-

 国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子
国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子

 二级病原微生物安全实验室
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
谷禾健康
产前抑郁
在围产期健康管理中,产前抑郁(antenatal depression, AND)正在受到越来越多关注。相比产后抑郁,产前抑郁过去长期处于相对被低估的位置,但它的发生率并不低。
既往荟萃分析显示,无论以抑郁症状还是临床诊断作为标准,产前抑郁的全球患病率均接近20%。
从临床结局来看,产前抑郁已被证明与多种不良母婴结局相关,包括流产风险增加、早产、低出生体重等。
近几年,研究视角开始进一步延伸到儿童早期发育。越来越多研究关注母亲孕期抑郁症状对子代神经发育的影响,例如婴儿认知、语言、运动、问题解决能力和社会行为等发育维度。
但目前相关结果并不完全一致。一个重要原因可能是不同研究关注的抑郁暴露时间窗口不同。过去不少研究集中在孕晚期,而对孕早期、孕中期,或整个孕期持续性抑郁症状的影响关注不足。
但胎儿脑发育贯穿整个孕期。只看孕晚期,可能会低估早期情绪风险对子代发育的影响。
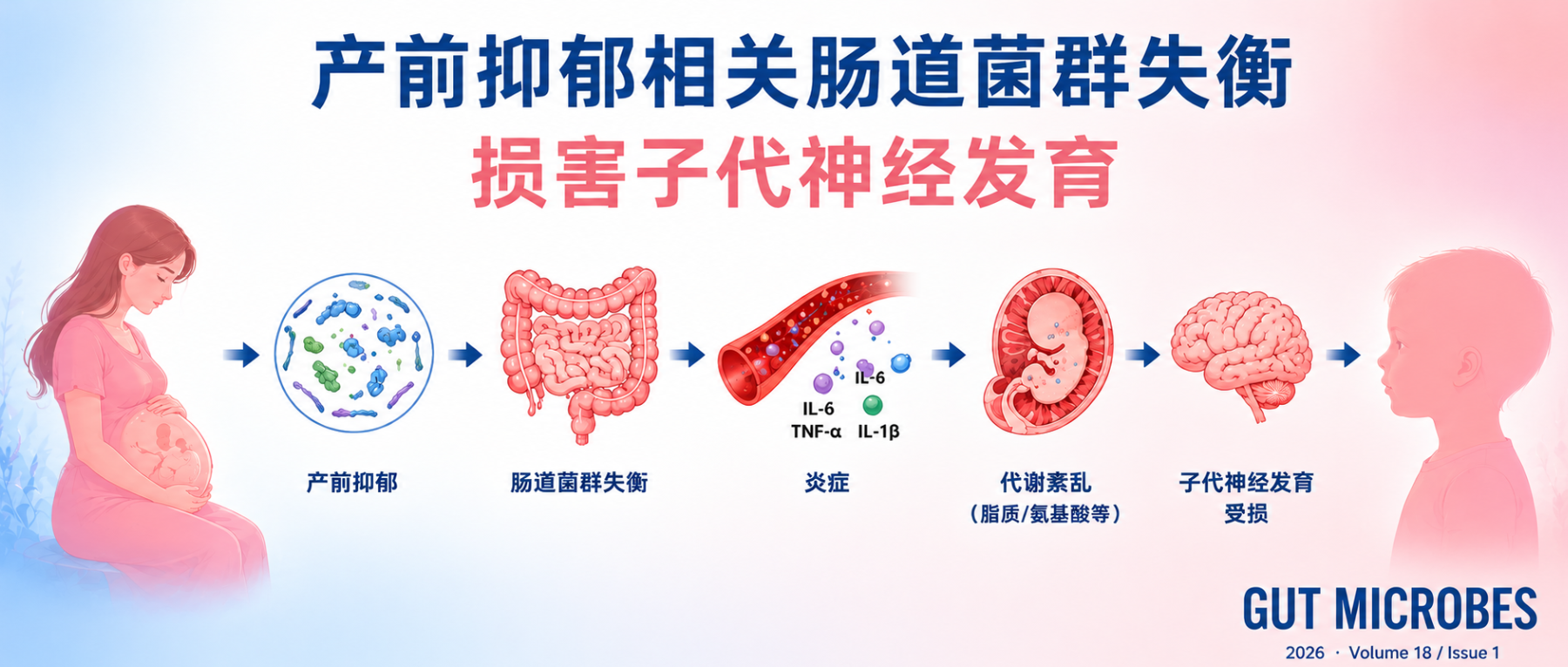
更关键的是,产前抑郁究竟如何影响子代神经发育,目前仍缺乏清晰机制。过去的解释更多集中在 HPA 轴、皮质醇、炎症反应、胎盘功能和母体代谢状态等方面。而近年来,肠道菌群逐渐成为一个新的机制切入点。
已有研究显示,抑郁和焦虑状态可改变肠道菌群组成;孕期焦虑、抑郁症状也与菌群多样性下降及特定菌群丰度变化有关。但这些研究多为横断面或小样本研究,较少追踪整个孕期菌群动态,也难以回答菌群变化是否真正参与了子代神经发育异常。
这里就出现了一个很关键的问题:
产前抑郁是否会通过改变母体肠道菌群,影响胎儿脑发育和出生后的神经认知表现?
近期,来自上海复旦大学妇产科医院和上海交通大学的研究团队发表在《Gut Microbes》的一篇研究,正是从这个问题切入。
研究人员结合多中心前瞻性孕妇队列、嵌套病例对照菌群分析、无菌小鼠粪菌移植实验以及丁酸补充实验,系统验证了孕期抑郁症状与婴儿神经发育受损之间的关系。
产前抑郁与婴儿早期神经发育受损相关,主要表现为婴儿问题解决和社交能力的下降,而且孕早、中、晚期的抑郁都会导致这一问题。机制上可能是通过母体肠道菌群失衡,尤其是产丁酸菌减少,导致肠屏障受损、炎症增强及胎儿脑代谢异常,从而影响神经发育。对母亲和婴儿的菌群检测显示,母婴之间肠道菌群传递较少。在小鼠实验中,粪菌移植实验表明,仅母体抑郁相关肠道菌群即可导致后代学习记忆能力下降。而丁酸补充可以改善炎症、代谢异常及后代认知表现。
对于围产期健康管理而言,这意味着孕期情绪风险背后,可能存在值得更早关注的微生态信号。
这项研究的设计比较完整,先在人群中确认关联,再通过菌群分析寻找机制线索,最后用动物实验验证因果可能。
◆ 人群队列:追踪孕期抑郁与婴儿发育
研究首先纳入 2053 名孕妇,在孕早期、孕中期和孕晚期分别使用 EPDS 量表评估产前抑郁症状,并在婴儿 3、6、12 月龄时随访神经发育表现。
随访重点包括:
注:EPDS量表是目前全球公认且广泛使用的围产期抑郁筛查工具。
◆ 菌群分析:寻找可能的机制线索
随后,研究者从队列中开展嵌套病例–对照分析,匹配出 135 名产前抑郁风险孕妇和 135 名对照孕妇,并对 504 份孕期母体粪便样本进行 16S rRNA 测序,追踪孕早、中、晚期肠道菌群变化。
在菌群分析上,研究除了关注哪些菌升高或降低,进一步将相关菌群归纳为共丰度模块,并结合功能预测,观察这些菌群变化可能对应哪些代谢通路异常。
同时,研究还收集了 290 份婴儿粪便样本,用于判断产前抑郁是否会影响婴儿出生后的菌群建立,以及母婴菌群传递是否参与神经发育异常。
◆ 动物实验:验证菌群是否具有因果作用
在人群研究之外,研究者进一步使用无菌小鼠进行粪菌移植(FMT):将产前抑郁风险女性和健康女性的菌群分别移植给无菌孕鼠,观察:
◆ 干预验证:丁酸是否是关键线索
最后,研究加入丁酸补充实验,用来验证产丁酸菌减少是否可能是关键干预点。
编辑
这样的设计超越了单纯的相关性分析,进一步挖掘了菌群是否参与其中及相关机制,以及是否存在可干预的代谢节点。
【 队列研究 】
在2053名女性的队列研究中,326名(15.9%)至少有一次EPDS ≥10 在妊娠期间(AND高危组),1634人(79.6%)没有(对照组)。
【 婴儿随访 】
对1岁以下的婴儿进行神经发育评估。AND高危组婴儿在3个月和6个月时的问题解决任务得分较低(P = 0.039,0.040)。
编辑
◐ 母亲孕期抑郁越明显,婴儿问题解决能力越弱
尤其是在 3 月龄时,EPDS 评分较高的母亲,其婴儿在问题解决能力和社交能力方面得分更低。
研究者还使用重复测量 GEE 模型进行分析,结果同样显示:无论是在孕早期、孕中期还是孕晚期,母亲 EPDS 评分越高,婴儿的问题解决能力越差。
◑ 嵌套病例–对照研究,再次验证
与对照组相比,存在产前抑郁症状的母亲,其婴儿在 3 月龄时问题解决能力更低,在 6 月龄时社交能力也更弱。
编辑
从这部分结果看,产前抑郁症状与婴儿早期神经发育表现下降存在关联,尤其集中在问题解决能力和社交能力等维度。
更重要的是,这种关联并不只出现在孕晚期。孕早期、孕中期和孕晚期 EPDS 评分升高,都可能与婴儿后续问题解决能力下降有关。也正因为如此,研究下一步需要回答:这种影响是否只是情绪状态本身带来的结果,还是与孕期母体肠道菌群变化有关?
在人群队列中,研究已经看到产前抑郁症状与婴儿早期神经发育表现下降有关。接下来,研究者进一步想了解:这种关联背后,母体肠道菌群是否发生了可识别的变化?
先区分“抑郁阶段”和“非抑郁阶段”
经过质量控制,504份来自不同孕期的母体粪便样本可供分析,覆盖孕早期、孕中期和孕晚期。
这里研究者并没有简单把孕妇分成“抑郁组”和“非抑郁组”,而是根据采样当时的 EPDS 评分,把样本进一步分为三类(下图d)。
编辑
这个分组很重要,可以帮助判断,菌群变化到底只出现在EPDS升高的阶段,还是在产前抑郁风险人群中具有一定持续性。
校正孕期阶段后,菌群差异仍然存在
孕期本身会影响肠道菌群。孕早期、孕中期和孕晚期,母体激素、免疫和代谢状态都在变化,菌群结构也会随之调整。
因此,研究人员采用PERMANOVA 分析评估了宿主因素在孕期对母体肠道菌群的影响。
结果显示,妊娠期是最显著的影响因素,其次为 产前抑郁症状。
编辑
具体来看,孕早期与孕中期之间的菌群差异显著(P < 0.05),孕早期与孕晚期之间的菌群差异更显著(P < 0.001),但孕中期与孕晚期之间差异不显著。
这说明,研究产前抑郁相关菌群变化时,必须先排除孕期自然变化的影响。
在校正妊娠阶段后,PERMANOVA 分析显示,AND-D 组与对照组之间(FDR P = 0.019),以及 AND-N 组与对照组之间(FDR P = 0.010)均存在显著差异。
编辑
分孕期来看:
也就是说,产前抑郁相关菌群改变并不只局限在某一次 EPDS≥10的抑郁阶段,即使某一阶段评分暂时低于阈值(也就是说非抑郁状态),菌群结构仍然可能与健康对照不同。
这提示一个关键信号:产前抑郁相关的母体菌群变化,更像是一种持续性的微生态特征,而不是短期情绪波动带来的偶然变化。
核心变化:产丁酸菌减少和代谢通路紊乱
接下来,研究者进一步分析到底是哪些菌群发生了改变。
他们将所有微生物群划分为 50 个共丰度菌群模块 CAGs,这种分析方式不是只看单个菌属升高或降低,而是观察一组具有相似变化趋势和潜在功能关联的菌群模块。
在校正孕前BMI、教育水平、孕周、产妇年龄、妊娠糖尿病、抗生素、益生菌、GBS感染等因素后,研究最终识别出 15个显著改变的CAGs。
编辑
其中,AND-D 和 AND-N 组中多个产丁酸菌相关模块持续下降,包括Eubacterium、Ruminococcaceae、Faecalibacterium、Oscillospiraceae 等。
这些菌群与短链脂肪酸,尤其是丁酸的产生密切相关。丁酸不仅是肠上皮细胞的重要能量来源,也参与维持肠屏障完整性、调节炎症反应和免疫稳态。
换句话说,产前抑郁相关菌群失衡的核心,指向了一个更具体的功能问题:产丁酸能力下降。
关于一些重要产丁酸菌属详见谷禾之前的文章:
肠道核心菌属——优/真杆菌属(Eubacterium),你为什么要关心它?
肠道核心菌属——普拉梭菌(F. Prausnitzii),预防炎症的下一代益生菌
肠道重要菌属——颤螺菌属 (Oscillospira),它为什么如此神秘?
与此同时,以 Streptococcus 为主的CAG36在产前抑郁相关样本中升高。
结合既往研究中Streptococcus与炎症、抑郁状态之间的关联,这一变化也提示母体肠道微生态可能向更促炎的方向偏移。
功能层面:脂肪酸和氨基酸代谢通路异常
菌群组成变化之外,研究还进一步做了功能预测。
功能预测还显示,抑郁阶段和非抑郁阶段都存在多条代谢通路异常,包括:
编辑
多不饱和脂肪酸与胎儿脑发育、神经细胞膜结构和炎症调节有关;氨基酸代谢则与神经递质合成、抗氧化能力和基础代谢密切相关。
这部分结果进一步说明:产前抑郁相关菌群失衡,可能不仅是菌群结构变化,还涉及母体肠道代谢功能的改变。
但这还只是母体菌群层面的发现。下一步更关键的问题是:这些菌群变化,是否真的和婴儿后续神经发育表现有关?
在明确产前抑郁孕妇存在持续性母体菌群失衡之后,研究进一步探讨了:这些孕期菌群变化是否会影响婴儿神经发育。
婴儿自身菌群:并非主要解释因素
研究者首先分析了婴儿出生后的肠道菌群,共纳入 290份婴儿粪便样本进行 16S rRNA 测序。
结果显示,影响婴儿肠道菌群组成的主要因素是采样时间、分娩方式和喂养方式,其中采样时间的影响最大。
编辑
这说明婴儿出生后肠道菌群本身处于快速建立和变化阶段,且明显受到出生方式、喂养方式等因素影响。
母亲是否存在产前抑郁症状,并没有显著影响婴儿出生后的菌群整体组成。
编辑
这表明婴儿自身菌群的波动很大,很难作为母亲情绪与婴儿神经发育之间关联的中介因素。
母婴菌群垂直传递证据有限
分析母婴共享的 ASVs 后发现,整体共享数量不多。只有一个Clostridium sensu stricto 1 相关 ASV 在产前抑郁风险组中共享率较高(P=0.034),并与婴儿神经发育评分存在一定关联趋势。但该 ASV 在极少数样本中出现,不足以解释整体神经发育差异。
编辑
由此可见,母婴菌群垂直传递并不是本研究中主要的机制线索。
孕期母体菌群更关键
研究者在校正母亲年龄、孕前 BMI、孕周、教育水平、妊娠糖尿病、抗生素和益生菌使用等因素后发现:
编辑
也就是说,这些菌群模块丰度越低,婴儿在认知、问题解决、个人-社会能力等方面的表现可能越弱。
编辑
婴儿出生后的菌群不构成主要中介。母婴垂直传递有限,婴儿肠道菌群受时间、分娩方式和喂养方式影响更大。
孕期母体菌群失衡可能是核心机制。特别是产丁酸菌相关模块下降和代谢通路功能扰动,与婴儿早期认知和社交能力存在正相关。
产前抑郁可能通过影响母体肠道菌群代谢环境,改变胎儿脑发育微环境,从而在婴儿早期表现为认知和社交能力下降。
为什么要做FMT验证?
因为孕期情绪状态、饮食、睡眠、家庭环境、遗传背景、妊娠并发症等因素,都可能同时影响母体菌群和婴儿发育结果。
因此,需要可以尽量把“菌群因素”从复杂的人群背景中单独提取出来。
FMT模型怎么做?
研究者从产前抑郁风险女性和健康对照女性中选择粪菌供体,将其菌群分别移植给无菌雌鼠,之后让小鼠妊娠,并观察孕鼠和后代的变化。
编辑
动物实验主要分为三组:
• Control_M组:接受健康对照女性菌群;
• AND_M组:接受产前抑郁风险女性菌群;
• AND + butyr_M组:接受产前抑郁风险女性菌群,同时在孕期补充丁酸。
编辑
FMT模型是否成功建立?
从菌群组成看,移植后小鼠的肠道菌群能够较好地反映对应供体来源。
注:E18.5,指胚胎第18.5天,接近小鼠足月分娩前阶段。
编辑
这说明FMT模型基本建立成功,后续观察到的母体和子代差异,可以进一步用于评估AND相关菌群的潜在作用。
产前抑郁相关菌群影响了后代学习记忆表现
研究者在后代小鼠12周龄时进行 Morris水迷宫实验,用于评估学习和空间记忆能力。
编辑
结果显示,与Control_M组相比,AND_M组后代在水迷宫测试中表现出明显的认知受损特征:
编辑
这些结果说明,接受产前抑郁风险女性菌群的孕鼠,其后代在学习记忆能力上出现下降。
值得注意的是,开放场实验中,AND_M组后代在活动距离、中心区停留时间、进入中心区次数等指标上没有显著差异。这提示AND相关菌群在该模型中更明显影响的是认知和学习记忆相关行为,而不是广泛诱导焦虑或抑郁样行为。
总的来说,在无菌小鼠FMT实验中,研究者进一步看到:仅仅移植AND相关母体菌群,就可以诱导后代出现认知行为异常。
这说明,产前抑郁相关菌群不只是一个伴随现象,而可能是参与子代神经发育异常的重要中介因素。
在人群研究中,产前抑郁症状、母体菌群失衡和婴儿神经发育下降之间存在关联。而在无菌小鼠实验中,研究者进一步看到,仅仅移植AND相关母体菌群,就可以诱导后代出现认知行为异常。
这说明,产前抑郁相关菌群不只是一个伴随现象,而可能是参与子代神经发育异常的重要中介因素。既然产前抑郁相关菌群本身能够影响后代认知表现,下一步就需要了解:它是通过什么路径影响胎儿脑发育的?
从动物实验和多组学结果来看,机制主要集中在几条连续线索上:
编辑
母体肠道:紧密连接蛋白下降,肠屏障受损
研究者首先观察孕鼠肠道屏障状态。结果显示,接受 AND 相关菌群的孕鼠,结肠组织中多个紧密连接蛋白表达下降,包括 claudin-1、ZO-1 和 ZO-2(下图a–d)。
编辑
这些蛋白是维持肠上皮屏障完整性的关键结构。它们下降,说明肠道屏障稳定性变差,肠腔内微生物相关成分或炎症刺激物更容易影响母体系统环境。
也就是说,AND 相关菌群进一步影响了母体肠道屏障功能。
母体和胎儿脑炎症信号同步增强
在肠屏障受损的同时,研究者还检测到母体炎症水平升高。
与对照组相比,AND_M 组孕鼠血清中的 TNF-α 和 IL-1β 水平均升高(下图e、f)。
编辑
这提示 AND 相关菌群可能诱导母体系统性低度炎症反应。
更关键的是,这种炎症信号并没有停留在母体端。胎儿小鼠脑组织中同样出现炎症相关变化:ELISA 结果显示,AND_M 组胎儿小鼠脑内 TNF-α 和 IL-1β 水平升高(下图j、k)。
编辑
转录组分析也支持这一点。AND_M 组胎儿小鼠脑中多条炎症相关通路被激活,包括 TNF 信号通路、IL-17 信号通路、Toll-like receptor 通路、NOD-like receptor 通路、趋化因子和细胞因子相互作用通路等(下图a、c)。
编辑
这说明,AND 相关菌群可能通过“肠屏障受损—母体炎症增强—胎儿脑炎症微环境改变”这一方式,参与胎儿脑发育异常。
胎儿脑代谢:PUFA 和氨基酸代谢异常
除了炎症,研究者还对胎儿小鼠脑组织做了代谢组分析。
结果显示,AND_M 组胎儿小鼠脑内代谢谱与对照组明显不同(下图g)。其中,一个突出的变化是多不饱和脂肪酸(PUFA)相关代谢通路下降,包括亚油酸、α-亚麻酸、花生四烯酸等相关代谢路径(下图k)。
编辑
具体代谢物上,AND_M 组胎儿脑中 anandamide、PC(14:0/20:4) 等脂质相关分子下降(下图h、i)。这些分子与神经细胞膜结构、突触功能、神经炎症调节都有关系。
编辑
同时,氨基酸相关代谢也受到影响。例如 L-alanine 水平下降(图j),并伴随半胱氨酸、甲硫氨酸、牛磺酸等相关代谢路径异常。
胎儿脑组织:突触和脂质代谢相关基因变化
转录组结果进一步补充了这一机制链。
在 AND_M 组胎儿脑中,脂质代谢相关通路和突触相关通路下调,包括谷氨酸能突触、5-羟色胺能突触等相关信号(图c)。
编辑
同时,关键基因如 Pla2g4b、Pla2g4d 表达下降(图e、f)。
编辑
综合来看,AND相关菌群可能通过多个相互关联的环节影响胎儿脑发育:
这些变化共同指向一个结果:胎儿脑发育所依赖的免疫和代谢环境被扰动,最终可能表现为后代认知功能下降。
既然前面的菌群结果反复指向产丁酸菌减少,而动物模型中也观察到肠屏障和炎症异常,那么如果补充丁酸,是否能够部分逆转这些异常?
研究者进一步设置了 AND + butyr_M 组:在接受 AND 相关菌群移植的基础上,于小鼠孕期补充丁酸,观察能否逆转部分异常表型。
丁酸补充提高了胎盘丁酸水平
结果显示,与 AND_M 组相比,AND + butyr_M 组胎盘丁酸水平明显升高。
编辑
这说明丁酸补充改变了模型中的丁酸供应状态,也为后续观察行为、炎症和代谢变化提供了基础。
后代认知行为得到部分改善
在 Morris 水迷宫实验中,丁酸补充组后代的认知表现有所改善。
与 AND_M 组相比,AND + butyr_M 组后代的小鼠学习记忆表现有所改善:
编辑
说明,丁酸补充可以部分缓解 AND 相关菌群导致的学习记忆能力下降。不过需要注意,这里是部分改善,并不是完全逆转所有行为异常。
胎儿脑代谢异常,出现恢复趋势
小鼠代谢组结果进一步显示,丁酸补充后,AND_M 组中被下调的部分 PUFA 代谢和氨基酸代谢通路出现恢复趋势。
编辑
PUFA 代谢与神经细胞膜结构、突触功能和炎症调节有关;氨基酸代谢则与神经递质、抗氧化能力和脑发育基础代谢密切相关。因此,这一结果提示,丁酸可能通过改善胎儿脑内脂质和氨基酸代谢环境,参与神经发育保护。
炎症信号和肠屏障损伤也有所缓解
小鼠实验的转录组结果显示,丁酸补充后,小鼠胎儿脑内部分炎症相关基因表达下降,例如 ccl21b、cd14、ms4a2 等;同时,脂质代谢相关基因 Pla2g4b 表达升高。
ELISA 结果也显示,AND + butyr_M 组胎儿脑内TNF-α 和 IL-1β 水平下降。
丁酸补充的影响并不只体现在胎儿脑代谢上。
在小鼠母体端,丁酸补充还部分恢复了结肠紧密连接蛋白表达,包括 claudin-1、ZO-1、ZO-2,并降低了母体血清中的炎症因子水平。
编辑
这些结果提示,产丁酸菌减少及丁酸不足,可能是 AND 相关菌群影响子代神经发育的重要机制节点。
更准确地说,产前抑郁相关菌群可能通过削弱产丁酸能力,影响母体肠屏障和炎症状态,并进一步扰动胎儿脑内代谢环境;而丁酸补充能够部分缓解这些变化。
但这仍然是动物实验结果,不能直接推导为临床建议。对于临床和健康管理场景,更稳妥的理解是:围产期微生态干预可以重点关注产丁酸菌、膳食纤维摄入、短链脂肪酸代谢能力和肠屏障状态。
过去在围产期健康管理中,情绪问题、肠道问题、营养问题和婴儿发育问题,往往是分开处理的。
这项研究提示,这些问题之间可能存在更深的连接。产前抑郁症状并不只是一个心理状态,它可能伴随母体肠道菌群重构,尤其是产丁酸菌减少,并进一步影响肠屏障、炎症反应、胎儿脑内多不饱和脂肪酸和氨基酸代谢。
也就是说,孕期情绪风险背后,可能有一部分生理和微生态基础可以被更早观察到。
围产期情绪管理可以前移,并做风险分层
这项研究显示,孕早期、孕中期和孕晚期的EPDS评分升高,都可能与婴儿后续神经发育表现下降有关。
临床产前抑郁筛查不应只集中在孕晚期或产后。孕早期建档时,就应该开始关注情绪风险。
同样是情绪风险,有些孕妇可能主要受压力、睡眠、家庭支持等因素影响;也有一部分孕妇,可能同时伴随肠道微生态、炎症和营养代谢方面的问题。
产丁酸菌,值得成为孕期微生态评估中的重点观察方向。在该研究中,产丁酸菌Eubacterium、Faecalibacterium、Ruminococcaceae、Oscillospiraceae 等相关菌群模块在产前抑郁风险样本中下降,并且与婴儿神经发育评分存在关联。
编辑
<来源:谷禾健康肠道菌群检测数据库>
实际评估中,核心菌属中这些产丁酸菌以及丁酸盐、丙酸盐、乙酸盐等短链脂肪酸指标,都可以作为观察产丁酸能力和肠道代谢支持状态的线索。
如果这类指标已经显示出不足,即使孕妇还没有明显情绪问题,也可以提示其肠道生态和代谢环境可能存在潜在风险。后续管理就可以更早关注她的饮食结构、膳食纤维摄入、肠道症状、睡眠状态以及炎症相关风险,而不是等到情绪或消化问题明显后才处理。
编辑
<来源:谷禾健康肠道菌群检测数据库>
这类信息不一定直接指向某种疾病,但可以帮助管理者更早识别需要重点关注的人群。但筛查只是第一步,还可以对情绪风险孕妇做进一步分层。
如果孕妇已经出现EPDS评分升高或明显情绪风险,肠道菌群检测可以辅助判别:这个情绪风险背后,是否同时存在肠道微生态和代谢环境异常。
若同时观察到产丁酸菌减少、短链脂肪酸能力不足、肠屏障压力或炎症相关指标异常,那么管理重点除了在心理支持上,也需要同步考虑饮食结构、膳食纤维摄入、肠道症状改善、睡眠和炎症风险管理。
编辑
<来源:谷禾健康肠道菌群检测数据库>
面向孕期健康的转化思路
肠道菌群检测适合嵌入临床已有的孕期评估流程,作为情绪筛查、产科评估和营养评估之外的补充信息,帮助医生判断是否需要进一步联合营养、肠道或生活方式干预。
在健康管理中,把菌群检测放进孕期服务流程中,比如孕早期建立肠道菌群基线状态,孕中晚期结合饮食、睡眠、肠道症状和菌群变化做再评估。
作为产品开发来说,这篇研究提供了一个更具体的思路。有转化价值的方向,可能是把产品功能落到产丁酸能力、短链脂肪酸生成、肠屏障支持和营养代谢这些环节上,设计检测、干预方案。
编辑
未来的围产期健康管理,会越来越强调前置识别。
不是等到婴儿发育问题出现后再回头寻找原因,也不是等到孕妇症状明显后才开始处理,而是在孕期就尽量识别那些可能影响母体状态和胎儿发育环境的信号。
情绪状态是信号,肠道菌群也是信号,营养和炎症状态同样是信号。
这篇研究让我们看到,这些信号之间可能存在联系。肠道菌群不是全部答案,但它可以成为理解孕期母体状态、进行风险分层和制定管理策略的重要入口。
主要参考文献
Zhou, Fangyue, et al. “Antenatal depressive symptoms impair offspring neurodevelopment by inducing maternal gut microbiota dysbiosis during pregnancy.” Gut Microbes 18.1 (2026): 2672188.

谷禾健康
传统治疗的局限
目前焦虑障碍的一线治疗方案主要包括药物治疗(如SSRIs、SNRIs)和认知行为治疗(CBT)。但它们的局限性也同时也存在:例如起效慢(通常需要2-4周)、有效率有限、存在副作用、停药后复发风险较高。
正是因为这些局限,才迫切地需要寻找新的治疗靶点和干预策略。而微生物-肠-脑轴的发现,为焦虑障碍的防治开辟了一个充满希望的新方向。
在深入讨论肠道菌群与焦虑的关系之前,我们先快速了解一下微生物-肠-脑轴:
微生物-肠-脑轴
这是一个连接肠道微生物、肠道和大脑的复杂通信网络,允许肠道微生物与中枢神经系统之间进行双向的信息交流。
简单来说:大脑的活动可以影响肠道菌群的组成,反过来,肠道菌群的变化也可以影响大脑的功能和行为。
这个通信网络主要通过四大通路实现:
了解了肠道和大脑之间存在密切的联系,接下来看看在真实的焦虑患者中,他们的肠道菌群真的发生了改变吗?如果有,具体是哪些细菌发生了变化?
在比较了焦虑障碍患者和健康对照者的粪便菌群组成,发现:
广泛性焦虑障碍 · 菌群特征
Chen等人在《Journal of Affective Disorders》上发表了一项针对36名广泛性焦虑障碍(GAD)患者的研究。研究结果表明,GAD患者的肠道菌群与健康对照者存在显著差异。
编辑
α多样性↓↓
α多样性也就是菌群的丰富度和均匀度:GAD患者的菌群丰富度显著降低,OTU(可操作分类单元)数量明显少于健康对照。这意味着,焦虑患者的肠道菌群多样性更低,而菌群多样性通常被认为是肠道健康的重要标志。
厚壁菌门↓↓ 拟杆菌门和梭杆菌门↑↑ 产丁酸菌↓↓
Faecalibacterium、Eubacterium rectale、Sutterella、Lachnospira、Butyricicoccus等产丁酸的有益菌属在健康对照中富集,而在GAD患者中减少。
Ruminococcus gnavus、Fusobacterium等潜在的促炎菌属在GAD患者中显著增加。
doi.org/10.1016/j.jad.2019.08.014
社交焦虑障碍 · 菌群特征
社交焦虑障碍(SAD)是另一种常见的焦虑障碍,但关于SAD患者菌群特征的研究相对较少。直到2023年,爱尔兰科克大学的Butler团队在《Translational Psychiatry》上发表了一项重要研究,首次采用全基因组鸟枪法测序技术对31名SAD患者和18名性别、年龄匹配的健康对照者进行了研究。
编辑
菌群特征:Anaeromassillibacillus↑↑
研究再次证实了焦虑患者存在独特的菌群特征:
在整体菌群组成(β多样性)上,SAD组与对照组存在显著差异;
编辑
doi.org/10.1038/s41398-023-02325-5
菌群功能:天冬氨酸降解I ↑↑
除了菌群组成的差异,这项研究还首次发现了SAD患者菌群功能的改变。在功能水平上,SAD患者的肠道代谢模块”天冬氨酸降解I“显著升高。天冬氨酸是一种重要神经递质前体,其代谢通路的异常可能通过改变神经递质水平影响大脑功能。
肠道屏障受损
研究人员还有一个重要的发现,SAD患者的肠道屏障完整性受损。肠道屏障受损意味着肠道通透性增加,细菌成分和代谢产物更容易进入血液循环,引发免疫激活和炎症反应,进而影响中枢神经系统功能。
跨研究特征:不同精神疾病共有的菌群标记
越来越多的研究表明,很多精神疾病,包括抑郁症、双相情感障碍、精神分裂症、自闭症等,都存在不同程度的菌群失调。而且,不同疾病之间似乎存在一些共同的菌群特征。
例如,2023年一项纳入了1054名参与者的队列研究发现,产丁酸菌的减少是多种精神疾病共有的特征,包括抑郁症和焦虑症。产丁酸菌的减少与炎症水平升高、神经递质合成异常密切相关。
编辑
doi:10.1001/jamapsychiatry.2021.2573
这些跨研究的共同特征提示我们:菌群失调可能不是某一种精神疾病的特有表现,而是多种精神疾病共享的潜在病理机制。这也解释了为什么焦虑和抑郁常常共病——它们可能有着共同的生物学基础,而肠道菌群失调就是其中之一。
可能有人觉得,相关性不等于因果性,这是科学研究中最重要的原则之一。横断面研究虽然清晰地展示了焦虑障碍与肠道菌群失调之间的关联,但无法确定因果方向:
孟德尔随机化:遗传学的自然实验
孟德尔随机化被称为自然的随机对照试验,是目前在观察性研究中确立因果关系最有力的方法之一。
它的基本原理很简单:根据孟德尔遗传定律,我们的基因是在出生时随机分配的,不受后天环境因素的影响。如果某个基因变异与我们感兴趣的暴露因素(如某种细菌的丰度)相关,而这个基因变异又与结局(如焦虑障碍)相关,那么我们就可以推断暴露因素与结局之间存在因果关系。
突破性发现:9种与焦虑因果相关的细菌属
2024年,上海交通大学的Li等人在《BMC Psychiatry》杂志上发表了一项具有里程碑意义的孟德尔随机化研究,首次在人类中系统地评估了肠道菌群与焦虑障碍之间的因果关系。
这项研究整合了多个大型GWAS数据集,共纳入了18,340名参与者的全基因组数据,分析了数百种细菌属的丰度与焦虑障碍风险之间的关系。通过严格的敏感性分析和多方法验证,研究人员最终确定了9种与焦虑障碍存在因果关联的细菌属。
编辑
这些发现通过了多种敏感性分析的验证,包括MR-Egger截距分析(检测水平多效性)、MR-PRESSO分析(检测异常值)和留一法分析(验证结果的稳定性),所有分析都得到了一致的结果。而且所有工具变量的F统计量均超过10。
这些发现意味着什么?
这项研究第一次在人类中通过遗传学方法确立了特定肠道菌群与焦虑障碍之间的因果关系。
仔细观察这些细菌的特点,会发现一些重要的规律:
4种保护菌属中有3种都是已知的产丁酸菌
Blautia、Butyricicoccus都是产短链脂肪酸菌(详细认识这些菌可以关注谷禾健康以往的文章),它们通过发酵膳食纤维产生丁酸。而丁酸是一种具有多重神经保护和抗焦虑作用的关键代谢产物。
其次,Parasutterella这个菌属在社交焦虑障碍的研究中也出现过——它在健康对照者中富集,而在SAD患者中减少。不同的研究方法(横断面研究和孟德尔随机化)得到了一致的结果,进一步增强了结论的可靠性。
如果多种风险菌同时增加、多种保护菌同时减少,累积的效应就会非常显著。而且,菌群是可以调节的——通过饮食、益生菌等方式增加保护菌的丰度,就有可能实实在在地降低焦虑风险。
现在我们已经知道,特定的肠道菌群确实可以因果性地影响焦虑风险。那么,这些肠道菌群,究竟是如何跨越肠-脑屏障,影响我们遥远的大脑的呢?
