-

 CNAS L23010
CNAS L23010

 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业 二级病原微生物安全实验室
二级病原微生物安全实验室- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业 二级病原微生物安全实验室
革兰氏阳性和阴性菌
在日常生活中,我们常常会看到药物或抗菌产品适应症会这样写到,对革兰氏阳性菌有效,对革兰氏阴性菌敏感,或者说对革兰氏阴性菌有效,对革兰氏阳性菌无效。可能很多人不是很清楚或搞不懂二者的区别。
本文主要介绍革兰氏阳性和阴性菌,它们的区别,代表性菌种以及针对用药等。
1884年,细菌学家Hans Christian Gram发明了革兰氏染色法来鉴别区分细菌。这种技术将细菌分成两大类,即革兰氏阳性菌(G+)和革兰氏阴性菌(G-)。区分主要是这两类细菌细胞壁成分不同,因而着色也不同所致。
这两类细菌的生理结构,疾病原因以及抗菌作用不一,因此,区分病原菌是革兰氏阳性菌,还是阴性菌,在临床确定感染和选择用药方面意义重大。
革兰氏阴性菌致病多由于患者有基础疾病或者体质比较差,肠道细菌感染引起的腹泻多是由肠道菌群中的革兰氏阴性菌所致,如大肠杆菌、沙门氏菌、志贺氏菌,布氏杆菌等,治疗这类细菌感染,一般使用三代头孢菌素以及喹诺酮类抗生素。注意大多数革兰氏阴性菌对青霉素耐药或不敏感。
大多数化脓性球菌都属于革兰氏阳性菌,它们能产生外毒素使人致病,常见的菌种有葡萄球菌、链球菌、肺炎双球菌、李斯特菌、炭疽杆菌、白喉杆菌、破伤风杆菌等。尤其在人体肠道内,革兰氏阳性致病菌致病几率更大。大多数由革兰氏阳性菌引起的感染可以用相当少量的抗生素治疗。青霉素、氯唑西林和红霉素足以覆盖 90% 的革兰氏阳性感染。
此外,某些广谱抗生素对革兰氏阳性菌和革兰氏阴性菌都有抗菌作用,如氨苄青霉素、庆大霉素、土霉素、磷霉素及环丙沙星等,但是作用效果可能不是最优。此外,磺胺类药物也属于广谱抑菌药物。
临床应用时,如果对细菌感染比较明确,尽量使用窄谱抗菌药物,如不太明确,可选用广谱抗菌药物。因此,临床治疗疾病时,首先要对药物的作用与用途要详细了解,然后再根据感染类型或诊断结果合理选择药物,这样才能取得最佳治疗效果。否则,药物选择不当,将会出现无效或越治越重的结果。
革兰氏阴性菌和革兰氏阳性菌之间的主要区别在于肽聚糖层的厚度和外部脂质膜的存在与否。
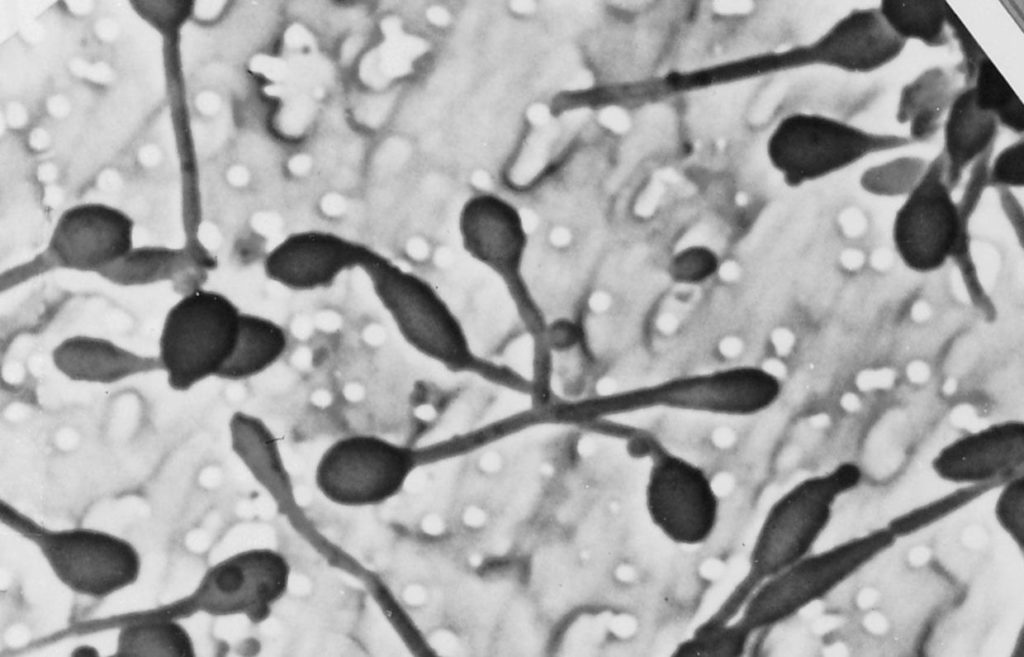
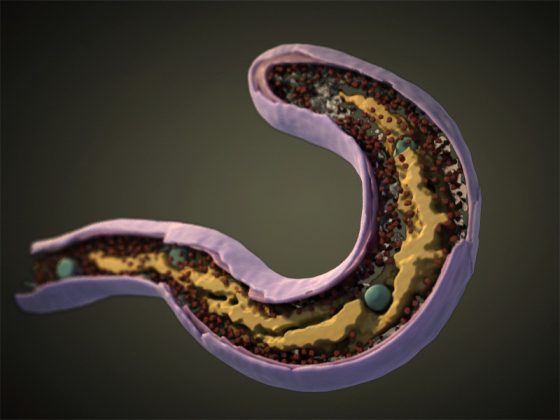
不同细菌的革兰氏染色
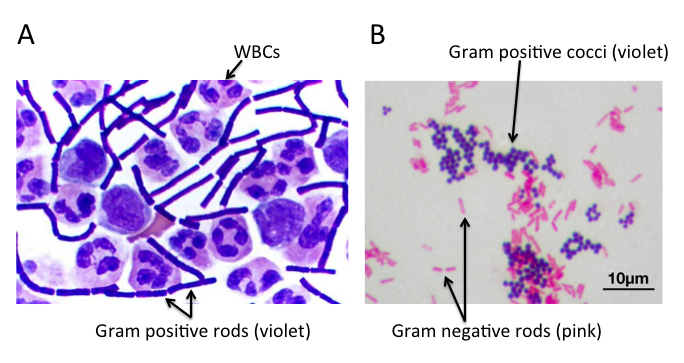
A) 脑脊液样本中的杆状革兰氏阳性炭疽杆菌(导致炭疽),也含有白细胞。B) 革兰氏阳性金黄色葡萄球菌(紫色或紫色)和革兰氏阴性杆菌大肠杆菌(粉红色)的革兰氏染色,它们是最常用的革兰氏染色参考细菌。参考来源:Wikipedia commons 和 tmedweb)
// 缺乏外膜,更容易受抗生素影响
革兰氏阳性菌的细胞壁含有肽聚糖、脂质、磷糖醛酸和磷壁酸。这种结构成分不同于由肽聚糖和外膜(由脂质、蛋白质和脂多糖组成)组成的革兰氏阴性细菌细胞壁。尽管革兰氏阳性菌具有较厚的肽聚糖层,但它们比革兰氏阴性菌更容易受到某些靶向细胞壁的抗生素的影响,因为它们缺乏外膜。
// 常见的致病菌
最常见的革兰氏阳性细菌包括葡萄球菌、链球菌、芽孢杆菌、梭状芽孢杆菌、李斯特菌、棒状杆菌等。这些革兰氏阳性菌的代表性物种是致病的,并可能引起多种疾病。
// 可用于治疗的抗生素
青霉素是影响革兰氏阳性菌的主要抗生素之一。
红霉素是另一种用于治疗革兰氏阳性细菌感染的强效抗生素。红霉素属于一类称为大环内酯类的抗生素,与阿奇霉素和克拉霉素同属一类。它通常用于对青霉素过敏的人。
甲氧苄啶/磺胺甲恶唑,克林霉素,克林霉素,强力霉素,万古霉素也可以用于特定的革兰氏阳性菌的感染。
// 层层抵抗更难杀死:细胞壁更硬,可改变外膜
革兰氏阴性菌有坚硬的保护外壳。它们的肽聚糖层比革兰氏阳性杆菌薄得多。
当它们的细胞壁受到干扰时,革兰氏阴性细菌会释放内毒素,症状更糟。同时,大多数抗生素为了接近它们的目标,必须通过外膜。例如,亲水性抗生素通过孔蛋白。革兰氏阴性菌可以通过改变它们的疏水特性或通过孔蛋白的突变来改变它们的外膜。这对这些细菌细胞产生了抵抗力。
// 暗藏玄机:比革兰氏阳性菌更危险
与革兰氏阳性菌相比,革兰氏阴性菌作为疾病生物体更危险,因为存在覆盖外膜的荚膜或粘液层。通过这种方式,微生物可以隐藏表面抗原,这个抗原可以触发人体免疫反应。
革兰氏阴性菌是一组臭名昭著的细菌,可导致多种疾病,包括肺炎、脑膜炎、淋病、细菌性痢疾、霍乱、胃炎等。在重症监护病房 (ICU)的患者,处于发病和死亡的高风险中,更容易遇见这类细菌,因此它们在医院具有重要的临床意义。
// 可用于治疗的抗生素
已经开发了许多不同种类的抗生素来杀死革兰氏阴性菌,例如头孢菌素、叶酸拮抗剂、哌拉西林-他唑巴坦、脲青霉素、内酰胺-β-内酰胺酶抑制剂、碳青霉烯类和喹诺酮类。它们是专门针对革兰氏阴性细菌而开发的,不过有时也对某些革兰氏阳性细菌有效。
革兰氏阳性菌与革兰氏阴性菌区别总结

参考来源:microbenotes
以上是革兰氏阳性菌和阴性菌的主要区别,接下来我们针对革兰氏阳/阴性菌,从细菌特征,形状表征,细胞结构等方面,进行更详细的介绍。
革兰氏阳性菌的定义是基于它们在革兰氏染色中用酒精短暂洗涤后保留结晶紫染料的能力。革兰氏阳性菌呈紫色。
这些细菌具有非常独特的特征,可以将其与其他类型的细菌区分开来。这些包括:
在革兰氏染色过程中,革兰氏阴性细菌在用酒精清洗后将失去结晶紫染料的颜色,并吸收反染物藏红花的粉红色/红色。
几乎在地球上的每个生活区域都可能发现革兰氏阴性细菌。
尽管大多数细菌是通过革兰氏染色染料进行区分的,但显微镜下的观察揭示了更多可用于定义和表征这些细菌的特征。
根据形状的定义,革兰氏阳性菌可分为两类:
革兰氏阳性细菌形成的其他特殊形状包括:
革兰氏阴性菌细胞的显微镜观察范围从杆状到芽孢杆菌,从球菌到螺旋状,螺旋状是最常见的形状。然而,有些表现出特殊的形状,如球杆菌、四分体、栅栏、毛状体等。例如:
革兰氏阳性菌的细胞壁含有肽聚糖、脂质、磷糖醛酸和磷壁酸。这种结构成分不同于由肽聚糖和外膜(由脂质、蛋白质和脂多糖组成)组成的革兰氏阴性细菌细胞壁。因此,革兰氏阳性菌的细胞壁很厚,并吸收了革兰氏染色的结晶紫染料。因此,显微镜下的革兰氏阳性菌呈紫色。
革兰氏阳性细菌细胞壁的结构特征
革兰氏阳性细菌具有由肽聚糖组成的厚的多层细胞壁(因为它含有肽和糖的混合物)。由于肽聚糖存在于大多数细菌中,但不存在于哺乳动物细胞中,因此它是抗菌药物的良好靶标(例如细胞壁合成抑制剂,包括青霉素、头孢菌素和万古霉素)。
这些抗生素会干扰转肽酶(也称为青霉素结合蛋白或PBP 的活性) 在细胞壁中催化相邻聚糖链之间的交联。
细胞壁还含有磷壁酸纤维,有助于细菌附着在宿主细胞膜(例如粘膜细胞)上,释放后会引起感染性休克,类似于革兰氏阴性菌释放的内毒素 (LPS) 产生的感染。
β-内酰胺酶(青霉素酶)是由细菌产生的酶家族,可水解 β-内酰胺抗生素(其中许多也是细菌来源)的四原子 β-内酯环,从而使其抗菌性能失活。
细菌细胞膜还可以包含ABC 外排泵这可能导致抗生素耐药性和对那些具有细胞内作用机制的药物(例如 DNA 促旋酶抑制剂或蛋白质合成抑制剂)的多药耐药性 (MDR)。
结晶紫染料附着在革兰氏阳性菌细胞壁的厚肽聚糖层上,在光学显微镜下观察时将它们染成紫色或紫色。
肽聚糖
它也被称为胞壁质(murein),占细菌细胞壁含量的 90%。
——维持形状,并保持细胞壁强度和弹性
它是一种优质聚合物,由两种相同的糖衍生物(N-乙酰氨基葡糖和 N-乙酰胞壁酸)以及 L-氨基酸链和蛋白质中很少发现的三种不同 D-氨基酸(即 D-谷氨酸、D-丙氨酸和内消旋二氨基庚二酸)组成,可保护细胞壁免受肽酶的攻击。
D-氨基酸和L-氨基酸连接到N-乙酰壁酸,L-氨基酸特别是赖氨酸可以取代中二氨基丙烯酸。
肽聚糖亚基的这种相互连接使肽聚糖具有很强的维持细菌形状和完整性的能力,并具有弹性和延展性。
肽聚糖也具有渗透性,允许分子进出细菌细胞。
——肽聚糖的生物合成
抑制细菌细胞壁肽聚糖层的合成是许多抗菌药物的分子靶点,包括 β-内酰胺类抗生素(青霉素、头孢菌素、碳青霉烯类和单环内酰胺类)和糖肽类抗生素(万古霉素和其他较新的类似物)。
这些药物的两个主要分子靶标是转肽酶,也称为青霉素结合蛋白 (PBP),因为它们与青霉素结合,而糖基转移酶 (GT) 可被万古霉素等糖肽抑制。
PBPs 有许多亚型,给定的细菌菌株可以表达多种 PBPs。这些 PBP 在其生理特性和与抗生素相互作用的敏感性方面可能有所不同。取决于 PBP 亚型,糖基转移酶可以作为单独的酶或作为与转肽酶相关的二聚体存在。
革兰氏阳性细胞壁生物合成
图片来源自(Wikipedia Commons 和 tmedweb)
细菌细胞壁由重复的 N-乙酰氨基葡萄糖 (NAG) 和 N-乙酰胞壁酸 (NAM) 亚基链组成。NAM 亚基附有短肽链。
肽链的组成因细菌而异,但近端的丙氨酸通常是 L-Ala,而远端的两个通常是 D-Ala。也与青霉素结合的细胞壁转肽酶(青霉素结合蛋白:PBPs)在肽侧链之间形成键,并从肽侧链之一排出末端 D-丙氨酸。
一旦形成交联,PBP 就会从壁上解离。用糖基转移酶 (GT)分离酶结构域NAM 和 NAG 残基之间的活性形成联系。一些高分子量 PBP(例如 PBP2)是含有转肽酶和糖基转移酶结构域的酶复合物。
磷壁酸纤维存在于革兰氏阳性菌的细胞壁中,由磷酸甘油或磷酸核糖醇的聚合物组成。它们参与细菌与黏膜细胞的附着,可诱发感染性休克,类似于革兰氏阴性菌释放的 LPS(内毒素)。
磷壁酸
这是由甘油共聚物组成的加固墙。
磷壁酸是水溶性的,占细菌细胞壁总干重的 50%。
它要么直接与肽聚糖共价连接,要么与细胞膜(脂磷壁酸)连接。通过 6-羟基 N-乙酰胞壁酸与肽聚糖直接相连。
带负电荷,延伸到肽聚糖表面,使细菌细胞壁带负电荷。
它还有助于维持细胞壁的结构。
它在革兰氏阴性菌中完全不存在。
脂质
它们在肽聚糖下方有一层薄薄的脂质,大约 2-5%,其作用是锚定细菌细胞壁。
细胞壁
——细胞壁非常复杂
结合细胞外膜的主要作用,加上一层肽聚糖,其功能特性复杂,这里是对细胞壁及其功能部分的描述。
革兰氏阴性细菌的细胞壁具有一层2-7nm的肽聚糖薄层和7-8nm厚的外膜。
——周质空间较大
显微镜下,细胞膜和细胞壁之间有一个空间,称为由周质组成的周质空间。在革兰氏阴性菌和革兰氏阳性菌中均能发现,但在革兰氏阴性菌中,周质空间较大。
革兰氏阴性细菌细胞壁
在结构上,革兰氏阴性细胞壁由细胞膜外部的两层组成:一层薄薄的肽聚糖(太薄而无法吸收大量甲基紫染色)和一层外膜(革兰氏阴性细菌独有),通常含有促进小(<700 Da)亲水分子(例如糖、氨基酸和维生素)扩散的孔蛋白。
许多抗生素(例如许多青霉素和头孢菌素)也可以通过孔蛋白扩散到达它们的作用部位。
然而,万古霉素 (1449 Da) 的质量太大,无法透过孔蛋白到达其作用部位,这使其对革兰氏阴性细菌无效。因此,外膜为革兰氏阴性菌提供了对某些抗生素的固有“内在抗性”,可以通过改变孔蛋白的表达水平或改变孔蛋白的孔特性以降低抗生素的渗透性来进一步修饰。
革兰氏阴性菌的外膜还含有脂多糖 (LPS)或内毒素,可被细菌排出,引起宿主强大的免疫反应。
周质空间
革兰氏阴性菌的周质空间由几种蛋白质组成,这些蛋白质有助于获取营养,例如攻击核酸和磷酸化分子的水解酶,以及积极协助将物质运输到细菌细胞中的结合蛋白。周质空间还具有合成肽聚糖和修饰可能对细胞造成伤害的有毒元素的酶。
肽聚糖
革兰氏阴性细菌细胞壁有一层薄的肽聚糖层,位于质膜上方,约占细胞干重的5%。厚度不超过4纳米,一些细菌如大肠杆菌只有2纳米厚的肽聚糖。
外膜和脂多糖
革兰氏阴性菌还有第二层脂质双层,位于肽聚糖层的外部。这种外膜通过布劳恩脂蛋白与肽聚糖相连。外膜和肽聚糖之间的紧密连接是维持外膜作为许多有毒分子和抗生素的不渗透屏障所必需的。
外膜上的粘附位点也加强了革兰氏阴性细胞壁,这些粘附位点在允许细胞接触和膜融合方面发挥作用。物质通过这些粘附位点进入细胞。
革兰氏阴性菌结构

图源:Jeff Dahl,wikipedia
外膜主要由脂多糖 (LPS)组成,脂多糖是由脂质和碳水化合物组成的大型复杂分子。脂多糖由3个单元组成:脂质A、核心多糖和O侧链。
脂质A由两种氨基葡萄糖糖衍生物组成,每个衍生物含有三种脂肪酸和焦磷酸盐,脂多糖的任何剩余部分都会伸出膜表面。
O侧链也称为O抗原,是从核心向外延伸的链。它由导致细菌菌株之间变异的糖组成。这些 O 抗原也负责细菌逃避抗体反应。
// 外膜及其脂多糖的作用
——脂多糖负责保护细胞壁免受外部攻击
LPS 带有负电荷,使电池表面带负电荷。因此,这稳定了膜结构。
脂质A是脂多糖的有毒成分,因此它起到内毒素的作用。
——防毒素进入,防成分丢失
外膜及其脂多糖有助于防止抗生素、胆汁盐和其他有毒元素进入并破坏细胞。
外膜由孔蛋白组成,使其具有渗透性,允许小分子(如葡萄糖)进入。维生素 B12 等较大的分子通过特定的载体运输穿过外膜。
外膜还有助于防止成分丢失,特别是来自周质空间。
下表描述了主要的革兰氏阳性致病菌,它们的基本形态特征以及它们在人类中引起的疾病。
已知革兰氏阴性菌是正常菌群,部分会导致严重的人类感染,从社区获得性感染到医院感染。
革兰氏阴性菌外膜的结构是其众多显着特征之一。脂多糖 (LPS) 存在于膜的外叶上,其脂质 A 部分用作内毒素。
// 革兰氏阴性菌感染:严重时可危及生命
如果由于某种原因,革兰氏阴性细菌能够到达动物的循环系统,脂多糖将激活免疫系统,并触发先天免疫反应,产生细胞因子和激素调节剂。这会引起炎症,并可能导致毒性反应,从而导致发烧、呼吸急促和低血压。这就是已知革兰氏阴性细菌会导致危及生命的休克的原因。
内毒素休克的一些症状:
发烧和发冷或体温下降、发炎、皮疹、呼吸急促、心率加快、低血压、多器官衰竭等。
下表给出了一些革兰氏阴性致病细菌的例子以及它们在人类宿主中引起疾病和感染时表现出的临床特征。

如上表所述,已知革兰氏阳性细菌会引起多种感染,如果不及时和适当地治疗和管理,可能对人类造成灾难性的影响。
革兰氏阳性杆菌感染用抗生素治疗。青霉素、氯唑西林和红霉素可治疗 90% 以上的革兰氏阳性菌。
常见的革兰氏阳性抗菌药物及作用机制

然而,抗生素耐药性正在成为革兰氏阳性感染的一个严重问题。研究人员正在开发新的药物来帮助解决这个问题。只有在绝对需要时才应使用抗生素。需要严格遵循感染控制标准,以防止抗生素耐药性感染的发展和传播。
由于它们的外膜,这些细菌对溶菌酶和青霉素具有抗性。这是因为存在保护内膜和细胞壁的外壁。
在周质空间(两个细胞膜之间的区域)中也发现了分解或改变抗生素的酶。用于治疗革兰氏阴性菌感染的治疗方法包括羧基、氨基和脲基青霉素。为了对抗可以消化这些药物的酶,有时将它们与β-内酰胺酶抑制剂结合使用。β-内酰胺酶是一种存在于周质中的酶。
针对细菌病原体的抗菌剂被称为抗生素。这些抗生素启动针对细菌细胞的阻断或抑制机制,以诱导细菌细胞增殖和复制。
用于对抗革兰氏阴性菌的抗生素示例

