-

 CNAS L23010
CNAS L23010

 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业 二级病原微生物安全实验室
二级病原微生物安全实验室- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业 二级病原微生物安全实验室
谷禾健康
自身免疫性疾病,包括炎症性肠病、多发性硬化和类风湿性关节炎,具有不同的临床表现,但肠道微生物群紊乱和肠道屏障功能障碍的潜在模式相同。但宿主因素的个体差异使得统一的方法不太可能。
我们往往会有这样的错觉:“自然”的东西一定是对健康有益的。我们是不是只要吃最原始的食物就能恢复健康?
事实上,优化人类健康的途径并不像照搬我们祖先的饮食或增加我们对微生物的接触那样简单。归根结底,过去和现在的环境是根本不同的,也许是不可逆转的。
因此,针对微生物群的饮食干预的目标不应是回到祖先的状态,而是操纵微生物群,优化宿主健康,直接适应日益工业化的世界。
本文介绍了关于饮食-局部炎症中的微生物群相互作用、肠道微生物群失衡和宿主免疫失调的知识。通过了解并结合个别饮食成分对微生物代谢输出和宿主生理的影响,研究了基于饮食的自身免疫性疾病预防和治疗的潜力。讨论了针对肠道微生物群的工具,如粪便微生物群移植、益生菌和正交小生境工程等。这些方法强调了在自身免疫性疾病不断增加的情况下,利用饮食精确操纵肠道微生物组的途径。
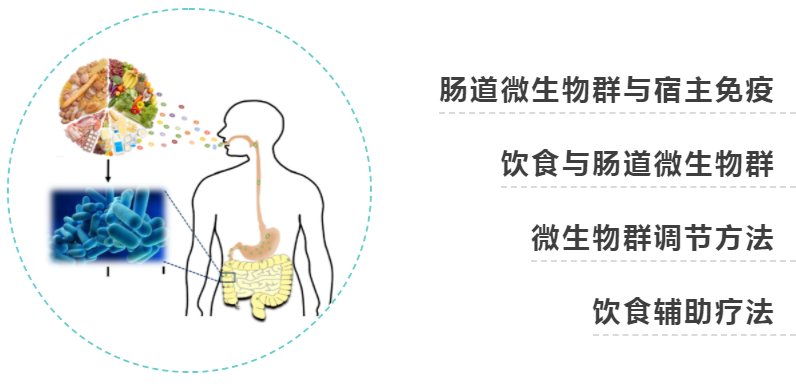
在肠道中,对微生物威胁作出适当反应的免疫平衡行为,对共生微生物和自身抗原的耐受,在生命早期尤其重要,这是定植微生物群和宿主免疫系统相互作用的窗口,引发促炎或抗炎倾向,可能对终身健康产生影响。
★ 婴儿期影响菌群的因素
看过我们文章的都知道,早期生活因素,如剖腹产、配方奶粉喂养和抗生素的使用,可能会干扰微生物群生长过程。
婴儿期母乳中的人乳低聚糖有助于双歧杆菌的定植,双歧杆菌在早期肠道微生物群中占主导地位,产生岩藻糖、乙酸盐、丙酮酸盐和1,2-丙二醇,通过交叉喂养支持微生物群的扩增,有助于对共生细菌的免疫耐受。
人类通常在6个月大时开始转变为固体食物饮食,自此菌群显著扩大,产生更多数量和种类的代谢物(例如,丁酸盐,它促进结肠粘液屏障的成熟,并阻止具有高致病潜力的细菌的生长)。
★ 抗性淀粉缓解系统性红斑狼疮
在一项涉及149名健康志愿者的研究中,脂多糖通过诱导TLR4耐受来抑制先天免疫过程。微生物抗原对TLR的不当刺激也可能影响自身免疫疾病的进展,并为其修复提供线索,如系统性红斑狼疮过度表达TLR7的小鼠模型中,通过添加抗性淀粉来改变微生物代谢产物和分类组成,狼疮进展得到缓解。
★ 微生物变化引起的免疫反应
在自身免疫性疾病患者中也发现了由微生物引起的适应性免疫反应失调。哺乳动物肠道中的共生微生物群,尤其是梭菌群IV和XIVa,它们将膳食纤维代谢成丁酸盐以诱导Treg细胞发育,与小鼠模型中的结肠炎严重程度呈负相关。
通常,自身免疫性疾病患者表现出产生耐受性IL-10的CD25 FOXP3 T细胞的丰度降低,自身反应性效应性T细胞亚群(如TH1细胞和TH17细胞)的丰度增加,从而使Treg细胞与效应性T细胞的比率偏离稳态水平。这些改变可能通过过度产生促炎细胞因子(如TNF和IL-17)进一步促进肠道通透性的增加,这些促炎细胞因子可调节紧密连接蛋白的表达。
自身免疫疾病共同的潜在驱动因素鼓励通过饮食干预或微生物群调节策略(下文讨论)来进行肠道微生物群工程,以将肠道微生物群转变为功能多样的“健康”状态。
饮食可直接影响肠道微生物群,调节其组成或代谢输出,从而可能促进疾病或形成稳态。
膳食碳水化合物发酵
在一项针对34名健康人参与者的每日抽样研究中,通过多次24小时食品召回评估,饮食占微生物组分变化的44%,另外三分之一的变化与性别、BMI和年龄等因素有关。
在分析的主要食物组中,谷物和水果纤维与微生物成分的相关性最强。
纤维与疾病之间的联系

益生元和后生元
国际益生菌和益生元科学协会定义,益生元是一种基质,具有被宿主微生物选择性利用及由此产生健康效益的特征。
★不同参数(如数量,溶解度等)影响纤维效果
使用一种灵长类小鼠模型,发现缺乏纤维的肠道微生物群以宿主粘蛋白聚糖为能源,导致肠道粘液屏障减少,并增加对肠道病原菌Citrobacter rodentium的易感性。然而,14种不同纯化益生元或其他高溶解度多糖(而非复杂的植物纤维)的混合物在喂食小鼠时无法阻止结肠粘液屏障的降解。
