-

 国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子
国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子

 二级病原微生物安全实验室
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
谷禾健康
产前抑郁
在围产期健康管理中,产前抑郁(antenatal depression, AND)正在受到越来越多关注。相比产后抑郁,产前抑郁过去长期处于相对被低估的位置,但它的发生率并不低。
既往荟萃分析显示,无论以抑郁症状还是临床诊断作为标准,产前抑郁的全球患病率均接近20%。
从临床结局来看,产前抑郁已被证明与多种不良母婴结局相关,包括流产风险增加、早产、低出生体重等。
近几年,研究视角开始进一步延伸到儿童早期发育。越来越多研究关注母亲孕期抑郁症状对子代神经发育的影响,例如婴儿认知、语言、运动、问题解决能力和社会行为等发育维度。
但目前相关结果并不完全一致。一个重要原因可能是不同研究关注的抑郁暴露时间窗口不同。过去不少研究集中在孕晚期,而对孕早期、孕中期,或整个孕期持续性抑郁症状的影响关注不足。
但胎儿脑发育贯穿整个孕期。只看孕晚期,可能会低估早期情绪风险对子代发育的影响。
更关键的是,产前抑郁究竟如何影响子代神经发育,目前仍缺乏清晰机制。过去的解释更多集中在 HPA 轴、皮质醇、炎症反应、胎盘功能和母体代谢状态等方面。而近年来,肠道菌群逐渐成为一个新的机制切入点。
已有研究显示,抑郁和焦虑状态可改变肠道菌群组成;孕期焦虑、抑郁症状也与菌群多样性下降及特定菌群丰度变化有关。但这些研究多为横断面或小样本研究,较少追踪整个孕期菌群动态,也难以回答菌群变化是否真正参与了子代神经发育异常。
这里就出现了一个很关键的问题:
产前抑郁是否会通过改变母体肠道菌群,影响胎儿脑发育和出生后的神经认知表现?
近期,来自上海复旦大学妇产科医院和上海交通大学的研究团队发表在《Gut Microbes》的一篇研究,正是从这个问题切入。
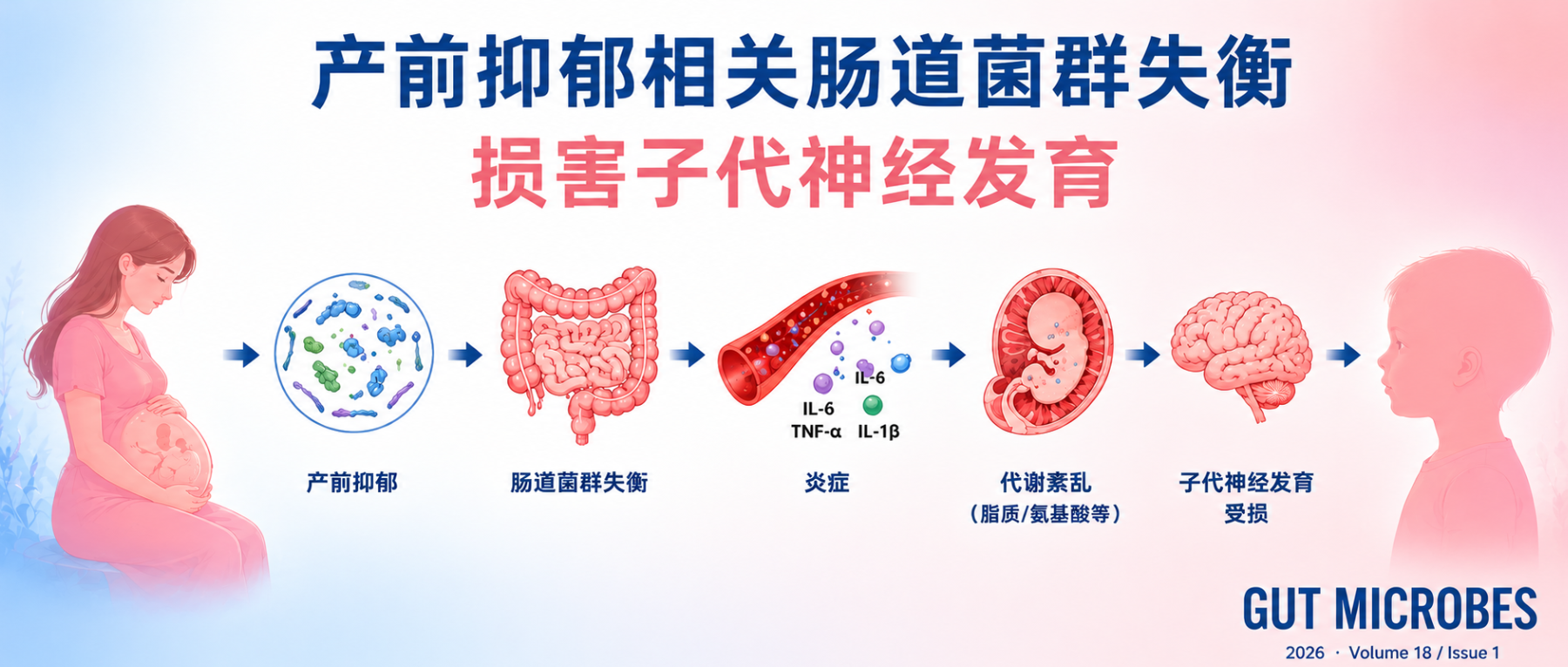
研究人员结合多中心前瞻性孕妇队列、嵌套病例对照菌群分析、无菌小鼠粪菌移植实验以及丁酸补充实验,系统验证了孕期抑郁症状与婴儿神经发育受损之间的关系。
产前抑郁与婴儿早期神经发育受损相关,主要表现为婴儿问题解决和社交能力的下降,而且孕早、中、晚期的抑郁都会导致这一问题。机制上可能是通过母体肠道菌群失衡,尤其是产丁酸菌减少,导致肠屏障受损、炎症增强及胎儿脑代谢异常,从而影响神经发育。对母亲和婴儿的菌群检测显示,母婴之间肠道菌群传递较少。在小鼠实验中,粪菌移植实验表明,仅母体抑郁相关肠道菌群即可导致后代学习记忆能力下降。而丁酸补充可以改善炎症、代谢异常及后代认知表现。
对于围产期健康管理而言,这意味着孕期情绪风险背后,可能存在值得更早关注的微生态信号。
这项研究的设计比较完整,先在人群中确认关联,再通过菌群分析寻找机制线索,最后用动物实验验证因果可能。
◆ 人群队列:追踪孕期抑郁与婴儿发育
研究首先纳入 2053 名孕妇,在孕早期、孕中期和孕晚期分别使用 EPDS 量表评估产前抑郁症状,并在婴儿 3、6、12 月龄时随访神经发育表现。
随访重点包括:
注:EPDS量表是目前全球公认且广泛使用的围产期抑郁筛查工具。
◆ 菌群分析:寻找可能的机制线索
随后,研究者从队列中开展嵌套病例–对照分析,匹配出 135 名产前抑郁风险孕妇和 135 名对照孕妇,并对 504 份孕期母体粪便样本进行 16S rRNA 测序,追踪孕早、中、晚期肠道菌群变化。
在菌群分析上,研究除了关注哪些菌升高或降低,进一步将相关菌群归纳为共丰度模块,并结合功能预测,观察这些菌群变化可能对应哪些代谢通路异常。
同时,研究还收集了 290 份婴儿粪便样本,用于判断产前抑郁是否会影响婴儿出生后的菌群建立,以及母婴菌群传递是否参与神经发育异常。
◆ 动物实验:验证菌群是否具有因果作用
在人群研究之外,研究者进一步使用无菌小鼠进行粪菌移植(FMT):将产前抑郁风险女性和健康女性的菌群分别移植给无菌孕鼠,观察:
◆ 干预验证:丁酸是否是关键线索
最后,研究加入丁酸补充实验,用来验证产丁酸菌减少是否可能是关键干预点。
编辑
这样的设计超越了单纯的相关性分析,进一步挖掘了菌群是否参与其中及相关机制,以及是否存在可干预的代谢节点。
【 队列研究 】
在2053名女性的队列研究中,326名(15.9%)至少有一次EPDS ≥10 在妊娠期间(AND高危组),1634人(79.6%)没有(对照组)。
【 婴儿随访 】
对1岁以下的婴儿进行神经发育评估。AND高危组婴儿在3个月和6个月时的问题解决任务得分较低(P = 0.039,0.040)。
编辑
◐ 母亲孕期抑郁越明显,婴儿问题解决能力越弱
尤其是在 3 月龄时,EPDS 评分较高的母亲,其婴儿在问题解决能力和社交能力方面得分更低。
研究者还使用重复测量 GEE 模型进行分析,结果同样显示:无论是在孕早期、孕中期还是孕晚期,母亲 EPDS 评分越高,婴儿的问题解决能力越差。
◑ 嵌套病例–对照研究,再次验证
与对照组相比,存在产前抑郁症状的母亲,其婴儿在 3 月龄时问题解决能力更低,在 6 月龄时社交能力也更弱。
编辑
从这部分结果看,产前抑郁症状与婴儿早期神经发育表现下降存在关联,尤其集中在问题解决能力和社交能力等维度。
更重要的是,这种关联并不只出现在孕晚期。孕早期、孕中期和孕晚期 EPDS 评分升高,都可能与婴儿后续问题解决能力下降有关。也正因为如此,研究下一步需要回答:这种影响是否只是情绪状态本身带来的结果,还是与孕期母体肠道菌群变化有关?
在人群队列中,研究已经看到产前抑郁症状与婴儿早期神经发育表现下降有关。接下来,研究者进一步想了解:这种关联背后,母体肠道菌群是否发生了可识别的变化?
先区分“抑郁阶段”和“非抑郁阶段”
经过质量控制,504份来自不同孕期的母体粪便样本可供分析,覆盖孕早期、孕中期和孕晚期。
这里研究者并没有简单把孕妇分成“抑郁组”和“非抑郁组”,而是根据采样当时的 EPDS 评分,把样本进一步分为三类(下图d)。
编辑
这个分组很重要,可以帮助判断,菌群变化到底只出现在EPDS升高的阶段,还是在产前抑郁风险人群中具有一定持续性。
校正孕期阶段后,菌群差异仍然存在
孕期本身会影响肠道菌群。孕早期、孕中期和孕晚期,母体激素、免疫和代谢状态都在变化,菌群结构也会随之调整。
因此,研究人员采用PERMANOVA 分析评估了宿主因素在孕期对母体肠道菌群的影响。
结果显示,妊娠期是最显著的影响因素,其次为 产前抑郁症状。
编辑
具体来看,孕早期与孕中期之间的菌群差异显著(P < 0.05),孕早期与孕晚期之间的菌群差异更显著(P < 0.001),但孕中期与孕晚期之间差异不显著。
这说明,研究产前抑郁相关菌群变化时,必须先排除孕期自然变化的影响。
在校正妊娠阶段后,PERMANOVA 分析显示,AND-D 组与对照组之间(FDR P = 0.019),以及 AND-N 组与对照组之间(FDR P = 0.010)均存在显著差异。
编辑
分孕期来看:
也就是说,产前抑郁相关菌群改变并不只局限在某一次 EPDS≥10的抑郁阶段,即使某一阶段评分暂时低于阈值(也就是说非抑郁状态),菌群结构仍然可能与健康对照不同。
这提示一个关键信号:产前抑郁相关的母体菌群变化,更像是一种持续性的微生态特征,而不是短期情绪波动带来的偶然变化。
核心变化:产丁酸菌减少和代谢通路紊乱
接下来,研究者进一步分析到底是哪些菌群发生了改变。
他们将所有微生物群划分为 50 个共丰度菌群模块 CAGs,这种分析方式不是只看单个菌属升高或降低,而是观察一组具有相似变化趋势和潜在功能关联的菌群模块。
在校正孕前BMI、教育水平、孕周、产妇年龄、妊娠糖尿病、抗生素、益生菌、GBS感染等因素后,研究最终识别出 15个显著改变的CAGs。
编辑
其中,AND-D 和 AND-N 组中多个产丁酸菌相关模块持续下降,包括Eubacterium、Ruminococcaceae、Faecalibacterium、Oscillospiraceae 等。
这些菌群与短链脂肪酸,尤其是丁酸的产生密切相关。丁酸不仅是肠上皮细胞的重要能量来源,也参与维持肠屏障完整性、调节炎症反应和免疫稳态。
换句话说,产前抑郁相关菌群失衡的核心,指向了一个更具体的功能问题:产丁酸能力下降。
关于一些重要产丁酸菌属详见谷禾之前的文章:
肠道核心菌属——优/真杆菌属(Eubacterium),你为什么要关心它?
肠道核心菌属——普拉梭菌(F. Prausnitzii),预防炎症的下一代益生菌
肠道重要菌属——颤螺菌属 (Oscillospira),它为什么如此神秘?
与此同时,以 Streptococcus 为主的CAG36在产前抑郁相关样本中升高。
结合既往研究中Streptococcus与炎症、抑郁状态之间的关联,这一变化也提示母体肠道微生态可能向更促炎的方向偏移。
功能层面:脂肪酸和氨基酸代谢通路异常
菌群组成变化之外,研究还进一步做了功能预测。
功能预测还显示,抑郁阶段和非抑郁阶段都存在多条代谢通路异常,包括:
编辑
多不饱和脂肪酸与胎儿脑发育、神经细胞膜结构和炎症调节有关;氨基酸代谢则与神经递质合成、抗氧化能力和基础代谢密切相关。
这部分结果进一步说明:产前抑郁相关菌群失衡,可能不仅是菌群结构变化,还涉及母体肠道代谢功能的改变。
但这还只是母体菌群层面的发现。下一步更关键的问题是:这些菌群变化,是否真的和婴儿后续神经发育表现有关?
在明确产前抑郁孕妇存在持续性母体菌群失衡之后,研究进一步探讨了:这些孕期菌群变化是否会影响婴儿神经发育。
婴儿自身菌群:并非主要解释因素
研究者首先分析了婴儿出生后的肠道菌群,共纳入 290份婴儿粪便样本进行 16S rRNA 测序。
结果显示,影响婴儿肠道菌群组成的主要因素是采样时间、分娩方式和喂养方式,其中采样时间的影响最大。
编辑
这说明婴儿出生后肠道菌群本身处于快速建立和变化阶段,且明显受到出生方式、喂养方式等因素影响。
母亲是否存在产前抑郁症状,并没有显著影响婴儿出生后的菌群整体组成。
编辑
这表明婴儿自身菌群的波动很大,很难作为母亲情绪与婴儿神经发育之间关联的中介因素。
母婴菌群垂直传递证据有限
分析母婴共享的 ASVs 后发现,整体共享数量不多。只有一个Clostridium sensu stricto 1 相关 ASV 在产前抑郁风险组中共享率较高(P=0.034),并与婴儿神经发育评分存在一定关联趋势。但该 ASV 在极少数样本中出现,不足以解释整体神经发育差异。
编辑
由此可见,母婴菌群垂直传递并不是本研究中主要的机制线索。
孕期母体菌群更关键
研究者在校正母亲年龄、孕前 BMI、孕周、教育水平、妊娠糖尿病、抗生素和益生菌使用等因素后发现:
编辑
也就是说,这些菌群模块丰度越低,婴儿在认知、问题解决、个人-社会能力等方面的表现可能越弱。
编辑
婴儿出生后的菌群不构成主要中介。母婴垂直传递有限,婴儿肠道菌群受时间、分娩方式和喂养方式影响更大。
孕期母体菌群失衡可能是核心机制。特别是产丁酸菌相关模块下降和代谢通路功能扰动,与婴儿早期认知和社交能力存在正相关。
产前抑郁可能通过影响母体肠道菌群代谢环境,改变胎儿脑发育微环境,从而在婴儿早期表现为认知和社交能力下降。
为什么要做FMT验证?
因为孕期情绪状态、饮食、睡眠、家庭环境、遗传背景、妊娠并发症等因素,都可能同时影响母体菌群和婴儿发育结果。
因此,需要可以尽量把“菌群因素”从复杂的人群背景中单独提取出来。
FMT模型怎么做?
研究者从产前抑郁风险女性和健康对照女性中选择粪菌供体,将其菌群分别移植给无菌雌鼠,之后让小鼠妊娠,并观察孕鼠和后代的变化。
编辑
动物实验主要分为三组:
• Control_M组:接受健康对照女性菌群;
• AND_M组:接受产前抑郁风险女性菌群;
• AND + butyr_M组:接受产前抑郁风险女性菌群,同时在孕期补充丁酸。
编辑
FMT模型是否成功建立?
从菌群组成看,移植后小鼠的肠道菌群能够较好地反映对应供体来源。
注:E18.5,指胚胎第18.5天,接近小鼠足月分娩前阶段。
编辑
这说明FMT模型基本建立成功,后续观察到的母体和子代差异,可以进一步用于评估AND相关菌群的潜在作用。
产前抑郁相关菌群影响了后代学习记忆表现
研究者在后代小鼠12周龄时进行 Morris水迷宫实验,用于评估学习和空间记忆能力。
编辑
结果显示,与Control_M组相比,AND_M组后代在水迷宫测试中表现出明显的认知受损特征:
编辑
这些结果说明,接受产前抑郁风险女性菌群的孕鼠,其后代在学习记忆能力上出现下降。
值得注意的是,开放场实验中,AND_M组后代在活动距离、中心区停留时间、进入中心区次数等指标上没有显著差异。这提示AND相关菌群在该模型中更明显影响的是认知和学习记忆相关行为,而不是广泛诱导焦虑或抑郁样行为。
总的来说,在无菌小鼠FMT实验中,研究者进一步看到:仅仅移植AND相关母体菌群,就可以诱导后代出现认知行为异常。
这说明,产前抑郁相关菌群不只是一个伴随现象,而可能是参与子代神经发育异常的重要中介因素。
在人群研究中,产前抑郁症状、母体菌群失衡和婴儿神经发育下降之间存在关联。而在无菌小鼠实验中,研究者进一步看到,仅仅移植AND相关母体菌群,就可以诱导后代出现认知行为异常。
这说明,产前抑郁相关菌群不只是一个伴随现象,而可能是参与子代神经发育异常的重要中介因素。既然产前抑郁相关菌群本身能够影响后代认知表现,下一步就需要了解:它是通过什么路径影响胎儿脑发育的?
从动物实验和多组学结果来看,机制主要集中在几条连续线索上:
编辑
母体肠道:紧密连接蛋白下降,肠屏障受损
研究者首先观察孕鼠肠道屏障状态。结果显示,接受 AND 相关菌群的孕鼠,结肠组织中多个紧密连接蛋白表达下降,包括 claudin-1、ZO-1 和 ZO-2(下图a–d)。
编辑
这些蛋白是维持肠上皮屏障完整性的关键结构。它们下降,说明肠道屏障稳定性变差,肠腔内微生物相关成分或炎症刺激物更容易影响母体系统环境。
也就是说,AND 相关菌群进一步影响了母体肠道屏障功能。
母体和胎儿脑炎症信号同步增强
在肠屏障受损的同时,研究者还检测到母体炎症水平升高。
与对照组相比,AND_M 组孕鼠血清中的 TNF-α 和 IL-1β 水平均升高(下图e、f)。
编辑
这提示 AND 相关菌群可能诱导母体系统性低度炎症反应。
更关键的是,这种炎症信号并没有停留在母体端。胎儿小鼠脑组织中同样出现炎症相关变化:ELISA 结果显示,AND_M 组胎儿小鼠脑内 TNF-α 和 IL-1β 水平升高(下图j、k)。
编辑
转录组分析也支持这一点。AND_M 组胎儿小鼠脑中多条炎症相关通路被激活,包括 TNF 信号通路、IL-17 信号通路、Toll-like receptor 通路、NOD-like receptor 通路、趋化因子和细胞因子相互作用通路等(下图a、c)。
编辑
这说明,AND 相关菌群可能通过“肠屏障受损—母体炎症增强—胎儿脑炎症微环境改变”这一方式,参与胎儿脑发育异常。
胎儿脑代谢:PUFA 和氨基酸代谢异常
除了炎症,研究者还对胎儿小鼠脑组织做了代谢组分析。
结果显示,AND_M 组胎儿小鼠脑内代谢谱与对照组明显不同(下图g)。其中,一个突出的变化是多不饱和脂肪酸(PUFA)相关代谢通路下降,包括亚油酸、α-亚麻酸、花生四烯酸等相关代谢路径(下图k)。
编辑
具体代谢物上,AND_M 组胎儿脑中 anandamide、PC(14:0/20:4) 等脂质相关分子下降(下图h、i)。这些分子与神经细胞膜结构、突触功能、神经炎症调节都有关系。
编辑
同时,氨基酸相关代谢也受到影响。例如 L-alanine 水平下降(图j),并伴随半胱氨酸、甲硫氨酸、牛磺酸等相关代谢路径异常。
胎儿脑组织:突触和脂质代谢相关基因变化
转录组结果进一步补充了这一机制链。
在 AND_M 组胎儿脑中,脂质代谢相关通路和突触相关通路下调,包括谷氨酸能突触、5-羟色胺能突触等相关信号(图c)。
编辑
同时,关键基因如 Pla2g4b、Pla2g4d 表达下降(图e、f)。
编辑
综合来看,AND相关菌群可能通过多个相互关联的环节影响胎儿脑发育:
这些变化共同指向一个结果:胎儿脑发育所依赖的免疫和代谢环境被扰动,最终可能表现为后代认知功能下降。
既然前面的菌群结果反复指向产丁酸菌减少,而动物模型中也观察到肠屏障和炎症异常,那么如果补充丁酸,是否能够部分逆转这些异常?
研究者进一步设置了 AND + butyr_M 组:在接受 AND 相关菌群移植的基础上,于小鼠孕期补充丁酸,观察能否逆转部分异常表型。
丁酸补充提高了胎盘丁酸水平
结果显示,与 AND_M 组相比,AND + butyr_M 组胎盘丁酸水平明显升高。
编辑
这说明丁酸补充改变了模型中的丁酸供应状态,也为后续观察行为、炎症和代谢变化提供了基础。
后代认知行为得到部分改善
在 Morris 水迷宫实验中,丁酸补充组后代的认知表现有所改善。
与 AND_M 组相比,AND + butyr_M 组后代的小鼠学习记忆表现有所改善:
编辑
说明,丁酸补充可以部分缓解 AND 相关菌群导致的学习记忆能力下降。不过需要注意,这里是部分改善,并不是完全逆转所有行为异常。
胎儿脑代谢异常,出现恢复趋势
小鼠代谢组结果进一步显示,丁酸补充后,AND_M 组中被下调的部分 PUFA 代谢和氨基酸代谢通路出现恢复趋势。
编辑
PUFA 代谢与神经细胞膜结构、突触功能和炎症调节有关;氨基酸代谢则与神经递质、抗氧化能力和脑发育基础代谢密切相关。因此,这一结果提示,丁酸可能通过改善胎儿脑内脂质和氨基酸代谢环境,参与神经发育保护。
炎症信号和肠屏障损伤也有所缓解
小鼠实验的转录组结果显示,丁酸补充后,小鼠胎儿脑内部分炎症相关基因表达下降,例如 ccl21b、cd14、ms4a2 等;同时,脂质代谢相关基因 Pla2g4b 表达升高。
ELISA 结果也显示,AND + butyr_M 组胎儿脑内TNF-α 和 IL-1β 水平下降。
丁酸补充的影响并不只体现在胎儿脑代谢上。
在小鼠母体端,丁酸补充还部分恢复了结肠紧密连接蛋白表达,包括 claudin-1、ZO-1、ZO-2,并降低了母体血清中的炎症因子水平。
编辑
这些结果提示,产丁酸菌减少及丁酸不足,可能是 AND 相关菌群影响子代神经发育的重要机制节点。
更准确地说,产前抑郁相关菌群可能通过削弱产丁酸能力,影响母体肠屏障和炎症状态,并进一步扰动胎儿脑内代谢环境;而丁酸补充能够部分缓解这些变化。
但这仍然是动物实验结果,不能直接推导为临床建议。对于临床和健康管理场景,更稳妥的理解是:围产期微生态干预可以重点关注产丁酸菌、膳食纤维摄入、短链脂肪酸代谢能力和肠屏障状态。
过去在围产期健康管理中,情绪问题、肠道问题、营养问题和婴儿发育问题,往往是分开处理的。
这项研究提示,这些问题之间可能存在更深的连接。产前抑郁症状并不只是一个心理状态,它可能伴随母体肠道菌群重构,尤其是产丁酸菌减少,并进一步影响肠屏障、炎症反应、胎儿脑内多不饱和脂肪酸和氨基酸代谢。
也就是说,孕期情绪风险背后,可能有一部分生理和微生态基础可以被更早观察到。
围产期情绪管理可以前移,并做风险分层
这项研究显示,孕早期、孕中期和孕晚期的EPDS评分升高,都可能与婴儿后续神经发育表现下降有关。
临床产前抑郁筛查不应只集中在孕晚期或产后。孕早期建档时,就应该开始关注情绪风险。
同样是情绪风险,有些孕妇可能主要受压力、睡眠、家庭支持等因素影响;也有一部分孕妇,可能同时伴随肠道微生态、炎症和营养代谢方面的问题。
产丁酸菌,值得成为孕期微生态评估中的重点观察方向。在该研究中,产丁酸菌Eubacterium、Faecalibacterium、Ruminococcaceae、Oscillospiraceae 等相关菌群模块在产前抑郁风险样本中下降,并且与婴儿神经发育评分存在关联。
编辑
<来源:谷禾健康肠道菌群检测数据库>
实际评估中,核心菌属中这些产丁酸菌以及丁酸盐、丙酸盐、乙酸盐等短链脂肪酸指标,都可以作为观察产丁酸能力和肠道代谢支持状态的线索。
如果这类指标已经显示出不足,即使孕妇还没有明显情绪问题,也可以提示其肠道生态和代谢环境可能存在潜在风险。后续管理就可以更早关注她的饮食结构、膳食纤维摄入、肠道症状、睡眠状态以及炎症相关风险,而不是等到情绪或消化问题明显后才处理。
编辑
<来源:谷禾健康肠道菌群检测数据库>
这类信息不一定直接指向某种疾病,但可以帮助管理者更早识别需要重点关注的人群。但筛查只是第一步,还可以对情绪风险孕妇做进一步分层。
如果孕妇已经出现EPDS评分升高或明显情绪风险,肠道菌群检测可以辅助判别:这个情绪风险背后,是否同时存在肠道微生态和代谢环境异常。
若同时观察到产丁酸菌减少、短链脂肪酸能力不足、肠屏障压力或炎症相关指标异常,那么管理重点除了在心理支持上,也需要同步考虑饮食结构、膳食纤维摄入、肠道症状改善、睡眠和炎症风险管理。
编辑
<来源:谷禾健康肠道菌群检测数据库>
面向孕期健康的转化思路
肠道菌群检测适合嵌入临床已有的孕期评估流程,作为情绪筛查、产科评估和营养评估之外的补充信息,帮助医生判断是否需要进一步联合营养、肠道或生活方式干预。
在健康管理中,把菌群检测放进孕期服务流程中,比如孕早期建立肠道菌群基线状态,孕中晚期结合饮食、睡眠、肠道症状和菌群变化做再评估。
作为产品开发来说,这篇研究提供了一个更具体的思路。有转化价值的方向,可能是把产品功能落到产丁酸能力、短链脂肪酸生成、肠屏障支持和营养代谢这些环节上,设计检测、干预方案。
编辑
未来的围产期健康管理,会越来越强调前置识别。
不是等到婴儿发育问题出现后再回头寻找原因,也不是等到孕妇症状明显后才开始处理,而是在孕期就尽量识别那些可能影响母体状态和胎儿发育环境的信号。
情绪状态是信号,肠道菌群也是信号,营养和炎症状态同样是信号。
这篇研究让我们看到,这些信号之间可能存在联系。肠道菌群不是全部答案,但它可以成为理解孕期母体状态、进行风险分层和制定管理策略的重要入口。
主要参考文献
Zhou, Fangyue, et al. “Antenatal depressive symptoms impair offspring neurodevelopment by inducing maternal gut microbiota dysbiosis during pregnancy.” Gut Microbes 18.1 (2026): 2672188.
谷禾健康
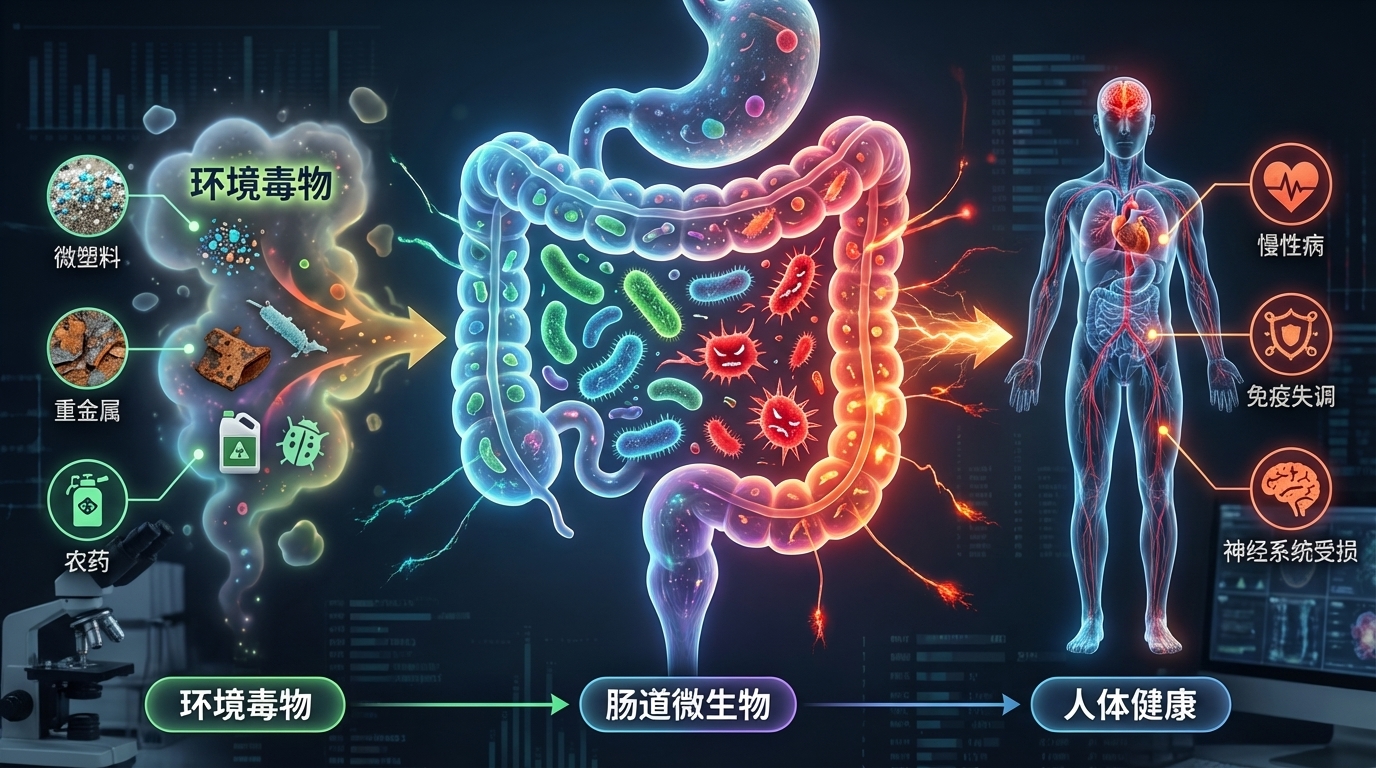
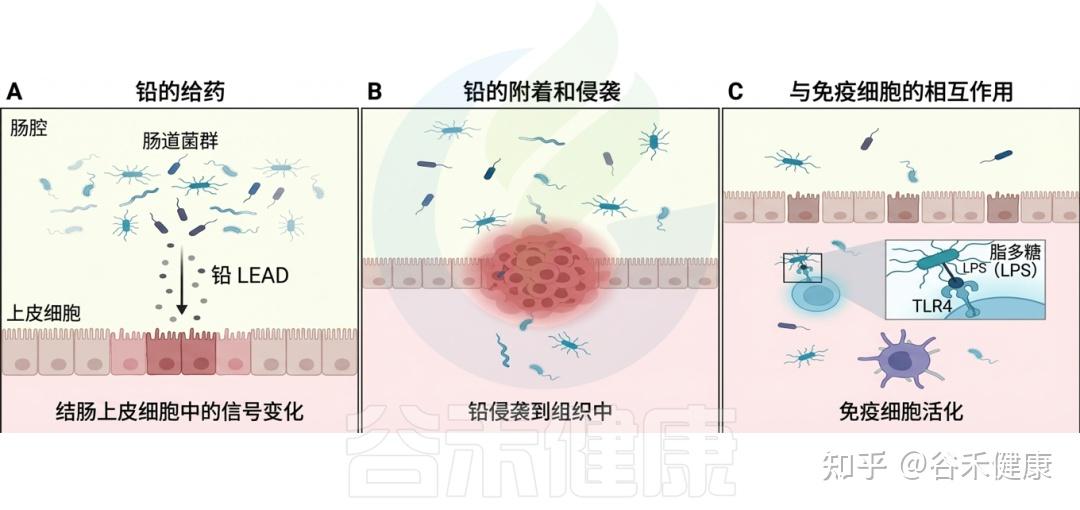
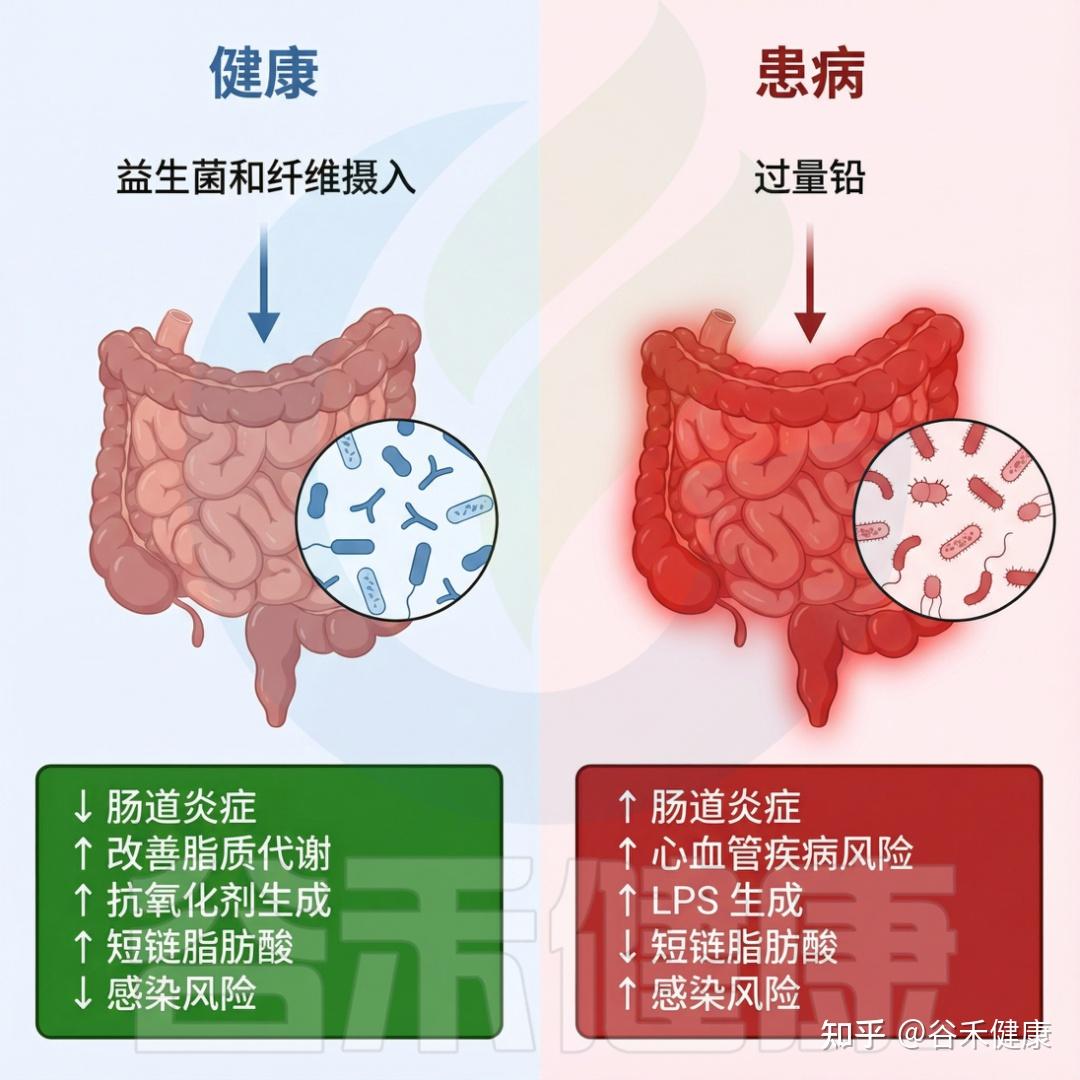
在现代社会,环境毒物已成为影响人类健康且尚未得到充分研究的风险因素。这些有害物质,从工业化学品和污染物到日常家居用品,均可能通过复杂的生物机制对健康产生深远影响。尤其令人关注的是,研究发现这些环境毒物不仅直接影响人体,还对肠道微生物群这个复杂的生态系统造成显著干扰。
肠道微生物群是我们消化道内的重要组成部分,对调节免疫系统、代谢功能和神经活动等方面起着关键作用。例如,肠道菌群能够通过产生短链脂肪酸(SCFAs)来调节免疫反应,增强肠道屏障的功能,从而降低慢性炎症的风险。然而,若这种微妙的平衡被扰动,将可能引发一系列健康问题,包括慢性炎症、肥胖、糖尿病、癌症和神经退行性疾病等。
研究表明,环境毒物的暴露(如铅、镉、农药和邻苯二甲酸盐等)会导致肠道微生物的组成和功能改变。例如,铅暴露与有益细菌数量的减少相关,导致乳球菌和阿克曼菌减少,而脱硫弧菌和大肠杆菌等机会性致病菌则显著增多。此外,还发现环境毒物影响下维生素和胆汁酸的浓度失衡,以及一氧化氮生成的增加也随之发生。这直接影响宿主的代谢和免疫功能。
相反,肠道微生物组也能影响环境毒物的毒性。例如,某些肠道细菌能够代谢并转化环境中的化学物质,从而减轻它们对宿主的损害。一项研究指出,某些益生菌如植物乳杆菌能够帮助降解体内的有机污染物,减少其生物积累和毒性。此外,肠道微生物能通过直接与环境毒物相互作用、影响其生物利用度,间接改变其对健康的影响。
本文通过综合各领域的研究成果,探讨环境毒物如何通过肠道微生物组影响人类健康,尽量基于研究结论去揭示环境毒物与肠道微生物组之间的复杂关系。本文还将探讨如何通过调整饮食与生活方式来调控肠道微生物反应,从而缓解环境毒物的潜在危害。
我们希望本文不仅能为学术界提供有价值的见解,还能激发公众对环境健康问题的关注,共同应对这一全球性挑战。
▸ 什么是环境毒物?
环境毒物是指广泛存在于空气、水、土壤、食品和日常用品中的有害化学或金属物质,如重金属(铅、汞、镉、砷)、农药、工业有机污染物、增塑剂(邻苯二甲酸盐)等。
它们可通过饮食、饮水、吸入或皮肤接触进入人体,扰乱机体免疫、代谢和神经等系统,进而增加多种慢性疾病甚至癌症的风险。
多种引起毒性的环境毒物
★ 剂量和胃肠道对物质毒性有重要影响
帕拉塞尔苏斯(Paracelsus,1493-1541)提出的名言“仅剂量决定某物不是毒药”奠定了当代毒理学的核心原则,强调评估物质毒性必须关注剂量。
他另一句“生命中最重要的事情发生在消化道的开始和结束之间”,突出了消化系统在毒理学中的重要性。由此引出对环境毒物与胃肠道微生物群相互作用的研究,强调剂量对毒性的关键影响,以及肠道内数万亿微生物对人类健康与疾病的重要作用。
▸ 胃肠道微生物与环境毒物
胃肠系统是人体微生物最密集的区域,细菌量从胃的101–103 CFU/mL到结肠的1011–1012 CFU/mL不等,已鉴定出数千种细菌,显示肠道微生物群高度复杂,以细菌为主,古细菌、真菌和原生动物为次要成分。
微生物群参与胃肠道的形成与稳态,并与免疫和神经系统协同,其与摄入外源性物质存在密切且重要的相互作用。
▸ 环境毒物扰动肠道微生物群
研究表明,环境毒物暴露会显著扰动肠道微生物群。如镉暴露与微生物多样性降低及特定菌群丰度变化相关;汞暴露同样可改变菌群构成,并可能促进其整体毒性作用。
由于肠道微生物群在营养代谢、免疫调节及抵御病原体方面发挥关键作用,这一脆弱生态系统的失衡将损害上述功能,从而危及宿主健康。
微生物组与环境毒物的相互作用
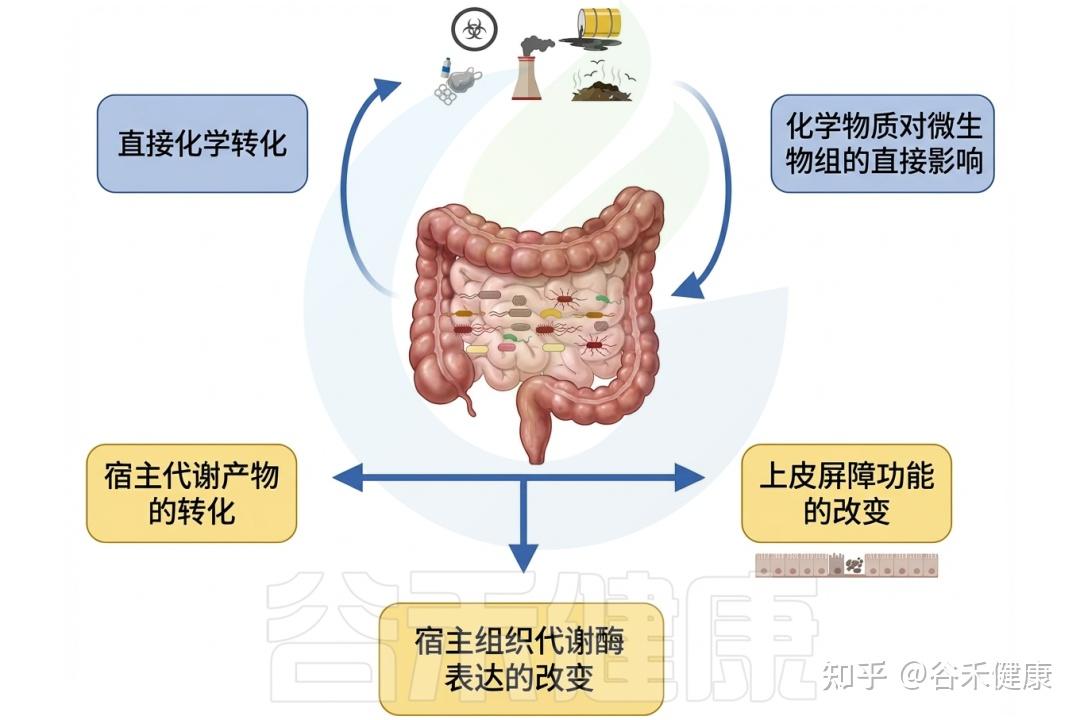
▸ 肠道微生物影响环境毒物毒性
相反,肠道微生物群可以通过各种机制影响环境毒物的毒性。微生物可以改变环境毒物的化学形态,影响其吸收和生物利用度。
此外,某些细菌具有解毒或隔离环境毒物的能力,可能减轻它们对宿主的有害影响。肠道微生物组参与调节炎症和免疫反应,使环境毒物与宿主毒性之间的关系进一步复杂化。
肠道微生物群影响环境毒物的生物利用度和毒性
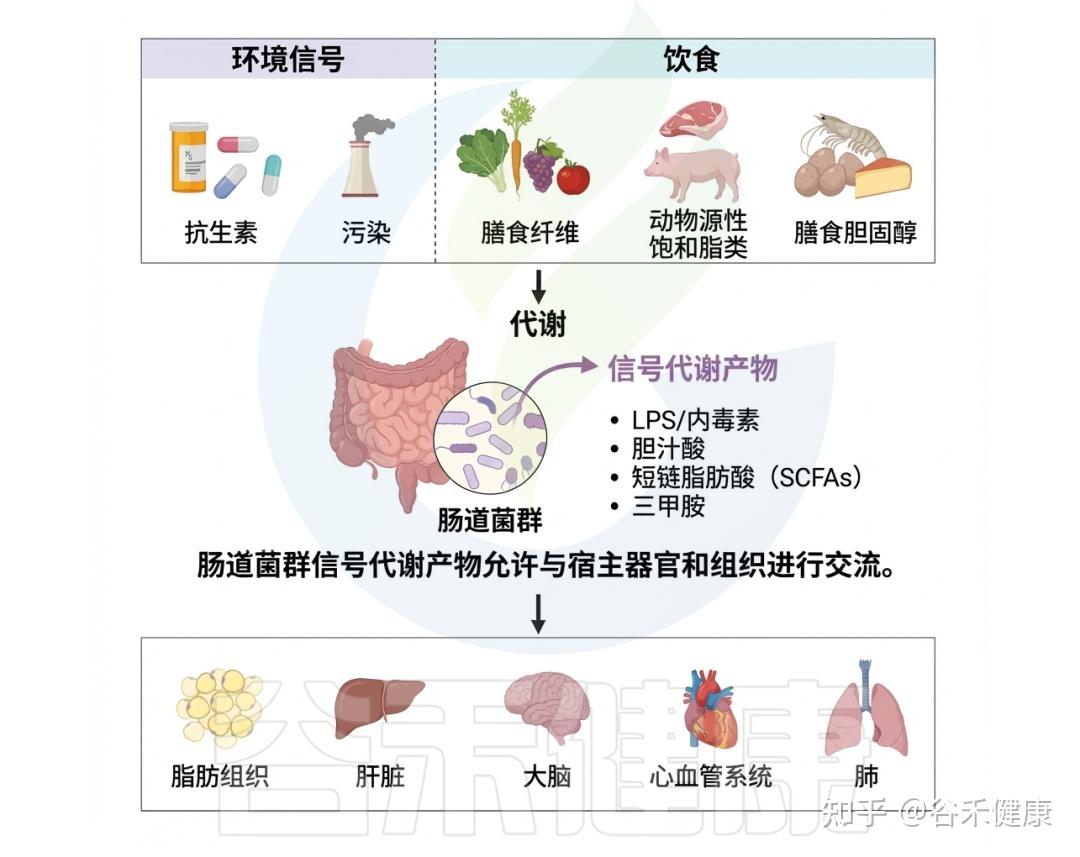
深入理解肠道微生物与环境毒物之间的复杂互作,是全面认识环境暴露相关健康结局的关键。环境毒物与肠道微生物组之间的精细关系突显了环境毒理学的多维性。随着对肠道微生物组认识的不断加深,其在调控环境毒物健康效应中的核心作用日益清晰。
环境毒物对肠道微生物群及人体的影响
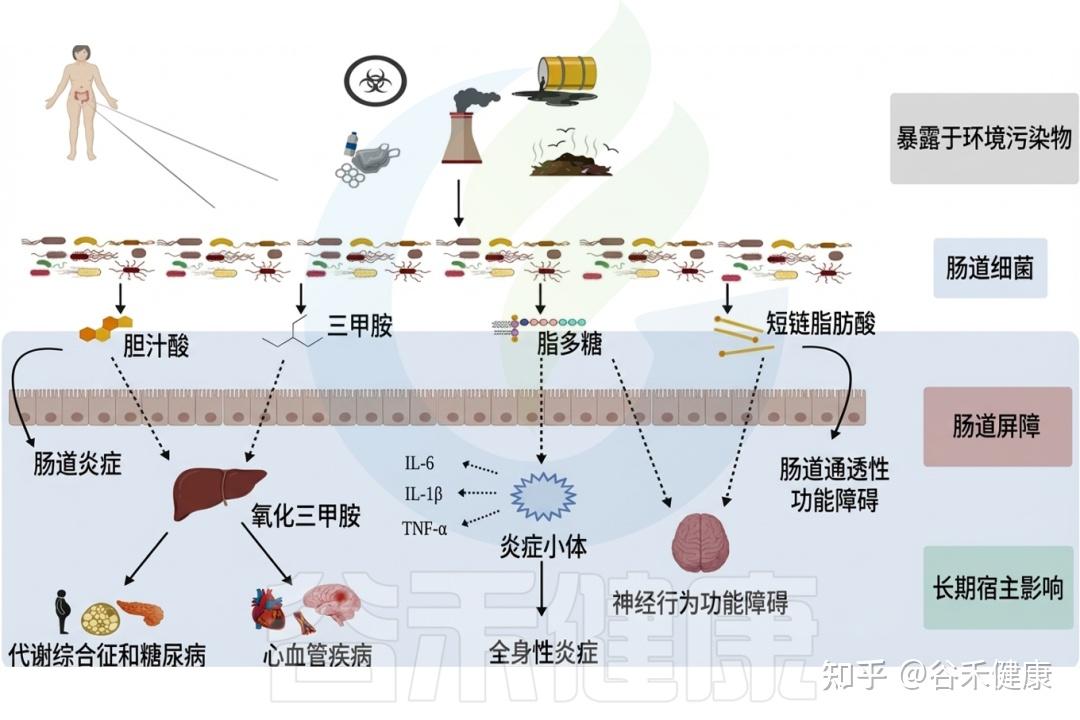
环境毒物影响肠道微生物组,反过来微生物组也调节环境毒物毒性,这一双向作用已成为快速发展的研究热点,对人类健康和环境管理具有重要意义。只有全面阐明环境污染物与胃肠道微生物群的相互作用,才能制定有效策略减轻健康损害,维护个体与生态系统的整体福祉。下面我们将详细具体地阐述日常生活环境中常见有毒物质与肠道微生物群的相互作用及其对人体健康的影响。
▸ 邻苯二甲酸盐(PAE):来源、健康影响
▸ 什么邻苯二甲酸盐(PAE)?
直接说“邻苯二甲酸盐”(PAE),很多人可能并不了解;但如果说它是一种用来让塑料更柔软、有弹性、不易脆裂的添加剂,这样可能就容易理解多了。
▸ 食品、日用品、玩具、医疗用品中都有PAE
人类接触邻苯二甲酸盐(PAE)可分为直接和间接两类。直接接触指直接使用PAE或含PAE的产品;间接接触则是PAE通过浸出或渗透污染环境或其他产品。研究显示,截至2017年,全球PAE年产量已超过600万吨,其年消耗量持续增加,人群体液样本中PAE代谢物检出频率亦在升高。
食品是人类暴露PAE的重要来源之一,已在酒精饮料、食用油、乳制品、饮用水、肉类、家禽和蔬菜等中检测到。其脂溶性、低熔点、高沸点及易溶于乙醇等特性,促进其由接触材料迁移至食物并被摄入。
在儿童用品中,塑料玩具、牙胶和餐具因价廉耐用而被广泛使用,部分含有邻苯二甲酸酯(PAEs)的产品可能导致儿童通过手口接触和含吮行为摄入该类毒物。
在医疗领域,邻苯二甲酸二(2-乙基己基)酯(DEHP) 常作为医疗器械增塑剂,为留置类器械提供柔韧性。因其不与塑料形成化学键,使用中易析出并在体内累积。药物制剂中部分杂质来源于增塑剂的浸出或迁移。片剂和胶囊通常包裹聚合物包衣,以调控其在胃肠道内释放的时间和部位。
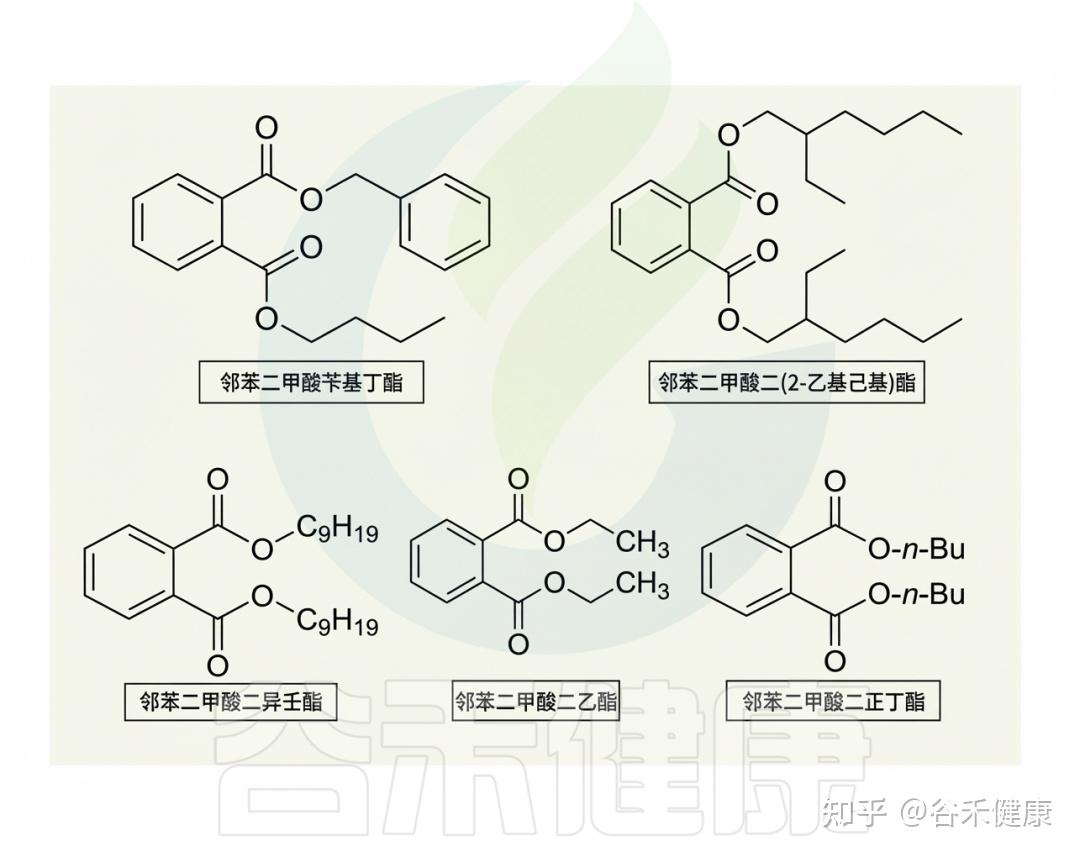
最常见的邻苯二甲酸盐
多种日常用品,如清洁剂、指甲油、香水和个人护理品中普遍含有PAE,邻苯二甲酸二乙酯(DEP)常作香精定香剂。研究表明,环境PAE可沉积于棉和聚酯纤维,且棉中浓度更高,这些纺织品主要经皮肤致暴露。
此外,PAE还可由家用产品、服装、建材和汽车部件等释放至空气,通过吸入进入人体。
▸ PAE暴露与多种健康问题有关
目前,PAE暴露已被关联于多种健康问题,包括内分泌与生殖功能紊乱、性早熟、子宫内膜异位症、性发育异常、不孕不育、胎儿发育异常、乳腺癌和皮肤癌、肥胖、2型糖尿病、注意力缺陷多动障碍、自闭症谱系障碍,以及心、肝、肾毒性、哮喘和过敏等。
邻苯二甲酸盐对人体健康造成的影响
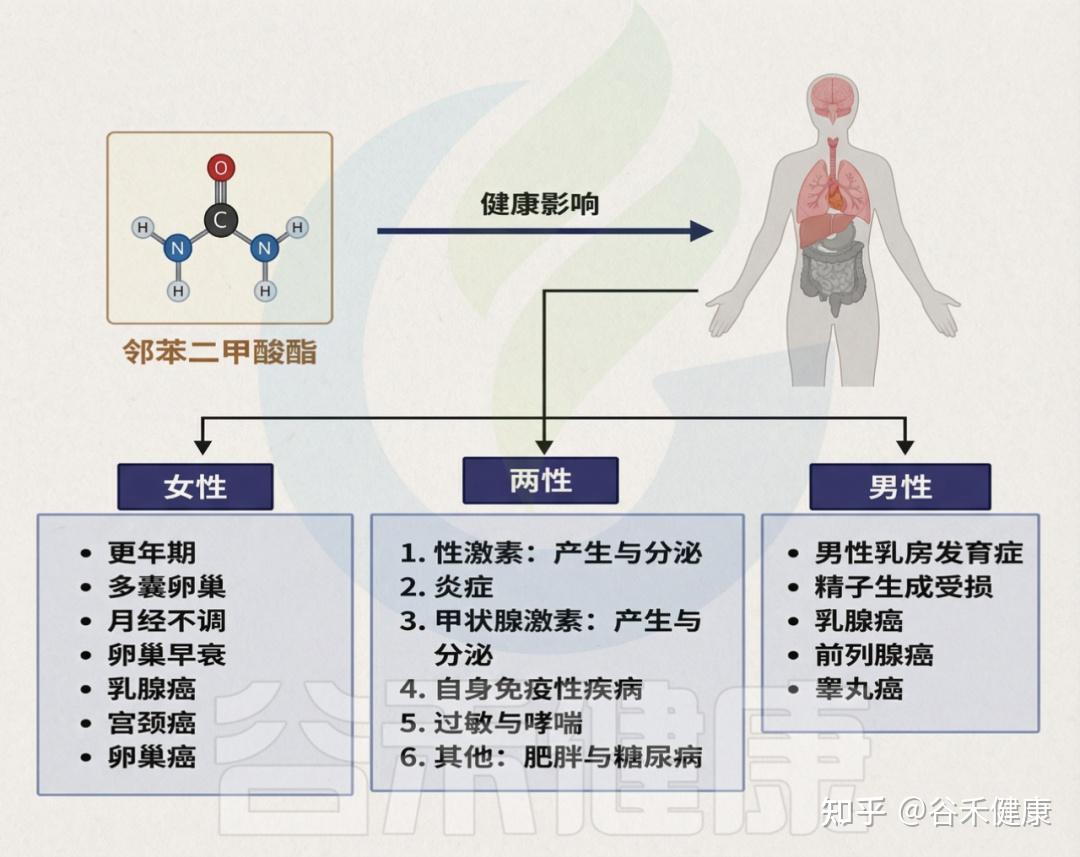
需强调的是,儿童与成人对PAE的反应可能不同,儿童因频繁手口行为而更易摄入,暴露风险更高。
▸ 邻苯二甲酸盐(PAE)与肠道微生物的互作
邻苯二甲酸盐(PAE)因广泛存在及其潜在健康危害而备受关注。近期研究逐步揭示PAE暴露与肠道菌群(GM)之间的复杂联系,表明其可影响微生物组成及宿主健康。
▸ 暴露于PAE后,乳杆菌和双歧杆菌减少
暴露于常见邻苯二甲酸盐(PAE)化合物邻苯二甲酸二(2-乙基己基)酯(DEHP) 后,多种关键菌种水平明显变化,其中乳杆菌和双歧杆菌等有益菌显著减少,导致整体菌群多样性和丰度发生重塑。
PAE经摄入或吸收后在体内代谢,产生可干扰微生物生长和代谢的活性产物;同时,PAE还可能损伤微生物细胞结构或关键代谢过程,打破菌群平衡。
▸ PAE可能营造导致菌群失衡的环境
新证据显示,PAE暴露可引发肠道炎症并削弱黏膜屏障,从而营造有利于菌群失衡的环境。人群流行病学研究亦支持这一关联:在成人队列中发现,尿中PAE代谢物水平与肠道微生物多样性及丰度变化之间存在显著相关。
这些结果突显PAE相关菌群生态失调在实际人群中的重要性,并提示其对公共卫生干预具有潜在影响。PAE导致的肠道微生物群改变不仅破坏肠道微生态,还影响宿主整体健康和疾病易感性。肠道微生物群在维持代谢稳态、调节免疫功能及保障肠屏障完整性等方面至关重要;因此,PAE 诱导的菌群失衡可能增加代谢综合征、胃肠道疾病、免疫失调及神经发育异常等多种健康风险。
▸ PAE还通过营养可用性和免疫影响肠道菌群
此外,PAE还可通过改变微生物生长与代谢所需营养和底物的可及性来影响肠道微生物群,干扰肠腔中膳食成分的吸收和利用,并通过调节肠上皮营养转运与代谢相关基因的表达,改变宿主—微生物相互作用。营养供给和宿主信号的变化共同塑造肠道微生物群的结构与功能。
PAE还可通过调节宿主免疫反应间接影响菌群。已有研究将PAE暴露与免疫失调联系起来,如炎症水平升高和免疫细胞功能异常,这会改变肠道微环境和菌群构成。肠道炎症有利于有害菌繁殖并抑制有益菌生长。PAE还与破坏肠上皮屏障完整性相关,导致通透性增加及微生物产物大量进入血液,即“肠漏”现象,从而诱发系统性免疫激活和炎症,进一步影响肠道微生物群的组成和功能。
▸ 肠道菌群调节PAE的代谢和生物效应
邻苯二甲酸盐(PAE)与肠道微生物群之间存在双向关系:PAE可改变肠道微生物群的组成和活性,而肠道微生物群又能调节PAE的代谢和生物效应。
研究发现,肠道细菌具有多种酶,可通过水解、还原和氧化等过程分解PAE。由于肠道微生物群的组成和代谢活性在个体间差异显著,PAE的代谢效率及对其不良作用的易感性亦随之不同。部分菌群可将 PAE 转化为毒性较低或更易排出的代谢物,有助于解毒并降低健康风险;而特定菌种则可能将PAE转化为更具毒性的代谢物,放大其对宿主的危害。
此外,肠道微生物群还能调节机体对 PAE 暴露的免疫和炎症反应,从而影响 PAE 相关疾病的发生风险。例如,肠道细菌可通过与宿主免疫系统互作,改变 PAE 诱导的炎症程度。研究表明,在炎症性肠病(IBD)等慢性炎症性疾病中,菌群失调引起的持续炎症,可进一步加剧 PAE 对肠道健康与功能的负面影响。
▸ 邻苯二甲酸盐(PAE)对肠道健康的影响
▸ 破坏肠道屏障功能
邻苯二甲酸盐(PAE)通过影响紧密连接蛋白破坏肠上皮完整性。PAE 可干扰这些蛋白的表达和组装,使紧密连接减弱、屏障通透性升高。多项研究发现,暴露于PAE会降低关键屏障蛋白水平,导致屏障功能下降。
此外,PAE 可在肠上皮诱导氧化应激(OS)和炎症,进一步削弱屏障功能。其通过提高活性氧(ROS)水平、抑制抗氧化酶活性,导致脂质、蛋白质和 DNA 氧化损伤,并破坏紧密连接结构。
PAE 还能诱导肠黏膜炎症,增加屏障负担,可激活巨噬细胞、树突状细胞等免疫细胞,促进 IL-6、TNF-α、IL-1β 等促炎因子及趋化因子释放,加重炎症并损伤屏障。
同时,PAE 可改变肠道微生物组组成与功能,诱发菌群失调,导致通透性升高和屏障障碍。其暴露会破坏菌群平衡,减少短链脂肪酸产生,部分菌还可将 PAE 代谢为有害或促炎产物,进一步加重屏障损伤。
▸ 邻苯二甲酸盐的促炎作用
PAE与肠道炎症密切相关,多项研究表明其长期暴露可促发并加重消化道炎症。PAE通过激活巨噬细胞、树突状细胞和淋巴细胞,诱导 IL-6、TNF-α、IL-1β、IL-8 等促炎因子和趋化因子产生,招募更多免疫细胞并加剧肠壁损伤和炎症,同时打破促炎与抗炎信号平衡,放大炎症反应。
PAE还能在肠黏膜诱导氧化应激,进一步推动炎症和组织损伤。其通过增加活性氧生成、抑制抗氧化酶活性,导致脂质、蛋白质和 DNA 氧化损伤,并激活炎症通路,放大肠道炎症级联反应。动物研究证实,PAE 暴露可显著引起肠道炎症。
▸ PAE可诱发炎症性肠病、肠易激综合征等疾病
PAE可通过破坏肠道屏障、诱导炎症和干扰肠道微生物群,整体损害肠道健康,并促进炎症性肠病(IBD)、肠易激综合征(IBS)和结直肠癌(CRC)等疾病的发生与进展。
慢性肠炎是炎症性肠病(IBD)核心病理环节,并与结直肠癌风险升高相关。动物研究表明,PAE能导致结肠炎症和氧化应激,产生类似结肠炎的病理改变并增加CRC易感性,提示 PAE 相关炎症可能通过组织破坏、免疫失衡及形成促癌微环境,加速肠道疾病进程。
此外,PAE 对肠道微生物群的干扰也是影响肠道疾病的重要因素。菌群组成和多样性改变(菌群失调)与 IBD、IBS、CRC 等密切相关。PAE 暴露可打乱菌群平衡,减少有益菌、增加有害菌,改变宿主—微生物互作、免疫稳态和代谢途径,从而共同促进肠道相关疾病的发生与恶化。
小结
邻苯二甲酸盐(PAE)广泛存在于消费品中,经摄入或吸收后代谢为具生物活性的产物,可显著改变肠道菌群组成与功能,降低多样性并引发菌群失调。
PAE 对肠道的危害不仅包括破坏屏障和诱导炎症,还在炎症性肠病、肠易激综合征和结直肠癌等疾病的发生与进展中发挥重要作用。通过纵向和机制研究加深对邻苯二甲酸盐–肠道微生物群关系的理解,将为降低PAE暴露相关健康风险提供依据,亦是维护肠道健康和应对环境暴露的关键。
3
农药和肠道微生物群
伴随全球人口增长,对高产与优质农产品的需求推动农药使用急剧增加,以满足粮食供给并追求经济效益。然而,农药的不受控和不合理施用对农产品和整个生态系统构成潜在毒性威胁。由于农药在土壤中的高持久性及其在大气和水体中的残留,非靶标生物(尤其是动物和人类)通过摄入受污染的食物和水而面临更高的农药暴露风险。
大量研究表明,农药暴露与癌症、哮喘、激素紊乱和过敏等疾病相关;同时,多种农药的联合作用可诱发肠道菌群失调,改变微生物组成与功能以及宿主代谢表型。
▸ 肠道菌群失调可作为农药暴露的生物标志物
肠道微生物群的多样性对维持宿主正常生理功能至关重要。最新研究表明,农药暴露会显著影响肠道微生物群,菌群失调可作为评估农药毒性的潜在生物标志物。
▸ 农药导致肠道微生物代谢改变
精确评估特定农药引起的菌群失调及肠道毒性,有助于理解微生物群对污染物暴露的响应机制,并挖掘诊断相关疾病的生物标志物。例如,在二嗪农(一种广谱、高效的有机磷杀虫剂)暴露下,肠道微生物相关信号通路发生改变,导致代谢物谱重塑,并以性别依赖的方式触发神经毒性。这类由微生物产生的神经毒性代谢物可用作二嗪农相关肠道毒性的生物标志物。
各类农药对肠道微生物群的影响
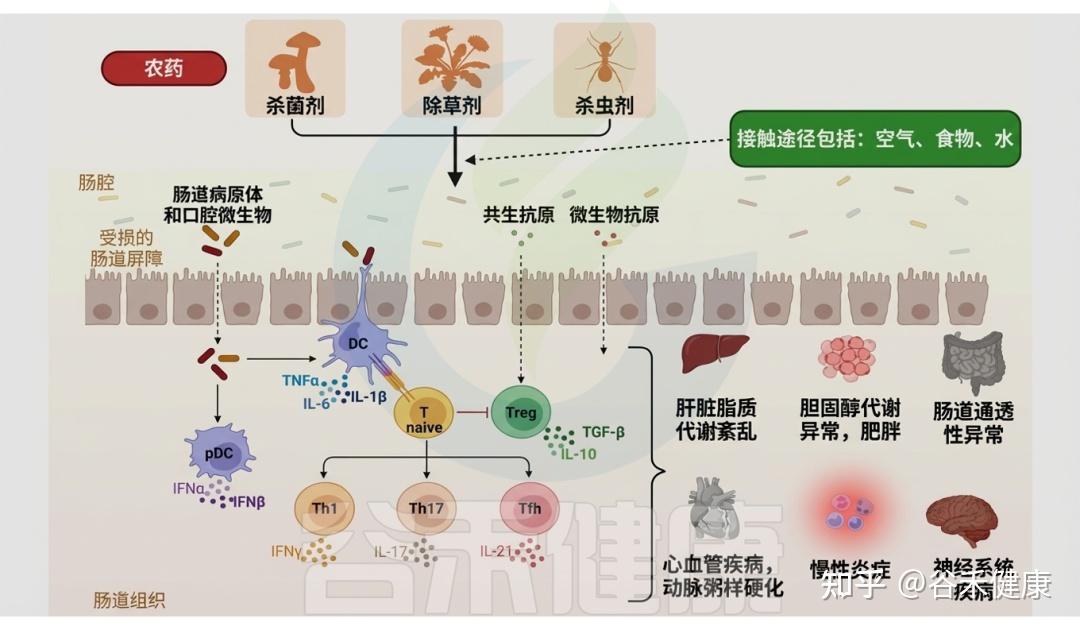
DOI:10.1201/9781003489221-6
农药毒理学风险评估依赖明确且可靠的终点。需要筛选合适的生物标志物,限定其使用条件,界定诊断阈值和异常范围,建立作用机制,并明确其在何种场景下最具适用性。
注:欧洲食品安全局已指出,微生物多样性在环境污染物毒性和健康风险评估中具有重要意义,但仍需进一步界定方法学的局限性、研究空白、纳入/排除标准以及数据处理与解释的标准化要求。
▸ 农药相关毒性还受饮食、年龄、胃肠状况影响
肠道菌群对宿主健康的影响及其在农药相关毒性中的作用极为复杂,还受到饮食、年龄、遗传背景、胃肠动力、地理位置和抗生素使用等多重因素干扰。
要全面理解菌群毒性与宿主健康之间的关系,仍需大量系统研究。针对农药暴露对肠道菌群和宿主健康影响的文献中普遍存在方法学不足:多数研究缺乏体内、体外实验流程及分析手段的标准化与优化。标准化是确保实验可重复性和结果可比性的关键步骤;体外发现也应在体内模型中得到验证。
此外,农药毒理实验的设计必须能准确测定化学残留量及其生物学效应,从而构建可靠的剂量–反应关系,并最终界定慢性暴露水平及其健康风险。
小结
除已阐明的分子通路外,多种细胞过程也可能受到农药毒性的影响,肠道微生物组成与农药毒性之间的关联仍需在人群中深入验证。近期研究已提示子宫内农药污染及其跨代效应,多代暴露人群的肠道微生物组更易发生改变。在农业生态系统中,农民和一线工人因高水平残留及职业暴露而面临严重健康风险;同时,农药在食物链中的渗透污染也亟需持续监测。
因此,有必要系统评估农药污染引起的肠道菌群失调对公众健康的长期影响,并制定有效的干预与缓解策略。对新型和现用农药的风险评估应在透明、统一的框架下进行,系统研究其使用与对人群和环境影响之间的关系。同时,必须为相关实验所得数据的解读制定明确的标准和指南,以规范各类农药风险研究与监管实践。
4
有机污染物和肠道微生物群
近年来,石化、塑料、合成纤维、焦化、印染、印刷等行业的快速发展,导致环境中高浓度有机污染物(OPs)广泛存在。
▸ 常见的有机污染物
有机污染物种类繁多,对环境和人类健康均构成潜在威胁。其来源广泛,包括农用化学品和药品、个人护理品、非法药物、工业污染物、阻燃剂、增塑剂、多环芳烃(PAHs)、卤代多环芳烃、二噁英及其他持久性有机污染物(POPs)残留等。它们通过多种人类活动进入环境,如污水处理厂排放、废弃物无序排放、前体物质降解、海滩塑料碎片累积、工业生产及热处理过程等。
常见的有机污染物类别及示例
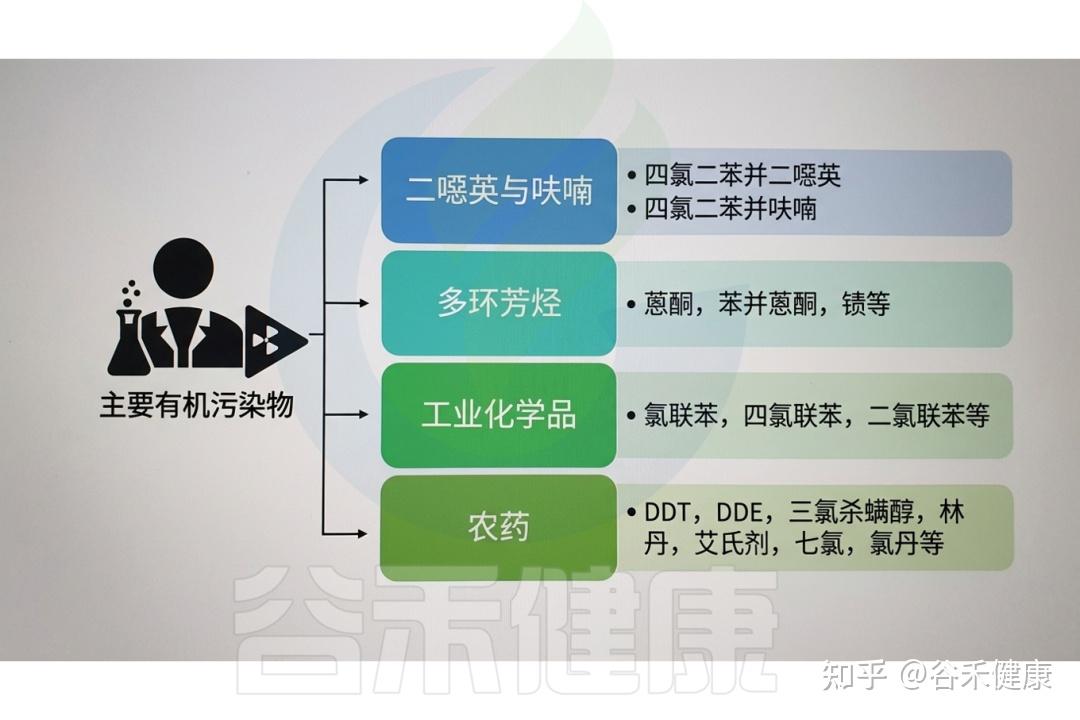
DOI:10.1201/9781003489221-7
常见的有机污染物(OPs)包括以下物质:
农药:用于农业、家庭和公共卫生等场景的害虫防治,包括杀虫剂、除草剂、杀菌剂和杀鼠剂。
溶剂:广泛用于清洗、脱脂及化学反应的有机溶剂,如苯、甲苯、二甲苯及三氯乙烯(TCE)等氯代溶剂。
药物:经人畜排泄或不当处置进入环境的药物化合物,包括抗生素、激素、止痛药及其他处方药。
多氯联苯(PCB):一类曾广泛用于电气设备、液压油和工业过程的合成有机物,属于具有生物累积性的持久性有机污染物。
二噁英:在废物焚烧及含氯化学品生产等工业过程中产生的剧毒副产物,具有高度持久性和生物累积潜力。
▸ 有机污染物与肠道微生物群的互作
▸ 有机污染物引发菌群失调、炎症
研究表明,有机污染物(OP)可扰乱肠道微生物群的平衡和功能,引发菌群失调、炎症,并增加多种疾病风险。
常用除草剂草甘膦(GLP)是其中一例,农产品中常可检出。GLP 暴露会降低动物和人群中的肠道微生物群多样性。研究发现,接触 GLP 会减少双歧杆菌、乳杆菌等有益菌,同时增加梭菌等潜在有害菌。
此外,多氯联苯(PCB)和多溴联苯醚(PBDE)等工业化学品也被证明与肠道微生物群变化相关,PBDE 暴露可导致变形菌增加、拟杆菌减少,引发菌群失调和炎症。
生态失调会导致各种不良的健康结果
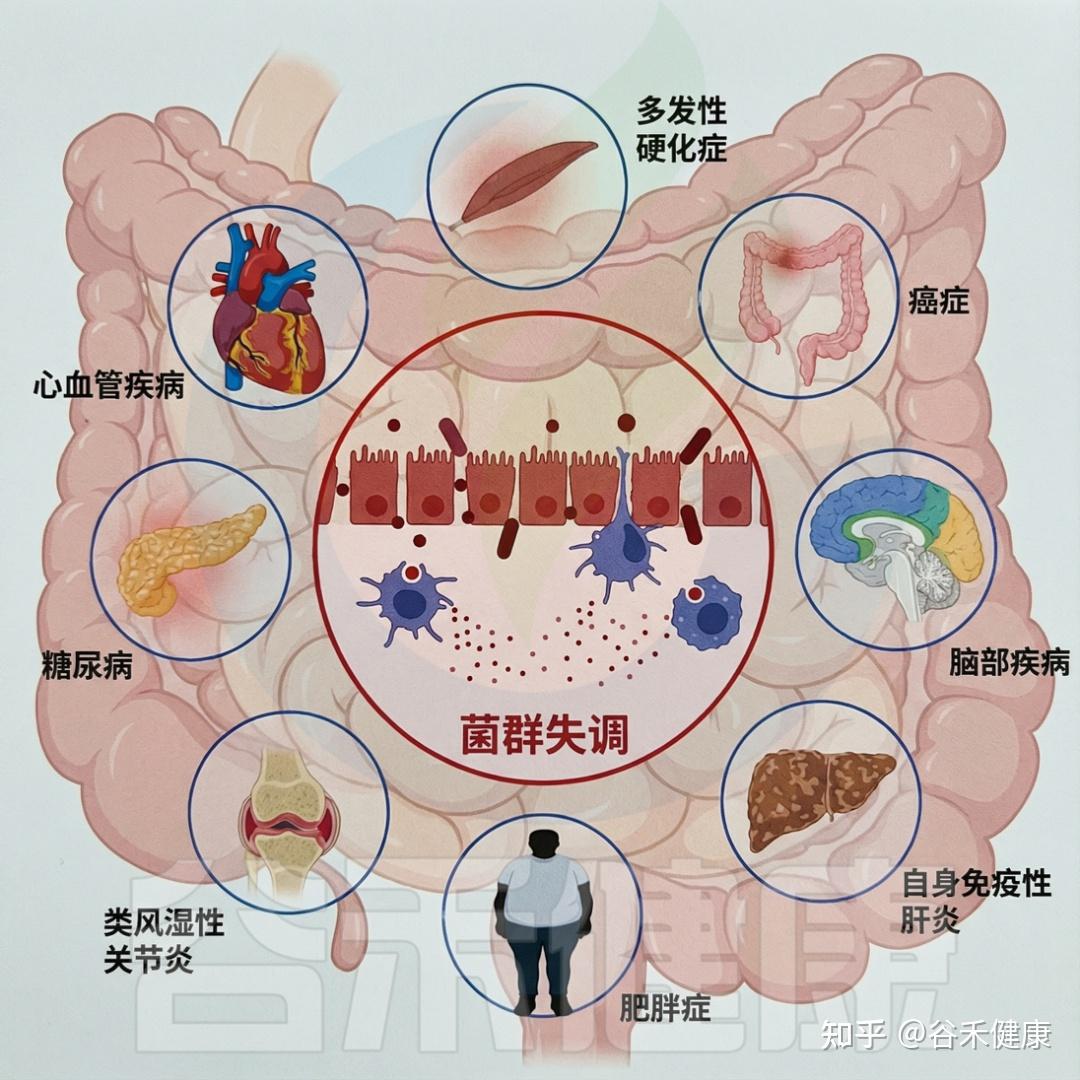
DOI:10.1201/9781003489221-7
▸ 有机污染物损害肠屏障、诱发肠炎
有机污染物(OP)还可能损害肠屏障完整性,屏障受损后,微生物产物和毒素更易进入血液,引起炎症,并促进炎症性肠病(IBD)和代谢综合征(MS)等疾病的发生与进展。
同时,OP 还能干扰宿主–微生物互作,打破有益菌与致病菌平衡。空气污染及烧烤、熏制食品中的多环芳烃(PAH)可改变肠道菌群对胆汁酸的代谢,引发失调和炎症。
有机磷杀虫剂与短链脂肪酸(SCFA)异常相关,而 SCFA 对肠道和免疫功能至关重要。长期 OP 暴露可致轻度慢性肠炎,促炎因子和免疫活性升高,进一步加剧菌群失调。研究还发现,多氯二苯并二恶英(PCDD)、多氯二苯并呋喃(PCDF)等工业污染物同样可诱发肠炎并改变特定菌群数量。
▸ 肠道微生物群能改变有机污染物毒性
另一方面,肠道微生物群也可通过多种酶促途径调节有机污染物的毒性。肠道微生物能够将有机磷转化为性质不同的代谢物,其毒性可能升高或降低。
