-

 国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子
国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子

 二级病原微生物安全实验室
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业

谷禾健康
导 语:关于抗体,免疫和微生物
肠道微生物群的组成因个体而异,肠道微生物群中的特定细菌类群可能是某些肠内外疾病的独特有效驱动因素。对人类而言,精准且优先识别影响疾病易感性和严重性的细菌仍然是一项重大挑战。
很多时候我们想知道:
微生物群是如何选择与我们定居下来的?
它们如何表现出成分多样性和功能稳定性?
它们如何适应我们生活方式的变化?
为什么有的人肠道菌群容易失调,有益菌不易定植?
想要通过一篇文章来破译这个话题比较困难,但是以上问题很关键一个原因是与我们身体的免疫抗体和免疫耐受性有关。
接下来的主题我们将持续关注和讨论免疫,抗体以及微生物之间的对话及其在人体健康或疾病中的作用。
人类复杂的饮食结构让胃肠道环境对免疫系统提出了巨大的挑战。经典的耐受机制受到来自饮食和共生微生物群的大部分无害外来抗原以及偶尔有害病原体的复杂和动态混合物的挑战。
免疫系统既要有效阻挡和消灭病原入侵,又要识别和保留肠道中的良性细菌,使得肠道达到动态平衡。这两年国际权威期刊《Cell》 《Science》《Nature》等主刊和子刊研究成果不断揭开了控制肠道菌群这一机制的“重要开关”——免疫球蛋白(IgA)。一致认为IgA是帮助肠道细菌“定居”肠道和保持物种多样性的关键,也是抵御微生物群和病原体的第一道防线。
免疫球蛋白 (Ig),也称为抗体,通过识别、结合和消除特定的细菌、真菌和病毒抗原,参与体内外来颗粒的清除和中和,人体内的五种免疫球蛋白 Ig :
IgA、IgD 、IgE、IgG 和IgM;
其中IgA最初发现于50年前,是机体黏膜分泌最多的抗体之一,负责防御病原体入侵。
人类每天生成3-5g IgA抗体,约占人体抗体总表达量的75%。一旦缺乏免疫球蛋白A,炎症微生物将繁殖,共生微生物将无法在肠道中长久寄居。IgA抗体能以多种方式来影响细菌的适应度,比如细菌的运动性被限制,或者其阻碍细菌所需的营养尤其糖类的摄入,或改变菌群基因的转录表达等。
IgA 在控制人类肠道微生物群组成方面起着关键且非冗余的作用。IgA 缺乏或过量均与疾病的发生发展有重要关系。
低水平会增加过敏、感染和自身免疫性疾病的风险,也意味着肠道屏障和/或肠道微生物群失衡。IgA 缺陷的常见自身免疫病包括类风湿关节炎、狼疮、乳糜泻或炎性肠病。
而高 IgA 可能是慢性感染和低度炎症以及代谢综合征的标志。目前比较关注的是IgA肾病,尽管 IgA肾病是一种影响肾脏的疾病,但它的起源也与肠道微生物群 – 肾轴密切相关。分泌型 IgA (SIgA) 与肠道中的这些微生物结合,从而调节它们的定植、增殖和潜在致病性。
一般遗传性的IgA缺乏症无法治愈。免疫疗法不能治疗它。但可以采取措施降低患病或感染的风险,其中包括生病或感染时服用抗生素。此外,预防也很重要,通过经常洗手和远离人群来限制细菌和疾病的传播。尤其在感冒和流感季节,甚至可以提前接种疫苗。
IgA 的测试一般用血液或脑脊液,我们通过长期的粪便检测和研发,构建了通过粪便检测来间接评估IgA 水平,对于判别感染,炎症,菌群失调等有一定的辅助作用。
本文将基于最新研究进展,全面讨论 IgA 尤其分泌型IgA (SIgA) 的来源及其结构形式,缺乏或过量的原因和症状,以及IgA如何决定微生物群的反应性,以维持宿主和细菌都受益的多样化生态位。IgA 与微生物群在生理、感染和疾病方面的密切相互作用以及生理条件下潜在的 IgA 和微生物群依赖性机制。另外基于目前许多病原体已经进化出阻止IgA 介导的清除的机制,探讨针对 IgA 和/或微生物群的疗法的发展。
本文预览:
01- 免疫球蛋白A(IgA)
02- IgA 功能
03- IgA 对微生物群的反应
04- SIgA:宿主-微生物群动态互作中的盟友
05- SIgA 介导的肠道病原体清除和稳态特性
06- IgA-微生物轴的缺陷导致病理疾病
07- 低IgA和选择性IgA缺乏:原因和症状及改善措施
08- 高IgA水平的原因和改善
09- IgA 的治疗潜力
-正文-
免疫球蛋白 A (IgA) 是一种由浆细胞(效应B细胞)产生的抗体,可保护身体免受感染。
注解:浆细胞(plasma cell),又称效应B细胞,常见于消化管和呼吸道固有膜的结缔组织内。浆细胞来源于B细胞。
浆细胞具有合成、贮存抗体即免疫球蛋白的功能,参与体液免疫反应。
正常机体浆细胞存在在脾、淋巴结的相应组织结构处,浆细胞生成抗体并将它们释放到周围的组织液中,最后汇集成静脉出淋巴结。所以抗体先进入组织液,而不是直接分布在血浆中。
抗体是指机体由于抗原的刺激而产生的具有保护作用的蛋白质,可结合并中和细菌和病毒等病原体。
在人体中,每天产生的 IgA 比所有其他抗体的总和还要多。
▸ 分泌型IgA
IgA 由浆细胞产生后被转运到粘膜细胞分泌的液体中。这种 IgA 称为分泌型 IgA(SIgA)。分泌型是IgA 是的优势抗体,主要在以下粘膜组织中产生:
IgA 也是血液中第二丰度的抗体(仅次于 IgG)。
循环 IgA 是单体形式;
分泌型 IgA 是二聚体(以两个连接的 IgA 单体的形式)。
效应位点包括B细胞和浆细胞,主要产生(>90%)IgA。这种保护性体液反应是全身最有效的免疫球蛋白途径,每天产生大量的免疫球蛋白A。在固有层有淋巴循环通过组织连接到肠系膜淋巴结,是免疫反应开始的部位。
每个抗体由四种较小的蛋白质组成。其中两种蛋白质称为轻链,另外两种称为重链。抗体通常根据它们所含的轻链和重链蛋白质类型进行分类。
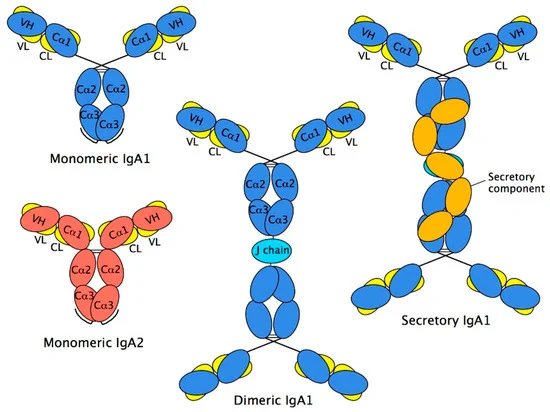
IgA 的分子大小为 160 kDa,血液中的 IgA 以单体形式出现,而身体分泌物中的 IgA 以二聚体或多聚体形式出现。
在分泌物中,除了 κ 或 λ L 链和 IgA 重链 α 外,IgA 还含有另外两条多肽链——分泌成分(SC)和 J 链(连接链)。
分泌链有助于外分泌 IgA 的胞吞作用并稳定 IgA 以防止蛋白水解降解。
人类表达两种IgA 亚型(小鼠只表达一种),称为 IgA1 和 IgA2。基因序列分析已证实 IgA 在所有类别的哺乳动物和鸟类中都存在。
存在显着的物种差异
大多数哺乳动物具有单一的 IgA 同种型。与人类相似的 IgA1 和 IgA2 亚类仅存在于相关的灵长类动物中,包括黑猩猩、大猩猩和长臂猿。
人类IgA1和IgA2的结构差异
就结构差异而言,人 IgA1 显示出一个延伸的铰链区,该铰链区由 IgA2 中不存在的两个 8 个氨基酸长的重复序列(每个轻链一个)组成。人类IgA1和IgA2的区别在于IgA1分子的铰链区域有13个不同氨基酸,导致其对细菌蛋白酶的敏感性增加,这个扩展的铰链区域赋予IgA1一个T型结构,有利于远程抗原识别。
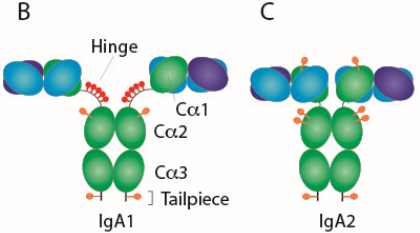

IgA1和IgA2的CH1和CH2结构域中都有高度的N-糖基化,糖约占其含量的6%。IgA1在铰链区含有由N-乙酰半乳糖胺、半乳糖和唾液酸组成的额外O联聚糖。
在黏膜部位,IgA常以二聚体IgA(dIgA)形式存在。
J链的存在是IgA在上皮细胞间的转运及其在黏膜表面分泌的先决条件。如下图:
聚合物Ig受体(pIgR)在上皮细胞的基底外侧极上表达,与J链结合,将IgA释放到管腔中,如SIgA。在此过程中,称为分泌成分(SC)的pIgR外结构域仍然以共价方式附着在IgA上。重N-糖基化SC稳定IgA并防止被蛋白酶快速水解,从而保护IgA在消化道中不被降解。
在粘膜表面发现的主要分子形式,称为分泌型 IgA (SIgA),是二聚体,尽管也存在一些较高分子量的物质,包括三聚体和四聚体。这里两个子类的相对比例更接近;平均分布约为 40% IgA1 和 60% IgA2,但这取决于取样的特定粘膜部位。
值得注意的是,IgA1铰链区的糖组成不均匀。有报道称异常糖基化参与了IgA肾病的发病。
肠道中 IgA2 转换的一般是由肠上皮细胞介导的,肠上皮细胞在通过 Toll 样受体感知共生细菌后分泌 APRIL。因此,局部肠道中不依赖 T 细胞的 IgA 产生对于确保宿主和微生物群之间适当的粘膜免疫至关重要。
注释:Toll样受体(Toll-likereceptors,TLR)是参与非特异性免疫(天然免疫)的一类重要蛋白质分子,也是连接非特异性免疫和特异性免疫的桥梁。
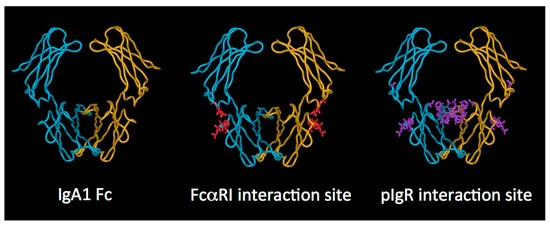
IgA与各种宿主受体相互作用,包括pIgR、转铁蛋白受体(CD71)、asialoglycoprotein receptor、dectin-1、Fcα/μR、DC-SIGN和FcαRI(CD89)。这些相互作用是通过抗体的FC段聚糖,或辅助分子(如J链或SC)结合来介导的。
IgA可以和一些细菌蛋白结合,如A群链球菌的M蛋白,B群链球菌的β抗原等。

doi.org/10.3390/antib8040057
小鼠和人的 IgA差异:
小鼠只有一个 IgA 亚类,并且可能使用替代受体,例如 Fcα/μR、转铁蛋白受体 (CD71) 和 pIgR,因为它们缺乏 FcαRI(别名CD89),这是主要的人体中的 IgA 受体。其他显着差异包括:
小鼠而非人类在脾脏中表现出 IgA 浆细胞的 B-1 B 细胞前体;
小鼠的体细胞超突变率是人类的三倍;
与人类相比,小鼠的产生中心或组织边界不明确
由于肝脏 pIgR 将血清 IgA 转运到胆汁中,然后进入肠腔,小鼠的全身 IgA 水平较低。
以下是 IgA 的近似值,具体取决于年龄:
根据大多数检测数据的说法,60-400 mg/dL 是成人的正常范围。
正常 IgA 水平会略有不同,具体取决于用于测试的实验室或机构。IgA 随着儿童的年龄和体重增长而逐渐增加,直到在成人中达到稳定水平。男性的血 IgA 水平高于女性。
总结:IgA 是免疫防御和耐受的重要参与者
低水平IgA 通常意味着免疫系统被削弱。低 IgA 水平和 IgA 缺乏是不一样的。大多数实验室认为任何低于 60 mg/dL 的 IgA 值都较低,但只有低于 7 mg/dL 的值才意味着缺乏。症状取决于根本原因。
慢性压力、睡眠不佳、疲惫和某些遗传疾病都可能导致 IgA 水平低或缺乏。虽然需要更多的研究,但一些研究也将低 IgA 与感染、过敏、自身免疫和肠道疾病联系起来。
