-

 国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子
国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子

 二级病原微生物安全实验室
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
谷禾健康
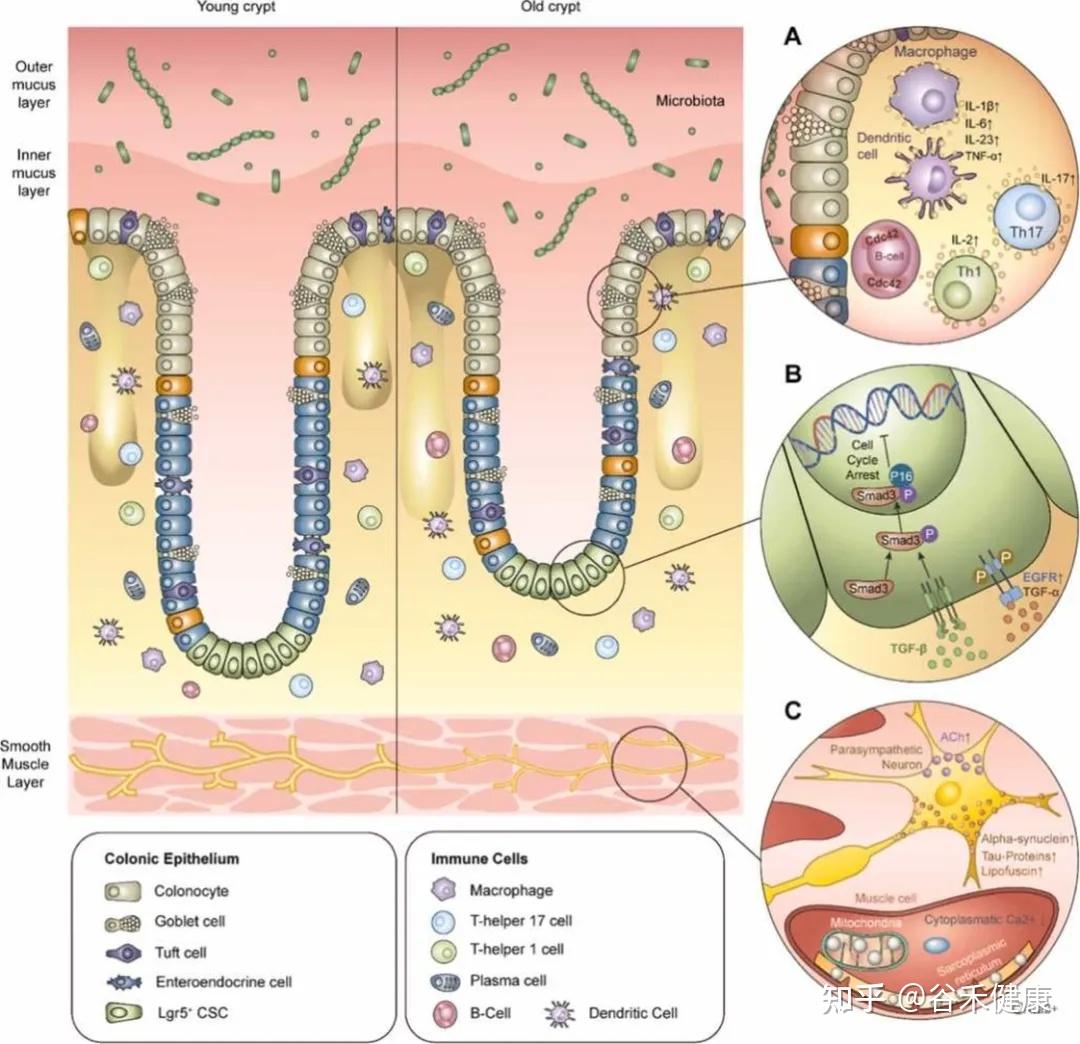
结肠,作为我们消化系统中的关键一环,却是最容易受到衰老影响的器官之一。随着年龄的增长,结肠的结构和功能发生了显著变化,这些变化会导致肠道屏障变得脆弱,进而影响我们的健康。
研究表明,衰老导致结肠黏膜上皮更新速度减缓,隐窝干细胞活性降低,紧密连接蛋白(如occludin、claudin、ZO-1)的表达异常。这些变化使得细胞通透性增加,黏液层变薄,黏液屏障的完整性下降。同时,免疫系统也出现了免疫衰老现象,降低了对有害菌的防御能力。
这一切都可能导致低度炎症状态,表现为内毒素易位和系统性炎症标志物的升高。而在微生态方面,老年人的肠道菌群多样性下降,产短链脂肪酸菌减少,潜在的促炎菌却相对增加,形成了一个恶性循环。
近年来,益生菌作为调节微生态和修复肠道屏障的干预策略,在老年相关便秘、抗生素相关腹泻、感染性腹泻、炎症性肠病辅助治疗和肠易激综合征等多个领域积累了丰富的临床证据。然而,益生菌的效果并非一概而论,它们的保护作用高度依赖于特定菌株,并受到个体差异的影响。
注:不同实验体系中的挑战因素(如大肠杆菌、TNF-α、LPS、氧化应激)、观测指标(如TEER、FITC-dextran通透性、紧密连接蛋白定位、炎症因子谱)和给药形式(活菌、热灭活菌、上清、菌体裂解物)都可能导致不同结论。因此,实际应用中仍需逐株验证其作用机制和可重复性,并明确其“预防型”、“抗炎型”或“修复型”的屏障支持作用。
本文将探讨衰老对结肠的影响,重点围绕益生菌是否能够改善肠屏障功能的临床证据,从结局指标和证据强度等方面进行梳理,尤其关注现有的随机对照试验,并总结不同菌株的具体功效及适应证,提出应用建议和研究展望。
01
衰老和结肠老化
随着年龄增长,身体各个器官以不同的速度衰老。这些变化跨度极大,可能只是轻微的外貌改变,也可能是严重的功能障碍,进而导致疾病、生活质量降低和死亡率增加。
脆弱的消化道:对衰老尤为敏感
在全球范围内,65岁及以上人群(本文定义的“老年人”)中消化系统疾病发病率显著上升。衰老波及整个消化系统,程度不一,其中结肠似乎最容易受累。常见的相关疾病包括憩室病、结直肠癌、显微镜下结肠炎及慢性便秘,都是老年人常见的问题。
结肠不同部位老化不一样,研究有挑战
虽然这些疾病发病率很高,但科学界直到最近才开始重视人类结肠衰老的分子机制研究。研究老化的结肠特别具有挑战性。因为结肠像一个复杂的工厂,不同区段(升结肠、降结肠等)的结构和功能差异巨大,导致它们老化的方式也不尽相同。
此外,结肠包含多种细胞,每种细胞的老化路径都不同,再加上肠道菌群和局部免疫系统等外部因素的干扰,这让研究变得更加困难。
下面我们将从微观层面详细拆解:
上皮:脆弱的第一道防线
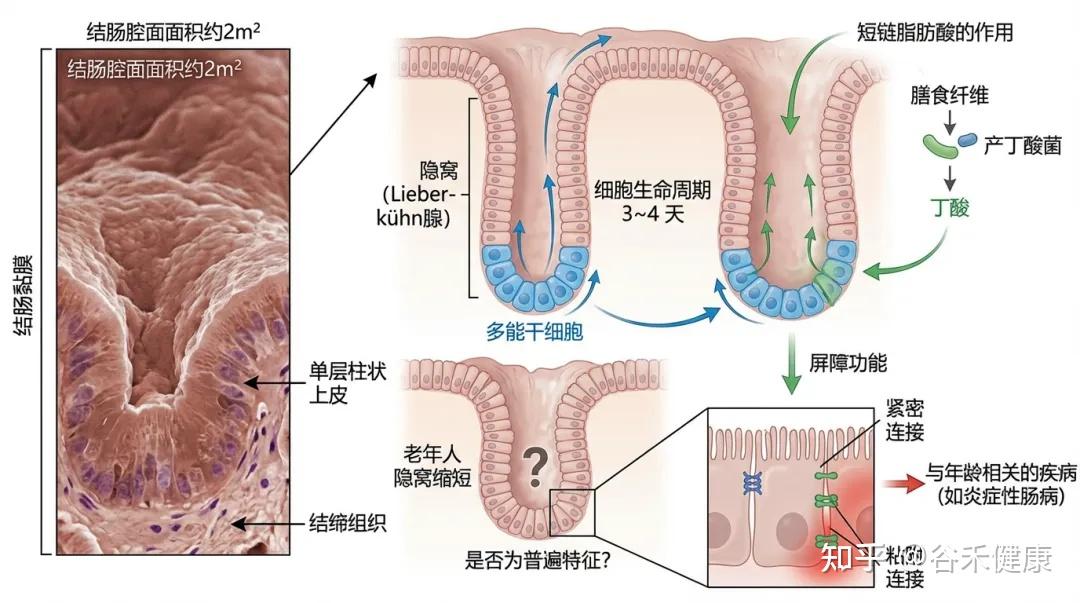
结肠的腔面面积约为2平方米,由结肠黏膜覆盖,黏膜由单层柱状上皮和结缔组织构成,上皮层向结缔组织内凹陷形成隐窝(Lieberkühn 腺)。
结肠的隐窝中有多能干细胞,负责快速更新肠细胞。这些细胞在3~4天内完成生命周期,最终脱落到腔内。(读过我们文章的可能了解,这个干细胞的更新需要短链脂肪酸,尤其丁酸,所以缺乏膳食纤维以及产丁酸菌会影响结肠隐窝底部的干细胞更新)
隐窝结构的老化
研究发现,老年人的隐窝可能会缩短,但是否这是结肠老化的普遍特征仍需进一步验证。
结肠上皮必须提供强大的屏障功能,允许选择性吸收水分、电解质和短链脂肪酸。屏障功能由紧密连接和粘附连接维持,任何功能障碍都可能导致与年龄相关的疾病加重,例如炎症性肠病(前面谷禾的文章也讲了很多关于肠道屏障的文章,其中上皮的构成紧密连接起到关键作用,相关阅读:什么是肠道屏障,影响因素,监测评估以及如何治疗干预 )。
研究显示,老年灵长类动物(非人)的结肠中,紧密连接蛋白显著减少,而在不同解剖区域的变化也存在差异。这表明,结肠的老化特征可能因部位而异。
杯状细胞的变化:为何重要?
衰老还影响结肠上皮的分泌成分。研究发现,老年小鼠的杯状细胞数量显著下降,伴随分泌功能受损。这些变化与慢性低度炎症及氧化应激有关,导致结肠屏障功能减弱(有兴趣可以翻翻以前关于肠道屏障的文章,影响因素以及干预等)。
在像结肠上皮这样的高复制组织中,组织的老化与结肠干细胞的老化密切相关,因为干细胞对于维持上皮的稳态至关重要。
干细胞老化的内外因素是什么?
影响结肠干细胞老化的因素可分为内因和外因。内因包括干细胞的基本特性和突变率,外因则涉及复杂的信号通路。例如,老年大鼠表现出表皮生长因子受体(EGFR)的基础活化增加,这可能与对TGF-α的敏感性增加有关。
Wnt信号在衰老中的作用:如何影响干细胞功能?
Wnt信号通路在维持干细胞功能中起着关键作用。研究发现,衰老小鼠的肠道干细胞中Wnt3水平降低,添加Wnt3可以恢复其形成类器官的能力。这表明Wnt信号在肠道衰老中扮演着重要角色。
内分泌细胞的功能:为何不可忽视?
尽管内分泌细胞在结肠上皮中所占比例不到1%,但它们在调节结肠运动和分泌中发挥着重要作用。老年小鼠的研究显示,肠内分泌细胞的数量显著增加,可能是对年龄相关受体抗性的补偿。
老年小鼠与人类的比较:有什么不同?
在人类乙状结肠中,尽管感觉神经密度降低,肠嗜铬细胞的数量却保持稳定。这表明老年结肠可能存在代偿性神经免疫适应。
表观遗传重塑与衰老:如何影响癌症风险?
衰老是结直肠癌的主要风险因素,结肠粘膜的表观遗传重塑可能是生理衰老与肿瘤发生之间的重要联系。研究发现,正常结肠粘膜中存在年龄相关的甲基化变化,这些变化可能导致癌症的发生。
表观遗传时钟的研究:如何评估衰老?
通过分析334份结肠粘膜样本,研究发现不同的表观遗传时钟与实际年龄之间存在强烈相关性。这些发现表明,功能失调而非加速的表观遗传衰老可能促进肿瘤易感性。
衰老结肠中的表观遗传漂移不仅反映了被动的分子磨损,更是一个塑造组织脆弱性的主动过程。受区域和环境因素影响的表观遗传衰老失调,可能是生理衰老与结直肠肿瘤发生之间的关键机制。
小 结
随着年龄的增长,身体各个器官就像老旧的机器,以不同的速度磨损,消化系统尤其敏感。65岁以上的人群中,结肠问题频繁出现,比如憩室病和结直肠癌,成为老年人常见的健康隐患。研究发现,结肠的不同部分衰老方式各异,内部细胞和微生物的互动让问题更加复杂。结肠上皮的脆弱性与干细胞老化和慢性炎症有关,这不仅影响消化,还可能增加癌症风险。了解这些变化,有助于减缓衰老带来的影响,提升生活质量。
在我们身体的消化系统中,结肠扮演着至关重要的角色。它的推进运动依赖于神经元与平滑肌细胞的精密协调。然而,随着年龄的增长,这种协调性逐渐受到影响,结肠的功能也随之下降。这一过程涉及复杂的生物机制,值得我们深入探讨。
神经网络的角色
结肠的神经网络是自主神经系统的重要组成部分,主要分布在以下两个区域:
这些神经元通过信号传递来调节肠道的运动和分泌,确保消化过程的顺利进行。
衰老对神经元的影响
研究发现,结肠中的神经元随着年龄的增长表现出不同的退行性变化:
-数量减少
胆碱能神经元(兴奋性)数量显著下降,而氮氧化酶神经元(抑制性)数量变化较小。
-形态变化
肠神经元的形态变化高度异质,且结肠周径的增加可能导致神经元密度的稀释效应。
这些变化意味着,尽管神经元的数量在减少,但观察到的密度下降可能并不完全反映真实的细胞损失。
在对13名成人尸检样本的分析中(<35岁 n=6;>65岁 n=7),研究者发现老年人肌间神经元总体减少。此外,另一项对16名患者(33-99岁)降结肠和乙状结肠的研究显示,Hu阳性和ChAT阳性神经元选择性丧失,而nNOS阳性神经元保持相对稳定。
有趣的是,最大规模的研究发现,肌间神经丛中的神经元总数随衰老并无显著变化。但在老年个体的升结肠中,ACh阳性神经细胞体数量却增加,这可能是由于轴突功能障碍导致神经递质在细胞体中积累。
神经支配的变化
结肠不仅受到内在神经元的支配,还受到来自交感神经和副交感神经的广泛外在神经支配:
随着年龄的增长,迷走神经和盆腔副交感神经的结构也经历了退化,影响了肠道的正常功能。
神经营养信号与细胞衰老
衰老过程还显著影响神经营养信号通路。研究表明,GDNF、RET和NOS1的表达在衰老过程中发生变化,提示营养支持的减少可能导致神经元维持能力的下降。此外,慢性衰老标志物p16的表达在老年患者的升结肠中增加,而在降结肠中未见类似变化。
胶质细胞的变化
肠道胶质细胞(EGCs)同样随着年龄的增长经历了结构重塑。尽管SOX10阳性肠道胶质细胞体的数量保持稳定,但S100阳性胶质细胞突起的密度显著下降。这种变化可能影响神经与胶质细胞之间的交流,从而损害神经肌肉协调能力。
小 结
随着年龄的增长,结肠的神经元和平滑肌细胞发生了明显变化。研究发现,兴奋性神经元数量减少,而抑制性神经元相对保留。神经元密度的下降可能是因为结肠周围的组织变大了。此外,衰老还影响了神经营养信号,导致神经元的维持能力下降。肠道胶质细胞也经历了结构变化,虽然数量保持不变,但功能可能受到影响。这些变化共同导致结肠的运动能力和整体健康下降。
结肠平滑肌细胞的作用是什么?
结肠中的平滑肌细胞(SMC)负责产生推进运动,并通过基线张力维持器官的结构完整性。收缩可以通过神经递质(乙酰胆碱,ACh)的化学方式或通过机电耦合来启动。在这两种情况下,收缩最终由钙离子从细胞外间隙和肌浆网流入细胞质来介导。这种钙内流激活肌球蛋白轻链激酶(MLC 激酶),它磷酸化肌球蛋白轻链(MLC),从而触发肌肉收缩。
年龄增长如何影响ACh的反应?
随着年龄的增长,ACh的作用似乎发生了变化。对老年大鼠和狒狒的多项研究表明,来自老年动物的结肠平滑肌细胞对ACh的收缩反应减少,或在接触ACh拮抗剂时放松受损。
在老年个体中,从升结肠分离的肌肉条对电场刺激表现出增强的放松反应,而降结肠的肌肉条则没有表现出类似的变化。
这些发现表明,衰老可能在结肠内引发区域特异性的神经肌肉功能改变,其中抑制通路在近端部分似乎被选择性增强。由于抑制性肠神经元的数量和功能能力在衰老过程中保持稳定,因此推测局部的胆碱能功能障碍可能是观察到的放松增强的根源,且这种障碍仅限于老化的升结肠。
钙稳态变化与胆碱能功能障碍:有什么联系?
关于结肠平滑肌细胞中钙稳态的年龄相关变化也有证据。在从老年大鼠的远端结肠提取的细胞中,细胞质钙水平减少,而钙则在线粒体和肌浆网中积累。
总体而言,观察到的结肠中的年龄相关变化似乎与其他器官中的平滑肌细胞变化相呼应。结肠平滑肌细胞是否存在特定于结肠的与衰老相关的变化仍需进一步研究。
小 结
结肠平滑肌细胞的主要功能是帮助推动食物并保持结肠的形状。随着年龄的增长,这些细胞对一种叫乙酰胆碱的神经递质的反应变得不如以前强,尤其是在升结肠部分,它们更容易放松。这种变化与细胞内钙的平衡失调有关,表明衰老可能导致结肠的神经和肌肉功能发生特定区域的变化。
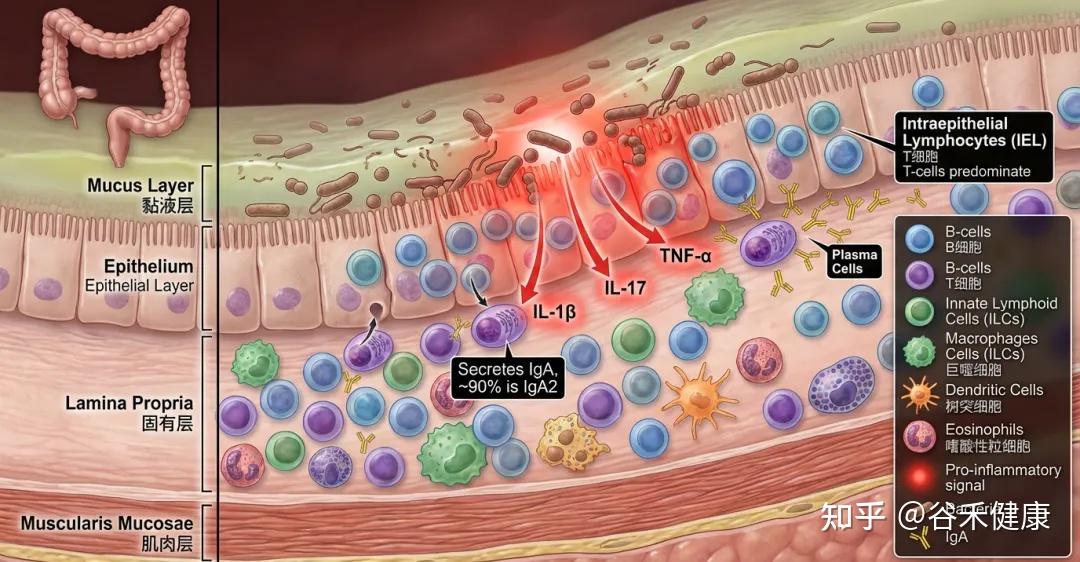
结肠里的免疫细胞有哪些?
结肠里的免疫细胞很多,主要集中在肠壁表面上皮细胞的下方(称为固有层),包括:
在上皮层内部还有一类“上皮内淋巴细胞”(IEL),以T细胞为主。结肠里“浆细胞”也很多,它们主要分泌IgA抗体(对肠道微生物的选择性定植非常关键),用来在肠道表面“温和地管住细菌”。其中大约90%是IgA2这一类型。
老年人的免疫系统:更易出现慢性炎症
老年人更容易出现“低度、长期”的炎症状态。衰老常见的特征之一是全身长期的低水平炎症,被称为“炎症衰老”。在结肠里也能看到类似现象:免疫系统更容易处在被激活状态,例如一些促炎信号分子增加(如IL-1β、IL-6、IL-17、TNF-α等),B细胞和T细胞也更活跃。
一种可能机制:黏液“隔离层”变弱,细菌更靠近肠壁,引发炎症。
黏液层的保护作用:为什么会变弱?
在衰老小鼠中,细菌更容易钻进这层本该相对干净的内层黏液,从而触发免疫反应和炎症。
进一步的链条可能是:衰老相关地出现一种小RNA(miR-124–3p)表达升高,它会抑制一种关键酶(T合成酶),这会影响MUC2这种黏液主要成分的“糖基化”(可以理解为:黏液蛋白需要正确“挂糖”才能形成坚固有效的保护层)。
免疫细胞的局部差异:老年小鼠的观察结果
研究人员用单细胞技术,对比了年轻和老年小鼠的结肠免疫细胞发现:老年小鼠结肠里“免疫细胞大类的比例”整体看起来变化不大。但如果把结肠按不同部位拆开看,就会发现明显的“局部差异”。
具体来说:
在盲肠和结肠远端(更靠近直肠的一段),老年小鼠的 T 细胞更容易处于被激活的状态,提示局部免疫反应更“紧绷”。
在结肠近端(更靠近小肠的一段),老年小鼠的 B 细胞和浆细胞更多。浆细胞是负责大量分泌抗体的细胞。
Cdc42基因的变化:与衰老有什么关系?
还有一个值得注意的发现:老年小鼠的 B 细胞里,Cdc42 这种基因/蛋白的表达更高。Cdc42 与干细胞衰老有关(包括造血干细胞和肠道干细胞),在人类老年人的血细胞中也发现会升高。这提示结肠局部免疫系统的老化变化,至少有一部分和血液中观察到的免疫老化现象是同方向的。
免疫系统衰老通常表现为免疫能力下降:更容易感染、肿瘤风险上升,也更可能出现免疫紊乱(包括自身免疫问题)。
研究结果显示,老年小鼠结肠免疫系统存在局部差异,反映出免疫功能的复杂变化及其对健康的影响。免疫能力下降导致肠道屏障脆弱,可能引发健康问题。因此,探索益生菌等干预手段以调节肠道功能、改善免疫状态和增强屏障功能显得尤为重要。接下来,我们将深入探讨益生菌对肠道健康的调节作用。
益生菌指的是“在适当剂量和适当组合下,能为所给生物体(如人类)带来健康益处的活微生物”。这些细菌长期被证明通过促进粘液层的形成、分泌抗菌因子、竞争性地粘附肠道上皮细胞以及增加紧密连接形成来调控肠道上皮功能。
益生菌通过多种机制调节肠道屏障功能,主要包括以下几个方面:
增强紧密连接结构
益生菌可通过上调紧密连接蛋白的表达来增强机械屏障功能。例如,研究发现某些乳杆菌和双歧杆菌菌株可增加occludin、claudins和ZO-1的表达,从而加固紧密连接结构,降低肠道通透性。
抑制炎症反应
慢性炎症是导致肠道屏障功能障碍的主要原因之一,而益生菌可通过多种途径抑制炎症反应:
特别值得一提的是,益生菌还可通过激活过氧化物酶体增殖物激活受体γ(PPARγ)来发挥抗炎作用。PPARγ是一种核受体,可调节炎症反应和细胞分化,某些益生菌菌株可通过激活PPARγ来保护肠道屏障功能。
调节肌球蛋白轻链激酶(MLCK)通路
肌球蛋白轻链激酶(MLCK)是调节紧密连接通透性的关键酶。炎症因子可通过激活MLCK,导致肌球蛋白轻链磷酸化,进而破坏紧密连接结构。益生菌可通过抑制MLCK表达或活性来保护肠道屏障。
例如,罗伊氏乳杆菌可通过下调MLCK表达,减轻肠毒素诱导的肠道屏障功能障碍。这一机制在预防和治疗感染性腹泻中具有重要意义。
激活模式识别受体
益生菌可通过与肠道上皮细胞表面的模式识别受体(PRRs)相互作用来调节肠道屏障功能。其中,Toll样受体2(TLR-2)是研究较多的受体之一。
研究发现,某些益生菌菌株可通过激活TLR-2,触发下游信号通路,促进紧密连接蛋白表达和抗炎因子产生。例如,双歧双歧杆菌BB1可通过TLR-2/p38 MAPK通路增强肠道屏障功能,同时抑制NF-κB激活,发挥双重保护作用。
调节肠道菌群平衡
益生菌作为肠道生物屏障的重要组成部分,可通过以下方式调节肠道菌群平衡:
肠道菌群平衡的恢复可减少有害菌对肠道屏障的破坏,同时增加有益菌的保护作用,形成良性循环。
改善消化功能
益生菌可促进食物消化和营养吸收,缓解多种消化问题:
调节免疫功能
肠道是人体最大的免疫器官,益生菌可通过调节肠道免疫细胞活性来增强全身免疫功能:
这些作用使益生菌在预防感染、过敏和自身免疫性疾病方面具有潜力。
影响代谢和体重管理
近年来研究发现,益生菌还可能通过”肠-脑轴”和代谢调节影响体重和代谢健康:
虽然这一领域的研究仍在深入,但已有证据表明某些益生菌菌株可能有助于预防肥胖和代谢综合征。接下来,我们将具体探讨哪些益生菌对肠道健康和代谢调节具有积极影响,以及它们如何在改善整体健康方面发挥作用。
乳杆菌属是发酵食品生产中最广泛使用的益生菌之一,这些食品来源包括动物(例如牛奶和肉类)和植物(例如蔬菜和谷物)。
乳杆菌属包括一大类异质的革兰氏阳性兼性厌氧菌,包括嗜酸乳杆菌(L.acidophilus) 、鼠李糖乳杆菌(L.rhamnosus)、发酵酵母菌(L.fermentum)、干酪乳杆菌(L.casei)、植物乳杆菌(L. plantarum)、瑞士乳杆菌(L. helveticus)、罗伊氏乳杆菌(L. reuteri)
除了在食物发酵中的作用外,乳杆菌属还因物种、宿主年龄或肠道位置不同而异,是人类和动物消化系统中健康的组成部分。
动物研究和临床前结果表明 ,乳杆菌可能有助于预防和治疗多种胃肠道疾病。这些疾病包括肠道感染、抗生素相关腹泻、早产儿 NEC、炎症性肠病、结直肠癌和肠易激综合征。
多项研究证实 ,乳杆菌在消化系统疾病管理中发挥重要作用,通过体外和体内维持上皮屏障完整性。该功能的多种机制包括调节细胞骨架、诱导粘液生成以及紧密连接蛋白的磷酸化,这些都会增强紧密连接功能和免疫反应,同时预防上皮细胞凋亡。
以下总结了乳杆菌的不同菌株及其在肠道健康中的作用。
✧ Lactobacillus amylophilus D14
研究发现,把Lactobacillus amylophilus D14 加到健康状态的肠道细胞(Caco-2)上时,细胞之间的紧密连接通透性不会发生变化,也就是不会让屏障变得更松或更紧。
但如果这些细胞先被致病菌(肠源性大肠杆菌 K88 或鼠伤寒沙门氏菌 SL1344)弄伤,D14 就能起到保护作用,它可以减少关键连接蛋白(ZO-1、claudin-1、E-cadherin)的受损,从而帮助维持紧密连接和屏障功能。其原因可能是 D14 抑制了细胞内的 ERK 信号通路活化(ERK 的磷酸化降低),并减少炎症因子 IL-8 的释放,进而减轻炎症和屏障破坏。
✧ 嗜酸乳杆菌 (Lactobacillus acidophilus)
嗜酸乳杆菌 (Lactobacillus acidophilus) 表面有一种特殊的“外衣蛋白”,叫表面层蛋白(Slp)。
嗜酸乳杆菌的外衣蛋白有什么作用?
研究发现,这种蛋白对肠道有多方面的保护作用,比如:
抢先占住肠道黏膜上的“附着位点”,让有害菌不容易黏上去,帮助肠道细胞减少不必要的凋亡(细胞过早死亡),降低脂多糖(LPS,常见于某些细菌成分)引发的炎症反应。
Slp如何增强肠道屏障?
在细胞实验中,研究人员把Slp作用在Caco-2细胞(常用来模拟肠道上皮的细胞模型)上,发现它能让肠道屏障更牢固,表现为:
注:TER可以理解为用电学方法测试上皮细胞层“漏不漏”。TER越高,说明细胞之间的连接越紧密、屏障越强;TER越低,说明屏障受损、变漏了。
不同菌株的效果差异明显
一项研究比较了20多种益生菌(不同物种和不同“菌株”)对肠道屏障的影响,重点看它们能不能把肠道上皮细胞之间的紧密连接变得更牢。
结果发现嗜酸乳杆菌整体上能明显增强这种封口功能,但同一种细菌的不同菌株效果差很多。
LA1:效果最好,增强幅度接近翻倍
LA2:只有大约一半的提升
LA3:几乎没有影响
这说明:益生菌不能只看“是什么菌”,还要看“是哪一株”。
LA1的保护机制是什么?
进一步研究显示,LA1之所以有效,是因为它能被肠道免疫系统表面的识别开关(TLR2与TLR1/6组成的受体组合)识别并触发信号,从而加强屏障。用小鼠实验也看到:在正常小鼠中,LA1可以减轻一种用DSS诱导的结肠炎;但在缺少TLR2的敲除小鼠里,这种保护作用就消失了,说明LA1的效果离不开TLR2这条通路。
注:TLR-2 指 Toll 样受体 2(Toll-like receptor 2),是一种先天免疫的模式识别受体(PRR),常表达在肠上皮细胞和免疫细胞表面,用来识别微生物成分并触发信号通路。
不同细胞中的信号通路有何不同?
最近的研究发现,同一种乳酸菌成分(LA1)在不同细胞里“走的通路”不一样,所以产生的效果也不一样。
在免疫细胞里LA1会通过TLR-2这类警报器启动经典的炎症信号通路(依赖MyD88),进而激活NF‑κB,让免疫细胞更容易进入应战状态。这属于比较符合预期的免疫激活反应。
在肠道上皮细胞里LA1同样需要TLR‑2,但不走MyD88这条路,而是改走PI3K通路。结果不是“点火”,而是“刹车”——它能抵消TNF‑α这种炎症因子造成的肠道紧密连接变松、通透性升高的问题,从而降低NF‑κB和MLCK等会破坏屏障的分子活性,帮助肠道屏障更稳。
其他乳杆菌的表现如何?
L. acidophilus W37:让TER提高约15%,说明屏障更“紧”。
L. brevis、L. casei:对TER没明显影响。
注:Caco‑2 是来源于人结肠腺癌的上皮细胞系。它常被用来在体外建立“肠上皮屏障模型”,因为在 Transwell 等培养条件下会分化成类似小肠吸收上皮的单层细胞,形成紧密连接,能模拟肠屏障的通透性变化。
常用指标:TER/TEER(跨上皮电阻)和 TER/TEER,以及分子通量(如葡聚糖/荧光葡聚糖)。
有研究发现,嗜酸乳杆菌分泌的一些代谢产物,能让肠道细胞之间的“缝隙”变得更紧:细胞屏障的紧密度指标(TER)会小幅上升(约25%),而一些大分子(比如葡聚糖)从细胞间漏过去的量会减少。进一步看机制,这种乳杆菌的“培养上清液”(相当于它在培养液里释放的各种物质)能抑制炎症信号通路NF-κB的激活,并把紧密连接相关蛋白(如occludin、claudin-1)的状态恢复到更正常,从而防止炎症因子IL‑1β导致的肠道屏障“变漏”。
热灭活的嗜酸乳杆菌还有用吗?
有报告指出,即使把嗜酸乳杆菌加热杀死(热灭活),它本身以及它的上清液也能在另一种肠道细胞模型(HT‑29)里减轻阿司匹林造成的屏障损伤,主要和调节紧密连接蛋白ZO‑1的表达与位置有关。
Lcr35对肠道屏障的保护作用如何?
为了研究另一株乳杆菌Lcr35对肠道屏障的保护作用,研究人员用Caco‑2细胞在体外搭了一个“肠道屏障模型”,并在细胞下方放入免疫细胞(PBMC)模拟免疫环境。然后用沙门氏菌的LPS(会引发炎症的细菌成分)去刺激细胞,模拟感染带来的炎症损伤。结果是:只用LPS会让屏障变差;但在LPS刺激后再加入Lcr35,并培养到48小时,屏障紧密度(TER)明显回升,比仅LPS组高了87%。这提示Lcr35可能帮助修复被LPS破坏的肠道屏障、减少“肠漏”。
Lcr35如何影响炎症反应?
此外,Lcr35 还能明显减少一种炎症信号分子 IL-8 在“肠上皮细胞 + 免疫细胞(PBMC)”共培养体系中向基底侧(相当于体内朝向血液的一侧)释放,说明它有助于降低炎症反应。
干酪乳杆菌(Lactobacillus casei)
最近有研究专门看了干酪乳杆菌(Lactobacillus casei,简称 L. casei)对“肠道屏障受损”的保护作用,并探讨是否和肥大细胞有关。
结果发现:和对照相比,干酪乳杆菌能让猪肠上皮细胞的“电阻值(TER)”提高约 60%(电阻越高通常表示屏障越紧密),同时让大分子(葡聚糖)的渗漏减少 80% 以上。面对会引发腹泻的致病菌产肠毒素性大肠杆菌 K88(ETEC K88)时,干酪乳杆菌能减轻肠屏障变漏的情况,关键在于它能阻止紧密连接相关蛋白(occludin、claudin-1、ZO-1)以及免疫识别相关受体(TLR-2、TLR-4)的表达被病原体“压低”。
L. casei DN-114 001
更早的研究也发现,L. casei DN-114 001 的“菌体裂解物”(把菌体打碎后的成分)可以在小鼠 DSS 诱导的肠炎模型中起到保护作用,带来健康收益。L. casei 能抑制 DSS 导致的肠道渗漏增加,这与它降低炎症因子 TNF-α 和 IFN-γ、并改变肠道菌群组成有关。
还有研究发现,L. casei DN-114 001 在 T84 肠上皮细胞中也能对抗肠致病性大肠杆菌(EPEC)感染带来的类似屏障损伤。
L. casei CRL 431:减少感染损伤
比较了不同乳杆菌对抗沙门氏菌感染的效果,发现:
在他们测试的菌株里,只有 L. casei CRL 431 能明显起到保护作用:表现为肠屏障功能更好、局部炎症更低,从而减少感染损伤。
有研究认为,起保护作用的是活菌,不是培养上清液
活的 L. casei 可以防止 TNF-α 和 IFN-γ 导致的 TER 下降。机制上,它可能通过激活 MAPK/PI3K/Akt 这类细胞信号通路、提高 TLR-2 表达,并保护 Caco-2 细胞中 ZO-1 蛋白的水平,从而避免炎症因子引起的通透性升高。
✧ 植物乳杆菌(Lactobacillus plantarum)
有研究发现,植物乳杆菌(Lactobacillus plantarum)以及它产生的一些成分,可能通过修补肠道屏障来减轻肠道炎症(比如溃疡性结肠炎模型中的结肠炎)。如果肠道屏障损伤,肠道通透性升高,细菌或毒素更容易“漏过去”,炎症就会加重。
下面是这些研究想说明的要点:
研究提示这种保护作用和 STAT3 这个细胞信号“开关”有关:如果在细胞实验里把 STAT3 敲低,再用 EPS116 处理,ZO-1 和 occludin 就上不去,肠道通透性反而变高。说明 EPS116 需要通过 STAT3 才能更好地“加固屏障”。
植物乳杆菌可在感染情况下保护肠道屏障
在人正常结肠上皮细胞模型(NCM460)中,植物乳杆菌能减轻 ETEC(产肠毒素大肠杆菌)感染对屏障的破坏。还有研究显示它能增强 NK 细胞产生 IL-22(一种有助于黏膜修复和屏障保护的免疫因子),即使在 ETEC 感染时也能帮助维持紧密连接屏障。
不同菌株对屏障电阻提升明显
屏障电阻(TER)可以粗略理解为细胞单层的密封程度指标:TER 越高,说明屏障越紧密、渗漏越少。
L. plantarum DSM 2648:让 Caco-2 细胞 TER 比对照提高约 235%;还能显著抵消致病性大肠杆菌(EPEC)对 TER 的破坏,并减少 EPEC 黏附。
L. rhamnosus HN001:也能提高 TER(约 148%)。
L. plantarum MB452:在 10 小时内让 TER 随剂量增加,最高比对照高约 60%;同时提高 ZO-1 和 occludin 的表达。
植物乳杆菌 HY7714 可能通过关掉炎症通路,保护紧密连接
在TNF-α(强促炎因子)处理的 Caco-2 单层模型里,HY7714 能保护 ZO-1、claudin-1、occludin 不被破坏。它还抑制一些与炎症和屏障破坏相关的信号(如 NF-κB、MLCK 等)被 TNF-α 诱导升高,提示其可能通过降低炎症信号来维持屏障完整。
植物乳杆菌 LR002 改善小鼠溃疡性结肠炎
修屏障 + 抗炎 + 调菌群。在 DSS 结肠炎/UC 模型中,LR002 能恢复紧密连接蛋白(ZO-1、Occludin、Claudin-3)的水平。同时降低促炎因子、减少氧化应激;调节肠道菌群,并让短链脂肪酸(SCFA,肠道有益代谢物)水平回升。
机制上可能与激活PPARγ信号、抑制 MAPK/NF-κB 炎症通路有关。
这些研究总体指向同一件事——某些植物乳杆菌菌株或其代谢产物,可能通过增强紧密连接蛋白、降低炎症信号、改善菌群与代谢物,来减少“肠漏”和肠道炎症;不同菌株差别很大;人体研究里未必能直接看到“阻止肠漏”的效果,但在组织/细胞与动物模型中,某些植物乳杆菌确实能增强紧密连接、减轻炎症、提高屏障相关指标。
扩展阅读:
客观认识植物乳杆菌 (L. plantarum) 及其健康益处
✧ 鼠李糖乳杆菌(Lactobacillus rhamnosus)
鼠李糖乳杆菌 LGG 在儿童克罗恩病中的初步研究
较早的研究把一种常见益生菌——鼠李糖乳杆菌 LGG——用于儿童克罗恩病(CD)。结果发现:肠漏明显改善(用尿液中的双糖通透性试验来测),疾病活动度也下降了。但可惜的是,这种改善在随访到 24 周时没有持续下去。
特定菌株 CNCM I-3690 的保护机制
后来又有研究指出:鼠李糖乳杆菌的某个特定菌株 CNCM I-3690,在小鼠炎症模型里能保护肠道屏障。它可以阻止炎症因子 TNF-α 让肠上皮细胞“缝隙变大”(表现为电阻下降),还会影响杯状细胞和黏液层(相当于让肠道表面的保护胶层更稳)。
小鼠的结肠基因表达分析还显示,这个菌株可能通过抑制某些体内蛋白酶,来增强一批与健康肠道屏障相关的基因活动,比如与营养吸收、细胞更新和防护功能有关的基因。
体外实验支持 LGG 的保护作用
很多体外实验(细胞/类器官实验)也支持:活的 LGG 对紧密连接有保护作用,能让屏障在炎症或病原体攻击时更不容易被破坏。
用 TNF-α 和 IFN-γ 模拟炎症攻击 Caco-2 细胞时,活 LGG 可以减轻屏障受损,可能与抑制 NF-κB、ERK1/2 等炎症通路有关;而一些其他乳杆菌即使用更高浓度,也未必能达到同样效果。
在人肠道类器官实验中,活 LGG 能防止 IFN-γ 把关键的紧密连接蛋白压下去。
在“麦醇溶蛋白”(与小麦相关蛋白)诱导屏障变漏的模型中,LGG 能把屏障功能拉回去;不仅活菌有效,热杀死的菌体和它的培养上清(菌分泌/代谢产物的混合物)在某些实验里也显示出一定恢复效果。
在细菌感染中的保护作用
细胞先用活 LGG 处理后,再感染肠出血性大肠杆菌(EHEC O157:H),可以减轻屏障电阻下降、通透性升高,并阻止紧密连接蛋白(如 claudin-1、ZO-1)被“挪位/打散”。但热灭活 LGG 在这类实验里往往效果不明显,提示“活菌状态”可能很关键。
其他不同的 L. rhamnosus 菌株也各有差异
有的本身对屏障指标影响不大,但在大肠杆菌攻击时能起到保护;有的能抑制 TNF-α 造成的损伤并减少 IL‑8(炎症信号)上升,但它的培养上清却未必有效。
总结来说,不少实验提示某些鼠李糖乳杆菌(尤其是 LGG 及特定菌株)可能通过稳定紧密连接、维持黏液层、调节炎症信号等方式,帮助肠道不那么容易漏。但不同菌株、活菌/灭活、以及人体长期效果差异很大;早期儿童克罗恩病研究里也出现了短期改善、长期不一定维持的情况。
✧ 发酵乳杆菌(Lactobacillus fermentum)
发酵乳杆菌 AGR1487
和其他益生菌菌株不一样,发酵乳杆菌 L. fermentum 的 AGR1487 这一株在肠上皮细胞模型(Caco-2)里会削弱屏障:给得越多,细胞层的电阻(TER,常用来衡量屏障是否紧密)下降得越明显,说明细胞之间的封条变松了。
但有趣的是,如果不把细菌本身加进去,而是加入它培养后留下的上清液(可以理解为细菌分泌物/代谢产物的混合液),反而会让TER比对照培养基提高约34%,也就是屏障变得更紧。
AGR1485的不同表现
相比之下,同一物种的另一株 AGR1485(无论是细菌本体还是上清液)对TER基本没影响。
另外,AGR1487 还会让甘露醇更容易穿过细胞单层(甘露醇通量增加),这通常意味着屏障通透性变高、漏得更多;而 AGR1485 不会造成这种变化。
这意味着即使是同一种乳杆菌,不同菌株也可能作用完全不同。AGR1487 会以不利于紧密连接屏障的方式改变细胞内相关基因和蛋白(包括微管蛋白及其相关蛋白)的水平,最终导致肠道屏障功能变差。
✧ 罗伊氏乳杆菌(Limosilactobacillus reuteri)
最近一些研究发现,某些罗伊氏乳杆菌(Limosilactobacillus reuteri)可能有助于加固肠道屏障、降低肠漏风险。
高脂饮食小鼠模型中的积极效果
在高脂饮食小鼠中的发现,研究人员从人类母乳中分离到一株罗伊氏乳杆菌(L. reuteri FN041)。把它给高脂饮食(HFD)的小鼠后,发现小鼠原本因高脂饮食而更漏的肠道(肠通透性升高)得到了缓解。
与此同时,一些与炎症相关的物质(如LPS、TNF-α、IL-16)水平下降。肠道屏障的关键“封条蛋白”(紧密连接蛋白,如occludin、ZO-1、claudin-6、claudin-7)在高脂饮食下通常会减少,而FN041能阻止这种下降。研究者认为,这种益处与肠道菌群产生的短链脂肪酸变化有关(研究中提示与短链脂肪酸减少相关)。
对细胞层渗漏的抑制作用
在仔猪肠道细胞+致病菌模型中的发现,使用从健康断奶仔猪粪便中分离的罗伊氏乳杆菌,先用它“预处理”肠上皮细胞(IPEC-1),再让细胞感染致病性大肠杆菌ETEC K88。
结果发现:预处理能减少细胞屏障被破坏、降低大分子(如葡聚糖)穿过细胞层的渗漏。但如果只用罗伊氏乳杆菌而不感染细菌,效果不明显或较弱。机制上,这种保护与维持紧密连接蛋白(尤其ZO-1、occludin)有关,并且可能通过MLCK相关通路来实现。也有研究观察到:单独用罗伊氏乳杆菌处理时,细胞层的渗漏指标甚至可降低约25%。
不同菌株的效果略有差异
处理IPEC-J2细胞10小时后,细胞层的电阻(TER,数值越高通常代表屏障越紧)只小幅上升约8%,并伴随claudin-1、occludin、ZO-1轻度增加。更重要的是,在LPS(内毒素)刺激下,这株菌的活菌或培养上清都能阻止紧密连接蛋白被下调,同时抑制炎症因子(TNF-α、IL-6)过度升高。
它也能让TER上升约10%,并减少ETEC导致的屏障变漏。
研究者还发现另一种益生菌詹氏乳杆菌(Lactobacillus jensenii)也能在ETEC攻击时起保护作用,但在没有感染压力时,单独使用对屏障影响不明显。它们的保护效应可能与提升热休克蛋白HSP-27以及增加ZO-1有关,意味着这个菌株是发挥感染压力下的保护作用,而不是预防或维持作用。
扩展阅读:
认识罗伊氏乳杆菌(Lactobacillus reuteri)
✧ 唾液乳杆菌 (Lactobacillus salivarius)
L. salivarius SMXD51处理Caco-2细胞24小时后,可使跨上皮电阻(TER)提高约20%;同时还能对抗铜绿假单胞菌PAO1引起的TER下降,起到保护肠道屏障的作用。这种益生与保护效果被认为与上调F-肌动蛋白细胞骨架有关。
进一步比较了不同L. salivarius菌株对过氧化氢(H₂O₂)造成的屏障损伤的保护能力。在检测的33株菌中,UCC118和CCUG38008能通过依赖ERK1/2磷酸化的途径,减轻H₂O₂导致的紧密连接(TJ)蛋白(ZO-1、occludin、claudin-1和JAM)的降解与错位;而无效菌株AH43324则没有这种作用。
在迄今为止报道的各种益生菌细菌中,Bifidobacterium spp.是研究最广泛和使用最广泛的益生菌细菌之一。双歧杆菌属属于放线菌门,目前包括80个(亚)种,它们分布在不同的生态位,包括人类的胃肠道,口腔等。
双歧杆菌属是最早定植人类肠道的细菌,与母乳衍生的低聚糖有关。因此,它们占母乳喂养婴儿肠道中微生物占比很高。B. breve、B. bifidum、B. longum、Bifidobacterium infantis是婴儿阶段常见的检测到的细菌,其中B. bifidum是最突出的物种,其次是B. breve、B. longum、B. infantis。
随着年龄的增长,双歧杆菌的总体下降,但在整个成年期保持相对稳定,并在老年时再次下降。成人肠道中常见的物种包括B. adolescentis和B. catenulatum,其次是B. longum 和 B. bifidum。
需要注意的是,并不存在绝对的“婴儿专属种”和“成人专属种”,只是不同年龄段更常见的种类不同。
为什么双歧杆菌被当作益生菌?
双歧杆菌因为被认为较安全(很多菌株具有GRAS地位,意思是通常被认为是安全的),而且与多种健康益处相关,所以被广泛做成益生菌产品。研究中常提到的潜在作用包括:
因为双歧杆菌本来就是人体肠道的常住居民,不少研究认为它对于维持肠道上皮屏障完整性很关键。
一些研究用16S rRNA(常见的菌群测序方法)分析溃疡性结肠炎患者的肠道菌群时发现:双歧杆菌明显减少,尤其是B. bifidum减少更突出。这提示双歧杆菌(特别是B. bifidum)可能与溃疡性结肠炎的发生发展有关,也可能成为一种有参考价值的微生物标志物。
双歧杆菌怎么加固肠道屏障?
在细胞实验中(如Caco-2、HT-29、T-84等肠上皮细胞模型),用某些双歧杆菌先处理细胞,能够减轻多种因素造成的紧密连接损伤。
机制上常见的解释包括:
一个具体例子:B. bifidum BB1菌株
研究发现,B. bifidum 的某个特定菌株(BB1)能以菌株特异性的方式把肠上皮细胞单层的屏障变得更紧(用TER指标衡量,最高可增加50%–80%,TER越高通常表示屏障越紧)。
保护肠道的机制
有研究用DSS(一种常用来在小鼠体内制造结肠炎/肠损伤的化学物质)做肠道损伤模型,发现一株双歧杆菌 B. bifidum FL-228.1 的预防效果最明显。它之所以能保护肠道,主要做了几件事:
不同双歧杆菌的保护效果
其他动物研究也发现,不同双歧杆菌在多种胃肠道疾病模型中都可能对肠屏障有帮助,例如:
NEC(新生儿坏死性小肠结肠炎)模型:B. infantis能减轻肠通透性升高(不那么漏),并更好地维持 occludin、claudin-4 等紧密连接蛋白在细胞连接处的正常分布,从而降低 NEC 发生率。
另一种 NEC 大鼠模型:B. bifidum 也被证明能改善肠道完整性。
DNBS 诱导的轻度肠炎模型:B. animalissubsp. lactis 能把肠道通透性拉回正常水平,机制包括让多种紧密连接蛋白(尤其 claudin-4)恢复正常,同时帮助结肠局部免疫细胞(Th1/Th2)维持更平衡的状态。
DSS 结肠炎模型:不同菌株效果不一样。有研究显示 双歧杆菌的某个菌株(CCM 7952)能维持紧密连接蛋白表达、降低血液中“肠漏”指标(如 FITC-葡聚糖),疾病严重度也更低;但同物种的另一个菌株(Bl 372)却做不到,说明“菌株差异”很关键。
酒精相关肝病与双歧杆菌的菌株特异性
在酒精相关肝病(长期饮酒)方面,很多研究认为:长期饮酒会导致肠道菌群和代谢物失衡,使肠屏障受损、通透性上升;这样细菌内毒素 LPS 更容易进入门静脉循环,激活 TLR4-NF-κB 通路,引发更多炎症因子。
体外实验也发现,双歧杆菌可以在 LPS 处理的肠上皮细胞模型中降低炎症因子(如 IL-6、TNF-α),并上调/稳定紧密连接蛋白(occludin、claudin-3、ZO-1),从而减少细胞间渗漏。
特别需要强调的是:双歧杆菌的作用高度菌株特异性。同属甚至同种的不同菌株,效果可能完全不同,甚至在免疫反应上方向相反。因此,不能因为某一株有效,就推断所有双歧杆菌都有效。
一些更间接的保护方式
B. infantis 的“无菌条件培养上清”(可以理解为它产生的可溶性代谢物混合物)能抑制 NF-κB 炎症通路,减少 TNF-α 等炎症因子,并阻止炎症因子诱导的肠通透性升高。
B. bifidum 通过酶作用产生成分(如半乳寡糖)后,可能减少沙门氏菌对肠道的黏附和侵袭(相当于降低致病菌“粘上去、钻进去”的机会)。
最后,虽然证据很多,说明双歧杆菌对维持肠屏障有益,但分子机制为什么会这样、到底哪条通路最关键,仍然没有完全弄清。
未来研究的挑战与方向
目前有以下主要难点:
1)菌株差异太大,很难用一套机制解释所有双歧杆菌;
2)它们和人体细胞、以及肠道里其他微生物之间相互作用非常复杂;
3)双歧杆菌不太容易做遗传改造,导致研究工具受限。
未来如果有更好的分子生物学工具、更聚焦的研究,才更有希望把双歧杆菌如何与人体互作并发挥作用的关键机制讲清楚。
扩展阅读:
其他益生菌
✧ 布拉氏酵母菌 (Saccharomyces boulardii)
对肠道屏障的保护作用
较早的研究表明,布拉氏酵母菌(S. boulardii)保护T84单层中EPEC诱导的TER降低和甘露醇通量增加;然而,布拉氏酵母菌不改变健康T84细胞中的TJ屏障功能。
布拉氏酵母菌的保护作用是通过ZO-1的保存和ERK1/2信号通路的抑制介导的。
临床效果
布拉氏酵母菌在第三个月末与未接受益生菌治疗的患者相比,CD患者的肠通透性降低33%。
此外,S. boulardii CNCM I-745已显示出保护免受病原体诱导的TJ屏障功能障碍。
影响上皮屏障完整性的作用
布拉氏酵母菌共孵育有助于维持上皮屏障完整性,而预孵育是显著减少IL-8分泌所必需的。布拉氏酵母菌通过阻断MLC磷酸化并抑制T84感染细胞中的NF-κB和MAPK信号通路来保护屏障功能并抑制EHEC诱导的炎症。
对 Shigella flexneri 感染的影响
另一项研究使用人肠上皮的体外和体内模型检查了布拉氏酵母菌对Shigella flexneri感染的影响。在感染期间,布拉氏酵母菌通过上调TJ蛋白ZO-2并减少ERK、Jun N-末端激酶(JNK)和NF-κB信号通路的激活来增强屏障完整性。
抗炎作用的验证
无细胞布拉氏酵母菌上清液重现了这些抗炎作用。布拉氏酵母菌的抗炎作用在人胎儿肠异种移植模型中得到证实,其中酵母菌株减轻S. flexneri造成的损伤和炎症,但不能预防感染。
✧ E. coli Nissle 1917 (EcN)
只有活的 EcN 益生菌(以及它培养后的上清液,也就是细菌分泌物所在的液体)才能明显增强肠道上皮细胞的“紧密连接”屏障;把细菌热杀死后就几乎没效果。
细胞模型中的屏障增强
在 HT-29 和 Caco-2 细胞模型里,活菌或上清液能让细胞层的电阻(TER,代表屏障紧密程度)提高约 40%–75%,同时让甘露醇这种小分子“漏过去”的程度降低约 40%。
TcpC蛋白的作用机制
研究认为关键原因是 EcN 产生的一种蛋白 TcpC,它会启动细胞内的一些信号通路(如 PKCζ 和 ERK1/2 的磷酸化),并促进紧密连接相关蛋白(如 claudin-14)增加,从而把细胞之间的“缝”收紧。
EcN上清液对肠道屏障的保护作用
EcN 的上清液能防止化疗药 5-FU 导致的肠上皮细胞(IEC-6)屏障变差,表现为 TER 不那么容易下降。
在脓毒症相关的炎症条件下,EcN 上清液可以在体内和体外保护紧密连接屏障:它通过抑制 NF-κB 相关通路,减少会“拉开细胞缝隙”的 MLCK-P-MLC 信号被激活。并且在 TNF-α 和 IFN-γ 这类炎症因子一起存在时,EcN 上清液仍能提高 TER,从而减轻屏障功能障碍。
外膜囊泡的保护机制
EcN 释放的外膜囊泡(OMV,可理解为细菌分泌的“微小包裹”)和可溶性因子,能在 EPEC 致病菌感染时保护 T-84 和 Caco-2 细胞的紧密连接屏障,这种保护与 occludin 和 claudin-14 等紧密连接蛋白不被破坏有关。
在 DSS 诱导的结肠炎小鼠模型中,EcN 还能防止 DSS 造成的肠道通透性升高(减少“肠漏”),这与紧密连接支架蛋白 ZO-1 的表达得以维持有关。
✧ 枯草芽孢杆菌 (Bacillus subtilis)
比较了三种不同的枯草芽孢杆菌(B. subtilis)菌株对肠道上皮细胞模型(Caco-2)屏障的影响。结果发现:只有其中一株(B. subtilis 29784)能让细胞屏障更“紧”(电阻TER提高约50%),另外两株要么没效果,要么反而让屏障变差。
同时,B. subtilis 29784之所以能增强屏障,和细胞里几种“把细胞缝起来”的关键紧密连接蛋白增加有关,包括ZO-1、occludin和claudin-1。
在抗炎方面,不同菌株的效果也不一样(有明显“菌株差异”):某些B. subtilis菌株能抑制IL‑1β引发的NF‑κB信号通路激活,从而显著减少炎症相关物质IL‑8的产生,并降低IL‑1β刺激下iNOS蛋白水平的上升。
在坏死性小肠结肠炎(NEC)的细胞和动物模型中研究了两种益生菌(L. plantarum 和 L. rhamnosus)对肠道紧密连接的影响。
细胞实验结果显示屏障增强
在细胞实验里:他们把这两种菌加到 Caco-2 肠上皮细胞表面 5 小时。
结果显示:两种菌都能让细胞层更不漏(TER 升高、葡聚糖更不容易穿过去),说明屏障更紧。
当用 LPS 或 EGTA 这类物质去破坏屏障时,提前用这两种菌处理能减轻损伤,可能与调节 ZO-1(一种关键紧密连接蛋白)有关。
其中 L. rhamnosus 的保护效果看起来比 L. plantarum 更强。
动物模型中益生菌效果不一致
但在大鼠 NEC 模型里(用阪崎克罗诺杆菌感染诱导):直接给乳杆菌组合反而观察到肠道通透性增加(更漏),这与细胞实验相反。
不过如果在感染前先用这组益生菌预处理,通透性并不会增加,而且还能减少肠道损伤。
作者因此提醒:益生菌并不一定永远是“有益无害”,在某些条件下可能也会刺激或伤害肠上皮,所以临床使用特定菌株需要谨慎。
BWI多菌株益生菌的保护作用
有研究发现,一个叫 BWI 的多菌株益生菌混合物(含 8 种活菌 + 1 种热处理菌株成分)对肠屏障更稳:在 Caco-2 与免疫细胞(THP-1)的共培养模型中,它能维持 occludin(紧密连接蛋白)水平,并激活 AMPK 通路(有助于紧密连接组装),从而保护屏障完整性。
在 LPS 引发的炎症环境下,它还能抑制 NFκB 通路,降低促炎因子相关基因表达。
TER 测量也支持:BWI 能随剂量增加而更有效地防止通透性升高(减少“渗漏”)。
B. bifidum + 嗜热链球菌
一项双盲研究显示:含 B. bifidum 和嗜热链球菌(S. thermophilus) 的商业益生菌配方,能显著减少婴儿的抗生素相关性腹泻。
L. rhamnosus + L. acidophilus + L. helveticus
有研究把L. rhamnosus、L. acidophilus、L. helveticus混合给健康志愿者:当用吲哚美辛(NSAID)人为造成胃肠屏障受损时,活菌能明显减轻“胃”的通透性升高,但对“肠”的通透性影响不明显,说明作用可能因部位不同而不同。
L. helveticus R0052 + B. longum R0175
动物实验中,L. helveticus R0052 与 B. longum R0175联用,能保护心肌梗死后大鼠出现的肠道紧密连接通透性升高;但对健康对照大鼠的屏障并没有明显提升。研究者推测机制可能与抑制NF-κB炎症信号、激活抗凋亡通路以及TLR-2相关调节有关。
很多实验提示改善肠上皮屏障往往需要“活的菌”,因为它们要通过黏附、与上皮细胞直接互动等方式来触发紧密连接的调整;但在个别情况下,细菌产生的特定产物(如胞外多糖)也可能提供类似保护。不同菌株、不同部位、以及是否处在炎症/损伤状态下,都会影响效果。
肠道内壁的上皮细胞之间有一套紧密连接蛋白,像拉链/密封条一样把细胞缝隙关紧,防止细菌、毒素和未消化的大分子随意穿过去,同时又让营养正常吸收。这道屏障一旦变漏,就很容易引发或加重多种胃肠道问题,比如炎症性肠病、肠易激综合征、坏死性小肠结肠炎以及一些病原体导致的肠炎。
越来越多的细胞实验、动物研究和部分临床研究提示:某些益生菌可以帮助维持或修复这道封口带,它们可能通过以下方式保护肠屏障:
而且这些效果往往非常看菌株——同一种类的益生菌,不同菌株效果可能差很多;有时还取决于益生菌是不是活的、它分泌的代谢物、或它表面的成分。另外,它在上皮细胞和免疫细胞里起作用的方式也可能不一样。
不过,我们目前对“益生菌到底是怎样在分子层面修好TJ屏障”的理解还不够完整。
研究结果之所以经常不一致,常见原因包括:
未来更需要做的是:明确哪个菌株通过哪条通路,对哪类人群最有效,并结合肠道菌群检测的结果,分析不同菌群的组成和功能,以指导益生菌的选择。同时,应将剂量、给药方式、疗程标准化,再用设计良好的临床试验去验证这些策略的有效性。
总体来说,益生菌作为一种思路合理的干预手段,确实有潜力帮助加强肠道屏障、缓解与肠漏相关的胃肠疾病,但要实现可靠应用,还需要更扎实的机制研究和更严格的临床证据,免疫功能低下人群使用活菌也应更谨慎。
注:本账号发表的内容仅是用于信息的分享,在采取任何预防、治疗措施之前,请先咨询临床医生。
主要参考文献
Hofmann C, Geiger H. Aging of the Colon – A Mechanistic View. Mech Ageing Dev. 2025 Dec 11:112143.
Baidoo N, Rasis E, Tahiri D, Sanger GJ. The impact of ageing on the structural compositions of the mucosa of human colon. Tissue Cell. 2025 Dec;97:103090.
Abahussin HM, Alotaibi MS, Alhazzaa OA, Alotaibi AG, Alsaab SM, Aljawini NA, Alawad AO. Exploring the intricate link between gut microbiota dysbiosis and the aging process: implications for age-related diseases. Gut Pathog. 2025 Dec 14.
Elfers K, Sehnert AS, Wagner A, Zwirner U, Linge H, Kulik U, Poehnert D, Winny M, Gundert B, Aselmann H, Mazzuoli-Weber G. Functional and Structural Investigation of Myenteric Neurons in the Human Colon. Gastro Hep Adv. 2024 Aug 24;4(1):100537.
谷禾健康
糖尿病,一个日益严峻的全球健康挑战,其影响远不止血糖升高那么简单。在血糖波动的背后,看不见的慢性低度炎症,正悄然侵蚀着患者的健康,加速并发症的到来。
当我们谈论炎症时,脑海中浮现的可能是皮肤割伤后的红肿热痛,那是身体对损伤的快速、有益的防御反应,属于急性炎症;然而还有一种更隐蔽、更持久的炎症形式——慢性低度炎症,它没有明显症状,却像温水煮青蛙般长期损害我们的身体,被认为是心脏病、癌症以及糖尿病等诸多慢性疾病的共同根源。
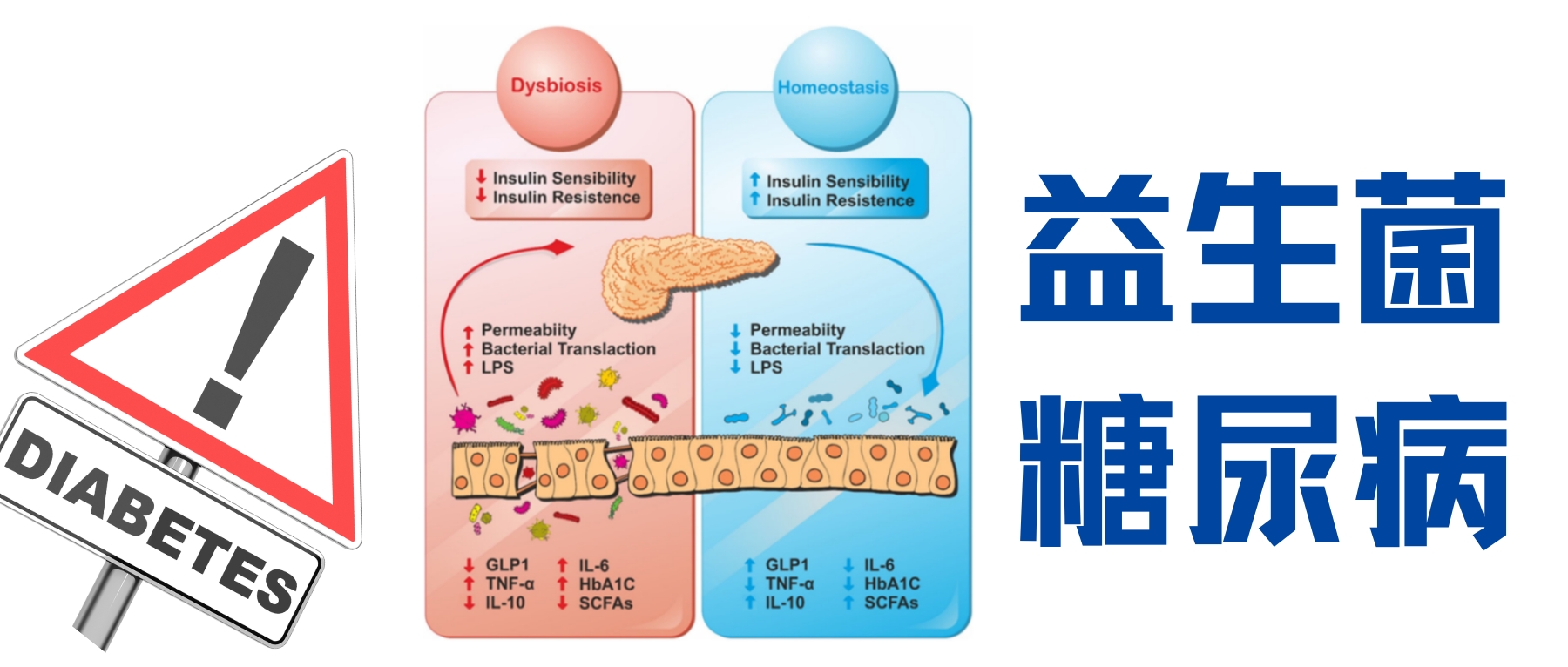
这场炎症风暴的源头可能隐藏在我们体内最复杂的微生态系统——肠道菌群中。研究发现,糖尿病患者常存在菌群失衡:有益菌(如双歧杆菌、乳杆菌)减少,潜在致病菌增多,这些改变会导致肠道通透性增加(肠漏),使脂多糖(LPS)等微生物成分进入血液,通过 Toll 样受体激活引发全身性炎症。形成“菌群失调→肠漏→炎症→胰岛素抵抗”的恶性循环。
能否通过调节肠道菌群来阻断这一恶性循环?益生菌,作为定植于肠道的友好菌群,为我们提供了新的解题思路。在代谢性疾病中常见的益生菌属包括双歧杆菌属、乳杆菌属等通过发酵膳食纤维产生短链脂肪酸(SCFAs)——如丁酸(增强肠道屏障、直接抗炎)、丙酸(调节肝脏糖代谢)、乙酸(参与全身代谢),这些短链脂肪酸具有抗炎和改善胰岛素敏感性的作用。
但益生菌的抗炎效果是否真能得到科学验证?不同菌株、剂量的干预差异何在?这些问题亟待更权威的证据解答。
近日,一篇荟萃分析发表在《Frontiers in Cellular and Infection Microbiology》上,该研究系统整合了46项随机对照试验,覆盖3580名糖尿病患者,旨在揭示益生菌及其代谢产物对糖尿病患者体内炎症标志物的真实影响。
这项研究不仅回答了“益生菌是否有效”的简单疑问,更深入探究了“哪种方案更优”、“需要多长时间见效”以及“其作用机制是否与短链脂肪酸直接相关”等核心问题。为临床实践提供了可靠的循证医学指导,也为未来精准干预策略的制定奠定了坚实的科学基础。
在糖尿病患者体内,慢性炎症状态尤为普遍。它不仅是疾病的结果,更是推动疾病恶化的帮凶。
炎症因子,作为炎症反应中的信使分子,在其中扮演了关键角色,就像一支特殊的信号部队,负责传递指令,协调免疫系统的行动。炎症因子主要分为两大类:
促炎因子
点燃和助长炎症的分子,如肿瘤坏死因子-α (TNF-α)、白细胞介素-6 (IL-6) 和 白细胞介素-1β (IL-1β)。
在糖尿病中,这些因子水平的升高会干扰胰岛素信号传导,加剧胰岛素抵抗,损害血管内皮功能,为心血管并发症埋下祸根。
抗炎因子
它们是负责对抗炎症和修复的分子,其中最著名的是白细胞介素-10 (IL-10)。它能够抑制促炎因子的产生,帮助炎症反应消退,维持免疫系统的平衡。
还有一个重要的炎症标志物——C反应蛋白 (CRP)。它由肝脏在炎症刺激下产生,是临床上常用于衡量全身性炎症水平的指标之一。在糖尿病患者中,CRP水平的持续轻度升高,预示着心血管事件的风险增加。
研究表明,在明显的临床症状出现之前,这些炎症因子的水平可能已经悄然改变。而最早发出预警信号的,可能就是我们的肠道菌群。
健康的肠道菌群平衡(共生)对维持新陈代谢和免疫稳态至关重要。然而,在糖尿病患者中,这种平衡常常被打破,出现所谓的菌群失调——有益菌(如双歧杆菌、乳酸杆菌)减少,而一些潜在的致病菌增多。
菌群失调会导致肠道屏障功能受损,即肠漏。这使得细菌的某些成分(如脂多糖LPS)更容易进入血液,触发全身性的炎症反应。因此,通过调节肠道菌群来恢复平衡,成为控制糖尿病相关炎症的一个极具前景的策略。
益生菌,即“摄入足够数量时能对宿主健康产生有益作用的活的微生物”,正是实现这一目标的有力工具。常见的用于改善代谢疾病的益生菌属包括乳杆菌属、双歧杆菌属等。
益生菌发挥作用的一个关键机制,是通过发酵我们无法消化的膳食纤维(如菊粉、低聚果糖等益生元),产生一系列代谢产物如短链脂肪酸 (SCFAs) 。其中最重要的三种是:
这些短链脂肪酸不仅在肠道局部发挥作用,还能进入血液循环,对全身的免疫细胞和代谢器官产生深远影响,被认为是连接肠道菌群与宿主免疫的关键信使。
那么,补充益生菌是否真的能增加短链脂肪酸,是否能有效降低糖尿病患者的炎症水平呢?
为了解决更深入了解以上问题,近日一项大规模的系统回顾和荟萃分析,严格遵循PRISMA指南,从全球六大数据库中检索了从2000年1月至2024年3月的所有相关研究。
经过层层筛选,最终纳入了46项符合标准的随机对照试验,共涉及3580名1型或2型糖尿病患者。这些研究评估了口服益生菌或合生元(合生元是指益生菌+益生元)对炎症标志物(CRP, IL-6, TNF-α, IL-10)和短链脂肪酸水平的影响。
(对炎症标志物和菌群代谢物的影响)
经过对46项研究数据的严谨分析,研究者们得出了相关结果。以下我们将详细解读益生菌/合生元对各项关键指标的影响。
C-反应蛋白(CRP)——显著下降
在34项报告了CRP数据的研究中,合并分析结果显示,与安慰剂组相比,益生菌/合生元干预显著降低了CRP水平(SMD = –0.54;95% CI:–0.72 至–0.35;p < 0.0001),异质性为中度(I² = 64%),这是一个中等强度的、具有临床意义的改善。
更深入的亚组分析揭示了几个关键因素:
干预时长是关键
干预时间≥8周的研究显示出更强的CRP降低效果(SMD = –0.66),而短于8周的干预效果较弱(SMD = –0.31)。这表明,调节肠道菌群并影响全身免疫系统需要一定的时间。
“1+1 > 2”的合生元效应
使用合生元(益生菌+益生元如菊粉或低聚果糖)的干预比单独使用益生菌效果更优(SMD = –0.71 vs. –0.45)。这印证了益生元为益生菌提供食物,从而增强其功能和代谢产物生成的理论。
研究者发现,体内丁酸和丙酸水平的升高与CRP的下降显著相关。这首次在如此大规模的人群证据中,将益生菌的代谢产物(SCFAs)与全身炎症标志物的改善直接联系起来,揭示了其作用的可能机制。
白介素-6(IL-6):关键促炎因子下降
IL-6是驱动胰岛素抵抗和血管功能障碍的关键促炎因子。在29项报告了IL-6数据的研究中,益生菌/合生元干预同样显示出显著的降低效果(SMD = –0.41)。
亚组分析发现,干预时长≥12周时效果更为明显(SMD = –0.51),进一步证实了长期干预的必要性。
此外,分析还提示了菌株的特异性,含有干酪乳杆菌 (Lactobacillus casei) 和 短双歧杆菌(Bifidobacterium breve) 的制剂在降低IL-6方面表现尤为突出。
多变量荟萃回归模型再次证实,即使在校正了年龄和BMI等混杂因素后,丁酸和丙酸水平的升高仍然是IL-6降低的独立预测因素。这强有力地支持了肠道来源的短链脂肪酸通过抑制细胞因子来缓解糖尿病中代谢应激的假说。
益生菌来源的SCFAs降低IL-6的机制通路
doi.org/10.3389/fcimb.2025.1677671
TNF-α:胰岛素抵抗元凶之一被抑制
TNF-α在脂肪组织炎症和胰岛素抵抗中扮演着核心角色。26项RCTs的数据显示,益生菌干预显著降低了TNF-α水平(SMD = –0.48)。分析发现了一些有趣的差异:
对2型糖尿病更有效
在2型糖尿病患者中,TNF-α的降低效果(SMD = –0.52)比1型糖尿病患者(SMD = –0.36)更显著。这可能因为2型糖尿病通常与更严重的代谢综合征和胰岛素抵抗相关,为益生菌提供了更大的改善空间。
多菌株的协同作用
使用多菌株混合制剂的干预效果优于单菌株制剂(SMD = –0.50 vs. –0.28),这提示不同菌株之间可能存在功能互补和协同增效的作用。
相关性分析再次给出了强有力的证据:
在那些同时测量了丁酸水平的研究中,丁酸水平的变化与TNF-α的变化呈现出极强的负相关(r = –0.72)。这意味着丁酸水平越高,TNF-α的下降越明显。
益生菌衍生的丁酸盐在糖尿病中调节TNF-α和炎症信号通路
doi.org/10.3389/fcimb.2025.1677671
IL-10:抗炎主力军增强
与促炎因子下降相反,抗炎因子IL-10的水平在干预后出现了显著升高(SMD = +0.38),这表明免疫系统正向着一个更平衡、更具耐受性的状态转变。在18项研究中,这一发现在统计学上非常稳健。
合生元再次显示出其优越性,其提升IL-10的效果最为强劲(SMD = +0.49)。与IL-10升高最相关的菌株包括植物乳杆菌 (Lactobacillus plantarum) 和 长双歧杆菌 (Bifidobacterium longum)。
荟萃回归分析发现,丙酸浓度的升高是IL-10增加的强预测因子。这为SCFAs不仅能抑制促炎因子,还能增强抗炎因子提供了证据,共同促进了免疫稳态的恢复。
微生物代谢物变化
21项研究直接定量了SCFAs,为上述所有发现提供了直接的生化基础。合并分析显示,干预后:
这些代谢物的增加在使用合生元和多菌株配方的研究中最为明显,特别是那些包含长双歧杆菌 (B.longum)、鼠李糖乳杆菌 (L. rhamnosus) 和 植物乳杆菌 (L.plantarum) 的制剂。
这些发现无可辩驳地证明,益生菌/合生元干预确实能够改变肠道菌群的代谢功能,增加有益的短链脂肪酸产量。而这些短链脂肪酸,正是连接肠道与全身免疫系统,发挥抗炎作用的关键介质。
稳健性于偏倚性
为了确保结论的可靠性,研究者还进行了一系列严格的敏感性分析和发表偏倚评估。结果显示,即使剔除单个研究或质量较低的研究,总体结论依然不变,表明结果非常稳健。
虽然在CRP的分析中检测到轻微的发表偏倚(倾向于发表阳性结果的研究),但经过校正后,其显著的抗炎效果依然存在。这大大增强了本研究结论的可信度。
doi.org/10.3389/fcimb.2025.1677671
这项荟萃分析的意义不仅在于学术层面,更在于为临床实践和个人健康管理提供了清晰的指导。它系统地证明了益生菌和合生元作为辅助疗法,在管理与糖尿病相关的代谢性炎症方面的巨大潜力。
研究结果反复指向短链脂肪酸的核心作用。科学界目前认为,短链脂肪酸主要通过以下途径发挥抗炎作用:
这些机制共同解释了在这项荟萃分析中,短链脂肪酸水平的升高与炎症标志物的改善呈现出如此强烈的相关性。
实践建议:如何选择和使用益生菌?
基于这项研究的发现,我们可以总结出一些给糖尿病患者的实用建议,以最大化益生菌的抗炎效益:
优先选择合生元或多菌株产品
研究明确指出,含有益生元(如菊粉、低聚果糖)的合生元产品,以及包含多种菌株(特别是乳杆菌属和双歧杆菌属)的混合产品,效果优于单一菌株的益生菌。寻找包含如干酪乳杆菌、植物乳杆菌、短双歧杆菌、长双歧杆菌等经研究验证的菌株。
保证足够的使用剂量和时长
为了达到有效的抗炎效果,建议的剂量通常在每天 109~1010 CFU(菌落形成单位)。更重要的是,干预需要足够长的时间,至少应持续 8-12周,甚至更长,才能观察到显著的免疫调节效果。
关注饮食协同
益生菌的效果离不开“燃料”。在补充益生菌的同时,增加膳食纤维的摄入(如全谷物、豆类、蔬菜、水果),可以为益生菌提供充足的食物,促进短链脂肪酸的产生,达到事半功倍的效果。
对于那些已经存在明显代谢性炎症(如CRP/IL-6升高)、中心性肥胖或非酒精性脂肪肝(NAFLD)的糖尿病患者,益生菌干预可能带来更大的获益。
作为辅助而非替代疗法
益生菌是一种辅助管理策略,不能完全替代标准的降糖药物和生活方式管理,益生菌可以从源头改善,通过调节肠道微生态,靶向慢性炎症,缓解糖尿病,且减少并发症的风险。
益生菌与常规治疗相结合,共同控制疾病。益生菌创造的低炎症、更健康的肠道环境,可能有助于提高身体对降糖药物的反应,并优化健康饮食的营养吸收效果。这种协同作用,最终可能带来比单一策略好得多的整体管理效果。
肠道菌群与糖尿病炎症的深度关联,已从基础研究走向临床转化的关键节点。益生菌通过调节短链脂肪酸代谢实现抗炎降糖,但“菌株选择、剂量优化、疗效预测”等核心问题,始终依赖对个体菌群特征的精准洞察。
益生菌通过重塑肠道微生态、修复肠道屏障、抑制慢性炎症,为2型糖尿病的辅助管理开辟了充满希望的新途径。它从源头上干预了由菌群失调驱动的代谢紊乱,展现出巨大的应用潜力。
从盲目补充到精准调养
肠道菌群检测将成为个性化健康管理的起点。通过检测,你能清晰了解自身菌群结构(如双歧杆菌、乳杆菌等有益菌丰度)、短链脂肪酸代谢能力及炎症风险,从而告别试错式补充,精准匹配最适合的菌株与饮食方案(如搭配益生元增强效果),让益生菌的抗炎潜力得到最大释放。
从单一产品到生态协同
人群研究数据(如干预≥8周效果提升、合生元方案优于单一益生菌)揭示,肠道菌群调节需“检测-干预-跟踪”的闭环支持。
主要参考文献
Xie Y, Zheng Y, Jiang F, Cai X. Meta-analytical insight on probiotic metabolites and inflammatory markers in diabetes. Front Cell Infect Microbiol. 2025 Sep 22;15:1677671.