研究显示,肠菌产生的酶可将多环芳烃分解为毒性较低或更易清除的形式。例如,部分梭菌属菌株可将致癌多环芳烃苯并[a]芘(BaP)转化为危害较小的羟基化代谢物;肠道微生物群来源的细胞色素 P450 单加氧酶可氧化多环芳烃,生成具有不同生物效应的代谢物。
对于以高毒性和持久性著称的多氯联苯 (PCB),肠道微生物群能通过还原脱氯途径促进其代谢,即逐步去除分子上的氯原子。厌氧菌 Dehalococcoidesspp. 已被证实可催化 PCB 脱氯,将高度氯化同系物转化为毒性较低或近乎无毒的产物。类似的微生物过程会影响 PCB 在机体内的生物利用度和实际危害。
▸ 假单胞菌、芽孢杆菌等可减轻有机污染物毒性
肠道细菌还可通过水解与解毒过程分解有机污染物(OP)。假单胞菌(Pseudomonas)、芽孢杆菌(Bacillus)等可产生有机磷水解酶,切断 OP 分子中的磷酸酯键,促进其在胃肠道内的解毒和清除,从而减轻对宿主的毒性。
此外,三氯乙烯(TCE)、四氯乙烯(PCE)等常见于受污染地下水和土壤的氯代溶剂,也可被肠道微生物群通过还原脱氯及其他酶促反应代谢。Dehalococcoides spp. 含有还原性脱卤酶,可去除 TCE 和 PCE 中的氯原子,将其转化为毒性更低的乙烯和乙烷。此类微生物转化过程对环境中氯代溶剂的自然衰减至关重要。
▸ 有机污染物对肠道健康的影响
大量研究表明,不同有机污染物可干扰肠屏障,损害胃肠道健康。
▸ 削弱肠道屏障,上调炎症因子
双酚 A (BPA) 暴露可通过改变紧密连接蛋白的表达和分布削弱肠屏障,而紧密连接蛋白对维持上皮完整性至关重要。
多氯联苯 (PCB)暴露则可诱导氧化应激和肠壁炎症,破坏屏障功能。进一步证实,PCB 同系物混合暴露会提高小鼠肠通透性,并伴随上皮促炎细胞因子增加及 NF‑κB 通路激活。
多种广泛存在于工业和消费品中的化学物质也被证明会影响肠屏障功能。全氟辛烷磺酸(PFOS)暴露会提高小鼠肠通透性,并伴随紧密连接蛋白减少和炎症因子上调。邻苯二甲酸二(2‑乙基己基)酯(DEHP)暴露会增加大鼠肠通透性,与 occludin 和 ZO‑1 表达下降及肠黏膜活性氧升高相关。
▸ 有机污染物会诱发或加重胃肠道疾病
有机污染物(OP)可通过干扰多种胃肠生理过程诱发或加重胃肠道疾病,并参与炎症性肠病(IBD)、肠易激综合征(IBS)和结直肠癌(CRC)的发生发展。
其主要机制包括影响肠道屏障、免疫调节及炎症。双酚A(BPA)暴露会破坏肠屏障并诱导肠黏膜炎症;动物实验表明,多氯联苯(PCB)可增加肠通透性和免疫细胞浸润,从而加剧肠炎并可能触发 IBD。OP 还可通过影响肠蠕动、内脏高敏和免疫反应与 IBS 相关:草甘膦 (GLP)暴露会扰乱肠道微生物群并在动物中诱导内脏高敏;全氟化合物(PFC)则与肠蠕动改变和内脏疼痛有关,提示其可能参与IBS发病。
▸ 调节肠道微生物以解毒有机污染物
肠道微生物群参与药物、环境污染物和膳食成分的代谢与清除,在解毒过程中至关重要。其与外源物之间的复杂相互作用依赖多种微生物酶和代谢途径,对维持宿主健康具有关键意义。
已经提出了几种方法来调整肠道微生物群以增强其解毒能力,包括饮食改变以及益生菌和益生元的使用。
▸ 纤维、多酚和发酵食品的摄入增强肠道菌群的解毒作用
一种策略是通过饮食干预促进有利于解毒的有益肠道菌群。研究表明,膳食纤维、多酚和发酵食品有助于维持肠道微生物群平衡并增强解毒功能。
高纤维、低脂、以植物性食物为主的饮食可促使肠道微生物群朝有利方向改变,如增加产丁酸菌、减少潜在有害菌,并与炎症标志物下降和代谢健康改善相关,从而可能提升机体排毒能力。
富含多酚的食物同样可以调节肠道微生物群并提高解毒潜力。研究发现,补充富含多酚的葡萄提取物可显著增加双歧杆菌和乳杆菌等有益菌,并改善代谢指标和氧化应激标志物,提示解毒能力增强。
▸ 补充益生菌和益生元改善代谢解毒能力
益生菌是活微生物,足量摄入可带来健康益处,而益生元是不可消化的膳食纤维,专门促进肠道有益细菌的生长。研究显示,益生菌补充剂对异生素代谢和解毒有积极影响。
研究发现,给大鼠补充益生菌植物乳杆菌可通过增加粪便排泄帮助去除镉和铅等重金属。另外,添加益生元也能改变肠道微生物组成并增强解毒能力。例如,菊粉作为益生元纤维的添加使益生菌增多,潜在有害菌减少,进而改善代谢指标和炎症标志物,解毒能力提升。
益生菌的干预显示出积极作用
DOI:10.1201/9781003489221-7
▸ 粪菌移植促进解毒和改善健康
此外,粪便微生物群移植(FMT)是一种新兴方法,用于调整肠道微生物组并提高解毒能力。FMT 通过将健康供体的粪便转移到受者体内来恢复健康的微生物组成。研究探索了 FMT 在促进解毒和改善健康方面的潜力。
FMT 也能改变肠道微生物组成,并缓解炎症性肠病(IBD)患者的症状,显示其在疾病状态下促进解毒的前景。
▸ 微生物衍生的酶和代谢物作为潜在治疗剂
微生物衍生的酶和代谢物作为潜在治疗剂也展现了改善解毒的潜力。微生物产生的酶,如葡萄糖醛酸酶和硫酸酯酶,对分解结合代谢物和帮助清除体内异生物质至关重要。
研究表明,添加微生物葡萄糖醛酸酶能促进癌症患者的新陈代谢并增强化疗效果。此外,短链脂肪酸及次级胆汁酸等微生物代谢物可影响宿主的代谢和免疫功能,从而影响解毒过程。例如,短链脂肪酸显著调控肝脏内药物代谢酶的表达与功能,有助于改善解毒和药物清除。
小结
有机污染物(OP)和肠道微生物群之间的复杂关系突显了环境毒理学和人类健康的重要性。该领域的研究揭示了 OP 破坏肠道微生物群和影响宿主生理机能的不同方式。这些机制包括微生物组成的变化、肠道屏障功能的紊乱、炎症反应的触发以及解毒过程的调整。 OP 会加重肠道炎症,破坏代谢平衡,并通过影响肠道微生物群在慢性疾病的发展中发挥作用。
研究有机污染物和肠道微生物群是一个结合了环境毒理学、微生物学和人类健康的动态领域。通过了解背后的机制及其对健康和环境可持续性的影响,研究人员可以帮助创建新方法来降低有机污染物风险并保障公众健康。这一领域的进一步探索对于解决有机污染物带来的复杂问题并加深我们对其对人类健康和环境影响的理解至关重要。
5
铅与肠道微生物群
重金属(HM)如汞、铅、镉在环境中广泛存在,并因其毒性而备受关注。铅(Pb)是一个非必需重金属,常见的环境污染物,对人类健康构成重大风险。
▸ 铅的存在情况及危害
研究表明,城市土壤中的铅浓度通常高于环境背景值,范围在100至1000毫克/千克。一项研究发现,经济地位较低和中等的国家儿童的平均血铅含量(0.84 µg/dl)更高,其中埃及为6 µg/dl、印度为5.46 µg/dl、中国为3.71 µg/dl。即使是极低水平(2 µg/dl)的铅暴露也对健康构成威胁,因此需进一步减少铅暴露。
▸ 铅会对神经、消化、免疫等产生危害
铅接触对社会影响深远,可能导致多种疾病,特别是对儿童的神经系统和认知发展造成严重危害。铅中毒可引发神经、血液、心脏、消化、免疫和肾脏等系统的问题。
学术研究表明,铅暴露显著提高死亡率。与其他重金属一样,铅的毒性与其对靶器官的影响密切相关。研究发现,铅可降低免疫反应、增加氧化应激和炎症,并导致金属稳态失调。此外,铅还可能干扰激素功能。尽管铅危害已知,但其中毒机制仍需进一步研究。
▸ 铅对肠道微生物群的改变
▸ 短期接触铅就会显著影响肠道菌群,代谢表达下降
短期接触铅可以直接影响肠道菌群。在一项初步研究中,口服铅两周的成年大鼠肠道微生物群中乳糖阴性大肠杆菌显著增加。
斑马鱼在暴露于30 µg/L铅7天后,其肠道微生物群发生显著变化,包括厚壁菌门和拟杆菌门显著增加,梭杆菌门和变形菌门减少,以及微生物丰度和种类的变化。
其他肝脏代谢组学研究发现与葡萄糖、脂质、氨基酸和核苷酸代谢相关的41种代谢物变化。通过饮水接触500 mg/kg铅14天后,糖酵解和脂质代谢相关基因(如Gk、Aco、Acc1、Fas、Apo和Dgat)的表达持续下降。
铅暴露对肠道微生物群的影响
DOI:10.1201/9781003489221-9
▸ 厚壁菌与拟杆菌比例上升
相较于急性铅暴露,长期铅接触较为少见,但会改变细菌的丰富度和多样性,可能导致肠道微生物群失调。
接触铅可能导致厚壁菌门与拟杆菌门的相对比例(F/B比)上升,厚壁菌门的优势显著增加,而拟杆菌门则相对减少。研究表明,高F/B比与肥胖和脂质代谢疾病密切相关。
▸ 铅接触减少了乳球菌、肠杆菌、阿克曼菌等
早期接触铅影响了六个分类群:乳球菌属、肠杆菌属和柄杆菌目(Caulobacterales)减少,而脱硫弧菌科、Barnesiella和梭状芽胞杆菌XIVb增加。脱硫弧菌将胆碱转化为三甲胺(TMA),后者在肝脏氧化为TMA N-氧化物(TMAO),与结肠癌和心血管疾病相关。
同年发现,在同一浓度和时间下,小鼠出现肾脏和肝脏结构与功能的改变以及认知能力下降,Lachnospiraceae_NK4A136_group显著增加,螺杆菌种群减少。
铅暴露后,微生物组的变化在前四周更为明显,研究人员推测那时肠道菌群处于铅敏感阶段。铅接触显著减少多种微生物,同时长期铅接触加重胃痛,降低瘤胃球菌科的比例,可能加重胃肠不适,且阿克曼氏菌显著下调,已与结肠炎相关。这些变化与肠道炎症和其他胃肠疾病密切相关,例如某些颤杆菌属(Oscillibacter)的戊酸可能与胃肠问题负相关,而某些毛梭菌(Lachnoclostridium)则促进次级胆汁酸的产生。
健康微生物群与受铅暴露影响的患病微生物群
DOI:10.1201/9781003489221-9
▸ 铅对肠道微生物群相关代谢的改变
▸ 维生素E和胆汁酸浓度失衡
研究人员采用多组学技术分析了暴露于10 ppm铅4周和13周的C57 BL/6小鼠的微生物组和代谢特征,结果显示铅暴露显著改变了微生物的代谢过程,包括能量代谢、氧化应激、维生素E及胆汁酸的变化以及防御/解毒机制。
共鉴定出1314种显著变化的分子特征,其中维生素E和胆汁酸浓度失衡。暴露于铅的小鼠胆固醇及其相关物质显著减少。
▸ 增加一氧化氮生成
在正常与异常胃肠道功能中,一氧化氮(NO)起到关键作用。铅暴露还可能引起编码铜亚硝酸还原酶的基因过度表达,从而增加肠道菌群中一氧化氮(NO)的生成。
核磁共振 (1NMR) 研究显示,饮用水中添加0.1 mg/L铅15周后,小鼠盲肠中15种代谢物水平显著变化,如4-胍基丁酸、胆碱和谷氨酸等增加,而β-半乳糖、苯丙氨酸等则减少。这些变化可能影响三羧酸循环、氨基酸代谢和宿主的能量代谢。
▸ 脂质代谢紊乱
有趣的是,铅处理后的小鼠肝脏显示出脂质代谢基因的剂量依赖性上调,包括参与新脂肪酸合成和转运的基因。暴露于铅八周的小鼠粪便中短链脂肪酸如丙酸、丁酸和醋酸盐的浓度显著降低,这可能与脂质代谢紊乱相关。
这些发现与早期研究一致,表明铅接触可能导致肠道微生物群及其代谢产物发生失调,从而影响宿主的代谢过程。
▸ 铅对肠道屏障的影响
铅暴露显著影响了肠道的结构和屏障功能。扫描电子显微镜显示,小肠表面绒毛不均匀,并出现大片退行性病变,有时缺乏肠上皮细胞微绒毛。
▸ 肠道屏障和通透性显著受损
铅几乎完全保留了与肠道屏障功能相关的ZO-1、Foxp3和Foxo4基因,而与炎症相关的基因(如Il1b、Tnf和Ifng)及抗炎基因(如Tgfb和Il-10)的表达显著下降。
在人类细胞模型中,进一步研究显示跨上皮电阻降低。口服高剂量(1 g/L)铅八周后,小鼠的肠道屏障和通透性显著受损,紧密连接(TJ)相关mRNA(ZO-1、claudin-1和occludin)表达下调,瘤胃球菌增多,Turicibacter减少。
▸ 紧密连接受损,微绒毛收缩
研究还发现,铅暴露使小鼠结肠和小肠中ZO-1、ZO-2、claudin-1和occludin基因表达显著降低。在铅处理后,小鼠的紧密连接mRNA表达进一步下调,尽管其肠道菌群已受损。
此外,长期接触铅对日本鹌鹑盲肠的组织学产生重大影响,导致粘膜脱落、利伯库恩腺损伤和淋巴细胞扩张。透射电子显微镜分析显示超微结构缺陷,包括细胞核固缩、线粒体空泡化和微绒毛收缩。盲肠形状的改变与IFN-γ和IL-2表达水平降低,以及TNF-α、NF-κB和IL-6水平升高相关,并且与粪杆菌和双歧杆菌的减少,以及拟杆菌的增加有关。
▸ 益生菌作为铅对抗治疗策略
铅是重要的有毒金属污染物之一,因此关于使用益生菌作为治疗干预以减少铅中毒的研究不断增多。
▸ 屎肠球菌Pb12能够吸收铅,减少生物体内的积累
研究首先测试了两种屎肠球菌株(Enterococcus faecium)EF031和M74在去除水中铅的能力。据初步研究,粪肠球菌能够有效捕获铅。屎肠球菌 Pb12 的变种是去除环境中铅 (Pb) 的有效菌株。这些细菌能够从鱼类胃肠系统中吸收铅,从而减少组织和器官中铅的生物累积。
▸ 罗伊氏乳杆菌也具有铅去除能力
研究发现,罗伊氏乳杆菌(Lactobacillus reuteri)P16表现出强大的胆汁和酸抵抗力,以及>15%的铅去除率。喂食P16(108 CFU/g)六周,结果显示可改善生长性能,减少组织铅积累,减轻氧化应激等影响。
从发酵食品中分离出的耐铅菌株肠膜状明串珠菌(Leuconostoc mesenteroides)显著降低肝酶水平并恢复部分雄性生殖功能。给暴露于铅的小鼠喂食特定肠道微生物群,结果显示铅去除和紧密连接蛋白表达增加。
▸植物乳杆菌螯合铅并减少损伤
植物乳杆菌CCFM8661能螯合铅并保护小鼠免受氧化应激。该菌在后续研究中显著降低鱼类的血红细胞核异常,提高铅引起的免疫状态。
综上所述,益生菌可能预防铅中毒,并通过改变肠肝循环、增加胆汁流量及谷胱甘肽的产生,促进胆汁和粪便中铅的排泄。使用FXR激动剂时,这种效果显著减弱。膳食补充剂中的益生菌及其他成分也可能提高抗铅效果,并帮助恢复铅暴露小鼠的记忆和学习能力。
小结与未来展望
近期研究显示,胃肠道中的铅对肠道有直接或间接影响。铅中毒损害肠上皮、紧密连接、免疫功能并引发炎症,导致大分子渗透增加。此外,铅暴露可能导致肠道微生物群失调,破坏肠道屏障,影响微生物多样性和代谢物,导致内毒素血症。
一些研究显示益生菌可能有效解毒和治疗铅中毒,但大多数研究关注铅对微生物种群的整体影响,而非特定菌株。因此,需要更深入的宏基因组和代谢组研究,以了解重要细菌对肠道和其他器官的影响。铅对消化道和其他器官的毒性尚不明确,特别是其对脂多糖和其他微生物副产物的影响,还需探讨益生菌减少铅暴露的机制。
▸ 汞污染及其各种来源
汞是全球普遍存在的污染物,存在多种形式,包括元素汞(Hg)和无机汞(Hg+和Hg2+)。
▸ 食物中的汞污染
食品中的汞污染是一个严重问题,尤其是在工业活动造成环境污染的地区。许多人喜欢食用鱼,而鱼类可能通过生物累积过程储存汞。
此外,作为全球许多人主食的大米,可能是在汞污染的土壤中种植,从而导致汞在谷物中积累。
▸ 药物、环境中的汞污染
含汞药物也增加了汞暴露的风险,尽管低于食品中的汞。过去,牙科汞合金、抗菌霜及其他药物中均含有汞。尽管由于其毒性,药品中使用的汞量有所减少,但它在某些补充和替代医疗中仍然是重要的接触源。
此外,空气中汞沉积在土壤中,一些细菌将其转化为甲基汞,植物吸收后使汞进入食物链,这使得汞更容易从土壤迁移到植物,并最终传递到更高级别的动物中。
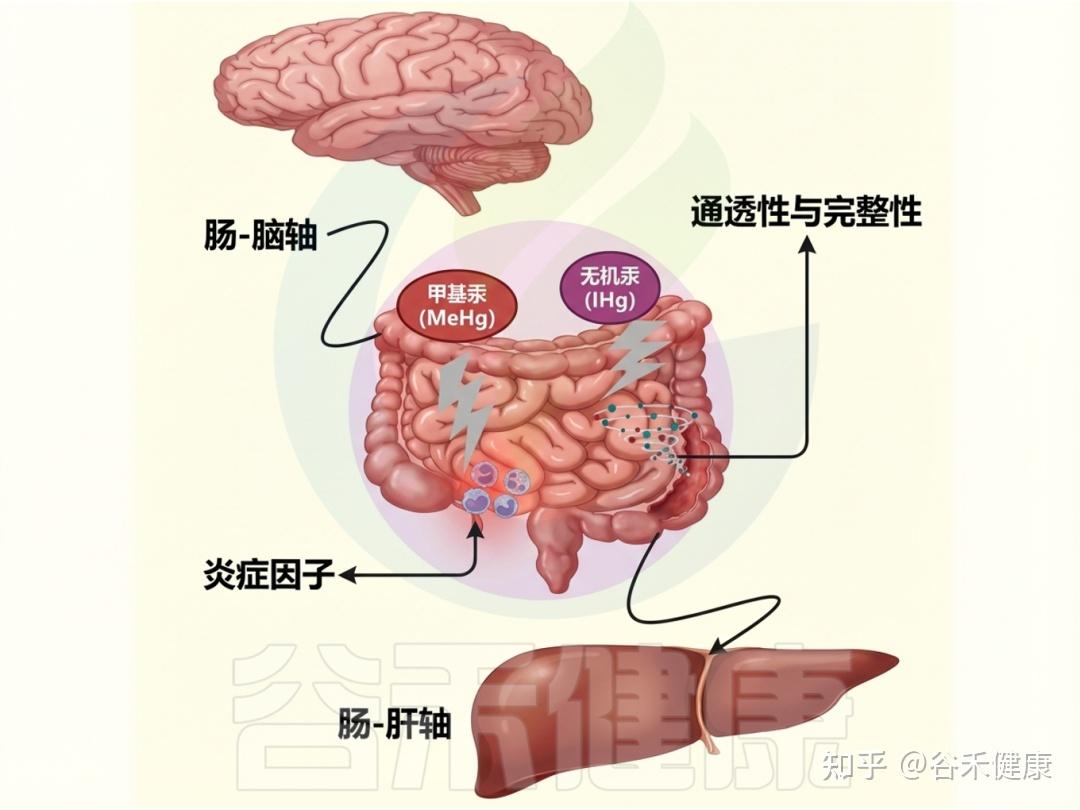
▸ 汞的毒性及危害
汞是已知的环境毒素之一,对人类健康产生复杂的负面影响,尤其是对肝脏、肾脏和中枢神经系统。它的毒性源于双重作用,表现为氧化应激增加和对特定器官系统的集中攻击。
汞对肠道屏障、微生物紊乱等的影响
▸ 氧化损伤:汞损害身体的主要方式
汞对身体有害的主要方式之一是引起氧化应激。当元素汞和无机汞相互作用时,会产生活性氧(ROS),如超氧自由基和过氧化氢。当这些活性氧水平过高时,将压垮人体的抗氧化防御系统,导致脂质、蛋白质和DNA的氧化损伤。
汞造成的氧化损伤广泛影响细胞结构,导致膜不稳定和细胞功能障碍。蛋白质的氧化改变了多种生理过程,而氧化应激诱导的DNA损伤则可能引起突变。
▸ 器官特异性毒性:汞的不同目标
汞对某些器官也有独特的影响。汞中毒影响的主要器官是肝脏、肾脏和中枢神经系统。
肝脏作为解毒中心,汞在进入循环系统后通常积聚于此。肝脏尝试清除汞,导致氧化应激增加,长期接触可能损害肝功能并增加肝毒性风险。
肾脏在过滤血液中的汞时,可能会受到损害。无机汞在肾脏组织中的积累会导致肾毒性,干扰其过滤和重吸收功能,进而损害肾功能。
汞的神经毒性特别影响中枢神经系统,因其能穿过血脑屏障。甲基汞的神经毒性尤为明显,影响整体神经健康、运动能力和认知功能。与汞接触相关的神经系统症状部分源于神经递质途径的破坏及炎症激活。
▸ 肠道微生物群影响汞的作用
▸ 微生物基因簇影响汞转化
某些基因簇(如hgcAB和mer操纵子)决定了肠道微生物群在汞转化中的作用。hgcAB基因簇促进无机汞(IHg)转化为甲基汞(MeHg),而mer操纵子(包括merA和merB基因)则将MeHg去甲基化还原回IHg。
肠道中汞生物转化的复杂过程
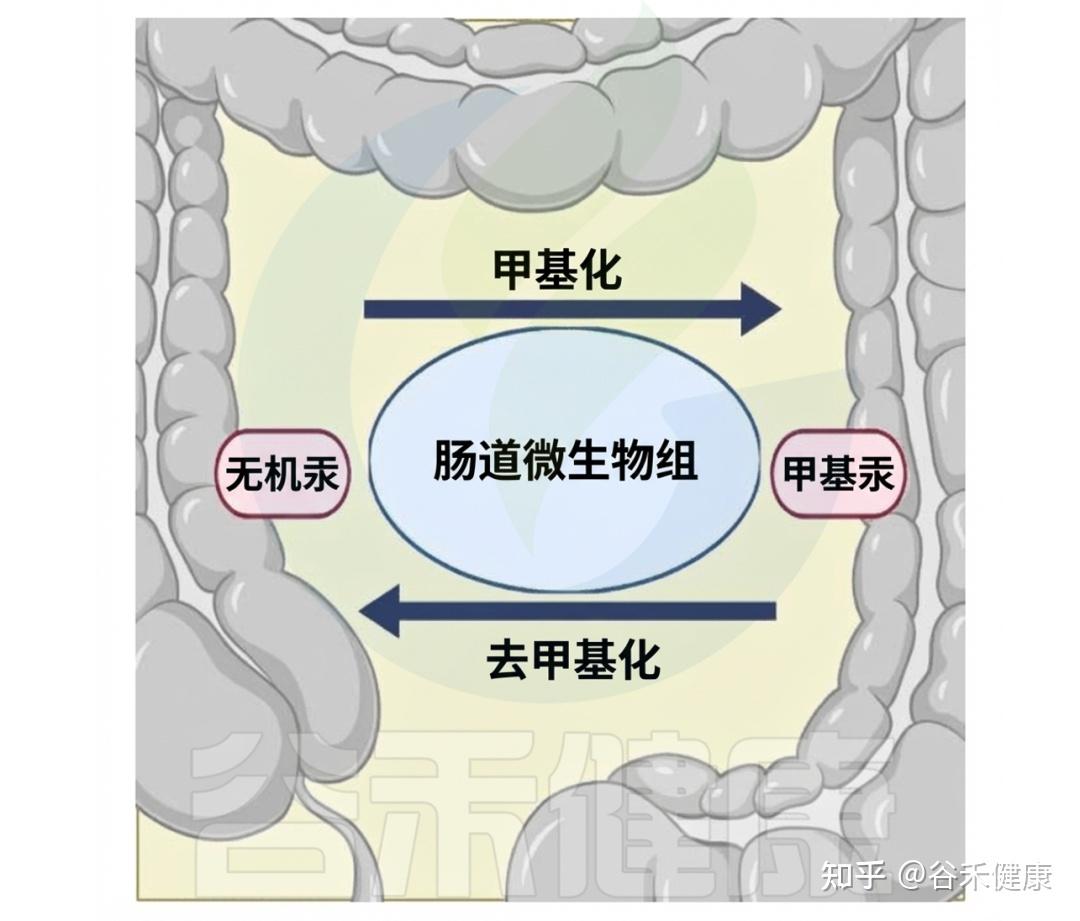
这些基因簇作为微生物工具,使肠道微生物能够参与汞的复杂变化,其丰度因接触汞而增加,显示出微生物群落的适应性和在肠道汞循环中的重要性。
▸ 影响汞的毒性和生物利用度
肠道微生物群在汞代谢中不仅限于甲基化和去甲基化,还积极改变汞的化学形态,影响生物体的命运。与汞暴露相关的毒理学后果受到微生物群调节汞形态能力的影响。同时,新的研究揭示了汞引起的肠道功能障碍与肠道菌群之间的复杂相互作用,为健康后果提供了新见解。
汞与肠道微生物群的最新研究
▸ 肠道菌群有助于减少汞的吸收和解毒
最新研究发现,肠道细菌在汞的解毒与生物转化中发挥关键作用,可能影响体内的汞形态和生物蓄积。有研究指出可利用人胃中的细菌减少对有害金属如汞的吸收,但两者之间的关系仍需进一步探讨。
益生菌可表达一种酶,将易吸收的汞形式转化为难以吸收的形式,为减少人类接触汞提供了可能的策略,尤其对于弱势群体。此外,研究中还显示,改变肠道微生物群(如通过抗生素或粪便微生物群移植)可以减轻汞暴露引起的肝脏损伤,并展示了汞暴露如何影响肠道微生物的多样性和构成,导致肝脏铁死亡(一种受控细胞死亡过程)。
▸ 汞的接触改变了肠道微生物群的组成
此外,汞暴露显著影响肠道菌群。一项研究显示,湿地植被的存在促进了汞的甲基化,而无植被沉积物中甲基汞的生成与微生物组变化有关。研究还表明,肠道微生物群的去甲基化能力可能受到汞暴露水平和来源的影响,且某些细菌如脱硫弧菌和产甲烷菌可能对肠道甲基汞去甲基化有所贡献。
重金属如汞的接触改变了肠道微生物群的组成,病原菌如气单胞菌(Aeromonas)数量显著增加。这表明健康、肠道微生物群以及重金属暴露之间可能存在密切联系。
坦桑尼亚的一项研究探讨了补充益生菌的酸奶是否能降低幼儿和孕妇的重金属,尤其是汞的含量,结果表明,益生菌酸奶能够防止汞含量上升,从而证明其减少重金属暴露负面影响的潜力。
这些发现强调了肠道微生物群与重金属(如汞)之间复杂而相互的关系,以及益生菌和其他疗法可能减轻重金属暴露负面后果的前景。
镉是一种有毒重金属,来源于自然和人为因素,其渗透到环境中对人类健康构成潜在威胁。
▸ 镉的来源和健康危害
作为一种广泛存在的环境污染物,这种重金属普遍存在于沉积物、溪流水和土壤中。中国同样面临镉暴露问题,在大米中检测到较高的镉含量。
▸ 镉会导致糖尿病、心血管疾病、慢性肾病等
镉与多种严重健康问题相关,包括癌症、糖尿病、心血管疾病、慢性肾病、骨质疏松、肝病和不良的神经发育结果。
研究估计,高浓度镉暴露可导致死亡率增加17%,突显了解决这一环境挑战的紧迫性。历史上,对镉影响健康的研究主要关注与接触相关的有害结果,包括金属动态失衡、氧化应激、炎症、内质网应激和基因组不稳定等问题。
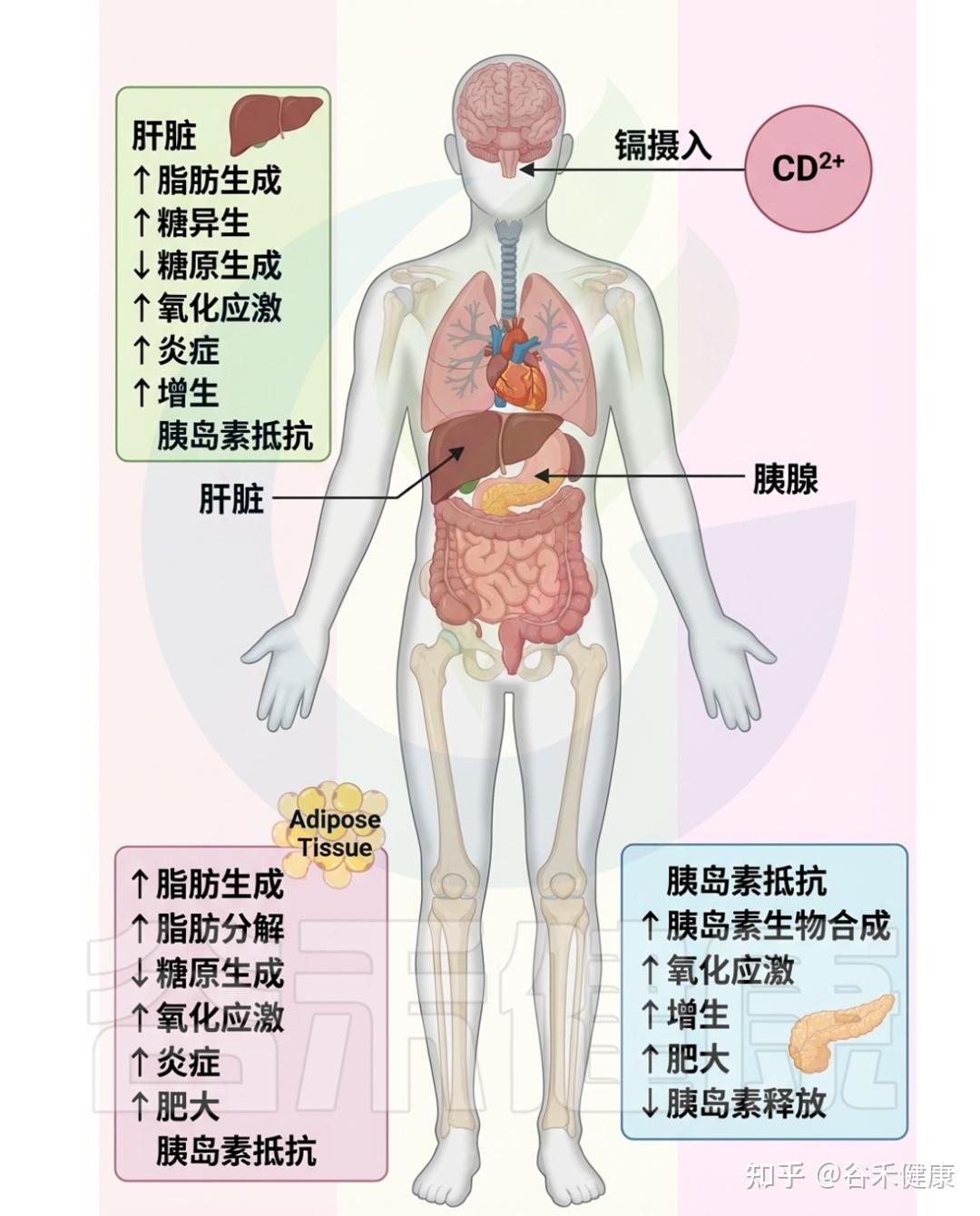
镉毒性的影响
▸ 导致锌、铜等微量元素的失衡
在细胞水平上,镉破坏金属稳态,导致锌和铜等必需微量元素的失衡,主要通过与这些金属竞争结合运输蛋白而实现。这种不平衡损害细胞功能,并引发各种健康问题。
▸ 损害脂质、蛋白质和DNA等细胞成分
从分子层面来看,镉诱发氧化应激,表现为活性氧(ROS)产生与机体对其的清除能力之间的不平衡。镉刺激超氧自由基和过氧化氢等的生成,损害脂质、蛋白质和DNA等细胞成分,导致细胞功能障碍,并激活与炎症和细胞凋亡相关的信号通路。此外,镉还会引发内质网应激,破坏其蛋白质折叠能力。
▸ 增加遗传突变风险,干扰正常内分泌
镉还与基因组不稳定性相关,影响细胞遗传物质的完整性,增加DNA损伤和突变的风险,从而可能导致癌症的发生。此外,镉还充当内分泌干扰物,模仿激素作用并扰乱正常内分泌功能,尤其在生殖和发育过程中影响显著。
总的来说,镉毒性对细胞和分子的影响表现为金属稳态破坏、氧化应激、内质网应激、基因组不稳定性和内分泌干扰。这些复杂机制共同导致与镉暴露相关的各种不良健康结果,反映出其多方面的毒理学影响。
▸ 镉与肠道微生物群的互作
镉的毒性不仅源于直接的细胞相互作用,还涉及肠道微生物群的复杂生态系统。镉一旦摄入,便穿过胃肠道,首先与粘膜内层的多种微生物群接触,可能与产生粘蛋白的杯状细胞相互作用。这些相互作用可能损害肠道屏障的完整性,促进镉向肠道深层的转移。
▸ 镉会诱导氧化应激扰乱肠道微生物平衡
镉诱导的氧化应激能扰乱肠道微生物群的平衡,促进某些细菌生长并抑制其他细菌繁殖。它还倾向于与酶和关键蛋白质的巯基结合,这种结合影响微生物的结构与功能,从而可能中断重要的代谢途径和营养循环。
▸ 镉还会通过肠道微生物群影响免疫功能
镉的影响还延伸至肠道微生物组的免疫调节功能,微生物群与免疫系统的相互作用对维持对病原体的有效反应至关重要。镉引起的微生物组成变化可能触发免疫反应,导致炎症加剧肠道疾病。因此,深入理解镉与肠道微生物组成的复杂相互作用非常重要。
▸ 镉暴露导致乳杆菌、梭状芽胞杆菌等减少
对不同年龄实验动物的研究发现,短期和长期镉暴露都会显著影响肠道微生物组的生理。暴露于镉42天的成年大鼠消化道中的需氧和厌氧乳酸菌均显著减少。镉在属和科水平上引起了肠道微生物群的特殊变化,例如,在饮用水中接触镉45天的小鼠中,
蜡样芽孢杆菌、乳杆菌、梭状芽胞杆菌和大肠杆菌数量显著减少,并呈剂量依赖性。而八周镉暴露则以剂量依赖的方式增加小鼠盲肠中放线菌的相对丰度,但对拟杆菌门和厚壁菌门的影响未达到统计显著性。
在盲肠菌群的科水平上,Coriobacteriaceae和Lactobacillaceae的相对数量以剂量依赖性方式增加,而Lachnospiraceae的百分比下降。尽管在镉暴露的小鼠中,粪便微生物的变化不那么明显,但Odoribacter和Alistipes属的相对丰度显著增加。
注:镉引起的微生物群变化在初期四周更为明显。由于无菌动物不易表现出促炎反应,观察到的NF-kB激活和促炎细胞因子的产生至少部分依赖于镉诱导的肠道微生物群改变。
▸ 镉还影响短链脂肪酸和细菌蛋白的表达
从三周开始,增加剂量的镉显著减少了肠道细菌数量及厚壁菌门与拟杆菌门的比例。乳杆菌的显著降低仅在三周后出现,而双歧杆菌的数量在一周后便开始下降。同时,TNF-α合成增加,伴随与短链脂肪酸(SCFA)相关的基因变化。
研究还表明,镉暴露降低了粪便中的短链脂肪酸含量。除了影响肠道生物多样性,短期镉治疗也对肠道微生物组功能产生影响。研究显示,接触镉后15分钟内,细菌蛋白质组发生显著变化,表明对镉暴露的反应可能与肠道微生物群中蛋白质表达的独特变化有关。
早期体外研究也表明,镉影响整体基因表达,尤其是在能量代谢、转运、细胞周期和结合蛋白基因的表达上,镉暴露后0、5、15和25分钟,相关基因的表达均有所上升,可能涉及脂多糖的生产或运输。
长期低剂量镉(Cd)暴露的影响
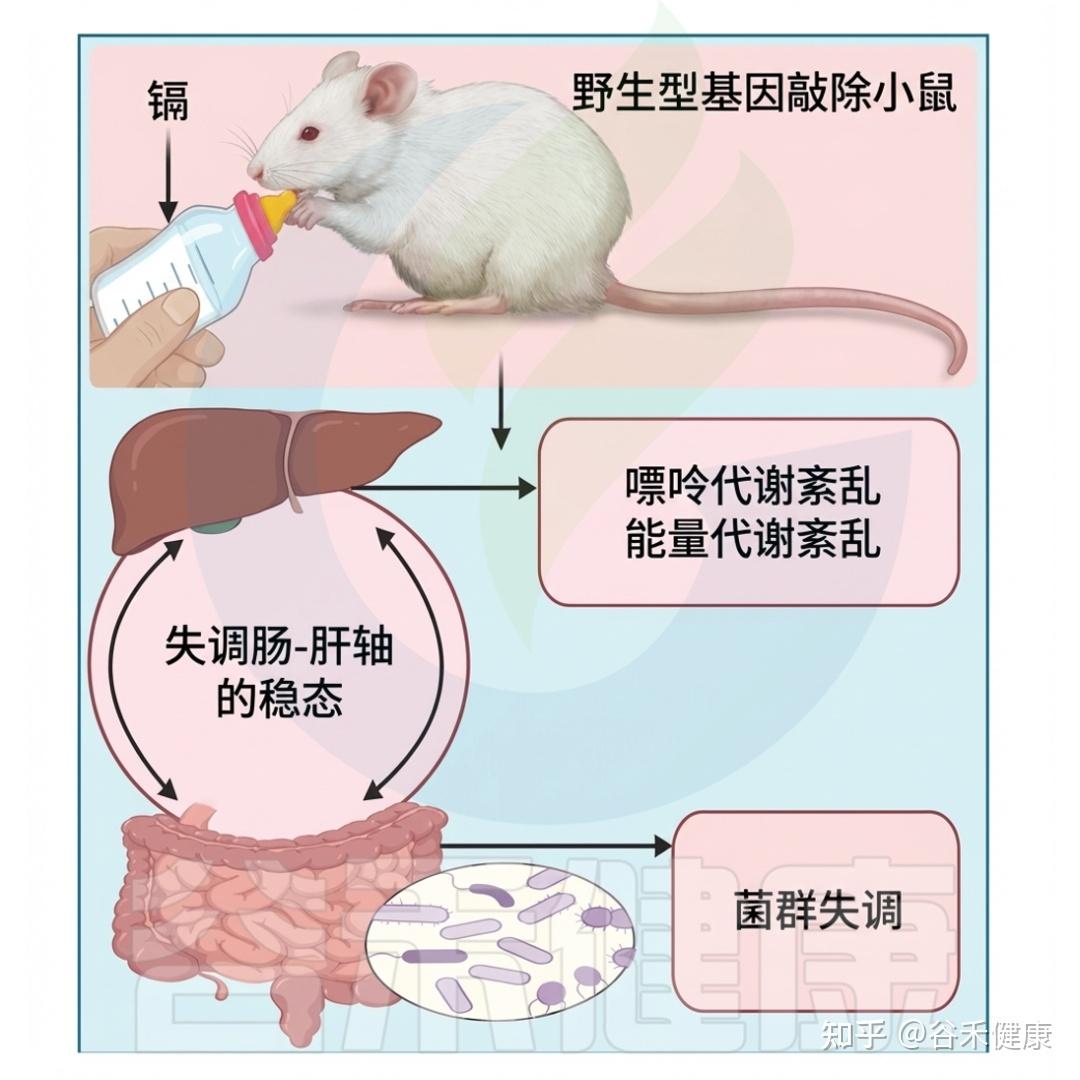
这些发现说明,镉几乎立即影响肠道微生物群。婴儿期低剂量镉暴露也显著改变肠道微生物群的数量和种类。在八周龄接触镉的雄性小鼠中,拟杆菌数量显著增加,而厚壁菌相应减少,双歧杆菌和普雷沃菌的数量下降幅度最大。
此外,接触镉的动物体内的粘液鞘氨醇单胞菌(Sphingomonas)数量明显增加,并与较高的肥胖程度相关。另一项研究显示,暴露于镉的五周大雄性小鼠的盲肠厚壁菌门显著降低,而拟杆菌门和g-变形菌门未显著下降。这些变化可能与肝脏能量平衡基因的改变有关,进而影响脂肪酸的生产和运输。
总体上,由于拟杆菌门丰度的增加及厚壁菌门数量的减少,多项研究表明这二者的比率显著上升。有关镉暴露对特定细菌群体影响的不一致性可能与不同的暴露方案(剂量、持续时间、种类)有关。
▸ 镉与肠道通透性和炎症
▸ 镉破坏肠道屏障,提高肠道通透性
研究表明,镉暴露显著降低肠上皮细胞活力,并损害紧密连接,影响细胞旁通透性。后续研究发现,HT-29细胞单层中紧密连接蛋白分布不均,镉暴露小鼠的结肠和空肠中ZO-1、ZO-2、occludin和claudin-1的mRNA表达显著减少。这些变化与肠道通透性提高相关,导致血液中LPS水平升高。
在Caco-2细胞培养中,镉显著破坏紧密连接,造成细胞间连接损伤,并上调与氧化应激、热休克蛋白和胱天蛋白酶3相关的基因。这验证了Caco-2细胞旁通透性显著增加的研究,伴随HSP70水平升高和坏死病变的存在。此外,镉暴露还导致小鼠血清脂多糖结合蛋白水平升高,且呈剂量依赖性。
▸ 提高炎症细胞因子水平,促使肠道炎症
长期接触镉改变肠道组织学,显示绒毛增厚变短,且伴有融合和坏死区域。肠道乳酸菌数量减少,TNF-α、IFN-γ、IL-1β和IL-17水平升高,导致肠道形态改变。肠系膜淋巴结呈现镉诱导的变化,包括细胞增殖和结构变化。
Caco-2细胞模型表明,镉通过I-kBa降解诱导NF-kB激活,促使肠道炎症反应。镉和脂多糖的联合暴露显著增强了肝损伤,相较于单独暴露,氧化、亚硝化应激和炎症反应加剧。共同暴露也影响一氧化氮的生成,并且镉会替代必需金属(Cu、Zn)。
▸ 高镉暴露还会导致内毒素血症
值得注意的是,高镉暴露与代谢综合征的频率显著相关。研究表明,镉可阻碍细胞中的脂多糖信号传导,抑制巨噬细胞中NF-kB通路,降低对感染因子的抵抗力。
镉诱导的氧化应激可能导致小鼠巨噬细胞反应性失调。在患有结肠炎的小鼠中,镉的接触显示促炎细胞因子表达的剂量依赖性减少。迄今的研究表明,镉可能通过增加肠道通透性来诱发内毒素血症,并提高脂多糖水平。然而,脂多糖与镉之间的关系可能表现出拮抗和协同作用,具体取决于剂量和作用机制。
▸ 益生菌作为镉暴露的治疗策略
大量研究表明,益生菌在镉中毒中发挥显著保护作用,可以缓解肠道菌群失调。
▸ 植物乳杆菌和菊粉等有助于减轻镉引起的损伤
含有凝结芽孢杆菌、植物乳杆菌和菊粉的膳食已被证实能减轻镉引起的肝脏和肾脏损伤。具体而言,富含益生菌的饮食显著减少了镉在肝脏和肾脏中的积累,并降低了血液中ALT、AST、肌酐和尿素的水平。
此外,接受益生菌治疗的镉暴露大鼠中,乳酸菌丰度增加,与观察到的健康改善一致。在暴露于镉的HT-29结肠细胞中,植物乳杆菌显著减弱了镉诱导的炎症反应,减少了TNF-α、IL-1β、IL-6和IL-8的产生。
植物乳杆菌治疗还减少了镉暴露大鼠的内毒素血症、肠道通透性和紧密连接异常,并增强了肾脏和肝脏的抗氧化反应,降低了镉造成的损伤。这些结果促使研究人员认为,与抗氧化疗法相比,益生菌可能是治疗镉中毒的更有效方法。
▸ 益生菌降低了镉的毒性并恢复肠道微生态
值得注意的是,尽管腹腔注射植物乳杆菌对接触镉的动物未能减少器官中镉的积累,但却增强了抗氧化活性并保护组织。含有鼠李糖乳杆菌、嗜酸乳杆菌和长双歧杆菌的益生菌可有效逆转镉诱导的乳酸杆菌和双歧杆菌数量减少,从而显著降低镉的遗传毒性,并减少肝细胞DNA损伤。
此外,益生菌通过与结肠中的金属离子结合,降低其生物可及性,从而对汞暴露产生预防作用,研究发现血液金属浓度下降、粪便中镉水平和乳杆菌含量增加之间存在显著相关性。
在体外模型中,Cd(NO3)2与开菲尔乳杆菌的预孵育显著降低了其对HepG2细胞的毒性。研究表明,益生菌在预防镉中毒方面主要针对镉引起的紧密连接破坏和肠道炎症,此外,添加锌补充剂可能增强益生菌的保护作用。
砷是全球最危险的环境污染物之一,广泛存在于自然环境中,主要来源于地表侵蚀。它与食物、饮水以及空气中土壤和颗粒密切相关,人类活动(尤其是砷采矿和工业使用)进一步加剧了这种污染。
砷化合物对大多数生物构成威胁,长期接触低水平砷也与多种疾病相关。国际癌症研究机构(IARC)已将砷及其无机化合物列为一级致癌物,与皮肤癌、肺癌和膀胱癌密切相关,并且暴露于砷与代谢紊乱、皮肤损伤、糖尿病、心血管疾病等非癌性疾病也有关。
砷存在于多个化学形式中,分别具有不同的毒性和生物可及性,其中As(III)和As(V)是饮用水中最常见的二价和五价形式。
▸ 肠道微生物群对砷的作用
肠道微生物可代谢砷,从而改变其在宿主中的毒性。相反,如果砷导致特定微生物的死亡,则可能削弱它们的保护作用。细菌砷加工的研究主要集中于研究微生物对砷有害影响的“抵抗力”,例如其导致细胞死亡的能力。
▸ 部分肠道细菌具有抗性基因或能编码代谢砷的酶
Ars操纵子是抗性基因簇,最初在大肠杆菌和金黄色葡萄球菌的质粒中发现,后来在多种医学相关病原体中得到验证,如耶尔森氏菌、单核细胞增生李斯特菌和空肠弯曲菌。Ars操纵子还在人类肠道共生体中被鉴定,如枯草芽孢杆菌和普通拟杆菌。
细菌编码的酶能够通过氧化、还原、甲基化和去甲基化过程改变无机和有机砷,这些过程与特定底物相关联。细菌代谢物可能对胃肠道中砷化合物的转化有显著影响。
砷与肠道微生物群间的相互作用
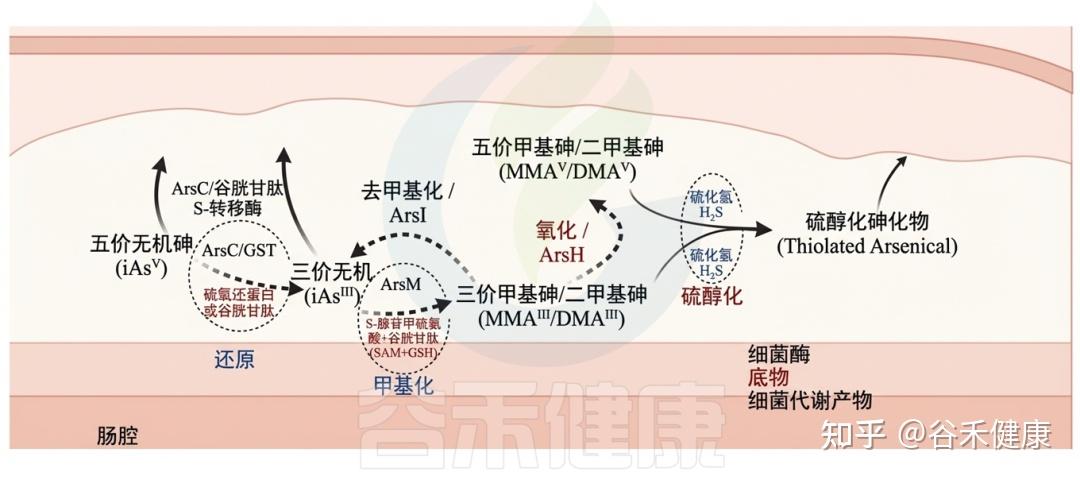
▸ 肠道微生物群能够改变砷暴露水平
最近的研究表明,大鼠盲肠中的大肠杆菌能够代谢二甲基亚砷酸盐(DMAsV),生成三甲基亚砷酸盐(TMAVO)和一种未确定的砷化合物。研究者还考察了微生物组成员在实验室中的砷吸收。小鼠的微生物组在酶促还原和甲基化无机砷(iAs)的能力方面表现出强大效果。
两项独立研究显示,在模拟人类肠道环境中,微生物群可减少无机砷(iAsV)的浓度并进行甲基化,生成毒性较低的砷化合物。即便在高压灭菌实验对照中,iAsV也仍能转化为iAsIII,这表明非酶机制可能参与此转化。
研究发现,给小鼠服用抗生素会显著减少其粪便中砷的排泄,同时增加其肝脏和肺组织中的砷积累。研究人员推测,抗生素暴露降低了肠道微生物的生物量,从而影响了砷的排泄。其他实验表明,无机砷(iAs)会被吸附到革兰氏阳性细菌的胞外聚合物上,而革兰氏阴性细菌则不然,这可能说明微生物对砷的积累是通过该过程实现的。
▸ 肠道微生物群可降低砷毒性
微生物组的“表型”与宿主内甲基化砷与无机砷比例的变化相关。然而,鲜有研究明确证实微生物组的修饰、变化或缺失与宿主健康之间的直接联系。研究人员使用已人源化且无菌的AS3MT-KO小鼠进行实验,旨在评估微生物组的异质性如何影响疾病结果,特别是死亡率。
研究发现,只有少数细菌始终表现出积极作用(即延长存活时间),这些细菌为人类肠道中一些常见物种。例如,经黏液真杆菌属(Blautia)的两个成员,以及来自毛螺菌科、瘤胃球菌属和粪杆菌科的细菌,与不同供体的生存有强相关性。
▸ 宿主基因决定肠道微生物表型对砷代谢的影响
大量证据表明,个体对砷暴露引发疾病的易感性差异显著,这通常与体内对砷的处理方式不同有关。在此环境中,肠道微生物群对砷代谢及多脏器的全身反应起着直接作用,表明微生物群特征可能影响砷化合物的代谢范围。
然而,宿主遗传学与肠道微生物组如何结合影响砷的生物转化机制尚不明确。我们采用综合方法,将16S rRNA基因测序与HPLC-ICP-MS砷形态分析结合,发现IL-10基因的缺失会显著改变肠道微生物组的组成,从而对砷代谢产生重要影响。
然而,证据表明,保留微生物组的小鼠组比无菌或接受抗生素治疗的小鼠组在砷引起的死亡时间上显著延长。因此,微生物组具有保护宿主免受砷毒影响的潜力。这些发现可能为开发砷中毒的创新治疗和预防方法提供基础。
▸ 砷对肠道微生物群的影响
研究发现砷能够改变肠道微生物群的组成,但其对微生物组功能的影响,尤其是在环境相关剂量下,尚未充分评估。使用16S rRNA测序和宏基因组测序,检查了在100 ppb砷处理下小鼠肠道微生物组的组成和功能。
▸ 影响碳水化合物代谢和维生素合成
结果显示,砷暴露不仅改变了微生物群的组成特征,还导致了α和β多样性的变化。宏基因组分析表明,碳水化合物代谢相关基因的表达显著变化,特别是在丙酮酸发酵、脂肪酸合成和淀粉利用方面。
此外,砷暴露小鼠的肠道微生物群中,脂多糖合成和压力响应基因的表达显著增加,与维生素(如叶酸、维生素B12、B6和K2)合成相关的基因含量也更高。同时,砷治疗后,与药物耐药性和接合转座子蛋白相关的基因表达显著上升。
▸ 破坏胆汁酸的代谢平衡
让 C57BL/6 小鼠的饮用水中砷浓度达到百万分之一 (ppm),目的是检查砷暴露对胆汁酸平衡的影响。研究结果表明,砷暴露导致初级胆汁酸(特别是那些未结合的胆汁酸)发生明显变化,并持续降低血清和肝脏中的次级胆汁酸。
拟杆菌门与厚壁菌门的比例与血流中胆汁酸的浓度相关。目前的研究提供的证据表明,砷暴露引起的肠道微生物群失衡可能会导致受砷影响的胆汁酸调节的破坏。
最终结果显示,即使在适宜的环境砷水平下,仍会破坏肠道微生物组的整体结构并显著改变一些关键的细菌功能途径。接触100 ppb砷会扰乱小鼠肠道微生物组的典型组成和功能,这对环境具有重要意义。能量代谢基因的表达发生了变化,与脂多糖生成、氧化应激和DNA修复相关的基因广泛上调。
此外,砷暴露还导致与接合转座子蛋白、外排多药元件及维生素合成相关的基因表达增加。这些发现为理解砷对肠道微生物组影响提供了新的见解,尤其是在环境相关的剂量下。
环境毒物无处不在, 如果积累到上限,不仅直接挑战人体的解毒器官,更在悄无声息中重塑着我们体内的“隐形生态”——肠道菌群。科学研究已经证实,环境污染物与肠道微生物之间存在着复杂的双向互作:过量的毒物可以破坏菌群平衡,诱发代谢与免疫紊乱;而健康的肠道菌群和肠道屏障则是人体抵御外源毒素的第一道防线,甚至能协助降解部分有害物质。
面对这一挑战,我们既无需过度恐慌,也不能掉以轻心。未来的健康管理将不再局限于对疾病的被动治疗,而是转向对环境暴露风险的主动评估。
这需要我们在三个维度上通过跨学科的协同努力实现突破:
首先是强化内在防线,通过回归自然饮食,利用富含膳食纤维和植物化学物的全谷物、蔬果来“喂养”体内的有益菌等自然疗法,激发菌群自身的解毒潜能;其次是落实精准医疗,结合基因组与代谢组数据,为不同体质的人群定制个体化的健康管理方案;最后是筑牢社会屏障,推动更严格的环境卫生政策,重点保护儿童及弱势群体。
本文仅供科研交流。
主要参考文献
Fuloria, Neeraj Kumar, Shivkanya Fuloria, Mahendran Sekar, Kamal Narain, and Sokindra Kumar. 2025. “Introduction to Environmental Toxicants.” In Gut Microbiome and Environmental Toxicants. 1st ed. CRC Press.https://www.taylorfrancis.com/chapters/edit/10.1201/9781003489221-1.
Campana AM, Laue HE, Shen Y, Shrubsole MJ, Baccarelli AA. Assessing the role of the gut microbiome at the interface between environmental chemical exposures and human health: Current knowledge and challenges. Environ Pollut. 2022 Dec 15;315:120380.
R.M. Abdel-Megeed, Probiotics: a promising generation of heavy metal detoxification, Biological Trace Element Research, 199 (2021) 2406–2413.
K.S. Alharbi, W.H. Almalki, M. Albratty, A.M. Meraya, A. Najmi, G. Vyas, S.K. Singh, K. Dua, G. Gupta, The therapeutic role of nutraceuticals targeting the Nrf2/HO-1 signaling pathway in liver cancer, Journal of Food Biochemistry, 46 (2022) e14357.
A.Giuliani, M. Zuccarini, A. Cichelli, H. Khan, M. Reale, Critical review on the presence of phthalates in food and evidence of their biological impact, International Journal of Environmental Research and Public Health, 17 (2020) 5655.
Y.Wang, H. Qian, Phthalates and their impacts on human health, Healthcare, 9 (2021) 603.
C.Tropini, How the physical environment shapes the microbiota, mSystems, 6 (2021) e0067521.
G.McCallum, C.Tropini, The gut microbiota and its biogeography, Nat Rev Microbiol, 22 (2024) 105–118.
H.R. Safford, K. Shapiro, H.N. Bischel, Wastewater analysis can be a powerful public health tool—if it’s done sensibly, Proceedings of the National Academy of Sciences, 119 (2022) e2119600119.
B.D. Anderson, J.E. Bisanz, Challenges and opportunities of strain diversity in gut microbiome research, Frontiers in Microbiology, 14 (2023) 1117122.
D.O’Connor, D. Hou, J. Ye, Y. Zhang, Y.S. Ok, Y. Song, F. Coulon, T. Peng, L. Tian, Lead-based paint remains a major public health concern: a critical review of global production, trade, use, exposure, health risk, and implications, Environment International, 121 (2018) 85–101.
T.E. Adolph, L. Mayr, F. Grabherr, J. Schwärzler, H. Tilg, Pancreas-microbiota cross talk in health and disease, Annual Review of Nutrition, 39 (2019) 249–266.
Y. Pan, Y. Song, Broken thermometer in hand: mercury elevation caused by improper debridement, Clinical toxicology, 60 (2022) 654–656.
F.U. Haider, C. Liqun, J.A. Coulter, S.A. Cheema, J. Wu, R. Zhang, M. Wenjun, M. Farooq, Cadmium toxicity in plants: impacts and remediation strategies, Ecotoxicology and Environmental Safety, 211 (2021) 111887.
Q.Gu, C.Wang, Q.Xiao, Z.Chen, Y.Han, Melatonin confers plant cadmium tolerance: an update, International Journal of Molecular Sciences, 22 (2021) 11704.
B.Lv, H.L. Yang, Y.D. Peng, J. Wang, Z. Zeng, N. Li, Y.E. Tang, Z. Wang, Q.S. Song, Cadmium exposurealters expression of protective enzymes and protein processing genes in venom glands of the wolf spider Pardosa pseudoannulata, Environmental Pollution, 268 (2021) 115847.
Martinez-Guryn K, Leone V, Chang EB. Regional diversity of the gastrointestinal microbiome. Cell Host & Microbe. 2019 Sep 11;26(3):314–24.
Chi L, Xue J, Tu P, Lai Y, Ru H, Lu K. Gut microbiome disruption altered the biotransformation and liver toxicity of arsenic in mice. Archives of Toxicology. 2019 Jan 21;93:25–35.