经过二十年的研究,科学家们已经基本阐明了菌群影响大脑的六大核心机制:HPA轴调节、免疫调节、色氨酸代谢、短链脂肪酸信号、内源性大麻素系统,以及海马神经发生。
HPA轴调节:应激反应的总开关
下丘脑-垂体-肾上腺轴(HPA轴)是人体对应激反应的核心调控系统,也被称为应激反应的”总开关”。当我们面临压力时,下丘脑会分泌促肾上腺皮质激素释放激素(CRH),刺激垂体分泌促肾上腺皮质激素(ACTH),ACTH再作用于肾上腺皮质,促进糖皮质激素(在人类中是皮质醇)的合成和释放。
编辑
大量的临床前研究表明,肠道菌群在HPA轴的早期发育和持续调节中发挥着关键作用。
无菌小鼠表现出过度的HPA轴反应——面对应激时,它们的皮质酮水平比正常小鼠高出两倍以上,表现出明显的焦虑样行为。而如果在生命早期给无菌小鼠定植正常的肠道菌群,就能逆转这种异常。
这说明,肠道菌群在生命早期为HPA轴的发育设定了基线。如果生命早期菌群失调,就可能导致HPA轴的调定点过高,使得个体在一生中都对应激更加敏感,更容易患上焦虑等应激相关疾病。
免疫调节:炎症是情绪的隐形推手
第二条重要的通路是免疫调节。肠道是人体最大的免疫器官,约70%的免疫细胞位于肠道相关淋巴组织中。肠道菌群是免疫系统最重要的训练者和调节者——它们帮助免疫系统区分”自己”和”异己”,维持免疫激活和免疫耐受之间的微妙平衡。
编辑
当肠道菌群失调时,这种平衡就会被打破,主要表现为两个方面:
这两个因素共同作用,会导致低度慢性炎症。与急性炎症不同,低度慢性炎症没有红、肿、热、痛等明显症状,而是表现为血液循环中促炎细胞因子(如IL-6、TNF-α、IL-1β等)水平的轻度升高。
这种低度慢性炎症正是连接肠道菌群与焦虑障碍的关键桥梁。
越来越多研究表明,焦虑障碍患者的外周血和脑脊液中,促炎细胞因子水平显著升高。
荟萃分析显示,焦虑患者的IL-6和TNF-α水平明显高于健康对照。而且,炎症水平与焦虑症状的严重程度呈正相关。
色氨酸代谢:5-羟色胺的前体争夺战
色氨酸是一种人体必需氨基酸,我们只能从食物中获取。色氨酸之所以重要,是因为它是5-羟色胺(血清素)的前体物质——而5-羟色胺是大脑中最重要的神经递质之一,与情绪调节、睡眠、食欲、冲动控制等多种功能密切相关。
肠道菌群能够影响大脑5-羟色胺水平的关键
人体中90%以上的5-羟色胺是在肠道中合成的,而不是在大脑中。虽然肠道中的5-羟色胺无法穿过血脑屏障进入大脑,但它可以通过多种途径间接影响大脑功能。更重要的是,大脑中5-羟色胺的合成还依赖于从血液中摄取的色氨酸。
进入人体的色氨酸有三条主要的代谢途径
编辑
对于焦虑障碍来说,最重要的是犬尿氨酸途径。多项研究已经证实,焦虑障碍患者的色氨酸代谢存在异常,犬尿氨酸途径过度激活。而Butler等人2025年的研究更是直接证明了社交焦虑障碍患者的犬尿氨酸代谢产物存在改变。
短链脂肪酸:细菌的神奇代谢产物
短链脂肪酸(SCFAs)是肠道细菌发酵难以消化的碳水化合物(膳食纤维)产生的主要代谢产物,主要包括乙酸、丙酸、丁酸三种。
短链脂肪酸可能是目前研究最充分的菌群-肠-脑轴信号分子,也是我们理解菌群如何影响大脑的关键钥匙。它们在肠脑交流中发挥着多重作用:
增强肠道屏障完整性
短链脂肪酸是结肠上皮细胞的主要能量来源,提供约70%的能量。它们可以促进肠道上皮细胞的增殖和分化,增强紧密连接蛋白的表达,维持肠道屏障的完整性,减少肠漏和后续的炎症反应。
调节血脑屏障通透性
短链脂肪酸不仅能保护肠道屏障,还能调节血脑屏障的通透性。研究表明,无菌小鼠的血脑屏障通透性增加,而补充SCFAs可以恢复血脑屏障的完整性。这意味着SCFAs可以保护大脑免受循环中神经毒素的侵害。
免疫调节作用
短链脂肪酸具有强大的抗炎作用。它们可以通过激活G蛋白偶联受体(GPR41、GPR43、GPR109A)和抑制组蛋白去乙酰化酶(HDAC)等途径,调节免疫细胞的分化和功能,抑制促炎细胞因子的产生。
调节HPA轴反应
SCFAs可以直接作用于肾上腺,调节糖皮质激素的合成,也可以通过迷走神经间接调节HPA轴的活性。研究表明,补充乙酸可以降低小鼠的应激反应,减少焦虑样行为。
直接的神经调节作用
SCFAs还可以直接作用于神经系统。它们可以刺激肠神经系统的感觉神经元,通过迷走神经向大脑传递信号;它们可以穿过血脑屏障,直接作用于大脑中的神经元和胶质细胞;它们还可以调节神经营养因子(如BDNF)的表达,促进神经发生和突触可塑性。
编辑
在这三种主要的SCFAs中,丁酸的研究最为充分,也被认为是最重要的有益代谢产物之一。有意思的是,几乎所有的保护菌属都是丁酸的主要产生菌。这完美地解释了为什么这些细菌可以降低焦虑风险——它们通过产生丁酸,发挥多重神经保护和抗焦虑作用。
海马神经发生:大脑可塑性的菌群调控
很长一段时间里,科学界普遍认为成年人类的大脑不会产生新的神经元。但近年来的研究已经证实,在大脑的两个特定区域——海马的齿状回和侧脑室的室管膜下区——神经发生可以持续终生。
海马是大脑中与学习记忆、情绪调节、应激反应密切相关的关键脑区
成年海马神经发生在这些功能中发挥着重要作用。越来越多的证据表明,海马神经发生减少是抑郁症和焦虑障碍的重要病理机制之一,而抗抑郁药和抗焦虑药的疗效也与促进海马神经发生有关。
肠道菌群是成年海马神经发生的重要调控者
多项研究表明,无菌小鼠的海马神经发生显著减少,而菌群定植可以恢复神经发生水平。益生菌干预、粪菌移植和高膳食纤维饮食(通过增加SCFAs产生)都可以促进海马神经发生,增加BDNF(脑源性神经营养因子)的表达。
BDNF是大脑中最重要的神经营养因子之一
它可以促进神经元的存活、增殖、分化和突触可塑性。BDNF水平降低在焦虑和抑郁的发病机制中扮演着核心角色,而几乎所有有效的抗抑郁治疗都会增加BDNF的表达。
几大通路的协同作用网络
这里,我们要强调的是,这六大通路并不是独立工作的,而是相互交织、相互影响,形成一个复杂的调控网络。例如:
这是一个恶性循环:任何一个环节的异常都会波及其他环节,最终导致整个系统的失衡。
但反过来,这也意味着我们有多个干预靶点:改善任何一个环节,都可能打破恶性循环,推动整个系统向健康的方向发展。例如,增加膳食纤维摄入→SCFAs增加→肠道屏障改善→炎症减少→HPA轴恢复正常→焦虑症状缓解。
可能大家更关心的问题是:我们能否通过主动调节肠道菌群来改善焦虑症状?
目前,基于微生物组的焦虑干预策略已经成为精神医学领域最热门的研究且干预逐步显现成效的方向之一。
益生菌:从补充细菌到改善情绪
在过去的二十年里,益生菌在情绪调节方面的作用得到了大量研究的验证:
健康人群的研究表明,益生菌可以降低自我报告的焦虑和抑郁水平,改善情绪认知加工;
在临床诊断的焦虑障碍患者中,益生菌干预可以显著降低汉密尔顿焦虑量表(HAMA)评分,同时降低血清炎症标志物水平;
在社交焦虑障碍患者中,益生菌干预可以改善社交焦虑症状,同时调节大脑边缘系统的活动;
在抑郁症患者中,益生菌作为辅助治疗可以显著增强抗抑郁药的疗效,降低复发风险。
需要特别注意的是,益生菌的作用具有高度的菌株特异性——不是所有的益生菌都有抗焦虑效果,也不是一种益生菌对所有人都有效。乳酸杆菌和双歧杆菌是两类已被用于焦虑治疗研究的细菌。
目前证据最充分的抗焦虑益生菌菌株主要集中在以下几个属:
乳杆菌属
鼠李糖乳杆菌
一项近期研究报告称,在一项包含 70 人的双盲随机对照试验(DB-RCT)中,含有鼠李糖乳杆菌及其他菌株的益生菌胶囊减少了焦虑。
一项动物研究中,据报道,补充鼠李糖乳杆菌可减少焦虑和抑郁行为,甚至可能改变小鼠大脑中 GABA 相关基因的活性。
瑞士乳杆菌
一项近期动物研究中,据报道,补充瑞士乳杆菌可减少大鼠与焦虑相关的行为。
相关研究还报道称,补充瑞士乳杆菌可保护小鼠免受西式饮食对焦虑行为造成的负面影响。
植物乳杆菌
一项包含 84 名肠易激综合征(IBS)患者的双盲随机对照试验(DB-RCT)中,含有植物乳杆菌的益生菌补充剂降低了与肠道相关的焦虑。
一项动物研究中,据报道,长期摄入植物乳杆菌可提高小鼠体内多巴胺和血清素水平,并减少焦虑样行为。另有报道称,它还能降低促炎性细胞因子,并提高经历早期生活压力的小鼠体内抗炎性细胞因子水平。
双歧杆菌属
长双歧杆菌
长双歧杆菌可减轻感染性结肠炎小鼠的焦虑样行为,这些作用似乎是通过迷走神经通路介导的。
一项人体研究中,据报道,连续 30 天每日服用瑞士乳杆菌和长双歧杆菌,可降低 25 名受试者的焦虑、敌意以及与压力相关的激素皮质醇水平。
产丁酸菌属
国际益生菌和益生元科学协会(ISAPP)推荐的益生菌剂量范围是每天至少摄入109-1010 CFU。需要坚持服用至少4-8周,因为菌群的改变需要时间。
益生元:喂养好细菌
如果说益生菌是直接补充好细菌,那么益生元就是喂养好细菌的食物。
益生元是人体无法消化的碳水化合物,但可以被肠道中的有益菌选择性发酵利用,促进它们的生长和代谢活性。
最常见的益生元包括:低聚果糖(FOS)、低聚半乳糖(GOS)、菊粉(Inulin)、抗性淀粉(Resistant Starch)、母乳低聚糖(HMOs)。
多项临床研究证实了益生元的抗焦虑作用。
例如,2015年的一项研究发现,健康志愿者连续3周服用低聚半乳糖(GOS)后,对消极情绪刺激的注意力偏向显著降低,这一效应与抗焦虑药的效果类似。
另一项研究发现,菊粉型果聚糖可以显著降低抑郁症患者的抑郁和焦虑评分,同时增加粪便中双歧杆菌的丰度。
值得注意的是,益生菌和益生元常常联合使用,这种组合被称为合生元。合生元可以同时发挥补充有益菌和喂养有益菌的双重作用,效果往往优于单独使用益生菌或益生元。
饮食调节
饮食是偏生活方式的菌群干预。饮食是塑造肠道菌群组成和功能的最重要的环境因素,其影响远远超过我们大多数人的想象。
研究表明,饮食的改变可以在短短几天内改变肠道菌群的组成。而且,饮食对菌群的影响是长期的——长期的饮食习惯会塑造出相对稳定的菌群特征。因此,通过调整饮食来改善菌群,进而改善情绪,是一种经济、安全、可持续的干预手段。
地中海饮食——情绪友好型饮食
地中海饮食泛指地中海沿岸国家的传统饮食模式,其核心特点包括:丰富的植物性食物(蔬菜、水果、全谷物、豆类、坚果)、橄榄油作为主要的脂肪来源、适量的鱼类和家禽、少量的红肉、适量的乳制品(主要是发酵乳制品,如酸奶、奶酪)、少量的红酒(随餐饮用)。
大量的流行病学研究表明,坚持地中海饮食与更低的抑郁和焦虑风险显著相关。
2017年著名的SMILES随机对照试验更是直接证明了饮食干预对抑郁症的治疗效果,饮食干预组有32%的患者达到了临床缓解标准,而对照组只有8%。
那么,地中海饮食为什么能改善情绪呢?
肠道菌群在其中发挥了关键的中介作用。
基于目前的科学证据,我们给出以下改善情绪的饮食建议:
应该多吃的食物
– 各种颜色的蔬菜(每天至少500克)
– 新鲜水果(每天200-350克)
– 全谷物(燕麦、糙米、藜麦、全麦面包等)
– 豆类(黄豆、黑豆、红豆、鹰嘴豆、扁豆等)
– 坚果和种子(核桃、杏仁、亚麻籽、奇亚籽等,每天一小把)
– 富含脂肪的鱼类(三文鱼、鲭鱼、沙丁鱼、凤尾鱼,每周2-3次)
– 发酵食品(无糖酸奶、开菲尔、泡菜、纳豆等)
– 橄榄油(作为主要的烹饪用油)
– 香料(姜黄、生姜、大蒜、肉桂等)
应该少吃的食物
– 加工食品(薯片、饼干、方便面、火腿肠等)
– 含糖饮料(可乐、果汁饮料、奶茶等)
– 精制碳水化合物(白米饭、白面包、面条、糕点等)
– 油炸食品和高脂肪食物
– 加工肉类(培根、香肠、火腿等)
– 过量的酒精和咖啡因
西式饮食模式与认知功能下降和情绪症状风险增加密切相关,长期高脂高糖低纤维的饮食会显著降低菌群多样性,减少产丁酸菌,促进有害菌生长,导致肠漏和慢性炎症。
植物提取物
一些植物提取物、氨基酸、脂肪酸和矿物质也被研究用于缓解焦虑。不过,这类干预的证据强度差异很大,可以作为辅助性的情绪调节工具。
有较多小型人体研究支持的
★ Kava(卡瓦)
卡瓦是一种生长在南太平洋岛屿的植物,通常作为茶或补充品食用。在几项人体研究中,卡瓦被报道可改善焦虑、紧张、不安、失眠、睡眠质量和情绪状态。
在动物中,卡瓦的抗焦虑效果被报道相当强大,甚至可能与苯二氮卓类药物(一类常用治疗焦虑症的药物)的效果相媲美。
一项对多种草本抗焦虑药进行比较的研究得出结论,卡瓦在减轻焦虑方面可能比大多数其他草本补充剂更有效。
机制上,Kava 可能增强 GABA-A 受体活性,并影响去甲肾上腺素摄取,也就是说“降低神经兴奋性、增强抑制性神经传递”。
注:需要特别注意的是,Kava 曾被报道与肝毒性相关,尤其可能与其他药物或肝脏负担叠加,因此有肝病、饮酒较多或正在服药的人不应自行使用。
★ 薰衣草
人体研究中,薰衣草被报道可改善焦虑和睡眠质量。它不一定是直接治疗焦虑障碍,而可能是通过改善紧张感和睡眠质量,间接降低焦虑体验。
在动物研究中,薰衣草精油中的几种成分(芳樟醇、龙脑)被报道能略微增加抑制性神经递质GABA的活性,这可能是其所谓减缓焦虑效果的基础。
编辑
★ 银杏叶
银杏是一种药用植物,广泛应用于中医,用于抵消与年龄相关的认知衰退,以及其他传统用途。
一项针对107名人类的研究,每天补充240或480毫克银杏叶连续4周,被报告显著减轻焦虑症状(相比无效安慰剂治疗)。
银杏提取物也被报道能减轻老年患者的焦虑、抑郁和痴呆症状。
一些动物研究的早期证据表明银杏叶可能在大鼠中表现出某些抗焦虑特性。
注:需要注意的是,银杏可能影响凝血功能,正在使用抗凝药、抗血小板药、准备手术或有出血风险的人群,不应自行使用。
★ 洋甘菊
洋甘菊中含有黄酮类成分芹菜素。洋甘菊更接近温和的神经镇静和压力调节方向。
两项分别为57名和61名参与者的人体研究报告,洋甘菊提取物可降低轻度至中度的焦虑、抑郁和压力。
据报道,服用标准补充剂量的着皮生素能减轻焦虑,同时不引起类似镇静的副作用。
一些早期证据表明,芹菜素可能通过与GABA受体结合来增强GABA通讯(类似于苯二氮卓类药物的作用方式)。有报道称,它还可能抑制谷氨酸家族的NMDA受体。
注:需要区分的是,标准化洋甘菊提取物和日常饮用的洋甘菊茶并不等同,茶饮中的有效成分剂量通常更低,不能简单理解为“喝洋甘菊茶就能治疗焦虑”。
★ 柠檬香蜂草
柠檬香蜂草传统上常作为镇静类草本茶饮使用。
根据多项人体用户研究,300~1600毫克的柠檬香蜂草提取物补充剂在20名、18名、20名和20名参与者的人体研究中被报告能改善焦虑、压力、失眠、记忆、情绪和认知处理。
其主要活性成分之一是迷迭香酸,可能通过抑制 GABA 分解相关酶,间接提高 GABA 水平。由于 GABA 是大脑中最重要的抑制性神经递质之一,这一机制可以解释它为何可能带来放松和抗焦虑效果。
编辑
★ 紫色西番莲
紫色西番莲是一种原产于美国东南部的开花藤本植物。
一项针对18名广泛性焦虑症患者的早期研究结果表明,紫西番莲可能有助于减轻焦虑,类似于苯二氮卓类药物(一种常用于治疗焦虑及其他精神疾病的药物)。
另一项研究报告称,手术前补充紫色西番莲能减轻30名手术患者的焦虑。
一项纳入 30 人的双盲随机对照试验(DB-RCT),单次服用 5,000 毫克紫花西番莲可降低公众演讲后的心率;而连续 6 天服用 500 毫克,则被报告可在公众演讲任务期间降低血压。
机制上,动物研究提示它可能影响海马区 GABA-A 和 GABA-C 通路。整体来看,可以理解为“降低应激反应和生理唤醒”的辅助工具,但现有人体样本量仍偏小。
有初步人体研究但证据仍不足的
► 南非醉茄 / 印度人参
南非醉茄可能不是直接作用于某一个焦虑症状,而是通过降低慢性压力负荷来改善焦虑状态。
据报道,51名更年期综合征女性的研究中,南非醉茄可以减少情绪波动和焦虑,一些研究人员甚至提出,南非醉茄提取物可能有助于在传统的精神病学实践中治疗焦虑症。
动物研究还提示,它的抗焦虑作用可能与劳拉西泮类似。
其他研究报告称,在治疗焦虑和社交隔离诱导行为时,其作用类似于情绪稳定剂。
编辑
► 茶氨酸 / 绿茶
L-茶氨酸是一种存在于白茶、红茶和绿茶中的氨基酸,因其镇静效果广受推崇。
一些早期研究也支持这一点。例如,在两项分别包含 12 名和 14 名参与者的研究中,受试者在完成脑力任务后服用 L-茶氨酸。研究报告称,L-茶氨酸有助于缓解压力症状,同时还能降低心率和血压。
一项针对20名精神分裂症患者的研究(DB-RCT)报告每日补充L-茶氨酸能减轻焦虑,20名健康个体中也显示有促进放松的效果。
动物研究提示,L-茶氨酸可能提高 GABA、多巴胺和 5-羟色胺等神经递质水平。
► 积雪草
积雪草(Centella asiatica)是一种生长在南亚湿地的植物,传统上既被用作食用蔬菜,也被用作药用草本。
在一项安慰剂对照研究中,积雪草被报告可抑制 40 名健康成年人中的惊跳反应。
一些大鼠研究报告称,积雪草及其主要成分积雪草酸(asiatic acid)可减少与焦虑相关的行为。
一项大鼠研究的早期证据提示,积雪草可能通过激活谷氨酸脱羧酶发挥作用。该酶能够提高大脑中的 GABA 水平。
注:证据有限但机制上值得关注,人体焦虑研究仍较少。
► 黄芩类草本
它们含有具有生物活性的黄酮类化合物,这类成分被认为具有较强的抗氧化作用。
一项小规模人体研究显示,与安慰剂相比,美洲黄芩被报告可减轻 19 名成年人的焦虑。
一些动物研究证据提示,补充中国黄芩可能有助于减少压力诱发的焦虑样行为,并降低血液中的皮质酮水平。
虽然这类草本中含有许多值得研究的不同化合物,但汉黄芩素(wogonin)被认为是中国黄芩中的主要活性黄酮。有研究报告称,它可减少小鼠的焦虑样行为,并增强 GABA-A 受体活性。
编辑
► 缬草
缬草(Valeriana officinalis)是一种开花植物,历史上曾用于香水制作,如今常以茶饮或根部提取物的形式作为补充剂使用。
根据一项初步人体研究,在 33 名健康个体中,每天两次补充 500 mg 缬草、持续 60 天,被报告可减轻焦虑、压力和抑郁。
一些动物研究也报告称,缬草可能有助于减少焦虑样行为。
机制上,缬草可能增强 GABA-A 信号,因此更接近“睡眠—镇静—焦虑缓解”路径。
注:由于它可能带来嗜睡,也可能与镇静催眠药、酒精或抗焦虑药叠加,不建议自行混用。
补充剂
► 5-羟色氨(5-HTP)
5-HTP 是一种天然存在的氨基酸。大脑会利用它来合成血清素,而血清素是一种神经递质,通常被认为与情绪调节密切相关。
一项针对 15 名年轻成年人的人体研究中,受试者因单恋而出现焦虑,连续 6 周补充 5-HTP 后,他们的焦虑有所减轻,这可能与血液中 BDNF 和血清素水平升高有关。
另一项人类研究报告称,仅一次200毫克的5-HTP剂量即可部分缓解24名惊恐障碍患者的惊恐发作。
注:需要强调的是,5-HTP 可能与 SSRIs、SNRIs、MAOIs 等抗抑郁药发生相互作用,如正在服用精神科药物请遵医嘱。
► 镁
镁与神经兴奋性、NMDA 受体、HPA 轴和肌肉紧张都有关。
一些病例研究报告称,开始补充镁后一周内,抑郁、焦虑和睡眠问题有所改善。
注:这些结果只基于少数个体使用者,仍需要在健康人群中开展更大规模的临床试验,才能确认这些效果。
一些研究人员认为,镁可能具有抗焦虑作用,这或许与它能够降低 HPA 轴的活性有关。HPA 轴是大脑中参与压力调节的重要系统之一。此外,镁还可能通过抑制 NMDA 受体发挥作用。
► 鱼油 / Omega-3 脂肪酸
Omega-3 脂肪酸主要包括 EPA 和 DHA,与炎症、神经可塑性和细胞膜功能密切相关。
在一项包含 49 人的研究中,Omega-3 脂肪酸水平较低与社交焦虑水平较高有关。
在一项针对 34 名健康人的研究中,每天补充 2.5 克 Omega-3 被报告可减轻焦虑症状,同时降低IL-6水平。IL-6 是一种重要的促炎性细胞因子。
另一项研究报告称,补充 3 克 Omega-3 可减轻 11 名物质滥用者的焦虑。
一项大鼠研究中,DHA(一种 Omega-3 脂肪酸)被报告可降低 TNF-α、IL-6 和一氧化氮合酶 的水平。这可能说明,DHA 的部分作用或许与减轻炎症有关。
其他动物研究报告称,EPA(另一种 Omega-3 脂肪酸)可能有助于增加海马体神经发生。
其他干预手段
除了上述主要方法,还有一些新兴的干预手段也在研究之中,包括粪菌移植(将健康捐赠者的粪便菌群移植到患者肠道中)、后生元(益生菌的代谢产物或灭活菌体成分)、噬菌体疗法(利用噬菌体精准清除有害菌)、菌群定向培养(从患者自身肠道中分离有益菌,体外扩增后回输)等。这些技术目前大多还处于研究阶段,但未来可能会成为重要的治疗手段。
基于的研究探索和临床实践,不同人群的菌群组成、肠脑轴功能以及对干预的反应存在较大的差异。因此,基于微生物组的焦虑干预需要个性化。
儿童与青少年:发育中的菌群-肠脑轴
儿童和青少年时期是大脑发育的关键时期,也是肠道菌群建立和稳定的关键时期。研究表明,生命早期的菌群失调(如剖腹产、配方奶喂养、抗生素使用等)与儿童和青少年时期的焦虑、抑郁、自闭症、注意力缺陷多动障碍(ADHD)等精神疾病风险升高密切相关。
对于儿童和青少年的焦虑干预,菌群-肠脑轴具有特别重要的意义:预防优先、早期干预效果更好、安全性更高。当然,儿童和青少年的菌群干预也需要特别注意安全性和适宜性,剂量需要根据年龄调整,饮食干预需要考虑生长发育需求。
老年人:菌群衰退与情绪健康
随着年龄的增长,肠道菌群会发生一系列特征性的改变,被称为”菌群衰老”:菌群多样性显著降低、有益菌(如双歧杆菌属、产丁酸菌)丰度显著减少、潜在致病菌丰度增加、菌群的代谢功能下降,短链脂肪酸产生减少。
菌群衰老与老年人的多种健康问题密切相关,包括免疫功能下降、炎症水平升高、认知功能衰退、虚弱等。同时,也与老年人的焦虑和抑郁风险升高密切相关。65岁以上老年人中,焦虑障碍的患病率也在上升。
对于老年人的焦虑干预,重点包括:增加膳食纤维和发酵食品的摄入、适当补充益生菌、规律运动、社交活动、避免不必要的药物使用。
共病躯体疾病患者:特殊考虑
焦虑障碍常常与多种躯体疾病共病,尤其是胃肠道疾病、自身免疫病、代谢疾病等。
肠易激综合征(IBS)
一定比例IBS患者伴有焦虑或抑郁症状,而焦虑反过来又会加重IBS症状,形成恶性循环。菌群干预对IBS患者具有双重益处——既可以改善胃肠道症状,又可以改善焦虑情绪。
炎症性肠病(IBD)
IBD患者的焦虑和抑郁患病率高于健康人。这些患者存在严重的菌群失调,改善菌群不仅有助于控制肠道炎症,也有助于改善情绪症状。
肥胖和代谢综合征
肥胖和代谢综合征患者的焦虑和抑郁风险显著高于正常体重人群。同时,肥胖人群通常伴有显著的菌群失调。对于肥胖伴焦虑的患者,菌群干预同样具有双重益处。
性别差异:为什么女性更容易焦虑?
焦虑障碍存在显著的性别差异——女性的患病率是男性的2倍左右。这一差异的原因是多方面的,包括激素、社会心理因素、遗传等。近年来的研究发现,肠道菌群的性别差异可能也是其中一个重要的原因。
肠道菌群中含有大量的β-葡萄糖醛酸酶,可以将结合态的雌激素重新转化为活性态的游离雌激素,增加体内的雌激素水平。这一过程被称为“雌激素的肠肝循环”。菌群失调时,β-葡萄糖醛酸酶活性改变,会影响雌激素的正常代谢,导致雌激素水平异常。
编辑
雌激素可以通过多种途径影响大脑功能和情绪:它可以调节5-羟色胺、多巴胺等神经递质的合成和代谢,可以调节HPA轴的活性,可以影响神经可塑性和炎症反应。因此,菌群失调导致的雌激素代谢异常,可能是女性更容易患上焦虑和抑郁的重要原因之一。
这也解释了为什么女性在一些特殊的生理时期,如经前期、产后、更年期,更容易出现情绪问题。对于女性的焦虑干预,需要特别考虑激素与菌群的相互作用。
尽管微生物-肠脑轴的研究已经取得了很大的进步,但是这个领域仍然非常年轻,还有很多未知的问题和挑战等待我们去解决。目前大多数研究的样本量还不够大,方法异质性较高,因果推断仍然存在困难,动物到人的转化也存在鸿沟。
客观地说,基于菌群的焦虑干预距离真正成为常规临床治疗手段,可能还需要几年甚至更长的时间。不过,这并不意味着我们现在什么都做不了。恰恰相反,基于现有的证据已经足够支持我们在日常生活中做出一些改变,来改善我们的菌群和情绪。
最后,想给给读者一些实用建议
基于目前的科学证据,我们提出以下几点实用建议:
1
优先调整饮食
这是最重要、最有效、也最安全的干预手段。饮食调整不需要完美,只需要开始,哪怕只是每天多吃一份蔬菜,或者每周多吃一把坚果,都会对你的菌群和健康产生积极的影响。
2
谨慎选择益生菌
如果你想尝试益生菌,请注意以下几点:
– 选择有临床研究证据支持的菌株,而不是随便买。
-基于自己的肠道菌群构成和菌群状态选择合适的益生菌。
– 注意活菌数量,一般建议每天至少摄入10⁹-10¹⁰ CFU
– 选择靠谱的品牌,注意产品的保质期和储存条件
– 坚持服用4-8周,菌群的改变需要时间
– 如果服用后出现严重的胃肠道不适,应该停止使用并咨询医生
需要特别提醒的是:益生菌不是万能药,不能替代正规的心理治疗和药物治疗。如果你患有严重的焦虑障碍,一定要首先寻求专业精神科医生和心理医生的帮助。
3
改善生活方式
除了饮食,很多其他的生活方式因素也会影响肠道菌群和情绪:
– 规律运动:运动可以显著增加菌群多样性,促进有益菌生长,同时本身就是有效的抗焦虑手段;
– 充足睡眠:睡眠不足会严重破坏菌群平衡,增加炎症水平,加重焦虑症状;
– 压力管理:长期压力会导致菌群失调,而菌群失调又会加重压力反应,形成恶性循环。学习冥想、深呼吸、正念等压力管理技巧,打破这个恶性循环;
– 接触自然:多去户外,接触土壤和植物,可以增加环境微生物暴露,改善菌群多样性;
– 避免不必要的抗生素:抗生素会严重破坏肠道菌群,除非医生认为确实必要,否则不要随意使用。
4
关注自己的肠道感受
注意观察什么食物让你感觉舒服,什么食物让你感觉不舒服。每个人的菌群都是独特的,没有一种饮食适合所有人。学会倾听你身体的声音,找到最适合你自己的饮食和生活方式。
5
寻求专业帮助
如果有严重的胃肠道问题或者情绪问题,要寻求专业医生或专业的检查,包括消化、精神、营养等相关的检查和咨询等。菌群干预可以作为辅助治疗手段,但不能替代正规的医学治疗。
肠道菌群——心理健康的新边疆
一个多世纪前,Sigmund Freud 创立并发展精神分析学时,人们开始尝试从童年经验、潜意识冲突和内在心理世界中理解精神痛苦。
约七十年前,第一代抗抑郁药的出现,又让我们看到神经递质、神经环路和大脑生物学在情绪障碍中的重要作用。
今天,随着微生物—肠—脑轴研究的不断深入,我们对焦虑、抑郁等精神健康问题有了更加立体的认识:情绪并不只是“大脑内部”的事件,它也与免疫、代谢、内分泌、肠道屏障以及数以万亿计的肠道微生物密切相关。
这不是对过去理论的简单否定,而是一次重要的拓展。我们越来越清楚地看到,大脑并不是孤立运行的指挥中心,而是与全身各系统持续对话的复杂网络。精神健康也不应被简单归为心理问题或大脑问题,而应被理解为心理、神经、免疫、代谢、环境和生活方式共同作用的结果。肠道菌群正是这个复杂系统中一个重要而活跃的参与者。
编辑
微生物—肠—脑轴的研究,为焦虑、抑郁和压力相关问题的预防与干预提供了新的思路。合理饮食、充足睡眠、规律运动、压力管理,以及在专业指导下使用益生菌、益生元或其他微生态干预方式,未来都可能成为精神健康综合管理的一部分。但我们也需要保持科学的谨慎:肠道菌群不是解释一切的万能答案,菌群干预也不能替代规范的心理治疗、药物治疗和医学评估。
对行业而言,肠道菌群研究最大的价值,不在于制造新的概念热潮,而在于推动我们以更系统、更精准、更个体化的方式理解人体健康。未来真正有价值的微生态产品和干预方案,必须建立在高质量临床证据、精准的检测结果、清晰作用机制和真实人群获益之上。
注:本账号内容仅供学习和交流,不构成任何形式的医疗建议。
主要参考文献
Xiong RG, Li J, Cheng J, Zhou DD, Wu SX, Huang SY, Saimaiti A, Yang ZJ, Gan RY, Li HB. The Role of Gut Microbiota in Anxiety, Depression, and Other Mental Disorders as Well as the Protective Effects of Dietary Components. Nutrients. 2023 Jul 23;15(14):3258.
Butler, M.I., Bastiaanssen, T.F.S., Long-Smith, C. et al. The gut microbiome in social anxiety disorder: evidence of altered composition and function. Transl Psychiatry 13, 95 (2023).
Butler, M.I., Kittel-Schneider, S., Wagner-Skacel, J. et al. The Gut Microbiome in Anxiety Disorders. Curr Psychiatry Rep 27, 347–361 (2025).
Ding, H. “Gut microbiota and anxiety: an exploration of key findings. J Depress Anxiety.” 2017
Lan, J., Wang, J., Huang, S. et al. Neuropeptide SP protects against colitis and linked anxiety-like behavior through the putative roles of gut microbiota and metabolite inositol. Nat Commun 17, 295 (2026).
Shen H, Wang SY, Zhao YY, Zhou JL, Zhao J and Zhu WK (2026) Brain-gut-microbiota axis: a review on the bidirectional regulatory mechanisms between gut microbiota and brain and their disease interactions. Front. Microbiol. 17:1768891.
Kumar, Akash, et al. “Gut microbiota in anxiety and depression: unveiling the relationships and management options.” Pharmaceuticals 16.4 (2023): 565.
karger.com/books/book/382/chapter/5564732/Is-Anxiety-Associated-with-the-Gut-Microbiota
selfhacked.com/blog/natural-treatments-anxiety/
Cao, Y., Cheng, Y., Pan, W. et al. Gut microbiota variations in depression and anxiety: a systematic review. BMC Psychiatry 25, 443 (2025).
Li, Jianbing, et al. “Association between gut microbiota and anxiety disorders: a bidirectional two-sample mendelian randomization study.” BMC psychiatry 24.1 (2024): 398.
谷禾健康
当一个孩子长期“吃了也不长”、身高曲线逐渐往下掉,大部分家长可能首先会想到是不是缺钙、缺锌,或者是不是肠胃吸收不好。事实上,儿童发育迟缓是一个比“矮一点”更复杂的问题。
儿童发育迟缓(stunting)诊断标准为年龄别身高(Height-for-age, HFA)低于同年龄、同性别儿童生长曲线中位数减去2个标准差(-2SD)。它不是简单的“个子小”,而是生命早期长期线性生长受限的表现。
传统营养支持仍是基础,但越来越多研究提示,营养不足、反复感染、环境性肠功能障碍(EED)、肠道菌群成熟延迟、肠屏障受损和宿主生长信号改变,可能共同参与儿童发育迟缓的形成与持续。
本文基于重点梳理儿童发育迟缓与肠道菌群的证据。探讨菌群如何通过菌群成熟、短链脂肪酸、胆汁酸、色氨酸、B族维生素、核苷酸、免疫和IGF-1/生长轴参与儿童生长,梳理列出发育迟缓儿童中观察到的菌群成熟延迟、发育轨迹异常。同时讨论母乳/辅食、微量营养素、铁剂、WASH、益生菌、HMO/GOS 等干预方向及其边界。
目录
儿童发育迟缓通常按世界卫生组织发布的《儿童发育标准》(Child Growth Standards)定义:年龄别身高低于同年龄、同性别儿童生长曲线中位数减去2个标准差(-2SD)。这里的关键是“年龄别”和“Z评分”。是把孩子的身长/身高放在同年龄、同性别标准曲线中判断。
发育迟缓与消瘦不同。消瘦更多反映近期体重相对于身长/身高不足,常与急性食物不足、急性疾病或快速体重下降有关。发育迟缓则更偏向长期线性生长受限,常反映生命早期较长时间的营养、感染和环境压力。
研究专家同时提醒不能把发育迟缓简单等同于遗传性矮小。父母身高、出生体重、孕周、青春发育、内分泌疾病和慢性疾病都需要综合考虑。因此,判断儿童发育迟缓需要连续生长曲线,而不是一次测量。
为什么前 1000 天特别重要?
从受孕到2岁左右的前 1000 天,是营养、免疫、肠道菌群和神经发育高度敏感的窗口。这个阶段,孩子从宫内营养转向母乳,再逐渐进入辅食和家庭饮食。每一步变化都在塑造肠道生态。
出生后0–3岁,肠道菌群通常从早期以 Bifidobacterium 等利用母乳低聚糖的细菌为主,逐渐走向更复杂、更成人样的厌氧发酵网络。这个过程受到分娩方式、母乳、辅食、感染、抗生素和家庭环境影响。
编辑
如果孩子在这个窗口长期处于营养不足、反复感染或不良卫生环境中,菌群成熟、肠屏障修复和身体生长信号都可能受到影响。它们不是单一因果关系,而是相互牵连的网络。
为什么发育迟缓值得重视?
发育迟缓不仅意味着身高落后。大量研究将早期线性生长不良与感染风险、认知发展、学习能力、成年健康和社会经济结局联系起来。这种关系并不意味着每一个发育迟缓儿童都会出现相同结局,但提示早期生长轨迹需要认真评估。
编辑
联合国儿童基金会、世界卫生组织和世界银行集团共同开展的儿童营养不良估计项目/报告指出:发育迟缓并非一句“孩子晚长”就能概括。它意味着儿童在关键生长窗口长期偏离正常线性生长轨迹,需综合评估营养、疾病、环境和发育状况。
儿童发育迟缓更像是长期叠加的“生长压力”:胎儿期营养不足、早产或低出生体重,出生后母乳或辅食质量不足,反复腹泻或无症状肠道感染,卫生环境差,家庭食物不安全,医疗可及性不足。这些因素叠加在一起,最终影响线性生长。
风险可能从出生前开始
胎儿生长受限、早产、低出生体重与后续线性生长相关。母亲营养不良、贫血、感染、都会改变胎儿和婴幼儿早期生长基础。
编辑
饮食质量:不只是吃饱
儿童维持正常线性生长,不只需要热量,还需要足够优质蛋白、脂肪、必需脂肪酸、铁、锌、维生素A、维生素D、维生素B12、叶酸、碘等营养素。长期只吃稀粥、汤泡饭、甜饮或零食,即便“看起来吃了很多”,也可能能量密度和营养密度都不足。
WHO 2023 年 6–23 月龄婴幼儿辅食指南强调,辅食应及时添加,并逐渐提高频次、质地、食物多样性和动物源性食物摄入。这些基础建议仍是所有菌群导向干预的前提。
感染:会影响消化吸收
反复腹泻、肠道病原体、寄生虫或无症状感染,会让孩子出现食欲下降、吸收不良和炎症消耗。多国出生队列研究表明,儿童生命前2年的腹泻等疾病负担、肠道病原体感染/暴露以及膳食摄入状况,与线性生长、体重增长等生长指标之间存在复杂的相互关系。
这也是为什么有些孩子“补了营养”但仍然恢复有限。营养进入肠道后,还需要健康的肠屏障、足够消化吸收能力和较低炎症负担才能被身体用于生长。
如果把儿童生长看作一棵树,食物是土壤和水分,激素和骨骼是枝干,肠道则像根系所在的土层。肠道菌群不是“让孩子长高”的开关,但它可能参与营养代谢、屏障维护、免疫训练、感染抵抗和生长信号调节或复杂网络中的调节节点。
儿童肠道菌群不是静态的:需要“按年龄长大”
健康儿童的肠道菌群会随年龄、饮食和环境逐步成熟。在谷禾跨年龄和地理人群研究中发现,人类肠道菌群在生命早期变化最快,随后逐渐向成人样结构过渡。这说明儿童菌群不能简单拿成人标准判断。
• 婴幼儿时期肠道菌群变化很快
婴儿早期,母乳喂养儿童常见 Bifidobacterium 优势。Bifidobacterium longum subsp. infantis 等能够利用母乳低聚糖(HMO),产生乳酸和乙酸,并可能帮助抑制部分肠杆菌科扩张。
进入辅食期后,膳食底物从乳糖和 HMO 扩展到淀粉、植物多糖、蛋白质和脂肪。此时,Bacteroides、Prevotella、Ruminococcus、Faecalibacterium、Blautia、Ruminococcaceae 和 Lachnospiraceae 等复杂厌氧发酵网络逐渐建立。这一过程与食物多样性密切相关。
肠道微生物群发育受喂养方式、出生方式、地理位置和环境暴露等多因素影响。研究显示,母乳喂养与较高水平的 Bifidobacterium密切相关,停止母乳喂养则会加速菌群成熟;阴道分娩婴儿肠道中 Bacteroides 水平也较高。尽管存在这些共性,不同个体的菌群发育轨迹仍受生活方式和环境因素影响而各有差异。
编辑
doi: 10.3233/NHA-170030
如何衡量菌群成熟度?