谷禾健康
自闭症谱系障碍(ASD)是一种神经发育障碍,越来越多的研究表明它与“微生物群-肠道-大脑轴”的功能失调有关,但其中起因果作用的微生物介质和分子机制仍不清楚。
2021年,谷禾团队参与构建并分析包含上千名中国儿童的大规模ASD队列,证实ASD儿童的肠道菌群发育早期即发生偏离,该研究成果发表于《Gut》期刊上。
近日,刘星吟教授研究团队基于之前发表的ASD队列研究,发现患有ASD的儿童体内乳酸杆菌属(Lactobacillus)的减少与更严重的肠胃道症状和社交缺陷相关。孕期母体免疫激活(MIA)已被确定为ASD的一个关键环境风险因素。在MIA诱导的ASD小鼠模型中,补充植物乳杆菌(Lactobacillus plantarum)或其衍生物苯甲酸钠(NaB),能够减轻自闭症小鼠的肠道菌群失调,改善其社交行为缺陷、谷氨酸-谷氨酰胺水平以及神经元活动异常。
单细胞RNA测序显示,苯甲酸钠(NaB)恢复了自闭症小鼠星形胶质细胞中Cxcl16等基因的表达,这与神经元和星形胶质细胞之间的谷氨酸代谢活动有关。该研究成果近日发表《Molecular Psychiatry》。
该研究构建了一个完整和严谨的证据链,从大规模人群的菌群分层观察,到代谢物鉴定,再到动物模型的因果验证和分子机制探索。它成功地回答了“是什么”(菌群差异)、“产生了什么”(苯甲酸钠)、以及“如何起作用”(调节星形胶质细胞等方面)的完整科学问题,是肠-脑轴领域从关联性研究向因果机制研究转变的典范。
本文将带大家深入了解这项研究及其对自闭症儿童未来基于菌群治疗干预的潜在价值。
为了实现上述目标,研究团队设计了严谨的多层次研究方案,并在方法学上展现出显著的创新性。
队列设计与分层策略
该研究利用了前期已建立的大规模队列,包括773名ASD儿童和448名年龄、性别匹配的正常发育(NT)儿童。
研究者基于16S rRNA测序数据,检测了每位受试者肠道中乳杆菌的相对丰度。他们定义了一个阈值(例如,相对丰度 > 0),将773名ASD儿童分为两个亚组:
这一分层策略在于,它将宏观的“ASD菌群失调”问题,转化为一个更具体、更可控的比较生物学问题,即比较NT、ASD-L和ASD-WL三组之间的差异,更精准评估乳杆菌在ASD肠道微生态中的独特作用。
多组学分析与网络构建
研究团队采用了多种先进的分析方法来揭示菌群结构和功能的改变:
PM2RA微生物网络分析
采用自主研发的PM2RA分析框架,构建并比较了三组(NT, ASD-L, ASD-WL)的微生物互作网络,揭示了乳杆菌作为对网络稳定性的影响。
宏基因组功能分析
在明确了网络结构差异后,研究进入功能层面。利用PICRUSt2等工具,分析了与肠道菌群相关的肠-脑功能模块(GBMs)和肠道代谢模块(GMMs)的活性变化,锁定了与神经递质和能量代谢相关的差异通路。
注:GBMs特指那些与神经递质合成、代谢(如多巴胺、GABA、色氨酸代谢)相关的通路,是连接肠道与大脑功能的桥梁。
靶向代谢组学验证
在功能预测的基础上,采用靶向气质联用(GC-MS)技术,直接定量检测了粪便和脑组织样本中关键代谢物(如苯甲酸钠)的浓度,实现了从“功能预测”到“实测验证”。
动物模型的功能验证
为了探究因果关系和分子机制,研究团队使用了母体免疫激活(MIA)小鼠模型。
注:该模型通过在母鼠孕期注射Poly(I:C)来模拟病毒感染,其后代会表现出类似ASD的核心行为缺陷。
益生菌与代谢物干预
对MIA模型小鼠分别灌胃植物乳杆菌(L. plantarum)或其代谢产物苯甲酸钠,并评估其对肠道菌群、肠道通透性及ASD样行为(社交缺陷、重复刻板行为)的改善效果。
单细胞转录组测序 (scRNA-seq)
对不同处理组小鼠的海马组织进行10x Genomics单细胞测序,以细胞类型特异性的方式解析苯甲酸钠对大脑基因表达谱的影响,从而锁定关键的细胞亚群(星形胶质细胞)和靶基因(Cxcl16)。
机制探索与功能验证
在证明苯甲酸钠能够改善行为后,研究者进一步探索其作用机制。他们检测了苯甲酸钠是否能穿过血脑屏障,并分析了其对大脑特定脑区(如下丘脑、前额叶皮层)中与社交行为密切相关的基因表达的影响。
这一系列从人体观察到动物模型验证,再到细胞、分子及表观遗传机制的深入探索,为揭示苯甲酸钠在肠-脑轴中的关键作用提供了强有力的证据。
该研究的核心发现层层递进,描绘了从菌群定植到行为改善的完整路径。
与没有乳杆菌定植的ASD儿童(ASD-WL)相比,ASD-L组携带有益共生菌(如双歧杆菌)的可能性更高,携带机会性致病菌(如Odoribacter)的可能性更低。
1
人群发现:乳杆菌缺失加剧ASD儿童肠道生态失调与功能异常
研究的第一个重要发现是,在ASD儿童群体中,乳杆菌的定植与否,是决定其肠道微生态网络结构和功能的关键变量。通过PM2RA网络分析,研究者发现:
菌群结构失调
与健康对照组(NT)相比,ASD-WL组(无乳杆菌)的微生物网络表现出最严重的失调,其网络复杂性和稳定性均显著降低,节点间的连接变得稀疏和脆弱。这表明缺乏乳杆菌的肠道生态系统更为混乱和不稳定。
改变的微生物群落网络:
ASD-WL与NT 和 ASD-L与NT
红色节点代表ASD组中上调的微生物群,绿色节点代表自闭症谱系障碍组中下调的微生物群。
ASD-L、ASD-WL和NT的微生物网络改变程度
ASD-L组(有乳杆菌)的网络结构则呈现出一种介于NT组和ASD-WL组之间的“中间态”或“恢复态”,乳杆菌的定植可能部分“纠正”或“缓冲”了ASD状态下的严重生态失调,使其网络结构向健康状态靠近。
代谢功能异常
功能层面的分析(GBM和GMM)进一步揭示了这种结构差异背后的功能后果。
在ASD-WL组中,多个与神经系统功能密切相关的肠-脑模块(GBM)出现显著异常。
更重要的是,这种代谢功能的失衡与ASD核心症状的严重程度(如ADOS评分)以及胃肠道不适(如便秘)显著相关。
而在ASD-L组中,这些与神经递质和胃肠道功能相关的代谢通路则与NT组无显著差异。
这些结果首次在大型队列中证实,乳杆菌的缺失与ASD儿童更严重的神经-胃肠道共病表型紧密相连,其机制可能在于调节了整个菌群的代谢产出。
乳杆菌定植和GMM及其与肠道不适的关系
这些发现的意义在于,它超越了简单地将乳杆菌视为一个有益菌的层面,而是揭示了乳杆菌作为网络核心,能够重塑整个生态系统的结构。这为理解益生菌如何发挥作用提供了全新的网络视角。
2
动物模型验证:植物乳杆菌及其关键代谢产物苯甲酸钠可逆转ASD样行为
为了验证人群中的关联发现并探究因果关系,研究者在MIA诱导的ASD小鼠模型中进行了干预实验。
首先,他们给MIA小鼠灌胃了在人群研究中富集的植物乳杆菌(L. plantarum)。结果显示:
菌群与代谢重塑
植物乳杆菌干预改变了小鼠的肠道菌群组成,能显著逆转MIA模型引起的肠道菌群结构失调。
功能通路富集分析显示,植物乳杆菌恢复了MIA小鼠中异常的氨基酸代谢通路。
行为改善
这种肠道层面的改善直接转化为了行为上的获益。植物乳杆菌干预显著改善了MIA小鼠的重复刻板行为(图G)和社交缺陷(图H,I)。
那么,植物乳杆菌是通过什么物质来发挥作用的呢?
研究团队通过代谢组学分析其培养上清液,发现其中富含一种神经活性代谢物——苯甲酸。随后,他们直接给MIA小鼠灌胃其更稳定、易于使用的盐式——苯甲酸钠(NaB),发现苯甲酸钠同样能够显著改善MIA小鼠的社交新颖性偏好和重复刻板行为(下图I, J)。
研究发现,粪便中苯甲酸钠的水平与ASD的核心症状严重程度(通过ADOS和ABC量表评估)呈现显著的负相关。也就是说,苯甲酸钠水平越高的ASD儿童,其社交障碍和重复刻板行为的症状越轻。
这一结果有力地证明:
苯甲酸钠是介导植物乳杆菌改善ASD样行为的关键效应分子之一。
3
机制解析(一):苯甲酸钠修复肠道屏障、穿透血脑屏障并纠正脑内神经病理
为了厘清苯甲酸钠的作用路径,研究者首先考察了它对肠-脑连接的影响。研究发现,苯甲酸钠通过两个关键步骤连接肠道和大脑:
MIA小鼠表现出肠道通透性显著增高,即“肠漏”现象,这可能导致肠道内有害物质进入血液。
而苯甲酸钠(NaB)干预能够有效降低MIA小鼠异常增高的肠道通透性,修复受损的肠道屏障功能,从源头上减少了潜在的负面影响。
作为一种小分子代谢物,苯甲酸钠(NaB)能够成功穿过血脑屏障,并在脑组织中有效富集。
这一发现至关重要,它打通了从肠道到大脑的直接物理通路,证实了肠道微生物代谢产物可以直接进入中枢神经系统发挥作用。
▶▷
进入大脑后,苯甲酸钠对MIA小鼠的神经病理状态产生了广泛的纠正作用。MIA小鼠表现出典型的神经病理特征,包括:
而补充植物乳杆菌或其代谢产物苯甲酸钠,均能显著缓解这些神经病理学上的异常现象,使大脑从一种“应激”和“失衡”的状态中恢复过来。
3
机制解析(二):苯甲酸钠通过调控星形胶质细胞Cxcl16,恢复谷氨酸稳态
为了找到苯甲酸钠在大脑中发挥作用的具体靶点,研究者利用单细胞RNA测序技术,高精度地解析了苯甲酸钠对海马区不同细胞类型的影响。结果清晰地指向了一类关键的非神经元细胞——星形胶质细胞。在MIA小鼠中,这类细胞的基因表达谱发生紊乱,而苯甲酸钠处理能够特异性地“纠正”这种紊乱。在众多被纠正的基因中,Cxcl16基因的变化最为显著。
与人群研究中观察到的谷氨酸代谢通路异常相呼应,MIA小鼠脑组织中的兴奋性神经递质谷氨酸(Glu)和谷氨酰胺(Gln)水平异常升高,提示大脑处于过度兴奋状态。而植物乳杆菌和苯甲酸钠干预均能显著降低其水平,使其恢复至正常。
星形胶质细胞正是调控脑内谷氨酸循环的关键。
一系列精巧的实验证实了Cxcl16的核心地位:在MIA小鼠海马的星形胶质细胞中特异性地过表达Cxcl16,可以直接恢复其社交缺陷和异常的神经元活动模式。
星形胶质细胞中的Cxcl16是介导肠道代谢物苯甲酸钠改善社交行为的关键分子。
3
机制解析(三):苯甲酸钠通过H3K27表观遗传修饰诱导Cxcl16表达
最后,来自肠道的代谢物苯甲酸钠,是如何在大脑中调控Cxcl16基因表达的?研究揭示了一个全新的表观遗传学通路。
研究发现,苯甲酸钠(NaB)可以作为底物,参与一种新型的组蛋白翻译后修饰——组蛋白H3第27位赖氨酸的苯甲酰化(H3K27bz)。
苯甲酸钠处理显著提升了星形胶质细胞内H3K27bz的水平,并且这种修饰特异性地富集在Cxcl16基因的调控区域,从而打开了该基因的转录。
这一发现首次将肠道微生物代谢物与宿主大脑中的新型表观遗传修饰直接联系起来,为理解肠-脑轴的调控深度提供了开创性见解。
科 学 意 义
提出ASD分层干预的新范式
该研究强调了ASD的异质性,并提出基于微生物特征(如乳杆菌定植)进行分层的精准医疗概念。这提示我们,未来的干预措施可能不适用于所有ASD患者,而是需要为具有特定肠道微生物特征个性化定制。“ASD-L”和“ASD-WL”的分组,可能代表了两种不同的病理生理亚型,需要不同的干预策略。
发现内源性神经调节剂
苯甲酸钠作为一种由特定菌群结构产生的、能够调节大脑关键社交通路的内源性分子,为我们理解共生微生物如何塑造宿主行为提供了全新的视角。未来可以挖掘更多能够调节宿主神经系统的神经活性代谢物(Neuroactive Metabolites)。
局 限 性
研究虽有重大突破,但仍有一些问题有待未来研究深入探讨:MIA小鼠未能完全模拟ASD;需在其他遗传模型验证NaB效果,评估其高浓度安全性;NaB仅部分改改善ASD行为,该益生菌产生的其他代谢物(如吲哚-3-乳酸)可能也发挥了协同作用;其作用机制和Cxcl16染色质可及性等都需进一步研究。
实 际 应 用
开发新型益生菌或合生元
针对缺乏乳酸杆菌的ASD-WL亚群,开发能够高效定植并产生苯甲酸钠的下一代益生菌或合生元产品,将成为极具前景的治疗策略。
苯甲酸钠作为潜在治疗药物
苯甲酸钠本身是一种FDA批准的食品防腐剂和临床药物,其安全性已得到广泛验证。这意味着将其重新定位为治疗ASD社交缺陷的药物,其临床转化路径可能相对较短。
诊断与分层标志物
肠道菌群健康检测,可以测粪便中的乳杆菌定植情况,有望成为筛选适合接受特定微生物疗法或苯甲酸钠疗法的ASD儿童的生物标志物,从而实现精准干预。
参考文献:
Li T, Chen J, Mi K, Lei J, Ning Z, Lu L, Su Z, Zhu Q, Lin M, Wang Y, Wang J, Xu R, Khaib L, Knight R, Hu Z, Liu X. Probiotics derived sodium benzoate improves social behavior of offspring exposed in the maternal immune activation through regulation of histone lysine benzoylation in astrocytes. Mol Psychiatry. 2025 Aug 25.
Lou M, Cao A, Jin C, Mi K, Xiong X, Zeng Z, Pan X, Qie J, Qiu S, Niu Y, Liang H, Liu Y, Chen L, Liu Z, Zhao Q, Qiu X, Jin Y, Sheng X, Hu Z, Jin G, Liu J, Liu X, Wang Y. Deviated and early unsustainable stunted development of gut microbiota in children with autism spectrum disorder. Gut. 2022 Aug;71(8):1588-1599.
谷禾健康
在过去十年中,临床微生物组研究的格局发生了巨大变化。通过利用体内和体外实验、多组学方法和计算生物学,发现了许多详细的作用机制和微生物关联指标,并确定了在许多疾病和治疗方式中改变微生物组的有效方法。
尽管微生物组领域的临床转化仍处于早期阶段,其基础性发现正稳步为医疗领域带来变革性应用奠定基础。重要的是,微生物组的相关发现从基础研究到转化的速度已超过许多其他重要的新兴生物医学进展。例如,癌症免疫疗法的相关发现是在第一种获批疗法面世前50年取得的,而癌症免疫疗法在过去十年极大地革新了癌症治疗。
同样,推动mRNA疫苗发展的尿嘧啶修饰创新发生在首次人体试验前15年(若非新冠疫情的紧迫需求,这一过程本会耗时更长)。临床微生物组研究在过去20年取得了实质性进展,正从相关性走向因果性和干预,并已催生首批FDA批准的疗法。
驱动微生物-微生物及微生物-宿主相互作用的具体功能机制正被越来越精准地揭示,这为药物靶点和患者群体的选择带来了改进。
这些进展,加之监管环境的日益完善,正在切实降低基于微生物组的治疗和诊断的临床转化风险。
本文介绍和讨论微生物组研究成果如何推进临床实践的关键研究示例。讨论将微生物组数据整合到常规临床实践、精准医疗以及个性化医疗保健和营养方面的主要挑战和机遇的前瞻性观点。
过去几年微生物如火如荼的研究通过有效使用微生物培养和临床前建模以及宿主和微生物基因组学、转录组学、蛋白质组学和代谢组学以及临床前和临床观察或干预试验中的高级生物信息学分析,阐明了微生物组在调节宿主免疫、代谢甚至行为中的作用。
但是将微生物组整合到临床实践中需要以下一项或多项组合:
这项机制、指标和调节因素原则可用于评估转化医学的进展,并明确在任何特定临床实践推广应用中所面临的挑战。
微生物组通过特定的生物化学和物理相互作用对宿主产生影响,这些作用主要通过免疫、神经和内分泌系统以及代谢合作与拮抗来发挥。可以通过研究这些相互作用,确定特定的药物靶点,从而阻断或增强微生物功能,进而促进健康结果。
统计学和机器学习方法可用于基于关联性识别潜在的微生物因素,这些因素可以预测或诊断健康或疾病,从而能够开发微生物群健康指数。
比较幸运的是,现在已经可以指出一些能够诊断疾病或指示治疗成功的微生物指标,但定义一个“健康的”微生物组仍然很困难。这是因为与健康或疾病相关的参考范围会受到诸如地理位置、年龄、饮食、生活方式和遗传等无数混杂因素的影响。
饮食、益生元、噬菌体、益生菌、活生物治疗剂、药物(包括但不限于抗微生物剂)和生活方式改变已成功用于通过微生物调节解决症状和改善宿主-微生物协同作用,尽管成功的个体间差异很大。未来使用考虑个体微生物组和现实生活方式干预的个性化微生物组干预将有助于缩小研究与实践之间的差距。
这三种模式(机制、度量和调节因素)中的每一种都有助于一定程度的临床实施,因此并非所有模式都适用于特定应用。自过去 5 年的研究示例,这些示例为这三种模式提供了令人信服的示例,其中一些已经成熟,可以临床采用。
这些如何(或可以)转化为临床实践,包括开发基于微生物组的诊断、治疗和营养策略。
接下来本文将按照身体部位组织分开讲述。
阴道微生物组如何构建天然防御屏障?
以乳杆菌属(Lactobacillus)为主的阴道微生物组创造了一个酸性环境,防止感染并支持平衡的微生物生态系统。
同一种微生物“亦敌亦友”
解码微生物群中的菌种甚至菌株水平基因型和表型变异,有助于识别致病潜力的差异。
从母体阴道到婴儿肠道的免疫保护
乳酸杆菌属的菌种和菌株会附着在阴道上皮细胞上,帮助维持3.5~4.5 之间的 pH 值,从而防止与细菌性阴道病 (BV) 和性传播疾病相关的病原体过度生长。
它们还可以跨代传播到新生儿的肠道微生物组,可遗传菌株在人类抗原呈递细胞上表现出免疫抑制表型。因此,确定成功传播且在调节炎症和粘附上皮方面更有效的乳酸杆菌种类和菌株,尤其是在不同种族和社会文化背景的女性中,应该是提高活体生物治疗药物和预防策略的疗效和特异性的研究重点。
结合临床观察和指标综合判别诊疗阴道感染,因为由于菌株的普遍性而变得复杂,这些菌株可能具有共生或感染作用,具体取决于环境。
同一种菌株在不同环境下——不同的行为特征
例如,加德纳菌属(Gardnerella)历来被标记为致病性,但已被证明在某些人中是良性的,其丰度会随着月经周期的变化和避孕药的使用而波动,进一步说明了微生物与宿主关系的动态性质。
菌株亚型:疾病风险的预测指标
使用阴道微生物指标需要对微生物组的地理、种族和时间动态进行广泛评估,以明确哪些分类群或菌株与健康或疾病相关。
B组链球菌(GBS)的研究为我们提供了理解微生物精细分类的重要案例。这种与新生儿脑膜炎相关的常见病原体,通过其VIIb型分泌系统的特征分析,已被识别出四种不同亚型,这些亚型可用于直接治疗和减少传播,尤其是免疫系统尚未完全发育的新生儿。
抗生素过度使用的隐忧
目前在美国,每预防一例新生儿 GBS 败血症,就有 200 名母亲及其婴儿接受产前高剂量抗生素预防。更好地定义微生物风险可以减少不必要的抗菌药物给药,而且鉴于早期抗菌药物给药、微生物组扰动和儿童慢性病发展之间的关系,这一点至关重要,以减少过度医疗干预。
阴道菌群治疗的创新前沿
许多临床疗法旨在通过调节 pH 值或雌激素或通过益生菌来重建主要的乳酸产生菌株。
——阴道菌群移植
在一项针对 5 名患者的研究中,该移植导致 BV 消退,缓解持续至少2年。
阴道微生物组移植也在单个案例研究中被探索为一种不孕症治疗方法,通过调节阴道 pH 值来提高精子活性,从而提高妊娠成功率。
阴道菌群干预:从益生菌到噬菌体技术
基于阴道的益生菌乳酸菌也用于诱导酸性 pH 值以降低 BV 风险或治疗疾病,但噬菌体对引入物种的捕食可能会使治疗复杂化。噬菌体在临床上也可用于治疗BV,公司现在正在将噬菌体和 CRISPR 方法相结合来治疗顽固性大肠杆菌尿路感染。
目前,越来越有机会利用从大规模阴道微生物组研究中获得的知识来推进精确的阴道微生物组干预,例如下面的ISALA 项目,这些研究将微生物群落分析与更深入地了解激素、生殖阶段、免疫反应(例如,GBS 特异性抗包膜抗体)、环境暴露、饮食、行为和个人卫生产品如何改变阴道微生物组。

