-

 国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子
国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子

 二级病原微生物安全实验室
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
谷禾健康
编辑
在现代快节奏的生活中,消化系统不适已成为困扰全球数亿人的常见问题。其中,腹胀和便秘是最为普遍的胃肠道症状。它们不仅带来身体上的沉重感、疼痛和不适,更深刻影响着患者的心理健康和生活质量。许多患者在经历了长期的腹胀和排便困难后,最终被诊断为肠易激综合征(Irritable Bowel Syndrome, IBS),这是一种以腹痛伴随排便习惯改变为特征的功能性肠病。
过去,这类问题常被简单归因于“消化不良”或“胃肠动力不足”。随着神经胃肠病学和微生物组学的发展,人们逐渐认识到其背后是复杂的脑-肠-微生物轴失调。
研究表明,腹胀和便秘并非孤立的局部问题,而是涉及中枢神经系统、肠神经系统(ENS)、肠道内分泌激素、免疫炎症反应以及数以万亿计的肠道微生物的系统性失调。
本文将基于最新的科学研究和临床指南(如罗马IV标准),深入探讨腹胀与便秘的定义、高发人群、病理生理机制,特别是肠道菌群和肠道激素在其中扮演的关键角色,并为您提供一套涵盖饮食、微生态调节、生活方式及中医药的综合干预建议。
腹胀是一个常见但复杂的症状,原因多种多样,从饮食和压力到肠道微生物群失衡或医疗状况。本文解释了胃肠科医生通常会如何应对腹胀,胃肠专家会诊时应期待的情况,以及消化健康评估如何识别腹部不适背后的模式。帮助判断何时采取基础干预,何时需要进一步检查。
▸ 什么是腹胀和便秘?
▸ 腹胀的定义
腹胀是腹部胀气或膨胀感的统称。普通人吃饭后偶尔会有轻度腹胀,这是正常生理现象。只有频繁发作或持续存在的腹胀才需要关注。
▸ 便秘的定义
便秘主要表现为排便次数减少、粪便干硬及排便困难。一般以每周排便少于3次,并伴有排便费力或不畅作为诊断标准。
注:每个人排便习惯存在差异,有人每日1–2次,也有人2–3天一次。只要规律且粪便不干硬,均不属于便秘。
下面将从临床与医学角度进一步解析腹胀和便秘的定义及其亚型。
▸ 临床与医学定义
(主观感受与客观体征)
在消化系统临床中,腹胀是最常见且困扰患者的症状之一。医学上需区分“Bloating”和“Distension”两个概念。根据罗马IV标准及国际疾病分类(ICD-11),其区别主要如下:
▸主观腹胀(Bloating):指患者主观感受到的腹部饱胀感、紧绷感或气体堵胀感。这是一种感觉,可能伴随也可能不伴随腹部的实际增大。
▸客观腹部膨隆(Distension):指客观上可测量到的腹围增加,通常是肉眼可见的腹部向外突出。
二者可单独或同时存在。普通人群中,主观腹胀患病率约19%,伴客观腹围增加约9%;而在肠易激综合征(IBS)等功能性肠病患者中可高达66%–90%。
★ 腹胀是多系统的复杂失调
罗马IV标准将功能性腹胀和腹部膨隆定义为独立的“肠-脑互动异常”疾病。其诊断要求为:反复出现腹胀或腹部膨隆,平均每周至少1天,症状在诊断前已持续6个月,且近3个月符合标准,并排除IBS、功能性便秘等其他功能性胃肠病。该区分提示腹胀并非单一器质性问题,而是涉及神经、肌肉、微生物及免疫系统的复杂失调。
▸ 腹胀的亚型
腹胀可以分为功能性腹胀和器质性腹胀:
▸功能性腹胀:最常见,无明显的解剖学或生化异常。其发生多与内脏高敏感性、胃肠动力异常、肠道菌群失调及腹膈肌协同失调有关。
▸器质性腹胀:由明确的疾病引起,如肠梗阻、腹水(肝硬化、心衰引起)、胃轻瘫、乳糜泻、卵巢肿瘤等。在重症医学科中,腹胀甚至可能是危及生命的腹腔间隔室综合征(ACS)的先兆。
相比之下,功能性腹胀缺乏常规临床评估可解释的器质性原因。
▸ 腹胀与腹部肥胖的区别
许多患者,尤其是女性,常将功能性腹胀导致的腹围增大误认为“发胖”或“腹部肥胖”,但两者本质上截然不同:
▸基础原因不同:腹部肥胖是由于皮下脂肪或内脏脂肪的长期过度蓄积;而腹胀引起的腹围增大,多由肠道气体、滞留粪便或腹壁肌张力异常导致腹腔内容物前移所致。
▸时间节律不同:肥胖引起的腹围增大相对稳定,短时间内变化不明显;而功能性腹胀呈明显昼夜节律,常表现为“晨轻暮重”:晨起腹部平坦,随进食逐渐加重,至傍晚或夜间达峰,经休息或排气排便后缓解。
▸伴随症状不同:腹胀常伴有肠鸣音亢进、频繁嗳气、排气增多、腹痛或排便习惯改变(如便秘);而单纯的腹部肥胖通常不伴随这些消化道急性症状。
1
总体患病率与风险因素
腹胀和便秘是全球常见的健康问题。流行病学调查显示,全球约有15%-18%的普通人群每周至少经历一次腹胀。在中国,成人慢性便秘的患病率约为4.0%~10.0%,且随着年龄的增长而显著升高,80岁以上老年人的患病率甚至可达38.0%。
两者常相伴发生。在便秘型肠易激综合征(IBS-C)和功能性便秘(FC)患者中,超过90%伴有明显腹胀。
▸ 共同风险因素
这两种症状的共同风险因素包括:
–不良的饮食习惯(如高脂、低纤维饮食,高肉类摄入);
–缺乏体力活动或运动;
–不规律的排便习惯、
–精神心理压力(焦虑、抑郁、睡眠质量差);
–年龄、疾病因素等。
编辑
2
女性更易患腹胀和便秘
需要注意的是,腹胀和便秘的性别差异极为显著,女性患病率通常为男性的1.5~2倍以上。
女性之所以成为腹胀和便秘的高发群体,性激素(雌激素和孕激素)的周期性波动和生命阶段性衰退起到了决定性作用。特别是在围绝经期和围产期,女性的胃肠道面临着巨大的生理挑战。
▸ 围绝经期女性的“肠道危机”
围绝经期是指女性绝经前后的一段过渡时期,其核心特征是卵巢功能衰退,雌激素和孕激素水平剧烈波动并最终显著下降。研究表明,绝经后女性患便秘和IBS的症状严重程度显著高于绝经前女性。其机制主要包括:
1.胃肠动力减弱:雌激素受体(ERα、ERβ)及G蛋白偶联受体(GPER/GPR30)广泛存在于肠神经系统和肌间神经丛。雌激素通过一氧化氮(NO)通路调节平滑肌收缩与舒张;其水平下降会减慢胃肠蠕动、延迟胃排空并延长结肠传输时间,从而引发便秘和腹胀。
2.肠道微生态失衡:肠道菌群与雌激素相互调节。肠内“雌激素代谢菌群”通过分泌β-葡萄糖醛酸酶,将结合态雌激素转为游离态,促进肠肝循环重吸收。围绝经期雌激素下降会降低菌群多样性,减少有益菌、增加有害菌,既降低雌激素利用率,又增强发酵产气,加重腹胀。
3.内脏敏感性增加:雌激素对中枢神经系统的痛觉调节具有保护作用;其缺乏会降低内脏痛觉阈值,使正常肠气或轻度扩张被放大为明显的腹胀和腹痛。
▸ 围产期女性的“双重压迫”
在怀孕期间及产后(围产期),女性同样极易发生便秘和腹胀,患病率可高达40%-52%。其原因包括:
1.激素风暴:孕期孕激素显著升高,其平滑肌松弛作用虽可防止子宫过早收缩,但也降低胃肠道平滑肌张力、减弱肠蠕动,使食物残渣滞留时间延长、水分过度吸收,从而导致粪便干结和气体滞留。
2.机械性压迫:随着胎儿的生长,不断增大的子宫会直接压迫直肠和乙状结肠,形成物理性梗阻,阻碍粪便和气体的排出。
3.产后盆底肌受损:分娩(尤其阴道分娩)可损伤盆底肌及阴部神经,导致排便协同失调,排便时盆底肌反而收缩,进而引起排便困难和继发性腹胀。
1
饮食不当(最常见)
饮食不当是腹胀的首要原因,占比超过60%。具体常见情况主要有以下几种:
▸ 吃太多容易产气的食物
豆类、洋葱、大蒜、韭菜、萝卜、卷心菜、甘蓝这些食物含有较多不易消化的碳水化合物。它们进入结肠后被细菌发酵产生大量气体,易引发腹胀。
许多人饮奶后腹胀,多为乳糖不耐受所致,中国人约70%存在不同程度乳糖不耐受。
▸ 吃饭太快
吃饭速度快会吞入大量空气。这些空气进入胃肠道,加重腹胀感。边吃边说话更容易吞入空气。
▸ 吃太多高脂高糖食物
高脂肪高糖食物减慢胃排空速度,食物在胃内停留时间延长,发酵产气增加。
可能导致腹胀发生的因素
编辑
2
功能性消化不良
功能性消化不良占腹胀原因的15%~20%。这种病无明显胃肠结构异常,主要表现为餐后饱胀、早饱和上腹胀痛。
其发病与胃动力异常及内脏高敏感有关,约半数患者合并幽门螺杆菌感染,根除后部分症状可缓解。
3
本身便秘
便秘患者粪便长时间积聚在结肠内,细菌发酵粪便产生大量气体。便秘本身就会引发严重腹胀。很多便秘患者第一主诉就是腹胀。
4
肠易激综合征
肠易激综合征是一种常见功能性肠病。腹胀是肠易激综合征便秘型最常见症状,大约80%患者有不同程度腹胀。腹胀症状在排便后通常会减轻。
肠易激综合征中的微生物群改变
编辑
Almonajjed MB,et al.Medicina (Kaunas).2025
5
肠道菌群失调
抗生素使用、饮食改变和年龄增长可改变肠道菌群组成;产气菌过度生长会增加气体生成,导致持续腹胀。
▸ 肠道菌群失调会加重腹胀和消化功能障碍
肠道菌群失调还意味着微生物群落失衡。当产气细菌数量超过其他细菌,或有益菌数量被耗尽时,发酵平衡会倾斜。
肠道微生物群的失调会导致中枢和消化生理过程的改变,包括运动性和免疫功能,这些都会加重腹胀和消化功能障碍。
▸ 腹胀可能与菌群失调相关的迹象:
6
胃酸过低(低盐酸)
这里小编要特别强调,胃酸不足是传统医学中常被忽视的根本原因之一。胃酸(HCl)对蛋白质分解、食物杀菌、启动消化级联反应及维持酶活性所需酸性环境至关重要;其不足时,食物尤其蛋白质在进入小肠前难以充分消化。
▸ 低胃酸可致小肠细菌过度生长,从而引起腹胀
文献综述显示,胃酸不足时未消化食物在胃肠道发酵,可能诱发小肠细菌过度生长(SIBO),并促进幽门螺杆菌等进一步增殖。
SIBO与腹胀、吸收不良和腹泻有关,直接与胃酸减少及随后的上小肠菌群失衡相关。关于低盐碱和细菌过度生长的研究发现,在低酸条件下,小肠中的细菌过度生长会导致本应用于重要代谢功能的蛋白质分解,从而产生消化和系统下游的影响。
▸ 腹胀可能与胃酸过低有关的迹象:
7
肠道蠕动缓慢
肠道内存在“清洁运转周期”,对腹胀的发生至关重要。该专业名词为迁移运动复合体(MMC):MMC 是空腹期周期性反复出现的肠蠕动模式,可在餐间“清扫”残留食物残渣、细菌和其他残留物。
缺乏正常 MMC 活性与胃轻瘫、肠道假性阻塞及小肠细菌过度生长有关,从而引起显著腹胀。研究显示,慢传输型便秘患者常出现 MMC 异常(如缺失或延长周期,以及关键阶段蠕动减退)。MMC 受损时,食物停滞并发酵,平时会被清除的菌群得以定植和繁殖,进而导致腹胀、便秘和 SIBO。
▸ 腹胀可能与肠道蠕动缓慢有关的迹象:
8
小肠细菌过度生长(SIBO)
小肠细菌会分解碳水并生成气体及短链脂肪酸;当细菌过多并“吞食”时,气体与其他副产物就会显著增多。它们还会消耗蛋白质和维生素B12,以及脂肪消化所需的胆汁盐。
因此,小肠细菌过度生长(SIBO)导致的腹胀往往特别令人困惑:常见的“健康食物”反而会诱发发作。益生元纤维、洋葱、大蒜、苹果、豆类和发酵食品会为细菌提供养分;若这些细菌在小肠而非结肠内大量繁殖,就会在不合适的时间、地点与剂量下发生发酵,从而引起症状。
SIBO 往往并非独立疾病,而是多因素共同导致:胃酸受损使细菌得以在小肠存活,MMC 功能障碍无法有效清除,并叠加结构性问题形成“藏匿口袋”。
▸ 腹胀可能与 SIBO 相关的迹象:
9
消化酶不足
消化酶是分解食物的关键:淀粉酶处理碳水,蛋白酶处理蛋白质,脂肪酶负责脂肪。若酶分泌不足,食物会以较大且未充分消化的形式进入肠道,随后被细菌分解,从而引发腹胀。
▸ 最常见的是外分泌胰酶缺乏,导致脂肪吸收差
临床上最公认的情况是外分泌胰功能不全(EPI)。EPI的核心特征是外分泌胰酶缺乏,而脂肪吸收不良是最典型表现,常伴腹部不适和腹胀。
不过,腹胀不一定要达到“明显缺乏酶”的程度。亚临床功能不足(常与胃酸过低有关,因胃酸参与触发酶释放的信号级联反应)、压力或衰老,也可能导致碳水与脂肪分解不良。
▸ 腹胀可能与消化酶不足相关的迹象:
10
慢性压力与神经系统失调
这一点常被低估:若你曾在恐惧或压力时感到“胃部下沉”,甚至出现消化仿佛完全停止的体验,你可能已亲身感受到肠-脑轴。
▸ 压力或心情不好会影响食物消化
研究表明,压力会抑制迷走神经,进而损害胃肠道及微生物群,并参与肠易激综合征、炎症性肠病等胃肠疾病的发生发展。
肠易激综合征病理生理学要素
编辑
doi: 10.3390/jcm12072558.
迷走神经是肠道与大脑之间的重要通路。处于交感神经优势时,消化会被“忽略”:胃酸减少、酶分泌下降、肠蠕动减缓,且 MMC 节律受影响。清单中的这些环节都可能因长期且未缓解的压力而加重。
此外,迷走神经对肠道功能具有关键的中枢调控作用;压力可通过该通路直接影响消化动力、分泌与免疫功能。
▸ 腹胀可能与压力相关的迹象:
11
激素波动
这一原因仅限于女性,在功能和传统环境中都被严重低估。
▸ 月经周期可能会影响女性消化和吸收
一项关于孕酮与胃肠蠕动的综述指出:孕酮会减慢胃肠蠕动,并以“剂量依赖”方式影响胃排空,从而促成许多女性在黄体期出现消化迟缓和腹胀。
在月经周期中,卵巢激素的波动会调节胃肠道的收缩力、运输、分泌、内脏敏感性和免疫功能;月经期间雌激素水平下降可能尤其增加腹痛、改变排便习惯并引发腹胀。
此外,研究显示肠道通透性与月经周期存在关联:黄体期孕酮相关的变化可能影响肠道通透性、细菌易位以及乳房肿胀、腹胀等生理症状的严重程度。
▸ 临床实际情况:
▸ 腹胀可能与激素有关的迹象:
12
胃肠道器质性疾病
▸ 慢性胃炎、胃下垂
慢性胃炎患者胃动力减弱,食物排空减慢,容易腹胀。
胃下垂患者胃排空能力明显下降,站立后腹胀加重。
▸ 幽门梗阻
幽门是胃通向十二指肠的出口。溃疡或肿瘤造成幽门梗阻,食物和气体无法顺利通过,会引发上腹部严重腹胀。
▸ 肠梗阻
肿瘤、粪块、粘连性肠梗阻都能阻挡肠道内容物通过。梗阻近端肠道积气积液,腹胀非常明显,同时伴随停止排气排便。
▸ 腹腔肿瘤
卵巢癌、结肠癌、胃癌增长到一定体积,占据腹腔空间,产生持续腹胀。
13
全身性疾病
▸ 糖尿病
糖尿病患者常合并胃肠自主神经病变,胃排空减慢,肠蠕动减慢,引发腹胀便秘。
▸ 甲状腺功能减退
甲状腺激素分泌不足,全身代谢减慢,胃肠道蠕动也减慢,容易腹胀便秘。
▸ 慢性肾功能不全
尿毒症毒素影响胃肠道平滑肌功能,引发腹胀食欲下降。
▸ 心力衰竭
右心衰竭患者腹腔脏器淤血,胃肠道水肿,蠕动减慢,易出现腹胀食欲减退。
1
饮食结构不合理(最常见)
当代饮食越来越精细,膳食纤维普遍摄入不足。纤维能增加粪便体积、促进肠道蠕动;纤维不足则粪便量小、对肠道刺激不够,因而更容易便秘。
中国居民膳食纤维推荐摄入量是每天25~30克。实际调查显示,国人平均摄入量只有10~15克,远达不到推荐标准。
▸ 具体饮食相关因素:
– 吃太多精加工米面,全谷物摄入太少;
– 蔬菜水果吃得不够;
– 喝水太少,粪便水分被过度吸收,变得干硬;
– 吃太多肉类,膳食纤维严重不足。
可能导致便秘发生的因素
编辑
2
不良生活习惯
▸ 缺乏运动
长期久坐不动,肠道蠕动减慢。很多办公室工作者每天坐着超过8小时,便秘发生率明显高于体力劳动者。
▸ 排便习惯不良
很多人因为工作紧张,有便意时憋着不排便。长期反复这样,直肠对粪便刺激敏感性降低,便意减退,引发便秘。
▸ 如厕习惯不好
许多人上厕所玩手机,排便时间延长,容易诱发盆底肌功能紊乱,加重便秘。
▸ 长期精神紧张压力大
压力会通过脑肠轴影响肠道蠕动,进而影响便秘。
3
年龄增长
随着年龄增长,胃肠道蠕动减弱,腹肌与盆底肌力量下降,排便动力随之减弱,因此老年人便秘的发生率明显高于年轻人。60岁以上老年人的便秘发生率约为年轻人的2–3倍。
4
药物副作用
很多常用药物会引起便秘,这是临床便秘重要原因之一。
▸ 常见引起便秘的药物:
5
肠道菌群紊乱
越来越多研究证实,肠道菌群失调在便秘发病中起到重要作用。具体哪些细菌和便秘有关,我们后面专门详细说。
肠道微生物群可能影响便秘的发病机制
编辑
doi: 10.1093/gastro/goab035.
