-

 国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子
国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子

 二级病原微生物安全实验室
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业

谷禾健康
肠道菌群检测临床版(16S rRNA测序),作为谷禾健康最早推向市场、历经十余年打磨的经典产品,凭借其成本效益、高效性与成熟度,为临床端肠道微生态评估以及诸多科研项目基线建立,提供了坚实、可靠且极具价值的数据基础(注:仅用于菌群科学研究和辅助参考,不直接用于临床诊断 )。

临床版对报告版式进行简化,以更符合临床检验的形式呈现分析结果,便于临床医生快速查看和判断异常。
临床版是主要面向临床和医疗机构的版本,主要用于临床科室已有明确症状或诊断,需要对肠道菌群进行进一步分析以为临床提供辅助判断。
该版本减少了基本介绍和文字说明,并对部分指标的异常判定范围和计算方式进行调整,更加适应临床需求。
报告内容截图:


谷禾健康

随着“它经济”的蓬勃发展和“科学养宠”理念的深入人心,宠物已成为家庭的重要成员。然而,面对“毛孩子”们无法言说的病痛,如反复腹泻、顽固皮肤病、食欲不振、呕吐等,传统兽医诊断往往面临挑战。
从宠物医院的实际经营来看,慢性疾病正成为他们面临的核心挑战。慢性肾病、老年痴呆、精神类疾病等病症不仅治疗费用昂贵,而且现有手段往往无法覆盖,特别是小型诊所更是心有余而力不足。
在与许多B端合作伙伴的深度交流中我们发现
宠物腹泻,肾病以及其他疾病等正在增加
后期医疗费用高昂让宠物主人无能为力
异常行为严重影响生活质量和主人养宠体验
情感难舍却不得不放弃…
因此,迫切需要一种更加
科学、经济、精准的健康管理方式
既能降低医疗成本
又能提供个性化的健康方案
还能避免过度医疗
这就需要我们从根本上
重新思考宠物健康管理的方法论
从”治疗导向“转向”预防导向“
✲✲✲✲✲✲✲✲✲✲✲✲✲✲✲✲✲✲✲✲✲✲✲✲✲✲✲✲✲✲
谷禾凭借在人类健康领域积累的深厚微生物组学经验,战略性延伸至动物医学领域,推出宠物菌群精准检测服务。
我们致力于解码宠物肠道微生态的奥秘,为兽医临床、宠物营养和家庭养护提供科学依据,开启宠物健康管理的精准化新时代。

从人类微生态到宠物微生态,不是简单的复制,而是技术能力的升维应用。人类肠道微生态的复杂性研究为我们提供了强大的算法基础和数据分析能力,以及多年来在宠物菌群科研中的持续投入,这些经验在宠物领域的应用中展现出了独特的技术优势。
研究表明,宠物肠道菌群不仅影响消化吸收功能,更与免疫调节、神经系统、皮肤健康等多个生理系统密切相关,成为宠物整体健康状况的重要晴雨表。
谷禾正有序推进构建涵盖不同品种、年龄、健康状况犬猫的肠道菌群数据库,结合最新的机器学习算法,实现对宠物肠道微生态健康状况和营养进行精准评估(注:仅用于菌群科学研究和辅助参考,不直接用于临床诊断 )。
宠物菌群报告展示采用更温馨活泼的配色,通过可视化图表和情感化设计,让复杂数据更直观,帮助主人轻松了解爱宠健康。
谷禾宠物菌群检测报告中包括菌群评估(整体指标)、肠道基础功能评估(屏障功能、炎症水平、代谢状态)、菌群代谢物评估(短链脂肪酸等)、炎症免疫评估(促炎、抗炎等指标)、营养饮食评估(维生素、微量元素)等。
…
…
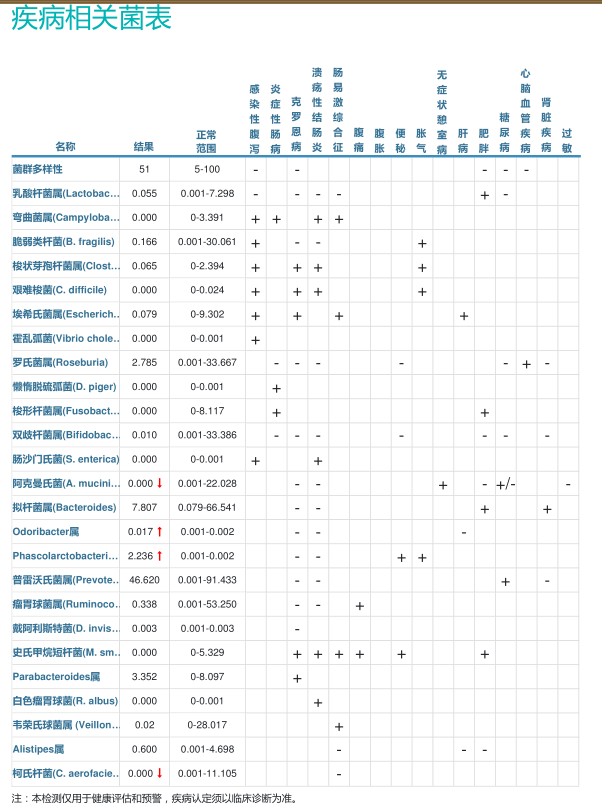
症状相关菌群分析,包括腹泻、呕吐、过敏等。
…
菌群代谢物评估(短链脂肪酸等)。
…
…
从多维度全面评估宠物肠道健康状况,为宠物主人提供科学的健康管理依据和个性化调理建议。