谷禾健康
肠道屏障是将宿主与外界隔离的主要防御,具有多项关键生理功能,包括营养消化、吸收以及防止潜在有害的膳食抗原和致病微生物的侵害。然而,饮食、药物、昼夜节律紊乱、年龄、肠道微生物群、微生物代谢物和遗传易感性等多种因素都可能破坏肠道屏障。这种破坏可能导致细菌易位,进而引发肠肝和全身性炎症。
目前,肠道屏障受损已被认为与多种疾病的发病机制有关,包括炎症性肠病(克罗恩病和溃疡性结肠炎)、肠易激综合征和结直肠癌等肠道疾病。此外,肝病(如代谢功能障碍相关脂肪性肝病、酒精性肝病和原发性硬化性胆管炎)和全身性代谢疾病(如糖尿病和肥胖)也与肠道屏障受损有关。
然而,目前大多数临床数据仅显示相关性,尚无法明确肠道屏障损伤是这些疾病的原因还是结果。目前,全世界药监督管理局尚未批准专门用于修复肠道屏障损伤的药物。现有疗法主要侧重于疾病的预防和管理,并严重依赖免疫抑制剂来控制炎症。
不幸的是,持续的屏障损伤和延迟愈合会降低这些治疗的疗效,并可能导致治疗耐受甚至复发。因此,开发直接靶向肠道上皮屏障的治疗策略至关重要。
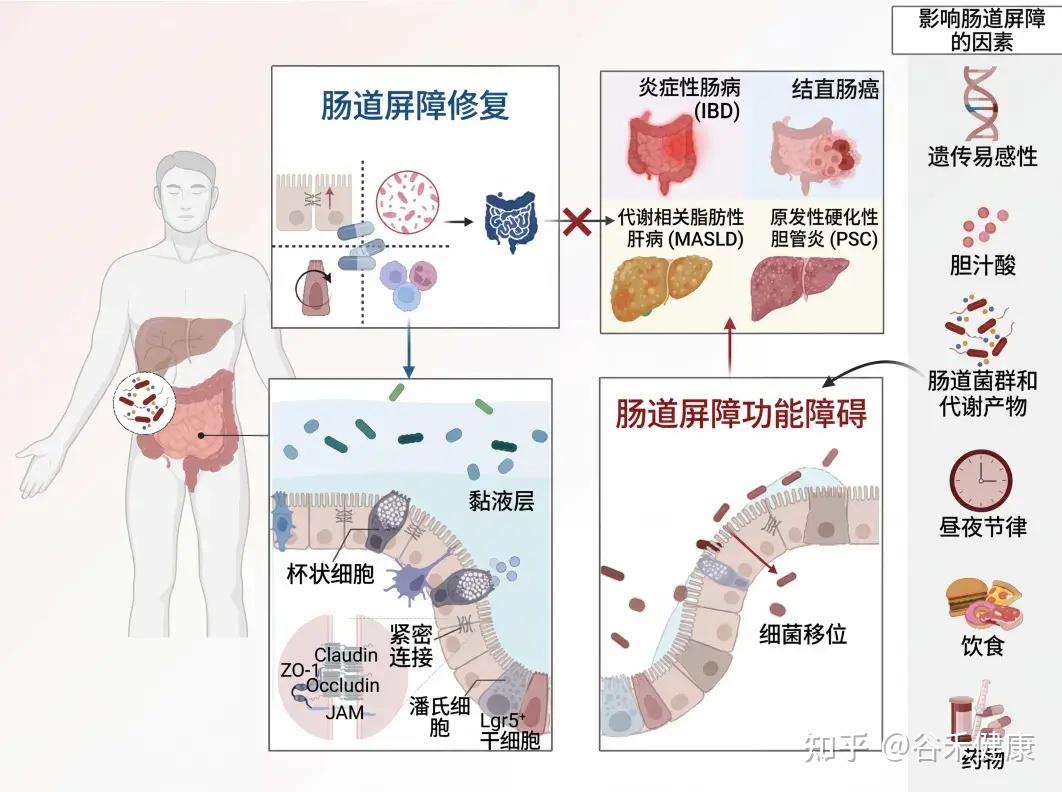
本文介绍肠道屏障的基本功能,生理结构和功能、影响其完整性的内外因素,重点介绍一些潜在的治疗策略,旨在恢复屏障完整性,改善和优化疾病管理。
肠道是一个独特的器官,在食物消化、营养吸收、动态的宿主与环境互动以及维持身体稳态方面发挥着关键作用。
为了保护宿主免受外部威胁,例如长期暴露于膳食抗原和病原微生物,肠道上皮细胞(IECs)形成了多种类型的屏障,包括机械屏障、富含共生微生物的粘液层,以及由免疫细胞及其活性物质组成的免疫屏障(下图)。