在 2014 年 Nature 研究中提出微生物群年龄 Z 评分(microbiota-for-age Z score, MAZ),用于评估儿童肠道菌群成熟度是否匹配实际年龄。研究团队基于健康马拉维儿童粪便菌群数据建立“菌群年龄”预测模型,并发现严重急性营养不良(SAM)儿童的菌群成熟度落后于同龄健康儿童,呈现发育不成熟或“低龄化”特征。
注:标准营养治疗可在短期内改善部分菌群特征,但治疗结束后,菌群成熟度可能再次下降,提示菌群成熟延迟可能参与营养不良的持续或复发。
谷禾健康基于 10 多年的肠道菌群检测实践,也构建了人体年龄预测方法及介质(GUHEAge 模型)。该模型通过可靠样本筛选降低噪声诱导关联的影响,并考虑不同生命阶段菌群发育速率的差异。尤其在婴幼儿期,肠道菌群经历快速定植、演替和成熟,短期内组成和功能潜力变化显著,因此模型在年龄尺度和样本分层上采用更精细的时间窗口,以捕捉早期菌群快速成熟的动态特征。
• 发育迟缓儿童的菌群年龄滞后现象
发育迟缓儿童的肠道菌群呈现出明显的“年龄滞后”特征,这种现象反映在菌群的多样性、组成结构和功能等多个方面。研究表明,这些儿童的肠道菌群发育水平往往落后于其实际年龄,这种滞后可能是导致发育迟缓的重要因素之一。
之前发表在《Gut》上的针对自闭症儿童菌群发育的研究显示,自闭症儿童的菌群发育要滞后于健康儿童。
编辑
在菌群组成方面,发育迟缓儿童表现出显著的特征性改变。首先是有益菌群的明显减少,特别是双歧杆菌和乳杆菌的含量显著低于同龄健康儿童。这些菌群对维持肠道健康、促进营养物质吸收和调节免疫功能具有重要作用。同时,研究还发现这些儿童的肠道中普氏菌(Prevotella)和粪杆菌(F.prausnitzii)等核心菌菌含量也明显降低。
编辑
doi.org/10.1136/gutjnl-2021-325115
但是需要注意,不管是MAZ 研究工具还是谷禾肠道擦年龄评估模型,都不能看作是临床诊断项目,不能把粪便菌群检测中的“菌群年龄”直接等同于发育迟缓诊断。
更应该理解为菌群成熟度比“某个菌多一点或少一点”更适合解释儿童营养不良的长期性。
早期菌群发育滞后对身体多系统产生影响
肠道菌群发育滞后会对婴幼儿的多个系统产生深远影响。
•在神经系统方面:可能导致认知发育迟缓、语言发育延迟、注意力不集中,甚至增加自闭症谱系障碍(ASD)的风险;
•在免疫系统方面:常见表现包括反复呼吸道感染、特应性皮炎、食物过敏,以及哮喘等过敏性疾病的发生率升高;
•在代谢系统方面:可能引起生长发育迟缓、体重增长异常、微量元素吸收不良等问题;在消化系统方面,则可能出现腹泻、便秘、肠痉挛等功能性胃肠道疾病。
特别是在出生后最初1000天这个关键期内,肠道菌群的发育状况对婴幼儿的长期健康具有决定性影响。
因此,规范的肠道菌群检测可作为婴幼儿发育评估的重要参考,内容包括微生物多样性指数、关键菌群(如双歧杆菌、乳酸菌、粪杆菌等)丰度及潜在致病菌监测。
建议在以下关键时间点进行检测:出生后1-2个月、辅食添加期(4-6个月)、断奶期(12个月左右)以及2岁左右。对于高危人群,如早产儿、剖宫产婴儿、有过敏家族史或自身免疫性疾病家族史的婴幼儿,需要更频繁的监测。
这种检测不仅能够及早发现发育风险(如自闭症倾向、过敏风险、免疫功能异常等),还能为个性化干预方案的制定提供科学依据。特别是对于已经出现发育迟缓、免疫功能低下、消化吸收障碍等问题的婴幼儿,定期的肠道菌群检测对评估干预效果和及时调整治疗方案具有重要的临床指导意义。
菌群如何影响能量和营养代谢?
肠道菌群可以分解人体自身难以消化的复杂碳水化合物,产生短链脂肪酸( SCFAs),包括乙酸盐、丙酸盐和丁酸盐。丁酸盐可为结肠上皮细胞供能,乙酸盐和丙酸盐可参与能量代谢和肝脏代谢调节。
• 产短链脂肪酸菌群会影响能量代谢
在发育迟缓或营养不良儿童中,Faecalibacterium、Blautia、Bifidobacterium、Roseburia、Megasphaera、部分 Ruminococcaceae 和 Lachnospiraceae 等与 SCFA 生成相关的菌群,在若干研究中呈减少趋势。这可能意味着膳食底物和微生物交叉喂养网络不足。
但是要注意一个误区:看到丁酸盐菌减少,并不等于“补丁酸盐菌就能长高”。SCFA 网络需要多样化食物底物、适龄辅食、低炎症环境和成熟菌群共同支持。单一菌株或单一代谢物补充不能替代基础营养。
B族维生素、核苷酸与氨基酸
多项微生物组研究提示,发育迟缓相关差异不只体现在菌的构成变化,还体现在功能通路。目前已经了解到到B 族维生素生物合成、核苷酸代谢、碳水化合物利用、氨基酸代谢等功能通路,可能与儿童生长指标相关。
B 族维生素参与能量代谢、核酸合成和神经发育。核苷酸与肠上皮更新、免疫细胞增殖和组织修复有关。氨基酸代谢则与蛋白质合成、免疫功能和生长轴支持相关。
但是菌群报告里功能通路来自微生物功能预测,不一定等同于血液或肠腔代谢物实测,更多反馈相关菌群功能潜力和膳食摄入情况。
肠屏障与黏膜免疫:为什么“低度炎症”会消耗生长资源?
肠道屏障是儿童将食物转化为生长材料的关键界面。健康屏障既促进营养吸收,又限制病原体和微生物产物入侵;一旦受损,脂多糖(LPS)等微生物产物可能激活Toll样受体4(TLR4)和 NF-κB 炎症通路。
在分子层面,LPS、鞭毛蛋白等病原相关分子模式可激活先天免疫信号;在细胞层面,巨噬细胞、树突状细胞和 T 细胞可能释放促炎因子;在组织层面,紧密连接、黏液层和绒毛结构可能受损;在系统层面,慢性炎症会增加能量消耗,并可能抑制生长支持信号。
短链脂肪酸(SCFA),尤其是丁酸盐,可能支持上皮细胞供能、紧密连接和调节性 T 细胞功能;色氨酸代谢物如吲哚类化合物,可能通过芳香烃受体(AhR)和 IL-22 相关通路促进黏膜修复。但这些机制在儿童发育迟缓中的直接证据仍有限,更适合作为机制假说,而非临床结论。
不能只看粪便菌群,还要结合小肠、胆汁酸和脂质吸收
粪便样本主要反映结肠生态,而很多营养吸收发生在小肠。发育迟缓相关研究提示,十二指肠可能出现“口腔样菌群”过度代表,如 Streptococcus、Veillonella、Haemophilus、Neisseria、Rothia、Actinomyces 等。这被称为消化道生态位“隔室化解除”。
• 小肠菌群异常可能影响胆汁酸代谢和脂质吸收
胆汁酸不仅帮助脂肪吸收,也通过 FXR 和 TGR5 等受体参与肠屏障、代谢和免疫调节。若小肠菌群、胆汁酸转化和局部炎症同时异常,儿童可能出现“吃了但利用不足”的状态。
不过,十二指肠取样具有侵入性,健康儿童对照难获得,样本量通常有限。因此这类证据很有机制价值,但不能用于家庭检测或普通商业菌群报告解释。
首先要强调的是发育迟缓儿童的菌群差异,不能用“好菌少、坏菌多”简单概括。更符合证据的说法是:若干研究观察到菌群成熟延迟、条件致病菌富集、丁酸盐产生菌减少、特定营养代谢异常和功能通路改变。但是这些的评估需要足够大的检测数据能涵盖不同地区、年龄、饮食和用药等差异。
菌群成熟延迟:发育迟缓儿童的肠道“年龄”可能偏小
如前面所述,严重急性营养不良(SAM)儿童存在菌群成熟延迟,营养治疗后可部分改善但恢复不完全。这一发现后来成为理解儿童营养不良菌群的核心框架。
成熟延迟并非指某种菌不应出现,而是整个菌群网络尚未达到同龄儿童常见的功能状态。例如,断奶后儿童通常需要发展更强的复杂碳水发酵、厌氧交叉喂养、短链脂肪酸生成及维生素/核苷酸代谢能力;若这些功能不足,可能影响辅食营养的充分利用。
基于这一思路,相关研究不再仅关注补充热量,而是通过设计食物底物,促进与健康成熟轨迹相关的菌群和血浆蛋白网络。
Bifidobacterium:早期重要,但不能简单理解为越多越好
双歧杆菌(Bifidobacterium)是婴儿早期备受关注的菌属。母乳喂养婴儿中,Bifidobacterium 可利用 HMO 产生乳酸和乙酸,并可能抑制肠杆菌科扩张。
在营养不良或发育迟缓相关研究中,Bifidobacterium 的解读需按年龄分层。婴儿早期不足,可能提示母乳利用、HMO 代谢或早期定植抵抗不足;但随着辅食添加和断奶,其相对丰度下降也可能属于正常成熟过程。
因此,不能简单认为 “Bifidobacterium 越多越好”。其意义取决于年龄、喂养方式、HMO 暴露、菌株功能及整体菌群成熟阶段。
条件致病菌富集:Escherichia/Shigella、Campylobacter 与炎症生态位
多项研究发现,发育迟缓或营养不良儿童中 Proteobacteria/Pseudomonadota、肠杆菌科、Escherichia/Shigella、Campylobacter等菌群富集。这些细菌常与病原负担、脂多糖、肠道炎症及定植抵抗不足有关。
Escherichia/Shigella 富集可能提示肠道炎症生态位,其脂多糖(LPS)和毒力因子可激活先天免疫,增加肠道炎症和通透性。Campylobacter 感染或定植在中低收入国家儿童中较常见,与腹泻及无症状感染负担相关。
但解读时需谨慎:粪便中某一菌属升高,并不能证明其单独导致发育迟缓。它可能是卫生暴露、饮食不足、抗生素使用、炎症或环境性肠功能障碍(EED)的结果,也可能参与维持炎症。
口腔样菌群和十二指肠隔室化解除
近年研究表明,小肠,尤其是十二指肠生态,可能比粪便样本更接近吸收障碍的发生现场。发育迟缓儿童的十二指肠中,常可见 Streptococcus、Veillonella、Haemophilus、Neisseria、Rothia、Actinomyces 等口腔相关菌过度富集。
这一现象可称为“隔室化解除”,即原本多见于口腔或上消化道的细菌在小肠异常增多,可能与胃酸、胆汁酸、肠蠕动、局部免疫及肠屏障异常有关。
这些口腔样菌群可能影响脂质吸收、胆汁酸信号和小肠局部炎症。若儿童本身饮食质量不足,再叠加小肠吸收效率下降,线性生长可能更易受影响。
注:但目前十二指肠取样不适合普通筛查,粪便商业菌群检测也不能替代小肠生态评估。
丁酸盐产生菌减少:Faecalibacterium、Blautia、Megasphaera等
若干发育迟缓研究观察到丁酸盐或 SCFA 相关菌减少,如 Faecalibacterium、Blautia、Megasphaera,以及 Ruminococcaceae、Lachnospiraceae 的部分成员。这些菌参与复杂碳水降解、乳酸利用、丁酸盐生成和微生物交叉喂养。
Faecalibacterium prausnitzii 常被视为重要丁酸盐产生菌,并与抗炎环境相关;Blautia 参与碳水发酵和 SCFA 生成;Megasphaera 可利用乳酸并参与交叉喂养网络。它们减少可能提示儿童肠道发酵网络发育不足。
丁酸盐产生菌减少可能源于饮食底物不足、感染、炎症、抗生素使用或菌群成熟延迟。改善这一网络应优先从适龄辅食、膳食多样性、感染控制和基础营养入手。
Prevotella、Ruminococcus 相关成熟轨迹
一些与健康成熟轨迹相关的菌群受到关注,如 Prevotella copri、Faecalibacterium prausnitzii、Bifidobacterium longum、Bacteroides thetaiotaomicron 等。不同研究显示,它们与食物底物利用、复杂多糖发酵或菌群成熟网络有关。
Prevotella 常与植物性多糖和高纤维饮食相关,但其健康意义取决于个体肠道环境,不能简单归为“好菌”或“坏菌”。在 MDCF 语境中,研究重点是特定食物组合如何促进与年龄适宜成熟相关的菌群网络,而非单纯提高某一菌属丰度。
Ruminococcus、Clostridium symbiosum 等曾在动物模型或定义菌群研究中被讨论为促生长候选菌,但仍属于转化研究线索,不能据此直接用于儿童补菌。
Desulfovibrio、噬菌体和病毒组:仍处于探索阶段
2024 年系统综述提出了一些探索性线索,包括 Desulfovibrio 等硫酸盐还原菌的潜在变化,以及噬菌体、病毒组与肠道菌群失衡和儿童生长发育异常的可能关联。但这些发现多来自少量研究或关联性观察,仍需更多纵向和机制研究验证。
Desulfovibrio 可产生硫化氢,理论上可能影响黏膜环境,但其在儿童发育迟缓中的因果证据仍不足。
噬菌体与细菌群落的共同变化,可能影响菌群稳定性和功能基因转移。目前这些方向更适合作为未来研究议题,不宜用于临床解释或家庭干预依据。
功能通路异常:从菌名走向“菌群能做什么”
从报告角度,功能通路比菌名清单更重要。发育迟缓相关研究反复提示,碳水化合物降解、B 族维生素、核苷酸、氨基酸、SCFA、胆汁酸和色氨酸代谢可能与儿童生长相关。
其中,B 族维生素和核苷酸通路与细胞增殖、肠道修复和能量代谢有关;SCFA 通路与屏障和免疫调节有关;胆汁酸和脂质吸收通路与小肠生态有关。
这些通路共同解释了为什么“吃进去”与“长出来”之间还隔着肠道功能这一层。
环境性肠功能障碍(Environmental Enteric Dysfunction, EED)是理解儿童发育迟缓的重要概念,常见于低资源、高病原暴露环境,表现为小肠慢性炎症、屏障受损、通透性升高、吸收不良及系统性炎症负担增加。
EED 的价值在于串联多个关键环节:卫生环境差使儿童反复接触病原体;病原暴露和菌群失衡诱发小肠炎症;屏障与绒毛结构受损降低营养吸收效率;慢性炎症进一步可能影响 IGF-1 和生长轴。
1
分子—细胞—组织—系统:EED 的机制层级
•在分子水平:LPS 等病原相关分子模式可能激活 TLR4/NF-κB 炎症通路。
•在细胞水平:巨噬细胞、树突状细胞和 T 细胞参与免疫激活,促炎因子释放增加。
•在组织水平:小肠绒毛结构、隐窝增生、紧密连接和黏膜修复可能受到影响。吸收面积下降时,即便食物进入肠道,也未必能高效转化为身体生长所需营养。
•在系统水平:儿童身体需要把更多能量用于免疫应答和组织修复。营养不足与炎症负担叠加时,线性生长支持可能下降。
2
EED 指标评估
研究中常用乳果糖/甘露醇试验、粪便 MPO、AAT、neopterin、calprotectin、宿主粪便 mRNA 等指标评估 EED 或肠道炎症。但 EED 缺乏统一常规临床金标准,这些指标多用于研究或专科评估。
因此,家庭不能靠商业菌群检测或单个粪便指标判断孩子是否存在 EED。家庭层面更应关注特征信号:持续腹泻、便血、贫血、反复感染、体重下降、身高增长速率下降等。
3
为什么单一干预常常不够?
研究提醒我们,改善水和卫生行为本身很重要,但在复杂低资源环境中,单独卫生未必足以显著改善线性生长。这可能因为病原暴露来源复杂,包括食物、动物粪便、土壤、家庭拥挤和社区环境。
更合理的策略是综合干预:基础营养、感染控制、安全饮水、食品卫生、疫苗接种、社会支持和儿童保健同时推进。
以下内容用于解释不同干预方向的证据边界,不是治疗方案。铁剂、锌剂、维生素A/D、益生菌、抗生素、特殊医学用途配方食品、RUTF/RUSF 或微生物群导向互补食品,都应根据儿童年龄、营养状态、感染风险和医生/营养师评估决定。
干预前先评估:孩子到底慢在哪里?
对于“生长轨迹偏慢”的儿童,第一步不是购买益生菌或微量元素套餐,而是先判断:是否确有低于同年龄、同性别儿童生长曲线中位数减去2个标准差(-2SD),是否合并消瘦、低体重或水肿,是否存在慢性腹泻、贫血、反复感染、早产或低出生体重等情况。
临床检查应根据病史、生长曲线和体征选择,如 CBC、铁蛋白联合 CRP、维生素 D、甲状腺功能、大便病原、寄生虫或慢性腹泻相关评估,而非对所有儿童套用统一套餐。
母乳:早期菌群的第一生态底物
母乳不仅提供能量、蛋白质和免疫因子,也提供母乳低聚糖(HMO)。HMO 不能被婴儿直接完全消化,却能为 Bifidobacterium 等菌提供底物。唾液酸化乳低聚糖可能通过菌群依赖方式支持生长。
6月龄内支持母乳喂养,母乳不足时接受专业哺乳指导,必要时用合适配方补充。
辅食:喂孩子,也是在喂菌群
约 6 月龄后,辅食成为儿童生长和菌群成熟的重要驱动因素。WHO 2023 指南强调,6–23 月龄儿童应逐步获得足够频次、合适质地、多样化食物和动物源性食物。
高质量辅食不仅提供能量和蛋白质,也提供复杂碳水、膳食纤维、铁、锌、维生素和脂肪,共同支持 Bacteroides、Prevotella、Faecalibacterium、Blautia 等成熟菌群网络建立。
家庭层面应重视“营养密度”:主食不应只依赖稀粥,还应搭配蛋、奶、鱼肉、禽肉、瘦肉、豆制品、深色蔬果和适量油脂。
微量营养素:补对,而不是越多越好
铁、锌、维生素A、维生素D、维生素B12、叶酸、碘等均与儿童生长、免疫和神经发育相关。存在缺乏时,规范补充有助于恢复正常生长轨迹;但盲目、长期、大剂量叠加补剂并不一定更好。
锌适用于急性腹泻等明确场景;维生素A缺乏地区或高风险儿童应按公共卫生或临床规范补充。维生素D 关系骨骼发育,B12和叶酸关系造血与神经发育,碘则关系甲状腺功能和生长。
编辑
关键在于“评估后补充”。挑食但无明确缺乏的孩子,与贫血、慢性腹泻且生长曲线下降的孩子,可能需要完全不同的管理路径。
铁剂:最需要谨慎解释的营养干预
铁缺乏性贫血会影响食欲、免疫、认知和生长,规范补铁是必要治疗。但在高感染负担环境中,铁可能被部分肠道病原菌利用,并与肠道炎症风险相关。
因此,不能简单表述为“铁会喂养坏菌,所以不要补铁”。更准确的说法是:缺铁性贫血需要规范补铁;对感染负担较高或慢性腹泻儿童,应由专业人员结合血红蛋白、铁蛋白、CRP、感染风险和膳食情况综合评估。
如需补铁,还应重视饮食配合。含铁辅食、动物源性食物和富含维生素 C 的食物有助于铁摄入;茶和咖啡不宜与儿童正餐同用。
与感染控制:卫生不是万能,但仍是基础
腹泻期间应继续喂养、及时补液,必要时按医嘱补锌。若出现长期反复腹泻、便血、发热、体重下降或慢性腹胀,应就医排查病原、寄生虫、吸收不良或慢性肠病。
卫生方面,安全饮水、洗手、食物充分加热、餐具清洁、妥善处理粪便,并减少儿童接触动物粪便和污染土壤,仍是降低感染暴露的基础。需强调的是,线性生长依赖营养、感染控制和社会环境的综合改善。
抗生素:用得准,而不是完全不用
抗生素可挽救细菌感染儿童,但不应作为“调菌”工具。病毒感冒、非细菌性腹泻或无明确适应证时,不应自行使用抗生素。
生命早期反复或不必要的抗生素暴露可能扰乱菌群成熟,减少双歧杆菌和厌氧共生菌,增加腹泻和耐药风险;但真正的细菌感染仍需及时、规范治疗。
益生菌:有证据的场景,不等于治疗发育迟缓
部分益生菌在急性感染性腹泻等场景中有证据支持,但这属于腹泻管理证据,并不等同于益生菌能改善发育迟缓。其效果高度依赖菌株、剂量、年龄、基础疾病、营养状态和原有菌群。
应尽量具体到菌株,而非笼统称“益生菌”。例如,Bifidobacterium infantis EVC001 常用于讨论早期 HMO 利用和双歧杆菌定植,但不能泛化为所有 Bifidobacterium 产品都能改善生长。
早产儿、SAM、免疫低下、重症或有中心静脉导管的儿童,使用活菌制剂需在医疗监督下进行。报告中不应建议家庭自行长期、高剂量使用益生菌。
益生元、HMO/GOS:儿童证据仍不足
益生元和 HMO/GOS 的核心思路是提供底物,支持特定菌群代谢。例如 GOS 可能支持 Bifidobacterium 和 SCFA 相关通路,并在部分铁补充研究中被讨论为缓解不良菌群变化的策略。
但儿童发育迟缓干预证据不足。HMO 是母乳重要成分,并不意味着补充 HMO 产品即可治疗发育迟缓。GOS 也不能替代均衡辅食和临床营养管理。
MDCF:最有前景的菌群导向营养策略之一
微生物群导向互补食品(Microbiota-Directed Complementary Food, MDCF)是该领域重要前沿。它并非简单增加食物种类,而是基于儿童菌群成熟轨迹,设计可促进健康菌群网络的食物组合。
2021 年 NEJM 研究纳入 123 名孟加拉国 12–18 月龄中度急性营养不良(MAM)儿童,比较 MDCF-2 与常规补充食品 RUSF。结果显示,MDCF-2 在体重相关指标(如 WLZ、WAZ)变化率、部分生长相关血浆蛋白和菌群特征方面更有利。2024 年 eBioMedicine 对同一人群的两年随访进一步提示,MDCF-2 组线性生长相关结局更好。这些研究为“通过食物底物支持菌群成熟,促进营养恢复”提供了重要临床线索。
但需明确,MDCF-2 是特定研究配方,适用于孟加拉国 12–18 月龄 MAM 儿童,并在研究团队监测和食品安全条件下使用,不能简化为“花生、香蕉、大豆随意混合”,也不能作为所有发育迟缓儿童的家庭处方。
因此,发育迟缓干预的第一步不是购买菌群产品,而是专业评估、基础营养、感染控制和卫生改善。益生菌和 HMO/GOS 是重要研究方向,但必须明确适用人群、配方、剂量和安全边界。
对家庭而言,最重要的不是寻找一种“神奇菌”,而是稳定做好生长监测、基础营养、感染预防和及时就医。对临床和公共卫生而言,重点是分层评估和多部门支持。
1
家庭可做什么?
•0–6月龄:优先支持母乳喂养,关注尿量、体重增长、喂养姿势和奶量。不要用米汤、果汁或成人食物替代奶。若体重增长不足、喂养困难或尿量减少,应就医评估。
•6–23 月龄:约 6 月龄及时添加辅食,逐步提高质地和频次。每餐尽量包括主食、蛋/奶/鱼肉/豆类、蔬果和适量油脂。避免长期稀粥、汤水或零食替代正餐。
•2岁以上:维持规律三餐和多样化饮食,减少甜饮和高糖零食。关注身高增长速率,而不是只看单次身高。
2
哪些情况需要就医?
出现以下情况应及时就医:
1.HAZ/LAZ < -2 或生长曲线持续明显下滑;
2.体重下降、消瘦、MUAC 异常或水肿;
3.持续或反复腹泻、便血、长期呕吐或明显腹胀;
4.贫血、反复发热或感染、慢性咳嗽、食欲显著下降;
5.发育倒退、神经发育迟缓或青春发育异常。
早产、低出生体重、免疫缺陷、慢性肠病、肝肾病、内分泌疾病或遗传病史儿童,也应尽早接受专业评估。
3
公共卫生需要做什么?
公共卫生策略应整合食物可及性、母乳和辅食教育、强化食品、免疫接种、感染控制和社会支持等。
儿童发育迟缓并不只是“吃得少”,也不能简单归因于“菌群不好”。它往往是营养质量不足、反复感染、卫生条件差、环境性肠功能障碍(EED)、肠道菌群发育不成熟、肠屏障受损、免疫炎症持续激活和生长轴调控异常等多因素长期共同作用的结果。
肠道菌群研究为理解“为什么吃进去了,孩子却不一定长起来”提供了新视角。前沿研究提示,未来营养干预可能不仅要“喂养孩子”,也要“喂养菌群”:通过支持放线杆菌、拟杆菌、链球菌、双歧杆菌、Prevotella 属及多种年龄相关关键菌群的发育,改善生长和整体健康结局。
不过,这一领域仍有许多问题待解:哪些微生物是真正关键的干预靶点?相关干预通过何种机制发挥作用?能否据此发展出更个体化的营养策略,用于应对营养不良和发育迟缓?这些仍需进一步研究。
从临床到公共卫生,这些发现都具有重要意义。医务人员可据此设计更有针对性的营养支持方案,促进脆弱儿童肠道微生物群健康发育;若将相关干预纳入国家营养项目和社区行动,也有望带来更持久的健康收益,帮助打破营养不良与贫困的恶性循环。
但目前最可靠、最基础的策略,仍是专业评估、合理营养、感染控制、卫生改善,以及持续的家庭和社会支持。
医学免责声明:本文为科普与综述性资料,不构成个体化医疗建议。
主要参考文献:
Subramanian S, Huq S, Yatsunenko T, Haque R, Mahfuz M, Alam MA, Benezra A, DeStefano J, Meier MF, Muegge BD, Barratt MJ, VanArendonk LG, Zhang Q, Province MA, Petri WA Jr, Ahmed T, Gordon JI. Persistent gut microbiota immaturity in malnourished Bangladeshi children. Nature. 2014 Jun 19;510(7505):417-21.
WHO Multicentre Growth Reference Study Group. WHO Child Growth Standards based on length/height, weight and age. Acta Paediatr Suppl. 2006 Apr;450:76-85.
WHO Guideline for complementary feeding of infants and young children 6–23 months of age [Internet]. Geneva: World Health Organization; 2023.
Guideline: Updates on the Management of Severe Acute Malnutrition in Infants and Children. Geneva: World Health Organization; 2013.
WHO guideline on the prevention and management of wasting and nutritional oedema (acute malnutrition) in infants and children under 5 years. Geneva: World Health Organization; 2023.
Bhutta ZA, Das JK, Rizvi A, Gaffey MF, Walker N, Horton S, Webb P, Lartey A, Black RE; Lancet Nutrition Interventions Review Group, the Maternal and Child Nutrition Study Group. Evidence-based interventions for improvement of maternal and child nutrition: what can be done and at what cost? Lancet. 2013 Aug 3;382(9890):452-477.
谷禾健康
在多组学研究中,重复测量数据是一个宝藏。
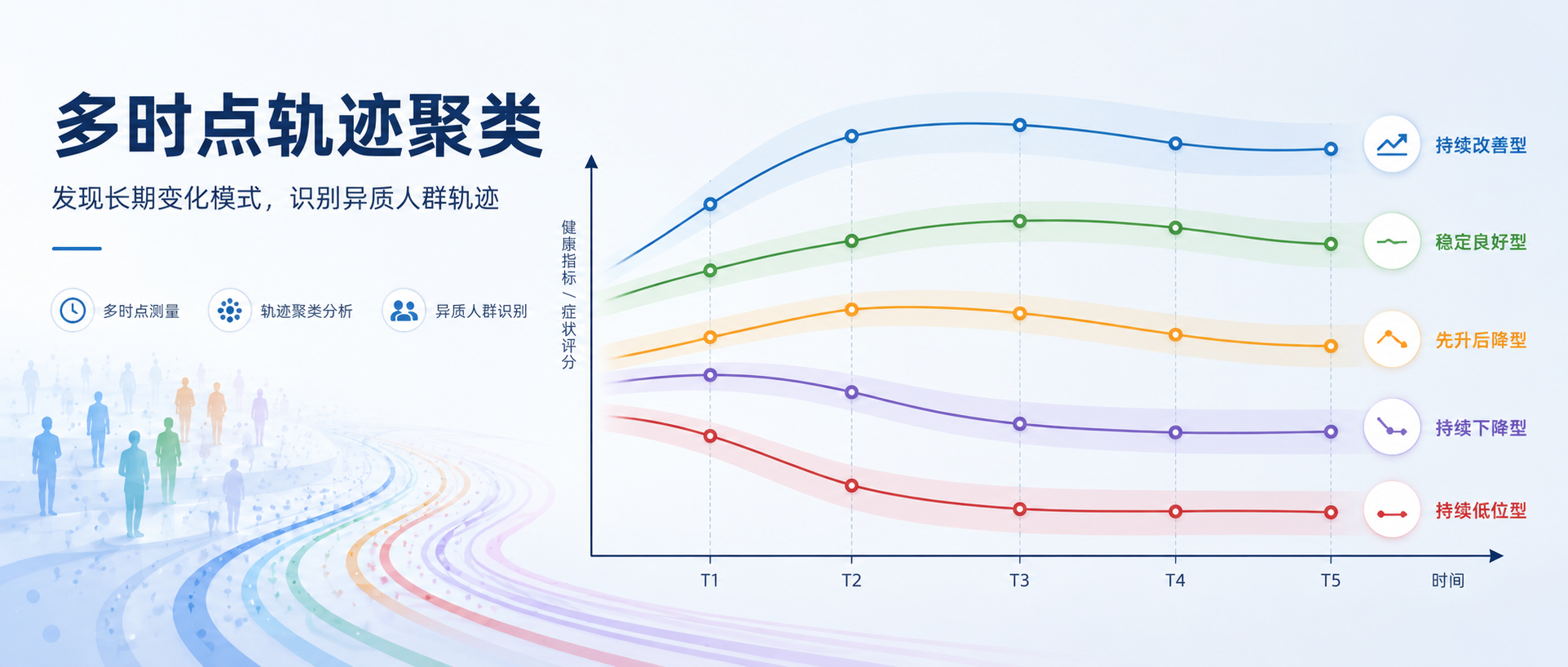
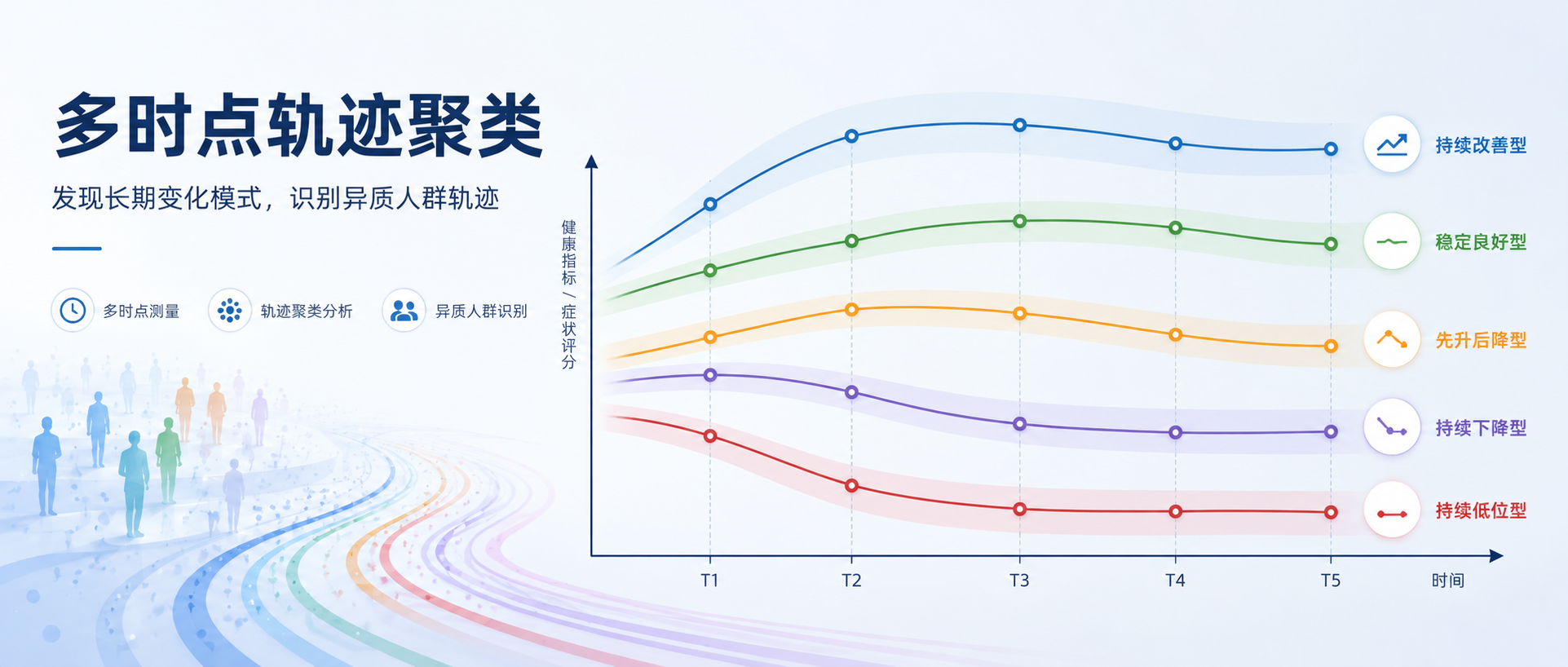
相比于传统的单时间点静态对比,追踪同一个体在多个时间点的动态变化,能够帮我们看到更细的异质性:有的人快速上升,有的人缓慢恢复,有的人看似稳定,但其实只是在绝对水平上分层。
近期正在整理一些开源数据,发现一份代谢组学数据,它包含 5 个时间点的重复测量信息,队列是在接受新辅助化疗的晚期直肠癌患者。面对这份数据,我们产生了一个问题:
能否利用这些连续时间点,为每个重要代谢物绘制出个体的“动态响应轨迹”?进一步通过轨迹聚类,发现具有不同响应特征的亚群?
为了回答这个问题,我们进行了探索性数据分析,建立了一套工作流:
原始代谢组矩阵→ log1p 转换→ ICC + 时间效应 + 变化幅度 + 斜率异质性筛选→ 基线中心化→ latrend 单变量轨迹聚类→ BIC / APPA / OCC / Relative Entropy / 完整病例 ARI 敏感性分析
这篇文章就以这次实战为例,讲一讲多时点重复测量数据如何做轨迹聚类。
原始数据中共有 557 个代谢物。
直觉上,我们似乎可以把每个代谢物都送进轨迹聚类模型里,然后让模型自己找模式。但这样做通常会带来三个问题:
所以,在聚类前,我们先做了一轮多维度过滤,筛出真正具有纵向动态潜力的候选分子。
代谢组丰度通常右偏明显,并且存在极端值。第一步先对原始丰度做 log 转换。
library(data.table)
# x 为某个代谢物的原始丰度向量
if (all(x >= 0, na.rm = TRUE)) {
y_log <- log1p(x)
} else {
# 如果存在负值,先整体平移再取 log
y_log <- log(x – min(x, na.rm = TRUE) + 1)
}
这里需要注意:log1p 不是“消除”右偏和异方差,而是减弱右偏、降低极端值影响、改善方差稳定性。
有些代谢物看起来有差异,但差异主要来自“这个人一直高、那个人一直低”。这种情况下,轨迹聚类很容易分成“高水平组”和“低水平组”,而不是我们真正关心的“变化模式组”。
因此,我们计算了每个代谢物的组内相关系数(Intraclass Correlation Coefficient, ICC):
ICC = between-subject variance / total variance
如果 ICC 很高,说明变异主要来自稳定的个体间差异。在这次分析中,我们将筛选阈值定义在 ICC < 0.70 ,优先保留个体内随时间变化占比更高的代谢物。
R 代码示意:
library(lme4)
fit_icc <- lmer(y_log ~ 1 + (1 | subject_id), data = df, REML = TRUE)
vc <- as.data.frame(VarCorr(fit_icc))
var_between <- vc$vcov[vc$grp == “subject_id”]
var_resid <- vc$vcov[vc$grp == “Residual”]
icc <- var_between / (var_between + var_resid)
接下来,我们使用线性混合模型(LMM)检验每个代谢物是否存在整体时间效应。
比较两个模型:
null model: y_log ~ 1 + (1 | subject_id)
time model: y_log ~ time + (1 | subject_id)
R 代码示意:
library(lme4)
m0 <- lmer(y_log ~ 1 + (1 | subject_id), data = df, REML = FALSE)
m1 <- lmer(y_log ~ factor(timepoint) + (1 | subject_id), data = df, REML = FALSE)
lrt <- anova(m0, m1)
p_time <- lrt$`Pr(>Chisq)`[2]
对所有代谢物的 p 值做 BH-FDR 校正:
screen_table[, time_FDR := p.adjust(p_time, method = "BH")]
筛选阈值:
time_FDR < 0.05
统计显著性不等于变化幅度有意义。
因此,我们又计算了每个代谢物在不同时间点的平均 log 丰度范围:
logFC_range = max(mean log abundance across timepoints)
– min(mean log abundance across timepoints)
R 代码示意:
fc_range <- df[, .(mean_y = mean(y_log, na.rm = TRUE)), by = timepoint][
, max(mean_y, na.rm = TRUE) – min(mean_y, na.rm = TRUE)
]
保留 logFC_range 位于全体代谢物前 25% 的分子,即:
logFC_range ≥ 75% 分位数
最后,我们希望候选分子不仅有整体时间变化,还要有个体间变化趋势差异。
做法是:对每个个体单独拟合一条简单线性回归,提取 slope,然后计算 slope 的标准差。
get_subject_slope <- function(d) {
if (sum(!is.na(d$y_log)) < 2) return(NA_real_)
coef(lm(y_log ~ time_num, data = d))[“time_num”]
}
slopes <- df[, .(slope = get_subject_slope(.SD)), by = subject_id]
slope_sd_OLS <- sd(slopes$slope, na.rm = TRUE)
筛选阈值:
slope_sd_OLS ≥ 75% 分位数
在这份数据中,筛选结果如下:
原始总代谢物:557 个
ICC < 0.70 且 time FDR < 0.05:331 个
严格满足 ICC、time FDR、logFC_range ≥ q75、slope_sd_OLS ≥ q75:63 个
这 63 个候选分子 就是最终进入轨迹聚类分析的代谢物。
在纵向数据中,一个常见问题是:模型可能按照个体的绝对基线水平分组。
比如某个代谢物,A 组从头到尾都高,B 组从头到尾都低。图上看起来分成了两条线,但这未必说明它们有不同的动态响应模式。
为了减少这种基线水平驱动的分组,在进入 latrend 前做了基线中心化:
y_bc,it = log1p(y_it) - log1p(y_i,Baseline)
其中:
i 表示第 i 个个体;t 表示时间点;Baseline 表示该个体自己的基线时间点。R 代码示意:
library(data.table)
setDT(df)
setorder(df, subject_id, time_num)
# df 包含 subject_id, timepoint, time_num, raw_value
# 先 log1p
if (all(df$raw_value >= 0, na.rm = TRUE)) {
df[, y_log := log1p(raw_value)]
} else {
df[, y_log := log(raw_value – min(raw_value, na.rm = TRUE) + 1)]
}
# 提取每个个体的 baseline log 值
baseline_values <- df[timepoint == “Baseline”, .(
baseline_log = y_log[1]
), by = subject_id]
# 合并回长表,并计算 baseline-centered value
df <- merge(df, baseline_values, by = “subject_id”, all.x = TRUE)
df[, y_bc := y_log – baseline_log]
基线中心化后的 y_bc 可以理解为:
log-scale change from baseline ≈ 相对于个体自身基线的 log fold-change
它的优点是:
当然,它也有代价:如果一个代谢物的生物学意义主要体现在绝对水平高低,那么 基线中心化可能会削弱分类信号。
所以这一步的目的不是让所有模型都变好,而是让聚类问题从“谁高谁低”转向“谁怎么变化”。
筛选出候选分子并完成基线中心化后,就可以进入轨迹聚类分析了。
这里我们使用 R 包 latrend,并通过其统一接口调用底层的 lcmm 模型。
对每个代谢物,分别拟合:
GBTM: Group-Based Trajectory ModelGMM: Growth Mixture Model
候选类别数:
K = 2, 3, 4
时间函数:
quadratic polynomial: poly(time, 2, raw = TRUE)
下面是一段可复用的核心代码框架。
library(latrend)
library(lcmm)
library(data.table)
make_methods <- function(K) {
K <- as.integer(K)
gbtm <- lcMethodLcmmGBTM(
fixed = y_bc ~ poly(time_num, 2, raw = TRUE),
mixture = ~ poly(time_num, 2, raw = TRUE),
id = “subject_id”,
time = “time_num”,
nClusters = K
)
gmm <- lcMethodLcmmGMM(
fixed = y_bc ~ poly(time_num, 2, raw = TRUE),
mixture = ~ poly(time_num, 2, raw = TRUE),
random = ~ time_num,
id = “subject_id”,
time = “time_num”,
nClusters = K
)
methods <- list(gbtm, gmm)
names(methods) <- c(paste0(“GBTM_K”, K), paste0(“GMM_K”, K))
methods
}
methods <- unlist(lapply(2:4, make_methods), recursive = FALSE)
models <- latrendBatch(
methods,
data = df[!is.na(y_bc)],
cartesian = FALSE,
errorHandling = “pass”
)
轨迹图很直观,但不能只凭肉眼判断。
在潜类别轨迹模型里,最容易犯的错误是:看到几条线分开了,就以为“分型成立”。实际上,轨迹聚类至少要同时回答三个问题:
1. 模型整体分得清不清?
