-

 CNAS L23010
CNAS L23010

 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业 二级病原微生物安全实验室
二级病原微生物安全实验室- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业 二级病原微生物安全实验室
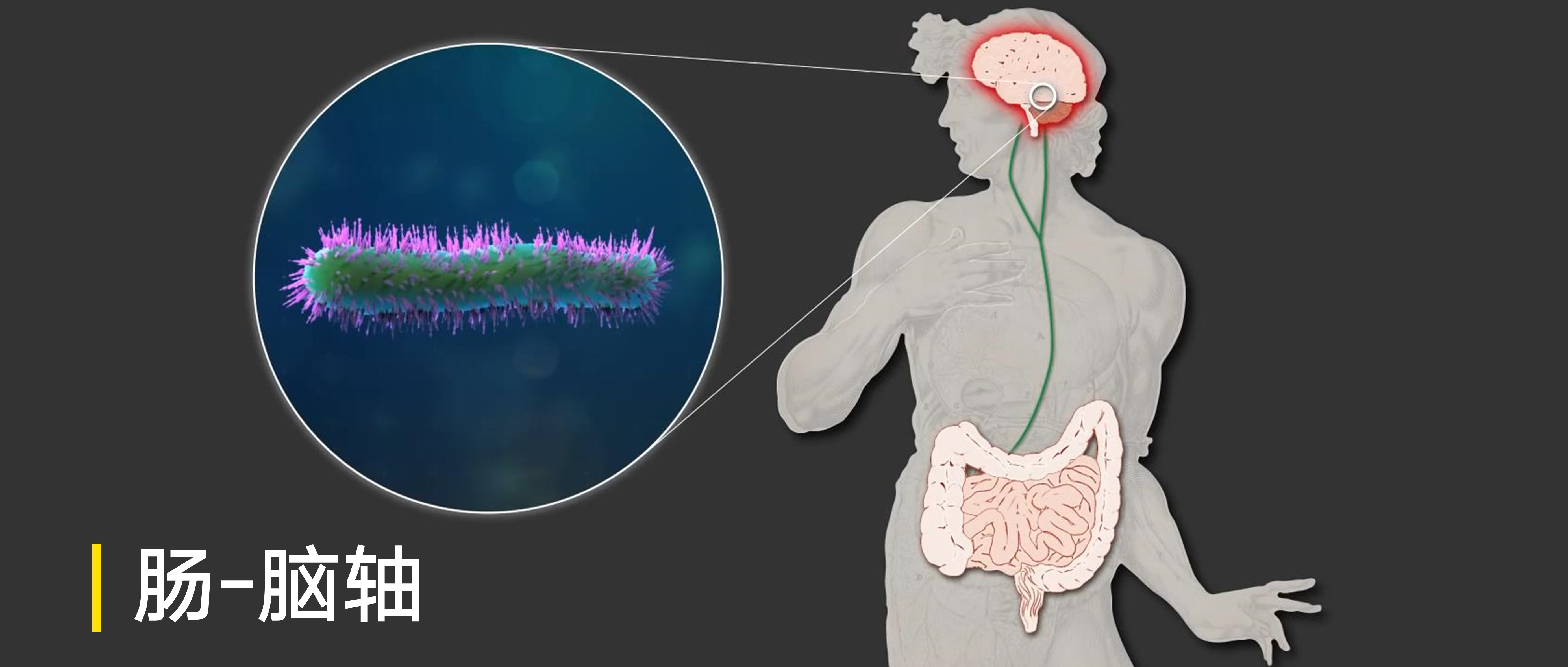
原创: 谷禾健康
说话晚、读书读不进、每天不开心不全是我们的错,还可能是肠道菌群有问题。肠道菌群不仅影响消化吸收,还影响神经系统。近年来,科学家们研究发现,肠道细菌可能在神经发育,焦虑和抑郁症的诱发过程中,甚至很多中枢神经系统疾病中,都起着重要的作用。
肠道菌群究竟是什么
首先,我们了解下肠道菌群。
肠道菌群包含居住在胃肠道中的大约100万亿微生物的集体基因组,我们肠道细菌的基因库包含比人类基因组多150倍的独特基因。
在人体定植的许多微生物群落中,肠道菌群正在成为影响宿主健康状况的主要参与者。肠道菌群的组成是在宿主发育早期建立的,并且可以在一生中经历无数的变化。越来越多的证据表明,肠道微生物组与中枢神经系统(CNS)相通 。
肠道菌群和我们的发育同步
是伴随和影响我们一生的伙伴
