-

 国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子
国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子

 二级病原微生物安全实验室
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
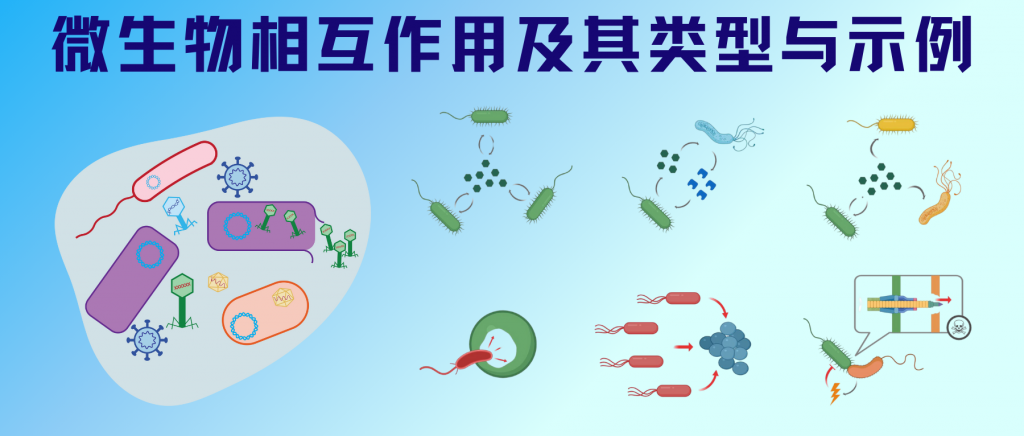
在自然界中,微生物之间的相互作用构成了一个复杂而精密的网络系统。这些相互作用不仅存在于任何生物群落中,更在全球生物地球化学循环中扮演着不可或缺的角色。从互惠共生的地衣,到肠道中的营养互补,从土壤中的捕食关系,到水体中的共营养现象,微生物间的相互作用无处不在、形式多样。
随着微生物学研究的深入,科学家们逐渐认识到这些相互作用的复杂性和重要性。这些互动可能是互惠互利的,如互惠共生和原始合作;也可能是单向获益的,如共生关系;甚至可能是竞争或对抗性的,如偏害共生和寄生关系。
微生物系统中最常见的合作互动是互惠互利的。根据两个种群和其中一个种群是否从关联中受益,或者一个或两个种群都受到负面影响,对两个种群之间的交互作用进行分类。

来源:microbenotes
理解这些相互作用不仅有助于我们更好地认识微生物世界的运作机制,也为解决环境保护、疾病防控等实际问题提供了新的思路和方法。
本文将系统地探讨微生物之间的各种相互作用类型,深入分析它们的特征、机制及其在生态系统中的重要作用,以期为相关领域的研究和应用提供参考。
共生是指相互作用中的每个生物体从关联中受益的关系。这是一种强制性的关系,其中互惠论者和宿主在代谢上相互依赖。
这种互惠关系是非常具体的,其中“协会”的一个成员不能被另一个物种取代。
共生关系要求相互作用的生物体之间有密切的物理接触。
共生关系允许生物体存在于任何一个物种都无法单独占据的栖息地中。
生物体之间的共生关系使它们能够作为一个单一的生物体行事。
共生的例子:
地衣
地衣是互惠共生的极好例子。
它们是特定真菌和某些藻类属的复合体。因为藻类生物是光合自养生物,所以真菌直接从藻类伙伴那里获得有机碳,反过来真菌保护藻类免受极端条件的影响,并为藻类提供水和矿物质。
地衣生长非常缓慢,但能够在不允许其他生物生长的栖息地聚集。大多数地衣耐高温和干燥。
草履虫-小球藻