高 IgA 通常表示慢性感染或炎症,尽管多种疾病可以提高其水平。在成人中,大多数实验室认为高于 300 mg/dL 的值很高。高水平不会引起任何症状。症状取决于根本原因和健康状况,应由医生进行评估。
如果出现大量感染,尤其是鼻窦、肺、胃或肠道感染,医生可能会要求进行免疫球蛋白A检测,尤其有下列情况:
一般还需要同时监测的其它指标:
检测样本有血液,脑脊液,还有肠道菌群也可以辅助。
要讲清楚IgA 的生产分泌,需要先了解下肠道相关淋巴组织,肠道实际上是最大的外周免疫器官。
肠道相关淋巴组织
人体肠道拥有人体内最大的淋巴组织,肠相关淋巴组织遍布整个肠道,覆盖面积约为260–300平方米,是粘膜相关淋巴组织的一个组成部分,它在免疫系统中起作用。
肠道相关淋巴组织 (GALT) 包括一系列的多泡结构,比如扁桃体、阑尾、结肠和盲肠斑片,Peyer’s斑片(派尔斑,PPs)、盲肠斑、以及一些称为孤立淋巴滤泡 (ILFs) 的较小的单个滤泡结构,这些组织遍布胃肠道,它们与肠系膜淋巴结(mesenteric lymph nodes, mLN)一起帮助保护宿主免受感染。
注释:
肠系膜淋巴结(mLN)指的是分布在肠系膜上的淋巴结,而且需要强调的是当患者出现肠系膜淋巴结炎的时候,患者会出现腹部疼痛、腹胀、发热等临床症状。
潘氏斑(或聚集性淋巴结节)是有组织的淋巴滤泡,以17世纪瑞士解剖学家约翰康拉德派尔的名字命名。
它们是肠道相关淋巴组织的重要组成部分,通常见于人类小肠的最下部,主要见于远端空肠和回肠,也可见于十二指肠。研究发现潘氏斑或能通过增加不同B细胞亚群的数量和效应功能来感知、增强并传递益生菌信号,从而就会导致IgA的产生、肠道菌群的改变以及保护机体抵御炎症的发生。
肠道相关淋巴组织是 IgA 诱导的主要部位
这些包括派尔斑 (PP)、肠系膜淋巴结(mLN)、孤立淋巴滤泡(ILF) 和盲肠斑。初始 B 细胞在 GALT 中分化和成熟;在该组织中,PPs 被认为是 IgA 的主要诱导位点。
在 PPs 中,微生物抗原通过滤泡相关上皮 (FAE) 上的 M 细胞转运到位于 FAE 下方的上皮下圆顶 (SED) 区域的树突状细胞 (DC),以启动 IgA 的产生。
肠道中的 IgA 是通过与许多免疫细胞和上皮细胞合作产生的
此外,在小肠固有层中发现了最大的 IgA + 浆细胞(PC )种群,而结肠固有层仅含有少量种群。
在唾液腺、肺以及哺乳期的肠外组织如肝脏、骨髓、乳腺中可检测到额外的少量 IgA 。
人类 IgA亚型显示出不同的表达模式,IgA1 在血清中占主导地位,而 IgA2 在远端肠道中占主导地位。
小肠中的 Peyer 斑和固有层分别是 T 细胞依赖性和独立稳态 IgA 产生的中心枢纽。然而,应该注意的是,据报道,结肠表现出不依赖于 T 细胞的 IgA 类转换重组,但仅在有组织的淋巴滤泡存在时。
无论如何,树突状细胞是主要的抗原呈递细胞,它们可以识别 T 细胞依赖性抗原和微生物分子模式,从而分别在派尔氏斑或固有层中刺激肠道 IgA 的产生。
此外,提到的肿瘤坏死因子(TNF)配体(CD40L、BAFF 和 APRIL)和细胞因子(TGFβ),其他研究也显示了其他环境因素,如视黄酸(RA)和其他白细胞介素(例如,IL-5、IL-6、IL-21 ),是 IgA 合成协同所需的。这些表明肠道中的 IgA 是通过与许多免疫细胞和上皮细胞合作产生的。
对肠道免疫系统至关重要的小分子是通过饮食和微生物提供的
例如,视黄酸参与表达醛脱氢酶的 DC 的功能,因此对于 IgA 的产生以及调节性 T 细胞 (Treg) 极化至关重要。这种分子被认为来源于饮食中的维生素 A,维生素 A 缺乏确实会降低小肠 IgA 的产生 。
膳食抗原也可能影响 PPs 和 ILFs 中 IgA 的产生,从而特别影响小肠 IgA 的产生。
另一方面,膳食纤维被肠道微生物群发酵,产生短链脂肪酸(SCFAs),对肠道免疫发挥各种作用。
SCFA 为B 细胞提供燃料以促进 IgA 的产生并通过 SCFA 受体 (GPR43) 以及组蛋白脱乙酰酶抑制活性激活 DC,以支持 IgA 类别转换。脂多糖和鞭毛蛋白等微生物成分可刺激 DC 和 CD4 T 细胞中的 toll 样受体,从而促进 IgA 的产生。
其他研究发现某些细胞因子,如 IL-21,在存在微生物抗原的情况下会增加 IgA 的产生。由于 T 细胞非依赖性 IgA 是响应内源性微生物群而产生的,因此 IgA 被确定为为相对非特异性和多反应性抗体。
虽然肠道是 IgA 合成的主要位点,但 IgA +PC (浆细胞)也存在于许多肠外组织中,包括骨髓 (BM) 和泌乳乳腺 (LMG)。而且 BM IgA + PC 可能是大多数血清 IgA 抗体的来源,这些抗体的特异性已通过用血清染色粪便细菌和 IgA-seq 来分析确定。
这些实验表明,血清 IgA 抗体通常会针对与肠道 IgA 靶向的微生物群相似的亚群起反应。
值得注意的是,血清 IgA 显着地与变形杆菌菌群结合,这些微生物在肠道中的相对丰度可能会影响 BM IgA +的大小PC 响应。
对从 BM IgAs 克隆的 mAb 的分析表明,这些单克隆抗体包括许多与变形杆菌菌群结合存在多反应特异性。
然而,与肠道 IgA 库相比,几乎所有 BM IgA 都是通过T细胞依赖反应产生的,这可能是因为诱导整合素等分子需要 T 细胞衍生的信号α4β1 和趋化因子受体(如 CXCR4)促进迁移和归巢至骨髓。
虽然在未怀孕女性的乳腺中发现了很少的 IgA + PC,但在怀孕和产后哺乳期间这些细胞大量积累,在哺乳停止后逐渐减弱。这些细胞可能会分泌在母乳中发现的高滴度 IgAs,这也是提倡母乳喂养的重要原因。
许多研究已经描述了 IgA 在粘膜防御肠道病原菌的背景下的功能,例如通过免疫排斥清除它们、限制它们的运动性和通过束缚来抑制它们的生长。
此外,IgA一直被描述为对抗黏膜表面病原体的第一个障碍,可以凝集细菌、干扰细菌运动、中和细菌毒素、抑制细菌对上皮的黏附,从而防止病原体传播到循环系统。
IgA对多种胃肠病原体如鼠伤寒沙门菌、志贺菌、艰难梭菌和一些病毒(仙台病毒、人类免疫缺陷病毒、流感病毒、SARS-CoV-2等)发挥中和作用。
病原体对 IgA 功能的规避
在系统发育和多样性分析的基础上,IgA-FcαRI 相互作用被认为是病原体和人类之间进化军备竞赛的焦点。IgA 上相互作用的中心位点,为了结合 FcαRI 而被保留下来,已经受到某些病原体产生的 IgA 结合蛋白进化的压力。
这些 IgA 结合蛋白已经进化为与同一位点相互作用,从而颠覆 IgA 反应,并推动了一个迭代的选择过程,在这个过程中,哺乳动物和病原体蛋白都在继续进化,试图“智取”另一个。事实上,靶向 FcαRI 相互作用位点只是病原微生物用来规避 IgA 保护能力的策略之一。不同 IgA 靶向机制的存在,以及这些机制似乎在不同生物体中独立出现的事实,表明它们通过允许更容易的粘膜定植和传播为微生物提供显着益处。
IgA 的保护能力也可能因许多重要病原细菌产生的蛋白水解酶的作用而受到损害。这些蛋白酶都在 IgA 的铰链区切割。除了少数例外,它们特异性地作用于 IgA1 的延伸铰链区,并且不切割 IgA2。
此类 IgA1 蛋白酶由引起口腔感染的细菌(如血链球菌、缓症链球菌和口腔链球菌)和生殖道感染的细菌(如淋病奈瑟菌)产生,这表明它们为细菌提供了优势来获得立足于黏膜表面。有人提出 IgA1 蛋白酶可用作治疗选择,以降解 IgA 肾病中异常糖基化 IgA1 的致病性免疫复合物,这是肾脏疾病的常见原因。
早期免疫——在每一个成人身上,都有曾经的孩童时候的印记;在每一个孩子身上,都有将来的成人的缩影,免疫也一样。
从逻辑上讲,IgA结合作为致病条件下潜在促炎性分类群的标记物的可能性应该是:IgA在建立肠道微生物与其宿主的早期生命平衡中很重要。
乳汁成分对微生物群的发展有着深远的影响
例如乳铁蛋白和乳低聚糖等其他乳蛋白会影响早期肠道中微生物群的营养和竞争。
缺乏针对特定细菌表位的IgA已被证明增加了肠内先天免疫系统激活的转录证据。母乳中分泌的抗体保护幼年哺乳动物免受自身粘膜免疫系统的过早刺激,并对其获得的微生物群的组成产生长期影响。
聚合免疫球蛋白保护早期粘膜,防止菌群转移
在某些情况下,机制可以从某些分类群的代谢能力推断,例如代谢特定糖类,或抗体中和病原体的潜力。然而,实验表明,通过这种转运机制主动分泌的聚合免疫球蛋白(IgA和IgM)可以形成幼犬获得的长期肠道微生物群。
也就是说,主动分泌的聚合免疫球蛋白的保护作用,不仅能够在其自身固有和适应性免疫机制发展之前保护脆弱的早期粘膜,包括防止肠道微生物穿透上皮防御系统转移到肠系膜淋巴结,同时也形成了长期组成的微生物群,这些微生物群落先后在早期肠道中繁殖。
IgA抗体是人类免疫系统中最常见的抗体,由黏膜中的专门细胞分泌,在人类免疫球蛋白中占三分之二。
机体产生的大多数IgA抗体都是针对肠道菌群中的良性细菌的。如果没有这种免疫保护,这些微生物也可能对健康产生不利影响,并引起肠内外疾病。
IgA抗体能在多个层面上特异性地限制良性细菌的适应度,这或许会促进免疫系统能够更好地微调肠道中的微生物平衡,如今研究人员成功阐明,免疫系统能识别并特异性地限制这些细菌的生长。
一个有用的起点是哺乳动物肠道表面IgA功能的多样性。这些措施包括中和毒素和病毒,阻止过度活的细菌粘附或易位,清除不需要的大分子结构在上皮表面,并定向采集管腔抗原。
研究人员进行了三年的研究,他们成功地以高度的精确度追踪了无菌小鼠肠道中的体外和体内效应。发现该抗体在几个方面影响细菌的适应性。例如,细菌的迁移率受到限制,或者它们阻碍了细菌代谢的糖结构单元的摄取。该效应取决于被特异性识别的表面成分。
研究支持抗体在肠道中的作用是介导耐受性的观点。肠道内的耐受性可被视为基于对其微生物群成员的免疫识别的静态稳态。
产生过量 IgA 的细菌可能比产生低至中等量 IgA 的细菌面临更大的竞争劣势。这需要长期的‘居民’不断地调节免疫显性决定因素,这可能为在肠道生态系统中观察到的异常水平的菌群多样性提供了一种解释。
研究表明适应性免疫系统是这些表面结构多样化的驱动力一致,其有益结果是促进肠道共生体和宿主之间的非炎症关系。
对细菌的先天免疫反应,包括 NO 的产生,在无脊椎动物和脊椎动物中是高度保守的。
适应性免疫系统在脊椎动物中的一个关键进化作用可能是适应更复杂的微生物群落,即使存在病原体定植和/或自身免疫易感性的额外风险。