抗生素耐药性是现在世界上的一个主要临床问题。
耐多药细菌在人群中变得越来越普遍,如果不进行有效治疗,这种感染可能会导致肾功能衰竭、败血症,甚至死亡。
微生物以多种方式抑制临床治疗中使用的许多抗菌剂。这些包括改变药物结合位点的方法,改变药物构象的方法,改变膜通透性的方法,可以导致耐药机制失活。
例如革兰氏阴性菌中有两层膜,外膜和内膜。脂多糖被认为是一种非常强的免疫反应诱导剂,它具有三个重要成分:脂质 A、亲水性多糖、抗原 O 的疏水域。
疏水域在细胞膜的外部表达。它是疏水成分脂质a,它负责内毒素作用。LPS 在细菌中是可变的,并且由于遗传变异,一些细菌只产生一种不被 Toll 样受体识别的弱抗原。然而,有大量的革兰氏阴性菌团体可能会引起这样的反应。免疫系统也被一些 toll 样受体 4 (TLR4) 激活,这些受体存在于与免疫系统有关的众多细胞中,如巨噬细胞、单核细胞、中性粒细胞和树突状细胞。
由 LPS 和 TLR4 受体介导的先天免疫反应的激活导致反应增强,产生细胞因子、趋化因子和干扰素等。
免疫系统的反应取决于感染过程的严重程度以及侵袭性细菌中 LPS 的结构,这与菌的毒力有关。因此,虽然一些细菌(如大肠杆菌)可以诱导免疫系统,但其他细菌(如幽门螺杆菌)仅具有弱抗原性。
// 对付耐药性细菌新思路:根据电荷相互作用原理设计新药
2017 年,伊利诺伊大学化学教授和当前研究的合著者Paul Hergenrother(ACPP 负责人/MMG)在《自然》杂志上报告说 ,发现的一个关键是,如果向它们添加带正电荷的基团,例如胺,一些抗生素可以使用特定的膜孔穿透革兰氏阴性细菌的细胞膜。
这项工作表明,抗生素上带正电荷的胺基与细菌孔内的负电荷有良好的相互作用。这些吸引力使带有胺基的抗生素以一种更有利于能量的方式排列,因为它穿过收缩区的孔的最狭窄部分。不含胺的抗生素面临更高的能量屏障去通过孔隙。
这或许意味着未来可以设计新药(或修改旧药),以攻击和杀死对抗生素治疗具有耐药性的微生物。
问
革兰氏阳性细胞和革兰氏阴性细胞之间的三个区别是什么?
答
革兰氏阳性菌有一层厚的肽聚糖作为它们的细胞壁,而革兰氏阴性菌有一层薄薄的肽聚糖和外膜。
革兰氏阴性菌有脂多糖(LPS),而革兰氏阳性菌没有。
一些革兰氏阳性细菌含有霉菌酸,它会在细胞壁上形成一层蜡质层。
问
什么是革兰氏阳性感染?
答
由革兰氏阳性菌引起的感染,如耐甲氧西林金黄色葡萄球菌 (MRSA)、耐万古霉素肠球菌 (VRE) 和艰难梭菌是常见的多重耐药菌感染。
问
革兰氏阳性菌更容易治疗吗?
答
革兰氏阳性细菌,即那些具有肽聚糖外层的物种,更容易被杀死——它们的厚肽聚糖层很容易吸收抗生素和清洁产物。因此,某些容易杀死革兰氏阳性菌的洗涤剂不会破坏革兰氏阴性菌。
问
肠道内很多革兰氏阳性菌致病吗?
答
是的,人体肠胃道的大多数革兰氏阳性菌都是条件致病菌。包括微球菌、肠球菌、金黄色葡萄球菌、表皮葡萄球菌、腐生葡萄球菌、肺炎链球菌、草绿色链球菌、酿脓链球菌、无乳链球菌、破伤风梭菌、肉毒杆菌、产气荚膜梭菌、产气荚膜梭菌、艰难梭菌, 单核细胞增生李斯特菌等。
问
革兰氏阳性菌在哪里发现?
答
根据革兰氏阳性菌种,它们可以在人类的土壤、水生沉积物、灰尘、皮肤、口腔、肠道或生殖道中找到。
问
革兰氏阳性球菌危险吗?
答
革兰氏阳性菌可能是球菌或杆菌。这些称为常驻菌群的细菌通常不会引起疾病。革兰氏阳性杆菌引起某些感染,包括:炭疽。
问
革兰氏阳性菌对抗生素的抵抗力更强吗?
答
不是。与革兰氏阳性菌相比,革兰氏阴性菌对多种抗生素的耐药性更强。由于它们的外膜,它们对抗生素的抵抗力更强。
革兰氏阳性细菌更容易被杀死,因为它们的厚肽聚糖层很容易吸收抗生素和清洁剂。另一方面,革兰氏阴性细菌具有薄的肽聚糖层,不会吸收周围的任何异物。
问
革兰氏阳性菌有内毒素吗?
答
不会。内毒素与革兰氏阳性菌无关。这些细菌没有内毒素,因为它们没有外膜。另一方面,革兰氏阴性细菌会产生内毒素。
这些内毒素是革兰氏阴性细菌细胞外膜的一部分,只有当细胞裂解或细菌死亡时才会释放出来。内毒素是形成革兰氏阴性菌细胞壁结构的热稳定性脂多糖-蛋白质复合物。
问
为什么革兰氏阳性菌对抗生素更敏感?
答
尽管革兰氏阳性菌具有较厚的肽聚糖层,但它们比革兰氏阴性菌更容易受到某些靶向抗生素的细胞壁的影响,因为它们缺乏外膜。
大多数抗生素为了接近它们的目标,必须通过外膜。例如,亲水性抗生素通过孔蛋白。因此,革兰氏阴性菌可以通过改变它们的疏水特性或通过孔蛋白的突变来改变它们的外膜。这对这些细菌细胞产生了抵抗力。革兰氏阳性菌缺乏这一因素,因此革兰氏阴性菌对抗生素的抵抗力比它们强。
问
革兰氏阳性菌是否致病?
答
是的,大多数革兰氏阳性菌都是致病菌。致病性革兰氏阳性菌的实例包括微球菌、肠球菌、金黄色葡萄球菌、表皮葡萄球菌、腐生葡萄球菌、肺炎链球菌、草绿色链球菌、酿脓链球菌、无乳链球菌、破伤风梭菌、肉毒杆菌、产气荚膜梭菌、产气荚膜梭菌、艰难梭菌, 单核细胞增生李斯特菌等。
问
革兰氏阳性菌引起的常见感染有哪些?
答
炭疽、白喉、腹泻、脑膜炎、恶心、皮肤感染、尿路感染。
问
哪种抗生素对革兰氏阳性菌有效?
答
对革兰氏阳性菌有效的抗生素是青霉素、氯唑西林和红霉素,几乎覆盖了 90% 的革兰氏阳性菌感染。其他还有万古霉素、甲氧苄啶/磺胺甲恶唑,克林霉素,克林霉素等。
问
革兰氏阴性菌引起的人类常见疾病有哪些?
答
革兰氏阴性菌会在医疗机构中引起感染,包括肺炎、血流感染、伤口或手术部位感染以及脑膜炎。此外还有霍乱、鼠疫、伤寒、脑膜炎和尿路感染是人类常见的细菌性疾病。
问
为什么革兰氏阴性菌比革兰氏阳性菌更有害?
答
革兰氏阴性菌细胞壁坚硬,不易对抗生素敏感,在抗生素作用下会释放内毒素。
问
什么会杀死革兰氏阴性菌?
答
这些抗生素包括头孢菌素类(头孢曲松-头孢噻肟、头孢他啶等)、氟喹诺酮类药物(环丙沙星、左氧氟沙星)、氨基糖苷类(庆大霉素、阿米卡星)等
问
革兰氏阴性菌的分泌系统是什么?
答
革兰氏阴性菌具有广泛封闭的分泌系统,可以转移微小分子、DNA、氨基酸、蛋白质。
问
革兰氏阴性菌从哪来?
答
革兰氏阴性细菌随处可见,几乎遍布地球上所有支持生命的环境。革兰氏阴性菌包括模式生物大肠杆菌,以及许多致病菌,如铜绿假单胞菌、淋病奈瑟菌、沙眼衣原体和鼠疫耶尔森菌。
问
如何自然去除革兰氏阴性菌?
答
天然抗生素。一些天然抗生素是大蒜、蜂蜜、卷心菜、葡萄柚籽提取物、生苹果醋、特级初榨椰子油、发酵食品等。
免责声明:本公众号内容仅作交流参考,不作为诊断及医疗依据。主要参考文献:
Oliveira J, Reygaert WC. Gram Negative Bacteria. 2022 Mar 26. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2022 Jan–. PMID: 30855801.
Carroll K.C., & Hobden J.A., & Miller S, & Morse S.A., & Mietzner T.A., & Detrick B, & Mitchell T.G., & McKerrow J.H., & Sakanari J.A.(Eds.), (2019). Jawetz, Melnick, & Adelberg’s Medical Microbiology, 27e. McGraw Hill.
Acheson DWK (2015): Patient information: Food poisoning (foodborne illness) (Beyond the Basics). In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/1/16
Apicella M (2015): Treatment and prevention of meningococcal infection. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/1/16
Baum SG (2016): Mycoplasma pneumoniae infection in adults. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/2/16
Crowe SE (2016): Bacteriology and epidemiology of Helicobacter pylori infection. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/1/16.
Crowe SE (2016b): Treatment regimens for Helicobacter pylori. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/1/16.
File TM (2016): Treatment of community-acquired pneumonia in adults in the outpatient setting. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/1/16
Ghanem KG (2016): Clinical manifestations and diagnosis of Neisseria gonorrhoeae infection in adults and adolescents. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/1/16
Hicks CB, Clement M (2016): Syphilis: Treatment and monitoring. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/2/16
Kanafani ZA, Kanj SS (2014): Acinetobacter infection: Epidemiology, microbiology, pathogenesis, clinical features, and diagnosis. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/10/16
Kanafani ZA, Kanj SS (2016): Acinetobacter infection: Treatment and prevention. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/10/16
Kelly CP, Lamont JT (2015): Clostridium difficile in adults: Treatment. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/2/16
Koulenti D et al (2009): Spectrum of practice in the diagnosis of nosocomial pneumonia in patients requiring mechanical ventilation in European intensive care units. Critical Care Med 37(9):2360-2369. doi: 10.1097/CCM.0b013e3181a037ac
Lamont JT (2016): Clostridium difficile in adults: Epidemiology, microbiology, and pathophysiology. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/2/16
Li X-Z, Nikaido H (2004): Efflux-mediated drug resistance in bacteria. Drugs. 64(2):159–204.
Lowy FD (2016): Methicillin-resistant Staphylococcus aureus (MRSA) in adults: Treatment of skin and soft tissue infections. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/1/16
Pegram PS, Stone SM (2016): Botulism. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/1/16
Riley LW (2015): Natural history, microbiology, and pathogenesis of tuberculosis. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/2/16
Sauvage E et al (2008): The penicillin-binding proteins: structure and role in peptidoglycan biosynthesis. FEMS Microbiol Rev 32:234–258. DOI:10.1111/j.1574-6976.2008.00105.x
Southwick F (2008): Infectious Diseases. A Clinical Short Course. McGraw Hill/Lange.
Sterling TR (2016): Treatment of pulmonary tuberculosis in HIV-uninfected adults. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/2/16
Stevens DL, Bryant A (2015): Group A streptococcus: Virulence factors and pathogenic mechanisms. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/1/16
Stevens DL (2016): Group A streptococcal (Streptococcus pyogenes) bacteremia in adults. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/1/16
Swygard H et al (2016): Treatment of uncomplicated gonococcal infections. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/1/16
Wanke CA (2015): Pathogenic Escherichia coli. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 7/28/16
Wanke CA (2015b): Travelers’ diarrhea: Clinical manifestations, diagnosis, and treatment. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/1/16
Yeh S (2015): Microbiology, epidemiology and treatment of Haemophilus influenzae. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/1/16
Sagar Aryal,Gram-Positive Vs Gram-Negative Bacteria- 31 Differences With Examples. Microbenotes.2022,January 9
Jawertz M., Alderbergs., Medical Microbiology 28th Edition.
Prescott M. L., Microbiology. 5th Edition
Lippincott Microbiology in review: 3rd edition
Faith Mokobi,Gram-Positive Bacteria- Cell Wall, Examples, Diseases, Antibiotics,2021, April 15
Faith Mokobi,Gram-Negative Bacteria- Cell Wall, Examples, Diseases, Antibiotics ,2021 ,April 15

