-

 国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子
国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子

 二级病原微生物安全实验室
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业

谷禾健康
现在越来越多的人出现血脂不正常、腰围增加、血糖略高,这背后的隐藏风险,可能正在悄悄累积。
在全球范围内,代谢综合征已成为最受关注的健康挑战之一,患病率仍呈逐年上升趋势。它不仅显著增加心血管疾病、2型糖尿病及脂肪肝的风险,也对医疗资源和生活质量带来巨大压力。更让人警惕的是,这种代谢异常往往早期无明显症状,很多人直到发现血脂异常或血糖升高才意识到问题。
医学界和科学界也一直都在不断探索代谢综合征的发病机制和新的干预途径。传统的观点主要关注遗传因素、生活方式以及宿主自身的代谢紊乱,但近年来多个证据同时表明,代谢综合征和血脂异常通常同时存在——肠道菌群是将它们联系起来的关键因素之一。
肠道菌群影响食物的消化方式、胆汁酸的处理以及炎症信号在全身的释放。当肠道生态系统失衡时,新陈代谢可能转向胰岛素抵抗和脂质处理改变——这会导致甘油三酯升高、高密度脂蛋白(俗称好胆固醇)降低,有时还会增加 LDL 颗粒。
相关机制包括短链脂肪酸、胆汁酸、TMAO等。例如肠道细菌将初级胆汁酸转化为调控能量平衡、葡萄糖代谢及胆固醇处理的次级胆汁酸,通过FXR和TGR5受体发挥作用。某些微生物模式减少有益胆汁酸转化,降低胆固醇清除效率,进一步加剧脂质调节异常。同时,代谢综合征与低度炎症相关,菌群失衡可增加肠道通透性,使细菌成分激活免疫通路,加重胰岛素抵抗和血脂异常,形成一个循环,可能延续血脂异常。
本文将从多个层面深入探讨肠道微生物群如何影响血脂代谢,以及这些微生物学的改变如何最终导致血脂异常的发生。首先介绍代谢综合征的流行病学现状和临床特征,然后聚焦于血脂异常在代谢综合征中的核心地位,接着详细阐述肠道微生物群影响血脂代谢的五大核心机制,总结代谢综合征患者特征性的微生物组模式改变,最后讨论为什么肠道菌群检测对于代谢综合征的防控具有重要意义,以及这一领域未来的发展方向和临床转化前景。
代谢综合征是什么?
代谢综合征(英文名为Metabolic Syndrome,经常被简称为MetS),它是一组代谢紊乱症候群的集合。
根据国际糖尿病联盟(IDF)及多学会联合发布的统一诊断标准,代谢综合征的诊断需要满足以下五项中的三项或更多:
从这个诊断标准我们可以看出,血脂异常本身就是代谢综合征的核心诊断指标之一,这也说明了血脂异常在代谢综合征中的重要地位。
代谢综合征的全球发病率持续上升
全球成人代谢综合征患病率约为20%–25%(多项流行病学研究估计),这个比例意味着每四个成年人中就有一个患病,患者总数已经超过了10亿人。
更值得警惕的是,在过去数十年中呈显著上升趋势。这种快速增长的背后,是全球范围内肥胖症的流行。肥胖,特别是腹型肥胖,正是代谢综合征最重要的危险因素之一。
主要风险因素
代谢综合征的发生发展是一个复杂的过程,是遗传因素与环境因素长期相互作用的结果。我们可以将这些风险因素分为不可控和可控两大类。
不可控的风险因素:
年 龄
年龄是一个无法改变的因素,然而,它能帮助我们识别高风险人群。随着年龄增长,患代谢综合征的风险显著增加。
具体而言,随着年龄的增长,肌肉量会逐渐减少,而肌肉是消耗葡萄糖的关键组织。肌肉量的减少直接导致胰岛素敏感性下降。与此同时,随着年龄的增长,内脏脂肪的积累也会加重代谢负担。
遗 传
遗传易感性也是一个重要的不可控因素。研究已经发现了数十个与代谢综合征相关的遗传位点,这些位点每个单独的效应虽然不大,但累积起来可以显著影响患病风险。
如果父母或兄弟姐妹中有人患有代谢综合征、2型糖尿病或心血管疾病,患病几率会显著增加。
不过,遗传易感性并不意味着必然患病,健康的生活方式仍然能够有效地预防疾病的发生。
性 别
性别我们前面已经提到了,男性患病风险高于绝经前女性,这主要是性激素的保护作用在起作用。
女性在绝经前由于雌激素对脂质代谢、脂肪分布和血管功能具有一定保护作用,风险相对较低;但绝经后,随着雌激素水平下降,内脏脂肪增加、LDL-C升高、胰岛素敏感性下降,代谢综合征发生风险会明显上升。
可控的风险因素:
饮食——关键变量
饮食不是吃得多或者少的问题,是能量摄入、营养结构、脂肪类型、碳水质量、膳食纤维水平和饮食节律共同决定了代谢后果。
——饮食结构
富含饱和脂肪、反式脂肪、添加糖和精制碳水化合物的饮食模式,会促进能量过剩、内脏脂肪堆积、肝脏脂质合成增加和胰岛素抵抗。长期如此,机体更容易出现中心性肥胖、甘油三酯升高、脂肪肝、血糖异常等代谢综合征相关表现。
膳食纤维、全谷物、豆类、蔬菜水果、多酚类食物和优质不饱和脂肪酸摄入不足,也会削弱代谢保护作用。尤其是膳食纤维不足,会减少肠道菌群可利用底物,影响短链脂肪酸生成、肠屏障功能和炎症调节。
——脂肪摄入问题
需要强调的是,脂肪本身并不是一概有害。适量膳食脂肪对细胞结构、脂溶性维生素吸收、饱腹感和能量平衡都有重要作用。需要关注的是脂肪来源和脂肪酸类型。
从肠道微生态角度看,饮食也是塑造肠道菌群结构和功能的主要外部因素。长期高脂、高糖、低纤维饮食可能降低产短链脂肪酸菌,影响胆汁酸代谢和肠道屏障功能,进一步加重代谢异常。
运 动
缺乏运动是另一个重要的可控危险因素。出门坐车,上楼电梯,办公室久坐,回家躺着看电视,每天的体力活动量比五十年前减少了很多。规律的运动不仅可以帮助我们控制体重,还能提高骨骼肌胰岛素敏感性,增加葡萄糖转运蛋白GLUT4表达,促进糖原合成,提高脂肪酸氧化能力,并降低内脏脂肪含量。
对于代谢综合征的人群,很多研究已经提示,即便体重下降幅度有限,规律有氧运动和抗阻训练也能改善胰岛素抵抗、降低甘油三酯、提高HDL-C,并改善血压和炎症状态。
世界卫生组织推荐成年人每周至少进行150分钟中等强度的有氧运动,但是调查显示,仅约20%–30%的成年人达到推荐水平。
吸烟 饮酒
吸烟和过量饮酒也会显著增加代谢综合征的患病风险。
烟草中的尼古丁会引起血管收缩,升高血压,还会影响胰岛素信号传导,降低胰岛素敏感性。长期吸烟的人,患代谢综合征的风险比不吸烟者高出许多。
过量饮酒(如每日酒精摄入超过20–30 g),影响肝脏的脂代谢,还会提供大量的空热量,容易导致肥胖。研究发现,每天酒精摄入量超过25克,代谢综合征患病风险大大增加。
压力 睡眠
长期精神压力也是一个容易被忽视的危险因素。现代社会生活节奏快,竞争压力大,很多人长期处于慢性应激状态。
慢性压力会导致体内皮质醇水平持续升高,皮质醇是一种糖皮质激素,长期高水平的皮质醇会促进内脏脂肪堆积,升高血糖和血压,还会打乱正常的饮食节律,导致暴饮暴食。这些都会增加代谢综合征的发病风险。
睡眠不足和昼夜节律紊乱同样重要。睡眠时间不足、睡眠质量下降、夜间进食、轮班工作等因素,都会影响胰岛素敏感性、食欲激素、交感神经活性和炎症状态。临床上,睡眠呼吸暂停也常与中心性肥胖、胰岛素抵抗、高血压和血脂异常共同出现。
胰 岛 素 抵 抗
生物标志物层面的中间危险因素。胰岛素抵抗被认为是代谢综合征最核心的病理生理基础。
所谓胰岛素抵抗,就是指身体对胰岛素的反应变得不敏感,胰岛素不能有效地发挥降低血糖、调节脂代谢的作用。身体需要分泌更多的胰岛素才能维持血糖正常,长期高胰岛素血症会进一步加重代谢紊乱。
其 他
慢性低度炎症也是一个重要的中间危险因素,代谢综合征患者体内的促炎细胞因子水平普遍升高,这些炎症因子会干扰胰岛素信号传导,损伤血管内皮功能,促进动脉粥样硬化的发生发展。
脂肪组织功能异常同样关键,当脂肪细胞储存过多的脂肪时,脂肪细胞会发生肥大、缺氧,释放出大量的游离脂肪酸和炎症因子,进入血液循环,影响肝脏、肌肉、胰腺等多个器官的代谢功能。
伴随症状与疾病进展
代谢综合征自然病程呈现出渐进性发展的特点,从正常代谢到明显的代谢综合征,往往需要几年甚至十几年的时间。了解这个发展过程,有助于我们理解在不同阶段应该采取什么样的应对策略。
★ 早期阶段:指标轻度异常,但干预价值最高
在疾病的早期阶段(也就是常说的前驱期),这个阶段往往没有明显的临床症状,很多人体检的时候才会发现一些轻微的异常。
例如:腰围逐渐增加,甘油三酯轻度升高,HDL-C略低,血压处于正常高值,空腹血糖轻度偏高,或者提示脂肪肝。
这个阶段很容易被忽视。因为患者可能没有明显不适,也没有达到糖尿病、高血压或严重高脂血症的诊断标准。但从代谢机制看,胰岛素抵抗、脂肪组织功能异常和慢性炎症可能已经开始出现。
实际上,这个阶段正是干预效果最好的时期,如果能够及时改变生活方式,完全可以逆转病情,阻止它进一步发展。
■ 中期阶段:多项代谢异常叠加,进入明确风险状态
如果在早期阶段没有及时干预,病情就会进展到中期阶段。这个阶段已经出现了明确的代谢异常,达到了代谢综合征的诊断标准。
出现明确高血压(≥130/85 mmHg)、部分患者到糖尿病前期(空腹血糖也开始升高,往往超过5.6 mmol/L)或2型糖尿病、明显高甘油三酯血症、低HDL-C、脂肪肝也常见(约50%–70%的患者合并非酒精性脂肪肝)、睡眠呼吸暂停(尤其是在严重肥胖的患者中,超过一半的人睡觉打呼噜,憋气,频繁缺氧)等问题。
这一阶段的特点是多个系统开始同时受累,相互影响,形成循环。
例如,脂肪肝会进一步加重肝脏胰岛素抵抗,促进VLDL分泌;高甘油三酯和低HDL-C会增加动脉粥样硬化风险;睡眠呼吸暂停会通过夜间缺氧和交感神经激活,加重血压和糖脂代谢异常。
到了中期阶段,虽然已经有了多种代谢异常,但是如果能够积极干预,仍然可以很好地控制病情,延缓甚至阻止并发症的发生。很多人通过积极的生活方式干预,显著改善代谢状态,减少药物的使用。
▲ 晚期阶段:出现器质性损伤和慢病并发症
如果病情继续发展,就会进入晚期阶段,这个阶段已经出现了严重的并发症,往往伴随难以完全逆转的器质性损伤。
例如:2型糖尿病并发症、动脉粥样硬化性心血管疾病、肝病、肾病等。
大量的队列研究已经证实了代谢综合征对健康的严重危害。
一项荟萃分析纳入了超过一百万人的随访数据,结果显示,代谢综合征患者发生心血管事件的风险是正常人群的约2-3倍,发生2型糖尿病的风险更是增加了5倍。
还有研究发现,代谢综合征不仅增加心血管疾病和糖尿病的风险,与多种癌症风险增加相关(证据在不同癌种中强度不一),包括乳腺癌、结肠癌、胰腺癌等。这可能与慢性低度炎症、胰岛素抵抗、性激素水平改变等多种机制有关。
此外,代谢综合征还会增加认知功能下降和痴呆的风险,对老年人的健康构成更大的威胁,而且证据仍在不断积累。
以上自然病程我们可以看出,代谢综合征实际上是肝脏、脂肪组织、肠道、免疫系统和血管系统共同作用后的结果。代谢综合征的预防和干预越早越好。
也正因为如此,下一部分就有必要把血脂异常单独拎出来讨论:为什么在代谢综合征中,血脂异常不仅是诊断指标,更是心血管风险、肠-肝代谢轴和菌群干预价值的关键入口。
血脂异常在代谢综合征中不仅仅是五项诊断指标之一,它实际上承载着代谢状态、胰岛素敏感性、炎症水平和心血管风险的综合信息。理解血脂异常的特点和机制,有助于制定更精准的风险分层和干预策略。
血脂异常在代谢综合征中的核心地位
在代谢综合征的诊断标准中,高甘油三酯和低高密度脂蛋白胆固醇(HDL-C)就已经占据两项核心指标。血脂异常不仅是代谢综合征的重要诊断依据,还能够反映整体代谢紊乱的严重程度及其潜在风险,同时也是连接其他代谢紊乱与心血管终点事件的关键环节。
临床流行病学显示:
血脂异常不仅是代谢综合征的结果,也可能在代谢综合征的发生发展中起到推动作用。
血脂异常与心血管风险:独立且显著
研究证实:
胰岛素抵抗:血脂异常的驱动力
这要从代谢综合征的病理生理机制说起。代谢综合征的核心是胰岛素抵抗,胰岛素抵抗不仅会影响血糖代谢,还会直接影响脂肪组织的脂解过程。
当胰岛素抵抗发生时,脂肪组织对胰岛素的抗脂解作用不敏感,导致脂肪组织持续释放大量的游离脂肪酸进入血液循环。这些游离脂肪酸被肝脏摄取之后,就会刺激肝脏合成更多的甘油三酯和极低密度脂蛋白,导致血脂异常的发生。同时,胰岛素抵抗还会影响脂蛋白脂酶的活性,影响甘油三酯的清除,进一步加重高甘油三酯血症。
从这个角度我们可以看出,是胰岛素抵抗的重要代谢表现之一,它反映了全身代谢紊乱的严重程度。因此,血脂异常的检测和管理,应该成为代谢综合征防控的核心环节之一。
代谢综合征——脂质三联征
代谢综合征血脂异常具有高度特征性,被称为脂质三联征或致动脉粥样硬化血脂谱。
注:因为这种血脂谱具有很强的致动脉粥样硬化作用,心血管危害远超单纯的高胆固醇血症。
具体来说,代谢综合征的脂质三联征包括三个主要特点:
1、高甘油三酯血症
大部分代谢综合征患者会出现空腹甘油三酯(TG)升高,一般诊断标准是空腹甘油三酯≥1.7 mmol/L。
甘油三酯升高是代谢综合征血脂异常最常见的表现,这和我们前面提到的游离脂肪酸(FFA)释放增加、肝脏合成极低密度脂蛋白(VLDL)增加密切相关。
高甘油三酯血症不仅仅本身是一个危险因素,更重要的是,高甘油三酯血症往往伴随着其他脂蛋白的异常改变,共同导致动脉粥样硬化的发生发展。
2、低高密度脂蛋白胆固醇血症
代谢综合征患者大多出现高密度脂蛋白胆固醇降低。诊断标准:
男性<1.0 mmol/L,女性<1.3 mmol/L。
高密度脂蛋白胆固醇被我们称为”好胆固醇“,高密度脂蛋白胆固醇降低,这种保护作用就会减弱,动脉粥样硬化的风险就会增加。
注:高密度脂蛋白胆固醇负责将胆固醇从周围组织转运回肝脏进行代谢和排泄,这个过程叫做胆固醇逆向转运,它具有抗动脉粥样硬化的作用。
在代谢综合征患者中,高密度脂蛋白胆固醇降低往往伴随着甘油三酯升高同时出现,这两者之间存在密切的代谢联系。当甘油三酯升高时,胆固醇酯转运蛋白活性增加,会促进高密度脂蛋白中的胆固醇转移到极低密度脂蛋白和低密度脂蛋白中,导致高密度脂蛋白胆固醇水平降低。
3、小而密低密度脂蛋白比例升高
很多人都知道低密度脂蛋白胆固醇是”坏胆固醇“,但其实低密度脂蛋白并不是均匀的,它还可以根据颗粒大小分为不同的亚型:
其中小而密的低密度脂蛋白致动脉粥样硬化作用远远强于大而轻的低密度脂蛋白。
原因是:小而密LDL颗粒更易氧化、穿透血管内皮并形成泡沫细胞,即使总LDL-C正常,也显著增加心血管事件风险。
这种协同效应使代谢综合征患者的血脂异常风险复杂而显著
多项大型临床研究已经证实,在部分研究中,TG/HDL比值或非HDL胆固醇等指标对心血管风险具有更好的预测能力。
对于代谢综合征患者来说,我们不能仅仅关注总胆固醇和低密度脂蛋白胆固醇,还需要关注甘油三酯和高密度脂蛋白胆固醇,在有条件情况下可进一步评估(如ApoB或LDL颗粒特征)。
传统干预策略
目前临床上对于代谢综合征相关的血脂异常,主要的干预策略包括生活方式干预和药物治疗两大类。这两种策略都取得了一定的效果,但是也存在明显的局限性,也需要寻求新的干预方式。
生活方式干预是基础
包括控制饮食、增加运动、减轻体重、戒烟限酒等。健康的生活方式确实可以改善血脂异常,也可以改善胰岛素敏感性。
但是现实问题在于,生活方式干预的长期依从性普遍较低(多项研究显示仅有少部分患者能够长期坚持)。大多数人在坚持几个月之后都会逐渐回到原来的不良生活习惯。这其中有很多原因,比如工作忙、压力大、社交应酬多、难以抵制美食的诱惑等等。所以,仅仅依靠生活方式干预,很难长期有效地控制血脂异常。
药物治疗是目前临床上控制血脂异常的主要手段
——高胆固醇血症
首选的药物是他汀类药物。
大量的临床试验证实,他汀可以抑制胆固醇合成,显著降低低密度脂蛋白胆固醇,减少心血管事件。
但是,他汀类药物也存在局限性。对甘油三酯升高和低HDL-C的改善相对有限,尤其是对于高甘油三酯血症和低高密度脂蛋白胆固醇血症,他汀的效果有限。还有少数患者不能耐受他汀,服用他汀之后会出现肌肉疼痛、乏力,甚至肝功能异常,这些不良反应导致患者不得不停药。
——高甘油三酯血症
临床上常用的药物是贝特类药物。
贝特可以有效降低甘油三酯,升高高密度脂蛋白胆固醇。
但是贝特类药物也存在问题,它的主要副作用是会增加他汀的肌肉不良反应,与部分他汀联合使用时可能增加肌病风险。而且,贝特类药物虽然可以改善血脂谱,但是对于减少心血管事件的获益证据并不像他汀那么充分。
除了他汀和贝特,还有烟酸、依折麦布、PCSK9抑制剂等其他调脂药物,但是这些药物也各有各的局限性。
更深层次的问题在于,主要针对血脂异常这一表型进行干预,而对上游代谢紊乱(如胰岛素抵抗)的直接作用有限。
代谢综合征相关的血脂异常,根源在于胰岛素抵抗、脂肪组织功能异常、肠道菌群紊乱等深层次的问题,传统药物并没有针对这些根本原因进行干预,所以往往需要长期甚至终身服药,停药之后血脂很容易再次升高。
这种现状促使科学界不断寻找新的治疗靶点,而肠道微生物组正是目前看来最具前景的方向之一。
近二十年来的研究已经证实,肠道微生物组在脂质代谢中发挥着非常重要的作用,很多代谢综合征患者的血脂异常,都和肠道微生物组的异常改变密切相关。通过调节肠道微生物组来改善血脂异常,有望从根本上解决问题,为代谢综合征的治疗提供全新的思路。
在血脂代谢领域,大量的研究已经明确证实,肠道微生物群通过多种不同的机制影响宿主的脂质吸收、合成、转运和代谢,这些机制共同作用,最终影响血脂水平。目前学术界已经比较明确的核心机制主要有五个方面:
10.1007/S11154-019-09512-0
机制一:短链脂肪酸(SCFA)的调控作用
短链脂肪酸是肠道微生物群对宿主代谢产生影响的重要信号分子,也是目前研究最为深入、证据最充分的机制之一。
肠道细菌可以产生各种各样的糖苷水解酶,能够分解膳食纤维,将它们发酵产生短链脂肪酸。
所以说,膳食纤维对健康的好处,很大一部分是通过短链脂肪酸实现的。
短链脂肪酸主要通过三条途径调控血脂代谢。
途径一 | 脂肪组织:抑制脂解与炎症
第一条途径是通过G蛋白偶联受体介导的信号通路,抑制脂肪组织的脂肪分解。
在肠道上皮细胞、免疫细胞、脂肪细胞上都表达着两种特殊的G蛋白偶联受体,叫做GPR41和GPR43,这两种受体的配体是短链脂肪酸。
当短链脂肪酸和这些受体结合之后,会激活细胞内的信号通路,产生一系列生理效应。其中一个很重要的效应就是抑制脂肪组织中的脂肪分解。
——抑制脂肪分解,改善血脂
脂肪分解是脂肪细胞把储存的甘油三酯分解成游离脂肪酸释放到血液中,这个过程如果过度活跃,就会导致血液循环中游离脂肪酸增加,过多的游离脂肪酸被肝脏摄取之后,就会刺激肝脏合成更多的甘油三酯和极低密度脂蛋白,导致血脂升高。
而短链脂肪酸通过激活GPR41/43受体,可以抑制这个过度活跃的脂解过程,减少游离脂肪酸的释放,从而减轻肝脏的脂代谢负担,降低血脂水平。
——抗炎,改善胰岛素敏感性
肥胖和代谢综合征患者的脂肪组织中会有大量的巨噬细胞浸润,这些巨噬细胞会释放促炎细胞因子,这些促炎细胞因子会进一步加重胰岛素抵抗,促进脂肪分解。
短链脂肪酸通过激活GPR43受体,可以抑制巨噬细胞的促炎激活,减少促炎细胞因子的释放,改善脂肪组织的慢性炎症,从而改善胰岛素敏感性,间接改善脂代谢。
途径二 | 肝脏:调节胆固醇与脂肪酸合成
短链脂肪酸产生后,通过肠道吸收,经门静脉进入肝脏,其中丙酸的作用最为显著。
——丙酸直接抑制关键酶
研究发现,丙酸可以直接抑制肝脏胆固醇合成的关键限速酶——HMG-CoA还原酶(3-羟基-3-甲基戊二酰辅酶A还原酶),这一酶也是他汀类药物主要靶点。
通过抑制HMG-CoA还原酶,丙酸能够减少肝脏内源性胆固醇合成,从而降低血液中的胆固醇水平。
动物实验研究证实,给高脂饮食喂养的小鼠补充丙酸,可以显著降低血清总胆固醇和低密度脂蛋白胆固醇水平。
编辑
——丙酸调控脂肪酸合成与甘油三酯降低
研究发现,丙酸可以抑制肝脏中脂肪酸合成关键酶ACC和FAS的表达,减少从头合成脂肪酸,从而减少肝脏甘油三酯的合成,降低血清甘油三酯水平。
途径三 | 肠道激素:调节胰岛素敏感性与食欲
第三条途径通过调节肠道激素分泌,改善胰岛素敏感性,减少食欲。
肠道中的L细胞能够分泌一类重要的肠道激素:
这两种肠道激素都有很重要的代谢调节作用:
研究发现,短链脂肪酸能够刺激肠道L细胞分泌更多的GLP-1和PYY,从而发挥这些有益的代谢效应。胰岛素敏感性改善之后,脂代谢也会随之改善,游离脂肪酸释放减少,肝脏合成甘油三酯也会减少。
食欲减少有助于控制体重,减轻肥胖,进一步改善代谢。
机制二:影响胆汁酸代谢
胆汁酸是肝脏合成的重要分子,主要生理功能是帮助膳食脂肪的消化吸收。肝脏合成的胆汁酸储存在胆囊中,吃饭后胆囊收缩,将胆汁酸排入肠道帮助脂肪乳化和吸收。大约95%的胆汁酸在回肠末端被重新吸收,通过门静脉回流至肝脏,形成肠-肝循环。
过去人们认为胆汁酸仅仅是一种消化液,但近二十年来的研究表明,它同时也是重要的信号分子,能够激活核受体和G蛋白偶联受体,调控糖脂代谢,对全身代谢具有深远影响。
而肠道微生物群在胆汁酸代谢中发挥关键作用,能够通过多种酶改变胆汁酸组成和结构,进而影响其信号功能,最终调控宿主脂质代谢。
1、肠道菌群通过胆汁盐水解酶对胆汁酸解偶联
肝脏合成的胆汁酸,在分泌到胆汁中的时候,会和甘氨酸或者牛磺酸结合,形成结合型胆汁酸。结合型胆汁酸才能发挥正常的生理功能。
——胆汁酸的解偶联过程
当胆汁酸进入肠道之后,肠道中的很多细菌,主要是乳杆菌、双歧杆菌这些有益菌,能够产生一种叫做胆汁盐水解酶(简称BSH)的酶,这种酶能够把结合型胆汁酸中的甘氨酸或者牛磺酸水解下来,变成游离型胆汁酸。
——解偶联之后的游离型胆汁酸和结合型胆汁酸有什么不同?
主要的区别在于它们在肠道的重吸收效率。
游离型胆汁酸更容易被肠道被动重吸收,但是也有一部分会随着粪便排出体外。
如果BSH活性发生改变,就会影响胆汁酸的肠肝循环效率,改变胆汁酸的返回量,从而影响肝脏中胆汁酸的合成。
2、肠道菌群通过FXR信号调控肝脏脂蛋白合成
FXR(法尼醇X受体)是一种核受体,它主要在肝脏和肠道表达,它的配体就是胆汁酸。
研究发现,肠道微生物改变导致胆汁酸组成异常,会过度激活肠道FXR,FXR激活之后会通过循环中的FGF15/19(成纤维细胞生长因子15/19)信号,影响肝脏的脂代谢,促进VLDL合成,导致高甘油三酯血症。
3、通过TGR5受体激活增加能量消耗
TGR5是一种G蛋白偶联受体,它在棕色脂肪组织中高表达。
研究发现,代谢综合征患者往往次级胆汁酸合成减少,TGR5激活不足,能量消耗减少,这也会促进肥胖和血脂异常的发生。
机制三:内毒素血症与慢性低度炎症
内毒素血症和慢性低度炎症是连接肠道菌群紊乱与代谢综合征的一个核心机制,这个机制也是最早被发现和研究比较透彻的机制。
革兰氏阴性细菌的细胞壁外面有一种成分叫做脂多糖,简称LPS,也称为内毒素。
<向上滑动阅览>
还有研究发现,通过饮食干预改善肠道菌群,降低内毒素血症,可以显著改善血脂异常,进一步证实了内毒素血症在血脂异常中的介导作用。
机制四:胆碱代谢与TMAO途径
每天我们从膳食中摄入胆碱,尤其在鸡蛋、动物肝脏、红肉等食物中含量较高。摄入的胆碱大部分被吸收利用,但一部分进入大肠,由肠道细菌代谢。特定菌群产生的胆碱三甲胺裂解酶能将胆碱转化为三甲胺(TMA),随后通过门静脉进入肝脏。肝脏中的FMO3(黄素单加氧酶3)将TMA氧化为TMAO。
TMAO影响胆固醇和血脂代谢主要通过以下途径。
1、抑制胆固醇逆向转运
首先是抑制胆固醇逆向转运,减少胆固醇外流。前面提到过,胆固醇逆向转运是把胆固醇从周围组织包括动脉粥样斑块转运回肝脏代谢排泄的过程,这个过程是对抗动脉粥样硬化的重要保护机制。这个过程中,ABCA1和ABCG1这两个转运蛋白发挥着关键作用,它们位于巨噬细胞表面,能够把细胞内多余的胆固醇转运出来,交给高密度脂蛋白。
研究提示TMAO可能通过下调巨噬细胞ABCA1和ABCG1的表达,抑制胆固醇外流,从而影响胆固醇逆向转运,促进动脉粥样硬化的发生。
2、促进泡沫细胞形成
第二条途径主流假说是通过促进泡沫细胞形成。泡沫细胞是动脉粥样硬化斑块中最早出现的细胞成分,它是巨噬细胞吞噬了大量的氧化低密度脂蛋白之后形成的。
部分研究显示TMAO能够上调巨噬细胞表面清道夫受体SR-A1的表达,SR-A1能够识别并吞噬氧化低密度脂蛋白,SR-A1表达增加之后,巨噬细胞吞噬更多的氧化低密度脂蛋白,更容易形成泡沫细胞。泡沫细胞大量形成,可能是动脉粥样硬化发生发展的关键步骤。
3、抑制胆汁酸合成
胆固醇7α-羟化酶,也就是CYP7A1,是肝脏合成胆汁酸的关键限速酶。
研究显示TMAO能够抑制肝脏CYP7A1的表达,导致胆汁酸合成减少。胆固醇是合成胆汁酸的原料,胆汁酸合成减少,意味着胆固醇消耗减少,更多的胆固醇留在体内,同时,胆汁酸排泄减少也会影响胆固醇的平衡,导致血液中胆固醇水平升高。
<向上滑动阅览>
从干预的角度来看,如果我们能够抑制肠道细菌产生TMAO,就能够降低血浆TMAO水平,从而降低心血管风险。
目前已经有一些研究在探索这个方向,比如使用特异性的小分子抑制剂抑制胆碱三甲胺裂解酶的活性,或者使用噬菌体特异性地清除产TMAO的细菌,这些研究都取得了一些初步的成果,未来有可能开发出新的降脂和抗动脉粥样硬化药物。
机制五:膳食脂肪吸收调控
除了我们前面介绍的这四个机制,肠道菌群还能够直接影响肠道上皮细胞对膳食脂肪的吸收效率,以及乳糜微粒的形成和分泌过程,这个直接的调控作用对血脂水平也有着重要的影响。
脂肪吸收
吃进去的脂肪,需要经过一系列的步骤才能被身体吸收利用。
肠道菌群和脂质通过菌群代谢物、胆汁酸和免疫细胞吸收
编辑
脂蛋白脂酶不仅存在于脂肪组织和肌肉组织的血管内皮表面,在肠道上皮细胞的表面也有表达,它能够帮助分解乳糜微粒中的甘油三酯,促进脂肪酸的吸收。
研究发现,肠道菌群代谢产生的一些短链脂肪酸和其他代谢产物,能够影响肠道上皮细胞脂蛋白脂酶的活性,如果有害菌过度生长,产生的一些有害代谢产物会增加脂蛋白脂酶的活性,导致更多的膳食脂肪被吸收,过多的脂肪吸收之后,会导致血液中甘油三酯升高,也会增加脂肪堆积。
肠道菌群还能够调节肠道通透性,影响乳糜微粒进入循环。肠道菌群紊乱会损伤肠道屏障,增加肠道通透性,不仅LPS和细菌内毒素更容易进入血液循环,乳糜微粒进入循环也会增加,更多的膳食脂肪被吸收进入血液,导致血脂升高。同时,肠道通透性增加还会引起慢性低度炎症,加重胰岛素抵抗和血脂异常,形成恶性循环。
近年来,大规模肠道微生物组组学研究揭示了代谢综合征患者特有的菌群模式,尤其在伴血脂异常的人群中表现明显。这些特征不仅是疾病结果,还可能参与代谢综合征的发生和发展。
α多样性显著降低
α多样性是衡量肠道微生物群落结构的一个常用指标,它反映了肠道中物种的丰富度(也就是有多少种不同的细菌)和均匀度(也就是不同细菌的丰度分布是否均匀)。
大量的研究一致发现,代谢综合征患者的肠道菌群α多样性显著降低。
在代谢综合征伴血脂异常中,肠道微生物群的变化通常包括整体多样性下降以及有益菌的缺失。
产短链脂肪酸菌减少
多样性下降通常伴随产短链脂肪酸菌减少,如丁酸生成菌减少,使肠道调节胰岛素敏感性、脂质代谢以及炎症信号的能力下降,调节影响甘油三酯和 HDL 的炎症信号的能力减弱。
同时,多样性丧失常伴胆汁酸代谢紊乱和肠道屏障功能受损,导致低度全身性炎症增加,从而强化血脂异常和胰岛素抵抗的循环。
关键有益菌属/菌种的特征性变化
属和种的水平,代谢综合征患者也呈现出特征性的变化,一些有益菌降低,一些有害菌升高,这些变化共同推动了血脂异常的发生发展。
⭐ 有益菌显著减少
✦Faecalibacterium prausnitzii(普拉梭菌)
【产丁酸,抗炎,改善肠屏障,丰度下降 → 丁酸减少 → 炎症增加 → 胰岛素抵抗和血脂异常加重】
普拉梭菌是人体肠道中丰度最高的产丁酸菌之一,它能够发酵膳食纤维产生大量的丁酸,丁酸不仅是肠道上皮细胞的主要能量来源,还具有抗炎作用,能够改善肠道屏障功能,减少炎症反应。
大量研究一致发现,代谢综合征患者中普拉梭菌的丰度降低。普拉梭菌减少,丁酸产生减少,肠道屏障功能受损,炎症增加,这些都会加重胰岛素抵抗和血脂异常。
✦Roseburia (罗氏菌属)
【产SCFA,丰度下降 → SCFA生成减少 → 代谢保护作用减弱】
罗斯氏菌属也是非常重要的产短链脂肪酸的细菌,它也能够发酵膳食纤维产生大量的丁酸和乙酸。罗斯氏菌减少,同样会导致短链脂肪酸产生减少,对代谢的保护作用减弱。
✦ Akkermansia muciniphila(嗜黏蛋白阿克曼菌,常简称它为Akk菌)
【增强肠屏障、改善胰岛素敏感性,降低甘油三酯】
Akk菌是一种专门降解肠道黏液层中黏蛋白的细菌,它对维护肠道屏障功能非常重要。
研究发现,Akk菌能够增强肠道屏障完整性,减少内毒素血症,还能够改善胰岛素敏感性,降低体重和血脂。
动物实验研究发现,给肥胖小鼠补充Akk菌,能够降低血清甘油三酯,改善胰岛素抵抗,减轻体重,效果非常显著。
✦ Christensenellaceae(克里斯滕森菌科)
【丰度与瘦体型及代谢健康正相关,代谢综合征患者显著减少】
这个科的细菌在健康人肠道中普遍存在。
多项研究发现,克里斯滕森菌科的丰度和代谢健康指数呈强正相关,和BMI、甘油三酯呈负相关。
代谢综合征患者中克里斯滕森菌科的丰度显著降低。有意思的是,研究还发现克里斯滕森菌科的丰度和瘦体型相关,携带克里斯滕森菌科多的人,不容易发胖,代谢也更健康。
其它还有:
⭐ 有害菌显著增加
✦Escherichia/Shigella(大肠杆菌/志贺氏菌属)
【LPS产生增加 → 内毒素血症 → 慢性炎症】
这些细菌属于变形菌门,是革兰氏阴性条件致病菌,它们的细胞壁含有大量的LPS。
代谢综合征患者中这些细菌的丰度显著升高,它们产生大量的LPS,更容易导致内毒素血症和慢性炎症。
✦ Desulfovibrio(脱硫弧菌属)
【高浓度硫化氢损伤肠屏障 → 炎症 → 代谢紊乱】
脱硫弧菌是一种硫酸盐还原菌,它能够还原硫酸盐产生硫化氢。硫化氢在低浓度的时候有一定的生理作用,但是高浓度的硫化氢对肠道上皮细胞有毒性作用,能够损伤肠道屏障,增加肠道通透性,还能够抑制线粒体呼吸,影响肠道上皮细胞的能量代谢,还会加重炎症,促进代谢紊乱。
研究发现,代谢综合征患者中脱硫弧菌的丰度显著升高,而且脱硫弧菌丰度和血压、甘油三酯呈正相关。
✦Clostridium clusters(一些梭菌属的细菌)
【影响胆汁酸代谢 → 血脂异常】
特定的一些梭菌集群和胆汁酸代谢紊乱相关,它们能够改变胆汁酸的代谢,影响胆汁酸的组成和信号功能,进而影响血脂代谢。这些梭菌过度生长,会导致胆汁酸代谢重编程,促进血脂异常的发生。
其它还有:
功能基因谱的特征变化
代谢综合征患者的微生物群功能能力下降,表现为:
这些功能性变化揭示了代谢综合征肠道微生态对血脂异常、胰岛素抵抗及慢性低度炎症的上游贡献。理解这些特征性变化,可以为精准干预提供明确靶点。
通用干预的局限性
常规药物在代谢综合征合并血脂异常的患者中疗效有时不理想,因为无法考虑影响甘油三酯、HDL 和全身炎症的特定肠道-胆汁酸-代谢物信号回路。
热量控制难以修复肠道生态
许多标准的健康饮食方法强调热量控制或食物限制(广泛限制),但它们可能无法可靠地恢复驱动胆汁酸受体信号(FXR/TGR5)和肝脂调控的肠道生态系统功能。
如果一个人的肠道菌群纤维发酵能力低且多样性减少,仅仅添加一些纤维或做出模糊的饮食改进,通常无法有效改善肠道胆汁酸转化模式,也难以调节VLDL生成或提升胰岛素敏感性。
低度慢性炎症难以消除
即便进行了饮食调整或生活方式干预,低度慢性炎症在代谢综合征患者体内仍可能持续存在。
这主要与短链脂肪酸产量下降和肠道屏障受损有关,导致内毒素(如LPS)进入循环,维持炎症。持续的炎症环境不仅影响脂质代谢,还强化胰岛素抵抗,使甘油三酯升高、HDL下降的血脂异常难以改善。
通用干预很少针对屏障完整性和 SCFA 生成的根本驱动因素(如持续可发酵的纤维数量/品种、富含多酚的植物摄入量,以及对发酵或益生元食品的个体耐受性)。如果不解决这些功能缺陷,脂质水平的改善可能会趋于平稳,因为炎症和胰岛素抵抗环境持续促进动脉粥样状、富含 ApoB 的脂蛋白谱。
个体差异决定效果
最重要的是一刀切的建议往往忽视了微生物生态和饮食反应中的个体间差异。
两个人可能都吃得更好,但他们的基线菌群组成、胆汁酸库、运输时间和代谢反应可能导致截然不同的结果。帮助一个人的微生物组产生 SCFA 或改善胆汁酸信号的机制,对另一个人可能影响不大,甚至因不耐受而加重症状。
如果没有基于功能标志物(如纤维摄入质量、炎症状态、血糖反应以及特定益生元/发酵策略耐受性)进行个性化调整,通用方案就不太可能在甘油三酯/HDL 和心血管代谢风险方面产生持久改善。
风险分层与早期预警
代谢综合征的发生发展是一个漫长的过程,从正常代谢到明确的代谢综合征,往往需要几年甚至十几年的时间。
肠道菌群变化:早于临床症状的预警信号
传统的危险因素,比如BMI、腰围、血压、血糖、血脂,这些都是在代谢异常已经发生之后才能发现异常。
而肠道菌群紊乱往往早于临床症状和生化异常的出现,也就是说,还没有出现血糖升高、血压升高、明确的血脂异常的时候,肠道菌群已经发生了改变。
风险分层与预测模型
基于这个特点,肠道菌群特征可以用于代谢综合征的风险分层和早期预警。
多项研究以及谷禾健康已经构建了基于肠道菌群特征的代谢综合征预测模型,这些模型的预测性能如何呢?
研究发现,基于肠道菌群特征构建的预测模型,对代谢综合征的曲线下面积(AUC)可以达到0.83-0.89,这个预测精度优于传统的危险因素,比如BMI、年龄这些指标。如果把肠道菌群特征和传统危险因素结合起来,预测精度还能进一步提高。
这个发现有什么临床意义呢?
它意味着我们可以通过检测肠道菌群,在疾病的早期阶段,甚至在还没有出现生化异常的时候,就识别出代谢综合征的高风险人群。
提前干预与疾病分期监测
研究发现,在糖耐量正常人群中,肠道微生物变化可比血糖异常提前3–5年被检测到。
也就是说,如果现在血糖、血脂都还正常,但是肠道菌群已经出现了特征性的改变,那么未来几年发生代谢综合征的风险就会显著升高,这个时候就可以提前进行干预,通过改变饮食、增加运动、补充益生菌益生元等方法,调整肠道菌群,阻止或者延缓代谢综合征的发生。这就是真正的上医医未病,预防重于治疗。
除了预测发病风险,肠道菌群变化还能够反映代谢综合征的疾病分期和严重程度。
研究发现,随着代谢综合征从早期到中期再到晚期,肠道菌群的紊乱程度也逐渐加重,α多样性逐渐降低,特征性改变也越来越明显。
因此我们可以通过检测肠道菌群,对疾病进展情况进行监测,了解疾病处于什么阶段,帮助我们制定更合适的干预策略。
个性化干预方案制定
现在医学界越来越强调精准医学,也就是根据每个人不同的特征,制定个性化的干预方案,而不是千人一方。肠道菌群检测就是实现个性化干预非常重要的一个工具,因为不同的人肠道菌群特征不一样,适合的干预方法也不一样。
例如,产短链脂肪酸菌缺乏。
如果肠道中产丁酸、产乙酸的有益菌明显减少,那么适合补充益生元,比如菊粉、低聚果糖这些益生元,它们能够选择性地促进这些有益菌的生长,增加短链脂肪酸产生,改善血脂代谢。
例如,Akkermansia muciniphila缺乏。
如果Akk菌丰度明显降低,那么可以补充特定的益生菌,或者吃一些能够促进Akk菌生长的食物,比如石榴、蔓越莓这些富含多酚的食物,能够促进Akk菌生长,改善肠道屏障功能,降低内毒素血症,改善血脂。
治疗反应预测与监测
评估药物干预效果
肠道菌群特征不仅仅能够帮助我们制定干预方案,还能够预测药物和生活方式干预的效果,通过肠道菌群特征预测对某种治疗方法的反应好不好,这对于临床治疗决策有辅助作用。
例如他汀类药物,他汀是临床上最常用的降脂药物,但是我们前面提到过,部分患者对他汀反应不佳,所以谷禾也一直在和临床推进合作看有没有办法在用药之前就预测哪些人反应好,哪些人反应不好呢?
研究发现,肠道菌群特征确实能够预测他汀的降脂疗效,特定的菌群特征。
评估减肥效果
减重,无论是通过生活方式减重还是减重手术,不同的人效果差异很大。
研究发现,基线肠道菌群中普雷沃氏菌/拟杆菌比例高的患者,减重干预的效果更好,体重减轻更多,血脂改善也更明显。另一方面,基线Akk菌丰度可以作为减重效果的预测因子,如果AKK菌丰度比较低,那么可能需要更强化的干预,或者先补充Akk菌,提高减重效果。
动态监测肠道菌群评估
在干预过程中,还可以通过动态监测肠道菌群来评估干预效果。干预之后,如果肠道菌群多样性恢复了,有益菌丰度增加了,有害菌丰度减少了,这些都可以作为代谢改善的早期生物标志物。
这些变化往往出现在血脂、血糖这些生化指标改善之前,所以我们可以通过监测菌群变化,早期了解干预是否有效,如果干预没有改善菌群,帮忙用户及时调整干预方案,避免无效治疗。
并发症风险评估
特定的肠道微生物标志物和代谢综合征并发症的风险相关,通过检测这些标志物,可以辅助评估并发症的发生风险,帮助我们进行风险分层和预防。
举例:和并发症风险有可能相关的指标:
次级胆汁酸升高
次级胆汁酸是肠道细菌代谢初级胆汁酸产生的,粪便次级胆汁酸升高和非酒精性脂肪肝的发生风险密切相关,如果有代谢综合征,同时粪便次级胆汁酸升高,那么可能需要提前进行筛查和干预。
<来源:谷禾健康肠道菌群检测数据库>
产短链脂肪酸菌减少
短链脂肪酸对代谢有保护作用,产短链脂肪酸菌减少,代谢性疾病进展的风险升高。所以检测发现产短链脂肪酸菌明显减少,那么需要更积极的生活方式干预,必要的时候补充益生元或者益生菌,增加短链脂肪酸产生,降低糖尿病等代谢疾病风险。
通过检测这些微生物标志物,我们可以更精准地评估不同并发症的风险,然后进行针对性的预防和干预。
一般支持策略
代谢综合征伴血脂异常越来越多地与肠道微生物组调节胆汁酸、炎症色调和代谢信号传递有关。一个核心支持策略是通过增加来自多样、最少加工的植物性食物(如豆类、全谷物、水果、蔬菜)的膳食纤维来改善微生物生态。
膳食纤维
膳食纤维为有益发酵菌提供能量,促进短链脂肪酸生成,如丁酸。丁酸可强化肠道屏障,减少循环内毒素(LPS),降低低度慢性炎症,从而改善胰岛素敏感性,减轻甘油三酯升高和HDL下降。可选益生元来源包括大蒜、洋葱、韭葱、芦笋、菊苣根/菊粉、青香蕉等,同时限制高度加工食品。
支持胆汁酸信号(天然食物)
由于肠道微生物也会调节胆汁酸,支持正常胆汁酸信号传递也是关键方法。某些细菌群落产生的二级胆汁酸可以与 FXR 和 TGR5 等受体相互作用,这些受体会影响肝脂的合成、葡萄糖处理和能量平衡。富含植物多酚和其他微生物群可及化合物的饮食可以促进更有利的细菌转变,并支持胆汁酸池,更好地支持代谢调节。
对于某些人来说,添加天然食物可能进一步增强微生物多样性和代谢反应性。每天通过多种植物(浆果、柑橘、石榴、绿叶蔬菜、十字花科蔬菜、特级初榨橄榄油、可可)增加多酚,以改善胆汁酸信号和微生物多样性。
长期饮食模式(减少加工食品)
持续减少超加工食品和过量添加糖的摄入,有助于防止菌群失调引发的动脉粥样性血脂模式(甘油三酯升高、HDL降低、非HDL/LDL升高)。
限制添加糖和精制淀粉(含糖饮料、甜食、白面包/糕点)的摄入,以减少甘油三酯负担,并限制微生物组驱动的炎症信号。
全食物饮食、逐步增加纤维摄入、改善胰岛素敏感性的生活方式,可长期强化肠-肝代谢轴,改善炎症标志物,帮助血脂向更健康的心血管风险水平转变。
如果能耐受,可以考虑发酵食品(如酸奶/克菲尔加活菌、酸菜、泡菜、天贝),以增强有益微生物并减少低度炎症。
健康脂肪与运动
选择健康脂肪而非饱和脂肪/反式脂肪(橄榄油、坚果、种子、牛油果;若耐受时可食用脂肪鱼类),以改善动脉粥样形成的血脂水平。
优先进行规律的体育活动(有氧+抗阻训练),减少久坐时间,以改善胰岛素敏感性和降低甘油三酯。
睡眠、压力管理
确保充足睡眠并管理压力(例如7-9小时睡眠、正念/呼吸/锻炼),以减少炎症,避免加重胰岛素抵抗和脂质指标。
个体化监测原则
由于胆汁酸代谢和肠道代谢物因人而异,应通过饮食和生活方式调整改善肠道功能,而尽量不要直接用补充剂替代饮食。益生菌或合生菌选择需依据有代谢或抗炎证据的菌株,并考虑剂量及个体反应。
对血脂异常患者,应避免在未监测的情况下过度干预,必要时结合血脂、血糖及炎症指标评估(如可用的CRP、血浆甘油三酯监测),尤其当存在他汀类药物或胆汁酸干预时。
复测/监测
复测的重要性和时间安排
对于伴有血脂异常的代谢综合征患者,在进行干预后进行复测是评估效果的重要环节。常见干预包括增加膳食纤维、提升植物多样性、摄入富含多酚的食物,有时还包括适量发酵食品。
一般建议干预8–12周后复测代谢和血脂标志物,以捕捉甘油三酯、HDL及非HDL/LDL的趋势性变化。这段时间足以让肠道代谢产物(如SCFA)和胆汁酸信号发生适应性改变,从而更明显地反映临床效果。
如果患者同时在进行降脂或胰岛素敏感性优化治疗,复测时间可由临床医生根据具体情况调整,但通常仍建议在同一个8–12周周期内进行安全性和有效性评估。
复测如何指导干预?
复测不仅评估血脂变化,还可用于验证干预模式是否改善肠道介导的炎症和胆汁酸代谢。需要注意的是,复测可以结合其他临床实验室指标解读结果。
最终目标是通过动态监测,确认肠道驱动的代谢物变化与甘油三酯、HDL及整体心血管代谢风险持续改善之间的相关性,而非依赖单次检测数据。这样可以为后续干预优化和个性化调整提供科学依据。
新兴治疗策略
近年来,针对代谢综合征和血脂异常的微生物干预研究呈现多条前沿方向,涵盖从益生菌到后生元的多种策略。
下一代益生菌
传统的益生菌主要是乳杆菌、双歧杆菌这些,而下一代益生菌是从健康人肠道中分离出来的,对代谢有明确益处的特定菌种,目前研究最多的就是Akkermansia muciniphila、Eubacterium hallii。
Akk菌我们前面多次介绍过,它能够改善肠道屏障,降低内毒素血症,改善胰岛素抵抗和血脂。
目前Akk菌已经进入了II期临床试验,初步结果显示,补充Akk菌能够显著降低代谢综合征患者的体重指数和甘油三酯,改善胰岛素抵抗,而且安全性很好,没有明显的不良反应。
Eubacterium hallii 是一种产丁酸菌,它也能够改善糖脂代谢,目前也在进行临床试验。相信在未来几年,这些下一代益生菌就会上市,给代谢综合征患者带来新的治疗选择。
粪菌移植(FMT)
粪便菌群移植就是把健康供体的肠道菌群移植到患者肠道内,重建患者正常的肠道菌群。
目前已经完成了一些小规模的研究,结果显示,健康供体FMT能够改善代谢综合征患者的胰岛素敏感性和血脂谱,部分患者改善效果能够维持三个月以上。
当然,目前FMT还存在一些问题,比如长期效果还不明确,供体菌群的质量控制,安全性等问题都还需要进一步研究。
未来通过把多种明确的有益菌按比例混合,代替完整的粪便,以解决供体质量控制和安全性问题,提升临床可行性,这个方向也是很有前景的。
噬菌体靶向调控
噬菌体干预通过特异性攻击目标有害细菌(如产TMAO菌),精准清除有害菌而不影响其他有益菌。相比广谱抗生素,这种方法副作用更小,精确性高。
动物实验显示,噬菌体可有效降低产TMAO菌群水平,减少血浆TMAO并降低动脉粥样硬化风险。
这个方向未来很可能开发出全新的治疗方法,精准地调节肠道菌群,改善血脂异常。
后生元干预
后生元不是活菌,而是细菌产生的有益代谢产物,或者细菌本身的一些成分,比如短链脂肪酸、特定的胆汁酸、细菌细胞壁成分等等。
后生元和活菌相比,有很多优势:
多个后生元产品已进入临床试验,未来几年有望面市。
除了这四个主要方向,还有很多其他的策略在研究之中,比如精准化营养饮食干预、益生菌+益生元的合生元、膳食益生元促进特定有益菌生长等等,这些策略都在进行研究,未来会有更多的干预方法可供选择。
整合医学模式
未来代谢综合征的管理,预测会走向多组学整合的精准医学模式,也就是整合多个层面的信息,为每个患者制定最精准的预防和治疗方案。
具体来说,整合主要包括四个层面:
基因组学:评估遗传易感性
每个人的遗传背景不同,对代谢综合征的易感性和药物反应也存在差异。通过基因组学检测,可以了解个体携带的易感位点、患病风险以及对特定药物的反应,为精准干预提供基础。
微生物组学:分析肠道菌群特征
肠道菌群是代谢综合征干预的重要对象。通过分析肠道菌群组成与功能基因,可识别有益菌减少、有害菌增加或功能异常的情况。结合这些信息,可以制定个性化干预方案,包括膳食、益生菌/益生元以及生活方式调整。
代谢组学:反映代谢状态
代谢组学通过检测血液、粪便等样本的代谢谱,如血脂、血糖、TMAO、胆汁酸和短链脂肪酸水平,直观反映个体代谢状态,同时也可评估肠道菌群功能。与宏基因组学结合,可以更全面地了解代谢综合征的发病机制和个体差异。
生活方式组学:定制可执行方案
每个人的饮食、运动、睡眠和压力水平不同。生活方式组学通过收集这些数据,将干预措施与个人习惯匹配,从而制定切实可行、易于长期坚持的方案,提升干预效果。
多组学整合的价值
通过整合基因组学、微生物组学、代谢组学和生活方式组学的数据,可以对代谢综合征风险进行精准评估,制定最适合个体的干预策略,实现精准预防和个性化治疗。这将是未来代谢综合征管理的发展方向。
代谢综合征伴血脂异常的发生与发展,不仅涉及传统的血脂、血糖和体重因素,更深层次地与肠道微生物群调控的胆汁酸代谢、炎症状态及代谢信号传递密切相关。本文梳理了肠道微生物群通过多个不同的机制影响宿主的脂质代谢:
这五个机制共同作用,最终影响血脂水平,促进血脂异常的发生发展。
从临床应用的角度来看,肠道菌群检测能够帮助我们进行风险分层和早期预警,制定个性化的干预方案,预测治疗反应,评估并发症风险。这些应用正在逐渐从研究走向临床,未来肠道菌群检测很可能会成为代谢综合征患者常规评估的一部分。
尽管当前仍存在科学问题和技术挑战,如微生物-宿主交互作用、长期干预安全性及微生物组-药物相互作用,但随着多组学整合和大规模临床研究的推进,这一领域有望实现从基础研究到临床实践的快速转化。
未来需要更大规模的前瞻性队列研究,来进一步明确肠道菌群改变和代谢综合征发生发展的因果关系,还需要设计严谨的大规模临床试验,来验证各种肠道菌群干预方法的长期有效性和安全性。
但是无论如何,这个领域的发展方向是非常明确的,随着研究的不断深入,我们对肠道微生物组在代谢综合征中作用的理解会越来越深入,干预方法也会越来越成熟。
未来已来,对于代谢综合征高风险人群,以及已经患有代谢综合征的患者而言,肠道微生物组有望成为代谢综合征管理的重要抓手,了解自己的肠道菌群特征,不仅仅是了解当下的健康状态,更是把握未来健康主动权的重要一步。
希望更多的人能够了解肠道微生物组与代谢健康的关系,重视肠道健康,通过健康的饮食和生活方式,维护良好的肠道菌群平衡,远离代谢综合征,远离心血管疾病和糖尿病,享受健康美好的生活。
注:本账号内容仅供学习和交流,不构成任何形式的医疗建议。
主要参考文献:
Mohammadi F, Rudkowska I. Dietary Lipids, Gut Microbiota, and Their Metabolites: Insights from Recent Studies. Nutrients. 2025 Feb 11;17(4):639.
Ko CW, Qu J, Black DD, Tso P. Regulation of intestinal lipid metabolism: current concepts and relevance to disease. Nat Rev Gastroenterol Hepatol. 2020 Mar;17(3):169-183.
Gildner TE. Links between metabolic syndrome and the microbiome. Evol Med Public Health. 2020 Feb 19;2020(1):45-46.
Singh A, Verma A, Ashraf S, Sarfraz Sheikh D, Irfan H, Riaz R, Venjhraj F, Meghwar S, Kumar R, Tariq MD, Hamza HM, Ahsan A, Satapathy P. Role of gut microbiota in the pathogenesis of metabolic syndrome: an updated comprehensive review from mechanisms to clinical implications. Ann Med Surg (Lond). 2025 Jul 29;87(9):5851-5861.
Gut Microbiome and Metabolic Syndrome: How Dyslipidemia Is Influenced. innerbuddies
Zeng L, Yu B, Zeng P, Duoji Z, Zuo H, Lian J, Yang T, Dai Y, Feng Y, Yu P, Yang J, Yang S, Dou Q. Mediation effect and metabolic pathways of gut microbiota in the associations between lifestyles and dyslipidemia. NPJ Biofilms Microbiomes. 2025 May 28;11(1):90. doi: 10.1038/s41522-025-00721-x.
Yin J, Zhang M, Jiang W, Huang X, Yin Y. Microbial Interactions with Intestinal Lipid Digestion and Absorption: Emerging Targets for Metabolic Disorders. Research (Wash D C). 2025 Sep 19;8:0904.
Schoeler M, Caesar R. Dietary lipids, gut microbiota and lipid metabolism. Rev Endocr Metab Disord. 2019 Dec;20(4):461-472.