适应性免疫系统的主要作用是通过选择性地对刺激先天系统的细菌产生免疫反应来维持与肠道微生物群的“联系”。这种安排允许宿主检测新的细菌系统发育型,并忽略它以前遇到过的那些(记忆)的存在,结果才能更大的多样性,而不会牺牲先天免疫系统在维持粘膜屏障方面的基本保护作用。在这个过程中几种抗体,尤其IgA、IgM以及IgG发挥关键性配合作用。
炎症性肠病 (IBD) 至少在一定程度上可以被视为未能对常驻肠道微生物群产生有效的适应性免疫反应:这种反应通常会阻止微生物抗原呈递给 T特定于社区成员的细胞。与该模型一致,TCRα-/-小鼠对肠道微生物的抗体反应在发生结肠炎时从多克隆变为寡克隆。此外,将 Ig 被动转移到该模型中可以改善疾病。
因此,不仅仅是寻找富含 IBD 相关抗体库的微生物靶标,确定库中存在“漏洞”的位置可能会提供信息,并可以提出基于主动或被动免疫的新治疗策略。
天然存在的全身性 IgA 主要是调节免疫的,与微生物几乎没有直接接触,部分原因是血液的无菌环境。先前的研究已经证明血清 IgA 能够通过抑制补体系统有效消除抗原而不会提醒宿主免疫系统。这使得血清 IgA 在清除体内抗原物质时充当“沉默的恐慌按钮”。
然而,当单体、非抗原携带 IgA1 与髓系 IgA Fc相互作用时受体、FcαRI,然后是含有 Src 同源区 2 结构域的磷酸酶 1 (SHP-1) 以 ERK 依赖性方式募集到名为抑制性免疫受体酪氨酸激活基序 (ITAM) 的对接位点 。
注释:FcR是一类能够和免疫球蛋白(Ig)重链羧基末端的功能区Fc段特异结合的细胞表面蛋白,在抗体依赖性免疫应答过程中至关重要。不同类型的细胞可以表达不同类型的FcR,不同结构类型的Ig也和不同类型的FcR结合,从而诱导后续的不同类型的免疫反应。按FcR所结合的Ig种类不同可将其分为五类,即IgG(FcγR)、IgE(FcεR)、IgA(FcαR)、IgM(FcμR)和IgD(FcδR)。
当 FcαRI 和 ITAM 与周围的脂筏共定位时,它们的复合物形成称为 ITAMi 的抑制体簇,由此产生的下游磷酸化受损会阻断免疫反应。
脂筏:膜脂双层内含有特殊脂质及蛋白质的微区。
与循环中 IgA 介导的免疫耐受相比,来自固有层的二聚体 IgA2 可以作为 SIgA 转移到肠腔中,并将其自身锚定在外粘膜表面上,与肠道细菌强有力地相互作用,以获得适当的免疫微生物群稳定性。这共同确立了血清和粘膜 IgA 在稳态条件下对免疫功能的重要参与。
血清和分泌型 IgA 的结构和功能
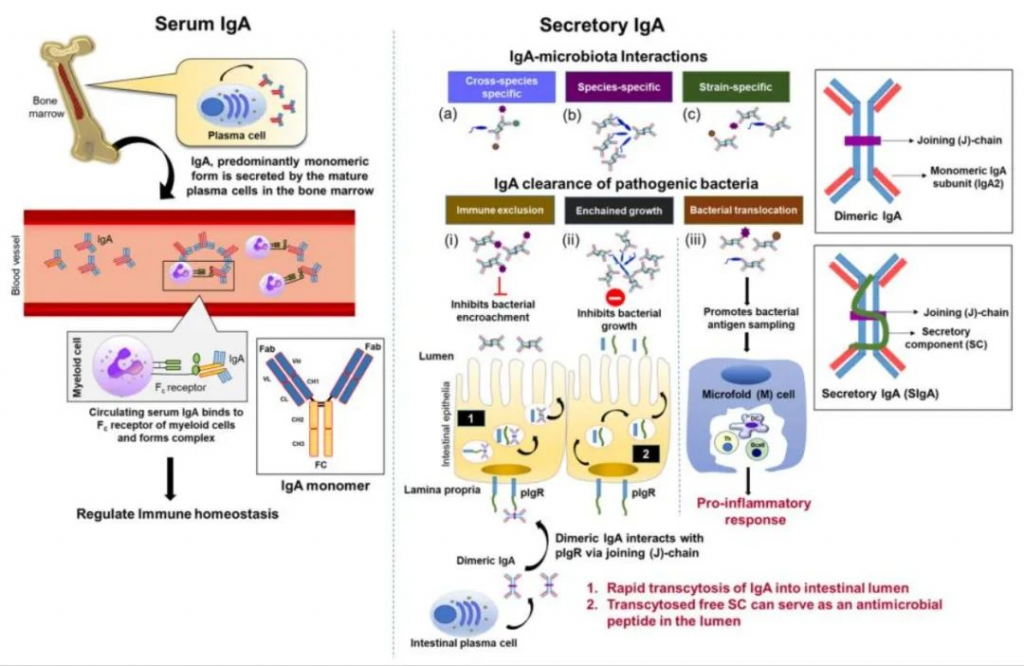

Abokor et al., Microorganisms. 2021
在左栏中,IgA,主要是单体 IgA,由骨髓中的成熟浆细胞分泌并进入体循环。循环血清 IgA 与位于骨髓细胞上的跨膜 Fc 受体形成免疫复合物,以诱导维持免疫稳态所必需的下游效应信号。
在右栏中,肠浆细胞通过两个 IgA 单体与连接 (J) 链的二价连接产生二聚体 IgA。J 链与位于肠上皮基底外侧表面的聚合 IgA 受体 (pIgR) 的分泌成分 (SC) 结合。IgA 以分泌型 IgA (SIgA) 的形式迅速转入肠腔。游离 SC 也被转胞吞到管腔中并用作抗菌肽。
与肠道微生物群相互作用,a ) 对各种细菌物种具有跨物种(多反应性)反应性,( b ) 物种特异性反应性或 ( c ) 菌株特异性反应性。对于病原体去除,SIgA 可以 ( i ) 与细菌结合并凝集,从而阻碍微生物附着和侵入宿主肠上皮细胞,这一过程称为免疫排斥,( ii ) 通过链式生长防止细菌结合以限制细菌增殖,以及 ( iii ) ) 加速细菌通过微折叠 (M) 细胞转移到派尔斑块中,以便常驻树突细胞 (DC) 进行抗原采样。
在局部细菌传播的情况下,当 SIgA 的前线防御不足时,二聚体 IgA2 通过与驻留的 Fcα/μR +滤泡树突细胞交联并募集 Fcα/μR +中性粒细胞来调理抗原。当交联时,Src 激酶 Lyn 磷酸化相关 ITAM 中的酪氨酸,这促进激酶/生长因子的募集,这些激酶/生长因子刺激与吞噬作用、呼吸爆发和炎性细胞因子分泌相关的免疫细胞。
同时,分泌白三烯 B4 (LTB4) 作为趋化信号,将更多的中性粒细胞募集到感染部位,从而形成正反馈回路以消除入侵的病原体。如果细菌感染和传播严重到足以到达门静脉循环,血清 IgA 会调理抗原,与枯否细胞(常驻肝巨噬细胞)交联,并诱导促炎反应。
注:白三烯B4是一种与炎症反应有关的白三烯类物质。它由响应炎症介质的白细胞产生,让白细胞活化并依附在内皮上,允许其穿过组织。
枯否细胞,被誉为人体肝脏内有一忠诚“卫士”,守护监视着进出肝脏这一人体化工厂的所有物体,以防机体受到任何外来侵害,它的名字叫枯否细胞(Kupffer Cells)。枯否细胞是定居于肝内的一种巨噬细胞,是我们人体内最大的固有巨噬细胞群,约占固有巨噬细胞总数的 80%~90%。
枯否细胞具有吞噬、免疫调节与监视、分泌等作用。生理条件下,枯否细胞不仅能非特异的吞噬和清除血流中的细菌、异物等抗原性物质,而且还具有特异性的免疫应答、抗肿瘤免疫、内毒素解毒、抗感染、调节微循环等方面的作用。病理条件下,枯否细胞可以释放肿瘤坏死因子、转化生长因子、干扰素、白介素、氧自由基及一氧化氮等多种炎性介质。
治疗策略——靶向自身抗体糖基化
值得注意的是,天然二聚体 IgA2 和单体 IgA1 的本身抗炎和促炎效应功能分别归因于它们不同的糖基化谱。两种抗体都含有几个N-糖基化位点,但只有 IgA1 具有多个O-糖基化位点,因此每个聚糖具有更多的末端唾液酸。
据报道,通过神经氨酸酶处理的去唾液酸化增加了 IgA1 的促炎能力,这反映了 IgA2由于循环糖基化 IgA 可促进各种自身免疫性疾病的进展,因此靶向自身抗体糖基化可能是一种潜在的治疗策略。
总而言之,如果我们将身体视为抵御病原体入侵者的堡垒,那么 SIgA 与肠道上皮细胞一起充当“封锁墙”,作为中和微生物不可或缺的第一道防线。当细菌侵入 SIgA 并突破黏膜层时,二聚体 IgA 充当第二道天然黏膜免疫防御,然后血清 IgA 和枯否细胞的协同作用成为消灭病原体的第三道也是最后一道防线。
IgA 对微生物群的反应通过T细胞依赖 (TD) 和非依赖(TI) 途径发生。
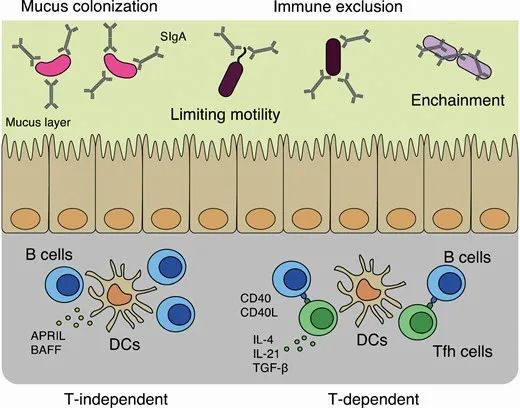

microbiology
SIgA抑制致病菌,结合有益菌
越来越多的证据表明,通过 T 细胞依赖性途径产生的亲和力成熟和特异性 SIgA 会影响肠道微生物群,特别是致病菌,以影响其运动性,将它们排除在肠道表面(免疫排斥)或通过束缚来抑制它们的生长。相比之下,IgAs 还可以与有益微生物结合,并将它们固定在粘液层中(粘液定植)。
IgA 促进拟杆菌在肠道内定植
拟杆菌属是肠道共生微生物群的一个核心菌属,一些报告表明,IgA 可以促进拟杆菌在肠道内定植。例如,脆弱拟杆菌可以以依赖于表面荚膜多糖的方式粘附在肠上皮细胞上,而 SIgA 在功能上参与了这一过程。SIgA 介导肠道定植Bacteroides thetaiotaomicron通过改变参与多糖利用的基因的表达。这些研究表明 SIgA 还可以直接改变拟杆菌属的功能以促进在肠道中的定植。
肠道环境影响IgA-微生物之间的互动
尽管许多研究都集中在 IgA 与肠道微生物群的相互作用中的特征,但同样重要的是要注意某些肠道环境也可能影响微生物的基因表达和功能,从而影响 IgA 微生物互动。SIgA在营养不良的情况下不太可能覆盖肠道乳酸杆菌。他们认为,某些乳酸杆菌菌株可以在营养不良期间进行代谢适应,从而逃避 SIgA 的识别。
稳态 IgA 反应的机制
在全身免疫的经典模型中,TI 反应发生在对多价抗原(例如细菌多糖)的反应中,并且涉及快速的细胞分化,几乎没有体细胞超突变。
相比之下,TD 反应通常针对蛋白质抗原,并涉及基于与 CD4+ T 滤泡辅助细胞 (Tfh) 的同源相互作用在抗体生发中心中进行的 SHM 和亲和力选择的迭代轮次。
然而,稳态粘膜 IgA 反应与这些过程相似的程度仍不清楚,并且有几条证据表明不同的机制和调节。
注:CD4并不是一种细胞是一种蛋白质“标签”,存在于在某些免疫细胞,如巨噬细胞、T细胞和单核细胞表面。具有CD4“标签”的细胞被称为CD4细胞。
CD4 +T细胞通常是作为是免疫系统中的“辅助”角色,它们大多数情况并不能直接中和感染,而是引导并触发机体对感染源的免疫应答,类似于免疫系统的“哨兵”。在CD启动免疫应答后,后再由CD8+T细胞来扮演“行刑官”的角色,清除感染源。
首先,尚未证实稳态 IgA 抗体对单个微生物抗原的特异性和高亲和力识别。相反,对单克隆抗体 (mAb) 的研究表明,IgA 衍生的抗体通常具有多反应性,并且与许多微生物抗原(包括脂多糖、DNA、鞭毛蛋白和荚膜多糖)具有低亲和力结合。
Peyer 斑块中 IgA 选择的机制
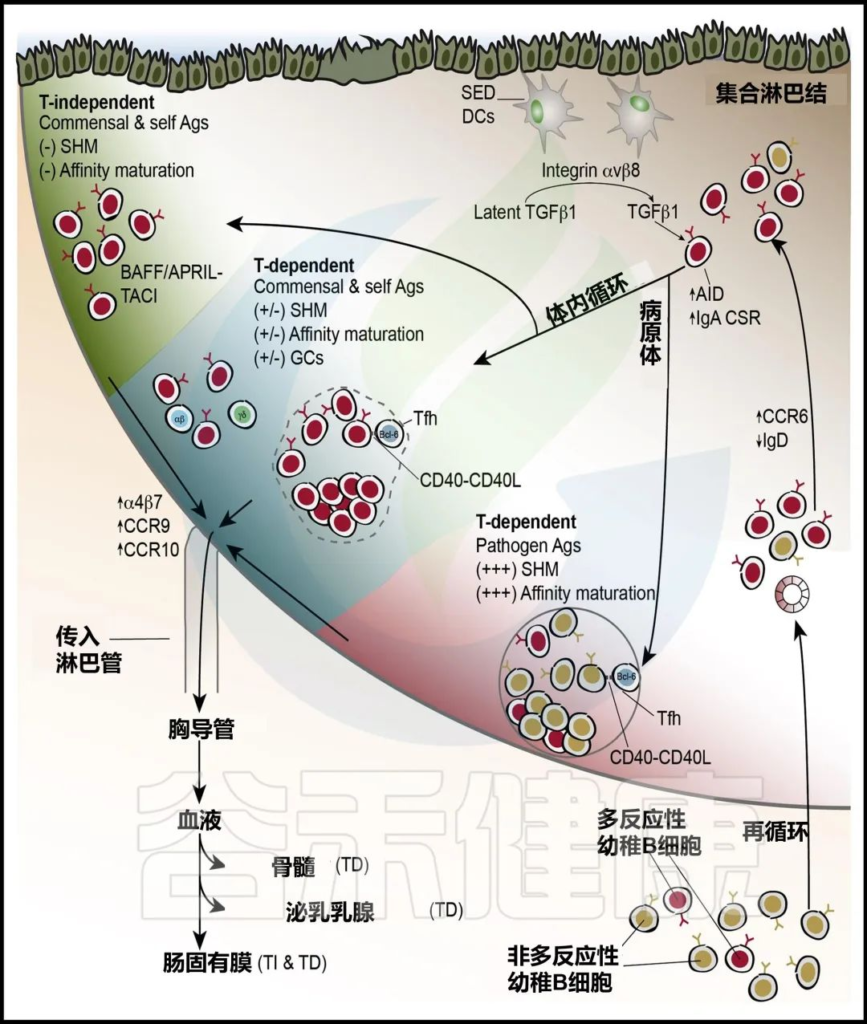
doi.org/10.1016/j.immuni.2018.08.011
此外,从幼稚 B 细胞或流感特异性反应克隆的随机多反应性 mAb 与体内涂有 IgA 的相同微生物群亚群结合。由病原体引发的聚糖反应性但非多反应性抗体通常也与共生细菌发生交叉反应。
总之,这些数据表明抗体多反应性和相关的自身反应性可能是 IgA 选择的主要驱动因素,并支持 IgA 多反应性能够与多种细菌表面分子低亲和力结合的模型。
在我们的一生中,微生物群塑造了先天和适应性免疫系统,在生命的前三年是细菌定植的最大变异性的关键时间点。我们已经讨论过不依赖 T 细胞的 SIgA 合成是通过微生物刺激来促进的,这可以被认为是在宿主和微生物群之间创造共生环境的有目的行为。
在以下部分中,我们将描述产生不同 SIgA 反应类型的机制以及这些 SIgA 亚群如何识别其清除的细菌目标。
SIgA 对肠道微生物群有选择性反应
SIgA 与微生物群相互作用以维持体内平衡,其稳态特性在很大程度上取决于抗体对各种微生物群落的特异性。据估计,在人类肠道中,单个细菌被近 19,000 个 SIgA 分子包被,而在小鼠中,这个数字增加到大约 60,000 个分子,用于包被 SIgA 的细菌。
三类SIgA 反应
这些抗体-微生物群的相互作用可以根据 SIgA 反应性明显分为三类:
(i) 跨物种
(ii) 物种特异性
(iii) 菌种特异性反应性 (如下图)
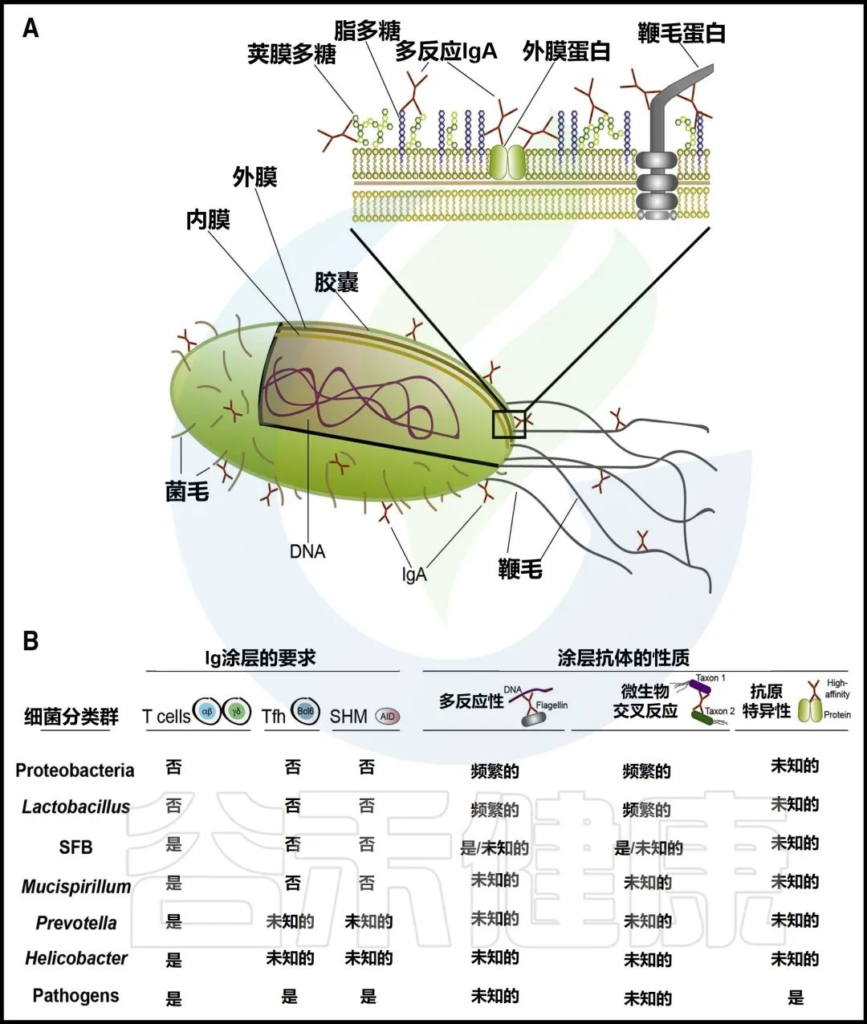
doi.org/10.1016/j.immuni.2018.08.011
跨物种反应性SIgA 是指具有结合各种不同种类细菌能力的 IgA 抗体,并且通常具有多反应性,因为它们能够结合结构上不同的抗原(例如 LPS、CpG)。