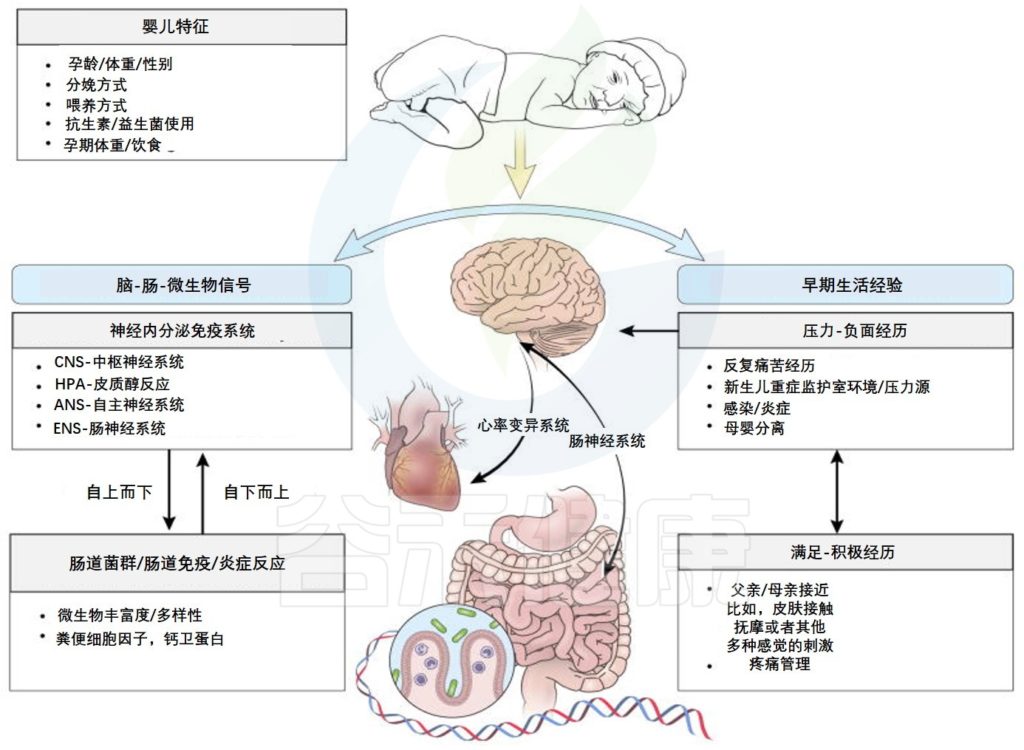
从怀孕时期就影响我们的神经发育
神经发育过程
经研究发现,神经发育的主要过程与母体和新生儿肠道微生物的变化一致

肠-脑轴参与婴儿早期神经发育与感受
接下来我们看看胃肠道是怎么构成的
胃肠道的构成
胃肠道(GI)长5米,上皮表面积约32平方米。它是身体70-80%免疫细胞的家园,超过1亿个神经元,以及多达100,000个外在神经末梢。
微生物组包含多达40万亿个细胞和至少数百种不同的物种。胃肠道的一个重要功能是感知和响应外部线索。
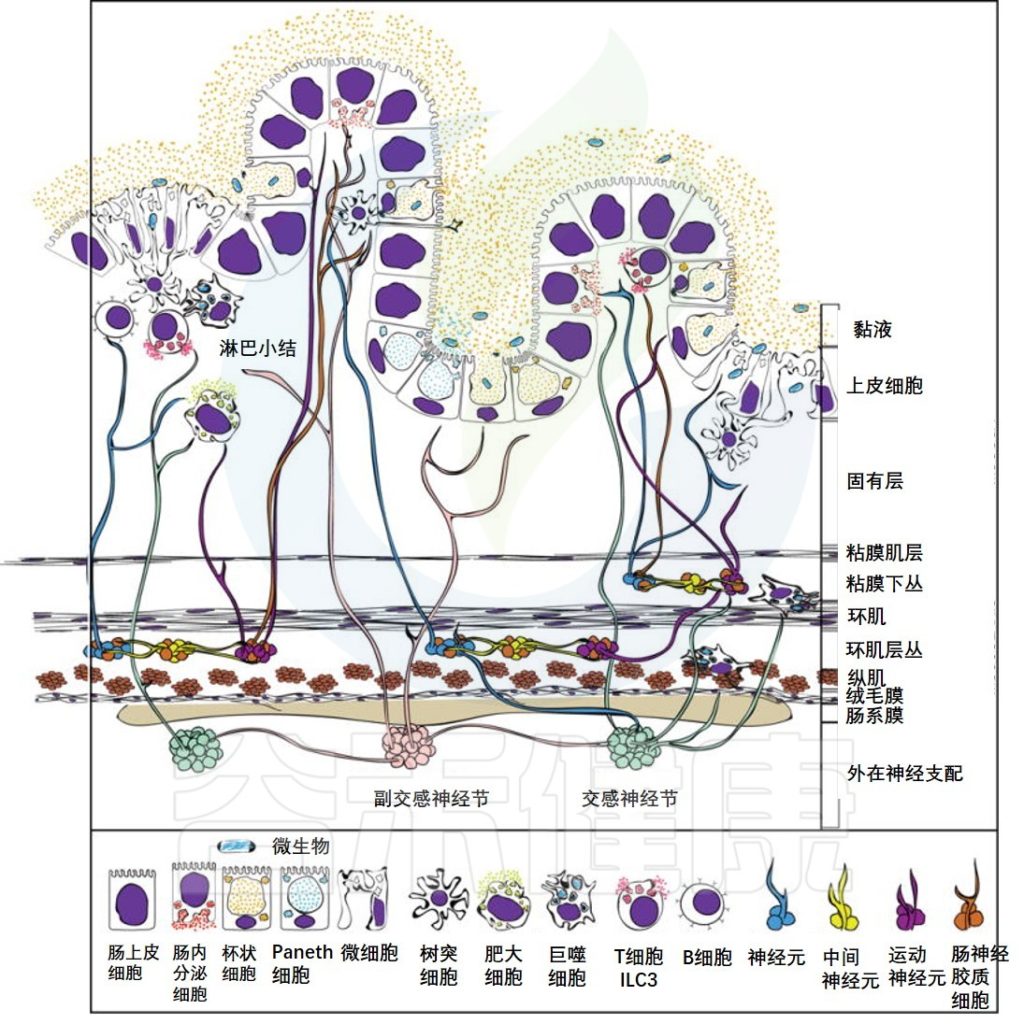
胃肠道由不同的横截面隔室组成。外在的,交感神经和副交感神经纤维通过肠系膜进入胃肠道并且可以延伸遍及肠组织的所有层。各种免疫细胞驻留在肌层,但在固有层中也非常丰富,特别是在Peyer氏斑块和淋巴滤泡中。
这些免疫细胞也与神经元和神经胶质细胞紧密相连。这里显示的上皮由5种不同的细胞类型组成,包括吸收肠上皮细胞,肠内分泌细胞,杯状细胞,潘氏细胞,微细胞。
胃肠道(GI)对于营养物质的吸收,粘膜和全身免疫反应的诱导以及健康的肠道微生物群的维持是必不可少的。
你会发现不只是消化的细胞
还有大量的神经元和免疫细胞
以及海量的肠道菌群
为什么有句话说
“肠胃不好顺带着心情和脑子也不好了”?