谷禾健康
微生物群代表宿主肠道中存在的整个微生物群。肠道内细菌界的“贫富差距”非常大,和人类社会创造的大部分的财富都流向少部分人口的现实类似,只有少数几十种的细菌分布在近乎90%的人群中。换句话说,大部分细菌都只能在特定的环境中生存,只有少数细菌适应能力超强,这可能也是我们需要重点关注的对象。
如果把不同细菌品种看作互相竞争的国家,那么细菌界的“超级大国”就属拟杆菌门和厚壁菌门了。当然它们都不是单独某一种细菌,而是一大类细菌的统称。
然而近年来随着患有肠内外疾病的人群越来庞大,变形菌门也逐渐被关注和研究,变形菌门是含有最丰富细菌的门,麾下包括多种“著名的”病原菌,如大肠杆菌、幽门螺杆菌、克雷伯氏菌、沙门氏菌、志贺氏菌、绿脓杆菌、霍乱弧菌、空肠弯曲菌、鼠疫杆菌、脑膜炎双球菌、淋球菌等,让其备受关注。
事实上,越来越多的数据将变形菌确定为疾病的可能微生物特征。目前主要证据涉及代谢紊乱和炎症甚至癌症。然而,最近的研究表明,在哮喘和慢性阻塞性肺病等肺部疾病中也有作用,有些疾病中变形菌不受控制扩张导致疾病易感和发生。
变形菌(proteobacteria)是细菌中最大、种类最多的一个门,它们在系统发育、生态和致病方面具有广泛的重要性。所有变形菌都是革兰氏阴性菌,外膜主要由脂多糖组成。
图源:esacademic
变形菌门主要是由核糖体RNA序列定义的,名称取自希腊神话中能够变形的神普罗透斯(这同时也是变形菌门中变形杆菌属的名字),因为该门细菌具有极为多样的形状,代谢特征等。
△ 形状:杆状和球菌、弯曲的、螺旋状的、环状的、丝状的和带鞘的细菌都有。
△ 新陈代谢:新陈代谢类型也多种多样,一系列代谢特征包括化学自养(从无机化合物的氧化中获取能量)、化学有机营养(从有机化合物的氧化中获取能量)和光养(从光中获取能量)。
△ 氧气利用:从严格厌氧菌和严格需氧菌到兼性厌氧菌和微需氧菌株的都有,但是大多数变形菌门的成员是兼性厌氧菌。
△ 运动:许多使用鞭毛移动,但有些不能移动或依赖细菌滑动,而一些细菌是不运动的。
△ 生态分布:变形菌门的成员具有极大的可变形态和多才多艺的生理学,这使它们在各种生态位中生存具有竞争优势。已观察到变形菌在不同生境中无处不在。
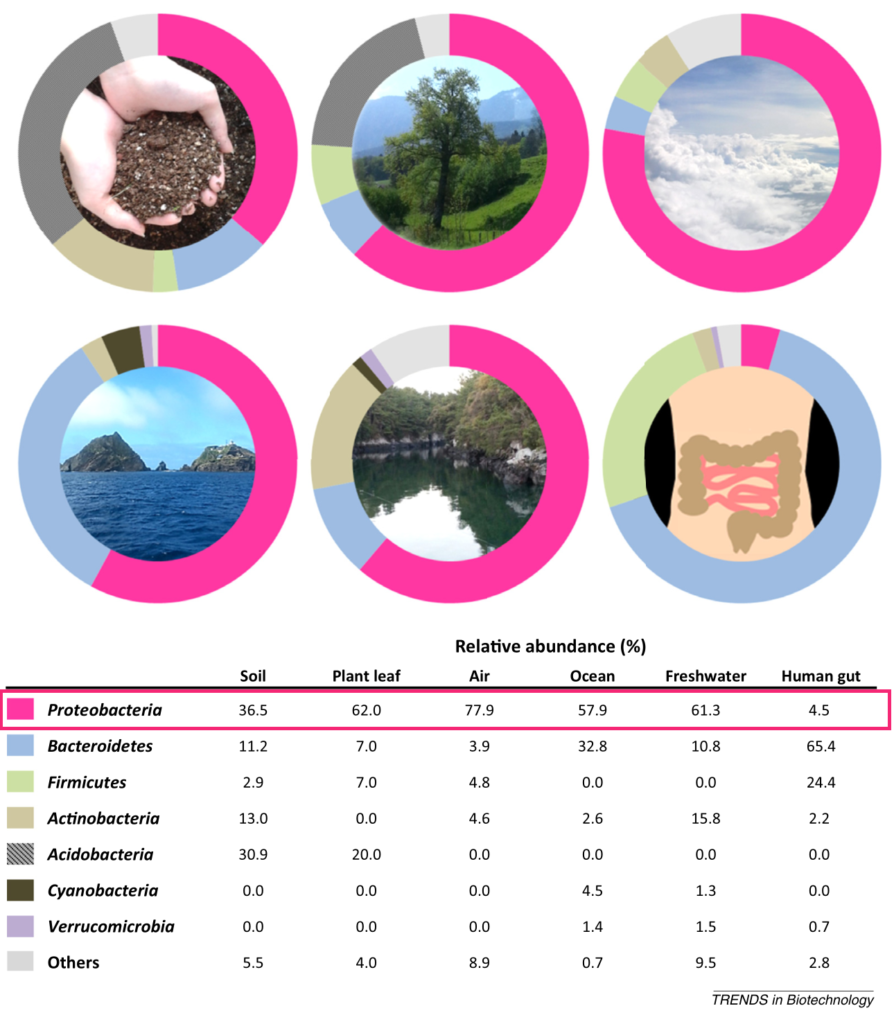
Shin NR, et al., Trends Biotechnol. 2015
植物 、海水、淡水 ,空气,以及人和动物的身体部位,包括肠道、口腔、皮肤、阴道。尽管存在研究间差异,但健康人口腔微生物群的变形菌相对丰度最高(17.2-36.8%),其次是皮肤(6.8-30.0%)、胃肠道(2.5-4.6%)和阴道(2.3%)。
在系统发育学上,变形菌是根据小核糖体亚单位RNA基因(16S rRNA)的测序定义的。这是一个巨大的革兰氏阴性原核生物门,原线粒体起源于此。
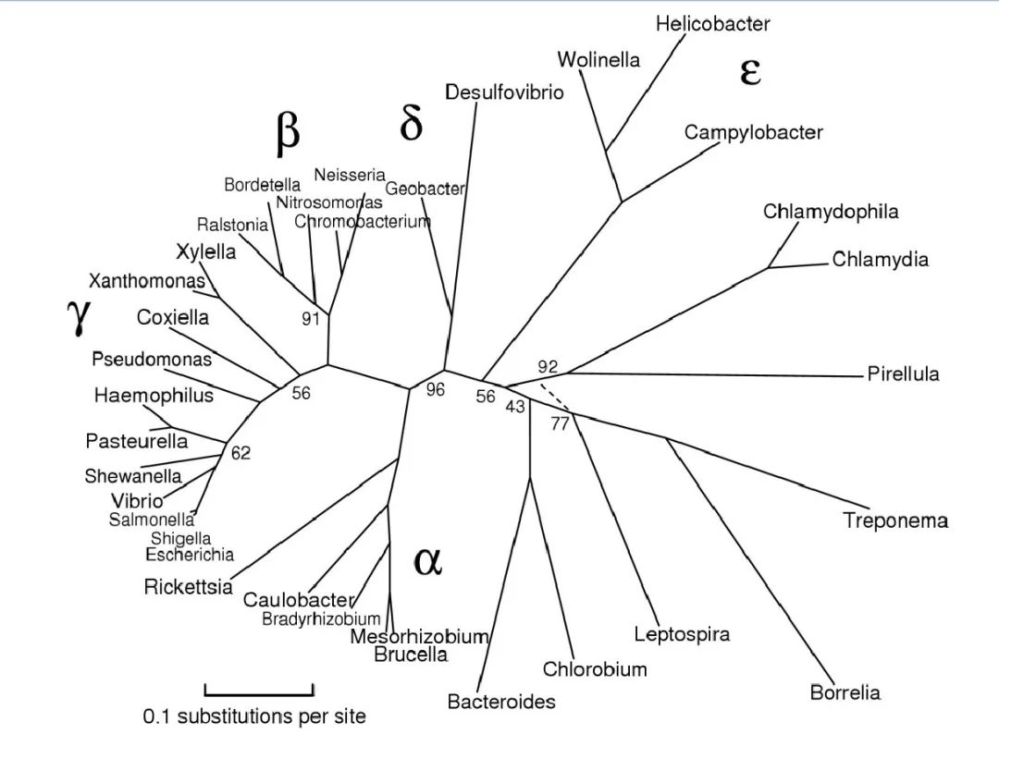
图片来源:Maria Lane,eportfolio
该门主要分为以下几大类:
最初,变形菌包括 α、β、γ 和 δ 四个亚类。ε变形菌 和 δ变形菌 通常被认为是最古老的变形菌群,因为它们包括利用硫化合物进行能量代谢的专性厌氧菌。
α变形菌(Alpha-proteobacteria)
第一类变形菌是α-变形菌。这一类的统一特征是它们是寡营养生物,能够生活在低营养环境中,如深海沉积物、冰川或深层地下土壤。同时α-变形菌是多样化的细菌分支之一,在生活方式、地理分布和基因组大小方面表现出极大的差异。
在 α-变形菌 中有两个重要分类群,衣原体和立克次体,它们是专性细胞内病原体,这意味着它们的部分生命周期必须发生宿主细胞内。由于它们无法合成自己的三磷酸腺苷 (ATP),因此,量需求依赖宿主于细胞。
立克次体属是人类很多严重疾病的病原体。例如,布鲁氏菌属、埃立克体属和立克次氏体。立克次氏杆菌会导致落基山斑疹热,这是一种威胁生命的脑膜炎(包裹大脑的膜发炎)。R. rickettsii 感染蜱,并可以通过被感染的蜱叮咬传播给人类。此外,布鲁氏菌科(Brucellaceae)和巴尔通氏菌科(Bartonellaceae)的细菌是人类病原体。
α-变形菌 还包括固氮细菌,例如固氮螺菌属和根瘤菌属。这两种细菌都使用一种称为固氮酶途径的复杂酶途径将大气中的氮 (N2) 转化为氨 (NH3)。此外,α变形菌还包括硝化细菌。这种类型的细菌将氨和铵 (NH4+) 还原为硝酸盐 (NO3–)。乙酸杆菌属和葡糖杆菌属的变形菌可用于生产乙酸。
β变形菌(Beta-proteobacteria)
与依靠最少量营养物质生存的 Alpha-proteobacteria 不同,Beta-proteobacteria 类是富营养生物,这意味着它们需要大量的有机营养物质。
Beta-proteobacteria 通常在需氧和厌氧区域之间生长(例如,在哺乳动物的肠道中)。一些属包括作为人类病原体的物种,能够引起严重的,甚至可能危及生命的疾病。例如,奈瑟球菌属包括淋病奈瑟菌( STI淋病的病原体)和脑膜炎奈瑟菌(细菌性脑膜炎的病原体)
β变形菌中的亚硝化单胞菌可以将亚硝酸盐还原为亚硝酸盐 (NO2–)。同时,硫杆菌属物种是将硫化氢 (H2S) 和元素硫氧化成硫酸盐 (SO42-) 的细菌,以及用于污水处理的菌胶团(Zoogloea)和Sphaerotilis 。
γ变形菌(Gamma-proteobacteria)
最多样化的革兰氏阴性细菌是γ-变形菌,它包括许多人类病原体。包括几个医学和科学上重要的细菌群,例如肠杆菌科、弧菌科和假单胞菌科。
此外,许多重要的病原体属于这一类,例如:
Richard B. Frankel
△ 铜绿假单胞菌
一个庞大而多样的科,假单胞菌科,包括假单胞菌属。铜绿假单胞菌在该属内,它是一种病原体,可以造成身体不同部位的各种感染。铜绿假单胞菌是一种严格需氧、不发酵、高度运动的细菌。
它通常可能造成伤口和烧伤感染,也可能是慢性尿路感染的原因,并且可能是囊性纤维化患者或机械呼吸机患者呼吸道感染的重要原因。
铜绿假单胞菌感染通常难以治疗,因为该细菌对许多抗生素具有抗性,并且具有形成生物膜的非凡能力。
△ 肠杆菌科
肠杆菌科是属于γ-变形菌 的一大类肠道细菌。它们是兼性厌氧菌,能够发酵碳水化合物。在这个家族中,微生物学家认识到两个不同的类别。
第一类,大肠杆菌,以其原型细菌种类大肠杆菌命名。大肠菌能够完全发酵乳糖(即产生酸和气体)。
第二类,非大肠杆菌,要么不能发酵乳糖,要么不能完全发酵(产生酸或气体,但两者不能同时产生)。
非大肠杆菌包括一些值得注意的人类病原体,例如沙门氏菌属,志贺氏菌,鼠疫耶尔森氏菌。
δ 变形菌(Delta-proteobacteria)
δ-变形菌(Delta-proteobacteria )包括基本好氧的形成子实体的粘细菌和严格厌氧的一些种类,如脱硫球菌属(Desulfococcus)、脱硫线菌属(Desulfonema)、硫酸盐还原菌(脱硫弧菌属(Desulfovibrio)、脱硫菌属(Desulfobacter)、和硫还原菌(如除硫单胞菌属Desulfuromonas),以及具有其它生理特征的厌氧细菌,如还原三价铁的Geobacter和互营菌属(Syntrophus)。
△ 蛭弧菌属:
δ-变形菌还包括蛭弧菌属,Bdellovibrio侵入宿主细菌的细胞,将自身定位在周质中,即质膜和细胞壁之间的空间,以宿主的蛋白质和多糖为食。这种感染对宿主细胞是致命的。
△粘细菌:
粘细菌(“粘液细菌”)是一组主要生活在土壤中并以不溶性有机物质为食的细菌。与其他细菌相比,粘细菌具有非常大的基因组,例如 9-1000 万个核苷酸。
Sorangium cellulosum 拥有最大的已知(截至 2008 年)细菌基因组,有 1300 万个核苷酸。
粘细菌产生许多在生物医学和工业上有用的化学品,例如抗生素。他们将这些化学物质输出到细胞外。
ε变形菌(Epsilon-proteobacteria )
ε-变形菌(Epsilon-proteobacteria) 是革兰氏阴性微需氧细菌(意味着它们在其环境中只需要少量氧气)。多数是弯曲或螺旋形的细菌,如沃林氏菌属(Wolinella)、螺杆菌属(Helicobacter)和弯曲菌属(Campylobacter)。它们都生活在动物或人的消化道中,为共生菌(沃林氏菌在牛中)或致病菌(螺杆菌在胃中或弯曲菌在十二指肠中)。
△ 弯曲杆菌:
变形菌门Epsilon-proteobacteria 中的两个临床相关属是弯曲杆菌属和螺杆菌属,它们都包括人类病原体。
弯曲杆菌可引起食物中毒,表现为严重的肠炎(小肠发炎)。这种由空肠弯曲杆菌引起的疾病在发达国家相当普遍,通常是因为食用了受污染的家禽产品。鸡通常携带空肠弯曲杆菌在胃肠道和粪便中,它们的肉在加工过程中可能会受到污染。
△螺杆菌:
螺杆菌是ε-变形菌的一个属,具有特征性的螺旋形状。它们最初被认为是弯曲杆菌属的成员,但自 1989 年以来,它们独立为自己的属。
螺杆菌属属于ε-变形菌,弯曲杆菌目,螺杆菌科,已经有超过 35 种。已经发现一些菌生活在上胃肠道的内壁,以及哺乳动物和一些鸟类的肝脏中。
该属中最广为人知的物种是幽门螺杆菌,它感染多达 50% 的人口。这种细菌的某些菌株对人类具有致病性,因为它与消化性溃疡、慢性胃炎、十二指肠炎和胃癌密切相关。它也作为该属的模式种。
幽门螺杆菌在胃的高酸性环境中存活的能力有些不同寻常。它产生脲酶和其他酶来改变其环境以降低其酸性。
幽门螺杆菌也有它存在的意义,可能抑制引起结核的细菌(结核分枝杆菌),预防哮喘,克罗恩病,食管反流,腹泻病以及食道癌。
❥ 识别微生物编码的基因,与特征相关联
栖息在哺乳动物肠道中的微生物编码了大量的蛋白质,这些蛋白质有助于广泛的生物功能,从调节免疫系统到参与新陈代谢。
我们从这些微生物中识别蛋白质编码基因并将基因水平与疾病、药物功效或副作用以及其他宿主特征相关联。
例如,与传统的高纤维农业饮食相关的人类肠道微生物群编码了参与纤维素和木聚糖水解的基因家族,而这些基因家族在吃典型西方饮食的人群(年龄匹配)中不存在。
一般编码适应肠道环境所必需的功能的微生物有很强的选择性,在不同宿主中具有大量冗余的基因库。然而,目前的研究和临床很容易忽略健康人类微生物组之间基因丰度的生理意义差异。
❥ 较少丰度的变形菌门,才是是跨宿主丰度变异性最大的基因的主要来源
人体肠道通常由拟杆菌门和厚壁菌门主宰,这些门内的进化枝(尤其是拟杆菌属、普氏菌属和瘤胃球菌科)是最常用于将个体聚集成“肠型”,因为它们解释了最多的分类变异。Bacteroidetes 与 Firmicutes 的比率也被推定为疾病或健康的潜在生物标志物。
有人提出,人类肠道微生物组中可能存在少量“肠型”,每一种都具有不同的分类组成。因此,虽然拟杆菌门和厚壁菌门可能对宿主之间的分类变异贡献最大,但变形菌门的丰度可能会捕获更多的功能变异。
与先前确定的肠型标记分类群相比,变形菌门的水平和可能的 Euryarchaeota 更好地解释了肠道微生物基因功能的人与人之间的差异。
在肠型研究中遗漏了这些不太丰富的门,可能是因为肠型是通过倾向于对高丰度分类群进行更多加权的方法鉴定的,并且肠型是从分类学而非功能数据中鉴定的。这对解释人类肠道微生物群的分类数据具有重要意义。
例如,变形菌门的过度生长与代谢综合征和炎症性肠病有关。通过 TLR5 敲除小鼠测试的肠道炎症关联到变形菌门(超过拟杆菌门和厚壁菌门),并且一些变形杆菌可以在这种背景下诱发结肠炎,可能导致反馈循环。因此,可变基因家族对解释人类肠道微生物群的分类数据具有重要意义。
备注:肠道受体蛋白TLR5参与积极地塑造新生小鼠肠道微生物群落的长期组成,敲除的Toll样受体(TLR5),是免疫系统识别鞭毛细菌(比如变形菌和梭状芽孢杆菌)的关键受体,缺乏它则机体可能不会在感知到细菌鞭毛时对细菌产生免疫应答。
肠道相关微生物群落组成的变化与许多人类疾病有关,但驱动这种不平衡(生态失调)的机制尚不完全清楚。
在肠道菌群失调期间观察到的最一致和最强大的生态模式是属于变形菌门的兼性厌氧细菌的扩张。
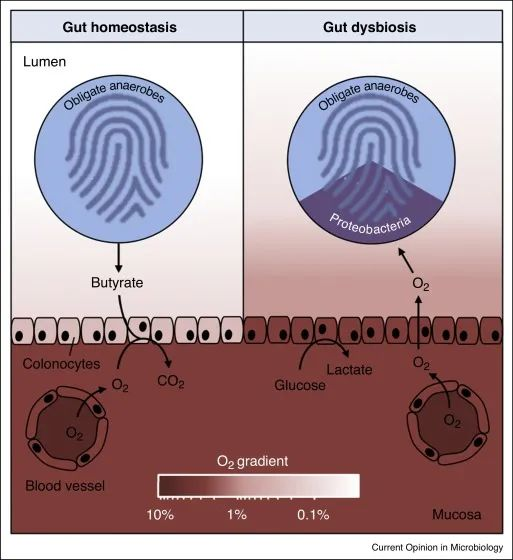
变形菌的菌群失调是上皮功能障碍的微生物特征
在肠道稳态期间(左),微生物群衍生的丁酸盐的 β 氧化导致上皮缺氧,从而维持大肠腔内的厌氧状态。反过来,腔内厌氧症导致肠道微生物群内专性厌氧菌占主导地位。
备注:丁酸(Butyrate acid,BA),俗称酪酸,是构成脂肪的一种脂肪酸,含有4个碳原子又称短链脂肪酸。人体的丁酸部分来自于食物中丁酸的吸收,主要的来自结肠厌氧菌的发酵产生。人体结肠产生的短链脂肪酸丁酸占比大部分)。
在肠道菌群失调期间(右),表面结肠细胞通过无氧糖酵解获得能量,从而导致上皮氧合增加,这种上皮功能障碍破坏了管腔中的厌氧菌,从而通过有氧呼吸推动兼性厌氧变形菌的扩张。
健康结肠的厌氧菌导致肠道微生物群的组成以专性厌氧菌为主,而菌群失调通常与兼性厌氧变形菌的丰度持续增加有关,这表明厌氧菌的破坏。
结肠上皮是缺氧的,但肠道炎症或抗生素治疗会增加结肠中的上皮氧合,从而破坏厌氧作用,通过有氧呼吸驱动兼性厌氧变形菌的菌群失调。
肠沙门氏菌(S. enterica)是一种食源性病原体,属于肠杆菌科,变形菌门,可引起小鼠结肠炎。在肠道沙门菌S. enterica诱导的结肠炎期间,肠腔内的氧气可用性增加,这表明结肠中病原体的氧气呼吸依赖性大量繁殖以及随之而来的专性厌氧梭状芽胞杆菌的丰度下降。
同样,结肠隐窝增生由鼠肠道病原体柠檬酸杆菌(肠杆菌科,变形菌门)引发,可提高肠腔内的氧气利用率,从而通过有氧呼吸推动变形菌病原体扩张。
这些观察结果表明,变形菌的菌群失调是上皮功能障碍的潜在诊断微生物特征,建议将变形菌负荷作为生态失调和疾病的潜在诊断标准,所以在谷禾即将更新的肠道菌群检测报告中,我们会加入变形菌门丰度和参考范围这一指标。
大肠中专性厌氧菌的优势可能是宿主环境的氧气限制严重的结果,这反过来又对用于营养物质的分解代谢途径产生重要影响。
避免被上消化道中的宿主酶降解的复合碳水化合物,可以被大肠中的专性厌氧细菌水解并发酵成更小的化合物。专性厌氧菌最终将许多发酵产物转化为短链脂肪酸,其中乙酸盐、丙酸盐和丁酸盐是最丰富的产物。宿主吸收了大约 95-99% 的微生物产生的短链脂肪酸,它到达血流以影响免疫发育。因此,大肠中专性厌氧菌的优势确保了维持肠道稳态的代谢物的产生。
变形菌是平衡的肠道相关微生物群落中的一个次要成分。然而,由遗传易感性、化学物质或肠道病原体感染引起的肠道炎症会导致小鼠模型中变形杆菌的管腔扩张不受控制。
同样,在患有严重肠道炎症的人类中,包括炎症性肠病、结直肠癌或坏死性小肠结肠炎的患者中观察到变形杆菌的丰度增加。此外,在包括肠易激综合征和代谢综合征在内的低水平肠道炎症条件下观察到大量变形菌。
肠道炎症增加了替代电子受体的可用性,这些电子受体通过厌氧呼吸支持兼性厌氧细菌的生长。肠道炎症过程中产生的活性氧可以将内源性硫化合物氧化为连四硫酸盐,这是一种电子受体,通过连四硫酸盐呼吸作用在鼠结肠中驱动类似肠沙门氏菌和Yersinia enterocolitica(一种属于肠杆菌科,变形菌门的病原体)的管腔扩张 。
一氧化氮由宿主酶产生化学诱导的结肠炎或由遗传易感性引发的结肠炎期间的诱导型一氧化氮合酶(iNOS) 。一氧化氮在肠腔内分解成硝酸盐,从而通过硝酸盐呼吸支持生长,从而增加小鼠结肠中共生大肠杆菌的丰度。类似,宿主衍生的硝酸盐的呼吸有助于在 S. enterica 诱导的小鼠结肠炎期间腔内病原体扩张。
有趣的是,即使在没有明显肠道炎症的情况下,例如在抗生素治疗期间,呼吸电子受体也有助于细菌群落从专性厌氧菌转变为兼性厌氧菌。为了支持这一观点,用链霉素治疗小鼠可将盲肠中的氧化还原电位提高到接近需氧培养液的水平。链霉素治疗通过硝酸盐呼吸和氧气呼吸的结合增加结肠中共生大肠杆菌或致病性肠杆菌的生长。
其他类似研究的结论也表明,氧气,单独或与其他呼吸电子受体结合,是广泛的胃肠道失衡中肠道菌群失调的常见驱动因素。因此,为了开发新的预防或治疗策略,必须了解在肠道菌群失调期间呼吸电子受体的可用性如何升高。
基于这些观察,有人提出变形菌的扩增是肠道菌群失调的微生物特征,而氧气、用药,遗传易感,肠炎驱动了变形菌的扩张,反过来加剧疾病的进展。
宿主遗传因素和外在环境因素,如饮食和生活环境,不断影响肠道微生物群的分类和功能组成。鉴于具有高度稳定性的平衡肠道微生物群与宿主的免疫系统具有共生相互作用,能够抑制变形杆菌失控的扩张,肠道中变形杆菌的大量繁殖可以反映肠道微生物群落结构的不稳定;这种不稳定的结构可以在非疾病状态下观察到(例如,新生儿期 和胃绕道手术后和疾病状态例如,代谢紊乱和肠道炎症)。
Shin NR, et al., Trends Biotechnol. 2015
在新生儿胃肠道的初始定植期间,兼性厌氧变形菌使肠道生态位有利于专性厌氧菌的定植;后者很快被专性厌氧的厚壁菌门和拟杆菌门所取代,它们在健康成年人的肠道微生物群中占主导地位。胃绕道手术导致的胃肠道重排可以改变 pH、胆汁流量和肠道激素,所有这些因素都会影响变形杆菌的丰度。
新生儿肠道中的变形菌
新生儿肠道中的微生物群备受关注,因为它不仅反映了细菌群落的脆弱结构,而且反映了哺乳动物肠道微生物群的真正起源。新生儿肠道中的细菌群落由于其快速的时间变化而不稳定。然而,这种脆弱性与更重要的肠道菌群定植有关,例如严格的厌氧菌。
具体来说,由于新生儿肠道中的氧气丰富,生命第一周的微生物群经常以兼性厌氧菌为主,主要是变形菌属(例如,埃希氏菌属、克雷伯氏菌属和肠杆菌属)。这些兼性厌氧菌通过消耗氧气、改变 pH 值、降低氧化还原电位并产生二氧化碳和营养物质,使栖息地适合严格的厌氧菌定殖。
因此,可以推测变形杆菌在为新生儿肠道准备好接受严格厌氧菌的连续定植方面发挥了作用,这些厌氧菌在健康成人的肠道中含量丰富。
最近对母体胎盘微生物组的一项研究描述了共生细菌群落的存在,其中大肠杆菌的丰度最高。尽管关于胎盘微生物群的活力和起源存在争议,但在母体胎盘中发现的这些有趣的细菌群落与来自母体羊水和新生儿胎粪的细菌群落重叠。
因此,新生儿肠道中的变形菌可能通过胎儿在子宫内吞咽羊水从母体胎盘传播。有趣的是,妊娠后期孕妇肠道中变形菌的比例增加。这意味着母亲微生物群中的这种特定细菌群转移到了新生儿身上。
在新生儿肠胃道中观察到的变形杆菌定植生长的持续时间很可能在母体控制之下。事实上,新生儿微生物群会受到各种母体因素的影响,例如分娩方式、饮食和怀孕期间接触抗生素。
最重要的是,新生儿肠道中变形菌的丰度受喂养类型的影响,这些细菌在配方奶喂养的婴儿中的频率更高,但在母乳喂养的婴儿中很少见。
人乳寡糖 和分泌型 IgA 的产生参与在最初的肠道定植过程中选择性抑制变形菌。因此,越来越多的人认为,及时减少变形菌的丰度是初始微生物定植的正常部分,而这种定植模式的紊乱与新生儿疾病的风险增加有关。
肠道中微生物群和宿主细胞之间的相互作用对于免疫系统的形成和调节至关重要,由于肠腔内有大量外源性抗原,免疫系统必须严格调节其反应以维持与共生菌的共生关系。共生体传递一种信号,诱导宿主免疫的耐受性反应。因此,宿主可以区分有益的本土微生物和有害病原体,并建立健康的微生物群。
变形杆菌的主要分类及其与IBD的关系
Mukhopadhya I, et al., Nat Rev Gastroenterol Hepatol. 2012
为了防止对共生细菌的炎症反应,肠道内的免疫细胞,如单核吞噬细胞(巨噬细胞和树突状细胞)和 CD4 + T 细胞,对微生物刺激反应迟钝或表现出共生反应。
同时,黏膜免疫系统负责清除病原体,这一过程需要积极的促炎信号级联反应。因此,不适当的免疫反应会破坏肠道稳态,引发生态失调,并导致局部和全身炎症和代谢功能障碍。
这种慢性进行性肠道炎症的状态在临床上被诊断为炎症性肠病 (IBD),其中包括溃疡性结肠炎 (UC) 和克罗恩病 (CD)。IBD 的确切病因仍然无法获得,但新出现的证据表明,肠道微生物群成为了这种疾病的主要嫌疑。
许多研究报告了动物和人类各种炎症持续条件下微生物群组成的改变。在这种情况下,通常发现变形菌在疾病中增加,变形菌在肠道炎症中的作用已在各种结肠炎小鼠模型中得到解决,与疾病呈正相关。
例如,使用易发炎症的小鼠模型,即鞭毛蛋白受体 TLR5 缺陷小鼠 (T5KO),发现,进展为结肠炎的小鼠表现出明确的微生物群特征,其特征是变形菌的水平增加,尤其是大肠杆菌属。并且一些作者已将其确定为微生物群不稳定性的潜在标志物,因此易诱发疾病发作。
与变形杆菌属大量繁殖的同时,结肠炎Tlr5-/- 小鼠表现出杂乱无章的结肠粘液层,与非结肠炎Tlr5-/- 同胞相比,感染性病原体的清除延迟。
这些结果表明,短暂不稳定的肠道微生物群,尤其是以变形菌为主的群落,会使遗传易感的小鼠易患慢性结肠炎。
先天免疫反应失调推动变形杆菌生长的假设这反过来又会促进肠道炎症,这一点得到了其他小鼠模型研究的支持,这些小鼠模型具有影响适应性免疫的突变,白细胞介素 (IL)-10 是对本地微生物群产生免疫耐受所需的主要免疫调节细胞因子。
IL-10 缺陷小鼠由于对肠道菌群不耐受而表现出自发性结肠炎。随着结肠炎症的发生和发展,在定植常规微生物群或缺乏特定病原体的微生物群的 IL-10-/- 小鼠中,变形杆菌和大肠杆菌比野生型小鼠多。
在另一项对 IL-10 缺陷小鼠的研究中,富含饱和乳脂的饮食扰乱了肠道微生物群,导致亚硫酸盐还原Delta-proteobacteriumBilophila wadsworthia 大量繁殖。这种病原菌在 IL-10 -/-小鼠中诱导促炎性黏膜免疫反应并促进自发性结肠炎的发生率和严重程度;它还在喂食高乳脂饮食的野生型小鼠中促进葡聚糖硫酸钠 (DSS) 诱导的结肠炎。
除了对结肠炎的易感性与肠道变形菌的相对丰度之间存在正相关性之外,对先天性和适应性免疫系统均缺陷的小鼠的研究提供了支持变形菌在肠道炎症中的致病作用的证据。
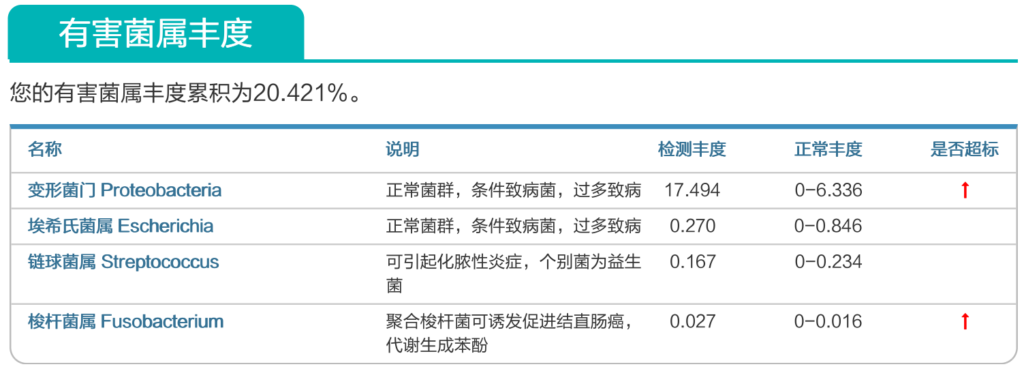
来源:谷禾健康肠道菌群数据库
谷禾健康肠道菌群检测大数据也显示,在炎症性肠病,结直肠癌等患者的肠道菌群检测报告中,85%以上的患者显示变形菌门超标或多项变形菌门病原菌超标或占比丰度偏高。
在最近的一项研究中重现了,结肠炎中变形杆菌的显着扩增,该研究比较了患有活动性结肠炎的 TRUC 小鼠的肠道微生物组与因庆大霉素、甲硝唑或抗肿瘤坏死因子 (TNF)-α 治疗而缓解的小鼠的肠道微生物组。
值得注意的是,从 TRUC 小鼠的粪便中分离出的两种肠杆菌科细菌(肺炎克雷伯菌和奇异变形杆菌)即使在没有任何遗传免疫缺陷的受体小鼠中也足以引发结肠炎。
然而,这两种微生物的致结肠潜力并未在无菌 TRUC 小鼠中复制,这表明结肠炎的发病机制需要其他共生成员。口服伤寒杆菌,另一种富含 TRUC 小鼠的变形菌,也会在非结肠炎 TRUC 小鼠中引发结肠炎,这些小鼠具有大量的促炎细胞因子(例如,TNF-α)。
遗传易患结肠炎的小鼠的生态失调与人类 IBD 特别相关,因为与 IBD 相关的风险等位基因或多态性与先天性和适应性免疫成分有关。与小鼠研究相似,两项人类研究表明,与健康受试者相比,IBD 患者肠道微生物群落的特点是微生物多样性低、变形菌门(尤其是肠杆菌科)的产物以及厚壁菌门的减少。
一项人类队列研究发现,核苷酸结合寡聚化结构域 (NOD)-2 风险等位基因剂量与 IBD 患者肠道标本中肠杆菌科的相对丰度呈正相关。
在 UC 患者中,与炎症的中度和轻度阶段相比,在严重阶段观察到的变形杆菌水平显着升高。
在新发 CD 的初治儿科患者和非 IBD 对照受试者之间,回肠和直肠活检(但不在粪便样本中)的粘膜相关微生物组存在明显差异。变形菌的相对丰度增加,包括肠杆菌科、巴氏杆菌科和奈瑟菌科,将 CD 相关细菌群落与健康对照组区分开来。与慢性炎症一致,伴随变形杆菌属优势的肠道微生物群落改变不仅见于传染性病原菌或原生动物寄生虫引起的急性炎症,而且见于实验性和人类结肠炎相关的结肠直肠癌。
最有趣的生物体,通过一个孤立的病例报告与 IBD 有关,该病例报告一名感染这种细菌的小男孩在放射成像上出现回肠增厚,这是克罗恩病的典型表现。
血清学研究表明,与健康对照相比,克罗恩病患者的大肠杆菌抗体数量增加。具体地说,已发现37-55 % 的克罗恩病患者、2-11% 的溃疡性结肠炎患者和 <5% 对照组患者的百分比。
此外,克罗恩病患者中这些抗体的存在与更严重的表型相关,其特征是小肠受累、疾病进展频繁、病程更长和对手术的需求更大,这表明它们可以用作克罗恩病的预后标志物。
饮食被认为是塑造肠道微生物结构的最关键的环境因素之一。
△ 肥胖:丰富的变形菌为特征
累积证据表明,人类和啮齿动物的健康和肥胖个体的肠道微生物群的分类和功能组成存在差异。
此外,肥胖表型通过粪便移植的传播能力表明肠道微生物群落的改变,作为主要触发因素,是因果关系而不是结果。
肠道微生物群的分类组成失衡,称为生态失调,在代谢紊乱中得到充分证明,并被视为厚壁菌门相对于拟杆菌门的相对丰度增加(F:B 比率)。尽管一致的研究结果普遍支持这一概念,但代谢紊乱期间的生态失调通常包括变形菌的患病率增加。
例如,一项对儿童肠道微生物群的研究发现,与低脂肪、高纤维饮食儿童相比,食用高热量、高脂肪、低纤维饮食的欧洲儿童中的变形杆菌数量更多。
这种差异揭示了肠道微生物群落对非洲儿童饮食的适应性,这可以提高他们从难消化的多糖中获取能量的能力。此外,一些导致有害代谢影响的因素,例如食用无热量的人造甜味剂和乳化剂(通常用作加工食品中的添加剂),也会损害血糖控制并诱发变形杆菌繁殖。
特别是,人造甜味剂介导的肠杆菌科和Delta-proteobacteria类相对丰度的升高与 2 型糖尿病 (T2DM) 患者的结果一致,表明葡萄糖稳态和肠道变形菌之间存在联系。相比之下,证明变形菌的丰度与糖尿病表型呈负相关,挑战代谢疾病患者中高丰度变形菌的概念。
为支持代谢紊乱与变形菌属的扩张之间的关系,变形杆菌属的致肥胖潜力已在无菌小鼠的单关联研究中被确定。
在对一名病态肥胖志愿者进行的减肥试验中,肠杆菌科的相对丰度逐渐减少,假设肠杆菌在代谢恶化中具有致病作用。用从肥胖的人类肠道中分离出来的阴沟肠杆菌B29对无菌小鼠进行单菌定植足以诱导肥胖和胰岛素抵抗。
这一发现支持了这样一个假设,即以丰富的变形菌为特征的不稳定的肠道微生物群落可能代表代谢紊乱的主动特征,而不是被动后果。
△ 营养不良儿童:变形菌成为优势菌
营养不良会导致其他健康问题,例如消瘦和夸希奥科病。在发展中国家,营养不良是威胁 5 岁以下儿童生命的疾病。
营养不良的主要病因是在孕期或产后头 3 年由于大量营养素缺乏和微量营养素缺乏导致的慢性能量负平衡。
然而,最近的研究表明,孟加拉国和马拉维营养不良儿童的肠道微生物群落结构和基因含量与营养良好的儿童不同。在这些研究中,在营养不良的儿童中普遍观察到变形菌的优势和肠道微生物群的低多样性,并被认为是肠道微生物群成熟的障碍。
此外,最近的一项研究揭示了肠杆菌科细菌与营养不良下的肠道黏膜免疫球蛋白 A (IgA) 反应之间存在机制上的相互关系,这会引发肠病并中断黏膜免疫的发展和健康微生物群的组装。
鉴于生态失调驱动的选择压力似乎干扰了微生物群的稳定性,变形菌随后借此机会增加了它们的适应性。微生物群落在异常代谢条件下的不稳定性已被解释为对定植的抵抗力受损。
当接种来自肥胖人类供体的培养细菌(“肥胖受体小鼠”)的无菌小鼠与携带来自瘦肉供体的细菌物种(低脂肪、高纤维饮食)的小鼠共同饲养时,它们被瘦肉有效定殖供体来源的细菌菌株及其肥胖表型得到改善。相比之下,瘦小鼠没有被来自肥胖小鼠的外源或外源细菌菌株定殖。
这一发现表明,生态失调的特点是传播能力减弱和对定植的抵抗力。鉴于 kwashiorkor 儿童的肠道微生物不成熟且富含肠道病原体营养不良被认为与对殖民化的抵抗力有缺陷有关。
总的来说,这一间接证据导致了这样一种观点,即肠道变形菌的扩张反映了宿主的能量不平衡和不稳定的微生物群。有趣的是,在非疾病状态下,如新生儿期和胃绕道手术后也观察到肠道微生物群落的不稳定结构和高丰度的变形菌。
与大多数细菌一样,在细胞外环境中对变形菌的初步识别是通过病原体识别受体 (PRRs) 发生的,PRRs 识别微生物相关分子模式 (MAMPs)——一个包括病原体相关分子模式 (PAMPs) 和危险相关分子模式的统称分子模式(DAMP)。
这些信号受体可分为三个家族:
尽管至关重要的是,只有 TLR 家族参与识别肠细胞表面的细菌配体。
存在于变形菌细胞表面的主要 MAMP 是脂多糖 (LPS) 和鞭毛蛋白,它们分别被 TLR4 和 TLR5 识别。其他参与细菌识别的TLR包括检测细菌脂蛋白的TLR2和检测未甲基化 CpG DNA 的细胞内受体 TLR9。
LPS 的产生和鞭毛组装是在原核生物中观察到的两个最动态的过程,这些结构组成的巨大差异反映在不同变形菌家族成员中观察到的先天免疫反应的强度和方向上。例如,弯曲杆菌和螺杆菌属LPS 与大肠杆菌LPS 的不同之处在于具有更长的酰基链和增加的链连接和脂质 A 磷酸基团的修饰。
在许多病原生物体(例如百日咳杆菌和幽门螺杆菌)中观察到脂质 A 锚中的一个或两个磷酸基团丢失,并且已被证明可提供对抗菌肽的抗性。
参与细菌识别的 TLR 的遗传变异与 IBD 相关。2010 年发表的一项荟萃分析表明,TLR4 Asp299Gly 和 Thr399Ile 变体都赋予白人患克罗恩病和溃疡性结肠炎的统计学显着风险。有趣的是,这两种变体都位于 LPS 结合域内 TLR4 的胞外域,并且被认为会影响蛋白质的二级结构。
这些功能变体的存在已被证明会影响 LPS 反应性,并使个体更容易受到革兰氏阴性菌的感染。证据还表明,这些遗传变异的存在可能会影响基础免疫状态。
因此,有理由推测,在 TLR4 基因变异的携带者中,在营养不良事件之前或期间发生的免疫反应改变,可能足以驱动 IBD 发生不可挽回的免疫反应改变。TLR9 中的遗传变异也与 IBD 易感性增加有关。证据不如TLR4那样令人信服,尽管这一警告可能反映了 TLR9 处理来自所有细菌的配体而 TLR4 反映革兰氏阴性菌易感性的事实。
变形菌门是肠道菌群中四个主要门(厚壁菌门、拟杆菌门、变形菌门和放线菌门)中最不稳定变化最快的门。变形菌门作为一线反应者,对环境因素(如饮食)反应敏感。
总的来说,迄今为止的许多研究都支持这样一个概念,即肠道中大量变形菌反映了生态失调或不稳定的肠道微生物群落结构。除了外源性肠致病性变形杆菌外,健康的哺乳动物肠道还含有数种属于该门的共生细菌,作为其天然肠道菌群。
这些细菌在比例较小时似乎是良性的,而在某些肠道环境下,它们会变成可引发炎症反应甚至代谢障碍。
然而,肠道中变形菌的长期富集可能代表不平衡的不稳定微生物群落结构或宿主的疾病状态。因此,时间顺序监测,而不是横断面研究,可能是根据肠道中变形菌的比例确定疾病风险的更好方法。
在健康肠道中,免疫系统严格调节其反应以维持与共生菌的共生关系。这种可能性表明存在正反馈循环。环境或宿主因素(例如低纤维饮食和急性或慢性炎症)破坏体内平衡,具有选择性并导致肠道内大量变形菌的生态失调。由于宿主无法保持共生的变形菌而导致变形菌的不受控制的扩张,在一小部分和微生物群落对定植的抵抗力降低的情况下,可以进一步促进炎症或外源性病原体的入侵。
因此,切断反馈回路的策略可能包括优化肠道微生物群和宿主之间的伙伴关系。鉴于大多数研究已经在与宿主生理学相关的背景下描述了微生物群落状态,因此对于未来的炎症和代谢干预治疗,首先需要判别变形菌的丰度以及是其是否不受控制扩张,另外需要确定变形杆菌大量繁殖的原因以开发有效的治疗方法。
主要参考文献:
Rizzatti G, Lopetuso LR, Gibiino G, Binda C, Gasbarrini A. Proteobacteria: A Common Factor in Human Diseases. Biomed Res Int. 2017;2017:9351507. doi: 10.1155/2017/9351507. Epub 2017 Nov 2. PMID: 29230419; PMCID: PMC5688358.
Mukhopadhya I, Hansen R, El-Omar EM, Hold GL. IBD-what role do Proteobacteria play? Nat Rev Gastroenterol Hepatol. 2012 Feb 21;9(4):219-30. doi: 10.1038/nrgastro.2012.14. PMID: 22349170.
Litvak Y, Byndloss MX, Tsolis RM, Bäumler AJ. Dysbiotic Proteobacteria expansion: a microbial signature of epithelial dysfunction. Curr Opin Microbiol. 2017 Oct;39:1-6. doi: 10.1016/j.mib.2017.07.003. Epub 2017 Aug 4. PMID: 28783509.
Shin NR, Whon TW, Bae JW. Proteobacteria: microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015 Sep;33(9):496-503. doi: 10.1016/j.tibtech.2015.06.011. Epub 2015 Jul 22. PMID: 26210164.
Shin NR, Whon TW, Bae JW. Proteobacteria: microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015 Sep;33(9):496-503. doi: 10.1016/j.tibtech.2015.06.011. Epub 2015 Jul 22. PMID: 26210164.
Rigottier-Gois L. Dysbiosis in inflammatory bowel diseases: the oxygen hypothesis. ISME J. 2013 Jul;7(7):1256-61. doi: 10.1038/ismej.2013.80. Epub 2013 May 16. PMID: 23677008; PMCID: PMC3695303.
谷禾健康