然而,最近发现 SIgA 体细胞超突变,而不是多反应性,赋予跨物种结合和高微生物群反应性。跨物种反应性 SIgA 在浆细胞分化之前先天在所有幼稚 B 细胞亚群中出现,并与广泛的微生物群亚群结合,其中包括变形菌门中的大多数成员,但这些 SIgA 在很大程度上缺乏与主要分类群拟杆菌门和厚壁菌门的结合。
这是意料之中的,因为如前所述,只有 7% 的肠道 SIgA 具有跨物种反应性,而大多数 IgA 是抗原特异性的。然而,尽管数量较少,但跨物种反应性 SIgA 在维持微生物群多样性方面和清除病原菌方面发挥着重要作用。
物种特异性反应性SIgA 是指仅与肠道中存在的不同细菌物种结合的 IgA 抗体。虽然尚不清楚 IgA 如何区分不同的细菌物种,但人们普遍认为细菌表面碳水化合物部分在 IgA 跨分类物种的选择性中发挥重要作用。
在用Bacteroides thetaiotaomicron (B. theta) 单克隆化的无菌小鼠中,诱导了高度特异性的肠道 SIgA 反应,而与其他密切相关的拟杆菌的交叉反应性较低。
还确定了抗B.theta IgA 靶向 B.theta 上的蛋白质多糖利用基因座 (PUL),这表明细菌果聚糖是物种特异性 SIgA 选择性所需的潜在表位。
另一项具有可逆体内无菌定植模型的研究进一步证实了物种特异性 SIgA 的高精度。
在这个模型中,无菌小鼠接受了营养缺陷型大肠杆菌 K-12突变体的三重突变体(称为 HA107 菌株)的诱导,但由于该菌株不能分裂也不能在体内持续存在,因此小鼠在体内恢复到无菌状态。72 小时,再次暴露于大肠杆菌HA107 后,无菌小鼠对该菌株产生了明显的黏膜 SIgA 反应,而首次暴露于鼠伤寒沙门氏菌(Salmonella typhimurium)在预处理的大肠杆菌HA107 无菌小鼠中没有引起物种特异性 SIgA 反应。
与此一致,经过大肠杆菌HA107 预处理的无菌小鼠后来被确定为缺乏大肠杆菌的微生物群定植,但缺乏具有大肠杆菌结合能力的SIgA,尽管总 IgA 产量没有减少。该证据表明,物种存在与否可能是 SIgA 物种特异性的先决条件。
有趣的是,对 B 细胞对肠道微生物群反应的功能分析检测到了针对普氏菌的 SIgA 抗体。
在一个人类队列中的健康个体的血浆和粪便中,这些人的肠道微生物群中显然缺乏普雷沃氏菌(Prevotella)。研究人员将这种现象归因于在某些时候之前接触过细菌的个体,并且在从系统清除后很长时间可能会产生针对普雷沃氏菌的抗体,他们认为这与之前对其他菌的其他研究一致。
菌株特异性 SIgAs 是对细菌物种内的各种遗传变异或亚型具有选择性的 IgAs。
最近证明,用卵形拟杆菌单菌定殖的小鼠引起强烈的粘膜 SIgA 反应。然而,已确定某些卵形双歧杆菌变体在诱导结肠 IgA 分泌浆细胞方面比其他变体更有效,导致这些特定亚型具有高IgA 分类—卵形双歧杆菌。
同样,观察到特定的双歧杆菌菌株能够在体外诱导大量的 IgA,而其他菌株只能微弱地诱导 IgA。此外,B.theta型菌株 VPI-5482 的单殖化引发了一种菌株特异性 IgA 库,被 B.theta VPI-5482 上发现的荚膜多糖吸引,而不是其他B.theta菌株。
同样,与突变株相比,IgA在体外极化上皮细胞单层上阻止了野生型伤寒杆菌的细菌粘附和侵袭,因为 IgA 识别野生型伤寒杆菌上的特定碳水化合物表位。这些发现共同突出了能够被 SIgA 识别的特定表位的多样性,并表明 IgA 反应利用大量的多反应库来广泛性结合分类众多的微生物群亚群。
SIgA 是 IgA 的主要形式,在保护宿主免受病原体侵害和塑造肠道微生物群组成以促进宿主-微生物群稳态方面发挥着动态作用。
与单体 IgA 相比,聚合 SIgA 对 Fc受体的激活效果较差,无法获得足够的下游效应信号。出于这个原因,SIgA 开发了几种机制,利用其交联能力和肠道环境来有效消除病原体。
SIgA 介导的微生物中和的第一个机制是称为免疫排斥的过程,其旨在以逐步方式拦截微生物进入肠上皮:
(i) 抗体凝集和交联
(ii) 粘液中的病原体截留
(iii) 通过蠕动去除
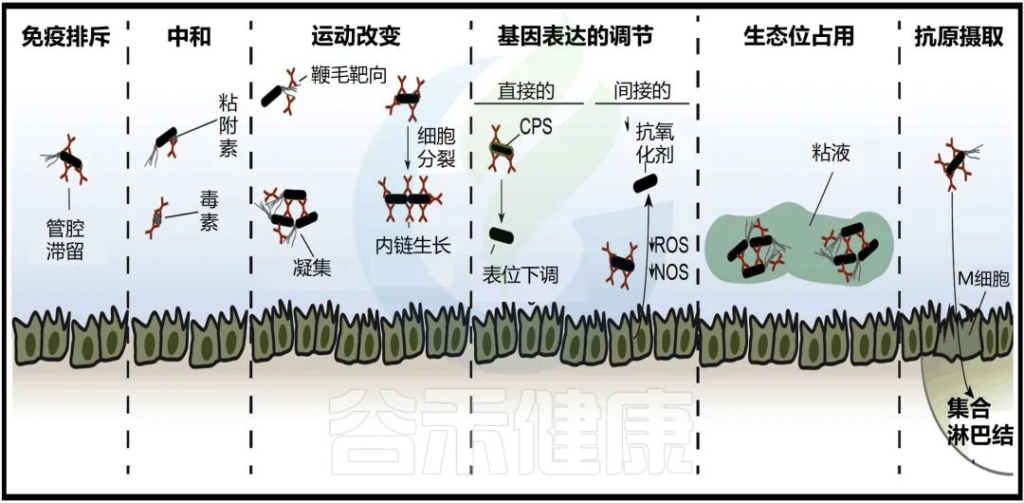
doi.org/10.1016/j.immuni.2018.08.011
在这方面,SIgA 更像是一道“阻挡墙”,以抑制微生物从管腔转移到血液中。例如,SIgA 可以预防口服接种鼠伤寒沙门氏菌的小鼠的全身感染,但 SIgA 无法预防相同细菌腹腔攻击后的菌血症和全身感染。
后来的研究结果表明,SIgA 的免疫排斥是在粘膜内特别指定的。另一项体内研究证实,志贺氏菌的免疫排斥需要 IgA 分泌糖基化残基来辅助定位抗体分子和最佳预防粘膜感染。
此外,IgA 介导的免疫排斥不仅发生于病原体细菌,也发生在共生真菌中。最近发现,SIgA 还可以靶向负责菌丝粘附和宿主细胞侵袭的细胞表面粘附蛋白,以防止白色念珠菌的附着和随后的人类感染。
免疫排斥的一个限制是它仅在高病原体密度下有效,这与典型感染不同。
出于这个原因,最近提出了另一种被称为“链式生长”的 SIgA 介导的病原体消除机制,在较低的病原体数量下相对有效。SIgA 介导的链式生长通过链式和分离细菌质粒供体和受体克隆来防止接合质粒转移。
然而,链式生长是有限的,尽管受到限制,但聚集的病原体生长可能会发生,直到获得一定的大小,然后破裂形成由密切相关的细菌组成的亚群。
链式生长的一个缺点是它对快速生长的细菌最有效。事实上,研究人员指出,具有高生长速率的细菌在子细菌之间的联系破裂之前复制,并产生更大的簇,而具有较慢生长和复制速率的细菌很可能更早遭受簇断裂并在复制时逃脱 SIgA 链式生长。
除了上述两种机制之外,SIgA 还具有一种称为“涂层”的独特功能,可增加派尔斑块中的细菌易位,从而无意中改善了常驻树突细胞的抗原采样和激活。例如,发现 SIgA 包被的福氏志贺菌(S. flexneri)被迅速转胞吞入派尔氏斑并被树突细胞内化,而未包被的S. flexneri无法穿透肠上皮。
据报道,SIgA 涂层对小肠中的共生细菌占优势,这反过来又有助于菌群定植,而不含 IgA 的细菌大多是结肠的固有细菌。
重要的是,尽管有足够的 SIgA 可用于覆盖几乎整个微生物群,但 SIgA 的这种“包被”作用似乎受到高度“监控”,因为 <5% 的 SIgA 被用于细菌包被,这可以推断是为维持与肠道微生物群共生的基本机制,而在疾病条件下,SIgA 涂层变得更加普遍。比较特别的是,IgG 和 IgM 几乎没有包裹厌氧菌的能力,更突显了 SIgA 是针对肠道抗原的主要反应性免疫成分。
已发现 SIgA 通过重塑肠道微生物群组成以促进肠道共生体生长和抑制病原菌增殖,对维持肠道稳态具有很大影响。
例如,专门覆盖粘液共生体 B.theta 的 SIgA已被证明可以上调一组临时命名为粘液相关功能因子 (MAFF) 的基因,这些基因的功能是确保厚壁菌门细菌之间的共生,并且还可以提供针对结肠炎的化学保护物质。
此外,SIgA通过共生定植因子 ( ccf ) 对共生脆弱拟杆菌的黏膜定植和单菌株稳定性至关重要。脆弱拟杆菌介导的荚膜多糖上调以吸引 IgA 结合。有趣的是,脆弱拟杆菌具有内切糖苷酶活性,因此可以利用在 SIgA 上重度修饰的复杂 N-聚糖进行必要的共生细菌生长,这最终有助于该细菌在特定的粘膜生态位内定植。这也是我们很多的粪便检测实践中发现脆弱拟杆菌的丰度占比较高。
此外,粘膜 IgA 通过限制共生真菌增殖来维持微生物群的稳态,因为已观察到 SIgA 对人类粪便中发现的光滑念珠菌、白色念珠菌、酿酒酵母和热带念珠菌具有反应性 。
SIgA 还促进出生时肠道微生物群中的健康生物多样性。研究发现母乳会转移某些细菌,例如链球菌属和Veillonella dispar,这导致了后代微生物群的整体变化 。
以上证据表明 IgA 对于微生物粘附到上皮表面至关重要,并且可以同时去除这些细菌以进行清除以保持适当的多样性。此外,抗体具有多种结合亲和力,可以覆盖肠道微生物群的大部分,目的是维持而不是消除微生物的多样性。
宿主 SIgA 和细菌之间的这种相互作用促进了肠道中额外的宿主免疫反应,从而产生了一个共生的调节回路来维持肠道稳态。
有趣的是,在厚壁菌门中属于簇 IV 和 XIVa 的梭状芽胞杆菌被观察到是多样化 IgA 生产所必需的 Foxp3 + T 细胞的有效诱导剂。反过来,依赖于 T 细胞的 IgA 对共生体Akkermansia muciniphilia的反应提供了针对肠道感染的“旁观者保护”,以进一步促进肠道健康和维持体内平衡。
尽管 SIgA 的肠道稳态功能仍然很微妙,但 IgA 反应的程度以及分泌黏膜 IgA 抗体支持其功能意义。尽管尚不清楚 IgA 抗体是否可能对 IgA 靶向微生物产生有利或有害的影响,但 IgA 包被的共生体的组成性存在表明任何有害影响通常不足以引起消除。
事实上,IgA 与细菌荚膜多糖的结合可能会被一些微生物群抑制以允许粘液层附着,从而防止竞争物种的生态位入侵。
肠道微生物群的组成和活性可以受到环境和宿主衍生因素的调节。其中,免疫球蛋白 A (IgA) 对胃肠道共生菌的稳态功能最近已在IgA 缺乏症患者中得到证实。
IgA 通过聚糖和脂多糖 (LPS) 相互作用调节肠道菌群组成,与 IgA 同源抗原识别无关
具体而言,IgA通过聚糖-聚糖相互作用(IgA-聚糖和 LPS)与细菌共生亚群如多形拟杆菌Bacteroides thetaiotaomicron ( B. theta )、B. vulgatus、B. fragilis和Parabacteroides distasonis 等结合。此外,IgA 增强了结肠外膜中细菌与宿主粘液和饮食衍生多糖的结合。
粘液相关的B. theta特别容易受到 IgA 涂层的影响
研究人员发现,暴露于 IgA 会导致B. theta上调两种未发现的蛋白质——称为粘液相关功能因子 (MAFF) C 和 D——它们在结构上与其他肠道细菌共享,并增强了B. theta的代谢活性。
在小鼠和人类的结肠粘液中,B. theta 诱导 MAFF 系统表达,这取决于与梭状芽孢杆菌等厚壁菌门成员的相互作用。同样,B. theta 产生的代谢物可诱导梭菌属成员的扩增,从而驱动更多样化的微生物群。
这些发现强调,IgA 驱动的B. theta MAFF 表达是调节网络的结果,不仅涉及肠道微生物群组成,还涉及其在粘膜生态位内的遗传结构和潜在功能。
在化学诱发结肠炎的小鼠模型中,接种了野生型 B.theta 的小鼠结肠在用硫酸葡聚糖钠治疗 10 天后恢复。相比之下,接种了缺乏 MAFF 蛋白的B.theta菌株的小鼠的结肠仍然显示出损伤和肠道炎症的迹象。这些结果表明,MAFF 诱导的肠道菌群增强了结肠上皮细胞的增殖和再生,同时也增加了肠道菌群的多样性。