谷禾健康
牙龈卟啉单胞菌(Porphyromonas gingivalis)是一种革兰氏阴性、绝对厌氧的杆状细菌,是生活在口腔内的700多种细菌中的一员,常定植于牙龈下菌斑,并且与牙周病理密切相关。
发表在《微生物学前沿》杂志上的一篇综述指出,数百种口腔细菌通常在相对平衡的环境中共存,但环境一旦改变,部分细菌可能占据优势,从而引发龋齿或牙龈疾病。
牙龈卟啉单胞菌(P.gingivalis)之所以能在口腔微生态失衡中“占据上风”,与其独特的生存策略密切相关:一方面依赖铁/血红素获取形成代谢优势;另一方面通过多种毒力因子(如脂多糖、牙龈蛋白酶、菌毛黏附结构以及外膜囊泡等)实现黏附侵袭、营养掠夺与免疫调控。
致病过程中,P.gingivalis可破坏上皮屏障与结缔组织稳态,诱导并放大炎症反应,同时干扰补体与吞噬清除等先天免疫环节,进而促使炎症持续化,最终表现为牙周附着丧失与牙槽骨吸收等典型病理改变。
更值得关注的是,牙周局部的炎症与细菌相关分子并不局限于口腔。牙龈出血、牙周袋溃疡面等为细菌或其组分进入体内提供了“通道”,它们可通过血液循环(以及吞咽/吸入等途径)到达远处组织,触发系统性炎症反应或加重既有疾病风险。近年来的研究不断提示,牙周炎与多种全身疾病之间存在流行病学与机制层面的联系,而P. gingivalis作为“关键病原体”可能在其中扮演重要角色,例如与心血管系统疾病风险增加、以及多种慢性炎症相关疾病(糖尿病、阿尔兹海默病、溃疡性结肠炎、类风湿性关节炎)的发生发展相互关联。
本文将围绕P.gingivalis的核心致病机制、其从口腔到全身的潜在健康影响及相关证据,并对干预策略进行概述与讨论。
♥
牙龈卟啉单胞菌(Porphyromonas gingivalis)是一种革兰氏阴性、严格厌氧的杆状细菌,主要定植于龈下(牙龈线以下)牙菌斑,是慢性牙周炎的主要致病菌。它是牙周病的重要致病菌之一:即使丰度较低,也能影响口腔菌群与宿主反应,诱发菌群失调和炎症。
1
细胞形态与基本特征
•革兰氏染色:革兰氏阴性。
•形态:短杆状细菌(无鞭毛,不运动)。
•氧气需求:专性厌氧菌——生长需要无氧环境。
•菌落形态:菌落多为光滑、隆起,直径约1–3mm,因沉积血红素(卟啉铁)复合物而常由边缘向中心逐渐变黑(产黑色素)。
•其他特征:不形成孢子。但细胞表面具有丰富的荚膜、多种蛋白质和脂多糖(LPS),并表达粗细两类菌毛(FimA主要菌毛、Mfa1次要菌毛)等结构。
•分类:属于拟杆菌门(Bacteroidota)卟啉单胞菌属(Porphyromonas)。
2
生长环境与营养需求
•定植:主要定植在龈下牙菌斑和牙周袋;也可在舌背、扁桃体隐窝等厌氧微环境中检出。
•环境偏好:温和条件(约37 °C、pH≈7),与人体口腔环境相似。
•主要营养来源:对铁和血红素依赖性极强,通过 HmuY 等血红素获取系统摄取血红素。
宿主蛋白/肽:降解血清白蛋白、血红蛋白、转铁蛋白及其他蛋白质,以获得碳、氮和铁。
3
人群分布
作为常驻口腔微生物群的一员,牙龈卟啉单胞菌(P.gingivalis)在健康个体与患病个体中均存在。
健康个体:P.gingivalis在健康者龈下菌斑中可检出,但丰度较低;检出率约为 10%–25%,随检测方法和人群而异。
牙周炎患者:约69%–79%的牙周炎患者可检出该菌;多项流行病学研究表明,其在慢性和侵袭性牙周炎中的检出率或菌量显著高于口腔健康者。
注:最新高分辨率测序表明,健康者与患者均可携带该菌;但高毒力株(如 W83/W50)在中重度牙周炎中显著富集,低毒或无毒力株(如 ATCC33277/381)则更普遍见于各类样本。
患病率因以下而异:
•国家和种族;
•年龄和口腔护理的可及性;
•诊断方法(培养、PCR、qPCR)。
4
代谢能力
典型的蛋白/肽发酵型厌氧菌:不能利用大多数糖类作为主要碳源;偏好利用多肽与氨基酸,尤其依赖宿主组织蛋白(胶原、纤维连接蛋白、血红蛋白等)及其他细菌代谢产物。
具有多种蛋白水解酶,可:
分解宿主组织与血浆蛋白获取营养;
释放铁、血红素等必需因子;
同时改变局部炎症和凝血环境。
例如Gingipains是由牙龈卟啉单胞菌产生的一类关键毒力因子,属于半胱氨酸蛋白酶(cysteine proteases),在牙周病及多种系统性疾病中发挥核心作用。
RgpA/RgpB——精氨酸特异性
Kgp——赖氨酸特异性
其能:
–降解宿主的结构蛋白和免疫蛋白(如白蛋白、补体组分、免疫球蛋白、细胞因子);
–释放肽和氨基酸以获取营养;
-从血红蛋白和其他血蛋白中释放血红素以获取铁;
-对组织破坏和免疫调节有重大影响。
其他营养获取酶还有二肽基肽酶IV(DPPIV):从宿主蛋白/肽中切割二肽,有助于肽的利用和毒力。
牙龈卟啉单胞菌具有专门的血红素/铁获取系统(如Hmu和Hus系统)用于捕捉和内化血红素:
首先将氧血红蛋白转化为更易受影响的高铁血红蛋白;
其次完全蛋白水解血蛋白以释放游离血红素;
最后通过特定的外膜受体和运输系统结合并引入血红素。
5
耐药性
P.gingivalis对甲硝唑、阿莫西林、四环素等常用抗菌药总体仍以敏感为主,但临床分离株中β-内酰胺类和大环内酯类耐药呈上升趋势。
注:生物膜状态可显著增强耐药表型与耐受性;“耐药并非单一基因决定,而是生态位与生理状态的产物”。
鉴于其关键性质和生物膜生活方式,机械清创加上局部/口腔卫生措施仍是基础,抗生素作为辅助而非单独治疗。
6
与其他菌群的互作
即使相对丰度较低,P.gingivalis也可通过调控宿主免疫并改变微环境,驱动群落菌群失衡。
P.gingivalis与Fusobacterium nucleatum、Prevotella intermedia、Treponema denticola等协同形成生物膜:其 gingipains 和几类外膜蛋白对于多种菌种生物膜形成至关重要。
与其他牙周病原体共聚并协同:
•齿垢密螺旋体(Treponema denticola)——形成具有增强毒力和稳定性的混合物种生物膜;
•与早期定殖者如格氏链球菌(Streptococcus gordonii)相互作用,利用特异性粘附蛋白-受体相互作用整合至现有斑块。
牙龈卟啉单胞菌(Porphyromonas gingivalis)与Tannerella forsythia和Treponema denticola可形成一种“红色复合体”。
菌群失调与生态变化:
通过降解宿主蛋白和调节炎症,牙龈卟啉单胞菌丰富了以下环境中的栖息地:
•血红素和肽,支持蛋白水解性厌氧菌生长。
•炎症性渗出物,有利于更具致病性的群体。
这导致形成促炎、破坏组织的微生物群。
口腔外的相互作用:
•肠道微生物群相互作用:实验研究表明,P.gingivalis可以定殖或通过肠道,改变肠道微生物组成和短链脂肪酸代谢,可能促成全身炎症和代谢变化。
•系统微生物组-免疫轴:由P. gingivalis引起的口腔菌群失调,是炎症介质和微生物产物的慢性来源,影响全身免疫力,并与其他黏膜微生物组(如肠道、气道)相互作用。
相互作用表:
向上滑动阅览
增强:
Bacteroidales
Bacteroides
Odoribacter
Peptococcaceae
抑制:
Bifidobacterium
Coriobacteriales
Adlercreutzia
Collinsella
Porphyromonas
Prevotella
Clostridium
Clostridiales incertae sedis
Clostridiales Family XIII. Incertae Sedis
Blautia
Coprococcus
Dorea
Lachnospiraceae
Ruminococcaceae
Ruminococcus
Dialister
Campylobacteraceae
Erysipelotrichaceae
被抑制:
Bifidobacterium
Coriobacteriales
Adlercreutzia
Collinsella
Bacteroidales
Bacteroides
Porphyromonadaceae
Odoribacter
Parabacteroides
Porphyromonas
Prevotella
Rikenellaceae
Alistipes
Turicibacter
Streptococcus
Clostridiales
Catabacteriaceae
Clostridium
Clostridiales incertae sedis
Peptoniphilus
Clostridiales Family XIII. Incertae Sedis
Lachnospiraceae
Blautia
Lachnospiraceae
Coprococcus
Dorea
Eubacterium
Lachnobacterium
Lachnospira
Roseburia
Lachnospiraceae
Peptococcaceae
Ruminococcaceae
Ruminiclostridium
Acetivibrio
Eubacterium
Faecalibacterium
Oscillospira
Ruminococcus
Acidaminococcus
Dialister
Phascolarctobacterium
Veillonella
Rubrivivax
Alcaligenaceae
Oxalobacter
Bilophila
Desulfovibrio
Campylobacteraceae
Enterobacteriaceae
Escherichia
Erysipelotrichaceae
Erysipelotrichaceae
Holdemania
Akkermansia
♥
研究发现,牙龈卟啉单胞菌(Porphyromonas gingivalis)是导致慢性牙周炎的主要病因。牙周炎是一种感染驱动的炎症性疾病,其特征是牙龈炎症和骨质流失。
注:全球有5.38亿人患有严重牙周病,其中2.76亿人失去了牙齿。随着人口增长和老龄化,这一数字预计还会上升。
牙龈卟啉单胞菌作为牙周关键病原体概述
Curtis MA,et al.J Periodontal Res.2025
◮ 牙周炎特征包括:
•牙龈发红、肿胀、流血;
•牙齿与牙龈之间的附着脱落;
•牙周韧带和牙槽骨的逐渐破坏;
•最终牙齿的活动性与脱落。
下面让我们一起进一步了解牙龈卟啉单胞菌的毒力因子、致病机制以及致病过程,重点梳理其主要毒力因子如何发挥作用,并从整体上把握其致病机制与致病过程的关键环节。
主要毒力因子
◮ 细胞表面与黏附相关因子
①菌毛(FimA主要菌毛+Mfa1次级菌毛+其他类型 )
•介导对牙面、唾液蛋白、ECM(纤维连接蛋白、层粘连蛋白)及其他细菌(如Streptococcus gordonii)的黏附与共聚集。
•不同 fimA 型别(I–V、Ib)在侵犯性、炎症诱导和骨吸收潜能上显著不同;侵袭力强的菌株(如 W83)往往携带特定 fimA 型别,并与 rgpB 基因型存在关联。
•对上皮细胞和成骨细胞的初始侵袭非常重要,但对后续抑制成骨分化和矿化作用不是必需的。
②荚膜
•多糖荚膜可抗吞噬、抗补体杀伤,包被脂多糖和表面蛋白,减弱模式识别。
•有荚膜菌株在牙周炎、菌血症模型中毒力更高,与全身炎症、心血管损伤相关。
③脂多糖
•P.gingivalis的脂多糖结构高度可塑,在 TLR2/TLR4 之间切换,可诱导低反应型或偏向 Th2/Th17 的炎症,避免有效清除。
•LPS 还影响骨代谢(促进 RANKL 表达、抑制成骨),并可在远处组织(血管、脑)诱导炎症与内皮损伤。
④外膜蛋白与血红素捕获蛋白
•HmuY、HemR、HBP35、HbR、OMP26/83 等:血红素/血红蛋白受体,是铁利用的核心。
注:HmuY蛋白是P.gingivalis特有的铁调控血红素结合蛋白,主要位于菌体表面,并可作为可溶性脂蛋白或外膜囊泡释放至环境中。
•LptO:参与 LPS 去酰化及 A‑LPS 与 CTD 蛋白分泌协调,是表面蛋白正确展示和 OMVs 形成的关键。
牙龈卟啉单胞菌的主要致病因子
编辑
doi: 10.1016/j.jmb.2021.166836.
◮ 分泌系统与外排结构
①IX 型分泌系统(T9SS)
•负责分泌并锚定一大批 CTD 蛋白,包括 gingipains、部分黏附蛋白、血红素获取蛋白等。
②外膜囊泡
•富集 gingipains、LPS、PPAD、HBP35 等毒力因子;
•体积小、易扩散,可穿透黏液和组织,既在局部牙周组织中扩散毒力,也可进入血流,参与远处器官(血管、肝、脑)的炎症和损伤。
P.gingivalis OMVs中的毒力因子及相关效应
编辑
doi.org/10.3390/app15126847
◮ 蛋白水解酶与代谢相关因子
①Gingipains(三种半胱氨酸蛋白酶)
•RgpA、RgpB(精氨酸特异)和 Kgp(赖氨酸特异),是最核心毒力因子:
•营养获取:大规模降解胶原、纤维蛋白原、白蛋白、免疫球蛋白等,释放多肽与氨基酸,同时从血红蛋白、血红素蛋白、铁蛋白中夺取 hemin/铁。
•免疫调控/免疫逃逸:切割 C3、C4、C5 生成或耗竭补体组分;剪切 CD4、CD8、CD14、TREM‑1 等免疫受体;激活或失活细胞因子(IL‑1β、IL‑6、TNF‑α、IL‑8)。
•组织破坏与骨吸收:降解基质金属蛋白酶抑制因子、激活前体 MMP;促进 RANKL 表达,诱导破骨细胞生成。
②二肽基肽酶 IV(DPPIV)及其他外肽酶
•分解宿主多肽、细菌信号肽,提供营养并调节细胞因子谱。
•DPPIV 活性与生物膜形成和牙周炎严重度正相关。
③氧化应激应答系统
•多种抗氧化酶(超氧化物歧化酶、过氧化氢酶样因子等)与调控网络,使其在充满活性氧的炎症牙周袋内仍能存活。
•抗氧化能力本身也被视为毒力决定因素之一。
致病机制:从“关键病原体”视角理解
◮ 关键病原体与多菌种协同
P.gingivalis在菌斑中的绝对丰度一般不高,却能通过毒力因子重塑宿主免疫与微环境。
主要通过:
•破坏上皮屏障(菌毛、LPS、SerB 等效应蛋白);
•调节补体–TLR 交叉通路(C5aR–TLR2 失衡、CR3 介导“伪吞噬”);
•提供丰富蛋白底物与使整个群落从“相对共生”转向“蛋白分解‑致炎”的菌群失衡(dysbiosis)。
•与“红色复合体”伙伴 T. denticola、T. forsythia 以及 Fusobacterium nucleatum、Streptococcus gordonii 等协同,相互黏附、交叉供养(代谢产物共享);共同构建耐药、耐清除的复杂生物膜。
◮ 免疫逃逸与免疫重编程
①先天免疫层面
•多种脂A异构体改变 TLR2/TLR4 信号,诱导偏低IL‑12产生和异常NF‑κB激活,削弱Th1清除反应。
•趋化因子瘫痪:通过降解 IL‑8 等趋化因子,削弱中性粒细胞募集。
②获得性免疫层面
•Protease 切割 CD4/CD8/TCR 及共刺激分子,抑制 T 细胞活化;
•PPAD 诱导瓜氨酸化新表位,驱动异常 B/T 细胞反应;
•在某些模型中,可减少调节性 T 细胞或诱导“致病性 Treg”亚群。
③细胞侵袭与细胞内生存
•通过菌毛、胶原黏附蛋白、SerB 等进入上皮细胞、内皮细胞、成骨细胞、巨噬细胞;
•高度利用宿主自噬途径作为“运输高速公路”,在自噬体/溶酶体中长期存活;
•改变宿主细胞骨架与紧密连接,破坏屏障并利于自身传播。
◮ 铁依赖与代谢优势
•严格依赖氯化血红素(Hemin)/铁生长,通过:
表面 hemin 受体(HmuR / HemR / HBP35 / IhtB 等);
Gingipain 和 DPPIV 等蛋白酶自宿主血红蛋白、hemopexin、触珠蛋白、转铁蛋白中释放 hemin;
IX 型分泌系统分泌血素样蛋白(HmuY)和 OMVs 携带 hemin。
•在铁缺环境(健康牙周袋)下具有明显优势,可储存表面黑色素,兼具营养库与抗氧化“缓冲层”的功能。
◮ 组织破坏与骨代谢失衡
本质是由 失衡炎症 + 破骨/成骨比失衡 驱动:
•Gingipain、LPS 及 OMVs 激活 RANKL‑RANK 轴,上调 RANKL、下调 OPG;
•诱导多种炎症因子(TNF‑α、IL‑1β、IL‑6、IL‑17),促进破骨细胞分化;
•降解骨基质与 ECM,并抑制成骨细胞功能和矿化,导致牙槽骨吸收与附着丧失。
致病过程(以牙周炎为主线)
一般可将牙龈卟啉单胞菌(Porphyromonas gingivalis)相关牙周炎发展分为四个阶段:
阶段1:定植与生物膜初建
①环境准备
•牙面先由需氧/兼性革兰阳性球菌(链球菌等)形成早期生物膜;
•牙龈炎形成后,龈沟加深、氧压下降,蛋白质/铁丰富,为P.gingivalis和其它厌氧菌提供生态位。
②进入与黏附
•依靠 fimbriae、Mfa1、HagA/HagB 等凝集素结合唾液受体、ECM、早期定植菌表面蛋白(如 S. gordonii GAPDH);
•借助脂多糖、外膜蛋白与宿主受体(整合素、TLR、CXCR4 等)相互作用加强黏附。
阶段2:微生态重塑与免疫失衡
①生物膜成熟与菌群失衡
•与T. denticola,T. forsythia, F. nucleatum 等协同,构成“红色复合体”和复杂多菌种生物膜;
•Gingipains/DPPIV 分解唾液蛋白与炎性渗出液,为整个群落提供氨基酸与肽类碳氮源。
②免疫重编程
•脂多糖结构变化、补体–TLR 信号交叉、趋化因子瘫痪 → 局部防御减弱而炎症持续;
•中性粒细胞、大量炎症细胞浸润但杀菌效率低,形成典型的 “低效炎症+高组织破坏” 状态。
阶段3:组织破坏与牙槽骨吸收
①软组织破坏
•Gingipains 和其他蛋白酶广泛降解胶原、纤维连接蛋白、基底膜成分;
•SerB 等效应蛋白通过胞内信号干扰上皮屏障、诱导凋亡或坏死。
②骨破坏与牙周袋形成
•炎症介质(PGE₂、IL‑1β、TNF‑α、IL‑6、IL‑17)+ RANKL/OPG 比值上升 → 破骨细胞活化;
•牙槽骨逐渐吸收,临床表现为出血、附着丧失、牙齿松动。
③持续慢性感染
P.gingivalis能侵入牙周组织细胞(上皮、成纤维细胞、内皮细胞、成骨细胞)及局部淋巴结,在细胞内长期存活,形成难以彻底清除的“储备库”。
阶段4:扩散与系统性影响
①外膜囊泡入血
•咀嚼、刷牙、牙周治疗等操作可造成P.gingivalis进入循环系统或其他组织。
•外膜囊泡(OMVs)更易进入血流并到达远处器官。
②系统炎症与靶器官损伤
•在动脉斑块、胰腺、肝脏、胎盘、脑组织中均可检出P.gingivalis DNA/抗原或 gingipains;
•通过上述机制参与心血管疾病、糖尿病、类风湿关节炎、神经退行性病变等的发生与进展。
牙龈卟啉单胞菌作为牙周炎发生与进展中的关键致病菌,即使在菌群中相对丰度不高,也能凭借多样的毒力因子与免疫调控能力,改变龈下微环境并推动菌群失衡,从而放大炎症反应并加速牙周组织破坏。
现有研究还提示其不同毒力株在病变人群中呈现差异性富集,且在生物膜状态下更易表现出更强的耐受与耐药表型,增加临床控制难度。因此,围绕牙龈卟啉单胞菌的精准检测、分型评估与靶向干预,对于牙周炎的早期预警、个体化治疗与长期维持具有重要意义。
案例分享
基本情况(主要诉求):口臭,牙周病,胃食管反流。
检测结果要点,如下图:
编辑
从图中可以看出,检测者口腔中的相关病原菌水平出现严重超标的情况,其中以牙龈卟啉单胞菌(P.gingivalis)最为突出,其检出量相较参考范围高出约39倍,提示该菌丰度异常升高。
编辑
编辑
•与牙周病(核心主线)
P.gingivalis是经典牙周致病菌之一。报告中牙周炎高风险与P.gingivalis显著超标一致,提示口腔内可能存在更活跃的炎症微生态环境与牙龈下菌斑问题。
•与口臭(症状表现)
口臭常与牙周袋、舌苔及厌氧菌代谢产生的口腔挥发性硫化物(VSC)增多相关。该报告在“口臭”模块中同时出现P.gingivalis明显超标及多种口臭相关菌升高,形成“菌群—代谢产物—异味”的同向线索:牙周炎越活跃、厌氧菌负荷越高,口臭越容易反复。
•与胃食管反流
从“过程”角度看:反流可改变口腔环境(酸暴露、口干、菌斑生态位变化),使牙周相关厌氧菌更容易在不利环境下获得优势;反过来,牙周感染灶与炎症负荷升高也可能让上消化道不适更难“稳定下来”。因此,报告提示的P.gingivalis高丰度更像是一个“口腔慢性炎症与微生态失衡”的信号,可能与胃食管反流形成双向叠加的困扰体验。
由此可进一步得知,口腔微生物状态是否健康不仅影响口腔本身,也与全身健康状况存在密切关联。
♥
越来越多证据表明,牙龈卟啉单胞菌(P.gingivalis)与多种全身性疾病相关。牙周炎不仅是牙齿脱落的重要原因,还与心血管疾病、糖尿病、阿尔茨海默病及类风湿性关节炎等疾病有关。
牙龈卟啉单胞菌传播的机制
口腔病原体牙龈卟啉单胞菌除致牙周炎外,还与多种全身疾病相关。其由口腔播散至其他器官主要有四种途径:
(1)分泌蛋白水解酶降解细胞—基质黏附分子,破坏黏膜屏障并促其侵入;
(2)经转胞作用进入细胞并跨层转移,逐步深入组织;
(3)被吞噬细胞摄取后随血流转运并在血中释放;(4)附着菌丝并随其穿透黏膜,进入更深结构。
牙龈卟啉单胞菌入侵全身的策略
编辑
doi: 10.3390/pathogens9110944.
1
心血管疾病
牙周炎患者发生冠心病的风险更高
研究显示,患有牙周炎的人患冠心病的风险显著高于无牙周炎者。2023年的一项荟萃分析证实,牙周病与心血管不良结局(如心肌梗死和中风)在不同人群和性别中存在一致关联。
编辑
同样,研究报告称,刷牙、清创或刮治等操作可能促使口腔病原体及其毒力因子进入血流。另有证据显示,可在动脉粥样硬化斑块中检出牙龈卟啉单胞菌,且牙周细菌在血液或血管病变内的原位存在与动脉瘤风险升高相关。
最新研究还表明,牙周病与冠状动脉疾病、中风及心力衰竭风险显著增加相关,且在男女中均一致观察到。
牙龈卟啉单胞菌会促进动脉粥样硬化发展
文献指出牙周炎可能影响心血管疾病发展的两种主要机制:直接机制涉及细菌侵入血管,而间接机制则通过激活宿主过度的炎症反应,进而影响系统性疾病的发生。
P.gingivalis诱导的加速动脉粥样硬化刺激
编辑
doi: 10.3390/pathogens9110944.
细菌抗原与宿主细胞的相互作用可以引发局部和全身免疫反应。牙龈卟啉单胞菌已被证明会侵入内皮细胞,诱导功能障碍,并触发促动脉粥样硬化反应,促进动脉粥样硬化病灶的发展和进展。
2
糖尿病
大量文献已证实糖尿病与牙周病存在生物学关联,且牙周炎可影响包括糖尿病在内的多种全身性疾病的发病机制。
牙周炎损害血糖控制、促进糖尿病
牙周炎与糖尿病呈双向交互关系:糖尿病促进牙周炎的发生和进展,而牙周炎则损害血糖控制,并促成糖尿病并发症的发展。糖尿病患者通常表现出比非糖尿病患者更严重的临床牙周炎症表现,牙周破坏的严重程度与全身性并发症的存在、血糖控制程度及糖尿病类型相关。
牙周感染的微生物活动是牙周炎与糖尿病双向关联的重要基础之一。牙周炎的高炎症状态可显著增加活性氧(ROS),并与高级糖基化终产物(AGEs)形成复杂互作;AGEs在高血糖与氧化条件下积累,进一步加剧氧化应激与炎症,损害牙周及全身健康。
牙周细菌诱发的感染产生脂多糖(LPS),促使炎症细胞因子升高并募集免疫细胞,从而加重糖尿病;而糖尿病的高血糖环境又利于致病菌生长,推动菌群失调。
炎症性细胞因子增多还会削弱血糖控制。牙周组织破坏始于牙龈龈沟液中升高的细菌激活炎症细胞因子,继而导致结缔组织与牙槽骨分解。
牙周炎与糖尿病之间的双向关系
编辑
doi.org/10.3390/app15126847
最新研究表明,糖尿病控制不佳的人群患牙周炎的风险显著更高,强调了有效血糖管理在牙周健康中的重要性。相反,牙周炎与糖尿病患者代谢控制恶化有关,突显了需要综合管理策略以同时应对这两种疾病。
3
阿尔茨海默病
阿尔茨海默病(AD)是一种累及全球数百万人的神经退行性疾病,随着寿命延长与人口老龄化,其患病率快速上升。有研究人员发现,牙周病暴露与AD风险增加1.7倍相关。
注:AD以神经元进行性丧失为特征,导致记忆、语言与认知功能逐步衰退,晚期可致死亡(阿尔茨海默病协会,2016)。
牙周病原体通过血流可进入大脑并释放毒素
牙龈卟啉单胞菌通路至大脑
编辑
Huang Z,et al.Front Immunol.2025
科学证据表明,阿尔茨海默病/痴呆患者常伴口腔健康受损,因而更易发生慢性口腔疾病。牙周病原体可侵入牙周袋上皮并进入血流,在循环中释放内毒素和外毒素,将感染扩散至包括大脑在内的多个部位;在AD患者脑组织中已检出大量脂多糖(LPS)。
牙周病(PD)与AD的相互作用可能通过多环节过程诱发神经元退变:口腔菌群失调促使病原体增殖并侵入牙周组织,继而入血;病原体及其毒性分子(如LPS)与小胶质细胞的Toll样受体2/4(TLR2/4)结合,诱导细胞因子和炎症介质释放。
炎症反应进一步促进神经元产生淀粉样前体蛋白(APP),并通过激活β、γ分泌酶增加淀粉样蛋白β(Aβ)分泌,尤其是Aβ42单体和sAPPβ,导致细胞外沉积及AICD生成;Aβ随后聚集为寡聚体、原纤维/纤维,最终形成淀粉样斑块。
AD患者在脑脊液和口腔中检测到牙龈卟啉单胞菌
编辑
doi: 10.1126/sciadv.aau3333.
牙龈卟啉单胞菌是Aβ斑块形成的重要风险因素
这些斑块可被小胶质细胞膜上的髓系细胞触发受体2(TREM2)识别,进而引发进一步的炎症反应并促进Aβ生成。与此同时,功能失调的神经元中tau蛋白磷酸化(p-tau)增加,形成神经纤维缠结。慢性牙周炎(CP)及牙龈卟啉单胞菌感染已被认为是Aβ斑块、痴呆和阿尔茨海默病发生的重要风险因素。
有研究人员提出,牙龈卟啉单胞菌通过分泌牙龈蛋白酶(gingipains)介导神经损伤,参与AD发病机制;其研究在AD大脑中检测到显著的牙龈蛋白酶免疫反应性(IR),并在生前确诊AD患者的大脑与脑脊液(CSF)中鉴定出P.gingivalis DNA,提示其DNA或可作为鉴别诊断标志。
研究指出,过度神经炎症可导致突触蛋白减少与记忆丧失,并对牙周炎中P.gingivalis-LPS介导的炎症通路提供机制性解释。近期研究进一步表明,牙周病原体可通过调控小胶质细胞激活加重神经炎症,可能加速AD的神经退行过程。
此外,纵向队列研究发现,有效牙周治疗与AD高风险人群认知衰退减缓相关,提示口腔健康干预或有助于AD管理。
4
其他神经退行性疾病
除阿尔茨海默病外,研究还发现牙龈卟啉单胞菌与帕金森病等其他神经退行性疾病相关。
牙龈卟啉单胞菌与神经退行性疾病
编辑
Huang Z,et al.Front Immunol.2025
牙周炎与帕金森病(PD)风险相关
流行病学与机制线索提示P. gingivalis可能通过氧化应激与神经炎症影响帕金森病(PD)发生发展。
在PD患者血液中检测到 P.gingivalis 的主要毒力因子(如 gingipain R1、P.g-LPS)的报道,并有研究提示 gingipains 可能在黑质相关区域出现沉积。
相较 AD,P. gingivalis 通过“氧化应激直接驱动 PD”的直接证据仍不足,但其脂多糖诱导线粒体功能障碍与神经炎症的结果与 PD 病理环节相契合。
多发性硬化(MS)
直接发表证据有限;但动物模型中口腔感染P.gingivalis 可加重自身免疫性脑脊髓炎样病理,提示外周炎症/感染可放大神经炎症与脱髓鞘过程。
5
溃疡性结肠炎
编辑
研究发现,溃疡性结肠炎(UC)患者较健康对照牙周炎发生率更高、牙周袋更深、缺牙更多。
口服P.gingivalis会加重溃疡性结肠炎(小鼠)
设计:C57BL/6 小鼠连续 40 天每周 2 次灌胃(生理盐水或活菌 P. gingivalis W83 等),后 10 天给予 DSS 诱导结肠炎。
结果:与单纯 DSS 相比,P. gingivalis 灌胃组出现更严重的结肠炎表型:
–DAI(疾病活动指数)更高;
–结肠更短
–组织学损伤更重(深溃疡、上皮/腺体脱落、基底膜破碎、炎性细胞浸润等)。
牙龈卟啉单胞菌会加重溃疡性结肠炎
编辑
doi: 10.1038/s41368-021-00136-2.
P.gingivalis可能加重已有炎症,并非诱导发病
研究讨论引用“口源菌到达肠道”的模型:口腔炎症(牙周炎)增加口源致病菌负荷;肠道已有炎症会破坏菌群稳态,使口源菌更易在肠道定植并取代常驻菌;两者协同使口源菌更可能通过胃酸屏障并在肠道扩增,从而加剧既有肠炎(更倾向于“加重已有炎症”,而非诱发健康肠道发病)。
P.gingivalis 部分通过诱导异常免疫反应、提高 Th17/Treg 比值,从而加重 UC 炎症。
6
类风湿性关节炎
类风湿性关节炎与牙周病共享多种病理机制,包括相似的炎症细胞参与、血清及局部细胞因子谱的平行变化,以及基质金属蛋白酶(MMPs)等组织降解介质的激活。两者亦均以破骨细胞介导的骨吸收为特征,造成显著骨损伤。
牙龈卟啉单胞菌能表达PAD,这在类风湿关节炎中起重要作用
类风湿关节炎(RA)发病机制的关键环节是瓜氨酸化,即由肽基精氨酸去亚胺酶(PAD)介导的翻译后修饰,将精氨酸转化为瓜氨酸。
注:PAD由多种免疫细胞(如T/B淋巴细胞、中性粒细胞、单核细胞和巨噬细胞)表达。
过度瓜氨酸化可形成自身抗原,诱导自身抗体生成,尤其是抗环瓜氨酸肽(anti-CCP)抗体,这是RA发病的核心。瓜氨酸化蛋白的积累进一步放大自身免疫反应,加重炎症与组织破坏。
迄今牙龈卟啉单胞菌是唯一已知可表达肽基精氨酸脱亚胺酶(PPAD)的微生物,该细菌酶与人类PAD同源。RA患者口腔中P.gingivalis的存在与抗CCP抗体高度相关,提示其可能在RA发病中具有关键作用。
假设牙龈卟啉单胞菌促进了瓜氨酸化蛋白在口腔内的积累,通过异常免疫反应激活产生自身抗体。特别是,类风湿关节炎患者对瓜氨酸化蛋白的免疫耐受降低可能促进这些自身抗体的生成,进而加重疾病的系统性炎症负担。
此外,瓜氨酸化蛋白的产生升高与更严重的疾病表现有关,包括关节侵蚀的快速进展。因此,抗CCP抗体的存在已被证明与类风湿性关节炎患者肺泡骨吸收增加相关,进一步强调了牙周病与类风湿关节炎严重程度之间的相互关联。
近期研究通过识别口腔微生物组中可能与牙龈卟啉单胞菌协同作用,加重类风湿关节炎的瓜氨酸化和系统性自身免疫,进一步扩大了这一视野。
7
慢性阻塞性肺病
慢性阻塞性肺病(COPD)是一组以气道气流受限为特征的疾病,包括慢性支气管炎和肺气肿。
牙周炎与慢性阻塞性肺病正相关
研究显示,牙周炎与COPD正相关,牙周治疗可改善肺功能并降低急性加重频率。基于16S rDNA的宏基因组分析在重症急性COPD患者气管抽吸物中检出P.gingivalis,提示其可能参与发病。
唾液中与该菌相关的蛋白酶等酶可改变黏膜表面黏附受体并破坏/降解唾液膜,从而促进呼吸道病原体在呼吸上皮定殖。因此,牙龈卟啉单胞菌可能通过促进呼吸道病原体定植推动COPD进展。
牙龈卟啉单胞菌在COPD发展过程中的影响
编辑
doi: 10.1038/s41368-025-00397-1.
8
不良妊娠结局
不良妊娠结局(APO)是指妊娠、分娩或产后发生的孕产妇、胎儿或新生儿并发症。流行病学证据一致表明牙周炎与APO正相关。
P.gingivalis的DNA可在胎盘、脐带中检出
牙龈卟啉单胞菌(P.gingivalis)的DNA和抗原也可在胎盘、脐带及羊水中检出,其在胎盘或脐带中的存在与早产(PB)、子痫前期及妊娠期高血压等并发症相关,提示其可通过侵入并损伤子宫-胎盘组织参与APO。
多种啮齿动物模型进一步证实,P.gingivalis感染可诱发子宫-胎盘病变(如子宫内膜动脉炎、轻度绒毛膜羊膜炎及伴胎盘结构障碍的子宫胎盘出血),并导致胎儿生长受限(FGR)、低出生体重(LBW)和早产等不良结局。
P.gingivalis通过免疫失衡影响FGR与早产
在孕鼠模型中,感染P.gingivalis可触发胎儿特异性胎盘免疫反应,提高胎盘Th1/Th2细胞因子比值(IFN-γ、IL-2、IL-12升高,IL-4、IL-10降低),与胎儿生长受限(FGR)相关;同时可增强母体免疫反应并加重FGR,表现为母体血清TNF-α升高、IL-10降低。此外,P. gingivalis LPS可通过诱导胎盘半乳糖凝集素-3并上调TNF-α、IL-8和环氧合酶2等炎症因子,直接或间接诱发早产。
体外研究显示,该菌可侵入人绒毛外滋养层细胞,诱导G1期阻滞与凋亡,可能涉及ERK1/2及DNA损伤反应通路;p53/p38与JNK介导的HSP27/p21通路等复杂信号网络亦可能参与其中。此外还提出其可通过侵入与免疫逃逸在子宫-胎盘组织持续存活,促发多微生物失调并与共生菌协同增加母胎界面微生物负荷,上调CRP、戊曲素3等急性期蛋白,并伴随活性氧过量、胎儿肾上腺皮质激素增加及胎儿应激等改变。
♥
鉴于牙龈卟啉单胞菌(Porphyromonas gingivalis)与牙周炎及多种疾病之间存在密切联系,对该菌进行控制或干预,被视为一种切实可行的疾病管理策略,并为相关疾病的防治提供新的切入点。
▸ 经典临床策略:控制菌斑与靶向抗菌
•机械清创 + 抗生素/抗菌剂:龈下刮治联合阿莫西林、甲硝唑、阿奇霉素等可降低P.gingivalis负荷,但临床获益并非总与菌量下降成正比,提示疾病是多因素的。
•氯己定及光动力疗法:可减少P.gingivalis计数和炎症,被认为是有效辅助方法。
▸ 日常居家护理:减少牙菌斑
•刷牙:每天两次用含氟牙膏刷牙和牙缝清洁(牙线、牙间刷)会破坏牙龈卟啉单胞菌定殖所需的牙菌斑基质。
•使用漱口水:白藜芦醇漱口水在与牙龈卟啉单胞菌相关病例中,配合使用时显著改善了口臭和临床指标。
▸ 饮食:减少P.gingivalis定植
•多酚(绿茶、蔓越莓、其他植物来源):绿茶、多酚和蔓越莓含有原花青素,可以抑制牙龈卟啉单胞菌的致病因子,如牙龈炎和生物膜形成。
•地中海风格、富含油酸的饮食:在老年小鼠中,富含油酸(典型橄榄油)的饮食相比富含棕榈酸的饮食,减少了牙龈卟啉单胞感染的全身性炎症后果,并减轻了骨质流失。支持采用地中海饮食(橄榄油、坚果、蔬菜、鱼类)来缓冲系统性影响。
•天然多酚/植物化学物:姜黄素、白藜芦醇、槲皮素等可抑制生长、黏附和生物膜,并显著下调黏附素和gingipain等毒力基因。
▸ 益生菌与疫苗
•Lactobacillus casei DS31:体外实验中,DS31细菌或游离上清液显著抑制了牙龈卟啉单胞菌生物膜的形成,并帮助消除了现有的生物膜;在牙周炎模型中,DS31缓解了炎症和组织损伤。
•其他乳杆菌和双歧杆菌菌株:系统综述显示,多种乳杆菌菌株可以减少P.gingivalis数量或干扰其粘连,尽管临床数据仍有限;双歧杆菌附着于龈下生物膜,并在体外减少牙龈卟啉单胞菌的生长。不过不同乳杆菌株在抑制P.gingivalis和炎症上具有菌株特异性。
•疫苗与免疫增强:以P.gingivalis纤维蛋白、外膜蛋白、热休克蛋白及gingipain为抗原的黏膜疫苗在动物实验中能阻断P.gingivalis介导的破坏性免疫反应,被视为长期预防的重要方向。
▸ 抑毒力与生物膜的创新策略
•纳米递药与植物外泌体:
抗体修饰脂质体包载人参皂苷Rh2可定向清除P.gingivalis,维持口腔菌群稳态。
姜来源外泌体样纳米颗粒被P. gingivalis特异摄取,可干扰多种毒力因子并在小鼠中减轻牙周炎。
•群体感应/酶学“抗毒力”药物:
AHL内酯酶Est816通过“干扰群体感应”显著破坏P.gingivalis生物膜、下调毒力基因并降低炎症,无明显细胞毒性。
针对P.gingivalis谷氨酰环化酶(PgQC)的小分子S‑0636可显著降低gingipain活性和上皮侵袭性,且不杀菌、不扰乱共生菌,也未见耐药产生。
直接gingipain抑制剂在阿尔茨海默病模型中可减少脑内P. gingivalis负荷、Aβ生成和神经炎症,提示有望降低其全身危害。
♥
基于上述认识,如何检测自身P.gingivalis的水平与变化,并据此进行针对性的调控或干预,以降低或减轻其带来的危害,已逐渐成为当前的重要研究方向与关注重点。现有策略主要分四类:
一、口腔卫生管理与牙周基础治疗以降低生物负荷;
二、必要时联合抗生素控制感染;
三、开发靶向关键毒力因子(尤其是gingipains)的小分子抑制剂以削弱致病力;
四、探索基于特定抗原结构的免疫预防与疫苗方案。
主要参考文献
How KY, Song KP, Chan KG. Porphyromonas gingivalis: An Overview of Periodontopathic Pathogen below the Gum Line. Front Microbiol. 2016 Feb 9;7:53.
Mysak J, Podzimek S, Sommerova P, Lyuya-Mi Y, Bartova J, Janatova T, Prochazkova J, Duskova J. Porphyromonas gingivalis: major periodontopathic pathogen overview. J Immunol Res. 2014;2014:476068.
Xu W, Zhou W, Wang H, Liang S. Roles of Porphyromonas gingivalis and its virulence factors in periodontitis. Adv Protein Chem Struct Biol. 2020;120:45-84.
Curtis MA, Garnett JA, Darveau RP. The Keystone-Pathogen Hypothesis Updated: The Role of Porphyromonas gingivalis in Periodontitis. J Periodontal Res. 2025 Oct 31.
Lunar Silva I, Cascales E. Molecular Strategies Underlying Porphyromonas gingivalis Virulence. J Mol Biol. 2021 Apr 2;433(7):166836.
Lv C, Shi K, Guo Z, Luo P, Wu Z, Yu P. Porphyromonas gingivalis-derived outer membrane vesicles travel from oral to distant organs via blood circulation in a time-dependent manner. BMC Oral Health. 2026 Feb 27.
Mei F, Xie M, Huang X, Long Y, Lu X, Wang X, Chen L. Porphyromonas gingivalis and Its Systemic Impact: Current Status. Pathogens. 2020 Nov 13;9(11):944.
Murugaiyan V, Utreja S, Hovey KM, Sun Y, LaMonte MJ, Wactawski-Wende J, Diaz PI, Buck MJ. Defining Porphyromonas gingivalis strains associated with periodontal disease. Sci Rep. 2024 Mar 14;14(1):6222.
Dominy SS, Lynch C, Ermini F, Benedyk M, Marczyk A, Konradi A, Nguyen M, Haditsch U, Raha D, Griffin C, Holsinger LJ, Arastu-Kapur S, Kaba S, Lee A, Ryder MI, Potempa B, Mydel P, Hellvard A, Adamowicz K, Hasturk H, Walker GD, Reynolds EC, Faull RLM, Curtis MA, Dragunow M, Potempa J. Porphyromonas gingivalis in Alzheimer’s disease brains: Evidence for disease causation and treatment with small-molecule inhibitors. Sci Adv. 2019 Jan 23;5(1):eaau3333.
Zhang L, Tian H, Ma Y, Xu J, Guo C, Wang Z, Ma J. Periodontitis pathogen Porphyromonas gingivalis promotes chronic obstructive pulmonary disease via affecting neutrophils chemotaxis and function. Int J Oral Sci. 2026 Jan 9;18(1):4.
Olczak T, Simpson W, Liu X, Genco CA. Iron and heme utilization in Porphyromonas gingivalis. FEMS Microbiol Rev. 2005 Jan;29(1):119-44.
Huang Z, Hao M, Shi N, Wang X, Yuan L, Yuan H, Wang X. Porphyromonas gingivalis: a potential trigger of neurodegenerative disease. Front Immunol. 2025 Feb 14;16:1482033.
Zhao X, Liu J, Zhang C, Yu N, Lu Z, Zhang S, Li Y, Li Q, Liu J, Liu D, Pan Y. Porphyromonas gingivalis exacerbates ulcerative colitis via Porphyromonas gingivalis peptidylarginine deiminase. Int J Oral Sci. 2021 Sep 30;13(1):31.