随着测序技术的进步,阴道菌群检测已从传统的培养法发展为利用高通量测序技术,使得菌群的构成分析更加全面。这些先进检测手段使临床医生能够获取阴道微生物组的完整图谱,包括多样性指数、优势菌种构成以及潜在致病菌的存在。
结合人工智能算法,这些检测数据可转化为个体化风险评估和治疗建议,将传统的”一刀切”治疗方案转变为基于个体微生物特征的精准干预策略。
扩展阅读:
胃肠道(GI)是宿主-微生物组-环境相互作用研究最广泛的生态位之一。这种串扰的结果往往超越了肠道,以系统的方式调节健康和疾病。
新生儿肠道微生物组
新生儿肠道微生物组如何形成?
新生儿出生时,肠道微生物主要来自母亲的阴道和肠道菌群。自然分娩的婴儿通过产道接触母体微生物,而剖腹产婴儿则缺少这一过程,导致其肠道菌群与自然分娩婴儿不同。

对这些队列的后续分析还确定了来自父亲微生物组的贡献。
这些发现表明上述两种方法可能有助于改善剖腹产婴儿的肠道健康,但要有效地将这些方法转化为临床应用,还需要:
母乳喂养如何塑造婴儿肠道菌群?
婴儿肠道微生物群的改变也可能来自营养和母乳的获取。
母乳含有多种成分,对婴儿肠道微生物组的建立与调节至关重要:
这些机制为新生儿重症监护病房营养计划以及富含关键母乳低聚糖、益生菌菌株、抗菌肽和 IgA 的合成牛奶替代品的开发提供信息,以模拟天然母乳的效果。母乳的动态成分(随时间变化)是复杂生物系统,现有合成配方难以完全模拟其效果。
◐ 纯母乳喂养的婴儿肠道菌群发展更规律
◑ 前3个月断奶使用配方奶粉会破坏这一发育过程,可能导致微生物群过早成熟,增加哮喘等疾病风险

CHILD 队列最近的一项研究使用因果模型来识别婴儿肠道微生物组中的生物体(如活泼瘤胃球菌Ruminococcus gnavus)和功能(如色氨酸代谢),这些生物体在机制上将纯母乳喂养与预防哮喘联系起来,为临床转化提供了具体目标。
关于活泼瘤胃球菌详见我们之前的文章:活泼瘤胃球菌(Ruminococcus gnavus)——多种疾病风险的潜在标志物
色氨酸代谢详见:色氨酸代谢与肠内外健康稳态
益生菌在新生儿健康中的作用与风险
近期几项研究已经确定了针对新生儿胃肠道微生物组的临床相关治疗的作用机制。
一个典型例子是益生菌——婴儿长双歧杆菌亚种(Bifidobacterium longum subsp. infantis):
——争议案例
虽然B. infantis在改善营养不良方面显示出潜力,但在特定人群(如早产儿)中使用时需要格外谨慎,可能存在安全隐患。
最近发生了一起与Evivo产品(含有B.infantis EV001和中链甘油三酯[MCT]油)相关的早产儿死亡事件。基于此事件,美国食品药品监督管理局(FDA)向临床医生发出警告信,提醒在新生儿重症监护病房(NICU)使用益生菌的潜在风险。

——循证医学
大型临床试验支持益生菌在早产儿中的使用是安全且有益的,可以降低健康并发症的风险,比如坏死性小肠结肠炎。
具体来说,2023年的一项涉及106个试验、共25,840名早产儿的荟萃分析发现,益生菌(包括婴儿双歧杆菌菌株)与全因死亡率的显著降低相关。特别是多菌株益生菌能够将死亡率降低高达31%。
这些研究结果表明,在早产儿护理中适当使用益生菌可能是一种有效的预防性干预措施,有助于改善早产儿的健康结果和生存率,但需要多项评估,评估人群适用性,避免过度依赖单一菌株。
成人肠道微生物组
成人肠道微生物组通过多种途径影响健康:
微生物组紊乱与疾病
当肠道微生物组遭到破坏,微生物之间及微生物与宿主之间的功能性相互作用扰乱时,可能导致疾病发生,例如:
这类功能紊乱可以通过基于微生物组的干预措施进行纠正,正如口服微生物组疗法VOWST(原名SER-109)在三期随机对照试验中所证明的那样。
微生物组与药物互作
肠道微生物组与环境(包括饮食)之间的相互作用十分复杂,并且与个体的微生物代谢功能能力密切相关,这对临床转化和潜在患者群体干预措施的安全性有重要影响。这种异质性可能影响微生物与药物的相互作用,进而影响药物疗效。
而通过给予次级胆汁酸熊去氧胆酸作为治疗手段可以逆转这一过程。

为什么药物对人效果不一?探索药物-微生物群相互作用对效果的影响
癌症、炎症和肠道微生物组
肠道微生物组如何影响免疫治疗的效果?
黑色素瘤:多样性低→免疫疗法成功率低
产丁酸菌高、益生菌高→免疫疗法成功率高
肠道微生物多样性低已被证明是晚期黑色素瘤免疫治疗失败的可靠预测指标。
在一项包含175名接受免疫治疗的黑色素瘤患者的队列研究中,肠道相关的产丁酸菌(Agathobaculum butyriciproducens)和益生菌干酪乳杆菌(Lactobacillus gasseri)的丰度可预测12个月无进展生存期的增加和增强的抗癌免疫力,而普雷沃氏菌属(Prevotella copri)则与更短的无进展生存期和炎症相关。
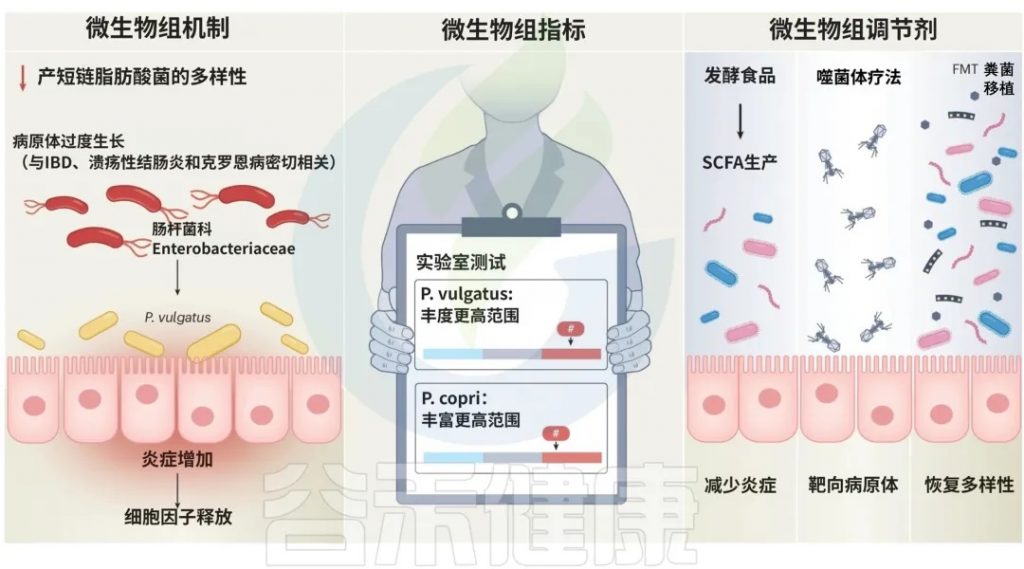
炎症性肠病:与P. copri 、Enterobacteriaceae有关
P. copri 已被强调为一种特别依赖环境的细菌,与正面和负面的健康结果都有关联。实际上,P. copri 菌株的比例以及更普遍地说,肠杆菌科(Enterobacteriaceae)细菌,已被密切联系到炎症性肠病(IBD)的发展和进展,包括溃疡性结肠炎(UC)和克罗恩病。溃疡性结肠炎患者可以根据粪便中源自Phocaeicola vulgatus 菌产生的蛋白酶丰度进行预后评估,这些蛋白酶通过破坏上皮屏障并导致炎症而与疾病严重程度相关。
如何通过靶向干预改善癌症治疗响应?
——噬菌体疗法
有针对性地消除促发疾病的菌株可改善患者预后,例如,一种针对肺炎克雷伯氏菌(Klebsiella pneumoniae)特定分支的噬菌体混合物在炎症性肠病患者体内富集,在临床前模型中能够抑制肠道炎症,并在一项I期人体试验中被证明是可行且安全的。
全面认识——肺炎克雷伯菌 (Klebsiella pneumoniae)
PHAGE-2研究将益生菌(动物双歧杆菌Bifidobacterium animalis subsp. lactis BL04)与一组针对大肠杆菌(E. coli)的噬菌体混合物结合,用于改善68名健康成人的胃肠道健康。这项随机对照试验发现,这种组合能显著减少胃肠炎症和结肠疼痛,改善粪便稠度,并增加乳酸杆菌的丰度,同时不影响整体肠道菌群多样性。
doi.org/10.1038/s41591-025-03615-9
来自动物研究和小型人类队列的有力证据表明,饮食对微生物组的调节可以影响癌症治疗中患者对免疫检查点阻断(ICB)的反应。一项研究发现,在128名接受ICB治疗的患者中,更高的膳食纤维摄入改善了无进展生存期,而益生菌削弱了这种影响,这在临床前模型中得到了证实。
——粪菌移植 (FMT)
也在一项包含16名患者的试验中进行了测试,其中从对黑色素瘤的抗PD-1治疗有成功结果的个体中选择捐赠者,为那些同一治疗失败的个体进行移植。来自响应者的FMT结合抗PD-1治疗在15名可评估患者中的6名显示临床益处,增加了CD8+ T细胞活化并减少了表达白细胞介素-8(IL-8)的髓样细胞。
——广谱抗生素
当在嵌合抗原受体T细胞输注前使用时,与较差的ICB结果相关,无进展生存期和总体生存期降低。
——饮食
饮食作为调节因素也被探索用于其他癌症类型。试验证明,食用海军豆增加了肠道微生物多样性和哌啶酸浓度,并降低了血清吲哚衍生物,这逆转了在肥胖和结直肠肿瘤患者中观察到的特征。
这表明通过使用饮食、FMT或益生菌来修改微生物组,并减少抗生素介导的修改,可能在癌症预防和治疗中起关键作用。
注:海军豆原产于美洲,椭圆形略扁,豆粒呈白色,是美国海军中水手的主食。
肠道微生物组和心脏代谢健康
肠道微生物如何促进肥胖?
——Megamonas rupellensis降解肌醇、脂质吸收增强
研究表明肠道微生物组的某些特征有助于促进肥胖。其中一种机制涉及Megamonas rupellensis主动降解肠道结肠细胞脂质膜中的肌醇,这减少了肌醇衍生的对脂肪酸摄取通路的抑制,从而增加脂质吸收。
肌醇缺乏还与胰岛素抵抗和多囊卵巢综合征相关,这表明该机制可能有更广泛的作用。
——微生物酶与能量提取
其他促肥胖的机制还包括微生物糖苷水解酶的作用。微生物产生的糖苷水解酶可以分解人体无法消化的复杂碳水化合物,增加宿主可利用的能量摄入。这种额外的能量提取能力是微生物组促进肥胖的另一重要机制。
为什么有些人节食减肥不成功?
在节食后未能减重人群中革兰氏阴性厌氧菌生长率的降低。
而那些节食后能够减重的人群中,短链脂肪酸的产生增加,胰高糖素样肽1(GLP-1)分泌增加。
肠道微生物群的某些指标,例如肠道相关的Prevotella的丰度,可以预测个体在膳食干预后减重的成功率。
体重管理的微观关键:脂肪组织、肠道菌群与肥胖的关联
微生物如何影响我们的食欲?
有趣的是,P. vulgatus能够产生泛酸,促进GLP-1的分泌。
肠道细菌产生谷氨酸也被认为可通过结肠嗜铬细胞增加食欲,这些细胞通过迷走神经向大脑发送信号。这展示了微生物组如何通过”肠-脑轴”调节我们的进食行为。
携带与支链氨基酸合成相关的生物合成功能的P. copri亚群也是2型糖尿病的诊断标志。
微生物组对肥胖影响的最新研究分析
与健康体重相关的菌——Christensenellaceae
人体研究表明,克里斯滕森菌科(Christensenellaceae)与低体重指数(BMI)和其他健康指标如低甘油三酯水平相关。基于这些观察,克里斯滕森菌科被提议作为治疗肥胖的候选新型活体生物治疗剂。
这些微生物与体重之间的关联可能是菌株特异性的。
对7,190个健康成人肠道宏基因组的分析发现,细菌基因组中的特定单核苷酸多态性与BMI相关,尽管在忽略单核苷酸多态性时,相同细菌种类的相对丰度与BMI并无关联。
意外的健康标志物:芽囊原虫
最近芽囊原虫属(Blastocystis)在肠道微生物组中的存在也被证明与多种健康指标相关:
注:芽囊原虫是一种多样性极高的单细胞真核生物,目前已被分类为至少28个亚型(subtypes, STs),其中至少有8个亚型在人类中发现。尽管早期的宏基因组研究已探讨了芽囊原虫的流行情况,但需要更大规模和多样化的队列研究来建立肠道芽囊原虫、营养和心脏代谢健康结果之间的关系。
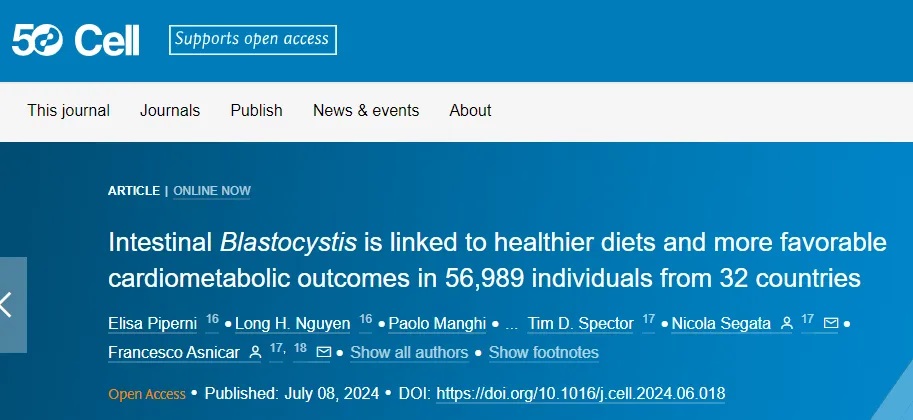
通过外部验证,研究显示芽囊原虫携带率与体脂、炎症性肠病(IBD)、结直肠癌(CRC)和糖尿病等疾病呈负相关。
总的来说,该研究揭示了芽囊原虫作为健康生活方式标志物的潜在有益作用,并提出了其可能解释个体对饮食反应差异的机制。重要的是,当代医疗实践经常试图从肠道中清除这种生物。
粪菌移植、药物干预治疗肥胖
关于微生物修饰策略治疗肥胖的研究主要集中在肠道菌群在调节饮食或手术对体重增减影响方面的作用。
粪菌移植(FMT)治疗肥胖的潜力产生了相互矛盾的结果,这可能是由于持续的饮食选择压力、微生物组的生态稳定性或相关的免疫反应,使得将微生物组修改为非致肥状态变得困难。
虽然一些动物模型的证据表明,以GLP-1为靶点的减肥药物(如索马鲁肽和替尔泊肽)也会改变微生物组,但几乎没有证据表明这些药物与微生物组有因果关系,导致体重减轻增强或停药后体重减轻时间延长。
微生物组定向辅食的突破
基于微生物组的饮食干预也已被证实可用于治疗营养不良。在一项涉及123名营养不良儿童的随机研究中,一种旨在促进微生物组介导体重增长的微生物组定向辅食(MDCF-2)能够调节肠道微生物群,并改善生长和健康结局。
——关键菌群 P. copri、F.prausnitzii
MDCF-2 的成功部分归因于普雷沃氏菌(P. copri)中碳水化合物酶代谢活性的变化,以及由普拉梭菌(Faecalibacterium prausnitzii)产生的N-酰基酰胺具有的促进食欲作用。
广泛存在于人群的双面使者——Prevotella copri与疾病和健康
肠道核心菌属——普拉梭菌(Faecalibacterium Prausnitzii),预防炎症的下一代益生菌
微生物组如何预测心脏代谢健康?
利用微生物组预测饮食和特定营养素对心脏代谢健康影响的兴趣日益增长。例如,在一项涵盖2500名非洲裔人群的队列研究中,微生物组的α-多样性及特定微生物特征均与高血压、高血糖、肥胖和糖尿病相关联。
甜味剂案例:其影响依赖基线菌群
在最早的实例之一中,非营养性甜味剂糖精对葡萄糖耐量产生了有害影响(最初在小鼠中发现)。在人类中,这种影响被发现依赖于补充前的微生物组,这种关系最近也适用于其他非营养性甜味剂。
个性化饮食方案:微生物组指导的精准营养学
更广泛地说,2015年一项研究开发出一种机器学习算法,将人体数据和微生物组数据整合,从而预测个体对任何食物项目的餐后葡萄糖反应。
基于这种方法,23位糖尿病参与者的微生物组数据被用来预测“个体化餐后靶向”饮食,在改善葡萄糖指标和代谢健康参数方面显著优于地中海饮食,在一项随机试点研究中,61%的受试者实现了糖尿病缓解。
一项更大规模的随机对照试验(n = 347)开发了一套基于粪便微生物组和患者健康数据的“个性化饮食方案”,在包括甘油三酯、糖化血红蛋白(HbA1c)、体重和腰围等心脏代谢健康标志物方面显示出显著改善。
间歇性禁食 & 肠道菌群 & 心血管代谢疾病
基于微生物组的个性化营养干预可能是未来营养医学的主要发展方向,有望革新我们对慢性代谢疾病的预防和管理方法。
微生物组-神经免疫轴:心血管疾病的预防和治疗希望
饮食-肠道微生物群对心血管疾病的相互作用
口腔微生物群可能成为喉咙甚至肺部微生物的来源,因此是健康监测和干预的重要部位。
口腔菌群在肠道定植引发疾病
有趣的是,口腔微生物在肠道中的高水平通常与不良健康结局相关。例如,口腔菌具核梭杆菌(Fusobacterium nucleatum)在健康个体的胃肠道中很少有显著丰度,但当肠道微生态被破坏时,该菌种会在结肠定植,并通过其表面蛋白FAP2与结直肠癌肿瘤中高度表达的一种糖蛋白特异性结合,优先定位于结直肠腺癌。一旦与肿瘤共定位,具核梭杆菌可能通过激活癌细胞增殖加剧CRC的恶性程度,这一过程可能与其产生的甲酸促进芳香烃受体蛋白1(AHR1)信号通路中的TLR4激活有关。
一些研究表明,结直肠癌患者口腔和远端胃肠道肿瘤中发现的具核梭杆菌菌株是相同的,而另一些研究则认为结直肠肿瘤中的具核梭杆菌菌株与口腔菌株不同。这些机制为阻断肿瘤形成提供了多种机会,无论是通过噬菌体或CRISPR技术有针对性地去除该菌种,还是抑制促进转移的微生物代谢机制。
最近的研究表明,口腔中的微生物生物膜能够促进抗生素耐药性和远端炎症的发生,增加牙周病的风险,并降低抗菌治疗的效果。通过刷牙和使用抗菌牙产品来破坏这些生物膜可能是有益的,但也可能破坏口腔中的氮循环,从而降低血液中的一氧化氮水平,导致血管收缩和血压升高。
口腔微生物组还可用于疾病诊断
例如,口腔微生物组与特发性肺纤维化有关。口腔微生物多样性较低与特发性肺纤维化患者的肺功能和生存率较高相关,链球菌(Streptococcus )被确定为与较好预后相关的关键物种。用抗生素复方新诺明(磺胺类抗菌药)治疗患者会导致变异链球菌(Streptococcus mutans)的富集,该菌被认为与疾病进展恶化相关,表明使用抗生素还存在其他未预料到的代价。
其他研究显示,口腔微生态失衡和拟杆菌门的变化可通过对淀粉样β斑块形成的特定微生物驱动影响,成为阿尔茨海默病的风险因素,这或许可以通过治疗手段加以解决。
肠-肺轴:远程控制如何影响肺部健康?
新兴研究正在揭示共生微生物影响肺部疾病的机制,包括直接作用于肺部和通过肠-肺轴的远程作用。
例如,临床研究表明,肠道微生物不仅能产生亚油酸,还能将其代谢为12,13-二羟基-9Z,15Z-十八碳烯酸,这种物质通过巨噬细胞效应功能、B细胞相互作用及表观遗传修饰,引发全身炎症,增强对过敏原的炎症反应,包括在肺部,从而加剧哮喘疾病。
慢性阻塞性肺疾病:肺内微生物的保护作用
——乳酸杆菌:肺部健康的守护者
关于慢性阻塞性肺疾病的一项研究表明,患病者肺内气道相关乳酸杆菌种类的缺失或活性降低与炎症、细胞凋亡及纤维化组织损伤有关。
——健康肺部的微生物代谢机制
而在健康对照组中,痰液内乳酸杆菌的丰富度与色氨酸代谢增加及吲哚-3-乙酸合成相关,这种代谢物通过AHR1介导的IL-22通路减轻了肺上皮的中性粒细胞炎症。
微生物靶向治疗——补充代谢物、菌群移植、噬菌体疗法
在小鼠模型中,无论是直接补充吲哚-3-乙酸,还是通过定植活乳酸杆菌于肺组织,都可改善肺功能并减少炎症,突出显示了以微生物—宿主相互作用为靶点的潜在治疗途径。
通过针对性移除引发疾病并发症的致病菌,或许也能实现对肺部微生物群的调控,例如,应用噬菌体疗法治疗囊性纤维化患者的下呼吸道感染
呼吸道病毒感染后,为何会引发细菌性肺炎?气道和肠道微生物组改变是关键
皮肤是一个独特的界面,在这里宿主免疫细胞与多样的外源性微生物群落之间持续交流。
表皮葡萄球菌:抗菌抗炎
凝固酶阴性的葡萄球菌,尤其是表皮葡萄球菌,是调节皮肤免疫的关键参与者,其机制已被充分研究。这些细菌产生的产物具有抗菌、抗炎甚至抗肿瘤的作用,从而增强了先天和适应性免疫反应。
皮肤微生物的保护机制:脂质代谢与免疫调节
例如,皮肤细菌代谢皮脂生成短链脂肪酸,可抑制组蛋白去乙酰化酶的活性,而表皮葡萄球菌产生的脂磷壁酸可激活TLR通路,阻断与皮肤组织损伤相关的炎症,促进伤口愈合,并有助于这些有益微生物的生存。
这也提示特定皮肤共生菌可能成为候选治疗剂,其应用在近期临床试验中已展现出良好前景。
微生物疗法:皮肤疾病治疗的创新途径
例如,将人表皮葡萄球菌涂敷于皮肤可减少金黄色葡萄球菌的存活,从而改善特应性皮炎患者的病情。
类似,具有类似抗金黄色葡萄球菌活性和免疫激活潜力的粘液玫瑰单胞菌(Roseomonas mucosa) 和线状透明颤菌(Vitreoscilla filiformis)也被证明可以改善特应性皮炎的疾病结局。
这些研究成果推动了皮肤微生物疗法从实验室到临床应用的快速转化,为患者提供了新的治疗选择。
微生物组标志物的诊断价值
人类皮肤微生物组研究正促进可预测或诊断多种疾病状态的指标的开发。
噬菌体疗法革新皮肤感染治疗
噬菌体疗法早于抗菌治疗出现,并且从20世纪初到今天一直用于治疗皮肤、口腔、肺部和胃肠道的感染。许多公司曾成立以推广这些疗法,其中一些至今仍以健康、美容或制药公司的形式存在。
噬菌体疗法目前在治疗系统性细菌感染方面具有很高的转化潜力。
一项回顾性研究对2008年至2022年间来自12个国家35家医院的难治性感染病例进行了个体化噬菌体疗法的评估,结果发现:
这项研究不仅强调了抗生素的协同作用,也突出了噬菌体耐药性的出现,强调今后在临床试验中需要采用个体化的方法来应对这一问题。
随着我们对皮肤微生物组的理解不断深入,未来可能出现更加个性化的皮肤健康管理方案:
随着微生物组研究的不断深入,基于微生物组的新型预防和诊疗手段正在逐步推动临床实践的变革,其中肠道菌群检测作为关键工具正在帮助科学家突破研究瓶颈。
但仍然存在许多挑战。
确定可以由医生测量、用于诊断疾病或作为治疗成功预后的特定微生物特征范围,依然是一项艰巨的挑战。
造成这一现象的主要原因包括:个体之间及种群间在菌株水平上的功能变异、微生物组成和结构存在显著的地理差异、缺乏纵向分析以解释特征丰度的时间波动从而证明其真实存在或缺失,以及缺乏对微生物特征的定量测量。
目前已经比较可靠的可以预测微生物组如何影响我们对特定影响因素(包括生活方式、营养、药物和时间等)的生理反应(除少数明显的例子外如使用抗生素或精准噬菌体疗法治疗特定感染)。
随着人体微生物组的研究越来越深入,科学家们不再只是发现微生物组与疾病之间的关联,而是开始探索它们之间的因果关系,研究微生物之间的相互作用,以及它们如何与人体互动,这使得基于微生物组的治疗和诊断方法的临床转化风险显著降低,帮助更好地选择药物靶点和确定适合治疗的患者群体。
未来的研究将进一步完善目前的理解,为利用人体微生物组力量的新型诊断、预防和治疗方法铺平道路。期待有足够的后端干预或精准药物等来实现对微生物组的精准调控以改善健康或提供治疗效果。
主要参考文献
Gilbert JA, Azad MB, Bäckhed F, Blaser MJ, Byndloss M, Chiu CY, Chu H, Dugas LR, Elinav E, Gibbons SM, Gilbert KE, Henn MR, Ishaq SL, Ley RE, Lynch SV, Segal E, Spector TD, Strandwitz P, Suez J, Tropini C, Whiteson K, Knight R. Clinical translation of microbiome research. Nat Med. 2025 Apr;31(4):1099-1113.
Schmartz, G. P. et al. Decoding the diagnostic and therapeutic potential of microbiota using pan-body pan-disease microbiomics. Nat. Commun. 15, 8261 (2024).
Piperni, E. et al. Intestinal Blastocystis is linked to healthier diets and more favorable cardiometabolic outcomes in 56,989 individuals from 32 countries. Cell 187, 4554–4570 (2024).
Lev-Sagie, A. et al. Vaginal microbiome transplantation in women with intractable bacterial vaginosis. Nat. Med. 25, 1500–1504
(2019) fda.gov/media/82945/download
Benoit, P. et al. Seven-year performance of a clinical metagenomic next-generation sequencing test for diagnosis of central nervous system infections. Nat. Med. 30, 3522–3533 (2024).