肠道又是如何联系到大脑
从而影响神经系统的呢?
经过大量的研究,我们逐步揭开了肠道菌群与大脑之间的联系。
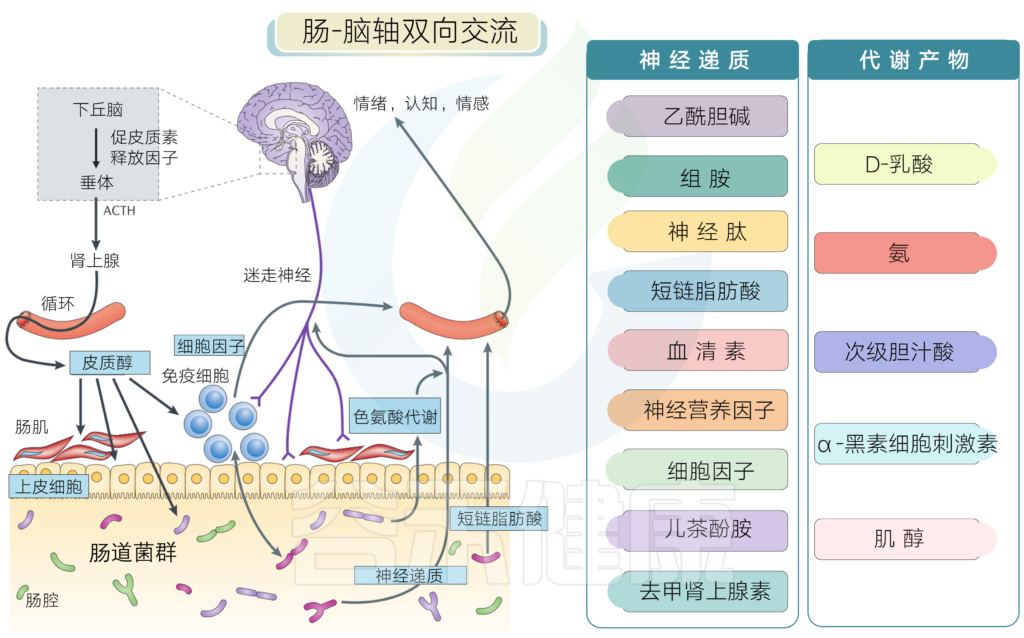
肠-脑轴
人体肠道微生物组以多种方式影响人类大脑健康:
(1)结构性细菌成分如脂多糖为先天免疫系统提供低级强直性刺激。由细菌生态失调,小肠细菌过度生长或肠渗透性增加引起的过度刺激可能产生全身和/或中枢神经系统炎症。
(2)细菌蛋白质可能与人类抗原交叉反应,刺激适应性免疫系统的功能失调反应。
(3)细菌酶可产生神经毒性代谢物,如D-乳酸和氨,甚至有益的代谢物如短链脂肪酸也可能发挥神经毒性。
(4)肠道微生物可以与人类产生相同的激素和神经递质。这些激素的细菌受体影响微生物的生长和毒力。
(5)肠道细菌直接刺激肠神经系统的传入神经元,通过迷走神经向大脑发送信号。通过这些不同的机制,肠道微生物塑造了睡眠和下丘脑 – 垂体 – 肾上腺轴的应激反应的结构。它们影响记忆,情绪和认知,并且在临床和治疗上与一系列疾病相关,包括酗酒,慢性疲劳综合症,纤维肌痛和不安腿综合征。
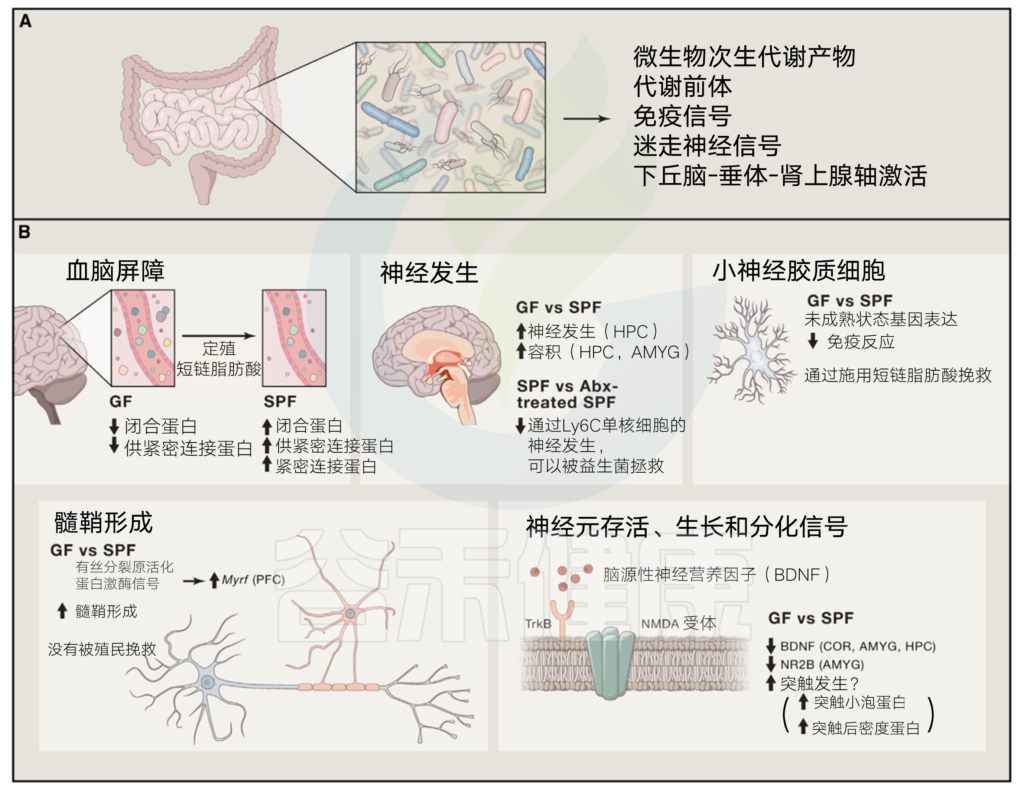
在健康和疾病的背景下,多个途径引导微生物组 – 肠 – 脑轴的向下和向上方向。
(A)向下,CNS通过影响营养的可用性饱食信号肽,影响肠功能和神经通路的内分泌物来控制肠道微生物组成。
皮质醇的HPA轴释放调节肠道运动和完整性。
免疫途径(细胞,细胞因子和sIgAs)可以开启,用以响应肠道功能的改变。内分泌和神经通路还可以调节来自特化肠上皮细胞的分泌,包括潘氏细胞,肠内分泌细胞和杯状细胞。它们的分泌产物影响菌群的存活和居住环境。
(B)向上,肠道微生物组通过神经(通过微生物组直接激活神经元),内分泌(例如5-羟色胺的肠内分泌细胞释放),代谢(神经活性分子的菌群合成)和免疫(CNS浸润免疫细胞和全身炎症)途径来控制CNS活动。
菌群在健康状态(神经发育)和疾病(一系列神经免疫和神经精神疾病)状态影响CNS。肠腔菌群,其产物由APC取样,附着上皮的SFB(肠内节丝状菌)介导外周免疫培养。