谷禾健康
肌醇(Inositol,i-Inositol或myo-Inositol)
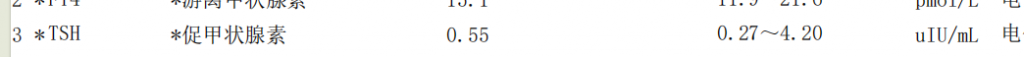
肌醇,它不仅是构成细胞膜的基础材料,更是一个连接新陈代谢、细胞信号和肠道微生态的核心枢纽。在其九种立体异构体中,主要以其特定形式(myo-inositol)参与广泛的生理过程,包括构成细胞膜磷脂、介导胰岛素信号、调节渗透压、基因表达等。
近年来,研究人员发现,肌醇代谢的紊乱正日益被视为多种疾病的促成因素,如代谢综合征、2型糖尿病、多囊卵巢综合征、神经退行性疾病如阿尔茨海默病乃至皮肤健康。
近期证据同时凸显,肠道菌群在塑造肌醇的生物可利用性和生物转化中扮演着决定性角色。有益的共生菌群(如Anaerostipes)能将膳食肌醇转化为促进健康的短链脂肪酸,而与肥胖相关的微生物则会过度降解肌醇,从而促进脂质吸收和脂肪堆积。
本文将一起整理讨论关于肌醇的以下方面知识:
1)肌醇:从化学结构到生理功能
2)肌醇缺乏或过量的影响
3)调控其合成、分解代谢与利用的机制
4)肌醇与肠道菌群的相互作用
5)影响肌醇代谢的因素
6)肌醇与代谢性疾病
7)肌醇与神经精神健康
8)肌醇与皮肤健康
9)诊断指标与临床相关性
10)恢复代谢稳态的循证营养与治疗策略
注:肌醇(Inositol) 是一个总称,它有多种异构体(分子式相同但结构不同)。
肌醇(myo-inositol) 是这些异构体中最常见、在自然界中分布最广、生物活性也最重要的一种形式。我们通常所说的“肌醇”,在文中没有特别指明的情况下,往往就是指这种形式。
游离myo-Ins 就是指不与其他分子结合、以独立形态存在的肌醇。
关于肌醇的研究发展
在过去二十年里,我们对肌醇的认识发生了深刻的转变。肌醇已从一个仅仅被视为“类维生素”的化合物,转变为一个深度参与葡萄糖和脂质调节的多功能信号分子。尽管有时被称为“维生素B8”,但肌醇并非必需维生素,因为人体自身就能以葡萄糖为原料(通过葡萄糖-6-磷酸经肌醇-3-磷酸合酶催化),实现内源性合成。
肌醇的重要性体现在其作为细胞信号的核心角色上。肌醇及其立体异构体D-手性肌醇(D-chiro-inositol, DCI)一起,充当着响应胰岛素的“第二信使”,负责传达葡萄糖摄取、糖原合成和脂肪代谢等关键指令。持续的研究发现,在糖尿病和胰岛素抵抗的状态下,这两种信使水平会显著下降。
与此同时,一个更前沿的发现是,肠道菌群对我们体内的肌醇水平有着深刻影响。肠道菌群不仅通过代谢膳食肌醇,还能将其转化为短链脂肪酸(SCFAs)。因此,理解肌醇、宿主代谢、肠道微生物群之间的这种三方互动,为个体化营养和精准医学提供了全新的视角。
肌醇的化学性质与合成
结构与立体化学
肌醇是一种结构类似于葡萄糖的天然多羟基环己六醇(C₆H₁₂O₆),形成一个稳定的六元环,每个碳原子上均带有一个羟基。
正是这些羟基在空间中朝向的不同,造就了肌醇的多种立体异构体。在其九种可能的立体异构体中,肌醇(myo-inositol)在哺乳动物组织中占比超过99%。每一种异构体都因其独特的空间结构,而具备了不同的生化活性和信号传导能力。
肌醇九种异构体的结构
生物合成途径
人体能够自行合成肌醇,内源性生物合成主要发生在肾脏、肝脏和大脑,这个过程始于我们最熟悉的能量物质——葡萄糖,通过一个两步酶促转化过程完成:
该途径受到葡萄糖可利用性、胰岛素水平和渗透压应激的严密调控,确保按需生产。
膳食来源
肌醇以游离myo‑Ins 和其结合形式植酸(肌醇六磷酸, IP6)两种形式存在,后者在以下食物中含量丰富:
加工过程(如浸泡、发酵)会显著影响其生物可及性,因为植酸酶能将植酸降解为可吸收的形式。
肌醇的生理功能
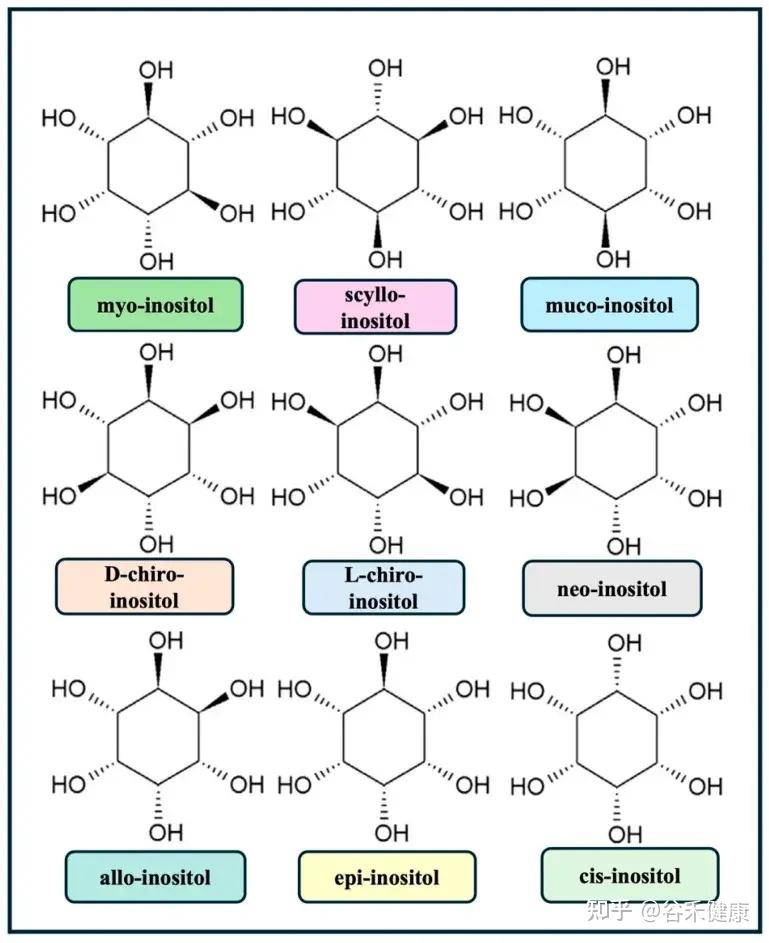
肌醇:细胞信号转导
肌醇是细胞信号传导通路中的关键一环。在细胞膜上,它的衍生物磷脂酰肌醇(如 PI(4,5)P₂)就像是为关键蛋白预留的停靠位点。
当胰岛素或激素(第一信使)敲响细胞的大门时,它自己不进去,而是派 IP₃(第二信使)在细胞内部跑腿传达命令。IP₃ 的一个重要任务就是去打开钙离子的阀门,而钙离子就像是启动细胞各种功能的电火花。
如果这个信号系统失灵会出现胰岛素抵抗,也就是说你吃了东西,胰岛素(第一信使)发出了指令,但细胞内部(第二信使)没反应过来,导致血糖降不下去,人会饭后特别困、容易饿、精力不济,并且脂肪更容易堆积。
渗透压调节:细胞的脱水危机与肌醇的救援
为了回收水分,我们的肾脏会创造一个盐分极高的“高渗环境”,普通细胞在这种环境下会像腌菜一样脱水。
肌醇(Myo-Ins)此时便作为一种有机渗透调节物,被 SMIT1 等转运蛋白大量泵入肾脏细胞内,提升细胞内部浓度,以对抗外界的高渗应激。这相当于给细胞喝足了保护液,防止其在极端环境下功能失常。
肌醇与脂肪肝:从源头阻断脂肪堆积
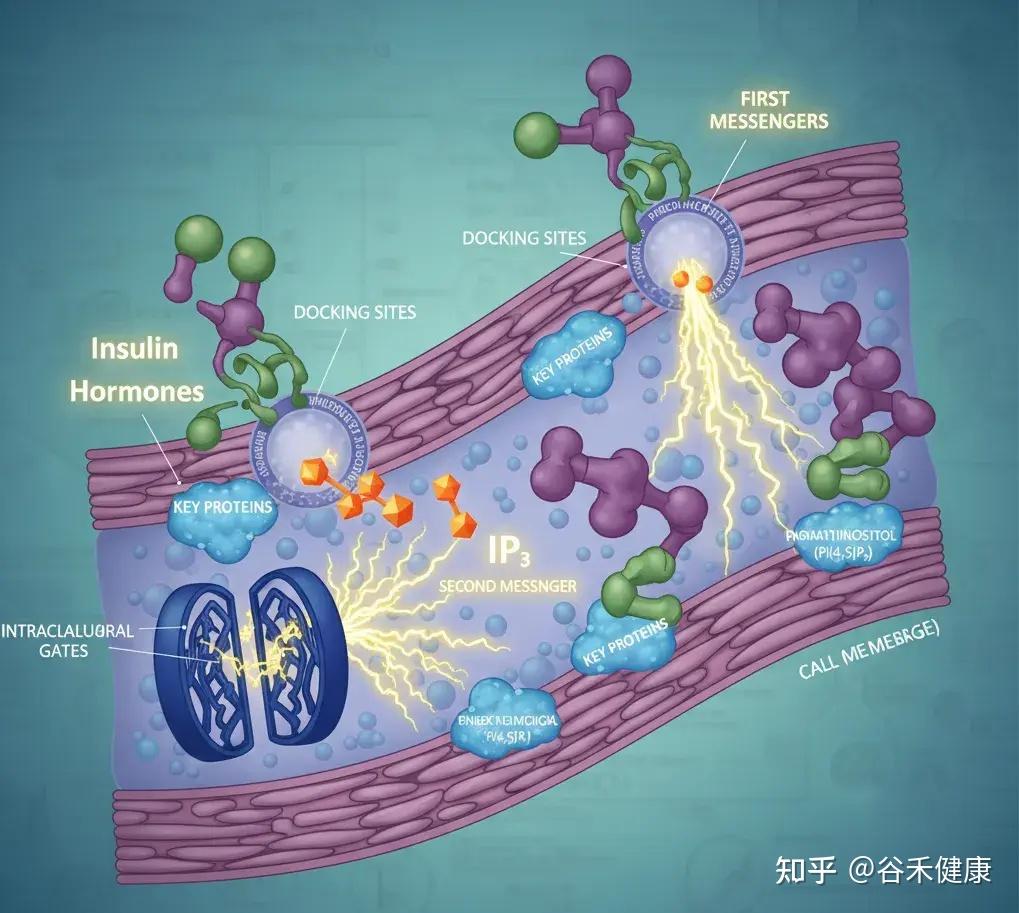
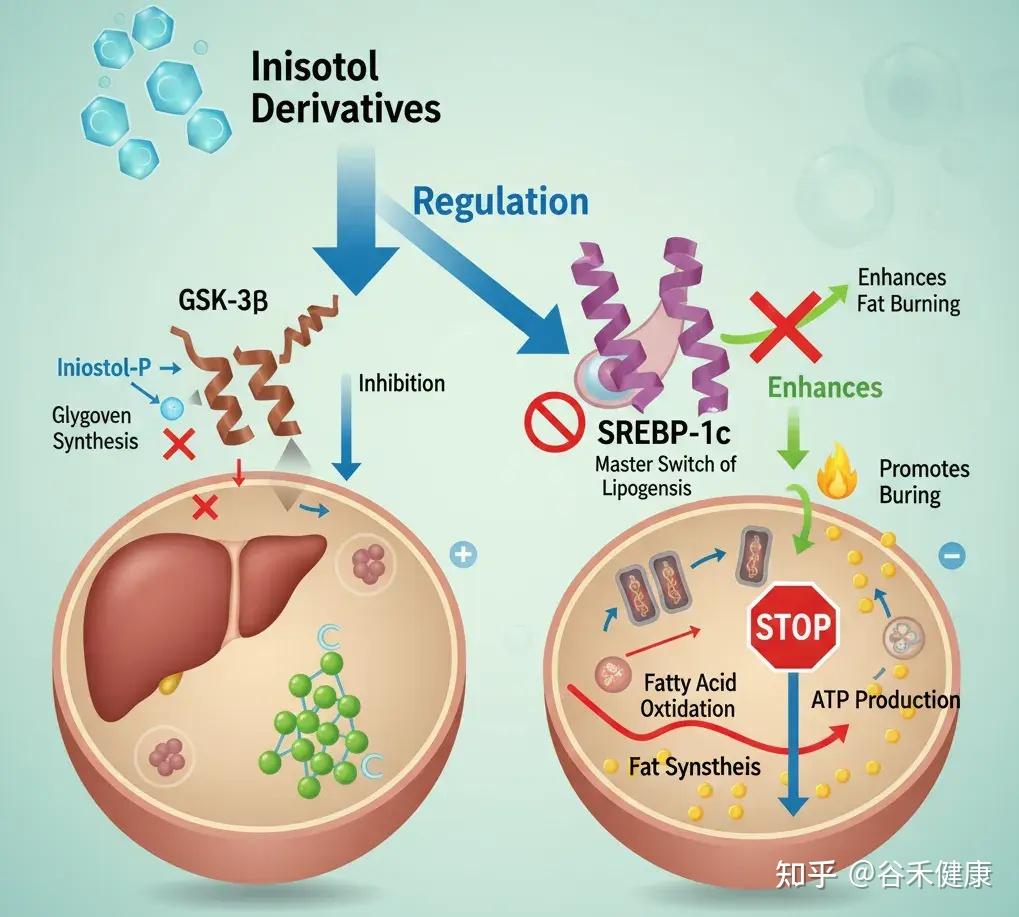
肌醇衍生物能精准调控肝脏和脂肪细胞内的关键酶,如糖原合酶激酶-3β(GSK-3β)和固醇调节元件结合蛋白-1c(SREBP-1c)。
例如,它能抑制 SREBP-1c 这个脂肪生成总开关的活性。当 SREBP-1c 过于活跃时,肝脏就会不停地合成脂肪。同时,肌醇还能增强脂肪酸氧化,也就是促进脂肪的燃烧。
当这个开关失控时,SREBP-1c 持续开启,会导致肝脏甘油三酯堆积,在体检报告上就表现为非酒精性脂肪肝。反映在体型上,就是腹部脂肪顽固、向心性肥胖。
肌醇:备孕路上的好助手
在女性体内,myo-肌醇调节卵泡刺激素(FSH),确保卵巢能准确接收指令,促进卵母细胞的正常成熟,这解释了为何补充肌醇能有效改善多囊患者的排卵状况和月经周期。
肌醇在男性精浆中的浓度非常高,它对精子的正常生理功能起着关键作用。补充肌醇,特别是肌醇(Myo-inositol),已被证明可以显著改善精子的活力、数量和形态。主要通过增强线粒体功能、提供能量和抗氧化作用。这使肌醇成为治疗男性不育的有效营养方案。
肌醇:如何稳定你的情绪?
在中枢神经系统中,肌醇衍生物参与磷脂酰肌醇循环,这与血清素受体活性、情绪稳定和神经保护有关。血清素是决定我们情绪的“快乐激素”。如果这个循环不畅,大脑就无法有效利用血清素,就会出现情绪波动大、容易焦虑、烦躁或感到莫名的情绪低落等问题。
平衡总是一体两面的。如果说缺乏肌醇会打破身体的稳定,那么,过量的肌醇就一定更好吗?接下来,我们就将深入探讨肌醇在体内的失衡状态——即缺乏与过量,分别会带来怎样的病理生理学后果。
缺乏机制
肌醇耗竭的产生途径包括:
临床表现
虽然不存在“经典的肌醇缺乏症”,但经验证据表明,组织中myo‑Ins水平低下与以下情况有关:
潜在毒性与过量
高达每日12克的剂量通常是安全的,仅有轻微的胃肠道不适报告。
当口服补充剂量很大时(通常指每日超过12克),肌醇可能无法被小肠完全吸收。未被吸收的肌醇在肠道内会产生渗透效应,像吸水海绵一样将水分吸入肠腔,从而导致轻微的胃肠道不适,如腹胀、排气和腹泻。
极高浓度可能扰乱渗透压平衡,尤其是在肾功能不全的患者中。
肌醇在人体内的转化与胰岛素信号
这里要介绍肌醇的两种异构体形式:
肌醇(Myo-Ins)和D-手性肌醇(DCI)。
生理转化(Myo ➜ DCI)
Myo‑Ins通过一种胰岛素依赖性的表异构酶(epimerase)转化为D-手性肌醇(DCI)。
myo/DCI的比例在不同组织中有所不同,例如:
这种差异化分布支持了组织特异性的胰岛素作用。
然而,在胰岛素抵抗的病理状态下,如在糖尿病和多囊卵巢综合征(PCOS)中,这种转化机制会受损。
胰岛素抵抗中的失衡
表异构酶活性的不足,导致myo‑Ins耗竭和DCI生成不足,损害了糖原合成和细胞葡萄糖摄取。
研究表明,补充肌醇(myo‑Ins)能有效恢复两种异构体的平衡,而单独补充DCI,则可能破坏生理比例,反而达不到理想的治疗效果。
肌醇在胰岛素信号传导中的作用机制
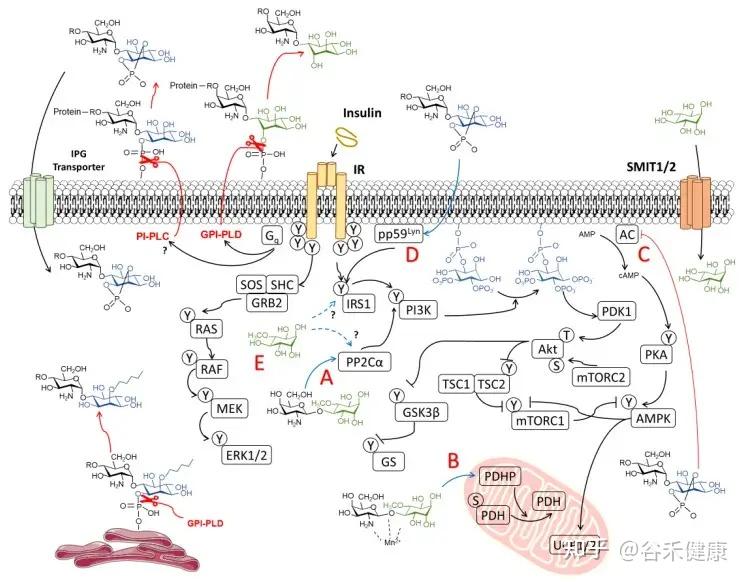
肠道菌群对肌醇的降解:诱发肥胖的关键机制
肠道菌群:肌醇代谢的隐形参与者
新兴证据正在引领我们对宿主-微生物协同代谢的理解。肠道菌群可以直接代谢膳食myo-肌醇,从而影响宿主的能量平衡。
Anaerostipes属等菌种能将肌醇转化为丙酸和乙酸,这两种主要的短链脂肪酸参与抑制肝脏糖异生和改善胰岛素敏感性。
相反,与肥胖相关的菌种(例如,高脂饮食下富集的某些厚壁菌门成员)表现出降解肌醇的活性,耗尽肠腔内的myo-Ins,并增强肠道对脂质的吸收。
肌醇缺乏,如何导致肥胖?
最新研究揭示了一个事实:肠道微生物可以降解肌醇,而这种降解与肥胖密切相关。
研究人员在肥胖人群中发现了一种名为Megamonas rupellensis(巨单胞菌代表物种)的细菌显著富集。这种细菌具有强大的肌醇降解能力,其降解途径(PWY-7237)及相关基因(iolG、iolI、iolE、iolD、iolB)在肥胖个体中丰度和表达水平均显著升高。
肌醇的抗肥胖作用
在正常情况下,肌醇能够通过下调几个基因表达,有效抑制小肠对脂肪的吸收。
肌醇下调的是三个关键脂肪酸转运蛋白基因:
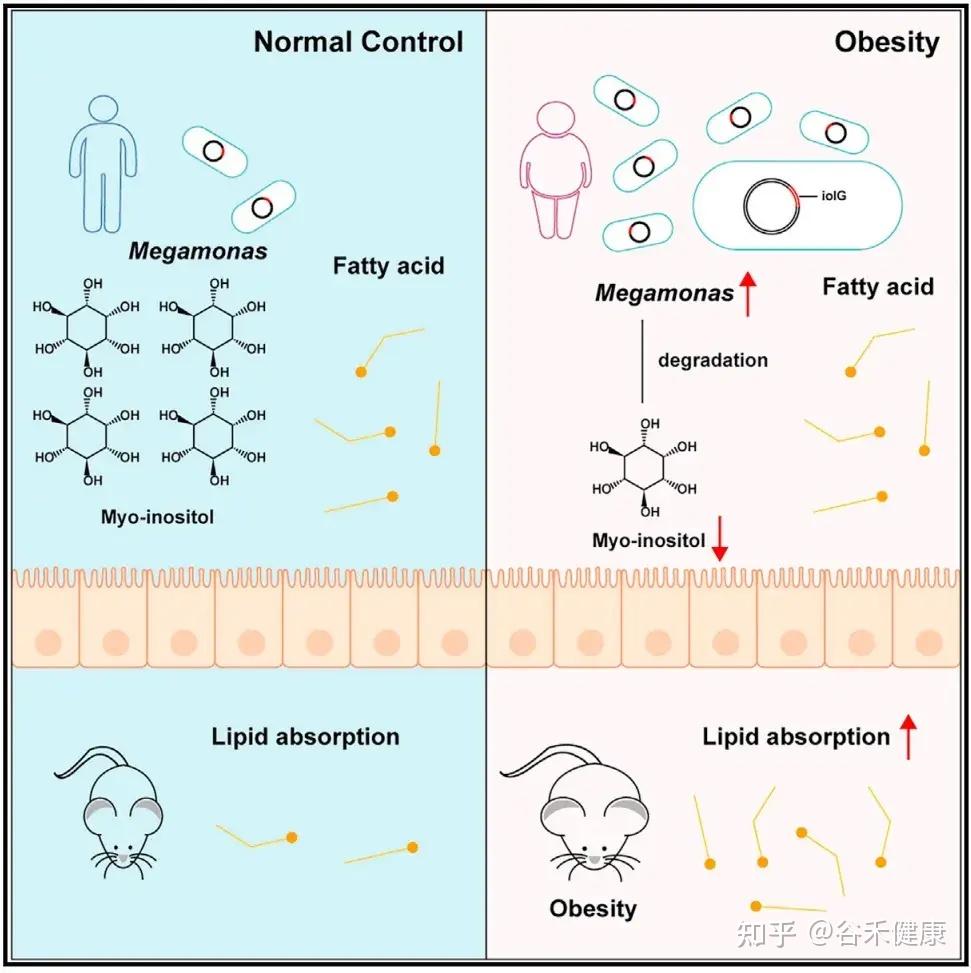
菌群降解肌醇的后果
当M. rupellensis 定植在小鼠肠道后,它会大量消耗肌醇,导致肠道内的myo-肌醇水平几乎耗尽。这直接削弱了肌醇抑制脂肪吸收的功能,最终引发一系列肥胖表型:肠道脂质吸收增加、体重与脂肪增加、肝脏脂肪堆积以及血糖升高等。
研究人员进一步进行了一项巧妙的实验:
仅将肌醇降解的第一个关键酶基因(iolG)转入原本不能降解肌醇的大肠杆菌中,就足以复现上述肥胖现象:体重增加、脂肪组织增多、肝脏脂肪堆积。这证明了:
肌醇降解本身(而非其他细菌因素),是导致肥胖的直接原因。
人体肠道内微生物肌醇及植酸降解推导途径
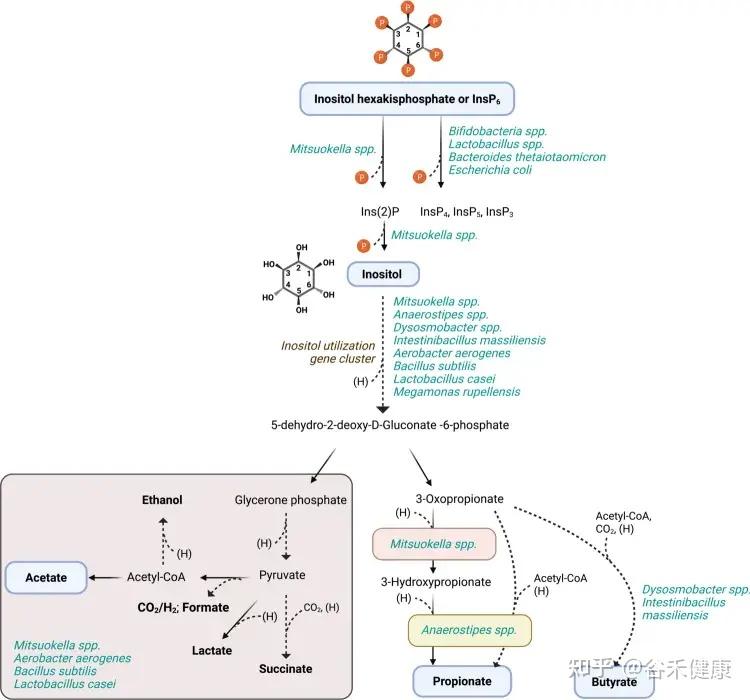
肠道菌群的另一面:合成有益的肌醇脂质
并非所有细菌都是反派。一些共生菌,特别是拟杆菌属,实际上能利用肌醇来合成对自身和宿主都有益的——肌醇脂质。
例如,人类肠道中的常驻菌多形拟杆菌(Bacteroides thetaiotaomicron)拥有完整的肌醇脂质合成基因簇。这些脂质能够帮助细菌抵抗宿主的抗菌肽,从而更好地在肠道中定植。
这体现在功能保护和结构支撑两方面:
然而,这个过程的影响远不止于细菌自身。研究发现,一旦细菌失去合成肌醇脂质的能力,会直接扰乱宿主的基因表达,特别是那些与碳水化合物降解和能量合成相关的关键代谢通路,这表明菌群的这一行为与宿主的能量平衡息息相关。
更重要的是,这背后还隐藏着一个精巧的生态循环系统。
我们摄入的食物中含有植酸(比如小麦、糙米、藜麦、玉米、花生、大豆、芝麻等),肠道中广泛存在的植酸酶,可以降解食物中的植酸,释放出肌醇。这些被释放的肌醇并不会被浪费,随后可被拟杆菌等细菌重新利用,作为原料去合成上述的肌醇脂质。
这一发现具有重要的临床意义:
不仅揭示了这种有益的合成能力在肠道菌群中远比我们想象的更为普遍,也展示了一个复杂的“宿主-微生物-肌醇”循环系统。
肌醇的降脂效果,要看肠道菌群的脸色
肌醇的代谢调控能力还体现在一个更宏观的战场上——直接对抗由现代不良饮食习惯(尤其是高糖饮食)引发的代谢紊乱。一个关键的动物研究,为我们深入揭示了肌醇是如何与我们的肠道菌群联手,共同调控肝脏脂肪代谢的。
高蔗糖饮食的影响
研究证实,高蔗糖饮食可触发一系列级联负面效应,涵盖肝脏代谢、基因转录及肠道微生态。
核心指标:肝脏甘油三酯水平显著升高(这是通往脂肪肝的快车道)。
伴随变化:肝脏总脂质、胆固醇含量及肝脏重量均呈现上升趋势,表明脂肪变性的发生。
上游通路激活:果糖代谢关键酶酮己糖激酶(KHK)与糖酵解关键酶丙酮酸激酶(PKLR)的表达被显著上调,为脂肪酸合成提供了充足的底物。
新生脂肪合成全面上调:涉及脂肪生成的全套关键基因表达均上调,包括:
核心转录因子激活:脂肪合成的主调控因子 SREBP1 与 ChREBP 的mRNA水平显著升高,从转录层面驱动了整个脂肪生成程序。
注:SREBP1(固醇调节元件结合蛋白1);
ChREBP(碳水化合物反应元件结合蛋白)。
肠道微生物组失调:
α-多样性显著降低:菌群的物种丰富度和系统发育多样性下降,生态系统稳定性减弱。
β-多样性显著变化:菌群的整体结构发生重组,偏离了健康状态。
特定菌属丰度改变:
削弱了有益菌(如Lactobacillus, Bifidobacterium),打击了负责肠道屏障的AKK菌,纵容了可能引发炎症或与代谢病相关的菌群(如 Desulfovibrio)的过度生长。
在这一片混乱中,肌醇的补充扮演了力挽狂澜的角色。
肌醇补充的干预效果
肌醇补充显著逆转了肝脏中甘油三酯和胆固醇的积累。
此作用具有选择性,因为肝脏的磷脂水平未发生变化,表明干预主要靶向异常的脂质沉积。
肌醇显著抑制了脂肪生成通路中的关键限速酶(尤其是FAS)的表达。
更重要的是,它显著降低了主转录因子 SREBP1 和 ChREBP 的mRNA水平,从源头上削弱了脂肪生成的转录驱动力。
同时,它减弱了糖酵解相关酶的上调趋势,减少了流向脂肪合成的碳水化合物底物。
肠道菌群:关键中介及潜在生物标志物
该研究最关键的发现之一,在于揭示了肌醇并非在孤军奋战,它的降脂效果,高度依赖肠道菌群这个关键盟友。
研究建立了一个重要的关联模型——乳杆菌属丰度的降低与拟杆菌属丰度的增加,这两项指标与血浆脂质升高及肝脏甘油三酯累积存在显著的正相关性。
这一结论提示,特定的肠道微生物特征可以作为评估宿主-微生物共代谢中肌醇代谢效率的有效生物标志物,为未来临床评估和干预提供了新的视角。
事实上,肌醇在体内的整个代谢网络,都受到一系列内在和外在因素的精密调控,任何一个环节的改变都可能影响其最终的生物学效应。接下来,我们将探讨哪些因素在影响着肌醇代谢。
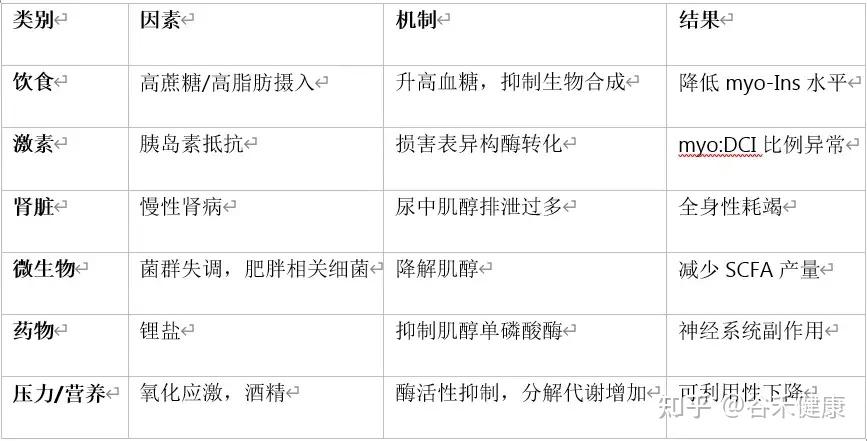
肌醇的体内平衡受多种因素调控,失衡会导致功能性缺乏。以下是五个主要影响因素:
高糖与高脂饮食
这是最核心的干扰因素,通过四种方式产生负面影响:
– 竞争抑制
葡萄糖与肌醇结构相似,当血糖水平过高时,葡萄糖会抢占细胞为肌醇准备的转运通道,导致细胞内的肌醇供应不足。
– 加速流失
高血糖导致糖尿时,会顺带让大量肌醇从尿液中流失,造成体内总量下降。
– 抑制合成
长期高糖饮食会抑制体内合成肌醇的关键酶活性,减少内源性供应。
– 扰乱菌群
高糖高脂环境促使肠道中有害菌增殖,这些菌会过度消耗肌醇,或阻止其向有益的短链脂肪酸转化。
胰岛素抵抗
胰岛素抵抗既是肌醇失衡的结果,也是其恶化的推手。
– 转化障碍
胰岛素信号失灵,身体将“基础版”的肌醇(myo-肌醇)转化为其重要异构体“D-手性肌醇”(DCI)的能力会下降,影响糖原合成等功能。
– 组织失衡
在某些组织(如卵巢)中,身体为了代偿胰岛素抵抗而分泌更多胰岛素,造成“高胰岛素血症”。这反而会过度消耗myo-肌醇,导致两种异构体比例失调,这也是多囊卵巢综合征(PCOS)的核心病理机制之一。
肠道菌群失调
肠道菌群扮演着“双刃剑”的角色。
有益菌:像 Anaerostipes 属的细菌,能将肌醇转化为丙酸、乙酸等有益的短链脂肪酸,帮助改善整体代谢。
有害菌:与肥胖相关的菌群则会直接吞噬肌醇,减少其在体内的可用量,并可能促进脂肪吸收,进一步加剧代谢问题。
肾脏功能障碍
肾脏是回收肌醇、防止其流失的关键器官。在健康状态下,肾脏会重吸收超过99%被过滤的肌醇。然而,当患有慢性肾病或糖尿病肾病时,肾脏的重吸收功能受损,会导致大量肌醇随尿液排出,引起全身性的耗竭。
药物与生活方式
药物影响:例如,用于治疗双相情感障碍的锂盐,会抑制肌醇在细胞内的循环再生,导致功能性缺乏。
生活方式:长期饮酒和慢性压力会持续增加肌醇的代谢消耗,同时干扰其在肝脏和神经系统中的正常功能。
以上我们了解到肌醇的体内平衡,易受到现代生活方式和普遍健康问题的干扰。这种平衡的打破,绝非一个简单的营养素缺乏问题,它构成了多种复杂疾病发生与发展的核心病理生理基础。
多囊卵巢综合征
多囊卵巢综合征(PCOS)是育龄女性最常见的内分泌与代谢紊乱疾病。虽然其典型症状如月经不调、高雄激素、卵巢多囊样改变——都指向生殖系统,但其病理根源远不止于此。PCOS的本质,是一种与胰岛素抵抗及代偿性高胰岛素血症密切相关的全身性代谢失调。
在这一背景下,肌醇作为一种关键营养素,其代谢紊乱被发现是PCOS病程中的核心环节。这引出了一个重要概念:PCOS患者普遍存在一种“功能性肌醇耗竭”。这并非指吃得不够,而是细胞层面无法有效利用肌醇,尤其是其两种关键异构体——肌醇(myo-Ins)和D-手性肌醇(DCI)。
两种肌醇,两大职责
在健康人体内,胰岛素如同“总指挥”,精确调控着两种肌醇的转化与功能。它们分工明确:
肌醇失衡:与PCOS有重要关联
在PCOS患者中,普遍存在的胰岛素抵抗破坏了这个过程。
细胞对胰岛素不敏感(即胰岛素抵抗),导致将 myo-Ins 转化为 DCI 的酶活性不足。这使得细胞无法有效利用葡萄糖,进一步加剧高血糖和高胰岛素血症。
矛盾的是,卵巢对胰岛素依然高度敏感。全身循环中的高水平胰岛素在卵巢过度激活了转化酶,导致卵巢内的 myo-Ins 被大量、过早地消耗,转化为 DCI。
这就造成了卵巢微环境中灾难性的后果:myo-Ins 严重相对缺乏,而 DCI 严重相对过量。
肌醇失衡如何破坏生育力?
这种卵巢内的肌醇失衡直接导致了PCOS的两大核心生殖障碍:
作为FSH信号的关键信使,myo-Ins的耗竭损害了卵泡对FSH的反应。卵泡在募集后无法发育成优势卵泡,最终发育停滞,形成了一串“小囊泡”——即卵巢的多囊样改变。同时,myo-Ins的缺乏也直接影响卵母细胞的质量,导致受孕困难和流产风险增加。
卵巢内过量的DCI会反过来刺激卵泡膜细胞,使其对胰岛素和促黄体生成素(LH)的信号反应过度,从而过量合成雄激素。
治疗思路:补充肌醇,打破不良循环
理解了肌醇原因,那么外源性补充肌醇的逻辑便十分清晰了。其目的正是为了纠正卵巢内的营养失衡,打破恶性循环。
临床研究发现,单独补充大剂量DCI效果不佳甚至可能有害。因为它无法解决卵巢中真正耗竭的myo-Ins,反而可能加剧失衡比例。
直接喂养处于“饥饿”状态的卵巢,恢复其对FSH信号的敏感性,促进卵泡正常发育与排卵。
从根源改善:在全身层面改善胰岛素敏感性,降低血清胰岛素水平,从而减轻对卵巢转化酶的过度刺激。
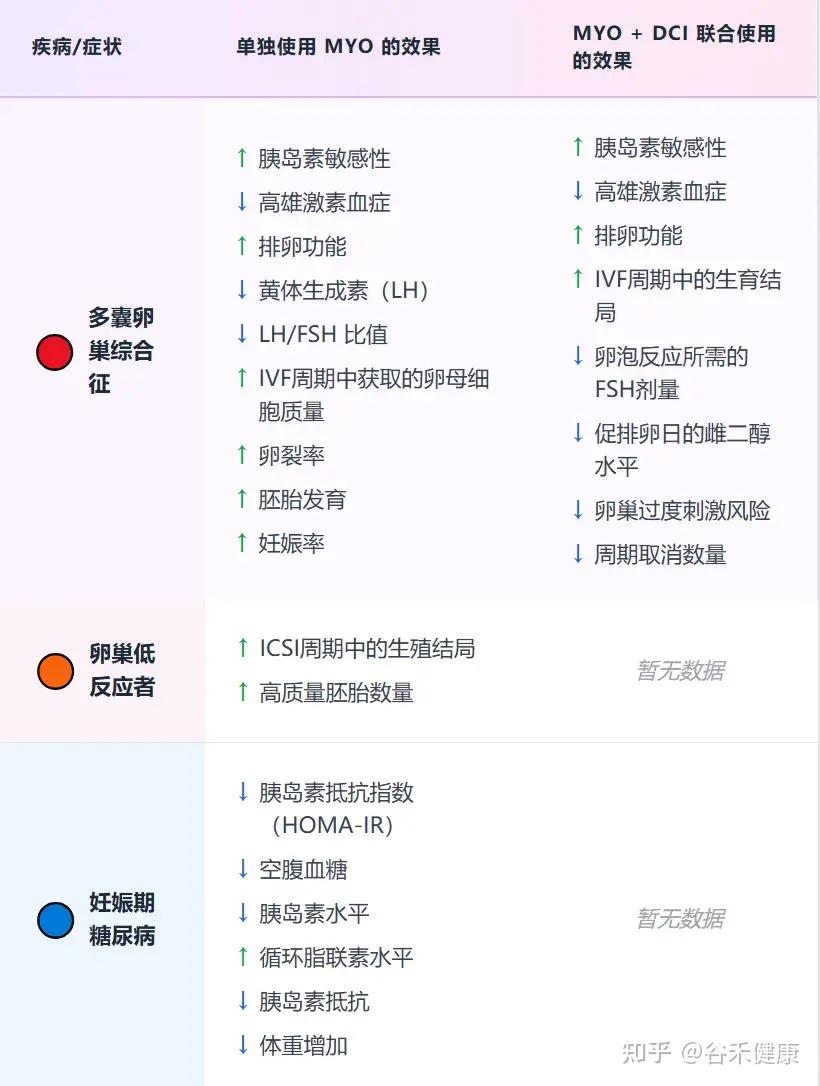
目前,大量高质量的临床研究证实,补充肌醇,特别是以生理比例40:1混合的myo-Ins和DCI,能够显著改善PCOS患者的内分泌和代谢指标,包括:
糖尿病
在糖尿病及其前驱状态(胰岛素抵抗)的复杂病理中,肌醇代谢的紊乱扮演着一个核心角色,它既是后果又是驱动因素,形成了一个自我强化的恶性循环。
高血糖如何耗尽肌醇?
高血糖通过“入口”和“出口”两个维度,造成了体内肌醇的严重耗竭。
葡萄糖与肌醇的化学结构高度相似,它们共享细胞膜上的部分转运通道,尤其是钠/myo-肌醇协同转运体(SMIT)。
当血糖持续升高时,血液中过量的葡萄糖霸占转运通道。这种竞争性抑制作用,极大地阻碍了细胞对肌醇的有效吸收。这就导致了一个矛盾的现象:即使血液中的肌醇浓度尚可,真正需要它的细胞(如神经、肝脏、肾脏细胞)内部却已处于饥饿状态,即功能性耗竭。
高血糖对肾脏也构成了巨大压力。当血糖超过肾脏的处理极限时,多余的葡萄糖会随尿液排出(即糖尿)。由于转运通道被葡萄糖占据,本应被肾脏回收的肌醇也同步受损,大量地随尿液流失。这直接削减了身体的肌醇总储备,是糖尿病患者体内肌醇水平显著偏低的关键原因。
肌醇耗竭如何反噬代谢?
当肌醇耗竭发生后,它会对糖尿病的病理产生关键的反噬作用,让情况变得更糟。
肌醇是胰岛素信号通路中不可或缺的第二信使。可以把它想象成信号传导链条上的一个关键齿轮。当细胞内肌醇水平下降,这个齿轮就脱落了,导致信号中断,加剧胰岛素抵抗。
也就是说,胰岛素虽然与细胞受体结合了,但信号无法有效向下游传递(无法有效激活下游的PI3K/Akt等关键激酶)。这使得负责将葡萄糖运入细胞的“大门”(GLUT4转运蛋白)无法正常打开,细胞摄取和利用葡萄糖的能力大幅下降。这直接加重了胰岛素抵抗。
恶性循环的闭环
至此,一个恶性循环正式形成:
高血糖 ➔ 抑制肌醇吸收 & 加速其流失 ➔ 细胞内肌醇耗竭 ➔ 胰岛素信号链中断 ➔ 胰岛素抵抗加剧 ➔ 血糖进一步升高
这个循环一旦启动,便会不断自我强化,使血糖控制变得愈发困难。
连锁反应:损害远不止于血糖
肌醇的缺乏会间接促进肝脏的脂肪生成,并导致血脂异常,从而显著增加糖尿病患者并发脂肪肝和心血管疾病的风险。
肌醇对维持神经细胞的渗透压平衡和正常信号传导至关重要。其长期耗竭是导致糖尿病周围神经病变(如手脚麻木、刺痛感)的核心原因之一。
因此,肌醇在糖尿病中的角色是一个从宏观器官功能(肾脏流失)到微观细胞信号(胰岛素抵抗)的系统性障碍。理解这一恶性循环,为通过补充肌醇来干预和改善糖尿病代谢状态提供了重要的理论依据。
脂质代谢(血脂异常)
代谢综合征(MetS)是一组以腹型肥胖、高血压、高血糖和血脂异常为核心特征的病理状态集合,其共同的病理生理基础是胰岛素抵抗。
研究表明,肌醇之所以能在改善血脂方面大有可为,是因为它能通过至少三条关键路径,协同作战,调控脂质代谢。

路径一:直击源头——为肝脏的脂肪工厂刹车
肝脏是全身的脂质代谢中心。在胰岛素抵抗和高糖负荷的状态下,肝脏的“新生脂肪合成”功能会失控,如同一个被过度激活的工厂,不停地将碳水化合物(糖)转化为脂肪。
肌醇的作用,就是为这个失控的工厂精准刹车。它通过抑制 SREBP-1c 和 ChREBP 这两个控制脂肪合成的总开关,直接从源头上减少了肝脏的脂肪产量。这不仅能降低血液中的甘油三酯(TG)水平,也是改善非酒精性脂肪肝(NAFLD)的关键一步。
路径二:切断原料——修复外周脂肪组织的失灵开关
胰岛素抵抗不仅会引发高血糖,还会导致另一个严重问题:脂肪组织中一个叫“激素敏感性脂肪酶(HSL)”的开关失灵了。
这些泛滥的原料被肝脏大量吸收,进一步加剧了甘油三酯和“坏胆固醇”(VLDL,极低密度脂蛋白)的生产。肌醇通过修复细胞内的胰岛素信号通路(如PI3K/Akt通路),重新恢复了对HSL的抑制作用。这相当于切断了肝脏过度生产脂肪的原料供应。
这个过程不仅能降低血脂,还能减少脂肪在肝脏、肌肉等不该堆积之处的沉积,从而进一步改善全身的胰岛素敏感性,形成一个宝贵的良性循环。
路径三:另辟蹊径——优化肠道菌群(肠-肝轴)
肌醇还通过一条全新的路径——调节肠道菌群,来间接实现降脂效果。
它扮演着益生元的角色,为特定的有益菌(如Anaerostipes属)提供食物。这些有益菌在代谢肌醇后,会产生丙酸和乙酸等短链脂肪酸。这些有益物质通过肠道进入肝脏,发挥重要作用,例如,丙酸可以抑制肝脏合成胆固醇。
更重要的是,一个健康的肠道菌群能够强化肠道屏障,防止脂多糖(LPS)等内毒素进入血液,从而减轻体内的低度炎症状态,而这种炎症正是驱动胰岛素抵抗和肝脏脂肪合成的关键推手。因此,肌醇通过优化肠道微生态,利用肠-肝轴,间接抑制了炎症和脂质生成。
其他潜在机制
除了上述三大路径,研究还提示肌醇可能通过以下方式发挥作用:
正是这种在多个层面、通过多种通路协同作用的综合效应,使肌醇成为一种极具潜力的天然营养干预策略,用以改善代谢综合征患者的血脂谱和整体健康。
肌醇是中枢神经系统中浓度第二高的有机小分子(仅次于谷氨酸),其重要性远不止于一个简单的结构单元。
它在维持神经精神健康中扮演着两大关键角色:既是细胞信号传导的基石,也是神经元功能的稳定器。肌醇代谢的紊乱,已与抑郁症、焦虑症、双相情感障碍等多种疾病的病理生理学紧密相连。其核心作用机制围绕着两个主要方面:
➷ 肌醇——信号传导的基石
肌醇是大脑最重要的信号传导系统,磷脂酰肌醇(PI)信号通路的绝对基石。
可以这样理解:几乎所有与情绪、认知相关的神经递质,包括血清素(5-HT)、去甲肾上腺素、多巴胺等,都好比是信使。但信使发出的信号要被细胞内部听到并执行,就需要一个高效的扩音系统,而PI通路就是这个系统。
细胞信号传导过程 – PI通路
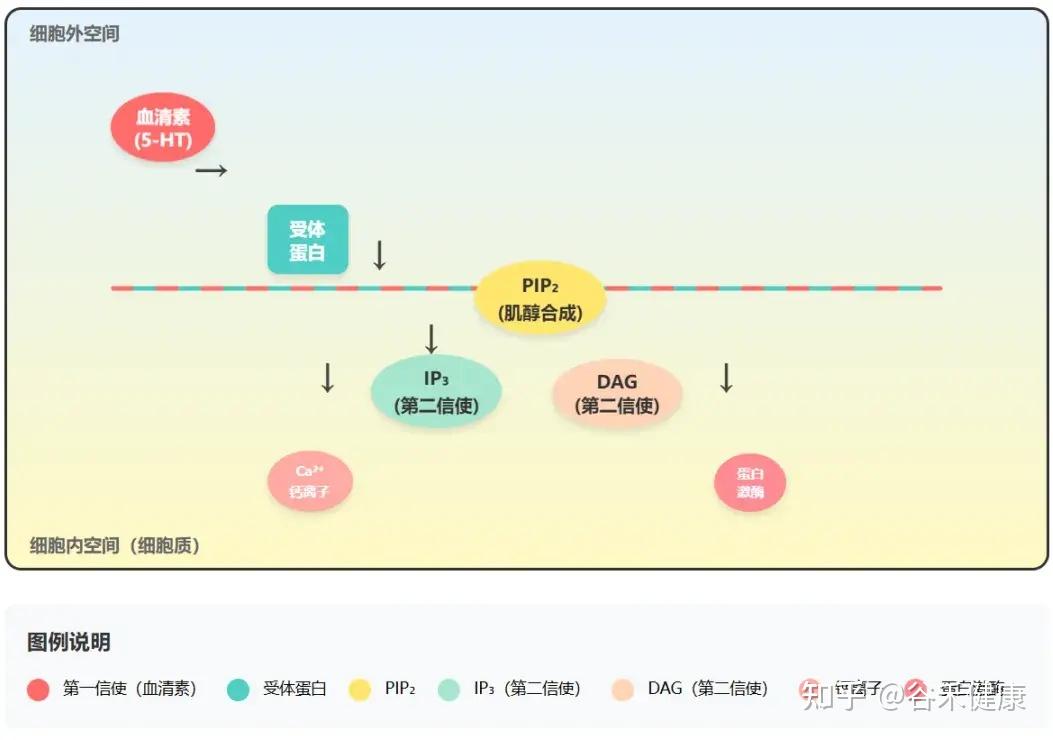
1.
“信使”(如血清素)与细胞外的受体结合。
2.
激活PI通路,将细胞膜上的PIP₂(由肌醇合成)分解为两个关键的“第二信使”——IP₃(肌醇三磷酸)和DAG(二酰甘油)。
3.
这两个第二信使在细胞内触发一系列反应,如IP₃调动钙离子、DAG激活蛋白激酶等,最终将“信使”的指令转化为实际的神经元活动。
因此,脑内充足的肌醇,是确保血清素等“快乐激素”的信号能被有效放大和传递的根本前提。
研究人员已反复在抑郁症和焦虑症患者的脑脊液中观察到,肌醇水平的显著降低。这强烈暗示,肌醇水平下降导致PI信号通路功率减弱,使得神经递质的信号无法被有效传导,从而引发情绪低落、快感缺失和焦虑。
这也解释了为何在多项临床试验中,补充大剂量肌醇(每日高达12-18克)显示出与标准抗抑郁药相当的疗效,它直接为这个“信号放大系统”补充了燃料。
➷ 肌醇——神经元功能的稳定器
大脑的细胞环境并非一成不变,脱水、高血糖等状态都可能引起渗透压的剧烈变化,威胁神经元的生存。
肌醇此时扮演着“渗透压稳定器”或“细胞海绵”的角色。它能在细胞内累积或释放,灵活调节细胞内外的水分平衡,防止神经元因过度肿胀或皱缩而受损。这种保护功能对于维持神经元的结构完整性和正常电生理活动至关重要。
来自临床与疾病的佐证
双相情感障碍的锂盐治疗
治疗双相情感障碍的经典药物——锂盐,其作用机制就从反面印证了肌醇的核心地位。锂盐通过抑制肌醇的再循环【锂盐抑制肌醇单磷酸酶(IMPase)和肌醇多磷酸-1-磷酸酶(IPPase)】,导致细胞内PI信号通路的信号减弱。这种信号抑制是锂盐平复躁狂心境的关键,这从反面印证了肌醇信号通路在情绪调节中的核心地位,但也可能因此产生副作用。
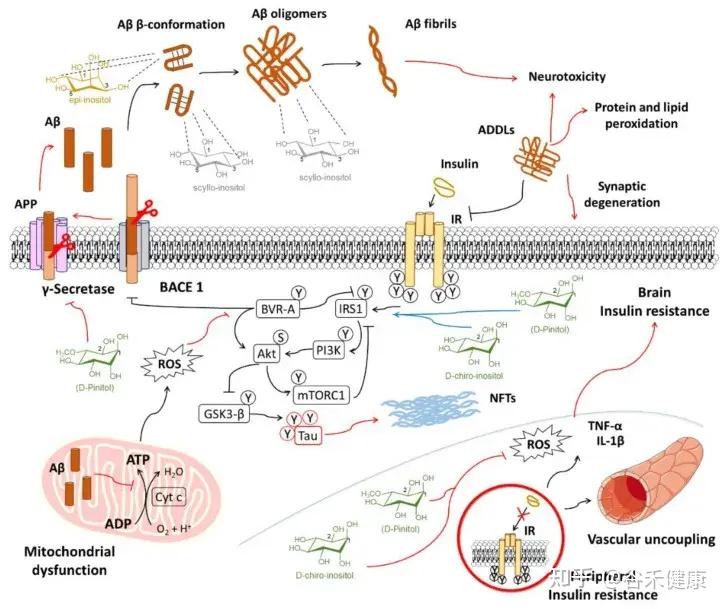
阿尔茨海默病的关联
研究发现,肌醇有助于预防脑内胰岛素信号传导的早期损伤和血管功能障碍。这些功能障碍会促进氧化应激和炎症,最终在阿尔茨海默病的背景下,助长淀粉样蛋白的病理改变。
肌醇在导致阿尔茨海默病进展的机制中相互作用的示意图
综上所述,肌醇并非直接充当神经递质,而是作为神经信号传递系统的燃料和细胞环境的稳定器。当大脑的肌醇供应不足或代谢受阻时,整个神经系统的效率都会大打折扣,引发情绪和认知问题。
因此,维持充足的肌醇水平,是通过恢复最基础的细胞功能来支持整体神经精神状态的健康。这也使其成为现代营养精神病学中备受关注的干预策略之一。
尽管肌醇在皮肤健康领域的知名度远不及在代谢和神经科学中的应用,但它作为细胞功能的基础分子,通过调节细胞信号、脂质代谢和炎症,对维持皮肤的整体稳态发挥着不可或缺的幕后作用。
肌醇对皮肤的影响,主要通过以下三个层面实现:
1
强化皮肤屏障,构筑坚固屏障
肌醇是构成细胞膜关键成分——磷脂酰肌醇(PI)的核心原料。一个健康的细胞膜,是维持皮肤细胞功能和构建坚固皮肤屏障的基础。
可以把皮肤屏障想象成一道砖墙:角质细胞是砖块,细胞间的脂质(神经酰胺、胆固醇等)是水泥。肌醇参与的PI信号通路,深刻影响着砖块也就是角质细胞的增殖、分化和凋亡。
2
调节皮脂与炎症,从根源控痘
肌醇通过其在脂质代谢和胰岛素信号中的核心作用,直接影响着皮脂腺的活动——这与痤疮(也就是痘痘)的形成密切相关。
痤疮的形成与皮脂分泌过多、毛囊口角化异常、痤疮丙酸杆菌增殖和炎症反应四大因素相关。
这种状态会刺激皮脂腺过度活跃,分泌过多油脂,同时促进角化,为痤疮的爆发创造了土壤。
肌醇,特别是myo-肌醇,扮演着代谢调节器的角色:
这与它在治疗多囊卵巢综合征(常伴有痤疮的疾病)中观察到的效果一致:通过改善内部代谢,皮肤状况也随之显著好转。
3
改善全身代谢
皮肤是身体内部健康状况的一面镜子。代谢综合征(由胰岛素抵抗、血脂异常和慢性炎症引起)常常伴随着典型的皮肤问题,如黑棘皮病(皮肤皱褶处天鹅绒般的色素沉着)、皮赘以及更差的伤口愈合能力。
肌醇通过系统性地改善糖脂代谢、降低全身炎症水平,能够从根本上扭转这些病理状态,从而改善甚至消除这些皮肤表现。一个代谢平衡的身体,其皮肤的血液循环、营养供应和自我修复能力都会得到提升,自然会显得更有光泽和弹性。
综上肌醇并非一种传统意义上的护肤维生素,而是由内而外地改善,确保了皮肤细胞能够正常沟通、有序更新,并帮助控制那些由内部代谢失衡所驱动的皮肤问题(如过度出油和炎症)。因此,维持充足的肌醇水平,是实现由内而外的持久皮肤健康的重要一环。
评估体内的肌醇状态,通常需要结合临床表现和实验室检测。可以从以下几个方面,判断是否存在肌醇代谢紊乱的可能性。
症状信号
当患者出现以下看似不相关、但背后可能都与肌醇代谢紊乱有关的特定情况时,应引起警惕:
代谢:
尽管努力控制饮食,但血糖水平依然居高不下或难以控制。
肝脏:
体重正常,甚至偏瘦,但体检却发现存在脂肪肝。
生殖:
育龄女性出现无排卵或月经周期严重不规律(提示多囊卵巢综合征的可能)。
精神:
患有抑郁症或焦虑症,但对常规的血清素类药物(如SSRI)反应不佳。
生物标志物
使用气相色谱-质谱(GC-MS)或液相色谱-质谱(LC-MS)定量检测血清、尿液或组织中的myo‑Ins可提供直接测量。
这是最直接、最准确的评估方法。
辅助指标
胰岛素抵抗评估
检测空腹血清胰岛素和计算HOMA-IR指数,是评估胰岛素抵抗严重程度的核心指标。
血脂与肝功能检测
检查肝脏中的甘油三酯和胆固醇水平,以评估与肌醇代谢密切相关的脂肪肝状况。
肠道微生态分析
通过粪便样本分析,评估肠道菌群的构成,以及肌醇在肠道内的代谢产物,短链脂肪酸的水平,从而了解肠道在肌醇代谢通路中扮演的角色。
综合这些临床症状和辅助指标,可以为判断是否存在肌醇功能性缺乏以及是否需要进行营养干预提供有力的证据。
一般情况下, 以下建议可以帮助补充肌醇。
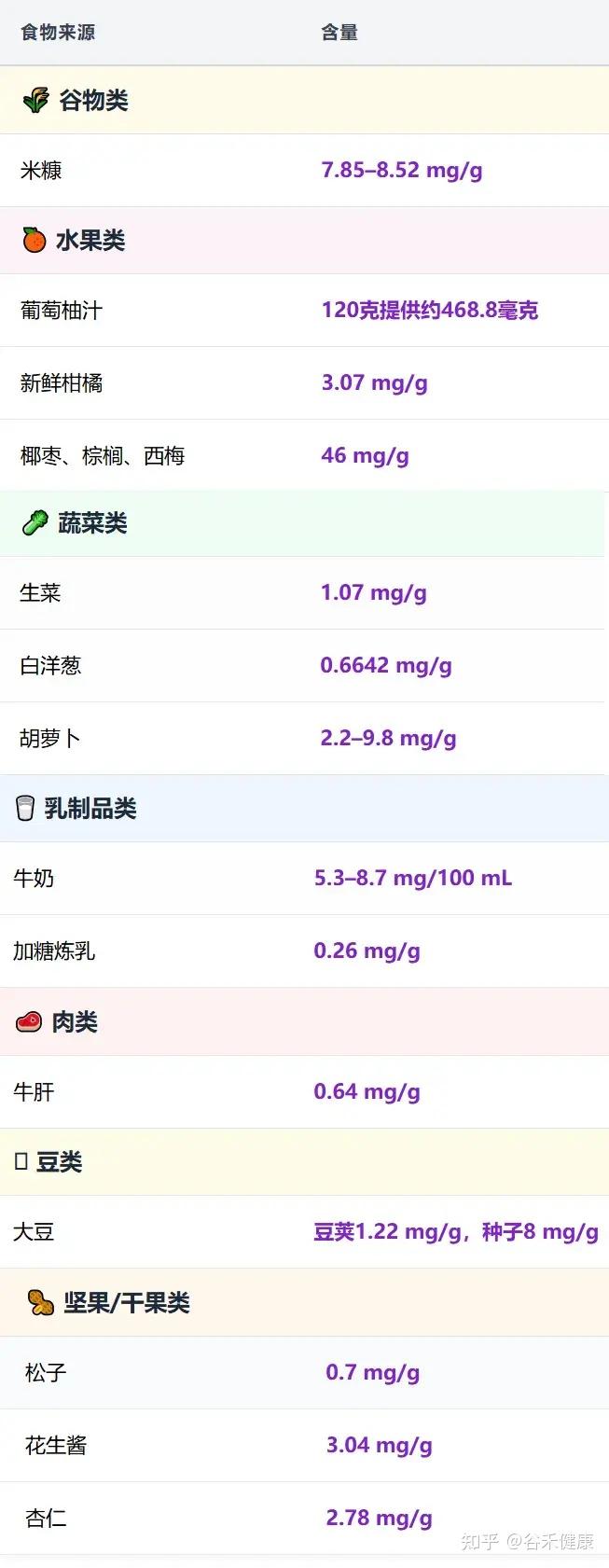
饮 食
富含天然肌醇来源的食物——全谷物、豆类、柑橘类水果、坚果、动物肝脏,可提供游离和结合形式的肌醇。发酵食物(如酸面包、酸奶发酵)能提高其生物利用度。
肌醇可以在以下食物来源中找到:
注:新鲜水果和蔬菜中的肌醇含量,远高于干燥或罐装水果和蔬菜。
磷脂酰肌醇也存在于卵磷脂中,因此以下所有富含卵磷脂的食物都是很好的来源:
注:动物来源通常比植物来源提供更大的卵磷脂来源。
D-手性肌醇(用作抗氧化剂,与肌醇结合用于多囊卵巢综合征)的最佳来源是荞麦。
肌醇食物来源含量表
补 充 剂 量
针对不同需求剂量有些差异
高剂量应用于神经与精神健康
对于抑郁症、焦虑症、强迫症及饮食失调等精神健康问题,临床研究倾向于使用每日12克甚至更多的剂量。研究人员认为需要如此高的剂量才能有效穿过血脑屏障,显著提升大脑中的肌醇与血清素水平,从而发挥稳定情绪的作用。
低剂量应用于代谢与生殖健康
相比之下,在处理代谢和生殖相关问题时,采用的剂量要温和得多。对于改善生育能力、调理多囊卵巢综合征、辅助减重以及提升胰岛素敏感性,每日2~4克的肌醇(Myo-inositol)已被证实是行之有效的。
在治疗PCOS时,一个关键的策略是将肌醇(Myo-inositol)与D-手性肌醇(D-chiro-inositol, DCI)联合使用,并遵循40:1的生理比例。一个典型的例子是:每日服用4克的肌醇,同时配以100毫克的D-手性肌醇,这种组合被证实能最有效地恢复代谢和卵巢功能。
也有单独的D-手性肌醇,每天500毫克~1.2克的剂量,改善激素状况并恢复排卵。每天1克改善了接受控制性卵巢过度刺激的女性的卵子质量。
注:联合补充剂可能比单独使用肌醇效果更好。
对于妊娠期糖尿病,每天服用1.75克肌醇和250毫克DCI,可以降低血糖和母婴健康风险。
补充形式
补充剂的形态对吸收率有显著影响。
研究表明,软胶囊的生物利用度远高于粉末,仅需约30%的剂量即可达到同等效果。例如,一项针对经前情绪障碍的研究发现,每日3.6克的软胶囊与12克的粉末效果相当。
增强效果的协同组合
为了最大化肌醇在生殖和代谢方面的益处,常常推荐将其与其他营养素协同使用。临床实践表明,将肌醇与叶酸或Omega-3脂肪酸联合补充,可以起到1+1>2的效果,进一步增强其对生殖和代谢健康的积极影响。
MYO或MYO + DCI给药对生育治疗和健康妊娠的影响
生 活 方 式
规律运动:有氧运动能增强体内将myo-肌醇转化为DCI的关键酶的活性。
优质睡眠:充足的睡眠有助于减少压力激素(皮质醇)对肌醇的降解。
限制不良习惯:减少精制糖的摄入和限制饮酒,能有效稳定血液中的肌醇水平。
调 节 肠 道 菌 群
益生元纤维(如菊粉、抗性淀粉)和益生菌(如鼠李糖乳杆菌、婴儿双歧杆菌)有助于培养有益的微生物群落,这些菌群能产生短链脂肪酸,支持肌醇介导的代谢健康。
根据肌醇缺乏机制定制
根据五大缺乏机制,可以针对性的策略:
1- 针对「膳食摄入不足」
策略:重点增加全谷物、豆类和坚果的摄入。如果饮食难以保证,应考虑直接补充。
2- 针对「内源合成障碍」(如基因缺陷)
风险人群:携带 ISYNA1 基因变异者;有唐氏综合征家族史的备孕人群。
策略:必须依赖外源性补充,且可能需要比常规更高的剂量。
3- 针对「肾脏过度清除」(如糖尿病状态)
风险人群:高血糖、糖尿病、肥胖及高血压患者。
策略:
-严格控制血糖是重中之重。
-补充 myo-肌醇以弥补流失。
-定期监测肾功能。
4- 针对「肠道吸收障碍或竞争」
A. 葡萄糖竞争
挑战:与高糖食物同服会降低吸收。
策略:选择在空腹或两餐之间服用肌醇。
B. 高剂量DCI的竞争
挑战:单独补充高剂量DCI会反过来抑制myo-肌醇的吸收。
策略:严格遵守40:1或更高的 myo-Ins/DCI 比例,避免使用高剂量DCI的单一配方。
C. 肠道菌群失调
策略:采用组合方案,myo-肌醇 (2-4g/天) + α-乳白蛋白 (作为益生元, 2-3g/天) + 乳杆菌/双歧杆菌益生菌,并配合高纤维饮食。
5- 针对「药物诱导的消耗」
A. 影响中枢神经的药物(锂盐、丙戊酸等)
策略:进行外周补充以缓解副作用。此时配比可能需要调整为 80:1 (myo:DCI)。需定期监测多尿、皮肤问题等代谢指标。
B. 影响肠道菌群的药物(PPI、他汀类、泻药等)
策略:联合补充益生元和益生菌,必要时临时增加myo-肌醇的补充。
特殊人群的注意事项
在为以下人群制定方案时,应格外谨慎,并从低剂量开始:
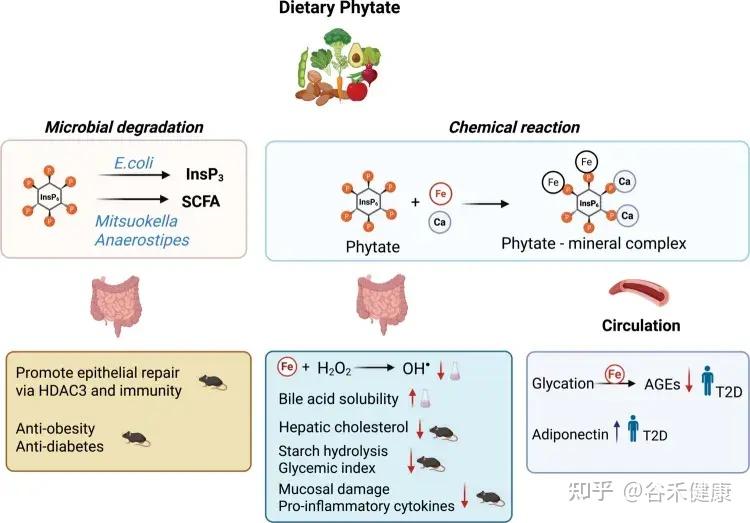
基于体外和体内模型以及人体研究肌醇肌醇六磷酸盐对健康影响
肌醇的研究正进入一个新纪元,其重要性已远远超越了传统的代谢调节范畴,延伸至癌症、神经保护和肠道健康等前沿领域,进一步巩固了它作为连接人体与微生物生态的核心分子的地位。
癌症代谢的新武器
研究发现,肌醇及其衍生物(如IP6)通过调节细胞内关键的 PI3K/AKT 信号通路并抑制肿瘤血管的生成,表现出显著的抗肿瘤增殖特性。这为开发基于营养的癌症辅助治疗策略提供了新思路。
神经保护的希望之星
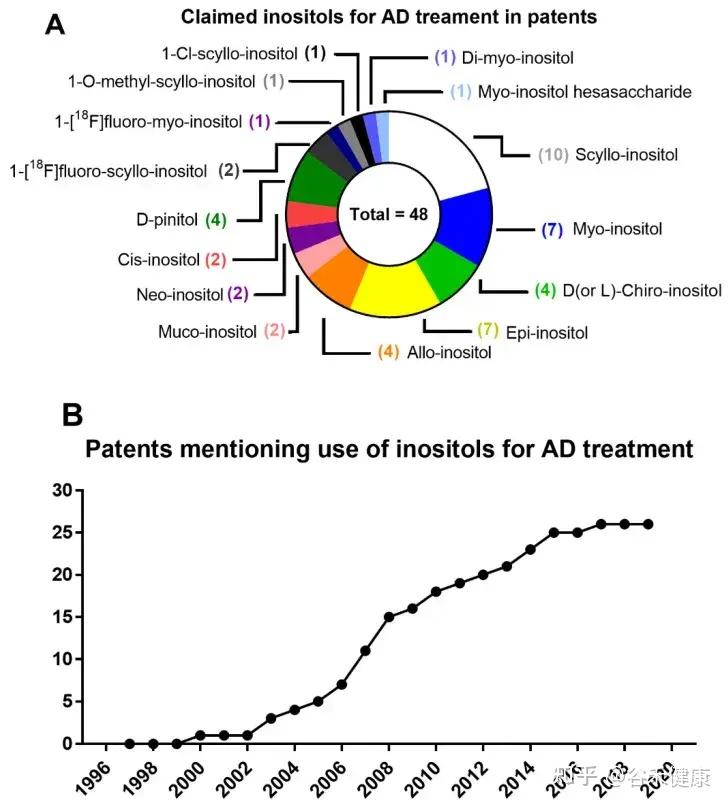
在阿尔茨海默病等神经退行性疾病的研究中,肌醇显示出巨大潜力。它被证实可以减少β-淀粉样蛋白的聚集和脑内的氧化应激,这是阿尔茨海默病的两大核心病理过程。事实上,已有大量专利被授予,用于探索和应用肌醇来治疗阿尔茨海默病,突显了其在该领域的应用前景。
提及和要求使用肌醇治疗阿尔茨海默的专利数量
肠-肝轴的协同调控
最新研究揭示,将肌醇与牛磺酸联合使用,能够有效重编程肝脏的基因表达,不仅能防止因高糖饮食诱导的脂肪肝,还能显著改善肠道内有益的短链脂肪酸。
这些发现共同描绘了:肌醇是一个卓越的整合分子,它能够巧妙地协调宿主自身的代谢节律与体内庞大的微生物生态系统。
肌醇深刻影响着全身的能量平衡与健康信号。过去,我们补充肌醇,期望它能在体内某个需要的地方恰好生效。现在,我们知道肌醇的效能并非孤立存在,它也同时取决于我们的肠道菌群。
基于此,“先养菌,再补剂”可能会成为新的黄金法则。我们可以先通过饮食、生活方式、益生元或益生菌来改造肠道环境,为肌醇补充铺平道路。未来可能将有更深入的研究,去识别究竟是哪些微生物基因簇在主导肌醇的转化;也许我们会找到那些由肌醇衍生出的、尚未被发现的新型健康信使。
主要参考文献:
Lepore E, Lauretta R, Bianchini M, Mormando M, Di Lorenzo C, Unfer V. Inositols Depletion and Resistance: Principal Mechanisms and Therapeutic Strategies. Int J Mol Sci. 2021 Jun 24;22(13):6796.
Jessen H, Bui TPN. Metabolism of inositol derivatives by the gut microbiome. Gut Microbes. 2025 Dec 31;17(1):2564765.
López-Gambero AJ, Sanjuan C, Serrano-Castro PJ, Suárez J, Rodríguez de Fonseca F. The Biomedical Uses of Inositols: A Nutraceutical Approach to Metabolic Dysfunction in Aging and Neurodegenerative Diseases. Biomedicines. 2020 Aug 20;8(9):295.
Wu C, Yang F, Zhong H, Hong J, Lin H, Zong M, Ren H, Zhao S, Chen Y, Shi Z, Wang X, Shen J, Wang Q, Ni M, Chen B, Cai Z, Zhang M, Cao Z, Wu K, Gao A, Li J, Liu C, Xiao M, Li Y, Shi J, Zhang Y, Xu X, Gu W, Bi Y, Ning G, Wang W, Wang J, Liu R. Obesity-enriched gut microbe degrades myo-inositol and promotes lipid absorption. Cell Host Microbe. 2024 Aug 14;32(8):1301-1314.e9.
Fazio A, Rosenstiel P. Language of a Long-Term Relationship: Bacterial Inositols and the Intestinal Epithelium. Cell Metab. 2020 Oct 6;32(4):509-511.
Caputo M, Bona E, Leone I, Samà MT, Nuzzo A, Ferrero A, Aimaretti G, Marzullo P, Prodam F. Inositols and metabolic disorders: From farm to bedside. J Tradit Complement Med. 2020 Mar 24;10(3):252-259.
Bui TPN, Mannerås-Holm L, Puschmann R, Wu H, Troise AD, Nijsse B, Boeren S, Bäckhed F, Fiedler D, deVos WM. Conversion of dietary inositol into propionate and acetate by commensal Anaerostipes associates with host health. Nat Commun. 2021 Aug 10;12(1):4798.
Lentini G, Querqui A, Monti N, Bizzarri M. PCOS and Inositols – Advances and Lessons We are Learning. A Narrative Review. Drug Des Devel Ther. 2025 May 21;19:4183-4199.
Min M, Afzal N, Maloh J, Dulai AS, Ahmad N, Pinzauti D, Sivamani RK. Prospective Comparative Study of an Oral Synbiotic and a Myoinositol-Based Herbal Supplement in Modifying Hormone Levels and the Gut Microbiome in Non-cystic Acne. Dermatol Ther (Heidelb). 2025 Jun;15(6):1331-1350.
Vitacolonna E, Masulli M, Palmisano L, Stuppia L, Franzago M. Inositols, Probiotics, and Gestational Diabetes: Clinical and Epigenetic Aspects. Nutrients. 2022 Apr 8;14(8):1543.
Placidi M, Casoli G, Tatone C, Di Emidio G, Bevilacqua A. Myo-Inositol and Its Derivatives: Their Roles in the Challenges of Infertility. Biology (Basel). 2024 Nov 16;13(11):936.