谷禾健康

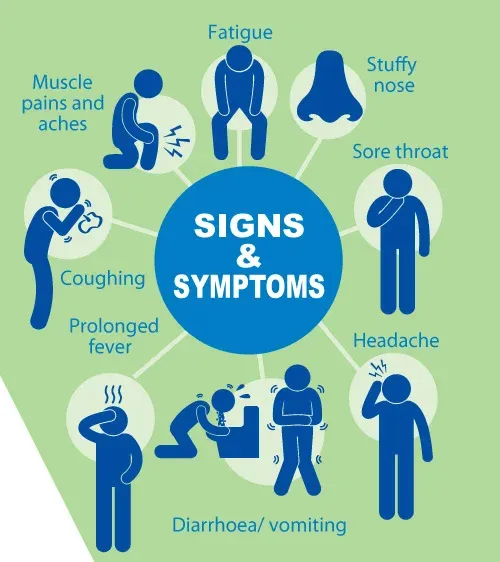
如果近期出现鼻子、喉咙、肺部相关症状,如咽喉痛、发烧(体温迅速升高通常超过38.5℃)、咳嗽、流鼻涕、鼻塞、乏力、发冷、浑身疼痛等情况,很有可能是甲流。
根据中国疾病预防控制中心2025年第1周流感监测周报显示(截至2025年1月5日),本周南、北方省份流感病毒检测阳性率继续上升,甲型H1N1流感占比99.4%。
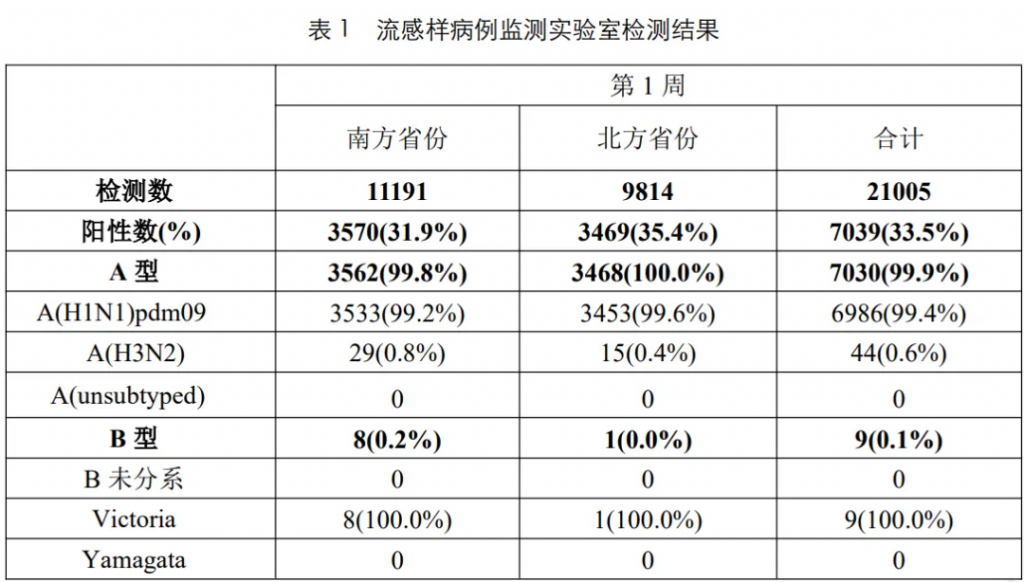
来源:Chinacdc
甲型流感是一种高度传染性的呼吸道感染疾病,其传播速度快、感染范围广,给公共卫生带来了巨大挑战,甲流每年导致300万-500万例严重感染和25万-50万例死亡。甲型流感主要通过咳嗽、打喷嚏或说话时的飞沫传播,症状可持续几天到几周。
近年来的研究发现,甲流感染可能会破坏肠道微生物群的平衡,而肠道生态失调可能会影响甲流患者的疾病进展。
调节肠道菌群可以通过肠-肺轴促进抗甲流病毒的免疫效应。这包括在感染的早期阶段促进甲流病毒清除,并在后期阶段减少炎症损伤。此外,肠道菌群还可以通过维持肠道屏障功能、产生具有保护作用的代谢产物(如短链脂肪酸、氨基酸、胆汁酸)等,促进宿主的免疫反应。
本文我们主要来详细了解一下甲流的症状、原因、潜伏期、危险因素、感染过程,包括肠道菌群及其代谢产物与甲流的关系,以及通过调节肠道菌群及其代谢产物来防治流感的潜在策略,包括益生菌、植物化学物质、中药等干预措施,这为甲流的预防和治疗提供了新的策略。
流感病毒有四种类型:甲(A)、乙(B)、丙(C)、丁(D)。甲型和乙型流感病毒几乎每个冬天都可能会引起季节性疾病流行。
甲流病毒是一种包膜病毒,包膜含有两种糖蛋白,一种叫血细胞凝集素(HA),另一种叫神经氨酸酶(NA)。
注:有18种不同的血凝素亚型和11种不同的神经氨酸酶亚型(分别为H1至H18和N1至N11)。
这两个可以排列组合形成不同的流感亚种,比如说H1N1,就是血细胞凝集素的1型和神经氨酸酶的1型组合。目前经常在人群中传播的甲型流感病毒亚型包括H1N1和H3N2。
甲型流感主要影响呼吸系统,包括鼻子、喉咙、肺。症状可能会突然出现,可能包括:
严重的症状
在严重病例中,甲流引起一系列症状,可能较少见:
图源:mypositiveparenting
并非所有流感都出现呼吸道症状
医生提醒,呼吸道症状是流感的典型症状,但以恶心、呕吐为主的消化道症状患者也大有人在。此外,还有部分患者会出现反应迟钝、嗜睡、躁动等精神系统症状。以上这些症状可能合并出现,也可单独出现,容易给人误导,导致没有及时就诊,延误流感的治疗。
当病毒感染上呼吸道(鼻窦,口腔和喉咙)或下呼吸道(肺部)的组织时,可能会发生甲型流感。病毒可以在这些组织内复制,从而引起症状。
这种流感具有高度传染性。你可以通过咳嗽、打喷嚏或在他人附近说话将病毒传染给他人。
甲流病毒通常在每个流感季节迅速变异。免疫系统可能无法立即抵抗新形式的病毒,即使以前得过流感,你也可能会有几天的症状。
如果被感染,通常会在接触后一到四天(潜伏期)出现流感症状,平均 2 天。
流感可以持续几天到两周。
发烧和身体疼痛等症状可能会突然出现,但通常比其他症状消失得更快。咳嗽或流鼻涕会持续更长时间。
普通感冒由鼻病毒等多种病原体导致,传染性弱,发病没有明显的季节特征,症状表现为鼻塞、流涕、喷嚏、咳嗽等,一般不发热或仅有低热,没有全身性症状,5-7天即可自愈,很少有并发症出现。
甲流传染性强,容易大范围流行,发病季节特征明显。患者表现为高烧、头痛、乏力,并伴有全身肌肉酸痛等。甲流有一定自限性,但容易引起其他并发症,包括肺炎、中耳炎、心肌炎、脑膜炎等严重并发症。
甲流和乙流之间有几个区别:
其他两种流感:
任何人都可能患上甲型流感。某些人出现严重症状和并发症的风险更大,例如:
大多数甲流病例在几周内消退,甲流可能更严重,并导致一些人的并发症,并发症可能包括:
耳部感染:对甲型流感的免疫反应会引起炎症,这会影响并导致咽鼓管中的液体积聚。这根管子把喉咙后部和中耳连接起来。这种液体会吸引可能导致耳部感染的细菌。尤其在儿童中较为常见。
鼻窦感染:甲型流感可能会导致鼻窦充血和炎症,从而吸引可能导致感染的细菌。
脑炎:免疫反应可能导致这种大脑炎症。甲流病毒可以通过血液或神经系统进入脑部,引起脑组织的炎症和损伤。
脑膜炎:表现为头痛、呕吐、意识障碍等症状。
心肌炎: 流感病毒可以直接攻击心肌细胞,引起心肌受损,表现为心悸、胸闷、胸痛、乏力等症状。
肌肉炎症:对流感过度活跃的免疫反应可导致肌肉炎症。症状包括疼痛、触痛和肌肉无力。
肺炎:甲型流感会削弱免疫系统,使其更容易受到细菌的感染,导致肺部感染。表现为高热、咳嗽加剧、呼吸困难等,严重时可导致呼吸衰竭。
继发性细菌性肺炎:它通常发生在病程的后期,在急性疾病观察到一段时间的改善之后,症状和体征是典型的细菌性肺炎。继发性细菌性肺炎比原发性病毒性肺炎更常见。
脓毒症:这是一种严重的、危及生命的感染反应。如果细菌进入血液就会发生。如果不治疗,败血症会导致器官损伤和死亡。
急性呼吸窘迫综合征(ARDS):在重症病例中可能出现,导致严重的呼吸困难。
这些并发症在免疫力低下的人群中更为常见和严重,如老年人、儿童、孕妇以及患有慢性疾病的人。因此,及时就医和治疗对于预防并发症的发生至关重要。
病毒入侵
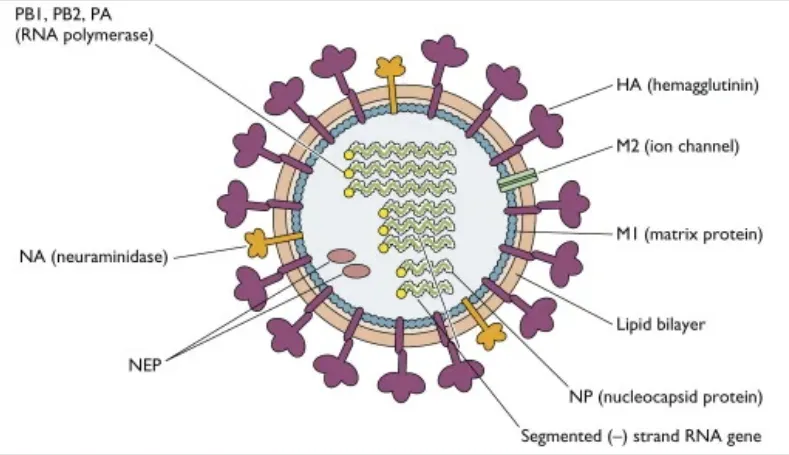
首先,甲流病毒主要通过呼吸道进入人体。
甲流病毒通过其表面的血凝素(HA)蛋白与呼吸道上皮细胞表面的唾液酸受体结合,从而进入宿主细胞。
病毒复制
病毒进入细胞后,其基因组RNA在细胞核内进行复制,并指导宿主细胞合成病毒蛋白。新合成的病毒颗粒在细胞内组装后,通过细胞膜释放到外界,感染其他细胞。
免疫反应
当甲型流感病毒入侵人体后,免疫系统会迅速识别病毒的病原相关分子模式(PAMPs),激活先天性免疫应答。
巨噬细胞、树突状细胞等免疫细胞被激活,开始分泌多种细胞因子,包括IL-1、IL-6、TNF-α和干扰素等。这些细胞因子一方面协调免疫细胞的募集和活化,增强机体的抗病毒能力;另一方面作用于下丘脑体温调节中枢,导致体温升高,同时还可引起全身性炎症反应,造成肌肉关节疼痛、乏力等症状。
症状形成机制
发热:发热是由于病毒感染触发免疫系统释放的细胞因子(如IL-1、IL-6等)作用于下丘脑温度调节中枢,使体温设定点上调所致,这种发热反应实际上有利于抑制病毒复制并增强免疫功能。
咳嗽:病毒直接感染呼吸道上皮细胞,导致细胞损伤、炎症反应和气道分泌物增多的结果,刺激气道感受器引起咳嗽反射。
全身症状:如肌肉酸痛、乏力、头痛等,主要是由细胞因子介导的系统性炎症反应引起,这些炎症介质可以作用于全身多个器官系统,影响神经-内分泌网络,导致患者出现全身不适。症状的严重程度往往与机体免疫反应的强度直接相关。
病毒变异特点
抗原漂移:点突变导致的小幅变异
抗原转变:基因重配导致的大幅变异
变异使病毒逃避免疫系统识别,是季节性流感和流感大流行的重要原因。
在传统认知中,我们往往将甲流视为一种单纯的呼吸道感染,但随着研究的深入,研究人员发现这种认识可能过于简单化。近年来,越来越多的研究表明,人体的免疫系统是一个高度统一的网络,其中肠道菌群作为人体最大的微生态系统,在调节全身免疫反应中扮演着关键角色。
肠道菌群可以通过调节免疫细胞的分布和功能,影响宿主对流感病毒的免疫反应,显著影响呼吸道感染的疾病进程和预后。此外,肠道菌群的代谢产物,如短链脂肪酸,也被发现能够调节免疫系统,从而可能对甲流的发病机制产生间接影响。接下来,我们进一步探讨肠道菌群是如何参与和调节甲流感染的过程。
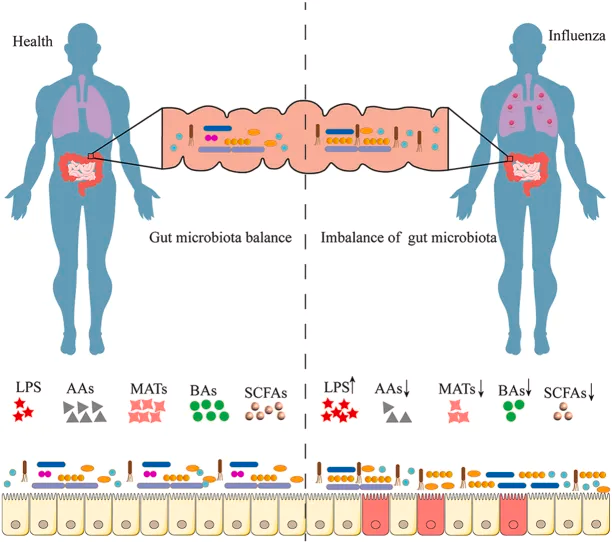
研究表明,肠道微生物群的丰富性和多样性响应于甲流感染发生了显著变化,包括拟杆菌门中细菌数量的增加和厚壁菌门中细菌数量的减少。同样,另一项研究表明,流感感染改变了肠道微生物群,促进肠道特异性厌氧菌的消耗和变形菌门的富集,导致肠道生态失调。
doi: 10.1016/j.heliyon.2024.e37661
▸甲流病毒感染
肠道菌群多样性增加,同时下列菌群增殖较为明显:
而一些有益菌的丰度则降低:
在健康个体中,肠道微生物群以平衡、稳定的方式在肠道中定植,与肠道粘膜屏障和抑菌物质一起维持肠道微生态。 一个平衡良好的肠道菌群能够增强机体的免疫功能,从而降低感染甲型流感的可能性。以下是几个关键机制:
调节免疫系统
在流感病毒感染后,CCR9 + CD4 + T细胞(其是源自肺的效应细胞)被募集到小肠,在小肠中它们分泌干扰素-γ(IFN-γ)。这导致肠道微生物群的不平衡,促进了小肠中的Th17细胞极化。最终导致IL-17 A分泌,其介导免疫损伤。接下来,我们从先天免疫和适应性免疫两方面详细了解肠道菌群对甲流的影响。
先天免疫
肠道微生物主要通过两种方式影响流感病毒感染的先天免疫:
首先,肠道菌群能够影响一种叫做天然淋巴细胞(ILCs)的免疫细胞。这些ILCs细胞在我们的肺部可以对抗病毒,而肠道菌群就像是一个“信号员”,通过释放一些信号物质,告诉肺部的ILCs该怎么做。比如,肠道菌群产生的短链脂肪酸(SCFAs)可以调节肺部ILCs的数量和功能,帮助我们更好地对抗流感病毒。
此外,肠道菌群还能够影响我们的身体识别病毒的能力。它们通过激活一些特殊的受体(比如Toll样受体和RIG-I样受体),帮助我们的身体更快地发现病毒并启动免疫反应。一旦有病毒入侵,就能迅速发出警报,让免疫细胞及时赶到“战场”去消灭病毒。
适应性免疫
肠道微生物群对甲型流感的适应性免疫影响主要体现在三个方面:
1- CD4+T细胞的调节
肠道微生物及其代谢产物可调节CD4+T细胞的分化。
在感染早期,主要促进Th1和Th17细胞分化,有助于病毒清除,乳酸杆菌通过参与色氨酸代谢和产生芳烃受体配体,促进IL-22表达,影响Th17分化;梭菌属通过诱导TGF-β释放,促进结肠Treg细胞分化。
短链脂肪酸(SCFA)通过多种机制促进Treg细胞分化,帮助控制炎症。
抗生素破坏菌群会显著降低CD4+T细胞数量,影响病毒清除。
2- 细胞毒性T淋巴细胞(CTL)的影响
CTL是消灭病毒感染的关键角色,而肠道微生物群能够通过调节树突状细胞(DCs)的活动来增强CTL的细胞毒性作用。
抗生素导致的菌群破坏会降低CTL的免疫应答。
3- 参与调节流感疫苗的免疫效果,促进流感特异性抗体的产生
预防流感的最佳方法是在流行期前接种流感疫苗,以产生特异性抗体。近期的临床试验表明,益生菌制剂可以有效增强流感疫苗的保护效力及提高个体抗体滴度。
然而,动物实验显示,接种前使用抗生素会降低疫苗诱导的抗体滴度及其中和能力,而在无菌小鼠中,恢复肠道植物群能恢复疫苗保护效力。此外,短链脂肪酸(SCFA)的不足会影响B细胞向特异性抗体产生浆细胞的分化。
维护肠道屏障功能
感染甲型流感病毒后,肠黏膜屏障功能障碍与感染的严重程度密切相关。
肠黏膜屏障由肠上皮细胞构成,分泌多种免疫因子并传递细菌抗原,在维持肠道菌群与宿主的共生关系中发挥着重要作用,并作为维持肠道菌群稳定性和生态平衡的控制开关。
流感病毒感染通过破坏肠上皮细胞中紧密连接蛋白和粘附蛋白的功能来影响肠道屏障,这些蛋白调节肠道屏障功能,阻止肠腔内大分子(如细菌和毒素)进入血液。
这些蛋白表达的降低可导致屏障功能受损,从而增加甲型流感病毒的疾病进展和危重症的发生。研究发现,小鼠感染H1N1导致肺和结肠中紧密连接蛋白的表达显著降低。这种减少导致这些器官的屏障结构受损,从而允许肠道细菌易位。随后,通过体液循环发生继发性细菌感染。
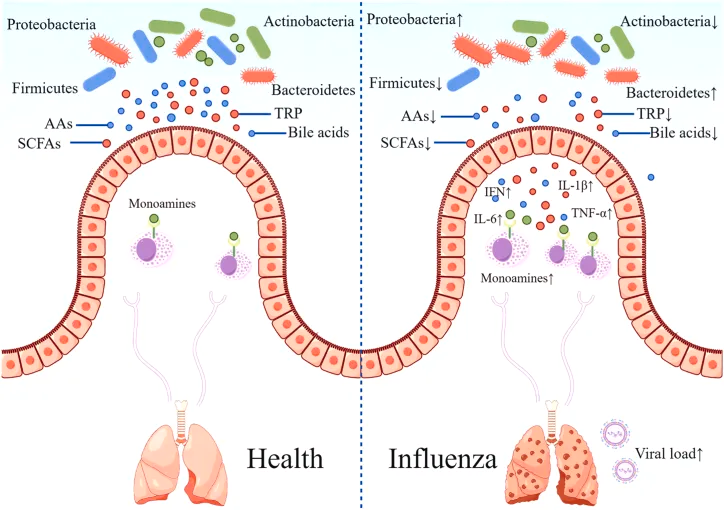
产生有益代谢产物
短链脂肪酸
短链脂肪酸包括乙酸盐、丙酸盐和丁酸盐,SCFAs可以通过调整流感病毒感染宿主的肠道菌群结构,如增加双歧杆菌等有益菌的数量,增强免疫适应性等来抵抗流感病毒。
短链脂肪酸能够增强肠道上皮细胞的免疫功能,促进免疫细胞的增殖和分化,同时具有抗炎作用,能够减轻流感病毒感染引起的炎症反应。
在流感期间补充短链脂肪酸,可以减轻肠道病原体如鼠伤寒沙门氏菌的移位,并提高合并感染的存活率。
肠道菌群及其代谢产物对正常人和流感患者的影响
doi: 10.1016/j.heliyon.2024.e37661
氨基酸
支链氨基酸被认为是身体健康的“潜在生物标志物”,在蛋白质合成、分泌和激素(如胰岛素和生长激素)释放中发挥重要作用。 研究表明,庆大霉素治疗后,肠道微生物群紊乱诱导流感小鼠体内支链氨基酸水平升高,从而抑制CD11b + Ly6G +细胞的发育,增加CD8+T细胞水平,导致流感病毒感染程度增加。
色氨酸是必需氨基酸,其缺乏会损害肠道免疫力,通过改善宿主先天免疫应答,降低流感期间的发病率和死亡率,补充色氨酸可能是预防流感的有效策略。
脱氨基酪氨酸(DAT),一种Clostridium orbiscindens菌产生的代谢物,可以通过提高I型干扰素(IFN)的水平,来增强身体对流感病毒的抵抗力,从而减少流感感染小鼠的体重减轻和死亡率。
胆汁酸
初级胆汁酸在肝脏产生,分泌到肠道中,约95%通过肠-肝循环被重吸收。
肠道菌群衍生的代谢产物脱氧胆酸(DCA)通过TGR5受体信号通路,抑制病毒复制,抑制嗜中性粒细胞依赖性组织损伤,增加宿主对流感病毒的抵抗力。
牛磺胆酸是一种胆汁酸。牛磺胆酸钠水合物(STH)抑制流感病毒复制,降低流感病毒RNA、互补RNA和mRNA水平,并对多种流感病毒株(H5N6、H5N1、H1N1、H3N2)表现出广谱抗病毒活性。 此外,STH降低流感病毒感染小鼠中炎症因子(TNF-α、IL-1β、IL-6)的表达,减轻其临床症状,抑制体重减轻,并降低其死亡率,这归因于STH通过抑制NF-B途径的激活而在流感抗性中的作用。
鹅去氧胆酸对甲型流感病毒(H5N1、H9N2和H1N1)具有抑制作用,它能够通过阻断病毒RNA复合物的核输出来抑制甲型流感病毒的复制。
以上我们了解到健康的肠道菌群通过多重免疫调控机制能够增强机体对甲型流感病毒的抵抗力,包括增强免疫系统的警戒能力、提升适应性免疫防护、调节炎症反应等,但在实际临床工作中,准确及时的诊断仍然是控制流感传播和开展针对性治疗的关键。接下来我们来看在临床中,医生可能借助的实验室检测手段。
甲型流感与其他病毒感染(如COVID-19)的症状类似。诊断测试包括:
★ 病毒分离和鉴定
从患者的呼吸道标本中分离出甲型流感病毒,并通过电镜或免疫荧光等方法进行鉴定。
优点:是诊断甲流的金标准。
缺点:操作复杂且耗时较长,一般不作为常规诊断方法
★ 甲型流感病毒核酸检测
通过聚合酶链式反应(PCR)等技术检测病毒的核酸。采集患者的呼吸道标本(如鼻咽拭子、口咽拭子等),进行核酸提取和扩增。
优点:准确率高,是确诊甲流的常用方法。
缺点:需要专业的实验室设备和技术
★ 快速抗原检测
通常使用检测试剂条,从鼻子或喉咙采集组织样本后检测病毒的存在,最快可在15分钟内显示结果,类似于新冠抗原检测。
优点:速度快;
缺点:敏感性和特异性相对较低。
★ 血常规检查
通过检测血液中的白细胞水平等指标,辅助诊断甲流。
优点:可以提供一些间接的诊断信息。
缺点:不能直接确诊甲流
★ 胸部X光
拍摄肺部图像以排除细菌感染或肺炎。
优点:可以排除其他肺部疾病。
缺点:影像学表现无特异性,不能单独用于确诊甲流。
利福昔明通过调节肠道菌群改善甲流病毒感染引起的肺屏障损伤
IAV引起了显着的体重减轻,并破坏了肺和肠的结构。16 S rRNA和代谢组学分析结果表明,感染甲流的小鼠粪便样品中益生菌Lachnoclosdium、Ruminococcocceae_UCG-013和色氨酸代谢产物的水平显著降低。
相比之下,补充50mg/kg利福昔明可以逆转这些变化,包括促进肺屏障的修复,增加粪便中Muribaculum、Papillibacter和色氨酸相关代谢物的含量。此外,利福昔明治疗增加了ILC3细胞数量、IL-22水平以及肺中RORγ和STAT-3蛋白的表达,减轻对肠屏障的损伤,同时增强小肠中AHR、IDO-1和紧密连接蛋白的表达。
在病原体清除方面,用冻干鼠李糖乳杆菌GG和加氏乳杆菌TMC0356进行口服预处理显著降低了PR8感染后6天小鼠肺部的病毒载量。
此外,单独口服煮沸的植物乳杆菌06CC2、植物乳杆菌DK119、热灭活b240和副干酪乳杆菌也降低了甲流病毒感染后BALF中的病毒载量。
口服加氏乳杆菌SBT2055可在感染后5天降低PR8感染小鼠肺部的病毒载量,并上调抗病毒基因Mx1和Oas1a的表达。益生菌治疗降低肺部病毒载量可能与促进对甲流病毒的先天免疫和适应性免疫有关。
煮沸的植物乳杆菌06CC2在感染早期(第二天)增加了抗病毒效应分子IFN-a、IFN-γ、IL-12的水平和NK细胞的活性。
口服植物乳杆菌DK119可增加BALF中IL-12和IFN-γ水平。
副干酪乳杆菌增加了甲流感染后肺组织中树突状细胞的募集。
植物乳杆菌0111口服预处理在H9N2感染后7天上调ISG转录,同时提高脾脏中CD3+CD4+TNF-α+T淋巴细胞百分比和CD3+CD8+TNF-β+T淋巴细胞百分比,增强对病毒的适应性免疫。
在感染的晚期,一些益生菌还可以抑制炎症损伤,促进组织修复。鼠李糖乳杆菌M21可减少甲流感染小鼠肺部的炎症损伤,并增加肺部裂解物中的IFN-γ和IL-2水平。
A. muciniphila可降低H7N9感染小鼠中促炎因子IL-1β和IL-6的水平,并增加炎症调节因子IL-10、IFN-β和IFN-γ的水平。
与益生菌粘膜乳杆菌L. mucosae1025和短双歧杆菌CCFM1026联合治疗可提高甲流感染小鼠盲肠粪便中的丁酸水平,并减轻肺组织中的炎症浸润。
一些口服益生菌可以双向作用,在甲流感染的早期阶段创造一个有利于病毒清除的炎症环境,并在后期抑制过度的炎症激活。
口服Bacteroides dorei在感染早期(第3天)更快地增加了1型干扰素的表达,降低了肺部的病毒载量,而在感染后期(第7天),它降低了1型IFN和其他促炎因子的水平,这有利于组织修复。与此同时,Bacteroides dorei治疗也改变了肠道菌群组成,增加了拟杆菌、普氏菌和乳杆菌,减少了大肠杆菌、志贺氏菌和副拟杆菌。
除此之外,益生菌制剂还可以通过促进甲流特异性抗体的产生来提高流感疫苗的有效性。
植物乳杆菌16和鼠李糖乳杆菌P118共同口服,改善了宿主对甲型流感病毒感染的防御。
研究人员利用C57BL/6和BALB/c小鼠模型的研究结果表明,单独施用植物乳杆菌16(L.plantarum 16)或鼠李糖乳杆菌P118(L.rhamnosus P118)并不能提供足够的流感防护。然而,植物乳杆菌16和鼠李糖乳杆菌P118的联合给药显著降低了呼吸道和肺部的病毒滴度,从而显著缓解了临床症状,改善了预后,降低了死亡率。
这种作用的机制涉及通过共同施用植物乳杆菌16和鼠李糖乳杆菌P118来调节宿主肠道微生物群和代谢,导致厚壁菌门富集和苯丙氨酸相关代谢增强,最终导致抗病毒免疫反应增强。值得注意的是,研究人员发现循环代谢分子2-羟基肉桂酸在抗击流感中起着重要作用。数据表明,植物乳杆菌16和鼠李糖乳杆菌P118这两种细菌或2-羟基肉桂酸在预防流感方面具有潜在的效用。
膳食中添加益生元短链低聚半乳糖和长链低聚果糖会增加小鼠流感疫苗特异性T辅助细胞1反应和mLN中特异性B细胞活化,导致IgG1和IgG2A水平升高。
β-葡聚糖重编程中性粒细胞以促进对甲型流感病毒的疾病耐受
最新研究表明,β-葡聚糖通过重编程造血干细胞来产生具有”调节”功能的中性粒细胞,这些特殊的中性粒细胞具有线粒体氧化代谢特征并能产生IL-10。这种机制通过RoRγt+ T细胞促进中性粒细胞向肺部组织募集,提高了机体对甲流病毒感染的疾病耐受能力,有助于维持肺部组织的完整性,从而降低发病率和死亡率,而这种保护作用并不依赖于抑制病毒载量。
益生元改善肠道菌群,延长流感疫苗抗体效果
接受肠内营养治疗的60岁以上老年人在标准肠内营养配方治疗的基础上,接受了双歧杆菌生长刺激素和半乳糖低聚糖治疗14周,并在治疗后第4周接种了三价季节性流感疫苗(A/H1N1、A/H3N2和B)。在第0、4、6、8和12周测量抗体滴度。结果显示,两组疫苗免疫应答率无差异,但实验组肠道内双歧杆菌数量明显高于对照组,实验组A/H1N1抗体滴度维持时间长于对照组。这表明益生元可能会增加肠道中益生菌的数量,从而维持抗体滴度。
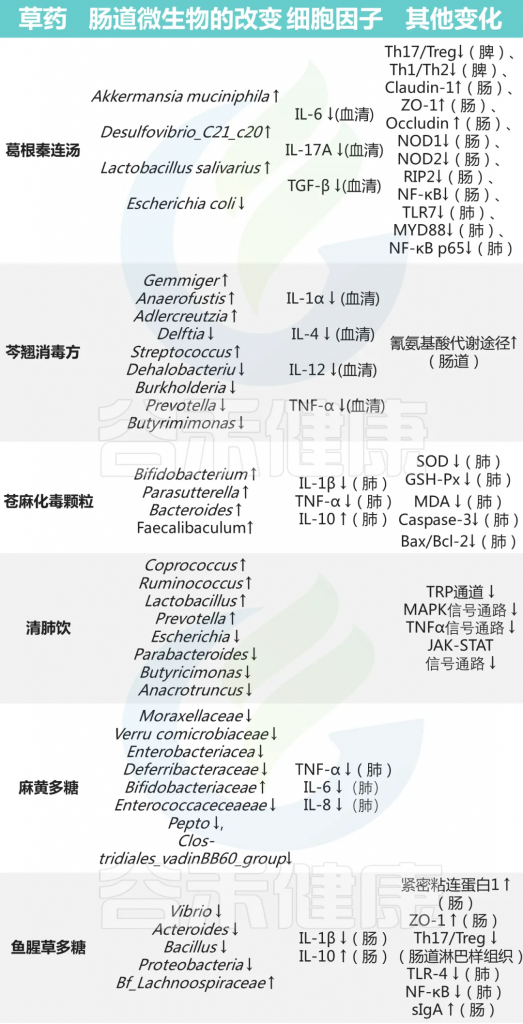
中药在临床应用已有一千多年的历史,最近研究表明,中药在减轻甲流患者肺部炎症、改善临床症状、缩短治疗时间、促进康复等方面具有功效。
感染肺部后,甲流病毒可以改变肠道微生物群的组成和代谢,导致肠粘膜屏障受损、免疫功能受损和炎症因子水平升高。尽管如此,中药干预具有调节肠道微生物群、恢复体内平衡、保持肠粘膜屏障完整性、增强免疫功能和调节炎症反应的潜力。
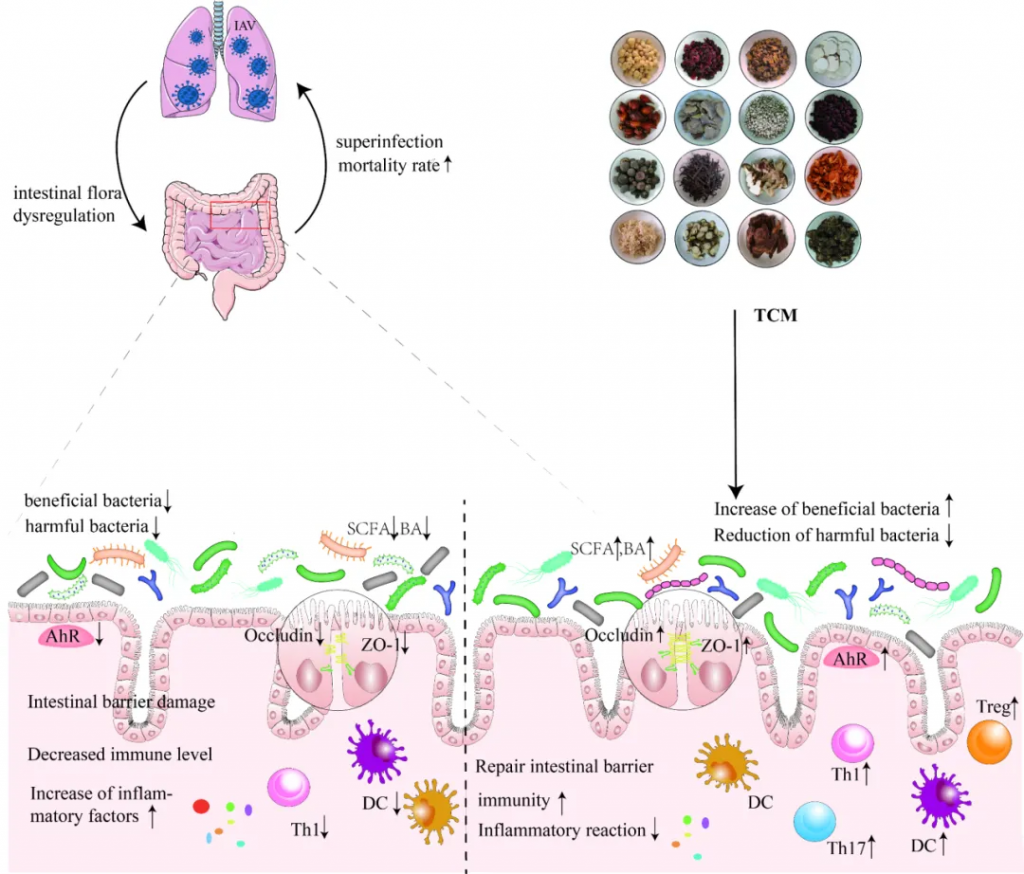
doi: 10.1186/s12985-023-02228-3.
宣肺败毒汤调节肠道微生物群多样性,并与拟杆菌、志贺氏菌、Eubacterium nodatum、Turicibacter、Clostridium sensu stricto 1 的变化呈正相关,而这些变化与 TNF-α 水平相关。
升麻素苷(Prim-O-glucosylcimifugin,POG)是中药防风的提取物,可以通过上调紧密连接蛋白Occludin、Claudin-3和ZO-1的表达水平来调节肠道菌群结构并修复肠道免疫屏障。
999 小儿感冒颗粒可以缓解H1N1感染小鼠的体重减轻,降低IL-6和IL-1β等炎性细胞因子的水平。 降低肺指数和病理损伤,通过维持结肠杯状细胞的数量来保护肠道屏障,降低结肠组织中IL-17 A的表达。
大黄中蒽醌类成分不仅增加了大鼠肠道中一些益生菌和产短链脂肪酸菌的丰度,而且通过上调ZO-1和闭塞素的表达水平来增强肠道屏障功能,从而抑制炎症。
其他中药对菌群及免疫的影响
编辑
doi: 10.3389/fimmu.2023.1147724
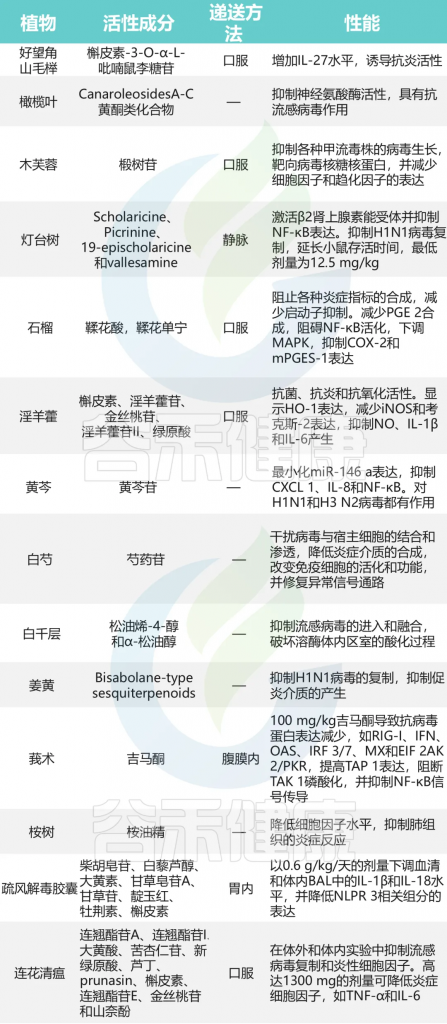
黄芪多糖具有免疫增强作用,有望成为甲流疫苗的新型佐剂
黄芪多糖组表现出更高的免疫球蛋白G(IgG)、IgG 1和IgG 3水平,以及中和抗体水平。此外,它增加了CD 8+细胞的频率,以增强对致死性感染的抵抗力。
感染后第14天,高剂量黄芪多糖组的存活率(71.40%)高于血凝素组(14.28%),体重恢复更快。
黄芪多糖还能改善肺泡损伤和肠道结构紊乱。上调肠组织紧密连接蛋白Occludin和Claudin-1的表达水平,降低血清TNF-α的表达水平。此外,Colidextribacter、消化球菌科和瘤胃球菌科的群体是病毒感染后黄芪多糖组中的优势肠道微生物群。
doi: 10.1002/ptr.8334