肠道微生物组成,菌群内的特定菌株,益生菌处理,菌群衍生产物和其他因素构成微生物组研究的范围。
由肠道微生物及其产物
直接或间接驱动的基本发育过程
细分来看,肠神经系统(ENS)就是肠道的大脑,同时还是联系着肠道外部(微生物群,代谢物和营养物)和内部(免疫细胞和基质细胞)微环境。
肠神经系统
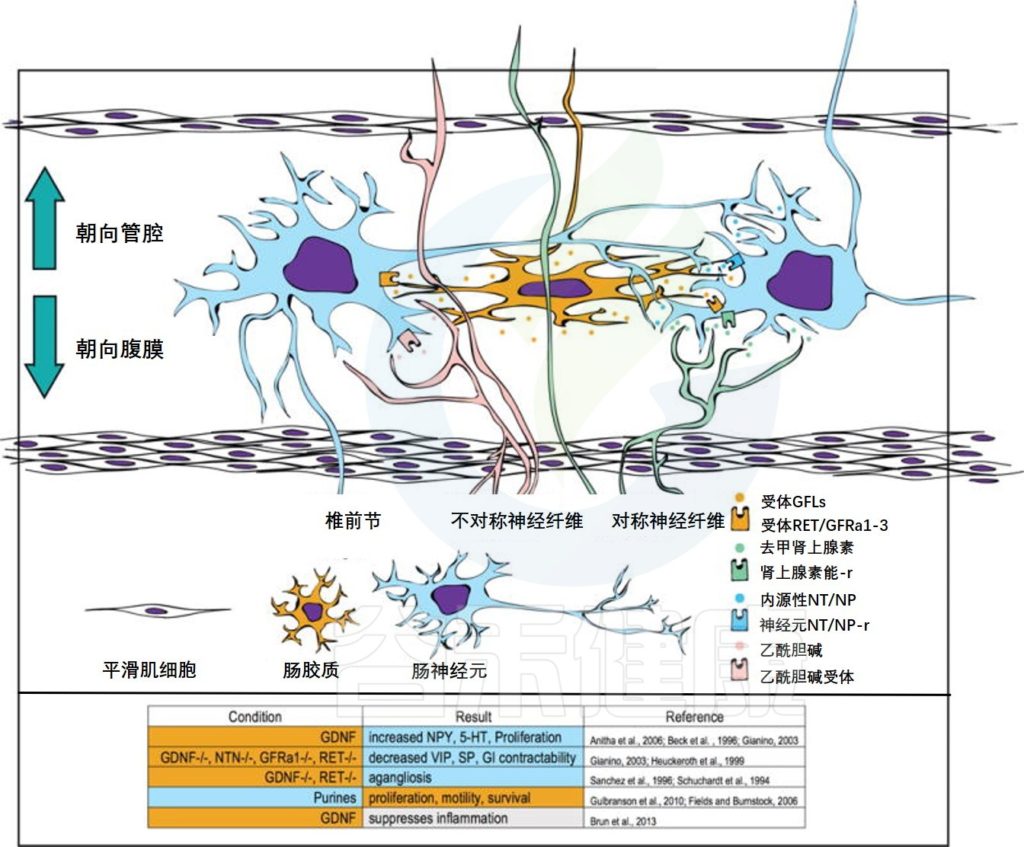
胃肠生理学的关键方面由肠神经系统(ENS)控制。ENS由神经元和神经胶质细胞组成。
肠神经元位于粘膜下或肌间神经丛中。两个丛都位于两个肌肉层之间。副交感神经纤维释放乙酰胆碱,交感神经释放去甲肾上腺素。这些外在神经纤维可以支配肠神经元,但也与平滑肌,固有层和上皮细胞相关。肠神经元可以相互支配或延伸到固有层,特定的肠道真菌(IFAN)可以突触到交感神经节。
肠神经胶质细胞产生和释放神经营养因子,与肠神经元结合,并延伸到整个粘膜。左列和中间列用颜色编码,分别代表产生特定条件的细胞和分子以及从这些特定条件产生的结果。
肠神经元与胶质细胞的连接
此外肠道菌群还通过释放不同物质和干预免疫系统最终影响血脑屏障和中枢神经系统(CNS)产生联系。
肠道用于收集营养和能量,防止有害的毒素和病原体,并清除废物,它是一个高度动态的环境,受到蠕动活动的周期性波动的影响。这些功能主要受两个肠神经系统(ENS)和驻留在肠道内的亿万共生细菌调节和控制。斑马鱼研究实验表明ENS调节肠道微生物群落成员身份以维持肠道健康。通过施用代表性抗炎细菌菌株或恢复ENS功能来预防ENS突变体中的炎症。
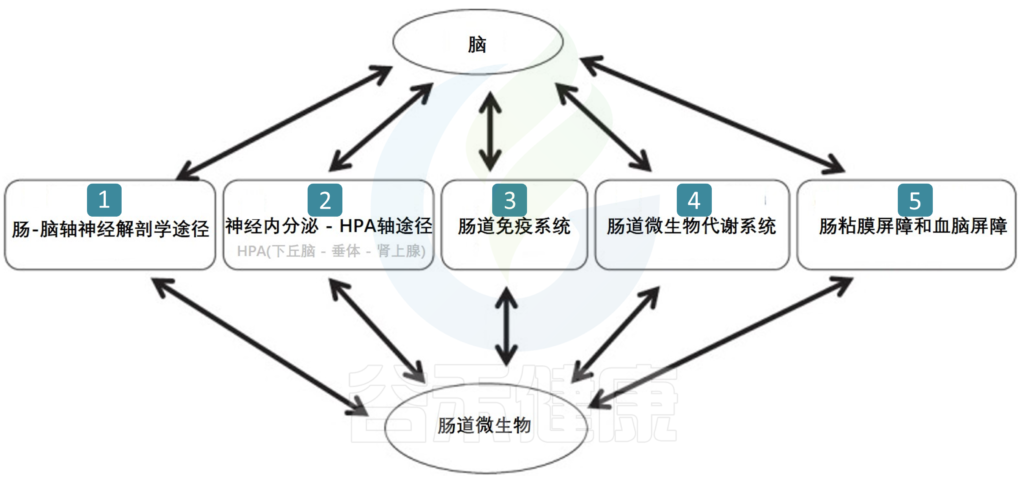
肠-脑之间通讯途径
肠道微生物群与大脑之间可能存在五种通信途径,包括肠道神经网络,神经内分泌 – HPA轴,肠道免疫系统,肠道菌群合成的一些神经递质和神经调节因子,以及包括肠粘膜屏障和血脑屏障在内的屏障。在这个通信网络中,大脑影响肠道运动,感知和分泌功能,来自肠道的内脏信号也影响大脑功能。