2. 每个亚群内部分类把握度够不够?
3. 换一批更严格的完整病例样本后,分型是否还能复现?
因此,我们综合使用了以下指标:
编辑
下面逐个解释这些指标。
group_sizes 指的是最终模型中每个潜在轨迹类别包含的个体数。例如:
group_sizes: A:90; B:75
意思是模型把个体分成 A、B 两类,其中 A 类 90 人,B 类 75 人。
min_group_n 则是所有类别中人数最少的那一类:
min_group_n = min(number of subjects in each trajectory class)
为什么要看这个?
因为潜类别模型有时会切出一个人数极少的小类。这个小类可能看起来很“特别”,但也可能只是模型为了追求更低 BIC 而切出来的噪声亚群。
在本次工作流中,我们对 165 例全样本主模型优选 设定:
min_group_n ≥ 20
也就是说,在主分析中,每个轨迹亚群至少要有 20 个个体,才进入优先考虑范围。
但需要注意:96 例完整病例分析的定位是敏感性分析。它主要用于检验“同样的模型类型和类别数,在完整 5 时间点样本中是否能得到相似分型”,因此我们在 96 例模型中报告实际的 group_sizes、APPA、OCC 和 ARI,但没有再次强制使用 min_group_n ≥ 20 作为模型入选门槛。
R 代码示意:
assignments <- trajectoryAssignments(final_model)
class_sizes <- table(assignments)
group_sizes <- paste(names(class_sizes), as.integer(class_sizes), sep = “:”, collapse = “;”)
min_group_n <- min(as.integer(class_sizes))
group_sizes
min_group_n
APPA,全称是 Average Posterior Probability of Assignment,即平均后验分配概率。
它衡量的是:
被分到某一类的个体,平均有多大概率真的属于这一类。
举个例子,如果 A 组的 APPA = 0.92,意思是:模型把这些人分到 A 组时,平均后验概率达到 0.92,分类把握度较高。
理论上,所有类别的 APPA 都应该达到:
APPA ≥ 0.70
如果某一类 APPA 很低,说明这个亚群边界模糊,很多个体其实处在“可分可不分”的状态。
在文章结果中,我们展示的是:
min APPA
也就是所有类别中最差的那个 APPA:
min APPA = min(APPA across trajectory classes)
为什么展示最小值?因为平均值可能掩盖问题。一个模型可能大组 APPA 很高,小组 APPA 很低,平均看起来还不错,但小组其实并不可靠。
OCC,全称是 Odds of Correct Classification,即正确分类几率比。
相比 APPA,OCC 的优势是:它考虑了类别样本量不均衡的影响。
直观地说,OCC 衡量的是:
模型把个体分到某一类的正确分类几率,相比随机分配提高了多少倍。
常见经验标准是:
OCC > 2.0:通常可认为分类优于随机分配OCC > 5.0:更严格,说明错分风险更低
本次分析中,我们在模型优选时采用了更严格的标准:
所有类别 OCC > 5
和 APPA 一样,文章中展示的是:
min OCC
也就是所有类别中最差的那个 OCC。这样可以避免某个边缘亚群分类质量很差,却被整体结果掩盖。
APPA 和 OCC 可以从 lcmm 模型的后验概率表中手动计算。
library(data.table)
calc_quality <- function(model, model_name) {
# latrend 模型底层存放的是 lcmm 对象
lc <- slot(model, “model”)
# pprob 包含:个体 ID、最终分类 class、以及 prob1/prob2/… 后验概率
pp <- as.data.table(lc$pprob)
n_total <- nrow(pp)
class_quality <- rbindlist(lapply(sort(unique(pp$class)), function(k) {
prob_col <- paste0(“prob”, k)
# 该类别人数
n_k <- sum(pp$class == k)
# 该类别占比
pi_k <- n_k / n_total
# APPA:被分到第 k 类个体,其第 k 类后验概率的平均值
appa <- mean(pp[[prob_col]][pp$class == k], na.rm = TRUE)
# OCC:正确分类 odds 相对于随机分配 odds 的提升倍数
occ <- (appa / (1 – appa)) / (pi_k / (1 – pi_k))
data.table(
model = model_name,
class = k,
n = n_k,
proportion = pi_k,
APPA = appa,
OCC = occ
)
}))
# 文章中展示的是最保守的 min APPA / min OCC
summary_quality <- class_quality[, .(
group_sizes = paste0(“class”, class, “:”, n, collapse = “;”),
min_group_n = min(n),
min_APPA = min(APPA, na.rm = TRUE),
min_OCC = min(OCC, na.rm = TRUE)
)]
list(
by_class = class_quality,
summary = summary_quality
)
}
相对熵(Relative Entropy,也常写作 Entropy)是一个全局分类不确定性指标。
它的取值范围通常在:
0 到 1
解释上可以这样理解:
Relative Entropy 越接近 1:类别之间边界越清晰,后验概率越集中Relative Entropy 越接近 0:类别之间重叠越严重,分类越不确定
在轨迹模型评估中,常见经验判断是:
Relative Entropy ≥ 0.80:分类边界较清晰,是比较理想的结果0.60–0.80:中等清晰,需要结合 APPA/OCC/ARI 判断< 0.60:整体分类边界模糊,需要谨慎解释
在 latrend 中,可以直接使用 metric() 提取相对熵:
library(latrend)
relative_entropy <- metric(final_model, “relativeEntropy”)
relative_entropy
如果希望自己从后验概率表中复核,也可以根据 posterior probability 手动计算。下面给出一个常见计算方式:
calc_relative_entropy <- function(model) {
lc <- slot(model, “model”)
pp <- as.data.table(lc$pprob)
# 提取 prob1, prob2, …, probK
prob_cols <- grep(“^prob[0-9]+$”, names(pp), value = TRUE)
P <- as.matrix(pp[, ..prob_cols])
n <- nrow(P)
K <- ncol(P)
# 避免 log(0)
P[P <= 0] <- .Machine$double.eps
# classification entropy
entropy_raw <- -sum(P * log(P))
# relative entropy, normalized to 0-1
relative_entropy <- 1 – entropy_raw / (n * log(K))
relative_entropy
}
relative_entropy_manual <- calc_relative_entropy(final_model)
relative_entropy_manual
这也是为什么在实战结果中,除了 APPA/OCC,我们还展示了每个模型的 relative entropy。
完整病例敏感性分析比较的是:
全样本模型:165 个个体,允许部分时间点缺失完整病例模型:96 个个体,均具有 5 个时间点
核心问题是:全样本模型和完整病例模型是否给出了相似的个体分型?
这里使用 Adjusted Rand Index(ARI) 比较两套聚类标签。
ARI 的取值通常可以这样理解:
ARI ≥ 0.90:高度稳健与可重复0.40–0.90:存在一定一致性,但需要结合图形和其他指标判断ARI < 0.40:分类存在高度不稳定性或随机性,需要警惕ARI 接近 0:一致性接近随机水平ARI < 0:比随机一致性还差
R 代码示意:
library(mclust)
library(data.table)
# all_assign: 165 例全样本模型的分类结果
# complete_assign: 96 例完整病例模型的分类结果
# 两个表都至少包含 id 和 traj_group 两列
all_assign <- fread(“final_assignments_all_165_baseline_centered.csv”)
complete_assign <- fread(“final_assignments_complete_96_baseline_centered.csv”)
# 只比较两套结果共有的 96 个完整病例个体
ari_dt <- merge(
all_assign,
complete_assign,
by = “id”,
suffixes = c(“_all165”, “_complete96”)
)
ari <- adjustedRandIndex(
ari_dt$traj_group_all165,
ari_dt$traj_group_complete96
)
ari
这里还有一个技术细节:潜类别模型可能发生 label switching,也就是 A/B/C 标签名称在不同模型里被置换。
ARI 本身不受标签名称置换影响,因此适合用于比较两套聚类结果的一致性;但如果要并排画图展示,就需要根据最大重叠关系,把完整病例模型的 A/B/C 标签重标到全样本模型的语义上。
我们的实际判断顺序是这样的。
第一步:先看全局 Relative Entropy。
Relative Entropy 相当于先给模型做一个“全身检查”:
Entropy ≥ 0.80:整体分类边界清晰,可以进入下一步局部检查Entropy < 0.60:整体边界模糊,即使图形好看,也要谨慎解释0.60–0.80:中间地带,需要结合 APPA、OCC、ARI 和轨迹形态综合判断
第二步:再看分亚群的 APPA 和 OCC。
在整体边界还可以的前提下,进一步排查每个亚群:
每个亚群 APPA 是否 ≥ 0.70?每个亚群 OCC 是否至少 > 2.0?如果采用更严格标准,是否能达到 OCC > 5.0?
这一步是为了防止某些“人数很少、边界很模糊”的亚群被整体指标掩盖。
第三步:看 group_sizes。
即使 APPA/OCC 达标,如果某个类别只有几个人,也不适合作为稳定亚型解释。本次设置了较保守的:
min_group_n ≥ 20
第四步:最后看 complete-case ARI。
如果全样本模型看起来很好,但 96 例完整病例模型给出完全不同的划分,那说明结果可能受到缺失时间点或样本组成影响。
因此,ARI 是最后的复现性检查:
ARI ≥ 0.90:非常稳健ARI < 0.40:高度不稳定,需要作为警示案例
一句话总结:
Relative Entropy 看整体边界,APPA/OCC 看每个亚群的分类把握度,group_sizes 看亚群是否太小,ARI 看全样本和完整病例之间是否可重复。四者结合,才比单纯看轨迹图可靠得多。
从下面的实战结果开始,每个代谢物都并排展示两张图:左侧为全样本 165 例模型,右侧为完整 5 时间点的 96 例敏感性分析模型。这样可以同时观察主分析轨迹形态和完整病例中的稳定性。
全样本模型,165 例:
final_model: GMM_K2
group_sizes: A:90; B:75
min APPA: 0.850
min OCC: 5.199
relative entropy: 0.550
完整病例模型,96 例,均有 5 个时间点:
complete_model: GMM_K2
group_sizes: A:42; B:54
min APPA: 0.866
min OCC: 5.026
relative entropy: 0.584
全样本 vs 完整病例标签一致性:
complete-case ARI: 0.558
编辑
-敏感性校验
全样本模型与完整病例模型的 ARI 为 0.558,提示两套分型之间存在中等一致性,但未达到高度稳健水平。
因此,Phenylalanine 的两类轨迹结构具有一定可重复性,但仍可能受到样本完整性或缺失时间点的影响。
-结果说明
在基线中心化后,Phenylalanine 的全样本轨迹可见两类相对变化模式。两组在时间轴上的均值曲线存在差异,提示该代谢物的动态变化并非完全由个体基线水平驱动。
APPA和OCC均达到预设标准,但Relative entropy < 0.60,提示全局分类边界并不十分清晰。该结果更适合作为探索性发现,而非直接作为稳定亚型结论。
全样本模型,165 例:
final_model: GBTM_K2
group_sizes: A:42; B:123
min APPA: 0.951
min OCC: 8.485
relative entropy: 0.842
完整病例模型,96 例,均有 5 个时间点:
complete_model: GBTM_K2
group_sizes: A:20; B:76
min APPA: 0.915
min OCC: 10.351
relative entropy: 0.856
全样本 vs 完整病例标签一致性:
complete-case ARI: 1.000
编辑
-敏感性校验
全样本模型与完整病例模型的 ARI 为 1.000,说明两套模型在共有个体上的分型完全一致。该结果支持该代谢物轨迹分型具有较高稳健性,受缺失时间点影响较小。
-结果解释
12alpha-Hydroxy-3-oxochola-4,6-dienoate 的全样本轨迹显示出两类较明确的相对变化模式。完整病例图中,两类轨迹的主要形态与全样本结果保持一致,说明该分型结构在样本限制后仍然可见。
APPA和OCC均达到预设标准,Relative entropy > 0.80,提示类别边界清晰。该代谢物是较适合后续分析的候选对象。
其分型同时具备清晰的视觉分离、较高的分类纯度、较好的全局 entropy 以及完整病例复现性,可优先用于后续与疗效、病理反应或代谢通路变化的关联分析。
全样本模型,165 例:
final_model: GMM_K3
group_sizes: A:122; B:21; C:22
min APPA: 0.871
min OCC: 11.440
relative entropy: 0.888
完整病例模型,96 例,均有 5 个时间点:
complete_model: GMM_K3
原始标签 group_sizes: A:17; B:64; C:15
按全样本语义重标后 group_sizes: A:64; B:15; C:17
min APPA: 0.891
min OCC: 27.438
relative entropy: 0.906
全样本 vs 完整病例标签一致性:
complete-case ARI: 0.966
编辑
-敏感性校验
全样本模型与完整病例模型的 ARI 为 0.966,说明在共有个体中,三类分型具有高度一致性。
需要强调的是,ARI 支持分型结构可复现,但不能抵消完整病例模型中小类样本量不足的问题。
-结果解释
N-Acetylisoniazid 的全样本轨迹呈现三类相对变化模式,而非简单的二分类结构。三类轨迹形态在完整病例中仍可观察到。
APPA和OCC均达到预设标准,Relative entropy > 0.80,提示三类结构具有较高分类确定性和较清晰的全局边界。在后续临床关联分析中,应特别注意小亚群样本量对效应估计和置信区间稳定性的影响。
全样本模型,165 例:
final_model: GBTM_K2
group_sizes: A:82; B:83
min APPA: 0.880
min OCC: 7.275
relative entropy: 0.651
完整病例模型,96 例,均有 5 个时间点:
complete_model: GBTM_K2
group_sizes: A:50; B:46
min APPA: 0.872
min OCC: 7.396
relative entropy: 0.669
全样本 vs 完整病例标签一致性:
complete-case ARI: 0.958
编辑
-敏感性校验
全样本模型与完整病例模型的 ARI 为 0.958,提示两套分型在共有个体中具有高度一致性。结合均衡 group sizes,该代谢物的二分类结果具有较好的稳健性。
-结果解释
5-Hydroxyectoine 的基线中心化轨迹可分为两类,且两类在全样本和完整病例图中均呈现相似的分离趋势。相较于极端不均衡分组,该代谢物的两类轨迹在人群规模上更接近。
APPA和OCC均达到预设标准,Relative entropy在0.60–0.80之间,说明各类别后验分配质量达标,但全局边界清晰度处于中等水平。算是适合后续比较分析的候选分子。
其优势在于两类样本量均衡、分类质量达标、完整病例复现性较好;但 Relative entropy 未超过 0.80,因此仍应避免将其解读为边界极其清晰的分型。
全样本模型,165 例:
final_model: GMM_K2
group_sizes: A:39; B:126
min APPA: 0.794
min OCC: 2.041
relative entropy: 0.519
完整病例模型,96 例,均有 5 个时间点:
complete_model: GMM_K2
group_sizes: A:43; B:53
min APPA: 0.846
min OCC: 5.493
relative entropy: 0.527
全样本 vs 完整病例标签一致性:
complete-case ARI: 0.309
编辑
-敏感性校验
全样本模型与完整病例模型的 ARI 为 0.309,低于 0.40,提示两套分型一致性较差,存在明显不稳定性。
-结果解释
Flavone 的轨迹图在全样本和完整病例中均可见两类曲线,单纯从图形上看,该代谢物似乎可以被划分为两类,但视觉分离并不能直接证明分型可靠。
全样本 min APPA 为 0.794,达到 APPA ≥0.70 的最低要求;但 min OCC 为 2.041,仅接近宽松阈值,未达到本次主分析优先采用的 OCC >5 标准,Relative entropy < 0.60,提示整体分类边界不清晰。
96 例完整病例模型的 min APPA 为 0.846,min OCC 为 5.493,Relative entropy < 0.60,说明完整病例模型内部虽然局部指标改善,全局分类边界仍不清晰。
本次分析使用的是单变量轨迹聚类。也就是说,每次只针对一个代谢物建模。
在真实的前瞻性临床研究中,还可以继续往下做:
例如,将某个代谢物的轨迹亚型作为核心暴露变量,进一步分析它与疗效、预后或不良事件之间的关系。
可选模型包括:
Cox proportional hazards modellogistic regressionlinear mixed modelGEE
如果有年龄、性别、BMI、疾病分期、用药、生活方式等元数据,可以在后续关联模型中进行校正。
单个代谢物的轨迹有时噪声较大。可以先通过 WGCNA、通路富集或 PCA 得到代谢模块得分,再对模块得分做轨迹聚类。
这种思路可能更接近系统生物学层面的动态异质性。
如果目标是联合多个代谢物的动态变化,可以尝试 Joint / Multivariate LCMM 或其他多变量纵向聚类方法。
谷禾健康
科研绘图不仅要展示真实严谨的数据,更要讲究排版的美观与可读性。
今天小编来分享一个兼具美感和实用性的R包——vbracket,它打破常规,将组间差异的统计学显著性括号直接集成到了图例的垂直结构中,助你的文章插图提升到顶刊发表级的水准。
vbracket?vbracket 是专门为 ggplot2 开发的一个高度定制化的图例增强包。
它的核心设计理念是:既然图例已经垂直排列了各个分组的名称和颜色,为什么不直接在图例的分组之间拉起垂直括号,来展示它们的两两差异呢?
vbracket 做法编辑
(注:图例右侧的垂直括号展示了 WT/Dox vs CH3+5 等组间的显著性差异)
vbracket 高阶美化编辑
vbracket 官方示例效果展示
编辑
vbracket 官方示例效果展示
下面我们以代谢物数据差异分析为例,展示如何快速上手 vbracket。
目前该包托管在 GitHub,可以通过 devtools 快速安装:
# 如果未安装 devtools,请先执行 install.packages("devtools")devtools::install_github("h20gg702/vbracket")
研究问题:益生菌干预前后,短链脂肪酸(SCFAs)的变化是否不同于安慰剂组?
这里的重点不是简单比较 0d 或 56d 某一个时间点两组有没有差异,而是两组从干预前到干预后的“变化幅度”是否显著不同:
每个受试者先计算:Δ = 56d 相对丰度 - 0d 相对丰度然后比较:Probiotic 组 Δ vs Placebo 组 Δ
suppressPackageStartupMessages({ library(ggplot2) library(vbracket)})metab_file <- file.path(base_dir,"scfa_metabolomics_filtered_by_subject_id_freq_gt1_samples.csv")meta_file <- file.path(base_dir,"filtered_metainfo_subject_id_freq_gt1_rows.csv")metab <- read.csv(metab_file, check.names =FALSE, stringsAsFactors =FALSE)meta <- read.csv(meta_file, check.names =FALSE, stringsAsFactors =FALSE)meta$ghname <- trimws(as.character(meta$ghname))meta$subject_id <- trimws(as.character(meta$subject_id))meta$time_point <- trimws(as.character(meta$time_point))meta$day <- ifelse(meta$time_point =="0d",0, ifelse(meta$time_point =="56d",56,NA_real_))meta$Group <- ifelse(meta$`研究分组` =="安慰剂干预","Placebo", ifelse(meta$`研究分组` =="益生菌干预","Probiotic", as.character(meta$`研究分组`)))meta$Group <- factor(meta$Group, levels = c("Placebo","Probiotic"))sample_cols <- setdiff(colnames(metab),"Metabolite")sample_cols <- sample_cols[sample_cols %in% meta$ghname]long_list <- lapply(seq_len(nrow(metab)),function(i){ data.frame( Metabolite = metab$Metabolite[i], ghname = sample_cols, intensity = as.numeric(metab[i, sample_cols]), stringsAsFactors =FALSE, check.names =FALSE)})long <- do.call(rbind, long_list)plot_df <- merge( long, meta[, c("ghname","subject_id","研究分组","Group","time_point","day")], by ="ghname", all.x =TRUE)
到这里,我们就得到了适合 ggplot2 绘图和统计分析的长格式数据。
一部分人的做法是直接在 0d 和 56d 分别比较两组,这样回答的是:
某个时间点,两组是否不同?
但如果你的问题是“干预前后变化是否不同”,更合适的做法是先在每个受试者内部计算变化量:
Δ56d-0d = intensity_56d - intensity_0d
脚本中使用 reshape() 将同一个 subject_id 的 0d 和 56d 拉成一行,然后计算 Δ:
wide <- reshape( sub[, c("Metabolite","subject_id","Group","day","intensity")], idvar = c("Metabolite","subject_id","Group"), timevar ="day", direction ="wide")wide$delta_56d_minus_0d <- wide$intensity.56- wide$intensity.0wide <- wide[!is.na(wide$delta_56d_minus_0d),]
随后,对每个代谢物分别比较两组的变化量:
delta_placebo <- wide$delta_56d_minus_0d[wide$Group =="Placebo"]delta_probiotic <- wide$delta_56d_minus_0d[wide$Group =="Probiotic"]pval <- t.test(delta_placebo, delta_probiotic)$p.value
也就是说,这里的统计检验是:
Placebo 组的 Δ56d-0d vs Probiotic 组的 Δ56d-0d
将 p 值转换成常见的显著性标记:
p_to_star <-function(p){if(is.na(p)) return("NA")if(p <0.001) return("***")if(p <0.01) return("**")if(p <0.05) return("*")"ns"}
最终,每个短链脂肪酸都会得到一个变化量比较结果,例如:
Δ56d-0d: nsΔ56d-0d: *
这个标签不是某个时间点的横向差异,而是干预前后变化幅度的组间差异。
vbracket展示显著性结果绘图部分保持经典的论文风格:
首先定义颜色:
colors <- c("Placebo"="#00AA00","Probiotic"="#FF4400")groups <- c("Placebo","Probiotic")
然后把变化量检验得到的显著性结果写进 vbracket 的比较结构中:
vb_label <- paste0("Δ56d-0d: ", stat_m$significance)comparisons <- add_bracket_comparisons( groups1 ="Placebo", groups2 ="Probiotic", labels = vb_label)
最后绘图:
p <- ggplot(sum_m, aes(x = day, y = mean, color = Group, group = Group))+ geom_line(linewidth =1.2)+ geom_point(size =3)+ geom_errorbar(aes(ymin = mean - sem, ymax = mean + sem), width =4, linewidth =0.8)+ scale_color_manual(values = colors)+ scale_x_continuous( breaks = c(0,56), limits = c(-3,59), labels = c("0d","56d"))+ labs( title = met, x ="Time point", y ="Relative Abundance")+ theme_classic(base_size =14)+ theme( plot.title = element_text(hjust =0.5, size =16, face ="bold"), legend.position ="none")+ legend_bracket( labels = groups, colors = colors, comparisons = comparisons, position ="topleft", output_width =6, output_height =4, width =0.36, text_size =9, sig_size =8)ggsave(file_png, p, width =6, height =4, dpi =300)
编辑
从图中可以直观看出,尽管起点不同,但益生菌干预显著促进了丙酸的产生
为了让大家在排版时少走弯路,在使用 vbracket 时有以下两点需要特别注意:
由于图例中文字宽度、括号间距的计算是基于物理绝对单位(如毫米或英寸)进行的,因此在 legend_bracket() 中设置的 output_width 和 output_height,必须与你最终用 ggsave() 保存时的宽高完全一致!否则,导出的 PDF 或 PNG 中括号和文字可能会发生错位。
在编写 legend_bracket(labels = ...) 时,填写的组名顺序要与你在 add_bracket_comparisons() 里使用的组名完全一致,且顺序应符合逻辑(通常是从上到下对应的实验组顺序)。
vbracket 包非常适合处理多时间点折线图+显著性标记。如果你也有类似的场景,一起用起来吧!
谷禾健康
川崎病 (KD) 是一种急性、免疫介导的血管炎,也是儿童获得性心脏病的主要原因。其最早由日本儿科医生川崎富作于1967年首次报告,此后全球六十多个国家相继发现病例。尽管临床诊治水平不断进步,但川崎病的病因至今仍未完全阐明。它不像普通感染那样有明确的病原体,也不像自身免疫病那样有清晰的自身抗体——它更像是一个由遗传易感性、环境触发因素和免疫异常反应共同编织的复杂谜团。而正是这种复杂性,使得川崎病在诊断、治疗和预后管理上始终面临诸多挑战。
更令人忧心的是,川崎病对儿童心脏的危害常被低估。若不及时治疗,约25%的患儿会出现冠状动脉异常,包括冠状动脉扩张和冠状动脉瘤,严重者可导致心肌缺血甚至心肌梗死。即使急性期症状消退,曾经出现冠状动脉损伤的患儿仍需长期甚至终身的心脏随访。这意味着,幼年一次发病,可能对其一生的心血管健康造成深远影响。
近年来,一个长期被忽视却逐渐浮出水面的关键角色——肠道菌群,正为理解川崎病发病机制提供新视角。研究显示,川崎病患儿急性期常出现明显菌群失衡:产短链脂肪酸的有益菌减少,条件致病菌增多,肠道屏障受损,微生物产物入血并放大全身炎症反应。更重要的是,这种失衡不仅与疾病进展相关,还可能影响IVIG治疗反应和冠状动脉损伤风险。肠道菌群正从“旁观者”转变为川崎病发生发展的参与者和潜在干预靶点。
本文将系统梳理川崎病从基础认知到前沿探索的完整知识脉络,首先初步介绍了川崎病的定义、病理特征、流行病学特点及风险因素,帮助读者建立基础认知。
其次整理了现有传统的川崎病治疗方法,包括静脉注射免疫球蛋白(IVIg)联合阿司匹林、糖皮质激素、IVIG抵抗补救方案及冠状动脉损伤后的抗栓治疗,呈现现有治疗的优势与局限。
然后揭示了患者肠道菌群失调的特征,短链脂肪酸不足、肠屏障受损、微生物产物易位与免疫炎症放大之间的关联,揭示其潜在致病机制。
最后,肠道微生物组作为川崎病的治疗靶点,将展望以微生态重塑为核心的干预新方向,从益生菌补充、益生元干预、粪菌移植到饮食调节,逐一分析各类策略的作用机制、现有证据及临床转化前景,为川崎病的精准治疗开辟新的思路。
什么是川崎病?
川崎病是一种主要发生在儿童(尤其是5岁以下)的急性全身性血管炎。可以把它理解为:孩子的免疫系统“被错误激活”,去攻击自身血管(尤其是中等大小动脉)的一种炎症性疾病。
编辑
★其中最关键的一点是:
它最容易损伤“冠状动脉”(给心脏供血的血管),这也是为什么川崎病被认为是:儿童中最常见的获得性心脏病原因之一。
注:川崎病(KD)最早由日本儿科医生川崎富作于1967年发表报告。1970年,在对10例诊断为KD后突发性心脏死亡的尸检后,这些心脏后遗症被记录下来。日本以外首次报告的病例出现在1970年代初的夏威夷;此后,全球60多个国家相继报告了KD病例。
川崎病有哪些症状及危害?
川崎病的诊断目前主要依赖临床表现,其经典特征可以归纳为“一加五”模式。
✅ 必备条件
“一”是指必备条件——患者主要表现为持续5天以上的高热(体温一般在39~40℃),且抗生素治疗无效。这是所有川崎病患儿共同的核心表现。
✅ 常见五大表现(满足其中4条即可高度怀疑)
1.双侧结膜充血:眼睛红,但没有分泌物。
2.口腔和嘴唇改变:
-嘴唇干裂、发红;
-“草莓舌”(舌头红、乳头突起)。
3.皮疹:全身性、多样性皮疹。
4.四肢改变:
-手脚红肿;
-恢复期出现指尖脱皮。
5.颈部淋巴结肿大。
与川崎病相关的变化
编辑
doi: 10.1161/ATVBAHA.124.321201.
✅ 川崎症在不同阶段表现出不同症状
川崎症按病程可为四期,不同阶段会有不同的症状:
•Ⅰ期,急性期:通常病程≤10天,临床表现为发热、结膜充血、皮疹、口腔病变、淋巴结肿大。
•Ⅱ期,亚急性期:病程在12~28天,表现为烦躁、脱皮,心脏表现如心包积液、房室瓣返流、心力衰竭、冠状动脉瘤形成等。
•Ⅲ期,恢复早期:病程在第28~45天,上述各种临床表现逐渐消失。
•Ⅳ期,恢复晚期:数月~数年,患儿大多已恢复正常,部分伴有心脏病变,如心电图异常、心律失常、心脏扩大,甚至缺血性心肌病、心功能不全。
✅ 川崎症的长期危害也很大
然而,川崎病真正的威胁远不止于这些表面症状。最核心、最严重的危害在于冠状动脉损伤。如果不及时治疗,约25%的患儿会出现冠状动脉异常,包括冠状动脉扩张和冠状动脉瘤(CAA)。冠状动脉瘤一旦形成,可能导致心肌缺血,甚至引发心肌梗死。
是的,儿童同样可能发生心梗——并显著增加猝死风险。
除此之外,由于川崎病本质上是一种全身性血管炎,它还可能波及心脏本身(引起心肌炎、心包炎)、消化系统(导致腹痛、呕吐),以及少见的神经系统损伤。
更值得警惕的是,即使急性期症状消退,曾经出现冠状动脉损伤的患儿仍需要长期、甚至终身的心脏随访,以监测潜在的心血管并发症。
川崎病的流行病学
川崎病是一种全球分布但发病率差异极大的儿童血管炎。近年来呈现:
川崎病有非常明显的流行病学特征,包括以下几点:
(1)地区差异(显著特征)
川崎病在不同地区发病率差异极大(可相差数倍):
• 东亚地区(高发区):持续上升、并存在流行高峰。
• 北美与欧洲(中低发区):亚裔儿童发病率显著更高。
• 其他地区:非洲、南美报道较少,全球总体发病趋势正在被越来越多识别。
(2)人群分布特征
• 年龄分布(极其典型):80–90%发生于5岁以下儿童,高峰年龄为6个月~4岁。(<6个月少见(可能受母体抗体保护),5岁后也明显减少)
• 性别差异:男孩发病率高于女孩,比例约为1.5:1。
• 种族/遗传背景:高风险人群为日本人、韩国人、中国人等亚洲人及亚裔后代(无论居住地)。说明遗传易感性在发病中起核心作用。
(3)时间分布特征
• 明显的季节变化:高发季节为冬季和早春;晚夏和秋季病例较低。
• 流行波动:周期性流行高峰,类似病毒传播的“波浪式模式”。
川崎病的风险因素
川崎病的本质是免疫介导的全身性血管炎,其核心病理过程是免疫系统对冠状动脉等血管壁的异常攻击。这些特征并不是孤立的,它们共同指向一个核心结论:“感染触发 + 遗传易感 + 免疫异常”三者共同作用的结果。导致疾病发生的风险因素主要有以下几种:
(1)遗传易感性(最强风险因素)
该疾病存在一定的家族聚集现象,在部分家庭中可见多名成员先后发病。同时已发现多个相关的易感基因位点(SNP),提示遗传因素在发病中具有重要作用:
(2)年龄因素
高风险:6个月–5岁。
可能原因:免疫系统尚未成熟、初次暴露病原体。
(3)性别因素
男性风险更高
可能原因:
(4)感染暴露
可能相关:
(5)种族与地理因素
日本、韩国、中国等亚洲人群风险较高,即使迁移仍保持高风险。
(6)季节与环境因素
冬春高发
可能相关因素:
(7)肠道菌群失衡(新兴风险因素)
研究发现,川崎病患者中:
注:肠道菌群与川崎病的关联将在后文进一步详细阐述。
(8)免疫状态异常
包括:
这些因素都可能会影响川崎病的发生。
本章小结
综上所述,川崎病是一种以儿童为主的免疫介导性全身血管炎,核心在于对冠状动脉的损伤,严重时可导致冠状动脉瘤、心肌缺血甚至心肌梗死,是儿童获得性心脏病的重要原因之一。
该病具有明显的年龄集中性(多见于5岁以下)、性别差异(男多于女)、种族倾向(亚裔高发)及季节性与流行波动特点。综合现有研究,川崎病的发生可归因于感染触发、遗传易感与免疫异常三者共同作用,其中遗传背景、年龄、性别、感染暴露及环境因素等均为重要风险因素,近年来肠道菌群失衡也被认为可能参与其发病过程。
整体而言,川崎病是一种多因素共同驱动的复杂免疫相关疾病,其早期识别与干预对于预防严重心血管疾病后果至关重要。
川崎病作为一种病因尚未完全明确的儿童急性系统性血管炎,其核心特征是异常激活的免疫反应及由此引发的血管内皮损伤。然而,一个长期未被充分重视却逐渐浮出水面的关键问题是:这一过度炎症反应”究竟由何触发,又如何被持续放大?
近年来,随着检测技术和微生态研究的发展,肠道菌群逐渐被认为是连接环境因素与宿主免疫反应的重要桥梁。越来越多证据表明,川崎病患儿存在显著的肠道菌群失衡,这种改变不仅体现在菌群组成的重塑,更涉及代谢功能紊乱及免疫调控异常,并与疾病的炎症程度、冠状动脉损伤以及IVIG治疗反应密切相关。因此,肠道菌群很可能并非单纯的“旁观者”,而是在川崎病的发生、发展乃至预后过程中发挥着主动而复杂的调控作用。
肠道微生物组与川崎病之间的潜在关联
编辑
doi: 10.1080/17460913.2025.2469432.
基于此,深入探讨肠道菌群在川崎病中的具体作用机制,对于理解其发病本质并寻找新的干预靶点具有重要意义。
川崎病中的肠道微生物失调
急性期川崎病(KD)患儿普遍存在肠道菌群α-多样性(丰富度与均匀度)显著降低及菌群结构失衡,典型表现为”产短链脂肪酸有益菌减少,条件致病菌过度增殖“的双重偏移。
编辑
doi: 10.1161/ATVBAHA.124.321201.
(1)产丁酸盐的菌属普遍降低:
• Faecalibacterium(特别是 F. prausnitzii):肠道中最主要的产丁酸盐菌之一;许多研究均证实其在川崎病(KD)急性期显著下调。其减少导致抗炎能力下降、肠道屏障受损。
• Ruminococcus 与 Roseburia:均为重要的产丁酸菌,负责维持紧密连接蛋白表达并调节Treg/Th17平衡;Ruminococcus在急性期减少,非急性期可回升。
• Clostridium(能产SCFA的梭菌簇)和Blautia:同样被发现在急性期丰度降低。
(2)经典的益生菌和保护性菌群减少:
• Bifidobacterium(双歧杆菌)和 Lactobacillus(乳杆菌):这两种最典型的益生菌在急性期丰度下降,这削弱了肠道黏膜屏障的保护功能。
• Akkermansia muciniphila(阿克曼氏菌):显著减少,该菌负责分解肠道黏液层,它的减少意味着黏液层的更新和维护可能受阻,屏障功能受损。
• Bacteroides(拟杆菌属):整体丰度下降,特别是其中的 Bacteroides thetaiotaomicron、B.uniformis 和 B. ovatus 等产SCFA的菌种。
注:值得注意的是,B. thetaiotaomicron 的减少被发现与IVIG耐药及心脏并发症风险增加相关。
川崎病肠道微生物组组成及关键代谢产物
编辑
doi: 10.1007/s00431-026-07008-z.
