-

 CNAS L23010
CNAS L23010

 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业 二级病原微生物安全实验室
二级病原微生物安全实验室- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业 二级病原微生物安全实验室
谷禾健康
在开始本文之前,首先祝祖国母亲生日快乐!
是祖国的繁荣昌盛
成就我们无限可能
目光所致皆为华夏
五星闪耀皆为信仰


-正文-
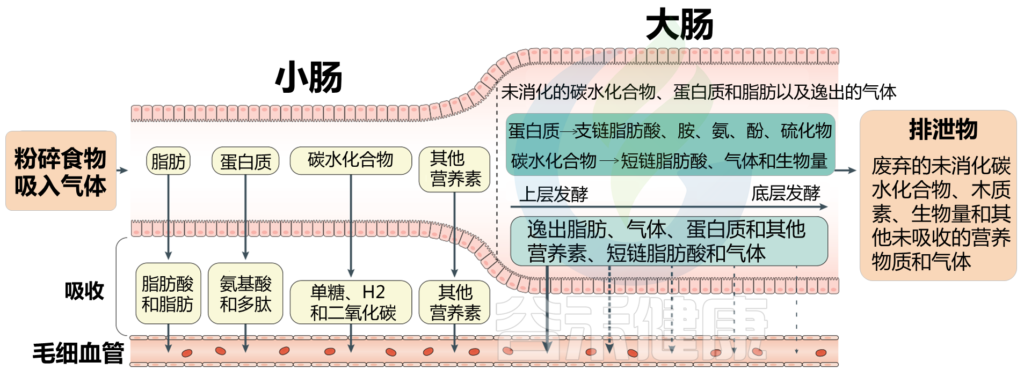
我们知道肠道微生物群影响健康。然而肠道的内部工作机制较为复杂,不仅有各种微生物,也包括它们产生的气体,比如二氧化碳、氢、甲烷和硫化氢以及各种微量气体,都是由肠道内的化学相互作用和微生物群产生的。
分析这些肠道气体及其对饮食变化的反应可以揭示肠道微生物群的产物和功能及其对人类健康的影响。更好地理解肠道内产生气体的复杂相互作用将提高我们预防、诊断、治疗和监测许多胃肠道疾病的能力。
本文主要概述了肠道气体的生理学及其在胃肠道疾病中的作用,并讨论了目前对肠道主要气体成分、其产生来源及其与饮食摄入的关系的认识。
了解肠道气体的生理学,无论是由微生物发酵还是化学活动产生的,是了解其对人类健康影响的第一步。
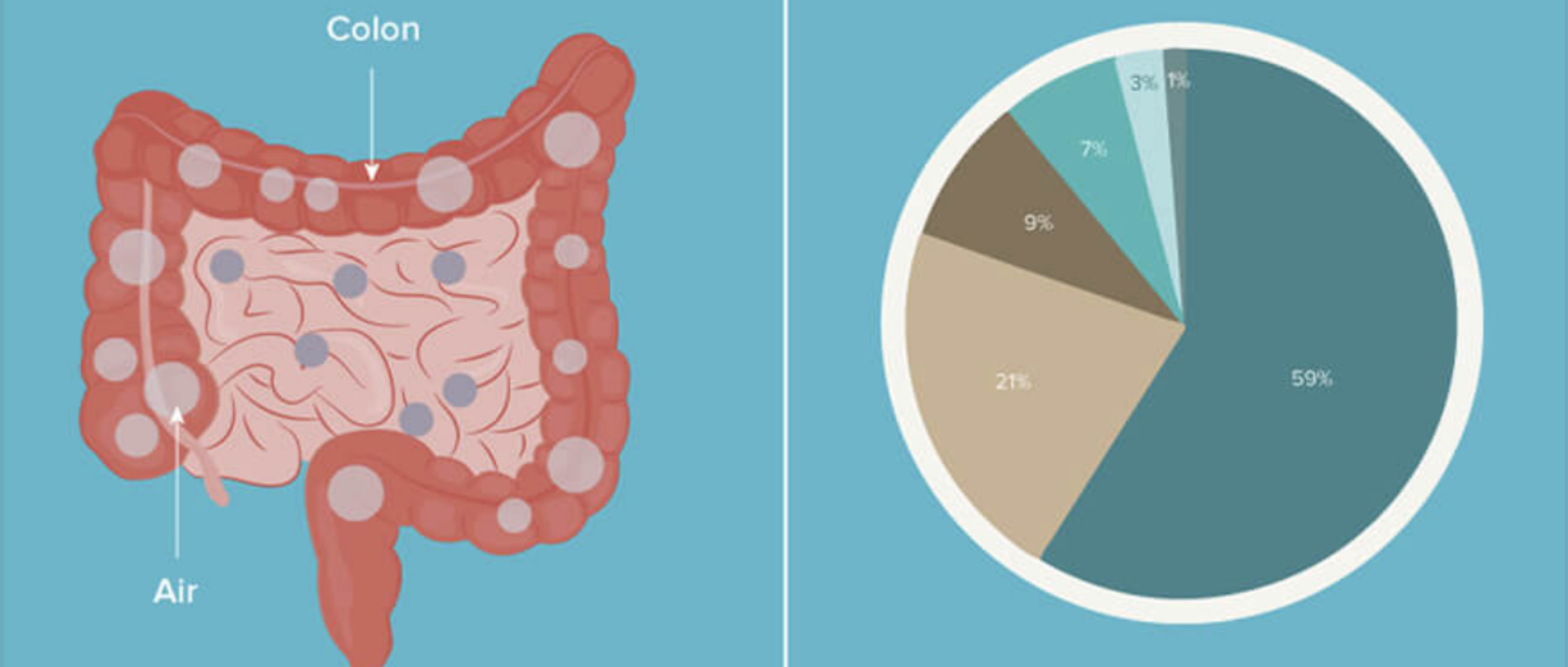
气体和食物沿着肠道的运输
» 气体成分
主要的肠道气体:
氮气(N2)、氧气(O2)、二氧化碳(CO2)、氢气(H2)、甲烷(CH4)
其他气体(以微量浓度存在):
硫化氢(H2S)、一氧化氮(NO)和含硫化合物等
» 肠道气体产生与分布
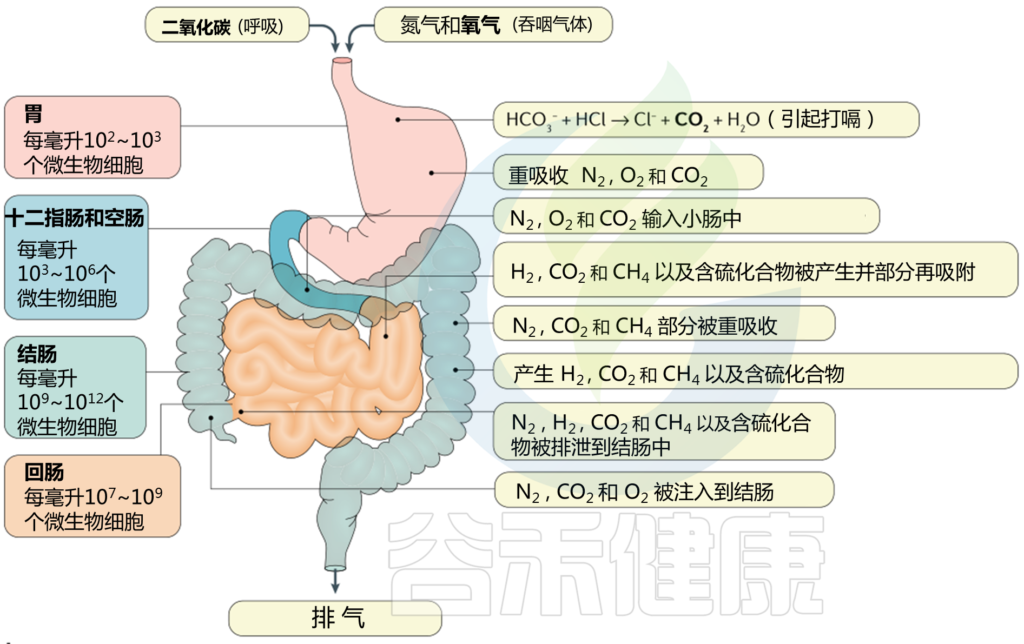
肠道气体可溶解在肠道的液体成分中,或以气体形式保留在顶空气体中。气体可能通过吞食进入肠道,也可能通过内部化学反应或细菌发酵产生。
在肠道中,氧气(完全)和N2(大部分)来自吞食的空气(尽管少量N2是通过结肠中蛋白质的微生物反硝化作用产生的)。由于N2在通过肠道期间未被吸收或代谢,因此其水平在整个胃肠道内保持合理稳定。
相反,肠道在整个长度内变得越来越无氧,随着管腔微生物数量的增加,氧气浓度逐步降低;因此,结肠中的氧气浓度非常低。(见下图 ↓↓)
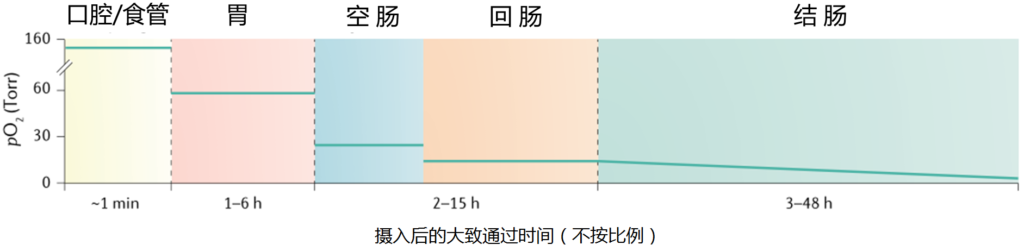
胃和小肠中的二氧化碳部分是通过化学反应产生的,虽然一些吞咽的呼吸性二氧化碳确实会与胃中的气体成分混合。
在近端小肠中,碳酸氢盐(HCO3−), 由胰腺分泌,与十二指肠胃食糜中的盐酸(HCl)发生化学反应,使管腔内pH值从酸性(~3)增加到接近中性(~7.4),并产生CO2.