细 菌
单一细菌,是一种微观的单细胞微生物,生活在地球上的几乎每个角落,从深海喷口到地表以下再到人类的消化道,都存在于其中。
结核分枝杆菌
图源:Centers for Disease Control and Prevention
革兰氏阳性结核分枝杆菌细菌(结核病的原因)的扫描电子显微照片
细菌细胞
细菌缺乏与膜结合的细胞核和其他内部结构,因此被称为原核生物。原核生物是地球上占主导地位的生物,在地球历史上大约有四分之三的时间都存在,并且已经适应了几乎所有可用的生态栖息地。
作为一个群体,它们表现出极其多样化的代谢能力,几乎可以使用任何有机化合物和一些无机化合物作为食物来源。一些细菌可以引起人类、动物或植物的疾病,但大多数是无害的,是有益的生态因子,其代谢活动维持更高的生命形式。
其他细菌是植物和无脊椎动物的共生体,它们对宿主起着重要的作用,如固氮和纤维素降解。没有原核生物,土壤就不会肥沃,死去的有机物腐烂的速度也会慢得多。一些细菌被广泛用于食品、化学品和抗生素的制备。对不同细菌群之间关系的研究,不断为地球生命起源和进化机制提供新的见解。
细菌——原核生物
地球上所有的生物都是由两种基本类型的细胞中的一种组成:一种是真核细胞,其遗传物质被包裹在核膜内;另一种是原核细胞,其遗传物质不与细胞的其他部分分离。传统上,所有的原核细胞都被称为细菌,被归为原核生物界。但是,它们的分类为Monera,在分类学上与其他王国(植物界,动物界,真菌和原生生物)相当,这低估了原核细胞相对于真核细胞表现出的显着遗传和代谢多样性。
1970年代后期,美国微生物学家卡尔·沃斯(Carl Woese)率先在分类上进行了重大变革,将所有生物分为真核生物、细菌(原名真细菌)和古细菌(原名古细菌)三个领域,以反映三条古老的进化路线。原核生物以前被称为细菌,然后被分为两个领域,细菌和古细菌。细菌和古细菌在表面上是相似的;例如,它们没有细胞内的细胞器,它们有环状DNA。但是,它们在本质上是截然不同的,它们的分离是基于其古老而又独立的进化谱系的遗传证据,以及其化学和生理学的根本差异。 这两个原核域的成员彼此之间的区别与它们与真核细胞中的区别一样。
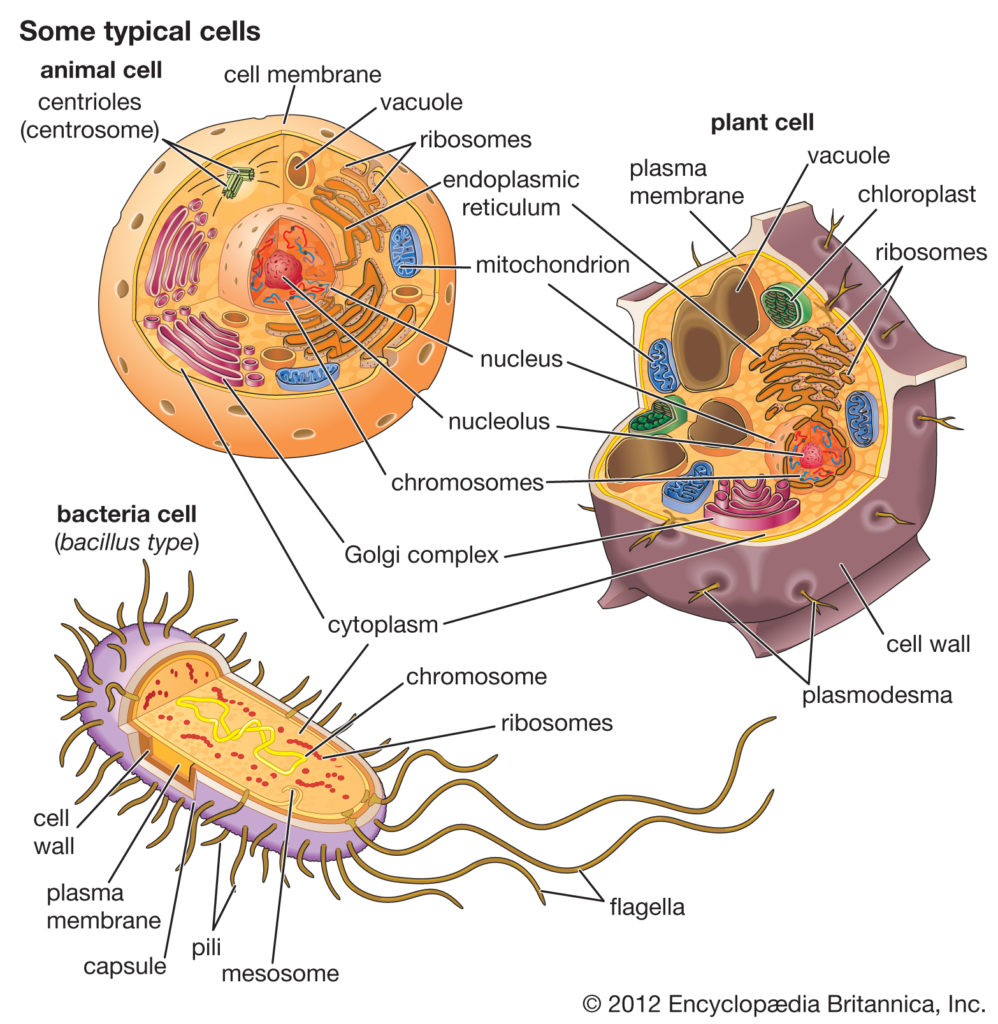
细菌,动物和植物细胞的比较
细菌细胞在几个方面不同于动物细胞和植物细胞。一个根本的区别是细菌细胞缺乏动物细胞和植物细胞中都存在的细胞内细胞器,例如线粒体,叶绿体和细胞核。
原核细胞(即细菌和古细菌)与构成其他生命形式的真核细胞有根本的不同。原核细胞的定义比真核细胞要简单得多。最明显的简化是缺乏细胞内的细胞器,这是真核细胞的特征。所有由细胞器执行的活动也发生在细菌中,但它们不是由专门的结构执行的。此外,原核细胞通常比真核细胞小得多。细菌体积小,设计简单,代谢能力强,使它们能够迅速生长和分裂,并在几乎任何环境中生存和繁衍。
杆菌型细菌细胞
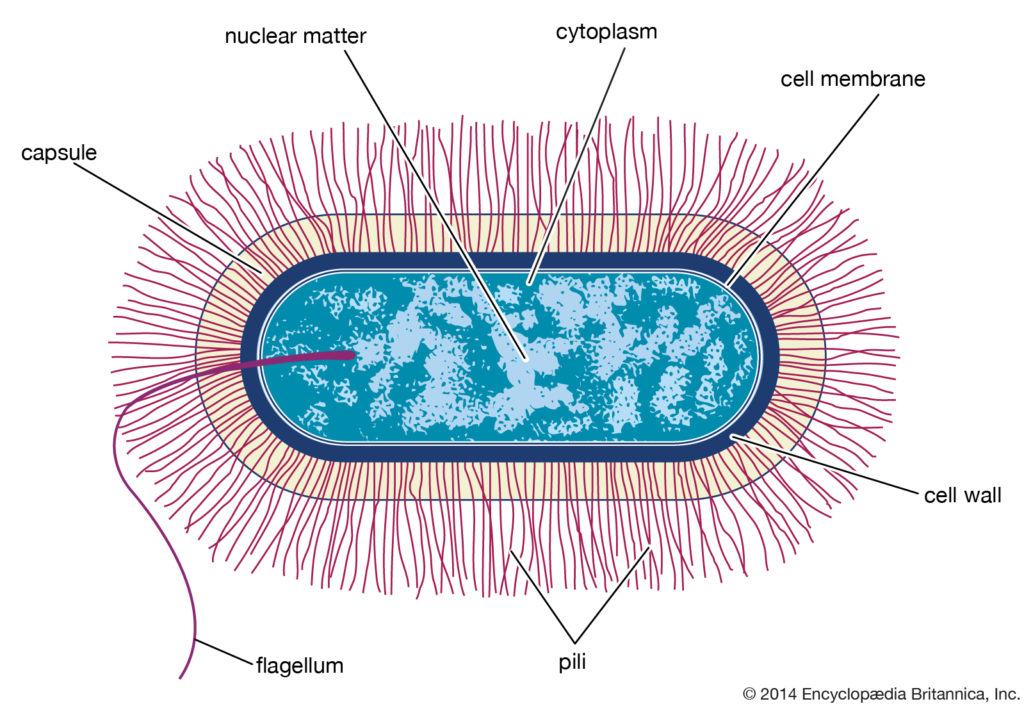
典型的芽孢杆菌属细菌细胞的结构示意图
原核和真核细胞在许多其他方面有所不同,包括脂质组成,关键代谢酶的结构,对抗生素和毒素的反应以及遗传信息的表达机制。 真核生物包含多个线性染色体,这些染色体的基因比编码蛋白质合成所需的基因大得多。 遗传信息的核糖核酸(RNA)副本(脱氧核糖核酸或DNA)的大部分被丢弃,剩余的信使RNA(mRNA)在被翻译成蛋白质之前已被充分修饰。相反,细菌具有一个包含所有遗传信息的环形染色体,它们的mRNA是其基因的精确副本,不会被修饰。
细菌结构的多样性
尽管细菌细胞在结构上比真核细胞小得多,也简单得多,但细菌是一个在大小、形状、生境和代谢上都有差异的极其多样化的有机体群体。许多关于细菌的知识来自对致病细菌的研究,这些细菌比许多自由生活的细菌更容易在纯培养中分离出来,也更容易被研究。必须注意的是,许多自由生活的细菌与适应作为动物寄生虫或共生体生活的细菌有很大的不同。因此,关于细菌的组成或结构没有绝对的规则,任何一般的说法都会许多例外。
单个细菌可以呈现三种基本形状之一:球形(球菌)、棒状(芽孢杆菌)或弧形(弧菌、螺旋体或螺旋体) 。在细菌的实际形状中可以看到相当大的变化,细胞可以在一维中拉伸或压缩。细胞分裂后不分离的细菌形成有助于鉴定的特征性簇。
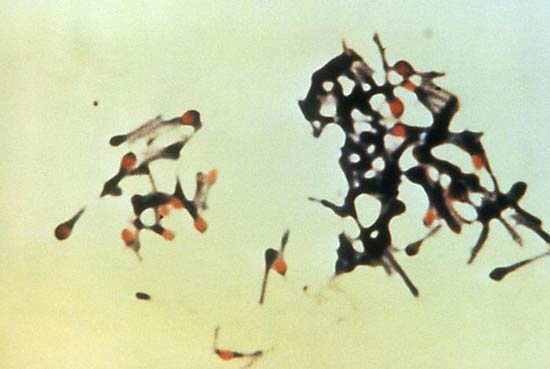
例如,有些球菌主要成对出现,包括肺炎链球菌Streptococcus pneumoniae(一种引起细菌性大叶性肺炎的肺炎球菌)和淋病奈瑟菌Neisseria gonorrhoeae(一种引起性传播疾病淋病的淋病球菌)。大多数链球菌类似于一长串珠子,而葡萄球菌则形成随机的团块(“葡萄球菌”这个名字来源于希腊语staphyle,意思是“葡萄簇”)。
此外,有些球菌呈方形或立方形。杆状杆菌通常单独出现,但有些菌株形成长链,如棒状杆菌的杆状杆菌,通常以任意角度彼此相连。有些杆菌的末端是尖的,而有些杆菌的末端是方的,有些杆菌的杆弯曲成逗号形状。这些弯曲的杆状病毒通常被称为弧菌,包括霍乱弧菌,它能引起霍乱。
其他形状的细菌包括弯曲和弯曲的螺旋形螺线管和螺旋形螺线管,螺旋形螺线管类似于开瓶器,其中细胞体包裹在称为轴向细丝的中央纤维周围。
变形链球菌
图源:David M. Phillips/Visuals Unlimited
变形链球菌细菌是球形(球菌)细菌的一个例子。这种细菌通常会聚集成对和短链。
细菌是最小的生命体。一种细菌的平均大小,如杆状大肠杆菌,是人类和动物肠道的“正常居民”,长约2微米(μm;百万分之一米),直径0.5μm,金黄色葡萄球菌的球形细胞直径可达1μm。
少数细菌种类甚至更小,如肺炎支原体是最小的细菌之一,其宽度约为0.1至0.25μm,长度约为1至1.5μm;百日咳的病原体为棒状百日咳杆菌,其直径为0.2至0.5μm,长度为0.5至1μm;梅毒病原体螺旋形梅毒螺旋体直径仅0.1~0.2μm,长度6~15μm。蓝藻聚球藻平均直径约0.5~1.6μm。有些细菌相对较大,例如固氮菌,其直径为2至5μm或更大;无色菌,其最小宽度为5μm,最大长度为100μm,视物种而定。用肉眼可以看到巨大的细菌,例如平均直径为750μm的Thiomargarita namibiensis和长度在30到600μm之间的杆状Epulopiscium fishelsoni。
细菌是单细胞微生物,因此通常不会组织起来。 每个细菌的生长和分裂都独立于其他任何细菌,尽管常会发现细菌的聚集体,有时包含不同物种的成员。许多细菌可以形成称为生物膜的聚集结构。
生物膜中的生物通常表现出与处于个体状态或浮游状态的同一生物基本不同的特性。 聚集到生物膜中的细菌可以传达有关种群大小和代谢状态的信息。 这种类型的通信称为群体感应,通过产生称为自动诱导剂或信息素的小分子来进行操作。
群体感应分子(最常见的是肽或酰化高丝氨酸内酯(AHL;特殊的信号化学物质))的浓度与生物膜中相同或不同物种的细菌数量有关,有助于协调生物膜的行为。
革兰氏染色
细菌如此之小,直到1677年才被首次发现,当时荷兰科学家安东尼·列文虎克借助原始显微镜(在设计上与现代放大镜比现代显微镜更相似)在各种物质中看到了微生物,其中一些能放大200倍以上。现在细菌通常在能放大1000倍以上的光学显微镜下进行检查;然而,只有借助更强大的透射电子显微镜才能观察到其内部结构的细节。除非使用特殊的合适的显微镜,否则细菌必须用有色染料染色,以使它们从背景中脱颖而出。
肺炎克雷伯菌
图源:AW Rakosy /Encyclopædia
从肺炎患者的肺脓肿中分离出的革兰氏阴性杆菌肺炎克雷伯菌。
对细菌最有用的染色反应之一叫革兰氏染色,由医生汉斯·克里斯蒂安·格兰姆(Hans Christian Gram)于1884年开发。
悬浮液中的细菌通过短暂加热固定在玻片上,然后暴露在两种染料中,这些染料会在每个细胞内结合形成大的蓝色染料复合物。 当用酒精溶液冲洗载玻片时,革兰氏阳性菌会保留蓝色,而革兰氏阴性菌会失去蓝色。然后用一种较弱的粉红色染料将玻片染色,这种粉红色的染料会使革兰氏阴性菌变成粉红色,而革兰氏阳性菌则保持蓝色。革兰氏染色剂对细菌细胞表面结构的差异起反应,当在电子显微镜下观察细胞时,这种差异是显而易见的。
金黄色葡萄球菌
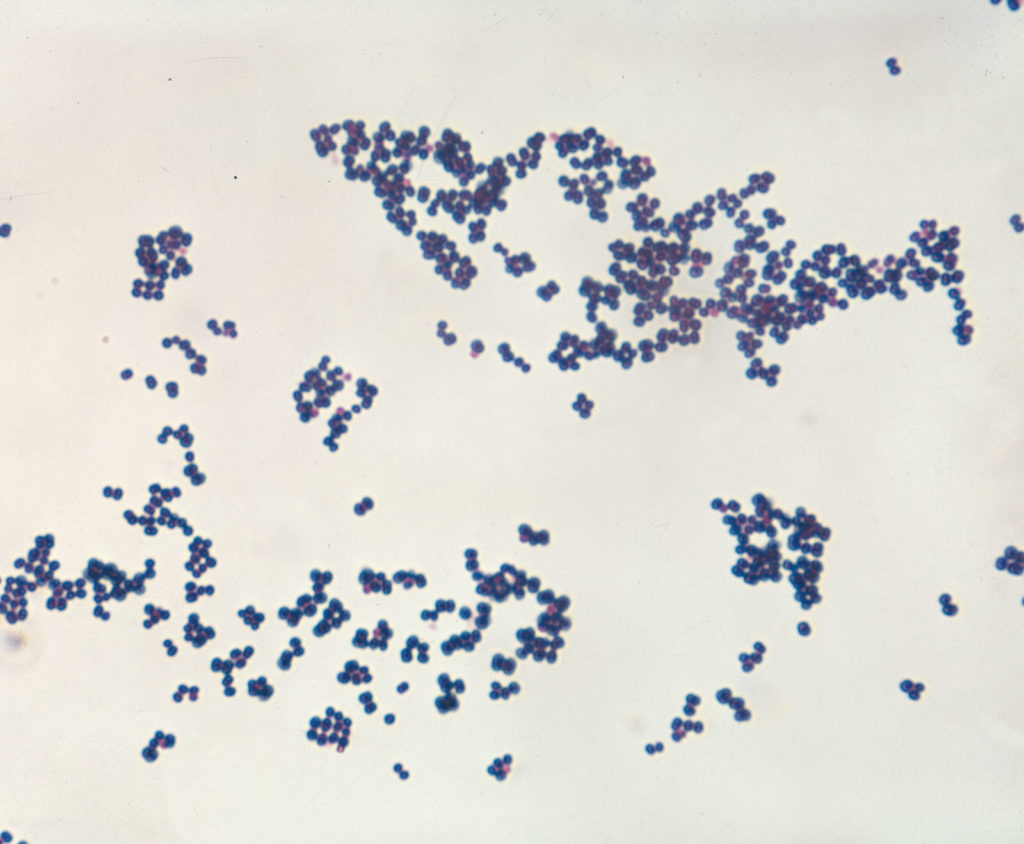
图源:AW Rakosy /Encyclopædia
实验室培养中的革兰氏阳性球菌金黄色葡萄球菌
细胞包膜
细菌细胞表面(或包膜)的结构变化很大,它在细胞的性质和能力中起着核心作用。所有细胞的一个特征是细胞质膜,它将细胞内部与外部环境分开,调节营养物质的流动,维持适当的细胞内环境,并防止细胞内容物的丢失。细胞质膜执行许多必要的细胞功能,包括能量产生、蛋白质分泌、染色体分离和有效的营养物质主动运输。它是一种典型的由蛋白质和脂类组成的单位膜,与包围所有真核细胞的膜基本相似。在电子显微照片中,它是由脂质和蛋白质组成的三层结构,完全包围细胞质。
水螺螺旋藻的肽聚糖层
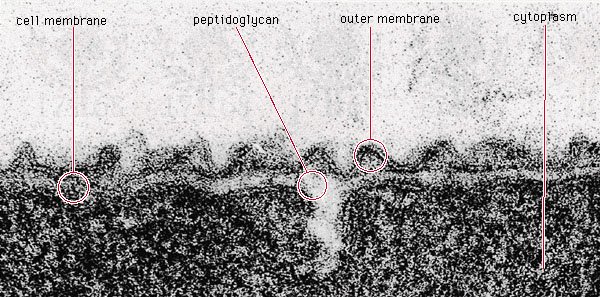
图源:AW Rakosy /Encyclopædia
革兰氏阴性细菌蛇形藻(Aquaspirillum serpens)具有薄的肽聚糖层,位于细胞膜和外膜之间。
肽聚糖只存在于细菌中(没有细胞壁的细菌除外,如支原体)。肽聚糖是两个重复糖(N-乙酰氨基葡萄糖和N-乙酰壁酸)的长链聚合物,其中相邻的糖链通过肽桥相互连接,从而提供刚性稳定性。
肽桥的性质在不同种类的细菌之间有很大差异,但一般由四种氨基酸组成:L-丙氨酸与D-谷氨酸相连,在革兰氏阴性细菌中与二氨基丙酸相连,或在革兰氏阳性细菌中与赖氨酸、L-鸟氨酸或二氨基丙酸相连,最后与D-丙氨酸相连。
在革兰氏阴性菌中,肽桥将一条链上的D-丙氨酸连接到另一条链上的二氨基丙酸。在革兰氏阳性细菌中,可以有一个额外的肽链延伸交叉连接的范围;例如,在金黄色葡萄球菌中有一个额外的五个甘氨酸桥。
肽聚糖的合成是许多有用的抗菌剂的目标,包括阻断肽桥交联的β-内酰胺类抗生素(如青霉素)。动物合成的一些蛋白质作为天然抗菌防御因子攻击细菌的细胞壁。例如,一种名为溶菌酶的酶分解肽聚糖分子的主链糖链。这些药物中的任何一种都会削弱细胞壁并破坏细菌。
在革兰氏阳性细菌中,细胞壁主要由一层厚的肽聚糖网状结构组成,该网状结构与被称为磷壁酸(来自希腊单词teichos,意思是“壁”)的其他聚合物以及一些蛋白质或脂质交织在一起。相反,革兰氏阴性菌有一个复杂的细胞壁,由多层组成,其中一个外膜层位于一个薄肽聚糖层的顶部。这种外膜由磷脂和脂多糖组成,磷脂是一种含有磷酸盐分子的复合脂质,脂多糖是一种复合脂质,通过脂质末端锚定在细胞外膜上,并有一长链糖从细胞延伸到培养基中。
脂多糖,通常被称为内毒素,对动物和人类是有毒的;它们在血液中的存在会导致发烧、休克,甚至死亡。
对于大多数革兰氏阴性细菌来说,外膜形成了一道屏障,阻碍了许多对细菌有害的化学物质的通过,例如通常溶解细胞膜的染料和洗涤剂。对油溶性化合物的不渗透性在其他生物膜中是不存在的,这是由于膜中存在脂多糖和外膜蛋白的不寻常特性造成的。作为外膜抵抗恶劣环境的能力的证据,一些革兰氏阴性细菌在浮油、喷气燃料箱、酸性矿井排水甚至是瓶装消毒剂中生长良好。
古细菌的表面结构与细菌明显不同。它们没有肽聚糖;相反,它们的膜脂是由支链类异戊二烯通过乙醚键与甘油相连组成的。一些古菌的壁材料与肽聚糖相似,只是与氨基酸桥相连的特定糖不是壁酸而是塔罗糖胺酸。许多其他古细菌使用蛋白质作为其细胞壁的基本组成部分,有些则缺乏坚硬的细胞壁。
胶囊和粘液层
许多细菌细胞以荚膜或粘液层的形式分泌一些细胞外物质。黏液层松散地与细菌结合在一起,很容易被洗掉,而胶囊则紧紧地附着在细菌上,并有明确的边界。通过将细胞置于印度墨水的悬浮液中,在光学显微镜下可以看到胶囊。胶囊排除了墨汁,在细菌细胞周围出现了清晰的光晕。
胶囊通常是单糖(多糖)的聚合物,尽管炭疽杆菌的胶囊是由聚谷氨酸制成的。大多数胶囊都是亲水的,可以通过防止水分流失帮助细菌避免干燥(脱水)。胶囊可以保护细菌细胞免受白细胞的吞噬和破坏。虽然逃避吞噬作用的确切机制尚不清楚,但可能是因为胶囊使细菌表面成分更光滑,帮助细菌逃避吞噬细胞的吞噬。肺炎链球菌中存在的胶囊是导致肺炎的最重要因素。肺炎链球菌的突变株失去了形成荚膜的能力,很容易被白细胞吸收,不会引起疾病。在许多其他种类的细菌中也发现了毒力和荚膜形成的联系。
醋酸钙不动杆菌
图源:T.J. Beveridge
这些细菌周围的荚膜材料(钙不动杆菌)显示在印度墨水的悬浮液中,并通过光学显微镜(放大约2500倍)观察。
胞外多糖材料的包膜层可以将许多细菌包裹成一个生物膜,并具有多种功能。引起龋齿的变形链球菌Streptococcus mutans,会分解食物中的蔗糖,并利用其中一种糖类来构建它的荚膜,使之紧紧地附着在牙齿上。被困在胶囊中的细菌利用其他糖来促进新陈代谢,并产生一种强酸(乳酸),攻击牙釉质。当铜绿假单胞菌定植于囊性纤维化患者的肺部时,它会产生一种厚的海藻酸囊膜聚合物,从而导致根除细菌的困难。Zoogloea属的细菌分泌纤维素纤维,将细菌缠绕成漂浮在液体表面的絮状物,使细菌暴露在空气中,这是该属新陈代谢的需要。一些杆状细菌,如Sphaerotilus,分泌长而复杂的管状鞘,这些鞘包围了大量的细菌。这些细菌和许多其他环境细菌的外壳会被铁或锰氧化物包裹。
变形链球菌
图源:Kateryna Kon / Shutterstock
变形链球菌是一种在口腔中发现的细菌,可导致蛀牙。
鞭毛、菌毛
许多细菌是运动的,能够在液体介质中游动,或在固体表面滑动或聚集。游动和成群的细菌拥有鞭毛,鞭毛是运动所需的细胞外附属物。
鞭毛是由单一类型的蛋白质组成的长而螺旋的细丝,位于杆状细胞的末端,如霍乱弧菌或铜绿假单胞菌,或遍布细胞表面,如大肠杆菌。鞭毛可以在革兰氏阳性和革兰氏阴性杆菌上找到,但在球菌上很少见,并且被困在螺旋体的轴向丝中。
鞭毛在其基部附着在细胞膜的基体上。在膜上产生的原动力被用来转动鞭毛丝,就像涡轮一样,由氢离子流通过基体进入细胞。当鞭毛以逆时针方向旋转时,细菌细胞以直线游动;顺时针方向旋转导致以相反方向游动,或者,如果每个细胞有多个鞭毛,则随机翻滚。趋化性使细菌能够调整自己的游泳行为,使其能够感觉到并向不断增加的引诱性化学物质或远离排斥性化学物质的方向迁移。
细菌不仅能够游向或滑向更有利的环境,而且它们还有附属物,使它们能够粘附在表面,防止被流动的液体冲走。有些细菌,如大肠杆菌和淋病奈瑟菌,会产生笔直、坚硬的尖刺状突起,称为菌毛,它们从细菌表面延伸出来,附着在这些菌株的其他细胞上的特定糖上,肠上皮细胞或泌尿道上皮细胞。菌毛只存在于革兰氏阴性菌中。某些菌毛是用来让一个细菌识别和坚持另一个在有性交配过程中称为接合。许多水生细菌产生一种酸性粘多糖固着物,使它们能够紧紧地附着在岩石或其他表面。
细胞质
虽然细菌在表面结构上有很大的不同,但它们的内部内容物非常相似,显示出相对较少的结构特征。
所有细胞的遗传信息都存在于DNA超长分子的含氮碱基序列中。与真核细胞中的DNA不同的是,细菌细胞中的DNA并没有被隔离在膜结合的细胞器中,而是呈长螺旋状分布在细胞质中。
在许多细菌中,DNA是以单个环状染色体的形式存在的,尽管有些细菌可能含有两条染色体,而且在某些情况下DNA是线性的而不是环状的。可变数量的较小的,通常是圆形的(虽然有时是线性的)DNA分子,称为质粒,可以携带辅助信息。
DNA中的碱基序列已经为数百种细菌所确定。细菌染色体的DNA含量从生殖支原体的580000碱基对到大肠杆菌的4700000碱基对,再到黄色粘球菌的9450000碱基对。粘液杆菌纤维素(Sorangium cellulosum),拥有最大的细菌基因组之一,包含超过1300万个碱基对。大肠杆菌染色体的长度,如果从细胞中取出并拉伸到最大限度,约为1.2毫米,考虑到细胞的长度约为0.001毫米,这蛮惊人的。
与所有生物体一样,细菌DNA含有四种含氮碱基腺嘌呤(A)、胞嘧啶(C)、鸟嘌呤(G)和胸腺嘧啶(T)。 双链DNA分子碱基配对的规则要求腺嘌呤和胸腺嘧啶碱基的数目相等,胞嘧啶和鸟嘌呤碱基的数目也相等。
G和C碱基对的数量与A和T碱基对的数量之间的关系是生物体内进化和适应性遗传变化的重要指标。
G + C的比例或摩尔比可以用G + C除以所有碱的总和(A + T + G + C)乘以100%来测量。生物体之间G + C比率的变化程度可能是相当大的。在动植物中,G + C的比例约为50%。 在原核生物中,G + C的比例范围更广,从大多数支原体的约25%到大肠杆菌中的约50%,到微球菌,放线菌和结实的粘菌中的近75%。但是,单个属中某个物种内的G + C含量非常相似。
细菌的细胞质包含高浓度的酶,代谢产物和盐。 另外,细胞的蛋白质是在分散在整个细胞质中的核糖体上制造的。 细菌核糖体与真核细胞中的核糖体不同之处在于它们更小,具有更少的成分(由三种类型的核糖体RNA和55种蛋白质组成,而真核生物中则由四种类型的rRNA和78种蛋白质组成)并且与作用于真核核糖体的抗生素相比,它们受到不同抗生素的抑制。
细菌细胞质中有许多包涵体或颗粒。这些物体从不被膜封闭,而是作为储存容器。糖原是葡萄糖的聚合物,是碳水化合物和能量的储备。 Volutin或变色颗粒含有聚合磷酸盐,是无机磷酸盐和能量的储存形式。许多细菌具有含有聚β-羟基丁酸聚合物酯或相关化合物的脂滴。这与真核生物不同,真核生物利用脂滴储存甘油三酯。在细菌中,储存颗粒是在有利的生长条件下产生的,并在营养物质从培养基中耗尽后被消耗掉。 许多水生细菌产生气体空泡(或叫气囊),这是一种蛋白质结合的结构,含有空气,其中包含空气,并使细菌能够调节其浮力。 细菌也可以有内部的膜结构,作为细胞质膜的外生物形成。
细菌的生物类型
致病细菌不断地与宿主的免疫系统搏斗,这一事实可能解释了属于同一物种但可通过血清学试验加以区分的不同菌株或类型的细菌数量之多,令人困惑。微生物学家通常通过细胞表面的特定分子来识别细菌,这些分子是用特定的抗体检测出来的。抗体是一种血清蛋白,在免疫反应中与外来分子(抗原)紧密结合,以清除或破坏抗原。抗体具有显着的特异性,蛋白质中甚至一个氨基酸的取代都可能阻止该蛋白质被抗体识别。
对于许多细菌种类,有成千上万种不同的菌株(称为血清变异体,用于血清学变异体),它们主要或仅在其脂多糖、鞭毛或荚膜的抗原特性上彼此不同。例如,肠内细菌的不同血清型,如大肠杆菌和沙门氏菌,常常被发现与栖息在不同宿主动物或引起不同疾病的能力有关。这些众多血清型的形成反映了细菌对免疫系统强烈的防御行为作出有效反应的能力。
生殖过程
二分裂法
大多数原核生物通过二分裂过程进行繁殖,细胞体积不断增大,直至分裂成两半,产生两个完全相同的子细胞。每个子细胞可以继续以与父细胞相同的速度生长。为了实现这一过程,细胞必须在其整个表面生长,直到细胞分裂时,在细胞中部的分裂隔膜处形成一个新的半球形极。
在革兰氏阳性细菌中,隔膜沿着细胞的中点从质膜向内生长;在革兰氏阴性细菌中,细胞壁更具弹性,当侧壁向内挤压时形成分裂隔膜,将细胞一分为二。为了使细胞分裂成两半,肽聚糖的结构必须在半球形帽中不同于在细胞壁的笔直部分,并且不同的壁交联酶必须在隔膜处比在其他地方更活跃。
出芽生殖
一群环境细菌通过出芽繁殖。在这个过程中,在母细胞的一端或丝上形成一个小的芽,称为前列腺癌。随着生长的进行,母细胞的大小保持不变,但芽会扩大。当芽的大小和母细胞差不多时,它就会分离。这种繁殖方式类似于萌芽真菌,如啤酒酵母(Saccharomyces cerevisiae)。分裂和出芽的一个区别是,在后者中,母细胞通常具有与后代不同的特性。在某些巴氏杆菌菌株(Pasteuria strains)中,子芽有鞭毛,并能运动,而母细胞没有鞭毛,但有长的菌毛,但在芽对面的末端具有长的菌毛和固定的附肢。在浮游生物中发现的相关浮萍菌在芽对面的末端具有长的纤维状茎。 在Hyphomicrobium中,菌丝(prostheca)从细胞的一端长出,芽从prostheca的尖端长出,与母细胞相距相对较长的距离。
多形生丝单胞菌
图源:Biological Photo Service
在萌芽的多形性hyphomonas细菌中,芽长在称为prostheca的细丝末端。
孢子繁殖
许多环境细菌能够产生稳定的休眠或休眠形式,作为其生命周期的一个分支,以增强其在不利条件下的生存能力。这些过程不是细胞生命周期的一个强制性阶段,而是一种中断。这种休眠形式被称为内生孢子、胞囊或异胞囊(主要见于蓝藻),这取决于孢子形成的方法,而孢子形成的方法因细菌群而异。
在许多属的细菌中都发现了形成内生孢子的能力,这些细菌主要是革兰氏阳性菌,包括需氧芽孢杆菌(aerobic rod Bacillus),微需氧菌芽孢杆菌(microaerophilic rod Sporolactobacillus),厌氧菌梭状芽孢杆菌(anaerobic rods Clostridium)和脱硫芽孢杆菌(Desulfotomaculum),球菌孢子菌(coccus Sporosarcina)和丝状嗜热放线菌(filamentous Thermoactinomyces)。
孢子的形成是对营养缺乏的反应。 因此,在营养物质可用之前,内生孢子不具有代谢活性,这时它们能够从孢子分化为营养细胞。 在孢子形成过程中,每个细菌细胞内部仅形成一个孢子。孢子的形成始于细菌染色体拷贝周围细胞质膜的内陷,从而使较小细胞的内容物与母细胞分离。
母细胞的膜吞噬了其细胞质内的较小细胞,有效地提供了两个同心的单元膜来保护正在发育的孢子。 薄的孢子膜和厚的肽聚糖皮质位于两个单元膜之间。 皮质外部形成坚硬的孢子皮,包围整个孢子结构。孢子皮具有类似于角蛋白的特性,能够抵抗热,干燥(脱水),冷冻,化学药品和辐射的致死作用。 内生孢子抵抗这些有害物质的能力可能来自于孢子内部极低的水含量。 甲基肌球菌属中的甲烷氧化细菌(Methylosinus)也会产生耐干燥性的孢子,称为外孢子。
包囊是由固氮菌(Azotobacter), 蛭弧菌(Bdellovibrio,蛭包囊)和粘球菌(Myxococcus,粘孢子)的休眠成员产生的厚壁结构。它们对干燥和其他有害条件有抵抗力,但程度低于内生孢子。在固氮固氮菌的包囊中,细胞分裂后形成一层厚厚的多层细胞壁和外壳,包围着静息细胞。
丝状放线菌产生两类生殖孢子:分生孢子,是在地上或基质菌丝体上形成的多个孢子链;孢子囊孢子,是在称为孢子囊的特殊囊中形成的。
遗传信息交流
细菌在其生命周期中没有一个专有性生殖阶段,但它们可以非常活跃地交换遗传信息。DNA中携带的遗传信息可以从一个细胞转移到另一个细胞;然而,这不是真正的交换,因为只有一个’同伴’接收到新的信息。
此外,转移的DNA量通常只是染色体的一小部分。发生这种情况有几种机制。在转化过程中,细菌吸收漂浮在培养基中的游离DNA片段。为了有效地吸收DNA,细菌细胞必须处于一种有能力的状态,这是由细菌结合DNA自由片段的能力来定义的,并且仅在有限数量的细菌中自然形成,例如嗜血杆菌(Haemophilus)、奈瑟菌(Neisseria)、链球菌(Streptococcus)和芽孢杆菌(Bacillus)。在实验室条件下,例如通过暴露于氯化钙(CaCl2)溶液,可以使许多其他细菌,包括大肠杆菌,变得人为地具有竞争力。转化是重组DNA技术中的一个主要工具,因为来自一个生物体的DNA片段可以被另一个生物体摄取,从而使第二个生物体获得新的特性。
细菌之间交换遗传物质
图源:Encyclopædia
细菌DNA可以通过结合和转导过程从一个细胞传递到另一个细胞。
转导是指通过一种称为噬菌体的细菌感染病毒将DNA从一种细菌转移到另一种细菌。转导是在细菌之间转移DNA的有效手段,因为封闭在噬菌体中的DNA受到保护,免受物理降解和环境中酶的攻击,并通过噬菌体直接注入细胞中。 然而,通过转导的广泛基因转移具有有限的意义,因为将细菌DNA包装到病毒中效率低下,并且噬菌体通常在它们可感染的细菌种类范围内受到高度限制。 因此,通过转导进行种间转移的DNA很少。
图源:Encyclopædia
接合是通过质粒(非染色体DNA分子)介导的细胞间直接接触的DNA转移。接合质粒编码一种极其有效的机制,它介导自己从供体细胞到受体细胞的转移。因为仅供体细胞包含结合质粒,所以该过程在一个方向上发生。在革兰氏阴性细菌中,供体细胞产生特定的质粒编码菌毛,称为性菌毛,该菌毛将供体细胞附着到受体细胞上。 连接后,两个细胞将直接接触,并形成一个偶联桥,DNA通过该桥从供体转移到受体。 许多接合质粒可以在大量不同的革兰氏阴性细菌之间转移,并在其中繁殖。 质粒的大小不等,从几千个到超过100,000个碱基对。 后者有时被称为巨质粒。
巴氏杀菌如何保护食物
图源:Encyclopædia
细菌可能是致病的(引起疾病),并可能产生毒素,使食物变质并引起食物中毒。细菌引起的某些疾病包括肉毒杆菌中毒,伤寒和肺炎。
细菌的种群增长
细菌培养物的生长被定义为一个群体中细菌数量的增加,而不是单个细胞大小的增加。细菌种群的生长是以几何或指数方式发生的:每一个分裂周期(世代),一个细胞产生2个细胞,然后是4个细胞,然后是8个细胞,然后是16个细胞,然后是32个细胞,依此类推。生成一代所需的时间,即生成时间(G),可通过以下公式计算:
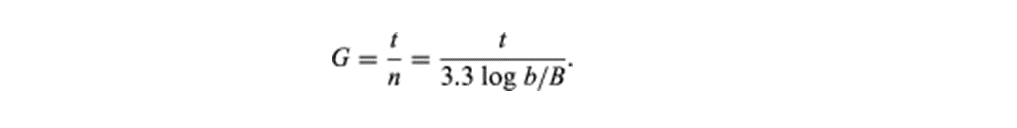
在公式中,B是观察开始时出现的细菌数量,B是时间段t后出现的细菌数量,n是世代数。这种关系表明,平均产生时间是恒定的,细菌数量增加的速度与任何给定时间的细菌数量成正比。这种关系只在种群以指数方式增长的时期有效,称为对数增长期。因此,显示细菌培养物生长的图形被绘制为细胞数的对数。
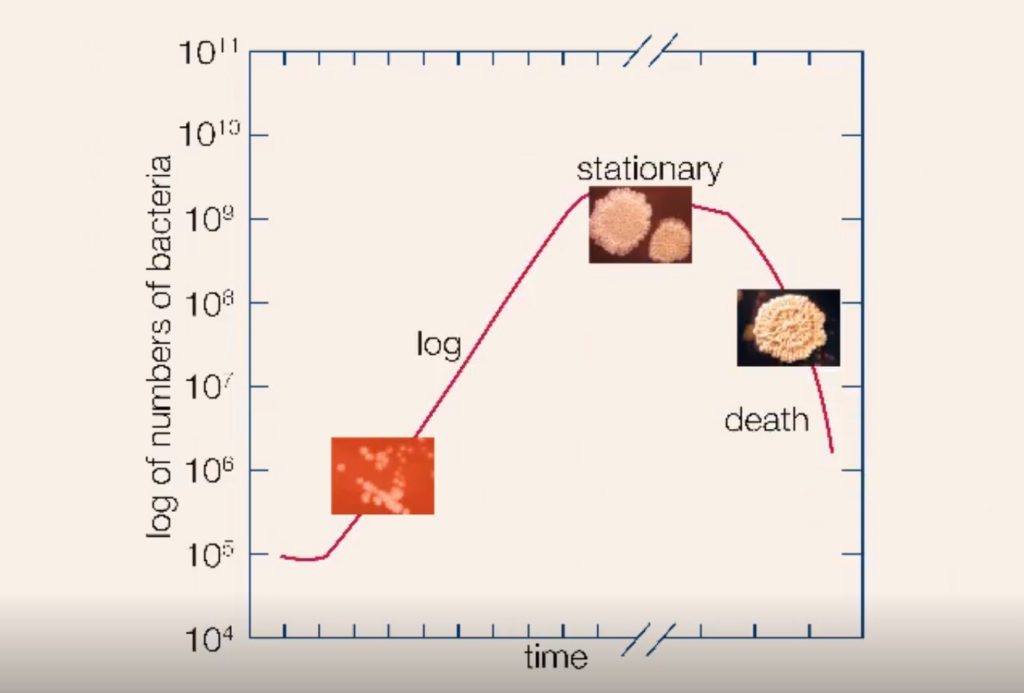
枯草芽孢杆菌生长周期
图源:AW Rakosy /Encyclopædia
细菌菌落通过生长的四个阶段进行:滞后阶段,对数阶段,静止阶段和死亡阶段。
细菌的产生时间因细菌而异,受许多环境条件和细菌种类的性质所控制。例如,生长最快的细菌之一产气荚膜梭状芽胞杆菌的最佳生成时间约为10分钟。 大肠杆菌每20分钟可以翻一倍; 生长缓慢的结核分枝杆菌的产生时间为12至16小时。 一些研究人员建议,某些生活在地球表面深处的细菌种群可能以极慢的速度生长,每隔几千年才繁殖一次。 “生长培养基” 例如摄入食物的组成是控制生长速率的主要因素。 当培养基提供更好的能源和细胞原本必须自己制造的更多生物合成中间体时,生长速率会增加到最大。
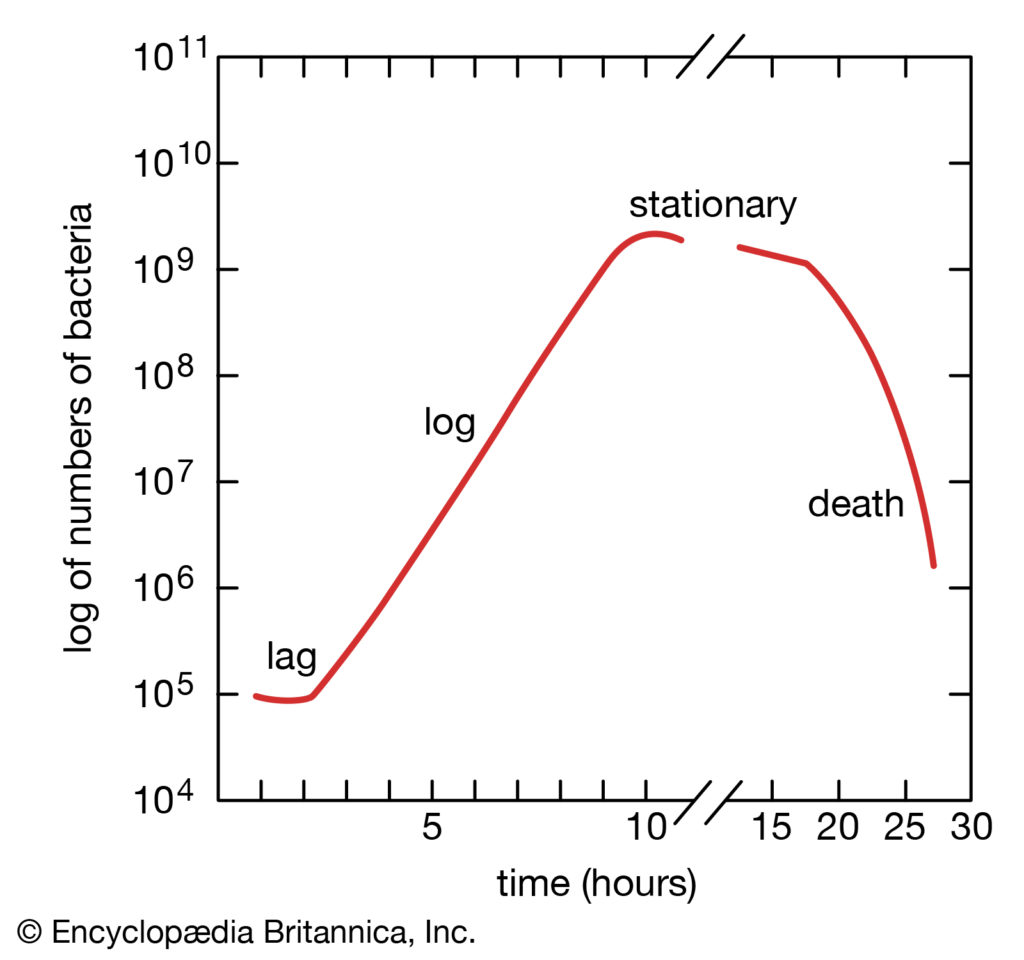
细菌生长曲线
广义细菌生长曲线显示了细菌菌落生长的各个阶段。
当细菌被放置在一个能提供其生长所需的所有营养物质的培养基中时,菌群表现出四个生长阶段,这四个阶段代表了典型的细菌生长曲线。一旦接种到新的培养基中,细菌不会立即繁殖,种群大小保持不变。在这一时期,称为滞后期,细胞代谢活跃,只增加细胞大小。它们还在新的环境条件下合成细胞分裂和种群增长所需的酶和因子。
然后,种群进入对数阶段,细胞数量以对数方式增加,每一代细胞与前一代细胞在相同的时间间隔内发生,从而使每一代细胞的成分均衡增加。对数阶段一直持续到营养物质耗尽或有毒物质积累,此时细胞生长速度减慢,一些细胞可能开始死亡。在最佳条件下,对数末期某些细菌的最大种群密度可达每毫升100亿至300亿个细胞。
枯草芽孢杆菌
图源:AW Rakosy /Encyclopædia
枯草芽孢杆菌的细菌菌落在37℃下进入生长的对数期培养18-24小时后,(98.6°F;放大约6倍)
细菌生长的对数阶段之后是稳定阶段,在这个阶段,细菌种群的大小保持不变,即使一些细胞继续分裂,而另一些细胞开始死亡。稳定期之后是死亡期,在这一阶段,群体中细胞的死亡超过了新细胞的形成。死亡阶段开始前的时间长短取决于物种和培养介质。即使在缺乏营养的情况下,细菌也不一定会死亡,并且它们可以长期存活。
大自然中的分布
原核生物在地球表面无处不在。 从极地冰到冒泡的温泉,从山顶到海底,从动植物到森林土壤,都可以找到它们。 有些细菌可以在接近冰点的温度下在土壤或水中生长,而另一些细菌则可以在接近沸腾(100°C 的温度下繁殖。 每种细菌都适合生活在特定的环境环境中,无论是海洋表面,泥浆沉积物,土壤还是其他生物的表面。 空气中的细菌含量很低,但是当灰尘被悬浮时含量会升高。 在未受污染的自然水体中,细菌数可以是每毫升数千。 在肥沃的土壤中,细菌数可以达到每克数百万。 在粪便中,细菌数量可能超过每克数十亿。