此外,富含纯化可溶性纤维(例如菊粉、低聚果糖或果胶)的食物可导致先前存在微生物群紊乱的小鼠肝细胞癌。这些发现表明,为了发挥最佳效果,可以考虑其他参数,如天然食品中天然膳食纤维或原纤维浓缩物的数量、溶解度和复杂分支形式。
在炎症性疾病的背景下,继续研究确定的纤维类型的影响,并使用高质量的随机试验,对于提供可操作的、经证实的工具来利用饮食对自身免疫性疾病的微生物群介导的影响至关重要。
★ 纤维补充不是越多越好
在一项对39名健康志愿者进行的为期17周的随机前瞻性研究中,高纤维饮食改变了CAZyme谱;然而,丁酸水平在干预后没有显著变化。相反,摄入的纤维量与粪便总碳水化合物密切相关,这表明参与者的肠道微生物群无法完全降解饮食中的纤维。这些发现提醒人们,如果纤维降解途径饱和,盲目增加纤维摄入可能不会转化为增加短链脂肪酸的生成。
★ 发病率——微生物群比单纯饮食更容易预测
在一项前瞻性队列研究中,对170776名妇女进行了26年的随访,结果表明,水果和蔬菜纤维与克罗恩病发病率呈负相关,但奇怪的是,其他纤维来源或溃疡性结肠炎发病率并非如此。另一项跟踪401326名参与者7年的研究发现IBD发病率与纤维摄入之间没有关联。这些发现提示,潜在微生物群的差异,特别是降解纤维生成短链脂肪酸的能力,可能比单纯饮食更好地预测疾病发生率。
★ 疾病发生前,短链脂肪酸生成力已受损
为此,对127例溃疡性结肠炎患者进行的研究表明,与健康对照组相比,患者体内关键产丁酸菌Roseburia hominis,Faecalibacterium prausnitzii的丰度降低,且这些细菌的丰度与疾病活动性呈负相关,这一发现在结肠粘膜刷毛细菌的靶向分析中得到了独立证实。总之,这些观察结果表明,在溃疡性结肠炎患者中,微生物短链脂肪酸生成受损可能先于疾病发生。
鉴于这些联系以及短链脂肪酸在维持免疫稳态和肠道屏障功能中的作用,人们有兴趣利用微生物衍生代谢物的有益宿主调节作用,包括“后生元”。例如,由主要降解物(如双歧杆菌)发酵的膳食纤维产生乙酸盐和其他中间产物,然后通过定义的结肠细菌亚群将其转化为丁酸酯。
丁酸盐
在小鼠和人类细胞系中,丁酸不足可导致产生粘蛋白的杯状细胞从β-氧化转变为厌氧糖酵解,从而对杯状细胞分化和构成结肠粘液层大部分的糖蛋白MUC2的合成产生负面影响。这种代谢变化还增加了氧气和硝酸盐的利用率,从而形成了一个正反馈回路,增加的氧气不利于专性厌氧菌,即短链脂肪酸的主要产生者。此外,丁酸盐可与G蛋白偶联受体(例如,GPR41、GPR43和GPR109A)结合,下调炎性细胞因子转录并促进Treg细胞分化。
丙酸盐
丙酸盐,另一种重要的短链脂肪酸,参与肝脏的糖异生,促进饱腹感和降低胆固醇。在一项概念证明研究中,143例多发性硬化症患者在补充丙酸和现有疗法后获得缓解,功能性Treg细胞增加,TH1和TH17细胞减少。尽管这种补充中的微生物组调节机制尚不清楚,但应答患者肠道微生物组的转录组学特征与与Treg诱导正相关的基因相对应,而与Treg细胞诱导负相关的基因被抑制,在无反应的患者中观察到相反的趋势。

Wolter M, et al., Nat Rev Gastroenterol Hepatol. 2021
排除或消除饮食
通过微生物组优化短链脂肪酸的生产可能是一种有希望的联合治疗策略;然而,评估宿主和微生物生物标志物(疾病状态的可测量的生化或生理指标)的迭代过程必须适应疾病状态,因为在疾病活跃期间,许多IBD患者在食用高纤维食物后报告腹胀和过量气体。
虽然在疾病活跃期或外科手术期间禁用复合碳水化合物,但它们仍可能在预防或延长缓解期方面发挥作用。各种排除或消除饮食,如专门为IBD患者提供的专用肠内营养(EEN)或部分肠内营养(PEN),不应完全忽视纤维,而应利用疾病活跃或缓解状态下即将进行的研究。例如,EEN和PEN是流行的治疗性饮食,它们缺乏纤维,能够诱导80-90%的克罗恩病患者病情缓解。
在所有患者中,EEN与一致的分类转变无关,但它似乎通过改变结肠微生物群的代谢输出,使其发酵成蛋白质而不是碳水化合物,从而增加克罗恩病的缓解率。这种功能转变对疾病结果的机制性影响尚未完全阐明;目前尚不清楚以下哪一项会缓解:排除纤维、缺乏另一种饮食成分、细菌密度或多样性降低,或这些因素的某种组合。
或者,通过肠道生态系统重建观察到的间歇性禁食对多发性硬化症进展的保护作用,也可能为EEN治疗的成功提供见解。
用于治疗的饮食
自身免疫方案饮食:基于古饮食,该方案强调食用营养丰富的新鲜或发酵食品,并消除可能引发炎症的食物组。在维持阶段,特定食物的逐渐重新引入因个体食物耐受性而异。