机械屏障:由紧密连接(TJs)形成的紧密排列的IECs层构成,确保肠道的结构完整性,调节肠道通透性,并控制水和大分子的运输。
粘液层:由杯状细胞分泌的粘蛋白组成,为共生细菌提供栖息地和营养。其独特的结构特征限制了病原体的渗透,进一步增强了TJs的物理隔离功能。
干细胞微环境:位于肠腺隐窝底部,由隐窝驻留肠道干细胞(ISCs)、间充质细胞、免疫细胞以及肠道分泌细胞(如杯状细胞和潘氏细胞)组成。这个微环境具有高度增殖性,因此负责组织更新和肠道屏障修复。它还介导抗原吞噬并释放抗菌肽(AMPs),通过清除潜在病原体来维持内部稳态。
这些元素共同构建了物理和生化屏障,保护宿主并调节内部和外部环境之间的交流。这种复杂的平衡对于胃肠系统的正常运作和维持身体整体平衡至关重要。
1
粘液层组成与免疫监测
粘液层的物理与生化屏障
粘液层是胃肠道的第一道防线,主要由90%-95%的水、1%-2%的脂质和1%-5%的粘蛋白组成。粘蛋白通过糖基转移酶高度糖基化,含有50%-80%的碳水化合物(w/w)。
粘蛋白聚糖多样而复杂的结构,为粘液相关细菌(如 R.torques、A.muciniphila、B.bifidum和R.gnavus)提供了理想的定植位点和营养来源。
动态防御:细菌即是住客,也是建筑师
MUC2是胃肠道中表达的主要粘蛋白。在细菌暴露后,杯状细胞通过meprin β介导的裂解机制分泌MUC2,形成保护性粘蛋白层。该结构允许共生微生物在外粘液层定植,并利用其多糖降解酶从粘蛋白O-聚糖中获取营养能量。这种宿主与微生物的相互作用,有助于调节近端结肠微生物群的结构和转录。
别让细菌吃肠壁:低纤维饮食的代价
研究表明,低纤维饮食会促进微生物群降解宿主粘蛋白,导致粘液层变薄,从而削弱屏障功能。Muc2缺陷小鼠表现出结肠组织学损伤增加、细菌易位至肝脏增多以及肠道紧密连接蛋白显著减少。此外,粘蛋白O-糖基化紊乱导致的粘液屏障完整性和功能受损与代谢疾病的发病机制有关。
粘液层的水龙头:谁在掌控肠道保护液的释放?
最近的研究发现,Gasdermin D (GSDMD) 是一种参与细胞凋亡的成孔效应蛋白,它通过scinderin介导的F-肌动蛋白解聚,促进钙依赖性胞吐作用,从而调节杯状细胞的粘蛋白分泌。GSDMD缺陷会破坏粘液屏障,使病原体粘附到上皮细胞,导致肠道疾病的发生。
严防细菌偷渡:一套保护肝脏的精密免疫系统
粘液层凭借其独特的粘弹性,能够有效滞留并扩散来自潘氏细胞和杯状细胞的抗菌物质及免疫细胞因子,形成化学屏障。
-抗菌肽 (AMPs) 与IgA的协同作用
潘氏细胞产生隐匿防御素、抗菌素和溶菌酶等AMPs。这些AMPs大量存在于肠道上皮表面,能够直接清除有害微生物。在新生非肥胖糖尿病小鼠模型中,生态失调导致的结肠AMPs缺乏会导致1型糖尿病中的胰腺自身免疫。
AMPs与微生物特异性免疫球蛋白IgA协调作用,在维持屏障稳定性和抑制炎症中发挥关键作用。
-IL-17和IL-22的调控
它们的调节受T辅助17(Th17)细胞和III型固有淋巴细胞(ILC3)产生的IL-17和IL-22的影响。
ILC3依赖于树突状细胞(DC)相关的Mincle信号通路,该通路与酪氨酸激酶偶联的C型凝集素受体有关。在Mincle缺失或酪氨酸激酶受损的情况下,肠道再生胰岛衍生III-γ(RegIIIγ)和IgA的合成会减少,从而导致肠道微生物群移位,进而引发肝脏炎症和脂质代谢失调。
这些发现表明粘液层完整性在维持肠肝稳态中的关键作用。
2
上皮连接:构筑肠壁防线的灰浆与砖块
微绒毛:不仅仅是吸收养分的地毯
功能:肠道上皮细胞顶端的微绒毛密集排列成刷状缘。它们既是营养吸收的高效界面(扩大表面积),又是阻止细菌附着的第一道物理防线。
脆弱性:在克罗恩病中,这些绒毛会变短、基因表达混乱。
破坏机制:就像拆除帐篷的支柱一样,肠出血性大肠杆菌的毒力因子通过CDK1-Formin信号轴,攻击支撑微绒毛的骨架蛋白(ACT-5),导致微绒毛坍塌消失,引发严重腹泻、出血性结肠炎等。
紧密连接(TJs):细胞间的拉链
在微绒毛下方,紧密连接(TJs)像拉链一样把相邻细胞的细胞膜紧紧锁死。
核心作用:这种吻合结构封堵了细胞间的空隙,相当于门控功能——只允许特定的物质通过,严防细菌和有害大分子渗透。
关键零件:谁在控制拉链的松紧?
病菌如何撬开防御?
紧密连接的稳定性高度依赖于细胞骨架的支撑,这成为了病原体的攻击目标:
3
动态防御机制:从干细胞再生到免疫感知
隐窝深处的生命源泉:肠道干细胞 (ISCs)
肠道屏障之所以能抵御消化道内持续的磨损与危害,归功于其惊人的自我更新能力。
核心机制: 位于肠道隐窝底部的肠道干细胞是这一过程的“总工程师”。它们通过持续增殖,不断分化补充受损的肠道上皮细胞(IECs),维持着组织的修复与动态平衡。
谁在调控修复?信号通路、压力与衰老
ISCs 的功能受到微环境信号的精密调控,同时也易受外部因素干扰:
-修复的加速器
当肠道受损时,IL-1R1 信号 和 Wnt 激动剂 RSPO3 会协同作用,强力促进 ISCs 的修复功能,加速伤口愈合。
-心理压力:让干细胞电量耗尽
心理压力不仅仅是情绪问题,它能产生实实在在的生理毒性。
心理压力会导致 ISCs 内部的 线粒体能量代谢 受损,进而干扰细胞分化。这种微观层面的能量危机削弱了宏观的肠道屏障,这科学地解释了为何精神疾病患者常伴有肠道问题。
-衰老的阻滞剂
随着年龄增长,ISCs 的数量和活性会下降,导致屏障完整性受损及菌群失衡。
关键原因:维持 ISCs 活性的关键信号——Wnt 信号通路随衰老而减弱,导致干细胞枯竭。
簇状细胞:不仅是免疫哨兵
簇状细胞是肠道屏障中一种重要的分泌型肠上皮细胞,与潘氏细胞和杯状细胞共同发挥作用。它们通过分泌IL-25来抵御病原体感染,激活2型免疫并清除病原体。簇状细胞还能感知病原体代谢物,并通过G蛋白偶联信号通路产生PGD2,从而刺激杯状细胞分泌粘液并促进自身增殖,进一步增强抗菌防御。此外,在肠道损伤时,簇状细胞可以充当储备肠道干细胞,协助屏障修复。
肠道屏障的健康受多种因素影响,包括遗传、饮食(如西式饮食)、药物(抗生素等)、疾病、生活习惯(昼夜节律)、心理状态(压力)和生理过程(衰老)。这些因素共同作用,可导致肠道微生物失衡,进而引发全身性代谢紊乱。这些代谢紊乱又会加剧炎症反应,进一步损害肠道屏障,形成一个恶性循环,持续破坏肠道健康。
下表总结了影响肠道和肝脏疾病的因素及机制与肠道屏障功能障碍相关的研究

1
遗传易感性
基因组关联研究(GWAS)的进展显著提升了疾病易感基因的识别和相关生物学通路的理解,这对临床转化具有重要价值。这些研究分析基因组中的遗传变异,以探索基因型和表型之间的关系。
免疫刹车失灵:IL-10 信号通路的遗传缺陷
尽管基因突变与肠道屏障功能障碍之间的直接联系有限,但GWAS已识别出许多与IBD(炎症性肠病)发展相关的基因座。例如,IL10基因突变是首批被发现能诱发IBD的突变之一。IL10基因敲除小鼠会自发发展结肠炎并增加肠道微生物易位。IL10受体(IL10R)的突变(由IL10RA和IL10RB基因编码的IL10R1和IL10R2蛋白组成)已与早发性小肠结肠炎相关联。这些突变损害了IL10诱导的信号传导,这可能增加TNF-α和其他促炎细胞因子的分泌,从而加剧炎症并削弱肠道耐受性。
离子转运障碍:SLC26A3 与物理屏障的松动
GWAS还显示SLC26A3基因(编码DRA蛋白,一种肠道氯离子转运蛋白)的显著下调。该基因在人类基因组中有害IBD变异中排名在前1%。SLC26A3表达的降低显著增加结肠旁细胞通透性,降低紧密连接(TJ)和黏附连接(AJ)蛋白的表达,从而增加对IBD的易感性。
黏液防线的溃败:ST6GALNAC1 与糖基化异常
糖蛋白组学分析揭示,先天性IBD(炎症性肠病)患者可能携带ST6GALNAC1(ST6)基因突变,该基因编码一种对维持黏液屏障稳态至关重要的唾液酸转移酶。该基因的突变会导致肠道黏膜层厚度减少,并破坏其保护功能。阐明这些调控机制对于理解先天性IBD的发病机制至关重要。此外,屏障功能还受到遗传易感性和环境因素相互作用的影响。DNA甲基转移酶编码基因DNMT3A的突变与IBD风险增加相关,已被证明能减少杯状细胞数量,缩短黏附连接(AJ)复合体,并增加肠道通透性。这些改变增加了对结肠炎的易感性,并阻碍了上皮再生和修复过程。
2
胆汁酸
胆汁酸(BAs)不仅是消化和脂质吸收的关键,还扮演着调节全身代谢和免疫的激素角色。它们通过激活FXR和TGR5等受体发挥作用。
修复的动力(次级胆汁酸)
最近研究发现,胆汁酸在维持肠道上皮屏障方面至关重要,能刺激Lgr5+肠道干细胞(ISCs)的自我更新,从而促进肠道修复。
修复的阻力(过量初级胆汁酸)
然而,过量的初级胆汁酸(如胆酸)会通过抑制脂肪酸氧化来减缓ISC的增殖,进而影响屏障修复。另一方面,TGR5在肠道代谢稳态中作用显著,其被次级胆汁酸(脱氧胆酸)激活后,会促进ISCs中YAP1和SRC等因子的转录,从而有效驱动肠道上皮的再生。
孕烷X受体 (PXR):抗炎与修复的化学感应器
孕烷X受体(PXR)是调节肠道上皮屏障稳态的关键核受体,它通过调控外源性物质代谢和先天免疫发挥作用。
溃疡性结肠炎患者中PXR及其靶基因的下调表明,PXR功能受损可能导致肠道损伤后屏障修复缺陷。PXR缺陷会加剧肠道上皮功能障碍,并增加对肠道损伤的易感性。
PXR的激活(例如通过石胆酸LCA)通过抑制NF-κB通路和炎症因子释放,以及维持紧密连接的完整性来发挥保护作用。
维生素 D 受体 (VDR):从抗菌防御到防癌屏障
胆汁酸也是维生素D受体(VDR)的内源性配体,对肠道稳态具有保护作用。VDR通过调节上皮分化和增强紧密连接表达来维护肠道屏障完整性,并支持潘氏细胞的抗菌功能。然而,在炎症性肠病(IBD)和结直肠癌(CRC)中,VDR的保护机制受损,其信号通路的失调与CRC的加速进展和不良预后密切相关。VDR激活不仅直接抑制肿瘤细胞增殖,还能通过增强黏膜屏障完整性来限制肿瘤进展,从而抑制CRC。
3
肠道微生物群及衍生代谢物
肠道微生物群通常与宿主共生,并被肠道上皮屏障限制在黏液层外。但有些微生物能突破屏障,损害胃肠道健康。抗生素虽看似能保护肠道,但实际上可能削弱屏障功能,增加新发炎症性肠病(IBD)的风险,这已在大量的研究中得到证实。与此相反,良好的肠道微生物结构对肠道屏障修复有积极作用。研究发现,肠道微生物群能通过激活巨噬细胞信号促进结肠上皮前体细胞生长,从而帮助修复受损的肠道上皮屏障。
色氨酸代谢物:PXR 与 AhR
肠道微生物群通过产生多种代谢物(如次级胆汁酸和色氨酸衍生物)来维护肠道屏障的完整性和整体健康。在IBD患者中,色氨酸水平显著降低,其代谢改变对疾病进程和预后有重要影响。
路径一:芳烃受体 (AhR) 的激活
色氨酸衍生物中的吲哚类物质是芳烃受体(AhR)的天然配体,AhR的激活能调节免疫细胞(如Tregs和Th17)及其细胞因子(特别是IL-22)的产生,从而促进肠道屏障的保护、修复和稳态。
路径二:孕烷 X 受体 (PXR) 的增强
除了AhR途径,这些代谢物还能通过孕烷X受体(PXR)增强屏障。例如,吲哚-3-丙酸(IPA)作为内源性PXR激动剂,通过TLR4信号通路减轻肠道通透性和炎症。PXR激活通过稳定紧密连接蛋白ZO-1、抑制MLCK表达和JNK1/2磷酸化来保护肠道屏障。
综上所述,色氨酸衍生的吲哚代谢物通过同时激活PXR和AhR信号通路,对肠道上皮发挥双重保护作用。
反面:犬尿氨酸 (Kyn) 途径
色氨酸代谢的犬尿氨酸(Kyn)途径与IBD进展密切相关,其限速酶IDO在炎症下促进Kyn途径,IDO的抑制或缺陷可减轻肠道炎症并增强屏障功能。
短链脂肪酸:能源与信号的结合
短链脂肪酸(SCFAs),包括丁酸、乙酸和丙酸,是肠道微生物分解膳食纤维等产生的关键代谢物,对肠道屏障功能至关重要。它们通过激活G蛋白偶联受体(如GPR43或FFAR2)来增强肠道屏障,并保护免疫细胞免受损伤,从而有助于预防结直肠癌(CRC)。
丁酸抑制组蛋白脱乙酰酶(HDACs)以调节基因转录,并通过调控紧密连接处的蛋白来促进屏障修复。GPCR信号和HDAC共同抑制维护上皮屏障完整性。此外,SCFAs还能刺激杯状细胞分泌黏蛋白,进而促进黏蛋白降解菌生长,通过消耗黏蛋白促进肠道干细胞分化,确保肠道上皮的再生能力。
4
昼夜节律
昼夜节律,常被称为生物钟,是代谢稳态的内部控制系统,旨在同步诸如光暗周期等周期性环境信号。该系统通过转录、转录后和翻译后修饰,在特定时间协调基因表达,以实现最佳代谢适应。
“什么时候吃”可能比“吃什么”更重要
多项研究表明,昼夜节律紊乱与代谢疾病之间存在密切关联。具体而言,不规律的进食时间会增加小肠暴露于膳食抗原和微生物刺激的风险,从而加重肠上皮细胞(IECs)主要组织相容性复合体II(MHC II)的负担。这种紊乱会削弱肠道微生态的调节功能,并减少IL-10的分泌。
神经免疫回路的失调:VIP 与 ILC3 的博弈
昼夜节律还会影响肠道中的神经免疫回路,这些回路受进食行为激活,并在饮食不规律时可能导致病理变化。食物摄入会触发肠道神经元分泌血管活性肠肽(VIP),该肽会上调与脂质吸收和转运相关的蛋白质。同时,VIP会降低IECs中的AMP水平,并减少ILC3产生的IL22。这种饮食节律的紊乱有助于病原体的肠道定植,尤其是在神经免疫回路的屏障功能受损时。
微生物振荡器:细菌也有生物钟
肠道微生物群自身也表现出丰度和功能的昼夜波动,被称为微生物振荡器,它们通过微生物代谢物或自身抗原影响宿主昼夜节律。肠道微生物群与宿主生物钟之间的这种相互作用显著影响屏障完整性和先天免疫反应。例如,短链脂肪酸通过抑制组蛋白脱乙酰酶(HDAC)活性,有助于调节小肠的昼夜节律相移。
肠道中的分节丝状细菌(SFB)驱动着与宿主节律同步的节律性ILC3回路振荡,从而通过时间依赖性地表达抗菌肽(AMPs)来介导感染抵抗力的昼夜变化。
5
饮食
高脂与快餐:胆汁酸的黑化与致癌风险
高脂肪和高糖、高加工饮食,与现代社会代谢疾病患病率的增加密切相关。饮食成分通过改变肠道微生物群以及主动参与宿主生理过程的次级代谢产物的产生,显著影响肠道微环境。
研究表明,现代快餐和西方饮食引起的肠道微生物群变化会提高脱氧胆酸(DCA)水平,这通过激活肠道FXR和I型干扰素(IFN)信号通路损害潘氏细胞。
动物研究进一步揭示,高脂肪饮食(HFD)在不同的结肠癌模型中,包括偶氮甲烷(AOM)-葡聚糖硫酸钠(DSS)诱导模型以及Apc突变诱导的自发模型,都会加剧肠道屏障损伤。HFD受损的肠道屏障允许更多病原体和衍生代谢物穿透上皮,从而加速结直肠癌(CRC)的发展。
糖衣炮弹:果糖与高血糖的双重打击
除了高脂肪摄入,过量的膳食果糖摄入也会损害上皮屏障。果糖水平升高会增加循环内毒素,从而激活巨噬细胞上的Toll样受体4(TLR4),引发全身性炎症反应。同时,葡萄糖代谢既对代谢综合征具有治疗潜力,也是肠道屏障功能的关键协调者。
在瘦素缺乏(ob/ob)和瘦素受体(LepR)缺乏(db/db)的2型糖尿病(T2DM)小鼠模型中,高血糖通过诱导肠上皮细胞(IECs)中葡萄糖转运蛋白2(GLUT2)依赖性转录重编程来破坏肠道屏障,这随后损害了紧密连接(TJ)和黏着连接(AJ)结构的完整性。
富含果糖的食物

6
药物
抗生素:精准打击与地毯式搜捕
抗菌素的使用对肠道微生物群落的丰度和结构有着深远的影响,进而影响肠道屏障的完整性。
例如,利福昔明-α (Rifaximin-α)常用来治疗小肠细菌过度生长:这种抗生素可以减少破坏粘膜的细菌,增加肠道内TNF-α和IL-17的水平,从而增强对病原体的抵抗力。它还能通过增加回肠中的乳杆菌水平来改善应激引起的肠道屏障功能障碍。
广谱抗生素会降低肠道微生物多样性,导致免疫失调,并增加感染的易感性。在健康成年小鼠中,广谱抗生素治疗会导致菌群失调,肠道上皮紧密连接(TJ)的完整性受损,表现为ZO-1表达减少和NLRP3炎症小体激活。
非抗生素药物:阿司匹林的隐形副作用
非抗生素药物也可能损害肠道屏障,例如阿司匹林,广泛用作消炎镇痛药,但会引起胃肠道损伤。
它会激活肠道FXR信号,并减少戈氏副拟杆菌(Parabacteroides goldsteinii)的数量。
戈氏副拟杆菌产生7-酮-LCA,这种物质能抑制肠道干细胞(ISC)的干性,从而减缓肠道屏障的修复。
化疗药物:再生能力的丧失
化疗药物:化疗是导致肠道屏障损伤的另一个主要原因。例如,5-氟尿嘧啶会加速黏膜细胞的死亡,而非再生。因此,超过40%的化疗患者会出现胃肠道损伤,表现为腹泻、便秘和消化不良等症状。
免疫检查点抑制剂:免疫激活的附带损伤
免疫检查点抑制剂(ICIs):靶向PD-1的ICIs彻底改变了抗肿瘤治疗。尽管这些抗体通过阻断PD-1通路来重建正常的免疫反应,但它们经常引起胃肠道毒性。
PD-1信号被发现对调节结肠淋巴组织诱导(LTi)细胞(ILC3的一个特定亚群,对维持免疫稳态至关重要)至关重要。
PD-1信号的缺失会导致LTi细胞中脂肪酸氧化过度激活,并反馈抑制LTi细胞的激活和IL-22的产生。这种失衡导致生态失调、肠道屏障损伤,并增加肠炎的易感性。
功能性测试:探针分子与 LMR 比值
乳果糖/甘露醇比值 (LMR) 是评估肠道通透性的重要生物标志物。
注:乳果糖(大分子)的尿液排泄量反映“肠漏”程度和上皮损伤。甘露醇(小分子)主要通过跨细胞途径吸收,代表非特异性上皮转运。
通过口服探针分子后检测尿液排泄来评估肠道通透性,这是最常用和直接的方法。
常用的探针分子包括不易代谢、吸收差的糖类(如甘露醇、鼠李糖、三氯蔗糖、乳果糖)或低分子量聚乙二醇(PEG)和乙二胺四乙酸(EDTA)。
为了避免饮食干扰,这些探针常使用同位素标记。
乳果糖-甘露醇测试 (LMT) 是临床广泛使用的评估方法。
在动物模型中,常用FITC标记的葡聚糖或铬-51标记的EDTA (51Cr-EDTA) 吸收到体循环的量来评估肠道屏障完整性。
其他方法:生物标志物的间接分析和多组学分析平台也是新兴的评估手段。
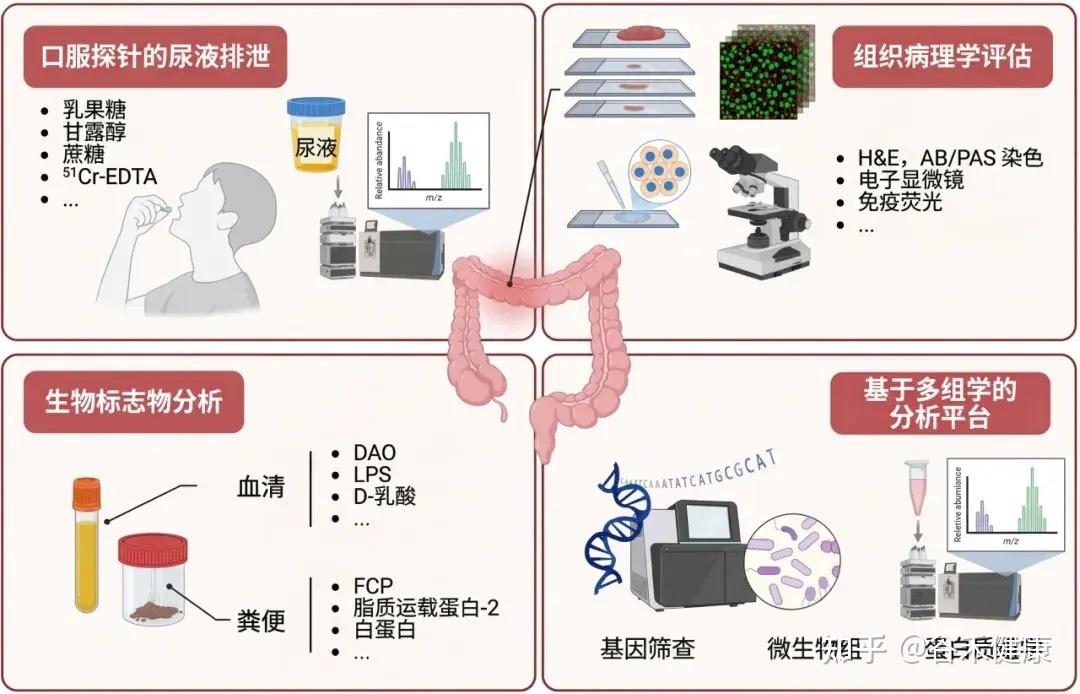
生物标志物:血液与粪便的线索
血清蛋白质组学和基因筛查技术:用于早期检测屏障功能障碍,并支持对其潜在原因的机制研究。
这些间接方法通过检测血液、粪便或分子水平上的特定物质,提供肠道屏障健康状况的重要信息。
组织病理学:屏障损伤的可视证据
内窥镜活检标本的组织病理学评估可以详细检查上皮绒毛形态和评估肠道屏障完整性。可以使用黏蛋白特异性阿尔辛蓝/过碘酸-希夫 (AB/PAS) 染色来评估黏液层,使用电子显微镜进行超微结构分析,以及使用免疫荧光技术精确定位和量化紧密连接 (TJ) 蛋白的表达,从而系统地分析黏膜成分的结构改变。
这些方法共同为评估肠道屏障完整性提供了全面的视角。
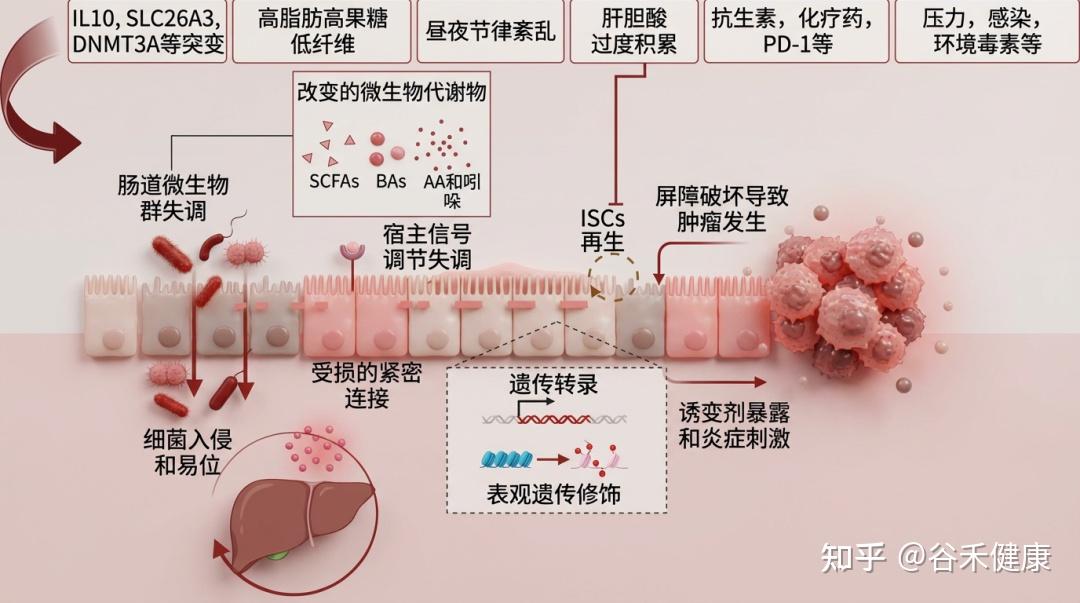
肠道上皮屏障的失调对多种肠道和肝脏疾病的发展和进展具有重要意义。肠道上皮的紧密连接功能障碍增加了通透性,促进细菌及其致病因子穿越屏障进入全身循环及其他组织。这种入侵会引发全身性炎症,破坏代谢稳态,而代谢稳态是肠道和肝脏疾病发生和进展的关键事件。虽然尚不确定肠道屏障的损伤是主要驱动因素还是偶然的病理特征,但越来越多的证据表明,肠道屏障的完整性在这些疾病的病因中起着关键作用。
1
炎症性肠病与结直肠癌
IBD 的病理核心:炎症因子的破坏机制
炎症性肠病 (IBD),包括克罗恩病 (CD) 和溃疡性结肠炎 (UC) 的特征是慢性胃肠道炎症。上皮屏障功能障碍是导致 IBD 发生和发展的关键因素。
临床研究表明,UC 和 CD 患者的肠道紧密连接 (TJ) 结构发生改变,导致肠道通透性增加。此外,黏膜免疫激活会引起TNF-α、IL-13、IL-17等细胞因子的产生。
屏障损伤是“因”而非“果”
进一步研究表明,由于 TJ 功能障碍和细胞骨架变化,细胞旁通透性增加可能在疾病发生前就已出现,例如 CD 患者亲属中早期肠屏障异常以及肠易激综合征 (IBS) 的较高患病率。同样,大规模队列研究已经发现 CD 诊断前三年内的屏障障碍标志物。这些发现支持屏障功能障碍是疾病进展的主要原因,而不是次要效应。
致癌风险:暴露于诱变环境
上皮屏障缺陷会使肠道内壁暴露于诱变化合物或长期炎症刺激,这可能通过氧化应激等机制启动并促进结直肠肿瘤的发生。在早期和晚期结直肠癌中,各种屏障相关蛋白的表达显著降低,这促进了微生物移位,引发炎症,并进一步加速肿瘤生长。然而,结直肠癌发生发展过程中上皮屏障成分异常表达的确切机制仍不清楚。
关键分子机制:GOLM1 与 NDRG2
事实上,实验模型表明,上皮屏障受损的小鼠更容易患结肠炎和炎症驱动的肿瘤。例如:
-GOLM1 缺失(细胞分化紊乱)
高尔基体膜蛋白 1 (GOLM1) 的缺失会过度激活 Notch 通路,从而破坏肠道稳态,改变肠上皮细胞 (IEC) 的分化和分泌细胞谱,并减少杯状细胞的数量,所有这些都会增加肠道通透性和促肿瘤炎症。机械屏障在预防结直肠癌发展中也至关重要。
-NDRG2(肿瘤抑制基因)
肿瘤抑制基因 N-myc 下游调节基因 2 (NDRG2) 已被证明可以增强 E3 连接酶 FBXO11 和 E-钙粘蛋白抑制剂 Snail 之间的相互作用,促进 Snail 泛素化。E-钙粘蛋白的这种稳定作用增强了黏附连接,从而限制了结肠炎相关肿瘤的发展。
2
慢性肝病
慢性肝病,包括 MASLD、ALD 和 PSC,常伴有肠道屏障受损和肠肝轴的失调。胃肠道物质,如营养素、分泌因子和微生物代谢物, 通过门静脉进入肝脏,并被代谢用于全身利用。这一过程使肝脏对病原体相关分子模式(PAMPs)极为敏感,成为肠道物质的主要靶点。
代谢功能障碍相关脂肪性肝病
代谢功能障碍相关脂肪性肝病 (MASLD, Metabolic dysfunction-associated steatotic liver disease):以前被称为NAFLD(非酒精性脂肪性肝病)。它是一种与代谢功能障碍相关的肝脏疾病,通常与肥胖、2型糖尿病、高血压和血脂异常有关。主要特征是肝脏中脂肪过度积聚,但与大量饮酒无关。
MASLD中,内毒素血症常见,提示肠屏障与肝炎症相关。DSS诱导小鼠肠屏障损伤加剧肝损伤。Il-10敲除鼠中,西方饮食+DSS处理降低BA水平,抑制肝脏FXR信号,加重MASLD。
肠道菌群失调是MASH早期驱动因素,宏基因组研究表明MASLD患者肝病发作前已存在肠道菌群失调。MASH患者粪外囊泡可能通过nmMLCK机制降低TJ表达,加剧肠屏障功能障碍。恢复肠屏障可逆转结肠炎引起的肝脂代谢失衡,特别是通过调节次级BA-TGR5/mTOR/氧化磷酸化通路。
酒精性肝病
酒精性肝病 (ALD, Alcoholic Liver Disease):是由于长期过度饮酒引起的肝脏损伤。 它可以表现为脂肪肝、酒精性肝炎和肝硬化等不同阶段。
酒精暴露破坏上皮连接,诱导肠屏障分解。ALDH2缺乏加剧酒精引起的肠道TJ/AJ蛋白降解,肠道ALDH2可能是酒精诱导的肠-肝轴损伤靶点。慢性酒精摄入减少肠道cDCs,导致AMP产量下降,保护性A. muciniphila丰度下降,AJs破坏,最终通过IL-12-IFNγ信号通路引起肝损伤。
原发性硬化性胆管炎 (PSC)
原发性硬化性胆管炎 (PSC, Primary Sclerosing Cholangitis):是一种慢性胆汁淤积性肝病,其特征是肝内外胆管的炎症和瘢痕形成。 最终可能导致胆管狭窄、肝硬化和肝功能衰竭。
PSC常与溃疡性结肠炎共病。PSC患者肠道微生物群与健康个体不同。门静脉中存在微生物提示肠屏障破坏,可能导致细菌易位。肺炎克雷伯菌等致病菌可通过接触依赖性细胞凋亡诱导上皮孔形成,加剧炎症和肝胆损伤。BDL诱导胆汁肝病小鼠模型中,肠道屏障破坏独立于菌群失调发生。敲除CHOP可缓解BDL小鼠肠屏障损伤、ISC干性丧失以及肝脏炎症和纤维化。
肠道屏障受损可刺激PSC中的保护性负反馈回路,LPS激活NF-κB通路抑制肝细胞BA代谢,减缓胆汁性肝病进展。抗生素或泛半胱天冬酶抑制剂能减弱菌群失调诱导的内毒素易位或NLRP3炎症小体激活,有望治疗PSC。
免疫靶向疗法,包括抗炎药、免疫抑制剂和生物制剂,已被证明在控制多种肠道疾病(如炎症性肠病)炎症方面有效。然而,这些治疗并不能直接解决肠道屏障的根本损伤。不幸的是,长期使用这些药物甚至可能增加感染、耐药性和疾病复发的风险,以及恶性肿瘤和死亡等严重不良反应。
确实,虽然减少炎症至关重要,但促进上皮屏障的愈合和恢复对于肠道疾病,尤其是炎症性肠病(IBD)的长期缓解至关重要。目前,没有临床批准的疗法专门针对肠道屏障的修复和维护。鉴于屏障功能障碍与肠道和肝脏疾病发病机制的密切联系,及时修复和恢复上皮屏障是一种有前景的治疗策略。与其仅关注症状管理,提升上皮愈合和健康完整性的治疗方法,可以减缓肠道和肝脏疾病的进展。
通过加强屏障,这类疗法还可能减少细菌易位、炎症以及肝损伤的加重。

1
紧密连接蛋白调节
靶向 MLCK:紧密连接稳态的核心调控
肠道屏障的维持在很大程度上依赖于多种 TJ 蛋白。肌球蛋白轻链激酶(MLCK)被视为开发炎症性肠病障碍疗法的有力候选目标,其在调节 TJs 及其分解中发挥核心作用,通过调节紧密连接(TJs)及其分解来保护屏障功能免受免疫诱导损伤。
对 MLCK 的缺失或抑制可以有效保护屏障功能免受免疫诱导损伤,但这些干预无法预防晚期结肠炎,后者涉及细胞凋亡和黏膜损伤,这些过程与 MLCK 介导的 TJ 分解无关。这些发现反映了 MLCK 抑制剂的治疗局限性。目前的 MLCK 抑制剂对上皮和平滑肌的催化域缺乏特异性,因此可能出现毒性副作用。
新机制发现:TCPTP 的双重保护
紧密连接 (TJ) 通过调节闭合蛋白 (occludin) 和封闭蛋白 (claudin) 来维持肠道完整性,而闭合蛋白和封闭蛋白是两种关键蛋白。
一项研究发现,上皮细胞中的蛋白酪氨酸磷酸酶 (TCPTP) 通过以下两种方式保护肠道屏障:
后者可以防止闭合蛋白 (occludin) 错位并降低上皮细胞的通透性。
目前尚无专门针对 TJ 蛋白的药物进入临床试验阶段,因此,需要在此领域进行深入研究和开发。
2
肠道微生物群的生态调控
益生菌疗法:从天然菌株到工程改造
益生菌已被发现通过多种机制调节屏障功能。例如,罗伊氏乳杆菌(Lactobacillus reuteri)激活Wnt/β-catenin信号通路,诱导Paneth细胞分化和AMP(抗菌肽)分泌,同时刺激Lgr5+肠道干细胞(ISCs)增殖以促进上皮修复。
此外,罗氏菌属(Roseburia)的鞭毛蛋白与Toll样受体5(TLR5)结合,可以上调occludin和MUC2,从而改善肠道屏障完整性,并增加IL-22和REG3γ水平,进一步调节肠道生态。
嗜黏蛋白阿克曼氏菌(AKK菌,Akkermansia muciniphila)也通过调节ISC程序来支持肠道上皮的修复、增殖、分化和稳定。因此,靶向肠道微生物失调成为恢复受损屏障功能的一种有前景的治疗策略。
最新的进展包括使用生物纳米材料包裹过表达人工酶的基因工程益生菌,这是一种用于黏膜修复和炎症治疗的新方法。长双歧杆菌(Bifidobacterium longum)经过修饰以表达过氧化氢酶和超氧化物歧化酶,表现出增强的肠道定植能力、强大的抗炎活性,促进肠道屏障重塑,并调节微生物平衡。这些进展有望减少传统抗炎药物的不良反应。
胆汁酸信号:FXR 激动剂的多重获益
胆汁酸在宿主与肠道菌群的交流以及肠道菌群的构成调节中起着关键作用。胆汁酸合成失调与多种疾病的发生密切相关。激活法尼酯X受体(FXR)能有效缓解胆汁酸过量带来的危害,它通过抑制胆固醇代谢、促进肝细胞将胆汁酸转运出去,从而减少胆汁淤积性肝损伤。
研究显示,FXR激动剂奥贝胆酸(OCA)能够:
此外,OCA治疗还能通过稳定内皮细胞内的β-连环蛋白来防止肠道血管屏障受损。
色氨酸代谢:AHR 通路与 IDO 的平衡
-AHR:屏障完整性的总开关
AHR(芳香烃受体)是一种色氨酸代谢物的受体,对维持肠道屏障的完整性至关重要。研究发现,Ahr−/−小鼠(即缺乏AHR的小鼠)的上皮屏障功能明显丧失,表现为肠道机械屏障受损以及细胞无法正常分化。缺乏AHR配体的小鼠也会出现类似的症状。
进一步研究表明,来自食物的AHR配体可以通过促进细胞内的锌离子(Zn2+)信号传导,从而提高紧密连接(TJ)蛋白的表达,有效预防损害肠道屏障功能的疾病。
-AHR 激动剂的治疗潜力:以尿石素 A 为例
其他AHR激动剂,例如微生物代谢产物尿石素A(UroA),也显示出保护肠道屏障功能的潜力。UroA具有抗炎作用,并且可以通过激活AhR–NrF2依赖性通路来上调TJ蛋白的表达,从而促进肠道屏障的修复。
这些研究表明,激活肠道AHR通路对于治疗酒精性肝病(ALD)具有重要意义,因为ALD与TJ屏障的丧失密切相关。因此,AHR可能成为修复肠道屏障的一个有价值的治疗靶点,尤其是在与肠道屏障功能障碍相关的慢性疾病中。
-IDO 抑制剂:代谢平衡与潜在毒性
正向:抑制 IDO -> 阻断色氨酸向犬尿氨酸转化 -> 迫使色氨酸转化为吲哚
抑制IDO(一种将色氨酸转化为犬尿氨酸的酶)可能有助于治疗肠道屏障功能障碍。IDO抑制剂通过促进色氨酸转化为吲哚,增加AHR配体的生成,从而改善肠道完整性。
虽然IDO抑制剂在癌症治疗中显示出潜力,但它们在治疗肠道屏障损伤方面的应用仍需更多研究。Indoximod是一种IDO抑制剂,已被证明能有效维持细胞间的紧密连接,并显著减轻DSS诱导的结肠损伤。
反向:过度抑制 IDO -> 减少犬尿酸生成 -> GPR35 失去激活 -> 削弱屏障保护
需要注意的是,IDO抑制也可能导致不良的肠道毒性反应。完全阻断犬尿氨酸通路可能会适得其反,损害上皮屏障的完整性。
研究表明,化疗药物引起的肠道损伤会激活色氨酸-犬尿氨酸-犬尿酸通路,增加肠道内犬尿酸的生成。犬尿酸随后激活GPR35,从而增强肠道完整性。这意味着,过度抑制犬尿氨酸通路可能会降低犬尿酸水平,削弱GPR35介导的保护作用,并可能延缓屏障修复。
综上所述,色氨酸代谢在肠道健康中扮演着复杂的角色。因此,在开发治疗肠道屏障功能障碍的新方法时,需要进行更深入的研究。
临床挑战:个体化差异
虽然大量研究表明直接调节肠道微生物群可以改善肠道屏障功能并缓解肠肝疾病,但基于微生物的疗法在临床应用上仍然受到限制。
肠道微生物群产生的有益代谢物,如吲哚-3-乳酸(ILA),其增强肠道屏障的特性取决于宿主特异性的微生物群调节。临床疗效主要取决于患者的初始肠道微生物群组成,这会影响微生物的定植情况。
此外,宿主的遗传变异也是治疗结果的关键因素。例如,携带CARD9风险基因的炎症性肠病(IBD)患者,其膳食色氨酸向芳香烃受体(AHR)激活代谢物的微生物转化能力受损。这些发现解释了微生物靶向疗法中个体差异显著的分子机制。
益生菌作为疾病状态下耗尽的共生微生物,需要稳定的宿主微环境才能定植和发挥作用。如果事先不恢复受损的肠道生态系统,微生物干预疗法往往无法建立持久的微生物平衡或实现有意义的临床结果。因此,未来的治疗策略应采用结合微生物调节和微环境恢复的双靶点方法。
3
肠道干细胞再生
Lgr5+ 肠道干细胞 (ISCs):修复的原动力
肠上皮细胞的更新、修复和再生在很大程度上依赖于位于隐窝底部的Lgr5+肠道干细胞(ISCs)。
Lgr5作为ISCs的特异性标记,编码一种受体,该受体能够响应Wnt等信号,从而触发ISCs重编程为上皮细胞谱系。ISC的活性受到细胞外信号和旁分泌信号的精密调控,以维持肠道稳态,并在损伤发生时启动适应性分化,从而保障肠道的基本生理功能。
调控胆汁酸水平,恢复干细胞活力
胆汁酸在介导肠肝轴通讯中扮演着关键角色,并整合了调控ISC功能的饮食和代谢信号。近期研究提示,通过减少病理条件下肠道内过量的胆汁酸累积,或可为治疗炎症性肠病(IBD)相关的肠道损伤提供新的干预策略。
例如,FXR激动剂治疗通过抑制肝脏中CYP8B1的表达,进而降低肠肝胆汁酸水平,从而减轻胆汁酸对Lgr5+ ISCs的相关毒性。上述发现提示,基于跨器官代谢调节靶向ISCs有望成为治疗IBD的新型目标。
4
免疫调节
传统抗炎疗法的局限性
传统的抗炎疗法,包括皮质类固醇、5-氨基水杨酸制剂以及新型TNF-α单克隆抗体,一直是炎症性肠病(IBD)和其他免疫介导疾病的标准治疗方法。
虽然这些疗法能有效减轻炎症并缓解症状,但它们在实现长期愈合方面往往力有不逮,尤其是在肠道屏障方面。许多患者随着时间推移出现疗效丧失和疾病复发。例如,尽管抗IL-6抗体疗法在临床试验中对中度至重度克罗恩病(CD)或溃疡性结肠炎(UC)有效,但在一些患者中仍持续发生脓肿和肠穿孔等严重不良反应。
这些疗法通常未能解决根本的上皮功能障碍问题,而这正是维持慢性肠道炎症的关键因素。
新兴策略:直接靶向屏障修复
为了解决这些局限性,人们对靶向更直接参与上皮屏障修复的免疫通路越来越感兴趣。IL-10是一种有效的抗炎和组织再生细胞因子,最近开发的IL-10制剂旨在通过靶向黏膜屏障来增强治疗效果。
另一个有前景的靶点是IL-22,其受体IL-22R在上皮细胞上表达,这使得IL-22和IL-22R成为旨在恢复上皮完整性和增强黏膜愈合疗法的潜在候选者。
IL-22通过刺激AMPs(抗菌肽)和粘蛋白的产生以及ISC(肠道干细胞)的再生,在维持肠道稳态中发挥关键作用。因此,IL-22被认为是屏障修复的潜在治疗方法。
IL-22 激动剂的临床与代谢获益
IL-22融合蛋白激动剂Efmarodocokin alfa (UTTR1147A) 目前正在研究中,用于活动性溃疡性结肠炎(UC)和克罗恩病(CD)的治疗(NCT02749630)。
临床试验表明,UTTR1147A在UC患者和健康个体中都能激活IL-22R信号通路,并改善UC相关的菌群失调。
此外,肠道中IL-22信号的特异性激活可以在代谢紊乱模型中以微生物依赖的方式增强肝脏和全身葡萄糖和脂质代谢稳态。此外,IL-22还对MASLD(代谢功能障碍相关脂肪性肝病)、ALD(酒精性肝病)和饮食诱导的肥胖表现出积极作用。
当外源性给药时,IL-22通过其在肠上皮细胞(IECs)而非肝细胞上的受体发挥治疗作用,然后激活STAT3并抑制WNT–β-catenin信号传导以减少吸收性肠上皮细胞的数量。
然而,IL-22在肠道屏障维持中的作用仍存在争议,人们担心可能产生致病性的免疫调节作用,例如在结肠组织中介导CXCR2+中性粒细胞的趋化作用,以及增加对IL-23单克隆抗体Ustekinumab的抵抗力。这些观察结果表明,IL-22靶向疗法可能并非总能达到预期的疗效,需要进一步研究以更好地了解IL-22激活的全部影响和潜在副作用。
5
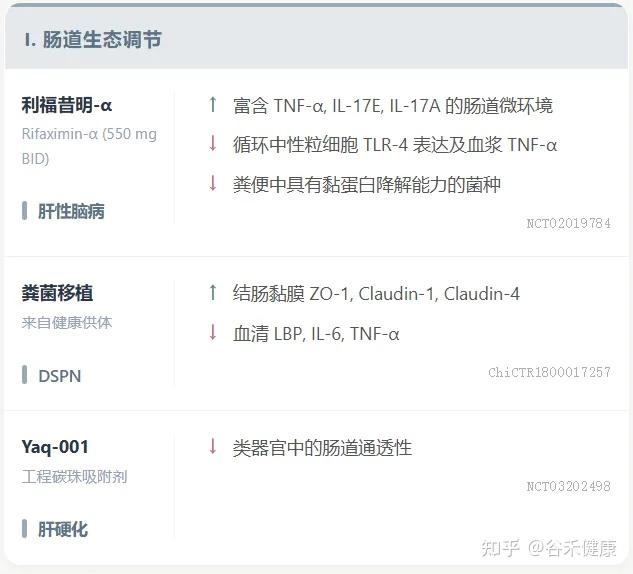
肠道屏障功能增强相关的临床试验进展
前面介绍了目前针对肠道屏障完整性的多种创新疗法,包括药物、微生物疗法、吸附剂、饮食干预和工程益生菌,并同时也探讨了这些疗法在临床验证中的进展和未来面临的挑战。
其实核心要点如下:
创新疗法:多项创新疗法正在进行临床验证,以靶向肠道屏障完整性。
小分子创新 / 微生物疗法
ISM5411:一种新型肠道限制性选择性脯氨酰羟化酶结构域(PHD)抑制剂,通过AI平台开发,已完成I期临床试验。其在肠黏膜修复和免疫调节方面具有双重机制,在IBD模型中显示出显著疗效。
利福昔明-α (Rifaximin-α):通过上调粪便中的TNF-α和IL-17E来调节肠道微环境,增强抗菌防御,有效促进肠道屏屏障修复。
ZED1227:在乳糜泻中,作为转谷氨酰胺酶2抑制剂,显著改善十二指肠黏膜结构,减少上皮内淋巴细胞浸润,通过抑制免疫原性谷蛋白肽中谷氨酰胺残基的脱酰胺化来防止T细胞活化和黏膜损伤。
粪便菌群移植(FMT):健康供体FMT在恢复糖尿病远端对称性多发性神经病变(DSPN)患者肠道屏障功能和减轻全身炎症方面显示出治疗潜力。
工程益生菌:在恢复肠道屏障完整性和维持黏膜稳态方面具有显著治疗潜力,目前研究重点是结合其屏障增强和免疫调节作用,但仍在临床前阶段。
吸附剂:非吸收性、肠道限制性工程化碳珠吸附剂Yaq-001通过改善肠道屏障功能障碍和减少全身内毒素负荷,在肝硬化中显示出临床疗效。