doi: 10.2147/IJGM.S361001
维生素C
维生素 C 具有重要的抗炎、免疫调节、抗氧化、抗血栓和抗病毒特性。诺贝尔奖获得者莱纳斯·鲍林从随机对照试验中得出结论,维生素 C 可以预防和缓解感冒。
一项针对儿童上呼吸道感染的荟萃分析发现,维生素C的补充能够减少感染持续时间约1.6天。
口服维生素 C(2-8 克/天)可以减少呼吸道感染的发生率和持续时间。
英国安慰剂对照试验由 168 名志愿者组成,他们在 60 天的冬季期间随机接受安慰剂或维生素 C(每天 2 × 500 毫克)。
维生素 C 组的感冒次数较少(37次 vs.50次,p=0.05)
病毒感染的“感冒”天数更少(85次 vs.178次,p=0.03)
严重症状持续时间较短(1.8天 vs.3.1天,p=0.03)。
试验期间患过两次感冒的参与者人数显着减少(维生素 C 组为 2/84,安慰剂组为 16/84;p=0.04)
维生素D
维生素D缺乏会影响呼吸道感染的发生和哮喘的恶化。
维生素D和肠道微生物组以多种类似的方式影响呼吸道疾病中的免疫系统。它们之间可能存在一些相互作用和/或协同作用。
肠道微生物组可以改变肠道维生素D代谢,1,25(OH)2D (维生素D的活性形式)水平较高的人更有可能拥有有利的肠道微生物群,尤其是更多的产丁酸菌。
维生素D->抗病毒感染->降低哮喘发生和恶化的风险
一项涉及不同国家儿童的队列研究发现,补充维生素D 与 RV感染呈负相关。此外,患有下呼吸道感染的儿童的平均维生素 D 水平明显低于对照组。下呼吸道感染的发生率和严重程度也与维生素 D 水平相关。
维生素D可以双向调节肠道微生物群
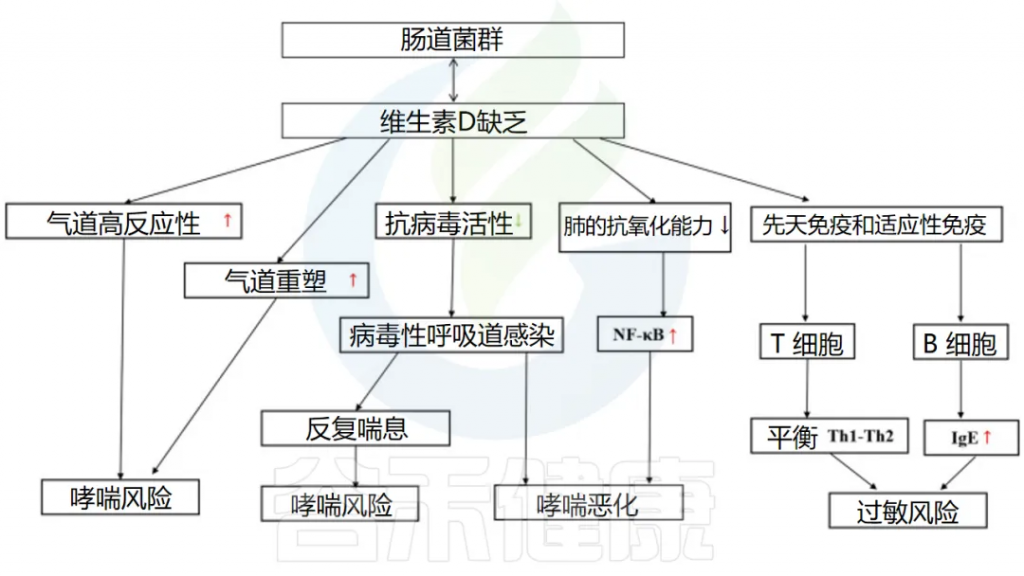
doi: 10.3389/fmicb.2023.1219942
维生素D通过多种途径影响哮喘的发生
因此,保持足够的维生素D水平对于预防和管理哮喘以及呼吸道感染至关重要。
研究表明,甲流病毒主要破坏气道上皮紧密连接,导致急性呼吸窘迫综合征,且胃肠道症状常与甲流病毒感染同时发生,提示肠-肺轴参与了宿主对甲流病毒的反应。在感染甲流病毒的患者和小鼠中发现SCFA(尤其是乙酸盐)显著减少。
在FMT后的小鼠中,这些条件被逆转,减少了肺部炎症损伤,并证实乙酸盐可能是肠-肺轴的重要介质。进一步的实验表明,乙酸盐激活了GPR 43,恢复了一些甲流病毒诱导的气道上皮屏障功能,并降低了TNF-α、IL-6、IL-1β水平。FMT能够有效调节流感感染小鼠肠道菌群结构紊乱。
粪菌移植通过增强I型干扰素通路改善甲流防御
在病毒感染中,I型干扰素(IFNs)非常重要,它们是病原体和宿主之间的重要调节因子。I型干扰素是抵御病毒感染的第一道防线。通过使用感染了甲流病毒的小鼠模型,研究人员发现IFN-κb是最早对H9N2感染做出反应的I型干扰素之一,并且IFN-κb能够有效地抑制多种流感病毒在培养的人类肺细胞中的复制。因此,阻断IFN-κ特异性途径可以作为预防和治疗甲流的依据。
I型干扰素途径对身体很重要。适当调节它既能抵御病毒感染,又能避免免疫或病理对身体造成过度损伤。比如降低IFNAR1水平可抑制流感病毒早期繁殖。
I型干扰素主要会激活一些干扰素刺激基因(ISGs)的表达。ISGs编码的蛋白质具有多种抗病毒功能。抗生素的使用会导致肺间质细胞中ISGs的减少,从而使身体更容易感染流感病毒。
然而,粪菌移植(FMT)可以逆转这一情况,增加肺间质中的I型干扰素信号通路和I型干扰素驱动的抗病毒状态,从而增强对流感病毒感染的防护。
在流感季节很容易感染甲型流感,因为它传染性很强。预防策略可以保护你免受流感或减少并发症的风险。一些常见的预防方法包括:
肠道微生物群是一个复杂而广泛的群落,显著影响各种生物系统,包括免疫系统和代谢功能。最近的研究越来越多地强调肠道微生物群与流感之间的潜在联系,表明肠道微生物群可能影响流感病毒的致病性,从而提供了一种新的治疗靶点。
总之,肠道微生物群表现出对流感的治疗潜力,这可以通过调节肠道微生态来解决。未来的研究应侧重于确定肠道微生物群中的关键菌株、特定代谢物和免疫调节机制,以精确靶向微生物群干预,预防和治疗流感和其他呼吸道病毒感染。
在此基础上,菌群组合的优化研究显得尤为重要。比如说通过研究植物乳杆菌16和鼠李糖乳杆菌P118等益生菌的协同作用,探索不同菌群配比对免疫调节的影响,从而制定个体化的菌群干预方案。
可以进行新型保护性代谢物的筛选与鉴定,深入研究短链脂肪酸、胆汁酸等代谢产物的作用机制,并结合代谢组学分析推进临床应用。
开展一系列临床转化研究,包括建立菌群检测作为辅助诊断的标准化体系,开展益生菌制剂的临床试验,并根据不同患者特点制定个性化预防和治疗方案。同时,预防策略的优化也需要重点关注,特别是探索疫苗免疫效果与肠道菌群的关系,建立预防性菌群调节方案,并为高危人群制定个性化预防策略。
这些研究方向相互关联、相互促进。分子机制的研究为菌群组合优化提供理论指导,代谢产物研究则为临床应用提供新靶点,而临床转化研究的结果又能反馈指导基础研究的方向。通过多维度、多层次的研究,将更全面地理解肠道菌群与甲型流感的关系,为开发新型防治策略提供坚实的科学依据。
主要参考文献
Khan N, Tran KA, Chevre R, Locher V, Richter M, Sun S, Sadeghi M, Pernet E, Herrero-Cervera A, Grant A, Saif A, Downey J, Kaufmann E, Khader SA, Joubert P, Barreiro LB, Yipp BG, Soehnlein O, Divangahi M. β-Glucan reprograms neutrophils to promote disease tolerance against influenza A virus. Nat Immunol. 2025 Jan 8.
Luo C, Yang Y, Jiang C, Lv A, Zuo W, Ye Y, Ke J. Influenza and the gut microbiota: A hidden therapeutic link. Heliyon. 2024 Sep 10;10(18):e37661.
Ma L, Ji L, Wang T, Zhai Z, Su P, Zhang Y, Wang Y, Zhao W, Wu Z, Yu H, Zhao H. Research progress on the mechanism of traditional Chinese medicine regulating intestinal microbiota to combat influenza a virus infection. Virol J. 2023 Nov 13;20(1):260.
Liu X, Wang Q. Effect of natural products on host cell autophagy induced by Influenza A virus infection. Front Cell Infect Microbiol. 2024 Sep 30;14:1460604.
Ho JSS, Ping TL, Paudel KR, El Sherkawi T, De Rubis G, Yeung S, Hansbro PM, Oliver BGG, Chellappan DK, Sin KP, Dua K. Exploring Bioactive Phytomedicines for Advancing Pulmonary Infection Management: Insights and Future Prospects. Phytother Res. 2024 Dec;38(12):5840-5872.
Ou G, Xu H, Wu J, Wang S, Chen Y, Deng L, Chen X. The gut-lung axis in influenza A: the role of gut microbiota in immune balance. Front Immunol. 2023 Oct 20;14:1147724.
Gounder AP, Boon ACM. Influenza Pathogenesis: The Effect of Host Factors on Severity of Disease. J Immunol. 2019 Jan 15;202(2):341-350.
Zhang Y, Chen Y, Xia J, Li L, Chang L, Luo H, Ping J, Qiao W, Su J. Rifaximin ameliorates influenza A virus infection-induced lung barrier damage by regulating gut microbiota. Appl Microbiol Biotechnol. 2024 Sep 19;108(1):469.
Wan C, Yan S, Lu R, Zhu C, Yang Y, Wu X, Yu Z, Jiang M, Peng W, Song W, Wu H, Fang B, He Y. Astragalus Polysaccharide improves immunogenicity of influenza vaccine as well as modulate gut microbiota in BALB/c mice. Microb Pathog. 2024 Oct;195:106893.

谷禾健康

随着人们健康意识的提高,”益生菌”早已不是什么陌生的名词。目前人们比较关心的问题是:
实际上,自从2002年联合国粮农组织和世界卫生组织——益生菌是当足量补充时,对宿主健康有益的,活的微生物。其中强调了3点,只有对宿主能发挥显著有效作用,能够确保安全性,存活在食品中能保持有效的菌数。
虽然我国益生菌行业起步较晚,但发展较为迅速, 尤其是近年来一些益生菌已被卫生部批准作为新资源食品。广泛应用于乳制品行业,同时也逐步被应用于其他功能性食品,如固体饮料,果汁、冰淇淋、糖果、巧克力、 泡菜等。常见益生菌主要指两大类乳酸菌群:一类为双歧杆菌;常见的有婴儿双歧杆菌、长双歧杆菌、短双歧杆菌和动物双歧杆菌。另一类为乳酸杆菌,常见的有罗伊氏乳杆菌,植物乳杆菌,鼠李糖乳杆菌和嗜酸乳杆菌。
但是菌种具有特异性,不同的菌种具有特定的健康益处,没有一种菌种是万能的。由于主要的益生菌是通过补充或膳食进入肠道,属于肠道菌群的一员。而肠道菌群被认为是影响人体健康的重要因素之一,其与代谢、免疫、神经系统等多方面的健康密切相关。
因此,益生菌作为调节肠道菌群的重要工具,已被广泛应用于功能性食品和临床干预。然而,由于个体间肠道菌群的组成和功能存在显著差异,传统的“一刀切”益生菌干预模式在某些情况下可能效果有限,甚至可能引发不良反应。
本文将探讨个性化益生菌选择的必要性、益生菌定植影响的内外部因素,以及益生菌补充不适当或者过量的案例。
人体内外栖息着大量微生物,这些微生物与健康存在密切的关系。有的能够促进健康,有的则会引起疾病;有的能促进消化、有的则会造成感染;有的能增强免疫系统,有的则会产生毒素破坏器官和组织。同样是细菌,可以被分为“有益的”细菌和“有害的”细菌。
联合国粮食及农业组织和世界卫生组织将益生菌定义为“活的微生物,当摄入足够量时,会给宿主带来健康益处”(又名友好细菌)。通俗地讲,益生菌就是“对人体有益的细菌”。
近几十年来,益生菌因其对人类健康的有利影响而逐渐被纳入我们的饮食中,成为越来越多人的健康选择。益生菌可能对身体产生多种积极影响,而不同种类的益生菌可能在作用机制和效果上有所不同。这些影响包括:
No.1
维持肠道微生态平衡
◆ 抑制有害菌定植
益生菌可以通过竞争肠道黏膜表面的结合位点,阻止有害菌的黏附与定植。例如,双歧杆菌和嗜酸乳杆菌等益生菌能够分泌抗菌物质,如乳酸、乙酸和细菌素等,抑制病原菌的生长。
益生菌还可以调节肠道的pH值,创造不利于有害菌生长的环境。
◆ 促进有益菌生长
益生菌可以为其他有益菌提供营养支持,促进其生长繁殖。例如,某些益生菌可以分解膳食纤维产生短链脂肪酸,为双歧杆菌等有益菌提供能量来源,从而促进它们的生长。
益生菌还可以通过信号传导等方式,调节肠道菌群的组成和结构,促进有益菌的生长和优势地位的确立。
No.2
改善消化系统功能
◆ 缓解便秘
益生菌可以增加肠道内的水分含量,使粪便变得柔软,易于排出。益生菌还可以促进肠道蠕动,加快粪便在肠道内的传输速度。例如,乳酸菌可以通过刺激肠道神经系统,促进肠道平滑肌的收缩,从而增强肠道蠕动。
◆ 减轻腹泻
对于感染性腹泻,益生菌可以通过调节肠道菌群,增强肠道免疫力,抑制病原菌的生长,从而减轻腹泻症状。例如,布拉氏酵母菌可以在肠道内定植,分泌抗菌物质,抑制致病性大肠杆菌等病原菌的生长,减轻腹泻症状。
对于抗生素相关性腹泻,益生菌可以预防和治疗由于抗生素使用导致的肠道菌群失调引起的腹泻。
◆ 促进营养物质的吸收
益生菌可以产生多种消化酶,帮助人体消化和吸收食物中的营养物质。例如,乳酸菌可以产生乳糖酶,帮助人体消化乳糖;双歧杆菌可以产生多种维生素,如维生素B1、B2、B6、B12等,以及短链脂肪酸等有益物质,促进肠道对这些营养物质的吸收。
◆ 改善肠道屏障
益生菌通过促进肠道上皮细胞修复,减少肠漏综合征的发生。益生菌还可以调节肠道黏膜的通透性,改善肠道屏障功能,减少营养物质的流失。
doi.org/10.1016/j.tifs.2023.06.013.
No.3
增强免疫功能
◆ 调节肠道免疫
益生菌可以刺激肠道黏膜免疫系统,增强肠道局部免疫力。例如,双歧杆菌和嗜酸乳杆菌等益生菌可以激活肠道黏膜中的免疫细胞,如巨噬细胞、T细胞等,促进免疫细胞的增殖和分化。
益生菌还可以调节肠道免疫细胞分泌的细胞因子,维持肠道免疫平衡。例如,某些益生菌可以促进抗炎细胞因子的分泌,抑制促炎细胞因子的产生,从而减轻肠道炎症反应。
◆ 全身免疫调节
益生菌可以通过肠道黏膜免疫系统与全身免疫系统的相互作用,调节全身免疫功能。例如,可以刺激肠道黏膜中的免疫细胞产生免疫调节因子,如白细胞介素-10(IL-10)等,这些因子可以进入血液循环,调节全身免疫系统的功能。
益生菌还可以增强机体的非特异性免疫,如提高吞噬细胞的活性、增强自然杀伤细胞的活性等,从而提高机体的抵抗力。
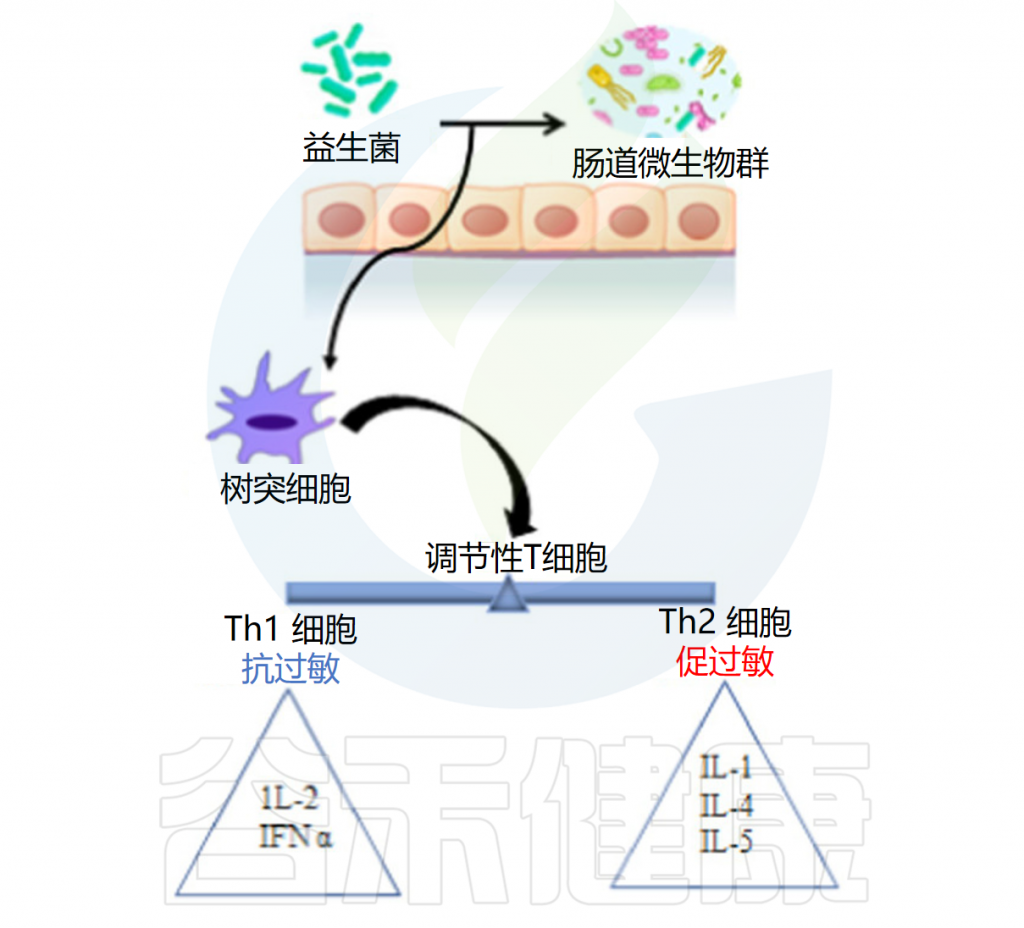
◆ 抗过敏作用
益生菌抗过敏作用的机制是通过抑制Th2免疫反应和有利于Th1细胞反应来增强辅助性T细胞(Th1/Th2)免疫平衡。益生菌调节树突状细胞的功能,而树突状细胞又具有外周Tregs的能力。Tregs控制过量的免疫反应并维持Th1和Th2细胞之间的平衡。
例如乳杆菌刺激调节性T细胞,调节性T细胞通过产生免疫抑制细胞因子和调节IgE、IgA和IgG的产生在平衡免疫反应中起着至关重要的作用。
口服罗伊氏乳杆菌有助于恢复结肠菌群的恶化特征并减轻过敏性腹泻。研究结果证实了罗伊氏乳杆菌的抗过敏活性是通过调节肠道菌群和增强耐受性免疫反应来促进的。它还增加了肥大细胞的活化,增强了血清免疫球蛋白E(IgE)的产生,抑制了辅助性 T 细胞因子1和2的产生,下调了GATA3的表达,并增加了TGF-b 、 IL-10和Foxp3的表达。
益生菌的抗过敏作用
Latif A,et al.Front Microbiol.2023
No.4
益生菌与病毒感染
益生菌在几种关键病毒感染中也发挥作用,包括 SARS-CoV-2感染、流感感染、病毒性肝炎、人类免疫缺陷病毒(HIV)、人瘤病毒(HPV)。
◆ 减轻新冠病毒感染患者症状
先前的研究表明,益生菌治疗呼吸道感染的给药具有有益效果,这可能是由于它们的免疫调节和抗炎作用。
在新型冠状病毒大流行期间还研究了益生菌菌株与 新型冠状病毒(SARS-CoV-2)相关潜在治疗作用的临床试验,在大多数研究中,使用的益生菌由乳酸杆菌和双歧杆菌菌株组成,因为这些细菌已被推广为潜在的免疫调节剂。
一项研究报告称,接受益生菌和酶补充剂的患者身体和精神疲劳减轻。在另一项随机安慰对照试验中,接受测试的SARS-CoV-2 患者的胃肠道症状似乎有所改善,并且与接受安慰剂的患者相比,接受益生菌的患者发生医院获得性腹泻的频率较低。
◆ 降低流感的易感性
人类流感病毒主要附着在上下呼吸道的上皮细胞内并复制。在流感感染的啮齿动物模型中使用了益生菌菌株副干酪乳杆菌CNCM I-1518,并报道摄入该菌株降低了对流感感染的易感性,减少了炎症细胞浸润到肺部,并增加了病毒清除率。
对啮齿类动物的其他研究表明,微生物组通过诱导流感病毒复制的早期难治性环境,从而减少流感病毒的早期感染,在介导肺组织中的IFN特征中发挥重要作用。抗生素诱导的啮齿动物生态失调可降低受体7(TLR7)和NF-kB mRNA的表达,导致抗病毒免疫力受损,而使用双歧杆菌和乳酸菌的益生菌菌株可逆转。
◆ 对肝炎具有一定的治疗作用
乙型肝炎(HBV)和丙型肝炎(HCV)感染是全球性的健康问题,病毒与宿主免疫系统之间的致病性相互作用可导致肝损伤,并可能导致肝硬化和肝细胞癌。
据报道,在体外细胞模型中,青春双歧杆菌SPM0212的细胞提取物抑制乙型肝炎(HBV),其抗病毒机制与Mx-GTP酶途径有关。Mx GTP酶通路是干扰素介导的抗病毒反应中的四种主要效应通路之一。
No.5
改善某些疾病
◆ 辅助癌症治疗
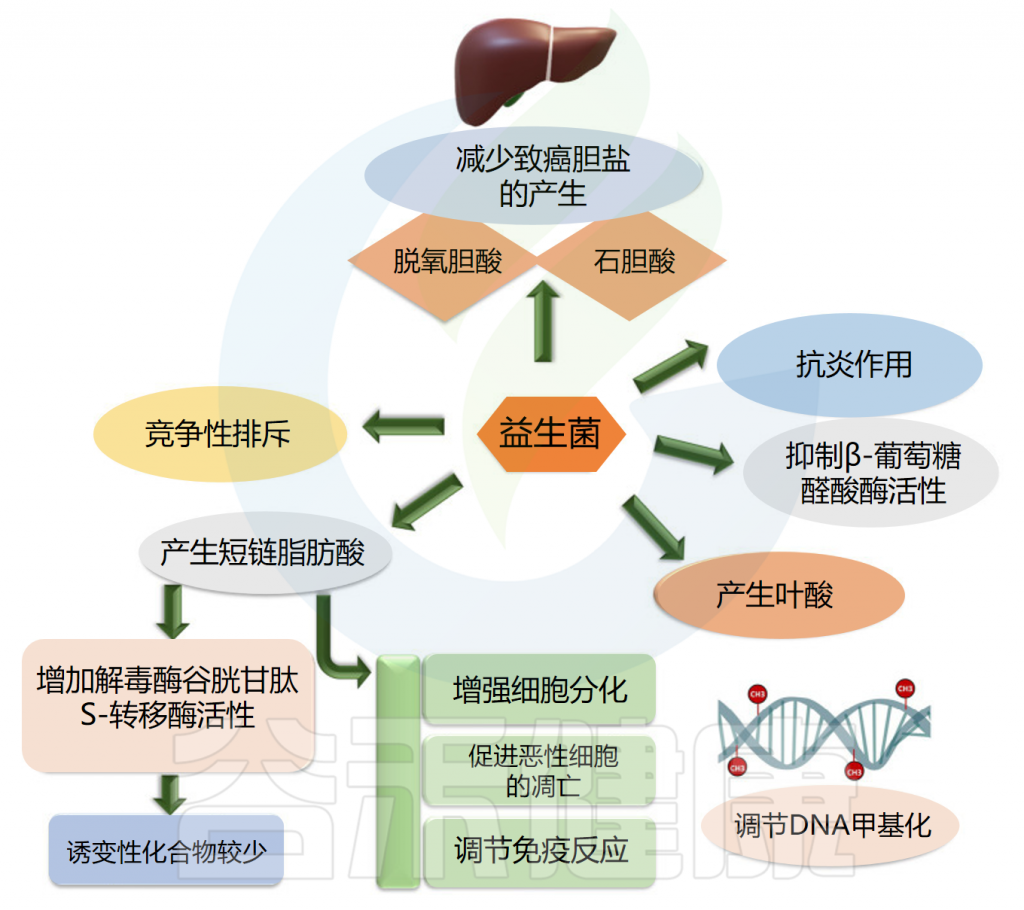
益生菌可用作癌症治疗的佐剂,因为它们具有调节肠道菌群和增强局部和系统免疫力的潜力。它们可以防止可移植或化学诱导的肿瘤的发生、进展和转移。在抑制肠癌和肠外癌方面都可以观察到益生菌的作用。
益生菌使用不同的途径来对抗癌症。益生菌抑制β葡萄糖醛酸酶活性,产生叶酸,最终调节DNA甲基化模式,保护基因组的完整性,产生短链脂肪酸,增强癌细胞的细胞分化和凋亡,排除可能导致癌症发展的慢性炎症病原体。
此外,益生菌通过竞争性排斥抑制病原体、减少致癌胆盐的产生、结合致癌物和诱变剂、下调细胞增殖的NF-kappa B依赖性基因产物和细胞存活率。益生菌还上调TNF相关的凋亡诱导配体。
益生菌的癌症抑制活性
Latif A,et al.Front Microbiol.2023
◆ 减轻炎症性肠病、肠易激综合征
益生菌在调节导致肠道相关疾病的微生物失调方面起着至关重要的作用。肠易激综合征(IBS)、乳糜泻和其他肠道疾病与肠道有益细菌的缺乏有关。通过荟萃分析评估益生菌对肠道炎症的治疗效果,得出结论:益生菌治疗可防止溃疡性结肠炎复发,其疗效与常用药物美沙拉嗪相当。然而,它们对溃疡性结肠炎的影响是特定于菌株的。
这些益生菌菌株的作用机制可能不同,但一般限于三种主要机制:抑制肠道细胞上的病原菌,通过诱导屏障细胞产生粘液或抗菌肽来增强肠道细胞的物理屏障,或通过改变自然杀伤活性来改变粘膜免疫系统, 核因子 kappa-B(NFkB)途径,并诱导T细胞凋亡,导致肠道抗炎细胞因子(IL-10、TFG-B)的产生增加以及促炎细胞因子(TNF-a、IFN-y、IL-8)的产生减少。
研究表明,益生菌可能对治疗与肠易激综合征(IBS)相关的症状有效,从而改善胀气、腹胀、食欲、排便频率和营养。一项研究表明,益生菌治疗IBS的可能机制是通过上调粘蛋白2(MUC2)基因表达来诱导人肠道细胞分泌粘蛋白,并在相同的粘附部位竞争、排除和置换病原体,防止病原菌的入侵。嗜酸乳杆菌CL1285、干酪乳杆菌LBC80R和鼠李糖乳杆菌CLR2已被确定为缓解IBS症状的潜在治疗方法。
◆ 降胆固醇,减少心血管疾病
益生菌可以用作降低血液胆固醇水平的有效工具,它们可以直接或间接地降低体内的胆固醇水平。直接机制包括尿酸、乳糖、乳清酸和乳清蛋白等低胆固醇血症因子抑制胆固醇的从头合成,以及以三种方式减少肠道对膳食胆固醇的吸收——同化、结合和降解。
益生菌降低胆固醇的间接机制是通过胆盐水解酶生产使胆盐(共轭糖脱氧胆酸和牛磺脱氧胆酸)解离胆盐。解离的胆盐通过肠道的重吸收较少,从而抑制胆汁的肠肝循环和粪便中的排泄增加。
益生菌对一些疾病的影响和相关机制
DOI:10.1002/fbe2.12078.
No.6
改善心理健康
◆ 通过肠脑轴联系
益生菌通过肠道与大脑之间的联系,可能对情绪和心理健康产生积极影响。益生菌可以通过调节肠道菌群,影响肠道神经系统和内分泌系统,进而影响大脑的功能和情绪。例如,某些益生菌可以产生神经递质如γ- 氨基丁酸(GABA)等,这些神经递质可以通过肠-脑轴传递到大脑,调节情绪和行为。
◆ 缓解焦虑和抑郁
一些研究表明,益生菌可以缓解焦虑和抑郁症状。例如,在动物实验中,补充益生菌可以改善焦虑和抑郁样行为;在临床研究中,益生菌也被发现可以改善焦虑和抑郁患者的症状。
No.7
促进女性健康
◆ 维持阴道健康
益生菌可以帮助维持阴道内的微生物平衡,通过竞争作用,阻止有害菌在阴道黏膜表面的黏附、定植与生长,预防阴道感染和尿路感染。
◆ 提高HPV清除率
在人乳头瘤病毒(HPV)感染的背景下对益生菌进行了研究。一项前瞻性研究纳入了54例 HPV 感染者,这些女性被诊断为低级别鳞状上皮内病变,研究发现,每天饮用益生菌饮料后,使用益生菌的患者中有29%清除了HPV,而对照组这一比例为19%
与短期服用益生菌相比,长期添加阴道益生菌在减少细胞学异常和提高HPV清除率方面具有更强的能力。然而,无论治疗方法或持续时间如何,益生菌的功能支持和保护性阴道微生物群的最终恢复似乎都是成功对抗 HPV 感染的关键要素。
◆ 改善妊娠健康
一些研究表明,益生菌可能对孕妇和胎儿的健康有益。健康的阴道微生物群与妊娠结果密切相关。益生菌的使用可能有助于降低早产、妊娠高血压等并发症的风险。
No.8
改善皮肤健康
◆ 促进皮肤修复、预防感染
益生菌可以帮助维持皮肤表面的微生物平衡,抑制有害细菌的生长,同时有助于加速皮肤伤口的愈合过程,减少痤疮和其他皮肤感染的风险。
体外研究表明,某些菌株,如唾液乳杆菌LS03、乳球菌和唾液链球菌,会产生抑制痤疮杆菌生长的细菌素。此外,由于抗菌活性,青春芽孢杆菌SPM0308被证明可有效控制痤疮丙酸杆菌和金黄色葡萄球菌的生长。
◆ 改善皮肤屏障功能
益生菌有助于增强皮肤的屏障功能,防止水分流失和外部刺激物的侵入,保持皮肤的水分和健康;减少皮肤敏感性和炎症反应。
◆ 抗氧化和抗衰老
益生菌可能通过减少自由基对皮肤的损伤;促进胶原蛋白生成,改善皮肤弹性,帮助减缓皮肤衰老过程。
除此之外,益生菌可能还存在着许多潜在的健康益处,这些益处尚未被人们完全认知和发现,可能在不同的生理和病理状态下发挥着重要的作用,影响着我们的整体健康和疾病预防。深入研究益生菌的多种功能将有助于揭示它们在健康管理中的潜力。
!
益生菌补充越多越好吗?
既然益生菌有如此多的健康益处,是否所有人都可以补充益生菌,且补充越多越好呢?
对于这个问题而言,我们要明确的是,补充益生菌的目的是为了通过改善菌群,从而达到更健康的状态。
◆ 并非每个人都需要补充益生菌
并不是每个人都需要补充益生菌,也并不是每个人都适合补充益生菌,所以首先第一步是判断:
是否真的需要补充益生菌?
目前已有研究的,与菌群相关的疾病包括:
-消化系统(腹泻、腹胀、便秘等);
-神经系统(自闭症、精神障碍等);
-免疫系统(哮喘等);
-代谢系统(肥胖、糖尿病等);
-心血管疾病(高血压、中风等);
-癌症(胰腺癌、结肠癌等);
假如你本身非常健康,同时也一直遵循着健康的饮食和生活方式,那就不需要补充。此外如果在你身上出现的症状和菌群完全没有关联,补充益生菌可能起不到太大的效果。
◆ 益生菌也不是补充越多越好
那有些人会觉得,我出现了与菌群相关的症状,那我就大量补充益生菌,把有益菌的丰度变得非常高。
这种做法也是不正确的
益生菌会与宿主常驻菌群相互作用,有些益生菌可以与宿主菌群形成共生关系,促进彼此的生长和代谢;而有些则可能导致竞争或拮抗作用。
过量补充益生菌可能破坏原有的菌群结构,导致生态失调或菌群结构过于单一,从而引起不健康症状。此外,对于免疫功能低下的人群,过量或不当的益生菌补充可能引发感染或其他健康问题。
谷禾的检测中就曾遇到过量补充益生菌的案例,下面让我们一起来看下:
①双歧杆菌和乳杆菌过高,菌群多样性非常低
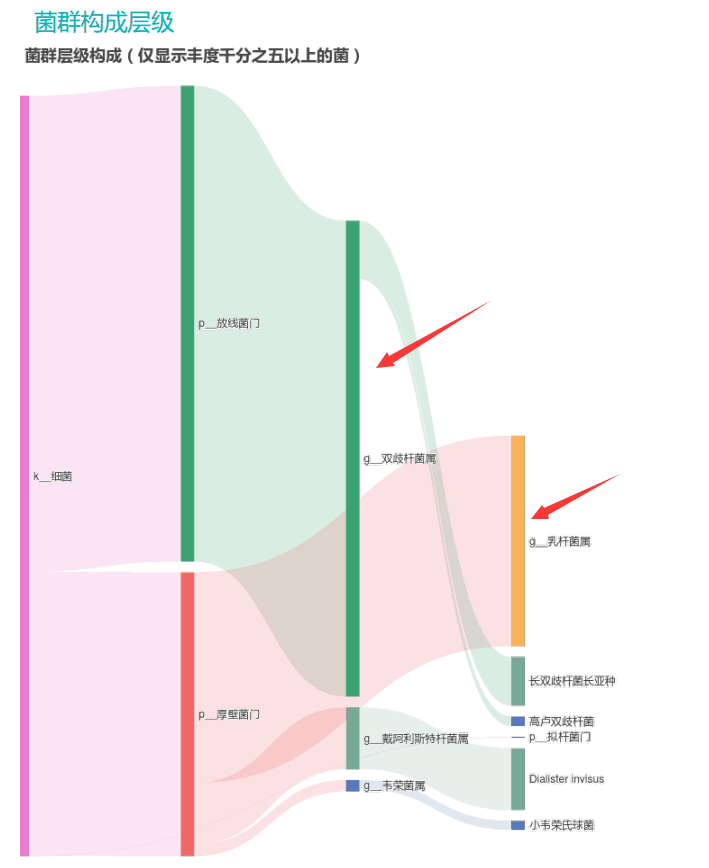
从菌群构成层级和核心菌属构成表中可以看到,该患者的双歧杆菌属和乳杆菌属严重超标。
其的有益菌水平非常高,但是反观菌群多样性却很低,菌群数量明显低于正常人群。并且由于其菌群构成不平衡,导致肠道产气过多,而次级胆汁酸、短链脂肪酸、吲哚等需要由其他菌株产生的代谢物严重缺乏,最终导致了身体的不健康状态。
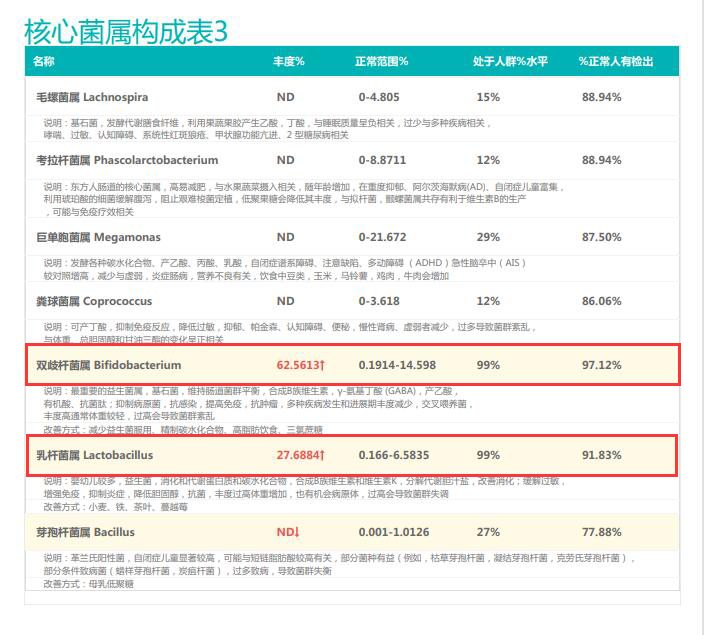
②双歧杆菌过高,核心菌属缺乏
观察另一个案例报告发现,尽管双歧杆菌含量较高,但许多核心菌属缺乏,且菌群多样性较低。因此导致了肠道产气和炎症水平较高,一些菌群代谢产物如脂多糖过量,而短链脂肪酸则存在缺乏。
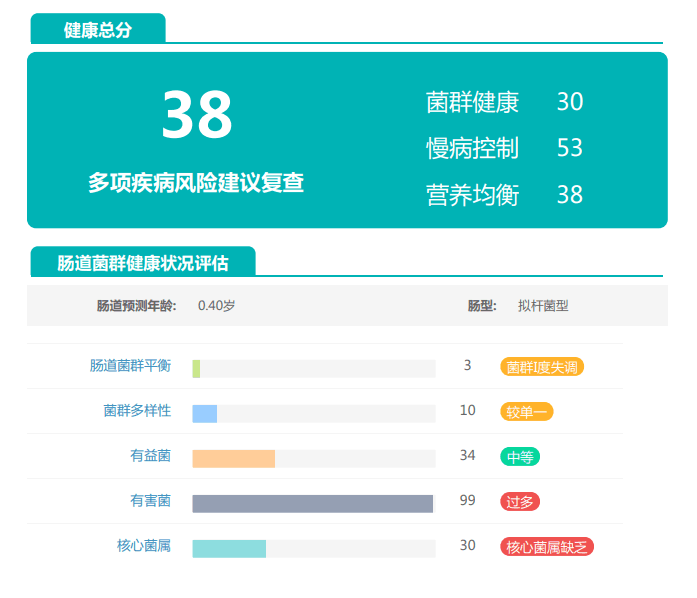
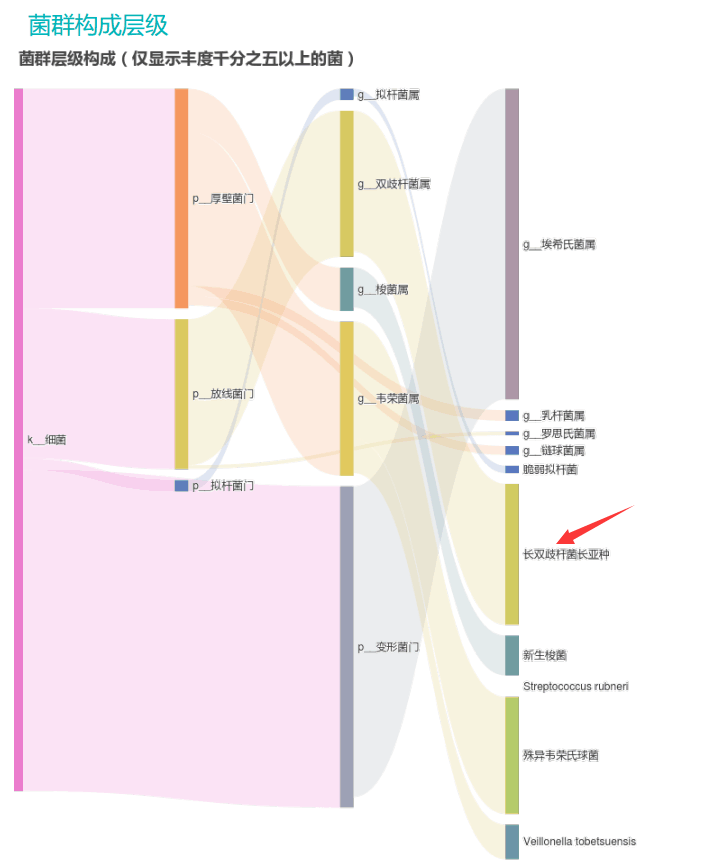
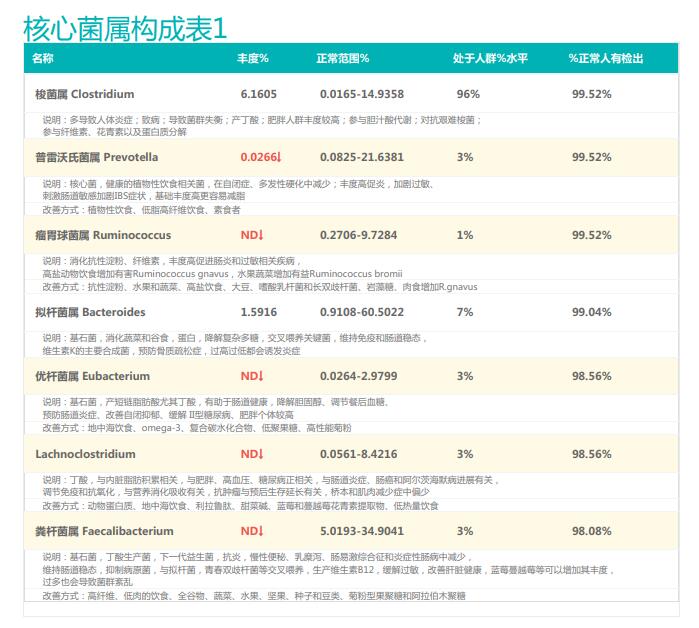
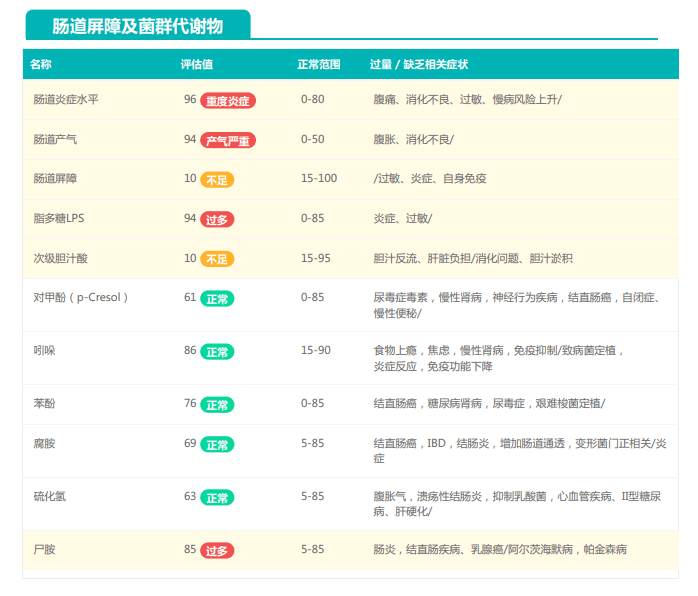