全部或部分肠内营养(EEN或PEN): 一种液态元素(氨基酸型)、半元素(寡肽)或聚合物(全蛋白基)配方饮食,不含固体食物(EEN)或限制摄入(PEN)1-3个月,以减少食物抗原暴露,促进肠道愈合。
克罗恩病排除饮食(CDED):为克罗恩病患者量身定制的缓解饮食,这种全食物饮食与PEN饮食一起使用,以减少暴露于可能对宿主免疫、肠道屏障功能和/或微生物群产生负面影响的饮食成分。
特殊碳水化合物饮食(SCD):最初是为治疗腹腔疾病而开发的,这种饮食通常用于帮助治疗IBD,允许大多数水果、一些蔬菜、坚果、肉类和鸡蛋,同时避免所有谷物、食糖、食品添加剂和大多数乳制品。
低FODMAP饮食:这种饮食强调限制可发酵低聚糖、双糖、单糖和多元醇(FODMAP),并分阶段进行——消除、重新引入和维持——以确定引发疾病症状的食物,并促进个性化饮食的发展。
生酮饮食:这种低碳水化合物、高脂肪的饮食会导致酮症,酮症是一种代谢状态,在这种状态下,脂肪(而不是葡萄糖)会被降解以产生能量。酮体的产生可以消耗双歧杆菌并降低促炎性T辅助细胞17的水平。
通过考虑这些消除饮食成功的微生物组介导机制,我们也许能够改进饮食干预措施。这种适应可能对提高患者依从性和持续缓解很重要,因为EEN配方饮食很难长期维持,甚至在与抗生素联用时可能对克罗恩病患者有害。
诱导缓解后,如克罗恩病排除饮食中所建议,可能建议选择性地用耐受性良好的纤维混合物补充EEN。在对78名克罗恩病儿童进行的随机对照试验中,CDED导致变形菌门降低,炎症和病情缓解的患者比例高于单独用EEN的患者。
为了支持这一发现,对患有活动性结肠炎的Il10−/−小鼠给予含明确纤维混合物的EEN,可降低疾病活动度和结肠炎。这一发现还伴随着固有层中CD4 FOXP3 Treg 细胞群的增加,盲肠中总短链脂肪酸水平的增加,以及屏障功能恢复的各种其他迹象,促使人们考虑在活动性炎症患者中是否建议使用特定纤维。
尽管如此,干预的重点不应局限于碳水化合物。许多饮食和饮食成分以微生物群依赖的方式共同影响宿主健康。
西方饮食(其特点是大量摄入脂肪、蛋白质、糖、盐和加工食品)与伴随的导致自身免疫疾病发展的微生物变化之间的联系越来越明显,但要解开设计治疗性饮食的机制并不那么简单。研究表明,在影响肠道微生物群改变的因素中,饮食是一种潜在的强有力的工具,可以直接改变微生物群,使其达到健康状态。
人体宿主、肠道菌群和饮食的时间趋势
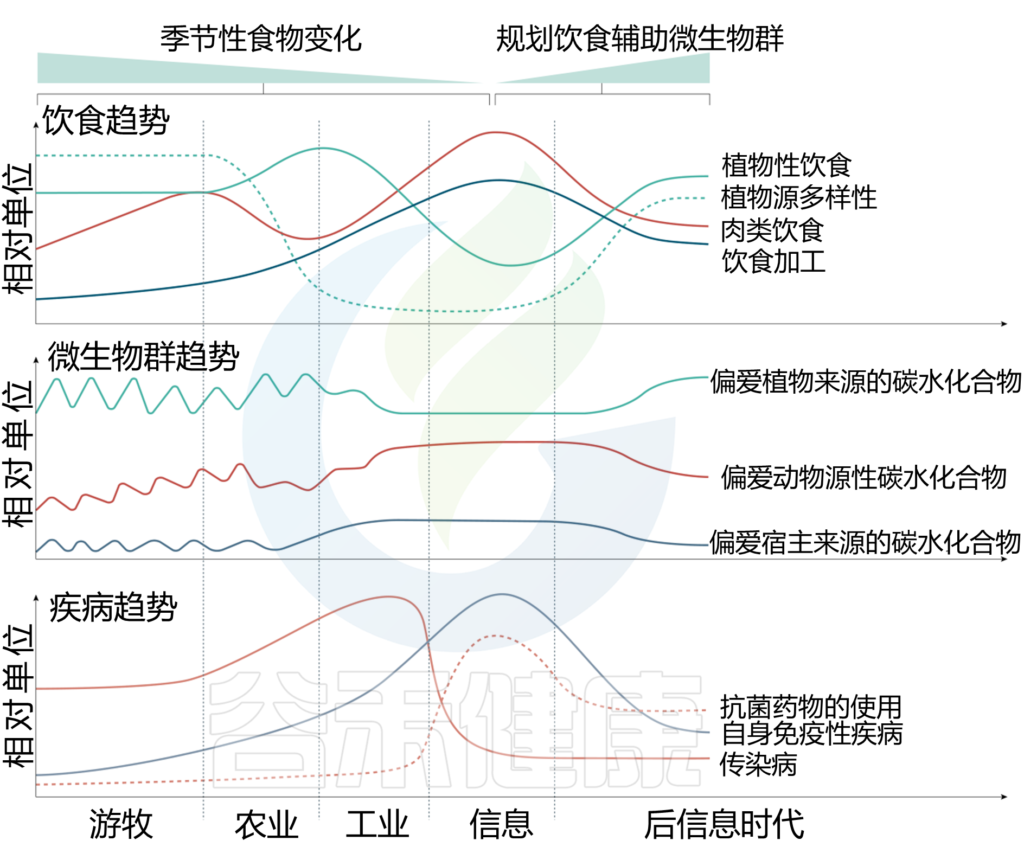
Wolter M, et al., Nat Rev Gastroenterol Hepatol. 2021
在游牧时代,人类饮食主要受季节性影响,相应的肠道菌群也存在相当明显的季节性变化。
当进入农业社会后,随着粮食种植,季节性变化逐渐减少,植物性饮食增加,但食物多样性在减少,相应的疾病以传染性疾病为主。
进入工业化之后,肉类饮食增加,加工食物快速增加,伴随的是自体免疫疾病的开始上升。进入信息时代后,随着抗菌药物的大量使用,自身免疫疾病快速增加并成为主要疾病类型,同时植物性饮食快速下降,肉类消费和加工食品达到最高。
我们希望通过进一步认知菌群和饮食的关系,有效规划饮食从而辅助微生物群的改善,减少抗菌药物使用,增加植物性饮食并改善摄入的多样性,减少加工食品的比例,最终减少自身免疫性疾病的发生。
蛋白质腐败
膳食蛋白质是另一种关键的大量营养素,也可以进行细菌发酵,产生影响宿主功能和细菌组成的关键代谢产物。
在消化过程中,膳食蛋白质首先被宿主酶水解成肽,游离氨基酸在小肠中被吸收。未被吸收的肽和氨基酸可到达结肠,在结肠中由共生微生物发酵,根据体外和体内测量,估计产生17–38%的结肠短链脂肪酸。肠道微生物群的蛋白质降解为宿主提供必需的游离氨基酸,支链脂肪酸(BCFA)可作为结肠细胞的替代能源。
然而,这一过程可能对局部肠道环境有害,因为有毒的代谢副产物如氨、硫化氢和酚的释放,增加了小鼠上皮细胞的通透性,降低了体外细胞活力。
★ 不同来源蛋白质作用不同
在IBD小鼠模型中,动物源性蛋白质通过促炎单核细胞加重结肠炎,而富含植物蛋白的饮食或无菌小鼠中未观察到这种情况,这表明动物蛋白的微生物代谢物导致过度炎症。
因此,将蛋白质类型或来源(例如,麸质或酪蛋白)个性化为个体患者的微生物群组成对于限制炎症代谢物的产生至关重要。
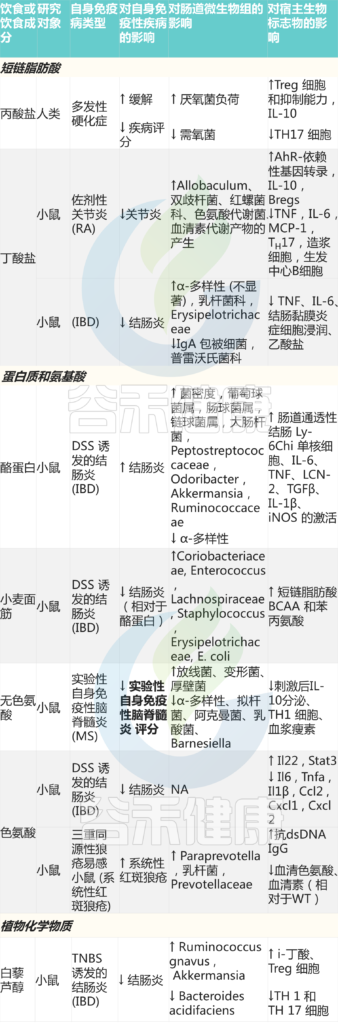
Wolter M, et al., Nat Rev Gastroenterol Hepatol. 2021
重要的是要考虑不同的蛋白质来源如大豆可能不容易在小肠中降解,因此增加的底物到达结肠,在那里它可以发酵。