在远端小肠和结肠中,肠道气体的产生主要来自微生物群。
氢气和二氧化碳:大多是在碳水化合物发酵过程中释放的(在较小程度上是内源性蛋白质和膳食蛋白质造成)
甲烷:是由大肠内古菌代谢二氧化碳和氢气产生的
硫化氢、含硫微量气体:是在蛋白质发酵过程中产生的,由硫还原菌还原硫酸盐和亚硫酸盐。
个体之间气体产生模式的巨大差异,并且与肠道微生物群的性质和可用底物有关。产气的底物主要是饮食,但粘液和糖蛋白的内源性分泌也起作用。
» 腔内微生物群和肠道气体
肠道微生物群是小肠和结肠产气的来源,其组成在个体内部和个体之间存在相当大的多样性。
虽然肠道微生物群的丰度比率差异很大,但五门涵盖了每人约160种的绝大多数,其中大多数属于拟杆菌门和厚壁菌门。
许多因素影响肠道微生物群的组成。由于功能和微环境的变化,胃肠道中的位置决定了存在的微生物的数量和类型。
胃中的微生物含量较低,且大多限制在每毫升约103个微生物细胞,而微生物数量沿肠道长度增加,在结肠中达到每毫升1010–1012个微生物细胞。
宿主因素,尤其是早期接触,在决定个体中发现的独特细菌组合方面起着关键作用。由于两岁后肠道菌群组成更加稳定,所以两岁前后的气体分布有很大的不同。所产生的气体的体积和分布在很大程度上取决于细菌和基质的丰度。
在健康人的胃肠道中,小肠近端产生的气体含量最低,而回肠和结肠的气体含量明显增加。
尽管微生物成分多样,但肠道微生物群总体具有类似于器官的关键功能特征。膳食碳水化合物的微生物发酵主要导致气体和短链脂肪酸的形成,后者直接影响宿主的健康。肠道微生物群还产生一系列具有不同功能的酶,包括复杂饮食基质的降解,从而影响气体分布。不同人群有不同情况,下一章节了解下病理生理学中的肠道气体。
阐明胃肠道症状和肠道气体之间的精确关系是未来生成气体特征库的关键,这些特征库可用于预防、诊断和监测胃肠道疾病。
◢ 胃肠道症状
胃肠道气体通常被认为是引起多种胃肠道症状的原因,这些症状可能发生在健康人或胃肠道疾病患者身上。
过度打嗝可能是由于非自愿吸入食道的空气排出(胃上打嗝)或吞咽的空气或气体饮料排出(胃打嗝)。
排气(胀气)是正常现象,但如果(或被认为)过多和/或恶臭,则可能是一个问题。过度胀气是吃了富含不易消化但可高度发酵的碳水化合物的食物的症状之一。
然而,一般来说,腹胀伴或不伴腹痛与产气过量无关,而是内脏敏感性改变的表现,或者在某些个体中,通过蠕动推进清除腔中气体的能力。
例如,在健康人和IBS患者中,服用乳果糖(一种不可消化的双糖)与结肠产气(如呼吸中H2水平显著增加所示)和腹胀有关,但只有IBS患者出现疼痛和腹胀症状。在摄入菊粉(一种高度发酵的益生元)后也有类似的发现,之后MRI显示的气态结肠扩张与IBS患者的疼痛有关,但在健康人中没有。
总的来说,排气(放P)的一些原因如下:
滞留在结肠中的未消化食物残渣会增加结肠细菌的发酵,从而产生气体。
吞咽空气的无意识习惯会在胃中积聚,但最终会以胀气的形式逃逸。肌肉发生变化后,空气会被排入胃中,这些变化允许吸入空气,并且试图通过打嗝排出空气的尝试无效。
这种胃酸过少的情况会抑制消化运动并引发消化紊乱,从而增加细菌发酵。在胃酸过低的情况下,未消化的食物颗粒会被某些细菌代谢,从而导致腹胀和排气。
食物在消化系统中停留的时间越长,细菌发酵和肠道气体产生过多的风险就越大。
产生的气体取决于肠道微生物群的健康状况。存在于消化系统中的支持性细菌和有害细菌的组成因人而异。这种微生物组组成会容易产生过多的气体。
肠道气体也可能对肠道产生直接影响,除了通过所谓的气体传递素效应增加肠道容积的作用外。例如,CH4在减缓胃肠运动中的作用或H2S在调节运动和内脏敏感性中的作用已被记录。
◢ 胃肠道疾病
微生物群和气体分子在胃肠道炎症和功能性疾病的发病机制中也具有重要作用。肠道缺氧和气态条件的调节会影响肠道微生物群及其功能。这种方法有可能改变短链脂肪酸的产生,短链脂肪酸具有多种稳态、抗炎、抗伤害和抗致癌作用。
粘膜通透性决定胃肠道内容物和体液交换的效率,并受食物分解和加工的影响。
在肠道中,氧化还原相关的气体物质是由化学反应、酶反应和细菌活动产生的,而这些反应反过来又改变了肠道粘膜的微生物生态系统。由于肠道的粘膜表面被各种微生物定植,异常的粘膜定植可导致局部粘膜和免疫系统的改变,进而导致胃肠道疾病的发病机制。
摄入可能改变微生物群落结构的膳食成分,单独或添加益生元和益生菌,可以潜在地逆转黏膜定殖肠道菌群的病理变化。
此外,肠粘膜的健康对热应激高度敏感,热应激可改变粘膜血流。血流量越高,通过粘膜溶解到血液中的气体越多,从而能够通过血流量更有效地清除肠道气体。通过使用提高气体渗透性的膳食添加剂,如肉桂,可以改善粘膜完整性。
IBD
与健康对照组相比,IBD患者的H2S、NO和VOCs(如氨、胺、戊烷、乙烷、丙烷、辛烯、甲基己烷和癸烯)水平有所升高。
已证明,饮食中的氨基酸组成和蛋白质消化率在改变IBD患者粪便中的VOC分布方面具有关键作用。此外,H2S通过其对平滑肌的作用参与结肠顺应性的调节,并与结肠伤害性感受和IBD有关。