由此可见,单一有益菌的过高可能导致菌群多样性较差和菌群失衡,这并不健康。因此,在补充益生菌的同时整体菌群多样性和核心菌属水平同样不容忽视。
★ 肠道菌群检测有助于益生菌的辅助判别和干预
利用好肠道菌群检测这个工具,有助于益生菌干预的辅助判别:
辅助干预:可以进行肠道菌群检测查看体内菌群比例,如发现某些菌群过高,可选择相应的益生菌来抑制致病菌的大量增殖,有针对性地施用益生菌;
辅助判别:在服用益生菌前后均进行肠道菌群检测,通过菌群变化情况判断益生菌服用效果。
益生菌虽然被认为是有益的,但并不一定在所有情况下都能产生良好的效果。实际上,有许多因素会影响益生菌的功效,这些因素可以分为外部因素和内部因素。这一节我们主要讲影响益生菌功效的外部因素。
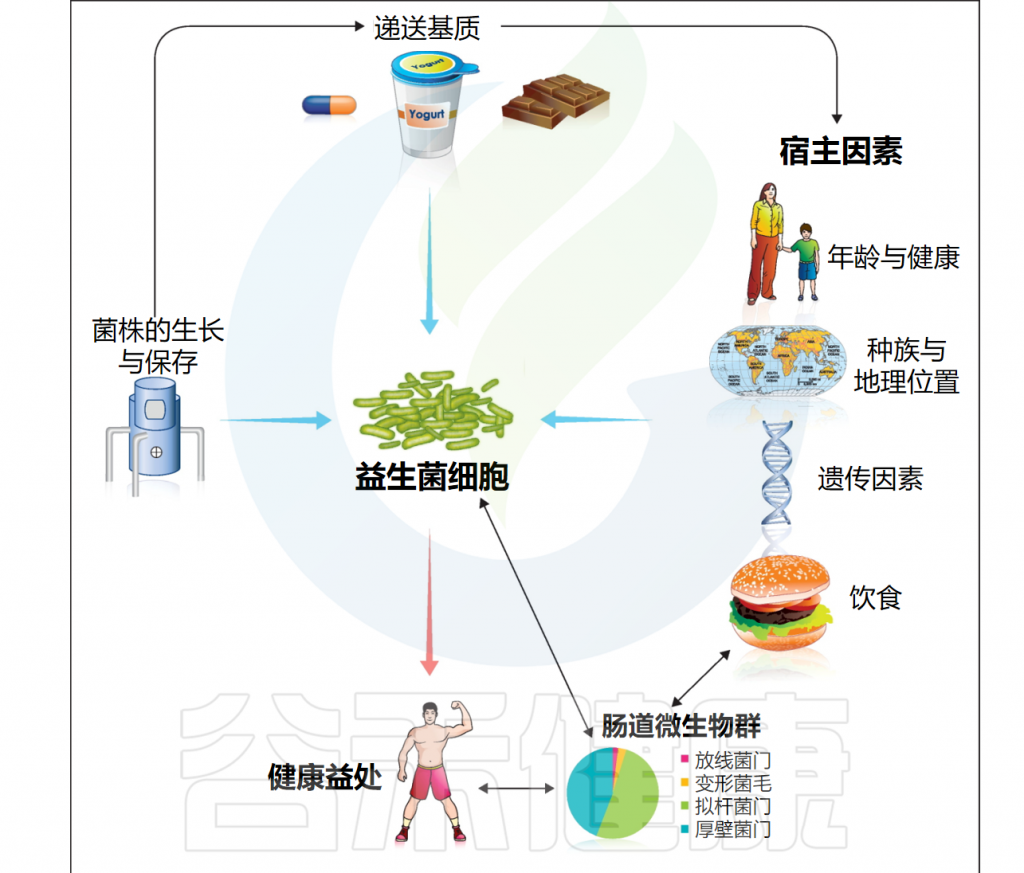
doi: 10.1016/j.copbio.2012.10.002.
益生菌在生产加工和储存过程中的存活率受多种因素影响,包括温度、pH值、氧气含量、水分、盐浓度、糖等营养资源和其他化合物(包括过氧化氢、细菌素、人工香料和色素)的存在。
这些因素可以显著改变益生菌在体外的生存和活性,影响其的生长速率和总细胞量,也会对益生菌在消费品和消化道中的存活产生下游影响。
此外,发酵条件、包装材料、食物基质、给药方式、干预持续时间、菌株特异性等参数也起着影响。
1
益生菌的生长条件
◆ 生长条件会影响效应分子的表达
益生菌的生长条件会影响宿主-益生菌和病原体-益生菌相互作用的效应分子的表达。例如,小鼠脾细胞对嗜酸乳杆菌L-92的反应产生的IL-12、IFN-γ、IL-4和IL-10的量因细菌是在培养基中生长还是在没有pH控制的情况下生长而不同。
◆ 生长条件会影响抵抗病原体的能力
生长条件也会影响益生菌细胞预防人类病原体感染的能力。这表现为在补充不饱和脂肪酸中生长后,约氏乳杆菌NCC533对沙门氏菌粘附和感染的抑制作用增强。
益生菌的免疫调节也是生长阶段依赖性的,这可以通过人外周血单核细胞对指数期和稳定期野生型和突变型植物乳杆菌WCFS1的IL-10和IL-12的差异表达来说明。
这些生长期效应得到了以下发现的支持:摄入指数期植物乳杆菌WCFS1激活了人十二指肠的细胞分裂和生长,而静止期和热杀伤细胞诱导了NF-κB免疫反应途径。
◆ 加工和保存影响益生菌的耐受水平
大量研究表明,加工和保存会显著影响益生菌的应激耐受水平,益生菌功能也会随培养加工步骤的不同而变化。
在益生菌制造和储存过程中以及胃肠道中更好的生长和存活能力,对酸、胆汁和胃肠道酶的保护,与肠上皮的粘附,抗菌特性和抗生素耐药性可以被认为是维持益生菌功效的重要因素。
2
益生菌的选择和特异性
◆ 多样化的益生菌一般效果更好
多菌株益生菌比单菌株益生菌在降低空腹血糖和胰岛素抵抗指数方面效果更好。益生菌产品中包含的菌株种类越多,其在体外条件下抑制病原菌的能力越强。多样化的菌株组合可以提供更好的抗菌效果。
◆ 不同的菌株效果各异
不同的益生菌菌株具有不同的功能特性。益生菌的功效是高度针对菌株的。同一物种的不同菌株对健康的影响不同。例如,嗜酸乳杆菌CL1285、干酪乳杆菌LBC80R和鼠李糖乳杆菌 CLR2,已显示出预防抗生素相关腹泻的功效,而其他菌株则没有显示出相同的益处。
◆ 活菌数量和施用速度
益生菌产品中的活菌数量是影响效果的重要因素。一般来说,活菌数量越高,可能发挥的作用越大。但这也不是绝对的,还需要考虑菌株的活性和生存能力。
产品在储存和使用过程中,活菌数量可能会逐渐减少。因此,选择具有较高初始活菌数量且能够在储存和消化过程中保持一定活性的产品更为重要。
◆ 疾病特异性
益生菌也是疾病特异性的。相同的益生菌菌株或菌株混合物可能对一种疾病有效,但对其他疾病类型无效。
益生菌使用的适应症多种多样,从预防疾病(例如阴道炎、旅行者腹泻、败血症、特应性皮炎)或预防疾病标准疗法的副作用(如幽门螺杆菌的治疗或化疗),到急性疾病的治疗(如艰难梭菌感染、急性儿童或成人腹泻、便秘)或慢性疾病的治疗(如炎症性肠病、肠易激综合征或肥胖)。
益生菌菌株的有效性可能因其用于治疗的疾病而异。例如,鼠李糖乳杆菌GG和布拉酵母菌CNCM I-745 在治疗特定胃肠道疾病方面显示出显著的疗效,而对其他疾病可能没有效果。
◆ 孢子益生菌具有更好的生存能力和稳定性
孢子益生菌指的是能使用蛋白质、矿物质和其他可用的化合物形成保护性内生孢子,使它们免受恶劣环境影响的一部分益生菌。
孢子益生菌通常属于芽孢杆菌属,以下是一些在临床环境中的常见孢子益生菌物种:
•印度芽孢杆菌(Bacillus indicus) HU36
•凝结芽孢杆菌(Bacillus coagulans) SC-208
•克劳氏芽孢杆菌(Bacillus clausii) SC-109
•枯草芽孢杆菌(Bacillus subtilis) HU58
•地衣样芽孢杆菌(Bacillus lichenformis) SL-307
•巨型芽孢杆菌(acillus megaterium) EM144
孢子益生菌相比于普通益生菌,具有更好的生存能力: 孢子益生菌能够通过胃酸和消化液而不会被破坏。其他益生菌,如乳酸菌和双歧杆菌则没有。并且孢子益生菌通常不需要冷藏,保质期更长。
临床试验和研究中报告了孢子益生菌的一些健康益处,包括:
•减少血液中的毒素
•降低胆固醇
•降低炎症
•改善免疫反应
•新陈代谢增加
•维生素合成
•抗氧化剂生产
3
益生菌依附的食物基质(载体)
益生菌依附的食物基质会显著影响其功效。不同的食物基质可以提供不同的营养成分和环境条件,从而影响益生菌的生长、存活和定植能力。
食品,特别是乳制品被认为是将益生菌输送到人体胃肠道的理想载体。目前,益生菌主要掺入乳制品中,如奶酪、酸奶、冰淇淋和其他乳制品甜点。尽管其他基质(例如果汁)的使用也越来越多。
◆ 食物基质会影响益生菌的粘附、细胞大小等
食物配方可能对益生菌产生直接影响,例如提供针对胃酸的特定营养物质的理化屏障。这些基质效应因细菌种类而异。
食物基质还会影响肠道递送部位的功能性状表达。尽管相关研究有限,但鼠李糖乳杆菌GG在酸奶中孵育后对Caco-2细胞的粘附性较在冰淇淋中增强,且其结合能力与产品储存时间相关。在食物输送基质中孵育可能还会导致其他生理差异。例如,与实验室培养基相比,干酪乳杆菌在牛奶中冷藏孵育后,细胞大小显著减少(约0.1μm),并诱导肠道中与功能相关的特定细胞表面和代谢蛋白的表达。
干酪乳杆菌BL23在不同培养基中的细胞形态

doi: 10.1016/j.copbio.2012.10.002.
(a)含有2%乳糖;(b)UHT牛奶。平均而言,在牛奶中培养的干酪乳杆菌BL23明显短于在MRS中培养的细胞
◆ 不同的食物基质益生菌活力水平存在差异
不同的酸奶在保质期内表现出不同的益生菌活力水平。观察到,在60天的保质期内,全套酸奶中的链球菌水平在6°C下下降了约1个对数周期。虽然它们在相同条件下在脱脂酸奶中仍然存活甚至繁殖。当全套酸奶与加热牛奶和糖的浓缩产品一起生产时,它们的存活率下降了1.5个对数周期。
与含有混合浆果或百香果的酸奶相比,普通酸奶在保质期内保持更高水平的嗜酸乳杆菌的能力显著。有趣的是,含有芒果或草莓的酸奶比普通酸奶含有更高水平的嗜酸乳杆菌,这表明了各种水果混合物的不同性质(如pH值)对酸奶中益生菌活力的影响。
不同的营养成分,如果汁中的维生素水平,也可能对生存能力产生影响。据报道,抗坏血酸强化可以提高酸奶中嗜酸乳杆菌的存活率,尽管这对双歧杆菌没有任何影响。抗坏血酸的氧清除作用是可能有助于提高益生菌存活率的原因之一。
◆ 蔬菜、谷物的微结构保护益生菌或促进其生长
尽管目前大多数益生菌食品主要以乳制品为基础,但由于许多人的乳糖不耐症和发酵乳制品中不利的胆固醇含量等问题,此外,对素食益生菌产品的需求不断增加。这导致了从各种食物中开发出新的益生菌产品,包括水果、蔬菜、豆类和谷物产品。
麦芽、小麦和大麦提取物对提高嗜酸乳杆菌、罗伊氏乳杆菌和植物乳杆菌的胆汁耐受性有积极影响。然而,麦芽培养基已经显示出比小麦或大麦培养基更好的支持嗜酸乳杆菌、发酵乳杆菌、罗伊氏乳杆菌和植物乳杆菌的生长。将植物乳杆菌(L.plantarum)固定在麦芽和大麦纤维中似乎对胃肠道耐受性起主要作用。
此外据报道,在储存期间以及体外和体内胃肠道条件下,副干酪乳杆菌、植物乳杆菌和一些其他益生菌在食用橄榄和朝鲜蓟中的存活率较高。这种高活力是由这些蔬菜的微结构引起的,粗糙可能会在酸性环境中保护益生菌,或存在益生元物质,并由于蔬菜中营养物质的释放而对生存产生积极影响。因此,不仅食品的化学成分,而且它们的物理结构对它们所具有的益生菌功效也很重要。
因此,可以通过调整和优化携带益生菌的食物产品的配方,以提高其疗效和功能。这种操纵不仅可以增强益生菌的存活率,还能改善其在肠道内的定植能力,从而更有效地发挥其健康益处。
◆ 微胶囊化或益生元封装提高了存活率
研究发现,用人工酶包裹的长双歧杆菌可以增加益生菌在肠道中的定植时间,增强其抗炎作用。另一项研究报告称,益生元封装的益生菌在小鼠结肠癌病变周围特异性富集,有效抑制结肠癌。
微胶囊化的乳双歧杆菌在胃液中表现出更高的存活率,其保质期内的存活率显著高于游离细胞。此外,微胶囊化还提高了冻干酸奶在4°C和21°C下储存6个月后的益生菌活力。这是因为微胶囊化通过在封装材料中保护细胞,减少了细胞损伤和损失。
4
干预持续时间

一项研究通过系统综述和荟萃分析的方法,评估了益生菌补充对2型糖尿病患者的血糖、血脂、血压和炎症控制的影响。研究发现干预持续时间的不同会影响益生菌补充的效果。
◆ 长期的干预可能具有更好的定植和改善效果
干预时间超过8周的益生菌补充在改善血糖控制、胰岛素抵抗、血脂水平、血压和炎症因子方面表现出更显著的效果。这表明较长的干预时间可能更有助于益生菌发挥其潜在的健康益处。
空腹血糖(FBS):干预时间超过8周的研究显示,FBS显著降低。
糖化血红蛋白(HbA1c):干预时间超过8周的研究在整体分析中显示出更显著的改善。
胰岛素抵抗:干预时间超过8周的研究显示,胰岛素抵抗指数(HOMA-IR)显著降低。
总胆固醇和甘油三酯:干预时间超过8周的研究显示,总胆固醇(TC)和甘油三酯(TG)显著降低。
血压:收缩压和舒张压都显著降低。
炎症因子:干预时间超过8周的研究显示,肿瘤坏死因子(TNF-α)显著降低。
上一节讨论了影响益生菌效果的环境因素,本节将重点介绍宿主因素。益生菌的效果受到多种宿主因素的影响,这些因素决定了益生菌在肠道内的生存、定植和功能。

DOI:10.1002/fbe2.12078.
个体的营养饮食习惯、肠道微生物组成、免疫系统状态、年龄、健康状况和药物使用等均可能显著影响益生菌的活性和功效。
1
宿主营养、饮食习惯
宿主的饮食习惯和营养成分可能对益生菌在体内获得生长和生存所需的营养能力产生重大影响。

Rytter H,et al.Gut Microbes.2023
◆ 不同饮食习惯下的益生菌活性和效果存在差异
对于单个菌株,无菌小鼠盲肠中植物乳杆菌(L.plantarum)WCFS1的整体转录组和定植水平在喂食标准食物或西式饮食(41%的卡路里来自脂肪)的小鼠之间有所不同,并且在活性水平和与宿主相互作用的代谢和细胞表面修饰途径方面存在差异。L. plantarum 299v 在人结肠和回肠中表达的适应与 L. plantarum WCFS1 在喂食西方饮食的小鼠中表达的性状最相似。
饮食也有助于植物乳杆菌WCFS1的持续存在和与免疫调节作用相关的细胞产物的差异表达。对饮食条件的反应能力可能会调节益生菌对人类病原体的保护作用。例如双歧杆菌菌株具有将糖发酵成醋酸盐的能力增强,醋酸盐是一种保护无菌小鼠免受大肠杆菌侵害的产品。
◆ 特殊饮食成分可以影响益生菌的免疫反应
饮食对益生菌性能的影响也可能与个别饮食成分有关。饮用水中的乳糖补充剂促进了嗜热链球菌LMD9在无菌大鼠体内的肠道定植,嗜热链球菌的乳酸产生与上皮细胞中单羧酸转运蛋白和细胞周期停滞蛋白的诱导有关。
罗伊氏乳杆菌从组氨酸中产生的组胺也被证明可以诱导上皮细胞对抑制促炎细胞因子 TNF 产生的反应。益生菌在消化道中将膳食成分转化为对免疫反应途径至关重要的生物活性形式。
2
宿主生理特征
益生菌临床试验中反应者与无反应者的一致发现表明,宿主的生理特征会影响益生菌的效果。年龄、遗传学、种族、性别差异和健康状况都可能单独或共同影响益生菌对健康的作用。
◆ 年龄<55岁的人群可能更利于益生菌定植
上述研究益生菌补充对2型糖尿病患者的影响,表明年龄≤55岁的人在血糖(FBS)和胰岛素抵抗(HOMA-IR)方面对益生菌补充剂的反应更强。
◆ 益生菌效果存在性别差异
一项研究调查了益生菌对肠道微生物群的性别依赖性影响,发现补充益生菌后,与男性相比,女性的促炎肠道微生物显著减少。此外,外周免疫细胞分析显示,益生菌降低了男性树突状细胞和CD14单核细胞的比例,但女性没有。这表明益生菌对肠道微生物群的调节存在性别特异性反应。
另一项研究表明,补充益生菌后,与男性运动员相比,女性运动员的厚壁菌门比例很高 。哥德堡大学进行的一项有趣的研究报告了补充植物乳杆菌后男性和女性患者沙门氏菌感染临床症状的差异。
◆ 体重指数影响益生菌效果
不同体重指数(BMI)人群的肠道环境存在差异,例如肠道pH值、胆汁酸浓度和肠道通透性等。肥胖者的肠道菌群多样性通常较低,且一些特定的细菌种类相对丰度发生改变。益生菌需要在适宜的肠道环境中生存和发挥功能,这些因素可能会影响益生菌的定植和生存。
然而有研究发现,当BMI≥30时,益生菌补充对改善血糖控制、胰岛素抵抗、血脂水平、血压和炎症因子的效果更显著,而当BMI<30时,效果则不明显。但这可能是由于基数较高,使得改善的变化更加明显。
◆ 地理区域或种族
来自东亚国家的受试者在血糖控制和胰岛素抵抗(HOMA-IR)方面的改善效果显著优于来自西方国家的受试者。
3
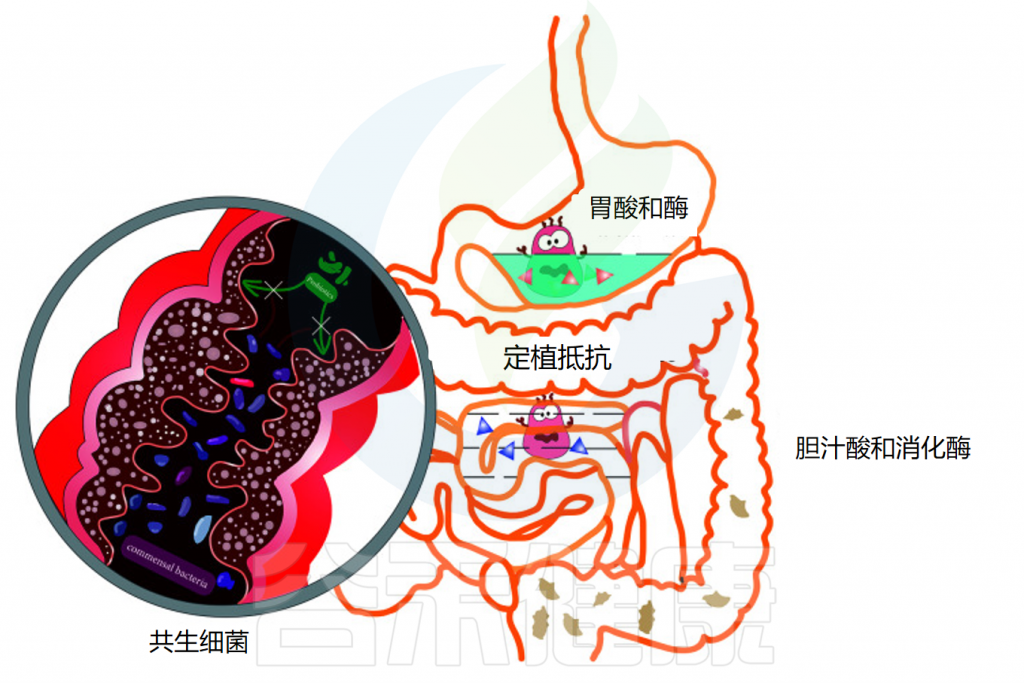
消化道的生理结构
胃肠道的成功定植是益生菌能够发挥足够的宿主相互作用以赋予健康益处的关键因素。口服后,益生菌经过口腔,通过胃,到达小肠和结肠。在这一过程中,消化系统的结构与蠕动及消化酶、氧气水平、pH变化、胆汁等都会影响益生菌的定植和效果。
影响益生菌在胃肠道运输过程中活力的因素
doi: 10.3389/fcimb.2021.609722.
◆ 唾液对益生菌的影响微乎其微
当摄入益生菌时,它们首先会接触到口腔中的唾液。对多种乳酸杆菌、片球菌和双歧杆菌菌株的体外研究表明,与对照组相比,当暴露于唾液时,细胞计数没有显著损失。虽然益生菌在口服后通过口腔的运输和它们与唾液的接触是短暂的,但唾液对益生菌存活率的影响似乎很小。
◆ 胃酸对大多数细菌都是极其致命的
通过食道后,益生菌到达胃部,在那里它们暴露在酸性胃液中。酸性环境对大多数细菌极为致命,尤其是对不耐酸的细菌,这会导致细菌细胞质pH值降低。通过胃的运输需要5分钟到2小时,长时间暴露在酸性环境中对益生菌来说是一个巨大的挑战。
此外,胃中存在的其他不良条件,包括离子强度、酶活性(胃蛋白酶)和机械搅动,已被证明对益生菌的生存能力有影响。例如,长双歧杆菌和短双歧杆菌的活细胞在一小时内在模拟胃液中变得检测不到。因此,在经过胃的时候可能有一部分的益生菌已经失活或失去作用。
◆ 小肠中的胆汁酸和消化酶影响益生菌生存
益生菌通过幽门后,会到达小肠,那里有丰富的胰液和胆汁。在肠液的中和作用下,小肠中的pH值约为 6.0-7.0,比胃液温和得多。然而,胆汁酸和消化酶(包括脂肪酶、蛋白酶和淀粉酶)也可以通过细胞膜破坏和DNA损伤来影响益生菌的活力。
体外研究表明,唾液乳杆菌Li01和戊糖片球菌Li05在模拟肠液中的活力降低。
◆ 增强在胃肠道中存活率的技术
为了增强益生菌对胃肠道中胃液和胆汁的耐受性,益生菌可以涂上保护壳,这种技术称为微胶囊化。近年来,在提高存活率和保证通过基于微胶囊的方法到达结肠的足够数量的活益生菌方面取得了重大进展。
◆ 结肠中的共生细菌可能产生定植抵抗
结肠具有最大的细菌密度,益生菌在这可能会遇到共生细菌的定植抗性。益生菌必须与宿主常驻微生物群竞争营养物质和粘附位点,才能在结肠粘膜定植和增殖。
并且由于定植抗性,大多数益生菌在口服给药后和停止食用后不久随粪便从结肠排出,因此无法检测到益生菌。产生定植抵抗的机制在下面的部分中详细说明。
拓展:益生菌的结构与定植
益生菌可以编码许多细胞表面因子,这些因子与粘附粘蛋白或肠上皮细胞有关。
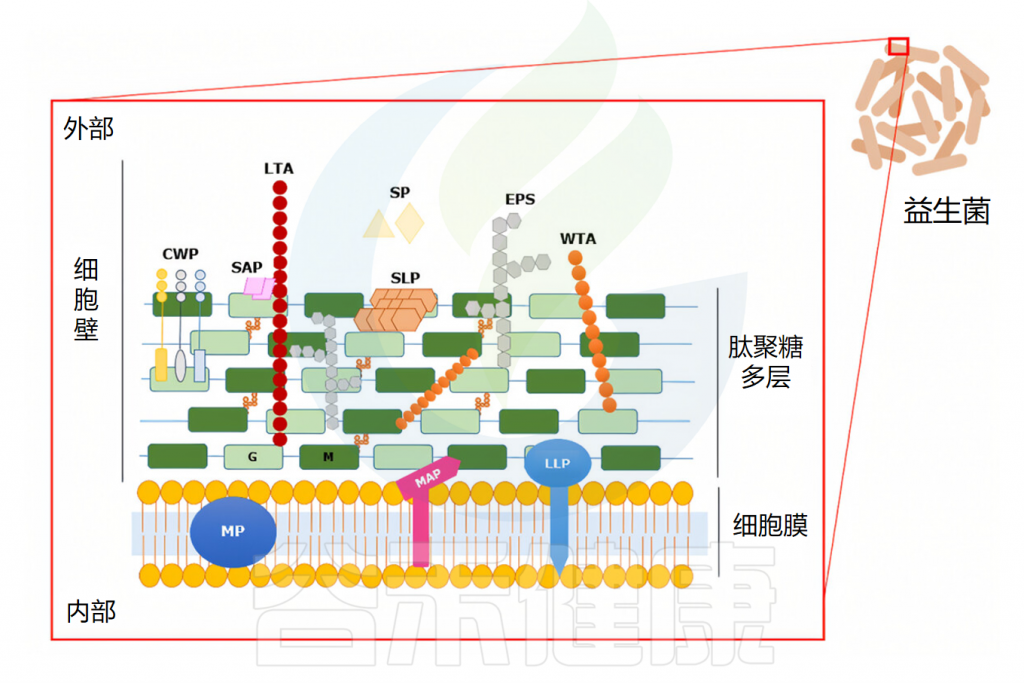
DOI:10.1002/fbe2.12078.
胞外多糖
胞外多糖可能促进细菌和肠道细胞之间的相互作用,从初始粘附机制到永久粘附机制。例如,长双歧杆菌NB667 含有与人肠道粘液的粘附直接相关的胞外多糖。益生菌产生的胞外多糖可能因物种而异,这些变化可能会影响相应细菌的粘附能力,因为它们可能需要不同的结合机制。
菌毛或鞭毛
菌毛或鞭毛是存在于特定革兰氏阳性菌和革兰氏阴性菌上的细胞表面的细长蛋白质突起。已经证明鼠李糖乳杆菌GG 中的菌毛在确保有效粘附人肠上皮细胞系方面起着至关重要的作用。
蛋白质成分
蛋白质成分已被证实在乳酸杆菌菌株的粘附促进因子中起关键作用。存在于乳酸杆菌CNCM-I-3698和法氏乳杆菌CNCM-I-3699表面的蛋白质成分对于它们与人肠道上皮细胞的粘附至关重要。
在乳酸杆菌的粘附过程中,表层蛋白和表面相关蛋白,包括粘液结合蛋白、胶原蛋白和纤连蛋白以及月光蛋白,通过与宿主受体相互作用而发挥关键作用,宿主受体在肠上皮定植中表现出不同的功能作用。
DOI:10.1002/fbe2.12078.
附着力
细菌粘附到粘膜的过程包括可逆和稳定的阶段。最初,益生菌通过非特异性物理接触与粘膜结合,建立可逆和弱的物理结合。随后,随着粘附素(通常是锚定在细胞表面的蛋白质)和互补受体之间的特异性相互作用,益生菌与粘液或肠上皮细胞(IEC)建立了稳定的结合,从而成功地定植了胃肠道。
4
肠道微生物群
常驻肠道微生物群对消化道益生菌功能的影响也不应被低估。肠道微生物群落的结构可能影响了对益生菌的反应,可能是通过改变益生菌在肠道中的生理状态和活性。
◆ 不同的菌群可能对益生菌“允许”定植或“抵抗”
同样是补充益生菌,有些人身上可以看到非常明显的效果,而有些人的效果则不明显,这是为什么呢?
一项有趣的研究,志愿者被分为两组,“允许型”和“抵抗型”。在允许组的人的肠道粘膜中益生菌菌株显著增加,而在抵抗组的人的肠道中没有检测到益生菌。
不同的肠道菌群对益生菌的定植反应复杂,有些适宜定植,而另一些则强烈抵抗。这些机制对益生菌的合理选择和干预效果至关重要。
◆ 定植抗性会严重影响补充益生菌的效果
人体自身正常的肠道微生物群形成稳定的细菌群落,抵抗外来细菌的入侵和病原体的扩张。这种现象被称为“定殖抵抗”。定植抗性机制可分为两大类:直接机制和间接机制。
直接定植抗性是指严格通过与肠道微生物群相关的因素限制外源性微生物定植,独立于与宿主的任何相互作用。间接定植抗性取决于宿主衍生因子,包括抗菌肽的产生、上皮屏障的维持以及通过与宿主相互作用调节胆汁酸浓度。
例如,细菌素是在革兰氏阳性菌或革兰氏阴性菌的核糖体中合成的蛋白质化合物,并且能够抑制密切相关的物种或利用相似营养物质或生态位的物种。
益生菌受到共生肠道微生物群施加的定植抗性的不利影响。一些研究表明,人类摄入的益生菌在局限于给药期间和之后不久在通过粪便排出。定植抵抗可能是限制益生菌长期影响的重要原因之一。
将由11种益生菌菌株组成的组合施用到成年、雄性无特异性病原体小鼠和无菌小鼠中。在指定的时间点分析粪便样本,然后在补充后第28天解剖胃肠道。发现在无菌小鼠中观察到的细菌活菌计数显着更高。对结果的一种解释可能是,与无菌小鼠相比,益生菌在常规小鼠中遇到更高程度的粘膜定植抵抗。
5
抗生素或药物的使用
我们知道,抗生素可以在杀死致病菌的同时,也会误伤体内的有益菌,破坏原有的菌群平衡状态。因此,其使用也可能会影响益生菌的效果。
◆ 抗生素和抗炎药会改变肠道环境,影响益生菌定植
抗生素的使用可能改变肠道环境,使得益生菌难以在肠道内定植。肠道黏膜屏障可能受损,影响益生菌的黏附和生存。
例如,抗生素可能导致肠道黏液层变薄,减少益生菌与肠道黏膜的接触机会。此外,抗生素还可能改变肠道pH值、胆汁酸浓度等,这些因素都可能影响益生菌的生长和定植。
例如非甾体类抗炎药(如布洛芬、阿司匹林等)可能引起胃肠道黏膜损伤,破坏肠道屏障功能。质子泵抑制剂(如奥美拉唑、兰索拉唑等)通过抑制胃酸分泌来治疗胃酸相关疾病。然而,胃酸减少可能会改变肠道菌群的组成,影响益生菌的生长和定植。
研究人员发现在服用抗生素4周后,接受11株益生菌治疗的人的肠道微生物群需要更长的时间才能恢复正常。
胃肠道成功定植是益生菌发挥功效并带来健康益处的关键。考虑到影响益生菌定植的众多外部和宿主因素,那么,究竟什么样的肠道更适合益生菌定植呢?
1
平衡和健康的菌群组成
菌群都具有高度个体特异性,也就是人和人之间的菌群相差较大。益生菌更有可能定植于具有平衡和健康微生物群的肠道。特定有益菌的存在和肠道环境的整体健康状况在益生菌的成功定植中起着至关重要的作用。
就婴儿而言,影响初始菌群定植的因素包括:胎龄(足月or早产)、分娩方式(自然分娩or剖腹产)、喂养方式(母乳or配方奶)、药物使用(抗菌药物)、生长环境(城市or农村)等。
就成年人而言,年龄、饮食、生活方式、疾病状况、药物使用、环境等多因素都会影响肠道菌群多样性,这在前已经详细阐述。这些所有叠加后会形成一个具有高度特异性的个体菌群,构成的整体菌群环境会影响到其他新的菌群的定植。
根据外来菌群和常驻菌群的反应结果进行分类。有四种可能:
“抵抗”(外来益生菌消失,所有常驻物种保持)
“增强”(外来益生菌维持,所有常驻物种保持)
“破坏”(外来益生菌消失,一些常驻物种也消失)
“位移”(外来益生菌维持,一些常驻物种消失)
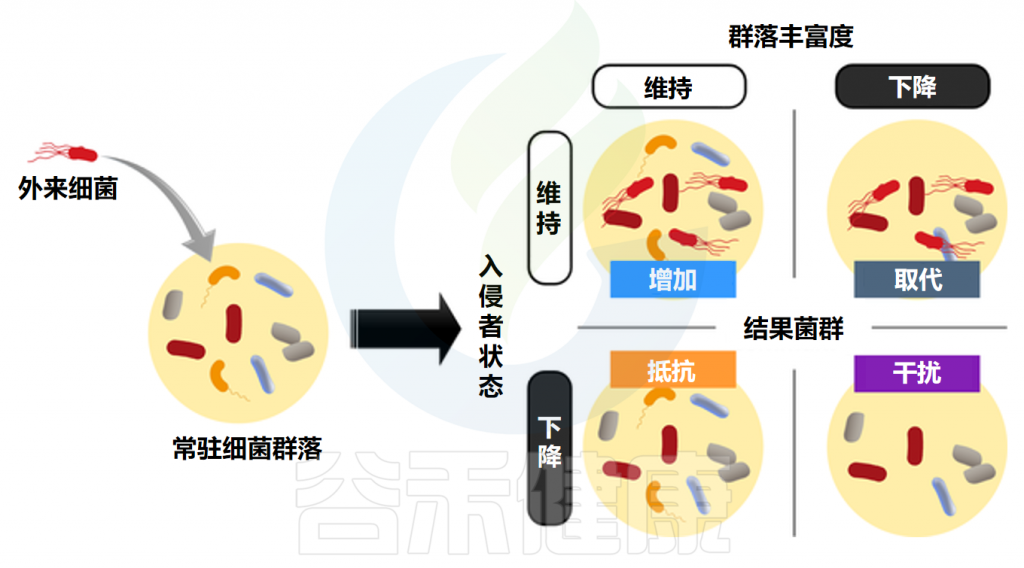
当常驻菌群创造的环境有利于外来菌群,施用益生菌就会更成功。常驻菌群本身的结构特征也比较重要。益生菌带来的健康益处可能不仅是由单独补充的益生菌菌株引起的,而是与常驻肠道微生物群相互作用的结果。
2
健康的肠道
益生菌更有可能在粘膜完整且炎症水平低的肠道中定植。益生菌有助于维持粘膜完整性和调节免疫反应,这也可以进一步支持它们的定植。
◆ 正常的肠道pH值
不同的益生菌对pH值有不同的适应范围,但一般来说,偏酸性的肠道环境(pH值约为5.5-7.0)更有利于益生菌的生长和定植。
例如,乳酸菌等益生菌在酸性环境下活性较高,而过高或过低的pH值会影响益生菌的存活。
◆ 完整的肠道黏膜屏障
健康的肠道黏膜可以为益生菌提供附着位点,同时阻挡有害菌和毒素的入侵。如果肠道黏膜受损,益生菌的定植可能会受到影响。例如,炎症性肠病患者的肠道黏膜屏障功能受损,益生菌的定植难度可能会增加。
◆ 正常的肠道蠕动
适度的肠道蠕动有助于益生菌在肠道内的分布和定植。过慢的肠道蠕动可能导致益生菌在局部堆积,而过快的肠道蠕动则可能使益生菌来不及定植就被排出体外。
3
适宜的饮食和生活方式
◆ 富含膳食纤维等益生元的饮食
膳食纤维可以被肠道中的有益菌发酵,产生短链脂肪酸等物质,为益生菌的生长提供营养支持。例如,摄入足够的蔬菜、水果、全谷物等富含膳食纤维的食物,可以促进益生菌的定植。
益生元是一种可溶于水的可溶性纤维,可以作为益生菌的“食物”。
益生元可以通过小肠到达结肠,然后有益细菌会把它消化或发酵,形成乙酸盐、丙酸盐和丁酸盐等代谢物。最广泛认可的益生元包括低聚果糖 (FOS)、菊粉和低聚半乳糖(GOS)等。
◆ 规律的生活习惯
良好的睡眠、适度的运动和减少压力等生活方式因素有助于维持肠道的正常功能,从而为益生菌的定植创造有利条件。例如,长期熬夜、过度劳累和精神压力过大可能会影响肠道菌群的平衡,降低益生菌的定植能力。
4
根据症状选择适合自身的益生菌
益生菌每个属内都有很多种和菌株,益生菌的作用因这些种和菌株而不同。哪怕同属于乳酸杆菌,菌株不同效果也不同,比如:嗜酸乳杆菌有助于血压、胆固醇、过敏和消化;鼠李糖乳杆菌可以帮助降低血糖、免疫功能和胆固醇;瑞士乳杆菌可以降低血压、改善睡眠质量和骨骼健康。
这里提供一些常见的益生菌及目前已知的功能作为参考。