饮食干预
临床研究证实,膳食干预在多种胃肠道疾病中有效。
谷氨酰胺补充剂:显著恢复肠道通透性并缓解感染后肠易激综合征的腹泻 (NCT01414244)。
在肠易激综合征-腹泻、代谢紊乱和轻中度克罗恩病中均显示出益处。

未来挑战
尽管这些进展显示了靶向屏障修复策略的广阔前景,但仍需要在人体试验中全面评估长期安全性和有效性,以推进临床应用。
肠道屏障功能障碍是多种胃肠和肠外疾病(如IBD、MASLD)的关键因素。传统免疫抑制疗法虽能改善症状,但无法根治屏障问题,且副作用明显。新型疗法应结合屏障修复策略,如干细胞再生、微生物疗法、胆汁酸调节、TJ调节剂等。
肠道屏障功能障碍与疾病互为因果,受遗传、环境等因素影响。屏障破坏可引发炎症和多器官功能障碍,导致MASH、IBD、CRC等。同时,疾病微环境反过来又损害屏障,形成恶性循环。
免疫抑制疗法可能抑制黏膜愈合,加剧微生物失衡。需深入研究肠道细胞间通讯,以确定有效治疗靶点。
独立于免疫抑制的屏障防御和修复是治疗肝肠疾病的重要目标。个性化治疗策略可能更有效。增强肠道屏障完整性在MASLD预防和逆转中潜力巨大。未来人体研究至关重要。
恢复肠道屏障是治疗肝肠疾病的重要机遇。新生物技术、再生医学和微生物组研究有望重塑胃肠道治疗格局。
主要参考文献:
Zhang Y, Liu Y, Liang X, Wen Y, Zhao J, He Y, Xie Q, Xie C. Intestinal barrier in chronic gut and liver diseases: Pathogenesis and therapeutic targets. Acta Pharm Sin B. 2025 Nov;15(11):5515-5536.
Macura B, Kiecka A, Szczepanik M. Intestinal permeability disturbances: causes, diseases and therapy. Clin Exp Med. 2024 Sep 28;24(1):232.
Farré R, Vicario M. Abnormal Barrier Function in Gastrointestinal Disorders. Handb Exp Pharmacol. 2017;239:193-217.
Brandl C, Bucci L, Schett G, Zaiss MM. Crossing the barriers: Revisiting the gut feeling in rheumatoid arthritis. Eur J Immunol. 2021;51(4):798–810.
Ramakrishna BS. Role of the gut microbiota in human nutrition and metabolism. J Gastroenterol Hepatol. 2013;28(Suppl 4):9–17.
Bäumler AJ, Sperandio V. Interactions between the microbiota and pathogenic bacteria in the gut. Nature. 2016;535(7610):85–93

谷禾健康

分娩标志着孩子和母亲的一个相当大的变化,对于母亲来说,分娩是一个转折点,过去我们常常将关注点集中在宝宝的成长发育上,却往往忽视了产后妈妈的身体恢复,如激素急剧变化、在营养补充和维持身材之间难以取舍、代谢异常、睡眠减少等困扰。
更隐秘的危机在精神领域悄然蔓延。世卫组织2023年的报告显示,全球每5位产妇就有1位陷入产后抑郁的泥沼,这些被激素风暴与育儿压力双重撕扯的母亲,往往在”为母则刚”的社会期待中被迫沉默。当一位母亲在产后面临身体不适时,对孩子的照料、家庭的和睦乃至整个社会都带来深远影响。
分娩后,女性体内发生着一系列显著的激素变化:雌激素和孕激素的急剧下降,催产素的波动,皮质醇等应激激素的起伏。激素变化会直接影响肠道菌群的组成和功能,而这些变化又与产后女性的代谢状态、食欲调节和营养吸收密切相关。
一方面,产妇需要充足的营养支持身体恢复和维持母乳喂养;另一方面,也希望逐渐恢复孕前体重。在这种情况下,找到产后营养补充与健康体重管理之间的平衡点变得尤为重要。理解肠道菌群与营养的双向互动关系,可以制定更有效的产后恢复策略,这一点在产后体重管理中尤为重要。
而当产后激素波动导致肠道菌群失衡时,不仅会影响营养物质的吸收和能量代谢,还可能通过炎症因子和神经递质的变化影响情绪,与产后抑郁等心理问题密切相关。
针对产后抑郁倾向的女性,菌群检测可与传统量表评估相结合,构建多模态评估体系,推动产后心理健康管理从单纯症状评估向生物标志物预警转变,提供更精准的个性化干预方案,有效支持产后女性的全面康复过程。
本文将重点探讨肠道菌群在产后女性健康中的关键作用,以及如何通过调节肠道菌群,为这群被忽视的重要人群提供更全面的关怀与支持。
疼痛修复期的身心挑战
产后女性经历着人类生物学中剧烈的生理重构,其变化涵盖内分泌、代谢、免疫等多个系统,并与肠道菌群形成复杂的双向调控网络。
生理层面——连锁效应
“肚子阵阵拧着疼”
很多女性在产后2-3天,下腹持续出现类似痛经的阵发性绞痛,哺乳时疼痛加剧,甚至需要咬牙忍耐。
子宫通过强烈收缩恢复孕前大小,宫底每日下降1-2厘米,约6周完成复旧。肌纤维缩短压迫血管止血。伴随这一过程,子宫内膜脱落形成恶露,初期为血性(持续3-4天),逐渐转为浆液性(持续约10天),最后为白色恶露(持续3周),全程4-6周。
哺乳时婴儿吸吮刺激催产素分泌,进一步强化宫缩。这种疼痛虽难熬,却是子宫复旧的积极信号。
“打个喷嚏就漏尿”
咳嗽、大笑时突然漏尿,尴尬又无奈。孕期增大的子宫压迫盆底肌长达数月,分娩时肌肉和神经被过度拉伸甚至断裂。松弛的盆底肌无法有效支撑尿道,导致腹压增加时尿液失控。
本文后面章节有介绍盆底肌修复的相关干预措施。
心理层面——从甜蜜到崩溃
“前一秒笑着哄娃,下一秒突然崩溃大哭”
分娩后24小时内,雌激素水平暴跌至孕前1/10,直接影响大脑5-羟色胺分泌,导致情绪调节功能紊乱。同时,睡眠剥夺(平均每天仅4小时碎片化睡眠)加剧前额叶皮层代谢异常。
“镜子里的我像个陌生人”——身材焦虑爆发
妊娠纹、松垮的肚皮、下垂的乳房引发强烈外貌焦虑。产后体内脂联素水平持续偏低,导致脂肪代谢减缓,而镜像神经元对体像改变的过度关注会激活杏仁核恐惧回路。
以上是产后康复涉及身体的方方面面,产后生理层面的变化是产后康复研究与实践的核心关注点。这些变化涉及多个系统,包括泌尿系统、体重管理、疼痛感知以及心理健康等方面,需要综合考虑各系统的代谢机制。
产后康复对于保障母婴健康、提升产妇生活质量具有关键意义。接下来们将聚焦于产后营养与体重管理这一关键领域,探讨如何通过科学的饮食策略来支持身体恢复并维持健康体重。
产后女性面临着双重挑战:一方面需要充足的营养来恢复体力、促进伤口愈合并支持母乳喂养;另一方面又期望逐渐恢复孕前体态。
产后超重一方面源于孕前超重或孕期增重过多,另一方面与产后体重滞留有关。
什么是产后体重滞留?
产后体重滞留(Postpartum Weight Retention, PPWR)是女性产后普遍面临的代谢失衡问题,定义为产后一年内体重较孕前增加≥5kg的状态。
主要以产后42天(产后6~8周)、产后6个月和产后12个月作为产后体重监测时点。
能量代谢重构与激素波动
脂肪储备与能量失衡
孕期母体脂肪储备量增加约3-4 kg,主要分布于腹部和臀部,用于支持胎儿发育及哺乳。
产后能量消耗模式改变:哺乳期基础代谢率较孕前提高15%-20%,但若能量摄入(尤其是高脂高糖饮食)超过哺乳消耗(约650 kcal/天),脂肪持续堆积。
激素与代谢调节紊乱
雌激素骤降:产后雌激素水平下降50%-70%,导致脂蛋白酶活性降低,脂肪分解受阻。
胰岛素抵抗:妊娠期胎盘分泌的激素(如胎盘催乳素)诱导胰岛素抵抗,产后恢复期若糖代谢未正常化,易引发脂肪合成亢进。
初始体重减轻幅度
分娩后,产妇体重平均减少约5-10斤(胎儿、胎盘、羊水等重量),但仍有60%-80%的增重需通过后续干预消除。
2013年中国不同产后时间和地区的乳母体重滞留量比较
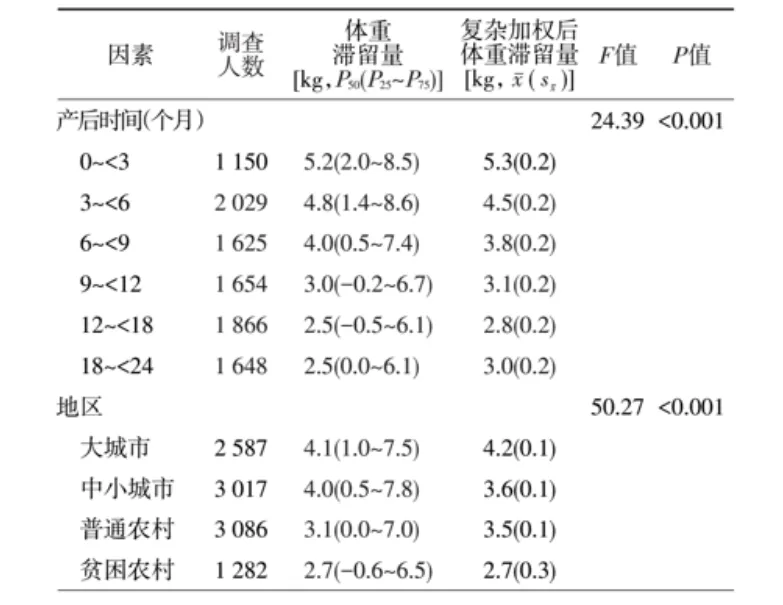
产后女性的肠道菌群变化
门水平
厚壁菌门(Firmicutes)是产后早期(<6周)和晚期(6周至1年)的优势菌门,部分研究还发现放线菌门(Actinobacteria)在产后期间的比例显著增加。
属水平
拟杆菌属在产后早期具有主导地位,也有研究发现Clostridiales在分娩后和产后4-6周内占优势。研究中常见的丰富菌属还包括:
为什么产后有些人很容易瘦,有些不容易?
产后肠道菌群组成的变化不仅仅是微生物学现象,更是体重调节与代谢健康的关键因素。肠道微生物组直接参与能量提取、内分泌代谢与脂肪分解过程,充当产后女性代谢调节网络中的核心枢纽。
当然,体重也会涉及其他方面因素,这里我们从肠道菌群的角度来分析:
★ 基线菌群组成不同
每个产妇初始肠道菌群组成存在个体差异;不同菌群组成可影响从相同食物中提取的能量总量,具有特定微生物组特征的人在相同热量摄入条件下,体重变化轨迹可能完全不同。
那些具有”瘦菌群”特征的产妇(如双歧杆菌、阿克曼氏菌丰度高,F/B比值低)更容易恢复体重;菌群多样性高的产后女性通常代谢恢复更为理想。
妊娠期遗留的菌群变化
妊娠期激素水平和代谢变化会改变肠道菌群,这些变化可能在产后持续存在,影响能量提取效率和代谢调节;菌群变化的恢复速度存在个体差异,检测肠道菌群的变化可以了解其恢复程度。
菌群对能量平衡、激素的调节
基于菌群特征,有些产妇从相同食物中获取更多能量,肠道菌群影响短链脂肪酸产生,进而影响脂肪燃烧和储存。
短链脂肪酸通过多种机制影响脂肪组织代谢:
特定菌群可促进脂肪组织褐变,增加能量消耗。
某些肠道菌如Dysosmobacter welbionis产生的12,13-diHOME能促进脂肪酸转运至棕色脂肪组织,增加能量消耗。
阿克曼氏菌被证明能增加褐变、脂肪酸氧化和棕色脂肪组织(BAT)活性,这一转化对能量消耗和代谢健康至关重要。
肠道菌群还通过影响胆汁酸代谢,间接影响激素水平,菌群可能影响产后激素敏感性,进而影响脂肪分解和糖代谢。
肠道屏障与炎症
肠道屏障功能受损时,细菌内毒素LPS可进入循环系统,激活炎症途径;LPS暴露改变脂肪因子分泌,与脂肪组织中巨噬细胞积累和促炎状态相关。
总之,产后体重恢复的个体差异在很大程度上取决于肠道菌群的组成与代谢功能,这些因素通过影响能量提取、脂肪储存、激素调节和炎症状态等多种机制,共同决定了产后女性体重恢复的难易程度和速度。了解和调节个人肠道菌群可能是未来产后体重管理的关键策略。
基于这一认识,产后营养补充不再是简单的能量与宏量营养素摄入,而应转向”营养-微生物-代谢”的整体观念。
产后营养
合理的产后营养不仅关乎体重管理,更是整体康复过程的基石。适当的营养摄入能够提升免疫功能,加速产后恢复,调节激素平衡,甚至对缓解产后抑郁情绪也有积极作用。
产后体重管理的重要时间窗口
产后1年内是体重恢复的关键时期,产后6个月左右恢复到孕前体重的女性,其后续10年超重的风险会较低。
因此,理解产后身体的特殊需求,并据此制定个性化的饮食策略,是帮助产后妈妈顺利度过产后恢复期的重要支持手段。
首先,产后的营养需求有什么特点?
产后女性对食物的吸收利用率往往有所变化,激素波动会影响消化系统功能,影响营养素吸收。
◆ 能量需求增加哺乳期每天额外需要约500卡路里能量以支持乳汁分泌。
《美国膳食指南(2020-2025)》建议,与哺乳期的孕前需求相比,哺乳期妇女在产后前6个月额外摄入330卡路里,6-12个月额外摄入400卡路里。
我国传统月子餐文化源远流长,”坐月子”被视为产后恢复的关键。传统观念认为产后女性处于特殊”虚弱”状态,需通过特定食物”补”回元气。传统月子餐通常以高能量、高蛋白、温热食物为主,如猪蹄、鸡汤、鲫鱼汤等。这些食物含有丰富蛋白质和胶原蛋白,有助于伤口愈合和组织修复。
然而,传统月子餐也存在局限性:过度强调高油高糖食物,食用红糖、米酒等”温补”食物可能导致血糖波动;单一食谱导致营养素摄入不均衡等。
◆ 蛋白质需求提高 乳汁分泌和身体组织修复都需要优质蛋白质支持。膳食蛋白质不仅是宿主营养的来源,也是肠道微生物的关键营养底物。
肠道菌群在蛋白质代谢中发挥着重要作用,特定菌群如拟杆菌属、梭状芽胞杆菌属和乳酸杆菌属能有效分解未消化的蛋白质,生成短链脂肪酸和多种氨基酸。产后如果肠道菌群紊乱,可能导致菌群分解能力下降,从而引起蛋白质消化不良,影响营养吸收效率。
适量优质蛋白摄入可促进双歧杆菌和乳杆菌等有益菌生长,形成良性循环,而过高或过低的蛋白质摄入都可能导致菌群失衡,影响产后恢复。此外,一些肠道菌群还能从头合成赖氨酸等必需氨基酸,并将其整合到宿主蛋白质中,为产后女性提供额外的营养支持。
◆ 微量营养素需求钙、铁、锌等微量元素与多种维生素的需求量也显著增加,比如铁、钙流失和骨质疏松风险上升,微量营养素缺乏对产后女性健康恢复造成显著影响。单纯补充有时并不有效,甚至可能产生不良副作用(如补铁引起的消化问题)。
肠道菌群在微量营养素代谢中扮演着关键角色,它们能够合成、转化和影响多种微量营养素的吸收与利用。
例如,特定的乳杆菌如Latilactobacillus sakei能产生叶酸(维生素B9),帮助改善宿主叶酸状态,同时增加短链脂肪酸产生;研究发现,维生素A的摄入与肠道微生物多样性呈正相关,特别是在孕期和产后,类胡萝卜素摄入充足的女性肠道微生物α多样性更高。
铁是许多细菌生长必需的营养素,但铁补充不当可能导致条件致病菌增殖,产生肠道不适。同时,某些菌群可提高微量营养素的生物利用度,如乳酸乳球菌N8和布拉酵母菌可将发酵食品中的核黄素和叶酸浓度提高40%-90%。
在这其中,肠道菌群与微量营养素之间存在重要的双向相互作用,在营养补充策略中需要加以考虑。
母乳喂养
美国儿科学会 (American Academy of Pediatrics) 推荐前 6 个月进行纯母乳喂养,然后继续母乳喂养并辅食至少 2 年。哺乳是一种能量密集型状态,理论上可以引起净负平衡,从而使产后体重减轻。
《美国膳食指南(2020-2025)》建议,与哺乳期的孕前需求相比,哺乳期妇女在产后前6个月额外摄入330卡路里,6-12个月额外摄入400卡路里。有稳定乳汁且想减重的女性可每天减少500卡路里摄入(相当于哺乳消耗的能量),但每天至少保持1800卡路里。这样可以每周减重1磅,4个月后可恢复孕前体重而不影响婴儿生长。
在哺乳期,坚持均衡的饮食,包括所有类别的常量营养素,对于维持母乳的成分和产量至关重要。一些流行的几种时尚饮食(例如,极低碳水化合物、旧石器时代、极低脂肪)可能会导致哺乳期妇女营养缺乏,应该避免。
膳食计划应强调营养丰富的全食物,如水果、蔬菜、豆类、全谷物和健康脂肪。选择纯素饮食的患者应评估并补充叶酸、维生素B12、铁、锌、胆碱、碘和EPA/DHA。
运动干预
剖宫产后,建议早期下床活动作为促进剖宫产后恢复(ERAC)方案的一部分。早期行走可缩短肠功能恢复时间、血栓形成风险和产后住院时间。
NICE指南:建议所有产后女性在6-8周后进行体重管理。

分期渐进:产后0-6周以低强度恢复性训练为主(如凯格尔运动、腹式呼吸),6周后逐步增加有氧与抗阻训练强度。
产后2天起:产褥期保健操(每1-2天增加1节,每节8-16次)。
产后6周后:逐步引入有氧运动(如散步、瑜伽),每周≥150分钟中等强度运动,分4-5次完成。
力量训练:针对核心肌群,改善基础代谢率。
为了安全起见,避免过早负重或高强度运动,运动强度以心率<140次/分钟为宜。
个体化:剖宫产者需延迟腹部加压训练至伤口愈合(通常8-12周),重点关注多裂肌与膈肌协同。
有效运动模式:
高强度间歇训练(HIIT)
RCT显示,每周3次、每次20分钟的HIIT(如自行车+深蹲组合)在12周内显著降低体脂率(-3.2% vs对照组-1.1%),且不影响母乳产量。
抗阻-有氧联合训练
对比静态核心训练与瑞士球训练,发现两者均能改善腰围(平均减少4.3cm)和内脏脂肪面积,但需结合饮食控制才能实现体重下降。
注:以上需结合自身状况,避免盲目锻炼。
中药调理
中药调理通过使用具有健脾祛湿功效的中药材,改善体内湿气过重。产后体虚、气血不足时,可在医生指导下服用补血益气类中药。
产后体重管理与全面康复是产后女性身体健康的一个方面,心理健康同样不容忽视,而肠道菌群失衡不仅影响代谢和体重,也直接参与情绪调节。产后激素波动引起的菌群变化,可能形成生理-心理-微生物的复杂互动网络。
当产后女性面临身体恢复压力的同时,还可能经历产后抑郁的困扰。这种情绪障碍影响产后妈妈健康的同时,也有可能传递给婴儿,影响下一代发育。因此,我们需要将产后照护视角从单纯的体重恢复拓展至情绪健康领域,探索肠道菌群在产后抑郁中的关键作用。
神经系统
产后抑郁症(PPD)是女性分娩后的常见疾病,症状包括情绪低落、易怒、睡眠障碍,严重时还会出现自杀或自杀意念。与产后抑郁相关的自杀是产后妇女死亡的主要原因,仅次于其他原因。
产后抑郁的发生率因地理位置而异,在经济发达国家为6.9%~12.9%,在经济欠发达或发展中国家超过20%。
近年来,社会和生活压力的增加与产后抑郁发病率的上升有关,这对患者的生活质量和身心健康产生了负面影响。病理机制很复杂,目前还不完全清楚。大多数学者认为,产后抑郁的发展可能是多因素的,受遗传、生理、心理、社会和家庭因素的影响。

一项荟萃分析显示,以下危险因素与产后抑郁相关:
• 妊娠糖尿病(OR = 2.71, 95%CI 1.78-4.14, I2 = 0.0%)
• 妊娠期抑郁(OR = 2.40, 95%CI 1.96-2.93, I2 = 96.7%)
• 孕妇生男孩 (OR = 1.62;95% CI 1.28-2.05;I2 = 0.0%)
• 妊娠期抑郁病史 (OR = 4.82,95%CI 1.32-17.54,I2 = 74.9%)
• 抑郁病史 (OR = 3.09,95%CI 1.62-5.93,I2 = 86.5%)
• 分娩时硬膜外麻醉 (OR = .81,95%CI .13-4.87,I2 = 90.1%)
产后抑郁症不仅影响母亲的身心健康和家庭关系,还通过多种途径传递给宝宝,影响宝宝的大脑发育。
抑郁状态下的妈妈往往缺乏与宝宝的积极互动,减少了皮肤接触和情感交流,这也会影响婴儿的神经系统发育。
可能通过喂养习惯的改变和照料质量下降影响宝宝,也可能通过母乳微生物群影响宝宝的肠道菌群定植,进而影响宝宝的认知发展和情绪调节能力。
产后一个月的这段时期对婴儿肠道菌群建立至关重要,而这正是大多数婴儿完全依赖母乳的阶段。一项研究发现,产后母亲压力与母乳微生物组成之间存在关联,研究发现,高压力组母亲的乳汁中链球菌、Gemella、Veillonella含量显著降低,而葡萄球菌、棒状菌和不动杆菌的含量升高,这些部分是条件致病菌。母亲的压力状态会影响乳汁的微生物组成,这可能会进一步影响婴儿肠道菌群的定植和后续的健康发展。
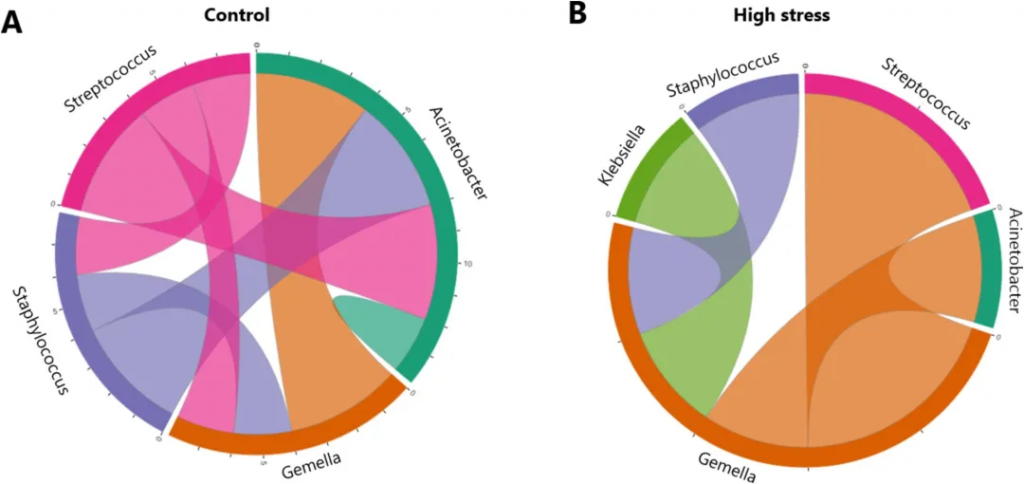
doi.org/10.1016/j.bbi.2024.11.020
链球菌和Veillonella的减少可能通过影响短链脂肪酸(SCFAs)的产生影响大脑发育,短链脂肪酸可穿过血脑屏障,对神经发育至关重要。
这种情况如果得不到及时干预,宝宝发育问题可能让产后妈妈的情绪雪上加霜,陷入恶性循环,导致抑郁症状加重,而对宝宝的影响也将长时间持续。
这种早期的微生物群失衡可能对宝宝的神经发育产生长期影响。研究发现,早期肠道菌群不健康与儿童期注意力缺陷、认知障碍、情绪调节问题甚至自闭症谱系障碍风险增加相关。由于大脑发育的关键期正是生命最初几年,这种影响可能会一直持续到青少年甚至成年期,形成跨代际的健康问题传递。
因此,针对产后抑郁的早期识别和干预至关重要。
产后抑郁治疗的困境
目前产后抑郁的治疗通常包括抗抑郁药、心理干预和激素补充。然而,这些方法经常出现副作用,表现出不一致的长期有效性,并可能在母乳中积聚,可能影响婴儿,从而限制其临床效用。
***
科学界开始逐渐认识到产后期女性不仅经历着激素的剧烈波动,也同时面临着肠道微生物生态的显著变化。这种变化并非独立存在,而是通过”肠-脑轴“与产后情绪密切相关。研究表明,肠道微生物通过产生多种神经递质和生物活性物质,如血清素(5-HT)、γ-氨基丁酸(GABA)、多巴胺等,直接影响中枢神经系统功能。
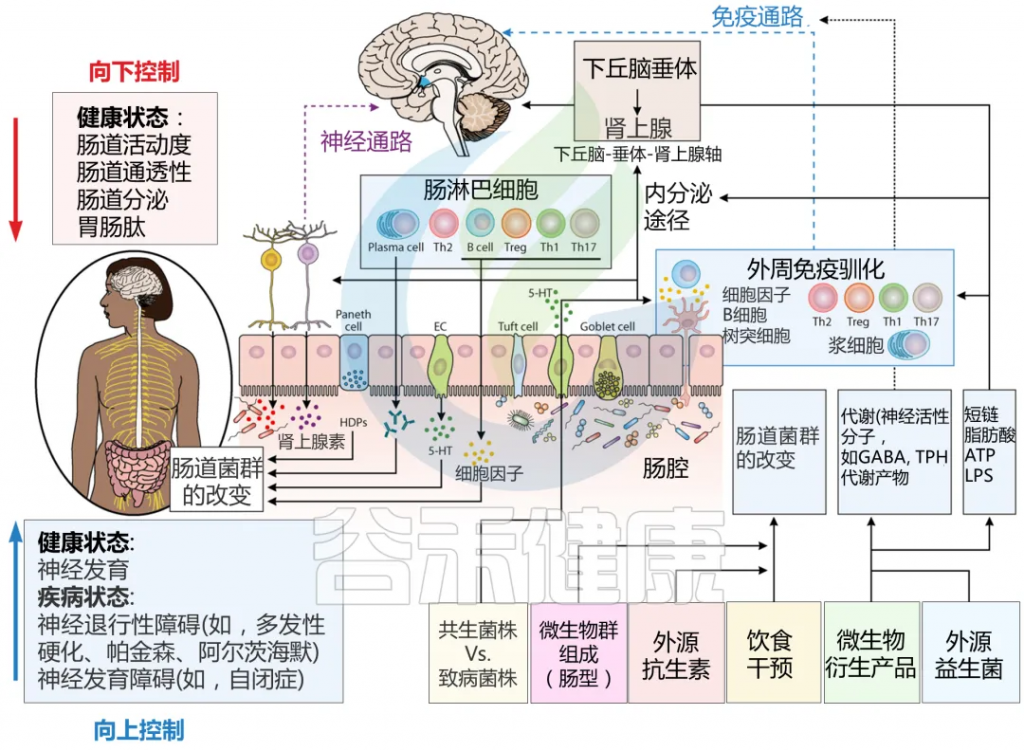
Sorboni SG, et al.,Clin Microbiol Rev. 2022
关于肠-脑轴详见我们之前的文章:
特别值得注意的是,血清素的合成受到肠道微生物群的显著调节,而血清素水平的异常与抑郁症状密切相关。肠道微生物还通过产生短链脂肪酸等代谢物,调节免疫系统功能,改善肠道屏障完整性,减少炎症反应。
这些发现为我们打开了理解产后抑郁的新视角——肠道微生物群的失衡可能是连接激素波动、免疫功能改变和情绪障碍的关键环节。
一项孟德尔随机化分析研究表明,产后抑郁患者的厚壁菌门与拟杆菌门之比(F/B)显著增加,这种菌群失衡与促炎状态相关。
α-变形杆菌属(属于变形杆菌门)、Clostridialesvadin BB60(属于拟杆菌属)和韦荣球菌科(属于厚壁菌门)与抑郁症风险的发展有关。
菌群“守护者”与“破坏者”的博弈
韦荣菌科(Veillonellaceae)通过代谢短链脂肪酸和脂多糖,增强肠道通透性,并激活Toll样受体通路,促进炎症因子释放,进而影响神经可塑性,这可能解释其在抑郁风险中的双重作用。
Clostridiales vadin BB60的保护作用可能通过调节F/B比值实现,与5-HT水平呈正相关,可能通过维持色氨酸代谢稳态发挥保护效应,但其具体机制需结合其对色氨酸代谢的影响进一步阐明。
RuminococcaceaeUCG011(保护性菌群)丰度下降会直接削弱其对5-HT的正向调节作用,5-羟色胺是一种与积极情绪相关的神经递质。
在产后抑郁大鼠模型中,该菌属的减少导致5-羟色胺合成酶(TPH)表达降低,从而引起5-羟色胺水平下降,可能导致情绪和行为调节障碍。
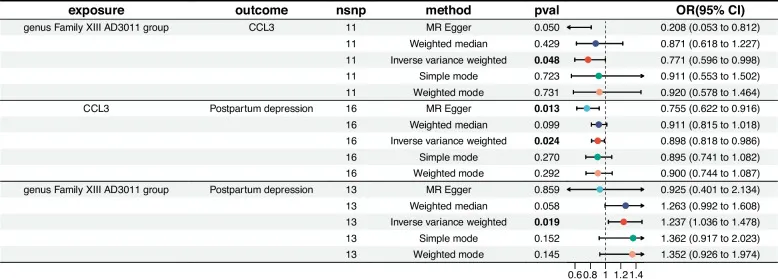
doi: 10.1186/s12884-025-07304-w
炎症——连接肠道菌群与产后抑郁的另一关键
血清趋化因子CCL5(OR=0.878)和CCL3(OR=0.898)水平升高与产后抑郁风险降低显著相关,但其作用机制独立于Family XIII AD3011属的致病效应。
动物实验证实,LPS诱导的神经炎症可导致海马神经元损伤和下丘脑-垂体-肾上腺(HPA)轴过度激活,最终引发皮质醇水平持续升高及突触可塑性受损。
通过对炎症介质的调节,可能能够降低产后抑郁的风险。这一理论为寻找新的干预手段提供了可能的方向。例如,益生菌或其他饮食干预策略被认为能够改善肠道菌群组成,并且有潜力减轻与产后相关的情绪障碍。
目前临床主要通过量表筛查进行初步判断,例如爱丁堡产后抑郁量表(EPDS),然而,量表评估依赖主观感受,可能存在漏诊或延迟干预的风险。近年来科研领域开始关注肠道菌群等相对客观生物标志物与产后抑郁的关联。
纵向研究表明,不同抑郁症状强度的孕产妇肠道菌群存在差异。
产后早期:在产后住院期间(产后1-3天)
中重度抑郁症中Enterobacteriaceae高
中重度抑郁症状组(MG组) Enterobacteriaceae细菌相对丰度显著高于轻度/无抑郁症状组(4.9%vs1.0%,p<0.05)。
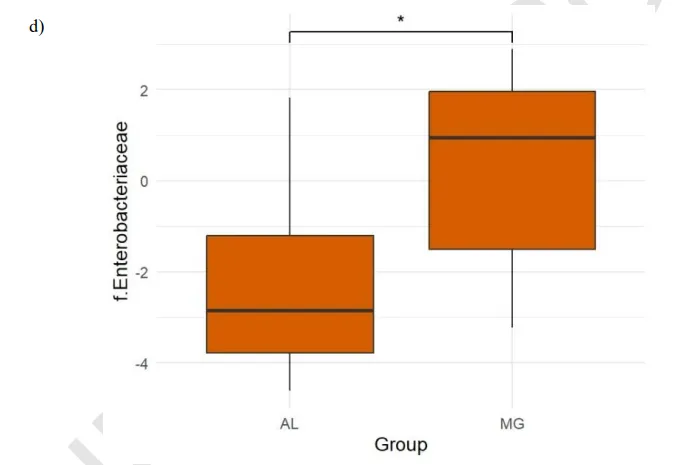
doi.org/10.47626/1516-4446-2024-3721
肠杆菌科细菌由革兰氏阴性菌组成,这些细菌可以转移到循环和淋巴系统,促进免疫反应的增加和炎性细胞因子的释放,如IL-1、IL-6等,这些细胞因子可以穿过血脑屏障并导致神经炎症,这是抑郁症女性的常见情况。
双歧杆菌属增加与抑郁症状评分(EPDS)降低存在相关趋势(p=0.2, cor= -0.19);Enterobacteriaceae与双歧杆菌属之间存在负相关趋势(cor:-0.14, p=0.2)。在接受精神治疗的中重度抑郁组中,双歧杆菌属的丰度随着时间的推移而增加。轻度/无症状组产后Faecalibacterium属丰度较高。
这项研究突显了将肠杆菌科(Enterobacteriaceae)视为妊娠晚期和产后恢复期间母体抑郁症状发作的合理生物标志物或促成因素的重要性。
识别出具有潜在预防或治疗意义的生物标志物,将有助于早期干预和个体化的治疗方案制定,最终减轻产后抑郁对母婴健康的影响。
临床应用思路
像上述研究证据,可以构建多模态评估体系:
通用益生菌、益生元等干预方式
编辑
——直接干预:产后母亲补充益生菌
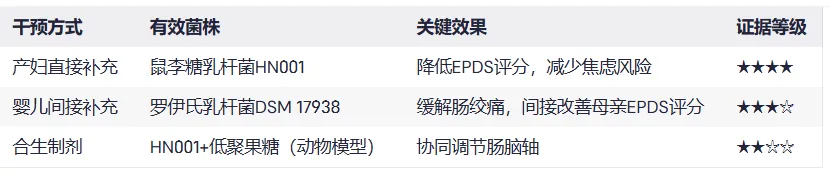
鼠李糖乳杆菌HN001
针对438名孕妇的双盲试验显示,补充HN001组产后6个月抑郁评分(EPDS)降低(Slykerman et al., 2017)。结果显示,焦虑评分(STAI-6)降低34%,抑郁风险减少50%(p=0.037)。
复合菌株干预
超重孕妇补充鼠李糖乳杆菌HN001+动物双歧杆菌420,产后12个月抑郁症状减轻(Hulkkonen et al., 2021)。
局限性:需联合鱼油使用,单独益生菌效果未达统计学显著。
doi: 10.3390/healthcare10060970
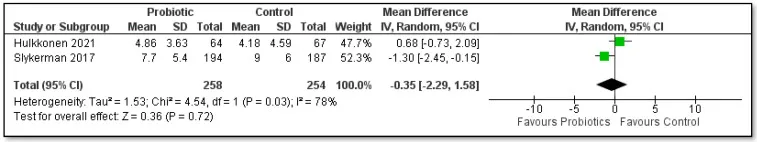
——间接干预:婴儿补充益生菌
罗伊氏乳杆菌DSM 17938
42名肠绞痛婴儿补充该菌株28天,母亲EPDS评分较安慰剂组降低2.3分(p<0.01)(Mi et al., 2015)。减少婴儿哭闹时长,缓解母亲育儿压力。
doi: 10.3390/healthcare10060970
个性化干预
基于上述纵向研究,如果产后女性情绪波动较大,量表评分趋于临界值,肠道菌群检测报告中有上述特征,肠杆菌较高,双歧杆菌较低,则可以针对性补充双歧杆菌来平衡肠道微生态,或者补充相关益生元为肠道有益菌提供养分,或者针对肠杆菌较高的情况进行饮食干预,吃西兰花、菠菜、胡萝卜等,具体情况因人而异。
有时候并不是简单地通过一个菌的高低来判别,而是要看个人肠道生态平衡状态,谷禾肠道菌群检测报告提供了更为精准的个体化干预方案。报告通常包含肠道菌群多样性评估、关键菌种分析以及代谢功能预测,尤其可以关注与情绪调节相关的微生物组成变化,如果偏高或者偏低,则有相关饮食、生活方式等干预措施,为产后女性提供个性化的调整建议。
产后女性可以在传统治疗方法之外,获得一种安全有效的补充干预途径,帮助她们更好地度过产后适应期,降低产后抑郁的风险。
维生素D
维生素D通过调节下丘脑-垂体-肾上腺轴(HPA轴)和促进神经递质合成发挥抗抑郁作用。
伊朗一项病例对照研究发现,维生素 D 水平低的妇女在分娩后 6-8 周患抑郁症的风险更大。
一项荟萃分析研究表明,低水平的维生素 D(低于 50 nmol/l)与产后抑郁的风险增加 3.67 倍相关。
从肠道菌群的角度,补充维生素D增加肠道菌群多样性,普氏菌属丰度增加,而嗜血杆菌和韦荣氏球菌丰度降低;促进双歧杆菌等益生菌生长,这些菌群参与血清素合成。维持肠道屏障完整性,减少促炎因子释放,调节免疫系统,改善Th17/Treg平衡。
一项研究发现,在怀孕的最后 2 周和分娩后的前 8 周每天服用 2,000 单位维生素 D 可以降低患产后抑郁的风险。
一项临床研究还发现,服用 2000 IU/d 的维生素 D 可以减轻抑郁症状。
B族维生素
维生素 B1 (硫胺素)
硫胺素是一种水溶性维生素,不会储存在体内,必须每天从牛肉、鸡肉、谷物、坚果和豆类等来源摄入。孕妇和哺乳期妇女每日需求量为 1.4 毫克。它对神经递质合成和能量产生至关重要,其缺乏会导致脑干变化和抑郁症状。补充硫胺素可以改善这些症状。
维生素 B2 (核黄素)
核黄素常见于乳制品、瘦肉、肝脏、绿叶蔬菜和酵母中,一项队列研究表明,适量摄入维生素 B2 有助于降低患产后抑郁的风险。
维生素 B6 (吡哆醇)
维生素 B6 或吡哆醇可以来自肉类、家禽、鱼、蛋、白土豆和淀粉类蔬菜。这种维生素对于血清素和多巴胺等必需神经递质的生物合成至关重要,这些神经递质可以改善情绪和认知功能。
抑郁症患者的同型半胱氨酸水平通常较高,而叶酸和维生素 B6 水平较低。同型半胱氨酸是一种非必需的硫氨基酸,对血管有毒并导致氧化应激和细胞死亡。 一项研究表明,服用维生素 B6 可以独立降低产后抑郁的发病率。
维生素 B12
血液中维生素B 的正常水平在147.6~664.2 pmol/L 之间。VB12 缺乏导致同型半胱氨酸水平增加以及叶酸和 VB12 效率降低,因为它被困在 5-甲基四氢叶酸循环中,而不是四氢叶酸的活性形式。
叶酸和VB12 在中枢神经系统的几种代谢途径中起着至关重要的作用。血清同型半胱氨酸水平升高表明VB12 和叶酸缺乏,见于抑郁症患者。
也有一项系统评价发现血清VB12、叶酸、铁蛋白水平与产后抑郁风险之间没有关联。
补充VB12 和叶酸可能在一定程度上提供保护作用,降低这些疾病的发病风险。
关于B族维生素和肠道菌群的关联,在我们之前的文章中有详细阐述:
铁
红肉、肝脏、种子、豆类和牡蛎等食物是铁的丰富来源。各种研究表明,与其他母亲相比,患有贫血的女性往往会经历更严重的产后抑郁。妊娠期贫血的诊断和治疗有助于降低患产后抑郁的风险。此外产后低铁蛋白水平也会导致产后抑郁。
中国一项涉及 1,592 名女性的队列研究表明,从怀孕期间开始摄入铁超过 6 个月有助于降低分娩后 6 个月和 12 个月患抑郁症的风险。
然而,另一项研究发现母体血清铁与产后抑郁症状之间没有关联。
一项对 649121 样本进行的 10 年高质量队列研究表明,在分娩后第一年,贫血和抑郁症之间存在显著关联。
维生素 C
维生素 C 在防止氧化应激方面起着至关重要的作用。它还充当一组酶的辅助因子,这些酶负责调节和合成整个身体的重要功能。
一项研究表明,高剂量的维生素 C(每天 3 克)可以降低 PPD 的严重程度。然而,另一项研究发现,压缩的橙皮滴剂对 PPD 或焦虑没有任何影响。
锌
锌是一种稀有金属离子,是人体中仅次于钙的第二大元素。它在免疫和内分泌系统中都起着至关重要的作用。锌负责刺激和抑制中枢神经系统中的化学介质。
根据一项全面的荟萃分析,抑郁症女性的血锌水平(1.85 μmoll/L)低于非抑郁症女性。
此外,一项系统评价发现,低锌水平与产后抑郁风险较高有关。然而,几项研究表明血清锌水平与产后忧郁综合征之间没有相关性。
此外,一项临床试验发现,锌对减轻产后焦虑和抑郁症状没有显着影响。
必需脂肪酸
必需不饱和脂肪酸,也称为多不饱和脂肪酸 (PUFA),分为两大类:n-6 和 n-3。
Omega-3 是一种亚麻酸,来自三个脂肪组,即 α-亚麻酸(ALA)、二十碳五烯酸(EPA)、二十二碳六烯酸(DHA)。n-3 多不饱和脂肪酸摄入量与产后抑郁之间的显著正相关。
研究表明,低水平的 omega-3 脂肪酸与产后抑郁高风险之间存在联系。其中,DHA 的作用更为明显。怀孕和哺乳自然会降低 DHA 水平。
然而,DHA 和产后抑郁之间的联系并不一致。在一项干预研究中,即使血清水平较低的女性每天摄入 200 毫克 DHA,产后前 4 个月内产后抑郁的患病率也没有变化。
一些研究发现鱼类摄入量或 omega-3 补充剂与产后抑郁之间没有相关性。一般来说,系统评价无法在omega-3脂肪酸和产后抑郁之间建立有效的关系。然而,素食中产后抑郁的患病率高于杂食性饮食。
碳水化合物
碳水化合物在能量产生中起着至关重要的作用,会影响情绪。研究表明,怀孕会导致胰岛素抵抗,这会使女性更容易患抑郁症和葡萄糖耐量受损。
一项队列研究发现,患有妊娠糖尿病的女性在怀孕和产后患抑郁的风险明显更高。此外,据推测,分娩后胰岛素水平降低,会降低血清素水平并导致抑郁症。血糖水平的波动也会导致炎症和脂肪因子标志物的分泌,例如白细胞介素IL-6,这些标志物与产后抑郁有关。
一项队列研究发现血糖指数与产后抑郁之间没有关联。
最后,低碳水化合物饮食往往会增加患抑郁症的风险,因为它们会减少大脑中血清素的产生。然而,富含碳水化合物的食物通过触发色氨酸的产生来促进幸福感。
临床实践中,产后女性补充维生素可以结合肠道菌群检测报告,针对个体差异制定个性化补充方案,以达到改善肠道菌群结构、提升情绪状态的最佳效果。
整合检测与干预
基线评估:产后进行首次肠道菌群检测与心理健康评估,建立个体基准数据。
分层干预:
动态监测:3个月左右复查肠道菌群和情绪状态,及时调整干预方案。
通过”检测-干预-评估-调整”的模式,产后女性可以获得高度个性化的维生素补充方案,靶向调节肠道菌群生态,有效支持情绪调节系统,实现身心健康的协同优化。
产后便秘、高血糖
产后便秘在新妈妈中较为常见,据相关研究显示,产后便秘的发生率在不同地区和人群中有所差异,但总体上可达到 20% – 40%。顺产和剖宫产的产妇均可能出现便秘问题,但剖宫产产妇由于手术和卧床休息等因素,便秘的发生率相对更高。长期便秘还可能增加患肠道疾病的风险。
分娩过程中抗生素使用、应激反应以及生理变化都会扰乱原有的肠道菌群平衡。特别是剖宫产女性,抗生素使用可能导致肠道菌群短期内显著改变,抗生素引起的生态失调会导致胆碱能神经元和整体肌间神经元减少,影响肠道蠕动。
产后盆底肌肉松弱或损伤会影响排便功能,而肠道运动减慢进一步导致粪便在肠道停留时间延长。关于盆底肌康复在下一章节介绍。
产后女性饮食习惯改变(如纤维摄入不足)、活动减少、水分摄入不足等因素,直接影响肠道菌群组成。
干预措施
饮食
增加膳食纤维的摄入是缓解产后便秘的重要方法。产妇应多吃蔬菜、水果、全麦面包等富含膳食纤维的食物,以增加粪便体积,促进肠道蠕动。同时,保持充足的水分摄入也很重要,每天至少饮用1500-2000毫升水,有助于软化粪便,使其更容易排出。
生活方式
适当运动可以促进肠道蠕动,缓解便秘症状。产后身体条件允许的情况下,新妈妈可以进行一些简单的运动,如散步、产后瑜伽等。
每天坚持 30 分钟左右的运动,有助于增强腹肌力量,促进排便。
定时排便训练
晨起或餐后2小时内尝试排便,利用生理性肠蠕动高峰(胃结肠反射)建立规律。
药物辅助
在医生指导下使用开塞露等药物,但不宜长期使用,以免形成依赖。乳果糖口服溶液也可在医生指导下使用,增加大便体积,缓解便秘。
中药如黄连上清丸、枳实导滞丸等,具有健脾消食、通腑泻下的作用,适用于产后气血虚弱所致的便秘。
腹部按摩
顺时针方向轻柔地按压下腹部,每次持续5-10分钟,有助于促进肠胃蠕动,缓解便秘症状。
以上是普适性的干预方式,我们也了解到,产后便秘的发生与肠道菌群失调密切相关,每个人肠道菌群不一样,便秘的原因也可能不一样。通过专业的肠道菌群检测,可以了解产后女性肠道微生物组成的具体情况,比如产丁酸菌的丰度、双歧杆菌和乳杆菌等有益菌的数量,以及潜在致病菌如艰难梭菌的水平。
如果是菌群紊乱的原因,那么结合菌群检测结果进行精准干预效果会更为显著。例如,对双歧杆菌缺乏的产妇可重点补充特定双歧杆菌株;对产丁酸菌不足者,可增加膳食纤维和低聚果糖等益生元摄入。
谷禾肠道菌群检测报告有便秘相关菌群过多或过少的提示,根据个人菌群报告可以有针对性地干预。
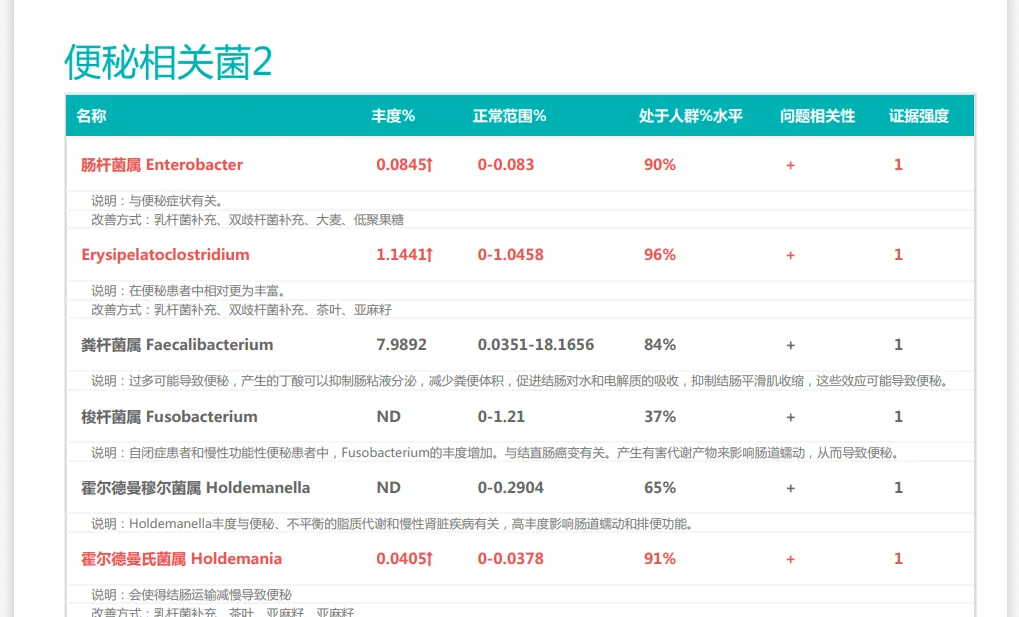
<来源:谷禾健康肠道菌群检测数据库>
这种个性化的微生物干预策略,不仅能更有效缓解便秘症状,还能从根本上改善肠道微生态环境,促进产后整体健康恢复。
一小部分女性产后出现血糖调节能力异常的状况,更有甚者这些女性可能会在产后5-10年内发展为2型糖尿病。
产后高血糖的常见表现
餐后异常疲劳感、口渴增加、排尿频繁、视力模糊、伤口愈合缓慢,以及无法解释的体重变化。
为什么产后会出现血糖波动?
分娩后胎盘激素急剧下降,理论上有利于胰岛素敏感性恢复,但实际情况更为复杂。
产后激素变化剧烈,催乳素和皮质醇水平波动可直接影响血糖;激素变化也会改变肠道菌群组成。
研究表明妊娠期糖尿病患者的肠道菌群失调在产后8个月仍然存在,拟杆菌减少,厚壁菌增加,这种菌群失衡直接影响肠道产生的短链脂肪酸水平,进一步影响胰岛素敏感性。
睡眠不足和作息紊乱会增加应激激素分泌,干扰胰岛素功能;这些因素也会改变肠道微生态环境,导致肠道通透性增加,使细菌代谢产物和内毒素进入血液循环,引发低度炎症反应。
产后恢复期能量消耗模式变化,哺乳消耗增加而活动量可能减少;
产后子宫收缩、伤口愈合和组织修复过程会导致代谢状态波动。同时,身体组成变化如脂肪分布调整也会影响葡萄糖代谢。这些生理变化通过改变肠道微环境pH值、氧气含量和营养物质分布,进一步影响肠道微生物组成。
这些因素共同作用,使产后血糖波动大且不可预测,特别是在产后头三个月。
产后阶段是女性身体快速恢复和调整的关键期,尤其对于妊娠期糖尿病的新妈妈,这一时期的营养管理更为重要,直接关系到恢复质量和未来健康风险。
肠道菌群检测帮助早期风险预警,辅助干预
相比传统血糖检测,菌群变化可能是代谢异常的更早期信号,为高风险人群提供预防性干预的时间窗口。
通过肠道菌群检测识别个体特异性的菌群失衡模式,结合临床指标(如炎症因子IL-6、皮质醇水平)建立个人”菌群-代谢”关联模型。
跟踪干预措施对肠道菌群的调节效果,为调整干预方案提供客观依据。
相关产后康复中心或临床营养或健康管理机构,可以根据菌群结构设计低血糖指数的膳食模式,针对特定菌群不足,选择相应的益生元食物。基于个体菌群特性,调整蛋白质、脂肪和碳水化合物的比例。
这种微生态层面的干预不仅能帮助产妇更有效地管理血糖,还能从根本上改善代谢健康,降低未来发展为2型糖尿病的风险。
盆底肌损伤
产后盆底肌损伤是许多产妇面临的常见问题。盆底肌损伤会引发尿失禁、漏尿、性生活质量下降、盆腔器官脱垂、衰老等问题。研究表明,产后妇女盆底功能异常发生率为50-64%。
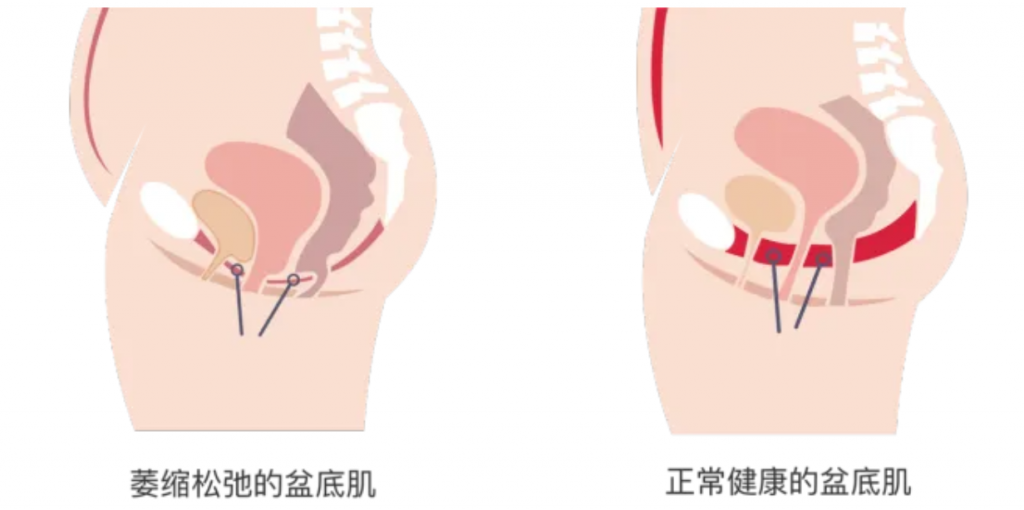
什么是盆底肌?
盆底肌是指位于骨盆底部的一组肌肉和筋膜组织,像一张“吊网”一样支撑着盆腔内的器官,包括膀胱、子宫、阴道和直肠等。盆底肌在维持盆腔器官正常位置、控制排尿和排便、支持性功能以及保持盆腔稳定等方面发挥着重要作用。
编辑
盆底肌损伤的原因
分娩压力:分娩过程中,胎儿通过产道时会对盆底肌肉施加巨大压力,导致肌肉损伤或拉伸,同时阴部神经牵拉导致神经传导功能下降,引发肌肉失用性萎缩。
激素变化:妊娠期孕激素和松弛素水平升高,抑制胶原蛋白合成,降低盆底肌的承托能力;产后雌激素骤降则延缓肌肉修复进程。
体重增加:怀孕期间体重的增加也会对盆底肌肉造成额外负担。
产后盆底肌修复
盆底肌训练(PFMT)
盆底肌训练是预防和治疗产后尿失禁的有效方法。训练通常包括收缩和放松盆底肌肉的动作,如凯格尔运动(Kegel exercises),并建议在专业理疗师或护理护士的指导下进行。
具体做法是找到盆底肌肉,在排尿时尝试中断尿流,然后收紧并上提盆底肌,保持5秒钟,然后放松10秒钟,每天进行3次,每次10-15分钟。
注:美国妇产科医生 Dr. Arnold Kegel 于 1948 年创建了盆底肌锻炼法,简称 Kegel 法,即指导患者做提肛运动。通过自主的、反复的盆底肌肉群的收缩和舒张,增强支持尿道、膀胱、子宫和直肠等脏器的盆底肌张力,恢复盆底肌功能。
生物反馈疗法
生物反馈疗法通过监测盆底肌肉的活动,让产妇直观了解自身肌肉收缩情况,帮助患者正确执行训练动作,提高康复效果。
电刺激疗法
电刺激疗法则通过特定频率和强度的电流刺激盆底肌肉或腹部肌肉,促进肌肉收缩,增强肌力。
一般来说,一周 3-5 次治疗,一疗程 10 次,每次 20-30 分钟,2-3 疗程。
但电刺激治疗需要专业设备和技术支持,且治疗过程可能会给产妇带来不适感。
中医调理
补肾固涩类方剂:如四神丸、金匮肾气丸等,用于调节内分泌系统,缓解压力性尿失禁。
健脾利水类中药:适用于气血虚弱导致的尿失禁及漏尿。
针灸疗法:通过在特定穴位如百会穴、关元穴等进行针刺,促进血液循环,帮助盆底肌修复。
手术治疗
对于严重的盆底肌损伤,如盆腔器官脱垂,严重尿失禁,影响日常生活,可能需要手术治疗。
尿道中段悬吊术(TVT/TOT):治愈率达85%-90%,但5年内复发率约10%-15%,且可能引发膀胱穿孔。
其他
使用盆底康复仪进行训练,可以帮助患者更有效地进行盆底肌锻炼。
针对性训练如腹式呼吸、脚趾叩击等也有助于增强盆底肌肉的力量。
多模式康复方法
多模式康复方法通过综合多种手段,以达到最佳康复效果。
Wang 等人的研究中,48 名产后腰骶骨盆疼痛女性参与随机对照试验,干预组接受为期 12 周的盆底肌训练结合腰背部肌肉神经肌肉电刺激,对照组仅接受神经肌肉电刺激。结果显示,干预组在 6 周和 12 周时疼痛、功能障碍和生活质量的身体领域均有显著改善。
进一步的研究由 Bhat 等人完成,比较了结合性唤起的凯格尔训练与单独凯格尔训练对产后盆底肌力和性功能的提升效果,涉及性活跃的初产阴道分娩女性。结果显示,结合性唤起的组别在每月评估中盆底肌力和性功能均有显著提升。这进一步证实了多模式康复方法的有效性,为产后康复提供了更多选择和可能性。
总的来说,骨盆底训练成为最常见的方法,躯干稳定练习和电刺激也强调恢复骨盆和核心力量。此外,创新技术,如脊椎按摩疗法,针灸等也是不断发展的产后护理方式。
这些产后康复方法不仅关乎生理恢复,更与产妇心理健康密切相关。研究表明,身体不适与心理健康之间存在双向互动关系。当产后女性遭受尿失禁、便秘或持续疼痛等困扰时,往往伴随着自尊心降低和社交回避倾向。
随着症状改善,产后女性对身体恢复的信心增强,减轻对永久性损伤的焦虑;其次,运动本身促进内啡肽分泌,天然调节情绪;运动通过影响肠道菌群组成间接改善了与情绪调节相关的神经递质平衡。
每一位母亲的产后康复都应被视为身心一体的整体康复过程。专业机构在设计产后康复方案时,如果超越传统的身体功能修复视角,整合肠道菌群调节的微生态方法,则能建立更全面、更有效的产后健康支持体系,帮助每位妈妈在迎接新生命的同时,也能守护自己的身心健康。
产后女性经历剧烈的生理重构,从激素波动到代谢变化,这些变化直接影响肠道微生态,而肠道菌群失衡又反过来影响产后恢复质量,形成复杂的双向调控网络。正是这种复杂的生理重构过程,使得产后康复面临独特挑战。
肠道菌群作为人体最大的微生态系统,不仅参与营养物质代谢和免疫调节,还通过肠-脑轴直接影响情绪和认知功能,是连接产后各种健康问题的核心纽带。通过解析产后女性独特的菌群变化特征,可以更精准地把握康复关键,从根本上促进身体各系统协调恢复。
目前,在临床应用中,肠道菌群检测不仅可辅助诊断产后抑郁、便秘和血糖异常等问题,更能通过”菌群-疾病”关联模型指导个性化治疗方案设计,减少药物依赖,缩短治疗周期。
月子中心可基于菌群检测结果定制营养餐,调节双歧杆菌、产丁酸菌等关键菌群,同时通过环境菌群优化降低感染风险,为母婴健康奠定微生态基础。
对健身机构而言,菌群检测结合训练能显著提升燃脂效率,使腰围减少更显著。健康管理机构通过菌群监测预警代谢风险,实施早期干预,避免产后5-10年内发展为2型糖尿病的风险。
未来,肠道菌群检测将与数字技术深度融合,推动产后康复服务从”经验型”向”精准型”转变。菌群检测+AI分析将实现健康异常早期预警;功能性菌株筛选与应用将催生新一代产后康复产品;母婴菌群联动管理将形成全新的家庭健康解决方案。
临床-科研-产业-政策四维联动机制,可以共同构建如”产后菌群健康分级评估指南”,制定产后菌群健康指导标准,推动产后菌群药物和营养品开发,为产后康复提供科学指导。
注:本账号内容仅供学习和交流,不构成任何形式的医疗建议。
主要参考文献
Mota AS, Sparvoli LG, Vanzele PAR, Naspolini NF, Tobaruela EC, Yoshizaki CT, Francisco RPV, Oliveira AMSS, Galletta MAK, Tess VLC, Taddei CR. Longitudinal Gut Microbiota Composition During Perinatal period in Women with Different Intensities of Depressive Symptoms. Braz J Psychiatry. 2024 Sep 21.
Wallace CJ, Audet MC. Diet and the gut microbiota-immune axis in the context of perinatal mental health: Protocol for a prospective cohort study. Womens Health (Lond). 2024 Jan-Dec;20:17455057241277072.
Wallace CJ, Audet MC. Diet and the gut microbiota-immune axis in the context of perinatal mental health: Protocol for a prospective cohort study. Womens Health (Lond). 2024 Jan-Dec;20:17455057241277072.
Macphail MG, Juul S, Wollny K, Negre JY, Metcalfe A, Chaput KH, Butalia S, Nerenberg KA. Nutrition Interventions for Lowering Cardiovascular Risk After Hypertensive Disorders of Pregnancy: A Systematic Review. CJC Open. 2023 Nov 4;6(2Part B):195-204.
Trifkovič KČ, Mičetić-Turk D, Kmetec S, Strauss M, Dahlen HG, Foster JP, Fijan S. Efficacy of Direct or Indirect Use of Probiotics for the Improvement of Maternal Depression during Pregnancy and in the Postnatal Period: A Systematic Review and Meta-Analysis. Healthcare (Basel). 2022 May 24;10(6):970.
Strobel KM, Juul SE, Hendrixson DT. Maternal Nutritional Status and the Microbiome across the Pregnancy and the Post-Partum Period. Microorganisms. 2023 Jun 13;11(6):1569.
Weerasuriya W, Saunders JE, Markel L, Ho TTB, Xu K, Lemas DJ, Groer MW, Louis-Jacques AF. Maternal gut microbiota in the postpartum Period: A Systematic review. Eur J Obstet Gynecol Reprod Biol. 2023 Jun;285:130-147.
Ghaedrahmati M, Alipour Z. The Association between Post-Partum Depression and Nutrition and Dietary Patterns: Systematic Review. Iran J Nurs Midwifery Res. 2024 Jul 2;29(3):280-289.
Neumann CJ, Pausan M-R, Haid V, Weiss E-C, Kolovetsiou-Kreiner V, Amtmann B, Winkler P, Mahnert A, Jantscher-Krenn E, Moissl-Eichinger C. The dynamics of the female microbiome: unveiling abrupt changes of microbial domains across body sites from prepartum to postpartum phases. Microbiol Spectr. 2024 Jun 25;12(8):e0014724.
Dellapiana, G., Nguyen, Q.T. & Naqvi, M. Navigating Postpartum Weight Loss: Evidence and Interventions. Curr Obstet Gynecol Rep 13, 207–212 (2024).
Ionescu RF, Enache RM, Cretoiu SM, Gaspar BS. Gut Microbiome Changes in Gestational Diabetes. Int J Mol Sci. 2022 Oct 25;23(21):12839.
Hasain Z, Mokhtar NM, Kamaruddin NA, Mohamed Ismail NA, Razalli NH, Gnanou JV, Raja Ali RA. Gut Microbiota and Gestational Diabetes Mellitus: A Review of Host-Gut Microbiota Interactions and Their Therapeutic Potential. Front Cell Infect Microbiol. 2020 May 15;10:188.
Hasain Z, Mokhtar NM, Kamaruddin NA, Mohamed Ismail NA, Razalli NH, Gnanou JV, Raja Ali RA. Gut Microbiota and Gestational Diabetes Mellitus: A Review of Host-Gut Microbiota Interactions and Their Therapeutic Potential. Front Cell Infect Microbiol. 2020 May 15;10:188.
Ohene-Agyei P, Iqbal A, Harding JE, Crowther CA, Lin L. Postnatal care after gestational diabetes – a systematic review of clinical practice guidelines. BMC Pregnancy Childbirth. 2024 Nov 4;24(1):720.
Xu H, Liu R. Comprehensive management of gestational diabetes mellitus: practical efficacy of exercise therapy and sustained intervention strategies. Front Endocrinol (Lausanne). 2024 Oct 3;15:1347754.
Gunabalasingam S, Kyrka A, Hopkins L, Lebrett R, Dyer E, Forde R, Heslehurst N, Meek CL, Schoenaker DAJM, Flynn AC, White SL. Interventions in women with type 2 diabetes mellitus in the pre-pregnancy, pregnancy and postpartum periods to optimise care and health outcomes: A systematic review. Diabet Med. 2025 Jan;42(1):e15474.
Juncker HG, Jakobsen RR, Naninck EFG, Davids M, Herrema H, van Goudoever JB, de Rooij SR, Korosi A. Maternal stress in the early postpartum period is associated with alterations in human milk microbiome composition. Brain Behav Immun. 2025 Feb;124:74-84.
谷禾健康
让食物成为你的药物,让药物成为你的食物
随着现代生活节奏的加快,疲劳、压力、不均衡的饮食,都在悄悄侵蚀着我们的健康。多糖,这些来自植物、真菌乃至海洋生物的天然赠礼,正以其独特的方式,为我们提供了一种全新的健康支持。
多糖由单糖通过糖苷键结合而成的高分子碳水化合物。研究表明,多糖具有多种生物活性,包括调节免疫功能、降低血脂和血糖、抗癌、抗病毒、抗肥胖、抗精神病、抗氧化、抗炎、抗凝血、止吐、抗辐射等作用。因此,多糖已成为最重要的天然成分之一,并引起了世界上越来越多研究者的关注。
虽然部分多糖如淀粉可以在人体胃和小肠中被消化吸收,但仍有许多特殊结构的多糖不能在这两个部位分解。对于那些不能被宿主消化的多糖,它们可以进入结肠,而结肠是大多数肠道菌群居住的地方。在结肠中,多糖可以与肠道菌群相互作用,从而发挥营养或药理作用。
多糖与肠道菌群之间相互作用可以影响健康,同时也通过肠道菌群的代谢作用,转化为有益的代谢产物,如短链脂肪酸,这些产物对维持肠道屏障的完整性、调节免疫反应发挥着至关重要的作用,甚至扩展到了全身的健康状况,可以影响我们的精力水平、情绪状态,对疾病的抵抗力等方方面面。
本文将深入探讨多糖,了解其在人体内的消化过程、与肠道菌群的相互作用,以及它们如何通过调节肠道菌群代谢物影响健康,如短链脂肪酸、三甲胺、色氨酸,还讨论了多糖在疾病预防和治疗中的应用,包括它们在改善代谢性疾病、炎症性肠病、缓解疲劳、改善肿瘤,神经系统疾病等方面的潜在效果。这为靶向肠道菌群开发新型的营养补充剂和药物提供了新的思路。

