-

 国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子
国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子

 二级病原微生物安全实验室
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
谷禾健康
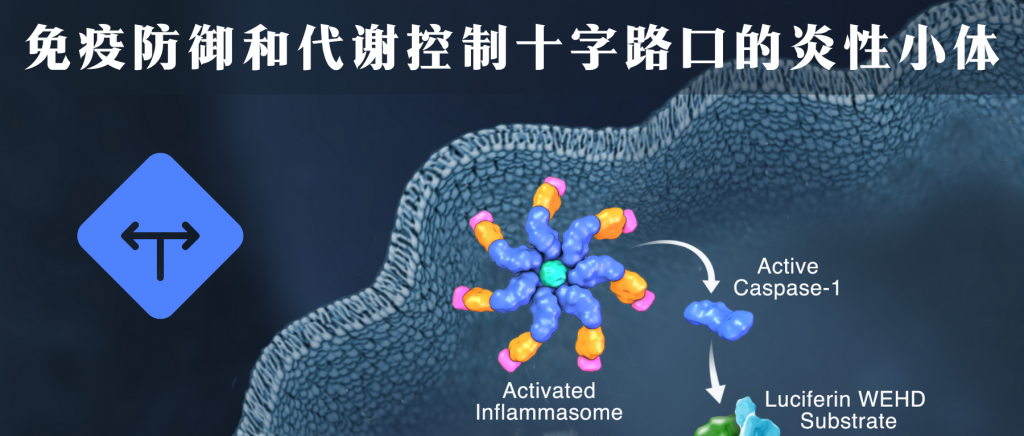
人体的肠道粘膜内层形成物理屏障和免疫防御系统,以防止微生物入侵。当身体受到感染或细胞遭受损伤时,免疫系统会启动炎症反应来应对这些情况。炎症是对感染和组织损伤的一种急性反应,以限制对身体的伤害,这种反应是身体自然的防御机制,旨在清除病原体并修复受损细胞或组织。
先天免疫系统包含多种种系编码的模式识别受体(PRRs),这些受体可以检测由细胞损伤或组织损伤产生的微生物抗原,称为病原体相关分子模式(PAMPs)或损伤关联分子模式(DAMPs)。
这些PRRs包括位于细胞膜上的Toll样受体(TLRs)和C型凝集素受体(CLRs),以及细胞内PRRs如RIG样受体。NOD样受体(NLRs)是其他可以识别来源于病原体和受损细胞的分子模式的PRRs。
炎性小体的概念在2002年首次被提出,它是细胞内多种蛋白质组成的复合体,处在免疫防御和代谢控制十字路口,是细胞完整性的守护者并调控各种关键细胞功能。主要介导宿主对微生物感染和细胞损伤的免疫反应,在骨髓细胞中产生,是免疫系统的重要组成部分。
在感知外界病原体或损伤后,传递信号给免疫系统,启动炎症。它们形成大型多蛋白信号传导平台来裂解和激活caspase-1,这是一种主要的炎症途径。活性caspase-1可以将非活性形式的促炎细胞因子白细胞介素1β(IL-1β)和IL-18裂解为活性形式来调节炎症和宿主防御反应,这些相关细胞因子介导针对感染的多种局部和全身免疫反应,包括诱导发热、白细胞迁移至损伤或感染部位,以及Th1、Th2和Th17反应的激活和极化。
此外,炎性小体激活与细胞焦亡有关,细胞焦亡又称细胞炎性坏死,是一种程序性细胞死亡。在对抗感染中发挥重要作用。研究发现其参与宿主防御鼠伤寒沙门菌(Salmonella typhimurium)、土拉热弗郎西丝菌(Francisella tularensis)和炭疽芽孢杆菌(Bacillus anthracis)。
各种内源性和外源性刺激已被证明可以激活炎性小体。由于大量微生物寄居在粘膜表面,维持人体和微生物群之间的稳定需要与炎性小体的共生相互作用。

炎性小体主要充当复杂的传感器,使宿主能够区分有益细菌和有害细菌,但它们也充当宿主与其肠道微生物群之间沟通的介质。肠腔的环境状态持续影响宿主反应,导致通过产生IL-1β或IL-18产生特定信号,进而调节肠道微生物群。
随后,调节的微生物群可以通过短链脂肪酸和胆汁酸衍生物等微生物副产物增强宿主反应。但是炎性小体的激活需要受到严格调节,以限制异常激活和对宿主细胞的损害。如果失调,可能会导致不同的疾病。包括自身免疫疾病、癌症、胃肠道(GI)疾病和炎症性疾病。
因此,炎性小体对于协调体内精确的相互作用是不可或缺的。通过针对与炎症小体信号传导相关的结构来研究炎症小体活性的适当调节和治疗干预,可能是一个有前途的研究领域。
本文旨在使人们更清晰地认识炎性小体,讨论了不同炎性小体在人体内的功能、外来细菌、病毒等病原体入侵时,炎性小体在抵抗感染的作用。此外,炎性小体的失调或异常激活可能与人体一些疾病相关。炎性小体和微生物群的相互作用影响着人类健康。
✦
炎性小体(inflammasome)是由多种蛋白质组成的复合体,也称炎症小体,是免疫系统的重要组成部分,此概念由于2002年首次提出。
炎性小体可识别多种炎症诱导的刺激,包括病原体相关分子模式(PAMPs)和损伤相关分子模式(DAMPs),并控制重要的促炎细胞因子如白细胞介素-1β(IL-1β)和IL-18的产生。
▼
炎性小体的功能
炎症小体是细胞内多聚蛋白复合物,是细胞完整性的守护者并控制各种关键细胞功能的完整性。具体来说,炎症小体的功能包括以下几个方面:
▸ 参与免疫反应,加工IL-1β和IL-18
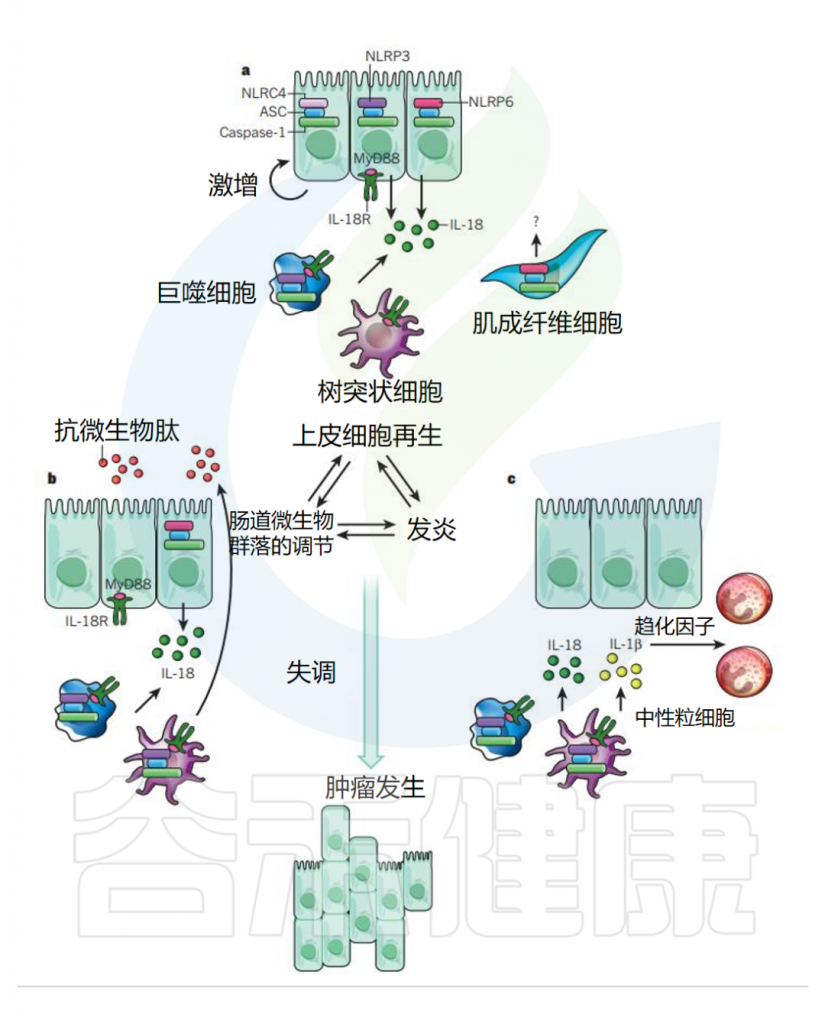
在体内,炎性小体已被证明参与抗微生物先天免疫反应。在这方面研究最广泛的炎症小体是NLRP3炎性小体,它参与了抗菌、病毒、真菌和寄生虫的免疫反应。
caspase-1的激活受炎症小体调节,caspase-1的激活会导致IL-1β和IL-18的加工。
在非经典途径中,小鼠体内caspase-11(人类直系同源物包括caspase4和5)的裂解会激活NLRP3炎症小体,该炎症小体在维持肠道免疫稳态中发挥着关键作用。
doi: 10.1038/nature10759.
需要注意的是,在急性炎症中,炎症小体的激活有助于去除死细胞并启动组织修复。然而,在慢性炎症中,炎症小体的持续激活是有害的,因为它会损伤组织。
▸ 促进细胞焦亡
此外,炎症小体激活与细胞焦亡有关。细胞焦亡是一种程序性细胞死亡,表现为细胞不断胀大直至细胞膜破裂,导致细胞内容物的释放进而激活强烈的炎症反应,是机体一种重要的天然免疫反应,在抗感染中发挥重要作用。
细胞焦亡最常发生在细胞内病原体感染时,并且可能形成抗菌反应的一部分。焦亡是caspase-1依赖性的,并且独立于促凋亡caspase发生。
注:尽管细胞焦亡被认为是程序性细胞死亡的一种形式,但它与细胞凋亡所呈现的免疫沉默细胞死亡不同。焦亡伴随着质膜破裂、水流入、细胞肿胀、渗透溶解和促炎细胞内容物的释放。焦亡还伴随着DNA裂解和核浓缩,这与细胞凋亡的DNA阶梯特征不同,因为核完整性并未受到损害。
细胞焦亡的调控尚不明确;然而,细胞焦亡的程度似乎随着炎症体刺激的增加而增加。细胞焦亡是否在产生更活跃的炎症小体途径(下文讨论)的遗传性自身炎症疾病中发挥病理作用尚待确定。
▸ 与炎症性疾病相关,调节肠道稳态
炎性小体激活需要严格调节,以限制异常激活和对宿主细胞的损害。炎症小体活性失调与多种炎症性疾病有关,包括自身免疫、癌症和胃肠道疾病。
尽管炎症小体主要充当复杂的传感器,使宿主能够区分有益细菌和有害细菌,但它们也充当宿主与其肠道微生物群之间沟通的介质。
肠腔的环境状态持续影响宿主反应,导致通过产生IL-1β或IL-18产生特定信号,进而调节肠道微生物群。
随后,调节的微生物群可以通过短链脂肪酸和胆汁酸衍生物等微生物副产物增强宿主反应。因此,炎症小体对于协调体内精确的相互作用是不可或缺的。在这方面,通过针对与炎症小体信号传导相关的结构来研究炎症小体活性的适当调节和治疗干预,可能是一个有前途的研究领域。
炎症小体调节微生物感染和自身炎症性疾病期间的炎症
doi: 10.1038/nature10759.
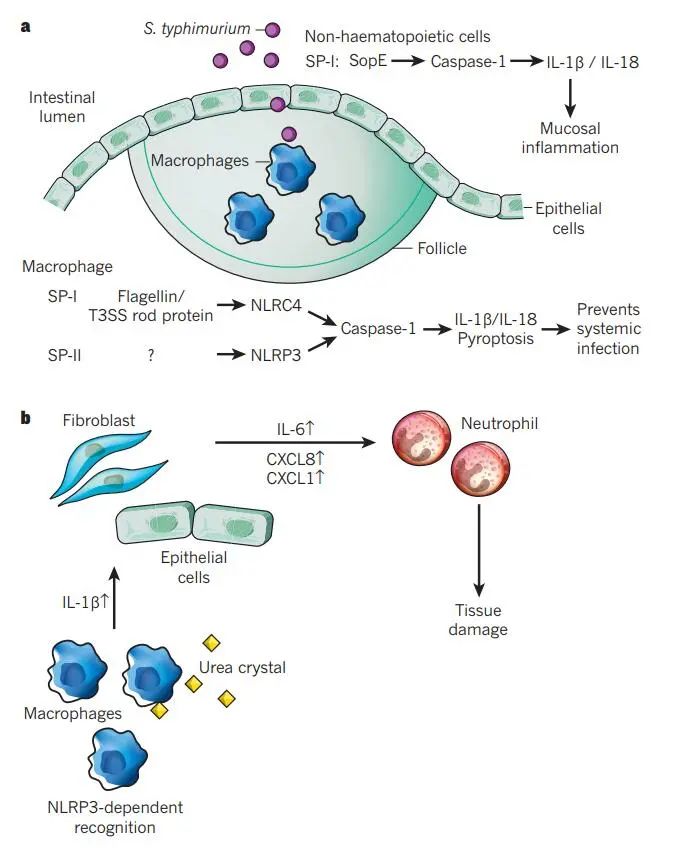
a.鼠伤寒沙门氏菌通过肠上皮屏障进入宿主。M细胞是一种特殊的上皮细胞类型,分布在Peyer’s斑块上,特别参与沙门氏菌的胞吞作用和Peyer’s斑块中巨噬细胞的感染。炎症小体和caspase-1参与了几种细胞类型和感染的几个步骤。将细菌效应蛋白SopE注射到上皮细胞中,通过涉及GTPase Rac1的过程诱导caspase-1独立于NLRP3和NLRC4的激活。由此产生的粘膜炎症依赖于非造血细胞产生的IL-1β和IL-18。在巨噬细胞感染时,细菌蛋白鞭毛蛋白和PrgJ (T3SS的一部分)通过NLRC4被感知。这导致caspase-1的激活,导致IL-1β/IL-18加工和焦亡,从而限制全身感染。NLRP3通过识别未知信号参与这些过程。
b.巨噬细胞吞噬尿酸钠(MSU)晶体诱导nlrp3依赖性caspase-1激活和IL-1β释放,刺激非造血细胞产生IL-6和趋化因子(CXCL1和CXCL8),吸引中性粒细胞。然后,活化的中性粒细胞引起组织损伤。治疗性阻断人IL-1β可改善痛风的炎症发作。
▸ 和自噬途径的相互调节
自噬是一种细胞保护过程,细胞通过该过程将受损的蛋白质、细胞器或病原体隔离在双膜室(自噬体)中,靶向这种细胞材料在溶酶体中降解,并回收组成分子。
自噬发生在正常生理条件下,但可以通过细胞应激(如饥饿、促炎信号传导(例如 IFNγ)或细菌感染)上调。
最近的报告揭示了炎症小体和自噬途径之间复杂的相互作用。在没有污染配体的情况下,用TLR4激动剂脂多糖处理不会诱导野生型巨噬细胞中的炎症小体活化。然而,通过自噬调节因子Atg16L1或 Atg7 的基因消融来阻断自噬,可以实现LPS依赖性炎症小体激活,这表明自噬通常会对抗LPS引起的炎症小体激活。
另一项研究发现,炎症小体对自噬有负向调节作用,这使炎症小体和自噬之间的联系更加复杂。
!