6
功能性疾病
▸ 肠易激综合征便秘型
肠易激综合征便秘型,除了便秘还伴随腹痛腹胀,排便后腹痛腹胀减轻。
▸ 结直肠肿瘤
肿瘤长大堵塞肠腔,造成排便困难。老年人突然发生便秘,一定要首先排除肿瘤可能。
▸ 直肠前突/盆底肌功能障碍
这是女性出口梗阻型便秘常见原因,多次生育女性更容易发生。
▸ 糖尿病
糖尿病神经病变影响肠道蠕动,很多糖尿病患者合并便秘。
▸ 甲状腺功能减退
甲状腺激素不足,全身代谢减慢,肠道蠕动也减慢。
▸ 神经系统疾病
中风、帕金森病、多发性硬化都能影响神经控制,引发便秘。
1
内脏高敏与腹膈肌协同失调
功能性腹胀的发生机制极其复杂,目前认为主要由以下几个核心环节构成:
▸ 内脏高敏感性
首先是内脏高敏感性。这是肠易激综合征(IBS) 和功能性腹胀的关键机制:患者的肠道会对正常生理刺激(如少量气体、食物残渣或正常蠕动)产生过度敏感。
其原因可能包括肠道黏膜的低度炎症、肠道通透性增加(“肠漏”),使潜在有害物质刺激黏膜下神经丛;信号经脊髓传入大脑后,被大脑皮层(如前扣带回、岛叶)错误地放大解读为胀痛感。
▸ 腹膈肌协同失调
其次是腹膈肌协同失调,它解释了为什么腹胀患者的腹围会明显增大。
健康情况下,肠道内容物增多(气体或食物)会触发协调反射:膈肌向上松弛、腹壁肌肉收缩,从而增加腹腔容积,避免腹部向前突出。
但在腹胀患者中,该反射出现异常:进食或肠腔轻度扩张时,膈肌反而出现矛盾性收缩(向下移动),同时前腹壁肌肉异常松弛。这种内脏-躯体反射失调会使腹腔内容物被向下、向前挤压,从而造成可见的腹部膨隆。
2
腹胀与便秘同行的疾病机理与致病因素
腹胀与便秘常常是“难兄难弟”,两者同发的概率很高。在慢传输型便秘和便秘型肠易激综合征(IBS-C)中,二者病理机制相互促进,形成恶性循环。
▸ 粪便滞留与过度发酵
当结肠蠕动减弱(慢传输)或排便出口受阻(排便障碍)时,粪便在肠内停留时间变长。水分被过度吸收使粪便干硬;更关键的是,滞留的食物残渣为肠道菌群提供底物,促使细菌长时间发酵,产生大量气体(如氢气、甲烷、二氧化碳)。
▸ 肠腔扩张与动力抑制
气体增多与粪便堆积会造成肠腔扩张。一方面直接引起腹部胀满;另一方面可触发抑制性反射,进一步减弱肠蠕动,加重便秘。
此外,部分气体(如甲烷)还可能具有神经肌肉毒性,直接抑制肠道平滑肌收缩,减慢肠道传输。
3
肠道激素与神经递质在胃肠动力中的作用
胃肠道不仅负责消化,也被认为是人体最大的内分泌器官。肠道激素与神经递质共同调控胃肠动力、感觉和分泌,起到“指挥官”的作用。
肠道微生物群对神经递质的调节
编辑
•血清素(5-羟色胺):人体约95%的血清素(5-HT)存在于胃肠道,主要由肠嗜铬细胞(EC细胞)合成并释放。5-HT是“脑-肠轴”连接的重要关键分子:当食物或气体刺激肠壁时,EC细胞释放5-HT,作用于肠神经系统的5-HT3与5-HT4受体。
5-HT4受体激活可促进乙酰胆碱释放,增强肠蠕动与分泌;5-HT3受体则参与疼痛信号向中枢传递。便秘患者常见5-HT通路异常(如合成减少、受体表达下调),导致肠道动力不足;而IBS患者则多见异常释放,引发内脏高敏与腹痛。
•胃动素:由小肠M细胞分泌,主要在空腹(消化间期)释放;它激活MMC的第III相,也就是“肠道清道夫”运动,将未消化的食物残渣和细菌推向大肠。若胃动素分泌不足或受体不敏感,便会导致胃排空延迟和小肠细菌过度生长(SIBO),进而引起腹胀。
•胆囊收缩素:主要由小肠I细胞分泌。进食脂肪和蛋白质后,CCK释放增多,促进胆囊收缩并刺激胰酶分泌;同时它显著抑制胃排空、减慢结肠传输。IBS患者可能对CCK更敏感,从而出现进食后夸张的动力抑制与腹胀。
•多巴胺:尽管多巴胺常被视为中枢“快乐递质”,但肠道同样可合成大量多巴胺。肠道多巴胺受体(如D2受体)被激活时,通常会抑制胃肠动力。因此,帕金森病患者往往伴随顽固便秘和腹胀(中枢多巴胺不足,同时外周受体敏感性可能发生改变)。
4
肠道产气机制及其对症状的放大效应
正常成年人每天肠道排气量约为500-1500毫升。肠道气体主要来源于吞咽的空气和肠道细菌的发酵。未被小肠完全吸收的碳水化合物(如膳食纤维、低聚糖、抗性淀粉)进入结肠后,被肠道菌群发酵,产生氢气(H2)、二氧化碳(CO2)以及微量的甲烷(CH4)和硫化氢(H2S)。
编辑
不同菌群代谢产生的气体对肠道功能影响迥异。例如,甲烷(CH4)不仅是一种温室气体,在人体内它被证明是一种神经递质调节剂。
产甲烷菌(如史氏甲烷短杆菌Methanobrevibacter smithii)消耗氢气生成甲烷。高浓度的甲烷能够直接抑制回肠和结肠的平滑肌收缩,显著延长肠道传输时间。临床研究证实,呼气试验中甲烷阳性的患者,其便秘的严重程度和腹胀频率远高于阴性患者。此外,硫化氢(H2S)虽然量少,但具有高度毒性,不仅产生恶臭,还可能损害肠道上皮细胞的能量代谢,加剧内脏高敏。
近年来研究已证实:肠道菌群在便秘与腹胀的发生发展中起关键作用。
便秘患者粪便菌群α多样性通常降低,厚壁菌门/拟杆菌门比例升高。
1
产气菌过度生长易导致腹胀
产气菌过度生长会使肠道内产生的气体增多,从而引发腹胀等不适症状。
–Klebsiella pneumoniae(肺炎克雷伯菌)
克雷伯菌过度生长会产生大量气体,引发明显腹胀。很多小肠细菌过度生长患者,克雷伯菌丰度显著升高。
–Streptococcus属某些物种
链球菌发酵碳水化合物产生大量气体,增加腹胀风险。
-Enterobacteriaceae科整体
肠杆菌科多数细菌发酵碳水化合物产气,丰度升高后气体产生增加。
人类肠道微生物组中主要的产气微生物
编辑
doi.org/10.1016/j.jff.2022.105367
2
为什么产气菌多会腹胀便秘?
产气菌过度增殖会消耗可发酵碳水化合物,生成大量氢气、二氧化碳和甲烷;气体积聚引起腹胀。与此同时,甲烷可减慢肠蠕动、延长结肠传输时间,从而加重便秘,因此不少便秘患者常同时出现腹胀。
细菌发酵中H2、H2S、CH4和CO2的生化途径
编辑
doi.org/10.1016/j.jff.2022.105367
3
哪些细菌增多易导致腹胀?
–Desulfovibrio物种
硫酸盐还原菌代谢产生硫化氢,高浓度硫化氢对肠道平滑肌有毒性作用,抑制蠕动。研究发现,便秘患者硫酸盐还原菌丰度高于健康人。
-黏液降解菌过度生长
Akkermansia muciniphila(嗜黏蛋白阿克曼菌)专门降解肠道黏液层。适量阿克曼菌对健康有益,过度生长会破坏黏液屏障,引发肠道炎症,影响蠕动功能。
微生物群和肠道屏障完整性
编辑
4
哪些细菌减少易导致便秘腹胀?
-产丁酸菌减少
丁酸盐是结肠上皮细胞主要能量来源,同时能够促进肠道蠕动,维持肠道正常节律。
主要产丁酸细菌包括:Faecalibacterium prausnitzii(普拉梭菌)、Roseburia属物种(如Roseburia inulinivorans)、Eubacterium rectale(直肠真杆菌)、Butyricicoccus pullicaecorum。
这些细菌丰度降低,丁酸产生减少,结肠上皮能量不足,蠕动功能减弱,容易便秘。
–Bifidobacterium(双歧杆菌)减少
双歧杆菌能够调节肠道蠕动,降低肠道pH值,促进有益菌生长。很多研究证实,便秘患者双歧杆菌丰度显著低于健康人。
–Lactobacillus(乳杆菌)某些物种减少
Lactobacillus reuteri(罗伊氏乳杆菌)能够改善肠道动力,减少炎症反应。补充罗伊氏乳杆菌能够缩短结肠传输时间。干酪乳杆菌也能调节肠道运动功能,增加排便频率。
近年来多个大型队列研究证实了肠道菌群和便秘腹胀的关系。随机双盲安慰剂对照试验证实:
补充 Bifidobacterium lactis能够增加每周排便频率,改善粪便性状;
补充Lactobacillus reuteri 缩短结肠传输时间15-20%;
益生元菊粉能够增加粪便中双歧杆菌数量,增加排便频率。
212种肠道微生物群与便秘的因果关联
编辑
doi: 10.3389/fmicb.2024.1438778.
5
针对特定细菌的干预方法
根据肠道菌群检测结果,不同菌群异常采用不同干预策略。
▸ 产气菌过度生长(克雷伯菌、肠杆菌科丰度升高)
饮食干预—减少精制碳水化合物摄入:产气菌喜欢利用容易发酵的简单糖产生气体。减少精制糖就是减少底物。
增加膳食纤维,特别是不可发酵纤维:不可发酵纤维能够促进肠蠕动,减少细菌过度生长。不可发酵纤维主要存在于全谷物外壳、蔬菜的纤维素部分。
低FODMAP饮食:低FODMAP饮食减少可发酵寡糖和单糖,减少气体产生。对小肠细菌过度生长引发的腹胀效果很好。
需要注意,低FODMAP饮食不建议长期使用,一般使用4-6周,症状缓解后需要逐步恢复高纤维饮食,避免长期低纤维饮食对菌群造成不良影响。
益生菌:补充产丁酸益生菌可通过竞争占位抑制产气菌生长;如布拉氏酵母菌、嗜酸乳杆菌等还能减少有害菌过度繁殖,改善肠道菌群环境。
抗生素:确诊为小肠细菌过度生长后,可在医生指导下短期使用抗生素(如利福昔明),通常疗程约1–2周;不建议自行用药,也不推荐长期使用抗生素。
▸ 产丁酸菌减少
增加膳食纤维摄入:尤其是菊粉、低聚果糖等益生元,可选择性促进产丁酸菌生长。常见富含益生元的食物包括:菊苣、芦笋、洋葱、大蒜、香蕉、燕麦。
多吃全谷物:全谷物因膳食纤维更丰富,可促进产丁酸菌生长;抗性淀粉同样能促进其增殖,例如冷却后的米饭和青香蕉都富含抗性淀粉。
直接补充:可以补充含有产丁酸菌的益生菌产品;或者补充丁酸栓剂或丁酸衍生物,但是注意直肠局部生产的丁酸效果更好。
▸ 双歧杆菌、乳杆菌减少
多吃含有低聚糖的食物:比如大豆、洋葱,能够促进双歧杆菌生长。
发酵乳制品:比如酸奶,含有活的乳酸菌,能够直接补充,注意选择无糖原味酸奶,避免额外添加过多糖分
直接补充:直接口服含有双歧杆菌或乳杆菌的益生菌制剂;选择明确标注菌株的产品,不同菌株功能不同。比如罗伊氏乳杆菌DSM17938明确研究证实对便秘有效,购买时可以关注菌株编号。
▸ Parabacteroides distasonis 低丰度
目前没有专门针对性食物。研究发现,蘑菇多糖能够促进P. distasonis 生长。克罗恩病患者食用蘑菇提取物后,P. distasonis 丰度升高,症状改善。可以尝试增加蘑菇摄入,每天吃100-200克新鲜蘑菇,有助于促进生长。
胃肠科医生在评估腹胀时通常会先进行细致而全面的检查:明确症状特点,识别风险,找出最可能引发不适的机制。尽管不同医生的具体方案会有所不同,但常见流程可能包括:
⑴ 询问临床病史
起效时间、发作频率及具体时段(如饭后、早晚),并评估可能诱因。需记录饮食习惯、纤维摄入、碳酸饮料,以及乳糖或果糖的暴露情况,和近期抗生素使用史。
抗生素使用可能产生的关联
编辑
同时梳理排便习惯(便秘、腹泻、腹痛急迫、排便不尽)及伴随症状(烧心、恶心、打嗝、疼痛),是否受月经周期影响,体重变化、疲劳与压力。
药物和补充剂使用情况(如二甲双胍、阿卡博糖、GLP-1 受体激动剂、阿片类药物、制酸剂、益生菌)。
⑵ 体格检查
进行腹部体检,观察是否存在压痛、腹胀、肠鸣音异常,以及器官肥大或肿块;同时评估是否有液体积聚(腹水)或疝气迹象。有时还会进行直肠检查,以了解粪便负担、张力及盆底相关问题。
⑶ 腹部不适的诊断与风险评估
症状出现的年龄、家族史(乳糜泻、炎症性肠病、结直肠癌)、旅行史,以及近期感染或手术史(如胆囊切除术后可能出现胆汁酸相关腹泻)。
⑷ 初步测试(如有指示)
基础化验常包括:全血细胞计数、电解质检测、炎症标志物(如C反应蛋白)、肠道菌群检测、甲状腺功能及乳糜泻相关血清学。
必要时可做粪便检测,如粪便钙卫蛋白用于评估炎症,或粪便病原检测以排除感染。呼气测试可评估乳糖不耐受或果糖吸收不良;而针对小肠细菌过度生长(SIBO),有时会采用乳果糖或葡萄糖呼气测试。
⑸ 询问临床病史
若出现警报症状或检查结果需要进一步评估,可能会进行上消化道内镜(EGD)、结肠镜、腹部超声或其他影像学检查,以排查结构性或炎症性疾病;怀疑胃轻瘫时,可考虑做胃排空检查。
通过整合这些数据点,胃肠科医生会制定计划,可能包括饮食调整、肠道习惯优化、微生物调节策略或治疗特定疾病的方案。重要的是,这很少是一个放之四海而皆准的流程。相反,它是迭代的,并由你个人的反应随时间引导。
!
重点:为什么这个诊疗对肠道健康很重要
腹胀不仅仅是烦恼,它常提示消化系统失衡。对部分人而言,可能是快速发酵的碳水化合物刺激了敏感肠道;对另一些人,缓慢排便与便秘会导致气体积聚;还有人则可能与微生物失衡或炎症有关。
由于肠道与免疫、新陈代谢、神经系统和微生物群紧密相连,理解腹胀有助于打开更广泛的消化健康视角。
积极处理持续腹胀可缓解不适,提升生活质量、精力与社交功能,也能避免反复试验带来的“走弯路”,减少误诊或延误评估的风险,防止症状在无意中加重。
若腹胀伴随非自愿体重减轻、便血、持续呕吐、吞咽困难、发热、贫血,或50岁后新出现的症状等警示信号,应尽早重视并进行及时、详细的评估。
▸ 腹胀并非单独出现,常见伴随症状包括:
长期且失控的腹胀会显著降低生活质量,并因饮食受限导致营养不良,进一步加剧肠—脑反馈循环。腹胀本身虽只是症状而非疾病,但可能提示肠易激综合征、功能性腹胀/膨胀、小肠细菌过度生长(SIBO)、乳糜泻、碳水化合物不耐受、胆汁酸吸收不良,或在特定情况下提示更严重的潜在病理。因此,当症状持续时,进行结构化评估并咨询胃肠专科尤为重要。
1
个体对腹胀原因的差异与不确定性
▸ 症状相似但可能由个性化的不同机制引起
每个人的消化系统都不完全相同。遗传、早期暴露、饮食、压力、激素、药物以及微生物群都会影响肠道对日常食物的反应:同样一顿饭,有人舒服,有人却胀气。
即使症状看起来相似,也可能由不同机制引起。例如,夜间腹胀的两个人可能都表现出相近外观:其中一人可能结肠排出缓慢、以产甲烷的菌群为主;另一人则可能主要由压力和快速发酵的碳水化合物导致内脏过敏。
2
仅根据症状本身难以确定根本原因
▸ 症状相似但可能由于不同病因
腹部腹胀表面上可能很相似:无论成因是气体过度产生、气体处理能力受损、肌肉协调障碍、炎症、酶缺乏还是微生物群失衡,都会出现类似表现。
肠易激综合征、功能性消化不良、乳糖不耐受和小肠细菌过度生长(SIBO)等多种疾病中也常有重叠症状。缺少具体背景时,同样的症状可能对应不同病因,甚至导致误诊。
案例一:许多人误以为所有腹胀都源于豆类和十字花科蔬菜,因此完全回避。但真正的主要诱因往往是严重便秘。相较于减少健康食物,改善排便频率与规律更能缓解腹胀。
案例二:有人会把腹胀归咎于麸质。检测已排除乳糜泻;反而,果聚糖和多元醇可能是个体的主要触发因素。通过结构化的重新引入,可更清楚界定耐受水平,从而保留更多饮食多样性。
案例三:有人在使用抗生素后反复腹胀与腹泻,怀疑乳糖不耐受;但呼气测试及症状出现时间提示,更可能是感染后IBS,且可能伴随SIBO。治疗重点应放在促进胃肠动力与调整微生物平衡。
这些例子说明了清晰诊断框架的价值。它减少了不必要的限制,将精力集中在最重要的地方,并为个性化策略打开了大门——尤其是在结合测试和细致跟进的客观见解时。
鉴于腹胀与便秘发病机制复杂,单一治疗常难以见效。现代医学通常建议采用多层次、多靶点的综合干预,包括饮食调整、微生态治疗、生活方式干预以及中西医结合治疗。
1
饮食干预:低FODMAP饮食与膳食纤维的精准应用
▸ 低FODMAP饮食法是针对腹胀的有效干预
对伴有明显腹胀的 IBS 患者而言,低 FODMAP 饮食是目前国际公认的一线干预方案。FODMAP 指“可发酵的寡糖、二糖、单糖和多元醇”。这类短链碳水在小肠中难以充分吸收,进入结肠后易被细菌快速发酵产气,同时具高渗透性,把水分吸入肠腔,从而引起腹胀并加重腹泻。
低FODMAP饮食分为三个阶段:
1.排除期(2-6周):严格去除所有高FODMAP食物(如小麦、洋葱、大蒜、苹果、西瓜、牛奶、豆类及人工甜味剂),以迅速缓解症状并建立消化基线。
2.重新引入期(6-8周):症状控制后,对各类FODMAP食物进行逐一、少量重引,评估耐受情况,明确具体“触发食物”。
3.个性化维持期:依据测试结果制定长期饮食方案,仅避免已证实不耐受的食物,以兼顾营养均衡与肠道菌群多样性。
▸ 膳食纤维的精准补充
膳食纤维是预防和治疗便秘的关键,但若患者同时伴有腹胀,补充需格外谨慎。不可溶性纤维(如麦麸)虽能增加粪便体积,却可能带来较强摩擦且发酵快,容易加重腹胀;因此更推荐可溶性、非黏性或低发酵性纤维。
例如,猕猴桃(每天2个)已在多项临床试验中证实可显著提高排便频率、软化粪便,并因其特异性酶(如猕猴桃碱)和低发酵特性,通常不易引起腹胀,患者满意度高。
西梅(每天100g)富含山梨糖醇和多酚,具有天然的渗透性通便作用;此外,部分水解瓜尔豆胶(PHGG)和洋车前子壳粉也属优选纤维补充剂,可形成凝胶润滑肠道,产气量较低。
2
微生态调节:益生菌与益生元的循证应用
纠正肠道微生态失衡是治本之策。微生态制剂(益生菌、益生元、合生元)通过竞争性抑制有害菌、修复肠黏膜屏障、调节免疫和神经递质分泌,有效改善腹胀和便秘。
▸ 精准益生菌株的选择
益生菌的效果具有显著的菌株特异性。对IBS相关腹胀与腹痛,长双歧杆菌35624®(Bifidobacterium longum 35624)和格氏乳杆菌LA806循证证实可显著降低IBS严重程度评分(IBS-SSS),缓解内脏高敏。
而对慢性便秘,乳双歧杆菌HN019和动物双歧杆菌BB-12可缩短肠道传输时间并提高排便频率。
▸ 益生元协同作用
益生元(如低聚果糖、菊粉)是益生菌的“食物”。对便秘患者而言,合生元(益生菌+益生元)的联合使用常可产生“1+1>2”的效果。需注意的是,部分益生元含FODMAP,摄入过量可能在早期加重腹胀,应从小剂量开始,逐步增加。
3
生活方式与心理行为干预
生活方式的调整与良好的精神状态,不仅是治疗方案的起点,也是所有治疗能够有效开展的基础。
▸ 日常保持一定的运动
避免久坐:每坐1小时起身活动2-3分钟,走动并做轻度伸展即可促进肠蠕动。
规律运动有助于改善便秘。每周约150分钟的中等强度有氧运动(如快走、慢跑)可机械性刺激胃肠蠕动、缩短结肠传输时间。太极拳与瑜伽等特定运动(如膝胸动作)配合腹式呼吸与腹部内脏按摩,既能促进排气,也有助于调节自主神经、降低交感兴奋,缓解压力。
推荐运动项目:
-快走或慢跑:每天30分钟,促进全身血液循环,增加肠蠕动;
-瑜伽:很多扭转体式能够按摩腹部肠道,促进蠕动;
-游泳:全身运动,促进代谢,改善肠道功能;
-腹肌锻炼:适当锻炼腹肌,增加排便动力,老年人特别需要注意腹肌锻炼。
▸ 压力管理
长期压力增大,通过脑肠轴抑制肠道蠕动。你可以尝试:
-每天10-15分钟深呼吸冥想;
– 规律作息,保证每天7-8小时睡眠;
– 找到适合自己的放松方式,比如听音乐、养花、散步。
▸ 环境调整
-旅行环境改变容易便秘,提前做好准备,带够膳食纤维食物,不要完全改变饮食习惯。
–长期精神紧张工作环境,试着拆分任务减少压力,定时起来活动。
▸ 心里干预
鉴于“脑-肠轴”的紧密关联,心理干预对难治性腹胀和便秘尤为关键。认知行为疗法(CBT)、肠道导向催眠疗法和正念冥想可降低焦虑与抑郁水平,纠正大脑对内脏痛觉信号的异常处理,从而从根本上缓解症状。
4
中医药干预:中草药、针灸与内外同治
中医药在治疗功能性胃肠病方面具有独特的“整体观念”和“辨证论治”优势,强调多靶点、双向调节。
▸ 中草药的独特机制
大黄:大黄是治疗便秘的中医经典药。现代研究认为其具有“双向调节”机制:蒽醌类成分(如番泻苷)在肠道菌群作用下转化为大黄酸,可刺激肠神经丛,促进肠液分泌与蠕动,从而通便泻下;而其中的鞣质类成分具收敛作用,可抑制肠道水通道蛋白(AQPs)表达,起到止泻作用。凭借这种动态平衡,大黄在腹泻-便秘交替型IBS的治疗中更具优势。
生姜与薄荷油:生姜含姜辣素和姜烯酚,具有促胃动力、抗炎与抗氧化作用,可加速胃排空,缓解上腹胀和恶心。薄荷油(主要成分薄荷醇)可作为天然的平滑肌钙离子通道阻滞剂,解除肠道痉挛,缓解IBS患者的腹痛和腹胀,因而被多国指南推荐为一线用药。
经典方剂:如白术、白芍、防风、陈皮用于肝郁脾虚型IBS,能抑肝扶脾、解痉止痛;木香、枳壳、乌药则能行气导滞,显著改善便秘型IBS患者的腹胀和排便困难。
▸ 中医外治法:针灸与穴位贴敷
针灸刺激特定穴位(如天枢、足三里、大肠俞、上巨虚),可调节自主神经平衡,促使胃肠激素(如胃动素、VIP)分泌趋于正常,实现肠道动力的双向调节。
穴位贴敷(如用生姜泥或健脾温肾中药贴敷神阙穴/肚脐)则借助药物透皮吸收与经络传导直达病所。临床研究显示,生姜热敷脐部可有效预防并缓解卧床患者(如急性心肌梗死者)的腹胀与便秘,且无全身性副作用。
1
微生物组检测在腹胀便秘背景下能揭示的信息
对于频繁或持续腹胀的人群,粪便微生物组检测报告可提供如下参考:
• 微生物多样性: 慢性胃肠症状患者有时会观察到较低的多样性,但这并非腹胀特有。这可以指导逐渐谨慎地重新引入不同纤维。
• 相对丰度: 甲烷相关信号升高可能与便秘为主的模式相符。某些有益类群(如双歧杆菌)含量偏低,可能反映了饮食、抗生素史或其他因素。
• 发酵潜力: 分类模式可能指示其碳水化合物发酵能力极强;如果与敏感性结合,这有助于塑造饮食试验。
• 潜在失衡的标志: 部分特征显示其易产生炎症或硫化物,因此应密切关注诱因和耐受性。
微生物组检测多以粪便样本为主,用于表征微生物多样性及细菌(有时也包括其他生物)的相对丰度。不同方法差异明显:不少检测采用16S rRNA基因测序分析细菌类群;也有部分采用散弹式宏基因组学,捕捉更广泛的微生物并推断功能通路。结果可能呈现多样性指标、特定群体潜在过度生长,以及与胀气、便秘或腹泻等症状相关的模式(往往为松散关联)。
在16S检测方面,数据库更庞大、模型构建时可纳入的因素更多,因而敏感性与稳定性通常更强。宏基因组则能识别真菌、毒力基因及部分代谢相关基因,从而更全面了解微生物并推断其功能。
需要注意的是,微生物组检测不能用于诊断SIBO或排除其他医疗状况;但当症状持续、且需要反复调整管理方案时,它可帮助进行个体化规划。
2
哪些时候可以考虑微生物组检测?