肠道气体由化学或肠道微生物群产生,根据其类型,可能对肠道内环境稳定产生一系列影响。
当胰腺中含有HCO3–时,上消化道会产生CO2 与胃中的盐酸反应,大部分被吸收和/或排出。每次摄入食物后,在胃中或肠道其他部分(未被微生物群缓冲)都会产生CO2。
此外,CO2是远端小肠和结肠中碳水化合物细菌发酵过程中产生的主要气体。
二氧化碳是一种惰性气体,因此,除了通过体积相关效应进行机械刺激外,可能没有其他特殊作用。与其他气体相比,它被被动吸收到循环中的速度相对较快,然后通过呼气排出。
细菌发酵是肠道产生H2的主要来源,产氢气的菌:
罗氏菌属Roseburia,瘤胃球菌属Ruminococcus,真杆菌属Eubacterium,梭状芽孢杆菌属,拟杆菌属
当肠道厌氧菌使用未完全氧化的营养基质(如丙酮酸盐)作为末端电子受体,部分减少它们以产生能量时,就会发生发酵;还原的吡啶和黄嘌呤核苷酸被再氧化以维持氧化还原平衡(下图)。
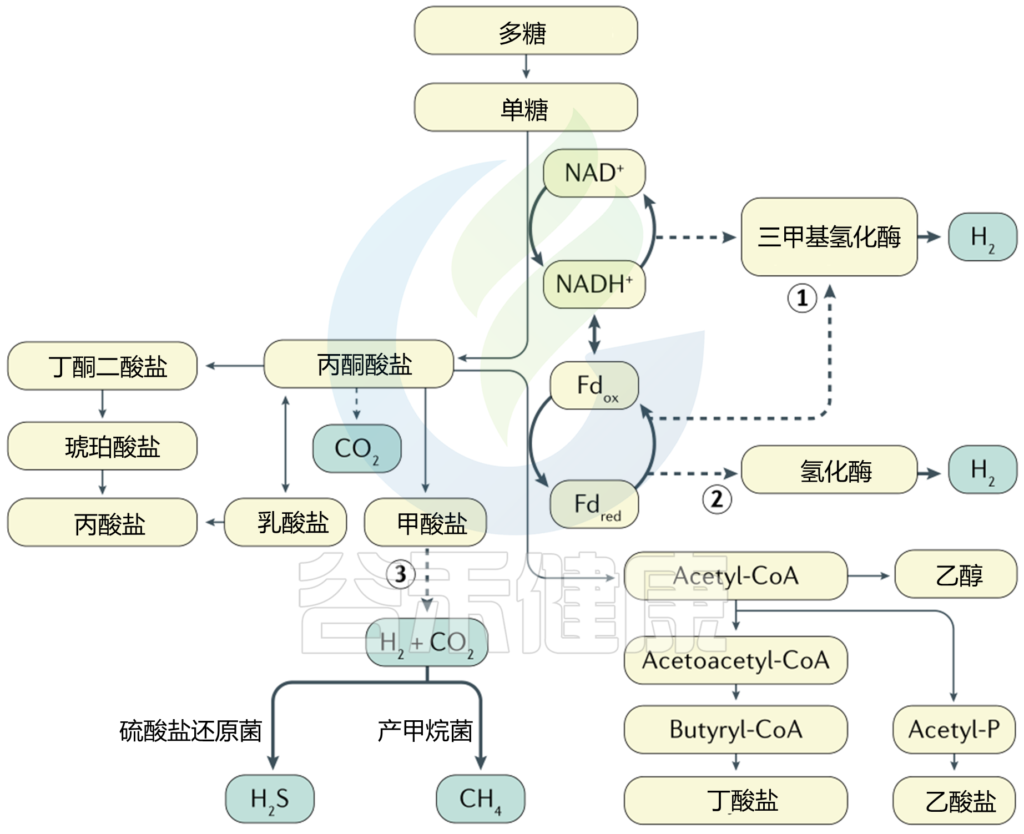
细菌产氢的其他途径包括丙酮酸裂解成甲酸盐或丙酮酸通过丙酮酸合成酶(如丙酮酸-铁氧还蛋白氧化还原酶)和氢化酶的活性生成。
氢气的产生通过三个主要机制进行调节。
1. H2分压的增加在热力学上限制了进一步发酵
2. H2的产生是通过粘膜吸收部分排泄来平衡的,然后通过肺排出体外。
3. 三类主要的氢营养(利用H2)微生物将H2转化为其他化学物质,如CH4(通过产甲烷古菌)、H2S(通过硫酸盐还原菌)、乙酸盐(通过还原性丙酮)。
▲ 微生物将氢气转化为甲烷
Methanobrevibacter smithii是人类肠道微生物组中数量最多的产甲烷菌,在粪便中的含量高达 109 CFU/g. 研究表明,在结肠样本中发现产甲烷菌的比例范围从 < 103到超过 109 CFU/g粪便。
最近,发现了一种新的甲烷形成途径,它使用细菌铁固氮酶从 CO2 中产生甲烷。纯铁固氮酶存在于多种微生物中,可能通过向厌氧代谢CH4的古菌和好氧嗜甲烷菌提供CH4,作为碳源和能量源,在调节微生物群落结构中发挥重要作用。
▲ 微生物将氢气转化为硫化物
在结肠中,产甲烷菌并不是唯一能够利用 H2的微生物群。硫酸盐还原菌能够还原硫酸盐作为其末端电子受体,形成硫化氢气体(H2S 或 HS–)。这种生化途径能够利用来自众多电子供体的还原当量,包括:乳酸、丙酮酸、乙醇、甲酸、短链脂肪酸和氨基酸。与产甲烷类似,硫酸盐还原为硫化氢气体是通过电子传递链进行的。
在脱硫弧菌属中发现了对 H2具有最高活性和亲和力的硫酸盐还原菌,其含量为104 –1011 CFU/g 湿重。
由于肠道微生物群中的硫酸盐还原菌和产甲烷菌竞争相同的 H2 ,决定性因素是用于硫化氢生产的硫酸盐的可用性。然而,从能量上讲,硫化氢生成是比甲烷生成更有利的反应。人类粪便样本的筛选试验表明,患者通常携带产甲烷菌或硫酸盐还原菌,但在H2丰富的情况下,有报告称两者都存在并具有活性。
在小鼠模型中比较产甲烷菌和H2S生成的体内研究证实,当产甲烷菌和H2S生成菌都存在时,并且在可用硫酸盐丰富的地方,H2S生成占主导地位,可行的产甲烷菌低于检测限值。通过减少可用硫酸盐的数量,甲烷生成已被证明是可回收的,这表明H2S生产和甲烷生成之间存在直接的反向联系,并证实硫酸盐还原菌在利用氢气方面胜过甲烷生成菌。
▲ 微生物将氢气转化为乙酸盐
利用肠道微生物发酵产生的H2的第三种机制是通过产乙酸作用,这是细菌利用分子H2作为还原当量来源将CO2还原为乙酸盐的途径。
WLP是一种多步骤酶促途径,利用八种还原当量和两种CO2形成醋酸盐(下图)。整个反应产生的能量甚至低于产甲烷或硫化物生成,ΔG′=−95kJ/mol. 在乙酰辅酶A最终底物水平磷酸化为乙酸盐期间,ATP由ADP生成。
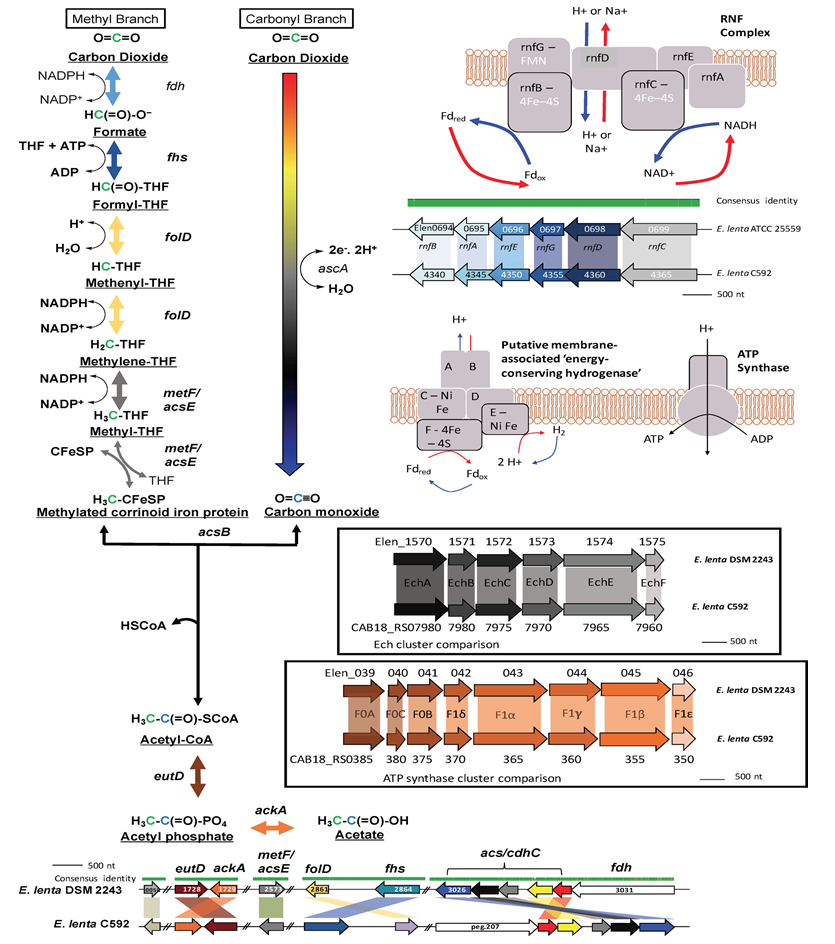
然而,最近有人提出,它可能是一种再生氧化吡啶核苷酸和Fd以维持细胞内氧化还原平衡的方法,而不是作为能量产生途径的乙酸生成。许多产乙酸细菌存在于电子受体稀缺的厌氧环境中,产乙酸细菌通过WLP将CO2用作电子受体的能力使其具有竞争优势。此外,大多数丙酮能够使用许多不同的电子受体和电子供体,使它们能够很好地适应其环境的能量和氧化还原要求。
微生物群产生的H2可通过饮食调节。例如,难以消化的膳食碳水化合物的发酵通常会增加H2的产生(在下一章节详述)。H2与其他气体的比率也可能因膳食发酵基质的可用性而不同。例如,纤维素和玉米麸皮可增加H2产量,但不影响CH4产量。
当产甲烷古细菌使用H2或甲酸盐作为电子供体还原CO2或甲醇时,会产生管腔CH4.
▌结肠产甲烷菌丰度的个体间差异很大
在人类胃肠道中,史密斯产甲烷短杆菌(Methanobrevibacter smithii)是负责将CO2和H2转化为甲烷的主要产甲烷菌。在呼吸中检测到甲烷导致将个体分类为“甲烷生产者”或“甲烷非生产者”。然而,由于在许多甲烷非产生者的粪便中仍发现产甲烷菌,这种分类似乎是人为的,甲烷的检测可能与呼吸中是否产生足够水平的甲烷有关。每克粪便中存在浓度>1×108菌落形成单位(CFU)的产甲烷菌已被认为是导致呼吸中甲烷可检测水平的必要条件。