炎性小体是好是坏
“抛开剂量谈毒性都是耍流氓”——炎症小体过少或过于活跃对健康都是不利的。炎症小体的活动是需要严格控制的,不能随意抑制和刺激,以避免产生过多的炎性细胞因子导致细胞死亡,伤及自身。
所以正常情况下,炎症小体,特别是NLRP3的表达在许多细胞中相对较低,需要诱导去引发信号。
先天免疫系统包含多种编码的模式识别受体(PRR),可检测微生物抗原,称为病原体相关分子模式(PAMP)或损伤相关分子模式(DAMP),由细胞或组织损伤产生。
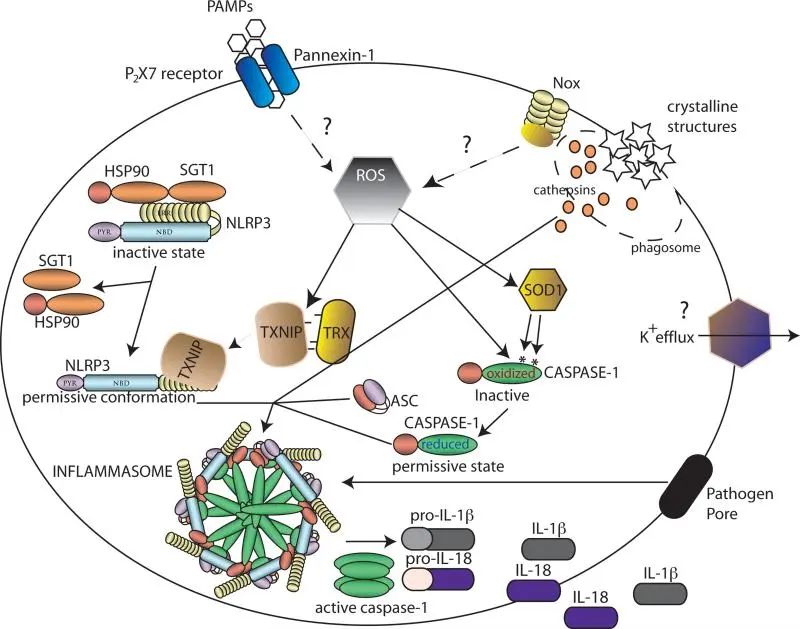
炎症小体如何被激活尚不清楚。由于激活炎症小体的PAMP、DAMP和病原体具有不同的性质,因此可能存在多种途径。
炎性小体在感知到PAMPs和DAMPs的结构多样性后进行组装。已经提出了几个模型来解释这些信号是如何被感知的,包括基于一般细胞应激识别的模型(图a和b)或基于激活信号的直接和间接识别的模型(图c-e)。
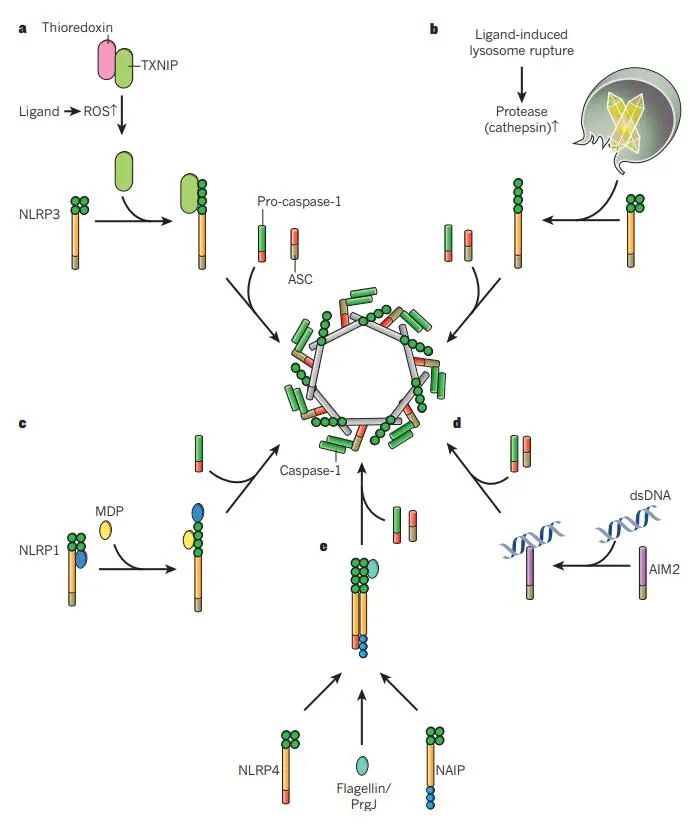
doi: 10.1038/nature10759.
(a) NLRP3感知细胞内(可能由线粒体)直接或间接由NLRP3炎症小体激活剂产生的活性氧(ROS)。硫氧还蛋白和硫氧还蛋白相互作用蛋白(TXNIP)的复合物可以感知ROS的增加,从而导致该复合物的解离。
随后,TXNIP与NLRP3结合导致NLRP3的激活,ASC和前caspase-1的募集,以及活性炎性体复合物的形成。
(b) 溶酶体失稳后,NLRP3被激活。特定晶体和颗粒结构的吞噬可导致溶酶体不稳定和溶酶体内容物(包括蛋白酶)的释放。这些蛋白酶可导致负调节因子的蛋白水解失活或NLRP3正调节因子的蛋白水解激活,导致炎性小体组装。
(c, d) NLRP1和AIM2直接感知配体。特异性配体(muramyl二肽(MDP)和双链DNA (dsDNA))的直接结合可导致NLRP1和AIM2的构象改变,导致炎性小体活化。
(e) NLRP1炎症小体的形成不依赖于ASC。NAIP蛋白感知细菌蛋白,导致NLRC4的募集和NLRC4炎症小体的组装。
此外,构成信号的限制因子如pro-IL1β和-IL18的转录上调是炎症小体激活的先决条件。除了编码原细胞因子的基因的诱导转录之外,NLRP3转录的激活也由NF-κB 激活剂(例如TLR配体)诱导。
doi: 10.1146/annurev-immunol-031210-101405.
大部分炎性小体主要由受体蛋白(NLR或ALR家族的成员),衔接蛋白ASC和效应蛋白caspase组成。
炎性小体作为一种重要的细胞结构,它在炎症和免疫反应中发挥着关键作用。不同类型的炎性小体在形态和功能上都有所不同,对于了解炎症过程和治疗炎症性疾病具有重要意义。下面将探讨不同类型的炎性小体的特点和对肠道微生物群的作用机制。
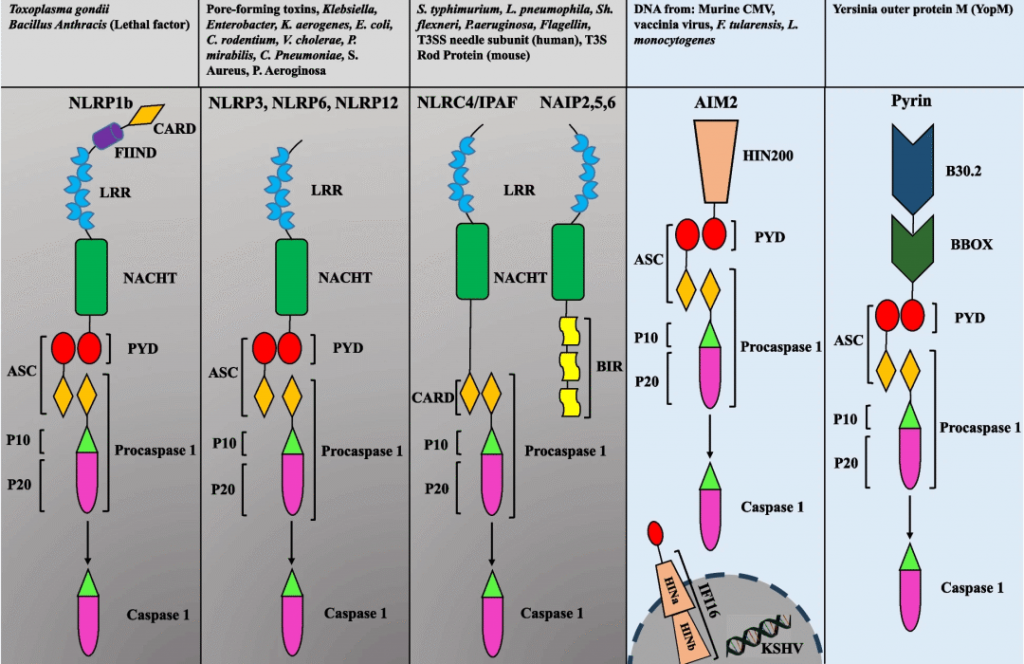
Manshouri S,et al.Cell Commun Signal.2024
NLRP1炎症小体
NLRP1是第一个报道的形成炎症小体的分子,对caspase-1、caspase-5和ASC的需求最低。
NLRP1在结构上与其他NLR的不同之处在于其额外的C端延伸,由具有未知功能的结构域和CARD结构域组成。
NLRP1炎症小体在小鼠和人类中是不同的。小鼠NLRP1炎症小体由Nlrp1a、b和c的三个旁系同源物组成,其中包含NR100结构域,而不是人类中看到的PYD。
▸ NLRP1会影响产生丁酸盐的菌群
研究发现NLRP1炎症小体可以影响肠道微生物群。Nlrp1缺陷的小鼠表现出产生丁酸盐的细菌数量增加。丁酸盐已被证明通过促进肠道屏障的功能(例如粘液产生和紧密连接)对炎症性肠病(IBD)具有有益作用。
NLRP1炎症小体可能通过减少肠道微生物群的丁酸盐产生而对IBD产生负面影响。IBD有两种主要的临床形式,包括克罗恩病和溃疡性结肠炎。短链脂肪酸(SCFA)是由有益肠道细菌通过高纤维饮食发酵产生的。这些SCFA在减少炎症、调节免疫功能和防止过度活跃的免疫反应方面发挥着至关重要的作用,从而减缓IBD的临床进展。
NLRP3炎症小体
NOD样受体蛋白3(NLRP3)炎性小体包括N末端的热蛋白结构域(PYD)、中央NACHT结构域(包括带有核苷酸三磷酸腺苷/三磷酸鸟苷 (ATP/GTPase) P 环的七个基序和Walker A 和 B 结合位点),以及C端的9个富含亮氨酸的重复序列 (LRR)。
▸ 与其他炎症小体相比,NLRP3炎症小体需要两个信号
信号1(启动)由微生物分子或内源性细胞因子或 PRR(例如 TLR)的激活提供,导致经典和非经典 NLRP3 炎性体成分的转录上调。
它由NLRP3和pro-IL-1β的转录上调以及非转录机制组成,例如N端 PYD 内残基的去磷酸化、PYD 和 NACHT 结构域之间关键丝氨酸残基的磷酸化和 NLRP3 去泛素化。
Caspase-8和FAS相关死亡结构域蛋白(FADD)通过调节NF-kB通路介导此步骤。Lys-63 特异性去泛素酶 BRCC36 (BRCC3) 和 IL-1 受体相关激酶 1 (IRAK1) 调节 NLRP3 的激活。
信号2(激活)由PAMP或DAMP、成孔毒素、K +外流、溶酶体破坏、线粒体活性氧产生、心磷脂重新定位到线粒体外膜以及氧化线粒体DNA的释放提供,然后Cl -流出。
NLRP3炎症小体的经典和非经典激活机制
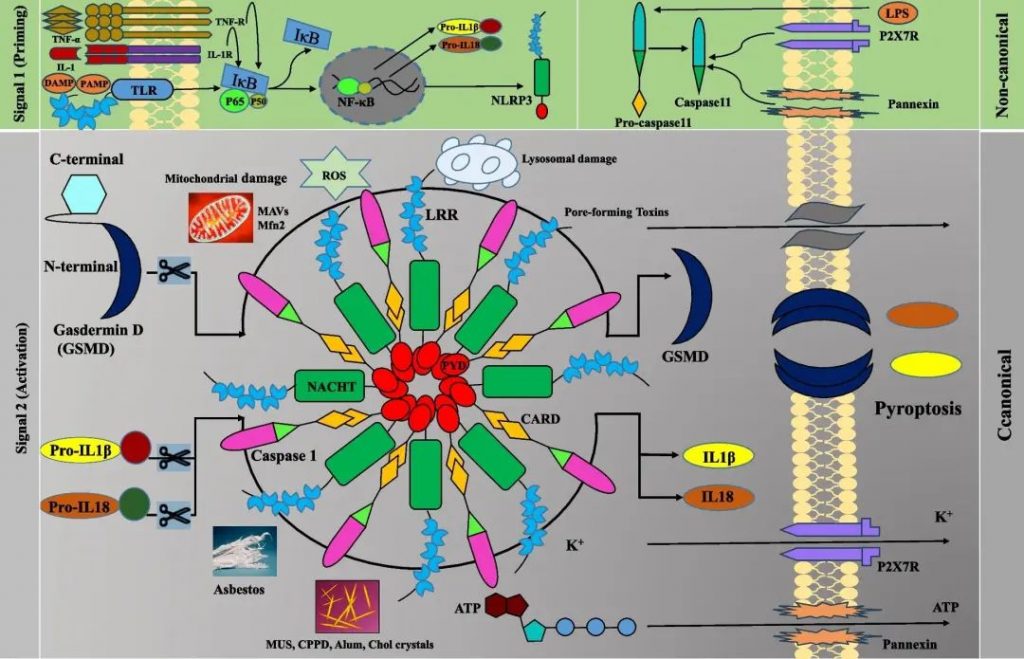
Manshouri S,et al.Cell Commun Signal.2024
经典途径涉及TLR信号传导,通过NF-κB途径诱导IL-1β、IL-18和NLRP3的转录。非规范途径涉及脂多糖等刺激,需要caspase-11来激活caspase-1。
▸ 微生物与NLRP3炎症小体的相互作用
一些共生肠道微生物可能会激活肠粘膜巨噬细胞中的NLRP3炎性小体。据报道,奇异变形杆菌(Proteus mirabilis)可能通过产生溶血素成为 NLRP3 激活剂。肠杆菌(Enterobacter)和克雷伯菌属(Klebsiella spp)在口腔定植可能会触发NLRP3炎症小体。存在于小鼠口腔中的产气克雷伯菌(K.aerogenes)通过巨噬细胞分泌IL-1β导致牙周炎。
研究还报道了肠道微生物群异常积累对年龄相关性心房颤动的因果影响,表明微生物群-肠道屏障-心房NLRP3炎性体相互作用可能作为治疗年龄相关性心律失常的潜在靶点。
在临床前研究中,研究人员发现金黄色葡萄球菌将线粒体与吞噬体隔离开,以逃避杀菌活性氧。这种巨噬细胞死亡的逃避依赖于NLRP3炎性体。使用小干扰RNA(siRNA)靶向NLRP3可以改善感染金黄色葡萄球菌的小鼠的细菌清除率。研究人员还发现,NLRP3 抑制和电子传递链复合物 II 抑制相结合,可以提高对人类单核细胞中金黄色葡萄球菌的杀伤力。
最近,证明了NLRP3炎性体在接触香烟烟雾后被激活,从而为肺部造成铜绿假单胞菌引起的急性损伤做好准备。这项研究表明,靶向NLRP3炎性体可能是治疗香烟烟雾引起的肺损伤的潜在治疗方法。
NLRP6炎症小体
NLRP6,也称为 PYPAF5,被描述为大多数免疫细胞中NF-κB和caspase-1表达的调节剂。这种蛋白质存在于肠上皮细胞中。研究表明,NLRP6对于调节肠道微生物组的组成和功能至关重要。
▸ 调节肠道微生物的组成和功能
NLRP6通过炎症小体依赖性和炎症小体独立途径、结肠炎相关肿瘤发生和杯状细胞中的粘液分泌来协调宿主与肠道病毒和细菌感染的相互作用。肠上皮细胞中的NLRP6缺陷与IL-18产生和caspase-1激活中断有关。NLRP6缺陷小鼠表现出普氏菌科和 TM7 的生长,以及乳杆菌和厚壁菌门的减少。
这些不平衡会引发结肠炎和肠道自发炎症。微生物群相关代谢物,例如牛磺酸和肠道共生细菌,可以激活NLRP6炎性体产生抗菌肽。杯状细胞可以通过 TLR-Myd88 信号传导激活 NLRP6 炎症小体,从而产生muc2。革兰氏阳性病原体产生脂磷壁酸,通过ASC募集激活NLRP6炎症小体,导致全身感染。应激诱导的促肾上腺皮质激素释放激素(CRH)抑制NLRP6炎症小体的激活,后者会导致肠道炎症和肠道微生物组的改变。
▸ NLRP6缺陷会导致胃肠道生态失调
NLRP6 缺陷的小鼠容易发生炎症。NLRP6 炎症小体在正常条件下是自我抑制的。脂磷壁酸和双链 RNA (dsRNA) 可以直接与 NLRP6 结合,从而产生可能的构象变化,以帮助液-液相分离(LLPS),这是炎症小体组装所必需的早期步骤。
此外,脂多糖可以直接与NLRP6结合,这可能导致LLPS的形成。然后,它与ASC相互作用激活 caspase-1或 caspase-11,从而激活 GSDMD 并导致质膜中孔的形成以及促炎细胞因子和细胞内内容物的释放。如果与ASC的相互作用不形成 NLRP6 炎症小体,则 LLPS 中的 NLRP6 通过诱导干扰素 (IFN) 和 IFN 刺激基因,诱导替代的炎症小体独立途径。
总而言之,在强烈的炎症反应具有破坏性的情况下,NLRP6可能通过 TLR 轴发挥保护作用,而其作用对于维持肠道稳态是必要的。NLRP6炎症小体与胃肠道的稳态有关。先前的研究表明NLRP6失调可能导致胃肠道生态失调。
此外,NLRP6炎症小体刺激抗菌肽(AMP)的表达,包括血管生成素-4(Ang4)。一些微生物代谢物,包括牛磺酸、精胺和组胺,似乎可以诱导NLRP6依赖性IL-18和AMP的产生。