并非所有腹胀患者都需要做微生物组检测。很多人通过日常调整并在必要时进行简短的临床评估即可改善。但若符合以下情况,补充相关信息可能会更有帮助:
在这些情况下,结合临床评估与个性化肠道微生物组报告的仔细审查,有助于指导有针对性的营养选择和微生物支持策略。目标不是“治疗检测结果”,而是将数据与症状、饮食和生活方式整合成一个连贯的计划。
3
决策支持:微生物组检测何时有意义?
当信息能改变您下一步的行动时,测试最有价值。
• 持续的不确定性: 你尝试过第一线策略(比如慢吃、减少碳酸饮料、优化排便、基础碳水化合物-乳糖试验),但都没有明确的答案。
• 反应不一: 有些改变能稍微改善,但症状会反复出现;目前尚不清楚是否应该增加纤维、更换纤维类型,还是继续避免。
• 便秘为主症状: 甲烷相关模式的证据支持关注运输时间和特定微生物方法。
• 抗生素后或感染后转变: 了解多样性和相对丰度有助于重新引入阶段并支持恢复。
• 复杂的饮食或限制: 客观的洞察有助于战略性地重建饮食多样性,避免不必要的回避。
相反,如果你的腹胀是轻微的,明显与暴饮暴食或汽水有关,并且通过基本调整就能缓解,检测效果可能不会太大。同样,如果出现明显症状,医疗评估优先于任何择期检测。
4
胃肠专科就诊咨询的帮助
胃肠专家咨询将对您的症状模式与可能诱发机制进行评估和对应。典型的就诊和随访可能包括:
• 明确目标:哪些结果最重要——是减少胀气、减少不适日、更明确的诱因,还是改善肠道规律?
• 症状日记/饮食记录: 在预约前后进行简短记录有助于识别时间模式(例如,与每日进食节律或便秘相关的下午-傍晚胀痛)。
• 选择性检测:如前所述,可能会根据风险、病史和检查安排针对性的化验或微生物测试,呼气测试等。
• 管理试验:指导性饮食调整(例如,FODMAP 轻量法并重新引入)、肠道优化(补水、可溶性纤维或渗透剂治疗便秘),或在适当情况下支持肠道蠕动。
• 微生物组及辅助见解:可选的微生物组检测可用于制定下一步,尤其是在反应部分或非典型时。
• 后续与迭代: 在规定的间隔后回顾进展,根据反应调整,避免长时间不必要的限制。
这一流程强调协作与灵活调整,而不只是追逐症状。它要把机制理清:气体来源可能与肠黏膜受损有关,饮食可能导致发酵增加并影响肠道蠕动,微生物失衡则可能与炎症或结构性问题相关。明确这些因素后,腹胀治疗方案便能更有针对性地选择。
5
管理腹胀的循证策略
管理与干预通常不依赖单一因素。相反,结合符合个体生理特点与偏好的多项调整往往效果更佳。潜在组成包括:
• 饮食行为:吃得更慢;彻底咀嚼;均匀间隔的用餐;限制碳酸饮料的使用;避免过量口香糖或硬糖,以免增加吸入空气。
• 靶向饮食试验:短暂、结构化的低 FODMAP 阶段,随后逐步重新引入以确定个人耐受性;如果呼气测试或病史支持不耐受,则要求乳糖或果糖限制;在工艺早期关注糖醇(山梨醇、甘露醇)和高度可发酵的纤维。
• 纤维:如果便秘,应优先选择可溶性纤维(例如部分水解的瓜尔胶、车前子)和充足的水分摄入。高不可溶性纤维可能会加重某些人的症状。
• 粪便调节:对于慢性便秘,建议在临床医生指导下使用循证渗透剂,必要时进行盆底评估,并定期活动以支持肠道蠕动。
• 肠道-大脑支持:减压、膈肌呼吸以及认知行为或肠道导向催眠疗法可以降低内脏敏感度,改善部分患者的症状控制。
• 其他:薄荷油肠涂胶囊有一定证据可能缓解与IBS相关的不适。西甲硅油在某些情况下或可帮助减轻胀气。益生菌可作为试验性选择,但效果会因菌株和个体差异而不同;最好结合临床与微生物组背景来选择合适的菌株。
• 医疗治疗:如果怀疑并确诊 SIBO,有时会与促进肠道蠕动的同时使用靶向抗生素或草药方案;如果发现胆汁酸腹泻,胆汁酸结合剂可能有帮助;如果存在胃排空延迟,可以考虑促动力学策略。
需要强调的是,这并非“一刀切”治疗。关键在于把治疗选择与驱动你症状的机制对齐,并持续、系统地重新评估。
腹胀与便秘,看似是日常生活中微不足道的“小毛病”,实则是人体内部复杂生态系统失衡的警报。从肠道内数以万亿计的微生物,到穿梭于神经突触间的5-羟色胺;从女性生命周期中起伏的雌激素,到内脏高敏、腹膈肌协同失调,从肠道激素的紊乱,再到产甲烷菌等微生态的失衡,每一个环节都相互交织,互为因果。大脑与肠道之间无声的对话,每一个环节的微小偏差都可能演变为令人痛苦的临床症状。
面对这种复杂的疾病网络,仅靠单一的泻药或促动力药物往往只能“治标”,且长期使用可能带来依赖与副作用。未来治疗方向应走向更精准、更综合。
通过低FODMAP饮食减少发酵底物,结合特定益生菌株与益生元重塑微生态;再辅以运动与心理干预,打破脑-肠恶性循环。同时充分发挥中医药多靶点、双向调节的优势,才能真正从“缓解症状”迈向“恢复稳态”的根本转变。
随着微生物组学测序技术普及以及人工智能在医疗数据分析中的应用,我们有望实现真正的精准医疗:基于个人肠道菌群图谱、基因型与代谢特征,制定专属的益生菌组合、饮食方案和药物干预策略。
对广大受困扰的患者,尤其处于特殊生理阶段的女性而言,理解疾病机制是战胜疾病的第一步。保持良好心态,科学调整饮食,合理补充膳食纤维并选择合适的益生菌;必要时及时咨询消化科与心理科医生。通过重建肠道微生态与脑-肠轴的平衡,才能重新获得轻松顺畅的健康生活。
注:本账号内容仅供学习和交流,不构成任何形式的医疗建议。
主要参考文献
中华医学会消化病学分会胃肠动力学组,功能性胃肠病协作组,食管疾病协作组.中国慢性便秘临床诊断与治疗规范[J].中华消化杂志,2024, 44(10): 649-658. DOI:10.3760/cma.j.cn311367-20240930-00373.
Napolitano M, Fasulo E, Ungaro F, Massimino L, Sinagra E, Danese S, Mandarino FV. Gut Dysbiosis in Irritable Bowel Syndrome: A Narrative Review on Correlation with Disease Subtypes and Novel Therapeutic Implications. Microorganisms. 2023 Sep 22;11(10):2369.
Zhang S, Wang R, Li D, Zhao L, Zhu L. Role of gut microbiota in functional constipation. Gastroenterol Rep (Oxf). 2021 Aug 6;9(5):392-401.
Almonajjed MB, Wardeh M, Atlagh A, Ismaiel A, Popa SL, Rusu F, Dumitrascu DL. Impact of Microbiota on Irritable Bowel Syndrome Pathogenesis and Management: A Narrative Review. Medicina (Kaunas). 2025 Jan 13;61(1):109.
Feng C, Gao G, Wu K and Weng X (2024) Causal relationship between gut microbiota and constipation: a bidirectional Mendelian randomization study. Front. Microbiol. 15:1438778.
Shaikh SD, Sun N, Canakis A, Park WY, Weber HC. Irritable Bowel Syndrome and the Gut Microbiome: A Comprehensive Review. J Clin Med. 2023 Mar 28;12(7):2558.
Mamieva Z, Poluektova E, Svistushkin V, Sobolev V, Shifrin O, Guarner F, Ivashkin V. Antibiotics, gut microbiota, and irritable bowel syndrome: What are the relations? World J Gastroenterol. 2022 Mar 28;28(12):1204-1219.
Bercik P. The Brain-Gut-Microbiome Axis and Irritable Bowel Syndrome. Gastroenterol Hepatol (N Y). 2020 Jun;16(6):322-324.
Ohkusa T, Koido S, Nishikawa Y, Sato N. Gut Microbiota and Chronic Constipation: A Review and Update. Front Med (Lausanne). 2019 Feb 12;6:19.

谷禾健康

抗生素可有效治疗细菌感染,但其广泛使用引发了抗生素耐药性,抗生素耐药性是个全球性的大问题,这与人类疾病密切相关。这里存在着一个被我们长期忽视的关键因素:肠道耐药组。
当我们在医学上使用抗生素时,肠道里的微生物数量和基因组成可能会发生变化,这样就增加了抗生素耐药性在肠道中积累的风险。
肠道耐药组(gut resistome)是指肠道微生物群中存在的所有抗生素耐药基因(ARG)的集合,可以从自然环境和食物摄入中获得,并且可以随着食物中抗生素和其他抗菌物质诱导的选择压力进一步积累。
这些耐药基因不只是让细菌对抗生素产生抵抗力,更可能通过影响肠道微生物平衡、改变代谢功能、触发炎症反应,与多种慢性疾病建立起令人担忧的联系,包括代谢紊乱、心血管疾病、肝病、神经系统疾病等。
当我们服用口服抗生素时,药物直接穿过消化系统,破坏肠道微生物的平衡,促进抗生素耐药基因的出现和扩散。更令人忧虑的是,这些耐药基因可以通过食物链、水源和环境污染从自然环境进入人体,并在肠道微生物间通过水平基因转移快速传播。
肠道耐药组的富集不仅加剧了抗生素耐药性,还导致了多重耐药性感染的出现。这些感染构成了重大的公共卫生挑战,因为它们更难治疗,通常需要更昂贵的疗法,这些疗法可能具有更大的副作用且效果较差。
肠道耐药组已成为连接抗生素使用、耐药危机和慢性疾病的关键环节。
已知膳食成分会影响肠道微生物组的发生率、活性和多样性,例如,研究表明,地中海饮食(大量摄入完整的、未加工的植物性食物、橄榄油和乳制品;适量的家禽和鱼类;以及最少食用红肉)与粪杆菌属和短链脂肪酸的增加有关,它们具有抗炎作用。相比之下,西方饮食(高热量,富含动物蛋白、饱和脂肪、单糖和超加工食品,而纤维、水果和蔬菜含量低)与 Blautia、拟杆菌属、瘤胃球菌属物种的丰度较高有关,这与代谢紊乱和慢性炎症的风险增加有关。
在此基础上,饮食不仅会影响有益细菌,而且还有可能改变肠道中抗生素耐药细菌的存在。到目前为止,已发现食品中的生物活性常量营养素、植物化学物质和益生菌可以改变肠道中的抗生素耐药细菌,为调节肠道耐药组的功能性食品打开了新的研究窗口。
随着科学进步,肠道菌群检测技术已能够识别耐药基因的存在和丰度,使个体ARG状况的评估成为可能,为个性化的肠道耐药组管理提供客观依据。
本文强调了肠道耐药组与人类疾病(如代谢紊乱、心血管疾病、肝病和神经系统疾病)之间的联系,包括抗生素作用机制和耐药性的发展,还讨论了饮食习惯和饮食成分(包括生物活性宏量营养素、植物化学物质和益生菌)通过增强抗生素疗效和潜在降低耐药性,对肠道抗药组的组成产生影响。
强调了对靶向肠道耐药组的功能性食品的新兴趋势,以及越来越关注具有调节抗生素耐药性潜力的生物活性植物化学物质。
靶向肠道耐药组的功能性食品,正从概念走向临床,未来,通过“膳食-菌群-肠道耐药组”三位一体的精准干预,有望将耐药危机防控窗口前移至日常饮食,从功能性食品的创新到临床营养的精准化应用,为全球公共卫生提供可持续解决方案。
★
随着抗生素在现代医学和农业中的广泛应用,一个隐形的健康危机正在悄然形成——肠道抗生素耐药基因组(ARG)的扩张。
ARG 可以从亲本细菌(垂直基因转移)遗传,也可以通过水平基因转移从其他微生物获得,水平基因转移是遗传物质在细菌之间移动的过程,甚至跨物种移动。
这些耐药基因不仅可以使细菌对抗生素产生抵抗力,更可能通过影响肠道微生物平衡、改变代谢功能、触发炎症反应,与多种慢性疾病建立起令人担忧的联系。
给药方法会显著影响肠道微生物组和耐药组
静脉注射(注射到血液中)或局部(直接涂抹在皮肤上)的抗生素通常与肠道微生物组的直接接触最少,因此对肠道耐药组的影响较小。
然而,口服抗生素是最常见的治疗形式,它通过消化系统,直接影响肠道微生物群。这种相互作用会破坏肠道微生物的平衡,并促进抗生素耐药基因(ARG)的出现。
Zhang等人强调了这一差异,表明小鼠口服四环素或阿莫西林会迅速增加肠道中ARG的存在,而静脉注射则会延迟或减少这种作用。这种差异与药物的排泄途径有关:氨苄西林主要通过肾脏排泄,限制了静脉注射后的肠道暴露,而四环素通过肾脏和胆汁排出,即使在静脉注射后也会暴露肠道。这些发现强调了口服抗生素在塑造肠道抗药性中的重要作用。
此外,抗生素的广泛使用,无论是直接在人类身上还是间接在动物身上,都会导致抗生素耐药性在自然环境和生物中的积累,从而促进人类肠道耐药性的扩大。
严重的健康问题与肠道耐药组有关,包括代谢紊乱、心血管疾病、肝病、神经系统疾病(下图)。
肠道耐药组对人体健康的影响
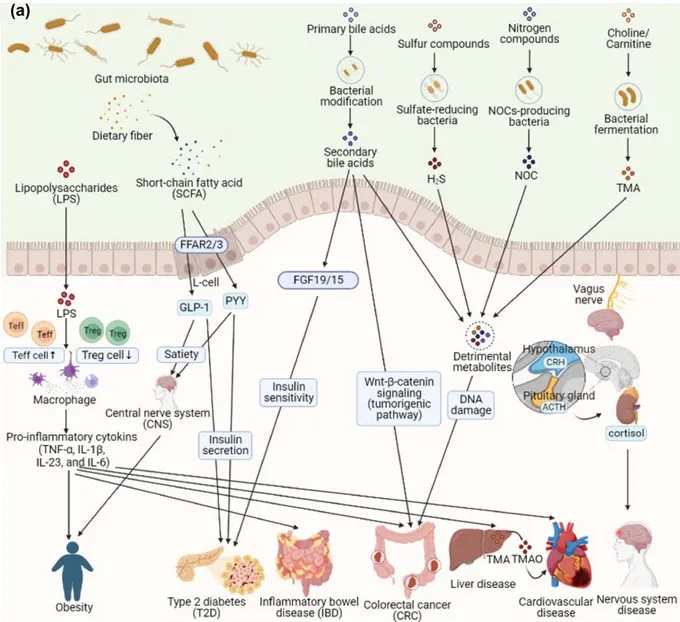
doi:10.1111/1541-4337.70143
a) 肠道微生物群在健康和疾病中的机制。短链脂肪酸(SCFA)来源于膳食游离脂肪酸受体(FFAR2/3),通过释放肽YY(PYY)和胰高血糖素样肽1(GLP-1)等肠道激素来促进饱腹感,同时增强胰岛素分泌和敏感性,导致肥胖和2型糖尿病(T2D)。来自革兰氏阴性菌的脂多糖(LPSs)通过激活巨噬细胞和释放促炎细胞因子(TNF-α、IL-1β、IL-6)触发全身炎症,导致肥胖、2型糖尿病、炎症性肠病(IBD)、癌症(CRC)、心血管疾病和肝脏疾病。次生胆汁酸、三甲胺-N-氧化物(TMAO)、硫化氢(H2S)和含氮有机化合物(NOC)会导致胆汁酸信号失调、氧化应激和DNA损伤,从而导致CRC等慢性疾病。此外,肠道和中枢神经系统(通过迷走神经)以及肠道衍生代谢物(如三甲胺(TMA)和TMAO)之间的相互作用会影响神经和心血管健康。
下面我们来看具体与肠道耐药组相关的疾病。
抗生素耐药性通过破坏肠道微生物群及其相关代谢途径,在包括肥胖、2型糖尿病(T2D)、炎症性肠病(IBD)和结直肠癌(CRC) 在内的代谢紊乱中发挥着至关重要的作用。肠道耐药组是肠道微生物组的一部分,也与这些疾病的发展有关。
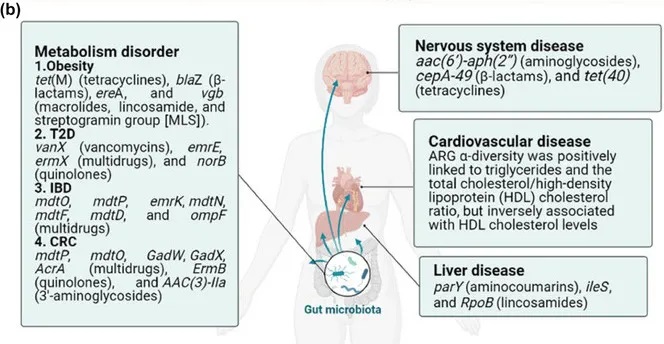
doi:10.1111/1541-4337.70143