(3)潜在致病菌和条件致病菌大量增殖:
与有益菌减少相对应,多种条件致病菌在川崎病急性期呈现过度富集,可能通过超抗原、毒力因子或促炎代谢产物加剧全身炎症:
• Enterococcus(肠球菌属,特别是E.faecalis):在几乎所有研究中都显示急性期显著增高,并且其丰度与体内炎症标志物白细胞介素-6(IL-6)的水平呈正相关。此外,它还被认为与IVIG耐药有关。
• Streptococcus(链球菌属):其多个菌种在急性期丰度升高,它们可能通过产生超抗原,非特异性地过度激活T细胞,从而引爆全身性的免疫风暴。
• Staphylococcus(葡萄球菌属)、Helicobacter(螺杆菌属)、Acinetobacter(不动杆菌属)的丰度也在急性期有所增加,共同加剧了炎症反应。
• Fusobacteria(梭杆菌属)和 Shigella(志贺氏菌属)也被发现与川崎病的发生有关。
肠道菌群相关代谢物的变化及影响
川崎病(KD)急性期最关键的代谢物改变是短链脂肪酸(尤其是丁酸盐)显著下降,其他代谢轴亦有变化。
(1)短链脂肪酸(SCFA)减少
变化趋势: 实测显示KD患儿粪便丁酸盐浓度显著降低,乙酸盐、丙酸盐、乳酸盐在部分研究中相近或轻度下降;动物模型(CAWS诱导)亦示三种SCFA均减少。
丁酸盐是结肠上皮细胞主要能量源,可上调Occludin/Claudin等紧密连接蛋白;促进CD4⁺FoxP3⁺ Treg分化、抑制Th17分化;抑制NLRP3炎症小体及NF-κB/MAPK通路。
川崎病中缺失丁酸盐会导致:
•Treg发育受阻 → Th17/Treg失衡偏向促炎方向;
•肠道通透性升高 → 微生物产物(LPS等)入血;
•巨噬细胞MKP-1下调 → MAPK通路持续活化 → TNF-α、IL-1β、IL-6升高 → 冠脉血管炎。
(2)其他潜在代谢变化
•LPS(脂多糖)易位增加: 因”肠漏”发生,革兰阴性菌LPS进入循环 → 经TLR4-MyD88激活下游炎症通路。
•胆汁酸代谢改变: 部分研究示胆酸代谢通路差异,阿司匹林可通过P. goldsteinii / 7-keto-LCA影响肠上皮修复,但KD中直接数据有限。
血清免疫球蛋白下降: KD急性期肠道来源IgA、IgG水平下降,提示肠黏膜免疫功能障碍,削弱对共生菌/致病菌的管控。
肠道菌群影响川崎病的具体机制
根据现有文献,肠道菌群影响川崎病的具体机制可概括如下:
①菌群失调与短链脂肪酸匮乏
遗传易感 + 早期环境(感染触发、抗生素暴露、喂养方式)→ 急性期川崎病特有菌群失调 → 产 SCFA 菌(Faecalibacterium、Roseburia 等)锐减 → 粪便丁酸盐等 SCFA ↓
②肠道屏障破坏
SCFA(尤其丁酸盐)不足 → 结肠上皮细胞能量危机 + 紧密连接蛋白(Occludin、Claudins、ZO-1)表达下调 + 黏蛋白层变薄(Akkermansia 少)→ 肠上皮通透性增加。
③微生物产物易位与免疫激活
肠道内条件致病菌超抗原(链球菌/金葡菌来源)或 脂多糖(LPS)穿越受损屏障入血/淋巴;
LPS → TLR4 → NF-κB/NLRP3 炎症小体激活;
超抗原 → 非特异性 TCR 交联(Vβ2⁺ T 细胞克隆扩增)→ 强烈T细胞活化。
肠道微生物群失调与川崎病的联系
编辑
doi.org/10.1038/s41592-025-02923-9
④系统性免疫放大:
在上述刺激下,机体进入持续的高炎症状态:
•IL-6、TNF-α、IL-1β、IFN-γ持续升高;
•中性粒细胞、单核/巨噬细胞被大量激活。
同时:
•炎症细胞从肠道相关淋巴组织(GALT)迁移至外周;
•形成全身免疫放大效应。
⑤血管内皮损伤
炎症因子作用下:
•ICAM-1、VCAM-1表达上调;
•内皮通透性增加;
促进炎症细胞黏附与浸润及内皮细胞激活。
此外,中性粒细胞早期浸润、巨噬细胞、T细胞持续参与,会导致中小动脉全层炎症。
传统川崎病疗法与肠道菌群间的联系
(1)静脉注射免疫球蛋白能帮助菌群复衡
多数研究显示 IVIG (静脉注射免疫球蛋白)治疗后,有益菌(Bifidobacterium、Lactobacillus、Roseburia、Bacteroides vulgatus)部分恢复,条件致病菌(Finegoldia、Enterococcus)下降,SCFA 合成通路上调——提示 IVIG 可能间接帮助菌群复衡,或菌群稳态影响 IVIG 疗效(Bacteroides thetaiotaomicron 低、E. faecalis 高与 IVIG 耐药相关)。
(2)阿司匹林也会影响菌群丰度和短链脂肪酸
尚无 KD 患儿直接数据;其他模型显示阿司匹林可改变菌群丰度(升 高Bifidobacterium、Lactobacillus、降低Fusobacterium、Enterococcus),在临床前研究中,阿司匹林给药降低了厚壁菌门与拟杆菌门 (F/B) 的比率并增加了特异性 SCFA 。
但长期/高剂量可致轻微胃肠黏膜损伤,推测在 KD 中可能参与微调菌群—需进一步验证。
(3)糖皮质激素会降低菌群多样性
动物/其他病种研究显示长期糖皮质激素(GC)降低菌群多样性、改变短链脂肪酸产生菌;但川崎病中GC作为补救/辅助治疗与菌群交互尚不明确。
(4)抗生素暴露可能增加川崎病风险
回顾性研究显示生命早期(<6~12 月)抗生素使用与 KD 发病风险增加相关——抗生素破坏婴幼儿菌群建立,可能导致 SCFA 产生菌定植不足,构成 KD 易感背景。
本章小结
川崎病表现出典型的肠道菌群失调特征,即短链脂肪酸(SCFA)产生菌减少而条件致病菌增加,进而导致丁酸盐等抗炎代谢物不足,肠道屏障受损及通透性升高,促进微生物产物入血,并诱导Th17/Treg免疫失衡,最终放大全身炎症反应并加重冠状动脉损伤。
另一方面,标准治疗手段,尤其是静脉注射免疫球蛋白(IVIG),在一定程度上可促进肠道菌群结构的恢复与重建,而早期不合理或过度使用抗生素则可能进一步推动易感性菌群的异常偏移,加剧菌群失衡状态。
川崎病作为一种以全身性血管炎为特征的儿童疾病,其治疗策略始终围绕炎症控制与心血管保护展开。随着临床研究的不断深入,已逐步形成较为规范的传统治疗体系,并在降低严重并发症方面取得显著成效。
然而,在实际临床应用中,不同患儿的治疗反应及预后仍存在一定差异,提示该领域仍具有进一步优化与发展的空间。基于此,有必要对当前传统治疗方法及其特点进行系统梳理与分析。
1
静脉注射免疫球蛋白(IVIg)
适用对象:所有确诊或高度疑似川崎病患儿,争取在发热 ≤10 天内治疗。
川崎病最核心、最关键治疗,所有指南一致推荐。
✔优点:
•显著降低冠状动脉损伤:从15–25%下降至3–5%;
•起效快:多数患儿在24–48小时内退热;
•广谱免疫调节作用:抑制炎症因子(IL-1、TNF-α等)、调节T细胞、B细胞功能;
•安全性总体较好(临床经验丰富)。
✘局限性与缺点:
•约10%–20%患者无反应(IVIg-resistant),此类患者冠脉瘤风险显著升高;
•输注相关不良反应:发热、寒战、皮疹、过敏性休克(罕见);快速输注增加风险。
•作用机制尚未完全阐明(推测为 Fc 受体阻断、细胞因子中和、病原体特异性抗体)。
•费用高、资源依赖强:成本较高、依赖血制品供应。
2
阿司匹林(Aspirin)
阿司匹林是治疗川崎病儿童的关键药物。中剂量阿司匹林(30~50 mg/kg/天)在急性期发挥抗炎作用,而小剂量阿司匹林(3~5 mg/kg/天)在亚急性期发挥抗血小板作用,从而预防冠状动脉血栓形成。
✔优点:
•抗炎(高剂量)及抗血小板(低剂量)双重作用,符合 KD 急性期炎症 + 高凝状态病理生理;
•使用方便、成本低;
•在预防血栓方面作用明确。
✘局限性与缺点:
•高剂量时可能引起胃肠道刺激、Reye 综合征风险(尤其在病毒感染如流感/水痘时需暂停);
•出血风险(尤其联用其他抗凝药时);需根据肝功能、尿酸调整;
•不同国家对高剂量持续时间(至退热48h vs 至病程第14天)尚无统一标准。
3
糖皮质激素—联合辅助治疗
适用对象:经Kobayashi或Egami评分判定的IVIg 耐药高危患者,非高危者不常规推荐。
✔优点:
•强效抗炎作用;
•对IVIG抵抗患者有效;
•可缩短发热时间(部分研究支持)。
✘局限性与缺点:
•对冠状动脉保护作用不一致;
•可能存在副作用:包括免疫抑制、血糖升高、感染风险增加;
•使用时机存在争议:是早期联合用?还是仅用于难治?
4
IVIG抵抗病例的治疗
定义:首剂静脉注射免疫球蛋白(IVIg)(2g/kg)后≥36h 仍发热(≥38℃)且 CRP 未降。
(1)第2剂(或第3剂)IVIg
用法:再次给予2g/kg IVIg,品牌通常不变。
(2)静脉甲泼尼龙脉冲
用法:30mg/kg/日×1–3天(多在二次 IVIg 失败后),可续以口服激素逐渐减量。
(3)英夫利昔单抗(Infliximab,抗TNF-α)
用法:单次5mg/kg 静滴(通常在 1 剂 IVIg 无反应且考虑二次 IVIg 替代/联用时)。
(4)环孢素 A(Cyclosporine A)
用法:4–6 mg/kg/日分两次,根据血药浓度调整(谷值 60–200 ng/mL 或按中心方案),用至退热+CRP 正常,最长 2–3 周。
(5)低剂量甲氨蝶呤(极少用)
用法:10 mg/m² 每周口服,至 CRP 正常。
5
冠状动脉损伤后的抗栓治疗
本章小结
传统川崎病治疗以静脉注射免疫球蛋白(IVIG)联合阿司匹林为核心,该方案显著降低了冠状动脉并发症的发生率,是当前临床标准治疗。然而,该治疗体系仍存在一定局限性,包括约10%–20%的IVIG抵抗、对冠状动脉损伤预防不完全、治疗缺乏靶向性以及个体化管理不足等问题。
对于难治性病例,虽可采用重复IVIG、糖皮质激素及其他免疫调节治疗,但其疗效与长期获益仍存在争议。此外,抗血栓治疗及长期随访虽有助于降低严重心血管事件风险,但也增加了治疗复杂性与患者负担。总体而言,传统治疗虽显著改善预后,但仍有进一步优化和精准化发展的空间。
随着肠道微生物组在川崎病发生发展中的关键调控作用逐渐被揭示,其已从“相关因素”转变为具有潜在干预价值的治疗靶点。
当前研究表明,肠道菌群不仅参与炎症放大和免疫失衡过程,还可能影响静脉注射免疫球蛋白(IVIg)治疗反应及冠状动脉病变风险。因此,围绕微生态重塑的干预策略,正逐渐成为川崎病治疗领域的研究热点。这类策略旨在恢复菌群稳态、重建肠屏障功能并调节免疫反应,从而为传统治疗提供有益补充,并为改善预后开辟新的方向。
肠道微生物组的川崎病治疗干预措施
编辑
doi: 10.1007/s00431-026-07008-z.
益生菌帮助改善川崎病
益生菌对于调节肠道微生物群、提高整体免疫力、增加有益细菌丰度以及减轻与许多疾病相关的症状至关重要。
✅ 补充A.muciniphila减轻川崎病血管炎严重程度
在临床前模型中,研究已证实口服补充巴氏灭菌或活性的 A. muciniphila及 F. prausnitzii可减轻川崎病小鼠血管炎严重程度;类似地,口服巴氏灭菌的 A. muciniphila 或其外膜蛋白 Amuc_1100 亦具有保护作用。
其潜在机制可能与增强肠道紧密连接表达、降低全身免疫反应及增厚肠道黏液层有关。
✅ 补充丁酸梭菌降低炎症因子减轻冠状动脉病变
短链脂肪酸的产生源于膳食成分与肠腔内肠道微生物群之间复杂的相互作用。已证明在川崎病小鼠模型中给予丁酸梭菌可显著提高产生 SCFA 的细菌的流行率,并通过降低炎症标志物 IL-1β 和 IL-6 的水平来减轻冠状动脉病变 (CAL)的发展。
这为益生菌(丁酸梭菌)及其代谢产物补充剂(SCFA)调节川崎病患者肠道微生态提供了新的见解。
产生短链脂肪酸细菌对心血管疾病的影响
编辑
doi: 10.1161/ATVBAHA.124.321201.
多摄入益生元
益生元是不可消化的膳食纤维,可选择性刺激有益肠道微生物群的增殖并促进短链脂肪酸的合成。在成人临床研究中,一项临床试验表明,富含乙酰化和丁酰化淀粉的饮食可通过增加产生 SCFA 的细菌数量并提高循环乙酸盐和丁酸盐水平来显著降低高血压患者的血压。
✅ 低聚果糖可能成为免疫增强补充剂
低聚果糖 (FOS) 等益生元也观察到了类似的效果,它可以增强 SCFA 的产生、抑制病原菌并帮助维持肠道微生物稳态。通过益生元改变肠道微生物群可能是诱导保护性免疫反应的一种有前途的方法。在临床前研究中,在健康小鼠的饮食中添加果胶寡糖(POS)可以增强免疫球蛋白的产生,这与肠道细菌和醋酸盐的增加相关。这些发现表明,FOS 和 POS 可以作为川崎病患者潜在的免疫增强补充剂。
然而,益生元对川崎病的影响研究有限,有研究提出补充益生元可以改善川崎病患者的肠道免疫功能,从而提高治疗效果和疾病预后。
粪菌移植
✅ FMT可能在川崎病的治疗中发挥特定的作用
粪菌移植(FMT)有望直接重建紊乱的肠道微生态结构,将“有益菌减少、条件致病菌增加”的失衡状态恢复为相对健康的菌群组成,从而纠正川崎病急性期特征性的菌群紊乱。通过增强短链脂肪酸(SCFA)的合成,特别是丁酸盐的合成来降低肠道通透性,从而减轻疾病的严重程度,这有助于保持上皮屏障的完整性。
此外,由于肠道微生物群在调控系统免疫中具有核心作用,FMT还可能通过整体性免疫重塑,减轻血管炎症反应,从而在一定程度上降低冠状动脉损伤风险。
尽管如此,肠道微生物组在川崎病发病机制中的作用仍处于研究的初步阶段,FMT治疗急性期KD患者可能还不成熟,其治疗效果还不稳定。
饮食调节
饮食会影响肠道菌群的组成,进而影响全身疾病。
✅ 多酚和类胡萝卜素对川崎病有积极作用
β-胡萝卜素和虾青素等膳食类胡萝卜素已被证明可以促进肠道免疫系统的成熟并增加 IgA 的产生,这对肠道健康至关重要。此外,大豆异黄酮可以影响 Fcγ 受体功能,这对于 KD 的发病机制及其对 IVIG 治疗的反应具有重要意义。
饮食中的多酚和类胡萝卜素可能对川崎病具有积极作用,而高脂饮食则未显示出类似益处。
肠道微生物组作为川崎病的重要调控因素,正逐渐成为具有潜力的治疗靶点。以微生态重塑为核心的干预策略,包括益生菌、益生元、粪菌移植及饮食调节等,均可通过恢复菌群稳态、促进短链脂肪酸生成、增强肠道屏障功能及调节免疫平衡,从不同层面减轻炎症反应并可能降低冠状动脉损伤风险。
A. muciniphila、F. prausnitzii及丁酸梭菌等益生菌,以及低聚果糖、果胶寡糖等益生元均显示出一定的保护作用;粪菌移植则在重建整体菌群结构方面具有潜在优势;而富含多酚和类胡萝卜素的饮食亦可能带来益处。
然而,目前相关研究仍以临床前和探索性研究为主,证据尚不充分,未来仍需更多高质量临床研究以验证其安全性与有效性。
越来越多证据提示,川崎病并不只是血管和免疫系统“出了问题”,肠道菌群失衡、短链脂肪酸减少、肠屏障受损以及微生物代谢产物入血,可能共同参与了炎症放大和血管损伤过程。更重要的是,肠道微生态状态还可能影响IVIG治疗反应及冠状动脉病变风险,使其成为疾病进展的重要参与者。
当然,目前关于肠道微生物群与川崎病关系的研究仍处于探索阶段。无论是益生菌、益生元、饮食调节,还是粪菌移植等微生态干预策略,虽然展现出一定潜力,但距离真正进入标准化临床应用仍需更多高质量研究验证其有效性、安全性和适用人群。
不过可以确定的是,肠道微生物组正在为川崎病研究打开一条新的路径。未来,随着菌群检测、代谢组学和个体化医学的发展,我们或许能够更早识别高风险患儿,更精准预测治疗反应,并在传统治疗基础上,通过调节肠道微生态进一步改善疾病预后。
从“治炎症”到“调节微生态”,川崎病的治疗思路正在被重新拓展。肠道中的一个个微生物,也许正隐藏着改变儿童心血管健康结局的重要答案。
注:本账号发表的内容仅是用于信息的分享,在采取任何预防、治疗措施之前,请先咨询临床医生。
主要参考文献
Mohsenzadeh A, Mohammadi A, Mohsenzadeh H, Kamali K, Moradi M, Ebrahimi N, Sheikhy M, Zabet AH, Mirheidari H, Bavari S, Elahi R. The gut microbiota and kawasaki disease: exploring the role of microbial dysbiosis and metabolites in pathogenesis and therapeutics. Eur J Pediatr. 2026 May 1;185(5):332.
Yang Q, Kang Y, Tang W, Li M, Zhao C. Interplay of gut microbiota in Kawasaki disease: role of gut microbiota and potential treatment strategies. Future Microbiol. 2025 Mar;20(4):357-369.
Jena PK, Arditi M, Noval Rivas M. Gut Microbiota Alterations in Patients With Kawasaki Disease. Arterioscler Thromb Vasc Biol. 2025 Mar;45(3):345-358.
Chen H, Liu H, Qiao L, Liu Y, Yu D, Wang Z, Wang T, Li W. The characteristics of the gut microbiota in patients with Kawasaki disease: a systematic review. Front Microbiol. 2025 Dec 18;16:1715478.
Chen J, Yue Y, Wang L, Deng Z, Yuan Y, Zhao M, Yuan Z, Tan C, Cao Y. Altered gut microbiota correlated with systemic inflammation in children with Kawasaki disease. Sci Rep. 2020 Sep 3;10(1):14525.
Shulman ST, Rowley AH. Kawasaki disease: insights into pathogenesis and approaches to treatment. Nat Rev Rheumatol. 2015 Aug;11(8):475-82.
Zeng Q, Zeng R, Ye J. Alteration of the oral and gut microbiota in patients with Kawasaki disease. PeerJ. 2023 Jul 10;11:e15662.
Han L, Liu X, Lan Y, Hua Y, Fan Z, Li Y. Metagenomic analysis demonstrates distinct changes in the gut microbiome of Kawasaki diseases children. Front Immunol. 2024 Jul 22;15:1416185.
Rife E, Gedalia A. Kawasaki Disease: an Update. Curr Rheumatol Rep. 2020 Sep 13;22(10):75.
Yang Q, Tang W, Ren J, Li M, Zhao C. Unveiling the gut-heart potential connection: microbiota’s role in kawasaki disease and coronary artery lesions. Front Cell Infect Microbiol. 2025 May 29;15:1560083.
McCrindle BW, Rowley AH, Newburger JW, Burns JC, Bolger AF, Gewitz M, Baker AL, Jackson MA, Takahashi M, Shah PB, Kobayashi T, Wu MH, Saji TT, Pahl E; American Heart Association Rheumatic Fever, Endocarditis, and Kawasaki Disease Committee of the Council on Cardiovascular Disease in the Young; Council on Cardiovascular and Stroke Nursing; Council on Cardiovascular Surgery and Anesthesia; and Council on Epidemiology and Prevention. Diagnosis, Treatment, and Long-Term Management of Kawasaki Disease: A Scientific Statement for Health Professionals From the American Heart Association. Circulation. 2017 Apr 25;135(17):e927-e999. doi: 10.1161/CIR.0000000000000484. Epub 2017 Mar 29. Erratum in: Circulation. 2019 Jul 30;140(5):e181-e184.
Dietz SM, van Stijn D, Burgner D, Levin M, Kuipers IM, Hutten BA, Kuijpers TW. Dissecting Kawasaki disease: a state-of-the-art review. Eur J Pediatr. 2017 Aug;176(8):995-1009.
Rowley AH, Shulman ST. The Epidemiology and Pathogenesis of Kawasaki Disease. Front Pediatr. 2018 Dec 11;6:374.
Yim D, Curtis N, Cheung M, Burgner D. Update on Kawasaki disease: epidemiology, aetiology and pathogenesis. J Paediatr Child Health. 2013 Sep;49(9):704-8.

谷禾健康
人体肠道内栖息着超百万亿个微生物,它们如同看不见的器官,深刻左右消化、免疫、代谢甚至精神健康;土壤、水体、动植物体表的微生物群落,更是维系地球生态循环的核心单元。近20年,依托高通量测序技术,微生物组研究迎来爆发,科研工作者最核心的目标之一,就是找到和疾病、环境、饮食等表型相关的关键微生物,为疾病标志物筛选、益生菌开发、生态修复提供依据。
然而,微生物组数据分析面临三大核心挑战:
•第一,稀疏性: 微生物组数据中存在大量零值。一个零可能意味着某种微生物真的不存在,也可能只是因为含量太低而未被检测到。传统方法往往用任意小数值填补零值,但这会扭曲统计结果,导致假阳性或假阴性。
•第二,组成性: 微生物组数据通常以相对丰度(百分比)的形式报告,所有物种的丰度加起来必须等于100%。这意味着,如果某一种微生物的真实数量增加了,其他微生物的相对比例就会被动下降——即使它们的绝对数量没有发生改变。
•第三,异方差性: 不同微生物的变异程度差异巨大,丰度高的物种波动大,丰度低的物种波动小,这使得传统的线性模型假设难以满足。
微生物测序数据自带的缺陷,让科研成果陷入偏差:大量测序结果出现零值、数据受样本总测序量约束只能得到相对比例,传统统计工具很难分清 “细菌是真的消失了” 还是 “测序没检测到”、“某菌变多” 还是 “别的菌变少挤占占比”。
2026年1月,哈佛大学公共卫生学院 Huttenhower 教授团队在顶级期刊《Nature Methods》上推出的新一代微生物组多变量关联分析工具MaAsLin3。它能够精准区分微生物”是否存在”与”数量多少”这两种截然不同的变化模式,旨在解决微生物组数据中的稀疏性、组成性和高维度挑战,能够更精准地识别微生物特征(如物种、基因)与临床或环境元数据之间的关联 。
MaAsLin 3 作为新一代微生物组多变量关联分析R包工具,相比前代及同类工具(如 ALDEx2, ANCOM-BC2),主要实现了以下三大技术突破:
双重建模(丰度 + 存在率)
传统方法通常只关注相对丰度的变化。MaAsLin 3 采用“跨栏模型”策略,将分析分为两步:
√存在率(Prevalence):使用逻辑回归分析微生物的“存在/缺失”状态。
√丰度(Abundance):使用线性模型分析非零数据的数量变化。
这种分离使得研究能发现仅由“定植状态改变”驱动的关联(例如某菌在疾病组中完全消失,而非仅仅减少),这在炎症性肠病(IBD)等研究中已被证实非常普遍 。
MaAsLin 3能够实现丰度和流行率建模,精度更高
编辑
doi.org/10.1038/s41592-025-02923-9
组成性校正
(Compositionality Correction)
针对测序数据固有的组成性偏差,MaAsLin 3 提供了两种校正路径:
√实验性校正:支持使用 spike-in(外源内参)或 qPCR 等绝对定量数据。
√计算性校正:引入中位数系数比较法。通过检验特定特征的系数是否显著偏离群落中所有特征系数的中位数,从而推断绝对丰度的变化,无需额外的实验数据即可缓解相对丰度带来的假阳性 。
扩展的分析能力
支持更复杂的现代流行病学设计,包括混合效应模型(处理纵向/重复测量数据)、有序预测变量检验以及特征特异性协变量(如宏转录组中的 DNA 丰度作为 RNA 表达的协变量)。
在多项模拟数据和真实数据集(如 HMP2 IBD 队列)的基准测试中,MaAsLin 3 表现出显著优势:
1
准确性
在样本量 >50 时,其 F1 分数优于或持平于主流工具。
2
高精度与高召回率的平衡
MaAsLin 3 在保持高精确度(Precision)的同时,随着样本量增加(>100),其召回率(Recall)显著优于 ALDEx2,并逼近 ANCOM-BC2。
3
稳健性
在模拟仅含测序零值(Sequencing zeros)或包含生物学零值的情况下,MaAsLin 3 均表现出较高的稳健性。
MaAsLin 3的默认模型组件提升了比简单回归模型
更准确的水平
编辑
doi.org/10.1038/s41592-025-02923-9
!
揭示新生物学发现:
在炎症性肠病(IBD)数据分析中,MaAsLin 3 发现 77% 的显著关联是由存在率变化驱动的,而非传统的丰度变化。例如,它首次揭示了 Dysosmobacter welbionis 在 IBD 患者中主要是检出率降低,而非丰度降低。
在炎症性肠病(IBD)研究中
的新发现
研究团队应用 MaAsLin 3 重新分析了 HMP2 炎症性肠病(IBD)多组学数据时发现:
√存在率的差异: 在识别出的 372 个显著关联中,77% 与存在率(Prevalence)有关,且 89% 为负相关。这意味着 IBD 相关的菌群失调更多表现为共生菌的“丢失”,而非仅仅是丰度降低。
√新生物标志物: 首次发现 Dysosmobacter welbionis 在 IBD 患者中显著“消失”,存在率下降。这是一个潜在的有益菌,MaAsLin 2 未能区分这一差别。
√年龄特异性:明确区分了成人和儿童 IBD 患者的特异性菌群变化,仅发现极少数(如 Klebsiella pneumoniae)在两个年龄段均与生态失调相关。
MaAsLin 3应用于HMP2 IBDMDB验证并扩展了以往肠道微生物群与IBD的关联
编辑
doi.org/10.1038/s41592-025-02923-9
★官方教程:
1
安装要求
•R 版本:建议 R >= 4.4 。
•安装命令:
#通过 BiocManager 安装稳定版MaAsLin 3:
if (!require(“BiocManager”, quietly = TRUE))
install.packages(“BiocManager”)
BiocManager::install(“remotes”)
BiocManager::install(“biobakery/maaslin3”)
for (lib in c(‘maaslin3’, ‘dplyr’, ‘ggplot2’, ‘knitr’, ‘kableExtra’)) {
suppressPackageStartupMessages(require(lib, character.only = TRUE))
}
2
基本运行示例
taxa_table_name <- system.file(“extdata”, “HMP2_taxonomy.tsv”, package = “maaslin3”)
taxa_table <- read.csv(taxa_table_name, sep = ‘\t’, row.names = 1)
metadata_name <- system.file(“extdata”, “HMP2_metadata.tsv”, package = “maaslin3”)
metadata <- read.csv(metadata_name, sep = ‘\t’, row.names = 1)
#分类变量因子化(用于IBD与健康对照比较):
metadata$diagnosis <-
factor(metadata$diagnosis, levels = c(‘nonIBD’, ‘UC’, ‘CD’))
metadata$dysbiosis_state <-
factor(metadata$dysbiosis_state, levels = c(‘none’, ‘dysbiosis_UC’, ‘dysbiosis_CD’))
metadata$antibiotics <-
factor(metadata$antibiotics, levels = c(‘No’, ‘Yes’))
output <- “maaslin3_results”
set.seed(1)
fit_out <- maaslin3(
input_data = taxa_table,
input_metadata = metadata,
output = ‘hmp2_output’,
formula = ‘~ diagnosis + dysbiosis_state + antibiotics + age + reads’,
normalization = ‘TSS’, # 总丰度标准化
transform = ‘LOG’, # 对数转换
augment = TRUE, # 启用数据增强(解决逻辑回归线性可分问题)
standardize = TRUE, # 连续变量Z-score标准化
max_significance = 0.1, # FDR阈值设为0.1
median_comparison_abundance = TRUE, # 丰度系数与中位数比较(应对组成性效应)
median_comparison_prevalence = FALSE,# 存在率系数与0比较
max_pngs = 250, # 最大可视化结果数
cores = 1 # 单核运行
)
3
结果解读
•significant_results.tsv:筛选后的显著关联列表,包含 coef(效应量)、qval_individual(个体 FDR)和 qval_joint(联合 FDR)。
•summary_plot.pdf:热图和系数排序图,直观展示主要驱动变量。
•association_plots/:每个显著关联的详细散点图或箱线图 。
#适用场景,MaAsLin 3 特别适用于:
•需要区分微生物是“消失/出现”还是“增多/减少”的研究。
•拥有复杂设计(如配对样本、时间序列、多因素交互)的数据集。
总结
MaAsLin3的出现,标志着微生物组关联分析从”模糊关联”迈向”精准提取“的重要一步。MaAsLin 3 为广大科研人员提供了一个更严谨的工具箱,特别是针对大样本量、复杂临床设计或关注低丰度关键物种的研究项目,它能有效减少因数据稀疏性导致的假阴性,并提供更具生物学解释力的结果。
参考文献:Nickols, W.A., Kuntz, T., Shen, J. et al. MaAsLin 3: refining and extending generalized multivariable linear models for meta-omic association discovery. Nat Methods 23, 554–564 (2026). https://doi.org/10.1038/s41592-025-02923-9.
谷禾健康
编辑
在人类肠道这个高度复杂的“微型生态系统”中,除了广为人知的细菌之外,还栖息着一类常被忽视却至关重要的微生物——古菌。
其中,Methanobrevibacter smithii(史氏甲烷短杆菌)作为人体肠道中最主要的产甲烷古菌,具有短杆/卵圆形、肽聚糖样细胞壁、丰富黏附蛋白等特征。它能通过消耗发酵细菌如Prevotella产生的氢气并生成甲烷,不仅维持了肠道微生态的代谢平衡,还在能量回收、菌群互作以及宿主健康中发挥着关键作用。
近年来,随着宏基因组学和代谢研究的推进,M.smithii 已从“沉默的共生者”转变为研究热点。该菌自生命早期定植并伴随终生,通过与细菌及宿主的多重互作调控能量代谢与疾病风险。在不同疾病中,其可能表现为“保护性减少”或“风险性增加”。目前,其在便秘型肠易激综合征、肥胖、炎症性疾病及结直肠癌中的潜在作用正逐步被揭示。
Methanobrevibacter smithii(史氏甲烷短杆菌)是人类肠道中最主要的产甲烷古菌,也是目前研究最深入的人体“古菌共生者”之一,其在能量代谢、肠道微生态和多种疾病中扮演重要角色。多项研究从形态学、基因组学、代谢特征和生态位等方面刻画了其“基本特性”。
1
分类学
史氏甲烷短杆菌(M. smithii)属于广古菌门、甲烷杆菌科 (Methanobacteriaceae),是人类肠道最优势的古菌代表,约占肠道古菌的 >90%。
2
细胞形态与结构
▸ 形态
编辑
▸ 细胞壁结构(关键差异)
▸ 特殊结构
提示:这些特征使其在免疫识别、抗性、进化上都不同于细菌。
3
生长环境
▸ 基本生理条件
为典型中温、专性厌氧、氢营养型产甲烷菌。
4
代谢能力
▸ 核心特征
其核心代谢途径是:
CO2+4H2→CH4+2H2O
•即利用H₂(由发酵细菌如 Bacteroides, Clostridium产生)还原 CO₂ 生成甲烷。
• 不能利用糖类、蛋白质等常见有机碳源,完全依赖 氢营养型代谢。
M. smithii产甲烷途径
编辑
doi: 10.1016/j.heliyon.2024.e36742.
也可利用:
▸ 清除氢气
这点极其关键,因为:
▸ 对短链脂肪酸的间接调控
通过“氢转移”
5
耐药性
▸ 对大部分抗生素耐药
古菌对多数抗细菌抗生素不敏感,如β-内酰胺类(如青霉素、头孢菌素,因缺乏细菌细胞壁合成靶点)、大环内酯类、氨基糖苷类、喹诺酮类。
▸ 有效药物
莫匹罗星(特异性抑制古菌异亮氨酰-tRNA合成酶)、部分硝基咪唑类(如甲硝唑,部分敏感,但易产生耐药)。利福昔明 + 新霉素(IMO常用)、甲烷抑制剂(研究中)。
!
提示
常规肠道菌群检测(细菌培养)可能会漏检,需特异性古菌PCR或宏基因组测序。
Methanobrevibacter smithii(史氏甲烷短杆菌)是人体内分布最广泛且最具代表性的产甲烷古菌,主要定植于胃肠道,尤其在结肠中丰度最高。
除肠道外,M. smithii 亦可在口腔、呼吸道及个别体液或组织样本中被检测到,提示其具有一定的跨部位分布潜力。
M.smithii在人类中的定植
编辑
doi: 10.1016/j.heliyon.2024.e36742.
▸ 胃肠道
目前关于M. smithii在肠道中的认识主要来源于粪便研究。
▸ 胃和小肠的环境不太利于M. smithii定植
实际上,胃、小肠和大肠在pH、氧分压、菌群组成及黏液厚度等方面存在显著差异,从而影响其定植。胃内强酸性环境(pH 2–3.5)、较高氧分压、低菌密度及薄黏液层均不利于M. smithii生长。
小肠中,随着从十二指肠到回肠pH升高(6至7.5)及氧分压下降,兼性厌氧菌(如肠杆菌科和链球菌科)逐渐占优势,而并非M. smithii的理想生态位。
▸ 结肠和直肠更适合M.smithii 定植
相比之下,结肠和直肠具备近中性pH(6.5–7)、强厌氧环境及较厚黏液层,更适合M. smithii定植,并支持其与拟杆菌科和瘤胃球菌科形成稳定的代谢网络。
▸ 口腔
M. smithii已在牙周病相关口腔样本中被分离鉴定。在对54份口腔样本进行气相色谱分析后,有20份检测到甲烷,其中3份通过间接免疫荧光和电子显微镜确认存在M. smithii。
▸ 吸烟者口腔中更容易存在M.smithii
对200份口腔样本(包括52名吸烟者和148名非吸烟者)的研究显示,32%样本检测到M. smithii 16S rRNA基因,其中吸烟者占73%。进一步通过针对mcrA基因的FISH检测,在阳性样本中有39.58%被确认,其中部分为M.smithii与M.oralis的共感染。对PCR阳性样本的培养共获得46例生长,其中包括一例同时分离出M. smithii的吸烟者口腔样本。
总体而言,这些结果表明吸烟与口腔中M. smithii的存在显著相关,提示其可能影响口腔甲烷古菌群落结构。
▸ 下呼吸道
M. smithii亦可在下呼吸道样本中检测到,包括痰液、支气管肺泡灌洗液和支气管抽吸液。通过PCR和RT-PCR扩增古菌及M. smithii的16S rRNA基因,在部分痰液和支气管肺泡灌洗样本中确认其存在。
进一步采用FISH结合共聚焦显微镜进行形态学分析,仅在一份支气管肺泡灌洗样本中观察到M.smithii,而痰液样本未检出阳性信号。
▸ 母乳
通过改进的DNA检测方法及抗氧化培养技术,研究在3份初乳和5份乳汁样本中成功分离出M. smithii。该菌分别通过培养和qPCR在38%(5/13)和29%(37/127)的母乳样本中被检测到,其存在与母体体重指数等代谢特征相关。
既往研究亦表明,母乳中含有多种源自母体肠道的微生物,且母亲与母乳喂养婴儿的肠道菌群具有显著重叠,提示包括M. smithii在内的肠道微生物可能经母乳传播。
总体而言,这些证据表明母乳喂养有助于关键共生菌的垂直传播,并在出生早期促进婴儿肠道微生物群的建立。
▸ M. smithii分布的时间和人群差异
根据文献数据显示,M.smithii在出生后1–2天即可在新生儿胃液和粪便中发现,可能通过母乳或母体来源实现早期定植;并且其在不同年龄、饮食结构及健康状态下,丰度呈动态变化。
▸ 不同地理环境和饮食会影响M. smithii丰度
中国 5 地区 792 名成人研究发现,地理位置是决定肠道古菌组成的主因,其次为城市化、饮食和民族;农村人群粪便中M. smithii 更丰富,饮用酥油奶茶与 M. smithii 升高相关。
而城市化会降低古菌多样性及古菌/细菌比值,M.smithii 减少伴随血清胆碱酯酶水平升高,与现代非传染性疾病上升趋势相呼应。
需要说明的是,M. smithii在有氧环境中的活动,如口腔、血液和母乳,取决于它找到无氧生态位或有利于其生存和潜在代谢活动的条件。例如,在口腔内,牙菌斑可以创造微环境,使M.smithii得以生存。在血液中,其存在通常表明异常状况,虽然可以检测到,但可能并非积极参与正常生理过程,而是参与病理状态。母乳中M.smithii的存在可能是短暂的,也可能是母体微生物群转移的一部分。
▸ 极高龄老人中M.smithii也比较丰富
编辑
影响因子:2.989
在19–109岁人群中,极高龄老人的高甲烷表型与 M. smithii 丰度增加相关,百岁老人的甲烷菌组成更接近年轻人,尤其表现为Methanobrevibacter smithii的回升。研究还揭示两种关键古菌(M.smithii与Ca.M.intestini)在不同年龄阶段的网络作用发生转变,并与产丁酸菌形成复杂共生关系,可能通过代偿机制维持肠道代谢与健康,从而与健康老龄化及长寿相关。
不同人群M.smithii的差异
编辑
▸ 与其它微生物的互作
M. smithii被揭示与细菌、真菌和病毒以及宿主均存在互作关系,构成了一个广泛的网络。
M. smithii及其微生物互作网络的图示
编辑
doi: 10.1016/j.heliyon.2024.e36742.