NLRP12炎症小体
NLRP12,也称为Nalp12和Pypaf-7,与ASC和胱天蛋白酶1形成炎症小体,使IL-1β成熟。它是最早与衔接蛋白ASC共定位并相互作用形成炎症小体的NLR之一。人类基因组中NLRP12编码序列的突变与IL-1介导的炎症性疾病有关。
▸ 识别鼠疫耶尔森氏菌等病原菌
尽管我们对NLRP12在健康和疾病中的作用的了解有限,但最近的数据表明,NLRP12对于识别鼠疫耶尔森氏菌(鼠疫病原体)至关重要。
NLRP12在巨噬细胞感染鼠疫杆菌后控制caspase-1裂解以及IL-1β和IL-18分泌。然而,NLRP12可以抑制骨髓源性巨噬细胞产生IL-12,并负向调节宿主对流产布鲁氏菌的防御。
NLRP12的确切配体目前未知;然而,它的激活需要一个功能正常的T3S系统。这表明细菌毒力因子进入宿主细胞质可能是直接激活NLRP12或改变宿主信号通路所必需的。无论激活机制如何,NLRP12驱动的IL-18分泌和相关的IFN-γ产生在小鼠抵抗鼠疫耶尔森氏菌感染方面发挥着关键作用。NLRP12缺陷小鼠在感染后表现出更高的死亡率和细菌载量。
▸ 抑制肠道炎症和肿瘤
除了形成炎症小体之外,NLRP12还通过负向调节NF-kB信号传导来抑制肠道炎症和肿瘤发生。几项独立研究表明,NLRP12 在生化检测、结肠癌和结肠炎模型中对经典和非经典NF-κB信号传导有负向调节作用。
NLRP12被认为在鼠伤寒沙门氏菌感染期间独立于炎症小体抑制宿主防御,因为Nlrp12缺陷小鼠比WT对照对鼠伤寒沙门氏菌感染具有更强的抵抗力,并且炎症细胞因子水平较低。
NLRP12在造血细胞中发挥抑制肿瘤发生的作用,但它不是造血细胞,而是非造血细胞,这对于限制肿瘤数量至关重要。尽管如此,两项研究都表明NLRP12在控制结肠炎症反应中发挥着重要作用。
NLRC4炎症小体
NLRC4(以前称为IPAF,Card12)在感染各种革兰氏阴性菌后可形成炎症小体,如鼠伤寒沙门菌(S.typhimurium)、嗜肺军团菌、福氏志贺菌和铜绿假单胞菌。NLRC4在髓系中表达,其基本作用是防止细菌入侵。
与其他炎症小体不同,NLRC4与另一种NLR蛋白NAIP结合被激活,NAIP作为NLRC4激活剂的受体。
▸ 有效抵御铜绿假单胞菌和伤寒沙门氏菌
几项研究表明,NLRC4在宿主防御有鞭毛的铜绿假单胞菌方面是有效的。在缺乏NLRC4或胱天蛋白酶-1激活的情况下,含有军团菌的吞噬体不能与溶酶体融合。相反,鞭毛蛋白突变的军团菌不能激活巨噬细胞中的胱天蛋白酶1。
鼠伤寒杆菌可以激活NLRC4和NLRP3,这导致ASC的形成和胱天蛋白酶向炎症小体的募集。NLRC4炎症小体感知PrgJ,一种III型分泌系统(T3SS)的成分,并通过CARD-CARD与胱天蛋白酶-1的相互作用启动炎症小体组装。
▸ 与婴儿期肠炎伴自体炎症相关
研究人员证明, NLRC4的功能获得性突变与一种极其罕见的疾病有关,这种疾病称为婴儿期肠炎伴自体炎症(AIFEC)。这种疾病的特点是巨噬细胞激活和胃肠道严重炎症。常驻肠道单核吞噬细胞(iMP),例如树突状细胞和巨噬细胞,可以对抗肠道病原微生物,同时保持对共生微生物的耐受性。
由于胃肠道的免疫细胞主要与许多共生微生物发生反应,它们应用多种机制来限制针对胃肠道共生微生物的不受控制的免疫反应。iMP中的NLRC-4激活后,分泌IL-1β诱导内皮细胞中粘附分子的表达。这些粘附分子促进中性粒细胞募集到肠粘膜和外来微生物的摄入。
PYHIN炎症小体
另一类与NLR不同的炎症小体已被鉴定为PYHIN家族。PYHIN是由四个人类基因(AIM2、IFI16、MNDA和IFIX)和13个小鼠基因组成的家族,并包含一个PYD和一个或两个HIN-200DNA 结合域。
AIM2和IFI16已被证明可形成caspase-1激活炎症小体。与NLR不同,AIM2和IFI16在这两种情况下都直接与其配体dsDNA结合。ASC是招募pro-caspase-1所必需的,因为AIM2和IFI16缺乏 CARD。
▸ AIM2对于肠道微生物群稳态非常重要
在感染过程中,AIM2感知来自鼠巨细胞病毒、牛痘病毒、土拉弗朗西斯菌和单核细胞增生李斯特菌的 DNA 。
AIM2炎症小体的一个功能是调节肠道微生物群。研究表明,AIM2炎症小体的激活导致肠道中IL-18和AMP的产生。Aim2缺陷小鼠的IL-18和AMP(例如REG3c和REG3b)减少。AIM2 的缺乏会导致肠道菌群失调,从而增加对结肠炎的易感性。
同时研究发现, Aim2缺陷小鼠粪便中肠杆菌科成员(例如大肠杆菌)的数量比普通小鼠高数百倍。当新杀弗朗西丝菌(F.novicida)(一种胞质病原体)从液泡逃逸到细胞质时,AIM2炎性小体就会受到刺激。缺乏逃离液泡的关键基因的F. novicida突变体无法触发AIM2炎症小体。
与NLRP6类似,AIM2炎性小体对于维持肠道微生物稳态至关重要。在胃肠道中,未经治疗的Aim2缺陷小鼠显示,Akkermansia muciniphila和 Anaeroplasma的数量较高,而双歧杆菌、普雷沃菌、Anaerostipes和Paraprevotella的数量较低。
Pyrin炎症小体
Pyrin是一种高分子量(86kDa)蛋白质,主要存在于免疫细胞中,包括中性粒细胞、单核细胞和树突状细胞。
与其他免疫传感器不同,pyrin通过细胞骨架重塑而不是微生物化合物来检测细菌毒力。
Pyrin在识别病原体对RhoA GTPase的失活修饰后,以ASC依赖性方式介导caspase-1炎症小体组装。小鼠pyrin有两个功能性磷酸化位点:Ser-205 和Ser-241,它们通过与14-3-3蛋白结合而使pyrin 失活。当毒素刺激或细菌感染时,导致Rho修饰,Ser-205和Ser-241去磷酸化,导致14-3-3解离。该级联导致吡啶激活并形成寡聚吡啶-ASC炎性体复合物。
▸ 在维持肠道稳态中发挥作用
尽管关于肠道微生物群产生的特定pyrin炎症小体激活剂的知识很少,但最近的研究揭示了它们在维持肠道稳态中的作用。在一项使用小鼠结肠炎的研究中,pyrin炎症小体信号可防止生物失调,促进肠道屏障完整性,并改善结肠炎症和肿瘤发生。
最近一项使用全基因组合并CRISPR筛选技术的研究中,两种胆汁酸类似物(BAA485和BAA473)被鉴定为在髓系和IEC系中诱导pyrin炎症小体信号传导的特异性配体。由于肠道细菌是胆汁酸代谢的丰富来源,类似的微生物组衍生的pyrin炎症小体激活配体可能有助于调节肠道稳态。
总体而言,pyrin炎症小体为与细胞骨架结合的先天免疫成分提供了新的范例,为细胞免疫的结构调节提供了新的机制。
✦
炎症小体正在成为宿主针对微生物病原体反应的关键调节因子。当微生物侵入组织或引起细胞损伤时,这些胞质多蛋白复合物会招募并激活半胱氨酸蛋白酶caspase-1。
炎症小体激活的caspase-1通过将促炎细胞因子IL-1β和IL-18裂解为其生物活性形式并将高迁移率族蛋白B1(HMGB1)释放到细胞外环境中来诱导炎症。此外,炎症小体通过称为细胞焦亡的炎症细胞死亡程序来对抗细菌复制并清除受感染的免疫细胞。
跟着谷禾一起来深入了解炎症小体在宿主与微生物相互作用中所扮演的重要角色,探讨其对免疫调节和疾病防御的影响,以及在维持体内微生物稳态和免疫平衡方面的关键功能。
炎症小体的组装和caspase-1的激活在病原体特异性方式下发生,尽管不同的炎症小体在感染过程中可能具有相似的作用。
▸ Nlrp1b炎症小体
Nlrp1b炎症小体识别细胞质中存在的炭疽芽孢杆菌致死毒素,Nlrp1b基因突变被确定为炭疽致死毒素诱导巨噬细胞死亡的关键易感位点。值得注意的是,Nlrp1b炎性体诱导的细胞焦亡赋予体内对炭疽芽孢杆菌孢子感染的抵抗力,突显了细胞焦亡对于宿主防御病原体的重要性。
▸ Nlrp3炎症小体
NLR家族成员Nlrp3的激活包括一个两步过程,需要用TLR和NLR配体启动以增强NF-κB驱动的Nlrp3转录,然后将巨噬细胞暴露于微生物毒素和离子载体(例如尼日利亚菌素和蓖麻毒素)或内源性毒素。
在巨噬细胞分别感染细菌、病毒和真菌病原体如金黄色葡萄球菌、肺炎链球菌、流感病毒和白色念珠菌的过程中,可以结合Nlrp3启动和激活步骤。类似于Nlrp1b炎症小体在炭疽杆菌感染中的作用,Nlrp3炎症小体激活缺陷使小鼠对念珠菌病高度敏感。
▸ Nlrc4炎症小体
Nlrc4炎性体可检测沙门氏菌(Salmonella)、假单胞菌(Pseudomonas)、军团菌(Legionella)和志贺氏菌(Shigella spp.)的III型和IV型细菌分泌系统的细菌鞭毛蛋白和基体杆成分。
除了分泌IL-1β和IL-18之外,最近还确定诱导焦亡细胞死亡是一种关键的体内机制,Nlrc4炎性体通过该机制清除表达鞭毛蛋白的细菌,例如嗜肺军团菌和伯克霍尔德菌。
人们认为细胞焦亡使细胞内细菌暴露于细胞外免疫监视,从而使它们被抗菌肽、免疫球蛋白和补体系统破坏,并被中性粒细胞和其他免疫细胞摄取。
▸ AIM2炎症小体
最后,AIM2响应土拉热弗朗西斯菌(F.tularensis)、单核细胞增多性李斯特菌和某些DNA病毒(例如CMV和痘苗病毒)以诱导caspase-1激活。caspase-1缺陷型小鼠对土拉菌病(土拉菌病的病原体)感染的敏感性增加,说明AIM2炎性小体在宿主对微生物病原体的防御反应中发挥着关键作用。
细菌和病毒效应物对炎症小体途径的调节
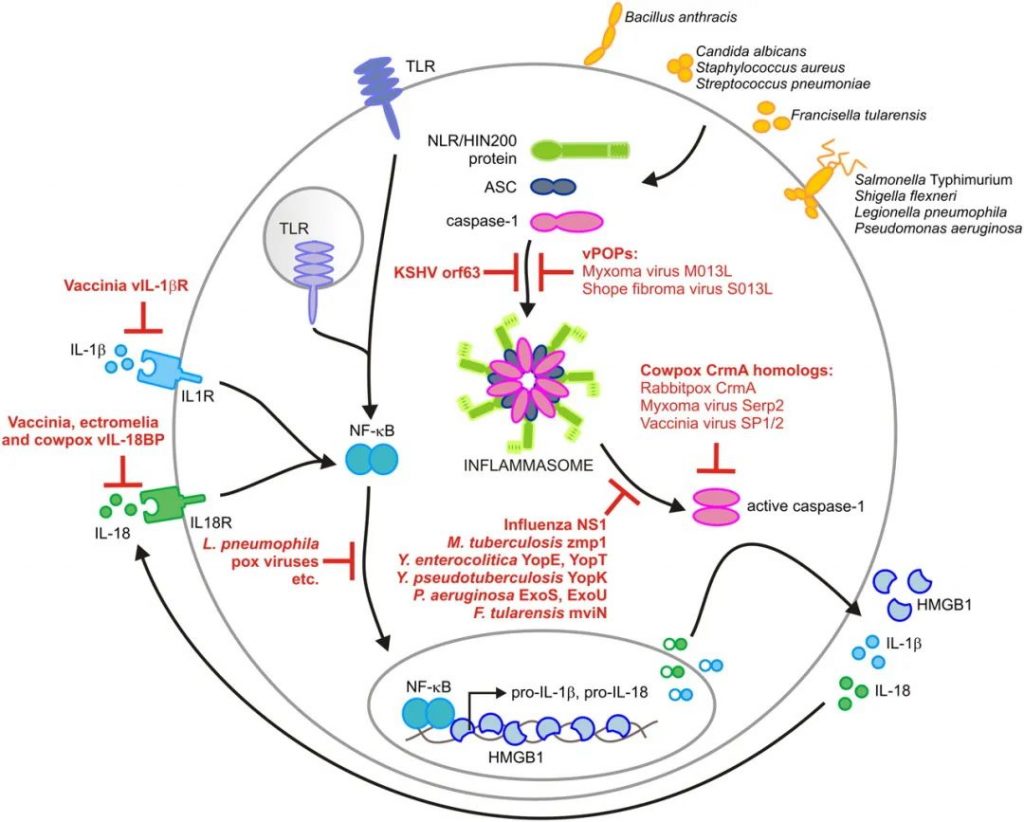
doi: 10.4049/jimmunol.1100229.