本文目录
01 多糖
02 多糖调节肠道菌群的组成
为什么多糖可以调节肠道菌群的组成?
多糖促进或抑制肠道微生物群
多糖分子量、糖苷键影响其细菌调节活性
03 肠道微生物将多糖代谢为短链脂肪酸
短链脂肪酸的生物学效应
人体内的多糖代谢
多糖补充与短链脂肪酸的生成
04 多糖调节其他肠道微菌群代谢物
三甲胺和氧化三甲胺(TMAO)
色氨酸及其代谢产物
胆汁酸、脂多糖、胃肠道气体
05 多糖调节肠道菌群修复肠道屏障
06 多糖通过肠道菌群改善疾病
2型糖尿病、非酒精性脂肪肝、肥胖、高血脂症
炎症性肠病、其他肠胃疾病、肿瘤
疲劳、神经系统疾病(认知障碍、抑郁等)
07 部分多糖营养与菌群调节
路易波士茶多糖、地黄多糖、五指毛桃根多糖
大蒜多糖、槐耳多糖、黄芩多糖、枸杞多糖
岩藻多糖、桑叶多糖、沙棘多糖、蘑菇多糖
08 结语
糖,这个小小的分子,是能量的源泉,是细胞的加油站。除了我们熟知的葡萄糖以外,还有一种叫做多糖,由许多糖分子手拉手组成,从植物的根茎到海洋生物的细胞壁,它们以复杂多样的形态存在。
根据糖单元的数量,碳水化合物可分为几类:
多糖是由多个单糖分子通过糖苷键连接而成的聚合物,属于高分子碳水化合物。它们广泛存在于自然界中,包括植物、真菌和海藻等生物体内。根据其来源和结构特性,多糖具有多种生物活性,如免疫调节、抗氧化、抗肿瘤等。
多糖是由10个以上相同或不同的单糖通过α或β糖苷键连接而成的大分子化合物,分子量从几万到数百万。
多糖的空间构象非常复杂,具有一级、二级、三级和四级结构。研究表明,多糖的活性与其结构密切相关。此外,通过分子修饰,如乙酰化、硫酸化、羧甲基化、硒化、磷酸化和磺化等可显著提高多糖的生物活性。
根据来源和结构的不同,多糖可以分为天然多糖和合成多糖,其中天然多糖又可根据其在自然界中的分布分为植物多糖、动物多糖、微生物多糖等。
例如,透明质酸和硫酸软骨素属于动物多糖,而纤维素、淀粉和糖原是常见的植物多糖。
淀粉
由大量葡萄糖分子通过α-1,4-糖苷键和α-1,6-糖苷键连接而成,形成直链淀粉和支链淀粉两种结构。广泛存在于谷物(如大米、小麦、玉米)、薯类(如土豆、红薯)等食物中。在人体消化过程中,被淀粉酶逐步分解为葡萄糖,为身体提供能量。
纤维素
由葡萄糖分子通过β-1,4-糖苷键连接而成,形成长而直的链状结构。是植物细胞壁的主要成分,在蔬菜(如芹菜、菠菜)、水果(如苹果)中含量丰富。由于人体缺乏分解β-1,4-糖苷键的酶,纤维素难以被人体消化吸收,但对促进肠道蠕动、预防便秘等具有重要作用。
果胶
是一种复杂的多糖,由半乳糖醛酸等组成。常见于水果(如柑橘、苹果)中。在食品工业中,常用于制作果酱、果冻等,增加其黏稠度和稳定性。
尽管功能性糖因其在健康和疾病预防中的潜在作用而受到越来越多的关注,但它们的天然可用性相对较小。这意味着,为了充分利用这些有益的分子,需要采用特定的提取方法来增加它们的可获得性,多糖提取常用的方法有热水提取、酸提取、碱提取和酶水解等。
近年来,一些新的方法,如超声波提取、微波提取、超滤、高压电场法、超临界流体萃取、亚临界水萃取等也用于多糖的提取。这些方法不仅能够提高多糖的提取率,还能够在一定程度上保护多糖的结构完整性,从而保留其生物活性。
通常,人体分泌的消化酶只能分解几种多糖,而纤维等许多多糖不能被吸收和直接使用。因此,多糖可以通过小肠进入结肠,这是大多数肠道细菌居住的地方,然后与肠道微生物群相互作用。
细菌在肠道中通过发酵降解多糖
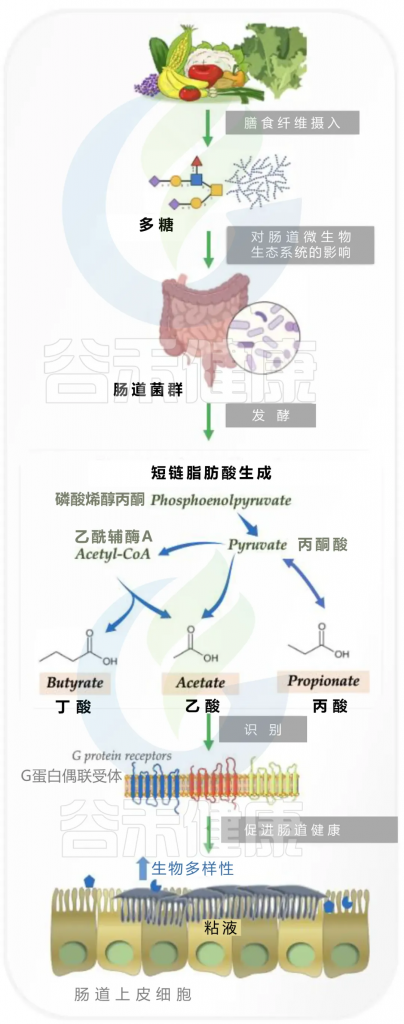
doi.org/10.3390/nu14194116
多糖转化为短链脂肪酸
首先与多糖能够发酵生成酸性的短链脂肪酸密切相关。未电离的短链脂肪酸能够穿过细菌细胞膜,对肠道细菌产生一系列影响,例如改变DNA合成和氨基酸摄取。
短链脂肪酸对肠道细菌的影响
例如,短链脂肪酸能够通过调节侵袭基因的表达,抑制沙门氏菌(一种常见的食源性病原体)的生长。因此,短链脂肪酸的增加可以改变肠道菌群的功能,进而影响其组成。
肠道pH值的变化
短链脂肪酸的增加还会导致肠道pH值下降,影响细菌的适应能力。每种细菌都有其适宜的pH范围,不同细菌在特定pH条件下的适应能力各异。
多糖的降解产物作为能量来源
多糖对肠道菌群组成的调节作用还与其降解产物有关。一些肠道细菌能够利用多糖的降解产物作为碳源和能量来源,而另一些细菌则不能。
综上所述,多糖通过影响肠道菌群的代谢功能,直接调节了肠道菌群的组成。这些发现为我们理解多糖如何通过肠道菌群影响宿主健康提供了新的视角。
多糖的益生元效应
具有选择性刺激有益微生物生长能力的多糖被称为益生元,例如果聚糖,它能丰富乳酸菌和/或双歧杆菌。这些有益细菌的减少与糖耐量受损密切相关。一些多糖能够丰富这些细菌,因此可以用来改善糖尿病表型。
多糖抑制病原菌
病原菌在胃肠道中的定植和增加会导致一系列疾病,而抑制它们可以控制疾病的严重程度。例如,致病性 Sutterella、Desulfovibrionaceae、Streptococcaceae 、Clostridium 的比例较高与肥胖发展呈正相关。
多糖的双向调节作用
疾病的发生是一个复杂的过程,涉及不同的微生物,包括有害细菌和有益细菌。例如:
多糖能够对肠道微生物发挥双向调节作用,即促进有益细菌的同时抑制有害细菌。
例如,口服灰树花多糖(GFHP)对非酒精性脂肪肝病的积极作用与调节肠道菌群有关。
在分子量方面,不同分子量的魔芋葡聚糖(KGM)对2型糖尿病(T2DM)大鼠的降血糖作用研究表明,中等分子量的KGM显著增加了Muribaculaceae,减少了Romboutsia和Klebsiella,但高分子量和低分子量的KGM对这些细菌的影响不显著。
灵芝的低分子量多糖(<10 kDa)具有更好的发酵和更高的产气能力,刺激肠道细菌快速生长。另一方面,高分子量多糖(>100 kDa)更难被肠道细菌发酵,并且在肠道中的停留时间更长,导致对肠道微生物群的影响更长。
高分子量的黄芪多糖具有一定的生物活性,但其相对分子量较大,溶解性差,生物利用度低,限制了其功效的发挥。低分子量的黄芪多糖具有较好的水溶性,能够在更大程度上刺激巨噬细胞摄取中性红、NK细胞增殖,发挥免疫活性。
多糖由各种通过糖苷键连接的单糖组成,糖苷键的类型和位置导致肠道微生物群的选择性发酵存在差异。
多糖的单糖组成越复杂,调节细菌的活性越强
一项关于龙眼多糖和燕麦多糖的研究表明,龙眼多糖能显著促进干酪乳杆菌、嗜酸乳杆菌、植物乳杆菌、粪肠球菌的增殖,但燕麦多糖的作用并不明显。原因是龙眼多糖由葡萄糖、甘露糖和阿拉伯糖组成,而燕麦多糖的单糖主要是葡萄糖。
短链脂肪酸(SCFA)是一组含有少于六个碳的脂肪酸,包括甲酸盐、乙酸盐、丙酸盐、丁酸盐、戊酸盐。
乙酸盐、丙酸盐和丁酸盐是肠道中的主要SCFA,约占所有SCFA的95%,三者的比例约为3:1:1。
作为肠道微生物群和宿主的重要能量来源,短链脂肪酸通过不同的作用模式在健康和疾病中发挥着重要作用。
作用机制
与GPRs的相互作用
抗炎作用
免疫调节
HDAC抑制作用
丁酸盐的作用
与疾病的关系
多糖对SCFA的调节及其对靶标的影响
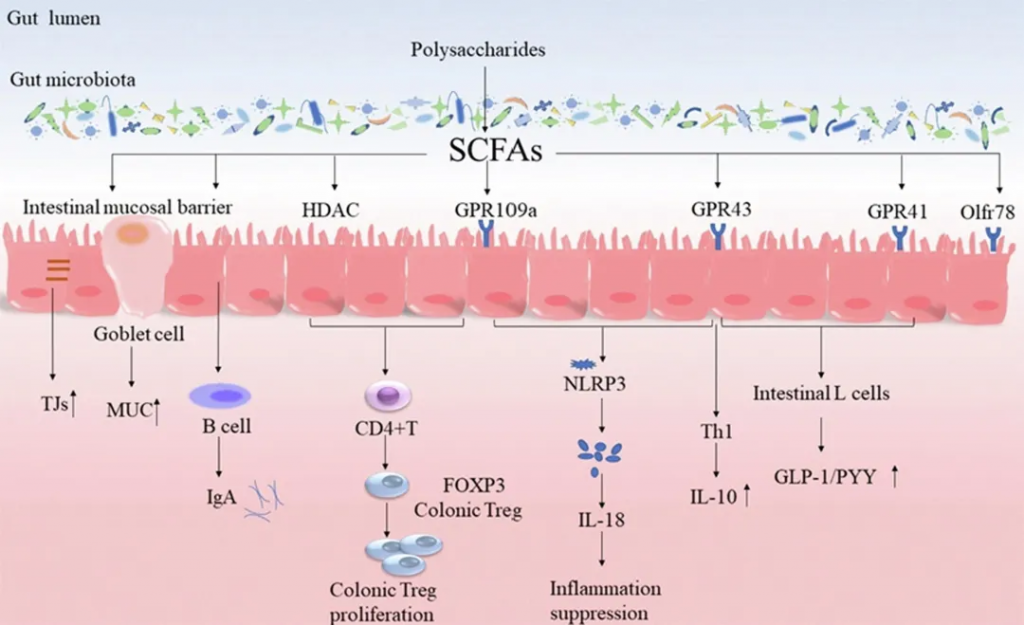
doi.org/10.1016/j.foodres.2022.111653
人体内消化酶的局限
在人体消化系统中,我们自身分泌的消化酶往往难以分解复杂的多糖。这些多糖分子,因其结构复杂,通常在我们体内无法被有效代谢。
肠道菌群的代谢作用
我们的肠道菌群拥有破解这些复杂多糖的秘密武器——一系列的酶,统称为碳水化合物酶(CAZymes)。这些酶能够分解多糖,将其转化为对人体健康有益的短链脂肪酸。
碳水化合物酶的种类
肠道菌群中的“专家”与“通才”
在肠道菌群中,拟杆菌门和厚壁菌门是编码CAZymes的两大主力。
多糖的初步降解
在属的水平上,多糖最初可以由某些微生物降解,例如双歧杆菌属、真杆菌属、梭菌属、罗氏菌属(Roseburia spp.)。
SCFAs的生成途径
尽管人体自身无法分解复杂的多糖,但我们的肠道菌群却具备了这一能力,它们通过一系列特殊的酶,将多糖转化为对人体健康有益的短链脂肪酸。
多糖对SCFAs生成的促进作用
饮食补充多糖可以为产生SCFAs的细菌提供有利的生长环境,从而促进SCFAs的生成。例如,沙棘多糖(CCPP)通过调节肠道菌群和SCFAs,能够缓解2型糖尿病。
沙棘多糖调节肠道菌群和SCFAs,缓解2型糖尿病
枸杞多糖调节肠道菌群,提高SCFAs
多糖结构对短链脂肪酸生成的影响
不同的多糖因其分子结构的不同,对SCFAs的调节作用也不尽相同。
多糖的疗效与SCFAs的非直接关联
尽管大多数多糖可以被代谢成SCFAs,但它们的疗效并不一定与SCFAs直接相关。
多糖通过调节肠道菌群和促进SCFAs的产生,对健康具有多方面的益处。然而,多糖的结构与它们对SCFAs生成的调节作用之间的关系仍需进一步研究。此外,多糖的疗效可能不仅限于SCFAs的产生,还可能涉及肠道菌群产生的其他分子。
在肠道微生物群的作用下,脂质和蛋白质等饮食成分可以代谢为一系列代谢产物,如三甲胺-N-氧化物(TMAO)、色氨酸、脂多糖(LPS)等。此外,肠道微生物群还可以与宿主合成和释放的化合物相互作用。例如,肠道微生物群可以将初级胆汁酸(BA)转化为次级胆汁酸。由于肠道微生物群的组成与肠道微生物群代谢物的产生直接相关,因此可以推断,除了SCFAs,多糖还可以调节其他肠道微生物群代谢产物,如BA。
三甲胺和TMAO的代谢过程
在肠道中,饮食中的四胺类物质如胆碱、L-肉碱和卵磷脂(来自红肉、鸡蛋、鱼、海鲜)首先被微生物胆碱三甲胺裂解酶分解成三甲胺(TMA)。随后,TMA被吸收进入门脉循环,并运输到肝脏,在黄素单加氧酶1和黄素单加氧酶3的作用下转化为三甲胺-N-氧化物(TMAO)。
TMAO的潜在危害
值得注意的是,TMAO是一种潜在的有害代谢产物。多项研究指出,TMAO水平的增加与多种疾病的发生和发展呈正相关。例如,TMAO可以直接增强血小板的反应性,促进血栓形成,并通过激活核因子κB(NF-κB)和NLRP3炎症体影响血管炎症。
多糖对TMA和TMAO代谢的影响
最近的研究表明,TMA和TMAO在理解多糖的作用机制中扮演重要角色。例如:
不同多糖对肠道菌群的调节作用
不同的多糖可以通过调节相同的肠道细菌来降低TMA和TMAO的代谢,例如变形菌门。研究表明,变形菌门负责TMA的转化,增加变形菌门可能导致TMA增加。然而,也有研究表明TMAO与变形菌门呈负相关,这表明多糖调节变形菌门与TMA产生的效应之间的关系需要进一步研究。
多糖通过调节肠道菌群,影响TMA和TMAO的代谢,从而可能对人体健康产生积极影响。然而,多糖与TMAO之间的关系复杂,需要更多的研究来阐明这些相互作用的确切机制。
色氨酸代谢的重要性
色氨酸(Trp)是人体必需的氨基酸。它的代谢可以分为内源性代谢和细菌性代谢。内源性代谢主要通过犬尿氨酸途径(KP)和5-羟色氨酸途径进行,前者产生犬尿氨酸(KYN)、犬尿酸(KA)、烟酸、黄嘌呤酸等,后者转化为5-羟色氨酸(5-HT)和褪黑素。
色氨酸代谢物的生理功能
增强免疫:色氨酸可加强免疫力,减少炎症。
神经保护:KA作为谷氨酸受体拮抗剂,具有神经保护和抗惊厥作用,还能调节能量代谢。
情绪调节:5-HT作为神经递质,可调节情绪、肠道通透性和肠道蠕动。
肠道菌群在色氨酸代谢中的作用
肠道细菌代谢色氨酸产生吲哚及其衍生物,如吲哚丙酸、吲哚乙酸等,这些物质可以缓解炎症,促进肠道上皮屏障功能。肠道菌群的色氨酸代谢异常与肠易激综合症、代谢综合症和结肠癌等疾病有关。例如,结肠癌患者常伴有色氨酸水平下降和KP代谢物水平升高。
多糖影响肠道微生物色氨酸代谢,从而改善疾病
吲哚是硫酸吲哚酚的前体,是一种蛋白结合尿毒症毒素,是心血管疾病的危险因素。对于患有终末期肾病心血管疾病的患者,吲哚水平升高,患者粪便中产吲哚细菌丰富。
总的来说,多糖可以通过调节肠道菌群来改变色氨酸代谢,最常见的是增加乳杆菌和阿克曼菌,从而缓解疾病。
胆汁酸的生物合成与功能
胆汁酸(BAs)是一类由肝脏产生的特殊类固醇分子,经过肠道菌群转化。肝脏中存在两种BA生物合成途径:
CA和CDCA是体内的主要胆汁酸。经过肠道菌群的改造,CA转化为脱氧胆酸(DCA),CDCA转化为鹅去氧胆酸(LCA)。
胆汁酸受体及其作用
胆汁酸受体包括细胞表面受体和细胞内受体。细胞表面受体包括TGR5,细胞内受体包括法尼酰X受体FXR、孕烷X受体、维生素D3受体(VDR)和组成型雄烷受体。胆汁酸通过激活相应的受体调节脂质、葡萄糖和能量代谢。例如,TGR5和VDR的激活导致GLP-1和FGF19分泌,GLP-1可以改善胰岛素敏感性,FGF19可以通过抑制脂肪生成减少肝脏脂肪变性。
多糖对胆汁酸代谢的调节作用
近年来的研究表明,多糖可以通过恢复胆汁酸的代谢来缓解疾病。
多糖对胆汁酸代谢影响的总结
多糖通过调节肠道菌群的组成,特别是Bacteroides、Lactobacillus、Clostridium、Ruminococcus、Bifidobacteria,影响胆汁酸的代谢。
某些多糖如岩藻聚糖和灰树花多糖减少了Clostridium的水平,这与文献报道的促进胆汁酸转化的作用似乎矛盾,需要进一步研究确认这些肠道细菌与多糖之间的关系。
脂多糖的危害
脂多糖(LPS)是由革兰氏阴性细菌(如大肠杆菌)产生的内毒素。持续暴露于LPS或LPS异常增加,可通过减少肠道上皮细胞活性、降低肠道细胞增殖、抑制肠道细胞迁移和诱导肠道细胞凋亡等方式,导致肠道损伤。
LPS的转移还能损伤肠道,并可能通过与多种受体的相互作用,如LPS结合蛋白、簇分化14、髓样分化2和Toll样受体4,引发糖尿病、非酒精性脂肪肝病、肥胖、动脉粥样硬化等一系列疾病。
多糖对LPS产生菌的抑制作用
许多多糖能够抑制产生LPS的细菌。例如,在链脲佐素(STZ)诱导的糖尿病肾病模型中:
多糖调节肠道菌群的矛盾效应
尽管多糖可以通过调节肠道细菌来抑制LPS,但对特定细菌的调节作用可能存在矛盾。例如,作为LPS产生菌的拟杆菌门,在多糖处理后的水平变化并不一致。有研究表明,黄精多糖和蝉花多糖增加了拟杆菌门的水平,而竹荪多糖却降低了它。这些研究表明,多糖对肠道菌群的调节效应有时可能相互矛盾,需要进一步研究以确认结果。
胃肠道气体的生成
胃肠道内通过细菌发酵食物,会产生一系列气体,包括氢气(H2)、甲烷(CH4)、二氧化碳(CO2)、硫化氢(H2S)和一氧化氮(NO)。这些气体在胃肠道中发挥着调节作用,例如影响结肠蠕动、神经通讯、血管功能和免疫反应等。
气体产生的部位和作用
CO2 主要在胃中产生,而其他气体如 H2、CH4、CO2 和 H2S 主要在小肠和结肠中产生。
这些气体对人体健康至关重要,它们可以调节肠道功能,影响营养物质的吸收和疾病的发生。
多糖对气体产生的调节
尽管多糖对 H2、CH4 和 CO2 的产生有明显影响,但关于多糖结构与气体产生之间具体关系的研究会相对较少。需要更多的研究来明确这些关系,以及多糖如何通过影响肠道菌群来调节气体的产生。
肠道菌群是一个复杂的微生物群落,具有显著的组成和功能多样性。不同的微生物可以介导相同或不同的代谢物的产生,相同的微生物也有助于不同代谢物的生产。
例如,持续的研究表明:
拟杆菌门(特别是Bacteroides thetaiotaomicron、Bacteroides fragilis)、厚壁菌门(如Clostridiaceae、Erysipelotrichia)、以及变形菌门可以促进TMA的产生。
放线菌门(如Bifidobacteria)、厚壁菌门(如Lactobacillus、Clostridium、Peptostreptococcus)、拟杆菌门(如Bacteroides)可以促进色氨酸(Trp)的转化。
双歧杆菌、乳酸菌、梭菌、Peptostreptococcus、拟杆菌也有助于次级胆汁酸(BAs)的产生。
因此,就像肠道菌群组成的调节一样,多糖对特定肠道菌群代谢物功能的调节作用不是孤立的。
肠道是我们抵御外界有害物质和病原体侵袭的第一道防线。它由多个层次的子屏障构成:
生物屏障:由肠道细菌和病毒组成;
化学屏障:包含免疫球蛋白A(IgA)、抗菌肽(AMPs)和粘液(MUC);
物理屏障:由肠道上皮细胞构成;
免疫屏障:含有T细胞、B细胞、巨噬细胞和树突细胞等免疫细胞。
这些子屏障协同工作,限制病原体与肠道的接触,维持肠道稳定。肠道屏障的损伤与多种疾病正相关,包括肠易激综合症(IBS)、代谢综合征、过敏、肝脏炎症等。
肠道菌群及其代谢物可以直接或间接影响肠道屏障:
越来越多的证据表明,多糖通过调节肠道菌群对肠道屏障有益。正常的肠道菌群组成可以通过竞争性排除,通过消耗营养源和占据附着位点,作为抵御外界病原体的屏障。
多糖→ 调节肠道菌群→ 修复肠道屏障
肠道菌群可以刺激宿主产生抗菌化合物,如IgA和AMPs,这些是化学屏障的关键组成部分。
例如,菊粉型果聚糖可以促进乳杆菌的丰度和IgA的分泌。在DSS诱导的溃疡性结肠炎小鼠模型中,金银花多糖通过增加双歧杆菌和乳杆菌,增加了分泌型IgA含量,从而调节肠道屏障。
在DSS诱导的结肠炎小鼠中,海蜇皮多糖增加Akkermansia,Akkermansia muciniphila作为粘液的降解者,可以增强肠道屏障的完整性,减少炎症。同时海蜇皮多糖增加结肠中TJs和MUC2的表达,保护了肠道屏障。
多糖→ 短链脂肪酸→ 修复肠道屏障
短链脂肪酸和胆汁酸等肠道菌群代谢物在调节肠道屏障功能中也扮演重要角色。
多糖→ 色氨酸和胆汁酸代谢→ 修复肠道屏障
这些研究表明,多糖可以通过调节肠道菌群及其代谢物,维护肠道屏障的完整性。
代谢性疾病包括一组因碳水化合物、脂质和蛋白质代谢错误而导致的疾病。2 型糖尿病 (T2DM)、非酒精性脂肪性肝病 (NAFLD) 和肥胖是常见的代谢性疾病。
生活方式干预、全身药物治疗和外科手术等多种方法被用于预防和治疗代谢性疾病。尽管代谢性疾病的药物治疗取得了最新进展,但潜在的不良反应仍然是关键挑战。
使用天然物质的药物治疗被认为是改善代谢疾病的一种有前途且可行的方法。
doi.org/10.1016/j.biopha.2023.114538
多糖通过多种机制在治疗2型糖尿病方面表现出良好的效果,比如:
肠道菌群在代谢紊乱,特别是 2 型糖尿病的发展中起着重要作用。
◤车前子多糖 对 STZ 诱导的 2 型糖尿病大鼠有抗糖尿病作用,这可能与其调节肠道菌群和增加短链脂肪酸水平有关。车前子多糖可显著增加糖尿病大鼠粪便中Bacteroides vulgatus、发酵乳杆菌、Prevotella loescheii、Bacteroides vulgates等结肠细菌的多样性和丰度,以及短链脂肪酸的浓度。
◤桑果多糖 可以丰富糖尿病小鼠的功能菌并调节微生物多样性。具体而言,该多糖显著富集了一些有益细菌(拟杆菌目、乳杆菌属、Allobaculum、拟杆菌属、阿克曼菌属),同时减少了一些致病菌(葡萄球菌、棒状杆菌属、Jeotgalicoccus、Aerococcus、Enterococcus、Facklamia)。
◤罗布麻叶的两种富含多糖的提取物改善了糖尿病小鼠的肠道微生物群失调,包括增加了Odoribacter、Anaeroplasma、Parasutterella、Muribaculum的丰度,并降低了肠球菌属、克雷伯菌属、Aerococcus的丰度。这可能有助于它们的抗糖尿病作用。
◤菊粉补充增加了双歧杆菌的丰度并增强了肠道屏障的完整性,这与2型糖尿病呈负相关。
◤青钱柳叶中分离的多糖通过增加 SCFAs 含量和有益的肠道细菌瘤胃球菌科来减轻 HFD/STZ 诱导的 2 型糖尿病大鼠的糖尿病症状。
◤苦瓜中的天然多糖通过增加 SCFAs 含量和Prevotella loescheii、Lactococcus laudensis 的丰度来改善 HFD/STZ 诱导的 2 型糖尿病大鼠的高血糖、高脂血症、高胰岛素血症。
◤天然南瓜多糖通过增加阿克曼氏菌和减少丹毒丝菌科(Erysipelotrichaceae)来显示出对 HFD/STZ 诱导的 2 型糖尿病的降血糖作用。此外,南瓜多糖还能增加 2 型糖尿病模型中肠道短链脂肪酸的产生。
◤灵芝多糖(GLP)通过恢复HFD/STZ诱导的肠道微生物群失调,特别是通过增加Blautia、拟杆菌、Dehalobacterium、Parabacteroides,以及减少有害的肠道细菌Aerococcus、Corynebacterium、Ruminococcus、Proteus,显示出抗糖尿病作用。
◤薏苡仁多糖通过降低厚壁菌门/拟杆菌门的比例和增加SCFAs的含量,在HFD/STZ诱导的T2DM小鼠模型中表现出降血糖活性。
◤葡甘聚糖作为铁皮石斛、芦荟和魔芋的天然多糖,通过增加厚壁菌门的丰度和减少拟杆菌门、变形杆菌的丰度,改善HFD/STZ喂养大鼠的T2DM代谢紊乱。
多糖对改善NAFLD具有有益作用,比如:
多糖可以改善肠道菌群失调并保护非酒精性脂肪性肝动物的肠道屏障完整性
◤枸杞多糖结合有氧运动通过改善肠道菌群失调改善 NAFLD,包括调节肠道菌群的丰度和多样性,增加微生物代谢产物 SCFA 的水平,减少变形菌和厚壁菌门/拟杆菌门的比例。厚壁菌门和拟杆菌门是参与宿主代谢和脂肪积累的关键细菌。
◤麦冬多糖可以通过调节肠-肝轴显著保护 NAFLD。具体来说,这种多糖显著降低了一些有害细菌的相对丰度,包括乳球菌、肠杆菌、Turicibacter、Clostridium- sensu-stricto -1、Tyzzerella、Oscillibacter,并增加一些有益菌的相对丰度,如Alistipes、Ruminiclostridium、Rikenella。这种多糖还显著增加了两种产SCFAs菌( Butyricimonas、Roseburia )的丰度以及乙酸和戊酸的水平,从而改善了炎症反应和肝脏脂质代谢。
◤灰树花杂多糖可通过调节肠道菌群来改善高脂饮食诱导的NAFLD,包括显著增加Allobaculum、拟杆菌属和双歧杆菌属丰度,减少Acetatifactor、Alistipes、Flavonifractor、Paraprevotella、Oscillibacter的丰度。
◤黄芪多糖可减轻HFD喂养小鼠的NAFLD,丰富了脱硫弧菌属,尤其是作为SCFAs、乙酸的产生者的Desulfovibrio vulgaris,减轻肝脂肪变性。
◤诺尼果多糖来源于辣木,通过促进短链脂肪酸的产生缓解HFD喂养小鼠的NAFLD,并通过改善肠道微生物群的多样性和组成逆转HFD诱导的肠道微生态失调。
◤核桃青皮多糖通过提高肠微生物群(包括普氏菌科、Allobaculum)的SCFAs含量和丰度,预防HFD喂养大鼠的肥胖和NAFLD。
◤从贻贝中提取的贻贝多糖,α-D-葡聚糖(MPA)可保护HFD喂养的大鼠的NAFLD,补充MPA可逆转HFD抑制的微生物微生态失调和SCFAs。
◤海带可溶性多糖通过降低厚壁菌门/拟杆菌门的比例,促进Verrucomirobia和丙酸盐产生菌拟杆菌和阿克曼菌,减轻高脂饮食喂养小鼠的NAFLD。
多糖通过多种机制表现出良好的抗肥胖作用,作用机制如:
◤枸杞多糖补充剂可降低厚壁菌门与拟杆菌门的比例,增加产短链脂肪酸菌,如Lacticigenium、Butyricicoccus、Lachnospiraceae_NK4A136_group数量,从而改善肥胖小鼠的肠道菌群失调。
◤桑叶多糖治疗可调节肥胖小鼠肠道菌群的组成和功能,这与增加Allobaculum、Parabacteroides、Porphyromonadaceae、Butyricimonas、Ruminococcus 的水平有关。
◤黄精多糖调节HFD喂养的肥胖大鼠的肠道微生物群结构,包括降低梭菌、肠球菌、Coprobacillus、乳球菌、Sutterella的相对丰度。
◤沙蒿多糖给药8周显著上调了屏障完整性的结肠基因,并通过增加有益细菌(双歧杆菌和Olsenella)和抑制有害细菌(Mucispirillum和幽门螺杆菌)改善了肥胖小鼠的肠道微生物微生态失调。同时,它显著富集了与促进SCFAs产生相关的碳水化合物代谢,同时显著抑制了与肥胖和肠道微生态失调相关的氨基酸代谢。
◤从海带中提取的天然多糖可通过使肠道菌群正常化来缓解小鼠HFD引起的肥胖,特别是通过增加拟杆菌目和Rikenellaceae的丰度。
◤从杏鲍菇中分离出的蘑菇多糖通过增加产生 SCFA 的肠道细菌Anaerostipes和Clostridium 的数量,在高脂饮食喂养的小鼠中表现出抗肥胖作用。
◤茶树菇多糖对HFD诱导的小鼠脂肪堆积和减肥的影响,发现脱硫弧菌减少,副拟杆菌增加,从而显著降低肥胖相关的TNF-α 和 IL-6 的水平。
◤从苦瓜中获得的多糖通过增加有益细菌(如放线菌、Coprococcus、乳酸杆菌)和减少有害细菌(变形菌和幽门螺杆菌)来改善HFD诱导的小鼠肥胖。
◤日本刺参的硫酸多糖通过富集益生菌Akkermansia、减少携带内毒素的变形杆菌和提高SCFAs含量来预防HFD诱导的小鼠肥胖。
◤ 破壁灵芝孢子多糖能逆转 HFD 喂养小鼠中许多细菌的相对丰度,特别是一些潜在的益生菌,包括Allobaculum、双歧杆菌,这与抗肥胖呈正相关。双歧杆菌、乳杆菌和阿克曼菌可促进SCFAs的产生,并抑制梭菌科、脱硫弧菌和肠球菌的丰度,这将有助于减少体重和脂质积累。
总的来说,多糖可通过作用于多个环节、调控多个疾病相关靶点来改善这三种代谢性疾病。
doi.org/10.3389/fmicb.2022.859206
高脂血症是指脂质代谢紊乱,其特征是甘油三酯 (TG)、总胆固醇 (TC) 和低密度脂蛋白浓度升高,同时高密度脂蛋白水平降低。
◤果胶多糖(高支链 RG-I,531.5 kDa)显著改善了 HFD 引起的脂质代谢异常,TG、TC、LDL-C 和游离脂肪酸水平降低。它还通过增加Roseburia、Clostridium等产生 SCFA 的细菌的数量来恢复肠道菌群失衡。
◤裙带菜多糖 ( Undaria pinnatifida )修复了高脂饮食引起的肠道微生物群改变,特别是Prevotellaceae_UCG-001,发现这与脂质代谢紊乱有关。
◤鲍鱼性腺多糖显著增加了丁酸水平,丁酸是一种重要的短链脂肪酸,它通过GPR依赖性途径抑制脂质相关基因的表达。
◤龙须菜多糖调节拟杆菌、瘤胃球菌_1和乳酸杆菌的相对丰度来增强胆固醇向BAs的转化。在遗传水平上,有人认为BA代谢的调节主要涉及CYP39A1和CYP7B1。
炎症性肠病 (IBD) 包括溃疡性结肠炎 (UC) 和克罗恩病 (CD),其特点是胃肠道持续炎症。IBD 的症状包括腹泻、腹胀、腹痛、便血、体重减轻和不适。
◤银耳多糖(TPs)通过多途径调节肠道菌群及其代谢物,改善了DSS诱导的溃疡性结肠炎。TPs可以增加Lactobacillus的丰度,从而改善色氨酸的分解代谢。这导致黄嘌呤酸、KA和吲哚衍生物(如5-羟吲哚、5-羟吲哚-3-乙酸、5-羟吲哚乙酰酸)的增加。
TPs还可以增加Romboutsia的水平,促进DCA的产生。因此,TPs可以通过影响色氨酸代谢和胆汁酸代谢来保护小鼠免受结肠炎的侵害。
◤金针菇多糖已被证明可以通过控制结肠微生物失调、增加短链脂肪酸和抑制 TLR4-NF-κB 信号通路来缓解结肠炎。能促进益生菌的生长,抑制致病菌的生长,恢复肠道稳态,缓解IBD症状。
◤竹荪多糖由59.84%的葡萄糖、23.55%的甘露糖和12.95%的半乳糖组成,已被证明可以通过增加粘蛋白和紧密连接蛋白的表达,抑制有害细菌(如γ-变形菌、变形菌、拟杆菌科、拟杆菌科和肠杆菌科)并增强有益细菌(如嗜酸乳杆菌)来改善肠道菌群组成和肠道屏障功能。
◤坛紫菜多糖通过上调紧密连接蛋白,增加粘液层及其分泌,调节肠道微生物群落,富集有益细菌,如拟杆菌、Muribaculum和乳酸杆菌,从而减轻DSS诱导的结肠损伤,从而改善结肠粘膜屏障的完整性。
◤白术多糖可以缓解在DSS诱导的溃疡性结肠炎小鼠模型炎症。白术多糖可以增加Butybacterium、Lactobacillus,同时减少Actinomyces、Akkermansia、Faecalibaculum、Verrucomicrobia、Bifidobacterium等。
肠道菌群的变化逆转了DSS引起的短链脂肪酸的减少以及色氨酸和色氨酸相关代谢物5-羟基-N-甲酰基犬尿氨酸和吲哚-3-乙酸的减少。白术多糖还剂量依赖性地逆转了LCA、DCA、缬氨酸、亮氨酸等的异常变化。
天然植物多糖治疗IBD的机制
doi.org/10.1016/j.ijbiomac.2023.126799
◤甘草多糖GPS上调乳杆菌科、S24–7、Turicibacteraceae、Verrucomicrobiaceae和双歧杆菌科的丰度,下调脱硫弧菌科、瘤胃球菌科、毛螺菌科、肠杆菌科、丹毒丝菌科的丰度。GPS能促进乳杆菌、拟杆菌和产SCFAs菌的生长繁殖,起到减轻炎症、升高IL-10水平、抑制TLR4活化、降低血浆LPS水平的作用,从而保护肠道免受LPS诱导的炎症。
◤何首乌多糖(TSG)的给药显著增加了厚壁菌门和拟杆菌门的相对丰度,同时也降低了幽门螺杆菌和拟杆菌门的属水平,改善了肠道菌群,起到治疗IBD的作用。

