-

 国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子
国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子

 二级病原微生物安全实验室
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
谷禾健康
食源性微生物是影响食品安全的主要致病菌,因人类食用被致病菌或其毒素污染的食品(主要是动物制品)而引发全球范围的人类疾病。其中多数微生物具有人畜共患的特性,对公共卫生和经济领域产生重大影响。
细菌是全球三分之二人类食源性疾病的病原体。例如,金黄色葡萄球菌、沙门氏菌属、弯曲菌属、单核细胞增生李斯特菌和大肠杆菌是引发食源性疾病和死亡的主要人畜共患细菌性病原体,与食用受污染产品有关。
近年来,食源性病原体已成为全球重要的公共卫生问题,其对健康(显著的发病率和死亡率)和经济的影响日益受到重视。不同报告显示,每年全球约有6亿人(全球人口的十分之一)因食用受污染的食物而患病。由于未被发现或未被报告的爆发,食源性疾病的统计数据有所增加。
目前正值夏季,本文介绍食源性细菌性病原体的背景,并回顾常见的主要食源性人畜共患细菌性病原体。动物是许多食源性人畜共患细菌性病原体的主要储存宿主,动物源性食品是主要的传播载体。肉类、乳制品和蛋类是人类暴露于人畜共患细菌的主要途径。
由于人口快速增长,城市化,人均收入提高,全球化以及消费者习惯的改变(偏爱高蛋白饮食),肉类、牛奶和鸡蛋等动物产品的消费量有所增加。这种情况导致对动物源性食物的需求量大增,并导致集约化动物生产和产品加工,尤其是大规模生产和产品在全球范围内的流通。在此期间,从农场到餐桌的任何环节都可能出现有缺陷的加工实践,从而增加了污染和食源性病原体传播的机会。
人类肉源性疾病进展
doi: 10.3389/fpubh.2022.1045599.
★ 食品在不同阶段都可能受到污染
食品可能在食品链的不同阶段受到污染,包括生产、加工、分销、制备或最终消费过程中。食品被污染的风险很大程度上取决于食品处理者的健康状况、个人卫生、知识以及食品卫生的实践。
肉类供应链每一步都有污染风险
doi: 10.3389/fpubh.2022.1045599.
根据世界卫生组织(WHO)的定义,食源性疾病是指因食用食物或水引起的具有传染性或毒性的疾病。中毒(病原体产生的毒素引起食物中毒)、感染(摄入含有病原体的食物)和毒素感染(在人体肠道中生长时产生毒素)是食源性疾病的三种类型。
★ 动物食品是食源性感染的主要载体
大多数病原体具有人畜共患病的起源,动物源性食品被认为是食源性感染的主要载体。食用动物(牛、鸡、猪、鸭等)是许多食源性病原体的主要储存库。动物产品(肉、奶、蛋、鱼等)及其产品由于病原体含量、天然毒素、掺杂物和其他可能的污染物而具有高风险,当增加动物源性食品的消费时,人类患食源性疾病的风险也在增加。
编辑
doi: 10.3389/fpubh.2022.1045599.
图A描述了自1961年以来选定国家的个人肉类消费量。事实证明,人均收入较高的国家比人均收入较低的国家消费更多的肉类。世界每年生产近3.4亿公吨肉类,是50年前的三倍。图1B所示,到2050年,肉类消费量估计将增加至4.6亿-5.7亿公吨。病原性疾病,如狼疮性皮肤病,在主要畜牧国家越来越常见,对全球肉类和肉类产品的供应构成重大威胁。
在引起食源性中毒的细菌中,一些细菌在发病频率或疾病严重性方面尤为重要。各种细菌(包括革兰氏阳性和革兰氏阴性细菌)会产生引起食源性中毒的毒素,导致的症状从胃肠紊乱到瘫痪甚至死亡。据报道,革兰氏阴性细菌约占细菌性食源性疾病病例的69%。
虽然目前已确定有31种病原体可导致食源性疾病,但包括金黄色葡萄球菌(S.aureus)、非伤寒沙门氏菌、弯曲杆菌属、单核细胞增生李斯特菌(L.monocytogenes)和大肠杆菌(E.coli)在内的细菌性病原体,是全球食源性疾病和死亡的常见原因。下面我们就为大家一一介绍这些常见的食源性细菌病原体。
★ 沙门氏菌是最常见的食源性病原体
沙门氏菌广泛分布于自然界,是人类和动物的主要致病菌。它们是食源性疾病爆发中最常见的细菌来源,每年在全球范围内导致约9380万例食源性疾病和15.5万例死亡。
沙门氏菌已被认为是食源性疾病的主要原因,也是一个严重的公共卫生问题,人们越来越关注包括工业化国家在内的抗菌素耐药菌株的出现和传播。人类和动物的耐药沙门氏菌感染是普遍关注的问题,尤其是在发展中国家。
除了给人类和动物带来的发病率和死亡率成本外,贸易限制和丢弃受污染的食物也是该细菌造成的重要社会经济问题。
1
病原学特征 (Etiology)
分类: 沙门氏菌属包含超过2500种已识别的血清型,分属于肠道沙门氏菌 (Salmonella enterica) 和邦戈沙门氏菌 (Salmonella bongori) 两个种。其中,肠道沙门氏菌是人畜共患的主要病原体,又可细分为六个亚种。
常见致病菌株: 超过150种血清型可引起食源性沙门氏菌病,但鼠伤寒沙门氏菌 (S.typhimurium) 和肠炎沙门氏菌 (S.enteritidis) 是最常见的两种。
基本特性: 这类细菌属于肠杆菌科,是革兰氏阴性、兼性厌氧、不形成孢子、呈直杆状的细菌。在显微镜下或普通营养培养基上,它们与大肠杆菌无法区分。
2
流行病学特征 (Epidemiology)
全球公共卫生问题:沙门氏菌是全球主要的公共卫生问题之一,无论在发展中国家还是发达国家,它都是最常见的食源性疾病之一。
主要宿主与传染源
动物携带:动物是沙门氏菌食源性疾病的主要宿主,其在动物中的普遍携带是重要的流行病学因素。
自然栖息地:沙门氏菌的主要栖息地是动物(如农场动物、鸟类、爬行动物、昆虫)和人类的肠道。
导致暴发的因素:食用生或不安全的食物、交叉污染、不当的食品储存、不良的个人卫生习惯、食品冷却和再加热不充分,以及制备与食用间隔时间过长等,都是促使人类沙门氏菌病暴发的因素。
环境耐受性:它们能在典型的餐饮冷藏温度下存活,并在温度滥用(即不当的温度控制)的情况下繁殖。
高风险原因:不卫生的生活条件、人畜混居、以及食用生或未煮熟的动物源性食品,都增加了感染风险。
3
传播途径 (Transmission)
主要途径: 食源性传播被认为是沙门氏菌感染的主要原因。动物源性食品及其制品是沙门氏菌传播给人类的最常见载体。
其他途径: 传播也通过摄入被动物粪便污染的水、食物以及被污染的食品加工设备而发生。
主要污染食品: 禽类、猪、牛及其产品(如肉、蛋、奶)是导致人类沙门氏菌病暴发的最常见食品来源。尤其是鸡肉产品(包括鸡蛋)被公认为是沙门氏菌的重要宿主。
非伤寒沙门氏菌(NTS)的宿主和传播
doi: 10.3389/fpubh.2022.1045599.
污染过程
屠宰环节: 受污染的动物产品通常源于受感染的动物本身,或在屠宰过程中胴体或可食用器官被污染。
蛋类污染: 蛋壳和蛋内容物可能在母鸡生殖系统内形成时或从环境中(如粪便接触)被污染。
粪便污染: 胴体的粪便或肠道污染是人类食源性感染的主要来源。
特定菌株: 最近出现的多重耐药性鼠伤寒沙门氏菌DT104型,主要通过摄入受污染的牛肉传播。
4
致病机理 (Pathogenesis)
影响因素: 致病性受菌株毒力、感染剂量、感染途径和宿主易感性等因素影响。毒力质粒、毒素、菌毛和鞭毛等毒力因子有助于建立感染。
入侵过程
靶细胞: 微皱褶细胞(M细胞)是沙门氏菌致病性的靶细胞。
机制: 细菌首先利用菌毛(粘附结构)粘附于肠道上皮细胞,然后侵入上皮细胞引发肠胃炎。这一复杂的入侵过程由多种染色体基因产物介导。
宿主反应: 细菌入侵后,会诱导靶细胞膜起皱,将细菌内化到膜结合的液泡中。这个过程会引发一系列反应,包括中性粒细胞的募集和迁移、上皮细胞分泌细胞因子以及液体和电解质的分泌。
定植与生存: 肠道正常菌群可以抵抗沙门氏菌定植,因此口服抗生素会促进感染的建立。沙门氏菌能够避开胃部的宿主防御,到达肠道进行感染。
5
临床症状 (Symptoms)
潜伏期: 潜伏期为12至72小时。
临床表现严重程度不一,从自限性肠胃炎到败血症不等,这主要取决于宿主的易感性和菌株的毒力。
消化道症状: 通常为胃肠道症状,包括恶心、呕吐、腹部绞痛以及水样、绿色、有恶臭的腹泻或带粘液的血性腹泻。
其他症状: 还包括头痛、虚脱、疲劳(肌肉无力)和中度发烧。
预后与并发症: 该病通常是自限性的,不需要特殊治疗。但在幼儿、老人和免疫功能低下者中可能导致严重并发症。反应性关节炎、镰状细胞性贫血和骨髓炎等并发症在普通人群中也较为常见。
6
检测方法 (Detection)
常规方法: 通常的程序是使用选择性增菌培养基(如沙门氏菌-志贺氏菌琼脂)进行细菌的培养和分离。
血清分型: 血清分型是区分沙门氏菌菌株的公认方法,是公共卫生的重要工具。
快速方法: 市售的快速检测方法包括新型选择性培养基、改良的传统程序、免疫学检测法和核酸检测法。
免疫学检测: 包括ELISA、乳胶凝集试验、免疫扩散和免疫层析法(试纸条)。
分子检测: 主要技术是直接杂交(DNA探针)和扩增(PCR)方法。
7
预防与控制 (Prevention and Control)
综合策略:需要建立生物安全和生物防护措施,改进食品加工方法以及制备和储存实践。减毒的DNA重组活沙门氏菌疫苗,结合对动物、饲料和动物食品的综合控制策略,将有助于减少沙门氏菌病。
食品安全实践:应采取安全的食品制备方法,包括彻底煮熟、重新加热食物、牛奶的巴氏消毒(煮沸)、充分冷藏,并避免宠物和其他动物进入食品处理区域。
高危人群建议:建议易感人群避免食用未煮熟的肉和家禽、生牛奶、生鸡蛋以及含有生鸡蛋的食物。
二次污染控制:通过清洁消毒、人员卫生和适当的加工来防止二次污染。
金黄色葡萄球菌是全球最常见的食源性病原体之一,其高发生率仅次于沙门氏菌。
它可以作为一种共生菌存在于健康人和动物的皮肤、鼻子和粘膜上。然而,它也是一种众所周知的机会性食源性病原体,可引起多种严重程度不一的感染性疾病,在人和动物中引起广泛的疾病。
★ 金黄色葡萄球菌是食源性中毒的重要原因
金黄色葡萄球菌在人类消费品中的存在对食品工业至关重要,因为某些菌株是食源性中毒的原因。它们是导致食品腐败、降低食品安全和保质期以及引起食源性中毒的原因。
金黄色葡萄球菌是因食用被葡萄球菌肠毒素污染的食物而导致食物中毒的主要原因。
并且由于与多重耐药性相关的死亡率不断上升,它引起了公众的关注。抗生素的广泛使用以及细菌快速发展和获得抗菌素耐药性的能力促进了耐药菌株的出现,例如耐甲氧西林金黄色葡萄球菌(MRSA)。
注:MRSA最早于20世纪60年代初被报道,现在被认为是全球主要的医院获得性病原体。MRSA是人和兽医医学中常见的病原体。
1
病原学特征 (Etiology)
基本分类:金黄色葡萄球菌(S.aureus)是一种革兰氏阳性、过氧化氢酶阳性、凝固酶阳性、氧化酶通常为阴性的兼性厌氧球菌。
形态与生化特性: 这种细菌细胞呈球形,不运动,常形成葡萄状的菌落。它可以通过产生金色菌落、凝固酶、发酵甘露醇和海藻糖以及产生耐热核酸酶等特性,与其他葡萄球菌区分开来。
生存能力: S.aureus对环境有极强的适应能力,可在广泛的温度(7°C至48°C)、pH值(4.2至9.3)和高盐浓度(最高15% NaCl)下生长。这使其能够在多种食品中存活,尤其是在加工过程中需要较多人工操作的食品(如奶酪等发酵食品)。
2
流行病学特征 (Epidemiology)
重要性日益增加:由于其导致的感染增多(特别是MRSA菌株),在医学中的重要性,以及近年来发现其与动物相关的克隆谱系和人畜共患的潜力,该微生物在动物中的流行病学研究备受关注。
食源性疾病:S.aureus是全球食源性细菌性中毒的主要原因之一,也是全球报告的最常见的食源性疾病之一。
宿主与传播
人类携带:大约50%的健康人群在鼻腔、喉咙和皮肤中携带这种细菌。
动物来源:患有乳腺炎的奶牛是生牛奶中S.aureus的常见来源。该细菌广泛存在于多种宿主中,包括人类和猪、牛、山羊、鸡、鸭等食用动物。
污染途径:食品污染可能直接来自受感染的食用动物,也可能是由于食品生产、零售和储存过程中的卫生条件不佳所致。
耐甲氧西林金黄色葡萄球菌的风险因素:
免疫抑制
血液透析
外周灌注不良
高龄
长期住院
抗生素治疗不当
体内留置装置(如导管)
需要胰岛素治疗的糖尿病
褥疮溃疡等
3
传播途径 (Transmission)
主要来源:S.aureus主要通过受污染的动物源性食品传播。细菌有潜力在加工、制备、包装、切碎和储存过程中污染动物产品,从而进入食物链。
常见载体:污染可见于多种食品,如畜禽产品、海鲜以及烘焙产品。潮湿的、富含淀粉和蛋白质的食物特别容易被葡萄球菌肠毒素污染。牛奶、乳制品、猪肉、牛肉、羊肉、禽肉和鸡蛋是导致葡萄球菌食物中毒的常见载体。生肉也是S.aureus存活和耐药菌株在社区传播的良好媒介。
人为污染:携带S.aureus的食品处理者,其身体或手套也可能污染食物。
4
致病机理 (Pathogenesis)
毒力因子:S.aureus拥有多种毒力因子,这些因子单独或共同作用可导致严重感染。它能表达多种分泌性和细胞表面相关的毒力因子,以促进其粘附于宿主细胞外基质、破坏宿主细胞并对抗免疫系统。
关键致病物质:其产生的胞外活性物质被认为是致病的关键,包括凝固酶、溶血素、核酸酶、酸性磷酸酶、脂肪酶、蛋白酶、纤维蛋白溶酶、肠毒素和中毒性休克综合征毒素。
产生条件:如果食物在室温下存放一段时间,食物中的细菌就能产生毒素。
毒素稳定性:肠毒素能抵抗蛋白水解酶,且耐热。因此,即使食物中的S.aureus细菌已被杀死,毒素仍可能存在。
种类与作用:已分类出23种不同的葡萄球菌肠毒素(SEs)和SE样毒素。肠毒素会刺激中枢神经系统的呕吐中枢,并抑制小肠对水和钠的吸收,从而引起急性肠胃炎症状。
5
临床症状 (Symptoms)
疾病谱:该细菌可引起多种疾病,症状从简单的皮肤感染到更严重甚至危及生命的感染,如败血症、坏死性筋膜炎、感染性心内膜炎、坏死性肺炎和中毒性休克综合征。
潜伏期:潜伏期很短,通常在食用被污染食物后2-4小时内出现临床症状。
典型症状:特征为恶心、呕吐,体温多为偏低,伴有寒战、头痛和腹部绞痛,可有或无腹泻,但通常不发烧。最常见的症状是腹部绞痛、恶心和呕吐。
高危人群:在儿童和老年人等易感人群中,偶尔可能导致死亡。
MRSA感染症状:MRSA或多重耐药S.aureus菌株可引起院内感染,导致快速进展且可能致命的疾病,如威胁生命的肺炎、坏死性筋膜炎、心内膜炎、骨髓炎、严重败血症以及中毒性休克综合征等毒素病。
6
检测方法 (Detection)
常规检测
培养基: 使用一系列选择性和诊断性培养基(如甘露醇盐琼脂)来检测和计数食品中的葡萄球菌。
鉴定标准: 鉴定致病性葡萄球菌最广泛接受的标准是其产生凝固酶的能力。标准方法包括选择性增菌、分离具有典型形态的菌落,并通过微生物学和生化方法确认。
细菌检测:实时荧光定量PCR (RT-PCR) 和定量PCR越来越多地用于临床实验室,以快速检测和鉴定MRSA菌株。快速乳胶凝集试验也是一种常见的实验室测试。
肠毒素检测:检测食品中肠毒素最重要的方法是酶联免疫吸附试验(ELISA)。其他血清学测试如凝集试验和凝胶扩散法也可用于检测。近年来,核酸杂交、PCR和荧光免疫分析等分子生物学方法已成为最流行的检测手段。
7
预防与控制 (Prevention and Control)
基本原则: 葡萄球菌无处不在,无法从环境中根除。预防的关键在于切断其各种传播途径。
预防措施
食品安全: 彻底煮熟食物、防止污染和交叉污染、控制关键控制点是有效的预防方法。
公众教育: 提高公众关于安全处理肉类的意识以及其他公共卫生干预措施是预防疫情的基石。
控制方案
卫生实践: 改善医护人员和食品处理者的个人卫生习惯。
环境控制: 对设备、表面、衣物进行消毒。
医疗相关: 明智地使用抗生素,并实施筛查计划。对MRSA患者所在的区域应使用消毒剂进行彻底清洁。
弯曲杆菌属(Campylobacter)是全球人类细菌性食物传播腹泻病的主要原因,主要源于动物源性食品的污染。弯曲杆菌属可以在大多数温血动物和家禽中定殖。
弯曲菌病的人畜共患性质使其在全球范围内具有重要的临床和经济意义。它们导致了15%的食源性疾病相关住院和6%的食源性疾病相关死亡,据估计,每年有4亿例病例是由弯曲菌感染引起的。
1
病原学特征 (Etiology)
命名与形态:“Campylobacter”一词源于希腊语,意为“弯曲的杆菌”。这类细菌属于弯曲杆菌科,是微小(0.2–0.8μm×0.5–5μm)的革兰氏阴性、呈弯曲或螺旋状的微需氧菌。它们具有独特的“飞镖式”运动能力,过氧化氢酶和氧化酶呈阳性。当两个或多个细菌细胞聚集时,会形成“S”形或海鸥翅膀样的“V”形。
分类:弯曲杆菌属目前包含25个种和8个亚种。
主要致病菌:在众多弯曲杆菌中,空肠弯曲杆菌(C.jejuni)和结肠弯曲杆菌(C.coli),是重要的食源性病原体。其中,空肠弯曲杆菌是最常报告的种类(占80%至90%),其次是结肠弯曲杆菌(占5%至10%)。
2
流行病学特征 (Epidemiology)
动物宿主:它们广泛分布于自然界,被认为是人畜共患病原体,存在于众多动物宿主体内,尤其是鸟类。它们可以定植在野生和家养鸟类及哺乳动物(包括所有食用动物)的消化道中,也可存在于动物和人类的生殖器官、肠道和口腔中。
食品来源:散装牛奶、牛的组织标本和生碎牛肉中都可能发现这种细菌,但鸡蛋通常不被认为是该菌的重要来源。
暴发特点:弯曲杆菌病的暴发通常是散发性的,虽然不常导致死亡,但可能引起继发性并发症。
人类感染风险:旅行是导致人类感染弯曲杆菌病的最重要风险因素,其次是食用未煮熟的鸡肉、环境暴露以及与农场动物直接接触。
3
传播途径 (Transmission)
主要途径:人类感染弯曲杆菌的主要传播途径是处理、制备和食用受污染的食物,特别是禽肉产品。
其他途径:食品处理者在制备过程中对即食食品的交叉污染,以及与动物的直接接触也被确定为传播途径,或接触受污染的设备、水传播。
弯曲杆菌的储存库和传播
doi: 10.3389/fpubh.2022.1045599.
主要食品来源:感染主要来自食用受污染的家禽、牛肉、猪肉或其他动物肉类、肉制品、生(未经巴氏消毒)牛奶或奶酪等乳制品。
4
致病机理 (Pathogenesis)
关键毒力因子:细菌的运动能力、粘液定植、上皮细胞入侵、毒素产生、附着、内化和易位在疾病发展中起着重要作用。
入侵过程
运动与附着:鞭毛使细菌能够到达肠道内的附着位点。病原体对肠道上皮的附着对于定植和增加细菌毒素的分泌至关重要。
细胞入侵:空肠弯曲杆菌在被摄入后,会在肠道中繁殖并损害粘膜上皮,侵入上皮细胞和固有层内的细胞,导致自限性腹泻和腹痛。腹泻病可能由其产生的一种不耐热毒素引起。
防御机制:弯曲杆菌能产生超氧化物歧化酶,这种酶可以分解超氧自由基,是细菌对抗氧化损伤的主要防御机制之一。
5
临床症状 (Symptoms)
潜伏期:潜伏期为3至5天。
主要症状:人类弯曲杆菌病的特征是水样或血性腹泻、腹痛、痉挛、发烧、不适和呕吐。它也是“旅行者腹泻”的主要病原体。
高危人群: 这对幼儿尤其危险,因为腹泻会导致脱水和钠、蛋白质等营养物质的流失。在幼儿(<1岁)和免疫功能低下的患者中,可能会出现中毒性巨结肠、脱水和败血症。
感染后并发症:格林-巴利综合征(GBS)是最主要的感染后并发症,通常在感染后2-4周出现。其特征是周围神经的多发性神经炎,可能导致持续数周的肢体短期或长期麻痹。
其他并发症还可能包括脑膜炎、尿路感染和短期反应性关节炎。
6
检测方法 (Detection)
传统方法:传统上,鉴定方法包括使用选择性培养基(如Preston琼脂、木炭-头孢哌酮-脱氧胆酸盐琼脂等)结合生化测试。
分子技术:近年来,PCR已越来越多地应用于弯曲杆菌的检测和鉴定。
免疫学方法:作为替代方法,也可以使用免疫测定法等多种技术。
7
预防与控制 (Prevention and Control)
养殖场控制: 控制取决于畜舍的卫生设施和卫生状况,以减少动物环境中细菌的数量。
加工厂控制: 在肉类加工厂,可以通过实施HACCP体系,包括清洗、处理和冷冻胴体,来减少和控制细菌数量。
食品制备:提高餐馆和家庭厨房的食品处理技能将减少细菌的传播。 将家禽等生肉充分烹饪至内部温度达到82°C将能消灭该细菌。
新型干预: 精油、益生元、益生菌、细菌素、噬菌体和免疫措施在控制弯曲杆菌方面也具有重要作用。
它是一种主要的食源性人畜共患细菌,对公共卫生构成严重威胁,主要通过食用受污染的动物源性食品传播。单核细胞增生李斯特菌作为食源性病原体的出现可以追溯到1980年,当时发生了很多与食用受污染食品有关的李斯特菌病爆发和散发病例。
★ 李斯特菌感染病例死亡率较高
单核细胞增生李斯特菌是全球控制和监测机构一直试图控制的最具毒力的病原体之一,其病例死亡率约为30%,远高于其他常见的食源性病原体。
此外,食品中存在单核细胞增生李斯特菌还会造成重要的经济后果,例如产品从消费者市场撤回以及涉案产品销量下降。由于李斯特菌属无处不在的特性,以及它们在包括pH、温度和盐度在内的广泛环境压力下生存的独特能力,它们被认为是重要的食源性病原体。
1
病原学特征 (Etiology)
基本特性:李斯特菌属的生物是嗜冷、革兰氏阳性、可运动、兼性厌氧、不形成孢子的杆状细菌。
分类:李斯特菌属目前包含10个种,包括单核细胞增生李斯特菌 (L.monocytogenes)、伊万诺夫李斯特菌 (L.ivanovii) 等。
主要致病菌:尽管李斯特菌属有多个种,但由人类李斯特菌病引起的病例几乎全部是由单核细胞增生李斯特菌(L.monocytogenes)引起的。它是人类和动物的主要病原体。
生存能力:该细菌具有极强的环境适应能力,能够在冷藏条件、低pH值和高盐浓度下存活。它是一种兼性胞内菌,可在0°C至45°C的温度和4.4至9.4的pH值范围内生长。
2
流行病学特征 (Epidemiology)
地理分布:单核细胞增生李斯特菌在世界范围内广泛存在。李斯特菌病在全球以散发或流行的形式出现。
环境分布:李斯特菌属细菌无处不在,广泛分布于正常环境中。它们最常见于被土壤和水污染的蔬菜等生食,以及生的动物产品中。
主要污染食品:单核细胞增生李斯特菌经常从动物源性食品中分离出来,如即食肉制品、碎牛肉、香肠、鱼和鱼制品、牛奶以及经过巴氏消毒的乳制品(如软奶酪和冰淇淋)。
注:尽管巴氏杀菌可以消灭生奶中的单核细胞增生李斯特菌,但并不能消除乳制品后续被污染的风险。
职业与高危人群:人类可能因职业原因从动物源感染李斯特菌病,特别是农民、屠夫、家禽工人和兽医。
侵袭性李斯特菌病的主要风险人群是免疫功能低下者,如孕妇、未出生或新生婴儿、器官移植接受者、癌症和艾滋病患者以及老年人。
3
传播途径 (Transmission)
主要途径: 人类感染的最常见途径是食用被单核细胞增生李斯特菌污染的动物源性食品。
高危人群:孕妇、新生儿、老年人和免疫功能低下的人群感染风险更高。
单核细胞增生李斯特菌引起的宿主、传播和疾病
doi: 10.3389/fpubh.2022.1045599.
4
致病机理 (Pathogenesis)
独特毒力:李斯特菌拥有独特的毒力因子,使其能够侵入宿主、逃避免疫细胞并引起感染。
入侵过程
附着:L.monocytogenes 表面有D-半乳糖残基,可以附着到宿主细胞(通常是肠道粘膜的M细胞和派尔集合淋巴结)上的D-半乳糖受体。
穿透:附着后,细菌可以穿过肠道膜进入血液,成为血源性(败血性)病原体。
胞内生长:进入宿主的单核细胞、巨噬细胞或多形核白细胞后,细菌可以在其中生长。
跨越屏障: 作为一种胞内菌,它能够感染多种类型的细胞,并能穿过肠道、血脑和胎盘等生物屏障,引发感染。
5
临床症状 (Symptoms)
典型症状: L.monocytogenes通常表现为典型的“食物中毒”症状,包括流感样症状(如发烧、疲劳)和胃肠道症状(恶心、呕吐和腹泻)。
持续时间:李斯特菌病的症状通常持续7-10天。
严重感染:在高危人群中,它可能导致严重的、危及生命的感染,如败血症、脑膜炎、自然流产、死产或胎儿感染。
6
检测方法 (Detection)
传统方法:方法包括在选择性培养基中进行增菌,随后在琼脂平板上进行划线培养,并进行各种物种鉴定测试。
注:Christie–Atkinson–Munch–Peterson (CAMP) 试验有助于识别李斯特菌属的种类。
分子方法:由于其准确、灵敏和特异,PCR和RT-PCR等分子方法越来越多地用于从食品中鉴定L.monocytogenes。
免疫学方法:使用单克隆抗体的ELISA已被开发用于鉴定食品中的李斯特菌。
7
预防与控制 (Prevention and Control)
卫生措施
表面消毒:对食品接触表面进行有效的卫生处理可以预防李斯特菌病。
规范操作: 良好的卫生习惯、良好生产规范和卫生设施是操作程序中最合适的策略。
高危人群防护: 建议易感人群(孕妇、老年人和免疫抑制者)避免食用未经巴氏消毒的乳制品以降低风险。
法规与生产控制
法规:标准化的法律法规和对肉制品生产的控制应是保护食品免受L.monocytogenes污染的根本途径。
生产链控制:在食品生产链的不同阶段,防止细菌在加工厂传播是保护食品的最重要方法之一。
管理体系:实施HACCP方法并建立有效的关键控制点,可以显著降低许多加工食品中李斯特菌的污染水平。各国采用的冰淇淋/冷冻甜点巴氏杀菌标准对于减少李斯特菌病具有重要意义。
大肠杆菌是众多可进入动物源食品的病原微生物之一,被认为是粪便、土壤和水污染的可靠指标。一种新兴的克隆差异型大肠杆菌最早于1982年被确定为重要的食源性人畜共患病原体,当时它与美国爆发的严重血性腹泻有关,而这种腹泻的源头被追溯到食用未煮熟的汉堡包。
大多数大肠杆菌是动物和人类胃肠道(回肠下段和大肠)的正常居民,而另一些则对人类具有致病性。大肠杆菌具有人畜共患性质,构成公共卫生危害。产志贺毒素大肠杆菌与全球范围内多起危及生命的食源性疫情相关。
1
病原学特征 (Etiology)
基本特性:大肠杆菌是一种属于肠杆菌科的革兰氏阴性杆状细菌,长度可达3微米,能发酵葡萄糖和其他糖类,有周生鞭毛使其能够运动,并常有菌毛。
致病性分组:根据其毒力特征,可分为五个致病性组:肠道集聚性大肠杆菌(EAEC)、肠道出血性大肠杆菌(EHEC)、肠道侵袭性大肠杆菌(EIEC)、肠道致病性大肠杆菌(EPEC)和肠道产毒性大肠杆菌(ETEC)。
主要致病血清型:O157:H7是最著名的血清型之一,其致病型可引起人类食源性感染。它是一种广为人知的产志贺毒素细菌,也是主要的食源性和人畜共患病原体。
2
流行病学特征 (Epidemiology)
全球公共卫生问题:大肠杆菌O157:H7是最重要的食源性病原体之一,在世界各地报道日益增多,是导致全球暴发的新兴食源性病原体。据估计,仅在美国,每年就有约74000例病例和61例死亡归因于该菌。
主要宿主:牛是大肠杆菌O157:H7的主要宿主。其次是绵羊和山羊。一些研究报告称,小型反刍动物通过粪便排菌也是人类感染的来源之一。此外,也从马、狗和鹿中分离出该菌。
主要传染源:未煮熟的牛肉、受污染的碎牛肉以及其他牛源性食品被确定为暴发中的主要感染来源。牛奶和乳制品也是其来源之一。
促发因素:大肠杆菌O157:H7感染发病率上升的主要促成因素包括饮食习惯的改变、大规模餐饮、复杂且漫长的食品供应链(国际流动增加)以及不良的卫生习惯。
近期趋势:然而,最近关于该菌在牛肉和乳制品中出现水平的报告有增加的趋势。近年来已开展了一些研究来评估其在肉类和奶类等动物源性食品中的出现情况。
3
传播途径 (Transmission)
主要途径:人类感染大肠杆菌O157:H7最常见的传播方式是食用受污染的食物和水。它也可以通过人与人直接接触以及偶尔的职业暴露传播。
大肠杆菌O157:H7的储存库和传播
doi: 10.3389/fpubh.2022.1045599.
污染过程
屠宰环节:在加工厂的屠宰过程中,通过皮肤到胴体或粪便到胴体的转移导致胴体污染,这是人类感染的主要风险因素。
后续环节:交叉污染也可能在加工厂的后续加工、零售市场的牛肉分销和储存过程中发生。
4
致病机理 (Pathogenesis)
关键毒力因子:细菌的致病机制包括毒力因子、志贺样毒素和黏附因子。
黏附与损伤
黏附素基因(Intimin gene):该基因负责细菌与肠道细胞的紧密黏附,导致出现黏附/清除(A/E)病变,并擦除肠细胞刷状缘的微绒毛。
细胞结构改变:细菌黏附后会破坏微绒毛,并改变肠细胞的细胞骨架结构。
志贺毒素(Shiga toxin):志贺毒素的产生是导致出血性腹泻和溶血性尿毒综合征的核心。O157:H7菌株产生志贺毒素1(stx-1)和志贺毒素2(stx-2)。毒素会刺激细胞分泌并杀死结肠上皮细胞。
5
临床症状 (Symptoms)
潜伏期:潜伏期为2至10天。
主要症状:感染后出现腹泻、腹痛、呕吐。最初可能是伴有腹部绞痛的腹泻,几天内可能转为严重的血性腹泻,通常没有发烧。
严重并发症:可能导致出血性结肠炎、伴有急性肾衰竭的溶血性尿毒综合征(HUS)和血栓性血小板减少性紫癜(TTP)。
6
检测方法 (Detection)
金标准:大肠杆菌O157:H7的培养分离仍然是鉴定的金标准,并辅以生化测试。
选择性培养基:添加了头孢克肟和亚碲酸钾的山梨醇-麦康凯琼脂(SMAC)是分离O157:H7最灵敏和鉴别性的培养基之一。O157:H7不发酵山梨醇,因此在该培养基上会形成无色菌落。
免疫学方法:多种免疫测定技术和血清学技术可用于特异性诊断。
分子方法:PCR-ELISA、多种qPCR方法、环介导等温扩增(LAMP)等是检测志贺毒素的主要有效方法。
7
预防与控制 (Prevention and Control)
通用原则:预防由大肠杆菌引起的食源性疾病的方法与其他食源性细菌病相同,但由于其对幼儿的严重后果,需要特别注意。
养殖场策略:在养殖阶段采取干预措施,如使用益生菌、疫苗、抗菌剂、氯酸钠和噬菌体,以增加畜群对感染的抵抗力。
加工干预:开发了多种干预技术,如皮肤和胴体清洗、使用抗菌剂、蒸汽巴氏杀菌、伽马射线辐照等。
HACCP体系:在加工环节实施HACCP方法,该方法包括对产品进行微生物测试。
食品安全计划:证据表明,食品处理人员培训、食品经营场所检查以及旨在推广正确食品处理和制备技术的教育项目,是减少公众暴露于食源性病原体的有效组成部分。
人畜共患疾病是一个严重的公共卫生问题。大多数人类传染病都是由动物引起的。这些病原体不仅在动物中引起疾病,而且还对人类健康构成严重威胁。本文详细介绍了五种主要的食源性细菌性病原体——非伤寒沙门氏菌、金黄色葡萄球菌、弯曲杆菌、单核细胞增生李斯特菌和大肠杆菌O157:H7。这些病原体具有以下共同特点:
主要特征与危害:
-广泛分布于自然界,以动物为主要宿主;
-具有人畜共患特性,对公共卫生构成严重威胁;
-主要通过受污染的动物源性食品传播;
-可引起从轻微胃肠道症状到危及生命的严重并发症。
许多人畜共患病目前已得到控制,但我们对许多疾病的认识仍存在差距,特别是对疾病分布、病原学、病原体、宿主、媒介生物学、动力学、传播周期、易感因素和危险因素的认识。宿主、病原体和环境之间的平衡可能在任何时候受到人类活动和自然活动的干扰,从而引起人畜共患病的发生。
除前述针对不同病原菌(沙门氏菌、弯曲杆菌等)的传统培养、血清分型、免疫学检测及分子诊断等常规方法外,高通量测序技术正逐步成为重要补充工具。
以16S rRNA测序、宏基因组测序为代表的新型技术,无需依赖传统培养流程,可直接对粪便、食品或环境样本中的微生物群落进行全面解析。
未来预防与控制建议:
1.建立”从农场到餐桌”的全链条防控体系
-在养殖阶段加强生物安全措施,使用益生菌、疫苗等预防手段;
-在屠宰加工环节严格实施HACCP体系,加强胴体清洗和消毒;
-在零售和消费环节加强食品安全教育和监管。
2.强化多部门协作的”一体化健康”理念
-建立兽医、公共卫生、食品安全等部门的协调机制;
-完善动物疫病监测预警系统;
-加强抗菌药物合理使用,遏制耐药菌株传播。
3.推进科技创新与检测技术发展
-开发更快速、准确的分子检测方法;
-研发新型疫苗和生物防控技术;
-利用人工智能等技术提升风险预测能力。
4.加强公众教育与意识提升
-普及食品安全知识,提高消费者自我保护能力;
-加强食品从业人员培训;
-建立食品安全社会共治格局。
5.完善法律法规与标准体系
-制定和完善食品安全法律法规;
-建立严格的食品安全标准和检测体系;
-加大对违法行为的处罚力度。
面对人畜共患病的复杂挑战,我们需要以科学的态度、系统的思维和协作的精神,构建覆盖全社会、全过程的食品安全防控网络。消化道病原菌直接关系到我们的饮食和健康,尤其夏季食物易腐败或受污染,其检测更为重要。只有通过持续的努力和创新,才能有效预防和控制食源性人畜共患病的传播,保障人类健康和食品安全。
主要参考文献
Ali S, Alsayeqh AF. Review of major meat-borne zoonotic bacterial pathogens. Front Public Health. 2022 Dec 15;10:1045599.
Abebe E, Gugsa G, Ahmed M. Review on Major Food-Borne Zoonotic Bacterial Pathogens. J Trop Med. 2020 Jun 29;2020:4674235.
Rahman MT, Sobur MA, Islam MS, Ievy S, Hossain MJ, El Zowalaty ME, Rahman AT, Ashour HM. Zoonotic Diseases: Etiology, Impact, and Control. Microorganisms. 2020 Sep 12;8(9):1405.
Ed-Dra A, Giarratana F, White AP, Yue M. Editorial: Zoonotic bacteria: genomic evolution, antimicrobial resistance, pathogenicity, and prevention strategies. Front Vet Sci. 2024 Mar 8;11:1390732.
Ed-Dra A, Filali FR, Khayi S, Oulghazi S, Bouchrif B, El Allaoui A, Ouhmidou B, Moumni M. Antimicrobial Resistance, Virulence Genes, and Genetic Diversity of Salmonella enterica Isolated from Sausages. Eur J Microbiol Immunol (Bp). 2019 Apr 9;9(2):56-61.
Feng Y, Pan H, Zheng B, Li F, Teng L, Jiang Z, Feng M, Zhou X, Peng X, Xu X, Wang H, Wu B, Xiao Y, Baker S, Zhao G, Yue M. An integrated nationwide genomics study reveals transmission modes of typhoid fever in China. mBio. 2023 Oct 31;14(5):e0133323.
Mughini-Gras L, Kooh P, Augustin JC, David J, Fravalo P, Guillier L, Jourdan-Da-Silva N, Thébault A, Sanaa M, Watier L; Anses Working Group on Source Attribution of Foodborne Diseases. Source Attribution of Foodborne Diseases: Potentialities, Hurdles, and Future Expectations. Front Microbiol. 2018 Sep 3;9:1983.
Dhama K, Rajagunalan S, Chakraborty S, Verma AK, Kumar A, Tiwari R, Kapoor S. Food-borne pathogens of animal origin-diagnosis, prevention, control and their zoonotic significance: a review. Pak J Biol Sci. 2013 Oct 15;16(20):1076-85.