▸ 产氢气细菌能促进M. smithii的丰度
M. smithii与肠道细菌之间存在复杂的代谢互作。Bacteroides、Prevotella、Ruminococcus和Faecalibacterium等菌通过发酵产生氢气,为M.smithii进行产甲烷提供关键底物。
编辑
Villa R,et al.Trends in Microbiology.2025
在体外研究中,B. thetaiotaomicron产生的氢气和发酵产物可支持M. smithii生长。矛盾的是,这些发酵副产物的积累会扰乱B. thetaiotaomicron的代谢分泌模式,阻碍M. smithii的生长。
相比之下,Christensenella minuta产生氢气的能力更强,对M. smithii的代谢支持更为显著。进一步的体外培养实验表明,M. smithii可抑制硫酸盐还原菌(如脱硫弧菌科)的生长;当其活性被抑制时,这类菌群则显著增加,提示二者在氢气利用上存在竞争关系。
▸ M. smithii可增强部分真菌代谢酶活性
目前尚无关于M. smithii与人类真菌相互作用的研究,但其与瘤胃真菌的关系已有较多报道。M.smithii与Neocallimastix frontalis和Piromonas communis共培养时,可降低真菌对离子载体抗生素(如monensin和lasalocid)的敏感性,但在香豆素或对香豆酸存在时未表现出保护作用。
此外,与N. frontalis、P. communis和Sphaeromonas communis共培养可增强真菌代谢酶活性,促进木聚糖和纤维素发酵,增加乙酸、木糖和己糖的释放;与N. frontalis、Piromyces和Caecomyces共培养则可改善黑麦草及木质化细胞壁的降解。
相反,稻草发酵提取物中的土曲霉可抑制M. smithii生长及其甲烷生成,其机制可能与真菌分泌的洛伐他汀抑制参与甲烷生成细胞膜合成的关键酶(HMG-CoA还原酶)有关。
▸ M.smithii可能会被噬菌体感染
人类病毒组由多种栖息于人体内的病毒组成,参与调控健康、疾病及微生物间相互作用。在消化道中,病毒以尾部双链DNA噬菌体(Caudoviricetes)为主。最新生物信息学研究在多个人源M. smithii基因组中发现了嵌入的Caudoviricetes相关序列,提示该古菌可能受到此类噬菌体感染。
▸ 与人体的相互作用
▸ 影响肠道动力
甲烷在肠道中可能通过多种机制减缓肠蠕动,其作用途径尚未完全明确。现有研究推测,这一过程可能涉及对肠道平滑肌活动的直接影响,同时通过调节肠神经系统的信号传导改变蠕动节律,此外还可能通过气体信号分子的调控发挥作用。这可能导致便秘的发生。
▸ 影响能量代谢
甲烷相关微生物可通过多种途径影响宿主代谢,包括提高能量的获取与利用效率,同时改变短链脂肪酸(SCFA)的组成谱,并进一步调节脂质代谢过程。
▸ M.smithii会影响免疫反应
在人类中,对暴露于M. stadtmanae、M. smithii和M. luminyens的外周血单核细胞(PBMC)及单核细胞来源树突细胞的研究显示,这些古菌可诱导TNF-α和IL-1β等促炎细胞因子释放,并干扰moDC中抗菌肽基因的表达。然而,在Caco-2/BBe肠上皮细胞中未观察到类似反应,提示其识别主要发生于免疫细胞。
此外,小鼠吸入M. smithii可使骨髓来源树突细胞积累增加约10倍。由M. smithii脂质构建的“古菌体”在增强免疫应答方面优于传统脂质体。
Methanobrevibacter smithii是人类肠道中最常见的产甲烷古菌之一,在维持肠道微生态稳态与宿主代谢中发挥重要作用。然而,M. smithii的过度增殖或功能失衡也与多种疾病相关。
1
便秘为主的肠易激综合征中增加 ↑↑↑
▸ 最明确
编辑
影响因子:3.141
▸ M.smithii可能通过抑制肠道蠕动导致便秘
研究显示,肠易激综合征(IBS)患者的M. smithii数量显著高于健康对照,且在便秘型(IBS-C)中最为显著;其丰度与排便频率呈负相关,提示其可能通过抑制肠道蠕动促进便秘。
▸ M.smithii产生的甲烷还可能导致腹胀
此外,M. smithii负荷与呼气甲烷水平及腹胀症状密切相关,支持其在IBS发病机制中的重要作用。
2
肥胖患者中丰度变化
▸ 存在争议
编辑
影响因子:3.514
有研究发现,Methanobrevibacter smithii(史氏甲烷短杆菌) 在肥胖个体中呈现出丰度显著增加的趋势,并通过独特的“代谢协同”机制促进宿主能量吸收和体重增加。
其具体机制包括:
▸ 氢清除效应与能量增效
肠道发酵会产生大量氢气(H₂),积累的 H₂ 会抑制细菌的进一步发酵。而M.smithii利用H₂还原CO₂生成甲烷(CH₄),清除了肠道内的H₂。
H₂降低后,发酵细菌的代谢活性被“解锁”,能更高效地将复杂多糖分解为短链脂肪酸(SCFAs)。使得宿主可吸收的 SCFAs(能量来源)总量增加,导致“能量收获”过度。
▸ 部分新研究发现肥胖者中M.smithii减少
但也有一些新研究发现M.smithii在肥胖患者中丰度呈现下降趋势(基于meta分析结果的综合观察),并且与“瘦相关菌群”表现出共现关系,
因此,现在普遍共识认为M.smithii与肥胖之间的关系具有明显的情境依赖性特征。
3
在高注意力人群中更为丰富 ↑↑↑
编辑
影响因子:7.74
▸ M.smithii丰度较高与更好的注意力及认知相关
一项研究探讨了肠道古菌(尤其是产甲烷菌Methanobrevibacter smithii)与认知功能之间的关系,发现其丰度越高与更好的注意力、执行功能和认知灵活性显著相关,并在两个独立人群队列(n=125;n=942)中得到验证。
高丰度M. smithii个体具有独特的肠道菌群结构及代谢特征,表现为丁酸盐、组氨酸、苯丙氨酸和胆汁酸代谢增强,同时伴随更低炎症及改善的代谢状态。
粪菌移植实验进一步证明,这种微生物生态可改善小鼠认知灵活性和体重,提示古菌通过“肠-肌-脑轴”调控代谢与认知,具有潜在治疗价值。
4
炎症性肠病患者中减少 ↓↓↓
编辑
影响因子:2.914
▸ M.smithii对炎症性肠病具有一定的诊断价值
研究了肠道古菌Methanobrevibacter smithii在炎症性肠病(IBD)中的变化及其作为生物标志物的潜力。
研究发现,IBD患者粪便中M. smithii的丰度显著低于健康人群,提示其与疾病发生密切相关;此外,在疾病缓解期该菌数量高于活动期,说明其可能参与疾病状态调节;ROC分析进一步表明,M. smithii具有一定的诊断价值,可能作为区分健康肠道与IBD的重要微生物标志物。
5
结直肠癌患者中增加 ↑↑↑
编辑
影响因子:12.121
▸ M.smithii可与具核梭杆菌形成互利代谢关系
一项研究通过整合19项临床宏基因组数据(共1882例样本)系统分析人类肠道古菌与疾病的关系,发现古菌与多种疾病存在关联,其中结直肠癌(CRC)患者中Methanobrevibacter smithii显著富集。
进一步通过代谢建模与体外共培养实验揭示,M.smithii可与CRC相关细菌(如具核梭杆菌)形成互利代谢关系,增强细菌生长并改变代谢产物;代谢组学分析表明古菌参与产生具有肿瘤调控作用的代谢物,首次从机制上证明肠道古菌通过“营养级控制”调节细菌群落并影响肿瘤微环境。
6
严重急性营养不良患者中耗竭 ↓↓↓
M. smithii被认为是在因饮食中断和营养不足导致严重急性营养不良(SAM)的非洲儿童肠道中缺失的关键微生物。对SAM儿童粪便的PCR检测显示,该菌显著且不可逆耗竭,仅在4.2%的病例中检出,而对照组为40.9%。
▸ M.smithii影响能量获取可能是导致营养不良的原因
其在营养不良中的作用可能与肠道微生物群的能量获取能力相关。相反,研究也发现其在肥胖人群中更为丰富,提示其可能促进能量提取并加重过度营养相关疾病。因此,其在营养不良中的影响还需结合与其他肠道微生物的相互作用及对营养吸收的调节来理解。
7
M.smithii与肠外感染
M.smithii还可能在一定情况下引发肠道以外部位的感染。
▸ 心内膜炎
在发热患者,尤其是感染性心内膜炎患者的血液中已检测并分离出M. smithii。在5594份厌氧血培养中,有27例(0.5%)细菌阳性且M. smithii PCR阳性。
进一步分析3例相关样本发现:在一次血培养中,通过自体荧光和电子显微镜检测到与金黄色葡萄球菌共存的M. smithii;同时,从两例患者血液中分离并测序获得与S. mutans和S. epidermidis相关的菌株。
▸ 尿路感染
在383份尿标本中,有9%检测到M. smithii。此外,感染尿液样本中53%、18%和3%的M. smithii分别与大肠杆菌、肺炎克雷伯菌和肠杆菌属相关。
▸ 细菌性阴道炎
在细菌性阴道炎(BV)患者的厌氧阴道液中检测到产甲烷活性。通过富集、氯化物色谱及间接免疫荧光分析,鉴定出两个接近M. smithii PS和ALI株的分离株。进一步研究显示,在33例阴道样本中有32例经16S rRNA RT-PCR检测为M. smithii阳性,并在厌氧SAB培养基中分离出9株,同时FISH在2例样本中证实其存在。
▸ 鼻窦炎
在116份来自难治性鼻窦炎患者的手术标本中检测产甲烷菌,PCR共检出12个阳性样本,涉及9名患者(10.3%)。进一步通过FISH和扩增子测序分析发现,其中4名患者存在M. smithii。
▸ 肌肉脓肿
M.smithii从一名41岁患者的椎旁肌肉脓肿中分离获得。测序证实其古菌16S rRNA和mcrA基因,并在SAB培养基联合抗生素培养下表现出对阿莫西林-克拉维酸的体外耐药性及对甲硝唑的敏感性。
▸ 脑脓肿
有一项研究在脑脓肿样本中检测到M. smithii,在18例标本中仅1例经RT-PCR呈阳性。
▸ 牙脓肿
在牙龈下牙菌斑样本中检测并培养出M.smithii。另在37°C厌氧条件下培养4个月,从一例样本中成功分离出M. smithii。
Methanobrevibacter smithii作为人体肠道中最主要的产甲烷古菌之一,在肠道微生态系统中扮演着不可替代的关键角色。它以严格厌氧和氢营养型代谢为主要特征,通过利用氢气并产生甲烷,维持微生物代谢的平衡,从而促进膳食底物的进一步分解与短链脂肪酸的生成,最终影响宿主的能量获取效率。因此,M. smithii 并非传统意义上的“产能微生物”,而更应被视为调控肠道代谢网络运行的重要“生态调节者”。
在健康与疾病状态中,M. smithii的作用主要体现在其对微生物功能与代谢通路的调控,而非单纯的丰度变化。例如,在帕金森病中,其丰度并无显著改变,但却深度参与以谷氨酸为核心的代谢网络重编程;在多发性硬化中,其丰度对治疗具有响应性,可能反映肠道微生态的恢复状态。
此外,通过与其他微生物的共营养作用,M. smithii 还可间接影响致病菌的生长及多种疾病过程,并在肥胖、炎症性肠病及其他慢性疾病中表现出一定的相关性。这些证据共同表明,其对人体健康的影响具有“间接性、系统性和功能导向性”的特点。
M. smithii有望成为连接肠道微生态与宿主代谢、免疫及神经功能的重要桥梁。在未来研究中,其功能状态(如代谢活性和通路参与)可能适合作为疾病生物标志物。同时,围绕其氢代谢和共营养机制的调控,有望发展出新的微生态干预策略,用于代谢性疾病、神经系统疾病以及个体化营养管理。此外,其在肠-脑轴中的潜在作用也为探索微生物参与神经退行性疾病提供了新的方向。
主要参考文献
Malat I, Drancourt M, Grine G. Methanobrevibacter smithii cell variants in human physiology and pathology: A review. Heliyon. 2024 Sep 6;10(18):e36742.
Villa R, Martin-Gallausiaux C, Gribaldo S …
Methanobrevibacter smithii Trends in Microbiology, 2025; 34, 217-218
Malat I, Bossi V, Drancourt M, Grine G, Ruimy R. Methanobrevibacter smithii strain U29 whole genome sequence delineates M. smithii intermediate cell variants. BMC Res Notes. 2025 Oct 1;18(1):414.
Kholwadwala AS, Quigley EMM. Small Intestinal Bacterial Overgrowth and Intestinal Methanogen Overgrowth: Are They Overdiagnosed? Acta Gastroenterol Latinoam. 2025 Dec 23;55(4):276-291.
Fumagalli A, Castells-Nobau A, Trivedi D, Garre-Olmo J, Puig J, Ramos R, Ramió-Torrentà L, Pérez-Brocal V, Moya A, Swann J, Martin-Garcia E, Maldonado R, Fernández-Real JM, Mayneris-Perxachs J. Archaea methanogens are associated with cognitive performance through the shaping of gut microbiota, butyrate and histidine metabolism. Gut Microbes. 2025 Dec;17(1):2455506.
Hu X, Yu C, He Y, Zhu S, Wang S, Xu Z, You S, Jiao Y, Liu SL, Bao H. Integrative metagenomic analysis reveals distinct gut microbial signatures related to obesity. BMC Microbiol. 2024 Apr 5;24(1):119.
Zhou Y, Tao E. The role of intestinal gases in pediatric functional constipation: a narrative review of pathophysiology and emerging therapeutics. Front Nutr. 2026 Jan 2;12:1694831.
Ghoshal U, Shukla R, Srivastava D, Ghoshal UC. Irritable Bowel Syndrome, Particularly the Constipation-Predominant Form, Involves an Increase in Methanobrevibacter smithii, Which Is Associated with Higher Methane Production. Gut Liver. 2016 Nov 15;10(6):932-938.
Catlett JL, Carr S, Cashman M, Smith MD, Walter M, Sakkaff Z, Kelley C, Pierobon M, Cohen MB, Buan NR. Metabolic Synergy between Human Symbionts Bacteroides and Methanobrevibacter. Microbiol Spectr. 2022 Jun 29;10(3):e0106722.
Dirks B, Davis TL, Carnero EA, Corbin KD, Smith SR, Rittmann BE, Krajmalnik-Brown R. Methanogenesis associated with altered microbial production of short-chain fatty acids and human-host metabolizable energy. ISME J. 2025 Jan 2;19(1):wraf103.
Conway de Macario E, Macario AJ. Methanogenic archaea in health and disease: a novel paradigm of microbial pathogenesis. Int J Med Microbiol. 2009 Feb;299(2):99-108.
Mohammadzadeh R, Mahnert A, Shinde T, Kumpitsch C, Weinberger V, Schmidt H, Moissl-Eichinger C. Age-related dynamics of predominant methanogenic archaea in the human gut microbiome. BMC Microbiol. 2025 Apr 4;25(1):193.
Mohammadzadeh R, Mahnert A, Zurabishvili T, Wink L, Kumpitsch C, Habisch H, Sprengel J, Filek K, Mertelj P, Pernitsch D, Hingerl K, Durdevic M, Gorkiewicz G, Diener C, Loy A, Kolb D, Trautwein C, Madl T, Moissl-Eichinger C. Cross-domain metabolic interactions link Methanobrevibacter smithii to colorectal cancer microbial ecosystems. Nat Commun. 2026 Feb 20;17(1):2979.
Baquero DP, Borrel G, Gazi A, Martin-Gallausiaux C, Cvirkaite-Krupovic V, Commere PH, Pende N, Tachon S, Sartori-Rupp A, Douché T, Matondo M, Gribaldo S, Krupovic M. Biogenesis of DNA-carrying extracellular vesicles by the dominant human gut methanogenic archaeon. Nat Commun. 2025 Jun 3;16(1):5093.
Samuel BS, Hansen EE, Manchester JK, Coutinho PM, Henrissat B, Fulton R, Latreille P, Kim K, Wilson RK, Gordon JI. Genomic and metabolic adaptations of Methanobrevibacter smithii to the human gut. Proc Natl Acad Sci U S A. 2007 Jun 19;104(25):10643-8.
谷禾健康
环境污染
环境污染已成为普遍的全球健康威胁,但其影响远远超出了直接的器官毒性。越来越多的证据表明,肠道微生物群是污染物诱发生理功能障碍的核心介导者。本文主要探讨和关注包括空气污染物、重金属、持久性有机污染物和新兴污染物如何扰动微生物组成、代谢活性及宿主-微生物信号传递的最新进展。
污染物暴露会改变微生物来源的代谢产物,如短链脂肪酸、胆汁酸和色氨酸衍生物,从而损害肠道屏障的完整性和免疫稳态。这些微生物群驱动的干扰会引发氧化应激、慢性炎症和神经内分泌失调,导致代谢紊乱、免疫失衡、神经毒性和致癌。
从众多的文献得出的机制上看,氧化还原失衡、TLR4/NF-κB和NLRP3通路的激活以及 AhR信号失调是连接环境暴露于疾病的关键交叉点。
本文整合170多篇发表的研究成果,包括流行病学调查、动物实验、体外研究和多组学研究。通过系统总结各种污染物如何影响微生物组成和代谢功能,以及微生物菌群失调如何影响宿主的新陈代谢、免疫和神经生理,构建一个机制框架,描述“环境暴露→微生物群破坏→宿主功能障碍→疾病发展”的级联反应。
编辑
还讨论了这一新兴领域面临的关键挑战,包括对复杂污染物混合物进行更精确评估、因果关系的识别,以及微生物与宿主之间分子相互作用的阐明。
希望以深化对环境污染和肠道健康影响的理解,指导针对微生物群的疾病预防和环境风险缓解策略的发展。
▸ 肠道菌群
人类肠道中蕴藏着庞大且多样的微生物群落,随着个体的成长,饮食和环境等会因内部或外部干扰而波动。肠道微生物群承担四项对人体健康至关重要的基本功能:代谢调节、免疫调节、屏障保护和神经行为交流。
编辑
doi.org/10.3389/fmicb.2026.1737229
▸ 污染物
在现代社会中,环境污染已成为对人类健康的隐蔽而普遍的威胁。环境中存在各种污染物,主要包括空气污染物、重金属、持久性有机污染物(POPs)和新兴污染物。
编辑
doi.org/10.3389/fmicb.2026.1737229
▸ 主要环境污染物及其进入人体的途径
环境污染物种类繁多,来源复杂,可通过不同途径进入人体,并影响多个器官系统。
常见的环境污染包括空气污染、水污染、土壤污染,以及放射性污染、光污染和噪音污染等。其中,空气污染物、重金属、持久性有机污染物和新兴污染物与人体健康关系尤为密切。
★ 环境污染物既直接损伤器官,也能通过干扰肠道菌群影响健康
近年来,越来越多研究发现,这些污染物不仅会直接损伤肺、肝、肾、神经系统和胃肠道等器官,还可能通过改变肠道微生物群影响人体健康。
肠道微生物群参与营养代谢、免疫调节和肠道屏障维持,因此一旦菌群平衡被破坏,可能进一步引发炎症、代谢紊乱和多种系统性疾病。
㈠空气污染物
空气污染物包括多种气体和颗粒物,例如:
其中,PM2.5、臭氧和二氧化氮是最受关注的空气污染物。它们在全球分布并不均匀,通常在工业化程度高、人口密集、交通繁忙的地区浓度较高。其主要来源包括工业排放、汽车尾气和生物质燃烧。
空气污染物可以在大气中悬浮较长时间,并随气流进行远距离运输。人体接触空气污染物最主要的途径是吸入。污染物进入呼吸道和肺部后,可引起局部炎症和氧化应激;部分污染物或其诱导的炎症信号还可能通过血液循环影响全身器官,包括肝脏、胃肠道和肠道微生物群。
因此,空气污染的影响并不局限于肺部。它还可能通过肺—肠轴改变肠道微生态,破坏肠道屏障,并进一步影响免疫和代谢功能。
㈡重金属
重金属也是重要的环境污染物,常见类型包括铅(Pb)、汞(Hg)、镉(Cd)和砷(As)。
这些重金属广泛存在于土壤、水体和沉积物中,主要来源包括采矿、冶炼、工业生产、污水排放,以及农业中含金属化肥和农药的使用。由于重金属不能被自然降解,它们会长期残留在环境中,并通过食物链逐渐富集。
人类接触重金属的主要途径包括:
重金属进入人体后,可能损伤肝脏、肾脏、神经系统和免疫系统。同时,它们也可能改变肠道菌群组成,影响肠道屏障功能,并通过微生物代谢改变加重毒性效应。
㈢持久性有机污染物(POPs)
持久性有机污染物,简称 POPs,是一类难以降解、容易在脂肪组织中积累的有毒化学物质。常见POPs包括多氯联苯(PCBs)、二噁英和有机氯农药等。
这些化合物曾被广泛用于工业和农业生产。虽然许多 POPs 目前已经被禁止或限制使用,但由于它们化学性质稳定、脂溶性强,仍然可以长期存在于全球生态系统中。
POPs还具有远距离迁移能力。它们可以随着大气环流、水流和食物链传播到远离污染源的地区,因此具有全球性污染特征。
人体接触 POPs 的主要途径是饮食摄入,尤其是摄入脂肪含量较高的食物,如肉类、乳制品和部分水产品。此外,人体也可能通过吸入污染空气或皮肤接触而暴露于 POPs。
POPs 进入人体后,可影响内分泌、免疫、代谢和神经系统。近年来研究还发现,POPs 可能通过改变肠道微生物群,进一步影响胆汁酸代谢、炎症反应和能量代谢。
㈣新兴污染物
近年来,新兴污染物逐渐受到关注,典型代表包括微塑料和药品及个人护理用品污染物(PPCPs)。
微塑料主要来源于塑料垃圾的分解、工业生产过程以及日常塑料制品的磨损。它们已经广泛存在于海洋、河流、湖泊、土壤甚至饮用水中。
PPCPs 包括药物、化妆品、防晒剂、清洁用品和个人护理产品中的化学成分。这些物质主要通过生活污水、医院废水和工业排放进入环境。
人类可能通过以下途径接触这些新兴污染物:
与传统污染物相比,新兴污染物的使用历史相对较短,环境行为和健康效应仍在研究中。目前对其长期影响,特别是对肠道微生物群的影响,证据仍然有限。因此,新兴污染物的健康风险评估仍需要更多长期研究支持。
主要环境污染物类别、代表性来源及暴露途径
编辑
doi.org/10.3389/fmicb.2026.1737229
★ 环境污染物暴露的共同特点
不同污染物的来源和进入人体的途径有所不同,但它们有一个共同特点:人体往往不是一次性接触单一污染物,而是长期、低剂量、同时接触多种污染物。
这种长期低剂量混合暴露更接近真实生活情境。例如,一个人可能同时通过空气吸入PM2.5,通过饮食摄入POPs或重金属,又通过饮用水接触微塑料和药物残留。多种污染物共同作用时,可能产生更复杂的健康影响。
▸ 肠道菌群在污染物健康效应中的作用
肠道微生物群被污染物扰乱后,可能通过以下方式影响健康:
因此,污染物对人体的影响并不一定只是“污染物直接损伤器官”这么简单。很多情况下,污染物可能先改变肠道微生态,再通过微生物代谢产物、免疫信号和炎症反应影响全身健康。
★ 肠道微生物群作为环境毒理学的新靶点
传统毒理学研究主要关注污染物对重要器官的直接损伤,例如肺、肝、肾和神经系统损害。但近年来,越来越多证据表明,肠道微生物群也是环境污染物作用的重要靶点。
这些发现重塑了环境毒理学的概念框架,表明肠道微生物群不仅是被动旁观者,更是污染物诱发生理变化的主动介导者。这一范式转变提出了一个关键的科学问题:环境污染物是否通过扰动肠道微生物群,诱发宿主生理功能障碍和疾病?
编辑
doi.org/10.3389/fmicb.2026.1737229
为了明确污染物选择标准,我们重点关注了四大类:空气污染物、重金属、持久性有机污染物(POPs)和新兴污染物。这些基于其全球流行率、对肠道微生物群的充分记录影响,以及世界卫生组织(WHO)、美国环保署(USEPA)和国家环境机构等监管机构日益关注。
每个类别都包含具有大量肠道微生物群调节证据的代表性化合物,可以提供一个全面且聚焦的污染物-微生物群-宿主相互作用跨化学类别的概述。
代表性污染物对肠道微生物群的影响及相关健康结果
编辑
doi.org/10.3389/fmicb.2026.1737229
▸ 空气污染物及其对肠道微生物群的影响
空气污染是全球重要的公共卫生问题。它的成分非常复杂,既包括空气中的颗粒物,也包括多种气体污染物。其中,最受关注的包括细颗粒物(PM2.5)、臭氧(O₃)和氮氧化物(NOx)。
过去,人们更多关注空气污染对呼吸系统和心血管系统的危害。近年来,越来越多研究发现,空气污染还可能通过 “肺—肠轴” 影响肠道微生物群。也就是说,污染物被吸入肺部后,不仅会引起局部炎症,还可能通过血液循环、免疫反应和代谢改变影响肠道环境,最终导致肠道菌群失调,并增加多种系统性疾病的风险。
①细颗粒物(PM2.5)
PM2.5是指空气动力学直径小于或等于2.5微米的细小颗粒物。由于颗粒很小,它们可以被吸入肺部深处,甚至到达肺泡。PM2.5 还具有较大的表面积,容易吸附重金属、多环芳烃等有毒物质,因此可能携带多种有害成分进入人体。
吸入后,PM2.5 可能通过血液或淋巴系统进入身体其他部位,包括肠道,并改变肠道微生物的组成。流行病学研究和动物实验均显示,PM2.5 暴露与肠道菌群变化有关,而且这种影响可能与暴露浓度和暴露时间有关。
在人群研究中,长期暴露于高 PM2.5 环境的人群,尤其是居住在交通繁忙或工业区附近者,其肠道微生物α多样性显著下降。
注:α多样性反映菌群的丰富度和均衡性,其降低意味着物种减少、生态稳定性减弱。
PM2.5的影响亦可发生于生命早期。一项纳入168852名母亲的队列研究显示,孕期较高 PM2.5 暴露与早产风险增加相关,提示空气污染不仅危害孕妇健康,也可能影响胎儿发育及妊娠结局。
动物实验进一步支持上述发现。例如,小鼠长期暴露于约 70.9 ± 26.8 μg/m³ 的浓缩颗粒物后,出现肠道屏障受损及菌群结构改变,可能与 TLR2/5–MyD88–NLRP3 信号通路激活有关,该通路与免疫炎症反应密切相关。
另一研究中,小鼠暴露于 198.5 μg/m³ PM2.5 后,先出现短暂体重下降,随后发生代偿性增加,同时伴随肠道微环境及代谢通路显著变化,表明 PM2.5 可同时影响菌群与能量代谢。
此外,通过气溶胶浓缩系统对雄性 C57BL/6J 小鼠进行 PM2.5 暴露,并结合抗生素清除菌群及粪菌移植实验发现,PM2.5 引起的葡萄糖代谢异常与肠道菌群失衡密切相关。
进一步分析表明,短链脂肪酸(SCFAs),尤其是乙酸盐,是关键代谢环节。补充乙酸盐可部分缓解代谢异常,提示肠道菌群可能是 PM2.5 影响葡萄糖代谢的重要中介。
②臭氧(O₃)和氮氧化物(NOx)
臭氧(O₃)由三个氧原子组成,具有强氧化性。平流层臭氧可阻挡紫外线,而近地面臭氧则是重要空气污染物,近年来已成为中国及其他快速发展地区的突出环境问题。
臭氧对肠道微生物群的影响主要通过氧化应激和全身炎症间接实现。氧化应激指体内氧化物质过多,超出清除能力,从而引发细胞损伤和炎症。
研究显示,将健康大鼠及心血管疾病易感大鼠暴露于不同浓度臭氧4小时后,其肺和肾出现不同程度炎症反应。尽管未直接观察到肠道损伤,但全身炎症已被激活,提示臭氧可能通过免疫与氧化应激通路间接改变肠道微环境和菌群。
氮氧化物(NOx),主要包括一氧化氮(NO)和二氧化氮(NO₂),是交通污染的重要来源。研究发现,交通相关空气污染与肠道菌群变化及空腹血糖升高有关。
具体而言,污染暴露与拟杆菌科(Bacteroidaceae)丰度下降、瘤胃球菌科(Ruminococcaceae)丰度升高相关;前者减少、后者增加均与空腹血糖升高相关。
路径分析显示,这两类菌群变化可解释交通污染与空腹血糖升高约 24%–29% 的关联,表明空气污染可能部分通过改变肠道菌群影响宿主的葡萄糖代谢。
总体来看,空气污染不仅会损害肺部和心血管系统,也可能通过肺—肠轴影响肠道微生物群。PM2.5、臭氧和氮氧化物等污染物可以通过炎症反应、氧化应激、肠道屏障损伤和代谢通路改变,导致肠道菌群失衡。
这些菌群变化又可能进一步影响葡萄糖代谢、免疫状态和全身健康。因此,肠道微生物群可能是连接空气污染暴露与代谢性疾病、炎症性疾病等健康风险之间的重要桥梁。
▸ 重金属对肠道微生物群的影响
重金属是一类常见的环境污染物。它们有两个重要特点:不容易被降解,并且可能在生物体内不断积累。常见的重金属包括砷、镉、铅、汞等。人类接触重金属的主要途径包括食用受污染的食物、饮用受污染的水,以及职业或生活环境中的长期接触。
长期暴露可带来多种健康风险,如增加癌症发生、造成DNA损伤、干扰免疫功能,甚至引发不可逆的组织损害。
近年来研究表明,重金属除直接损伤器官外,还可通过影响肠道微生物群和肠道屏障危害健康。其在肠道内会对菌群产生选择压力,促使耐受菌增殖,并通过诱导氧化应激、改变基因表达,破坏肠道微生态平衡。
不同重金属的毒性机制各异,对肠道菌群及人体健康的影响也存在差别。
①砷:可改变肠道菌群,并影响全身代谢
砷可以以不同形式存在,主要包括无机砷和有机砷。其中,无机砷,如三价砷和五价砷,毒性通常更强。对人类来说,受污染的饮用水是砷暴露的重要来源。在一些地区,例如孟加拉国以及中国西部部分地区,地下水中的砷含量可能超过安全限值。
流行病学研究表明,高砷暴露与肠道菌群结构改变相关。例如,生活在砷污染地区的儿童肠道中变形菌门丰度升高,同时富集与毒力和抗生素耐药相关的功能基因。
注:变形菌门包含多种机会致病菌或病原菌,如大肠杆菌、幽门螺杆菌、克雷伯氏菌、沙门氏菌、志贺氏菌、铜绿假单胞菌、霍乱弧菌和空肠弯曲菌等,其增多通常提示肠道微生态失衡。
此外,从这些儿童体内分离的大肠杆菌携带砷抗性基因且表达增强,表明长期砷暴露可促使肠道细菌获得或强化抗砷能力。
动物实验亦支持这一结论:小鼠经饮水长期暴露于不同浓度砷后,出现肝损伤及菌群失调;暴露数周的无机砷亦可显著改变肠道微生物结构。
代谢组学分析进一步发现,砷暴露会引起血液、肝脏和粪便中多种代谢物异常,其中多与肠道微生物相关,提示砷不仅改变菌群组成,还影响其代谢功能,并扰乱全身代谢平衡。
②镉:可能通过“肠—脑轴”影响认知功能
镉和铅是环境中较常见的重金属污染物。它们的肠道毒性机制有一定相似性:都可能诱发氧化应激,破坏肠道微生态平衡,并促进某些耐受重金属的细菌增殖。
镉主要通过饮食进入人体,尤其来源于受污染土壤中种植的农作物,如稻米等主食。
动物研究表明,长期摄入含镉饮水可导致肠道菌群失调并影响神经功能。例如,小鼠连续 9 周暴露于一定浓度镉后出现认知损伤;同时伴随肠道屏障受损、炎症因子变化、海马相关基因表达异常,以及肠源性神经活性代谢物紊乱。
这些结果提示,镉的神经毒性不仅源于对大脑的直接作用,也可能通过肠—脑轴间接实现,其中肠道菌群和屏障功能是关键中介。
此外,研究还发现镉暴露可抑制呼吸代谢和免疫反应,诱导氧化应激,并改变能量代谢相关基因表达,可能反映机体对长期镉胁迫的适应性反应。
③铅:可能促进耐重金属细菌增加
铅的暴露途径与镉相似,主要来自受污染的食物、水及环境或职业接触。
一项纳入696名参与者的研究发现,尿铅水平与肠道菌群多样性变化相关:尿铅较高者肠道中变形菌门及伯克氏菌目等细菌丰度增加。
这表明铅可能通过改变肠道微生态,促进耐重金属的机会性细菌生长,进而削弱菌群稳定性,并增加炎症或感染风险。
④汞:可能损伤肠道屏障,并通过食物链累积
汞可在环境中转化为毒性更强的甲基汞,并在食物链中逐级富集。人类主要通过食用鱼类,尤其是处于食物链高位的大型掠食性鱼类而暴露。
鱼类实验表明,膳食甲基汞未必导致急性死亡,但在体内累积至一定水平后,会显著影响其生长、繁殖和行为;野外研究亦提示,低剂量暴露也可能影响种群稳定。
在人类和哺乳动物中,无机汞和甲基汞均具有神经毒性和免疫毒性。相较之下,其对肠道上皮和菌群的直接影响研究较少。
细胞实验显示,肠上皮细胞暴露于无机汞或甲基汞后,氧化还原平衡被破坏,通透性增加,紧密连接受损,即肠道屏障变“更漏”。
因此,肠道屏障损伤可能是汞引发全身毒性的重要途径之一。
★ 肠道微生物也会“改造”重金属
重金属进入肠道后并非保持不变,肠道微生物可通过酶系统和氧化还原反应参与其转化。
例如,部分微生物可对砷和汞进行甲基化或去甲基化,改变其化学形态、毒性及吸收程度。硫酸盐还原菌可将无机汞转化为更易吸收且神经毒性更强的甲基汞;而其他微生物过程则可能将其转化为更难吸收或更易排出的形式,从而降低毒性。
此外,铬、硒等元素也可被微生物改变价态,不同价态对应不同毒性和健康风险。
这些转化受微生物基因调控,如编码金属还原酶、氧化酶和转运蛋白的基因;在长期暴露下,这些基因可能富集,使菌群逐步适应重金属压力。
因此,肠道微生物对重金属的转化具有“双刃剑”效应:既可能降低毒性,也可能生成更易吸收、危害更大的形式。
▸ 持久性有机污染物对肠道微生物群的影响
持久性有机污染物,简称 POPs,是一类很难在环境中降解的有毒化学物质。它们通常容易溶解在脂肪中,因此会在动物和人体内长期积累。POPs还可以随着空气、水流和食物链传播到很远的地方。
常见的 POPs 包括:
这些物质可以通过食物链不断富集。例如,小鱼摄入污染物后被大鱼吃掉,大鱼又被人类或其他动物食用,污染物浓度就会一级一级升高。因此,POPs 不仅会影响生态系统,也可能危害人体健康。
近年来,肠道微生物群逐渐受到关注。研究发现,肠道微生物并不只是被 POPs 影响的“受害者”,它们还可能参与污染物的代谢,并影响宿主对污染物的反应。也就是说,POPs 会改变肠道菌群,而肠道菌群的变化又可能进一步影响人体的代谢、免疫功能,甚至神经行为。
①PCBs和TCDD:通过 AhR 影响肠道和免疫
PCBs 和 TCDD 可激活芳基烃受体(AhR),这一细胞内“环境感应器”参与外源化学物代谢及免疫调控。
AhR 被激活后,宿主细胞与肠道微生物的基因表达均可能改变,打破菌群平衡,导致失调并损害肠道功能。
尽管PCBs已被管控多年,但因其高度稳定,仍广泛存在。动物研究发现,母体暴露于 PCB-126 会使成年后代肠道菌群的丰富度和多样性下降,并改变特定菌群组成,从而可能增加其晚年慢性疾病风险,这种影响并不完全依赖于饮食或运动。
机制上,PCB 通过激活 AhR 上调炎症因子(如 IL-6、TNF-α)表达,使肠道处于长期低度炎症状态,抑制有益菌生长并加剧菌群失衡。
②TCDD:影响胆汁酸代谢和肠道-肝轴
TCDD 是一类高毒性的二噁英,可持续激活 AhR,并影响肠道菌群及宿主代谢。
小鼠研究表明,TCDD 暴露会改变胆汁酸代谢,并可能诱发非酒精性脂肪肝(NAFLD)。同时,肠道菌群发生变化,如乳酸杆菌增多、次级胆汁酸水平升高。
宏基因组分析进一步发现,与胆汁酸合成相关的微生物基因也发生改变,提示 TCDD 可能通过“肠道-肝轴”影响肝脏功能,即菌群变化影响胆汁酸代谢,进而作用于肝脏。
另一项研究显示,母体在妊娠和哺乳期暴露于 TCDD,高剂量会导致母体及后代菌群失调、色氨酸代谢紊乱,并增加潜在致病菌。
但也有研究发现,低剂量 TCDD 在某些情况下反而减少后代肠道致病菌,提示其对肠道微生物的影响具有剂量依赖性,甚至可能产生相反效应,并具有跨代影响。
③DDT和PBDEs:也会扰乱肠道菌群
DDT 是曾广泛使用的有机氯农药,虽已在多国被限制或禁用,但因其高度稳定,仍可在土壤、水体和生物体内检测到。
PBDEs 是用于塑料、纺织品和电子产品的阻燃剂,亦属常见持久性有机污染物,其结构和毒理作用与 PCBs 部分相似。
研究表明,DDT 和 PBDEs 均可改变肠道微生物群,如减少有益菌、增加与炎症或代谢紊乱相关的菌群。但其影响受暴露剂量、时间、物种、饮食及个体原有菌群状态等多因素调控,因此具有明显的情境依赖性。
有研究发现,围产期暴露 PBDE-47 会干扰后代肠道菌群的发育、组成及代谢;而孕期和哺乳期补充罗伊氏乳杆菌(Lactobacillus reuteri)可在性别依赖性下缓解这些影响,恢复菌群多样性,并改善体重和神经行为表现。
总体来看,POPs 不只是直接损害器官和组织,它们还会通过改变肠道微生物群影响健康。PCBs、TCDD、DDT 和 PBDEs 等污染物都可能破坏肠道菌群平衡,并进一步影响炎症反应、胆汁酸代谢、色氨酸代谢以及肠道-肝轴功能。
换句话说,肠道微生物群可能是连接环境污染物暴露与慢性疾病风险之间的重要桥梁。
编辑
doi.org/10.3389/fmicb.2026.1737229
▸ 新兴污染物及其对肠道微生物群的影响
随着环境检测技术进步,许多以往难以发现的污染物逐渐进入公众视野,被称为新兴污染物,如微塑料、纳米塑料、抗生素耐药基因以及药品和个人护理品残留等,已受到广泛关注。
这类污染物通常具有难降解、分布广等特点,并可能干扰人体与肠道微生物的平衡,可通过食物、饮水及日常环境进入人体。
进入体内后,它们可能通过破坏肠道屏障、释放有害物质或促进耐药基因传播等途径影响肠道健康。因此,其对人体健康的潜在影响已成为环境毒理学的重要研究方向。
①微塑料与纳米塑料
微塑料指直径约1微米至5毫米的塑料颗粒,纳米塑料则小于1微米,来源包括塑料制品在环境中的风化破碎及工业生产过程。
这类颗粒已广泛存在于水体、土壤、空气和食物链中。研究表明,它们可通过物理损伤、化学毒性和载体效应三种机制影响肠道微生物:既可能直接刺激或破坏肠道,也可释放有害添加剂,或作为“载体”携带其他污染物进入体内。