总之,共生类拟杆菌的最佳体内代谢能力取决于 IgA-多糖相互作用。IgA 对肠道稳态的作用取决于不同微生物群之间的相互作用,并且由 MAFF 微生物基因的表达调节。
正如前面部分所述,IgA 由于其结构意义、分泌、糖基化、定位和受体相互作用而在免疫系统中发挥着重要作用以及 IgA 强烈影响各种共生微生物的微生物群组成及其相关基因表达的方式。
重要的是,持续的研究强调微生物群组成是越来越多疾病的牵连病因,包括肠胃病(如结肠炎,结直肠癌)、肾病、过敏、哮喘,精神类疾病和代谢疾病甚至人类寿命。由于 IgA 生物学功能障碍也可导致多种疾病,因此了解 IgA -微生物群轴的缺陷可以帮助预防和治疗各种疾病的发展。
高和低 IgA 水平都与死亡风险增加有关。不过,需要更多的研究才能得出任何结论。
与死亡率的关联
在 4,255 名越战老兵中,较高的 IgA 水平死于传染病的风险增加 2 倍有关。
在 90 – 99 岁的人群中,较高的 IgA 水平与较高的死亡率相关。
同样,在 8 岁儿童中,高 IgA 水平(>400 mg/dl)与较高的死亡率相关。IgA 水平与男性癌症死亡率有关。
较高的IgA与癌症(特别是非肺癌)和呼吸系统疾病死亡风险的降低有关。
严重的 IgA 缺乏与诊断后前 10 至 15 年内较高的死亡率相关。
高和低 IgA 水平可能有许多潜在原因。因此,引发异常 IgA 水平的健康状况比 IgA 水平本身更可能影响一个人的死亡风险。
选择性 IgA 缺乏症 (SIgAD)
无法检测到的血清 IgA 含量低于 7 mg/dL。人类 IgA 缺乏症的临床表现被称为选择性 IgA 缺乏症 (SIgAD) 。SIgAD 是最常见的原发性免疫缺陷,其余 Ig 水平正常。20-30% 的 SIgAD 患者患有自身免疫性疾病。研究表明 SIgAD 患者的一级亲属有 10% 的自身免疫性疾病发生率,是一般人群中估计的 5% 的两倍。
重要的是要注意,即使 B 细胞或 IgA 不存在,肠上皮细胞也可以启动其他保护性防御,例如诱导干扰素诱导的免疫反应途径,但前提是存在微生物群。
关于人类 SIgAD 是否与肠道微生物生态学的重大变化有关,存在相互矛盾的报道。
有趣的是,最近的一项研究表明,SIgAD 患者的体循环中存在代偿性 IgG 反应,其中 IgG 对共生体具有抗菌特性。另一项宏基因组研究表明,SIgAD 患者的微生物多样性降低,但富含大肠杆菌等机会性细菌。
常见可变免疫缺陷 (CVID)
除 SIgAD 外,还有一种称为常见可变免疫缺陷 (CVID) 的疾病,原因是抗体产生无效,主要是 IgG 和 IgA,这是由于普遍的 B 细胞缺陷。
Omenn 综合征 (OS)
由亚型 RAG 突变引起的 Omenn 综合征 (OS),会在不经意间导致 IgA 缺乏。CVID 和 OS 患者感染细菌感染的风险自然更高,但他们也可能有非感染性自身免疫并发症,例如炎症性肠病和肠病。尽管对该主题的研究很少,但暗示不稳定的肠道微生物确实在 CVID 和 OS 患者的自身免疫反应中起作用。
炎症性肠病 (IBD) 是整个胃肠道剧烈炎症的结果,每年估计有 70,000 例新的 IBD 病例被诊断出,全球有 680 万例。微生物群在 IBD 中起着不可或缺的作用,但其作用是原因、结果还是相关性是 IBD 研究中的一个持续问题。
细菌诱导IBD -> IBD -> 菌群失调 -> IBD 加重
一方面,某些细菌可能是导致 IBD 进展中必不可少的炎症过程升级的抗原刺激物。事实上,无菌小鼠表明微生物群是自发性结肠炎的诱导物和侵袭物。
另一方面,当研究缺乏上皮特异性极化分选因子衔接蛋白 (AP)-1B 的小鼠的自发性结肠炎时,发现 IBD 的易感性引发了微生物群组成向生态失调的不利变化,其中大量减少硫和产生乳酸的细菌可能解释了加重的结肠炎。
其他小鼠研究支持某些个体细菌作为前结肠炎,包括人类衍生的产生丁酸盐的菌株Anaerostipes hadrus BPB5 、吃粘蛋白的AKK菌 、肺炎克雷伯菌和奇异变形杆菌等。这与观察到IBD 患者肠杆菌科和变形菌门增加的人类研究相关。
这些变化同时被认为是人类 IBD 的生物标志物和治疗相关的靶点,以减少微生物群加剧 IBD。
虽然这些研究表明微生物群组成差异只是炎症的并发症,但值得注意的是转化乙酸丁酸的Roseburia的减少。最近发现,在健康对照组中,在 IBD 之前并在 IBD 期间持续存在,这表明微生物群参与了疾病本身的病因学。
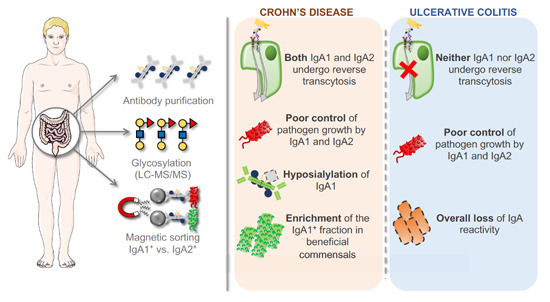
doi.org/10.15252/emmm.202115386
鉴于其先前确定的影响肠道微生物群组成的能力,SIgA 在微生物群-IBD 关系中非常重要,其中 IBD 患者可能对共生真菌和细菌的粘膜耐受性功能障碍。
例如,pIgR 基因敲除小鼠由于 SIgA 转运缺陷和微生物群不稳定而更容易患结肠炎。关于真菌,几种念珠菌属与 IBD 病理学有关,特定的念珠菌菌丝形态与 IBD 严重程度增加有关。此外,观察到 SIgA 靶向病原真菌中的粘附和菌丝细胞,以防止结肠炎期间念珠菌相关的损伤。
关于细菌,缺乏诱导型共刺激配体 (ICOSL) 的小鼠对 IBD 自发敏感,其 IgA 减少,并且对来自毛螺菌科粘液相关细菌的鞭毛蛋白的抗原识别受损。
此外,激活转录因子 3 (ATF3) 缺陷的小鼠表现出肠道菌群失调,这有利于促炎性普氏菌的丰度,并显示肠道中 T FH细胞发育受损,导致 SIgA 产生显着降低。这种反应在缺乏先天效应蛋白、骨髓分化初级反应 88 (MyD88) 的小鼠中是相似的,它通过诱导 CD4 + T 细胞和调节性 T 细胞在调节 IgA 对肠道微生物群的反应中起关键作用。
IgA 对 IBD 和肠道菌群失调的反应需要 MyD88 信号传导
MyD88 缺陷型小鼠的结肠炎严重程度加重,肠道菌群失调突出,分段丝状细菌过多和细菌负荷增加,表明 IgA 对 IBD 和肠道菌群失调的反应需要 MyD88 信号传导。相比之下,甲基化控制的 J 蛋白(一种线粒体内膜蛋白)的缺失导致 IBD 相关细菌Ruminococcus gnavus 大量繁殖,但令人惊讶地增加了 SIgA 水平。
升高的SIgA可能是一种补偿反应,准备对抗病原体
同样,吲哚胺 2,3-双加氧酶 (IDO) 敲除小鼠对柠檬酸杆菌具有较高的 SIgA 基础水平,并且对柠檬酸杆菌诱导的结肠炎具有抗性。最后两项研究表明,在某些情况下,升高的 SIgA 可能是一种补偿反应,以确保共生清除并创造一个准备好对抗病原体的环境。
IBD 患者的粪便中含有更多的 SIgA 涂层细菌
最近的一项研究表明,与对照组相比,IBD 患者的粪便中含有更多的 SIgA 涂层细菌。因此,IgA-Seq 已被用于分析 SIgA 包被的细菌,从而识别 IBD 相关微生物。例如,通过 IgA-Seq 选择 SIgA 包被的细菌,从 IBD 患者中分离和厌氧培养这些微生物,然后在无菌小鼠中定殖,进行了一项具有里程碑意义的研究。
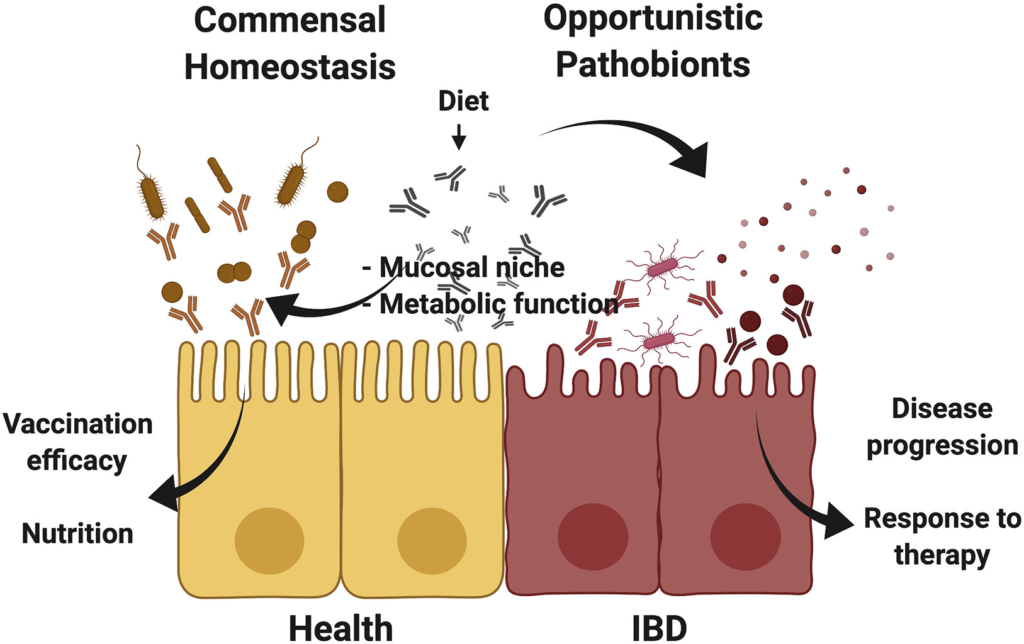
doi.org/10.1016/j.chom.2020.12.014
SIgA 涂层作为免疫介导的降低肠道细菌负荷的目标
除了作为生物标志物外,SIgA 涂层似乎还可以作为免疫介导的降低肠道细菌负荷的目标。因为 SIgA 高的小鼠品系 CBA/CaJ (CBA) 对葡聚糖硫酸钠 (DSS) 诱导的急性结肠炎具有抗性,因为它继承了增加的 SIgA 涂层和减少的粪便细菌负荷,而 SIgA 低的小鼠品系C57BL/6 (B6) 易患结肠炎。
令人印象深刻的是,最近的一份报告发现了一部分克罗恩病患者在含有核苷酸结合寡聚结构域(NOD2)发生突变,携带抗原的 SIgA 逆行转运到派尔斑块中增加。作者在 NOD2 缺陷小鼠中证实了这一观察结果,这支持了粘膜炎症增加可能是由于过度活跃的 SIgA 逆行转运的概念。
注:核苷酸结合寡聚结构域(NOD2)肽聚糖(细菌细胞壁)的产物,是一种位于上皮细胞和免疫细胞内的胞质 ,能够感知胞壁酰二肽。这种胞质对于病原体入侵和几种炎症性疾病期间的免疫反应至关重要,从而调节粘膜细菌定植。
一般来说,这一证据表明,人类的高 IgA 反应可以预防结肠炎,因此,消除或抑制 SIgA 包被的细菌是潜在治疗的途径。
研究发现与同种型对照相比,对 IL-10 缺陷小鼠进行丁酸钠治疗可减少 SIgA 涂层细菌的数量,同时增加肠道生物多样性。
利用IgA靶向和抑制有害菌
作为一种更直接的方法,最近开发了一种工程化 IgA 克隆 W27,它靶向和抑制有害共生细菌,但不抑制有益细菌,从而在几种小鼠模型中预防结肠炎和提高肠道微生物群多样性。
另一个需要考虑的来源是母乳衍生的 SIgA。证明其早期暴露可改善葡聚糖硫酸钠诱导的上皮损伤。研究这一观察结果是否可能是由于表达 RORγ 的调节性 T 细胞在多代传递中的免疫调节是很有趣的。针对 IgA 微生物群轴的持续研究肯定会推进治疗 IBD 的临床困境。
坏死性小肠结肠炎(NEC)是易感婴儿最严重、最常见的肠道疾病。早产儿小肠结肠炎的风险在中度至极低体重的新生儿中增加,后者婴儿的死亡率估计为 20-30% 。
坏死性小肠结肠炎进展的促成因素集中在新生儿期肠道不成熟和不适当的微生物定植。
NEC 通常在出生后 8-10 天出现,此时肠道被变形菌和厚壁菌门的兼性厌氧菌定植。当早产儿进入新生儿重症监护室 (NICU) 时立即使用抗生素可能会干扰适当的细菌定植,因此会导致坏死性小肠结肠炎。实质上,细菌侵入肠壁引起局部感染,随后是上皮损伤、潘氏细胞耗竭、屏障功能受损、炎症、坏死、菌血症和内毒素血症。
在潘氏细胞破坏坏死性小肠结肠炎啮齿动物模型中,添加配方奶会加剧肠道损伤,而与肠道微生物菌群失调无关。这一结果强调了早产儿喂养需要谨慎。
母乳降低坏死性小肠结肠炎发病率,与IgA有关
相比之下,发现母乳显着降低坏死性小肠结肠炎发病率,这表明母乳中的抗菌成分负责保护。
最近的一项开创性研究发现母乳是出生后第一个月 SIgA 的主要来源,当时 IgA 涂层有助于减少肠杆菌科细菌并保护小鼠免受坏死性小肠结肠炎。
这一发现进一步得到了以下观察结果的支持:暴露于母乳的 IgA 缺陷幼崽仍然易患坏死性小肠结肠炎。值得注意的是,γ-变形杆菌特异性 IgA 负责从未成熟微生物群到成熟微生物群的转变,而 IgA 缺乏会导致 γ-变形杆菌大量繁殖。
回顾到成熟微生物群的最终转变是从 γ-变形菌到梭状芽孢杆菌,可以假设梭状芽孢杆菌耗竭可能是坏死性小肠结肠炎婴儿微生物群成熟停滞的指标。