细菌、病毒和真菌病原体感染巨噬细胞和树突状细胞会诱导炎性体复合物的组装。这些多蛋白复合物驱动诱导caspase-1的激活,从而使IL-1β、IL-18和高迁移率族蛋白B1(HMGB1)释放到细胞外。
鉴于炎症小体在控制微生物病原体复制和传播中的重要性,细菌进化出一套机制来对抗炎症小体组装并干扰caspase-1效应机制的诱导也就不足为奇了。
▸ Yop蛋白等毒力因子抑制IL-1β分泌
例如,肠道病原性小肠结肠炎耶尔森氏菌通过专用的III型分泌系统将称为Yop蛋白的毒力因子注入宿主细胞胞浆中。在这些效应蛋白中,YopE和YopT抑制caspase-1激活以及随后成熟IL-1β的分泌。
这些Yop蛋白是Rho GTP酶和Rho介导的过程(例如细胞骨架重组和吞噬作用)的负调节因子。
尽管目前尚不清楚Yop效应蛋白和细胞骨架过程如何干扰炎症小体信号传导,但显性失活蛋白和化学抑制剂导致Rho GTPase Rac1失活表明该Rho GTPase在caspase-1激活和IL-1β分泌中发挥着关键作用。
假结核耶尔森氏菌使用名为YopK的第三种效应蛋白来掩盖细菌III型分泌系统并阻止其被Nlrp3和Nlrc4炎性体识别。这导致宿主巨噬细胞中的细菌存活率增加,说明炎症小体在控制侵入性耶尔森氏菌细胞内增殖中的重要性。
▸ 毒力因子外酶抑制caspase-1激活
表达毒力因子外酶(Exo)U的铜绿假单胞菌分离株使用不同的策略来抑制人类吞噬细胞中caspase-1的激活。这种革兰氏阴性病原体编码一种名为ExoU的具有磷脂酶A2活性的酶,可抑制Nlrc4炎性体驱动的受感染巨噬细胞分泌IL-1β和IL-18。
ExoS是另一种假单胞菌毒力因子,可干扰炎症小体诱导的IL-1β产生。该效应蛋白通过涉及其ADP-核糖基转移酶活性的不完全表征过程抑制caspase-1激活。
▸ 干扰炎性小体的信号传导
嗜肺军团杆菌(L.pneumophila)代表了革兰氏阴性病原体如何干扰炎症小体信号传导的另一个例子。Nlrc4炎性小体限制体外培养的巨噬细胞和受感染小鼠肺部的军团菌生长。该病原体干扰炎性小体接头ASC的转录上调,以保护其在人单核细胞中的增殖。
F.tularensis利用假定的脂质II翻转酶mviN来抑制AIM2炎症小体的激活。mviN突变株感染小鼠,由于AIM2炎性体介导的IL-1β分泌增强和巨噬细胞焦亡,导致体内毒力受损。
革兰氏阳性病原体(例如结核分枝杆菌)也已进化出干扰炎性体功能的机制。被称为Zmp1的推定Zn2+金属蛋白酶对Nlrc4炎性体激活和IL-1β分泌的抑制使这种人类结核病病原体在骨髓细胞中增殖。
炎症小体信号传导的调节并不局限于细菌病原体。事实上,病毒提供了一些最具特征的机制,通过这些机制影响炎症小体。
▸ 牛痘病毒编码的丝氨酸蛋白酶抑制剂抑制 Caspase-1
牛痘病毒细胞因子反应修饰物A(CrmA)及其在牛痘病毒中的同源物直接靶向影响Caspase-1的酶活性。
CrmA和其他丝氨酸蛋白酶抑制剂在增强痘病毒毒力方面的重要性通过以下观察得到证实:CrmA的缺失会减弱BALB/c和C57BL/6小鼠鼻内和颅内感染的毒力。同样,感染缺乏CrmA同源Serp2的粘液瘤病毒突变体的兔子的病毒滴度显著降低。
此外,正痘病毒、痘苗病毒和副痘病毒产生可溶性IL-18结合蛋白,可防止细胞因子诱导的IL-18受体激活。因此,正痘病毒通过丝氨酸蛋白酶抑制剂和清道夫受体分别对caspase-1活性和下游炎症体效应子的联合抑制作用来增加毒力。
注:清道夫受体是吞噬细胞表面的一组异质性分子
▸ 病毒诱饵蛋白对炎症小体组装的抑制作用
除了直接靶向caspase-1的酶活性并干扰IL-1和IL-18受体的连接外,病毒还部署了阻止炎症小体组装的分子。
卡波西肉瘤相关疱疹病毒(KSHV)Orf63的作用很好地说明了这一点,Orf63是一种病毒Nlrp1同源物。KSHV Orf63与人Nlrp1和Nlrp3相互作用,以阻止其各自炎症小体的组装以及随后的caspase-1依赖性先天免疫反应。
KSHV Orf63表达的转录下调降低了病毒复制率,这是由于Nlrp1和Nlrp3介导的IL-1β分泌增强以及KSHV感染的人单核细胞和293T细胞中焦亡诱导的结果。
吡啶结构域蛋白(POP),例如粘液瘤病毒M013L和纤维瘤病毒S013L,代表了病毒诱饵蛋白抑制炎性体的另一个例子。由于宿主炎症反应增加和病毒复制减弱,缺乏编码M013L基因的粘液瘤病毒突变体的病毒血症显著减少,从而强调了粘液瘤病期间病毒POP的重要性。
此外,人类CARD蛋白ICEBERG、COP、INCA和CASP12 S被认为通过与caspase-1前结构域中的CARD基序的同型CARD相互作用来清除caspase-1,从而干扰炎症小体组装。然而,与病毒POP不同,人类CARD-only蛋白的病毒对应物仍有待鉴定。
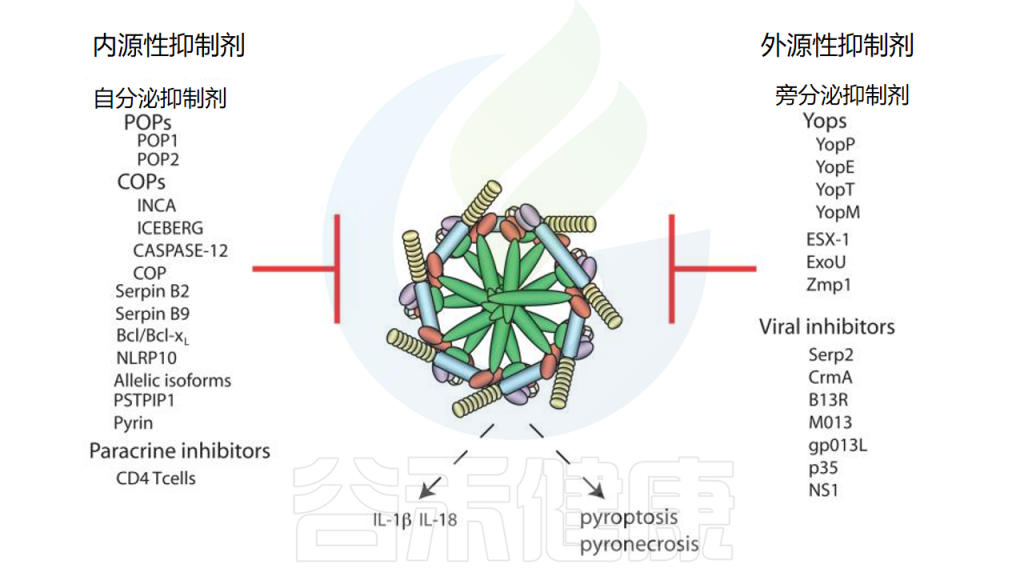
炎症小体的抑制
doi: 10.1146/annurev-immunol-031210-101405.
▸ 流感病毒抑制炎症小体信号传导
有趣的是,流感病毒使用与上述正痘病毒无关的机制来阻止caspase-1的激活并干扰炎症小体信号传导。人类流感A/PR/8/34(H1N1)的突变病毒,其中流感NS1基因被删除,触发受感染宿主细胞分泌显著增加的IL-1β和IL-18水平,并未能阻止巨噬细胞中caspase-1的成熟。
这些突变病毒在体外被减毒,但caspase-1依赖性和非依赖性机制在多大程度上促成了这种表型尚不清楚。流感NS1驱动的caspase-1激活抑制似乎仅依赖于NS1的N-末端RNA结合/二聚化结构域,而羧基末端效应结构域对于抑制IL-1β和IL-18分泌是可有可无的。对流感病毒NS1抑制胱天蛋白酶-1激活的分子机制的进一步分析可能揭示病毒靶向炎症小体的有趣的新机制。
我们的观点
总体而言,宿主-病原体相互作用本质上是动态的。病毒利用了人体的基因,并利用它们来规避免疫系统。细菌也进化出了复杂的机制。病原体对炎症小体的特异性靶向强调了其在先天免疫中的重要性。
✦
炎症小体是一种在人体免疫系统中起关键作用的多蛋白复合物,其异常活化或抑制与多种炎症性疾病的发生和发展密切相关。炎症小体在调节炎症反应、细胞焦亡和免疫调节中发挥重要作用。
但由于炎症小体成分的遗传突变以及调节缺陷而导致的不适当的炎症小体反应与多种人类疾病有关。研究表明,炎症小体的功能异常与自身免疫疾病、感染性疾病和肿瘤等的发生有关。
1
在肠道炎症和肿瘤发生中的作用
结直肠癌是一种常见的恶性肿瘤,发病率和死亡率在全球范围内居高不下。据统计,结直肠癌是全球第三常见的癌症,也是溃疡性结肠炎和克罗恩病等炎症性肠病的主要并发症。炎症性肠病和结直肠癌通常与炎症细胞因子的过度产生有关。
IL-1α/β、IL-6和TNF-α等炎症细胞因子在炎症促进的肿瘤发生中发挥重要作用。基于炎症小体在IL-1β加工中的关键作用,研究了NLRP3炎症小体在结肠炎和结肠炎相关癌症(CAC)中的作用。
▸ 炎症小体在控制肠道稳态和预防肿瘤中起作用
多个研究小组意外地发现炎症小体的成分在控制肠道稳态和预防肿瘤发生方面发挥着保护作用。
NLRP3、ASC或caspase-1缺陷的小鼠更容易患结肠炎和CAC。这种表型与局部和全身IL-1β和IL-18分泌减少有关。数据表明,NLRP3负责防止肠道炎症和肿瘤发生增加。
此外,观察到caspase-1对DSS诱导的结肠炎具有类似的保护作用。Casp1−/−小鼠在DSS治疗后表现出肠道炎症和NF-κB激活增强以及组织修复受损。
▸ IL-18可能介导了炎症小体对肠道的保护作用
IL-18是肠道稳态和炎症所必需的。研究发现,IL-18信号传导可防止DSS诱导的结肠炎和DSS+氧化偶氮甲烷诱导的CAC动物模型中的组织损伤。此外,外源性IL-18使Casp1−/−小鼠免受结肠炎诱发的体重减轻影响。因此,IL-18似乎负责NLRP3炎性体介导的针对肠道炎症、组织损伤和肿瘤发生的保护作用。
▸ NLRP3炎症小体在化疗抗肿瘤反应中起作用
还研究了NLRP3炎症小体在肿瘤发生中的作用。研究指出,NLRP3炎症小体对化疗诱导的抗肿瘤反应是必不可少的。从机制上看,化疗诱导的垂死肿瘤细胞释放的ATP激活了NLRP3炎症小体,进而通过IL-1β的分泌进一步激活产生IFN-γ的CTL。
考虑到用于激活NLRP3炎症小体的外源性ATP浓度远高于化疗诱导的垂死肿瘤细胞释放的ATP浓度(mM与μM),其他内源性NLRP3激活剂,如尿酸,可能在化疗期间释放,从而激活体内的NLRP3炎性小体。
2
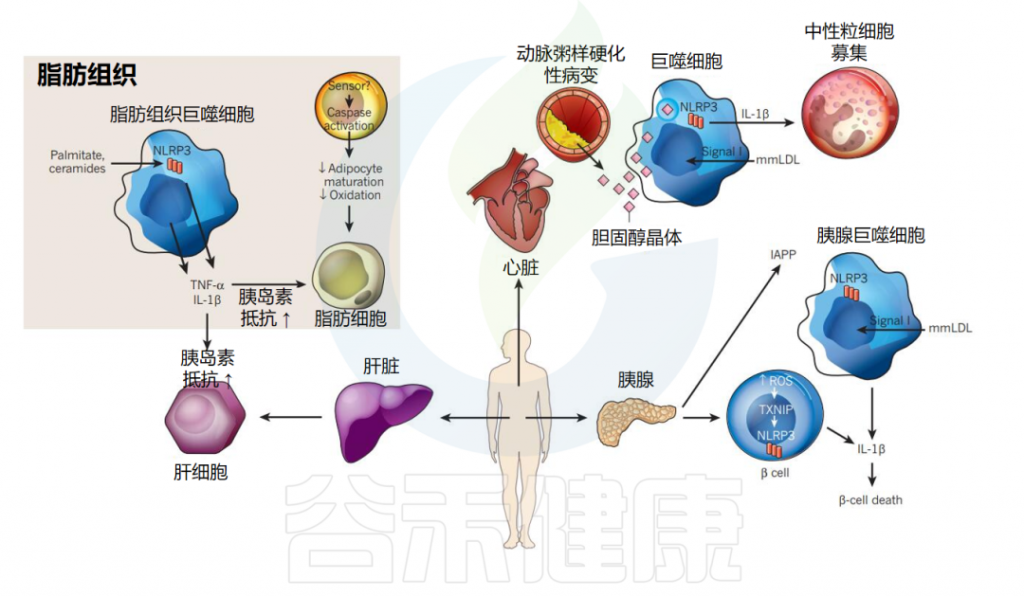
NLRP3炎症小体和代谢紊乱
近几十年来,肥胖、2型糖尿病、动脉粥样硬化等代谢性疾病的发病率急剧上升,严重威胁人类健康。
▸ 肥胖患者的促炎细胞因子上调
在过去的十年中,人们已经清楚地认识到慢性炎症是代谢紊乱的一个关键预测因素。例如,肥胖与细胞因子产生的上调和炎症信号通路的激活有关。
肥胖状态下,脂肪组织中的炎症小体活化会导致促炎细胞因子的过度分泌,如IL-1β和IL-18等。这些促炎细胞因子的释放会引发炎症反应,进而影响胰岛素信号传导、葡萄糖代谢和脂质代谢,加剧肥胖相关的代谢紊乱。
▸ 炎症小体分泌IL-1β破坏胰岛素调节
NLRP3炎症小体在2型糖尿病(T2D)中扮演着重要角色。它作为代谢应激传感器,在治疗T2D的临床试验中加强了IL-1β受体拮抗作用。
IL-1β升高是发生T2D的危险因素,并通过拮抗胰岛素信号传导导致胰岛素抵抗。IL-1β还介导胰岛中长期高血糖(糖毒性)的毒性作用,导致β细胞破坏并调节葡萄糖诱导的胰岛素分泌。
最近的一项研究描述了慢性高血糖期间小鼠胰岛中IL-1β的分泌情况:高细胞外葡萄糖通过NLRP3炎症小体触发IL-1β分泌。
此外,NLRP3结合蛋白TXNIP作为胰腺β细胞死亡和外周葡萄糖摄取失败的介质,与T2D密切相关。
炎性小体在代谢综合征中的作用
doi: 10.1038/nature10759.