草履虫(原生动物)可以在其细胞质中寄生小球藻(藻类)。
藻类小球藻为原生动物伙伴提供有机体碳和 O2,反过来原生动物提供CO2和其他生长因子。
只要有足够的光线,草履虫中小球藻的存在有助于原生动物在厌氧条件下存活。
在这种关联中,一种生物体的生长要么依赖于另一种生物体提供的基质,要么由另一种生物体提供的基质来改善。
在互生中,两种相关的生物体都可以从彼此中受益。
在这个互生理论例子中:
例如:Lactobacillus arobinosus、Enterococcus faecalis(粪肠球菌)。
在最小培养基中,两种菌必须共同生长,单独都无法生存。
这是一个典型的互补型营养共生关系。
在这种关系中,一个协会中的有机体彼此互惠互利。
这种互动类似于互惠互利,但原始合作中生物之间的关系并不像互惠互利那样是强制性的。
原始合作是一种生物间的互利关系,其中参与的生物体都能从这种关系中获益。
与互利共生的区别:
举例说明:
脱硫弧菌(Desulfovibrio)和Chromatium的关系,这是碳循环和硫循环之间的原始合作关系。
两种细菌通过各自的代谢活动互相促进,但并非完全依赖。
此外,参与生物地球化学循环的细菌,固氮菌与纤维素分解菌(如Cellulomonas)的互动。
两者通过各自的代谢活动互相促进生长,但都能独立存在。
在这种关系中,协会中的一个生物体(共生体)受益,而协会的另一个生物体(宿主)既没有受益也没有受到伤害。
它是一个单向的关联,如果共生体与宿主分离,它就可以存活。
共生的例子:
a. 人类肠道中的非致病性大肠杆菌:大肠杆菌是一种兼性厌氧菌,它使用氧气并降低肠道中的 O2 浓度,从而为拟杆菌等专性厌氧菌创造合适的环境。大肠杆菌是不受拟杆菌影响的宿主。
b. 黄杆菌(Flavobacterium)(宿主)和嗜肺军团菌(Legionella pneumophila)(共生):黄杆菌排泄嗜肺军团菌使用的胱氨酸,并在水生栖息地中存活。
c. 韦荣氏球菌和链球菌利用其他婴儿肠道细菌的碳水化合物发酵的最终产物(如乳酸)产生丙酸,形成重要的营养链。
d. 硝化作用中亚硝化单胞菌(Nitrosomonas)(宿主)和 硝化杆菌(Nitrobacter)(共生菌)的结合 :亚硝化单胞菌将氨氧化成亚硝酸盐,最后,硝化杆菌使用亚硝酸盐获取能量并将其氧化成硝酸盐。