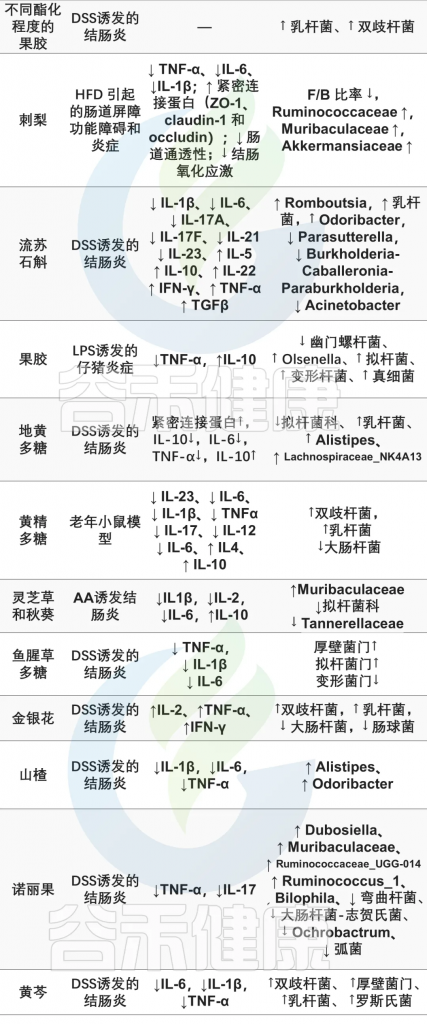
doi.org/10.3390/nu15153321
◤乳果糖通过重塑肠道菌群组成和代谢物,改善了由洛哌丁胺引起的便秘小鼠模型中的肠道水和盐代谢。具体来说,乳果糖上调了Bacteroides的丰度,并显著降低了厚壁菌门和Verrucomicrobia的水平。
此外,乳果糖减少了胆汁酸(包括CA、DCA等)、粪便中高浓度的吲哚(高浓度吲哚对细胞有毒)并增加了丙酸。
◤西洋参多糖(WQP) 可增强大鼠肠道结构的恢复,降低炎性细胞因子水平,改善短链脂肪酸 (SCFA) 水平,促进肠道菌群和肠黏膜屏障的恢复,并减轻盐酸林可霉素引起的腹泻和菌群失调等抗生素相关副作用。
◤葛根多糖( PPL )可缓解抗生素相关性腹泻引起的结肠病理改变和肠道菌群失调。
◤紫菜中提取的多糖(Nemacystus decipiens)能显著缓解小鼠抗生素相关性腹泻(AAD)的症状,并且显著增加了 Muribaculum、Lactobacillus 和 Bifidobacterium 的丰度,降低Enterobacter 、Clostridioides 的丰度。
◤茯苓多糖(PCP)通过恢复7种肠道菌菌缓解了抗生素相关性腹泻小鼠的症状,包括:Parabacteroides distasonis、Akkermansia muciniphila、Clostridium saccharolyticum、Ruminococcus gnavus、Lactobacillus salivarius、Salmonella enterica、Mucispirillum schaedleri.
适当调节免疫反应可以降低炎症反应引起的病原体入侵的风险。
结直肠癌
◤灵芝多糖在缓解结直肠癌症状方面比瓜尔胶更有效,因为它们能增加Akkermansia、结肠长度,并下调直肠癌相关基因。灵芝多糖通过动态调节肠道菌群和宿主免疫反应,已证明具有预防和治疗癌症的功能。
灵芝多糖通过调节乳酸杆菌、双歧杆菌等有益菌的相对丰度,诱导SCFAs的产生,改善肠道屏障损伤,抑制TLR4/MyD88/NF-κB信号通路,从而降低结肠炎和致癌风险。
◤绞股蓝与灵芝多糖联合使用显著提高了SCFAs产生菌的丰度,提高了丁酸和异丁酸水平,抑制了硫酸盐还原菌的丰度。
乳腺癌
◤来自灵芝破壁孢子(分子量为 3659 Da)的多糖可作为乳腺癌治疗的天然佐剂,增加细胞毒性 T 细胞和辅助性 T 细胞的数量。
灵芝孢子提取物(ESG)重塑了4T1荷瘤小鼠的肠道菌群:厚壁菌门和变形菌门的相对丰度增加,放线菌、拟杆菌门和蓝藻的相对丰度降低。
◤灵芝多糖联合紫杉醇对4T1乳腺荷瘤小鼠有抗肿瘤作用。联合治疗能显著富集拟杆菌、瘤胃球菌等5个菌属,降低脱硫弧菌和Odoribacter的丰度,平衡肠道菌群,抑制肿瘤代谢。
疲劳是一种普遍的不适感,表现为极度疲倦和力竭,通常在生理、病理或心理失衡时出现。体力劳动、心理压力、高原缺氧和长期疾病都可能引发疲劳。疲劳不仅影响日常生活,还可能导致内分泌、免疫、代谢等系统功能受损,甚至与癌症、糖尿病等严重疾病相关。此外,疲劳还与焦虑、抑郁和神经系统疾病有关。
近年来,天然多糖因其在缓解运动性疲劳中的潜在效果和较少的副作用而受到关注。研究表明,肌肉功能与肠道菌群的多样性和组成密切相关,而天然多糖如决明子、灵芝、枸杞和冬虫夏草等可通过不同机制发挥抗疲劳作用。
doi.org/10.3390/foods12163083
多糖抗疲劳机制如下:
抗疲劳多糖干预后肠道菌群的变化
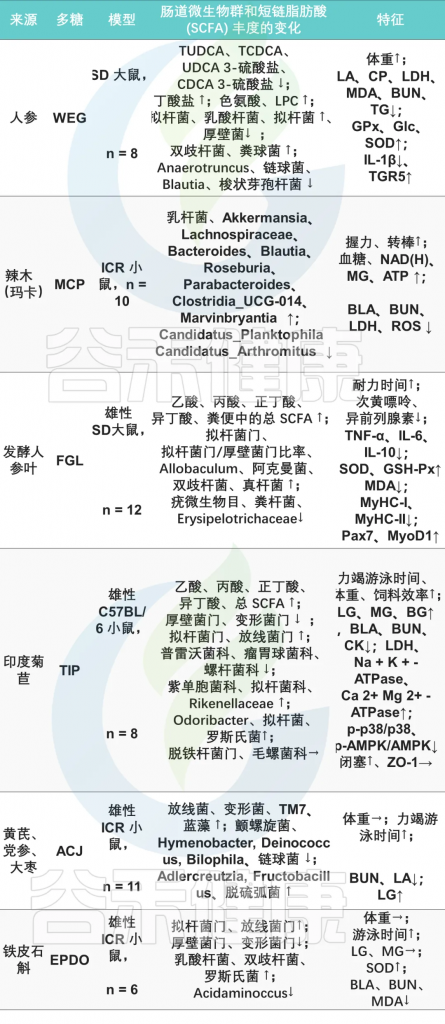
doi.org/10.3390/foods12163083
肠道-肌肉轴是肌肉与消化道之间的双向沟通,微生物可以通过微生物-肠道-肌肉轴作用于全身的肌肉。肠道微生物在膳食多糖的作用下,产生一些代谢产物(短链脂肪酸等),有些代谢产物会直接穿过肠道上皮细胞,通过血液循环直接或间接作用于肌肉组织和细胞,引起细胞发生生理生化反应,对疲劳产生一定的影响。
补充膳食多糖通过作用于肠道菌群及其代谢产物,间接激活AMPK/PGC-1α、PI3K/AKT、NF-κB、Nrf2/Keap1信号通路,调节能量代谢,降低炎症水平,增强线粒体功能和抗氧化能力,进一步维持肌肉质量和功能,从而缓解疲劳。
扩展阅读:
◤银杏叶中的一种水溶性多糖(GPS)可减轻压力引起的抑郁症并逆转肠道菌群失调。GPS 治疗可以缓解压力引起的血清素阳性和多巴胺阳性细胞密度降低。GPS 逆转了与抑郁相关的肠道菌群失调,并增加了乳杆菌的丰度,而乳杆菌已被证明是缓解抑郁的途径。
◤从秋葵中提取的多糖,发现它对抑郁小鼠的肠道菌群有明显的恢复作用,表现为厚壁菌门比例上调,拟杆菌门和放线菌门相对比例下调。这种调节有助于强化肠黏膜屏障,维持肠道免疫系统正常功能,减少肠道炎症反应,对抗抑郁有效,抑郁症小鼠的抑郁症状有所改善。用秋葵多糖治疗的小鼠体内的SCFAs显著增加,而SCFAs作为重要的通讯介质,对抗抑郁障碍有积极的影响。
◤接受金针菇多糖 (FVP)治疗的小鼠的肠道微生物组成发生显著改变,放线菌、丹毒菌和拟杆菌的丰度增加,梭菌的丰度降低,并且接受FVP治疗的小鼠的学习和记忆能力得到改善。
◤肉苁蓉多糖可以通过恢复小鼠模型中D-半乳糖诱导的衰老引起的肠道菌群稳态来抑制氧化应激和外周炎症,从而改善小鼠的认知功能。
◤从黄芪中提取的一种多糖已被证明可以通过改变糖尿病小鼠的肠道菌群来改善认知障碍。
扩展阅读:
以下是关于一些多糖的详细介绍,包括其功效,与肠道菌群的关联等,更深入地了解多糖在人体中的重要作用。
路易波士茶是什么?
路易波士茶(Rooibos)又名Aspalathus linearis,中文也有译作“路易博士茶”,取自原产于南非的一种豆科植物的茎叶。虽然带有一个茶字,但路易波士茶并不是传统意义上的茶叶。
路易波士茶因不含咖啡因、单宁含量低而受到南非人的喜爱,并在全球范围内进行商业化种植和销售。2014年,中国卫生和计划生育委员会批准路易波士茶作为新的食品原料,丰富了中国的食品和药物资源。
路易波士茶具有良好的抗氧化、抗过敏、解痉和降血糖作用。也可以预防心血管疾病、神经退行性疾病、各种癌症、骨质疏松症等。
路易波士茶多糖
一项研究从路易波士茶中分离得到均一酸性多糖(ALPs) ,水溶性多糖ALP由β-糖苷键连接,含有吡喃糖环,主要由岩藻糖、鼠李糖、阿拉伯糖和半乳糖组成。
结合RT-PCR结果推测,ALP可能通过降低Cyp2e1和Keap1的mRNA表达,增加Nrf2和HO-1的mRNA表达,激活Cyp2e1/Keap1-Nrf2-HO-1信号通路,调控下游抗氧化酶活性和炎症因子表达,减轻氧化应激损伤和炎症反应造成的损伤,从而改善急性酒精性肝损伤。
路易波士茶多糖对肠道菌群的影响
多样性
急性酒精性肝病模型对照组(MC)的物种多样性和丰富度显著降低(p≤0.05)。高剂量和低剂量ALP处理组的物种多样性和丰富度有所增加,其中高剂量组的增加更为显著。
干预后改善的菌群
ALP 干预后疣微菌丰度显著升高(p≤0.01),而脱硫杆菌丰度及F / B值均降低,但差异不显著。
肠道菌群中乳酸杆菌科的丰度与肝脏 AST 和 ALT 水平呈负相关。小鼠ALP干预后,乳酸杆菌科的丰度显著增加(p ≤ 0.05),而Rikenellaceae的丰度显著降低(p ≤ 0.05)。
ALP 显著改善了小鼠急性酒精性肝损伤中Alloprevotella和Alistipes丰度显著降低的情况( p ≤ 0.05)。
“肠-肝轴”途径
对属级别排名前20位的菌种进行了 Spearman 相关性分析。
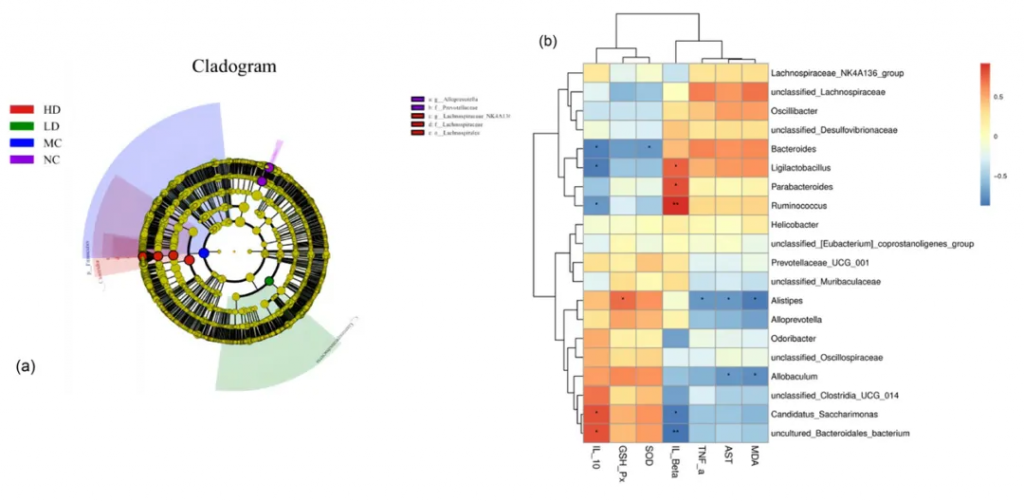
推测ALP可能通过“肠-肝轴”通路促进肠道菌群与抗氧化酶及炎症因子相互作用,从而减轻酒精性肝损伤,而上述菌群与酒精损伤标志物、抗氧化酶及炎症因子的表达均密切相关。
酒精及其代谢产物会影响肠道菌群的平衡和组成,导致肠道功能受损。这反过来又会影响肝脏健康,而肝脏健康与肠道菌群稳态密切相关。肠道和肝脏之间的相互作用被称为“肠-肝轴”通路。
地黄是玄参科地黄属植物,在我国拥有久远的药用历史,作为滋阴补肾的传统中药,也被《神农本草经》列为上品。
多糖是地黄中的主要活性成分之一。地黄多糖具有免疫调节、抗肿瘤、抗氧化、抗衰老等多种生物活性。
迄今为止,从地黄中分离纯化了20多种多糖,主要由阿拉伯糖、鼠李糖、半乳糖、葡萄糖、甘露糖、木糖、岩藻糖和半乳糖酸组成。
地黄多糖能增加DSS诱发小鼠的体质量指数和结肠长度、降低DAI评分,改善组织病理学损伤。同时,地黄多糖能阻断NF-κB信号通路,降低细胞内促炎因子表达,减轻炎症,增加紧密连接蛋白表达,维持肠道上皮屏障。
地黄多糖可能在肠道微生物作用下发酵转化为SCFAs,增加肠道中乙酸、丙酸和丁酸的含量,起到缓解IBD的作用。
拟杆菌属、乳酸杆菌属、Alistipes是导致DSS结肠炎组肠道微生物组失衡的关键细菌类型,而补充地黄多糖可以逆转这种有害变化。
五指毛桃,又叫粗叶榕(Ficus hirta Vahl),常被用作滋补品的草药成分,以其丰富的多糖含量和生物活性而闻名。
一项研究发现,FHVP-3 对肠道微生物群产生影响:
下列菌群富集:
FHVP-3 抑制了下列机会性致病菌属的丰度:
作为可发酵底物,FHVP-3 还增加了短链脂肪酸的浓度,包括乙酸盐、丙酸盐和丁酸盐。FHVP-3 对脂多糖 (LPS) 诱导的 RAW 264.7 巨噬细胞表现出显着的抗氧化活性和显着的抗炎作用。
doi.org/10.1021/acsfoodscitech.3c00626
大蒜多糖 (GP) 占大蒜干重的 75% 以上。它们的特征是具有 2,1- β – d -Fruf 主链和 2,6- β – d -Fruf 分支的果聚糖。
研究表明,大蒜多糖在调节肠道微生物群方面发挥着作用,但它们是否具有维持肠道健康的全面功能并可作为有效的益生元仍不清楚。
为了探索这一点,通过管饲法给昆明小鼠施用不同剂量的大蒜多糖(1.25-5.0g/kg 体重)和菊粉(作为阳性对照),并评估它们对肠道上皮、化学和生物屏障的影响。还使用洛哌丁胺建立了便秘模型,以研究大蒜多糖对缓解便秘的潜在影响。
施用大蒜多糖显著上调昆明小鼠小肠组织中紧密连接蛋白和粘蛋白的表达。大蒜多糖提高了盲肠丁酸含量,降低了脱硫杆菌的丰度,并降低了厚壁菌门与拟杆菌门的比例(F/B)。大蒜多糖还促进了 Bacteroides acidifaciens、Clostridium saccharogumia的生长。
Tax4Fun 功能预测表明,大蒜多糖具有预防人类疾病的潜力,可降低胰岛素抵抗、传染病和耐药性的风险。
大蒜多糖还通过增强小肠转运、软化粪便稠度、加速排便和促进兴奋性神经递质的释放,在缓解洛哌丁胺引起的便秘症状方面表现出有益作用。
多年来,槐耳 (Trametes robiniophila Murr) 一直被用于药物治疗。槐耳含有多种成分,包括多糖、蛋白质、酮和生物碱,其中蛋白聚糖和多糖是主要的生物活性成分。
槐耳提取物具有免疫调节活性,并可通过激活自噬、抑制铁死亡、抑制内质网应激等过程对细胞发挥保护作用。研究表明,槐耳水提取物可通过抑制NLRP3炎症囊泡活化,减轻肠道屏障损伤和炎症反应,并抑制DSS和氧化偶氮甲烷 (AOM) 联合诱导的结肠肿瘤形成。
可缓解 DSS 引起的肠道菌群紊乱
一项小鼠研究显示,槐耳多糖干预显著逆转了 DSS 引起的Muribaculaceae_unclassified、Anaerotruncus、Ruminococcaceae_unclassified丰度的下降以及Escherichia-Shigella丰度的增加( p < 0.05)。
其中,Muribaculaceae_unclassified是健康人中发现的肠道微生物,参与丁酸代谢和色氨酸代谢,可产生对人体有益的短链脂肪酸。
相关性分析,Muribaculaceae_unclassified与结肠长度、SOD 和 T-AOC 呈正相关,而与 DAI 评分以及炎症和氧化指标呈负相关。
Anaerotruncus与结肠长度、SOD 和 T-AOC 呈正相关,但与炎症标志物 LPS、MDA 和 MPO 呈负相关。
黄芩的根通常用作药物,用于清热利湿、泻火解毒。多糖是黄芩的最重要成分之一。
一种来自黄芩的多糖通过抑制 NF-κB 信号传导和NLRP3 炎症小体活化来改善溃疡性结肠炎。在多糖的分离和纯化过程中,研究人员还获得了另一种名为 SP2-1 的均质多糖。SP2-1由甘露糖、核糖、鼠李糖、葡萄糖醛酸、葡萄糖、木糖、阿拉伯糖和岩藻糖组成。
研究人员发现其对肠道菌群紊乱、肠道屏障改善以及短链脂肪酸产生影响。
在UC患者中,SP2-1显著抑制了促炎性细胞因子IL-6,IL-1β和TNF-α。
溃疡性结肠炎患者的屏障完整性被破坏,TJ 蛋白的表达发生改变,SP2-1增加小鼠TJ蛋白的表达,修复肠道屏障。
SP2-1对肠道菌群的影响
SP2-1组的粪便微生物群多样性明显高于DSS组。
临床上,溃疡性结肠炎患者的双歧杆菌和乳酸杆菌的丰度降低。与模型组相比,SP2-1 组的双歧杆菌、乳酸杆菌和Roseburia的水平提高。
而拟杆菌和葡萄球菌的种群受到抑制。肠道菌群中存在过量的拟杆菌和葡萄球菌对肠道免疫系统有害。
Roseburia 通过调节调节性 T 细胞的发育和分化、增加抗炎细胞因子的分泌和抑制促炎细胞因子的产生来缓解UC。
枸杞多糖(简称LBPs)是从枸杞中提取的一类多糖物质。枸杞是一种多年生灌木,属于茄科,枸杞多糖因其多样的药理活性和生理功能而受到越来越多的关注。
枸杞多糖 (LBPs)作为最重要的生物活性分子,可通过肠道微生物参与有益作用,包括调节代谢、降血糖、神经保护、抗衰老、保护各种器官免受氧化应激相关疾病的侵害。
枸杞多糖对肠道菌群的影响
癌症
一般来说,抗癌化疗药物除了会诱导癌细胞凋亡外,还会对肠道菌群产生不利影响,主要表现在肠绒毛缩短,乳酸杆菌和肠球菌丰度下降,节段丝状菌丰度增加。而枸杞多糖治疗可通过增加有益菌相对丰度来改善肠道环境和免疫功能,逆转环磷酰胺引起的有害菌(瘤胃拟杆菌科、Longibraceae、脱硫弧菌和厌氧拟杆菌科)相对丰度的增加。
还发现毛螺菌科、瘤胃菌科、脱铁菌科、脱硫弧菌科、Aneoplasmataceae与细胞因子IL-2、IL-6、IL-1β、TNF-α、IFN呈负相关。因此,主要肠道菌群的相对丰度可能与免疫调节有关。
厚壁菌门与拟杆菌门(F/B)比例的变化与许多疾病状态有关,它被视为菌群失调的重要指标,有助于了解肝脏和代谢疾病的发展。枸杞多糖可降低高脂饮食大鼠的 F/B 比,表明补充枸杞多糖有助于调节肠道菌群失调。
doi.org/10.1080/10408398.2022.2128037
神经系统
枸杞多糖可通过调节肠道菌群-肠-脑轴的神经免疫通路,对中枢神经系统产生多方面的保护作用。枸杞多糖可改善菌群失调、肠道屏障受损等问题,并通过抑制细胞凋亡、促进自噬等机制发挥神经保护效应。
肝脏
枸杞多糖能够影响NAFLD患者的肠道菌群组成、肠道屏障及肝脏炎症。
代谢(肥胖、糖尿病)
肥胖个体的研究中,肠道内F/B比例较高,因此推测肠道内F/B比例与肥胖呈显著正相关。
后续研究发现,LBPs可通过调节肠道菌群组成和短链脂肪酸生成来改善肥胖。
枸杞多糖可作为2型糖尿病的潜在辅助药物。
LBPs能调节肠道菌群,激活大鼠肠黏膜TLR2+上皮细胞γδT细胞,增强肠道屏障功能,改善糖尿病。此外,LBPs能明显降低血浆中促炎性细胞因子IL-1β、IL-6、IL-17A和TNF-α,而抗炎性细胞因子IL-10水平在糖尿病大鼠中有所升高。
哮喘
枸杞多糖还可以通过直接或间接地改变肠道菌群,参与炎症介质的调控,从而改善肺功能和过敏性哮喘症状。
肠道菌群测序分析显示,LBPs能够促进哮喘小鼠肠道中乳酸杆菌和双歧杆菌增加,并降低厚壁菌门和放线菌水平,通过肠道介导缓解哮喘。
岩藻糖(Fucose),参与构成肠上皮细胞(IEC)顶端表达的聚糖,并介导肠道中的许多生物过程,尤其是宿主-微生物相互作用。
释放的岩藻糖可被微生物用作膳食聚糖、能量来源或合成结构蛋白。肠道中岩藻糖的变化影响微生物群的定植。
岩藻多糖
岩藻多糖是一种含有岩藻糖和硫酸基团的多糖,可改善糖尿病肾病。
一项小鼠研究发现,岩藻多糖可显著改善肾小球滤过率高滤过和肾纤维化,其机制与短链脂肪酸产生菌富集、增加盲肠内乙酸浓度、提高肾脏ATP水平以及改善线粒体功能障碍有关。此外,岩藻多糖还可通过抑制MAPKs通路来改善肾脏炎症和纤维化。总之,岩藻多糖可通过改善线粒体氧化应激和抑制MAPKs通路,靶向肠道菌群-线粒体轴,改善早期糖尿病肾病。
桑叶的药用功能最早在2000多年前的汉代被发现,并记载于《神农本草经》。明代李时珍在《本草纲目》中对桑叶的药用功效有更详细的描述,包括活血化瘀、祛风、清热解毒等功能。桑叶已被列入国家卫生健康委员会公布的食药同源资源名单。
桑叶多糖(Mulberry Leaves Polysaccharides,MLPs)是从桑树(Morus alba L.)叶片中提取的一种植物多糖。它们是桑叶中主要的活性成分之一,由多种单糖组成,主要包括木糖、阿拉伯糖、果糖、半乳糖、葡萄糖、甘露糖等。
桑叶多糖对人体的影响
桑叶多糖具有多种生物学活性,包括降低血糖、抗氧化、免疫调节、抗肿瘤、抗菌、抗凝和调节肠道菌群等。这些活性使得MLPs在医药和食品领域具有广泛的应用前景。并且安全、有效、低毒、副作用小。
doi.org/10.1016/j.ijbiomac.2023.128669
桑叶多糖对肠道菌群的影响
桑叶多糖通过调节肠道菌群的平衡,进而对人体的健康产生积极的影响。以下是桑叶多糖影响的肠道菌群及其变化情况:

桑叶多糖能够调节短链脂肪酸和肠道菌群的相对丰度,降低真细菌与过敏性细菌的比例,从而改善肠道屏障功能。
沙棘果实在藏族食品和药物中已有数千年的传统。沙棘多糖 (SP) 是沙棘果实中的主要功能成分之一。
对高脂饮食诱导的肥胖小鼠:沙棘多糖治疗提高了 p-AMPKα 和 PPARα 蛋白的表达,刺激了小鼠肝脏中 ACC1 的磷酸化,并抑制了 FAS、PPARγ 和 CD36 的蛋白表达。
沙棘多糖上调Muribaculaceae_unclassified、双歧杆菌、Rikenellaceae_RC9_gut_group、Alistipes、Bacteroides的比例,并下调Lactobacillus、 Firmicutes_unclassified 、Dubosiella Bilophila、 Streptococcus 的比例,重组了HFD诱导的肥胖小鼠的肠道微生物群。
此外,粪便中的微生物代谢物短链脂肪酸 (SCFAs) 的产生也有所增加。此外,相关性分析结果表明沙棘多糖的肥胖改善作用与粪便中的SCFAs水平高度相关。因此,沙棘多糖对肝脏脂质代谢的调节可能是由于肠道微生物群的变化和SCFAs产生量的增加。这些结果表明,沙棘多糖可以通过调节肠-肝轴发挥改善肥胖的潜在营养保健作用。
蘑菇多糖是一类存在于蘑菇中的生物活性多糖,它们包括但不限于几丁质、甘露聚糖、半乳糖聚糖、木聚糖、葡聚糖、云芝多糖、灵芝多糖、半纤维素。这些多糖在蘑菇细胞壁中含量丰富,赋予蘑菇独特的结构和生物活性。
蘑菇多糖的功效
蘑菇多糖对人体具有多种潜在的健康益处。它们可以增强免疫系统、具有抗肿瘤活性、调节肠道菌群、抗氧化、抗糖尿病、抗衰老作用。
蘑菇多糖对肠道菌群的影响
促进益生菌生长
蘑菇多糖通过选择性地促进益生菌的生长,增强肠道健康。例如,灵芝和茯苓中的多糖被发现可以增加有益细菌的数量,这些细菌可以对抗肥胖、产生短链脂肪酸和乳酸。香菇中的多糖也显示出对嗜酸乳杆菌(Lactobacillus acidophilus)有促进作用。
抑制病原菌
蘑菇多糖能够通过增强肠道屏障功能和促进益生菌的生长来间接抑制病原菌。双孢蘑菇中的多糖已被证明可以限制大肠杆菌的生长。
增强肠道屏障功能
蘑菇多糖通过增强肠道上皮细胞的功能,提高肠道屏障的完整性,减少有害物质的渗透。云芝(Trametes versicolor)中的多糖肽PSK和PSP能够调节肠道菌群,增加有益菌双歧杆菌和乳杆菌的数量,同时减少有害菌如梭状芽孢杆菌和金黄色葡萄球菌。在降低腹泻、艰难梭菌感染、炎症性肠病等方面发挥作用。
调节免疫反应
蘑菇多糖通过激活肠道相关淋巴组织,增强机体的免疫反应。灵芝多糖能刺激和增加免疫细胞如自然杀伤细胞、T淋巴细胞和巨噬细胞的数量。
产生短链脂肪酸
蘑菇多糖在肠道发酵过程中产生短链脂肪酸,这些物质对维持肠道健康和调节宿主代谢具有重要作用。蚝菇(Pleurotus ostreatus)中的β-葡聚糖衍生物能够诱导前列腺癌细胞的凋亡,并且显示出免疫调节、巨噬细胞激活、抗肿瘤和免疫刺激活性。

多糖的多样性和复杂性使其在人体内的作用千变万化,它们能够通过与肠道菌群的互动,从调节免疫功能到改善代谢性疾病等。
然而,利用天然多糖通过肠道菌群治疗疾病仍存在一些限制和挑战。对肠道菌群和多糖之间相互作用的全面了解需要进一步研究,由于大多数研究都是在动物身上进行的,因此开展研究多糖-微生物组-疾病相互作用的临床试验并实现临床转化至关重要。
幸运的是,随着生命科学领域新兴技术的发展,我们有了更多的工具来揭示这些复杂问题。高通量测序技术、多组学技术、人工智能和大数据分析的交叉融合,为研究多糖和肠道菌群的相互作用提供了强大的技术支持,推动了这一领域的快速发展。
此外,多糖与肠道菌群之间的相互作用不仅揭示了多糖的生物活性,也突显了肠道菌群对健康的重要贡献。多糖与肠道菌群的相互作用为我们提供了一个全新的视角,了解个体的肠道菌群组成,不仅有助于我们理解自身的健康状况,更为个性化的营养和健康管理提供了科学依据。肠道菌群检测可以揭示个体对多糖等营养成分的响应差异,从而为制定个性化的饮食和治疗计划提供指导。
注:本账号内容仅作交流参考,不作为诊断及医疗依据。
主要参考文献
Xue H, Mei CF, Wang FY, Tang XD. Relationship among Chinese herb polysaccharide (CHP), gut microbiota, and chronic diarrhea and impact of CHP on chronic diarrhea. Food Sci Nutr. 2023 Aug 6;11(10):5837-5855.
Xu X, Wang L, Zhang K, Zhang Y, Fan G. Managing metabolic diseases: The roles and therapeutic prospects of herb-derived polysaccharides. Biomed Pharmacother. 2023 May;161:114538
Zhang D, Liu J, Cheng H, Wang H, Tan Y, Feng W, Peng C. Interactions between polysaccharides and gut microbiota: A metabolomic and microbial review. Food Res Int. 2022 Oct;160:111653.
Chen R, Zhou X, Deng Q, Yang M, Li S, Zhang Q, Sun Y, Chen H. Extraction, structural characterization and biological activities of polysaccharides from mulberry leaves: A review. Int J Biol Macromol. 2024 Feb;257(Pt 2):128669.
Lan Y, Sun Q, Ma Z, Peng J, Zhang M, Wang C, Zhang X, Yan X, Chang L, Hou X, Qiao R, Mulati A, Zhou Y, Zhang Q, Liu Z, Liu X. Seabuckthorn polysaccharide ameliorates high-fat diet-induced obesity by gut microbiota-SCFAs-liver axis. Food Funct. 2022 Mar 7;13(5):2925-2937.
Feng Y, Song Y, Zhou J, Duan Y, Kong T, Ma H, Zhang H. Recent progress of Lycium barbarum polysaccharides on intestinal microbiota, microbial metabolites and health: a review. Crit Rev Food Sci Nutr. 2024;64(10):2917-2940.
Cui L, Guan X, Ding W, Luo Y, Wang W, Bu W, Song J, Tan X, Sun E, Ning Q, Liu G, Jia X, Feng L. Scutellaria baicalensis Georgi polysaccharide ameliorates DSS-induced ulcerative colitis by improving intestinal barrier function and modulating gut microbiota. Int J Biol Macromol. 2021 Jan 1;166:1035-1045.
Tang YF, Xie WY, Wu HY, Guo HX, Wei FH, Ren WZ, Gao W, Yuan B. Huaier Polysaccharide Alleviates Dextran Sulphate Sodium Salt-Induced Colitis by Inhibiting Inflammation and Oxidative Stress, Maintaining the Intestinal Barrier, and Modulating Gut Microbiota. Nutrients. 2024 Apr 30;16(9):1368.
Zhao Q, Jiang Y, Zhao Q, Patrick Manzi H, Su L, Liu D, Huang X, Long D, Tang Z, Zhang Y. The benefits of edible mushroom polysaccharides for health and their influence on gut microbiota: a review. Front Nutr. 2023 Jul 6;10:1213010.
Álvarez-Mercado AI, Plaza-Diaz J. Dietary Polysaccharides as Modulators of the Gut Microbiota Ecosystem: An Update on Their Impact on Health. Nutrients. 2022 Oct 3;14(19):4116.
Tang M, Cheng L, Liu Y, Wu Z, Zhang X, Luo S. Plant Polysaccharides Modulate Immune Function via the Gut Microbiome and May Have Potential in COVID-19 Therapy. Molecules. 2022 Apr 26;27(9):2773.
Sun CY, Zheng ZL, Chen CW, Lu BW, Liu D. Targeting Gut Microbiota With Natural Polysaccharides: Effective Interventions Against High-Fat Diet-Induced Metabolic Diseases. Front Microbiol. 2022 Mar 15;13:859206.
Gan L, Wang J, Guo Y. Polysaccharides influence human health via microbiota-dependent and -independent pathways. Front Nutr. 2022 Nov 9;9:1030063.
Chen J, Gao Y, Zhang Y, Wang M. Research progress in the treatment of inflammatory bowel disease with natural polysaccharides and related structure-activity relationships. Food Funct. 2024 Jun 4;15(11):5680-5702.
Chen P , Hei M , Kong L , Liu Y , Yang Y , Mu H , Zhang X , Zhao S , Duan J . One water-soluble polysaccharide from Ginkgo biloba leaves with antidepressant activities via modulation of the gut microbiome. Food Funct. 2019 Dec 11;10(12):8161-8171.
Wang, A.; Liu, Y.; Zeng, S.; Liu, Y.; Li, W.; Wu, D.; Wu, X.; Zou, L.; Chen, H. Dietary Plant Polysaccharides for Cancer Prevention: Role of Immune Cells and Gut Microbiota, Challenges and Perspectives. Nutrients 2023, 15, 3019.
Zhou, Y.; Chu, Z.; Luo, Y.; Yang, F.; Cao, F.; Luo, F.; Lin, Q. Dietary Polysaccharides Exert Anti-Fatigue Functions via the Gut-Muscle Axis: Advances and Prospectives. Foods 2023, 12, 3083
Shen, Y.; Song, M.; Wu, S.; Zhao, H.; Zhang, Y. Plant-Based Dietary Fibers and Polysaccharides as Modulators of Gut Microbiota in Intestinal and Lung Inflammation: Current State and Challenges. Nutrients 2023, 15, 3321
Lv H, Jia H, Cai W, Cao R, Xue C, Dong N. Rehmannia glutinosa polysaccharides attenuates colitis via reshaping gut microbiota and short-chain fatty acid production. J Sci Food Agric. 2023 Jun;103(8):3926-3938.
谷禾健康
曾经关节痛只是老年人身上常见的病,没想到这事竟然发生在不到30岁的小白身上。
有一天,小白突然感觉手腕使不上劲儿,以为只是平时电脑鼠标用多了,贴个膏药就好了…
但某一天,手指关节又开始疼痛并且肿起来了,可小白认为只是哪里撞到了没当回事儿…
直到有一天晚上,小白的膝关节突然觉得疼到不行,连翻身都不容易,第二天走路都困难,她终于决定去医院看看。
在经历各项指标检查后,医生告诉她,是类风湿性关节炎…… 这对小白来说犹如晴天霹雳……