谷禾健康

葡萄球菌(Staphylococcus)属于厚壁菌门,是一类革兰氏阳性球菌,因常呈葡萄串状聚集而得名。已经鉴定出大约89种葡萄球菌及亚种。金黄色葡萄球菌(Staphylococcus aureus)为该菌属的模式菌种。
葡萄球菌是兼性厌氧菌,但在有氧条件下生长更快、更丰富,金黄色葡萄球菌亚种和解糖葡萄球菌厌氧除外。
它们可在18°C至40°C、pH4.5~9.8范围内生长,最适温度37°C,最适pH7.4。葡萄球菌可以在高盐(15%氯化钠)和高胆汁浓度(40%胆汁)的环境中生存,展现出强大的适应性。
葡萄球菌分布非常广泛,普遍存在于人类和其他温血动物皮肤、皮肤腺体和粘膜上。但也能在肠道和会阴区域定植。根据谷禾数据库和相关文献统计,葡萄球菌肠道定植的发生率约为20%。过量的葡萄球菌定植于肠道会导致肠道菌群平衡受到破坏,另外由于其具有产毒素的能力,还可能引发肠道炎症、并影响肠道屏障功能。
葡萄球菌以呼吸作用或发酵为主,利用碳水化合物或氨基酸作为碳源和能量来源。大多数菌种发酵葡萄糖的主要产物是乳酸,而在有氧条件下则产生乙酸和CO2。
大多数葡萄球菌为非致病菌,少数可导致疾病,一般为凝固酶阳性葡萄球菌,例如金黄色葡萄球菌,是最常见的化脓性球菌,是医院交叉感染的重要来源。因为它能够产生一种称为凝固酶的物质;当分泌时,凝固酶会导致血液凝结,从而使细菌能够在组织中持续存在并导致脓肿的发展。是导致一系列感染的原因,从轻微的皮肤刺激到肺炎和败血症等危及生命的疾病。
但有一些葡萄球菌也可作为共生菌生活在人体体表或是肠道,了解葡萄球菌的独特结构特征、分布、致病机制及耐药性等对其在人类健康与疾病中的作用至关重要。让我们一起深入认识它。
▸ 发现历史
葡萄球菌(Staphylococcus)是柯赫(1878年)、巴斯德(1880年)和奥格斯顿(1881年)从脓液中发现的,由F. J. Rosenbach于1884年首次纯培养并详细研究。
从金黄色葡萄球菌的细胞壁分离出的蛋白质A可与免疫球蛋白(主要为IgG)进行特异性结合,这是于1959年发现的,现已被应用于各种免疫反应。
▸ 细胞形态
葡萄球菌(Staphylococcus)属于厚壁菌门下的芽孢杆菌目-葡萄球菌科,是一类革兰氏阳性球菌,因常呈葡萄串状聚集而得名。
葡萄球菌细胞呈球形,直径为0.5-1.5μm,单独出现、成对出现、以四分体形式出现或以短链形式出现(3-4个细胞),其特征是在多个平面分裂形成不规则的葡萄状簇。
注:这种簇状结构将葡萄球菌与链球菌区分开来,链球菌通常呈链状生长。
金黄色葡萄球菌10000倍放大倍率下的图像
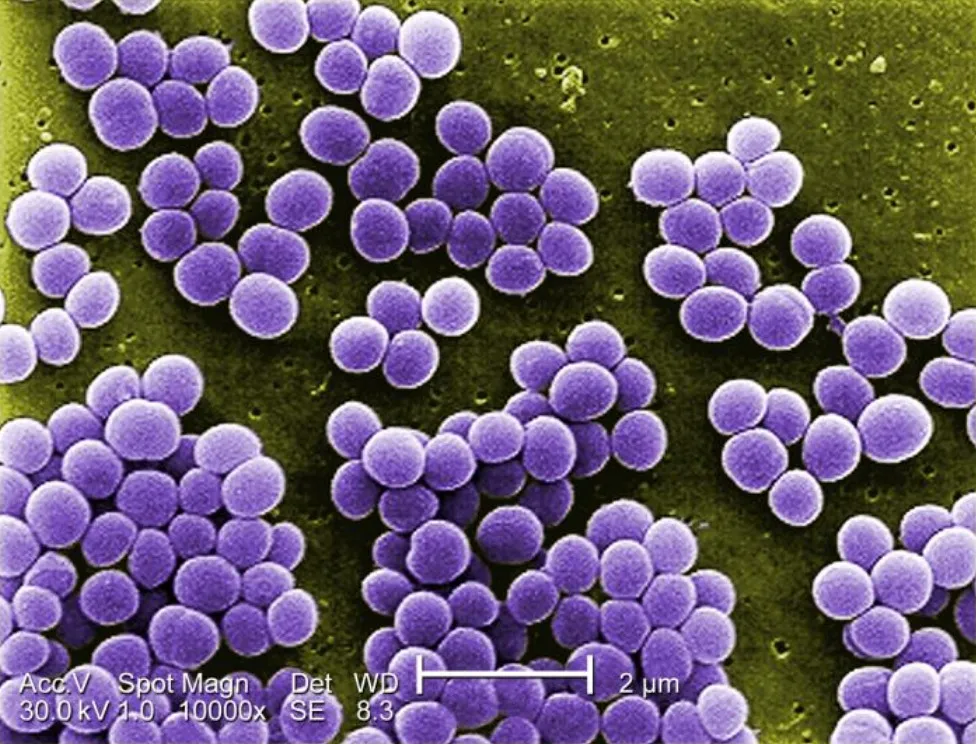
来源:https://www.lecturio.com/concepts/staphylococcus/
葡萄球菌不具备鞭毛或纤毛,因此通常是不运动的,也不形成内生孢子。但葡萄球菌具有厚的、富含肽聚糖的细胞壁,肽聚糖层为细胞提供结构支持,并在维持细胞形状和完整性方面起着重要作用。
固体琼脂上的葡萄球菌菌落通常是圆形、光滑且略微凸起的。颜色可以从白色或奶油色到金黄色不等,具体取决于菌株。
来源:britannica
特别注意,一些葡萄球菌可能会产生围绕细胞壁的多糖保护层,这种细胞外涂层可以保护细菌免受宿主免疫防御,并与毒力增加有关。
▸ 生长代谢
葡萄球菌是兼性厌氧菌,但在有氧条件下生长更快、更丰富,金黄色葡萄球菌亚种和解糖葡萄球菌厌氧除外。
它们可在18°C至40°C、pH4.5~9.8范围内生长,最适温度37°C,最适pH7.4。因此人体的环境非常适宜其定植。此外,葡萄球菌可以在高盐(15%氯化钠)和高胆汁浓度(40%胆汁)的环境中生存,展现出强大的适应性。
一些种类以呼吸作用或发酵为主,利用碳水化合物或氨基酸作为碳源和能量来源。大多数菌种发酵葡萄糖的主要产物是乳酸,而在有氧条件下则产生乙酸和CO2。过氧化氢酶也呈阳性。
营养需求是可变的。大多数物种需要有机氮源,即某些氨基酸和B族维生素。其他的可以在(NH4)2SO4作为底物氮的唯一来源条件下生长。某些物种可能需要尿嘧啶或可发酵的碳源进行厌氧生长。
注:金黄色葡萄球菌能够产生类胡萝卜素,使得菌落呈现从深橙色到浅黄色的特征。此外,它们产生接触酶,并且能够分解各种蛋白质和脂质,如血红蛋白、纤维蛋白、卵白、酪朊、明胶以及脂类,释放出脂肪酸。
▸ 分类
据报道,已经鉴定出大约89种葡萄球菌和亚种。金黄色葡萄球菌(Staphylococcus aureus)为该菌属的模式菌种。
▸ 凝固酶阳性葡萄球菌
根据其血浆凝固酶活性一般可分为两类:凝固酶阳性葡萄球菌和皮肤上常见的凝固酶阴性葡萄球菌。例如金黄色葡萄球菌是一种致病性的物种,属于凝固酶阳性组。此外,凝固酶检测呈阳性的葡萄球菌属物种还包括中间葡萄球菌(S.intermedius)、S.delphini、S.schleiferi subsp和S.hyicus。
注:血浆凝固酶是能使含有枸椽酸钠或肝素抗凝剂的人或兔血浆发生凝固的酶类物质,致病菌株多能产生,常作为鉴别葡萄球菌有无致病性的重要标志。
对人类具有重要意义的金黄色葡萄球菌是伤口感染、疖子和其他人类皮肤感染的主要病原体,是食物中毒的最常见原因之一。金黄色葡萄球菌还会导致脑膜炎、肺炎、尿路感染和乳腺炎。此外,局部葡萄球菌感染可导致中毒性休克综合征,这是一种与毒素从感染部位释放到血液中有关的疾病。
中间葡萄球菌(S.intermedius)是一种狗的机会性病原体,可引起外耳道炎、脓皮病、脓肿、生殖道感染、乳腺炎和化脓性伤口。
S.hyicus被认为是猪感染性渗出性表皮炎和感染性多关节炎、牛和马皮肤病变、家禽和牛骨髓炎的病原体,偶尔与牛的乳腺炎有关。
▸ 凝固酶阴性葡萄球菌
其他葡萄球菌基本都是凝固酶阴性,凝固酶阴性葡萄球菌是人体菌群的重要组成部分。
在凝固酶阴性葡萄球菌中,表皮葡萄球菌(S.epidermidis)是与院内疾病最相关的物种,表皮葡萄球菌是一种轻微的病原体,在抵抗力较低的人群中具有致病性。近年来发现表皮葡萄球菌与菌血症、自体和人工瓣膜心内膜炎、骨髓炎、脓性关节炎、纵隔炎、永久性起搏器感染、血管移植物、脑脊液分流、假体、骨科和泌尿关节以及泌尿道感染(包括尿道炎和肾盂肾炎)有关。
其他凝固酶阴性物种可能也与人类和动物的感染有关。溶血葡萄球菌(S.haemolyticus)与自体瓣膜心内膜炎、脓毒症、腹膜炎和尿路感染有关,有时与伤口、骨骼和关节感染有关。
S.caprae已经发现了感染性心内膜炎、菌血症和尿路感染的病例。
除此之外,其他葡萄球菌见附录。
葡萄球菌种型菌株关系树
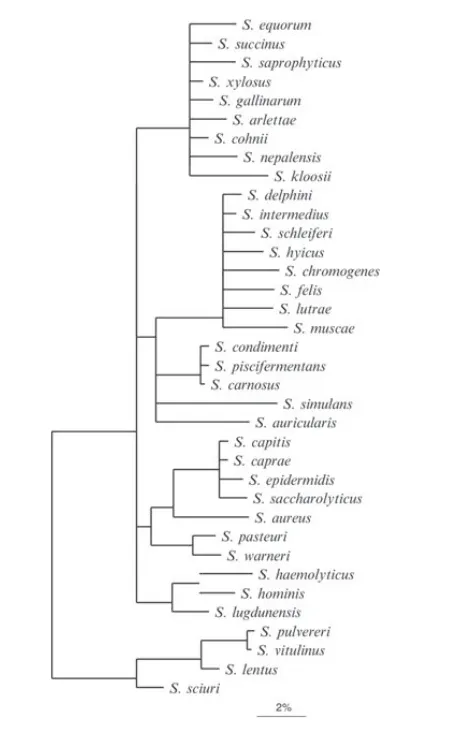
DOI: 10.1002/9781118960608.gbm00569.
▸ 葡萄球菌的分布
▸ 人和动物来源
葡萄球菌物种普遍存在于人类和其他温血动物皮肤、皮肤腺体和粘膜上。它们可能在皮肤上作为常驻细菌或瞬态细菌被发现。
注:常驻细菌是宿主固有的,并主要通过已有细菌的繁殖来增加数量。瞬态细菌来源于外源,主要存在于暴露的皮肤上,很容易被洗掉。
金黄色葡萄球菌可以在许多灵长类动物中发现,尽管偶尔也可以在不同的家畜或鸟类身上发现。在人类中,金黄色葡萄球菌偏爱于鼻腔前部,尤其是在成人中,约30%的人类鼻子和其他粘膜中存在金黄色葡萄球菌。
表皮葡萄球菌是人类皮肤上最常见和持久存在的葡萄球菌。它广泛分布于体表,在前鼻孔、腋窝、腹股沟、会阴区和趾蹼等湿润且营养丰富部位数量最多。偶尔也可在家畜等其他宿主上发现,可能源自人类传播。
人葡萄球菌(Staphylococcus hominis)也普遍存在于人体皮肤上。在大汗腺较多的皮肤部位(如腋窝、腹股沟和会阴区),其种群大小通常次于表皮葡萄球菌。它比其他物种更能成功定居于四肢等干燥皮肤区域。
溶血葡萄球菌与人葡萄球菌有许多共同的栖息地,但通常发现的较少,有些个体可能携带大量溶血葡萄球菌。
头葡萄球菌(Staphylococcus capitis)在青春期后大量繁殖于人类头皮,并存在于成人前额、面部、眉毛和外耳道等部位。耳葡萄球菌(Staphylococcus auricularis)是成人外耳道的主要菌种之一,对该生态位有强烈偏好。
从家禽和山羊中分离出了Staphylococcus arlettae,从马中分离出了Staphylococcus equorum,从家禽中分离出了Staphylococcus gallinarum。
中间葡萄球菌(Staphylococcus intermedius)是家养狗中存在的主要种类;猫葡萄球菌(Staphylococcus felis)是家猫的主要种类之一;
猪葡萄球菌(Staphylococcus hyicus)和产色葡萄球菌(Staphylococcus chromogenes)主要存在于猪、牛和马等家养有蹄类动物身上。Staphylococcus lentus已从家养绵羊和山羊中大量分离出来,偶尔来自其他家畜,是兔唾液中存在的细菌。
▸ 环境和食物来源
葡萄球菌零星地从各种环境来源中分离出来,如土壤、沙滩、海水、淡水、植物表面和饲料、肉类、乳制品,以及炊具、器具、家具、衣服、地毯、纸币的表面,以及各种居住地区的灰尘和空气。
金黄色葡萄球菌已被证实是食物中毒的主要病原体。其他葡萄球菌可能参与某些发酵食品的生产。
发酵肉中的葡萄球菌主要是肉葡萄球菌(Staphylococcus carnosus),50多年来,肉葡萄球菌一直被单独使用或与乳酸菌联合使用,作为生产发酵香肠的发酵剂。在干香肠的成熟过程中,肉葡萄球菌发挥了几种理想的功能。
从酱油醪中分离到调料葡萄球菌(Staphylococcus condimenti);以及从表面成熟的奶酪中提取到Staphylococcus fleurettii,从发酵虾和鱼中提取到的Staphylococcus piscifermentans。
金黄色葡萄球菌和木糖葡萄球菌(Staphylococcus xylosus)可以在仅含无机氮源的环境中生长,因此可能比其他葡萄球菌分布更广泛。这些物种已从沙滩、天然水域、沼泽草和植物产品中分离出来。
▸ 基因组信息
葡萄球菌基因组大小约为2-3 Mbp。目前已完全测序八种菌株(七种金黄色葡萄球菌和一种表皮葡萄球菌),其基因组大小分别为2.82-2.9Mbp和2.5Mbp。
两种菌株均含有1个质粒和3个致病岛。将引起急性感染的金黄色葡萄球菌N315的基因组与引起慢性感染的表皮葡萄球菌RP62A的基因组进行比较。
与金黄色葡萄球菌相比,表皮葡萄球菌毒力基因较少
在金黄色葡萄球菌N315的125个毒力基因中,仅22个(18%)在表皮葡萄球菌RP62A中有同源基因,反映了两者致病潜力的差异。这解释了为何金黄色葡萄球菌是侵袭性病原体,而表皮葡萄球菌则相对无害。
在金黄色葡萄球菌的40个毒素基因中,表皮葡萄球菌仅有3个同源基因。这与表皮葡萄球菌毒素产生减少的观察结果一致,也与其感染严重程度较低相符。此外,与金黄色葡萄球菌相比,表皮葡萄球菌的外泌酶和粘附素等毒力因子数量也明显减少。
▸ 危害与致病性
虽然大部分葡萄球菌可以在人体共存,但一些葡萄球菌物种对人体具有危害,其中最具代表性的是金黄色葡萄球菌(Staphylococcus aureus)。
• 葡萄球菌可能产生毒素
葡萄球菌,尤其是金黄色葡萄球菌,会产生多种毒素,导致其致病性。这些毒素会在感染宿主时引起一系列症状和疾病。葡萄球菌毒素包括:
α-毒素:这种溶细胞毒素会损害细胞膜,并可能导致各种宿主细胞(如红细胞和白细胞)以及感染部位的其他细胞裂解(破裂)。它还可能导致组织破坏。
β-毒素:β-毒素是一种影响细胞膜的鞘磷脂酶。它在红细胞的破坏中发挥作用,并可能导致脓肿的形成。
δ-毒素:δ-毒素是一种小的疏水性肽,可在宿主细胞膜上形成孔。它会损害红细胞并导致细胞裂解。
γ-毒素:γ毒素是一种溶血素,可损害红细胞并促进感染扩散。
去角质毒素:金黄色葡萄球菌会产生剥落毒素,在葡萄球菌烫伤皮肤综合征等情况下会导致水泡和皮肤脱皮。这些毒素会破坏表皮的完整性。
肠毒素:葡萄球菌肠毒素(SEs)`是导致食物中毒的原因。当它们在受污染的食物中产生并摄入时,它们会导致恶心、呕吐、腹泻和腹部绞痛等症状。
中毒性休克综合征毒素(TSST-1):这种毒素与中毒性休克综合征(TSS)有关,可导致快速而严重的疾病,并伴有高烧、皮疹、低血压和器官衰竭等症状。
杀白细胞素(PVL):PVL是一种针对白细胞的毒素,可导致金黄色葡萄球菌引起的皮肤和软组织感染的严重程度。
葡萄球菌超抗原:这些毒素通过激活大量T细胞来刺激夸张的免疫反应。它们是葡萄球菌感染中一些强烈炎症的原因。
• 葡萄球菌的致病种及其危害
金黄色葡萄球菌是多种感染的罪魁祸首。在20世纪50年代末和60年代初,金黄色葡萄球菌作为住院患者的一种医院病原菌,引起了相当大的发病率和死亡率。
该物种引起的主要人类感染包括局部皮肤感染(疖、痈、脓疱疮)、中毒性表皮坏死松解(烫伤皮肤综合征)、肺炎、骨髓炎、急性心内膜炎、心肌炎、心包炎、小肠结肠炎、乳腺炎、膀胱炎、前列腺炎、宫颈炎、脑炎、脑膜炎、菌血症、败血症、中毒性休克综合征,以及肌肉、皮肤、泌尿生殖道、中枢神经系统和各种腹内器官的脓肿。此外,葡萄球菌肠毒素与食物中毒有关。
注:金黄色葡萄球菌也能在多种其他哺乳动物和鸟类中产生感染。我们会在后文详细介绍金黄色葡萄球菌的致病过程和机制。
中间葡萄球菌是犬的一种严重机会致病菌,可引起外耳炎、脓肿、生殖道感染、乳腺炎和脓性伤口感染。Staphylococcus hyicus被认为是猪的感染性渗出性表皮炎(油猪病)和脓毒性多发性关节炎、牛和马的皮肤病变、家禽和牛骨髓炎的病原体,偶尔与牛的乳腺炎有关。Staphylococcus delphini与海豚的化脓性皮肤病变有关。
尽管凝固酶阴性葡萄球菌是人体正常菌群的组成部分,但过去二十年来,其在医院感染中越来越多的被发现。这类感染的增加与假体和留置装置的广泛使用以及免疫功能低下患者数量的增长密切相关。
在凝固酶阴性葡萄球菌中,表皮葡萄球菌是最常与疾病相关的物种。它似乎具有最大的致病潜力和适应多样性。该物种涉及菌血症、原生和人工瓣膜心内膜炎、骨髓炎、化脓性关节炎、持续动态透析期间的腹膜炎、永久性起搏器感染、血管移植、脑脊液分流、假关节和各种骨科装置,以及包括尿道炎和肾盂肾炎在内的尿路感染。
最近发表了关于表皮葡萄球菌和其他凝固酶阴性物种引起的人类感染综述。医院内的耐甲氧西林表皮葡萄球菌(MRSE)菌株在20世纪80年代成为一个严重的临床问题,特别是在植入人工心脏瓣膜或接受其他形式心脏手术的患者中。表皮葡萄球菌有时也与牛的乳腺炎有关。
一些其他凝血酶阴性的葡萄球菌也可能与人类或动物的感染有关。溶血葡萄球菌是人类临床感染中常见的,它与先天性瓣膜心内膜炎、败血症、腹膜炎和尿路感染有关,偶尔与伤口、骨和关节感染有关。
Staphylococcus caprae与感染性心内膜炎、菌血症和尿路感染病例有关。Staphylococcus lugdunensis与天然和人工瓣膜心内膜炎、败血症、脑脓肿、慢性骨关节炎以及软组织、骨、腹膜液和导管感染有关,尤其是在患有基础疾病的患者中。
Staphylococcus schleiferi与人骨膜炎、菌血症、伤口感染以及与颅骨引流管和颈静脉导管相关的感染有关。腐生葡萄球菌(Staphylococcus saprophyticus)是人类尿路感染中一种重要的机会性病原体,尤其是在年轻、性活跃的女性中。它被认为是这些患者中第二常见的尿路感染原因,如急性膀胱炎或肾盂肾炎。该物种偶尔也会从伤口感染和败血症中分离出来。
▸ 耐药性
葡萄球菌对各种抗生素的敏感性或耐药性,可能因菌株而异,并且由于耐药菌株的出现,可能会随着时间的推移而变化。以下是金黄色葡萄球菌抗生素敏感性的概述:
• 一部分菌株耐甲氧西林
一些金黄色葡萄球菌菌株对甲氧西林和其他β-内酰胺类抗生素敏感,包括青霉素和头孢菌素。它们通常可以用一系列抗生素治疗,例如萘夫西林、苯唑西林和万古霉素。
但耐甲氧西林金黄色葡萄球菌(MRSA)菌株已对甲氧西林和许多其他β-内酰胺类抗生素产生耐药性。它们通常对多种抗生素具有耐药性,因此难以治疗。MRSA菌株通常对万古霉素、利奈唑胺、达托霉素等抗生素和头孢洛林等较新药物敏感。
耐甲氧西林金黄色葡萄球菌(MRSA)菌株出现于20世纪80年代,是医院的一个主要临床和流行病学问题。
• 一部分菌株耐万古霉素
金黄色葡萄球菌已显示出对多种抗生素产生耐药性的显著能力,部分原因是获得耐药基因和存在可移动遗传元件,例如质粒。
一些金黄色葡萄球菌菌株对万古霉素的敏感性降低,万古霉素是治疗耐甲氧西林金黄色葡萄球菌(MRSA)的关键抗生素。在极少数情况下,报道了万古霉素-中间金黄色葡萄球菌(VISA)和耐万古霉素金黄色葡萄球菌(VRSA)菌株,使治疗更具挑战性。
除了β-内酰胺类和万古霉素耐药外,一些金黄色葡萄球菌菌株还对其他类别的抗生素产生了耐药性,例如大环内酯类、四环素类和氨基糖苷类抗生素。这导致了多重耐药金黄色葡萄球菌菌株的出现。
下面通过葡萄球菌属中的典型和最引人重视的金黄色葡萄球菌和表皮葡萄球菌,来说明葡萄球菌对人体存在的危害和可能的益处,有助于更深刻地认识葡萄球菌。
金黄色葡萄球菌(Staphylococcus aureus)是最臭名昭著、分布最广的细菌性病原体之一,每年在全球范围内造成难以估计的皮肤感染,以及可能数十万至数百万例更严重的侵袭性感染。
它还是肺炎和其他呼吸道感染、手术部位、假体关节、心血管感染以及院内菌血症的主要病原体。
其他金黄色葡萄球菌感染,如中重度皮肤感染,包括疖、脓肿和伤口感染,通常不会危及生命,但可能伴有明显的发病率和疼痛。由于发病频率高,它们构成了相当大的公共卫生负担。最后,金黄色葡萄球菌也与特应性皮炎的发展有关。
注:2012年的一项综述估计,金黄色葡萄球菌菌血症的发病率为每年20至50例/10万人,其中10%至30%的患者将死于感染。在2017年的一项最新研究中,据报道,美国每年因金黄色葡萄球菌菌血症死亡的人数为2万人。金黄色葡萄球菌菌血症导致的死亡人数比获得性免疫缺陷综合征(艾滋病)、结核病和病毒性肝炎加起来造成的死亡人数还要多。
▸ 感染的来源
金黄色葡萄球菌感染通常源于无症状定植,或更罕见地通过污染物或他人传播,尤其在医院中。鼻孔是其主要定植部位,除肠道外,金黄色葡萄球菌也可定植于多种皮肤部位。
• 通过破损皮肤感染
金黄色葡萄球菌感染依赖于细菌突破上皮屏障。例如,皮肤感染可由轻微划伤发展并可能变为侵袭性。此外,金黄色葡萄球菌可通过α毒素激活含金属蛋白酶结构域的蛋白10(ADAM10),裂解E-钙粘蛋白,破坏粘附连接并损害肌动蛋白细胞骨架。
α毒素在金黄色葡萄球菌感染中的作用
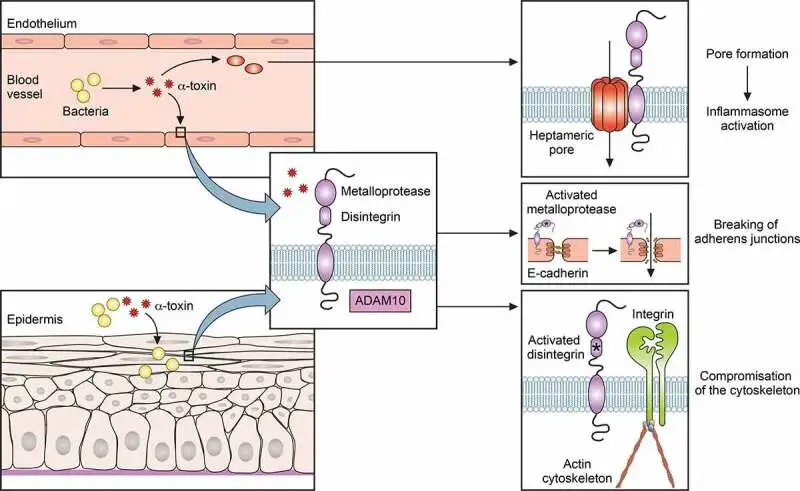
doi: 10.1080/21505594.2021.1878688.
• 通过医疗器械感染
留置医疗器械的污染是医院中常见的感染途径。其主要机制是金黄色葡萄球菌可在器械插入后迅速粘附于塑料材料及其基质分子上,并形成生物膜。
• 通过食物摄入
食物中毒是急性金黄色葡萄球菌感染的一种特殊情况,由摄入含葡萄球菌肠毒素(SE)的受污染食物引起。SE属于超抗原毒素,可非特异性激活T细胞,导致多克隆T细胞活化和大量细胞因子释放,引发过度免疫反应。肠道金黄色葡萄球菌定植可能是其扩散至其他上皮部位的储存库,与其他感染形式存在一定关联。
最后,金黄色葡萄球菌还可作为机会性病原体,利用其他病原体或易感因素引发感染。例如,在流感等病毒感染导致的肺部感染中,继发性金黄色葡萄球菌感染常成为最终致死原因。
▸ 感染的过程
金黄色葡萄球菌全身感染通常始于细菌突破皮肤屏障或从留置医疗器械上的生物膜传播。在血液中,细菌可通过溶细胞毒素攻击并消除中性粒细胞,或在其中存活以实现全身分布。
经过肝脏时,细菌需面对库普弗细胞(Kupffer cell)的吞噬作用,这一阶段是全身感染的关键阶段。一旦存活,细菌可通过血流进一步扩散,依靠MSCRAMM表面蛋白附着并侵入组织细胞。
脓肿的形成受多种细菌因子影响,包括特定表面蛋白、毒素和外酶。
金黄色葡萄球菌的全身感染

doi: 10.1080/21505594.2021.1878688.
• 避免吞噬细胞的杀伤
金黄色葡萄球菌会通过许多机制避免被中性粒细胞消除,包括:
1)抑制中性粒细胞从血流外渗到组织中、中性粒细胞活化和趋化性;
2)通过聚集、保护性表面结构和生物膜形成抑制吞噬作用;
3)抑制调理素作用;
4)抑制中性粒细胞杀伤机制;
5)通过溶细胞毒素直接消除中性粒细胞或触发细胞凋亡。
• 感染的恶化
一旦形成脓肿,除了大量白细胞浸润外,细菌增殖也随之而来。此外,高细菌密度意味着营养物质变得稀缺。金黄色葡萄球菌会产生一系列溶细胞素,可以裂解细胞和酶以消化释放的营养大分子。
除了杀白细胞素对白细胞的溶细胞特性外,一些金黄色葡萄球菌溶细胞素协同作用,实现极强的溶血。
表皮葡萄球菌(S.epidermidis)是迄今为止研究最多的凝固酶阴性葡萄球菌(CoNS)成员。表皮葡萄球菌可从所有皮肤微环境中分离,包括干燥、湿润、皮脂腺和足部。
一项宏基因组学研究显示,不同皮肤微环境和个体间的健康皮肤表皮葡萄球菌分离株具有高度的时空多样性。这些群落在高选择压力下,通过质粒和噬菌体多次水平基因转移,适应并维持其特定的皮肤生态位。
★ 表皮葡萄球菌的多样性非常重要
表皮葡萄球菌菌株水平多样性及其对整体皮肤健康影响的一个机制例子是辅助基因调节(agr)群体感应系统。
表皮葡萄球菌的agr调控着一小部分潜在毒力因子的产生,如蛋白酶、脂肪酶和免疫调节酚溶性调节素(psm),而agr系统的保留对于皮肤定植是必要的。重要的是,每个表皮葡萄球菌菌株都是单一的agr型。虽然大多数个体主要由单一表皮葡萄球菌agr型定殖,但在特定皮肤部位的非显性agr型的小亚群也很常见。
某些表皮葡萄球菌agr类型以及其他凝固酶阴性葡萄球菌物种可以产生抑制非同源的表皮葡萄球菌agr信号传导的小肽。这一观察结果表明,agr异质性与总凝固酶阴性葡萄球菌多样性可能是促进稳态的重要因素。
由于agr活性受细菌密度控制,皮肤上表皮葡萄球菌的绝对数量低也可能导致稳态条件下agr活性低。
▸ 表皮葡萄球菌皮肤定植的益处
表皮葡萄球菌及其作为有益皮肤共生菌的作用受到了广泛关注。
• 激活先天免疫通路,增强抗菌作用
一些表皮葡萄球菌激活了人类角质形成细胞中不同的先天免疫信号通路,以增强抗菌肽(AMP)介导的对金黄色葡萄球菌的杀伤作用,尽管这种增强所必需的分泌因子尚未阐明。
表皮葡萄球菌酚溶性调节蛋白(PSM)是一种小的、两性的α-螺旋肽,在正常表皮和毛囊中大量产生。PSM与宿主AMPs协同作用,增强病原菌化脓性链球菌的杀伤作用。
• 减轻炎症反应
一些表皮葡萄球菌菌株还可以抑制金黄色葡萄球菌诱导的中性粒细胞募集和促炎细胞因子的产生,这可能对更严重的皮肤感染有潜在的保护作用。
在小鼠皮肤损伤模型中,表皮葡萄球菌脂质胆酸和脂肽LP78都通过toll样受体(TLR)-3依赖机制减轻炎症反应,加速伤口愈合。
• 有助于适应性免疫的发育和启动
除了调节对皮肤感染或损伤的先天免疫反应外,表皮葡萄球菌的定植还有助于适应性免疫系统的发育和启动。对共生小鼠的研究表明,表皮葡萄球菌的皮肤定植对于效应T细胞的发育和功能,以及粘膜相关不变T细胞(MAIT细胞)的早期定位和启动是必要的。
注:MAIT细胞是介导宿主-共生串扰不同模式的非经典皮肤免疫信号传导的重要组成部分。
皮肤也是最大的效应T细胞亚群库之一,人们越来越认识到这些组织识别淋巴细胞和表皮葡萄球菌等定植体之间串扰的深度和复杂性。
综上所述,表皮葡萄球菌在启动先天和适应性防御病原体和促进体内平衡方面无疑是重要的。未来可能会揭示其他凝固酶阴性葡萄球菌,与表皮葡萄球菌一起,对皮肤发育和健康的共同作用。
▸ 表皮葡萄球菌的潜在危害
虽然被广泛认为是一种丰富的皮肤共生体,但新出现的证据表明,表皮葡萄球菌特定菌株在皮肤上的定植实际上可能在某些条件下对宿主有害。完整的皮肤对病原体和共生生物都是一个强大的屏障,但是通过基因突变或物理破坏这种屏障,可以极大地改变表皮葡萄球菌的行为,从良性到致病性。
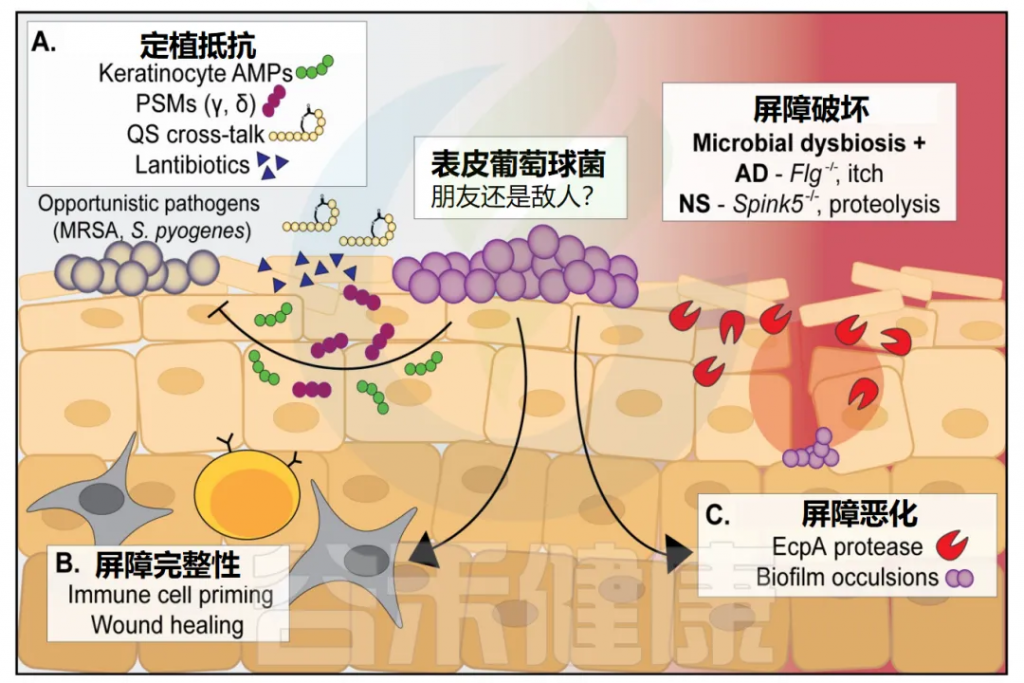
• 表皮葡萄球菌只有在皮肤完整时具有抗菌能力
表皮葡萄球菌预处理的小鼠皮肤只有在屏障完好无损时才对金黄色葡萄球菌的攻击有抵抗力,而在细菌接种前被物理破坏时则没有。
• 可能加剧特应性皮炎
在特应性皮炎(AD)中,患者通常在病灶部位高度定植金黄色葡萄球菌,这种细菌“繁殖”与疾病严重程度呈正相关。纵向宏基因组学研究表明,一些AD患者在病变部位可以被表皮葡萄球菌而不是金黄色葡萄球菌高度定植。据推测,这种生长可能同样与疾病严重程度相关;然而,关于表皮葡萄球菌介导的AD屏障恶化的机制基础的研究很少。
最近,半胱氨酸蛋白酶EcpA被确定为表皮葡萄球菌诱导的特应性皮炎屏障降解的关键介质。EcpA存在于所有表皮葡萄球菌菌株中,但似乎仅由一个亚群表达。EcpA降解多种皮肤屏障成分,包括LL-37和粘粒素-1,并在AD小鼠模型中显著增加炎症和屏障功能障碍。
最后,表皮葡萄球菌形成生物膜的倾向也可能加剧特应性皮炎(AD),因为在AD病变部位的一些汗腺中已经发现了金黄色葡萄球菌和表皮葡萄球菌的炎症生物膜群落。
• 恶化内瑟顿综合征
除了特应性皮炎,表皮葡萄球菌的过度扩张和EcpA的产生也与内瑟顿综合征(NS)的恶化有关,内瑟顿综合征是一种皮肤疾病,以丝氨酸蛋白酶活性高水平为特征,由spink5基因突变引起。
重要的是,EcpA的产生受表皮葡萄球菌agr群体感应系统的调节。这表明表皮葡萄球菌可能通过生态失调和agr信号去抑制加重特应性皮炎和内瑟顿综合征,其中抑制型表皮葡萄球菌或共生菌的减少是关键因素。
在这两种皮肤病中,EcpA和其他毒力因子的表达增强,再加上遗传或环境屏障的破坏,将为表皮葡萄球菌的扩张和恶化提供理想的环境。
除了金黄色葡萄球菌和表皮葡萄球菌外,还存在一些可能致病的葡萄球菌物种或是一些可以在人体共生的葡萄球菌。
1
溶血葡萄球菌
溶血葡萄球菌(Staphylococcus haemolyticus)是人类皮肤微生物群的组成部分。它已成为医院感染的一个重要原因,在医院和医务人员中普遍存在。
与其他凝固酶阴性葡萄球菌相比,溶血葡萄球菌分离株,特别是那些导致医院获得性感染的葡萄球菌,表现出更高水平的抗生素耐药性。大量证据表明,溶血葡萄球菌具有将耐药基因传播给其他葡萄球菌的能力。
它在免疫功能低下的人群中尤其普遍,并与严重感染有关,包括脑膜炎、心内膜炎、假体关节感染、菌血症、败血症、腹膜炎和中耳炎。
建立生物膜的能力是致病性溶血葡萄球菌分离株的一个显著特征,溶血葡萄球菌分泌多种促进细菌粘附和侵袭的因子,以及生物膜的形成,如肠毒素、溶血素和纤维连接蛋白结合蛋白。
2
沃氏葡萄球菌
沃氏葡萄球菌(Staphylococcus warneri)是一种凝固酶阴性的机会致病菌,已知可引起多种感染,特别是在留置医疗器械的患者中。
沃氏葡萄球菌是皮肤菌群的成分,特别集中在头部,颈部,手臂和大腿。作为一种机会致病菌,沃氏葡萄球菌的重要性正在上升。
从这些感染中分离的大多数沃氏葡萄球菌菌株对β-内酰胺类抗菌药物耐药。其毒力与粘附素、酶、细胞外毒素、荚膜、铁摄取系统、毒力调节因子、生物膜形成及侵入和损伤上皮细胞的能力有关。目前尚无法完全解析其致病机制,包括逃避宿主免疫系统并持续存在的策略。
3
头葡萄球菌
头葡萄球菌(Staphylococcus capitis)于1975年首次从人体皮肤中分离出来。在特定菌株中发现的蛋白质对生物膜的形成、粘附性和毒力至关重要。
一项研究发现,头葡萄球菌TE8菌株含有14种促进粘附的粘附素,从而促进其在人体皮肤上的定植。根据另一项研究,在从患者胃中分离出的所有凝固酶阴性葡萄球菌中,S.capitis分离株表现出最强的尿素酶活性。它们建立生物膜的能力是S.capitis物种的主要致病因素。
4
人葡萄球菌
人葡萄球菌(Staphylococcus hominis)是凝固酶阴性葡萄球菌中第三大常见病原体,可在新生儿和免疫抑制的成人血液中引起机会性感染。近年来,由人葡萄球菌引起的菌血症、败血症、眼内炎和心内膜炎的报告病例数量显著增加。
人葡萄球菌对多种抗菌剂具有耐药性,给治疗带来挑战。其耐药性源于获得mecA基因,该基因通过改变青霉素结合蛋白,降低了对青霉素等β-内酰胺类抗生素的亲和力。此外还发现了耐甲氧西林人型葡萄球菌(MRSHo)的出现。
尽管对几种抗生素具有耐药性,人葡萄球菌仍然对包括头孢他林、四环素、氨基糖苷类、头孢菌素、糖肽类和脂糖肽类(如万古霉素),以及其他抗菌剂如达托霉素的治疗敏感。
葡萄球菌属还有几十种,在此就不一一列举了。
附录:其他葡萄球菌
Staphylococcus agnetis
Staphylococcus argensis
Staphylococcus argenteus
Staphylococcus arlettae
Staphylococcus aureus
Staphylococcus auricularis
Staphylococcus capitis
Staphylococcus caprae
Staphylococcus carnosus
Staphylococcus chromogenes
Staphylococcus cohnii
Staphylococcus condimenti
Staphylococcus delphini
Staphylococcus devriesei
Staphylococcus epidermidis
Staphylococcus equorum
Staphylococcus faecalis
Staphylococcus felis
Staphylococcus fleurettii
Staphylococcus gallinarum
Staphylococcus haemolyticus
Staphylococcus hominis
Staphylococcus hyicus
Staphylococcus intermedius
Staphylococcus kloosii
Staphylococcus leei
Staphylococcus lentus
Staphylococcus lugdunensis
Staphylococcus lutrae
Staphylococcus lyticans
Staphylococcus massiliensis
Staphylococcus microti
Staphylococcus muscae
Staphylococcus nepalensis
Staphylococcus pasteuri
Staphylococcus petrasii
Staphylococcus pettenkoferi
Staphylococcus piscifermentans
Staphylococcus pseudintermedius
Staphylococcus pseudolugdunensis
Staphylococcus rostri
Staphylococcus saccharolyticus
Staphylococcus saprophyticus
Staphylococcus schleiferi
Staphylococcus schweitzeri
Staphylococcus sciuri
Staphylococcus simiae
Staphylococcus simulans
Staphylococcus sp.
Staphylococcus sp. 3348O2
Staphylococcus sp. C9I2
Staphylococcus sp. WB18-16
Staphylococcus sp.C10c
Staphylococcus sp.FZ-6
Staphylococcus stepanovicii
Staphylococcus succinus
Staphylococcus vitulinus
Staphylococcus warneri
Staphylococcus xylosus
environmental samples
unclassified Staphylococcus
主要参考文献
Britannica, T. Editors of Encyclopaedia (2024, May 3). staphylococcus. Encyclopedia Britannica.
Ahmad-Mansour, N., Loubet, P., Pouget, C., Dunyach-Remy, C., Sotto, A., Lavigne, J. P., & Molle, V. (2021). Staphylococcus aureus Toxins: An Update on Their Pathogenic Properties and Potential Treatments. Toxins, 13(10), 677.
Karl-Heinz Schleifer, Technische Universität München, Lehrstuhl für Mikrobiologie, Am Hochanger 4, Freising D-85350,Germany Julia A. Bell, Michigan State University, Food Safety and Toxicology Center, East Lansing, MI 48824, USA
Paharik AE, Horswill AR. The Staphylococcal Biofilm: Adhesins, Regulation, and Host Response. Microbiol Spectr. 2016 Apr;4(2):10.1128/microbiolspec.VMBF-0022-2015.
Brown MM, Horswill AR (2020) Staphylococcus epidermidis—Skin friend or foe? PLoS Pathog 16(11): e1009026.
Cheung GYC, Bae JS, Otto M. Pathogenicity and virulence of Staphylococcus aureus. Virulence. 2021 Dec;12(1):547-569.
Heath, V., Cloutman-Green, E., Watkin, S., Karlikowska, M., Ready, D., Hatcher, J., Pearce-Smith, N., Brown, C., & Demirjian, A. (2023). Staphylococcus capitis: Review of Its Role in Infections and Outbreaks. Antibiotics (Basel, Switzerland), 12(4), 669.
Becker, K., Heilmann, C., & Peters, G. (2014). Coagulase-negative staphylococci. Clinical microbiology reviews, 27(4), 870–926.
medicallabscientist.org/staphylococcus/
biologyinsights.com/understanding-staphylococcus-types-diagnosis-and-resistance/
lifeder.com/staphylococcus

谷禾健康
随着冬季到来,天气逐渐转冷,大部分呼吸道病原体在外界存活时间延长,同时人体的免疫力下降,呼吸道感染性疾病进入高发季节。
2024年第48周(11月25日—12月1日),对全国哨点医院(不含港澳台)采集的门急诊流感样病例和住院严重急性呼吸道感染病例的呼吸道样本,开展了多种呼吸道病原体检测,检测结果如下表所示:
呼吸道病原体检测阳性率(%)
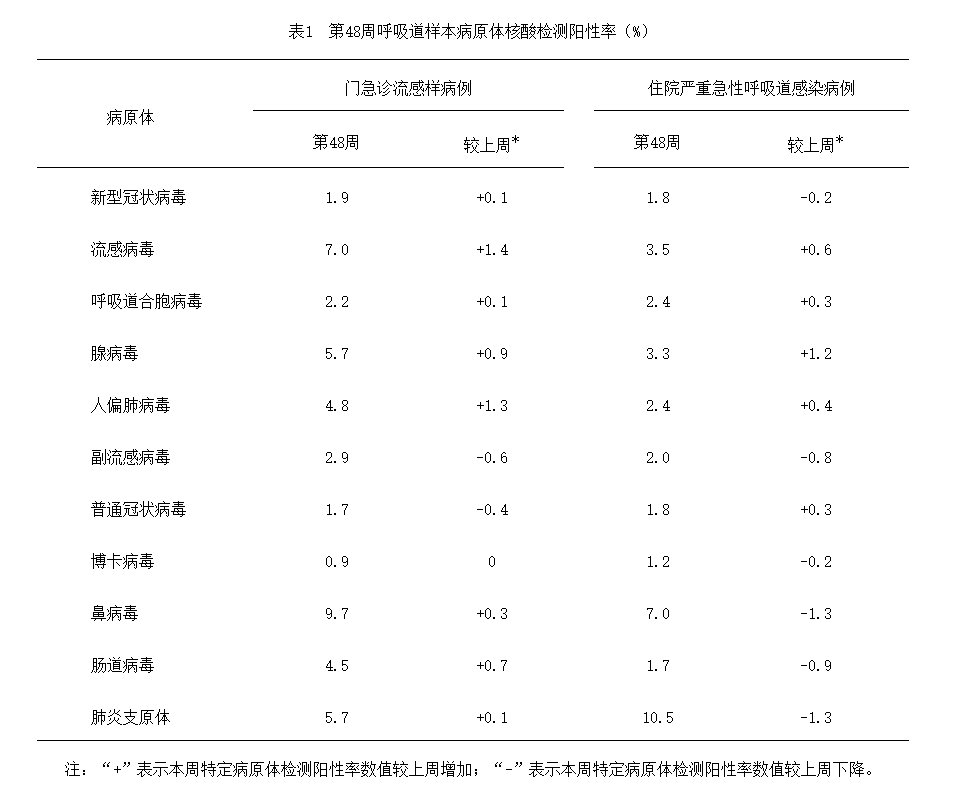
来源:中国疾病预防控制中心
呼吸道主要病原体检测阳性率区域差异