▸ NLRP3炎症小体在动脉粥样硬化中起重要作用
最近的研究还表明NLRP3炎症小体在动脉粥样硬化中发挥着核心作用。动脉粥样硬化是一种慢性炎症性疾病,其特征是动脉粥样硬化病变中炎性成分的积累和免疫细胞的募集。
观察到在早期饮食引起的动脉粥样硬化病变中存在微小胆固醇晶体的沉积,这与巨噬细胞的募集有关。体外生成的胆固醇晶体在脂多糖引发的人外周血单核细胞和小鼠巨噬细胞中诱导NLRP3/ASC炎性体激活和caspase-1/IL-1β/IL-18裂解。
此外,使用骨髓嵌合体的体内实验表明,骨髓细胞来源的NLRP3、ASC和IL-1β/β在动脉粥样硬化病变的发展中发挥着关键作用。由于活性氧(ROS)还促进动脉粥样硬化的发展,因此测试ROS在胆固醇晶体诱导的NLRP3炎性体激活中的参与将很有意义。
3
炎症小体和适应性免疫
NLRP3炎性小体除了在先天免疫反应中的促炎作用外,最近的研究强烈表明NLRP3炎性体介导的细胞因子(IL-1β和IL-18)在形成适应性免疫反应中发挥着重要作用。
▸ IL-1β调节早期TH17细胞分化
据报道,IL-1β信号传导可调节早期Th(辅助性T)17细胞分化,并在实验性变态反应性脑脊髓炎(EAE)诱导中发挥重要作用。
从机制上讲,IL-1β信号传导诱导IRF4和RORγt的表达,这是参与Th17分化的两个重要转录因子。IL-1β 还被证明可以与IL-23协同作用,诱导产生IL-17的 γδ T 细胞的发育,从而促进EAE的发展。因此,人类Th17细胞的分化需要IL-1β的存在。
▸ IL-18协同其他细胞因子影响T细胞的反应
与IL-1β相反,IL-18信号在Th细胞分化中的作用取决于其他协同细胞因子。例如,IL-18与IL-12的协同作用诱导产生IFN-γ的Th1细胞,而IL-18与IL-2的组合增强了IL-13(一种Th2细胞因子)的产生。
通过与IL-23协同作用,IL-18扩增极化Th17细胞产生的IL-17。因此,与IL-1β相比,IL-18在形成适应性T细胞反应方面表现出更灵活的功能,这可以解释IL-1β和IL-18在某些疾病模型(如肠道炎症和2型糖尿病)中的不同功能。
基于IL-1β和IL-18在T细胞分化和自身免疫性疾病中的作用,几个研究了炎症小体在T细胞介导的疾病中的作用。数据表明NLRP3在加剧EAE发展中发挥着重要作用。这是由于抗原呈递巨噬细胞和DC需要NLRP3才能最佳地激活初始T细胞形成Th1和Th17效应细胞。
总之,NLRP3炎症小体介导IL-1β和IL-18的产生,IL-1β和IL-18与其他炎症细胞因子配合调节T效应细胞的产生并影响疾病进展。这些研究将炎症小体的作用扩展到适应性免疫的调节。
4
炎症小体和痛风
痛风是一种自身炎症性疾病,其特征是严重的关节炎症,导致关节病和相当大的疼痛。痛风与代谢紊乱密切相关,导致血尿酸水平升高(高尿酸血症)和 尿酸盐(MSU)晶体在关节中沉积。
▸ 尿酸盐是NLRP3炎症小体的有效激活剂
最近的研究阐明了尿酸盐(MSU)依赖性关节炎症的潜在机制。MSU在体外是NLRP3炎症小体的有效激活剂,并且MSU依赖性中性粒细胞募集在体内依赖于ASC衔接子、caspase-1和IL-1R。
IL-1β拮抗剂在临床试验中的成功支持了炎症小体调节的IL-1β在人类痛风和密切相关的假痛风中的致病作用。
5
炎症小体和肝损伤
尽管NLRP3炎性体在DSS诱导的结肠炎期间的组织损伤中发挥保护作用,但对乙酰氨基酚(APAP)诱导的肝损伤动物模型的研究表明,NLRP3炎性体会放大免疫反应并加剧肝损伤。
▸ NLRP3可能是造成肝损伤的关键介质
APAP治疗通过有毒代谢中间产物诱导肝毒性,导致肝细胞死亡。最近的一项研究发现TLR9和NLRP3炎症小体是APAP诱导的肝损伤和炎症的关键介质。TLR9检测APAP诱导的肝细胞死亡后释放的内源DNA,并上调pro-IL-1β和pro-IL-18的产生,这些物质进一步被NLRP3炎性小体裂解。
因此,TLR9和NLRP3炎症小体在APAP诱导的肝损伤和炎症过程中发挥着作用。然而,NLRP3炎症小体的刺激尚未得到充分研究。基于APAP治疗诱导的急性和强烈的细胞死亡,从死亡细胞释放的尿酸、ATP、线粒体或透明质酸可能会激活TLR9或激活巨噬细胞中的NLRP3炎症小体。
此外,AIM2最近被鉴定为介导caspase-1激活和IL-1β/IL-18加工的胞质DNA传感器。需要进一步的研究来测试AIM2在肝损伤和其他涉及广泛细胞死亡的疾病模型(如脓毒症)中的作用。
▸ 拓展:其他炎症小体可能影响的疾病
NLRP3以外的NLRP突变与人类疾病相关。NLRP12突变与一种名为FACS2的类似FCAS的发热综合征有关。
研究人员认为,这些患者的NLRP12突变可能破坏了该蛋白的NF-κB抑制活性。然而,考虑到NLRP12和NLRP3之间的高度同源性以及FCAS和FCAS2患者症状的相似性,这些患者的炎症小体活性可能存在失调。
NLRP1的突变与白癜风等多种自身免疫性疾病有关。最近,NLRP2突变被发现与一例家族性Beckwith-Wiedmann综合征有关,这是一种导致胎儿过度生长和印记障碍的疾病。
此外,NLRP7突变与家族性和复发性葡萄胎有关,这是一种异常妊娠状态,胎盘绒毛退化,受精卵无法存活。这些基因突变参与的疾病机制以及炎症小体途径的潜在参与仍有待进一步阐明。
检测炎症小体激活的方法
激活炎症小体具有多个重要特征,包括ASC斑点的形成、促炎性细胞死亡、具有生物活性IL-1β/IL-18细胞因子的分泌以及HMGB1的表达。一般可以通过以下几种方法检测:
1.使用RT-qPCR检测NF-κB诱导的pro-IL-1β和NLRP3是否上调;
2.使用荧光显微镜或流式细胞术监测细胞系中ASC斑点的形成;
3.使用Western blot检测caspase-1的裂解或pro-IL-1β/IL-18的成熟;
4.使用ELISA测定IL-1β、IL-18或HMGB1的释放;
5.使用乳酸脱氢酶(LDH)测定或碘化丙啶(PI)染色法检测细胞焦亡;
6.使用检测IL-1β、IL-18分泌的报告基因功能细胞系。
以上方法各有利弊,可以适当的结合这些方法来检测炎症小体的激活。
✦
越来越清楚的是,炎症小体激活caspase-1以多种方式有助于保护宿主免受入侵微生物的反应。例如通过分泌IL-1β和IL-18诱导炎症,介导HMGB1等“警报素”的释放,并触发受感染宿主细胞的焦亡以消除微生物病原体。
炎症小体与肠道微生物群之间的相互作用在维持肠道稳态和调节免疫反应中也发挥着至关重要的作用。然而炎症小体的功能具有两面性(有害与有益)。这种相互作用的失调可能导致各种胃肠道疾病的发生。因此,必须对其进行严格监管,以限制异常激活和对宿主细胞的损害。
NLRP6炎性小体缺乏被证明与促进自身炎症的微生物群的扩张有关,如普氏菌科(Prevotellaceae)。炎症小体可以感知微生物群成员或群落,调节组织修复和再生,以及在稳态和炎症状态下协调粘膜免疫反应。
在之前的研究中发现,炎症小体缺乏和肠道菌群的改变都与人类代谢综合征(如肥胖和动脉粥样硬化)的发展倾向有关。炎症小体对肠道菌群的调节是否会影响体重、代谢和炎症,预计将成为该领域的主要研究方向。
主要参考文献
Manshouri S, Seif F, Kamali M, Bahar MA, Mashayekh A, Molatefi R. The interaction of inflammasomes and gut microbiota: novel therapeutic insights. Cell Commun Signal. 2024 Apr 2;22(1):209.
Liang Z, Damianou A, Di Daniel E, Kessler BM. Inflammasome activation controlled by the interplay between post-translational modifications: emerging drug target opportunities. Cell Commun Signal. 2021;19:1–12.
Sim J, Park J, Moon J-S, Lim J. Dysregulation of inflammasome activation in glioma. Cell Commun Signal. 2023;21(1):239.
Pellegrini C, Antonioli L, Lopez-Castejon G, Blandizzi C, Fornai M. Canonical and non-canonical activation of NLRP3 inflammasome at the crossroad between immune tolerance and intestinal inflammation. Front Immunol. 2017;8:36.
Davis BK, Wen H, Ting JP. The inflammasome NLRs in immunity, inflammation, and associated diseases. Annu Rev Immunol. 2011;29:707-35.
Carriere J, Dorfleutner A, Stehlik C. NLRP7: From inflammasome regulation to human disease. Immunology. 2021 Aug;163(4):363-376.
Lamkanfi M, Dixit VM. Modulation of inflammasome pathways by bacterial and viral pathogens. J Immunol. 2011 Jul 15;187(2):597-602.
Strowig T, Henao-Mejia J, Elinav E, Flavell R. Inflammasomes in health and disease. Nature. 2012 Jan 18;481(7381):278-86.
Próchnicki T, Latz E. Inflammasomes on the Crossroads of Innate Immune Recognition and Metabolic Control. Cell Metab. 2017 Jul 5;26(1):71-93. doi: 10.1016/j.cmet.2017.06.018. PMID: 28683296.

谷禾健康
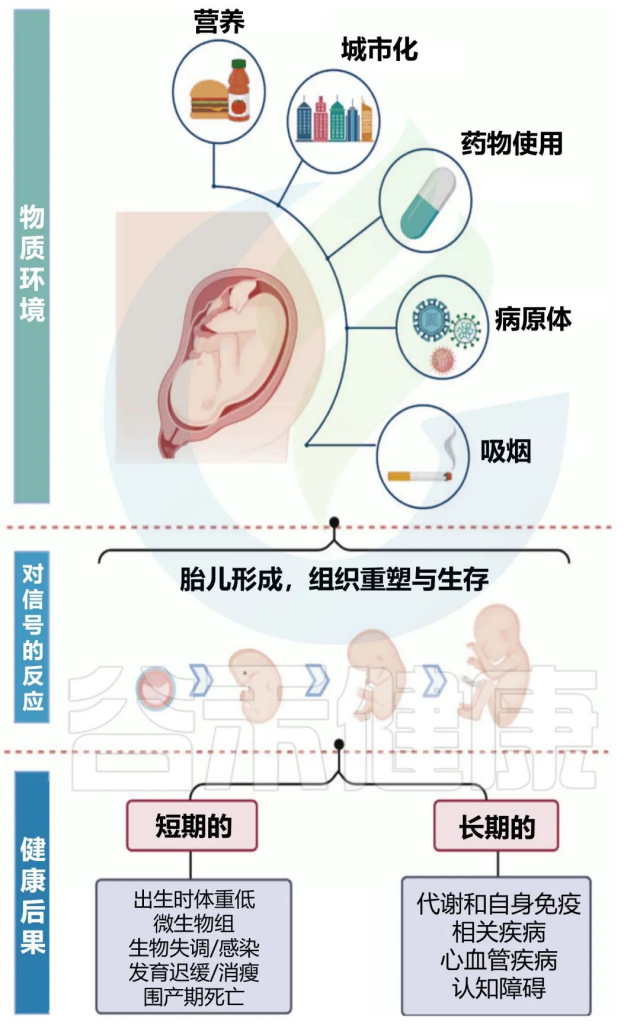
怀孕的母亲与体内的胎儿是息息相关的。由于婴儿接触母体微生物群,母亲和孩子之间的微生物联系在怀孕期间形成。而宿主与微生物群的联系在出生后成熟,并进化成为个体生命中最重要的共生关系之一,对响应营养和环境刺激的稳态调节至关重要。


最近,有报道称微生物群在子宫内发育,并在整个妊娠期间改变。这一发现表明,胎儿很可能通过胎盘早期暴露于母体微生物群。
✦多种因素共同作用婴儿的微生物群
婴儿的基因组成、分娩方式、抗生素使用、喂养类型、疾病状况和其他环境暴露可能影响新生儿微生物群的建立和成熟。
在子宫内和早期生活中的营养和环境暴露是形成健康肠道微生物群的关键因素。强大的微生物群有助于粘膜免疫系统的发育和成熟,有助于维生素合成和营养吸收,并在中枢神经系统的生化信号中发挥作用。
健康与疾病的发展起源

Mua B,et al.Comprehensive Gut Microbiota.2022
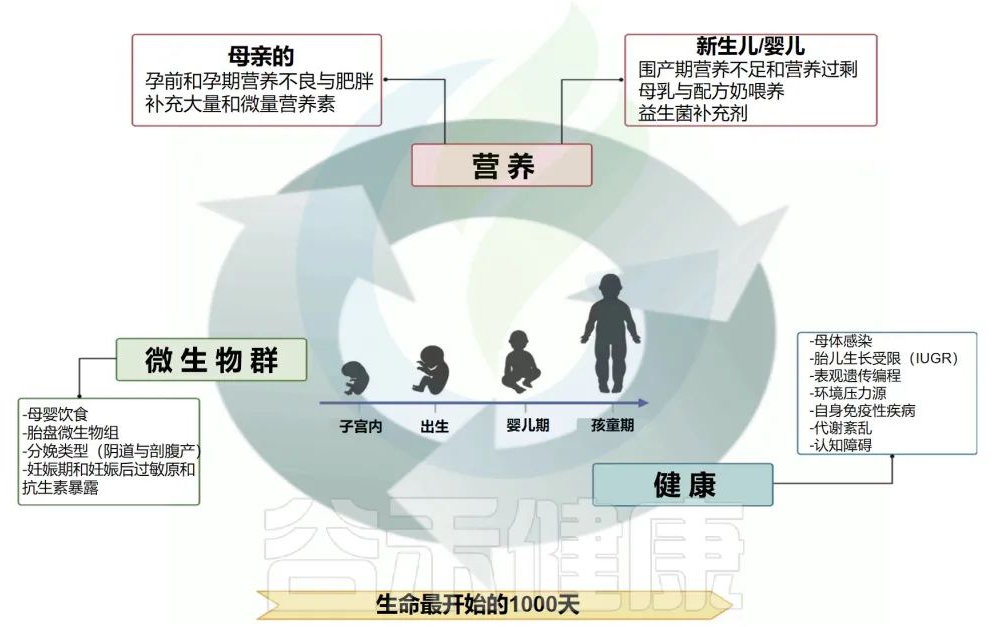
生命的前1000天,是发育可塑性窗口。这是一个快速成长和表观遗传调控的关键时期。
✦微生物群改变与许多疾病有关