细菌在有机分解中的作用是从垃圾填埋场和水中去除不需要的生物材料的过程的一部分。
细菌是其栖息生态系统中的重要成员。尽管它们的体积很小,但它们的数量之多意味着它们的新陈代谢在外部环境中的元素转化中起着巨大的作用,有时有益,有时有害。可能每一种自然产生的物质,以及许多人工合成的物质,都能被某些种类的细菌降解(代谢)。奶牛最大的胃——瘤胃是一个发酵室,细菌在其中消化草料和饲料中的纤维素,将其转化为脂肪酸和氨基酸,这是奶牛使用的基本营养素,也是奶牛产奶的基础。污水或堆肥堆中的有机废物被细菌转化为适合植物新陈代谢的营养物质,或转化为气态甲烷(CH4)和二氧化碳。包括动植物在内的所有有机物质的残余物最终通过细菌和其他微生物的活动转化为土壤和气体,从而可供进一步生长。
许多细菌生活在溪流和其他水源中,它们在水样中的低种群密度并不一定表明水不适合饮用。然而,含有大肠杆菌等细菌的水,是人类和动物肠道的正常居民,表明污水或粪便物质最近污染了该水源。这类大肠菌群本身可能是病原体(致病生物),它们的存在表明其他不太容易检测到的细菌和病毒病原体也可能存在。净水厂中使用的沉淀、过滤和氯化程序旨在去除这些以及任何其他可能存在于人类饮用水中的微生物和传染源。水中微生物分解有机物质消耗氧气(生化需氧量),导致氧气水平下降,这对接收废水的溪流和湖泊中的水生生物非常有害。污水处理的一个目标是在有机物排放到水系统之前尽可能地氧化它们,从而降低废水的生化需氧量。为此,污水消化池和曝气装置专门利用细菌的代谢能力。
土壤细菌通过转化土壤的各种物质、腐殖质和矿物质,在影响生物化学变化方面极为活跃。对生命至关重要的元素,如碳、氮和硫,被细菌从无机气态化合物转化为可供植物和动物使用的形式。
细菌还将动植物代谢的最终产物转化为细菌和其他微生物可以利用的形式。氮循环可以说明细菌在影响各种化学变化中的作用。氮在自然界中以几种氧化状态存在,如硝酸盐、亚硝酸盐、二氮气体、几种氮氧化物、氨和有机胺(含有一种或多种取代烃的氨化合物)。固氮是将大气中的二氮气体转化为可供生物利用的形式。一些固氮细菌,如固氮菌、巴氏梭菌和肺炎克雷伯菌,是自由生活的,而根瘤菌则与豆科植物密切相关。
土壤中的根瘤菌生物识别并侵入其特定植物宿主的根毛,进入植物组织,并形成根瘤。 这个过程导致细菌失去许多自由活动特性。 它们变得依赖于植物所提供的碳,并且作为碳的交换,它们将氮气转化为氨,氨被植物用于其蛋白质的合成和生长。 另外,当硝酸盐用作电子受体时,许多细菌可以将硝酸盐转化为胺,以合成细胞材料,或将其转化为氨。反硝化细菌将硝酸盐转化为氮气。 氨或有机胺向硝酸盐的转化是通过需氧生物硝化单胞菌和硝化细菌的联合活动来完成的,它们利用氨作为电子供体。
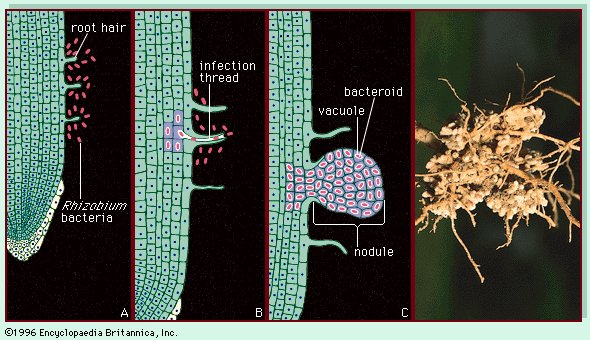
固氮细菌 (右)豌豆植物(Pisum sativum)的根部,其根瘤内藏有固氮细菌(Rhizobium)。(左)根瘤是根瘤菌与植物根毛之间共生关系的结果。细菌识别根毛并开始分裂(A),通过感染线(B)进入根部,使细菌进入根细胞,然后分裂成结节(C)
在碳循环中,二氧化碳被植物和自养原核生物转化为细胞物质,有机碳通过异养生物返回大气。 微生物分解的主要分解产物是二氧化碳,它是由呼吸需氧生物而形成的。
甲烷是碳代谢的另一种气体最终产物,是全球碳循环中相对次要的组成部分,但在当地情况下具有重要意义,是供人类使用的可再生能源。甲烷的产生是由高度专业化和专性厌氧产甲烷原核生物进行的,它们都是古细菌。产甲烷菌利用二氧化碳作为末端电子受体,从氢气(H2)中接收电子。其他一些物质可以被这些生物转化为甲烷,包括甲醇、甲酸、乙酸和甲胺。
尽管产甲烷菌可利用的物质范围极窄,但在许多有机物质(包括纤维素、淀粉、蛋白质、氨基酸、脂肪、醇和大多数其他底物)的厌氧分解过程中,甲烷的生成非常普遍。从这些物质中生成甲烷需要其他厌氧菌将这些物质降解为醋酸盐或二氧化碳和氢气,然后由产甲烷菌(methanogens)使用。产甲烷菌通过去除甲烷代谢活动中产生的氢气来支持混合物中其他厌氧菌的生长。消耗氢气会刺激其他细菌的新陈代谢。
尽管产甲烷菌的代谢能力非常有限,而且对氧非常敏感,但它们在地球上很普遍。大量的甲烷是在厌氧环境中产生的,比如沼泽和沼泽,但是土壤和反刍动物也会产生大量的甲烷。大气中至少80%的甲烷是由产甲烷菌作用产生的,其余的则是从煤层或天然气井中释放出来的。
食物中的细菌
健康奶牛的牛奶最初含有很少的细菌,这些细菌主要来自奶牛的皮肤和处理牛奶的程序。牛奶是许多细菌的极好生长介质,除非牛奶经过适当加工,否则细菌的数量会迅速增加。如果存在致病菌,细菌生长会破坏牛奶,甚至对健康造成严重危害。可以从被感染的牛传播的疾病包括结核病(结核分枝杆菌,Mycobacterium tuberculosis),不发烧的发热(流产的布鲁氏菌Brucella abortus)和发热的病原体(柯氏杆菌Coxiella burnetii)。
此外,伤寒可通过受感染的牛奶处理者的牛奶传播。 巴氏杀菌程序将牛奶的温度升高30分钟至63°C(145°F)或15秒升高至71°C(160°F),这可以杀死可能存在的任何病原菌,尽管这些程序不能杀死所有微生物。
某些细菌将牛奶转化成有用的乳制品,如酪乳、酸奶和奶酪。商业培养的酪乳是从接种了乳酸菌(Lactococcus)发酵剂(通常是乳酸乳杆菌或乳酪乳杆菌)的牛奶中制备的。酸奶和其他发酵乳制品是用不同的细菌培养物以类似的方式生产的。许多奶酪也是通过细菌的作用制成的。产酸细菌如乳乳酸杆菌在牛奶中的生长会使酪蛋白沉淀成凝乳。除去水分和加盐后,凝乳在其他微生物的作用下成熟。不同的细菌赋予食物不同的风味和特性;例如,干酪乳杆菌、嗜热链球菌和舍曼丙酸杆菌的混合物负责瑞士奶酪的成熟,并产生其特有的味道和大气泡。其他类型的细菌长期以来被用于制备和保存通过细菌发酵生产的各种食品,包括腌制产品、泡菜和橄榄。
在食物中传播的许多致病菌的毒素在摄入时会引起食物中毒。其中包括一种由金黄色葡萄球菌(Staphylococcus aureus)产生的毒素,这种毒素能引起迅速、严重但有限的胃肠道不适,或者肉毒梭菌(Clostridium botulinum)的毒素,这种毒素通常是致命的。在密封前未完全煮熟的罐装非酸性食品中可能产生肉毒杆菌毒素。肉毒梭菌形成耐热孢子,可以发芽为营养细菌细胞,在厌氧环境中茁壮成长,这有利于产生其极强的毒素。其他食源性感染实际上是由受感染的食物处理者传播的,包括伤寒、沙门氏菌病(沙门氏菌属)和志贺氏菌病(痢疾志贺氏菌属)。