肠道微生物群与脑之间可能存在的五种通讯途径
神经递质和代谢产物
很多肠道菌群能代谢产生大量神经递质及其类似物,此外肠道菌群的部分代谢物质也会通过免疫系统影响神经系统。
连接菌群和大脑的
还离不开一个重要通道
——血脑屏障
血脑屏障越来越多的证据表明,菌群与中枢神经系统(CNS)相互作用,并可以调节其许多功能。这种相互作用的一种机制是在血脑屏障(BBB)的水平上。
细菌可以直接将因子释放到体循环中或可以转移到血液中。一旦进入血液,微生物组及其因子可以改变外周免疫细胞,促进与BBB的相互作用,并最终与神经血管单元的其他元素相互作用。
在菌群影响下从外围部位释放的细菌及其因子或细胞因子和其他免疫活性物质可穿过BBB,改变BBB完整性,改变BBB转运率,或诱导屏障细胞释放神经免疫物质。
由菌群代谢产物,例如短链脂肪酸,可穿过BBB以影响脑功能。通过这些和其他机制,微生物组-BBB相互作用可以影响疾病的进程。

Logsdon et al, Experimental Biology and Medicine, 2018
图中1层
菌群与全身免疫细胞相通,可影响血脑屏障(BBB)和CNS功能。肠腔不断暴露于来自外部环境的细菌。肠上皮屏障的破坏可允许肠道菌群不受调节的移动进入固有层。
图中2层
细菌可以渗透GALT(肠道相关的淋巴组织)和血腔,它们与各种免疫细胞相互作用,包括T细胞。
图中3层
某些细菌可以刺激效应型T细胞分化。调节性T细胞测量在GALT,血液和脑脊液中局部菌群的变化可促进T细胞脑浸润。
图中4层
循环细菌可以上调炎性细胞因子水平,影响BBB完整性并促进神经炎症。LPS(脂多糖)由细菌因子产生,并且可以作用于内皮TLR(Toll样受体)以促进神经炎症和CNS疾病。
图中5层
细菌代谢物可以上调紧密连接蛋白并改善BBB完整性。
图中6层
代谢物也可穿过BBB以影响神经胶质细胞和神经炎症。微生物组对周细胞的作用仍不清楚。
那么肠道菌群如果发生变化,
会带来哪些神经系统疾病和问题呢?