b) 抗生素抗性基因对疾病的影响。特定的抗生素抗性基因(ARG)与代谢紊乱、神经系统疾病和肝病有关。ARGα多样性与心脏代谢风险增加有关。
肥胖
在肥胖人群中,肠道生态失调通过改变短链脂肪酸和脂多糖(LPS)影响能量代谢和慢性炎症(图a)。短链脂肪酸通过刺激GLP-1和PYY等激素的释放来调节饱腹感,这些激素作用于下丘脑。此外,肠道细菌的不平衡,特别是革兰氏阴性菌如变形杆菌的丰度增加,会导致LPS水平升高,并通过增加促炎细胞因子(TNF-α、IL-1β、IL-6)的产生引发全身炎症。
一项研究中,与富营养化参与者相比,肥胖个体中梭杆菌、肠球菌、大肠杆菌的丰度更高,同时ARG水平升高,如tet(M)(四环素类)、blaZ(β-内酰胺类)、ereA和vgb(大环内酯类、林可酰胺类、链菌素组[MLS])。这表明ARG与革兰氏阴性菌之间的关系,是由拟杆菌门和变形杆菌门之间的不平衡所驱动的。
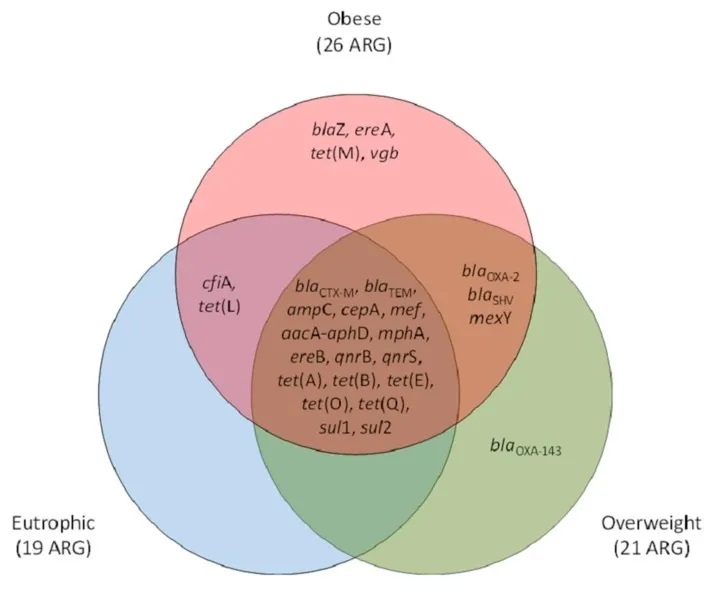
doi: 10.3390/genes10050349
2型糖尿病
在2型糖尿病中,肠道微生物失调,包括抵抗体的变化,导致SCFA产生减少,损害PYY和GLP-1的分泌,这对胰岛素调节和胰高血糖素水平至关重要(图a)。
胆汁酸代谢的改变刺激成纤维细胞生长因子19/15(FGF19/15)的释放,进一步促进胰岛素敏感性和葡萄糖耐量。
ARG丰度的增加,包括Vancomycin_vanX、Multidrug_emrE、MLS_ermX、喹诺酮_norB,与2型糖尿病风险的增加有关,耐药组的变化发生在肠道微生物群变化之前(图b)。
在IBD中,能够产生短链脂肪酸的细菌数量减少,这会削弱短链脂肪酸的抗炎作用。短链脂肪酸通常有助于Treg和效应性T细胞(Teff)的分化。由于这种减少,炎症和肠道上皮细胞损伤会加剧,从而为有害细菌的滋生创造条件,并导致慢性炎症的持续。大肠杆菌中携带的抗生素抗性基因,如mdtO、mdtP、emrK、mdtN、mdtF、mdtD、ompF,变得更加普遍。这些ARGs通过破坏肠道屏障功能,增加肠道的通透性,进而促进炎症反应(如图b所示)。
此外,ARGs和细菌的毒力因子可以通过水平基因转移在微生物群落中传播,进一步加重IBD中的炎症状况。
在结直肠癌中,肠道菌群失衡会通过减少粘液层厚度和增加肠道通透性来促进癌症的发生和发展(图a)。
有害细菌过度生长会导致脂多糖(LPS)和其他有毒代谢物的释放,比如次生胆汁酸、三甲胺-N-氧化物(TMAO)、硫化氢(H₂S)和含氮有机化合物,这些物质会引发低度炎症并激活致癌信号。

上述研究发现,结直肠癌患者体内大肠杆菌、柠檬酸杆菌、不动杆菌水平升高,同时存在耐药基因如mdtP、mdtO、GadW、GadX、AcrA(多药耐药)、ErmB(喹诺酮类耐药)和AAC(3)‐IIa(3′-氨基糖苷类耐药)等耐药基因水平升高(图b)。这些抗生素抗性基因通过加剧肠道菌群失衡、炎症反应和肠道屏障功能障碍,进一步推动CRC的发展,突出了肠道耐药组在代谢性疾病发病机制中的作用。
总体而言,新出现的证据表明,肠道耐药组不仅反映了微生物失衡,而且通过抗生素耐药性的传播积极推动代谢紊乱。ARG在肠道微生物群中的存在和传播会破坏关键的生理过程,如能量代谢、免疫调节和肠道屏障的完整性。通过营造一个有利于致病菌及其毒力因子的环境,耐药组会放大炎症,改变代谢信号通路,并促进生态失调。
肠道微生物组通过炎症反应和TMAO代谢机制参与心血管疾病的发病机制,包括心力衰竭和冠状动脉疾病(图a)。
心血管疾病患者体内产丁酸菌减少,这会引发局部炎症,加剧肠道菌群失衡,并导致肠道屏障功能受损。这些患者体内TMAO水平的升高,会干扰胆固醇运输、促进泡沫细胞形成和诱导血小板聚集,从而可能导致急性冠状动脉综合征,引发动脉粥样硬化的发展。
此外,较高的肠道抗生素抗性基因α多样性与心脏代谢风险增加有关。
一项研究发现,较高的肠道ARGα多样性指数与甘油三酯和总胆固醇/高密度脂蛋白胆固醇比值(TC/HDL)呈正相关,但与高密度脂蛋白胆固醇(HDL)水平呈负相关。
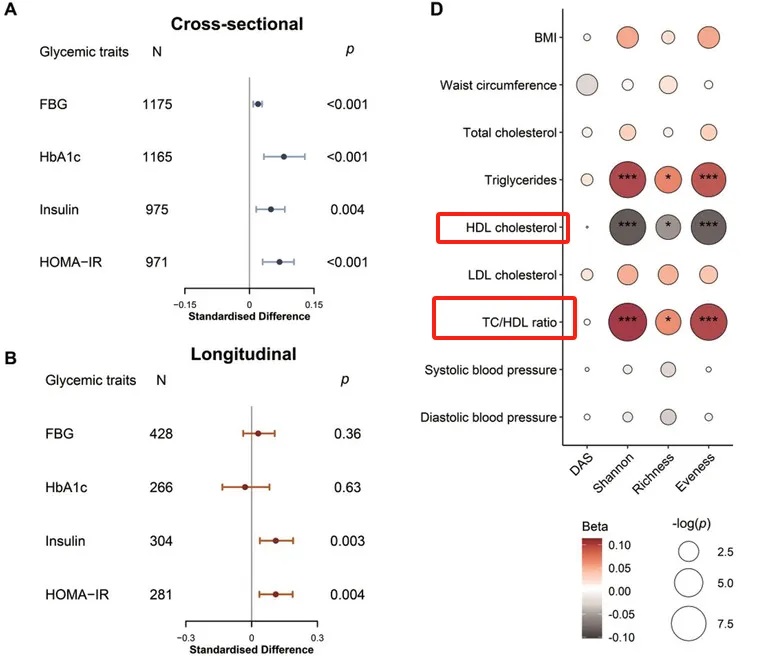
doi: 10.1002/advs.202104965
这表明ARG在扰乱脂质和葡萄糖代谢方面发挥作用,并可能通过血脂异常和胰岛素抵抗增加心血管疾病的风险。
肠道微生物群通过引发炎症反应,显著影响肝脏的正常功能(图a)。
当肠道微生物组因抗生素耐药性发生变化时,肠道的通透性会增加,使细菌及其产物(如脂多糖LPS)能够通过肝肠循环进入肝脏。LPS进入肝脏后,会引发急性炎症反应,导致肝细胞受损,进而可能引发多种肝脏疾病,如非酒精性脂肪肝、脂肪性肝炎、酒精性肝病、肝癌,甚至肝性脑病。
肠道耐药组的变化与肝脏疾病的发生密切相关。研究发现,肝硬化患者的肠道耐药组中,与肠杆菌科、链球菌、肠球菌和不动杆菌属等病原菌相关的耐药基因丰度更高。
此外,这些患者还表现出对β-内酰胺酶、大环内酯类、喹诺酮类、糖肽类、磷霉素、四环素类抗生素的更大耐药性。肝性脑病患者则表现出更高水平的氨基香豆素耐药基因parY和林可酰胺耐药基因(ileS和RpoB)。这些抗性组的变化会破坏肠道与肝脏之间的正常相互作用,以及胃肠道的免疫反应,随着疾病的进展,这种破坏会进一步恶化。
肠道微生物组在通过肠道-大脑轴塑造神经系统的发育和功能方面起着关键作用。迷走神经传递神经元、内分泌和免疫信号,而下丘脑通过释放促肾上腺皮质激素释放激素(CRH)来调节应激反应,导致皮质醇的产生,从而影响肠道的上皮屏障和免疫系统。
肠道耐药组的改变也与神经发育障碍有关。
一项研究发现,患有自闭症的儿童具有更高丰度的屎肠球菌aac(6′)-aph(2′)基因、Megasphaera elsdenii 的cepA‐49和tet(40)基因以及脆弱拟杆菌的cepA-49基因。

研究还发现,健康个体和自闭症患者在对大环内酯类抗生素耐药性的基因方面存在显著差异,包括肠球菌和未培养细菌的氨基糖苷类耐药基因、拟杆菌属和Acidaminococcus的β-内酰胺抗性基因,以及来自Megasphaera和Alistipes的四环素抗性基因。因此,肠道微生物群平衡和抗性基因的破坏可能会降低神经代谢潜力,影响大脑功能所需的神经递质和代谢物的产生。
通常用抗生素治疗的疾病与扩大的肠道耐药性有关,这表明曾经有接触抗生素史对疾病相关菌株的ARG获得产生了相当大的选择压力。
未来的研究需要更好地了解肠道耐药性与疾病之间的相互作用及其机制。将多基因风险评分与肠道宏基因组风险模型相结合可以提高对常见慢性病的预测能力,为疾病预测和预防提供更全面的方法。
通过上述研究,我们看到肠道耐药组对人体健康的多层面影响,是如何超越简单的感染问题而演变成一个系统性的健康风险。然而,要有效应对这一挑战,我们需要深入了解:这些耐药基因是如何产生的?它们又是如何从环境进入人体并在肠道微生物间传播的?…
★
抗生素耐药性并非凭空出现,而是有其明确的生物学机制和传播途径,下面章节我们来详细了解这一过程中的关键环节,包括垂直遗传、水平基因转移等核心机制,以及食物链、水源和环境污染在耐药基因传播中的角色。
在这里重点介绍针对细菌的抗生素,介绍它们的作用机制,并讨论抗生素耐药性如何在肠道中发展和积累。
将治疗细菌性疾病的抗生素引入临床应用是 20 世纪最大的医学突破。抗生素的抗菌机制可细分为五类:
抗生素作用机制和抗生素耐药性
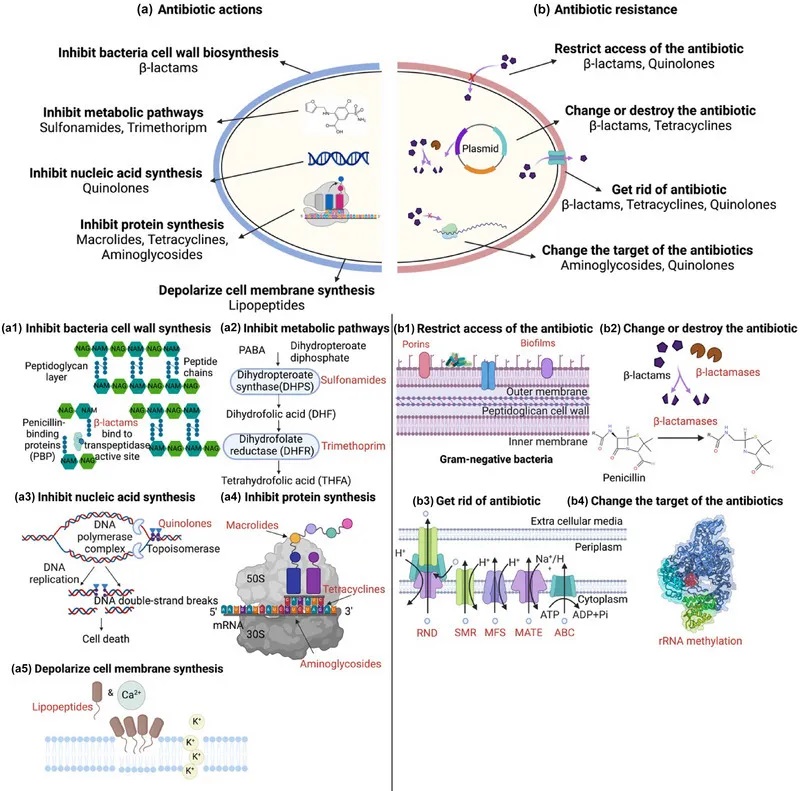
doi:10.1111/1541-4337.70143
a)抗生素作用:抗生素针对关键的细菌过程来抑制生长和存活。
b)抗生素耐药性:细菌通过各种策略产生耐药性。
下面我们先来看一下抗生素的作用。
阻碍细菌细胞壁的合成——结构性破坏
β-内酰胺类抗生素,可以伪装成细菌细胞壁合成所需的关键分子。这类抗生素会:
• 模仿细菌细胞壁合成的天然底物(d-丙氨酸)
• 结构相似性使抗生素能够与DD转肽酶结合(d-丙氨酸是DD转肽酶的底物),防止DD转肽酶与其天然底物进一步结合
• 破坏肽聚糖层的交联,肽聚糖框架减弱
• 最终导致细胞壁结构损坏,细胞死亡
典型的β-内酰胺类抗生素包括:青霉素类、头孢菌素类、碳青霉烯类、单巴坦类。
抑制细菌代谢途径——叶酸合成阻断
磺胺类药物抑制二氢蝶酸酯合酶,这是叶酸生产中必不可少的酶。这种抑制阻断了DNA和RNA生产所必需的核苷酸的合成,阻止了细菌复制并导致细胞死亡。
最常用的磺胺类抗生素:磺胺甲氧基嘧啶、磺胺二甲嘧啶、磺胺甲恶唑、磺胺嘧啶。
协同用药:甲氧苄啶(靶向二氢叶酸还原酶,DHFR)与磺胺类联用,双重阻断叶酸代谢链。
抑制核酸生物合成所需的细菌RNA聚合酶
作用靶点:细菌RNA聚合酶β亚基
机制:阻断转录起始→抑制mRNA合成→细胞分裂停滞
代表药物:利福霉素类(如利福平)
作用靶点:DNA旋转酶(拓扑异构酶II/IV)
机制:
与 DNA 和旋转酶结合→阻止DNA超螺旋/松弛
捕获DNA上的拓扑异构酶→抑制核酸合成→DNA双链断裂→抑制细菌的生长
代表药物:喹诺酮类(诺氟沙星、环丙沙星、氧氟沙星)
阻断蛋白质合成
大环内酯类(红霉素、克拉霉素、阿奇霉素)
与50S核糖体亚基上的23S rRNA产生的结合肽基转移酶中心结合→抑制核糖体沿mRNA的运动→多肽链延伸终止→细菌蛋白质合成的抑制→细菌细胞死亡
四环素类(四环素、土霉素、多西环素)
靶点:与30S核糖体亚基结合
阻断氨酰-tRNA与A位点结合→抑制氨基酸掺入生长的多肽链
氨基糖苷类(卡那霉素、链霉素、庆大霉素)
靶点:30S核糖体16S rRNA上的 A位点
诱导mRNA误读→多肽链提前终止→破坏蛋白质合成完整性→细胞死亡
破坏细胞膜完整性
脂肽类抗生素(达托霉素)
结构特征:含脂尾的肽核
钙离子依赖型寡聚→达托霉素分子嵌入细胞膜形成孔道→膜电位去极化→胞内成分泄漏→细胞裂解
不同种类抗生素作用的方式

Uddin TM, et al., J Infect Public Health. 2021
细菌抗生素耐药性是微生物数千年进化的自然现象。这种固有的抗生素耐药性也被称为固有耐药性。
除了固有耐药性外,过度使用抗生素还会导致抗生素耐药性的积累。细菌采用各种策略来抵抗抗生素,例如减少抗生素的摄取,修饰或灭活抗生素,主动将其从细胞中排出,或改变抗生素的靶标以阻止其作用。
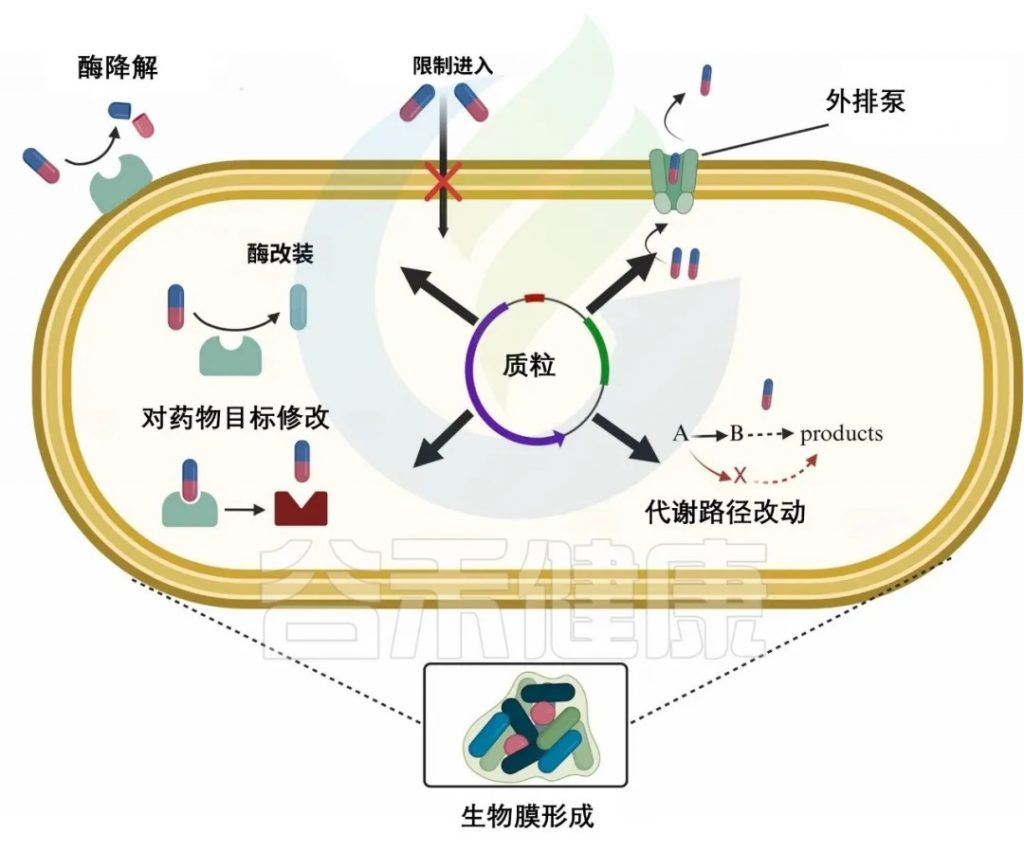
doi.org/10.1016/j.glmedi.2024.100081
细菌耐药核心机制如下:
1. 细菌改变外膜疏水性、孔蛋白突变或形成生物膜,阻止抗生素渗透,导致耐药性
革兰氏阴性菌有一层外膜,可以作为屏障,阻止抗生素(β-内酰胺类、喹诺酮类)进入细胞。大多数抗生素需要穿透外脂膜才能到达目标。疏水性药物如大环内酯类(红霉素)可以通过扩散穿过膜,而亲水性抗生素如β-内酰胺类需要通过孔蛋白(图b1)。
外膜的变化,如疏水性的改变或孔蛋白的突变,可能导致革兰氏阴性菌产生抗生素耐药性。一些细菌可以产生大量的细胞外聚合物,包括胞外多糖、蛋白质、细胞外DNA和脂质。这些物质可以在生物膜内的细菌细胞周围形成物理屏障,防止抗生素直接到达细菌细胞。这减少了抗生素的直接暴露,导致抗生素疗效明显降低。
例如,金黄色葡萄球菌菌株产生的生物膜可以显著降低苯唑西林、万古霉素和头孢噻肟的渗透性。
2. 细菌产生分解抗生素分子的酶,对抗生素修饰或降解,使其无效
β-内酰胺酶
细菌产生β内酰胺酶来降解β内酰胺类,这些酶通过不可逆地打开β-内酰胺环使其失活,从而使其无法与青霉素结合蛋白结合,使其无效。
氨基糖苷修饰酶
如AAC(3)乙酰转移酶修饰庆大霉素,降低其与核糖体结合能力,从而降低其疗效。
3. 细菌外排泵,以能量依赖性方式从细菌细胞中去除抗生素,使细菌在高浓度抗生素中存活
细菌药物外排泵可分为五个家族——ABC、SMR、MATE、MFS、RND。
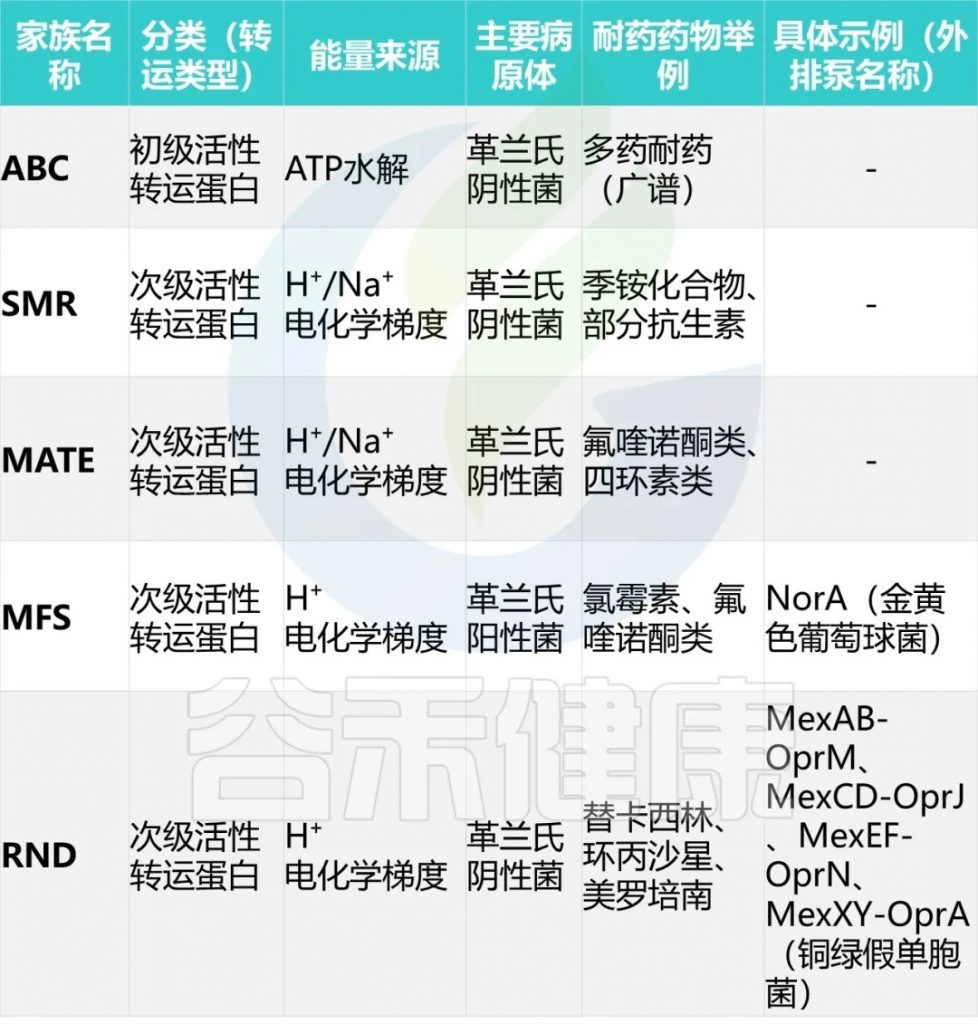
4. 改变抗生素的靶点结合位点,降低结合效率
16S rRNA甲基转移酶改变氨基糖苷类核糖体结合位点的构象,从而降低氨基糖苷类对核糖体的亲和力,使抗生素更难有效结合。
氨基糖苷修饰酶可分为两类,甲基化G1405和A1408。
G1405甲基化:改变了核糖体RNA在这些4,6-二取代2-脱氧链霉素(DOS)氨基糖苷类关键结合位点的结构,如卡那霉素、庆大霉素、妥布霉素、阿米卡星。
A1408甲基化(NpmA酶):可对4,5和4,6二取代的2-DOS氨基糖苷类产生耐药性。
随着分子技术的进步,了解抗生素耐药性背后的机制变得越来越复杂。其中,基因测序已成为一种强大的工具,能够根据遗传数据预测抗性表型。这允许更精确地跟踪耐药性演变,并为有针对性的干预措施提供信息,以防止耐药菌株的传播。
此外,细菌采用多种策略来确保其存活,即使是少量耐药菌株也可能在胃肠道的复杂微生物环境中持续存在,从而导致耐药细菌种群的选择和存活。
了解分子耐药机制对于减轻抗生素耐药性和揭示人体肠道中耐药性积累的动态是必要的。
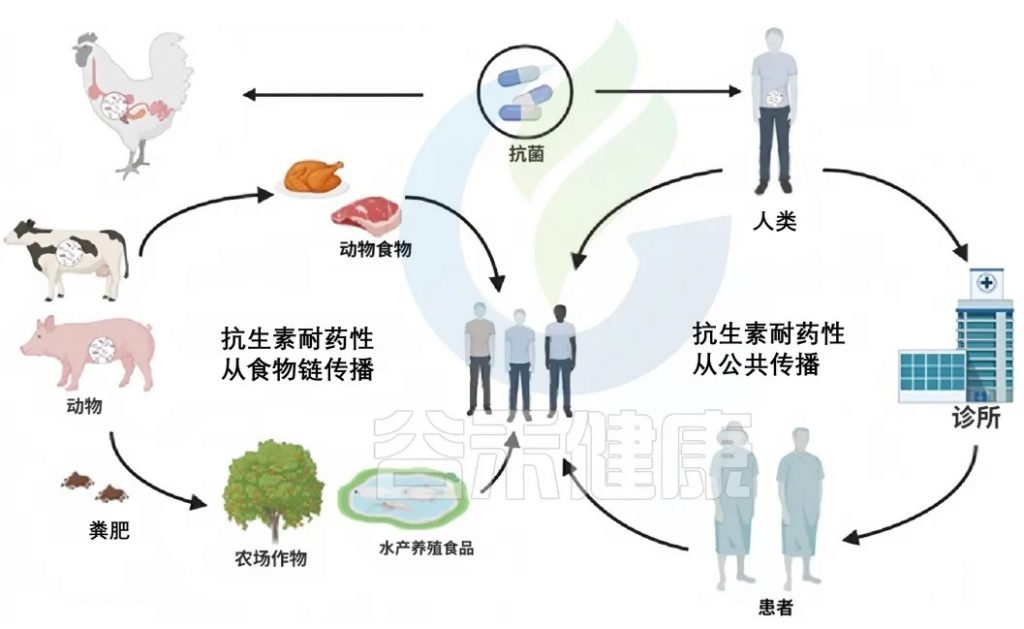
抗生素耐药性发生在自然环境,如土壤、水生态系统、动物粪便,在社区环境中也有,如医院污水、农业实践中。当人类食用食物中的抗生素耐药细菌时,抗生素耐药性可以通过食物链从自然环境传播给人类,比如,通过受污染的食物和直接的公共接触传播给人类。
环境中抗生素耐药性的累积
doi:10.1111/1541-4337.70143
在自然环境中,不同的生物和基质(如动物、土壤和水)之间存在联系。这种相互联系促进了抗生素耐药性在各种环境因素中的传播。土壤、水和沙子是抗性细菌和遗传元素可以持续存在并相互作用的水库。
整个自然界就像一张巨大的“互联网络”——动物、土壤、水甚至沙子之间都有看不见的“网线”连接。这些“网线”让抗药细菌和它们的耐药基因能在环境中四处“串门”。
抗生素耐药性从环境传播到植物、动物和昆虫涉及多种机制。农业实践,包括在畜牧业中使用抗生素和将处理过的动物粪便施用到田地里,将抗性基因引入土壤。水源可能通过农田径流,特别是那些用牲畜粪便施肥的农田径流,被抗生素抗性细菌和遗传元素污染。水生生态系统中的昆虫和其他生物可以成为抗性基因的载体,进一步在环境中传播抗生素抗性。
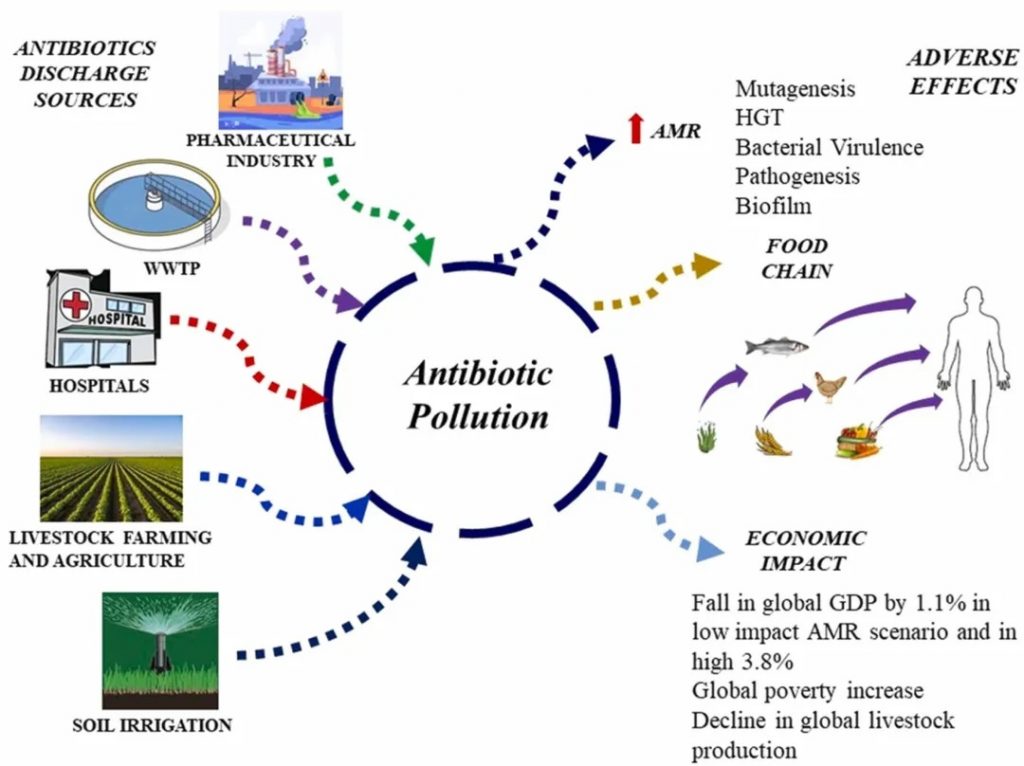
doi.org/10.1016/j.hazl.2024.100105
人类活动对抗生素排放到环境中起着重要作用。这通过多种途径发生,包括城市和医院废物、工业制造、农业径流和垃圾填埋场渗滤液。因此,抗生素耐药性可能通过各种途径进入人体肠道。人类可能通过受污染的食物、水和与环境的直接接触(如家庭、社区或医疗保健互动)接触到抗生素抗性细菌和遗传因素。
宠物狗和猫可携带多种多重耐药细菌,包括耐甲氧西林金黄色葡萄球菌和耐β-内酰胺类抗生素的肠杆菌科细菌。