来源:中国疾病预防控制中心
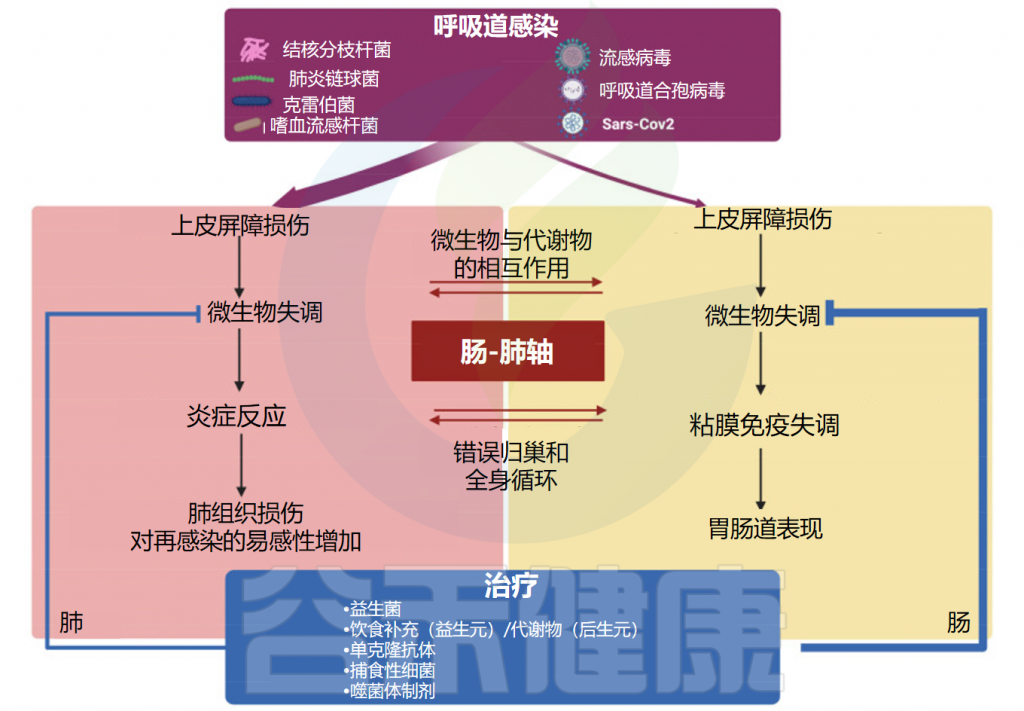
呼吸道感染是最常见的传染病类别,也是全球发病率和死亡率的主要原因之一。2019年冬天爆发新型冠状病毒(SARS-CoV-2)就对人类产生了深远且持续的影响,影响了健康、日常生活、经济和心理等多个方面。
呼吸道是一个复杂的器官系统,分为不同的区域;上呼吸道包括鼻腔、咽和喉,而下呼吸道则由传导气道(气管和支气管)、小气道(细支气管)和呼吸区(肺泡)组成。曾经被认为是无菌器官的健康肺现在被描述为拥有自己的特定微生物种群——肺微生物群。
肺微生物群被认为是不断吸入和消除的细菌的短暂定居点。这些共生细菌作用于免疫系统,诱导保护性反应并防止病原体入侵和定植。同时,它们通过生产抗菌物质直接抑制病原体的生长。共生菌与常驻上皮细胞和免疫细胞之间的持续互作支持肺稳态。
肺部微生物群被视为“肺部健康状况的镜子”:多项研究表明,在肺部疾病发生期间,肺部细菌组成和肺部环境会发生巨大变化。另一方面,肺与肠道进行连续的双向串扰,并且任一部位微生物群组成的改变都会导致远端疾病的发展和进展。
在本文中,我们将重点关注宿主-微生物群相互作用在健康和最常见的呼吸道细菌(结核分枝杆菌、肺炎链球菌、肺炎克雷伯菌和流感嗜血杆菌)和病毒(流感病毒、呼吸道合胞病毒和严重急性呼吸系统综合症冠状病毒)感染中的作用,描述肺微生物群在生理防御以及疾病环境中的变化。
最后,本文总结了基于微生物群的治疗方法在呼吸道感染中的作用,包括新型微生物基础疗法(益生菌和膳食补充剂)以及微生物靶向疗法(抗菌单克隆抗体和噬菌体),有助于重塑常驻微生物群落并恢复呼吸道健康。
doi: 10.3390/ijms25074051.
肺部微生物群是指存在于肺部的微生物群落,包括细菌、病毒、真菌和其他微生物。尽管肺部长期被认为是无菌的,但近年来的研究表明,健康的肺部也存在复杂的微生物群,其组成和功能对维持肺部健康和免疫平衡至关重要。
★ 肺部相比肠道微生物较少,但也存在
肺部环境通常不适合细菌群落的发育,导致与肠道环境相比,细菌复制率相对较低,生物量也较低(103-105VS1011-1012个细菌/克组织)。
健康个体的上呼吸道(URT)微生物群更复杂一些,其中口腔共生菌普遍存在。相比之下,下呼吸道(LRT)的微生物群生物量较低,其组成受上呼吸道微吸入的影响,同时因粘膜纤毛活动和先天免疫功能的作用,局部微生物复制受到限制。
大量研究表明,健康的肺部含有拟杆菌门(Bacteroidetes)和厚壁菌门(Firmicutes)的微生物群,其中普雷沃氏菌(Prevotella)、链球菌(Streptococcus)、奈瑟菌(Neisseria)、嗜血杆菌属(Haemophilus)、梭杆菌属(Fusobacterium)和Veilonella是最丰富的属。
16S rRNA 分析确定了健康URT和LRT之间微生物群的空间差异。从口腔到下肺不断发现与链球菌属的细菌重叠。然而,葡萄球菌通常栖息在上呼吸道,而普雷沃氏菌属和韦荣氏菌主要栖息在下呼吸道。
★ 呼吸道微生物定植在出生后就开始
呼吸道定植在出生后24小时内立即开始。在健康状态下,来自链球菌属(Streptococcus)、葡萄球菌属(Staphylococcus)、普雷沃氏菌属(Prevotella)、莫拉氏菌属(Moraxella)、嗜血杆菌属(Haemophilus)、乳杆菌属(Lactobacillus)、棒状杆菌属(Corynebacterium)和Dolosigranulum的物种代表了生命早期呼吸道的初始定植者,有助于平衡和动态的微生物群落。
从新生儿到成熟期,呼吸道中的细菌数量逐渐增加,多样性和功能能力受到分娩方式、母乳喂养、抗生素使用和烟雾暴露等因素的影响。动物模型研究显示,出生后前两周内细菌量明显增加,菌群从γ‐变形菌纲和厚壁菌门逐步转向拟杆菌门。
★ 微生物可以通过空气吸入和扩散进肺部
微生物迁移是通过吸入空气中的细菌、沿粘膜表面直接扩散发生的,后者被认为是健康受试者中占主导地位和普遍存在的途径。事实上,下呼吸道的细菌群落在很大程度上类似于口腔微生物群的组成。
与微生物群落具有高度抗性的高生物量粘膜(即口腔、肠道)的微生物相比,肺微生物组的动态性质可能是一个重要的独特特性。
★ 影响肺部微生物群的因素
影响肺微生物组成的因素主要有以下几个方面:
1.宿主因素
-年龄
-性别
-体重
-遗传因素
-健康状况(基础疾病)
2.环境因素
-空气质量
-吸烟
-职业暴露
-居住环境
3.生活方式因素
-饮食
-口腔卫生
-运动
-抗生素使用
4.其他因素
-季节变化
-医院环境暴露
-抗生素的使用
-疫苗接种
此外还有局部微生物竞争、宿主上皮细胞相互作用和免疫细胞活化等。术语“菌群失调”表示健康肺微生物组的三个决定因素(微生物迁移、微生物消除和繁殖率)平衡的变化,肺部菌群失调与许多不良生物学事件有关,并参与呼吸系统疾病的发生和发展。
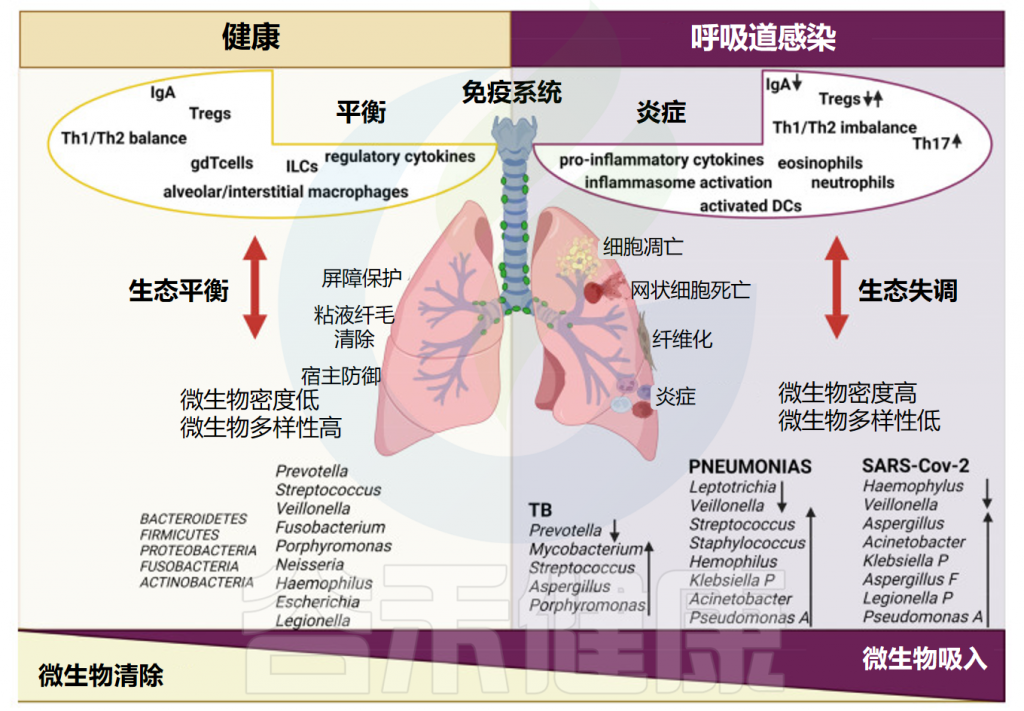
★ 肺部微生物群的变化与疾病存在相关性
比较患病肺和健康肺发现,肺部微生物组成存在显著差异,疾病状况与细菌多样性的丧失有关,或者与一小群分类群的优势有关。微生物菌群失调是各种肺部疾病的特征,其中微生物多样性减少可能与疾病进展有关。
肺微生物组的生长条件在病理过程中发生了显著变化,导致适合受伤气道的疾病和患者的微生物群落。例如肺部富集的口腔厌氧菌群(如Prevotella和Veillonella)与炎症增强和免疫反应改变导致的感染易感性增加有关。
此外,肺微生物群成分的改变对某些个体造成更大的感染风险。肺微生物群的改变与多种肺部疾病的恶化有关,不同的细胞免疫反应与暴露于各种肺部微生物有关。例如在慢性肺部炎症中,来自病理性人支气管肺泡系统的假单胞菌(Pseudomonas)和乳杆菌(Lactobacillus)的富集与Th17型反应的增强相关。一些致病体还会诱导严重的不依赖性Toll样受体2的气道炎症和肺部免疫病理学。
呼吸道微生物组成影响人类和小鼠的炎症反应,调控肺γδ T细胞产生IL-17及肺泡巨噬细胞的分化,并影响抵抗呼吸道感染的能力。副流感嗜血杆菌通过TLR4激活促炎反应,阻碍皮质类固醇通路,诱导炎症性Th2途径,最终导致支气管反应。
健康和呼吸道感染期间的肺微生物群和免疫
doi: 10.3390/ijms25074051.
肺部感染包括病原体的获得、传播和侵入下呼吸道。
肺炎是肺实质的炎症。在病因学上,它被归类为社区获得性肺炎(先前健康的个体的感染)或医院获得性肺炎(住院个体在入院后48小时内的感染)。革兰氏阳性肺炎链球菌(Streptococcus pneumoniae)是大多数社区获得性肺炎病例的原因,也是2岁以下儿童、老年人和免疫功能低下个体患病的主要原因。
★ 不同人群易感的病原体有所差异
由流感嗜血杆菌和肺炎克雷伯菌引起的肺炎易在50岁以上的慢性阻塞性肺病或酗酒患者中传播,而由肺炎支原体和肺炎衣原体引起的肺炎则在儿童中更为广泛。由呼吸道合胞病毒(RSV)和腺病毒引起的病毒性肺炎在健康人群中很少见,而由流感病毒引起的肺炎仍然是老年人和基础疾病患者高死亡率的原因。
在成人中,肺炎链球菌(S.Pneumonia)、乳杆菌(Lactobacilli)和罗斯氏菌属(Rothia)的优势与肺炎有关。新生儿中肺炎链球菌、流感嗜血杆菌或莫拉菌的定植与患支气管炎的风险较高有关。在流感患者中还发现假单胞菌增多。
主要病原体检测阳性率年龄组差异

来源:中国疾病预防控制中心
★ 呼吸道病原体感染导致肺部疾病的发展
人体研究表明,不同菌株的早期定植与发生呼吸道感染的风险增加有关。因此,病原体感染和微生物相互作用都可能影响疾病的进程。从生态学上讲,感染的特点是微生物负荷增加和群落多样性减少,以及宿主炎症和组织损伤增加。上呼吸道微生物组的改变是导致肺炎的细菌感染发生的原因。
最近的一项研究描述了严重肺部感染中病毒复制的显著增加,以及轻度和重度疾病患者之间微生物相互作用的差异,尤其是与常见病原菌之间的关联。流感导致的高死亡风险主要归因于继发性细菌感染。病毒通过多种机制增加了宿主对细菌定植的脆弱性。
注:鼻内接种H1N1病毒以模拟人类流感的小鼠微生物组、肺转录组和代谢组显示出显著变化,这些变化在恢复期持续存在。这表明感染对微环境的稳态产生了长期影响,有利于潜在病原体。
结核病
结核分枝杆菌(Mtb)是导致肺结核(TB)疾病中肺部肺炎的原因。在小鼠和人类身上进行的研究表明,肺部微生物组在抵抗结核分枝杆菌感染方面发挥作用。
结核病患者肺部存在特殊的菌群富集
一些细菌菌株可能与结核病的发病、复发和治疗失败有关。例如假单胞菌的丰度与治疗失败的风险增加有关。与健康对照组相比,感染结核分枝杆菌的个体的微生物群多样性降低,并且经常表现出链球菌和假单胞菌的富集。
在结核病中,肺部和整个下呼吸道具有特殊的微生物特征:结核分枝杆菌、金黄色葡萄球菌和乳酸克鲁维酵母(Kluyveromyces lactis)在123名结核病患者的支气管肺泡灌洗液 (BALF)中高度富集,而副流感嗜血杆菌在未感染的肺部富集。
使用鼻咽拭子样本的研究表明,结核分枝杆菌感染显著改变了微生物群组成:与健康对照组相比,结核病患者的变形杆菌、γ-变形杆菌、假单胞菌和莫拉菌科增加,而芽孢杆菌和毛螺菌属减少。
肺泡微生物多样性降低可能由炎症环境引起
与健康对照组相比,结核病患者的肺泡微生物群多样性降低(链球菌和梭杆菌减少,结核分枝杆菌丰度增加)。这些变化可能是由炎症环境决定的,因为结核分枝杆菌可以释放毒力因子,从而抑制巨噬细胞的反应。
在另一项研究中,链球菌在结核病中显著增加,而结核病中的Th1反应可能是由奈瑟菌(Neisseria)和嗜血杆菌(Haemophilus)引发的。
新冠肺炎
2019年,一种新型冠状病毒严重急性呼吸系统综合症2(SARS-CoV-2)引起了新型冠状病毒病(COVID-19)大流行。疾病严重程度和死亡率因年龄及合并症而异,其中许多与肠道和肺部微生物改变有关。这表明菌群失调可以在一定程度上预测COVID-19的严重程度。
新冠病毒感染者肺部微生物群失调
一般来说,与健康或病情较轻的COVID-19个体相比,危重症COVID-19患者的肺部物种多样性降低,细菌负荷增加。不良的临床结果与支原体的下呼吸道富集有关。
一项研究描述了来自重症COVID-19患者的支气管肺泡灌洗液(BALF)样本,假单胞菌丰度显著更高,而来自COVID-19肺炎阴性患者的 BALF 的特征是流感嗜血杆菌和韦荣氏球菌等的富集。
据报道,20名已故 COVID-19 患者的肺组织微生物组以不动杆菌(Acinetobacter)为主,不动杆菌通常与引起肺炎的肺部感染有关。
在上呼吸道和下呼吸道中,宿主免疫系统对潜在的有害物质做出反应,并将它们与自身成分、外来非危险物质和有益共生微生物群区分开来。
◆ 气道上皮产生粘蛋白形成抵御病原体的屏障
气道上皮是与当地微生物群相互作用的场所,也是生物物理保护屏障。产生粘液的气道上皮分泌细胞是肺先天免疫系统的关键要素。一些数据表明,呼吸道微生物群调节塑造粘液产生的上皮屏障。具体来说,在小鼠模型中,证明了粘蛋白与肺部感染反应之间存在联系。
◆ 产生lgA阻止病原体的吸附
除粘蛋白外,气道上皮细胞还提供抗原特异性分泌型IgA屏障,能够保护气道表面。分泌型IgA的作用是阻止病原体的吸附,中和它们在粘膜上皮的毒性产物,介导感染上皮细胞中病毒的消除,并促进对病原体的杀伤。
多项研究证明,IgA在防御呼吸道感染中发挥重要作用:IgA缺陷小鼠对牛分枝杆菌卡介苗(BCG)鼻内感染的易感性增加。鼻内给药中和IgA后,接种流感病毒(IV)或呼肠孤病毒导致小鼠模型中肺部病毒滴度显著降低,肺炎严重程度减轻。值得注意的是,静脉注射抗原特异性聚合物IgA可通过血清中的IgA分泌保护小鼠免受流感感染。
◆ 分泌抗菌肽来对抗病原体
局部呼吸道还分泌称为抗菌肽(AMP)的保护性介质,包括溶菌酶、乳铁蛋白、脂质运载蛋白、过氧化物酶、氨肽酶、集合蛋白(表面活性剂蛋白A和表面活性剂蛋白D)、甘露聚糖结合凝集素(MBL)、cathelicidins和β-防御素。
值得注意的是,β-防御素和cathelicidins具有抗菌和免疫调节作用,并参与塑造微生物群组成。事实上,炎症反应的直接抗菌活性和免疫调节是由微生物群的成员触发的。
免疫细胞在呼吸道病原体感染中至关重要,是机体防御系统的核心。接下来,我们一起来了解一下不同免疫细胞与病原微生物感染之间的相互作用。
▸ 巨噬细胞
肺中存在不同类型的先天免疫细胞:肺泡巨噬细胞(AMs)是最具代表性的细胞,似乎是几种呼吸道感染发病机制的核心,包括结核分枝杆菌(MTB)、肺炎链球菌、鼻病毒、流感病毒(IV)和呼吸道合胞病毒(RSV)。
◆ 肺巨噬细胞失调影响呼吸道感染的死亡率和并发症
AMs启动白细胞募集,并使用几种病原体特异性机制直接消除病原体,例如分泌促炎细胞因子/趋化因子(IL-6、IL-8或CXCL10),启动I型IFN信号传导,增强模式识别受体的表达,以及抑制病毒基因组的核输出。
并发症发生率和死亡率的增加与肺泡巨噬细胞耗竭有关,无论是在实验过程中还是在自然过程中,呼吸道病毒感染都是相关的。此外还发现,重症COVID-19的一个一致特征是肺巨噬细胞失调。
▸ 树突状细胞
肺部的树突状细胞(DC)在防止呼吸道感染方面发挥作用,对病原体产生强大的适应性免疫反应。树突状细胞可以启动抗病毒CD8细胞毒性T细胞反应,导致病毒清除,还可以控制炎症反应的水平。
◆ 树突状细胞可作为继发性肺炎的生物标志物
炎症期间和之后的树突状细胞(DC)改变可用作继发性肺炎易感性的生物标志物,以及有希望的治疗靶点,以改善患者的预后。在结核分枝杆菌(MTB)感染中,DC在将先天免疫反应转变为适应性免疫方面发挥着多因素作用。值得注意的是,树突状细胞在防御SARS-CoV-2感染方面也发挥着关键作用。
▸ 黏膜相关不变T细胞
黏膜相关不变T(MAIT)细胞是非常规T细胞的一个亚群,对微生物感染进行免疫监视和免疫反应。
◆ 进行免疫监视和免疫反应
不同的研究表明,MAIT 细胞通过识别和杀死细菌感染的细胞(包括树突状细胞和肺上皮细胞)在结核分枝杆菌感染的免疫控制中发挥作用。
此外,MAIT细胞在人和小鼠中都需要细胞因子来响应结核分枝杆菌抗原,这可能是由于它们在感染部位募集。一项新的研究证实,急性SARS-CoV-2感染患者循环中的MAIT被激活,但频率降低,而它们在致命性COVID-19患者的肺部蓄积。
▸ 自然杀伤T细胞
不变自然杀伤T(iNKT)细胞在控制共生菌(包括机会性病原微生物群)中发挥作用,反过来,微生物群调节iNKT细胞。
在原发性呼吸道合胞病毒(RSV)感染期间,肺iNKT细胞的激活除了导致肺嗜酸性粒细胞增多和纤维化外,还导致抗病毒CD8 T淋巴细胞反应和病毒清除。
◆ 活动性结核病患者存在iNKT细胞缺乏
人类iNKT细胞缺乏可能是活动性/急性结核病发展的基础:与潜伏性结核病患者相比,活动性结核病患者的外周iNKT细胞较少,并且通过活动性结核病的治疗可以重建正常的iNKT细胞频率。据报道,重症 COVID-19 肺炎患者 iNKT 细胞减少,表明该亚群可能作为疾病严重程度的生物标志物发挥作用。
▸ γδT细胞
肺驻留γδT细胞是粘膜上皮屏障的主要T细胞成分,对维持肺稳态和影响多种肺部疾病的进展至关重要。γδT细胞是肺炎克雷伯菌感染中 IL-17A 的主要来源,也是宿主对急性铜绿假单胞菌肺部感染的早期免疫防御。
◆ γδT细胞对肺部感染进行早期免疫防御
在肺炎链球菌肺部感染期间,观察到活化的γδ T细胞数量显著增加。γδ T 细胞对冠状病毒感染的反应仍在研究中,之前关于SARS-CoV-2感染的报道显示对感染的靶标单核细胞系具有很强的溶细胞活性。
▸ 调节性T细胞
调节性T(Treg)细胞对于肺对空气传播的过敏原的免疫耐受以及减少对自身和非自身抗原的危险免疫反应至关重要。Treg耗竭通过抗原致敏在衣原体肺炎感染的发病机制中发挥作用。
◆ 调节性T细胞对肺炎链球菌肺炎具有保护作用
此外,Tregs通过与TGF-β通路相关的机制对肺炎链球菌引起的肺炎具有保护作用。其他研究使用呼吸道合胞病毒和甲型流感病毒的小鼠模型表明,Treg细胞的耗竭可能导致CD8+ T细胞亚群的迁移延迟。
最近的研究显示,COVID-19患者的Tregs数量显著减少,导致Treg/Th17比值失衡,这与呼吸衰竭风险相关。
此外,呼吸道微生物群的成员通过TLR反复暴露于病原体相关分子模式(PAMPs)和损伤相关分子模式(DAMPs),诱导树突状细胞和肺泡巨噬细胞的模式识别受体(PRR)耐受性。总之,呼吸道微生物群与气道上皮细胞和吞噬细胞形成正反馈回路,以实现免疫耐受并避免失控的炎症反应。
呼吸道中的宿主-微生物相互作用主要发生在粘膜部位。常驻微生物可以局部或全身引发免疫细胞(如上皮细胞、树突状细胞和中性粒细胞)的反应。这些物质能够进入循环并到达其他器官。
◆ 呼吸道微生物群与免疫和呼吸健康密切相关
呼吸道微生物组为宿主免疫系统提供重要信号,这些信号对免疫训练、器官发生和免疫耐受的维持至关重要。观察结果表明,在生命早期存在一个关键时期,正确的微生物群感知对免疫成熟和呼吸健康至关重要。
动物模型研究了微生物组在影响和管理宿主免疫系统中的作用,以及免疫系统在塑造微生物组中的作用。
例如,在小鼠模型中,拟杆菌门的成员相比流感嗜血杆菌减少了炎症、中性粒细胞募集和TLR2介导的细胞因子产生。
鼻内接种金黄色葡萄球菌导致单核细胞募集到肺部,并分化为抑制肺泡巨噬细胞的IV诱导炎症反应。
尽管肺炎链球菌被视为病原体,但它也是上呼吸道的一种共生菌。在小鼠鼻腔中同时定植肺炎链球菌和流感嗜血杆菌创造了炎症环境,导致C-X-C基序趋化因子配体2和中性粒细胞的高水平募集。协同反应依赖于肺炎链球菌产生的溶细胞毒素,表明其存在调节了对流感嗜血杆菌的免疫反应。遭受流感嗜血杆菌攻击的小鼠表现出严重的肺部疾病,显示出明显的中性粒细胞通路和高浓度的促炎细胞因子。
然而,如果小鼠在吸入共生普雷沃氏菌后进行预处理,炎症会显著减少,且不存在组织病理。腺病毒诱导的记忆肺泡巨噬细胞在小鼠急性鼻内感染中表现出主要组织相容性复合体II类(MHCII)表达增加,以及与宿主防御、趋化性、抗原呈递和糖酵解代谢相关的基因转录上调。
在呼吸道病原体引起的菌群失调期间,共生细菌受到干扰,致病菌可同时出现在肺和肠道中,因此可能导致两个部位的组织损伤。
肠道和肺之间的密切生理和病理联系主要依赖于宿主-微生物的串扰。事实上,肺和肠道细菌的成员可以通过血流成分和代谢物直接交换,从而促进这两个部位的健康或疾病。
肠-肺轴主要相互作用的示意图
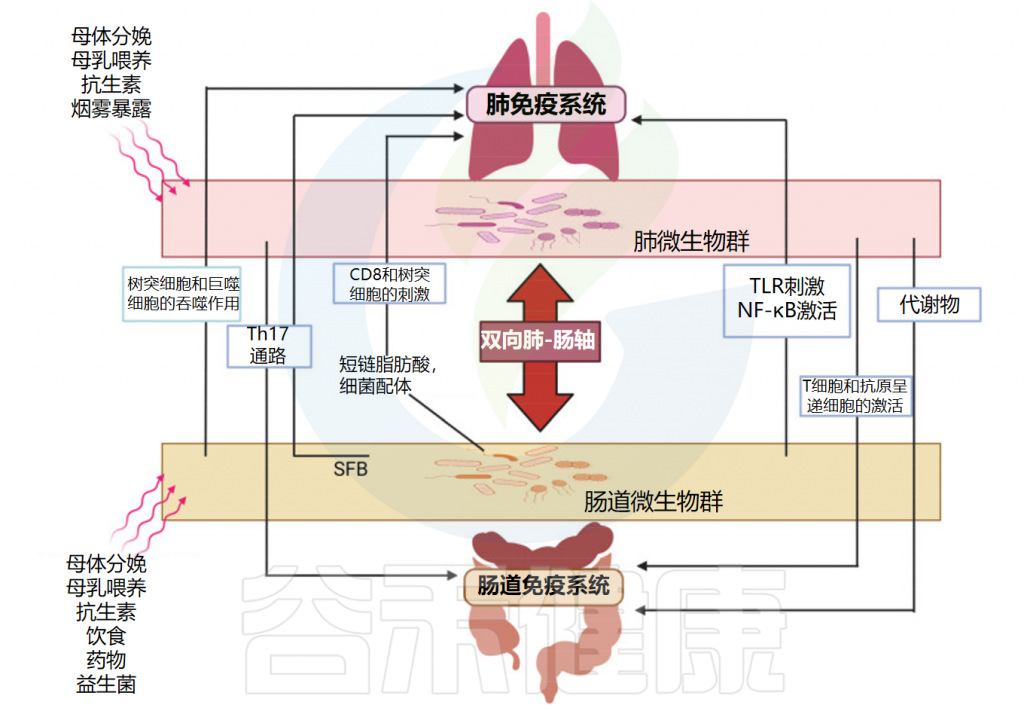
doi: 10.3390/ijms25074051.
呼吸道感染对肠道菌群的影响
研究发现,肺微生物群的改变调节肠道微生物群落,从而影响肠道信号传导。已知结核分枝杆菌感染会导致免疫系统失调,从而使肠道微生物组改变。
◆ 结核病患者肠道微生物群发生显著改变

在一项研究中,比较了成年结核病患者与健康对照的肠道微生物组,发现厚壁菌门、变形菌门和疣微菌门减少,而放线菌门、拟杆菌门和梭杆菌门增加。
另一项研究分析了新发和复发性结核病患者,报道了拟杆菌门、普雷沃氏菌属和毛螺菌属的减少,而放线菌门和变形菌门的富集。
最后,在一组受影响的儿童中,观察到放线菌门和厚壁菌门的减少,而包括拟杆菌属、双歧杆菌属、Dorea属、粪杆菌属、瘤胃球菌属和普氏粪杆菌(F.prausnitzii)属、肠球菌属和普氏菌属的数量增加。
◆ 结核病患者的微生物代谢也变得不平衡
结核病患者可能导致微生物代谢物的产生不平衡,例如短链脂肪酸(SCFA),这可能会重置肺微生物组和通过“肠-肺轴”的免疫反应。这些发现也可能解释了结核分枝杆菌在胃肠道中的定植和肺结核患者发生肠结核的原因。普氏粪杆菌(F.prausnitzii)还被描述具有抗炎作用,可抵御胃肠道疾病。
很少有研究分析人类呼吸道病毒感染期间发生的肠道微生物群改变的性质。在流感期间,尽管肠道中明显不存在病毒,但一些患者仍会出现类似胃肠炎的症状。
◆ 流感病毒感染患者的肠道菌群也存在失衡
一项针对H7N9病毒感染患者的研究显示,拟杆菌门(包括经黏液真杆菌属(Blautia)、罗氏菌属(Roseburia)和瘤胃球菌(Ruminococcus))减少,但厚壁菌门和变形菌门以及属(包括埃希氏菌(Escherichia)、梭菌属(Clostridium))增加。
另一项在流感亚型H1N1患者中进行的研究报告了放线菌门和厚壁菌门的减少,以及包括Dorea、粪杆菌、瘤胃球菌、链球菌在内的属,放线菌科和微球菌科的富集。
在最近的一篇综述中,11项不同的研究报告了与健康对照相比,确诊或疑似呼吸道感染(RTI)患者的肠道微生物组改变。总之,患者的肠道微生物组改变始终是多样化的,包括一些菌的枯竭和富集。
◆ 新冠病毒感染可在肠道中检测到病毒RNA
与甲型流感病毒(IAV)和呼吸道合胞病毒(RSV)的情况相反,在SARS-CoV-2(新型冠状病毒)感染期间,即使病毒RNA不再存在于呼吸道中,也可以在肠道中检测到病毒RNA,从而指出消化道是病毒复制和活性的潜在部位。
◆ 产丁酸抗炎菌减少,机会性病原体增多
SARS-CoV-2感染患者的肠道菌群失调与COVID-19疾病进展和严重程度以及COVID-19后综合征相关。它的特点是双歧杆菌和粪杆菌等抗炎菌减少,丁酸盐生产者丰度降低,例如瘤胃球菌科和毛螺菌科的几个属。
相反,还存在炎症相关微生物群(包括链球菌和放线菌)的富集,以及机会性细菌病原体的过度生长。
最近的一篇论文也表明,SARS-CoV-2感染会导致小鼠肠道微生物组失调,以及潘氏细胞和杯状细胞以及屏障通透性标志物的改变。同样,从96名 COVID-19患者收集的微生物组样本显示,抗菌素耐药物种在内的机会性病原菌属大量繁殖,这种肠道菌群失调与肠道细菌的继发性血流感染有关。
◆ 通过促炎因子和食物摄入减少影响肠道菌群
肺部感染的细胞免疫反应会导致食欲不振,从而改变肠道微生物组和代谢。研究发现,消除CD8+细胞可以阻止食物摄入量的减少并逆转肠道微生物群的变化,这可能是通过分泌TNF-α实现的。实际上,在呼吸道合胞病毒感染期间,中和这种细胞因子可以减少体重减轻并缓解肠道微生物群的扰动。
总的来说,炎性细胞因子的释放和食物摄入量减少是急性病毒性呼吸道感染影响肠道微生物群的可能机制。另一种机制涉及浸润的CD4+ T细胞或全身 IFN释放,它改变了上皮细胞的代谢,导致营养物质的积累,肠腔的微生物为此竞争。这些发现可能在呼吸道病毒感染期间的肠道菌群失调和胃肠道疾病中发挥作用。
肠道微生物群对呼吸道感染的影响
◆ 炎症性肠病患者通常存在肺部受损
肠道和肺之间的重要联系主要体现在炎症性肠病(IBD)患者的肺部受累范围广泛且严重,从亚临床改变到明显的慢性炎症性肺病。气道受累的症状最常见于有长期IBD病史的患者,这与肠道发育不良或全身性不当免疫反应有关。
◆ 肠道菌群失调与呼吸道感染易感性增加相关
除了慢性疾病,肠道菌群失调还与呼吸道感染的易感性增加有关。临床观察强调了健康肠道微生物群在预防病毒呼吸道感染方面的重要性。在肾移植受者和同种异体造血干细胞移植(HSCT)患者中,产丁酸盐的肠道细菌减少与病毒呼吸道感染的风险和发生率增加相关。
在COVID-19(新冠病毒感染)患者中也发现了肠道菌群失调和肠道代谢产物的变化,这与炎症反应和疾病并发症相关。值得注意的是,将COVID-19患者的粪便移植到无菌小鼠体内会导致肺部炎症,并在多重耐药肺炎克雷伯菌感染期间导致更糟糕的结果,这表明微生物群可能直接导致疾病后遗症。
此外,肠道菌群还可以调节血管紧张素转换酶2(ACE2)受体的结肠表达。这些证据可能有助于解释肠道菌群失调患者(如老年人、免疫功能低下患者和有其他合并症的患者)疾病易感性和胃肠道症状的增加。
◆ 肠道微生物及其代谢物可以调节肺部免疫反应
在几种细菌和病毒呼吸道感染的实验模型中,肠道共生菌的缺失或通过抗生素耗竭导致微生物传播、炎症、器官损伤和死亡率增加。这些影响大多与肠道微生物群塑造全身免疫的能力有关。由肠道微生物及其代谢物(如SCFAs)触发的免疫细胞和细胞因子可以到达体循环,并调节肺部、健康和疾病中的免疫和炎症反应。
此外,据报道,炎症性肠病患者的肠道淋巴细胞缺乏组织特异性;这可以解释IBD患者肠外器官存在炎症的原因。菌群失调介导的炎症也会导致粪便钙卫蛋白、血浆C反应蛋白、IL-6和IL-8的循环水平升高,这可能导致肺部感染期间的并发症发生。
肠道菌群失调也可能通过减少营养吸收和能量可用性来影响肺部感染的结果,这反过来会干扰患者产生有效免疫反应的能力。
关于肠道微生物群如何影响呼吸道感染的机制,动物研究提供了更多见解。
在抗生素处理的小鼠中,腹膜巨噬细胞中IFN-γRI、MHC-I、CD86和CD40分子的表达在对病毒感染的早期反应中减弱,这表明肠道微生物群在病毒在宿主体内复制之前就向先天免疫反应发出信号。
Th1、IgA和巨噬细胞对呼吸道病毒感染的反应建立取决于肠道微生物。直肠TLR刺激为IL-1β和IL-18分泌提供信号,恢复了抗生素处理小鼠的肺CD4+和CD8+ T细胞对感染的反应。
最近报道了一种常见细菌种类,即分段丝状细菌(SFB)在肠道中的定植,重新编程了肺泡巨噬细胞,使其增殖、补体产生和吞噬作用增强,从而增强了对流感病毒、呼吸道合胞病毒和SARS-CoV-2的保护。
总体而言,这些发现证实了肠道微生物刺激在先天抗病毒免疫反应激活中的重要性。在感染模型中,抗生素消耗肠道微生物群增加了结核分枝杆菌的负担和传播。菌群失调特别降低了肺树突状细胞中先天受体和巨噬细胞诱导的C型凝集素的表达,导致对幼稚T细胞的刺激功能受损,从而减少了感染小鼠的效应细胞和记忆T细胞数量。
◆ 肺部重叠感染可能受肠道微生物群的影响
肺部重叠感染可能是肺部原发感染诱导的肠道微生物组成改变的进一步后果。对继发性细菌感染的易感性增加,特别是由肺炎链球菌和金黄色葡萄球菌诱导的细菌感染,经常发生在患有呼吸道病毒感染的儿童和老年人中,导致并发症和死亡。
流感期间的肠道菌群失调通过改变短链脂肪酸的产生导致肺炎链球菌重叠感染。在接受甲型流感病毒条件微生物群的小鼠中,乙酸盐的产生减少,改变了肺泡巨噬细胞的杀菌活性,降低了肺对继发性肺炎链球菌感染的防御能力,并促进了重叠感染小鼠的死亡。肠道疾病也可能导致重症 COVID-19 患者伴随或继发性细菌感染。
因此,局部肺部或肠道菌群失调,改变了微生物间相互作用的动力学以及微生物代谢,可能会增强潜在致病细菌物种的增殖。
揭示肺和肠道之间的复杂相互作用有助于更好地了解共生微生物群作为各种呼吸道传染病的治疗靶点。
使用益生菌(微生物)、有利于其生长的产品(如益生元)或微生物代谢物(如后生元)的给药可以通过与病原微生物的直接竞争、改善上皮屏障功能或免疫调节,在呼吸系统疾病期间提供宿主保护。
病毒感染下的治疗
◆ 使用益生菌能够减少流感风险和发生率
在一项系统评价中,评估了益生菌在大规模健康受试者中预防病毒性呼吸道感染(RTI)的疗效。乳杆菌(Lactobacillus)是最常用的益生菌,其次是双歧杆菌(Bifidobacterium)和乳球菌(Lactococcus)。
大多数研究表明,益生菌的使用与病毒RTI的风险和发生率降低相关。然而,在临床表现、病毒载量和免疫学结局的改善方面尚无共识。益生菌鼠李糖乳杆菌和短乳杆菌也与流感感染发生率的减少有关。
在新型冠状病毒(COVID-19)背景下,一项针对SARS-CoV-2诱发肺炎的ICU患者的回顾性研究显示,与单独使用标准治疗相比,使用乳杆菌、双歧杆菌和链球菌属的益生菌混合物治疗与死亡率降低相关。
◆ 益生菌增强了免疫反应,有助于提高生存率
几份报告描述了口服益生菌对病毒性呼吸道感染结果的影响,也提供了机制见解。在感染流感病毒或呼吸道合胞病毒之前服用益生菌,如乳杆菌、双歧杆菌、肠球菌或乳球菌,可缓解症状并提高生存率。肺部和鼻腔冲洗液中的病毒载量也有一定程度的降低。
从机制上讲,益生菌可以通过参与免疫细胞并诱导特定的细胞因子/趋化因子产生来引发针对病毒呼吸道感染的保护性反应,尽管其作用似乎是高度菌株特异性的。
研究表明,益生菌给药后,自然杀伤细胞活性增加,浸润性巨噬细胞和中性粒细胞减少,支气管肺泡灌洗液中的病毒特异性IgA/G滴度增加。粘膜乳杆菌抑制呼吸道合胞病毒复制并降低血炎细胞(如粒细胞和单核细胞)的比例。
用副流感病毒对用益生菌治疗的小鼠肺淋巴细胞进行离体刺激,导致IFN-α和IFN-β的高度表达。病毒感染期间IL-10的上调和IL-6的相应减少也由益生菌给药引发。除了这些保护作用外,经鼻给药鼠李糖乳杆菌GG和嗜酸乳杆菌L-92还增加了IL-1β和单核细胞趋化蛋白1细胞因子以及趋化因子eotaxin和M-CSF的水平。
注意:尽管益生菌具有令人满意的安全性,但它们的使用可能与体弱人群感染或发病率的较高风险有关。因此,人们对使用非活微生物越来越感兴趣。动物研究证明了热灭活益生菌在呼吸道感染中的有益作用,尽管它们的全球影响似乎仅次于活益生菌。
◆ 通过饮食补充益生元提高了抗病毒感染的能力
肠道微生物群可以通过产生短链脂肪酸来调节免疫反应。摄入微生物可利用的膳食纤维(益生元),促进特定共生微生物的多样性和活性的增加,导致对微生物代谢物产生不同影响,进而对宿主对感染的反应产生不同影响。
高纤维(可发酵菊粉)饮食通过两种互补机制传达对流感的保护。小鼠表现出增强的选择性激活巨噬细胞的骨髓生成,这些巨噬细胞促进CXCL1介导的中性粒细胞募集到气道的能力有限,从而导致感染期间组织免疫病理学受限。
同时,饮食衍生的短链脂肪酸刺激CD8+ T细胞抗病毒活性。这种作用是由丁酸盐通过游离脂肪酸受体(FFAR)介导的。同样,高纤维(可发酵果胶)饮食通过刺激肺上皮细胞中的I型IFN 反应来防止呼吸道感染,这种保护是由乙酸盐通过GPR43介导的。
此外,有研究报道,流感感染期间口服醋酸盐补充剂以FFAR2依赖性方式加强了肺部对继发性肺炎球菌感染的防御能力,并降低了重叠感染小鼠的致死结局。同样,鼻内醋酸盐增加了鼻病毒感染期间的干扰素依赖性反应,降低了肺病毒载量。
有趣的是,一项评估益生元(低聚半乳糖和聚葡萄糖)预防新生儿病毒性呼吸道感染功效的试验表明,与益生菌相比,益生元显示出更好的有益效果,这可能与对预先存在的有益细菌生长的直接刺激作用有关。
细菌感染下的治疗
通过调节微生物群对肺部细菌感染又有怎样的效果,让我们接着往下看。大多数研究分析了益生菌在预防和治疗ICU患者肺部感染中的疗效。
◆ 口服益生菌降低了细菌性肺部感染的概率
口服或口咽给予干酪乳杆菌和鼠李糖乳杆菌,可以减少铜绿假单胞菌或相关病原体对下呼吸道的定植和感染。一项研究观察到,在脓毒症患者中,使用短双歧杆菌、副干酪乳杆菌和低聚半乳糖的混合物后,呼吸机相关性肺炎(VAP)的发生率降低。
对感染肺炎链球菌的小鼠研究表明,口服不同的益生菌,如乳杆菌属和链球菌属的菌株,可增加对感染的抵抗力,降低肺细菌负荷,提高存活率。
◆ 鼠李糖乳杆菌等益生菌增强了肺部的免疫反应
鼠李糖乳杆菌的使用通过增加 Foxp3 + Treg 和减少促炎IL-6来增强抗炎反应。在气管内给予其他乳杆菌菌株后,在感染的小鼠中也观察到这种抗炎特征。
活的或灭活的益生菌长双歧杆菌 51A的给药通过增强肺泡巨噬细胞中活性氧的产生和减少促炎性TNF-α和IL-6来刺激肺炎克雷伯菌的肺清除。然而,只有活的益生菌诱导了IL-10水平的同时升高,主要是由醋酸盐介导的。
经鼻或口服接种能有效激活Nod2受体的细菌菌株(分别为脆弱乳杆菌、金黄色葡萄球菌和表皮葡萄球菌,或罗伊氏乳杆菌、粪肠球菌和解黄酮梭菌)可保护小鼠免受肺炎链球菌或肺炎克雷伯菌感染。
补充乳杆菌还可以恢复肺部树突状细胞介导的抗结核分枝杆菌免疫。在感染结核分枝杆菌的小鼠中,口服A.muciniphila或A.mucinihila介导的棕榈烯酸通过表观遗传抑制肿瘤坏死因子,强烈抑制了结核病感染。
呼吸道益生菌
虽然大多数用作益生菌的共生菌起源于肠道,但已经尝试使用呼吸道共生菌。特别是,在婴儿小鼠中鼻内施用Corynebacterium pseudodiphteriticum能够改善呼吸道合胞病毒原发感染和继发性肺炎链球菌重叠感染的特征,降低病原体负荷和肺损伤。据我们所知,迄今为止还没有人类研究评估过呼吸益生菌(即滴注或雾化到下呼吸道的活菌群)的潜力。
在动物模型中,细菌B.bacteriovorus和M.aeruginosavorus的鼻内给药大大降低了呼吸道肺炎克雷伯菌的负担。它们随后被宿主先天免疫机制消除,没有产生不良影响,这预示了它们可能用于治疗人类细菌性肺炎。
◆ 单克隆抗体特异性对抗细菌病原体
使用单克隆抗体(mAb)选择性消耗机会性细菌病原体,通过靶向和灭活细菌及其毒力因子或毒素,成为对抗传染病的一种有前景的方法。
这些抗体因其靶标特异性,不会对本身微生物群产生不良影响,且不太可能导致广泛耐药性。此外,多价mAb经过工程改造,能够发挥多种抗菌作用,包括灭活毒力因子、促进补体沉积和激活先天免疫。
Gremubamab(MEDI3902;AstraZeneca)是一种双特异性人IgG1 mAb,能选择性结合铜绿假单胞菌毒力因子,用于预防高危患者的医院肺炎。该单克隆抗体促进中性粒细胞清除细菌,并防止其附着在气道上皮细胞。Gremubamab的预防性和治疗性给药在急性铜绿假单胞菌肺炎中表现出高度保护作用。
单克隆IgM抗体帕诺巴单抗(AR-101,Aerumab;Aridis Pharmaceuticals)正在临床开发中,用于治疗医院获得性肺炎中的铜绿假单胞菌。帕诺巴单抗的给药减少了急性铜绿假单胞菌感染小鼠模型中的细菌负荷,并减轻了肺部炎症。
另外两种抗体已被检测用于预防和治疗金黄色葡萄球菌肺炎。舒拉妥珠单抗(MEDI4893;Aridis Pharmaceuticals)特异性结合并灭活金黄色葡萄球菌的成孔α-毒素,这是一种关键的毒力因子。在针对金黄色葡萄球菌定植的机械通气ICU患者的临床试验中,单克隆抗体显著缩短了部分患者的肺炎、住院时间和ICU住院时间。
◆ 噬菌体及其内溶素有助于治疗细菌感染
噬菌体疗法在治疗细菌病原体的呼吸道感染方面受到了临床关注,因为它能够在不影响宿主微生物群的情况下选择性地消除目标细菌,与抗生素相容性好,免疫原性低。
噬菌体制剂在治疗铜绿假单胞菌、肺炎克雷伯菌、鲍曼不动杆菌或大肠杆菌方面显示出很强的临床疗效,目前正在临床上评估用于治疗人类呼吸道感染。四种针对铜绿假单胞菌呼吸道感染的专性裂解噬菌体混合物被用于成功治疗肺炎和肺气肿患者。
噬菌体衍生的内溶素或溶酶在对抗细菌感染方面可能优于全噬菌体制剂,因为其细胞壁靶标的保守性使得耐药性难以产生。在鼻咽定植小鼠模型中,噬菌体内溶素的给药降低了肺炎链球菌的滴度,并有效保护小鼠免受致命性肺炎球菌肺炎或铜绿假单胞菌感染,显示出其在预防和治疗呼吸道细菌感染中的潜力。
呼吸道感染是最常见的病毒或细菌来源的传染病之一,造成严重社会和经济负担。研究发现呼吸道感染会导致肺部微生物群的多样性和组成发生变化,进而影响宿主与病原体的相互作用、炎症信号和免疫细胞因子的产生,最终影响疾病的进展和结果。
肺和肠道之间的重要性和复杂的串扰以及肠-肺轴与呼吸健康的紧密联系越来越受到认可,特别强调了肺-肠轴在调节炎症和免疫反应中的双向通信作用。然而,对涉及肠-肺轴的机制的理解,特别是在呼吸道感染的情况下才刚刚开始。需要更好地了解微生物组、呼吸道粘膜和潜在免疫途径之间的相互作用。
迄今为止,大多数用作益生菌的共生菌都来自胃肠道。由于肠道微生物群在影响免疫系统方面起关键作用,它会影响局部和全身(肺部)对病原体的反应。因此,改变肠道微生物组的微生物群靶向疗法(如益生菌、益生元和噬菌体疗法)已被证明对急性和慢性呼吸系统疾病都有益处。
然而,最近的研究表明,与肠道微生物群落相比,肺微生物群与肺免疫变化的相关性始终更密切。此外,肺共生菌的实验性调节直接和持续地改变了肺免疫反应,表明它们可能具有作为局部应用益生菌预防和管理呼吸道感染的潜力。
值得注意的是,致病细菌的抗生素耐药问题。下呼吸道感染是多重耐药感染导致死亡的主要原因,通常与金黄色葡萄球菌、肺炎克雷伯菌、肺炎链球菌、鲍曼不动杆菌和铜绿假单胞菌等重点病原体相关。同时,新变种的出现使呼吸道病毒感染愈加难以控制。
未来的研究应进一步揭示肺和肠道微生物群在健康和疾病状态下的具体机制,特别是它们如何通过代谢产物和免疫细胞影响呼吸道的免疫稳态。此外,需要更多的临床数据来验证这些微生物组干预措施在改善呼吸道感染预后方面的有效性。通过深入理解肺-肠轴的复杂相互作用,有望开发出新的预防和治疗策略,以减轻呼吸道感染的负担并改善患者的整体健康状况。
主要参考文献
Marrella V, Nicchiotti F, Cassani B. Microbiota and Immunity during Respiratory Infections: Lung and Gut Affair. Int J Mol Sci. 2024 Apr 5;25(7):4051. doi: 10.3390/ijms25074051.
Wypych T.P., Wickramasinghe L.C., Marsland B.J. The influence of the microbiome on respiratory health. Nat. Immunol. 2019;20:1279–1290.
Mathieu E., Escribano-Vazquez U., Descamps D., Cherbuy C., Langella P., Riffault S., Remot A., Thomas M. Paradigms of Lung Microbiota Functions in Health and Disease, Particularly, in Asthma. Front. Physiol. 2018;9:1168.
Sommariva M., Le Noci V., Bianchi F., Camelliti S., Balsari A., Tagliabue E., Sfondrini L. The lung microbiota: Role in maintaining pulmonary immune homeostasis and its implications in cancer development and therapy. Cell. Mol. Life Sci. 2020;77:2739–2749.
Natalini J.G., Singh S., Segal L.N. The dynamic lung microbiome in health and disease. Nat. Rev. Microbiol. 2023;21:222–235.
Wu B.G., Segal L.N. The Lung Microbiome and Its Role in Pneumonia. Clin. Chest Med. 2018;39:677–689.
Yagi K., Huffnagle G.B., Lukacs N.W., Asai N. The Lung Microbiome during Health and Disease. Int. J. Mol. Sci. 2021;22:10872.
Belizário J., Garay-Malpartida M., Faintuch J. Lung microbiome and origins of the respiratory diseases. Curr. Res. Immunol. 2023;4:100065.