★ 特定蛋白质对微生物组和代谢产物的影响各不相同
因为并非所有蛋白质都同样适合发酵,也并非所有细菌都配备了适当的酶途径。例如:
• 尽管所有氨基酸都经过氨释放脱氨作用,但只有支链氨基酸(缬氨酸、亮氨酸和异亮氨酸)产生支链脂肪酸;
• 硫酸化氨基酸有利于硫酸盐还原菌的增殖以提供硫化氢,这一过程在溃疡性结肠炎患者中升高;
• 根据使用人类微生物组数据的代谢模型,肠杆菌属和大肠杆菌属优先发酵芳香氨基酸以产生酚类物质。例如,色氨酸是一种基本的芳香族氨基酸,主要由宿主酶代谢;然而,它也可以通过具有良好特征的吲哚途径被肠道微生物群代谢。其代谢产物为宿主芳基烃受体提供配体,降低促炎细胞因子和趋化因子的表达,促进IL-22的产生,IL-22刺激上皮细胞释放调节小鼠微生物组生长的抗菌肽。
与这些免疫代谢过程一致,饮食中补充色氨酸或产生色氨酸的乳酸杆菌菌株可缓解各种小鼠模型中的实验性结肠炎。相反,饮食色氨酸可能通过增加循环自身反应性CD4+T细胞数量并促进其通过血脑屏障迁移而加剧实验性自身免疫性脑脊髓炎(EAE),EAE是多发性硬化症的临床前小鼠模型。
因此,虽然膳食色氨酸已被提议作为缓解IBD患者结肠炎的治疗策略,但其微生物群介导的作用可能是器官特异性和疾病特异性的。
膳食脂肪代谢
除了作为肠道微生物的潜在营养源外,膳食脂肪还通过增加宿主胆汁酸的产生来改变微生物的组成,这可以促进具有高致病潜力菌的生长,如沃氏嗜胆菌Bilophila wadsworthia。
★ 脂肪改变微生物群,调节此生胆汁酸
由膳食脂肪引起的肠道微生物群组成改变也可以调节微生物来源的次生胆汁酸的丰度。例如,食用富含不饱和脂肪的核桃,会在健康成年人的肠道中富集粪杆菌、玫瑰杆菌和梭菌,从而减少微生物来源的、促炎症的次生胆汁酸。
★ 高脂饮食也可通过增加LPS暴露影响宿主健康
已经在小鼠中证明,高脂肪饮食也可以通过增加LPS暴露影响宿主健康,LPS可诱导宿主促炎性TLR4信号通路,导致代谢性内毒素血症的低度全身炎症。考虑到在IBD患者中观察到的LPS水平与疾病严重程度之间的强烈相关性,以及LPS与T1DM之间已证实的联系,这种代谢性内毒素血症可能对自身免疫活动的开始产生深远的影响。
自身免疫疾病和稳态中的饮食-肠道微生物-宿主轴

Wolter M, et al., Nat Rev Gastroenterol Hepatol. 2021
LPS暴露升高与疾病相关的详细机制仍有待充分阐明;然而,基于对小鼠的研究,它可能是多因素的,包括:革兰氏阴性杆菌的丰度增加,这是一种具有更强免疫原性的改变的LPS来源;抑制LPS诱导炎症的细菌减少,如双歧杆菌;以及宿主因素,如由于脂肪或微生物活动的直接影响而增加的肠道通透性。
★ 特定类型的脂肪对于精确设计非常重要
未定义的高脂肪饮食通常用于人类微生物群研究,然而,特定类型的脂肪对肠道微生物群和宿主健康结果有重大影响。尽管西方高脂肪饮食支持有害的微生物转移,但对于定义的脂肪类型,如ω-3多不饱和脂肪酸或复杂的脂肪源,如核桃,存在许多有益微生物变化的例子。
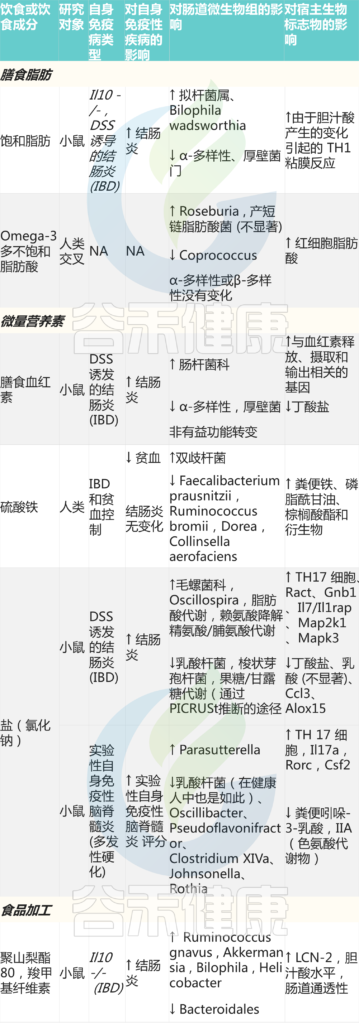
Wolter M, et al., Nat Rev Gastroenterol Hepatol. 2021
此外,在一项没有评估临床参数的研究中,多发性硬化症患者在服用生酮饮食(低碳水化合物高脂肪)6个月后,表现出包括直肠真杆菌Eubacterium rectale 和Faecalibacterium prausnitzii 在内的细菌水平恢复,这被认为反映了适当的发酵能力。
与右旋糖酐硫酸钠(DSS)中的低碳水化合物饮食相比,生酮饮食以类似的微生物组依赖方式减轻了结肠炎-诱导结肠炎小鼠模型。此外,在生酮饮食中产生酮体,特别是β-羟基丁酸盐,可消耗双歧杆菌并降低人类的TH17细胞水平,在自身免疫性癫痫儿童中观察到类似的生酮饮食诱导TH17细胞减少。
这些发现挑战了高脂肪饮食作为促炎症饮食的全面分类,但强调了需要进一步研究特定饮食脂肪如何调节肠道微生物组或宿主-微生物组的相互作用(例如,通过直接改变免疫系统,然后反馈给肠道微生物组,反之亦然)。分析特定类型的脂肪及其对微生物组的影响是使用膳食脂肪精确设计微生物组的关键。
食品加工
食品加工通常随着人类历史的发展而增加,包括一系列可能是热加工(例如烹饪或膨化)、机械加工(例如研磨、切片或搅打)或化学加工(例如氢化或乳化),或者涉及添加剂的使用。这些操作通常用于延长保质期或改善外观或味道。
食品加工也可能使小肠中的营养物质消化饱和,对结肠微生物群的资源可用性影响不明确。