这些耐抗生素微生物可能通过直接接触、体外寄生虫和气溶胶从宠物传给人类。特别是弱势群体,包括免疫功能低下的老年人和婴儿,可能面临更大的风险。
详见我们之前的文章:宠物猫可能塑造人类肠道中的抗生素耐药性和益生菌
ARG在自然环境、动物和人类中的发生、积累和传播依赖于垂直遗传和水平基因转移(下图)。在细菌的生长和繁殖过程中,基因突变使它们对特定抗生素产生耐药性,它们的后代将继承突变基因(下图a)。携带突变基因的细菌,更有可能存活,并且可以世代垂直传播。随着时间的推移,细菌种群将由具有耐药基因的细菌主导,这种选择性优势使耐药细菌得以增殖,有助于抗生素耐药性的扩大。
ARG具有高流动性,可以通过水平基因转移传播,包括四种机制:基因转化、基因转导、基因接合、膜囊泡融合(下图b)。
通过垂直遗传、水平基因转移以及影响这些机制的因素在肠道中积累抗生素耐药性
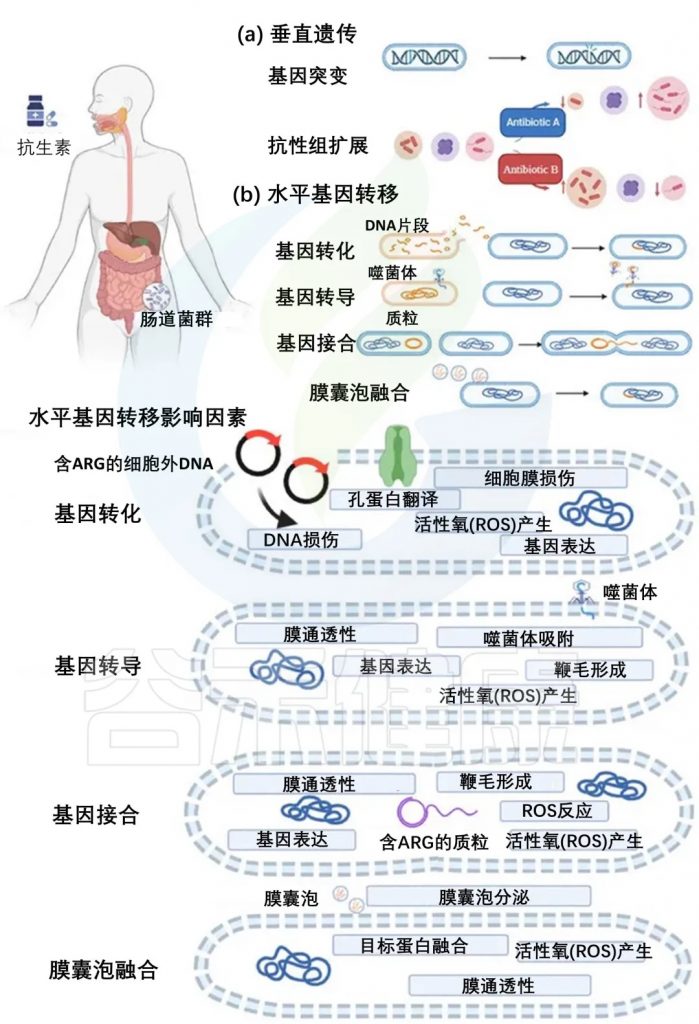
doi:10.1111/1541-4337.70143
a) 垂直遗传:抗生素耐药性可以通过基因突变和耐药体扩增进行垂直遗传。
b) 水平基因转移及其影响因素:
水平基因转移发生在四种途径中:
抗生素可以影响细菌中的水平基因转移,特别是在ARG的情况下。它们创造了一个有利于携带抗性基因的细菌存活和增殖的选择性环境。除了抗生素选择压力外,各种非抗生素因素,包括抗菌剂和环境条件,都会影响ARG的水平转移。
在大多数细菌中,获取抗性基因的主要和最有效的方法涉及使用可移动的遗传元件,如质粒作为基因交换的载体。在多种细菌中观察到水平基因转移,如大肠杆菌和肺炎克雷伯氏菌,使耐药基因能够在细菌种群中快速有效地传播。一般来说,ARG的水平转移会加剧肠道中抗生素耐药性的传播。
总体而言,垂直遗传和水平基因转移是导致ARG在自然环境、农场动物和最终人类肠道中发生和传播的两条主要途径。值得注意的是,ARGs的水平基因转移是决定其传播的关键因素。
饮食干预在抑制ARGs的水平基因转移中起着重要作用。已经发现,一些膳食植物化学物质,如酚类化合物、硫化物、萜类化合物、生物碱,有可能诱导ROS的产生,导致细菌细胞膜不可逆的损伤,生物膜活性降低,毒力基因表达下调,从而抑制水平基因转移。下面章节将进一步讨论食物成分在控制ARG中的作用。
★
通过饮食成分改变肠道耐药性是一个新兴的研究领域,人们越来越关注食物选择如何影响肠道中的抗生素抗性细菌。例如,发酵食品中含有大量微生物,乳酸菌和凝固酶阴性葡萄球菌被确定为四环素、青霉素、氯霉素和大环内酯类ARG的携带者。这表明食用发酵食品可能会直接影响肠道耐药性。
食品中存在的许多天然分子,如酚类、萜类和酚酰胺,具有抗菌能力,能够调节肠道微生物群。然而,关于食物成分对肠道抗生素耐药细菌影响的研究有限,这是一个新兴的研究领域。
这里总结了对大量人群的营养研究,以了解饮食习惯对肠道耐药性的潜在影响,并强调了选定类别的饮食成分对抗生素耐药细菌的调节作用。
长期的饮食习惯会影响肠道微生物群和肠道抵抗力。
Stege等人(2022)选择了149名荷兰个体,他们的饮食习惯分为四个不同的群体:杂食者、鱼素食者、素食者、纯素食者。研究发现长期的饮食习惯并没有显著影响肠道微生物组的关键组成和多样性。在前10个最丰富的ARG中没有观察到显著差异。然而,两个ARG[lsa(C)和tet(L)]在杂食者和鱼食者之间显示出不同的丰度。
此外,宏基因组分析显示,与鱼素者相比,tet(X)在杂食动物的抗性体中更为丰富。
在来自三种不同饮食习惯(杂食性、蛋乳素食主义、严格素食主义)的58名志愿者的肠道抵抗力中观察到了类似的结果。
在所有习惯组中,最常见的耐药基因是针对四环素类抗生素[tet(A)、tet(B)、tet(M)、tets(O)、tet(Q)]、β-内酰胺类(blaTEM、blaSHV、mef)和MLS耐药基因[erm(B)、erm(C)],其次是磺胺类(sul1、sul2)和氨基糖苷类耐药基因(aacA-aphD)。
blaCTX‐M仅在卵乳素食组的样本中发现,而tet(E) 仅在杂食组的样本中检测到。有趣的是,这些经常检测到的ARG也常见于农场动物粪便中,这表明饮食习惯和环境ARG之间存在潜在联系。
虽然将饮食与肠道抗药性联系起来的确切机制尚不清楚,但饮食干预可能提供一种有前景的方法来减轻抗生素耐药性的负担。
Oliver等人(2022)研究了290名健康成年人,以探索饮食、肠道微生物组和抗生素耐药性之间的关系,并观察到高膳食纤维摄入量的个体ARG水平较低。
低ARG人群在肠道中显示出更多梭菌科的专性厌氧菌,而高ARG人群的链球菌科和肠杆菌科水平更高。此外,低ARG个体摄入的蛋白质较少,尤其是来自牛肉和猪肉的蛋白质。
该研究表明,增加纤维摄入量可能会促进肠道微生物组环境,有利于专性厌氧菌的生长,同时限制兼性厌氧菌,从而可能降低肠道中抗生素耐药性的发生率。
这是因为由专性厌氧菌发酵的膳食纤维会产生短链脂肪酸,降低肠道环境的pH值,使其对专性厌氧杆菌有利,但对兼性厌氧菌不利。
特定的饮食成分通过各种机制在调节肠道微生物群方面发挥着至关重要的作用。生物活性常量营养素,如凝集素、乳铁蛋白和多不饱和脂肪酸(PUFA),来源于碳水化合物、蛋白质和脂质,主要因其营养作用而被认可,但它们也具有生物活性,通过破坏营养可用性和细菌膜完整性来抑制抗生素细菌。
多酚等植物化学物质通过破坏细胞功能、干扰代谢过程或抑制酶活性表现出抗菌特性。
益生菌,如乳杆菌和双歧杆菌,通过竞争性排斥、分泌抗菌化合物和免疫调节来抑制病原体。
有趣的是,其中一些化合物还具有选择性抑制抗生素耐药细菌的能力,通过靶向其脆弱性,如损害生物膜形成、抑制耐药酶和破坏外排泵。
研究这些化合物如何特异性靶向细菌,包括抗生素耐药菌株,对于制定控制或减少肠道致病菌群的饮食策略至关重要。这有助于提供了一种非抗生素方法来管理肠道健康,并有助于遏制抗生素耐药性的传播。
生物活性常量营养素
常量营养素包括碳水化合物、蛋白质、脂肪,是人类饮食中支持生命和调节各种生理功能的重要组成部分。虽然它们的主要作用是为身体提供能量和构建块,但某些生物活性常量营养素也具有固有的抗菌特性。
这些特性可能有助于控制抗生素耐药细菌的生长,减轻其对肠道微生物组的影响,从而可能提供一种对抗抗生素耐药性的替代策略。
碳水化合物
不仅是能量的来源,而且具有生物活性,可以提高抗生素的有效性。
来自紫色鞘豆(Dioclea violacea)种子的甘露糖结合凝集素已被证明可以提高抗生素的疗效。关于什么是凝集素可以详见我们之前的文章:什么是凝集素,食物中的凝集素如何影响肠漏和自身免疫
Santos等人(2023)证明,从紫色鞘豆中提取的凝集素作为佐剂,通过碳水化合物识别增加细菌膜附近氨基糖苷类的生物利用度,并促进抗生素进入细菌细胞质,从而增强新霉素对多药耐药金黄色葡萄球菌和铜绿假单胞菌的活性。
蛋白质
如母乳和牛乳中的乳铁蛋白,也表现出抗菌和抗菌膜特性。
Avery等人(2021)报告称,人和牛乳铁蛋白都可以显著抑制测试的多重耐药鲍曼不动杆菌菌株中生物膜的形成,而人乳铁蛋白的最低抑菌浓度(MIC)低于牛乳铁蛋白,表明其效力更高。除了乳铁蛋白,来自植物的抗菌肽(AMP)在对抗抗生素耐药细菌方面也显示出了希望。
Heymich等人(2021)从鹰嘴豆中鉴定出21种AMP候选物,其中两种对耐甲氧西林的金黄色葡萄球菌ATCC 3300具有杀菌活性。这些肽与细菌膜相互作用,通过膜破裂导致细胞裂解,这与它们的杀菌作用直接相关。
膳食脂肪
特别是PUDA,也可以提高某些抗生素的有效性。
Zang等人(2021)发现,花生四烯酸(AA)和二十二碳六烯酸(DHA)等PUFA可增强氨基糖苷类药物对多重耐药鲍曼不动杆菌的疗效。
AA和DHA都通过细胞外隔离减轻了粘菌素的抗菌作用,使其难以穿透细菌的亲水性包膜和脂多糖屏障,并上调了外排系统基因表达(adeABC和adeIJK),从而通过将抗生素排出细胞来增加细菌的抗药性。
上述研究强调了生物活性常量营养素在增强抗生素有效性和影响耐药机制方面的作用。
果胶等碳水化合物可以通过与抗生素相互作用来提高抗生素的疗效,而乳铁蛋白等蛋白质则具有强大的抗菌和抗菌膜特性。然而,AA和DHA等膳食脂肪的作用更为微妙,因为它们都可以通过激活外排泵来增强抗生素作用并上调细菌耐药性机制。
总之,生物活性常量营养素不仅可以作为能量来源,还可以在对抗抗生素耐药细菌方面提供巨大的潜力。虽然它们在调节细菌耐药机制方面的作用很复杂,但这些发现强调了进一步研究饮食成分补充或增强传统抗生素治疗潜力的重要性。通过利用这些天然化合物,可能能够开发出新的策略,来缓解抗生素耐药性日益增长的挑战。

doi:10.1111/1541-4337.70143
植物化学物质
植物化学物质来源于植物性食品,主要包括多酚、含硫化合物、萜类、生物碱。这些化合物对抗生素耐药细菌具有抗菌活性,其中一些会破坏细菌膜结构,另一些则通过抑制外排泵来增强抗生素的有效性,从而防止耐药性的发展。
★ ■ 多酚类
柿子单宁——直接抗菌作用
一种经过充分研究的多酚,即来自青涩柿子的柿子单宁,已显示出对耐甲氧西林金黄色葡萄球菌的抑制作用。柿子单宁抑制细胞增殖,降低膜电位和细胞内ATP浓度,破坏全细胞蛋白质完整性,并诱导细胞S期细胞周期阻滞。
EGCG、原花青素——协同作用、增强抗生素疗效
Parvez等人(2019)报道了表没食子儿茶素没食子酸酯(EGCG)与庆大霉素联合使用时具有明显的协同作用,增强了对金黄色葡萄球菌和大肠杆菌多药耐药菌株的抗生素疗效。这是因为EGCG可以破坏细菌细胞膜,阻碍DNA超螺旋,增加细胞通透性,最终导致细菌死亡,并减轻耐药性的发展。
同样,从美国蔓越莓(Vaccinium macrocarpon L.)中提取的原花青素与β-内酰胺类抗生素显示出显著的协同作用。
Gallique等人(2021)报告称,原花青素抑制β-内酰胺酶,并选择性增强苯唑西林和羧苄青霉素对耐甲氧西林葡萄球菌的有效性,可能是通过调节葡萄球菌中变异转肽酶PBP2a的表达。PBP2a的低表达降低了细菌抵抗β-内酰胺类抗生素的能力,使细菌更容易受到抗生素的影响。
扩展阅读:
★ ■ 含硫化合物
大蒜素衍生物——释放硫化物破坏致病生物膜
例如,Xu等人(2018)将从大蒜鳞茎中分离出的大蒜素转化为纳米级硫化铁。这些纳米颗粒可以释放杀菌的有机硫化物(Polysulfanes),有效抑制生物膜形成和耐药的金黄色葡萄球菌。
异硫氰酸盐——抑制生物膜代谢活性
同样,来源于旱金莲和辣根的异硫氰酸酯对铜绿假单胞菌等抗生素耐药细菌具有抗菌潜力,特别是在生物膜抑制方面。当与美罗培南联合使用时,异硫氰酸酯通过抑制铜绿假单胞菌生物膜的代谢活性来增强抗生素疗效。
皂苷——破坏细胞壁结构,改变膜通透性
此外,da Silva等人(2024年)从Sarcomphalus joazeiro中提取了六种皂苷——红枣皂苷B、红枣皂苷III、酸枣仁苷、红枣皂苷IV、红枣皂苷II和红枣皂苷I,以及三种皂苷衍生物。当与庆大霉素和诺氟沙星联合使用时,富含皂苷的组分显示出增强作用,增加了它们对多药耐药铜绿假单胞菌和金黄色葡萄球菌的活性。皂苷与细菌细胞壁相互作用,导致结构损伤,改变膜通透性,并诱导膜破裂。
★ ■ 萜类药物
1,8-桉叶醇——增加膜通透性、诱导氧化应激导致成分泄漏
Moo等人(2021)发现,1,8-桉叶醇对碳青霉烯类抗生素耐药肺炎克雷伯菌具有抗菌作用。它增加了细菌表面电荷,增强了外膜通透性,并诱导了氧化应激,导致膜损伤、细胞内成分(如核酸、蛋白质和脂质)泄漏,最终导致细胞死亡。
芳樟酯——产生ROS、引发脂质过氧化
另一种萜烯,邻氨基苯甲酸芳樟酯(LNA),存在于薰衣草和百里香等植物中,对碳青霉烯类耐药肺炎克雷伯菌具有杀菌活性。
Yang等人(2021)证明,当LNA与美罗培南联合使用时,会降低MIC,使细菌更容易感染。LNA的抗菌作用归因于其通过产生ROS诱导氧化应激的能力,ROS可能进一步引发脂质过氧化并对细菌膜造成损伤。
萜类化合物——细菌外排泵的重要抑制剂
Oluwatuyi等人(2004)在迷迭香中鉴定出几种萜类化合物,如鼠尾草酸、鼠尾草酚和12-甲氧基-反式鼠尾草酸。这些化合物已被证明可以通过抑制NorA外排泵来增强红霉素对多药耐药金黄色葡萄球菌的活性,从而降低细菌排出抗生素的能力并增加其细胞内浓度。
★ ■ 生物碱
原小檗碱类生物碱——抑制H+-ATP酶泵,阻断ATP合成
Guefack等人(2022)从绿花恩南番茄(Enantia chlorantha)中鉴定出的原小蘖碱类生物碱(protoberberine alkaloids)(哥伦比亚胺、假哥伦比亚胺、药根碱、巴马汀、4,13-二羟基-3,9,10-三甲氧基原小檗碱和13-羟基-2,3,9,10-四甲氧基原黄连素)可以有效抑制多药耐药细菌的生长,包括大肠杆菌、产气肠杆菌、肺炎克雷伯菌、普罗维登斯氏菌(Providencia stuartii)、金黄色葡萄球菌,MIC值低于100µg/mL。重要的是,其中一种名为哥伦比亚胺的生物碱抑制了H+-ATP泵,阻碍了细胞ATP的产生。
苦参碱——抑制Mex泵,增强氟喹诺酮敏感性
来自苦豆子种子的苦参碱与环丙沙星对铜绿假单胞菌的抗生素耐药菌株显示出协同作用。总生物碱显示出抑制生物膜形成和降低Mex泵活性的作用,从而增强细菌对氟喹诺酮类药物的敏感性。
研究人员从Erythroxylum revolutum Mart.中分离出生物碱,包括6-(2′-甲基丁酰氧基)-3-羟基托烷和6-丁酰氧基-3-羟基托巴烷,可增强诺氟沙星和红霉素对多药耐药金黄色葡萄球菌的疗效。这种作用可能与托烷生物碱削弱耐多药金黄色葡萄球菌中ABC多药转运蛋白的能力有关。
总之,植物化学物质通过破坏细菌膜、抑制外排泵和阻碍细菌生物膜的形成来发挥抗菌能力。当与抗生素结合时,一些植物化学物质表现出协同作用,可以提高抗生素的有效性并改变抗生素的耐药性,表明有可能管理肠道中的抗生素耐药细菌并影响肠道耐药组。
未来,有必要研究这些植物化学物质在人体肠道中的生物利用度、剂量和长期影响,这可能会导致更有针对性的饮食干预。深化我们对特定植物化学物质如何选择性调节肠道抗药性的理解,对于开发创新方法来减轻抗药性基因的传播和改善人类健康结果至关重要。

doi:10.1111/1541-4337.70143
益生菌
益生菌是具有健康益处的活微生物,通常被掺入酸奶和奶酪等发酵食品中。然而,这些产品通常只有在用未经巴氏消毒的牛奶制成时才含有益生菌,因为巴氏消毒可以显著减少或消除活的益生菌菌株。益生菌可以通过调节肠道微环境、增强免疫功能和抑制过敏反应来影响肠道中的抗生素耐药细菌。
★ ■ 直接抗菌效应
某些益生菌菌株已显示出对耐药病原体的抗菌活性。
乳杆菌类——破坏细胞膜完整性,与抗生素协同抑制有害菌
例如,嗜酸乳杆菌NCFM和鼠李糖乳杆菌GG通过破坏细菌细胞膜的完整性对耐甲氧西林金黄色葡萄球菌表现出抗菌作用。
益生菌胶囊(嗜酸乳杆菌CL1285、干酪乳酸杆菌LBC80R、鼠李糖乳杆菌CLR2)与妥布霉素联合使用时,通过限制病原体定植和阻碍生物膜的形成,对耐甲氧西林的金黄色葡萄球菌和铜绿假单胞菌表现出协同作用。