▌CH4的产生也受饮食基质的数量和类型的影响
例如,在健康人群中,饮食中含有大量不易吸收和消化的可发酵短链碳水化合物(称为FODMAPs),导致CH4生成量显著减少,H2量相应增加(通过呼吸试验检测到)。这种效应在IBS患者中并不显著,IBS患者的气体总量减少与症状的产生高度相关。此外,摄入木聚糖和果胶会增加CH4的呼吸水平,而乳果糖则不会。
由于不同地区不同人群的肠道微生物群和饮食模式差异很大,因此被归类为CH4产生者的人口比例因种族而异也就不足为奇了。例如,一半的以色列成年人口和高比例的北非和中东个人是CH4生产商,而在西方人口中,只有三分之一的个人是CH4生产商。
在CH4生产者队列中,CH4产量随着年龄的增长而增加。事实上,在3岁之前没有检测到呼吸CH4,但从近0%增加到6%。以色列3-4岁年龄组为4%,8-14岁年龄组为18%,成人约为50%.
▌ CH4作为一种气体传递素也具有生理效应
事实上,CH4减缓结肠运动,与便秘有关。此外,由于CH4的存在将其“母体”分子(H2和CO2)占据的体积减少了约20%,因此减少了体积效应。CH4还抑制胃肠运动,这一点可以从它与慢传输便秘的密切关系中看出。
在临床环境中,诊断试验中呼气中CH4谱的价值比H2更具争议性,但在诊断试验碳水化合物吸收不良(例如,IBS患者)时,当个体对不可消化的碳水化合物没有产生可测量的H2呼气水平时,CH4谱在诊断试验中具有作用。因此,CH4测量通常与H2分析一起使用,以提高诊断的整体准确性。
硫化氢 (H2S)
★ 硫化氢的产生
人体内硫化氢的生物生成主要通过两条途径进行:内源性专门酶和作为肠道微生物群内微生物代谢途径的终产物或中间产物。
◥ 途径一:来自微生物群的硫化氢
肠道微生物组代谢半胱氨酸和蛋氨酸,以产生含硫结构、代谢物、信号分子和细胞能量学。硫酸盐还原细菌利用硫代硫酸盐生成硫化氢。在结肠腔内,几种细菌能够将Cys转化为H2S。
许多菌群(梭杆菌属、梭菌属、大肠杆菌属、沙门氏菌属、克雷伯氏菌属、链球菌属、脱硫弧菌属、肠杆菌属)通过半胱氨酸脱硫酶将Cys转化为H2S、氨和丙酮酸。
特别是,大肠杆菌和鼠伤寒沙门氏菌缺乏将蛋氨酸转化为半胱氨酸的酶。因此,这些物种不能使用蛋氨酸作为生长的来源。
相比之下,梭杆菌至少有四个基因编码催化H2S产生的不同酶。
微生物群的存在不仅与某些肠道(结肠和盲肠)中的游离H2S浓度较高有关,而且与血浆中的游离H2S浓度较高有关。
微生物群的存在对CSE活性和Cys生物利用度有影响
一种可能的解释是,细菌产物泄漏到血液中,并诱导产生H2S的酶。另一种可能性是,在常规小鼠的血液和组织中发现的H2S生物等效物的一部分是由肠腔中的硫代谢细菌产生的。此外,个人饮食的特点和组成可能会对肠道中的硫铝酸盐浓度产生影响。
◥ 途径二:内源性产生H2S的酶
哺乳动物内源性产生硫化氢由三种酶进行:
CBS、CSE、MST
注:CBS 胱硫醚β合酶;CSE 胱硫氨酸γ-裂解酶;
MST 3-巯基丙酮酸硫转移酶
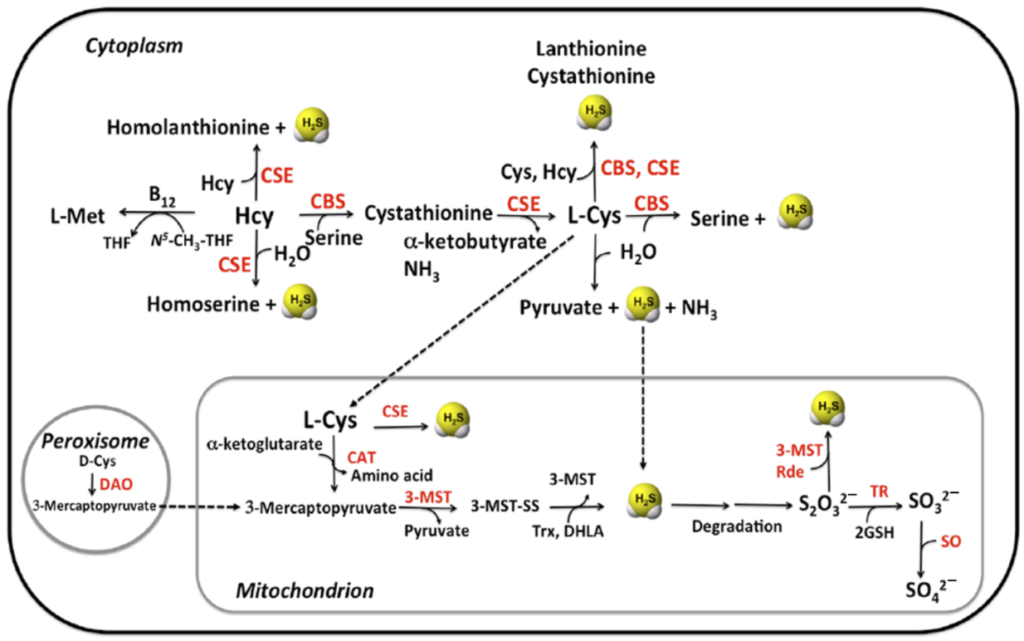
CBS和CSE是转硫途径中依赖吡哆醛5’磷酸(PLP-)的酶,可将同型半胱氨酸转化为半胱氨酸。细胞中所有可用的同型半胱氨酸都来源于蛋氨酸分解代谢。
CBS催化丝氨酸(Ser)被同型半胱氨酸取代形成半胱硫氨酸。在下游,CSE在α,γ-消除反应中催化半胱硫氨酸转化为半胱氨酸,α-酮丁酸和氨。CBS和CSE参与多种副反应,产生信号分子硫化氢(H2S)作为副产物。在CBS和CSE催化的反应中,L-Cys和L-Hcy生成胱硫醚都会生成H2S。此外,CSE还分别催化底物组合Cys/Cys和Hcy/Hcy生成的产物镧硫氨酸和高镧硫氨酸,这两种底物组合都会生成H2S。硫醚代谢物镧硫氨酸和高镧硫氨酸被认为是CBS缺乏患者产生硫化氢的生物标志物。
第三种PLP依赖性途径涉及具有半胱氨酸转氨酶(CAT)活性的天冬氨酸转氨酶(AAT)。与PLP非依赖性酶3-巯基丙酮酸硫转移酶(MST)结合,可生成硫酸盐硫,还原后可释放硫化氢。
一旦进入循环,H2S主要通过肾脏以游离或结合硫酸盐的形式排出。
◥ 其他途径
红细胞还通过无机和有机多硫化物产生硫化氢据估计,红细胞在恒定浓度为170µmol/(L cells)/min的时候可产生H2S。人类红细胞通过葡萄糖支持、巯基和谷胱甘肽依赖反应将大蒜衍生的有机多硫化物转化为H2S。
线粒体酶半胱氨酰tRNA合成酶(CARS2)在催化Cys转化为Cys-per/多硫化物物种中起主要作用。
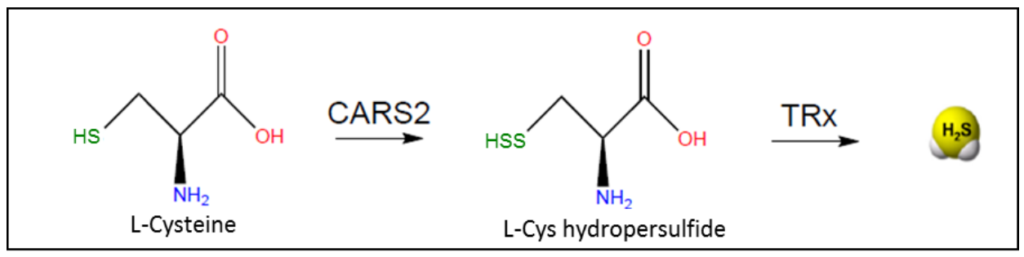
用NADPH作为电子源,硫氧还蛋白和GSH酶机制可有效地还原Cys/多硫化物。因此,这些非标准酶途径可能有助于细胞产生大量H2S。