谷禾健康
在肠道菌群检测系列产品成熟之后,谷禾健康将深耕多年的微生物组学技术平台,延伸至关乎女性全生命周期健康的另一核心领域——阴道微生态。
还包括子宫颈沙眼衣原体、HPV、HSV、EB病毒、巨细胞病毒等。
谷禾阴道菌群检测报告引入科学前沿的菌群状态分型(CST)概念,将复杂的菌群构成归纳为几种易于理解的健康状态类型。
例如,以卷曲乳杆菌为主的CST-I型代表健康的稳定状态,而以加德纳菌等多种厌氧菌为主的CST-IV型则与细菌性阴道病高度相关。这为临床判断和干预效果评估提供了科学支持。
谷禾阴道菌群检测报告中包括阴道菌群总体评估、CST分型、致病菌表(细菌性阴道病,需氧菌性阴道炎,外阴念珠菌病等)、列出异常菌群及相关说明,菌群详细构成等(注:仅用于菌群科学研究和辅助参考,不直接用于临床诊断)。
…
…
…
…
阴道菌群检测让我们能够更全面地了解阴道微生物组的组成及其变化,以及它是如何随着时间的推移或对各种因素(如环境、激素变化、性活动和抗生素使用等)的反应而变化的。
注:本产品可辅助评估和筛查,不用作临床诊断。
05

谷禾健康
谷禾16S+tNGS技术结合了超多重PCR和高通量测序的优势,旨在提供比传统16S rRNA测序和宏基因组测序更优、更全面的病原体及耐药基因检测方案。

传统16S + 病原体精准分型 = 全新升级
它在保留16S报告的基础上
以接近16S的成本和周期
用靶向测序技术在原16S的基础上
增加了125 种消化道病原体的检测
还包括耐药基因和毒力基因等
如幽门螺杆菌、大肠杆菌、艰难梭菌分型
弥补了传统16S无法检测
非细菌/古菌病原体的不足
这是一款突破传统16S检测瓶颈而生的产品
值得一提的是,谷禾在tNGS技术的研发道路上并非一帆风顺。这项看似成熟的技术,在不同应用场景下却面临着截然不同的挑战难度。
tNGS对血液和上呼吸道样本检测较简单,因其主要含病原体。但在消化道特别是粪便样本检测时,技术难度大幅增加。
经过无数次的实验优化和迭代
我们最终突破了这一技术瓶颈
为大家带来真正可靠的
消化道病原体检测解决方案
谷禾16S+tNGS产品特点
弥补16S检测技术边界
实现靶向病原体精准检测
谷禾16S+tNGS报告内容涵盖所有16S报告的内容,即包括健康总分评估、慢病风险预警、肠道屏障及代谢物、神经递质分析、个性化营养评估等。此外还包括常见消化道病原体(注:仅用于菌群科学研究和辅助参考,不直接用于临床诊断)。
例如:
细菌病原体
…
病毒
真菌、寄生虫、其他病原体
…
毒力基因
…
耐药基因
…

也包括相关病原微生物的解释
…
例如,通过检测幽门螺杆菌毒力基因组合,可判断是否需立即治疗,避免对弱毒株患者的不必要抗生素使用。强毒株感染会损伤胃黏膜,增加胃炎和溃疡风险,早期预警能在胃黏膜不可逆损伤前提供治疗窗口期。
注: 由于该技术是检测粪便中的幽门螺杆菌,当浓度低于检测下限(50 copies/mL)时,可能出现假阴性结果。因此,对于临床症状明显但检测结果为阴性的患者,建议结合其他检测方法。

开始写作或按/来选择区块