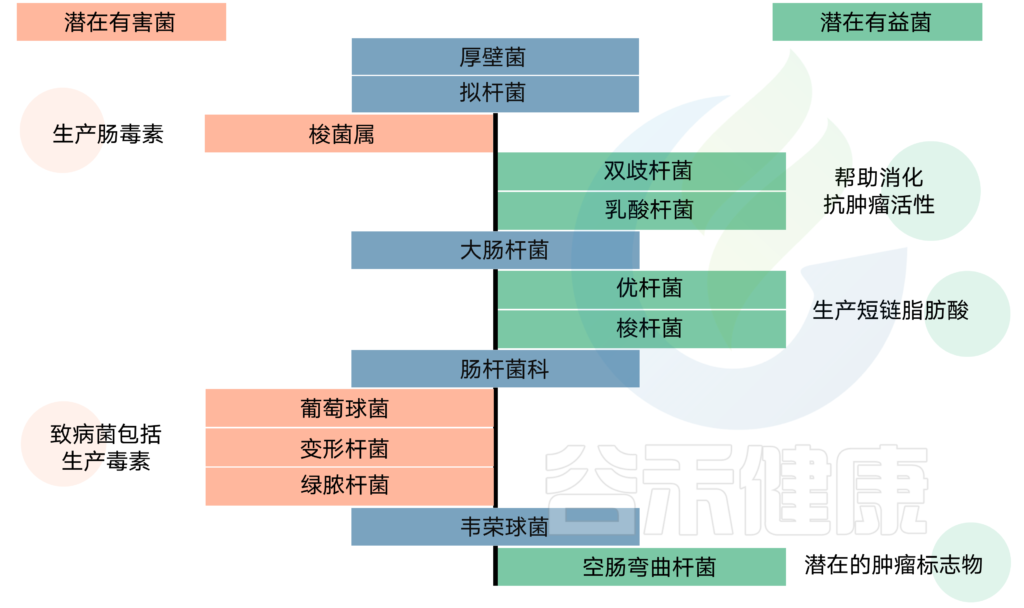
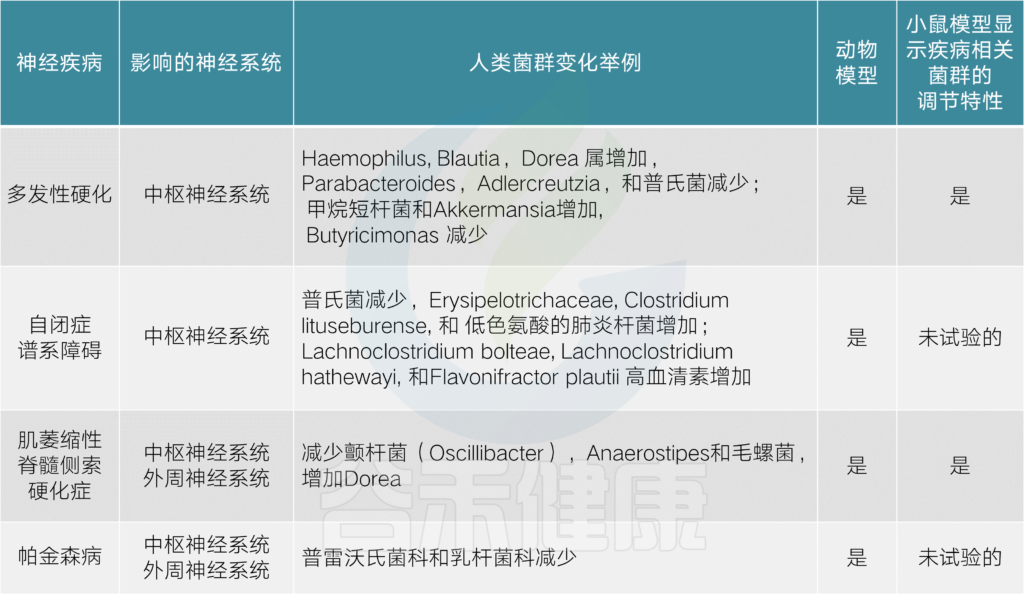
神经系统疾病的相关菌
目前已经发现一些与神经系统疾病(包括多发性硬化症,自闭症,帕金森病等)研究相关的菌。研究发现,这些疾病患者的某些菌群的数量明显发生变化,具体如下表:
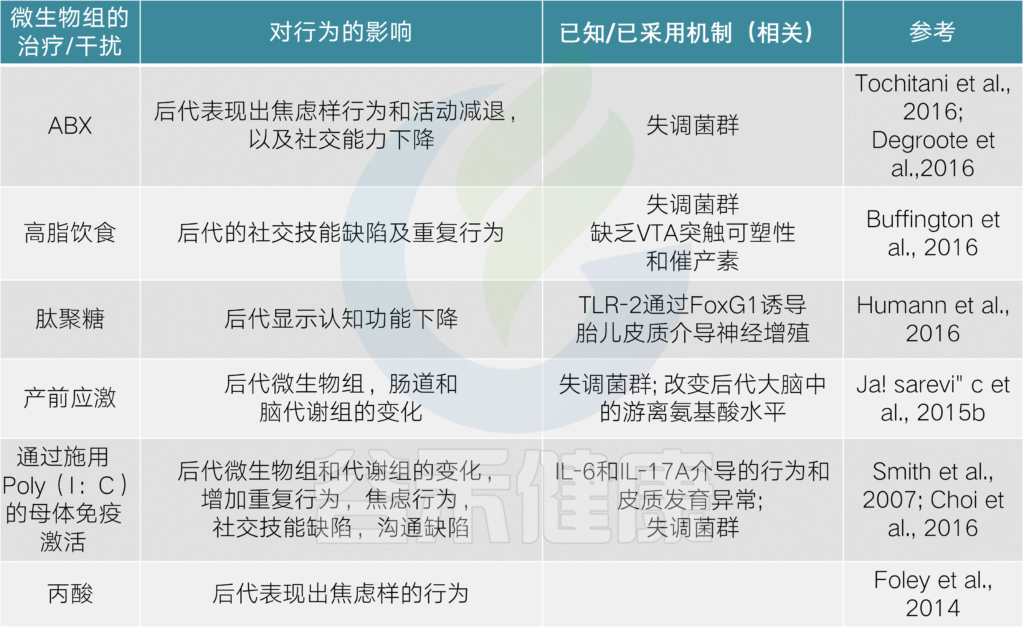
菌群干预或异常会导致的问题
目前有越来越多的证据表明,肠道微生物群在指导和促进大脑发育过程中发挥着重要作用,对健康具有长期的影响。
菌群和菌群产物的扰动
会影响小鼠模型和人类的行为结果
产前效果
产后效果




菌群如何影响中枢神经系统疾病

免疫介导的CNS疾病
多发性硬化症
多发性硬化症(MS)是由针对中枢神经组织的自身反应性免疫攻击介导的慢性CNS脱髓鞘疾病。这是通过研究患者和使用称为实验性自身免疫性脑脊髓炎(EAE)的MS动物模型来实现的。
如在一系列研究中观察到的,用单一细菌或细菌混合物口服治疗可调节EAE。益生菌动物双歧杆菌减少了大鼠EAE模型中症状的持续时间。
乳酸杆菌(包括LcS),单独施用或与其他双歧杆菌属菌株组合施用,倾向于通过相互调节促炎细胞因子和抗炎细胞因子反应来缓解小鼠EAE症状。
脆弱拟杆菌和乳酸片球菌(菌株R037)的益生菌治疗也显着降低了小鼠对EAE的易感性。
视神经脊髓炎
视神经脊髓炎(NMO),是一种CNS自身免疫疾病,其特征在于视神经和脊髓的免疫介导的脱髓鞘。
研究发现水通道蛋白血清阳性的NMO和NMO谱系疾病患者血清对胃肠道的抗原(最常见的饮食蛋白)抗体水平高于健康对照组,暗示NMO患者微生物群组成和免疫状态的改变。
格林 – 巴利综合征
格林 – 巴利综合征(GBS)是一种周围神经系统的自身免疫性疾病。
空肠弯曲杆菌在家禽中发现的肠道共生物种是由食物污染引起的人类肠炎的主要原因。研究表明弯曲杆菌肠炎患者的GBS风险高。
此外,弯曲杆菌与几种GBS的病理形式有关。不同的弯曲杆菌菌株以及宿主因子在GBS发育过程中形成自身反应性免疫反应中起重要作用。
因此,空肠弯曲杆菌代表了一种介导神经自身免疫的肠道相关病原体。
其他免疫介导的疾病
脑膜炎是CNS保护膜的炎症。病毒或细菌感染可能导致脑膜炎。据报道,成年肠道共生大肠杆菌 K1能够通过母体转移给新生儿引起脑膜炎。
慢性疲劳综合征(CFS),也称为肌痛性脑脊髓炎(ME),目前尚不清楚病因。据推测,共生细菌的转运升高可能是某些CFS患者疾病活动的原因。
非免疫介导的CNS疾病
自闭症
自闭症谱系障碍(ASD)是一系列发育性神经行为障碍,其特征是社交互动和沟通受损。新出现的数据表明肠道微生物组与ASD之间存在联系,可能是直接因果关系,也可能是间接的非典型摄食和营养模式的结果。
肠道微生物群的破坏可能促进产生神经毒素的细菌的过度定殖,从而导致自闭症症状。据报道,在自闭症儿童的粪便样本中存在的梭菌属下的物种数量更多,Bacteroidetes和Firmicutes门的不平衡也表现在自闭症儿童身上。
此外,其他肠道共生物的水平改变,包括双歧杆菌,乳酸杆菌,Sutterella,普氏菌和Ruminococcus属以及Alcaligenaceae家族,与自闭症相关。
肠道微生物组介导的新陈代谢也会影响自闭症。
抑郁症
抑郁症是由神经精神障碍或免疫失调导致的情绪障碍的主要形式。益生菌治疗已经显示出抑制动物抑郁症模型的功效。乳杆菌属下的物种特别表征为抗抑郁剂,包含鼠李糖乳杆菌和瑞士乳杆菌菌株的益生菌混合物通过使皮质酮水平正常化来改善母体分离诱导的抑郁。
类似地,鼠李糖乳杆菌菌株JB-1通过以迷走神经依赖性方式调节皮质酮和GABA受体来减少抑郁相关行为。
双歧杆菌的种类也是有效的抗抑郁药。
如大鼠强迫游泳试验(FST)和母体分离模型所示,Bifidobacterium infantis减轻了抑郁症。涉及的机制包括促炎细胞因子的减弱,色氨酸代谢的调节和CNS神经递质。
此外,含有高水平多不饱和脂肪酸(PUFA)n-3的饮食配方通过与瑞士乳杆菌和长双歧杆菌相似的机制减弱大鼠MI后抑郁症。
焦虑和压力
焦虑和压力是具有神经,内分泌和免疫学基础的情绪障碍的常见形式。暴露于诸如化学,生物或环境刺激的压力因素可引发压力和焦虑反应,其涉及激活HPA轴(下丘脑-垂体-肾上腺轴)。如前所述,焦虑和压力的共病已经在剧烈和轻微的肠功能障碍类型中被感知,强调了肠 – 脑信号如神经递质和免疫因子的作用。
与具有正常肠道微生物群的SPF小鼠相比,GF小鼠显示出增加的运动活性和减少的焦虑。这种行为表型与GF小鼠的CNS中更高水平的神经递质和降低的突触长期增强相关。
