω-3 PUFA 和 MUFA 则具有积极作用,例如增加乳酸杆菌和双歧杆菌属。由于膳食脂肪的代谢需要氧气,因此脂肪不太可能成为厌氧肠道细菌的能量来源。然而,较高的脂肪摄入量可能会取代饮食中的膳食纤维和碳水化合物;因此,对肠道微生物群的影响可能是由于饮食替代或较低的碳水化合物底物可用性,而不是较高的脂肪摄入量。膳食脂肪可能对细胞膜有杀菌作用,导致细菌丰度降低。此外,膳食脂肪可以诱导细菌增加胆汁酸代谢,以应对增加的脂肪。
一项研究旨在分析鱼和红花油的混合物对接受小肠肠造口术的早产儿(n = 16)肠道微生物群的影响(即在腹壁上开一个开口,以便在出生后进行肠内喂养)因炎症性疾病而需要进行腹部手术,例如坏死性小肠结肠炎 (NEC),与标准护理相比这是早产儿的常见病症。
与标准营养疗法相比,干预措施持续 9 周,结果增加了微生物α 多样性,降低了潜在致病菌(肠杆菌科、梭状芽胞杆菌)的丰度,并丰富了碳水化合物代谢的预测基因功能。
总之,这些研究表明,ω-3 LCPUFA 干预可能会改善婴儿肠道微生物群多样性、组成和预测功能,特别是患 NEC 的早产儿,但母亲在怀孕期间摄入 LCPUFA 不会影响婴儿肠道微生物组。
蛋白质
膳食蛋白质由形成肽的氨基酸 (AA) 组成,肽是组织的基本组成部分。氨基酸对于骨骼肌功能、生长、健康、发育和生存至关重要,可以为宿主和肠道微生物群提供能量,而肠道微生物群可以改变宿主对氨基酸的生物利用度。
膳食蛋白质的消化从胃开始,蛋白酶将蛋白质切割成大肽。小肠中的肽酶分解这些肽,以便随后由肠上皮细胞和腔细菌进行转运和分解代谢。
增加蛋白质摄入量通常会导致结肠消化率降低和可发酵底物增加。细菌将游离氨基酸转化为多肽有助于肠道中氨基酸的代谢和生物利用度,而 L-谷氨酰胺等氨基酸的可用性可以调节必需和非必需氨基酸的小肠细菌代谢。
结肠细菌,特别是拟杆菌、梭状芽胞杆菌和大肠菌,将摄入的蛋白质和来自宿主酶、粘蛋白和脱落的肠细胞的内源蛋白质转化,产生副产物,例如较短的肽、AA、脂肪酸(例如 SCFA)和气体例如氨和硫化氢。
整个肠道的蛋白水解活性各不相同:例如,体外实验发现,与回肠微生物群相比,结肠微生物群可以更有效地降解结肠中的牛血清白蛋白,具体取决于 pH 水平、碳水化合物可用性和肠道模型保留时间。
▸ 较高的蛋白质摄入量:促进潜在致病菌
较高的蛋白质摄入量(例如西方饮食中的蛋白质)被认为会增加 pH 值,从而减少双歧杆菌等严格厌氧菌的生长。并促进蛋白质发酵,潜在致病性兼性厌氧细菌。
这些细菌,例如大肠杆菌和沙门氏菌,可以通过形成降解产物(例如三甲胺氧化物以及芳香族和支链氨基酸)来破坏肠道屏障和免疫系统;这些副产品与胰岛素抵抗和 2 型糖尿病有关。除了摄入水平之外,蛋白质来源、浓度和 AA 组成也会影响肠道微生物群的组成和功能。
▸ 蛋白质来源会导致微生物组成和功能的变化
由于植物细胞壁的原因,豆类等植物蛋白不能被宿主酶完全消化,导致它们作为微生物发酵的益生元运输到结肠。
难解析出植物蛋白来源中的蛋白质、纤维或其他化学物质的个体效应,但总的来说,这些蛋白质来源往往与细菌组成的有益变化有关,包括乳酸菌的生长和更大的微生物多样性。
进行了两项试验,利用 V4 区的 16S 测序来检查植物性蛋白质来源(包括豇豆、黄豌豆和普通豆)对肠道微生物群的影响。
一项试验比较了烤豇豆粉(25% 蛋白质,21% 纤维)、烤普通豆粉(25% 蛋白质,28% 纤维)和挤压煮熟的玉米大豆混合 (CSB) 面粉(13% 蛋白质,8% 纤维) ,6 个月大的儿童每天服用 6 个月,豇豆导致双歧杆菌属的比例更高。
与普通豆和 CSB 相比, 9 至 12 个月期间普雷沃氏菌丰度较低,6 至 9 个月期间埃希氏菌/志贺氏菌丰度较低;这是出乎意料的,因为与普通豆相比,豇豆的纤维含量较低。
另一项针对儿童(6-10 岁)的研究检查了四种不同剂量的补充微量营养素的豆类蛋白质(黄豌豆)(即 6、8、10 或 12 克),连续 1 个月每天食用两次,在厚壁菌门和拟杆菌门中发现了 9 个有区别的类群,其中许多与植物多糖发酵和 SCFA 生产有关——对应于肠道微生物组的变化。
三个类群(普雷沃菌属、颤旋菌属、普氏镰刀菌)与儿童线性生长的增加显着相关,并且所有类群的相对丰度发生变化,特别是在最低蛋白质剂量组(6 g)和最高蛋白质剂量组(12 g)。这些结果强调了豆类对肠道微生物群的潜在剂量反应效应,并确定了与功能结果(例如儿童线性生长)相关的分类群以进一步研究。
与植物蛋白相比,动物来源的蛋白质在近端肠道更容易消化,因此运输到结肠的蛋白质较少,导致结肠微生物群调节减少,并可能抑制病原体。
维生素A
维生素A是一类具有全反式视黄醇生物活性的类视黄醇化合物,包括视黄醛(视黄醛)、视黄酸、视黄酯(主要是棕榈酸视黄酯)和类胡萝卜素化合物,如β-胡萝卜素、α-胡萝卜素和β-胡萝卜素。
▸ 维生素 A 的功能
维生素 A 对于维持肠道屏障的完整性和受损粘膜上皮的再生至关重要,特别是肠上皮细胞的正常增殖和产生粘液的杯状细胞分化,这会影响肠道微生物群的定植和粘附。
膳食维生素 A 可以作为植物来源的维生素 A 原(类胡萝卜素)或动物来源的预制维生素 A(视黄酯)形式食用。
▸ 维生素 A在体内的加工
消化从咀嚼开始,包括通过胆汁盐和胰脂肪酶的胃作用,胆汁盐和胰脂肪酶在肠腔中从膳食脂质形成混合胶束以供吸收。虽然预先形成的维生素 A 来源更容易被宿主肠细胞吸收以运输至储存或循环,但维生素 A 原类胡萝卜素必须进一步加工才能转化为类视黄醇并被宿主使用。
▸ β-胡萝卜素的吸收
根据食物基质的不同,大约 5-50% 的 β-胡萝卜素被吸收。例如,膳食脂质的存在会增加小肠中类胡萝卜素的吸收;如果β-胡萝卜素被纤维吸附,它可以绕过上肠的吸收并被转运到结肠。结肠微生物群可以消化纤维,释放β-胡萝卜素供结肠细胞吸收。在结肠粘膜中发现了类胡萝卜素,主要是在升结肠中。
一项研究发现,大肠杆菌表达一种与 β-胡萝卜素单加氧酶 1 同源的耐盐酶,使大肠杆菌能够积累 β-胡萝卜素;然而,没有证据表明这种β-胡萝卜素被裂解形成视网膜以供进一步使用。
▸ 维生素A与普通拟杆菌:此消彼长
在一项针对小鼠的研究中,普通拟杆菌(Bacteroides vulgatus)先前被证明是一种生长歧视类群,随维生素 A 补充而丰度降低,并因维生素 A 缺乏而丰度增加,可能与胆汁酸代谢的改变有关。作者报告说,这些变化是由于普通双歧杆菌视黄醇流出系统的破坏以及随后对视黄醇和胆汁辅助敏感性的影响造成的。
▸ 肠道共生菌与视黄酸代谢
另一项针对小鼠的研究发现,粘膜树突状细胞 (DC) 对肠道共生婴儿双歧杆菌进行摄取,导致其从维生素 A 向视黄酸的转化增加,并且具有更高数量的具有耐受性的粘膜 DC,例如抑制 1 型 T 辅助细胞(Th1) 和 Th17 细胞。这种效果也随着宿主视黄酸状态的不同而变化。然而,还需要对人类进行进一步的研究,因为啮齿类动物的类胡萝卜素代谢效率比人类高得多,并且没有其他动物模型能够完全代表人类类胡萝卜素代谢。
最近的一项研究检查了补充维生素 A 对新生儿肠道微生物群的影响,结果发现对细菌组成(包括大双歧杆菌属)有积极的以及性别特异性的影响。
与安慰剂相比,入组时单剂量 50,000 国际单位剂量不会改变α-多样性,但在 6-15 周龄时对雄性产生双歧效应,但对雌性没有影响;作者推测,这可能有助于解释本研究中男性死亡率低于女性的原因。
维生素E
维生素 E 主要通过 α-生育酚活性发挥抗氧化剂的作用。
α-生育酚可来源于坚果、植物油、乳制品、奶酪和鸡蛋,也可以作为酯化 α-生育酚的补充剂形式食用。
作为自由基清除剂,α-生育酚可破坏细胞膜和血浆脂蛋白中的自由基链,以维持 LCPUFA 的完整性。维生素E与膳食脂质一起被吸收到肠道细胞中,融入乳糜微粒中,分泌到淋巴系统中,并转运到肝脏,然后被极低密度脂蛋白吸收并分泌到与α-生育酚转移蛋白结合的血液中; 然而,其吸收和贩运的许多细节基本上不为人所知。
经过额外的代谢步骤后,未结合的维生素 E 异构体通过粪便和尿液排出体外。维生素 E 与铁一起,与氧化电位降低有关;因此,维生素 E 被认为可以改善铁相关炎症的影响,改善肠道微生物群的结果。
一项针对美国缺铁 6 个月大婴儿的研究发现,每天两次服用 18 毫克维生素 E 和 15 毫克元素铁,持续 2 个月,并不会导致细菌多样性的变化,但确实会导致相对细菌多样性的变化出现差异。随着时间的推移,维生素 E 与铁的结合导致拟杆菌门(尤其是拟杆菌科)数量减少,而厚壁菌门(尤其是毛螺菌科)和罗氏菌属(Roseburia)数量增加。Roseburia产生丁酸盐,可以刺激结肠血流。
铁
铁是人体内最丰富的微量营养素,对于人体的氧运输、氧化还原反应、新陈代谢和电子传递链机制以及许多细菌的新陈代谢和毒力功能至关重要。
膳食血红素铁 (10%) 和非血红素铁 (90%) 的摄取和吸收存在不同的机制。牛肉是血红素铁的来源;美国的一项对测序研究使用牛肉作为干预措施。从 16S测序数据来看,以牛肉作为补充食品的群体中双歧杆菌属没有下降。与基线相比,4 周后细菌种类丰富度更高;4 个月后,放线菌和 XIVa 族梭菌(丁酸盐生产者)的丰度增加,并且与铁强化婴儿谷物相比,拟杆菌门较低。
差异可能是由于肉类中铁的生物利用度较高,以及未吸收的铁进入结肠,以及铁强化谷物类中存在植酸盐。
▸ 铁的生物利用度
铁的生物利用度随着植酸盐和多酚的存在而降低,而抗坏血酸(维生素 C)则提高铁的生物利用度。胃酸和蛋白水解可能会从食物基质中释放非血红素铁,需要更多的消化才能释放血红素和铁蛋白。一些细菌可以通过特殊机制获取食物中的铁源,例如解没食子链球菌或路邓葡萄球菌降解多酚单宁酸。
营养干预措施对肠道菌群影响的潜在机制
doi.org/10.1146/annurev-nutr-021020-025755
▸ 未吸收的铁被认为会刺激肠道中细菌病原体的生长和毒力
例如大肠杆菌,而宿主铁的状态会影响细菌免疫防御和宿主炎症反应,包括乳杆菌科和伯氏疏螺旋体在内的一些细菌不依赖铁生长,而是利用锰。
链球菌属可以使用铁或锰,具体取决于可用性。大肠杆菌使用 Feo 摄取系统来摄取二价铁,而三价铁在摄取之前首先被细胞外还原酶还原为二价铁,或者被摄取为柠檬酸铁或与细菌铁载体结合。
▸ 感染期间可用铁量的变化向毒力基因发送信号
低铁会导致铁载体抑制和毒素上调,而较高的铁可用性会诱导细菌粘附到肠上皮细胞,如肠沙门氏菌的情况鼠伤寒血清型。缺铁还会减少肠道细菌(包括Roseburia 、直肠真杆菌和梭状芽胞杆菌IV 族成员)等产生的短链脂肪酸。
在易缺铁的人群中,婴儿和幼儿最常需要口服铁剂治疗,例如补充剂,尽管他们的肠道微生物群仍在成熟。
▸ 铁干预措施对腹泻:负面影响
多项研究表明铁干预措施对腹泻等胃肠道症状有负面影响以及肠道微生物群的变化,例如较高的肠杆菌科和较低的乳杆菌科和双歧杆菌属。
▸ 铁补充剂,取决于基线铁或贫血状态
为了帮助抵消铁的这些影响,儿童随机试验检查了铁与维生素 E 等抗氧化剂结合使用的效果或冻干覆盆子作为抗氧化剂来源。例如,在加拿大一项研究中,与仅接受铁强化谷物的婴儿相比,接受含冻干覆盆子的电解铁强化米粉的婴儿在2 -4周后肠道微生物组的丰富度和多样性有所增加。
然而,接受冻干覆盆子的组活性氧并没有显着减少,与胃肠道中过量铁相关的炎症过程的引发剂;这可能是由于冻干覆盆子的消耗量很小,或者与新鲜覆盆子中的抗氧化剂含量相比可能较低,而新鲜覆盆子没有被任何群体食用。
在未来的研究中,考虑其他抗氧化剂来源(例如新鲜浆果或提取物)和不同剂量的这些抗氧化剂来源,以及对大量贫血婴儿的研究或铁充足,将有助于建立铁和抗氧化剂之间潜在的相互作用。
▸ 使用铁剂联合干预的随机试验
包括叶酸, 锌和多种微量营养素(MMN),无论是片剂的形式还是作为微量营养素粉(MNP)的一部分。铁的形式包括硫酸亚铁、富马酸亚铁、NaFeEDTA 和电解铁,它们可能对儿童肠道微生物群产生不同的影响;其他变量包括给药剂量、基线铁和贫血状况、抗氧化剂和其他微量营养素等辅助干预措施以及研究环境。
大多数检查 MNP 的研究都是在肯尼亚针对开始添加辅食的 6 个月大婴儿进行的;MNP 与玉米粥一起服用 3 至 4 个月。MNP 的微量营养素成分各不相同,但都含有维生素 A、叶酸和维生素 C,并且除了一种还含有 B 族维生素、维生素 D、铜、碘、硒和锌。
根据靶向 qRT-PCR 结果,MNP ( MixMe ) 加上 2.5 mg NaFeEDTA 4 个月,导致大肠杆菌/志贺氏菌丰度增加,以及肠杆菌科:双歧杆菌属比例增加。与单独使用相同MNP产生的丰度相比,10个月大时的比例增加。
在一项比较 MNP(Sprinkles)加 12.5 毫克富马酸亚铁与单独使用相同 MNP 的试验中,以及当含铁组组合并与仅使用 MNP 的组进行比较时,也观察到了类似的结果。
在另一项试验中,与单独使用相同的 MNP 相比,MNP 中的铁含量(2.5 mg NaFeEDTA 和 2.5 mg富马酸亚铁)对微生物组组成有显着影响,包括较低丰度的有益菌(如乳杆菌属和双歧杆菌属)以及较高比例的梭菌目,而拟杆菌门在各组之间保持相似。在一项辅助研究中,抗生素治疗并没有抵消铁的作用,铁仍然导致肠杆菌科细菌的增加。
最后,在另一项试验中,与单独的 MNP 相比,含铁的 MNP(12.5 毫克富马酸亚铁)在多样性或相对丰度方面没有组间差异。然而,接受含铁 MNP 的组梭状芽胞杆菌增加,双歧杆菌减少,而仅接受 MNP 的组大肠杆菌减少。
尽管 MNP 配方和 16S 测序区域存在差异,但这些研究一致发现铁会导致有益细菌减少和可能有害细菌增加,这为铁对肠道微生物组的影响提供了多种证据。需要进一步的研究来检查铁在其他环境中对肠道微生物组的影响。
▸ 铁强化和补充剂:对肠道菌群的影响
在科特迪瓦 6-14 岁儿童中(n = 60),铁强化饼干(20 毫克电解铁)不会导致微生物多样性发生变化,肠杆菌科细菌增加,乳杆菌属细菌减少,并且双歧杆菌属没有差异。6个月后这些发现与之前对婴儿进行的研究一致。
在南非进行的另一项针对 6-11 岁儿童的研究中,与安慰剂组(两组均服用维生素 C)相比,饮用含 50 毫克硫酸亚铁的饮料会导致组内相对含量存在差异;然而,尽管铁剂量较高,但没有发现组间差异。这些发现可能是由于肠道病原体负担和卫生设施的差异造成的,包括优质水和卫生设施、饮食质量和感染负担。
▸ 存在于全食物基质中的铁可能有利于铁的吸收
如LNS,不会产生与铁相关的负面影响,例如增加潜在致病菌、炎症和腹泻等胃肠道疾病症状。
LNS由植物油、花生、奶粉、糖、维生素和矿物质组成,食用量约为54g小袋,或在少量的情况下,食用量为20g(~6g脂肪),以帮助解决大量营养素和能量不足问题,并满足微量营养素需求。
已经进行了三项随机试验来检验直接给予婴儿 LNS 的影响或在怀孕期间给予母亲,然后给予婴儿。在同一队列的两项研究中,6 个月大的儿童接受基于牛奶蛋白的 LNS、基于大豆蛋白的 LNS、CSB 或 12 个月内不接受干预。通过 16S rRNA V4 区域测序评估肠道微生物群,双歧杆菌属的定植率或计数没有差异。与未接受营养补充剂的对照组相比,接受 LNS 和 CSB 补充剂的组中发现了金黄色葡萄球菌或其他肠道细菌。
在另一项试验中,将 SQ-LNS(20 克铁)给予孕妇(妊娠 <29 周),然后给予 6 至 18 个月大的婴儿;对照组包括给怀孕期间和哺乳期前 6 个月的母亲服用含有 20 毫克铁的 MMN,或者给怀孕期间的母亲服用标准护理铁和叶酸(含有 60 毫克铁)加上安慰剂哺乳期的前 6 个月。
12 个月时,与任一对照组相比,SQ-LNS 组的婴儿肠道微生物α 多样性和均匀度更高,而β 多样性没有差异或观察年龄微生物群 z 评分 (MAZ)。
然而,由于干预组之间干预的类型、数量、接受者和给药方案存在差异(只有 SQ-LNS 组中的婴儿接受直接补充),因此很难确定任何一种特定的营养素或干预类型可能会造成这些影响。总之,这些发现表明,怀孕和哺乳期间补充铁可能不会影响新生儿和婴儿的微生物组。
锌
微量矿物质锌在体内发挥催化、结构和调节作用,包括肠道健康和免疫健康,并已广泛用于治疗严重腹泻。
锌的吸收主要发生在人体的小肠中,受到食品中含锌物质是否存在以及食品加工方法的影响。例如,植物性锌来源(如种子、根和块茎)中的植酸盐会抑制锌的吸收,而铁等其他微量营养素会竞争性地抑制锌的吸收,而谷物的发酵可以提高锌的吸收。
此外,由于存在组氨酸等氨基酸增强锌的溶解度,动物源产品中的锌更容易被利用。过量的锌,包括未吸收的膳食锌和内源性锌,以受控机制通过粪便排出体外,以维持体内平衡。由于体内没有锌的储存,低锌饮食后可能会迅速出现锌缺乏症。
▸ 共生菌和病原菌在肠道中的定植和功能可能受到锌的调节
锌对于许多细菌(例如毒力因子)至关重要,并因其抗菌作用而用于动物生产。
在大肠杆菌中,锌受到严格调控,并在 300 多种蛋白质中发挥结构和催化作用,并且许多细菌拥有重金属外排系统或进行特定重金属质粒的基因转移。乳酸杆菌等细菌对膳食锌的抗性也不同。
关于锌和肠道微生物群的研究主要在动物模型中进行;这些研究的作者发现了细菌组成的差异,特别是乳杆菌属、梭菌属和肠杆菌科。
一项针对患有继发于肺炎的抗生素相关性腹泻的 2-36 个月大儿童的研究,检查了锌补充剂与益生菌的作用,为期 14 天,与单独益生菌相比,双歧杆菌属的计数没有显着差异。与基线相比,每个干预组中大肠杆菌减少。
另一项针对 6 个月大儿童食用铁和锌强化或仅铁强化谷物的研究发现,锌可以抵消铁强化对肠道微生物群的潜在不利影响。
上面的证据表明肠道微生物组在生命早期生长中的作用,为儿童营养不良的营养和药物治疗提供了新的目标。
目前预防发育迟缓和消瘦的营养策略并不能完全解决短期和长期生长缺陷或相关的临床结果。6-24 个月龄小剂量脂质营养补充剂可将发育迟缓发生率降低12%,将严重发育迟缓发生率降低 17%。
使用即用型治疗食品 (RUTF) 对 SAM 进行社区管理可显着改善营养恢复;然而,复杂的 SAM 后仍然存在高死亡率和长期生长缺陷。
总的来说,目前的治疗方法通过抗生素解决感染负担,并通过单糖、脂质和微量营养素恢复宿主营养;然而,这些治疗方法并不专门针对肠道微生物组,而肠道微生物组可能有助于改善生长。
事实上,目前对复杂 SAM 的建议(包括在所有情况下进行抗生素治疗)可能会损害肠道微生物组的恢复。而越来越多的试验报告了针对微生物群的干预措施对儿童营养不良的临床有益效果。
益生菌
少数益生菌试验报告了在营养不良负担较高的环境中益生菌对儿童生长的不同影响。对 795 名 SAM 儿童进行的一项大型随机临床试验发现,尽管观察到门诊死亡率有降低的趋势,但多物种益生菌和益生元联合治疗对营养恢复、死亡率或相关临床症状没有影响。较小规模的试验表明,其他益生菌种类对 SAM 和其他形式的营养不良具有潜在益处。
长双歧杆菌亚种由于婴儿益生菌与资源匮乏环境中的生长呈正相关,因此它具有作为促进生长的潜在益生菌的巨大潜力。在孟加拉国开展的一项 SAM 儿童试验,在临床稳定和急性期管理后,将 62 名参与者随机分为婴儿双歧杆菌益生菌(婴儿双歧杆菌 EVC001)、益生菌和纯化 HMO(乳-N-新四糖)或安慰剂,为期4周在医院。
在开始治疗 8 周后的研究终点,益生菌组的年龄别体重 Z 评分 (WAZ) 和 MUAC 显着更高。鼠李糖乳杆菌GG (LGG) 和动物双歧杆菌亚种组合的试验。SAM 治疗期间的乳酸菌和 LGG 也证明了感染发生率和门诊腹泻减少的证据。
由于患有复杂 SAM 的儿童的临床不稳定和高死亡率,特定益生菌用作 SAM 标准治疗的潜力将在很大程度上取决于成本、可持续性和安全性。
此外,需要合理选择益生菌,以确定那些能够适当定植于营养不良儿童受损肠道的益生菌,并针对可以改善生长恢复的特定途径。
来自动物研究的进一步数据可能有助于为未来合适益生菌的临床试验提供信息,例如针对内分泌生长途径的益生菌。
即食治疗性食品(RUTF)
针对肠道微生物群的改良营养疗法有潜力成为儿童营养不良的可持续且经济有效的疗法。SAM 的证据表明,RUTF 喂养后微生物组成熟度的恢复是短暂的,并且往往在治疗后 3-4 个月恢复到不成熟状态。
因此,针对 SAM 恢复中肠道微生物组和预防发育迟缓的改良补充食品可能有助于促进生长。
一项试点试验评估了在 SAM 治疗期间向 F75 和 F100 治疗奶中添加可微生物发酵的碳水化合物菊粉或豇豆粉,发现与对照组相比,对营养恢复没有改善作用;然而,补充营养可以防止住院治疗期间物种多样性的暂时丧失,这部分是由抗生素治疗引起的。
豇豆辅助喂养可以显着减少有发育迟缓风险的儿童的一些 LAZ 缺陷;然而,它仅引起肠道微生物组组成的适度变化,包括双歧杆菌增加和大肠杆菌/志贺氏菌减少,表明对生长的有益影响可能与肠道微生物组无关。
来自孟加拉国的一个队列的研究,与标准 RUTF 疗法相比,微生物群导向的补充食品 (MDCF) 对 MAM 恢复儿童生长的影响显着增强。通过合理设计由当地可用的、文化上可接受的食物组成的补充食品组合 (MDCF-2),该组合可促进人源化动物模型中早期生命肠道微生物组的成熟。
这些有希望的数据表明,与目前的疗法相比,针对微生物群的补充食品可以在更大程度上促进儿童营养不良后的生长恢复。从长远来看,这些小但显着更大的生长改善是否与持续的生长改善、认知益处或感染和慢性病风险的降低相对应,目前尚不清楚。
来自高收入环境的临床试验越来越多的证据支持针对微生物群的治疗在各种感染、胃肠道和代谢疾病中的功效。
由于世界上超过五分之一的儿童发育迟缓或消瘦,旨在进一步表征导致儿童营养不良的微生物介导的病理生理途径的研究对于为更好的治疗提供信息至关重要。
儿童营养不良涉及许多生理系统的紊乱,包括代谢、免疫和内分泌系统,其中许多系统与肠道微生物组密切相关,因此可能适合针对微生物组的干预措施。
未来潜在的微生物组靶向疗法将补充针对宿主营养需求和感染负担的现有疗法。此外,必须根据国家或地区的具体要求调整这些干预措施,以制定具有成本效益且本国或本地区上可接受的可持续干预措施。
Tips
在这篇文章中综合了营养干预试验的现有证据,这些试验测试了多种饮食干预措施对婴儿和儿童微生物组相关结果的影响,并讨论了来自实验室和动物研究的支持证据,以及来自人类研究的观察数据,旨在指导未来的研究并增强可解释性,为饮食相关政策提供信息。
可比性较差,因为纳入的研究在干预类型和持续时间上有所不同,是在有或没有既往疾病的不同儿科人群中进行的,并且是在可能影响基线微生物组成的各种环境中进行的。
虽然调查婴儿微生物群的研究发现,母亲在怀孕期间的饮食变化几乎没有影响儿童肠道微生物群。但是通过单独检查研究而不是包括整个全球背景,很难将研究偏差与真实的生物现象分开。可以使用 Qiita 等工具尝试进行此类分析,前提是研究数据与符合标准的样本元数据是公开的,以促进未来分析中的跨研究可比性。
在进行 16S rRNA 测序的 19 项研究中,只有两项对 16S V4 区域进行测序的试验可用于 Qiita 的进一步分析。作者将这两个试验人群的肠道微生物群与 2012 年具有里程碑意义的微生物组队列研究的亚组进行了比较。
在 2012 年的研究中发现,马拉维出生的婴儿和美国出生的婴儿之间存在显着的微生物分离,但在纳入最近的两项试验时,缺乏数据来深入研究营养干预措施的效果与2012年的研究对比。
尽管微生物组生物信息学取得了很大进展,并且研究的人群类型和健康状况的广度不断扩大,但在营养和肠道微生物组领域,特别是儿童,仍需要解决许多研究空白和问题。
为了解决基于发现的差距,未来的研究人员将需要:
a)识别饮食细分成分响应微生物群并确定它们是否影响宿主的生理结果;
b)与最适合现实世界应用的特定膳食成分相比,了解全食干预措施的作用;
c)提高复制性和再现性,例如在不同人群中使用相同的饮食干预措施,以及在同一人群中测试不同的饮食干预措施;
d)确定食品加工和准备的作用,了解熟食或生食如何影响肠道微生物群(烹饪的影响最近已被研究,但仍是一个研究不足的领域);
e)考虑到依从性的差异,设计和使用饮食干预措施,这需要大量的队列和长时间的随访;
f)将研究结果和序列数据整合到一个数据库中,用于对类似处理的序列进行汇总分析并了解全局现象,同时最大限度地减少研究偏差。
介入/治疗差距
未来的研究应解决以下五个介入和治疗差距。
首先,需要解决稳定性和可塑性问题。虽然短期饮食变化可以改变肠道微生物群,但长期饮食模式与更稳定的微生物群构象相关,并且很难改变。仍有待确定的是哪种饮食模式具有最大或最小的可塑性,以及这种可塑性是否也取决于基线微生物种群和过去的饮食模式。
第二个差距是干预研究中针对不同人群的营养素和食物研究不足。尽管本次综述确定了几项微量营养素粉 (MNP) 试验,而关于补充个别维生素、矿物质和天然食品的试验相对较少,但仍然需要在中国以及其他国家重复研究中对 MNP 进行调查。
第三,应进行纵向、长期的人体研究。需要对 2-18 岁婴儿、青春期、成年早期和晚年等人群进行长期喂养研究,以了解衰老微生物群的动态。
第四,解决精准营养健康问题。
第五,未来健康建议应考虑大局。建议食物和饮食(生活方式)改变需要全面的方法和来自营养师和营养科学家、心理学家、医生和生物统计学家的多方面团队的支持。
小编寄语
中国在医疗和队列研究方面的发展仍有待提升。正如首都医科大学附属北京天坛医院院长王拥军教授所指出的:“中国目前还缺乏具有全球影响力的标志性队列研究,尤其是能够改变临床指南的里程碑试验”。
举例来说,美国的弗明汉心脏研究和英国的UK Biobank人群队列都对临床研究产生了重大影响。弗明汉心脏研究自1948年开始,最初只有5200名参与者,但经过70年的发展,参与者数量增加到15000人,产生了3698篇文章。这项研究改变了几乎所有心血管疾病的临床危险因素的认知。
另外,英国的UK Biobank人群队列从2006年开始策划,至今已发表超过3000篇文章。该队列成为世界上其他国家难以逾越的临床队列研究之一。
因此,谷禾也愿意和期待与各大医疗机构合作构建大型研究队列,尤其是关于青少年儿童的。通过对这些队列进行全面多组学以及生活营养指标的调查分析,将为我国医疗研究带来新的机遇和突破,为改善人们的健康水平做出重要贡献。
主要参考文献
Jones HJ, Bourke CD, Swann JR, Robertson RC. Malnourished Microbes: Host-Microbiome Interactions in Child Undernutrition. Annu Rev Nutr. 2023 Aug 21;43:327-353.
Acosta A, De Burga R, Chavez C, Flores J, Olortegui M, et al. 2017. Relationship between growth and illness, enteropathogens and dietary intakes in the first 2 years of life: findings from the MAL-ED birth cohort study. BMJ Glob. Health 2(4):e000370
Alves da Silva AV, de Castro Oliveira SB, Di Rienzi SC, Brown-Steinke K, Dehan LM, et al. 2019. Murine methyl donor deficiency impairs early growth in association with dysmorphic small intestinal crypts and reduced gut microbial community diversity. Curr. Dev. Nutr. 3(1):nzy070
Mehta S, Huey SL, McDonald D, Knight R, Finkelstein JL. Nutritional Interventions and the Gut Microbiome in Children. Annu Rev Nutr. 2021 Oct 11;41:479-510.