血液中也显示H2S的非酶生成,对前体代谢物半胱氨酸具有特异性。该反应对全身基础H2S浓度的影响很小,但在以释放铁升高为特征的病理条件下(如溶血性和出血性疾病)可能是有害的。
★ 硫化氢的分解
硫化氢的分解代谢主要通过线粒体中的氧化途径发生,同时产生ATP。H2S氧化发生在多种哺乳动物组织中,包括肝脏、肾脏、心脏和结肠细胞。
硫化氢的氧化途径
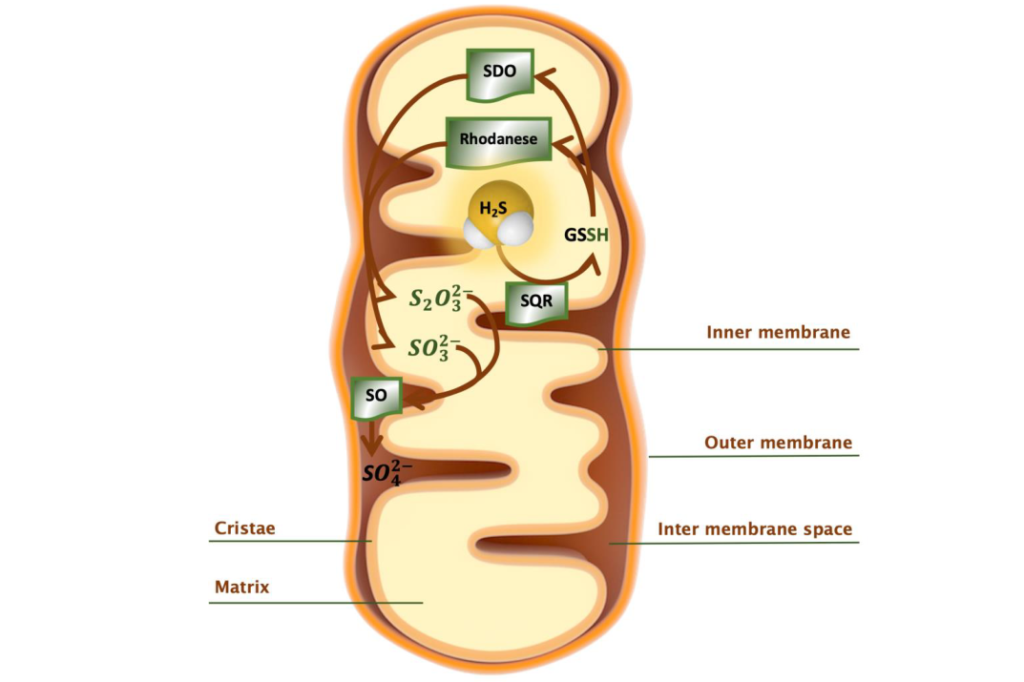
在线粒体基质中,SQR将H2S氧化为GSSG,并将衍生电子转移到呼吸链,从而减少辅酶Q并驱动ATP合成。GSSG通过SDO进一步氧化形成亚硫酸盐。或者,硫氰酸酯催化GSSG,形成硫代硫酸盐阴离子。下一步,产物亚硫酸盐和硫代硫酸盐阴离子被亚硫酸盐氧化酶氧化为硫酸盐。
SQR=硫化物醌氧化还原酶,SDO=过硫化物双加氧酶,
SO=亚硫酸盐氧化酶, GSSH=谷胱甘肽过硫化物
★ 与硫化氢有关的饮食影响
研究表明,在饮食中缺乏维生素B6(一种CSE和CBS辅助因子)可使粪便中的硫化氢减少50%。饮食中维生素B6的缺乏显著降低了粪便中硫化氢的水平,可能是由于抑制了结肠组织中硫化氢的酶合成。
在六周缺乏维生素B6的饮食后,粪便中的硫化氢水平恢复到了与对照组相同的水平。这表明,通过增加硫酸盐还原菌活性,无菌小鼠肠道中硫化氢的生成向非酶途径转移。
营养食品多酚(浆果提取物)可将H2S氧化为有效的抗氧化多硫化物,从而引发细胞保护作用。该机制表明,多酚会自动氧化为相应的半醌类物质,然后与H2S反应生成巯基,最终形成多硫化物和硫代硫酸盐。
★ 硫化氢对肠道的影响:双刃剑
H2S在低浓度下具有保护作用,但在高浓度下具有有害作用。这一悖论反映在:
一方面,H2S作为一种气体递质,可能有助于调节肠道的细胞过程,如炎症、运动、上皮分泌和痛觉。
H2S可用于治疗结肠炎,起到减少失调和帮助粘液层重建的作用。
而另一方面,H2S是一种有毒气体,在高浓度下被视为健康危害。
它对人体组织具有潜在毒性,尤其是与NO一起存在时,由于其损害β-氧化和脂质及蛋白质合成的综合作用,但它具有重要的细胞信号特性。
★ 硫化氢对心血管的影响
一项实验检测了结肠中H2S可用性的增加对大鼠血流动力学的影响。Na2S(一种H2S供体)结肠内给药可产生强效、持久的降压作用,其持续时间是此前报道的肠外输注后的数倍(>90分钟)。
有趣的是,高血压大鼠的动脉血压比正常大鼠下降更明显。此外,新霉素组大鼠硫代硫酸盐和磺胺硫含量显著降低,对Na2S的降压反应有增强趋势。这些数据表明,肠道来源的H2S可能产生全身效应,结肠H2S稳态的变化可能与高血压有关。
肠道细菌产生的硫化氢在血压控制中发挥作用
该研究中低血压的作用最可能是由于外周血管舒张和心率降低。相反,小肠血流的局部变化不是H2S依赖性低血压的可能原因。结肠内H2S供体的血流动力学影响伴随着门静脉而非外周血H2S氧化产物水平的升高。因此,肠道H2S的全身效应似乎是由一些肝脏依赖机制或结肠H2S对肠道神经系统的影响产生的。
硫化氢(H2S)对心血管作用的推测途径
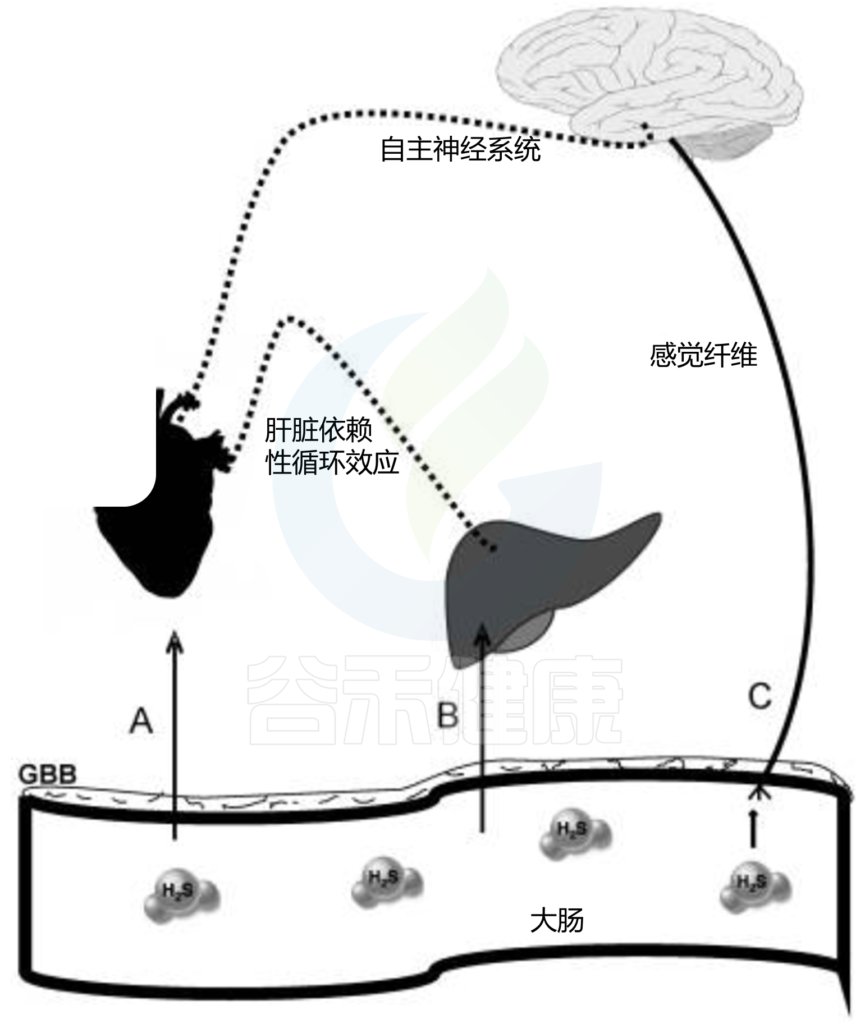
(A) H2S穿过肠道-血液屏障(GBB),绕过肝脏(直肠丛),并靶向心脏和血管。
(B) H2S穿过GBB,影响与循环系统稳态相关的肝功能。
(C) H2S刺激肠神经系统的感觉纤维,该纤维投射到大脑中枢,通过自主神经系统控制循环系统。
一氧化氮是研究最多的生物递质之一。
目前有明确证据表明,肠道细菌通过厌氧反硝化作用也会产生NO,溃疡性结肠炎患者的NO水平升高。
首先是硝酸盐亚硝酸盐NO途径。硝酸盐被共生口腔细菌还原为亚硝酸盐,肠道细菌通过非酶酸性还原或亚硝酸盐还原酶进一步还原亚硝酸盐。最后,肠粘膜表达NO合酶,该合酶合成NO将l-精氨酸转化为l-瓜氨酸。NO的循环效应主要是通过各种组织的酶促生成来评估的。
研究还发现,益生菌乳酸杆菌和双歧杆菌通过降低肠道pH值,增加非酶亚硝酸盐还原,在肠道NO生成中发挥作用。相比之下,脱硫弧菌将NO转化为硝酸盐。
许多研究表明内源性产生的NO广泛参与自主神经系统和肠道神经系统,以及其在胃肠道稳态中的功能。
NO在动脉和静脉中形成,有助于控制血流。证据表明NO作为炎症介质促进胃肠平滑肌的非肾上腺素能非胆碱能抑制。
NO也可能在肠道中作为神经递质或神经调节剂发挥作用,而内源性NO可能介导胃粘膜保护和调节胃粘膜血流。刺激NO生成的药物已被用于预防急性粘膜损伤,虽然目前还不知道这种治疗策略的进展。
许多研究表明一氧化碳在循环系统中所起的作用的重要性。与 NO 和 H2S 类似,已发现 CO 具有血管舒张和心脏保护作用。
CO 是在血红素加氧酶 (H2O) 催化的反应中产生的。诱导型 H2O (HO-1) 和组成型 H2O (HO-2) 主要用于哺乳动物组织中的内源性 CO 产生。在胃肠系统中,CO 可能由表达 HO-1 的肠粘膜产生。此外,据报道,肠道微生物群(大肠杆菌)也表达 HO 同源物并在小鼠中诱导 HO-1 的结肠表达。肠道源性 CO 对体循环的可能影响仍有待阐明。