后来的研究证实了GF条件下焦虑样行为的减少,这可以通过其他神经化学变化来解释,例如神经递质受体减少和色氨酸代谢增加。因此推测肠道微生物组调节HPA轴的设定点。
有益的益生菌可以改善焦虑。乳杆菌属和双歧杆菌属的特定种类具有抗焦虑作用。用长双歧杆菌,婴儿双歧杆菌,瑞氏乳杆菌或鼠李糖乳杆菌的某些菌株进行益生菌处理单独或联合使用,在动物焦虑模型中归一化行为表型。
Lactobacillus farciminis还抑制了应激诱导的肠道泄漏并减弱了HPA轴应激反应。
由瑞士乳杆菌和长双歧杆菌组成的益生菌制剂显示出在大鼠中的抗焦虑样活性和对健康人受试者的有益心理作用。
痛
通过益生菌调节微生物组可以减轻由对刺激的外周神经反应和对CNS的信号转导引起的伤害性疼痛。在乳杆菌属的种中可见抗伤害感受作用。
罗伊氏乳杆菌还减轻正常大鼠CRD诱导的内脏疼痛。
L. paracasei使抗生素扰动小鼠的CRD内脏超敏反应正常化。
嗜酸乳杆菌通过诱导阿片样物质和大麻素受体在肠道疼痛中产生镇痛作用。
此外,两项研究支持IBS背景下特定婴儿双歧杆菌菌株的抗伤害感受作用。
其他神经精神症状
微生物组与其他神经精神疾病有关,其中经常发生基于免疫和非免疫的病因的混合物。GF动物表现出缺陷的记忆和认知能力。用小鼠菌群重新定殖GF小鼠可以增强或减少探索行为。海马脑源性神经营养因子水平与探索行为正相关。
益生菌能够改善感染引起的记忆功能障碍和糖尿病引起的认知缺陷。肠道微生物组的膳食改变也调节了小鼠的认知和学习行为。
总而言之,肠道菌群的研究对于CNS疾病相关的诊断,预后和治疗都有很大的意义。
主要参考文献
Sharon G et al. (2017) The Central NervousSystem and the Gut Microbiome. Cell. 167: 915–932.
Cong et al. (2016) Early life experience and gut microbiome: the Brain-Gut-Microbiota signaling system. Adv Neonatal Care. 15(5): 314–323.
Rolig et al. (2017). The enteric nervous system promotes intestinal health by constraining microbiota composition. PLoS Biol. 15(2): e2000689.
Logsdon AF et al. (2018) Gut reactions: How the blood–brain barrier connects the microbiome and the brain. Exp Biol Med. 243(2): 159–165.
Wang HX and Wang YP (2016). Gut Microbiota-brain Axis Chin Med J (Engl). 129(19): 2373–2380.
Vighi G et al. (2008) Allergy and the gastrointestinal system. Clin Exp Immunol.153:3–6
Hooper LV et al. (2012) Interactions between the microbiota and the immune system. Science. 336:1268–1273
Mowat AM. (2003) Anatomical basis of tolerance and immunity to intestinal antigens. Nat Rev Immunol. 3:331–341.
Bryan B. Yoo et al. (2018) The Enteric Network: Interactions between the Immune and Nervous Systems of the Gut. Immunity. 46(6): 910–926.
Johansson MEV et al. (2016) Immunological aspects of intestinal mucus and mucins. Nat Rev Immunol. 16:639–649.
Gerbe F et al. (2016) Intestinal epithelial tuft cells initiate type 2 mucosal immunity to helminth parasites. Nature. 529:226–230.