其毒性与粒径、表面特性及塑料类型密切相关;通常粒径越小,与组织和微生物的相互作用越强,潜在影响也越大。
√物理影响:小颗粒可能更容易穿过肠道屏障
相比尺寸较大的塑料颗粒,纳米塑料由于体积更小,通常表现出更强的生物活性。动物实验显示,摄入纳米级塑料后,对肠道和造血系统造成的影响可能比大颗粒塑料更明显。
研究发现,小鼠口服微塑料或纳米塑料后可出现肠道菌群失衡,并伴随造血系统异常,提示肠道微生物变化可能影响全身健康。
机制上,纳米塑料可穿过肠上皮紧密连接进入黏膜层并直接接触微生物,导致细菌膜受损、通透性增加,甚至引发部分菌体死亡。
此外,小鼠长期口服约2微米的聚氯乙烯微塑料会出现肠道损伤,并伴随菌群结构和代谢物改变,提示即使低剂量、长期摄入也可能对肠道健康构成风险。
√化学效应:塑料中的添加剂可能释放出来
塑料并非单一材料,生产中常加入增塑剂、稳定剂、阻燃剂等以改善性能。这些添加剂进入人体后可从塑料颗粒中释放,产生额外毒性。
例如,常见增塑剂邻苯二甲酸酯(如 DEHP)长期暴露可导致胆固醇代谢紊乱。研究表明,DEHP 还能改变肠道微生物组成,影响胆汁酸代谢,并通过肠—肝轴干扰肝脏胆汁酸合成。
这表明,微塑料相关化学物质不仅影响肠道,还可能进一步影响全身代谢。
此外,不同塑料的效应存在差异,如聚苯乙烯较聚乙烯或 PET 更易吸附有毒物质,因而可能对肠道微生物产生更强干扰。
√载体效应:微塑料可能“搭载”其他污染物进入肠道
微塑料和纳米塑料还有一个特殊问题:它们表面积较大,容易吸附环境中的其他污染物,例如重金属、持久性有机污染物以及塑料添加剂等。因此,它们可能像“污染物载体”一样,把其他有害物质一起带入人体肠道。
研究表明,微塑料可吸附邻苯二甲酸酯并将其带入小鼠肠道,其在肠道中的累积程度与对微塑料的吸附能力相关,即吸附性越强,越易随微塑料进入并滞留。
在一项联合暴露实验中,小鼠连续 30 天接触被 DEHP 污染的微塑料后,出现肠道通透性升高、炎症增强及菌群结构改变,部分受影响菌群与能量代谢和免疫功能密切相关;同时,与氧化应激和炎症相关的基因表达也发生变化。
这些结果表明,实际环境暴露多为多种污染物共同作用,微塑料可与其他化学物协同影响,产生更复杂甚至更强的健康效应。
②抗生素耐药基因(ARGs)
抗生素耐药基因(ARGs)是赋予细菌抗药性的遗传信息,可随可移动遗传元件在细菌间传播,近年来被视为重要的新型环境污染物。
ARGs 广泛存在于土壤、水体和动物粪便中,并可通过受污染的食物或饮水进入人体肠道。进入体内后,这些基因可通过“水平基因转移”在细菌间扩散,使原本不具耐药性的菌获得抗生素抵抗能力;一旦转移至致病或机会致病菌,可能增加耐药感染风险。
在谷禾健康检测报告的抗生素耐药风险评估中,部分用户即使近期并未使用抗生素,仍检出较高的耐药风险。这可能与抗生素耐药基因在环境中的污染及其传播有关。
编辑
<来源:谷禾健康肠道菌群检测数据库>
目前研究主要聚焦两个问题:一是 ARGs 在肠道中的传播效率,二是其对肠道微生物群组成的影响。
在肠道内,细菌可通过转导(病毒介导)和接合(直接接触)等方式交换遗传信息,促进共生菌与潜在致病菌之间的基因传播。随着生物信息学技术的发展,研究人员已能更准确识别耐药基因的宿主及其在群落中的传播频率,为制定干预策略、遏制耐药基因扩散提供了依据。
③药品和个人护理产品(PPCPs)
药品和个人护理产品(PPCPs)包括抗生素、止痛药、抗抑郁药、激素、防腐剂、紫外线滤镜和化妆品等,正成为重要的环境污染物。它们主要通过废水排放和农业径流进入环境,且难以在传统处理系统中完全降解,从而对水生生物和人类造成长期、低剂量暴露风险。
越来越多证据表明,PPCPs 即使在环境相关浓度下也可扰乱肠道微生物群的组成、多样性和功能。例如,常见抗菌剂三氯生可降低Lachnospiraceae 和 Ruminococcaceae 的丰度,同时增加促炎的变形菌门;在小鼠中,长期暴露还会改变微生物代谢通路,损伤上皮屏障并加重结肠炎。
非抗生素类 PPCP,如卡马西平和氟西汀,也能调控与神经递质代谢和应激反应相关的微生物基因表达。斑马鱼研究显示,氟西汀暴露可增加放线菌丰度并改变氨基酸生物合成通路,可能影响神经行为。此外,多种 PPCP(如 NSAIDs 和紫外线滤镜)联合暴露的人群研究发现,胆汁酸代谢改变和短链脂肪酸(SCFA)下降,提示其与炎症和代谢综合征相关。
尽管已有进展,关于长期效应、混合暴露及个体差异仍存在明显不足。目前多数功能研究集中于啮齿动物或水生模型,缺乏人群高分辨率纵向数据。未来需结合宏基因组、代谢组和转录组等多组学与环境暴露评估,以阐明其与肠道菌群失调的因果关系。
总体而言,污染物对肠道微生物群的影响具有复杂性和动态性:短期暴露多为可逆改变,而长期暴露(尤其在动物中)可导致持续性改变。但在人类中的长期证据仍有限,有待进一步纵向与干预研究验证。
污染物引发的肠道微生物群改变通常源于直接与间接机制的共同作用。直接作用包括氧化应激、膜损伤、基因毒性及群体感应干扰,常见于重金属、抗生素和纳米颗粒。
间接作用则通过诱导宿主的代谢、内分泌和免疫反应改变微生物组成。例如,全氟化合物可通过影响胆汁酸代谢和免疫状态重塑肠道菌群。这些过程涉及氧化应激反应、芳香烃受体(AhR)信号激活、肠上皮屏障受损及黏膜炎症增强等通路,不仅驱动菌群变化,也可能进一步影响全身健康。
▸ 氧化应激在微生物群破坏中的核心中介作用
氧化应激是连接环境污染暴露、肠道菌群失衡与宿主肠道损伤的关键机制。重金属、微/纳米塑料、持久性有机污染物、农药及电离辐射等进入机体后,主要通过两条途径升高活性氧(ROS):一是受损线粒体电子传递链产生的线粒体 ROS,二是细胞膜 NADPH 氧化酶系统(NOX/DUOX)生成的 ROS。
√ 污染物干扰线粒体导致氧化失衡
当污染物干扰线粒体氧化磷酸化时,会增加复合物 I 和 III 的电子泄漏,降低膜电位并产生过量 ROS,进而引发脂质过氧化、DNA 损伤和细胞凋亡。与此同时,NOX1、NOX2 和 DUOX2 虽参与应激和免疫防御,但长期过度激活会削弱肠道屏障并促进慢性炎症。
因此,线粒体与 NOX/DUOX 来源的 ROS 共同导致肠道氧化还原失衡,不仅损伤宿主细胞,也进一步改变肠道微生态结构与功能。
编辑
doi.org/10.3389/fmicb.2026.1737229
上面示意图展示了污染物促进肠道氧化应激的两条主要途径:
(1)污染物损伤线粒体呼吸链,导致线粒体 ROS 过量产生;
(2)污染物激活肠上皮细胞表面的 NOX/DUOX 酶系统,进一步促进 ROS 生成。
这两类 ROS 来源可相互作用并产生放大效应,加剧肠道氧化还原失衡。持续升高的 ROS 会损伤紧密连接、削弱黏液层并扰乱微生态,促进炎症发生。
√ 过量活性氧会削弱肠上皮的防御功能
过量活性氧(ROS)还会降低肠上皮防御功能,增加通透性,使脂多糖(LPS)等微生物产物更易穿过屏障进入体内,形成“肠漏”,进而促进促炎菌增殖、抑制有益菌,导致菌群失调。
在分子层面,ROS 可激活 MLCK/肌球蛋白及 NF-κB/MAPK 通路,下调闭塞蛋白、ZO-1 等关键屏障蛋白并增强炎症因子释放,形成“氧化应激—屏障损伤—菌群失调—炎症”的恶性循环。
同时,肠道微生物也能调控 ROS 水平。共生菌产生的乳酸及相关信号可激活上皮细胞 NOX,促进 ROS 生成并调节干细胞更新:生理水平有助于维持稳态,过量则可能引发异常增生和菌群紊乱,体现宿主—微生物相互作用对肠道生态的影响。
√ ROS还扰动代谢平衡
短链脂肪酸(SCFA),尤其丁酸,可通过G蛋白偶联受体和 Nrf2/ARE 通路减轻氧化应激、增强屏障功能;而菌群失调导致 SCFA 减少时,抗氧化与屏障保护能力随之下降,使肠道更易受损。
因此,在颗粒物、辐射、农药等环境暴露下,ROS 介导的屏障破坏与菌群失调可能成为系统性炎症和代谢紊乱的起点,并通过肠—肝轴、肠—脑轴影响全身健康,为污染暴露与疾病之间的关联提供了重要机制解释。
▸ AhR 作为连接环境感测与肠道免疫的集成枢纽
芳香烃受体(AhR)信号通路是环境污染物毒理作用的关键枢纽,连接外源物代谢与免疫调控。AhR 在胞质中通常与热休克蛋白 90(HSP90)和 X 相关蛋白 2(XAP2)等伴侣蛋白形成复合体;当内源或外源配体结合后,复合体构象改变、伴侣蛋白解离,促使配体-AhR 复合体转位入核。
AhR连接污染物、肠道菌群与免疫机制
编辑
doi.org/10.3389/fmicb.2026.1737229
许多环境污染物,如 TCDD、二噁英、多氯联苯(PCBs)和多环芳烃(PAHs),都可以作为 AhR 的外源性配体,激活肠上皮细胞和免疫细胞中的 AhR 信号通路。
AhR 被配体激活后由胞质转位入核,与核转运子 ARNT 形成异二聚体,并结合芳香烃响应元件(AHRE),调控靶基因表达,从而影响外源物代谢、免疫反应、肠道屏障及微生物群组成。
√ AhR 过度激活与肠道炎症失衡
AhR 信号具有明显的“双向调节”作用。不同来源的配体会产生不同的生物学效应。外源性有毒污染物,如 TCDD、二噁英、PCBs 和 PAHs,通常会导致 AhR 持续、过度激活。
这种异常激活可上调多种代谢酶,例如 CYP1A1、CYP1B1 和 ALDH3A1,同时促进炎症因子 IL-6、TNF-α 和 IL-1β 的释放。长期来看,这会增强氧化应激,破坏肠道黏膜平衡,并推动慢性肠道炎症和菌群失调的发生。
√ 内源性AhR配体的保护作用及其免疫调控机制
相比之下,内源性或肠道微生物来源的 AhR 配体通常发挥保护作用。例如,肠道菌群代谢色氨酸后产生的吲哚类代谢物,如吲哚-3-醛(I3Ald)和吲哚-3-乙酸(IAA),可以温和、短暂地激活 AhR。这种生理性 AhR 激活有助于促进 IL-22 分泌,增强上皮修复能力,维持黏液层和杯状细胞功能,并支持肠道免疫耐受和黏膜稳态。
此外,AhR信号还与氧化应激和炎症通路密切交互。AhR 的激活可以影响 NF-κB 信号通路和 NLRP3 炎症小体活性,从而参与调节炎症因子的产生。同时,它也会影响短链脂肪酸(SCFA)水平、杯状细胞功能以及肠道微生物群组成。因此,AhR 可以被看作是整合“污染物暴露—微生物代谢—免疫炎症—氧化应激”信号的关键节点。
总体来看,适度的 AhR 激活有助于维持肠道屏障完整性、促进组织修复并保持免疫平衡;而污染物诱导的持续 AhR 过度激活,则会打破这种平衡,使肠道微环境从稳态转向氧化应激、慢性炎症和菌群失调。
▸ 肠道屏障完整性的破坏
肠道屏障是连接宿主与外界的首道防线,由黏液层、上皮紧密连接和免疫成分协同维持稳态。环境污染物通过诱导氧化应激、炎症因子过度产生及直接损伤上皮蛋白,破坏屏障结构,增加通透性并引发菌群失调。
编辑
doi.org/10.3389/fmicb.2026.1737229
环境污染物可通过多种方式损伤肠道屏障。主要机制包括:
(1)减少杯状细胞分泌黏蛋白 MUC2,使黏液层变薄;
(2)通过 ROS 相关氧化应激信号,下调或破坏紧密连接蛋白,如 occludin、claudin 和 ZO-1;
(3)激活 MLCK 和 PKC 等信号通路,引起细胞骨架收缩。
这些变化会使肠上皮细胞之间的连接变得松散,增加肠道通透性,从而使肠腔内的细菌、脂多糖(LPS)和其他毒性产物更容易穿过肠屏障,进入黏膜甚至全身循环,最终促进慢性炎症和肠道菌群失调。
√ 环境污染物诱导的肠道屏障损伤与炎症级联
肠道黏液层是保护上皮的第一道防线,兼具物理与生化屏障功能,可阻止病原体和毒素接触上皮细胞。污染物如双酚 A(BPA)可通过诱导线粒体功能障碍和氧化应激,抑制杯状细胞分泌黏蛋白 2(MUC2),使黏液层变薄,降低对细菌和毒素的防御能力。
MUC2 表达受 p53 等转录因子调控,其下降会显著削弱黏膜保护。研究表明,黏液屏障受损是溃疡性结肠炎等炎症性肠病的早期特征之一,可促进微生物易位和炎症发生。
同时,污染物还会破坏上皮细胞间的紧密连接。PM2.5 及镉、铅等重金属可通过氧化应激下调 occludin、claudin 和 ZO-1 等蛋白,增加肠道通透性;MLCK 和 PKC 的激活则引发细胞骨架收缩,扩大细胞间隙,加重“肠漏”。
持续的屏障损伤使 LPS 等微生物产物进入血液,与免疫细胞表面的 TLR4 结合,激活 NF-κB 炎症通路,进而引发内毒素血症、代谢紊乱和免疫失衡,形成恶性循环。
因此,肠道屏障既是污染物损伤的直接靶点,也是连接污染暴露、氧化应激、菌群失调与全身炎症的关键环节,其破坏会打破宿主与微生物平衡,并推动局部炎症向全身性病变发展。
▸ 免疫炎症反应的扩增与持续性
在氧化应激和屏障破坏后,环境污染物会引发一连串免疫激活,可能从急性黏膜炎症进展为慢性系统性失调。持续暴露维持细胞因子的产生,招募免疫细胞,改变肠道微生物群,形成自我强化的炎症和氧化损伤循环。
正反馈循环推动污染物诱导的慢性炎症
编辑
doi.org/10.3389/fmicb.2026.1737229
√ 污染物驱动的免疫失衡与炎症放大机制
炎症早期,模式识别受体尤其是 TLR4 识别 LPS 或污染物信号,激活 MyD88–NF-κB 和 MAPK 通路,促进 IL-6、TNF-α、IL-1β 等产生;短期有利于防御,但持续激活会转为慢性炎症。重金属和多环芳烃(PAHs)还可激活 NLRP3 炎症小体,促进 IL-1β、IL-18 过度释放,放大炎症。
ROS 还促进巨噬细胞向促炎 M1 型极化并抑制 M2 型,导致炎症加重;同时,污染物促进 Th17 分化和 IL-17A 释放,抑制 Treg 及 IL-10,削弱抗炎调节。
此外,污染物引起菌群失调,减少产短链脂肪酸(SCFAs)的有益菌,降低丁酸盐水平,削弱其对 HDAC 和 NF-κB 炎症通路的抑制作用,使炎症更难终止。
√ AhR配体缺失与炎症正反馈循环
同时,能代谢色氨酸的共生菌减少,导致 AhR 配体(如吲哚-3-醛)生成下降,削弱 IL-22 介导的黏膜修复,使受损屏障更难恢复并加剧炎症。
因此,污染物、ROS、LPS、炎症因子与菌群失调形成正反馈循环:污染物诱导 ROS,ROS 破坏屏障;屏障受损促进 LPS 移位并激活免疫;免疫反应持续释放炎症因子;炎症与污染物进一步扰乱菌群,减少 SCFAs 等保护性代谢物;这些变化又加重屏障损伤和氧化应激,最终推动炎症长期化。
在全身层面,进入血液循环的 LPS 和炎症细胞因子还可以通过肠-肝轴和肠-脑轴影响远端器官。例如,它们可激活肝脏中的库普弗细胞和中枢神经系统中的小胶质细胞,进而参与非酒精性脂肪肝、神经炎症和代谢综合征等肠外疾病的发生。这说明肠道免疫激活不仅是局部炎症反应,也是污染物导致全身毒性效应的重要机制。
编辑
doi.org/10.3389/fmicb.2026.1737229
小结
总体来看,污染物诱导的免疫反应并不是单一、短暂的过程,而是会通过 ROS 过量产生、炎症信号激活、肠道屏障损伤和菌群失调等多个环节相互放大,使急性上皮炎症逐渐发展为慢性、系统性的炎症状态。
理解这一正反馈机制,有助于开发抗炎治疗以及靶向肠道菌群的干预策略,从而减轻污染物相关慢性疾病的发生和发展。
过去十年中,越来越多研究表明,环境污染物不仅直接损伤特定器官,还可以通过扰乱肠道菌群,引发全身性毒性反应。这种影响可进一步改变宿主代谢、免疫稳态和疾病易感性。然而,目前该领域仍存在一些关键问题尚未解决。这在一定程度上与传统毒理学长期采用“器官中心”研究模式有关,即更多关注污染物对肝脏、肺、肾脏或神经系统等单一器官的直接损伤,而对肠道菌群介导的全身效应认识不足。
污染物、菌群失调与疾病之间的因果关系仍不清楚
目前多数研究主要停留在相关性分析层面,即观察污染物暴露后哪些菌群发生变化,以及这些变化是否与炎症、代谢紊乱或组织损伤相关。但这类研究往往难以回答一个关键问题:菌群失调究竟是污染物毒性的原因,还是毒性结果的一部分?
换句话说,污染物可能直接损伤宿主组织,也可能先改变肠道菌群,再通过菌群代谢物、免疫调节和屏障功能改变引发疾病。目前很多研究尚未有效区分这两类机制。
此外,宿主自身因素也会影响污染物毒性和菌群反应,包括年龄、性别、饮食结构、遗传背景和基础健康状态等。例如,高纤维饮食可能通过促进短链脂肪酸产生,缓解污染物导致的菌群功能下降;而衰老个体的肠道菌群恢复能力较弱,可能更容易受到污染物影响。免疫调节或解毒代谢相关基因的差异,也可能导致个体对污染物暴露的敏感性不同。
因此,未来研究应更多采用多污染物共暴露模型,并纳入宿主个体差异因素,以更接近真实环境暴露情境。同时,暴露组学、微生物组学和计算建模的结合,有助于解析复杂暴露条件下污染物与肠道菌群之间的网络关系。
主要参考文献:
Ma W, Xiong X, Tian Z, Li L and Huang Y (2026) Environmental pollutants and the gut microbiota: mechanistic links from exposure to systemic disease. Front. Microbiol. 17:1737229.
AgarwalM.HoffmanJ.Ngo TenlepS. Y.SantarossaS.PearsonK. J.SitarikA. R.et al. (2023). Maternal polychlorinated biphenyl 126 (PCB 126) exposure modulates offspring gut microbiota irrespective of diet and exercise. Reprod. Toxicol. 118:108384.
Aravind KumarJ.KrithigaT.SathishS.RenitaA. A.PrabuD.LokeshS.et al. (2022). Persistent organic pollutants in water resources: fate, occurrence, characterization and risk analysis. Sci. Total Environ. 831:154808.
AldereteT. L.JonesR. B.ChenZ.KimJ. S.HabreR.LurmannF.et al. (2018). Exposure to traffic-related air pollution and the composition of the gut microbiota in overweight and obese adolescents. Environ. Res.161, 472–478
AlexanderC.RietschelE. T. (2001). Bacterial lipopolysaccharides and innate immunity. J. Endotoxin Res. 7, 167–202
AndersenO. (2016). A characterisation of low-grade inflammation and metabolic complications in HIV-infected patients. Dan. Med. J. 63:B5291.

谷禾健康
现在越来越多的人出现血脂不正常、腰围增加、血糖略高,这背后的隐藏风险,可能正在悄悄累积。
在全球范围内,代谢综合征已成为最受关注的健康挑战之一,患病率仍呈逐年上升趋势。它不仅显著增加心血管疾病、2型糖尿病及脂肪肝的风险,也对医疗资源和生活质量带来巨大压力。更让人警惕的是,这种代谢异常往往早期无明显症状,很多人直到发现血脂异常或血糖升高才意识到问题。
医学界和科学界也一直都在不断探索代谢综合征的发病机制和新的干预途径。传统的观点主要关注遗传因素、生活方式以及宿主自身的代谢紊乱,但近年来多个证据同时表明,代谢综合征和血脂异常通常同时存在——肠道菌群是将它们联系起来的关键因素之一。
肠道菌群影响食物的消化方式、胆汁酸的处理以及炎症信号在全身的释放。当肠道生态系统失衡时,新陈代谢可能转向胰岛素抵抗和脂质处理改变——这会导致甘油三酯升高、高密度脂蛋白(俗称好胆固醇)降低,有时还会增加 LDL 颗粒。
相关机制包括短链脂肪酸、胆汁酸、TMAO等。例如肠道细菌将初级胆汁酸转化为调控能量平衡、葡萄糖代谢及胆固醇处理的次级胆汁酸,通过FXR和TGR5受体发挥作用。某些微生物模式减少有益胆汁酸转化,降低胆固醇清除效率,进一步加剧脂质调节异常。同时,代谢综合征与低度炎症相关,菌群失衡可增加肠道通透性,使细菌成分激活免疫通路,加重胰岛素抵抗和血脂异常,形成一个循环,可能延续血脂异常。
本文将从多个层面深入探讨肠道微生物群如何影响血脂代谢,以及这些微生物学的改变如何最终导致血脂异常的发生。首先介绍代谢综合征的流行病学现状和临床特征,然后聚焦于血脂异常在代谢综合征中的核心地位,接着详细阐述肠道微生物群影响血脂代谢的五大核心机制,总结代谢综合征患者特征性的微生物组模式改变,最后讨论为什么肠道菌群检测对于代谢综合征的防控具有重要意义,以及这一领域未来的发展方向和临床转化前景。
代谢综合征是什么?
代谢综合征(英文名为Metabolic Syndrome,经常被简称为MetS),它是一组代谢紊乱症候群的集合。
根据国际糖尿病联盟(IDF)及多学会联合发布的统一诊断标准,代谢综合征的诊断需要满足以下五项中的三项或更多:
从这个诊断标准我们可以看出,血脂异常本身就是代谢综合征的核心诊断指标之一,这也说明了血脂异常在代谢综合征中的重要地位。
代谢综合征的全球发病率持续上升
全球成人代谢综合征患病率约为20%–25%(多项流行病学研究估计),这个比例意味着每四个成年人中就有一个患病,患者总数已经超过了10亿人。
更值得警惕的是,在过去数十年中呈显著上升趋势。这种快速增长的背后,是全球范围内肥胖症的流行。肥胖,特别是腹型肥胖,正是代谢综合征最重要的危险因素之一。
主要风险因素
代谢综合征的发生发展是一个复杂的过程,是遗传因素与环境因素长期相互作用的结果。我们可以将这些风险因素分为不可控和可控两大类。
不可控的风险因素:
年 龄
年龄是一个无法改变的因素,然而,它能帮助我们识别高风险人群。随着年龄增长,患代谢综合征的风险显著增加。
具体而言,随着年龄的增长,肌肉量会逐渐减少,而肌肉是消耗葡萄糖的关键组织。肌肉量的减少直接导致胰岛素敏感性下降。与此同时,随着年龄的增长,内脏脂肪的积累也会加重代谢负担。
遗 传
遗传易感性也是一个重要的不可控因素。研究已经发现了数十个与代谢综合征相关的遗传位点,这些位点每个单独的效应虽然不大,但累积起来可以显著影响患病风险。
如果父母或兄弟姐妹中有人患有代谢综合征、2型糖尿病或心血管疾病,患病几率会显著增加。
不过,遗传易感性并不意味着必然患病,健康的生活方式仍然能够有效地预防疾病的发生。
性 别
性别我们前面已经提到了,男性患病风险高于绝经前女性,这主要是性激素的保护作用在起作用。
女性在绝经前由于雌激素对脂质代谢、脂肪分布和血管功能具有一定保护作用,风险相对较低;但绝经后,随着雌激素水平下降,内脏脂肪增加、LDL-C升高、胰岛素敏感性下降,代谢综合征发生风险会明显上升。
可控的风险因素:
饮食——关键变量
饮食不是吃得多或者少的问题,是能量摄入、营养结构、脂肪类型、碳水质量、膳食纤维水平和饮食节律共同决定了代谢后果。
——饮食结构
富含饱和脂肪、反式脂肪、添加糖和精制碳水化合物的饮食模式,会促进能量过剩、内脏脂肪堆积、肝脏脂质合成增加和胰岛素抵抗。长期如此,机体更容易出现中心性肥胖、甘油三酯升高、脂肪肝、血糖异常等代谢综合征相关表现。
膳食纤维、全谷物、豆类、蔬菜水果、多酚类食物和优质不饱和脂肪酸摄入不足,也会削弱代谢保护作用。尤其是膳食纤维不足,会减少肠道菌群可利用底物,影响短链脂肪酸生成、肠屏障功能和炎症调节。
——脂肪摄入问题
需要强调的是,脂肪本身并不是一概有害。适量膳食脂肪对细胞结构、脂溶性维生素吸收、饱腹感和能量平衡都有重要作用。需要关注的是脂肪来源和脂肪酸类型。
从肠道微生态角度看,饮食也是塑造肠道菌群结构和功能的主要外部因素。长期高脂、高糖、低纤维饮食可能降低产短链脂肪酸菌,影响胆汁酸代谢和肠道屏障功能,进一步加重代谢异常。
运 动
缺乏运动是另一个重要的可控危险因素。出门坐车,上楼电梯,办公室久坐,回家躺着看电视,每天的体力活动量比五十年前减少了很多。规律的运动不仅可以帮助我们控制体重,还能提高骨骼肌胰岛素敏感性,增加葡萄糖转运蛋白GLUT4表达,促进糖原合成,提高脂肪酸氧化能力,并降低内脏脂肪含量。
对于代谢综合征的人群,很多研究已经提示,即便体重下降幅度有限,规律有氧运动和抗阻训练也能改善胰岛素抵抗、降低甘油三酯、提高HDL-C,并改善血压和炎症状态。
世界卫生组织推荐成年人每周至少进行150分钟中等强度的有氧运动,但是调查显示,仅约20%–30%的成年人达到推荐水平。
吸烟 饮酒
吸烟和过量饮酒也会显著增加代谢综合征的患病风险。
烟草中的尼古丁会引起血管收缩,升高血压,还会影响胰岛素信号传导,降低胰岛素敏感性。长期吸烟的人,患代谢综合征的风险比不吸烟者高出许多。
过量饮酒(如每日酒精摄入超过20–30 g),影响肝脏的脂代谢,还会提供大量的空热量,容易导致肥胖。研究发现,每天酒精摄入量超过25克,代谢综合征患病风险大大增加。
压力 睡眠
长期精神压力也是一个容易被忽视的危险因素。现代社会生活节奏快,竞争压力大,很多人长期处于慢性应激状态。
慢性压力会导致体内皮质醇水平持续升高,皮质醇是一种糖皮质激素,长期高水平的皮质醇会促进内脏脂肪堆积,升高血糖和血压,还会打乱正常的饮食节律,导致暴饮暴食。这些都会增加代谢综合征的发病风险。
睡眠不足和昼夜节律紊乱同样重要。睡眠时间不足、睡眠质量下降、夜间进食、轮班工作等因素,都会影响胰岛素敏感性、食欲激素、交感神经活性和炎症状态。临床上,睡眠呼吸暂停也常与中心性肥胖、胰岛素抵抗、高血压和血脂异常共同出现。
胰 岛 素 抵 抗
生物标志物层面的中间危险因素。胰岛素抵抗被认为是代谢综合征最核心的病理生理基础。
所谓胰岛素抵抗,就是指身体对胰岛素的反应变得不敏感,胰岛素不能有效地发挥降低血糖、调节脂代谢的作用。身体需要分泌更多的胰岛素才能维持血糖正常,长期高胰岛素血症会进一步加重代谢紊乱。
其 他
慢性低度炎症也是一个重要的中间危险因素,代谢综合征患者体内的促炎细胞因子水平普遍升高,这些炎症因子会干扰胰岛素信号传导,损伤血管内皮功能,促进动脉粥样硬化的发生发展。
脂肪组织功能异常同样关键,当脂肪细胞储存过多的脂肪时,脂肪细胞会发生肥大、缺氧,释放出大量的游离脂肪酸和炎症因子,进入血液循环,影响肝脏、肌肉、胰腺等多个器官的代谢功能。
伴随症状与疾病进展
代谢综合征自然病程呈现出渐进性发展的特点,从正常代谢到明显的代谢综合征,往往需要几年甚至十几年的时间。了解这个发展过程,有助于我们理解在不同阶段应该采取什么样的应对策略。
★ 早期阶段:指标轻度异常,但干预价值最高
在疾病的早期阶段(也就是常说的前驱期),这个阶段往往没有明显的临床症状,很多人体检的时候才会发现一些轻微的异常。
例如:腰围逐渐增加,甘油三酯轻度升高,HDL-C略低,血压处于正常高值,空腹血糖轻度偏高,或者提示脂肪肝。
这个阶段很容易被忽视。因为患者可能没有明显不适,也没有达到糖尿病、高血压或严重高脂血症的诊断标准。但从代谢机制看,胰岛素抵抗、脂肪组织功能异常和慢性炎症可能已经开始出现。
实际上,这个阶段正是干预效果最好的时期,如果能够及时改变生活方式,完全可以逆转病情,阻止它进一步发展。
■ 中期阶段:多项代谢异常叠加,进入明确风险状态
如果在早期阶段没有及时干预,病情就会进展到中期阶段。这个阶段已经出现了明确的代谢异常,达到了代谢综合征的诊断标准。
出现明确高血压(≥130/85 mmHg)、部分患者到糖尿病前期(空腹血糖也开始升高,往往超过5.6 mmol/L)或2型糖尿病、明显高甘油三酯血症、低HDL-C、脂肪肝也常见(约50%–70%的患者合并非酒精性脂肪肝)、睡眠呼吸暂停(尤其是在严重肥胖的患者中,超过一半的人睡觉打呼噜,憋气,频繁缺氧)等问题。
这一阶段的特点是多个系统开始同时受累,相互影响,形成循环。
例如,脂肪肝会进一步加重肝脏胰岛素抵抗,促进VLDL分泌;高甘油三酯和低HDL-C会增加动脉粥样硬化风险;睡眠呼吸暂停会通过夜间缺氧和交感神经激活,加重血压和糖脂代谢异常。
到了中期阶段,虽然已经有了多种代谢异常,但是如果能够积极干预,仍然可以很好地控制病情,延缓甚至阻止并发症的发生。很多人通过积极的生活方式干预,显著改善代谢状态,减少药物的使用。
▲ 晚期阶段:出现器质性损伤和慢病并发症
如果病情继续发展,就会进入晚期阶段,这个阶段已经出现了严重的并发症,往往伴随难以完全逆转的器质性损伤。
例如:2型糖尿病并发症、动脉粥样硬化性心血管疾病、肝病、肾病等。
大量的队列研究已经证实了代谢综合征对健康的严重危害。
一项荟萃分析纳入了超过一百万人的随访数据,结果显示,代谢综合征患者发生心血管事件的风险是正常人群的约2-3倍,发生2型糖尿病的风险更是增加了5倍。
还有研究发现,代谢综合征不仅增加心血管疾病和糖尿病的风险,与多种癌症风险增加相关(证据在不同癌种中强度不一),包括乳腺癌、结肠癌、胰腺癌等。这可能与慢性低度炎症、胰岛素抵抗、性激素水平改变等多种机制有关。
此外,代谢综合征还会增加认知功能下降和痴呆的风险,对老年人的健康构成更大的威胁,而且证据仍在不断积累。
以上自然病程我们可以看出,代谢综合征实际上是肝脏、脂肪组织、肠道、免疫系统和血管系统共同作用后的结果。代谢综合征的预防和干预越早越好。
也正因为如此,下一部分就有必要把血脂异常单独拎出来讨论:为什么在代谢综合征中,血脂异常不仅是诊断指标,更是心血管风险、肠-肝代谢轴和菌群干预价值的关键入口。
血脂异常在代谢综合征中不仅仅是五项诊断指标之一,它实际上承载着代谢状态、胰岛素敏感性、炎症水平和心血管风险的综合信息。理解血脂异常的特点和机制,有助于制定更精准的风险分层和干预策略。
血脂异常在代谢综合征中的核心地位
在代谢综合征的诊断标准中,高甘油三酯和低高密度脂蛋白胆固醇(HDL-C)就已经占据两项核心指标。血脂异常不仅是代谢综合征的重要诊断依据,还能够反映整体代谢紊乱的严重程度及其潜在风险,同时也是连接其他代谢紊乱与心血管终点事件的关键环节。
临床流行病学显示:
血脂异常不仅是代谢综合征的结果,也可能在代谢综合征的发生发展中起到推动作用。
血脂异常与心血管风险:独立且显著
研究证实:
胰岛素抵抗:血脂异常的驱动力
这要从代谢综合征的病理生理机制说起。代谢综合征的核心是胰岛素抵抗,胰岛素抵抗不仅会影响血糖代谢,还会直接影响脂肪组织的脂解过程。
当胰岛素抵抗发生时,脂肪组织对胰岛素的抗脂解作用不敏感,导致脂肪组织持续释放大量的游离脂肪酸进入血液循环。这些游离脂肪酸被肝脏摄取之后,就会刺激肝脏合成更多的甘油三酯和极低密度脂蛋白,导致血脂异常的发生。同时,胰岛素抵抗还会影响脂蛋白脂酶的活性,影响甘油三酯的清除,进一步加重高甘油三酯血症。
从这个角度我们可以看出,是胰岛素抵抗的重要代谢表现之一,它反映了全身代谢紊乱的严重程度。因此,血脂异常的检测和管理,应该成为代谢综合征防控的核心环节之一。
代谢综合征——脂质三联征
代谢综合征血脂异常具有高度特征性,被称为脂质三联征或致动脉粥样硬化血脂谱。
注:因为这种血脂谱具有很强的致动脉粥样硬化作用,心血管危害远超单纯的高胆固醇血症。
具体来说,代谢综合征的脂质三联征包括三个主要特点:
1、高甘油三酯血症
大部分代谢综合征患者会出现空腹甘油三酯(TG)升高,一般诊断标准是空腹甘油三酯≥1.7 mmol/L。
甘油三酯升高是代谢综合征血脂异常最常见的表现,这和我们前面提到的游离脂肪酸(FFA)释放增加、肝脏合成极低密度脂蛋白(VLDL)增加密切相关。
高甘油三酯血症不仅仅本身是一个危险因素,更重要的是,高甘油三酯血症往往伴随着其他脂蛋白的异常改变,共同导致动脉粥样硬化的发生发展。
2、低高密度脂蛋白胆固醇血症
代谢综合征患者大多出现高密度脂蛋白胆固醇降低。诊断标准:
男性<1.0 mmol/L,女性<1.3 mmol/L。
高密度脂蛋白胆固醇被我们称为”好胆固醇“,高密度脂蛋白胆固醇降低,这种保护作用就会减弱,动脉粥样硬化的风险就会增加。
注:高密度脂蛋白胆固醇负责将胆固醇从周围组织转运回肝脏进行代谢和排泄,这个过程叫做胆固醇逆向转运,它具有抗动脉粥样硬化的作用。
在代谢综合征患者中,高密度脂蛋白胆固醇降低往往伴随着甘油三酯升高同时出现,这两者之间存在密切的代谢联系。当甘油三酯升高时,胆固醇酯转运蛋白活性增加,会促进高密度脂蛋白中的胆固醇转移到极低密度脂蛋白和低密度脂蛋白中,导致高密度脂蛋白胆固醇水平降低。
3、小而密低密度脂蛋白比例升高
很多人都知道低密度脂蛋白胆固醇是”坏胆固醇“,但其实低密度脂蛋白并不是均匀的,它还可以根据颗粒大小分为不同的亚型:
其中小而密的低密度脂蛋白致动脉粥样硬化作用远远强于大而轻的低密度脂蛋白。
原因是:小而密LDL颗粒更易氧化、穿透血管内皮并形成泡沫细胞,即使总LDL-C正常,也显著增加心血管事件风险。
这种协同效应使代谢综合征患者的血脂异常风险复杂而显著
多项大型临床研究已经证实,在部分研究中,TG/HDL比值或非HDL胆固醇等指标对心血管风险具有更好的预测能力。
对于代谢综合征患者来说,我们不能仅仅关注总胆固醇和低密度脂蛋白胆固醇,还需要关注甘油三酯和高密度脂蛋白胆固醇,在有条件情况下可进一步评估(如ApoB或LDL颗粒特征)。
传统干预策略
目前临床上对于代谢综合征相关的血脂异常,主要的干预策略包括生活方式干预和药物治疗两大类。这两种策略都取得了一定的效果,但是也存在明显的局限性,也需要寻求新的干预方式。
生活方式干预是基础
包括控制饮食、增加运动、减轻体重、戒烟限酒等。健康的生活方式确实可以改善血脂异常,也可以改善胰岛素敏感性。
但是现实问题在于,生活方式干预的长期依从性普遍较低(多项研究显示仅有少部分患者能够长期坚持)。大多数人在坚持几个月之后都会逐渐回到原来的不良生活习惯。这其中有很多原因,比如工作忙、压力大、社交应酬多、难以抵制美食的诱惑等等。所以,仅仅依靠生活方式干预,很难长期有效地控制血脂异常。
药物治疗是目前临床上控制血脂异常的主要手段
——高胆固醇血症
首选的药物是他汀类药物。
大量的临床试验证实,他汀可以抑制胆固醇合成,显著降低低密度脂蛋白胆固醇,减少心血管事件。
但是,他汀类药物也存在局限性。对甘油三酯升高和低HDL-C的改善相对有限,尤其是对于高甘油三酯血症和低高密度脂蛋白胆固醇血症,他汀的效果有限。还有少数患者不能耐受他汀,服用他汀之后会出现肌肉疼痛、乏力,甚至肝功能异常,这些不良反应导致患者不得不停药。
——高甘油三酯血症
临床上常用的药物是贝特类药物。
贝特可以有效降低甘油三酯,升高高密度脂蛋白胆固醇。
但是贝特类药物也存在问题,它的主要副作用是会增加他汀的肌肉不良反应,与部分他汀联合使用时可能增加肌病风险。而且,贝特类药物虽然可以改善血脂谱,但是对于减少心血管事件的获益证据并不像他汀那么充分。
除了他汀和贝特,还有烟酸、依折麦布、PCSK9抑制剂等其他调脂药物,但是这些药物也各有各的局限性。
更深层次的问题在于,主要针对血脂异常这一表型进行干预,而对上游代谢紊乱(如胰岛素抵抗)的直接作用有限。
代谢综合征相关的血脂异常,根源在于胰岛素抵抗、脂肪组织功能异常、肠道菌群紊乱等深层次的问题,传统药物并没有针对这些根本原因进行干预,所以往往需要长期甚至终身服药,停药之后血脂很容易再次升高。
这种现状促使科学界不断寻找新的治疗靶点,而肠道微生物组正是目前看来最具前景的方向之一。
近二十年来的研究已经证实,肠道微生物组在脂质代谢中发挥着非常重要的作用,很多代谢综合征患者的血脂异常,都和肠道微生物组的异常改变密切相关。通过调节肠道微生物组来改善血脂异常,有望从根本上解决问题,为代谢综合征的治疗提供全新的思路。
在血脂代谢领域,大量的研究已经明确证实,肠道微生物群通过多种不同的机制影响宿主的脂质吸收、合成、转运和代谢,这些机制共同作用,最终影响血脂水平。目前学术界已经比较明确的核心机制主要有五个方面:
10.1007/S11154-019-09512-0
机制一:短链脂肪酸(SCFA)的调控作用
短链脂肪酸是肠道微生物群对宿主代谢产生影响的重要信号分子,也是目前研究最为深入、证据最充分的机制之一。
肠道细菌可以产生各种各样的糖苷水解酶,能够分解膳食纤维,将它们发酵产生短链脂肪酸。
所以说,膳食纤维对健康的好处,很大一部分是通过短链脂肪酸实现的。
短链脂肪酸主要通过三条途径调控血脂代谢。
途径一 | 脂肪组织:抑制脂解与炎症
第一条途径是通过G蛋白偶联受体介导的信号通路,抑制脂肪组织的脂肪分解。
在肠道上皮细胞、免疫细胞、脂肪细胞上都表达着两种特殊的G蛋白偶联受体,叫做GPR41和GPR43,这两种受体的配体是短链脂肪酸。
当短链脂肪酸和这些受体结合之后,会激活细胞内的信号通路,产生一系列生理效应。其中一个很重要的效应就是抑制脂肪组织中的脂肪分解。
——抑制脂肪分解,改善血脂
脂肪分解是脂肪细胞把储存的甘油三酯分解成游离脂肪酸释放到血液中,这个过程如果过度活跃,就会导致血液循环中游离脂肪酸增加,过多的游离脂肪酸被肝脏摄取之后,就会刺激肝脏合成更多的甘油三酯和极低密度脂蛋白,导致血脂升高。
而短链脂肪酸通过激活GPR41/43受体,可以抑制这个过度活跃的脂解过程,减少游离脂肪酸的释放,从而减轻肝脏的脂代谢负担,降低血脂水平。
——抗炎,改善胰岛素敏感性
肥胖和代谢综合征患者的脂肪组织中会有大量的巨噬细胞浸润,这些巨噬细胞会释放促炎细胞因子,这些促炎细胞因子会进一步加重胰岛素抵抗,促进脂肪分解。
短链脂肪酸通过激活GPR43受体,可以抑制巨噬细胞的促炎激活,减少促炎细胞因子的释放,改善脂肪组织的慢性炎症,从而改善胰岛素敏感性,间接改善脂代谢。
途径二 | 肝脏:调节胆固醇与脂肪酸合成
短链脂肪酸产生后,通过肠道吸收,经门静脉进入肝脏,其中丙酸的作用最为显著。
——丙酸直接抑制关键酶
研究发现,丙酸可以直接抑制肝脏胆固醇合成的关键限速酶——HMG-CoA还原酶(3-羟基-3-甲基戊二酰辅酶A还原酶),这一酶也是他汀类药物主要靶点。
通过抑制HMG-CoA还原酶,丙酸能够减少肝脏内源性胆固醇合成,从而降低血液中的胆固醇水平。
动物实验研究证实,给高脂饮食喂养的小鼠补充丙酸,可以显著降低血清总胆固醇和低密度脂蛋白胆固醇水平。
编辑
——丙酸调控脂肪酸合成与甘油三酯降低
研究发现,丙酸可以抑制肝脏中脂肪酸合成关键酶ACC和FAS的表达,减少从头合成脂肪酸,从而减少肝脏甘油三酯的合成,降低血清甘油三酯水平。
途径三 | 肠道激素:调节胰岛素敏感性与食欲
第三条途径通过调节肠道激素分泌,改善胰岛素敏感性,减少食欲。
肠道中的L细胞能够分泌一类重要的肠道激素:
这两种肠道激素都有很重要的代谢调节作用:
研究发现,短链脂肪酸能够刺激肠道L细胞分泌更多的GLP-1和PYY,从而发挥这些有益的代谢效应。胰岛素敏感性改善之后,脂代谢也会随之改善,游离脂肪酸释放减少,肝脏合成甘油三酯也会减少。
食欲减少有助于控制体重,减轻肥胖,进一步改善代谢。
机制二:影响胆汁酸代谢
胆汁酸是肝脏合成的重要分子,主要生理功能是帮助膳食脂肪的消化吸收。肝脏合成的胆汁酸储存在胆囊中,吃饭后胆囊收缩,将胆汁酸排入肠道帮助脂肪乳化和吸收。大约95%的胆汁酸在回肠末端被重新吸收,通过门静脉回流至肝脏,形成肠-肝循环。
过去人们认为胆汁酸仅仅是一种消化液,但近二十年来的研究表明,它同时也是重要的信号分子,能够激活核受体和G蛋白偶联受体,调控糖脂代谢,对全身代谢具有深远影响。
而肠道微生物群在胆汁酸代谢中发挥关键作用,能够通过多种酶改变胆汁酸组成和结构,进而影响其信号功能,最终调控宿主脂质代谢。
1、肠道菌群通过胆汁盐水解酶对胆汁酸解偶联
肝脏合成的胆汁酸,在分泌到胆汁中的时候,会和甘氨酸或者牛磺酸结合,形成结合型胆汁酸。结合型胆汁酸才能发挥正常的生理功能。
——胆汁酸的解偶联过程
当胆汁酸进入肠道之后,肠道中的很多细菌,主要是乳杆菌、双歧杆菌这些有益菌,能够产生一种叫做胆汁盐水解酶(简称BSH)的酶,这种酶能够把结合型胆汁酸中的甘氨酸或者牛磺酸水解下来,变成游离型胆汁酸。
——解偶联之后的游离型胆汁酸和结合型胆汁酸有什么不同?