因此,对于未来的研究来说,了解母体 IgA 的供应是否与成熟微生物群相关,即NEC 保护婴儿中梭状芽胞杆菌的恢复,这一点很重要。
上述证据强调了坏死性小肠结肠炎中提高 IgA 和调节 IgA 相关微生物群的治疗潜力。
然而,在坏死性小肠结肠炎中靶向 IgA 并不一定是新的方法,1988 年的一项研究发现口服 IgA-IgG 补充剂可有效预防早产儿坏死性小肠结肠炎。
无论如何,促进和/或维持 IgA 水平的治疗方法可能会挽救生命。考虑到患有炎症性肠病的母亲通过母乳水平转移给婴儿的 IgA 可用性较低,其中,母亲和孩子都可能需要补充 IgA。
然而,通过最近的一项研究,必须意识到 IgA“过度涂层”的可能性。产前压力增加了后代微生物群中的 IgA 涂层,并以性别依赖的方式加重了坏死性小肠结肠炎。因此,其他微生物方法(即益生菌)也可用于治疗坏死性小肠结肠炎,特别是乳杆菌属。
结直肠癌 (CRC) 是癌症相关死亡的第二大常见原因,而 IBD 恰好是 CRC 发展的主要前兆。与任何癌症一样,早期发现对于降低死亡率至关重要。
IgA 作为一种重要的结直肠癌筛查工具
几项研究调查了自身反应性抗体,特别强调 IgA 作为一种重要的 CRC 筛查工具。特别是,与肿瘤相关抗原癌胚抗原 (CEA) 反应的 IgA 已成为 CRC 患者的重要特征。
最近用水凝胶生物芯片证明,在诊断中结合抗 CEA 和抗聚糖抗体可提供更好的预测值。特别值得注意的是,在用重组 CEA 对 CRC 患者进行免疫时,发现 IgA 抗 CEA 抗体对肿瘤细胞具有细胞毒性并提高了患者的存活率。对某些细菌(如具核梭杆菌和产毒素的艰难梭菌)特异的 IgA也已被证明具有高度特异性和敏感性的诊断价值。
虽然 IBD 患者 IgA 不足,但最近的文献可能表明 IgA 的缺乏实际上可能是预防侵袭性 CRC 的一种保护机制。
在Apc Min/+ CRC 小鼠模型中,确定了肿瘤微环境中 IgA +淋巴细胞的扩增。这与 CRC 患者晚期肿瘤中浆细胞的优势相匹配,其中 B 细胞亚群 IgA + IGLC2 +与预后不良有关。
值得注意的是,前 B 样细胞可能在 CRC 发展的早期阶段具有抗肿瘤功能 ,但是当它们在晚期 CRC 中分化为浆细胞时,这可能会变得不那么有效。
IgA 维持微生物群稳态
具体而言,通过研究研究表明与宿主故意限制 IgA 可用性本身的想法相反,IgA 迁移到结肠肿瘤细胞的缺乏,理论上可以促进支持致癌生长的促炎环境。研究表明,缺乏 IL-33 的小鼠 IgA 水平显着降低、菌群失调、结肠炎和最终发生 CRC ,这支持了 IgA 是维持微生物群稳态以预防肠道疾病所必需的传统观念。
肠道微生物组和 IgA 与 CRC 之间的关系相对未描述,未来的研究可能会揭示筛选和治疗这种致命癌症的新方法。一个值得的研究方向是了解 IgA 涂层对 CRC 相关细菌(如具核梭菌和脆弱拟杆菌)在疾病进展中的作用。可能还有许多其他肠道微生物可能在 CRC 中发挥作用,需要进一步研究。
中枢神经系统(CNS)的复杂疾病是由遗传和环境因素共同引起的。人体研究和动物模型表明,宿主体内的共生微生物可以影响中枢神经系统疾病(下图)。
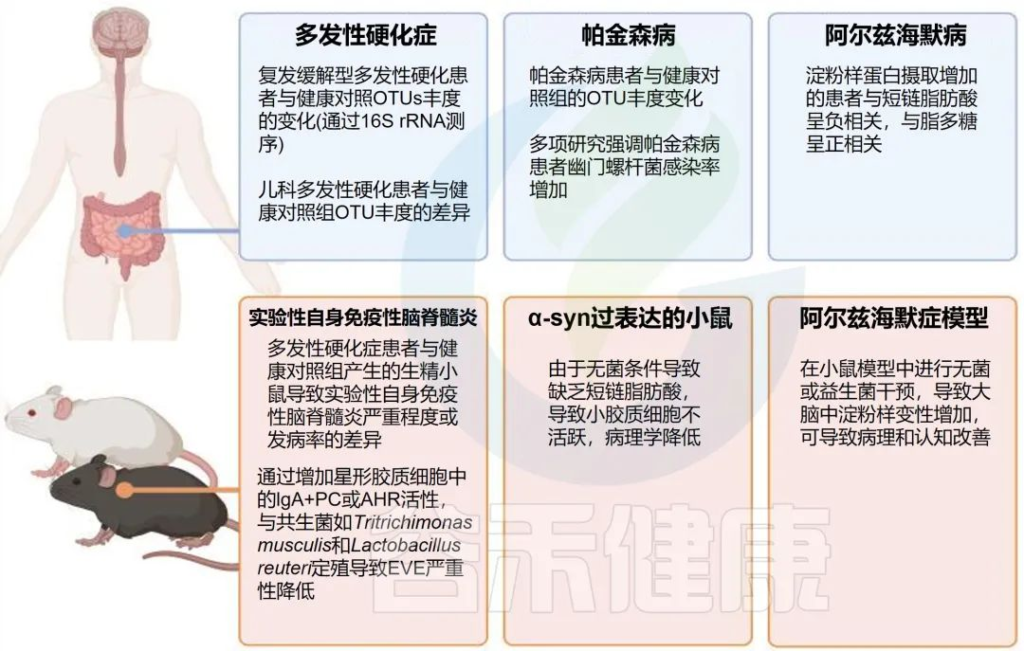
doi: 10.3389/fimmu.2021.742173
有两个关键宿主因素,肠道 IgA 和衰老,它们对微生物组的形成具有深远的影响。且而且宿主因素(IgA 和衰老)之间的相互作用会塑造微生物组,进而影响中枢神经系统疾病背景下的淋巴细胞和神经胶质细胞行为。
肠道微生物组与中枢神经系统神经炎症和神经退行性变之间的假定联系:
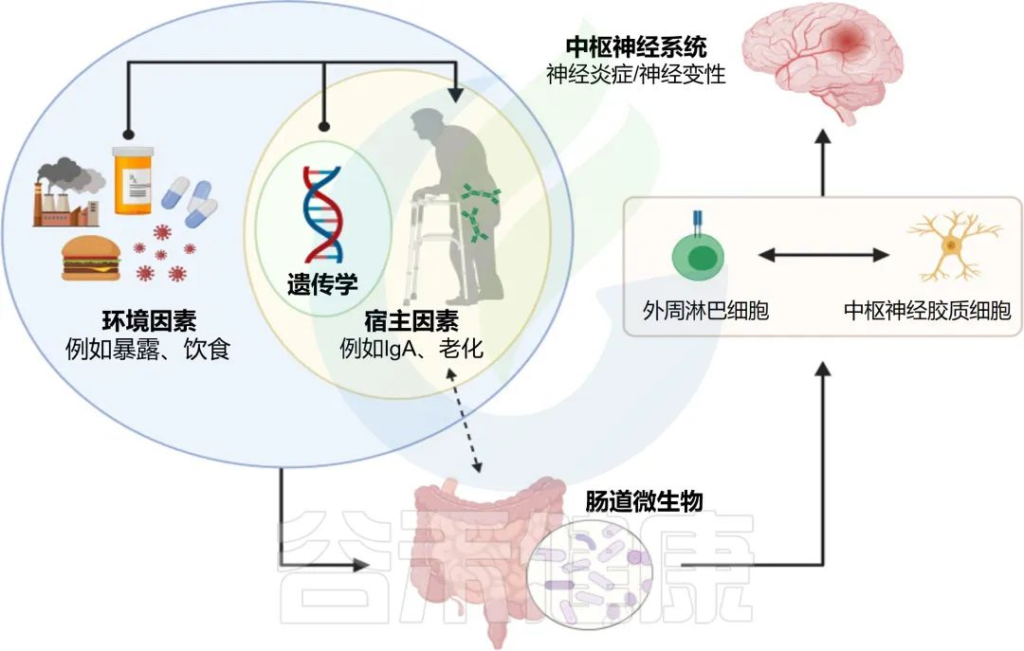
doi: 10.3389/fimmu.2021.742173
实验性自身免疫性脑脊髓炎
虽然 IgA + ASCs (抗体分泌细胞)可以在体内平衡期间归巢到硬脑膜 ,但在稳态中枢神经系统中不存在克隆扩增的 IgA,并且仅在炎症期间出现。在实验性自身免疫性脑脊髓炎期间,SILP 中的 IgA + ASC 显着减少。
此外,在中枢神经系统疾病中发现过继转移的肠道来源的 IgA + ASC 对小鼠来源的肠道细菌有反应,并显示通过在实验性自身免疫性脑脊髓炎(EAE)的慢性阶段产生 IL-10 来减轻神经炎症。
过量的 IgA + ASC 能够减少 T 细胞产生的 GM-CSF,这是一种促进神经炎症的重要细胞因子。毛滴虫( T.mu ) 是一种啮齿类共生动物,可促进 IgA 的产生。T.mu +小鼠的 EAE 发生率和严重程度以及脊髓炎症和脱髓鞘减少。T.mu +小鼠还表现出血清和粪便 IgA 水平升高,肠道、骨髓和大脑中 IgA + ASCs 的频率增加。
虽然以上强调了动物模型的主要发现,但也有早期证据表明微生物群驱动的 IgA 反应在人类疾病中的重要性。
多发性硬化:肠道菌群 IgA降低,脑脊液 IgA 相应升高
由 IgA-seq 鉴定的细菌在多发性硬化患者与健康对照中的表达差异很大。按疾病活动分层,与缓解患者相比,复发的多发性硬化患者粪便样本中 IgA 结合的肠道细菌百分比降低,脑脊液 IgA 相应升高。
中枢神经系统浸润的 IgA + B 细胞对肠道微生物抗原具有特异性,表明在复发期间产生 IgA 的细胞从肠道迁移。
多发性硬化患者的脑脊液中IgA也升高。重要的是,共生特异性 IgA +在多发性硬化患者的炎性病变中观察到 ASCs。然而,这种现象可能不是 IgA 独有的,因为已发现多发性硬化患者脑脊液中的 IgG 对多发性硬化相关的肠道细菌裂解物具有反应性。这些细菌反应性 IgG 在疾病中的影响尚未完全阐明。
最后,虽然 IgA + ASC 现在已经在发炎的 EAE 和 多发性硬化中进行了描述,但现在人们认识到这些细胞在体内平衡中起着重要作用。具体而言,已在健康小鼠和人类的软脑膜中检测到肠道共生特异性 IgA + ASC,但在无菌小鼠中不存在。这些细胞可能在硬脑膜窦附近维持屏障完整性;然而,它们也可能有助于中枢神经系统的静止。
总而言之,除了在塑造微生物组方面广受赞赏的作用外,产生 IgA 的 ASC 在健康和多发性硬化/EAE 中枢神经系统中同样发挥着重要作用。
IgA肾病(IgAN)
1968 年首次发现IgA 肾病(IgAN,别名Berger 病)描述了由于在肾脏中形成炎性免疫复合物而导致半乳糖缺乏的 IgA1 在肾小球系膜中沉积和随后的肾小球肾炎。
IgAN 是全世界最常见的原发性肾小球肾炎,其临床特征通常是无症状血尿和进行性肾病。
最近的估计表明,大约四分之一的 IgAN 患者在 20 年内最终发展为终末期肾病,因此死亡风险增加。
IgAN 背后的病因似乎始于固有层中肠激活 B 细胞和抗体分泌细胞 (ASC) 的扩张。
小鼠和人类研究均表明,APRIL 或高同源性 BAFF 的转基因表达会导致IgA1 铰链区的异常O-糖基化和 IgA1 产生的高反应性。当 IgA + ASC 离开次级淋巴组织进入循环时,它们可以进一步分化为长寿命的 IgA +浆细胞。半乳糖缺乏的 IgA1 的系膜沉积物会过度激活补体系统 和/或与 IgG 自身抗体复合,这共同导致促炎反应和肾损伤。
尽管 IgAN 是一种影响肾脏的疾病,但它的起源也与肠道微生物群—肾轴密切相关。2011 年发现四个新的 IgAN 基因座富含与“用于 IgA 产生的肠道免疫网络”相关的 KEGG 途径,包括与粘膜免疫即局部病原体多样性的强正相关。
IgA 血管炎的发病机制
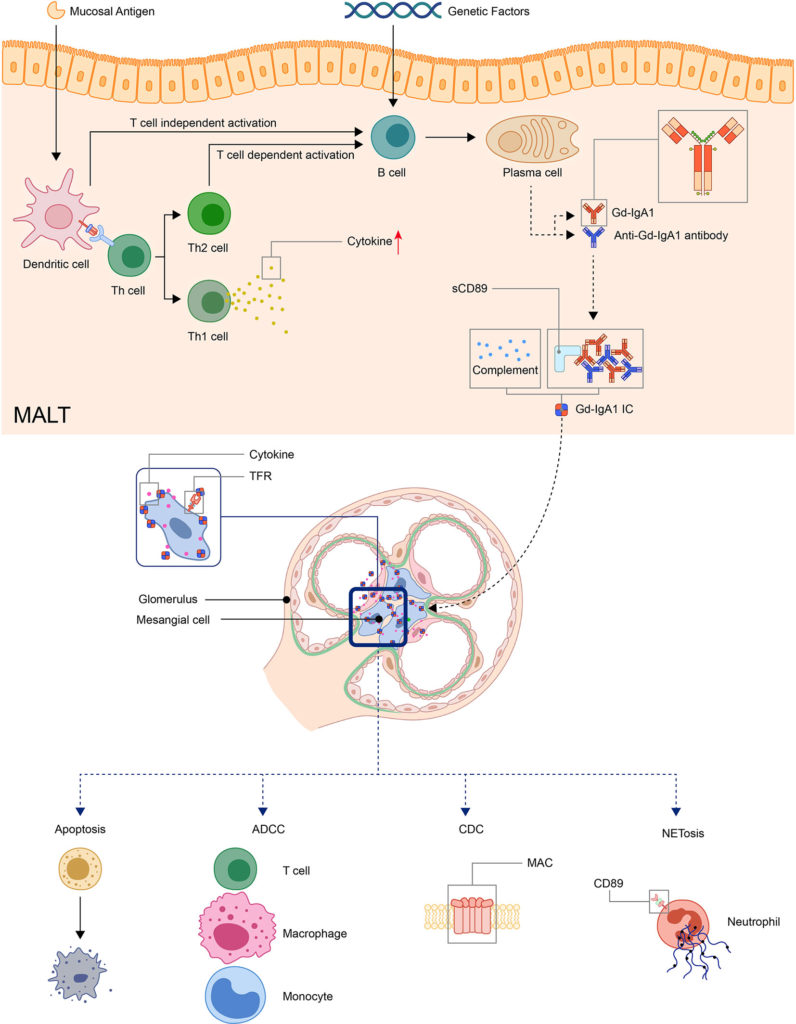
doi.org/10.3389/fimmu.2021.771619
黏膜抗原可以通过T细胞依赖或独立的方式激活MALT中的B细胞。后者通过 TLR 途径激活 B 细胞。在遗传因素的作用下,活化的 B 细胞变成浆细胞并产生 Gd-IgA1。Gd-IgA1 和抗 Gd-IgA1 自身抗体与其他成分(包括 sCD89 或补体)一起形成循环免疫复合物。然后,免疫复合物沉积在器官中并激活炎症反应。在肾脏中,免疫复合物可以通过 TfR 激活系膜细胞,导致肾细胞凋亡和炎症细胞的募集。
最近,在 2021 年发现了几个与 IgAN 中微生物组成变化相关的微生物组数量性状基因座,例如,Dialister和Bacilli的丰度降低,但Lachnobacterium的丰度增加。IgAN 患者的关键细菌有显着的分类学改变,包括埃希氏菌、志贺氏菌水平升高和罗氏菌属、毛螺菌科、梭菌属水平降低。
有趣的是,在 IgAN 患者中,某些肠道代谢物(如短链脂肪酸)与其细菌产生物同时显着减少。
值得注意的是,对人源化小鼠进行抗生素治疗足以显着降低 IgAN 的病理生理特征,包括 IgA1 系膜沉积、免疫复合物和肾小球炎症。因此,肠道微生物群似乎是产生粘膜来源的肾毒性 IgA1 的重要因素,但需要更多的研究来进一步确定 IgAN 中的肠道微生物特征。
值得注意的是,微生物蛋白酶可以从肾小球中去除 IgA 免疫复合物 ,这表明有机会以依赖微生物群的方式治疗性解决 IgAN。
近日研究发现:IgAN患者志贺氏菌属的显着扩增
该研究选取127名IgA肾病(IgAN)患者和127名对照,随机分为发现和验证队列,对 77 名患者的亚组进行了前瞻性随访,以进一步剖析 6 个月免疫抑制治疗后肠道菌群变化与治疗反应之间的关系。
结果发现,治疗前,α-多样性降低(Shannon,P = 0.03),变形菌门-γ变形菌纲-肠杆菌目-肠杆菌科-埃希氏杆菌-志贺氏菌属显著扩增,这种情况在免疫抑制治疗 6 个月后达到临床缓解的患者中逆转。
IgAN患者肠道菌群失调的标志,以埃希氏菌-志贺氏菌属的显着扩增为特征,可作为 IgAN 有前景的诊断生物标志物和治疗靶点。
IgA 血管炎(IgAV)
在 IgAN 患者中经常同时发现的是 IgA 血管炎 (IgAV),也称为过敏性紫癜,一种 IgA 沉积在血管中导致炎症的疾病。关于 IgAV 和 IgAN 是否是同一疾病在不同组织中的两种临床表现,一直存在争议。两种疾病的病原学概念基本相同,但在症状和流行病学方面存在细微差别。
与 IgAN 主要发生在成人和血尿是第一个临床指标相比,IgAV 患者在儿科人群中更常见,他们表现出症状:
IgAV 患者的多样性显着下降,但IgAV 患者的梭杆菌数量增加。IgAV 患者与Dialister (梭状芽胞杆菌家族的前一个成员)和更丰富的大肠杆菌科的志贺氏菌呈负相关。
有趣的是,其他血管炎病例,如川崎病,也表现出更多的梭杆菌,而 Behçet 综合征患者的丁酸生成细菌较低,例如Roseburia和Clostridia.