这一发展时期允许通过多种因素调节妊娠结局和微生物组,包括母亲营养、药物滥用、吸烟、城市化、抗生素暴露以及婴儿的喂养类型。胎儿期对此类外部压力源的敏感性也会持续影响到成年的微生物组的组成和功能。
在这个早期时间点,微生物群多样性的改变还与许多疾病有关,包括但不限于慢性代谢紊乱、自身免疫性疾病和神经发育障碍。
本文总结了孕期和产后母婴微生物组的生理及病理变化。此外,我们将讲述常见的母体和营养因素对发育的影响及其在成年期疾病发病中的作用。
本文主要从以下几个方面讲述
●妊娠期间微生物组的变化
●影响婴儿微生物群的因素
●母体微生物对后代免疫的影响
●微生物对儿童疾病风险的影响
几十年来,人们对妊娠相关的激素和代谢变化进行了广泛研究,而对妊娠期间发生的微生物组成变化的研究则较少。
✦母体微生物群发生显著变化
母体微生物群在怀孕过程中发生了显著变化,其特征是微生物群落内丰度和多样性或α多样性降低,微生物群落间丰度或β多样性增加。
这些变化受育龄妇女营养、胎龄、遗传、种族和环境因素的差异影响。此外,孕期母体微生物群组成与母体饮食、孕前体重和孕期体重增加相关,并表明肠道、阴道、口腔和胎盘微生物群可能会根据母体生活方式和孕期环境暴露而发生变化。
孕期母体微生物组成变化
Mua B,et al.Comprehensive Gut Microbiota.2022
健康怀孕的特点是不同部位微生物组丰度和多样性的增加和减少。口腔、胎盘、肠道和阴道微生物组的改变与妊娠炎症和整体健康有关。
细菌的种类(放线菌属、变形菌属、厚壁菌门和拟杆菌门)分别按颜色(绿色、红色、蓝色和黄色)区分。微生物群落多样性在文本中用黑色箭头表示,它们各自的增加和减少。
在整个孕期,肠道微生物组对于营养素获取、免疫重塑和抵抗病原体至关重要。对人类微生物组的研究发现,肠道微生物组的组成在怀孕过程中发生变化,并与生理和代谢变化相关。
✦妊娠期间肠道微生物改变
妊娠前期体重指数较高或患有妊娠期糖尿病的女性在妊娠晚期发现β多样性微生物群落水平升高,且在产后持续存在。还发现,在适当的营养干预后,妊娠期糖尿病患者表现出α多样性增加。
这些发现表明,怀孕是一个动态过程,根据女性怀孕前和怀孕期间的身体状态(即体重和激素水平)、心理压力和环境因素,改变了微生物组特征;所有这些都会在怀孕期间影响肠道微生物组的组成。
•怀孕前三个月与正常妇女相似
例如,在怀孕的前三个月,母体肠道微生物组的组成与健康的非孕妇相似,其中厚壁菌(Firmicutes)比拟杆菌(Bacteroides)占优势。
•三个月后产丁酸的细菌减少
在头三个月后,母体肠道微生物群中产生丁酸的细菌(如普拉梭菌)减少,双歧杆菌(Bifidobacteria)、变形杆菌(Proteobacteria)和乳酸产生菌增加。
反过来,肠道微生物组的改变可能会对妊娠疾病的发生率、胎儿状况、妊娠结果和胎儿免疫发育产生重大影响。
✦激素变化影响细菌生长
•双歧杆菌增加
妊娠相关的激素变化也与宿主激素引起的微生物组组成改变有关。向雌性小鼠补充了孕激素,并确定双歧杆菌(Bifidobacteria)丰度增加,孕激素促进了孕晚期双歧杆菌的生长。
此外,母体胃肠粘膜免疫反应的变化,以及妊娠期间代谢激素的变化,可能引发低级促炎状态,从而诱导肠道通透性,促进葡萄糖从肠道上皮向内腔扩散。这可能会导致体重增加,进而改变肠道微生物组组成。
总的来说,最近的研究结果表明,整个孕期肠道微生物组组成发生了显著变化,与初始体重和饮食、体重增加、炎症水平和代谢参数有关。
✦怀孕对免疫性疾病存在有益影响
有趣的是,怀孕对某些自身免疫性疾病,包括炎症性肠病的病程有有益的影响。
炎症性肠病包括一组广泛的疾病,这些疾病具有潜在的免疫介导的失调和各种肠道炎症状态。
炎症性肠病的两种主要形式为溃疡性结肠炎(UC)和克罗恩病(CD)。
肠道微生物组的组成对炎症性肠病的临床过程很重要,在大多数炎症性肠病病例中,由于肠道失调,细菌多样性降低。
注:炎症性肠病与肠道菌群的关联在谷禾前不久的文章中有具体描述。
详见:肠道菌群失调与炎症性肠病的关联
•怀孕期间炎症性肠病改善
新出现的证据表明,在怀孕期间和怀孕后,炎症性肠病的总体病程有所改善。雌激素和孕酮通过积极调节肠道上皮,对疾病活动有有益影响。
这些激素缓解了压力,减少了促炎细胞因子的产生,刺激了伤口愈合,并增加了上皮屏障功能。
注意:妊娠对炎症性肠病的积极影响可能取决于患者的特征,例如潜在的遗传风险因素、妊娠前或妊娠期间持续的疾病严重程度、环境因素(即饮食、药物、酒精)以及随后对微生物组组成的影响。
阴道微生物组由许多细菌物种控制,包括乳杆菌属(Lactobacillus)以及梭状芽孢杆菌目(Clostridium)、拟杆菌(Bacteroidales)和放线菌目(Actinomycetales)的成员。
✦阴道微生物组在母婴健康中有重要作用
•抑制细菌和病毒
例如,乳杆菌属(Lactobacillaceae)的不同成员可以增强阴道健康的各种特征。这些产乳酸细菌在维持低pH值(<4.5)方面发挥了作用,从而形成了抵抗病原体入侵的屏障。它们还分泌可以抑制泌尿生殖道细菌和病毒感染的代谢物。
•产生细菌素,杀死病原体
此外,除了降低阴道pH值和增加阴道分泌物以防止致病菌进入子宫并引起感染外,乳酸杆菌还会产生细菌素,杀死入侵的病原体。
在母体阴道道中发现的某些细菌种类也可能具有抗菌特性,有助于预防细菌性阴道病和淋病奈瑟菌感染。
✦怀孕后阴道微生物多样性降低
在阴道分娩期间,新生婴儿接触阴道微生物群,这对婴儿微生物群的定植至关重要。重要的是,与未怀孕妇女相比,随着妊娠进展,母体阴道微生物组发生变化,总体细菌多样性降低,不过乳酸杆菌(Lactobacillales)、梭状芽孢杆菌(Clostridiales)、拟杆菌(Bacteroidales)和放线菌目(Actinomycetales)的优势增加。
最近证实,随着乳酸杆菌的增加,母体阴道微生物组的多样性降低,支原体和脲原体的含量降低,这两者都与早产和出生体重低等负面结果相关。
•微生物变化可能是为了保护胎儿
在孕妇体内观察到的阴道微生物组变化可能是保护胎儿健康和在出生时提供特定微生物种类的适应性反应的一部分,因为健康妊娠相关微生物组的变化可能会使母亲容易产生负面健康结果。
母体阴道微生物组组成取决于胎龄和种族。有趣的是,怀孕后期的母体微生物群落与未怀孕状态的相似。
注:也有研究整个怀孕期间没有发现阴道微生物组有任何显著变化。然而,他们发现产后阴道微生物群落与分娩后长达一年的肠道微生物群落更为相似。在他们的研究中,分娩后检测到从乳酸杆菌到各种厌氧细菌的转变,包括Peptoniphilus、Prevotella和Anaerococcus。
✦细菌性阴道病
通常,当阴道群落内的乳酸杆菌浓度改变时,会发生生物失调。阴道失调的特点是乳酸杆菌的丰度低,厌氧微生物增多。
细菌性阴道病(BV)是一种由阴道中自然发现的细菌过度生长引起的阴道炎症,是妊娠期最常见的一种生殖道感染。细菌性阴道病是一种由阴道环境变化引起的病症,它允许机会致病菌在阴道区域定居。怀孕期间的细菌性阴道病对母亲和孩子都是健康风险。
机会致病菌——正常菌群与宿主之间、正常菌群之间,通过营养竞争、代谢产物的相互制约等因素,维持着良好的生存平衡。在一定条件下这种平衡关系被打破,原来不致病的正常菌群中的细菌可成为致病菌,称这类细菌为机会性致病菌,也称条件致病菌。
•细菌性阴道病有更高的早产风险
一些初步研究表明,患有细菌性阴道病的孕妇存在更高的早产风险,这突出了怀孕期间阴道微生物组的重要性。
✦影响细菌性阴道病的因素
多年来,营养被认为是细菌性阴道病发展的一个因素。报告称,妊娠期缺铁和维生素D与细菌性阴道病风险增加有关。
扩展阅读:
人与菌对铁的竞争吸收 | 塑造并控制肠道潜在病原菌的生长
维生素D与肠道菌群的互作
•富含纤维的饮食发病率降低
在一项平行研究中,维生素A、维生素C、维生素E和维生素b-胡萝卜素的血清浓度较低与细菌性阴道病相关。最近的一项发现表明,富含纤维的饮食与细菌性阴道病发病率降低相关。
•超重和肥胖发病率更高
此外,一项流行率研究得出结论,超重和肥胖女性的细菌性阴道病发病率更高。这些发现强调了饮食对阴道微生物组组成的影响,进而强调了微生物组对怀孕期间阴道健康的重要性。
▸ 口腔含有仅次于肠道的第二大的微生物组,里面居住着700多种不同的细菌,包括链球菌(Streptococcus)、乳酸杆菌、葡萄球菌(Staphylococcus)和棒状杆菌。
微生物定植于牙齿的硬表面和口腔粘膜的软组织,通常以生物膜的形式存在。
注:微生物组研究的一个局限性是在没有交叉污染(即肠道或肺部)的情况下对微生物组进行采样。然而,由于口腔微生物组采样的简便性,它已成为迄今为止研究最深入的微生物组。
口腔微生物组在促进健康或疾病进展中起着关键作用。它对于维持口腔稳态、保护口腔和预防疾病发展至关重要。
✦怀孕期间口腔微生物发生变化
•总活菌数显著增加
在怀孕的不同阶段,口腔微生物组的组成会发生变化。在对非怀孕妇女口腔中存在的七种常见细菌种类的丰度进行比较时,妊娠早期、妊娠中期和妊娠晚期发现,与非怀孕妇女相比,怀孕期间的总活菌数显著增加。与其他妊娠阶段相比,妊娠早期存活微生物计数最高。
•妊娠中期致病菌水平升高
与未怀孕妇女相比,妊娠早期和中期组牙龈卟啉单胞菌(Porphyromonas)和放线聚集杆菌(均为致病菌)的水平升高。
牙龈卟啉单胞菌是一种非酵解糖的革兰氏阴性厌氧球杆菌,是研究广泛且证据充足的重要牙周致病菌之一。
最后,念珠菌(Candida)水平在妊娠中期和末期显著升高。有趣的是,雌激素与念珠菌感染有关,并可能解释怀孕期间念珠菌的丰富。
尽管口腔微生物组已被广泛研究,但口腔微生物组与饮食和妊娠结合仍是未来研究的领域。
▸ 胎盘是孕期母亲和胎儿之间的主要营养和激素调节界面。它在感知和引导母体营养信号到胎儿方面发挥着重要作用。
✦胎盘连接母体和胎儿的物质交换
胎盘促进氧气和生长因子向胎儿的交换,并将废物从胎儿转移到母体循环系统。
虽然长期以来认为胎儿和胎盘是无菌环境,不过这是一个有争议的话题。
最近有证据表明,在分娩前通过胎盘进行微生物交换。健康孕妇的脐带血、羊水和胎盘中检测到微生物颗粒。然而,这种细菌DNA是否是细菌从母亲血液中定植或易位的证据尚不清楚。
•细菌可能由母体经胎盘进入胎儿
一些理论表明,细菌可能由免疫细胞通过血流和淋巴进入胎盘,并通过胎盘进入胎儿一侧,以帮助启动胎儿免疫系统。事实上,在健康新生儿的胎盘组织中检测到几种细菌种类,包括放线菌属(双歧杆菌、丙酸杆菌、链霉菌、红球菌)、厚壁菌属(乳杆菌)、拟杆菌属(拟杆菌)和变形杆菌属(大肠杆菌、肠杆菌)。
此外,在通过剖宫产分娩的健康母亲的所有29例胎盘活检中发现了乳酸杆菌和双歧杆菌(Bifidobacterium)。
✦胎盘中的微生物组暂不明确
尽管如此,最近一项更有力的研究表明,没有证据表明人类胎盘内存在微生物组,尽管它可能含有潜在的病原体。他们证明,胎盘样本中的大多数阳性细菌信号是出生或实验室污染的结果。
然而,在同一项研究中,该小组在临产前采集的约5%的样本中发现了无乳链球菌(S.agalactiae)的证据。由于无乳链球菌是新生儿败血症的主要原因,胎盘可能是败血症新生儿感染的起始部位。
注:新生儿的败血症和早产也归因于母体口腔感染,如牙周病。此外,胎盘中发现的细菌门与母体口腔中发现的菌门最为相似。尽管似乎有一些证据表明母亲和胎儿之间存在胎盘微生物组交换,但对分娩期间或分娩后以及实验室中污染的担忧仍然是科学界争论的话题。
母亲的健康状况和微生物群状况对儿童微生物群的定植和发展有着深远的影响。同样,在出生时和婴儿早期建立健康的肠道微生物群对儿童免疫系统的发育和成熟至关重要。
由于共生微生物群在肠道免疫中起着关键作用,任何改变其组成的环境或宿主因素都可能导致负面健康结果。我们在此罗列了一些影响婴儿微生物群定植和发育的主要因素:母体营养、分娩方式、妊娠期和围产期抗生素的使用、喂养类型和胎盘微生物群。
什么是营养不良?