★ 生 v.s. 熟 (作用不同)
烹饪是最早的食品加工实例之一,对营养物质的消化率有着重要影响,Carmoty等人通过对小鼠和人类的控制喂养研究证明了这一点。他们发现,与煮熟的块茎相比,食用富含淀粉的生块茎会破坏肠道微生物群,降低细菌的丰度,并上调异源代谢的微生物途径。这些发现指向了挖掘生食物中具有治疗用途的成分的可能性,因为微生物的外生代谢可能对宿主有益或有毒,或者它可以修改药物,这取决于肠道微生物组成。
★ 不同烹饪、保存方式影响微生物组
另一项研究发现,将全麦面粉置于蒸煮-冷冻循环中会显著增加抗性淀粉的百分比,从而导致体外发酵过程中丙酸盐和双歧杆菌水平的增加。由于这些温度循环而增加的营养物质的生物利用度或化学变化很少是饮食设计中的因素,但鉴于生、熟和冷冻食品可能以不同方式影响微生物组的发现,这些过程可能需要更仔细地考虑。
★ 添加剂——损害紧密连接完整性和增加肠道通透性
在过去的一个世纪里,使用添加剂进行的有针对性的食品改性(通常用于保存、颜色或质地改性或口味增强)大幅增加。这是通过损害紧密连接完整性和增加肠道通透性来部分解释自身免疫相关上升的原因。
事实上,避免加工食品在许多用于调节自身免疫疾病进展的排除或消除饮食中是常见的,包括AIP饮食、特定碳水化合物饮食和低发酵低聚糖、双糖、单糖和多元醇(FODMAP)饮食。
Wolter M, et al., Nat Rev Gastroenterol Hepatol. 2021
★ 乳化剂—— 利于促炎微生物群生长
在具有结肠炎遗传易感性的小鼠模型中,暴露于两种常见乳化剂,羧甲基纤维素和聚山梨酯80,增加了粘液层的通透性,增加了细菌对粘膜的粘附性,并有利于更具促炎性的微生物群。
★ 误将乳化剂当膳食纤维
羧甲基纤维素在化学上来源于纤维素,在结构上被认为是一种纤维,因此,它有助于增加欧洲营养标签上的膳食纤维含量。这可能会误导那些打算根据标签值增加膳食纤维摄入量的消费者。监管机构对此类添加剂的安全性评估也没有纳入潜在的微生物群介导的健康影响,可能是因为这一研究相对较新。然而,在动物模型和患者团体的研究表明,食品添加剂可能比现行规定更具破坏性,强调需要考虑它们对微生物和宿主的影响。
总的来说,越来越多研究强调了碳水化合物、脂肪、蛋白质、植物化学物质、微量营养素和食品添加剂对微生物组成和代谢产物的影响。随着这些年测序技术的爆炸式发展,我们能够更深入地了解肠道内的复杂菌群及其在健康和疾病中的作用,但有针对性的个性化营养措施却跟不上。需要适应性、个性化的研究方法和工具来量身定制饮食计划或确定潜在的微生物目标,以缓解自身免疫性疾病患者的病情。
由于在诊断为各种自身免疫性疾病的患者中观察到微生物多样性较低,因此丰富多样性是一种普遍建议的策略,有可能最大限度地提高人类肠道微生物组的健康效益。
然而,将“健康”微生物群等同于高度多样化微生物群这样的过度简化,可能掩盖了潜在的生物标志物或来自疾病潜在驱动因素的信号。布鲁索的警告:不要依赖多样性指标,在没有探索潜在机制的情况下,在疾病背景下将一个微生物群标记为“失调”。
基于最大化有益微生物生理学(微生物已实现的代谢或物理特性)和最小化有害影响的方法可以避免这种陷阱,前提是可以定义适当的生理目标。为此,机器学习方法是在自身免疫疾病背景下利用现有宏基因组和代谢组学数据的强大工具。这些目标还可以提供基于遗传、环境因素和存在于个体中的潜在微生物群的个性化干预的机会。
在患者队列和动物模型中的现有研究支持使用定制饮食或特定饮食成分来调节各种疾病环境中的肠道微生物群和宿主反应。

Wolter M, et al., Nat Rev Gastroenterol Hepatol. 2021
许多宿主因素,包括基因和预先存在的肠道微生物群组成,可以影响个体对饮食的反应。鉴于越来越多的证据表明,与饮食或治疗相关的个性化肠道微生物群反应,我们必须积极将更动态的方法纳入当前的自身免疫性疾病治疗方案中,利用疾病或缓解状态的潜在机制知识,最大限度地提高现有饮食疗法在特定个体中的成功率。例如,在个性化营养学中,个体可能会跟踪饮食,通过纳入相关的协变量,如疾病状态、治疗方案、微生物组和其他生物标记物,预测性建模工具可用于建议食用或避免食用特定结果的食物。
肠道微生物组工程工具的个性化应用方法
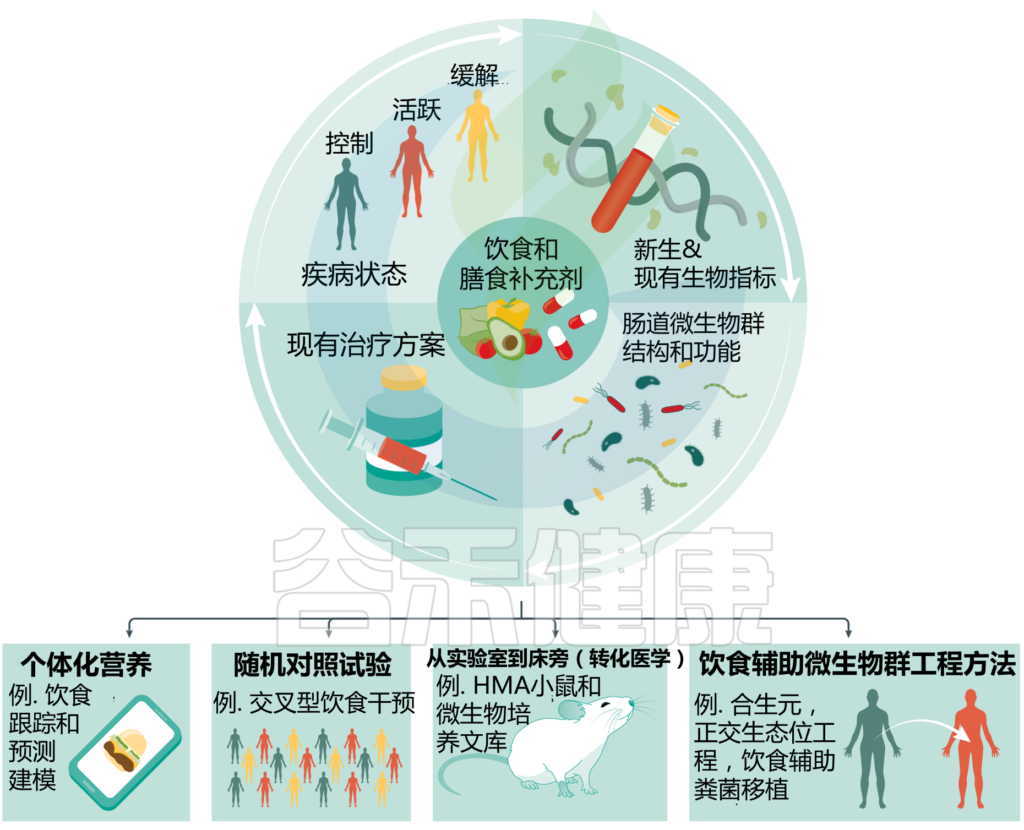
Wolter M, et al., Nat Rev Gastroenterol Hepatol. 2021
★ 设计定制饮食改变血糖水平