谷禾健康
《血糖,微生物,胰岛素》
2型糖尿病(T2D)是一种全球性慢性代谢性疾病,目前已影响超过 4.6亿人,其社会经济负担巨大,迫切需要寻求有效的治疗方法。
根据中华医学会糖尿病学分会发布的《中国2型糖尿病防治指南》,我国2型糖尿病的管理路径清晰而规范:以生活方式干预为基石,以二甲双胍为一线首选,后续根据情况采取阶梯式的强化治疗。
然而,尽管指南明确,临床上仍然面临治疗反应个体差异大这一核心难题,这导致我国仍有近半数患者血糖控制不达标。为何同样的“金标准”方案,效果却天差地别?
多项研究发现,2型糖尿病的肠道菌群组成与健康人存在明显差异,菌群失衡可能通过影响短链脂肪酸合成、胆汁酸代谢及胰岛素敏感性等途径促进代谢紊乱。
甚至连二甲双胍这些药物的降糖效应,竟有相当一部分是依赖其对患者肠道微生物群的重塑和调节。
本文将介绍当前有关肠道微生物群调控葡萄糖稳态的潜在机制,并重点探讨近年来通过靶向肠道菌群改善糖代谢的研究进展。其中包括粪菌移植、益生元、益生菌、合生元与外源性代谢调节物质等多种策略在动物模型中的应用及其转化潜力。这些非侵入性干预手段有望为2型糖尿病带来新的、有效的治疗或至少是辅助选择。
要揭示肠道菌群与个体糖尿病的关系,需要谨慎和长期的队列研究
肠道菌群:一个多因素互动的复杂网络
然而,事情并非绝对“好细菌”与“坏细菌”的简单对抗。肠道菌群是一个复杂的生态系统,这个复杂的网络包括:
所有这些因素交织在一起,共同决定了我们的代谢健康状况。
因此,要真正揭示肠道菌群在糖代谢中的作用机制,就需要做到:
这类研究不仅能为糖尿病的早期预测和个体化干预提供坚实证据,也将为利用肠道菌群进行精准营养调控和代谢重塑奠定科学基础。
谷禾已经通过14,846例2型糖尿病患者(包括妊娠糖尿病)构建了早期糖尿病预测模型,但该模型仍面临若干挑战。
未来谷禾会继续推进研究结合代谢组,药理学等多组学数据,并通过多中心、纵向的设计来控制药物等潜在混杂因素,以实现更高精度、更具普适性的肠道菌群早期糖尿病预测模型。
通过深入探索这些微生物是通过哪些机制影响血糖平衡和代谢功能的,也许会为糖尿病治疗带来新的突破。
肠道屏障受损:免疫与糖代谢的微妙联系
肠道屏障是先天性免疫系统的重要组成部分,它的主要工作就是筑起一道坚固的墙,将肠道内的细菌、病原体及其代谢产物牢牢地限制在肠道内,防止进入血液循环。
▸当肠道屏障出现漏洞
然而,高脂饮食(HFD)会破坏肠道微生物群平调,诱发肠漏,这使得细菌内毒素,特别是革兰阴性菌产生的脂多糖(LPS),更容易进入血液循环,形成“代谢性内毒素血症”。
▸炎症的触发机制
—警报响起
泄漏到血液中的LPS会激活免疫细胞表面的Toll样受体4 (TLR4)。
—信号传导
TLR4被激活后,会启动一个信号级联反应,导致细胞核内的 NF-κB激活。NF-κB是调控炎症反应的总开关。
—促炎因子
被激活的NF-κB会命令细胞大量生产和释放促炎细胞因子,如肿瘤坏死因子-α (TNF-α)、白介素-6(IL-6)等。在全身造成低度慢性炎症。
值得注意的是,近期研究发现,不同菌种来源的LPS在影响葡萄糖代谢、炎症反应及肠屏障完整性方面存在差异,说明仅以LPS总量评价肠漏程度可能并不准确。
▸ 慢性炎症:导致胰岛素抵抗的元凶
这种炎症状态是导致代谢紊乱的关键因素之一,例如,TNF-α会直接干扰肌肉细胞对胰岛素的响应,阻碍血糖的正常利用,最终引发胰岛素抵抗、高血糖及高胰岛素血症。
▸免疫系统与菌群的双向互动
免疫系统和肠道菌群的关系并非单向的,而是一个复杂的双向互动。
关键免疫细胞:Th17细胞
这是一种特殊的T细胞,它在正常情况下分泌IL-17和IL-22等因子,对于维持肠道屏障的完整性和抵御病原体至关重要。可以看作是城墙的守护者。
➦ 菌群 → 免疫系统
证据1(生酮饮食):生酮饮食会减少肠道中Th17细胞的数量。但如果给无菌小鼠移植了采用生酮饮食的人的粪菌,小鼠的Th17细胞也会减少。而补充双歧杆菌则可以恢复Th17细胞的水平。
证据2(高糖高脂饮食):高糖高脂饮食会减少那些能够诱导Th17细胞发育的特定细菌。如果给高糖高脂饮食喂养的小鼠补充这些特定的细菌,它们的Th17细胞水平得以恢复,血糖也得到改善。
➦ 免疫系统 → 菌群
证据(过继转移):将Th17细胞直接移植到肥胖小鼠体内,这些小鼠的肠道菌群结构发生了有益的改变,葡萄糖耐量也得到了改善。
doi.org/10.1146/annurev-physiol-051524-094728
►▷
不良饮食 → 菌群失调 → 肠漏 → LPS进入血液 → 激活免疫系统(TLR4)→ 慢性炎症(TNF-α)→ 胰岛素抵抗。
菌群的改变会影响关键免疫细胞(如Th17)的数量,而免疫系统的状态反过来又能重塑肠道菌群的构成。
这种菌群与肠道免疫之间错综复杂的相互作用,是理解代谢性疾病病理生理的核心,也为未来的治疗提供了新的靶点。
短链脂肪酸:菌群送给你的控糖礼物
短链脂肪酸(SCFAs)包括丁酸、丙酸、乙酸,它们是由肠道细菌通过碳水化合物发酵产生。
它们不仅仅是代谢产物,更是重要的信号分子,是肠道菌群与我们身体对话的语言。
▸ 短链脂肪酸如何调控血糖?
▸ 动物研究,直接补充SCFA改善血糖
多项动物实验表明,外源性补充SCFAs可改善能量代谢与血糖稳态,表现为体重下降、葡萄糖耐受性提升及胰岛素抵抗减轻。
▸ 人体研究,效果不一致
例如,口服丁酸仅在健康受试者中改善血糖控制,而对代谢综合征患者无显著效应,提示代谢紊乱本身,可能就损害了人体对SCFA信号的响应能力。
▸ 口服补充为何会失效?
口服补充SCFA是不符合生理规律的。
在自然状态下,绝大多数SCFA是在远端肠道(结肠)中由细菌产生的。
口服的缺陷:口服的SCFAs在到达结肠之前,可能早已在胃或小肠中被吸收或代谢掉,无法到达它们应该发挥主要作用的地方。
有力的证据: 一项巧妙的研究解决了这个问题。科学家设计了一种“菊粉-丙酸酯”,这种特殊结构可以保护丙酸不被提前吸收,精准地将其运送到结肠释放。结果发现,这种精准投放的丙酸,确实能够有效改善超重和肥胖人群的能量代谢和胰岛素敏感性。
这说明,SCFA在哪里释放,比释放了多少,可能更为重要。
▸ 为何病人的粪便里SCFA反而更高?
库存 ≠ 产量
一些研究发现,代谢疾病患者粪便中的SCFA含量反而更高。这似乎与“SCFA是有益的”这一观点相矛盾?
合理解释:粪便中的SCFA含量,仅仅是未被肠道吸收和利用的“剩余物资”。
▸ 肠道菌群-短链脂肪酸-肠道激素轴
短链脂肪酸通过与肠内分泌细胞(EECs)上的受体结合,刺激激素的释放,比如说,GLP-1(胰高糖素样肽-1)、PYY(肽YY)、GIP(胃抑制肽)、CCK(胆囊收缩素)等。GLP-1与GIP为主要的“促胰素”,能够在摄食后迅速加强胰岛素分泌反应,是控制餐后血糖的关键。
doi.org/10.1146/annurev-physiol-051524-094728
近年来的新发现进一步揭示,微生物群可通过调控宿主营养感知机制影响肝脏糖异生,形成由GLP-1介导的“肠–脑–肝”信号轴。
►▷
由于肠道菌群与上皮层密切接触,它可能通过改变肠内分泌细胞的营养感知机制,并利用短链脂肪酸及其他微生物代谢物作为信号分子,调控肠激素分泌和葡萄糖平衡。这种微生物与宿主的相互作用网络为解析肠源信号调控代谢疾病提供了新的理论框架。
菌群把胆汁酸变成影响血糖的信号
肠道是一个高度复杂的生态系统,如同一个动态的生物反应器,在这个反应器里,无数的微生物与我们吃进去的食物、以及我们身体自己分泌的物质(如胆汁)发生反应,生成了大量独特的代谢产物。
关于肠源性代谢产物我们前面已经讨论过短链脂肪酸,现在焦点转向另一位主角:胆汁酸。
★ 胆 汁 酸
▸ 初级胆汁酸
出生地: 肝脏。 原料: 胆固醇。
初始形态:在肝脏合成后,它们会与牛磺酸(主要在小鼠中)或甘氨酸(主要在人类中)结合,形成结合型初级胆汁酸。
主要工作:进入肠道,帮助我们消化和吸收脂肪。
▸ 次级胆汁酸
一旦初级胆汁酸完成了消化任务并进入肠道后段,肠道菌群就接管了,对胆汁酸进行两步关键的改造:
这种从初级到次级的转变至关重要,因为不同形态的胆汁酸,会像不同的钥匙一样,激活不同的细胞受体“锁”,从而触发完全不同的生理效应。
doi.org/10.1146/annurev-physiol-051524-094728
▸ 两大关键受体:TGR5 与 FXR
次级胆汁酸主要通过激活两个著名的受体来发挥信号作用:
—— TGR5受体 (明确益处)
激活机制:菌群将初级胆汁酸(如鹅去氧胆酸)转化为次级胆汁酸(如石胆酸),后者是TGR5受体的强效激动剂。
明确的益处:激活肠道细胞上的TGR5受体,能够促进GLP-1的分泌,从而改善高脂饮食动物的血糖稳态。
—— FXR受体 (更具争议)
FXR受体则要复杂得多,它的作用似乎取决于它在哪个器官被激活。
在肝脏中(作用清晰):
激活肝脏的FXR受体,可以改善健康和糖尿病小鼠的胰岛素敏感性。
在肠道中(作用矛盾):
一些研究报告称,使用激动剂激活肠道FXR能改善胰岛素抵抗和葡萄糖耐量。
而另一些研究(包括我们之前讨论的产乳酸菌的研究)则发现,使用拮抗剂抑制肠道FXR,或者直接把肠道FXR基因敲除掉,反而能改善血糖稳态。
这说明,FXR在肠道中的作用远非简单的“开”或“关”,而是受到多种因素调控的复杂网络。
★ 其他代谢产物
这些肠源性代谢产物通过影响能量代谢、免疫反应以及宿主信号通路,构成肠道微生物–代谢–宿主疾病之间的重要生物学纽带。
既然我们已经明确,肠道菌群是糖尿病发生发展的关键一环,那么一个问题随之而来:我们目前广泛使用的那些经典降糖疗法,比如二甲双胍等,它们的疗效背后,是否也隐藏着菌群的秘密?下面我们就来看看,常见的治疗方式是如何通过影响肠道菌群来帮助我们控制血糖的。
二甲双胍
二甲双胍是最常见的降糖药之一。它降低血糖的传统机制是减少肝脏制造葡萄糖的能力。但近些年,人们发现——它在肠道里也发挥着巨大作用。
研究发现,口服二甲双胍比注射更能有效降糖,提示它在消化道内可能有额外的作用通路。科学家进一步发现,二甲双胍能改变肠道菌群结构:
更有趣的是,如果把“吃过二甲双胍的小鼠”的小肠菌群移植到其他糖尿病老鼠体内,这些受体小鼠体内的营养感知通路得以恢复,进而抑制了肝脏的葡萄糖生成——也就是说,药效可以通过菌群“传递”出来。
这说明,二甲双胍不仅是“肝药”,更是“肠药”。它同时通过肠道菌群、胆汁酸,以及“肠–脑–肝”信号网络,共同调控血糖代谢。
GLP-1受体激动剂
GLP-1受体激动剂(胰高糖素样肽-1受体激动剂,GLP-1RAs)是一类降糖药物,GLP-1受体激动剂通过多靶点机制发挥作用:
▸肠道菌群对GLP-1的双重影响
促进分泌:肠道菌群能够促进肠道内GLP-1激素的分泌,这是一种有益的代谢效应。
限制作用:分泌的GLP-1在体内极易被二肽基肽酶-IV(DPP-IV)快速降解,导致其对血糖和食欲的系统性调节作用受限,仅产生局部、短暂的代谢效应。
注:DPP-IV 是 Dipeptidyl Peptidase-IV 的缩写,中文名称为 二肽基肽酶-4。它是一种丝氨酸蛋白酶(serine protease),广泛存在于人体的多种组织中,包括肠道上皮、肝脏、肾脏、免疫细胞以及血浆中。
因此,肠道菌群在介导GLP-1药物系统性效应中的具体角色仍不明确。
▸动物与人体研究中的差异
关于GLP-1RAs对肠道菌群的影响,现有研究结果存在不一致。
动物研究:在高脂饮食或糖尿病动物模型中,GLP-1RAs治疗能够增加肠道菌群的多样性,并提升有益菌(如Akkermansia muciniphila、乳酸杆菌)的丰度。
🧍♀️人类研究:也有观察到双歧杆菌数量增加。但大多数人类临床研究报告称,在接受GLP-1RAs治疗后,患者的肠道菌群组成并无显著变化。
▸给药途径:可能是造成差异的关键因素之一
大部分GLP-1RAs均经皮下注射给药,因此菌群改变或许是药物通过影响肠道生理功能(如延缓胃排空、降低肠蠕动等)所致的间接效应,而非药物直接作用于菌群。
目前,虽然口服剂型司美格鲁肽已获批准,但其对肠道菌群的直接影响尚缺乏系统性评估。
▸新视角:来自肠道菌群的DPP-IV
近期的一项关键研究为理解这一复杂关系提供了重要的新视角:
部分肠道微生物自身能够产生并分泌DPP-IV酶。这种细菌源性的DPP-IV同样会降解宿主的GLP-1,从而削弱其生理活性。
临床关联: 研究发现,在对DPP-IV抑制剂药物西他列汀反应不佳的个体中,其肠道微生物的DPP-IV活性显著更高。
干预潜力: 基于此,研究人员已鉴定出一种能特异性抑制微生物DPP-IV的抑制剂,并在动物模型中证实其能改善葡萄糖耐受性,并降低粪便DPP-IV活性。
►▷
这一发现为2型糖尿病(T2D)的治疗开辟了新思路。未来可能开发出一种联合干预策略,即同时靶向宿主和菌群来源的DPP-IV。这种策略有望更有效地保护内源性GLP-1,从而实现更优的血糖控制效果。
减重手术
两种常见的减重手术——Roux-en-Y胃旁路术(RYGB)和 袖状胃切除术(VSG),在胃肠道生理结构上的改变机制不同,但它们在治疗肥胖和糖尿病方面的效果及益处相当。
起初,人们认为手术后的机械性限制,也就说胃容量变小减少食物摄入,是体重下降与代谢改善的主要原因。然而,后续研究发现,减重手术会引发一系列深远的胃肠道生理适应,包括肠道激素分泌、胆汁酸代谢及 肠道微生物群的显著变化,这些因素彼此关联,共同促进体重下降与血糖耐受性改善。
▸ 肠道菌群的核心作用与证据
——菌群持久且独立的变化
研究表明,减肥手术后,肠道菌群会发生显著且持久的变化(至少维持十年)。这种改变似乎是独立于体重减轻或热量限制本身。
证据:与仅节食减肥个体相比,RYGB手术患者的菌群多样性增加;并且,RYGB手术的肥胖大鼠菌群,与体重相当的假手术组大鼠(未真正手术但体重通过节食控制)的菌群显著不同。
——因果关系的探索 (动物、人类)
动物实验:将接受了RYGB手术小鼠的肠道菌群移植到无菌小鼠体内,能够复制体重下降的效果,这强烈暗示了由手术介导的菌群改变在驱动体重减轻中可能具有部分因果作用。
人类研究:然而,在人类研究中结果更为复杂。将术后一年且体重下降30%的供体的菌群移植给代谢综合征患者,并未能改善其葡萄糖稳态。但值得注意的是,这种移植确实阻止了受体胰岛素敏感性的进一步恶化(相比之下,接受其他代谢综合征患者菌群移植的对照组则出现了恶化)。
这说明,肠道微生物群在RYGB的代谢改善中虽起到一定作用,但更可能是与其他机制协同发挥效应,而非单一决定因素。
▸ 菌群发挥作用的关键途径
近期的证据揭示了菌群影响代谢的具体机制,主要涉及以下两个方面:
——胆汁酸信号通路
垂直袖状胃切除术的降糖效果依赖于肠道菌群,并且需要功能正常的FXR和TGR5受体(两者均为胆汁酸受体)。
手术后的菌群改变会增加肠道中胆汁酸转运体的表达,从而促进石胆酸 (lithocholic acid) 的吸收并将其运输至肝脏。
在肝脏,石胆酸被转化为CA7S,这种物质可以直接激活TGR5受体,从而增加GLP-1的分泌,最终改善血糖控制。
——短链脂肪酸信号通路
减肥手术后,肥胖个体循环系统中的丁酸和丙酸水平显著增加,而乙酸水平则降低。
这些短链脂肪酸的变化,同样被认为有助于改善体重和葡萄糖稳态。
减肥手术带来的全面健康益处(体重减轻和葡萄糖稳态改善)不能归因于任何单一机制。它很可能是多种因素,包括肠道菌群的深远影响、胆汁酸信号的改变和短链脂肪酸的产生等共同作用的结果。
总结来说,不同的减重手术(如胃旁路术和袖状胃切除术)虽然操作方式差别很大,但减重与控糖效果相近。关键并非只是吃得少、吸收少,而是对肠道—代谢轴的系统性重启。
肠道菌群、胆汁酸、激素信号和神经代谢网络共同协作,把机体从易胖、胰岛素抵抗的状态,切换回更健康、更稳定的能量代谢模式。
以上这些发现让我们意识到,我们可能一直在间接利用菌群的力量。这自然而然地将我们引向了糖尿病治疗的下一个前沿:与其依赖药物对菌群的附带效应,不如直接将肠道菌群作为干预的核心。接下来,我们将探索那些专门为此设计的创新途径。
FMT 是目前临床上应用最广泛且成功率最高的治疗方法之一,特别是在治疗艰难梭菌感染以及其他多种胃肠道疾病中表现突出。近年来,FMT 也被探索作为一种新型的神经系统、免疫系统及代谢性疾病(包括肥胖与2型糖尿病)替代疗法。
▸ 动物研究中,FMT显著成效
糖尿病小鼠实验:持续为糖尿病模型小鼠(db/db小鼠)移植健康人类的粪便菌群,能够改善它们的葡萄糖耐量和新陈代谢。
运动与饮食实验:从健康饮食并坚持运动的小鼠身上获取菌群,并将其移植给久坐不动的高脂饮食小鼠,可以改善后者的葡萄糖耐量并减少其脂肪堆积。
肠道内容物移植:将健康大鼠的小肠内容物移植到高脂饮食的大鼠体内,可以迅速改善其对葡萄糖的耐受能力,并减少葡萄糖的生成。
动物研究普遍表明,移植健康的肠道菌群能够有效改善代谢问题。
▸ 🧍♀️人体临床研究的:效果有限且短暂
关键研究案例: 在两项人类研究中,患有代谢综合征的男性受试者,接受了来自瘦型的健康捐赠者的FMT。
短期效果(6周后): 受试者的胰岛素介导的葡萄糖摄取能力得到改善,糖化血红蛋白(HbA1c)水平也下降了。这是一个积极的信号。
长期效果(18周后):然而,之前观察到的有益效果不再显著。
▸ 为什么在人类身上效果不佳?
▸ 未来的改进方向
为了让FMT成为治疗代谢疾病的可行方案,需要更精细化的策略:
优化方案:需要调整FMT的时间、重复和剂量。
联合疗法:将FMT与生活方式干预(如改变饮食、补充膳食纤维)相结合。
近期的两项临床试验证明,当FMT与饮食改变或膳食纤维补充相结合时,这种联合策略确实能改善肥胖或2型糖尿病患者的代谢结果。
个性化FMT:类似于个性化医疗,未来可能需要发展“个性化粪菌移植”。即根据每个患者的具体情况,精心挑选或设计最适合他们的菌群来进行移植。
谷禾也会根据肠道菌群检测报告为合作方提供相关指标,便于更好地筛选与匹配。
多项研究表明,在代谢性疾病(包括肥胖与2型糖尿病)的动物和人类模型中,补充益生菌可在急性及长期阶段均显著改善多项代谢指标。
在这里,介绍用产乳酸菌、产丁酸菌和特定物种Akkermansia muciniphila治疗2型糖尿病和其他代谢疾病状态的效果。
AKK菌|Akkermansia muciniphila
Akkermansia muciniphila(简称AKK菌)是一种革兰氏阴性菌,具有降解黏蛋白的能力,主要定植于肠道黏液层,并可在粪便样本中检测到。
肠道粘液层的主要成分是粘蛋白(Mucins),粘蛋白是一种富含糖基的结构蛋白,是肠道黏液层的重要组成部分,这也是Akk菌的“主食”。
AKK菌与健康的关系:绝大多数证据指向有益
大量研究发现,在啮齿动物与人类中,肥胖或2型糖尿病患者体内的Akk菌丰度都显著偏低;当其丰度减少时,常伴随炎症水平上升、肝脏脂肪变性以及胰岛素抗性增强。
有一项宏基因组学研究报告指出,AKK菌的增加与2型糖尿病相关,理由是它降解粘蛋白会破坏肠道粘液层的完整性。
更多的研究表明,AKK菌的丰度与更厚的粘液层和更低的肠道通透性(即更少的“肠漏”)正相关。它似乎能通过一种尚不完全明确的机制,刺激肠道中负责生产粘蛋白的杯状细胞数量增加,从而实现“越吃越有”的良性循环。
Akk菌如何发挥作用?
Akk菌的益处主要通过其菌体上的特定成分和它分泌的蛋白质来实现。
Akk菌外膜上的一种叫 Amuc_1100 的蛋白质。
作用过程: 这种蛋白质可以激活肠道细胞上的TLR2受体,进而调节一系列负责细胞连接的紧密连接蛋白(如Claudin 3,Occludin等)的表达。
最终效果: 这大大增强了肠道细胞间的连接,加固了肠道屏障。屏障加固后,肠道中的有害物质(如内毒素LPS)就难以泄漏到血液中,从而减轻了全身的低度炎症,最终改善胰岛素抵抗和血糖控制。
最近,科学家发现Akk菌还能分泌一种名为 P9 的新型蛋白质。
作用效果: 在小鼠实验中,P9被证实能够促进GLP-1(一种重要的降糖激素)的分泌,从而改善葡萄糖耐量。
研究发现,活的Akk菌和巴氏杀菌的Akk菌都有效,但高温彻底灭活(Heat-killed/Autoclaved)的Akk菌则无效。
原因在于:巴氏杀菌的温度(约70°C)足以杀死细菌,但不会破坏关键蛋白Amuc_1100的结构和活性。而更高的灭菌温度则会使其变性失效。这为开发安全的菌剂产品提供了重要依据。
🧍♀️人体临床证据
Akk菌的益处已经在人体上得到初步验证。
安全性:早期的临床研究证实,无论是活菌还是巴氏杀菌的Akk菌,对人体都是安全且耐受性良好的。
有效性: 在一项针对代谢综合征患者的研究中,与安慰剂组相比,服用巴氏杀菌Akk菌的患者出现了显著的积极变化:
►▷
综合来看,上述动物与早期临床研究一致显示:
AKK菌的干预可重建肠道屏障完整性、降低代谢炎症并改善血糖稳态,提示该菌株具有成为治疗肥胖与2型糖尿病的潜在候选益生菌的巨大应用前景。
产丁酸菌
产丁酸菌并非单一菌种,而是一个庞大的功能性菌群。产丁酸菌是一类能够在厌氧环境下、通过发酵多种底物产生丁酸的肠道共生菌群。
★ Faecalibacterium prausnitzii
F. prausnitzii 是一种严格厌氧、产丁酸的共生菌,在小鼠与人类结肠中含量丰富。
为什么它如此重要?(关联性证据)
研究发现,2型糖尿病患者体内 F. prausnitzii 的丰度显著降低,而在接受减重手术后,该菌丰度显著增加,且与炎症标志物水平呈显著负相关。
这一系列发现表明,F. prausnitzii 可能通过减少肠道炎症、增强肠屏障完整性、改善代谢性内毒症,从而发挥潜在益生作用。
它如何发挥作用?
在结肠炎小鼠模型中,给予 F. prausnitzii 或其上清液,可显著增加肠上皮紧密连接蛋白表达,并降低炎症性细胞因子水平。
一个关键的发现是:单独使用丁酸,其抗炎效果远不如用F. prausnitzii的培养上清液。这说明,F. prausnitzii的益处并不仅仅来自于丁酸,它还分泌其他抗炎的有益物质。
研究人员后来确实鉴定出了一种由F. prausnitzii产生的“微生物抗炎分子”(microbial anti-inflammatory molecule, MAM),这种分子能够修复糖尿病小鼠的肠道屏障功能,并上调紧密连接蛋白表达。
给予F.prausnitzii可以改善啮齿动物的葡萄糖稳态,并且对2型糖尿病患者进行GLP-1RA治疗可以增加粪便中F.prausnilzii的丰度,这与空腹血糖呈负相关。
虽然目前尚无直接临床试验验证F. prausnitzii 在血糖调节中的疗效,但近期一项研究已证实,其长期补充在人体中是安全且可耐受的,并已启动相关的临床试验以进一步评估其作为下一代益生菌的潜力。
★ Anaerobutyricum soehngenii
这个菌它最初是在一项粪菌移植临床试验中引起关注的。研究者观察到,Anaerobutyricum属增加与受试者胰岛素敏感性改善密切相关。
动物实验:给糖尿病小鼠补充 A. soehngenii 可显著提高粪便中丁酸及次级胆汁酸水平,增强胰岛素敏感性,提升能量消耗。
🧍♀️人体临床试验:两项针对代谢综合征人群的临床试验表明,A. soehngenii 的口服或十二指肠输注均安全且耐受良好;单次十二指肠灌注实验显著提升GLP-1水平,并改善胰岛素分泌与敏感性。
►▷
无论是F. prausnitzii还是A. soehngenii,它们的益处是多方面的,不仅仅是生产丁酸,还包括分泌其他抗炎分子和调节宿主激素(如GLP-1)。这些产丁酸菌的研究都凸显了它们作为治疗代谢疾病的新型疗法的巨大潜力。
产乳酸菌
产乳酸菌是一类耐酸的革兰氏阳性菌,通常不具运动性,主要通过发酵碳水化合物产生乳酸作为主要代谢产物。乳酸是厌氧呼吸中的重要代谢物,既是宿主能量代谢中关键的中间产物,也可作为底物被其他肠道菌群利用以产短链脂肪酸。
明星成员: 其中最著名、研究最广泛、应用最普遍的就是乳杆菌属和双歧杆菌属的成员。
动物研究,效果显著
在啮齿类动物模型中,补充多种乳酸生成菌可显著带来以下代谢益处:
在多项研究中,使用以下菌株治疗,显著改善小鼠的血糖控制:
乳杆菌属
双歧杆菌属
核心作用机制:巧妙抑制FXR信号,促进GLP-1分泌
产乳酸菌改善血糖的机制相当精妙,其中一个核心通路与胆汁酸受体FXR有关:
1
分泌“胆盐水解酶” (BSH)
许多乳杆菌能产生一种叫做“胆盐水解酶”的工具。
2
分解胆汁酸
胆盐水解酶这个工具可以将“结合型胆汁酸”分解为“游离型胆汁酸”。
3
抑制肠道FXR活性
“游离型胆汁酸”激活肠道FXR受体的能力较弱,因此,这一过程的最终结果是降低了肠道FXR信号的整体活性。
4
解放GLP-1
关键点来了,肠道中的FXR受体被激活时,会抑制GLP-1(一种重要的降糖激素)的分泌。因此,当产乳酸菌抑制了FXR后,就相当于解除了对GLP-1的束缚,从而促进了GLP-1的释放,改善了血糖控制。
双重协同作用:抑制FXR还能增强短链脂肪酸诱导GLP-1分泌的效果。同时,某些产乳酸菌自身也能促进丁酸等短链脂肪酸的产生。这种 抑制FXR + 增加短链脂肪酸的协同作用,可能共同放大了GLP-1的分泌。
🧍♀️ 人体临床研究,好坏参半
与动物实验的普遍成功形成鲜明对比,产乳酸菌在人体临床试验中的表现,结果好坏参半:
☺一项研究显示,包含多种产乳酸菌的复合益生菌产品,相比安慰剂,能够降低2型糖尿病患者的糖化血红蛋白(HbA1c)和空腹血糖。
☺另一项研究发现,单独使用植物乳杆菌也能降低餐后血糖和HbA1c。
☹ 然而,与安慰剂对照组相比,补充罗伊氏乳杆菌、嗜酸乳杆菌或乳双歧杆菌并没有显著改善健康或糖尿病患者的血糖控制或胰岛素敏感性。
☺一个值得注意的细节是,在一项研究中,乳双歧杆菌(B. lactis)虽然没有改善代谢综合征患者的指标,但成功维持了他们的胰岛素敏感性,阻止了其进一步恶化。
鉴于在糖尿病患者体内,许多产乳酸菌(尤其是双歧杆菌)的数量有所减少,那么通过“缺啥补啥”的思路来恢复它们的水平,理论上应是一种有前景的治疗方法。
然而,现有的人体临床证据有限,乳酸菌对改善代谢性疾病的作用不足,因为每个人的饮食、基线肠道菌群和疾病表现都千差万别(即高度异质性),想用标准化的益生菌对所有人都产生效果,是不容易的。因此,益生元等可以增加整体微生物群的多样性和丰富性,而不是单一细菌,这可能会为更广泛的人群提供更有效的治疗选择。
2型糖尿病的治疗策略
doi.org/10.1146/annurev-physiol-051524-094728
益生元被定义为:“能够被选择性发酵,并导致胃肠道微生物群组成和/或活性产生特定变化,从而对宿主健康带来益处的成分。”
简单来说,一种物质要被称为“益生元”,必须满足苛刻的条件:
基于这些标准,许多食物或成分都可以被归入益生元的范畴。在众多候选物质中,当前研究最为集中、并在糖尿病治疗中表现出潜在疗效的包括几类新型功能性益生元。
菊粉型果聚糖(ITFs)
菊粉型果聚糖(ITFs)——主要包括菊粉(inulin)、低聚果糖(OFS)和果寡糖(FOS),它们本质上是一类由果糖分子链组成的不可消化性碳水化合物。能够被肠道细菌发酵利用,从而促进宿主健康。
它们如何发挥作用?—— 多途径、多靶点的系统工程
核心作用:精准“施肥”,优化菌群
机制一:放大GLP-1信号,控制血糖
这是菊粉型果聚糖改善血糖的核心途径。
证据: 益生菌(特别是双歧杆菌)利用菊粉型果聚糖作为底物,发酵产生短链脂肪酸,如乙酸和丁酸。而短链脂肪酸是已知的肠道L细胞分泌GLP-1的强效刺激剂。
铁证: 在GLP-1受体被基因敲除的小鼠中,或者在使用了GLP-1受体拮抗剂的小鼠中,菊粉型果聚糖带来的所有降糖益处都完全消失了。这证明了GLP-1通路在其中的决定性作用。
菊粉型果聚糖也能显著提高人和动物的餐后GLP-1水平。
机制二:激活GLP-2信号,修复肠道屏障
GLP-2可以看作是GLP-1的兄弟,它主要负责维持和修复肠道屏障的完整性。
研究发现,补充菊粉型果聚糖能够增加内源性GLP-2的分泌。如果在小鼠中阻断GLP-2受体,那么菊粉型果聚糖带来的修复肠道屏障、抗炎等益处也大部分会消失。
机制三:调节内源性大麻素系统
这是一个更深层的机制。菊粉型果聚糖诱导的菌群变化(或AKK菌的增加)能够调节肠道的内源性大麻素信号系统(endocannabinoid),增加2-棕榈酰甘油(2-palmitoylglycerol)、2-油酰甘油(2-oleoylglycerol)和2-花生四烯酰甘油(2-arachidonoylglycerol);降低花生酰胺(anandamide)水平;进而增强肠道屏障功能,减少LPS泄漏。
🧍♀️ 人体临床证据
虽然不是所有研究都报告了积极结果,但大量的临床试验已经证实了菊粉型果聚糖在人体中的益处:
►▷
菊粉类果聚糖代表了一种非常有前景的、非侵入性的2型糖尿病治疗方案。它不像直接补充益生菌那样面临能否存活和定植的问题,而是通过为体内已有的有益菌(如Akk菌和双歧杆菌)提供精准的养料,激发一连串有益的生理反应——核心是驱动GLP-1和GLP-2的分泌,最终达到控制血糖和修复肠道屏障的双重目的。
其他膳食纤维
首先,一个基本共识是:增加膳食纤维的摄入量,能显著降低患上肥胖和2型糖尿病的风险。
膳食纤维主要分为两大类:
🧬 植物来源的主要可溶性纤维包括:
然而,研究发现,不同可溶性纤维的效果差异很大,这取决于纤维的用量、食物来源、补充方式(是直接吃食物还是吃提纯的纤维补充剂)等因素。
几种可溶性纤维的表现:
🧪 β-葡聚糖—— 全能选手
来源: 主要存在于燕麦和大麦中。
一项对比研究发现,在高脂饮食中添加富含β-葡聚糖的大麦粉,能增加肠道丁酸、改善血糖。但如果直接添加提纯的β-葡聚糖纤维,同样能改善代谢。这表明β-葡聚糖本身就是强效的功能成分。
⁎ 作用机制
无论是动物还是人体研究,都强有力地支持β-葡聚糖作为一种益生元,在改善血糖和治疗代谢疾病方面具有巨大潜力。
🌽 抗性淀粉—— 瞒过小肠的特工
抗性淀粉它本质上是淀粉,但由于其特殊的结构,能抵抗小肠的消化,完整地到达结肠,成为微生物的食物。
来源:冷却的米饭、土豆、未完全成熟的香蕉等。
⁎ 作用机制(与β-葡聚糖类似):
🧍♀️ 人体证据
在患有代谢综合征和2型糖尿病的人群中,补充抗性淀粉(特别是2型抗性淀粉,如高直链玉米淀粉)能够改善胰岛素敏感性并降低空腹血糖,同时也能观察到短链脂肪酸产量的增加。
一个有趣的发现:补充高直链玉米淀粉会特异性地增加双歧杆菌的丰度,而双歧杆菌正是一种擅长发酵抗性淀粉的细菌。
🍎 果胶 —— 机制独特
来源: 广泛存在于水果中,如苹果、柑橘等。
⁎ 作用机制(与其他不同):
►▷
简而言之,膳食纤维是“好菌的食物,也是代谢的调节器”:当你吃更多含纤维的天然食物(燕麦、豆类、全谷、蔬果),肠道菌群就会发酵产出丁酸等代谢产物,能修复肠道、提升GLP-1分泌、降低炎症、帮助降糖。β-葡聚糖与抗性淀粉尤其被视为下一代代谢健康促进纤维,未来可能成为糖尿病膳食疗法的核心成分。
合生元是一种同时包含益生菌和益生元的产品。
这个设计的初衷是,通过提供益生菌最喜欢吃的益生元,来帮助它在复杂的肠道环境中更好地存活、定植并发挥作用。
鉴于益生菌能产生短链脂肪酸,而益生元是产生短链脂肪酸的代谢前体物质(原料),那么将两者结合,应能够协同增加短链脂肪酸的产生,从而对代谢健康产生比单独使用任何一种都更强大的益处。
🧬 合生元的常见组合
双歧杆菌/乳杆菌 + 低聚糖/膳食纤维
这种组合的设计理念是:由益生元提供底物,使益生菌能够在肠道定植并活跃代谢,从而增强肠道生态修复与宿主代谢调节效应。
并未实现“1+1 > 2”的突破
动物与人体研究均表明,补充益生元或益生菌本身即可带来代谢性益处,合生元干预能够改善肠道菌群生态与增加粪便SCFA含量。然而,在大多数研究中,联合应用(合生元)并未显示出明显优于单独使用益生元或益生菌的效果。
为什么这个看似完美的策略效果不理想?
研究人员认为是由于缺乏精准匹配,也就是说大多数研究在设计合生元产品时,只是想当然地将一种常见的益生菌和一种常见的益生元组合在一起,但未能首先验证,这个益生菌是否真的喜欢吃、并且能高效利用配给它的那个益生元。
这并不意味着合生元的理念是错误的,而是说明我们的执行方式需要更加科学和严谨。
改进:在进行昂贵且复杂的人体临床试验之前,必须增加一个关键的验证步骤:
►▷
合生元的概念本身极具潜力,但需确保益生菌和益生元是真正的天作之合,这种“1+1>2”的协同效应才会在临床上有更好的效果。这要求我们对菌株和底物的特性有更深入的了解和更严格的前期验证。
外源化合物(Xenobiotics)被定义为:“在机体内非天然存在的外来化学物质。”这个范畴非常广泛,从化妆品、药物到膳食补充剂中的成分都可能属于外源物。
前文已提到二甲双胍(metformin)与GLP-1受体激动剂(GLP-1RA)的降糖作用同样被认为与肠道微生物介导机制相关,而这些药物本质上也属于外源化合物的范畴。
虽然很多外源物对健康有害,但我们这里聚焦的是那些有益的外源物,重点分析两种备受关注的有益外源物:多酚和小檗碱。
多 酚 类 化 合 物 (Polyphenols)
多酚是一大类存在于植物中的化合物,如白藜芦醇、类黄酮等。
多酚类化合物是植物来源的次级代谢物,主要包括:
这些分子既能调节肠道微生物群结构,也能改善2型糖尿病的多种代谢指标。
💊白藜芦醇(Resveratrol)
白藜芦醇是多酚中研究最为深入的代表性化合物之一。
来源: 葡萄皮、红酒、花生等。
它可显著改善胰岛素敏感性、葡萄糖稳态、血脂水平、高血压等。但它的生物利用度很低,口服后能进入血液循环的量很少,而且大部分还和蛋白质绑定,无法发挥活性。
那它是如何起作用的?
答案指向了肠道。
口服有效。 多项研究发现,口服白藜芦醇可以改善小鼠的血糖,但腹腔注射(绕过了肠道)则无效。这有力地证明了它的作用点在肠道内。
口服补充剂也有益地改变了啮齿动物的肠道微生物组,降低了厚壁菌门/拟杆菌的比例,增加了Akk菌、双歧杆菌和乳杆菌的丰度,这与改善葡萄糖稳态和炎症标志物有关。
最终证据——粪菌移植: 将服用过白藜芦醇的小鼠的粪菌移植给普通高脂饮食的小鼠,后者也出现了血糖改善的效果。
🧍♀️ 人体临床研究
尽管研究尚少,但已有临床试验发现,补充白藜芦醇可以增加代谢综合征男性体内的AKK菌,并轻微改善血糖。
💊 黄酮类化合物(Flavonoids)
除白藜芦醇外,黄酮类化合物也能增加肠道有益菌数量,并改善葡萄糖耐受性、抑制炎症反应、增强肠屏障功能。
来源: 广泛存在于各种水果、蔬菜、茶中。
动物实验研究
在高脂饮食诱导的肥胖或糖尿病小鼠模型中:
蔓越莓提取物可提高胰岛素敏感性、降低HOMA-IR与循环内毒素(LPS)水平;同时伴随AKK菌丰度显著上升与肠屏障完整性增强。
蓝莓原花青素含有丰富的花青素与原花青素,同样可改善葡萄糖耐受;增加AKK菌含量;并增厚胃肠道黏液层。
苹果原花青素处理高脂饮食小鼠时,也观察到一致的结果:肠道屏障功能及AKK菌丰度同步提升。
这些研究揭示不同植物来源的黄酮类多酚均通过促进有益菌的富集和黏液层重塑,实现改善肠屏障与系统代谢功能的作用。
🧍♀️ 人体及体外研究证据
尽管人群中关于单独考察黄酮类化合物作用的临床研究仍然有限,但已有体外模拟与部分干预研究提供了支持性证据:
红酒葡萄提取物或红茶多酚在人体肠道模拟模型中均能增加AKK菌的丰度。
可可黄烷醇则在健康人群中可显著增加双歧杆菌和乳杆菌的数量,表明其对肠道菌群结构也具有积极调节作用。
综上,黄酮类化合物以其独特的双重功能而受到关注:
这些作用共同促进了葡萄糖代谢与胰岛素敏感性的改善,为其作为代谢性疾病营养干预的潜在候选物提供了有力依据。
小 檗 碱
来源: 黄连、黄柏等中草药的主要活性成分。
小檗碱可以直接作用于肝细胞,激活AMPK(一个关键的能量代谢开关),从而改善血糖(增强胰岛素敏感性与葡萄糖稳态控制)。但它的口服生物利用度同样很低。
小檗碱是否进入循环系统,依赖肠道菌群
小檗碱只有在被肠道菌群代谢为更易吸收的衍生物后,其药效才能被充分发挥。
——肠道菌群:加工
肠道菌群能将小檗碱转化为更容易被人体吸收的形式,帮助它进入血液循环。
例如,某些肠道细菌可将小檗碱还原为二氢小檗碱,这种形式在肠上皮中的吸收效率更高,随后可重新氧化为小檗碱并在全身循环中发挥作用。
——肠道菌群:效应放大器
动物实验表明,口服(而非注射)小檗碱能增加肠道产丁酸菌的数量和丁酸产量,并改善空腹血糖。这说明小檗碱的另一部分功效是通过调节菌群-丁酸实现的。
临床研究与争议
动物研究:在动物模型中,小檗碱改善血糖的机制似乎与增加GLP-1/GLP-2、增加次级胆汁酸(DCA)从而激活TGR5受体、增加SCFA产量等有关。这是一个非常清晰、正面的通路。
人体研究的反转:迄今为止唯一一项相关的糖尿病人体研究却得出了矛盾的结果:小檗碱使血液中次级胆汁酸和有益的产短链脂肪酸菌的显著下降了,但同时它确实又显著降低了患者的糖化血红蛋白。
这凸显了小檗碱作用机制的复杂性。小檗碱确实是一种有效的降糖物质,但它究竟是如何通过菌群在人体内发挥作用的,远比我们想象的要复杂,目前仍是一个充满争议和未解之谜,其肠道依赖机制亟需更多临床研究验证来阐明。
►▷
小檗碱就像一个“需要肠道助手的药”:它自己难以被吸收,但一旦被肠道菌群“加工”成可利用的形式,便能帮助调节血糖、减少炎症、修复肠屏障。不过,因为每个人肠道菌群不同,它在不同人身上可能表现出截然不同的效果。
尽管大量动物研究已明确证实,肠道菌群在血糖稳态调控中扮演着核心角色,但将这些发现直接转化为对人类有效的通用疗法却挑战重重。其根本原因在于人类肠道菌群的复杂性与高度个体化。
这种个体差异正是许多干预措施(如益生元或益生菌)效果因人而异的关键。
益生元:当人们吃下富含β-葡聚糖的特制大麦面包后,只有一部分人的代谢状况得到了改善。科学家检查后发现,这些“有效者”在干预开始时,体内就存在着“无效者”所缺乏的特定肠道细菌。是这些细菌帮助他们更好地利用了膳食纤维。
益生菌:吃下益生菌,不代表它们就能在你的肠道里安家落户,一项研究发现,11种常见益生菌能否成功在肠道定植,取决于每个人已有的菌群构成。
既然每个人都是不同的,那么治疗方案也必须因人而异,兼顾菌群个体差异与动态变化。这个“个性化”会是什么样子呢?
比如,个性化饮食算法,这个算法不仅分析一个人吃了什么,更整合了这个人肠道菌群数据、血液指标等多种参数。这样不仅可以预测这个人吃下特定食物后的血糖反应,更能反过来为他量身定制一套可以最大程度平稳餐后血糖的个性化饮食方案。
在糖尿病前期人群中,通过整合基线菌群特征,机器学习模型,也能预测哪些患者能通过什么样的运动有效改善血糖,而哪些患者则收效甚微。
简而言之,要让基于微生物群的疗法真正在人体中奏效,我们需要摆脱“一刀切”的思路,而是把每个人的肠道菌群当作独特的生态系统。未来的糖尿病干预,可能需要“定制化的微生物处方”:先读取你的菌群模式,再精准投喂适合的菌与食物,让微生物群自己帮你稳糖、抗炎、调代谢。
深入理解并善用
每个人的独有菌群信息
将是我们开启
代谢疾病精准治疗
新时代大门的钥匙
注:本账号内容仅作交流参考,不作为诊断及医疗依据。
主要参考文献
Weninger, Savanna N., Andrew Manley, and Frank A. Duca. “Managing Glucose Homeostasis Through the Gut Microbiome.” Annual Review of Physiology 88 (2025).
Howard, Elizabeth J., Tony KT Lam, and Frank A. Duca. “The gut microbiome: connecting diet, glucose homeostasis, and disease.” Annual review of medicine 73 (2022): 469-481.
Cho, Hyoung-Soo, et al. “Structure of gut microbial glycolipid modulates host inflammatory response.” Cell 188.19 (2025): 5295-5312.
Sun, Hanxiao, et al. “GLP‐1 receptor agonists alleviate colonic inflammation by modulating intestinal microbiota and the function of group 3 innate lymphoid cells.” Immunology 172.3 (2024): 451-468.
Su, Lili, et al. “Health improvements of type 2 diabetic patients through diet and diet plus fecal microbiota transplantation.” Scientific reports 12.1 (2022): 1152.
Anhê, Fernando F., et al. “Metabolic endotoxemia is dictated by the type of lipopolysaccharide.” Cell reports 36.11 (2021).
Baroni, Irene, et al. “Probiotics and synbiotics for glycemic control in diabetes: A systematic review and meta-analysis of randomized controlled trials.” Clinical Nutrition 43.4 (2024): 1041-1061.
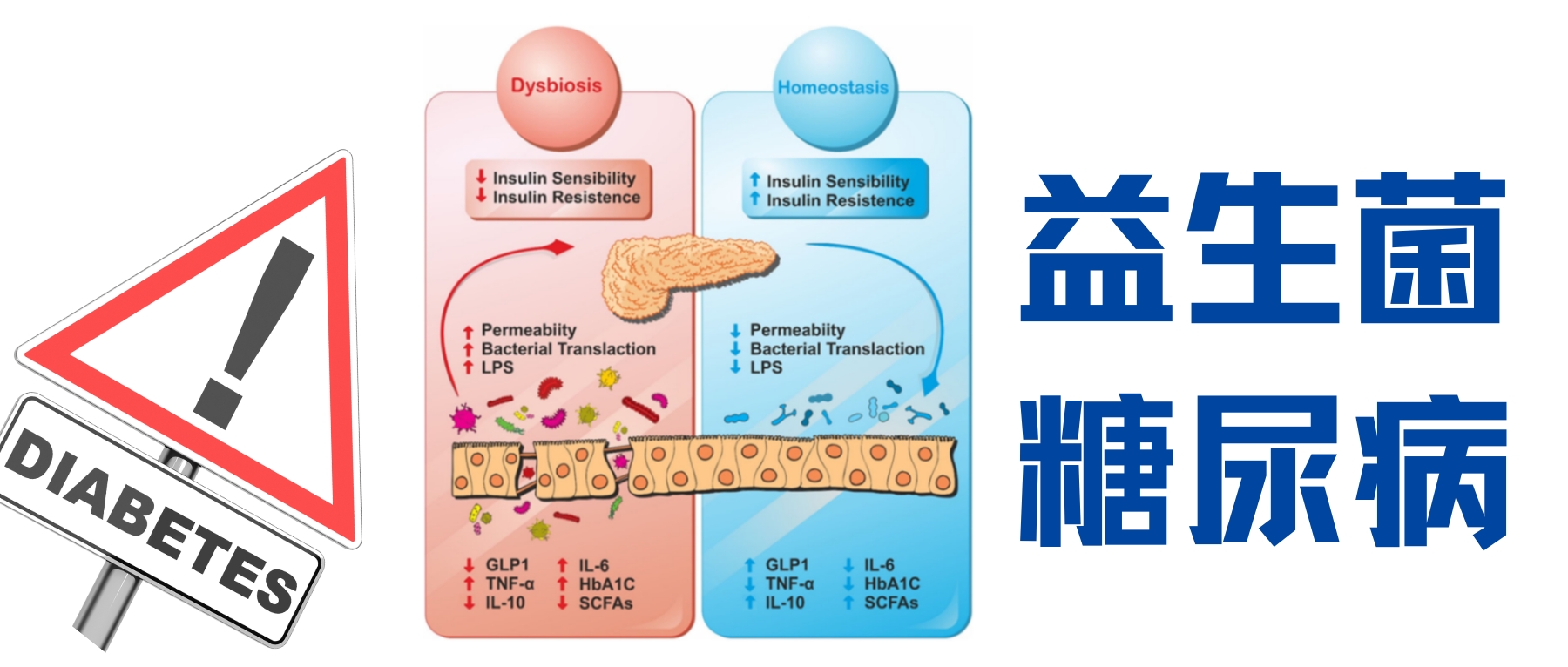
谷禾健康
糖尿病,一个日益严峻的全球健康挑战,其影响远不止血糖升高那么简单。在血糖波动的背后,看不见的慢性低度炎症,正悄然侵蚀着患者的健康,加速并发症的到来。
当我们谈论炎症时,脑海中浮现的可能是皮肤割伤后的红肿热痛,那是身体对损伤的快速、有益的防御反应,属于急性炎症;然而还有一种更隐蔽、更持久的炎症形式——慢性低度炎症,它没有明显症状,却像温水煮青蛙般长期损害我们的身体,被认为是心脏病、癌症以及糖尿病等诸多慢性疾病的共同根源。
这场炎症风暴的源头可能隐藏在我们体内最复杂的微生态系统——肠道菌群中。研究发现,糖尿病患者常存在菌群失衡:有益菌(如双歧杆菌、乳杆菌)减少,潜在致病菌增多,这些改变会导致肠道通透性增加(肠漏),使脂多糖(LPS)等微生物成分进入血液,通过 Toll 样受体激活引发全身性炎症。形成“菌群失调→肠漏→炎症→胰岛素抵抗”的恶性循环。
能否通过调节肠道菌群来阻断这一恶性循环?益生菌,作为定植于肠道的友好菌群,为我们提供了新的解题思路。在代谢性疾病中常见的益生菌属包括双歧杆菌属、乳杆菌属等通过发酵膳食纤维产生短链脂肪酸(SCFAs)——如丁酸(增强肠道屏障、直接抗炎)、丙酸(调节肝脏糖代谢)、乙酸(参与全身代谢),这些短链脂肪酸具有抗炎和改善胰岛素敏感性的作用。
但益生菌的抗炎效果是否真能得到科学验证?不同菌株、剂量的干预差异何在?这些问题亟待更权威的证据解答。
近日,一篇荟萃分析发表在《Frontiers in Cellular and Infection Microbiology》上,该研究系统整合了46项随机对照试验,覆盖3580名糖尿病患者,旨在揭示益生菌及其代谢产物对糖尿病患者体内炎症标志物的真实影响。
这项研究不仅回答了“益生菌是否有效”的简单疑问,更深入探究了“哪种方案更优”、“需要多长时间见效”以及“其作用机制是否与短链脂肪酸直接相关”等核心问题。为临床实践提供了可靠的循证医学指导,也为未来精准干预策略的制定奠定了坚实的科学基础。
在糖尿病患者体内,慢性炎症状态尤为普遍。它不仅是疾病的结果,更是推动疾病恶化的帮凶。
炎症因子,作为炎症反应中的信使分子,在其中扮演了关键角色,就像一支特殊的信号部队,负责传递指令,协调免疫系统的行动。炎症因子主要分为两大类:
促炎因子
点燃和助长炎症的分子,如肿瘤坏死因子-α (TNF-α)、白细胞介素-6 (IL-6) 和 白细胞介素-1β (IL-1β)。
在糖尿病中,这些因子水平的升高会干扰胰岛素信号传导,加剧胰岛素抵抗,损害血管内皮功能,为心血管并发症埋下祸根。
抗炎因子
它们是负责对抗炎症和修复的分子,其中最著名的是白细胞介素-10 (IL-10)。它能够抑制促炎因子的产生,帮助炎症反应消退,维持免疫系统的平衡。
还有一个重要的炎症标志物——C反应蛋白 (CRP)。它由肝脏在炎症刺激下产生,是临床上常用于衡量全身性炎症水平的指标之一。在糖尿病患者中,CRP水平的持续轻度升高,预示着心血管事件的风险增加。
研究表明,在明显的临床症状出现之前,这些炎症因子的水平可能已经悄然改变。而最早发出预警信号的,可能就是我们的肠道菌群。
健康的肠道菌群平衡(共生)对维持新陈代谢和免疫稳态至关重要。然而,在糖尿病患者中,这种平衡常常被打破,出现所谓的菌群失调——有益菌(如双歧杆菌、乳酸杆菌)减少,而一些潜在的致病菌增多。