Amadi B, Fagbemi AO, Kelly P, Mwiya M, Torrente F, et al. 2009. Reduced production of sulfated glycosaminoglycans occurs in Zambian children with kwashiorkor but not marasmus. Am. J. Clin. Nutr. 89(2):592–600
Amadi B, Zyambo K, Chandwe K, Besa E, Mulenga C, et al. 2021. Adaptation of the small intestine to microbial enteropathogens in Zambian children with stunting. Nat. Microbiol. 6(4):445–54
Ansaldo E, Farley TK, Belkaid Y. 2021. Control of immunity by the microbiota. Annu. Rev. Immunol. 39:449–79
Attia S, Versloot CJ, Voskuijl W, van Vliet SJ, Di Giovanni V, et al. 2016. Mortality in children with complicated severe acute malnutrition is related to intestinal and systemic inflammation: an observational cohort study. Am. J. Clin. Nutr. 104:1441–49
Barratt MJ, Nuzhat S, Ahsan K, Frese SA, Arzamasov AA, et al. 2022. Bifidobacterium infantis treatment promotes weight gain in Bangladeshi infants with severe acute malnutrition. Sci. Transl. Med. 14(640):eabk1107
Bartelt LA, Bolick DT, Mayneris-Perxachs J, Kolling GL, Medlock GL, et al. 2017. Cross-modulation of pathogen-specific pathways enhances malnutrition during enteric co-infection with Giardia lamblia and enteroaggregative Escherichia coli. PLOS Pathog. 13(7):e1006471
Bartz S, Mody A, Hornik C, Bain J, Muehlbauer M, et al. 2014. Severe acute malnutrition in childhood: hormonal and metabolic status at presentation, response to treatment, and predictors of mortality. J. Clin. Endocrinol. Metab. 99(6):2128–37
Bhattacharjee A, Burr AHP, Overacre-Delgoffe AE, Tometich JT, Yang D, et al. 2021. Environmental enteric dysfunction induces regulatory T cells that inhibit local CD4+ T cell responses and impair oral vaccine efficacy. Immunity 54(8):1745–57.e7
Black RE, Victora CG, Walker SP, Bhutta ZA, Christian P, et al. 2013. Maternal and child undernutrition and overweight in low-income and middle-income countries. Lancet 382(9890):427–51
Bourdon C, Lelijveld N, Thompson D, Dalvi PS, Gonzales GB, et al. 2019. Metabolomics in plasma of Malawian children 7 years after surviving severe acute malnutrition: “ChroSAM” a cohort study. EBioMedicine 45:464–72
Aakko J, Grzeskowiak L, Asukas T, Paivansade E, Lehto KM, et al. 2017. Lipid-based nutrient supplements do not affect gut Bifidobacterium microbiota in Malawian infants: a randomized trial. J. Pediatr. Gastroenterol. Nutr. 64:610–15
Aly H, Said RN, Wali IE, Elwakkad A, Soliman Y, et al. 2017. Medically graded honey supplementation formula to preterm infants as a prebiotic: a randomized controlled trial. J. Pediatr. Gastroenterol. Nutr. 64:966–70
Amarri S, Benatti F, Callegari ML, Shahkhalili Y, Chauffard F, et al. 2006. Changes of gut microbiota and immune markers during the complementary feeding period in healthy breast-fed infants. J. Pediatr. Gastroenterol. Nutr. 42:488–95
Andrews SC, Robinson AK, Rodriguez-Quinones F. 2003. Bacterial iron homeostasis. FEMS Microbiol. Rev. 27:215–37
Arimond M, Zeilani M, Jungjohann S, Brown KH, Ashorn P, et al. 2015. Considerations in developing lipid-based nutrient supplements for prevention of undernutrition: experience from the International Lipid-Based Nutrient Supplements (iLiNS) Project. Matern. Child Nutr. 11:31–61
Arrieta MC, Stiemsma LT, Amenyogbe N, Brown EM, Finlay B. 2014. The intestinal microbiome in early life: health and disease. Front. Immunol. 5:427
Black RE, Victora CG, Walker SP, Bhutta ZA, Christian P, de Onis M, Ezzati M, Grantham-McGregor S, Katz J, Martorell R, Uauy R; Maternal and Child Nutrition Study Group. Maternal and child undernutrition and overweight in low-income and middle-income countries. Lancet. 2013 Aug 3;382(9890):427-451.
谷禾健康
偏头痛
偏头痛是一种常见且使人衰弱的神经系统疾病,也是世界经济的重大负担。大约14%的成年人患有偏头痛。其特点是反复出现一侧或两侧严重头痛(抽痛),并伴有独特的相关症状,包括畏光、恐声、恶心、呕吐等。
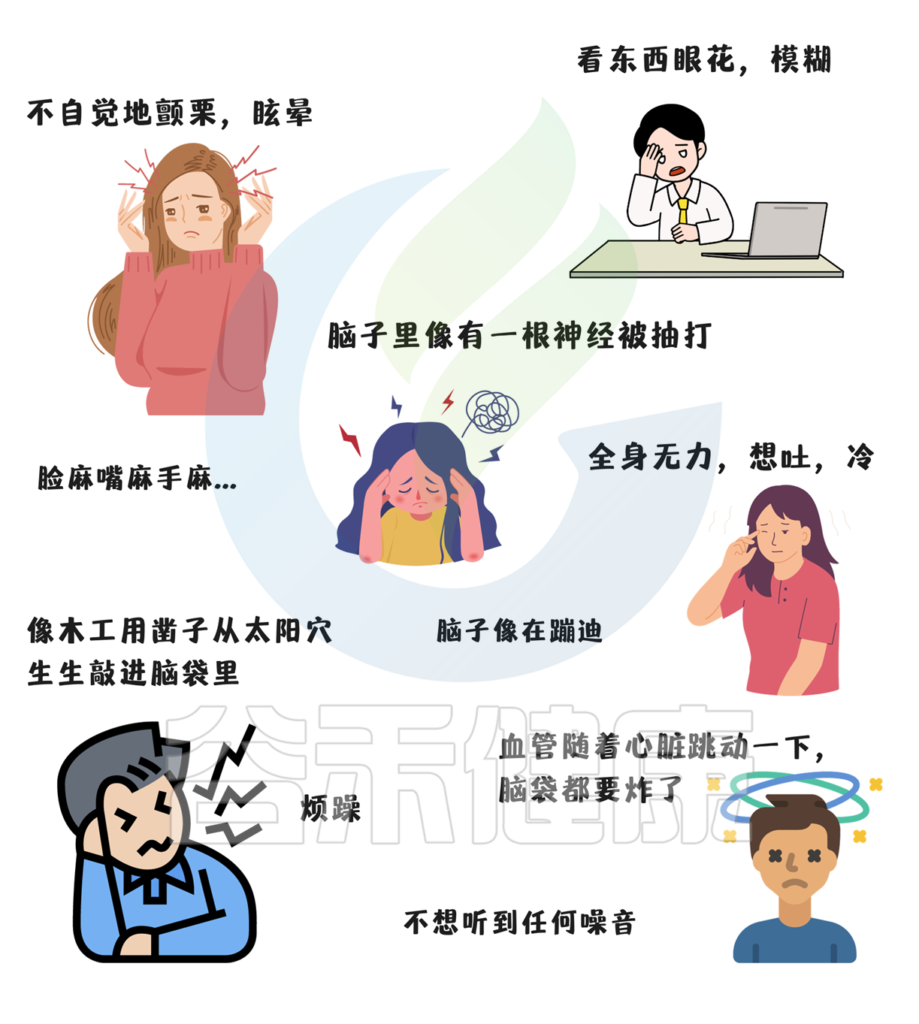
偏头痛在儿童晚期/青春期早期发病,在 40 多岁的人群中发病率最高,此后发病率下降。在所有年龄段中,女性偏头痛的发病率始终高于男性。
潜在危险因素包括压力、饮食、急性药物过量、激素波动、睡眠、环境、天气等。
对偏头痛根本原因的研究已经跨越了几个世纪,涉及外周和中枢机制等,但其根本机制仍未完全了解。已经确定三叉神经系统的激活和敏化在偏头痛发作期间至关重要。
胃肠道系统和中枢神经系统之间存在双向关系。多项研究表明偏头痛与某些胃肠道疾病有关,例如幽门螺杆菌感染、肠易激综合征(IBS)、胃轻瘫、乳糜泻等。
越来越多的证据表明微生物介导的肠-脑串扰可能导致偏头痛的发病机制。在偏头痛患者中,肠道微生物群的组成发生了改变,包括肠道菌群多样性下降,产丁酸菌减少,梭状芽孢杆菌增加等。
偏头痛患者肠道和大脑之间的相互作用,可能受到多种因素的影响,如炎症介质(IL-1β、IL-6、IL-8 和 TNF-α)、肠道微生物群、肠道菌群代谢产物、血清素途径、谷氨酸途径、神经肽(包括 CGRP、P物质、PACAP、NPY)等。
本文主要介绍了偏头痛相关症状、发病人群、四个阶段,基于偏头痛的发病机制,阐述肠道微生物群如何通过各个途径参与偏头痛的病理生理学,并探讨了潜在的干预策略。

很多人在听到偏头痛这个词的时候,会想到严重的头痛,但头痛并不直接等于偏头痛,头痛只是偏头痛的一种症状。
偏头痛是一种常见的神经系统疾病,会引起多种症状,最明显的是头部一侧的搏动性头痛,伴随着头部不适感和其他一些症状。
偏头痛是原发性头痛,这意味着它不是由其他疾病引起的。偏头痛的发作通常会对患者的日常生活和工作造成较大的干扰,那么偏头痛发作时,具体会有什么表现?
偏头痛可能会有以下症状:
日常体力活动、运动,甚至咳嗽或打喷嚏都会加剧头痛。
偏头痛最常发生在早晨,尤其是刚醒来时。有些人会在可预测的时间出现偏头痛,例如月经前或一周紧张工作后的周末。许多人在偏头痛后感到疲惫或虚弱,但在两次发作之间通常没有症状。
偏头痛的频率可以是每年一次、每周一次、或介于两者之间的任意时间。
每月发生两到四次偏头痛是最常见的。
未经治疗的发作持续 4~72 小时。
大多数偏头痛持续约 4 小时,但严重的可能会持续 3 天以上。
偏头痛的两种主要类型是:
先兆偏头痛
以前称为经典偏头痛,包括视觉障碍和其他神经系统症状,这些症状在实际头痛前约10-60分钟出现,通常持续不超过一小时。个人可能会暂时失去部分或全部视力。这种先兆可能会在没有头痛的情况下出现,并且随时可能发作。
其他典型症状包括:
说话困难;身体一侧感觉异常、麻木或肌肉无力;手或脸有刺痛感,意识混乱。头痛之前可能会出现恶心、食欲不振以及对光、声音或噪音的敏感性增加。
无先兆偏头痛
或普通偏头痛是更常见的偏头痛形式。症状包括毫无征兆地出现头痛,通常在头部的一侧感觉到,并伴有恶心、意识模糊、视力模糊、情绪变化、疲劳以及对光、声音或噪音的敏感性增加。
其他类型包括:
经期偏头痛
这些通常发生在经期开始前 2 天,并持续到经期后 3 天。可能还会在每月其他时间出现其他类型的偏头痛,但月经前后的偏头痛通常没有先兆。
沉默偏头痛
会出现前兆症状,但没有头痛。事实上,前兆通常是这种类型偏头痛的主要警告信号。但可能还会出现恶心和其他偏头痛症状。它通常只持续约20-30分钟。
前庭偏头痛
有平衡问题、眩晕、恶心、呕吐,伴或不伴头痛。这种情况通常发生在有晕车病史的人身上。
腹部偏头痛
可能会导致胃痛、恶心和呕吐。它经常发生在儿童身上,随着时间的推移可能会变成典型的偏头痛。
偏瘫偏头痛
身体一侧出现短暂瘫痪 (偏瘫)或无力。可能还会感到麻木、头晕或视力变化。这些症状也可能是中风的征兆,因此请立即寻求医疗帮助。
眼部偏头痛
也称为眼部或视网膜偏头痛。它会导致一只眼睛短暂、部分或完全丧失视力,并伴有眼睛后面的钝痛,这种疼痛可能会扩散到头部的其他部位。请立即就医检查这些症状。
带脑干先兆的偏头痛
头痛之前可能会出现头晕、意识混乱或失去平衡。疼痛可能会影响头后部。这些症状通常突然出现,并可能伴有说话困难、耳鸣和呕吐。这种类型的偏头痛与激素变化密切相关,主要影响年轻的成年女性。请立即就医检查这些症状。
持续偏头痛
这种严重类型的偏头痛可持续超过 72 小时。疼痛和恶心非常严重需要去医院。有时,药物或停药可能会导致它们出现。
眼肌麻痹性偏头痛
这会导致眼睛周围疼痛,包括周围肌肉麻痹。这是医疗紧急情况,因为这些症状也可能是由眼睛后面的神经受压或动脉瘤引起的。其他症状包括眼睑下垂、复视或其他视力变化。
活动性偏头痛的高峰发病年龄为 40 岁左右,其中 35%左右的女性和15%左右的男性受到影响。
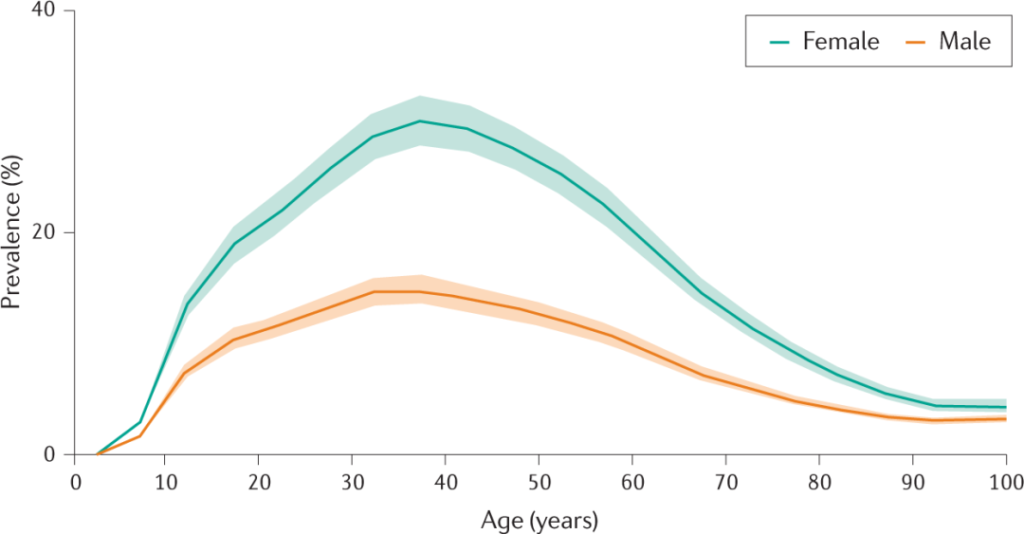
doi: 10.1038/s41572-021-00328-4.
一些常见的偏头痛诱因包括:
许多女性注意到她们在月经期间、怀孕期间或排卵期间会感到头痛。症状也可能与更年期、使用激素避孕或激素替代疗法有关。
当感到压力时,大脑会释放化学物质,这些化学物质会导致血管变化,从而可能导致偏头痛。
某些食物和饮料,例如陈年奶酪、酒精、硝酸盐(意大利辣香肠、热狗和午餐肉中的硝酸盐)、味精等食品添加剂,高盐和加工食物,可能对部分人造成影响。
摄入过多或未达到习惯的摄入量都会导致头痛。咖啡因本身可以治疗急性偏头痛。
风暴锋、气压变化、强风或海拔变化都可能引发偏头痛。
大声的噪音、明亮的灯光和强烈的气味都会引发偏头痛。
口服避孕药、血管扩张剂可以扩张血管,从而引发这些症状。
这包括体育锻炼和性生活等。
当睡眠过多或不足时,可能会感到头痛。
不良习惯包括不吃饭(或推迟进餐),吸烟,酗酒等,还有晕车也都可能引发偏头痛。
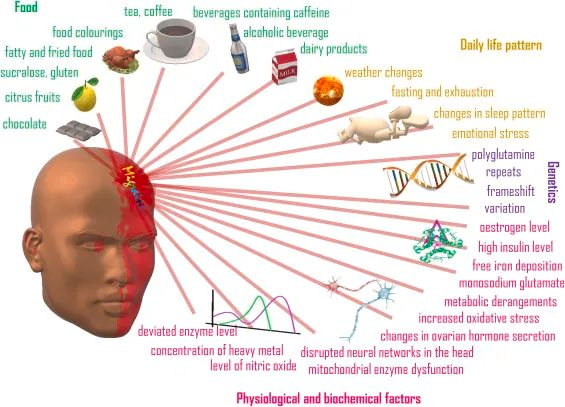
doi.org/10.1016/j.biopha.2021.111557
前驱症状阶段
有时称为头痛前阶段,此阶段的特点是在偏头痛出现前数小时或数天出现,特点是没有疼痛的症状。这些包括情绪波动、对食物的渴望、颈部僵硬、严重口渴、腹胀、便秘或腹泻、注意力不集中、排尿增多等。
先兆阶段
先兆是指偏头痛之前或期间发生的感觉障碍。这些障碍会影响一个人的视力、触觉或言语,但并不是每个偏头痛患者都会出现视觉异常期。例子包括:
视力模糊、看到黑点、波浪线、闪光或不存在的东西、渐渐扩大的盲点、手臂麻木、手臂和腿部有沉重感、口齿不清或语句混乱。
头痛阶段
这是通常出现疼痛的阶段,疼痛程度可能从轻微到无法忍受。
偏头痛通常以钝痛开始,然后发展为抽痛,有时候是钻孔感。也可能出现颈部疼痛、僵硬、鼻塞、恶心、呕吐、失眠等。
体力活动以及接触光、声音和气味可能会加剧疼痛。然而,有些人在发作偏头痛时并不会出现头痛。
恢复阶段
最后阶段是疼痛消退时。在此阶段,人们可能会感到疲惫、困惑或总体不适。
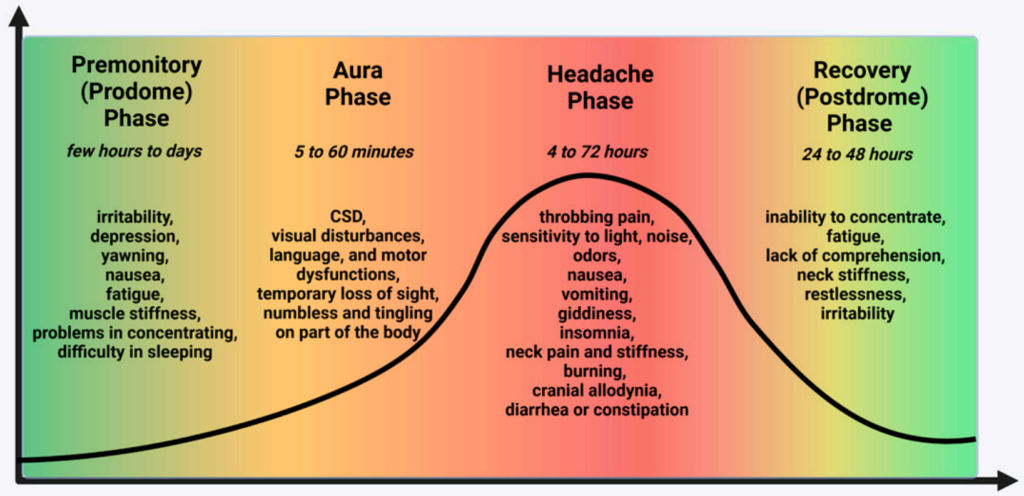
doi.org/10.3390/neurolint15030073
简单来说,在偏头痛发作之前几天,可能会感到疲劳,情绪变化,打哈欠,出现睡眠问题,恶心等情况,对光和声音敏感,这些预警信号与大脑中某个特殊的区域——下丘脑有关。下丘脑通常控制体内激素平衡,昼夜节律等,偏头痛发作之前下丘脑很活跃。
偏头痛还有常见的症状,视觉变化,刺痛感,言语困难等,这些感觉与细胞膜电位变化相关,导致脑活动和血流变化在大脑中扩散,电位变化快速在大脑表面传播,它传向哪里,哪里就出问题(*>﹏<*) …当它侵入视觉皮层,可能出现视觉盲点。
在头痛阶段,三叉神经起到关键作用,三叉神经会传递来自脸部皮肤,头皮,覆盖大脑皮层的血管和膜层的触觉,温度等,一旦被激活,三叉神经会传递痛觉信号,当偏头痛发作时,这个痛觉通路会变得敏感化,也就是说能触发痛觉的阈值下降了,那么平时不太会引起疼痛的也变得容易痛了,比如说,咳嗽,弯腰,一些强光,噪声等都会引起疼痛。
具体来说,关于偏头痛的发病机制,目前主要有以下几种理论。
I) 血管学说
该学说认为由血管舒缩功能障碍引起。基于先兆期间大脑血管收缩引起的三叉神经过度兴奋,随后产生炎症神经肽释放引起血管舒张,导致偏头痛。
II)神经学说 (皮层扩散性抑制)
该学说认为偏头痛是原发性神经功能紊乱疾病。其中通过皮层-三叉神经通路,大脑皮层的过度神经兴奋(异常的神经放电)会增加细胞外谷氨酸、钙离子、钠离子和钾离子的释放,从而引发去极化作用,造成皮质扩散性抑制(CSD)。
皮质扩散抑制是什么意思呢?
它其实是一种缓慢传播的大脑活动改变波,涉及神经、血管功能的巨大变化,皮质扩散抑制可能是偏头痛先兆的原因。这些慢电波可以激活负责感知大脑周围覆盖物(“脑膜”)疼痛的神经,并改变血管功能,这两者都可能导致偏头痛。
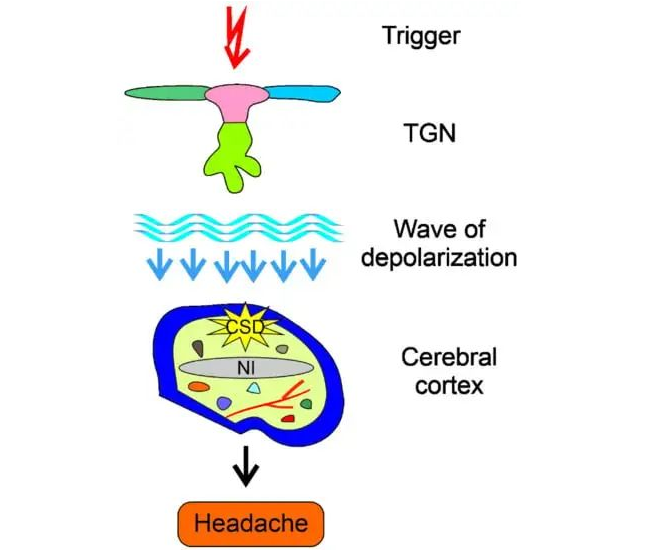
doi.org/10.3390/ijms221810134
III)三叉神经血管学说 (近年流传较广)
三叉神经感觉纤维的激活(传递疼痛信号),导致脑膜血管(硬脑膜)发生痛性神经源性炎症,此过程通过炎症神经肽(CGRP、SP、NK-A)的释放介导,这表明CGRP释放与神经源性硬脑膜血管舒张有关,这可能在偏头痛的发生中很重要。
注:关于CGRP在第四章节会详细介绍。
简单来说,激活三叉神经血管通路:
激活三叉神经元 → 释放P物质、CGRP等血管活性神经肽 → 血管扩张和促炎分子(组胺、缓激肽)的释放 → 神经源性炎症 → 激活脑膜痛觉受体
偏头痛中的 CGRP 和三叉血管系统
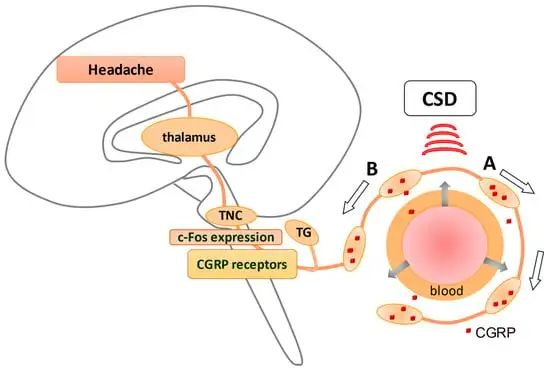
doi.org/10.3390/ijms22168929
三叉血管系统是一组感知面部和大脑覆盖层疼痛的神经细胞。三叉血管系统的反复激活会导致神经系统过敏和持续疼痛。
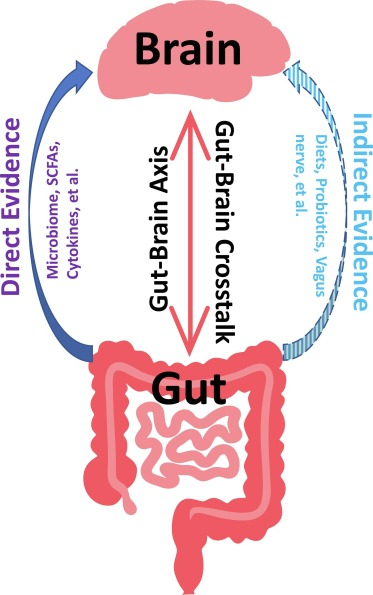
IV)肠脑轴(GBA)理论
肠-脑轴作为肠道功能和维护的协调系统,将大脑的情绪中心与肠外周机制连接起来,包括肠反射、肠道通透性、免疫反应和肠内分泌信号传导等。
肠道微生物群组成的改变可能会影响肠脑轴并引起炎症,从而可能导致偏头痛。
通过肠-脑轴影响偏头痛
doi.org/10.1111/ene.15934
众所周知,睡眠不足和压力是原发性头痛障碍的常见诱因,不良饮食会增加偏头痛的严重程度和频率。
a) 上述因素导致肠道微生物组多样性和组成的改变,导致肠道细菌产生的短链脂肪酸减少,LPS转移到系统循环中。
b) 此外,这些因子激活免疫细胞,产生的促炎细胞因子也进入系统循环。
c) 这反过来又促进了肠道微生态失调,增加了肠道上皮的通透性,并将分子转移到系统循环中。
肠道微生物组的功能障碍,会导致与头痛病理生理学有关的必需神经递质GABA和血清素的耗竭。这反过来又改变了迷走神经功能。
而系统性促炎细胞因子和LPS,改变了HPA轴的功能,并激活了孤束核,孤束核介导了大脑中的疼痛处理途径,包括前扣带、岛叶皮层、运动皮层和感觉皮层,导致疼痛感知的改变。
神经炎症刺激三叉神经纤维释放CGRP,并驱动皮层扩散性抑郁(CSD),改变兴奋性和抑制性传递,激活三叉神经-颈复合体,加剧头痛严重程度并介导头痛频率。
接下来,我们主要就以上肠脑轴理论展开讨论一下,这其中涉及到的肠道菌群及其代谢产物与偏头痛有什么样的关联。
偏头痛常伴随一些胃肠道症状,包括恶心、呕吐、腹泻、便秘和消化不良。一些研究表明,肠道微生物群失调,可能会下调免疫系统功能,改变宿主代谢,破坏肠-脑轴,导致代谢和神经疾病的发生。胃肠道中的炎症和神经免疫调节可能在偏头痛的发病机制中发挥重要作用。
偏头痛患者的肠道微生物群的物种多样性(α多样性)和代谢功能降低,产丁酸菌减少。
而梭状芽孢杆菌增加,如:
硝酸盐已被确定为一种普遍的偏头痛诱因。
与没有偏头痛的人相比,在偏头痛患者的口腔和粪便样本中观察到能够减少硝酸盐、亚硝酸盐和一氧化氮的细菌丰度更高,如嗜血杆菌属和Rothia属。
编辑
doi.org/10.3390/microorganisms11082083
特殊年龄段的偏头痛研究队列:
儿童和青少年
偏头痛在儿童和青少年中表现出多种形式,估计全世界儿童和青少年的总患病率为 10%。儿童双侧偏头痛更为常见,且持续时间较短。偏头痛儿童会出现多种胃肠道症状,包括腹痛、恶心、呕吐、腹泻、便秘。
在381名 7-18 岁儿童的队列中(40 名患有偏头痛,341 名没有偏头痛,平均年龄 11.5 岁),在偏头痛儿童和健康对照儿童有明显的菌群差异。
与健康对照相比,偏头痛儿童下列菌群升高:
老年患者

一项针对108位老年女性(54 名偏头痛患者、 54 名健康受试者)的宏基因组关联研究发现偏头痛患者的细菌特征如下:
厚壁菌门,尤其是梭菌属显著增加。
在健康对照中有更多有益菌,例如:
普氏粪杆菌、青春双歧杆菌、史密斯甲烷杆菌。
同时,该研究发现犬尿氨酸降解、谷氨酸降解和γ-氨基丁酸(GABA)合成途径较多,提示神经传递和代谢可能发生转变。
接下来我们看这些肠道菌群代谢产物会怎样参与到偏头痛的发病机制中。
谷氨酸和γ-氨基丁酸(GABA)等都参与疼痛感知。肠道中的这些神经递质参与了几种信号通路,这些信号通路除了调节疼痛外,还调节促炎细胞因子的释放。
★ 谷 氨 酸
谷氨酸作为一种兴奋性神经递质,通过不同的作用,包括皮质扩散抑制、中枢敏化和刺激三叉血管系统,在偏头痛病理生理学中发挥作用。
研究证实,各种细菌菌株,包括在环境中发现的或用于食品发酵的菌株,都能够产生谷氨酸。
偏头痛患者谷氨酸水平升高
由于谷氨酸可以沿着三叉神经血管通路对伤害性神经元产生刺激作用,因此它可能在偏头痛和偏头痛相关的中枢敏化的病理生理学中至关重要。这一理论得到了进一步的证明,偏头痛患者的血液谷氨酸水平在间歇期和发作期都有升高,在血浆和脑脊液中均升高。
有研究表明,血浆谷氨酸水平可能是慢性偏头痛和阵发性偏头痛的潜在指标。

此外,有证据表明谷氨酸在皮质扩散性抑制中起主要作用,皮质扩散性抑制被假设为偏头痛先兆的生理底物。基于这些,谷氨酸能神经传递可能是偏头痛和微生物组之间的联系。
扩展阅读:
★ 血 清 素
血清素存在于大脑、肠道和血液的组织中,其中肠道含有~95%的人体总血清素。血清素能神经元存在于中缝背核和整个疼痛通路中。
它在体内有很多作用,例如在神经细胞之间发送信号并帮助身体控制血管收缩的方式。它还在控制情绪、睡眠、饮食和消化方面发挥作用。
血清素从血小板释放到血浆中可能与偏头痛先兆期的病理生理学有关。偏头痛发作期间储存在血小板中的血清素被耗尽;血清素血浆水平降低,而相应代谢物羟基吲哚乙酸 (5-HIAA) 水平升高。
血清素既是血管扩张剂又是血管收缩剂,可作为伤害性疼痛的调节剂
偏头痛患者经常报告说,呕吐后头痛就会停止,呕吐会刺激肠道蠕动并提高血液血清素水平。而血清素的合成与肠道菌群有关,例如肠球菌、链球菌、大肠杆菌等。
血清素可能具有促伤害感受和抗伤害感受特性,具体取决于亚型
现在估计至少有七种不同的受体家族(5-HT1-7),其中许多具有亚型,偏头痛的发生涉及不止一种亚型。由于色氨酸是血清素的前体并且可以穿过血脑屏障,因此大脑中血清素的量取决于色氨酸的水平。
接下来我们看看关于色氨酸的代谢。
★ 色 氨 酸
色氨酸在人体内以三种主要途径代谢:血清素(5-HT),犬尿氨酸(l-kyn)和微生物群相关的吲哚途径。
一些报告表明,增加膳食色氨酸的摄入量可预防偏头痛的发生或减轻其胃肠道相关症状,如恶心、呕吐以及畏光。
色氨酸的代谢途径在五个肠道门中最有效:放线菌门、厚壁菌门、拟杆菌门、变形菌门、梭杆菌门。梭菌属、Burkholderia、链霉菌属、假单胞菌属、芽孢杆菌属5个属也具有较大的肠道内色氨酸代谢潜力。
色氨酸代谢的犬尿氨酸途径,可能对偏头痛具有治疗潜力
色氨酸代谢产物可能作用于谷氨酸能系统,该系统参与疼痛传递、中枢敏化和皮质扩散性抑制,谷氨酸能离子型和代谢型受体参与偏头痛的发病机制,KYNA(犬尿喹啉酸)是NMDA受体的竞争性拮抗剂,其抑制作用可防止谷氨酸诱导的兴奋性毒性,因此有理由推测色氨酸代谢的犬尿氨酸途径可能对偏头痛具有治疗潜力。
注:NMDA受体是离子型谷氨酸受体的一个亚型
犬尿氨酸途径是色氨酸转化的主要途径,因为它在正常情况下约占色氨酸代谢的95%,并且与神经系统疾病的行为和认知症状有关。
犬尿氨酸(l-kyn)途径(KP)将l-色氨酸转化为几种神经活性化合物。犬尿氨酸和喹啉酸的产生,耗尽了作为血清素前体的色氨酸和色胺的供应。

doi.org/10.3390/ijms221810134
犬尿氨酸衍生物具有有效的镇痛作用,其机制包括抑制CGRP。犬尿氨酸途径产物是芳烃受体(AhR)的重要配体,其激活与胃肠道炎症和偏头痛的发病机制有关。
AhR、TLR 和 MyD888 激活可能强调犬尿氨酸参与偏头痛和功能性胃肠道疾病。
扩展阅读:
★ 短 链 脂 肪 酸
短链脂肪酸 (SCFA) 是胃肠道细菌的代谢产物,对葡萄糖稳态、饱腹感、抗炎作用和大脑信号传导有许多局部和全身影响。
通过调节免疫细胞和细胞因子→影响神经炎症
如短链脂肪酸的施用可以减少硝酸甘油诱导引起的痛觉过敏,并导致肠道促炎细胞因子如 TNF-α 和 IL-1β 的释放减少。
丙酸可以保护血脑屏障免受氧化应激
一些菌群如:普通拟杆菌、B.uniformis、Alistipes putredinis、Prevotella copri、Roseburia inulinivorans、Veilonella、Akkermansia mucinophila等,具有很高的丙酸盐生产能力。
丁酸和丙酸等短链脂肪酸通过迷走神经,穿过血脑屏障,然后激活可能改变多巴胺能和血清素能信号传导的受体。
神经保护特性:如丁酸盐
丁酸盐影响肠嗜铬细胞释放血清素,前面我们已经了解到血清素在偏头痛发病中的重要性。
与产丁酸相关的菌群,例如:
快速和极端的饮食变化直接影响肠道微生物群,影响肠道微生物群的β多样性。在高脂肪饮食中添加益生元(可发酵纤维)可以恢复产丁酸菌和双歧杆菌水平,这也突出了饮食对肠道微生物群组成的关键影响。
丁酸盐刺激齿状回的细胞增殖和分化,并增强脑源性神经营养因子(BDNF)和胶质源性神经营养因子(GDNF)的表达。丁酸盐还通过抑制 TNF-α 的合成而在大脑中显示出抗炎作用,TNF-α 是由内毒素脂多糖 (LPS) 通过抑制NF-κB 诱导的。
下面我们来看看炎症因子在偏头痛发病中扮演怎样的角色。
炎症介质
在炎症性疾病和偏头痛中都观察到炎症免疫反应增强。促炎性免疫反应的主要触发因素是由于肠道通透性增加而导致脂多糖进入循环,那么,炎症反应可能出现在身体的各个部位,当三叉神经上伤害感受器的激活时,偏头痛就出现了。
研究表明偏头痛发作期间,下列炎症因子升高:
这些炎症因子是如何在偏头痛发病中发挥作用的呢?