世界卫生组织将营养不良定义为“人体能量或营养摄入的不平衡(不足和过量)”。营养不足和营养过剩都被认为是营养不良的表现形式;然而,过度摄入某些特定营养素通常被称为肥胖。
营养不良包括缺乏基本的宏观和微观营养素,发育迟缓、消瘦和体重不足。营养过剩包括超重和肥胖,其特征是营养消耗过量和体质指数超标。这两种形式的营养不良都可能导致非传染性饮食相关疾病的发展,如心脏病、中风、糖尿病甚至是癌症。
✦营养不良的影响
•增加了疾病和死亡率
营养不良是一个全球性的问题,它影响着每个国家的人民,最脆弱的人群是中低收入国家的五岁以下儿童和育龄妇女。营养不良增加了母亲和儿童疾病和死亡的可能性,是五岁以下儿童死亡的主要原因,占所有死亡人数的近一半(45%)。
•导致肠道微生物失调
营养不足和营养过剩都会导致屏障功能和肠道完整性失调,从而导致生物失调。如前所述,在怀孕期间,健康且平衡的微生物组保护母亲和胎儿免受机会病原体的侵害,产生妊娠发育所需的营养物质,并将营养物质吸收到血液中,以促进胎儿健康成长。
当微生物组因营养不良而受损时,这些有益功能就会丧失,导致母体肠道炎症水平升高。孕期营养不良还与围产期并发症、流产和早产风险增加、出生体重低以及婴儿后期心血管疾病、中风、糖尿病和癌症等非传染性饮食相关疾病的发生有关。
✦母体营养对微生物群影响的一些研究
大多数关于母体营养在促进婴儿健康微生物群中的作用研究都是在动物模型中进行的。
怀孕期间高脂饮食与微生物组成的差异相关
与正常饮食对照组相比,热量限制的怀孕小鼠的细菌组成和肠道微生物组的多样性没有显著差异。
然而,与正常饮食的雌性小鼠相比,怀孕前和怀孕期间喂养高脂肪饮食的雌性老鼠在怀孕后期肠道微生物群发生了变化。
营养与婴儿的生长结果相关
检查了营养不良婴儿的母乳,发现与健康婴儿母亲的母乳相比,唾液酸化的母乳低聚糖(HMO)的含量显著降低。
在小鼠和小猪后代的饮食中添加唾液酸化的母乳低聚糖可以改善婴儿的生长结果。不过目前只有少数研究关注了孕期母体营养在人类中的作用。
素食导致罗氏菌和毛螺菌丰度增加
另一项研究指出,素食导致微生物中罗氏菌属(Roseburia)和毛螺菌科(Lachnospiraceae)的丰度增加,而与杂食性饮食相比,未检测到a多样性的差异。高脂肪、低纤维摄入孕妇的细菌密度较低。
此外,在怀孕早期,不饱和脂肪酸的高摄入量也与微生物密度降低相关。一项队列研究表明,早产母亲在怀孕期间维生素D和视黄醇摄入量较高,与微生物a多样性降低和促炎性及变形杆菌丰度增加相关。
视黄醇又称为维生素A,对于人体的多项生命活动都具有非常重要的作用。
反过来,维生素E的摄入与促炎性的减少相关。在超重孕妇中,膳食纤维和多不饱和脂肪酸导致微生物群丰富度更高,血清中zonulin蛋白水平降低。
zonulin是一种不利调节肠道紧密连接通透性的蛋白质。
✦营养不良导致微生物缺乏
一项纵向比较研究报告称,与健康对照组相比,严重急性营养不良儿童(由食物和水不安全以及母乳喂养不足引起)存在肠道微生物组缺乏和发育不成熟。
含有严重急性营养不良微生物群的小鼠也被发现具有较高水平的无害梭状芽孢杆菌(Clostridium)和沃氏嗜胆菌属(Bilophila wadsworthia),这两种细菌分别与免疫受损患者和炎症性肠综合征有关。
产妇产后时期微生物群的变化显著。母亲的微生物群在出生一个月后仍未恢复到基线水平。产后的特点是激素发生剧烈变化,包括孕酮和雌激素水平大幅下降。孕期不饱和脂肪酸消耗量的增加与分娩后变形杆菌(Proteobacteria)和厚壁菌(Firmicutes)的丰度相对于其他门的减少有关。
注:由于只有少量关于产后期的研究,因此需要进一步调查,以更好地了解这一阶段的微生物群。
✦母体营养不良会影响营养输送
•营养不良易导致宫内生长受限
母亲营养不良也会影响胎盘的营养输送和大小,并与炎症途径增加导致的宫内生长受限(IUGR)相关。
低出生体重与宫内生长受限相关,这涉及胎盘结构、形态受损和胎盘血管系统异常发育,从而损害胎盘营养物质的运输。在这些条件下,发育中的胎儿表现出较低的代谢率和较慢的生长率,以弥补营养不足。因此,胎儿的生长潜力低于其胎龄的正常水平。
•非传染性疾病风险增加
妊娠期间的母体营养限制也可能以男性特有的方式影响妊娠后半期胎儿脑内大麻素系统的发育,这在母体营养限制的狒狒模型中提供了证据。宫内生长受限可能导致非传染性疾病的风险增加,如代谢综合征、胰岛素抵抗易感性、2型糖尿病和成年期心血管疾病。
注意:除了营养不足,胎盘功能不全还可能由母亲年龄、吸烟和吸毒、高海拔妊娠、胎盘血流异常(子痫前期)、感染、炎症和胎儿先天性疾病引起。
// 建议
为了使母亲和婴儿拥有更好的健康,需要关注孕期和产后的营养摄入,尽量保证饮食均衡且有营养,不缺乏也不过量。
✦分娩类型对婴儿早期微生物群有影响
分娩类型对婴儿早期微生物群的定植有影响。由于暴露于阴道、粪便和皮肤微生物群,大多数早期微生物定植发生在自然分娩期间。
•阴道分娩
阴道分娩期间,兼性厌氧菌(如大肠杆菌、葡萄球菌和链球菌)定植于新生儿肠道,并允许严格的厌氧菌(例如拟杆菌和双歧杆菌)增殖。
•剖腹产分娩
相比之下,剖腹产出生的婴儿改变了微生物组,这可能会影响以后的健康。剖腹产婴儿的双歧杆菌和拟杆菌定植延迟,艰难梭菌水平较高。
基于丙酸杆菌、棒状杆菌和链球菌(Streptococcus)在剖腹产婴儿中增加,新生儿肠道微生物组存在显著差异,这与皮肤微生物组相似,而在阴道产婴儿中观察到乳酸杆菌和普雷沃氏菌(Prevotella)水平增加,这与母体阴道微生物组相似。
✦出生地也会影响婴儿早期微生物
有趣的是,婴儿出生的地点也会影响婴儿早期微生物群的定植。发现,与家庭分娩相比,在医院阴道分娩的婴儿中梭状芽孢杆菌(Clostridium)和肠杆菌(Enterobacteriaceae)的水平更高。
通过对771名足月医院出生的婴儿和母亲进行纵向抽样和宏基因组分析,在剖腹产婴儿和阴道分娩时使用母体抗生素预防或未在新生儿期母乳喂养的婴儿中,拟杆菌的母体传播受到干扰,以及肠球菌、肠杆菌和克雷伯氏菌等机会性病原体的高度定植,观察到的影响持续到出生后一年。
// 建议
阴道分娩相较于剖腹产分娩微生物群更健康。作为一种在新生儿中引入阴道微生物群的方法,阴道接种是一种很有前途的方法,可以恢复剖腹产婴儿的微生物组成。
该方法需要对新生儿进行擦拭,新生儿通常缺乏双歧杆菌和拟杆菌,这些细菌在出生时存在于母亲的阴道微生物组中。使用这种方法,已经被证明剖腹产婴儿的微生物组组成部分得到恢复。
改变早期微生物组的另一个主要因素是妊娠期和围产期使用抗生素。怀孕期间的抗生素治疗与新生儿第一次粪便中细菌多样性的减少以及肠道中乳酸杆菌和双歧杆菌的丰度降低有关。
✦使用抗生素导致多样性降低
在生命早期使用抗生素也会导致物种和菌株水平上的微生物多样性降低,某些有益物种如梭状芽孢杆菌减少,抗生素抗性细菌如肠球菌的数量增加。抗生素暴露对后代微生物组更深远的影响将在后面继续讲述。
// 建议
由于抗生素的使用,婴儿的微生物多样性会降低,还会对健康造成一定影响。因此建议不是必要情况,尽量减少抗生素的使用。
✦母乳促进免疫发育
出生后,儿童微生物群发育的最重要因素是母乳喂养。母亲母乳是细菌、抗原、营养素和生物活性物质的唯一来源,所有这些都能促进婴儿免疫系统发育、新陈代谢和肠粘膜屏障功能。
•母亲的饮食影响母乳组成
母亲的饮食是母乳组成的重要因素。最近证明,母亲的饮食显著改变了母乳低聚糖的组成,地理位置和体重也在一定程度上影响母乳的组成。
一项全面系统综述得出结论,母亲饮食中脂肪酸、脂溶性维生素、维生素B1和维生素C都与母乳成分有关。
注:虽然强烈建议母乳喂养,但有时这是不可能的或不充分的,只能用配方奶粉代替。婴儿配方奶粉试图模仿母乳,通常以补充维生素、铁、脂肪酸和益生菌的大豆或牛奶等替代品为基础。
•母乳喂养下的主要菌群
尽管母乳成分因母亲健康状况、饮食、分娩方式和哺乳时间而异,但母乳主要由几个微生物属(葡萄球菌、链球菌、丙酸杆菌、鞘氨醇杆菌、双歧杆菌和乳酸杆菌)构成,并导致双歧杆菌、拟杆菌的早期定植,以及大肠杆菌和艰难梭菌的水平较低。
•配方奶喂养下的主要菌群
相比之下,配方奶喂养与母乳喂养婴儿相比,艰难梭菌(C. difficile)、脆弱拟杆菌(Bacteroides fragilis)和大肠杆菌(E. coli)的定植率更高,其肠道微生物组与更典型的成人模式相似。
✦母乳对于婴儿十分重要
母乳不仅作为细菌来源,还含有细菌抗原和其他生物活性物质,可刺激免疫系统的发育和成熟。最近,使用宏基因组方法发现,母乳喂养的样本中与免疫、代谢和生物合成活动相关的基因的共表达相比配方奶粉喂养的样本增加。
•促进免疫系统发育和调整
母乳中含有益生母乳低聚糖,可促进双歧杆菌生长,进而在免疫系统的发育和微调以及抑制致病物种生长方面发挥重要作用。这些研究证明了母乳喂养在早期婴儿微生物群发育和成熟中的重要性。
// 建议
条件允许的话,最好还是用母乳喂养婴儿(不过母乳喂养的前提需要保证母亲是健康的)。如果不行的话也应选用成分相似的配方奶进行喂养。
在婴儿期建立健康的肠道微生物群对于儿童日后免疫力的发育和成熟至关重要。孕期母体肠道微生物组可能通过细菌代谢物或免疫球蛋白直接或间接影响胎儿免疫发育。
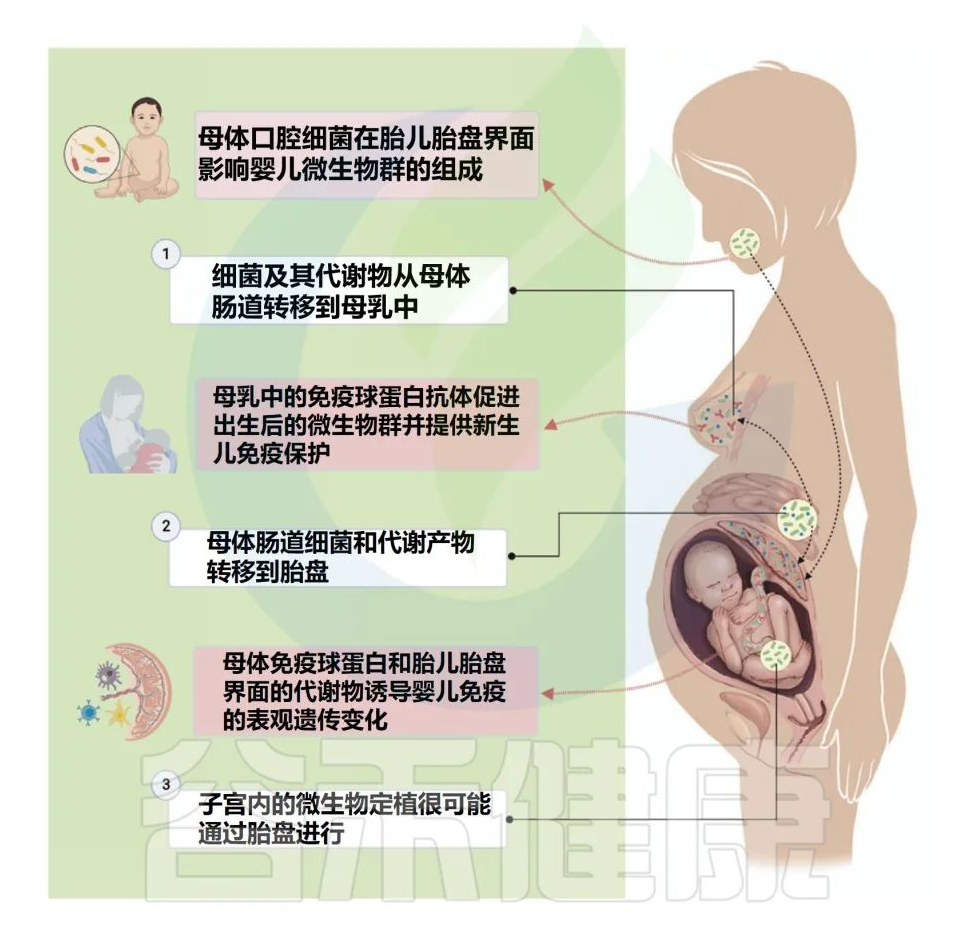
肠-乳腺通路是肠道细菌在怀孕期间通过肠系膜淋巴网络从母体胃肠道向乳腺的生理易位机制,可能有助于后代的免疫发展。
尽管确切的机制尚待确定,但研究表明母体微生物组与胎儿免疫之间存在联系。
产前母子之间的微生物联系
Mua B,et al.Comprehensive Gut Microbiota.2022
怀孕期间,母婴界面存在定向微生物群影响。所谓的肠乳途径是怀孕期间肠道细菌通过肠系膜淋巴网络易位的潜在机制,被认为通过表观遗传变化诱导后代的免疫发育。
母体肠道细菌和代谢产物从母体胃肠道输送到乳腺,通过肠道-母乳轴影响产后肠道定植和免疫。母体的口腔和胎儿胎盘串扰会导致子宫内共生肠道细菌的早期定植,从而获得终身免疫。
怀孕期间,母体肠道中产生的代谢物(如短链脂肪酸)会转移到乳腺,在乳腺中通过母乳喂养传递给婴儿。
对微生物和代谢物的动物和人类研究为孕期微生物易位及其对胎儿免疫的影响提供了证据。例如,怀孕小鼠在妊娠晚期口服乳酸菌菌株乳酸乳球菌和唾液乳球菌,然后在小鼠的母乳和组织中检测到。
肠道微生物组产生代谢产物,在调节宿主免疫、生理和能量产生中发挥关键作用。
✦有助于婴儿生理发育
由于短链脂肪酸与人类微生物群、免疫和神经内分泌系统的相互作用,短链脂肪酸在人体稳态中发挥着关键作用,并可能有助于婴儿的神经、代谢和免疫发育。
母乳中由细菌或肠道人类细胞产生的代谢物,通过促进或抑制细菌生长的机制,帮助调节乳汁和婴儿微生物组组成。
将具有特应性表型的母亲与非特应性母亲的乳汁成分进行了比较,发现特应性妈妈的乙酸和丁酸水平明显较低,与婴儿体重指数呈负相关。
母亲特应性状态定义为有过敏症状,如特应性皮炎、哮喘和食物过敏等。同样,母乳激素瘦素和胰岛素水平的变化与婴儿肠道微生物多样性和通透性的差异有关。
✦调节宿主免疫
•母体视黄酸提高后代免疫力
母体视黄酸作为胎儿3型先天性淋巴细胞的调节因子,并通过诱导小鼠胎儿发育过程中的次级淋巴器官来提高后代的免疫力。
视黄酸——又称维A酸,是体内维生素A的代谢中间产物,主要影响骨的生长和促进上皮细胞增生、分化、角质溶解等代谢作用。
3型先天淋巴细胞(ILC3)具有抗原呈递作用,可以激活T细胞反应。
•短链脂肪酸调节肠道免疫,防止致病性感染
微生物短链脂肪酸(包括丁酸、乙酸盐和丙酸盐)影响肠道免疫,调节结肠调节性T细胞(一种免疫T细胞的特殊亚群),增强口腔对食物的耐受性,并防止肠道致病性感染。
•对哮喘有积极影响
同样,妊娠期间的细菌代谢产物和母体饮食通过补充纤维和乙酸盐增强调节性T细胞的丰度和功能,对哮喘的发展产生积极影响,这在人类哮喘小鼠模型研究中得到了证明。
细菌代谢产物在母体-胎儿之间的联系是母亲和社会普遍关注的一个话题,因此,应在进一步的实验和综合研究中加以解决。
在妊娠期间,母体免疫球蛋白(IgG)通过胎盘转移至胎儿,诱导出生前的被动体液免疫和出生后的先天免疫,以防止感染。
免疫球蛋白G(IgG)是血清中免疫球蛋白的主成分,约占血清中免疫球蛋白总含量的75%。
✦母体免疫球蛋白影响胎儿免疫力
母乳中的IgG抗体会影响产后微生物群和哺乳期的免疫力。母亲的母乳抗体也可以通过与细菌结合的胎盘运输,从而影响胎儿在子宫内的免疫规划。
母体IgG抗体在胎盘的合胞体滋养层细胞内主动运输并与新生儿Fc受体(FcRn)结合。一旦结合,IgG被包装到内体中,并保护其不被降解,直到其达到胎儿循环。
✦母体健康会影响免疫球蛋白
孕期母体健康状况也是免疫球蛋白转移至胎儿的关键因素。妊娠期高血糖降低了母亲母乳和血液中的免疫球蛋白丰度,并降低了IgG通过胎盘向胎儿的转移。
注:跨胎盘免疫调节也可能由其他细菌成分介导,如脂多糖、细胞因子和激素。
微生物的组成因身体部位而异,并在不同个体内部和个体之间形成了不同微生物群落的独特环境。
✦肠道是微生物最丰富的区域
胃肠道内发现了最丰富的微生物多样性和丰度,主要以厚壁菌门(Firmicutes)、拟杆菌门(Bacteroidetes)、放线菌门(Actinobacteria)和变形菌门(Proteobacteria)的兼性厌氧菌为特征。
✦微生物变化时会产生一系列影响
宿主-微生物关系复杂,当发生变化时,会对免疫、代谢和神经系统的稳态产生负面影响,导致对环境过敏原和营养刺激过敏。
这些影响主要通过免疫、神经、胃肠和内分泌调节肠-脑轴(一种调节体内稳态的双向神经体液系统)的组成部分对子宫中的后代进行表观遗传编程。
表观遗传机制是细胞分化的关键调节因子,通过在生物和病理条件下将环境线索印记到细胞和组织的生理学中来发挥作用。
注:与健康变化相关的所有暴露量的测量被称为暴露量,从怀孕开始就涉及环境和职业暴露。
在妊娠期间,DNA甲基化和组蛋白修饰打开和关闭基因,允许不同的细胞类型分化并获得胎儿暴露体的表观遗传记忆,这将在个体的整个生命周期中保存。
✦环境因素影响表观遗传
孕期母亲的生活方式、饮食、年龄、压力、感染和其他环境因素会影响母亲和胎儿的表观遗传途径,导致后代的遗传、代谢和神经发育改变。
在生命的前1000天,表观遗传调节和可塑性、胎儿器官发育和成熟与疾病易感性的印记一起被编程。如果外部因素不健康,可能会发生表观遗传变化,从而改变基因表达,增加成年后患病的风险。
后代的表观遗传调控是通过母亲在肠-脑轴中的饮食成分进行的。这些成分包括叶酸和营养素,它们在母体肠道中代谢和吸收,从而帮助维持健康的肠道微生物群落。
✦肥胖和正常体重的母亲微生物组成不同
怀孕肥胖母亲的微生物群组成与正常体重母亲的明显不同,其特征是葡萄球菌(Staphylococcus)和肠杆菌(Enterobacteria)水平升高,拟杆菌和双歧杆菌水平降低。