菌群失调会导致肠道屏障功能受损,即肠漏。这使得细菌的某些成分(如脂多糖LPS)更容易进入血液,触发全身性的炎症反应。因此,通过调节肠道菌群来恢复平衡,成为控制糖尿病相关炎症的一个极具前景的策略。
益生菌,即“摄入足够数量时能对宿主健康产生有益作用的活的微生物”,正是实现这一目标的有力工具。常见的用于改善代谢疾病的益生菌属包括乳杆菌属、双歧杆菌属等。
益生菌发挥作用的一个关键机制,是通过发酵我们无法消化的膳食纤维(如菊粉、低聚果糖等益生元),产生一系列代谢产物如短链脂肪酸 (SCFAs) 。其中最重要的三种是:
这些短链脂肪酸不仅在肠道局部发挥作用,还能进入血液循环,对全身的免疫细胞和代谢器官产生深远影响,被认为是连接肠道菌群与宿主免疫的关键信使。
那么,补充益生菌是否真的能增加短链脂肪酸,是否能有效降低糖尿病患者的炎症水平呢?
为了解决更深入了解以上问题,近日一项大规模的系统回顾和荟萃分析,严格遵循PRISMA指南,从全球六大数据库中检索了从2000年1月至2024年3月的所有相关研究。
经过层层筛选,最终纳入了46项符合标准的随机对照试验,共涉及3580名1型或2型糖尿病患者。这些研究评估了口服益生菌或合生元(合生元是指益生菌+益生元)对炎症标志物(CRP, IL-6, TNF-α, IL-10)和短链脂肪酸水平的影响。
(对炎症标志物和菌群代谢物的影响)
经过对46项研究数据的严谨分析,研究者们得出了相关结果。以下我们将详细解读益生菌/合生元对各项关键指标的影响。
C-反应蛋白(CRP)——显著下降
在34项报告了CRP数据的研究中,合并分析结果显示,与安慰剂组相比,益生菌/合生元干预显著降低了CRP水平(SMD = –0.54;95% CI:–0.72 至–0.35;p < 0.0001),异质性为中度(I² = 64%),这是一个中等强度的、具有临床意义的改善。
更深入的亚组分析揭示了几个关键因素:
干预时长是关键
干预时间≥8周的研究显示出更强的CRP降低效果(SMD = –0.66),而短于8周的干预效果较弱(SMD = –0.31)。这表明,调节肠道菌群并影响全身免疫系统需要一定的时间。
“1+1 > 2”的合生元效应
使用合生元(益生菌+益生元如菊粉或低聚果糖)的干预比单独使用益生菌效果更优(SMD = –0.71 vs. –0.45)。这印证了益生元为益生菌提供食物,从而增强其功能和代谢产物生成的理论。
研究者发现,体内丁酸和丙酸水平的升高与CRP的下降显著相关。这首次在如此大规模的人群证据中,将益生菌的代谢产物(SCFAs)与全身炎症标志物的改善直接联系起来,揭示了其作用的可能机制。
白介素-6(IL-6):关键促炎因子下降
IL-6是驱动胰岛素抵抗和血管功能障碍的关键促炎因子。在29项报告了IL-6数据的研究中,益生菌/合生元干预同样显示出显著的降低效果(SMD = –0.41)。
亚组分析发现,干预时长≥12周时效果更为明显(SMD = –0.51),进一步证实了长期干预的必要性。
此外,分析还提示了菌株的特异性,含有干酪乳杆菌 (Lactobacillus casei) 和 短双歧杆菌(Bifidobacterium breve) 的制剂在降低IL-6方面表现尤为突出。
多变量荟萃回归模型再次证实,即使在校正了年龄和BMI等混杂因素后,丁酸和丙酸水平的升高仍然是IL-6降低的独立预测因素。这强有力地支持了肠道来源的短链脂肪酸通过抑制细胞因子来缓解糖尿病中代谢应激的假说。
益生菌来源的SCFAs降低IL-6的机制通路
doi.org/10.3389/fcimb.2025.1677671
TNF-α:胰岛素抵抗元凶之一被抑制
TNF-α在脂肪组织炎症和胰岛素抵抗中扮演着核心角色。26项RCTs的数据显示,益生菌干预显著降低了TNF-α水平(SMD = –0.48)。分析发现了一些有趣的差异:
对2型糖尿病更有效
在2型糖尿病患者中,TNF-α的降低效果(SMD = –0.52)比1型糖尿病患者(SMD = –0.36)更显著。这可能因为2型糖尿病通常与更严重的代谢综合征和胰岛素抵抗相关,为益生菌提供了更大的改善空间。
多菌株的协同作用
使用多菌株混合制剂的干预效果优于单菌株制剂(SMD = –0.50 vs. –0.28),这提示不同菌株之间可能存在功能互补和协同增效的作用。
相关性分析再次给出了强有力的证据:
在那些同时测量了丁酸水平的研究中,丁酸水平的变化与TNF-α的变化呈现出极强的负相关(r = –0.72)。这意味着丁酸水平越高,TNF-α的下降越明显。
益生菌衍生的丁酸盐在糖尿病中调节TNF-α和炎症信号通路
doi.org/10.3389/fcimb.2025.1677671
IL-10:抗炎主力军增强
与促炎因子下降相反,抗炎因子IL-10的水平在干预后出现了显著升高(SMD = +0.38),这表明免疫系统正向着一个更平衡、更具耐受性的状态转变。在18项研究中,这一发现在统计学上非常稳健。
合生元再次显示出其优越性,其提升IL-10的效果最为强劲(SMD = +0.49)。与IL-10升高最相关的菌株包括植物乳杆菌 (Lactobacillus plantarum) 和 长双歧杆菌 (Bifidobacterium longum)。
荟萃回归分析发现,丙酸浓度的升高是IL-10增加的强预测因子。这为SCFAs不仅能抑制促炎因子,还能增强抗炎因子提供了证据,共同促进了免疫稳态的恢复。
微生物代谢物变化
21项研究直接定量了SCFAs,为上述所有发现提供了直接的生化基础。合并分析显示,干预后:
这些代谢物的增加在使用合生元和多菌株配方的研究中最为明显,特别是那些包含长双歧杆菌 (B.longum)、鼠李糖乳杆菌 (L. rhamnosus) 和 植物乳杆菌 (L.plantarum) 的制剂。
这些发现无可辩驳地证明,益生菌/合生元干预确实能够改变肠道菌群的代谢功能,增加有益的短链脂肪酸产量。而这些短链脂肪酸,正是连接肠道与全身免疫系统,发挥抗炎作用的关键介质。
稳健性于偏倚性
为了确保结论的可靠性,研究者还进行了一系列严格的敏感性分析和发表偏倚评估。结果显示,即使剔除单个研究或质量较低的研究,总体结论依然不变,表明结果非常稳健。
虽然在CRP的分析中检测到轻微的发表偏倚(倾向于发表阳性结果的研究),但经过校正后,其显著的抗炎效果依然存在。这大大增强了本研究结论的可信度。
doi.org/10.3389/fcimb.2025.1677671
这项荟萃分析的意义不仅在于学术层面,更在于为临床实践和个人健康管理提供了清晰的指导。它系统地证明了益生菌和合生元作为辅助疗法,在管理与糖尿病相关的代谢性炎症方面的巨大潜力。
研究结果反复指向短链脂肪酸的核心作用。科学界目前认为,短链脂肪酸主要通过以下途径发挥抗炎作用:
这些机制共同解释了在这项荟萃分析中,短链脂肪酸水平的升高与炎症标志物的改善呈现出如此强烈的相关性。
实践建议:如何选择和使用益生菌?
基于这项研究的发现,我们可以总结出一些给糖尿病患者的实用建议,以最大化益生菌的抗炎效益:
优先选择合生元或多菌株产品
研究明确指出,含有益生元(如菊粉、低聚果糖)的合生元产品,以及包含多种菌株(特别是乳杆菌属和双歧杆菌属)的混合产品,效果优于单一菌株的益生菌。寻找包含如干酪乳杆菌、植物乳杆菌、短双歧杆菌、长双歧杆菌等经研究验证的菌株。
保证足够的使用剂量和时长
为了达到有效的抗炎效果,建议的剂量通常在每天 109~1010 CFU(菌落形成单位)。更重要的是,干预需要足够长的时间,至少应持续 8-12周,甚至更长,才能观察到显著的免疫调节效果。
关注饮食协同
益生菌的效果离不开“燃料”。在补充益生菌的同时,增加膳食纤维的摄入(如全谷物、豆类、蔬菜、水果),可以为益生菌提供充足的食物,促进短链脂肪酸的产生,达到事半功倍的效果。
对于那些已经存在明显代谢性炎症(如CRP/IL-6升高)、中心性肥胖或非酒精性脂肪肝(NAFLD)的糖尿病患者,益生菌干预可能带来更大的获益。
作为辅助而非替代疗法
益生菌是一种辅助管理策略,不能完全替代标准的降糖药物和生活方式管理,益生菌可以从源头改善,通过调节肠道微生态,靶向慢性炎症,缓解糖尿病,且减少并发症的风险。
益生菌与常规治疗相结合,共同控制疾病。益生菌创造的低炎症、更健康的肠道环境,可能有助于提高身体对降糖药物的反应,并优化健康饮食的营养吸收效果。这种协同作用,最终可能带来比单一策略好得多的整体管理效果。
肠道菌群与糖尿病炎症的深度关联,已从基础研究走向临床转化的关键节点。益生菌通过调节短链脂肪酸代谢实现抗炎降糖,但“菌株选择、剂量优化、疗效预测”等核心问题,始终依赖对个体菌群特征的精准洞察。
益生菌通过重塑肠道微生态、修复肠道屏障、抑制慢性炎症,为2型糖尿病的辅助管理开辟了充满希望的新途径。它从源头上干预了由菌群失调驱动的代谢紊乱,展现出巨大的应用潜力。
从盲目补充到精准调养
肠道菌群检测将成为个性化健康管理的起点。通过检测,你能清晰了解自身菌群结构(如双歧杆菌、乳杆菌等有益菌丰度)、短链脂肪酸代谢能力及炎症风险,从而告别试错式补充,精准匹配最适合的菌株与饮食方案(如搭配益生元增强效果),让益生菌的抗炎潜力得到最大释放。
从单一产品到生态协同
人群研究数据(如干预≥8周效果提升、合生元方案优于单一益生菌)揭示,肠道菌群调节需“检测-干预-跟踪”的闭环支持。
主要参考文献
Xie Y, Zheng Y, Jiang F, Cai X. Meta-analytical insight on probiotic metabolites and inflammatory markers in diabetes. Front Cell Infect Microbiol. 2025 Sep 22;15:1677671.
谷禾健康
糖尿病是一种复杂的多系统代谢紊乱,其特征是高血糖,它还会导致并发症,降低生活质量并增加死亡率。糖尿病病理生理学包括β细胞、脂肪组织、骨骼肌和肝脏功能障碍。
1型糖尿病(T1D)是由免疫介导的β细胞破坏引起的。更常见的2型糖尿病(T2D)是一种异质性疾病,其特征是不同程度的β细胞功能障碍与胰岛素抵抗共同作用。
肥胖和2型糖尿病之间的密切关联涉及由中枢神经系统调节的通路,这些通路控制食物摄入和能量消耗,并整合来自外周器官和环境的输入。
患糖尿病或其并发症的风险代表了遗传易感性和环境因素之间的相互作用,包括营养食品的可用性和其他健康的社会决定因素。
糖尿病已经困扰人类数千年。从对这一疾病的“早期”描述到现代,目前人们对全球日益增多的糖尿病病人的流行病学、病理生理学、并发症和治疗选择的理解已经大大加深。
在过去的 50 年里,我们共同目睹了有关1型糖尿病和2型糖尿病病理生理学的知识爆炸式增长,而这种知识正在彻底改变糖尿病的治疗和预防方法。因此,任何关于糖尿病病理生理学和治疗进展的观点都不可能详尽无遗,或完全统一。
本文根据顶级期刊《Cell》近期发布的关于近50年有关糖尿病研究进展总结内容,与大家分享。
◮ 关于糖尿病的基础知识
糖尿病是由于胰岛素不足以刺激生理性葡萄糖处理,从而促进能量在脂肪组织、肌肉和肝脏中的储存而发展起来的。
糖尿病的表现范围从几乎完全的胰岛素缺乏症,如1型糖尿病(T1D)中出现的情况,到在胰岛素抵抗的情况下胰岛素相对缺乏的情况——2型糖尿病(T2D)的特征。
虽然糖尿病的诊断是基于测量血糖或糖化血红蛋白,但这种疾病应该被视为一个与多种合并症相关的多系统疾病。
◮ 糖尿病的大致分类
•1型糖尿病:由免疫介导的胰岛细胞破坏引起;
•2型糖尿病:与胰岛素抵抗和相对胰岛细胞功能不全相关;
•特定单基因疾病、药物毒性或胰腺功能不全引起的糖尿病综合征;
•妊娠期糖尿病
◮ 糖尿病的患病情况
2型糖尿病患者人数最多,1型糖尿病则不到所有病例的5%。2021年,全球糖尿病的患病率估计为6.1%,相当于5.29亿人,某些地区的患病率甚至高达12.3%。2型糖尿病占96%的病例,其中超过50%与肥胖有关。
糖尿病流行的趋势令人担忧,预计到2050年,将有13.1亿人患糖尿病。其他分析指出,2021年全球患病率已经超过10%。此外,2021年估计还有4.64亿人有糖耐量受损,2.98亿人有空腹血糖受损,总体上代表了糖尿病前期。
◮ 糖尿病大大增加了患其他病的风险
糖尿病大大增加了由心血管疾病和肾脏病造成的全因死亡率,且导致多种其他病症,包括失明、肢体丧失、慢性疼痛和残疾。
尽管升高的循环葡萄糖是任何原因引起的糖尿病的特征,但2型糖尿病是一种异质性疾病,不同人群亚群的结果存在差异。
鉴于肥胖与2型糖尿病之间的关联,有人认为,只要更加注重改善营养、增加身体活动和减少肥胖的政策,这种负担的大部分是可以预防的。然而,糖尿病的异质性表明,预防和治疗策略最好量身定制,以最大限度地发挥其在特定人群中的功效。
◮ 1型糖尿病概况
1型糖尿病(T1D)占所有糖尿病病例的5%–10%,由自身免疫介导的胰腺β细胞破坏导致。
胰岛素的发现使1型糖尿病从曾经的致命诊断变为可管理的慢性病。尽管取得了管理进步和新疗法的问世,只有约20%的患者能实现最佳血糖控制,且预期寿命较短。
◮ 遗传研究和疾病进展
全基因组关联研究(GWAS)鉴定了60多个与1型糖尿病相关的基因位点,显示出该病的高度遗传性。
对自然史的研究通过新生儿筛查和自身抗体筛查提供了对环境因素、疾病触发因素、胰岛自身免疫轨迹及代谢和免疫表型的见解。
重要观察之一是综合分析显示,有两种或更多胰岛自身抗体的人在15年内发展为临床1型糖尿病的风险超过80%。
◮ 疾病分期
1期T1D:存在两种或两种以上自身抗体
2期T1D:多种自身抗体阳性和血糖紊乱
3期T1D:明显的高血糖症
历史背景
在上世纪末,关于2型糖尿病的遗传学理解主要集中在少数几个基因座上,这些基因座与环境因素共同影响个体患病风险。
现代研究进展
随着全基因组关联研究(GWAS)的发展,现在已发现数百种与2型糖尿病相关的遗传变异。这些变异大多对患病风险的影响很小。
变异的作用机制
大多数与2型糖尿病相关的变异并不直接改变蛋白质功能,而是通过影响非编码基因组序列中的调控元件来改变基因表达的丰度。这些调控元件在特定的细胞类型和发育时间点发挥作用。
挑战与机遇
当前的挑战是将这些调控信号与具体的效应基因(转录本)关联起来,以了解其对糖尿病风险的影响。识别这些基因有助于理解糖尿病的机制,并可能发现新的治疗靶点。
单细胞多组学数据的应用
单细胞分辨率多组学数据集的出现为研究人员提供了工具,以更好地将遗传变异与其效应基因关联起来。这些数据结合高通量细胞表型分析,有助于评估变异相关的基因表达改变与疾病的关系。
临床应用的局限性
尽管这些研究提供了对糖尿病潜在病理生理学的生物学洞察,但目前尚无直接的精准诊断或治疗方法。
多样性研究的需求
大多数遗传学研究是在欧洲人群中进行的,因此需要在更广泛和多样化的人群中开展类似研究,以避免因缺乏多样性导致的健康差异,尤其是在糖尿病护理方面。
未来研究方向
研究人员对将心脏代谢和血糖特征的基因数据与2型糖尿病风险的基因数据结合起来很感兴趣。这些共享信号可能揭示潜在的组织和机制,通过这些机制,变异体会影响糖尿病或其并发症的风险。
聚类分析
通过不同聚类方法,研究人员正在识别2型糖尿病中常见的缺陷过程(如胰岛素作用、β细胞功能、血脂异常),并评估其临床效用。
20世纪90年代初,通过识别单基因糖尿病的突变,揭示了葡萄糖代谢与胰岛素分泌偶联机制的重要性。这些研究表明,关键的糖酵解酶如葡萄糖激酶的功能丧失突变影响胰岛素分泌。
转录因子HNF1A/HNF4A对内分泌胰腺的发育和维持至关重要,其发现为进一步研究胰腺和内分泌细胞发育的重要步骤奠定了基础。
基因变异的影响
罕见的完全渗透性突变在生命早期表现为糖尿病,而影响较小的等位基因也会增加2型糖尿病的风险。
胰腺功能的复杂性
研究发现外分泌和内分泌胰腺之间存在意想不到的联系。罕见的消化酶基因突变和与糖尿病相关的常见变异揭示了胰腺疾病(如胰腺炎和囊性纤维化)与内分泌细胞功能障碍之间的联系。
对β细胞功能受损的各种方式的机制理解得益于人类基因发现(下图)。
胰腺β细胞因环境和遗传因素受损的潜在方式示意图
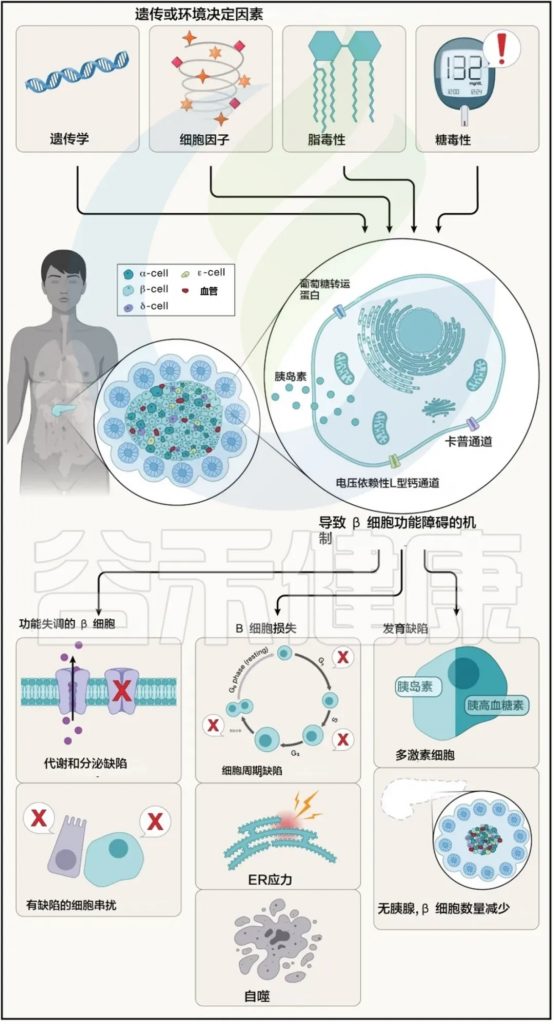
Abel ED,et al.Cell.2024
提升β细胞功能的兴趣
由于β细胞在维持正常葡萄糖耐受性中的关键作用,提高“功能性β细胞质量”被视为治疗1型和2型糖尿病的重要策略。
潜在治疗靶点
人类遗传学研究支持K-ATP通道和葡萄糖激酶为改善胰岛素分泌的潜在靶点。
SLC30A8基因中的特定蛋白质变体(ZnT8的锌转运蛋白)主要在胰腺β细胞中表达,并被证明可保护个体免受2型糖尿病的侵袭,因此相关药物的开发在进行中。
GLP-1受体(GLP-1R)
虽然在GWAS证实其作为治疗靶点的有效性之前人们就已经对它进行了广泛研究,但人类遗传学为其在降低血糖和改善心脏代谢健康方面的作用提供了有力证据。
GLP-1R激动剂不仅有效控制血糖,还具有减肥和降低心血管死亡率的积极作用。
开发新疗法的挑战
GLP-1类药物的多重成功显示了从多个角度治疗疾病生物学的巨大潜力,但也增加了专注于改善β细胞功能新疗法开发的难度。
社会健康决定因素(SDoH)指的是影响健康的社会条件,包括经济、教育、食品安全、医疗保健等因素。它们对包括2型糖尿病在内的多种疾病的发病和进展有深远影响。
SDoH通过复杂的途径影响2型糖尿病的发展,从生活方式行为到分子机制(例如HPA轴激活、微生物群失调)。粮食不安全和空气污染等具体因素已经被证明与糖尿病有关,表明需要多层次的干预策略。
粮食和营养不安全
粮食安全被定义为“所有人在任何时候都能获得足够的食物以过上积极和健康的生活”。
营养安全进一步扩展为“能够公平、稳定地获取、负担得起和利用促进健康、预防和治疗疾病的食品和饮料的状态”,包括饮食质量和社会健康决定因素(SDoH)。
粮食不安全的影响:粮食不安全导致水果和蔬菜摄入减少,精制碳水化合物、饱和脂肪和添加糖摄入增加,整体饮食质量下降。长期不良饮食习惯导致氧化应激、炎症和脂肪组织扩张。
粮食不安全与增加的炎症、生物荷负和肠道微生物多样性降低有关。社会经济压力则加剧了这种状态,影响补偿行为,并激活炎症途径。
空气污染
空气污染是重大环境健康风险,社会经济地位较低的人群受影响更大。关键污染物包括臭氧 (O₃)、二氧化氮 (NO₂) 和各种颗粒物 (PM),如PM2.5。
对糖尿病的影响:空气污染与胰岛素抵抗、血糖紊乱、高脂血症和2型糖尿病的发病率增加有关。观察性研究与孟德尔随机化研究支持其因果关系。
机制:初级启动途径:包括氧化应激、淋巴细胞活化、组织内DAMP产生及直接影响胰腺β细胞等。
次级效应途径:涉及全身炎症、神经内分泌失调、肝脂肪变性以及肠道微生物群失调等。
共同的病理生理影响:无论是营养不良还是空气污染,均通过氧化应激、炎症反应、HPA轴激活等机制影响糖尿病的发展。
粮食不安全和空气污染等因素可能在从胚胎到婴儿期影响后代未来的糖尿病风险。饥荒和空气污染对母体的营养和污染暴露都有证据表明会影响后代的代谢健康。
研究和政策发展:需跨学科团队的合作来深度研究SDoH的病理生理影响,指导新的干预策略和政策制定,以提高糖尿病预防和管理的公平性。
总之,粮食不安全和空气污染通过复杂的生物和环境机制显著影响2型糖尿病的进程。理解这些机制将有助于开发更有效的公共卫生政策和临床干预措施。
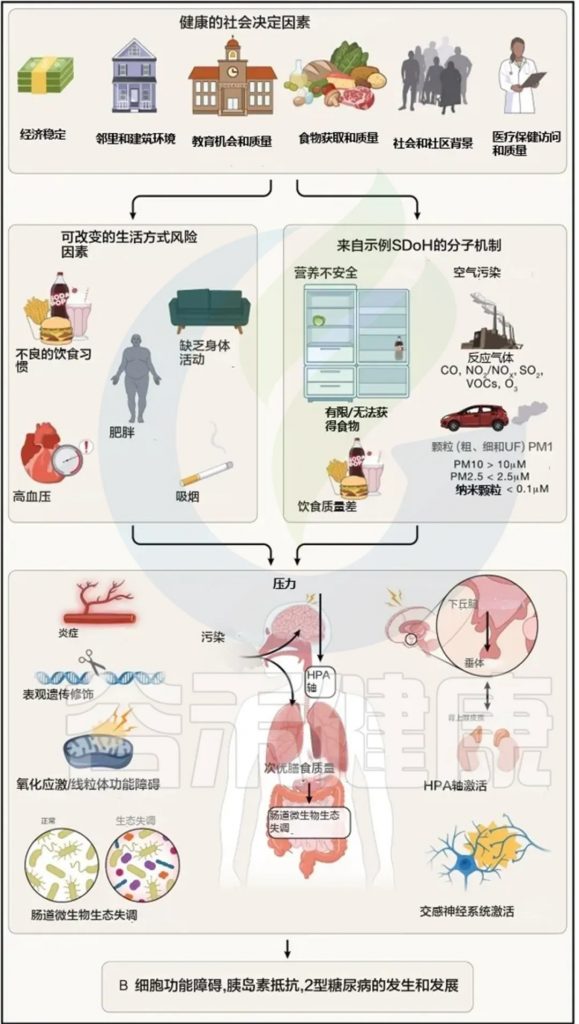
Abel ED,et al.Cell.2024
大量临床前数据强调了大脑在控制体重以及易感人群中胰岛素抵抗和肥胖发展中的重要作用。尤其中枢神经系统(CNS)在控制血糖和2型糖尿病病理生理学中的重要性。
◮ 重要性与作用
大量临床前数据显示,大脑在体重控制、胰岛素抵抗(IR)、肥胖及2型糖尿病发病机制中扮演关键角色。其中肥胖是2型糖尿病的重要风险因素。
食欲和能量消耗的调节对于肥胖的发生至关重要,受中枢系统全面调控。
肠道激素如生长素释放肽和瘦素,以及肠道微生物群的变化,在调节过程中起重要作用。
已在下丘脑和脑干中对这些调控神经回路的了解取得显著进展。
临床研究
人类研究重点关注中枢及自主神经系统如何整合体重调节及葡萄糖代谢,尤其在维持循环葡萄糖浓度中的作用。
动物模型研究
动物模型研究提供了许多见解,显示迷走神经及中枢神经系统回路对营养感知的重要性。
研究表明脂肪或糖摄入与多巴胺释放及暴饮暴食行为之间存在关联。
大脑与外周之间相互作用的示意图
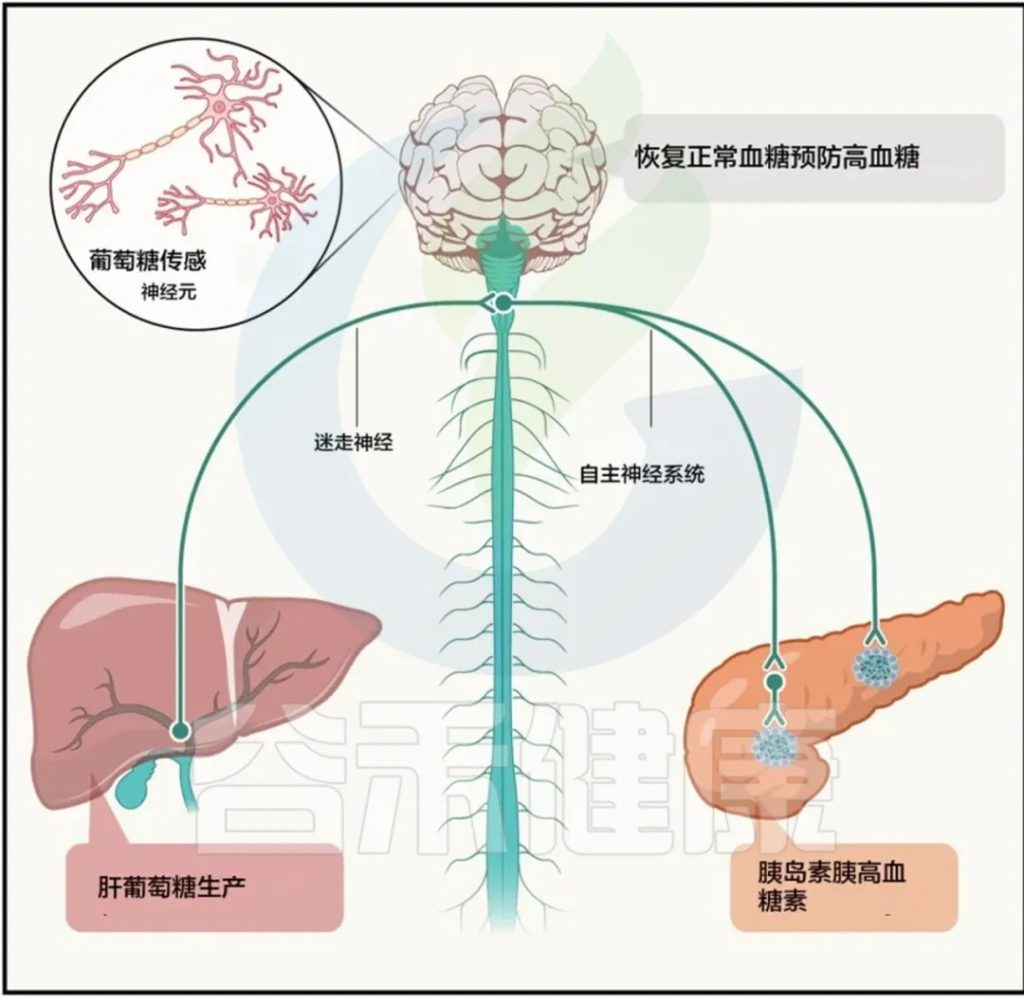
Abel ED,et al.Cell.2024
注:这些见解大多来自动物模型研究
为了更好地科普中枢神经系统(CNS)在糖尿病和代谢调节中的作用,可以从以下几个方面进一步理解:
基本概念
中枢神经系统(CNS)不仅仅是控制思维和行为的中心,它还在调节身体的代谢功能中发挥重要作用。特别是在血糖调节方面,CNS通过调控肝脏的葡萄糖生成和分解来维持血糖的稳定。
机制
下丘脑是CNS中一个关键区域,它通过神经信号影响胰腺的功能,调节胰岛素的分泌。胰岛素是降低血糖的主要激素,因此下丘脑的调控对于维持正常血糖水平至关重要。
胰岛素的作用
胰岛素不仅在外周组织中发挥作用,它在大脑中也有重要功能。研究表明,胰岛素可以通过鼻内给药直接作用于大脑,影响胰岛素的分泌和全身的胰岛素敏感性。
个体差异
不同个体对大脑胰岛素的反应存在差异,这种差异可能影响到胰岛素的分泌和代谢功能。这种差异也与肥胖和胰岛素抵抗(IR)有关。
FGF的潜力
成纤维细胞生长因子(FGF)在动物实验中显示出通过作用于大脑来缓解糖尿病的潜力。这种方法不依赖于体重减轻,而是直接改善葡萄糖代谢。
药物的CNS作用
一些用于治疗2型糖尿病的药物,如GLP-1受体激动剂,通过中枢神经系统发挥作用,影响食欲、体重和胰岛素分泌。
关联性
2型糖尿病与认知功能受损和神经退行性疾病(如阿尔茨海默病)之间存在关联。研究发现,糖尿病患者的大脑胰岛素反应性可能受损,这与认知功能下降有关。
潜在治疗
鼻内胰岛素和GLP-1受体激动剂在临床研究中显示出神经保护作用,可能降低痴呆症的发病率。
更具脑渗透性的药物
开发能够更好地穿透血脑屏障的药物,可能增强其神经保护作用,同时减少不良反应。
共同途径的探索
由于糖尿病和神经退行性疾病的发病率随着年龄增长而增加,研究共同的病理机制可能带来同时治疗这两种疾病的新方法。
胰岛素抵抗(IR)
当胰岛素无法有效地降低血糖时,称为胰岛素抵抗。这种情况不仅影响糖代谢,还导致脂肪组织释放更多的游离脂肪酸(FFA)进入血液。
脂解作用增加
在IR条件下,脂肪组织经历脂肪分解增加,产生更多的FFA。这些游离脂肪酸流入血液并扩散到其他组织,如肝脏和骨骼肌,进一步导致这些组织的发展胰岛素抵抗。
二酰甘油的作用
二酰甘油(DAG)是一种重要的信号分子。研究表明,在胰岛素抵抗的状态下,血浆中DAG水平升高。DAG可以与细胞膜上的蛋白激酶Cε (PKCε)结合,从而使胰岛素受体发生抑制性磷酸化。这一过程会限制胰岛素受体的激酶活性,削弱胰岛素的信号传导,导致细胞对胰岛素的反应降低。
特异性结合:DAG的不同立体异构体对作用靶点的结合亲和力不同,其中sn-1,2 DAG与PKCε结合最强,定位在质膜上,而其他异构体如sn-1,3 DAG和sn-2,3 DAG倾向于定位在脂滴和内质网(ER)。
神经酰胺的作用
神经酰胺是另一种导致胰岛素抵抗的脂质。高FFA水平促进长链神经酰胺(如C16和C18)的产生。神经酰胺通过激活特定的激酶,如蛋白激酶C ζ (PKCζ)和蛋白磷酸酶2A (PP2A),抑制关键信号分子Akt的磷酸化。这种抑制作用破坏了正常的胰岛素信号传导。
选择性影响:神经酰胺特异性地促进胰岛素抵抗,这是因为其他类似脂类(如二氢神经酰胺和鞘磷脂)未能在实验模型中表现出同样的效果。
异位脂质沉积
骨骼肌和肝脏中DAG和神经酰胺的积累会加剧胰岛素抵抗。这些组织的『选择性胰岛素抵抗』意味着通常应该增加葡萄糖吸收的组织变得不再敏感,反而可能继续将葡萄糖转化为脂质储存。
加速糖尿病的进展
当骨骼肌和肝脏对胰岛素的敏感性降低时,会加速糖尿病的进展。这些脂质代谢产物的积累不仅影响胰岛素信号传导,还可能引发脂质毒性,损伤细胞功能并加重代谢紊乱。
以上细节描述展示了脂质介质如何在分子水平上影响胰岛素信号通路,从而推动全身性胰岛素抵抗的发展。这些见解为开发针对性更强的干预措施和治疗策略提供了依据,以改善代谢健康并减缓或逆转糖尿病。
下面是肥胖和脂肪组织通过多种机制驱动胰岛素抵抗(IR)的详细总结:
1.脂肪酸羟基脂肪酸酯与胰岛素敏感性
脂肪酸羟基脂肪酸酯(FAHFA)的角色:FAHFA是一类由两个脂肪酸通过酯键结合形成的复合脂质。研究表明,FAHFA可以改善胰岛素敏感性,其水平在肥胖状态下和2型糖尿病中会降低。
机制:FAHFA通过与某些代谢重要组织中的G蛋白偶联受体结合,起到调节小鼠胰岛素敏感性、脂肪生成和能量消耗的作用。此作用途径表明FAHFA可能通过影响受体来调控广泛的代谢过程。
2.脂肪甘油三酯脂肪酶与FAHFA
脂肪甘油三酯脂肪酶(ATGL)的双重角色:ATGL是一种负责脂肪分解的酶,被认为可能在FAHFA代谢中也发挥作用。
ATGL可能充当FAHFA的合酶,通过改变ATGL功能,提供了FAHFA与2型糖尿病中的脂肪分解之间的潜在联系。调节ATGL功能的变化可能影响FAHFA的合成和分解。
3.前馈循环与全身性胰岛素抵抗
前馈循环:肥胖状态下,脂肪组织中多种信号分子的变化(包括FAHFA的减少)会驱动一种前馈的代谢循环。这种循环会进一步加剧全身胰岛素抵抗。
注释:代谢生理学中,”前馈循环” (feed-forward loop) 指的是一种生物学过程,其中某一信号或变化会增强或放大某一响应,而这种响应又会使初始信号或变化更加显著。上文中前馈循环是指肥胖状态下脂肪组织中多种信号分子的变化(例如 FAHFA 的减少)导致的一系列代谢事件。这些变化不仅直接影响机体的代谢状况,还通过促进某些过程或信号的放大,进一步加剧全身胰岛素抵抗。
具体来说,脂肪组织中信号分子的变化作为初始刺激,会引发代谢反应或变化,这些代谢反应可能通过各自的信号机制,反过来进一步促进初始信号或加大初始变化。如果这种循环不被打断,就可能导致长期的代谢失调,比如全身性的胰岛素抵抗,从而增加糖尿病等代谢疾病的风险。
4. 脂肪组织在全身性胰岛素抵抗中的作用
脂肪组织重塑:肥胖引起的脂肪组织重塑是全身性胰岛素抵抗、炎症以及葡萄糖和脂质代谢异常的关键驱动因素。
5. 高血糖与胰岛素抵抗的恶性循环
高血糖的影响:高血糖本身会通过增加己糖胺生物合成途径的通量,加剧胰岛素抵抗。这一途径生成尿苷二磷酸-N-乙酰葡萄糖胺 (UDP-GlcNAc),是N和O连接糖基化的前体。
O连接糖基化:包括Akt在内的胰岛素信号蛋白的O连接糖基化会进一步诱导胰岛素抵抗,形成恶性循环。
在胰岛素抵抗状态下,脂肪细胞的葡萄糖摄取功能受损。这种情况与脂肪细胞释放的胰岛素敏感因子和复合脂质的变化有关,这些变化会影响其他组织对胰岛素的响应。
肝脏和骨骼肌是胰岛素作用的主要目标组织,在胰岛素抵抗状态下,其糖代谢功能受损,尤其是肝脏的葡萄糖输出增加和骨骼肌的葡萄糖摄取减少。这些机制是糖尿病患者血糖水平升高的主要驱动因素。
糖原合成减少
在胰岛素抵抗的早期阶段,骨骼肌中胰岛素介导的糖原合成减少。这是因为胰岛素信号传导受损,影响了糖原合成酶的活性。
GLUT4易位受损
胰岛素通常促进葡萄糖转运蛋白4(GLUT4)从胞内储存池转移到质膜,以增加葡萄糖摄取。然而,在胰岛素抵抗状态下,这一过程受损,导致葡萄糖摄取减少。
葡萄糖氧化减少
由于胰岛素信号传导的缺陷,骨骼肌中葡萄糖的氧化能力降低,这进一步影响能量代谢。
长期影响
随着时间推移,骨骼肌胰岛素抵抗会导致肌肉萎缩、运动能力下降以及线粒体功能和质量的下降。
肝糖输出(HGP)增加
肝脏胰岛素抵抗的主要表现是肝糖输出增加。这是因为胰岛素对糖异生基因的抑制作用受损,导致肝脏中糖异生增加。
脂质积累
胰岛素抵抗还促进肝脏中脂质的积累,进一步影响肝脏功能。
组织间通讯的理解
未来的研究需要深入了解不同组织间的通讯机制,包括外泌体等新型介质在内的作用。
脂质动员机制
研究控制组织和细胞器之间脂质动员的机制,以及未表征脂质的功能,将有助于揭示2型糖尿病中的脂质失调。
复杂病理生理学的探索
进一步探索这些途径在持续胰岛素抵抗中的复杂病理生理学影响,将有助于开发新的治疗策略,改善代谢稳态。
综上所述,理解骨骼肌和肝脏的胰岛素抵抗机制及其与脂肪组织的相互作用,对于揭示2型糖尿病的发病机制和寻找新的治疗靶点至关重要。
近年来,越来越多的研究揭示了2型糖尿病与代谢功能障碍相关脂肪肝病(MASLD)之间的紧密联系,除了影响血糖稳态外,还对心血管并发症以及其他健康问题有显著的影响。
病理生理学背景
糖尿病患者常伴有肝脏代谢功能障碍,这是血糖稳态受损和糖尿病心血管并发症的主要因素之一。肥胖引发这些患者肝脏葡萄糖和脂质的异常增加。
MASLD的演变
MASLD从单纯脂肪变性(最初是可逆且普遍的状态)到代谢功能障碍相关脂肪性肝炎(MASH),可以进展至纤维化,这在严重情况下是主要的死亡原因。
下面是对2型糖尿病和MASLD相互作用的总结:
1.多重打击假说
这一理论解释不同患者发展严重并发症的原因,重点在于脂质蓄积后非实质细胞(NPC)的异常激活。对于早期疾病,GLP-1 基药物疗法可能有效,但对晚期纤维化无明显效果。
2.过量从头脂肪生成(DNL)
过量从头脂肪生成(DNL)在2型糖尿病患者中明显增加。健康人群中,胰岛素通过Akt信号通路抑制肝糖输出(HGP),同时激活脂肪生成的促进元素,如SREBP-1c。但在胰岛素抵抗状态下,FoxO1抑制失效增加葡萄糖生成,然而胰岛素仍然能推动DNL。
尽管胰岛素信号转导的某些功能失效,DNL却仍然活跃。这可能由于糖尿病患者CHREBP诱导Akt磷酸化调控机制存在异常,尤其是PHLPP2磷酸酶水平降低,导致了过量DNL。
3.胰岛素信号转导的动力学变化
在慢性高胰岛素血症背景下,正常的胰岛素信号分叉模型可能发生变化,影响胰岛素在早期和晚期阶段的作用。
早期阶段:胰岛素的快速作用通过抑制FoxO1来减少葡萄糖生成。
晚期阶段:延长的Akt活性则促进了DNL和脂质合成,潜在机制包括细胞自主信号、自主脂肪信号以及肠道介导的信号。
4.双向风险与相互促进
2型糖尿病和MASLD不仅拥有共同的风险因素,还可能通过如上机制双向增加各自的风险。研究显示,肝脏脂肪过多与2型糖尿病高度相关,即便在调整了BMI和其他潜在混杂因素后,二者之间的联系依然显著。
5.临床与研究启示
由于代谢功能障碍相关脂肪肝病(MASLD)在肥胖率较高的人群中表现为一种日益增长的健康挑战,当前的研究重点在于理清组织间通讯的复杂网络、揭示潜在的细胞机制、探索治疗新策略以及更好地管理代谢性肝病。
通过全面的机制研究和临床分析,我们可以更好地理解2型糖尿病与MASLD的复杂交互机制,为治疗和预防这些相关病理状态带来新的希望。
在2型糖尿病与代谢功能障碍相关脂肪肝病(MASLD)的研究中,理解肝脏脂质堆积及其进展至肝脏病变的过程尤为重要。以下是详细汇总和对当前研究状况的总结:
1.肝脂质过量与疾病进展
影像学发现:许多2型糖尿病患者在影像学检查中展示了肝脂质过量。然而,哪些患者将发展为有临床意义的肝脏疾病,以及进展的时间和诱导因素,仍不明确。
遗传因素:基因组广泛关联研究(GWAS)揭示了一些与体重和肝脏脂质积累相关的常见风险等位基因(如FTO、PNPLA3、TM6SF2和APOB)。然而,这些基因标记在预测疾病进展为代谢功能障碍相关脂肪性肝炎(MASH)方面的效能有限。
2.脂质管理障碍
2型糖尿病和MASLD患者的肝细胞可能有储存脂质的倾向,但难以在不损伤细胞的情况下管理过量的脂质。这导致了细胞的炎症反应和纤维化。
3.肝细胞的信号贡献
肝细胞在疾病中的作用:研究表明肝细胞不仅在血糖调节中起作用,还通过分泌促进NPC浸润和活化的趋化性和纤维化细胞因子,直接参与肝脏炎症和纤维化过程。
Notch信号的再激活:这是2型糖尿病和MASLD患者肝细胞炎症的一部分,影响HGP和DNL的增加。
4.临床研究和治疗前景
治疗性干预:使用如GalNAc修饰的反义寡核苷酸或siRNA,可以靶向性调制肝细胞信号通路,有可能改善病情。
5.当前研究的主要方向和挑战
高胰岛素血症的作用:2型糖尿病患者IR导致的高胰岛素血症,与肝脏DNL正相关,可能是MASLD病理机制之一。现有研究正在探讨抑制胰岛素分泌的策略,但其在糖尿病患者中的适用性仍需验证。
其他激素和营养因素:胰高血糖素、果糖和胆固醇也可能对肝脏脂质生成有影响。此外,肝交感神经系统的参与是否在脂质生成功能中有重要贡献,需要进一步探索。
通过这些研究,肝脏的代谢调节能力、遗传和环境因素的交互作用构成了2型糖尿病和MASLD在病理进程中的复杂图景。理解这些机制对于开发有效的治疗方案和风险评估方法至关重要。
尽管最近取得了进展,但许多问题依然存在,包括一下几点。
1.高胰岛素血症和非激素因素的作用
2型糖尿病的胰岛素抵抗(IR)显示出代偿性高胰岛素血症。人类数据表明血浆胰岛素水平与肝脏DNL正相关,动物研究证实胰岛素作用时机不当可能导致MASLD。有趣的是,奥曲肽通过阻断胰岛素分泌降低了大鼠的DNL标记物和肝脏甘油三酯。
目前,这一概念正在非糖尿病个体中使用二氮嗪(NCT05729282)进行测试,其结果是否适用于T2D患者尚不明确。其他激素如胰高血糖素也通过促进糖原分解和降低肝脏脂质起作用。
此外,果糖和胆固醇可能影响肝脏脂质生产。最后,非营养和非激素的肝糖输出(HGP)决定因素,如交感神经对肝脏的影响及其对脂质生产的共调节作用,仍不清楚。
2.脂肪性肝炎的空间决定因素
组织异质性:肝脏作为一种异质性组织,由于不同区域的氧张力和营养状态,产生了肝内糖异生和脂肪生成功能的分区。这种分区影响了MASH的区域特异性,表现为中枢周围或门管周围类型,后者尤为常见于儿童患者。
区域性亚型:代谢综合征和2型糖尿病患者倾向于发生门管周围类型的疾病。这种区域亚型与代谢和肝脏病理的发展程度相关,但其机制尚不清楚。针对肝脏区域特定通路(例如Notch、法呢醇X受体[FXR]、甲状腺激素受体[TR]和过氧化物酶体增殖激活受体[PPAR])的治疗可能提供新的干预策略。
3.纤维化消退的途径
尽管对肝脏促炎和纤维化途径的理解有所增加,但对纤维化清除及肝细胞途径影响纤维化消退的研究较少。我们推测,所有肝细胞决定的“纤维化开启”信号都有相应的“纤维化关闭”信号。
系统研究回归途径对肝纤维化的影响,类似于动脉粥样硬化病变的解决,可能带来重要的影响。对于部分对肠促胰岛素治疗抵抗的T2D患者,新的治疗靶点可能特别有价值。
4.双向肝细胞-非实质细胞串扰
虽然肝细胞在脂质过载中的作用被强调,但非实质细胞(NPC)对肝细胞功能也有显著影响。例如,肝星状细胞不仅是肝细胞再生的决定因素,还是一个潜在的调节因子。在2型糖尿病中,免疫细胞增加从而改变胰岛素敏感性。
6.对脂质超载的遗传适应
近来的研究揭示了MASLD/MASH患者肝细胞中FOX01的体细胞突变,这些突变可能是一种保护机制,减轻脂质超载的损伤。这种基因适应可能说明MASLD与2型糖尿病的关联。
7.与心血管疾病的关系
肝脏脂质过多增加了肝病相关死亡率的风险,但与2型糖尿病患者类似,MASLD/MASH患者的主要死因是心血管疾病(CVD)。鉴于直接加速CVD的常见并发症,需在临床前动物模型和人类中进行进一步机制研究以揭示其潜在机制。
糖尿病是一种影响全身的慢性疾病,会对多种系统造成广泛且复杂的影响,显著增加患病率和死亡率。以下是对糖尿病相关代谢异常和其主要并发症的详细分析。
增高的风险:糖尿病患者患动脉粥样硬化性心血管疾病(ASCVD)的风险增加2至5倍。
其他风险因素:包括高血压、血脂异常、肥胖和代谢综合征。
糖尿病性心肌病:糖尿病导致特有的心脏结构和功能异常,不完全依赖于冠状动脉疾病(CAD),而与异常代谢环境直接相关。
综合风险:没有单一机制可以解释CVD风险的增加,糖尿病引起的多种风险因素复杂相互作用导致CVD。
机制:与内皮细胞(EC)和血管平滑肌细胞(VSMC)功能障碍有关。
糖尿病影响:特征是内皮功能障碍,包括一氧化氮合酶功能下降和NO缺乏。
动脉粥样硬化驱动因素:糖尿病通过激活髓系细胞和炎症细胞(如单核细胞、巨噬细胞)的生成,直接加剧动脉粥样硬化。
风险因子和生物标志物:LDL胆固醇、血脂异常、HbA1c、肾功能和身体活动为疾病预测因子。新型生物标志物(如线粒体代谢物)可能用于风险分层。
Abel ED,et al.Cell.2024
风险增加:糖尿病增加心力衰竭风险,这与冠状动脉疾病(CAD)风险增加无关。
机制:碳毒性(脂毒性和糖毒性)
氧化应激
线粒体生物能量学受损
线粒体解偶联
心肌兴奋-收缩偶联受损
促纤维化途径激活
选择性胰岛素抵抗引起的肥大性信号通路激活
人类研究证据:心脏移植研究显示,正常供体心脏在移植到糖尿病患者体内后数月内出现代谢异常。
观察到甘油三酯超载、神经酰胺等有毒脂质积聚、线粒体呼吸功能不全、氧化应激和炎症。
注:前驱糖尿病患者也观察到线粒体氧化缺陷,二甲双胍治疗可能减轻这些变化。
Abel ED,et al.Cell.2024
特征:糖尿病降低多种促血管生成因子的表达,导致血管生成信号通路紊乱。
影响:增加外周血管疾病和严重肢体缺血的风险。
机制:VEGF抵抗
一氧化氮信号受损
血管生成干细胞前体水平降低
周细胞丢失
微小RNA和长链非编码RNA的失调
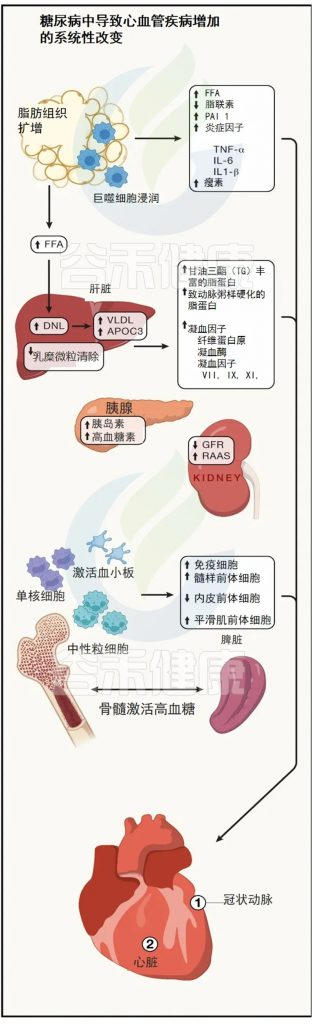
Abel ED,et al.Cell.2024
特征:白蛋白尿和估计肾小球滤过率(eGFR)降低。
患病率:约40%的糖尿病患者会发展为DKD。
肾血管功能障碍:肾小球高滤过是早期特征之一;
肾小管肾小球反馈机制导致肾小球传出动脉收缩。
肾小球和肾小管上皮细胞变化:
足细胞代谢改变,氧化应激增加。
RhoA/Rac1通路导致足突消失。
mTOR和生长因子信号改变导致足细胞增大。
足细胞丢失是疾病进展的关键步骤。
近端小管(PT)细胞:
高代谢活性,负责大量水和盐的重吸收。
初期细胞大小和数量增加,后期出现能量耗竭和细胞身份丧失。
线粒体损伤激活炎症途径。
小结
总的来说,糖尿病通过多种复杂的机制影响心血管系统和肾脏,涉及代谢异常、血管功能障碍、炎症反应和表观遗传变化等多个方面。
这些发现为理解糖尿病并发症的发生机制和开发新的治疗策略提供了重要的科学基础。
传统疗法的局限:早期治疗通常在T1D的3期开始,对于改变疾病进展的效果有限。
新进展:创新的分期系统为早期干预提供了框架,报告聚合抗CD3单克隆抗体teplizumab治疗延迟T1D进展的结果。这一研究促使FDA批准teplizumab作为首个T1D疾病改良疗法。
病毒传播与诊断挑战:目前缺乏统一的筛查指导方针,正测试多种自身抗体筛查策略。
胰岛素管理技术(如优化的药物动力学胰岛素、胰岛素泵和持续血糖监测设施)不断革新。
β细胞替代疗法:胰腺或供体胰岛移植显示出β细胞替代的可行性,但供体资源有限。
干细胞技术:通过多能干细胞分化得到的β细胞有望克服移植局限性,Vertex制药公司正在测试相关疗法(VX-880)。
异质性挑战:T2D患者的疾病表现由于遗传和环境因素而异,例如,非洲-加勒比和亚洲人群的T2D表现差异明显。
◮ 精准医疗的方法
临床分层工具:如BMI和eGFR用于HbA1c反应的预测,最新的研究表明此类因子可以指导选择更有效的治疗方法。
复杂数据整合:利用机器学习分类法如集群分析,识别糖尿病亚型,与特定并发症风险相关。
◮ 临床实施与研究挑战
标准化的必要性:研究需要一致的方法来评估精准治疗的优劣及其经济性。
证据差距:精准糖尿病治疗需要更多随机对照试验来证明其临床优势和成本效益。
精准医疗成功的例子:例如葡萄糖激酶突变的单基因糖尿病无需药物治疗,而HNF1A突变可用磺酰脲类药物有效管理。
综合来看,1型糖尿病的治疗正在向提前预防和细胞治疗方向发展,而2型糖尿病的精准医疗方法强调对不同人群亚型的特定管理。
随着研究的深入和技术的进步,这些策略有望显著改善糖尿病患者的治疗效果和生活质量。
在2型糖尿病的精准医疗和全球健康公平领域,取得了许多令人瞩目的成就,但仍面临资源可及性不均与种族多样性不足等挑战,并且新型疗法不断涌现,进一步推动病情管理和并发症预防。
◮ 资源限制与成本效益
2型糖尿病精准医疗是否能在不同资源环境中实施仍不明确,当前诊断和治疗的费用较高,尤其在中低收入国家。
如果精准医疗能提高治疗精度,减少资源浪费,则可能长期节约成本,但这些长期获益难以精准量化。
◮ 全球资源差异
许多中低收入国家患者难以获得基本药物,更复杂的精准医疗诊断尚难普及。简便的分层方法和决策支持工具可能在这些地区更具实用性。
糖尿病护理的差距往往影响社会经济地位低、少数民族群体。未经过周密计划和执行的精准医疗可能导致不平等加剧。
◮ 种群多样性的影响
研究多样性不足:大多数精准医疗研究在欧洲白人群体中进行,跨种族的应用性受限。需在不同种族背景下进行更多研究,以开发具有全球代表性的解决方案,适应不同遗传机制。
基因组研究局限:多样性缺乏的基因组研究可能导致偏见。例如,某些族群的2型糖尿病特征存在重大差异。开发跨祖先适用的基因风险评分,不同族群的参与和持续研究至关重要。
总体来看,精准医疗已经在2型糖尿病领域取得长足进步,但其广泛和公平实施受到资源、技术和种族多样性等多个因素的限制。未来的研究应致力于开发合适的工具和管理策略,以改善全球范围内的健康结果。
药物创新
GLP-1受体激动剂、DPP-4抑制剂和SGLT-2抑制剂在改善血糖及心血管、肾脏健康方面展现无与伦比的效果。
GLP-1RA的增强版本显示出非凡的减肥和心血管保护作用。SGLT-2抑制剂同样有助于不同心肾健康目标。
开发多激动剂如Tirzepatide(GIP受体-GLP-1R共激动剂),有效性超越单一疗法。此类疗法在减重和改善心血管风险方面效果显著,开创了治疗新方向。
基于GLP-1的治疗心脏代谢和神经退行性疾病
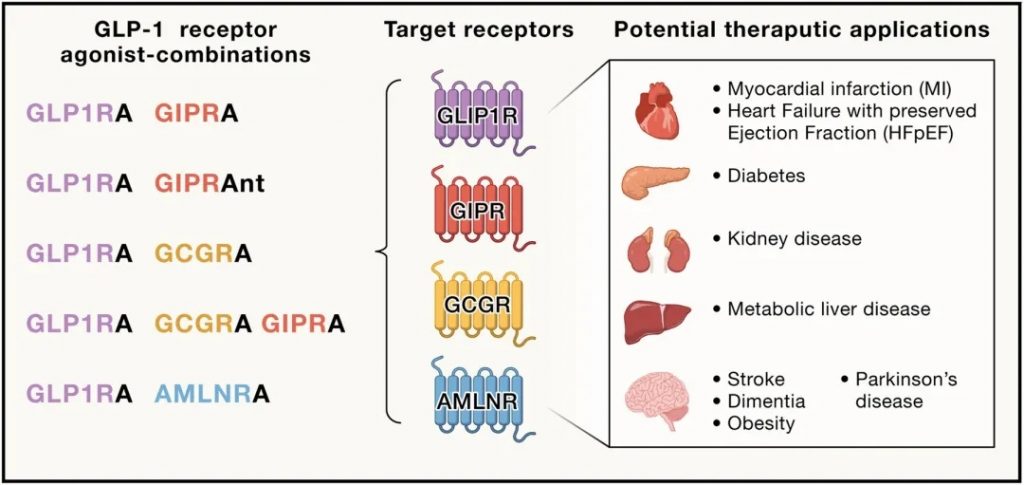
Abel ED,et al.Cell.2024
针对不同疾病机制的新疗法和联合疗法的临床试验持续进行,探索能够提供更高效、个性化处理方案的创新疗法。
多种基于GLP-1的联合疗法目前正处于临床后期开发阶段,期待它们在提高血糖控制和体重减轻效果的同时,继续展现心血管益处。这些治疗选项包括:
合成及联合应用疗法
-卡格列肽与索马鲁肽每周一次联合使用;
-胰高血糖素受体GLP-1R共激动剂如survodutide;
-GIP受体拮抗剂—GLP-1R激动剂抗体;
-三联胰高血糖素-GIP-GLP-1R多激动剂。
注:卡格列肽是一种肠促胰岛素激素,用于治疗2型糖尿病。
索马鲁肽,英文名称 “Semeglutide”,是一种新型的,长效的,每周皮下注射一次的 GLP-1 类似物,是基于利拉鲁肽基本结构而开发的长效剂型,其治疗2型糖尿病的效果更好。
新兴口服疗法
-包括danuglipron和orforglipron在内的小分子口服GLP-1R激动剂,以及ECC5004和GSBR-1290等其他小分子GLP-1RA。
-高剂量口服索马鲁肽(每天高达50毫克)与新配方吸收促进剂的应用。
这些创新药物不仅可能增强降糖和减重效果,还可能在外周动脉疾病、代谢性肝病、神经退行性疾病等领域拓展GLP-1药物的应用。
此外,对成瘾相关行为、遗传性肥胖、多囊卵巢综合征、1型糖尿病等多种适应症的研究也在进行之中。未来十年有望出现更为便捷和有效的GLP-1药物,这些药物的广泛适应症及安全性将通过更大规模临床试验的验证,为更多患者提供治疗选项。
随着糖尿病大流行的发展,我们对病理生理学以及治疗和预防方法的理解也呈指数级增长。目前的知识为提高识别增加β细胞功能障碍易感性标志物的特异性奠定了基础,特别是在肥胖环境中。
我们对大脑在体重调节中的作用以及脂肪组织分泌的新因子的理解可能有助于改进治疗或预防肥胖的方法。增加对肝功能障碍对胰岛素抵抗(IR)贡献的理解以及增加对代谢功能障碍相关肝病的理解代表了进一步研究的重要领域,以避免可能成为日益流行的肝功能衰竭。
心血管和肾脏疾病仍是糖尿病的主要死亡和发病因素,复杂的病理生理学涉及多器官系统和全身环境的交互作用。
糖尿病大流行由环境和社会因素推动,需综合管理。治疗进展为预防或治愈1型糖尿病和治疗2型糖尿病带来希望,不仅改善代谢稳态,还降低心肾疾病风险。
最后,随着我们了解和开发出识别导致糖尿病及其并发症的潜在异质性的工具,我们将为有针对性的治疗和预防策略奠定基础,以优化其在不同人群和医疗资源中的应用。
参考文献:
Abel ED, Gloyn AL, Evans-Molina C, Joseph JJ, Misra S, Pajvani UB, Simcox J, Susztak K, Drucker DJ. Diabetes mellitus-Progress and opportunities in the evolving epidemic. Cell. 2024 Jul 25;187(15):3789-3820.