图源:Encyclopædia
工业中的细菌
各种细菌的厌氧糖发酵反应会产生不同的最终产物。用酵母生产乙醇已经被酿酒工业开发了几千年,并被用于燃料生产。在醋的生产过程中,特定的细菌将酒精氧化成醋酸。其他的发酵过程可以制造出更有价值的产品。有机化合物,如丙酮、异丙醇和丁酸,是由各种梭状芽孢杆菌(Clostridium)在发酵过程中产生的,可以在工业规模上制备。在极端环境的生物中也发现了其他细菌产物和反应。 从嗜热细菌中分离出的酶引起了极大的兴趣,在这些酶中,由于可以在较高的温度下进行反应,因此可以以较高的速率进行反应。
来源:Encyclopædia
烃类氧化细菌和真菌对石油产品的微生物分解具有重要的生态学意义。石油的微生物分解是一个好氧过程,如果石油沉淀到底部的厌氧沉积物层(厌氧环境中的天然石油沉积物有数百万年的历史),那么这一过程就会被阻止。碳氢化合物氧化细菌附着在水面上漂浮的油滴上,它们的作用最终将油分解为二氧化碳。 将此类细菌及其生长因子喷洒到溢油上以提高非挥发性脂族和芳族烃的降解速度已成为一种常见的做法。
医学中的细菌
菌性疾病在人类历史上起着主导作用。 霍乱和鼠疫的广泛流行使世界上某些地区的人口减少了三分之一以上。细菌性肺炎可能是老年人死亡的主要原因。 也许有更多的军队被伤寒,痢疾和其他细菌感染击败,而不是被武力击败。 随着管道和卫生设备的现代发展,细菌疫苗的发展以及抗菌素的发现,细菌性疾病的发生率已降低。 然而,细菌没有继续作为传染因子消失,因为它们继续进化,产生了越来越强的毒株并获得了对许多抗生素的抗性。
尽管大多数细菌对地球上的生命都是有益的,甚至是生命所必需的,但少数细菌却对人类有害。 目前,没有一种古细菌被认为是病原体,但是包括人类在内的动物不断遭到大量细菌和各种细菌的轰击和居住。 与动物接触的大多数细菌都可以通过宿主的防御系统迅速消除。 口腔,肠道和皮肤被大量特定类型的细菌定殖,这些细菌适应这些栖息地的生活。 这些生物在正常情况下是无害的,并且仅在它们以某种方式穿过身体的屏障并引起感染时才变得危险。
一些细菌擅长入侵宿主,被称为病原体或疾病产生者。 一些病原体作用于人体的特定部位,例如脑膜炎球菌(Neisseria meningitidis),它会侵袭并刺激脑膜,大脑和脊髓周围的膜,并刺激脑膜。白喉细菌(Corynebacterium diphtheriae),最初感染喉咙; 以及在肠道中繁殖的霍乱细菌(霍乱弧菌),其产生的毒素导致该霍乱的大量腹泻。 其他能感染人类的细菌包括葡萄球菌(主要是金黄色葡萄球菌),它能感染皮肤引起疖子(疖子),血液引起败血症(血液中毒),心脏瓣膜引起心内膜炎,或骨头引起骨髓炎。
来源:Encyclopædia
人体中的细菌
图源:Encyclopædia
人体屏障(例如皮肤和肠道内壁)中的天然细菌在正常的人类生理过程中起着重要的作用。
侵入动物血液的病原菌可以利用多种机制中的任何一种来逃避宿主的免疫系统,包括形成长的脂多糖链,从而对一组血清免疫蛋白(称为补体)产生抵抗力,补体通常会抑制细菌的生长。细菌表面蛋白的致病性重组阻止动物产生的抗体识别病原体,在某些情况下使病原体能够在吞噬性白细胞中存活和生长。许多病原菌产生毒素,协助它们入侵宿主。这些毒素包括蛋白酶,分解组织蛋白质的酶,以及脂肪酶,分解脂质(脂肪)并通过破坏细胞膜来损伤细胞的酶。其他毒素通过在细胞膜上形成孔或通道来破坏细胞膜。一些毒素是修饰参与蛋白质合成或控制宿主细胞代谢的特定蛋白质的酶;例如白喉、霍乱和百日咳毒素。
补体激活途径
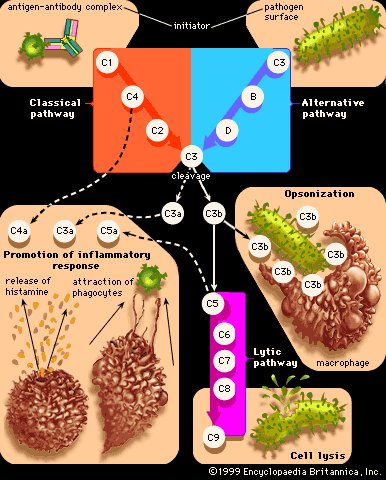
补体蛋白的主要功能是通过刺穿病原体的外膜(细胞裂解)或使它们对吞噬细胞(例如巨噬细胞)更具吸引力来帮助破坏病原体(这一过程称为调理作用)。一些补体成分还通过刺激细胞释放组胺和将吞噬细胞吸引到感染部位来促进炎症。
一些病原细菌在宿主体内形成区域,在这些区域中它们被封闭并不受免疫系统的保护,例如葡萄球菌形成的皮肤疖子和结核分枝杆菌形成的肺部空洞。脆弱拟杆菌(Bacteroides fragilis)大量存在于人体肠道中,但是一般不会对宿主造成任何困难。如果这种细菌通过损伤进入体内,细菌囊会刺激身体将细菌隔离成脓肿,从而减少细菌的传播。在许多情况下,细菌感染的症状实际上是免疫系统过度反应的结果,而不是细菌产生毒性因子的结果。
对抗致病性细菌感染的其他方法包括使用生物治疗剂或益生菌。这些无害的细菌会干扰病原菌的定殖。另一种方法是利用噬菌体,即杀死细菌的病毒,来治疗特定细菌病原体的感染。此外,20世纪80年代发展起来的重组DNA技术使细菌合成几乎任何蛋白质成为可能,大肠杆菌是这一过程中常见的宿主。重组DNA技术用于廉价、大规模生产极为稀缺和有价值的动物或人类蛋白质,如激素、凝血因子,甚至抗体。
细菌自地球生命史以来就已经存在。 在岩石中发现的细菌化石至少可以追溯到泥盆纪时期(4.192亿至3.589亿年前),并且令人信服的论点是自前寒武纪大约35亿年前就已经存在细菌。细菌至少在大约18亿年前的古元古代末期就已在地球上广泛传播,当时由于蓝细菌的作用,大气中出现了氧气。 因此,细菌有足够的时间来适应其环境,并产生了许多后代。
关于生命起源所涉及的原始祖先的性质,人们有相当多的猜测。有人认为原始细胞可能使用RNA作为其遗传物质,因为研究表明RNA分子可以具有多种催化功能。在这个时期,细菌和古菌很早就从它们共同的前体中分化出来。这两类原核生物倾向于栖息在不同类型的环境中,并以不同的速度产生新的物种。许多古菌喜欢高温生态位。古细菌树的一个主要分支仅由嗜热物种组成,而另一个主要分支中的许多产甲烷菌可以在高温下生长。相反,没有一个主要的真细菌分支仅仅由嗜热菌组成。细菌和古细菌都含有能够在高温下生长的成员,以及其他能够在低温下生长的物种。另一个显著的区别是细菌广泛地适应好氧条件,而许多古细菌是专性厌氧菌。没有古细菌是专性光合作用的。也许古细菌是一种较原始的生物体,对变化的环境条件的遗传反应受损。 适应新情况的能力有限可能会将古细菌限制在恶劣的环境中,与其他生命形式的竞争较少。
根据三域系统的生命之树
有机体必须不断进化或适应不断变化的环境,很明显,突变,即有机体DNA中核苷酸序列的变化,在所有有机体中不断发生。DNA序列的变化可能会导致该DNA片段编码的蛋白质的氨基酸序列发生变化。因此,改变后的蛋白质可能更适合或不太适合在当前条件下发挥功能。尽管DNA中可能发生的许多核苷酸变化对细胞的适应性没有影响,但如果核苷酸变化能促进细胞的生长,哪怕是很小的程度,那么突变型细胞就能够增加其在群体中的相对数量。然而,如果核苷酸的改变阻碍了细胞的生长,那么突变形式就会被其他细胞所取代而丧失。
在生物体之间传递遗传信息的能力是适应环境变化的一个主要因素。DNA交换是高等真核生物生命周期的重要组成部分,在所有真核生物中都有发生。基因交换也发生在整个细菌世界,虽然转移的DNA量很小,但这种转移可以发生在远缘生物之间。携带在质粒上的基因可以进入细菌染色体,成为细菌遗传的稳定部分。生物体通常拥有称为转座子的可移动遗传元件,可以重新排列染色体上任何基因的顺序和存在。转座子可能在帮助加快进化速度方面发挥作用。
细菌快速进化的例子很多。在20世纪40年代以前,抗生素还没有用于医疗实践。当抗生素最终投入使用时,大多数致病菌对它们很敏感。然而,从那时起,细菌对一种或多种抗生素的耐药性增加,以至于以前有效的抗生素对某些类型的细菌不再有用。大多数致病细菌对抗生素耐药的例子并不是由于突变改变了抗生素攻击的蛋白质,尽管这种机制可能发生。相反,抗生素抗性通常涉及细菌产生的酶,这些酶会改变抗生素并使之失活。抗生素耐药性传播的一个主要因素是可传播的质粒,它将药物灭活酶的基因从一个细菌携带到另一个细菌。尽管这些酶的基因的原始来源尚不清楚,但可移动的基因元件(转座子)可能在它们的出现中起了作用,也可能允许它们转移到其他细菌类型。
影响细菌生长的因素
细菌在最佳生长所需的条件上有很大的不同。就营养需求而言,所有细胞都需要碳、氮、硫、磷、多种无机盐(如钾、镁、钠、钙和铁)以及大量其他微量元素(如锌、铜、锰、硒、钨和钼)。碳是细菌最需要的元素,因为氢和氧可以从水中获得,这是细菌生长的先决条件。同时还需要一种能量来源来促进细菌的新陈代谢。组织细菌的一种方法是基于这些基本的营养需求:碳源和能源。
细胞可用于碳的来源有两种:无机化合物和有机化合物。使用无机化合物二氧化碳(CO2)作为碳源的生物称为自养生物。 需要碳的有机源(例如糖,蛋白质,脂肪或氨基酸)的细菌称为异养生物(或有机营养菌)。 许多异养生物,例如大肠杆菌或铜绿假单胞菌,都由简单的糖(例如葡萄糖)合成所有细胞成分,因为它们具有必要的生物合成途径。 其他异养生物失去了其中一些生物合成途径。 为了生长,它们要求其环境包含化学上完整的特定氨基酸,含氮碱或维生素。
铜绿假单胞菌
除了碳,细菌还需要能量,能量几乎总是通过电子从电子供体转移到电子受体来获得的。有三种基本的能源:光、无机化合物和有机化合物。光营养细菌利用光合作用从光能中产生三磷酸腺苷(ATP)形式的细胞能量。化学营养体从化学物质(有机和无机化合物)中获取能量;化学石营养体从与无机盐的反应中获取能量;化学异养体从有机化合物中获取碳和能量(能量源也可作为这些生物体的碳源)。
在大多数情况下,细胞能量是通过电子转移反应产生的,其中电子通过一种途径从电子或有机供体分子移动到受体分子,从而保留了电子在转移过程中释放出的能量,其形式为电子俘获 细胞可用于其化学或物理工作的物质。 从电子传输中捕获的主要能量形式是ATP。 分解有机分子以产生能量的代谢过程称为分解代谢反应。 相反,合成分子的代谢过程称为合成代谢反应。
许多细菌可以使用大量的化合物作为碳和能源,而其他细菌的代谢能力受到很大限制。尽管碳水化合物是真核生物的常见能源,但是由于大多数细菌不具有代谢这些通常复杂的分子所必需的酶,因此这些分子仅通过有限数量的细菌进行代谢。 相反,许多细菌都依赖于其他能源,例如氨基酸,脂肪或其他化合物。 对细菌重要的其他化合物包括磷酸盐,硫酸盐和氮。 由于许多细菌无法合成磷酸盐,因此在许多环境中,尤其是在水中,磷酸盐含量低可能是细菌生长的限制因素。 另一方面,大多数细菌可以将硫酸盐或硫化物转化为蛋白质合成所需的有机形式。 生命有机体从氨中吸收氮的能力是广泛存在的,细菌将其他形式的氮(例如土壤中的硝酸盐或大气中的二氧化氮转化为细胞物质的能力也有所不同。
有些细菌是专性寄生虫,只在活的宿主细胞内生长。例如,立克次体(Rickettsia)和衣原体(Chlamydia), 在真核细胞中生长,蛭弧菌(Bdellovibrio)在细菌细胞中生长。梅毒螺旋体(Treponema pallidum)很难(即使不是不可能)在培养中生长,可能是因为它需要低的氧张力和低的氧化还原水平,这是由于存在动物细胞而不是任何特定的营养素所致。因为有些细菌可能只以动物或植物寄生虫的形式生长,或者只在牛奶等营养丰富的来源中生长,所以它们在自然界中不可能以游离细菌的形式生长。许多来自自然环境的细菌与其他细菌联合存在,很难从该伙伴关系的其他成员中分离和培养。
对于不同的细菌类型来说,最适合细菌生长的物理条件有很大的不同。作为一个群体,细菌在不同环境中的生存能力在所有生物中表现出最大的差异。
氧 气
细菌之间最显著的区别之一是它们对大气氧(O2)的需求和反应。实际上,所有的真核生物都需要氧气才能茁壮成长,而许多种类的细菌可以在厌氧条件下生长。需要氧气才能生长的细菌称为专性需氧细菌。在大多数情况下,这些细菌需要氧气才能生长,因为它们产生能量和呼吸的方法依赖于电子向氧气的转移,而氧气是电子传递反应中的最终电子受体。
专性需氧菌包括枯草芽孢杆菌(Bacillus subtilis)、铜绿假单胞菌(Pseudomonas aeruginosa)、结核分枝杆菌(Mycobacterium tuberculosis)和氧化亚铁硫杆菌(cidithiobacillus ferrooxidans)。
只有在缺氧条件下才能生长的细菌,如梭状芽孢杆菌、类杆菌和产甲烷的古细菌(产甲烷菌)被称为专性厌氧菌,因为它们产生能量的代谢过程与氧气的消耗相无关。事实上,氧气的存在实际上会毒害它们的一些关键酶。有些细菌(肺炎链球菌)是微需氧或耐氧厌氧菌,因为它们在低浓度的氧气中生长得更好。在这些细菌中,氧气通常会刺激微小的代谢过程,从而增强能量产生的主要途径。兼性厌氧菌可以根据氧气的存在改变其代谢过程,在氧气存在时使用更有效的呼吸过程,在缺氧时使用效率较低的发酵过程。兼性厌氧菌包括大肠杆菌和金黄色葡萄球菌。
破伤风梭状芽胞杆菌
图源:Centers for Disease Control and Prevention
细菌对氧的反应不仅仅取决于它们的代谢需要。氧是一种非常活泼的分子,会形成一些有毒的副产物,如超氧物(O2-)、过氧化氢(H2O2)和羟基自由基(OH·)。有氧生物产生的酶可以使这些氧气产物解毒。
最常见的排毒酶是分解过氧化氢的过氧化氢酶和分解超氧化物的超氧化物歧化酶。 厌氧细菌通常不产生过氧化氢酶,其超氧化物歧化酶的水平随细胞对氧气的敏感性而成比例地变化。 许多厌氧菌对氧气过敏,短时间接触就会被杀死,而其他厌氧菌,包括大多数梭状芽孢杆菌,对氧的存在更具耐受性。这些酶的联合作用,以消除过氧化氢和超氧物是重要的,因为这些副产品与铁一起形成极为活跃的羟基自由基,这是能够杀死细胞。
温度
细菌已适应多种温度。 在低于约15°C(59°F)的温度下生长的细菌是嗜冷菌。细菌在低温下生长的能力并不意外,因为温带土壤的平均地下温度约为12°C(54°F),90%的海洋温度为5°C(41°F)或更低。专性嗜冷菌是从北极和南极海水和沉积物中分离出来的,其最适生长温度约为10℃(50℉),如果暴露在20℃(68℉)下,则无法存活。
大多数嗜冷细菌属于革兰氏阴性菌属假单胞菌属、黄杆菌属、无色杆菌属和产碱菌属。中温细菌是指在20至45°C(68至113°F)之间发生最佳生长的细菌,尽管它们通常可以在10至50°C(50至122°F)的温度下存活和生长。动物病原体一般是嗜中性的。
嗜热原核生物可以在高于60°C(140°F)的温度下生长。这些温度在腐烂的堆肥堆、温泉和海洋地热喷口中都会遇到。在温泉的径流中,在温度下降到70℃左右的源头附近发现了嗜热菌,如水热菌(生长的最适温度为70℃[158℉];最高温度为79℃[174℉])。
蓝藻合成球藻和光营养滑动细菌氯曲菌的厚席在径流较冷的部分发育。古生酸硫菌对酸性条件有很高的耐受性,允许在pH值约为1.0至6.0的范围内生长,最适温度为80°C(176°F)。许多细菌和古细菌适应于50至70°C(122至158°F)的温度范围,包括芽孢杆菌属、热放线菌属、甲烷杆菌属、甲基球菌属和硫菌属的一些成员。
最引人注目的是上世纪80年代中期在深海海底营养丰富、温度极高的热液喷口中发现了细菌和古细菌。焦菌属的古细菌在80到110°C(176到230°F)的温度范围内繁衍生息,在这种温度下,水只有在极高的压力下才能保持液态。
pH值
大多数细菌在中性pH值范围内(5-8之间)生长,尽管有些物种已经适应了酸性或碱性极端条件下的生活。 嗜酸细菌的一个例子是氧化铁曲霉。 当煤层通过采矿作业暴露于空气中时,黄铁矿硫化物沉积物会受到A.ferrooxidans的攻击而生成硫酸,从而将pH值降低至2.0甚至0.7。 但是,铁氧化农杆菌的耐酸性仅适用于硫酸,因为这些细菌在暴露于等浓度的其他酸(例如盐酸)时会死亡。许多细菌不能耐受酸性环境,特别是在厌氧条件下,结果,植物聚合物在酸性沼泽(pH值在3.7和5.5之间),松树林和湖泊中会缓慢降解。 与嗜酸菌相反,嗜碱菌能够在高达10至11的pH值的碱性浓度下生长。嗜碱菌已从土壤中分离出来,大多数是革兰氏阳性芽孢杆菌属。
盐和水
水是生命的基本需求。有些细菌喜欢咸的环境,因此被称为嗜盐菌。极端嗜盐菌,如嗜盐杆菌,在含盐量为20%至30%的条件下表现出最佳生长,如果盐含量降低,它们会溶解(破裂)。这种细菌存在于死海、盐水池中,有时也存在于咸鱼和兽皮上。中度嗜盐细菌在含盐量为5%至20%的条件下生长,在盐卤和泥浆中也有发现。
死海中的盐柱