双歧杆菌——促进四环素进入,破坏蛋白质合成
双歧杆菌会产生生物表面活性剂和乙酸盐,这有助于分离粘附的大肠杆菌并改变细胞内的阴离子组成。这可以增强四环素进入大肠杆菌,破坏蛋白质合成,最终导致协同杀死大肠杆菌。这些研究为益生菌作为功能性食品调节肠道抗菌药物耐药性的潜在应用铺平了道路。
★ ■ 发酵食品中的益生菌活性
除了这些单独的益生菌菌株外,酸奶和泡菜等发酵食品中发现的益生菌已被证明可以抑制抗生素耐药细菌的生长。
酸奶来源菌株抗菌物质:类细菌素肽或蛋白
从商业酸奶中分离出的石蜡乳杆菌对14种多药耐药细菌具有抗菌活性,包括大肠杆菌、铜绿假单胞菌、鲍曼不动杆菌、产气杆菌、奇异变形杆菌、肺炎克雷伯菌。该菌株产生的抗菌物质可能是一种细菌素样肽或蛋白质,对密切相关的细菌菌株具有活性,表明其在抑制致病性多药耐药细菌生长方面具有潜在作用。
泡菜来源菌株——抑制耐药菌、生物膜
Yi和Kim(2023)从泡菜中分离出益生菌乳酸菌(Pediococcus inopinatus K35),该菌能有效抑制多药耐药铜绿假单胞菌的生长和生物膜形成。这些研究共同强调了益生菌的巨大潜力,特别是那些来自发酵食品的益生菌,作为能够对抗抗生素耐药细菌的天然药物。

doi:10.1111/1541-4337.70143
然而,一个值得关注的问题是益生菌菌株可能携带ARG,ARG可以转移到致病菌,从而导致耐药性的传播。
例如,Selvin等人(2020)从膳食补充剂中分离出益生菌菌株,发现粪肠球菌和肠系膜芽孢杆菌对青霉素G有耐药性,嗜酸乳杆菌对氨苄青霉素有耐药性。
Montassier等人(2021)的一项研究表明,摄入市售益生菌补充剂(含有11种常见的益生菌菌株:嗜酸乳杆菌、干酪乳杆菌、副干酪乳酸杆菌、鼠李糖乳杆菌、植物乳杆菌、双歧杆菌、短双歧杆菌、长双歧杆菌亚种、婴儿长双歧杆菌、乳链球菌和嗜热链球菌)导致ARG丰度降低。
然而,益生菌也可以作为肠道耐药组扩展的储库,因为益生菌的摄入可以通过水平基因转移将携带ARG的菌株扩展到病原体,因为移动遗传元件含量(转座酶和整合酶)的检测与ARG丰度相关。
这些发现强调了在将益生菌纳入饮食时需要谨慎,特别是在商业益生菌补充剂的情况下。虽然益生菌可以在减少致病菌定植和增强抗菌活性方面发挥作用,但必须仔细评估它们作为ARG载体的作用。为了降低潜在风险,需要监测补充剂和功能性食品中使用的益生菌菌株中的ARG含量,确保它们不会导致耐药性的进一步传播。
总之,虽然益生菌作为功能性食品具有调节肠道耐药性和对抗抗生素耐药性的潜力,但它们储存和转移ARG的潜力需要更彻底的研究。未来的研究应侧重于鉴定ARG转移风险最小的益生菌菌株,并优化益生菌食品,以提高其治疗效果,同时限制对肠道耐药的不利影响。
以上我们已经了解了抗生素耐药性的发生机制、传播途径、饮食干预的方法和机制。在实施上述饮食干预策略的同时,如何客观评估其对肠道耐药组的影响至关重要。
随着测序技术的快速发展,肠道菌群检测已经能够识别肠道中的抗生素耐药基因,例如,谷禾宏基因组肠道菌群检测报告中,专门设有抗生素耐药基因分析板块,可检测β-内酰胺酶、氨基糖苷类、大环内酯类、喹诺酮类、四环素耐药基因等多种耐药机制的存在和丰度,这为评估饮食干预效果提供了客观依据。
<来源:谷禾宏基因组肠道菌群检测报告>
通过干预前后的对比检测,我们可以了解特定饮食成分对耐药基因表达的影响,从而不断优化干预方案,实现个性化的肠道耐药组管理。
例如,患者A因肺炎需要使用抗生素治疗,在用药前安排了肠道菌群检测。报告显示其肠道中存在高水平的β-内酰胺酶耐药基因,那么根据报告调整相关抗生素方案,避开容易产生耐药的β-内酰胺类抗生素,减少治疗期间耐药基因的扩增风险。
通过识别高丰度ARGs,预测治疗失败风险,避免可能造成更加严重抗生素耐药。
一名反复发生尿路感染的患者B,常规抗生素治疗无效。通过肠道菌群宏基因组检测,发现其肠道中携带高丰度的 blaCTX-M(广谱β-内酰胺酶基因) 和 mecA(甲氧西林耐药基因),提示对β-内酰胺类及甲氧西林类药物耐药。
根据检测结果,避免使用头孢类(靶向β-内酰胺酶敏感菌),改用磷霉素(对blaCTX-M无交叉耐药)或联合多黏菌素(针对mecA阳性菌)。补充蔓越莓原花青素(抑制β-内酰胺酶活性)或大蒜素纳米颗粒(破坏生物膜),增强抗生素渗透性。
识别关键耐药基因,指导临床选择非交叉耐药抗生素并辅以天然抗菌成分,从源头减少抗生素滥用导致的耐药性扩增。
一名2型糖尿病患者宏基因组检测显示,其肠道中 Vancomycin_vanX(万古霉素耐药基因) 和 Multidrug_emrE(多药耐药基因) 丰度升高,且与胰岛素抵抗指数呈正相关。
ARG丰度高的糖尿病患者血糖控制失败率相对较高,因此可以从这方面入手,增加苦参碱摄入(抑制Mex泵活性),联合二甲双胍改善胰岛素敏感性。
通过识别耐药基因丰度,可预测糖尿病血糖控制失败的风险,针对性干预,减少耐药基因对慢性疾病的协同恶化作用。
抗生素耐药性已经从单纯的医学挑战演变为一个多维度的健康问题,而肠道耐药组的研究为我们提供了新的思路应对这一挑战。
肠道菌群检测技术已能够识别耐药基因的存在和丰度,使个体ARG状况的评估成为可能,这不仅可以帮助临床医生了解患者的耐药风险,也为饮食干预提供了科学依据。
日常饮食的预防性策略
加富含多酚的食物(如绿茶、浆果、深色蔬菜)、含硫食物(如十字花科蔬菜、大蒜)及发酵食品(如酸奶、泡菜),通过天然成分抑制耐药基因传播。
控制高脂高糖饮食(如红肉、加工食品),避免促进肠杆菌科等耐药菌增殖。地中海饮食模式(富含纤维、橄榄油)可提升短链脂肪酸水平,抑制耐药菌生长。
通过将这些科学发现转化为具体的饮食策略和产品,结合肠道菌群检测技术,我们有望在不依赖新抗生素开发的情况下,通过日常饮食管理减轻抗生素耐药性的负担,为公共健康问题提供可持续的补充解决方案。
功能性食品的开发创新
开发含高浓度EGCG的绿茶提取物饮品,或添加原花青素的蔓越莓咀嚼片,辅助治疗泌尿系统耐药菌等相关感染。
利用大蒜素纳米颗粒或异硫氰酸盐制成肠道缓释胶囊,抑制生物膜形成并减少水平基因转移。
基因编辑乳杆菌(如表达抗菌肽的菌株),靶向清除肠道内携带ARGs的致病菌。
通过微胶囊化或脂质体包裹提高多酚类成分的肠道吸收率;结合AI预测模型,从天然产物库中筛选新型抗菌分子,加速功能性配方的迭代。
建立功能性食品中ARGs迁移风险的动态监测体系,避免益生菌载体成为耐药基因传播媒介。
功能性食品作为“可食用疗法”,通过精准调控肠道耐药组,为临床耐药危机提供了“非抗生素”解决方案。未来需融合合成生物学、纳米技术与营养学,推动第三代功能性食品的落地。
临床营养的精准化应用
本文中列举的一些研究表明,植物多酚(如柿子单宁、表没食子儿茶素没食子酸酯)、含硫化合物(如大蒜素)及萜类(如1,8-桉叶油醇)可通过破坏细菌生物膜、抑制外排泵或增强抗生素渗透性,降低耐药菌存活率。临床可开发含此类成分的功能性营养剂,辅助治疗耐药菌感染(如MRSA、铜绿假单胞菌),或用于术后耐药菌感染预防。
特定益生菌(如乳酸杆菌、双歧杆菌)可通过竞争排斥或分泌抗菌肽抑制耐药菌定植。临床中可针对术后感染或抗生素滥用患者,定制含益生菌的肠内营养配方,重建肠道菌群平衡。
乳铁蛋白、植物抗菌肽等可增强抗生素疗效。例如,乳铁蛋白与氨基糖苷类联用可显著抑制多重耐药鲍曼不动杆菌,适合用于重症患者的营养支持方案。
2型糖尿病患者中,苦参碱与二甲双胍联用可抑制肠道内产气荚膜梭菌的Mex泵活性,改善胰岛素敏感性。
结直肠癌患者化疗期间,添加大蒜素纳米颗粒的营养配方可减少肠道内携带mdtP基因的多重耐药大肠杆菌丰度,缓解化疗相关性腹泻。
然而,也需要建立植物化学物质与抗生素的相互作用数据库,避免协同毒性(如EGCG与环丙沙星联用可能加重肝损伤)。
同时可以开展剂量梯度临床试验,明确如多酚类成分的最佳治疗窗口。
联合微生物学、药理学与食品科学,开发兼具营养支持与耐药调控功能的特医食品。通过跨学科协作,特医食品不仅可成为感染患者的营养支持方案,也能作为耐药危机的一线防控工具。
未来,跨领域合作结合人工智能,有望加速功能性食品的迭代,在降低抗生素依赖的同时,重塑肠道健康生态,为应对耐药危机提供可持续路径,最终实现“以食为药”的精准健康管理。
主要参考文献:
Abbas A, Barkhouse A, Hackenberger D, Wright GD. Antibiotic resistance: A key microbial survival mechanism that threatens public health. Cell Host Microbe. 2024 Jun 12;32(6):837-851.
Liang Z, Liang Z, Hu HW, Howell K, Fang Z, Zhang P. Food substances alter gut resistome: Mechanisms, health impacts, and food components. Compr Rev Food Sci Food Saf. 2025 Mar;24(2):e70143.
Pramod Barathe, Kawaljeet Kaur, Sagar Reddy, Varsha Shriram, Vinay Kumar, Antibiotic pollution and associated antimicrobial resistance in the environment,Journal of Hazardous Materials Letters, Volume 5,2024.
Langford BJ, Soucy JR, Leung V, So M, Kwan ATH, Portnoff JS, Bertagnolio S, Raybardhan S, MacFadden DR, Daneman N. Antibiotic resistance associated with the COVID-19 pandemic: a systematic review and meta-analysis. Clin Microbiol Infect. 2023 Mar;29(3):302-309.
Laxminarayan R. The overlooked pandemic of antimicrobial resistance. Lancet. 2022 Feb 12;399(10325):606-607.
Sarmiento MRA, de Paula TO, Borges FM, Ferreira-Machado AB, Resende JA, Moreira APB, Dutra Luquetti SCP, Cesar DE, da Silva VL, Diniz CG. Obesity, Xenobiotic Intake and Antimicrobial-Resistance Genes in the Human Gastrointestinal Tract: A Comparative Study of Eutrophic, Overweight and Obese Individuals. Genes (Basel). 2019 May 7;10(5):349.

谷禾健康

春节假期期间,亲朋好友欢聚一堂,美食佳肴各种狂炫,然而当传统节日遭遇现代饮食文化,频繁的高脂高蛋白摄入、不规律的进食节奏,正不断冲击着肠道屏障的免疫调控网络,部分人群可能正被食物过敏困扰。
食物过敏(FA)是一个当今面临公共卫生问题。一个人可以在任何时候对任何食物过敏,但有引起过敏反应最普遍的食物包括:牛奶、鸡蛋、花生、坚果、大豆、小麦、鱼类和贝类。这些过敏反应范围从肿胀和荨麻疹到危及生命的过敏性休克。
更不乐观的是,食物过敏在当今全球工业化社会中的流行率呈现显著代际增长(一代比一代增强)。与此同时食物过敏率的上升与很多疾病的增加相呼应,包括肥胖、糖尿病、哮喘、自闭症和炎症性肠病(以及其他疾病)。我们如何解释这种代际变化?
食物过敏是过敏性疾病的一部分,被称为I型超敏反应,是由食物蛋白(抗原)通过肥大细胞和/或嗜碱性细胞表面上的免疫球蛋白同种型E(IgE)交联引起的,这与由食物成分引起的食物敏感性不同(例如,乳糖不耐症)。
与此同时,我们的肠道相关淋巴组织(GALT)每天都面临着相当大的挑战。它遇到的抗原估计来自1014微生物和每年超过约30公斤的食物蛋白质。这要求它必须将这些无害的抗原与潜在的病原体区分开来,并能同时引起适当的宿主免疫应答。
目前越来越明确的是,与先天免疫细胞和常驻微生物群密切相互作用的功能性上皮屏障对于建立和食物过敏以及口服耐受性至关重要。而口服耐受性的形成取决于年龄、遗传学、粘膜屏障状态、抗原的物理性质、暴露剂量和频率以及肠道微生物群。
是什么驱动了对食物抗原的过敏反应,近年来有关微生物群在调节抗原摄取和抗原呈递中的作用开始被解开。在4至6个月大的婴儿中早期引入致敏性食物已被证明有希望降低部分但并非所有儿童的IgE-食物过敏(IgE-FA)发病率。
IgE-FA尚无明确的治愈方法,金标准治疗是在发生过敏反应时避免使用对症治疗。幸运的是,疾病修饰疗法,包括免疫疗法,生物制剂等有希望。
本文汇总和依托这些文献结果,讨论了IgE-食物过敏在其自然史的背景下,发展的风险因素,免疫生物学,诊断和治疗,包括新兴疗法。同时,我们也将探讨肠道微生物群在食物过敏中的作用,以及如何通过调节肠道微生物群来预防和管理食物过敏。
食物过敏是一种由免疫球蛋白E(简称IgE)引起的过敏反应。这种病近几十年来越来越常见,可能会威胁生命。特别是对于那些难以完全避开的食物(比如牛奶或鸡蛋)过敏的人来说,生活质量会受到很大影响。
对牛奶、鸡蛋、小麦和大豆的过敏,随着年龄增长可能会慢慢好转。
对花生、坚果和海鲜的过敏,通常会伴随终身。
食物过敏常常和其他过敏性疾病一起出现,比如:
虽然理论上任何食物蛋白质都可能引起过敏,但最常见的是:
小麦过敏和乳糜泻是两种不同的病:
小麦过敏是对小麦蛋白的过敏反应;
但在乳糜泻中,部分水解的麦醇溶蛋白被小肠粘膜中的组织转氨酶脱酰胺。这种特异性和有序的脱酰胺作用增强了小肠固有层中CD4+T细胞对麦胶蛋白的识别,导致免疫应答。细胞因子的后续释放导致肠粘膜的组织学变化。
◆ 过敏疾病是如何发展的?
医生们发现,很多孩子的过敏疾病会按照一定规律逐步发展,这个过程被称为“过敏进行曲”。大约有30-50%的过敏儿童会经历这个过程。不过每个孩子的发展过程可能不太一样。
◆ 过敏疾病通常会按这个顺序出现:
湿疹(特应性皮炎):通常在出生后6个月内最先出现
食物过敏:在1岁前就可能出现
过敏性鼻炎和哮喘:在有湿疹的孩子中,差不多三分之二会在3岁前出现这些症状。
特别提醒:如果湿疹控制得不好,更容易得其他过敏病。
◆ 为什么会得食物过敏?
食物过敏的形成和很多因素有关:
个人因素:
严重的早期湿疹
父母有过敏病史
环境因素:
接触过敏物的时间
肠道中的菌群平衡情况
环境因素如何影响过敏?