在一项针对800名健康人的里程碑式研究中,Zeevi等人设计了一种机器学习算法,通过考虑与葡萄糖反应密切相关的个人健康指标来推荐定制饮食。将该模型扩展到100名个体的队列中,同样允许有针对性地降低血糖反应,这表明通过考虑一组个性化因素,可以简化此类饮食的设计。设计定制饮食以引起血糖水平预期变化的能力只是冰山一角;在将这项工作扩展到其他非代谢性疾病时,机制上的见解应该为生物标记物的选择提供信息,以制定个性化的饮食建议。
★ 自适应平台试验有望进一步治疗自身免疫性疾病
提高我们对饮食-微生物组在健康和疾病中相互作用的理解的方法可能涉及提供饮食或饮食成分的人类队列,以根据反应对患者进行分层,同时也纳入相关因素以帮助理解干预效果的差异。
为此,自适应平台试验有望进一步治疗自身免疫性疾病,因为在试验设计中建立了测试治疗方案以确定应答者亚群的迭代过程。然而,为了产生机理知识,对小鼠模型的研究对于验证或激发人类转化工作仍然是不可或缺的。这一策略将有助于深入了解饮食-微生物组-宿主轴,并确定新的干预选择。
★ 研究时要考虑空间异质性
出于实用性考虑,许多研究根据粪便样本中检测到的分类群推断肠道微生物群的组成,但宿主特征和胃肠道微生物生理学的细微变化可能与体内稳态密切相关。例如,居住在粘液或结肠隐窝中的细菌,如Akkermansia muciniphila或不动杆菌属,与多发性硬化症和病原体诱导的结肠炎有关,但它们在粪便衍生微生物组中的丰度可能低于在结肠粘膜中的实际丰度。然而,临床相关部位的微生物群调查,如结肠黏膜,需要侵入性活检,因此小鼠模型可以很容易地研究肠道微生物组成的空间异质性。
没有一种单一的方法足以理解疾病驱动因素并改善健康结果;必须利用所有相关的模式生物、系统(如电子、体外或体内)和方法(如还原论或生态学)。
有许多新兴技术旨在通过引入具有新的或失去的有益功能的细菌,或通过消除具有有害功能的细菌,使肠道微生物群达到功能健康状态。这些技术的一个子集,如粪菌移植(FMT)或正交小生境工程,显示出与特定饮食或饮食补充剂结合使用的高潜力,以解决其一些局限性或潜在放大其效果。将特定饮食添加到这些技术中增加了另一个可修改的变量,有可能提供更高程度的系统控制,并提高处理个性化反应的能力。
这里介绍和讨论了在自身免疫性疾病的背景下设计肠道微生物组的新方法,这些方法已经与饮食或饮食补充相结合进行了探索。
粪菌移植和饮食
尽管粪菌移植成功的机制尚未完全解决,但该程序有可能与特定饮食或膳食补充剂结合使用,更有效。
粪菌移植试图通过将健康供者的粪便物质转移到肠道微生物群紊乱的患者肠道,重新建立一种多样的、稳态的微生物群组成。关于粪菌移植在我们的很多文章里都有提到,此处不详述。
在这里我们将重点关注饮食在粪菌移植成功中的潜在作用。
鉴于饮食对微生物组的显著影响,接受移植的患者的营养习惯对移植成功和持久性的影响是一个潜在的重要话题。
尽管大部分未经探索,但有两种直观的方法:
1. 接受者的饮食可以设计为复制提供者的饮食,从而最大限度地提高供体微生物群的传递保真度
2. 接受者可以考虑一种支持潜在有益菌的饮食,如克罗恩病排除饮食或菊粉型果糖补充,以支持产短链脂肪酸的Roseburia 或Faecalibacterium
一项随机临床试验比较了20例活动性溃疡性结肠炎患者在补充或不补充果胶的情况下粪菌移植的有效性,表明补充果胶可能改善移植微生物群的持久性;然而,需要进一步的研究,包括足够数量的患者和合适的对照,以更好地了解饮食对提高粪菌移植成功率的潜力。
单独使用粪菌移植可能部分足以提供疾病缓解特性或诱导缓解,但在肺气肿小鼠模型中,给小鼠喂食高纤维饮食结合粪菌移植可对微生物组分和短链脂肪酸输出产生额外的有益影响。在肥胖小鼠模型中观察到,肠道微生物组的有益和疾病缓解作用可通过粪菌移植(例如,从食用富含植物化学物质饮食的供体)传播到食用高脂肪或高糖饮食的受体,这也提出了一个问题,即是否修改供体的饮食,应该进一步研究,而不仅仅是接受者的饮食。
最后,在供体肠道微生物群组成不清楚的情况下,设计支持粪菌移植饮食的能力可能会受到限制。因此,应考虑使用从健康捐献者粪便中提取的特定细菌培养物。
益生菌-饮食组合和合生元
独立于粪菌移植,对预先存在的肠道微生物群直接施用单一微生物菌株或菌株联合体,如益生菌制剂VSL#3,是IBD广泛探索的治疗选择。
在服用益生菌时,有两个主要考虑因素:
第一,可能是最重要的,是益生菌的微生物组成;
第二,选择益生菌所在食品的基质或胶囊。
无论是单一的益生菌物种还是一个群落的口服给药已经被广泛地探索。这种方法对实验性IBD的各种小鼠模型以及溃疡性结肠炎患者有效,但在克罗恩病患者中无效,克罗恩病需要长期缓解或积极诱导缓解。由于益生菌在治疗自身免疫性疾病,特别是IBD方面的应用和潜力已在其他文章中详细阐述。