当一个微生物种群产生对其他微生物种群具有抑制性的物质时,这种种群间关系称为偏害共生(Ammensalism 或 Antagonism)。
这是一种负向关系。
第一个产生抑制性物质的种群不受影响,或者可能会获得竞争并在栖息地中生存,而其他种群则受到抑制。这种化学抑制被称为抗生作用。
偏害共生的例子(amensalism):
竞争代表了两个微生物种群之间的负相关关系,其中两个种群的生存和生长都受到不利影响。
当两个种群使用相同的资源(例如相同的空间或相同的营养)时,就会发生竞争,因此,微生物种群的最大密度或增长率较低。
微生物种群争夺任何限制生长的资源,如碳源、氮源、磷、维生素、生长因子等。
竞争抑制了两个种群占据完全相同的生态位,因为一个将赢得竞争,另一个将被淘汰。
示例:
尾草履虫(Paramecium caudatum)和金草履虫(Paramecium aurelia)之间的竞争:当这两种原生动物被放在一起时,它们都以相同的菌群为食。由于竞争的结果,金草履虫比尾草履虫具有更好的生长速率。
在这种关系中,一个种群(寄生体)受益,并从受到伤害的协会中的其他种群(宿主)那里获得营养。
宿主-寄生体关系的特点是接触时间相对较长,这可能是物理的或代谢的。
一些寄生虫生活在宿主细胞之外,称为外寄生虫,而其他寄生虫生活在宿主细胞内,称为内寄生虫。
寄生的例子:
a. 病毒:病毒是一种专性细胞内寄生虫,表现出极强的宿主特异性。有许多病毒是细菌(噬菌体)、真菌、藻类、原生动物等的寄生虫。
b. 蛭弧菌(Bdellovibrio):蛭弧菌是许多革兰氏阴性菌的体外寄生虫。
当一种生物(捕食者)吞噬或攻击其他生物(猎物)时,这是一种普遍的现象。
捕食的例子:
未来的研究方向可能会更多地关注这些相互作用在分子水平的调控机制,特别是在肠道微生态系统中各种微生物之间的信号交流网络。
在实践层面,这些知识为微生物组的调控、疾病防治等领域提供了重要的理论指导。例如,通过理解肠道菌群中的共营养关系,我们可以更好地调节人体微生物组的平衡;通过了解偏害共生机制,可以开发新的抗菌策略。
随着研究技术的进步,特别是高通量测序和生物信息学等技术的发展,我们有望对微生物间的相互作用有更深入的认识,从而为人类的健康管理提供新的解决方案。
主要参考文献:
microbialinteraction.conferenceseries.com/2017/call-for-abstracts.php#collapse12