新出现的数据表明,在免疫系统发育过程中暴露于非微生物环境因素,包括空气过敏原、清洁剂、污染物和微塑料,可能会导致屏障功能障碍。霉菌、蟑螂和尘螨等空气过敏原含有促进上皮蛋白分解的蛋白酶,导致上皮渗透性增加,例如,清洁剂破坏角质层内的紧密连接和脂质依赖性相互作用,从而增加细胞旁渗透性。
◆ 食物过敏的风险因素:
对过敏原蛋白的反应会受到多种因素的影响,包括:
个人的敏感程度:有些人对过敏原特别敏感,而有些人反应较轻。
过敏原的特性:不同食物中的过敏原性质不同,反应也会有所差异。
食物加工方式:加工过程可能会改变过敏原的性质,比如加热可能降低或增强过敏性。
有的加工对某些过敏原有帮助,比如鸡蛋中的卵清蛋白和卵粘蛋白在加热后过敏原性下降。
但对某些过敏原(如花生蛋白)可能起反作用,加热会使其形成更稳定的抗性结构,增加过敏原性。
◆ 胃肠道屏障与过敏风险
婴幼儿风险更高: 由于他们的胃肠道屏障和免疫系统尚未成熟,食物过敏更容易发生。
胃酸不足: 服用抗酸剂可能降低胃酸浓度,使更多未被分解的蛋白质进入肠道,增加过敏风险。
肠道通透性增加: 肠道屏障功能受损可能导致更多过敏原进入免疫系统,增加致敏和过敏反应的可能性。
食物成分的相互作用:食物中的其他成分可能会影响过敏原的作用。
◆ 过敏反应的严重程度
敏感人群的过敏反应可能从轻微到严重不等,严重时甚至可能危及生命(如过敏性休克)。因此,避免接触过敏原是目前最有效的预防方法。
◆ 为什么食物过敏难以治疗?
目前还没有针对食物过敏的有效治疗方法,因为科学家对引发过敏的分子机制还没有完全弄清楚。因此,过敏患者最好的办法就是避免食用含有过敏原的食物。但问题是,只有在明确知道食物中含有过敏原时,才能做到有效避免。
◆ 食品标签的重要性
食品标签是帮助消费者了解食物中是否含有过敏原的重要手段。然而,确保全球食品标签的准确性并不容易,主要有以下几个挑战:
◆ 哪些食物最容易引发问题?
牛奶是最常见的未标注过敏原,其次是小麦和大豆。这些食物经常出现在食品召回事件中,很大程度因为它们被意外添加或未正确标注。
对于过敏患者来说,了解食品标签和避免过敏原是保护自己的关键。但由于标签错误、交叉污染等问题,完全避免过敏原仍然具有挑战性。
食物过敏原主要是一类特殊的蛋白质(糖蛋白),它们通常能溶于水,而且比较”顽固”-不容易被热、酸和消化酶破坏。
不过,不同的过敏原特性会有很大差异。有的需要糖分子的修饰才能发挥作用,有的则不需要。
过敏原的结构特点:
大小不一:从小分子到大分子都有。
结构多样:有的是单个分子,有的会几个分子组合在一起。
稳定性强:通常含有特殊的化学键(二硫键),使它们不容易被破坏。
结合能力:能与金属、脂肪等物质结合,这种结合会让过敏原更稳定。
不同食物的过敏原如果结构相似,可能会导致交叉过敏。
常见的交叉过敏组合:
过敏原的识别位点(表位):
直线型表位:像一串珠子一样排列的氨基酸片段
立体型表位:蛋白质折叠形成的特殊立体结构
一般认为直线型表位更重要,因为它们更稳定,不容易被破坏。
但在某些情况下,立体型表位也能在食物中保持稳定,发挥作用。
了解这些特性有助于预测哪些食物可能会引起交叉过敏;改进食物加工方法,降低过敏风险;开发更好的过敏预防和治疗方案。
致敏是指免疫系统首次接触到某种过敏原后,开始产生针对它的特异性抗体(IgE),但此时不会引发过敏症状。
致敏的主要途径:
小麦过敏是一种IgE介导的I型过敏反应,部分人群对小麦中的某些蛋白质成分较为敏感,即使在护肤品中接触也可能出现过敏症状。
如果你已知对小麦过敏,或者属于敏感肌肤,建议谨慎使用含有水解小麦蛋白的护肤品。
花生过敏者可能因皮肤接触花生而致敏,尤其是皮肤屏障受损的人。
胃肠道致敏(1类食物过敏): 最常见的方式,食物过敏原通过消化道进入身体,导致过敏反应,尤其常见于婴幼儿,因为他们的肠道屏障和免疫系统尚未完全发育。常见过敏原包括牛奶、鸡蛋和豆类。
吸入性致敏(2类食物过敏): 通过呼吸道接触到空气中的过敏原(如花粉、动物毛发等),随后对相关食物产生过敏。
皮肤接触致敏: 通过皮肤接触到过敏原而致敏。例如:
蜱叮咬致敏: 蜱叮咬可能导致对红肉的过敏反应,这是一种特殊的皮内致敏途径。
子宫内暴露: 胎儿可能通过胎盘接触到母体食物中的过敏原(如牛奶、鸡蛋和花生),但目前没有确凿证据表明母亲的饮食会直接影响婴儿的食物过敏风险。
B细胞的双重角色 | 引起过敏/预防过敏
什么是B细胞?
B细胞是我们身体里的一种免疫细胞,它最重要的工作就是产生抗体(免疫球蛋白)。在食物过敏中,B细胞主要产生一种叫IgE的抗体。
食物过敏发生的过程:
第一步: 识别“入侵者”
B细胞会产生IgE抗体,这些IgE能够识别食物中的特定蛋白质(过敏原)。
第二步: 设置“警报器”
IgE会附着在特殊的免疫细胞(肥大细胞和嗜碱性粒细胞)表面,这些细胞就像身体的“警报器”。
第三步: 过敏反应的触发
当吃到过敏食物时,IgE认出并抓住过敏原
“警报器”被触发,免疫细胞释放出化学物质,这些化学物质引起过敏症状(如皮疹、呼吸困难等)。
关于IgE的特点:
IgE在所有抗体中数量最少,但它在过敏反应中起着关键作用。
B细胞的变化过程:
B细胞可以通过特殊方式转变,开始产生IgE,这个转变需要其他免疫细胞(T细胞)的帮助,还需要特定的信号分子(IL-4和IL-13)的参与。
对花生过敏的研究发现,科学家从花生过敏患者的血液中找到了产生IgE的B细胞。这些B细胞寿命较短。
不同过敏患者的IgE抗体可能会以相似的方式识别花生中的过敏原,这些发现帮助我们更好地理解食物过敏的发生机制,为开发新的治疗方法提供线索,从而可能帮助预防或减轻过敏反应。
图 控制对过敏原反应的免疫过程
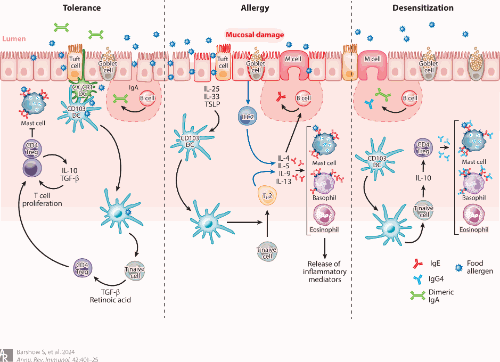
(左)耐受性:耐受性是一种主动免疫过程,在此过程中暴露于过敏原不会引发过敏性炎症。耐受性可涉及特异性抗原呈递细胞、TcB的诱导和活化以及B细胞同种型转换为伊加。
(中)过敏症:在过敏症中,上皮细胞功能障碍使抗原进入并触发alarmins IL-25、IL-33和TSLP的释放。幼稚T细胞分化成TH2细胞,其分泌促炎性TH2细胞因子IL-4、IL-5、IL-9和IL-13,其导致B细胞同种型类别转换为IgE。IgE通过高亲和力IgE受体(FcεR1)与肥大细胞和嗜碱性粒细胞结合,导致急性过敏性炎症。
(右)脱敏:过敏原免疫疗法的脱敏与IL-10的Treg产生相关。B细胞类别从IgE转变为IgG 4和伊加产生。IgG 4和伊加可通过分别与IgE竞争变应原结合位点和直接结合食物变应原而充当阻断抗体来抑制TH2适应性免疫应答。缩略语:DC,树突状细胞;TH2,2型辅助性T细胞; Treg,调节性T细胞; TSLP,胸腺基质淋巴细胞生成素。
不是所有抗体都会引起过敏;B细胞除了能产生引起过敏的IgE抗体,还能产生其他种类的保护性抗体,这些保护性抗体可以帮助预防食物过敏。
保护性抗体是如何工作的?
科学家通过小鼠实验发现粘膜中的IgA抗体能阻止过敏原被肠道吸收。
IgG抗体有两种保护作用:
直接捕获并中和过敏原;
通过FcγRIIb受体的作用抑制过敏反应。
此外,治疗食物过敏的新发现在口服免疫治疗(OIT)中观察到,保护性抗体IgG4会明显增加,IgG4与致敏抗体IgE的比值升高。这些变化与患者对食物的耐受性直接相关。
调节性B细胞的重要性
研究发现牛奶过敏患者与健康人对比,过敏患者的调节性B细胞明显减少,这些细胞产生的IL-10也较少。
IL-10的重要作用可以减少致敏抗体IgE的产生,促进保护性抗体IgG4的产生。
B细胞的信使功能:
B细胞也可通过IgE促进的抗原呈递作用作为T细胞的抗原呈递细胞(APC),其中IgE-过敏原复合物被低亲和力IgE受体CD 23(FcεRII)内化,后者在B细胞上表达。
值得注意的是,最近的一项研究表明,B细胞上CD 23的表面密度通过IgE促进的抗原呈递决定了过敏原特异性T细胞的活性,并与总血清IgE水平呈正相关。抗CD 23单克隆抗体鲁昔单抗通过靶向CD 23-IgE相互作用抑制IgE合成。
B细胞治疗方面潜力:
预防方面:
T细胞在食物过敏中的角色
1. TH2细胞:过敏反应的”指挥官”
研究发现,TH2细胞是引发食物过敏的核心细胞,它通过三种方式发挥作用:
2. 治疗的新发现
口服免疫治疗(OIT)的研究显示:能够抑制TH2细胞的活动,减少过敏相关物质的产生,帮助建立对食物的耐受性。
3. 新发现的重要T细胞类型
最新研究发现Tfh13细胞在食物过敏中的重要作用,动物实验证明,这种细胞对产生高效过敏抗体必不可少,能直接影响过敏反应的强度。
注:Tfh13(滤泡辅助性T细胞13)是一种特殊的免疫细胞亚型,属于CD4+ T细胞家族。它在免疫反应中起着重要作用,尤其是在过敏反应和IgE抗体的产生过程中。
此外,还有TH2A细胞,这是一种特殊的记忆性T细胞,该细胞只在过敏人群中发现,其表面有特定标记物(CD161、CRTH2、CD49d)。发现该细胞不仅与食物过敏有关,还与其他过敏病相关,例如,湿疹、过敏性鼻炎、哮喘和食管炎。
4. T细胞的共同识别标记
研究人员通过CD154标记发现:花生过敏患者的T细胞有特殊特征,17%的特定序列在不同花生过敏患者中相同。这说明花生过敏可能有共同的免疫机制。
5. 调节性T细胞(Treg):过敏的“刹车系统”
Treg细胞是免疫系统的调节者,有两种来源:
动物实验显示,牛奶过敏小鼠经过治疗后:Treg细胞数量增加,产生更多保护性物质(IL-10和TGF-β)。
人类研究发现,鸡蛋过敏儿童成功治疗后:特定类型Treg细胞增加。与此同时,花生过敏治疗后:Treg相关基因活性增加。
T细胞在诊断和治疗方面应用潜力:
肥大细胞:驻扎在“人体边防”的过敏触发器
主要在身体的”边界”位置:皮肤、呼吸道、消化道。
研究发现:对胃肠道过敏患儿的活检发现,他们的胃部和结肠中,肥大细胞数量比健康孩子多(通过CD117和类胰蛋白酶标记检测)。
临床观察:在食物过敏患者的肠道检查中发现,过敏相关物质明显增加:
特定条件下可能缓解过敏
小鼠实验发现,肥大细胞表面有个“刹车开关”(FcγRIIb),能接收IgG抗体的“停战指令”,抑制过敏反应,这可能是未来治疗的关键靶点。
嗜碱性粒细胞:血液中的哨兵
基本特征:在血液中数量很少(仅占白细胞的0.5-1%),但在过敏反应中作用重要。
临床应用:可以通过流式细胞术(BAT)检测,使用CD63和CD203c作为标记,帮助诊断食物过敏,监测治疗效果。
治疗相关发现:口服免疫治疗(OIT)研究显示,成功治疗后:
小鼠实验发现:在 IgE-FA 的小鼠模型中,嗜碱性粒细胞耗竭减轻了腹泻的发生率,临床过敏反应减弱。同一项研究表明,嗜碱性粒细胞来源的 IL-4 对于肥大细胞募集和/或增殖至关重要。
临床意义:可以通过检测这两种细胞,判断过敏严重程度,评估治疗效果。
治疗方向:可以破坏 IgE-FA 中肥大细胞和嗜碱性粒细胞反应性的新型干预措施,可能代表未来研究的有吸引力的治疗靶点。
抗原呈递细胞(APCs):免疫系统的“情报分析员”
主要包括三类细胞:
抗原呈递细胞是如何工作的?
➤ 发现并捕获食物过敏原 → 分解成小片段(肽段) → 用MHC II分子“举牌”展示
➤ 指挥CD4+ T细胞分化成不同细胞(TH1/TH2/Treg等)
不同派系的树突状细胞:过敏派 vs 和平派
◈ 过敏派(CD209+树突状细胞)
锁定目标:锁定致敏蛋白,如花生、坚果等。
煽风点火:释放信号诱导T细胞变身TH2(分泌IL-4/IL-5/IL-13)
研究证据:
◈ 和平派(CD103+树突状细胞)
维稳专家:在小鼠肠道训练T细胞成为Foxp3+ Treg(分泌IL-10),有助于预防过敏。
制衡机制:表达诱导性T细胞共刺激配体(ICOS-L),促进“调解员细胞”生成。
树突状细胞功能的其他重要考虑因素包括共刺激分子如 CD80 和 CD86 等表达,它们是 CD28 的天然配体;这些分子与 T 细胞表达的 CD28 的相互作用受损可能导致激活减少或缺失,并最终导致 T 细胞无反应。
花生口服免疫治疗降低了APC上CD86的表达,尽管这些变化可能是暂时的。
诊断方面:通过分析树突状细胞的亚型特征和活化状态,可评估个体过敏风险等级,并预测潜在的过敏反应发生概率。
治疗突破:为过敏治疗开辟了新路径。
1)精准调控不同功能亚型DC的比例平衡;
2)靶向干预DC表面关键分子(如共刺激分子)的表达;
3)特异性增强具有免疫调节功能的DC活性。
总之,免疫系统的多个部分协同作用。
正常耐受状态是B细胞生产保护性抗体(IgA),调节性T细胞(Treg)担任”指挥”角色,共同抑制过度反应,防止肥大细胞引发过敏症状。
过敏状态TH2细胞过度活跃,产生大量致敏物质,指导B细胞产生过敏抗体(IgE)。多种免疫细胞被激活,包括肥大细胞、嗜碱性粒细胞、嗜酸性粒细胞等。受损的肠道细胞会释放:IL-25、IL-33、TSLP,这些物质进一步加重过敏反应。
成功的脱敏治疗会带来以下变化:
T细胞方面,抑制致敏T细胞(TH2)、增加调节性T细胞;
B细胞方面,增加保护性抗体(IgA和IgG4)、减少致敏抗体(IgE)。
来自人类和动物研究的越来越多的证据支持肠道微生态失调在食物过敏发展中的关键作用。
基于16S rRNA测序的研究发现,与没有食物过敏的儿童相比,患有食物过敏的儿童具有独特的肠道微生物组结构。
表. 在有和没有FA的小儿患者之间主要的肠道微生物组差异(基于16S-rRNA测序的研究)
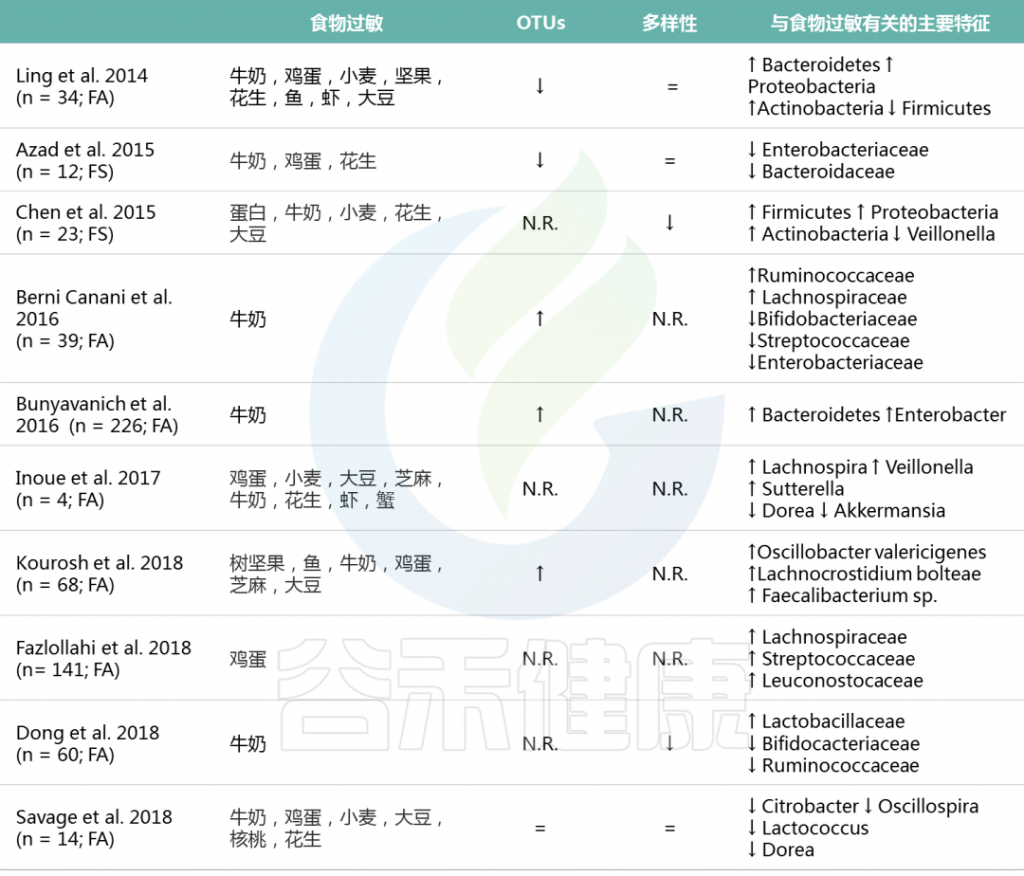
M.D.Costanzo et al, Int.J.Mol.Sci. 2020
FA:食物过敏; FS: 食物过敏; OTUs:操作分类单元; N.R.: 未报道过的; ↑ : 增加;↓: 减少; =: 不变.