IL-1β 是一种主要细胞因子,可驱动大脑和全身炎症,激活普遍存在的细胞表面受体 IL-1R1。
IL-1β/IL-1R1 轴激活一系列炎症分子,包括其他细胞因子和趋化因子。IL-1β 激活脑膜伤害感受器并使之敏感,是皮质扩散性抑制后三叉神经激活的关键介质。也就是说,IL-1β/IL-1R1 轴在外周TG和中枢神经系统的三叉血管通路中尤其发挥着至关重要的作用。
IL-1和IL-6 是由调节性B细胞响应微生物群的紊乱而释放的。
研究表明,肠道微生物群失调通过三叉神经伤害感受系统中的TNF-α水平上调,促进了类似偏头痛疼痛的持续性。
肠道菌群通过间接信号传导维持肠脑轴正常平衡的作用机制
doi: 10.1186/s10194-020-1078-9
IL-8的表达,涉及到神经肽,与P物质有关,P物质可以帮助调节免疫细胞及其产生的各种细胞因子。
接下来,我们来看偏头痛中重要的——神经肽。
★ CGRP(降钙素基因相关肽)
对于患有慢性偏头痛的患者,CGRP 水平在偏头痛发作期间持续升高,并且在两次发作之间继续升高。CGRP是我们反复提到的词,我们来看看它在偏头痛疾病中发挥什么样的作用。
CGRP,降钙素基因相关肽 ,一种由37个氨基酸组成的肽,可能通过胃肠道免疫细胞和肠道运动与肠道微生物有关。在偏头痛的情况下,它在三叉神经节、三叉神经尾核、杏仁核、丘脑和皮质中浓度很高。
CGRP有两种形式:
CGRP升高如何与偏头痛关联起来:
星形胶质细胞和神经元参与了CGRP合成释放过程的正反馈循环,并维持了增加的炎症和致敏状态。
CGRP 可以作为一种有价值的治疗标志物
目前,许多用于治疗和预防偏头痛药物的原理,就是针对这个CGRP或其受体的,抑制 CGRP 转录并减少其释放,从而缓解偏头痛。
★ P 物 质
P物质(substance P),是一种广泛分布于细神经纤维内的神经肽,主要作用是传递痛觉信息。
呕吐是最常见的胃肠道反应之一,受P物质的控制
在中枢神经系统中,延髓有面突区和孤束核的区域,这两个区域控制呕吐反射并含有高水平的P物质。P物质释放结合到NK1R上会触发呕吐反应。
P物质的一个功能是通过改变细胞信号通路作为神经递质和疼痛感知调节剂
有一个假说是P物质有助于突触后神经元对谷氨酸的敏感性,有助于将疼痛信号传递到大脑的体感区域。在许多其他中枢神经系统病理中通过神经源性炎症引起早期血脑屏障的破坏。
P物质的关键作用是双向调节免疫细胞及其产生的各种细胞因子
IL-1、IL-4和IFN-γ可以诱导巨噬细胞表达NK1R。诱导细胞因子释放会导致巨噬细胞和树突状细胞的招募,这有助于刺激IL-8的表达。P物质间接参与了免疫细胞的招募和迁移。P物质能刺激和诱导肥大细胞脱颗粒化。
★ PACAP
在与偏头痛病因机制相关的几个结构中发现了PACAP(垂体腺苷酸环化酶激活肽)的存在,包括硬脑膜、脑血管、三叉神经节、三叉神经原球和颈椎脊髓。
PACAP在神经调节、神经源性炎症和痛觉中起着重要作用。PACAP在感觉神经元炎症后上调。PACAP与CGRP在一些硬膜神经纤维中共同表达,也与疼痛的中枢敏化和情绪负荷有关。
PACAP对革兰氏阳性菌、革兰氏阴性菌和真菌具有抗菌活性,因此它可以直接影响膜的杀菌功能。有效的防御能力和较低的细菌耐药性使抗微生物神经肽成为一类有吸引力的新型抗生素。
★ NPY(神经肽Y)
NPY是去甲肾上腺素能系统功能的可能指标,通过脑循环调节影响脑血流量。值得一提的是,在偏头痛发作期检测到较高水平的NPY。
NPY已经在肠-脑轴的各个层面上被检测到。NPY通路也有助于改变胃肠道功能及其血流、免疫系统和炎症状态、疼痛、能量稳态、情绪、情绪以及行为和大脑认知功能等。
★ 胆囊收缩素 (CCK)
胆囊收缩素(CCK)由小肠粘膜内层(I 细胞)、哺乳动物大脑(例如皮质、丘脑、中脑边缘、导水管周围灰质和中脑)和脊髓的肠内分泌细胞合成。这种肽抑制胃排空和胃酸分泌,刺激胆囊收缩和胰腺分泌,并通过 CCK1 受体在大脑中激发饱腹感。
CCK在导水管周围灰质中产生,可能是偏头痛内源性疼痛信号系统及其水平升高的原因。CCK也存在于三叉神经节中,因此刺激三叉神经节会导致局部CCK增加。
CCK——肥胖和偏头痛之间的桥梁之一
肥胖患者偏头痛发作的频率和严重程度明显更高。偏头痛和肥胖之间联系的一种可能解释是,是高脂饮食引起的CCK分泌,因为十二指肠内游离脂肪酸会刺激CCK的分泌。
关于CCK可以详见我们之前肠道激素的文章:
当考虑到性激素时,我们知道偏头痛在女性中的发病率是男性的2-3倍,女性的发作时间更长、更严重,恢复时间也更长。
无先兆偏头痛的风险在月经前 2 天和月经期间最高,这也是生殖道微生物群多样性最低的时候。周期中期雌激素和孕酮水平高与无先兆偏头痛的风险较低以及生殖道内微生物群落的稳定性较高有关。
众所周知,雌激素会改变肠道内的紧密连接和粘蛋白保护,改变肠道通透性。
雌激素受体广泛表达于中枢神经系统,有证据表明雌激素调节痛觉传导,雌激素敲除模型在脊髓背角水平导致CGRP升高,从而改变了发起偏头痛攻击所需的阈值。临床上支持这一点的是黄体晚期偏头痛发作的可能性增加。虽然孕酮撤退似乎不会触发偏头痛发作,但它可能在疼痛敏感性和病程演化中起作用。
关于雌激素与肠道菌群详见我们之前的文章:
睾酮降低了对CSD的易感性,增加了血清素能调,并具有抗炎特性。较高水平的睾酮与肠道微生物群多样性增加有关,研究表明,患有偏头痛的男性存在雄激素缺乏和雌二醇水平增加。
因此,肠-脑轴可能介导性激素和头痛疾病之间的关系。
以上我们阐述了肠道菌群及其代谢产物等参与偏头痛发病机制,包括炎症介质、肠道微生物群、神经肽、血清素途径等。有证据表明偏头痛和胃肠道疾病之间存在共同的病理生理学,下面我们来了解一下它们之间的关联。
偏头痛和胃肠道合并症的病理生理学都很复杂,尚未完全阐明;然而,科学证据确实表明这些复杂疾病之间的病理生理学重叠。研究这种重叠有助于阐明常见的病理生理异常或生物学机制。
特别是,自主神经系统(ANS)被认为在偏头痛和胃肠道功能障碍之间的关联中起重要作用,因为它们的症状特征相似,包括恶心,呕吐,消化不良和胃轻瘫。
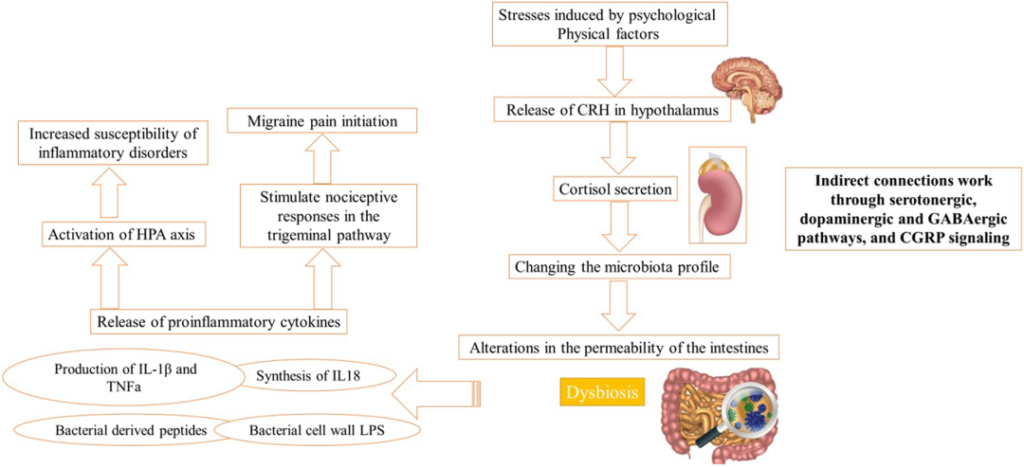
doi: 10.1111/head.14099
肠易激综合征(IBS)和炎症性肠病(IBD)的患者中,偏头痛的发生率增加。两者都是严重的肠道疾病,与肠道通透性增加和微生物引起的炎症有关。此外,中枢性、内脏性和热性皮肤过敏在IBS和偏头痛中很常见。患有长期和更频繁头痛的偏头痛患者更有可能被诊断为IBS。
偏头痛与IBS有一些共同的重要特征,这两种疾病都是慢性疼痛相关疾病,在女性中比男性更普遍,并且在纤维肌痛、慢性疲劳综合征、间质性膀胱炎、失眠、抑郁等方面与心理合并症有关。据估计,约60%的偏头痛患者患有异常性疼痛,除内脏超敏反应外,大多数IBS患者还报告异常性疼痛。
在IBS患者的粪便微生物群中评估出偏头痛的严重程度与肠道菌群失调之间存在强正相关。约一半没有先兆的年轻女性偏头痛患者患有肠道微生物群失调。
美国肠道计划队列研究发现,偏头痛与口腔微生物中具有还原硝酸盐、亚硝酸盐和一氧化氮功能活性的菌群水平呈正相关关系。
IBD是一种慢性复发-缓解炎症性疾病,由克罗恩病和溃疡性结肠炎组成。关于偏头痛和IBD之间可能关系的数据很少。
虽然机制不明确,IBD中存在的自身免疫性炎症反应、吸收不良、内皮功能障碍以及为该疾病开具的免疫抑制治疗可能参与了炎症性肠病和偏头痛之间的病理生理关联。
扩展阅读:
根据罗马IV标准,功能性消化不良分为餐后不适综合征(PDS)和上腹疼痛综合征(EPS)。
偏头痛是功能性消化不良和餐后症状患者的常见合并症。
扩展阅读:
基于人群的研究,个体主要是女性,最常见的症状(无论是持续性还是偶发性)是恶心和呕吐,但也可能包括腹痛、腹胀、体重减轻、餐后饱腹感和早饱感。
在患有特发性胃轻瘫和腹痛的个体中,腹痛严重的人比症状较轻的人更有可能患有重叠偏头痛。
在胃轻瘫中,观察到胃排空时间与偏头痛发作患者头痛、恶心和对光敏感的严重程度显著相关。多潘立酮是一种多巴胺受体拮抗剂,可用于治疗胃轻瘫,并已被证明在早期高剂量给药时可预防大多数偏头痛发作。另一种多巴胺受体拮抗剂甲氧氯普胺可用于治疗胃轻瘫和恶心,作为偏头痛的急性静脉注射治疗是有效的。
根据 5 项病例对照研究的荟萃分析结果,约 45% 的偏头痛患者患有幽门螺杆菌感染,而健康对照组的患病率估计约为 33% 。有证据表明根除幽门螺杆菌可能与缓解偏头痛症状有关。
幽门螺杆菌感染与持续的慢性炎症状态有关,这反过来可能导致炎症介质和血管活性化合物的产生增加。因此,偏头痛和幽门螺杆菌感染之间的关联机制可能包括诱导免疫、炎症和血管反应,以及随后免疫细胞、炎症和血管活性物质释放到胃粘膜中,最终可能导致大脑疼痛敏感结构的超敏反应。
扩展阅读:
正确认识幽门螺杆菌
乳糜泻具有多种神经系统表现,如癫痫、共济失调、小脑性共济失调、情绪障碍、脑炎、周围神经病变、神经肌肉疾病、痴呆、学习障碍、发育迟缓和偏头痛。
与没有这种疾病的人相比,被诊断患有乳糜泻(一种由谷蛋白肽引发的自身免疫性疾病)的人表现出更高的偏头痛发生率,据估计,大约 21-28% 的乳糜泻患者患有偏头痛。
相反,患有偏头痛的人乳糜泻的患病率更高。临床医生可能会考虑在存在此类改变的偏头痛受试者中寻找乳糜泻,特别是如果存在其他提示乳糜泻的症状。
偏头痛和乳糜泻之间的联系可归因于几种同时存在的机制,包括面筋引起的促炎细胞因子的激活、吸收不良导致的重要维生素和必需元素的缺乏、血管张力的紊乱、神经系统敏感性的增加、脑灌注不足、血管周围炎症等。
✔ 消除饮食
消除饮食需要先确定首先引发头痛的饮食成分,然后从常规饮食中排除这些成分。这是一种个人方法,个人可以识别引发或加剧头痛的成分。
如果某种食物在接触后 1 天内引起头痛,则可将其视为触发因素。大多数患者存在多种触发因素;因此,很难识别单一成分。况且有些食物含有不止一种成分,因此要完全识别出来并不简单。
✔ 综合饮食
富含ω-3和ω-6脂肪酸的低脂、高叶酸和生酮饮食对偏头痛患者有益。
改良的阿特金斯饮食提供神经保护,改善血清素能功能障碍,抑制神经炎症,改善线粒体功能。
生酮饮食会增加产生的酮体浓度,这对头痛患者有帮助。
低血糖饮食也能改善炎症状态。保持ω-6脂肪酸和ω-3摄入之间的微妙平衡可以减少炎症,维持血管张力,增强血小板功能。
偏头痛发作频率与坚持“健康”饮食模式(大量食用水果、蔬菜、鱼类、豆类、发酵食物等)成反比(P=0.04)。与坚持“西方”饮食方式(大量食用可乐、咸坚果、加工肉和快餐)成正比(P=0.02).
生姜、薄荷和辣椒都具有天然的止痛功效。生姜和薄荷有助于减少与偏头痛相关的恶心。
薰衣草可以减少血管炎症,通常是治疗焦虑、压力和压力引发的偏头痛有效的草药。
✔ 低钠饮食 / 高钠饮食
对脑脊液的分析显示,与对照组相比,偏头痛患者的钠含量较高,尤其是在头痛发作时。然而,低钠饮食的效果取决于许多因素,因此在考虑低钠饮食之前应详细评估其效果。
低钠饮食可以有效减少老年高血压患者头痛的发生。除了控制血压之外,低钠饮食还可以降低头痛的发生率。
相比之下,在没有高血压的低至正常体重指数的年轻女性人群中,高钠饮食有助于降低偏头痛发作的频率。
因此,低/高钠饮食应该根据患者群体进行调整。
低热量饮食 or 生酮饮食?
有研究认为,生酮饮食对偏头痛患者更有益,低热量饮食对偏头痛患者不利 (n = 108) 。在另一项研究中,生酮饮食显着改善了这些患者的头痛特征,并减少了发作频率和药物使用。停止生酮饮食后 2 个月观察到持续改善。
虽然看起来生酮饮食患者的头痛频率有所减少,但过渡期的头痛频率更严重。然而,经过3个月和6个月的随访后,低热量饮食组的头痛频率显著改善。
✔ 低血糖饮食(LGD)
通常从碳水化合物中获取总热量的 50-55% 是安全饮食。饮食转向低血糖指数碳水化合物已被证明可以增加短链脂肪酸水平。在某些情况下,LGD 是一种重要的替代方案,例如高脂血症、糖尿病、癫痫和体重管理。
罗马进行的一项队列研究中报告称,增加全麦面包(P = 0.04)和全麦面食(P = 0.004)的消费,以及减少白面包(P = .004)的消费,与偏头痛发作频率和每月使用药物救治的统计显著减少有关。
✔ 低脂饮食
一项研究调查了54名成年人减少脂肪饮食对偏头痛治疗的影响。患者被告知他们的脂肪消耗量将降至<20 g/天(12周)。发现头痛药物的发生率、严重程度和需求显著降低。
在一项为期 12 周的试验中,患有慢性偏头痛的成年人被随机分配接受高omega-3 / 低omega-6 饮食或低 omega-6 饮食。与采用低omega-6 饮食的偏头痛患者相比,采用高omega-3/低 omega-6 饮食的人的头痛改善程度更高。
可能与此相关:omega-6 和 omega-3 之间的平衡有助于炎症控制。omega-6 脂肪酸促进血管舒张。
谨慎选择下列食物
▸
味精在高浓度和溶解在液体中可能会引发偏头痛。并不是说引发每个患者的偏头痛发作,不能一概而论。
▸
咖啡因可能缓解偏头痛或者引发偏头痛。与阿司匹林和对乙酰氨基酚结合,咖啡因是一种高效的止痛药。咖啡因戒断会引发咖啡因使用者的偏头痛发作。需要注意自己的剂量。
咖啡因与偏头痛频率之间的联系没有定论。一些研究数据表明,不同的偏头痛和慢性日常头痛在咖啡因使用者中比不摄入这种物质的人更常见。也有研究认为偏头痛频率和咖啡因摄入之间没有关联。
▸
饮酒是偏头痛发作最常见的原因之一。酒精可以刺激三叉神经节中的脑膜伤害感受器,触发疼痛信号,然后在三叉神经脊核传递到丘脑核,最后传递到体感皮层。也可能涉及其他机制,例如血管舒张作用、脱水、毒性等。除了酒精,酒精饮料还含有某些化合物(酒精发酵的副产品),这些化合物都可能会引发偏头痛。
▸
许多研究提出了巧克力摄入与头痛之间的联系,但其确切的生理机制尚不清楚。可能是巧克力中的黄烷醇刺激内皮一氧化氮合酶(eNOS)活性,这可以通过增加一氧化氮(NO)的产生导致血管舒张。
也有研究认为巧克力含有许多维生素和矿物质(例如镁和核黄素),用于预防偏头痛。富含可可的饮食通过抑制CGRP的表达来预防三叉神经节神经元的炎症反应。
▸
酪胺(Tyramine),是一种衍生自酪氨酸的胺化合物,存在于各种食品中,包括陈奶酪、腌肉、熏鱼、啤酒、发酵食品和酵母提取物等。酪胺有可能通过促进去甲肾上腺素的释放并对α-肾上腺素受体产生激动作用而引发头痛。
▸
阿斯巴甜是一种人造甜味剂。几项研究表明,它会引起各种神经或行为症状;也会引起头痛,尤其是对于长期使用中等或高剂量(900–3000 mg/天)的人。
基于以上所述,识别和避免饮食中的偏头痛诱因至关重要,因为这有助于降低偏头痛的频率,让偏头痛患者能够控制让他们感到疲惫和无助的情况。
B族维生素
维生素B2对氧化代谢至关重要,可以降低偏头痛的风险。根据随机对照试验,服用维生素B2可以显著减少每月偏头痛天数1-3天。通过增加产短链脂肪酸菌丰度,维生素B2和其他B族维生素有助于对肠道微生物组产生正面影响。
扩展阅读:
维生素C
水溶性维生素C是最重要的非酶抗氧化剂之一,参与神经代谢。维生素C的膳食来源是水果和蔬菜。
维生素C可能调节偏头痛过程中的神经炎症和氧化应激活性的影响。
一项开放标签研究中,患者接受了含有120 mg粉红树皮提取物、60 mg维生素C和30 IU维生素E的胶囊(每粒胶囊)。在完成治疗期的患者中,得到了改善,包括头痛频率和头痛严重程度的降低。该研究人员还进行了一项开放标签研究,以检查维生素C(150 mg)和辐射松树皮提取物对偏头痛症状的影响。患者服用上述剂量的维生素C和辐射松树皮提取物3个月。治疗后,患者表现出显著改善:头痛频率和头痛严重程度减轻。
扩展阅读:
维生素D3
补充维生素D3会影响肠道微生物群的组成。为健康人补充8周维生素D3也显著减少了幽门螺杆菌的数量。几项研究报告称,血清维生素D水平可能与偏头痛/头痛风险增加有关。
偏头痛患者中这种维生素缺乏/不足的患病率可能更高。补充维生素D对偏头痛发作的强度和频率也有良好的影响。
镁
镁参与多种酶促反应,还维持细胞膜的平衡,影响其通透性并减少自发去极化的可能性。它影响外周和中枢神经系统的兴奋性和神经传导,因此在偏头痛病程中起着重要作用。
镁缺乏导致皮质扩散抑制或谷氨酸能神经传递异常是镁与偏头痛关系的一个可能机制。镁不足可能会影响 NMDA 受体阻断、钙通道、谷氨酸和 NO 活性,以及血清素受体的亲和力。
许多临床医生认为,以正确的剂量和形式补充镁可以成为预防偏头痛发作的治疗方法。补充镁也可以配合其他药物。
益生菌是偏头痛的另一种潜在疗法,它与饮食干预一样能够改变肠道微生物群。
在 NTG 诱导的偏头痛小鼠模型中,发现口服益生菌混合物显著抑制抗生素治疗引起的偏头痛样疼痛延长。
在一项随机双盲对照试验中观察到,与安慰剂对照相比,在 40 名发作性偏头痛患者和 39 名慢性偏头痛患者中,为期 8 周的益生菌试验显著降低了偏头痛的严重程度、频率和药物使用量。
其中含有14种益生菌菌株:
益生菌组偏头痛发作的平均频率显著降低(平均变化:-9.67 与 -0.22;p ≤ 0.001)。补充益生菌显著降低了严重程度(平均变化为-2.69;p≤0.001.),降低慢性偏头痛患者的发作持续时间(平均变化:-0.59;p≤0.034),减少每天服用药物数量(平均变化为-1.02;p<0.001)。
doi.org/10.3390/neurolint15030073
将芽孢杆菌、双歧杆菌、乳酸杆菌和链球菌菌株的混合物应用8-10周,可以降低发作性偏头痛患者的发作严重程度和频率,并降低慢性偏头痛患者的频率、严重程度、持续时间和每天服用的药物数量。
姜黄素
姜黄素是从姜黄根茎中提取的主要姜黄素,也是最著名的植物多酚之一。姜黄素具有许多有益的特性,如抗氧化、抗动脉粥样硬化、抗菌、免疫调节、抗衰等特性。
姜黄素可以显著抑制 IL-1β 和 TNF-α 表达、氧化应激和蛋白质积累。
对偏头痛治疗进行了一项研究,比较了单独使用舒米曲普坦(ST)和与姜黄素一起使用的效果,采用了硝酸甘油诱导的偏头痛大鼠模型。
姜黄素以下面两种形式静脉注射,因为姜黄素的肠道吸收较低:
a)酒精溶液(用生理盐水稀释)
b)脂质体的形式
当姜黄素以脂质体的形式进行注射时,其抗氧化作用更为显著。作者将所得结果归因于姜黄素能够清除羟基或过氧自由基、直接与过氧化物自由基相互作用以及抑制参与炎症性因子NF-κBs的活性。
偏头痛发病机制的一个考虑的机制涉及肿瘤坏死因子α(TNF-α)的参与,通过引发神经元超兴奋性、刺激伤害感受器和前列腺素的产生,从而引发神经炎症。
一项研究,旨在确定姜黄素和ω-3脂肪酸对TNF-α基因表达的协同作用。接受两种物质治疗的患者在血浆中显示出TNF-α mRNA水平的降低(反映了TNF-α表达的减少)。在只接受其中一种分析物的患者中,没有发现这种情况。
辅酶Q10
除了抗氧化特性外,辅酶Q10还表现出抗炎特性,参与嘧啶合成(作为辅因子),进而参与DNA复制和RNA修复过程。由于其潜在的积极作用,是许多神经系统疾病的膳食补充剂。
酶Q10被列为预防偏头痛最常用的治疗方法之一。给予偏头痛患者辅酶Q10会导致CGRP的降低。
儿童和青少年中研究了补充辅酶Q10与偏头痛发生之间的关系。患者每天服用100毫克辅酶Q10,为期4个月。结果显示,在主要结果方面,两组之间没有统计学上的显著差异。然而,与对照组相比,偏头痛患者的研究组头痛发作频率呈下降趋势。此外,在补充辅酶Q10的前四周,观察到发作性偏头痛患者头痛的严重程度降低。
在辅酶Q10的抗氧化特性中有以下方面,即超氧化物歧化酶活性的调节(导致细胞抗氧化防御的改善)、氧化应激OS的减少或增加谷胱甘肽还原形式(用作抗氧化剂)浓度的能力。上述机制可缓解偏头痛,降低偏头痛发作的频率,缩短偏头痛发作的持续时间。
银杏内酯B
银杏内酯B是从银杏叶中提取的一种草药成分。银杏叶提取物具有神经保护特性,对记忆有有益作用,也有抗氧化剂的某些特性。
一项为期六个月的开放标签多中心研究,证明了银杏内酯B对每月至少发作一次的偏头痛患者治疗先兆偏头痛的有效性。
槲皮素和银杏内酯B对减轻偏头痛症状有特殊作用。银杏叶提取物成分可降低氧化应激,抑制血小板活化因子,并影响谷氨酸能传递。
所有生物学机制都会增加神经保护作用,从而降低偏头痛的频率,减轻偏头痛,缓解先兆症状。
小白菊
小白菊(Tanacetum parthenium)属菊科植物,广泛分布于南美洲。其特征是具有抑制醛糖还原酶活性的高潜力,并显示出必要的抗氧化特性。
小白菊用于治疗多种疾病,不仅可以缓解偏头痛的症状,还可以缓解另一种来源的疼痛,如炎症、恶心和呕吐。
一项双盲、多中心、安慰剂对照研究,以确定小白菊稳定提取物(MIG-99)在降低偏头痛发作频率方面的有效性。最显著的效果记录为6.25mg剂量的MIG-99。三年后,同一作者发表了剂量为6.25 mg的MIG-99制剂的有效性和安全性结果。在服用该制剂的组中,偏头痛发作频率每月减少1.9次(安慰剂组每月减少1.3次)。
粪菌移植可以通过改变肠道菌群,帮助恢复肠道微生物平衡,使肠道微生物恢复更加有效和持久,理论上可减轻偏头痛发作和偏头痛严重程度,超过340项临床试验正在进行中。肠道菌群的改变可导致炎症介质和血清素途径的正常化,从而缓解偏头痛的频率和强度。
扩展阅读:
吴茱萸汤是一种临床用于治疗和预防偏头痛的配方,可anaerostipes和acidifaciens丰度,以恢复细菌的比例。由于5-HT合成与微生物群、功能和代谢紊乱相关,吴茱萸汤可以通过影响中枢和外周5-HT来缓解痛觉过敏。研究表明,在基因敲除小鼠和豚鼠中,5-HT1D受体具有抑制神经肽释放的作用,从而改变硬脑膜神经源性炎症反应。
良好的生活习惯
建立规律的睡眠时间。每天同一时间起床和上床睡觉,甚至周末也是如此。一天结束时放松一下。
减少吸烟。偏头痛患病率与每日吸烟的相关性为正相关(Spearman系数,rs=0.49)。
定期运动
身体活动期间,身体会释放某些化学物质来阻隔脑部疼痛信号。这些化学物质还有助于缓解焦虑和抑郁症,这两种状况都可能加剧偏头痛。
肥胖也会增加慢性头痛的风险。通过运动和饮食维持健康体重可能也有助于管理偏头痛。
值得尝试的有氧运动可能包括快走或慢跑、骑自行车、瑜伽、游泳等。
缓解压力
简化生活,明确管理时间,适当休息,可以选择接触大自然,做深呼吸(专注且缓慢),冥想,瑜伽等放松的方式,保持积极心态,享受乐趣,每天抽出至少 15 分钟做自己喜欢的事。
写偏头痛日记
记下偏头痛发作时周围的环境条件。写下感官刺激(明亮的灯光、响亮的音乐、奇怪的气味等)、压力的原因、饮食习惯和睡眠习惯。几次发作后,回顾一下日记,找出偏头痛发作前通常存在的常见情况。这些情况可能是“触发因素”,下次尽量避免。
当偏头痛发作时的缓解小方法:
关灯、静音
光线和声音会使偏头痛加剧,尽量在昏暗且安静的房间里放松,闭目休息,能睡就睡吧,让眼睛和大脑得到放松,帮助缓解头痛的不适感。
找一个安静的环境,避免嘈杂的声音,减轻对偏头痛的刺激。
如果不能马上找到这样的地方,可以考虑戴上太阳镜和耳塞。或者尝试舒缓的音乐。
试试温度疗法
热敷或冷敷头部或颈部。
按摩、放松
用双手的食指和中指,以小幅度打圈的方式轻轻按摩太阳穴、颈侧、颈后。
通过深呼吸放松身体和心理,减轻紧张和疼痛感。适量饮水,保持身体水分平衡,有助于缓解头痛。
偏头痛是由多种因素共同引起的,由遗传、内分泌、代谢和/或环境因素相互交织促成的。基于各种初始因素,广泛接受的观点是皮质扩散抑制假说和三叉神经血管假说。前者指出了大脑与神经炎症之间的相互关系,后者指出神经和血管是发生的关键因素。偏头痛复杂的机制和病理生理学仍在广泛研究中。
本文我们了解到,肠道微生物群通过参与肠-脑轴各个途径,肠道微生物群和大脑通过各种途径相互作用,包括免疫系统、色氨酸代谢、迷走神经和肠神经系统等,可能发挥着关键作用,或将作为偏头痛的有前途的生物标志物和治疗靶点。了解这种疾病的多系统联系,极大地扩展了对偏头痛机制的理解,并提供了更广泛的治疗蓝图。
偏头痛本身会产生一些诊断问题,这些问题可能会因合并症而加剧,包括功能性胃肠道疾病等。同时,复杂的疼痛通路,食物和药物代谢、免疫反应等造成的个体差异,会给在偏头痛的临床管理治疗过程中带来难度,目前药物并不能完全缓解症状或满足要求。
考虑以互补的方式将临床前和临床研究结合起来,同时包括科研人员、临床医生、健康管理以及其他相关机构的成员之间的相互交流、协作,在此过程中纳入肠道菌群健康检测,集结众多力量的合作,从预防偏头痛辅助判别其潜在风险,到偏头痛发作的疾病管理,再到预后减少发病次数等多个阶段,帮助降低偏头痛和偏头痛合并的各种胃肠道疾病的风险,改善偏头痛患者的生活质量,为他们带来更多希望。
注:本账号内容仅作交流参考,不作为诊断及医疗依据。
主要参考文献
Ferrari MD, Goadsby PJ, Burstein R, Kurth T, Ayata C, Charles A, Ashina M, van den Maagdenberg AMJM, Dodick DW. Migraine. Nat Rev Dis Primers. 2022 Jan 13;8(1):2.