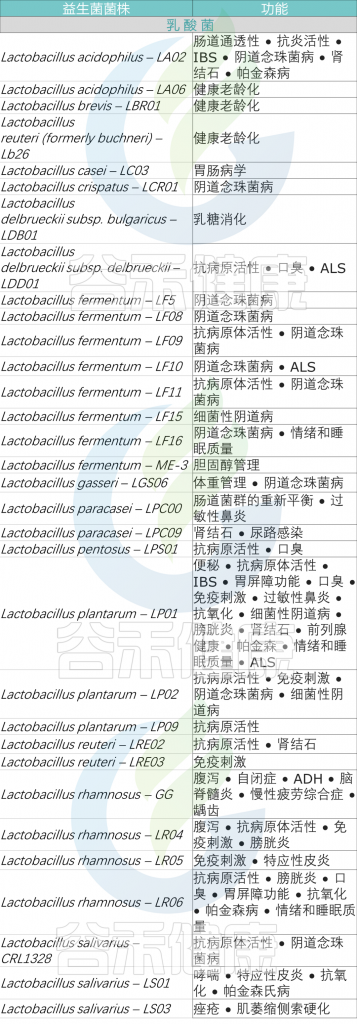

<来源:谷禾健康数据库>
不同益生菌菌株在不同疾病中的疗效存在显著差异,最终,在充分考虑益生菌菌株的特异性以及与特定疾病相关的特异性因素的情况下,挑选出一个最适合的益生菌进行补充,以便有效改善健康状况和促进身体的整体功能。
不能有效定植,是否意味着益生菌无效?
有时候,益生菌并不是完全依靠在肠道的定植来产生影响。就算不能定植,它也有其他很多种方式来发挥优势。包括以下几种方式:
(1)通过产生抗菌物质和与病原体竞争结合上皮细胞来阻断致病细菌的作用;
(2)通过增加屏障功能、粘液生成、存活和细胞保护反应来促进肠上皮细胞的稳态;
(3)通过增加先天免疫,如IgA和防御素的产生,上调抗炎细胞因子的产生,抑制促炎细胞因子的产生,确定必要和过度防御免疫之间的平衡;
(4)通过产生神经递质和迷走神经调节肠脑轴。
根据市场分析,益生菌产业呈现高态势增长,这背后离不开消费者需求的多元化发展以及企业的积极布局。一方面,人们对健康生活方式的追求,推动了益生菌产品从单一功能向个性化、多样化方向发展。针对不同人群和特定健康需求,企业纷纷推出定制化的益生菌组合,全方位满足消费者的健康诉求。另一方面,国内外研究机构对益生菌的探索不断深入,为企业的产品创新提供了扎实的科学基础。
目前谷禾与众多益生菌品牌以及科研院校合作,通过益生菌与其他健康元素(如益生元,维生素、矿物质、膳食纤维等)或干预手段(饮食生活方式,粪菌移植)的联合使用,有望产生”1+1>2″的协同效应,为消费者提供更加全面、更加个性化的健康解决方案。
注:本账号内容仅作交流参考,不作为诊断及医疗依据。
主要参考文献
Lau L Y J , Quek S Y .Probiotics: Health benefits, food application, and colonization in the human gastrointestinal tract[J].Food Bioengineering, 2024, 3(1).DOI:10.1002/fbe2.12078.
Latif A, Shehzad A, Niazi S, Zahid A, Ashraf W, Iqbal MW, Rehman A, Riaz T, Aadil RM, Khan IM, Özogul F, Rocha JM, Esatbeyoglu T, Korma SA. Probiotics: mechanism of action, health benefits and their application in food industries. Front Microbiol. 2023 Aug 17;14:1216674.
Maftei NM, Raileanu CR, Balta AA, Ambrose L, Boev M, Marin DB, Lisa EL. The Potential Impact of Probiotics on Human Health: An Update on Their Health-Promoting Properties. Microorganisms. 2024 Jan 23;12(2):234.
Rytter H, Combet E, Chassaing B. Probiotic: is diet part of the efficacy equation? Gut Microbes. 2023 Jan-Dec;15(1):2222438.
Marco ML, Tachon S. Environmental factors influencing the efficacy of probiotic bacteria. Curr Opin Biotechnol. 2013 Apr;24(2):207-13.
R.D.C.S. Ranadheera, S.K. Baines, M.C. Adams,
Importance of food in probiotic efficacy,Food Research International,Volume 43, Issue 1,
2010,Pages 1-7,ISSN 0963-9969,https://doi.org/10.1016/j.foodres.2009.09.009.
McFarland LV, Evans CT, Goldstein EJC. Strain-Specificity and Disease-Specificity of Probiotic Efficacy: A Systematic Review and Meta-Analysis. Front Med (Lausanne). 2018 May 7;5:124.
de Melo Pereira GV, de Oliveira Coelho B, Magalhães Júnior AI, Thomaz-Soccol V, Soccol CR. How to select a probiotic? A review and update of methods and criteria. Biotechnol Adv. 2018 Dec;36(8):2060-2076.
Ferreiro A, Dantas G, Ciorba MA. Insights into How Probiotics Colonize the Healthy Human Gut. Gastroenterology. 2019 Feb;156(3):820-822.
Han S, Lu Y, Xie J, Fei Y, Zheng G, Wang Z, Liu J, Lv L, Ling Z, Berglund B, Yao M, Li L. Probiotic Gastrointestinal Transit and Colonization After Oral Administration: A Long Journey. Front Cell Infect Microbiol. 2021 Mar 10;11:609722.
Teng Ma, Xin Shen, Xuan Shi, Hafiz Arbab Sakandar, Keyu Quan, Yalin Li, Hao Jin, Lai-Yu Kwok, Heping Zhang, Zhihong Sun,Targeting gut microbiota and metabolism as the major probiotic mechanism – An evidence-based review,Trends in Food Science & Technology,Volume 138,2023,Pages 178-198,ISSN 0924-2244,https://doi.org/10.1016/j.tifs.2023.06.013.
Zmora N, Zilberman-Schapira G, Suez J, Mor U, Dori-Bachash M, Bashiardes S, Kotler E, Zur M, Regev-Lehavi D, Brik RB, Federici S, Cohen Y, Linevsky R, Rothschild D, Moor AE, Ben-Moshe S, Harmelin A, Itzkovitz S, Maharshak N, Shibolet O, Shapiro H, Pevsner-Fischer M, Sharon I, Halpern Z, Segal E, Elinav E. Personalized Gut Mucosal Colonization Resistance to Empiric Probiotics Is Associated with Unique Host and Microbiome Features. Cell. 2018 Sep 6;174(6):1388-1405.e21.
Liang T, Xie X, Wu L, Li L, Yang L, Gao H, Deng Z, Zhang X, Chen X, Zhang J, Ding Y, Wu Q. Comparative analysis of the efficacies of probiotic supplementation and glucose-lowering drugs for the treatment of type 2 diabetes: A systematic review and meta-analysis. Front Nutr. 2022 Jul 18;9:825897.
Chandrasekaran P, Weiskirchen S, Weiskirchen R. Effects of Probiotics on Gut Microbiota: An Overview. Int J Mol Sci. 2024 May 30;25(11):6022.
Kothari D, Patel S, Kim SK. Probiotic supplements might not be universally-effective and safe: A review. Biomed Pharmacother. 2019 Mar;111:537-547

谷禾健康

益生菌被世界卫生组织定义为“当摄入足够量时,可为宿主带来健康益处的活微生物”。近年来,随着人们发现其可用于预防、减轻或治疗特定疾病以及改善健康,益生菌在食品和临床治疗中的应用越来越广泛。
大量研究表明,益生菌有助于维持肠道菌群的平衡,促进消化和吸收营养物质;可以改善皮肤状况,减少皮肤问题如湿疹、痤疮等的发生;对治疗肠道疾病如腹泻、便秘、肠炎等有积极作用;此外,益生菌还可以减少细菌性阴道病,预防婴儿特应性皮炎,减少口腔病原体和龋齿,减少常见上呼吸道感染的发病率和持续时间。益生菌和益生元也被用于预防代谢综合征的一些表现,包括超重、2型糖尿病和血脂异常等。
然而,益生菌也不是什么“灵丹妙药”,盲目地补充益生菌也是不好的。谷禾曾检测发现一些人过量补充益生菌,导致肠道微生物群极度不平衡,这也是不利于健康的。
并且一些研究已证实了益生菌导致的不良事件是存在的。国际益生菌和益生元科学协会召开了一次会议,涉及益生菌潜在的急性和长期风险、对弱势群体的风险、益生菌产品质量满足弱势群体需求的重要性以及与益生菌使用相关的不良事件报告。
益生菌理论上可能会导致五种潜在风险:
1.微生物群组成或功能的改变;
2.入侵其他组织导致感染;
3.有害的代谢活动;影响药物功能;
4.易感个体的过度免疫刺激;
5.抗生素耐药基因转移。
此外,有大量证据表明益生菌的有效性针对菌株和疾病具有特异性。益生菌的效果因使用的菌株、给药周期和持续时间而异。并且对干预措施的反应也可能会因人而异,他们的健康状况、年龄和肠道微生物组的组成,都会影响使用益生菌的效果。
鉴于益生菌的使用日益广泛,必须全面了解其风险和益处,重新审视益生菌的安全问题。在本文中,我们回顾了益生菌所带来的一些健康益处,并讨论了其使用过程中存在的一些风险以及特别需要注意的人群。尽管益生菌在大多数时候是安全的,但在某些患者群体中应谨慎使用,尤其是早产儿或免疫缺陷的新生儿。
Merenstein D,et al.Gut Microbes.2023
益生菌一词源自拉丁语“pro”,意思是“为了”,希腊语“biotic”意思是“生命”。益生菌被广泛认为是促进健康的微生物。
联合国粮食及农业组织(FAO)和世界卫生组织(WHO)将益生菌定义为“在摄入足够量时可为宿主带来健康益处的活微生物”。
此外,国际奥委会(IOC)表示:“益生菌是活的微生物,口服几周后可以增加肠道中有益细菌的数量。这些与肠道健康的一系列潜在益处以及免疫功能的调节有关”。与其他膳食补充剂相比,益生菌制剂是独一无二的,它含有足够数量的活的、有活力的、明确的微生物,可以提供有益的健康效果。
乳杆菌和双歧杆菌是最常用的益生菌,但酵母菌和一些大肠杆菌和芽孢杆菌也被使用。新型益生菌还包括最近被批准的丁酸梭菌。
唾液乳杆菌UCC118的电镜照片
World Gastroenterology Organisation.2017
★ 益生菌用于改善健康已有悠久历史
益生菌的原理可以追溯到100多年前,早在1857年,法国微生物学家巴斯德就已经在酸奶中发现了益生菌。1908 年,乳酸菌之父 Elie Metchnikoff 提出,改变我们体内的微生物群并用有用的微生物取代有害的微生物是可能的。
在过去的几十年里,益生菌领域的研究取得了长足的进步,由于益生菌产品对健康有益,益生菌产品的使用和接受度在全球范围内不断增加。
迄今为止,人类对于益生菌的研究依然处在一个世界热门课题阶段,可见益生菌对于人类健康事业的重要意义。
★ 益生菌的作用机制
益生菌通常在胃肠道发挥作用,可能影响肠道微生物群。益生菌可以以高度个性化的模式短暂定植于人类肠道粘膜,具体取决于基线微生物群、益生菌菌株和胃肠道区域。
益生菌影响微生物群和疾病进程
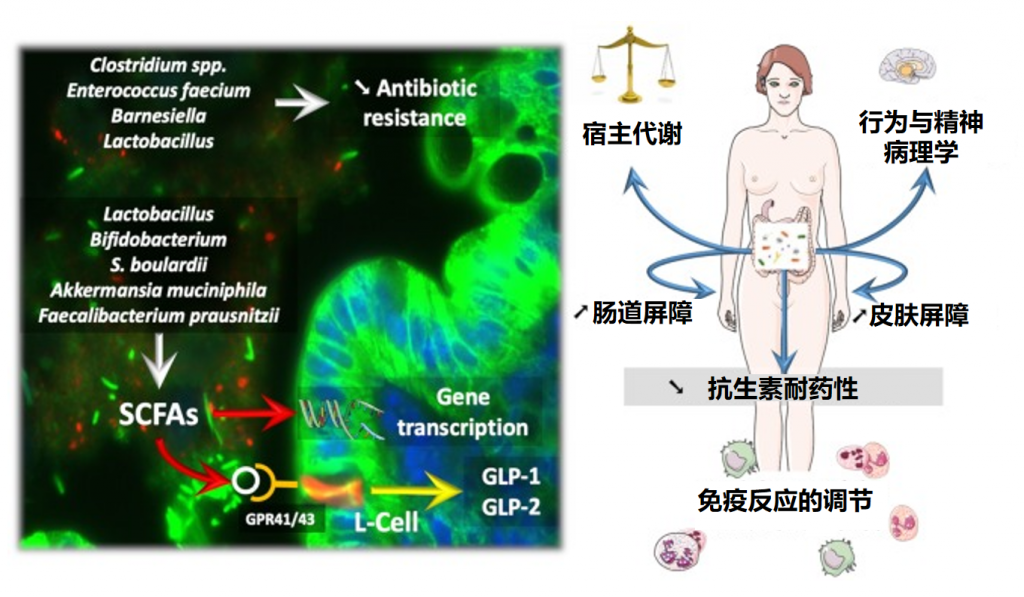
Wieërs G,et al.Front Cell Infect Microbiol.2020
益生菌还通过非特异性、物种特异性和菌株特异性机制发挥健康作用。常用益生菌补充剂的菌株、物种甚至属之间的非特异性机制差异很大。这些机制包括抑制胃肠道中病原微生物的生长(通过促进定植抵抗、改善肠道转运或帮助受干扰的微生物群正常化)、产生生物活性代谢物(例如短链脂肪酸)以及减少管腔结肠中的pH值。
物种特异性机制包括维生素合成、肠道屏障强化、胆汁盐代谢、酶活性和毒素中和。
菌株特异性机制非常罕见,并且仅被特定物种的少数菌株使用,包括细胞因子产生、免疫调节以及神经递质分子参与肠-脑轴通讯、对内分泌和神经系统的影响。通过所有这些机制,益生菌可能对人类健康和疾病产生广泛的影响。
微生物群和益生菌与宿主相互作用的机制
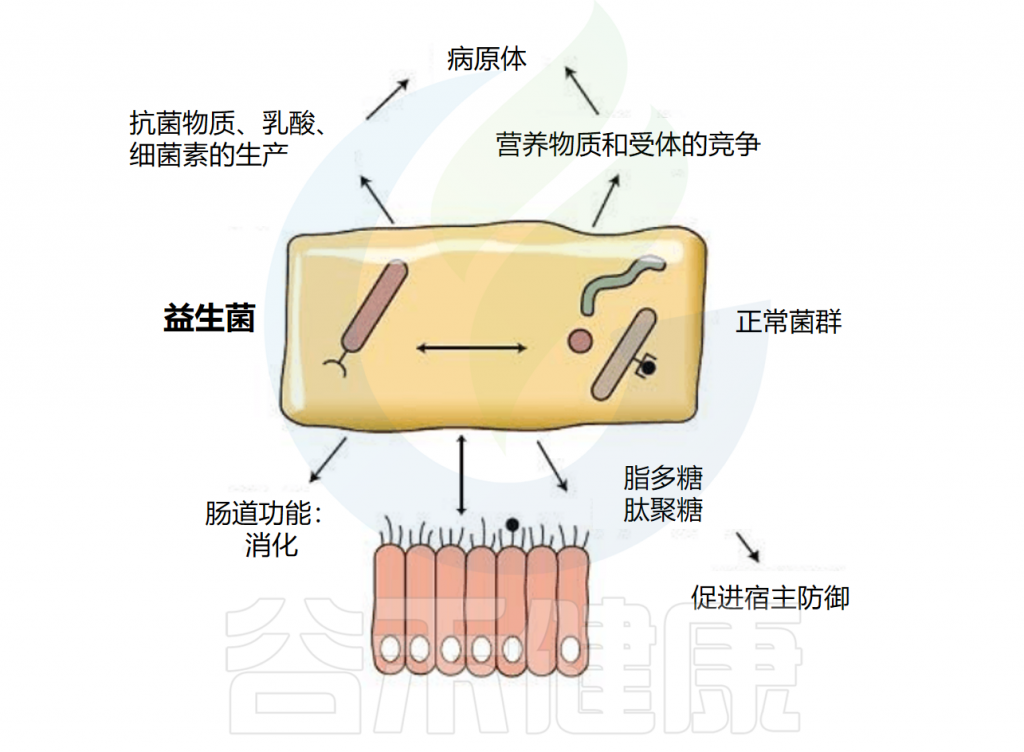
World Gastroenterology Organisation.2017
注:由于益生菌的作用可能特定于某些物种和菌株,因此其在临床或研究中使用的建议需要针对物种和菌株而定。此外,汇集不同类型益生菌研究的数据可能会导致关于其功效和安全性的误导性结论。
★ 益生菌的安全性也值得重视
尽管有潜在的好处,但益生菌对人类健康的有效性仍然存在不确定性,这主要是因为许多研究没有考虑到这些影响可能是针对菌株、剂量和条件的,并且对干预措施的反应可能会因人而异。独特的部分取决于他们的健康状况、年龄和肠道微生物组的组成。
重要的是,许多研究未能报告不良反应,或没有提供有关不良反应的具体细节,例如症状发生的时间、持续时间和严重程度。缺乏安全数据收集和报告可能会导致不良或意外的健康结果。
临床医生、研究人员和政策制定者已经提出了与益生菌安全性相关的担忧。这些问题可以广泛地分为与益生菌菌株、产品质量或益生菌管理相关的问题。
益生菌携带的抗生素抗性基因转移
一个令人担忧的问题是,益生菌携带的抗生素抗性基因可以转移到常驻的潜在病原体、宿主携带的其他微生物或环境微生物中,从而增加抗生素耐药性基因的生态库。
微生物污染物的存在
特别值得关注的是有害活微生物污染物的存在。由于益生菌被设计为活微生物形式施用,因此与经过有意灭菌过程的产品相比,被致病或潜在致病微生物污染的风险更大。最终产品的微生物污染以及过敏原或其他污染物的存在也令人担忧。
与益生菌产品配方安全相关的问题包括需要确定最终产品的纯度、效力(提供的活微生物的数量)和成分。此外,益生菌产品必须经过充分的测试(适合预期用途)以检测潜在污染物。
使用益生菌的方式也必须是安全的
给予或服用益生菌的方式也必须是安全的。安全给药包括对宿主的适当给药途径以及益生菌在现场的正确操作或制备。产品应以安全的剂量和最终配方交付。
例如,此类管理问题之一是一旦益生菌在现场打开并使用,医院环境和弱势患者可能会受到交叉污染。在医院病房内混合干燥的益生菌会导致静脉导管感染。
发酵食品
食用发酵食品是获取益生菌的一种重要方式,发酵食品是通过各种活微生物培养物的生长和代谢活动制成的。其中许多食物都含有丰富的活微生物和潜在有益微生物。
一些发酵食品,例如酵母面包和大多数商业泡菜,在发酵后进行加工,在食用时不含有活菌。而许多酸奶含有益生菌微生物,例如保加利亚乳杆菌(Lactobacillus bulgaricus)和嗜热链球菌(Streptococcus thermophilus)。
• 发酵食品中的部分活微生物无法到达远端肠道
用于制造许多发酵食品(包括酸奶)的活微生物通常在产品的整个保质期内都能很好地存活。然而,它们通常无法通过胃,并且可能无法抵抗小肠中水解酶和胆汁盐的降解,因此可能无法到达远端肠道。
然而,酸奶或其他食物中含有的部分益生菌菌株确实可以在肠道运输中存活下来。
含有活菌但不确定是否是益生菌微生物的发酵食品包括许多奶酪、泡菜(一种韩国发酵卷心菜)、普洱茶(一种发酵茶)、酸菜(发酵卷心菜)、味噌(一种发酵大豆酱),以及由发酵苹果糖制成的未经过滤的生苹果醋。
• 一些未发酵食品中也添加了微生物
某些未发酵食品,例如牛奶、果汁、冰沙、麦片、营养棒以及婴幼儿配方奶粉,都添加了微生物。这些食物是否是真正的益生菌取决于它们食用时所含的微生物水平、它们是否能在肠道运输中存活,以及它们的特定物种和菌株是否对健康有影响。
益生菌补充剂
益生菌也可以作为含有多种菌株和剂量的膳食补充剂(胶囊、粉末、液体和其他形式)提供。这些产品通常含有活微生物的混合培养物而不是单一菌株。
• 益生菌补充剂并不是含有的菌种数量越多带来的健康效果就越好
许多益生菌补充剂每剂含有1至100亿CFU,但有些产品含有高达500亿CFU或更多。然而,较高的CFU计数并不一定会改善产品的健康效果。
益生菌以菌落形成单位(CFU)为单位进行测量,它表示活细胞的数量。数量可以写在产品标签上,例如,1×10^9表示10亿CFU,或1×10^10表示100亿CFU。
由于益生菌必须活着食用才能对健康有益,而且益生菌可能在保质期内死亡,因此用户应寻找在产品保质期结束时(而不是在制造时)标有CFU数量的产品。
目前的法规仅要求制造商在益生菌产品的补充成分标签上列出微生物的总重量;该细胞团可由活微生物和死微生物组成,因此与产品中活微生物的数量无关。
益生菌的潜在健康益处是大量科学研究的焦点。本节介绍了使用益生菌在预防或治疗一些疾病中的作用,包括:特应性皮炎、小儿急性感染性腹泻、炎症性肠病、肠易激综合征、高胆固醇血症和肥胖。
1
特应性皮炎
特应性皮炎是最常见的湿疹形式,也是最常见的慢性炎症性皮肤病之一,影响全世界约15%至20%的儿童和1%至3%的成人。
许多益生菌研究评估了不同种类和菌株的细菌对预防特应性皮炎的作用,并且一些荟萃分析综合了这些研究的结果。
• 怀孕和婴儿早期接触益生菌可能降低儿童特应性皮炎的风险
研究和荟萃分析表明,在怀孕期间和婴儿早期接触益生菌可能会降低儿童患特应性皮炎的风险。例如,2018年的一项荟萃分析包括27项随机对照试验和一项对照队列研究,共有6907名婴儿和儿童在子宫内接触益生菌2周至7个月(通过母亲口服补充剂)或出生后2至13个月的婴儿口服给药。
在6个月至9岁之间,使用单一菌株或包括乳杆菌、双歧杆菌和丙酸杆菌菌株的混合物进行益生菌治疗可显著降低特应性皮炎的风险,从对照组的34.7%降至益生菌组的28.5%。
• 益生菌的治疗效果因菌株而异
此外,益生菌治疗的效果因益生菌菌株而异。例如,补充鼠李糖乳杆菌(Lactobacillus rhamnosus)或副干酪乳杆菌(Lactobacillus paracasei)可显著降低特应性皮炎的发病率,而补充罗伊氏乳杆菌或嗜酸乳杆菌则不会。
另一项荟萃分析包括8项随机临床试验,共有741名从出生到36个月大的参与者接受了乳酸菌或双歧杆菌治疗4至24周。结果表明,含有乳酸菌的益生菌可能会减轻婴儿和幼儿的特应性皮炎症状,但含有双歧杆菌的益生菌则不然。该治疗显著改善了患有中度至重度疾病的参与者的症状,但没有改善轻度疾病参与者的症状。
对2599名1至55岁参与者(大多数是儿童)使用单一益生菌和益生菌混合物治疗湿疹的39项随机对照试验进行了审查,发现益生菌治疗可能会略微降低 SCORAD评分。
SCORAD评分是一种用于评估湿疹严重程度的临床工具,它考虑了湿疹的病变严重程度、病变范围和瘙痒程度。
总体而言,现有证据表明,使用益生菌可能会降低患特应性皮炎的风险,并导致特应性皮炎SCORAD评分显著降低,但这些产品只能有限地缓解病情。
此外,益生菌的效果因使用的菌株、给药时间和患者年龄而异,因此很难提出建议。
2
小儿急性感染性腹泻
急性腹泻通常被定义为稀便或液状粪便或排便频率增加(通常24小时内至少3次)。急性腹泻可伴有发烧或呕吐,通常持续不超过7天。
• 使用益生菌缩短了腹泻的持续时间
对8014名参与者(主要是婴儿和儿童)进行了审查,发现单菌种和多菌种益生菌可显著缩短急性感染性腹泻的持续时间约25小时。这些补充剂还可以将腹泻持续4天或以上的风险降低59%,并且与未服用益生菌的患者相比,服用益生菌的患者第二天排便次数减少约1次。
对总共2444名参与者的11项随机临床试验的评估表明,鼠李糖乳杆菌(LGG)在治疗感染性腹泻方面最有效,每日剂量至少为10^10 CFU。布拉氏酵母菌(最常见的是10^9至10^10 CFU/天,持续5-10天)可减少腹泻持续时间和排便频率。
在这两项分析中,LGG和布拉氏酵母菌将急性感染性腹泻的持续时间缩短了大约1天。
然而,随后的两项临床试验发现,为期5天的LGG疗程(一项试验中每天两次单独服用1×10^10 CFU,另一项试验中每天两次总计4×10^9 CFU LGG 和瑞士乳杆菌R0052)没有效果。
然而,最近的研究表明,益生菌在急诊科可能并不有效,因为大多数急性感染性腹泻的发作都是自限性的,除了补液治疗外不需要其他治疗。
3
炎症性肠病
炎症性肠病(IBD)是一种慢性炎症疾病,包括溃疡性结肠炎和克罗恩病。IBD的确切原因尚不清楚,但可能是遗传因素和环境因素的结合,包括遗传改变和免疫系统功能障碍。IBD有多种治疗方法,包括口服类固醇和其他药物,但尚无治愈方法。
研究人员正在探索IBD患者的肠道微生物组是否发生改变,以及益生菌是否有助于控制炎症性肠病。研究得出了类似的结论——某些益生菌可能对溃疡性结肠炎有一定的有益作用,但对克罗恩病没有作用。
• 益生菌补充配合常规疗法可能使治疗效果更好
美国胃肠病学协会2020年的一项系统综述研究了益生菌在治疗胃肠道疾病中的作用。根据有限的证据,这些补充剂与常规疗法相结合可能会适度降低轻度至中度溃疡性结肠炎患者的疾病活动度。
但证据的局限性包括现有研究使用不同的患者群体、益生菌配方、治疗持续时间和伴随疗法。
2020年对865名溃疡性结肠炎参与者进行的14项研究也表明,益生菌可能有助于诱导缓解,并且益生菌与5-ASA(一种常用于治疗IBD的抗炎药物)联合使用可能优于单独使用5-ASA。
尽管益生菌可能对溃疡性结肠炎有一定的益处,但不常规使用。对于克罗恩病,还没有发现任何益处的证据。
还需要进行更多研究,包括强有力的随机对照试验,以确定哪些炎症性肠病患者可能受益于益生菌以及哪些益生菌菌株最有效。
4
肠易激综合症
肠易激综合征(IBS)是一种常见的胃肠道功能性疾病,其特征是反复出现腹部不适或疼痛、腹胀以及大便形式或频率的变化。尽管IBS的病因尚不完全清楚,但越来越多的证据表明肠道微生物群在其病理生理学和症状中具有潜在作用。
IBS患者体内含有丰富的促炎细菌,包括肠杆菌科细菌,而乳杆菌和双歧杆菌的数量通常也会相应减少。益生菌产品通常含有乳杆菌和双歧杆菌,因此有可能恢复一些缺失的微生物功能,从而帮助控制肠易激综合症症状。
• 益生菌对肠易激综合症患者具有一定积极作用
多项荟萃分析评估了益生菌对肠易激综合症患者的作用。大多数发现益生菌具有积极的、虽然有限的有益作用。例如,对总共2575名患者的23项 RCT 进行的荟萃分析发现,总体而言,益生菌可将IBS 症状持续或未改善的风险降低21%。
不同种类和菌株的益生菌对整体IBS症状、腹痛、腹胀和肠胃气胀具有有益作用,但研究质量较低。在此分析中,某些益生菌组合优于单个菌株,但没有特定组合优于其他组合。
对总共1793名IBS患者的15项RCT进行的第二次荟萃分析发现,治疗8至10周后,益生菌比安慰剂更能减轻总体症状和腹痛;在儿童中,这些补充剂还改善了粘膜屏障功能。
• 不同菌株对肠易激综合症的作用具有差异
不同菌株的益生菌对IBS的有益作用可能取决于所评估的IBS症状。在一项荟萃分析中,共有877名成人接受益生菌或安慰剂治疗4周至6个月,与安慰剂治疗相比,服用含有短双歧杆菌、长双歧杆菌或嗜酸乳杆菌的益生菌后疼痛评分显著改善。
相反,唾液链球菌嗜热亚种、动物双歧杆菌、婴儿双歧杆菌、干酪乳杆菌、植物乳杆菌、保加利亚乳杆菌和布拉酵母菌的益生菌无显著影响。
而使用含有短双歧杆菌、婴儿双歧杆菌、干酪乳杆菌或植物乳杆菌的益生菌可改善腹胀。使用所有测试的益生菌后,胀气现象有所减少。
总体而言,现有证据表明益生菌可能会减轻肠易激综合症的某些症状。然而,需要额外的高质量临床试验来确认所需的具体菌株、剂量和治疗持续时间,以及可以用益生菌有效治疗的肠易激综合症类型(例如腹泻或便秘)。
5
肥胖
肠道微生物群在从食物中提取营养和能量方面发挥着重要作用。研究表明,肠道微生物群不仅影响饮食中能量的使用,还影响宿主体内的能量消耗和储存。
评估益生菌对肥胖相关影响的临床试验结果不一致。例如,一项为期12周的临床试验,随机抽取210名年龄在35至60岁、内脏脂肪较多的健康成年人,每天饮用加氏乳杆菌的发酵奶。
• 益生菌对肥胖的影响可能并不显著
与对照组相比,接受10^7或10^6 CFU/g加氏乳杆菌牛奶的参与者的内脏脂肪面积(平均分别减少8.5%和8.2%)、体重指数、腰围和臀围以及身体脂肪量显著减少。
在另一项随机临床试验中,连续24周每日补充3.24x 10^8 CFU鼠李糖乳杆菌CGMCC1.3724,并在前12周进行能量限制饮食(比估计的卡路里需求少500kcal/天)并没有显著影响体重。然而,与安慰剂相比,77名女性参与者在12周(减轻1.8公斤)和24周(减轻2.6公斤),补充乳酸菌确实减轻了体重。
最近对957名超重或肥胖个体的15项随机对照试验进行的另一项系统回顾和荟萃分析发现,补充不同剂量和菌株的益生菌3至12周可导致体重(减少0.6公斤)、体重指数下降。比安慰剂低0.27kg/m2和脂肪百分比(低 0.6%)。然而,这些影响较小。
综上所述,这些结果表明,益生菌对体重和肥胖的影响可能取决于多种因素,包括益生菌菌株、剂量和持续时间以及使用者的某些特征,包括年龄、性别和基线体重。需要进行更多研究来了解益生菌对人体脂肪、体重和肥胖的潜在影响。
6
高胆固醇血症
血液中高水平的胆固醇或动脉壁中的胆固醇是心血管疾病的危险因素。低密度脂蛋白(LDL)将胆固醇携带至组织和动脉。LDL水平越高,患心血管疾病的风险就越大。高密度脂蛋白(HDL)将胆固醇从组织运送到肝脏并导致其排泄。高密度脂蛋白水平低会增加患心血管疾病的风险。
• 益生菌影响胆固醇水平的机制
研究人员研究了使用益生菌来改善血脂状况。它们对胆固醇浓度的影响机制包括通过增加胆固醇的分解代谢:
•影响胆盐水解酶活性,增加对新胆汁酸的需求,从而降低血清胆固醇水平;
•与小肠中的胆固醇结合,从而减少身体吸收的量;
•将胆固醇同化并掺入细菌中,从而降低血液中的胆固醇水平;
•乳杆菌和双歧杆菌产生短链脂肪酸,降低肝脏胆固醇合成并调节胆固醇代谢。
• 一些益生菌降低了总胆固醇和低密度脂蛋白水平
一项对30项随机对照试验的荟萃分析显示,与安慰剂组相比,接受益生菌治疗3至12周的受试者总胆固醇降低7.8mg/dL,低密度脂蛋白胆固醇浓度降低7.3mg/dL。
在亚组分析中,在持续8周或更长时间的研究中,以及在基线胆固醇水平高于240 mg/dL的参与者中,益生菌的益处略大。在三项以上研究中包括的菌株中,嗜酸乳杆菌、乳双歧杆菌以及植物乳杆菌的混合物与总胆固醇和低密度脂蛋白胆固醇浓度的显著降低有关,但瑞士乳杆菌和粪肠球菌则不然。
• 益生菌对高密度脂蛋白水平可能没有影响
在对602名胆固醇水平正常或高胆固醇成年人进行的11项随机对照试验中进行的一项小型荟萃分析显示,与安慰剂治疗组相比,接受益生菌治疗2至10周的患者总胆固醇水平降低了6.6mg/dL,低密度脂蛋白胆固醇水平降低了8.5mg/dL ,但益生菌治疗对高密度脂蛋白胆固醇水平没有显著影响。
另一项综述发现,使用含有多种菌株的益生菌可显著降低总胆固醇和低密度脂蛋白胆固醇水平(分别降低12.0和20.1mg/dL),而使用单一菌株的试验则没有这种效果。
总体而言,研究表明,组合使用多种益生菌菌株以及含有嗜酸乳杆菌和乳双歧杆菌的混合物或植物乳杆菌的益生菌可能会降低总胆固醇和低密度脂蛋白胆固醇水平。然而,需要更多的研究来证实这些发现。
尽管有潜在的好处,但益生菌人类健康的有效性仍然存在不确定性,这主要是因为许多研究没有考虑到这些影响可能是针对菌株、剂量和条件的,并且个人对干预措施的反应可能会因人而异。
独特的部分取决于他们的健康状况、年龄和肠道微生物的组成。在这节中我们会讲述当前使用益生菌还存在的一些安全隐患。
微生物群组成或功能改变
• 大多数口服益生菌不会长期定植
益生菌微生物的功效并不依赖于长期定殖于宿主的能力。长期定植通常意味着在停止给药后数周或数月仍可从宿主中检测到施用的微生物。
对于口服益生菌来说,这意味着微生物必须积极复制并且已经在宿主体内持续存在。迄今为止积累的关于从粪便中回收益生菌的证据表明,目前大多数益生菌不会定植。这几乎肯定是由于无法与常驻微生物群竞争造成的。
对于大多数人来说,仅仅遇到微生物并不会导致定植,但如果存在“开放生态位”,外来的微生物可能能够填补该生态位并进行定殖。
• 潜在风险有哪些?
有哪些安全隐患?在大多数情况下,来自常见共生微生物群的益生菌的低水平定植不会对宿主健康造成问题。即使没有明显毒力潜力的高水平微生物也不会对宿主健康产生负面影响。然而,可以想象的是,长期定植的益生菌所固有的暴露增加可能会带来以下风险:
(i) 益生菌可能会取代发挥重要功能的微生物;
(ii) 益生菌可能会对周围微生物群的结构和功能产生负面影响;
(iii) 如果正常的肠道屏障被破坏,益生菌可能会进入体循环,导致侵袭性感染。
• 潜在好处是什么?
有人可能会说,益生菌的长期存在(根据其定义可带来健康益处)可能是一种提供长期健康益处的有效方式。事实上,一种能够永久占据空缺生态位并提供缺失的代谢功能(例如代谢婴儿母乳寡糖的能力)的微生物,有助于宿主健康,可以代表一种优秀的益生菌候选者。
另一个考虑因素是,如果生态位注定要被占领,那么长期定殖的益生菌是否会比未知微生物更安全、更理想的占领者?这波新的菌株似乎更适合在人体中存在和生活,有可能实现独特和卓越的益处。例如,通过长期定植益生菌可以最好地实现赋予酶能力以补偿苯丙酮尿症等代谢紊乱。
• 建议:生物组分析有助于评估益生菌摄入后的健康作用
根据目前的理解,建议开发长期定植益生菌时,必须有一个明确的目标,即实现不易、可靠或经济上难以实现的效益,并权衡风险与这些明确定义的效益。建议仔细考虑确定哪些长期安全数据可能与宿主体内持续存在的益生菌菌株相关。应进行研究以确定有助于评估长期定植益生菌安全性的相关急性暴露试验和生物标志物。
生物组分析是评估基线微生物群组成和群落结构以及转录和代谢读数的过程。评估益生菌摄入前后的微生物组结果可能表明益生菌在塑造微生物组中的因果作用。
此类研究可能会为有关促进健康、引起安全问题或确定益生菌治疗反应者和非反应者特征的微生物机制和途径的假设提供信息。这种分析可能有助于解释或在一定程度上预测益生菌对特定宿主产生有益作用的能力。
抗生素抗性基因的转移
自从益生菌开始使用以来,人们担心抗生素耐药性可能从益生菌菌株转移到肠道微生物群中更多的致病菌。
例如许多乳杆菌菌株对万古霉素具有天然耐药性,这引发了人们对这种耐药性可能转移到更具致病性的生物体,特别是肠球菌和金黄色葡萄球菌的担忧。然而,乳杆菌的万古霉素抗性基因是存在于染色体的,因此不容易转移到其他物种。研究也尚未发现乳杆菌的万古霉素抗性基因转移到其他属。
• 评估益生菌中的抗生素抗性基因变得重要
评估益生菌基因组中现有抗生素抗性(AR)基因的风险需要基因型和表型方法。表型方法需要评估菌株表达的AR基因,通过最小抑制浓度技术确定。在某些情况下,AR 基因的表达是物种固有的,而不是由于可以获得的基因的表达。
常见益生菌种类的正常AR范围已经确定。然而,当开发信息未知的物种菌株(下一代菌株)时,需要进一步研究。
需要注意的是,即使不存在非内在AR基因的转移风险,也存在通过裸露DNA转化进行转移的可能性, 或通过噬菌体转导。
• 当考虑益生菌抗生素抗性基因的安全性时,应考虑几个问题:
(i)是与提供水平转移的可能机制的基因相关的基因;
(ii)如果转移至初始宿主,这些基因是否能够进行功能表达;
(iii)是该物种典型的抗生素耐药表型(内在耐药性);
(iv)耐药表型是否如此普遍,以至于益生菌不会对其在环境微生物中的存在产生实质性影响;
(v)是否对临床相关抗生素的耐药性。
这些问题必须作为严格分析的一部分来考虑,以平衡患者或消费者安全和公共卫生问题的利益。分析必须考虑益生菌是否被开发用于广泛分发给一般健康的消费者(食品或补充剂,其合理确定的无害性是一般标准)或限制分发给患者群体(药物)。
由于益生菌有可能加剧或减轻人类体内存在抗生素抗性基因库,因此可能需要采取具体情况具体分析的安全方法。
此外,基因修饰方法可用于改变或消除所关注的抗性元件。根据所采用的技术,该选项可能适用于食品应用的益生菌(例如质粒固化)或可通过药物途径的下一代益生菌。
侵袭性感染
在极少数情况下,益生菌可能会从胃肠道转移,导致侵袭性感染。
从心内膜炎、菌血症局部感染中分离出的菌株