•影响后代的健康
胃肠道改变扰乱了母体微生物群,影响了后代对代谢紊乱的敏感性,并产生长期影响。
例如,高脂饮食的母亲因脂多糖增加而导致的叶酸水平变化会影响肠道通透性和微生物组,进而影响婴儿的微生物定植,从而增加成年后的疾病风险。
动物模型为这一假设提供了进一步的支持
在妊娠期高脂饮食的狒狒模型中发现,高脂饮食会影响妊娠前和妊娠期间后代肝脏内源性大麻素成分的发育调节。胎儿大麻素系统的改变会增加成年后肝脏损伤和肥胖的风险。
综上所述,母体微生物组间接影响后代的代谢发育,因此可能是预防成年期胃肠道疾病的有效靶点。
✦母体代谢成分对后代的影响
•增加后代心血管病风险
很少有研究探讨孕期母体代谢成分通过表观遗传机制在后代免疫中发挥的调节作用。母亲饮食中长链多不饱和脂肪酸摄入量低会诱导血管生成因子基因的DNA甲基化,导致胎盘排列、血管调节发生改变,并增加后代晚年的心血管疾病风险。
通过胰岛素生长因子(IGF-1)启动子的组蛋白修饰,母体高血糖可能会增加成年后对胰岛素抵抗的敏感性。
•调节炎症反应
母亲补充膳食DNA甲基化物,如叶酸、维生素B12、胆碱、蛋氨酸和锌,可能有助于在胎儿细胞快速生长和增殖期间调节炎症反应。
胎儿发育过程中的炎症也可能诱导免疫系统过度激活并增加疾病风险。
•影响胎儿发育及免疫功能
母亲补充胆碱会影响DNA和组蛋白甲基化,从而改善成年期胎儿大脑发育和功能。维生素D是另一种必需的微量营养素,当缺乏时,会改变表观遗传途径,并通过胎盘中的DNA甲基化和组蛋白修饰导致炎症、先兆子痫和不良妊娠结局。
细菌代谢物和其他环境分子也可能通过母胎界面驱动的表观遗传调节影响胎儿免疫发育。研究发现,怀孕期间接触醋酸可以增强调节性T细胞的数量和功能,抑制后代的过敏性呼吸疾病,最有可能是因为FoxP3启动子乙酰化程度增加。
妊娠期间胃肠道成分的调节已被证明对儿童的免疫和代谢发育有强烈的影响,因此应在该领域的未来研究中加以解决。
✦母乳喂养对儿童健康有积极影响
•不易发生肠道感染
母乳喂养对儿童的健康结果有积极影响。头两年的亚最佳母乳喂养与死亡率增加相关。母乳喂养的婴儿较不容易发生肠道感染,从而导致慢性炎症和腹泻概率降低。
•母乳喂养降低儿童肥胖和2型糖尿病风险
然而,如果过早暂停母乳喂养,以引入配方食品或固体食品,则会使婴儿的肠道微生物组向成人样成分转变,并与儿童肥胖风险增加相关。
有证据表明,母乳喂养的婴儿青春期肥胖和2型糖尿病的风险降低。剖腹产也会破坏母亲和孩子之间的正常微生物群交换,这与儿童期和成年期乳糜泻、1型糖尿病和肥胖的风险增加有关。
其他研究人员报告称,孕前和孕期母亲超重、剖腹产和婴儿微生物群丰富的厚壁菌增加了儿童肥胖的风险。反过来,儿童肥胖可能会导致其他负面的短期和长期健康后果,包括低自尊、心血管问题以及成人肥胖和糖尿病。
✦孕期使用抗生素与儿童感染有关
孕期母亲使用抗生素是影响母亲肠道和母乳微生物群的另一个因素,并与儿童肥胖风险增加相关。最近,研究了孕期母亲抗生素治疗与生命后期感染风险之间的关系。他们发现,妊娠期抗生素治疗会增加四岁以下儿童患扁桃体炎的风险。
此外发现,孕期抗生素治疗与儿童因感染而住院的风险增加有关,并且在临近分娩时服用抗生素的风险更高。
小结
这些研究强调了一个事实,即母亲的微生物多样性在新生儿微生物组组成和健康的发展中至关重要。
早产儿长期接受抗生素治疗会增加其败血症、坏死性小肠结肠炎和死亡的风险。此外,生命早期的抗生素治疗会增加生命后期超重和肥胖的风险。
孕期和生命早期的抗生素治疗可还能会改变婴儿微生物组组成的发育,导致生命后期的负面健康结果。
在婴儿和儿童中,包括皮肤、呼吸道和食物过敏在内的自身免疫性疾病是全球关注的问题。研究发现,新生儿和婴儿微生物组参与肠道耐受性的调节和免疫系统的调节。
最近的研究集中于肠道微生物组在先天性和适应性免疫发展以及过敏性疾病发病中的作用。晚年过敏症的发展与新生儿和婴儿期微生物组发育和成熟的变化有关。这些情况也可能源于导致不同表型和症状的遗传和环境因素。
✦皮肤过敏
健康的人类皮肤微生物组主要由丙酸杆菌属(主要在皮脂部位丰富)、棒状杆菌属和葡萄球菌属(主要存在于潮湿的微环境中)以及马拉色菌属组成。
皮肤微生物组的组成受性别、年龄、种族、环境暴露和生活方式等因素的影响。婴儿最典型的过敏表现之一是湿疹或特应性皮炎,这是新生儿对过敏原的先天反应改变和皮肤微生物多样性受损的结果。
•患有特应性皮炎婴儿的菌群变化
患有急性和慢性特应性皮炎的婴儿的特征是表皮葡萄球菌(Staphylococcus)的数量减少,多于金黄色葡萄球菌,这可能通过下调表皮蛋白和促进皮肤蛋白酶而增加过敏原致敏的风险,从而导致皮肤屏障损伤。
•特应性皮炎受到微生物的影响与调节
新生儿表皮葡萄球菌的皮肤定植与toll样受体2诱导的抗菌信号和调节性T细胞介导的宿主免疫应答的调节和激活有关。
特应性皮炎的发展也受到肠道微生物群的影响。特应性皮炎患者的肠道微生物组中双歧杆菌(Bifidobacterium)的浓度较低,普拉梭菌(Faecalibacterium prausnitzii)的丰度较高,这与克罗恩病患者中这些细菌的比例较低以及抗炎粪便细菌代谢物有关。
然而,一项大型前瞻性研究表明,在儿童期患上特应性皮炎的婴儿中,普拉梭菌(Faecalibacterium)和毛螺菌属(Lachnobacterium)的比例明显较低。
尽管有些矛盾,但这些结果表明,微生物类群丰度的改变可能对特应性皮炎的发病起到保护作用。产前和产后补充口服双歧杆菌和乳酸杆菌可以诱导T细胞介导的反应改变,降低婴儿特应性皮炎的风险。
✦食物过敏
动物和人类研究表明,肠道微生物组可能与生活早期对饮食抗原的口服耐受、食物致敏和过敏结果密切相关。
•饮食过敏影响微生物多样性
与健康婴儿相比,对食物敏感的婴儿的α多样性微生物群和拟杆菌(Bacteroidetes)数量减少,厚壁菌(Firmicutes)丰度增加。人类这些特定结果的可能原因包括抗生素的直接和间接使用以及膳食纤维的消耗减少。
早期喂养的类型也可能导致不同的健康结果。食用牛奶配方的奶粉会增加总厌氧菌的数量,并增加患牛奶过敏的风险。与肠杆菌和双歧杆菌相比,患有牛奶过敏的婴儿的乳酸杆菌感染率更高。
此外,从婴儿期到儿童期,牛奶过敏儿童中厚壁菌(Firmicutes)和梭状芽孢杆菌(Clostridia)的丰度均升高。有趣的是,与对照组相比,坚果过敏儿童的肠道微生物组显示拟杆菌(Bacteroidetes)种类比例增加,微生物多样性降低。
同样,与健康对照组相比,鸡蛋过敏儿童的毛螺菌科(Lachnospiraceae)和瘤胃球菌科(Ruminococcaceae)浓度增加。
•细菌可能有助于解决食物过敏
添加Lactobacillus rhamnosus GG的牛奶配方有助于解决婴儿早期的牛奶过敏,最有可能是通过肠道微生物组中丁酸盐产量的增加。
L. rhamnosus GG还可能诱导花生过敏儿童的免疫变化和口服耐受性。口服微生物补充剂可能是食物过敏的一种潜在治疗策略,但需要进一步研究以阐明微生物菌株对饮食中过敏原的持续敏感性的影响。
✦呼吸道过敏
•肠道微生物影响哮喘
与其他过敏表现相比,出生第一个月的肠道微生物组成是与学龄期哮喘发作相关的最相关因素之一。
在婴儿早期和整个儿童期进行的观察和纵向研究表明,毛螺菌属(Lachnospira)、普拉梭菌(Faecalibacterium)和小杆菌属(Dialister)细菌类群的减少,与儿童期哮喘风险增加有关。
•其他影响哮喘的因素
然而,与婴儿微生物组相关的其他因素也可能在哮喘风险中发挥重要作用,包括接触抗生素、动物皮毛、阴道分娩与剖腹产以及母体孕龄。
抗生素增加了哮喘风险
抗生素的使用和出生前后的暴露与小鼠的气道超敏反应和反应性有关,并增加了人类患哮喘的风险。
母乳喂养可降低哮喘风险
婴儿期的喂养类型也是婴儿早期形成肠道微生物群的一个重要因素,当婴儿长时间母乳喂养时,在生命的头两年内发生哮喘的风险可以降低。
•微生物多样性紊乱易导致儿童后期哮喘
婴儿微生物组多样性的紊乱也是儿童后期哮喘发病的关键因素。粪便杆菌和双歧杆菌的多样性降低是有哮喘风险的婴儿肠道微生物组的特征。
这些婴儿的无菌粪水用于刺激成人外周血单核细胞(PBMC),这导致调节性Foxp3细胞减少,CD4+IL-4产生细胞增加。
新生儿肠道微生物组环境的改变通过调节CD4+T细胞功能和增殖影响儿童过敏性哮喘的易感性。
•肺部微生物影响呼吸健康
生命早期的肺部微生物多样性影响呼吸健康。婴儿出生头两周内的肺部微生物组促进树突状细胞中程序性死亡配体1(PDL1)的表达,这是调节性T细胞抑制过敏性呼吸道免疫反应所必需的。
在出生的头两个月,婴儿的气道被链球菌、莫拉克氏菌或嗜血杆菌定植,这与婴儿和儿童的严重下呼吸道感染和哮喘有关。变形杆菌和拟杆菌也与哮喘和其他免疫相关的加重有关。
如前所述,这些细菌类群的副产物和代谢物可能介导其保护作用。丁酸盐和其他短链脂肪酸作为膳食纤维的发酵产物,如普拉梭菌(Faecalibacterium prausnitzii),它们通过诱导Tregs促进上皮屏障通透性和抗炎作用,和粘膜B细胞产生IgA。
实验研究和人体临床试验还表明,通过补充肠道微生物组衍生的丁酸,可以降低变应原敏感性和哮喘的发病率。
不足之处
过敏性疾病发病和严重程度的生理机制仍在研究中,一些介入性研究表明,补充益生菌和益生元可以预防和下调此类综合征,然而,与哮喘相关的研究报告,补充和不补充益生药的婴儿之间没有显著差异,因此需要更多的研究来进一步检验这些影响。
生命的前1000天除了肠道微生物组的建立,还是神经健康规划和中枢神经系统后期正常运作的关键时期。
肠-脑轴由肠道和中枢神经系统以及神经和下丘脑-垂体-肾上腺轴(HPA轴)调节。因此,该轴的任何损伤,包括神经元损伤或死亡,都会导致神经变性,并可能导致神经变性疾病的发生和发展。
✦营养不良对认知障碍有影响
母婴营养不良通常与认知技能、入学准备和表现、不良生长结果、感染和死亡等方面的损害有关。由于母亲健康和营养不良以及婴儿护理和喂养不当而发育不良的儿童会出现身体和认知延迟。孕期母亲炎症和高脂饮食影响微生物群定植,增加成年期神经退行性疾病的风险。妊娠期以性别特异性的方式影响发育中胎儿大脑中的内源性大麻素途径。
✦孕期母亲的生活方式会导致神经发育障碍
因此,孕期母亲的生活方式和环境暴露可能导致神经发育障碍的发作,包括胚胎神经管缺陷、唐氏综合征、自闭症、雷特综合征和其他神经心理缺陷。
✦肠道微生物调节认知与行为
微生物群组成与认知和行为发育改变之间的双向相互作用已在肠-脑轴中得到确认,有证据支持这种相关性。
缺乏肠道微生物群的无菌小鼠表现出与焦虑相关的表型增加,神经递质水平和与神经可塑性有关的代谢物发生变化,从而表明肠道定植对大脑发育和功能有积极影响。包括双歧杆菌和乳酸杆菌在内的一些细菌菌株以及摄入的益生菌与调节行为变化有关。
注意
需要注意的是,当无菌小鼠在早期生活中重建了正常的微生物群,但在成年后没有重建时,行为模式和神经递质浓度正常化。
母乳喂养被发现可以促进大脑发育,并对婴儿的认知能力产生积极影响。一些因素,包括文化和社会信仰以及对母乳喂养的污名化、缺乏营养和教育、母亲压力和抑郁,都可能导致过早停止母乳喂养;所有这些都会对婴儿的肠道微生物群和早期和后期的认知发育产生负面影响。
生命早期营养、肠道微生物组和健康之间的相互关联在个体一生的稳态中起着关键作用。
生命最初营养、肠道微生物组和健康之间的关系

Mua B,et al.Comprehensive Gut Microbiota.2022
母亲和新生儿营养不良、喂养习惯和营养/益生菌补充对子宫、婴儿和儿童期微生物组的建立有很大影响。母体微生物组变化和过敏原/抗生素暴露也会影响儿童早期的微生物组组成。
微生物组组成和丰富性的变化会影响妊娠结局和产妇健康,并通过表观遗传编程使发育中的孩子在以后的生活中易患自身免疫、认知和代谢障碍。
生命的前1000天是一个“可塑性窗口”,在这个窗口中,后代的健康会根据母体和环境的影响进行变化。
由于饮食生活方式以及暴露于外来生物和其他环境压力源,代谢、免疫学和神经退行性疾病的患病率在全球范围内不断增加。
在微生物组水平上理解母亲和孩子之间的独特联系,有助于研究对孕期和婴儿期肠道微生物组的变化及其对母婴健康的直接和间接影响,使母亲和孩子在未来都拥有更健康的生活。
主要参考文献
Mua B , Nga B , Fjaa B , et al. The Gut Microbiota, Nutrition, and Long-Term Disease Risk: A Mother and Child Perspective[J]. Comprehensive Gut Microbiota, 2022:289-307.
Apostol, A.C., Jensen, K.D.C., Beaudin, A.E., 2020. Training the fetal immune system through maternal inflammationda layered hygiene hypothesis. Front. Immunol. 11, 1–14.
Cortes-Macías, E., Selma-Royo, M., García-Mantrana, I., et al., 2021. Maternal diet shapes the breast milk microbiota composition and diversity: impact of mode of delivery and antibiotic exposure. J. Nutr. 151 (2), 330–340.
Cunha, A.J.L.A., Santos, A.C., Medronho, R.A., Barros, H., 2020. Use of antibiotics during pregnancy is associated with infection in children at four years of age in Portugal. Acta Paediatr. 110 (6), 1911–1915.
Forgie, A.J., Drall, K.M., Bourque, S.L., et al., 2020. The impact of maternal and early life malnutrition on health: a diet-microbe perspective. BMC Med. 18 (1), 1–15.
Mesa, M.D., Loureiro, B., Iglesia, I., et al., 2020. The evolving microbiome from pregnancy to early infancy: a comprehensive review. Nutrients 12 (1), 1–21.