这暗示梭杆菌最有可能是血管炎中的病原菌,而短链脂肪酸丁酸盐则是一种有益的代谢物,在血管炎和肾病期间的可用性有限。
尽管如此,仍需要更大规模的临床研究来了解微生物群在肾病和血管炎中的相关性,并评估微生物群依赖性预后和 IgAN 和 IgAV 的治疗策略。
胆汁是一种黄绿色的分泌产物,负责乳化饮食中的脂质和脂溶性维生素。肝脏负责胆汁合成并将其运输到胆囊中进行储存。在人类中,每天大约有 5–50 µg/mL 的蛋白质以胆汁的形式从肝脏排出。
IgA 进入胆囊的机制
IgG 在肝胆汁中占优势,IgA 在胆囊胆汁中含量最高。具体来说,胆囊胆汁含有聚合 IgA、聚合 SIgA 和游离分泌成分 (SC)。IgA 进入胆囊的机制因物种而异。例如,在人类中,聚合 IgA 由沿肝胆树的相邻浆细胞产生,然后被胆管上皮细胞上表达的 SC-pIgR 复合物捕获,以分泌到胆汁中。
IgA 从循环到胆汁的转运是去除抗原的自然途径
胆汁中大量存在 SIgA 表明 IgA 在肝胆系统中具有重要的生物学功能。胆汁中的 IgA 有几个提议的功能 。1980 年代的大量研究得出结论,IgA 从循环到胆汁的转运是去除抗原的自然途径。将各种免疫球蛋白静脉注射到小鼠体内的放射性标记抗原显示 IgA,而不是 IgG 或 IgM,是抗原转运到胆汁中的主要抗体。
胆汁 IgA减轻继发性肝胆感染
除了预防原发性肝脏感染外,胆汁 IgA 的这一功能对于减轻肠道细菌和寄生虫感染的继发性肝胆感染至关重要。通过将杀死的大肠杆菌菌株注射到 Peyer 斑块中对大鼠进行免疫,产生胆道 IgA 特异性抗大肠杆菌可防止肝胆感染、胆管炎和全身性败血症。
肝胆疾病患者发生胆囊感染的风险更大,与IgA有关
必须注意的是,患有肝胆疾病(例如胆汁淤积、胆石症)的患者发生胆囊感染的风险更大,因为胆管上皮细胞的损伤会导致肝胆 IgA 清除受损和胆汁 IgA 回流到血液中。与此一致,SIgAD 患者自然更容易患肝胆疾病,例如原发性胆汁性肝硬化和胆囊感染 。有趣的是,胆汁液中 IgA 包被细菌的检测可能与肝胆感染患者的临床症状有关,例如发热和白细胞增多。
肝脏中的局部 IgA 具有微生物群反应性
没有太多研究调查肠道微生物群是否会影响肝胆感染中的 IgA。直到最近的研究表明,肝脏中的局部 IgA 被鉴定为具有微生物群反应性,并且来源于离开派尔斑的抗体分泌细胞(即浆母细胞)。
先前的研究还表明,胆结石患者表现出肠道菌群失调,大约 70% 的肠道细菌 OTU 在胆道中可检测到;然而,这是否会影响肝脏和胆汁 IgA尚不清楚。
胆道感染引起的胆汁微生物群变化
最近,人们对胆道感染引起的胆汁微生物群变化给予了更多的认识。与肠道微生物群平行,胆汁微生物群中的四个优势门是变形菌门,厚壁菌门、拟杆菌门和放线菌门。肝吸虫Opisthorchis felineus感染的存在导致胆汁微生物群的 β 多样性增加,螺旋体门的成员增加,克雷伯氏菌属、具合杆菌属、乳酸杆菌属、密螺旋体属、副流感嗜血杆菌和金黄色葡萄球菌的丰度增加。
肝吸虫感染后肠道菌群也发生变化
例如毛螺菌科、瘤胃球菌科和乳杆菌科增加,但紫单胞菌科、丹毒科和真杆菌科减少。
未来的研究应确定胆道和肠道微生物群之间的单一或同时变化是否会影响 IgA 功能,以及这是否可以确定肝胆感染的严重程度。
免疫球蛋白 A 是抵御有害微生物的第一道防线,但它也能维持免疫耐受性。正常的 IgA 水平可以平衡免疫系统。低水平会增加过敏和自身免疫的风险。
低 IgA 水平
较低的 IgA 可能意味着免疫力减弱。此外,最近的研究结果现在暗示它可能表明肠道屏障和/或肠道微生物群失衡。
在成人中,大多数实验室认为低于 60 mg/dL 的 IgA 值较低。
慢性压力、睡眠不足和疲惫会降低唾液中需要第一防御功能的 IgA。单独的血液 IgA 测试不会显示这种降低的免疫反应。
低IgA会增加过敏、感染和自身免疫性疾病的风险。
一些潜在的原因包括睡眠不足、慢性压力、肠道疾病、过敏、某些药物以及罕见的遗传疾病。
85% 到 90% 的 IgA 缺乏症患者没有症状。他们可能只表现出潜在感染或免疫疾病的症状。
IgA 缺乏的人更容易出现过敏症,包括哮喘、过敏性鼻炎/结膜炎、药物过敏或食物过敏等。
过敏性鼻炎患儿的 IgA 水平降低。较低的水平与更严重的症状相关。
母乳中的 IgA 可预防婴儿特应性皮炎。一项研究发现,在生命的第一年摄入母乳中的 IgA 可降低特应性皮炎的风险,直至 4 岁。
较高的血液 IgA 水平与 18 个月大的湿疹发生率较低有关。
婴儿中较高水平的 IgA 似乎与 4 岁时较少发生的过敏有关 。
唾液中的高 IgA 水平与儿童较少的过敏症状相关。
在对牛奶过敏的患者中,自然产生耐受性的患者血液 IgA 水平升高。
在严重哮喘患者的肺中发现了低 IgA 水平。较低的 IgA 水平与更严重的症状相关。
然而,在严重哮喘中,IgA 可能会加重现有炎症而不是促进耐受性。
此外,IgA 缺乏的人,痤疮和慢性自发性荨麻疹的患病率也较高。
如溶血性贫血、1 型糖尿病、类风湿性关节炎、甲状腺疾病和狼疮在 IgA 缺乏的人群中也更常见。
这种情况没有特定的治疗方法(例如用于其他免疫缺陷的免疫球蛋白替代品)。抗生素用于细菌感染患者,对预防严重并发症非常重要。
没有药物治疗的重度抑郁症患者在缓解状态(当他们没有症状时)血液 IgA 水平显着降低。
即使在健康人中,当参与者回忆起使他们沮丧的事件时,IgA 也会减少。
对 IgA 和自闭症的研究好坏参半。自闭症在 IgA 缺乏的受试者及其亲属中更为普遍。
一项研究表明,自闭症与低正常 IgA (<97 mg/dL) 之间存在关联。
然而,另一项研究并未发现自闭症患者的 IgA 水平较低 。
在小型研究中,低 IgA 水平与抑郁症和自闭症有关,但还需要更多的研究。
关于 IgA 和肥胖的研究也参差不齐,尚无定论。
与体重正常的人相比,肥胖的人血液中的 IgA 水平更高。
在同一项研究中,代谢综合征患者的血液 IgA 水平也较高。这包括甘油三酯升高、高血糖或高血压的人。
然而,其他研究发现肥胖人群的 IgA 水平正常,肥胖儿童的IgA水平甚至较低。
这种相互矛盾的研究可能是由于肥胖的原因并不单一。一些引发肥胖的潜在因素可能会增加 IgA 水平,而其他因素可能会降低IgA 水平。需要额外的研究来明确肥胖和IgA之间的联系。
感染风险
IgA 缺陷患者的一级亲属的自身免疫风险也增加(10%,而普通人群中估计为 5%)。
IgA 保护肺部和肠道免受有害微生物的侵害。IgA 缺乏症患者感染的风险更高,包括呼吸系统、肠道、关节和泌尿道感染 。
长时间运动会降低 IgA, 与超马拉松运动员的呼吸道感染增加有关。
选择性 IgA 缺乏
低 IgA 可能由选择性 IgA 缺乏引起,此时只有 IgA 水平低,而其他抗体(IgG、IgM、IgD 和 IgE)正常。
选择性 IgA 缺乏可以遗传或自发发生(由于感染、药物或未知原因)。导致人们缺乏 IgA 的遗传疾病在高达 1% 的人口中被发现。
IgA 缺乏是最常见的原发性免疫缺陷。白人的患病率较高,而亚洲人的患病率较低。
选择性 IgA 缺乏是指 IgA 水平 <7 mg/dL 但 IgM 和 IgG 正常。它可能是由遗传疾病和非遗传因素引起的。
85% 到 90% 的 IgA 缺乏症患者没有症状。有症状的人通常有呼吸道或肠道感染。
感染和乳糜泻
IgA 缺乏的人有发生肠道感染和疾病的倾向,例如吸收不良、乳糖不耐症、乳糜泻和溃疡性结肠炎。
患有选择性 IgA 缺乏症(无法检测到 IgA,但 IgG 和 IgM 水平正常)的患者患乳糜泻的风险要高出 10 – 20 倍。
多项研究报告了乳糜泻患者中 IgA 缺乏症的患病率增加,以及 IgA 缺乏症患者的乳糜泻患病率增加。
肠易激综合征和肠易激综合征
在一项针对 12 名 IBS 患者和 11 名健康对照者的初步研究中,IBS 患者的 IgA 产生细胞较少。虽然应该记住,这项研究非常小,需要更多的跟进。
IBD 与 IgA 缺乏有关。
在克罗恩病和溃疡性结肠炎患者中,肠道中 IgA 的产生减少了。
在多达 8% 的 IBD 患者中发现了低 IgA 。
然而,肠道中较高的 IgA 产量可能导致 IBD 。
涂有高水平 IgA 的细菌可能是 IBD 患者肠道炎症的原因。当移植到患有溃疡性结肠炎的小鼠体内时,涂有 IgA 的细菌会增加肠道炎症。
一些研究将 IgA 缺乏症与 IBD 和 IBS 联系起来,但研究仍无定论。
癌症风险
患有 IgA 缺乏症的人患癌症的风险会适度增加,尤其是肠癌。这项研究是作为一个队列进行的,并且在随访的第一年显示风险增加,这表明存在监测偏差。需要更多的研究来确定癌症风险的程度。
其他疾病
IgA 缺乏症患者与 1 型糖尿病(高达 10 倍)、狼疮、幼年特发性关节炎、IBD(克罗恩病和溃疡性结肠炎)、类风湿性关节炎和甲状腺疾病的相关性更高。
饮食因素
禁食
在 15 名肥胖受试者中,14 天的禁食增加了血液 IgA 水平。
间歇性禁食的小鼠具有更高的 IgA 水平,并且对感染的抵抗力更强。
高蛋白饮食
膳食蛋白质是分泌性IgA产生的主要驱动因素,高蛋白饮食可以通过肠道菌群分泌的细胞外囊泡或代谢产物琥珀酸盐激活Toll样受体4,增加细胞因子APRIL水平,从而促进分泌性IgA的水平增加。
蘑菇
在 24 名志愿者中,食用白蘑菇的人的 IgA 产量增加了。

编辑
白蘑菇增加了小鼠的 IgA 。
在灵芝中发现的化合物会增加小鼠的 IgA。
生活方式
减少慢性压力
持续数天、数周或数月的慢性压力会降低免疫反应。在中年和老年受试者中,慢性压力与较低的 IgA 相关。
牙科学生的自感压力与低IgA相关。
在母亲中,那些经历更多焦虑、抑郁、愤怒、疲劳和混乱的人的母乳 IgA 较低。
接受托儿服务的幼儿中,托儿质量较低的儿童的 IgA 水平较低。
管理压力可以帮助扭转 IgA 的减少。
在24名志愿者中,20分钟的放松显著增加了IgA的产生。此外,那些连续三周每天放松一次的人,其IgA水平比第一次放松的人增加幅度更大。
十分钟的放松使79名日本女性医务人员的IgA增加。
在14名乳腺癌患者中,那些参与放松的患者术后IgA水平较高。
音乐
听音乐提高了 87 名本科生的基线 IgA 水平。
在一组 66 名大学生中,接触背景音乐 30 分钟的人 IgA 水平升高。

参与音乐可能会产生更大的影响。
在 33 名受试者中,那些积极唱歌或演奏打击乐的人的 IgA 水平比只听音乐的人增加得更多。
另一项研究表明,在合唱团中唱歌会增加 IgA。
短期/中度运动,避免剧烈运动
IgA 水平会随着短期或适度运动而增加。这有助于降低呼吸道感染的风险。
与 10 名久坐不动的对照组相比,9 名受试者在休息时有规律的适度运动增加了 IgA。
几项研究表明,适度运动会增加老年人的 IgA。
在 45 名老年人中,每周一次的 60 分钟阻力和 60 分钟中等耐力训练在 12 个月后显着增加了 IgA。
IgA 水平会根据运动的强度和持续时间以及身体活动的类型而变化。短期和适度运动会增加IgA水平,而长时间运动会降低IgA水平。
在 155 名超级马拉松运动员中,IgA 水平在比赛后下降。
因此如果想要增加IgA水平要避免剧烈运动。
足够的睡眠
在一项对32名志愿者的研究中,在快动眼睡眠(REM睡眠)剥夺的四个晚上,IgA水平下降,但在完全睡眠剥夺后没有下降。即使在三个晚上的睡眠恢复后,IgA水平也没有恢复到基线水平。
一些药物可以阻止REM睡眠,并可能降低IgA水平。这些药物包括抗抑郁药和拟交感神经药(模拟肾上腺素和多巴胺作用并激活交感神经系统的药物)。
唾液IgA在睡眠中增加。在昼夜节律紊乱的小鼠中,唾液IgA在睡眠期间没有增加。研究表明,IgA的增加依赖于交感神经系统(战斗或逃跑)的激活,这与REM睡眠有关。
戒烟
一些研究表明,吸烟会降低 IgA 水平。
与不吸烟者相比,嚼烟者和吸烟者的 IgA 水平降低。此外,吸烟者的 IgA 水平明显低于咀嚼者。
在小鼠中,流感病毒感染前三个月的香烟烟雾暴露导致 IgA 水平降低和肺部炎症增加。
然而,一些研究表明吸烟者的 IgA 水平没有差异。
幽默感
使用幽默作为应对技巧的人具有更高的基线 IgA 水平。
看喜剧增加了 15 名大学生和 39 名女性的 IgA。
同样,与观看教育演示的 18 名同学相比,有趣的演示提高了 21 名五年级学生的 IgA 水平。
压力事件会降低 IgA 水平。在 40 名受试者中,具有幽默感的人不太可能因压力而降低 IgA。
光线充足
在一项针对 7 名女性的研究中,与暴露在昏暗光线下相比,白天暴露在强光下会增加 IgA 水平。
母乳喂养婴儿
婴儿通过母乳接受 IgA。然后,肠道内的IgA生成逐渐受到肠道微生物群的刺激。
几项研究表明,母乳喂养的婴儿的IgA水平较高。
益生菌
在一项包含 47 人的研究 (DB-RCT) 中,每天摄入三周的益生菌罗伊氏乳杆菌会增加 IgA 水平。
在 30 名运动员中,安慰剂组训练后 IgA 显着下降,但服用瑞士乳杆菌的运动员没有(DB-RCT)。
含有罗伊氏乳杆菌的口香糖显着增加唾液中的 IgA (DB-RCT) 。
益生菌增加了 40 名烧伤儿童的 IgA 水平并改善了恢复。
在一项针对 66 名孕妇的研究 (DB-RCT) 中,大剂量多菌株益生菌导致婴儿的 IgA 水平更高并改善了肠道功能。