表1中所有研究均调查了IgE介导的食物过敏。有趣的是,关于46例受非IgE介导的牛乳过敏(CMA)影响的患者的数据显示,与健康对照组相比,失调的菌群中拟杆菌Bacteroides (Bac 12) 和Alistipes丰富,IgE介导的CMA儿童的特征重叠,是从健康到IgE介导的CMA患者的拟杆菌逐渐增加。
在同一项研究中,患有非IgE介导的CMA的儿童的粪便中丁酸浓度明显低于健康人。
菌群失调先于食物过敏发作
来自人体研究的可用数据表明,菌群失调先于食物过敏发作。
Azad 等人研究发现,在婴儿早期肠道微生物组含量较低的情况下,肠杆菌科(Enterobacteriaceae) / 拟杆菌科(Bacteroidaceae)的比率增加和Ruminococcaceae的丰度较低与随后的食物敏感性相关,这表明早期肠道菌群失调对食物过敏的后续发展有促进作用。
此外,人类研究的可用数据表明,生命早期,尤其是生命的头6个月,微生物组构成与食物过敏的发展更为相关;
在出生后的前 6 个月内,在食物过敏儿童的肠道菌群中占比过高的菌如下:
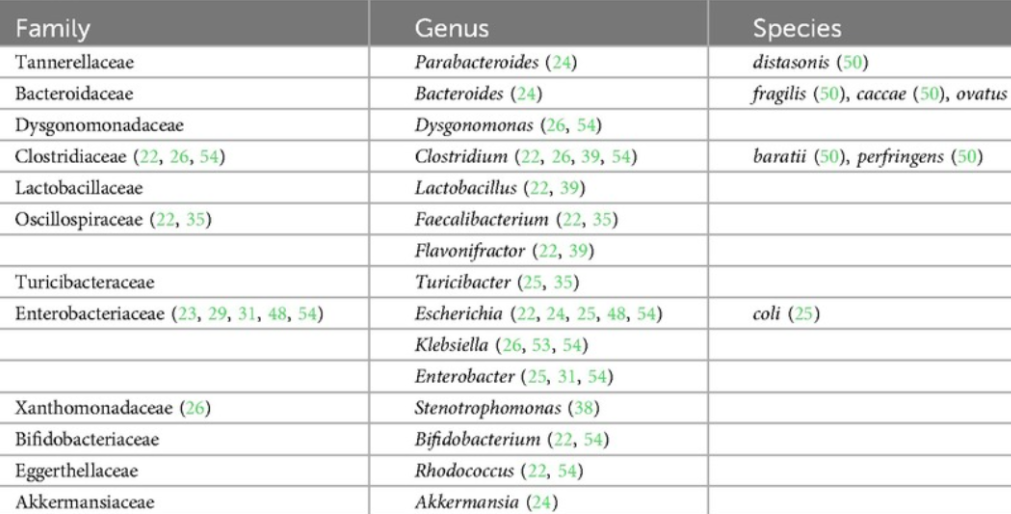
doi.org/10.3389/falgy.2024.1438252
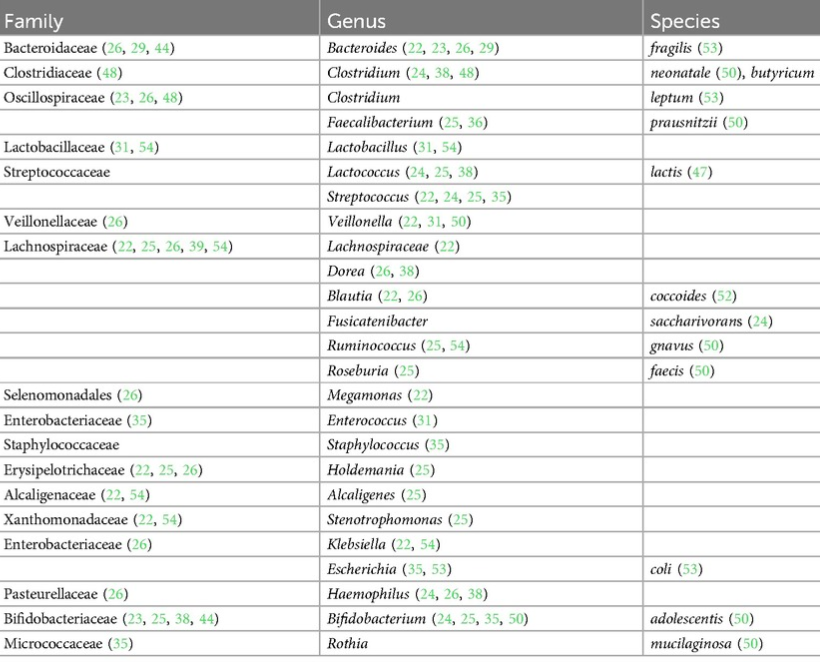
在出生后的前 6 个月内,在食物过敏儿童的肠道菌群中缺乏的菌如下:
doi.org/10.3389/falgy.2024.1438252
▲ 有害菌增加:
肠杆菌等有害菌释放脂多糖(LPS)等毒素:
▼ 有益菌减少:
双歧杆菌、乳杆菌等能分解食物纤维,产生短链脂肪酸(如丁酸),这类物质:
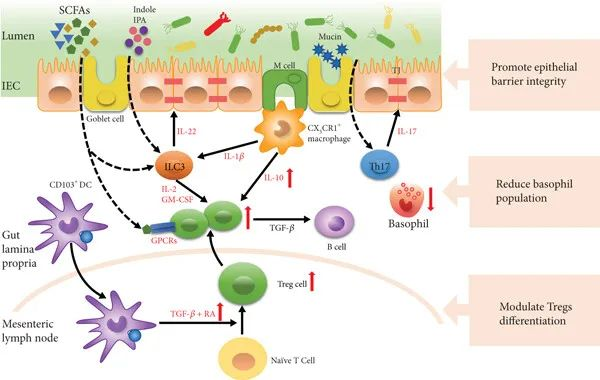
调节免疫耐受
肠道菌群通过调节Tregs细胞的分化和功能,增强对食物抗原的耐受性。例如,梭菌和双歧杆菌能够通过其代谢产物(如SCFAs)和信号分子(如IL-10、TGF-β)促进Tregs的分化和增殖,从而抑制过敏反应。
肠道菌群还通过调节树突状细胞(DCs)的功能,影响免疫反应。例如,健康的肠道菌群能够使DCs产生TGF-β和视黄酸(RA),从而诱导Tregs的分化。
共生菌介导免疫耐受
doi.org/10.1155/2023/9575410
增强肠道屏障功能
肠道菌群通过产生短链脂肪酸(如丁酸)和色氨酸代谢产物(如吲哚)来增强肠道上皮屏障的完整性。这些代谢产物能够促进黏液分泌和紧密连接蛋白的表达,从而防止食物抗原进入血液循环,减少过敏反应的发生。
例如,丁酸能够通过诱导IL-22的产生,增强肠道上皮细胞的屏障功能,减少过敏原的渗透。
调节过敏效应细胞
肠道菌群能够调节过敏效应细胞(如嗜碱性粒细胞)的数量和功能。例如,抗生素处理或无菌小鼠的嗜碱性粒细胞数量增加,而补充益生菌可以减少这些过敏效应细胞的数量,从而减轻过敏症状。
关于肠道菌群的干预措施,我们将在06章节详细阐述。
IgE-食物过敏的诊断依赖于临床病史和过敏试验的结合,如食物特异性IgE的存在。诊断指南和标准临床实践在国际上各不相同。这些差异主要是由于多种方法可用于评估IgE-FA;这些方法在灵敏度、特异性、成本、时间和可及性方面各不相同。准确的诊断对于防止不必要的食物避免和提供及时和适当的治疗建议至关重要。
金标准
双盲安慰剂对照食物激发试验(DBPCFC)是最可靠的确诊方法。由于12.9%的试验可能出现安慰剂反应,其准确性优于非盲法试验。
确定IgE对给定食物致敏的两种主要方法是:
体内皮肤点刺试验(SPT)
操作:将微量食物(新鲜的或更常见的作为提取物)刺入皮肤表层
判读标准:15-20分钟后,检查皮肤刺痛部位,若出现比阴性对照大3mm以上的红肿包块(因肥大细胞释放组胺所致)即为阳性。
编辑
血清特异性IgE(sIgE)
sIgE测定定量外周血中存在的过敏原特异性IgE的量。
传统标准:≥0.35 kU/L为阳性
新型检测:临界值可低至0.1 kU/L
注意:两项检测结果必须谨慎解释,均需结合症状分析,因可能出现”致敏但无过敏反应”的假阳性。
检测结果优化策略:
使用较大的SPT包块直径或较高的sIgE临界值可增加临床反应性的可能性,但这是以牺牲敏感性为代价的。
组合检测优势:
对1247例食物激发试验数据分析显示:
SPT+sIgE联合检测或sIgE/总IgE比值(sIgEr)
的阳性预测值显著优于单项检测。
应用价值:在临床试验等场景中可替代部分激发试验,但仍需更多验证。
现存挑战:许多具有临床相关IgE-FA的患者的SPT和/或sIgE值可能低于已发表的临界值,强调需要更稳健的诊断方法来取代口服食物激发试验。
食物过敏检测升级版:成分检测与表位检测有什么用?
除了常规的皮肤点刺、抽血验IgE,医生手里还有两把更精细的“放大镜”——成分检测和表位检测。它们能帮你区分“假警报”和“真危险”,甚至预测过敏反应有多严重。
成分检测:锁定过敏食物里的“关键蛋白质”
原理:
食物过敏其实是对其中某些蛋白质过敏。比如花生含有Ara h1、Ara h2等多种蛋白质,成分检测(CRD)可以单独测出你对哪一种蛋白质过敏。
实际用途:
区分“花粉过敏连带反应”和“真过敏”。
比如有人吃花生过敏,但检测发现只对Ara h8蛋白敏感,这通常是因为它和桦树花粉蛋白相似,属于“交叉反应”(可能症状轻微,比如口腔发痒)。
但如果对Ara h1/2/3敏感,则是真正的花生过敏,严重时可能引发全身反应。
预判过敏风险高低
研究证实:对花生中的Ara h2蛋白过敏的人,严重过敏风险更高;而另一种Ara h6蛋白(与Ara h2结构相似)阳性,也可能指向真实过敏。

表位检测: 看清IgE攻击的“精确靶点”
原理:如果把过敏蛋白比作一根长绳,表位就是绳子上被IgE抗体“咬住”的具体位置(分线性和构象两种结构)。表位检测能识别这些关键攻击点,帮助判断过敏是否会长期存在或容易引发严重反应。
在花生过敏的儿童和成人中,针对花生过敏的BBEA表位检测法(检测Ara h2蛋白的两个线性表位):灵敏度92%,特异性94%,准确度碾压常规检测:比皮肤点刺、普通IgE检测更准。
另一个相关的体液测试是 过敏原特异性IgG4
IgG4抗体通过充当阻断抗体而在IgE-FA中具有保护作用。一项研究发现,花生致敏但临床耐受或花生过敏并接受OIT治疗的儿童的血浆能够抑制花生诱导的肥大细胞活化。重要的是,当血浆中的IgG4耗尽时,肥大细胞的活化不能完全恢复。
但是目前需要进一步的研究来更全面地了解IgG4单独或与其他生物标志物结合在食物过敏诊断和/或预后中的作用。
编辑
诊断方法仍然主要限于研究环境,包括嗜碱性粒细胞活化试验(BAT)和肥大细胞活化试验(MAT)。
BAT与MAT的优劣对比:
BAT和MAT都是基于体外流式细胞术的实验室检测,用于测量过敏原刺激后活化标志物的表达。
BAT检测(嗜碱性粒细胞活化试验)
需要采集新鲜血液立即检测。
BAT在诊断IgE-食物过敏时的特异性高于SPT和sIgE水平,在花生过敏中的特异性为98%,灵敏度为75%。
但存在15%人群天生对检测无反应。
MAT检测(肥大细胞活化试验)
可用冷冻血清来激活培养的原代人血源性肥大细胞。
在花生过敏诊断中比BAT更准确。
重复检测结果高度一致(组内相关性:MAT 0.96 VS BAT0.43)。
新的诊断方法包括:
评估食物过敏原特异性B和T细胞
外周血中的过敏原特异性B和T细胞由于其稀缺性而历来难以分离。由于技术的进步,这些罕见的细胞现在可以通过几种方法来识别。
关于T细胞,这些方法包括:
a)用所选抗原或肽体外刺激PBMC,然后通过表达特异性活化标志物经由流式细胞术鉴定活化的T细胞;
b)用重组肽-MHC(pMHC)复合物离体染色T细胞。pMHC四聚体或多聚体技术也可用于鉴定变应原特异性B细胞。
多组学研究
包括基因组学、表观基因组学、转录组学、蛋白质组学、代谢组学、微生物组学和免疫组学在内的组学科学有望成为研究过敏性疾病(包括IgE-FA)的高通量方法。需要多组学方法来全面识别生物标志物,如基因,蛋白质和/或代谢产物,可以提高IgE-食物过敏诊断和预后的准确性。
此外,非常需要鉴定可预测IgE-食物过敏反应严重程度的生物标志物。特别令人感兴趣的是血小板活化因子乙酰水解酶(PAF-AH)。之前的研究已经证实,PAF-AH可以通过灭活PAF来预防过敏级联反应的进展。
肠道菌群检测
通过高通量测序技术获得菌群组成和功能数据,结合机器学习模型(如随机森林、神经网络)识别过敏相关特征。肠道菌群检测报告菌群失衡程度、关键菌群丰度及代谢物水平,提示过敏风险。
干预后复查菌群,观察过敏症状是否缓解,验证菌群与过敏的因果关系。
该方法是非侵入性的,且可以早期预警,因为很多时候菌群失衡可能早于过敏症状出现。通过个性化干预,指导益生菌、膳食调整等精准治疗。
肠道菌群检测可作为食物过敏的辅助诊断工具,为个性化治疗提供依据。可以结合临床传统方法(如食物激发试验)综合判断,通过连续检测评估过敏风险和干预效果,整合IgE检测、皮肤点刺试验和菌群数据,提高准确性。
1. 当前的食物过敏治疗方式
目前,食物过敏没有治愈方法,治疗的核心是避免接触过敏原。如果意外接触导致过敏反应,通常使用肾上腺素和抗组胺药进行处理。
由于食物过敏带来的严重健康和经济负担,科学界投入了大量资源研究新的治疗方法。
2. 预防的转变:从回避到早期接触
过去建议高过敏风险的婴儿(如父母有过敏史)避免接触过敏性食物,比如花生。
但后来研究发现,在婴儿早期引入过敏性食物(如花生)反而可以降低过敏风险。这项发现改变了过敏预防的建议,也激发了对免疫疗法的研究热情。
3. 免疫疗法的探索
免疫疗法的目标是通过逐步改变免疫系统的反应,让身体对过敏原“习惯”并耐受。方法包括:
其中,口服免疫疗法(OIT)效果最好,但也存在风险。比如,患者可能会出现轻微不适(如口痒、肚子痛),甚至更严重的过敏反应(如全身性过敏或嗜酸性食管炎)。
doi.org/10.1146/annurev-immunol-090122-043501
4. 辅助治疗:提高疗效,减少副作用
为了让免疫疗法更安全有效,科学家正在研究辅助治疗:
此外,新的治疗IgE-FA包括疫苗、纳米颗粒、Janus激酶(JAK)和布鲁顿的酪氨酸激酶抑制剂。
5. 基于微生物群的干预
研究发现,肠道中的某些细菌和它们的代谢产物(如丁酸盐)可以调节免疫系统,帮助身体更好地耐受过敏原。下列措施可以在调节肠道菌群的基础上干预食物过敏。
饮 食
增加膳食纤维
高纤维饮食(如全谷物、蔬菜、水果)可促进双歧杆菌、乳酸菌等有益菌增殖,增加短链脂肪酸的生成,抑制促过敏的Th2免疫反应。
举例:
菊粉(如洋葱、大蒜):增加双歧杆菌。
果胶(如苹果、柑橘):改变厚壁菌门与拟杆菌门的比例,增加血清和粪便中SCFA的含量,并通过损害树突状细胞功能来预防炎症的发展。
高纤维摄入通过重塑肠道微生物群和增加SCFA水平,特别是乙酸盐和丁酸盐水平,保护小鼠免受花生过敏。此外,这种保护作用取决于SCFA的受体GPR43和GRP109A,因为缺乏其中一种的小鼠表现出更严重的食物过敏。
减少高脂高蛋白饮食
过多动物蛋白和脂肪可能促进拟杆菌属等菌群占优,增加炎症风险,间接诱发过敏。建议适量摄入,平衡植物蛋白(如豆类)。
高脂饮食诱导的肠道菌群变化也被证实可以诱导食物过敏。
一项研究显示,高脂饮食通过增加Ruminococcaceae(梭菌目)、Desulfovibrionaceae(δ-变形菌纲)和Rikenellaceae(拟杆菌纲)的相对丰度,同时减少Muribaculaceae和Prevotellaceae的丰度来诱导食物过敏。
增加优质脂肪
ω-3脂肪酸,ω-3主要来自海洋来源(如鱼油、藻类)和某些植物(如亚麻籽):通过抑制树突状细胞(DCs)的激活、抑制CD4 T细胞的增殖和活化、减少促炎因子、降低IgE水平以及抑制肥大细胞脱颗粒,从而减轻食物过敏。
增加膳食多酚
多酚是次生植物代谢产物,富含水果、蔬菜、谷物、咖啡和茶。非结合多酚通过肠道菌群调节、紧密连接蛋白表达和 β-防御素分泌来缓解食物过敏。
可可多酚:
研究表明,富含可可多酚的饮食可以减少厚壁菌门和变形菌门,增加Tenericutes和蓝藻门(Cyanobacteria),从而抑制特定抗体和肠道IgA的合成,有助于减少过敏反应。
矢车菊素-3-O-葡萄糖苷(Cyanidin-3-O-glucoside):
通过增加有益菌(如乳酸杆菌和Odoribacter)和减少有害菌(如幽门螺杆菌和Turicibacter)来上调紧密连接蛋白的表达,促进分泌型IgA和β-防御素的分泌,并平衡Th1/Th2水平,有助于增强肠道屏障功能,减少过敏原的渗透,从而减轻食物过敏症状。
槲皮素(Quercetin):
槲皮素具有抗氧化、抗炎和抗癌活性。它可以通过与卵清蛋白(OVA)结合,减少OVA的致敏性,并降低特异性免疫球蛋白水平,从而减轻过敏反应。
咖啡酸和表儿茶素没食子酸酯(Epigallocatechin-3-gallate, EGCG):
与乳清蛋白结合后,可以减轻乳清蛋白引起的口服致敏反应,降低小鼠血清中特异性抗体和mMCP-1水平,并减少体外CD4 T细胞的增殖,有助于减少乳清蛋白引起的过敏反应。
益生菌
双歧杆菌(Bifidobacterium)
乳杆菌(Lactobacillus)
丁酸梭菌(Clostridium butyricum)
混合益生菌
注意:
益生元
通过特定益生元干预食物过敏的机制主要是调节肠道菌群,促进短链脂肪酸生成,抑制过敏相关炎症反应。以下为具体益生元及研究证据:
临床试验显示,补充该组合可显著降低高风险婴儿食物过敏发生率,可能与促进双歧杆菌生长有关。
2’-岩藻糖基乳糖(2’-FL)和乳糖-N-新四糖(LNnT):添加到配方奶粉中可缓解牛奶蛋白过敏(CMA),降低IgE水平和炎症因子(如IL-4、IL-6)。
乳-N-岩藻五糖III(LNFP III):摄入量高的婴儿CMA风险更低。
动物实验表明,母体补充可诱导免疫耐受,降低后代小麦过敏风险。
多不饱和脂肪酸(PUFA)等可能减少儿童食物致敏风险。
合生元
合生元是一种包含活菌(益生菌)和宿主微生物选择性利用的底物(益生元)的混合物,能够为宿主带来健康益处。
在一项多中心随机对照试验(RCT)中,研究了含有短双歧杆菌M-16V和低聚果糖(scGOS/lcFOS)的氨基酸配方(AAF-S)对IgE介导的牛奶过敏(CMA)婴儿的安全性。
结果显示,与普通氨基酸配方(AAF)相比,AAF-S在控制过敏症状和促进正常生长方面同样有效,但在减少感染、用药次数和住院率方面效果更好。具体数据如下:AAF-S组婴儿的感染发生率显著降低、使用的药物更少、住院率显著降低(8.8% vs. 20.2%,p = 0.036)。
粪便微生物组分析显示,AAF-S组婴儿的双歧杆菌丰度显著增加,而Eubacterium rectale和C. coccoides的丰度显著降低。
代谢产物变化:AAF-S组婴儿的粪便中乙酸和L-乳酸水平显著升高,粪便pH值显著降低。
粪菌移植
最近,一项小鼠模型研究表明,FMT抑制了特应性皮炎小鼠中OVA诱导的过敏反应。
除了恢复肠道微生物群外,FMT还降低了IgE水平;受监管的Tregs;肥大细胞、嗜酸性粒细胞和嗜碱性粒细胞减少;提高了SCFA的含量;恢复Th1/Th2平衡。
与其他方法相比,FMT是一种相对简单的治疗策略,可以改变人体肠道微生物群;然而,迄今为止,这项研究还不够充分,需要开展大量工作来提高我们对用FMT治疗食物过敏的理解。
中草药
中药因其低成本、高安全性以及高生物活性,近年来在食物过敏治疗中受到越来越多的关注。虽然传统中医中没有“食物过敏”的具体术语,但一些中药方剂在现代研究中显示出对食物过敏的潜在治疗效果。
食物过敏草药配方-2(FAHF-2):
来源:FAHF-2是从传统的“乌梅丸”(Wu Mei Wan)中开发而来的。
临床研究:在一项多中心、双盲、随机、二期临床试验中,68名12-45岁的食物过敏患者每天三次服用FAHF-2或安慰剂,持续六个月。结果显示,FAHF-2安全且耐受性良好。
免疫调节作用:FAHF-2能够显著抑制外周血单个核细胞(PBMCs)产生的IL-5,并增加IL-10和调节性T细胞(Tregs)的生成,显示出良好的免疫调节效果。
改进形式:由于FAHF-2的剂量较大,研究者开发了两种改进形式:丁醇纯化的FAHF-2(BF2)和乙酸乙酯及丁醇纯化的FAHF-2(EBF2)。在花生过敏的小鼠模型中,EBF2显示出很强的抑制IgE产生的能力,被认为是对花生过敏最有潜力的治疗药物。
尽管中药在治疗过敏性疾病中被广泛应用,但关于食物过敏的临床研究样本量小,且研究受限,需要进一步验证中药在食物过敏治疗中的效果。
从概念上讲,人类食物供应中的任何蛋白质都可能是潜在的过敏原,因此建立“无过敏原食品”体系不现实。由于不同个体的免疫系统对相同的过敏原可能会有不同的反应,因此很难对食物过敏做出普遍适用的定义。
现有诊断工具(如特异性IgE检测)可靠性有限,过敏原结构、加工方式及食物基质相互作用均影响致敏性。
引起个体过敏反应所需的最小量(阈值)也取决于个体的敏感性。大多数过敏性食物的阈值尚未确定,导致警示标签(如“可能含”)难以精准保护敏感人群。 需开发特异性强、灵敏度高的检测方法,并基于主要过敏食物(如牛奶)建立分级阈值标准——从鉴定关键过敏原及其表位入手,明确检测目标。
肠道菌群可能通过调节免疫耐受影响过敏风险。未来可探索菌群检测(如特定菌群丰度与过敏表型关联)作为辅助诊断工具,或通过益生菌等多种干预手段重塑菌群平衡,降低致敏性。
协同策略:
整合免疫学、微生物组学与食品加工技术,推动从“一刀切”警示转向动态风险评估,实现过敏防控的精准化与人性化。
注:本账号内容仅作交流参考,不作为诊断及医疗依据。
主要参考文献:
Iweala OI, Nagler CR. The Microbiome and Food Allergy. Annu Rev Immunol. 2019 Apr 26;37:377-403.
Januszkiewicz, Emilia, et al. “The importance of the gut microbiome in the development of allergic diseases.” Alergologia Polska-Polish Journal of Allergology 10.3 (2023): 202-209.
Venter, Carina. “Immunonutrition: Diet diversity, gut microbiome and prevention of allergic diseases.” Allergy, Asthma & Immunology Research 15.5 (2023): 545.
Jiachen Liang, Baodong Zheng, Yi Zhang, Hongliang Zeng, Food allergy and gut microbiota, Trends in Food Science & Technology, Volume 140, 2023, 104141, ISSN 0924-2244
Farnetano M, Carucci L, Coppola S, Oglio F, Masino A, Cozzolino M, Nocerino R, Berni Canani R. Gut microbiome features in pediatric food allergy: a scoping review. Front Allergy. 2024 Sep 25;5:1438252.