哺乳动物的肠道内由几种细菌和胃肠道组织形成大量的氨(NH3)。事实上,肠道菌群脲酶对尿素的降解(约7 g/天)是体内约50%的总氨氮的来源。人体肠道中NH3的产生速率为4-10 g/天。未结合的NH3或随粪便(~5 ~ 25 μg/g)排出,或被肠道菌群转化为氨基酸,或被GBB吸收。健康人血浆游离NH3浓度约为35 μmol/L。循环NH3可以在肝脏中转化为尿素或谷氨酰胺,也可以随尿液排出(2-3毫克/天)。
与高氨血症相关的肝脏疾病和相关的神经毒性作用已经得到了很好的描述,肝功能衰竭患者经常使用抗生素治疗,如新霉素,以清除肠道污染,减少细菌产生NH3。
一些证据表明,NH3可能会影响循环系统的控制。例如,有报道称NH3对离体大鼠心脏有正性肌力作用。此外,已有研究发现,健康成人吸入NH3可导致脑血管扩张,但不影响动脉血压。最后,肝衰竭患者血浆NH3水平升高。
以上是对几种主要的肠道气体的产生及其功能的阐述,下一章节我们来了解饮食对气体分布的影响。
迄今为止,习惯性饮食已被证明对肠道微生物群的组成有重大影响,因此,长期内对气体分布有重大影响。此外,已经证明,通过使用大幅度增加的饮食基质的短期饮食变化,肠道微生物生态系统的调节是可能的。
食物特别是其组成的大量营养素(膳食碳水化合物、蛋白质和脂肪)及其个体微量营养素——通过多种机制改变胃肠道分泌、吸收和运动,直接或间接地影响肠道。
这些效应可能是非特异性机械感受器反应的结果,继发于管腔拉伸。然而,这种机械感受器反应也可能是由食物的体积效应、管腔含水量(由于食物的渗透效应而增加)、食物中包含的气体(例如碳酸饮料)、吞食的空气(N2没有被吸收或代谢,有证据表明它主要向远端移动)或腔内产生气体(化学反应或细菌发酵)引起的。
摄入食物后肠道扩张也会导致内脏-躯体反射,这可能会改变前壁肌肉组织和膈肌的激活,并相应改变腹腔容积,可能导致胃肠道症状。然而,肠易激综合征发病机制中气体产量和容量增加的直接作用仍存在争议。
在与食物摄入相关的潜在气体来源中,只有从空气中吞下的N2和肠道发酵产生的气体可能会使肠道膨胀。虽然吞咽的空气在临床上很难调节,但在大多数IBS患者中,选择易发酵、缓慢吸收或难以消化的短链碳水化合物含量较低的食物会导致气体生成减少(如呼吸中H2排泄减少所示)和症状改善。
♨ 食物的选择影响排气
食物选择对每个肛门通过的肠道气体量有重大影响.例如,当在饮食中限制发酵底物时,健康个体和功能性肠道疾病患者每天的总排气量平均<300ml。当健康志愿者被喂食富含可发酵、不可消化基质(包括每天200克烤豆)的饮食时,这一体积增加到每天500–1500毫升。
食物的选择也会影响肠胃的气味,这通常与蛋白质发酵的程度以及随后产生的含硫微量气体有关。
因此,通过调节大量营养素摄入(膳食碳水化合物、蛋白质和脂肪)进行饮食操作可以改变肠道气体分布。下面详细了解这三大类营养物质。
膳食碳水化合物是不同长度和大小的碳基分子。在一般的西方饮食中,估计每天有40克的膳食碳水化合物在小肠中逃逸宿主酶消化和吸收,到达肠道的发酵部分。
☞ 小肠
在小肠中,只有单糖被直接吸收,吸收过程的效率因单糖而异。例如,葡萄糖和半乳糖被迅速吸收,而果糖被缓慢吸收,除非存在至少等摩尔浓度的葡萄糖。当过量的果糖负荷过大、通过小肠的速度过快或小肠吸收糖的能力减弱时,就会发生果糖吸收不良,这是一种生理上的正常事件。从淀粉酶介导的淀粉消化过程中释放的膳食双糖(如乳糖和蔗糖)和低聚糖需要在吸收前通过刷状缘酶消化。因此,当这些酶的活性较低时,它们的吸收就会失败,如乳糖酶或蔗糖酶-异麦芽糖酶缺乏。
其他低聚糖(如低聚果糖和低聚半乳糖)和非淀粉多糖由于小肠中缺乏合适的水解酶而不能消化,因此根本不能被吸收。
☞ 结肠
未被吸收的碳水化合物到达富含细菌的结肠,在那里可以发酵。一般来说,短链碳水化合物的发酵速度比长链碳水化合物(即聚合度(DP)大于10的碳水化合物)更快,在较短的时间内产生更多的气体。
非淀粉多糖的溶解度和发酵性差异很大。碳水化合物结构的这种异质性,以及由此导致的小肠对碳水化合物的处理和在结肠中的命运,导致了三个功能概念:FODMAP、纤维和益生元概念。
✔ FODMAP
FODMAP是短链(DP<10)、缓慢吸收或难以消化的碳水化合物,易于发酵。在肠道中,FODMAP具有渗透活性,导致小肠管腔含水量增加,并在近端结肠中发酵,产生气体。因此,由于过量含水量或气体产生的体积效应,摄入FODMAP与管腔扩张有关。
当内脏过敏出现时,摄入FODMAP可能会出现腹痛、腹胀和排便习惯改变等症状。过量摄入FODMAPs可促进H2的生成而不是CH4,减少FODMAPs可促进CH4的生成而不是H2,可能与H2的生成速率有关。减少FODMAPs的饮食摄入现在是缓解此类症状的关键循证饮食疗法。
✔ 纤维
纤维有许多定义,但通常包括DP大于2的吸收不良或难以消化的碳水化合物,因此包括许多FODMAP。