谷禾健康

日常生活中大部分人都可能碰到过食用了不卫生或过期变质食物以及水源后,出现过恶心、腹痛,并伴有腹泻、食欲不振、呕吐等症状?如果有就要当心,这可能是病原体感染引起的胃肠道炎症。
但是,即使食用了同样食物水源,不同人表现不一样。有的人表现无严重症状,有的人却症状严重,那么是什么因素导致病原体进入肠道后定植或无法定植,什么因素影响其进一步的感染?

胃肠道感染,也称作胃肠炎,是指病毒、细菌或其他病原微生物侵入消化道,引起的一系列消化系统症状的疾病。
这些微生物主要是通过食物或水进入人体后,在胃肠内繁殖并产生毒素,导致黏膜损伤和免疫细胞浸润,进而引发一系列病理生理变化。胃肠道感染的症状包括腹泻、腹痛、恶心呕吐以及食欲减退等。严重时还可能出现脱水、电解质紊乱甚至死亡。
有人可能会问:我们胃肠道中有那么多的微生物,为什么稍微一点外来的微生物就会让我们生病呢?的确,人体肠道内栖息着大量微生物,包括数万亿细菌,统称为肠道微生物群。微生物群的各种细菌成员在多个复杂层面上参与合作和竞争的生理网络。在过去10年中,测序技术领域的进步极大地增进了我们对共生微生物群影响的各种生理和病理过程的理解。
越来越多的人类疾病,如炎症性肠病、2型糖尿病、肥胖、心血管疾病、过敏和结直肠癌等被发现与微生物群组成的改变有关。此外,关于健康个体中人类微生物群的组成、其随时间和不同人之间的差异以及环境因素(即饮食)和遗传背景如何塑造微生物群的也正在变得更加清晰。
正常、健康的肠道微生物群可以在肠道中产生不利于肠道病原体定植的条件,这被称为定植抗性。肠道微生物群通过多种机制提供针对感染的保护,包括抗菌物质的分泌、营养竞争、上皮屏障完整性的支持、噬菌体和免疫激活。这些机制共同有助于抵抗外源微生物的定植。
当微生物群稳态受到干扰时,定植抗性可能会暂时受到破坏,病原体就有机会生长到高水平。这种破坏可能是由接触抗生素、饮食变化、益生菌和药物的应用以及各种疾病引起的。定植抗性的减弱会促进内在病原体的定植或增加对感染的易感性。
病原体扩张的后果之一是引发宿主炎症反应和病原体介导的疾病。同时在肠道炎症患者中,通常会观察到微生物群组成异常,称为“生态失调”。这通常的特征是兼性厌氧细菌(例如肠杆菌科、芽孢杆菌)的相对丰度增加,同时,拟杆菌属等专性厌氧细菌的减少。不过到目前为止,尚不清楚生态失调是肠道炎症的原因还是结果,但我们有充分的理由证明两者之间存在显著的相关性。
除了肠道微生物外,宿主营养通过影响宿主免疫系统和病原体资源的可用性,进而影响感染结果。从宿主角度看,维持活跃的免疫反应需耗费大量精力。营养状况的降低可能削弱宿主抵抗感染的能力,并在其他条件不变的情况下增加病原体的危害。
另一方面,从病原体角度看,宿主营养的数量(食物量)和质量(特定营养成分)的变化可能影响感染期间病原体生长资源的可用性和类型。在此情况下,生长较快的病原体或更高的病原体适应性/负载量可能导致更大的毒力。
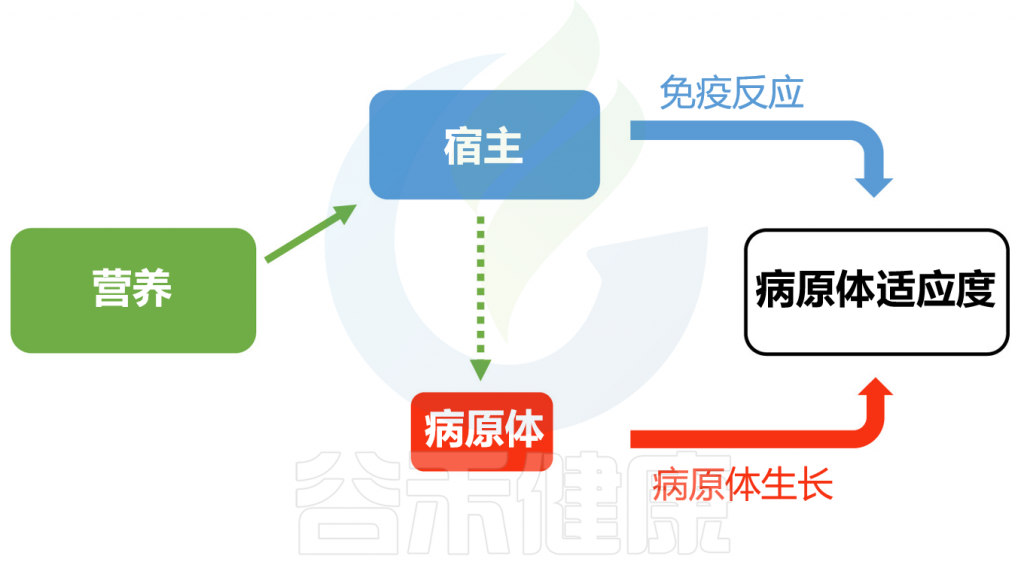
Pike VL,et al.Proc Biol Sci.2019
在本文中,我们将讲述健康人体肠道微生物的抗感染作用;讨论了营养与感染的关联。此外,本文重点关注肠道感染中炎症与微生物群的双向作用、发炎肠道中肠道病原体和肠道微生物群的竞争,以及最后提出了通过营养与微生物作用有助于预防和治疗感染的方法。
▸ 如何区分病原菌和共生菌
在感染医学中,共生菌和致病菌之间的区分仍然是一个重要标准,有益的共生现象和对立的致病性代表了细菌行为范围的两个极端,尽管很少有细菌微生物组成员表现出真正的致病行为。多细菌物种或菌株可以根据其环境背景和宿主生理状态动态改变其作为共生体或病原体的能力。
例如,微生物组失调和宿主免疫缺陷,可以将肠球菌(Enterococcus faecium)从一个几乎无害的肠道微生物组成员转变为血流感染的原因。
单个水平基因转移事件可以改变共生生活方式和致病生活方式之间的平衡,例如,当预测的编码毒素成为肠出血性大肠杆菌或皮肤定植和口咽定植的白喉杆菌的主要毒力因子时。这意味着,原本可能是相对无害的微生物,在获得某些特定基因后,其生物学性质和行为可能发生显著变化,从而增加了其致病潜力。这种基因的转移和表达可以使这些微生物在宿主体内表现出更强的侵袭性和病原性,从而对宿主健康构成更大的威胁。
扩展阅读:
在这篇文章中,我们将常规引起感染的外来细菌称为病原体,而不常引起感染的细菌称为共生菌,越来越多的证据表明,一些共生细菌对人类健康至关重要。
▸ 稳定的肠道共生菌对健康至关重要
而健康的肠道内有一个稳定的微生物群落,可以抵抗外来细菌和病原体的侵袭,这就是所谓的“定植抵抗”,这个概念至少从上世纪50年代就被认识到了。
不成熟的微生物群落(比如婴儿的微生物群落)或者受到抗生素或者饮食破坏的微生物群落可能会失去这种保护作用。
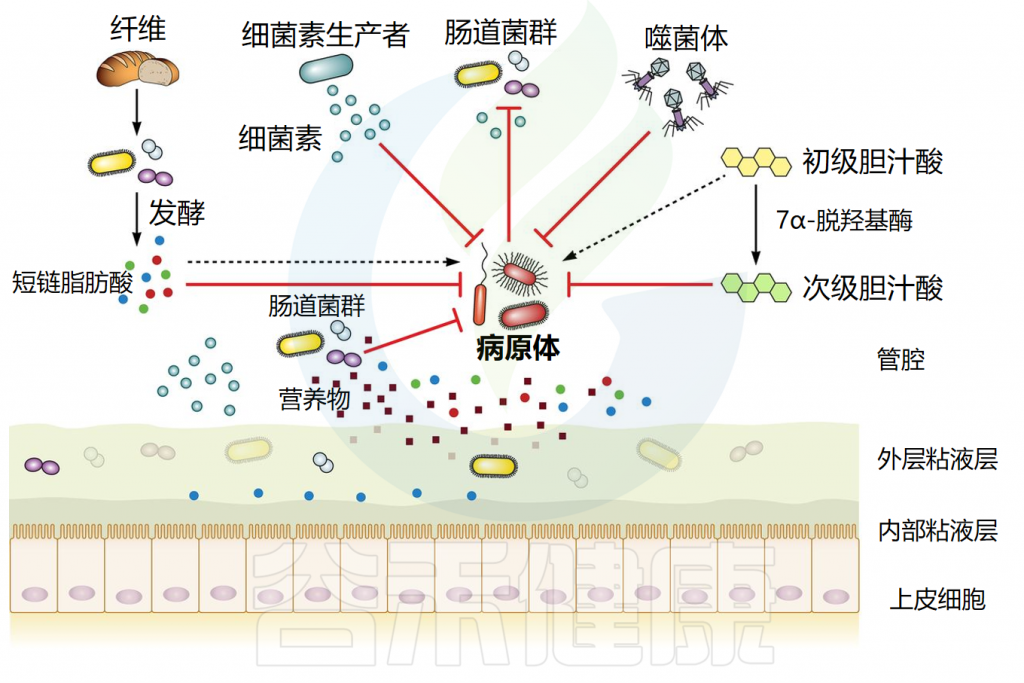
在健康个体中,共生稳定的肠道微生物群通过多种机制提供针对感染的保护,包括抗菌物质的分泌、营养竞争、上皮屏障完整性的支持、噬菌体和免疫激活。这些机制共同有助于抵抗外源微生物的定植。
肠道微生物介导的定植抵抗机制
Ducarmon QR,et al.Microbiol Mol Biol Rev.2019
肠道微生物群能产生各种具有抗菌作用的产物,包括短链脂肪酸、次级胆汁酸和细菌素。每一种都以特的方式抵抗外源微生物的定植。它们的作用机制如下所述。
// 短链脂肪酸影响细胞内pH值和代谢功能来抑制细菌生长
短链脂肪酸(SCFA)主要由细菌通过不易消化的碳水化合物发酵产生。三种主要的SCFA是乙酸盐、丙酸盐和丁酸盐,占总量的90%至95%。在稳态条件下,丁酸盐是肠上皮细胞的主要营养物质,并通过β-氧化进行代谢,可以维持肠道内的厌氧环境。
短链脂肪酸通过影响细胞内pH值和代谢功能来抑制细菌生长。研究表明,SCFA浓度与肠道不同区域的pH值呈负相关。在较低pH值下,短链脂肪酸以其非离子形式更为普遍,这些非离子酸可以穿过细菌膜,扩散到细胞质中。在细胞质内,它们解离,导致阴离子和质子积聚,从而降低细胞内pH值。
短链脂肪酸已被确认为抑制鼠伤寒沙门氏菌(Salmonella typhimurium)生长的关键因素,并且对致病性大肠杆菌和艰难梭菌具有抑制作用。这些SCFA主要由拟杆菌和梭菌等厌氧共生细菌产生,这些细菌是成年哺乳动物微生物群中的重要成员。
短链脂肪酸还可以影响病原体的毒力:例如,丙酸和丁酸可以抑制鼠伤寒沙门氏菌的毒力因子。短链脂肪酸还可以作用于宿主,降低氧气浓度,为病原体生长创造一个不利环境。
// 胆汁酸具有抗菌作用,减少病原体定植
胆汁酸是分泌到小肠中的两亲性胆固醇衍生分子。它们的主要功能是乳化脂肪和脂溶性维生素以供吸收,但它们也具有抗菌特性。
大部分结合的初级胆汁酸(50%至90%)在远端回肠中被重吸收,而其余部分可以在结肠中进行细菌代谢。一些细菌(主要是梭状芽胞杆菌)通过复杂的生化途径进行 7α-脱羟基作用,将解离的初级胆汁酸转化为两种主要的次级胆汁酸:脱氧胆酸和石胆酸。
一些胆汁酸对许多细菌具有杀菌作用,包括金黄色葡萄球菌、多形拟杆菌、艰难梭菌等。证据如下:
-其中一种细菌,C. scindens ,通过产生次级胆汁酸与对艰难梭菌的定植抗性相关。C.scindens能够保护小鼠免受艰难梭菌的侵害,并恢复次级胆汁酸水平。
-使用HT-29细胞系,石胆酸已被证明可以增强肠上皮中抗菌肽LL-37的转录。
-此外,初级胆汁酸鹅去氧胆酸通过激活小肠FXR受体,间接启动先天防御机制,具有保护作用。
// 细菌素通过多种机制发挥其抗菌作用
细菌素是由特定细菌物种产生的短有毒肽,能够抑制其他物种的定殖和生长。细菌素通过多种机制发挥其抗菌作用,主要方式包括直接杀死目标细菌或抑制其生长。这些机制具体如下:
-细胞膜破坏:许多细菌素可以通过插入到细菌的细胞膜中,形成孔洞。这些孔洞破坏了细胞膜的完整性,导致细胞内容物的泄漏,最终引起细菌细胞的死亡。
-干扰细胞壁合成:某些细菌素能够干扰细菌细胞壁的合成。例如,它们可能会抑制细胞壁的主要构建块(如肽聚糖)的形成,从而阻止细胞壁的正常建造和修复,导致细菌无法在分裂或生长过程中维持其结构的完整性。
-抑制核酸合成:直接或间接影响DNA和RNA的合成或功能。例如,它们可能会阻止核酸的复制或转录,或者干扰核酸的修复过程,从而抑制细菌的生长和繁殖。
-蛋白质合成抑制:细菌素还可能通过抑制蛋白质的合成来发挥作用。这通常是通过与细菌的核糖体结合,阻断蛋白质合成的关键步骤来实现的。
-代谢途径干扰:此外,一些细菌素可能会干扰细菌的代谢途径,如阻断能量产生的关键酶的活性,从而削弱细菌的生存能力。
细菌会在肠道中竞争营养物质,这对于同一物种的不同菌株尤为重要,因为它们通常需要相同的营养物。多项研究使用不同的大肠杆菌菌株已经证明了营养竞争在细菌定植抵抗中的重要性。以下是一些研究证据:
// 利用相同营养物的菌株竞争有助于抑制病原菌
本土大肠杆菌菌株与致病性大肠杆菌O157:H7 竞争氨基酸脯氨酸。在粪便悬浮液中,高脯氨酸利用率的菌株通过耗尽脯氨酸池抑制致病菌的生长,而添加脯氨酸可逆转此抑制效果,证实了菌株间的营养竞争。
除氨基酸外,不同的大肠杆菌菌株还利用肠粘液中的不同糖类。当小鼠肠道中存在两种大肠杆菌菌株共同利用与O157:H7相同的糖时,O157:H7无法在小鼠中定植。但若只有一种共生菌株存在,O157:H7则能成功定植,显示这两种共生体能共同耗尽所有致病菌所需的糖。
营养竞争还涉及微量营养素,如铁。已知鼠伤寒菌在感染过程中从发炎的肠道吸收大量铁。益生菌大肠杆菌Nissle能高效清除铁,单次给药可显著降低鼠伤寒菌的水平。对铁的竞争可能是大肠杆菌Nissle 能够减少鼠伤寒沙门氏菌在小鼠体内定植的另一种解释。
总之,这些研究表明,通过营养竞争吸收病原体所需的关键营养物质,可以有效地抵抗病原体定植。因此,未来的策略可能会集中在培养能在特定营养素上胜过病原体的益生菌菌株,这在肠道微生物群紊乱时尤为重要,如抗生素治疗期间及其后,因为这是外源细菌最易定植的时期。
// 对粘附位点的空间竞争可以预防感染
除了基于营养的功能性生态位外,细菌还必须竞争物理空间。有些物种更喜欢以内腔或外粘液层的食物为生,或更罕见的是上皮表面的食物。
与上皮细胞的密切身体接触是某些病原体生活方式的重要组成部分(例如弯曲杆菌、某些致病性大肠杆菌、鼠伤寒沙门氏菌),因此对粘附位点(通常是聚糖结构)的物理竞争有助于预防感染或病理。
定植抵抗的直接和间接机制
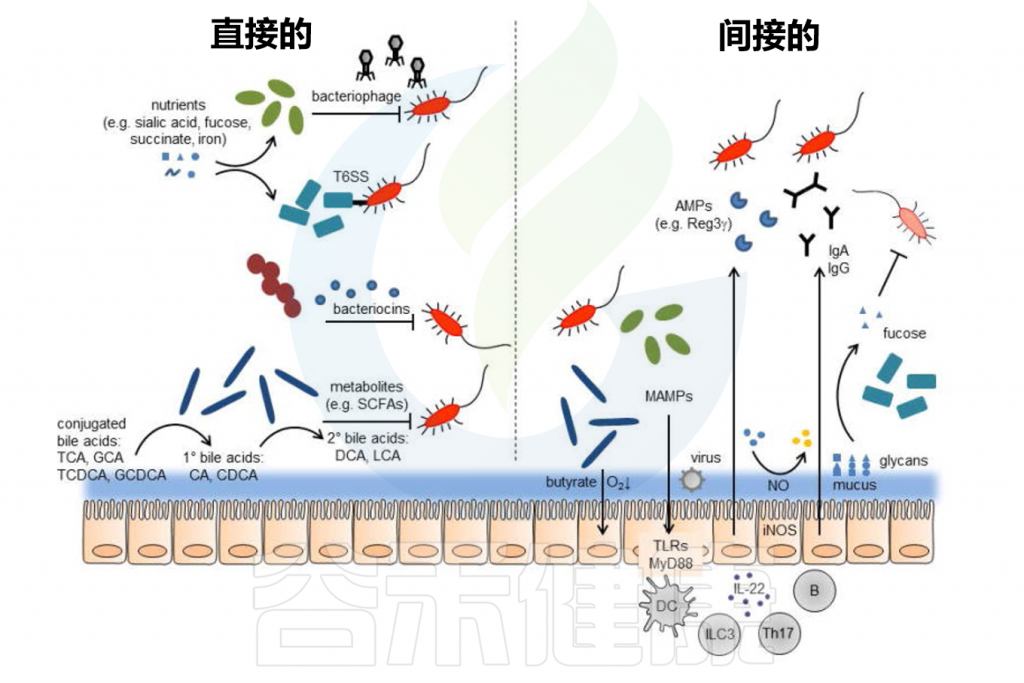
Pickard JM,et al.Immunol Rev.2017
肠道屏障由内外粘液层、上皮屏障及其相关的免疫屏障组成。内部粘液层不可穿透且牢固地附着在上皮层上,形成了一道防止细菌直接接触上皮层并引发潜在炎症的物理屏障。
// 缺乏可利用营养物质时粘液降解细菌会消耗外粘液层
共生肠道微生物居住在非附着的外粘液层中并代谢营养物。粘液层变薄会增加对病原体定植的敏感性,这可能由于西式饮食中缺乏微生物可利用的碳水化合物(MAC)引起。
当MAC稀缺时,粘液降解细菌如Akkermansia muciniphila和Bacteroides caccae会消耗外粘液层,使细菌更接近上皮层。宿主通过增加粘蛋白(MUC2)的表达来适应,但这种适应往往不足。
然而,内粘液层的损伤可以通过施用长双歧杆菌逆转,这可能因其刺激粘液生成。
// 肠道微生物对于维持粘液屏障的完整性非常重要
一方面,有益的微生物如乳酸菌和双歧杆菌可以通过产生短链脂肪酸等代谢产物来增强粘液层的保护作用,这些代谢产物能刺激粘液的产生和分泌,从而增强屏障功能。
另一方面,某些条件下如不健康的饮食习惯导致有益微生物减少,而粘液降解细菌的比例增加,可能会导致粘液层被过度消耗,粘液屏障变薄,从而使肠道更容易受到病原体的侵袭。
总之,粘液层是抵御外源微生物定植的首要屏障。研究已证明饮食是维持屏障正常功能的重要因素,这表明通过饮食干预或特定的益生元和益生菌可能成为未来的治疗选择。
目前,肠道微生物影响全身免疫反应的最佳机制可能是其对适应性免疫系统T细胞的影响。
// 肠道微生物影响T细胞的分化、影响炎症反应
研究表明,胃肠道微生物群可以影响T细胞群分化为辅助性T(Th)Th1、Th2和Th17细胞或具有调节表型的T细胞。具体来说,丁酸盐作为短链脂肪酸促进外周诱导的调节性T细胞的分化,并以这种方式能够抑制全身炎症的发展。
SCFA还能够重新编程细胞的代谢活动,从而诱导调节性B细胞并通过戊酸抑制Th17细胞的生成,这可能与炎症性肠病和自身免疫性疾病有关。
此外,微生物来源的ATP可以诱导Th17细胞的扩增,色氨酸分解产物可以导致上皮内CD4+ CD8αα+ T细胞的增加,细菌来源的多糖可以启动调节性T细胞。通过其诱导调节群体的能力,微生物组可以支持抑制炎症反应。
噬菌体是地球上最丰富的微生物,也在人类肠道中大量存在。噬菌体已被提议作为抗生素的潜在替代品,因其高度特异性,仅针对单一或少数细菌菌株,从而极大地减少对微生物群共生成员的影响。
// 使用噬菌体有效减少了霍乱弧菌感染
在实验中,使用小鼠和兔子的预防性噬菌体混合物可以有效控制霍乱弧菌感染。这种预防性混合物在体外能够杀死霍乱弧菌,减少其在小鼠肠道中的定植,并预防兔子的霍乱样腹泻。
研究表明,肠道中噬菌体的浓度是预防感染成功的关键因素,噬菌体的给药时间与霍乱弧菌接种的时间间隔与治疗效果密切相关。空肠弯曲杆菌在鸡体内的定植也通过噬菌体混合物显著降低。
// 噬菌体为肠道共生细菌提供竞争优势
噬菌体还可以提供共生体竞争优势。例如,粪肠球菌V583含有能够感染并杀死其他粪肠球菌菌株的噬菌体,为粪肠球菌V583创造了竞争优势。
噬菌体在排除特定肠道细菌方面发挥重要作用,对肠道健康具有潜在贡献。噬菌体的人类治疗应用尚未广泛进行,主要是因为缺乏足够的安全性和有效性证据。然而,最近的病例报告显示,噬菌体治疗在对抗多重耐药细菌方面展现出明显的潜力。
VI 型分泌系统(T6SS)是在一些革兰氏阴性细菌中发现的蛋白质易位复合物,其与一些噬菌体蛋白质具有相似的机制,具有注射毒素到邻近细胞的能力,这一功能使其在微生物间的竞争以及宿主与病原体间的相互作用中发挥关键作用。
T6SS的结构类似于细菌噬菌体的尾部,能够将效应蛋白直接转运到靶细胞中,这些效应蛋白能够破坏靶细胞的细胞结构或功能,从而抑制或杀死竞争对手。
T6SS 示意图
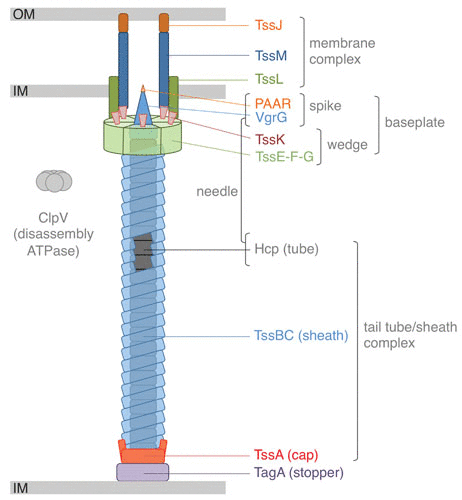
Cherrak Y,et al.Microbiol Spectr.2019
// T6SS通过注射毒素杀死或抑制病原细菌
在抵抗病原体感染方面,T6SS可以被视为一种防御机制。一些非病原性或共生细菌利用T6SS对抗侵入的病原细菌,通过直接向病原体注射毒素来抑制其生长或直接杀死这些病原体。
注:这种机制不仅限于细菌间的相互作用,也可能影响更高级生物的细胞,如真核宿主细胞,进而影响病原体的感染能力和宿主的免疫反应。
最近,在拟杆菌门成员中发现了一个新的T6SS蛋白家族,该门与厚壁菌门一起在哺乳动物肠道中占主导地位。多项研究表明,T6SS及其相关效应子和免疫蛋白的存在在小鼠肠道内拟杆菌属物种之间的竞争中发挥着重要作用。重要的是,T6SS 介导的竞争是接触依赖性的,可以涉及效应子和免疫蛋白的多种组合,并且可以具有比其他杀伤机制更广泛的目标范围。
// T6SS能够调节宿主的免疫系统
此外,T6SS还能够调节宿主的免疫系统。一些研究表明,T6SS可以通过调控炎症反应和影响免疫细胞的活性来影响宿主的免疫环境。例如,T6SS可以影响巨噬细胞的吞噬作用和炎症因子的释放,从而调节宿主的免疫反应。
因此,T6SS不仅是细菌间相互作用的武器,也是细菌与宿主互作的重要因素,对于维持微生物群落的平衡、抵抗病原体侵袭以及调节宿主免疫反应具有重要作用。
通过进一步研究T6SS的具体机制和作用,可以为开发新的抗感染策略提供理论基础和潜在靶点。
营养与感染之间存在密切的相互关系。首先,营养直接影响人体免疫系统的发展。此外,营养状况还会影响感染的发生,包括胃肠道感染、食物中毒、肠道疾病以及其他全身性传染病。
营养与感染的关系可分为以下几种:
(1)营养对人体免疫系统发育的影响;
(2)营养对感染(如胃肠道感染)、食物中毒、肠道疾病(如微生物性腹泻)和全身感染性疾病(如布鲁氏菌病、伤寒)发生的影响;
(3)营养不良与感染的关系;
(4)严重联合免疫缺陷患者的营养;
(5)暴饮暴食与感染的关系。
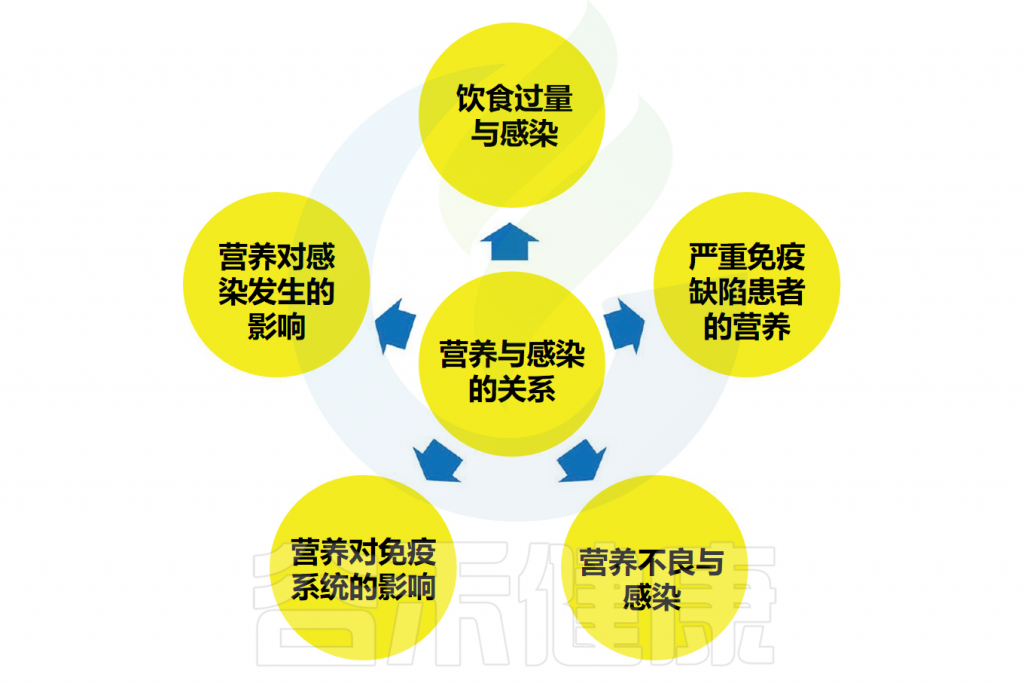
// 营养对免疫系统的发育至关重要
营养从胚胎阶段起就对人体免疫系统的发育产生影响。怀孕期间,尤其是前三个月,如果母亲摄入足够的蛋白质、维生素和矿物质,胚胎组织将得到良好的发育。胎儿营养不良会对免疫系统的正常发育造成不利影响,如果免疫系统在这一关键时期未能有效发展,将来对抗病原体的能力会受到影响。
// 营养不良抵抗病原体的能力会受到影响
母乳喂养是建立强健免疫系统的关键步骤。未经母乳喂养的营养不良婴儿,容易因缺乏蛋白质和维生素而易感染疾病,并且对疫苗的反应也不佳。因此,良好的营养是提高人体对环境病原体防御能力的基石。
广泛的研究已经证实了营养在增强抵抗感染能力方面的重要作用。例如,研究显示,相比仅接种结核疫苗的儿童,饮食充足的儿童患结核病的风险更低。同时保证营养健康和接种疫苗的人,患结核病的风险显著减少。
进行了一项研究来显示儿童感染与营养不良之间的关系。结果显示,营养不良的儿童经常出现腹泻。这些儿童的腹泻发生率和严重程度较高。结果表明,营养不良程度与腹泻风险之间存在直接关系。另一项研究评估了婴儿从出生到24个月大期间腹泻的影响。结果显示,腹泻发生率每增加 5%,生长障碍就会增加(约16%)。
营养不良和菌血症风险之间存在可变关系,营养不良儿童更有可能患革兰氏阴性菌血症。在坦桑尼亚进行的一项血培养研究结果表明,假单胞菌属、肠杆菌属和金黄色葡萄球菌的患病率很高。
溶组织内阿米巴寄生虫引起的阿米巴病与营养不良儿童腹泻的发生率和严重程度增加之间存在密切关系。据观察,溶组织内阿米巴感染可导致发展中国家儿童腹泻的发生率为2%–10%。全球每年由阿米巴病引起的死亡率和感染率估计分别约为100000例和5000万例。分泌性抗体(免疫球蛋白A)可促进针对溶组织阿米巴腹泻的免疫保护。营养不良儿童中IgA分泌细胞的减少会增加溶组织内阿米巴感染的发生率。
// 受污染的食物和水会导致人体发生感染
食物是人体的主要能量来源,但如果受到微生物污染,可能会引发消化系统疾病和食物中毒等问题。在不同季节,人们可能会遭遇不同程度的食物中毒。尤其在炎热天气中,肠道疾病的发生率较高。
// 受霍乱弧菌污染的食物会使人感染霍乱
其中霍乱是最危险的一种,霍乱是由一种叫做霍乱弧菌(Vibrio cholerae)的细菌引起的急性肠道传染病。这种病主要通过饮用或食用被霍乱弧菌污染的水和食物传播。人们在卫生条件差、饮用水处理不当或个人卫生习惯不佳的环境中更容易感染霍乱。
// 布鲁氏菌病
此外,通过水和食物传播的微生物污染可能导致儿童严重腹泻和传染病。布鲁氏菌病是通过摄入受布鲁氏菌(Brucella)污染的食物和水引起的传染病,这是人类和动物之间的常见疾病。
布鲁氏菌可感染绵羊、山羊、牛、猪和狗,人类则可能通过食用未经高温消毒的牛奶或未煮透的感染动物肉类而感染。密切接触受感染动物的分泌物也可能将病菌传播给人类。
// 其他可能通过食物导致人体的感染
阿米巴原虫病主要通过摄入受污染的水和食物引起,而弓形虫病则是通过食用生或未煮熟的肉类传播。当寄生虫包囊进入人体胃部,胃酸作用下释放,通过肠粘膜传入肝脏和淋巴结。
蛲虫感染是一种通过食物传播的寄生虫病,在全球范围内普遍存在,儿童的感染率高于成人。其他如带虫病和钩虫感染也常由不良营养引起。
注:由于病原体可能具有抗生素耐药性,这给治疗带来了挑战。
1968年,世界卫生组织报告就指出,感染可能会加剧人类营养不良,这种现象被称为协同作用。感染通常会对人的营养状态产生负面影响,从而导致营养不良,反过来又可能加重感染。
// 感染期间代谢和吸收功能会被削弱
多种因素可能削弱人体对抗感染的能力并引发营养不良。例如,厌食可能导致营养不良,从而削弱免疫系统。某些传统行为也会加剧营养不良和感染问题,如在一些文化中,发烧或腹泻的人被禁止进食。肠道感染可能导致肠道吸收功能降低,蛋白质、碳水化合物和脂肪的吸收量可能分别减少43%、42%和72%,具体数值取决于感染类型。
感染期间,如蛋白质丢失等代谢损伤会增加膳食蛋白质的需求。脂质和碳水化合物的代谢紊乱(如脂肪酸自身代谢紊乱、酮体和甘油三酯的变化)在各种感染中可见。此外,感染期间,通过糖异生途径,氨基酸可转化为葡萄糖。在呼吸道感染期间,血液中的维生素A浓度可能降低,肝炎、急性扁桃体炎、类风湿性关节炎等疾病也会降低血清维生素A浓度。
// 感染时体内许多营养素过低
感染时,血液中维生素C浓度降低,而尿液中维生素C浓度增加,即使是接种过麻疹和天花疫苗的人也会出现。感染后,体内维生素B2(核黄素)含量会减少,这种减少具有统计学意义。
铁是感染期间体内减少的另一种营养素,铁的有效利用有助于降低病原体活性并治疗感染。锌和铜是感染时浓度可能降低的其他元素,研究显示感染中铜和锌的平衡呈负值,尽管血液中铜浓度可能增加,但感染者体内铜水平可能显著降低。
有人说,营养不良会增加感染的风险,那我尽量多吃行不行,其实这也是不可取的。
// 暴饮暴食会降低抗感染能力
暴饮暴食可能会对人体的免疫系统造成负面影响,从而降低抗感染的能力。当人体摄入过量的食物,尤其是高脂肪、高糖的食物时,可能会导致炎症反应增强,肠道微生物失衡,这些都可能削弱身体的免疫功能。
因此,暴饮暴食不仅对健康有害,还可能增加感染的风险,包括消化系统的感染如胃肠炎等。合理的饮食习惯和适量的食物摄入对维护免疫系统和预防感染至关重要。
此外,肥胖的人有患糖尿病的潜力,所有糖尿病人都对感染敏感。一些研究还表明,超重的人患呼吸道感染的风险更大。
炎症和感染之间存在密切的关系,炎症通常是身体对感染的一种防御反应。当病原体如细菌、病毒或寄生虫侵入人体时,免疫系统会激活,产生炎症反应以抵抗感染。这种反应包括血液中的白细胞和其他免疫分子到达感染部位,以消灭病原体。
人类肠道微生物群的组成表现出很大的个体差异。而感染和肠道炎症可导致肠道微生物群组成发生改变,称为生态失调。
关于微生物群如何参与肠道炎症发病机制,人们提出了不同的理论:
(1)导致粘膜屏障功能缺陷(例如粘液层、先天杀伤、抗菌肽)的突变涉及共生细菌的过度易位和促炎信号的触发;
(2)宿主免疫调节异常会引起针对内在共生菌的过度免疫反应;
(3)不明病原体的存在会导致疾病的诱发;
(4)失调的微生物群,其特征是“有益”和“潜在有害”共生细菌之间的不平衡,是疾病的触发因素或驱动因素。
慢性结肠炎或肠道病原体感染过程中的炎症可能会破坏正常的微生物群组成,诱发生态失调,并有利于病原体和共生菌的过度生长,并增加毒力潜力。因此,菌群失调不仅被认为是肠道炎症的原因,而且也是肠道炎症的结果。
炎症相关的菌群失调有哪些特点?越来越多的证据表明,感染病原体导致肠道微生物群落的扰动可能会助长丰度较低的有害细菌大量繁殖,从而进一步加剧肠道炎症。
这些研究使用分子、独立于培养的技术,如16S rRNA 基因测序和宏基因组学。
事实上,远端肠道的生态失调通常以特定专性厌氧革兰氏阳性菌(如瘤胃球菌科、毛螺菌科)的丰度降低,同时兼性厌氧菌(如肠球菌和链球菌)以及革兰氏阴性变形菌(特别是肠杆菌科成员)也随之增加。
已提出将人类肠道中丰富的丁酸盐生产菌——普氏栖粪杆菌(F. prausnitzii)的减少作为活动性疾病的微生物标志物。
// 肠杆菌的大量增加可能是肠道炎症的一个重要标志物
肠杆菌科(Enterobacteriaceae)是革兰氏阴性兼性厌氧菌的一个大家族,由于它们对从上皮屏障扩散的氧气具有相对较高的耐受性,因此通常位于靠近肠道上皮的位置。事实上,肠杆菌科仅占远端肠道微生物群落的一小部分,大约为0.1%。然而,它们在各种肠道炎症环境中过度生长,例如炎症性肠病、乳糜泻和结肠癌。
肠道炎症是由病原体感染后产生的,炎症引起的环境和营养变化可能赋予肠杆菌科细菌生长优势。例如在克罗恩病(CD)和溃疡性结肠炎(UC)(炎症性肠病的两种主要形式)患者中观察到肠杆菌科细菌(包括粘附性侵袭性大肠杆菌)的患病率增加。
// 许多肠道病原体利用炎症环境来促进自身生长
许多肠道病原体能够利用炎症反应来谋取自身利益。在健康的肠道中,复杂的厌氧微生物群有效地阻止主要人类肠道病原体的定植和感染。这种“定植抵抗”在肠道炎症存在时会减弱,从而使病原体过度生长。
一个典型的例子是啮齿类柠檬酸杆菌(Citrobacter rodentium),一种引起鼠类传染性结肠增生的病原体;空肠弯曲杆菌;以及引起人类小肠结肠炎的鼠伤寒沙门菌(S. Typhimurium)。
许多研究已经表明,其他病原体如肺炎克雷伯菌、奇异变形杆菌、霍乱弧菌、艰难梭菌和肠球菌属也可能从肠道炎症环境中获益。因此,肠道中的炎症环境能够改变肠道病原体及其相关共生物种的存活、附着或生长情况。
细菌转录组的分析揭示了炎症对微生物功能及其在炎症环境中生存能力的影响。
// 病原菌粘附能力的增强使之持续存在并诱导肠道炎症
粘附性侵袭性大肠杆菌(AIEC)能够通过常见的1型菌毛粘附素FimH粘附到回肠肠上皮细胞,并识别在克罗恩病(CD)回肠上皮细胞上异常表达的癌胚抗原相关细胞粘附分子6(CEACAM6)。
最近的DNA序列分析表明,AIEC菌株主要表达带有新近氨基酸突变的FimH。这些新突变显著增强了AIEC对表达CEACAM的肠上皮细胞的粘附能力,使其能在遗传易感宿主中持续存在并诱导肠道炎症。
// 在不同炎症条件下表达特定的蛋白来促进生存
在另一项研究中,分析了从IBD和非IBD儿科患者中分离的粘膜相关大肠杆菌菌株的FimH基因突变模式,发现了每种大肠杆菌特征(即状态、门群和粘附类别)的不同FimH突变模式。溃疡性结肠炎(UC)患者的大肠杆菌菌株显示出突变数量增加,而克罗恩病(CD)患者的分离株显示出突变率增加,但出现了不同的FimH突变。
这些结果表明,在特定的选择压力下,FimH蛋白会发生选择性氨基酸突变以维持细菌的生存,并暗示大肠杆菌在不同炎症条件下(如CD和UC)采用独特的生存策略。
因此,可以想象,炎症环境可能会选择表达特定的微生物蛋白,从而促进细菌存活并进一步维持炎症。
许多文献已经提出了导致发炎肠道中一些有害细菌大量繁殖的机制,包括营养变化、粘蛋白利用、抗菌剂的产生、无氧/有氧呼吸和金属利用。我们这里主要以肠道炎症中的标志物种肠杆菌(Enterobacteriaceae)为例来介绍。
// 炎症下的营养变化使病原菌更具竞争优势
居住在远端肠道的微生物群落竞争有限的食源性碳水化合物或宿主粘液源性聚糖。因此,饮食对肠道微生物群的组成起着关键作用,饮食的变化可能导致肠道微生物群落结构的扰动。
在健康肠道中,专性厌氧梭菌和拟杆菌仍然保持对兼性厌氧肠杆菌科的主导地位。在正常情况下,梭状芽胞杆菌和拟杆菌利用糖苷水解酶分解复杂的碳水化合物,并通过结合蛋白质增加其表面的碳水化合物浓度,最终通过主动转运系统将碳水化合物转运穿过梭状芽胞杆菌的细胞质膜和拟杆菌的外膜。相比之下,肠杆菌科因缺乏糖苷水解酶,其分解复杂碳水化合物的能力较差,只能通过外膜扩散通道被动运输寡糖。
因此,在健康状态下肠杆菌科细菌在与专性厌氧菌竞争高能营养物质时处于劣势,这种竞争性生长劣势解释了健康远端肠道中梭状芽胞杆菌和拟杆菌对肠杆菌科细菌的优势。
而在炎症过程中,肠上皮损伤导致死亡上皮细胞脱落增加,增强了上皮细胞膜衍生磷脂(如磷脂酰胆碱和磷脂酰乙醇胺)的可用性。特别是,乙醇胺可被变形菌门中的某些细菌及病原菌(如沙门氏菌和假单胞菌)作为唯一的碳或氮源利用。这种利用乙醇胺的能力有助于这些细菌在肠道中成功定植并维持其致病机制。
// 粘蛋白的分解促进发炎肠道中病原菌的定植
肠杆菌科细菌在发炎肠道中扩张的另一个机制是利用粘蛋白。粘液层覆盖肠上皮,分为两层;外层可移动,被细菌定殖,通常限制共生菌的定殖,而内层牢固地附着在上皮上,基本上无细菌。
分泌的凝胶形成粘蛋白MUC2是人结肠粘液的主要成分。值得注意的是,MUC2缺陷小鼠表现出细菌对表面上皮的粘附增强、肠道通透性增加以及发生自发性或DSS诱导的结肠炎和结直肠癌的易感性增加。
最近的一篇论文强调了粘蛋白衍生的唾液酸在DSS治疗诱导小鼠肠道炎症期间促进肠杆菌科细菌扩张的作用。唾液酸是粘蛋白中的主要碳水化合物之一,可以被无法从头合成这些糖的细菌(如大肠杆菌)摄取,并掺入细菌荚膜和脂寡糖中。
此外,鼠伤寒沙门氏菌和艰难梭菌在肠道内扩张期间,采用了一种常见策略来分解代谢微生物群释放的粘膜糖,如岩藻糖和唾液酸。这些观察结果表明,唾液酸的分解代谢可能为发炎肠道中肠杆菌科细菌的生长提供优势。
// 肠杆菌产生大肠杆菌素来抑制其他细菌
肠杆菌科细菌还可以通过产生抗菌分子来战胜其他细菌,更利于自身在肠道中的繁殖。例如,大肠杆菌素是由某些大肠杆菌菌株产生的细菌素,对系统发育上的近亲致命。在发炎肠道中,大肠杆菌素Ib(col1B)的产生赋予鼠伤寒沙门氏菌相对于敏感大肠杆菌菌株的竞争优势。
col1B的表达受到低铁可用性和SOS反应的正向调节,这两种情况通常由中性粒细胞募集和氧化应激诱导的DNA损伤在发炎肠道中引发。
因此,肠道中的炎症环境似乎创造了一种有利的条件,可以增强大肠杆菌素的作用,大肠杆菌素作为适应因素,为肠杆菌科细菌的大量繁殖提供竞争性生长优势。
// 肠杆菌科的呼吸灵活性使其在炎症条件下更好地生长
导致肠杆菌科细菌在发炎的远端肠道中扩张的进一步机制是宿主在这种基本上厌氧的环境中诱导的生长条件的变化。
炎症期间血流量和血红蛋白升高而产生的高水平氧气,可以为兼性厌氧菌(如肠杆菌科)提供比专性厌氧菌(如梭菌属和拟杆菌属)更有利的生长优势。肠道炎症期间产生的新呼吸电子受体可能通过无氧呼吸(包括硝酸盐呼吸)支持细菌生长。
已经证明硝酸盐是作为宿主炎症反应的副产物而产生的。这种源自宿主的硝酸盐的富集可以为肠杆菌科(例如大肠杆菌和鼠伤寒沙门氏菌)带来适应性优势,因为编码硝酸还原酶的基因存在于大多数肠杆菌科细菌中,但在属于梭菌属和拟杆菌属的专性厌氧菌中基本上不存在。
由宿主炎症反应产生的活性氧(ROS)可以与内源性硫化合物(即硫代硫酸盐)反应,产生一种新的呼吸电子受体,称为四硫酸盐。这种新产生的电子受体为伤寒沙门氏菌提供了选择性生长优势,而不是发炎肠道中竞争的发酵肠道微生物。这些观察结果表明,病原体可以利用宿主反应来超越肠道微生物群。
肠杆菌科的呼吸灵活性使它们能够对肠道内不同的氧气供应量做出反应。例如,在没有氧气的情况下,大肠杆菌可以使用硝酸盐、亚硝酸盐、三甲胺-N-氧化物(TMAO)、二甲基亚砜(DMSO)和富马酸盐作为电子受体,而在氧气存在下,大肠杆菌表达使用氧作为电子受体的末端氧化酶。
此外,链霉素治疗导致小鼠体内产生丁酸的共生梭状芽孢杆菌的耗竭,导致丁酸水平降低,上皮氧合升高,伤寒沙门氏菌有氧繁殖。鼠类柠檬酸杆菌(C.rodentium)使用III型分泌系统(T3SS)促进小鼠结肠隐窝增生,这反过来增加了表面上皮的氧化,并促进了C.rodentim在结肠中的有氧扩张。
鼠类柠檬酸杆菌(C.rodentium)感染是常用的模型之一。
// 许多病原体进化出了高亲和力的金属摄取机制
肠杆菌科细菌在肠道炎症中大量繁殖的另一机制是金属获取。铁是宿主和病原菌重要的营养物质,大部分储存在细胞内,使得病原体难以获取。
然而,为了克服这种铁限制,许多病原体已进化出高亲和力的铁摄取机制,与宿主的限制性铁环境竞争。这些机制包括释放铁螯合铁载体、血红素获取系统和转铁蛋白/乳铁蛋白受体。
例如,大肠杆菌能产生肠杆菌素,一种儿茶酚铁载体,有效抑制中性粒细胞的杀菌髓过氧化物酶,使大肠杆菌在发炎肠道中具有明显的生存优势。因此,大肠杆菌释放的铁载体既能获取铁,也能抵御宿主源性氧化应激。
此外,粘附性侵袭性大肠杆菌(AIEC)生长依赖铁,chuA(血红素铁获取)的存在与其在巨噬细胞内持续存在的能力相关。这些结果支持铁载体需氧菌素增强巨噬细胞内存活及AIEC NRG857c (O83:H1)在小鼠肠道中的定植,该菌株是从CD患者回肠分离的AIEC临床菌株。
总体来看,这些研究突显了铁获取在促进发炎肠道中毒性更强的肠杆菌科细菌扩张的关键作用。肠杆菌科细菌也进化出了获取其他金属(如锌和锰)的策略,以利于它们在发炎肠道中的生长。
!
注意
肠道炎症通常会促进毒性更强的肠杆菌科细菌的出现,这些细菌已进化出多种策略来逃避宿主免疫反应、战胜共生细菌,并在发炎的肠道中茁壮成长。
众所周知,营养对肠道微生物群和免疫系统具有显著影响,在健康与疾病的发展中扮演着关键角色。例如,西方饮食通过诱导骨髓祖细胞的表观遗传和转录重编程,与炎症反应的增强有关,这直接影响了多种疾病的发展。
增强对肠道微生物群、宿主反应及其他微生物间关系的理解,为通过营养调节这三者之间的互动提供了可能,帮助维护肠道稳态和抵抗感染。
应考虑到不同的饮食成分,如矿物质、碳水化合物、维生素、脂质和蛋白质,它们具有特定的功能特性,能够以直接或通过微生物组间接的方式影响宿主与病原体的相互作用。在这些营养素之间建立机制联系为影响健康提供了多种可能。
因此,饮食干预应视为一种调节感染风险、预防病原微生物入侵、减轻感染严重程度及支持感染治疗的重要手段。尽管已知多种营养化合物对宿主微生物组和免疫系统有影响,但膳食纤维、益生元和益生菌仍然是研究的热点。
益生元是不易消化的食物成分,通过选择性刺激结肠中一种或有限数量的细菌的生长或活性,对宿主产生有益影响。典型的益生元是人乳低聚糖、菊粉、低聚果糖和低聚半乳糖。
膳食纤维不是典型的益生元,但具有益生元特性,例如,β-葡聚糖、阿拉伯木聚糖、果胶和抗性淀粉。益生元和特定的膳食纤维通过充当发酵底物促进肠道中有益细菌的生长,同时通过生态位排除抑制病原体的生长。
// 益生元与膳食纤维的代谢产物具有抗菌活性有助于预防胃肠道感染
主要发酵产物是短链脂肪酸,如上所述,它对免疫系统具有重大影响,因此可以抑制感染的发展。除短链脂肪酸外,益生元和膳食纤维还可以通过排除和抗菌活性直接预防胃肠道感染。
// 与免疫细胞作用预防感染、并减轻感染后的炎症
此外,益生元和膳食纤维与上皮细胞和免疫细胞的直接相互作用也有助于预防感染。β-葡聚糖和阿拉伯木聚糖等膳食纤维已被证明可以激活CLR dectin-1,这是一种参与诱导训练免疫的重要受体,可增强针对继发感染的免疫反应。
人乳低聚糖、阿拉伯木聚糖和果胶还与Toll 样受体 (TLR)相互作用,从而提高树突状细胞(DC)的功效,通过肠上皮细胞诱导耐受性DC,并保护胃肠道免受过度的TLR信号传导影响,而且还支持解决胃肠道感染后的炎症。
饮食改变与肠道炎症相关的肠道微生物群
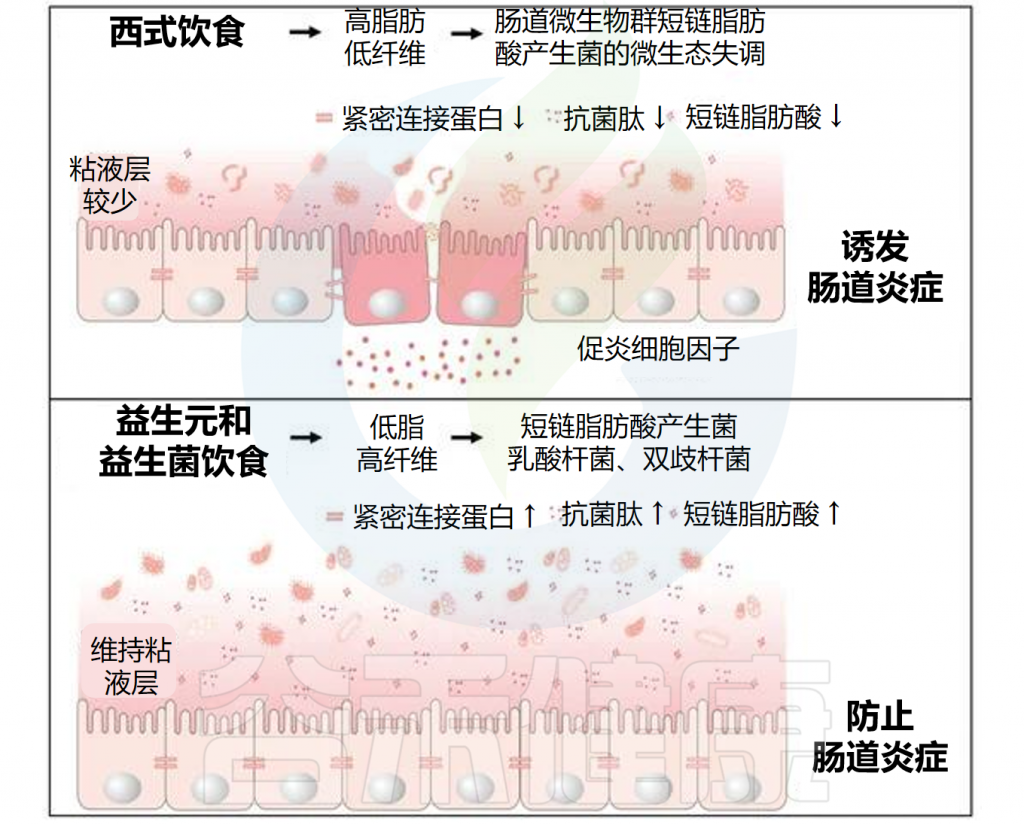
Lobionda S,et al.Microorganisms.2019
益生菌是活细菌,当摄入足够量时,可为宿主提供健康益处。使用益生菌的基本原理主要是基于它们改变肠道微生物群的能力,支持共生菌的生长而不是致病菌的生长。它们能够通过提供短链脂肪酸、维生素和其他通过粘蛋白降解产生的食物来源等代谢物来刺激常驻细菌的生长。
在一项关于益生菌治疗急性腹泻功效的研究中,益生菌通过缩短急性感染性腹泻的持续时间和减少平均排便次数具有明显的积极作用。
许多研究探讨了益生菌在预防和治疗感染方面的潜在作用,包括以下几种机制:
// 竞争性抑制
益生菌可以与病原菌竞争肠道黏膜的结合位点。通过占据这些位点,益生菌阻止病原菌的附着和进一步的入侵。此外,益生菌还能竞争肠道内的营养资源,限制病原菌的生长和繁殖。
// 产生抗菌物质,营造病原菌生存不利的环境
许多益生菌能产生抗菌物质,如细菌素、过氧化氢等。这些物质能直接杀死或抑制病原菌的生长,帮助清除肠道中的感染。
乳酸菌(LAB)可以通过产生抗菌肽(例如细菌素)来防止病原体入侵,抗菌肽可以通过在细菌细胞壁上形成孔并抑制细胞壁合成来消灭病原菌。此外,益生菌通过产生乳酸和乙酸来降低pH值,创造酸性环境,具有抗菌作用,不利于细菌病原体的生长。
// 增强肠道屏障功能
肠道完整性的损害可以解释胃肠道感染的发展。因此,益生菌增强肠道屏障的能力可能会对感染提供一些保护。
据报道,乳酸杆菌通过调节参与紧密连接信号传导的基因表达来改善肠道屏障功能。VSL3(益生元和益生菌的混合物)治疗可促进MUC2的表达和粘液分泌,有助于增强肠道屏障。这种屏障的增强有助于减少病原体的侵袭和内毒素的吸收,从而减轻感染的严重程度。
乳杆菌(尤其是L.reuteri)和动物双歧杆菌亚群,通过乳酸产生促进粘液粘附的蛋白质,称为粘液结合蛋白(MUBs),增强益生菌与宿主之间的相互作用。
// 调节免疫系统
宿主免疫系统和细胞因子谱的调节可能是益生菌有益的主要机制之一。益生菌和宿主免疫系统之间的相互作用在微生物相关分子模式中可见,包括细胞壁成分,如多糖、肽聚糖、脂蛋白和脂磷壁酸,这些成分可被上皮或宿主免疫细胞中表达的模式识别受体 (PRR) 识别。
Toll样受体2(TLR2)是一种跨膜受体,可在多种细胞类型(包括小胶质细胞、单核细胞、巨噬细胞和树突状细胞)中表达,在先天免疫反应中起着重要作用
益生菌还可以刺激肠道免疫系统产生特定的抗体(如IgA),这些抗体能够中和病原体,减少其感染性。
// 调节炎症反应
益生菌能够调节宿主的炎症反应,减少由过度免疫反应引起的组织损伤。它们通过产生抗炎细胞因子(如IL-10)和降低促炎细胞因子(如TNF-α)的水平来实现这一点。
除了单独使用益生元和益生菌外,还有将益生元和益生菌组合成合生元混合物的营养概念。考虑到上述临床研究的局限性,合生元已显示出临床成功,因此有望成为未来的治疗选择。
!
通过营养治疗感染需要注意的
与药物化合物的靶向作用相比,饮食治疗可能更加多因素。个体对营养化合物的反应受到个体遗传特征的影响。例如,炎症基因(如IL-1B、IL-6和TNF-α)中的单核苷酸多态性(SNP)会导致不同的炎症反应,这可以部分解释观察到的对营养化合物反应性的差异。
此外,越来越多的证据表明,每个人的肠道微生物群都是独一无二的,这种多样性使得个体对营养和治疗感染的反应存在显著差异。
研究显示,肠道微生物的组成可以影响营养物质的吸收和代谢,同时也会影响宿主的免疫系统功能。因此,了解个体的肠道微生物组成对于制定个性化的营养和治疗策略至关重要。
通过针对特定肠道微生物群优化的营养干预,可以更有效地预防和治疗各种感染,提高治疗的精准性和效果。这一新兴领域的研究不仅有助于推动精准医疗的发展,同时也为临床实践提供了新的视角和方法。
肠道感染仍然在全世界范围内造成沉重的疾病和经济负担。如何更好的预防和治疗胃肠道感染成了许多百姓和临床工作者关心的问题。
大量研究表明营养不良与感染之间存在双向作用。总体而言,营养影响人体免疫系统发育,而某些因素会导致营养不良并削弱人体抵抗感染的能力。包括厌食、肠道吸收减少、代谢损伤、脂质和碳水化合物代谢紊乱、维生素、铁、锌、铜减少。需要注意的是暴饮暴食可能也会增加感染风险。
与此同时,肠道微生物群在抵抗肠道病原体定植、生长和促进宿主免疫系统成熟以及影响宿主代谢方面发挥着不可或缺的作用。一些人类疾病被确定与特定微生物群的改变具有相关性或因果关系。
越来越多的人类疾病被确定与特定微生物群的改变具有相关性或因果关系。肠道中的炎症免疫反应(炎症性肠病和病原体诱导的)也可以直接塑造微生物群的组成并引发菌群失调。这一发现具有深远的影响。它从根本上改变了我们对人类肠道炎症发病机制的理解。
肠道微生物群在帮助宿主应对全身感染方面可能还有许多其他功能尚未被发现。越来越多的证据表明肠道菌群失调与慢性肠道炎症相关疾病的表现有关,尤其是炎症性肠病。抑制肠道炎症、利用益生菌或调整营养变化(如益生元和金属)可能是未来限制肠杆菌等致病细菌大量繁殖的潜在方法。
主要参考文献
Stecher B. The Roles of Inflammation, Nutrient Availability and the Commensal Microbiota in Enteric Pathogen Infection. Microbiol Spectr. 2015 Jun;3(3).
Pike VL, Lythgoe KA, King KC. On the diverse and opposing effects of nutrition on pathogen virulence. Proc Biol Sci. 2019 Jul 10;286(1906):20191220.
Cristofori F, Dargenio VN, Dargenio C, Miniello VL, Barone M, Francavilla R. Anti-Inflammatory and Immunomodulatory Effects of Probiotics in Gut Inflammation: A Door to the Body. Front Immunol. 2021 Feb 26;12:578386.
Ducarmon QR, Zwittink RD, Hornung BVH, van Schaik W, Young VB, Kuijper EJ. Gut Microbiota and Colonization Resistance against Bacterial Enteric Infection. Microbiol Mol Biol Rev. 2019 Jun 5;83(3):e00007-19.
Wiertsema SP, van Bergenhenegouwen J, Garssen J, Knippels LMJ. The Interplay between the Gut Microbiome and the Immune System in the Context of Infectious Diseases throughout Life and the Role of Nutrition in Optimizing Treatment Strategies. Nutrients. 2021 Mar 9;13(3):886.
Lobionda S, Sittipo P, Kwon HY, Lee YK. The Role of Gut Microbiota in Intestinal Inflammation with Respect to Diet and Extrinsic Stressors. Microorganisms. 2019 Aug 19;7(8):271.
Farhadi, Sedigheh1; Ovchinnikov, Roman S.2,. The Relationship between Nutrition and Infectious Diseases: A Review. Biomedical and Biotechnology Research Journal (BBRJ) 2(3):p 168-172, Jul–Sep 2018.
Cherrak Y, Flaugnatti N, Durand E, Journet L, Cascales E. Structure and Activity of the Type VI Secretion System. Microbiol Spectr. 2019 Jul;7(4).
Pickard JM, Zeng MY, Caruso R, Núñez G. Gut microbiota: Role in pathogen colonization, immune responses, and inflammatory disease. Immunol Rev. 2017 Sep;279(1):70-89.