Khan J, Asoom LIA, Sunni AA, Rafique N, Latif R, Saif SA, Almandil NB, Almohazey D, AbdulAzeez S, Borgio JF. Genetics, pathophysiology, diagnosis, treatment, management, and prevention of migraine. Biomed Pharmacother. 2021 Jul;139:111557.
Arzani M, Jahromi SR, Ghorbani Z, Vahabizad F, Martelletti P, Ghaemi A, Sacco S, Togha M; School of Advanced Studies of the European Headache Federation (EHF-SAS). Gut-brain Axis and migraine headache: a comprehensive review. J Headache Pain. 2020 Feb 13;21(1):15.
Aurora SK, Shrewsbury SB, Ray S, Hindiyeh N, Nguyen L. A link between gastrointestinal disorders and migraine: Insights into the gut-brain connection. Headache. 2021 Apr;61(4):576-589.
Zhou Y, Pang M, Ma Y, Lu L, Zhang J, Wang P, Li Q, Yang F. Cellular and Molecular Roles of Immune Cells in the Gut-Brain Axis in Migraine. Mol Neurobiol. 2023 Sep 11.
He Q, Wang W, Xiong Y, Tao C, Ma L, Ma J, You C; International Headache Genetics Consortium. A causal effects of gut microbiota in the development of migraine. J Headache Pain. 2023 Jul 17;24(1):90.
Kim JH, Lee Y, Kwon YS, Sohn JH. Clinical Implications of the Association between Respiratory and Gastrointestinal Disorders in Migraine and Non-Migraine Headache Patients. J Clin Med. 2023 May 12;12(10):3434.
Cámara-Lemarroy CR, Rodriguez-Gutierrez R, Monreal-Robles R, Marfil-Rivera A. Gastrointestinal disorders associated with migraine: A comprehensive review. World J Gastroenterol. 2016 Sep 28;22(36):8149-60.
Slavin M, Li HA, Frankenfeld C, Cheskin LJ. What is Needed for Evidence-Based Dietary Recommendations for Migraine: A Call to Action for Nutrition and Microbiome Research. Headache. 2019 Oct;59(9):1566-1581.
Park CG, Chu MK. Interictal plasma glutamate levels are elevated in individuals with episodic and chronic migraine. Sci Rep. 2022 Apr 28;12(1):6921.
Sgro M, Ray J, Foster E, Mychasiuk R. Making migraine easier to stomach: the role of the gut-brain-immune axis in headache disorders. Eur J Neurol. 2023 Jun 17.
Do TP, Hougaard A, Dussor G, Brennan KC, Amin FM. Migraine attacks are of peripheral origin: the debate goes on. J Headache Pain. 2023 Jan 10;24(1):3.
Pleș, H.; Florian, I.-A.; Timis, T.-L.; Covache-Busuioc, R.-A.; Glavan, L.-A.; Dumitrascu, D.-I.; Popa, A.A.; Bordeianu, A.; Ciurea, A.V. Migraine: Advances in the Pathogenesis and Treatment. Neurol. Int. 2023, 15, 1052-1105.
Kappéter, Á.; Sipos, D.; Varga, A.; Vigvári, S.; Halda-Kiss, B.; Péterfi, Z. Migraine as a Disease Associated with Dysbiosis and Possible Therapy with Fecal Microbiota Transplantation. Microorganisms 2023, 11, 2083.
Yamanaka, G.; Suzuki, S.; Morishita, N.; Takeshita, M.; Kanou, K.; Takamatsu, T.; Suzuki, S.; Morichi, S.; Watanabe, Y.; Ishida, Y.; et al. Role of Neuroinflammation and Blood-Brain Barrier Permutability on Migraine. Int. J. Mol. Sci. 2021, 22, 8929.
Martami F, Togha M, Seifishahpar M, Ghorbani Z, Ansari H, Karimi T, Jahromi SR. The effects of a multispecies probiotic supplement on inflammatory markers and episodic and chronic migraine characteristics: A randomized double-blind controlled trial. Cephalalgia. 2019 Jun;39(7):841-853.
Fila, M.; Chojnacki, J.; Pawlowska, E.; Szczepanska, J.; Chojnacki, C.; Blasiak, J. Kynurenine Pathway of Tryptophan Metabolism in Migraine and Functional Gastrointestinal Disorders. Int. J. Mol. Sci. 2021, 22, 10134.
Goschorska, M.; Gutowska, I.; Baranowska-Bosiacka, I.; Barczak, K.; Chlubek, D. The Use of Antioxidants in the Treatment of Migraine. Antioxidants 2020, 9, 116.
Spekker, E.; Nagy-Grócz, G. All Roads Lead to the Gut: The Importance of the Microbiota and Diet in Migraine. Neurol. Int. 2023, 15, 1174-1190
谷禾健康
假单胞菌属(Pseudomonas)是最多样化和普遍存在的细菌属之一,其物种存在于沉积物、临床样本、植物(或植物根际)、患病动物、水、土壤、海洋、沙漠等,这反映在它们多变的代谢能力和广泛的适应环境的潜力上。
假单胞菌属是一类革兰氏阴性的好氧或微需氧的细菌,属于 Proteobacteria(变形菌门),代表种包括:
Pseudomonas aeruginosa(铜绿假单胞菌)
Pseudomonas putida (恶臭假单胞菌)
图源:CDC
假单胞菌属是机会性病原体,是全世界医院获得性感染的主要原因。它可以侵入多个器官系统,包括血液、肺、软组织(皮肤、肌肉和肌腱)、身体的其他部位。症状取决于感染部位。
与假单胞菌属感染相关包括免疫系统受损患者的感染和医疗器械相关感染等,如刚接受过手术的人和免疫功能低下的人,艾滋病或糖尿病患者。
假单胞菌属与人体健康相关,其中一些菌株可以引起感染和疾病,如铜绿假单胞菌(Pseudomonas aeruginosa),可以引起呼吸道、肠道、泌尿道和皮肤感染。
免疫抑制个体和住院患者的铜绿假单胞菌肠道携带率显着升高,因此感染和抗生素相关性腹泻的风险增加。
假单胞菌属与其他菌属如葡萄球菌、普雷沃氏菌属、不动杆菌属、芽孢杆菌属、肠杆菌属、肠球菌属等菌属有互作关系。
假单胞菌不被认为是肠道微生物组的典型成员,并且假单胞菌在肠道定植与发生肺部感染和死亡率的风险增加有关。
研究表明,假单胞菌与肠道屏障功能障碍和感染有关,可能导致败血症和多器官功能障碍综合征。此外,假单胞菌与炎症性肠病的发生也有关系,它在IBD患者的肠道中检测到丰度更高,而且它参与引起其他菌群的失衡会影响疾病的进展。
细菌有可能在人体内的不同部位之间转移,防止肠道定植或肠道到其它器官尤其肺部的传播,可能是预防重症患者假单胞菌感染的有效策略。
本文主要分享假单胞菌属(Pseudomonas)的生态,分类,致病因素以及与人体相关健康特性和可能的生活干预措施。
假单胞菌属(Pseudomonas),是革兰氏阴性、好氧菌属,于 1894 年被描述,是最多样化和普遍存在的细菌属之一。有些物种对人类、动物或植物具有致病性。
▼
假单胞菌属(Pseudomonas)属于假单胞菌科,包含200多个有效描述的物种。该属的成员表现出大量的代谢多样性,因此能够在广泛的生态位中定居。
它们易于体外培养,并且越来越多的假单胞菌菌株基因组序列可供使用,这使得该属成为科学研究的绝佳焦点。
Photograph by Kateryna Kon
研究最好的物种包括铜绿假单胞菌。直的或略微弯曲的棒状但非螺旋状,0.5–1.0 × 1.5–5.0 µm。由一根或数根极鞭毛运动;很少不动。在某些物种中,也可能形成短波长的侧鞭毛。有氧,具有严格的呼吸型代谢,以氧为末端电子受体;在某些情况下,硝酸盐可用作替代电子受体,从而允许厌氧生长。
大多数物种(不是全部)无法在酸性条件(pH 4.5 或更低)下生长。大多数物种不需要有机生长因子。氧化酶阳性或阴性。过氧化氢酶阳性。化学有机营养的。
▼
——分布广
假单胞菌属是变形菌门的一个庞大而多样的属。从北极苔原到热带雨林,从干旱的土壤到雨云,几乎在地球的每个角落都可以找到该属的成员。这种令人难以置信的环境适应性归功于假单胞菌非凡的新陈代谢多样性。
——野蛮生长
假单胞菌可以在 0 至 42 °C 的温度范围内生长,甚至可以在更极端的温度下存活。它们对营养的需求很少,可以利用多种碳源。尽管假单胞菌在有氧环境中生长最佳,但它们也可以利用氮进行厌氧呼吸。
它们可以作为自由生活的浮游细胞或作为生物膜群落的成员生活,并且具有将微生物信号和环境线索转化为特定生态位过程的非凡能力。
基因组分析表明,许多其他因素有助于假单胞菌属的多样性和适应性。除了赋予调节和代谢灵活性的常见基因的大量等位基因差异外,水平基因转移影响了致病性假单胞菌属的能力。
▼
第一个假单胞菌基因组测序于 2000 年,有 630 万个碱基对,是当时最大的细菌基因组测序。基因组包含大量参与分解代谢、运输、流出、运动和信号响应调节的基因。
事实上,铜绿假单胞菌基因组中超过 8% 的基因被认为参与了调控,这远远超过了在任何其他细菌基因组中观察到的百分比。很明显,假单胞菌成功的关键在于它可以表达其基因的可塑性,这是由多层监管复杂性提供的。

Sem Photograph by David M. Phillips
自 2000 年以来,已测序的 1000 多个假单胞菌基因组中的绝大多数都是铜绿假单胞菌的临床菌株。
总的来说,铜绿假单胞菌基因组的主要部分(约 4000 个基因)在所有菌株中都是保守的,代表了“核心基因组”。多达另外 20% 的基因驻留在共同代表“附属基因组”的基因组岛上。正是这个附属基因组赋予了铜绿假单胞菌的可塑性,并包含许多参与代谢、毒力和抗生素抗性的基因。
由于已在测序分离株的附属区域鉴定出大约 10 000 个独特基因,因此估计铜绿假单胞菌泛基因组可以接近甚至超过 100 000 个基因,这意味着假单胞菌这一物种的遗传库将远远超过人类。
该属的分类学多年来一直存在争议,因为许多最初包含在假单胞菌属中的细菌类群已被重新归类为其他属或来自不同类别的变形杆菌的物种。
可以根据16S rRNA、gyrB、rpoB和rpoD基因的序列分为三个谱系和13个群。
临床常见菌种主要包括:
• 铜绿假单胞菌(Pseudomonas aeruginosa)
• 斯氏假单胞菌(Pseudomonas stutzeri)
• 恶臭假单胞菌(Pseudomonas putida)
• 荧光假单胞菌(Pseudomonas fluorescens)
• 门多萨假单胞菌(Pseudomonas mendocina)
• 产碱假单胞菌(Pseudomonas alcaligenes)
• 假产碱假单胞菌(Pseudomonas pseudoalcaligenes)
其它菌种见附录一
致病性菌株见附录二
▼
详见附录三
此外,铁是细菌必需且稀缺的营养素,也是重要的信号因子。铜绿假单胞菌已经开发出多种铁吸收系统来隔离足够的铁以维持其生存,在毒力因子的释放和生物膜的形成中具有重要的调节作用。
▼
假单胞菌适应进化的特性对人类既有益又有害。
——有益
一方面,我们利用假单胞菌的力量进行生物修复和生物防治。例如,荧光假单胞菌和P. protegens已被证明在害虫控制和作物保护方面特别成功,它们被认为可以战胜和/或对抗植物病原体。
假单胞菌的分解代谢能力也被用于杀虫剂、重金属和碳氢化合物(例如石油泄漏)以及许多其他污染物的生物降解和/或解毒。
——有害
另一方面,某些种类的假单胞菌对植物和动物具有致病性,导致极难根除的感染。例如,铜绿假单胞菌是世界范围内医院获得性感染最常见的原因之一,并在使用消毒剂和抗生素后存活下来。
它可以在许多解剖部位引起感染,包括皮肤、大脑、眼睛、耳朵、泌尿道、肺部等。免疫抑制的个体,尤其是那些有过度烧伤、囊性纤维化或中性粒细胞减少症的个体,尤其处于危险之中。(这在后一章节会详细介绍)
▼
一般非特异性症状(许多疾病中常见的症状)包括:
假单胞菌感染的症状取决于它们发生在身体的哪个部位。
假单胞菌(Pseudomonas)可引起以下器官感染:
耳朵(外耳炎或游泳者的耳朵):
疼痛、水肿(肿胀)、压痛和分泌物
皮肤:
假单胞菌通过压疮、烧伤和手术伤口或从血流进入菌血症患者的皮肤,还可能引起毛囊炎等。
具体可表现为:脓肿(感染肿块)、皮疹、脓液、皮肤溃疡、坏死组织(皮肤变黑,表明组织正在死亡)。
骨骼或关节:
受影响区域行动不便、关节或背部疼痛、肿胀、疼痛或发热。
这些通常是由于静脉内 (IV) 药物使用或感染的手术伤口或受伤造成的伤口细菌传播到皮肤、骨骼和关节的结果。
伤口:
伤口有分泌物或结痂(通常是黄绿色),可能有甜味或难闻的气味。
消化道:
阑尾炎(由于治疗导致白细胞低的人发生肠道感染)腹痛、腹胀和腹泻的症状。
肺:
假单胞菌可能会导致严重的肺炎,通常发生在住院、已经生病和免疫功能低下的人身上。它可引起囊性纤维化患者的慢性肺部感染(遗传病),支气管扩张(气道永久性扩张),或慢性阻塞性肺病。
具体可表现为:呼吸急促、胸痛、咳痰(伴有白色、黄色或绿色粘液)和充血。
泌尿道:
排尿疼痛、尿液难闻或有甜味、尿急、尿液浑浊。
因此,假单胞菌感染的特征在于广泛的症状。
▼
紧急情况下:呼吸急促、心率加快、脉搏微弱、意识模糊、发烧、发抖或剧烈疼痛是败血性休克的症状,是对血流感染的全身反应。
在极少数情况下,心脏瓣膜可能会受到感染,尤其是在静脉注射药物的人群中。
脏东西的细菌可以进入血液,会传播到心脏并感染心脏瓣膜,这种情况称为心内膜炎。
使用静脉注射药物的人出现新的杂音(心音异常)可能会怀疑与假单胞菌相关的心内膜炎。
▼
未经治疗、治疗不足或复发的假单胞菌感染会导致更广泛的感染,例如败血症,这可能是致命的。
更可能出现严重并发症的高危人群包括:
值得注意的是,假单胞菌属内的抗生素耐药性正在增长。这些细菌正在寻找新的方法来避免死于以往最强大有效的抗生素。
假单胞菌属与人体健康相关,其中一些菌株可以引起感染和疾病,如Pseudomonas aeruginosa 绿脓杆菌,又称铜绿假单胞菌,可以引起呼吸道、泌尿道、皮肤感染。虽然不是正常口腔菌群的成员,但在口腔疾病和牙齿矫正治疗中可能会富集。
▼
假单胞菌属不被认为是肠道微生物群落的典型成员,肠道定植假单胞菌属与发展肺部感染和死亡风险增加有关。
假单胞菌属的过度生长可能会导致肠道炎症和系统性炎症反应。
假单胞菌属感染会引起机体的Th17细胞反应和系统性炎症反应,因此,干预Th17细胞免疫反应可能是缓解肺炎的有效方法。此外,假单胞菌属也可以通过产生GABA来影响蛋白质代谢和细胞稳态,从而影响宿主的摄食行为。
▼
假单胞菌属与其他菌属如大肠杆菌、沙门氏菌等有关,与肠道微生物群的平衡和健康密切相关。
假单胞菌属与其他菌属如葡萄球菌属和普雷沃氏菌属等,共同存在于囊性纤维化患者的肺部感染中。
假单胞菌属与其他菌属如不动杆菌属(Acinetobacter)等一起被发现在冷鲜乳中,产生脂肪酶和蛋白酶,参与乳制品和肉类的变质。
假单胞菌属与其他菌属如Bacillus、Enterobacter、Enterococcus等一起参与了肠-肺轴的微生物和免疫调节,影响呼吸道疾病的发生和发展。
▼
与假单胞菌属相关的疾病包括:免疫系统受损患者的感染和医疗器械相关感染等。免疫抑制个体和住院患者的铜绿假单胞菌肠道携带率显著升高,因此感染和抗生素相关性腹泻的风险增加。
研究表明,假单胞菌属与肠道屏障功能障碍和感染有关,可能导致败血症和多器官功能障碍综合征。假单胞菌属与肺部疾病也有关联,如支气管扩张症。
肺部疾病
研究发现,肺部假单胞菌属感染会影响肠道菌群,而通过调节肠道菌群可以影响肺部疾病的治疗效果。
肠道定植通常先于肺部感染,并且相同菌株经常在肠道和肺部中发现,表明肠道充当了假单胞菌属的储存库,可以传播到肺部和其他感染部位。
肠道微生物群的免疫调节对呼吸道疾病的发生有影响,肠道共生菌通过屏障效应提供菌群抵抗力,从而保护肠道生态环境不被改变。
以肺部假单胞菌、肠道拟杆菌和肠道酵母菌为特征的高肠-肺相互作用集群,与加重的恶化,整体支气管扩张严重相关。
炎症性肠病(IBD)
假单胞菌属与炎症性肠病的发生也有关系。
研究表明,IBD患者肠道中假单胞菌属的种类和数量与非IBD患者存在显著差异。
其中,克罗恩患者中假单胞菌属的检出率明显高于非IBD患者。此外,假单胞菌属的多样性在活动性IBD患者中也明显降低。这些结果表明假单胞菌属可能与IBD的发病机制有关。具体而言,假单胞菌属可能通过影响肠道菌群的平衡和多样性,导致肠道免疫系统的异常反应,从而引发IBD的发生。
因此,调节肠道菌群可能是预防和治疗肠道疾病的一种新策略。
▼
铜绿假单胞菌(Pseudomonas aeruginosa)是一种普遍存在的革兰氏阴性细菌,存在于许多不同的环境中,包括不同的生命来源,如动物和人类。
铜绿假单胞菌能够以最低的营养需求生存并耐受各种身体条件,这使得该生物体能够在社区和医院环境中持续存在。
图源:EYE OF SCIENCE / SCIENCE PHOTO LIBRARY
引发感染
铜绿假单胞菌是一种机会性病原体,这意味着它利用宿主防御系统中的一些漏洞来引发感染。
铜绿假单胞菌是世界范围内医院感染的罪魁祸首,其医学和兽医重要性日益增加,并且主要在免疫系统受损的患者中引起感染。
这种细菌几乎从不感染未受损的组织,但如果组织防御以某种方式受损,它就会发起攻击。
铜绿假单胞菌可能引起尿路感染、呼吸系统感染、皮炎、软组织感染、菌血症、骨和关节感染、胃肠道感染和各种全身感染,特别是在严重烧伤患者和免疫抑制的癌症和艾滋病患者中。
在免疫缺陷的人群中,如严重烧伤患者、人工呼吸机使用者和支气管扩张症患者。
在囊性纤维化患者中,铜绿假单胞菌是慢性肺部感染的主要来源,导致患者的发病率和死亡率增加。
铜绿假单胞菌感染会导致肺部炎症和肺损伤,引起细胞浸润和肺泡水肿。在肺移植患者中,铜绿假单胞菌的分离率更高,但与非囊性纤维化肺移植患者相比,死亡率没有增加。铜绿假单胞菌感染会导致肺血管通透性增加,从而引起肺水肿。
耐药性
铜绿假单胞菌显示出对抗生素的高内在耐药性,并具有获得新耐药机制的非凡能力。由于临床治疗选择有限,多重耐药 (MDR) 铜绿假单胞菌分离株的流行率不断上升是一个全球性健康问题。
对来自西班牙两个不同地区的儿童粪便样本中铜绿假单胞菌的发生率、抗菌素耐药性表型、毒力基因型和遗传谱系表征数据显示:从1,443 个粪便样本中,有72份分离分离出来铜绿假单胞菌,并检测到低水平的抗菌素耐药性:头孢他啶 (8%)、头孢吡肟 (7%)、氨曲南 (7%)、庆大霉素 (3%)、环丙沙星 (1%) 和亚胺培南 (1%);对美罗培南、阿米卡星、妥布霉素、左氧氟沙星和粘菌素敏感,此外还发现了四种多重耐药菌株。
定植
人类粪便样本中的定植率范围为 2.6% 至 24 %。关于铜绿假单胞菌分离株在健康人粪便中定殖的数据很少。
其他研究表明,肠易激综合征患者的小肠和粪便样本中铜绿假单胞菌的定植率很高。迄今为止,关于铜绿假单胞菌分离株在健康人粪便中的定植及其抗菌素耐药性和毒力特征的数据相对较少。
尽管肠道微生物群作为抵御肠道病原体的屏障,但铜绿假单胞菌通过产生一系列令人印象深刻的毒力因子,克服了肠道微生物群和先天免疫系统介导的定植抵抗。
铜绿假单胞菌凝集素 LecA 和 LecB 以及细胞外附属物,如鞭毛和菌毛,在细菌附着到宿主上起着重要作用,并与粘附有关。
此外,铜绿假单胞菌携带属于双伙伴分泌(TPS)家族(也称为Vb型分泌系统,T5bSS)的大型蛋白质系统,TPS系统主要由分泌的TpsA效应蛋白及其TpsB伙伴转运蛋白组成。TpsA1 (CdiA2, 573 kDa) 和 TpsA2 (CdiA1, 361 kDa) 通过接触依赖性生长抑制系统 (CDI) 以及粘附和生物膜形成促进细菌竞争。
因此 TPS 系统效应子被指定为对革兰氏菌有益的主要毒力决定因素阴性病原体。
▼
铜绿假单胞菌等的特殊能力引起如此多样化感染的假单胞菌属是因为它们能够产生名副其实的毒力因子库,包括毒素、生物膜、蛋白酶和溶血素。
考虑到铜绿假单胞菌的医学重要性,假单胞菌领域的大部分研究工作都致力于了解影响这些毒力因子释放的调节、生物合成和环境因素。
此外,它们还可以获得赋予进一步抗菌素耐药性的移动遗传元件;因此,可以鉴定出泛耐药菌株。
▼
假单胞菌感知和适应环境的能力导致了一系列活动,例如许多酶和其他生物分子的分泌。虽然这些毒素、降解酶和抗菌剂可以将假单胞菌的环境转变为理想的微生物生态位,但它们可能对植物和动物宿主造成严重后果。例如,绿脓素是一种蓝/绿色色素毒素,它赋予铜绿假单胞菌培养物特有的颜色,并作为一种抗菌剂可以杀死竞争性微生物。然而,它也会破坏真核细胞过程,这会对人类细胞产生不利影响。
▼
过去几年的工作提供的证据表明,群体感应是一种通用的调节机制,它允许细菌以种群密度依赖的方式发起统一、协调的反应,以完成即使不是不可能,也很难完成的任务。
群体感应系统广泛存在于假单胞菌中,人类机会性病原体铜绿假单胞菌,属于研究最广泛的细胞间通讯系统。在这种生物体中,群体感应非常复杂,由两个相互关联的 N-酰基高丝氨酸内酯 (AHL) 依赖性调节回路组成,这些调节回路由非 AHL 相关信号分子和许多调节剂进一步调制,这些调节剂在转录过程中起作用和转录后水平。这种遗传复杂性可能是导致铜绿假单胞菌具有巨大环境多样性的关键因素之一。
过去几年的工作表明,群体感应对于一系列毒力因子的表达以及铜绿假单胞菌中生物膜的形成至关重要,因此代表了设计用于治疗铜绿假单胞菌的新型药物的有吸引力的目标感染。
此外,细胞间通讯能力也在许多额外的假单胞菌中得到证实。铜绿假单胞菌,因此可以用于设计新型药物来治疗铜绿假单胞菌感染。
长时间住院、创伤ICU住院、消毒干预等与肠道假单胞菌过度生长有关,而TAP暴露与抗菌剂干预则与肠道假单胞菌过度生长有负相关。
注:TAP的暴露是指肠道上皮细胞上的TAP暴露在肠道腔中,从而使肠道内的微生物可以通过TAP进入肠道上皮细胞内部。一些研究表明,TAP的暴露与肠道微生物群失调和某些疾病(如炎症性肠病)有关。而抗菌剂干预可以抑制肠道内细菌的生长,从而减少TAP的暴露,因此与假单胞菌肠道过度生长有负相关。
对于免疫功能低下的患者,应特别注意水的纯度,有些水中可能含有假单胞菌等细菌。
研究发现用海洋益生元岩藻依聚糖补充饮食可以通过抑制分泌性毒力因子 (TpsA/CdiA) 与粘蛋白的相互作用并促进有益种群的生长,从而介导铜绿假单胞菌从肠道菌群失调和去定植中的早期恢复。
芪归饮是一种用于治疗铜绿假单胞菌感染的中药。在门和属水平上显著抑制脱铁杆菌和粘孢子菌的过度积累。鉴定了11种潜在的代谢产物,这些代谢产物在铜绿假单胞菌感染中异常表达,服用芪归饮显著逆转。
此外,部分研究表明以下物质也可以抑制肠道内的假单胞菌属。
假单胞菌属相比于其它细菌引起人们广泛的研究兴趣,该属不仅包括人类病原体铜绿假单胞菌,还包括一系列与植物致病性、生物修复和环境微生物学相关的其他重要物种。
假单胞菌被广泛用作模型原型细菌,用于研究宿主-病原体相互作用、细胞-细胞通讯系统、进化生物学、基因调控和代谢网络、分泌系统、抗生素(耐药性)、生物修复、生物膜、细菌基因组学以及与微生物学和分子生物学广泛相关的许多其他主题。
假单胞菌属因其复杂的信号和调节系统,特别是细胞密度依赖性群体感应而被广泛研究。未来只有了解复杂调节回路中涉及的机制,才能确定新型抗菌方法的可能目标。铜绿假单胞菌中的锌、铁稳态是替代抗菌策略的一个有希望的目标。
由于其复杂性和适应性,它存在于无数的环境和临床生态位中,以及它在不同层面(种内、种间和与其宿主)相互作用的能力,假单胞菌属很可能在未来许多年内吸引更多研究人员。我们期待未来有更好的解决假单胞菌属,尤其人类病原体铜绿假单胞菌的抗菌策略。
附录一:
其他菌种
附录二:
致病菌菌株
附录三:
可能参与的代谢

谷禾健康

感染是人类面临的健康威胁之一。各种病原体,如细菌、病毒、真菌、寄生虫等,存在于我们日常接触的环境、物品、食物等中。一些常见的感染病例包括感冒、流感、腹泻、组织器官或血液感染等,在全球范围内广泛传播。这些疾病的传播方式多样,可能通过空气、水源、接触和食物等途径传播。
感染的症状包括发热、头痛、咳嗽、呕吐、腹泻等,严重感染可能导致器官衰竭、败血症等并发症。感染的原因包括接触感染源、免疫力下降、不良生活习惯等,易感人群有免疫力低下者、老年人、慢性疾病患者等。
研究发现,肠道菌群与感染也有密切关系。肠道菌群有助于消化、吸收营养物质,同时还调节人体免疫系统的功能。但如果肠道菌群失衡,会导致多种疾病,包括感染。
本文我们来了解一下关于感染的种类,症状,致病途径,易感因素,免疫系统应对病原体的三大防线,肠道菌群失衡与免疫系统的关联,以及肠道菌群检测在临床感染案例中的应用。
▼
感染可以按多种不同方式分类。
按影响部位分为局部和全身的:
按影响持续时间分为急性和慢性的:
按病原体类型划分:
病毒、细菌、真菌、寄生虫等都是导致感染的不同类型的病原体。
► 细菌感染
细菌感染是由体内过量的有害细菌引起的。正常情况下,人体的免疫系统能够通过白细胞和吞噬细胞等机制对抗细菌感染。但如果人体的免疫系统出现异常或受到外界因素干扰,细菌就可能趁虚而入,导致细菌感染的发生。
常见的细菌感染包括:肺炎、腹泻、皮肤感染、脑膜炎、膀胱炎、中耳炎等。
► 真菌感染
真菌感染多为继发性感染,由机会致病性真菌引起,特别是深部真菌感染多是由于各种诱因使机体免疫功能显著下降所致。
真菌是一类与细菌和病毒不同的微生物,它们包括霉菌、酵母菌等,常见的真菌感染有白色念珠菌感染、曲霉菌感染等。
► 病毒感染
病毒感染与细菌感染不同,因为病毒本身并不能独立存在,只有当它侵入人体细胞后才能产生病毒性病变。当病毒进入细胞时,它会留下迫使细胞复制的遗传物质。
当细胞死亡时,它会释放新的病毒感染其他细胞。并非所有的病毒都会破坏细胞——有些病毒会改变细胞的功能。
病毒会导致多种疾病,包括普通感冒、流感、登革热、水痘、艾滋病等。
► 寄生虫感染
寄生虫是需要以其他生物为食才能生存的生物。有些寄生虫不会对人产生明显影响,而有些寄生虫会生长、复制并侵入器官系统。
常见的寄生虫感染有阿米巴病、血吸虫病、肠道蠕虫感染等。
▼
无论何时发生感染,身体的一线反应都是炎症性的。
炎症是身体抵御疾病的方式,同时促进受影响组织的愈合。炎症的主要症状:红、肿、热、痛、组织功能的暂时丧失。
在感染期间,由于潜在的炎症反应,人们通常会出现非特异性的全身症状。症状和严重程度可能因受影响的器官系统而异,但可能包括:
▼
► 病史采集
询问患者或病人家属发病情况、既往病史、服药史、过敏史、流行病学史等。
► 体格检查
体温、呼吸、心率、血压、身体部位肿胀、红肿、局域性压痛、淋巴结肿大等都是感染体征,可以作为诊断和判断感染病情严重程度的参考指标。
► 实验室检查(抽血、尿检等)
通过血液、尿液、呼吸道分泌物、伤口分泌物等样本的化验检查可以更直接地确定病原体的种类和数量,以及感染程度的严重程度。
► 影像学检查(做CT等)
如X射线、CT、MRI等技术,可以帮助医生了解感染的程度以及可能受影响的部位,从而确定感染的类型和严重程度。影像学检查为临床医生在明确诊断之前的经验性治疗提供线索。
► 快速诊断测试(做核酸等)
如血清学检测、ELISA试剂盒、PCR技术等,可以在短时间内快速准确地检测出患者血清中的病原体抗体、抗原以及核酸等,提供对感染病原体的敏感性和特异性的快速诊断。
▼
呼 吸 道
普通感冒的另一个名称是上呼吸道感染。鼻病毒是引起感冒的最常见病毒。其他通过空气传播的传染病也以这种方式感染。
感冒和流感通常不会直接影响肺部,但它们可能会导致继发性细菌感染——肺炎。
消 化 道
如果不小心吃了被细菌或病毒感染的食物、饮料或水、或是农产品在种植或运输过程中接触了污染物,或者过期变质的食物等,病原体也一并被吞入,感染胃部或肠道,有时表现为腹泻和/或呕吐。一个常见的例子是细菌性肠胃炎,也称为食物中毒。
泌尿生殖系统
病原体也可以通过泌尿系统进入人体,如尿路感染,或生殖系统,如性传播疾病。传染源可能停留在局部,也可能进入血液。例如,性传播疾病最常感染生殖器,而艾滋病毒通过体液携带,也可以通过唾液、精液或血液传播。
皮 肤 接 触
皮肤的功能之一是充当抵御感染的屏障。但如果有割伤、抓伤、虫咬或任何类型的开放性伤口,皮肤阻挡不住,病原体可能会进入血液。如:蜂窝组织炎、脓疱疮等都是通过皮肤接触开始的常见感染。
▼
无论来源如何,任何人都可能感染。但有些人更容易感染,或感染并发症的风险增加,例如:
以上我们可以看到,那些免疫系统功能低下的人更容易发生感染。那么当病原体入侵时,免疫系统正常运作会发生什么呢?
