-

 CNAS L23010
CNAS L23010

 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业 二级病原微生物安全实验室
二级病原微生物安全实验室- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业 二级病原微生物安全实验室
谷禾健康
尊敬的谷禾用户:
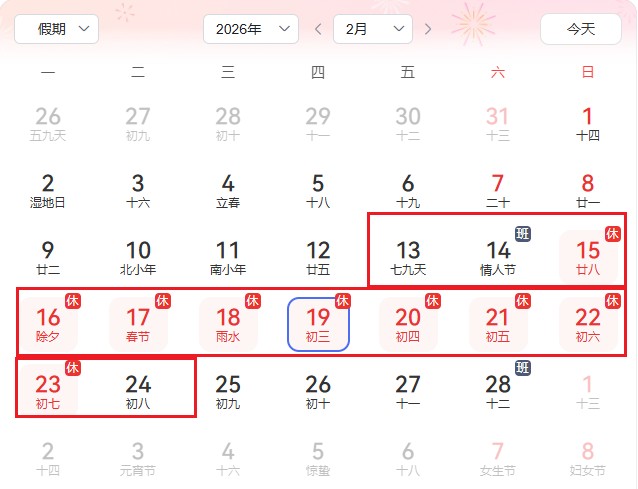
公司安排2026年2月13至2026年2月24日放假,年后初九开始恢复上班。12号(上午9点前)收到样本安排上机并出报告。
2月13号至24号暂停接收样本,节后初九开始上班处理样本。
杭州谷禾信息技术有限公司
2026年1月 20日