纤维在胃肠道松动中起着关键作用,特别是通过扩大细菌数量和在管腔内保留水分,以及通过增加管腔传输速度,引起结肠内容物膨胀。
然而,纤维在溶解度、发酵性和链长方面具有高度的异质性,因此它们对结肠生理学有着广泛的影响。

了解纤维在大肠中的发酵位置是了解如何在临床实践中使用它们的关键
纤维发酵的位置最终取决于其链长以及是否与其他类型的纤维一起施用。如果单独服用,容易发酵的低聚糖和高度发酵的长链纤维,如某些形式的抗性淀粉和燕麦麸皮,大部分在近端结肠中发酵。
相反,缓慢发酵的纤维,如麦麸纤维,在整个结肠内发酵。建议摄入缓慢可发酵或不可发酵的纤维(如不可发酵的可溶性纤维梧桐和甲基纤维素)以及易发酵的纤维(如高直链淀粉),以将纤维发酵的位置向远端移动,改善大肠各区域的发酵分布。
✔益生元
益生元被定义为“宿主微生物选择性利用的具有健康益处的底物”。目前,益生元主要局限于低聚果糖、低聚半乳糖、低聚木糖、菊糖和抗性淀粉,在较小程度上,因为大多数纤维导致细菌的非特异性增殖。益生元的代谢(发酵)主要释放气体。
因此,为了达到临床效果(如促进松弛、减少气体形成或改变肠道微生物群的相对组成),对膳食碳水化合物和肠道气体的调控需要考虑多个方面。
大部分膳食蛋白质被上消化道的内在酶分解并吸收,估计每天有12-18克剩余的未消化残余蛋白质到达结肠。
在结肠中,这些蛋白质的代谢通过两条主要途径进行。细菌可以分解结肠中的蛋白质,并利用产生的氨基酸生物合成结构蛋白质或其他细菌蛋白质。或者,蛋白质可以由细菌发酵,这一过程产生的代谢物比碳水化合物发酵产生的代谢物更加多样化。例如,在缺乏氧气的情况下,梭菌属和梭杆菌属(严格或兼性厌氧细菌)可以利用氨基酸进行发酵。该工艺的产出包括短链脂肪酸、支链脂肪酸(如异丁酸、2-甲基丁酸和异戊酸)、H2、CO2和氨以及少量H2S、甲硫醇、酚、醇和有机酸的生产。
暴露于膳食蛋白质会改变微生物组的多样性和组成。然而,增加蛋白质输送到肠道微生物群的主要影响似乎是改变参与氨基酸降解的微生物的代谢特征。
在蛋白质发酵产生的气体代谢物中,粪便中的氨浓度可能被用作有价值的蒸汽生物标记物,因为氨浓度会随着蛋白质摄入量的增加而增加(10–30 mmol/l)。
H2S也是蛋白质发酵的一个潜在有用的生物标志物,因为它的管腔浓度在食用富含肉类的食物的人身上增加,并且在体外新鲜排出的粪便中添加半胱氨酸会显著刺激H2S的产生。然而,了解蛋白质发酵对宿主生理学和病理生理学的影响仍处于早期阶段,进一步了解需要良好控制的人类干预研究。
由于多达7%的膳食脂肪在小肠中未被吸收,肠道微生物群暴露于相当数量的膳食脂质中。迄今为止,只有少数研究调查了膳食脂肪对人体肠道微生物群的影响,几乎没有一项研究关注气体的产生。
据报道,高脂肪饮食的摄入与人类粪便短链脂肪酸水平和双歧杆菌数量的大幅下降有关;然而,在这项研究中,碳水化合物的摄入没有得到控制。
在小鼠中,高脂肪饮食引起的微生物群变化似乎是可逆的,不太可能归因于肥胖表型本身。在小鼠中,高脂肪饮食也会引起肠道的低度炎症。这种变化可能会改变气体剖面,但这一假设需要未来的具体研究。
目前,对肠道气体分布及其随时间和整个肠道长度的变化的了解还较为肤浅,主要是因为大多数肠道气体测量方法要么是间接的,要么是高度侵入性的。了解这些气体分布将增强我们对食物和药物对人体影响的认识。肠道气体的评估可能是未来具有较高诊断和/或监测价值的疾病生物标记物。
通过肠道菌群检测,了解与气体产生相关菌的代谢,也可以侧面反映肠道气体的情况,在预防、诊断和干预疾病的所有阶段可将其正确利用作为参考。
1. 缓慢而有意识地进食
快速进食而不正确咀嚼食物会刺激消化系统并产生气体。
有意识地进食不仅可以进入放松状态,从而确保健康的消化,还可以:
2. 两餐之间多喝水
通过在两餐之间充分补水,确保食物通过消化道的适当运动。水合作用是清除体内废物的关键,因为它可以润滑纤维,使其膨胀,防止其在消化道中停滞,可以最大限度地减少发酵并减少气体形成。
虽然全天喝大量的水是必不可少的,但请尽量避免在进餐时喝水。与进食相结合,水会稀释为代谢食物而分泌的消化液,并可能导致问题。
3. 使用消除饮食
消除饮食可以帮助你深入了解哪些食物可能导致胀气症状。这个过程将使你从饮食中去除引发气体的常见食物罪魁祸首,包括:
4. 解决胃酸水平
胃酸在消化的各个阶段都是必不可少的。胃酸负责:
如果没有适当的胃酸水平,我们就无法对食物进行消毒、杀死不需要的微生物以及有效地消化和同化食物。这会给整个消化系统带来更多压力,并导致微生物发酵。
微生物发酵的天然副产物是气体。当我们不能有效地代谢硫时,就会放出非常“有味道”的气体。
5. 消化酶
消化酶可以帮助分解可以喂养细菌并产生气体的食物成分。这些消化酶可以以补充剂的形式存在,并且存在于许多未加工和发酵的食品和饮料中,例如苹果醋。
这些酶会分解糖类、核酸、氨基酸和脂肪酸,使它们更容易被人体吸收。例如,胀气和腹胀是胰酶缺乏的症状,胰酶可以得到多种消化酶的支持,例如淀粉酶、脂肪酶和蛋白酶 。