microbialinteraction.conferenceseries.com/2017/call-for-abstracts.php
Sagar Aryal. 2022. Microbial interaction and its types with examples.Edited By: Sagar Aryalmicrobenotes
谷禾健康
生态学的一个长期目标是捕获多种物种相互作用的多样性,生态系统中存在多种物种时产生的不可预测的影响。例如,动物具有多样的微生物群落,它们的组成是可变的,受随机过程控制,这会影响系统的整体行为。许多涉及复杂网络结构的生态研究通常集中于成对相互作用,忽略了涉及三个或更多的分类单元之间的高阶交互作用(higher-order interactions)带来的潜在影响。
高阶交互作用
高阶交互作用(higher-order interactions):高阶相互作用一直是遗传学领域中相对严格的研究对象,在遗传学领域中,它们被讨论为上位性,或基因和突变之间的非线性相互作用。尤其是高阶上位性,因为这些相互作用包含了理解和研究其他系统甚至微生物中的高阶相互作用的所有复杂性和挑战。
上位性(epistasis)是指某一基因受不同位点上别的基因抑制而不能表达的现象。现也将在群体遗传学和数量遗传学中非等位基因的遗传效应为非相加性时统称之为上位性。也就是位于不同座位上的基因间的非相加性相互作用。
与基因组不同,肠道微生物群是由单个实体或信息群(在这种情况下,是微生物群中的组成类群)之间的相互作用所定义的复杂系统。因此,科学家会预测微生物群中的生物群之间的更高阶相互作用可能是微生物群相关生物表型的基础。
例如一项研究了多物种相互作用对确定群落功能(即淀粉酶表达)的贡献。在存在更高阶的交互作用时,加性无效模型(没有交互作用)在预测社区功能方面的预测能力会降低。但是,通过将行为和人口动力学效应都计入其无效模型,高阶交互确实为社区功能提供了良好的预测。
最近发表的一篇关于此研究的文章,作者利用一种简单的数学方法——Hadamard-Walsh变换,研究了昆虫肠道菌群中的高阶相互作用。以此说明不同环境下肠道菌群之间的高阶相互作用是如何影响宿主表型的。希望能打破仅关注群落水平或两两互作关系的局限性,更多关注三阶、四阶以至多阶的分类群之间的互作关系。