类风湿性关节炎并不是专属于老年人,它可以发生在任何年龄段。其中高发年龄在40-60岁,而且女性是男性的2-3倍。
且类风湿性关节炎很“狡猾”,往往前期不容易被发现,且容易与其他疾病混淆。
但随着时间的推移,病情会逐渐恶化。未经及时发现、规范治疗的患者可能会出现关节骨质的破坏,最终导致关节畸形和功能丧失。
本文我们来详细了解一下类风湿性关节炎及其与肠道菌群之间的关联,以及针对类风湿性关节炎的建议。
类风湿性关节炎(RA)是一种自身免疫性疾病,可导致关节慢性炎症。
自身免疫性疾病是人体免疫系统攻击自身组织时产生的疾病。免疫系统包含一个由细胞和抗体组成的复杂组织,通常设计用于“寻找并消灭”身体的入侵的病原体。患有自身免疫性疾病的人血液中含有针对自身组织的抗体和免疫细胞,这些抗体和免疫细胞可能与炎症有关。
该疾病也可导致关节外炎症和其他器官损伤,可以影响身体的多个器官。
始于16岁以下人群的类风湿性关节炎称为幼年特发性关节炎或JIA。
疲劳、食欲不振、低热、肌肉和关节疼痛、关节发红、关节肿胀、关节压痛、关节热、关节畸形、类风湿结节、僵硬、失去关节活动范围、关节功能丧失、一瘸一拐……
类风湿关节炎患者以上症状都可能出现。
除此之外,类风湿关节炎患者也可能会伴有抑郁症、贫血、挫败感,社交退缩等情况。
类风湿关节炎可分为几个阶段(以下分类是由美国风湿病学会开发的分类系统。这个系统可以帮助医学专业人员根据软骨、韧带和骨骼对类风湿性关节炎的严重程度进行分类)
第一阶段(早期RA)
虽然可能有骨质变薄的迹象,但在X光片上没有发现损伤
第二阶段(中度进行)
在X光片上,关节周围骨质变薄的证据,有或没有轻微的骨质损伤
可能有轻微的软骨损伤
关节活动可能受到限制;未观察到关节畸形
邻近肌肉萎缩
可能存在关节周围软组织异常
第三阶段(严重进展)
在X光片上,关节周围有软骨和骨损伤以及骨变薄的证据
关节畸形,关节无永久性硬化或固定
广泛性肌肉萎缩
可能存在关节周围软组织异常
第四阶段(终末期进展)
在x光片上,关节周围有软骨和骨骼损伤和骨质疏松
关节畸形伴关节永久固定(称为强直)
广泛性肌肉萎缩
可能存在关节周围软组织异常
类风湿性关节炎患者的功能状态分类如下:
一级:完全能够进行日常生活的日常活动
二级:能够进行日常的自我护理和工作活动,但工作以外的活动有限(如运动、家务活)
三级:能够进行日常的自我护理活动,但工作和其他活动都有限
四级:无法进行日常自我护理、工作和其他活动
具体症状:
早期类风湿性关节炎的症状可能很轻微。可能是疼痛和关节僵硬,尤其是在早晨。
类风湿性关节炎患者的手部症状包括难以完成简单的日常生活任务,如转动门把手和打开罐子。
脚的小关节也会受累,导致行走疼痛,尤其是在早上起床后。
当只有一个关节受累时,关节炎可以模拟由其他形式的关节炎(如痛风或关节感染)引起的关节炎症。
在发作期间,关节通常会变得红肿、温热、疼痛。这是因为关节(滑膜)的衬里组织发炎,导致产生过多的关节液(滑液)。滑膜也因炎症(滑膜炎)而增厚。
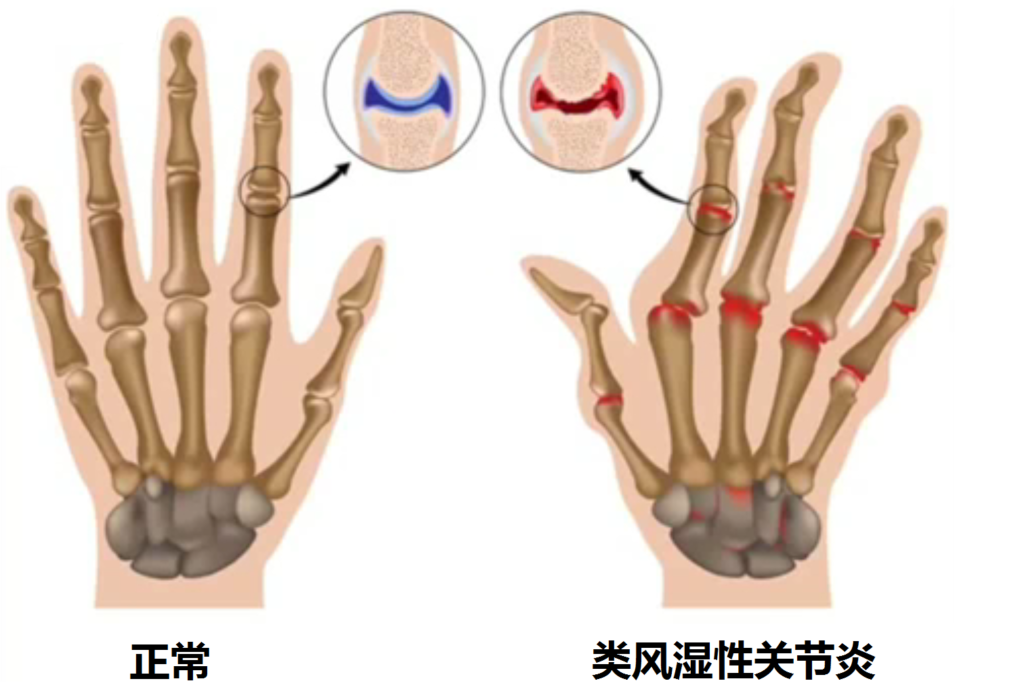
慢性炎症可导致身体组织受损,包括软骨和骨骼。这会导致软骨丧失、骨骼和肌肉的侵蚀和虚弱,导致关节畸形、运动范围丧失、功能丧失。
罕见类风湿性关节炎甚至会影响负责收紧声带以改变声调的关节,即环杓关节。当关节发炎时,会导致声音嘶哑。
类风湿性关节炎患儿的症状包括跛行、易怒、哭泣和食欲不振。
类风湿性关节炎是一种多因素疾病,宿主和环境之间存在复杂的相互作用,决定了疾病易感性、持久性和严重程度的总体风险。
发生类风湿关节炎的危险因素一般可分为宿主相关因素和环境相关因素(下图)。
Romão VC, et al., Front Med (Lausanne). 2021
与类风湿关节炎发生相关的宿主因素可进一步归类为遗传因素,表观遗传,激素、生殖和神经内分泌,共病宿主因素。环境风险因素包括吸烟和其他空气接触,微生物群和感染因子,饮食,社会经济因素。
类风湿关节炎不是关节的问题吗,怎么会和肠道有关联?
在20世纪90年代早期,医生已经检测到关节炎患者的微观肠道炎症,从而发现了胃肠道炎症和脊椎关节炎(SpA)之间的联系。
肠道失调的存在可能导致SpA中肠道炎症的发生,于是科学家想到类似的机制也可能在RA中起作用。
肠道炎症
对一部分早期RA患者进行的研究表明,几乎所有患者都存在亚临床肠道炎症。这些患者肠道炎症的特征是浸润性单核细胞、T细胞、B细胞和CD68+巨噬细胞数量增加,以及淋巴滤泡的存在。这些组织学发现表明,早期类风湿关节炎患者的肠道发生慢性炎症过程,这与类风湿关节炎是一种全身性疾病的概念一致。
为了支持这一观点,来自韩国的一项基于人群的研究强调了IBD和RA之间的显著相关性(OR3.31). 这些数据强化了IBD和RA之间潜在共同致病机制的观点,并强调了肠道在RA发病中的潜在作用。
肠道屏障功能改变
小肠的主要功能是从食物中吸收营养和矿物质。虽然暴露于外界的身体相当大的表面积,但肠上皮细胞形成了一个动态的物理屏障,通过细胞旁途径严格控制抗原运输。
在存在炎症的情况下,肠道屏障功能的改变以及伴随的肠道通透性和细菌易位的增加可促进遗传易感性宿主的IBD和自身免疫。
肠道屏障破坏机制
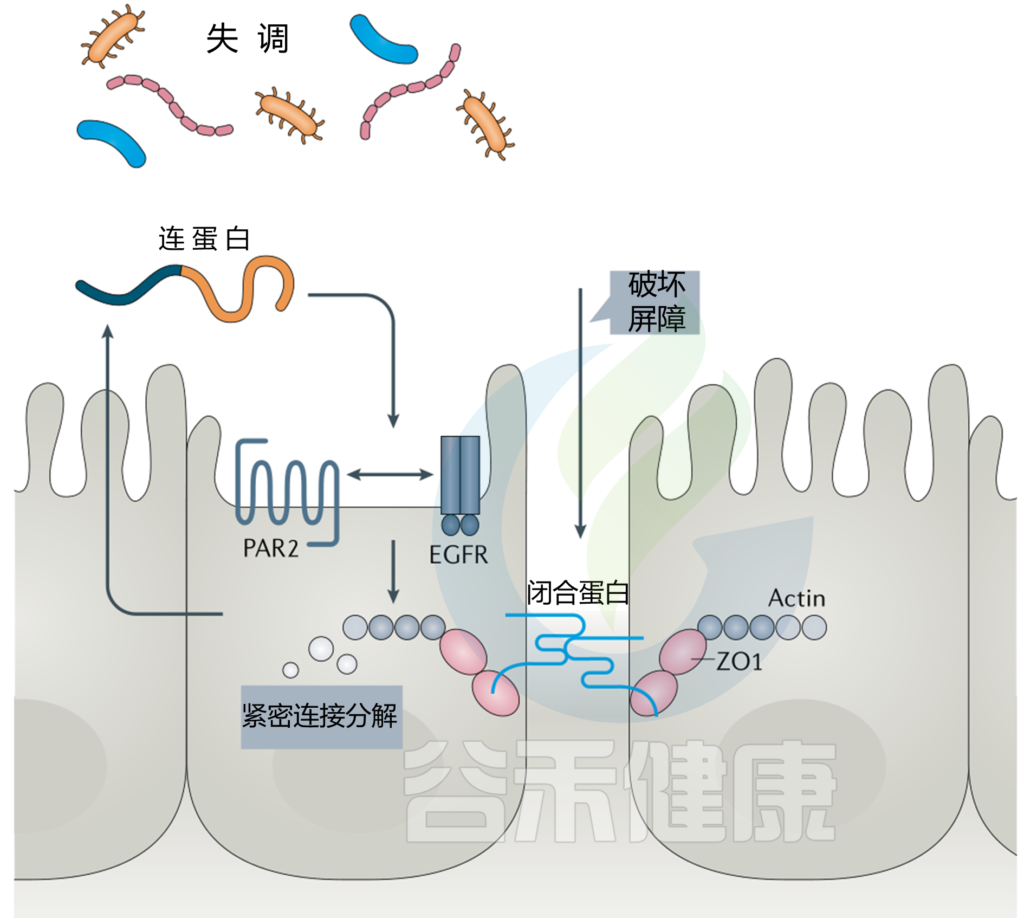
Zaiss MM, et al.,Nat Rev Rheumatol. 2021
肠道屏障破坏的主要机制似乎是通过增加连蛋白(zonulin)的产生,zonulin是肠上皮细胞紧密连接完整性的主要调节因子。
上皮细胞间紧密连接是调节肠屏障细胞旁转运的关键结构。zonulin浓度的增加与一种可能导致免疫介导疾病的机制有关。
Zonulin分泌依赖于衔接蛋白MYD88,通过导致蛋白质ZO1和闭塞素从紧密连接复合体中分解,从而降低肠屏障功能。
zonulin家族的肽可能是RA患者肠道失调与肠道炎症和屏障功能降低之间的良好候选物。
有趣的是,两项研究调查了无麸质纯素饮食(预计可降低zonulin浓度)与纯素饮食对RA的影响,发现接受无麸质饮食的患者炎症标志物有显著改善。事实上,之前的报道表明,麦胶蛋白(谷蛋白的一种成分)诱导zonulin的释放,zonulin通过与趋化因子受体CXCR3结合,增加小肠的通透性,并放大小肠的损伤。
此外,在RA发病前以及新发RA患者(同时伴有紧密连接蛋白下调)的肠道中,zonulin家族肽的表达增加,以及细菌可能影响zonulin表达。该结果支持RA患者肠道生态失调和肠道通透性改变之间的潜在联系。
肠道通透性改变
目前在已确诊的类风湿性关节炎患者和在类风湿性关节炎前期的患者中已观察到肠道通透性改变的迹象。
在一组法国女性患者中,在没有明确胃肠道疾病的情况下有慢性腹泻史与RA发病风险增加相关(HR 1.70),这表明在RA发病之前,肠道内稳态紊乱可能存在多年。
也就是说,隐匿的慢性改变可能先于关节炎的发病,在临床环境中可能诊断不足。
与无活动性关节疾病的RA患者相比,活动性疾病的RA患者肠道通透性异常。然而,在其中一些研究中,活动性RA患者肠道通透性改变的存在与非甾体抗炎药的摄入相混淆,非甾体抗炎药也影响肠道通透性。因此很难判断RA对肠道通透性的影响。
总的来说,目前的数据表明,肠道通透性的改变可能是RA固有的,并可能受到肠道失调的调节,然后因使用NSAIDs(非甾体抗炎药)而恶化。
目前可用的有限数据并不完全支持RA患者存在肠道炎症和肠道屏障破坏,需要进一步的研究来明确类风湿关节炎亚临床肠道炎症的确切患病率。还需要进一步的研究来确定出现肠道炎症症状的患者的疾病表型以及决定肠道受累的致病因素,如环境、遗传背景、性别和年龄等。
上述数据表明,肠道炎症以及屏障功能的降低(这两种情况都在RA患者中观察到)促进了临床RA的发病。屏障功能紊乱、肠道炎症和关节炎之间的联系可能由两种非相互排斥的途径介导。
接下来简要讨论这两种途径。
类风湿性关节炎的肠-关节轴
Zaiss MM, et al.,Nat Rev Rheumatol. 2021
自身抗体的产生
在临床RA发病前数年,血清中发现了大量的自身反应性IgA抗体。鉴于粘膜表面是IgA抗体产生的主要部位,这些结果表明与RA相关的自身抗体最初可能在粘膜表面(如肠道)产生。
一项2020年队列研究表明,早在RA诊断前14年和检测到IgG类风湿因子增加前几年,IgA类风湿因子浓度与配对对照组显著不同。此外,IgA ACPAs的出现发生在临床RA发病前6年,提示该疾病临床前阶段的粘膜过程有助于向临床RA的发病过渡。RA患者的肠组织样本也显示识别食物抗原的IgA和IgM抗体浓度增加。
总的来说,这些数据指向粘膜表面,特别是肠道,作为对外部刺激(包括食物源性抗原和易感个体自身抗原)产生抗体的场所。
促炎免疫细胞可进入全身部位和关节
肠道屏障功能的改变和肠道通透性的增加与肠道组织中的炎性免疫细胞可能进入关节有关。
一些对人类的研究支持细胞从肠道运输到关节的一般观点。例如,肠道激活的B细胞有效地粘附在肠道和滑膜的高内皮微静脉上,但不粘附在外周淋巴结的高内皮微静脉上,这表明来自肠道的免疫细胞可能进入关节。
此外,在SpA患者的关节和肠道中发现了相同的T细胞克隆,RA患者的滑膜中含有表达肠道归巢受体αEβ7整合素的T细胞。
在类风湿性关节炎患者中,几种粘膜来源的固有免疫细胞被扩增和激活,支持肠-关节细胞轴的观点。
类风湿关节炎中微生物群、肠上皮和免疫细胞之间的相互作用

Zaiss MM, et al.,Nat Rev Rheumatol. 2021
1)病理生物通过趋化因子受体CXCR3和髓样分化初级反应88(MYD88)介导的信号,增加zonulin的产生,从而促进上皮细胞的炎症反应
2)Zonulin导致上皮紧密连接紊乱,从而增加粘膜下微生物和微生物产物(如ATP和游离脂肪酸受体2(FFAR2)激动剂)的外显率
3)树突状细胞对增加的ATP产生反应,使T细胞成为T辅助细胞17(TH17),产生IL-17。细菌源性FFAR2激动剂和其他细菌产物直接激活第3组固有淋巴细胞(ILC3s),从而诱导IL-22分泌。微生物还直接激活固有免疫细胞,如巨噬细胞和固有样细胞,如粘膜相关不变T(MAIT)细胞,触发一个放大级联反应,导致肠道炎症以及T细胞、巨噬细胞和ILC3s的局部激活和分化
4)TH17细胞、ILC3s和MAIT细胞可以迁移到血液中,增加了这些细胞将炎症转移到关节的可能性
RA 的临床表现源于各种因素之间的复杂相互作用,包括自身抗体和先天性和适应性免疫系统的信号转导途径。
在 RA 患者中,关节组织通常被 T 细胞、B 细胞和巨噬细胞等免疫细胞浸润,产生多种促炎细胞因子,促进炎症并最终导致组织破坏。
而在整个生命过程中,肠道菌群在免疫系统的诱导、教育和功能以及个体对自身抗原的反应中起着基础性作用。
在RA患者的肠道、口腔、肺中已证实存在菌群失调,为该疾病发病机制的粘膜基础假说提供了证据。由于黏膜被特定的微生物群所定植,这些部位的菌群失调可能改变局部的,甚至可能是全身的免疫反应,从而促成了RA的发病机制。
接下来的章节详细介绍类风湿性关节炎与菌群之间的关联。
RA 患者的肠道微生物群 (GM) 的组成和功能发生了改变,与健康对照相比,微生物多样性显著下降。
Faecalibacterium、Flavobacterium、双歧杆菌、拟杆菌、嗜血杆菌Haemophilus减少;乳酸杆菌、普雷沃氏菌、梭状芽胞杆菌增多。
类风湿性关节炎患者微生物群组成的统计显著变化
Zaiss MM, et al.,Nat Rev Rheumatol. 2021
许多研究中都提到了与类风湿性关节炎相关的一个重要菌——普雷沃氏菌属,因此有必要单独介绍一下。
普雷沃氏菌属
来自《柳叶刀》风湿病学的一项涵盖6776名受试者(包括 2686 名 RA 患者)的横截面研究,通过逻辑回归对 RA 多基因风险评分(PRS) 进行分析,结果表明普雷沃菌属与RA的遗传风险显著相关,普雷沃菌属还与RA高危受试者(RA患者的直系亲属)的临床前期症状相关。
普雷沃氏菌属可能通过介导 Th17 细胞的炎症反应与 RA 相关。在RA 发病前的临床前阶段,普雷沃菌的富集表明肠道菌群的失调在RA 的发展中发挥着主要作用。
RA患者(新发和慢性)要么对Pc-p27或P.copri产生IgA样抗体反应,与TH17细胞因子的产生和ACPA的发生相关,要么表现出与滑液中存在Prevotella DNA相关的抗Prevotella IgG抗体,Prevotella特异性TH1细胞反应和较低浓度的ACPA。
肠道菌群和免疫系统相互依存
肠道菌群可能调节肠道免疫耐受机制,因为它影响结肠调节性T细胞(Treg)的数量和功能。Treg通过分泌抗炎细胞因子抑制效应T细胞的不适当激活。
另一方面,粘膜免疫系统在发展和维持健康的肠道菌群中起着至关重要的作用。由于这种相互依存的关系,肠道菌群失调(肠道菌群的成分和功能改变)可能会影响宿主对类风湿性关节炎的易感性。
肠道菌群参与类风湿性关节炎机制
肠道上皮、免疫系统和共生细菌之间的串扰是启动全身炎症反应的关键。抗炎和促炎细胞因子之间的失衡,包括白细胞介素(IL)-1β、肿瘤坏死因子、干扰素(IFN)-γ、白细胞介素-6、白细胞介素-12和白细胞介素-17,在参与RA发病机制的炎症过程中起着核心作用。
为了支持肠道菌群成分变化在类风湿性关节炎发病和进展中起重要作用的假说,已经提出了肠道菌群与关节炎相关的几种机制。
这些包括调节宿主免疫系统(触发T细胞分化)、通过作用Toll样受体(TLR)或NOD样受体(NLR)激活抗原呈递细胞(APC)、通过酶促作用促进肽的瓜氨酸化、抗原模拟和增加肠粘膜通透性。关于对APCs TLRs表达的影响,这可能导致Th17/Treg细胞比率失衡,这种局部免疫反应可能导致系统性自身免疫。
因此,现有文献表明,在RA的临床前和临床阶段,肠道菌群可能促进或阻止自身免疫和炎症的扩展,并且肠道菌群可能影响这些阶段之间的转换。
类风湿性关节炎病理生理学和肠道微生物群影响其发病机制的拟议机制
Ferro M, et al ., Front Pharmacol. 2021
肠道菌群代谢产物参与调节免疫功能
某些肠道菌群代谢产生短链脂肪酸,特别是丁酸盐,可以增强肠道屏障功能,因为丁酸盐通过增加TJ成分闭塞带ZO-1、ZO-2和扣带回蛋白的表达来加强屏障。
短链脂肪酸也通过调节免疫细胞功能影响免疫反应和全身炎症。SCFA作为多种白细胞功能的调节因子,包括类二十烷酸、趋化因子和细胞因子(如TNF、IL-2、IL-6和IL-10)的产生,具有抗炎特性。
此外,肠道有益菌及其代谢产物可以在对肠道菌群的耐受性和对有害细菌定植、粘附和易位的抵抗力之间保持平衡。这些特性可能有助于纠正炎症性关节炎肠关节轴肠壁的高渗性。
益生菌调节免疫反应
特定的益生菌通过影响参与先天性和获得性免疫的不同细胞来调节免疫反应,例如上皮细胞和树突状细胞 (DC)、自然杀伤细胞 (NK)、巨噬细胞和淋巴细胞。
益生菌对类风湿关节炎患者全身炎症的影响机制
Ferro M, et al ., Front Pharmacol. 2021
一项随机试验的荟萃分析研究了乳酸杆菌作为单一菌种或与双歧杆菌菌种混合培养的效果,因此补充益生菌可降低血清IL-6水平。
另一项调查补充干酪乳杆菌(L.casei)治疗RA有效性的系统性综述和荟萃分析报告,该特定菌株的C-反应蛋白(CRP)显著降低。研究还报告,干酪乳杆菌可能有助于缓解RA症状,抑制正在接受疾病修饰抗风湿药物(DMARDs)治疗的个体的促炎细胞因子,这表明DMARDs和益生菌对关节炎具有积极的协同作用。
临床试验中益生菌对类风湿关节炎的作用

Ferro M, et al ., Front Pharmacol. 2021
目前,体外微生物细菌生长研究和动物试验表明,随着益生菌的启动,细胞因子(IL-6)减少。在类风湿关节炎中,IL-6是一种与关节破坏相关的关键细胞因子。类风湿关节炎患者益生菌利用率上升。益生菌补充剂有助于减轻肿胀,并建立一个具有最小负面影响的多样化微生物群。
总的来说,需要更多前瞻性系统评价,评估确定益生菌在类风湿性关节炎中的疗效,以及哪些种类的益生菌提供了最大的益处。
牙周炎是这些因素的主要因素,由口腔微生物群失调引起,并与 RA 风险增加有关。
这两种疾病的关联性体现在?
这两种疾病之间的关系是双向的(即 RA 患者也有更高的牙周炎可能性)。它们具有相似的遗传(例如,HLA-SE 等位基因)和环境(例如,吸烟、营养)风险因素并且两者都导致慢性炎症、骨侵蚀和组织破坏 。
重要的菌群:
牙龈卟啉单胞菌 P. gingivalis
该菌是牙周炎的主要原因,也是与 RA 相关的最重要的病原体 。
它的参与不仅是间接的,还是一种病因,类似于吸烟机制。
P. gingivalis 的独特之处在于它有自己的PAD,可导致细菌和宿主蛋白质的慢性瓜氨酸化,导致免疫耐受的破坏,ACPA的产生,最终通过分子模拟和/或表位扩散,导致RA。
注:PAD, peptidyl-arginine deiminase, 一种多肽瓜氨酸化酶
ACPA,Anti-citrullinated peptide antibodies,抗环瓜氨酸肽抗体。有诊断意义的生物标记物。有关节痛或者早期RA病人需要进行该抗体检查。
最近的一项研究进一步表明,即使在ACPA阳性高危人群的牙周健康部位,也存在富含牙龈假单胞菌的口腔菌群失调。
放线共生放线杆菌
Aggregatibacter actinomycetemcomitans
该菌通过中性粒细胞介导瓜氨酸化的不同机制直接与RA有关。
不过需要提醒的是,虽然这么多理论基础和证据都支持牙周炎与RA之间的联系,但更大的、基于人群的前瞻性研究要么还没能证明牙周炎与RA事件相关,要么其效应量很小(OR 1.16-1.17),且没有对吸烟等主要混杂因素进行调整。这是由于方法学和疾病定义问题造成的,还是实际上反映了牙周炎在RA风险中的较小作用尚不清楚。
怀孕如何影响类风湿性关节炎症状?
一般来说,大约 60% 的女性类风湿性关节炎患者在怀孕期间有所改善。不幸的是,怀孕期间关节炎症的减轻通常不会一直持续到分娩后。
另外,大约 20% 的女性在怀孕期间的症状可能会恶化。这种变化的确切机制仍在研究中。
类风湿关节炎对宝宝有什么影响?
RA 可能导致低出生体重婴儿。在新生儿中,3-5% 可能有出生缺陷。一些药物有可能导致出生缺陷。然而,在大多数情况下,只要采取了足够的预防措施,患有 RA 的孕妇都会有健康的妊娠和婴儿。
怀孕期间不要使用非甾体抗炎药,包括布洛芬、萘普生等,不应使用甲氨蝶呤或来氟米特。由于对胎儿的潜在风险,必须在怀孕前尽早停止使用。
当类风湿性关节炎在怀孕期间活动时,类固醇药物如强的松和强的松龙通常用于缓解关节炎症。这些药物不会对胎儿产生不利影响。
其他药物可能需要在受孕前停用,RA患者备孕期间应详细咨询医生。
类风湿性关节炎孕妇需要知道什么?
患有风湿性关节炎的女性在怀孕期间应特别注意以下事项:
类风湿性关节炎会增加妊娠并发症的几率吗?
管理不善的类风湿性关节炎女性早产(37 周前分娩)和低出生体重婴儿的风险较高。类风湿性关节炎还会增加高血压和通过尿液丢失蛋白质的可能性(子痫前期)。据报道,有中度至高度疾病活动的妇女进行剖腹产分娩。
精神或情绪压力已被证明会引发类风湿性关节炎复发。说到这里不得不提到一个,肠-脑轴。
在类风湿性关节炎中,外周炎症信号(如促炎细胞因子、激酶途径)通过自主神经系统进入中枢神经系统。
迷走神经是最重要的传出途径之一,它控制TNF和其他促炎细胞因子的产生。这可能是由于脾神经激活了脾CD4+T细胞(胆碱乙酰转移酶阳性,ChAT+)上的β-肾上腺素能受体,通过胆碱能信号抑制其他免疫细胞产生细胞因子。来自外周的炎症信号激活CNS驻留细胞,特别是小胶质细胞和星形胶质细胞,这被认为是类风湿性关节炎维持神经炎症状态的原因。
研究表明,类风湿性关节炎可能是由压力引起的。压力会增加血液中炎症介质的水平,引发免疫系统的炎症反应,释放细胞因子,从而引发类风湿性关节炎。压力越大,炎症就越严重。
* 细胞因子是一种化学物质,在炎症中发挥重要作用,可以增加一些患者风湿性关节炎的严重程度。
心态不好,雪上加霜
现实生活中的主要应激事件会导致类风湿性关节炎患者疾病控制不佳。
一项横断面研究,使用PRIME Registry患者登记时收集的数据进行。注:PRIME Registry是一个大型、独立、前瞻性观察队列,包括风湿科医生诊断为RA、SLE、PsA或AS的患者。
507例RA患者(平均年龄42.3±12.6岁,女性73.6%,病程80±22个月)的资料。36%的队列报告在过去一年中有严重的心理压力和创伤。未发现年龄、性别和婚姻状况的统计相关性,但发现受教育程度低(p=0.042)、病程较长(p=0.044)、DAS-28值较高(p<0.001)和其他类风湿性关节炎疾病活动标志物(SJC、TJC、ESR、患者整体健康)的统计相关性。
在多元逻辑回归分析中,前一年的主要心理压力和创伤与活动性疾病显著相关(DAS-28;OR 1.67,CI 1.17-2.4,p=0.005).
这也说明了压力管理可能是传统药物疗法治疗风湿性炎症疾病的重要辅助手段。
压力和类风湿关节炎的恶性循环
类风湿性关节炎会造成压力,影响心理健康,特别是当症状持续时间较长时。持续的关节疼痛和睡眠不足会形成恶性循环。每一种症状都会使其他症状恶化,并增加患者已经感到的压力。当病人因为压力而感到疲倦时,他们就不想锻炼了。
缺乏锻炼会引起疼痛,使人难以入睡。患者会对未来的残疾、怀孕或处理治疗的经济负担感到焦虑。这些事情只会增加更多的压力。
如何尽可能缓解压力及RA症状详见后面章节。
每5名类风湿性关节炎患者中,就有1名患有抑郁症。类风湿性关节炎容易增加抑郁风险,反过来,抑郁又会进一步加重类风湿性关节炎,导致更多的关节疼痛、功能下降(卧床的时间更长)和更多的就医次数。所有这些都会进一步影响患者的心理健康,导致更多的压力和抑郁。
在15–17%的抑郁症患者中发现RA共病,提示一种双向关系。
这项观察是在三项大型纵向队列研究发表之后进行的,这些研究发现抑郁症使患RA的风险增加28–65%.
有趣的是,其中一项研究表明,在抑郁症患者中使用抗抑郁药对RA的发展具有保护作用[HazardRatio(HR)0.74,95%CI 0.71–0.76]. 而另一项研究发现它与随后的血清阴性RA相关(HR 1.75,95%CI 1.32–2.32)。
对抑郁症发病机制的新见解,表明显著的全身炎症机制作为与RA相关的可能。
研究证实了精神分裂症对RA的发展具有显著的保护作用(OR 0.48-0.65).
最近的数据加强了遗传免疫学理论。研究表明,这两种情况之间存在SNP遗传负相关性,并在已确定的HLA风险基因中识别出多效性SNP,这些基因对RA和精神分裂症有差异(即,基于同一基因内的特定等位基因变体)。
尽管仍存在争议,但这些有趣的复杂遗传机制有助于解释一种特定疾病如何降低发展为另一种看似无关疾病的可能性。
类风湿性关节炎与神经精神共病显著增加疾病负担,恶化治疗反应和结果;另一方面,类风湿关节炎的抗炎治疗缓解了神经精神症状,如抑郁和焦虑。这可能证明肠-脑轴在RA中的重要性,这其中微生物群的作用正在被更多理解,并为讨论该疾病的进一步治疗靶点提供基础。
心血管疾病
RA 患者可能与心血管死亡风险增加有关,因为许多心脏结构都参与了病理过程,这可能导致动脉粥样硬化、动脉僵硬、冠状动脉炎、充血性心力衰竭、瓣膜病和纤维蛋白性心包炎。它可能包含高血压和血脂异常等疾病的预后标志物。
一项涉及 41,490 名患者的 14 项对照观察性研究的荟萃分析报告显示,与普通人群相比,RA 患者的 CVD 风险增加了 48% 以上。
此外,一项病例对照研究使用多普勒超声心动图技术评估了 47 名没有表现出心血管症状的 RA 患者的潜在心脏异常,并显示肺动脉高压和左心室舒张功能障碍的高发病率。
肾脏疾病
肾脏表现很少见,包括肾小球肾炎和间质性肾病,这与血管炎的存在相关,而神经系统并发症可能导致周围神经病变和脊髓型颈椎病。
贫血
RA 患者最常见的血液学异常是贫血,这是由于铁调素刺激抑制铁转运。此外,据报道,铁调素可能是 RA 中有价值的预后生物标志物。
其他还包括恶性肿瘤、中性粒细胞减少症、嗜酸性粒细胞增多症和血小板减少症等。
使用热敷或冷敷
热敷可促进血液流动,缓解僵硬。
泡热水澡或淋浴,在疼痛的关节上放一个加热垫,或者在温水池里游泳,温水会放松紧绷的关节,促进血液流动,缓解疼痛。为了安全起见,把温度保持在33-37摄氏度之间。泡20分钟的疗程似乎最有效。
冰袋可作用于发炎的关节。寒冷可以减轻肿胀和疼痛。拿一个冰袋敷在关节上,每次15分钟。
找到感觉舒适的适合自己的方式。
另外,热敷和冷敷可缓解运动前后症状的方式。

按摩可以缓解疼痛,缓解压力和焦虑。
服用止痛药
镇痛药有助于缓解关节疼痛。按照医生的规定按时服用。确保白天喝足够的水。
寻求认知行为疗法
心理治疗师使用认知行为疗法来帮助病人改变他们对可能导致焦虑的情况的看法。较低的压力水平也可能减轻症状。
饮食
类风湿性关节炎没有特殊的所谓的“饮食疗法”。应该说,健康饮食是治疗类风湿性关节炎药物的重要补充。
前面我们知道,肠道菌群对类风湿性关节炎有很大影响。而饮食可以改变肠道菌群。
鱼、谷物和蔬菜含量较高的饮食可降低患 RA 的风险,其原因不能确定是因为鱼、谷物和蔬菜的直接抗炎作用,还是因为肠道菌群的变化。西方饮食则会增加其风险。

鱼油中的 Omega-3 脂肪酸可能具有抗炎作用,因此鲱鱼、鲭鱼、鳟鱼、鲑鱼和金枪鱼等鱼类可能是健康饮食的一部分。如果选择服用鱼油补充剂,请咨询医生了解合适的剂量。试着每周吃几次,大概 85克-170克这类鱼。
姜黄素在膳食姜黄(咖喱中的一种成分)中的抗炎作用可能有助于减轻类风湿性关节炎的症状。一些针对风湿性关节炎患者的小型研究发现,高剂量的姜黄关键化合物药片与普通止痛药一样有效。
少吃加工食品。omega-3脂肪酸有助于缓解炎症,而omega-6脂肪酸可能起到相反的作用,玉米油、花生油和蔬菜油中都含有这种物质。很多零食中也有,如薯片、饼干和包装好的烘焙食品等,以上尽量少吃。囤一些更健康的东西,比如新鲜水果、生蔬菜、坚果等。
乳香属(Boswellia)。它的活性成分可以帮助缓解疼痛,改善关节活动。研究还表明,它可以减缓软骨流失。

绿茶可以减缓关节损伤。虽然没有强有力的科学证据证明这一点,但绿茶富含可以镇静炎症和缓解疼痛的营养成分,绿茶中的一些抗氧化剂可能会减缓软骨的破坏。
关于咖啡、茶和咖啡因与RA的关系各研究不一致。
一项荟萃分析包括五项研究(2项队列研究和3项病例对照研究)和134901名参与者,发现咖啡总摄入量(RR 2.43,95%可信区间1.06–5.55)增加了风险,而与茶叶摄入量无关。亚组分析调查队列研究,含咖啡因的无咖啡因咖啡、咖啡因剂量或血清阴性RA不显著,但与血清阳性RA存在同质中度相关性(RR 1.33,95%可信区间1.16–1.52)。
另一项大型前瞻性队列研究(n=76853)发现,饮用咖啡(含咖啡因或不含咖啡因)不会增加RA发病率,而摄入含咖啡因的茶会增加40%的风险(HR1.40,95%可信区间1.01–1.93)。这与荟萃分析和大多数以前的研究不一致,因此,咖啡、茶和RA之间的联系仍然模棱两可。
发酵食品可以成为益生菌载体,并且可能是 RA 患者补充益生菌的有希望的替代品。发酵食品可能会改变肠道有益菌的数量和类型,在对健康个体进行的研究中,有证据表明发酵食品在减少炎症生物标志物方面具有有益作用。含有活微生物的发酵食品有酸奶,开菲尔,味噌,泡菜,纳豆,豆豉等。

类风湿关节炎补充剂
钙和维生素D等补充剂用于预防类风湿性关节炎患者的骨质疏松症。
叶酸被用作补充,以防止甲氨蝶呤治疗类风湿性关节炎的副作用。服用甲氨蝶呤的类风湿关节炎患者应尽量减少或避免饮酒。
维生素C作为强大的抗氧化剂,可以抑制引发关节炎症的促炎因子。此外,维生素C还参与胶原蛋白的合成,而胶原蛋白是关节组织和骨骼中的主要蛋白质。
RA患者体内的维生素C(抗坏血酸)含量远低于正常水平。因此,在临床治疗上,RA患者会通过服用维生素C补充剂来维持自身的维生素C水平,达到缓解疼痛、保护软骨、缓解自身免疫反应的治疗效果。
积极锻炼
需要根据每个人的能力定制活动和锻炼计划。
对关节创伤较小的运动,包括瑜伽和太极,冥想,深呼吸,有助于保持灵活性和力量,还可以改善人们的总体幸福感。
适当的有规律的运动对于保持关节的灵活性和加强关节周围的肌肉非常重要。
太极轻柔、流畅的动作使其成为在不冲击关节的情况下保持锻炼的理想方式。研究表明,它可以帮你更轻松地四处走动,改善运动范围,改善低落情绪,减少压力和焦虑。

瑜伽结合了各种姿势与深呼吸、冥想。尽管关于瑜伽与类风湿关节炎的研究尚未证明它有效,但研究表明,这种练习可以提高手部力量并缓解疼痛。瑜伽还可以增加灵活性、减轻压力、增强肌肉、改善平衡。
游泳特别有帮助,因为它可以在关节压力最小的情况下进行锻炼。
此外,步行,爬楼梯,椭圆机等低强度运动也有帮助,可量力而行。
经培训过的治疗师可提供具体的运动指导,并能提供夹板支撑。例如,手腕和手指夹板有助于减少炎症和保持关节对齐。
手杖、马桶座升降器和罐子夹持器等装置可以协助日常生活活动。
任何运动都要记住,调整自己的节奏;选择适合自己的运动且不要过度;如果引起疼痛,立即停止。
针灸
针灸可以降低肿瘤坏死因子(TNF)等物质的水平,这些物质会导致关节肿胀。它还会引发天然止痛药的释放。当然还需要更多的研究来证明其缓解作用。
听舒缓的音乐,即兴唱跳
当患者无法回避消极的想法时,放一些自己喜欢的、欢快的音乐,随之哼唱或舞动。
享受生活
患者应该做他们喜欢做的事情。看一部电影、画画、散步、在花园里干活、点香熏、泡个澡。
芳香疗法
植物油芳香疗法,目前尚不清楚芳香疗法是否对风湿性关节炎有效,但有证据表明,它可以缓解疼痛和疲劳,并改善情绪。
寻找病友
如果患者发现难以应对压力和类风湿性关节炎,可以寻找病友,一起建立互相支持小组。
补充和替代疗法
自然疗法与传统疗法结合使用时,对某些人是有用的。例如,阿育吠陀疗法提供了一种治疗风湿性关节炎的整体方法。
类风湿性关节炎患者的肠道菌群发生了改变。肠道菌群的正常定植与免疫系统的发育和免疫耐受的形成密切相关,肠道菌群失衡会改变上皮和粘膜通透性,使机体丧失免疫耐受功能,致敏免疫细胞和抗原在关节处聚集,导致类风湿关节炎的发生。
但关于肠道菌群的失调到底是类风湿性关节炎的一个启动因素还是一个被动产物还没有定论。菌群失调对类风湿关节炎发生发展的确切关系,在类风湿关节炎发生发展中起关键作用的细菌特征的筛选等都是我们关心的问题。
本文信息仅供学习参考交流,不参与提供医疗建议。
主要参考文献
Ferro M, Charneca S, Dourado E, Guerreiro CS, Fonseca JE. Probiotic Supplementation for Rheumatoid Arthritis: A Promising Adjuvant Therapy in the Gut Microbiome Era. Front Pharmacol. 2021 Jul 23;12:711788. doi: 10.3389/fphar.2021.711788. PMID: 34366867; PMCID: PMC8346200.
Zaiss MM, Joyce Wu HJ, Mauro D, Schett G, Ciccia F. The gut-joint axis in rheumatoid arthritis. Nat Rev Rheumatol. 2021 Apr;17(4):224-237. doi: 10.1038/s41584-021-00585-3. Epub 2021 Mar 5. PMID: 33674813.
Romão VC, Fonseca JE. Etiology and Risk Factors for Rheumatoid Arthritis: A State-of-the-Art Review. Front Med (Lausanne). 2021 Nov 26;8:689698. doi: 10.3389/fmed.2021.689698. PMID: 34901047; PMCID: PMC8661097.
Radu AF, Bungau SG. Management of Rheumatoid Arthritis: An Overview. Cells. 2021 Oct 23;10(11):2857. doi: 10.3390/cells10112857. PMID: 34831081; PMCID: PMC8616326.
Wells PM, Adebayo AS, Bowyer RCE, Freidin MB, Finckh A, Strowig T, Lesker TR, Alpizar-Rodriguez D, Gilbert B, Kirkham B, Cope AP, Steves CJ, Williams FMK. Associations between gut microbiota and genetic risk for rheumatoid arthritis in the absence of disease: a cross-sectional study. Lancet Rheumatol. 2020 Jun 25;2(7):e418-e427. doi: 10.1016/S2665-9913(20)30064-3. PMID: 33345197; PMCID: PMC7729822.
Zhao Y, Cheng M, Zou L, Yin L, Zhong C, Zha Y, Zhu X, Zhang L, Ning K, Han J. Hidden link in gut-joint axis: gut microbes promote rheumatoid arthritis at early stage by enhancing ascorbate degradation. Gut. 2021 Jul 8:gutjnl-2021-325209. doi: 10.1136/gutjnl-2021-325209. Epub ahead of print. PMID: 34244347.
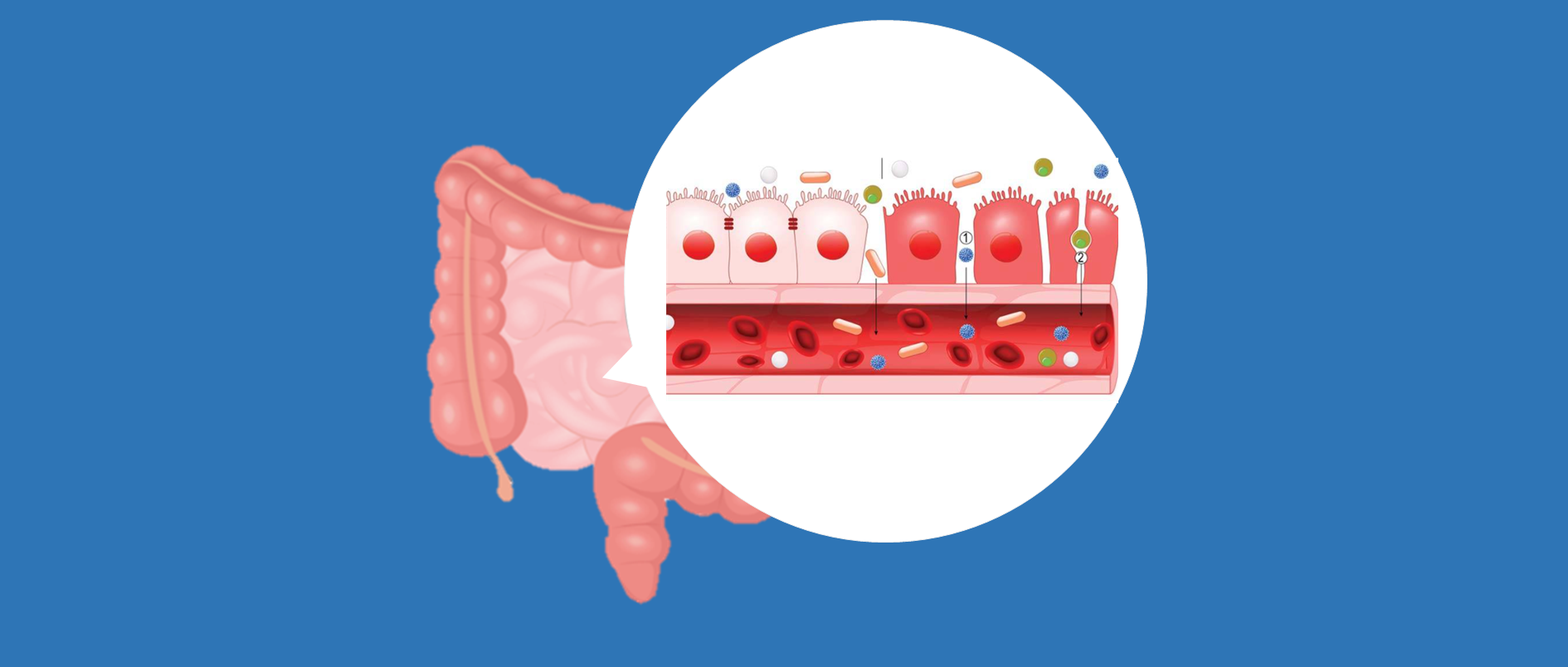
谷禾健康
出于消化和吸收的目的,哺乳动物已经形成了一个非常复杂且高度专业化的胃肠系统,由粘膜屏障维持。
越来越多的数据强调了肠道屏障和肠道通透性对健康和疾病的重要作用。然而,这些术语定义不明确,其评估存在争议,临床意义也没有明确确定。
肠漏症正在成为一种流行病。虽然它尚未被医学界广泛认可,但全球数亿人可能患有与肠漏综合征相关的症状和疾病。
本文讨论的是关于粘膜屏障及其在疾病预防和治疗中的作用的知识。
首先,定义相关术语“肠到屏障”、 “肠道通透性”、“肠漏”。
其次,描述了影响肠道通透性的肠道屏障的关键要素。这个屏障代表了一个巨大的粘膜表面,数十亿细菌面对着身体最大的免疫系统。一方面,完整的肠道屏障保护人体免受微生物和毒素的侵袭,另一方面,这个屏障必须打开以吸收必需的液体和营养。这两个看起来矛盾的目标是通过肠道屏障组成的复杂解剖和功能结构来实现统一的,这正是我们常听说的“肠道通透性”。
第三,描述了饮食和细菌对肠道通透性的调节。特别是,必须考虑潜在的屏障破坏因素,例如肠道灌注不足、感染和毒素,还必须考虑选择过量的营养物质、药物和其他生活方式因素。
第四,讨论肠道菌群对肠道通透性的互相影响,以及与通透性增加相关的特定疾病,例如危重病、炎症性肠病、乳糜泻、食物过敏、肠易激综合征,以及肥胖和代谢疾病。所有这些疾病都以炎症为特征,炎症可能由管腔成分易位到宿主中引发。
最后,讨论了评估肠道通透性的方法。手段差异很大,可能评估屏障的不同功能组件。
“肠道屏障”和“肠道通透性”描述了同一解剖结构的两个不同方面。
“肠道通透性”是一个术语,主要由电生理学家使用动物或人类的组织外植体在 Ussing 室中研究上皮通透性而形成。
注:Ussing Chamber(尤斯室,也叫尤斯灌流室)是研究跨上皮转运的工具,可用于包括离子转运、营养物质转运及药物转运等的研究。通过跨上皮转运的研究,可以了解上皮的离子通道机制、营养成分及药物透过上皮的吸收、影响上皮屏障功能以及通透性的因素等等。来源:百度百科
“肠道屏障”用来描述将内部环境与管腔环境分开的复杂结构。物理屏障包括由血管内皮、上皮细胞和粘液层组成。除了这个物理屏障之外,化学物质也参与了屏障功能。它们由消化分泌物、免疫分子、细胞产物如细胞因子、炎症介质和抗菌肽组成,主要由小肠隐窝中的潘氏细胞(Paneth cell)产生。
肠道屏障基本上将肠道内容物与身体分开。它由单层细胞(上皮细胞、传感细胞以及产生酶和神经递质的细胞)组成。这些细胞通过紧密连接(TJ)蛋白连接起来。
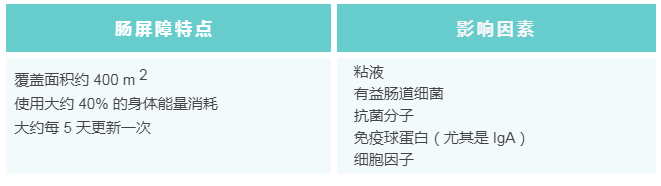
肠道屏障是一个动态实体,由多个元素组成,与各种刺激相互作用并做出反应。
首先,在管腔中,细菌和抗原被胆汁、胃酸和胰液降解,共生细菌通过产生抗微生物物质来抑制病原体的定植。
屏障的另一个要素是由未搅拌的水层、糖萼和粘液层组成的小环境,它们通过免疫球蛋白 A (IgA) 分泌以及糖萼和粘液提供的物理屏障防止细菌粘附。由顶端连接复合物连接的上皮细胞具有运输管腔内容物的能力,但它们也通过分泌氯化物和抗菌肽对有害刺激作出反应。
▃ 上皮层