谷禾健康

这篇文章总结了关于肠道微生物群(GM)在糖尿病中的作用的研究现状与结论,内容来自《Diabetes》、《Diabetes Care》和《Diabetologia》最近举办的一次国际专家论坛,该论坛在2023年的欧洲糖尿病研究协会年会上举行。论坛参与者包括在肠道微生物组和代谢领域的领军研究人员,其中既有临床医生,也有基础科学家。
讨论主要分为四个主题:流行病学、生理学和病理生理学、技术和方法以及临床应用。他们得出的结论如下:
肠道微生物群参与2型糖尿病的病理生理过程。微生物产生的代谢物与疾病有正面和负面的关联,而且最近通过人类的孟德尔随机化研究,发现了肠道微生物功能(如丁酸盐生产基因)与葡萄糖代谢之间的机制联系。
肠道微生物群的高度个体化特性是研究的一大障碍。需要大型队列和深度测序的方法来稳健地评估关联性和因果关系。
由于单次采样对于捕捉糖尿病个体内肠道微生物群的动态变化有些困难,建议未来需要要对个体多次重复测量的研究,就是要增加纵向队列的研究。
未来还需要大量研究来确定这些不断扩展的知识如何应用于糖尿病的诊断和治疗。新技术和改进的计算工具将对实现这一目标很重要。
研究小组承认,对肠道菌群在代谢疾病中的作用的理解存在许多差距,这并非糖尿病领域所独有,而是反映了更广泛的需求。开展更多控制良好的前瞻性和回顾性人体研究,并在机制上跟进模型系统研究,改进计算工具,将有利用将目前的微生物组数据应用于未来糖尿病的诊疗。
Perspectives
1)
肠道菌群与糖尿病之间的流行病学关联
肠道微生物群是人体最大且最复杂的微生物群落,它连接着我们的外部和内部环境。对肥胖症和心血管代谢疾病(包括2型糖尿病)中肠道微生物群进行流行病学研究的动机源于啮齿动物研究,这些研究证明了肠道微生物群、脂肪性和葡萄糖耐受性之间的联系。
在人类研究中,流行病学研究观察到肥胖症患者的微生物多样性降低,但通过对小型队列进行16S rRNA基因测序或全基因组宏基因组学分析的荟萃分析,并未发现普遍适用的与肥胖相关的肠道微生物特征(换句话说,微生物与疾病的研究需要大队列和大数据库)。
然而,一项对以色列和美国34057名个体进行深度测序全基因组宏基因组学的大规模肠道微生物研究显示,肠道微生物与表型之间存在一致的关联,并且基于微生物组数据训练的机器学习模型对体重指数(BMI)和糖化血红蛋白(HbA1c)的预测准确性可以在不同队列间复制。
通过对训练队列进行子采样,发现随着队列规模的增加,预测准确性也随之提高,约7500-10000个体的样本量可以获得可重复的结果(目前谷禾糖尿病模型使用了9989个样本队列)。这一发现强调了在人类肠道微生物横断面研究中使用大型队列(包含数百名个体)和深度测序组学的必要性,以充分代表人群中的个体异质性以及地区和人口统计学变异。
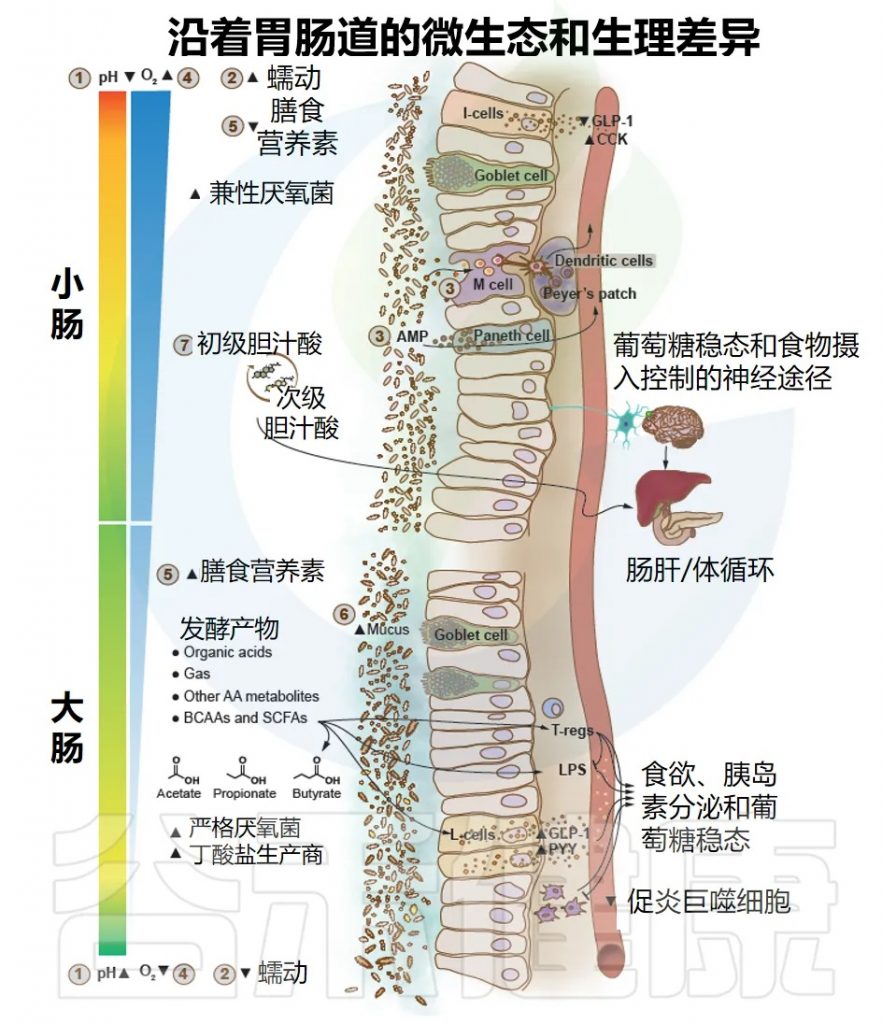
Byndloss M, et al.Diabetes Care.2024
胃肠道(GI)沿线的微生态和生理差异反映了各种环境因素的变化,这些变化依赖于物理、营养和生物学的宿主因素。这些因素导致肠道不同区域的细菌及其生理功能的适应和差异,通过可能影响葡萄糖代谢和糖尿病风险的多向交互产生效果。
以下是影响不同区域微生物数量和组成的主要因素:
pH值:从胃(pH1.0–4.4)到小肠(pH 5.5–7.0),然后更为渐进地到达结肠,在那里由于复杂碳水化合物(纤维)的微生物发酵,pH值可能再次下降(pH5.5)。在粪便中pH值再次升高(可达pH7.8)。
肠道蠕动:在小肠中,肠道通过时间较短,肠蠕动更为强烈,而在大肠中则不然。
肠道上皮细胞:小肠的泛素细胞分泌抗菌肽(AMPs),充当天生防御屏障以减少细菌定植;而M细胞从肠腔中捕获细菌。
氧气浓度:从小肠到大肠,氧气水平逐渐降低。
膳食营养:主要在小肠中由宿主酶消化和快速吸收蛋白质、脂肪和简单碳水化合物,因此限制了细菌对这些营养物质的获取;而部分未消化的膳食残留(例如复杂碳水化合物和部分水解的蛋白质/氨基酸)则积聚在大肠,为细菌提供营养。
宿主糖基:它是由杯状细胞产生的粘液层的一部分,在大肠中远比小肠厚,也是肠道细菌的营养来源,支持其生长。
胆汁酸:分泌至小肠,抑制和有利于某些参与其代谢和再循环的特定细菌的生长。
以上所有生物和非生物因素影响生态条件,使得细菌群体可以在向肠道最远端部分移动时更加密集(从十二指肠的每克102–104个细菌到回肠的107–109个,以及结肠的1011–1012个),并导致细菌组成的差异,其中兼性厌氧菌优先在小肠定殖,而严格厌氧菌在大肠内的微生物中占主导地位,包括丁酸盐生产者。
因此,为了在种族/民族、生活方式和其他人口特征差异之外识别疾病特异性的微生物特征,重要的是在大型人群中进行研究,并将来自不同研究的健康个体/对照参与者作为参考;这些方法已被证明可以提高疾病预测准确性。
2)
肠道微生物代谢与葡萄糖稳态的关联
多项观察性研究报告了肠道微生物群与2型糖尿病之间的关联。在全球流行病学研究中发现,2型糖尿病和糖耐量受损/空腹血糖异常患者的肠道微生物组成发生了一致的改变,这些改变在代谢综合征中也存在,主要特征是微生物多样性降低和产生短链脂肪酸(SCFA)丁酸盐的细菌丰度减少。一些研究还观察到机会性病原体的增加。
最近,粘液降解菌Ruminococcus gnavus被确定为代谢综合征多个特征的预测因子,包括低度炎症、腰围增大、血清甘油三酯升高、糖化血红蛋白升高和高密度脂蛋白胆固醇降低。然而,正如对不同疾病(包括胃肠道和代谢性疾病)肠道菌群改变的荟萃分析所指出的,这些特征中的几个并非疾病特异性,可能代表了一般的肠道菌群的失调。
与葡萄糖代谢和2型糖尿病相关的肠道微生物代谢物和信号分子,肠道微生物群的结构蛋白和分泌蛋白参与调节免疫反应和炎症,正如F. prausnitzii(普拉梭菌)分泌的蛋白质(微生物抗炎分子[MAM]),它能够抑制核因子-κB(NF-κB)通路。另一个例子是A. muciniphila(AKK菌)的外膜蛋白Amuc_1100,它能改善肠道屏障并降低炎症。
①短链脂肪酸
肠道菌群生成短链脂肪酸(SCFAs),SCFAs调节几个过程,包括肠道蠕动和pH值、肠道屏障免疫反应以及通过影响糖异生、胰岛素敏感性和胰岛素分泌的途径来调节系统代谢。
此外,SCFAs调节免疫细胞功能,增强肠道屏障,并刺激肠神经的信号传导。其中丁酸盐还为结肠细胞提供主要能量,并通过激活过氧化物酶体增殖物激活受体γ(PPARγ)增加结肠细胞的β-氧化(β-ox)。
然而,人类研究显示,2型糖尿病患者粪便或血液中不同SCFAs的水平存在广泛变异,这可能是由于方法学局限性造成的。支持SCFAs在调节葡萄糖代谢中作用的最强有力证据来自动物研究和最近一项使用孟德尔随机化统计方法的人类研究(参见下文)。
②胆汁酸
胆汁酸是两性分子,可介导膳食脂肪和脂溶性维生素的吸收。这些分子也被认为是调节脂质、葡萄糖和能量代谢的主要参与者。因此,在2型糖尿病和其他肥胖相关疾病中发现了胆汁酸池的改变,并被认为可能是2型糖尿病病理生理学的潜在贡献因素。
12α-羟基化胆汁酸水平的增加和6α-羟基化胆汁酸水平的降低与胰岛素抵抗有关,并在2型糖尿病患者中出现。胃旁路手术后观察到6α-羟基化胆汁酸水平增加,可预测2型糖尿病的缓解。
干预研究也调查了胆汁酸在人类代谢中的潜在重要性。在减肥手术后,系统性胆汁酸水平升高以及肠道信号激发GLP-1释放的现象得到了验证,尤其是餐后增加被认为特别重要。
然而,在一些进行过胆囊切除术的个体中发现胆汁酸反应过度,并与GLP-1和胰岛素反应的进一步增强相关联。在2型糖尿病患者中,已证明二甲双胍通过减少脆弱拟杆菌的丰度来改善葡萄糖代谢,这与肠道中甘牛胆酸水平的增加和法尼醇X受体(FXR)的抑制有关。
然而,对肠道微生物群与胆汁酸之间直接相互作用及其与2型糖尿病及相关疾病的发展和治疗的关联的理解仍然有限,还需要更多的人体研究来探索。
③色氨酸代谢物
肠道菌群色氨酸代谢物,如吲哚丙酸(IPA)和吲哚丙烯酸(IA),通过孕烷X受体(PXR)改善肠道屏障,并通过肠道免疫细胞上的芳香烃受体(AHR)进行信号传导,增加白介素-22(IL-22)的产生,从而调节免疫和代谢反应。
④其他代谢物
在血液中,IPA和IA还提供抗氧化和抗炎功能。研究还表明,3-吲哚丙酸(色氨酸的代谢产物)的循环水平升高与胰岛素分泌和敏感性改善以及2型糖尿病风险降低相关。
咪唑丙酸和支链氨基酸(BCAAs)通过激活雷帕霉素复合体1(mTORC1)的机制型靶标来损害胰岛素信号传导。
肠道菌群还产生乙醇,与脂肪肝病和胰岛素抵抗有关。
肠道微生物群产生的氨基酸代谢物也与2型糖尿病相关(前两个图)。研究表明,3-吲哚丙酸(色氨酸的代谢产物)的循环水平升高与胰岛素分泌和敏感性改善以及2型糖尿病风险降低相关。
此外,在存在胰岛素抵抗和2型糖尿病的个体中,血浆组氨酸代谢的细菌产物——咪唑丙酸水平增加。来自肠道菌群对芳香氨基酸代谢的这些代谢物和其他一些代谢物也与欧洲和美国独立队列中的心血管事件风险和死亡率有关。
肠道菌群还产生乙醇,与脂肪肝病和胰岛素抵抗有关。
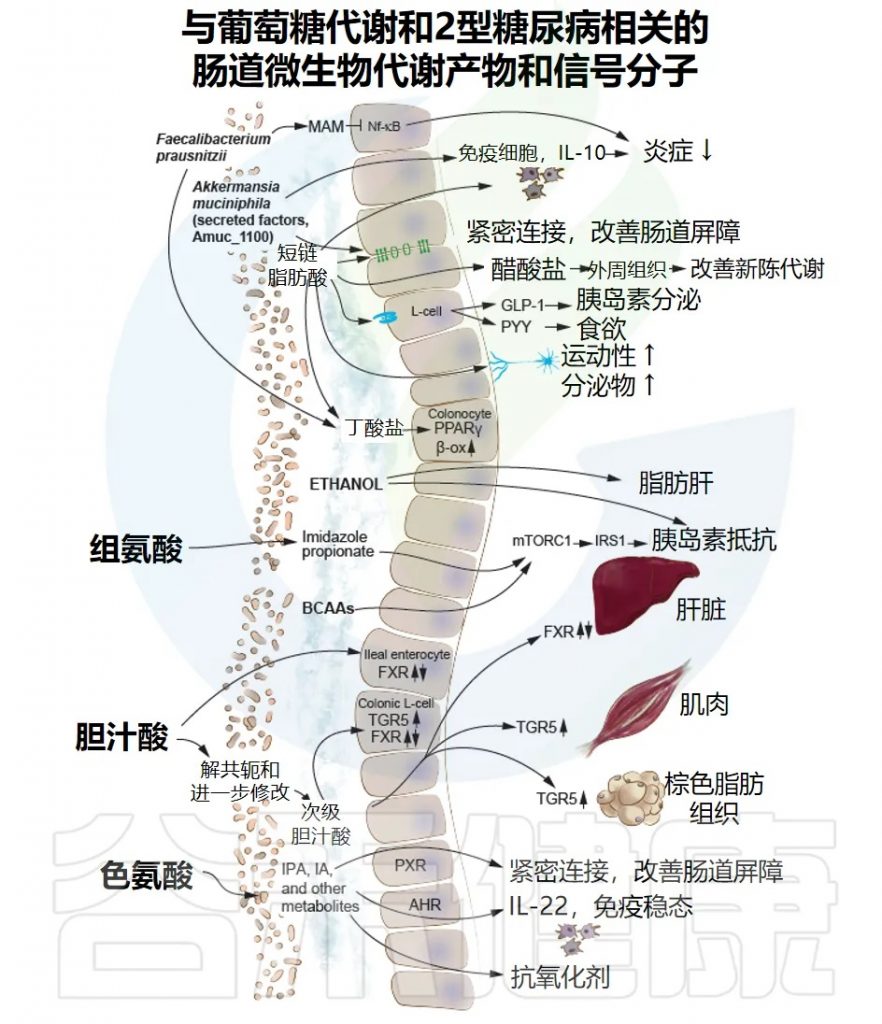
Byndloss M, et al.Diabetes Care.2024
3)
肠道菌群或其代谢产物和降糖药物治疗的关系
支持肠道微生物群在2型糖尿病中作用的证据已通过观察性和干预性研究得到加强。这些研究表明,二甲双胍使用者的肠道菌群中多种细菌物种的相对丰度发生了变化。
二甲双胍使用者肠道菌群发生显著变化
在多个涉及接受二甲双胍治疗个体的队列研究中,发现大肠杆菌(Escherichia coli)的相对丰度升高,而Intestinibacter bartlettii的丰度降低。
此外,最近一项大规模宏基因组研究发现,接受二甲双胍治疗的个体中Escherichia marmotae增加,而Romboutsia timonensis减少。
随机试验和对未使用药物的个体的研究为这些肠道菌群差异在2型糖尿病中的因果效应提供了支持。这些研究表明,肠道菌群组成的变化转化为丙酸盐和丁酸盐产生的增加以及胆汁酸池的调节,这可能介导了二甲双胍降血糖效应的一部分。
然而,肠道菌群也可能是约30%服用二甲双胍的个体经历短暂或持续肠道不适的原因(例如,通过一些大肠杆菌属种增加气体产生)。
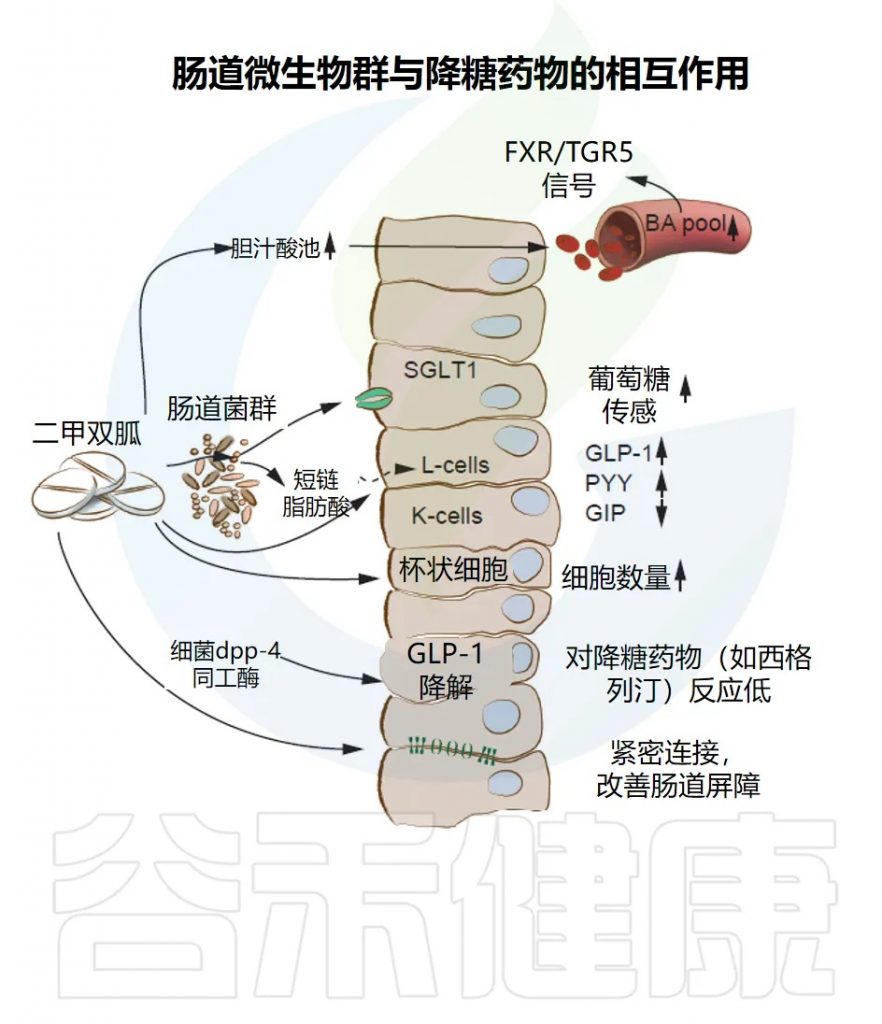
Byndloss M, et al.Diabetes Care.2024
肠道微生物代谢物参与二甲双胍的作用机制
肠道微生物代谢物参与二甲双胍的作用机制,包括通过胆汁酸受体FXR和TGR5进行胆汁酸信号传导,以及生产短链脂肪酸,这些都调节了内分泌细胞(K细胞和L细胞)中胰高血糖素样肽-1(GLP-1)、胃抑制多肽(GIP)和肽YY(PYY)等促胰岛素分泌激素的释放。
其他与二甲双胍作用相关的肠道菌群依赖机制包括通过钠-葡萄糖共转运蛋白1(SGLT1)改善葡萄糖感知和提高肠道屏障功能(例如,恢复紧密连接和增加产粘蛋白的杯状细胞)。
然而,肠道菌群通过表达二肽基肽酶-4(DPP-4)同工酶,可能降低GLP-1活性并影响降糖药物的疗效。
其他降糖药物与肠道微生物群的关联
关于其他口服降糖药物,研究表明二肽基肽酶4(DPP-4)抑制剂和α-葡萄糖苷酶抑制剂对肠道微生物群和微生物代谢物有影响,但钠-葡萄糖共转运蛋白2(SGLT2)抑制剂、噻唑烷二酮类和胰高血糖素样肽1(GLP-1)受体激动剂的影响较不明确。
迄今为止,大多数涉及SGLT2抑制剂的研究都是在小鼠模型中进行的,少数现有的人体研究结果存在矛盾,无法清楚地区分SGLT2抑制剂的效果与之前或同时使用二甲双胍治疗或同期生活方式改变的影响。
GLP-1受体激动剂可能通过激活上皮内淋巴细胞GLP-1受体产生抗炎作用,进而可能有助于调节肠道微生物群。尽管需要更多研究,但现有证据表明肠道菌群可能介导一些降糖治疗的益处,某些益生菌或益生元可能通过影响肠道微生物或其功能进一步改善这些药物的降糖效果。
需要进一步的干预和转化研究来确定药物诱导的肠道菌群变化是否在介导健康效应中起因果作用,并揭示潜在机制。
重要的是,肠道菌群还可能影响降糖药物的效果,例如通过表达人类DPP-4的同源物,这可能降低GLP-1的活性并影响葡萄糖代谢(下图)。由于细菌DPP-4同源物似乎对一些靶向人类DPP-4的药物具有耐药性,因此可能需要抑制细菌同工酶来改善当前药物的代谢反应。
4)
孟德尔随机化在阐明因果关系中的作用
肠道微生物群能够通过多种方式影响和与宿主健康互动,因果关系的方向通常是双向的甚至是多向的。肠道微生物群特征在不同层次(如群落、物种、途径、基因和代谢物)能影响宿主表型(例如改变肥胖的风险),同时,表型的发展(如肥胖)又可以反过来改变肠道微生物群。
孟德尔随机化(MR)是一种统计方法,利用与暴露相关的人类遗传变异体来区分疾病结果的因果效应、混杂、反向因果关系或其他因素导致的关联。为了应用MR研究肠道微生物与2型糖尿病的联系,所研究的肠道微生物特征需要受到一个或多个强烈的人类遗传变异体的影响,以便在工具变量分析中用作工具。
尽管已经有若干全基因组关联研究针对不同的肠道菌群特征(如肠道细菌分类群的相对丰度和人类粪便微生物代谢物)进行,但研究其在2型糖尿病中的因果作用的大型MR研究仍然有限,且尚未在重复研究中得到证实。
部分鉴定出与粪便短链脂肪酸(SCFA)水平相关的人类遗传变异体,并报告了肠道菌群丁酸盐生成潜力(即负责肠道丁酸盐生成的基因)与口服葡萄糖耐量测试中胰岛素反应改善之间的潜在因果联系。这些作者还发现异常的粪便丙酸水平与2型糖尿病风险增加之间存在因果关系。
另一项MR研究报告称,2型糖尿病和肾脏疾病提高了血浆中肠道菌群依赖的代谢物三甲胺氧化物(TMAO)的水平,并提出此前观察到的较高TMAO水平与心血管疾病风险升高的证据可能是由于混杂或反向因果关系,而非因果效应。
另一项近期研究则表明某些细菌属与2型糖尿病之间可能存在因果联系。考虑到MR(如多效性和与弱工具变量相关的问题)及肠道菌群研究(如方法学差异、个体间异质性和个体内可变性)的局限性,需要进行大型高质量研究,以评估使用MR的宿主遗传变异体模拟特定肠道菌群特征(无论是特定细菌物种、属还是代谢产物)的能力,以理解与2型糖尿病发病机制的因果关系。
Relationship
▸ 膳食纤维摄入量较少可能导致2型糖尿病
膳食纤维摄入量减少与2型糖尿病风险增加有关;因此,新的糖尿病管理饮食建议鼓励多食用加工程度低的植物性食物,如全谷物、蔬菜、全水果、豆类、坚果和种子。
饮食是肠道生态系统的驱动因素,微生物可及碳水化合物促进菌群多样性和短链脂肪酸生成,从而减少炎症并支持肠道屏障的维持。
就肠道菌群和葡萄糖代谢而言,增加纤维摄入量与不同菌种水平的增加有关,例如Prevotella copri(现已更名为Segatella copri)。研究还表明,纤维对 HbA 1c的有益作用可能是由特定的基线肠道菌群组成和纤维促进的短链脂肪酸产生菌的多样性介导的。
▸ 肠道微生物的特定成分会影响其宿主反应
然而,即使在控制良好的饮食干预中也会观察到不同的影响,而且鉴于肠道菌群的个体间差异很大,对肠道菌群的饮食反应是高度个性化的。
精准营养或“个性化”营养是一个不断发展的领域,它基于识别可用于设计饮食干预措施的个体特定反应预测特征。利用关于肠道菌群成分的个人数据以及血液生物标志物和饮食习惯等其他信息,机器学习方法已被用于预测标准化膳食后的血糖反应,其准确度高于其他预测方法。
这些研究表明,肠道微生物的特定成分会影响其宿主的特定反应(即在不同细菌存在的情况下,对饮食的反应不同)。因此,肠道菌群至少在一定程度上决定了人类代谢的异质性。肠道菌群可改变且代谢活性高,为更精确的生活方式干预和新疗法提供了可能性。
知识差距、挑战和机遇
目前已有数个大型的高质量参考基因组目录 ,它们为人类研究中肠道微生物的分类和功能表征提供了极大的便利。然而,这些数据库并非没有局限性。
对于流行病学分析而言,肠道菌群数据充满了挑战,包括个体间和个体内巨大的变异性、高维性(即观察到的肠道菌群特征的数量可能大于样本和受试者的数量)和稀疏性(即肠道菌群特征,如物种,仅在部分样本中检测到)。
在种群水平上,肠道菌群由数千个相互作用的物种组成,每个物种在不同宿主之间以及在同一宿主内随时间推移都具有遗传多样性;然而,常见的分析往往忽略了这种非独立性、微生物之间复杂的加性与相互作用效应,以及肠道菌群的可修改性和波动性。然而,最近的一些分析表明,不同细菌物种的个体内变异和对宿主生理的适应模式不同。
其他挑战与肠道微生物群可能影响和响应的大量表型和环境因素有关。大规模群体研究的重要性在人体遗传学中已被无可置疑地证明;大多数多基因性状已知受许多具有小效应的遗传变异影响,但这些变异可以汇总成具有临床重要性的强大多基因风险评分。
类似地,大规模宏基因组研究结果表明,单一的细菌物种可能与人类表型存在低效应大小的关联,或以低丰度存在。因此,需要大样本量以获得足够的统计能力和涵盖个体间变异性,以获得可复制的结果和高预测准确性。
为了更好地理解肠道菌群变异和动态对2型糖尿病的长期影响,前瞻性研究至关重要。在目前发表的少数前瞻性研究中,肠道菌群特征与中国地区多样人群中发生的2型糖尿病事件和西班牙一项临床试验的子集相关,这些研究均使用16S rRNA基因测序。
在一个拥有18年随访的大规模芬兰基于人群的队列研究中,肠道菌群特征也与2型糖尿病有关,使用的是浅层宏基因组测序。然而,在这些研究中,发生病例的数量有限,分析的分辨率也有限。
例如,Faecalibacterium prausnitzii是下一代益生菌最有希望的候选者之一,但也有其他有希望的候选者,如Akkermansia muciniphila和P. copri。关于F. prausnitzii,已在人体肠道中发现了几种潜在亚种,它们具有利用复杂多糖的不同功能潜力 。
与此观察一致,大型宏基因组学研究也发现了几种F. prausnitzii 亚种,但仅其中一部分亚种与 BMI 呈负相关。就P. copri 而言,已发现其与宿主代谢表型(如内脏脂肪和葡萄糖反应)呈正相关和负相关。
这些不一致的发现可以部分地用种内和种间多样性来解释。例如,这些差异可能将同一物种内的菌株定义为共生菌或致病菌,如脆弱拟杆菌和艰难梭菌的情况,这取决于该菌株是否编码毒力因子。
Perspectives
在进化过程中,哺乳动物必须适应充满微生物、病毒和真菌的世界。在无菌的宫内环境中,哺乳动物在出生期间和出生后立即暴露于潜在的有害微生物中。进化创造了实质性的屏障,包括胃肠道运输过程、免疫球蛋白A (IgA) 、粘液、上皮层、内皮屏障、淋巴结和肝脏,所有这些屏障阻止微生物易位进入体内,但为微生物生态系统创造了最佳储存库。上消化道中的微生物数量较少。同时,大肠内微生物密度和丰富度较高,从小肠到大肠,pH值和有氧/厌氧条件发生生理变化,大肠内为厌氧条件。
1)
微生物的基本功能
肠道微生物除了对哺乳动物造成潜在的有害威胁外,还为哺乳动物提供必需功能,包括训练免疫系统、保护其免受病原体的侵害(即抗定植)、代谢功能、营养物质供应(如维生素)、肠道蠕动和外来生物的解毒。
同时,微生物和宿主在小肠中竞争营养物质,微生物产生的常量营养素副产物被提供给宿主。
营养物质(即纤维)和哺乳动物代谢物(如葡萄糖醛酸苷、粘液多糖和胆汁酸)通过微生物代谢进行发酵或转化。微生物代谢以及微生物细胞的死亡和周转形成了外周血中微生物代谢物池,其中约30%的外周血代谢物与肠道菌群及其代谢物有关。
这些微生物代谢物被G蛋白偶联受体(GPCR) 或芳烃受体(AHR)等受体识别,或被哺乳动物酶(如 TMAO)进一步加工,通过表观遗传修饰调节哺乳动物基因表达,对代谢健康产生影响。
2)
不可消化纤维及其代谢物的作用
不可消化的碳水化合物是大肠中特定细菌的能量来源,这些细菌含有宿主所缺乏的酶,这些酶可以代谢这些纤维并促进SCFA的产生。大量研究表明,外源性短链脂肪酸(尤其是丙酸和丁酸)对糖尿病样表型的啮齿动物模型有益。然而,1型和2型糖尿病临床试验的证据尚不明确。
在结肠中,短链脂肪酸(SCFA)通过与GPCR和游离脂肪酸受体2和3结合激活肠内分泌细胞(EEC),诱导肠道肽(主要是GLP-1和肽YY)的释放。为了支持这一发现,啮齿动物和人类补充益生元可以改善葡萄糖耐量和胰岛素抵抗,并且与肠道肽水平升高有关 。
在一项研究中,高纤维饮食改善了2型糖尿病患者的葡萄糖耐量,这种效果与粪便丁酸水平和循环GLP-1水平升高有关。GLP-1通过增加胰岛素分泌、促进胰岛素敏感性和减少肝葡萄糖生成来调节葡萄糖稳态。
此外,短链脂肪酸(SCFA)对于维持整体肠道健康和肠道屏障至关重要,因为丁酸是结肠细胞的主要燃料来源。相反,丁酸减少会促使结肠细胞进行无氧糖酵解,从而增加上皮氧合,破坏结肠的厌氧环境。
3)
肠道菌群在肠道屏障功能中的作用
肠道微生物在肠道屏障功能中起着至关重要的作用。肠道屏障受损会导致肠漏,从而引发低度全身性炎症,这是肥胖和糖尿病的特征。尽管这些机制主要在实验模型中进行研究,但导致全身性炎症的一个潜在机制是来自革兰氏阴性细菌细胞包膜的脂多糖(LPS)内毒素增加,也称为代谢性内毒血症。
肠道菌群失调会导致促炎反应,影响葡萄糖稳态
脂多糖(LPS)可以作用于全身特定的病原体相关分子模式 (PAMP) — Toll 样受体 4 (TLR4) — 从而引发促炎免疫反应,从而对葡萄糖稳态产生负面影响。一系列研究表明A. muciniphila通过作用于 TLR4 和肠道屏障,在介导肠道微生物改变对全身炎症的一些影响方面发挥潜在作用;
然而,关于其在代谢疾病中介导葡萄糖代谢作用的证据较少 。然而,还需要更多的研究来确定TLR4以外的 PAMPs 对代谢物的感知是否与调节宿主-微生物串扰和人类肠道屏障完整性有关。
与此同时,在肥胖个体的肠道中观察到促炎性巨噬细胞的积聚、CD8αβ T 细胞浸润和 IgA+ 免疫细胞减少 ,导致胰岛素抵抗。肠道菌群调节策略可以减轻高热量饮食对肠道免疫的不利影响。例如,减少促炎性巨噬细胞的比例并增加 3 型先天淋巴细胞和调节性 T 细胞与改善葡萄糖代谢有关。
尽管如此,了解驱动肠道微生物与免疫相互作用的精确分子机制及其在人类中的应用也需要未来进行广泛的研究。
Progress
在微生物组领域,将现象学与实际生物学区分开来需要工具和方法来识别机制,以解析微生物组是否是代谢疾病的驱动因素或提供治疗机会。在这里,我们讨论了推动该领域发展的最有希望的技术发展。
模型系统
在比较研究肠道微生物与代谢疾病之间关系的模型系统时,必须同时考虑传统模型(例如无菌和无菌小鼠)和新兴技术(例如器官芯片和非鼠类GF模型,如斑马鱼和猪)。
无菌模型有利于研究肠道菌群与特定疾病的关联
无菌动物已被广泛用于研究人类肠道微生物在肥胖和糖尿病中的作用。这些动物出生时没有任何微生物群,可以在没有微生物组的情况下对干预措施进行研究。因此,我们可以深入了解微生物组是否对给定的生物过程必不可少。
通过将整个肠道菌群(通过供体粪便)或特定的分离菌株定植到无菌小鼠体内,建立无菌疾病模型。研究表明,将肥胖和2型糖尿病患者粪便中的微生物接种到无菌动物体内后,无菌动物成功复制了疾病表型,这为肠道菌群与代谢疾病的关联提供了证据。
此外,从患有病态肥胖和糖尿病的人的肠道中分离出一种过度生长的产生内毒素的细菌阴沟肠杆菌B29,它在无菌 C57BL/6J 小鼠中诱发了肥胖、脂肪肝和胰岛素抵抗,而这些小鼠对高脂饮食引起的代谢缺陷具有抵抗力。敲除B29菌株中产生内毒素的基因或C57BL /6J小鼠中的Tlr4基因可预防代谢缺陷,强调了特定肠道细菌和宿主反应在代谢疾病的发生和发展中的因果关系。
然而,某些关于肠道微生物小鼠的概念一直流传,而这些概念只是研究一种基因型的结果。例如,GF C57BL/6J 小鼠对饮食诱导的肥胖具有抗性,而 GF Swiss Webster 小鼠则不能;因此,由于大多数 GF 小鼠研究都使用 C57BL/6J 小鼠,因此 GF 小鼠通常必须比传统小鼠吃得更多才能维持体重,这已成为事实。这些模型对高脂饮食的不同反应凸显了遗传背景在研究结果中的重要性。
其他GF模型(如猪和斑马鱼)的存在补充了 GF 小鼠的使用。在开发与人类微生物群相关的无菌模型时,GF 猪和仔猪比小鼠能提供更多与人类相关的见解,尽管饲养它们所需的空间对于许多机构而言过于宽裕,或限制了研究只能使用少数动物。另一方面,无菌斑马鱼已被证明可用于研究肠道微生物和不同宿主细胞发育阶段 。
鱼体的透明度、在不同细菌存在下荧光标记和成像不同细胞类型的能力,以及饲养和繁殖斑马鱼的便利性,都有利于研究特定问题。这些模型不能完全复制人类生理学,但它们允许在严格控制的条件下进行纵向和侵入性采样,这在探究机制问题时非常重要。
器官芯片利于研究人体组织内的相互作用
器官芯片(如肠道芯片)提供了更多与人类相关的系统,因为它们可以直接从人体组织或血液来源的诱导性多能干细胞中获得,这些干细胞保留了宿主的遗传特征,因此,它们使我们可以在受控环境中研究复杂的人体组织和细胞相互作用。
最近的研究已经证明了在半厌氧环境中将微生物群接种到肠道芯片中的能力,目前许多研究小组正在测试在这些芯片上接种日益复杂的群落的效率。虽然肠道芯片模型缺少一些关键的细胞类型,如免疫细胞,但重大进展包括能够连接不同的器官芯片,如肠道芯片和神经元芯片,以模拟肠脑相互作用。利用器官芯片研究微生物组的创造性方法将继续涌现,并可能填补重要的空白,补充动物模型。
了解细菌基因和功能
对细菌全基因组进行测序和组装的能力是一种非常强大的方法,可用于识别菌株的谱系和相关性,以及识别可能与人类健康或疾病相关的特定细菌表型所涉及的假定途径。如果我们思考已从基因改造小鼠研究中阐明的人类疾病机制,就不难想象在细菌中进行同样研究可以获得的大量信息。敲除和操纵细菌基因的能力并不是什么新鲜事。
近80年的细菌遗传学研究已经阐明了病原体如何在肠道上皮中定植并分泌毒素从而导致霍乱等疾病,它们如何相互共享信息以适应不同的环境,以及营养选择如何驱动它们在宿主中的组成。大肠杆菌可以被认为是 C57BL/6 小鼠的细菌版本;其基因定义明确且易于改造,而且它已经成为在特定环境下测试改造效果的主力。然而,共生肠道细菌的多样性远不止大肠杆菌;因此,研究人员正在积极寻求对肠道j基因的更深入了解,例如使用拟杆菌和梭菌作为代表性生物 ,因为大量人类和小鼠研究已经证明了这些生物在健康和疾病中的重要作用。
人工智能等先进计算工具通过预测蛋白质的三维结构,为细菌基因组中未注释部分提供了新的见解,AlphaFold2 等技术极大地推进了这一任务。通过分析这些结构,研究人员可以根据它们的形状和结合位点推断出可能的功能。这些潜在作用可以通过生化和微生物学研究中的实验验证来证实。这些知识,特别是关于蛋白质如何影响代谢途径的知识,对于将微生物活动与糖尿病等健康状况联系起来至关重要,有助于深入了解疾病机制和潜在的治疗靶点。
无参考数据分析
目前,微生物组测序分析中依赖数据库的方法最关键的问题是它们在检测新型或研究不足的微生物方面存在局限性。当使用基于已充分表征的细菌的参考基因组的数据库分析微生物群落样本时,不匹配的序列会被忽略或错误分类。
这一过程导致对微生物生态系统的看法出现偏差,可能会遗漏在健康和疾病(包括糖尿病)中发挥重要作用的关键成分。因此,推进微生物组研究需要开发和使用能够发现和表征这些代表性不足的微生物实体的方法。
从宏基因组测序数据中从头组装基因组是微生物组研究中一种强大的方法,该方法涉及直接从测序读取构建基因组,而不依赖于参考数据库。
该方法使用先进的计算算法将样本中的 DNA 片段拼凑在一起,从而可以识别来自各种生物体的遗传物质,包括那些以前未测序或编目的生物体。通过组装这些基因组,研究人员可以发现新物种并揭示新的基因功能,从而大大扩展我们对微生物多样性及其在包括人体在内的各种环境中的潜在作用的理解。
对于16S测序来说,由于其经济和丰富的样本积累,对于构建疾病的大数据库来说存在很大的优势,虽然16S分辨率没有宏基因组高,但是通过方法和技术的弥补,可以大大提供16S的预测精准度。
例如谷禾健康结合宏基因组数据和已有的菌群构成数据,通过算法可以大大提高扩增子数据的物种分辨率。此外病毒、真菌、寄生虫和部分16S难区分的病原菌目前通过增加靶向引物进行tNGS的方式来实现,大大拓展了二代测序微生物多样性检测领域的涵盖范围。
这种方法在揭示微生物生命的全部范围方面特别有用,包括可能在健康和疾病中发挥关键作用的物种或基因。
了解小肠微生物群
小肠是营养吸收、肠肝循环和承受肠道激素刺激的主要部位;因此,深入了解人体这一部位的微生物功能至关重要。然而,我们对人类微生物组的大部分了解都基于粪便样本和结肠微生物群,因为即使使用现代内窥镜检查方法,获取小肠微生物群也很困难。
最近的进展使用了创新方法,例如可摄入的胶囊,它可以对整个胃肠道中的肠道物质进行取样。由于每个胶囊都由肠道中不同的 pH 值触发,因此这种方法可以提供肠道群落的微生物图谱。这些工具正在进一步完善和商业开发,以用于诊断和研究。但需要注意的是,样本采集到胶囊内后,微生物有可能继续生长,因此不能准确地代表原生微生物群落。
此外,这些胶囊和其他胶囊都是为空腹状态下的取样而开发的,因此对餐后反应的研究仍然有限,尽管这些反应对于全面了解微生物对葡萄糖代谢调节的贡献可能很重要。解决这些问题对于确保使用此类设备进行微生物组研究的可靠性和准确性至关重要。
同工酶和小分子筛选
微生物组研究中,同工酶和小分子筛选对于识别可以作为治疗目标的特定细菌产物至关重要。微生物同工酶是具有不同分子结构但催化与宿主酶相同反应的酶。筛选这些产物可以揭示微生物代谢的变化,这些变化可能会影响健康并可能干扰药物治疗,例如细菌 DPP-4 同工酶的情况。
小分子筛选专注于识别微生物产生的生物活性化合物 。这些化合物可对宿主的病理生理学产生显著影响。通过识别特定的同工酶和小分子,研究人员可以针对它们进行降解或增强,从而为糖尿病等疾病提供潜在的治疗策略。
Treatment
如上所述,目前尚未发现针对1型或2型糖尿病的诊断性和广义的粪便微生物群分类学特征。因此,未来的研究应转向在大规模前瞻性人群中进行菌种和菌株水平的研究,并在可能的情况下,重点研究胃肠道肠道微生物的功能分析,特别关注稳定同位素前体,以研究不同胃肠道区域重要微生物代谢物的生成和底物通量。
1
高纤维饮食和基于短链脂肪酸的治疗
关于基于肠道菌群的糖尿病疗法,高纤维饮食已被证明可有效控制1型和2型糖尿病患者的血糖水平并降低胰岛素抵抗。尽管膳食纤维通过肠道菌群直接起作用的方式仍有待证实,但这些试验强调了将肠道菌群调节策略纳入糖尿病干预试验的潜在重要性,特别是对于生产有益代谢物(如 SCFA)而言。
但是,如上所述,口服短链脂肪酸(丁酸盐)补充剂的干预试验并未显示在1型或2型糖尿病患者的血糖控制或其他糖尿病调节标志物上 ,可能是因为给药部位与内源性产生不同。对于其他短链脂肪酸,包括丙酸盐和乙酸盐,数据太少,无法得出关于对代谢调节的可能影响的任何结论。
2
传统益生菌和新一代益生菌
用于糖尿病的益生菌疗法可分为传统益生菌,特别是乳酸杆菌和双歧杆菌菌株,它们历来被用于发酵食品或补充剂中以促进健康,以及下一代益生菌,它们是最近被鉴定为人类肠道菌群成员的新菌种菌株。这些菌株与健康有关,但在疾病环境中存在会减少。
传统益生菌
关于传统益生菌,一项关于益生菌(双歧杆菌、乳酸杆菌和唾液链球菌菌株)的开放标签试验发现,益生菌对1型糖尿病患者的兄弟姐妹的1型糖尿病易感性和进展有益。
针对新发1型糖尿病的前瞻性随机对照试验 (RCT) 正在进行中(NCT03961854、NCT03961347、NCT04769037 和 NCT05767450),一项规模较小的试验显示,其对长期 1 型糖尿病仅有中等效果。
在2型糖尿病中,最近的一项荟萃分析描述了这些益生菌菌株在代谢控制和降低胰岛素抵抗方面具有一定功效。
新一代益生菌
关于下一代益生菌,在人体中产生的数据较少。例如,尽管特定菌株(例如Akkermansia)与健康的代谢表型相关,但使用A. muciniphila的 RCT 干预并未发现强烈的代谢影响。
这一发现可能是由于这些测试菌株与代谢综合征缺乏因果关系、通过胃后活力降低、剂量不足或引入人体肠道时缺乏定值。
由于小肠对1型和2型糖尿病的病理生理学都很重要,因此需要进一步分析1型和2型糖尿病患者的小肠微生物群,并研究确定的下一代益生菌菌株组合作为糖尿病的可能干预措施。然而,这种努力应该考虑细菌菌株的植入条件、对其他细菌成员的生态或功能依赖性、以及功能上的潜在冗余。
3
供体粪菌移植
在完成针对特定菌株组合的研究之前,供体粪菌移植(FMT)可能有助于了解调节肠道菌群的效果以及这种调节对糖尿病病理生理和潜在可逆性的影响。
最近发表了一项研究,探新鲜粪菌移植在维持残余β细胞功能和抑制新发1型糖尿病患者自身免疫方面的功效。其他针对 2 型糖尿病和胰岛素抵抗的研究显示,FMT 对胰岛素抵抗和非酒精性脂肪肝疾病有适度影响 ,而一项研究表明对这些参数没有影响。
此外,微囊供体 FMT 和纤维补充的联合干预显示出对葡萄糖代谢有益的影响,这表明干预设计不仅需要合成菌株联合体,还需要饮食支持(例如纤维来滋养菌株)。最后,评估生活方式干预后自体 FMT 是否有助于防止体重反弹的研究发现,饮食引起的低丰度细菌变化可能是维持减肥效果的原因,这可以指导更精准的干预措施,减少伦理负担,降低传播疾病的风险。
总体而言,与针对性菌株或代谢物的干预措施相比,供体 FMT 是一种更为分散的方法。此外,现有数据集中的粪便物质给药方式(胶囊与新鲜 FMT)、肠道 pH 值(例如由于抗酸药)和结肠转运时间存在差异,并且施用的粪便微生物群的数量似乎也会影响供体细菌菌株的植入。
关于糖尿病和基于肠道菌群的疗法的试验结果,使用随时间动态测量葡萄糖代谢(例如混合餐测试或连续血糖监测)可以更好地了解在 FMT 和施用确定的菌株组合期间肠道微生物、饮食和葡萄糖稳态之间的相互作用。
对小肠微生物组的研究仍不足,但对肠道微生物的新认识越来越多地将其与人类糖尿病联系起来。对人类 FMT 的干预研究已经能够从因果关系中分离出关联,并且确实显示出一些临床益处,虽然平均而言相对较小的治疗效果和伦理问题阻碍了这种治疗方法在糖尿病临床护理中的广泛实际应用。
因此,还需要在多民族队列中对肠道菌群和糖尿病之间的前瞻性关联进行更多研究。与此同时,还应探索合成的肠道菌群衍生的细菌菌株或群落和工程系统在糖尿病中靶向肠道输送已识别代谢物的治疗潜力。
Perspectives
在过去二十年中,肠道微生物的改变与糖尿病患者的异常葡萄糖代谢和脂肪变性有关。流行病学研究中的较大样本量现已开始显示肠道菌群与肥胖或2型糖尿病相关的人类代谢特征之间的相关性的大小和可能的一致性;然而,对于1型糖尿病,情况就不那么明朗了。
未来的研究应更严格地考虑糖尿病药物与种族和饮食摄入之间的相互作用。此外,近年来,人们对肠道菌群的功能有了更多的了解,而不仅仅是其成分,这些信息与之前关于特定代谢物(包括短链脂肪酸、支链氨基酸和胆汁酸)与肥胖和糖尿病之间联系的报告非常吻合。
关于肠道菌群成分,只有少数研究探讨了噬菌体和真菌的作用以及这些菌落与细菌菌株在糖尿病中的相互作用。显然,未来的研究还需要关注小肠微生物群的功能,以及开发足够的生物信息学流程和正确组装基因组。
我们还必须考虑到,迄今为止的大多数数据都是在小鼠研究中产生的,由于小鼠和人类在饮食、遗传和寿命方面存在巨大差异,其与人类糖尿病的相关性需要进一步证实。尽管如此,在人类糖尿病环境中对单一菌株和粪菌移植的人体干预研究表明,存在一系列临床代谢效应(与药物更一致的效果相比),但没有严重的副作用。
总之,经过近二十年的研究,我们仍然必须期待未来的努力,以阐明肠道微生物研究对人类糖尿病的临床诊断和治疗适用性。
主要参考文献
Caesar R. Pharmacologic and Nonpharmacologic Therapies for the Gut Microbiota in Type 2 Diabetes. Can J Diabetes. 2019 Apr;43(3):224-231.
Ridaura VK, Faith JJ, Rey FE et al (2013) Gut microbiota from twins discordant for obesity modulate metabolism in mice. Science 341:1241214.
Sze MA, Schloss PD (2016) Looking for a signal in the noise: revisiting obesity and the microbiome. mBio 7:e01018-16.
Duvallet C, Gibbons SM, Gurry T, Irizarry RA, Alm EJ (2017) Meta-analysis of gut microbiome studies identifies disease-specific and shared responses. Nat Commun 8:1784.
Pasolli E, Truong DT, Malik F, Waldron L, Segata N (2016) Machine learning meta-analysis of large metagenomic datasets: tools and biological insights. PLoS Comput Biol 12:e1004977.
Rothschild D, Leviatan S, Hanemann A, Cohen Y, Weissbrod O (2022) An atlas of robust microbiome associations with phenotypic traits based on large-scale cohorts from two continents. PLoS One 17:e0265756.