Howitt MR et al. (2016)Tuft cells, taste-chemosensory cells, orchestrate parasite type 2 immunity in the gut. Science. 351:1329–1333.
Shivani Ghaisas et al. (2016) Gut microbiome in health and disease: linking the microbiome-gut-brain axis and environmental factors in the pathogenesis of systemic and neurodegenerative diseases. Pharmacol Ther. 158: 52–62.
Yatsunenko T et al. (2012) Human gut microbiome viewed across age and geography. Nature. 486:222–227.
Dogra S et al. (2015) Dynamics of infant gut microbiota are influenced by delivery mode and gestational duration and are associated with subsequent adiposity. MBio. 6
Villaran RF et al. (2010) Ulcerative colitis exacerbates lipopolysaccharide-induced damage to the nigral dopaminergic system: potential risk factor in Parkinson’s disease. J Neurochem. 114:1687–1700.
Wang Y, Kasper LH. (2014) The role of microbiome in central nervous system disorders. Brain Behav Immun. 38: 1–12.
Mayer EA et al. (2017) The gut and its microbiome as related to central nervous system functioning and psychological wellbeing: Introduction to the Special Issue of Psychosomatic Medicine. Psychosom Med. 79(8): 844–846.
Cryan JF et al (2012). Mind-altering microorganisms: the impact of the gut microbiota on brain and behaviour. Nature reviews. Neuroscience. 13:701–712.
Belkaid Y, Naik S. (2013) Compartmentalized and systemic control of tissue immunity by commensals. Nature immunology. 14:646–653.
Knuesel I et al. (2014) Maternal immune activation and abnormalbrain development across CNS disorders. Nat Rev Neurol. 10:643–660.
Möhle L et al. (2016) Ly6Chi Monocytes Provide a Link betweenAntibiotic-Induced Changes in Gut Microbiota and Adult HippocampalNeurogenesis. Cell Rep. 15:1945–1956.
Matcovitch-Natan et al. (2016) Microglia development follows astepwise program to regulate brain homeostasis. Science
Desbonnet L. et al. (2015) Gut microbiota depletion from earlyadolescence in mice: Implications for brain and behaviour. Brain Behav Immun. 48:165–173.