图片来源:Peter Carmichael/ASPECT
细菌代谢
异养代谢
如上所述,异养(或有机营养)细菌需要有机分子来提供它们的碳和能量。产生能量的分解代谢反应可以有许多不同的类型,尽管它们都涉及电子转移反应,其中电子从一个分子到另一个分子的运动与产生ATP的能量捕获反应相耦合。一些异养细菌可以代谢糖或复杂的碳水化合物来产生能量。这些细菌必须产生许多特定的蛋白质,包括将多糖降解为其组成糖单位的酶,在细胞内积累糖的运输系统,以及将糖转化为代谢中间产物之一的酶,如葡萄糖-6-磷酸。糖酵解的Embden-Meyerhof途径和磷酸戊糖途径都存在于真核细胞中。有些细菌拥有Entner-Doudoroff途径,主要将葡萄糖转化为丙酮酸,还有一些途径可以用较少的酶催化步骤将葡萄糖转化为较小的化合物。
糖代谢通过发酵和呼吸两个不同的过程为细胞产生能量。发酵是在没有任何外部电子受体的情况下进行的厌氧过程。有机化合物,如糖或氨基酸,被分解成更小的有机分子,这些分子接受在能量源分解过程中释放的电子。
这些分解代谢反应包括几个步骤,直接形成ATP。当葡萄糖被分解成乳酸时,就像某些乳球菌和乳酸杆菌以及高等真核生物的肌肉细胞中发生的那样,每个葡萄糖分子只产生两个ATP分子,相当数量的葡萄糖必须被降解以提供足够的能量供细菌生长。由于有机分子在发酵过程中仅被部分氧化,发酵细菌的生长会产生大量的有机终产物,而消耗的每个葡萄糖分子的能量输出相对较小。
很少有细菌只产生乳酸,乳酸对细菌毒性很大,限制了菌落的生长。特定细菌利用多种额外的发酵途径分解葡萄糖;这些途径的特征性终产物有助于细菌的鉴定。这些最终产物通常比乳酸毒性小,或者是利用额外的代谢能量形成的。例如,大肠杆菌混合酸发酵的产物包括乳酸、琥珀酸、乙酸、甲酸、乙醇、二氧化碳和氢气。产气肠杆菌能产生大部分相同的发酵产物,以及大量的2,3-丁二醇,它是非酸性的,允许更多的细菌生长。
细胞可以从呼吸中获得相当多的能量,在这个过程中,糖分子的电子不是转移到另一个有机分子,而是转移到一个无机分子上。最常见的呼吸过程(有氧呼吸)使用氧气作为最终的电子受体。糖被完全分解成二氧化碳和水,每分子葡萄糖最多产生38个ATP分子。
电子通过电子传递链转移到氧中,电子传递链是位于细胞膜上的酶和辅助因子系统,其排列方式使电子沿着链的传递与质子(氢离子)穿过膜并离开细胞的运动相耦合。电子传输引起带正电荷的氢离子向细胞外移动,带负电荷的离子向细胞内移动。这种离子梯度导致外部介质的酸化和带电的质膜,其电荷为150至200毫伏。离子梯度的产生,包括质子动力(质子梯度),是所有生物能量产生和储存的一个共同方面。
质子的梯度被细胞直接用于许多过程,包括营养物质的主动运输和鞭毛的旋转。质子还可以通过一种被称为F1F0质子转位ATP酶的膜酶从细胞外部进入细胞质,该酶将质子运动与ATP合成结合,其过程与真核细胞线粒体中发生的过程相同。
能够利用呼吸作用的细菌每一个糖分子产生的能量比发酵细胞多得多,因为能量源的完全氧化(分解)可以完全提取所有可用的能量,如呼吸生物的ATP产量比发酵细菌的ATP产量大得多所示。呼吸有机体利用一定量的营养物质获得更高的细胞物质产量;它们也产生更少的有毒最终产品。然而,氧在水中的溶解度是有限的,需氧菌群的生长和存活与可用的氧供应量成正比。只有与空气接触的细菌才能获得连续的氧气供应,当细菌能够漂浮在暴露在空气中的表面上时,或者当细菌所在的介质被剧烈搅拌时,就会发生这种情况。
在厌氧条件下,呼吸也可以通过称为厌氧呼吸的过程发生,其中最终的电子受体是无机分子,如硝酸盐(NO3-)、亚硝酸盐(NO2-)、硫酸盐(SO42-)或二氧化碳(CO2)。在硫酸盐和二氧化碳的情况下,使用这些受体的细胞可获得的能量产量比氧气的呼吸低得多,但它们仍然比发酵可获得的能量产量高得多。一些细菌利用无机分子进行无氧呼吸的能力可能具有环境意义。大肠杆菌可以使用氧气,硝酸盐或亚硝酸盐作为电子受体,斯图氏假单胞菌在反硝化,硝酸盐转化为亚硝酸盐和氮气(N2)方面具有重要的全球意义。脱硫弧菌和脱硫单胞菌分别还原硫酸盐和元素硫,产生硫化物(S2-),而木醋杆菌和产甲烷古细菌,如嗜热甲烷杆菌,将二氧化碳还原为乙酸盐和甲烷。古细菌通常使用氢作为电子供体,使用二氧化碳作为电子受体以产生甲烷,或者使用硫酸盐作为电子受体以产生硫化物。
自养代谢
自养细菌以二氧化碳为碳源合成所有的细胞成分。从二氧化碳合成有机化合物的最常见途径是还原性戊糖磷酸(Calvin)循环,还原性三羧酸循环和乙酰辅酶A途径。 由美国生物化学家梅尔文·卡尔文(Melvin Calvin)阐明的卡尔文循环是这些途径中分布最广泛的途径,在植物,藻类,光合细菌和大多数需氧自养细菌中起作用。
卡尔文循环的关键步骤是核酮糖1,5-二磷酸与二氧化碳的反应,产生两个分子的3-磷酸甘油酯,葡萄糖的前体。这种循环在能量方面对细胞来说是极其昂贵的,因此一个甘油醛-3-磷酸分子的合成需要消耗九个ATP分子和六个电子供体分子的氧化,即烟酰胺腺嘌呤二核苷酸磷酸(NADPH)的还原形式。自养行为取决于细胞进行光合作用或有氧呼吸代谢的能力,这是唯一能够提供足够能量来维持碳固定的过程。
卡尔文循环
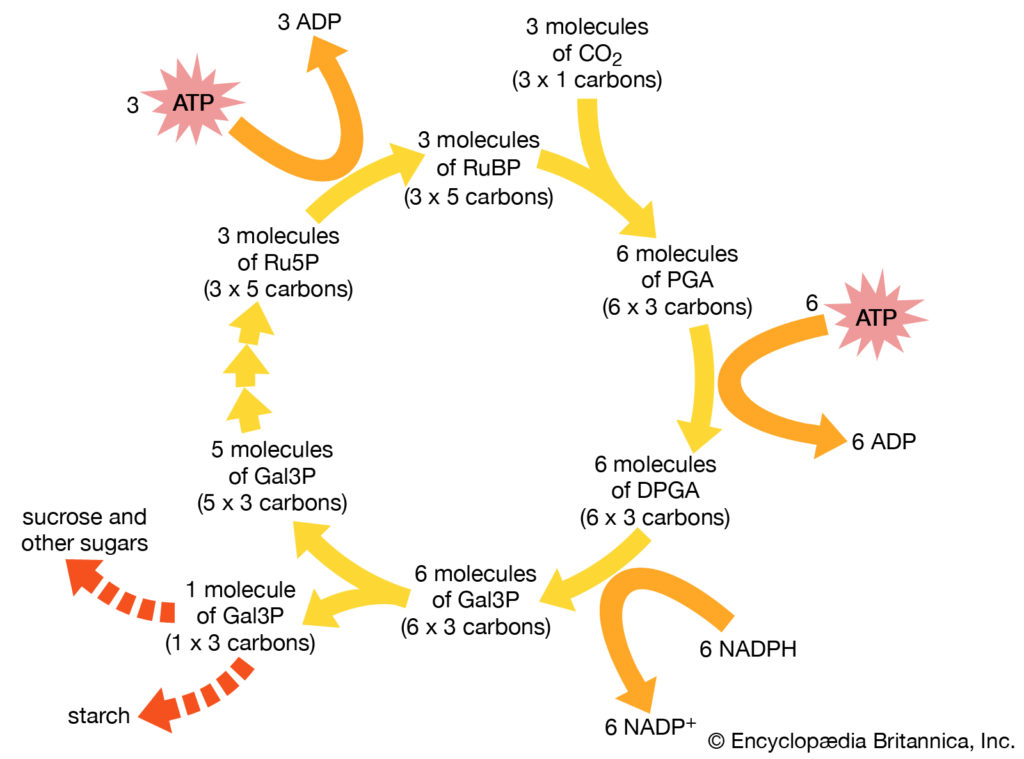
细菌使用加尔文循环来合成有机化合物。核糖1,5-二磷酸(RuBP)与二氧化碳的反应导致生成3-磷酸甘油酸酯(PGA)分子,该分子通过多次中间反应转化为一分子3-磷酸甘油醛(Gal3P)。然后可以将3-磷酸甘油醛转化为其他分子,包括糖或淀粉。
需氧非光合自养菌是指不仅以二氧化碳为唯一碳源,而且以氧为电子受体的无机化合物(电子供体)为能源的细菌。这些细菌在分类上是多样的,通常由它们使用的电子供体来定义。例如,欧洲亚硝基单胞菌将氨(NH4+)氧化为亚硝酸盐,而Nitrobacter winogradskyi将亚硝酸盐氧化为硝酸盐。 硫杆菌将硫代硫酸盐和元素硫氧化为硫酸盐,而氧化铁曲霉将亚铁离子氧化为三价铁。
这种不同的氧化能力使氧化亚铁杆菌能够耐受高浓度的许多不同离子,包括铁、铜、钴、镍和锌。所有这些类型的细菌似乎都是专性的岩性滋养菌,不能在很大程度上利用有机化合物。一氧化碳(CO)被寡糖carboxidovorans氧化成二氧化碳,而氢气(H2)被真核产碱杆菌(Alcaligenes eutrophus)氧化,在较小程度上也能被其他许多细菌氧化。
代谢能是由这些电子供体的氧化提供的,其方式基本上与呼吸异养生物相同,异养生物将电子从有机分子转移到氧气中。当电子沿着电子传递链传递到氧时,质子在细胞膜上产生梯度。这个梯度被用来产生ATP分子。岩石自养生物中存在的其他反应是用于从无机供体中去除电子和固定二氧化碳的反应。
光营养代谢
地球上的生命依靠光合作用将太阳能转化为细胞能量。光合作用的一般过程是利用叶绿素来吸收来自太阳的光能,并释放出一个能量更高的电子。这种电子通过电子传递链,通过形成质子梯度和伴随的ATP合成产生能量。电子最终返回叶绿素。这种循环反应路径可以满足电池的能量需求。
然而,为了使细胞生长,二氧化碳固定的卡尔文循环必须被激活,电子必须转移到辅助因子NADP以形成NADPH,这是循环运行所需要的大量的NADPH。因此,光营养细胞的生长需要一个电子源来取代生物合成反应中消耗的电子。
光合生物根据这些电子来源的性质分为两大类。其中一类包括高等植物、真核藻类和蓝藻(蓝绿藻);这些生物含有色素叶绿素a,在产生氧气的反应中以水为电子源。
据认为,到18亿年前,蓝藻的前身已经在全球产生了足够的氧气,开始允许更高形式的生命的发展。析氧光合作用需要两个独立的光吸收系统的作用,将水中电子的能量提高到足以转移到NADP的水平。因此,在这些生物体中存在两个不同的光反应中心,一个用于产氧反应,另一个用于能量产生的循环过程。
在蓝藻中,两个光反应中心都含有叶绿素a。它们的光合器官还含有其他光吸收色素,这些色素作为触角来捕获光能并将其转移到反应中心。蓝藻触角包括额外的叶绿素a分子和藻胆体,叶绿素a分子将能量转移到循环反应中心,藻胆体是一种蛋白质色素,它们吸收短、高能波长的光,并将能量传递到析氧反应中心。在几乎所有的蓝藻中,光合器都包含在一个广泛的细胞内扁平膜囊系统中,称为类囊体,其外表面布满规则排列的藻胆体颗粒。这种色素聚集体存在于类囊体表面的排列称为光系统。
其他光合细菌只含有一种不同色素的单一反应中心,称为细菌叶绿素,它能吸收长而低能波长的光。这些生物需要除水以外的电子供体,并且不释放氧气。绿色细菌(绿藻科)和紫色硫细菌(藻科)使用元素硫、硫化物、硫代硫酸盐或氢气作为电子供体,而紫色非硫细菌使用氢或有机基质中的电子。这些细菌需要厌氧条件才能进行光合作用。绿色细菌的光系统与高等植物的光系统Ⅰ有关,而紫色细菌的光系统Ⅱ则与光系统Ⅱ有关,后者提供了从细菌到植物的进化轨迹。
细菌的生物合成途径
许多原核生物能够将任何给定的碳源转化为生物合成的组成部分,例如氨基酸、嘌呤、嘧啶、脂类、糖和酶辅因子。在这些生物合成途径中,每种酶的数量和活性都受到严格的调节,因此细胞在任何时候只产生所需的任何化合物。
在进化过程中,一些细菌失去了编码某些生物合成反应的基因,因此可能需要营养补充。例如,支原体的DNA含量约为大肠杆菌的四分之一,它有许多营养需求,甚至丧失了形成细胞壁的能力。
支原体
图源:Centers for Disease Control and Prevention
革兰氏阴性人型支原体和在琼脂培养基上生长的常见机会性T株支原体的分离株。
细菌是肉眼看不到的单细胞微生物,是地球上最古老的生物之一,它们无处不在且数量惊人,难以逃避。由于我们看不到它们并且对它们的了解相对较少,因此细菌的世界对我们来说是神秘的,并且通常不为大众所重视。
人类每天使用细菌。细菌可用于保存食物,为农作物增添养分,制作奶酪和酸奶以及消除固体废物。这些用途有益于人类的日常生活。一些细菌在生物研究以及药物和疫苗的开发中非常有用。有的细菌帮助管理化粪池系统和清理漏油,或制作酸奶,奶酪和葡萄酒。人体中的细菌可以使身体受益,这取决于它们是细菌的类型。一些细菌有助于训练免疫系统,并有助于预防过敏。其他的则可以保护人体免受有害的致病细菌的侵害。细菌负责帮助我们消化食物,有些细菌会产生对人体非常有用的酶,激素和维生素。
细菌还会导致人患病或死亡。当人打喷嚏或咳嗽时,传染病(例如普通感冒和流感)可能会从细菌中传播。粪便中的细菌也会使人生病。如果一个人上完洗手间后没有正确洗手,他们可能会感染大肠杆菌和自己。这些特性使人类需要花更大精力来研究和重视细菌。
客观全面认识细菌,是我们正确管理和利用它们的第一步。
附录:
常见革兰氏阳性菌属:
Corynebacterium 棒杆菌属 , Propionibacterium 丙酸杆菌属, Alloiococcus 差异球菌属, Oscillospira 颤螺菌属, Enterococcus 肠球菌属, Dialister 戴阿利斯特杆菌属, Anaerostipes 丁酸弧菌属, Butyrivibrio 丁酸弧菌属, Mobiluncus 动弯杆菌属, Planomicrobium 动性杆菌属, Brevibacillus 短芽孢杆菌属, Actinomycetales 放线菌目, Actinomyces 放线菌属, Mycobacterium 分枝杆菌属, Finegoldia 芬戈尔德菌属, Faecalibacterium 粪杆菌属, Coprococcus 粪球菌属, Coprobacillus 粪芽孢菌属, Gordonia 戈登氏菌, Holdemania 霍尔德曼氏菌属, Rhodococcus 红球菌属, Arthrobacter 节杆菌属, Blautia 经黏液真杆菌属, Rubrobacter 红色杆菌属, Cetobacterium 鲸杆菌属, Catonella 卡托氏菌属, Lachnobacterium 毛螺旋菌属, Collinsella 柯林斯氏菌属, Nocardioides 类诺卡氏菌属, Paenibacillus 类芽孢杆菌属, Streptococcus 链球菌属, Ruminococcus 瘤胃球菌属, Roseburia 罗氏菌属, Rothia 罗氏菌属, Vagococcus 漫游球菌属, Lachnospira 毛螺菌属, Moryella 毛螺旋菌属, Oribacterium 毛螺旋菌属, Leuconostoc 明串珠菌属, Ureaplasma 尿枝原体属, Dermabacter 皮杆菌属, Staphylococcus 葡萄球菌属, Aerococcus 气球菌属, Carnobacterium 肉杆菌属, Lactobacillus 乳杆菌属, Lactococcus 乳球菌属, Slackia 史雷克氏菌属, Bifidobacterium 双歧杆菌属, Fusobacterium 梭杆菌属, Parvimonas 微单胞菌属, Weissella 魏斯氏菌属, Peptostreptococcus 消化链球菌属, Peptococcus 消化球菌属, Sporosarcina 芽孢八叠球菌属, Bacillus 芽孢杆菌属, Anaerococcus 厌氧球菌属, Alicyclobacillus 脂环酸杆菌属, Dactylosporangium 指孢囊菌属
常见革兰氏阴性菌属:
Escherichia 埃希氏菌属, Eikenella 艾肯氏菌属, Acidaminococcus 氨基酸球菌属, Hymenobacter 薄层菌属, Phenylobacterium 苯基杆菌属, Burkholderia 伯克霍尔德氏菌属, Acinetobacter 不动杆菌属, Filifactor 产丝菌属, Asticcacaulis 不粘柄菌属, Ochrobactrum 苍白杆菌属, Oxalobacter 草酸杆菌属, Flavobacterium 产黄菌属, Enterobacter 肠杆菌属, Comamonas 丛毛单胞菌属, Acetobacter 醋杆菌属, Brevundimonas 短波单胞菌属, Capnocytophaga 二氧化碳嗜纤维菌属, Delftia 代尔夫特菌属, Paracoccus 副球菌属, Arcobacter 弓形菌属, Devosia 德沃斯氏菌属, Mesorhizobium 根瘤菌属, Citrobacter 枸橼酸杆菌属, Azospirillum 固氮螺菌属, Stenotrophomonas 寡养单胞菌属, Marinobacter 海杆菌属, Rhodobacter 红细菌属, Rhodoplanes 红游动菌属, Porphyromonas 红棕色单胞菌属, Vibrio 弧菌属, Methylobacterium 甲基杆菌属, Pseudomonas 假单胞菌属, Succiniclasticum 解琥珀酸菌属, Chryseobacterium 金黄杆菌属, Kingella 金氏菌属, Megasphaera 巨球形菌属, Megamonas 巨单胞菌属, Klebsiella 克雷伯氏菌属, Lautropia 口动菌属, Aggregatibacter 凝聚杆菌属, Bradyrhizobium 慢生根瘤菌属, Agrobacterium 农杆菌属, Sphingomonas 鞘氨醇单胞菌属, Rikenella 理研菌属, Myroides 类香味菌属, Helicobacter 螺杆菌属, Chloronema 绿丝菌属, Erwinia 欧文氏菌属, Bilophila 嗜胆菌属, Treponema 密螺旋体属, Moraxella 莫拉氏菌属, Neisseria 奈瑟氏菌属, Sphingobacterium 鞘氨醇杆菌属, Bacteroides 拟杆菌属, Prevotella 普雷沃氏菌属, Sporocytophaga 生孢噬纤维菌属, Psychrobacter 嗜冷杆菌属, Hyphomicrobium 生丝微菌属, Prosthecobacter 突柄杆菌属, Cytophagales 嗜细胞菌目, Pectinatus 梳状菌属, Lysobacter 溶杆菌属, Sutterella 萨特氏菌属, Serratia 赛氏杆菌属, Cupriavidus 贪铜菌属, Leptotrichia 纤毛菌属, Haemophilus 嗜血杆菌属, Cytophaga 噬纤维菌属, Leptolyngbya 瘦鞘丝藻属, Campylobacter 弯曲杆菌属, Desulfobulbus 脱硫叶菌属, Enhydrobacter 水栖菌属, Pedomicrobium 土微菌属, Desulfovibrio 脱硫弧菌属, Veillonella 韦荣菌属, Shewanella 希瓦氏菌属, Gemmatimonas 牙单胞菌属, Selenomonas 月形单胞菌属, Phormidium 席蓝细菌属, Nitrospira 硝化螺菌属, Cardiobacterium 心杆菌属, Anaerovibrio 厌氧弧菌属, Halomonas 盐单胞菌属, Anaerobiospirillum 厌氧螺菌, Bdellovibrio 蛭弧菌属
以上信息整理来自谷禾健康,若需转载,请备注出处。
相关阅读:
主要参考资料:
Robert J. Kadner, Kara Rogers, Yamini Chauhan Swati, Yamini Chauhan, Swati Chopra, Aakanksha Gaur, Parul Jain, Robert Lewis, Gloria Lotha, Deepti Mahajan, Richard Pallardy, Chelsey Parrott-Sheffer, Marco Sampaolo, Veenu Setia, Shiveta Singh, Grace Young. Science Biology Archaea & Bacteria Bacteria. (1999-2020)
GEORGE M. GARRITY et al. (eds.), Bergey’s Manual of Systematic Bacteriology, 5 vol., 2nd ed. (2001–12)
MICHAEL T. MADIGAN et al., Brock Biology of Microorganisms, 14th ed. (2015)
JOANNE M. WILLEY, LINDA M. SHERWOOD, and CHRISTOPHER J. WOOLVERTON, Prescott’s Microbiology, 10th ed. (2015)
LUCY SHAPIRO and RICHARD LOSICK (eds.), Cell Biology of Bacteria (2011)
THOMAS D. BROCK, The Emergence of Bacterial Genetics (1990)
LARRY SNYDER et al., Molecular Genetics of Bacteria, 4th ed. (2013)
CLIVE EDWARDS (ed.), Microbiology of Extreme Environments (1990)
CHARLES GERDAY and NICOLAS GLANSDORFF (eds.), Physiology and Biochemistry of Extremophiles (2007)

谷禾健康

抗生素已经彻底改变了现代医学,但其有效性受到多重耐药细菌传播的威胁,而目前尚无有效的治疗方法。抗生素耐药性(AMR)通常由不必要的抗生素使用引起,是一种严重的全球健康和经济威胁。
特别值得关注的是,抗生素生产线不断减少,开发中的抗生素数量有限(数量和多样性都有限),以满足当前和预期的患者需求。此外,科学和经济挑战促使许多大型全球制药公司停止其抗生素开发计划,使得对新的感染控制方法的需求更加迫切。因此迫切需要用于治疗革兰氏阴性感染的新疗法。
最近一种潜在的方法是使用活的掠食性细菌。由于蛭弧菌Bdellovibrio bacteriovorus广泛存在于自然界中,而目前也尚没有与蛭弧菌相关的疾病报告,科学家认为有可能围绕它们建立一种安全无害的生物治疗手段,去抗击病原菌的感染。但毕竟蛭弧菌本身也是一种细菌,人和动物的免疫系统如果发现它们,会怎样对待这些“友军”?会增强对蛭弧菌的抵抗力吗?
本文讨论了支持掠食性细菌替代抗生素的可行性的基础科学。
蛭弧菌 Bdellovibrio bacteriovorus是寄生于其他细菌(也可无寄主而生存)并能导致其裂解的一类细菌。它虽然比通常的细菌小,能通过细菌滤器,有类似噬菌体的作用,但它不是病毒,确确实实是一类能”吃掉”细菌的细菌。
1962年首次发现于菜豆叶烧病假单胞菌体中,随后从土壤、污水中都分离到了这种细菌。根据其基本特性,命名为Bdellovibrio bacteriovorus。
其中,” Bdello”一词来自希腊字,是”水蛭”的意思,”vibrio”意为”弧菌”,而种名”bacteriovorus”是 “食细菌”的意思。“捕食”的对象正落在多出产致病菌的细菌类群——革兰氏阴性菌(如大肠杆菌、志贺氏菌等)中。

B. bacteriovorus通常会持续3-4 h消耗其革兰氏阴性细菌的猎物。这种掠夺性生命周期是一个复杂的过程,在分子水平上已经开始被理解。最初,B. bacteriovorus识别,附着并进入猎物细胞,对其进入口进行加固,穿越和重新密封。侵袭伴随着猎物细胞的圆形化,稳定的胶质细胞的形成以及猎物细胞的死亡。
同时,通过依次释放一组酶,B. bacteriovorus消化了猎物,并利用所形成的养分库长出了长丝。 单个细丝的同步分裂会产生奇数或偶数个子代细胞,每个子代细胞会产生鞭毛或滑行引擎(取决于条件),然后从死亡的猎物细胞中爆发并开始寻找新的猎物。 根据猎物细胞的大小,以及猎物细菌内部可用的养分,每个细胞平均释放4–6个B. bacteriovorus后代。 尽管被认为是专性的捕食性细菌,但B. bacteriovorus仍可转变为不依赖宿主的生活方式,可以在全培养基上的无菌生长。
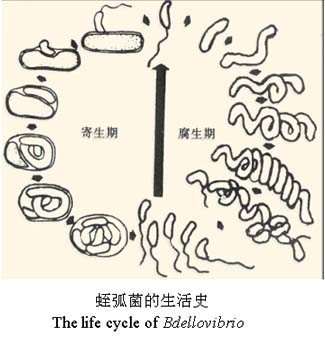
B. bacteriovorus生命周期和基因组的一些特征决定其成为一种潜在的抗革兰氏阴性细菌病原体的治疗剂。在捕食生命周期中,被捕食细胞在短时间内(<30分钟)被杀死,因此,被捕食者必须快速表达防御手段,以抵抗捕食,这是尚未见到的。
与某些抗生素不同,某些抗生素可引起一连串事件,导致细菌自溶和炎症分子释放,B. bacteriovorus细菌捕食不会导致猎物的最初溶解,因为在溶解之前,猎物的内容物是从稳定的蛭质体结构内消耗的。此外,没有单一的受体识别和附着猎物。在猎物入侵后,在数量和功能多样性方面,具有潜在遗传冗余的猎物破坏酶都出现了上调,这表明简单的猎物对B. bacteriovorus捕食的抵抗不太可能发生。
为了实现B. bacteriovorus的治疗潜力,必须在生物学相关系统中充分表征其对革兰氏阴性病原体的捕食性。 这种表征还必须同时解决诸如宿主反应,毒性,炎症,组织损伤或伤口愈合抑制等问题。
其中一些问题已通过体外细胞培养和体内动物模型解决,评估了掠食性细菌对免疫系统各个组成部分以及整个宿主的影响。 许多人类细胞系,包括角膜-上皮细胞,血液单核细胞,巨噬细胞,肾上皮细胞,肝上皮细胞和脾单核细胞,已经暴露于不同的捕食性细菌中,测定了不同暴露的时间范围、持续时间(2至24小时),下促炎和抗炎细胞因子水平。
已知可响应细菌外膜脂多糖(LPS)而被刺激的细胞因子,是宿主抵抗病原体所必需的。 这些研究表明,尽管B. bacteriovorus在免疫反应方面并不沉默,但暴露后产生的炎性细胞因子水平可忽略不计或很低。 B. bacteriovorus不像同时检测到革兰氏阴性病原体那样具有免疫刺激性,这可能部分是由于其独特的脂质A结构以及它拥有带鞘的鞭毛。
额外的细胞活力成像,细胞毒性测量以及暴露于掠食性细菌后对动物和人类培养细胞形态变化的评估初步表明,B. bacteriovorus对人细胞无毒,尽管还需要进行更多的研究。 Raghunathan及其同事使用人类巨噬细胞系(U937细胞)研大量掠食性细菌能够在细胞内存活长达24小时,从而确定了一段持久性和潜力捕食细胞内病原体的能力。
另外,通过在药理学抑制剂存在下进行吸收实验,证明了宿主肌动蛋白细胞骨架的作用及其在B. bacteriovorus吸收中的重排。B. bacteriovorus最终通过吞噬体途径被转运,这是由于它们靶向酸性液泡。 鼠巨噬细胞系中的类似观察结果支持了这一点,并且两者都是考虑给予B. bacteriovorus作为治疗剂的重要观察结果。 这些研究说明了B. bacteriovorus靶向细胞内病原体的潜力,而许多抗生素和其他生物控制剂(例如噬菌体)可能无法利用这些细菌。 尽管后一点正在积极研究中。 这些体外细胞实验以及下面描述的动物模型,是研究功效,缺乏毒性和潜在捕食者生物利用度的重要步骤。
早期体内宿主反应集中于B. bacteriovorus在吸热和放热脊椎动物肠道中的生存能力。研究了在鲶鱼(点状黄疸)、豹蛙(林蛙)、小鼠和兔子的肠道中实验投喂的B. bacteriovorus菌株MS7的生存能力和持久性,表明在接种后24-48小时内,B. bacteriovorus几乎没有恢复。通过饮用水给小鼠提供的3天B. bacteriovorus不能从肠道中完全恢复。同样,在注射后24小时左右,无论是使用无菌培养物,还是同时注射B. bacteriovorus,都没有或很少从兔回肠环中发现B. bacteriovorus。
B. bacteriovorus对动物没有致病性。这些研究已经扩展到包括评估宿主发病率、组织病理学、促炎和抗炎细胞因子水平、体内捕食性细菌传播以及长期评估大鼠、小鼠斑马鱼幼虫模型的一般健康状况。总的来说,在一系列动物模型中以及通过多种给药途径在体内给药已经证明,它们不会损害这些动物的健康。
事实证明,B. bacteriovorus对多种猎物具有体外功效,包括与肠道,口腔,伤口和眼部感染有关的细菌和生物制药。B. bacteriovorus成功地减少了实验室缓冲液和人血清中的病原体数量和生物膜中的猎物,而生物膜通常是抗生素治疗的重大阻碍。 重要的是,已经显示出许多具有多重耐药性的人类临床分离株容易被B. bacteriovorus捕食。 该清单包括许多需要新治疗的E(S)KAPE病原体,包括表达mcr-1的革兰氏阴性大肠菌素抗性分离株。 体外研究还调查了成功捕食所需的B. bacteriovorus与猎物的比率; 如果抗生素不能治疗感染,则作为将来的体内实验和临床的重要考虑因素。