下一章节我们来了解一下,默默帮助我们抵抗病原体的——免疫系统。
前面章节我们知道感染是微生物进入人体并造成伤害的结果。引起感染的生物体很多,包括病毒、细菌、真菌、寄生虫等。
当它们试图攻击的时候,免疫系统是如何运作的?
第一道防线——物理屏障
正常情况下,皮肤、呼吸道等组成的物理屏障在保护你,一般只有水分、气体等小分子物质进出,细菌、病毒难以入侵。
然而,也许你只是摔了一跤擦破了皮,也许只是水果刀不小心划伤了…皮肤作为免疫系统的第一道防线破了。
这时候细菌就开始争先恐后进入体内,不断繁殖,增强武力值。
第二道防线——先天性免疫防线
(广泛执行任务)
巨噬细胞——以吞制敌
巨噬细胞开始介入这场搏斗,它往往通过吞噬的方式以一敌百,不管三七二十一先吞下将其困在膜泡里,利用酶等物质分解杀敌,如果细菌还来不及生长繁殖就被吞完了,这场搏斗也就随之终止了。该过程可能引发局部的炎症,表现为红,肿,痛,热等。

但如果一直持续下去,巨噬细胞也可能累了,它开始发出紧急信号,召集后援团…
中性粒细胞——不分敌我直接干
中性粒细胞出现了,它离开了原本在血液中巡逻的路线,第一时间冲过来加入,武力值开始飙升,健康细胞也会被它误杀,它也会给细菌设置障碍,从而困住细菌将其干掉。
它们也会“适可而止、功成身退”,几天后自杀,不带来更多伤害。
如果这还不足以击退,发出的求救信号越来越多,当中枢神经检测到,下丘脑便会开始调节体温上升,也就是发烧。较高的体温可以抑制病原体的活性与繁殖。
以上就是人体免疫系统的第二道防线,也就是先天性免疫(非特异性免疫)。
如果第二道防线没有阻止入侵者呢?
最终对抗会发生在第三道防线,也就是后天性免疫。
第三道防线——后天性免疫防线
(形成记忆,长期保护)
后天性免疫系统是一个成长、学习和经验的累积,学习和记忆着病原体的模式,进而识别并产生特异性的免疫应答。具有极强的打击和防守能力,可以持续有效地保护我们的身体。

这时候,树突状细胞登场了。
树突状细胞——先天性免疫和后天性免疫之间的桥梁
树突状细胞开始手撕敌军,把病原体碎片暴露在自己的细胞膜上,带着病原体的信息去淋巴结,这里有很多T细胞待激活,当T细胞出生时会经过一个训练,之后会配备一个装备。
树突状细胞要寻找的正是可以和病原体表面特征蛋白结合的那个T细胞。
T细胞——等待激活,加入战斗
找到后该T细胞会被瞬间激活,然后开始分裂复制,并分化出三种T细胞,分别为:
B细胞——产生抗体,继续奋斗
当辅助T细胞过去找到B细胞识别结合之后,B细胞瞬间激活,大量复制,产生抗体。
抗体的出现意味着胜利的曙光,抗体表面有多个结合点,可以把多个病原体结合在一起,病原体捆在一起行动不便,失去活力。其他免疫细胞过来可以轻松把它们干掉。
抗体还可以拉着病原体连接杀伤细胞,将其消灭。
记忆T细胞和B细胞——学会对付,默默守护
在清除病原体后,分化出来的大量免疫细胞完成任务,会程序性自杀,防止继续占用资源。会有一小部分留下来,也就是记忆T细胞和记忆B细胞,等待病原体下次来的时候直接杀敌。
以上就是免疫系统的三大防线,它们的存在为人体提供了多重保护。正是因为它们的不懈努力,在感染发生后我们也会挺过来。
研究发现肠道菌群与人体免疫功能以及许多疾病的发生密切相关。
肠道菌群失衡如何与感染相关?肠道菌群与免疫系统之间存在怎样的关联?
下一章节我们继续来看。
肠道菌群中某些微生物和宿主的免疫系统之间存在相互作用,肠道菌群能够影响和调节上述第三大防线中B细胞和T细胞的活性,通过调节免疫细胞的行为,促进抗原递呈细胞的活化和作用,影响细胞因子及其受体的表达等。
肠道微生物代谢产物能够影响免疫平衡。例如,肠道菌群可以通过代谢产生短链脂肪酸、色氨酸胆汁酸等代谢产物,这些代谢产物能够通过激活特定的神经元和免疫细胞来调节免疫反应,从而维持肠道黏膜的免疫稳态。
短链脂肪酸可能具有抗炎和免疫调节作用,当然短链脂肪酸只是一部分,因为除了产生的酶和其他代谢物之外,细菌本身的成分,包括脂多糖、细胞囊碳水化合物和其他内毒素,也可能被释放并对宿主产生二次影响。
肠道微生物群衍生的代谢物丙酸盐(一种短链脂肪酸)水平降低可能导致 Th17/Treg 细胞分化的病理失衡。
一些菌群代谢物,如 4-甲基苯甲酸、3,4-二甲基苯甲酸、己酸和庚酸等,已被证明会加剧结肠上皮细胞的损伤,从而影响免疫。
在肠道菌群失调的情况下,某些有益菌可能会缺乏消化酶等必要的代谢产物和物质,使机体免疫系统无法及时有效地对病原体进行攻击。
许多致病菌与肠道屏障的特定元素相互作用。一些有益的菌群可以激活上皮细胞,加强它们的黏附能力和抗病原性,从而提高肠黏膜的免疫防御能力。
致病菌和抗生素的使用可能会通过增强黏液降解,或抑制正常的共生黏液生成触发器来扰乱肠道黏液层,从而破坏肠道屏障,也就是我们常说的“肠漏”。
肠道屏障的破坏也会诱发体内炎症:与微生物相关的代谢物离开肠道并进入血液。作为回应,身体会产生细胞因子和其他介质,有效地启动炎症反应。
肠道上皮组织内的细胞将细菌代谢物传递给免疫细胞,从而在局部和全身范围内促进炎症。这种情况的持续存在可能导致亚急性或慢性炎症,随后可能会导致炎症性肠病、糖尿病或心血管疾病等疾病的发展。
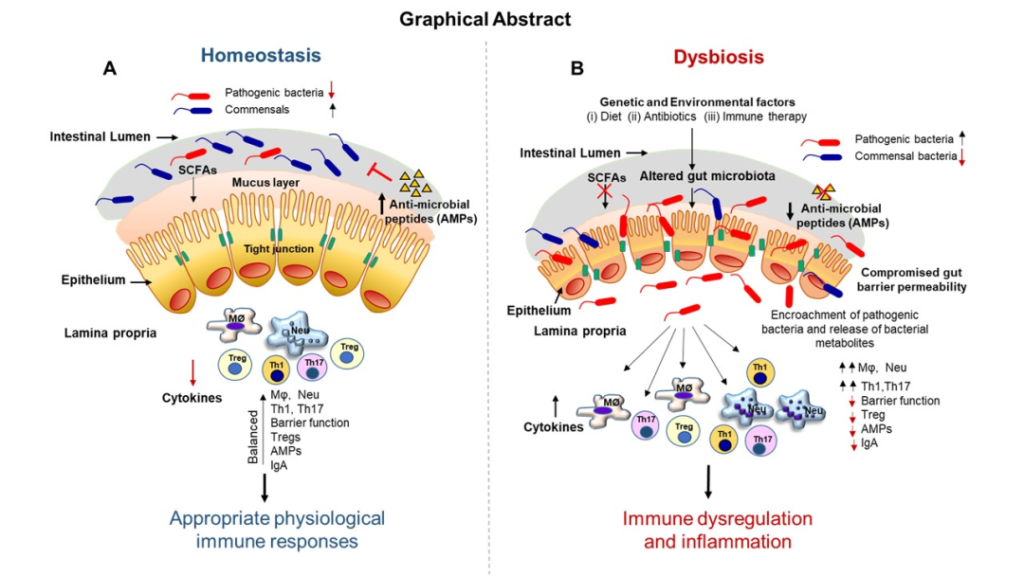
doi.org/10.3390/biomedicines11020294
C 反应蛋白 (CRP) 是一种下游炎症标志物,可以通过特定肠道微生物的抗炎代谢产物的作用下调。当巨噬细胞和 T 细胞分泌白细胞介素 IL-6 时,该分子随后以高水平释放到循环中。我们在肠道菌群检测报告中也可以用CRP来辅助判断炎症状况。
炎症标志物是复杂的双向调节机制的组成部分,并且参与多个重叠过程,与感染和体内稳态的维持有关。
由此,肠道菌群变化对于免疫功能强弱具有重要指示作用,那么我们可以结合肠道菌群检测中的实际案例,来了解更多关于肠道菌群和感染的关联。
在谷禾肠道菌群健康检测中,有许多关于感染的病例,我们选取一例来详细看一下。
年初,一名7岁女孩入院,经诊断后发现患有:
菌血症是一种严重的感染症状,通常由细菌或真菌等微生物感染引起。菌血症分为原发性和继发性。
只有大约20%病例是原发性菌血症,也就是说病原体最初进入血液系统并引起血流感染,例如创伤、手术、注射等导致血管受损直接感染。
其余大多数都是继发性菌血症,也就是在其他部位先感染,未能及时处理,致使病原菌进入血液系统引起继发性菌血症。
常见的菌血症类型包括葡萄球菌菌血症,肺炎链球菌菌血症,鲍曼不动杆菌菌血症,克雷伯菌菌血症,真菌菌血症等,都是相应的致病菌感染引起的。
入ICU后,出现腹胀问题,且愈发严重 ⇊
肠道菌群检测结果显示:
1. 患儿肠道菌群严重失调
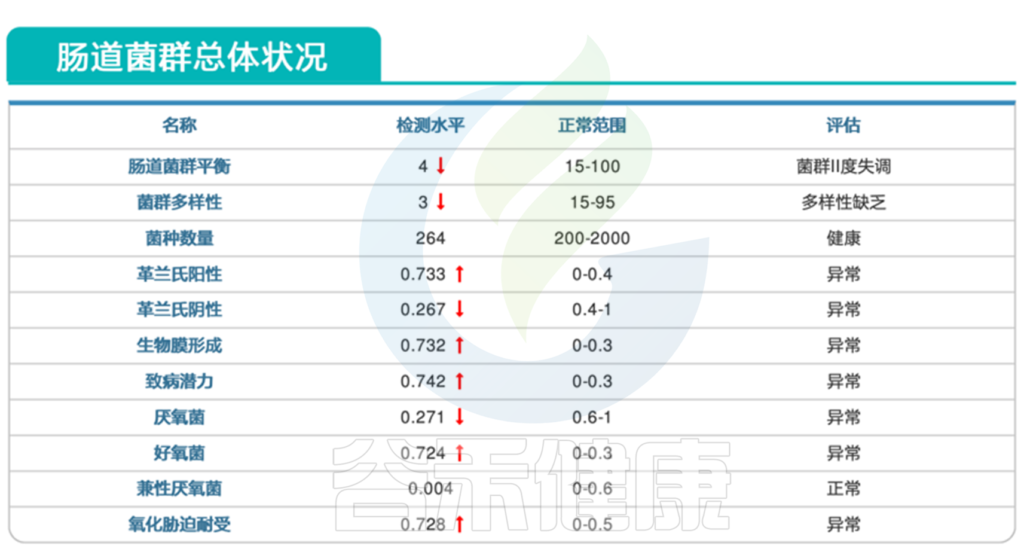
<来源:谷禾健康肠道菌群检测数据库>
显然,该患者的肠道菌群失调,多项指标均显示异常。
从报告的肠道菌群总体状况这部分,我们可以看到,菌群多样性下降,其中革兰氏阳性菌明显偏高,革兰氏阳性菌包括葡萄球菌,链球菌,梭菌等,可能与多种感染相关。同时报告中致病潜力这项指标也是大幅上升。
2. 存在艰难梭菌携带超标、酵母菌感染、不动杆菌超标严重(感染)情况
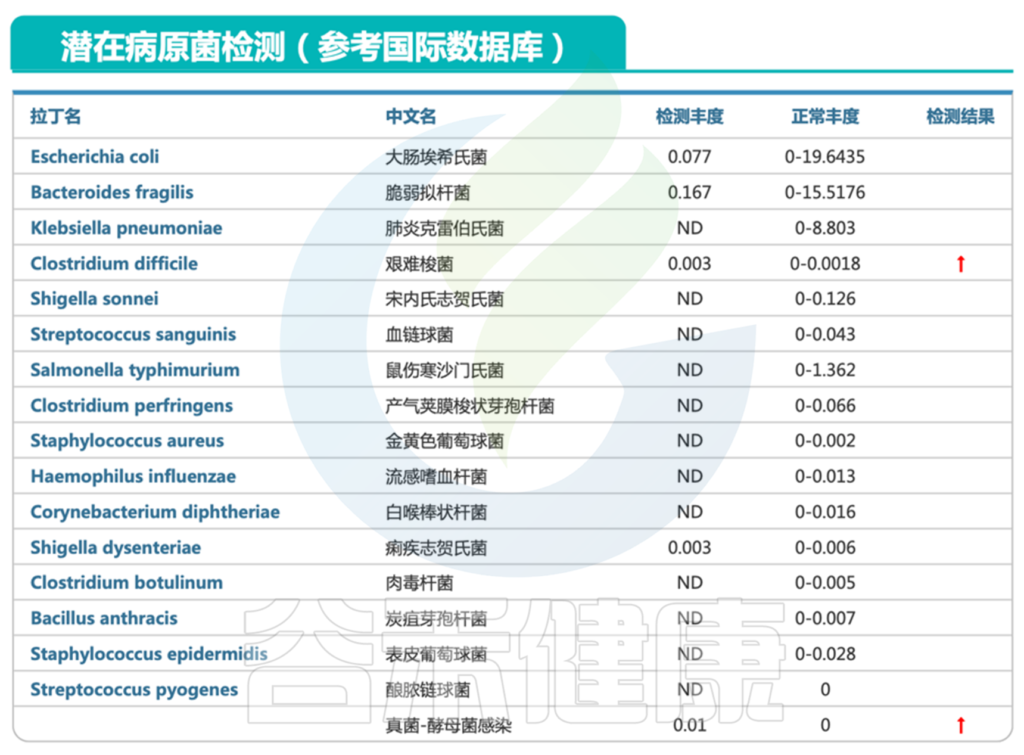
<来源:谷禾健康肠道菌群检测数据库>
艰难梭菌超标 ↑
艰难梭菌是一种常见的肠道病原菌。它可以引起广泛感染,症状最常见的是腹泻,伴随着腹部疼痛,刺痛,腹胀等不适,有些人还可能会出现发热,发冷,胃部压痛或疼痛,恶心呕吐的症状。
更严重的可能出现水样腹泻,心跳加快、体温升高、腹部绞痛、体重下降、腹部肿胀、肾功能衰竭等。
这与患儿的某些症状相符,从肠道菌群检测中可以发现艰难梭菌感染。
艰难梭菌感染与免疫功能有怎样的关联?
艰难梭菌毒素通过炎症小体和TLR4、TLR5和含有核苷酸结合寡聚结构域的蛋白1(NOD1)信号通路刺激先天免疫细胞,随后产生大量促炎细胞因子(如IL-12、IL-1β、IL-18、IFN-γ和TNFα)和趋化因子(MIP-1a、MIP-2、IL-8),导致粘膜通透性增加、肥大细胞脱颗粒、上皮细胞死亡和中性粒细胞浸润,引发免疫功能低下。
这与患者一开始诊断中中性粒细胞功能低下相符。
艰难梭菌感染通常与抗生素使用相关
通过某些抗生素,特别是克林霉素,可能会去除大量肠道中的有益细菌,使艰难梭菌得以繁殖,导致结肠炎和随后的腹泻。
真菌——酵母菌感染 ↑
报告中可以看到酵母菌超标,存在感染。这与入院时的粪便培养中的真菌感染相吻合。
真菌感染可能通过影响肠道微生物群和免疫反应,促进病原体的定殖和发展。
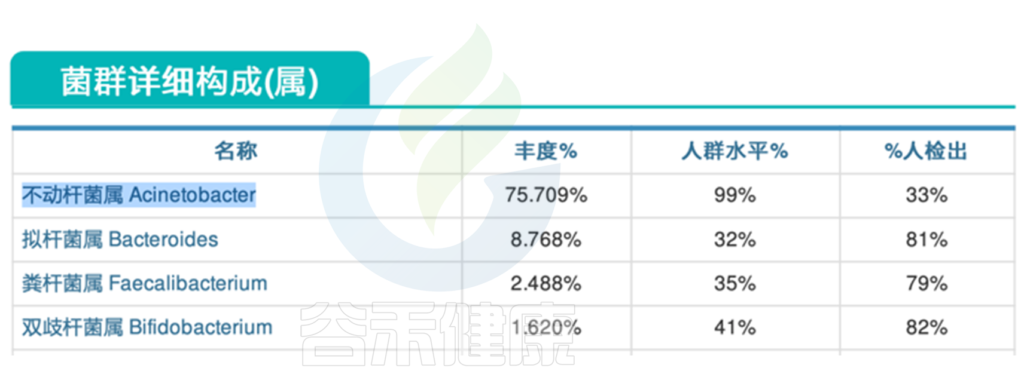
<来源:谷禾健康肠道菌群检测数据库>
不动杆菌属(75.709%)占比过多 ↑
从报告中的菌群详细构成这部分可以看到,不动杆菌属的丰度竟然超过75%,严重超标。
这种情况下,不动杆菌属严重挤压了其他肠道核心菌属的生存空间,有益菌生长受限,菌群多样性下降,肠道菌群整体失调。
不动杆菌属Acinetobacter是一类革兰氏阴性菌,该菌属于常见的医院感染病原体之一。不动杆菌属感染常见的病症包括呼吸道感染、泌尿道感染、创伤感染等,严重情况下还可能导致败血症。
不动杆菌属对常见抗生素经常会产生抗药性。这里不动杆菌属丰度较高可能与抗生素使用较多相关。
结合该患者的情况,一开始入院时的组合抗生素一周未见好转,可能与不动杆菌属占比过多相关,对许多常见的抗生素已经产生耐药性,且免疫功能低下,因此难以好转。
针对上述情况,临床考虑口服抗生素,针对消化道微生物进行抗感染治疗。
初步选定:口服利福昔明 + 甲硝唑治疗。2天未见明显改善。
后改为:口服万古霉素 + 阿莫西林治疗。当天腹胀症状明显缓解,3天治疗腹胀明显改善。
同时辅助康复新液+B族维生素+谷氨酰胺治疗。
用药两周后,患儿整体情况平稳,仍存在轻微腹胀⇊

肠道菌群复查提示菌群结构基本恢复正常。
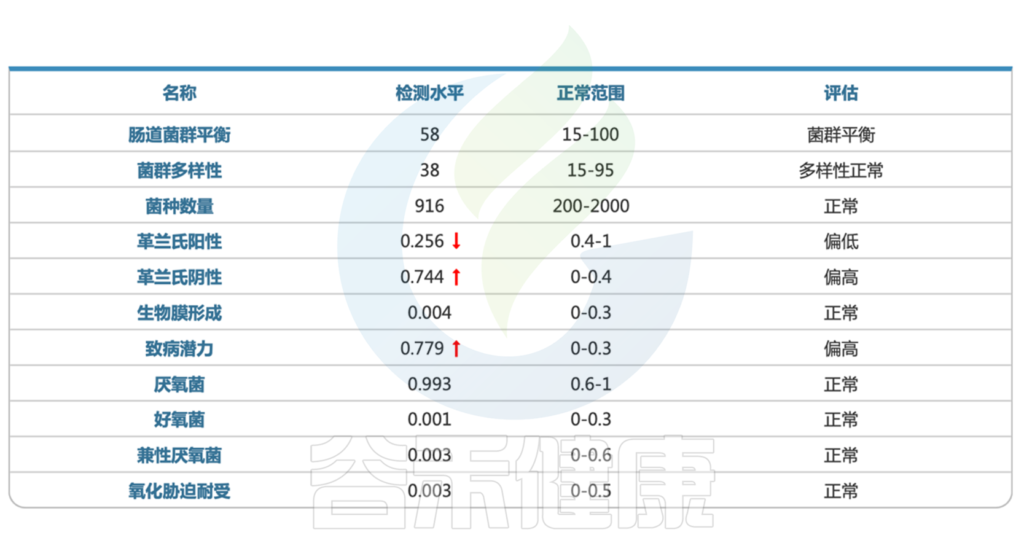
<来源:谷禾健康肠道菌群检测数据库>
一年后随访,该儿童身体已经好转,生活正常。
备注:临床中还有其它治疗药物等没有全部分享,主要分享了与菌群相关的治疗,仅做交流,不能作为治疗参考。
宿主感染与肠道菌群之间存在复杂的相互作用,免疫系统有两个分支,即先天性和适应性,它们协同工作以保护身体免受外部和内部威胁。这两个免疫系统分支都受到严格调节,肠道微生物群是其中的一个重要因素。
要揭开这其中的机制关联可能需要依靠营养学、流行病学、医学、免疫学和微生物学等多学科研究人员的共同努力。
不过从以上案例我们可以看到,肠道菌群健康检测在感染治疗中发挥重要价值。
辅助判别
在传统诊断中得知患者存在菌血症,真菌感染,但通过肠道菌群健康检测可以了解更多具体的感染信息,比如说艰难梭菌感染可能引起的腹胀等问题,不动杆菌属占比过大形成的抗生素耐药性。
除了致病菌超标等直观的信息之外,我们还可以判断整个肠道微生态的健康状况。肠道微生物群的失调可能导致肠道易感性增加,使得易感性疾病如艰难梭菌感染更容易发生。
辅助用药
针对这些具体信息可以使用药更加精确,而不是直接使用常见的组合抗生素。
用对抗生素对于恢复速度有很大提升,同时也避免了患者因使用不必要的抗生素产生耐药性。

谷禾健康

色氨酸(Tryptophan,简称 Try)是人体必需氨基酸,也是唯一含有吲哚结构的氨基酸,由食物尤其膳食蛋白质提供,是正常细胞稳态所必需的,是维持细胞生长和协调机体对环境和饮食线索的反应(其中色氨酸代谢物充当神经递质和信号分子)。
不同组织内的色氨酸代谢与许多生理功能有关:
在哺乳动物中,色氨酸是代谢物的生化前体,显著影响哺乳动物的生理机能,包括胃肠道功能、免疫力、新陈代谢和神经系统。色氨酸及其代谢物水平的失衡与广泛的人类病理学相关,包括抑郁症、精神分裂症、自身免疫、神经退化和癌症。同时它也是自身免疫、癌症、神经退行性或肠道疾病的一个非常有吸引力的治疗靶点。
本文将总结和讨论色氨酸及色氨酸代谢的生理和病理学作用,肠道中色氨酸代谢物的产生和调控、肠道菌群衍生的色氨酸代谢物在全身健康稳态中的作用、以及基于色氨酸代谢药物开发的巨大机遇和挑战。
/
/
▼
色氨酸是一种必需氨基酸,是体内许多重要分子的前体。
如果您读过我们很多文章,可能读到比较多是短链脂肪酸。短链脂肪酸(SCFA),尤其是丁酸盐,通常会在肠道菌群失调和慢性疾病的状态下耗尽。
但短链脂肪酸只是肠道代谢物的一大类之一。其他两类——色氨酸代谢物和胆汁酸,在维持肠道健康方面发挥着同样重要的作用。所以我们也将逐步关注和分享它们。
● 什么是色氨酸?
色氨酸是一种氨基酸——我们体内蛋白质的众多组成部分之一。
色氨酸是一种必需的芳香族氨基酸,由连接到吲哚基团 3 位的 β 碳组成。在 20 种常见的经典氨基酸中,色氨酸的分子量最大。
虽然色氨酸是蛋白质和细胞中含量最少的氨基酸,但它是大量微生物的生物合成前体和宿主代谢物。
大多数游离色氨酸通过犬尿氨酸 (Kyn) 途径 (KP) 或血清素途径降解为具有生物活性的化合物。
色氨酸分解代谢途径
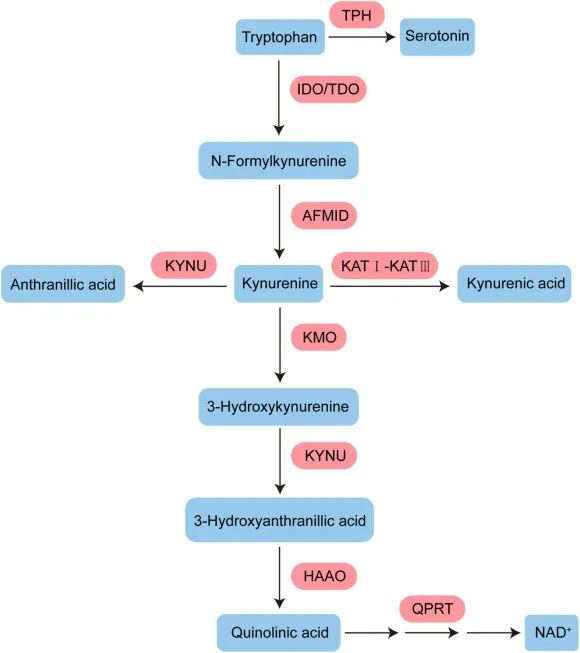
血清素途径产生血清素,可进一步转化为 N-乙酰血清素 (NAS) 和褪黑激素,后者对于昼夜节律调节和抗衰老至关重要。
肠道菌群对色氨酸的吸收很重要,限制和调节宿主细胞的使用。在此过程中生成吲哚衍生物,扩大了色氨酸分解代谢在不同组织器官中中的通讯作用。
色氨酸是体内许多其他化合物的重要前体,包括:
● 色氨酸 / 起源与生产
色氨酸是人体无法产生的必需氨基酸,必须通过饮食获取,主要来自动物或植物性蛋白质来源。
起源:酪蛋白分离
色氨酸是在 1900 年代初期从酪蛋白(一种在牛奶中发现的蛋白质)中分离出来后被发现的。几年后确定了它的分子结构。
释放:进入血液循环
小肠中膳食蛋白质的消化导致色氨酸的释放,色氨酸可以通过肠上皮细胞吸收并进入血液。色氨酸在血液中循环,主要与白蛋白结合,而在血液循环中只有 10-20% 的色氨酸是游离态。被吸收的色氨酸以其游离形式循环或与外周血流中的白蛋白结合。
据报道,健康献血者的总色氨酸平均血清水平为 73 ± 14.9 μmol/l 。
游离色氨酸的一个重要生理功能是对宿主蛋白质合成的贡献。
除了作为蛋白质合成的成分外,色氨酸还是生产多种重要生物活性物质的必需底物。例如,色氨酸是血清素合成(情绪相关)以及褪黑激素(睡眠相关)合成的底物。
全身和细胞色氨酸水平由食物摄入量、生物转化以及降解色氨酸的途径酶活性共同决定。
● 色氨酸的常见天然食物来源
乳制品、燕麦、香蕉、豆类、黑芝麻、李子干、金枪鱼、奶酪、面包、家禽、花生、黑巧克力、鱼肉、三文鱼、杏仁、南瓜和南瓜子等。
世界卫生组织将推荐的色氨酸摄入量设定为 4 毫克/千克/天,迄今为止,没有关于饮食中色氨酸过量的不良影响的报道。
注:含有色氨酸的食物对于制造激素血清素至关重要。但不应高估其影响。
一般来说,动物蛋白中的色氨酸含量往往高于植物蛋白。虽然色氨酸可以补充形式服用,但最好将其作为全食物中完整蛋白质的一部分。
● 为什么需要色氨酸?
色氨酸在体内的浓度是所有氨基酸中最低的,然而,色氨酸摄入量低与抑郁、焦虑、情绪低落、睡眠质量差、视觉认知能力下降以及学习和记忆受损有关。它还可能改变肠道微生物组并削弱肠道免疫力。
另一方面人们普遍认为色氨酸过量会导致困倦。比如在美国的感恩节食用大量火鸡,火鸡中的色氨酸含量很高,进食大餐会刺激胰岛素的产生,而胰岛素会清除血液中除色氨酸以外的所有氨基酸,会导致困倦。
/
/
▼
大量的数据表明色氨酸代谢的调节对环境条件很敏感,并且会影响生理和行为过程。
它因物种、细胞类型、诱导剂而异,并且可以通过组织之间的相互作用进行调节。
● 宿主色氨酸代谢
这里我们对色氨酸代谢先有个整体的认识:
色氨酸分解代谢主要两条通路:
▸犬尿氨酸通路占整体色氨酸降解的约95%
色氨酸 (TRP) 通过犬尿氨酸 (KYN) 通路 (KP) 的分解代谢,该通路占整体色氨酸降解的约 95%,形成主要最终产物 NAD+。
注:犬尿氨酸通路是炎症和免疫反应的重要参与者。
首先,色氨酸被转化为N-甲酰基-L-犬尿氨酸
该反应由三种限速酶之一催化:
注:这三种酶都是血红蛋白,并使用分子 O2作为共底物,这也使它们能够利用活性氧 (ROS) 并调节细胞内的氧化还原平衡。
IDO 和 TDO 酶在不同的组织中表达,暴露于不同的刺激物时被诱导,表明它们在健康和疾病中具有不同的功能。
TDO 在基础条件下催化色氨酸分解,而在免疫调节中具有关键作用的 IDO-1 受到多种刺激物的诱导和调节,例如炎症信号 。
进一步的,N-甲酰基-犬尿氨酸形式酰胺酶将 N-甲酰基-L-犬尿氨酸水解为 L-犬尿氨酸
成为三种具有不同氧化应激和器官毒性特性的替代代谢物:
最终在3-羟基邻氨基苯甲酸3,4-双加氧酶(3-HAAO)的催化下,进一步分解为喹啉酸、烟酰胺腺嘌呤二核苷酸(NAD+).
肝外色氨酸犬尿氨酸通路不提供所有必需的酶;因此,其中间代谢物及其特性在这些病症的发病机制和调节中变得至关重要(下图)。
注:在生理条件下,肝外通路仅占整体色氨酸降解的 5-10%.
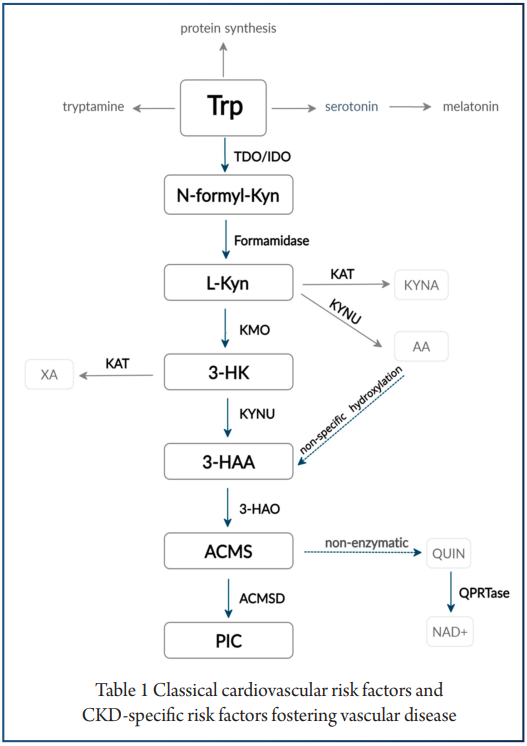
编辑
B 族维生素作为辅因子起着至关重要作用
KP 对B族维生素浓度的变化很敏感。维生素 B6(5′-磷酸吡哆醛,PLP)的活性形式影响犬尿氨酸酶 (KYNU) 和犬尿氨酸氨基转移酶 (KAT)。
维生素B6缺乏后,会影响色氨酸代谢。
▸ 大约不到5%的色氨酸会转化为5-羟色胺
通过色氨酸羟化酶(TPH)催化生成5-羟色胺(5-HT),也就是血清素。
doi.org/10.3389/fendo.2019.00158
以上是色氨酸代谢的两种主要途径。感染、压力和肠道菌群的变化都可以将色氨酸代谢从 5-HT的产生分流到犬尿氨酸途径,因此如抑郁症之类的病理变化,与人类的营养因素、压力和免疫功能有关。
简化的人类色氨酸的主要代谢途径
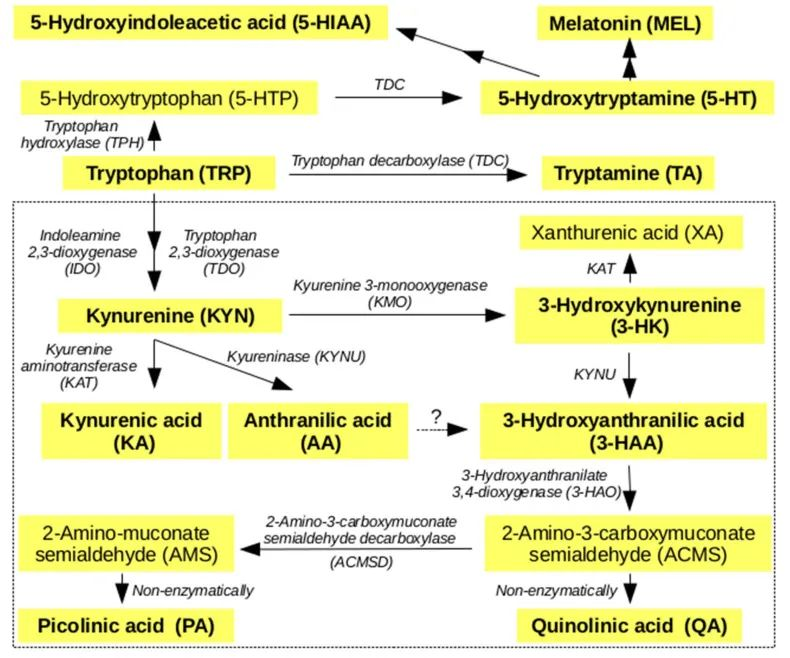
doi: 10.3390/metabo10050208.