谷禾健康

银屑病,又叫牛皮癣,会导致出现皮疹伴发痒的鳞状斑块,最常见于膝盖、肘部、躯干和头皮。通常呈周期性发展,发作数周或数月,然后消退一段时间,长期的发作和复发会给患者带来很大的痛苦和困扰,严重影响到生活质量和日常工作。银屑病影响着全球超过 6000 万成人和儿童。
银屑病是一种免疫介导的炎症性皮肤病,涉及遗传和环境因素之间复杂的相互作用。免疫学和遗传学研究确认IL-17和IL-23是银屑病发病机制中的关键驱动因子。然而,由于银屑病的反复发作和难治性质,目前还没有简单治愈方法。
在有遗传倾向的个体中,非遗传因素也是触发牛皮癣发病和复发所必需的,其中包括感染、皮肤和肠道微生物群失调、脂质代谢失调、性激素失调、精神疾病等,其他环境因素也可能诱发牛皮癣,例如皮肤创伤、不健康的生活方式和药物。
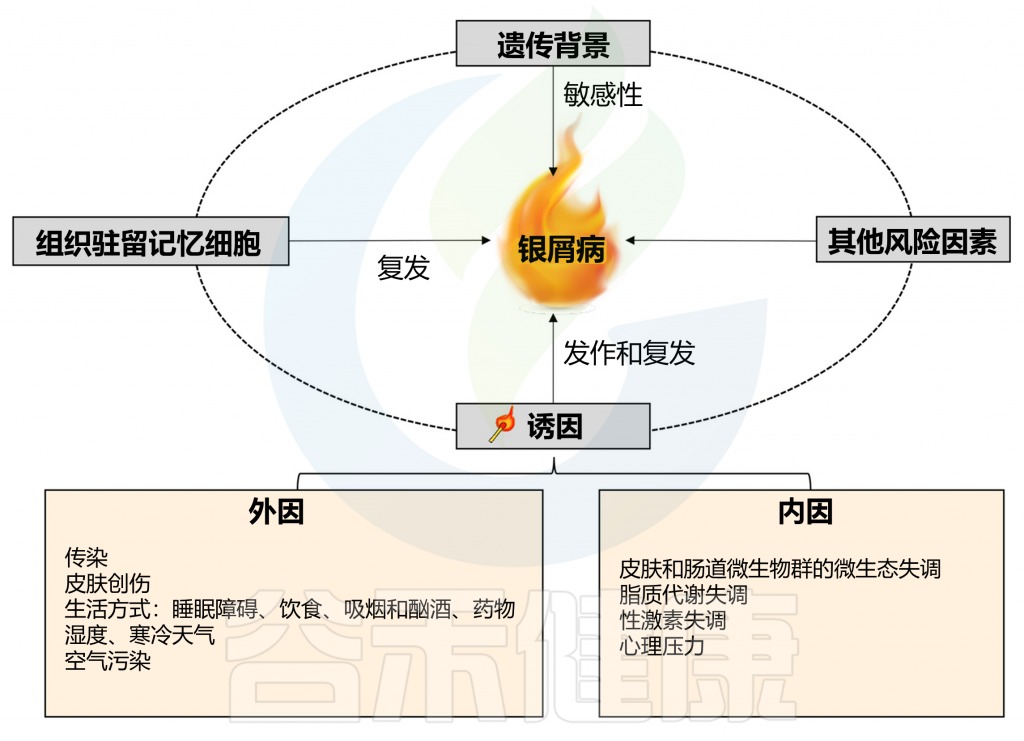
Liu S, et al., Cell Communication and Signaling, 2024
大量研究发现,银屑病不再被认为是仅影响皮肤的疾病,而是被视为一种全身性炎症性疾病,这与多种共病相关,包括结肠直肠癌、代谢综合征、肥胖、克罗恩病和溃疡性结肠炎等。微生物感染被认为是银屑病的重要病因,尤其是链球菌上呼吸道感染。
肠道和皮肤之间存在很强的双向联系,在肠道-皮肤轴中,皮肤炎症会影响肠道健康,而肠道微生物群也会影响皮肤健康,与健康对照相比,牛皮癣患者的肠道菌群失调,多样性降低,Firmicutes/Bacteroidetes(F/B)异常,大肠杆菌水平往往更高,Lachnospira、Akkermansia muciniphila、Blautia、Faecalibacerium减少,这种变化与人类结肠微生物群中丁酸盐代谢和产丁酸菌相关。丁酸与调节多种炎症因子有关,包括脂多糖、TNF-α、IL-10、IL-1β等。
扩展阅读:
其他菌群变化也与银屑病患者中与炎症相关标志物的程度相关联,特别是IL-2受体,其与Phascolarctobacterium呈正相关,与Dialister呈负相关。Phascolarctobacterium和Dialister的相对丰度可作为银屑病活动的预测指标。详见:
肠道核心菌属——考拉杆菌属(Phascolarctobacterium)
近日,来自华中科技大学同济医学院附属协和医院皮肤科的研究人员,总结了牛皮癣发病和复发的触发因素,发表在《Cell Communication and Signaling》上。

本文结合牛皮癣与肠道菌群的关联,更新了有关这些因素如何引发该疾病的潜在机制的当前证据。了解这些触发因素如何在银屑病的发病和复发中发挥作用,可以深入了解银屑病的发病机制,并更好地进行临床治疗。
银屑病是一种常见的红斑鳞屑性皮肤病,具有多种皮肤表现和全身参与,可影响任何皮肤部位,发生于任何年龄和任何地理区域。
在牛皮癣患者中,免疫系统错误地攻击了健康的皮肤细胞,导致皮肤细胞生长过快,形成皮肤表面的鳞屑。
银屑病的常见体征和症状包括:
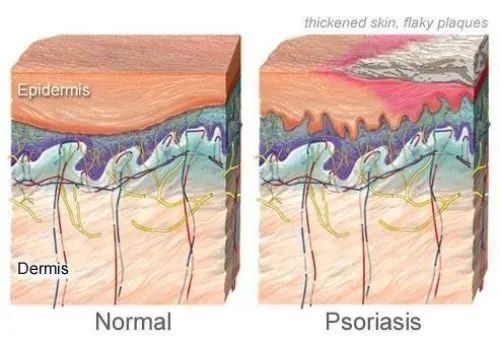
图源:WebMD
根据银屑病临床表现的不同,通常分为以下类型:
其他还包括:
牛皮癣和湿疹一样吗?
牛皮癣和湿疹是两种不同的皮肤病。这两种情况都会引起类似的症状,如皮肤变色、皮疹和瘙痒。
牛皮癣会传染吗?
牛皮癣不会传染。接触他人的牛皮癣皮疹不会感染牛皮癣。
牛皮癣是一种慢性炎症性皮肤病,世界卫生组织最近的报告指出,牛皮癣的患病率正在上升,在发达国家从 1.5% 到 5% 不等。
据统计,中国的牛皮癣发病率在1.3%-2.4%之间,约有1000万-2000万的患者。男性的发病年龄在30-39岁和60-69岁,而女性的发病时间大约要早十年,年轻人群体中发病率也有上升的趋势。
许多银屑病患者可能持续数年毫无症状,直至某种环境因素诱发这种疾病。常见的银屑病诱发因素包括:
银屑病关节炎
一般来说,银屑病越严重,患银屑病关节炎的风险就越高。2015 年欧洲一项研究发现,十分之一的银屑病患者患有未确诊的银屑病关节炎。未确诊的银屑病关节炎患者的生活质量受到很大影响。
心血管疾病
研究表明牛皮癣与心血管疾病之间存在联系,慢性炎症可能是心血管疾病和中风风险增加的原因,因为随着时间的推移,炎症会对动脉造成损害。
代谢综合征
根据一项研究,患有牛皮癣的人患代谢综合征的风险,是其他没有牛皮癣的人的六倍。
肥胖
研究表明,与一般人群中的其他人相比,牛皮癣患者更有可能肥胖。
炎症性肠病
牛皮癣和炎症性肠病 之间存在联系。《JAMA Dermatology》杂志 2018 年的系统综述和荟萃分析结果表明,牛皮癣与炎症性肠病相关。
抑郁症
抑郁症是银屑病的首要合并症。患有皮肤问题会导致抑郁。对于某些人来说,治疗牛皮癣可以帮助减轻抑郁症状并提高生活质量。
以上我们了解了关于银屑病的基本情况,接下来,我们从感染,微生物群失调,脂质代谢失调,心理压力和精神障碍,性激素失调,其他环境因素等方面来了解银屑病的相关触发机制。
多种微生物是银屑病的触发因素,见下表。
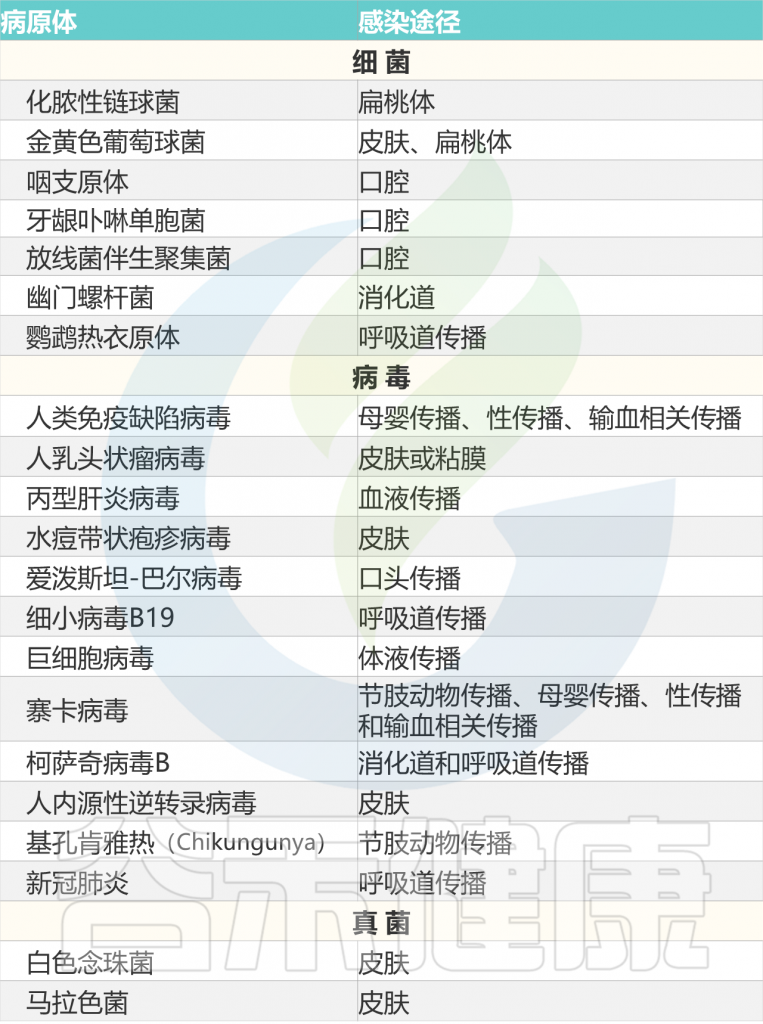
Liu S, et al., Cell Communication and Signaling, 2024
化脓性链球菌引起的扁桃体感染,可引发或加剧斑块状和滴状银屑病中的银屑病皮肤病变。
研究人员通过遗传关联研究将链球菌咽喉感染与银屑病联系起来,表明公认的银屑病风险等位基因HLA-C06:02是链球菌扁桃体炎的风险因素,而推断的银屑病风险单倍型HLA-C06:02/HLA-B*57:01是扁桃体炎最强的风险因素。
儿童银屑病与复发性扁桃体炎密切相关
一项临床队列研究还报道,年龄从10岁-11岁的儿童银屑病与复发性扁桃体炎密切相关。在银屑病患者的皮肤和扁桃体组织中发现了相同的T细胞克隆,提示在链球菌感染后的疾病中扁桃体内产生致病性T细胞。因此,建议将扁桃体切除术作为解决银屑病的干预措施,这可以减少循环T细胞的数量。
但仍应进行长期随访,验证扁桃体切除术的适应症和长期益处。同时,没有确凿的证据表明抗链球菌干预措施的有效性。有趣的是,肛周链球菌感染也会引发喉结型银屑病,但它不如喉咙感染常见。
化脓性链球菌喉咙感染和银屑病之间发病联系
链球菌扁桃体炎可能通过以下多种机制来引发牛皮癣:
a) 化脓性链球菌(SP)的M蛋白模仿人类 K17
CD8 + T细胞识别银屑病患者中链球菌M蛋白和人角蛋白17(K17)共享的表位,K17可以成为CD8的自身抗原和靶点 + 以HLA-C*06:02限制性模式浸润银屑病皮肤病变的T细胞。
b) SP超抗原激活IL-12的释放,然后促进CD4+T 细胞中皮肤归巢CLA的表达
皮肤寻求性皮肤淋巴细胞相关T细胞(CLA + T细胞)与化脓性链球菌的相互作用,为理解银屑病的免疫发病机制提供了新的概念。通过刺激IL-12的产生途径,化脓性链球菌超抗原诱导T细胞上皮肤特异性归巢受体(CLA抗原)的表达,并促进CLA+ T细胞向皮肤的迁移。
此外,在携带HLA-C*06:02的银屑病患者中,如果他们患有化脓性链球菌引起的扁桃体炎,其CLA+ T细胞和表皮细胞的培养中观察到高Th17反应。化脓性链球菌能够在斑块型和滴状银屑病中诱导循环CLA+ T细胞产生IL-17,这进一步在CLA+ T细胞迁移到皮肤后诱导银屑病自身抗原(如ADAMTS样蛋白5和LL-37)的产生。在体外银屑病模型中,化脓性链球菌提取物诱导CLA+ T细胞产生IL-9,从而上调IL-17A的产生。
c) 化脓性链球菌肽聚糖 (PG) 的辅助作用
含有PG的巨噬细胞与PG特异性CD4+密切接触银屑病病变中的T细胞,然后是PG特异性CD4 + T细胞以HLA-DR等位基因限制的方式增殖并产生IFN-γ。此外,PG固有识别的改变增强了T细胞对化脓性链球菌的反应,并诱导银屑病。
图2 感染和皮肤微生物失调可能诱发银屑病
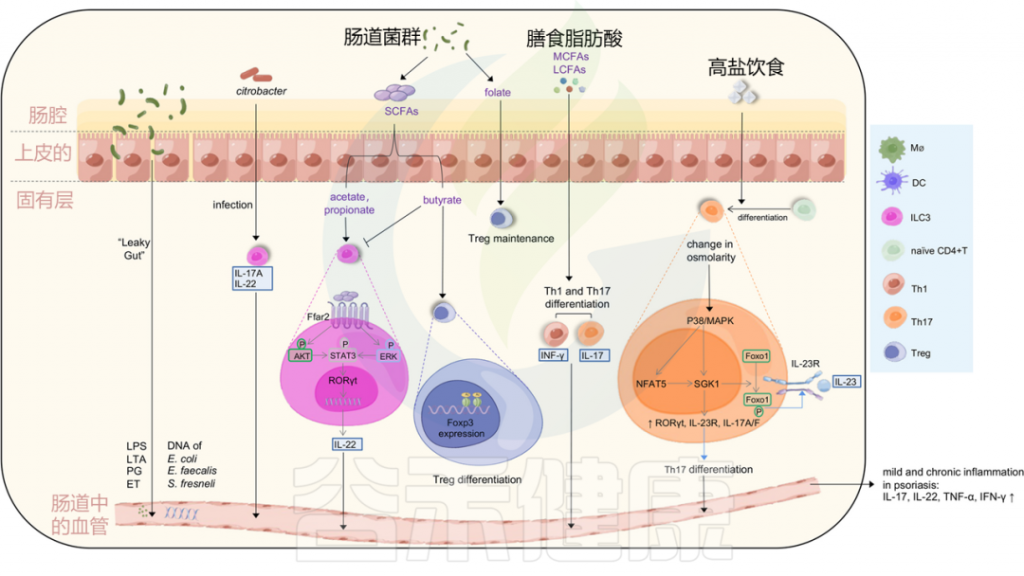
Liu S, et al., Cell Communication and Signaling, 2024
金黄色葡萄球菌在大约60%的银屑病患者的皮损和鼻孔中,而在5%至30%的健康个体中定植。
从22名银屑病患者中的11名患者的喉咙中分离出金黄色葡萄球菌。
一项研究显示,与健康对照组相比,银屑病受试者对超级抗原毒素的炎症皮肤反应增加,银屑病表皮中TNF-αmRNA水平增加。
然而,在银屑病病变中没有发现表达特异性T细胞受体Vβ的T细胞的选择性扩增,这是超抗原刺激的标志。这种T细胞依赖性反应可能是由于角质细胞中HLA-DR的高表达,增强了皮肤对超级抗原的炎症反应。
此外,银屑病的严重程度与葡萄球菌肠毒素的产生显著相关。
牙周炎患者患银屑病的风险较高
牙周炎期间,口腔微生物群可能影响银屑病的发展和恶化。一项涉及13项研究的荟萃分析显示,牙周炎患者患银屑病的风险高于对照组。
口腔微生物(尤其是咽支原体)在银屑病发展中的作用
一名患者最初患有滴状银屑病,后来发展为斑块型银屑病,其皮肤感染了咽支原体(Mycoplasma faucium),属于口腔软壁菌门,这种细菌出现在银屑病棘层角质形成细胞中,以及银屑病损害上层真皮的细胞外。
与对照组(骨关节炎)相比,在银屑病关节炎(PsA)患者的血清和滑液中也发现了更多种类和更高浓度的口腔细菌(牙龈卟啉单胞菌和Prevotella nigrescens)DNA。
牙龈病原体(尤其是牙龈卟啉单胞菌)在牙周炎与银屑病发病机制中的桥梁作用
牙龈卟啉单胞菌和放线菌聚集杆菌(Aggregatibacter actinomycetemcomitans)是与牙龈炎相关的病原体,可激活人CD14 + 单核细胞在体外增强Th17分化和IL-17产生。牙龈卟啉单胞菌蛋白酶可以通过降解其他关键细胞因子如IL-12,增强Th17系列反应,而髓样抗原呈递细胞(APCs)被触发产生与Th17相关的细胞因子IL-1β、IL-6、IL-23。然而,与健康受试者相比,牙周炎患者的牙龈组织中IL-17+细胞的频率增加,而在外周血中没有增加。
牛皮癣的皮肤炎症可以通过宿主的抗病毒免疫反应失调由病毒感染引发。
视黄酸诱导基因I (RIG-I)是病毒的主要细胞质传感器。通过激活RIG-I抗病毒信号传导,病毒感染可以在遗传易感个体中触发CD11c+树突细胞产生IL-23的表达,从而导致银屑病的发展。
HIV
与普通人群相比,人类免疫缺陷病毒(HIV)患者的银屑病标准化发病率更高。HIV可以直接作为超抗原或抗原呈递的共刺激因子触发银屑病,在HIV感染期间,活化的CD8 +T细胞产生更多的IFN-γ。神经肽物质P可以从HIV感染的免疫细胞中释放,然后调节炎症和免疫反应,刺激角质细胞的增殖。
HPV
人乳头瘤病毒(HPV)也与银屑病相关。一项针对66274名HPV感染患者的全国性人群队列研究显示,HPV感染后银屑病的患病率更高。
SARS-CoV2
严重急性呼吸综合征冠状病毒2型(SARS-CoV2)也被认为是银屑病恶化的原因。在严重急性呼吸系统综合征冠状病毒感染患者的血浆中炎性细胞因子水平升高,粒细胞集落刺激因子和TNF-α的浓度与疾病严重程度相关。
此外,一些接种新冠肺炎疫苗的患者患有慢性免疫介导的皮肤病,如牛皮癣,但皮肤反应通常轻微且自限性的。
NLRP1炎症小体
NLRP1是已鉴定的炎症小体形成模式识别受体(PRRs)之一,先天免疫系统可以通过该受体检测病原体。在正链RNA病毒感染过程中产生的长双链RNA(dsRNA),例如Semliki Forest病毒,可以结合并激活人类角质形成细胞中的NLRP1炎症小体。NLRP1炎症体在促进银屑病发病的过程中发挥作用,无论是通过增加对银屑病的易感性,还是通过促炎细胞因子(包括IL-1β和IL-18)的失调释放。
同样,NLRP1具有感知细菌病原体外毒素的能力,如铜绿假单胞菌分泌的外毒素A和白喉棒状杆菌分泌的白喉毒素,并诱导细胞死亡和IL-1β / IL-18分泌。
注:NLRP1,核苷酸结合域和富含亮氨酸重复pyrin结构域的蛋白质1(NLRP1),一种在人体免疫系统中发挥作用的蛋白质。
银屑病皮肤中的多种真菌已被鉴定为通过遗传易感个体的先天免疫系统激活银屑病。
白色念珠菌
来自白色念珠菌等微生物的超级抗原可能导致感染患者银屑病的恶化。接触白色念珠菌也可在银屑病皮肤中引发对IL-17的临床相关反应。银屑病CLA + T细胞/表皮细胞共培养物通过增加IL-9、IL-17A和IFN-γ的产生,对白色念珠菌提取物产生反应。
此外,皮肤白色念珠菌感染通过产生IL-17的CD4 + TRM诱导复发性银屑病。在小鼠模型中,CD4 + TRM在感染30天后成为IL-17的主要来源。
除白色念珠菌外,马拉色菌可能与头皮银屑病的恶化有关。
幽门螺杆菌
与对照组相比,幽门螺杆菌感染在中度和重度银屑病患者中显著增加,但在轻度银屑病患者中没有增加。在幽门螺杆菌感染的银屑病患者中,银屑病面积和严重程度指数(PASI)评分较高,银屑病相关细胞因子IL-1β、IL-6、IL-8和TNF-α的粘膜水平也较高。然而,2015年的一项发现表明,银屑病患者的幽门螺杆菌患病率没有比健康对照组增加。还需进一步研究。
目前,许多研究都致力于人类微生物组在银屑病发病机制中的作用,特别是皮肤和肠道微生物组之间的关系,即“肠道-皮肤轴”。
银屑病患者厚壁菌门增加
几位研究人员推测,银屑病可能与宿主皮肤微生物群的失调密切相关(图2)。与非病变皮肤和对照组相比,银屑病病变皮肤的细菌载量更高,但细菌多样性更低。厚壁菌门和放线菌门分别是银屑病患者和健康对照组中最常见的细菌门,病变皮肤中厚壁菌的增加和放线菌的相应减少是显著的。然而,另一项研究报道了银屑病病变中放线菌门和厚壁菌门都增加。这种差异可能是由于采样方法、皮肤部位、药物和分析方法的多样性。
棒状杆菌丰度与银屑病严重程度相关
根据新的证据,与未受影响和健康的皮肤相比,银屑病病变的棒状杆菌(Corynebacterium)浓度更高,Cutibacterium更低。棒状杆菌丰度与疾病严重程度相关,大多数种类的棒状杆菌在小鼠皮肤中诱导强烈的IL-23依赖性反应。在小鼠皮肤上涂抹假白喉棒状杆菌后,皮肤IL-1β蛋白水平升高,真皮中γδT17细胞增多。
银屑病耳朵皮肤显示葡萄球菌过度表达
较低丰度的表皮葡萄球菌和痤疮丙酸杆菌可能促进金黄色葡萄球菌在银屑病中的定植,这可以刺激小鼠模型中的Th17极化并引发IL-17介导的皮肤炎症。
为了解释银屑病的发病机制,提出了从屏障破坏到微生物群紊乱,再到病变加重的循环来解释银屑病的发病机制。
具有表皮屏障缺陷的小鼠具有增加的细菌载量和抗微生物肽(AMP)表达。在施用局部抗生素后,通过减少皮肤上的细菌负荷,以及减少IL-17和IL-22的产生,可以缓解小鼠的银屑病样表型。
除了皮肤微生物群的微生态失调外,紊乱的肠道微生物群也会影响银屑病的病理生理学(图3)。与健康对照组相比,银屑病患者的肠道微生物群在组成和功能潜力方面都发生了变化。银屑病患者的肠道微生物群特征显著紊乱,细菌多样性低,厚壁菌门/拟杆菌门(F/B)比率升高,并与PASI评分呈正相关。除了厚壁菌门和拟杆菌门外,银屑病患者和健康对照组16种属水平上的类群也存在显著差异。
银屑病患者肠道微生物的相关研究