在此讨论益生菌与膳食补充剂的结合以及它们的传递基质。
输送微生物的基质可以有不同的形式,最常见的是食品基质,如酸奶、谷物棒和果汁,或保护性乳液或生物聚合物中的封装。选择给药基质至关重要,因为益生菌使用的主要限制是通过胃肠道时的细菌活力和定植抗性,这是内源性微生物群的特性,可防止以前不存在于微生物群中的新细菌定植。
★ 营养利用度
由于营养限制是定植抵抗背后的核心机制之一,并且如前所述,基于饮食的营养利用度对肠道微生物群产生重大影响,提供益生菌的食物或饮料的选择对益生菌菌株的活力和持续植入有相当大的影响。
★ 环境压力因素
这一过程不仅是由于营养素的可用性,而且还由于环境压力因素,如生产和储存期间的pH值、氧气和温度,以及胃的高酸度和通过胃肠道时肠道中胆汁酸的存在。尽管研究最广泛的食品基质是乳制品,但人们一直在努力探索不同的食品基质,如谷物、水果、蔬菜和肉制品。
★ 合生元
益生元和益生菌可以组合成所谓的合生元,合生元可以定义为互补或协同。
互补合生元包括益生元和益生菌,它们相互独立地产生健康益处。
协同合生元旨在最大限度地发挥益生菌的有益功能。
协同设计背后的原理是:益生元可以提高益生菌抵抗环境压力的生存能力;益生菌确保肠道微生物群以有益的方式代谢益生元的能力;益生元可以通过开辟一种新的营养生态位,支持益生菌植入宿主微生物群。
虽然使用合生元的人类研究才刚刚开始出现,但某些组合已被发现是有效的,并用于治疗自身免疫性疾病。
在IBD的背景下,双歧杆菌和乳酸杆菌与车前草、低聚果糖和菊糖的结合显示出良好的前景。
类似地,添加菊粉的嗜酸乳杆菌、干酪乳杆菌和双歧双歧杆菌的混合物对类风湿性关节炎患者产生有益效果,乳酸杆菌产孢菌与麦芽糊精和低聚果糖的组合对T1DM患者有益。
另一种提高生存能力和定植的方法是将细菌包埋在保护性乳液或生物聚合物中。
这种封装不仅可以保护细菌免受环境危害,并有可能实现特定部位的释放,还可以通过将细菌封装在含有益生元的基质中,创造复杂的合生元结构;例如,海藻酸钠-菊糖-黄原胶或聚(乳酸-共-乙醇酸)-海藻酸钠胶囊。
正交小生境工程
正交小生境工程是合生元概念的一种演变,因为微生物-底物关系是利用细菌在宿主菌群中的持续植入。虽然合生元的益生元部分通常由普通营养素组成,但在正交小生境工程中,利用未使用或不常见的营养素,为细菌稳定植入宿主微生物群创造专属小生境。例如,降解卟啉(一种海藻多糖)的能力被用来稳定地将一种人类共生细菌移植到小鼠肠道中。
尽管目前的研究仅限于小鼠,但利用这些不常见的代谢途径,可以通过间歇性或持续的饮食补充精确控制植入的细菌。
Wolter M, et al., Nat Rev Gastroenterol Hepatol. 2021
虽然人类肠道微生物群降解海藻的能力比最初假设的更为普遍,但正交小生境工程在克服肠道微生物群中细菌持续植入的挑战方面有着巨大的希望,特别是考虑到利用的广泛潜在基因。
新的生态位,如琼脂糖、卟啉或卡拉胶降解,可用于帮助植入有益菌,这些细菌可以是野生型、原始菌株的改良版本,也可以是参与利用正交营养素的基因已转移的不同菌株。
★ 概念性应用
正交小生境工程的一个概念性应用是筛选工业化过程中丧失有益功能的细菌,这些细菌仍然与我们的现代生活方式相关,并将其重新引入人类肠道。这种细菌丢失的一个例子是经过充分研究的益生菌罗伊氏乳杆菌Lactobacillus reuteri,其对系统性红斑狼疮和多发性硬化症的免疫调节作用已经在小鼠模型中进行了研究。
另一个概念性应用是在重组细菌中引入罕见的营养利用途径,设计用于产生治疗性化合物,如用于IBD治疗的IL-10或防御素-5。使用不常见的营养素利用途径可以精确控制定植的剂量和持续时间,从而使治疗能够专门针对个别患者的需要进行调整。
然而,这种方法的长期可操作性可能是具有挑战性的,因为持续喂养会随着时间的推移降低植入细菌的活性。对这种现象的一种可能解释是,这种喂养可能会促进小生境分配,从而通过IgA结合将植入的细菌限制在结肠隐窝中,从而减少了与底物的接触。此外,还存在通过水平基因转移将这种特性传播给其他肠道细菌的风险,这可能导致特异性降低或反应完全改变。
这一研究领域涉及具有众多变量的高度复杂的相互作用,例如肠道微生物组内出现的相互作用,这些相互作用进一步受到宿主基因组、免疫系统和多种生活方式因素的影响,每一个都可能是不完整的特征。
因此,即将进行的研究应该具有双重目标,即识别一般和亚群体特异性的生物标志物,以提供最佳治疗,并解决拟议干预措施的各种应对措施背后的根本机制。