6. 益生菌
超过 500 种不同种类的细菌存在于人类肠道中,益生菌可用于补充健康细菌的浓度。益生菌的天然食物来源:椰子水开菲尔、康普茶、酸奶以及酸菜和泡菜等发酵蔬菜。
改善肠道菌群的平衡对于减少有害微生物的占有率至关重要,有害微生物会导致过多气体积累。
7. 天然植物类
茴香种子、豆蔻果实、肉桂、姜、芹菜种子、甘草、香芹籽、薄荷、橙皮
8. 镁
镁可能是最容易被忽视的矿物质之一,你可能需要它来改善消化并减少气体的产生和排出。镁对于支持 300 多种酶过程的活动至关重要,包括沿胃肠道作为肌肉松弛剂。
镁在健康的肠道蠕动中起着重要作用,通过刺激消化系统中的肌肉和通过镁移动发酵食物,有效地有助于减少气体积聚。
主要参考文献:
Tomasova L, Konopelski P, Ufnal M. Gut Bacteria and Hydrogen Sulfide: The New Old Players in Circulatory System Homeostasis. Molecules. 2016;21(11):1558. Published 2016 Nov 17.
Hylemon PB, Harris SC, Ridlon JM. Metabolism of hydrogen gases and bile acids in the gut microbiome. FEBS Lett. 2018 Jun;592(12):2070-2082.
Y ao, C. K., Muir, J. G. & Gibson, P . R. Review article: insights into colonic protein fermentation, its modulation and potential health implications. Aliment. Pharmacol. Ther. 43, 181–196 (2016).
Gottlieb, K., Wacher, V., Sliman, J. & Pimentel, M. Review article: inhibition of methanogenic archaea by statins as a targeted management strategy for constipation and related disorders. Aliment. Pharmacol. Ther. 43, 197–212 (2016)
Kalantar-Zadeh K, Berean KJ, Burgell RE, Muir JG, Gibson PR. Intestinal gases: influence on gut disorders and the role of dietary manipulations. Nat Rev Gastroenterol Hepatol. 2019 Dec;16(12):733-747.
Ong, A. M. L. et al. Diaphragmatic breathing reduces belching and proton pump inhibitor refractory gastroesophageal reflux symptoms. Clin. Gastroenterol. Hepatol. 16, 407–416.e2 (2018).
Blachier F, Beaumont M, Kim E. Cysteine-derived hydrogen sulfide and gut health: a matter of endogenous or bacterial origin. Curr Opin Clin Nutr Metab Care. 2019 Jan;22(1):68-75.
Myszkowska J, Derevenkov I, Makarov SV, Spiekerkoetter U, Hannibal L. Biosynthesis, Quantification and Genetic Diseases of the Smallest Signaling Thiol Metabolite: Hydrogen Sulfide. Antioxidants (Basel). 2021 Jul 1;10(7):1065.
Betesh AL, et al. Is achlorhydria a cause of iron deficiency anemia? Am J Clin Nutr. 2015 Jul; 102(1): 9-19
Szabo C. A timeline of hydrogen sulfide (H2S) research: From environmental toxin to biological mediator. Biochem. Pharmacol. 2018;149:5–19.