● 色氨酸微生物代谢
肠道微生物将未吸收的 L-色氨酸 代谢成几个分子,如吲哚衍生物 [indole-3-aldehyde (IAld)、indole-3-acetic acid (IAA)、indole-3-propionic acid (IPA)、indole-3-acetaldehyde (IAAld)、吲哚-3-乳酸 (ILA) 和吲哚丙烯酸],还有色胺和粪臭素。
最近表明,其中一些分子不仅由微生物群合成,而且还通过 L-氨基酸氧化酶 (IL-4I1) 的作用由肿瘤细胞合成,代谢 L -色氨酸 转化为吲哚-3-丙酮酸,随后转化为 IAA、IAld 和 ILA,从而以 AhR 依赖性方式逃避免疫系统、存活和肿瘤运动。
AhR 信号是免疫反应屏障位点的重要组成部分。它通过作用于上皮更新、屏障完整性和许多免疫细胞类型(如上皮内淋巴细胞、T 辅助 (Th)17 细胞、先天性淋巴样细胞、巨噬细胞树突状细胞和中。
肠道环境的细菌色氨酸代谢
由于不同的微生物拥有不同的催化酶,需要两种以上的细菌相互合作才能从色氨酸中产生一种代谢物。与动物内源性色氨酸代谢相对简单的背景不同,人类肠道环境在细菌色氨酸代谢方面相对复杂。
肠道菌群通过各种代谢途径产生多种色氨酸代谢产物,例如:
肠道中微生物群相关的色氨酸代谢
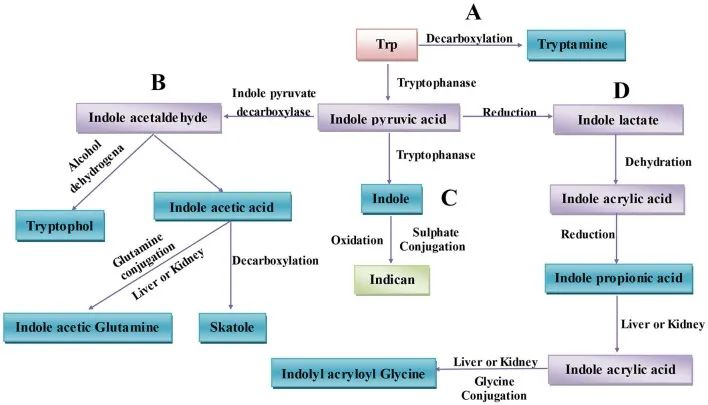
不同菌种可能存在相同代谢能力
比如:消化链球菌属的相同代谢功能可能是基于这些菌种拥有苯乳酸脱水酶基因簇,在下列菌群中也发现与它们产生IPA能力一致的同源基因簇:
不同菌种之间也存在一定代谢能力差异
比如:几种拟杆菌属和梭菌(Clostridium bartlettii)可以产生ILA和吲哚乙酸(IAA),而双歧杆菌属(Bifidobacterium spp.) 产生ILA 。
通过 5-HT、Kyn 和吲哚/AhR 途径的色氨酸代谢途径
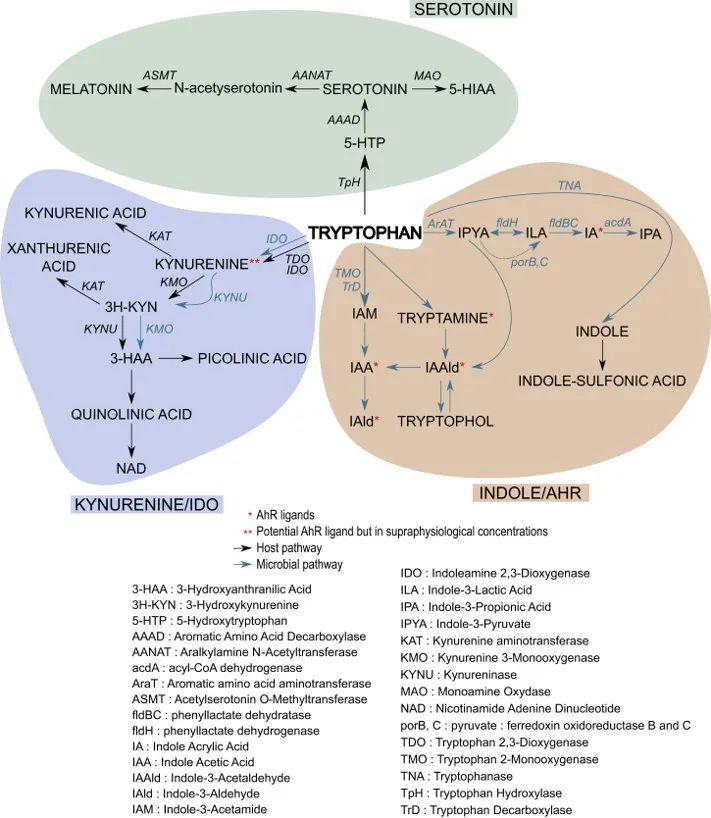
doi.org/10.1016/j.chom.2018.05.003
吲哚也是一种种间信号分子,能够控制细菌生理学的各个方面,例如抗生素抗性、孢子形成和生物膜形成。
在不产生吲哚的细菌中,吲哚及其衍生物显着抑制群体感应并调节毒力因子。然而,这些复杂现象在肠道生态系统中的重要性尚未得到具体解决。
微生物代谢的作用在肠道 AhR 活性中占主导地位。事实上,无菌或失调小鼠的肠道内容物缺乏 AhR 激动剂。只有少数共生物种能够产生 AhR 配体,例如Peptostreptococcus russellii罗氏消化链球菌和乳杆菌属已被表征,许多可能仍有待发现。
● 肠道色氨酸代谢平衡
虽然大部分色氨酸被小肠吸收,但其中一些会继续进入大肠,在那里它可以被微生物和宿主细胞作用。从上一小节的阐述,我们可以看到色氨酸在肠道内的三个主要归宿:
1) 吲哚/AhR 通路
肠道细菌直接将色氨酸转化为吲哚和相关分子。就像锁和钥匙一样,其中一些吲哚分子与整个肠道和其他器官细胞表面的芳烃受体 (AhR)结合。这会引发广泛的反应,促进肠道稳态。该途径的活性取决于饮食和微生物群的组成。
2) 犬尿氨酸(KP)通路
一些色氨酸被肠道上皮细胞和免疫细胞吸收,在那里它被 IDO1 酶转化为犬尿氨酸。犬尿氨酸可以进一步代谢为其他分子,例如具有神经毒性作用的喹啉酸。应激、炎症或感染会增加该通路的活性。
3) 血清素途径
色氨酸也被吸收到肠道肠内分泌细胞中,然后通过酶 TpH1转化为神经递质血清素。肠道中的血清素调节肠道运动、分泌和吸收,并在肠-脑信号传导中发挥作用。该通路的活性受禁食、饮食、肠道感染和某些微生物的影响。
当然,关键是平衡。在健康的肠道中,这三种途径是平衡的,从而产生最佳的肠道屏障功能、动力、免疫力和神经功能。
宿主生理学中肠道菌群控制下的综合色氨酸代谢
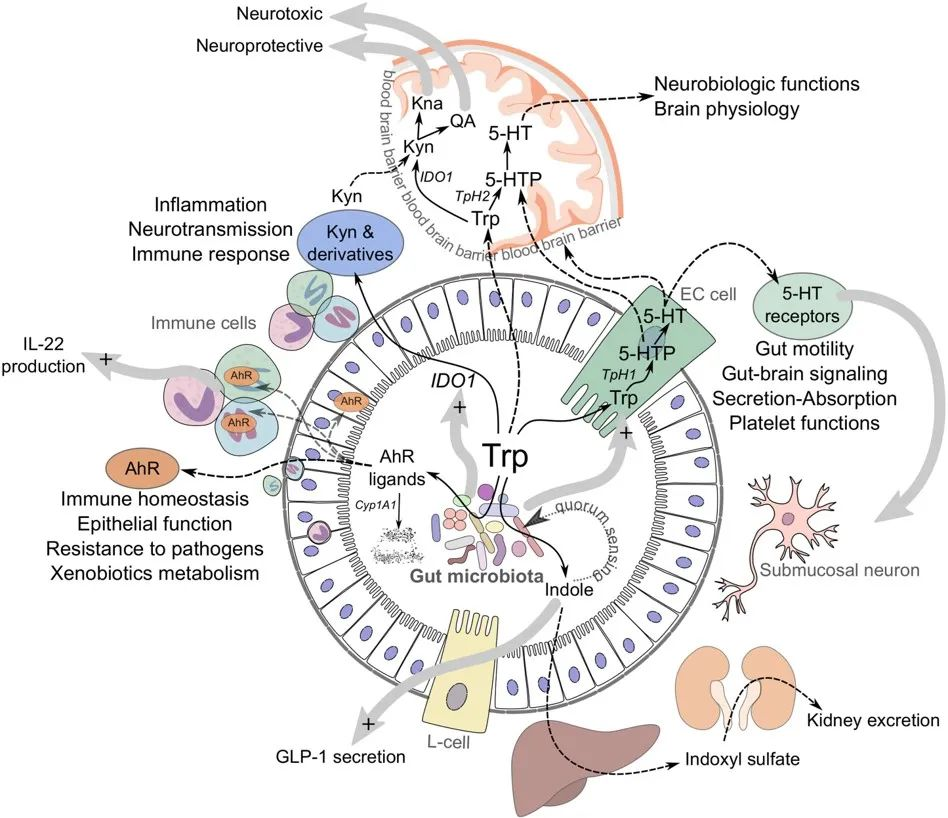
doi.org/10.1016/j.chom.2018.05.003
▼
在慢性疾病中,上述三种途径的平衡似乎出现了偏差,导致肠道功能受损和系统性影响。
/
/
▼
色氨酸及其代谢物水平的失衡与广泛的人类病理学相关,包括肠道疾病、抑郁症、精神分裂症、自身免疫、神经退化、癌症、心血管疾病、过敏、代谢综合征、肥胖、衰老等。
本章节我们对这些疾病中的色氨酸代谢先做个大致了解,后面章节会对各类疾病一一展开阐述。
由于许多 KP 代谢物具有神经活性,因此通常由炎症损伤引起的 KP 酶功能障碍可引发或促进中枢神经系统 (CNS) 疾病。
对于中枢神经系统疾病,人们越来越关注通过靶向特定 KP 酶来纠正 KP 代谢物变阻器的变化以实现净神经保护作用,以及色氨酸及其代谢物在调节肠道微生物组和大脑之间的相互作用中的作用。
肠道微生物组对膳食色氨酸吸收和代谢的影响也越来越受到关注,并且与中枢神经系统疾病以及肠易激综合征、胰腺炎和糖尿病具有潜在相关性。
降低维生素 B2 浓度会导致依赖于黄素腺嘌呤二核苷酸的犬尿氨酸 3-单加氧酶 (KMO) 的活性降低。B 族维生素,包括核黄素 (RBF) 和吡哆醇 ,在预防中风和中风后恢复中发挥作用。据报道,异常 KP 与神经系统疾病、癌症、心血管疾病和中风有关。
色氨酸分解代谢——涉及的关键器官
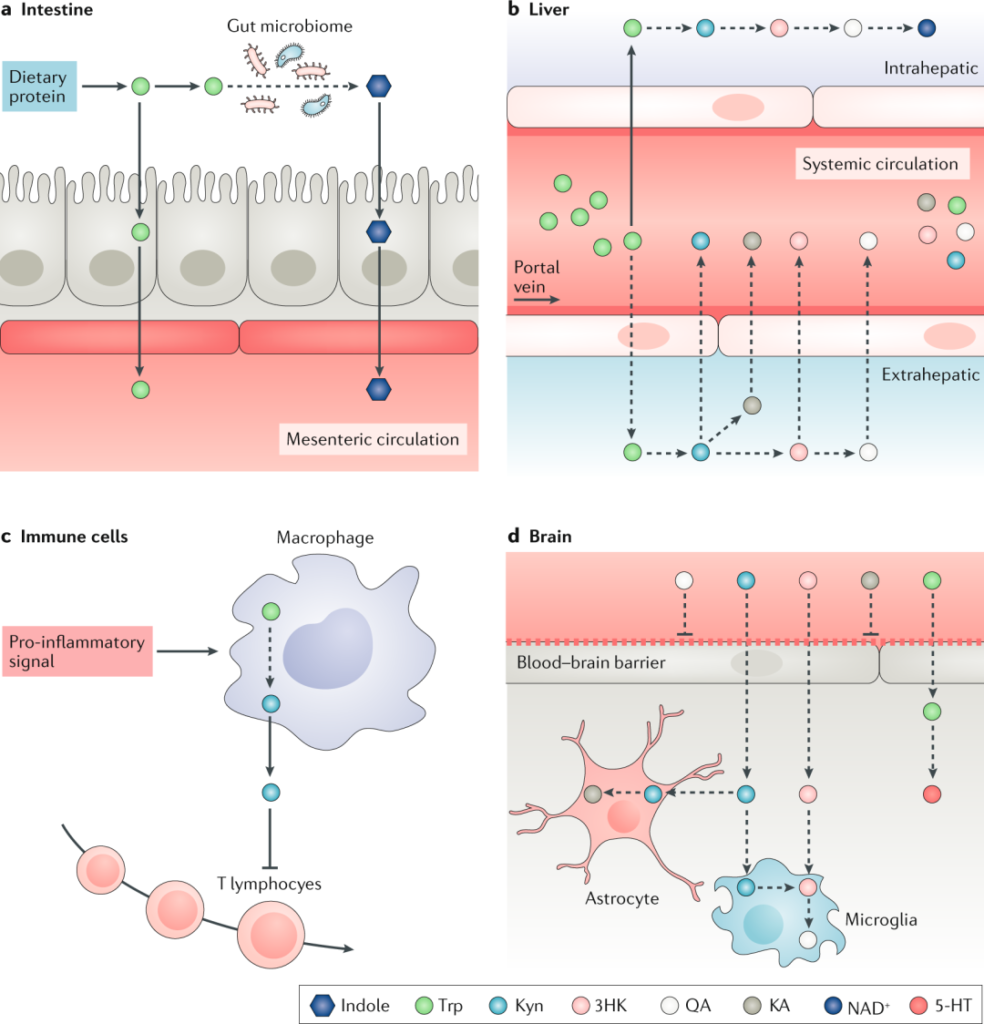
doi.org/10.1038/s41573-019-0016-5
a | 摄入膳食蛋白质后,肠上皮细胞将L-色氨酸转运穿过顶膜进入间质和肠系膜循环。或者,肠道微生物群合成色氨酸并将其代谢为吲哚并将其释放到体循环中。
b | 然后色氨进入肝脏,其中大部分被氧化为乙酰乙酰辅酶 A 并用于合成 NAD+。沿着犬尿氨酸 (Kyn) 途径 (KP) 代谢色氨酸 的肝外器官,包括肾脏、脾脏和免疫细胞,对 Kyn 和 KP 代谢物的循环水平贡献最大。
c | 在促炎性刺激后由骨髓细胞释放的 KP 代谢物抑制 T 细胞反应。
d | 色氨酸、Kyn 和 3-羟基犬尿氨酸 (3HK) 被转运穿过血脑屏障并被星形胶质细胞、小胶质细胞和神经元吸收。星形胶质细胞主要产生具有神经保护作用的犬尿酸 (KA),而小胶质细胞产生具有神经毒性的 KP 代谢物,例如喹啉酸 (QA)。
大约 5% 的色氨酸被代谢为血清素 (5-HT)、5-羟基吲哚乙酸 (5-HIAA)、褪黑激素 (MEL) 和色胺 (TA)。最近研究表明,5-HT除了在神经传递、血管收缩或血管舒张、止血控制和血小板功能中的作用外,还参与调节人体的能量平衡、食欲、肠道蠕动、免疫力、肝脏修复以及心血管和肺部生理学。
疾病中色氨酸代谢的扰动
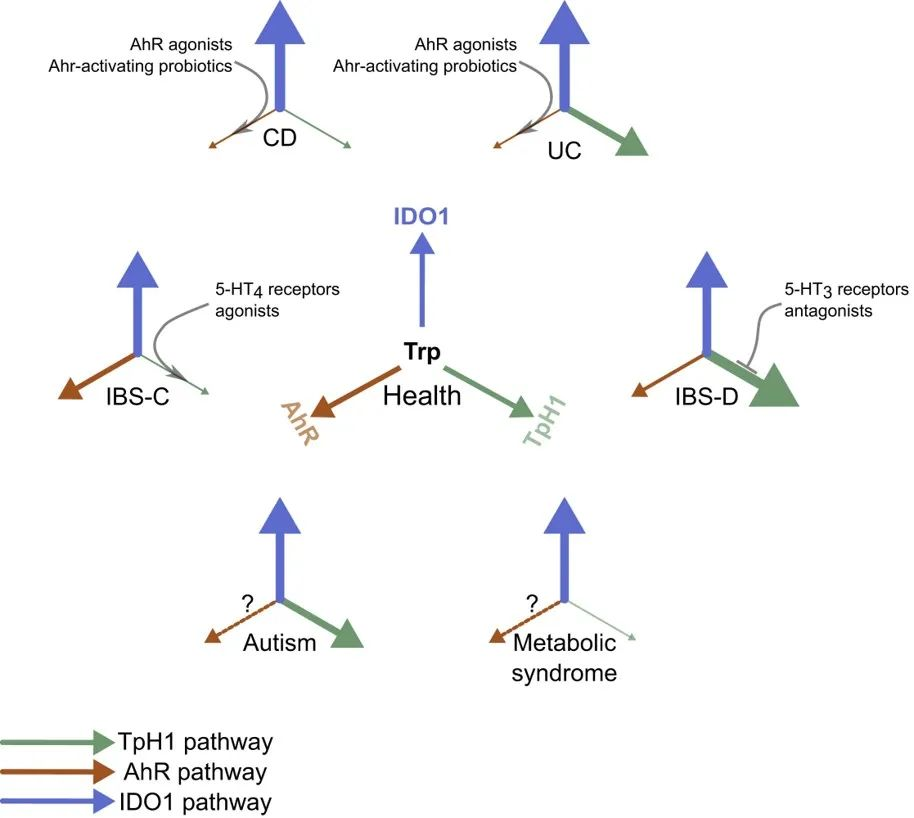
doi.org/10.1016/j.chom.2018.05.003
使用免疫组织化学监测 KP 代谢物的抗体的最新发展使得能够确定组织中 KP 代谢物的积累。
这些工具表明,犬尿氨酸通路在 IDO1 阳性癌症中积累,兴奋性毒性代谢物喹啉酸在脑肿瘤和神经退行性疾病的神经元中积累,而黄嘌呤酸 (XA) 是谷氨酸能突触传递的调节剂,定位于躯体和健康大脑中神经元的树突。
色氨酸代谢物与一系列疾病的联系导致人们在治疗上调节 KP 方面付出了巨大努力,特别是通过抑制所涉及的关键酶,包括 IDO1、TDO 和 KMO。
在癌症中,IDO1 和 TDO 的异常激活导致抗肿瘤免疫抑制。近年来 IDO1 抑制剂在癌症免疫治疗中得到了深入研究。
临床试验中有多种化合物,通常与免疫检查点抑制剂等其他药物联合使用。人们普遍预计领先的 IDO1 抑制剂将接近监管批准,但最近的 III 期试验终止引发了对该方法可行性的质疑,并强调需要更深入地了解 KP。
/
/
▼
● 神经退行性疾病中的色氨酸代谢
色氨酸代谢与多种神经退行性疾病有关,包括亨廷顿病 (HD)、阿尔茨海默病(AD)、肌萎缩侧索硬化 (ALS) 和帕金森病 (PD)。
尽管病理生理触发因素各不相同,但所有这些疾病的共同点是容易聚集的蛋白质引起神经元退化,从而导致细胞应激和有害的先天免疫反应。
基于人群的研究表明,就这些病理学特征而言,衰老和神经退行性疾病之间存在相当大的重叠,个体间差异很大。
虽然遗传和环境对色氨酸代谢的影响尚不完全清楚,但据信色氨酸代谢会导致衰老和神经退行性变,并且所涉及的机制即使不完全相同。这一观察得到了小鼠研究的支持,其中 TDO 的缺失已被证明会导致海马体和脑室下区的神经发生增强,可能抵消神经变性。
尽管生物标志物研究表明色氨酸代谢在神经退行性疾病患者中的活性不同,目前尚不清楚这是原发性倾向的结果还是神经变性或附带的先天免疫激活的结果。流行病学研究表明,KP 的激活与痴呆症风险增加有关。
然而,很难与生理老化明确区分。KP 对感染性和炎症性损伤的敏感性明显损害了其作为神经变性标志物的稳健性。另一方面,炎症对 KP 的激活可能在多发性硬化症等疾病中的神经炎症和神经变性之间建立联系。
由色氨酸代谢介导的神经变性的潜在机制包括:
➤ 阿尔茨海默氏病
色氨酸代谢物、肠道微生物和相关的神经炎症变化对阿尔茨海默病的病理生理学有显着影响。
阿尔茨海默患者的肠道微生物分类有显着差异,厚壁菌门和放线菌门减少,拟杆菌门增加。CSF 生物标志物升高与某些属的丰度相关,尤其是拟杆菌属和Blautia。
一项研究中,发现阿尔茨海默患者的循环色氨酸显著减少,犬尿氨酸/色氨酸比率升高,这反过来又与认知能力较差和促炎细胞因子升高相关。
几项临床前研究表明,在阿尔茨海默动物模型中具有保护作用。色氨酸代谢产物以芳基烃受体依赖的方式调节小胶质细胞和星形胶质细胞的活化。
此外,延缓阿尔茨海默进展的药物治疗的研究表明,肠道微生物和色氨酸代谢产物在阿尔茨海默的发展中可能发挥作用。
研究人员提供了犬尿氨酸代谢物在阿尔茨海默中的潜在毒性作用的间接证据,因为持续向小鼠腔内灌注犬尿氨酸会导致小鼠后代的学习和记忆缺陷。
吲哚途径代谢产物也可能介导阿尔茨海默病的发病。吲哚途径代谢物IPA在体外可抑制淀粉样蛋白-β诱导的神经毒性,并已被开发为治疗阿尔茨海默的神经保护剂。
➤ 帕金森病
帕金森病是一种进行性神经退行性疾病,其中α-突触核蛋白的聚集导致黑质神经毒性,导致多巴胺能神经传递不足。
大量数据表明肠道微生物组通过诱导炎性神经毒性参与帕金森发病机制。这些患者肠道拟杆菌属的丰度与运动症状严重程度和促炎性TNFα和IFNγ水平相关。尤其是疣微菌门(Verrucomicrobia)与循环中较高水平的IFNγ相关,说明了与IDO和色氨酸代谢产物的可能相互作用。
最近的研究还表明,帕金森病患者的色氨酸代谢紊乱,是潜在的治疗目标。帕金森患者血浆中3-HK显著升高,3-HANA降低。在这些患者中,犬尿酸显著降低,喹啉酸水平与疾病严重程度相关,表明色氨酸代谢产物在加重兴奋性毒性损伤中的潜在致病作用,尽管因果作用仍有待确定。
帕金森患者脑脊液和血浆中的犬尿氨酸/色氨酸比率升高,犬尿氨酸转氨酶活性降低。因此,犬尿酸合成类似物已成为治疗帕金森、亨廷顿病和阿尔茨海默病的神经保护药物。
➤ 其他神经退行性疾病
已知 NMDA 受体过度激活和随之而来的神经元兴奋性毒性在几种神经退行性疾病的发病机制中发挥作用。
——肌萎缩侧索硬化症
犬尿酸可能作为一种内源性神经保护剂发挥其拮抗 NMDA 受体过度激活的作用。临床数据表明了潜在的作用。晚期和延髓起病的肌萎缩侧索硬化症患者的 CSF 中 KA 水平显着升高。
——亨廷顿病
在亨廷顿病患者中,产生自由基的 3-HK 在早发性疾病中高度升高,同时纹状体和皮质喹啉酸也升高。然而,对于更晚期的疾病,这些浓度会降低。
虽然亨廷顿病患者的 CSF 中犬尿酸水平升高,对亨廷顿病大脑的尸检分析显示,与对照组相比,犬尿酸浓度降低,血清犬尿氨酸/色氨酸 比率升高。有趣的是,谷氨酰胺重复次数和疾病严重程度与循环色氨酸水平呈负相关。
亨廷顿病大鼠模型表明 3-HK 增强了神经兴奋性毒性,而自由基清除剂抑制了这种作用。
犬尿酸的合成类似物在原位产生神经保护和抗癫痫作用。鉴于IPA的抗氧化作用,研究人员还提出使用这种吲哚衍生物对亨廷顿病患者进行神经保护。
➤ 多发性硬化症
多发性硬化症是一种慢性、进行性和复发性中枢神经系统炎性脱髓鞘疾病。许多证据表明,这主要是由B和T细胞驱动的过程。最近,许多靶向B细胞和T细胞活化的药物被证明在预防复发方面具有临床疗效。
多发性硬化患者肠道菌群变化
多发性硬化患者粪便样本的微生物组分析显示,与对照组相比,多发性硬化患者中的Methanobrevibacter和Akkermansia增加,Butyricimonas 减少。
肠道微生物代谢产物参与多发性硬化发病
色氨酸代谢产物和I型IFN信号已显示在多发性硬化的实验性变态反应性脑脊髓炎(EAE)模型中激活星形胶质细胞AHR,从而抑制中枢神经系统炎症。
评估这种疾病中的犬尿氨酸途径的研究取得了有趣的结果。复发患者犬尿酸水平升高,而尸检样本显示犬尿氨酸转氨酶活性降低。
此外,喹啉酸可能诱导少突胶质细胞凋亡,导致脱髓鞘损伤。在其他EAE模型中,数据显示了有毒的犬尿氨酸代谢产物的集中聚集。
在这些模型中,肠道微生物也会影响中枢免疫,因为继发于微生物变化的免疫过度激活会加剧炎症损伤。
● 神经精神疾病中的色氨酸代谢
KP 的不平衡导致具有特定神经活性特性的代谢物过多,被认为是导致多种神经精神疾病的原因。
➤ 焦虑和抑郁
色氨酸代谢产物,尤其是血清素,与焦虑和抑郁的发病机制密切相关。
促进中枢5-羟色胺可用性的药物,特别是选择性5-羟色胺再摄取抑制剂(SSRIs)、MAO抑制剂(MAOIs)和三环类抗抑郁药(TCAs)已经彻底改变了这些疾病的治疗。
重度抑郁症、自杀倾向与喹啉酸水平升高相关
例如,重度抑郁症与 KP 的 3-羟基犬尿氨酸 (3HK) 分支下的新陈代谢增加有因果关系,导致大脑神经毒性喹啉酸水平高于神经保护性 KA。
同样,与 KA 和吡啶甲酸相比,喹啉酸水平升高也与自杀倾向相关。社会心理压力、感染或细胞因子治疗引起的免疫激活会导致抑郁症状。
系统性 IDO1 激活与抑郁症中 3HK 分支的激活有关
小鼠中的 IDO1 抑制或敲除可减轻抑郁样行为,细胞因子诱导的抑郁症易感性与 IDO1 基因的多态性有关。因此,系统性 IDO1 激活被认为与抑郁症中 3HK 分支的激活有关,但目前尚不清楚为什么 KA 和喹啉酸在对 IDO1 诱导的反应中没有同样上调。
随着对不同疾病过程中肠道微生物组组成的理解的增加,很明显,肠道微生物可能在这些疾病的起源和临床表型中发挥关键作用。
无菌小鼠表现出比常规饲养小鼠更焦虑的行为,这种行为在宿主断奶后不易随着微生物的重新繁殖而逆转,这表明肠道微生物组可能支持心理发育的关键时期。
一些益生菌在动物模型和人类中显示出减少焦虑和抑郁的功效。
补充色氨酸减少焦虑?仍然存在争议
瑞士乳杆菌R0052和长双歧杆菌R0175等物种的益生菌分别降低了小鼠和人类的焦虑、增强了情绪幸福感和抑郁症状。
患有抑郁症患者粪便微生物移植的小鼠表现出更严重的焦虑,这与更高的循环犬尿氨酸和犬尿氨酸/色氨酸比率有关。
有趣的是,已知慢性应激会增加循环色氨酸和皮质醇,由于糖皮质激素诱导的TDO表达增强,导致5-羟色胺代谢向犬尿氨酸及其代谢产物分流。
小鼠应激诱导的结果表明,外源性丁酸盐调节应激诱导的抑郁行为,降低海马血清素,增加海马脑源性神经营养因子(BDNF)。
肠道微生物群被抗生素耗尽的小鼠表现出类似焦虑的行为,循环的犬尿氨酸升高。在喂食高脂肪饮食的肥胖大鼠中,花青素可以防止神经炎症,并且循环色氨酸降低,犬尿酸增加。
➤ 精神分裂症
色氨酸向血清素的转换可能在精神分裂症的发病机制中受损,因为某些TPH1多态性增加了对精神分裂症和自杀的易感性。
精神分裂症患者 KA 的水平升高,与认知缺陷相关
精神分裂症和精神病似乎是由 NMDA 受体拮抗剂 KA 的形成增加引起的。在精神分裂症患者的死后大脑和脑脊髓液中测量到 KA 水平升高。KA 水平升高与在精神分裂症中观察到的认知缺陷相关、而KA 形成减少与认知功能改善相关。
同样,特别是在发育中的大脑中的神经炎症与精神分裂症的认知缺陷特征有关。KMO中的单核苷酸多态性基因与精神分裂症和双相情感障碍相关,这表明 KP 3HK分支下的流量减少可能使 Kyn 转向 KA 形成,KA 的积累与这些疾病有关。
脑脊液中低5-HIAA水平与自杀和攻击行为相关
由于精神分裂症患者皮质犬尿酸水平升高,犬尿氨酸代谢产物也可能起到致病作用。动物模型研究表明,色氨酸抑制攻击性行为,可能与增加中枢血清素的可用性有关。
小胶质细胞突触修剪过度激活
Sekar及其同事发表了一项具有里程碑意义的全基因组关联研究,该研究确定了与精神分裂症发病机制有关的基因位点,这涉及补体C4介导的小胶质细胞突触修剪过度激活。
肠道失调与免疫失调联系起来
鉴于肠道微生物组在介导中枢免疫中的既定作用,以及病例对照研究在精神分裂症患者中的优势,研究人员试图将肠道失调与免疫失调联系起来,导致大脑发育关键时期突触修剪过度活跃。
流行病学研究也支持这样的假设,即全身感染诱导的母体免疫激活是后代患精神分裂症的独立危险因素。
最近的一项研究表明,产前免疫暴露导致额叶皮质C4活性上调。母体微生物组向后代的垂直转移也可能导致持续的免疫功能障碍,增加突触过度修剪的风险。
需要进一步的机制研究来了解肠道微生物、色氨酸代谢产物和宿主免疫在精神分裂症和其他神经发育障碍发病机制中的相互作用。
➤ 自闭症
根据循环色氨酸的临床研究和排泄的犬尿氨酸代谢产物的检查,自闭症谱系障碍患者可能缺乏色氨酸。
自闭症患者来源的淋巴母细胞的代谢组学分析显示,当色氨酸是唯一可用的能量来源时,NADH生成减少,表明这些患者的喹啉酸降解途径可能受损。
某些微生物物种可能参与了自闭症的发病机制
几项评估自闭症患者肠道微生物丰度差异的研究将自闭症症状与Prevotella、Coprococcus、Veillonellaceae丰度较低联系起来。
脆弱拟杆菌(一种胰蛋白酶合成细菌),可能会降低自闭症患者的色氨酸可用性。非色氨酸衍生的微生物代谢产物也可能起到因果作用,一项观察自闭症小鼠母体免疫激活(MIA)模型中肠道微生物代谢产物的研究显示,微生物代谢产物4-乙基苯基硫酸盐增加了46倍,如果小鼠被脆弱拟杆菌定殖,则其正常化。
色氨酸代谢在肠道菌群-脑轴中的潜在作用
doi: 10.1093/advances/nmz127
通过各种方式(例如,抗生素和益生菌)对肠道微生物群组成和代谢的调控有助于在 5-羟色胺合成和色氨酸降解途径之间改变中枢色氨酸代谢,从而影响大脑功能和行为。
● 中枢神经系统疾病中靶向 KP 酶
尽管临床试验的重点(部分仍然是)补充或剥夺色氨酸或其代谢物以治疗神经精神疾病,但目前神经退行性和神经精神疾病药物开发的临床前工作主要集中在通过改变神经活性 KP 代谢物的变阻器抑制参与 喹啉酸或 KA 形成的酶。
从概念上讲,所有 KP 酶都代表潜在的治疗靶点,并且有几项研究调查了药理学抑制的影响。
例如,IDO1 抑制剂黄连碱已被证明可以减缓阿尔茨海默小鼠模型的认知障碍,尽管其对 IDO1 的特异性尚不清楚。
有趣的是,环氧合酶抑制通过抑制海马 TDO 表达,来防止类似阿尔茨海默模型中的行为下降。当使用 TDO 的药理学抑制剂时,观察到类似的神经保护作用。这些研究连同阿尔茨海默病和亨廷顿病患者中 KP 激活的证据, 表明抑制色氨酸降解中限速的第一个酶促步骤是一种潜在可行的治疗方法,可以抵消由淀粉样蛋白形成蛋白的积累引起的神经毒性。
尽管 IDO1 和 TDO 的抑制剂阻止了 KP 代谢物的产生,但这不会直接影响 KA/喹啉酸 喹啉酸变阻器,但会阻止两者的产生。这种治疗方法是可行的,因为它可以防止色氨酸的消耗,这可以减少在临床前模型中观察到的蛋白质毒性。
KAT 在辅助因子 pyridoxal-5-phosphate (PLP) 的帮助下催化 Kyn 转化为 KA。KATII 是哺乳动物大脑中最普遍的 KAT,并且正在寻求作为精神分裂症和认知障碍疾病的药物靶点。由于最近显示 KATII 也能催化 3HK 105形成 XA ,因此之前归因于 KA 的基于抑制 KATII 的效应也可能涉及 XA。
▸ KATII 抑制剂
KATII 的可逆抑制剂已经开发出来,包括 Kyn 类似物 (S)-4-(ethylsulfonyl)benzoylalalanine ( S -ESBA) ,它被证明可以降低大鼠大脑中的 KA 水平。
高效和选择性脑渗透不可逆抑制剂 PF-04859989 也报道了相同的抑制模式。然而,这些化合物都没有进入临床研究,这可能是由于它们与 KAT 同工酶和所有其他 PLP 依赖性酶所需的 PLP 辅因子发生不可逆相互作用而引起的毒性。
将 KATII 抑制剂推进临床试验的主要挑战包括由脑 KA 水平降低引起的潜在毒性、获得足够的效力和选择性以及 KATII 抑制剂效力的种间差异的发生。
▸ KMO 抑制剂
为了抑制 KP 的 喹啉酸分支和增加拮抗 KA 水平,KMO 抑制剂正在积极开发中。有关 KMO 晶体结构的信息有助于生成特异性更高的 KMO 抑制剂。众所周知的 KMO 抑制剂 Ro 61-8048已用于大量临床前研究,证明其作用范围从改善神经变性到减少大麻素滥用。
另一种广泛使用的工具化合物,UPF-648, 是一种不含氨基的 Kyn 类似物,在构象上受到环丙基环的限制。这种化合物,以及高效的恶唑烷酮 GSK180(在胰腺炎的背景下研究),是所谓的 I 型 KMO 抑制剂,它模仿 Kyn 并刺激有害的过氧化氢产生。
在一项基于结构的药物化学合作研究中,开发并评估了一种新的芳基嘧啶先导化合物 CHDI-340246,用于治疗 HD。然而,这种选择性 KMO 抑制剂的长期治疗并未显着改变 HD 小鼠模型的行为表型或自然进展,尽管它恢复了电生理学改变。
结构研究最近破译了 I 型和 II 型KMO抑制剂112、121之间的区别。II 型 KMO 抑制剂 GSK065 和 GSK366 显示出比 I 型 KMO 抑制剂更好的类药特性,因为它们具有皮摩尔亲和力、增加的停留时间和不产生过氧化物。
GSK065以GSK3335065(NCT03245619)的名称进入治疗胰腺炎的I期临床试验。有趣的是,KMO 抑制剂的外周给药足以影响 CNS KP 。然而,KMO 抑制剂是否需要穿透血脑屏障才能发挥作用,这仍然是一个有争议的问题。
最后,抑制初始限速 KP 酶 IDO1 和 TDO,它们分别在炎症条件或慢性社会心理压力下诱导,在神经退行性疾病和精神疾病中也可能值得探索。由于这些酶的抑制剂目前正在开发用于癌症治疗,因此可以使用多种化合物在临床环境中测试这些方法。
/
/
▼
● 感染中的色氨酸代谢
几条证据最近揭示了色氨酸代谢作为宿主-病原体相互作用和塑造宿主微生物群中免疫反应的重要调节因子的关键作用。
通过特定的色氨酸代谢酶,色氨酸代谢在细菌、病毒、真菌和寄生虫感染部位增加。通常以低基础水平表达,在抗原呈递细胞 (APC)中观察到 IDO1 增加,例如树突细胞 (DC) 和巨噬细胞,以响应多种微生物刺激,包括 Toll 样受体 (TLR) 配体(例如,脂多糖 (LPS) , CpG 寡核苷酸和聚肌胞苷酸 。
炎症刺激物诱导IDO1,IDO1 会耗尽色氨酸
此外,据报道,I 型和 II 型干扰素 、肿瘤坏死因子 (TNF)、前列腺素 和膜结合分子 等炎症刺激物可在特定APC类型中诱导IDO1。
在传染病中,IDO1 活性具有多效性,是一把双刃剑。实际上,IDO1 会耗尽色氨酸以饿死和重新编程营养缺陷型入侵者,同时有助于对在急性感染期间未清除的微生物产生 Kyn 依赖性免疫抑制状态或那些已经能够重新激活色氨酸生物合成的。
因此,已经表明色氨酸营养缺陷型病原体对 CD4 + T 细胞激活的巨噬细胞高度敏感。 在特定的环境条件下,色氨酸的微生物营养缺陷型可能会消失。
特定条件微生物重新获得合成必须氨基酸的能力
某些微生物可以在特定的胁迫条件下重新获得合成这种必需氨基酸的能力。此外,天然能够合成色氨酸的微生物群菌株可以在特定感染期间扩大,从而在色氨酸缺乏的条件下提供额外的这种必需氨基酸供应。
最近的研究结果表明,结核分枝杆菌等特定病原体可以在压力条件下重新获得合成色氨酸的能力,从而抵消 IDO1 饥饿驱动的抗菌作用。
此外,衣原体在由局部色氨酸剥夺引起的应激条件下进入非复制的持久状态。同样,IDO1 依赖性持久性已被记录在其他细菌物种中,包括肺炎积瘤。
除了调节病原体负荷外,通过 IDO1 活性进行的色氨酸代谢对于抑制最终阻止病原体根除的免疫病理也至关重要。
在这方面,最近对肠道微生物群的研究发现:
色氨酸代谢与通过充当特定 AHR 配体的微生物或细菌毒力因子在粘膜屏障表达的 AHR 激活之间存在重要联系。
值得注意的是,AHR +由于产生色氨酸代谢物(即吲哚 3 醛)的乳酸杆菌的选择性扩增,即使在 IDO1 缺乏的情况下,也会诱导产生IL-22 的第 3 组先天淋巴样细胞 (ILC3s)能够激活 AHR,从而在真菌感染模型中诱导保护性耐受状态。
TDO 在感染过程中的潜在作用
用 LPS 攻击的小鼠肝脏中 TDO 表达增加,而 TDO 缺陷小鼠更容易受到内毒素攻击。因此,在对弓形虫和金黄色葡萄球菌感染进行的体外研究中,已经报道了 TDO 依赖性抗菌和免疫调节作用。此外,代谢组分析揭示了原发性登革热感染患者 TDO 激活的变化。
因此,在宿主细胞中的三种不同色氨酸分解代谢酶中,IDO1 的影响已在几种临床前感染模型中得到解决。具体而言,据报道,IDO1 在体内抑制某些细胞内寄生虫和细菌的复制,例如弓形虫、衣原体和杜氏利什曼原虫。另一方面,弱 IDO1 抑制剂 1-甲基- L – Trp (L-1-MT) 增强了沙眼衣原体抗生素清除的功效,尽管可能涉及额外的 IDO1 独立机制。
IDO1活性还可以在体外抑制特定病毒的复制
例如人巨细胞病毒 (CMV)、2 型单纯疱疹病毒和痘苗病毒。然而,体内情况可能有所不同,因为病毒感染可能会诱导 IDO1 和 KP 逃避宿主免疫反应。