谷禾健康

目前越来越多营养概念诸如”低碳水化合物饮食”正在流行,然而,所有的碳水化合物都是不好的吗? 其实并非如此。
其中,抗性淀粉就是一种特殊的碳水化合物,它与我们通常所熟知的淀粉有区别。抗性淀粉之所以得名,是因为它能够抵抗人体消化酶的作用,逃离被吸收的命运,直接进入肠道。这种特性使得抗性淀粉拥有许多健康功效,它不仅能调节血糖,还能促进有益菌的生长,改善肠道功能,甚至还能增强饱腹感,帮助控制体重。
抗性淀粉天然存在于豆类(如扁豆、鹰嘴豆)、某些谷物(如大麦、燕麦)、块茎(如土豆、山药)等食物中。此外,各种食品加工和烹饪方法可以提高食品中的抗性淀粉含量,进一步使这种淀粉的潜在膳食来源多样化。
作为一种膳食纤维,抗性淀粉可以作为微生物发酵的底物,例如拟杆菌属,瘤胃球菌,双歧杆菌等可以发酵抗性淀粉。
一些肠道细菌促进抗性淀粉发酵产生短链脂肪酸。短链脂肪酸的主要功能是改善肠上皮屏障的完整性,缓解局部和全身炎症,产生全身效应,影响代谢健康、免疫功能,甚至可能影响大脑健康。
目前代谢性疾病负担正在增加,一些膳食可以通过调节肠道菌群干预疾病,其中抗性淀粉也很重要,它结合了传统和科学,促使研究人员和消费者重新审视传统饮食习惯并认识到其潜在的健康影响。
本文探讨了抗性淀粉和肠道微生物组之间复杂的相互作用,包括抗性淀粉影响微生物群落的机制、这些相互作用对健康的影响。此外,还讨论了富含抗性淀粉的饮食对肠道健康的影响。随着对肠道菌群在健康和疾病中的作用了解越来越多,强调从天然来源获取抗性淀粉的饮食将会不断增长,为营养干预提供新途径。
▼
什么是抗性淀粉?
抗性淀粉(RS)是一种不被人体消化酶分解的淀粉,被认为是膳食纤维。
淀粉作为葡萄糖的储存形式天然存在于植物中。食品中的淀粉可分为慢消化淀粉(SDS)、快速消化淀粉(RDS)或抗性淀粉(RS)。
一般淀粉在小肠中迅速分解并转化为葡萄糖,并迅速吸收到血液中,而抗性淀粉则抵抗正常消化,因此得名。它不会分解成葡萄糖,而是不受干扰地通过消化道,直到到达结肠。
首先,淀粉的基本结构基于两种不同的葡萄糖聚合物——直链淀粉和支链淀粉。
直链淀粉主要是由α-1,4-葡萄糖单元组成的线性分子,与支链淀粉的分支结构形成对比,支链淀粉还包含α-1,6键连接。
这两种组分的比例及它们在淀粉颗粒内的排列方式显著影响淀粉的可消化性。这些分子越密集、排列越紧密,消化酶就越难接近它们。
抗性淀粉(RS)是一种独特的膳食纤维,不是在小肠中消化,而是在大肠中发酵。不同食物类别(包括谷物、豆类、块茎和某些加工食品)中抗性淀粉的含量差异很大。
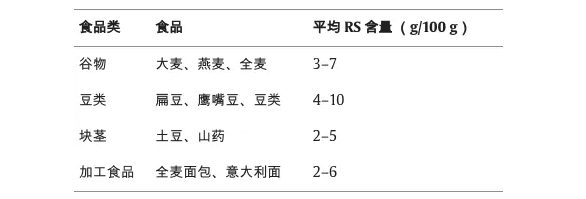
doi.org/10.1016/j.fochx.2024.101118
谷物,特别是大麦、燕麦和全麦等全谷类食物,是抗性淀粉的重要来源,尤其是在它们经过最低限度加工的情况下。比如,煮熟并冷却大米会增加其抗性淀粉含量,这是由于直链淀粉的逆淀粉化作用。
扁豆、鹰嘴豆等豆类也富含抗性淀粉。它们的抗性淀粉含量归因于它们的高直链淀粉和支链淀粉比率,这有助于它们缓慢的消化率。烹饪和冷却过程进一步增加了其抗性淀粉含量。
块茎,包括马铃薯和山药,含有抗性淀粉,尤其是在煮熟和冷却时,这是一个诱导淀粉回生的过程。这使得冷土豆沙拉等菜肴成为良好的抗性淀粉来源。
某些加工食品,特别是那些由全谷物制成或含有 抗性淀粉作为成分的食品,可能是重要的抗性淀粉来源。经过挤压烹饪等过程的全麦面包和意大利面保留了大量的抗性淀粉。
了解这些来源及其抗性淀粉含量对于饮食计划和营养优化至关重要。
在此基础上,抗性淀粉可以根据其起源和特性大致分为四种主要类型:
RS1型(RS1):由于食物基质和蛋白质外壳形成的保护屏障,这种形式的 RS 在物理上无法被酶接近。常见的来源包括全谷物和种子。
RS2型(RS2):RS2 的特点是其天然颗粒形式,主要存在于某些生食中。例子包括生土豆、青香蕉、高直链淀粉玉米。这些来源的高直链淀粉含量导致紧密堆积的颗粒结构,限制了酶的获取。
RS3型(RS3):也称为逆行淀粉,当某些食物煮熟然后冷却时会形成 RS3。这种冷却过程导致淀粉分子的重新排列和重结晶,进一步使它们对酶分解具有抵抗力。煮熟和冷却的土豆、意大利面和米饭等食物是 RS3 的主要来源。
RS4型(RS4):这种类型包括食品中天然不存在的化学改性淀粉,例如一些商业生产的面包和糕点。各种工业过程在淀粉分子中引入交联或取代,以增强其对消化的抵抗力。
部分分类有:
RS5型(RS5): 这是一个较新的类别。它需要通过加热和冷却含有特定脂质(例如脂肪或蜡)的淀粉类食品的过程产生的抗性淀粉。例如含有脂肪成分的面包或含有人工制造的淀粉-脂质复合物的食物。

虽然这些类别有助于讨论和研究,但许多现实世界的食物都含有抗性淀粉类型的混合物。此外,食品加工方法,储存条件和其他食品成分的存在等因素可以显着调节这些食品中的抗性淀粉含量。
抗性淀粉的潜在健康益处和生理影响主要来自其在大肠中的发酵。然而,这些益处的程度和特异性可能因抗性淀粉类型而异。例如,不同的抗性淀粉类型可能优先促进特定微生物物种的生长或导致挥发性脂肪酸的产生速率不同。
总之,抗性淀粉的生化结构和分类对于确定其与肠道微生物组的相互作用以及随后的健康结果至关重要。全面了解这些基础方面对于旨在利用抗性淀粉潜在益处的饮食干预的研究和应用至关重要。
保持血糖稳定
由于抗性淀粉消化缓慢,因此可以保持血糖水平稳定。这可以帮助减少餐后血糖峰值,这对糖尿病患者特别有益。
抗性淀粉具有第二餐的效果:根据一项小型研究的结果,早餐吃抗性淀粉可以降低午餐时的血糖。
2022年1月发表在《Frontiers in Nutrition》 的一篇评论指出,在饮食中添加抗性淀粉是一种简单的生活方式调整,可以帮助糖尿病管理。
促进心脏健康
2018年6月《Nutrition Research》发表的荟萃分析结果,抗性淀粉可以通过降低胆固醇水平有益于心脏健康。它还可以改善血糖控制,正如2017年《Nutrition Journal》上发表的一项针对超重成年人的小型研究所证明的那样,它通过促进肠道中健康细菌的生长来实现这一点,这些细菌产生具有有益作用的短链脂肪酸。
根据2022年3月发表在《国际分子科学杂志》上的一篇评论,短链脂肪酸有助于调节交感神经系统。抗性淀粉可能有助于治疗因神经系统过度活跃而加剧的心脏病,例如慢性心力衰竭、高血压和冠状动脉疾病。
减肥效果
抗性淀粉非常有饱腹感,可能会降低食欲。与其他碳水化合物相比,它的热量也较低,通过这些方式,抗性淀粉可能有助于减肥。
虽然早期研究表明抗性淀粉可能在减肥中发挥作用,但还需要进一步的研究来证实任何此类益处。2017 年《营养杂志》上发表的研究表明,每天吃 30 克抗性淀粉,持续六周,18 名超重成年人减少饥饿激素和无意识地吃零食,但不会改变身体成分。
支持肠道健康
抗性淀粉的作用类似于纤维,而纤维会被肠道中的健康细菌发酵。“这些有益的肠道细菌可以产生短链脂肪酸,这对肠道健康有帮助。
例如,短链脂肪酸可以帮助保持肠道内壁坚固,并有助于粘液产生和炎症,还可能有助于降低结直肠癌的风险。这在后面章节我们会详细阐述。
成人每天应摄入约15克抗性淀粉。然而,据估计大多数人每天的摄入可能不到这个量。
抗性淀粉食品含量表
(每100克食品的平均含量)
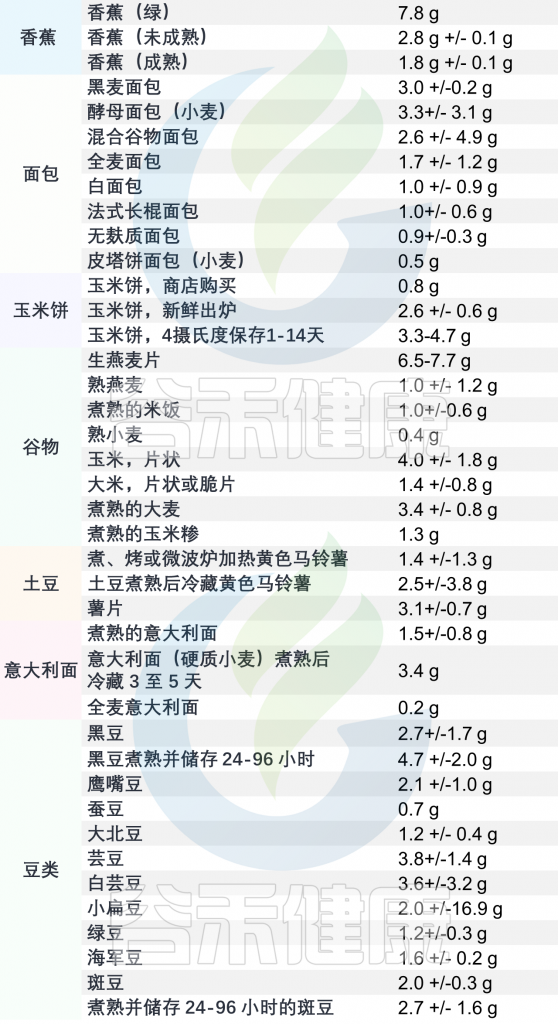
注:如果要增加摄入量时,请逐步增加,一下子吃太多可能会腹胀和胀气。
挤压烹饪,广泛用于生产即食谷类食品和零食,可以增加最终产品的抗性淀粉含量,取决于应用的条件,诸如含水量、螺杆速度和温度等参数可以进行调整以优化抗性淀粉的形成。
退火过程涉及水化淀粉颗粒而不使其明胶化,已发现这一过程可以增加一些谷物中的抗性淀粉含量。
发酵,是各种文化中历史悠久的烹饪和保鲜方法,具有提高抗性淀粉水平的固有能力。这一过程通常涉及有益细菌或酵母分解和发酵糖类,可以改变淀粉结构,使更多的淀粉对消化具有抗性。例如,将谷物发酵制成酸面包或某些传统非洲菜肴不仅赋予了独特的风味,还增加了它们的抗性淀粉含量。
虽然探索和应用这些方法可以显著增加抗性淀粉含量,但必须考虑更广泛的营养后果。并非所有增加抗性淀粉的方法都是普遍有益的。有些加工方法可能会剥夺食物的重要营养素,或引入不良化合物。
在增强抗性淀粉获得肠道健康益处,和确保食物整体营养价值保持完整之间取得平衡至关重要。
抗性淀粉(RS)与肠道微生物组之间错综复杂的相互作用主要发生在结肠,结肠是大多数未消化碳水化合物达到代谢目的的地方。
肠道微生物群发酵抗性淀粉
人类结肠内有着丰富多样、复杂的微生物群,对发酵未消化膳食成分,尤其是抗性淀粉,起着至关重要的作用。
抵达结肠后,抗性淀粉将被居住在肠道的微生物群体进行厌氧发酵。这一发酵过程导致了短链脂肪酸的产生,主要是乙酸、丙酸和丁酸,以及氢气、甲烷和二氧化碳等气体。
在短链脂肪酸中,丁酸在结肠健康中发挥着关键作用。作为结肠细胞的主要能源来源,丁酸还具有抗炎性能,加强结肠防御屏障,并潜在降低结肠癌的风险。此外,短链脂肪酸通过调节肠道pH值,有利于有益菌的生长,同时抑制致病菌株的增殖。
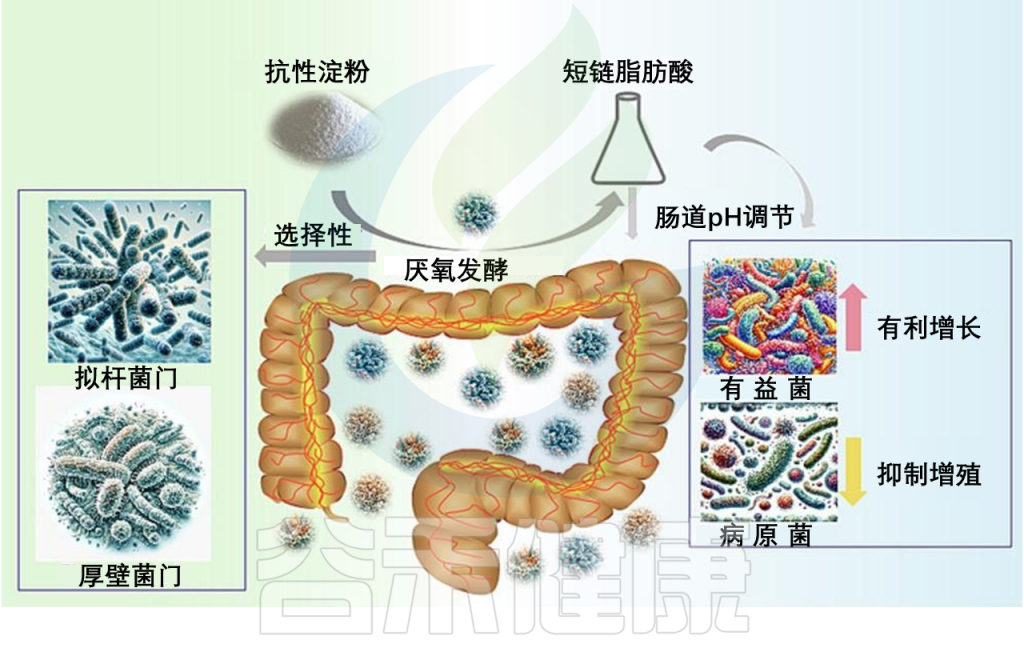
doi.org/10.1016/j.fochx.2024.101118
选择性发酵:抗性淀粉增多→有益菌随之增加
抗性淀粉发酵的另一个有趣的方面是其选择性。并非所有肠道微生物都能够有效发酵抗性淀粉;特定菌群,特别是来自拟杆菌门和厚壁菌门的细菌群,是主要的抗性淀粉发酵者。
这种选择性发酵会导致肠道微生物组成的变化。持续的抗性淀粉摄入可以促进这些抗性淀粉发酵细菌的增殖,使肠道富含有益微生物,进一步提高发酵效率和短链脂肪酸的产生。因此,抗性淀粉和肠道微生物群之间的动态相互作用有望进行有针对性的干预,有可能通过饮食策略调节肠道微生物组成和活性。
抗性淀粉为微生物发酵提供了底物,作为回报,肠道微生物会产生有益于宿主健康的代谢物。
前面我们了解了结肠中抗性淀粉发酵的机制,这里了解这一过程中的关键微生物参与者也很重要。
拟杆菌门
积极参与抗性淀粉发酵的主要群体属于拟杆菌门,尤其是拟杆菌属,拟杆菌的代谢能力使它们能够在各种复杂的碳水化合物中茁壮成长,包括抗性淀粉。它们的酶库有助于将抗性淀粉分解成更简单的单元,然后发酵以产生短链脂肪酸。
厚壁菌门
抗性淀粉发酵的另一个重要贡献者是厚壁菌门,尤其是瘤胃球菌属,瘤胃球菌是这一领域的关键物种,因其在启动抗性淀粉降解方面的无与伦比的效率。瘤胃球菌(R. bromii)进行的初步降解使抗性淀粉更易于其他微生物群进一步发酵。考虑到在摄入富含抗性淀粉饮食的个体中的统治地位,其重要性变得明显。此外,瘤胃球菌的丰度较高与改善的肠道健康状况相关,表明其潜在的保护作用。
放线菌门
虽然拟杆菌门和厚壁菌门脱颖而出,但另一个门放线菌门通过双歧杆菌属促进抗性淀粉发酵。双歧杆菌是备受推崇的益生菌,已知具有无数的健康益处。在抗性淀粉的背景下,双歧杆菌发酵它以产生短链脂肪酸,从而降低肠道 pH 值,从而创造不利于病原菌的环境。此外,抗性淀粉的双歧杆菌效应,即补充抗性淀粉导致双歧杆菌增加,已在各种研究中得到充分证明。
产甲烷古细菌
古菌,特别是产甲烷的Methanobrevibacter smithii,在抗性淀粉发酵领域也发挥作用。
M. smithii消耗其他微生物在抗性淀粉发酵过程中产生的氢气,将其转化为甲烷。这种氢气的去除至关重要,因为它防止了结肠中氢气的积累,否则可能会阻碍发酵过程。因此,M. smithii通过维持其他发酵者的适宜环境,间接支持抗性淀粉发酵过。
总而言之,结肠中的抗性淀粉发酵并不是归因于单个微生物分类群的孤立过程。这是一项涉及多个微生物群体的协同合作,每个微生物群体都为该过程及其健康益处做出了独特的贡献。
只有少数菌群如瘤胃球菌和青春双歧杆菌能够利用淀粉,这些细菌本身并不直接产生丁酸盐,而是依靠与其他肠道细菌的交叉喂养相互作用来产生丁酸盐。
他们的集体行动强调了肠道是一个代谢“器官”的概念,其中饮食成分,主要是抗性淀粉,以协调的方式代谢。
丁酸梭菌(Clostridium butyricum)是一种降解抗性淀粉的菌,能够在多种类型和来源的抗性淀粉上生长,在这个过程中产生大量的丁酸盐。它通过使用一种酶系统来实现这一点,该酶系统虽然是迄今为止在能够降解抗性淀粉的细菌中发现的最简单的酶系统,但在抗性淀粉的消化过程中表现出高度的协同作用和功能多样性。在一个生物体中,消化抗性淀粉和丁酸生产的结合有可能绕过交叉喂养网络的复杂性,而交叉喂养网络通常是抗性淀粉消耗过程中生产丁酸所必需的。
这种独特的性状组合表明,它可能作为一种与抗性淀粉协同使用的菌株,促进更广泛的丁酸反应,从而为更多人群解锁这种益生元的健康益处。
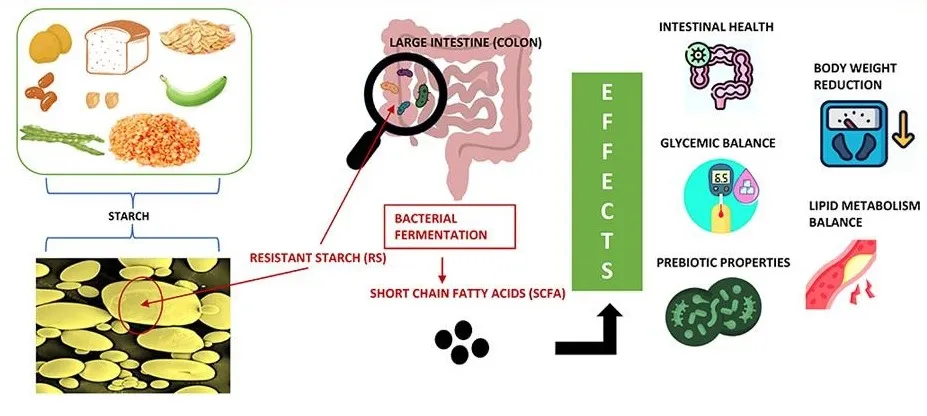
doi.org/10.1016/j.jff.2022.105094
我们了解到肠道微生物群领域及其与抗性淀粉的错综复杂互动是广阔而多层次的。抗性淀粉可以影响肠道微生物群的组成和功能,但同样明显的是,这些影响的程度和性质受到各种因素的调节。揭示这些因素可以增进我们对肠道健康复杂性的理解,并帮助更有效地定制膳食和治疗干预措施。
个体肠道微生物群的基线组成
一个主要决定因素是个体肠道微生物群的基线组成。每个人的肠道微生物群具有独特的特征,受到遗传、早期生活暴露、抗生素和饮食模式等因素的影响。引入膳食时,抗性淀粉可能会因个体肠道微生物的起始点而在个体之间产生不同效应。例如,基线拟杆菌水平较低的个体,在摄入抗性淀粉后,可能会比那些已经拥有更高丰度的个体出现这些细菌增加更显著。
不同类型的抗性淀粉会被特定菌群优先代谢
消费的抗性淀粉类型是另一个重要因素。根据其物理化学性质和来源,抗性淀粉有多种类型:RS1、RS2、RS3、RS4。每种类型可能会被特定微生物类群优先代谢。例如,瘤胃球菌对高直链淀粉玉米中的RS2表现出明显的偏好,而某些拟杆菌物种可能更青睐来自逆行淀粉的RS3。因此,膳食中包含的抗性淀粉类型可以引导微生物群变化的轨迹。
其他营养素的影响
抗性淀粉消费的膳食背景也不容忽视。其他膳食纤维、蛋白质、脂肪和微量营养素的存在可以影响抗性淀粉的可获得性和发酵性。例如,富含可溶性纤维的饮食可能通过促进有益菌(如乳杆菌)的生长来放大抗性淀粉的益生效应。相反,富含蛋白质的饮食可能会使一些结肠细菌转向蛋白质发酵,产生像氨之类的潜在有害化合物。
抗性淀粉摄入的持续时间
抗性淀粉摄入的持续时间也起着关键作用。初始引入抗性淀粉可能会导致微生物群组成的快速变化。然而,随着摄入时间的延长,微生物群可能会稳定下来,表明适应性。长期摄入抗性淀粉可以导致更具弹性和多样化的微生物群,这些微生物群更能抵抗干扰和潜在的菌群失调。
其他宿主相关因素
最后,宿主相关因素,如年龄、健康状况和遗传,调节着抗性淀粉与微生物群的相互作用。与微生物群的年龄相关变化、肠道传输时间的差异和酶活性可能会影响抗性淀粉在肠道中的发酵。同样,患有肠道紊乱症状如肠易激综合征(IBS)或炎症性肠病(IBD)的个体可能对抗性淀粉有不同反应,鉴于这些情况下肠道环境和微生物群的组成发生了改变。
总之,抗性淀粉与肠道微生物群之间的互动是一个受多种因素影响的动态过程。认识和理解这些因素对于个性化营养策略旨在利用抗性淀粉的肠道健康益处是至关重要的。这些见解呼唤着在营养和肠道健康领域采取更个性化的方法,更胜于一刀切的建议。
抗性淀粉已成为膳食的关键成分,其影响远远超出了其营养价值。抗性淀粉最重要的作用之一在于它能够维持肠道的屏障功能。这种错综复杂的粘膜细胞和细胞间连接是我们全身健康的关键,可防止病原体入侵并维持代谢平衡。鉴于肠道相关疾病的负担不断加重,了解抗性淀粉如何影响这一屏障可以为疾病预防和治疗干预提供关键的见解。
肠道屏障是一个动态和反应灵敏的系统,而不是一个静态的实体。在其核心,上皮细胞形成前线,作为对管腔环境的主要防御。
抗性淀粉促进粘蛋白分泌
抗性淀粉通过支持细胞更新和促进粘蛋白的分泌来增强上皮屏障,粘蛋白是一种糖蛋白,可润滑和保护上皮表面免受潜在病原体和研磨性食物颗粒的侵害。粘蛋白层不仅形成保护毯,还为共生细菌提供栖息地,有助于宿主和微生物群之间的双向关系。
抗性淀粉正向调节紧密连接蛋白
紧密连接蛋白,结合上皮细胞的微观结构,对于维持屏障完整性至关重要。这些蛋白质决定了屏障的渗透性,决定了哪些物质被允许通过,哪些物质仍然被排除在外。在“肠漏”的情况下,这些蛋白质会受到损害,导致肠道通透性增加。这种情况会允许不需要的物质(包括病原体和毒素)进入血液,引发全身炎症。研究表明,抗性淀粉正向调节这些蛋白质。抗性淀粉发酵产生短链脂肪酸,特别是丁酸盐,在上调紧密连接蛋白的表达、强化肠道屏障方面发挥作用。
抗性淀粉间接影响局部免疫反应
驻留在肠道粘膜内的免疫细胞为屏障的防御机制增加了另一层。在这里,抗性淀粉展示了其免疫调节能力。通过改变肠道微生物群组成,抗性淀粉间接影响局部免疫反应。它促进有益细菌的生长,进而与免疫细胞相互作用,指导它们的功能。这种串扰确保了潜在病原体的迅速消除,同时保持了对膳食抗原和共生微生物的耐受性。
抗性淀粉通过神经,免疫,血管等相互作用,间接提供保护
除了这些直接影响外,抗性淀粉诱导的肠道微生物群变化也会影响肠脑轴。肠道和中枢神经系统之间的这种双向沟通渠道对整体健康至关重要。肠道屏障功能的破坏与神经系统疾病有关,强调了抗性淀粉等膳食成分在神经保护中的重要性。
此外,肠道内的血管结构,包括血液和淋巴管,在屏障功能中发挥作用。它们确保营养吸收和免疫细胞运输。抗性淀粉通过其代谢物调节血管内皮屏障,优化营养吸收并确保有效的免疫监测。
总之,抗性淀粉与肠道屏障的细胞、免疫和血管成分错综复杂的相互作用,为应对环境挑战提供了强大的防御能力。拥抱抗性淀粉的治疗潜力可以重新定义面向胃肠道健康及其他方面的策略。
炎症是免疫系统对病原体、伤害或有害刺激发起的保护性反应,当放松管制时,可能会成为一把双刃剑。特别是在肠道内,持续的炎症会加剧从炎症性肠病发展到结直肠癌等多种疾病。人们的注意力已经转向可以调节炎症的饮食成分,其中抗性淀粉已成为一个至关重要的参与者。
抗性淀粉促进抗炎短链脂肪酸 (丁酸盐) 产生
与其他淀粉不同,抗性淀粉在小肠中抵抗消化,基本完好无损地到达结肠。一旦进入结肠,抗性淀粉就会充当某些有益肠道细菌的底物,导致短链脂肪酸的产生,主要是乙酸盐、丙酸盐和丁酸盐。值得注意的是,丁酸盐因其显着的抗炎作用而得到认可。丁酸盐通过抑制促炎细胞因子的产生来发挥作用,例如TNF-α和IL-6,它们在炎症传播中起着核心作用。
抗性淀粉影响免疫细胞分化
抗性淀粉发酵和随后的短链脂肪酸产生已被证明会影响免疫细胞分化,尤其是调节性 T 细胞(Tregs)。这些细胞在维持肠道免疫稳态方面起着不可或缺的作用。Tregs数量的增加与炎症减少有关,这证明了它们抑制异常免疫反应的能力。短链脂肪酸,特别是丙酸盐,影响幼稚T细胞分化为Tregs,确保肠道内平衡的免疫反应。
抗性淀粉影响肠道神经系统
肠道神经系统功能微妙,确保肠道蠕动和分泌,同时与免疫系统密切相互作用。肠道神经系统的破坏会导致肠道运动障碍,从而为细菌过度生长和炎症创造有利的环境。抗性淀粉通过其代谢物,尤其是丁酸盐,影响肠道神经系统功能。它有助于维持肠道神经元的健康和功能,随后促进肠道的定期肌肉收缩,最大限度地减少细菌停滞和炎症的机会。
抗性淀粉维持平衡的肠道pH值
此外,抗性淀粉可以通过调节肠道的 pH 值来影响肠道炎症。抗性淀粉发酵产生的短链脂肪酸导致结肠中的微酸性环境。这种酸度阻止了病原菌的生长,同时促进了有益共生细菌的增殖。这两个细菌群之间的平衡对于维持肠道健康至关重要,任何向致病性优势的转变,称为生态失调,都可能引发炎症。通过维持酸性 pH 值,抗性淀粉间接阻止炎症的发生和发展。
总之,抗性淀粉在调节肠道炎症中的复杂作用揭示了其潜在的治疗应用。它能够改变微生物组成,促进抗炎短链脂肪酸的产生,影响免疫细胞分化,并维持平衡的肠道pH值,这表明其在确保肠道稳态方面的多方面方法。随着肠道相关炎症的患病率不断上升,利用抗性淀粉的益处可以为提供预防和治疗潜力的新型饮食干预铺平道路。
肠道和免疫系统之间的界面是人体内最具活力的相互作用之一。我们整个免疫系统的近70%都存在于肠道内,随时准备对来自食物和病原体的各种抗原做出反应。正是在这种背景下,抗性淀粉等膳食成分占据了中心位置。抗性淀粉不仅仅是消化过程中的旁观者,它还以多种方式塑造和影响肠道的免疫反应。
抗性淀粉发酵产物,减少促炎细胞因子
抗性淀粉天然抵抗上消化道的消化,到达结肠基本保持不变。在结肠中,抗性淀粉由特定菌群发酵,导致短链脂肪酸的产生增加,主要是乙酸盐、丙酸盐和丁酸盐。除了作为结肠细胞的能量底物外,这些短链脂肪酸还调节各种免疫细胞功能。例如,短链脂肪酸可以降低炎性细胞因子的表达并增加抗炎介质,从而有效地抑制过度的免疫反应。特别是丁酸盐对中性粒细胞功能具有深远的影响,并减少炎症介质如TNF-α和IL-6的产生。
肠道相关淋巴组织 (GALT) 是免疫系统不可或缺的一部分,在维持肠道稳态方面起着至关重要的作用。在 GALT 内,树突状细胞不断对肠道的管腔内容物进行采样。这些细胞在遇到细菌代谢物(如抗性淀粉发酵产生的短链脂肪酸)时,其活性受到调节,导致调节性T细胞的产生增加,这些T细胞在控制炎症和自身免疫方面起着关键作用。此外,已经注意到短链脂肪酸对巨噬细胞的直接影响,观察到抗炎细胞因子产生增加和促炎细胞因子产生减少。
抗性淀粉增强屏障功能,减少LPS易位
抗性淀粉发酵产物会影响肠道屏障的完整性。维持肠上皮层连续性的紧密连接蛋白被短链脂肪酸上调,从而增强屏障功能,并减少细菌内毒素如脂多糖(LPS)进入体循环的易位。LPS易位减少导致内毒素血症相关免疫激活减少,有益于整体健康。
抗性淀粉促进有益菌生长,IgA升高
抗性淀粉作为一种益生元,选择性地滋养有益菌,进而积极调节免疫反应。例如,双歧杆菌和乳酸杆菌等有益细菌的富集通常与抗性淀粉消耗有关,与免疫球蛋白 A(IgA)的产生增强有关,IgA是粘膜防御中的一抗。升高的IgA水平在中和病原体和维持粘膜稳态方面起着关键作用。
从本质上讲,抗性淀粉与免疫系统之间的相互作用强调了饮食、微生物群和免疫力之间复杂的相互作用。通过其发酵产物和肠道微生物群的调节,抗性淀粉有可能成为调节免疫反应和维持肠道健康的重要膳食成分。它带来了全身益处,并为免疫调节的饮食策略开辟了途径。
抗性淀粉越来越被认为是一种重要的膳食成分,不仅因为它对肠道健康的直接影响,还因为它更广泛的代谢影响,特别是与代谢综合征有关。代谢综合征是一组疾病,包括血压升高、高血糖、腰部脂肪过多以及胆固醇或甘油三酯水平异常,会增加患心脏病、中风和糖尿病的风险。抗性淀粉减轻代谢综合征方面的潜力主要归因于其肠道微生物群发酵产物,如乙酸盐、丙酸盐和丁酸盐。
尤其是丁酸盐,在维持肠道屏障完整性方面发挥着重要作用,并具有抗炎特性,这对于对抗与代谢综合征相关的炎症过程至关重要。丁酸盐还通过增强结肠中的能量消耗和脂肪氧化,与改善胰岛素敏感性有关,胰岛素敏感性是代谢综合征的关键因素。此外,丙酸盐具有糖异生作用,有可能调节血糖水平,这对患有或有2型糖尿病风险的人至关重要。
此外,抗性淀粉在食欲调节中的作用也值得一提。随着短链脂肪酸的产生,它们会刺激厌食激素的释放,如肽YY(PYY)和胰高血糖素样肽-1(GLP-1),从而增加饱腹感并减少卡路里摄入量。这种食欲调节作用,加上对血脂和血压的潜在益处,使抗性淀粉消费成为预防或管理代谢综合征的有前途的策略。
体重管理和肥胖的全球挑战与饮食成分及其代谢结果有着内在的联系。肥胖的增加伴随着 2 型糖尿病、心血管疾病和几种癌症风险的增加,使其成为最重要的健康问题。从这个角度来看,抗性淀粉不仅作为一种膳食纤维脱颖而出,而且作为对抗肥胖症的潜在变革性膳食成分脱颖而出。
一些研究表明,抗性淀粉可能对体重管理有直接影响。一个主要机制是食物的热效应,这是消化和加工食物时消耗的能量的量度。抗性淀粉对立即消化具有抵抗力,往往会增加这种热效应,导致在大肠发酵过程中消耗更高的能量。这不仅有助于负能量平衡,还会影响脂肪储存并增强脂肪氧化,这对体重管理至关重要。此外,如前几节所述,抗性淀粉发酵导致短链脂肪酸的产生,短链脂肪酸通过释放 PYY 和 GLP-1 等激素在控制食欲方面发挥积极作用。调节食欲等同于减少热量摄入,这是体重管理的一个重要方面。
此外,抗性淀粉与改善肠道健康有关,这对肥胖有间接影响。健康的肠道生物群与更瘦的表型有关。当肠道微生物群发酵抗性淀粉时,它会导致微生物组成的变化,有利于与肥胖呈负相关的有益菌。
近日,一项针对 37 名超重或肥胖参与者的随机安慰剂对照交叉设计试验(ChiCTR-TTRCC-13003333) 中,研究人员测试了抗性淀粉作为膳食补充剂是否会影响肥胖相关的结果。
研究表明补充抗性淀粉 8 周有助于实现体重减轻(平均 -2.8 公斤)并改善体重超重个体的胰岛素抵抗。
补充青春双歧杆菌(一种与减轻研究参与者肥胖显著相关的物种)可以保护雄性小鼠免受饮食引起的肥胖。从机制上讲,抗性淀粉诱导的肠道微生物群变化会改变胆汁酸分布,通过恢复肠道屏障来减少炎症,并通过调节ANGPTL4抑制脂质吸收,提高脂肪组织对FGF21的敏感性。
次级胆汁酸,例如甘氨脱氧胆酸、脱氧胆酸、甘氨胆酸和牛磺脱氧胆酸,对于提高胰岛素敏感性和改善肝脂肪变性具有重要作用。胆盐水解酶负责次级胆汁酸的去偶联。补充抗性淀粉降低了胆盐水解酶的产生,增加了次级胆汁酸的水平。
抗性淀粉至少可以部分通过青春双歧杆菌促进体重减轻,并且肠道微生物群对于抗性淀粉的作用至关重要。
因此,持续食用富含抗性淀粉的食物可能会导致肠道环境不太容易使体重增加和肥胖。
糖尿病是一种以慢性高血糖为特征的代谢紊乱,是一个不断升级的全球健康问题,具有从个人健康恶化到国家经济负担的多方面影响。鉴于发病率不断上升,迫切需要饮食干预来缓解或可能逆转这种疾病的进展。抗性淀粉是一种引人注目的膳食成分,其多种代谢影响与糖尿病管理和血糖控制有关。
糖尿病管理的核心是调节餐后血糖和胰岛素反应。摄入抗性淀粉似乎有利地调节这些反应。与快速消化的淀粉不同,抗性淀粉不会直接导致餐后血糖峰值,因为它绕过了小肠的消化,它在大肠中的发酵会产生短链脂肪酸。特别是,丙酸盐可促进肝脏葡萄糖产生调节,降低餐后葡萄糖偏移的风险。此外,丁酸盐在促进胰高血糖素样肽-1(GLP-1)分泌中的作用。
注:GLP-1是一种增强胰岛素分泌和减少胰高血糖素释放的激素,协调血糖水平。
此外,长期服用抗性淀粉与提高胰岛素敏感性有关,这是 2 型糖尿病发病机制的关键因素。研究表明,食用富含抗性淀粉的饮食的胰岛素抵抗个体表现出胰岛素敏感性的显著改善。这种改善被认为与短链脂肪酸的抗炎特性有关,尤其是丁酸盐,以及它在减少氧化应激中的作用,氧化应激有助于胰岛素抵抗。此外,抗性淀粉培养有益肠道微生物群组成的能力对代谢健康有间接影响,进一步强调了其在糖尿病管理中的潜在作用。
莲子抗性淀粉抑制高脂血症大鼠中下列菌群的生长:
莲子抗性淀粉促进初级胆汁酸(CA、CDCA 、β-MCA)和次级胆汁酸(LCA、UDCA)的产生,降低高脂血症中TCA、DeHydro-LCA、isoLCA、LCA-3-S、THDCA的含量。
Blautia、norank_f__Muribaculaceae、norank_f__Eubacteria_coprostanoligenes_group与 DeHydro-LCA、isoLCA、TCA、LCA-3-S、 TCHO、TG和 LDL-C呈正相关。
莲子抗性淀粉通过调节肠道菌群并加速肝脏中胆固醇分解为胆汁酸来改善血脂水平。
doi.org/10.1016/j.foodchem.2022.134599
认知能力下降是衰老的常见后果。缺乏纤维和高饱和脂肪的饮食模式会引发促炎症途径和代谢功能障碍,从而加剧认知障碍。新的证据强调了富含纤维的饮食对神经认知的益处以及肠道-微生物-大脑信号传导的关键作用。
经过为期20周的饮食方案,包括含有5%重量比来自芸豆(PTB)、黑眼豆(BEP)、扁豆(LEN)、鹰嘴豆(CKP)或菊糖纤维(INU)的抗性淀粉的西式饮食(实验组),与不含抗性淀粉的西式饮食(对照组),发现抗性淀粉特别是来自扁豆的抗性淀粉,可以改善西式饮食引起的认知障碍。
从机理上看,抗性淀粉通过改善肠道菌群-代谢组,包括增加短链脂肪酸和降低支链氨基酸水平,从而改善神经认知功能评估。这种肠道菌群-代谢物-大脑信号级联抑制了神经炎症、细胞衰老和血清瘦素/胰岛素水平,同时通过改善肝功能增强脂质代谢。总的来说,数据证明了抗性淀粉的益生菌效应可通过调节肠-脑轴改善神经认知功能。
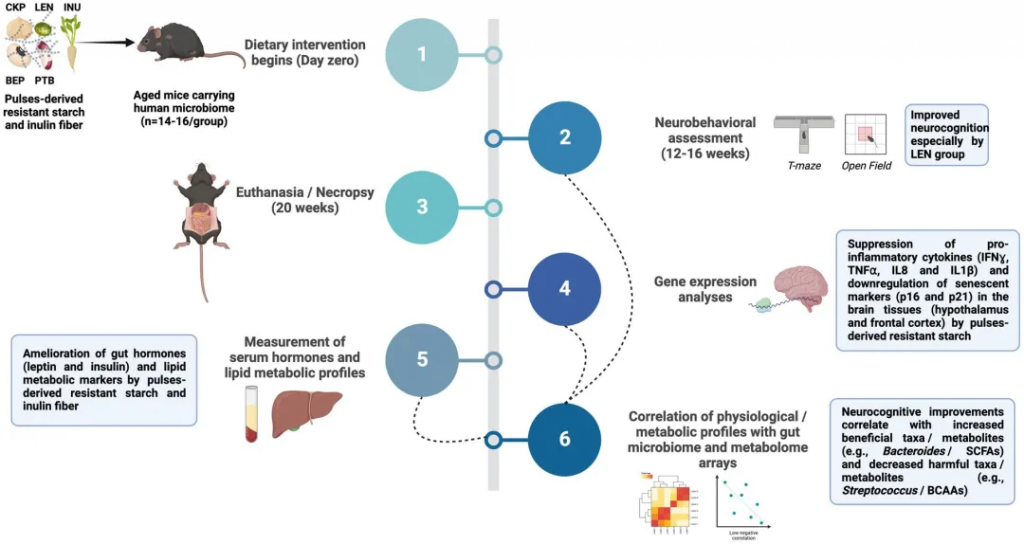
doi.org/10.3389/fnut.2024.1322201
溃疡性结肠炎是一种复杂的炎症性疾病,发病率不断上升。一项研究通过酶法分离方法从紫甘薯中获得抗性淀粉(PSPRS)。然后,研究了PSPRS的结构特性及其对葡聚糖硫酸钠诱导的结肠炎的保护功能。
结构表征结果表明,PSPRS的结晶度从CA型转变为A型,并且在酶水解过程中层状结构被完全破坏。与结肠炎小鼠相比,PSPRS给药以剂量依赖性方式显著改善病理表型和结肠炎症。
ELISA 结果表明,给予PSPRS的结肠炎小鼠表现出较高的 IL-10 和 IgA 水平,但较低的 TNF-α、IL-1β 和 IL-6 水平。同时,高剂量(300 mg/kg)的PSPRS显著增加了乙酸盐、丙酸盐和丁酸盐的产生。
16S rDNA高通量测序结果显示,PSPRS治疗组中厚壁菌门与拟杆菌门的比例以及潜在益生菌水平显著增加,如乳杆菌、Alloprevotella, 毛螺菌科_NK4A136_组、双歧杆菌。同时,高剂量 PSPRS 显著抑制了拟杆菌属、葡萄球菌属和阿克曼氏菌等有害细菌 (p < 0.05)。因此,PSPRS有潜力成为促进肠道健康、缓解溃疡性结肠炎的功能食品。
慢性肾病是与心血管疾病、发病率和死亡率风险增加相关的主要健康问题。最近的研究表明,慢性肾病的进展可能与肠道菌群的变化有关。抗性淀粉是一种膳食纤维,可以作为微生物发酵的底物。一些研究发现补充抗性淀粉可以改善慢性肾病患者的肠道菌群紊乱。
在 2022 年随机对照试验的荟萃分析中,发现补充抗性淀粉可以降低患有透析患者血液中硫酸吲哚酚、磷、IL-6和尿酸的水平。
抗性淀粉还对肠道环境产生有益影响,包括增加Ruminococcus bromide。
Ruminococcus brucei是厚壁菌门的主要成员之一,是一种主要的抗性淀粉发酵菌株。通过其针对抗性淀粉的特殊活性,Ruminococcus brucei从淀粉中释放能量以逃避宿主酶的消化。此外,摄入富含抗性淀粉的食物可以增加肠道短链脂肪酸水平,调节微生物代谢物,并改善葡萄糖稳态和胰岛素敏感性。
通过摄入抗性淀粉,胆固醇和甘油三酯降低,胰岛素敏感性提高,可以大大降低代谢综合征的发生率。慢性肾病患者也可能受益于更好的葡萄糖代谢、血脂水平和更好的体重管理。
近年来,富含抗性淀粉(RS)的食物因其多方面的健康益处而受到越来越多的关注,从肠道健康到调节全身新陈代谢。
益处
作为一种必需的膳食成分,抗性淀粉与典型的淀粉不同,主要是由于其对小肠消化的抵抗力,它基本上完好无损地到达大肠,在那里它作为微生物发酵的底物,产生有益的代谢物,如短链脂肪酸。
抗性淀粉中天然丰富的食物包括青香蕉、豆类、全谷物和某些类型的煮熟然后冷却的食物,如土豆和米饭。食用此类食物的好处之一是它们有可能增强血糖控制。富含抗性淀粉的食物具有较低的升糖指数,转化为较慢的餐后血糖升高。这一特性对患有糖尿病等代谢紊乱的人特别有益。此外,抗性淀粉发酵产生的短链脂肪酸,特别是丁酸盐,丁酸盐是结肠细胞的主要能量来源,并具有抗炎特性,使其对结肠健康不可或缺。
注意事项
抗性淀粉摄入量的快速和大量增加会导致胃肠道不适,包括腹胀、胀气和排便习惯改变。通常建议逐渐将富含抗性淀粉的食物引入饮食中,以使肠道微生物群有时间适应。此外,患有某些健康状况的人,例如患有肠易激综合征(IBS)或特定碳水化合物不耐受的人,应谨慎对待富含抗性淀粉的食物,并在专业指导下。抗性淀粉的发酵有时会加剧这些人的症状。
总之,虽然富含抗性淀粉的食物具有许多健康益处,尤其是在肠道健康和代谢调节方面,但个人应注意摄入饮食中的抗性淀粉的来源和数量。
最好取得平衡:优化健康益处,同时最大限度地减少潜在的不利影响。
抗性淀粉的作用超越了其个人益处,使其成为各种饮食模式和制度的关键成分。
地中海饮食
例如,当考虑因其保护心脏的益处而广受赞誉的地中海饮食时,我们发现豆类,抗性淀粉的天然来源,是其成分的基石。经常食用豆类,其丰富的抗性淀粉含量不仅有助于增强肠道健康,还有助于与这种饮食模式相关的心脏保护作用,因为抗性淀粉具有调节餐后血糖反应的潜力。
旧石器饮食
旧石器时代的饮食,俗称旧石器饮食,是另一个有趣的领域,其中抗性淀粉找到了相关性。对旧石器饮食的当代解释集中在块茎和某些根的消费上,当通过特定方法制备时,例如烹饪后冷却,这些根会富含抗性淀粉。这证实了我们的祖先可能已经消耗了大量的抗性淀粉,为他们的肠道微生物群提供了可发酵的底物。从理论上讲,肠道微生物与其宿主之间的共生关系已经共同进化了数千年,而抗性淀粉可能是推动这一进化过程的关键饮食元素。
低碳和生酮饮食
低碳水化合物和生酮饮食,在减肥和代谢健康方面很受欢迎,通常会限制淀粉的摄入。然而,将抗性淀粉整合到这些饮食中可以提供明显的优势。由于抗性淀粉不表现出与普通淀粉相同的消化率,因此它的加入不会显著提高血糖水平。这意味着这种饮食的人可以获得抗性淀粉的好处,例如增强肠道健康和饱腹感,而不会影响酮症或低碳水化合物方案的状态。从根本上说,抗性淀粉允许当代饮食方法之间的共生关系,重点是减肥或代谢益处,以及滋养肠道微生物群的古老进化重要性。
将抗性淀粉纳入饮食中不仅要承认其生理益处,还要了解其最佳摄入量并融入各种饮食中,以最大限度地发挥其潜力。随着越来越多的证据描绘抗性淀粉的多方面优势,从调节肠道微生物群到调节血糖水平,向更广泛的公众提供可操作的指南变得至关重要。
循序渐进
首先,身体可能需要一些时间来适应饮食中添加抗性淀粉。因此,需要循序渐进逐步添加。
添加少量抗性淀粉。例如,早餐中加入一根绿色香蕉,午餐中加入四分之一杯扁豆。
食物中获取
最好从食物中获取抗性淀粉,常见的食物补充包括:青香蕉、豆类(豌豆、扁豆)、全谷物(燕麦和大麦)、煮熟并冷却的米饭。
注:增加纤维摄入量时,要慢慢喝大量的水,以减少胃肠道副作用。

补充剂
补充性抗性淀粉通常以粉末形式服用,可混合到食品或饮料。如绿色香蕉粉、木薯粉等。
烹饪,可进一步提高抗性淀粉的含量
必须认识到并非所有抗性淀粉来源都是一样的。虽然豆类、全谷物和某些块茎天然富含抗性淀粉,但烹饪方法可以进一步调节其抗性淀粉含量。例如,烹饪然后冷却土豆或米饭等淀粉类食物可以增加其抗性淀粉含量,提供了一种直接的策略来提高饮食中的抗性淀粉水平,而无需任何剧烈变化。
抗性淀粉的推荐每日摄入量通常在 15-30 克之间。这可以通过多样化的饮食来实现,包括冷意大利面沙拉、隔夜燕麦或豆类菜肴等食物。
考虑个人的消化耐受性的重要性
抗性淀粉摄入量的突然激增会导致一些人的胃肠道不适。因此,谨慎的做法是在几周内逐渐增加抗性淀粉的摄入量,让肠道进行调整。此外,将富含抗性淀粉的食物与益生菌食物(如酸奶或开菲尔)相结合,可以产生协同效应,为有益的肠道细菌茁壮成长营造一个好的环境。与任何饮食改变一样,应根据个人口味、健康状况和饮食习惯定制抗性淀粉,确保方法既平衡又可持续。
抗性淀粉作为益生元可以调节肠道微生物群,对于肠道菌群失调相关的许多疾病包括炎症性肠病、肠易激综合征、结直肠癌等具有重要意义,为这些疾病辅助治疗的潜在用途奠定了基础。
抗性淀粉的另一个治疗意义在于其抗炎特性。慢性炎症在心血管疾病、2型糖尿病和某些癌症等多种疾病的发病和进展中发挥着关键作用。肠道微生物群发酵抗性淀粉会产生短链脂肪酸,尤其是具有抗炎作用的丁酸,可以抑制促炎细胞因子,使抗性淀粉成为减轻炎症引起的疾病的潜在候选者。
抗性淀粉也有望成为对抗肥胖和相关并发症的饮食策略。鉴于其调节饱腹感、提高胰岛素敏感性和调节脂质代谢的能力,抗性淀粉可以作为代谢综合征和相关病症的辅助或预防措施。例如,将抗性淀粉纳入饮食中可以改善糖耐量受损个体的餐后葡萄糖反应,强调其潜在的治疗相关性。
抗性淀粉在个性化营养方面的潜力也不容忽视,但仍需谨慎对待。剂量、持续时间和个体差异在决定抗性淀粉在任何治疗应用中的功效方面发挥着关键作用。
个体对抗性淀粉的反应可能存在很大差异。年龄、遗传、肠道微生物群组成和整体健康状况等因素会影响人们处理抗性淀粉摄入及益处。例如,摄入相同量抗性淀粉的两个人可能会表现出不同的餐后葡萄糖反应或结肠中不同的短链脂肪酸产生情况。此外,肠道微生物群将抗性淀粉发酵成有益代谢物的能力在个体之间可能有所不同,特别是在微生物群多样性高的人和微生物群较少的人之间差异较大。
这种个体差异突出了个性化营养方法的必要性。与其采取一刀切的抗性淀粉摄入建议,不如根据个人独特的代谢和菌群特征来定制饮食建议。随着深入探索个性化医疗时代,整合肠道菌群数据或许能为优化个人抗性淀粉摄入、获得健康效益提供更精准的建议。
主要参考文献:
Chen Z, Liang N, Zhang H, Li H, Guo J, Zhang Y, Chen Y, Wang Y, Shi N. Resistant starch and the gut microbiome: Exploring beneficial interactions and dietary impacts. Food Chem X. 2024 Jan 3;21:101118.
Li H, Zhang L, Li J, et al., Resistant starch intake facilitates weight loss in humans by reshaping the gut microbiota. Nat Metab. 2024 Mar;6(3):578-597.
Junejo SA, Flanagan BM, Zhang B, Dhital S. Starch structure and nutritional functionality – Past revelations and future prospects. Carbohydr Polym. 2022 Feb 1;277:118837.
Arp CG, Correa MJ, Ferrero C. Modified celluloses improve the proofing performance and quality of bread made with a high content of resistant starch. J Sci Food Agric. 2023 Apr;103(6):3041-3049.
Du X, Wu J, Gao C, Tan Q, Xu Y. Effects of Resistant Starch on Patients with Chronic Kidney Disease: A Systematic Review and Meta-Analysis. J Diabetes Res. 2022 Jul 18;2022:1861009.
Tekin T, Dincer E. Effect of resistant starch types as a prebiotic. Appl Microbiol Biotechnol. 2023 Feb;107(2-3):491-515.
Wang, Z.; Gao, M.; Kan, J.; Cheng, Q.; Chen, X.; Tang, C.; Chen, D.; Zong, S.; Jin, C. Resistant Starch from Purple Sweet Potatoes Alleviates Dextran Sulfate Sodium-Induced Colitis through Modulating the Homeostasis of the Gut Microbiota. Foods 2024, 13, 1028
Chen R, Zhang C, Xu F, Yu L, Tian F, Chen W, Zhai Q. Meta-analysis reveals gut microbiome and functional pathway alterations in response to resistant starch. Food Funct. 2023 Jun 6;14(11):5251-5263.
Kadyan S, Park G, Hochuli N, Miller K, Wang B, Nagpal R. Resistant starches from dietary pulses improve neurocognitive health via gut-microbiome-brain axis in aged mice. Front Nutr. 2024 Jan 24;11:1322201.
Pickens TL, Cockburn DW. Clostridium butyricum Prazmowski can degrade and utilize resistant starch via a set of synergistically acting enzymes. mSphere. 2024 Jan 30;9(1):e0056623.