doi.org/10.3390/ijms25041984
肠道屏障完整性与银屑病严重程度呈正相关
肠道脂肪酸结合蛋白(FABP)是肠道屏障完整性的生物标志物,其水平与银屑病的严重程度呈正相关。肠道微生物群失调可能通过减少粘液层的厚度、干扰肠道上皮细胞的增殖和代谢以及影响AMPs的产生来增加肠道通透性,也称为“肠漏”。
肠道微生物失调导致肠漏,促进细菌和抗原转移
肠道细菌可以通过DC通过上皮细胞之间的过程逃逸到血液中,而不影响紧密连接功能,或者通过覆盖在Peyer贴片上的微折叠细胞,将微生物产物呈递给APC。渗漏的肠道促进细菌移位,并允许外部抗原从肠腔进入血液和淋巴循环,从而驱动局部和全身免疫反应。
银屑病患者中细菌DNA易位与全身炎症的增加
与其他患者和健康对照组相比,斑块型银屑病患者血液样本中细菌DNA易位增加,主要是由肠道细菌引起的,包括大肠杆菌、粪肠球菌和弗氏志贺菌。细菌DNA易位的患者也表现出更高水平的全身炎症反应。
另一项研究还报道,在25%的斑块型银屑病患者的血液中观察到细菌DNA,与其他肠道类型的患者相比,肠型2(普雷沃氏菌占优势)的患者更容易发生细菌移位。这些微生物在被重新激活后可能会释放出高效的炎症因子,如脂多糖(LPS)和脂磷壁酸(LTA),这可能会导致宿主机体中的轻度和慢性炎症,银屑病患者会受此困扰。从肠道吸收的细菌内毒素(ET)和PGs也会加剧银屑病,银屑病的治疗已通过阻止其吸收或破坏内毒素来证明这一点。
短链脂肪酸促进Treg分化,调节免疫活性
微生物群可以通过肠道中的微生物代谢产物来改变免疫活性。短链脂肪酸(SCFAs)是肠道微生物组不可消化碳水化合物的主要发酵产物,主要包括乙酸盐、丁酸盐和丙酸盐。其中,丁酸盐可增强Foxp3基因座启动子区的组蛋白H3乙酰化,表明其有可能影响Treg细胞的分化。
微生物群对T细胞分化的影响:叶酸与色氨酸代谢途径
叶酸来自肠道微生物群和饮食,而饮食中的叶酸对维持Foxp3+Tregs有选择性作用。作为宿主色氨酸代谢途径之一,犬尿氨酸途径可以通过吲哚胺2,3双加氧酶将成熟的DC转化为耐受性DC,从而增强Tregs并抑制效应T细胞。这些结果可能表明,微生物群代谢产物通过触发肠道T细胞的分化,成为银屑病的非感染性风险因素。
固有淋巴细胞ILC3的调控:微生物代谢产物的双重作用
近年来,第3组固有淋巴细胞(ILC3)的功能受到了广泛关注。ILC3主要在肠道和皮肤中发现,并通过产生IL-17A和IL-22在银屑病中发挥致病作用。
当身体感染某些细胞外病原体(如柠檬酸杆菌)时,ILC3产生IL-22和/或IL-17,用于对抗病原体的粘膜免疫。ILC3的功能也可以受到微生物代谢产物的调节,如短链脂肪酸。丁酸盐可由厚壁菌门产生,而乙酸盐和丙酸盐主要由拟杆菌门产生。
图3 肠道微生物失调和饮食可能会在银屑病中诱导轻度和慢性炎症
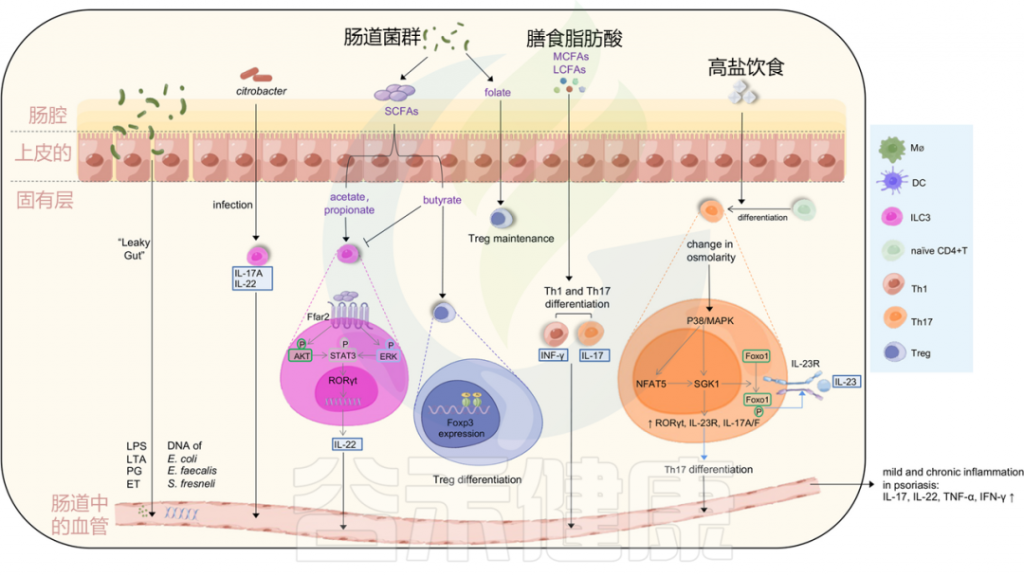
Liu S, et al., Cell Communication and Signaling, 2024
肠道通透性增加导致的“渗漏性肠病”可以诱导脂多糖(LPS)、脂蛋白(LTA)、外毒素(ET)和肽聚糖(PG)等强效炎症介质的释放,以及肠道细菌DNA进入血液。
柠檬酸杆菌感染可以刺激ILC3s产生IL-22和IL-17A,从而有助于黏膜免疫。ILC3s的功能也受到微生物代谢产物短链脂肪酸(醋酸、丁酸和丙酸)的影响。
乙酸和丙酸与结肠ILC3s上的FFAR2受体相互作用,激活AKT或ERK信号通路,并通过STAT3轴释放IL-22;
而丁酸则降低ILC3s的数量,同时,丁酸增强Treg细胞的分化,叶酸有助于维持Treg细胞,中长链脂肪酸支持Th1和Th17细胞的分化。
高盐饮食导致渗透压改变激活P38/MAPK通路,进而上调下游靶点NFAT5和SGK1,这反过来驱动转录因子RORγt、IL-23R、IL-17A和IL-17F的表达,导致Th17分化。上调的SGK1通过磷酸化Foxo1促进IL-23R的表达和稳定Th17分化。
益生菌的治疗已证明在改善银屑病方面具有潜在的益处,尽管尚未制定标准化的治疗方法。粪菌移植提供了另一种可能的治疗策略,因为它们通过允许健康粪便微生物组的“真生物”在受影响患者的肠道中重新定居来缓解自身免疫性疾病。未来更好地了解微生物群失调无疑将有助于缓解银屑病的治疗方法,我们在后面第八章也有具体的介绍。
许多研究已经表明肥胖和血脂异常与银屑病的关系,肥胖是银屑病的一个重要风险因素。而且,肥胖会加重已有的银屑病,这种自身免疫性疾病与肥胖之间的可能联系依赖于脂肪组织。
一般来说,脂肪组织可能通过肥大脂肪细胞分泌大量激素和细胞因子(也称为脂肪因子),包括IL-6、TNF-α和瘦素,在脂质代谢失调的患者中充当免疫器官,这些因子调节炎症途径和免疫细胞的活动。各种银屑病RNA-seq数据集表明,脂质代谢途径与银屑病的发病机制密切相关。
图4 将血脂异常与银屑病联系起来的潜在机制
Liu S, et al., Cell Communication and Signaling, 2024
最近的研究表明,与肥胖相关参数无关的饮食成分,可能在银屑病的恶化中发挥关键作用。一项使用小鼠银屑病模型的研究提供了证据,证明饮食中的游离脂肪酸(FFAs),特别是饱和脂肪酸(SFAs),是银屑病皮炎的关键放大器。
饱和脂肪酸:加重银屑病皮炎的可能潜在机制
循环中的饱和脂肪酸被转运到皮肤中,并诱导骨髓源性树突细胞产生各种促炎细胞因子,如IL-1β。这些促炎细胞因子随后促进角质形成细胞分泌趋化因子和炎性细胞因子,导致中性粒细胞和单核细胞募集到皮肤,并导致银屑病皮炎放大。
饱和脂肪酸还可以通过激活TLR2和TLR4,来调节单核细胞或巨噬细胞中的NALP3炎症小体和炎症小体介导的IL-1β分泌。饱和脂肪酸在银屑病皮炎恶化中的另一种机制是通过激活DC来促进Th1/Th17分化。
体内游离脂肪酸的增加可能通过支持表皮中TRM细胞的存活而促使银屑病的复发。此外,膳食游离脂肪酸、中链和长链脂肪酸(MCFA和LCFA)的成分指导肠形Th细胞分化,这与上述丁酸盐诱导的Treg分化相反(图3)。
多不饱和脂肪酸(PUFA)及其衍生的生物活性脂质介质
除饱和脂肪酸外,多不饱和脂肪酸(PUFA)和多不饱和脂肪酸衍生的生物活性脂质介质(LMs)也可介导银屑病的炎症反应。
其中,生物活性LMs来源于两种多不饱和脂肪酸,亚油酸(LA,n-6 PUFA)和α-亚麻酸(ALA,n-3 PUFA),它们分别因其在银屑病中的促银屑病和抗炎特性而闻名。
n-6
作为代表性的n-6 多不饱和脂肪酸衍生的LMs,LTB4有助于中性粒细胞趋化,PGE2有助于角质细胞增殖。
n-3
衍生自 n-3 多不饱和脂肪酸的专门的促分解脂质介质(SPMs),包括脂氧素(Lxs)、resolvins (Rvs)、保护蛋白(PD) 和Maresins (MaRs),在银屑病中具有抗炎和免疫调节功能。
一个研究小组专注于基于液相色谱-串联质谱(LC–MS/MS)分析鉴定人类银屑病中的生物活性LMs和SPMs。根据他们的研究,衍生自n-6 多不饱和脂肪酸的生物活性LMs在银屑病皮肤中丰富,同时resolving D1(RvD1)、resolving D5(RvD5)、保护蛋白D1(PD1)及其双加氧异构体10S,17S diHDHA(又名PDx),阿司匹林触发形式的脂氧素A4和脂氧素B4(AT-LXA4和AT-LXB4)可能是解决银屑病病理生理学中炎症反应的专门的促分解脂质介质。
脂质代谢与铁死亡(Ferroptosis)密切相关,铁死亡促进银屑病角质形成细胞引发炎症,这涉及一系列连续事件,即活性氧(ROS)的积累导致脂质过氧化并进一步诱导铁死亡。
PUFA促进银屑病角质形成细胞铁死亡和炎症的机制
在银屑病期间,脂质过氧化与Th22/Th17通路在单细胞水平上呈正相关,证明了脂质过氧化的增强。
角质形成细胞对铁死亡也具有时间和浓度依赖性。在角质形成细胞的脂质代谢中,循环中积累的PUFA被催化为关键底物PUFA-CoA,最终酯化为PUFA-PLs,当存在生物活性铁时,PUFA-PL-OH可以过氧化为PUFA-PL-OH。随后,PLOOH可以通过产生脂质羟基自由基和脂质过氧自由基使细胞对铁死亡敏感。
MUFA-CoA和Fer-1通过抑制脂质过氧化抑制银屑病铁死亡
相反,来自饱和脂肪酸的单不饱和脂肪酸(MUFA)的产物MUFA-CoA可以通过抑制PUFA-PLs的过氧化来减少脂质过氧化的可用底物,从而抑制铁死亡。
此外,各种研究表明,Ferrostatin-1(Fer-1)是一种有效的脂质过氧化抑制剂,可以抑制铁死亡并阻断银屑病中的炎症反应。
除了氧化应激外,脂质还可以启动内质网(ER)应激,这具有双向作用:
饱和游离脂肪酸通过诱导ROS和ER应激参与银屑病发病机制
在非脂肪细胞中,过量的饱和游离脂肪酸通过脂质代谢和信号通路诱导ROS和ER应激。随后的线粒体和ER功能障碍是导致终末细胞死亡的关键步骤。此外,长期的内质网应激可导致氧化应激,脂质诱导的ROS也可能间接触发内质网应激,尽管确切机制尚不清楚。
脂质抗原激活限制性T细胞促进银屑病炎症反应
脂质自身抗原在驱动血脂异常相关自身免疫性疾病中的作用也引起了关注。银屑病病变含有高水平的磷脂酶A2(PLA2),其参与新脂皮肤抗原的产生。在IFN-α的诱导下,细胞质PLA2组IVD(PLA2G4D)可以以外泌体的形式从银屑病肥大细胞中释放,并转移到邻近的表达CD1a的Langerhans细胞。然后,新脂质抗原被脂质特异性CD1a反应性T细胞识别,释放IL-22和IL-17A。
自身脂质反应性T细胞在高脂血症与银屑病关联中的角色
除了CD1a限制性T细胞外,其他CD1分子(如CD1b和CD1c)限制性T淋巴细胞也对自身脂质产生反应并诱导细胞因子的产生。
在一项研究中,通过皮肤病变中积累的磷脂和胆固醇的一些自身抗原直接激活了CD1b自身反应性HJ1 T细胞。在患有高脂血症血清的小鼠中,CD1b+树突细胞增加了IL-6的产生,以及HJ1 T细胞分泌IL-17A,表明高脂血症和银屑病之间的潜在联系可能在于自身脂质反应性T细胞。
心理压力与银屑病的发病、严重程度和复发之间可能存在相关性
在31-88%的患者报告压力是银屑病的诱因,在过去12个月内经历压力事件的受试者中,银屑病的发病率更高。
另一项病例对照研究利用Holmes和Rahe的社会适应评定量表来评估压力生活事件,得出结论,压力在银屑病的发展中起着重要作用,特别是在复发和延长方面。
然而,一项荟萃分析报告称,没有令人信服的证据表明应激事件与银屑病之间存在这种关联。因此,未来应在基于人群的研究中前瞻性地审查这种关系,利用标准化的压力测量工具,并结合额外的生理和生化压力标志物。
心理压力激活HPA轴促进银屑病发病
压力可能通过下丘脑-垂体-肾上腺(HPA)轴、免疫途径和外周神经系统在银屑病的病理生理学中发挥作用。(图5)
作为对心理压力的反应,下丘脑产生促肾上腺皮质素释放激素(CRH),进一步激活垂体促肾上腺皮质激素(ACTH)和肾上腺皮质醇的分泌。CRH能够抑制角质细胞的凋亡,这是银屑病的典型现象。
另一方面,CRH通过刺激血管内皮生长因子(VEGF)增强血管生成,并增加血管通透性,促进银屑病斑块中炎症细胞的渗透。肥大细胞(MC)也可以被CRH激活,然后释放细胞因子和趋化因子,包括IL-1、IL-6、IL-31、TNF和CXCL-8。
此外,压力刺激皮肤外周神经末梢神经肽的释放,导致MC激活后神经源性炎症的发展。这些神经肽包括神经降压素(NT)、物质P(SP)、神经生长因子(NGF)和垂体腺苷酸环化酶激活多肽(PACAP)。
重度抑郁、PTSD、焦虑
除压力外,重度抑郁障碍或创伤后应激障碍患者患银屑病的风险显著高于对照组。
甚至父母常见的精神障碍(焦虑和抑郁)与后代患银屑病风险之间的关系也已确定。
双相情感障碍
另一项研究报告称,一名患有双相情感障碍的女性随后发展为银屑病,并在每次躁狂发作期间银屑病病变加重。
自闭症
根据自闭症谱系障碍患者银屑病炎症的临床升高,自闭症小鼠模型中高水平全身炎症,表明自闭症与银屑病活动之间存在联系。此外,先前的研究已经证实,严重精神障碍和银屑病之间存在遗传重叠。
越来越多的研究阐明了性激素对皮肤的多种生物和免疫调节作用。银屑病的自然病程似乎受到妊娠、月经和更年期的调节,从而暗示女性激素诱导的机制可能参与调节皮肤炎症。此外,研究表明,与女性相比,男性银屑病的患病率和严重程度更高,尤其是在雌激素充足的年龄,这表明不同性激素对银屑病的调节作用不同。
雌激素对银屑病具有保护作用
研究发现,雌激素通过下调中性粒细胞和巨噬细胞产生的IL-1β,这一过程通过雌激素受体α和β(ERα和ERβ)介导,具有潜在的抗银屑病作用。
同样,一项体内研究表明,雌二醇通过调节中性粒细胞和巨噬细胞的功能,在咪喹莫特(IMQ)诱导的小鼠银屑病炎症中发挥保护作用。体外,17β-雌二醇阻断了10 kDa的IFN-γ/干扰素诱导蛋白(IP-10)的正反馈回路,支持银屑病中Th1介导的炎症。
雌激素在银屑病中也可能具有促炎特性
一些孕妇的银屑病症状有所改善,而另一些孕妇的症状则有所恶化。
一份病例报告表明,一名患者在接受三苯氧胺治疗(一种抗中风药物)后,银屑病症状得到缓解,但在围绝经期症状恶化。值得注意的是,与对照组相比,男性银屑病患者的血清雌二醇水平显著升高,这表明雌激素可能参与银屑病的发展。
此外,一项使用咪喹莫特诱导的银屑病模型的体内研究也表明,雌激素通过Erα诱导IL-23,在银屑病中发挥促炎作用。
总之,这些证据支持雌激素在特定情境下对银屑病有双重影响,这导致了偶尔出现的相互矛盾的观察结果。
孕激素在银屑病中具有保护作用
临床观察证明银屑病通常在妊娠期间改善或消退,但在分娩后再次出现。
一项病例对照研究确定了孕妇受影响体表面积的改善与雌二醇、雌三醇、雌激素与孕酮比例升高之间的相关性。一些研究人员已经证明,角质细胞通过在银屑病皮肤中表达孕酮受体(PR)而成为孕酮的靶标。此外,孕酮在妊娠期间诱导转录改变,这些改变富含与银屑病相关的基因。STAT1和STAT3显著下调,其下游靶点,包括IL-12β、OSM和CXCL10,受到影响。
睾酮
一些研究强调雄性激素在牛皮癣中的作用。他们表明,严重的牛皮癣与低血清睾酮水平有关。无论患者年龄(40 岁以上或以下)如何,总睾酮和游离睾酮与 PASI 之间均呈负相关。
睾酮可以作为一种免疫抑制因子,下调树突状细胞的活性,干扰B细胞的分化,减少促炎细胞因子的释放。此外,在动物模型中,睾酮可以通过降低 IL-17 的水平来充当肝脏炎症的抑制剂,IL-17 是一种已知在银屑病发展中发挥关键作用的白细胞介素。
雄激素剥夺疗法(ADT)在晚期前列腺癌中作为加剧或缓解银屑病因素
一份病例报告显示一名前列腺癌患者ADT后银屑病加重。相反,一项调查发现ADT与银屑病风险降低之间存在相关性。另一项研究显示,无论年龄组如何,总睾酮或游离睾酮与PASI之间存在显著的负相关。
催乳素:刺激角质形成细胞的增殖
催乳素是垂体前叶分泌的一种多肽激素,刺激角质形成细胞的增殖,充当血管生成的诱导剂,并促进 Th1 细胞浸润银屑病病变。
此外,催乳素在调节免疫反应方面具有复杂的作用。它可以作为体液和细胞免疫反应以及 Th1 和 Th17 反应的刺激剂。
催乳素可以刺激角质形成细胞的增殖。最近的一项荟萃分析支持了激素在银屑病发病机制中发挥重要作用,该分析评估了循环催乳素水平与银屑病严重程度之间的关系。该荟萃分析包括 12 项研究,总共涉及 446 名银屑病患者和 401 名健康受试者。银屑病患者的循环催乳素水平显著较高,并且与疾病严重程度呈正相关。
总之,目前探索性激素对银屑病影响的研究主要依赖于观察性研究,缺乏深入的机制探索。这些关于雌激素和雄激素的矛盾发现表明,需要更多高质量的证据来更好地理解性激素与银屑病发病机制之间的复杂联系。
一些触发因素的潜在机制无法归类为感染因素、皮肤微生物群失调、肠道微生物群失调或脂质代谢失调,如下图。
图5 与银屑病发生相关的其他诱因
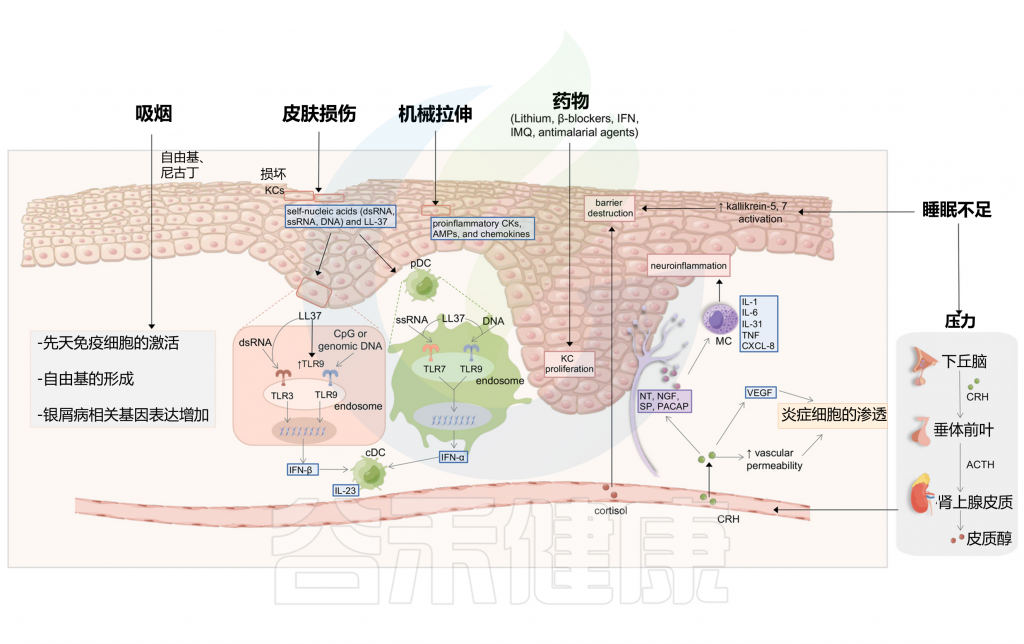
Liu S, et al., Cell Communication and Signaling, 2024
科布内氏现象与银屑病:皮肤损伤诱发疾病
皮肤创伤或压力可引发银屑病,称为科布内氏现象(Koebner phenomenon,KP)。
注:科布内氏现象,也称同形反应,是指在某些皮肤病患者健康皮肤区域受到损伤后出现的与原发皮肤病相同的皮损。
拔罐疗法与银屑病:传统治疗方法的争议
拔罐疗法作为一种传统的中医药,曾被用于治疗银屑病,但现在却存在争议,因为一些银屑病患者通过同形反应产生局部皮肤病变,而没有达到预期的治疗效果。
在银屑病患者中,拔罐治疗会导致拔罐部位出现同形反应,而Hijama(中东国家进行的一种湿拔罐)只会导致切口区域出现同形反应。
皮肤损伤激活角质细胞和免疫细胞参与银屑病发病
在皮肤损伤过程中,受损的角质细胞释放自身核酸,包括dsRNA、单链RNA(ssRNA)和DNA,并诱导LL-37的表达。
LL-37能够通过TLR7或TLR9在浆细胞样树突状细胞(pDC)中识别ssRNA或DNA,这最终导致IFN-α的分泌。LL-37暴露也可以通过DNA-LL-37复合物非依赖性机制或通过TLR3对dsRNA的识别诱导IFN-β的产生。对于前一种机制,LL-37增加TLR9的表达,从而促进TLR9配体的识别,如CpG或基因组DNA。浆细胞样树突状细胞的IFN-α和角质细胞的IFN-β促进常规树突状细胞的成熟。银屑病在创伤部位的复发归因于创伤部位TRM细胞的积聚和再激活。
机械拉伸与皮肤损伤:银屑病病变潜在触发因素
一份病例报告描述了一名患寻常型银屑病的妇女在插入组织扩张器后发展出新的银屑病病变。机械拉伸被怀疑会触发角质细胞释放ATP(5’-三磷酸腺苷),并随后产生Th17极化细胞因子,如前IL-1β和IL-6。被释放的ATP也可以激活表皮兰氏细胞。
在小鼠皮肤扩张模型中,观察到表皮过度增殖、皮肤屏障功能受损以及表皮角质细胞中银屑病相关细胞因子的上调。在人类角质细胞中,连续拉伸可以导致银屑病相关促炎细胞因子、抗微生物肽和趋化因子的产生。
除了拉伸,角质细胞的抓伤损伤可以通过细胞因子或趋化因子CCL20触发科布内氏现象,在较小程度上,这种触发作用与抓痕线数有关,也与CXCL8有关。
吸烟强度和持续时间与银屑病有关
与普通人群相比,银屑病患者中吸烟患病率更高,并揭示了吸烟开始和戒烟对银屑病的因果影响。吸烟强度和持续时间可能对银屑病的发病率具有剂量依赖性影响。作为银屑病发展的一个独立风险因素,吸烟对银屑病患者有许多负面影响,包括较高的PASI评分、指甲受累增加和心血管疾病的发展。
吸烟可能通过炎症、氧化和遗传机制引发银屑病
尼古丁通过释放炎性细胞因子刺激先天免疫细胞,如DC、巨噬细胞和角质细胞。此外,吸烟会引发自由基的形成,从而激活银屑病相关的蛋白质信号通路。
吸烟上调银屑病相关基因表达
在遗传学方面,吸烟上调银屑病相关基因的表达,包括HLA-C*06:02、HLA-DQA1*0201和CYP1A1。
烟草与银屑病:保护作用or风险因素?
最近的一项研究阐明了烟碱受体基因CHRNA5在银屑病的发展和发病机制中的作用。沉默的CHRNA5可以抑制人类角质细胞的增殖和迁移。有趣的是,吸烟也会增加普通人群患银屑病的风险,但吸烟在银屑病患者中表现出保护作用,这被称为“吸烟悖论”。然而,最近的一项孟德尔随机化研究涵盖了105912名具有生活方式因素、生物化学和基因型数据完整信息的个体,该研究表明吸烟是银屑病的一个独立风险因素,但不是银屑病的因果风险因素。
睡眠障碍:促炎因子显著增加,影响表皮屏障,加剧银屑病
睡眠障碍通常是银屑病的危险因素之一。一个具有全国代表性的基于人群的数据集表明,当发生阻塞性睡眠呼吸暂停时,银屑病和精神分裂症的风险增加。睡眠不足可能通过失眠的心理压力改变屏障稳态和角质层完整性。研究人员发现,银屑病小鼠在睡眠不足后,促炎细胞因子(IL-1β、IL-6和IL-12)显著增加,抗炎细胞因子(如IL-10)减少。
睡眠不足也促进了银屑病皮肤中kallikrein-5和kallikrein-7的活性,从而影响表皮屏障,导致银屑病的发展。此外,皮质醇在失眠等一些睡眠障碍中会增加。皮质醇刺激皮肤MC,破坏皮肤屏障功能,并上调促炎细胞因子,从而进一步加剧银屑病。
饮食因素在银屑病发病机制中的作用
73%的患者在坚持无麸质饮食三个月后出现临床改善,银屑病真皮中的Ki67淋巴细胞也减少了。
除麸质外,氯化钠(NaCl)摄入量的增加被认为对银屑病的发病机制有潜在影响(图3)。在高盐条件下,活化的p38/MAPK通路可以上调下游靶向活化T细胞核因子5(NFAT5)和血清/糖皮质激素调节激酶1(SGK1)。靶基因的上调可以驱动转录因子RORγt、IL-23R、IL-17A和IL-17F的表达,从而导致银屑病Th17细胞从初始CD4 + T细胞分化。SGK1对于通过Foxo1的磷酸化促进IL-23R表达和稳定Th17细胞分化至关重要。
银屑病与饮酒之间存在着复杂的多因素关系
银屑病患者的过度饮酒率高于普通人,酗酒会增加银屑病的严重程度并降低治疗效果。中重度银屑病患者的死亡风险也会增加,酒精是一个主要因素。然而,一项调查报告称,饮酒与银屑病的发展没有显著联系,孟德尔随机化研究也没有发现饮酒与银屑病之间的因果关系。目前还没有足够的证据来确定饮酒是否与银屑病的发病和复发有关。
许多药物都能引发银屑病,如锂、β受体阻滞剂、抗疟药、非甾体抗炎药、血管紧张素转换酶抑制剂、IFN、IMQ、特比萘芬、他汀类药物、贝特类药物和抗程序性细胞死亡蛋白1(PD-1)或抗程序性死亡配体1(PD-L1)抗体。在罕见病例中,TNF抑制剂也可能自相矛盾地诱发银屑病。
银屑病是锂的最常见皮肤不良反应
锂诱发和加重银屑病的发病率范围从3.4%至45%不等。
β受体阻滞剂治疗引发的角质细胞过度增殖及T细胞活性变化
当用β受体阻滞剂治疗时,负责促进细胞分化和抑制增殖的细胞内信使CAMP在表皮显示减少,最终导致角质细胞过度增殖。
此外,银屑病T细胞与对照组在蛋白酪氨酸磷酸化活性方面存在重要差异,蛋白酪氨酸激酶的诱导对包括淋巴细胞和角质细胞在内的细胞的激活和增殖至关重要。
抗PD-1/PD-L1诱导银屑病及潜在疗法
随着抗PD-1或PD-L1抗体在晚期恶性肿瘤治疗中的应用迅速增长,抗PD-1/PD-L1诱导的银屑病病例逐渐被报道。有研究者认为,PD-1的抑制通过加速表皮CD8 + 的浸润而促进皮肤炎症参与与角质细胞致病性串扰的T细胞。他们进一步证明了IL-6靶向疗法对抗PD-1/PD-L1诱导的银屑病的潜在疗效。
其他药物也通过不同的机制与银屑病有关
银屑病也可以由生物制剂引发
这被认为是一种自相矛盾的反应。一项研究显示,216例疑似TNF抑制剂诱导或加重的银屑病报告病例,英夫利昔单抗治疗银屑病的发生率更高,在克罗恩病和类风湿关节炎治疗的第一年最为普遍。对TNF抑制剂诱导的银屑病患者的回顾性分析也得出了一致的结果,表明英夫利昔单抗是主要的触发因子,而克罗恩病和类风湿关节炎是最常见的原发性疾病。这种自相矛盾的反应可能与通过抑制易感个体TNF活性而诱导的免疫改变有关。其发病机制也与IL-23/Th17轴参与TNF抑制的设定有关。
其中一个悬而未决的重大谜团是,在靶向TNF-α、IL-23和IL-17A/IL-17RA的生物制剂停用后,银屑病病变往往在相同的区域复发。目前最流行的观点是,现有生物制剂主要用于抑制致病性免疫细胞的活性,而不是完全消除它们。
地中海饮食(MeD),因为它具有抗炎作用。重点是增加蔬菜、橄榄油和豆类等富含营养和抗氧化剂的食物的消费,同时限制肉类和奶制品的摄入。
一项针对 3557 名银屑病患者的队列研究发现,与地中海饮食更一致的饮食模式与较低的银屑病严重程度相关。
一种方式是通过增加拟杆菌门细菌来降低F/B比例,从而帮助恢复短链脂肪酸的平衡,并促进丁酸盐的产生,这反过来又可以减轻肠道和全身炎症,对牛皮癣具有重要影响。
肠道中其他产丁酸菌包括Akkermansia和普拉梭菌,地中海饮食通过增加这些细菌也促进了短链脂肪酸的产生。
doi.org/10.1177/24755303241226626
食用特级初榨橄榄油含有高含量的多酚,已在健康患者中显示出能减少IFNy和IL7R的基因表达,表明减少银屑病患者炎症的潜在途径。
间歇性禁食可能为难以遵循低热量饮食的患者提供另一种策略。尽管关于这种饮食的研究仅限于斋月禁食,这增加了几个混杂因素,但总的来说有益处。以减重为重点的饮食改善可能的原因可能是由于腹部脂肪和脂肪率的减少,随后降低脂肪因子水平并减少银屑病进展所依赖的炎症。
血清脂肪因子之一的脂联素与PASI评分呈正相关,IL-22也是如此。肥胖加剧银屑病的机制因此可能是由于脂肪因子诱导Th-17相关细胞因子的增加。对于这些建议,应避免严重的饮食改变,以防止营养不良。低热量饮食和间歇性禁食可能推荐给有适当动机的患者。
无麸质饮食意味着避免摄入小麦、大麦和黑麦中的这种蛋白质,这对患者有益。银屑病患者食用无麸质饮食 3 个月后发表了积极结果,显著改善了PASI 评分。其他研究小组证实,一年的无麸质饮食可以改善抗麦胶蛋白肽 IgA 水平较高的群体的 PASI 评分。
高脂饮食破坏IL-23和胆汁酸信号通路的稳态,促进皮肤的γδT细胞浸润并增强其产生IL-17A的能力来诱发银屑病样皮炎。也会通过改变粘液屏障和肠道微生物群来加剧银屑病皮肤炎症,从而导致全身 IL-17 反应增强,从而加剧银屑病。
极低热量生酮饮食(VLCKD)可以显著减少炎症,是缓解肥胖银屑病患者症状的有效手段,可能与微生物群-肠道-皮肤轴有关,但无论短期还是长期都有一定的副作用。
鱼油已被加入到许多银屑病患者用来缓解症状的补充剂中。2014 年,对几项有关在银屑病患者中补充鱼油的研究进行荟萃分析发现,虽然一些研究显示了中等结果,但其他研究尚未发现任何相关性。
最近,在一大群个体(超过 25,000 名男女登记受试者)中,在他们的饮食中补充了维生素 D(2000 IU/天)和/或 omega-3 脂肪酸(1000 毫克/天),并且该组随访 5 年,记录了银屑病 与其他自身免疫性疾病的发病率。研究结果表明,在补充组中,包括银屑病在内的自身免疫性疾病的发病率降低了 22% 以上。
姜黄素是一种抗炎的天然化合物,口服后会在胃肠道中积聚,并可能通过调节肠道微生物群的微生物多样性和组成来发挥其调节作用。一项人类随机安慰剂对照试验,补充组每天接受6000 mg姜黄提取物,并在治疗开始时和8周后进行微生物群分析。大多数梭菌属、拟杆菌属、柠檬酸杆菌属、Cronobacter、肠杆菌属、肠球菌属、克雷伯菌属、拟副杆菌属和假单胞菌属在反应参与者中均匀增加。
产生丁酸的细菌数量和粪便丁酸水平增加,对银屑病患者口服补充姜黄素12周进行评估,结果PASI评分显著降低,IL-22血清水平降低。
在斑块型银屑病患者的局部光疗的同时口服姜黄素似乎可以诱导对治疗的更快、更渐进的治疗反应。考虑到这些炎症发病机制与银屑病受试者的发病机制相似,补充姜黄素可能代表了该病理管理的未来前景。
白藜芦醇是一种抗炎的非黄酮类多酚化合物。在小鼠实验中也发现了白藜芦醇对肠道菌群的有利效果。口服白藜芦醇能够增强肠道屏障功能,同时减少通透性和炎症。白藜芦醇治疗后肠道菌群拟杆菌属、Alistipes、Rikenella、Odoribacter、Parabacteroides和Alloprevotella分类的丰度水平增加,白藜芦醇治疗恢复了小鼠的菌群失调。
此外,白藜芦醇的给药,8周内每公斤400毫克白藜芦醇,增加了高脂饮食喂养的大鼠中产丁酸菌Blautia和Dorea。因此,白藜芦醇补充可能改善患者肠道微生物多样性。
槲皮素是一种植物黄酮醇,它可能存在于各种水果、蔬菜、叶子、种子和谷物中。以其抗炎、细胞保护和免疫抑制特性闻名。
最近的一项研究针对亚甲基丙烯酰亚胺诱导的小鼠,探讨了槲皮素补充(30、60和120 mg/kg)的效果,显示PASI评分显著降低,银屑病斑块改善。此外,槲皮素成功降低了小鼠血清中的TNF-α、IL-6和IL-17水平,加强了抗炎效果。
此外,研究表明口服补充槲皮素,减少了亚甲基丙烯酰亚胺诱导的小鼠类银屑病样皮炎,显著降低了角质形成细胞的增殖和异常分化,以及炎症细胞的浸润。口服槲皮素后,观察到IL-23/Th17轴上的细胞因子表达降低和Th17细胞反应减少。
其他中草药如雷公藤等,也对银屑病患者有用,改善了他们的临床参数,可作为辅助治疗。
回顾性分析显示,83.7% 的银屑病患者在接受唾液链球菌K-12治疗 24 周后,PASI 评分改善了 100%,并且随着治疗持续时间的延长,疗效持续改善。
一项随机对照试验发现,持续口服婴儿双歧杆菌35624(B.fantis)可显著改善银屑病的进展,并降低C反应蛋白和肿瘤坏死因子TNF-α的表达,显示出免疫调节作用。人体微生物群的作用不仅限于粘膜免疫系统,还延伸到全身免疫系统。
在明确单一益生菌的作用机制后,益生菌的组合应用以及相应的标准和方法仍需探索。不同的饮食和环境,可能会导致肠道微生物群的差异。
一项为期12周的开放标签、单中心临床试验评估了益生菌[印度芽孢杆菌(HU36)、枯草杆菌(HU58)、凝结芽孢杆菌(SC208)、草绿芽孢杆菌(SL307)和克劳氏芽孢杆菌(SC109)]和精确益生元(低聚果糖、低聚木糖和低聚半乳糖)对接受局部治疗的银屑病患者的有效性。接受抗银屑病局部治疗和益生菌及益生元补充的患者,在疾病活动性的测量中表现更好,包括银屑病面积和严重性指数、皮肤病生活质量指数、炎症标志物和皮肤厚度。肠道菌群发生了有利的变化,转变为抗炎的特征。
喂食富含菊粉的高纤维饮食(HFD)降低了咪喹莫特诱导的皮炎的增厚和总严重程度评分,并降低了表皮厚度、炎症浸润,包括Ly6G+中性粒细胞和表皮Ki67+增殖细胞。
富含菊粉的高纤维饮食增加了拟杆菌门和拟杆菌属的相对丰度,增加了血清和粪便中丙酸盐的浓度。膳食菊粉补充剂部分通过丙酸盐改善银屑病样皮炎,可能是一种很有前途的银屑病辅助治疗方法。
当然,何时使用益生菌、益生元以及针对不同个体的精准应用,还需结合肠道菌群检测进一步研究。
FMT是纠正肠道菌群失调、重建肠道微生态平衡的方法。近年来,这种疗法已用于治疗牛皮癣、系统性红斑狼疮、肠易激综合征、帕金森病疾病等。
FMT治疗的效果取决于不同的疾病、移植的形式和数量、给药途径以及所使用的供体。
在一项为期五周的干预性临床试验中,一名患有斑块型银屑病和IBS的受试者通过内窥镜和结肠镜接受了两次FMT。身体表面积、PASI评分、皮肤病学生活质量指数、肠道症状和血清TNF-α水平在干预后都有所改善,没有观察到不良反应。
然而,在周围性银屑病关节炎患者中,FMT在治疗活动性疾病方面并不那么有效。FMT可能在减轻银屑病患者的疾病严重程度方面带来一些益处,但需要更多的临床试验来证明这一点。
其他相关预防干预措施,皮肤护理等详见我们之前的文章:
以上我们了解了银屑病发病和复发的危险因素和潜在病理机制。银屑病的发展是复杂的,可能是由多种触发因素引起的。这些触发事件可能在不同条件下独立发生,或者表现出累积或协同效应。因此,很难确切地将这种疾病归因于特定的诱因。
鉴于银屑病是由遗传基础上的环境风险因素引发的,疾病的预防和管理值得应有的重视。根据这些明确的触发因素,可以制定银屑病风险评估和疾病管理指南,有助于轻症或非典型病例的早期诊断和银屑病的精准管理。例如,感染史(不仅是化脓性链球菌感染,还包括这里列出的其他感染)、肥胖和高血脂水平、过度的心理压力、吸烟、睡眠障碍、高盐饮食、服用特定药物史都应被视为银屑病的风险因素。
从患者的角度来看,消除这些风险因素对于他们个人对疾病的管理至关重要。结合肠道菌群检测报告查看相关菌群超标或缺乏情况例如链球菌,AKK菌,产丁酸菌等,以及整体菌群平衡情况,对于疾病预警有着重要作用。通过及时调整生活方式、饮食习惯和适当药物治疗,可以有效地降低银屑病的发病风险,提高患者的生活质量。
从治疗角度来看,患者可能受益于早期治疗,包括但不限于结合菌群的抗生素治疗、标准化益生菌补充、抗高脂血症治疗等方式。了解触发因素在银屑病发病机制中的作用,与肠道菌群的关联,也将为开发针对银屑病发病和复发期间触发机制的新疗法提供线索。
注:本账号内容仅作交流参考,不作为诊断及医疗依据。
主要参考文献
Zhu Q, Wu K, Yang Q, et al. Advances in psoriasis and gut microorganisms with co-metabolites[J]. Frontiers in Microbiology, 2023, 14: 1192543.
Liu S, He M, Jiang J, et al. Triggers for the onset and recurrence of psoriasis: a review and update[J]. Cell Communication and Signaling, 2024, 22(1): 108.
Kranyak A, Haran K, Smith P, et al. The Mediterranean Diet as a Potential Solution to the Gut Microbiome Dysbiosis in Psoriasis Patients[J]. Journal of Psoriasis and Psoriatic Arthritis®, 2024: 24755303241226626.
Ryguła I, Pikiewicz W, Grabarek B O, et al. The Role of the Gut Microbiome and Microbial Dysbiosis in Common Skin Diseases[J]. International Journal of Molecular Sciences, 2024, 25(4): 1984.
Buhaș, M.C.; Candrea, R.; Gavrilaș, L.I.; Miere, D.; Tătaru, A.; Boca, A.; Cătinean, A. Transforming Psoriasis Care: Probiotics and Prebiotics as Novel Therapeutic Approaches. Int. J. Mol. Sci. 2023, 24, 11225.
Constantin, C.; Surcel, M.; Munteanu, A.; Neagu, M. Insights into Nutritional Strategies in Psoriasis. Nutrients 2023, 15, 3528.
Tampa, M.; Mitran, M.I.; Mitran, C.I.; Matei, C.; Georgescu, S.R. Psoriasis: What Is New in Markers of Disease Severity? Medicina 2024, 60, 337.
Zang C, Liu J, Mao M, Zhu W, Chen W, Wei B. Causal Associations Between Gut Microbiota and Psoriasis: A Mendelian Randomization Study. Dermatol Ther (Heidelb). 2023 Oct;13(10):2331-2343.