一个由五个核心类群组成的肠道群落中。发现根据宿主特征,三向,四向和五向相互作用占所有可能病例的13-44%。然而,低阶相互作用(2对)仍然占系统中所有观察到的表型的至少一半。Sanchez-Gorostiage等人的工作研究了多物种相互作用对确定群落功能(即淀粉酶表达)的贡献。在存在更高阶的交互作用时,加性无效模型(没有交互作用)在预测社区功能方面的预测能力会降低。但是,通过将行为和人口动力学效应都计入其无效模型,高阶交互确实为社区功能提供了良好的预测。前述研究提供了可如何测量高阶相互作用的示例,并表明它们与理解微生物分类群如何影响某些表型有关。尽管多样性和主体互动的重要性很明显,但据我们所知,还没有研究试图明确区分环境环境中高阶互动的影响。
Hadamard-Walsh沃尔什-阿达玛变换使人们能够量化潜在相互作用的物体或地块之间相互作用的显著程度。它的主要输出是沃尔什系数(Walsh coefficient),该系数传达给定交互的大小(交互有多大)和符号(正交互或负方向)。
该方法以向量的形式实现表型(本研究中指宿主感染风险)值,然后将其重新格式化为Hadamard矩阵(然后通过对角线矩阵进行缩放)。输出的是与分类群之间相互作用的强度相对应的系数的集合。
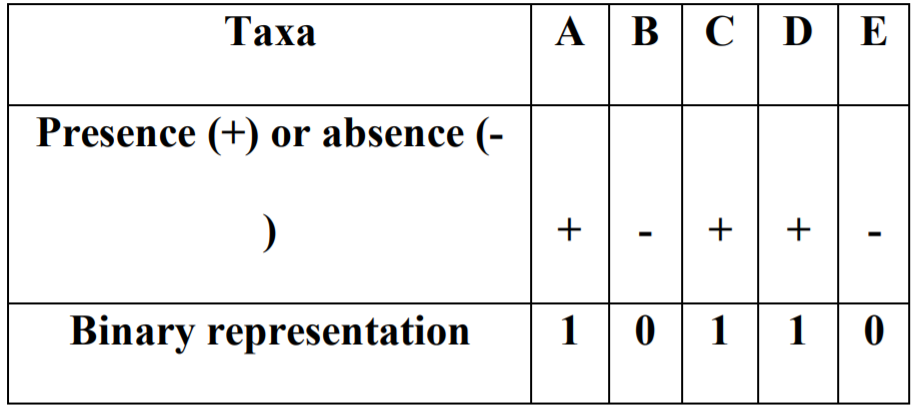
作者将沃尔什系数以二进制形式重新编码,例如: *B*DE 被编码为 01011
“*”表示不存在的分类群,在这个示例中存在BDE三个分类群,并将这三者之间的相互作用贴上“三阶相互作用”的标签,以此类推零阶即为*****或00000,五阶即为ABCD或11111。对于一个庞大的微生物群落,可以计算三阶相互作用的强度,并将它们与四阶相互作用的强度进行比较。
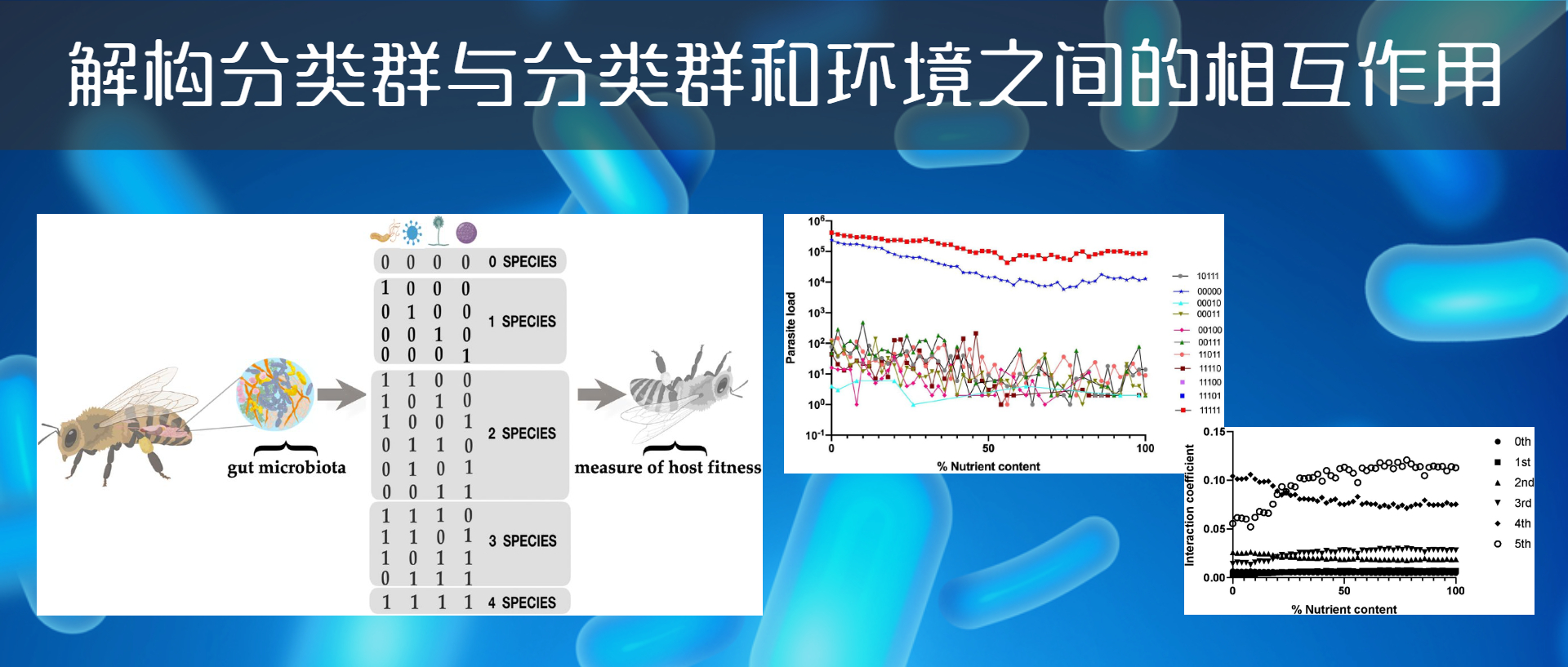
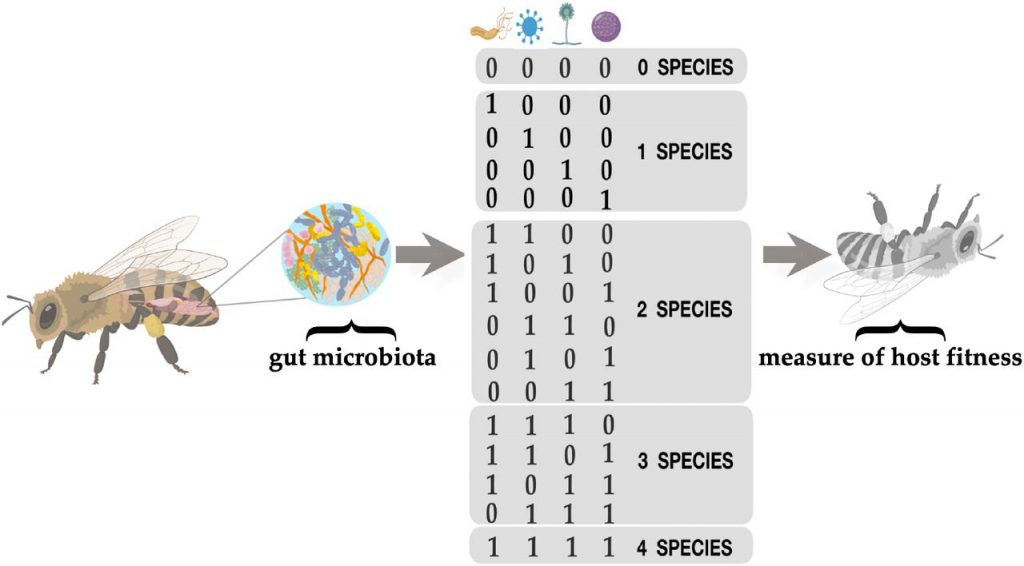
作者构建了一个昆虫肠道菌群的理论环境。用于研究饮食中不同品质的营养对宿主肠道菌群中寄生虫的易感性的影响。
利用从优质饲料(食物培养基中不含甲基纤维素)到低品质饲料(用甲基纤维素取代食物培养基的10%、20%、30%、40%、50%、60%、70%、80%、90%)的不同资源水平,对不同营养环境对鳞翅目害虫抗寄生性的影响进行了实证研究。
0%的饮食相当于极低质量的营养饮食,而100%的饮食相当于由昆虫的标准食物量组成的高质量饮食。