主要的区别在于它们在肠道的重吸收效率。
游离型胆汁酸更容易被肠道被动重吸收,但是也有一部分会随着粪便排出体外。
如果BSH活性发生改变,就会影响胆汁酸的肠肝循环效率,改变胆汁酸的返回量,从而影响肝脏中胆汁酸的合成。
2、肠道菌群通过FXR信号调控肝脏脂蛋白合成
FXR(法尼醇X受体)是一种核受体,它主要在肝脏和肠道表达,它的配体就是胆汁酸。
研究发现,肠道微生物改变导致胆汁酸组成异常,会过度激活肠道FXR,FXR激活之后会通过循环中的FGF15/19(成纤维细胞生长因子15/19)信号,影响肝脏的脂代谢,促进VLDL合成,导致高甘油三酯血症。
3、通过TGR5受体激活增加能量消耗
TGR5是一种G蛋白偶联受体,它在棕色脂肪组织中高表达。
研究发现,代谢综合征患者往往次级胆汁酸合成减少,TGR5激活不足,能量消耗减少,这也会促进肥胖和血脂异常的发生。
机制三:内毒素血症与慢性低度炎症
内毒素血症和慢性低度炎症是连接肠道菌群紊乱与代谢综合征的一个核心机制,这个机制也是最早被发现和研究比较透彻的机制。
革兰氏阴性细菌的细胞壁外面有一种成分叫做脂多糖,简称LPS,也称为内毒素。
<向上滑动阅览>
还有研究发现,通过饮食干预改善肠道菌群,降低内毒素血症,可以显著改善血脂异常,进一步证实了内毒素血症在血脂异常中的介导作用。
机制四:胆碱代谢与TMAO途径
每天我们从膳食中摄入胆碱,尤其在鸡蛋、动物肝脏、红肉等食物中含量较高。摄入的胆碱大部分被吸收利用,但一部分进入大肠,由肠道细菌代谢。特定菌群产生的胆碱三甲胺裂解酶能将胆碱转化为三甲胺(TMA),随后通过门静脉进入肝脏。肝脏中的FMO3(黄素单加氧酶3)将TMA氧化为TMAO。
TMAO影响胆固醇和血脂代谢主要通过以下途径。
1、抑制胆固醇逆向转运
首先是抑制胆固醇逆向转运,减少胆固醇外流。前面提到过,胆固醇逆向转运是把胆固醇从周围组织包括动脉粥样斑块转运回肝脏代谢排泄的过程,这个过程是对抗动脉粥样硬化的重要保护机制。这个过程中,ABCA1和ABCG1这两个转运蛋白发挥着关键作用,它们位于巨噬细胞表面,能够把细胞内多余的胆固醇转运出来,交给高密度脂蛋白。
研究提示TMAO可能通过下调巨噬细胞ABCA1和ABCG1的表达,抑制胆固醇外流,从而影响胆固醇逆向转运,促进动脉粥样硬化的发生。
2、促进泡沫细胞形成
第二条途径主流假说是通过促进泡沫细胞形成。泡沫细胞是动脉粥样硬化斑块中最早出现的细胞成分,它是巨噬细胞吞噬了大量的氧化低密度脂蛋白之后形成的。
部分研究显示TMAO能够上调巨噬细胞表面清道夫受体SR-A1的表达,SR-A1能够识别并吞噬氧化低密度脂蛋白,SR-A1表达增加之后,巨噬细胞吞噬更多的氧化低密度脂蛋白,更容易形成泡沫细胞。泡沫细胞大量形成,可能是动脉粥样硬化发生发展的关键步骤。
3、抑制胆汁酸合成
胆固醇7α-羟化酶,也就是CYP7A1,是肝脏合成胆汁酸的关键限速酶。
研究显示TMAO能够抑制肝脏CYP7A1的表达,导致胆汁酸合成减少。胆固醇是合成胆汁酸的原料,胆汁酸合成减少,意味着胆固醇消耗减少,更多的胆固醇留在体内,同时,胆汁酸排泄减少也会影响胆固醇的平衡,导致血液中胆固醇水平升高。
<向上滑动阅览>
从干预的角度来看,如果我们能够抑制肠道细菌产生TMAO,就能够降低血浆TMAO水平,从而降低心血管风险。
目前已经有一些研究在探索这个方向,比如使用特异性的小分子抑制剂抑制胆碱三甲胺裂解酶的活性,或者使用噬菌体特异性地清除产TMAO的细菌,这些研究都取得了一些初步的成果,未来有可能开发出新的降脂和抗动脉粥样硬化药物。
机制五:膳食脂肪吸收调控
除了我们前面介绍的这四个机制,肠道菌群还能够直接影响肠道上皮细胞对膳食脂肪的吸收效率,以及乳糜微粒的形成和分泌过程,这个直接的调控作用对血脂水平也有着重要的影响。
脂肪吸收
吃进去的脂肪,需要经过一系列的步骤才能被身体吸收利用。
肠道菌群和脂质通过菌群代谢物、胆汁酸和免疫细胞吸收
编辑
脂蛋白脂酶不仅存在于脂肪组织和肌肉组织的血管内皮表面,在肠道上皮细胞的表面也有表达,它能够帮助分解乳糜微粒中的甘油三酯,促进脂肪酸的吸收。
研究发现,肠道菌群代谢产生的一些短链脂肪酸和其他代谢产物,能够影响肠道上皮细胞脂蛋白脂酶的活性,如果有害菌过度生长,产生的一些有害代谢产物会增加脂蛋白脂酶的活性,导致更多的膳食脂肪被吸收,过多的脂肪吸收之后,会导致血液中甘油三酯升高,也会增加脂肪堆积。
肠道菌群还能够调节肠道通透性,影响乳糜微粒进入循环。肠道菌群紊乱会损伤肠道屏障,增加肠道通透性,不仅LPS和细菌内毒素更容易进入血液循环,乳糜微粒进入循环也会增加,更多的膳食脂肪被吸收进入血液,导致血脂升高。同时,肠道通透性增加还会引起慢性低度炎症,加重胰岛素抵抗和血脂异常,形成恶性循环。
近年来,大规模肠道微生物组组学研究揭示了代谢综合征患者特有的菌群模式,尤其在伴血脂异常的人群中表现明显。这些特征不仅是疾病结果,还可能参与代谢综合征的发生和发展。
α多样性显著降低
α多样性是衡量肠道微生物群落结构的一个常用指标,它反映了肠道中物种的丰富度(也就是有多少种不同的细菌)和均匀度(也就是不同细菌的丰度分布是否均匀)。
大量的研究一致发现,代谢综合征患者的肠道菌群α多样性显著降低。
在代谢综合征伴血脂异常中,肠道微生物群的变化通常包括整体多样性下降以及有益菌的缺失。
产短链脂肪酸菌减少
多样性下降通常伴随产短链脂肪酸菌减少,如丁酸生成菌减少,使肠道调节胰岛素敏感性、脂质代谢以及炎症信号的能力下降,调节影响甘油三酯和 HDL 的炎症信号的能力减弱。
同时,多样性丧失常伴胆汁酸代谢紊乱和肠道屏障功能受损,导致低度全身性炎症增加,从而强化血脂异常和胰岛素抵抗的循环。
关键有益菌属/菌种的特征性变化
属和种的水平,代谢综合征患者也呈现出特征性的变化,一些有益菌降低,一些有害菌升高,这些变化共同推动了血脂异常的发生发展。
⭐ 有益菌显著减少
✦Faecalibacterium prausnitzii(普拉梭菌)
【产丁酸,抗炎,改善肠屏障,丰度下降 → 丁酸减少 → 炎症增加 → 胰岛素抵抗和血脂异常加重】
普拉梭菌是人体肠道中丰度最高的产丁酸菌之一,它能够发酵膳食纤维产生大量的丁酸,丁酸不仅是肠道上皮细胞的主要能量来源,还具有抗炎作用,能够改善肠道屏障功能,减少炎症反应。
大量研究一致发现,代谢综合征患者中普拉梭菌的丰度降低。普拉梭菌减少,丁酸产生减少,肠道屏障功能受损,炎症增加,这些都会加重胰岛素抵抗和血脂异常。
✦Roseburia (罗氏菌属)
【产SCFA,丰度下降 → SCFA生成减少 → 代谢保护作用减弱】
罗斯氏菌属也是非常重要的产短链脂肪酸的细菌,它也能够发酵膳食纤维产生大量的丁酸和乙酸。罗斯氏菌减少,同样会导致短链脂肪酸产生减少,对代谢的保护作用减弱。
✦ Akkermansia muciniphila(嗜黏蛋白阿克曼菌,常简称它为Akk菌)
【增强肠屏障、改善胰岛素敏感性,降低甘油三酯】
Akk菌是一种专门降解肠道黏液层中黏蛋白的细菌,它对维护肠道屏障功能非常重要。
研究发现,Akk菌能够增强肠道屏障完整性,减少内毒素血症,还能够改善胰岛素敏感性,降低体重和血脂。
动物实验研究发现,给肥胖小鼠补充Akk菌,能够降低血清甘油三酯,改善胰岛素抵抗,减轻体重,效果非常显著。
✦ Christensenellaceae(克里斯滕森菌科)
【丰度与瘦体型及代谢健康正相关,代谢综合征患者显著减少】
这个科的细菌在健康人肠道中普遍存在。
多项研究发现,克里斯滕森菌科的丰度和代谢健康指数呈强正相关,和BMI、甘油三酯呈负相关。
代谢综合征患者中克里斯滕森菌科的丰度显著降低。有意思的是,研究还发现克里斯滕森菌科的丰度和瘦体型相关,携带克里斯滕森菌科多的人,不容易发胖,代谢也更健康。
其它还有:
⭐ 有害菌显著增加
✦Escherichia/Shigella(大肠杆菌/志贺氏菌属)
【LPS产生增加 → 内毒素血症 → 慢性炎症】
这些细菌属于变形菌门,是革兰氏阴性条件致病菌,它们的细胞壁含有大量的LPS。
代谢综合征患者中这些细菌的丰度显著升高,它们产生大量的LPS,更容易导致内毒素血症和慢性炎症。
✦ Desulfovibrio(脱硫弧菌属)
【高浓度硫化氢损伤肠屏障 → 炎症 → 代谢紊乱】
脱硫弧菌是一种硫酸盐还原菌,它能够还原硫酸盐产生硫化氢。硫化氢在低浓度的时候有一定的生理作用,但是高浓度的硫化氢对肠道上皮细胞有毒性作用,能够损伤肠道屏障,增加肠道通透性,还能够抑制线粒体呼吸,影响肠道上皮细胞的能量代谢,还会加重炎症,促进代谢紊乱。
研究发现,代谢综合征患者中脱硫弧菌的丰度显著升高,而且脱硫弧菌丰度和血压、甘油三酯呈正相关。
✦Clostridium clusters(一些梭菌属的细菌)
【影响胆汁酸代谢 → 血脂异常】
特定的一些梭菌集群和胆汁酸代谢紊乱相关,它们能够改变胆汁酸的代谢,影响胆汁酸的组成和信号功能,进而影响血脂代谢。这些梭菌过度生长,会导致胆汁酸代谢重编程,促进血脂异常的发生。
其它还有:
功能基因谱的特征变化
代谢综合征患者的微生物群功能能力下降,表现为:
这些功能性变化揭示了代谢综合征肠道微生态对血脂异常、胰岛素抵抗及慢性低度炎症的上游贡献。理解这些特征性变化,可以为精准干预提供明确靶点。
通用干预的局限性
常规药物在代谢综合征合并血脂异常的患者中疗效有时不理想,因为无法考虑影响甘油三酯、HDL 和全身炎症的特定肠道-胆汁酸-代谢物信号回路。
热量控制难以修复肠道生态
许多标准的健康饮食方法强调热量控制或食物限制(广泛限制),但它们可能无法可靠地恢复驱动胆汁酸受体信号(FXR/TGR5)和肝脂调控的肠道生态系统功能。
如果一个人的肠道菌群纤维发酵能力低且多样性减少,仅仅添加一些纤维或做出模糊的饮食改进,通常无法有效改善肠道胆汁酸转化模式,也难以调节VLDL生成或提升胰岛素敏感性。
低度慢性炎症难以消除
即便进行了饮食调整或生活方式干预,低度慢性炎症在代谢综合征患者体内仍可能持续存在。
这主要与短链脂肪酸产量下降和肠道屏障受损有关,导致内毒素(如LPS)进入循环,维持炎症。持续的炎症环境不仅影响脂质代谢,还强化胰岛素抵抗,使甘油三酯升高、HDL下降的血脂异常难以改善。
通用干预很少针对屏障完整性和 SCFA 生成的根本驱动因素(如持续可发酵的纤维数量/品种、富含多酚的植物摄入量,以及对发酵或益生元食品的个体耐受性)。如果不解决这些功能缺陷,脂质水平的改善可能会趋于平稳,因为炎症和胰岛素抵抗环境持续促进动脉粥样状、富含 ApoB 的脂蛋白谱。
个体差异决定效果
最重要的是一刀切的建议往往忽视了微生物生态和饮食反应中的个体间差异。
两个人可能都吃得更好,但他们的基线菌群组成、胆汁酸库、运输时间和代谢反应可能导致截然不同的结果。帮助一个人的微生物组产生 SCFA 或改善胆汁酸信号的机制,对另一个人可能影响不大,甚至因不耐受而加重症状。
如果没有基于功能标志物(如纤维摄入质量、炎症状态、血糖反应以及特定益生元/发酵策略耐受性)进行个性化调整,通用方案就不太可能在甘油三酯/HDL 和心血管代谢风险方面产生持久改善。
风险分层与早期预警
代谢综合征的发生发展是一个漫长的过程,从正常代谢到明确的代谢综合征,往往需要几年甚至十几年的时间。
肠道菌群变化:早于临床症状的预警信号
传统的危险因素,比如BMI、腰围、血压、血糖、血脂,这些都是在代谢异常已经发生之后才能发现异常。
而肠道菌群紊乱往往早于临床症状和生化异常的出现,也就是说,还没有出现血糖升高、血压升高、明确的血脂异常的时候,肠道菌群已经发生了改变。
风险分层与预测模型
基于这个特点,肠道菌群特征可以用于代谢综合征的风险分层和早期预警。
多项研究以及谷禾健康已经构建了基于肠道菌群特征的代谢综合征预测模型,这些模型的预测性能如何呢?
研究发现,基于肠道菌群特征构建的预测模型,对代谢综合征的曲线下面积(AUC)可以达到0.83-0.89,这个预测精度优于传统的危险因素,比如BMI、年龄这些指标。如果把肠道菌群特征和传统危险因素结合起来,预测精度还能进一步提高。
这个发现有什么临床意义呢?
它意味着我们可以通过检测肠道菌群,在疾病的早期阶段,甚至在还没有出现生化异常的时候,就识别出代谢综合征的高风险人群。
提前干预与疾病分期监测
研究发现,在糖耐量正常人群中,肠道微生物变化可比血糖异常提前3–5年被检测到。
也就是说,如果现在血糖、血脂都还正常,但是肠道菌群已经出现了特征性的改变,那么未来几年发生代谢综合征的风险就会显著升高,这个时候就可以提前进行干预,通过改变饮食、增加运动、补充益生菌益生元等方法,调整肠道菌群,阻止或者延缓代谢综合征的发生。这就是真正的上医医未病,预防重于治疗。
除了预测发病风险,肠道菌群变化还能够反映代谢综合征的疾病分期和严重程度。
研究发现,随着代谢综合征从早期到中期再到晚期,肠道菌群的紊乱程度也逐渐加重,α多样性逐渐降低,特征性改变也越来越明显。
因此我们可以通过检测肠道菌群,对疾病进展情况进行监测,了解疾病处于什么阶段,帮助我们制定更合适的干预策略。
个性化干预方案制定
现在医学界越来越强调精准医学,也就是根据每个人不同的特征,制定个性化的干预方案,而不是千人一方。肠道菌群检测就是实现个性化干预非常重要的一个工具,因为不同的人肠道菌群特征不一样,适合的干预方法也不一样。
例如,产短链脂肪酸菌缺乏。
如果肠道中产丁酸、产乙酸的有益菌明显减少,那么适合补充益生元,比如菊粉、低聚果糖这些益生元,它们能够选择性地促进这些有益菌的生长,增加短链脂肪酸产生,改善血脂代谢。
例如,Akkermansia muciniphila缺乏。
如果Akk菌丰度明显降低,那么可以补充特定的益生菌,或者吃一些能够促进Akk菌生长的食物,比如石榴、蔓越莓这些富含多酚的食物,能够促进Akk菌生长,改善肠道屏障功能,降低内毒素血症,改善血脂。
治疗反应预测与监测
评估药物干预效果
肠道菌群特征不仅仅能够帮助我们制定干预方案,还能够预测药物和生活方式干预的效果,通过肠道菌群特征预测对某种治疗方法的反应好不好,这对于临床治疗决策有辅助作用。
例如他汀类药物,他汀是临床上最常用的降脂药物,但是我们前面提到过,部分患者对他汀反应不佳,所以谷禾也一直在和临床推进合作看有没有办法在用药之前就预测哪些人反应好,哪些人反应不好呢?
研究发现,肠道菌群特征确实能够预测他汀的降脂疗效,特定的菌群特征。
评估减肥效果
减重,无论是通过生活方式减重还是减重手术,不同的人效果差异很大。
研究发现,基线肠道菌群中普雷沃氏菌/拟杆菌比例高的患者,减重干预的效果更好,体重减轻更多,血脂改善也更明显。另一方面,基线Akk菌丰度可以作为减重效果的预测因子,如果AKK菌丰度比较低,那么可能需要更强化的干预,或者先补充Akk菌,提高减重效果。
动态监测肠道菌群评估
在干预过程中,还可以通过动态监测肠道菌群来评估干预效果。干预之后,如果肠道菌群多样性恢复了,有益菌丰度增加了,有害菌丰度减少了,这些都可以作为代谢改善的早期生物标志物。
这些变化往往出现在血脂、血糖这些生化指标改善之前,所以我们可以通过监测菌群变化,早期了解干预是否有效,如果干预没有改善菌群,帮忙用户及时调整干预方案,避免无效治疗。
并发症风险评估
特定的肠道微生物标志物和代谢综合征并发症的风险相关,通过检测这些标志物,可以辅助评估并发症的发生风险,帮助我们进行风险分层和预防。
举例:和并发症风险有可能相关的指标:
次级胆汁酸升高
次级胆汁酸是肠道细菌代谢初级胆汁酸产生的,粪便次级胆汁酸升高和非酒精性脂肪肝的发生风险密切相关,如果有代谢综合征,同时粪便次级胆汁酸升高,那么可能需要提前进行筛查和干预。
<来源:谷禾健康肠道菌群检测数据库>
产短链脂肪酸菌减少
短链脂肪酸对代谢有保护作用,产短链脂肪酸菌减少,代谢性疾病进展的风险升高。所以检测发现产短链脂肪酸菌明显减少,那么需要更积极的生活方式干预,必要的时候补充益生元或者益生菌,增加短链脂肪酸产生,降低糖尿病等代谢疾病风险。
通过检测这些微生物标志物,我们可以更精准地评估不同并发症的风险,然后进行针对性的预防和干预。
一般支持策略
代谢综合征伴血脂异常越来越多地与肠道微生物组调节胆汁酸、炎症色调和代谢信号传递有关。一个核心支持策略是通过增加来自多样、最少加工的植物性食物(如豆类、全谷物、水果、蔬菜)的膳食纤维来改善微生物生态。
膳食纤维
膳食纤维为有益发酵菌提供能量,促进短链脂肪酸生成,如丁酸。丁酸可强化肠道屏障,减少循环内毒素(LPS),降低低度慢性炎症,从而改善胰岛素敏感性,减轻甘油三酯升高和HDL下降。可选益生元来源包括大蒜、洋葱、韭葱、芦笋、菊苣根/菊粉、青香蕉等,同时限制高度加工食品。
支持胆汁酸信号(天然食物)
由于肠道微生物也会调节胆汁酸,支持正常胆汁酸信号传递也是关键方法。某些细菌群落产生的二级胆汁酸可以与 FXR 和 TGR5 等受体相互作用,这些受体会影响肝脂的合成、葡萄糖处理和能量平衡。富含植物多酚和其他微生物群可及化合物的饮食可以促进更有利的细菌转变,并支持胆汁酸池,更好地支持代谢调节。
对于某些人来说,添加天然食物可能进一步增强微生物多样性和代谢反应性。每天通过多种植物(浆果、柑橘、石榴、绿叶蔬菜、十字花科蔬菜、特级初榨橄榄油、可可)增加多酚,以改善胆汁酸信号和微生物多样性。
长期饮食模式(减少加工食品)
持续减少超加工食品和过量添加糖的摄入,有助于防止菌群失调引发的动脉粥样性血脂模式(甘油三酯升高、HDL降低、非HDL/LDL升高)。
限制添加糖和精制淀粉(含糖饮料、甜食、白面包/糕点)的摄入,以减少甘油三酯负担,并限制微生物组驱动的炎症信号。
全食物饮食、逐步增加纤维摄入、改善胰岛素敏感性的生活方式,可长期强化肠-肝代谢轴,改善炎症标志物,帮助血脂向更健康的心血管风险水平转变。
如果能耐受,可以考虑发酵食品(如酸奶/克菲尔加活菌、酸菜、泡菜、天贝),以增强有益微生物并减少低度炎症。
健康脂肪与运动
选择健康脂肪而非饱和脂肪/反式脂肪(橄榄油、坚果、种子、牛油果;若耐受时可食用脂肪鱼类),以改善动脉粥样形成的血脂水平。
优先进行规律的体育活动(有氧+抗阻训练),减少久坐时间,以改善胰岛素敏感性和降低甘油三酯。
睡眠、压力管理
确保充足睡眠并管理压力(例如7-9小时睡眠、正念/呼吸/锻炼),以减少炎症,避免加重胰岛素抵抗和脂质指标。
个体化监测原则
由于胆汁酸代谢和肠道代谢物因人而异,应通过饮食和生活方式调整改善肠道功能,而尽量不要直接用补充剂替代饮食。益生菌或合生菌选择需依据有代谢或抗炎证据的菌株,并考虑剂量及个体反应。
对血脂异常患者,应避免在未监测的情况下过度干预,必要时结合血脂、血糖及炎症指标评估(如可用的CRP、血浆甘油三酯监测),尤其当存在他汀类药物或胆汁酸干预时。
复测/监测
复测的重要性和时间安排
对于伴有血脂异常的代谢综合征患者,在进行干预后进行复测是评估效果的重要环节。常见干预包括增加膳食纤维、提升植物多样性、摄入富含多酚的食物,有时还包括适量发酵食品。
一般建议干预8–12周后复测代谢和血脂标志物,以捕捉甘油三酯、HDL及非HDL/LDL的趋势性变化。这段时间足以让肠道代谢产物(如SCFA)和胆汁酸信号发生适应性改变,从而更明显地反映临床效果。
如果患者同时在进行降脂或胰岛素敏感性优化治疗,复测时间可由临床医生根据具体情况调整,但通常仍建议在同一个8–12周周期内进行安全性和有效性评估。
复测如何指导干预?
复测不仅评估血脂变化,还可用于验证干预模式是否改善肠道介导的炎症和胆汁酸代谢。需要注意的是,复测可以结合其他临床实验室指标解读结果。
最终目标是通过动态监测,确认肠道驱动的代谢物变化与甘油三酯、HDL及整体心血管代谢风险持续改善之间的相关性,而非依赖单次检测数据。这样可以为后续干预优化和个性化调整提供科学依据。
新兴治疗策略
近年来,针对代谢综合征和血脂异常的微生物干预研究呈现多条前沿方向,涵盖从益生菌到后生元的多种策略。
下一代益生菌
传统的益生菌主要是乳杆菌、双歧杆菌这些,而下一代益生菌是从健康人肠道中分离出来的,对代谢有明确益处的特定菌种,目前研究最多的就是Akkermansia muciniphila、Eubacterium hallii。
Akk菌我们前面多次介绍过,它能够改善肠道屏障,降低内毒素血症,改善胰岛素抵抗和血脂。
目前Akk菌已经进入了II期临床试验,初步结果显示,补充Akk菌能够显著降低代谢综合征患者的体重指数和甘油三酯,改善胰岛素抵抗,而且安全性很好,没有明显的不良反应。
Eubacterium hallii 是一种产丁酸菌,它也能够改善糖脂代谢,目前也在进行临床试验。相信在未来几年,这些下一代益生菌就会上市,给代谢综合征患者带来新的治疗选择。
粪菌移植(FMT)
粪便菌群移植就是把健康供体的肠道菌群移植到患者肠道内,重建患者正常的肠道菌群。
目前已经完成了一些小规模的研究,结果显示,健康供体FMT能够改善代谢综合征患者的胰岛素敏感性和血脂谱,部分患者改善效果能够维持三个月以上。
当然,目前FMT还存在一些问题,比如长期效果还不明确,供体菌群的质量控制,安全性等问题都还需要进一步研究。
未来通过把多种明确的有益菌按比例混合,代替完整的粪便,以解决供体质量控制和安全性问题,提升临床可行性,这个方向也是很有前景的。
噬菌体靶向调控
噬菌体干预通过特异性攻击目标有害细菌(如产TMAO菌),精准清除有害菌而不影响其他有益菌。相比广谱抗生素,这种方法副作用更小,精确性高。
动物实验显示,噬菌体可有效降低产TMAO菌群水平,减少血浆TMAO并降低动脉粥样硬化风险。
这个方向未来很可能开发出全新的治疗方法,精准地调节肠道菌群,改善血脂异常。
后生元干预
后生元不是活菌,而是细菌产生的有益代谢产物,或者细菌本身的一些成分,比如短链脂肪酸、特定的胆汁酸、细菌细胞壁成分等等。
后生元和活菌相比,有很多优势:
多个后生元产品已进入临床试验,未来几年有望面市。
除了这四个主要方向,还有很多其他的策略在研究之中,比如精准化营养饮食干预、益生菌+益生元的合生元、膳食益生元促进特定有益菌生长等等,这些策略都在进行研究,未来会有更多的干预方法可供选择。
整合医学模式
未来代谢综合征的管理,预测会走向多组学整合的精准医学模式,也就是整合多个层面的信息,为每个患者制定最精准的预防和治疗方案。
具体来说,整合主要包括四个层面:
基因组学:评估遗传易感性
每个人的遗传背景不同,对代谢综合征的易感性和药物反应也存在差异。通过基因组学检测,可以了解个体携带的易感位点、患病风险以及对特定药物的反应,为精准干预提供基础。
微生物组学:分析肠道菌群特征
肠道菌群是代谢综合征干预的重要对象。通过分析肠道菌群组成与功能基因,可识别有益菌减少、有害菌增加或功能异常的情况。结合这些信息,可以制定个性化干预方案,包括膳食、益生菌/益生元以及生活方式调整。
代谢组学:反映代谢状态
代谢组学通过检测血液、粪便等样本的代谢谱,如血脂、血糖、TMAO、胆汁酸和短链脂肪酸水平,直观反映个体代谢状态,同时也可评估肠道菌群功能。与宏基因组学结合,可以更全面地了解代谢综合征的发病机制和个体差异。
生活方式组学:定制可执行方案
每个人的饮食、运动、睡眠和压力水平不同。生活方式组学通过收集这些数据,将干预措施与个人习惯匹配,从而制定切实可行、易于长期坚持的方案,提升干预效果。
多组学整合的价值
通过整合基因组学、微生物组学、代谢组学和生活方式组学的数据,可以对代谢综合征风险进行精准评估,制定最适合个体的干预策略,实现精准预防和个性化治疗。这将是未来代谢综合征管理的发展方向。
代谢综合征伴血脂异常的发生与发展,不仅涉及传统的血脂、血糖和体重因素,更深层次地与肠道微生物群调控的胆汁酸代谢、炎症状态及代谢信号传递密切相关。本文梳理了肠道微生物群通过多个不同的机制影响宿主的脂质代谢:
这五个机制共同作用,最终影响血脂水平,促进血脂异常的发生发展。
从临床应用的角度来看,肠道菌群检测能够帮助我们进行风险分层和早期预警,制定个性化的干预方案,预测治疗反应,评估并发症风险。这些应用正在逐渐从研究走向临床,未来肠道菌群检测很可能会成为代谢综合征患者常规评估的一部分。
尽管当前仍存在科学问题和技术挑战,如微生物-宿主交互作用、长期干预安全性及微生物组-药物相互作用,但随着多组学整合和大规模临床研究的推进,这一领域有望实现从基础研究到临床实践的快速转化。
未来需要更大规模的前瞻性队列研究,来进一步明确肠道菌群改变和代谢综合征发生发展的因果关系,还需要设计严谨的大规模临床试验,来验证各种肠道菌群干预方法的长期有效性和安全性。
但是无论如何,这个领域的发展方向是非常明确的,随着研究的不断深入,我们对肠道微生物组在代谢综合征中作用的理解会越来越深入,干预方法也会越来越成熟。
未来已来,对于代谢综合征高风险人群,以及已经患有代谢综合征的患者而言,肠道微生物组有望成为代谢综合征管理的重要抓手,了解自己的肠道菌群特征,不仅仅是了解当下的健康状态,更是把握未来健康主动权的重要一步。
希望更多的人能够了解肠道微生物组与代谢健康的关系,重视肠道健康,通过健康的饮食和生活方式,维护良好的肠道菌群平衡,远离代谢综合征,远离心血管疾病和糖尿病,享受健康美好的生活。
注:本账号内容仅供学习和交流,不构成任何形式的医疗建议。
主要参考文献:
Mohammadi F, Rudkowska I. Dietary Lipids, Gut Microbiota, and Their Metabolites: Insights from Recent Studies. Nutrients. 2025 Feb 11;17(4):639.
Ko CW, Qu J, Black DD, Tso P. Regulation of intestinal lipid metabolism: current concepts and relevance to disease. Nat Rev Gastroenterol Hepatol. 2020 Mar;17(3):169-183.
Gildner TE. Links between metabolic syndrome and the microbiome. Evol Med Public Health. 2020 Feb 19;2020(1):45-46.
Singh A, Verma A, Ashraf S, Sarfraz Sheikh D, Irfan H, Riaz R, Venjhraj F, Meghwar S, Kumar R, Tariq MD, Hamza HM, Ahsan A, Satapathy P. Role of gut microbiota in the pathogenesis of metabolic syndrome: an updated comprehensive review from mechanisms to clinical implications. Ann Med Surg (Lond). 2025 Jul 29;87(9):5851-5861.
Gut Microbiome and Metabolic Syndrome: How Dyslipidemia Is Influenced. innerbuddies
Zeng L, Yu B, Zeng P, Duoji Z, Zuo H, Lian J, Yang T, Dai Y, Feng Y, Yu P, Yang J, Yang S, Dou Q. Mediation effect and metabolic pathways of gut microbiota in the associations between lifestyles and dyslipidemia. NPJ Biofilms Microbiomes. 2025 May 28;11(1):90. doi: 10.1038/s41522-025-00721-x.
Yin J, Zhang M, Jiang W, Huang X, Yin Y. Microbial Interactions with Intestinal Lipid Digestion and Absorption: Emerging Targets for Metabolic Disorders. Research (Wash D C). 2025 Sep 19;8:0904.
Schoeler M, Caesar R. Dietary lipids, gut microbiota and lipid metabolism. Rev Endocr Metab Disord. 2019 Dec;20(4):461-472.