由于它们具有诱导 Treg细胞的能力 ,因此 IDO1 消耗色氨酸并产生 Kyn 是抑制抗菌 TH17 和 TH1 驱动的炎症的重要手段。
因此,病原体可能会劫持 IDO1 的免疫抑制作用,并利用它们来促进自身的生命周期。在这方面,尿道致病性大肠杆菌(UPEC) 在泌尿道的上皮细胞中诱导 IDO1 ,并且色氨酸分解代谢的免疫反应减弱使得 UPEC 能够成功定植。
HIV-1 等病毒利用 IDO1 的免疫抑制活性建立 HIV 慢性感染
KP 活性的增加也与丙型肝炎病毒感染患者的进行性肝硬化有关。
同样,小鼠感染甲型流感/PR/8/34 (PR8) 会刺激肺部和肺引流纵隔淋巴结中 IDO1 活性的快速升高,导致发病率增加、恢复减慢和肺部效应 T 细胞反应降低,尽管在原发性甲型流感病毒感染期间,IDO1 诱导不会影响病毒清除。在其他情况下,例如在真菌感染中,IDO1 可用作建立共生或慢性感染的逃避机制。
● 在传染病中靶向 KP 酶
在选定的微生物物种中调节特定的色氨酸生物合成途径并靶向宿主细胞中的 IDO1-AHR-微生物群轴可能代表了抗生素开发或补充抗病毒疗法的新颖有吸引力的策略。有必要更全面地了解特定感染期间色氨酸分解代谢酶或下游酶的作用,以便了解旨在调节色氨酸分解代谢以根除病原体同时保持与微生物群平衡的疗法的效用。
基于以上总结的证据,可以假设特定的 IDO1 阻断剂可能会发现潜在的应用作为辅助疗法来提高抗病毒药物的疗效,但可能证明对真菌感染有害,其中色氨酸分解代谢主要通过 IDO1 作用于维持免疫稳态和保护性耐受。
然而,这种作用可能构成使用 IDO1 抑制剂作为抗肿瘤药物的潜在缺点(下面讲)。实际上,在使用 IDO1 通路调节剂 1-甲基-D -Trp (D-1-MT)对转移性实体瘤患者进行的 I 期试验中,感染是最常见的不良事件。
有趣的是,最近的一项研究表明,KYNU 的靶向抑制会影响铜绿假单胞菌基因表达和群体感应,这表明一种新的潜在抗毒策略。具体而言,与 Kyn 具有结构相似性的S-苯基-L-半胱氨酸亚砜可抑制对铜绿假单胞菌毒力至关重要的邻氨基苯甲酸盐的产生。
/
/
▼
● 自身免疫中的色氨酸代谢
自身免疫是未能发展出对自身的中枢(胸腺)耐受性和外周耐受性维持不足的结果。免疫区室中的色氨酸代谢主要由 IDO1 启动,它代表主要促炎刺激的靶基因。
在这方面,IDO1 介导的色氨酸降解可被视为调节过度活跃的免疫反应的关键反馈机制,这是自身免疫性疾病的标志。
IDO1 在发炎组织中转录激活抑制适应性免疫反应的作用已经从最初在胎盘中观察到的维持胎儿耐受性扩展到多种自身免疫性疾病。
尽管 IDO1 缺陷不会导致与 Treg细胞重要检查点缺陷相关的整体自身免疫表型,但它与更微妙的炎症表型相关。这种关联可能部分是由于与其他双加氧酶共享的酶功能冗余。越来越多的证据表明,人类自身免疫性疾病是由免疫和/或基质细胞未能上调 IDO1 以响应炎症刺激驱动的。
然而,与自身免疫相关的上调 IDO 的结构性缺陷的潜在原因尚未阐明。连锁分析将IDO1和IDO2基因的多态性分别与克罗恩病的严重程度和风险相关联。需要进一步的研究来确定上调组织中色氨酸代谢的本构或诱导缺陷是否会导致组织特异性自身免疫。
许多针对多发性硬化症、类风湿性关节炎、狼疮和自身免疫性糖尿病的自身免疫性疾病小鼠模型的研究已经证明色氨酸代谢在调节疾病活动中的相关性。
综上所述,这些研究表明 IDO1 在组织驻留的骨髓细胞中表达,并限制对自身抗原和炎症病理学的先天性和适应性免疫。
然而,自相矛盾的是,在自发性类风湿性关节炎的动物模型中,使用 D/L-1-MT 对 IDO1 的药理学抑制减轻了疾病的严重程度,这可能是自身反应性 B 细胞活化减少的结果。
这一发现说明了 IDO1 在自身免疫中的复杂免疫调节功能,这取决于细胞区室。例如,B 细胞中免疫抑制细胞因子 IL-10 的表达依赖于 IDO1,这表明 IDO1 不仅会触发免疫抑制机制,还会协调对炎症的复杂免疫调节反应。
重要的是,转录激活和蛋白质表达不一定转化为人类 B 细胞中描述的酶活性。 在这方面,需要更多的研究来阐明 IDO1 的非酶功能。
此外,需要谨慎看待使用 D-1-MT 得出关于 IDO1 功能的关键结论的研究,因为 D-1-MT 不会抑制 IDO1 并显示出相当大的脱靶效应,从而导致 p38 MAPK 通路的激活。
对自身免疫性疾病模型的研究表明,与 IDO1 相比,IDO2 充当自身免疫的促进者,特别是由于体液免疫反应的调节。由于致病性自身抗体和抗体分泌细胞的减少,IDO2 缺陷小鼠表现出关节炎症减少。尽管与人类疾病的相关性仍不清楚, 这些研究突出了双加氧酶对色氨酸代谢的复杂和高度分隔的调节。
尽管 IDO1 介导的免疫调节的主要作用被认为是由组织炎症的局部微环境中的活动驱动的,但在患有自身免疫性疾病的患者中观察到色氨酸代谢的全身激活。在干燥综合征患者中,血清中色氨酸的降解增加,并与循环 Treg细胞频率增加有关。
相比之下,在多发性硬化症中,血清中的 IDO1 活性与健康对照相比没有显着差异,但抗炎治疗降低了 IDO1 活性。由于全身性 IDO1 活性会受到大量非特异性和难以控制的刺激的影响,包括感染、压力和营养,尝试通过循环色氨酸代谢物监测组织特异性自身免疫将具有挑战性。然而,对血清中 KP 代谢组的更详细分析不仅揭示了 KP 在多发性硬化症患者中的激活,而且还将 KP 活性的程度与疾病严重程度相关联。因此,KP 活性可以作为能够指导多发性硬化症治疗的预测性生物标志物。
● 靶向自身免疫性疾病中的色氨酸代谢
在治疗上针对色氨酸代谢的努力主要集中在开发具有 Kyn 样特性的药物上。
曲尼司特
曲尼司特是一种具有 AHR 激动特性的 AA 衍生物,能够在多发性硬化症和类风湿性关节炎的临床前模型中诱导免疫耐受和改善疾病活动。然而,一项针对类风湿性关节炎患者的 II 期临床试验(NCT00882024)因肝毒性而终止。
拉喹莫德
拉喹莫德是一种喹啉甲酰胺,在开发治疗多发性硬化症时显示出与 KA 的结构相似性,它以 AHR 依赖性方式抑制多发性硬化症临床前模型中的自身反应性 T 细胞免疫和疾病活动。
在针对复发和进行性多发性硬化症患者的一系列 II/III 期临床试验中,拉喹莫德未达到预先指定的主要终点,包括减少复发率和残疾进展,因此被终止 (NCT01707992)。
AHR 配体改善自身免疫神经炎症
AHR 的特定内源性配体足够稳定,可以在临床前疾病模型中进行肠胃外给药。2-(1’H-indole-3′-carbonyl)-thiazole-4-carboxylic acid methyl ester (ITE) 通过以 AHR 依赖性方式诱导致耐受性 DC,在实验性自身免疫性脑脊髓炎 (EAE) 模型中诱导 Treg细胞并改善自身免疫性神经炎症。
AHR 激活配体也可以与自身抗原偶联,从而导致 APC 的特异性靶向,然后耐受性抑制自身反应性 T 细胞反应,从而抑制系统性自身免疫。
从概念上讲,色氨酸代谢也可以通过全身给药色氨酸来增强,色氨酸在口服灌胃后会迅速代谢成 Kyn。尽管这种方法导致 TH17 免疫力的不同抑制,但这并不转化为实验性自身免疫性神经炎症的改善。
阻断IDO1降解,维持外周耐受性
认识到蛋白酶体降解是调节自身免疫中色氨酸代谢的免疫抑制活性的重要机制后,另一种治疗途径是阻断IDO1降解,从而维持外周耐受性。
硼替佐米是一种批准用于治疗多发性骨髓瘤的蛋白酶体抑制剂,可防止 IDO1 降解并以 IDO1 依赖性方式在临床前动物模型中改善自身免疫性糖尿病。
基于 IDO1 感受态细胞的疗法
尽管间充质干细胞的过继转移已被证明可以独立于IDO抑制自身免疫性神经炎症的临床疾病活动,但 IDO1 已被证明在其他自身免疫性体内疾病模型中与间充质干细胞的免疫抑制特性密切相关。
增强或诱导宿主 IDO 表达的另一种方法是通过局部基因治疗。例如,腺病毒将 IDO1 递送至移植器官可诱导免疫耐受并防止大鼠发生移植排斥反应。
IDO2 直到最近才成为潜在的治疗靶点
迄今为止,还没有对 IDO2 具有足够特异性的小分子。在自身免疫性关节炎的临床前模型中,一种通过内化靶向 IDO2 的抗体通过抑制自身反应性 T 细胞和 B 细胞减轻了疾病。
新开发的 IDO2 特异性测定系统和基于计算结构的研究可能有助于开发对 IDO1 没有交叉反应的 IDO2 抑制剂。
/
/
▼
● 肠 病
最近研究强调了肠道色氨酸代谢的改变与肠道微生物的潜在联系。发现 IBD 患者微生物群产生的 AhR 配体减少,这是受遗传因素的影响。与健康受试者相比, 肠道组织中 AhR 的表达降低。
IBD还与宿主和肠道细菌色氨酸代谢物的改变有关。IBD 患者的犬尿氨酸和 KA 血浆水平升高,血浆色氨酸浓度降低。
几种特定的肠道细菌色氨酸代谢物也参与 IBD 的病理生理学
在患有 IBD 的狗中,被认为在肠道中具有抗炎功能的细菌色氨酸代谢物(吲哚乙酸盐和吲哚丙酸盐)显着减少。在 IBD 患者中,粪便中 IAA(肠道抗炎功能)水平降低,表明细菌色氨酸代谢减少可能是 IBD 的病因。
此外,在 IBD 患者中,可利用 α-L-岩藻糖苷酶从肠粘蛋白中切割末端岩藻糖残基的细菌数量显着减少,这与来自色氨酸的吲哚丙烯酸和吲哚-3-丙酸产量减少有关。
IBD 患者的 IDO1 活性更高
据报道,IBD 患者外周血和结肠细胞中的 IDO1 活性增加。在 IBD 中,增加的促炎细胞因子,包括 IFN-γ、IL-1 和 IL-6,已被建议诱导色氨酸分解代谢途径以降低血浆色氨酸水平,并增加色氨酸分解代谢物水平。
此外,UC 患者血清中的 IPA 降低。在肠道局部观察到 IDO1 的过度激活和免疫系统过渡激活。与非活动性 IBD 患者相比,活动性 IBD 患者的 IDO1 活性更高,并且色氨酸和 C 反应蛋白血清水平呈负相关,这一假设得到了支持,C 反应蛋白是一种常用的生物标志物,对炎症反应增加。
IBD 中 5-HT 通路激活的状态存在争议。限速酶TpH1的表达增加在克罗恩病中已报道。
小鼠研究表明AhR 缺乏会增加实验性结肠炎的严重程度
这种结肠炎是由 T 细胞转移或通过施用葡聚糖硫酸钠 (DSS) 以化学方式驱动的。在这些模型中,AhR 缺陷部分通过改变白细胞介素 (IL)-22 的产生来驱动结肠炎,白细胞介素 (IL)-22 是一种对肠道稳态具有众所周知影响的细胞因子。
缺乏 caspase 募集域 9 (Card9)(一种 IBD 易感基因)的小鼠的肠道菌群失调无法将色氨酸催化成 AhR 配体,导致 IL-22 释放减少并最终导致Card9的易感性更高-/-小鼠对 DSS 诱导的结肠炎 。
在人类中也发现了一些功能相关性,因为 AhR 的药理学激活减少了促炎细胞因子干扰素 (IFN) γ 的产生,并增加了 IBD 患者固有层单核细胞中 IL-22 的产生。
此外,正如在患有 UC 的人类中观察到的那样,IPA 和吲哚在 DSS 诱导的结肠炎小鼠血清中减少,另外的证据表明口服 IPA 在该模型系统中具有保护特性。
KP 的改变也可能在机制上参与 IBD 发病机制
IDO1 -/-小鼠更易患结肠炎,表明 IDO1 是肠道炎症的负调节剂。与 IDO1 缺乏相关的病理损伤部分是由于促炎细胞因子的激活和结肠中 CD4+ Foxp3+ 调节性 T 细胞数量的减少。然而,所涉及的确切机制和代谢产物仍然未知。
众所周知,Kyn 是一种 AhR 激动剂,但在肝癌细胞系中引发报告基因 AhR 活性所需的浓度让人怀疑它在生理条件下作为 AhR 激活剂的相关性,可能涉及导致抗炎代谢物(如 Kna)缺乏的下游代谢途径的改变,但这仍有待证明。在 IBD 的背景下,来自失调微生物群的异常信号可能是 KP 的驱动因素。
5-HT 加重肠道炎症
化学诱导的结肠炎的严重程度在TpH1 −/−小鼠和用 5-HT 合成抑制剂对氯苯丙氨酸处理的小鼠中减弱,表明 5-HT 加重肠道炎症。此外,删除 SERT 会导致 5-HT 可用性增加,从而导致实验性结肠炎恶化。
这些促炎作用可能部分是由DC上 5-HT 7受体的激活驱动的。然而,新的线索表明 5-HT 还通过作用于 5-HT 4 发挥抗炎作用对肠上皮细胞屏障功能产生积极影响。
总之,这些数据表明在 IBD 中观察到的色氨酸代谢改变可能在疾病发病机制中发挥积极作用。就这些微生物产生 AhR 激动剂的能力受损而言,微生物群的参与是显而易见的,但也可能解释了在生理条件下微生物群的直接影响下发生的 IDO 和 TpH1 的局部激活加剧。
● 肠易激综合症
IBS 的病因在很大程度上是未知的,但可能与色氨酸代谢受损有关。IBS 患者血清中 Kyn 升高,外周 IDO1 活性与 IBS 严重程度呈正相关。
IBS 与通过 KP 增加的色氨酸代谢有关
犬尿氨酸:色氨酸比率与 IBS 症状严重程度呈正相关,IFN-γ 激活和随后的色氨酸 IDO1 氧化可能是 IBS 的致病机制。
此外,血清素能系统的功能障碍与 IBS 的病理生理学有关。与急性色氨酸耗竭治疗相比,IBS 患者通过急性色氨酸增加治疗进行的 5-羟色胺能调节导致更严重的胃肠道症状。
肠道运动的改变是 IBS 的关键特征之一,与 5-HT 代谢障碍有关
与健康对照组相比,IBS 患者的直肠活检组织中发现 TpH1 和 SERT 表达水平降低。
此外,5-HT 结肠内容物在便秘型和腹泻型 IBS 中分别减少和增加。5-HT 的多效性与其受体的多样性有关,这些受体能够触发特定器官的特定功能。
在胃肠道中表达最多的5-HT 3和 5-HT 4亚型将 5-HT 与内脏伤害感受和运动障碍联系起来。5-HT 的作用已经被开发为治疗靶点,使用 5-HT 3受体拮抗剂和 5-HT 4受体激动剂分别显示出对腹泻和便秘为主的 IBS 的一些疗效。
然而,受肠道微生物群调节的中枢血清素作用紊乱也可能参与 IBS 发病机制。肠道菌群对 5-HT 产生和肠道运动的影响已在小鼠身上得到证实,并表明 IBS 发病机制部分与微生物群对 5-HT 产生的功能失调控制有关。
● 与年龄有关的胃肠功能障碍
高龄增加了个体对胃肠道功能障碍的易感性,这归因于与年龄相关的神经元丢失。
5-HT4 激动剂刺激发育中的肠道中的神经突生长和网络形成,也已被证明可以防止神经元凋亡和炎症诱导的轴突变性和自噬。
此外,5-HT4 受体激动作用可促进成人肠道神经发生。相应地,其他方面健康的高龄个体表现出循环色氨酸减少,可能会限制血清素的可用性。
/
/
▼
● 衰 老
衰老与肠道微生物群的变化有关,这通常与胃肠道的生理变化有关,同时免疫系统功能下降可能导致感染、营养不良和其他功能缺陷的风险增加。
老年人菌群特征
老年人的肠道微生物群通常以细菌多样性降低、优势菌种改变、有益微生物减少和兼性厌氧菌增加为特征,所有这些都表明与衰老相关的微生物变化具有潜在的不利影响。微生物群组成的变化与老年人的免疫衰老和炎症有关。
色氨酸代谢受衰老影响
色氨酸在诱导免疫耐受和维持肠道菌群方面起着至关重要的作用。
对老年人和年轻人肠道核心微生物组直系同源基因的分析表明,丰度增加的年龄相关基因参与了色氨酸代谢途径 (ko00380),这与在百岁老人血清中发现的与年龄相关的色氨酸浓度降低一致。研究表明,肠道微生物群对色氨酸消耗的潜在增加可能会影响宿主对色氨酸的生物利用度。
最近的一项研究报告了血清色氨酸水平降低与免疫激活增加之间的关系。还推测微生物群依赖性色氨酸减少会增强百岁老人的炎症。
食物传感信号通路调节寿命,与色氨酸关联
几种食物传感信号通路,包括胰岛素/胰岛素样生长因子 (IIS) 通路和哺乳动物雷帕霉素靶标 (mTOR) 通路,已被证明可以调节模式生物的寿命,并且已经提出了类似的关联对于KP途径。
在人类中,表示该通路活性的 Kyn:色氨酸 比率随着年龄的增长而增加。这种增加与 65 岁以上人群的虚弱有关,并预示着 90 多岁人群的死亡率。
KP 的活性与衰老之间存在因果关系
此外,对成人个体外周血中年龄相关基因表达变化的荟萃分析将 KYNU 酶(犬尿氨酸酶,色氨酸降解途径关键酶)鉴定为表达差异最大的基因之一。在对秀丽隐杆线虫的后续研究中,通过 RNA 干扰 (RNAi) 敲低 KYNU 比敲低任何其他差异表达基因所达到的寿命更长,这表明 KYNU 对衰老有重要贡献。
连同独立发现,线虫和黑腹果蝇中 TDO 活性的遗传减少, 导致 Trp:Kyn 比率显着增加,延长寿命,这些研究表明 KP 的活性与衰老之间存在因果关系。
Kyn/Trp 分流在炎症中的后果
色氨酸代谢向肝外 Kyn 产生的炎症相关分流,可能影响衰老过程中一系列器官中色氨酸代谢物的功能。
编辑
doi.org/10.3389/fimmu.2019.02565
与年龄相关的组织稳态下降会导致生理上的低度慢性炎症表型,称为炎症。我们假设色氨酸向Kyn途径代谢,以控制与年龄相关的炎症。色氨酸和Kyn代谢产物的相应紊乱可能与年龄相关疾病和寿命缩短有关。
色氨酸参与调节寿命机制
KP 调节衰老的机制尚不清楚。已经针对不同的无脊椎动物和脊椎动物模型描述了氨基酸(包括色氨酸)在调节寿命方面的作用。在大多数情况下,色氨酸可用性的降低或细胞摄取的阻断可延长寿命。然而,这种机制与 TDO 抑制(增加色氨酸)延长寿命的发现相悖,除非这与细胞摄取减少有关。
此外,用 Kyn 喂养果蝇会缩短寿命,这表明该通路下游的代谢物水平也可能参与寿命的调节。TDO 耗尽对秀丽隐杆线虫延长寿命的影响取决于 FOXO 转录因子 DAF-16,它是寿命调节通路的介质,例如驱动细胞防御通路表达的 IIS 通路,表明它具有保护细胞免受细胞侵害的作用伤害。
有趣的是,防止与年龄相关的蛋白质毒性(这也是由秀丽隐杆线虫中的 TDO 耗尽引起的)不依赖于 DAF-16,并且独立于 KP 63中的下游酶。这一观察结果表明,延长寿命的效果要么是这种保护的结果,要么是由一种独立的机制引起的。
由于 NAD +正在成为一种潜在的延长寿命分子,KP 的改变可能通过 NAD +产生延长寿命的效果。然而,无脊椎动物的寿命更长是 KP 活性降低的结果,而通过外部供应其他 NAD +前体来延长寿命则表明 KP 活性的增加也是有益的。
需要更多的研究来理解这些看似矛盾的发现。由于 IDO1 或 TDO 的敲除小鼠是可行的,这些模型对于进一步研究 KP 中的寿命调节机制和潜在治疗靶点可能很有价值。KP 调节的寿命延长效应可能源于一般健康益处,而不是疾病特异性效应。
● 代谢综合征和肥胖
在患有代谢综合征的人类患者中,据报道 IDO1 过度激活会导致血清 Kyn 水平升高以及 Kyn/Trp 比率与肥胖、代谢综合征、BMI 和血液甘油三酯之间的相关性。
肥胖患者IDO1 的局部激活
IDO1和 KP 下游酶(如犬尿氨酸酶 (KYNU)、犬尿氨酸氨基转移酶 (KAT) 和犬尿氨酸 3-单加氧酶 (KMO))的基因表达增加已在肥胖患者的脂肪组织中观察到,表明 IDO1 的局部激活。
然而,循环 5-HT 水平在代谢综合征中降低,并且与 BMI 和体脂呈负相关。
色氨酸转化产物吲哚衍生物起作用
微生物群通过色氨酸转化产生的几种吲哚衍生物可能在代谢综合征的发病机制中起作用。
吲哚本身已被证明可以刺激肠内分泌 L 细胞产生胰高血糖素样肽-1 (GLP-1),这是一种刺激胰腺 β 细胞分泌胰岛素的肠降血糖素。这种机制涉及快速抑制刺激 GLP-1 分泌的电压门控 K+ 通道,但受 ATP 合成抑制的长期影响控制,减少 GLP-1 分泌。
硫酸吲哚酚促炎和氧化作用,与心血管和肾病等相关
吲哚也在肝脏中被吸收并代谢为硫酸吲哚酚。在肾衰竭期间,这种代谢物会积累,其促炎和氧化作用与动脉粥样硬化、动脉硬化、充血性心力衰竭和其他心血管并发症的发病机制有关,这些并发症在慢性肾衰竭患者中尤为突出。
硫酸吲哚酚在肾功能正常的受试者中的作用仍有待确定。KP 也与动脉粥样硬化有关。在小鼠模型中,IDO1 缺乏症通过 IL-10 产生失调减少了动脉粥样硬化病变的发展,这是一种通过施用 Kna 逆转的表型。在人类中,高 Kna 水平与不稳定的斑块表型相关。
低度慢性炎症可能有助于 IDO1 激活
KP 的过度激活也可能参与低度炎症情况下胰岛素抵抗的发生,例如肥胖、抑郁、丙型肝炎病毒感染和心血管疾病。人体和实验数据表明,黄嘌呤酸和 KP 的其他产物对胰岛素的产生和释放以及对靶组织的影响具有有害影响。
大脑中产生的血清素会引起饱腹感,但在微生物群的直接影响下产生的肠道来源的 5-HT 不会穿过血脑屏障。然而,色氨酸和直接 5-HT 前体 5-HTP 确实可以穿过血脑屏障,从而间接调节中枢 5-HT 的产生和功能。
外周 5-HT 独立于任何中枢效应影响宿主代谢
肠道来源的 5-HT 能够诱导食欲减退和饱腹感,其水平在禁食期间增加并刺激脂肪组织中的脂肪分解和肝细胞中的糖异生,有利于血糖控制。随后,通过一种涉及产热棕色脂肪组织消耗更多能量的机制,给予高脂肪饮食的 TpH1 基因或化学消融小鼠可免于肥胖、胰岛素抵抗和非酒精性脂肪肝疾病 (NAFLD)。
然而,这些结果可能不适用于棕色脂肪组织含量低且随年龄增长而减少的成年人。此外,人类肥胖与外周 5-HT 减少有关,表明其在发病机制中的复杂作用。
已经使用小鼠模型研究了 AhR 在代谢综合征中的作用,但尚未得出明确的结论。这可能与 AhR 的多重作用有关,AhR 在参与代谢综合征发病机制的各种细胞类型(肠细胞、肝细胞和免疫细胞)中表达。
/
/
▼
● 癌症中的色氨酸代谢
多项证据表明色氨酸代谢在癌症中具有重要作用,通过抑制抗肿瘤免疫反应和增加癌细胞的恶性特性来促进肿瘤进展。
首先,色氨酸降解酶在多种癌症中表达
IDO1 在大约 58% 的人类肿瘤中表达,其表达与多种癌症的不良临床结果相关,包括黑色素瘤、妇科癌症、结肠癌和血液系统恶性肿瘤。
IDO1 表达要么作为一种反调节机制被诱导,以响应从肿瘤浸润性免疫细胞释放的细胞因子,要么它的表达通过肿瘤固有的致癌信号传导维持。
TDO 催化与 IDO1 相同的反应,在神经胶质瘤、黑色素瘤、卵巢癌、肝癌、乳腺癌、非小细胞肺癌、肾细胞癌和膀胱癌中表达,并已被证明可促进肿瘤进展。
其次,各类癌症患者中全身色氨酸水平降低
已在成人 T 细胞白血病、结直肠癌 、妇科癌症、恶性黑色素瘤、肺癌和恶性神经胶质瘤患者中测量到全身色氨酸水平降低。在患有这些癌症的患者的血液中很少观察到 KP 代谢物浓度升高,这可能表明肿瘤微环境中 Kyn 和下游代谢物的局部变化受到更多限制。
第三,色氨酸降解在调节 Treg细胞和癌症中的免疫细胞浸润中发挥作用
FOXP3 + T reg细胞与宫颈癌引流淋巴结中表达 IDO1 的 DC 直接接触,IDO1 表达与转移性胰腺导管腺癌患者CD4 + CD25 + FOXP3 + T reg细胞增加有关,急性髓性白血病 (AML) 33和非霍奇金淋巴瘤。
此外,IDO1 表达与 CD3 + T 细胞、CD8的低肿瘤浸润相关+ T 细胞和 CD3 +和 CD8 + T 细胞以及 CD57 +自然杀伤细胞分别存在于结直肠癌、卵巢癌和子宫内膜癌患者中。
最近的一项研究表明,肿瘤再生细胞将 Kyn 转移到 CD8 + T 细胞,这反过来又以 AHR 依赖性方式上调程序性细胞死亡蛋白 1 (PD-1)。总而言之,这些观察结果为色氨酸代谢在肿瘤细胞免疫逃逸中的作用提供了机制解释。
第四,色氨酸代谢物可以有效促进癌细胞的运动和转移
例如,体外研究表明,TDO 在胶质母细胞瘤或乳腺癌细胞中的表达可促进肿瘤细胞迁移和侵袭。类似地,IDO1 的过表达增强了肺癌细胞的运动性,而敲除则降低了运动性。
这种促迁移表型也反映在临床前模型中由色氨酸降解引起的转移形成促进。药理学 TDO 抑制减少了肺癌小鼠模型肺部肿瘤结节的数量。
植入小鼠体内的人肺癌细胞中的 IDO1 过表达增加了大脑、肝脏和骨骼中的转移形成,而 IDO1 缺乏减少了转移负担并提高了乳腺癌衍生肺转移小鼠模型的存活率。
此外,TDO-AHR 信号轴促进了对贴壁依赖性细胞从周围细胞外基质分离时发生的程序性细胞死亡的抵抗,这是转移的关键步骤。最后,肿瘤内 IDO1 表达已被证明与结直肠癌肝转移、肝细胞癌远处转移和子宫内膜癌淋巴结转移的频率相关。
第五,NAD+ 在癌症生物学中通过色氨酸 de novo 途径产生的作用
在小鼠中,色氨酸代谢受损导致肝脏中从头合成 NAD+ 受到抑制,从而通过 DNA 损伤促进肝肿瘤发生。
在人类神经胶质瘤中,从色氨酸重新产生的 NAD+ 赋予对放化疗诱导的氧化应激的抗性。有趣的是,胶质瘤细胞和小胶质细胞合作产生 NAD+。
此外,在人类癌细胞中,IDO1 与通过产生 NAD +改善 DNA 修复和介导对治疗的抗性有关,例如 PARP 抑制剂奥拉帕尼、γ-辐射和化疗剂顺铂。因此,抑制色氨安代谢也可能通过从头形成 NAD +来防止治疗耐药性;然而,根据 NAD +合成所必需的 KP 酶的表达,这种效应可能是组织特异性或细胞特异性的,因此需要进一步研究。
● 靶向癌症中的 IDO1 和 TDO
基于 IDO1 和 TDO 的肿瘤促进功能,已经研究了这些酶的小分子抑制剂用于癌症治疗。临床阶段 IDO1 抑制剂 epacadostat (INCB024360)、navoximod (NLG-919/GDC919)等化学结构已被公开。未公开结构的化合物KHK2455、LY3381916和MK-7162也作为IDO1抑制剂进入临床评估。
TDO 抑制剂(最初被开发为抗抑郁药以提高全身色氨酸水平,从而提高大脑血清素浓度)也正在探索用于癌症治疗,但尚未进入临床试验阶段。
此外,indoximod 正在临床试验中进行研究,但与 L-1-MT 237不同,它们不是 IDO1抑制剂及其作用机制,尽管它似乎与 IDO1 表达有关,但仍存在争议。
然而,IDO1 抑制的最大治疗潜力预计是它与其他疗法的联合使用,这一直是大多数 II 期和 III 期研究的重点。