谷禾健康

病毒-细菌合并或继发感染
引起呼吸道感染的病毒是导致全世界高发病率和死亡率的原因,数十年来通常发生在冬季。在冬天,空气干燥,那些可能含有病毒的飞沫可以在空气中停留更长时间,并可以进一步传播。此外人的免疫力在冬季的表现也会不佳。因为纤毛是上呼吸道中微小的毛发状突起,通常将异物从鼻子中移出,但在较低的温度和较低的湿度下,纤毛的功能无法有效发挥作用。
在过去 20 年中,世界经历了六次重大传染病爆发(SARS-CoV:2002-2004 年;H1N1 流感:2009-2010 年;中东呼吸综合征冠状病毒:2012-2020 年;埃博拉病毒:2013-2016 年;寨卡病毒: 2015-2016 年;SARS-CoV-2:2019 年至今),其中四种病毒(SARS-CoV、H1N1 流感、MERS-CoV、SARS-CoV-2)感染会导致呼吸道感染。
病毒感染(尤其是肺部病毒)的一项主要并发症是细菌在病毒感染的器官中定植,这与高发病率和死亡率有关。虽然继发细菌感染或重复感染很大程度上是原发病毒感染引起的免疫易感性的结果,但合并感染是同时发生的多种感染(病毒/细菌/酵母)。病毒大流行期间,合并感染、继发感染或“重复感染”很常见。1918 年西班牙流感大流行导致约 5000 万人死于细菌合并感染,其中许多人是由于细菌重复感染导致继发性肺炎。即使在抗生素时代,在 1957 年 H2N2 和 1968 年 H3N2 大流行中,超过一半的严重感染患者出现细菌并发症。而在 2009 年 H1N1 流感大流行期间,高达 34% 的死亡是细菌合并感染造成的。这些并发症在临床记录中常常被忽视。
因此,我们需要更好地了解病毒感染后继发细菌性肺炎的发病机制,以便在治疗这种破坏性并发症方面取得进展。
病毒后细菌感染的潜在机制很复杂,包括由病毒、细菌和宿主免疫系统之间的相互作用介导的多因素过程。过去十几年的研究表明,呼吸道病毒常常通过补充机会性细菌病原体的生长而促进继发性细菌感染。病毒感染从组织学和功能上损害呼吸道。
细胞损失、杯状细胞增生、粘液分泌改变、纤毛跳动频率降低、粘液纤毛清除功能不协调以及氧交换减少是病毒感染的特征。这些影响与不同的分子机制有关,通过这些机制,病毒在呼吸道中发生易感性,从而促进细菌感染。
同时病毒感染可能用来驱动急性加重的另一种机制是诱导血管舒张或紧密连接开放因子,这可能会增加浸润率。这也增加了过敏原与气道免疫细胞接触的机会。
呼吸道是一个复杂的器官系统,分为不同的区域;上呼吸道包括鼻腔、咽和喉,而下呼吸道则由传导气道(气管和支气管)、小气道(细支气管)和呼吸区(肺泡)组成。总的来说,这个巨大的表面积蕴藏着一系列不同的微生物群落,其中细菌负荷最高的位于上呼吸道。
呼吸道微生物组的发现始于 2008 年的人类微生物组计划,这要归功于使用 16S rRNA 基因的高通量测序的发展。从那时起,各种研究证明了上呼吸道(URT)和下呼吸道(LRT)中微生物组的存在及其对肺稳态的重要性。自过去十年以来,进行了大量研究来了解病毒感染对微生物组的影响。
各种 NGS 研究表明,呼吸道合胞病毒(Respiratory Syncytial Virus,RSV)和流感病毒通过直接或间接过程改变呼吸道微生物组,导致生态失调。对呼吸道病毒感染的人类样本进行的研究表明,从拟杆菌门到变形菌门的转变,包括许多革兰氏阴性致病菌。
迄今为止,URT的细菌定植被认为是原发性病毒感染后细菌侵入肺部的第一阶段。流感病毒诱导的信号,例如 ATP 的释放,导致生物膜中分散的细菌增加,与致病表型相关。
上呼吸道病毒感染后可出现细菌性肺炎,并使下呼吸道病毒感染复杂化。继发性细菌性肺炎是流感相关死亡的主要原因。
我们评估了以下假设:
i)呼吸道病毒通过改变上呼吸道(URT)的细菌群落结构来影响肺炎的病因;
ii)呼吸道病毒促进或抑制下呼吸道(LRT)的定植)由 URT 中的某些细菌种类引起。
通过对文献的系统回顾,发现病毒既改变了URT中的细菌群落,又促进了LRT中的细菌定植。
流感和细菌性肺炎是全世界传染病发病和死亡的主要原因。多种病毒、细菌和真菌病原体吸入肺部后可引起肺炎。几乎所有这些严重肺炎病例都用抗生素治疗,因为继发细菌感染会使下呼吸道病毒感染复杂化。因此,即使在确定仅由病毒病因引起的肺炎的病例中,病毒和细菌的细菌相互作用也可能发挥一定作用。
确定肺炎病因的挑战
即使在美国等定期进行肺炎监测的国家,大约 65-85% 的住院肺炎病例也没有记录微生物病原学信息。重症患者通常不被纳入监测范围,在患者临床出现时或检测之前,由于怀疑肺炎时需要快速使用抗生素治疗,致病途径上的微生物可能已被清除,并且很少对老年人进行尸检。为了最佳地确定病因,需要通过支气管肺泡灌洗直接取样,但由于易于收集、道德问题和成本,通常对血液、痰液和尿液进行致病因子检测。成人肺炎病例中仅 7%–13% 观察到菌血症,儿童肺炎病例中观察到菌血症为 1–5%,痰液可能被 URT 中的细菌污染,且很难从儿童身上获取,血液和尿液抗原检测需要进一步检测验证或仅限于成人,并且仅针对少数病原体(例如肺炎链球菌和军团菌)。尽管现代分子生物学技术使得对所有存在的细菌、病毒和真菌物种进行非针对性筛查成为可能,但仍然很难区分感染、定植或污染。需要继续努力开发更准确的方法来确定肺炎的病因,从而最大限度地发挥治疗和预防作用。
本文主要探讨了常见的病毒性呼吸道感染以及其可能引起的继发性细菌感染。比较了上呼吸道和下呼吸道微生物组的组成差异,并分析了病毒感染后细菌性肺炎发生的潜在机制。接下来着重讨论了肠道微生物组与呼吸道感染之间的相互作用,以及这种相互作用如何影响宿主的健康和疾病进程。通过这些分析,本文旨在为预防和干预病毒感染后的继发细菌感染提供重要的见解和警示。
病毒感染是一个常见的问题,尤其是在年轻人和老年人中。呼吸道是微生物暴露的主要门户,病毒感染可导致无症状、轻度和自限性或严重且有时致命的感染。尽管已经确定病毒特异性特性(例如寿命和复制动力学)会影响临床表现,但人们不太清楚为什么在感染同一病毒株的人群中可能会出现不同的感染结果。新出现的证据表明肺部和胃肠道微生物组的组成存在人际差异,特别是乳酸菌的成员属,作为定义呼吸道病毒感染结果的关键组成部分。
甲流病毒变异与疫苗挑战
甲型流感病毒 (IAV) 是一种由负义片段化 RNA 病毒引起的人类和动物高度传染性上呼吸道 (URT) 疾病。它被认为是一个主要的公共卫生问题,每年都会造成严重的疾病和经济负担。
注:负义片段化RNA病毒是指RNA病毒的基因组由负义片段组成。在这种病毒中,基因组的信息被存储在负义链上,而不是通常的正义链。负义链的信息需要通过反向转录过程转化为正义链,以便进行蛋白质合成。
频繁的核苷酸替换会导致甲流病毒颗粒表面的血凝素和神经氨酸酶糖蛋白发生变化(也称为抗原漂移),从而需要持续更新疫苗。在疫苗设计和病毒突变之间的这种进化“军备竞赛”下,平均每年导致 3-500 万例重症病例和高达 291,000 ~ 646,000 人死亡。
甲型H1N1流感病毒的传播
分段甲流病毒基因组的模块化结构允许与其他不同的甲流病毒进行遗传重配(抗原转移),从而导致能够引起大规模流行病或大流行的新型病毒的零星出现。过去,新的甲流病毒在未接触过的人群中传播曾引起过大流行,导致严重的发病率和死亡率,其中最引人注目的是 1918 年和 1919 年,当时的西班牙流感导致全球约 20 至 5000 万人死亡。
上呼吸道合并感染
对 1918 年大流行病尸检标本的回顾性分析揭示了由上呼吸道细菌引起的继发性重复感染的流行。然而,细菌合并感染在疾病预后中的作用不仅限于流行病;季节性流感流行期间细菌和病毒的双重感染通常会导致住院人数、重症和死亡人数增加。
微生物组与甲流病毒感染
尽管非患病个体的微生物组相对稳定,但甲流病毒感染已被证明会增加上呼吸道中存在的细菌类群的多样性。具体而言,甲流病毒可导致葡萄球菌属和拟杆菌属以及嗜血杆菌属、梭杆菌属和其他类群的相对丰度发生变化。
在急性或慢性疾病期间,由于局部上皮细胞的变化而导致微生物群的暂时紊乱也是感染的诱发因素。上呼吸道微生物群也可能在调节甲流病毒感染期间诱导的炎症反应中发挥有益作用。
此外,肠道微生物组组成在甲流病毒感染后可正向调节 Toll 样受体 7 信号通路。不过,甲流病毒复制和诱导先天免疫反应对人类或动物上呼吸道微生物组组成的影响,仍有待在社区范围内进行深入阐明和分析。
人类 URT 病毒感染期间假单胞菌大量繁殖
对感染者和健康患者菌群分析结果发现,在感染者队列中最常见的类群是γ变形菌,而在健康患者队列中最常见的类群是放线菌。除此之外,所有其他的分类单元都被归类到一个名为“其他”的附加分类单元中。在流感感染个体的所有样本中,假单胞菌是最丰富的分类群。系统发育推断将该分类单元可靠地定位为假单胞菌属,但无法进一步确定到物种水平。
其他数量较少的门包括拟杆菌门、厚壁菌门、放线菌门和变形菌门的一些其他科,如红杆菌科和巴斯德氏菌科(Gammaproteobacteria)和根瘤菌目的布鲁氏菌科(Alphaproteobacteria)。在对个体进行时间动态分析时,假单胞菌也被确认为主要的分类群。对于未感染的受试者,放线菌是最主要的分类群,而假单胞菌则是存在数量最少的分类群。这些结果在个体受试者的分析中也得到了验证。其他较少丰富的门包括疣微菌门和变形菌门、α变形菌门和ε变形菌门。
未感染 IAV(左)和感染 IAV(右)的人类受试者的综合分类分类
每列都是一个特定样本。该图总结了 >5% 的样本中存在的分类群在类水平上的相对分类丰度(参见下面的图例),按每个队列中最丰富的分类群按降序排序。γ 变形菌(假单胞菌,橙色)在感染患者中普遍存在(右),而放线菌在健康患者中最为丰富。
b在感染患者中观察到的 γ 变形菌目(左)和科(右)水平细分。
流感后细菌性肺炎的严重程度
流感后细菌性肺炎在季节性和大流行性流感病毒疾病相关的发病率和死亡率中发挥着重要作用。大流行预防和管理的一个重要部分是了解流感感染与继发细菌感染之间的关系。
在美国 2003-2004 年流感季节的一项全国调查中,传染病专家观察到成人和儿童患者细菌并发症的发生率分别为 1.6% 和 2%,我国的这个调查数据没有查到,应该也差不多。
——死亡人数
1918-1919 年流感大流行中的大多数死亡可能直接由常见上呼吸道细菌引起的继发性肺炎造成。肺组织样本表明,估计的 20-6000 万人死亡中,大部分是由于细菌重复感染,而不是病毒的直接影响。
——住院情况
在季节性流行期间,流感细菌合并感染与住院人数增加有关。入住 ICU 的四分之一的严重甲型流感感染患者出现细菌或病毒合并感染。系统回顾和荟萃分析显示,细菌合并感染的频率变化很大,范围从 2% 到 65%。
最常见的共感染细菌是:肺炎链球菌和金黄色葡萄球菌
这两种细菌分别占 35%(95% CI,14-56%)和 28%(95% CI,16-40%)。其他多种病原体,例如肺炎嗜血杆菌或肺炎克雷伯菌,引起了其余的感染。
有个研究在2010年至2018年的八个流感季节进行了一项回顾性、观察性研究。在209名流感相关肺炎入院患者中,41名(19.6%)被确定为社区获得性细菌合并感染,主要是沙门氏菌感染和金黄色葡萄球菌。这种现象在流感相关肺炎中经常观察到,但迄今为止尚未发现危险因素。
细菌合并感染可能预测严重程度,并且是院内死亡的独立危险因素。此外,金黄色葡萄球菌和流感的混合感染经常导致致命的协同作用。
呼吸道合胞病毒(Respiratory Syncytial Virus,RSV)是 1 岁以下儿童细支气管炎的最常见原因。RSV 还导致老年人和免疫功能低下的成年人发生急性下呼吸道感染。该疾病通常与同时或继发的细菌感染有关。
RSV 和细菌的共同感染
对医院内 RSV 感染患者进行的多项研究显示,RSV 感染患者与下呼吸道细菌合并感染有关,17.5% 至 44% 的患者 RSV 和细菌合并感染均呈阳性。
最常见的分离细菌是肺炎链球菌和流感嗜血杆菌。然而,最近的研究表明 RSV 和金黄色葡萄球菌之间存在最普遍的关联,尤其是耐甲氧西林金黄色葡萄球菌(MRSA) 。与单纯 RSV 感染相比,细菌合并感染与 RSV 感染通常与易感人群中更严重的疾病相关。
SARS-CoV-2是一种单股正链RNA病毒,属于β冠状病毒属。它通过飞沫传播和接触传播,感染人类的呼吸道,引起了全球范围内的疫情。
截至 发现3年内,SARS-CoV-2 已迅速成为全球主要病原体,COVID-19 大流行影响了全球超过 2 亿人,并导致超过 超过400万人死亡。
因新冠入院的患者细菌合并感染的频率较低
最近一项大型多中心前瞻性队列研究表明,在英国第一波大流行期间,经微生物学证实的细菌感染(主要是继发性感染)在因 COVID-19 入院的患者中很少见(低于 2.3%)。这一结果与之前的几项观察性研究和荟萃分析一致,这些研究和荟萃分析报告了因 COVID-19 入院的患者细菌合并感染的频率较低。
细菌合并感染在重症患者中更为常见
最近,对 3338 名 COVID-19 患者进行了荟萃分析,以评估细菌合并感染。3.5%的患者出现这种现象(95% CI 0.4-6.7%),但14.3%的患者也观察到继发细菌感染(95% CI 9.6-18.9%)。
一般来说,出现细菌感染的 COVID-19 患者比例为 6.9%(95% CI 4.3-9.5%)。然而,细菌合并感染在重症患者中更为常见(8.1%,95% CI 2.3-13.8%)。
在一般住院患者中观察到细菌合并感染的频率较低,可能不需要抗菌治疗。其他荟萃分析研究中也观察到类似的结果。
与单纯感染患者相比,社区获得性混合感染和医院获得性重复感染的 COVID-19 患者的预后最差。
常见的共感染细菌
主要检出肺炎链球菌、肺炎支原体、金黄色葡萄球菌、铜绿假单胞菌、大肠杆菌、流感嗜血杆菌,尤其是危重患者。
相比之下,其他研究声称重症 COVID-19 患者中存在细菌(和真菌)混合感染,包括鲍曼不动杆菌和肺炎克雷伯菌。
所有临床数据都表明,SARS-CoV-2感染者的细菌或真菌合并感染率低于流感病毒感染者。当然这也可能是由于漏报问题、抗生素的广泛使用或限制几种呼吸道病原体传播的控制措施的实施造成的。迄今为止,对 COVID-19 患者合并感染的重要性及其对发病机制的影响仍知之甚少。
继发性细菌感染及其在病毒感染期间的免疫反应列表

编辑

Manna S et al., J Infect Public Health. 2020,1397-1404.
吸道微生物组的多样性与健康关联
人类呼吸道的微生物群是多样且异质的,与多种疾病和表型相关。
上呼吸道栖息着丰富而复杂的微生物组,以健康个体的口腔共生类群为主。
相比之下,健康个体的下呼吸道微生物组(声带下方)的生物量通常相当低,其定义是通过上呼吸道微抽吸输入的竞争动态,以及通过粘液纤毛活动和先天免疫功能清除的竞争动态,可能存在一些有限的局部微生物复制。
因此,上呼吸道微生物组的变化不仅会影响局部微生物组与宿主的相互作用,还会影响下呼吸道的微生物组与宿主的相互作用。
微生物组变化与疾病状态的联系
在各种疾病状态下,通过增加进入下呼吸道、清除缺陷或下呼吸道微生物生长增加,维持低微生物量肺微生物组的平衡可能被破坏。
上呼吸道微生物群在晚期肺病、HIV 感染、血管炎和流感等情况下以及吸烟等暴露条件下会发生改变。
下呼吸道(肺)微生物组不仅在化脓性肺部疾病(如囊性纤维化和肺炎)中发生改变,而且在通常不被认为是微生物引起的肺部疾病,包括哮喘、慢性阻塞性肺病(COPD)和肺纤维化,以及机械暴露等肺部疾病中也发生改变。
微生物组与病毒感染的关系
上呼吸道微生物组的变化与病毒感染的易感性有关,特别是在流感暴露个体中,并且与 RSV 感染的疾病严重程度有关。例如,一项家庭传播研究发现链球菌属的丰度增加。病毒感染也可以改变细菌微生物组,从而导致潜在的双向相互作用。因此,人们对气道微生物组在 COVID-19 中的潜在作用的研究产生了浓厚的兴趣。
呼吸道微生物组对免疫系统的影响
呼吸道微生物群影响宿主健康的方式之一是通过局部和全身免疫系统的调节。如在 COVID-19 中的免疫反应与疾病进展和结果密切相关。
在健康方面,肺部免疫张力受到通过上呼吸道微抽吸获得的生理微生物群的调节,气道生态失调与炎症增加有关,例如哮喘或接触香烟烟雾。
重要的是,过去五年的研究表明,呼吸道微生物组在调节急性肺损伤患者或有急性肺损伤风险的患者的严重程度方面发挥着作用。肺微生物组负荷和组成与急性肺损伤患者的局部和全身炎症相关,并预测患有急性呼吸窘迫综合征(ARDS)的机械通气危重患者的临床结果。
创伤后,肺微生物组的组成与 ARDS 的发生有关,并且被认为可以介导吸烟对 ARDS 发生风险的影响 。因此,呼吸道微生物组与多种可导致严重肺损伤的情况下的局部和全身机制相关,并可能通过局部和全身机制影响结果。
成人上呼吸道的主要细菌属和微生物组种类
doi: 10.3390/biology9100318
在 COVID-19 中,已观察到免疫抑制和失控的炎症,并可能导致更严重的疾病,而保护性免疫涉及诱导体液和细胞介导的反应。感染可诱导多种促炎细胞因子,如 IL-1B、IL-6、TNF、IL1RA、CXCL10/IP10、MIP-1α 和 CCL2。几种 SARS CoV-2 蛋白能够通过延迟 I 型干扰素反应来抑制抗病毒免疫。
上面讲过上呼吸道和下呼吸道具有相似的微生物群落组成,尽管前者在健康宿主中的微生物密度要高得多。已知有几个因素会影响气道微生物组的组成,包括感染史、年龄、遗传和结构性肺疾病。
上呼吸道是一个相互连接的系统,由前鼻孔、鼻腔、鼻咽、鼻窦、咽鼓管、中耳腔、口腔、口咽和喉组成,每个系统都作为独特的生态位,拥有自己的微生物群落。
在健康成年人中,鼻腔中存在的细菌通常是与皮肤相关的细菌,主要是放线菌门(例如棒状杆菌属、丙酸杆菌属),其次是厚壁菌门(例如葡萄球菌属)和变形菌门。
口咽部含有厚壁菌门、变形菌门和拟杆菌门的成员,包括链球菌、奈瑟菌、嗜血杆菌和毛螺旋菌属。皮肤和口腔谱系以鼻咽为代表,例如链球菌、葡萄球菌、棒状杆菌和普雷沃菌。
包括肺炎链球菌、脑膜炎奈瑟菌和流感嗜血杆菌在内的有限数量的病原体是 URT 的共生细菌。
呼吸道微生物组的分布与动态
在健康个体中,LRT 中的微生物群落丰富度(即细菌类群总数)低于 URT 中的微生物群落丰富度(即细菌类群总数)。
与正常健康的肺部是无菌环境相反,可以使用测序技术来识别独特且有些动态的肺部微生物组,微抽吸是微生物从 URT 迁移到 LRT 的主要途径。
健康/慢性气道疾病中肺部微生物组的特征
健康肺部的主要门是拟杆菌门和厚壁菌门,其中主要包括普雷沃氏菌、韦荣球菌和链球菌。患有慢性气道疾病(例如囊性纤维化、慢性阻塞性肺病)的个体肺部细菌数量增加,并且某些物种的相对丰度存在差异。
微生物群落与呼吸道感染
由于内在或外在因素导致的气道清除受损导致可以利用这种生长机会的细菌种类的增殖。URT 的细菌定植通常被认为是侵袭性细菌感染发展的第一步,包括呼吸道病毒感染后的继发性细菌感染。下面将更详细地讨论细菌丰度、物种多样性以及影响后续感染免疫反应的因素。
呼吸道合并感染和重复感染涉及多种机制。长期以来,病毒感染对上皮屏障的影响被认为是细菌二重感染的主要原因。最近,多项研究表明,抗病毒免疫反应也在混合感染中发挥作用。
病毒对继发细菌感染的易感性机制
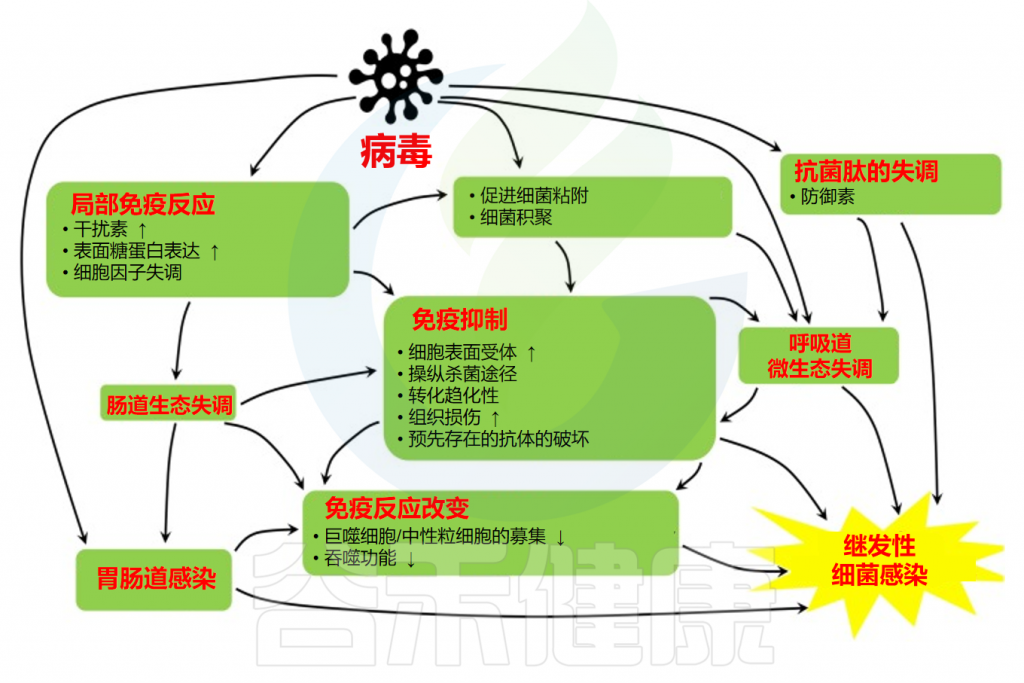
doi: 10.1016/j.jiph.2020.07.003
尽管在下文单独整理讲述,但是生理学和免疫学机制是同时存在且密切相关的。
病毒感染可破坏上皮完整性,导致紧密连接和细胞骨架功能障碍
上皮细胞在防止吸入病原体和颗粒物的入侵方面具有至关重要的作用。上皮细胞通过紧密连接组装成假复层结构,为病原体创造了不可渗透的屏障。SARS-CoV-2感染后,在气液界面培养的人原代上皮细胞(HAE)中观察到跨上皮阻力急剧下降和细胞形态改变,表明上皮完整性被破坏。
使用类似的体外模型对流感和 RSV 进行了类似的观察。在动物模型中还发现病毒感染后观察到的脱屑、纤毛缺失、免疫细胞浸润和坏死之间存在相关性。
单次病毒感染后经常观察到紧密连接和细胞骨架功能障碍。RSV 和流感病毒可以通过直接或间接靶向相关蛋白质(例如claudin、occludin 或ZO-1)来减少紧密连接,并诱导F-肌动蛋白细胞骨架重排,从而导致细胞形态改变。
呼吸道病毒引起的上皮损伤是导致继发感染的原因之一
原发性流感或RSV感染会引起上皮损伤,导致动物模型对金黄色葡萄球菌或肺炎链球菌的易感性更高。
病毒诱导的细胞凋亡也可能通过多种机制(例如 FasL/TRAIL 途径)导致上皮屏障的丧失。尽管没有经过专门研究,但一些研究表明病毒诱导的细胞凋亡/坏死与对细菌重复感染的较高易感性之间存在相关性,细胞碎片可改善细菌粘附和侵袭。
病毒感染→粘液分泌增加和纤毛损伤→破坏粘液纤毛清除机制
病毒引起的气道功能改变也是呼吸道合并感染的原因。上皮可以通过粘液纤毛清除来阻止感染,粘液纤毛清除重新组合了两种基本机制:
粘液纤毛清除机制受损→气道阻塞→为继发性感染创造条件
众所周知,上皮细胞在感染后会产生粘液,以减少流感、RSV 或 SARS-CoV-2 的感染。然而,这会导致气道阻塞,反映出在患者中观察到的肺活量下降。RSV 和流感病毒在感染后也会增加粘液相关蛋白,例如粘蛋白。
呼吸道病毒主要针对纤毛上皮细胞,诱导细胞死亡,从而导致纤毛损失。到目前为止,还没有研究明确关注合并感染期间粘液纤毛清除的分子机制。
RSV 和流感病毒增强病原体粘附
原发性病毒感染也会增加呼吸道中的细菌粘附。RSV 感染通过 G 糖蛋白与细菌成分的直接结合,增加肺炎链球菌对上皮细胞的粘附和毒力。
G 糖蛋白在感染后锚定在细胞膜上并充当细菌受体。转录组学研究表明,RSV 增加了 RSV 感染细胞表面的粘附分子,例如 CD47,导致肺炎链球菌粘附增加。
病毒蛋白质作为细菌受体
流感病毒还通过增加受感染细胞膜上的纤维蛋白原、糖蛋白修饰和唾液酸,来增强各种模型中金黄色葡萄球菌或肺炎链球菌的粘附。血小板激活因子受体 (PAF-R)也与细菌相互作用,促进重复感染。流感糖蛋白血凝素和神经氨酸酶也促进细菌粘附,充当或暴露细菌受体。
冠状病毒与特定细菌的重复感染
另一种人类冠状病毒 (HCoV-NL63) 增强了LLC-MK2 和 HAE 细胞中肺炎链球菌的重复感染,但对金黄色葡萄球菌、流感嗜血杆菌或铜绿假单胞菌等其他细菌没有增强作用。
重复感染加剧病理严重程度
流感和肺炎链球菌或金黄色葡萄球菌重复感染的各种小鼠模型在流感感染后 7 天接种细菌时表现出致命的协同作用,这表明重复感染在病毒感染后期的修复过程中增强了病理严重程度。
病毒感染干扰细胞修复机制
观察到修复细胞反应受损,尤其是巨噬细胞和上皮细胞的修复细胞反应受损,细胞再生减少和稳态信号通路改变。RSV 可以通过增加 MMP 或生长因子的产生来干扰修复机制,从而导致纤维化加剧。
细胞修复延迟与重复感染相关
流感病毒和肺炎链球菌重复感染小鼠模型的转录谱显示,48hpi 上皮细胞增殖和上皮修复增加。此外,观察到基因上调与疾病严重程度之间存在相关性,表明修复机制的改变与重复感染有关。最近证明,流感病毒诱导的 I 型和 III 型干扰素可延迟修复阶段的上皮细胞增殖。注意到 p53 激活和细胞周期改变导致细胞凋亡增加,导致细胞分化和生长抑制以及上皮修复延迟。
感染流感的Ifnlr1−/−小鼠在重复感染肺炎链球菌后具有更好的存活率,这表明病毒感染后诱导的IFN-λ延迟了上皮修复并导致继发性细菌感染。
共同感染的生理机制总结如下:
与细菌重复感染相关的生理机制的示意图
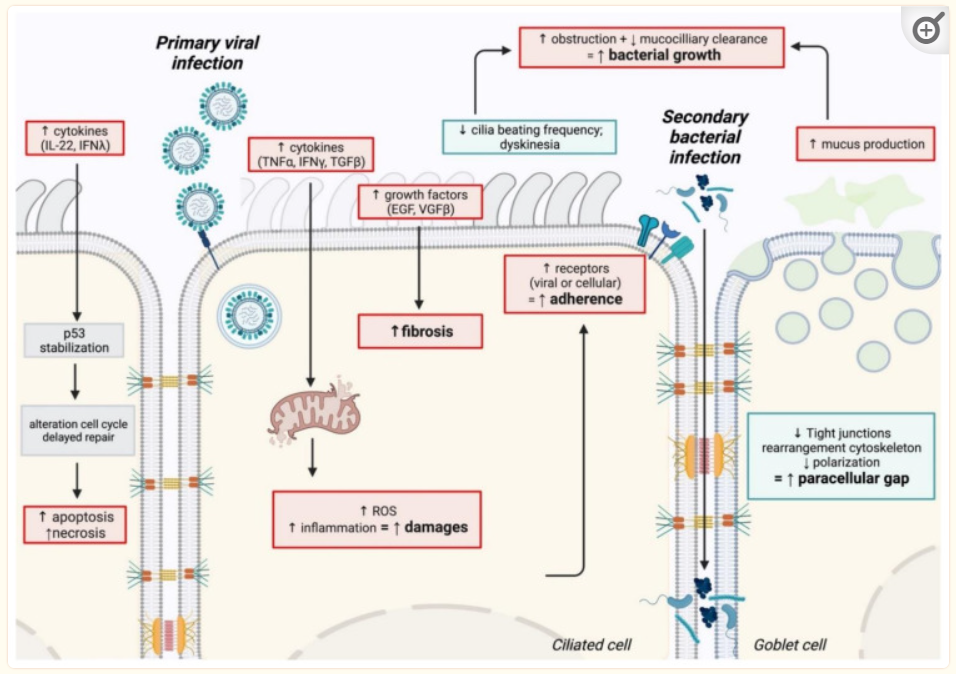
原发性病毒感染对上皮完整性和功能的影响(上皮损伤、修复延迟)有助于为继发性细菌感染的建立创造有利的环境。
呼吸道病毒通过URT进入人体,其一种可能的机制是改变上呼吸道的微生物组成,促进病原体的生长,并促进大量细菌随后进入。
横断面研究
病毒感染与细菌定植的互作
考虑到病毒对增强细菌对上皮细胞粘附的影响,对人类受试者以及动物模型的多项研究表明,病毒感染与潜在致病菌的定植增加有关,这也许并不奇怪。
使用 qPCR 检测感染或未感染甲型流感的成年患者中的特定细菌进行的比较分析显示,感染患者中分别有 12%、24% 和 32% 存在金黄色葡萄球菌、肺炎链球菌和流感嗜血杆菌,而 5% 的感染患者中共同存在金黄色葡萄球菌、肺炎链球菌和流感嗜血杆菌。在体外实验模型中,病毒感染会增加 URT 中各种细菌的定植率 ,包括肺炎链球菌和流感嗜血杆菌。
较高的致病菌负荷与呼吸道病毒合并感染,和继发性细菌感染风险增加相关
在儿童中,流感与鼻咽部肺炎链球菌滴度增加 15 倍相关。动物模型也同样证实,病毒感染,尤其是流感,会增加 URT 中的细菌定植率,从而增加继发细菌感染的风险。
在调整年龄和性别后,较高的肺炎球菌定植密度与呼吸道病毒合并感染和侵袭性肺炎球菌肺炎有关。
另一项比较患有和不患有肺炎的鼻咽细菌的病例对照研究也发现,鼻咽部肺炎链球菌(而非流感嗜血杆菌和卡他莫拉菌)的负荷与病毒合并感染和肺炎之间存在关联。
病毒感染可能会增强细菌的传播
在一项对小鼠定植肺炎链球菌并在 3 天后感染甲型流感病毒的研究中,只有当所有小鼠都感染流感并被流感中和抗体阻断时,才会发生肺炎链球菌传播。然而,虽然特定细菌可能在病毒感染期间获得竞争优势,但这并不普遍适用于所有细菌类群。最近一项针对患有和未患有呼吸道病毒感染的受试者的研究表明,与未感染的对照组相比,病毒感染受试者鼻咽样本中的总体细菌丰度较低。
急性病毒感染和细菌定植之间的关系是双向的
细菌携带或其配体可以增加或降低病毒感染率,从而对随后的宿主对病毒感染的免疫反应产生积极或消极的影响。接触肺炎链球菌可以增强呼吸道中的病毒复制。
与没有定植的患者相比,携带肺炎链球菌的患者更有可能经历随后的急性呼吸道疾病发作。此外,气道中存在的细菌可以调节宿主针对病毒感染的反应。
鼻咽共生体的存在,可以保护小鼠免受 RSV 诱导的气道高反应性
感染 RSV 的小鼠在接受抗生素介导的鼻咽部草绿色链球菌清除后,表现出炎症淋巴细胞数量和气道高反应性增加,以及调节性 T 细胞数量和转化生长因子-β 产生的减少。
特定细菌定植对免疫应答的影响
其他研究表明,金黄色葡萄球菌在上呼吸道的定植可通过将 2 型 CC 趋化因子受体+分化簇 (CD)11b +单核细胞子集募集到肺部并诱导 M2,从而显著降低小鼠中流感引起的急性肺损伤和死亡率。
病毒感染对微生物组多样性的影响不一致,受多因素影响
随着新一代 16S rRNA 测序的出现,基于微生物组的研究试图辨别病毒感染期间每个解剖生态位的细菌群落的整体变化模式,例如多样性的变化。多样性可以使用各种指数来评估,例如微生物组的独特物种总数(即丰富度)或考虑群落成员的丰富度和相对丰度均匀度的其他衡量标准(例如香农指数)。
当将病毒感染受试者与健康对照进行比较时,微生物组分析的结果并未显示出一致的多样性变化。考虑到采样对象的可变性、病毒感染的类型和严重程度、样本收集的类型和时间以及分析方法的差异。
多样性与严重程度有关
法国对因流感入院的儿童进行的一项研究表明,随着流感严重程度的增加,鼻咽微生物群的多样性也随之增加。
多样性与病毒感染的类型有关
我国最近一项针对 H7N9 禽流感患者的研究表明,与健康对照相比,H7N9 感染患者的口咽微生物组多样性显着增加,特别是继发细菌性肺炎的 H7N9 患者。
相反,法国的一项鼻咽样本研究和韩国的一项急性呼吸道病毒感染患者口咽样本研究均显示,与健康对照相比,病毒感染期间多样性指数有所下降 。
这两项研究的受试者范围从婴儿到 80 岁以上的成人,限制了有关年龄相关影响的结论。在接受鼻病毒实验性自我接种的健康志愿者中进行的纵向研究也未能证明 URT 微生物组多样性的显着变化,而对健康成年人注射 LAIV 疫苗导致病毒攻击后多样性指标的增加。
因此,与多样性降低被认为对宿主有害的其他疾病不同,病毒感染对多样性本身的影响是可变的,目前不被认为是并发症风险的良好指标,包括继发性细菌性肺炎。
微生物组测序研究还使研究人员能够同时识别多个细菌类群之间的丰度变化,而不仅仅是可以单独培养的细菌类群。这使得研究人员能够确定哪些细菌群在病毒感染期间一致变化,哪些细菌群相互竞争。
各种病毒感染中,微生物群的变化
法国最近一项基于宏基因组学的研究报告称,与健康对照相比,确诊呼吸道病毒感染受试者的鼻咽样本中金黄色葡萄球菌、肺炎链球菌、流感嗜血杆菌、卡他莫拉菌和肺炎克雷伯菌富集。
对感染和未感染 2009 年甲型 H1N1 流感大流行病毒感染的肺炎患者的口咽微生物组进行的检查显示,厚壁菌门(包括葡萄球菌和链球菌)和变形菌(主要是杏仁假单胞菌、荧光假单胞菌、假单胞菌UK4、鲍曼不动杆菌)——在流感患者中显着富集。
另一项针对 2009 年大流行 H1N1 流感感染患者的研究表明,尽管未包括正常对照,但携带大流行 H1N1 流感的患者上呼吸道(鼻和鼻咽样本)的主要门是放线菌门、厚壁菌门和变形菌门;然而,作者认为流感与变形菌的扩张有关,而变形菌在健康宿主中的数量通常较少。这些发现得到了另一组的支持,他们发现莫拉氏菌和肠杆菌属(被归类为变形菌门)是从大流行性 H1N1 流感患者的鼻咽样本中获得的最具代表性的细菌。然而,这些研究表明,受试者之间存在相当大的变异性,强调需要进行纵向研究来破译病毒感染后的变化。
特定病毒是否始终与某些细菌类群的富集相关
在澳大利亚原住民和非原住民儿童的鼻咽腔中,检测到 HRV 与肺炎链球菌、流感嗜血杆菌和卡他莫拉氏菌携带之间以及腺病毒与卡他莫拉氏菌之间存在正相关性。另一项研究通过 PCR 检测 20 种呼吸道病毒的存在以及儿童鼻咽部细菌携带的流行率,发现金黄色葡萄球菌定植与流感病毒之间存在很强的正相关性。
此外,肺炎链球菌定植与 HRV 和肠道病毒的存在呈正相关。流感嗜血杆菌与 HRV 和 RSV 呈正相关;卡他莫拉氏菌定植与冠状病毒和腺病毒呈正相关。
一项针对患有急性 RSV 或 HRV 呼吸道感染的婴儿进行的基于 16s rRNA 测序的研究报告称,患有 RSV 的婴儿的葡萄球菌丰度显著更高。
上呼吸道细菌群特征与宿主年龄相关
通过不依赖于培养物的焦磷酸测序对 57 名健康无症状个体和 59 名流感病毒、副流感、hRV、RSV、冠状病毒、腺病毒或偏肺病毒患者的 URT 细菌含量进行分析,揭示了六种不同的细菌谱,即链球菌+普雷沃氏菌+韦荣氏菌、链球菌+嗜血杆菌+奈瑟菌、链球菌、莫拉氏菌、嗜血杆菌和克雷伯氏菌。然而,这些特征与病毒类型无关,但与受试者的年龄有关。
病毒后细菌性肺炎成因复杂,需深入研究
鉴于许多人类研究本质上是横断面的,目前尚不清楚病毒后细菌性肺炎是否可能是病毒感染增强细菌定植或获得、定植细菌影响宿主对呼吸道病毒感染的易感性或两者结合的结果。另一个复杂的因素,特别是在病毒感染期间检查微生物组的横断面研究中,是组群控制不佳,而且考虑到可能影响呼吸道微生物组的变量数量(例如年龄、性别、口腔卫生和挖鼻孔习惯、基于医疗保健的就业状况、吸烟状况、药物使用、接触幼儿等。
病毒感染的潜在类型、感染发生后的采样时间点、感染的严重程度以及伴随的抗菌药物使用情况其他混杂因素。这可能是病毒感染患者微生物组研究中高度可变且有时不一致的观察结果的基础。
纵向研究
由于感染前采样困难,很少有临床研究比较患有急性病毒感染的其他健康个体感染前和感染后微生物组的基线。然而,现有的研究相对较少,可以深入了解细菌定植模式随时间的动态和稳定性,以及急性病毒感染引起的扰动是否以及如何改变这些模式。
在健康儿童中,鼻咽部微生物群的主要门是变形菌门、厚壁菌门、拟杆菌门、放线菌门和梭杆菌门,其中莫拉菌、嗜血杆菌、链球菌、黄杆菌、多洛西颗粒菌、棒状杆菌和奈瑟菌为主要属。
不同季节鼻咽微生物组变化的研究不一致
不同季节观察到鼻咽微生物组多样性的变化,秋季冬季以变形菌门和梭杆菌门为主,春季以拟杆菌门和厚壁菌门为主;这些差异与最近的抗生素和病毒合并感染无关。
然而,对 40 名哮喘儿童和青少年相隔 5.5-6.5 个月收集的两次鼻咽冲洗液进行的另一项分析表明,不同季节鼻咽微生物组多样性没有显着差异,尽管嗜血杆菌、莫拉氏菌、葡萄球菌和棒状杆菌的平均相对丰度在夏季和秋季样本之间以及不同年龄组之间存在显著差异。
此外,在 87.5% 的患者中,患者微生物丰度在不同时间点之间存在显着差异。对冬季和夏季无症状医疗人员的细菌和病毒载量的频率和季节变化进行的一项调查显示,在冬季接受测试的 100 名受试者中,34 人被至少一种细菌定植,11 人至少检测出一种病毒的阳性。
最常检测到的病原体是耐甲氧西林金黄色葡萄球菌(MRSA)、卡他莫拉菌和冠状病毒。相比之下,在夏季测试的 100 名受试者中,37 名受试者至少携带一种细菌(主要是 MRSA 和肺炎克雷伯菌),四名受试者检测出一种病毒呈阳性。
另一项对 31 个有小孩的家庭进行纵向鼻拭子采样的研究表明,鼻病毒感染与社区中肺炎链球菌感染的增加以及肺炎链球菌在家庭内传播的增加有关。
其他小组研究了将 hRV 实验接种到 URT(鼻孔)中的效果。这些研究报告称,随着时间的推移,鼻咽样本或咽拭子的总读数或主要门(例如放线菌门、厚壁菌门和变形菌门)没有显著变化。在口咽部,鼻病毒感染与副流感嗜血杆菌、微黄奈瑟菌(Neisseria subflava)相对丰度短暂增加的强烈趋势以及金黄色葡萄球菌相对丰度增加的微弱趋势相关。到 60 天时,这些细菌的丰度已恢复到基线。
鼻咽取样显示完全相反的结果,在 hRV 感染后的受试者中,嗜血杆菌和奈瑟菌属的相对丰度降低,但正常鼻共生丙酸杆菌增加。没有观察到葡萄球菌的差异。然而,这两项研究的受试者数量都很少,限制了检测随时间变化的能力。
与 URT 微生物组的研究相比,由于难以从肺部获取未污染的样本,因此对病毒感染后的 LRT 微生物组的研究相对较少。方便的样本(例如痰液)会受到口腔污染,但支气管镜样本是侵入性的,并且定期获取的成本很高。
此外,目前还不清楚,除了慢性肺病(例如慢性阻塞性肺病)患者之外,肺部微生物负荷是否足以对免疫反应和病毒感染期间继发细菌感染的风险产生强大影响。
来自流感感染小鼠模型的数据似乎表明,流感感染对肺部微生物组的细菌计数、多样性和组成仅产生适度的影响。
在 hRV 感染后患有慢性阻塞性肺疾病(COPD) 的受试者中,但在健康个体中则不然,基线时细菌负荷和细菌生长有所增加,特别是流感嗜血杆菌。
研究人员观察到,细菌的生长似乎源于现有的群落。将肺炎链球菌鼻内接种到预先感染流感病毒的小鼠体内,首先在鼻子中定植,几天后气管和肺部出现化脓性炎症。然而,在未感染的动物中没有观察到这种效应。这表明,在流感病毒感染者中,肺炎球菌感染可能从URT顺序发展到LRT。因此,一些流感感染者的肺部微生物组可能会因 URT 微生物群落的变化而发生变化。
微生物间相互作用
细菌中已经进化出竞争优势的机制,使它们能够在原本恶劣的宿主环境中生存。然而,种间竞争也可以通过捕获稀缺资源(例如铁)的能力或有针对性地杀死其他细菌(例如细菌素)来维持微生物群落的稳态,从而防止一种微生物主宰群落。
因此,急性病毒感染引起的免疫反应、病毒引起的宿主上皮表面的变化或病毒本身可能导致消除负责控制病原体的宿主共生体。例如,鼻孔中表皮葡萄球菌和痤疮丙酸杆菌的丰度与金黄色葡萄球菌携带呈负相关。
了解这些相互作用可能会为旨在减少流感流行或大流行期间致病菌定植的治疗干预措施创造新途径。
一组已被检查其在抑制金黄色葡萄球菌和肺炎链球菌鼻腔携带中的作用的共生菌是棒状杆菌属。日本的一项早期研究报告了将棒状杆菌菌株引入金黄色葡萄球菌持续携带者的健康成年医院工作人员的鼻孔中的效果,并成功根除 71% 的受试者,该机制似乎不依赖于细菌素。
相比之下,表皮葡萄球菌植入没有效果。所使用的表皮葡萄球菌菌株是否表达丝氨酸蛋白酶Esp(其抑制金黄色葡萄球菌生物膜形成和鼻定植)尚不清楚。
另一个小组的后续研究报告称,假白喉念珠菌抑制金黄色葡萄球菌生长,而Corynebacterium accolens和金黄色葡萄球菌似乎相互支持彼此的生长。相反,其他研究人员观察到棒状杆菌属。
在未鼻腔定植肺炎球菌的儿童中富集,并证明C. accolens通过表达脂肪酶来抑制体外肺炎链球菌生长,该脂肪酶从皮肤表面三酰甘油释放游离脂肪酸,从而抑制肺炎球菌生长。
因此,对共生体之间种间竞争的鉴定和机制了解,可能会带来新的见解,即病毒感染如何为致病生物赋予竞争优势,以及如何利用共生体采用的自然策略来恢复宿主微生物生态位的稳态。
有趣的是,最近的一项临床前研究使用RSV和肺炎链球菌双重感染的鼠模型,通过假白喉梭菌菌株进行鼻引发,以增强宿主对病毒感染的防御,从而增强对继发细菌攻击的清除并减少肺损伤。
感染病毒后的继发感染
感染病毒对细菌的直接影响可能促进病原体转变。宏基因组分析表明,与未感染患者中的相同细菌相比,pH1N1 相关的气道微生物群富含与细胞运动、转录调节、代谢和趋化反应相关的基因。这些数据表明,流感感染会扰乱呼吸道微生物组,导致产生包括免疫调节分子在内的次级代谢产物。病毒还会损害细菌生物膜的形成并破坏现有的生物膜。流感已被证明会通过下调与定植状态相关的基因表达和上调细菌素来影响肺炎链球菌转录组。因此,病毒对细菌转录模式的直接影响可能是定植细菌获得侵入潜力的一种机制,从而导致细菌重复感染。
在人体的生态位中,肠道微生物群落是研究最深入的,迄今为止已发表超过 20,000 篇论文。虽然病毒组和真菌组也在分析中,但大部分文献都集中在微生物组的细菌成分上,因此我们对肠道微生物组与宿主免疫和慢性疾病发病机制之间关系的大部分理解来自主要来自细菌群落的研究。
这里主要介绍肠道微生物组如何维持健康的宿主免疫环境的几个重要方面。
肠道微生物代谢产物与免疫
首先,肠道共生体产生的细菌代谢产物有助于维持完整的上皮完整性、调节性 T 细胞发育和相对抗炎的免疫状态。
特别是,短链脂肪酸(SCFA),例如乙酸盐、丙酸盐和丁酸盐,是大肠细菌发酵膳食纤维和碳水化合物的产物。短链脂肪酸可以:
同时,微生物代谢物对于促进肠道针对病原体的免疫反应至关重要,包括诱导 IL-18和防御素的分泌。因此,微生物组代谢产物对于粘膜屏障完整性和免疫稳态的适当调节至关重要。
此外,细菌群落的特定成员可以促进免疫系统的正常成熟和发育。虽然这仍然是一个正在深入研究的领域,但一个值得注意的例子是发现分段丝状细菌是肠粘膜 IgA 产生 和 Th17 细胞诱导的关键促进剂。
肠道微生物失衡与慢性疾病
菌群失调或微生物群组成的不平衡与多种慢病有关,其中许多疾病以慢性炎症或代谢异常为特征,包括炎症性肠病、心血管疾病、糖尿病等。因此,培养平衡的肠道微生物群多样性和组成对于促进健康和免疫稳态至关重要。
肠道微环境影响微生物动态变化
在健康期间,微生物群的组成受到每个解剖生态位特有的许多选择压力的控制,包括温度、营养可用性、pH、氧张力和局部免疫环境。由疾病、抗生素使用或饮食变化(例如饥饿)引起的肠道微环境的短期扰动可以改变肠道微生物群,并随后导致免疫反应的短暂改变。
因此,研究流感和其他呼吸道病毒是否改变胃肠道微生物组可能对病毒介导的抗菌免疫反应抑制具有机制意义。
肠道微生物群在免疫方面的这种有益作用不仅限于肠道区室,还延伸到全身区室和远处器官,例如肺部。
缺乏微生物群(即无菌)或口服抗生素(广谱或靶向抗生素,如新霉素、甲硝唑或万古霉素)治疗的小鼠对全身和呼吸道感染的反应受损。
在由例如甲型流感病毒(IAV)和呼吸道合胞病毒(RSV)引起的病毒性呼吸道感染的情况下,抗生素治疗对肠道微生物群生态的严重扰乱削弱了宿主的先天防御和适应性防御。
肠道微生物群对肺部产生I型干扰素的影响
众所周知,这些干扰素可控制病毒感染,包括SARS-CoV-2,COVID-19 的病原体。在这种情况下,脱氨基酪氨酸(源自类黄酮和氨基酸代谢)和短链脂肪酸等微生物代谢产物至关重要。
例如,研究表明,由专性梭状芽孢杆菌(Clostridium orbiscindens、对甲硝唑和万古霉素敏感)能够消化植物中黄酮类化合物,产生的脱氨基酪氨酸可以扩散到血液中,到达肺部并启动(先天)免疫系统,帮助身体抵抗流感病毒的感染。
脱氨基酪氨酸促进肺部 IFN 刺激基因的合成,而肺吞噬细胞在此机制中至关重要。在这种情况下,脱氨基酪氨酸通过 IFN-α/β 受体和信号转导器和转录激活剂增强 I 型 IFN 信号传导。例如,研究表明乙酸盐可以扩散到血液中,并激活基质细胞表达的GPR43。这种激活途径改善了 I 型干扰素反应并增加了干扰素刺激的基因表达。GPR43 促进 I 型 IFN 介导的流感感染保护作用的机制需要阐明。
除了微生物代谢物外,源自完整肠道微生物群的微生物膜成分(包括TLR配体)也在宿主防御病毒性呼吸道感染中发挥着关键作用。
事实上,局部和远端接种 Poly (I:C)(TLR3 激动剂)或肽聚糖(TLR2 激动剂)可以挽救抗生素治疗小鼠的免疫损伤。
值得注意的是,除了肠道微生物群发出的这些远端信号外,来自上呼吸道的局部微生物也调节下呼吸道免疫力,并在肺部防御病毒感染方面发挥作用。这些机制在很多研究中得到证实。
↑抗生素改变肠道菌群可能增加严重程度
肠道微生物群不仅影响先天免疫反应,还可以增强 CD8 + T 细胞效应器功能,这一过程也参与病毒(流感)清除。如上所述,用抗生素改变肠道微生物群会增加病毒性呼吸道感染(例如流感)的严重程度。
↓高纤维饮食刺激肠道菌群可能减轻严重程度
用高纤维饮食刺激微生物组会产生相反的效果。就流感而言,高纤维(可发酵菊粉)饮食通过两种不同的途径提供保护。膳食纤维的发酵导致骨髓造血功能改变,导致 IAV 感染小鼠肺中替代激活的巨噬细胞积聚。这些巨噬细胞产生较少的趋化因子(CXC 基序)配体 1,从而减少早期中性粒细胞浸润到气道中。
该途径极大地减少了过度的肺部炎症和损伤。该饮食还促进CD8+T细胞代谢,增强CD8+T细胞的效应功能,有效增强病毒清除。
保护作用需要 GPR41(也称为游离脂肪酸受体 3),口服 SCFA(丁酸盐)足以提供保护。在 RSV 感染期间也观察到了类似的保护作用。高纤维(可发酵果胶)饮食通过调节肺上皮细胞中的 I 型干扰素反应和增加肺部干扰素刺激基因的表达来防止 RSV 感染。保护作用需要 GPR43,口服乙酸盐足以提供保护。
肠道微生物群在病毒性呼吸道感染中的作用
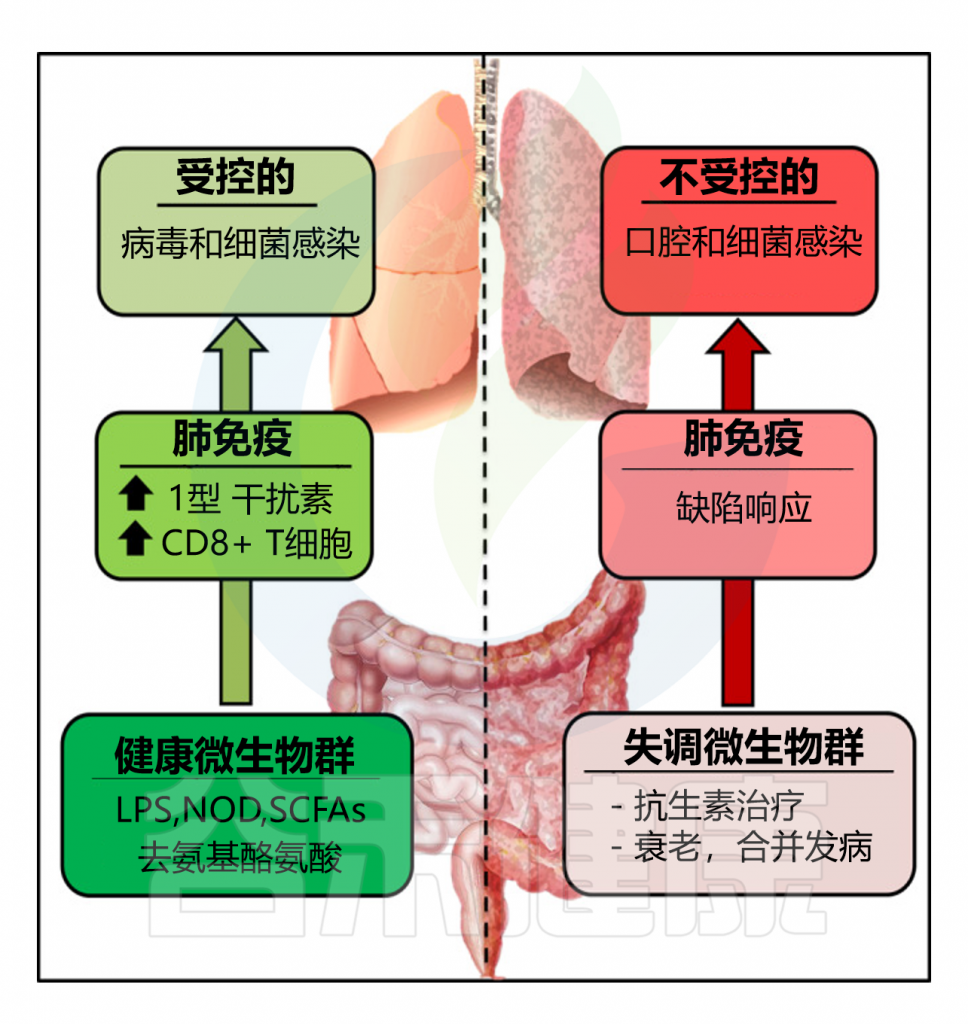
Sencio V, et al. 2021,14:296-304.
健康肠道微生物群释放的因子可以帮助肺部抵御病毒性呼吸道感染。相反,抗生素治疗会扰乱肠道内的生态平衡并改变肺部的防御能力。衰老和合并症也与肠道微生物群功能的改变和呼吸道感染的易感性增加有关。
在肺部发挥抗病毒作用的肠道共生细菌的性质仍然难以捉摸。如上所述,厌氧梭菌成员(脱氨基酪氨酸)和产短链脂肪酸菌对于肺部先天免疫系统的启动非常重要。最近,一项基于易感动物和耐药动物比较的研究表明,双歧杆菌属(假长双歧杆菌和动物双歧杆菌)的定植与流感感染小鼠的生存密切相关。很可能在不久的将来,其他共生成员将被识别出来,并将作为新的生物标志物来预测严重病毒性呼吸道感染患者的严重程度和死亡率。
因此,在稳定状态下,肠道微生物群中的常驻细菌可以自然地远程支持肺部对抗病毒性呼吸道感染。
研究表明,除了饮食模式,呼吸道病毒感染及其他压力因素也会影响肠道微生物群的组成。流感和RSV肺部感染的动物模型显示,即使肠道中没有检测到呼吸道病毒,肠道微生物组也会受到感染的显著影响。
流感病毒和肠道菌群
流感感染可导致轻度至重度肺炎、急性呼吸窘迫综合征 (ARDS)(临床上定义为急性呼吸衰竭)和多器官功能障碍。尽管肠道内明显没有病毒,但一些患者仍然出现类似胃肠炎的症状,如腹痛、恶心、呕吐、腹泻。这表明肺-肠轴在流感感染期间可能很重要。
根据可操作分类单位数量评估,细菌负荷和 α 多样性并未受到甲流病毒感染的强烈影响。在门水平上,观察到的变化很少。
H1N1 亚型 IAV 感染导致拟杆菌门/厚壁菌门比率下降。
H3N2 和 H1N1 亚型感染期间,还观察到疣微菌(主要由阿克曼氏菌属组成)增加。这些细菌会降解肠道粘液层,瘤胃球菌也是如此(这可能与感染期间粘液增加有关),其数量在 IAV 感染期间也会增加。IAV 感染后还观察到放线菌减少(主要是由于双歧杆菌科属减少)。
简而言之,大多数研究发现γ变形菌(潜在致病性大肠杆菌)增加,少数情况下发现芽孢杆菌类减少。微生物群分析还强调了厚壁菌门内部的变化。综上所述,感染过程中下列菌群数量相对较多:
总体而言,来自流感小鼠模型的数据表明,感染促进了潜在有害细菌种类的出现,例如γ变形菌门和粘液降解细菌的成员。相反,感染似乎会抑制促进健康的细菌(如乳酸杆菌、双歧杆菌和分段丝状细菌)的生长。
此外,小鼠体内 IAV 感染与分节丝状细菌(梭菌科)比例下降有关。这种细菌与肠上皮细胞密切相互作用,对于宿主抵抗啮齿类柠檬酸杆菌等肠道病原体具有重要作用。
流感感染→SFB及乳杆菌减少→肠道损伤
在流感感染的小鼠模型中,研究人员发现,虽然肠道内的细菌总数没有减少,但分段丝状菌(SFB)和乳杆菌/乳球菌的数量减少,同时肠杆菌科细菌增加。有趣的是,虽然 SFB 之前已被证明可以诱导 Th17 细胞 ,但感染流感的小鼠小肠和结肠中的 IL-17A 水平和 Th17 细胞数量增加,这似乎导致肠道损伤。
在这项研究中,流感感染前的抗生素治疗减轻了肠道损伤的程度,但没有改善肺部损伤,这表明肠道菌群失调导致局部炎症,而不是全身炎症。
活病毒感染→变形菌增加→结肠炎易感性增加
注射减毒流感活疫苗(LAIV),表明这些变化需要活病毒感染。变形菌的增加似乎是由 I 型干扰素 (IFN) 介导的,它不仅耗尽了厌氧菌,而且还增加了继发性沙门氏菌结肠炎的易感性。然而,热量限制也会导致变形菌门的相对丰度增加,以及拟杆菌门与厚壁菌门的比例增加,这增加了流感期间口服摄入量减少可能导致微生物组变化的可能性。
全身炎症信号介导肠道微生物变化
还表明,流感感染通过招募到肠道的肺源性 T 细胞产生的 II 型干扰素来改变肠道微生物群组成。因此,肠道微生物组的变化似乎不是由直接病毒效应引起的,而是由从肺部传播并触发肠道局部炎症反应的全身炎症信号引起的。
肠道微生物群可以调节肺部免疫活性
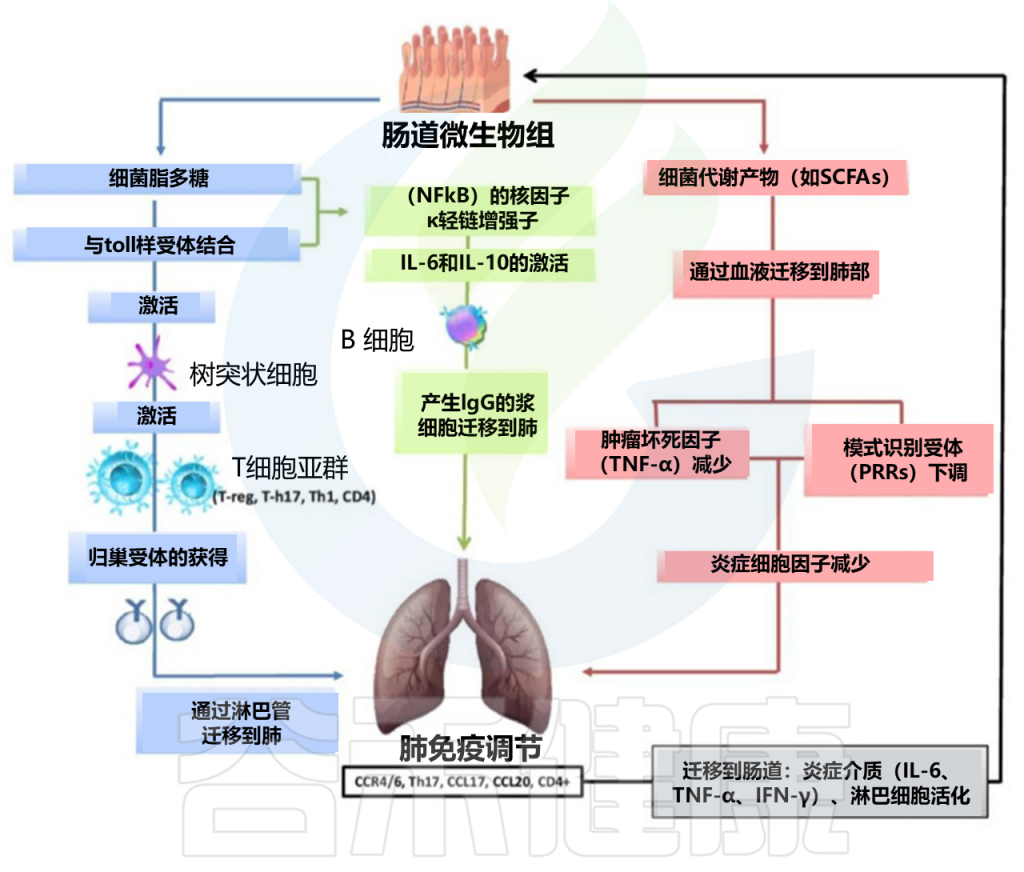
doi: 10.3390/biology9100318
肠道微生物群可以调节肺部免疫活性的双向假设:脂多糖 (LPS) 能够与肠粘膜上存在的TLR结合,从而激活树突状细胞,从而有利于各种 T 细胞的激活(T-reg、T-h17、Th-1、CD4);随后,获得角受体(CCR6、CCR9、CCR4、a4b7)并通过循环淋巴流迁移至肺部(CCR4/6Treg、CCR4/6Th17、CCR4/6Th1、CCR4/6CD4 +)。它还激活(IL-18、INF-y、TNF-a、TGF-b、IL-4、IL-1),这些细胞将进入循环(INF-y、TNF-a、IL-6)。
核因子 kappa-轻链增强子 (NFkB) 激活 IL-6、IL-10 以及 IgA 和 IgG 浆细胞 B 细胞的产生以及 IgG 向肺部的迁移。在肺部,CCL17、CCL20 增加,并且存在 CCR4/6、CD4+、CCR4/6 和 Th17。
细菌代谢物(例如,短链脂肪酸)通过血流迁移至肺部,导致模式识别受体(PRR)下调,从而减少炎症细胞因子(IL-1、IL-12、IL-18)、肿瘤坏死因子 α (TNF-α)、干扰素γ (IFNγ) 和粒细胞-巨噬细胞集落刺激因子 (GM-CSF)。
反过来,在疾病发生时,肺部会将炎症介质和淋巴细胞输送到肠道。
除了(不变)自然杀伤 T 细胞的配体之外,粘膜相关不变 T (MAIT) 细胞的配体合成也可能在病毒呼吸道感染期间发生改变。事实上,细菌共生体,包括属于拟杆菌门和变形菌门的物种,以及病原菌,可以产生维生素 B2 和维生素 B9 代谢物,已知这些代谢物可作为 MAIT 细胞的激动剂或拮抗剂。
关于 MAIT 细胞在粘膜免疫中的关键作用,在病毒呼吸道感染期间改变 MAIT 细胞配体的合成可能对疾病结果产生重要影响,这一假设需要进行研究。
呼吸道合胞病毒和肠道菌群
在小鼠模型中,RSV 导致感染后第 7 天肠道微生物群多样性(但不是丰度和 α 多样性)发生显着改变,拟杆菌门增加,厚壁菌门减少。
拟杆菌门的增加主要是由于拟杆菌科和S24-7科的增加,而厚壁菌门丰度的减少与毛螺菌科和乳杆菌科的减弱有关。RSV 感染对肠道微生物群代谢活动的影响很少被讨论。在一项研究中,RSV 感染主要增加脂质代谢,包括鞘脂、多不饱和脂肪酸和 SCFA 戊酸酯。
冠状病毒和肠道微生物群
一项研究通过 RNA 鸟枪法宏基因组学测序揭示,在患有高感染率的 COVID-19 患者的粪便样本中存在机会性细菌病原体。
相比之下,粪便样本中 SCFA 和色氨酸生产者含量丰富,具有低至无 SARS-CoV-2 感染性的特征。有趣的是,具有高 SARS-CoV-2 感染性的粪便具有更高的微生物组功能,用于核苷酸从头生物合成、氨基酸生物合成和糖酵解。
值得注意的是,在 COVID-19 患者中还描述了机会性真菌病原体(曲霉属和念珠菌属)的过度生长。有趣的是,流感 (H1N1) 和 SARS-CoV-2 患者之间的相似点和差异。核心肠道微生物特征和相关代谢物的发现可以作为潜在的诊断标志物。
急性病毒性呼吸道感染期间肠道微生物群发生变化的原因有多种:这些可能包括炎症细胞因子的释放和食物摄入量的减少。
感染病毒→食欲不振→肠道菌群紊乱
在小鼠中,感染 IAV 或 RSV 会导致体重大幅减轻,减轻幅度为初始体重的 10% 至 20%,并在感染后 5-7 天达到峰值。这种体重减轻主要是由于食欲不振。
众所周知,食物和卡路里摄入量的减少会扰乱肠道微生物群。配对喂养实验的结果清楚地表明,食物摄入量的迅速下降模拟了感染期间观察到的肠道微生物群的变化,即疣微菌门、α变形菌纲和副杆菌属的丰度增加,而毛螺菌科、瘤胃球菌属和乳杆菌属的丰度减少,尽管也观察到差异(梭菌目)。
纤维摄入量减少影响结肠上皮细胞代谢
纤维摄入量的减少一致,配对喂养的小鼠 SCFA 浓度较低。其中,丁酸盐为结肠上皮细胞提供能量,促进上皮氧消耗,从而导致肠腔缺氧。由于食物(纤维)摄入量减少而导致短链脂肪酸的下降可能会改变上皮细胞的代谢。
炎症细胞因子对肠道菌群的影响
病毒感染期间食欲不振的主要原因之一是炎症细胞因子的过量产生,包括肿瘤坏死因子α。
RSV 感染期间这种细胞因子的中和减少了体重减轻,并部分减弱了肠道微生物群的扰动。同样,在 RSV 感染期间消耗产生细胞因子的 CD8 + T 细胞可减少食欲不振并逆转肠道微生物群的变化。
I 型和 II 型干扰素是宿主抗病毒反应的重要组成部分——尤其是在流感期间。这些细胞因子强烈扰乱肠道微生物群。相对于野生型小鼠,I 型 IFN 受体缺陷的小鼠表现出变形菌(埃希氏菌属)繁殖减少,相反,分节丝状细菌的比例升高。
同样,IAV 感染期间 IFN-γ 的消耗恢复了分段丝状细菌(在 IFN-γ 活性小鼠中减少)、乳杆菌属(减少)和肠杆菌科(增强型)。因此,炎症细胞因子和食欲不振可能会导致病毒性呼吸道感染期间肠道菌群失调,至少在小鼠系统中是这样。
氧气梯度变化对肠道菌群的影响
其他机制也可能受到牵连。肠道炎症,例如由于浸润的 CD4 + T 细胞或全身性 IFN 释放而引起,已知会改变上皮细胞的代谢,从而导致一组新的营养物质的积累,而居住在肠腔中的微生物则需要这些营养物质竞争。随着氧气可用性的增加,这种现象似乎可以解释从专性厌氧菌到兼性厌氧菌(例如变形菌肠杆菌科)的转变。
缺氧是呼吸道病毒感染急性期的主要临床症状,包括 COVID-19 患者。它还与慢性肠道损伤有关。关于氧气在肠道稳态中的作用,包括微生物群的组成和功能,由于 SCFA 的可用性而造成的氧气梯度破坏,可能在呼吸道病毒感染期间肠道菌群失调和胃肠道疾病中发挥作用。呼吸道病毒感染期间肠道(上皮)免疫功能缺陷(抗菌肽产生减少)也可能参与生态失调。
病毒性呼吸道感染期间的“肠肺轴”
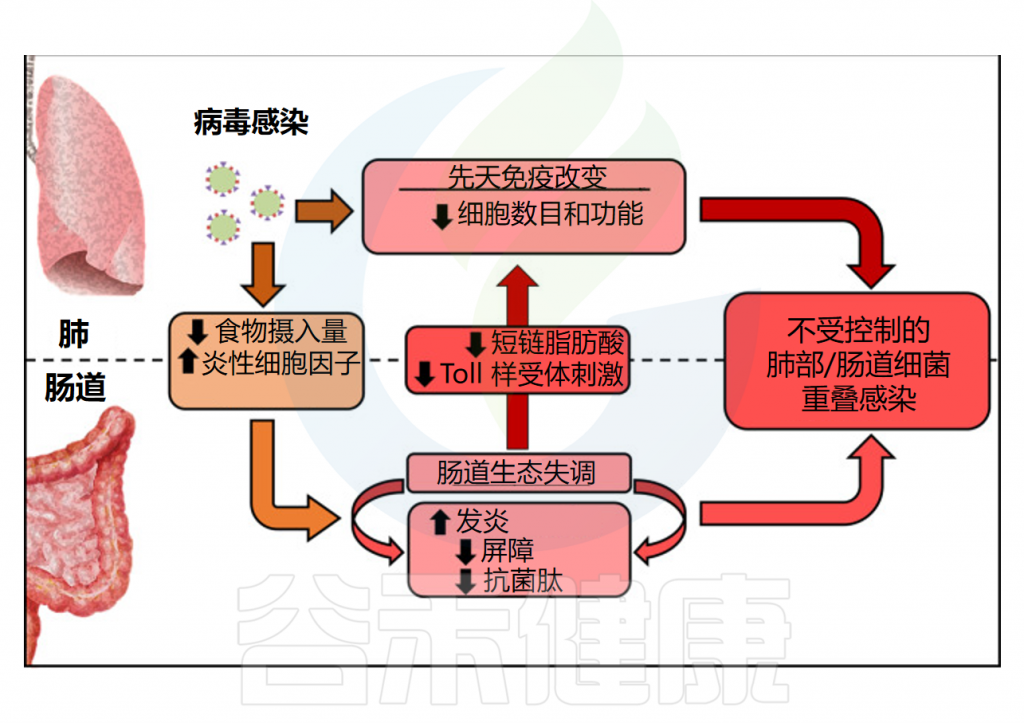
Sencio V, et al. 2021,14:296-304.
总的来说,病毒性呼吸道感染,例如流感,会改变肺部免疫和基质(上皮)细胞的功能,从而导致继发性细菌感染。与此同时,全身炎症细胞因子、食欲不振(纤维减少)、氧水平变化和上皮代谢改变会破坏肠道微生物群的组成和功能。
这些改变会导致肠道炎症、上皮屏障破坏和抗菌肽 (AMP) 产生减少。然后发生继发性肠道感染。上皮渗漏可能会增强细菌易位并引发全身炎症和器官功能障碍。病毒性呼吸道感染引起的菌群失调还会导致微生物相关分子模式的产生减少,包括 Toll 样受体 (TLR) 和核苷酸寡聚结构域 (NOD) 样激动剂以及短链脂肪酸等微生物代谢产物,从而降低抗菌肺部免疫力。因此,呼吸道病毒感染通过改变肠道稳态导致细菌重复感染。
肺和肠道的相互联系在 SARS-CoV-2 感染期间可能特别相关。
消化道可能是病毒复制和活动的场所
与 IAV 和 RSV 的情况相反,在冠状病毒感染(包括 SARS-CoV-2 感染)期间,在肠道中检测到病毒 RNA。大约一半的 COVID-19 患者的粪便中含有 SARS-CoV-2 RNA,即使呼吸道中不再发现它。
重要的是,在 COVID-19 患者的粪便样本中检测到传染性病毒,表明消化道可能是病毒复制和活动的场所。
同样,最近几项使用人类小肠类器官的研究表明 SARS-CoV-2 在肠细胞中复制。因此,局部病毒复制可能会扰乱当地生态系统,导致肠道微生物群组成和功能发生变化。
ACE2表达下调与肠道微生态变化
病毒刺突蛋白与细胞表面受体血管紧张素转换酶 II (ACE2) 的结合导致后者的表达下调。关于 ACE2 在维持肠道微生物生态中的关键作用(通过氨基酸运输故障、色氨酸缺乏和抗菌肽产量减少),SARS-CoV 期间缺乏可用的 ACE2 很可能-2感染在生态失调中起着至关重要的作用。
粪菌移植实验表明,从流感 (H7N9) 感染后幸存的小鼠中收集的肠道微生物群可以为受到 IAV 攻击的受体(未接触过的)小鼠提供保护。这种保护作用是否适用于其他 IAV 亚型和其他呼吸道病毒尚不清楚,值得未来研究。因此,呼吸道病毒感染引起的肠道微生物群组成和功能活动的改变可能会产生有益的影响。然而,如下所述,急性呼吸道病毒感染引起的菌群失调也会导致有害影响并参与感染的结果。
肠道微生物群对于维持上皮完整性和调节性 T 细胞的发育至关重要。这一关键作用主要取决于微生物群的代谢物。
在稳态期间,肠道发挥相对抗炎的免疫状态
肠道微生物群的扰动会导致多种肠道疾病,例如炎症性肠病、肠易激综合征、乳糜泻和结直肠癌。根据这些观察结果,研究人员调查了病毒性呼吸道感染期间观察到的肠道菌群失调对肠道稳态和功能可能产生的负面影响。
病毒性呼吸道感染对肠道的影响
如上所述,病毒性呼吸道感染,包括 IAV、RSV 和冠状病毒感染,可诱发胃肠炎样症状,如腹痛、恶心、呕吐、腹泻。
感染 IAV 的小鼠出现肠道损伤,包括结肠长度缩短(炎症标志物)、小肠粘膜层消失和轻度腹泻。同时,观察到炎症细胞因子和 IFN 刺激基因的 mRNA 表达增强。
肠道屏障的破坏
肠道的一个重要特征是其屏障功能;在健康条件下,这可以防止微生物成分的过度扩散。许多疾病与屏障功能破坏有关,从而导致细菌移位、全身炎症和休克。IAV 对肠道屏障特性的影响值得深入研究。就RSV而言,除了粪便中脂质运载蛋白2(肠道炎症标志物)水平升高之外,在小鼠模型中没有观察到明显结肠炎症的组织学证据。
肠道菌群改变,可能是炎症的结果,也可能是炎症的原因
人们可能会质疑,在病毒性呼吸道感染的情况下,微生物群的改变是否会引发肠道炎症,或者炎症是否会导致菌群失调。这是一个“先有鸡还是先有蛋”的情况。如上所述,炎症可能会影响肠道微生物群的组成。另一方面,失调的微生物群可能直接引发和维持肠道疾病。
事实上,IAV 感染期间的抗生素治疗(以限制肠杆菌科细菌的增加)可以减轻肠道炎症。此外,粪便移植实验表明,经历过 IAV 的微生物群触发了辅助 T17 细胞的局部积累,随后引起肠道炎症。
与肠杆菌科细菌一样,粘液降解细菌(例如瘤胃球菌和阿克曼氏菌)也可能是肠道炎症维持和/或放大的重要因素。事实上,已知这些细菌会侵蚀结肠粘膜并促进肠腔细菌与肠上皮之间的相互作用。反过来,这会导致炎症和屏障功能受损。
总体而言,病毒性呼吸道感染可能通过改变共生体平衡来影响肠道稳态。肠道疾病的性质以及感染对肠道生理学(例如,摄入的食物和液体的消化和吸收)、代谢和免疫稳态的影响仍有待详细分析。
病毒性呼吸道感染后肠道功能的改变可能会对健康产生直接和长期的影响。例如,肠道屏障对于维持体内平衡至关重要,任何扰动都可能导致细菌成分的系统性传播,从而导致有害的健康结果。
肠道微生物群通过直接微生物拮抗和刺激宿主效应反应(例如抗菌肽)来局部控制病原性感染至关重要。生态失调会增加发生肠道细菌感染的风险。动物模型实验表明,IAV 增强了肠沙门氏菌Thyphimurium 继发肠道感染的易感性。这种增强至少部分归因于 I 型干扰素以及相关的抗菌肽和炎症细胞因子释放的减少。
肠道微生物群多样性的减少也可能与继发性肠道感染有关;例如(在小鼠系统中),分段丝状细菌(已知在 T 辅助细胞 17 介导的免疫反应中很重要)的比例在 IAV 感染期间下降。
与此同时,SCFA 生产者比例的下降对于继发性肠道感染可能很重要。已知这些脂肪酸可以控制肠道微生物群的平衡,并防止肠道病原体(如粪肠球菌和鼠伤寒沙门氏菌)的发展。 IAV 感染期间口服补充 SCFA 是否可以降低继发肠道感染仍有待观察。
呼吸道病毒感染的主要并发症之一是继发细菌感染的易感性增加,主要是由肺炎链球菌、金黄色葡萄球菌和流感嗜血杆菌引起的继发细菌感染。这种并发症主要影响儿童和老年人,并导致流行病和大流行期间观察到的发病率和死亡率过高。动物模型实验表明,敏感性增强部分是由于抗菌先天免疫反应受损。
健康的肠道微生物群在肺部免疫中具有有益的生理作用,包括在控制肺炎链球菌或金黄色葡萄球菌等机会性细菌的发育方面发挥积极作用。
研究了流感感染期间肠道微生物群的扰动可能增加肺部细菌重复感染发生率的可能性。事实上,粪菌移植实验表明,IAV 条件下的微生物群损害了肺部对肺炎球菌感染的防御能力。
从机制上讲,乙酸盐(微生物群产生的主要短链脂肪酸)产量的下降影响了肺泡巨噬细胞的杀菌活性。在流感和肺炎球菌双重感染的情况下,补充乙酸盐可以降低细菌负荷,减少肺部疾病,并提高生存率。因此,流感期间肠道微生物群的变化(以及 SCFA 产生的减少)与细菌重复感染有关。该领域的进一步研究可能有助于定义预测标记(例如,系统性 SCFA)和/或开发针对细菌重复感染的治疗方法,例如通过益生元和/或益生菌利用肠道微生物群的力量。
值得注意的是,除了肠道微生物群之外,与急性病毒性呼吸道感染相关的呼吸道微生物组成和功能的变化也可能在细菌重复感染中发挥重要作用。这种局部生态失调可能会改变微生物间相互作用的动态,从而增强潜在致病细菌物种的增殖。局部微生物代谢输出的变化也可能降低肺部对继发细菌感染的防御能力。
总的来说,病毒性呼吸道感染(至少是流感)期间的肠道菌群失调会影响肺部的细菌重复感染。微生物变化是否会影响病毒性呼吸道感染期间的其他疾病结果,例如急性呼吸窘迫综合征、脓毒症和多器官功能障碍,仍然是一个悬而未决的问题。
流感和其他呼吸道病毒感染后常常导致细菌性肺炎,这种现象在历史和现在上已造成大量死亡。病毒感染能导致呼吸道结构和功能的损伤,改变呼吸道的微生物组,增加细菌的侵袭和定植机会,从而促进细菌性肺炎的发生。研究表明,病毒感染不仅影响上呼吸道(URT)的微生物群落结构,而且还可能促进下呼吸道(LRT)的细菌定植,从而增加了肺炎的风险。尽管在临床上普遍使用抗生素以防治细菌性并发症,但病毒与细菌之间的相互作用仍可能在病程中发挥作用。
呼吸道病毒感染后引发细菌性肺炎的机制是复杂的,涉及宿主免疫系统的变化、微生物组的改变以及病原体之间的相互作用等因素。以下是一些可能的机制:
1
免疫系统损伤
病毒感染,如流感病毒或冠状病毒感染,会损伤宿主的免疫系统,特别是呼吸道黏膜的屏障功能和局部免疫应答。这使得细菌更容易侵犯和定植在下呼吸道,从而导致细菌性肺炎。
2
微生物组改变
病毒感染可以改变呼吸道的微生物群落结构,这可能破坏了正常的微生物群平衡,使得某些致病细菌(如肺炎链球菌、葡萄球菌和流感嗜血杆菌)得以增殖和引起感染。
3
病原体协同作用
一些研究表明,病毒和细菌之间可能存在协同作用,病毒感染可能会促进细菌的黏附和侵袭能力,或者细菌可能利用病毒感染后宿主免疫应答的变化来促进自己的生长。
4
炎症反应
病毒感染引起的炎症反应可能会损害呼吸道的细胞,导致细胞死亡和组织损伤,这为细菌提供了更多的营养物质和一个更易于感染的环境。
5
抗生素使用
在治疗病毒感染的过程中,过度或不当使用抗生素可能会导致抗生素敏感的细菌被清除,而抗生素耐药的细菌得以生长,这可能增加了细菌性肺炎的风险。
6
肠道微生物组的改变
感染促进了潜在有害细菌种类的出现,例如γ变形菌门和粘液降解细菌的成员。相反,感染会抑制促进健康的细菌(如乳酸杆菌、双歧杆菌和分段丝状细菌)的生长。与此同时,全身炎症细胞因子、食欲不振(纤维减少)、氧水平变化和上皮代谢改变会破坏肠道微生物群的组成和功能。
7
宿主因素
宿主的年龄、基础疾病、免疫状态等因素也会影响病毒感染后细菌性肺炎的风险。
8
继发细菌感染
在病毒性疾病的紧急情况下,注意力最初集中在原发感染的临床管理上,但必须考虑患者在初次感染期间或之后发生的继发细菌感染。此外,混合感染(同时感染另一种病毒或细菌病原体)也可能发生,并最终导致相同的结果:患者患有由两种不同病原体引起的并发症。
未来诊断细胞性肺炎的项目通常需要综合临床表现、影像学检查以及微生物学检测。微生物学检测可能包括痰液培养、血液培养、支气管肺泡灌洗液(BAL)或粪便培养或分子检测。这些检测有助于确定引起肺炎的具体细菌种类,并为抗生素的选择提供指导。
在干预和管理方面,预防继发性感染策略包括:
改善免疫力
确保患者获得适当的营养和休息,以支持免疫系统的正常功能。对于易感人群,如慢性疾病患者,应考虑接种针对特定细菌的疫苗(如肺炎球菌疫苗)。
这些营养素在免疫健康中发挥作用:
β-胡萝卜素
β-胡萝卜素存在于植物性食品中,如红薯、菠菜、胡萝卜、芒果、西兰花和西红柿。
维生素 C
富含维生素 C 的食物,包括柑橘类水果、浆果、甜瓜、西红柿、青椒和西兰花。
维生素 D
维生素 D 存在于富含脂肪的鱼类和鸡蛋中。牛奶和 100% 添加维生素 D 的果汁也是很好的来源。
锌
锌更容易从牛肉和海鲜等食物中吸收,但也存在于植物性来源中,包括麦芽、豆类、坚果和豆腐。
益生菌
益生菌是促进健康的好细菌。发酵乳制品(例如酸奶)和发酵食品(例如开菲尔和泡菜)中有。
蛋白质
蛋白质来自动物和植物来源,包括牛奶、酸奶、鸡蛋、牛肉、鸡肉、海鲜、坚果、种子、豆类和扁豆。
抗病毒
对于治疗某些呼吸道病毒感染,如病毒感染,可以使用抗病毒药物来减少病毒复制,从而降低继发细菌性肺炎的风险。
维持微生物群平衡
使用益生菌和益生元可能有助于保持或恢复正常的微生物和落平衡。这可以通过补充或通过饮食调整来实现,例如增加富含纤维或益生菌的食物来促进有益菌的生长。
抗生素的审慎慎用
在病毒感染的患者中,不宜过分使用抗生素,以免造成不必要的抗生素抗性和抗生素失衡。只有在明显有细菌感染或经过微生物学验证的情况下,才应使用抗生素。
症状管理和支持治疗
针对呼吸道病毒感染的症状进行管理,如使用退热药、镇咳药和解痉药。同时,保持良好的水分和营养平衡,维生素B族,氨基酸等,必要时提供氧气支持。
监测和早期识别
对于高风险患者群体,应进行定期监测,以便于早期识别继发性细菌性肺炎的迹象,及时进行干预。
通过这些策略的综合应用,可以有效地管理呼吸道病毒感染后的气道和肠道微生物组的变化,并预防继发性肺炎的发生。然而,每个患者的具体情况可能不同,因此,治疗计划应根据个体的需求和病情进行个性化调整。
主要参考文献:
Hanada S, Pirzadeh M, Carver KY, Deng JC. Respiratory Viral Infection-Induced Microbiome Alterations and Secondary Bacterial Pneumonia. Front Immunol. 2018 Nov 16;9:2640.
Lee KH, Gordon A, Foxman B. The role of respiratory viruses in the etiology of bacterial pneumonia: An ecological perspective. Evol Med Public Health. 2016 Feb 15;2016(1):95-109.
Kaul D, Rathnasinghe R, Ferres M, Tan GS, Barrera A, Pickett BE, Methe BA, Das SR, Budnik I, Halpin RA, Wentworth D, Schmolke M, Mena I, Albrecht RA, Singh I, Nelson KE, García-Sastre A, Dupont CL, Medina RA. Microbiome disturbance and resilience dynamics of the upper respiratory tract during influenza A virus infection. Nat Commun. 2020 May 21;11(1):2537.
Sencio V, Machado MG, Trottein F. The lung-gut axis during viral respiratory infections: the impact of gut dysbiosis on secondary disease outcomes. Mucosal Immunol. 2021 Mar;14(2):296-304.
Manna S, Baindara P, Mandal SM. Molecular pathogenesis of secondary bacterial infection associated to viral infections including SARS-CoV-2. J Infect Public Health. 2020 Oct;13(10):1397-1404.
Santacroce L, Charitos IA, Ballini A, Inchingolo F, Luperto P, De Nitto E, Topi S. The Human Respiratory System and its Microbiome at a Glimpse. Biology (Basel). 2020 Oct 1;9(10):318.