谷禾健康

在研究肠道菌群或复杂微生物样本构成时,“门”(Phylum)是细菌分类的高级分类单位之一。
细菌分类依次为门纲目科属种亚种,最大的分类层面是门,以前写过人群肠道菌群构成主要是以拟杆菌门和厚壁菌门为主,大部分人群占比超90%(个别的变形菌门或放线菌门占比也很高),作为两大“细菌王国”,这两大王国有着不同的生态和代谢属性,又有着相似的营养和生存需求,它们如同天平的两端的砝码,不同的人有着不同倾斜,即使同一个人不同时期不同身体状态下天平的倾斜都不一样。
拟杆菌门被比喻为肠道微生物群落的“多面手”,有超过7000种不同的革兰氏阴性菌组成,主要来自拟杆菌属、副拟杆菌属、普氏菌属和卟啉单胞菌属。它们的脂多糖和鞭毛蛋白与细胞受体相互作用,并通过细胞因子合成增强免疫反应。
拟杆菌门作为多糖降解联盟的成员,尤其是拟杆菌属和普雷沃氏菌属,具有多种可以利用多糖的聚糖和糖苷酶。它们有助于分解膳食纤维和淀粉释放能量,并且它们可能是丙酸盐的主要来源。
拟杆菌门一些细菌的功能还包括胆汁酸的解离。肠道微生物群,特别是肠道拟杆菌,一定程度上也具有使初级胆汁酸解离和脱水并将其转化为人结肠中次级胆汁酸的能力。它们也参与蛋白质分解过程中有毒产物的释放。比如氨,组胺,硫化氢等,这些高浓度的产物会对肠黏膜产生毒性。
该群体的成员具有一些可能有助于抑制炎症的活动,但它们也有可能促进炎症,其中一些已知是机会病原体。
厚壁菌门细菌大多为革兰氏阳性菌,通过短链脂肪酸合成在宿主的营养和代谢中发挥关键作用。
通过其代谢产物,厚壁菌门细菌与其他组织和器官间接连接并调节饥饿和饱腹感。厚壁菌门由大量功能更加多样化的核心细菌组成。从代谢的角度来看,共生梭菌簇 XIVa 和 IV 通过产生短链脂肪酸在宿主和肠道稳态中发挥着重要作用,使肠道通透性正常化,参与脑肠轴调节。许多厚壁菌门的能力与宿主的体重有关。
F/B比率升高或降低被视为生态失调,前者通常与肥胖,代谢障碍相关,这可能与从食物中提取热量增加、脂肪沉积和脂肪生成、胰岛素敏感性受损有关;而后者则与炎症性肠病,抑郁,阿尔茨海默病等相关,这可能与短链脂肪酸尤其丁酸的生成减少,蛋白代谢物组胺,脂多糖累积等相关诱导的免疫炎症反应相关。
目前许多研究工作都集中在识别特定的微生物特征,特别是与肥胖、2型糖尿病和炎症相关的微生物特征。
我们从谷禾肠道菌群检测数据库中随机抽取5万例样本,统计的人群厚壁菌门/拟杆菌门(F/B)比率如下:
可以看出大部分人群的厚壁菌/拟杆菌比率在0.30-1.0之间(谷禾健康数据库随机抽样5万例样本),表明国人大部分人群肠道菌群的构成中拟杆菌占比多一点;同时抽样人群中1/10人厚壁菌/拟杆菌比率大于2.9,表明也有相当一部分人群厚壁菌门占比较高。
一般而言,从出生到成年,厚壁菌门/拟杆菌门的比例不断增加,并随着年龄的增长而进一步改变。该比率在婴儿、成人和老年人之间存在差异。它可能与生命不同阶段细菌谱的整体变化有关。
厚壁菌门/拟杆菌门比率是一个考虑高水平分类等级(即门)的指数,同一群体的受试者之间厚壁菌门和拟杆菌门的相对丰度差异很大。这可能是由于许多与生活方式相关的因素造成的,包括饮食、疾病状态、食品添加剂和污染物、抗生素消耗、体力活动等影响胃肠道微生物群组成的因素。
由于菌群是一个复杂的生态体系,门层面的简单划分只能提供一个最简单的线索,想要了解肠道菌群需要从最主要的核心菌属,常见的有害菌属以及大量功能菌属的构成来更加准确的分析,此外菌群的代谢通路也提供了不同角度的信息。
Wexler AG,et al.Nat Microbiol.2017
大多数健康成人微生物群仅由两个细菌门控制——革兰氏阳性厚壁菌门(许多属)和革兰氏阴性拟杆菌门(主要是拟杆菌属、另枝菌属、副拟杆菌属和普氏菌属)——它们共同构成了微生物中的大多数细菌分类群。
此外还有其他分类群包括变形菌门、放线菌门、梭杆菌门、疣微菌门、产甲烷古菌、真核生物(原生生物和真菌)和其他更短暂的定植者。
健康个体中发现的类群在家庭成员(例如父母及其子女)之间往往最相似,而在不同文化和地理空间(例如西方社会与非西方社会)之间则最不相似。
厚壁菌门(Phylum Firmicutes)是细菌的一个门,大多数具有革兰氏阳性细胞壁结构。然而,一些细菌,如巨球菌属、梳状菌属、月单胞菌属和嗜发酵菌属,具有多孔的假外膜,导致它们被染成革兰氏阴性。
科学家们曾经将厚壁菌门分类为包括所有革兰氏阳性细菌,但最近将它们定义为相关形式的核心组,称为低 G+C 组。与放线菌相反。它们有圆形细胞,称为球菌(单球菌),或杆状细胞(芽孢杆菌)。厚壁菌门缺乏脂多糖,而是向宿主呈现厚厚的肽聚糖层;研究人员已经发现了200多种不同类型的属于厚壁菌门的细菌。
★ 厚壁菌门中有许多有益的菌
厚壁菌门的成员包括许多有益的菌。比如乳酸菌是一种常见于酸奶和其他发酵乳制品中的益生菌,属于该门。这些微生物会产生醋酸盐(另一种促进健康的短链脂肪酸)以及乳酸和抗菌物质,以防止病原体定植。
又比如普氏栖粪杆菌(Faecalibacterium prausnitzii),又名:普拉梭菌,是人类肠道菌群中最重要的细菌之一,占健康人粪便样本中检测到的细菌总数的5-15%,是丁酸的重要生产者之一,具有抗炎作用,维持细菌酶的活性,保护消化系统免受肠道病原体的侵害。
★ 厚壁菌门中也存在一些致病菌
但是一些致病菌也属于该门。例如,产气荚膜梭菌是一种引起胃肠道感染的细菌。还有金黄色葡萄球菌,它是一些严重感染的常见原因。
丁酸盐是研究最深入的短链脂肪酸之一,是肠道上皮细胞营养的主要来源。丁酸盐的消耗与肠道屏障完整性受损有关。
注:研究支持,长寿、健康的老年人肠道内的丁酸细菌含量明显较高。
人类肠道包含一个非常密集的代谢活跃微生物群落。然而,只有有限数量的肠道细菌为丁酸生产者。人类肠道中大多数丁酸合成细菌分布于厚壁菌门。在厚壁菌门中,毛螺菌科、瘤胃球菌科、真杆菌科和梭状芽孢杆菌科是四个重要的丁酸合成科。
梭菌目的簇 IV、XIVa、XVI 和 I 包含大多数丁酸盐生产菌种。其中两个重要的物种,直肠真杆菌和普拉梭菌分别属于梭菌簇 XIVa 和梭菌簇 IV,占健康成人粪便样本中肠道菌群总数的12-14% 。此外,还有罗氏菌属,粪球菌属,瘤胃球菌属, Anaerostipes ,丁酸弧菌属和梭菌属。
分布在XIVa 簇中的Butyricicoccus pullicaecorum、Subdolicapsulu variabile、Anaaerotruncus colihominis和分布在 IV 簇中的Papillibacter cinnamivorans是其他产丁酸菌种。
据报道,属于韦荣氏球菌科、热厌氧杆菌科 III 的Megasphaera elsdenii和Caldocellum saccharolyticum也能合成丁酸盐。
最近,对人类样本的高通量宏基因组测序表明,Rawsonibactera saccharolyticus是瘤胃球菌科中的一个新物种,并且肠单胞菌AF211编码丁酰辅酶A:用于合成丁酸的乙酰辅酶A转移酶。
许多厚壁菌门细菌可以产生内生孢子,其耐干燥并且可以在极端条件下生存。孢子在细菌生长中起着重要的作用。细菌的孢子是一种休眠状态,可以在不利环境下存活,并在适宜条件下重新发芽成为活跃的细菌。这种休眠状态使得细菌能够在恶劣条件下存活并传播。
★ 内生孢子可以帮助细菌在恶劣条件下存活
内生孢子由细菌的DNA和部分细胞质组成,周围有一层非常坚韧的外层。它们可以在没有营养的情况下生存,并且能够抵抗紫外线、干燥、高温、极端冷冻和化学消毒剂。
厚壁菌门的代谢偏好
科学家发现,人类肠道微生物群虽然独特,但都可以按照几个分类划分下,称为肠型。第三种肠道型,被称为“谷物爱好者” ,通常以瘤胃球菌(厚壁菌门的成员)为主。
肠型往往可以追溯到长期的饮食习惯。顾名思义,具有“谷物爱好者”微生物组类型的人倾向于消耗更多全谷物中的抗性淀粉和膳食纤维。
✦ 高纤维饮食的人体内厚壁菌更丰富
高纤维饮食可以增加厚壁菌门的丰度并减少拟杆菌的丰度 ,从而增加肠道内短链脂肪酸的浓度。
而葡萄籽原花青素显著降低了肥胖大鼠肠道菌群中厚壁菌门的含量,提高了拟杆菌门的含量,显著降低了厚壁菌门与拟杆菌门比值。
✦厚壁菌偏向从膳食蛋白和循环尿素中获取氮
在厚壁菌门成员中,偏好尿素氮的属往往是菊粉的疯狂使用者,即使用菊粉和尿素合成自己的氨基酸。这包括一些脲酶阴性菌属,它们可能通过交叉喂养获得尿素氮。
此外,在厚壁菌中也看到了一些属更喜欢从膳食蛋白质中获得氮,而其他菌属更喜欢循环尿素。
静脉注射尿素以提高循环尿素浓度后,偏好尿素的厚壁菌以及阿克曼菌的丰度大幅增加。
膳食蛋白质和循环尿素是厚壁菌的主要氮原料,而分泌的宿主蛋白质为拟杆菌提供氮。
!
如果厚壁菌门过高意味着什么?
可能的原因:
– 不良的饮食习惯
– 生态失调
– 消化不良
– 胃酸过少
– 代谢疾病
可能的治疗方法和注意事项:
– 平衡共生细菌
– 当厚壁菌门含量较高时,考虑主要使用双歧杆菌益生菌和布拉氏酵母菌
– 乳杆菌属和芽孢杆菌属(益生菌中发现)可以提高厚壁菌门的数量
– 优化饮食结构:低脂肪饮食可能有助于厚壁菌门水平正常化
滋养肠道内厚壁菌门的食物
纤维和淀粉,是这些细菌的能量来源。富含水果、蔬菜、豆类和全谷物的饮食也是理想的。
研究表明,地中海饮食与更丰富的厚壁菌门有关,包括普拉梭菌。另一方面,富含动物蛋白、脂肪和糖的西方饮食会对有益的这些细菌产生负面影响。
如果想增加肠道中厚壁菌门的丰度,那么应该多吃高纤维食物。以下是一些可以添加到饮食中的食物,以支持您的肠道微生物组并促进厚壁菌门和丁酸盐的产生:
-苹果
-蒜
-大麦
-菊苣根
-燕麦
-亚麻籽
-洋葱
拟杆菌是肠道生态系统中非常成功的竞争者,表现出相当大的营养灵活性以及对宿主和肠道环境施加的压力做出反应的能力。很难权衡肠道拟杆菌对宿主是否产生负面或正面影响。
拟杆菌是复杂碳水化合物的主要降解者,迄今为止调查的所有生态系统中都普遍发现该属,在土壤以及人类和动物肠道中尤其占主导地位。大多数已测序的拟杆菌基因组编码大量多糖降解酶(CAZymes),表明代谢重点是聚糖降解。
CAZymes 在碳水化合物活性酶数据库中分为类别和家族,其中降解酶存在于糖苷水解酶(GH)、多糖裂解酶(PL)、碳水化合物酯酶(CE)和辅助活性酶 (AA) 类别中。
拟杆菌还通过与免疫系统的相互作用,在激活T细胞介导的反应中发挥作用,从而有助于宿主的整体健康。
✦ 防止特定炎症反应
基于多项研究,发现脆弱拟杆菌通过产生称为两性离子多糖的多糖来激活CD4+ T细胞。CD4+ T细胞的产生会刺激白细胞介素10的释放,从而防止特定的炎症反应和脓肿形成。
肠道免疫保护取决于拟杆菌、白细胞介素36信号和巨噬细胞的发育。拟杆菌的这种作用需要其保守的共生定植因子的多糖利用位点。
✦ 抑制胃肠道中的病原体
与胃肠道中发现的其他一些细菌相比,研究表明拟杆菌门的一些成员对各种抗生素和胆汁等物质具有耐药性,这给了他们竞争优势。
除了这种竞争优势之外,这些细菌还被证明在调节胃肠道中其他正常菌群和病原体的数量方面发挥着重要作用。
拟杆菌可以积极改善肠道环境,使其更适合自身和其他微生物。
✦降低肠道内氧水平从而促进厌氧菌生长
例如,许多编码细胞色素bd氧化酶,据推测该酶可以降低细胞内氧水平,进而降低肠道氧水平,从而允许严格厌氧菌生长,否则这些厌氧菌会被氧气的存在杀死。
这种耐受和降低氧气水平的能力可能有助于拟杆菌传播到新宿主,这可能是它们在哺乳动物中如此广泛传播的主要原因。
✦改变肠道的营养状况
它们还可以通过促进宿主的生理变化来诱导某些食物来源(例如岩藻糖基化糖蛋白)的产生,或通过从糖蛋白中释放岩藻糖和唾液酸残基来改变肠道的营养状况,这些残基可以被其他微生物消耗,包括病原体。
通过比较缺乏自身微生物的无菌动物和与单个拟杆菌属菌株和突变体单相关的无菌动物(出生时无菌但后来被微生物定殖),已经鉴定了许多这些拟杆菌属依赖性的肠道环境变化。
✦拟杆菌拥有多糖利用位点有助于代谢
尽管人们可能会想象肠道中的生命充满了微生物的过多食物选择,但在结肠(细菌密度最高的地方),简单、容易获得的糖大部分已经被消耗或吸收。其中的残留物是复杂的长链多糖,它们不能自由吸收,也不能被人类酶促消化。对于许多细菌来说,这些复杂的多糖同样难以处理,并且无法跨膜运输。
然而,早期研究证实拟杆菌是熟练的聚糖降解剂,具有识别和代谢十多种植物和宿主来源的多糖的不同寻常的能力。拟杆菌属物种通过基因簇来实现这一点,这似乎是拟杆菌特有的,称为多糖利用位点(PUL)。
多糖利用位点可以确定拟杆菌可以占据哪些代谢生态位,甚至可以确定它们在肠道内的生物地理位置。多糖利用位点被如此分类是因为它们的susC和susD的同系物,以及其他成分(糖苷水解酶、多糖裂解酶、糖基转移酶、碳水化合物酯酶),它们对于分解多种植物或宿主来源的聚糖很重要。
肠道细菌的其他谱系,包括革兰氏阳性放线菌,降解和输入膳食多糖的途径远不那么复杂,这在很大程度上是由于缺乏外膜屏障。例如,短双歧杆菌使用多结构域、细胞表面锚定酶(而不是一系列单独的酶),可以结合并降解聚合物,例如淀粉、支链淀粉和糖原。
拓展
比较基因组分析已在每个拟杆菌属中鉴定出数十种不同的多糖利用位点。例如,Bacteroides thetaiotaomicron是第一个对其基因组进行测序的拟杆菌属成员,将其 6.26 Mbp 基因组的近五分之一专用于88个不同的多糖利用位点。
拟杆菌不是同时表达所有多糖利用位点以捕获周围环境中可能存在的任何多糖,而是通过大量SusR样调节因子、混合双组分系统和涉及胞质外 σ/抗的跨包膜信号通路来严格调节其表达。
也许是由于在跨外膜运输之前需要部分降解长多糖链,B. thetaiotaomicron和其他肠道拟杆菌已经成功地将适当的多糖利用位点转录与其相应多糖分解中间体的丰度(而不是丰度)耦合起来。
这使得肠道细菌能够快速适应营养可用性的变化。糖分解物质形成甲酸盐、乙酸盐、乳酸盐、丙酸盐和琥珀酸盐作为主要发酵产物。拟杆菌可能是结肠中丙酸盐形成的主要贡献者。
由于多糖利用位点激活是肠道环境的产物,富含植物源性多糖和糖蛋白的饮食将导致多糖利用位点暂时上调,从而降解这些膳食营养素。然而,当这些底物稀缺时,例如在禁食期间或某些低纤维饮食期间(如高糖、高脂肪的西方饮食),亲糖拟杆菌会改变其转录谱以消耗宿主衍生的聚糖,这些暂时的转换可能是导致某些细菌在婴儿期最初在肠道定殖并在我们一生中持续存在的主要因素。
根据最近使用动物模型的研究,拟杆菌属可以在相对较短的宿主婴儿期主要依靠宿主聚糖生存,但当在多个宿主世代的时间内被迫以低纤维饮食为生时,它们最终可能会灭绝。
有的菌会通过破坏中性粒细胞趋化性等机制帮助逃避免疫系统(例如,伤寒沙门氏菌)和逃避宿主补体系统(例如肺炎克雷伯菌)。
相比之下,拟杆菌使用一种称为相变的过程,以开/关方式控制其荚膜多糖生物合成位点的表达。例如,脆弱拟杆菌从八个生物合成位点产生不同的荚膜多糖,其表达受到可逆启动子区域和反式位点抑制剂的严格调控,限制多个位点的同时表达,从而使脆弱拟杆菌的同基因群体共同表现出广泛的多样性。当脆弱拟杆菌漏入通常无菌的腹膜腔时,可能会发生腹膜炎。
注:绝大多数人类腹膜炎病例都是由脆弱拟杆菌引起的。
目前,拟杆菌门中研究的最充分的,包括拟杆菌属和普氏菌属以及卟啉单胞菌属。
✦ 卟啉单胞菌属可导致急性坏死溃疡性牙龈炎
卟啉单胞菌属的成员作为共生体或机会性病原体生活在人类口腔中。牙龈卟啉单胞菌引起临床上侵袭性牙龈炎,可导致急性坏死溃疡性牙龈炎或扩展到口腔和面部组织,这种情况称为坏疽性口炎或口腔癌。
✦ 普雷沃氏菌也是一种条件性致病菌
普氏菌属是一个大属,包括50多个不同的物种。大多数可以从口腔和肠道中分离出来。普雷沃氏菌属存在于人类中,帮助分解蛋白质和碳水化合物食物。也可作为条件致病菌,引起牙周和牙齿、肠道炎症、类风湿性关节炎、细菌性阴道炎等问题。
大型基因组计划,包括美国国立卫生研究院(NIH)资助的人类微生物组项目(http://nihroadmap.nih.gov/hmp/)和欧盟资助的 MetaHIT 项目 (http://www.metahit.eu)对50多种人类来源的拟杆菌门的拟杆菌和普氏菌分离株进行了测序。
基因组草图可从 Genbank 公开获得,并揭示这些物种拥有多种糖苷酶,这些糖苷酶具有预测的对抗植物和宿主来源的多糖的活性,以及维生素和辅因子合成的许多途径。
拟杆菌门的基因组大小相对较大,例如B.thetaiotaomicron 为 6.26 Mb。迄今为止在拟杆菌属中发现的16S rRNA基因的最高拷贝数为7个(在普通拟杆菌中)。
✦ 拟杆菌的组成和代谢受到饮食调节
在全基因组水平上,拟杆菌属与副拟杆菌属共享超过1000个蛋白质家族的核心,但与卟啉单胞菌属和普雷沃菌属共享较少数量的核心家族蛋白质家族。它们的组成和代谢活动在很大程度上受饮食调节。
拟杆菌与高脂肪和蛋白质的摄入量有关,而普雷沃氏菌与富含植物的饮食(高水平的复合碳水化合物和水果和蔬菜的摄入量)有关。因此,拟杆菌属和普雷沃氏菌属物种在人体内具有其特定的栖息地,因为它们大多数是拮抗的。
厚壁菌门与拟杆菌门之间的比率(F/B)与维持体内平衡有关,该比率的变化可能导致各种病理。
例如,特定厚壁菌门或拟杆菌门物种丰度的增加分别导致肥胖和肠道炎症。尽管人们非常关注 F/B 比率,但应该记住,这一比率可能会受到其他门的增加的影响,但是其他门的菌群失调增加或减少并不一定会改变 F/B 比率。
Stojanov S,et al.Microorganisms.2020
肥胖是一种复杂的、多因素的疾病,其原因包括宿主遗传背景、体力活动减少和食物摄入过多等多种因素。在过去的几十年里,肠道微生物群被认为是有利于脂肪储存、体重增加和胰岛素抵抗的额外因素。
世界范围内的研究正在不断寻找其致病因素。厚壁菌门和拟杆菌门的改变首先在肥胖动物和受试者中被描述,这些动物和受试者表现出厚壁菌门丰度增加,而拟杆菌门却减少了。
✦ F/B 比率较高时相同食物会吸收更多能量
当F与B的比例较高或体重增加和肥胖状态时,这两个细菌门影响我们体重的原因是它们调节我们吸收的脂肪量。想象一下两个人都消耗2000卡路里,但F与B的比例不同。F/B 比率较高的一种会吸收更多的卡路里,因此会比另一种增加更多的体重。因此,重点应该是平衡F与B的比例,一方面防止肥胖,另一方面预防炎症性肠病的发生。
当受试者接受热量限制饮食一年后,他们的拟杆菌门丰度增加,厚壁菌门/拟杆菌门比例正常化,同时体重减轻。这些研究得到了对喂食高脂肪或高纤维饮食的动物的研究的支持,这些研究显示厚壁菌门和拟杆菌门的丰度分别较高。
基于这些结果以及从肥胖动物和人类身上获得的其他结果,有人提出厚壁菌门比拟杆菌更能有效地从食物中提取能量,从而促进有效吸收热量和随后的体重增加。
肥胖双胞胎的微生物组富含编码营养转运蛋白的基因,而瘦双胞胎的微生物组富含编码与碳水化合物代谢相关的酶的基因。
最近发表的一项系统综述显示,大多数研究都支持 F/B 比率增加与肥胖之间的关系。
对肥胖和非肥胖日本受试者的粪便样本进行的一项研究显示,他们的 F/B 比率存在显著差异。厚壁菌门的百分比为 37.0 ± 9.1%(非肥胖者)和 40.8 ± 15.0%(肥胖者),而拟杆菌门的百分比为 44.0 ± 9.8%(非肥胖者)和 37.0 ± 14.0%(肥胖者)。
在61名乌克兰成年人中也报告了类似的结果,其中 F/B 比率与体重指数(BMI)显著相关。F/B 比率≥ 1 的个体超重的可能性比F/B 比率<1 的个体高出23%。
同样,卡塔尔受试者(37名肥胖者和36名瘦者)表现出肠道微生物群的改变;肥胖和瘦受试者的 F/B 比率分别为 2.25 ± 1.83 和 1.76 ± 0.58。在哈萨克族和比利时学童中,肥胖组的F/B比值显著高于对照组。
✦ 肥胖人群与正常人群拟杆菌存在差异
然而,两个拟杆菌属的存在差异:与肥胖组相比,正常组中拟杆菌属的高出20%,而肥胖组中普雷沃氏菌属比正常组高出19%。拟杆菌属和普雷沃氏菌属都属于拟杆菌门。
同一群体的受试者之间厚壁菌门和拟杆菌门的相对丰度差异很大。这可能是由于许多与生活方式相关的因素造成的,包括饮食、体力活动、食品添加剂和污染物、抗生素消耗等影响胃肠道微生物群组成的因素。
这种高度变异性可能解释了在比较正常体重和肥胖受试者之间的微生物群时观察到的矛盾结果,使得很难将厚壁菌门/拟杆菌门比率与确定健康状况联系起来。
重要的是要观察肠道微生物种群结构的整体组成,将其作为肥胖和肥胖相关病理(例如GSD)的指标,而不仅仅是厚壁菌门/拟杆菌门的比率。然而,在我们看来,可以修改这个比率,例如引入表征拟杆菌门底下的拟杆菌属或普氏菌属与梭状芽胞杆菌 XIVa 簇菌的比例。
!
F/B 比改变导致肥胖的争议
然而,与这些结果相反,许多研究没有观察到该参数的任何改变,甚至报告肥胖动物和人类中厚壁菌门/拟杆菌门的比例降低。
事实上,在大多数研究中,肥胖患者比瘦受试者表现出更少的细菌多样性,这表明在科、属或种水平上存在其他组成变化,这可能比厚壁菌门/拟杆菌门比率更相关。
✦肥胖增加可能与长期暴露脂多糖有关
关于这一点,代谢性内毒素血症的假说提出,肥胖增加和全身炎症的发展可能是由于长期暴露于脂多糖所致,脂多糖是一种源自革兰氏阴性细菌的促炎分子,它会通过肠腔进入血液。
这一假设与肥胖症中所报道的拟杆菌门丰度下降的情况并不相符,因为该门是肠道微生物群中革兰氏阴性菌的主要群体。
这种差异可以通过这样的事实来解释:来自拟杆菌门细菌的脂多糖的内毒素活性被认为低于来自其他革兰氏阴性细菌(例如属于变形菌门的细菌)的内毒素活性。有趣的是,在肥胖受试者或动物中也观察到变形菌门的增加,并且在无菌小鼠中施用变形菌门成员肠杆菌会导致这些动物出现肥胖和胰岛素抵抗。
✦ F/B比率增加与短链脂肪酸产生无关
另一方面,厚壁菌门/拟杆菌门比例的增加与肥胖个体中观察到的短链脂肪酸产生无关。事实上,据报道,拟杆菌门主要产生乙酸和丙酸,而厚壁菌门则产生更多的丁酸。丁酸盐被认为是一种促进健康的分子,因为它具有增加胰岛素敏感性、发挥抗炎活性、调节能量代谢和增加瘦素基因表达的能力。
结肠中的丙酸盐刺激L-肠内分泌细胞释放GLP-1和 PYY,从而抑制食欲。它还可能到达门静脉循环,主要被肝脏捕获,参与肝糖异生并减少参与脂肪酸和胆固醇从头合成的酶的表达。
乙酸也被吸收并到达全身循环和周围器官,包括脂肪组织、肌肉和大脑。在肝脏中,与丙酸盐相反,它刺激肝脏合成脂质,导致血脂异常。在大脑中,它激活副交感神经系统,分别促进胰腺和胃粘膜分泌胰岛素和生长素释放肽。
这些事件会导致脂肪储存和食欲增加,从而导致肥胖。根据这些结果,醋酸盐通常被认为更容易导致肥胖。肥胖个体中厚壁菌门/拟杆菌门比例的增加意味着这些受试者中丁酸盐的产量较高,丙酸盐和乙酸盐的产量较低,这一发现与这些短链脂肪酸各自的抗肥胖和致肥胖作用部分矛盾。一种解释是,肥胖个体中产生丁酸的细菌减少,并逐渐被属于同一门的其他细菌取代,导致结肠腔中丁酸的产生量降低。例如,葡萄球菌属和罗伊氏乳杆菌的丰度增加(均来自厚壁菌门)
肥胖状态还与A. muciniphila (疣微菌门)丰度较低有关,这是一种粘蛋白降解细菌,有助于稳定肠道屏障功能、分泌抗菌肽和控制炎症。
✦生理及病理状况都会影响肠道微生物群
另一方面,性别、年龄、宿主遗传学的差异。男性和女性的肠道微生物群存在差异,例如男性中拟杆菌-普雷沃氏菌群的水平较高,而女性中厚壁菌门的比例较高。然而,个别研究也发现女性中拟杆菌门的丰度最高。
此外,存在病理状况(糖尿病、心血管疾病、炎症性肠病等)。例如,Oddi括约肌松弛与胆管结石有关,可能是由于肠内容物回流增强改变了微环境所致。
治疗(抗生素、二甲双胍等)通过富集Akkermansia muciniphila以及几种产生短链脂肪酸的微生物群(丁酸弧菌、两歧双歧杆菌等)来改变肠道微生物群组成。
总之,这些导致厚壁菌门/拟杆菌门比率并不是与肥胖相关的微生物群失调的有力标志。
我们建议与肥胖相关的肠道微生物组研究不应该研究肥胖本身的分类标志物,而应侧重于确定用于将患者分层为亚组的分类标志物。引入微生物组患者分层将通过直接操纵患者微生物组来制定个性化治疗决策,从而改善肥胖的管理。
炎症性肠病代表小肠和大肠炎症反应引起的肠道疾病,由两种疾病组成:克罗恩病(CD)和溃疡性结肠炎(UC)。
溃疡性结肠炎是由结肠和直肠(从大肠末端开始到肛门结束的腔室)的长期炎症引起的,而克罗恩病是由结肠或回肠(小肠的最后部分)或两者的长期炎症引起的。
✦ 炎症性肠病患者厚壁菌门丰度下降
肠道微生物群会导致这些疾病。具体来说,当厚壁菌门减少或拟杆菌门相对于厚壁菌门增加时(两种情况下肠道菌群失调),就会导致慢性炎症和消化系统疾病,如克罗恩病和溃疡性结肠炎。
一项研究检查了克罗恩病和溃疡性结肠炎患者以及健康对照活检组织中的微生物多样性。观察到厚壁菌门丰度下降;更准确地说,克罗恩病患者中梭状芽胞杆菌类细菌减少,但溃疡性结肠炎患者或健康个体中则没有减少。
相反,与溃疡性结肠炎患者和健康个体相比,克罗恩病患者的拟杆菌丰度显著增加(分别为74.97%、64.31%和67.41%)。
✦肠道微生物变化与炎症性肠病严重程度相关
肠道微生物群的变化也与克罗恩病和溃疡性结肠炎的疾病活动度和严重程度相关。例如,与患有非活动性疾病的患者相比,患有活动性疾病的溃疡性结肠炎患者中厚壁菌门的含量较低。同样,与患有非侵袭性疾病的克罗恩病患者相比,患有侵袭性疾病的克罗恩病患者的厚壁菌门丰度显著降低。
具有抗炎作用的普氏粪杆菌减少
普氏粪杆菌(F. prausnitzii)是一种厚壁菌门物种,在炎症性肠病患者中减少,其减少与克罗恩病术后六个月复发相关。
普氏粪杆菌在体内和体外均表现出抗炎作用。在患有诱导性结肠炎的小鼠中,活普氏粪杆菌(或其上清液)降低了结肠炎的严重程度并纠正了生态失调。此外,普氏粪杆菌对血液单核细胞的体外刺激导致IL-10分泌增加以及IL-12和干扰素-γ(IFN-γ)分泌减少。
拟杆菌在炎症性肠病中增加
相比之下,拟杆菌在炎症性肠病中通常会增加,并与其进展和发展相关。对炎症性肠病患者和健康个体肠道炎症和非炎症区域的粘膜活检显示,炎症性肠病样本中厚壁菌门减少,拟杆菌门丰度增加,而肠杆菌科仅在克罗恩病患者中增加。活检还显示,与非发炎区域相比,发炎区域的细菌失调程度更高。
另一项对克罗恩病和溃疡性结肠炎患者进行粘膜活检和粪便菌群构成的研究证明了肠道微生物群和 F/B 比率的改变。克罗恩病和溃疡性结肠炎患者的F/B比值均较对照组显著降低。
此外,种、属和科的组成也发生了变化。与克罗恩病患者和对照组相比,溃疡性结肠炎患者中拟杆菌和乳酸菌的丰度较高;与对照组相比,溃疡性结肠炎患者中大肠杆菌的丰度也有所增加。相反,克罗恩病患者中球状梭菌减少。
✦ 不同拟杆菌种类与不同炎症性肠病程度相关
不同的拟杆菌与不同程度的炎症性肠病相关。在经过抗生素预处理且存在转化生长因子(TGF)βRII和IL-10R2信号传导缺陷的小鼠系中,在用普通拟杆菌和多形拟杆菌灌胃后观察到严重的溃疡性疾病。
与显示 F/B 比率对肥胖影响不大的研究类似,并不是在所有炎症性肠病病例中都观察到 F/B 比率降低。炎症性肠病患者粪便微生物分析显示,拟杆菌门、厚壁菌门和疣微菌门的丰度下降,而变形菌门、放线菌门和梭杆菌门的丰度增加。在48名克罗恩病患者中,粪便样本中的拟杆菌细菌丰度不到健康个体的10%,梭菌亚群 XIVab(厚壁菌门)也显著降低。
然而,大多数研究表明,拟杆菌因内毒素而表现出促炎特性,并影响细胞因子的产生,从而导致炎症性肠病。此外,厚壁菌门细菌具有抗炎作用,可以缓解炎症性肠病的进展。
2型糖尿病(T2DM)是一种代谢性疾病,其主要原因是与肥胖相关的胰岛素抵抗。然而,其他一些因素,如精神压力、感染和遗传倾向也可能导致糖尿病。
肥胖和糖尿病的特征都是慢性低度炎症状态,并伴有多种炎症介质(例如肿瘤坏死因子和白细胞介素)的异常表达和产生。
最近的研究基于大规模16S rRNA 基因测序、定量实时PCR (qPCR) 和荧光原位杂交 (FISH),显示了肠道微生物群组成与糖尿病等代谢疾病之间的关系。
有人提出,肠道微生物群引导增加肠道对单糖的摄取,并指示宿主增加肝脏产生与胰岛素抵抗相关的甘油三酯。
✦ 2型糖尿病患者中产丁酸厚壁菌门丰度降低
权威研究显示主要的丁酸盐合成菌厚壁菌门在2型糖尿病患者中显著减少。多项人体研究支持这样一个事实:与健康受试者相比,2型糖尿病受试者中合成丁酸的厚壁菌门丰度显著降低。
对这些患者的粪便微生物群分析显示,与正常耐糖受试者相比,新诊断或长期患有糖尿病的受试者中瘤胃球菌科和毛螺菌科等科的丰度有所下降。
还报告了罗氏菌属(Roseburia)和普拉梭菌(Faecalibacter prausnitzii)的丰度在健康对照中比2型或1型糖尿病患者更高。
对粪便样本进行深入的宏基因组学和宏蛋白质组学分析证实,在患有糖尿病前期 (Pre-DM) 或初治2型糖尿病 (TN-T2D) 的个体中,产生丁酸盐的菌含量显著降低。与正常耐糖个体相比, 患有糖尿病前期个体中的普拉梭菌的水平低于正常耐糖个体和初治2型糖尿病个体。
肠道微生物组可以反映整个身体的健康状况。厚壁菌门和拟杆菌门是结肠的主要门,可以影响与肥胖相关的疾病,这些疾病也是乳腺癌的危险因素。
✦ 乳腺癌患者F/B比值比健康人群低很多
因此,研究对乳腺癌患者和健康对照者的血清中提取细菌细胞外囊泡,使用微生物组测序分析厚壁菌门和拟杆菌门。分析乳腺癌的预后因素和血清学检测结果与 F/B 比值的相关性。发现乳腺癌患者的 F/B 比值比健康对照者低三倍。
此外,乳腺癌的危险因素,如空腹血糖,被发现与F/B比值有关。F/B比值可以作为乳腺癌的危险因素,并可能作为解释影响乳腺癌发生的潜在机制的线索之一。
健康对照者和乳腺癌患者中 F/B 的比率
An J,et al.J Clin Med.2023
(a)健康对照组和乳腺癌患者中厚壁菌门和拟杆菌门占总微生物群的比例;(b)健康对照者和乳腺癌患者的F/B比率;(c)F/B比与体重指数之间的关系;(d)F/B比与饮食习惯的关系(*p≤0.05,**** p≤0.0001)。
调查研究发现中国饮食中的动物脂肪和蛋白质含量高于印度饮食中的全谷物和植物性素食。而印度参与者微生物组中拟杆菌的比例几乎是中国参与者的四倍,分别为16.39%和4.27%(p=0.001)。
✦ 动物产品食用较少与拟杆菌数量较多有关
据推测,印度人体内拟杆菌门的丰度较高是由于他们对动物产品的消耗较少;表明饮食中动物产品含量较低与拟杆菌数量较多有关。
对欧洲儿童和布基纳法索儿童的微生物群的比较也显示了由于饮食习惯不同而导致的微生物群组成的差异,欧洲儿童的饮食富含动物蛋白、糖、淀粉和脂肪,纤维含量低。以素食为主的饮食主要由碳水化合物、纤维和非动物蛋白组成。欧洲儿童微生物组富含厚壁菌门和变形菌门,而放线菌门和拟杆菌门在非洲儿童中较多。
✦ 营养不良儿童的肠道微生物多样性较低
研究发现,与同一地区中高收入家庭的健康儿童相比,孟加拉国贫困社会经济地位家庭的营养不良儿童的肠道微生物群多样性较低,其特点是拟杆菌门的相对丰度较低,变形菌门占主导地位。
注:拟杆菌门的存在率较低众所周知,它们可以消化复杂的膳食材料,从而提高从各种食物中提取能量,而潜在致病性变形菌的较高含量可能有助于解释营养不良儿童的健康状况不佳。
✦ 增加纤维摄入量可以增加厚壁菌门的数量
一项研究将西方饮食的美国儿童与食用大米、面包和扁豆等植物性饮食的孟加拉国儿童进行了比较。在食用西方饮食的美国儿童中,厚壁菌门/拟杆菌门的比例减少了三倍。
另一项研究要求参与者增加纤维摄入量并避免西方减肥食品。研究表明厚壁菌门:拟杆菌门比率增加了。
通过分析三名健康受试者6-12个月的肠道宏蛋白质组,进一步证实了这些结果。在这项研究中,参与碳水化合物运输和代谢的蛋白质占检测到的蛋白质的10%以上,构成了所有测试对象中发现的核心宏蛋白质组的一部分。
糖酵解途径尤其引人注目,并鉴定出几种相关酶。在为每个门的每个COG功能类别分配光谱命中后,很明显,厚壁菌门和放线菌门负责活跃的碳水化合物代谢,而拟杆菌门则显示出更多的混合功能。
这些例子反映了将某些门与特定饮食广泛联系起来主要挑战是需要考虑涵盖整个微生物组的微生物的状态和相互作用动态。
✦ 当F/B比率增加时
即更多厚壁菌门,拟杆菌较少。或两者兼而有之,厚壁菌门增长更快。可能导致以下疾病:
-肥胖(BMI较高)
-高血压中的肠道菌群失调
-自闭症谱系障碍
-慢性疲劳综合症
-肠易激综合症
-干燥综合症
-类风湿性关节炎
✦当F/B比率下降时
即厚壁菌门较少,更多拟杆菌。或两者兼而有之,厚壁菌门减少得更快。可能导致以下疾病:
-系统性红斑狼疮
-乳糜泻
-克罗恩病
-老龄化
我们审查评估了饮食干预如何影响微生物组成的相关文章。总体而言得出以下建议供参考:
✦ 增加植物蛋白摄入,避免摄入过量动物蛋白
总体而言,蛋白质摄入量,尤其植物蛋白的摄入与肠道拟杆菌门,细菌多样性呈正相关。乳清和豌豆蛋白可以增加双歧杆菌和乳酸菌的数量,而乳清还可以减少脆弱拟杆菌和产气荚膜梭菌的数量,这两种细菌都是致病性的。
此外,豌豆蛋白被发现可以增加短链脂肪酸水平,短链脂肪酸被认为具有抗炎作用,并在粘膜完整性中发挥作用。然而,一项研究发现,高动物蛋白/低碳水化合物饮食会导致罗氏菌属和直肠真杆菌(这两种有益细菌)减少。动物蛋白可能含有大量脂肪,而脂肪也会影响肠道微生物群。
✦ 尽量摄入不饱和脂肪
当研究人员观察脂肪时,发现并不是所有的脂肪都是平等产生的。在一项研究中,小鼠被喂食猪油或鱼的脂肪。喂猪油的小鼠中拟杆菌属和嗜胆菌属(Bilophila)增加,而双歧杆菌和Adlercreutzia、乳酸菌(乳杆菌和链球菌)和疣微菌属(Akkermansia muciniphila)在鱼油喂养的小鼠中增加。
猪油喂养的小鼠还出现白色脂肪组织炎症和胰岛素敏感性受损。喂食鱼油的小鼠总体表现更好,研究人员得出结论,肠道微生物群促进了高饱和脂肪饮食的整体炎症反应。
✦ 高纤维的饮食可以减少炎症
碳水化合物,包括可消化的(淀粉和糖)和不可消化的(纤维),可能是研究最多的常量营养素,因为它们具有改变肠道微生物组的能力。
人类喂食大量葡萄糖、果糖和蔗糖,双歧杆菌水平增加,拟杆菌减少。相反,当喂食糖精(一种人工甜味剂)时,拟杆菌丰度增加,罗伊氏乳杆菌减少,微生物发生与天然糖完全相反的变化。
纤维被认为是一种不易消化的膳食成分,有时被称为益生元,因为它通过刺激有益肠道细菌的生长而对宿主有益。
来源包括大豆、菊粉、未精制的小麦和大麦、生燕麦和不可消化的低聚糖,例如果聚糖、聚葡萄糖、低聚果糖 (FOS)、低聚半乳糖 (GOS)、低聚木糖 (XOS) 和阿拉伯低聚糖 (AOS)。
低纤维饮食与较低的细菌丰度有关,而富含这些不易消化的碳水化合物的饮食会增加双歧杆菌和乳酸杆菌。较高的纤维摄入量导致促炎细胞因子IL-6和胰岛素抵抗减少。
✦ 发酵食品可以增加双歧杆菌和乳杆菌水平
除了常量营养素和纤维对肠道细菌的影响外,含有乳酸菌的发酵食品(例如发酵乳制品)也被发现可以增加双歧杆菌和乳酸杆菌的水平。
✦ 多酚具有抗氧化活性能够减少一些致病菌
最后但并非最不重要的是多酚,包括儿茶素、黄酮醇、黄酮、花青素、原花青素和酚酸(存在于水果、种子、蔬菜、茶、可可制品和葡萄酒等食品中),被发现可以增加拟杆菌,减少金黄色葡萄球菌、鼠伤寒沙门氏菌和梭菌等致病菌。
拟杆菌门的成员被假设可以降低细胞内的氧水平,从而有利于厌氧菌的生长,从而促进肠道平衡的维持,并且它们被认为是关键的聚糖降解细菌比厚壁菌门等其他类群更能代谢多酚。
多酚可以改变肠道环境,它们可以被肠道细菌分解代谢,将它们转化为多种化合物比原始化合物具有更强的抗氧化活性。
厚壁菌门/拟杆菌门(F/B比率)作为一个指标在很多关于肠道菌群的研究中被提及和表征,从疾病发生进展,到药物饮食干预,涉及肠道菌群的研究,首先会描述细菌门水平的变化,而人类或动物肠道细菌约90%都属于厚壁菌门与拟杆菌门,而它们的比率也被认为是肠道菌群动态平衡的标志。
拟杆菌门主要为革兰氏阴性菌,能激活T细胞介导的免疫反应,降解多糖保护肠道和其他病原菌,厚壁菌门主要为革兰氏阳性菌,尤其产丁酸的菌,但是更容易吸收能量。
但是肠道菌群受到抗生素,药物,年龄以及分析注释数据库等的影响,此外随着肠道菌群研究队列的扩大,以及对菌群构成与不同疾病状态和药物饮食等因素的相关性研究的深入,F/B比率作为一个相对粗放的指标已经无法准确反映复杂的肠道菌群状况。我们需要更加深入和全面的通过菌属甚至菌种的构成变化来解释和分析肠道菌群特征,并进而更准确的解释肠道菌群和健康及生活方式的关联。
主要参考文献
Stojanov S, Berlec A, Štrukelj B. The Influence of Probiotics on the Firmicutes/Bacteroidetes Ratio in the Treatment of Obesity and Inflammatory Bowel disease. Microorganisms. 2020 Nov 1;8(11):1715.
Mariat D, Firmesse O, Levenez F, Guimarăes V, Sokol H, Doré J, Corthier G, Furet JP. The Firmicutes/Bacteroidetes ratio of the human microbiota changes with age. BMC Microbiol. 2009 Jun 9;9:123.
Gui X, Yang Z, Li MD. Effect of Cigarette Smoke on Gut Microbiota: State of Knowledge. Front Physiol. 2021 Jun 17;12:673341.
Grigor’eva IN. Gallstone Disease, Obesity and the Firmicutes/Bacteroidetes Ratio as a Possible Biomarker of Gut Dysbiosis. J Pers Med. 2020 Dec 25;11(1):13.
Vaiserman A, Romanenko M, Piven L, Moseiko V, Lushchak O, Kryzhanovska N, Guryanov V, Koliada A. Differences in the gut Firmicutes to Bacteroidetes ratio across age groups in healthy Ukrainian population. BMC Microbiol. 2020 Jul 22;20(1):221.
Indiani CMDSP, Rizzardi KF, Castelo PM, Ferraz LFC, Darrieux M, Parisotto TM. Childhood Obesity and Firmicutes/Bacteroidetes Ratio in the Gut Microbiota: A Systematic Review. Child Obes. 2018 Nov/Dec;14(8):501-509.
Hidalgo-Cantabrana C, Delgado S, Ruiz L, Ruas-Madiedo P, Sánchez B, Margolles A. Bifidobacteria and Their Health-Promoting Effects. Microbiol Spectr. 2017 Jun;5(3).
An J, Kwon H, Kim YJ. The Firmicutes/Bacteroidetes Ratio as a Risk Factor of Breast Cancer. J Clin Med. 2023 Mar 13;12(6):2216.

谷禾健康

拟杆菌是革兰氏阴性、专性厌氧菌,在人类肠道微生物群中含量丰富,构成了肠道菌群中大部分比例。
Bacteroides uniformis (一般简称为 B.uniformis),翻译为单形拟杆菌或普通拟杆菌,是拟杆菌属的主要成员,在肠道中起着重要的作用。也被鉴定为阴道微生物组的组成部分。
Bacteroides uniformis是与人类粪便中异黄酮染料木黄酮降解相关的特定细菌物种,可以产生纤维素酶和淀粉酶,降解复杂多糖。
Bacteroides uniformis与肠炎、厌食、肥胖、糖尿病、腹胀、认知发育与情绪运动、多动症、慢性肾病、骨质疏松等相关,是可引起人类或动物疾病的物种,不太可能在人群中传播,并且存在足够的预防或治疗方法。
对单形拟杆菌进行深入研究,有助于我们更好地理解其在人体肠道中的功能,并为相关疾病的预防和治疗提供新的思路。
Bacteroides uniformis是一种革兰阴性细菌,呈杆状。它们通常具有弯曲或弯钩形态,并且没有芽胞。

图源:Microbe Canvas
生理特性
Bacteroides uniformis是一种厌氧菌,不能在氧气存在的环境中生长。它们通常能够利用多糖类物质作为碳源,如淀粉、纤维素和其他复杂碳水化合物。还可以产生一些酶,如纤维素酶和淀粉酶,用于降解消化这些复杂碳水化合物。
分布和生存环境
Bacteroides uniformis是人体肠道,阴道中常见的细菌之一,也可以在其他动物的肠道中找到。它们在肠道中扮演重要的生态角色,参与食物消化、营养吸收和免疫调节等过程。Bacteroides uniformis能够适应肠道的微生态环境,与其他肠道细菌相互作用。
功能和代谢
Bacteroides uniformis在肠道中具有多种功能和代谢活性。它们参与葡萄糖和其他碳水化合物的代谢,并产生短链脂肪酸(如丙酸和乙酸),这些酸有益于肠道健康。此外,Bacteroides uniformis还参与胆固醇代谢、氨基酸代谢和维生素合成等生物化学过程。
肥胖和肥胖相关疾病构成了世界范围内的重大公共卫生问题。除了传统的减肥药物外,下一代益生菌(NGP)似乎非常有希望作为潜在的肥胖预防和治疗药物。
主要的候选NGP已在肥胖和肥胖相关疾病的临床前模型中显示出前景,例如:
B.uniformis已被证明具有对抗肥胖和代谢紊乱的有益特性。在动物模型中, B. uniformis与减肥、降低血清胆固醇和甘油三酯水平以及改善肝脂肪变性有关。此外,它还与血清瘦素水平降低、空腹血糖浓度降低和糖耐量改善有关。
推测的机制包括调节肠道菌群和改善肠道菌群失调、改善肠道屏障功能、减少慢性低度炎症和调节肠道肽分泌。
一项对41名体重指数(BMI)正常的个体肠道菌群和内脏脂肪面积 (VFA)的研究显示,内脏脂肪面积高/低(VFA)两组的肠道菌群存在显著差异,在 L-VFA 组中,B. plebeius 和B.uniformis含量很高。此外,B.uniformis与血清 LDL-胆固醇水平呈负相关。
另一项对32名肥胖成年人(14 名接受了腹腔镜袖状胃切除术 (LSG))和30名基线健康对照者的肠道菌群以及身体指标的研究,并在术后 6 个月进行了随访。通过磁共振成像测量腹部内脏脂肪面积(VFA)和皮下脂肪面积(SFA)。腰围、臀围、腰臀比(WHR)和体重指数(BMI)被列为简单的肥胖参数。通过宏基因组测序分析肠道微生物组。在与腹部内脏脂肪面积(VFA)负相关的物种中,Eubacteria eligens相关性最强,其次是Clostridium citroniae、C. symbiosum、Bacteroides Uniformis、E. ventriosum、C. hathewayi等。
一项涉及 45 名儿童(6 至 12 岁)的横断面研究调查了肠道微生物群与儿童肥胖之间的关系。发现两个BMI组之间的多样性存在差异。这种差异包括真杆菌属(Eubacterium)、普拉梭菌(Faecalibacterium prausnitzii)、Dialister、Monoglobus pectinilyticus、Bifidobacterium pseudocatenulatum、Intestinibacter bartlettii、 Bacteroides intestinalis、Bacteroides uniformis (单形拟杆菌)和 史密斯甲烷杆菌(Methanobrevibacter smithii)。
一项针对减肥手术是否显著改善体重的研究显示单形拟杆菌的丰度可能影响手术后的效果。
术前属于拟杆菌门和厚壁菌门的成员丰度存在差异无应答组:
富含拟杆菌科、拟杆菌属、单形拟杆菌、Alistipes Finegoldii、Alistipes alistipes、Dorea formicigenerans 、瘤胃球菌。
应答组:
富含消化链球菌科、 Gemmiger、Gemiger formicilis、Barnesiella、Prevotellaceae 、 Prevotella。
线性判别分析 >2;p < 0.05。与有反应组相比,无反应组中普氏菌与拟杆菌的比率显着较低 (p = 0.048)。
神经性厌食症(Anorexia nervosa, AN)的特点是一种神秘的食物回避模式,导致严重的营养不良,这种疾病在青春期早期发展。近期,在《Nature Microbiology》上发表的文章研究了77名患有神经性厌食症的成年女性和70名被认为是健康体重的对照组。
利用宏基因组学,作者对微生物组的细菌和病毒成分进行了深入分析。细菌基因组中的结构变异,特别是Bacteroides uniformis(单形拟杆菌)中的结构变异,可能造成了硫胺素生产的破坏。
硫胺素(或称维生素B1)缺乏会影响人体健康,包括记忆丧失、焦虑、抑郁、易怒、失眠、食欲减退和胃肠道不适。这也可以解释在神经性厌食症患者中普遍观察到的硫胺素缺乏的现象。
认知发展
一项针对健康以色列阿拉伯儿童的横断面研究显示一些菌群分类学差异与儿童智商测试FSIQ评分显着正相关,包括Prevotella(普雷沃氏菌属)、Dialister属、Sutterella(萨特氏菌属)、Ruminococcus callidus和Bacteroides uniformis(单形拟杆菌)。
情绪相关
一项针对长期封闭环境中心理变化和肠道菌群的研究,鉴定出四种潜在的“精神抗生素”(单形拟杆菌Bacteroides Uniformis、Roseburia inulinivorans、Faecalibacter prausnitzii、Eubacterectale )。
根据宏基因组、宏蛋白质组和代谢组学分析,四种潜在的精神抗生素主要通过与神经系统功能相关的三种途径改善情绪:
首先,通过发酵膳食纤维,它们可能产生短链脂肪酸,例如丁酸和丙酸;
其次,它们可能调节天冬氨酸、谷氨酸、色氨酸等氨基酸代谢途径(例如,将谷氨酸转化为γ-氨基丁酸;将色氨酸转化为血清素、犬尿酸或色胺);
第三,它们可能调节其他途径,例如牛磺酸和皮质醇代谢。此外,动物实验的结果证实了这些潜在的精神抗生素对情绪的积极调节作用和机制。
运动相关
肠道微生物群可能有助于改善运动表现。日本庆应义塾大学先进生命科学研究所通过分析25名男性长跑运动员的比赛成绩后发现,运动员粪便中单形拟杆菌(Bacteroides uniformis)的丰度与其3000米比赛成绩间存在相关性。
在随后的一项随机、双盲、安慰剂对照研究中,研究人员让健康男性志愿者服用α-环糊精,以增加肠道中的单形拟杆菌总量。结果表明,补充α-环糊精可以提高人体的耐力运动能力。
此外,在小鼠实验中,食用单形拟杆菌显著延长了动物游泳的持续时间;动物盲肠短链脂肪酸浓度以及肝脏中与糖异生相关酶的基因表达均上升,肝糖原含量降低。该研究表明,单形拟杆菌可能通过促进肝脏内源性葡萄糖生成,增强个体的耐力运动能力。
妊娠糖尿病
妊娠合并糖尿病包括孕前糖尿病(PGDM)和妊娠期糖尿病(GDM)。 一项针对234份孕妇粪便样本的16S rRNA测序数据显示,随着妊娠的进展,妊娠糖尿病PGDM 组的肠道细菌种类数量有所减少。巨单胞菌、拟杆菌和 Roseburia Enteris 在 PGDM 组中富集,而 Bacteroides vulgatus、Faecalibacter prausnitzii、Eubacterectale、Bacteroides uniformis(单形拟杆菌)、Eubacteria eligens、Subdolilegum在对照组中比例更高,这些菌血糖与血糖特征呈负相关。
二型糖尿病
一项针对亚洲人的包括36个不同二型糖尿病T2DM研究(健康,n = 3378;T2DM,n = 551)的meta分析显示,T2DM 组中普通杆菌菌、单形拟杆菌和普氏粪杆菌较低。
另一项包括29名T2DM患者和28名健康对照者研究显示,Megamonas funiformis 和 Escherichia 的丰度在T2DM患者的肠道中显着增加。相反,Bacteroides stercoris、单形拟杆菌和Phascolarctobacterium faecium的丰度降低。
单形拟杆菌 (Bacteroides uniformis) 还可能调节动物肠道稳态,减轻小鼠结肠炎。一项研究使用B.uninis JCM5828 及其代谢物对患有葡聚糖硫酸钠盐 (DSS) 诱导的结肠炎的雌性 C57BL/6J小鼠进行治疗。
Bacteroides uniformis治疗显著减轻了结肠炎的进展并恢复了机械和免疫屏障蛋白的表达。此外,Bacteroides uniformis还增加了共生菌双歧杆菌和阴道乳杆菌的丰度,同时减少了致病性埃希氏菌、志贺氏菌的丰度,并调节了肠道胆汁酸代谢。
Bacteroides uniformis在很大程度上调节结肠组织中NF-κB和丝裂原激活蛋白激酶(MAPK)信号通路关键调节蛋白的表达以及TH17细胞的分化。然而,Bacteroides uniformis在体外不能直接抑制TH17细胞分化;它应该是通过参与胆汁酸代谢和调节关键代谢物(α-鼠胆酸、猪去氧胆酸和异石胆酸)来调节固有层的过程,从而调节肠道免疫反应。
此外在一项溃疡性结肠炎(UC)患者的菌群研究中,六种拟杆菌属表现出显着较低的相对丰度,十二种拟杆菌属被发现与至少一项疾病活动指标显着相关。五个物种( Alistipes putredinis、Bacteroides stercoris、Bacteroides Uniformis、Bacteroides rodentium、Parabacteroides merdae )的相对丰度与溃疡性结肠炎活性的四个指标中的三个相关。
12 个关键物种表达了与 UC 发病机制相关的5 个基因(TARP、C10ORF54、ITGAE、TNFSF9和LCN2 )。作为潜在生物标志物的关键物种的丧失可能会加剧 UC 活动。
比较功能性腹胀和膨胀(FABD)患者和12健康个体粪便微生物群的多样性和组成,并评估小肠细菌过度生长 (SIBO) 。结果显示FABD 组的微生物多样性显着低于对照组。与健康对照相比,FABD 患者中变形菌的比例显着升高,放线菌的比例显着降低。与健康对照相比,FABD患者中粪杆菌的比例显着升高,而SIBO(+)FABD患者中普氏菌和粪杆菌的比例也显着升高。与健康对照相比,FABD 患者的普氏粪杆菌 (Faecalibacter prausnitzii) 的丰度显着增加,但单形拟杆菌 (Bacteroides uniformis)和青春双歧杆菌数量明显减少。
乳糜泻患者肠道中的 Bacteroides distasonis、Bacteroides fragilis / Bacteroides thetaiotaomicron、Bacteroides Uniformis、Bacteroides ovatus比对照组更少(p<0.05)。
骨质疏松
一项研究中,与对照组相比,几个物种的丰度发生了变化,在骨质疏松组中发现了特定的功能途径,骨质疏松组中下列菌群的水平下降:
类风湿性关节炎
一项研究包括 76 名类风湿关节炎患者的粪便宏基因组和血浆代谢组的分期概况。
骨关节炎
骨关节炎是一种病因不明的慢性退行性疾病。一项研究针对骨关节炎患者(n = 32)和正常对照组(n = 57)肠道菌群检测,宏基因组测序显示,骨关节炎患者中Bacteroides stercoris、Bacteroides vulgatus、Bacteroides uniformis在物种水平上的丰度显著降低,大肠杆菌、肺炎克雷伯菌、Shigella flexneri、唾液链球菌在骨关节炎中显著增加。
特应性皮炎AD
使用 16S rRNA 测序分析了 93 名志愿者的粪便样本,其中包括 44 名 AD 患者和 49 名健康对照受试者,年龄为 6-22 岁。
卟啉单胞菌科、Blautia、Parabacteroides、Bacteroides ovatus、Bacteroides uniformis 和 Prevotella stercorea 的相对丰度显著较高(P < 0. 05) AD 患者高于健康对照者。与 AD 患者相比,健康对照受试者中梭状芽胞杆菌和粪疟原虫含量较高 (P < 0.05)。
儿童多动症
对11项儿童多动症研究的meta分析显示,拟杆菌属与多动和冲动水平相关。在物种水平上,多动症儿童Faecalibacterium丰度较低,但Bacteroides uniformis、肠球菌、双歧杆菌和Odoribacter增加,可能导致中枢神经系统多巴胺相关功能受损。
此外,多动症儿童肠道中Faecalibacterium丰度的降低可能导致炎症细胞因子的渗透性和交叉性更高。
一项调查多动症儿童及其饮食模式和肠道菌群的研究显示,在注意缺多动障碍(ADHD)组中,粪肠拟杆菌(B. coprocola)的相对丰度降低,而单形拟杆菌(B.uniformis)、卵形拟杆菌(B.ovatus)和粪类杆菌(S. stercoricanis)的相对丰度升高。
结直肠癌转移组
转移性结直肠癌组和非转移性结直肠癌组患者的肠道菌群存在差异。在属水平上,拟杆菌属菌种脆弱拟杆菌和单形拟杆菌组成在转移组中显着高于非转移组。
PD-1免疫疗法反应
肠道微生物群被认为是调节免疫检查点抑制剂治疗功效的关键因素。一项波兰黑色素瘤患者队列中PD-1疗法反应与基线肠道微生物组的研究显示,普氏菌和单形拟杆菌的丰度与反应有关,而无反应者的肠道微生物群富含普氏粪杆菌和脱硫弧菌以及一些未分类的厚壁菌门。
慢性肾病
一项台湾地区人群肠道菌群研究显示,在慢性肾病 (CKD)患者中发现的最丰富的属是拟杆菌属,在物种水平上,血液透析患者中的B. ovatus、B. caccae 、 B.uniformis表现出显著丰度,而腹膜透析患者中的Blautia表现出较高的丰度。
白塞病
在活动性白塞病BD患者其单形拟杆菌丰度显著高于其匹配的健康对照和处于非活动状态的疾病患者 ( p = 0.038)。
单形拟杆菌可以消化多种膳食纤维,作为一种代谢膳食纤维产生乙酸的肠道菌已有大量研究表明日常膳食纤维摄入会影响单形拟杆菌的丰度。
一项横断面研究中检查了全谷物和精制谷物、纤维和麸质的习惯摄入是否与肠道微生物群相关。对779 名参与者137 项食物频率的调查问卷和肠道菌群的分析显示。最高全谷物摄入量四分位与较高丰度的普通拟杆菌、普氏粪杆菌、Blautiaproducta和Rikenellaceae以及较低丰度的单形拟杆菌有关。
一项对不同特异性(低-低聚果糖、低至中-2 型抗性淀粉、中-果胶和高-不溶性 β-1,3-葡聚糖特异性)纤维的微生物发酵研究显示,高特异性不溶性β-葡聚糖促进目标细菌大量增加,Anaerostipes sp. 平均从0.3%增加到16.5%,单形拟杆菌平均为 2.5% 至 17.9%,这与这些细菌所在的每个微生物群落中相关代谢物(分别为丁酸盐和丙酸盐)比例的增加有关。
水溶性醋酸纤维素 (WSCA) 是一种合成纤维来源,可以导致拟杆菌属成员的丰度增加,尤其是单形拟杆菌。在四种具有代表性的人类肠道拟杆菌属中,只有单形拟杆菌在WSCA上的纯培养物中生长并活跃地产生乙酸。
一项对健康志愿者进行了为期 4周的营养干预分析天然单宁提取物混合物对肠道微生物变化的研究显示。从前两周起就可以观察到微生物群的组成和功能的变化,测序显示以下菌相对丰度增加,例如:
利用同位素标记发,发现食用菊粉富集的前三种细菌为单形拟杆菌(Bacteroides uniformis)、产酸拟杆菌(Bacteroides acidifaciens)和狄氏副拟杆菌(Parabacteroides distasonis)。
一项研究评估富含黄酮类化合物的蔓越莓多酚(CP)和龙舌兰(AG)(一种高支链龙舌兰衍生的新果聚糖)对C57BL6雄性小鼠的心脏代谢反应、肠道微生物群组成、代谢内毒素血症和粘膜免疫调节的影响,该雄性小鼠喂食致肥胖高脂肪高糖(HFHS)饮食9周。AG,单独或与CP(CP+AG)联合,主要刺激聚糖降解细菌:
与未经治疗的小鼠相比,接受AG的肥胖小鼠中聚糖降解细菌的增加与丁酸水平的显著增加一致。
总的来说,研究发现高特异性不溶性β-葡聚糖、天然单宁提取物混合物、菊粉、蔓越莓多酚和龙舌兰能增加Bacteroides uniformis的丰度。
主要参考文献:
Vallianou NG, Kounatidis D, Tsilingiris D, Panagopoulos F, Christodoulatos GS, Evangelopoulos A, Karampela I, Dalamaga M. The Role of Next-Generation Probiotics in Obesity and Obesity-Associated Disorders: Current Knowledge and Future Perspectives. Int J Mol Sci. 2023 Apr 4;24(7):6755.
Nie X, Chen J, Ma X, Ni Y, Shen Y, Yu H, Panagiotou G, Bao Y. A metagenome-wide association study of gut microbiome and visceral fat accumulation. Comput Struct Biotechnol J. 2020 Sep 20;18:2596-2609.
Murga-Garrido SM, Ulloa-Pérez EJ, Díaz-Benítez CE, Orbe-Orihuela YC, Cornejo-Granados F, Ochoa-Leyva A, Sanchez-Flores A, Cruz M, Castañeda-Márquez AC, Plett-Torres T, Burguete García AI, Lagunas-Martínez A. Virulence Factors of the Gut Microbiome Are Associated with BMI and Metabolic Blood Parameters in Children with Obesity. Microbiol Spectr. 2023 Feb 14;11(2):e0338222.
Lapidot Y, Maya M, Reshef L, Cohen D, Ornoy A, Gophna U, Muhsen K. Relationships of the gut microbiome with cognitive development among healthy school-age children. Front Pediatr. 2023 May 19;11:1198792.
Cheng M, Zhao Y, Cui Y, Zhong C, Zha Y, Li S, Cao G, Li M, Zhang L, Ning K, Han J. Stage-specific roles of microbial dysbiosis and metabolic disorders in rheumatoid arthritis. Ann Rheum Dis. 2022 Aug 19;81(12):1669–77.
Shirvani-Rad S, Ejtahed HS, Ettehad Marvasti F, Taghavi M, Sharifi F, Arzaghi SM, Larijani B. The Role of Gut Microbiota-Brain Axis in Pathophysiology of ADHD: A Systematic Review. J Atten Disord. 2022 Nov;26(13):1698-1710.
Wang LJ, Yang CY, Chou WJ, Lee MJ, Chou MC, Kuo HC, Yeh YM, Lee SY, Huang LH, Li SC. Gut microbiota and dietary patterns in children with attention-deficit/hyperactivity disorder. Eur Child Adolesc Psychiatry. 2020 Mar;29(3):287-297.
Hao Z, Meng C, Li L, Feng S, Zhu Y, Yang J, Han L, Sun L, Lv W, Figeys D, Liu H. Positive mood-related gut microbiota in a long-term closed environment: a multiomics study based on the “Lunar Palace 365” experiment. Microbiome. 2023 Apr 24;11(1):88.
Ren Y, Hao L, Liu J, Wang P, Ding Q, Chen C, Song Y. Alterations in the Gut Microbiota in Pregnant Women with Pregestational Type 2 Diabetes Mellitus. mSystems. 2023 Apr 27;8(2):e0114622.
Wu X, Park S. Fecal Bacterial Community and Metagenome Function in Asians with Type 2 Diabetes, According to Enterotypes. Biomedicines. 2022 Nov 21;10(11):2998.
Lv M, Li L, Li W, Yang F, Hu Q, Xiong D. Mechanism research on the interaction regulation of Escherichia and IFN-γ for the occurrence of T2DM. Ann Palliat Med. 2021 Oct;10(10):10391-10400.
Yan Y, Lei Y, Qu Y, Fan Z, Zhang T, Xu Y, Du Q, Brugger D, Chen Y, Zhang K, Zhang E. Bacteroides uniformis-induced perturbations in colonic microbiota and bile acid levels inhibit TH17 differentiation and ameliorate colitis developments. NPJ Biofilms Microbiomes. 2023 Aug 14;9(1):56.
Nomura K, Ishikawa D, Okahara K, Ito S, Haga K, Takahashi M, Arakawa A, Shibuya T, Osada T, Kuwahara-Arai K, Kirikae T, Nagahara A. Bacteroidetes Species Are Correlated with Disease Activity in Ulcerative Colitis. J Clin Med. 2021 Apr 17;10(8):1749.
Noh CK, Lee KJ. Fecal Microbiota Alterations and Small Intestinal Bacterial Overgrowth in Functional Abdominal Bloating/Distention. J Neurogastroenterol Motil. 2020 Sep 30;26(4):539-549.
Wang H, Zhang K, Wu L, Qin Q, He Y. Prediction of Pathogenic Factors in Dysbiotic Gut Microbiomes of Colorectal Cancer Patients Using Reverse Microbiomics. Front Oncol. 2022 Apr 27;12:882874.
Ye S, Yan F, Wang H, Mo X, Liu J, Zhang Y, Li H, Chen D. Diversity analysis of gut microbiota between healthy controls and those with atopic dermatitis in a Chinese population. J Dermatol. 2021 Feb;48(2):158-167.
Zhou P, Dai Z, Xie Y, Li T, Xu Z, Huang Y, Sun D, Zhou Y. Differences in tissue-associated bacteria between metastatic and non-metastatic colorectal cancer. Front Microbiol. 2023 Jun 9;14:1133607.
Shivani S, Kao CY, Chattopadhyay A, Chen JW, Lai LC, Lin WH, Lu TP, Huang IH, Tsai MH, Teng CH, Wu JJ, Hsieh YH, Wang MC, Chuang EY. Uremic Toxin-Producing Bacteroides Species Prevail in the Gut Microbiota of Taiwanese CKD Patients: An Analysis Using the New Taiwan Microbiome Baseline. Front Cell Infect Microbiol. 2022 Apr 26;12:726256.
Kim JC, Park MJ, Park S, Lee ES. Alteration of the Fecal but Not Salivary Microbiome in Patients with Behçet’s Disease According to Disease Activity Shift. Microorganisms. 2021 Jul 6;9(7):1449.
Um CY, Peters BA, Choi HS, Oberstein P, Beggs DB, Usyk M, Wu F, Hayes RB, Gapstur SM, McCullough ML, Ahn J. Grain, Gluten, and Dietary Fiber Intake Influence Gut Microbial Diversity: Data from the Food and Microbiome Longitudinal Investigation. Cancer Res Commun. 2023 Jan 11;3(1):43-53.
Cantu-Jungles TM, Bulut N, Chambry E, Ruthes A, Iacomini M, Keshavarzian A, Johnson TA, Hamaker BR. Dietary Fiber Hierarchical Specificity: the Missing Link for Predictable and Strong Shifts in Gut Bacterial Communities. mBio. 2021 Jun 29;12(3):e0102821.
Molino S, Lerma-Aguilera A, Jiménez-Hernández N, Rufián Henares JÁ, Francino MP. Evaluation of the Effects of a Short Supplementation With Tannins on the Gut Microbiota of Healthy Subjects. Front Microbiol. 2022 Apr 27;13:848611.