● 与免疫检查点抑制剂联合
IDO1 抑制剂的临床评估最先进的是它们与针对免疫系统检查点的单克隆抗体的组合,例如细胞毒性 T 淋巴细胞相关蛋白 4 (CTLA4)、PD-1 或其配体 (PD-L1),其中一些已被批准用于近年来治疗多种癌症的基础上部分患者出现前所未有的反应。
然而,由于相当大比例的患者无法从检查点抑制剂中获益,因此人们非常有兴趣确定缺乏治疗反应和治疗耐药性的分子基础,因为这些知识可能表明潜在的联合疗法可以改善反应。
有趣的是,在使用 PD-1 受体阻断剂 pembrolizumab 治疗期间,肉瘤患者的 Kyn:Trp 血浆比率增加,表明 IDO1 可能由免疫检查点封锁诱导。最有可能的是,这种 IDO1 的诱导,预计会抵消免疫检查点抑制的免疫刺激作用,是通过活化的 T 细胞产生的 IFNγ 介导的。
一项临床前研究表明,抑制 IDO1 会略微增强抗 CTLA4、抗 PD-1–PD-L1 和抗 GITR(糖皮质激素诱导的 TNFR 相关蛋白)疗法的疗效。
这些发现虽然不大,但引发了对 IDO1 抑制剂与免疫检查点抑制剂联合治疗的广泛临床研究。在 epacadostat 与 pembrolizumab 联合治疗的 I/II 期单臂试验获得令人鼓舞的数据后,在无法切除或转移性黑色素瘤患者中进行了 III 期试验。
尽管 ECHO-301 试验的阴性结果明显代表了 IDO1 抑制剂在癌症免疫治疗中的开发受挫,但它也激励人们利用临床试验来更多地了解 IDO1 抑制剂在癌症中的作用机制,以开发更复杂的生物标志物用于患者选择和治疗监测,并利用该途径中的新靶点,例如 AhR。
● 与免疫检查点抑制剂联合
目前已经计划在验证试验中继续研究 IDO1 抑制剂在联合免疫疗法中的潜力,包括不同于与 PD-1 和 PD-L1 拮抗剂组合的策略。
几项测试 IDO1 抑制剂 epacadostat 与抗肿瘤疫苗联合应用的临床试验正在进行中,可能会显示阻断 IDO1 是否会提高抗肿瘤疫苗接种的功效。这背后的基本原理是干扰素信号对 IDO1 的上调涉及多种免疫相关途径。例如,TLR 的激活通过干扰素诱导 IDO1 表达。
抑制抗肿瘤免疫反应的其他几种途径也与驱动色氨酸降解酶的肿瘤表达有关,包括 AhR 信号、TGFβ 信号和信号转导和转录激活因子 3 (STAT3)。
因此可以设想两种情况:
• 如果这些途径的抑制剂非常有效并且同时完全消除了色氨酸降解酶的表达,那么它们可能会使 IDO1 或 TDO 抑制剂在这种情况下变得可有可无。
• 相反,如果这些药物不能完全减轻 IDO1 和/或 TDO 的表达,它们可能与色氨酸代谢抑制剂协同作用。相比之下,其他治疗方法可能会诱导 IDO1 作为一种不良影响,这表明这些疗法与 IDO1 抑制剂的组合可能是有益的。
/
/
▼
前面概述了色氨酸及其在肠道中的三种命运。在这里主要谈论吲哚/AhR 通路,将深入探讨 AhR 信号的好处、为什么大多数人都缺乏这种途径、增加肠道中 AhR 活性的潜在策略,以及 AhR 可能被过度刺激的一些例外情况。
● 什么是AhR?
芳烃受体 (AhR)是一种转录因子——一种调节基因表达的蛋白质。结合并激活受体的分子称为激动剂。
AhR 的激动剂(即激活剂)主要分为三类分子:
AhR 最初因其在对二恶英和其他芳基碳氢化合物等环境毒素作出反应中的作用而被发现。这些污染物是 AhR 的非常强的激活剂。它们的结合增加了有助于促进其解毒的酶的表达。
肠道细菌,包括各种梭菌属、拟杆菌属、真杆菌属、乳杆菌属和双歧杆菌属,可以直接将色氨酸转化为称为吲哚的化合物,其中许多结合并激活 AhR。
近年来,膳食化合物也被证明可以激活 AhR。Indole-3-carbinol (I3C) 是一种源自十字花科蔬菜分解的化合物,可以结合并激活 AhR。
虽然环境污染物对 AhR 的慢性激活可能对健康产生负面影响,但肠道代谢物和膳食化合物对 AhR 的瞬时激活具有许多积极的下游影响。
● 肠道 AhR 激活的诸多好处
规律的、短暂的 AhR 信号在肠道和整体健康中起着许多重要作用。
1) 维持肠道屏障功能
AhR 刺激肠道中的先天免疫细胞产生细胞因子 IL-22,这是一种促进粘液产生和抗菌肽分泌的信号分子。在称为隐窝的肠道屏障口袋中,AhR 还支持干细胞增殖,这对于正常的肠道更新和修复至关重要。
2) 调节肠道菌群的组成
缺乏 AhR 刺激会导致促炎性肠杆菌科的扩张和产丁酸梭菌的减少,这是肠道菌群失调的常见特征。
3) 维持肠道免疫细胞群并减少炎症
AhR 支持肠道上皮细胞内足够数量的淋巴细胞。它还在将调节性 T 细胞引导至肠道并支持其抑制炎症的能力方面发挥关键作用。
4) 调节肠神经系统和肠蠕动
AhR 已被证明在调节蠕动方面发挥作用,蠕动是沿着胃肠道移动食物的肌肉收缩。AhR 也可能与损伤后肠神经的再生有关。
5) 防止念珠菌和其他肠道感染
AhR 激活通过支持 IL-22 信号传导在维持对酵母白色念珠菌和细菌病原体的定植抗性方面发挥重要作用。
6) 支持肺部的免疫防御
肠道 AhR 在防止其他粘膜表面(如肺)感染方面也起着重要作用。2019 年的一项研究发现,抗生素治疗后提高肠道中的 AhR 活性可显着减少肺部致病菌数量。
7) 促进健康的皮肤屏障功能
肠道 AhR 对于维持皮肤屏障的完整性至关重要。2016 年的一项研究发现,从饮食中去除 AhR 配体会损害皮肤屏障功能,而重新添加 AhR 激活剂吲哚-3-甲醇可挽救屏障缺陷,即使在老年小鼠中也是如此。
8) 激活解毒途径
AhR 在许多物质的解毒中发挥作用,包括多环芳烃、霉菌毒素、重金属和雌激素,激活整个身体的 解毒途径。
9) 保护肝肾功能
来自肠道色氨酸代谢的 AhR 信号也被证明可以预防非酒精性脂肪肝、酒精性肝损伤和肾纤维化。
10) 支持神经系统健康
星形胶质细胞中通过 AhR 发出的膳食色氨酸代谢物信号已被证明可以限制中枢神经系统的炎症。肠道 AhR 活性还促进成人神经发生,即新神经元的形成。
AhR 信号减少:许多慢性病的一个特征
在多种慢性疾病中观察到肠道 AhR 活性降低,包括炎症性肠病、肠易激综合征、结直肠癌、肥胖、代谢综合征、高血压、动脉粥样硬化、抑郁症、炎症性皮肤病、乳糜泻和多发性硬化症等疾病。
● 是什么导致 AhR 信号减少?
影响因素很多,包括:
改变的肠道微生物群组成通常无法产生已知可激活 AhR 的化合物,包括色氨酸衍生的吲哚和短链脂肪酸丁酸盐。
色氨酸(细菌吲哚形成的底物)的摄入量减少和/或植物性食物中 AhR 激动剂的摄入量减少都会减少 AhR 激动剂的总量。人造甜味剂的消费也被证明会减少 AhR 信号。
压力、炎症和某些类型的感染可以将色氨酸代谢从 AhR 转移到其他途径(后面我们会持续分享)。
解决这些根本原因始终是恢复 AhR 活动的第一步。
● 增加 AhR 活性的其他策略
以下是已知会暂时增加 AhR 活性的其他干预措施的总结。但是需要注意 AhR 激动剂具有物种特异性和组织特异性作用。它们的效果还取决于浓度,在存在多种化合物的情况下,它们甚至可能相互竞争——因此虽然这里列出了很多可能性,但“厨房水槽”方法并不一定是理想的。
希望在接下来的几年里,我们将看到更多的人体临床试验,以阐明这些疗法中的哪些可能对以 AhR 缺陷为特征的疾病状态最有帮助。以下信息不能视为医疗建议。
▸Indole-3-carbinol (I3C)
这种化合物由球芽甘蓝、卷心菜、西兰花、花椰菜和芥菜等十字花科蔬菜中的葡糖甘蓝素分解产生,是一种有效的 AhR 激活剂。
在动物模型中,I3C 已被证明可以诱导调节性 T 细胞的形成、抑制 Th17、保护粘液层、增加丁酸盐的产生、上调 PPAR-γ 并防止结肠炎。还对其潜在的抗癌和抗氧化作用进行了研究。虽然 I3C 或其衍生物二吲哚基甲烷 (DIM) 以补充剂形式提供,但较高剂量可能存在风险且人体研究有限,因此最好以整个食物形式食用 I3C。
注意:为了最大限度地提高膳食 I3C 的生物利用度,食用酸菜等生发酵形式的十字花科蔬菜,或在烹饪后加入芥末籽粉(含有黑芥子酶)。
▸丁酸盐
一种短链脂肪酸,是人肠上皮细胞中 AhR 的直接激活剂。在健康的肠道中,丁酸盐是由膳食纤维的发酵产生的,在较小程度上是蛋白质的发酵。它也以补充形式提供。一定情况下对结肠需要更有针对性。
▸尿石素 A
这种化合物由石榴、覆盆子和黑莓中的鞣花单宁分解产生,已被证明可以通过 AhR 增强肠道屏障功能。然而,估计只有 30-40% 的人拥有可以进行这种转化的细菌。尿石素 A也可以作为补充剂服用,并且作为食品成分已获得 FDA 公认的安全状态。
▸阳光照射
2019 年的一项研究发现,在小鼠身上,仅 15 分钟的 UV-B 照射就会诱导 AhR 靶基因在血液和外周组织(包括肠道)中的表达。
▸婴儿双歧杆菌(Bifidobacterium infantis)
该菌株在人乳低聚糖上生长后产生吲哚-3-乳酸,一种 AhR 的激活剂。它已在婴儿中得到充分研究,可用作婴儿益生菌。该菌株尚未在成人中进行研究。该配方确实含有大量的乳糖和残留量的大豆。
▸鼠李糖乳杆菌
某些乳杆菌菌株已被证明在色氨酸丰富时自然产生 AhR 激动剂。发现唯一已知可增加 AhR 活性且可商购的菌株是鼠李糖乳杆菌GG。
但是注意,不建议在使用抗生素期间或之后立即使用基于乳酸杆菌的益生菌,也不建议患有组胺不耐受/肥大细胞活化综合症的人服用。
▸Akkermansia muciniphila
至少在一项动物研究中,这种细菌或其外膜上的一种蛋白质增加了循环中的吲哚化合物并上调了 AhR 靶基因。
▸美沙拉嗪 (5-氨基水杨酸,5-ASA)
该药物是炎症性肠病 (IBD) 的一线治疗药物。这种药物上调 PPARgamma 和促细胞凋亡和抗增殖作用的能力。有趣的是,美沙拉嗪似乎也能激活 AhR。
▸咖啡
咖啡提取物,尤其是过滤较少的咖啡,如土耳其咖啡,已被证明可诱导肠上皮细胞中的 AhR 表达,并在啮齿动物模型中预防结肠炎。
▸萝卜硫素
虽然这种化合物的作用通常归因于 Nrf2 通路,但 AhR 似乎介导了它的许多保护作用。在喂食西方饮食的小鼠中,萝卜硫素增加了肠道中吲哚乙酸的产生,从而上调了 AhR 活性。
▸多酚
槲皮素、白藜芦醇和姜黄素都可以通过抑制控制 AhR 激动剂分解的 CYP1A1 酶来间接激活 AhR。其中,槲皮素在增强 AhR 信号传导方面最有效。
▸血清素
这种神经递质及其副产物 (5-HIAA) 也可以通过部分抑制 AhR 配体的清除来间接激活 AhR。这种效果取决于功能正常的血清素运输。
关于反馈调节而不是长期过度刺激 AhR 的重要性的说明:
这里不能忽略AhR 过度刺激的问题。虽然大多数慢性炎症性疾病都以 AhR 缺陷为特征,但在少数情况下,AhR 可能会过度激活,从而产生负面后果。这通常是由于污染物或霉菌暴露、严重病毒感染、或慢性肾病等引起的显着环境毒性。
这些条件的特点是持续的 AhR 激活,其对基因表达的影响与短暂的 AhR 激活截然不同。相反,由于某些解毒酶的上调,结合 AhR 的天然化合物在结合 AhR 时会被有效代谢。这种负反馈回路确保 AhR 信号是短暂的。
拥有足够的维生素,尤其B12 和叶酸可以防止慢性 AhR 过度刺激。尽管如此,在某些极端毒性或感染的情况下,短暂的 AhR 刺激是不提倡的。
/
/
▼
● 益 生 菌
益生菌,如属于乳杆菌属和双歧杆菌属的细菌,对色氨酸代谢产生有益影响。
益生菌促进血清素合成
一方面,益生菌,如乳杆菌和双歧杆菌中的物种,可以直接将色氨酸转化为血清素。
另一方面,一些益生菌乳杆菌菌株,如干酪乳杆菌327,可以通过增加TPH1表达间接促进结肠血清素合成。
益生菌与犬尿氨酸途径的调节密切相关
与血清5-羟色胺水平升高一致,大鼠口服约氏乳杆菌(Lactobacillus johnsonii) 无细胞上清液,也会导致血清中的犬尿氨酸水平降低,同时肠道IDO活性降低。口服约氏乳杆菌8周后,观察到人血清犬尿氨酸水平下降,色氨酸含量增加的明显趋势。
此前的一项研究还表明,大鼠服用益生菌婴儿双歧杆菌( Bifidobacteria infantis)会导致色氨酸水平升高,血液循环中的犬尿氨酸与色氨酸比率降低。
这些研究表明,一些属于乳杆菌和双歧杆菌的益生菌物种可能通过抑制犬尿氨酸途径改变宿主色氨酸代谢。
益生菌将色氨酸降解为吲哚化合物
此外,据报道,一些属于乳杆菌的细菌能够将色氨酸降解为吲哚化合物,如IAld、ILA和IAA。
向结肠炎易感小鼠口服3种色氨酸代谢乳杆菌菌株可促进微生物色氨酸新陈代谢芳基烃受体(Ah)依赖性信号传导,从而影响外周色氨酸的有效性。
尽管操纵肠道微生物群影响色氨酸代谢途径的机制尚未完全了解,但以肠道微生物群为靶点可能是调节色氨酸新陈代谢的一种有前途的方法。
● 抗 生 素
口服抗生素能够重塑肠道微生物群的组成和代谢。口服广谱抗生素会导致肠道微生物群耗竭,降低结肠血清素水平,进而延缓小鼠结肠运动。
正如结肠中关键合酶TPH1的下调所证明的,这项研究指出了共生微生物群在调节肠道血清素合成中的可能作用。
据报道,抗生素的微生物操纵会影响犬尿氨酸途径,因为抗生素诱导的微生物群耗竭会导致小鼠和猪的循环色氨酸可用性增加,并降低沿犬尿氨素途径的代谢。
此外,一些研究表明,抗生素诱导的肠道微生物改变也有利于猪体内的微生物色氨酸降解途径。随着循环色氨酸水平的增加,口服抗生素降低了空肠中色氨酸的可用性,并降低了猪大肠中的微生物色氨酸脱羧活性。
此外,口服抗生素会增加猪大肠中吲哚和吲哚化合物的含量。
有趣的是,最近的一项研究表明,回肠末端输注专门针对大肠微生物群的广谱抗生素会导致血液循环中的色氨酸水平降低,并增强微生物色氨酸降解,从而增加大肠中的吲哚水平。与之前的研究结果相反,该研究的发现表明,肠道微生物群在响应抗生素操作而调节色氨酸代谢方面发挥了独特的作用。
● 饮 食
饮食被认为是影响微生物色氨酸代谢的重要因素。
高脂肪饮食:抑制微生物从色氨酸向吲哚代谢物的转化,从而影响免疫调节
例如,最近的一项研究表明,高脂肪饮食会耗尽小鼠盲肠中的微生物代谢产物IAA和色胺,这表明在高脂肪饮食下,微生物色氨酸降解途径可以减弱。
高脂肪饮食增加了Alistipes和Bacteroides的丰度,同时减少了 Faecalibacterium。高脂肪饮食增加了致病菌属Alistipes,同时减少了有益菌 Parabacteroides distasonis ,导致小鼠肠道屏障功能受损。
高脂饮食显著增加小鼠肠道的IDO活性,促进色氨酸分解代谢为犬尿氨酸。
在暴露于高脂饮食的情况下,肠道微环境受到影响,随后抑制微生物从色氨酸向吲哚代谢物的转化,特别是吲哚-3-丙酸、吲哚-3-乳酸和吲哚乙酸盐。这些代谢物被认为是 AhR 激动剂,在免疫调节中起着关键作用。
配方奶:影响新生猪色氨酸代谢
母乳不仅是早期营养的唯一来源,而且有助于宿主肠道微生物群的成熟。有趣的是,之前的一项研究发现,配方奶引起的肠道微生物群的改变使新生猪结肠中的色氨酸代谢从血清素转变为色胺。
碳水化合物:影响色氨酸代谢速率
微生物色氨酸代谢的速率可能会受到管腔内营养物质(如碳水化合物)可用性变化的影响。
正如先前的体外研究所证明的,从仔猪粪便中分离出的一株利用色氨酸的细菌使用色氨酸进行细菌蛋白质合成,以可消化碳水化合物(葡萄糖)为底物,而不可消化碳水化合物(低聚果糖)是吲哚产生的底物。
此外,通过添加不可消化的碳水化合物,如低聚果糖和抗性淀粉,增加碳水化合物的可用性,促进碳水化合物代谢,从而增加短链脂肪酸的产量,同时减少色氨酸降解和仔猪大肠中的吲哚化合物。
事实证明,通过盲肠淀粉输注增加大肠碳水化合物的可用性可以抑制微生物色氨酸降解,从而导致大肠和血清中色氨酸水平的增加。
这些研究表明,增加碳水化合物的可用性抑制了肠道中的微生物色氨酸降解,这将进一步影响循环色氨酸库。
相比之下,增加碳水化合物的可用性促进了肠道血清素的合成,这与增加胃肠道传输有关,正如先前在口服多糖的小鼠中进行的一项研究所报告的那样。微生物短链脂肪酸的产生增强可以参与这一过程,因为它们已经被证明可以刺激结肠EC中的血清素释放。
腔内色氨酸可用性是影响微生物色氨酸代谢的另一个直接因素
IDO激活或饮食限制导致的宿主色氨酸耗竭可减少微生物增殖,尤其是乳酸杆菌中的细菌,据报道,其中一些细菌是利用色氨酸的细菌。通过饮食喂养选择性地恢复色氨酸水平导致乳杆菌的扩张,这进一步导致微生物色氨酸代谢的增强,IAld增加。
鉴于色氨酸也会被宿主直接吸收,肠道微生物群、管腔色氨酸可用性和宿主色氨酸代谢之间的复杂串扰需要进一步研究。
富含麦麸的饮食在调节色氨酸代谢物的合成和生物转化中的作用
富含麦麸的饮食有效地抑制了色氨酸向犬尿氨酸途径代谢物的转化,同时增加了褪黑激素和微生物分解代谢物,即吲哚-3-丙酸、吲哚-3-乙醛和 5-羟基-吲哚-3-乙酸。
麦麸增加了促进健康的细菌(例如,Akkermansia和Lactobacillus),它们与色氨酸衍生的吲哚类代谢物显著相关。
富含麦麸的饮食可有效调节与免疫功能相关的微生物转化和色氨酸合成(即增加 AhR 和 IL-22 的结肠表达),同时改善葡萄糖和脂质稳态,以及增加肠道健康促进菌的丰度。
/
/
色氨酸代谢调节炎症、肠道稳态和大脑功能等。色氨酸可用性和代谢的微生物调节对许多肠脑轴疾病具有重要意义,包括伴有精神疾病的胃肠道疾病,如IBS 、IBD,其他具有胃肠道功能障碍的中枢神经系统疾病,如自闭症等。
由于宿主色氨酸代谢直接或间接受肠道菌群调节,许多因素会影响肠道微生物群的组成和代谢,包括饮食、抗生素、益生菌等可以调节肠道微生物群,调节色氨酸的可用性,因此靶向肠道菌群干预是治疗肠脑轴疾病的有前途的方法。
色氨酸代谢可以作为与年龄相关的病理和寿命的调节剂。犬尿氨酸通路及其代谢产物可能成为预测衰老相关疾病的潜在风险标记物。
色氨酸主要经犬尿氨酸代谢,既促进肿瘤细胞固有的恶性特性,又限制肿瘤免疫,因此它是癌症免疫治疗的重要药物开发靶点。肿瘤中色氨酸代谢的改变常伴随色氨酸相关酶基因表达的异常,基于此,IDO抑制剂、TDO抑制剂及联合治疗被应用于大量的临床试验中。
随着这方面研究的不断深入,我们会持续关注和更新色氨酸代谢和全身健康稳态以及药物进展等。
主要参考文献
Le Floc’h N, Otten W, Merlot E. Tryptophan metabolism, from nutrition to potential therapeutic applications. Amino Acids. 2011 Nov;41(5):1195-205. doi: 10.1007/s00726-010-0752-7. Epub 2010 Sep 25. PMID: 20872026.
Gao K, Mu CL, Farzi A, Zhu WY. Tryptophan Metabolism: A Link Between the Gut Microbiota and Brain. Adv Nutr. 2020 May 1;11(3):709-723. doi: 10.1093/advances/nmz127. PMID: 31825083; PMCID: PMC7231603.
Sorgdrager FJH, Naudé PJW, Kema IP, Nollen EA, Deyn PP. Tryptophan Metabolism in Inflammaging: From Biomarker to Therapeutic Target. Front Immunol. 2019 Oct 30;10:2565. doi: 10.3389/fimmu.2019.02565. PMID: 31736978; PMCID: PMC6833926.
Richard DM, Dawes MA, Mathias CW, Acheson A, Hill-Kapturczak N, Dougherty DM. L-Tryptophan: Basic Metabolic Functions, Behavioral Research and Therapeutic Indications. Int J Tryptophan Res. 2009 Mar 23;2:45-60. doi: 10.4137/ijtr.s2129. PMID: 20651948; PMCID: PMC2908021.
Platten M, Nollen EAA, Röhrig UF, Fallarino F, Opitz CA. Tryptophan metabolism as a common therapeutic target in cancer, neurodegeneration and beyond. Nat Rev Drug Discov. 2019 May;18(5):379-401. doi: 10.1038/s41573-019-0016-5. PMID: 30760888.
Chen G, Zhou S, Chen Q, Liu M, Dong M, Hou J, Zhou B. Tryptophan-5-HT pathway disorder was uncovered in the olfactory bulb of a depression mice model by metabolomic analysis. Front Mol Neurosci. 2022 Oct 10;15:965697. doi: 10.3389/fnmol.2022.965697. PMID: 36299862; PMCID: PMC9589483.
Roth W, Zadeh K, Vekariya R, Ge Y, Mohamadzadeh M. Tryptophan Metabolism and Gut-Brain Homeostasis. Int J Mol Sci. 2021 Mar 15;22(6):2973. doi: 10.3390/ijms22062973. PMID: 33804088; PMCID: PMC8000752.
van der Goot AT, Nollen EA. Tryptophan metabolism: entering the field of aging and age-related pathologies. Trends Mol Med. 2013 Jun;19(6):336-44. doi: 10.1016/j.molmed.2013.02.007. Epub 2013 Apr 2. PMID: 23562344.
Yao K, Fang J, Yin YL, Feng ZM, Tang ZR, Wu G. Tryptophan metabolism in animals: important roles in nutrition and health. Front Biosci (Schol Ed). 2011 Jan 1;3(1):286-97. doi: 10.2741/s152. PMID: 21196377.
Höglund E, Øverli Ø, Winberg S. Tryptophan Metabolic Pathways and Brain Serotonergic Activity: A Comparative Review. Front Endocrinol (Lausanne). 2019 Apr 8;10:158. doi: 10.3389/fendo.2019.00158. PMID: 31024440; PMCID: PMC6463810.
Gibson EL. Tryptophan supplementation and serotonin function: genetic variations in behavioural effects. Proc Nutr Soc. 2018 May;77(2):174-188. doi: 10.1017/S0029665117004451. Epub 2018 Jan 25. PMID: 29368666.
Gostner JM, Becker K, Kofler H, Strasser B, Fuchs D. Tryptophan Metabolism in Allergic Disorders. Int Arch Allergy Immunol. 2016;169(4):203-15. doi: 10.1159/000445500. Epub 2016 May 4. PMID: 27161289; PMCID: PMC5433561.
Liu XH, Zhai XY. Role of tryptophan metabolism in cancers and therapeutic implications. Biochimie. 2021 Mar;182:131-139. doi: 10.1016/j.biochi.2021.01.005. Epub 2021 Jan 16. PMID: 33460767.
Yan T, Shi L, Liu T, Zhang X, Yang M, Peng W, Sun X, Yan L, Dai X, Yang X. Diet-rich in wheat bran modulates tryptophan metabolism and AhR/IL-22 signalling mediated metabolic health and gut dysbacteriosis: A novel prebiotic-like activity of wheat bran. Food Res Int. 2023 Jan;163:112179. doi: 10.1016/j.foodres.2022.112179. Epub 2022 Nov 19. PMID: 36596122.