数据集来自用于生成理论上的适应度地形(fitness landscapes,遗传学算法中的名词,指由一系列基因型组成的三维的地形图)的原始数据。
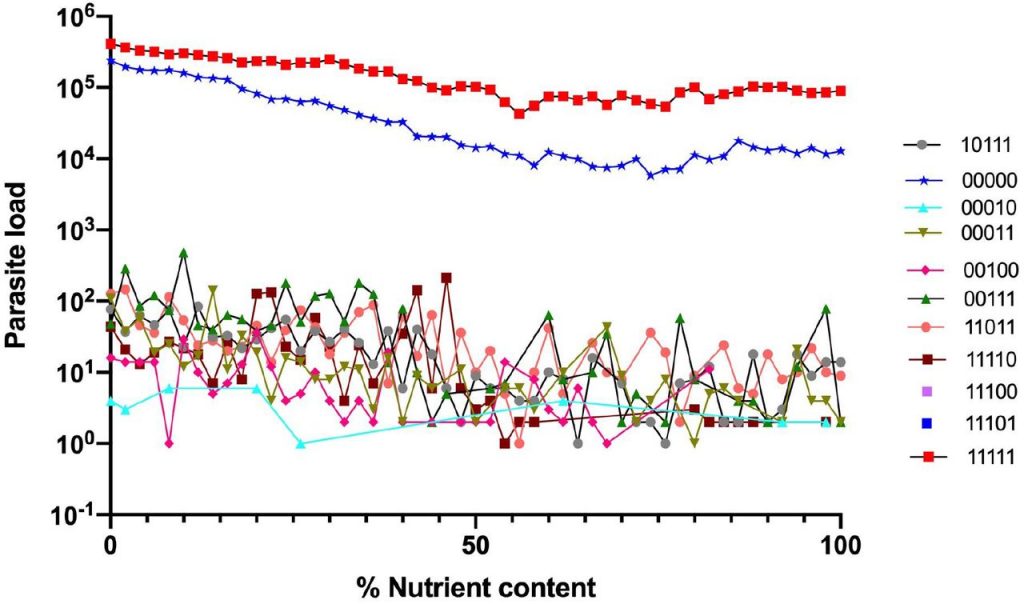
这些原始数据由5位字符串组成。这些数据集可用于研究群体遗传学的高级课题,比如高阶上位性。数据被定义为字符串(例如,01011或11001),每个字符串具有对应的“表现型”( phenotype)值。由此组成了含161个微生物群的昆虫肠道环境。如下图:

图一

图二
数据集是5个分类群(A-D)的存在和缺失(+或-)的组合。这种组合的可能性相当于四种不同昆虫环境中2的5次方=32个分类群的理论组合。
图一表示营养含量的消耗(x轴)与寄生虫含量(y轴)之间的反应关系。
结果显示与其他昆虫微生物区系组合相比,对应于00000(不含分类群) 和11111(存在各种分类群)的两个昆虫肠道最易感染寄生虫。相比之下所有其他昆虫微生物群组合(包括组合00100;11011;11101),感染的寄生虫含量显著较低。
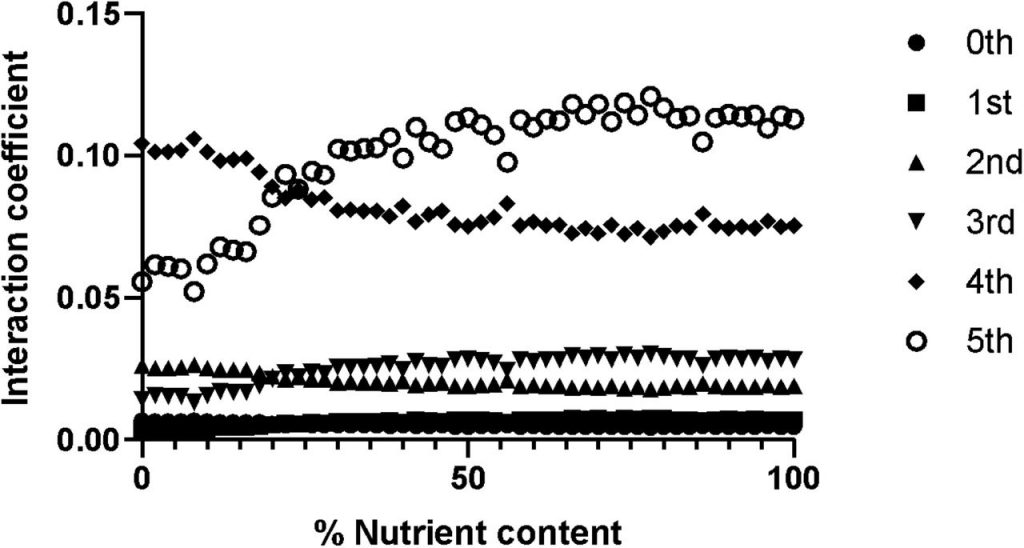
图二说明了阶级变化与相互作用强度是相关的,比如低阶的如零一二三,它们的相互作用强度也较低,相反,高阶的较高。以及营养含量对相互作用模式的影响,比如营养含量为20%时,可以明显看见五阶分类群的相互作用强度开始上升,四阶的下降。

图三

图四
上图结果表明,高阶微生物群的相互作用限制了寄生虫在营养环境中的入侵。肠道菌群的营养状况对宿主的健康起着重要作用。
使用高阶建模方法能够捕捉到来自复杂生态相互作用的丰富信息,将这些方法应用于实验数据的分析,将会推动微生物群动力学研究。
相关阅读:
Science|180种动物肠道菌群测序结果有哪些信息值得我们关注
参考文献
Deconstructing taxa x taxa x environment interactions in the microbiota: A theoretical examination
Senay Yitbarek, John Guittar, Sarah A. Knutie, C. Brandon Ogbunugafor
bioRxiv 647156; doi: https://doi.org/10.1101/647156