随着可用工具的完善,微生物领域可以从基于相关性的研究转向旨在揭示机制的研究,从而开发新的以微生物为重点的治疗方案。
“诸事利弊共存”
1. 在实现疾病中肠道微生物群的饮食驱动操作方面仍面临许多挑战。人体肠道微生物组的弹性是健康条件下的有利属性,这意味着将小鼠模型的发现转化为人类时,干预的效果可能不如预期的明显。
2. 肠道微生物组的相对稳定性可能会阻碍对自身免疫性疾病患者的组成或功能进行持久改变的努力,尤其是如果不解决稳定疾病状态的潜在驱动因素。虽然饮食是引起微生物变化的有效工具,但停止饮食通常会导致恢复到以前的微生物状态。因此,干预可能需要持续进行,这带来了确保遵守方案的额外挑战。通过将饮食干预与其他微生物组调节方法相结合,这种变化可能更稳定;然而,这一预测仍有待研究。
3. 对稳定群落的重复采样应减少典型微生物波动产生的背景噪音,从而揭示特定疾病的特征。通过了解基线微生物群,在单个患者中提出和测试有效治疗可能确实更容易。
因此,随着方法的发展,将有价值的机理知识从动物模型转化为人类或将其扩展到研究较少的疾病的研究对于完善饮食在微生物组工程中的应用至关重要。
微生物群驱动的屏障功能障碍是各种自身免疫疾病的基础,这一观点鼓励更密切地检查这些功能变化,以恢复有益的宿主-微生物群相互作用。
“精准——时代的召唤”
研究人员和临床医生必须考虑到这样一个事实,即许多宿主、环境甚至时间因素可能会在个体基础上决定“完美”的微生物组,并积极地将一种更动态、更量身定制的方法纳入当前的治疗方案。
通过实施精准医学方法和对驱动疾病的潜在机制的认识,可以提高现有治疗的成功率。我们正处于微生物组工程时代的前沿,但为了有效地实现这一转变,必须优先研究对饮食干预的个性化反应以及形成微生物组的精确机制。
主要参考文献:
Fugger, L., Jensen, L. T . & Rossjohn, J. Challenges, progress, and prospects of developing therapies to treat autoimmune diseases. Cell 181, 63–80 (2020).
Wolter M, Grant ET, Boudaud M, Steimle A, Pereira GV, Martens EC, Desai MS. Leveraging diet to engineer the gut microbiome. Nat Rev Gastroenterol Hepatol. 2021 Sep 27.
Scher, J. U., Nayak, R. R., Ubeda, C., T urnbaugh, P . J. & Abramson, S. B. Pharmacomicrobiomics in inflammatory arthritis: gut microbiome as modulator of therapeutic response. Nat. Rev. Rheumatol. 16, 282–292 (2020)
Johnson, A. J. et al. Daily sampling reveals personalized diet- microbiome associations in humans. Cell Host Microbe 25, 789–802.e5 (2019).
Horwat P, Kopeć S, Garczyk A, Kaliciak I, Staręga Z, Drogowski K, Mardas M, Stelmach-Mardas M. Influence of Enteral Nutrition on Gut Microbiota Composition in Patients with Crohn’s Disease: A Systematic Review. Nutrients. 2020 Aug 23;12(9):2551.
Wastyk HC, Fragiadakis GK, Perelman D, Dahan D, Merrill BD, Yu FB, Topf M, Gonzalez CG, Van Treuren W, Han S, Robinson JL, Elias JE, Sonnenburg ED, Gardner CD, Sonnenburg JL. Gut-microbiota-targeted diets modulate human immune status. Cell. 2021 Aug 5;184(16):4137-4153.