尽管实验性体外系统,尤其是实验室缓冲液或富含生长培养基的环境可能显示出掠食性功效,但它们不足以替代体内治疗的复杂性,而体内治疗必须考虑到宿主的免疫反应以及病原体的生存策略。 动物感染模型在评估B. bacteriovorus在体内捕食革兰氏阴性细菌的能力以及解决此类宿主对宿主的任何反应(以及由此产生的安全性)方面都发挥了作用,越来越多的证据表明,至少在动物中,以及将来将这种掠食性细菌用于治疗应用的可行性,将其扩展到人类。
一项对感染肠炎沙门氏菌p125109的鸡群实验表明。 在B. bacteriovorus处理后的三天内,与对照动物相比,经细菌性芽孢杆菌处理的禽类的盲肠中沙门氏菌数量显着降低。 此外,与对照动物的许多盲肠相比,经B. bacteriovorus处理的禽中盲肠的外观是正常的。
进一步的重要研究表明B. bacteriovorus能成功地在体内捕食病原体。 使用大鼠模型,Shatzkes及其同事证实B. bacteriovorus可以治疗肺炎克雷伯菌感染的大鼠,显示与对照组相比,细菌性芽孢杆菌治疗的动物的病原体负担明显减少。 另外一项通过向大鼠尾静脉注射引发疾病的败血病模型。 B. bacteriovorus不能显着减少感染。 这是首次将B. bacteriovorus直接施用到血流中,并且是评估掠食性细菌清除血流感染能力的重要一步。
斑马鱼幼虫的物理特征以及与人类的广泛基因组同源性包括良好的理解,充分发展的免疫系统和光学透明的性质,非常适合于创新的活荧光显微镜检查。
诺丁汉大学伊丽莎白·肖克特和伦敦帝国理工学院的塞尔吉·莫斯托维小组用模式生物斑马鱼做了研究。

接下来,研究者放心地开始了“以菌治菌”环节。他们用一种叫福氏志贺氏菌(Shigella flexneri)的病原菌来感染斑马鱼——所选取的菌株同时对链霉素与羧苄青霉素具有耐药性。他们向斑马鱼的后脑先接种了致死剂量的志贺氏菌,再注射蛭弧菌。由于这两种细菌被带上了不同颜色的荧光蛋白标记,研究者得以观察它们的数量和分布变化情况。他们发现,相比于对照组,注射了蛭弧菌的斑马鱼后脑内志贺氏菌大量减少,被感染的斑马鱼在72小时后的存活率也更高。在更高分辨率的显微观察中,研究人员也找到了蛭弧菌在斑马鱼体内和体外都能够侵染并杀死志贺氏菌的证据。
研究者认为,在抗击志贺氏菌感染这件事上,蛭弧菌和斑马鱼免疫系统其实某程度上达成了巧妙的“配合”:蛭弧菌对志贺氏菌的“捕食”开始得非常迅速,能够在感染初期控制住志贺氏菌繁殖的势头,帮免疫系统减轻应对的压力。而等到大批白细胞赶来时,蛭弧菌已经饱餐过一顿,收拾残余志贺氏菌的工作,免疫系统自己也能完成好。
1897年,日本细菌学家志贺洁发现了志贺氏菌(Shigella).
作为杆菌性痢疾的祸首,这类细菌每年导致约1.63亿严重痢疾病例,并夺走超过100万人的生命,可谓是最臭名昭著的病原菌之一。志贺氏菌是一类革兰氏阴性的杆状细菌,可以制造出能杀死细胞的志贺毒素。
在最近的一项研究中,Russo及其同事证明了B. bacteriovorus 可用于显着减少实验感染小鼠肺部的鼠疫杆菌(yersinia pestis)数量。 但是并非所有的体内B. bacteriovorus 施用都已成功地减少了病原体数量。 尽管在先前的研究中使用组织培养模型在体外取得了令人鼓舞的结果,但给予B. bacteriovorus 治疗经牛莫拉氏菌(Moraxella bovis )感染的牛犊是牛角膜结膜炎的病原体,未能导致角膜溃疡形成的显着改善。
从上面的大量研究可以看出,越来越多的证据表明Bdellovibrio sp持续非病原性且足够长以具有治疗活性,对免疫微生物群的不良影响极小,并且不成为正常宿主微生物群的一部分。 无论是成功的还是不成功的体内试验,如本文所述,对于评估哪些适应症,给药途径(局部给药还是静脉给药)和感染部位都至关重要,潜在的掠食性细菌给药将最有效地对其进行评估。
治疗后动物和人类感染的康复结果取决于多种因素的相互作用,例如患者的免疫系统和总体健康状况包括肠道菌群,病原体的性质以及治疗的类型和提供方式。 在将体外和体内细菌捕食视为宿主反应的独立实体时,未捕获革兰氏阴性病原体的细菌捕食与宿主反应之间的相互作用和协同作用。

该模型表明,B. bacteriovorus在非致病性条件下持续了足够长的时间,可以在实验性感染期间有效地捕食志贺氏菌。 在这项工作中,用显微镜观察了与宿主免疫系统细胞的相互作用。 此外,这项研究发现B. bacteriovorus的最大治疗益处来自细菌捕食和宿主免疫系统的协同作用。 给药后,掠食性细菌种群最终会通过与宿主免疫系统的相互作用而清除,实际上这种清除可能对(自我)限制治疗有益。
在考虑治疗掠食性细菌感染时,研究人员可以向噬菌体研究和临床界寻求动力,以应对从体外到临床环境的挑战。 尽管在一些国家已经使用了多年,但最近的一些引人注目的病例已证明在临床环境中成功施用了噬菌体鸡尾酒来治疗耐药性感染。
从表面上看,B. bacteriovorus和噬菌体具有许多潜在的生物防治剂特性。 它们都表现出掠食性或寄生性生命周期,它们是自我复制和自我限制的,仅在存在易感宿主或猎物时才持续存在,并且在裂解性噬菌体的情况下,两者都具有广泛相似的生命周期。 当将它们引入动物和人类体内时,它们似乎也几乎没有副作用。 但是,两者之间存在一些显着差异,这可能会限制和区别它们在某些适应症中的使用。
首先,与噬菌体相比,B. bacteriovorus无疑是“活的”并且具有代谢活性,噬菌体是惰性颗粒,直到它们通过特定受体与宿主建立接触。 这可能对B. bacteriovorus既有好处,也有缺点,因为它可能使用需要能量的过程,例如噬菌体无法获得的主动“定位”猎物。 但是,如果未发现猎物,则B. bacteriovorus不能进入休眠状态,并且会死亡。 相反,噬菌体可以在不消耗任何能量的情况下保持“生存”许多年,但无法主动寻找宿主。 他们必须依靠随机概率遇到宿主,并通过比B. bacteriovorus更快速地复制并且具有更大的爆发来补偿这种相对的低效率。
先前对伤寒沙门氏菌和大肠杆菌的研究表明,噬菌体并不总是消除其宿主的全部种群。B. bacteriovorus和噬菌体的复制都可能受到最低宿主/猎物阈值的限制,在此阈值之下,它们无法无限期地维持其种群。 然而,这可能在治疗上是有益的,因为通常不需要消灭病原体即可显着减轻或完全缓解疾病症状。
单个噬菌体的宿主范围通常限于一个菌种或一个或两个密切相关的菌种内的许多菌株,这使得其种群的维持比B. bacteriovorus更具挑战性。需要考虑宿主特异性和细菌对噬菌体耐药性的快速获得;与之相比,广泛的细菌捕食范围和缺乏掠食性细菌的简单抗药性机制使其具有优势。 有趣的是,最近有科学金工作强调了将掠食性细菌与噬菌体一起使用的组合能力。 噬菌体敏感性和Bdellovibrio捕食的独立动力学使得在某些条件下更大程度地杀死了大肠杆菌。
当被引入到吸热动物体内时,B. bacteriovorus和噬菌体都将面临挑战性条件。 与细菌噬菌杆菌相比,噬菌体在遇到苛刻的物理和化学条件(例如更高的温度和极端的pH值)时更有可能具有弹性。噬菌体可以穿过血脑屏障,但如果不进行故意修饰,可能无法在细胞内持续存在。 确实,以前的研究表明,当它们进入血液时,它们可以被网状内皮系统迅速隔离。 至少在某些情况下,可以通过在哺乳动物中连续传播噬菌体来重新分离能够在哺乳动物中长时间循环的噬菌体来应对这一问题。 众所周知,噬菌体能够通过普遍的和专门的转导在细菌之间转移DNA,这是一种不适用于B. bacteriovorus的风险。可以通过仔细筛选噬菌体基因组以除去具有整合能力的特定噬菌体和其他可能带有编码细菌毒素或其他毒力因子的基因的噬菌体,来减轻这种风险。
Bdellovibrio和噬菌体的治疗应用需要两者的工业生产。 每种噬菌体都面临着不同的挑战,因为噬菌体的产生数量更大且速度更快,但是它们的产生可能需要大量培养病原性宿主细菌。 相反,可以使用非病原性的Bdellovibrio宿主,例如大肠杆菌和恶臭假单胞菌,但是需要将它们与最终的治疗制剂有效地分开。 另外,维持Bdellovibrio制剂的生存力可能比噬菌体更具挑战性。 有趣的是,从生产这些生物防治剂的治疗制剂的技术挑战中可以学到什么教训,以及确定将它们一起用作治疗剂是否可以实现任何协同作用。
最近从事DARPA病原体捕食计划的一些研究人员,已经改变了研究领域,并证明了使用活的全食性B. bacteriovorus在体外和体内杀死广泛的抗药性(AMR)临床病原体的潜力。体内进一步支持其作为治疗手段的承诺。要被视为可靠的治疗选择,需要来自未来的人类试验的有力证据表明,这种治疗有效,至少在特定情况下,对患者没有(或最小)负面影响,并且与现有治疗相比具有一定优势。
研究人员需要证明其临床价值并衡量治疗效果,以说明对患者的益处。但现在是否拥有所需的所有信息? 如果没有,还需要解决什么?
根据体内动物模型,研究人员需要在人类身上进行安全性和有效性试验。动物模型是至关重要的,但也有局限性,迄今为止的协议管理捕食性细菌之前,或不久之后,病原体的兴趣。需要对更确定的感染进行调查,并支持确定捕食性细菌的剂量范围(以及可能的病原体数量)、剂量数量和治疗感染所需的时间表。也许令人惊讶的是,在感染和康复期间,对人体不同部位病原体实际数量的了解仍然是一个非常发展的领域。大规模捕食者生长和纯化方法的发展目前受到限制,需要扩大更大规模的安全性和有效性试验。感染治疗是时间关键,因此评估稳定性,长期储存和提供一个活的,掠夺性的积极治疗需要进一步的工作。
总之,用另一种细菌来治疗细菌感染似乎是反直觉的,离使用它们作为药物改善患者的生存状况还有一段时间。虽然蛭弧菌对其他病原的杀伤能力还有待验证,而且考虑到人体内大量必要有益的共生菌群,如何避免大量蛭弧菌“伤及无辜”也是一个值得关注的问题。需要进一步的工作来评估捕食性细菌从给药部位的传播,并确定暴露对宿主或其驻留生态位微生物群的长期影响。
但是我们相信,未来以细菌对抗细菌治疗某些感染会是一条后抗生素时代新的机遇。
相关阅读:
参考文献:
Laloux G. Shedding light on the cell biology of the predatory bacterium Bdellovibrio bacteriovorus . Front Microbiol 2020; 10:3136
Harding CJ, Huwiler SG, Somers H, Lambert C, Ray LJ et al. A lysozyme with altered substrate specificity facilitates prey cell exit by the periplasmic predator Bdellovibrio bacteriovorus. Nat Commun 2020; 11:4817
Ardal C, Balasegaram M, Laxminarayan R, McAdams D, Outterson K et al. Antibiotic development – economic, regulatory and societal challenges. Nat Rev Microbiol. 2019
Tacconelli E, Carrara E, Savoldi A, Harbarth S, Mendelson M et al. Discovery, research, and development of new antibiotics: the WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect Dis 2018; 18:318–327
Atterbury Robert J,Tyson Jess,Predatory bacteria as living antibiotics – where are we now?[J] .Microbiology (Reading), 2021, 167.
Laxminarayan R, Van Boeckel T, Frost I, Kariuki S, Khan EA et al. The Lancet infectious diseases Commission on antimicrobial resistance: 6 years later. Lancet Infect Dis 2020; 20:e51–e60
谷禾健康
细菌感染严重威胁人类健康。如果你去医院就医,医生怀疑你有感染,可能会要求进行革兰氏染色检查。该测试还可以帮助医生了解细菌是革兰氏阴性还是革兰氏阳性,从而选择合适的治疗方式。
然而你可能对于革兰氏阳性菌阴性菌这些概念并不熟悉,今天为大家讲解下,革兰氏阳性菌,革兰氏阴性菌的区别,革兰氏染色法,常见的革兰氏阳性菌类型以及相应的治疗。
首先,我们了解下,革兰氏阳性菌和阴性菌各自的特征。
革兰氏阳性菌特征
革兰氏阳性细菌的主要特征是它们的结构。一般有以下特点:
没有外膜。革兰氏阳性细菌没有外膜,但革兰氏阴性细菌有。
复杂的细胞壁。包围细胞质膜的细胞壁由肽聚糖、多糖、磷壁和蛋白质组成。它很容易吸收外来物质。
厚的肽聚糖层。在革兰氏阳性细菌中,肽聚糖有40到80层厚。
某些表面附属物。革兰氏阳性细菌可能有鞭毛,可以帮助它们移动。它们很少有被称为菌毛的毛发状结构。
革兰氏阴性菌特征
革兰氏阳性菌和革兰氏阴性菌具有不同的结构。通常,革兰氏阴性菌具有以下特征:
外脂质膜
肽聚糖薄层(2 – 3纳米)
通常不含磷壁酸
可以有鞭毛或毛
主要的区别是外脂膜。它很难渗透,这给了革兰氏阴性细菌额外的保护。革兰氏阳性细菌没有这种特征。
由于这种差异,革兰氏阴性细菌更难杀死。这意味着革兰氏阳性菌和革兰氏阴性菌需要不同的处理方法。
革兰氏阳性菌和阴性菌是怎么鉴定的呢?
——通过革兰氏染色法。
革兰氏染色法是一种根据细菌细胞壁进行细菌分类的方法。它使科学家能够确定一个有机体是革兰氏阳性还是革兰氏阴性。这项使用显微镜的测试是由汉斯·克里斯蒂安·克拉姆在1884年发明的。
在这个过程中,结晶紫染料被应用到细菌样本上。这种化学染料能将较厚的肽聚糖层染色。
在显微镜下,革兰氏阳性细菌呈现紫蓝色,因为它们厚厚的肽聚糖膜可以容纳染料。由于阳性结果,这种细菌被称为革兰氏阳性细菌。
革兰氏阴性菌染成粉红色。它们的肽聚糖层更薄,所以不会保留蓝色。测试结果为阴性。
革兰氏染色可在各种类型的标本上进行,包括:
血液、组织、粪便、尿液、痰液等。
医生将样本送到实验室进行革兰氏染色检测,从而了解是否是细菌导致了该症状,以及存在哪些类型的细菌。他们也可能会要求其他测试来帮助诊断疾病。这对于确定后续的治疗方案很有帮助。
根据不同的特性,革兰氏阳性菌可进一步分为以下几类:
革兰氏阳性球菌
革兰氏阳性球菌呈圆形或椭圆形。术语“球菌”的意思是球形,表明细菌通常是圆形的。
以下是革兰氏阳性球菌:
葡萄球菌属
葡萄球菌呈葡萄状丛生生长。正常情况下,它们存在于我们的皮肤和粘膜上,不会造成任何问题。但如果葡萄球菌进入人体,它们会引起严重的感染。
链球菌属
链球菌呈链状生长。这是因为细胞在分裂后并没有完全分离。
与葡萄球菌一样,链球菌通常存在于体内。它们通常存在于皮肤、口腔、肠道和生殖道。
链球菌可分为以下类别:
S. pyogenes (Group A)、S. agalactiae (Group B)、Enterococci (Group D)、S. viridans、S. pneumoniae
革兰氏阳性杆菌
当革兰氏阳性细菌呈棒状时,它们被称为杆菌。这些细菌大多在皮肤上发现,但有些会导致严重的疾病。
革兰氏阳性杆菌根据其产生孢子的能力被进一步分类。
有芽胞型:
芽孢杆菌和梭状芽孢杆菌可以形成孢子,这有助于细菌在高温等恶劣条件下生存。
这些杆菌根据它们对氧气的需要被细分。芽孢杆菌需要氧气才能生存(需氧),而梭状芽孢杆菌不需要氧气(厌氧)。
无芽胞型:
李斯特菌(Listeria )和棒状杆菌(Corynebacterium)不产生孢子。李斯特菌是厌氧菌,棒状杆菌是好氧菌。
许多革兰氏阳性菌都是致病菌。虽然有100多种致病性革兰氏阳性菌,但最显著的种类包括:
葡萄球菌
葡萄球菌通常是细菌感染的原因。大多数情况是由以下物种引起的。其他致病性葡萄球菌不太常见,很少导致疾病。
金黄色葡萄球菌
金黄色葡萄球菌是致病性最强的葡萄球菌。它导致了大多数葡萄球菌感染,包括:
皮肤感染,如蜂窝织炎和毛囊炎、感染性关节炎、脓肿、心内膜炎、细菌性肺炎、食物中毒、中毒性休克综合征、烫伤皮肤综合征、MRSA等。
表皮葡萄球菌
通常,表皮链球菌会在免疫系统薄弱或住院的人身上引起感染。会引起:
导尿管等医疗器械感染、菌血症、纵隔炎、手术部位感染、眼部角膜炎、眼内炎(内眼感染)等。
腐生葡萄球菌
通常见于生殖道和会阴。会引起: 无并发症的尿路感染(最常见)、尿道炎、前列腺炎、急性肾盂肾炎 、附睾炎等。
链球菌
链球菌也是常见的致病菌。以下是最常见的生物体。一般来说,其他链球菌群可引起食源性疾病,引起喉咙痛。
肺炎链球菌
肺炎链球菌是社区获得性肺炎最常见的病因。它还引起:红眼、鼻窦感染、脑膜炎等。
酿脓链球菌
会引起脓毒性咽喉炎、蜂窝织炎、咽炎、脓疱炎、猩红热、风湿热、坏死性筋膜炎、肾小球肾炎等。
无乳链球菌
通常会引起新生儿感染,包括:败血症、肺炎、脑膜炎、化脓性关节炎。
肠球菌
肠球菌主要在结肠中发现。它们会引起胆道和泌尿道感染。
芽孢杆菌
作为孢子形成细菌,杆菌产生孢子并释放毒素。大多数杆菌对人类没有致病性,但以下两种可导致严重的疾病。
炭疽杆菌(Bacillus anthracis )
炭疽杆菌孢子产生炭疽毒素,引起严重的疾病。人类可通过吸入或接触受感染的动物而感染炭疽。
根据炭疽热的传播方式,它可以引起各种症状。这些可能包括:恶心、呕吐、腹痛、咳血、高烧等。
蜡样芽胞杆菌(Bacillus cereus )
蜡样芽孢杆菌是一种孢子形成细菌,存在于土壤和一些食物中。它与吃未煮熟或重新加热的米饭而导致的疾病有关。会引起:腹泻、恶心、伤口感染、呼吸道感染、眼内炎等。
梭菌(Clostridium)
大约有30种梭状芽孢杆菌会引起人类疾病。像杆菌一样,这些细菌形成毒素,导致严重的情况。梭状芽孢杆菌通常与食源性疾病有关,但最令人担忧的细菌包括:
肉毒梭状芽胞杆菌(Clostridium botulinum )
肉毒杆菌的孢子产生肉毒杆菌毒素,这是对人类最危险的毒素。它会导致肉毒中毒,包括:
食源性肉毒中毒(最常见)、婴儿肉毒中毒、伤口肉毒中毒、吸入肉毒中毒等。
产气荚膜梭菌(Clostridium perfringens)
产气荚膜通常与肉类生产和加工有关。如果一个人吃了受污染的肉,他们可能会食物中毒。症状包括持续不到24小时的腹泻和腹部绞痛。
艰难梭菌(Clostridium difficile )
艰难梭菌通常影响住院的老年人。一般发生在服用抗生素之后,引起结肠炎、腹部绞痛、严重腹泻。
破伤风梭菌(Clostridium tetani)
破伤风梭菌孢子产生破伤风毒素,一种神经毒性物质。一般它会出现在土壤、灰尘和生锈的工具上,如果这种毒素引起感染,就叫做破伤风。这是个严重的医疗紧急事件。
单核细胞增多性李斯特氏菌(Listeria monocytogenes )
唯一致病性李斯特菌是单核增生杆菌。在健康人身上,它通常会引起轻微的食源性疾病症状。但在免疫系统较弱的人群中,这种细菌会导致危及生命的情况:脑膜炎、败血症、利斯特氏菌病。
白喉杆菌(Corynebacterium diphtheriae)
约有30种棒状杆菌与人类疾病有关。然而,这些微生物很少引起疾病,通常影响免疫系统受损的人。白喉是本组的主要致病菌。引起:白喉、咽炎、呼吸道感染、感染性关节炎、皮肤感染、骨髓炎、心内膜炎等。
治疗革兰氏阳性细菌引起的疾病时,最佳选择取决于:细菌类型、抗生素耐药性、细菌是否形成毒素等。常见的治疗方法包括:
盘尼西林
一种常见的用于多种感染的抗生素。主要是干扰细菌的肽聚糖层,从而杀死细菌。主要用于链球菌感染,包括:链球菌性喉炎、鼻窦感染、尿路感染、蜂窝织炎等。
糖肽类
糖肽类抗生素常用于治疗耐药细菌引起的严重感染。像青霉素一样,它们通过破坏细菌的细胞壁来发挥作用。可以治疗:耐多药肺炎、MRSA、结肠炎。
红霉素
红霉素属于大环内酯类抗生素,大环内酯类抗生素还包括更知名的阿奇霉素和克拉霉素。它是一种能阻止细菌生长的抗生素,对革兰氏阳性和革兰氏阴性细菌都有效。
通常,红霉素是给对青霉素过敏的人开的。治疗的情况如下:细菌性肺炎、红眼、咽喉炎、葡萄球菌性皮肤感染等。
液体疗法
在某些情况下,治疗可能包括液体疗法。它有助于补充身体的水分水平和防止脱水。一般来说,治疗毒素引起的疾病需要液体管理。
抗毒素
对于与毒素相关的疾病,如炭疽和肉毒中毒,治疗包括抗毒素。这种药通过靶向和清除体内毒素而起作用。合适的抗毒素取决于具体的毒素。此外,它还与其他疗法结合使用。
革兰氏染色测试可以帮助医生诊断疾病,从而选择最佳的治疗方式,如果能更快速准确地了解是由何种菌引起的疾病,将会为治疗提供捷径。
【参考文献】
Gillespie S, et al. (2012). Medical microbiology and infection at a glance. West Sussex, UK: Wiley-Blackwell.
Kulkarni AP, et al. (2019). Current perspectives on treatment of gram-positive infections in India: What is the way forward?
Mahony J, et al. (2015). Gram-positive phages: From isolation to application. Lausanne, Switzerland: Frontiers Media SA
Malanovic N, et al. (2016). Gram-positive bacterial cell envelopes: The impact on the activity of antimicrobial peptides.
Sizar O, et al. (2019). Gram positive bacteria.
Vazquez-Guillamet C, et al. (2014). Treatment of gram-positive infections in critically ill patients.