Doron S,et al.Clin Infect Dis.2015
• 一小部分接受益生菌给药的患者曾出现过感染
对1995年至2021年间儿童中与益生菌给药相关的败血症、菌血症和真菌血症进行了系统回顾。大多数符合感染临床定义的儿童年龄在两岁以下,并且有早产或留置静脉导管等诱发因素,94%的儿童通过抗菌治疗成功治愈。
最常报告的单一事件是菌血症,至少有33份报告称,食用益生菌布拉氏酵母菌的患者的血培养物中存在酿酒酵母或布拉氏酵母菌(这些微生物在微生物学上无法区分)。
至少有8例与乳杆菌相关的菌血症已被报道,包括嗜酸乳杆菌(Lactobacillus acidophilus)、干酪乳杆菌(Lactobacillus casei)和鼠李糖乳杆菌(Lactobacillus rhamnosus GG)。
已报道了9例明显脓毒症病例,与布拉氏酵母菌(S.boulardii)、鼠李糖乳杆菌(Lactobacillus rhamnosus GG)、枯草芽孢杆菌(Bacillus subtilis)、短双歧杆菌(Bifidobacterium breve)或组合益生菌有关。
乳杆菌和链球菌益生菌引起的心内膜炎事件也有报道。与鼠李糖乳杆菌相关的脓肿的发展也被报道过两次。
人类使用益生菌相关的细菌性感染病例
Boyle RJ,et al.Am J Clin Nutr.2006
• 在使用益生菌时,应考虑其感染风险和应对措施
由于多种原因,益生菌相关侵袭性感染的实际频率很难确定。大多数已发表的病例报告都发生在临床试验之外。并非所有临床实验室都会从血培养物中常规培养和鉴定益生菌,并且益生菌试验中的危害报告通常是不完整的。
尽管如此,在大多数对早产儿使用益生菌的试验中,经培养证明的侵袭性感染被作为主要或次要结果进行研究,荟萃分析表明,益生菌的使用不会增加这一弱势群体的败血症发生率。然而,益生菌相关脓毒症的真实发生率仍不确定。
我们认为,应在临床环境中认真监测侵袭性感染和菌血症,并在所有益生菌试验中充分报告。使用菌株水平分子技术,应将临床分离株与施用的益生菌进行比较。
补充益生菌微生物与侵入性临床分离株的分子匹配支持益生菌与全身感染之间的关联。在评估任何下一代益生菌的安全性时,应确定易位的可能性,并权衡风险与益处。
降解粘蛋白,影响肠道屏障
粘蛋白是一种蛋白质,是粘液的核心结构元素,粘液是整个消化道粘膜表面的重要组成部分。除其他作用外,粘蛋白还提供了保护宿主上皮免受微生物和化学入侵的屏障。
• 粘蛋白降解细菌过多可能不利于黏膜屏障稳定
当许多细菌将粘液用作碳源时,会发生一定程度的粘液降解。例如,Akkermansia muciniphila以粘蛋白为食,将其转化为短链脂肪酸。这些短链脂肪酸是胃肠道细胞的重要能量来源。
已鉴定出40多个与粘蛋白代谢相关的细菌基因。全基因组测序可用于鉴定与粘蛋白降解相关的细菌基因,并可通过评估细菌菌株在液体或含有粘蛋白作为唯一碳源的琼脂平板上生长的能力来体外分析其活性。
一些人认为,粘蛋白降解细菌的变化引起的生理平衡变化会导致人类疾病和感染,但据报道,这种特性在肠道微生物内受到非常严格的调节,并且受膳食多糖可用性的影响。
粘蛋白降解能力是益生菌综合安全性分析的一部分,因此,了解益生菌物种基因组中包含的粘蛋白降解基因的数量是益生菌菌株安全性整体审查的一部分,因为单独的粘蛋白降解潜力不应被视为毒力特征。
对药物功能的影响
• 微生物影响药物的代谢可能提高药效也可能产生毒性
肠道微生物群可以对药物的代谢产生直接和间接的影响,从而影响药效和毒性。例如,当大鼠接受抗生素治疗时,口服偶氮类药物Prontosil的大鼠中, 62%的总磺胺通过微生物偶氮还原酶的作用而减少,说明药物激活可以由微生物群介导。
微生物群也可以减少药物相关的毒性。一个众所周知的例子是葡萄糖醛酸化,结合物水解反应,通过 UDP-葡萄糖醛酸基转移酶将葡萄糖醛酸与底物连接成亲水性且带负电荷的葡萄糖醛酸苷。许多厌氧细菌可以诱导β-葡萄糖醛酸酶,这种酶能够解聚外源物质和之前通过葡萄糖醛酸化途径解毒的内源性化合物。这种解结合可以增强毒素、激素和各种药物的肠肝再循环以及局部致癌物的形成。因此,过量的β-葡萄糖醛酸酶可能会增加患结肠癌的风险。
然而,一定量的β-葡萄糖醛酸酶活性对于保证维生素D、甲状腺激素或雌激素等必需化合物的肠肝再循环非常重要。
• 益生菌影响药物的能力具有一定安全隐患
益生菌影响药物功能的能力可能会产生安全后果。毒物微生物组学或药物微生物组学是一门相对较新的学科,研究微生物群和外源化合物之间的相互作用。了解微生物群多样性、饮食和药物处置和反应之间的相互作用以及这如何影响未来的个性化医疗很重要。
过度的免疫刺激
研究还表明,肠道微生物群对于刺激正常免疫发育,特别是肠道相关淋巴组织的发育很重要。肠道微生物群的存在对于一系列免疫功能是必需的,包括抗体的产生、对食物抗原的口服耐受的发展和持续,以及淋巴滤泡内生发中心的形成。
由于益生菌已被证明会影响先天性和适应性免疫系统,包括对细胞因子分泌和树突状细胞功能的影响,因此人们担心益生菌可能会过度刺激某些个体的免疫反应。
• 新生儿和孕妇补充益生菌可能对免疫产生影响
肠道微生物群在正常免疫发育中的关键作用表明,旨在改变微生物群的操作可能具有显著的免疫调节作用。这些操作对宿主的长期影响很难预测,并且仍然有可能对免疫发育产生不利影响。
这在新生儿益生菌补充剂领域尤其重要,在该领域可能会实现微生物群的中长期改变或免疫反应的终生改变。
第二个可能面临不良免疫刺激风险增加的群体是孕妇。在怀孕期间,T细胞反应存在对Th2表型的偏向,这被认为对于维持胎儿活力很重要,因为Th1细胞因子与妊娠失败相关。
益生菌乳杆菌已被证明可以在体外抑制Th2细胞因子反应,并且在一些人体研究中发现可以增加Th1细胞因子干扰素γ的产生。这些影响可能不利于妊娠活力。
目前,实证研究很少支持益生菌导致免疫发育不良的假设,但这是一个值得进一步研究的领域。
尽管据报道益生菌对人群具有有益作用,但免疫功能低下的宿主由于抵御微生物入侵的能力降低,可能面临更高的不良事件风险。
益生菌在高危人群(例如免疫功能减弱/受损的个体、老年人、新生儿,特别是早产儿)中的安全性还有待研究。
√ 益生菌有时会变成机会性病原体
短期观察的证据表明,某些益生菌菌株可能在免疫功能低下、压力大、老龄或新生儿人群中充当机会病原体。
不良事件包括危及生命的肺炎、心内膜炎和败血症。一般来说,在弱势群体中,如果存在单一主要风险因素(例如免疫功能低下状态)或多个次要风险因素,则在使用益生菌时应谨慎。
然而,如果有令人信服的证据表明益生菌可以使一些弱势群体受益,则应考虑使用益生菌。根据现有数据,当对弱势目标人群施用益生菌时,需要进行额外监测。
益生菌对孕妇和哺乳期妇女的风险已得到审查。在100项妊娠期间服用益生菌的研究中,有28项报告了 不良事件。其中,只有11种报告的不良事件可能与治疗存在因果关系,包括胃肠道问题、恶心和头痛;但没有关于母亲或婴儿严重健康问题的报道。
一项研究报告称,施用鼠李糖乳杆菌GR-1和罗伊氏乳杆菌RC-14时,阴道分泌物风险增加,并且粪便稠度发生变化。
√ 一些疾病患者使用益生菌时应注意潜在风险
另一项研究结果显示,当对肥胖女性(先兆子痫风险增加)施用益生菌时,先兆子痫的风险增加(472名服用益生菌的女性中有31例先兆子痫,而安慰剂组中有17例)。尽管数据并不一定可靠,但我们仍然建议轻度至病态肥胖孕妇服用益生菌的同时监测先兆子痫的潜在风险。
而在非怀孕人群中,也有报道与使用益生菌相关的不良反应,包括胃肠道副作用,如腹部绞痛、恶心、软便、胀气和味觉障碍。
人们对一些人群中使用的某些益生菌的代谢活性表示担忧。D-乳酸性酸中毒可能发生在因手术改变肠道解剖结构的人身上,例如短肠或减肥手术,这是由常驻微生物的活动引起的。很少有公开报道将此与益生菌补充剂联系起来。由于缺乏早产儿数据,因此对该人群使用产生D-乳酸的益生菌持谨慎态度,尽管一项针对健康足月婴儿的对照试验表明,产生D-乳酸的罗伊氏乳杆菌DSM-17938不会导致酸中毒和含益生菌的婴儿配方奶粉与酸中毒无关。
√ 生命早期补充益生菌对发育可能有影响
鉴于在众多相互竞争的产前、围产期和产后因素中调节微生物群结构和功能的独特机会窗口,早产儿值得特别考虑。新生儿微生物群靶向疗法有可能影响宿主整个生命周期的生物学,可以通过在最有利于定殖的条件下引入异源微生物菌株,或者通过影响包括大脑在内的重要器官的早期发育轨迹。
迄今为止,几乎没有证据表明生命早期补充益生菌会对神经发育结果产生不利影响。在一项针对1099名极早产儿的后续研究中,接受益生菌治疗的存活婴儿与未接受益生菌治疗的婴儿相比,3-5岁时的主要神经发育结果没有差异。
有趣的是,接受益生菌治疗的儿童中耳聋的发生率较低,这不能归因于抗生素疗程数量或接受万古霉素或庆大霉素总天数的差异。
其他研究已经确定了生命早期微生物群的改变与肥胖之间的联系。一项针对美国333353名儿童的大型队列研究报告称,在生命的头两年内服用抗生素和抑酸药物与儿童后期的肥胖有关;这些改变微生物群的药物每增加一类,每增加30天的处方,这些关联就会得到加强。
现有的有限证据并不表明生命早期使用益生菌会增加儿童期不良后果的风险。没有足够的数据来确定围产期益生菌使用与成年期不良事件之间是否存在潜在关联。
因此,我们鼓励对围产期接受益生菌的早产儿进行至少2年的随访,以比较代谢、过敏、免疫和其他健康结果。
需谨慎使用益生菌的人群
然而使用益生菌时,一些特定人群需要格外注意:
• 免疫系统较弱的个体
如HIV感染者、正在接受化疗或免疫抑制治疗的患者,因为他们更容易受到感染。婴幼儿和老年人由于免疫系统不够成熟或衰弱,使用益生菌时也应谨慎。
• 患有消化系统问题的人
对于患有病毒性肠炎、细菌性胃肠炎等疾病的人群,为了避免益生菌的大量繁殖,导致病毒或细菌进一步增长,这类人群也需要慎重食用益生菌。
• 胃肠道屏障不完整的人
如消化道溃疡、胃肠道手术恢复期患者、胃出血等不建议服用,益生菌有随血液扩散至全身引起感染的风险。
• 身体过于虚弱者
因严重发热、感冒等导致的体能虚弱患者,摄入益生菌后,可能会加重身体负担,不利于身体恢复。
• 对益生菌成分过敏者
对益生菌成分过敏者吃了益生菌后,可能会出现皮肤起丘疹、腹泻次数增加、胃部不适等情况。
此外,怀孕和哺乳期妇女在使用益生菌前应咨询医生。总之,在使用益生菌补充剂前,最好先评估体内原有的肠道微生物群并咨询医疗专业人士是否需要额外补充益生菌。
益生菌的最佳剂量取决于菌株和产品。因此,建议患者使用益生菌的临床医生指定益生菌菌株、剂量和使用持续时间。
益生菌因其健康益处而被越来越多的消费者使用,并受到许多医疗保健专业人士的提倡。因此我们更需要了解使用益生菌的益处和一些潜在风险。
研究发现益生菌对于大部分健康的人来说是安全的,但由于存在感染的风险,因此对某些人群应谨慎使用。其中包括免疫功能低下的患者、早产儿、短肠综合征患者、使用中心静脉导管的患者以及心脏瓣膜疾病患者。
尽管关于益生菌的作用机制和施用还有很多东西有待了解,但很明显不同的菌株可以产生非常特定的效果。益生菌特异性取决于剂量、菌种、持续时间、给药途径和适应症等影响因素,此外还可能因施用对象的健康或疾病状况、不同年龄、肠道微生物组成而异。
因此,一个群体中一种益生菌菌株的临床试验结果不能自动推广到其他菌株或不同群体。需要进一步研究来探索益生菌的机制和对人体的作用。我们建议人们在使用益生菌之前了解自身具体的健康状况和微生物组成,以便能够“对症下药”。
主要参考文献
Merenstein D, Pot B, Leyer G, Ouwehand AC, Preidis GA, Elkins CA, Hill C, Lewis ZT, Shane AL, Zmora N, Petrova MI, Collado MC, Morelli L, Montoya GA, Szajewska H, Tancredi DJ, Sanders ME. Emerging issues in probiotic safety: 2023 perspectives. Gut Microbes. 2023 Jan-Dec;15(1):2185034.
Doron S, Snydman DR. Risk and safety of probiotics. Clin Infect Dis. 2015 May 15;60 Suppl 2(Suppl 2):S129-34.
Boyle RJ, Robins-Browne RM, Tang ML. Probiotic use in clinical practice: what are the risks? Am J Clin Nutr. 2006 Jun;83(6):1256-64; quiz 1446-7.
Sheyholislami H, Connor KL. Are Probiotics and Prebiotics Safe for Use during Pregnancy and Lactation? A Systematic Review and Meta-Analysis. Nutrients. 2021 Jul 13;13(7):2382.
Asha MZ, Khalil SFH. Efficacy and Safety of Probiotics, Prebiotics and Synbiotics in the Treatment of Irritable Bowel Syndrome: A systematic review and meta-analysis. Sultan Qaboos Univ Med J. 2020 Feb;20(1):e13-e24.
Jäger R, Mohr AE, Carpenter KC, Kerksick CM, Purpura M, Moussa A, Townsend JR, Lamprecht M, West NP, Black K, Gleeson M, Pyne DB, Wells SD, Arent SM, Smith-Ryan AE, Kreider RB, Campbell BI, Bannock L, Scheiman J, Wissent CJ, Pane M, Kalman DS, Pugh JN, Ter Haar JA, Antonio J. International Society of Sports Nutrition Position Stand: Probiotics. J Int Soc Sports Nutr. 2019 Dec 21;16(1):62.
Roe AL, Boyte ME, Elkins CA, Goldman VS, Heimbach J, Madden E, Oketch-Rabah H, Sanders ME, Sirois J, Smith A. Considerations for determining safety of probiotics: A USP perspective. Regul Toxicol Pharmacol. 2022 Dec;136:105266.

谷禾健康

在20世纪初,Elie Metchnikoff(著名生物学家,酸奶之父)发现了有益的肠道微生物,该微生物可使肠道健康正常化并延长寿命,后来被称为“益生菌”。益生菌是指“以适当的剂量给予宿主健康有益的活生物体” 。
益生菌如今越来越受欢迎,主要是因为它们对我们整体健康的重要性。益生菌主要存在于人体肠道内,通过维持肠道微生物平衡,在宿主体内发挥有益作用。
不久前发表于《自然•化学》的一项最新研究成果表明,“吃土”也能调节肠道微生物组和治疗肠炎。
究其原因是许多益生菌来源于土壤微生物 (SBO)。细菌在土壤中有着惊人的多样性和丰富性,它们在土壤生态系统中的作用方式与它们在肠道生态系统中的作用相似。
注:该研究中的“吃土”并不是直接食用土壤,而是“人造土”,通过人工合成的方法构建的土壤仿生材料。事实上,自然界土壤中的微生物与肠道菌群中的微生物并不完全相同,且土壤中可能存在有害物质,如重金属、农药等,因此,直接食用土壤可能会引起不必要的健康问题。
考虑土壤微生物的多样性和复杂性,为了让大家更好地了解来源于土壤微生物的益生菌,本文将介绍其中最常见的土壤益生菌及其作用。希望对未来益生菌的选择和恢复肠道微生态提供一定价值。
▼
土壤微生物一词涵盖了土壤中高度多样化的细菌(和其他生命形式),这些细菌在土壤中自然存在。在引入工业化和现代农业之前,我们每天都会定期接触这些细菌。近年来,一些土壤微生物已被分离并用作益生菌。
许多土壤微生物的一个关键特征是它们可以形成孢子。当条件不太有利时,土壤微生物会形成小孢子,这是细菌的一种休眠形式,具有坚硬的保护性外涂层。在这种形式下,细菌对热、酸和大多数抗生素具有高度抵抗力。
土壤微生物和肠道微生物组息息相关
doi: 10.3390/microorganisms7090287
▼
• 通过触发抗体来帮助调节免疫系统;
• 对抗肠易激综合征,调节肠道功能;
• 减轻炎症;
• 治疗腹泻、腹痛和腹胀;
• 帮助消化和营养吸收;
• 防止感染;
• 支持情绪和心理健康;
• 支持平衡肠道微生物群;
• 促进新陈代谢,加速脂肪流失;
• 缓解过敏症状
▼
以下是益生菌中最常用的八种土壤微生物:
Bacillus coagulans(凝结芽孢杆菌)
Bacillus subtilis (枯草芽孢杆菌)
Bacillus clausii(克劳氏芽孢杆菌)
Bacillus indicus (印度芽孢杆菌)
Bacillus licheniformis(地衣芽孢杆菌)
Enterococcus faecium (屎肠球菌)
Enterococcus faecalis(粪肠球菌)
Clostridium butyricum(丁酸梭菌)
注:印度芽孢杆菌是Bacillus属的微生物,原产地为中国。
在下文中介绍了八种来源于土壤微生物的益生菌在人类临床试验中的作用,以确定哪些菌株有较好的治疗前景。
01
凝结芽孢杆菌 (Bacillus coagulans)
凝结芽孢杆菌(Bacillus coagulans)是一种产生L-乳酸的革兰氏阳性细菌。最适生长温度为45-50℃,最适pH为6.6-7.0。它表现出乳酸杆菌科的许多典型特征,但与大多数乳酸菌不同,凝固芽孢杆菌可以形成孢子。
图源:JBMbio
六种凝结芽孢杆菌菌株在人体临床试验中得到充分研究:
✦六种不同菌株显示能够调节肠道微生物群、改善肠易激综合征
GBI-30, 6086
Bacillus coagulans GBI-30, 6086 (“Ganeden BC30”):通过至少10项人类随机对照试验,这是研究最深入的土壤菌株之一。GBI-30、6086已被证明可以改善肠易激综合征 、免疫功能并且可能有益地调节肠道微生物群。
体外研究还表明,它可能有助于果糖和乳糖的消化。
LBSC (DSM 17654)
Bacillus coagulans LBSC (DSM 17654):两项随机对照试验表明该菌株对肠易激综合征患者和急性腹泻或腹部不适患者有益。
MTCC 5856
MTCC 5856(“LactoSpore”):两项随机对照试验表明,该菌株可显著减轻肠易激综合征患者的腹胀、腹痛和抑郁症状。
SANK 70258
Bacillus coagulans SANK 70258 (“Lacris-S”):
一项随机对照试验和一项开放研究发现,该菌株可改善健康成人的大便频率,减少大便次数,且无不良影响。
最近一项使用人体肠道模型系统的研究发现,该菌株抑制了肠杆菌科中的促炎细菌,并增加了健康个体中产丁酸盐的毛螺菌科的丰度。
注:自1966年以来,该菌株已在日本用作食品成分。
SNZ 1969
Bacillus coagulans SNZ 1969:一项安慰剂随机对照试验发现,这种菌株显著改善了结肠转运时间并减少了轻度间歇性便秘人群的肠道不适。它也被证明对细菌性阴道病有效。
另一项使用SNZ 1969以及克劳氏芽孢杆菌和枯草芽孢杆菌的随机对照试验发现,胃肠道不适的成年人的嗳气、腹胀、疼痛和总体症状有所减轻。
Unique IS-2
Unique IS-2(“ProDURA”):四项人类随机对照试验发现该菌株对儿童和成人的细菌性阴道病和肠易激综合征均有效。该菌株还被证明可以在体外代谢果糖。
所有这六种菌株都作为食品成分进行了安全评估,并获得了食品药品监督管理局的“公认安全”(GRAS) 状态。
▸ 小结
这些凝结芽孢杆菌菌株似乎是安全的、耐受性良好的,并且可能对患有细菌性阴道病或肠易激综合征的患者特别有益。
枯草芽孢杆菌(Bacillus subtilis)是一种形成孢子的革兰氏阳性细菌,被认为是人类肠道的正常居民。
枯草芽孢杆菌天然存在于纳豆中,纳豆是一种由发酵大豆制成的日本传统食品。
在引入抗生素之前,枯草芽孢杆菌培养物就已在世界范围内广泛用作免疫刺激剂,以帮助治疗泌尿道和胃肠道疾病。
B.subtilis可以产生一种降解草酸盐的酶,也可能产生少量维生素K2。

图源:eol.org
三种枯草芽孢杆菌菌株已证明对人类有治疗作用,并已通过作为食品成分的安全性测试:
✦三种不同菌株显示能够改善人体健康状况
DE111
Bacillus subtilis DE111:在训练期间对运动员进行的两项随机对照试验发现枯草芽孢杆菌DE111具有良好的耐受性。女运动员的体脂率下降幅度更大;男性运动员发现血液中的炎症标志物减少,对身体机能没有影响。
其他安慰剂随机对照试验发现枯草芽孢杆菌DE111改善了肠道不规律,增加了抗炎免疫细胞群的活性,降低了空腹血糖并降低了健康成人的总胆固醇。
该菌株被食品药品监督管理局公认为安全。
CU1
Bacillus subtilis CU1(“LifeinU”):一项随机对照试验发现,为期10天的B. subtilis CU1补充剂耐受性良好,并改善了老年人的免疫功能(粪便和唾液 中的免疫球蛋白)。
它没有表现出任何抗生素耐药性,并被证明在体外不存在产毒活性。
MB40
Bacillus subtilis MB40 (“OPTI-BIOME”) :
一项随机对照试验发现,该菌株在健康成人中具有良好的耐受性,可降低男性受试者的腹胀强度和胃肠道症状,并适度改善总体健康状况。
该菌株被食品药品监督管理局公认为安全 。
在亚洲对另外两种菌株进行了深入研究:
R0179
Bacillus subtilis R0179(“Medilac-S”):
该菌株存在于亚洲益生菌制剂Medilac-S中,该制剂还含有Enterococcus faecium R0026。几项安慰剂随机对照试验发现,两种菌株的制剂减少了肠易激综合征相关的腹痛,并改善了结肠镜检查的肠道。
最近一项针对临床试验的荟萃分析得出结论,Medilac-S也可有效诱导溃疡性结肠炎的缓解。
两项随机对照试验还发现,分离的枯草芽孢杆菌R0179具有良好的耐受性,并在通过人体胃肠道后存活下来。
TO-A
Bacillus subtilis TO-A (“BIO-THREE”) :
这种枯草芽孢杆菌菌株与E. faecalis T-110和C. butyricum TO-A相结合,作为益生菌在亚洲得到了相当深入的研究。在随机对照试验中,这三种菌株已被证明可以减少手术患者的术后感染并缩短急性腹泻患儿的住院时间。
▸ 小结
通常研究的枯草芽孢杆菌菌株似乎是安全的、耐受性良好的,并且没有不良反应的报道。一些菌株似乎对改善免疫功能和肠道规律性特别有益。
克劳氏芽孢杆菌(Bacillus clausii)是一种形成孢子的革兰氏阳性细菌,以其对金黄色葡萄球菌和艰难梭菌的抗菌活性而闻名。它被认为是一种共生微生物,已从健康个体的小肠和粪便中回收。
只有两种菌株组合已在人类身上进行过研究:
✦两种不同菌株组合显示对过敏、肠易激综合征有益
Bacillus clausii OC、NR、SIN和T
自1958年以来,这四种菌株已被广泛研究并用作药物制剂Enterogermina(美菌纳)。这些菌株已被证明具有酸和胆汁抗性,并且在人类单次口服给药后可在粪便中恢复4-12天。
这些菌株可能对那些有过敏症、减少鼻塞和抗组胺药需求的人特别有益。
它也被证明可以减少幽门螺杆菌抗生素治疗期间的副作用。
一项针对40名诊断为“小肠细菌过度生长”患者的研究发现,服用美菌纳一个月可使 47%的患者的葡萄糖呼气试验结果恢复正常。
值得注意的是,这个比率与许多用于小肠细菌过度生长治疗的抗生素所见的正常化率相当。只有一名患者报告了便秘的副作用。
尽管美菌纳有许多记录在案的益处和总体安全性,但仍有零星的克劳氏芽孢杆菌败血症病例报告。大多数报告都是在老年人、重病患者或免疫功能低下的个体中使用益生菌。
Bacillus clausii UBBC-07
在一项针对印度急性腹泻儿童的安慰剂对照随机试验中,该菌株被证明可以改善粪便稠度并缩短腹泻持续时间。该菌株经过了毒理学研究,被确定可供人类安全食用。
▸ 小结
克劳氏芽孢杆菌已在许多随机、安慰剂对照试验中证明了疗效,并且可能对过敏或肠易激综合症患者特别有益。
脓毒症的个别病例报告表明,老年人、重病或免疫功能低下的人以及最近服用广谱抗生素的人可能禁忌使用它。
印度芽孢杆菌(Bacillus indicus)是一种形成孢子的细菌,因其能够有效刺激免疫系统并产生高水平的类胡萝卜素、维生素和喹啉而受到吹捧。
✦暂未有毒性或致病性的发现
体外和动物研究未能发现B.indicus HU36的任何潜在毒性或致病性,该菌株是益生菌中最常用的菌株,并且没有关于人类感染的报道。
▸ 小结
关于人类印度芽孢杆菌的科学研究,目前还缺乏人类证据。
地衣芽孢杆菌(Bacillus licheniformis)是一种革兰氏阳性、产芽孢的细菌,通常用于工业酶生产。
在所有基于土壤使用的益生菌中,地衣芽孢杆菌可能是最具争议的。
✦可能具有一定毒性
根据一些报道,地衣芽孢杆菌可能是一种机会性病原体,可导致免疫功能低下的宿主感染。
然而,其他报告得出的结论是,地衣芽孢杆菌的毒力非常低。地衣芽孢杆菌存在于韩国传统消费的许多发酵食品中,并且已从健康人类志愿者的胃肠道中分离出来,因此至少某些菌株似乎是共生的。
虽然已经出现了与大量该物种的分离相关的急性自限性胃肠炎病例, 对肠上皮细胞的毒性或直接影响尚未得到证实。很难确定这些报告病例中的物种(数量相当有限)是否积极参与了感染,或与一种不明病原体一起分离。
只有四项关于分离的地衣芽孢杆菌的人体研究,没有一项提供菌株信息,并且只有一项是随机和安慰剂对照的。
▸ 小结
虽然含有地衣芽孢杆菌的传统发酵食品几乎是安全的,但目前没有足够的证据支持大剂量补充地衣芽孢杆菌。最好谨慎行事,避免使用含有该物种的益生菌。
✦发酵碳水化合物
Enterococcus faecium是革兰氏阳性菌。虽然它不形成孢子,但它可以耐受多种环境条件。E. faecium是人类肠道的正常居民,发酵碳水化合物并产生乳酸作为副产品。
粪肠球菌的一些菌株可以在传统发酵食品中找到。
by Dennis Kunkel Microscopy
✦可能致病
然而,粪肠球菌也可能致病,引起新生儿脑膜炎和心内膜炎等疾病。这种细菌的许多菌株已经产生了抗生素抗性和毒力因子,使其能够聚集并形成生物膜。
在美国,80-90%的医疗器械相关感染可归因于耐抗生素粪肠球菌。因此,人们担心将其用作益生菌。
已经研究了许多不同的菌株,但只有两种菌株值得强调,它们已经在多项人体试验中进行了研究:
M-74
Enterococcus faecium M-74:该菌株有许多与之相关的“随机”临床试验,但每一个菌株都服用益生菌和50微克硒。
许多益生菌指出这些研究表明粪肠球菌可以降低胆固醇,但缺乏硒对照组意味着硒单独提供治疗效果是完全合理的。
R0026
Enterococcus faecium R0026 (“Medilac-S”):
该菌株存在于亚洲益生菌制剂Medilac-S中,该制剂还含有枯草芽孢杆菌R0179,并在国内得到了很好的研究。
如前面所述,多项随机对照试验发现,双菌株制剂可减少肠易激综合征相关的腹痛,并改善结肠镜检查的肠道准备,还可以有效诱导溃疡性结肠炎的缓解。
▸ 小结
除了Medilac-S中的菌株外,很少有精心设计的随机对照试验证明了粪肠球菌的功效。
鉴于其可能获得多重抗生素耐药性并引起感染。谨慎使用高剂量的分离粪肠球菌。
Enterococcus faecalis是一种革兰氏阳性菌。与Enterococcus faecium类似,它不能形成孢子,但对恶劣的环境条件相当耐受。
✦在肠道免疫中起重要作用
粪肠球菌是人类肠道的正常居民,存在于大约90-95%的人中。这种细菌通常是生命早期定植于人类胃肠道的首批微生物之一,在肠道免疫系统的发育中发挥着重要作用。
T-110
Enterococcus faecalis T-110:这种粪肠球菌菌株与枯草芽孢杆菌TO-A和丁酸梭菌TO-A结合使用作为益生菌(BIO-THREE)。这三种菌株一起可以减少手术患者的术后感染并缩短急性腹泻患儿的住院时间。
YM0831
Enterococcus faecalis YM0831:一项交叉研究发现,对健康人类受试者单次施用该菌株可显着改善血糖对蔗糖耐量试验的反应。
▸ 小结
除了BIO-THREE中的菌株外,很少有精心设计的随机对照试验证明了Enterococcus faecalis作为益生菌的功效,还需要更多的研究。
✦丁酸梭菌是肠道重要的菌属
丁酸梭菌(Clostridium butyricum)是人类胃肠道的天然居民。它是肠道中发酵膳食纤维并产生有益的短链脂肪酸丁酸盐的众多细菌之一。丁酸盐作为结肠上皮细胞的能量来源,有助于维持肠道屏障,具有抗炎作用,并可预防结肠癌。
doi.org/10.1007/s00535-015-1084-x
三种菌株已在人体中得到充分研究:
✦三种不同菌株显示能够调节肠道稳态、减轻肠炎
CBM 588
Clostridium butyricum CBM 588:这种丁酸梭菌菌株早在1963年就用作益生菌,用于一种名为Miyairisan的药物制剂中。它已经过安全性评估,不携带任何编码任何已知毒素或毒力因子的基因。
两项随机对照试验发现,该菌株可减少接受幽门螺杆菌根除治疗的患者的腹部症状和腹泻发生率的副作用。
另一项针对溃疡性结肠炎患者的安慰剂对照随机对照试验发现,接受MIYAIRI 588治疗的受试者中,患上肠结肠炎的人数较少。
尽管其影响在统计学上并不显著,而安慰剂组则服用乳糖,这会加重许多人的结肠炎。
TO-A
Clostridium butyricum TO-A:这种丁酸梭菌菌株与粪肠球菌T-110和枯草芽孢杆菌TO-A结合使用,作为益生菌在亚洲得到了广泛使用和深入研究。
A Tai Ning
该菌株包含在一种名为阿泰宁的产品中,最近在一项大规模、多中心、的随机对照试验中显示,该菌株对以腹泻为主的肠易激综合征具有显著益处。
该研究包括200名患者。4周后,与安慰剂组相比,益生菌组在整体肠易激综合征症状、大便频率和生活质量方面有显著改善。
几项研究发现丁酸梭菌降低了早产儿败血症和腹泻的发生率,并改善了抗原特异性免疫治疗。在动物研究中,丁酸梭菌已被证明可以改善肠道稳态并减轻结肠炎。
▸ 小结
丁酸梭菌是肠道重要的菌属,在随机试验中似乎是安全且耐受性良好的,并且帮助增加丁酸盐水平的同时,极少引起毒性。
➣土壤基益生菌的安全性仍存在争议,值得进一步关注
支持者声称它们是益生菌,可以使肠道功能正常化、帮助消化、有益地刺激免疫系统并帮助重新播种肠道微生物群。他们还称赞土壤微生物具有抵抗胃酸的能力并且不需要冷藏。
反对者认为,由于它们形成孢子的性质,它们会迅速增殖,与我们常驻的肠道微生物竞争,在某些情况下,它们甚至可能致病。由于它们的孢子形成能力和对大多数抗生素的天然抗性,意外的过度生长将很难治疗。
在随机、安慰剂对照的人体临床试验中,许多单独的土壤微生物菌株已被证明是有益的,几乎没有报告不良反应。
然而,其他菌株的临床证据有限或没有临床证据,可能会导致免疫系统受损的人感染。我们需要进行更细致的讨论——考虑每个特定物种、菌株和配方,而不是广泛地给土壤微生物贴上好或坏的标签。
基于土壤的微生物得到了很好的研究,并且已经在随机临床试验中显示出对一些症状有效,但仍需更多研究来确定其安全性和有效性。建议选择使用高质量和经过临床测试的菌株,在服用益生菌制剂前最好咨询医生的建议。
每个人对益生菌的反应都不同。反应将取决于服用的益生菌、剂量、肠道中已有的微生物以及肠道和免疫系统的健康状况。
可以通过微生物测序技术来检测使用益生菌前后的肠道菌群组成及相关健康状况。再选择针对自身特定健康状况的益生菌菌株。
同时也期待更多关于土壤微生物的单个菌株和配方在不同情况下影响肠道微生物群和肠道环境的研究。
注:本账号内容仅作交流参考,不作为诊断及医疗依据。
主要参考文献:
Mu Y, Cong Y. Bacillus coagulans and its applications in medicine. Benef Microbes. 2019 Jul 10;10(6):679-688. doi: 10.3920/BM2019.0016. Epub 2019 Jun 17. PMID: 31203635.
Jäger R, Purpura M, Farmer S, Cash HA, Keller D. Probiotic Bacillus coagulans GBI-30, 6086 Improves Protein Absorption and Utilization. Probiotics Antimicrob Proteins. 2018 Dec;10(4):611-615. doi: 10.1007/s12602-017-9354-y. PMID: 29196920; PMCID: PMC6208742.
Acosta-Rodríguez-Bueno CP, Abreu Y Abreu AT, Guarner F, Guno MJV, Pehlivanoğlu E, Perez M 3rd. Bacillus clausii for Gastrointestinal Disorders: A Narrative Literature Review. Adv Ther. 2022 Nov;39(11):4854-4874. doi: 10.1007/s12325-022-02285-0. Epub 2022 Aug 26. PMID: 36018495; PMCID: PMC9525334.
Ianiro G, Rizzatti G, Plomer M, Lopetuso L, Scaldaferri F, Franceschi F, Cammarota G, Gasbarrini A. Bacillus clausii for the Treatment of Acute Diarrhea in Children: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Nutrients. 2018 Aug 12;10(8):1074. doi: 10.3390/nu10081074. PMID: 30103531; PMCID: PMC6116021.
Kovács ÁT. Bacillus subtilis. Trends Microbiol. 2019 Aug;27(8):724-725. doi: 10.1016/j.tim.2019.03.008. Epub 2019 Apr 15. PMID: 31000489.
Marzorati M, Van den Abbeele P, Bubeck S, Bayne T, Krishnan K, Young A. Treatment with a spore-based probiotic containing five strains of Bacillus induced changes in the metabolic activity and community composition of the gut microbiota in a SHIME® model of the human gastrointestinal system. Food Res Int. 2021 Nov;149:110676. doi: 10.1016/j.foodres.2021.110676. Epub 2021 Aug 30. PMID: 34600678.
Kawarizadeh A, Pourmontaseri M, Farzaneh M, Hosseinzadeh S, Ghaemi M, Tabatabaei M, Pourmontaseri Z, Pirnia MM. Interleukin-8 gene expression and apoptosis induced by Salmonella Typhimurium in the presence of Bacillus probiotics in the epithelial cell. J Appl Microbiol. 2021 Jul;131(1):449-459. doi: 10.1111/jam.14898. Epub 2020 Dec 7. PMID: 33058340.
Muras A, Romero M, Mayer C, Otero A. Biotechnological applications of Bacillus licheniformis. Crit Rev Biotechnol. 2021 Jun;41(4):609-627. doi: 10.1080/07388551.2021.1873239. Epub 2021 Feb 16. PMID: 33593221.
Gudiña EJ, Teixeira JA. Bacillus licheniformis: The unexplored alternative for the anaerobic production of lipopeptide biosurfactants? Biotechnol Adv. 2022 Nov;60:108013. doi: 10.1016/j.biotechadv.2022.108013. Epub 2022 Jun 22. PMID: 35752271.
Gök ŞM, Türk Dağı H, Kara F, Arslan U, Fındık D. Klinik Örneklerden İzole Edilen Enterococcus faecium ve Enterococcus faecalis İzolatlarının Antibiyotik Direnci ve Virülans Faktörlerinin Araştırılması [Investigation of Antibiotic Resistance and Virulence Factors of Enterococcus faecium and Enterococcus faecalis Strains Isolated from Clinical Samples]. Mikrobiyol Bul. 2020 Jan;54(1):26-39. Turkish. doi: 10.5578/mb.68810. PMID: 32050876.
Stoeva MK, Garcia-So J, Justice N, Myers J, Tyagi S, Nemchek M, McMurdie PJ, Kolterman O, Eid J. Butyrate-producing human gut symbiont, Clostridium butyricum, and its role in health and disease. Gut Microbes. 2021 Jan-Dec;13(1):1-28. doi: 10.1080/19490976.2021.1907272. PMID: 33874858; PMCID: PMC8078720.
Chen D, Jin D, Huang S, Wu J, Xu M, Liu T, Dong W, Liu X, Wang S, Zhong W, Liu Y, Jiang R, Piao M, Wang B, Cao H. Clostridium butyricum, a butyrate-producing probiotic, inhibits intestinal tumor development through modulating Wnt signaling and gut microbiota. Cancer Lett. 2020 Jan 28;469:456-467. doi: 10.1016/j.canlet.2019.11.019. Epub 2019 Nov 14. PMID: 31734354.