每天摄入干酪乳杆菌会增加 14 名受试者的 IgA 水平。
在一项针对 98 名新生儿的研究中,双歧杆菌增加了低出生体重婴儿的 IgA 水平。
在一项针对 413 名婴儿的研究中,那些服用富含乳酸杆菌的配方奶粉的婴儿的 IgA 水平较高,与母乳喂养婴儿的水平相似。
益生元
益生元是有益细菌的食物。它们改善了肠道菌群。
益生元混合物增加了 45 名超重成人 (DB-RCT)的IgA 水平并改善了代谢参数(CRP、胰岛素、总胆固醇和甘油三酯) 。
在一项针对 187 名婴儿的研究 (DB-RCT) 中,仅喂食配方奶粉并给予益生元的婴儿的 IgA 水平高于接受安慰剂的婴儿。
雪莲果粉含有 50 – 70% 的益生元纤维,服用 18 周后,59 名学龄前儿童的 IgA 增加。
谷氨酰胺
一项对 13 项研究和 1,034 名患者的荟萃分析得出结论,谷氨酰胺增加了肠癌患者的 IgA 并减少了感染并发症。
在 13 名跑步者的高强度间歇训练期间,谷氨酰胺增加了鼻腔但不增加唾液 IgA。
小球藻
4 周的小球藻补充剂增加了 15 名男性的 IgA。
小球藻还增加了 26 名接受强化训练的受试者的静息 IgA。
小球藻增加了 18 名孕妇母乳中的 IgA 浓度。
人参
人参增强了小鼠肠道 IgA 的产生。
然而在其较高的剂量下,它也阻止了 IgA 的释放。
维生素A
分泌型 IgA 穿过黏膜的运输和释放需要维生素 A。
缺乏维生素 A 的大鼠和小鼠肠道中总 IgA 水平降低,但它们的血液 IgA 水平正常。
补充维生素 A 的女性母乳中的 IgA 水平较高。
总的来说,通过健康的饮食和生活方式,保持轻松的状态避免慢性压力、保证充足的睡眠和适度的运动、戒烟、保持良好的幽默感,必要的时候可以配合益生菌、益生元、谷氨酰胺、维生素 A 和间歇性禁食的生活方式可以增强免疫反应并增加 IgA.
研究表明,高 IgA 可能是慢性感染和低度炎症的标志。
没有与高 IgA 水平相关的症状。IgA 高的人可能只会出现感染或炎症性疾病的症状。这时候医生可能会进行其它的测试,以确定IgA高水平的真正原因。
高 IgA 通常表示慢性感染或炎症,尽管许多疾病可以提高其水平。高 IgA 不会引起症状。人们表现出潜在健康问题的症状。
当 IgA 在血管中积聚时发生IgA 血管炎;当 IgA 在肾脏中积聚时,就会发生IgA 肾病。两者都可能产生严重的健康后果。
在 IgA 血管炎中,IgA 沉积在引起炎症的小血管中。常见的症状是皮疹、关节疼痛和肿胀。
IgA 血管炎在儿童中更为常见,这种疾病通常会在数周内消退,无需治疗。在成人中,它可能更复杂、更持久,伴有更严重的肾脏疾病。
在 IgA 肾病中,IgA 复合物沉积在肾脏中。大约 20%-50% 的患者会出现进行性肾功能衰竭。
正常的 IgA 水平可保护身体免受感染、过敏和自身免疫。
IgA 水平是免疫健康的标志。如果没有症状不用太担心,低或高水平不一定表示有问题。
提高 IgA 水平不一定会直接导致免疫平衡的任何改善,但它可以用作免疫健康的生物标志物。
以下是改善免疫健康的补充方法列表,这些方法也被发现可以平衡高 IgA 水平。
尽管研究表明各种饮食和生活方式因素可能会降低 IgA 水平,但仍需要进行更多的大规模研究。
除了下面列出的因素外,请记住 IgA 是炎症的非特异性标志物。要告知医生以解决高 IgA 水平的根本原因,例如慢性炎症或感染。
有关降低炎症的补充方法,例如服用姜黄素或黑孜然油、锻炼或练习瑜伽,以及遵循地中海式饮食。
IgA 指向慢性炎症或感染。医生可能会建议采取措施来降低炎症并监测其他炎症标志物。
可能减少炎症的因素(通过降低高 IgA 来衡量)
可可
在动物研究中,可可降低了血液、肠道和唾液中的 IgA。
避免饮酒
血液 IgA 水平往往会随着饮酒而增加。在 460 人中,大量饮酒者的 IgA 水平很高。
白葡萄酒增加了 5 名男性的血液 IgA 。
然而,虽然血液 IgA 增加,但肠道 IgA 实际上可能会因酒精而减少。在小鼠中,酒精增加了总 IgA,但减少了肠道 IgA。酒精可能会减少 IgA 释放到肠道中。
禁食
避免禁食。在一项研究中,禁食增加了肥胖受试者的血液 IgA 水平。
愤怒管理
在 18 名有愤怒和抑郁症状的健康男性和女性中,容易表达愤怒的人中 IgA 升高。
容易表达愤怒的家庭虐待者的 IgA 水平较高。
愤怒管理技巧可以帮助人们学习如何控制愤怒。
雌激素水平
在 86 名女性中,雌二醇(主要雌激素)较高的女性 IgA 水平较高。
在细胞研究中,雌激素会增加 IgA 向粘液中的转运,从而减少细菌入侵。
虽然雌激素有助于女性保持健康和强大的免疫系统,但理论上过多的雌激素可能会过度提高 IgA。检查雌激素水平以确保激素平衡。
高 IgA 表示慢性炎症。建议采取措施来减轻炎症。一些补充方法包括瑜伽、草药补充剂(姜黄素)和遵循健康饮食。研究表明,可可、避免大量饮酒和避免禁食也可以减少炎症和降低 IgA 水平。
从微生物学的角度来看,关键问题仍然是关于如何靶向 IgA。从其独特的功能能力来看,有必要考虑IgA的临床应用。
免疫后血清和/或分泌物中通常会发现特异性 IgA 升高。虽然通过全身途径接种疫苗往往会产生血清反应,但通过鼻内或口服途径接种疫苗可以引发保护性粘膜反应。作为一个典型的例子,口服霍乱疫苗作为一种诱导保护性黏膜 IgA 反应的手段已得到广泛认可。
使用从捐献血浆中纯化的抗体的免疫球蛋白替代疗法已作为 IgA 缺乏症的常规治疗方法实施。
包括合成工程多价双特异性抗体 (BsAbs) 在内的新疗法已经开发出来,并在临床上被批准用于治疗各种癌症,例如急性淋巴细胞白血病和小细胞肺癌,但目前没有 BsAb 被批准,并且仅限于治疗 IgA 相关疾病。啮齿动物模型的阳性报告支持 BsAb 可有效缓解 IBD.
微生物群中的某些共生体(例如乳酸乳杆菌)已被证明是一种释放这些 BsAb 的传递系统,可用于改善 IgA 相关疾病,例如结肠炎。可以预期,基于 IgA 的 BsAb 与一线微生物群靶向治疗药物(即益生菌和益生元)相结合,将支持免疫疾病和传染病治疗的范式转变。
肠道黏膜 IgA 对微生物群的反应可以为疾病分类、评估肠道病原体感染/负担的影响,以及评估当前或新治疗干预措施提供有效性的方法。
以上我们基本了解IgA-微生物组轴的破坏如何导致结肠炎、结直肠癌和肾病等病理生理状况。此外,甚至在哮喘、食物过敏和肥胖症中都记录了 IgA 对肠道微生物群的反应改变,这进一步说明了在免疫疾病范围之外对 IgA 进行额外研究的必要性。
技术的最新进展促进了我们对各种类型的 SIgA 不同地调节肠道微生物群的理解。关于 IgA-微生物群相互作用的知识可能会为有效调节微生物群的新型基于 IgA 的疗法打开一扇大门。
主要参考文献:
Bunker JJ, Bendelac A. IgA Responses to Microbiota. Immunity. 2018 Aug 21;49(2):211-224.
Furusawa, Y. et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 504, 446–450 (2013).
Takeuchi, T. et al. Acetate differentially regulates IgA reactivity to commensal bacteria. Nature (2021). doi:10.1038/s41586-021-03727-5
Palm, N. W. et al. Immunoglobulin A coating identifies colitogenic bacteria in inflammatory bowel disease. Cell 158, 1000–1010 (2014).
Kau, A. L. et al. Functional characterization of IgA-targeted bacterial taxa from undernourished Malawian children that produce diet-dependent enteropathy. Sci. Transl. Med. 7, 276ra24 (2015).
Nakajima, A. et al. A soluble fiber diet increases Bacteroides fragilis group abundance and immunoglobulin a production in the gut. Appl. Environ. Microbiol. 86, e00405-20 (2020).
de Sousa-Pereira P, Woof JM. IgA: Structure, Function, and Developability. Antibodies (Basel). 2019 Dec 5;8(4):57.
Huus KE, Bauer KC, Brown EM, Bozorgmehr T, Woodward SE, Serapio-Palacios A, Boutin RCT, Petersen C, Finlay BB. Commensal Bacteria Modulate Immunoglobulin A Binding in Response to Host Nutrition. Cell Host Microbe. 2020 Jun 10;27(6):909-921.e5.
Moll JM, Myers PN, Zhang C, Eriksen C, Wolf J, Appelberg KS, Lindberg G, Bahl MI, Zhao H, Pan-Hammarström Q, Cai K, Jia H, Borte S, Nielsen HB, Kristiansen K, Brix S, Hammarström L. Gut Microbiota Perturbation in IgA Deficiency Is Influenced by IgA-Autoantibody Status. Gastroenterology. 2021 Jun;160(7):2423-2434.e5.
Bunker JJ, Erickson SA, Flynn TM, Henry C, Koval JC, Meisel M, Jabri B, Antonopoulos DA, Wilson PC, Bendelac A. Natural polyreactive IgA antibodies coat the intestinal microbiota. Science. 2017 Oct 20;358(6361):eaan6619.
Nakajima A, Vogelzang A, Maruya M, et al. IgA regulates the composition and metabolic function of gut microbiota by promoting symbiosis between bacteria. J Exp Med. 2018; 215(8):2019-34.
Abokor AA, McDaniel GH, Golonka RM, Campbell C, Brahmandam S, Yeoh BS, Joe B, Vijay-Kumar M, Saha P. Immunoglobulin A, an Active Liaison for Host-Microbiota Homeostasis. Microorganisms. 2021 Oct 8;9(10):2117.
Planer JD, Peng Y, Kau AL, Blanton LV, Ndao IM, Tarr PI, Warner BB, Gordon JI. Development of the gut microbiota and mucosal IgA responses in twins and gnotobiotic mice. Nature. 2016 Jun 9;534(7606):263-6. doi: 10.1038/nature17940. Epub 2016 May 25.
Moll JM, Myers PN, Zhang C, Eriksen C, Wolf J, Appelberg KS, Lindberg G, Bahl MI, Zhao H, Pan-Hammarström Q, Cai K, Jia H, Borte S, Nielsen HB, Kristiansen K, Brix S, Hammarström L. Gut Microbiota Perturbation in IgA Deficiency Is Influenced by IgA-Autoantibody Status. Gastroenterology. 2021 Jun;160(7):2423-2434.e5.
Pu A, Lee DSW, Isho B, Naouar I, Gommerman JL. The Impact of IgA and the Microbiota on CNS Disease. Front Immunol. 2021 Sep 15;12:742173.