谷禾健康
民以食为天,食以安为先。
随着夏季来临,我们最近检测的报告中发现以上食源性致病菌的检出率和超标率也在逐步上升。
尽管许多人可能没有意识到,近年来,食源性疾病的发病率逐年升高,食源性病原体正在引起大量疾病,对人类健康和经济产生重大影响,已成为全球范围内的重大公共卫生问题。
食源性疾病是一个全球性问题,影响所有年龄和背景的人,尤其是 5 岁以下的人和的65岁老人以及生活条件差的人。我国细菌性食源性疾病每年发病人数可达9411.7万人次,根据美国疾病控制与预防中心 (CDC) 的估计,美国每年也有4800 万例疾病、128000 例住院治疗和 3000 例死亡是由食源性细菌引起的。澳大利亚,每年约有五分之一的人患有这些疾病,住院人数超过 30,000 人。
致病菌广泛存在于各种食品中,食源性致病菌是食源性疾病的首要病因,由其引起的食源性疾病长期以来一直是食品安全的主要威胁,尤其在发展中国家,形势更严峻。它们会导致 200 多种疾病——从腹泻到癌症。在我国,每年因副溶血性弧菌导致急性腹泻665.5万人,导致急性胃肠炎病例估计为728.1万人。
最常见的致病细菌包括:蜡样芽胞杆菌,空肠弯曲杆菌,肉毒梭菌,产气荚膜梭菌,坂崎肠杆菌,埃希氏菌,李斯特菌,沙门氏菌属,志贺氏菌属,Staphylococccus黄色葡萄球菌,弧菌属等。
定 义
食物中毒:指食用了被有毒有害物质污染的食品或者食用了含有有毒有害物质的食品后出现的急性,亚急性以及其它食源性疾病。
目前一些-国家和国际组织以及很少使用食物中毒的概念,经常使用的是“食源性疾病”,食物中毒仅是食源性疾病的一部分,是一大类最常见最典型的食源性疾病
食源性疾病:世界卫生组织认为,凡是通过摄食进入人体的致病因素,是人体患感染性的或中毒性的疾病,都称之为食源性疾病。
我国《食品安全法》定义食源性疾病:指食品中致病因素进入人体引起的感染性,中毒性疾病以及其它疾病。
食物中毒的症状
细菌性
食物中毒的症状因人而异,有些人的病情会比其他人更严重。但是,一些常见的症状是:
不太常见的症状也包括:
保持水分和休息通常是治疗食物中毒的唯一方法,因为身体可以自然地清除污染物。但是,在某些情况下,需用抗生素来帮助身体抵抗感染,特别是在单核细胞增生李斯特菌中毒的情况下以及当易感受伤害的人受到影响时。
病毒性
与细菌不同,病毒不会在食物上生长和繁殖。相反,病毒细胞存在于食物中,当被食用时,它们开始接管人体细胞并感染它们。
常见的食源性疾病病毒类型是:
诺如病毒是导致大多数与食品污染有关的疾病的罪魁祸首 。

谷禾健康
http://v.qq.com/s/videoplus/457452244皮肤菌群如何导致痤疮、湿疹?如何维持皮肤健康?_腾讯视频

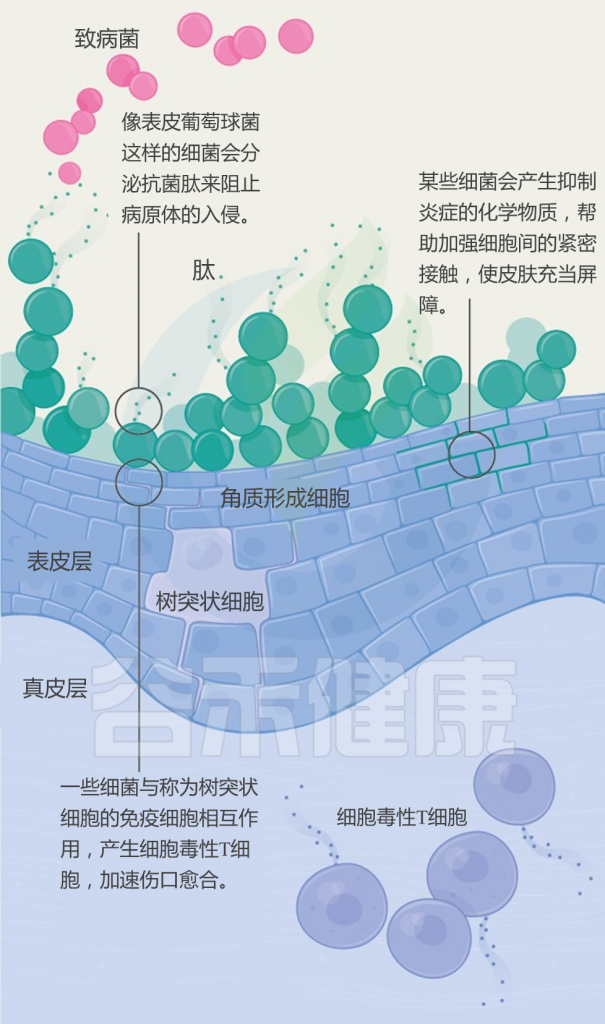
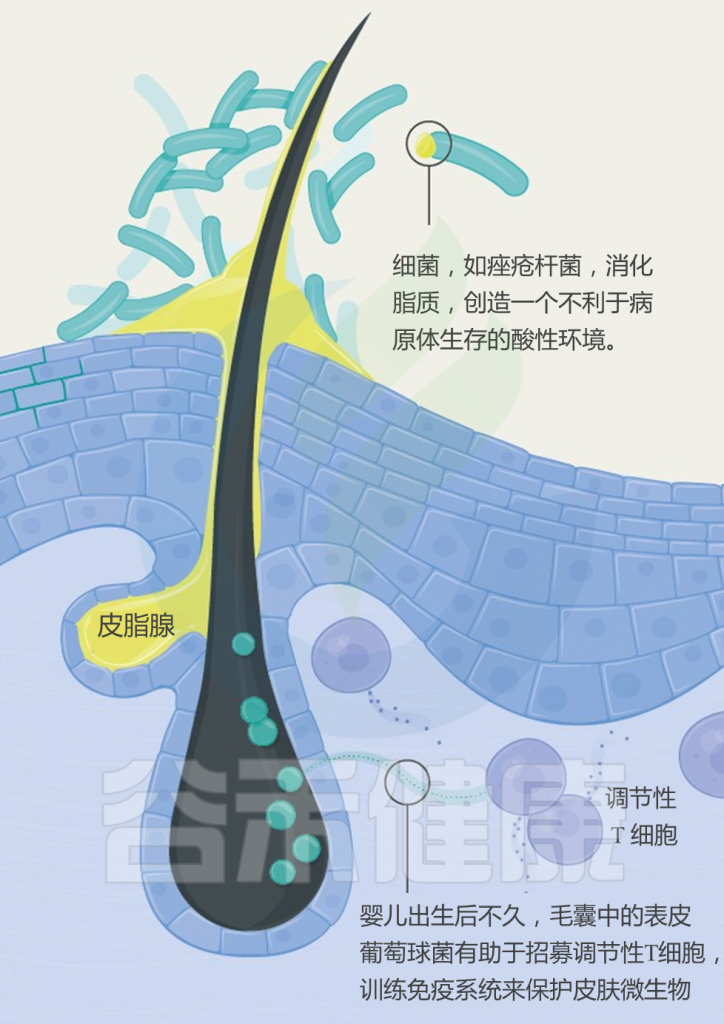
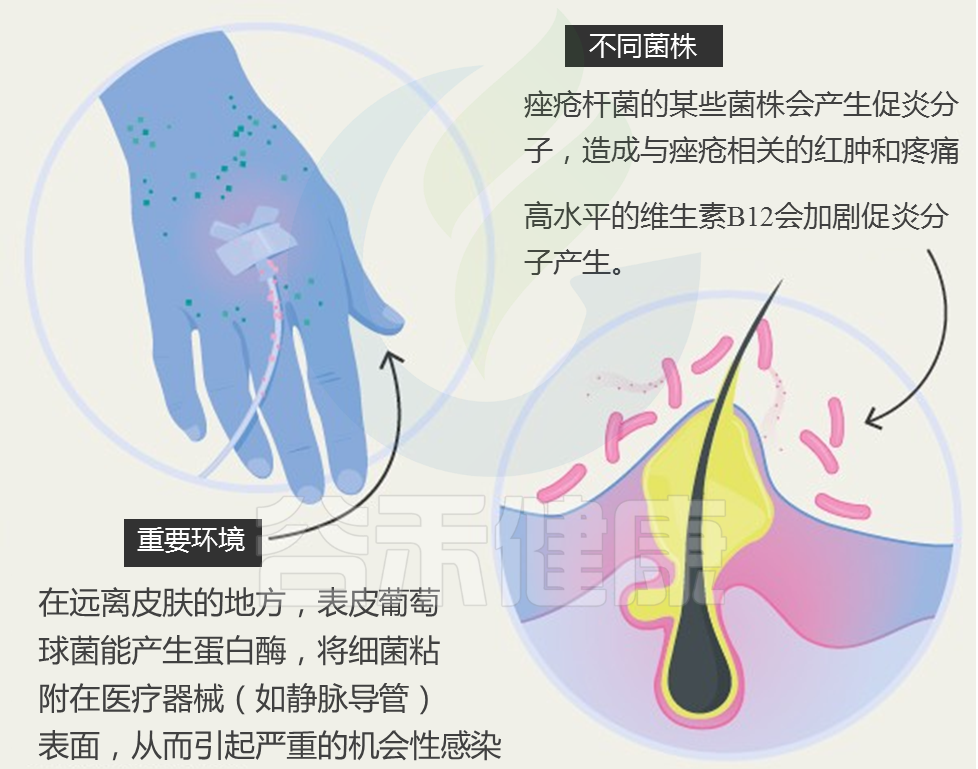



相关阅读: