-

 CNAS L23010
CNAS L23010

 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业 二级病原微生物安全实验室
二级病原微生物安全实验室- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业 二级病原微生物安全实验室

谷禾健康

抗生素的诞生彻底改写了人类对抗微生物感染的历史,从远古先民偶然利用霉变物质疗伤,到现代医学精准调控分子机制,抗生素通过抑制细菌细胞壁合成、干扰蛋白质生成等机制,成为治疗感染的核心武器。
它们按来源可分为天然(如青霉素)、半合成(如阿莫西林)和全合成(如喹诺酮类);按作用谱分为广谱(覆盖革兰氏阳性与阴性菌)和窄谱(针对特定菌种)。
然而,抗生素的过度使用(如农业促生长、医疗误用)导致耐药性急速发展。细菌通过β-内酰胺酶分解青霉素、修饰核糖体靶点逃避大环内酯类药物、外排泵排出喹诺酮类等策略,使药物失效。
耐药基因通过质粒、噬菌体在环境中扩散,甚至存在于未接触抗生素的偏远人群肠道中。通过评估个体肠道菌群中耐药基因的分布特点(如喹诺酮耐药基因的丰度),临床可优先选择对特定菌株敏感的抗生素,减少盲目使用广谱药物带来的生态风险。
本文谷禾通过查阅文献以及指南等,带大家一起深入理解抗生素的分类体系和作用机制,以及如何根据来源、作用谱和效果来认识不同的抗生素,及其针对不同微生物的使用策略探讨,在理解和允许的情况下结合感染或致病菌合理选择抗生素,并知晓其可能的副作用和耐药防控。
“抗生素”一词源自希腊语“anti”(反对)和“bios”(生命),指通过特异性作用于细菌靶点来抑制或杀灭微生物的有机分子。
天然抗生素最初由微生物(如真菌、细菌)产生,例如青霉素由青霉菌合成,链霉素来自链霉菌。现代抗生素的定义已扩展至包括全合成或半合成化合物,如磺胺类、喹诺酮类等。
抗生素的发现是医学史上的里程碑。

图源:tansuo
抗生素的现代角色抗生素在治疗细菌感染(如肺炎、尿路感染、皮肤感染)、预防术后感染及控制流行病中不可或缺。例如,β-内酰胺类抗生素(如青霉素)至今仍是许多感染的一线药物。
回顾抗生素的发展历程,这些微观世界的”分子武器”究竟是如何发挥作用的?它们为何能在细菌的生命过程中制造如此精准的”障碍”?
按来源分类
■ 天然抗生素
抗生素的天然来源涉及从生物成分中生产抗生素,其中包括微生物作为主要来源。
天然来源可能包括从细菌、真菌或放线菌中提取抗生素。
■ 半合成抗生素
这包括使用微生物生产从发酵中获得的物质,然后通过添加官能团进行化学修饰,从而产生变异。
例如:青霉素-氨苄西林、阿莫西林、甲氧西林。
■ 合成抗生素
属于这一类的抗生素是通过化学合成获得的。
示例:磺胺类药物、喹诺酮类药物。
按活性谱分类
■ 广谱抗生素
这些对革兰氏阳性菌和革兰氏阴性菌均具有活性。
例子包括:四环素类、氟喹诺酮类、“第三代”和“第四代”头孢菌素类。
■ 窄谱抗生素
它们的活性有限,主要仅对特定种类的微生物(革兰氏阳性或革兰氏阴性)有用。
例如,糖肽和杆菌肽仅对革兰氏阳性菌有效,而多粘菌素通常仅对革兰氏阴性菌有效。
氨基糖苷类和磺胺类药物仅对需氧微生物有效,而硝基咪唑通常仅对厌氧菌有效。
按抗菌活性分类
■ 抑菌剂
指那些阻止或抑制细菌生长的抗生素,也就是说细菌不会繁殖或产生,但它们不会杀死细菌。
例如:四环素、氯霉素
■ 杀菌剂
指根据所使用的抗生素,通过任何机制杀死细菌的抗生素。
示例:氨基糖苷类
基于行为模式的分类
细胞壁抑制剂:β-内酰胺、杆菌肽
细胞膜抑制剂:聚霉素、唑类抗真菌药物
蛋白质合成抑制剂
核酸合成抑制剂:喹诺酮类、利福平、呋喃妥因
叶酸合成抑制剂:甲氧苄啶、磺胺类

Uddin TM, et al., J Infect Public Health. 2021
按化学成分分类
■ β-内酰胺类抗生素
在引入后的20年里,青霉素是唯一一类β-内酰胺类抗生素。20世纪60年代中期,青霉素类抗生素与头孢菌素类抗生素并列,20世纪末,碳青霉烯类抗生素和单杆菌类抗生素并列。
这些抗生素具有相同的抗菌作用机制,并且都具有β-内酰胺环作为其结构的组成部分,但它们在其他特征上存在很大差异。
这些抗生素的作用机制是抑制细胞壁。
如:青霉素、头孢菌素、碳青霉烯类、单巴坦
■ 四环素类抗生素
这些是一类具有四个环核的抗生素,都是从土壤放线菌中获得的。
四环素最早是在20世纪40年代和50年代开发的,有几种至今仍在使用,如四环素本身、土霉素和金霉素。
多西环素和米诺环素是分别于1966年和1972年发现的更有效的半合成类似物,此后直到2005年引入替加环素(一种甘氨酰甘氨酸衍生物)才有了重大进展,替加环蛋白通常比其他四环素更有效,并对一些对该组早期成员产生耐药性的生物体保持活性。
四环素在实验室浓度下可能具有杀菌性,但体内安全浓度下仅为抑菌剂。它们通过结合敏感生物的30S核糖体来抑制蛋白质合成。
■ 氨基糖苷类
这些是一组天然和半合成的抗生素,具有与两个或多个氨基糖残基糖苷连接的多元氨基。
氨基糖苷类抗生素具有杀菌作用,在碱性条件下活性更强。它们通过干扰细菌蛋白质合成发挥作用,主要针对需氧革兰阴性杆菌,但抗菌谱各异。
氨基糖苷类的所有成员都具有杀菌作用,在碱性pH值下更具活性。
它们通过干扰细菌蛋白质合成来发挥作用。
它们主要对需氧革兰阴性杆菌有活性,但光谱不同。
如:链霉素、新霉素、卡那霉素、阿米卡星、庆大霉素

■ 多肽
低分子量阳离子多肽抗生素,均为强效杀菌剂,但因毒性较高,极少全身使用。
■ 糖肽
目前仅有的两种重要糖肽抗生素是万古霉素和替考拉宁。
它具有复杂的三环糖肽结构,其大分子量意味着它不能穿透大多数革兰氏阴性菌的外膜,因此它的使用实际上仅限于治疗需氧或厌氧革兰氏阳性菌的感染。
万古霉素在接近其最低抑菌浓度(MIC)的浓度下对大多数易感细菌具有杀菌作用,并且是细菌细胞壁肽聚糖合成的抑制剂,其位置与β-内酰胺类抗生素不同。
■ 大环内酯类
如: 红霉素、克拉霉素、阿奇霉素
大环内酯类抗生素是由12-16碳内酯环组成的大分子,通过糖苷键与氨基糖连接。
红霉素于1952年首次发现,至今仍是重要抗生素。它常用于治疗呼吸道、皮肤及软组织感染。大环内酯类药物通过抑制细菌蛋白质合成发挥抗菌作用,通常为抑菌剂,但在高浓度下可能具有杀菌活性。红霉素的抗菌活性具有pH依赖性,随pH值升高而增强,最佳pH值约为8.5,该特性在其他大环内酯类药物中也不同程度地存在。
■ 利福霉素
利福平口服吸收良好,蛋白结合率高(80-90%),能穿透细胞壁作用于细胞内病原体(如结核杆菌)。利福昔明因肠道吸收率低,主要用于局部感染。
如:利福平、利福巴林
■ 链霉素
普利司他霉素、维吉尼亚霉素
奎奴普汀/达福普汀是以30:70比例混合的两种链阳霉素类抗生素。它们源自链霉菌,经化学修饰后,通过抑制蛋白质合成发挥作用。
■ 抗真菌药物
灰黄霉素、多烯类
■ 合成药物
喹诺酮类、咪唑类、硝基呋喃衍生物
■ 氯霉素
氯霉素最初于1947年从委内瑞拉链霉菌中分离得到,后经化学合成。它是一种黄白色结晶固体,可溶于水。其抗菌活性可能源于其硝基苯结构。氯霉素的作用机制是抑制蛋白质合成。
■ 夫西地酸
夫西地酸是一种类似类固醇的杀菌抗生素,主要用于对抗葡萄球菌。
它对青霉素耐药的金黄色葡萄球菌菌株有活性,包括MRSA,可以与红霉素或克林霉素联合使用,用于严重的葡萄球菌感染。
■ 克林霉素
克林霉素是另一种对革兰氏阳性球菌具有显著抑菌活性的抗生素,尽管对革兰氏阴性球菌的活性较低,对肠杆菌的活性完全没有。
克林霉素在结构上与大环内酯类无关,但具有相似的作用机制,因此它们之间可能会发生交叉耐药性。
■ 林可霉素
它是克林霉素的前身,具有类似的抗菌和毒性特性,但效力较弱。
以上,我们已基本了解了抗生素的分类体系及其作用机制,这些分子武器本应是人类对抗微生物感染的坚固屏障。然而,当这些强大的武器被不当使用时,细菌迅速发展出层出不穷的耐药策略,将医学奇迹逐渐转变为临床危机。
全球健康的紧迫威胁
许多与人类疾病流行相关的细菌病原体在抗生素使用后已演变成多重耐药(MDR)形式。例如,多重耐药结核分枝杆菌(MDR M. tuberculosis)是发展中国家和工业化国家都发现的主要病原体,成为20世纪版本的古老病原体。
其他严重感染包括医院相关感染,如:
“超级细菌”指那些因多重突变而具有增强致病性和死亡率的微生物。

超级细菌的崛起:抗生素滥用的致命代价
近十年来,结核病的治疗遭遇了重大危机,广泛耐药结核菌(XDR-TB)和完全耐药结核菌(TDR-TB) 接连出现,这些“超级结核菌”对至少四种一线抗生素完全失效,且在全球快速传播。更令人担忧的是,像大肠杆菌、肺炎克雷伯菌等常见致病菌,也因抗生素滥用纷纷进化出耐药性。其中,青霉素类抗生素(如阿莫西林)和头孢类抗生素的过度使用,直接促使细菌发展出“β-内酰胺酶”这种分解抗生素的酶,让药物彻底失效。
医院里的危险分子也在增加:
某些超级细菌的爆发竟是抗生素滥用的直接后果。例如,艰难梭菌引发的严重腹泻,高毒力产毒素菌株已被确认,作为一种革兰氏阳性孢子形成菌,很容易通过医院人员、设备和气溶胶传播。它的重新突出被认为是医院广泛使用抗生素的结果,如广谱头孢菌素,新的青霉素和氟喹诺酮类药物,导致革兰氏阴性肠道菌群的显著消耗,从而提高了艰难梭菌。
在感染数量和后果方面,霍乱弧菌仍是头号杀手。亚洲和南美洲的疫情仍因缺乏有效监测系统难以控制。这些案例无不警示:抗生素的每一次误用,都在加速催生更危险的敌人。
耐药机制:细菌的“生存绝招”
细菌对抗生素的耐药性,本质上是它们演化出的保命技能。科学家发现,这些技能涉及基因突变、酶分解药物、改变自身结构等多种手段,甚至能通过“共享基因”在细菌间传播耐药能力。
以下是几种典型的耐药策略:
1
分解抗生素的“盾牌”——β-内酰胺酶
β-内酰胺酶是一种酶,能像剪刀一样切断青霉素、头孢类抗生素的结构(β-内酰胺环),让药物失效。β-内酰胺酶基因可能是分布最广泛的耐药基因。
这种酶的基因广泛存在于细菌的质粒(可移动的基因片段)上,甚至能在不同细菌间“快递传播”。
例如,新的超广谱β-内酰胺酶(CTX-M酶)最初在土壤中的Kluyvera菌里被发现,并于20世纪90年代出现在临床上,后来传播到致病的大肠杆菌中,导致头孢类抗生素失效。
2
改变“药物靶点”——大环内酯类耐药
红霉素等药物,广泛用于革兰氏阳性感染治疗。
通过结合细菌的核糖体(蛋白质工厂)来杀菌。而耐药菌通过修改核糖体的RNA或蛋白质结构,让药物无法锁定目标。
结果:即使更换新型大环内酯类药物,细菌仍能通过类似机制抵抗,形成“交叉耐药”。
3
升级“防御系统”——喹诺酮类耐药
当这类高效抗生素在1987年首次推出时,一些专家过于乐观地认为:”这种新型抗生素几乎不可能产生耐药性,因为细菌需要同时发生至少两种基因突变才能对它产生明显的抵抗力。”
然而,现实很快就打破了这些预测:
科学家感叹,永远别低估细菌的适应能力。
4
天生的“耐药基因”——内在耐药性
内在耐药性指细菌基因组中存在可能产生耐药表型的基因,即原始或准耐药性。不同属、种、菌株等表现出不同范围的抗生素反应表型。例如,某些肠道菌天生对磺胺类药物不敏感。
潜在威胁:这些基因平时潜伏,一旦被其他致病菌获取,可能引发新的耐药危机。
科学家们开发了一种方法来预测潜在的耐药基因。通过系统性地敲除细菌基因组中的每一个基因,观察哪些基因缺失会使细菌对抗生素变得更敏感。
逻辑很简单:如果缺少某个基因会使细菌更容易被抗生素杀死,那么这个基因过度表达可能会使细菌更具抗药性。
这种前瞻性研究已在多种细菌中进行:
5
环境中的“耐药基因库”
长期以来,人们知道可以通过在实验室中将环境细菌培养在含抗生素的培养基上来分离耐抗生素的细菌菌株。
产抗生素细菌的自我保护机制
对于那些能够产生抗生素的细菌(如放线菌),它们拥有耐药性并不令人惊讶。这是一种自我保护机制,细菌必须有能力抵抗自己生产的”武器”。在许多情况下,科学家们已经确定这种耐药机制是通过特定的酶来修饰抗生素,使其失去活性。
链霉菌:β-内酰胺酶的重要来源
链霉菌(Streptomycetes)能产生多种β-内酰胺酶(一种能破坏青霉素类抗生素的酶)。这很可能是如今临床上出现的一些β-内酰胺类抗生素耐药性的源头。同样,环境中的Kluyvera菌已被证实是CTX-M基因(一种重要的耐药基因)的起源。
多重耐药机制的进化蓝图
在产抗生素的细菌中,多重耐药机制十分常见,如四环素生产者Streptomyces rimosus就具有这种特性。根据生化和遗传相似性分析,这些天然存在的耐药机制预示了后来在致病细菌中发现的耐药机制。换句话说,致病菌的耐药策略很可能是从环境细菌那里”借鉴“而来的。
启示:耐药性为何难以控制?
对其他产抗生素细菌的进一步研究,如芽孢杆菌科、假单胞菌、蓝藻和多种放线菌科(这是一个已知能产生许多生物活性小分子的细菌群体),将有助于我们更全面地了解自然界中存在的耐药基因的本质和多样性。这些知识对于预测和应对未来可能出现的临床耐药问题至关重要。
当我们越来越理解这场微生物与药物之间的分子博弈,便不得不直面一个问题:如何在保持抗生素治疗效力的同时,为抗生素的未来开辟一条可持续发展的道路?下一章节我们来继续了解。
常见感染与抗生素选择
1. 尿路感染 (UTI)
无并发症尿路感染
– 首选药物:甲氧苄啶/磺胺甲噁唑(Bactrim Ds)、呋喃妥因(硝基呋喃类)
– 替代方案:头孢氨卡(Keflex,第一代头孢菌素)、阿莫西林/克拉维酸(Augmentin)
复杂性或上尿路感染(如肾盂肾炎)
– 首选药物:氟喹诺酮类(如环丙沙星Cipro、左氧氟沙星Levaquin)
– 替代方案:第三代头孢菌素(如头孢曲松)、氨基糖苷类(庆大霉素)
用药注意事项
– 磺胺类药物可能引起结晶尿,需充分水化
– 氟喹诺酮类因肌腱损伤和神经毒性风险,禁用于儿童及孕妇
– 氨基糖苷类雲监测肾功能及耳毒性
2. 肺 炎
社区获得性肺炎(CAP)
– 非重症患者:阿奇霉素(Zithromax,大环内酯类)或多西环素(Vibramycin,四环素类)
– 合并基础疾病或耐药风险:阿莫西林/克拉维酸(Augmentin)联合大环内酯类
医院获得性肺炎(HAP)
– 首选药物:第三代头孢菌素(如头孢他啶)或碳青烯类(如美罗培南)联合氟喹诺酮类
用药注意事项
– 大环内酯类(如阿奇霉素)可能延长QT间期,禁用于心律失常患者
– 碳青霉烯类需螫惕超广谱β-内酰胺酶(ESBL)耐药菌感染
3. 皮肤及软组织感染
轻度感染(如蜂窝织炎)
– 首选药物:头孢氨苄(Keflex)或双氯西林(耐酶青霉素)
疑似耐甲氧西林金黄色葡萄球菌(MRSA)感染
– 首选药物:复方磺胺甲噁唑(Bactrim DS)、克林霉素(Cleocin)
– 替代方案:万古霉素(糖肽类)或达托霉素(环脂肽类)
用药注意事项
– 需警惕艰难梭菌感染克林霉素可能引起伪膜性肠炎
– 万古霉素需根据肾功能调整剂量,监测血药浓度
4. 链球菌性咽炎 (Strep Throat)
首选药物
– 青霉素V钾(Penicillin VK)或阿莫西林(Amoxil)
青霉素过敏患者
– 阿奇霉素(Zithromax)或克拉霉素(Biaxin)
用药禁忌
– 四环素类(如多西环素)对链球菌活性低,不推荐作为一线治疗
– 氟喹诺酮类对链球菌无效
禁忌症与用药原则
病毒性感染的绝对禁忌
流感、普通感冒、急性病毒性鼻窦炎等无需抗生素治疗
滥用抗生素将增加耐药风险:60%的急性支气管炎为病毒性,仅需对症处理
特殊人群剂量调整
肝肾功能不全者:氨基糖苷类需减少剂量;磺胺类慎用于肾功能减退患者
孕妇与哺乳期妇女:禁用四环素类(影响胎儿骨骼发育)、氟喹诺酮类(致畸风险)
药物相互作用风险
大环内酯类(如红霉素)抑制CYP3A4酶,可能升高华法林、他汀类药物浓度
复方磺胺甲噁唑(Bactrim)增强磺脲类降糖药作用,需监测血糖
不合理用药的后果
耐药性加速:如氟喹诺酮类滥用导致铜绿假单胞菌外排泵基因突变频发
生态失衡:广谱抗生素(如第三代头孢菌素)破坏肠道菌群,增加艰难梭菌感染风险
经济负担:非必要使用增加医疗成本
抗生素的选择需基于病原体敏感性、感染严重程度及患者个体化因素。
临床实践中应严格区分细菌性与病毒性感染,优先使用窄谱抗生素。通过规范用药,可最大限度延缓耐药性蔓延。
通过肠道菌群检测识别抗生素耐药基因
宏基因组测序可获得细菌群体的全部基因组信息,通过序列比对和功能注释,可鉴定出各种已知和新型耐药基因,全面评估耐药基因的种类和数量。
例如,在人体肠道宏基因组中发现了大量β-内酰胺酶等耐药基因。
2021年一项研究显示,印度某社区爆发的碳青霉烯耐药肺炎克雷伯菌感染,其耐药基因与当地养殖场废水中分离的菌株高度同源,证实耐药基因通过水源传播至人体。
数据驱动的防控策略:从预警到干预
基于宏基因组分析的耐药风险评估,可制定精准防控措施:
对畜牧从业人员、医护人员等暴露于耐药基因环境的人群定期检测肠道菌群,早期发现耐药定植。
临床用药指导:建立区域耐药基因数据库,实时更新病原体耐药谱,帮助医生相对个性化地选择的窄谱抗生素(如用头孢他啶替代广谱碳青霉烯类药物)。
谷禾宏基因组肠道菌群检测报告中,就有抗生素耐药基因检测相关内容。
<来源:谷禾宏基因组肠道菌群健康检测数据库>
筛查应用:如对术前患者进行肠道耐药基因检测,若检出 vanA(万古霉素耐药基因),可优先选择替考拉宁或达托霉素替代万古霉素,降低术后感染风险。
用药方案优化:在复杂性尿路感染治疗中,若患者肠道中qnrS(氟喹诺酮耐药基因)丰度较高,应避免使用左氧氟沙星,转而选择磷霉素或呋喃妥因。
存在争议
支持证据
Cochrane综述表明,在抗生素后服用益生菌的儿童腹泻较少。
注:Cochrane综述是由国际非营利组织Cochrane协作网制作的系统性综述,被公认为循证医学领域的最高标准,旨在通过系统性方法评估医疗干预措施(如药物、手术、诊断测试)的有效性、安全性及适用性,为临床决策和政策制定提供可靠证据。
Cochrane对23项研究(3938名参与者)的综述调查了给予含有以下一种或多种的益生菌:
22/23项报告抗生素相关性腹泻发病率的试验结果显示,与活性、安慰剂或无治疗对照组相比,益生菌具有显著益处(益生菌组为8%,对照组为19%)。
报告副作用的16项试验(n=2455)均未记录任何可归因于益生菌的严重副作用,最常见的副作用是皮疹、恶心、胀气、肠胃气胀、腹胀、腹痛、呕吐、痰增、胸痛、便秘、味觉障碍和食欲低下。作者的结论是,益生菌对预防抗生素相关性腹泻具有保护作用。相对风险为0.46(95%CI 0.35至0.61),NNT为10。
作者认为,鼠李糖乳杆菌或布拉氏酵母菌的菌落形成单位为50亿~400亿个/天是最合适的选择。他们还评论,尽管在这些试验中,在其他健康儿童中没有观察到严重的不良事件,但在具有潜在风险因素(如中心静脉导管使用)的严重衰弱或免疫功能低下的儿童中观察到了严重的不良事件,并建议在进行进一步研究之前,应避免在有不良事件风险的儿科人群中使用益生菌。
反对观点
以色列魏茨曼科学研究所的科学家发现,当患者服用抗生素后,如果连续四周补充含11种菌株的复合益生菌,肠道菌群需要整整六个月才能恢复正常。尽管这些益生菌成功在肠道“安家落户”,但新引入的细菌和酵母菌株反而干扰了原有菌群的恢复。
对比发现:未服用益生菌的患者,停用抗生素后仅需三周,肠道菌群就自行恢复正常。
关键提示:
替代方案?
科学家正在探索其他恢复肠道的方法:
注:目前异体粪便移植目前是主流,但自体粪便移植可避免供受体不匹配的问题,部分研究仍处于早期阶段,需进一步验证长期安全性和有效性。
实用建议
如果你决定服用含有抗生素的益生菌,请在开始服用抗生素的同一天开始服用,但不要与抗生素完全同时服用。
选择高活性菌株,如鼠李糖乳杆菌GG、布拉氏酵母菌(每日5-400亿菌落单位)。
间隔服用时间,抗生素服用后2小时再服益生菌,避免活性被抑制。
抗生素的未来发展需在科学突破、临床实践与健康管理之间找到平衡点。面对耐药性危机,人类正通过多维度创新重塑感染治疗的格局。
1. 新型抗生素研发:从基因靶点到智能设计
宏基因组学驱动,通过分析环境样本(如深海沉积物、极端环境微生物)的宏基因组数据,挖掘新型抗生素合成基因簇。
合成生物学技术:利用CRISPR-Cas9基因编辑改造放线菌的代谢通路,定向生产改良抗生素。如改造链霉菌以高产泰斯巴汀(一种可靶向细胞壁脂质的广谱抗生素)。
噬菌体-抗生素联用,穿透生物膜杀灭耐药菌。例如,噬菌体搭载 β-内酰胺酶抑制剂,可恢复青霉素对产酶菌的活性。
缩短新药研发周期(从传统10-15年压缩至5-8年),降低开发成本,并为“无药可治”的超级细菌感染提供解决方案。
2. 精准医疗:从“一刀切”到个体化治疗
通过宏基因组测序分析患者肠道耐药基因谱(如 CTX-M、NDM-1),指导抗生素选择。例如,若检测到高丰度氟喹诺酮耐药基因(qnr),则优先选择磷霉素或呋喃妥因替代左氧氟沙星。
针对抗生素导致的菌群失衡,开发“益生菌-益生元-后生元”组合产品。例如,含 Akkermansia muciniphila 的微生态制剂可加速肠道屏障修复,减少艰难梭菌定植风险。
催生“感染精准医疗”细分市场,涵盖检测试剂、数据分析平台和微生态调节产品,预计2030年全球市场规模将突破520亿美元。
3. 耐药性防控:环境-医院-社区的联防联控
建立全球耐药基因数据库(如 ResistoMap),追踪耐药基因热点区域。例如,印度恒河流域因高浓度 blaNDM-1 基因被列为“耐药红色预警区”。
医院综合当地耐药数据、患者过敏史、肝肾功能、菌群检测报告等各类数据,个性化推荐窄谱抗生素。
设立“微生物学-药学-数据科学”交叉学科,开展耐药知识科普,提高公众对不当使用抗生素危害的认知,利用菌群检测手段辅助判别耐药情况,指导居民合理用药。
通过环境、医院、社区的协同联动,构建一个全方位的耐药性防控体系,减缓耐药菌的传播速度,保障公众健康。
抗生素的未来不仅是科学问题,更是社会命题。从基因编辑实验室到养殖场,从医院处方系统到家庭药箱,每个环节的革新都在重塑人类与微生物的关系。将技术创新、精准医疗和全球治理相结合,让抗生素这一“医学奇迹”持续守护生命,而非成为耐药危机的推手。
主要参考文献
Denyer S.P, Hodges N, Gorman S.P and Gilmore B.F. 2011. Hugo and Russell’s Pharmaceutical Microbiology. Eighth edition. A John Wiley & Sons, Ltd. Publication. Page no.169-193.
Antibiotics: Comprehensive Guide
Last updated: August 10, 2022 by Sagar Aryal
Davies J, Davies D. Origins and evolution of antibiotic resistance. Microbiol Mol Biol Rev. 2010 Sep;74(3):417-33.
Uddin TM, Chakraborty AJ, Khusro A, Zidan BRM, Mitra S, et al., Antibiotic resistance in microbes: History, mechanisms, therapeutic strategies and future prospects. J Infect Public Health. 2021 Dec;14(12):1750-1766.

谷禾健康

抗生素可有效治疗细菌感染,但其广泛使用引发了抗生素耐药性,抗生素耐药性是个全球性的大问题,这与人类疾病密切相关。这里存在着一个被我们长期忽视的关键因素:肠道耐药组。
当我们在医学上使用抗生素时,肠道里的微生物数量和基因组成可能会发生变化,这样就增加了抗生素耐药性在肠道中积累的风险。
肠道耐药组(gut resistome)是指肠道微生物群中存在的所有抗生素耐药基因(ARG)的集合,可以从自然环境和食物摄入中获得,并且可以随着食物中抗生素和其他抗菌物质诱导的选择压力进一步积累。
这些耐药基因不只是让细菌对抗生素产生抵抗力,更可能通过影响肠道微生物平衡、改变代谢功能、触发炎症反应,与多种慢性疾病建立起令人担忧的联系,包括代谢紊乱、心血管疾病、肝病、神经系统疾病等。
当我们服用口服抗生素时,药物直接穿过消化系统,破坏肠道微生物的平衡,促进抗生素耐药基因的出现和扩散。更令人忧虑的是,这些耐药基因可以通过食物链、水源和环境污染从自然环境进入人体,并在肠道微生物间通过水平基因转移快速传播。
肠道耐药组的富集不仅加剧了抗生素耐药性,还导致了多重耐药性感染的出现。这些感染构成了重大的公共卫生挑战,因为它们更难治疗,通常需要更昂贵的疗法,这些疗法可能具有更大的副作用且效果较差。
肠道耐药组已成为连接抗生素使用、耐药危机和慢性疾病的关键环节。
已知膳食成分会影响肠道微生物组的发生率、活性和多样性,例如,研究表明,地中海饮食(大量摄入完整的、未加工的植物性食物、橄榄油和乳制品;适量的家禽和鱼类;以及最少食用红肉)与粪杆菌属和短链脂肪酸的增加有关,它们具有抗炎作用。相比之下,西方饮食(高热量,富含动物蛋白、饱和脂肪、单糖和超加工食品,而纤维、水果和蔬菜含量低)与 Blautia、拟杆菌属、瘤胃球菌属物种的丰度较高有关,这与代谢紊乱和慢性炎症的风险增加有关。
在此基础上,饮食不仅会影响有益细菌,而且还有可能改变肠道中抗生素耐药细菌的存在。到目前为止,已发现食品中的生物活性常量营养素、植物化学物质和益生菌可以改变肠道中的抗生素耐药细菌,为调节肠道耐药组的功能性食品打开了新的研究窗口。
随着科学进步,肠道菌群检测技术已能够识别耐药基因的存在和丰度,使个体ARG状况的评估成为可能,为个性化的肠道耐药组管理提供客观依据。
本文强调了肠道耐药组与人类疾病(如代谢紊乱、心血管疾病、肝病和神经系统疾病)之间的联系,包括抗生素作用机制和耐药性的发展,还讨论了饮食习惯和饮食成分(包括生物活性宏量营养素、植物化学物质和益生菌)通过增强抗生素疗效和潜在降低耐药性,对肠道抗药组的组成产生影响。
强调了对靶向肠道耐药组的功能性食品的新兴趋势,以及越来越关注具有调节抗生素耐药性潜力的生物活性植物化学物质。
靶向肠道耐药组的功能性食品,正从概念走向临床,未来,通过“膳食-菌群-肠道耐药组”三位一体的精准干预,有望将耐药危机防控窗口前移至日常饮食,从功能性食品的创新到临床营养的精准化应用,为全球公共卫生提供可持续解决方案。
★
随着抗生素在现代医学和农业中的广泛应用,一个隐形的健康危机正在悄然形成——肠道抗生素耐药基因组(ARG)的扩张。
ARG 可以从亲本细菌(垂直基因转移)遗传,也可以通过水平基因转移从其他微生物获得,水平基因转移是遗传物质在细菌之间移动的过程,甚至跨物种移动。
这些耐药基因不仅可以使细菌对抗生素产生抵抗力,更可能通过影响肠道微生物平衡、改变代谢功能、触发炎症反应,与多种慢性疾病建立起令人担忧的联系。
给药方法会显著影响肠道微生物组和耐药组
静脉注射(注射到血液中)或局部(直接涂抹在皮肤上)的抗生素通常与肠道微生物组的直接接触最少,因此对肠道耐药组的影响较小。
然而,口服抗生素是最常见的治疗形式,它通过消化系统,直接影响肠道微生物群。这种相互作用会破坏肠道微生物的平衡,并促进抗生素耐药基因(ARG)的出现。
Zhang等人强调了这一差异,表明小鼠口服四环素或阿莫西林会迅速增加肠道中ARG的存在,而静脉注射则会延迟或减少这种作用。这种差异与药物的排泄途径有关:氨苄西林主要通过肾脏排泄,限制了静脉注射后的肠道暴露,而四环素通过肾脏和胆汁排出,即使在静脉注射后也会暴露肠道。这些发现强调了口服抗生素在塑造肠道抗药性中的重要作用。
此外,抗生素的广泛使用,无论是直接在人类身上还是间接在动物身上,都会导致抗生素耐药性在自然环境和生物中的积累,从而促进人类肠道耐药性的扩大。
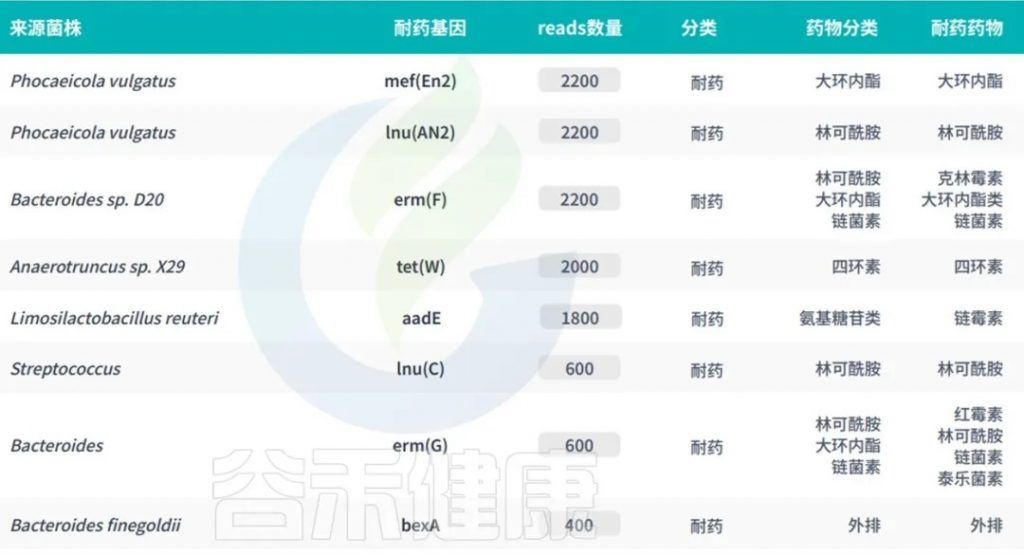
严重的健康问题与肠道耐药组有关,包括代谢紊乱、心血管疾病、肝病、神经系统疾病(下图)。
肠道耐药组对人体健康的影响
doi:10.1111/1541-4337.70143
a) 肠道微生物群在健康和疾病中的机制。短链脂肪酸(SCFA)来源于膳食游离脂肪酸受体(FFAR2/3),通过释放肽YY(PYY)和胰高血糖素样肽1(GLP-1)等肠道激素来促进饱腹感,同时增强胰岛素分泌和敏感性,导致肥胖和2型糖尿病(T2D)。来自革兰氏阴性菌的脂多糖(LPSs)通过激活巨噬细胞和释放促炎细胞因子(TNF-α、IL-1β、IL-6)触发全身炎症,导致肥胖、2型糖尿病、炎症性肠病(IBD)、癌症(CRC)、心血管疾病和肝脏疾病。次生胆汁酸、三甲胺-N-氧化物(TMAO)、硫化氢(H2S)和含氮有机化合物(NOC)会导致胆汁酸信号失调、氧化应激和DNA损伤,从而导致CRC等慢性疾病。此外,肠道和中枢神经系统(通过迷走神经)以及肠道衍生代谢物(如三甲胺(TMA)和TMAO)之间的相互作用会影响神经和心血管健康。
下面我们来看具体与肠道耐药组相关的疾病。
抗生素耐药性通过破坏肠道微生物群及其相关代谢途径,在包括肥胖、2型糖尿病(T2D)、炎症性肠病(IBD)和结直肠癌(CRC) 在内的代谢紊乱中发挥着至关重要的作用。肠道耐药组是肠道微生物组的一部分,也与这些疾病的发展有关。
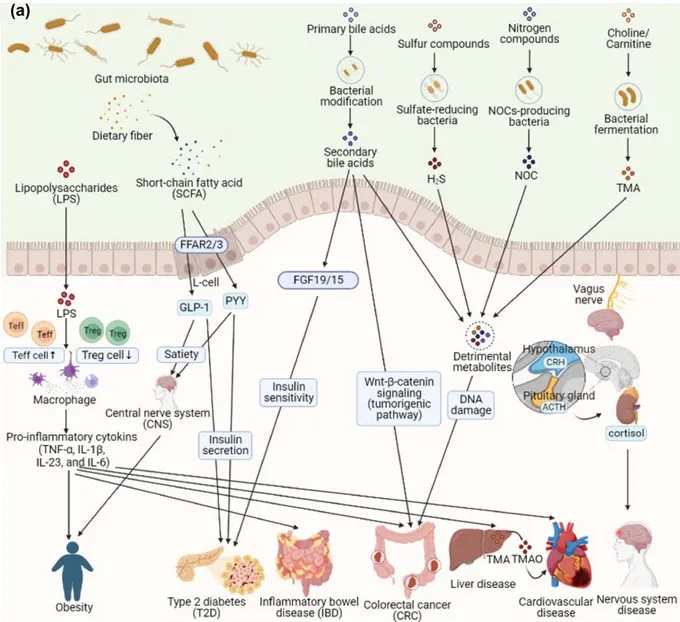
doi:10.1111/1541-4337.70143
b) 抗生素抗性基因对疾病的影响。特定的抗生素抗性基因(ARG)与代谢紊乱、神经系统疾病和肝病有关。ARGα多样性与心脏代谢风险增加有关。
肥胖
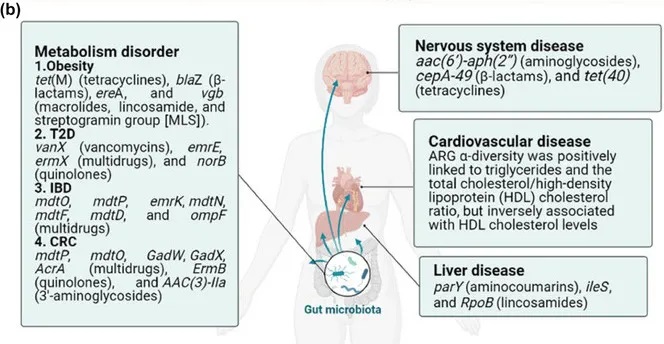
在肥胖人群中,肠道生态失调通过改变短链脂肪酸和脂多糖(LPS)影响能量代谢和慢性炎症(图a)。短链脂肪酸通过刺激GLP-1和PYY等激素的释放来调节饱腹感,这些激素作用于下丘脑。此外,肠道细菌的不平衡,特别是革兰氏阴性菌如变形杆菌的丰度增加,会导致LPS水平升高,并通过增加促炎细胞因子(TNF-α、IL-1β、IL-6)的产生引发全身炎症。
一项研究中,与富营养化参与者相比,肥胖个体中梭杆菌、肠球菌、大肠杆菌的丰度更高,同时ARG水平升高,如tet(M)(四环素类)、blaZ(β-内酰胺类)、ereA和vgb(大环内酯类、林可酰胺类、链菌素组[MLS])。这表明ARG与革兰氏阴性菌之间的关系,是由拟杆菌门和变形杆菌门之间的不平衡所驱动的。
doi: 10.3390/genes10050349
2型糖尿病
在2型糖尿病中,肠道微生物失调,包括抵抗体的变化,导致SCFA产生减少,损害PYY和GLP-1的分泌,这对胰岛素调节和胰高血糖素水平至关重要(图a)。
胆汁酸代谢的改变刺激成纤维细胞生长因子19/15(FGF19/15)的释放,进一步促进胰岛素敏感性和葡萄糖耐量。
ARG丰度的增加,包括Vancomycin_vanX、Multidrug_emrE、MLS_ermX、喹诺酮_norB,与2型糖尿病风险的增加有关,耐药组的变化发生在肠道微生物群变化之前(图b)。
在IBD中,能够产生短链脂肪酸的细菌数量减少,这会削弱短链脂肪酸的抗炎作用。短链脂肪酸通常有助于Treg和效应性T细胞(Teff)的分化。由于这种减少,炎症和肠道上皮细胞损伤会加剧,从而为有害细菌的滋生创造条件,并导致慢性炎症的持续。大肠杆菌中携带的抗生素抗性基因,如mdtO、mdtP、emrK、mdtN、mdtF、mdtD、ompF,变得更加普遍。这些ARGs通过破坏肠道屏障功能,增加肠道的通透性,进而促进炎症反应(如图b所示)。
此外,ARGs和细菌的毒力因子可以通过水平基因转移在微生物群落中传播,进一步加重IBD中的炎症状况。
在结直肠癌中,肠道菌群失衡会通过减少粘液层厚度和增加肠道通透性来促进癌症的发生和发展(图a)。
有害细菌过度生长会导致脂多糖(LPS)和其他有毒代谢物的释放,比如次生胆汁酸、三甲胺-N-氧化物(TMAO)、硫化氢(H₂S)和含氮有机化合物,这些物质会引发低度炎症并激活致癌信号。

上述研究发现,结直肠癌患者体内大肠杆菌、柠檬酸杆菌、不动杆菌水平升高,同时存在耐药基因如mdtP、mdtO、GadW、GadX、AcrA(多药耐药)、ErmB(喹诺酮类耐药)和AAC(3)‐IIa(3′-氨基糖苷类耐药)等耐药基因水平升高(图b)。这些抗生素抗性基因通过加剧肠道菌群失衡、炎症反应和肠道屏障功能障碍,进一步推动CRC的发展,突出了肠道耐药组在代谢性疾病发病机制中的作用。
总体而言,新出现的证据表明,肠道耐药组不仅反映了微生物失衡,而且通过抗生素耐药性的传播积极推动代谢紊乱。ARG在肠道微生物群中的存在和传播会破坏关键的生理过程,如能量代谢、免疫调节和肠道屏障的完整性。通过营造一个有利于致病菌及其毒力因子的环境,耐药组会放大炎症,改变代谢信号通路,并促进生态失调。
肠道微生物组通过炎症反应和TMAO代谢机制参与心血管疾病的发病机制,包括心力衰竭和冠状动脉疾病(图a)。
心血管疾病患者体内产丁酸菌减少,这会引发局部炎症,加剧肠道菌群失衡,并导致肠道屏障功能受损。这些患者体内TMAO水平的升高,会干扰胆固醇运输、促进泡沫细胞形成和诱导血小板聚集,从而可能导致急性冠状动脉综合征,引发动脉粥样硬化的发展。
此外,较高的肠道抗生素抗性基因α多样性与心脏代谢风险增加有关。
一项研究发现,较高的肠道ARGα多样性指数与甘油三酯和总胆固醇/高密度脂蛋白胆固醇比值(TC/HDL)呈正相关,但与高密度脂蛋白胆固醇(HDL)水平呈负相关。
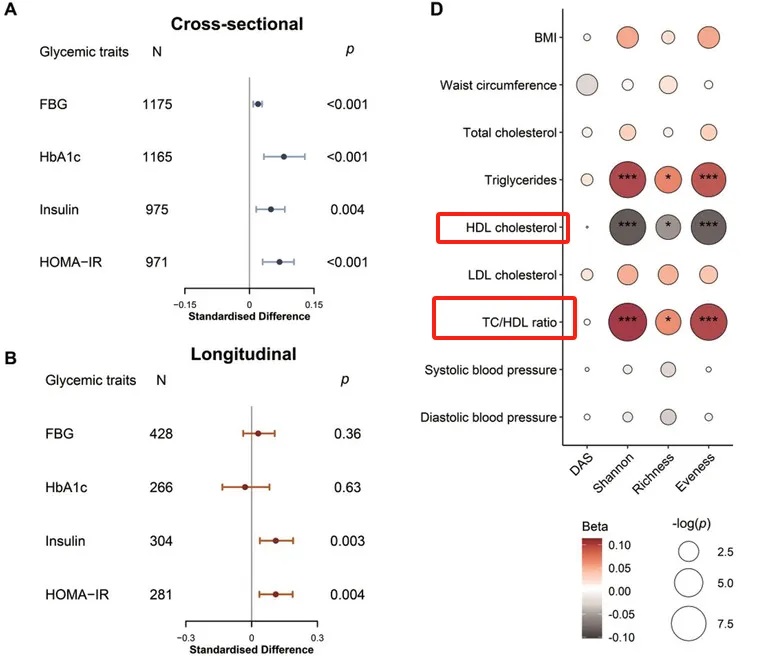
doi: 10.1002/advs.202104965
这表明ARG在扰乱脂质和葡萄糖代谢方面发挥作用,并可能通过血脂异常和胰岛素抵抗增加心血管疾病的风险。
肠道微生物群通过引发炎症反应,显著影响肝脏的正常功能(图a)。
当肠道微生物组因抗生素耐药性发生变化时,肠道的通透性会增加,使细菌及其产物(如脂多糖LPS)能够通过肝肠循环进入肝脏。LPS进入肝脏后,会引发急性炎症反应,导致肝细胞受损,进而可能引发多种肝脏疾病,如非酒精性脂肪肝、脂肪性肝炎、酒精性肝病、肝癌,甚至肝性脑病。
肠道耐药组的变化与肝脏疾病的发生密切相关。研究发现,肝硬化患者的肠道耐药组中,与肠杆菌科、链球菌、肠球菌和不动杆菌属等病原菌相关的耐药基因丰度更高。
此外,这些患者还表现出对β-内酰胺酶、大环内酯类、喹诺酮类、糖肽类、磷霉素、四环素类抗生素的更大耐药性。肝性脑病患者则表现出更高水平的氨基香豆素耐药基因parY和林可酰胺耐药基因(ileS和RpoB)。这些抗性组的变化会破坏肠道与肝脏之间的正常相互作用,以及胃肠道的免疫反应,随着疾病的进展,这种破坏会进一步恶化。
肠道微生物组在通过肠道-大脑轴塑造神经系统的发育和功能方面起着关键作用。迷走神经传递神经元、内分泌和免疫信号,而下丘脑通过释放促肾上腺皮质激素释放激素(CRH)来调节应激反应,导致皮质醇的产生,从而影响肠道的上皮屏障和免疫系统。
肠道耐药组的改变也与神经发育障碍有关。
一项研究发现,患有自闭症的儿童具有更高丰度的屎肠球菌aac(6′)-aph(2′)基因、Megasphaera elsdenii 的cepA‐49和tet(40)基因以及脆弱拟杆菌的cepA-49基因。

研究还发现,健康个体和自闭症患者在对大环内酯类抗生素耐药性的基因方面存在显著差异,包括肠球菌和未培养细菌的氨基糖苷类耐药基因、拟杆菌属和Acidaminococcus的β-内酰胺抗性基因,以及来自Megasphaera和Alistipes的四环素抗性基因。因此,肠道微生物群平衡和抗性基因的破坏可能会降低神经代谢潜力,影响大脑功能所需的神经递质和代谢物的产生。
通常用抗生素治疗的疾病与扩大的肠道耐药性有关,这表明曾经有接触抗生素史对疾病相关菌株的ARG获得产生了相当大的选择压力。
未来的研究需要更好地了解肠道耐药性与疾病之间的相互作用及其机制。将多基因风险评分与肠道宏基因组风险模型相结合可以提高对常见慢性病的预测能力,为疾病预测和预防提供更全面的方法。
通过上述研究,我们看到肠道耐药组对人体健康的多层面影响,是如何超越简单的感染问题而演变成一个系统性的健康风险。然而,要有效应对这一挑战,我们需要深入了解:这些耐药基因是如何产生的?它们又是如何从环境进入人体并在肠道微生物间传播的?…
★
抗生素耐药性并非凭空出现,而是有其明确的生物学机制和传播途径,下面章节我们来详细了解这一过程中的关键环节,包括垂直遗传、水平基因转移等核心机制,以及食物链、水源和环境污染在耐药基因传播中的角色。
在这里重点介绍针对细菌的抗生素,介绍它们的作用机制,并讨论抗生素耐药性如何在肠道中发展和积累。
将治疗细菌性疾病的抗生素引入临床应用是 20 世纪最大的医学突破。抗生素的抗菌机制可细分为五类:
抗生素作用机制和抗生素耐药性
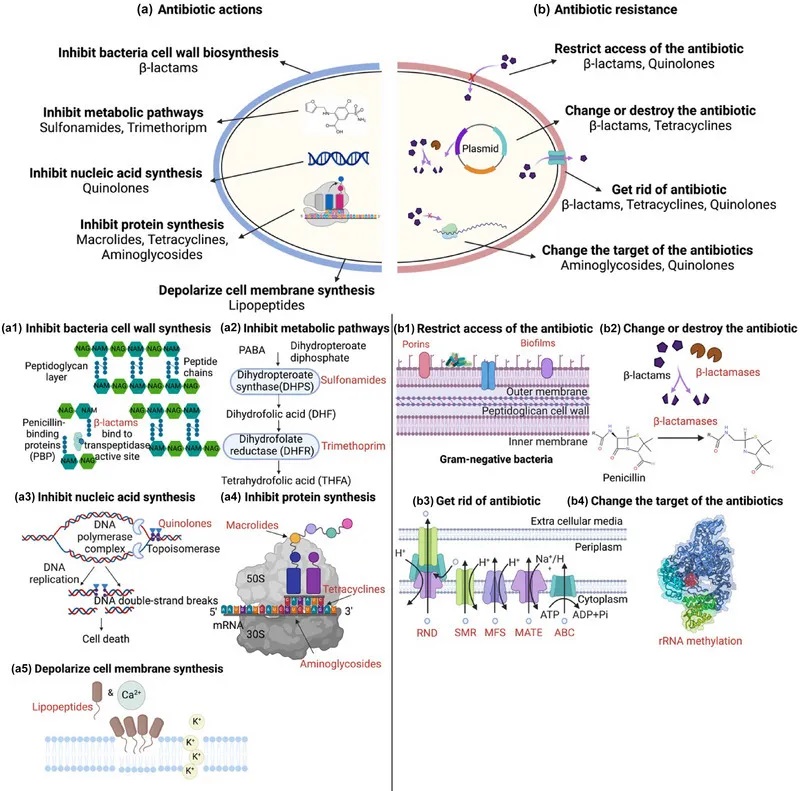
doi:10.1111/1541-4337.70143
a)抗生素作用:抗生素针对关键的细菌过程来抑制生长和存活。
b)抗生素耐药性:细菌通过各种策略产生耐药性。
下面我们先来看一下抗生素的作用。
阻碍细菌细胞壁的合成——结构性破坏
β-内酰胺类抗生素,可以伪装成细菌细胞壁合成所需的关键分子。这类抗生素会:
• 模仿细菌细胞壁合成的天然底物(d-丙氨酸)
• 结构相似性使抗生素能够与DD转肽酶结合(d-丙氨酸是DD转肽酶的底物),防止DD转肽酶与其天然底物进一步结合
• 破坏肽聚糖层的交联,肽聚糖框架减弱
• 最终导致细胞壁结构损坏,细胞死亡
典型的β-内酰胺类抗生素包括:青霉素类、头孢菌素类、碳青霉烯类、单巴坦类。
抑制细菌代谢途径——叶酸合成阻断
磺胺类药物抑制二氢蝶酸酯合酶,这是叶酸生产中必不可少的酶。这种抑制阻断了DNA和RNA生产所必需的核苷酸的合成,阻止了细菌复制并导致细胞死亡。
最常用的磺胺类抗生素:磺胺甲氧基嘧啶、磺胺二甲嘧啶、磺胺甲恶唑、磺胺嘧啶。
协同用药:甲氧苄啶(靶向二氢叶酸还原酶,DHFR)与磺胺类联用,双重阻断叶酸代谢链。
抑制核酸生物合成所需的细菌RNA聚合酶
作用靶点:细菌RNA聚合酶β亚基
机制:阻断转录起始→抑制mRNA合成→细胞分裂停滞
代表药物:利福霉素类(如利福平)
作用靶点:DNA旋转酶(拓扑异构酶II/IV)
机制:
与 DNA 和旋转酶结合→阻止DNA超螺旋/松弛
捕获DNA上的拓扑异构酶→抑制核酸合成→DNA双链断裂→抑制细菌的生长
代表药物:喹诺酮类(诺氟沙星、环丙沙星、氧氟沙星)
阻断蛋白质合成
大环内酯类(红霉素、克拉霉素、阿奇霉素)
与50S核糖体亚基上的23S rRNA产生的结合肽基转移酶中心结合→抑制核糖体沿mRNA的运动→多肽链延伸终止→细菌蛋白质合成的抑制→细菌细胞死亡
四环素类(四环素、土霉素、多西环素)
靶点:与30S核糖体亚基结合
阻断氨酰-tRNA与A位点结合→抑制氨基酸掺入生长的多肽链
氨基糖苷类(卡那霉素、链霉素、庆大霉素)
靶点:30S核糖体16S rRNA上的 A位点
诱导mRNA误读→多肽链提前终止→破坏蛋白质合成完整性→细胞死亡
破坏细胞膜完整性
脂肽类抗生素(达托霉素)
结构特征:含脂尾的肽核
钙离子依赖型寡聚→达托霉素分子嵌入细胞膜形成孔道→膜电位去极化→胞内成分泄漏→细胞裂解
不同种类抗生素作用的方式

Uddin TM, et al., J Infect Public Health. 2021
细菌抗生素耐药性是微生物数千年进化的自然现象。这种固有的抗生素耐药性也被称为固有耐药性。
除了固有耐药性外,过度使用抗生素还会导致抗生素耐药性的积累。细菌采用各种策略来抵抗抗生素,例如减少抗生素的摄取,修饰或灭活抗生素,主动将其从细胞中排出,或改变抗生素的靶标以阻止其作用。
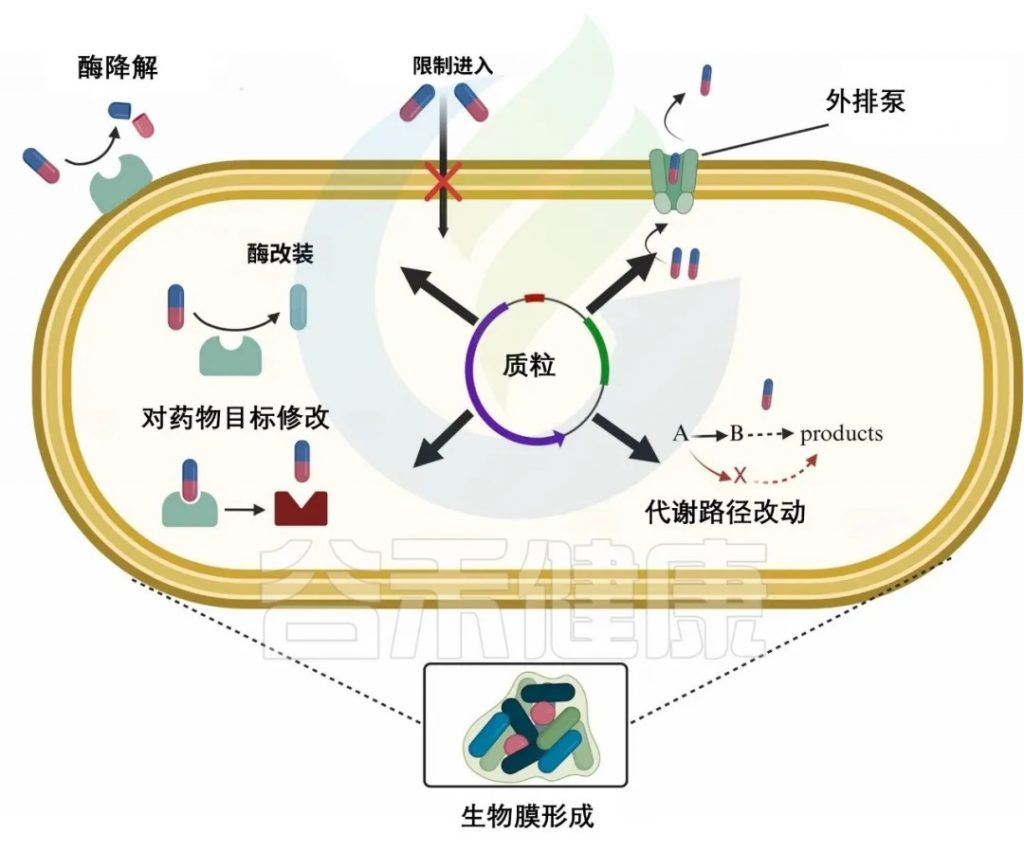
doi.org/10.1016/j.glmedi.2024.100081
细菌耐药核心机制如下:
1. 细菌改变外膜疏水性、孔蛋白突变或形成生物膜,阻止抗生素渗透,导致耐药性
革兰氏阴性菌有一层外膜,可以作为屏障,阻止抗生素(β-内酰胺类、喹诺酮类)进入细胞。大多数抗生素需要穿透外脂膜才能到达目标。疏水性药物如大环内酯类(红霉素)可以通过扩散穿过膜,而亲水性抗生素如β-内酰胺类需要通过孔蛋白(图b1)。
外膜的变化,如疏水性的改变或孔蛋白的突变,可能导致革兰氏阴性菌产生抗生素耐药性。一些细菌可以产生大量的细胞外聚合物,包括胞外多糖、蛋白质、细胞外DNA和脂质。这些物质可以在生物膜内的细菌细胞周围形成物理屏障,防止抗生素直接到达细菌细胞。这减少了抗生素的直接暴露,导致抗生素疗效明显降低。
例如,金黄色葡萄球菌菌株产生的生物膜可以显著降低苯唑西林、万古霉素和头孢噻肟的渗透性。
2. 细菌产生分解抗生素分子的酶,对抗生素修饰或降解,使其无效
β-内酰胺酶
细菌产生β内酰胺酶来降解β内酰胺类,这些酶通过不可逆地打开β-内酰胺环使其失活,从而使其无法与青霉素结合蛋白结合,使其无效。
氨基糖苷修饰酶
如AAC(3)乙酰转移酶修饰庆大霉素,降低其与核糖体结合能力,从而降低其疗效。
3. 细菌外排泵,以能量依赖性方式从细菌细胞中去除抗生素,使细菌在高浓度抗生素中存活
细菌药物外排泵可分为五个家族——ABC、SMR、MATE、MFS、RND。
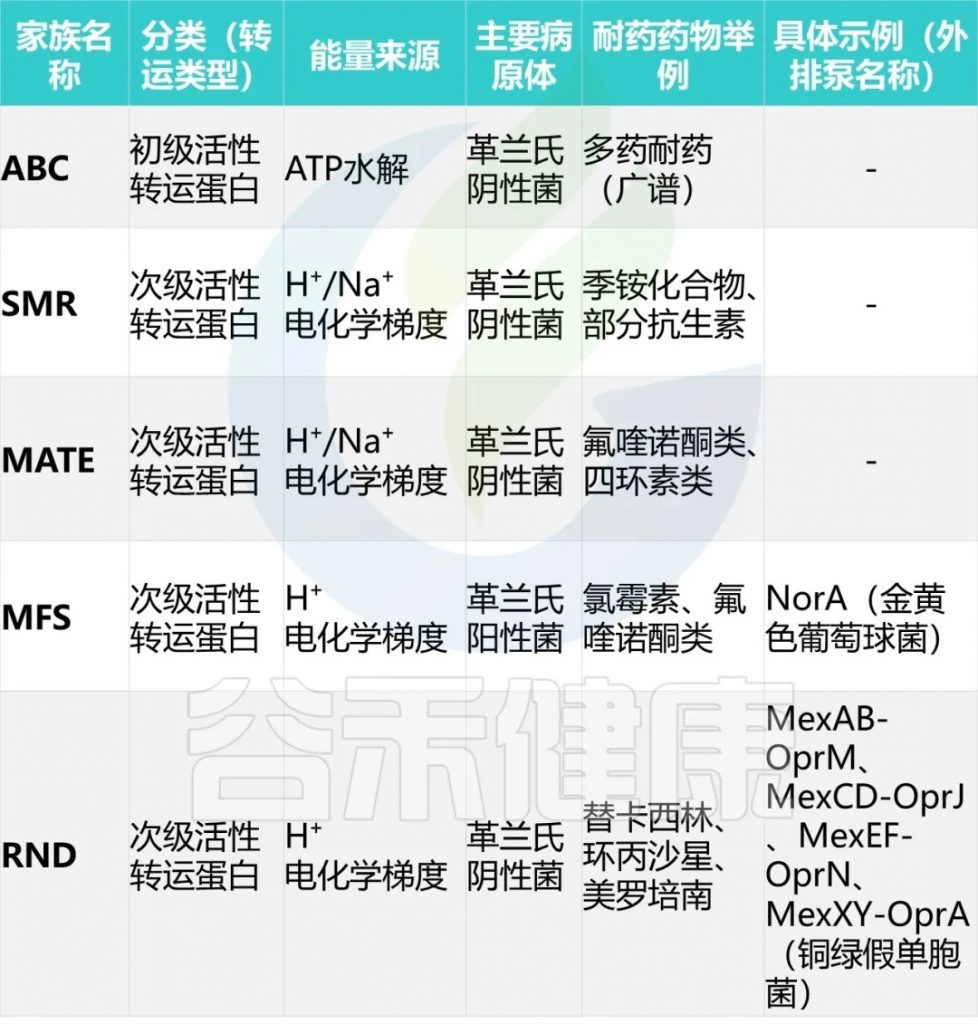
4. 改变抗生素的靶点结合位点,降低结合效率
16S rRNA甲基转移酶改变氨基糖苷类核糖体结合位点的构象,从而降低氨基糖苷类对核糖体的亲和力,使抗生素更难有效结合。
氨基糖苷修饰酶可分为两类,甲基化G1405和A1408。
G1405甲基化:改变了核糖体RNA在这些4,6-二取代2-脱氧链霉素(DOS)氨基糖苷类关键结合位点的结构,如卡那霉素、庆大霉素、妥布霉素、阿米卡星。
A1408甲基化(NpmA酶):可对4,5和4,6二取代的2-DOS氨基糖苷类产生耐药性。
随着分子技术的进步,了解抗生素耐药性背后的机制变得越来越复杂。其中,基因测序已成为一种强大的工具,能够根据遗传数据预测抗性表型。这允许更精确地跟踪耐药性演变,并为有针对性的干预措施提供信息,以防止耐药菌株的传播。
此外,细菌采用多种策略来确保其存活,即使是少量耐药菌株也可能在胃肠道的复杂微生物环境中持续存在,从而导致耐药细菌种群的选择和存活。
了解分子耐药机制对于减轻抗生素耐药性和揭示人体肠道中耐药性积累的动态是必要的。
抗生素耐药性发生在自然环境,如土壤、水生态系统、动物粪便,在社区环境中也有,如医院污水、农业实践中。当人类食用食物中的抗生素耐药细菌时,抗生素耐药性可以通过食物链从自然环境传播给人类,比如,通过受污染的食物和直接的公共接触传播给人类。
环境中抗生素耐药性的累积
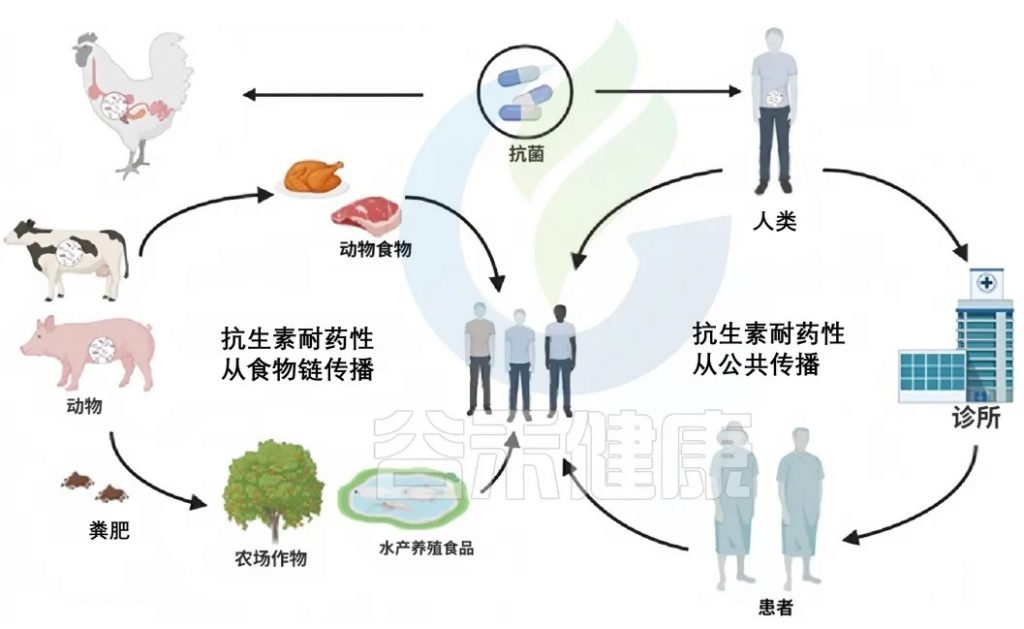
doi:10.1111/1541-4337.70143
在自然环境中,不同的生物和基质(如动物、土壤和水)之间存在联系。这种相互联系促进了抗生素耐药性在各种环境因素中的传播。土壤、水和沙子是抗性细菌和遗传元素可以持续存在并相互作用的水库。
整个自然界就像一张巨大的“互联网络”——动物、土壤、水甚至沙子之间都有看不见的“网线”连接。这些“网线”让抗药细菌和它们的耐药基因能在环境中四处“串门”。
抗生素耐药性从环境传播到植物、动物和昆虫涉及多种机制。农业实践,包括在畜牧业中使用抗生素和将处理过的动物粪便施用到田地里,将抗性基因引入土壤。水源可能通过农田径流,特别是那些用牲畜粪便施肥的农田径流,被抗生素抗性细菌和遗传元素污染。水生生态系统中的昆虫和其他生物可以成为抗性基因的载体,进一步在环境中传播抗生素抗性。
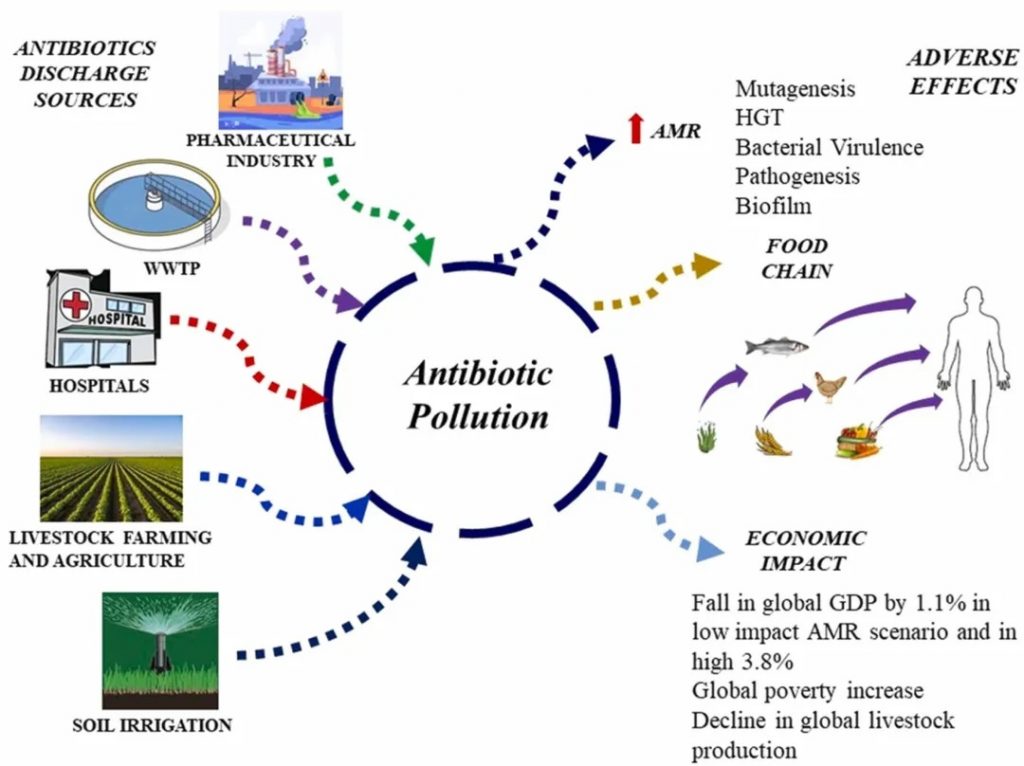
doi.org/10.1016/j.hazl.2024.100105
人类活动对抗生素排放到环境中起着重要作用。这通过多种途径发生,包括城市和医院废物、工业制造、农业径流和垃圾填埋场渗滤液。因此,抗生素耐药性可能通过各种途径进入人体肠道。人类可能通过受污染的食物、水和与环境的直接接触(如家庭、社区或医疗保健互动)接触到抗生素抗性细菌和遗传因素。
宠物狗和猫可携带多种多重耐药细菌,包括耐甲氧西林金黄色葡萄球菌和耐β-内酰胺类抗生素的肠杆菌科细菌。
这些耐抗生素微生物可能通过直接接触、体外寄生虫和气溶胶从宠物传给人类。特别是弱势群体,包括免疫功能低下的老年人和婴儿,可能面临更大的风险。
详见我们之前的文章:宠物猫可能塑造人类肠道中的抗生素耐药性和益生菌
ARG在自然环境、动物和人类中的发生、积累和传播依赖于垂直遗传和水平基因转移(下图)。在细菌的生长和繁殖过程中,基因突变使它们对特定抗生素产生耐药性,它们的后代将继承突变基因(下图a)。携带突变基因的细菌,更有可能存活,并且可以世代垂直传播。随着时间的推移,细菌种群将由具有耐药基因的细菌主导,这种选择性优势使耐药细菌得以增殖,有助于抗生素耐药性的扩大。
ARG具有高流动性,可以通过水平基因转移传播,包括四种机制:基因转化、基因转导、基因接合、膜囊泡融合(下图b)。
通过垂直遗传、水平基因转移以及影响这些机制的因素在肠道中积累抗生素耐药性
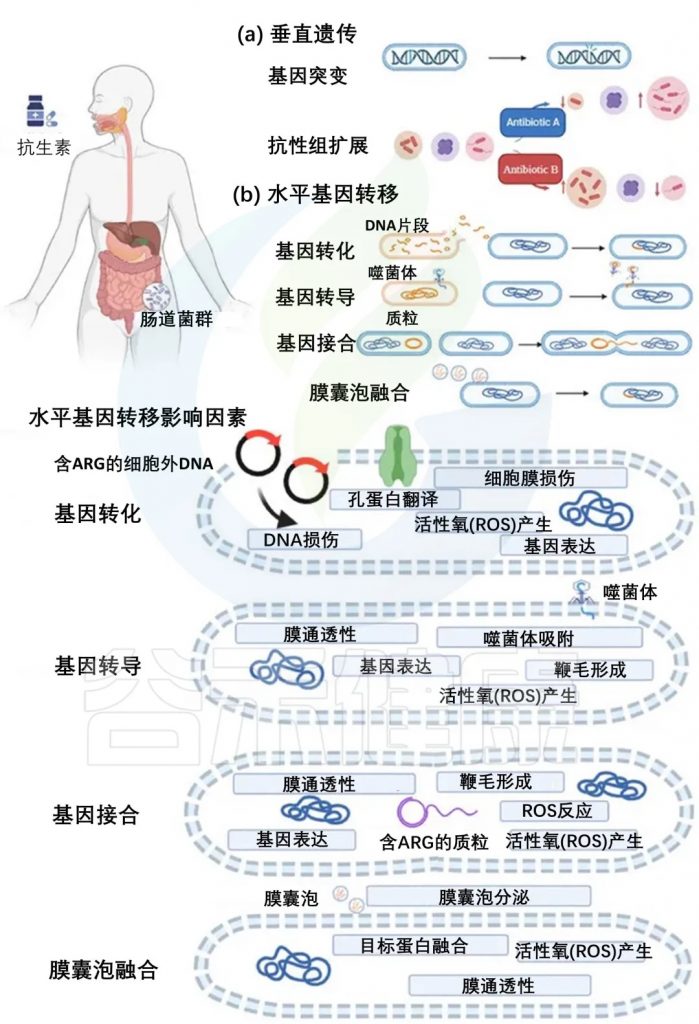
doi:10.1111/1541-4337.70143
a) 垂直遗传:抗生素耐药性可以通过基因突变和耐药体扩增进行垂直遗传。
b) 水平基因转移及其影响因素:
水平基因转移发生在四种途径中:
抗生素可以影响细菌中的水平基因转移,特别是在ARG的情况下。它们创造了一个有利于携带抗性基因的细菌存活和增殖的选择性环境。除了抗生素选择压力外,各种非抗生素因素,包括抗菌剂和环境条件,都会影响ARG的水平转移。
在大多数细菌中,获取抗性基因的主要和最有效的方法涉及使用可移动的遗传元件,如质粒作为基因交换的载体。在多种细菌中观察到水平基因转移,如大肠杆菌和肺炎克雷伯氏菌,使耐药基因能够在细菌种群中快速有效地传播。一般来说,ARG的水平转移会加剧肠道中抗生素耐药性的传播。
总体而言,垂直遗传和水平基因转移是导致ARG在自然环境、农场动物和最终人类肠道中发生和传播的两条主要途径。值得注意的是,ARGs的水平基因转移是决定其传播的关键因素。
饮食干预在抑制ARGs的水平基因转移中起着重要作用。已经发现,一些膳食植物化学物质,如酚类化合物、硫化物、萜类化合物、生物碱,有可能诱导ROS的产生,导致细菌细胞膜不可逆的损伤,生物膜活性降低,毒力基因表达下调,从而抑制水平基因转移。下面章节将进一步讨论食物成分在控制ARG中的作用。
★
通过饮食成分改变肠道耐药性是一个新兴的研究领域,人们越来越关注食物选择如何影响肠道中的抗生素抗性细菌。例如,发酵食品中含有大量微生物,乳酸菌和凝固酶阴性葡萄球菌被确定为四环素、青霉素、氯霉素和大环内酯类ARG的携带者。这表明食用发酵食品可能会直接影响肠道耐药性。
食品中存在的许多天然分子,如酚类、萜类和酚酰胺,具有抗菌能力,能够调节肠道微生物群。然而,关于食物成分对肠道抗生素耐药细菌影响的研究有限,这是一个新兴的研究领域。
这里总结了对大量人群的营养研究,以了解饮食习惯对肠道耐药性的潜在影响,并强调了选定类别的饮食成分对抗生素耐药细菌的调节作用。
长期的饮食习惯会影响肠道微生物群和肠道抵抗力。
Stege等人(2022)选择了149名荷兰个体,他们的饮食习惯分为四个不同的群体:杂食者、鱼素食者、素食者、纯素食者。研究发现长期的饮食习惯并没有显著影响肠道微生物组的关键组成和多样性。在前10个最丰富的ARG中没有观察到显著差异。然而,两个ARG[lsa(C)和tet(L)]在杂食者和鱼食者之间显示出不同的丰度。
此外,宏基因组分析显示,与鱼素者相比,tet(X)在杂食动物的抗性体中更为丰富。
在来自三种不同饮食习惯(杂食性、蛋乳素食主义、严格素食主义)的58名志愿者的肠道抵抗力中观察到了类似的结果。
在所有习惯组中,最常见的耐药基因是针对四环素类抗生素[tet(A)、tet(B)、tet(M)、tets(O)、tet(Q)]、β-内酰胺类(blaTEM、blaSHV、mef)和MLS耐药基因[erm(B)、erm(C)],其次是磺胺类(sul1、sul2)和氨基糖苷类耐药基因(aacA-aphD)。
blaCTX‐M仅在卵乳素食组的样本中发现,而tet(E) 仅在杂食组的样本中检测到。有趣的是,这些经常检测到的ARG也常见于农场动物粪便中,这表明饮食习惯和环境ARG之间存在潜在联系。
虽然将饮食与肠道抗药性联系起来的确切机制尚不清楚,但饮食干预可能提供一种有前景的方法来减轻抗生素耐药性的负担。
Oliver等人(2022)研究了290名健康成年人,以探索饮食、肠道微生物组和抗生素耐药性之间的关系,并观察到高膳食纤维摄入量的个体ARG水平较低。
低ARG人群在肠道中显示出更多梭菌科的专性厌氧菌,而高ARG人群的链球菌科和肠杆菌科水平更高。此外,低ARG个体摄入的蛋白质较少,尤其是来自牛肉和猪肉的蛋白质。
该研究表明,增加纤维摄入量可能会促进肠道微生物组环境,有利于专性厌氧菌的生长,同时限制兼性厌氧菌,从而可能降低肠道中抗生素耐药性的发生率。
这是因为由专性厌氧菌发酵的膳食纤维会产生短链脂肪酸,降低肠道环境的pH值,使其对专性厌氧杆菌有利,但对兼性厌氧菌不利。
特定的饮食成分通过各种机制在调节肠道微生物群方面发挥着至关重要的作用。生物活性常量营养素,如凝集素、乳铁蛋白和多不饱和脂肪酸(PUFA),来源于碳水化合物、蛋白质和脂质,主要因其营养作用而被认可,但它们也具有生物活性,通过破坏营养可用性和细菌膜完整性来抑制抗生素细菌。
多酚等植物化学物质通过破坏细胞功能、干扰代谢过程或抑制酶活性表现出抗菌特性。
益生菌,如乳杆菌和双歧杆菌,通过竞争性排斥、分泌抗菌化合物和免疫调节来抑制病原体。
有趣的是,其中一些化合物还具有选择性抑制抗生素耐药细菌的能力,通过靶向其脆弱性,如损害生物膜形成、抑制耐药酶和破坏外排泵。
研究这些化合物如何特异性靶向细菌,包括抗生素耐药菌株,对于制定控制或减少肠道致病菌群的饮食策略至关重要。这有助于提供了一种非抗生素方法来管理肠道健康,并有助于遏制抗生素耐药性的传播。
生物活性常量营养素
常量营养素包括碳水化合物、蛋白质、脂肪,是人类饮食中支持生命和调节各种生理功能的重要组成部分。虽然它们的主要作用是为身体提供能量和构建块,但某些生物活性常量营养素也具有固有的抗菌特性。
这些特性可能有助于控制抗生素耐药细菌的生长,减轻其对肠道微生物组的影响,从而可能提供一种对抗抗生素耐药性的替代策略。
碳水化合物
不仅是能量的来源,而且具有生物活性,可以提高抗生素的有效性。
来自紫色鞘豆(Dioclea violacea)种子的甘露糖结合凝集素已被证明可以提高抗生素的疗效。关于什么是凝集素可以详见我们之前的文章:什么是凝集素,食物中的凝集素如何影响肠漏和自身免疫
Santos等人(2023)证明,从紫色鞘豆中提取的凝集素作为佐剂,通过碳水化合物识别增加细菌膜附近氨基糖苷类的生物利用度,并促进抗生素进入细菌细胞质,从而增强新霉素对多药耐药金黄色葡萄球菌和铜绿假单胞菌的活性。
蛋白质
如母乳和牛乳中的乳铁蛋白,也表现出抗菌和抗菌膜特性。
Avery等人(2021)报告称,人和牛乳铁蛋白都可以显著抑制测试的多重耐药鲍曼不动杆菌菌株中生物膜的形成,而人乳铁蛋白的最低抑菌浓度(MIC)低于牛乳铁蛋白,表明其效力更高。除了乳铁蛋白,来自植物的抗菌肽(AMP)在对抗抗生素耐药细菌方面也显示出了希望。
Heymich等人(2021)从鹰嘴豆中鉴定出21种AMP候选物,其中两种对耐甲氧西林的金黄色葡萄球菌ATCC 3300具有杀菌活性。这些肽与细菌膜相互作用,通过膜破裂导致细胞裂解,这与它们的杀菌作用直接相关。
膳食脂肪
特别是PUDA,也可以提高某些抗生素的有效性。
Zang等人(2021)发现,花生四烯酸(AA)和二十二碳六烯酸(DHA)等PUFA可增强氨基糖苷类药物对多重耐药鲍曼不动杆菌的疗效。
AA和DHA都通过细胞外隔离减轻了粘菌素的抗菌作用,使其难以穿透细菌的亲水性包膜和脂多糖屏障,并上调了外排系统基因表达(adeABC和adeIJK),从而通过将抗生素排出细胞来增加细菌的抗药性。
上述研究强调了生物活性常量营养素在增强抗生素有效性和影响耐药机制方面的作用。
果胶等碳水化合物可以通过与抗生素相互作用来提高抗生素的疗效,而乳铁蛋白等蛋白质则具有强大的抗菌和抗菌膜特性。然而,AA和DHA等膳食脂肪的作用更为微妙,因为它们都可以通过激活外排泵来增强抗生素作用并上调细菌耐药性机制。
总之,生物活性常量营养素不仅可以作为能量来源,还可以在对抗抗生素耐药细菌方面提供巨大的潜力。虽然它们在调节细菌耐药机制方面的作用很复杂,但这些发现强调了进一步研究饮食成分补充或增强传统抗生素治疗潜力的重要性。通过利用这些天然化合物,可能能够开发出新的策略,来缓解抗生素耐药性日益增长的挑战。

doi:10.1111/1541-4337.70143
植物化学物质
植物化学物质来源于植物性食品,主要包括多酚、含硫化合物、萜类、生物碱。这些化合物对抗生素耐药细菌具有抗菌活性,其中一些会破坏细菌膜结构,另一些则通过抑制外排泵来增强抗生素的有效性,从而防止耐药性的发展。
★ ■ 多酚类
柿子单宁——直接抗菌作用
一种经过充分研究的多酚,即来自青涩柿子的柿子单宁,已显示出对耐甲氧西林金黄色葡萄球菌的抑制作用。柿子单宁抑制细胞增殖,降低膜电位和细胞内ATP浓度,破坏全细胞蛋白质完整性,并诱导细胞S期细胞周期阻滞。
EGCG、原花青素——协同作用、增强抗生素疗效
Parvez等人(2019)报道了表没食子儿茶素没食子酸酯(EGCG)与庆大霉素联合使用时具有明显的协同作用,增强了对金黄色葡萄球菌和大肠杆菌多药耐药菌株的抗生素疗效。这是因为EGCG可以破坏细菌细胞膜,阻碍DNA超螺旋,增加细胞通透性,最终导致细菌死亡,并减轻耐药性的发展。
同样,从美国蔓越莓(Vaccinium macrocarpon L.)中提取的原花青素与β-内酰胺类抗生素显示出显著的协同作用。
Gallique等人(2021)报告称,原花青素抑制β-内酰胺酶,并选择性增强苯唑西林和羧苄青霉素对耐甲氧西林葡萄球菌的有效性,可能是通过调节葡萄球菌中变异转肽酶PBP2a的表达。PBP2a的低表达降低了细菌抵抗β-内酰胺类抗生素的能力,使细菌更容易受到抗生素的影响。
扩展阅读:
★ ■ 含硫化合物
大蒜素衍生物——释放硫化物破坏致病生物膜
例如,Xu等人(2018)将从大蒜鳞茎中分离出的大蒜素转化为纳米级硫化铁。这些纳米颗粒可以释放杀菌的有机硫化物(Polysulfanes),有效抑制生物膜形成和耐药的金黄色葡萄球菌。
异硫氰酸盐——抑制生物膜代谢活性
同样,来源于旱金莲和辣根的异硫氰酸酯对铜绿假单胞菌等抗生素耐药细菌具有抗菌潜力,特别是在生物膜抑制方面。当与美罗培南联合使用时,异硫氰酸酯通过抑制铜绿假单胞菌生物膜的代谢活性来增强抗生素疗效。
皂苷——破坏细胞壁结构,改变膜通透性
此外,da Silva等人(2024年)从Sarcomphalus joazeiro中提取了六种皂苷——红枣皂苷B、红枣皂苷III、酸枣仁苷、红枣皂苷IV、红枣皂苷II和红枣皂苷I,以及三种皂苷衍生物。当与庆大霉素和诺氟沙星联合使用时,富含皂苷的组分显示出增强作用,增加了它们对多药耐药铜绿假单胞菌和金黄色葡萄球菌的活性。皂苷与细菌细胞壁相互作用,导致结构损伤,改变膜通透性,并诱导膜破裂。
★ ■ 萜类药物
1,8-桉叶醇——增加膜通透性、诱导氧化应激导致成分泄漏
Moo等人(2021)发现,1,8-桉叶醇对碳青霉烯类抗生素耐药肺炎克雷伯菌具有抗菌作用。它增加了细菌表面电荷,增强了外膜通透性,并诱导了氧化应激,导致膜损伤、细胞内成分(如核酸、蛋白质和脂质)泄漏,最终导致细胞死亡。
芳樟酯——产生ROS、引发脂质过氧化
另一种萜烯,邻氨基苯甲酸芳樟酯(LNA),存在于薰衣草和百里香等植物中,对碳青霉烯类耐药肺炎克雷伯菌具有杀菌活性。
Yang等人(2021)证明,当LNA与美罗培南联合使用时,会降低MIC,使细菌更容易感染。LNA的抗菌作用归因于其通过产生ROS诱导氧化应激的能力,ROS可能进一步引发脂质过氧化并对细菌膜造成损伤。
萜类化合物——细菌外排泵的重要抑制剂
Oluwatuyi等人(2004)在迷迭香中鉴定出几种萜类化合物,如鼠尾草酸、鼠尾草酚和12-甲氧基-反式鼠尾草酸。这些化合物已被证明可以通过抑制NorA外排泵来增强红霉素对多药耐药金黄色葡萄球菌的活性,从而降低细菌排出抗生素的能力并增加其细胞内浓度。
★ ■ 生物碱
原小檗碱类生物碱——抑制H+-ATP酶泵,阻断ATP合成
Guefack等人(2022)从绿花恩南番茄(Enantia chlorantha)中鉴定出的原小蘖碱类生物碱(protoberberine alkaloids)(哥伦比亚胺、假哥伦比亚胺、药根碱、巴马汀、4,13-二羟基-3,9,10-三甲氧基原小檗碱和13-羟基-2,3,9,10-四甲氧基原黄连素)可以有效抑制多药耐药细菌的生长,包括大肠杆菌、产气肠杆菌、肺炎克雷伯菌、普罗维登斯氏菌(Providencia stuartii)、金黄色葡萄球菌,MIC值低于100µg/mL。重要的是,其中一种名为哥伦比亚胺的生物碱抑制了H+-ATP泵,阻碍了细胞ATP的产生。
苦参碱——抑制Mex泵,增强氟喹诺酮敏感性
来自苦豆子种子的苦参碱与环丙沙星对铜绿假单胞菌的抗生素耐药菌株显示出协同作用。总生物碱显示出抑制生物膜形成和降低Mex泵活性的作用,从而增强细菌对氟喹诺酮类药物的敏感性。
研究人员从Erythroxylum revolutum Mart.中分离出生物碱,包括6-(2′-甲基丁酰氧基)-3-羟基托烷和6-丁酰氧基-3-羟基托巴烷,可增强诺氟沙星和红霉素对多药耐药金黄色葡萄球菌的疗效。这种作用可能与托烷生物碱削弱耐多药金黄色葡萄球菌中ABC多药转运蛋白的能力有关。
总之,植物化学物质通过破坏细菌膜、抑制外排泵和阻碍细菌生物膜的形成来发挥抗菌能力。当与抗生素结合时,一些植物化学物质表现出协同作用,可以提高抗生素的有效性并改变抗生素的耐药性,表明有可能管理肠道中的抗生素耐药细菌并影响肠道耐药组。
未来,有必要研究这些植物化学物质在人体肠道中的生物利用度、剂量和长期影响,这可能会导致更有针对性的饮食干预。深化我们对特定植物化学物质如何选择性调节肠道抗药性的理解,对于开发创新方法来减轻抗药性基因的传播和改善人类健康结果至关重要。

doi:10.1111/1541-4337.70143
益生菌
益生菌是具有健康益处的活微生物,通常被掺入酸奶和奶酪等发酵食品中。然而,这些产品通常只有在用未经巴氏消毒的牛奶制成时才含有益生菌,因为巴氏消毒可以显著减少或消除活的益生菌菌株。益生菌可以通过调节肠道微环境、增强免疫功能和抑制过敏反应来影响肠道中的抗生素耐药细菌。
★ ■ 直接抗菌效应
某些益生菌菌株已显示出对耐药病原体的抗菌活性。
乳杆菌类——破坏细胞膜完整性,与抗生素协同抑制有害菌
例如,嗜酸乳杆菌NCFM和鼠李糖乳杆菌GG通过破坏细菌细胞膜的完整性对耐甲氧西林金黄色葡萄球菌表现出抗菌作用。
益生菌胶囊(嗜酸乳杆菌CL1285、干酪乳酸杆菌LBC80R、鼠李糖乳杆菌CLR2)与妥布霉素联合使用时,通过限制病原体定植和阻碍生物膜的形成,对耐甲氧西林的金黄色葡萄球菌和铜绿假单胞菌表现出协同作用。
双歧杆菌——促进四环素进入,破坏蛋白质合成
双歧杆菌会产生生物表面活性剂和乙酸盐,这有助于分离粘附的大肠杆菌并改变细胞内的阴离子组成。这可以增强四环素进入大肠杆菌,破坏蛋白质合成,最终导致协同杀死大肠杆菌。这些研究为益生菌作为功能性食品调节肠道抗菌药物耐药性的潜在应用铺平了道路。
★ ■ 发酵食品中的益生菌活性
除了这些单独的益生菌菌株外,酸奶和泡菜等发酵食品中发现的益生菌已被证明可以抑制抗生素耐药细菌的生长。
酸奶来源菌株抗菌物质:类细菌素肽或蛋白
从商业酸奶中分离出的石蜡乳杆菌对14种多药耐药细菌具有抗菌活性,包括大肠杆菌、铜绿假单胞菌、鲍曼不动杆菌、产气杆菌、奇异变形杆菌、肺炎克雷伯菌。该菌株产生的抗菌物质可能是一种细菌素样肽或蛋白质,对密切相关的细菌菌株具有活性,表明其在抑制致病性多药耐药细菌生长方面具有潜在作用。
泡菜来源菌株——抑制耐药菌、生物膜
Yi和Kim(2023)从泡菜中分离出益生菌乳酸菌(Pediococcus inopinatus K35),该菌能有效抑制多药耐药铜绿假单胞菌的生长和生物膜形成。这些研究共同强调了益生菌的巨大潜力,特别是那些来自发酵食品的益生菌,作为能够对抗抗生素耐药细菌的天然药物。

doi:10.1111/1541-4337.70143
然而,一个值得关注的问题是益生菌菌株可能携带ARG,ARG可以转移到致病菌,从而导致耐药性的传播。
例如,Selvin等人(2020)从膳食补充剂中分离出益生菌菌株,发现粪肠球菌和肠系膜芽孢杆菌对青霉素G有耐药性,嗜酸乳杆菌对氨苄青霉素有耐药性。
Montassier等人(2021)的一项研究表明,摄入市售益生菌补充剂(含有11种常见的益生菌菌株:嗜酸乳杆菌、干酪乳杆菌、副干酪乳酸杆菌、鼠李糖乳杆菌、植物乳杆菌、双歧杆菌、短双歧杆菌、长双歧杆菌亚种、婴儿长双歧杆菌、乳链球菌和嗜热链球菌)导致ARG丰度降低。
然而,益生菌也可以作为肠道耐药组扩展的储库,因为益生菌的摄入可以通过水平基因转移将携带ARG的菌株扩展到病原体,因为移动遗传元件含量(转座酶和整合酶)的检测与ARG丰度相关。
这些发现强调了在将益生菌纳入饮食时需要谨慎,特别是在商业益生菌补充剂的情况下。虽然益生菌可以在减少致病菌定植和增强抗菌活性方面发挥作用,但必须仔细评估它们作为ARG载体的作用。为了降低潜在风险,需要监测补充剂和功能性食品中使用的益生菌菌株中的ARG含量,确保它们不会导致耐药性的进一步传播。
总之,虽然益生菌作为功能性食品具有调节肠道耐药性和对抗抗生素耐药性的潜力,但它们储存和转移ARG的潜力需要更彻底的研究。未来的研究应侧重于鉴定ARG转移风险最小的益生菌菌株,并优化益生菌食品,以提高其治疗效果,同时限制对肠道耐药的不利影响。
以上我们已经了解了抗生素耐药性的发生机制、传播途径、饮食干预的方法和机制。在实施上述饮食干预策略的同时,如何客观评估其对肠道耐药组的影响至关重要。
随着测序技术的快速发展,肠道菌群检测已经能够识别肠道中的抗生素耐药基因,例如,谷禾宏基因组肠道菌群检测报告中,专门设有抗生素耐药基因分析板块,可检测β-内酰胺酶、氨基糖苷类、大环内酯类、喹诺酮类、四环素耐药基因等多种耐药机制的存在和丰度,这为评估饮食干预效果提供了客观依据。
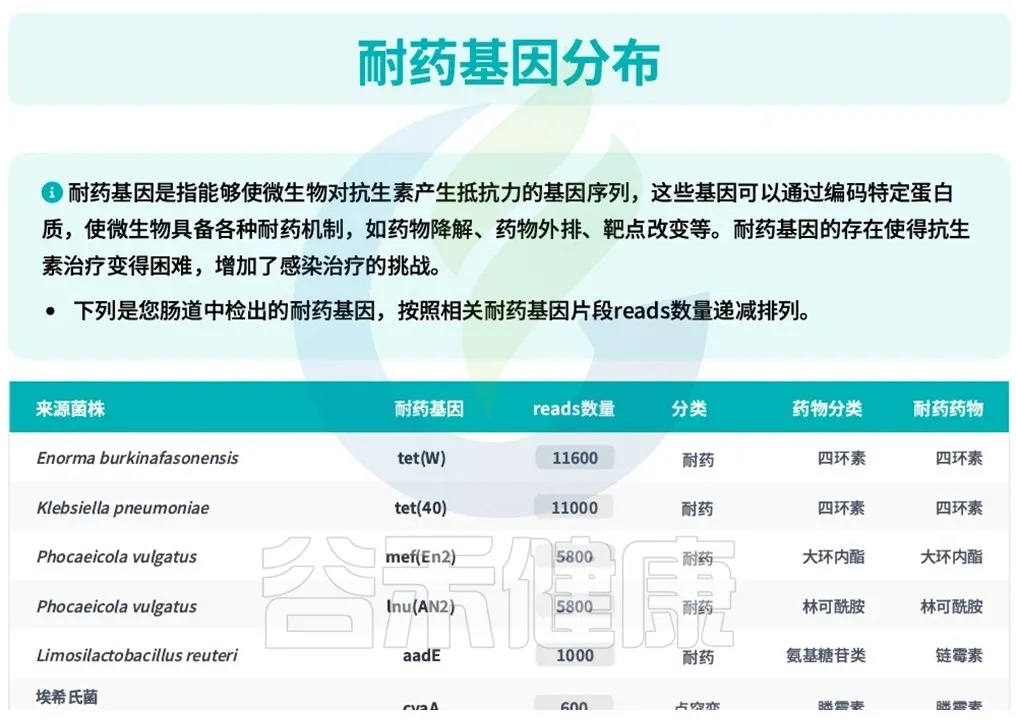
<来源:谷禾宏基因组肠道菌群检测报告>
通过干预前后的对比检测,我们可以了解特定饮食成分对耐药基因表达的影响,从而不断优化干预方案,实现个性化的肠道耐药组管理。
例如,患者A因肺炎需要使用抗生素治疗,在用药前安排了肠道菌群检测。报告显示其肠道中存在高水平的β-内酰胺酶耐药基因,那么根据报告调整相关抗生素方案,避开容易产生耐药的β-内酰胺类抗生素,减少治疗期间耐药基因的扩增风险。
通过识别高丰度ARGs,预测治疗失败风险,避免可能造成更加严重抗生素耐药。
一名反复发生尿路感染的患者B,常规抗生素治疗无效。通过肠道菌群宏基因组检测,发现其肠道中携带高丰度的 blaCTX-M(广谱β-内酰胺酶基因) 和 mecA(甲氧西林耐药基因),提示对β-内酰胺类及甲氧西林类药物耐药。
根据检测结果,避免使用头孢类(靶向β-内酰胺酶敏感菌),改用磷霉素(对blaCTX-M无交叉耐药)或联合多黏菌素(针对mecA阳性菌)。补充蔓越莓原花青素(抑制β-内酰胺酶活性)或大蒜素纳米颗粒(破坏生物膜),增强抗生素渗透性。
识别关键耐药基因,指导临床选择非交叉耐药抗生素并辅以天然抗菌成分,从源头减少抗生素滥用导致的耐药性扩增。
一名2型糖尿病患者宏基因组检测显示,其肠道中 Vancomycin_vanX(万古霉素耐药基因) 和 Multidrug_emrE(多药耐药基因) 丰度升高,且与胰岛素抵抗指数呈正相关。
ARG丰度高的糖尿病患者血糖控制失败率相对较高,因此可以从这方面入手,增加苦参碱摄入(抑制Mex泵活性),联合二甲双胍改善胰岛素敏感性。
通过识别耐药基因丰度,可预测糖尿病血糖控制失败的风险,针对性干预,减少耐药基因对慢性疾病的协同恶化作用。
抗生素耐药性已经从单纯的医学挑战演变为一个多维度的健康问题,而肠道耐药组的研究为我们提供了新的思路应对这一挑战。
肠道菌群检测技术已能够识别耐药基因的存在和丰度,使个体ARG状况的评估成为可能,这不仅可以帮助临床医生了解患者的耐药风险,也为饮食干预提供了科学依据。
日常饮食的预防性策略
加富含多酚的食物(如绿茶、浆果、深色蔬菜)、含硫食物(如十字花科蔬菜、大蒜)及发酵食品(如酸奶、泡菜),通过天然成分抑制耐药基因传播。
控制高脂高糖饮食(如红肉、加工食品),避免促进肠杆菌科等耐药菌增殖。地中海饮食模式(富含纤维、橄榄油)可提升短链脂肪酸水平,抑制耐药菌生长。
通过将这些科学发现转化为具体的饮食策略和产品,结合肠道菌群检测技术,我们有望在不依赖新抗生素开发的情况下,通过日常饮食管理减轻抗生素耐药性的负担,为公共健康问题提供可持续的补充解决方案。
功能性食品的开发创新
开发含高浓度EGCG的绿茶提取物饮品,或添加原花青素的蔓越莓咀嚼片,辅助治疗泌尿系统耐药菌等相关感染。
利用大蒜素纳米颗粒或异硫氰酸盐制成肠道缓释胶囊,抑制生物膜形成并减少水平基因转移。
基因编辑乳杆菌(如表达抗菌肽的菌株),靶向清除肠道内携带ARGs的致病菌。
通过微胶囊化或脂质体包裹提高多酚类成分的肠道吸收率;结合AI预测模型,从天然产物库中筛选新型抗菌分子,加速功能性配方的迭代。
建立功能性食品中ARGs迁移风险的动态监测体系,避免益生菌载体成为耐药基因传播媒介。
功能性食品作为“可食用疗法”,通过精准调控肠道耐药组,为临床耐药危机提供了“非抗生素”解决方案。未来需融合合成生物学、纳米技术与营养学,推动第三代功能性食品的落地。
临床营养的精准化应用
本文中列举的一些研究表明,植物多酚(如柿子单宁、表没食子儿茶素没食子酸酯)、含硫化合物(如大蒜素)及萜类(如1,8-桉叶油醇)可通过破坏细菌生物膜、抑制外排泵或增强抗生素渗透性,降低耐药菌存活率。临床可开发含此类成分的功能性营养剂,辅助治疗耐药菌感染(如MRSA、铜绿假单胞菌),或用于术后耐药菌感染预防。
特定益生菌(如乳酸杆菌、双歧杆菌)可通过竞争排斥或分泌抗菌肽抑制耐药菌定植。临床中可针对术后感染或抗生素滥用患者,定制含益生菌的肠内营养配方,重建肠道菌群平衡。
乳铁蛋白、植物抗菌肽等可增强抗生素疗效。例如,乳铁蛋白与氨基糖苷类联用可显著抑制多重耐药鲍曼不动杆菌,适合用于重症患者的营养支持方案。
2型糖尿病患者中,苦参碱与二甲双胍联用可抑制肠道内产气荚膜梭菌的Mex泵活性,改善胰岛素敏感性。
结直肠癌患者化疗期间,添加大蒜素纳米颗粒的营养配方可减少肠道内携带mdtP基因的多重耐药大肠杆菌丰度,缓解化疗相关性腹泻。
然而,也需要建立植物化学物质与抗生素的相互作用数据库,避免协同毒性(如EGCG与环丙沙星联用可能加重肝损伤)。
同时可以开展剂量梯度临床试验,明确如多酚类成分的最佳治疗窗口。
联合微生物学、药理学与食品科学,开发兼具营养支持与耐药调控功能的特医食品。通过跨学科协作,特医食品不仅可成为感染患者的营养支持方案,也能作为耐药危机的一线防控工具。
未来,跨领域合作结合人工智能,有望加速功能性食品的迭代,在降低抗生素依赖的同时,重塑肠道健康生态,为应对耐药危机提供可持续路径,最终实现“以食为药”的精准健康管理。
主要参考文献:
Abbas A, Barkhouse A, Hackenberger D, Wright GD. Antibiotic resistance: A key microbial survival mechanism that threatens public health. Cell Host Microbe. 2024 Jun 12;32(6):837-851.
Liang Z, Liang Z, Hu HW, Howell K, Fang Z, Zhang P. Food substances alter gut resistome: Mechanisms, health impacts, and food components. Compr Rev Food Sci Food Saf. 2025 Mar;24(2):e70143.
Pramod Barathe, Kawaljeet Kaur, Sagar Reddy, Varsha Shriram, Vinay Kumar, Antibiotic pollution and associated antimicrobial resistance in the environment,Journal of Hazardous Materials Letters, Volume 5,2024.
Langford BJ, Soucy JR, Leung V, So M, Kwan ATH, Portnoff JS, Bertagnolio S, Raybardhan S, MacFadden DR, Daneman N. Antibiotic resistance associated with the COVID-19 pandemic: a systematic review and meta-analysis. Clin Microbiol Infect. 2023 Mar;29(3):302-309.
Laxminarayan R. The overlooked pandemic of antimicrobial resistance. Lancet. 2022 Feb 12;399(10325):606-607.
Sarmiento MRA, de Paula TO, Borges FM, Ferreira-Machado AB, Resende JA, Moreira APB, Dutra Luquetti SCP, Cesar DE, da Silva VL, Diniz CG. Obesity, Xenobiotic Intake and Antimicrobial-Resistance Genes in the Human Gastrointestinal Tract: A Comparative Study of Eutrophic, Overweight and Obese Individuals. Genes (Basel). 2019 May 7;10(5):349.

谷禾健康

当地时间12月19日,微软联合创始人、亿万富翁比尔·盖茨发布了对来年的年度预测,称 2024 年将是一个“转折点”。
在这封长达 10 页的信中他展示了对人工智能领域的更多创新、婴儿营养不良问题的突破、气候变化谈判的进展等多方面的期待。
人工智能可以让世界变得更加公平。盖茨对人工智能的预测超越了去年的超声波预测,他断言人工智能的进步将广泛改善全球健康,同时促进创新。
人工智能可以帮助世界各地对抗抗生素耐药性疾病,识别高风险妊娠并估计孕龄等,他预计人工智能在开发新药、诊断疾病和增强教育资源方面将发挥至关重要的作用。
盖茨表示,世界在利用肠道微生物补充剂来解决儿童营养不良方面即将取得突破。
本文我们来看看盖茨眼中的儿童营养不良问题及解法,盖茨预计,随着对微生物组的深入研究,人们将能够开发出以微生物为基础的新型营养治疗方案,这将给解决全球儿童营养危机带来重要进展。
期待已久的营养不良问题的突破即将到来
在盖茨基金会,我们愿意下大赌注。我们知道每一次冒险都可能没有回报,但这没关系。我们的目标不仅仅是逐步取得进展。我们的目标是将我们的努力和资源投入到重大项目中,这些项目一旦成功,就能拯救和改善生命。
当你下一个大赌注时,你往往要等待很长时间才能看到它是否有回报。当你终于意识到它会成功时,那种感觉是难以置信的。我最兴奋的一次豪赌就接近了这一时刻:利用我们对肠道微生物群的了解来预防和治疗营养不良。
经常有人问我,如果只能解决一个问题,我会选择什么。我的答案不变:营养不良。这是世界上最严重的健康不平等现象,大约每四个儿童中就有一个受到影响。如果在生命的头两年得不到足够的营养,身体和智力都无法正常发育。通过解决营养不良问题,我们可以减少导致儿童死亡的最大因素之一。
营养不良的原因远比没有得到足够的食物要复杂得多。大约15年前,研究人员注意到脊髓灰质炎等口服儿童疫苗在营养不良率高的地区效果不佳,他们开始怀疑生活在肠道中的微生物群可能在其中发挥了作用。很明显,有什么东西妨碍了它们被正常吸收。
这种怀疑在2013年得到了证实,当时生物学家杰夫·戈登(Jeff Gordon)发表了一项有里程碑意义的研究,关于马拉维一对双胞胎婴儿的微生物群。微生物群不仅是健康的副产物,还是健康的决定因素。这是第一条重要线索,表明我们或许可以通过改变肠道微生物群来减少营养不良问题。
经常有人问我,如果只能解决一个问题,我会选择什么。我的答案不变:营养不良
在过去的十年里,我们对肠道微生物群的了解比之前的1000年还要多。我们发现,生活在肠道中的细菌可能处于功能失调状态,从而导致炎症,使人无法吸收营养。我们发现,如果及早干预,就能最大程度地改善肠道微生物群。
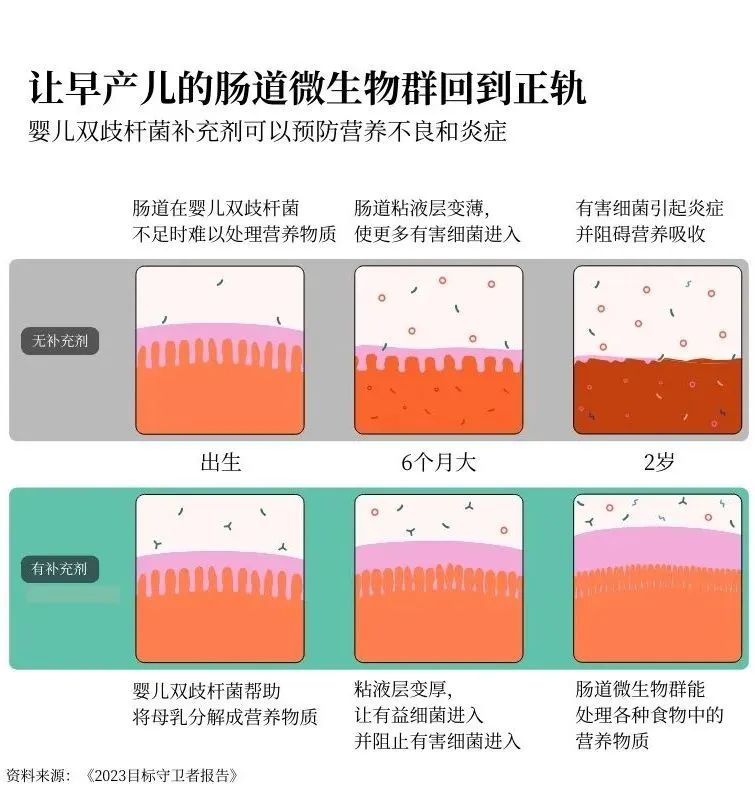
在人类发育过程中最先出现的肠道细菌之一叫做婴儿双歧杆菌。它能帮助将母乳中的糖分分解成人体生长所需的营养物质。反过来,母乳又为婴儿双歧杆菌和整个肠道微生物群提供食物。这是一个良性循环。但是,如果婴儿一开始没有足够的婴儿双歧杆菌,他们就可能无法从母乳中吸收足够的营养来支持其他必要的肠道细菌的生长。
要克服这种缺陷几乎是不可能的。你可以获得世界上所有富含营养的食物,但这并不管用。如果你的肠道生长路径过早被打乱,你可能永远无法吸收到所需的全部营养。
但是,如果我们能给高危婴儿服用婴儿双歧杆菌作为益生菌补充剂呢?我们能否及早干预,让他们正常发育?
这正是基金会合作伙伴多年来一直在研究的问题——我们终于找到了答案。我们正在对一种可添加到母乳中的婴儿双歧杆菌粉末补充剂进行三期临床。来自5个国家的16000名婴儿参与了试验,研究人员正在对每个婴儿进行跟踪,以确保益生菌既安全又有效。
目前的研究结果令人惊叹:通过给婴儿喂食这种益生菌,可以帮助他们的微生物群进入积极状态,从而使他们能够长大并充分发挥潜力。这对预防营养不良大有裨益。
在婴儿出生前就能改善肠道微生物群,这也让我感到兴奋。新的研究发现,婴儿的微生物群与母亲的微生物群息息相关。解决子宫内的炎症问题会给产妇、胎盘和发育中的胎儿带来更多益处。
如果我们能给准妈妈提供一种益生菌补充剂,让她的孩子从出生第一天起肠道就正常发育,那会怎么样呢?目前还不清楚这些活体生物药到底是什么样的,也不清楚如何给药,这还只是非常早期的研究。但研究表明,健康的微生物群可帮助婴儿在妊娠晚期每天增加5克体重。
在过去十年里,儿童健康领域发展速度超出了我一生中的预期。看到微生物群从一个完全看不见的东西变成解决世界上最大健康不平等之一的关键策略,真是令人惊叹。我迫不及待地期待在接下来一年里我们将学到多少新知识,以及我们将如何运用这些知识来拯救生命…
——比尔·盖茨
儿童营养不良问题的现状如何?带来什么问题?
儿童营养不良问题给全球健康带来巨大挑战,据世界卫生组织和联合国儿童基金会的数据统计,到 2020 年,估计有1.49亿 5岁以下儿童生长发育迟缓(年龄身高低),而 4500万儿童出现消瘦(WLZ 低)。
营养不良及其长期后遗症是该年龄段人群发病和死亡的主要原因。后遗症包括线性生长、免疫和代谢功能以及神经发育的持续损害——所有这些都对当前的干预措施有很大的抵抗力。
从比尔·盖茨的年度总结我们也可以看到,他十分重视肠道微生物对健康的影响,总的来说,微生物方面的研究给儿童健康带来了很大希望。
从肠道微生物组的角度入手,对营养不良儿童进行基于微生物群的相关干预措施,比尔及梅琳达·盖茨基金会已支持多项相关研究。我们来看近期得到该基金会支持的两篇研究文献。
在健康儿童中,肠道微生物组在分类和功能多样性方面稳步增长,直到3岁,其中最明显的变化发生在断奶期间。相比之下,营养不良儿童的肠道微生物组是年龄倒退的,即肠道微生物组与年幼儿童相似。
抗生素是严重急性营养不良的标准治疗方法,因为即使儿童没有出现明显的疾病,他们也可能因急性感染而突然恶化。
在马拉维和尼日尔,与安慰剂相比,短期(即7天)阿莫西林给药已被证明在降低全因死亡率、住院率、腹泻病和改善人体测量方面具有益处。
然而,抗生素治疗对严重急性营养不良儿童的潜在后果(如抗生素耐药性的发展和微生物组破坏)仍存在不确定性。
研究设计
研究人员想要确定 7 天的阿莫西林治疗,对接受严重急性营养不良治疗的儿童肠道微生物组和抗生素耐药组的急性和长期变化的影响。该成果发表在《Lancet Microbe》。

研究人员对尼日尔门诊治疗的严重急性营养不良儿童(6-59 个月)的阿莫西林随机、双盲、安慰剂对照试验 (NCT01613547) 进行了二次分析。从2013年9月23日至2014年2月3日从整个队列中随机选择了161名儿童(n = 2399)进行最初12周的随访。
根据人体测量结果从这161名儿童中选择了一个方便样本,2年后(2015年9月28日至10月27日)进行随访。儿童在基线、第 1 周、第 4 周、第 8 周、第 12 周以及 2 年随访队列中的第 104 周提供了粪便样本。研究人员进行了宏基因组测序,然后对粪便样本进行了微生物组和耐药组分析。38 名无严重急性营养不良的儿童和 6 名与原始队列基线年龄相匹配的严重急性营养不良儿童被用作参考对照。
研究结果
结果表明,营养不良的尼日尔儿童在接受抗生素治疗后,虽然发现克雷伯氏菌属、埃希氏菌属等增加,这些菌可能与更严重的感染相关,但是,微生物组和耐药组扰动的负面影响似乎是短暂的,在三周内完全消失。
进一步观察到阿莫西林治疗的一些意想不到的长期益处,包括改善长期微生物组丰富度,多样性和成熟度。
阿莫西林对长期微生物组成熟的影响
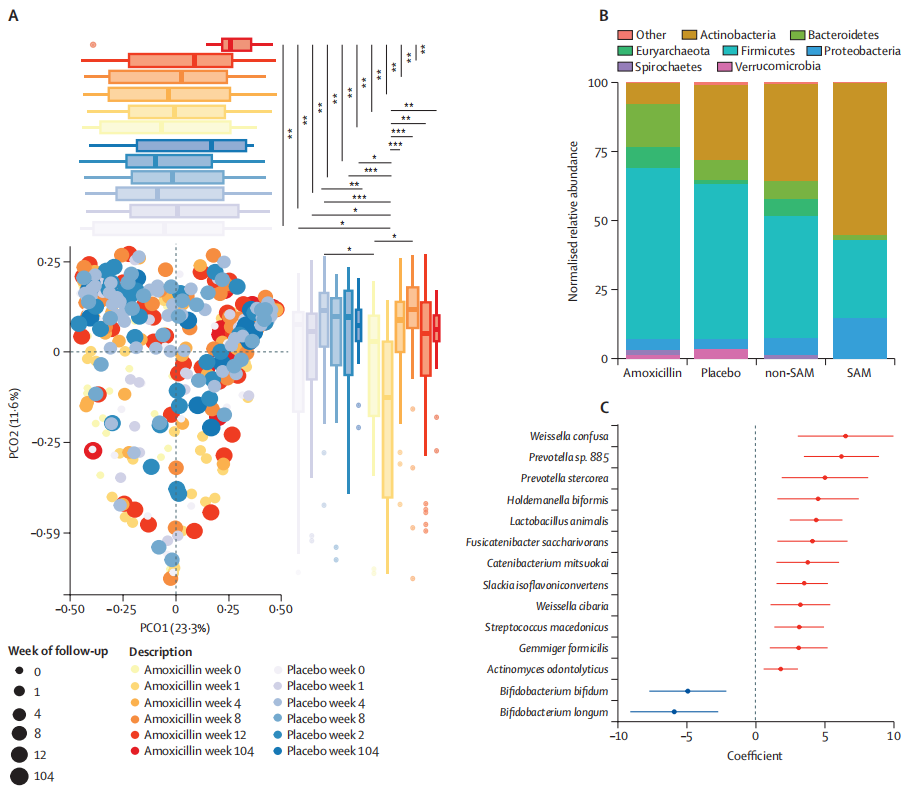
doi.org/10.1016/S2666-5247(23)00213-6
生命头三年肠道微生物组急剧成熟的一些关键驱动因素包括婴儿在出生期间和出生后接触母体微生物、加入牛奶、奶粉以及向固体食物的过渡。在营养不良的儿童中,肠道微生物组的成熟过程和生长都受到阻碍。但在治疗计划中加入抗生素改善了他们的身体测量和微生物组发育。
抗生素可能起到重置的作用,使有助于固体食物消化的微生物群在肠道生态系统中繁衍生息。
“
接受抗生素治疗的效果,对严重营养不良儿童来说算是个好消息,对于这一特定人群,抗生素治疗的益处似乎大于风险,但风险确实存在。研究人员警告说,儿童中已知的耐药细菌和耐药基因的增加不应被忽视。
营养干预措施,如即食治疗性食品,已被证明可以暂时改善肠道微生物组的成熟度和人体测量得分;然而,这种改善并不总是持续的。
在过去的5年里,合理设计的以微生物群为导向的治疗性食品在中度和重度急性营养不良儿童的微生物组恢复和人体测量得分方面显示出了希望。
接下来我们来看对于微生物群为导向的治疗性食品研究的最新进展。
生命头两年,肠道微生物群的发育需要与身体其他部分、其他器官系统的发育同步。当这个微生物群不完全形成时,仅仅增加热量是无法修复的。研究人员试图寻找特定的食物成分,以滋养健康的肠道微生物群,希望修复营养不良儿童肠道微生物群的功能失调。
使用传统食疗食品治疗可减少死亡,但并不能实质性改善营养不良的其他长期影响,包括新陈代谢、骨骼生长、免疫功能和大脑发育问题。
近日,来自华盛顿大学医学院的一项研究,已经确定了一种新型治疗食品中天然存在的关键生化成分,以及处理这些成分的重要细菌菌株。
该研究表明,识别这些成分以及作为其治疗靶标的关键促生长肠道细菌菌株,研究强调了了解细菌菌株如何加工特定食物成分的重要性,可以帮助指导当前食品配方的治疗,并可以在未来创造新的、更有效的配方。该研究成果于2023年12月13日发表在《Nature》杂志。
研究人员从试验参与者的粪便微生物群中重建了1000个细菌基因组(宏基因组组装基因组,简称MAG),确定了75个丰度与体重生长(体重长度Z评分变化,简称WLZ)正相关的MAGs。将MAG基因表达的变化表征为治疗类型和WLZ反应的函数,并定量MDCF-2和粪便中的碳水化合物结构。
结果表明,两个与WLZ呈正相关的普雷沃氏菌MAG是MDCF-2诱导的代谢途径表达的主要贡献者,这些代谢途径涉及利用MDCF-2的组分聚糖。
结果强调了微生物组反应的显著菌株特异性,并指出两种普氏菌菌株(MAG Bg0018 和 MAG Bg0019)是 MDCF-2 聚糖代谢和宿主体重生长反应的关键介质。
在生长相关细菌分类群代谢的MDCF中鉴定生物活性聚糖结构,有助于指导关于其在急性营养不良儿童中使用的建议,并有助于开发其他制剂。
在这项工作的基础上,世界卫生组织与比尔及梅琳达·盖茨基金会正在支持一项大型多站点临床试验,研究这种新的治疗性食品——MDCF-2,或微生物组导向的补充食品。
MDCF-2 治疗性食品包括哪些食物成分?
在 2021 年的临床试验中,MDCF-2 治疗性食品作为膳食补充剂提供,以提供儿童每日能量需求的约 20%。
Prevotella copri 两个菌株对MDCF-2干预改善营养不良的关键作用
研究人员发现:普雷沃氏菌Prevotella copri与儿童的生长发育呈正相关。
P. copri是拟杆菌门的成员,该门的成员含有一组叫多糖利用基因座(PUL)的基因,这些基因介导特定聚糖或一组聚糖的检测、导入和代谢。
它们在利用MDCF-2治疗食品中有益的生物活性碳水化合物结构的代谢途径中表现出活性的增加。
与接受传统治疗性食物的儿童相比,接受MDCF-2的儿童血液中支持肌肉骨骼生长和神经发育的某些蛋白质水平较高,与炎症有关的蛋白质水平较低。也表明,微生物群修复的作用远不止于肠道。
研究人员对这些儿童的粪便样本进行了广泛的基因组分析,了解对这些食物成分有反应的细菌,以及这些细菌对治疗的反应所具有和表达的代谢能力。事实证明,许多与儿童生长密切相关的细菌富含碳水化合物代谢的途径。
分析不同食物中的关键有效成分
在该研究中,通过质谱分析MDCF-2和RUSF两类食品,确定其组成成分。
MDCF-2、RUSF 及其食品成分中的多糖
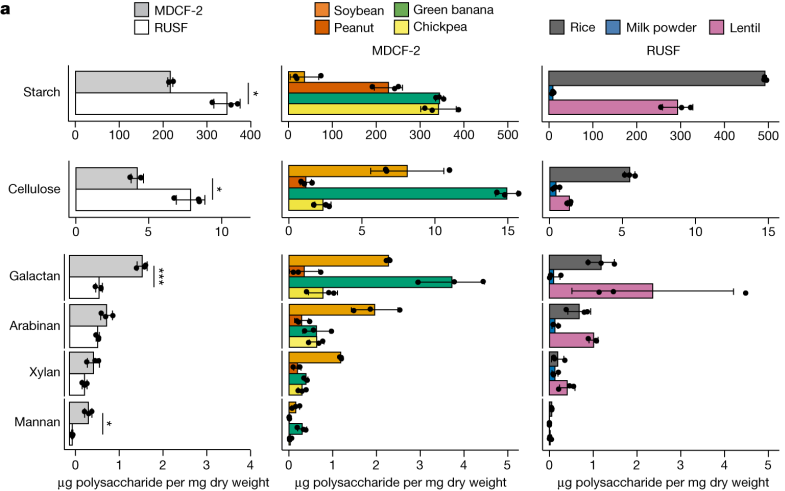
DOI: 10.1038/s41586-023-06838-3
对这些食物详细分析表明,MDCF-2食物中L-阿拉伯糖、D-木糖、L-岩藻糖、D-甘露糖和D-半乳糖醛酸 (GalA) 显著更丰富(P < 0.05)。
也就是说,MDCF-2含有更多的多糖,即半乳聚糖和甘露聚糖。而传统的治疗性食物RUSF含有更多的淀粉和纤维素。
MDCF-2 有效提高营养不良儿童生长
尽管MDCF-2的热量密度比高热量食物低15%,但与接受传统补充食物的儿童相比,接受MDCF-2治疗的儿童表现出更高的生长速度。
与MDCF-2治疗相关的生长增加与儿童微生物群中某些普氏菌菌株中存在的代谢途径的表达增加有关,这些途径与生物体利用MDCF-2中存在的多糖有关。
“来自食物频率问卷的信息表明,对MDCF-2反应最大的儿童也食用了更多的坚果和豆类作为他们日常饮食的一部分” ,研究人员表示,“这些坚果和豆类具有一些与MDCF-2相同的多糖。这表明可能有机会调整MDCF-2的成分和剂量,以进一步增强其治疗效果”。
营养不良儿童“微生物组修复”的一个定义是,重新平衡有益菌的表现和表达功能,使其呈现出更有利于健康微生物组-宿主共同发育的构型。
以这种方式将膳食聚糖和微生物代谢联系起来,为基于培养的计划提供了一个起点,这些计划旨在检索这些“效应”类群的分离物,用作潜在的益生菌制剂,或者如果与他们渴望的关键营养素相结合,修复对单独基于食物的干预反应不足紊乱的微生物群,提供合生元配方。
当然,P.copri菌株和MDCF-2聚糖之间的关系并不排除其他宏量或微量营养素对MDCF-2在增重方面优于RUSF的贡献。
总之,为改善微生物群的营养干预提供了一种新的见解。
这些和其他研究的结果有助于加深我们对微生物如何与人类细胞和器官合作,从而影响发育生物学的理解,帮助相关产业科研人员开发微生物组导向疗法的方法,帮助开发构建儿童营养健康的新策略,以确保在出生后的头几年形成健康的微生物组,尽可能规避儿童营养不良风险。
同时,随着人工智能技术及高通量测序等先进技术的不断发展,整合食物-微生物-人体交互作用研究,通过对婴幼儿的肠道菌群进行定期检测,可以帮助家长更好地监测儿童肠道菌群的变化,及时发现异常,采取相应的纠正措施,有利于促进儿童的身心健康发育。

谷禾健康

我们知道,微量营养素在人体健康中发挥着重要作用。然而,在全球范围内,约有3.4亿儿童患有多种微量营养素缺乏症,并伴有高致病性负担和耐多药细菌导致的死亡风险。
这些儿童经常用抗生素治疗营养不良相关疾病。可矛盾的是,由于潜在的微量营养素缺乏,他们的肠道微生物组可能已经为抗生素耐药性做好了准备…
通常,这些缺陷发生在生命早期前1000天,并可能带来即时和终身的健康缺陷。生命早期前1000 天是肠道微生物组成熟的关键时期,扰动会产生直接和长期的后果。
越来越多的研究让人认识到微生物与营养不良之间的密切关联。一些对单一微量营养素缺乏症的动物研究表明,诱导缺乏症后微生物特征发生了改变。例如,锌缺乏的雏鸡变形杆菌和厚壁菌门的丰度增加;妊娠、哺乳和断奶期间的维生素A缺乏症大鼠显示,其幼崽的微生物组成发生了显著变化,特别是厚壁菌门机会性病原体较高。
肠道环境中另一个容易被遗忘的元素是肠道抗性基因组(resistome),它是广泛表达于肠道微生物群栖息地内的共生菌和致病菌中的抗生素耐药基因(ARGs)和机制的完整集合。随着共生菌群的形成,抵抗基因组在婴儿期更加丰富,并在头几个月内高度动态变化。
抗生素耐药性可以是内在的,也可以是后天的。2019年的一份最新报告估计,5岁以下儿童中每5个死亡案例中就有1个是由于耐药细菌引起的。对营养不良微生物群组的理解仍然不完整,干预和治疗策略也可能被误导。
来自不列颠哥伦比亚大学的研究人员使用最近开发的小鼠模型,研究了多种微量营养素缺乏症,即维生素A、B12、叶酸、铁和锌,这是最常见的缺乏症,对成熟的肠道微生物组、结构和耐药性的影响。研究人员发现微量营养素缺乏会显著改变肠道菌群、真菌群、病毒组的组成,并且提供了以前未知的关于微量营养素如何塑造发育中的肠道抗性基因组的数据。

近日其研究成果发表在《Nature Microbiology》上,这为早期微量营养素缺乏症的长远后果提供了重要的见解。解决微量营养素缺乏问题,不仅仅是克服营养不良,也可能是抗击全球抗生素耐药性祸害的关键一步。我们一起来详细了解一下。
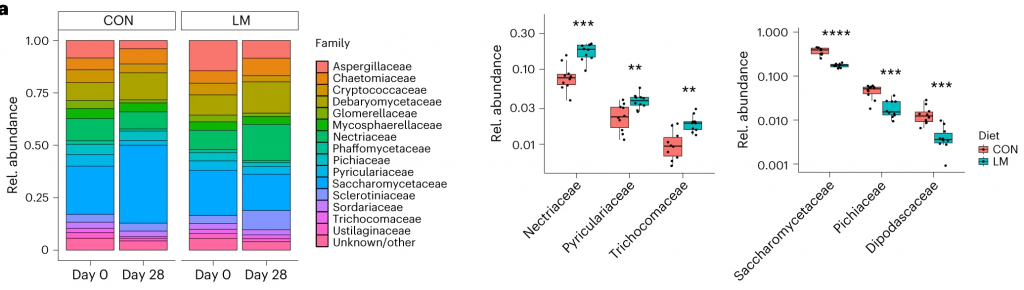
断奶C57BL/6N雄性小鼠接受等热量实验对照或多种微量营养素缺乏(即低锌、叶酸、铁、维生素A和B12)饮食28天。正如预期的那样,实验结束时,低微量营养素(LM)饮食的小鼠体重增加较少,发育迟缓和贫血。这里用鸟枪法宏基因组测序来表征饮食治疗前后的肠道微生物组。
第28天,在LM小鼠中观察到变形菌门、肠杆菌门和Tannerellaceae科的扩增,而CON组的厚壁菌门和丹毒科扩增。LM小鼠中,下列菌群的相对丰度较高:
而下列菌群丧失:
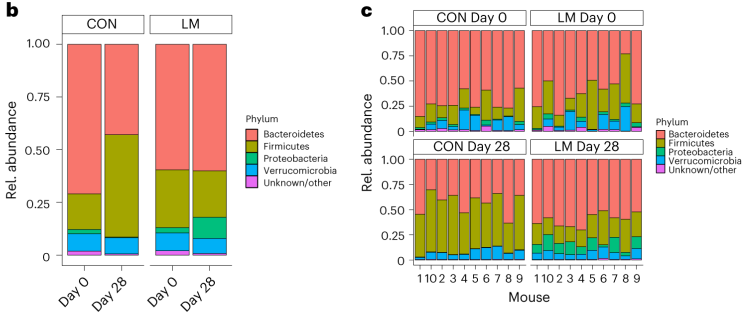
实验结束时,LM小鼠的α多样性较高(Shannon指数,P = 0.002);
PCA图显示,饮食处理后,两组之间存在明显的差异(p=0.001)
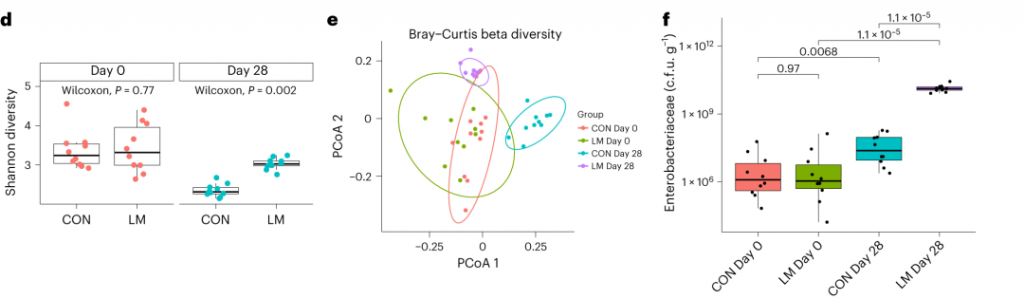
第28天,LM组中下列菌群增加:
下列菌群丰度降低:
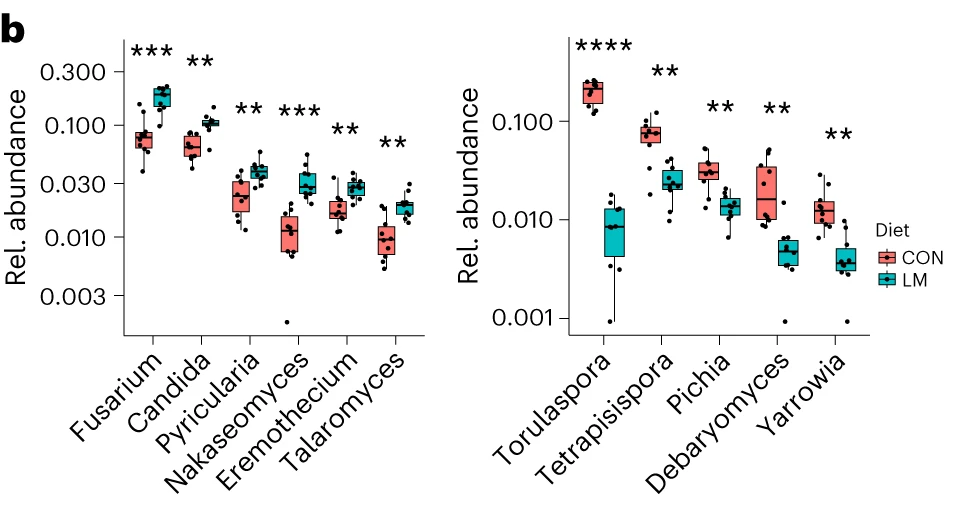
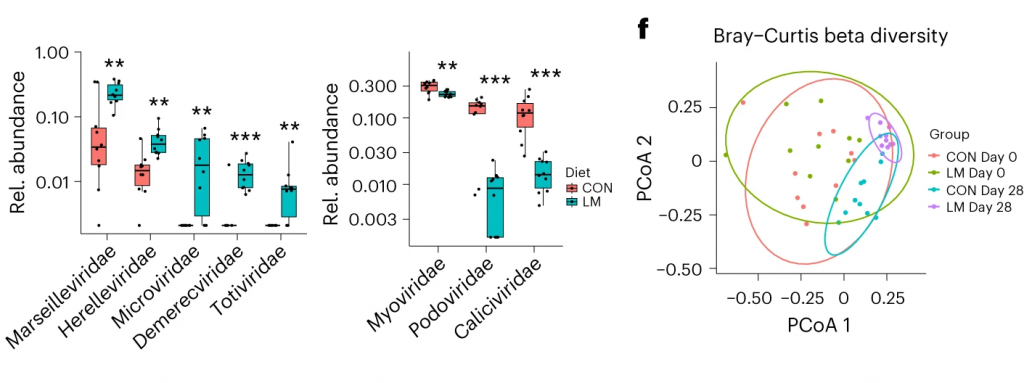
LM小鼠中,下列噬菌体丰度更高:
对照小鼠中,下列菌群的丰度更高:
LM组的β多样性与对照在第28天有显著差异(Bray-Curtis,PERMANOVA,P < 0.01)
发现了14种指示细菌:
在真菌组和噬菌体组中,LM小鼠中下列相对丰度显著增加:
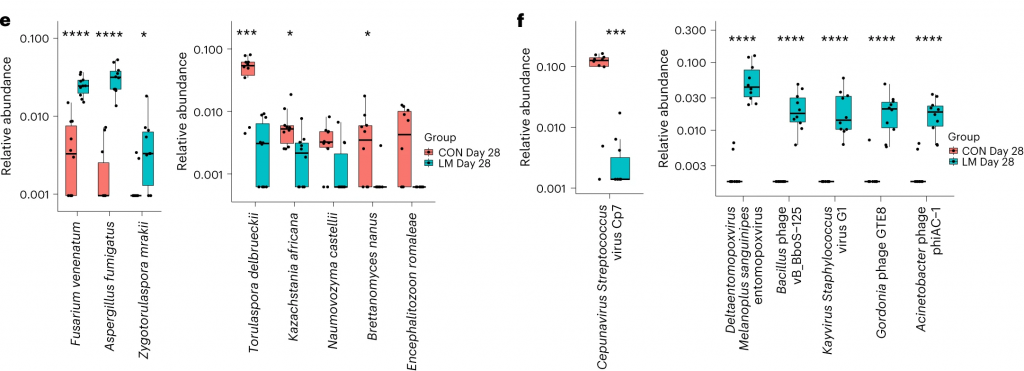
在营养不良的儿童中,机会性细菌的扩张(在不利情况下可能引发危害的微生物)是肠道的常见表型。因此,研究人员将模型中的机会性细菌与常见于营养不良儿童中的细菌进行表型比较。
第28天,LM小鼠中总机会性细菌的相对丰度总体增加(P < 0.0001)。11个分类单元中,有7个在第28天LM组的相对丰度显著增加:
有3个在LM组中丰度减少:
在LM的小鼠真菌群落中,观察到Candida dubliniensis 的相对丰度较高(P < 0.019)。
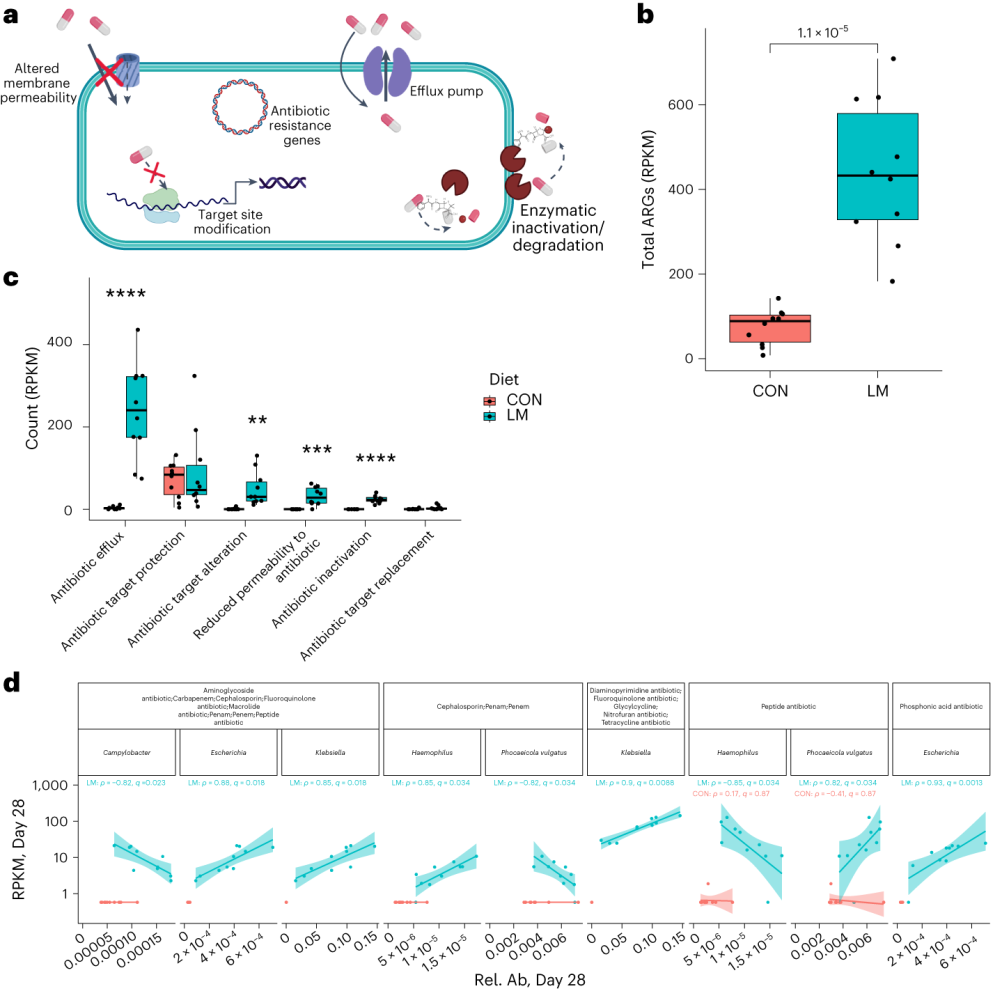
抗生素耐药性,无论是内在的还是后天的,都是由几个关键机制驱动的。
使用综合抗生素耐药性数据库(CARD)对肠道耐药性组进行分析显示,总ARGs的相对丰度增加(P = 1. × 10−5)。
随后的分析表明,抗生素外排和抗生素失活是改变的最常见的内在耐药性机制。
第28天LM组的ARG α多样性显著高于对照组(P = 0.0092)。
基于Bray–Curtis的相异性指标显示,低微量营养素饮食强烈影响了早期肠道抵抗。ARGs的计数显示,绝大多数(73%)ARGs仅在LM第28天组中流行≥30%。
将CARD数据库映射和药物分类分析显示,氟喹诺酮类/大环内酯类/青霉素类抗生素(Q < 0.0001)是LM处理小鼠中最常见的药物耐药机制改变,其次是磷酸酯类(即福氏霉素)(Q < 0.001),以及二氨基嘧啶类/氟喹诺酮类/甘氨环素类/硝呋环酮类/四环素类(Q < 0.001)。
▸ 接下来,研究人员提出是否有药物类别或耐药性机制与任何细菌门或机会性细菌相关?
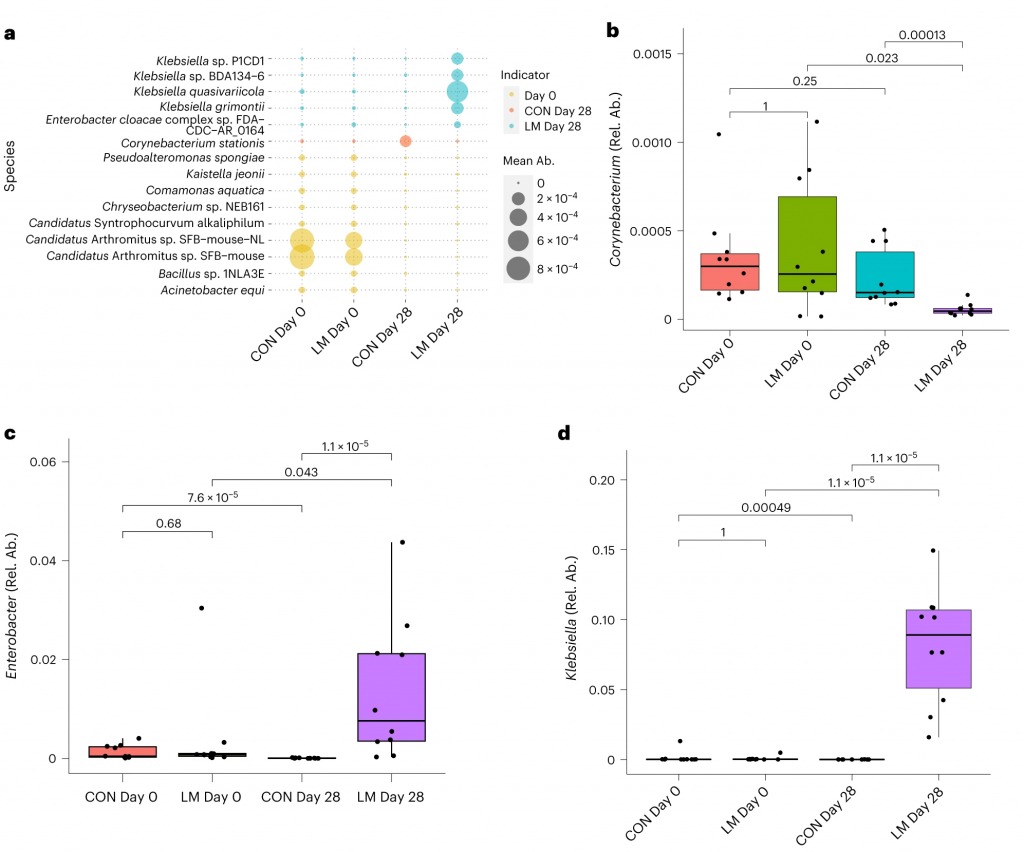
事实上在LM小鼠中,克雷伯氏菌、嗜血杆菌、埃希氏菌、弯曲杆菌、Phocaeicola vulgatus和几种药物类别之间存在正相关(Q < 0.05)。不出所料,克雷伯菌和大肠杆菌与ARGs药物类别的相关性比其他机会细菌更强。
在个体机会性细菌中,克雷伯菌和弯曲杆菌与外排机制相关,克雷伯菌,志贺菌,弯曲杆菌和埃希氏菌与抗生素失活相关。
肠球菌与外排机制相关,但仅在对照组中。
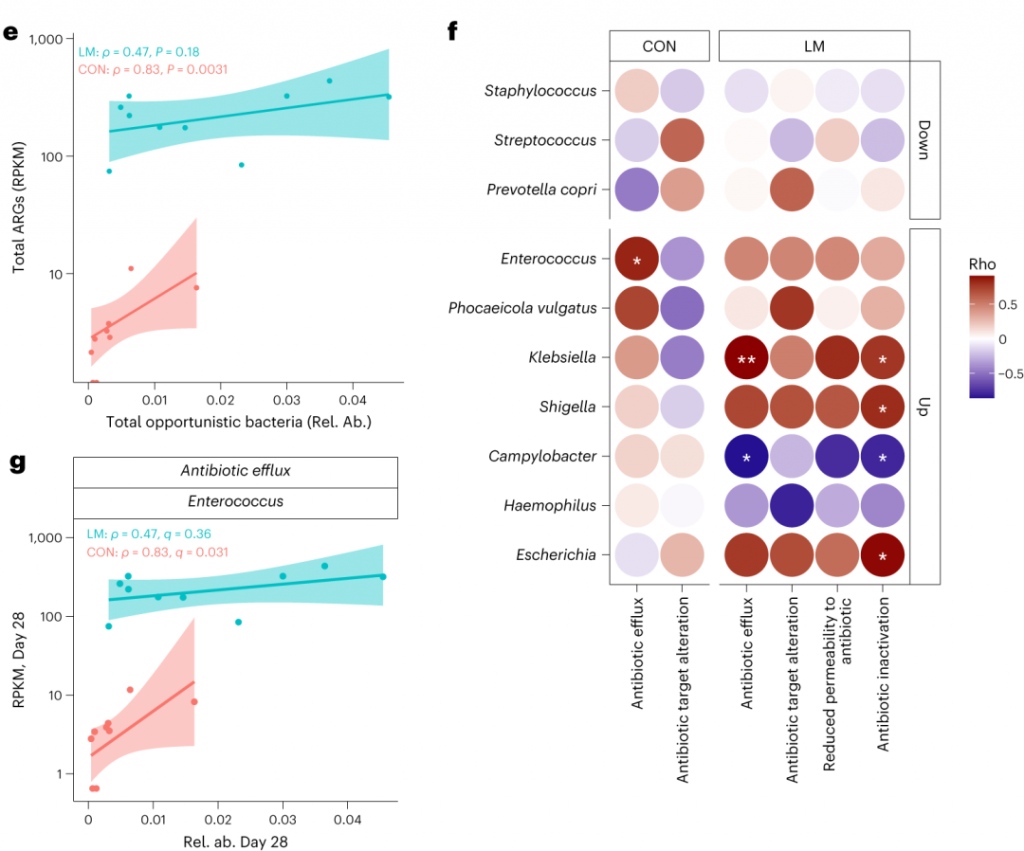
环境压力源,如营养缺乏,已被证明会触发细菌的压力反应,从而诱导抗生素耐药性机制作为生存策略。矛盾的是,细菌也可以产生活性氧(ROS),在致命压力的存在下自毁。
研究人员研究了LM喂养的小鼠的应激反应机制是否发生了改变,以及它们是否与个体ARGs相关。
事实上,在LM小鼠中发现编码氧化应激和氧化应激反应的基因更丰富,总ARGs与氧化应激之间具有很强的相关性(P = 0.0098)。
具体来说,氧化应激反应与下列之间存在强烈的相关性:
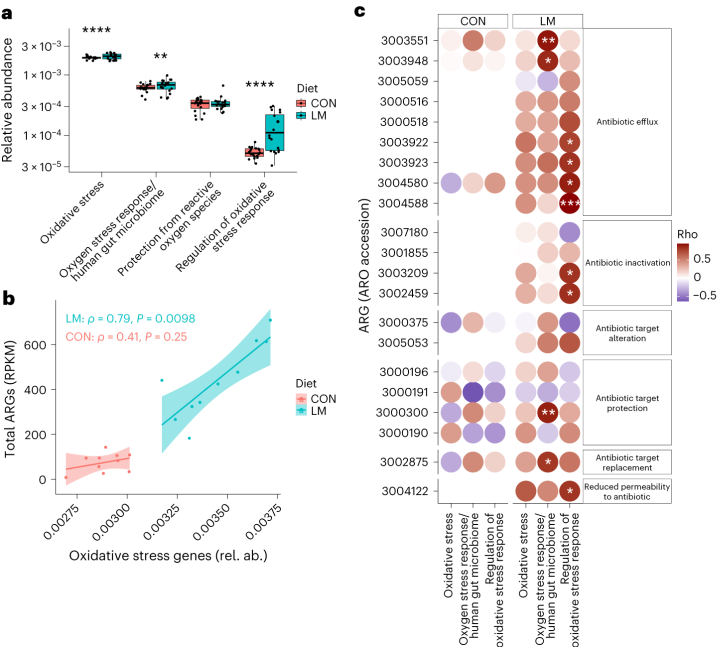
在对照小鼠中,总体氧化应激与嗜血杆菌之间存在强烈的相关性(p<0.05)。
在LM小鼠中,氧化应激反应的调节与克雷伯杆菌、志贺氏菌、弯曲杆菌、大肠杆菌、肠杆菌科菌和变形菌门菌的增加相关。
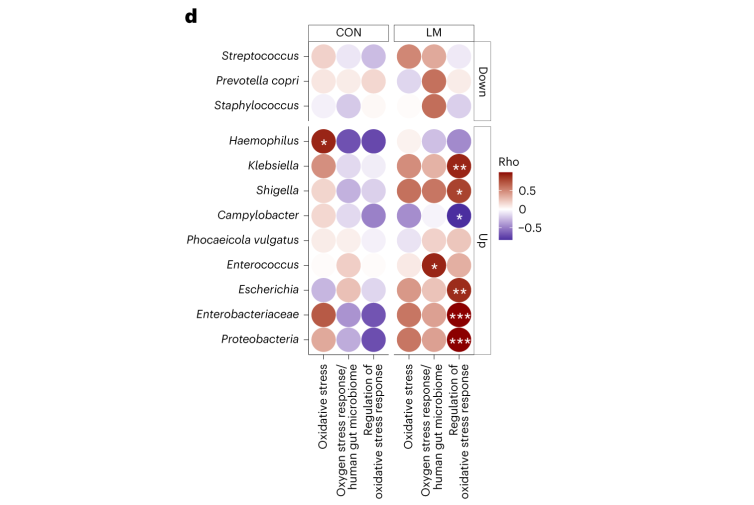
微量营养素缺乏是一项重要的公共卫生危机,同时也是一个容易忽视的研究领域。该研究基于先前的动物研究,强调了这些微量营养素缺乏如何影响发育中的肠道微生物组。此外还探索了耐药基因组。
低微量营养素 & 肠道微生物组
在低微量营养素饮食28天的小鼠体内,细菌、真菌和病毒/噬菌体粪便种群发生了变化 。由于微量营养素不足的影响,变形菌门细菌和机会性细菌,如克雷伯菌、大肠杆菌、志贺菌、衣原体、肠球菌、弯曲杆菌、肠杆菌、Phocaeicola vulgatus富集。
研究表明,饮食扰动确实会破坏肠道中正在发育的真菌生物群落和病毒组/噬菌体。还观察到LM小鼠体内病毒/噬菌体的富集,这表明了一个不成熟(即婴儿)表型和发育延迟。这一特征可能对细菌适应能力和宿主免疫反应产生影响。
真菌组在早期生命发育阶段受到饮食和其他环境因素的影响,可能对短期和终身健康产生影响。两个实验组的真菌携带量随着群落成熟而增加。然而,在第28天,低微量营养素饮食组的小鼠体内明显增加了致病真菌念珠菌,这是一种常见的侵袭性物种。
这些发现与一个在马拉维队列中的研究结果一致。该研究报道了在无锌情况下给予多微量营养素粉剂(MNP)后,马拉维队列的肠道微生物组中原生动物和真菌群落的增加。埃希氏菌-志贺菌也在12个月大的婴儿中富集,这些婴儿接受了补充或不补充锌的MNP,强调了微量营养素在形成营养不良儿童肠道群落组成中的作用。
低微量营养素 & 抗生素耐药性
抗生素耐药性是一项迫切的全球公共卫生威胁,肠道微生物组是耐药基因(抗性基因组)的重要来源。
一项先前的研究发现,健康婴儿的耐药基因组中存在预测超过30类抗生素耐药性的400多个基因。其中,167个基因对多个抗生素表现出表型耐药性。有趣的是,婴儿的耐药基因组中还包括对婴儿之前未接触过的多种抗生素的耐药性。这进一步强调了内在耐药基因组作为当前和新兴抗生素耐药性的病原库的重要性。
在本次研究中,发现低微量营养素饮食的小鼠中富集了参与内在抗生素耐药性机制的基因,这些基因与机会性细菌的存在具有关联性。例如,与氟喹诺酮和膦酸(如磷霉素)耐药相关的基因得到了富集。这对于常见患有多药耐药细菌引起的菌血症的营养不良儿童来说,具有重要的临床意义。
本次研究提供了重要发现,并强调了微量营养素缺乏与肠道抗药基因组之间的重要联系。这些结果表明,我们对营养不良微生物组的认识还不完整,需要进一步研究成熟的抗药基因组。
低微量营养素 & 肠道微生物组 & 耐药性
此外,还展示了微量营养素缺乏引起了肠道微生物组成的显著变化。这导致了变形菌门的扩增、机会性细菌和真菌病原体的增加,进一步促进了多种内在耐药基因的表达。
尤其是发现了细菌氧化应激与抗药基因组之间的显著相关性。确定了微量营养素缺乏引起的细菌应激在营养不良宿主中显著促进内在抗生素耐药性的作用。因此,未来解决营养不良和抗生素耐药性问题的策略应将内在耐药基因组纳入考虑,并与可持续发展目标(良好健康和福祉)相一致,最大程度造福于相关儿童。
本研究的一个局限性是初步的微生物组特征分析中使用了雄性小鼠(基于先前的研究表明雄性对营养不良比雌性更加敏感),研究人员计划在未来探索雌性小鼠。
另一个潜在的局限性是使用了多种微量营养素;然而,这种设计增强了模型,反映了实际应用中与儿童相符的多种微量营养素的复杂协同作用。一个主要的优势是将测序和生物学评估相结合,用于微生物组和抗生素耐药性的特征化。进一步探索数据集可能揭示共存微量营养素缺乏的功能和微生物动态。
未来可以调查肠道生理学以及抗药基因组在致病感染易感性中的作用。进一步探索真菌耐药基因组和该模型中内在抗真菌性能。然而,抗真菌耐药性数据库(MARDy)是目前唯一经过策划的抗真菌耐药性数据库,但更新不够频繁,限制了它的应用。
目前可用的抗生素耐药性分析数据库往往存在高估基因关联性的倾向。数据库的选择取决于研究人员想要回答的问题类型。
CARD数据库的一个局限性是它基于已发表的基因组数据进行策划,可能对经过深入研究的生物体比对少研究的生物体有偏见。不过它的全面列表、专家的定期更新和机器学习算法支持使其成为该领域中的宝贵工具。
编辑
抗生素耐药性通常归因于抗生素的过度使用和误用,但本研究表明,微量营养素缺乏的“隐性饥饿”是一个重要因素。需要进一步加强对营养不良与抗生素耐药性之间关系的研究,增进我们对这一领域的理解。还需要制定和实施综合性的干预策略,以解决儿童营养不良及其对健康的连锁反应,以及抗生素耐药性的双重挑战。总的来说,该研究为减少抗生素耐药性提供新的视角,为这方面的研究带来新的思路,并在全球范围内推动健康和可持续发展目标的实现。
参考文献:
Littlejohn PT, Metcalfe-Roach A, Cardenas Poire E, Holani R, Bar-Yoseph H, Fan YM, Woodward SE, Finlay BB. Multiple micronutrient deficiencies in early life cause multi-kingdom alterations in the gut microbiome and intrinsic antibiotic resistance genes in mice. Nat Microbiol. 2023 Nov 16.

谷禾健康
健康的肠道微生物群特点是氧气含量低,并且存在大型专性厌氧菌细菌群落,这些共生菌通过限制肠道病原体的扩张为宿主提供益处。
那么是什么导致肠道健康走下坡路,是什么原因引起肠道菌群失调,在科学界一直有2种观点:
■ 一种是“营养生态位假说”
大肠内多种微生物的共存要求群落中的每个成员,都能够比所有其他成员在少数有限资源条件下,能更快地生长。
■ 另外一种是“氧气假说”
氧气增加将中断或抑制厌氧菌群的生长繁殖,同时导致将赋予兼性厌氧菌甚至需氧菌生态选择优势,使它们变得更具竞争力并可能过度生长。相反,对氧气敏感并因其存在而受到伤害的专性厌氧菌将处于不利地位。
当我们有肠道炎症时会发生什么?
上皮细胞,即肠道内的细胞,开始发炎,它们开始将氧气泄漏到肠道中。当这种情况发生时,许多机会性病原体如大肠杆菌、沙门氏菌和肠球菌属(Enterococcus)等喜欢氧气的菌,可以在有氧条件下利用氧气作为电子受体代替其他代谢途径,从而获得更多的能量和生长优势。
在不同的慢性疾病中,常常观察到兼性厌氧菌的过度生长,这些微生物可以在氧气存在时使用它们,并且会“竞争”,消耗其他有益微生物所需的营养物质,包括一些对维持肠道屏障非常重要的产丁酸菌,如毛螺菌科、梭状芽孢杆菌、真杆菌属、罗氏菌属、普拉梭菌等,导致肠道菌群失衡,从而引起多种消化系统问题和慢性疾病。
人体需要氧气才能生存。几乎人体内的每个细胞都需要氧气。
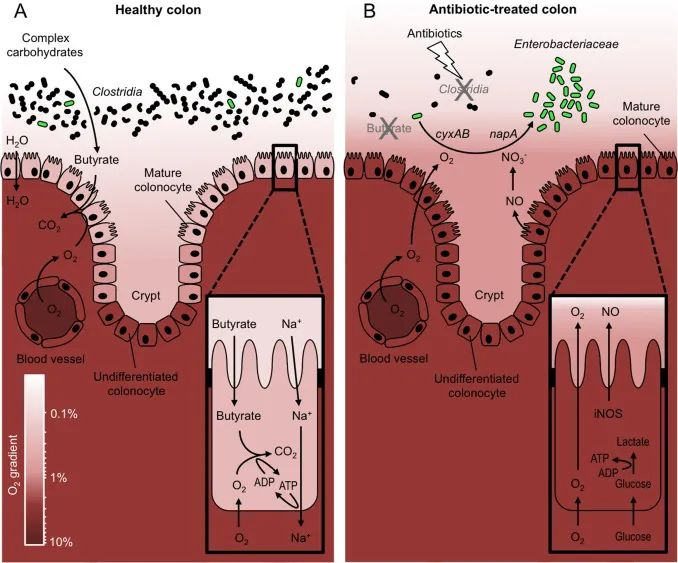
然而地球上大部分微生物却不能很好地耐受氧气,例如肠道的大多数微生物细胞在缺氧的环境中才可以茁壮成长。在健康状态下,结肠是一个低氧环境。如果肠道状态受到干扰,比如使用抗生素,发炎,氧气泄漏到肠道中,它会加剧细菌失衡和炎症,从而开始肠道生态失调、细胞处理能量饥饿和炎症的恶性循环中。
本文我们将重点讨论氧气与肠道菌群失调之间的联系,并分享总结氧气对肠道治疗的潜在影响。
# 1
健康的人类结肠含有估计超过30万亿个细菌细胞,这些细菌很重要,可以分解复杂的碳水化合物,蛋白质等,产生重要的肠道代谢物(如短链脂肪酸)等。然而这些细菌中大部分只能在缺氧的环境中生长和繁殖,属于专性厌氧菌。
婴儿时期,低氧环境利于专性厌氧菌定植
在婴儿的“经典”定植模式中,第一步涉及兼性厌氧菌,例如大肠杆菌(Escherichia coli)和肠球菌(Enterococcus)。
兼性厌氧菌在胃肠道的定植发生在,出生后立即发生氧化还原电位 (简称Eh) 高或呈阳性的肠道环境中。兼性厌氧菌的增加导致可用氧的消耗,从而产生一个氧气减少的环境,有利于出生后几天内专性厌氧菌的到来。然后,与氧气浓度相关的 Eh 降低到一个低水平,类似健康成人的肠道那样。
氧化还原状态通过创造抑制竞争者和潜在肠道病原体的生理环境来参与抵抗肠道定植。未经处理的小鼠大肠中的 Eh 约为 -200 mV。用链霉素处理后,脂肪酸水平下降,氧化还原态变为 +200 mV,同时沙门氏菌数量增加。
健康成人的肠道内:大量专性厌氧菌
在健康成人中,胃肠道远端的特点是低氧张力,通常含有大量专性厌氧菌群落:
大部分胃肠道的细菌在体外很难或还没有被培养出来,部分原因是它们的厌氧或对氧气的敏感性,及时培养出来的也需要长时间和费力的技术。
在过去的几十年中,健康人肠道微生物群的组成已经通过独立于培养的技术进行了探索,并通过测序分析进一步描述,在大人群水平的微生物群中鉴定出超过 20个门。两个主要的门,厚壁菌门和拟杆菌门,占肠道细菌的 90%,主要是专性厌氧菌。
在较低的分类学水平上,一个个体通常存在多达一千种。在个体间变异中,个体间共享的物种少于 60 种。这些物种代表了核心肠道微生物群。
肠道核心菌属
人体肠道菌群中的核心菌属是指在不同人群中普遍存在、数量相对较高、对人体健康具有重要作用的菌属。
目前对于核心菌属的定义还没有统一标准,但一般认为核心菌属应该具备以下特征:
1. 在不同人群中普遍存在:核心菌属应该在不同人群中都能够被检测到,而不是仅存在于某些特定人群中。
2. 数量相对较高:核心菌属应该在肠道菌群中占据较高的比例,而不是仅占据极小的比例。
3. 对人体健康具有重要作用:核心菌属应该对人体健康具有重要作用,如参与食物消化、维持肠道屏障功能、调节免疫系统等。
谷禾健康基于庞大的60万人肠道菌群检测样本数据,给出了人体肠道菌群核心菌属的标准:
在90%人群检出,人群平均丰度1%以上的菌属为核心菌属(双歧杆菌和乳杆菌作为益生菌相对丰度低于1%但是重要性很高)。
核心菌属及有益菌累加占总肠道菌群比例低于60%就可能出现肠道菌群紊乱。
# 2
现在,“肠道菌群失调”通常是指肠道微生物群的状态改变,一般与疾病有关。在肠道生态失调期间观察到的最一致和最强大的生态模式可能是属于变形菌门的兼性厌氧细菌的扩张。
变形菌门是人类肠道中常见的五种主要细菌门之一。它包括多种属,包括埃希氏菌属、志贺氏菌属、沙门氏菌属、螺杆菌属、弧菌属、耶尔森氏菌属、假单胞菌属、弯曲杆菌属和脱硫弧菌属,其中大多数被认为是机会性病原体。
关于变形菌门详见:
氧气的存在使变形菌门有了生存优势
在平衡的生态系统中,低丰度时无害的微生物,但当环境变得特别适合它们的生长时,它们会迅速扩散并引起问题。
导致变形杆菌快速扩张的一个环境因素是——氧气。大多数变形菌是兼性厌氧菌,这意味着它们可以在氧气存在的情况下生存和繁殖。这使它们在含氧环境中比有益的专性厌氧菌具有显着的竞争优势。
变形菌扩张的同时,产丁酸菌减少
值得注意的是,变形杆菌的扩张几乎总是伴随着产丁酸盐细菌丰度的减少。高变形菌和低产丁酸菌这种组合,是微生物群失调特征,并与许多慢性疾病有关,包括:炎症性肠病、肠易激综合症、结直肠癌、憩室炎、组胺不耐受、2 型糖尿病、肥胖等。
这个特征还表明,存在潜在的上皮功能障碍,这个后面会讲到。
# 3
专性厌氧菌产生的一种代谢物是丁酸盐。丁酸盐是这些细菌在结肠中代谢膳食纤维时产生的短链脂肪酸之一。我们之前写过丁酸盐对健康的好处,包括它能够减轻神经炎症、预防结肠癌和帮助维持肠道屏障功能。
丁酸盐给结肠细胞提供能量,该过程耗氧量大,由此导致的低氧对维持肠道稳态很重要
在健康的肠道中,丁酸盐提供结肠细胞所需能量的大约 70%。这些结肠细胞排列在结肠上,形成肠道屏障。被结肠细胞摄取后,丁酸盐和其他短链脂肪酸通过β 氧化途径在线粒体中分解。该过程使用大量氧气。事实证明,这种结肠细胞耗氧量对于维持肠道稳态非常重要。
2015 年,科罗拉多大学研究小组证明,丁酸盐的肠道代谢是维持结肠“生理性缺氧”所必需的。通过一系列实验,他们证明了丁酸盐以及较小程度上的短链脂肪酸丙酸盐和乙酸盐会耗尽结肠细胞中的氧气水平。低氧导致缺氧诱导因子 (HIF)的蛋白质稳定,它在细胞中充当一种“氧传感器”。
氧气含量高低如何影响HIF?
当氧气水平低时,HIF 会促进有助于协调肠道屏障保护的基因的表达。如果氧气水平升高,HIF 将不再稳定,这些肠道保护基因将不再表达。
抗生素是否会影响缺氧,从而影响肠道屏障?
研究人员仅使用三天广谱抗生素后,丁酸盐水平急剧下降,肠道含氧量上升,上皮缺氧状态消失。氧气传感器 HIF 不再稳定,肠道保护基因不再表达,导致肠道屏障功能丧失。
这不仅仅是因为缺乏纤维(纤维是生产丁酸盐的基质),抗生素治疗小鼠的肠道菌群,完全丧失了从膳食可发酵纤维中产生丁酸盐或其他短链脂肪酸的能力。不过好在他们又发现,补充丁酸盐能够挽救“生理性缺氧”,修复肠道屏障功能。
# 1
上皮细胞是排列在肠道壁上的细胞,是宿主与微生物通讯的主要界面。
正常代谢过程,正反馈循环维持稳态
当肠道健康且处于体内平衡状态时,结肠细胞主要通过利用大量氧气的过程代谢丁酸等脂肪酸。由此导致的肠道粘膜缺氧有助于维持以专性厌氧菌为主的肠道微生物群。
这些专性厌氧菌反过来通过将纤维发酵成短链脂肪酸(如丁酸盐)来促进健康,短链脂肪酸被结肠上皮细胞吸收。这种正反馈循环可维持肠道健康状态。
异常:结肠细胞从其他途径获取能量带来的问题
然而,当干扰使结肠上皮细胞的代谢远离脂肪酸的 β 氧化时,系统就会崩溃。能量匮乏的结肠细胞必须寻找其他能量来源,最终从血液中提取葡萄糖并将其发酵成乳酸,这个过程不使用氧气。由此产生的炎症也会导致硝酸盐的产生增加。由于没有其他地方可去,氧气、乳酸盐和硝酸盐会“泄漏”到肠道粘膜中。
那么氧气泄露到肠道粘膜后会发生什么呢?
这种变化有利于变形杆菌门中的病原体茁壮成长。例如沙门氏菌、克雷伯氏菌、柠檬酸杆菌和大肠杆菌,它们可以耐受氧气并依靠乳酸和硝酸盐。
同时,结肠的氧合作用抑制了专性厌氧菌的生长,包括非常重要的产丁酸菌。换句话说,“结肠细胞的新陈代谢充当肠道微生物群的控制开关,调节群落在稳态和失调之间的转变。”
兼性厌氧菌:有氧也可以生长,包含许多肠道病原体,引发炎症
健康的结肠还可能含有少量兼性厌氧菌,它们能够在有氧或无氧环境中生长和繁殖。兼性厌氧菌包括许多肠道病原体。健康肠道的低氧浓度和丰富的专性厌氧菌都会抑制这些兼性厌氧菌的生长。
所以一般来说,兼性厌氧菌往往更容易发炎,所以它们往往会给肠道带来压力,引起炎症、结肠上皮细胞缺氧,肠道通透性增加,并且还会直接使肠道上皮细胞发炎,然后导致肠道产生更多的氧气。
病原菌破坏肠道菌群,继续扩张,形成恶性循环
因此,这些病原体几乎可以破坏肠道新陈代谢,为自己提供生存所需的底物,开始扩张,然后形成了一个恶性循环。兼性厌氧菌大多数都属于变形菌门,特别是肠杆菌科在这种氧气泄漏增加的情况下最常见,如在IBD中那样,潜在有害的变形菌,尤其是肠杆菌科的大量繁殖,也就是增强炎症反应。一些粘附侵入性大肠杆菌 (AIEC) 在 IBD 发病机制中发挥重要作用(关于IBD中肠道菌群变化与氧气相关在后面章节有详细介绍)。
这些观察结果表明变形杆菌的生态失调扩张是肠道菌群失调和上皮功能障碍的潜在诊断微生物特征。
# 2
抗生素介导的肠道微生物群破坏,会增加感染沙门氏菌引起的胃肠炎的风险,它会增加患者肠道携带沙门氏菌血清型的持续时间,并且偶尔会导致复发。
使用一种特殊的染色技术,有研究人员证明,抗生素治疗增加了结肠细胞的氧合作用并导致肠道粘膜缺氧的丧失(也就是说进去了些氧气)。这种缺氧丧失导致沙门氏菌和其他兼性厌氧菌在氧气驱动下扩张。
链霉素 -> 肠道菌群破坏 -> 兼性厌氧菌扩张
链霉素介导的小鼠肠道菌群破坏,与兼性厌氧肠杆菌科(例如共生大肠杆菌)或致病性肠沙门氏菌不受控制的扩张有关。
注:实验室特别选择了链霉素,因为它在消除梭状芽孢杆菌方面特别有效,这样他们就可以研究丁酸盐消耗对结肠代谢的影响。链霉素通常不用于人体口服;然而,已知许多其他广谱抗生素会影响产生丁酸盐的细菌,因此 1-2 周的其他抗生素疗程可能也会通过相同的机制促使氧气泄漏到肠道中。
链霉素治疗 -> 耗尽产短链脂肪酸菌 -> 短链脂肪酸降低
链霉素治疗,优先耗尽肠道相关微生物群落中属于梭状芽胞杆菌纲(Clostridia)的细菌。链霉素处理后恢复定植抗性的尝试表明,转移梭状芽胞杆菌(Clostridia)在防止小鼠肠道内共生大肠杆菌扩张方面最有效。
梭状芽胞杆菌是肠道相关微生物群落中短链脂肪酸的重要生产菌。链霉素治疗与大肠中短链脂肪酸浓度的显着降低有关。由于短链脂肪酸可在体外阻碍大肠杆菌或肠道沙门氏菌(S. enterica)的生长,因此有人提出短链脂肪酸这些代谢物的存在,通过代谢排斥限制了大肠中肠杆菌科的生长。
也有人认为,短链脂肪酸的代谢排斥不足以解释问题
然而,有的研究者认为,短链脂肪酸的代谢排除,无法解释共生肠杆菌科在健康个体中保持低丰度,因为过多的抑制物会导致它们被消除,而过少的抑制物会导致它们的扩张,直到另一种资源变得有限。将肠杆菌(Enterobactericeae)维持在恒定的低水平需要持续保持一种精确的抑制剂浓度,这与体内发生的短链脂肪酸浓度波动不相容。
一个更强大的抗定植概念框架:营养生态位假说
它提出大肠内多种微生物物种的共存,要求群落中的每个成员都能够比所有其他成员在少数有限资源上更快地生长。
为了解释抗生素治疗后肠杆菌科不受控制的扩张,营养生态位假说预测,抗生素介导的肠道微生物群破坏,增加了通常限制肠杆菌科生长的关键资源的可用性。
因此,要了解定殖抗性,揭示此类有限资源的特性,并阐明抗生素处理后它们的丰度升高的原因至关重要。
抗生素介导的肠道菌群消耗,增加了唾液酸和岩藻糖的可用性
肠道相关微生物群落中的糖分解细菌可以从复杂的碳水化合物中释放出这些糖分,并随后消耗它们。链霉素治疗提高了唾液酸和岩藻糖的可用性,这有助于链霉素治疗小鼠肠腔扩张。
链霉素处理后在小鼠盲肠粘膜中观察到的另一个变化是,诱导型一氧化氮合酶(iNOS)的合成增加,一种宿主酶,可催化l-精氨酸产生一氧化氮 (NO)。
源自一氧化氮的活性氮物种可以氧化单糖,例如半乳糖和葡萄糖,分别产生葡糖二酸和半乳糖二酸,这在链霉素处理的小鼠的生长过程中赋予大肠杆菌和鼠伤寒沙门氏菌的管腔适应性优势。
总的来说,这些数据表明糖是肠道中的关键资源,但目前尚不清楚为什么肠杆菌科在这些有限资源上,比肠道微生物群的所有其他成员生长得更快。
最近的数据表明,肠杆菌科可以在可用碳源上生长得更快,因为抗生素介导的微生物群落破坏,通过改变宿主生理学提高了呼吸电子受体的可用性。
在这方面特别重要的一种微生物代谢物是短链脂肪酸丁酸盐。肠道微生物群通过乙酰辅酶 A 途径、戊二酸途径、4-氨基丁酸途径或赖氨酸途径产生丁酸,编码这些途径的大多数细菌都是梭状芽胞杆菌纲的成员。
丁酸盐是结肠细胞的重要能量来源。结肠的主要功能是通过吸收钠 (Na + )产生渗透梯度来吸收水分。Na +通过位于表面结肠细胞顶膜中的通道沿着电化学梯度扩散,然后被位于其基底外侧膜中的 Na +泵(Na + K + ATP 酶)主动挤出。

DOI: 10.1016/j.freeradbiomed.2016.09.022
表面结肠细胞激活 Na+ 所需的 ATP运输来自微生物群衍生的丁酸盐氧化成二氧化碳 (CO2)。这种新陈代谢消耗大量氧气,从而使表面结肠细胞缺氧,这对应于小于 7.6 mmHg(<1% 氧气)的氧分压 (pO2)。相比之下,含氧量正常的宿主组织的 pO2在 23 和 70 mmHg 之间(即含氧量在 3% 和 10% 之间)。
# 3
某些病原体也可能利用这种结肠细胞转换在肠道中获得竞争优势。如果你曾经因急性食物中毒而倒下,之后又出现肠道健康问题,这可以解释原因。
在前面中提到的某些沙门氏菌菌株(特别是肠沙门氏菌血清型鼠伤寒沙门氏菌,以下缩写为S. Tm)可以操纵宿主上皮细胞以促进肠道菌群失调。
S. Tm 是一种特别有毒的细菌,它会侵入宿主粘膜,引起严重的炎症。这种炎症导致产生丁酸盐的梭状芽胞杆菌耗尽,进一步增强了S. Tm 在肠道中增殖的能力。换句话说,这表明某些病原体可能会“破坏”肠道新陈代谢以增加自身健康,从而损害健康细菌。
值得注意的是,丁酸盐生产者的消耗似乎比抗生素治疗更缓慢,大约需要 1-3 周,但恢复速度也慢得多。感染后 4 周,梭菌的丰度仍比基线水平低两个半数量级。
由S. Tm 引起的炎症还导致活性氧和氮物质释放到肠道中,它们与单糖反应形成底物,选择性地喂养S. Tm 和肠杆菌科(变形杆菌门)内的其他微生物。
这不仅适用于S. Tm。在小鼠模型中证明,柠檬酸杆菌(Citrobacter)和空肠弯曲杆菌(Campylobacter jejuni)感染也能够引起宿主肠道炎症并驱动肠杆菌科过度生长。
空肠弯曲杆菌详见:细菌大盘点(二) | 葡萄球菌、沙门氏菌、弯曲杆菌
总的来说,这表明肠道感染可能有助于结肠的氧合作用并促进肠道生态失调的长期状态。因此,清除现有感染可能是恢复肠道上皮细胞正常代谢和健康肠道微生物群的关键步骤。

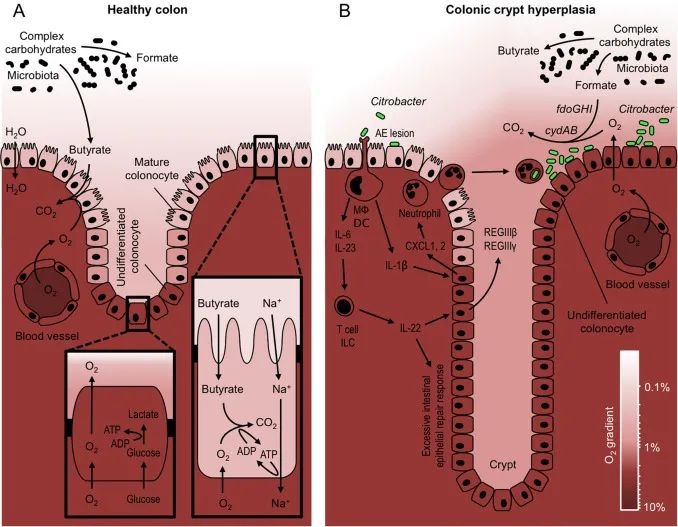
DOI: 10.1016/j.freeradbiomed.2016.09.022
(A) 专性厌氧菌降解复合碳水化合物产生发酵产物,如甲酸盐和丁酸盐。成熟的结肠细胞通过将丁酸氧化成二氧化碳 (CO2 ) 来产生能量 (ATP),这会导致粘膜表面缺氧。相反,位于隐窝中的未分化结肠细胞通过将葡萄糖发酵成乳酸来获取能量。
(B) 病原体C. rodentium ( Citrobacter ) 利用其毒力因子紧密附着在结肠细胞表面(AE 损伤)。树突状细胞 (DC) 和巨噬细胞 (MΦ) 引起的宿主反应导致产生作用于结肠细胞和淋巴细胞(T 细胞和 ILC)的细胞因子(IL-6、IL-23 和 IL-1β),从而诱导合成嗜中性粒细胞趋化因子(CXCL1, 2) 和 IL-22,可激活上皮增殖。
过度的上皮增殖导致隐窝伸长和粘膜表面未分化的含氧量正常的结肠细胞的积累。由此产生的氧气扩散到肠腔的增加,使用cydAB基因驱动C. rodentium的有氧扩张,编码高亲和力细胞色素bd氧化酶。由fdoGHI基因编码的氧依赖性甲酸脱氢酶使病原体能够消耗微生物群衍生的甲酸。
右下角显示的色标表示原理图着色方案表示的氧浓度。
#4
到目前为止,我们已经看到两个例子,其中丁酸盐耗尽导致肠道氧合和生态失调。鉴于丁酸盐的第一大来源来自膳食纤维,从理论上讲,低纤维饮食很可能可以通过相同的机制促进变形菌的扩张。
如果膳食纤维摄入量低,丁酸盐和其他短链脂肪酸的产生水平将无法满足结肠细胞的能量需求。结肠细胞将转向厌氧葡萄糖代谢。无氧代谢会消耗更少的氧气,并导致更多的氧气泄漏到肠道中。
虽然这种机制中的所有步骤并没有像低纤维饮食对抗生素和肠道感染的影响那样有序地得到证明,但一些研究确实将低纤维摄入量与较高水平的变形杆菌联系起来:
那么低纤维生酮饮食是否也如此?
正如之前讨论的,酮体乙酰乙酸和 β-羟基丁酸可以补充丁酸作为肠道上皮细胞的燃料来源。因此,低纤维生酮饮食不太可能激活这种机制来驱动肠道菌群失调。事实上,酮可能有助于恢复上皮缺氧。
#5
有趣的是,所有这些肠道菌群失调的驱动因素,包括抗生素、肠道感染和低纤维加工饮食,都与肠道炎症有关。
肠道炎症
2007 年,在动物模型中证明,肠道炎症本身足以破坏肠道微生物群并促进肠杆菌科的过度生长。通过敲除 IL-10,无论是接触葡聚糖硫酸钠(一种破坏肠道屏障完整性的化学物质),还是严重的遗传易感性,都能够导致肠道菌群失调。
其他更温和的炎症因子也可以促进这些炎症细菌的扩张。2015的一项研究表明,给小鼠喂食羧甲基纤维素和聚山梨醇酯 80(加工食品中常用的两种乳化剂),12 周会降低微生物多样性并导致粘膜相关变形菌增加。同样,发现给小鼠喂食无热量甜味剂阿斯巴甜 8 周会导致肠杆菌科细菌增多。
关于食品添加剂与肠道菌群详见:
你的焦虑可能与食品添加剂有关,警惕食品添加剂引起的微生物群变化
心理压力
压力还会促进炎症和肠道菌群失调。研究报道了在慢性社会心理压力的小鼠模型中变形杆菌的扩张。
食物不耐受
未被识别的食物不耐受也可能导致肠道炎症、结肠细胞代谢改变和肠道生态失调,尽管需要更多的研究来证实这一点。
以上,我们已经回顾了一些可能导致肠道缺氧和导致肠道菌群失调的因素。
在患有 IBD 的患者中观察到微生态失调。反复观察表明专性厌氧菌(F. prausnitzii)减少,兼性厌氧菌(肠杆菌科)增加,甚至出现需氧菌。加上远端肠道的正常厌氧肠道生理学,这些观察结果支持了细菌群落变化的假设,该变化是由氧气增加引起的,从而改变了胃肠道中的 Eh。
F. prausnitzii对氧气极其敏感,但仍可以看到粘附在氧气从上皮细胞扩散的肠道粘膜上。为了解释这个悖论,2012有研究报道说F. prausnitzii可以在低氧环境下生长。他们表明F. prausnitzii 利用黄素和硫醇的细胞外电子穿梭将电子转移到氧气。然而,存在于健康人体肠道中 的黄素和硫醇是普拉梭菌在氧气存在下存活所必需的,而在IBD患者的受损组织中可能并非如此。
IBD 患者中的肠道微生态失调
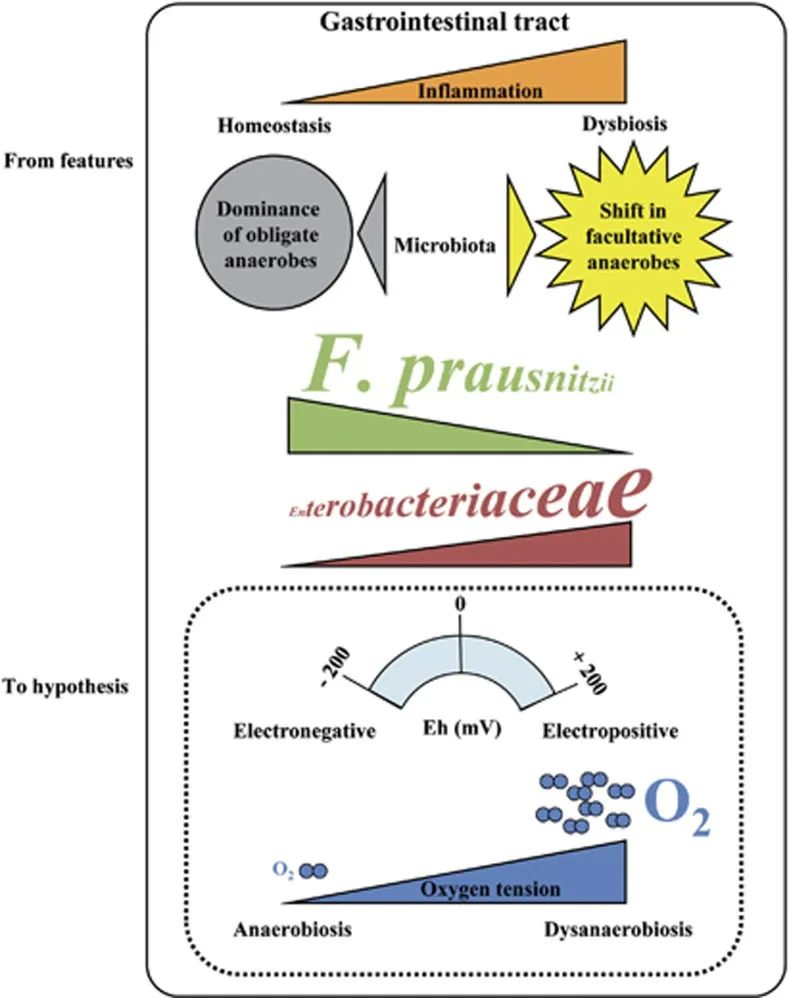

doi: 10.1038/ismej.2013.80
肠道微生物在肠道中的氧依赖性变化
研究在小肠移植和回肠造口术后监测微生物成分,肠道细菌群落从专性厌氧菌转变为以兼性厌氧菌为主的群落,并且肠杆菌科增加,就像 IBD 患者的微生物群一样。
在回肠造口关闭后,假设它提供了氧气入口,转变被逆转,专性厌氧菌再次占主导地位。
这些研究作者提出,氧气是菌群从专性厌氧菌转变为兼性厌氧菌或需氧菌的原因。
虽然肠杆菌科增加,但在大多数患者中未观察到炎症。与 IBD 菌群失调形成鲜明对比的是,乳酸杆菌也大量繁殖,后者很少见乳酸菌增加的报道。研究表明,乳酸杆菌及其细胞壁成分具有抗炎特性,这表明调节或补充乳酸杆菌可能会限制 IBD 患者的炎症反应。
菌群失调与炎症和氧气增加相关
在 IBD 中,氧气浓度的增加可能是由于血液进入胃肠道引起的,这种情况在慢性炎症期间经常观察到,这会促使携带氧气的血红蛋白在肠道细菌所在的肠粘膜和内腔中释放。小鼠模型表明,炎症可能导致微生物群失调,并促进肠杆菌科的过度生长。
一项初步研究评估了克罗恩病中抗肿瘤坏死因子-α 抗体治疗对粪便微生物群的组成和活性的影响,并报告了Faecalibacterium的适度增加, 这表明阻断炎症反应可能会促进这些菌增加。
氧气条件的变化也可能是由于炎症反应本身,导致肠道组织氧化爆发,例如,通过中性粒细胞释放活性氧。有趣的是,活性氧可以与内源性腔内硫化合物反应,形成肠道沙门氏菌使用的呼吸电子受体血清型 Typhimurium(一种好氧物种和属于肠杆菌科的肠道病原体),赋予沙门氏菌相对于专性厌氧菌的竞争性肠道微生物群的生长优势。
“
——丁酸盐有助于恢复肠道菌群
第一步只是确保有基本的健康。我们可以讨论各种先进的肠道干预措施,但最基础的永远都是要从源头上开始,比如说健康的饮食、锻炼、睡眠和压力管理等方式,如果饮食等很糟糕,那么再多肠道治疗,先进的疗法改变肠道和健康状况都不会持久。所以这是第一步。
健康的饮食对于不同人有不同的定义
是的。我们并不完全知道对肠道微生物组最好的饮食是什么,而且并没有一种所谓对肠道微生物组最好的饮食。每个人的微生物组都不同,大家知道,有些人可能对生酮饮食反应非常好,而有些人可能对包含更多植物性食物的饮食效果更好。
建议大家可以从自己身上进行健康探索,比如某一阶段进行某种健康的饮食或生活方式,坚持一段时间后进行肠道菌群检测,配合自身感受,更多角度更细微层次地去了解自己的身体状况。
假如说肠道真的发炎了,做一个短期的低过敏性饮食(比如自身免疫方案)可能是有益的,去除一些更容易引起炎症的食物,至少在一段时间内促进康复。减少炎症将从根本上阻止导致菌群失调的氧气泄漏。
如果变形菌过多和氧气失调的情况发生怎么办?它的典型症状是什么?
通过我们多年研究发现这个可以以很多不同的方式表现出来。基本上这种高变形杆菌和低产丁酸盐的微生物特征存在于各种不同的疾病中:炎症性肠病、肠易激综合症、结直肠癌,也可能是2型糖尿病、肥胖症、过敏以及牛皮癣等。
像这样情况的人通常倾向于腹泻或大便不成形?
不一定。在持续腹胀和胀气的患者身上看到了很多。这是看到的一个非常常见的问题,但它也可能因谷禾目前客户群体而有所偏差。
扩展阅读:“肚子像气球?”“好像怀孕?”——可能是腹胀惹的祸
建议补充丁酸盐可能会扭转这种生态失调状态?
是的,当然并不是单靠补充丁酸盐就一定能够解决这种生态失调。因此,本文概述了一些不同的因素,包括饮食和生活方式因素,以及可能有助于防止氧气泄漏到肠道并恢复体内平衡状态的补充因素,丁酸盐是其中的重要方式之一。
如果没有丁酸盐,氧气没有得到利用,就会溢出肠腔喂养病原体
基本上发生的氧气泄漏的部分原因是肠道内的上皮细胞缺乏能量。当极度缺乏能量时,如果它们没有丁酸盐,就不会代谢丁酸盐。所以从血液中提取葡萄糖,基本上发酵成乳酸,然后乳酸渗入肠道,除了通常通过丁酸代谢使用的氧气。所以实际上如没有丁酸盐存在,就意味着从血液流入上皮细胞的氧气没有得到利用。那就是它最终溢出到肠腔并喂养病原体的时候。
因此,从本质上讲,提供补充丁酸盐就是提供的是上皮细胞喜欢的能量。它们将使用更多的氧气作为丁酸代谢的一部分,并且基本上不需要进行厌氧葡萄糖发酵。这会产生乳酸,最终不会耗尽氧气,这意味着氧气和乳酸会渗入肠道,并喂养沙门氏菌、肠杆菌科和许多不同的变形菌。
抗生素治疗期间补充丁酸盐——雪中送碳
2019 年,在小鼠模型中证明,同时给予丁酸盐和抗生素可以减轻艰难梭菌引起的结肠炎。艰难梭菌是一种革兰氏阳性、形成孢子的细菌,是使用抗生素后肠道感染的常见原因。
在使用抗生素前一天开始补充丁酸盐,并在整个抗生素疗程和 5 天感染挑战期间持续使用。有趣的是,丁酸盐对艰难梭菌定植或毒素产生没有影响,但通过稳定 HIF-1 和增加肠道屏障的完整性,丁酸盐减少了肠道炎症和细菌穿过肠道屏障的运动。
研究人员继续测试另外两种提供丁酸盐的策略。在感染前后的三天内给予高剂量三丁酸甘油酯与丁酸盐具有同样的保护作用,就像在感染前服用抗生素后喂食高纤维饮食(含有高达25%的菊粉)一样。
显然,丁酸盐可以防止抗生素后病原体的扩张。但丁酸盐能否通过支持结肠细胞代谢来预防与抗生素相关的所有生态失调,这仍有待在对照研究中确定。
1
在分子生物学领域,过氧化物酶体增殖物激活受体 (PPAR) 是一组核受体蛋白,作为转录因子调节基因的表达。PPAR 在调节细胞分化、发育和代谢(碳水化合物、脂质、蛋白质)以及肿瘤发生中起着重要作用。
PPAR-γ:一个具体的结肠细胞代谢“开关”
如果说前面一直在抽象地谈论导致肠道菌群失调的结肠细胞代谢“开关”,那么这里来个具体的。研究人员已经确定了一种特殊的基因 PPAR-γ,它似乎可以调节这种转换。PPAR(过氧化物酶体增殖物激活受体的缩写)是一组与 DNA 结合以直接影响基因表达的蛋白质。PPAR-γ 在许多细胞中表达,但在脂肪(脂肪)组织和结肠中表达最高。
健康肠道:
丁酸盐 ⇔ PPAR-γ:互相促进,正向循环
在健康的肠道中,丁酸盐不仅可以为结肠细胞提供能量,还可以增强 PPAR-γ 的激活。这起到了一个正反馈循环的作用:PPAR-γ 激活了增加丁酸盐和其他脂肪酸代谢的基因。这会降低上皮细胞和肠粘膜中的氧气浓度,从而抑制致病性变形菌的生长,并促进有益的、产生丁酸盐的细菌的生长。
失调肠道:没有足够丁酸盐→ 较低的 PPAR-γ 表达→ 致病菌生长
然而,在失调的肠道中,没有足够的丁酸盐或其他底物来激活 PPAR-γ。较低的 PPAR-γ 表达导致 Nos2 的表达增加,Nos2 是编码诱导型一氧化氮合酶 (iNOS) 的基因,并且增加了硝酸盐释放到肠道中。这与厌氧糖酵解产生的乳酸和氧气一起促进了致病菌的生长。
PPAR-γ 激活对于维持肠道先天免疫也至关重要
2010 年发表在PNAS杂志上的一项研究通过一系列实验证明,PPAR-γ 有助于维持抗菌肽β-防御素的持续表达,后者调节结肠的微生物定植。缺乏 PPAR-γ 的小鼠表现出对白色念珠菌、脆弱拟杆菌、粪肠球菌和大肠杆菌的免疫防御缺陷。
分泌型 IgA 的产生也需要 PPAR-γ 。
IgA 详见:对抗病原菌,帮助共生菌定植的“重要开关”
2
多项研究表明,PPAR-γ 激活可能会预防或逆转与免疫激活相关的肠道菌群失调和组织损伤。例如,PPAR-γ 表达在炎症性肠病 (IBD) 中显着降低。
药物激活PPAR-γ :罗格列酮(不适合长期使用)
罗格列酮是一种与 PPAR-γ 结合并增加其活性的药物,在动物模型中已被证明在急性给药时可预防生态失调并减轻结肠炎的症状。虽然这种药物有时仍被用作抗糖尿病药物,但它有一些不良副作用,不适合长期使用。尽管如此,它证明了该途径对肠道微生物群产生重大变化的能力。
药物激活PPAR-γ :美沙拉嗪 (目前使用较多)
研究人员一直在寻找其他方法来刺激这一途径。另一种药物美沙拉嗪也可以激活 PPAR-γ,但程度适中。它在肠道中具有更多的局部作用,因此相对具有更少的全身副作用。这种药物现在被用作 IBD 的一线治疗药物。这种药物的抗炎作用是通过其上调 PPAR-γ 的能力介导的。此外,对照研究表明,美沙拉嗪处理可降低变形杆菌的丰度并增加粪杆菌和双歧杆菌的丰度。
药物激活PPAR-γ :丹参素冰片酯(激活程度小,但仍然提供显著保护作用)
北京的一组研究人员还发现丹参素冰片酯(DBZ)是一种 PPAR-γ 激动剂,具有预防或逆转肠道菌群失调的潜力。DBZ 是传统中药复方丹参中天然存在的两种化合物的合成版本。与罗格列酮和其他经典噻唑烷二酮类药物相比,DBZ 被发现对 PPAR-γ 的激活程度更小,但仍然能够在小鼠饮食模型中提供显着的保护作用,防止肠道菌群失调、肠屏障功能障碍、胰岛素抵抗和体重增加-诱发肥胖。
丁酸盐补充剂也可能参与激活
丁酸盐补充剂也被证明可以改变人类的肠道生态系统。一项针对 49 名 IBD 患者的前瞻性、随机、安慰剂对照研究发现,每天 1800 毫克的丁酸盐不仅可以减轻炎症并改善生活质量,而且还会增加产丁酸菌的数量。
经过两个月的补充,患有克罗恩氏病的人的丁酸球菌和Subdoligranulum丰度增加,而患有溃疡性结肠炎的人的毛螺菌科则大量增加。虽然研究人员没有直接测量 PPAR-γ,但考虑到产丁酸菌和炎症标志物的丰度发生了如此巨大的变化,这一途径可能参与其中。
▼
总而言之,这是一个非常有趣的研究领域,在未来几年无疑会受到更多关注。
《科学》杂志上的评论中写道:“结肠细胞的代谢重编程以恢复上皮细胞缺氧,代表了一种有前途的新治疗方法,可以在广泛的人类疾病中重新平衡结肠微生物群”。
换句话说,如果能够靶向结肠细胞的新陈代谢,我们就可以恢复肠道中的低氧环境,并有可能逆转肠道微生态失调。
谷禾也正积极开展研究合作,以确定丁酸盐和其他 PPAR-γ 激动剂是否可以预防抗生素引起的各种菌群失调。
3
以下是可能增加肠道 PPAR-γ 活性以支持肠道缺氧的干预措施的总结。可能有助于解决对其他治疗没有反应的肠道菌群失调疑难病例,尤其是那些以高变形杆菌和低丰度产丁酸菌为特征的肠道菌群失调。
首先应该关注的地方依然是日常能做到的,比如说健康的饮食,定期锻炼,充足的睡眠、阳光和健康的社交互动。
其他疗法包括:
美沙拉嗪 (5-ASA):该药物通常用作 IBD 的一线治疗药物。它的抗炎作用已被证明是通过其上调 PPAR-γ 的能力介导的。
丹参素冰片酯(DBZ):这种化合物来源于丹参酚和冰片,存在于中药复方丹参中。它上调 PPAR-γ,并已证明具有减轻微生态失调的潜力。注意:草药的来源和剂量应谨慎,最好是在草药方面经验丰富的医生的指导下。
丁酸盐:一种短链脂肪酸和 PPAR-γ 的有效刺激剂。即使是低浓度的丁酸盐也能将 PPAR-γ 蛋白表达增加 7 倍。
酮类: β-羟基丁酸和乙酰乙酸会激活肠上皮细胞中的 PPAR-γ,就像丁酸一样。生酮饮食已被证明可以在许多组织中上调 PPAR-γ,还可以为 β 氧化和上皮能量产生提供底物。希望能有更多支持这方面的研究。
禁食/热量限制:一项研究发现,在热量限制期间,交感神经系统激活需要肠道 PPAR-γ。然而,禁食或热量限制在多大程度上在肠道中诱导该通路仍不清楚。
运动:一个研究小组发现,在结肠炎模型和饮食诱导的肥胖模型中,自愿运动对肠道的保护作用是由运动增加肠道内源性糖皮质激素和上调 PPAR-γ 的能力介导的。
压力管理:压力会降低肠道中 PPAR-γ 的表达。
大麻素:大麻二酚 (CBD) 降低了溃疡性结肠炎患者直肠活检中的 iNOS 活性,这种作用是通过激活 PPAR-γ 介导的。
萝卜硫素:2008 年发现这种来自十字花科蔬菜的植物化学物质通过激活 PPAR-γ 增强先天免疫成分。
姜黄素:一项研究发现姜黄素通过激活 PPAR-γ 抑制小鼠化学诱导的结肠炎。达到这些效果所需的口服剂量尚不清楚。
其他草药:洋甘菊、当归、水飞蓟素、甘草根和柠檬香脂都是 PPAR-γ 的部分激活剂。
脂肪酸:共轭亚油酸 (CLA) 和 omega-3 脂肪酸 (DHA) 均可增强 PPAR-γ 的表达。
益生菌:对结肠细胞的体外研究表明,布拉酵母菌(Saccharomyces boulardii)具有增加 PPAR-γ 表达的能力。
益生元:对结肠细胞的体外研究表明,低聚糖 alpha3-唾液酸乳糖和 FOS 的抗炎作用是通过它们诱导 PPAR-γ 的能力介导的。
维生素 A:视黄酸是维生素 A 的一种形式,是 PPAR-γ 的激活和功能所必需的。维生素 A也称为“抗感染素”。
注:本账号内容仅作交流参考,不应将以上信息视为医疗建议。采取干预措施前请咨询医生,了解治疗是否适合。
主要参考文献
Bradley PH, Pollard KS. Proteobacteria explain significant functional variability in the human gut microbiome. Microbiome. 2017 Mar 23;5(1):36. doi: 10.1186/s40168-017-0244-z. PMID: 28330508; PMCID: PMC5363007.
Lucy Mailing. The oxygen-gut dysbiosis connection(What causes gut health to go downhill, and how to break the cycle of gut inflammation, dysbiosis, and epithelial energy starvation). 2019,11.14, lucymailing.
Rigottier-Gois L. Dysbiosis in inflammatory bowel diseases: the oxygen hypothesis. ISME J. 2013 Jul;7(7):1256-61. doi: 10.1038/ismej.2013.80. Epub 2013 May 16. PMID: 23677008; PMCID: PMC3695303.
Henson MA, Phalak P. Microbiota dysbiosis in inflammatory bowel diseases: in silico investigation of the oxygen hypothesis. BMC Syst Biol. 2017 Dec 28;11(1):145. doi: 10.1186/s12918-017-0522-1. PMID: 29282051; PMCID: PMC5745886.
Byndloss MX, Olsan EE, Rivera-Chávez F, Tiffany CR, Cevallos SA, Lokken KL, Torres TP, Byndloss AJ, Faber F, Gao Y, Litvak Y, Lopez CA, Xu G, Napoli E, Giulivi C, Tsolis RM, Revzin A, Lebrilla CB, Bäumler AJ. Microbiota-activated PPAR-γ signaling inhibits dysbiotic Enterobacteriaceae expansion. Science. 2017 Aug 11;357(6351):570-575. doi: 10.1126/science.aam9949. PMID: 28798125; PMCID: PMC5642957.
Roudsari NM, Lashgari NA, Zandi N, Pazoki B, Momtaz S, Sahebkar A, Abdolghaffari AH. PPARγ: A turning point for irritable bowel syndrome treatment. Life Sci. 2020 Sep 15;257:118103. doi: 10.1016/j.lfs.2020.118103. Epub 2020 Jul 15. PMID: 32681913.
What Is Dysbiosis? Written by WebMD Editorial Contributors. Medically Reviewed by Dan Brennan, MD on December 06, 2022
Yajie Wang, Yue Chen, Xiaoyu Zhang, Yangpeng Lu, Haixia Chen, New insights in intestinal oxidative stress damage and the health intervention effects of nutrients: A review, Journal of Functional Foods,2020. Volume 75,104248,
Shelton CD, Byndloss MX. Gut Epithelial Metabolism as a Key Driver of Intestinal Dysbiosis Associated with Noncommunicable Diseases. Infect Immun. 2020 Jun 22;88(7):e00939-19. doi: 10.1128/IAI.00939-19. PMID: 32122941; PMCID: PMC7309626.
谷禾健康
迄今为止最全面的全球抗菌素耐药性 (AMR) 研究发现,由耐药菌引起的感染是所有年龄段人群死亡的主要原因之一。
22年发表在《柳叶刀》杂志上的分析估计,2019 年有 495 万人死于细菌性 AMR 发挥作用的疾病。其中,127 万人死亡是 AMR 的直接结果——这意味着耐药性感染导致的死亡人数超过了艾滋病毒/艾滋病(864,000 人死亡)或疟疾(643,000 人死亡)。
细菌性 AMR 感染最常见的三个部位是胸部、血液和腹部——身体这些部位的感染占直接归因于 AMR 死亡的 78.8%。六种最致命的细菌病原体导致了将近四分之三的所有因耐药性导致的死亡。2019 年,仅耐抗生素大肠杆菌就导致约 20 万人死亡。
“AMR 确实是一个全球性问题,需要决策者和全球卫生界采取紧急行动,以避免可预防的死亡,”
对抗生素有耐药性的细菌比例正在上升。在一个抗生素使用变得如此普遍的世界里,耐药细菌比那些被药物杀死的细菌更胜一筹。
伤口敷料的微观纤维上的金黄色葡萄球菌
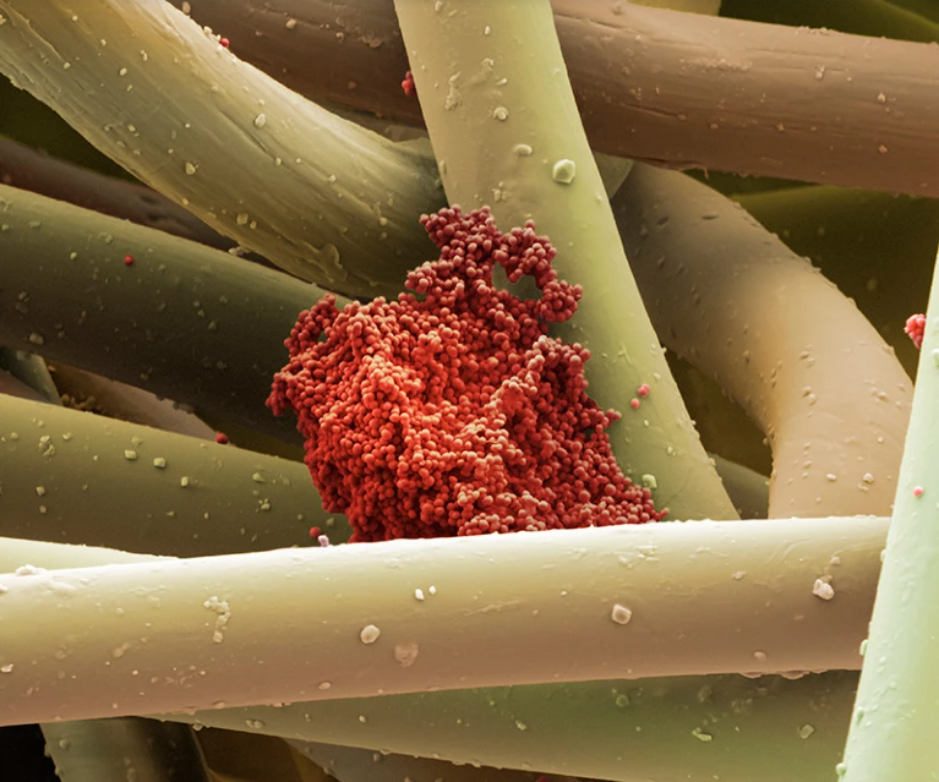
图片来源: Science Photo Library
预计到 2050 年,每年可能有多达 1000 万人死于抗菌素耐药性。如果任其发展,以前可以用几天抗生素治愈的感染可能变得无法治愈。
如果细菌感染无法控制会导致严重的健康问题,比如梅毒会引起失明和神经损伤。女性感染衣原体会导致不孕。梅毒可以传染给未出生的孩子,导致出生时患有这种疾病的婴儿流产、死产或大脑和器官受损。
淋病细菌(如图)对某些抗生素产生了耐药性
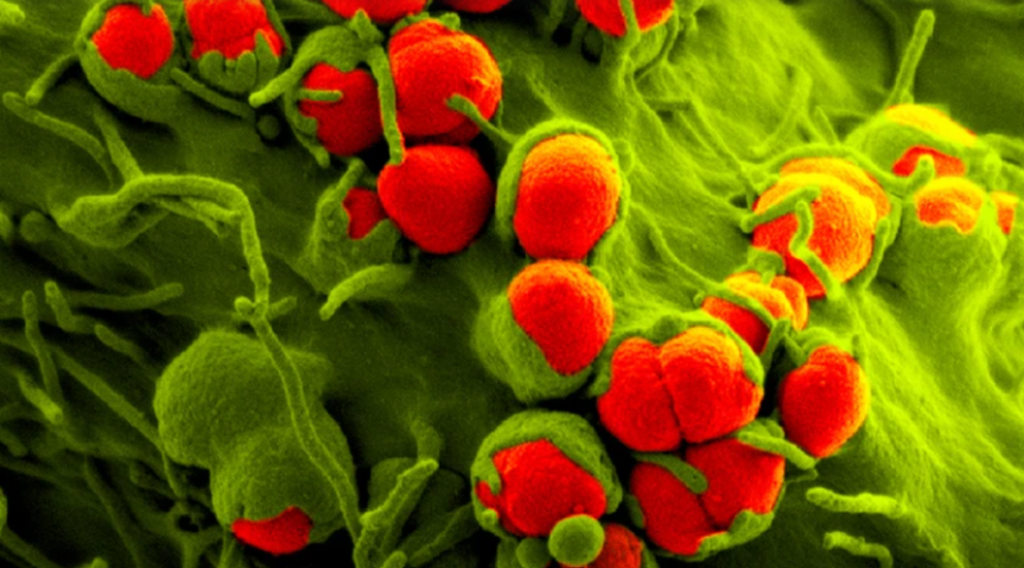
图片来源: SPL
数据显示,低收入国家的 AMR 相关死亡率最高。在 21 个 GBD 地理区域中,西撒哈拉以南非洲直接归因于 AMR 的死亡率最高,每 10 万人中有 27.3 人死亡。大洋洲最低,每 10 万人中有 6.5 人死亡。
低收入地区的耐药率和耐药菌感染数量均高于富裕国家。造成这种情况的原因包括环境卫生和卫生条件差、用于治疗的检测设施不足以及无法获得最新的抗生素和疫苗。
但是注意,许多低收入和中等收入国家的抗微生物药物耐药性数据很少,这表示实际的AMR比调查的数据更严重,这凸显了在这些地区大幅扩大实验室能力的必要性。
抗生素药物旨在消灭细菌,但它们不一定会杀死所有的细菌,它们只是将细菌减少到安全水平。但是不幸的是,这个过程选择留下了最强壮的细菌个体,然后可以通过基因传递产生耐药性。几十年后,就会有这么多种细菌进化出对一系列抗生素的抗性。
对抗生素产生耐药性的致病细菌的出现,往往归因于人类和牲畜过度和不合理使用抗生素。虽然这是问题背后的主要机制,但这不是唯一的因素。昆士兰大学的研究人员希望测试其他药物,即使是那些没有抗生素特性的药物,是否可以在超级细菌的出现中发挥作用。
最近昆士兰大学的研究人员已经将注意力集中在另一个潜在的抵抗力驱动因素上:抗抑郁药。通过研究在实验室中生长的细菌,已追踪到抗抑郁药如何引发耐药性。
“即使接触几天后,细菌也会产生耐药性,不仅针对一种抗生素,还针对多种抗生素”,布里斯班昆士兰大学澳大利亚水与环境生物技术中心的资深作者郭建华(英译)说。
早期线索
2014 年,郭开始对非抗生素药物对抗生素耐药性的可能贡献产生兴趣,因为他的实验室发现生活废水样本中循环的抗生素耐药基因比抗生素使用率更高的医院废水样本中循环的抗生素耐药基因更多。
郭的团队和其他团队还观察到,抗抑郁药——世界上使用最广泛的药物之一——可以杀死或阻碍某些细菌的生长。郭解释说,它们会引发“SOS 反应”,从而触发细胞防御机制,使细菌能够更好地在随后的抗生素治疗中存活下来。
在 2018 年的一篇论文中,该小组报告说,大肠杆菌在接触氟西汀后就对多种抗生素产生了耐药性,氟西汀通常以百忧解的形式出售。最新研究检查了6种此类药物中的5种其他抗抑郁药和13种抗生素,并调查了大肠杆菌的耐药性是如何产生的。
备注:百优解(盐酸氟西汀胶囊)是一种选择性血清素(5-羟色胺,5-HT)再吸收抑制剂(SSRI)型的抗忧郁药,其药物形态为盐酸氟西汀(Fluoxetinehydrochloride),商品名为“百优解”或“百忧解”(Prozac)。在临床上用于成人忧郁症、强迫症等。
在氧气充足的实验室条件下生长的细菌中,抗抑郁药会导致细胞产生活性氧:一种激活微生物防御机制的有毒分子。最重要的是,这激活了细菌的外排泵系统,这是许多细菌用来消除各种分子(包括抗生素)的通用排出系统。这可能解释了细菌如何在没有特定抗性基因的情况下抵抗抗生素。
在抗抑郁药存在的情况下,革兰氏阴性细菌大肠杆菌可以抵御抗生素
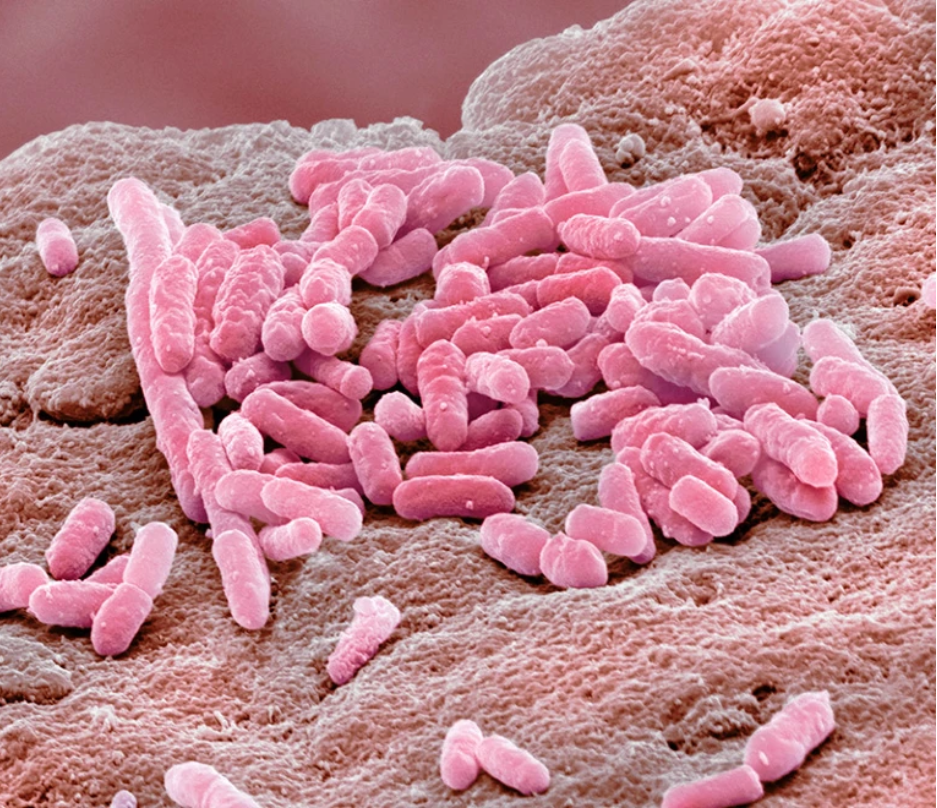
图片来源:Steve Gschmeissner/Science Photo Library
但是大肠杆菌暴露于抗抑郁药也导致微生物突变率增加,以及随后选择各种抗性基因。然而,在厌氧条件下生长的细菌中,活性氧水平要低得多,抗生素耐药性的发展要慢得多。
此外,至少一种抗抑郁药舍曲林(舍曲林是5-羟色氨再摄取抑制剂,是一种新型的抗抑郁剂,具有较强抗抑郁,抗焦虑作用,既可用于抑郁症,焦虑症、双相障碍的抑郁相,强迫障碍的治疗)促进了细菌细胞之间的基因转移,这一过程可以加速耐药性在人群中的传播。这种转移可以发生在不同类型的细菌之间,从而使耐药性在物种之间转移——包括从无害细菌到致病细菌。
越来越多的认可
英国剑桥大学研究微生物组-化学相互作用的 Kiran Patil 说,在过去五年中,人们越来越认识到许多针对人体细胞的非抗生素药物也会影响细菌并导致抗生素耐药性。
德国图宾根大学的 Lisa Maier 指出,研究药物与微生物组之间的相互作用,要了解抗抑郁药如何驱动抗生素耐药性,研究人员需要确定药物针对细菌中的哪些分子并评估药物对更广泛的临床相关细菌种类的影响。
2018 年,Maier 和她的同事调查了 835 种不针对微生物的药物,发现 24% 的药物抑制了至少一种人类肠道细菌菌株的生长。
Patil 和 Maier 表示,重要的是要收集证据来评估抗抑郁药在现实世界中对耐药性的影响,例如抗抑郁药是否正在推动抗生素耐药细菌的积累,尤其是致病细菌在人、动物或环境中的积累。
尽管在废水中发现了大量的抗抑郁药,但报告的水平往往低于郭的小组在大肠杆菌中看到显着效果的浓度。不过,预计在服用这些药物的人的大肠中会达到在这项研究中具有强烈作用的一些抗抑郁药的浓度。
后续研究
Maier 说,现在有几项研究将抗抑郁药和其他非抗生素药物与细菌的变化联系起来,并且初步研究已经给出了关于这些药物如何影响服用它们的人的微生物组的“初步提示”。
但在健康人类中,大肠杆菌主要存在于大肠中,那里的条件是厌氧的,这意味着论文中描述的过程可能不会以相同的速度发生在人身上,Maier 说,未来的研究应该使用细菌生长条件来模拟抗抑郁药可能起作用的部位。
郭说他的实验室现在正在研究给予抗抑郁药的小鼠的微生物组。早期未发表的数据表明,这些药物可以改变动物的肠道微生物群并促进基因转移。
但是郭和相关研究人员告诫人们不要根据这项研究停止服用抗抑郁药。“如果您患有抑郁症,则需要以最好的方式进行治疗,其次是关注肠道细菌”。
但是研究人员和制药公司需要量化非抗生素药物对抗生素耐药性的贡献。非抗生素药物是我们不应忽视的一个大问题。
寻找新抗生素的需求迫在眉睫,但许多科学家和政策制定者正在从其他角度解决抗菌素耐药性问题。《Nature》期刊详细地介绍了三种方法。
➤ 等离子清洗
富含化学不稳定形式的氧和氮(也称为自由基和活性物质)的等离子活化水被认为是潜在的新型消毒剂。“如果细菌被自由基淹没,它们最终就会死亡,” 澳大利亚阿德莱德大学的生物医学研究员 Katharina Richter 说。
Richter 和她的同事正在研究与未经治疗的伤口相比,血浆活化水清除感染耐甲氧西林金黄色葡萄球菌(MRSA) 的伤口的速度有多快。他们还将这项技术与通过静脉滴注施用抗生素进行比较。
她说,尚未公布的初步结果还不错。“治疗改善了伤口并比没有治疗更快地清除了感染,” 她说,“它不如抗生素治疗有效,但 Richter 说实验设计可能是罪魁祸首。将其与局部给予的抗生素进行比较会更公平。我们的下一项研究将有更好的控制。”
➤ 金属奇迹
细菌虽然在自然界中是单细胞,但确实聚集在一起并互相帮助以逃避药物和防腐剂。他们这样做的一种方法是形成生物膜——生活在它们自己制造的粘液环境中的细菌群。
生物膜保护居住在其中的单个细胞;据认为,大约 80% 的慢性人类感染是由生物膜引起的。金属元素镓会干扰细菌对铁的吸收,最终导致微生物缺乏营养。
正因为如此,含镓药物是一种正在探索的破坏生物膜的途径。英国曼彻斯特大学的科学家发现,镓化合物可以减少多达 87% 的细菌生长(JM Baker et al. Life Sci . 305 , 120794; 2022)。
这项工作建立在中国上海交通大学研究人员的研究结果之上,该研究表明镓可以有效溶解 MRSA 生物膜的结构,使细菌可以用常用抗生素剂量的十分之一杀死(W. Xia et al., ACS Infect. 2021, 2565–2582)。现在的研究重点是如何最好地输送镓以及输送多少剂量。
➤ 分子签证
抗生素应具有三个关键特性:溶解性,易于与细菌结合的能力,以及穿透细胞膜的能力。这使得设计一种新的抗生素成为一项艰巨的任务。
因此,一些研究人员并没有创造全新的抗生素,而是通过“大型数字图书馆”进行筛选,以预测哪些现有化合物可能已经具备所需的条件。
新加坡生物技术初创公司 BIOptimize 的计算化学家 Javad Deylami 说:“已经有既定的方法可以在几分钟内评估前两个特性,但渗透是这个难题中缺失的一块。”
Deylami 说,这意味着科学家们经常评估理论上可以很好地杀死细菌的化合物,但在进入细菌内部的第一个障碍上却达不到要求。“这就像他们到达边境但没有签证可以通过。”
Deylami 构建了一个细菌外细胞膜的计算机化版本,在上面进行模拟,测试分子穿透细胞膜的能力。他的模型确定了影响潜在药物通过或失败的力,从而使 Deylami 能够计算化合物的渗透性。
通过该程序运行已知的分子结构,Deylami 的团队能够在人工智能的帮助下了解化合物需要哪些“品质”来提高其渗透性。这些知识应该可以帮助他们在“庞大的图书馆”中寻找具有这些特性的现有药物。
主要参考文献
Wang, Y. et al. Proc. Natl Acad. Sci. USA 120, e2208344120 (2023).
Drew L. How antidepressants help bacteria resist antibiotics. Nature. 2023 Jan 24. doi: 10.1038/d41586-023-00186-y. Epub ahead of print. PMID: 36693968.
Antimicrobial Resistance Collaborators Lancet 399, 629–655 (2022).
Jin, M. et al. Environ. Int. 120, 421–430 (2018).
Maier, L. et al. Nature 555, 623–628 (2018).
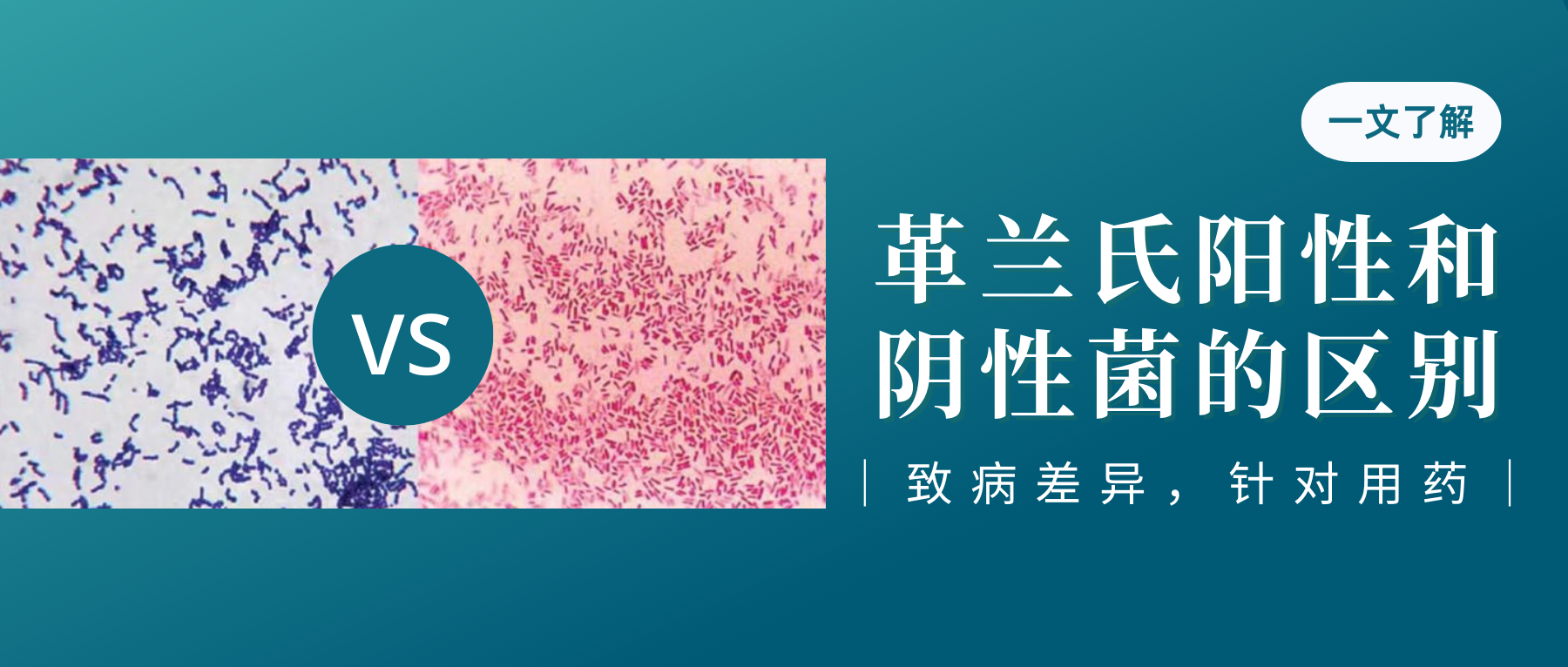
革兰氏阳性和阴性菌
在日常生活中,我们常常会看到药物或抗菌产品适应症会这样写到,对革兰氏阳性菌有效,对革兰氏阴性菌敏感,或者说对革兰氏阴性菌有效,对革兰氏阳性菌无效。可能很多人不是很清楚或搞不懂二者的区别。
本文主要介绍革兰氏阳性和阴性菌,它们的区别,代表性菌种以及针对用药等。
1884年,细菌学家Hans Christian Gram发明了革兰氏染色法来鉴别区分细菌。这种技术将细菌分成两大类,即革兰氏阳性菌(G+)和革兰氏阴性菌(G-)。区分主要是这两类细菌细胞壁成分不同,因而着色也不同所致。
这两类细菌的生理结构,疾病原因以及抗菌作用不一,因此,区分病原菌是革兰氏阳性菌,还是阴性菌,在临床确定感染和选择用药方面意义重大。
革兰氏阴性菌致病多由于患者有基础疾病或者体质比较差,肠道细菌感染引起的腹泻多是由肠道菌群中的革兰氏阴性菌所致,如大肠杆菌、沙门氏菌、志贺氏菌,布氏杆菌等,治疗这类细菌感染,一般使用三代头孢菌素以及喹诺酮类抗生素。注意大多数革兰氏阴性菌对青霉素耐药或不敏感。
大多数化脓性球菌都属于革兰氏阳性菌,它们能产生外毒素使人致病,常见的菌种有葡萄球菌、链球菌、肺炎双球菌、李斯特菌、炭疽杆菌、白喉杆菌、破伤风杆菌等。尤其在人体肠道内,革兰氏阳性致病菌致病几率更大。大多数由革兰氏阳性菌引起的感染可以用相当少量的抗生素治疗。青霉素、氯唑西林和红霉素足以覆盖 90% 的革兰氏阳性感染。
此外,某些广谱抗生素对革兰氏阳性菌和革兰氏阴性菌都有抗菌作用,如氨苄青霉素、庆大霉素、土霉素、磷霉素及环丙沙星等,但是作用效果可能不是最优。此外,磺胺类药物也属于广谱抑菌药物。
临床应用时,如果对细菌感染比较明确,尽量使用窄谱抗菌药物,如不太明确,可选用广谱抗菌药物。因此,临床治疗疾病时,首先要对药物的作用与用途要详细了解,然后再根据感染类型或诊断结果合理选择药物,这样才能取得最佳治疗效果。否则,药物选择不当,将会出现无效或越治越重的结果。
革兰氏阴性菌和革兰氏阳性菌之间的主要区别在于肽聚糖层的厚度和外部脂质膜的存在与否。
不同细菌的革兰氏染色
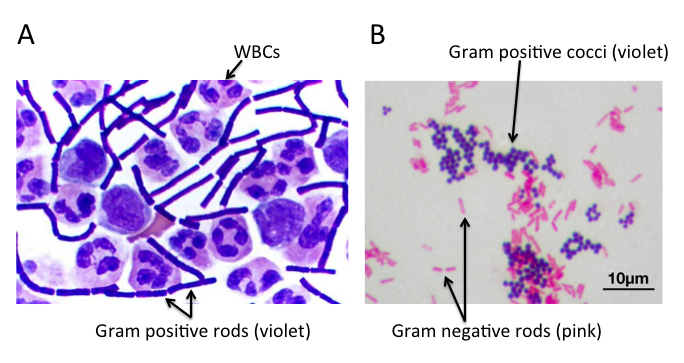
A) 脑脊液样本中的杆状革兰氏阳性炭疽杆菌(导致炭疽),也含有白细胞。B) 革兰氏阳性金黄色葡萄球菌(紫色或紫色)和革兰氏阴性杆菌大肠杆菌(粉红色)的革兰氏染色,它们是最常用的革兰氏染色参考细菌。参考来源:Wikipedia commons 和 tmedweb)
// 缺乏外膜,更容易受抗生素影响
革兰氏阳性菌的细胞壁含有肽聚糖、脂质、磷糖醛酸和磷壁酸。这种结构成分不同于由肽聚糖和外膜(由脂质、蛋白质和脂多糖组成)组成的革兰氏阴性细菌细胞壁。尽管革兰氏阳性菌具有较厚的肽聚糖层,但它们比革兰氏阴性菌更容易受到某些靶向细胞壁的抗生素的影响,因为它们缺乏外膜。
// 常见的致病菌
最常见的革兰氏阳性细菌包括葡萄球菌、链球菌、芽孢杆菌、梭状芽孢杆菌、李斯特菌、棒状杆菌等。这些革兰氏阳性菌的代表性物种是致病的,并可能引起多种疾病。
// 可用于治疗的抗生素
青霉素是影响革兰氏阳性菌的主要抗生素之一。
红霉素是另一种用于治疗革兰氏阳性细菌感染的强效抗生素。红霉素属于一类称为大环内酯类的抗生素,与阿奇霉素和克拉霉素同属一类。它通常用于对青霉素过敏的人。
甲氧苄啶/磺胺甲恶唑,克林霉素,克林霉素,强力霉素,万古霉素也可以用于特定的革兰氏阳性菌的感染。
// 层层抵抗更难杀死:细胞壁更硬,可改变外膜
革兰氏阴性菌有坚硬的保护外壳。它们的肽聚糖层比革兰氏阳性杆菌薄得多。
当它们的细胞壁受到干扰时,革兰氏阴性细菌会释放内毒素,症状更糟。同时,大多数抗生素为了接近它们的目标,必须通过外膜。例如,亲水性抗生素通过孔蛋白。革兰氏阴性菌可以通过改变它们的疏水特性或通过孔蛋白的突变来改变它们的外膜。这对这些细菌细胞产生了抵抗力。
// 暗藏玄机:比革兰氏阳性菌更危险
与革兰氏阳性菌相比,革兰氏阴性菌作为疾病生物体更危险,因为存在覆盖外膜的荚膜或粘液层。通过这种方式,微生物可以隐藏表面抗原,这个抗原可以触发人体免疫反应。
革兰氏阴性菌是一组臭名昭著的细菌,可导致多种疾病,包括肺炎、脑膜炎、淋病、细菌性痢疾、霍乱、胃炎等。在重症监护病房 (ICU)的患者,处于发病和死亡的高风险中,更容易遇见这类细菌,因此它们在医院具有重要的临床意义。
// 可用于治疗的抗生素
已经开发了许多不同种类的抗生素来杀死革兰氏阴性菌,例如头孢菌素、叶酸拮抗剂、哌拉西林-他唑巴坦、脲青霉素、内酰胺-β-内酰胺酶抑制剂、碳青霉烯类和喹诺酮类。它们是专门针对革兰氏阴性细菌而开发的,不过有时也对某些革兰氏阳性细菌有效。
革兰氏阳性菌与革兰氏阴性菌区别总结

参考来源:microbenotes
以上是革兰氏阳性菌和阴性菌的主要区别,接下来我们针对革兰氏阳/阴性菌,从细菌特征,形状表征,细胞结构等方面,进行更详细的介绍。
革兰氏阳性菌的定义是基于它们在革兰氏染色中用酒精短暂洗涤后保留结晶紫染料的能力。革兰氏阳性菌呈紫色。
这些细菌具有非常独特的特征,可以将其与其他类型的细菌区分开来。这些包括:
在革兰氏染色过程中,革兰氏阴性细菌在用酒精清洗后将失去结晶紫染料的颜色,并吸收反染物藏红花的粉红色/红色。
几乎在地球上的每个生活区域都可能发现革兰氏阴性细菌。
尽管大多数细菌是通过革兰氏染色染料进行区分的,但显微镜下的观察揭示了更多可用于定义和表征这些细菌的特征。
根据形状的定义,革兰氏阳性菌可分为两类:
革兰氏阳性细菌形成的其他特殊形状包括:
革兰氏阴性菌细胞的显微镜观察范围从杆状到芽孢杆菌,从球菌到螺旋状,螺旋状是最常见的形状。然而,有些表现出特殊的形状,如球杆菌、四分体、栅栏、毛状体等。例如:
革兰氏阳性菌的细胞壁含有肽聚糖、脂质、磷糖醛酸和磷壁酸。这种结构成分不同于由肽聚糖和外膜(由脂质、蛋白质和脂多糖组成)组成的革兰氏阴性细菌细胞壁。因此,革兰氏阳性菌的细胞壁很厚,并吸收了革兰氏染色的结晶紫染料。因此,显微镜下的革兰氏阳性菌呈紫色。
革兰氏阳性细菌细胞壁的结构特征
革兰氏阳性细菌具有由肽聚糖组成的厚的多层细胞壁(因为它含有肽和糖的混合物)。由于肽聚糖存在于大多数细菌中,但不存在于哺乳动物细胞中,因此它是抗菌药物的良好靶标(例如细胞壁合成抑制剂,包括青霉素、头孢菌素和万古霉素)。
这些抗生素会干扰转肽酶(也称为青霉素结合蛋白或PBP 的活性) 在细胞壁中催化相邻聚糖链之间的交联。
细胞壁还含有磷壁酸纤维,有助于细菌附着在宿主细胞膜(例如粘膜细胞)上,释放后会引起感染性休克,类似于革兰氏阴性菌释放的内毒素 (LPS) 产生的感染。
β-内酰胺酶(青霉素酶)是由细菌产生的酶家族,可水解 β-内酰胺抗生素(其中许多也是细菌来源)的四原子 β-内酯环,从而使其抗菌性能失活。
细菌细胞膜还可以包含ABC 外排泵这可能导致抗生素耐药性和对那些具有细胞内作用机制的药物(例如 DNA 促旋酶抑制剂或蛋白质合成抑制剂)的多药耐药性 (MDR)。
结晶紫染料附着在革兰氏阳性菌细胞壁的厚肽聚糖层上,在光学显微镜下观察时将它们染成紫色或紫色。
肽聚糖
它也被称为胞壁质(murein),占细菌细胞壁含量的 90%。
——维持形状,并保持细胞壁强度和弹性
它是一种优质聚合物,由两种相同的糖衍生物(N-乙酰氨基葡糖和 N-乙酰胞壁酸)以及 L-氨基酸链和蛋白质中很少发现的三种不同 D-氨基酸(即 D-谷氨酸、D-丙氨酸和内消旋二氨基庚二酸)组成,可保护细胞壁免受肽酶的攻击。
D-氨基酸和L-氨基酸连接到N-乙酰壁酸,L-氨基酸特别是赖氨酸可以取代中二氨基丙烯酸。
肽聚糖亚基的这种相互连接使肽聚糖具有很强的维持细菌形状和完整性的能力,并具有弹性和延展性。
肽聚糖也具有渗透性,允许分子进出细菌细胞。
——肽聚糖的生物合成
抑制细菌细胞壁肽聚糖层的合成是许多抗菌药物的分子靶点,包括 β-内酰胺类抗生素(青霉素、头孢菌素、碳青霉烯类和单环内酰胺类)和糖肽类抗生素(万古霉素和其他较新的类似物)。
这些药物的两个主要分子靶标是转肽酶,也称为青霉素结合蛋白 (PBP),因为它们与青霉素结合,而糖基转移酶 (GT) 可被万古霉素等糖肽抑制。
PBPs 有许多亚型,给定的细菌菌株可以表达多种 PBPs。这些 PBP 在其生理特性和与抗生素相互作用的敏感性方面可能有所不同。取决于 PBP 亚型,糖基转移酶可以作为单独的酶或作为与转肽酶相关的二聚体存在。
革兰氏阳性细胞壁生物合成
图片来源自(Wikipedia Commons 和 tmedweb)
细菌细胞壁由重复的 N-乙酰氨基葡萄糖 (NAG) 和 N-乙酰胞壁酸 (NAM) 亚基链组成。NAM 亚基附有短肽链。
肽链的组成因细菌而异,但近端的丙氨酸通常是 L-Ala,而远端的两个通常是 D-Ala。也与青霉素结合的细胞壁转肽酶(青霉素结合蛋白:PBPs)在肽侧链之间形成键,并从肽侧链之一排出末端 D-丙氨酸。
一旦形成交联,PBP 就会从壁上解离。用糖基转移酶 (GT)分离酶结构域NAM 和 NAG 残基之间的活性形成联系。一些高分子量 PBP(例如 PBP2)是含有转肽酶和糖基转移酶结构域的酶复合物。
磷壁酸纤维存在于革兰氏阳性菌的细胞壁中,由磷酸甘油或磷酸核糖醇的聚合物组成。它们参与细菌与黏膜细胞的附着,可诱发感染性休克,类似于革兰氏阴性菌释放的 LPS(内毒素)。
磷壁酸
这是由甘油共聚物组成的加固墙。
磷壁酸是水溶性的,占细菌细胞壁总干重的 50%。
它要么直接与肽聚糖共价连接,要么与细胞膜(脂磷壁酸)连接。通过 6-羟基 N-乙酰胞壁酸与肽聚糖直接相连。
带负电荷,延伸到肽聚糖表面,使细菌细胞壁带负电荷。
它还有助于维持细胞壁的结构。
它在革兰氏阴性菌中完全不存在。
脂质
它们在肽聚糖下方有一层薄薄的脂质,大约 2-5%,其作用是锚定细菌细胞壁。
细胞壁
——细胞壁非常复杂
结合细胞外膜的主要作用,加上一层肽聚糖,其功能特性复杂,这里是对细胞壁及其功能部分的描述。
革兰氏阴性细菌的细胞壁具有一层2-7nm的肽聚糖薄层和7-8nm厚的外膜。
——周质空间较大
显微镜下,细胞膜和细胞壁之间有一个空间,称为由周质组成的周质空间。在革兰氏阴性菌和革兰氏阳性菌中均能发现,但在革兰氏阴性菌中,周质空间较大。
革兰氏阴性细菌细胞壁
在结构上,革兰氏阴性细胞壁由细胞膜外部的两层组成:一层薄薄的肽聚糖(太薄而无法吸收大量甲基紫染色)和一层外膜(革兰氏阴性细菌独有),通常含有促进小(<700 Da)亲水分子(例如糖、氨基酸和维生素)扩散的孔蛋白。
许多抗生素(例如许多青霉素和头孢菌素)也可以通过孔蛋白扩散到达它们的作用部位。
然而,万古霉素 (1449 Da) 的质量太大,无法透过孔蛋白到达其作用部位,这使其对革兰氏阴性细菌无效。因此,外膜为革兰氏阴性菌提供了对某些抗生素的固有“内在抗性”,可以通过改变孔蛋白的表达水平或改变孔蛋白的孔特性以降低抗生素的渗透性来进一步修饰。
革兰氏阴性菌的外膜还含有脂多糖 (LPS)或内毒素,可被细菌排出,引起宿主强大的免疫反应。
周质空间
革兰氏阴性菌的周质空间由几种蛋白质组成,这些蛋白质有助于获取营养,例如攻击核酸和磷酸化分子的水解酶,以及积极协助将物质运输到细菌细胞中的结合蛋白。周质空间还具有合成肽聚糖和修饰可能对细胞造成伤害的有毒元素的酶。
肽聚糖
革兰氏阴性细菌细胞壁有一层薄的肽聚糖层,位于质膜上方,约占细胞干重的5%。厚度不超过4纳米,一些细菌如大肠杆菌只有2纳米厚的肽聚糖。
外膜和脂多糖
革兰氏阴性菌还有第二层脂质双层,位于肽聚糖层的外部。这种外膜通过布劳恩脂蛋白与肽聚糖相连。外膜和肽聚糖之间的紧密连接是维持外膜作为许多有毒分子和抗生素的不渗透屏障所必需的。
外膜上的粘附位点也加强了革兰氏阴性细胞壁,这些粘附位点在允许细胞接触和膜融合方面发挥作用。物质通过这些粘附位点进入细胞。
革兰氏阴性菌结构

图源:Jeff Dahl,wikipedia
外膜主要由脂多糖 (LPS)组成,脂多糖是由脂质和碳水化合物组成的大型复杂分子。脂多糖由3个单元组成:脂质A、核心多糖和O侧链。
脂质A由两种氨基葡萄糖糖衍生物组成,每个衍生物含有三种脂肪酸和焦磷酸盐,脂多糖的任何剩余部分都会伸出膜表面。
O侧链也称为O抗原,是从核心向外延伸的链。它由导致细菌菌株之间变异的糖组成。这些 O 抗原也负责细菌逃避抗体反应。
// 外膜及其脂多糖的作用
——脂多糖负责保护细胞壁免受外部攻击
LPS 带有负电荷,使电池表面带负电荷。因此,这稳定了膜结构。
脂质A是脂多糖的有毒成分,因此它起到内毒素的作用。
——防毒素进入,防成分丢失
外膜及其脂多糖有助于防止抗生素、胆汁盐和其他有毒元素进入并破坏细胞。
外膜由孔蛋白组成,使其具有渗透性,允许小分子(如葡萄糖)进入。维生素 B12 等较大的分子通过特定的载体运输穿过外膜。
外膜还有助于防止成分丢失,特别是来自周质空间。
下表描述了主要的革兰氏阳性致病菌,它们的基本形态特征以及它们在人类中引起的疾病。
已知革兰氏阴性菌是正常菌群,部分会导致严重的人类感染,从社区获得性感染到医院感染。
革兰氏阴性菌外膜的结构是其众多显着特征之一。脂多糖 (LPS) 存在于膜的外叶上,其脂质 A 部分用作内毒素。
// 革兰氏阴性菌感染:严重时可危及生命
如果由于某种原因,革兰氏阴性细菌能够到达动物的循环系统,脂多糖将激活免疫系统,并触发先天免疫反应,产生细胞因子和激素调节剂。这会引起炎症,并可能导致毒性反应,从而导致发烧、呼吸急促和低血压。这就是已知革兰氏阴性细菌会导致危及生命的休克的原因。
内毒素休克的一些症状:
发烧和发冷或体温下降、发炎、皮疹、呼吸急促、心率加快、低血压、多器官衰竭等。
下表给出了一些革兰氏阴性致病细菌的例子以及它们在人类宿主中引起疾病和感染时表现出的临床特征。

如上表所述,已知革兰氏阳性细菌会引起多种感染,如果不及时和适当地治疗和管理,可能对人类造成灾难性的影响。
革兰氏阳性杆菌感染用抗生素治疗。青霉素、氯唑西林和红霉素可治疗 90% 以上的革兰氏阳性菌。
常见的革兰氏阳性抗菌药物及作用机制

然而,抗生素耐药性正在成为革兰氏阳性感染的一个严重问题。研究人员正在开发新的药物来帮助解决这个问题。只有在绝对需要时才应使用抗生素。需要严格遵循感染控制标准,以防止抗生素耐药性感染的发展和传播。
由于它们的外膜,这些细菌对溶菌酶和青霉素具有抗性。这是因为存在保护内膜和细胞壁的外壁。
在周质空间(两个细胞膜之间的区域)中也发现了分解或改变抗生素的酶。用于治疗革兰氏阴性菌感染的治疗方法包括羧基、氨基和脲基青霉素。为了对抗可以消化这些药物的酶,有时将它们与β-内酰胺酶抑制剂结合使用。β-内酰胺酶是一种存在于周质中的酶。
针对细菌病原体的抗菌剂被称为抗生素。这些抗生素启动针对细菌细胞的阻断或抑制机制,以诱导细菌细胞增殖和复制。
用于对抗革兰氏阴性菌的抗生素示例

抗生素耐药性是现在世界上的一个主要临床问题。
耐多药细菌在人群中变得越来越普遍,如果不进行有效治疗,这种感染可能会导致肾功能衰竭、败血症,甚至死亡。
微生物以多种方式抑制临床治疗中使用的许多抗菌剂。这些包括改变药物结合位点的方法,改变药物构象的方法,改变膜通透性的方法,可以导致耐药机制失活。
例如革兰氏阴性菌中有两层膜,外膜和内膜。脂多糖被认为是一种非常强的免疫反应诱导剂,它具有三个重要成分:脂质 A、亲水性多糖、抗原 O 的疏水域。
疏水域在细胞膜的外部表达。它是疏水成分脂质a,它负责内毒素作用。LPS 在细菌中是可变的,并且由于遗传变异,一些细菌只产生一种不被 Toll 样受体识别的弱抗原。然而,有大量的革兰氏阴性菌团体可能会引起这样的反应。免疫系统也被一些 toll 样受体 4 (TLR4) 激活,这些受体存在于与免疫系统有关的众多细胞中,如巨噬细胞、单核细胞、中性粒细胞和树突状细胞。
由 LPS 和 TLR4 受体介导的先天免疫反应的激活导致反应增强,产生细胞因子、趋化因子和干扰素等。
免疫系统的反应取决于感染过程的严重程度以及侵袭性细菌中 LPS 的结构,这与菌的毒力有关。因此,虽然一些细菌(如大肠杆菌)可以诱导免疫系统,但其他细菌(如幽门螺杆菌)仅具有弱抗原性。
// 对付耐药性细菌新思路:根据电荷相互作用原理设计新药
2017 年,伊利诺伊大学化学教授和当前研究的合著者Paul Hergenrother(ACPP 负责人/MMG)在《自然》杂志上报告说 ,发现的一个关键是,如果向它们添加带正电荷的基团,例如胺,一些抗生素可以使用特定的膜孔穿透革兰氏阴性细菌的细胞膜。
这项工作表明,抗生素上带正电荷的胺基与细菌孔内的负电荷有良好的相互作用。这些吸引力使带有胺基的抗生素以一种更有利于能量的方式排列,因为它穿过收缩区的孔的最狭窄部分。不含胺的抗生素面临更高的能量屏障去通过孔隙。
这或许意味着未来可以设计新药(或修改旧药),以攻击和杀死对抗生素治疗具有耐药性的微生物。
问
革兰氏阳性细胞和革兰氏阴性细胞之间的三个区别是什么?
答
革兰氏阳性菌有一层厚的肽聚糖作为它们的细胞壁,而革兰氏阴性菌有一层薄薄的肽聚糖和外膜。
革兰氏阴性菌有脂多糖(LPS),而革兰氏阳性菌没有。
一些革兰氏阳性细菌含有霉菌酸,它会在细胞壁上形成一层蜡质层。
问
什么是革兰氏阳性感染?
答
由革兰氏阳性菌引起的感染,如耐甲氧西林金黄色葡萄球菌 (MRSA)、耐万古霉素肠球菌 (VRE) 和艰难梭菌是常见的多重耐药菌感染。
问
革兰氏阳性菌更容易治疗吗?
答
革兰氏阳性细菌,即那些具有肽聚糖外层的物种,更容易被杀死——它们的厚肽聚糖层很容易吸收抗生素和清洁产物。因此,某些容易杀死革兰氏阳性菌的洗涤剂不会破坏革兰氏阴性菌。
问
肠道内很多革兰氏阳性菌致病吗?
答
是的,人体肠胃道的大多数革兰氏阳性菌都是条件致病菌。包括微球菌、肠球菌、金黄色葡萄球菌、表皮葡萄球菌、腐生葡萄球菌、肺炎链球菌、草绿色链球菌、酿脓链球菌、无乳链球菌、破伤风梭菌、肉毒杆菌、产气荚膜梭菌、产气荚膜梭菌、艰难梭菌, 单核细胞增生李斯特菌等。
问
革兰氏阳性菌在哪里发现?
答
根据革兰氏阳性菌种,它们可以在人类的土壤、水生沉积物、灰尘、皮肤、口腔、肠道或生殖道中找到。
问
革兰氏阳性球菌危险吗?
答
革兰氏阳性菌可能是球菌或杆菌。这些称为常驻菌群的细菌通常不会引起疾病。革兰氏阳性杆菌引起某些感染,包括:炭疽。
问
革兰氏阳性菌对抗生素的抵抗力更强吗?
答
不是。与革兰氏阳性菌相比,革兰氏阴性菌对多种抗生素的耐药性更强。由于它们的外膜,它们对抗生素的抵抗力更强。
革兰氏阳性细菌更容易被杀死,因为它们的厚肽聚糖层很容易吸收抗生素和清洁剂。另一方面,革兰氏阴性细菌具有薄的肽聚糖层,不会吸收周围的任何异物。
问
革兰氏阳性菌有内毒素吗?
答
不会。内毒素与革兰氏阳性菌无关。这些细菌没有内毒素,因为它们没有外膜。另一方面,革兰氏阴性细菌会产生内毒素。
这些内毒素是革兰氏阴性细菌细胞外膜的一部分,只有当细胞裂解或细菌死亡时才会释放出来。内毒素是形成革兰氏阴性菌细胞壁结构的热稳定性脂多糖-蛋白质复合物。
问
为什么革兰氏阳性菌对抗生素更敏感?
答
尽管革兰氏阳性菌具有较厚的肽聚糖层,但它们比革兰氏阴性菌更容易受到某些靶向抗生素的细胞壁的影响,因为它们缺乏外膜。
大多数抗生素为了接近它们的目标,必须通过外膜。例如,亲水性抗生素通过孔蛋白。因此,革兰氏阴性菌可以通过改变它们的疏水特性或通过孔蛋白的突变来改变它们的外膜。这对这些细菌细胞产生了抵抗力。革兰氏阳性菌缺乏这一因素,因此革兰氏阴性菌对抗生素的抵抗力比它们强。
问
革兰氏阳性菌是否致病?
答
是的,大多数革兰氏阳性菌都是致病菌。致病性革兰氏阳性菌的实例包括微球菌、肠球菌、金黄色葡萄球菌、表皮葡萄球菌、腐生葡萄球菌、肺炎链球菌、草绿色链球菌、酿脓链球菌、无乳链球菌、破伤风梭菌、肉毒杆菌、产气荚膜梭菌、产气荚膜梭菌、艰难梭菌, 单核细胞增生李斯特菌等。
问
革兰氏阳性菌引起的常见感染有哪些?
答
炭疽、白喉、腹泻、脑膜炎、恶心、皮肤感染、尿路感染。
问
哪种抗生素对革兰氏阳性菌有效?
答
对革兰氏阳性菌有效的抗生素是青霉素、氯唑西林和红霉素,几乎覆盖了 90% 的革兰氏阳性菌感染。其他还有万古霉素、甲氧苄啶/磺胺甲恶唑,克林霉素,克林霉素等。
问
革兰氏阴性菌引起的人类常见疾病有哪些?
答
革兰氏阴性菌会在医疗机构中引起感染,包括肺炎、血流感染、伤口或手术部位感染以及脑膜炎。此外还有霍乱、鼠疫、伤寒、脑膜炎和尿路感染是人类常见的细菌性疾病。
问
为什么革兰氏阴性菌比革兰氏阳性菌更有害?
答
革兰氏阴性菌细胞壁坚硬,不易对抗生素敏感,在抗生素作用下会释放内毒素。
问
什么会杀死革兰氏阴性菌?
答
这些抗生素包括头孢菌素类(头孢曲松-头孢噻肟、头孢他啶等)、氟喹诺酮类药物(环丙沙星、左氧氟沙星)、氨基糖苷类(庆大霉素、阿米卡星)等
问
革兰氏阴性菌的分泌系统是什么?
答
革兰氏阴性菌具有广泛封闭的分泌系统,可以转移微小分子、DNA、氨基酸、蛋白质。
问
革兰氏阴性菌从哪来?
答
革兰氏阴性细菌随处可见,几乎遍布地球上所有支持生命的环境。革兰氏阴性菌包括模式生物大肠杆菌,以及许多致病菌,如铜绿假单胞菌、淋病奈瑟菌、沙眼衣原体和鼠疫耶尔森菌。
问
如何自然去除革兰氏阴性菌?
答
天然抗生素。一些天然抗生素是大蒜、蜂蜜、卷心菜、葡萄柚籽提取物、生苹果醋、特级初榨椰子油、发酵食品等。
免责声明:本公众号内容仅作交流参考,不作为诊断及医疗依据。主要参考文献:
Oliveira J, Reygaert WC. Gram Negative Bacteria. 2022 Mar 26. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2022 Jan–. PMID: 30855801.
Carroll K.C., & Hobden J.A., & Miller S, & Morse S.A., & Mietzner T.A., & Detrick B, & Mitchell T.G., & McKerrow J.H., & Sakanari J.A.(Eds.), (2019). Jawetz, Melnick, & Adelberg’s Medical Microbiology, 27e. McGraw Hill.
Acheson DWK (2015): Patient information: Food poisoning (foodborne illness) (Beyond the Basics). In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/1/16
Apicella M (2015): Treatment and prevention of meningococcal infection. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/1/16
Baum SG (2016): Mycoplasma pneumoniae infection in adults. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/2/16
Crowe SE (2016): Bacteriology and epidemiology of Helicobacter pylori infection. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/1/16.
Crowe SE (2016b): Treatment regimens for Helicobacter pylori. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/1/16.
File TM (2016): Treatment of community-acquired pneumonia in adults in the outpatient setting. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/1/16
Ghanem KG (2016): Clinical manifestations and diagnosis of Neisseria gonorrhoeae infection in adults and adolescents. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/1/16
Hicks CB, Clement M (2016): Syphilis: Treatment and monitoring. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/2/16
Kanafani ZA, Kanj SS (2014): Acinetobacter infection: Epidemiology, microbiology, pathogenesis, clinical features, and diagnosis. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/10/16
Kanafani ZA, Kanj SS (2016): Acinetobacter infection: Treatment and prevention. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/10/16
Kelly CP, Lamont JT (2015): Clostridium difficile in adults: Treatment. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/2/16
Koulenti D et al (2009): Spectrum of practice in the diagnosis of nosocomial pneumonia in patients requiring mechanical ventilation in European intensive care units. Critical Care Med 37(9):2360-2369. doi: 10.1097/CCM.0b013e3181a037ac
Lamont JT (2016): Clostridium difficile in adults: Epidemiology, microbiology, and pathophysiology. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/2/16
Li X-Z, Nikaido H (2004): Efflux-mediated drug resistance in bacteria. Drugs. 64(2):159–204.
Lowy FD (2016): Methicillin-resistant Staphylococcus aureus (MRSA) in adults: Treatment of skin and soft tissue infections. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/1/16
Pegram PS, Stone SM (2016): Botulism. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/1/16
Riley LW (2015): Natural history, microbiology, and pathogenesis of tuberculosis. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/2/16
Sauvage E et al (2008): The penicillin-binding proteins: structure and role in peptidoglycan biosynthesis. FEMS Microbiol Rev 32:234–258. DOI:10.1111/j.1574-6976.2008.00105.x
Southwick F (2008): Infectious Diseases. A Clinical Short Course. McGraw Hill/Lange.
Sterling TR (2016): Treatment of pulmonary tuberculosis in HIV-uninfected adults. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/2/16
Stevens DL, Bryant A (2015): Group A streptococcus: Virulence factors and pathogenic mechanisms. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/1/16
Stevens DL (2016): Group A streptococcal (Streptococcus pyogenes) bacteremia in adults. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/1/16
Swygard H et al (2016): Treatment of uncomplicated gonococcal infections. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/1/16
Wanke CA (2015): Pathogenic Escherichia coli. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 7/28/16
Wanke CA (2015b): Travelers’ diarrhea: Clinical manifestations, diagnosis, and treatment. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/1/16
Yeh S (2015): Microbiology, epidemiology and treatment of Haemophilus influenzae. In: UpToDate, Basow, DS (Ed), Waltham, MA. Cited 8/1/16
Sagar Aryal,Gram-Positive Vs Gram-Negative Bacteria- 31 Differences With Examples. Microbenotes.2022,January 9
Jawertz M., Alderbergs., Medical Microbiology 28th Edition.
Prescott M. L., Microbiology. 5th Edition
Lippincott Microbiology in review: 3rd edition
Faith Mokobi,Gram-Positive Bacteria- Cell Wall, Examples, Diseases, Antibiotics,2021, April 15
Faith Mokobi,Gram-Negative Bacteria- Cell Wall, Examples, Diseases, Antibiotics ,2021 ,April 15

谷禾健康
人的身体拥有数千种细菌,这些细菌在维持健康方面发挥着重要作用。当这些细菌失控繁殖并侵入身体的其他部位或将有害细菌引入身体的系统时,可能会发生细菌感染。
细菌感染的严重程度取决于所涉及的细菌类型和所感染的部位等。细菌最常感染肠道、皮肤和呼吸系统,包括肺、泌尿道和阴道。
目前应对细菌感染的主要治疗方法还是使用抗生素,但是抗生素管理,或改进抗生素的处方和使用方式,对于优化感染患者的治疗、保护患者免受伤害和对抗抗生素耐药性至关重要。
今天我们重点了解下什么是细菌感染,感染的症状、原因和类型,包括诊断测试和治疗方案,抗生素什么时候使用,什么时候不用,抗生素的耐药性如何产生以及使用抗生素的注意事项等。
了解什么是细菌感染以及如何治疗细菌感染,我们应该先了解细菌的概念。
来源:MedicineNet
地球上所有的生物都是由两种基本类型的细胞中的一种组成:
真核细胞,其遗传物质被包裹在核膜内;
原核细胞,其遗传物质与细胞的其他部分不分离。
传统上,所有的原核细胞都被称为细菌,被归为原核生物界。
1970年代后期,美国微生物学家卡尔·沃斯(Carl Woese)率先在分类上进行了重大变革,将所有生物分为真核生物、细菌(原名真细菌)和古细菌(原名古细菌)三个领域,以反映三条古老的进化路线。原核生物以前被称为细菌,然后被分为两个领域,细菌和古细菌。

published by Woese et al.
细菌和古细菌在表面上是相似的,例如,它们没有细胞内的细胞器,它们有环状DNA。但是,它们在本质上是截然不同的,它们的分离是基于其古老而又独立的进化谱系的遗传证据,以及其化学和生理学的根本差异。这两个原核域的成员彼此之间的区别与它们和真核细胞中的区别一样。
细菌细胞在几个方面不同于动物细胞和植物细胞。一个根本的区别是细菌细胞缺乏动物细胞和植物细胞中都存在的细胞内细胞器,例如线粒体,叶绿体和细胞核。细菌体积小,设计简单,代谢能力强,使它们能够迅速生长和分裂,并在几乎任何环境中生存和繁衍。
细菌,动物和植物细胞的比较

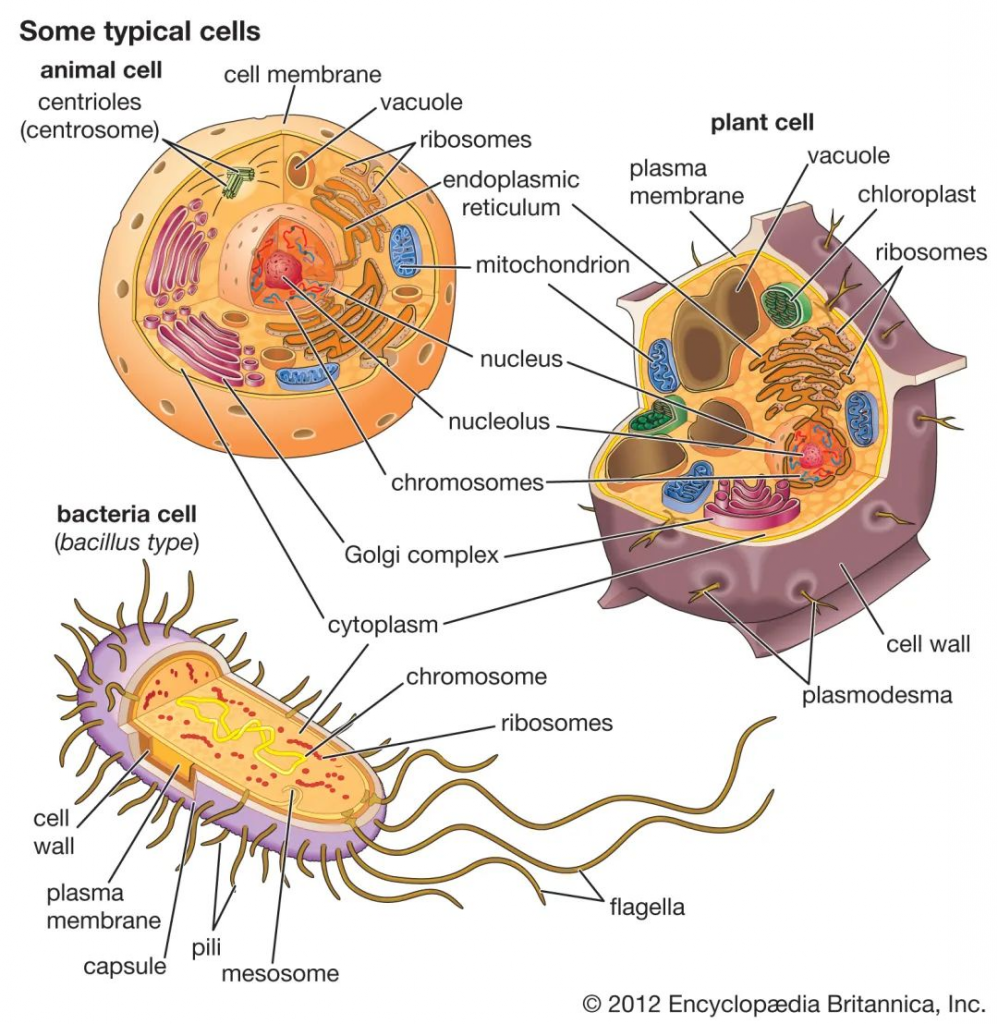
除此之外,原核和真核细胞在许多其他方面有所不同,包括脂质组成,关键代谢酶的结构,对抗生素和毒素的反应以及遗传信息的表达机制。
真核生物包含多个线性染色体,这些染色体的基因比编码蛋白质合成所需的基因大得多。遗传信息的核糖核酸(RNA)副本(脱氧核糖核酸或DNA)的大部分被丢弃,剩余的信使RNA(mRNA)在被翻译成蛋白质之前已被充分修饰。相反,细菌具有一个包含所有遗传信息的环形染色体,它们的mRNA是其基因的精确副本,不会被修饰。
细菌细胞在结构上比真核细胞小得多,也简单得多,但细菌是一个在大小、形状、生境和代谢上都有差异的极其多样化的有机体群体。
许多关于细菌的知识来自对致病细菌的研究,这些细菌比许多自由生活的细菌更容易在纯培养中分离出来,也更容易被研究。必须注意的是,许多自由生活的细菌与适应作为动物寄生虫或共生体生活的细菌有很大的不同。因此,关于细菌的组成或结构没有绝对的规则,任何说法都会有许多例外。
大多数细菌对地球上的生命都是有益的,甚至是生命所必需的,但少数细菌却对人类有害。目前,没有一种古细菌被认为是病原体,但是包括人类在内的动物不断遭到大量细菌的“轰击和居住”。
口腔,肠道和皮肤被大量特定类型的细菌定殖,这些细菌适应这些栖息地的生活。这些微生物在正常情况下是无害的,并且仅在它们以某种方式穿过身体的屏障并引起感染时才变得危险。
一些细菌擅长入侵宿主,被称为病原体或疾病产生者。一些病原体作用于人体的特定部位,例如:
在食物中传播的许多致病菌的毒素在摄入时会引起食物中毒。
由金黄色葡萄球菌(Staphylococcus aureus)产生的毒素,能引起迅速、严重但有限的胃肠道不适。
肉毒梭菌(Clostridium botulinum)的毒素,通常是致命的。在密封前未完全煮熟的罐装非酸性食品中可能产生肉毒杆菌毒素。肉毒梭菌形成耐热孢子,可以发芽为营养细菌细胞,在厌氧环境中茁壮成长,这有利于产生其极强的毒素。
其他食源性感染实际上是由受感染的食物处理者传播的,包括伤寒、沙门氏菌病(沙门氏菌属)和志贺氏菌病(痢疾志贺氏菌属)。
关于食物中毒相关细菌详见:正值夏季,警惕食源性疾病,常见的食物中毒的病原菌介绍
细菌感染是指病原菌侵入宿主组织。当有害细菌进入人体或伤口并繁殖,导致疾病、器官损伤、组织损伤或疾病时,就会发生细菌感染。细菌可以感染身体的任何部位。
细菌感染很常见,但它们并不完全相同。细菌有很多种,每种对身体都有不同的影响。广义上来讲,细菌感染也可以是由任何细菌引起的任何内部或外部疾病的广义术语。
细菌存在于与人体的各种关系中。它们在体表内定殖并提供益处,正常情况下细菌可以与人类身体共存不会产生有害的影响(在共生关系中)。但是有时候,通常无害地存在于我们体内或皮肤上,而不会造成任何异常问题的细菌会失控并导致感染。这种情况通常发生在我们的免疫系统不够强大,无法保持它们的平衡,或者我们的微生物区系的组成发生了一些变化,对某些细菌生存繁殖更有利。
细菌感染是由体内的坏细菌引起的。一些细菌感染可能危及生命。标准的医学治疗通常涉及一剂抗生素。细菌感染很容易与病毒感染混淆。但是他们并不一样,后面我们会单独解释。
当皮肤粘膜有破损或发生化脓性炎症时,细菌则容易侵入体内。
人体的免疫反应可分为非特异性免疫反应及特异性免疫反应两种,后者又可分为细胞免疫与体液免疫两方面。当机体免疫功能下降时,不能充分发挥其吞噬杀灭细菌的作用时,即使入侵的细菌量较少,致病力不强也能引起感染;条件致病菌所引起的医源性感染也逐渐增多。
人体免疫功能正常时,进入血中的细菌迅速被血中防御细胞如单核细胞、嗜中性粒细胞等所清除,而患肝硬变、糖尿病、血液病、结缔组织病等慢性病者,可因代谢紊乱、体液免疫及细胞免疫功能减低,易招致败血症发生;各种免疫抑制药物的使用、放射治疗亦是导致败血症发病率高的原因。广谱抗菌药物使用后,对药物敏感的细菌虽被抑制或杀灭,而一些耐药菌乘机繁殖,亦可酿成败血症。
细菌侵入人体后是否引起感染,除了与人的防御、免疫功能,还与细菌的毒力及数量有关。毒力强或数量多的致病菌进入机体,引起感染的可能性较大。
严重烧伤时,创面为细菌敞开门户,皮肤坏死、血浆渗出又为细菌繁殖提供了良好环境,故极易发生感染。
尿路、胆道、胃肠道、呼吸道粘膜受破坏后,若同时有内容物积滞、压力增高,细菌更易进入血中,保留导尿管、静脉等血管内留置导管、人工辅助呼吸时插管等,也使细菌易于侵入。
细菌是微观的,通常是单细胞生物,到处都可以找到(例如,在我们体内、皮肤上、空气中、水里、土壤中或我们吃的食物里)。细菌具有传染性,因此,它可以通过与患者的密切接触在人与人之间传播。接触受污染的表面、食物、水以及打喷嚏和咳嗽就是例子。
发生感染的几个条件:
此外,细菌还可以导致急性感染(快递治疗)和慢性感染(持续很长时间甚至终生),以及潜伏感染(最初可能没有任何明显迹象或症状但可以过段时间自然发展)。症状可以从轻度、中度到重度不等。在严重的情况下,导致数百万人死亡;例如,许多人死于黑死病或鼠疫。
细菌感染的常见特征:
常见的各部位细菌感染的症状

细菌性感冒
细菌性感冒(Bacterial cold)是指继发细菌感染引起的感冒。感冒在习惯上分为病毒性感冒和细菌性感冒。
病毒性感冒有:普通感冒、流行性感冒和病毒性咽炎等。其主要不同是致病因素不同,病毒性感冒是由于病毒所致,而细菌性感冒是由于细菌所致。
注:流行性感冒(流感通过病毒传播,不是细菌性感冒,注意区分),是由流感病毒引起的急性呼吸道传染病。
细菌的感染还取决于受感染细菌的类型,常见的细菌感染例如:
细菌感染具有高度传染性,因此需要特别注意避免通过洗手、打喷嚏和咳嗽时遮盖以及不共用杯子和饮料瓶来传播感染。
细菌有多种传播方式,包括:

当你忍不住咳嗽或打喷嚏时,用纸巾捂住口鼻或用手肘内侧。
在下述情况下需要洗手,至少洗20秒:
如果没有肥皂和水,可以使用酒精含量至少为 60% 的酒精类洗手液。
除此之外,其他需要注意的:
接种疫苗。如轮状病毒是一种导致婴儿和幼儿严重肠胃炎的病毒。所有婴儿都应在 6 周和 4 个月大时接种轮状病毒疫苗。
流感疫苗。当患上流感时,免疫系统会被削弱。这使细菌有机会侵入肺部并引起感染。每年注射一次流感疫苗可以防止感染肺部细菌。
肺炎球菌多糖疫苗。美国疾病控制与预防中心 (CDC) 推荐 65 岁及以上的人接种肺炎球菌多糖疫苗 (PPSV)。它也适用于免疫系统较弱或患肺炎风险较高的成年人。
治疗取决于感染的严重程度、患者的年龄、免疫系统有多强、是否存在任何并存症状以及现有的医疗条件。
治疗可能包括:
扩展阅读:
细菌素——对抗感染、保存食品、重塑肠道菌群
还在滥用抗生素?15种天然抗生素助你调节肠道菌群
这里我们主要来了解一下,用于细菌感染的药物——抗生素。
目前抗生素是对抗细菌感染的常用药物。它们通过破坏细菌细胞生长和增殖所需的过程来发挥作用。
抗生素根据抗菌谱可以分为:
广谱抗生素,中谱抗生素和窄谱抗生素。
✓ 广谱抗生素
广谱抗生素可对抗多种细菌。广谱抗生素可治疗革兰氏阳性菌和阴性菌,因此如果医生不确定有哪些细菌,他或她可能会开出其中一种抗生素。
阿莫西林、奥格门汀、头孢菌素(第 4 代和第 5 代)、四环素氨基糖苷类和氟喹诺酮类(环丙沙星)是广谱抗生素的例子。
✓ 中谱抗生素
中谱抗生素针对一组细菌。青霉素和杆菌肽是流行的中谱抗生素。
✓ 窄谱抗生素
窄谱抗生素用于治疗一种特定的细菌。多粘菌素属于这一小类抗生素。当确定患有哪种细菌感染时,治疗会更容易和更有效。
抗菌活性通常分为五种机制:
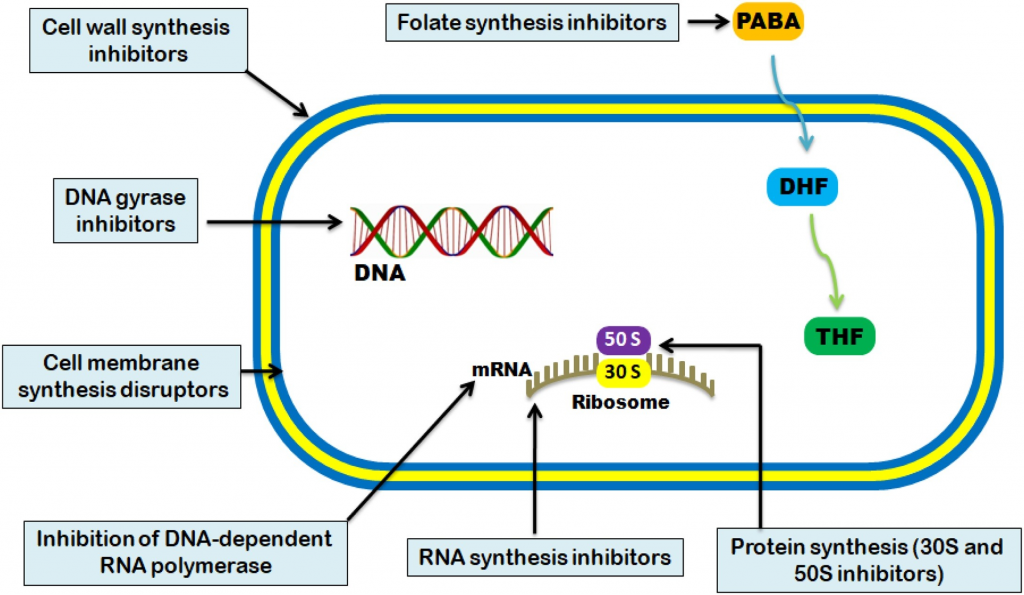

Uddin TM, et al., J Infect Public Health. 2021
不同种类抗生素作用的方式


Uddin TM, et al., J Infect Public Health. 2021
青霉素
最初的青霉素仅用于革兰阳性菌感染的治疗,由于金黄色葡萄球菌很快产生一种青霉素酶对青霉素产生了耐药,人们研制出了以甲氧西林、苯唑西林为代表的耐酶青霉素,用于产青霉素酶的葡萄球菌( 甲氧西林耐药者除外) 感染。
之后,以氨苄西林、阿莫西林为代表的广谱青霉素由仅对革兰阳性(G+) 菌有效扩展到对革兰阴性(G-) 菌( 主要为肠杆菌科细菌) 也具有抗菌活性。在广谱青霉素的基础上又发展为对铜绿假单胞菌等非发酵菌也具抗菌活性者,当前应用主要为脲基类青霉素,包括: 哌拉西林、阿洛西林、美洛西林。
代表药物: 阿莫西林(amoxicillin)
阿莫西林是一种抗细菌的青霉素抗生素。
阿莫西林用于治疗由细菌引起的多种不同类型的感染,例如扁桃体炎、支气管炎、肺炎以及耳、鼻、喉、皮肤或泌尿道感染。
阿莫西林有时还与另一种称为克拉霉素( Biaxin ) 的抗生素一起用于治疗由幽门螺杆菌感染引起的胃溃疡。这种组合有时与一种称为兰索拉唑(Prevacid)的胃酸减少剂一起使用。
四环素
四环素是一类抗生素,可用于治疗由易感微生物引起的感染,例如革兰氏阳性菌和革兰氏阴性菌、衣原体、支原体、原生动物或立克次体。
它们是在 1940 年代发现的,第一个四环素是从链霉菌中获得或衍生的。
四环素抑制微生物 RNA 中的蛋白质合成(一种重要的分子,作为 DNA 的信使)。它们主要是抑菌剂,这意味着它们可以防止细菌繁殖,但不一定会杀死它们。
四环素类药物现今应用的品种除四环素外,更常用有多西环素( 强力霉素) 和米诺环素( 二甲胺四环素) 。由于常见病原菌对本类药物耐药性普遍升高及其不良反应多见,当前本类药物临床应用已受到很大限制。
代表药物:强力霉素(doxycycline)
强力霉素是一种四环素抗生素,可以对抗体内的细菌。
强力霉素用于治疗许多不同的细菌感染,例如痤疮、尿路感染、肠道感染、呼吸道感染、眼部感染、淋病、衣原体、梅毒、牙周炎(牙龈疾病)等。
强力霉素也用于治疗由红斑痤疮引起的瑕疵、肿块和痤疮样病变。
某些形式的强力霉素用于预防疟疾、治疗炭疽或治疗由螨虫、蜱虫或虱子引起的感染。
什么是林可霉素衍生物?
林可霉素衍生物是一小类抗生素,可抑制细菌蛋白质的合成,而细菌蛋白质对细菌的生存至关重要。林可霉素衍生物保留用于治疗由肺炎球菌、葡萄球菌和链球菌的敏感菌株引起的感染。一种衍生物也可用于治疗恶性疟原虫(疟疾)。
林可霉素类包括林可霉素及克林霉素。克林霉素的体外抗菌活性优于林可霉素,主要可应用于敏感肺炎链球菌、其他链球菌属( 肠球菌属除外) 及甲氧西林敏感金葡菌所致的各种感染。该类药物对厌氧菌有良好的抗菌活性常与其他抗菌药物联合用于腹腔感染及盆腔感染,也由于其骨组织浓度较高适用于骨和关节的G + 菌感染。使用本类药物时,应注意假膜性肠炎的发生,如有可疑应及时停药。本类药物有神经肌肉阻滞作用,应避免与其他神经肌肉阻滞剂合用,应注意静脉滴注速度宜缓慢滴注,不可静脉推注。
代表药物:克林霉素(Clindamycin)
克林霉素是一种抗生素,可以对抗体内的细菌。
克林霉素用于治疗由细菌引起的严重感染。
服用此药前 如果对克林霉素或林可霉素过敏,则不应使用该药。
为确保克林霉素的安全,请告诉医生是否曾经有过:结肠炎、克罗恩病或其他肠道疾病; 湿疹或皮肤过敏反应; 肝病; 哮喘或对阿司匹林的严重过敏反应; 对黄色食用色素过敏。
什么是喹诺酮类药物?
喹诺酮类是一种抗生素。抗生素杀死或抑制细菌的生长。
有五种不同的喹诺酮类。此外,另一类抗生素,称为氟喹诺酮类,是从喹诺酮类通过氟修饰其结构衍生而来的。氟喹诺酮类抗生素可导致严重或致残的副作用,这些副作用可能不可逆。
喹诺酮类药物和氟喹诺酮类药物有许多共同点,但也有一些区别,例如它们对哪些生物有效。有些人互换使用喹诺酮和氟喹诺酮这两个词。
喹诺酮类和氟喹诺酮类对细菌产生的两种酶(拓扑异构酶 IV 和 DNA 促旋酶)的功能产生不利影响,因此它们不能再修复 DNA 或帮助其制造。
现今临床主要应用为氟喹诺酮类,常用有诺氟沙星、环丙沙星等主要用于单纯性下尿路感染或肠道感染。但应注意,现今国内尿路感染的主要病原菌大肠埃希菌中,耐药株已达半数以上。环丙沙星尚与β-内酰胺类抗生素联合用于治疗中、重度革兰阴性杆菌感染包括铜绿假单胞菌的感染。
近年来研制的新品种左氧氟沙星、莫西沙星等对肺炎链球菌、化脓性链球菌等G + 球菌的抗菌作用增强,对衣原体属、支原体属、军团菌等细胞内病原或厌氧菌的作用亦有增强,但对G-菌活性增强不明显,对铜绿假单胞菌的抗菌活性仍以环丙沙星为最强。左氧氟沙星、莫西沙星除可用于尿路感染或肠道感染外也可用于呼吸道感染,特别适合于肺炎链球菌( 包括耐青霉素肺炎链球菌) 、支原体、衣原体、军团菌等所致社区获得性肺炎的治疗,此外亦可用于皮肤软组织感染。
在治疗腹腔、胆道感染及盆腔感染时除莫西沙星外需与甲硝唑等抗厌氧菌药物合用。部分品种可与其他药物联合应用作为治疗耐药结核分枝杆菌和其他分枝杆菌感染的二线用药。
代表药物:环丙沙星(Ciprofloxacin)和左氧氟沙星(Levofloxacin)
环丙沙星和左氧氟沙星是一种氟喹诺酮抗生素,用于治疗不同类型的细菌感染。也用于治疗接触过炭疽或某些类型鼠疫的人。环丙沙星和左氧氟沙星应仅用于无法用更安全的抗生素治疗的感染。
左氧氟沙星、莫西沙星:对肺炎链球菌、A 组溶血性链球菌等革兰阳性球菌、衣原体属、支原体属、军团菌等细胞内病原或厌氧菌的作用强。
环丙沙星、左氧氟沙星:主要适用于肺炎克雷伯菌、肠杆菌属、假单胞菌属等革兰阴性杆菌所致的下呼吸道感染。
头孢霉素
头孢菌素是一大类从霉菌顶孢菌(以前称为头孢菌素)中提取的抗生素。头孢菌素具有杀菌作用(杀死细菌),其作用方式与青霉素相似。它们结合并阻断负责制造肽聚糖的酶的活性,肽聚糖是细菌细胞壁的重要组成部分。它们被称为广谱抗生素,因为它们对多种细菌有效。
自 1945 年发现第一个头孢菌素以来,科学家们一直在改进头孢菌素的结构,以使其对更广泛的细菌更有效。每次结构发生变化,都会产生新的“一代”头孢菌素。迄今为止,头孢菌素已有五代。所有头孢菌素均以 cef、ceph 或 kef 开头。请注意,该分类系统在不同国家/地区的使用并不一致。
第一代头孢菌素是指发现的第一组头孢菌素。它们的最佳活性是对抗革兰氏阳性细菌,如葡萄球菌和链球菌。它们对革兰氏阴性菌几乎没有活性。
第二代头孢菌素尚可应用于由流感嗜血杆菌、大肠埃希菌、奇异变形杆菌等中的敏感株所致的尿路感染、皮肤软组织感染、败血症、骨及关节感染和腹腔、盆腔等感染。但如用于腹腔感染和盆腔感染时需与抗厌氧菌药合用。头孢呋辛尚可用于脑膜炎球菌、流感嗜血杆菌所致脑膜炎的治疗,也可作为围术期预防用药。
第三代头孢菌素:对肠杆菌科细菌等革兰阴性杆菌具有强大抗菌作用,头孢他啶和头孢哌酮对铜绿假单胞菌亦具较强抗菌活性;注射品种有头孢噻肟、头孢曲松、头孢他啶、头孢哌酮等,口服品种有头孢克肟和头孢泊肟酯等,口服品种对铜绿假单胞菌均无作用。应注意的是,不是所有的三代头孢菌素都可用于非发酵菌( 如铜绿假单胞菌、不动杆菌等) 感染的治疗,除了头孢哌酮和头孢他啶;外,如头孢曲松和头孢噻肟不可以用于非发酵菌感染的治疗。
第四代头孢菌素:常用者为头孢吡肟,对肠杆菌科细菌作用与第三代头孢菌素大致相仿,对铜绿假单胞菌的作用与头孢他啶相仿,对革兰阳性球菌的作用较第三代头孢菌素略强。其不同于三代头孢菌素之处为对产头孢菌素酶( AmpC) 的肠杆菌属有效,其次是增强了对作用靶位细菌青霉素结合蛋白( PBP) 的亲和力以及其抗革兰阳性菌的抗菌活性亦较三代头孢菌素增强。
第五代(或下一代)头孢菌素头孢洛林( Teflaro ) 对耐甲氧西林金黄色葡萄球菌(MRSA)具有活性。Avycaz 含有β-内酰胺酶抑制剂avibactam。
代表药物:头孢氨苄(Cephalexin)
头孢氨苄是一种头孢菌素(SEF 一种低孢子)抗生素。它通过对抗体内的细菌起作用。
头孢氨苄用于治疗由细菌引起的感染,包括上呼吸道感染、耳部感染、皮肤感染、尿路感染和骨骼感染。
头孢氨苄用于治疗成人和至少 1 岁儿童的感染。
注意事项:
如果对头孢氨苄或类似抗生素(如Ceftin、Cefzil、Omnicef等)过敏,则不应使用该药。如果对任何药物(尤其是青霉素或其他抗生素)过敏,请告诉医生。
对任何药物(尤其是青霉素)过敏;
肝脏或肾脏疾病;要么肠道问题,例如结肠炎,请告诉医生。
头孢氨苄可以进入母乳。如果正在哺乳婴儿,请咨询医生。
β-内酰胺酶抑制剂
β-内酰胺酶抑制剂是一类阻断 β-内酰胺酶(也称为 β-内酰胺酶)活性,防止 β-内酰胺类抗生素降解的药物。本类药物适用于因产β-内酰胺酶的细菌感染,其抗菌谱主要依据原有抗生素的活性而酶抑制剂仅有对抗细菌产酶的作用,一般不增加抗菌活性。
β-内酰胺酶由以下细菌的某些菌株产生:拟杆菌属、肠球菌属、流感嗜血杆菌、卡他莫拉菌、淋病奈瑟菌和葡萄球菌属,无论是组成型还是暴露于抗微生物剂。
β-内酰胺酶裂解易感青霉素和头孢菌素的 β-内酰胺环,使抗生素失活。一些抗微生物药物(如头孢唑啉和氯唑西林)对某些 β-内酰胺酶具有天然耐药性。β-内酰胺类:阿莫西林、氨苄青霉素、哌拉西林和替卡西林,可以通过与β-内酰胺酶抑制剂组合来恢复和扩大其活性。
克拉维酸、舒巴坦和他唑巴坦都是β-内酰胺酶抑制剂。
大环内酯类
大环内酯类药物主要作为青霉素过敏患者的替代药物,用于治疗β-溶血性链球菌、肺炎链球菌中的敏感菌株所致的上、下呼吸道感染、敏感β-溶血性链球菌引起的猩红热及蜂窝织炎等感染。
大环内酯类是从红糖多孢菌(原名红链霉菌)中提取的一类抗生素,一种土壤传播的细菌。
红霉素是第一个发现的大环内酯类;其他大环内酯类包括阿奇霉素、克拉霉素和罗红霉素。
它们的作用主要是抑菌,但在高浓度时可能具有杀菌作用,或取决于微生物的类型。
代表药物:红霉素
红霉素为大环内酯类原型代表药物,由于其胃肠道反应大、口服吸收差使其疗效受到影响。当下在临床应用的主要为大环内酯类新品种罗红霉素、阿奇霉素和克拉霉素等,其他大环内酯类疗效多不如该三种药物。
罗红霉素主要是改善了其药代动力学,口服生物利用度明显提高、给药剂量减小、不良反应明显减少,但其抗菌谱没有明显的改善,是红霉素的替代药物。
氨基糖苷类
临床常用的氨基糖苷类药物主要有: 链霉素、卡那霉素、丁胺卡那霉素、庆大霉素、妥布霉素、奈替米星、依替米星。
其中链霉素当前主要用抗结核治疗,其次可用于治疗鼠疫及布鲁菌病; 庆大霉素、妥布霉素、奈替米星、依替米星肠杆菌科细菌和铜绿假单胞菌等G - 杆菌具强大抗菌活性,对葡萄球菌属亦有良好作用者。所有氨基糖苷类药物对肺炎链球菌、溶血性链球菌的抗菌作用均差。
链霉素、卡那霉素:对肠杆菌科和葡萄球菌属细菌有良好抗菌作用,但对铜绿假单胞菌无作用者。
庆大霉素、妥布霉素、奈替米星、阿米卡星、异帕米星、小诺米星、依替米星:对肠杆菌科细菌和铜绿假单胞菌等革兰阴性杆菌具强大抗菌活性,对葡萄球菌属亦有良好作用。
碳青霉烯类
碳青霉烯类药物是当下抗菌药物中抗菌谱最广的药物,对各种革兰氏阳性球菌、革兰氏阴性杆菌( 包括铜绿假单胞菌的非发酵菌) 和多数厌氧菌具强大抗菌活性,对大多数β-内酰胺酶高度稳定,但对甲氧西林耐药葡萄球菌和嗜麦芽窄食单胞菌等抗菌作用差。
当下在国内应用的碳青霉烯类抗生素有亚胺培南-西司他丁、美罗培南、帕尼培南-倍他米隆、比阿培南。厄他培南与上述品种不同对非发酵菌缺乏抗菌活性,而其他抗菌作用与上述药品基本相同,适合于治疗社区与医院早期特别是G-杆菌感染的重症患者。
碳青霉烯类对各种革兰阳性球菌、革兰阴性杆菌(包括铜绿假单胞菌、不动杆菌属)和多数厌氧菌具强大抗菌活性,对多数β-内酰胺酶高度稳定,但对甲氧西林耐药葡萄球菌和嗜麦芽窄食单胞菌等抗菌作用差。
现用于细菌感染的抗生素被认为是医学史上最重要的突破之一。不幸的是,细菌的适应性很强,抗生素的过度使用让许多人对抗生素产生了耐药性,造成了严重的问题,尤其是在医院的环境里。
抗生素耐药性是细菌抵抗抗生素杀伤力的能力。换句话说,以前治疗感染的抗生素没有效果了,或者说根本无法杀死细菌。
由于过度使用抗生素,抗生素耐药率持续上升,新的抗菌药物开发缓慢。感染耐药细菌可能导致住院时间更长、费用更高,并增加死于感染的风险。
抗生素耐药性是对全球健康的紧迫威胁,包括后来逐渐出现危险耐药细菌——“超级细菌”。
疾病控制和预防中心 (CDC)发布了对美国的前 18 种耐药性威胁的清单,至少包括:
注:以上这些并未全面列出所有耐药菌。抗生素耐药性模式不断演变,细菌可能并不总是对每位患者的选择抗生素表现出耐药性。在正常情况下,抗生素的选择应基于感染部位和由医疗保健专业人员评估的临床表现、培养/敏感性和其他所需的实验室结果、局部耐药性/敏感性模式以及患者的特定特征。
细菌通过以某种方式调整其结构或功能作为防御机制,从而对抗生素产生抗药性。
当细菌以某种方式发生变化时,细菌会抵抗药物。这种变化可以保护细菌免受药物的影响或限制药物接触细菌。或者这种变化可能导致细菌改变药物或破坏药物。
细菌可以用以下方式与抗生素抗争:
如果说以上列举的是抗生素产生耐药性的内因,那么除细菌本身外的因素也会导致抗生素耐药性的出现。
早前,医生有时会依赖不可靠或不准确的知识,开具抗生素“以防万一”,或在特定窄谱抗生素可能更合适时却开出广谱抗生素。这些情况加剧了选择压力并加速了抗菌素耐药性。
当医生不清楚是细菌或病毒加剧了感染时,他们可能会开抗生素。抗生素对病毒感染不起作用,可能会产生耐药性。
抗生素自我治疗(SMA)与药物使用不当的可能性有关,这会使患者面临药物不良反应的风险,掩盖潜在疾病的迹象,并在微生物中产生耐药性。
过度使用抗生素,尤其是在不正确治疗的情况下服用抗生素,会促进抗生素耐药性。根据疾病控制和预防中心的数据,人类使用大约三分之一的抗生素既不需要也不合适。
抗生素可以治疗由细菌引起的感染,但不能治疗由病毒引起的感染(病毒感染)。例如,抗生素是可以治疗由细菌引起的链球菌性咽喉炎。但对于大多数由病毒引起的喉咙痛,并不是正确的治疗方法。
每天都有成千上万的患者、工作人员和访客到达医院,每个人的衣服上和身体上/体内都有自己的一套微生物组。如果医院没有适当的程序和规程来帮助保持空间清洁,细菌就会传播。
抗生素被用作动物的生长补充剂和生长促进剂。在家畜中发现的耐抗生素细菌可能对人类致病,很容易通过食物链传播给人类,并通过动物粪便在生态系统中广泛传播。在人类中,这可能会导致复杂的、无法治疗的和长期的感染。
由于技术挑战、缺乏知识、对抗细菌生理学的重大困难,制药业对新抗生素的发明在很大程度上放缓了,当新抗生素普及时,耐药性的发展(在相对较短的时间内)几乎是不可避免的。
世界范围内抗生素的过度使用和滥用正在导致抗生素耐药性的全球医疗问题。可能会发生抗生素耐药性感染,在最坏的情况下,可能会没有有效的抗生素。这种情况在严重感染时可能会危及生命。
大多数病毒性疾病不需要特殊药物并且是“自限性”的,也就是说患者自身的免疫系统可以抵抗疾病。病毒性疾病的患者可以休息、多喝水并使用对症治疗。
由病毒引起的疾病包括:
有时,在复杂或长期的病毒感染中,细菌也可能侵入,并导致所谓的“继发感染”。在这些情况下,如果需要,可以使用抗生素。
在感染由病毒引起的疾病中,例如咳嗽、感冒或流感,患者不应要求医生开抗生素。抗生素不能治愈病毒感染,患者可能会因不必要的药物而产生副作用。医生可以用其他方法来帮助患有病毒性疾病的患者进行治疗。
如果感染没有得到正确治疗,抗生素耐药细菌也可能传播给其他人。
——有以下几个原因:
它可能不是治疗感染的正确抗生素
它可能已过时且无效
整个疗程可能没有足够的药物
如果新疾病是病毒感染,则不需要抗生素
一些疫苗可以预防细菌性疾病。抗病毒疫苗,例如流感疫苗或COVID-19 疫苗,可以帮助预防可能与继发性细菌感染(如严重的肺部肺炎)相关的原发性疾病。
在治疗的最初几天感觉好些了的时候,仍应完成整个抗生素疗程。
抗生素使用注意事项
✓ 按照指示使用抗生素
按照医生的指示治疗感染。医生会选择最能对抗导致感染的特定细菌的抗生素类型。目前市面上有许多不同种类的抗生素,大部分需要医生开出的非处方抗生素。
✓ 了解抗生素
有的抗生素需要进行皮试,比如青霉素,以防过敏,不同的抗生素杀菌的作用原理不尽相同,比如青霉素是通过能破坏细菌的细胞壁并在细菌细胞的繁殖期起杀菌作用的一类窄普抗生素,杀菌作用强,可用于敏感菌所致的严重感染,由于该类药物作用的靶位为细菌细胞壁,而人体的细胞没有细胞壁,因此毒性低,用于老年人,新生儿和孕妇时安全性相对较高。
✓ 确保清楚抗生素的使用方式
确保明确知道应该服用多少抗生素,以及何时服用。有些抗生素需要与食物一起服用,有些需要在晚上服用等。特别注意要用完整的病征疗程抗生素,有些人担心抗生素对身体有副作用,病情稍微好转就会停止服用或注射抗生素,但是这可能导致体内的致病细菌未完全清楚,再次发生感染,或引起形成抗生素抗药性,使得以后的感染治疗变得困难。
✓ 清洁伤口以防止细菌感染
通过立即正确清洁和包扎伤口来预防皮肤感染。适当的急救治疗对于帮助预防细菌感染至关重要,但不应尝试自己治疗严重的皮肉伤口。如果伤口很深、很宽或出血很多,应该立即寻求医疗帮助。
治疗伤口前先洗手。如果你用脏手治疗伤口,会增加细菌感染的机会。用温水和抗菌肥皂洗手 20 秒,然后擦干。如果有条件可以戴上手套。
细菌感染性疾病起病急、进展快,重症疑难患者病原细菌复杂,诊断及治疗困难,可危及生命。早期、准确地明确病原微生物对感染判定至关重要,有助于改善预后。
准确诊断细菌感染对于避免不必要的抗生素使用和集中适当的治疗至关重要。
细菌感染是细菌的存在与炎症或全身功能障碍的结合;因此,通常需要不止一种诊断方式进行确认,确定患者是否符合临床病例定义的病史和检查。
不同的病原微生物检测技术对感染的判定价值不同,临床上也会根据患者的病情,可能感染的部位,可供采集的标本等选择适宜的检测技术。
当前,病原微生物检测技术层出不穷,能识别的微生物种类越来越多,但如何正确利用和分析微生物检测结果仍是临床关注的难题。值得注意的是,感染首先是个临床诊断,所有的判定必须基于临床。
定植容易对感染判定产生混淆。正常生理状态下人体口腔、胃肠道、呼吸道及体表等部位均有细菌、真菌存在,患者无感染症状时称为定植菌。当定植菌的致病力改变或机体防御能力下降时,可大量繁殖或被带入机体深部引起感染成为致病菌。
临床医生应如何判断所获取的病原体为定植还是感染,是长久以来的难题,且未形成成熟统一的解决方案。
呼吸道定植菌的综合分析:
就呼吸道定植菌而言,既往认为可根据菌落数量或菌种拷贝数来判定定植与感染,但结果存在偏差。越来越多的学者认为,临床上判定是定植还是感染,需结合患者症状、体征及其他检查结果综合分析。
若患者存在与检测阳性结果相匹配的临床症状和体征,如发热、咳嗽、咳痰等,感染指标升高,肺部出现新发病灶,应考虑感染并给予相应的抗感染治疗。如仅培养阳性,患者无任何感染相关临床表现,则倾向于定植菌。
念珠菌定植的综合分析:
念珠菌可广泛定植于呼吸道、胃肠道及泌尿生殖道,正常屏障破坏、免疫功能受损或局部菌群失调时,定植的念珠菌可生长繁殖引起感染。痰和尿标本中分离出的念珠菌亦应根据临床症状和体征鉴别是定植还是感染,念珠菌血培养阳性时要高度警惕念珠菌血症。
此外,病原微生物检测结果阳性,并不意味着患者一定存在感染或感染一定由检出的病原体所致,需结合标本质量、采集部位、病史及其他检测结果综合分析。
不同部位标本检测出同一种病原体,其代表的临床意义可能并不相同。
以肺炎克雷伯氏菌为例,从肠道粪便样本中检出肺炎克雷伯氏菌,其中丰度占比超过1%的人群有3765例,占比28.2%【谷禾健康数据库】。但是当肺炎克雷伯菌进入血液并感染时,通常会发生细菌血症。症状可能包括发冷,发烧,发抖,肌肉酸痛,疲劳和嗜睡。当肺炎克雷伯菌感染肝脏时,会引起脓性病变,也被称为化脓性肝脓肿。这种类型的感染更常见于患有糖尿病一段时间或已经服用抗生素多年的人。
因病原微生物种类不同,微生物检测方法的选择也不相同;即便检测同一种微生物,因采集部位不同所选择的检测方法也有所差别。
如大肠埃希菌,血培养阳性时需考虑血流感染,怀疑致病性大肠埃希菌感染肠道时需特殊培养或者血清学、PCR,16s测序检测等。因普通大肠埃希菌可寄居在肠道中,粪便普通培养有大肠埃希菌生长并不能直接考虑大肠埃希菌肠道感染。
病原微生物检测技术快速发展,能识别的微生物种类越来越多,但感染首先是个临床诊断,所有的判定必须基于临床。随着检测技术革新带来的是临床辅助手段的进步和多样化,更快、更准确地判定感染一直都是病原学检测的挑战,不同方法各有优劣。当前,并没有一项技术可通过从标本中识别微生物来直接判定机体是否感染该病原体,甚至依靠检测的阴性结果来排除感染都很难实现。
在面对细菌感染诊疗过程中始终需要思考3个问题:患者是否存在感染?如果有感染,致病病原体是什么?如何治疗干预?
充分利用现有的病原微生物检测方法和其他实验室检查手段,综合判定,给出合适的抗菌治疗或其他综合治疗方法,是科学可持续的手段。
主要参考文献:
Uddin TM, Chakraborty AJ, Khusro A, Zidan BRM, Mitra S, Emran TB, Dhama K, Ripon MKH, Gajdács M, Sahibzada MUK, Hossain MJ, Koirala N. Antibiotic resistance in microbes: History, mechanisms, therapeutic strategies and future prospects. J Infect Public Health. 2021 Dec;14(12):1750-1766. doi: 10.1016/j.jiph.2021.10.020. Epub 2021 Oct 23. PMID: 34756812.
Cheung GYC, Bae JS, Otto M. Pathogenicity and virulence of Staphylococcus aureus. Virulence. 2021 Dec;12(1):547-569. doi: 10.1080/21505594.2021.1878688. PMID: 33522395; PMCID: PMC7872022.
Tshibangu-Kabamba E, Yamaoka Y. Helicobacter pylori infection and antibiotic resistance – from biology to clinical implications. Nat Rev Gastroenterol Hepatol. 2021 Sep;18(9):613-629. doi: 10.1038/s41575-021-00449-x. Epub 2021 May 17. PMID: 34002081.
BRM, Mitra S, Emran TB, Dhama K, Ripon MKH, Gajdács M, Sahibzada MUK, Hossain MJ, Koirala N. Antibiotic resistance in microbes: History, mechanisms, therapeutic strategies and future prospects. J Infect Public Health. 2021 Dec;14(12):1750-1766. doi: 10.1016/j.jiph.2021.10.020. Epub 2021 Oct 23. PMID: 34756812.
Carmen Fookes, BPharm, Bacterial Infection. January 13, 2020 Medicine
Spagnolo F, Trujillo M, Dennehy JJ. Why Do Antibiotics Exist? mBio. 2021 Dec 21;12(6):e0196621. doi: 10.1128/mBio.01966-21. Epub 2021 Dec 7. PMID: 34872345; PMCID: PMC8649755
Baquero F, Coque TM, Cantón R. Counteracting antibiotic resistance: breaking barriers among antibacterial strategies. Expert Opin Ther Targets. 2014 Aug;18(8):851-61. doi: 10.1517/14728222.2014.925881. Epub 2014 May 31. PMID: 24881465.
武洁, 王荃. 病原微生物检测在感染判定的意义[J]. 中国小儿急救医学, 2020, 27(3):6.
李丹鹤, 荣爱国, 马瑞芝,等. 病原微生物检验在抗感染经验治疗中的临床意义[J]. 医学理论与实践, 2019, 32(09):127-129.

谷禾健康
说起抗生素,可能会觉得有些抽象,那如果说阿莫西林、头孢、红霉素、阿奇霉素等等这些日常生活经常使用的药物,会不会有一些熟悉的感觉。当我们发现身体有一些小炎症(如头痛发热、喉咙痒疼、咳嗽)的时候,首先会想到要使用这类药物(抗生素) 进行消炎。
在现实生活中,抗生素被许多人当作是包治百病的妙药,一遇到头痛发热或喉痒咳嗽,首先想到的就是使用抗生素,而对滥用抗生素产生耐药性的危害却知之甚少。
世界卫生组织在1996年时就向全世界发出了警告,因为人们对抗生素的滥用,产生了一些严重的后果。其中之一就是导致了细菌的耐药性增强,其结果直接提高了人类对感染类疾病的治疗的难度,增加治疗所需的成本,甚至患病人群的死亡率剧增。
在人类和其他哺乳动物中,所有暴露于环境的表面都是无数古细菌、细菌、病毒和真核微生物的家园,这些定殖微生物被称为微生物群。
微生物群的定植为我们抵御病原微生物感染提供了最有效的屏障之一。微生物群通过启动免疫防御、通过代谢将病原体排除在其首选的生态位中以及通过直接的抗微生物拮抗作用来防止感染。微生物群的破坏,尤其是抗生素的破坏,是细菌病原体定植的主要风险因素。
研究表明,常见的革兰氏阴性菌感染包括由克雷伯氏菌、铜绿假单胞菌和大肠杆菌以及革兰阳性菌感染如艰难梭菌感染,最初的定植与治疗复发均与抗生素的使用和被破坏的肠道菌群有关。
例如,肺炎克雷伯菌是属于肠杆菌科的革兰氏阴性变形菌。它是包括人类在内的几种哺乳动物的胃肠道和上呼吸道共生的,估计人类携带率约为 3-8%。在住院患者中,胃肠道定植的发生率可能会显着升高(高达 38%)。在一部分宿主中,肺炎克雷伯菌会引起呼吸道、泌尿道、伤口和血流感染 。
胃肠道定植是肺炎克雷伯菌的关键步骤发病机制。住院时间和抗生素的使用会增加肺炎克雷伯菌的定植率。住院时间可能会增加患者通过医护人员和污染物接触细菌的可能性。抗生素的使用导致微生物群失调,为肺炎克雷伯菌提供了有利条件。抗生素耐药性的发展也放大了肺炎克雷伯菌的感染几率。
此外,革兰氏阳性菌感染中艰难梭菌感染可能是最广为人知且临床上最常见的感染。艰难梭菌感染的主要危险因素之一是之前的抗生素治疗 。感染的初始治疗通常依赖抗生素甲硝唑和万古霉素;然而,高达 35% 的患者会出现感染复发,并且与免疫抑制和进一步的抗生素治疗有关 。 此外,临床研究认为氟喹诺酮类、克林霉素和头孢菌素也是使患者对艰难梭菌易感的主要抗生素。
本文主要探讨抗生素究竟对人体肠道菌群有哪些影响,以及对人体健康的主要影响。
抗生素
抗生素是指由微生物(包括细菌、真菌、放线菌属) 或高等动植物在生活过程中所产生的具有抗病原体或其他活性的一类次级代谢产物,能干扰其他生活细胞发育功能的化学物质。简单来说就是在非常低浓度下对所有的生命物质有抑制和杀灭作用的药物。临床常用的抗生素有微生物培养液中的提取物以及用化学方法合成或半合成的化合物。
自人们发现抗生素的使用大大的降低了传染病导致的死亡率后,抗生素就开始被广泛的应用于疾病的治疗中,并在全球范围内彻底改变了传染病的治疗方法。在20世纪更是一度被称为 “预期寿命延长” 的一大促成因素。
抗生素的本质作用其实就是把感染我们的微生物消灭,清除掉病原体,控制住疾病的发展,最终得以将病症治疗康复。但因为人类、兽医学以及畜牧业中过度使用、误用等情况的发生,导致了当前出现了抗生素耐药性的危机。
抗生素的种类很多,目前国内在医学和兽医日常应用的抗生素不少于几十种。不同的抗生素对病菌的作用原理不尽相同:
1. 有的抗生素是干扰细菌的细胞壁的合成,使细菌因缺乏完整的细胞壁,抵挡不了水份的侵入,发生膨胀、破裂而死亡。
2. 有的抗生素是使细菌的细胞膜发生损伤,细菌因内部物质流失而死亡。
3. 有的抗生素能阻碍细菌的蛋白质合成,使细菌的繁殖终止。
4. 有的抗生素是通过改变细菌内部的代谢,影响它的脱氧核糖核酸的合成,使细菌(还有肿瘤细胞)不能重新复制新的细胞物质而死亡。
在日常生活以及医疗当中所指的抗生素主要是针对细菌、病毒微生物的药物,种类是相当多的,大概可以分成十余种大类。在临床上常见的更是有百余种之多。
在此简单的对其抗菌谱、化学结构、用途等方面进行分类。
◾ 按抗菌的范围(抗菌谱),分为:
◆ 广谱抗生素
指的是抗菌谱比较宽的药物,简单说就是能够抵抗大部分细菌的药物;
如. 氯霉素、金霉素、土霉素、四环素、甲砜霉素等
◆ 窄谱抗生素
是专门杀灭某一种或一类细菌的药物。
如. 青霉素G
◾ 按照化学结构,分为:
◆ 喹诺酮类抗生素
◆ β-内酰胺类抗生素
◆ 大环内酯类抗生素
◆ 氨基糖苷类抗生素
……
◾ 按照用途,分为:
◆ 抗细菌抗生素
◆ 抗真菌抗生素
◆ 抗肿瘤抗生素
◆ 抗病毒抗生素
◆ 畜用抗生素
◆ 农用抗生素
◆ 其他微生物药物(如麦角菌产生的具有药理活性的麦角碱类,有收缩子宫的作用) 等。
列举常用的各类抗生素及其作用、对应的病症等:
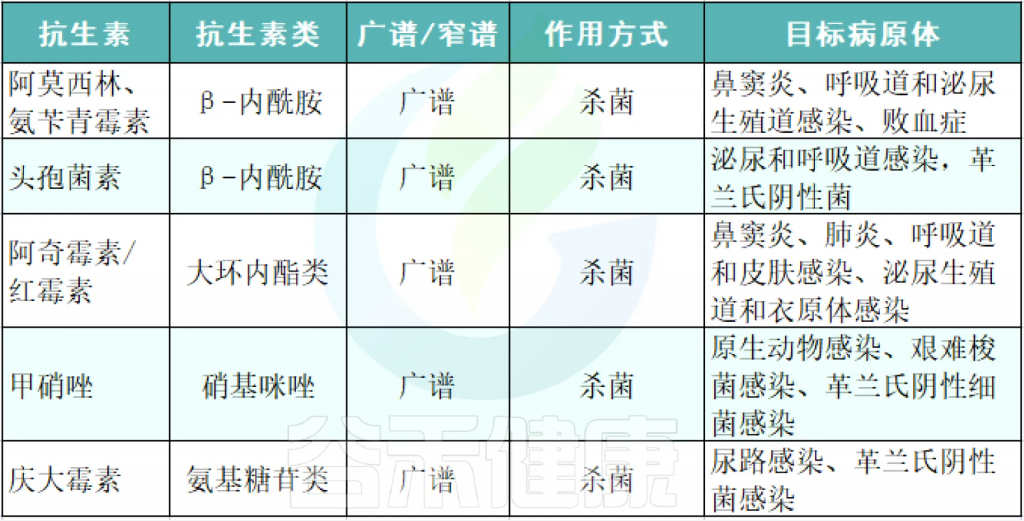

Patangia DV, et al., Microbiologyopen. 2022
肠道微生物群的组成
肠道微生物群对于宿主的健康起着重要的作用 —— 可以防止病原体的定植,调节肠道免疫,提供必需的营养物质、生物活性代谢物、参与能量稳态等。
肠道微生物群是由细菌、古细菌、微真核生物和病毒组成,且根据迄今为止的多项研究可以得出,平衡且稳定的微生物群组成和丰富的菌群多样性对于机体功能等至关重要。健康成人的肠道微生物群主要由厚壁菌门和拟杆菌门组成,其次是放线菌门、变形菌门和疣微菌门。人们普遍认为,肠道微生物群是在婴儿出生时就获得的,其母体是婴儿肠道微生物群最重要的来源,此后在肠道免疫力的发展中发挥着重要作用。婴儿肠道中菌群的多样性减少、微生物群组出现不平衡与肠道疾病和晚年易患的某些疾病有关。而健康婴儿肠道微生物群的建立以及其后续的持续发展是一个长久的过程,并受诸多因素的影响发生变化。
微生物群的变化随多种因素的影响而改变。其中对于婴儿来说,大部分的肠道菌群受其母体的影响最为严重,主要是与分娩方式、胎龄以及产后喂养习惯等方面有关;对于抗生素的使用同样是影响肠道菌群的一大常见因素。
◾ 分娩方式是影响婴儿肠道菌群的首要因素之一。通过对比发现,阴道分娩婴儿的微生物群明显更加的多样化,与其母亲的阴道微生物群相似度更高;而剖宫产出生的婴儿则因为被缺乏了部分相应的生产过程以及母体的接触,因此具有肠道微生物群仅与其母亲的皮肤和医院环境所类似。其中,
在剖宫产出生的婴儿中 ——
丰度增加的包含:
丙酸杆菌(Propionibacterium)
棒状杆菌(Corynebacterium)
葡萄球菌(Staphylococcus)
艰难梭菌(C.difficile)
链球菌(Streptococcus)
丰度降低的包含:
双歧杆菌(Bifidobacteria)
拟杆菌(Bacteroides)
在阴道分娩的婴儿中发现了 ——
乳酸杆菌(Lactobacillus)
普雷沃氏菌(Prevotella)
斯奈斯菌属(Sneathia spp.)
◾ 胎龄也是改变肠道菌群的因素。早产儿明显比足月新生儿的肠道菌群多样低,其中 ——
变形菌(Proteobacteria)的丰度较高;
专性厌氧菌(如. 双歧杆Bifidobacterium)、拟杆菌(Bacteroides)、Atopobium的水平较低。
◾ 喂养习惯是影响婴儿肠道菌群的另一个关键因素。由于母乳中存在大量的低聚糖(母乳低聚糖) 用来产生双歧杆菌(Bifidobacteria)。与配方奶喂养的婴儿相比:
母乳喂养的婴儿 ——
双歧杆菌的水平更高,且在断奶后的比例仍然很高;
检测出的菌群包含:
拟杆菌(Bacteroides)
链球菌(Streptococcus)
乳酸杆菌(Lactobacillus)
……
配方奶喂养的婴儿 ——
丰度较高的菌包含:
大肠杆菌(Escherichia coli)
艰难梭菌(C. difficile)
脆弱拟杆菌群(Bacteroides fragilis group)
乳酸杆菌(Lactobacilli)
以下,为各种抗生素对人体肠道微生物群影响:

Patangia DV, et al., Microbiologyopen. 2022
怀孕期间的阴道微生物群发生了特定的变化。有研究报道,与未怀孕女性的阴道微生物群相比,怀孕期间的阴道微生物群的变化重点表现在高细菌负荷、高乳酸菌丰度、低丰富度和菌群多样性的降低等。在整个怀孕期,表示机体健康的特有特征包括了细菌的负荷增加以及肠道微生物群组成发生的明显变化。在妊娠的前三个月至妊娠晚期,随着生理的变化,会出现诸如变形菌和放线菌总含量的明显增加以及其丰富度下降的现象。
抗生素所引起微生物组成的变化会对宿主的健康产生直接或间接的负面影响。
其中包括 ——
微生物多样性下降
微生物群功能属性发生变化
抗生素抗性菌株的形成和选择
这些变化会使宿主更容易感染艰难梭菌等病原体。
以下,是过度使用和滥用抗生素可能对宿主健康产生负面影响:

Patangia DV, et al., Microbiologyopen. 2022
◾ 母体&婴儿常使用的抗生素及原因 ——
◆ 怀孕期间 最常用的一些抗生素包含:
β-内酰胺类抗生素
磺胺类
甲氧苄啶
大环内酯类
林可酰胺
链霉素
其他规定的抗生素类
◆ 分娩期间 经常给母体服用抗生素,目的:
防止 B 群链球菌的传播
减少并预防子宫内膜感染的风险
以及预防伤口感染的产生
◆ 婴儿 最常用的抗生素包括:
阿莫西林
苄青霉素
头孢菌素
庆大霉素
万古霉素
克林霉素
阿奇霉素
……
这些抗生素适用于呼吸道和耳部的感染、支气管炎、咽炎和高温。
对于孕妇和幼儿来说要尤其注意,上肠道微生物群可能会因受到短期或低剂量抗生素的干扰,而对健康造成了长期深远的严重影响。希望人们可以警戒,不过度滥用抗生素。
◾ 广谱抗生素对肠道微生物群有害影响 ——
广谱抗生素会降低肠道微生物群的多样性,以杀死相关病原体,并根除有益微生物,其结果对宿主是有害的。
在西方国家,高达 35% 的女性在怀孕和分娩期间都会接触抗生素,并经常使用预防性抗生素来预防和治疗感染。女性在怀孕期间抗生素的使用就占了接触药物总量的 80%左右。
◾免疫性疾病
接触过产时抗生素预防(IAP) 的婴儿肠道菌群的多样性会发生改变。如果在儿童早期或发育阶段所接触到的微生物群种类较少或不足,则可能导致免疫相关疾病的产生。为此,世界卫生组织曾提出建议表示尽量不要在分娩后预防性的使用抗生素。
◆ 对于新生儿尤其是早产儿来说
由于其健康状况以及感染的高度敏感性和免疫力较低,导致经常性的要接受抗生素的相关治疗。
经常接触抗生素,会导致 ——
微生物群发生改变
增加患感染性疾病的可能性
常见如. 哮喘和过敏性致敏、NEC 和侵袭性真菌感染。
若延长使用抗生素进行治疗,则可能会 ——
增加发生迟发性败血症(主要由 B 组链球菌引起)
坏死性小肠结肠炎(NEC)
机体性死亡率增加的风险
◆ 在婴儿期使用抗生素,也会产生类似的影响。在哺乳期间,母体所使用抗生素会影响乳汁微生物群的变化,这又将会影响到婴儿肠道菌群的组成。抗生素等一些外在因素会对母体微生物群的多样性进行改变,从而直接或间接地影响到婴儿肠道微生物群的多样性、免疫力的增减以及疾病的发生。过早期的使用抗生素增加了儿童期患哮喘过敏、特应性皮炎和 IBD 等疾病的风险。
◆ 围产期(怀孕28 周到产后一周) 和围产期抗生素的使用会影响肠道菌群的定植以及婴儿的耐药性。
为了了解妊娠期间抗生素给药对后代的潜在影响,科学家们在白细胞介素 10(IL-10) 缺乏的结肠炎小鼠模型 (Miyoshi) 中研究了,在围产期给药头孢哌酮对母体及其后代微生物群随着给药时间的改变所带来的影响。暴露于头孢哌酮的后代在成年后显现出发生改变的肠道微生物群落,并且对自发性和化学性诱发的结肠炎的易感性增加。且用抗生素改变的微生物群落接种无菌怀孕小鼠后,观察到,改变的微生物群落被传播给缺乏 IL-10 的后代,此变化会导致结肠炎的显着增加。另一项研究表明,怀孕期间服用抗生素会导致出生前阴道微生物的组成发生变化;这可能会影响婴儿出生时所会接触的微生物组成。据报道,孕期母体抗生素摄入与微生物组成的改变有关,具体的改变趋势取决于抗生素类型。
对于抗生素会增加婴儿患哮喘和过敏的风险度、免疫学的改变和后代糖尿病的发生几率以及发育和认知功能的障碍等的相应结论,现阶段还存在争议和不确定性。
◾ 免疫发育
婴儿期,免疫系统就进行了对抗病原体的练习,而此时正是微生物定植的重要时期。这种共同生长发育的过程体系导致了对微生物定植的任何干扰都会影响免疫系统的成熟稳定。
通过对无菌小鼠的研究证实,如果肠道中没有微生物群则会影响肠道环境的生理结构和免疫特性,这些变化主要包括了粘液厚度和成分的改变、胃动力的降低、肠细胞和免疫细胞的非正常发育和功能缺失以及免疫系统发育不当。实验显示,抗生素治疗会减少结肠粘液层的厚度,从而增加小鼠(8-10 周龄) 病原体入侵和肠道炎症的风险。
另一项针对小鼠的研究报告称,抗生素诱导的微生物群改变将 TH1/TH2 平衡转变为 TH2 的主导免疫——伴随着淋巴细胞数量的减少,将导致特应性的发生。
在新生大鼠的相关治疗中观察到,微生物组成和基因成熟谱的改变,例如编码MHC 1b类和II类蛋白质和潘氏细胞产物(如防御素) 的基因下调,这可能会影响粘膜屏障的发育。一项研究表明,肠道菌群产生的某些分子与免疫系统的成熟发育有关。相关研究还发现了,用产生细菌多糖的脆弱拟杆菌来定殖无菌小鼠,可以纠正其T细胞的缺陷、改善 T(H)1/T(H)2 失衡同时促进淋巴器官的发育;然而在不产生多糖的突变体脆弱拟杆菌中并没有观察到该现象。
肠上皮细胞分泌抗菌肽受微环境中微生物群的调节。用常规或人类微生物群或特定益生菌物种或 LPS 定植的无菌小鼠表现出增加的抗微生物肽(如 REGIII-γ) 的产生,增强了先天免疫反应。一项针对小鼠的研究表明,产前抗生素不仅改变了新生小鼠的微生物群定植模式,而且还对影响其免疫反应的病毒感染的 CD8+ T 淋巴细胞的活性产生了负面影响。此外,还观察到,在更加严格的卫生环境下的新生小鼠更容易受到感染。另一种细菌多样性低的小鼠模型在抗生素治疗后有类似的结果产生,同时发现了 CD4+ T 淋巴细胞产生的细胞因子减少以及干扰素-γ 的产生减少。在一项涉及流感患者的研究中发现,感染滴度低并接受抗生素治疗的受试者的免疫反应较低,并会伴有微生物群的丢失以及免疫球蛋白(Ig) G1、IgA 和抗感染的次级胆汁酸水平的降低。这些研究证明了微生物群与宿主免疫反应之间的复杂关系,以及抗生素对其相互作用的影响,部分影响需要进一步相关研究证明。可能还会影响到使用抗生素疫苗治疗的有效性。
◾ 先天性异常
妊娠期间抗生素使用与新生儿和先天性异常之间存在关联。研究报告称,在怀孕期间接受大环内酯类药物治疗的母亲所生的婴儿患脑性麻痹、癫痫、心脏和生殖器畸形的风险增加,在孕早期服用这些药物的有害影响更大。
在少数病例中发现,在孕早期使用阿莫西林与婴儿的唇裂和腭裂的形成有关联,其有利证实了怀孕期间使用抗生素的不利影响。
妊娠前三个月使用磺胺类药物和呋喃妥因与小眼症、左心发育不全综合征、房间隔缺损和唇裂伴腭裂等出生缺陷也有关连;孕期使用甲氧苄啶-磺胺类药物与心血管畸形的高风险相关;可能是由于在子宫内器官发生和早期发育阶段对新生儿产生了影响。
虽然相关的病例还不能总结出确切的关系,但是对于妊娠期间抗生素的使用应该重新进行评估。
◾ 肥 胖
由于肠道微生物群在饮食代谢中的作用,肥胖与早期菌群的定植有着广泛的关联。早期使用抗生素对体重指数的增加、超重以及引起中心性肥胖症有潜在的关联。对此现象在男性中的影响大于女性,所以此类影响可能是具有性别特异性的。
在一项针对 96 人的研究中(其中抗生素组和对照组各 48 人),研究人员发现使用万古霉素和庆大霉素治疗的患者在感染性心内膜炎发作后的体重显着且持续增加。
在对动物的进一步研究后得出,如果早期使用低剂量或亚治疗浓度的抗生素会扰乱微生物组成,影响参与免疫和碳水化合物代谢基因的表达,并可能会破坏代谢的稳态,使宿主在后期更容易出现肥胖的症状。
婴儿期以及儿童期使用抗生素可能会与微生物组成以及代谢功能的改变、晚年的肥胖有关。
在功能水平上,无论是静态的还是灭活的抗生素,不同的类型都会对肠道微生物群产生不同的影响。抗生素可以影响原微生物群并导致耐药菌株的生长,这些菌株在使用治疗后可以持续保留数年。
◾对个体微生物影响
◆ 某种抑菌药物导致了革兰氏阴性菌增多,其影响了 ——
脂多糖(LPS)合成的基因数量的增加
杀菌药物与革兰氏阳性菌的增加有关
其现象还伴有内生孢子形成的基因过度表达
◆ 母体使用了产时抗生素是会影响到婴儿口腔微生物群,其中母体使用了抗生素治疗的婴儿中 —— 放线菌门、拟杆菌门、变形杆菌门更为丰富。氨苄青霉素在母体用于产时预防性药物对抗 B 组链球菌,其结果同样降低了婴儿双歧杆菌的水平含量。
◆ 产前暴露于抗生素的婴儿中拟杆菌和双歧杆菌有所降低,并且发现改变的微生物群类似于同期新生儿重症监护室中的耐药菌。许多相关的影响与产后服用抗生素观察到的结果相似。
◆ 接触 IAP 的婴儿特别是新生儿出生后的最初几周内 ——
放线菌、拟杆菌的比例较低
口腔变形杆菌属水平较高
双歧杆菌的水平较低
◆ 出生后3个月,使用抗生素的婴儿 ——
含量较低的菌属包含:
拟杆菌属
副杆菌属
高等肠球菌属
梭菌属
丰度较高的菌属包含:
而肠杆菌科
◆ 在早产儿中观察到——
双歧杆菌丰度下降
拟杆菌丰度下降
肠球菌丰度增加
早产儿短期和长期接触抗生素都会对其肠道菌群有所改变,包括细菌种群的多样性减少、肠道中物种丰富度降低、抗生素抗性基因增多等方面。
◾相关疾病的微生物
由于微生物群在宿主代谢和生理学中所起到的作用,大量研究提出了“缺失的微生物”(即.假设微生物失衡) 可能会与肥胖、糖尿病和哮喘等疾病具有一定的关联性。
通过微生物群的组成和饮食习惯等可控因素来控制来调控多因素条件,从而尽可能的防止疾病发生。
◆ 研究抗生素使用与肥胖之间的关联发现——
厚壁菌门与拟杆菌门的比例增加(不确定性)
短链脂肪酸(SCFA) 含量更高
◆ 重复性的使用广谱抗生素与糖尿病之间存在着联系。在易患糖尿病的动物中,部分原因是特异性抗原负荷的降低或耐受性 APC 的生长。微生物组成的变化与Ⅱ型糖尿病之间的关联更为明显,Ⅱ型糖尿病患者中产生丁酸盐的细菌水平有所下降。在研究了 121 名糖耐量正常、糖尿病前期以及新诊断为糖尿病的受试者得出,在糖尿病前期阶段肠道微生物组成受到调节,这可以作为糖尿病的一个状态发展标志。
抗生素所引起的微生物的定植变化与Ⅰ型糖尿病之间同样存在着关联性。对自身免疫性Ⅰ型糖尿病的新生非肥胖糖尿病小鼠,给予单独的广谱抗生素或与万古霉素的组合使用会发现——
大肠杆菌的种类增加
乳酸杆菌的种类增加
梭菌目减少
其微生物群发生了显着的变化
回肠和结肠固有层中IL-17 细胞产生大量减少
这会影响到宿主的防御机制。此外,葡萄糖稳态和胰岛素抵抗相关的代谢变化,也与糖尿病的患病有关。
◆ 抗生素导致抗生素相关性腹泻(AAD) ,克林霉素可导致微生物群发生改变,从而促进艰难梭菌等潜在病原体的定植,进而导致腹泻和结肠炎的发生。抗生素治疗导致 ——
α和β多样性下降
导致水平下降的包含:
血清素
色氨酸水解酶
次级胆汁酸
进一步影响肠道的蠕动和代谢问题。NEC 和 AAD 还与生命早期延长或预防性抗生素摄取有关。
◆ 许多抗生素通常用于牙科手术。这些抗生素可以增加口服耐药菌株的数量,还可以增加最低抑菌浓度,以及消除非致病菌株——这些菌株会导致全身感染和炎症。
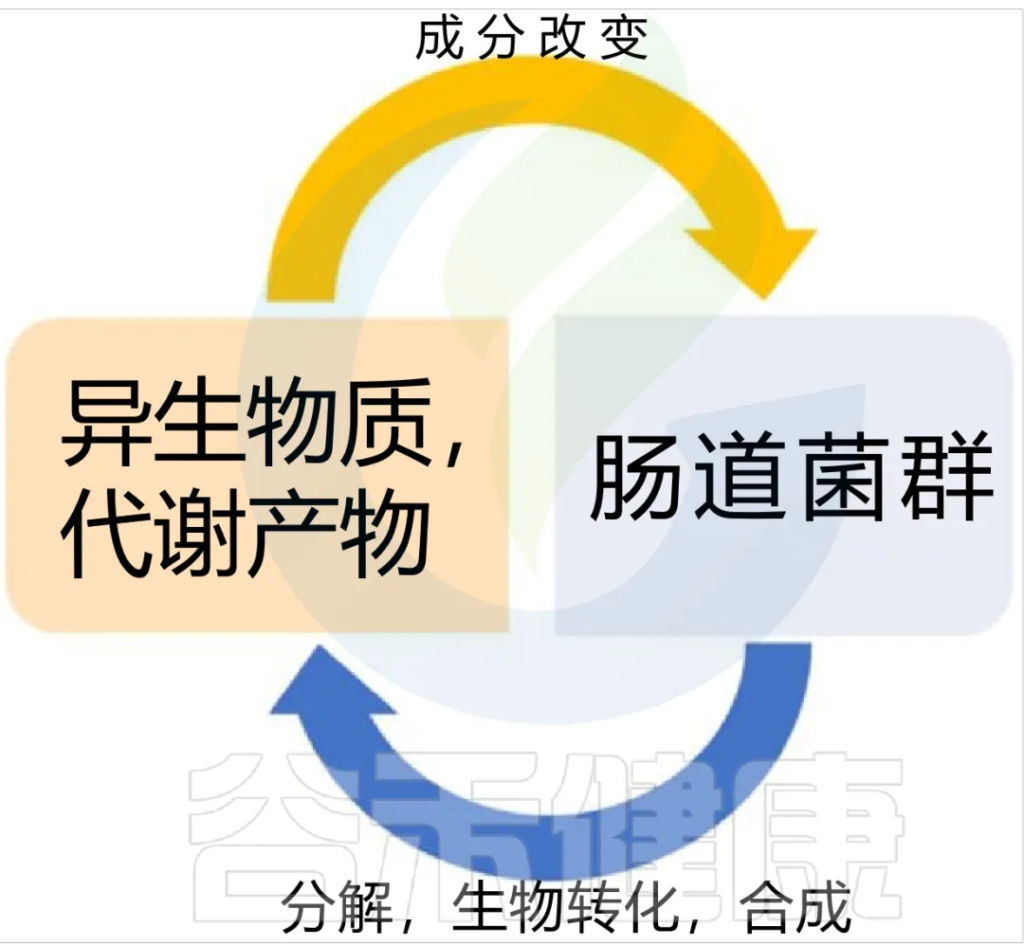
Patangia DV, et al., Microbiologyopen. 2022
◾代谢物
通过影响微生物群落的组成,抗生素还会改变微生物群的功能,从而改变产生的代谢物。
◆ 肠道微生物群负责产生许多必需代谢物,包括 SCFA 和氨基酸,所产生的丁酸盐和丙酸盐具有抗炎作用,促进调节性 T 细胞的产生和分化,在能量代谢中发挥作用。抗生素的摄取会导致微生物群中的蛋白质表达、能量代谢发生变化。
抗生素治疗后,分析代谢的变化——
氨基酸代谢相关代谢产物浓度较高;
氨基酸浓度降低;
丁酸盐和丙酸盐水平降低;
SCFA产量下降;
脂肪酸氧化分解代谢;
氨基酸生物合成发生下降;
影响激素的合成途径(各类细胞因子的基因表达上调);
在微生物群的恢复中,抗生素治疗改变了细菌代谢产物,其中包括——
SCFAs、氨基酸水平的降低;
以及胆汁酸和寡糖等前体增加;
提高胰岛素敏感性;
改变了全身葡萄糖代谢;
肝脏和回肠中参与葡萄糖和胆汁酸代谢的基因表达;
可能会导致屏障功能受损,其作为溃疡性结肠炎和沙门氏菌感染的致病因素。
◆ 抗生素对宿主代谢具有性别依赖性,已研究发现万古霉素和环丙沙星-甲硝唑治疗仅导致雌性小鼠厚壁菌门和 SCFAs 显着减少;两种抗生素暴露都显着降低了雌性小鼠结肠内容物中丙氨酸、支链氨基酸(亮氨酸、异亮氨酸和缬氨酸)和芳香族氨基酸的水平,但雄性小鼠中没有发现。
◾异生素的积累
肠道微生物群可以影响宿主的异生物质半衰期、到达靶受体的程度,以及宿主代谢异生物质的能力。
◆ 异生素(包括抗生素、重金属和环境化学品) 对肠道微生物组成的影响是周期性的,因微生物群是外源生物转化所必需的,所以异生素在到达其靶器官部位之前的代谢很大程度上取决于微生物群。
◆ 通过体内和体外研究都表明了,肠道微生物群参与了异生物质的生物转化,其中缺乏肠道微生物群会影响许多肝酶基因的表达。缺乏代谢特定化合物所必需的微生物会导致其在宿主中的累积,从而导致产生毒性。
对 SPF 和无菌小鼠使用环丙沙星后,显示 —— 肝脏 Cyp3a11 表达降低,这与肠道微生物群的改变有关。由于抗生素给药后产生 LCA 的细菌减少从而导致较的低石胆酸 (LCA) 水平,这可能是Cyp3a11 表达减少的原因。对人类的类似影响可导致多种 CYP3A4(Cyp3a11 的人体类似物) 依赖性药物的清除及减少。
◆ 使用抗生素对肠道微生物组成的改变,会产生的影响 ——
肝脏脂质积累水平增加
甘油三酯(TG) 水平升高
唾液酸和琥珀酸盐的水平增加
对沙门氏菌和艰难梭菌的易感性增加
导致某些仅由细菌产生的代谢物或维生素缺乏
导致铜 (Cu) 代谢发生变化
由于铜在一些功能中所发挥的作用,这可能会对免疫和肠道屏障产生影响。
◾细菌的相关影响
抗生素可以改变几个主要功能基因的转录,其中主要包括 —— 编码转运蛋白的基因;参与碳水化合物代谢的基因和蛋白质合成。
◆ 交叉喂养是肠道微生物群的一个重要特征,常见如 利用低聚果糖和淀粉产生乳酸和乙酸;而产生丁酸盐的厌氧菌不能利用低聚果糖和淀粉,而是依赖乳酸和乙酸作为生长底物。在许多细菌群中,通过交叉喂养间接促进了产丁酸盐物种的增殖和扩张。对此进行的相关研究发现万古霉素是一种革兰氏阳性靶向抗生素,可减少肠道中革兰氏阴性细菌的丰度,这可能是由于其细菌中的相互依赖性所导致的。相互依赖可能会因使用抗生素而受到干扰,从而导致某些代谢物/化合物的积累增加或缺乏。
◆ 细菌使用模式识别受体 (PRR) 通过产生胆汁酸、SCFA、脂肪酸、氨基酸、LPS、脂磷壁酸、鞭毛蛋白、CpG DNA 和肽聚糖的信号与其宿主相互作用,这些信号分子既可以作为其他细胞的能量来源,也可以通过 G 蛋白偶联受体和核受体家族、游离脂肪酸受体调节或调节免疫细胞(如单核细胞、巨噬细胞、T 细胞)的功能。
抗生素的使用导致这些细菌的减少,从而减少了 PRR,产生如 TLR 信号传导和先天防御的下游调节。对其研究发现,抗生素介导的丁酸盐产生细菌减少会导致通过细胞内的丁酸盐传感器的过氧化物酶体增殖物其激活受体 γ 的上皮信号传导减少。
同样,由于产生 IL-22 的淋巴样细胞的迁移中断,共生微生物群的抗生素破坏,增加了其肺炎的易感性;这种效应可以通过在出生时将共生微生物群转移给小鼠来逆转。共生菌还在防止结肠损伤和维持肠道稳态方面具有重要作用,共生体通过 TLR 诱导保护因子的释放,然而这些因子在缺乏共生细菌的抗生素治疗的小鼠中没有得到释放;因此,由于微生物群落组成的变化,抗生素可以影响复杂的宿主-微生物相互作用。
◾ 抗生素抗性基因
肠道已被确定为抗生素耐药性的中心,人体也是抗生素抗性基因(antibiotic resistance genes; ARGs)的储存库。在肠道中,细菌可以通过移动遗传元件将基因水平垂直传递给其他相关或不相关的细菌。
◆ 抗生素抗性基因存在于环境中,包括海洋和淡水体、土壤、冰川、食物链以及内部人类;除细菌外,病毒也是抗生素抗性基因的携带者。有些ARG的传播是历史遗留性的,例如在一些未受污染/未受污染的环境中;但其中大部分是因为人类广泛使用抗生素所传播的,除了用于治疗人类感染外,抗生素还被广泛用作动物体重增加的生长促进剂以及治疗和控制感染的使用,甚至应用于水产养殖。
其中一些抗生素与治疗人类感染的抗生素相同或结构相似,例如红霉素、庆大霉素、恩诺沙星、新霉素、链霉素。农业和水产养殖中抗生素抗性细菌的发展是一个严重的问题,因为这些细菌可以通过食物链进入人体,促进交叉耐药性,并降低传染性细菌对抗生素治疗的敏感性。
◆ 有研究发现来自偏远地区的人类中存在抗生素抗性基因,然而这些人对抗生素的接触使用非常有限;其中 —— 四环素、氨苄青霉素、甲氧苄啶/磺胺甲恶唑、链霉素和氯霉素等抗生素的高水平获得性耐药。
进而研究发现,尽管没有使用过抗生素,但健康的人体内也有抗生素抗性基因——对从未接触过抗生素的健康婴儿和儿童进行的研究表明,存在对 β-内酰胺类、氟喹诺酮类、四环素类、大环内酯类、磺胺类药物或多种药物产生耐药性的基因。
该研究发现其肠球菌属、葡萄球菌属、克雷伯氏菌属、链球菌属和大肠杆菌/志贺氏菌属是 ARGs 的主要携带者。
以下,列举出可用于 ARGs 转移的各种可移动遗传元件:
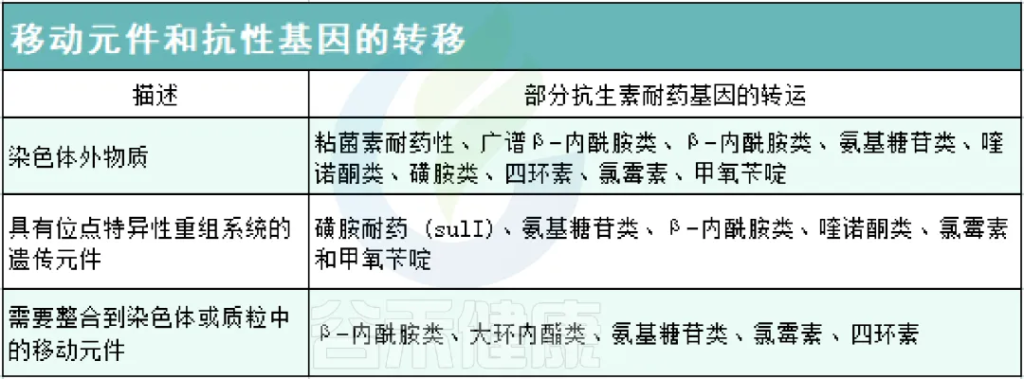
Patangia DV, et al., Microbiologyopen. 2022
抗生素可对宿主组织产生直接毒性作用,如 ——
线粒体损伤
抑制核糖体基因表达
哺乳动物细胞中的氧化组织损伤
该结论现今依旧存在一些争议。抗生素有时会使原本要治疗的病情变得恶化。许多 β-内酰胺类抗生素的杀菌作用会增加毒素的产生,例如 —— 从肠出血性大肠杆菌中释放的志贺毒素,使宿主更容易患溶血性尿毒症综合征。
抗生素还有可能直接影响宿主的代谢,无需微生物作为介质,同时会使目标病原体对抗生素的敏感性降低;宿主代谢物的这些变化大多发生在感染部位,其中包括高水平的 AMP,这会降低抗生素的功效并增加吞噬的活性。由于抗生素对免疫细胞呼吸活动的抑制作用,这可能会导致免疫功能的损伤。
以下,列举出可单独使用或在某些情况下与抗生素治疗联合使用的各种抗生素替代品:

Patangia DV, et al., Microbiologyopen. 2022
因为抗生素的使用会导致微生物组成发生变化,很可能会对宿主产生有害的影响。为减少其影响,使用一些方法在使用抗生素后或与抗生素一起进行调节使用,得以更快地恢复其微生物群。
抗生素的使用其主要问题之一就是对于细菌耐药性的产生。目前正在研究诸如噬菌体(噬菌体) 疗法和细菌素等替代物作为抗生素的替代品或抗生素的补充疗法,以克服耐药性问题。
◆ 噬菌体疗法在1900年代初首次被提出和使用,但由于抗生素的引入,噬菌体疗法在西方医学中逐渐被摒弃,但在国外有部分国家仍旧使用着。随着抗生素耐药性危机的日益严重,需要对噬菌体疗法进行重新审视。
如今已经证明噬菌体疗法可以成功地治疗呼吸道铜绿假单胞菌的感染。随着使用特异性噬菌体疗法,逐步实现了对多重耐药鲍曼不动杆菌患者的感染进行了清除和成功治疗;以及对细菌性前列腺炎、败血症和急性肾损伤患者的病原体实现了临床症状的减轻以及根除。
对96名患者的伤口和溃疡的实验中有 67 名在噬菌体治疗后愈合,其愈合与病原体的减少有关;还在使用工程化噬菌体治疗耐药性脓肿分枝杆菌中显示了囊性纤维化患者的临床改善。对其进行深入研究后,发现使用噬菌体溶解蛋白作为抗菌化合物的方法,可以使噬菌体成为抗生素的强大抗菌竞争者。
◆ 细菌素 —— 另一类潜在的抗生素替代品,是由细菌产生的核糖体产生的抗菌肽,由于特定的免疫机制,细菌本身对杀伤肽免疫。迄今为止,细菌素在食品工业中主要用作食品安全剂和防腐剂;在动物研究中,细菌素作为抗菌剂已逐渐展示出期望的结果。
在小鼠模型的研究中,成功使用绿脓杆菌治疗铜绿假单胞菌肺部感染,其疗效并高无任何副作用;在给予产乳链菌肽和片球菌素的乳酸乳球菌和乳酸乳酸菌菌株有助于减少肠道内耐万古霉素的肠球菌定植。
细菌素已成功应用于治疗和预防牛乳腺炎,且其疗效与抗生素相当。此外,对由葡萄球菌引起乳腺炎的8名哺乳期女性的研究中,已经证明了乳酸链球菌肽可有效的对其进行治疗。
◆ 单克隆抗体作为替代品或与抗生素结合使用,也可以解决抗生素耐药性的问题。单克隆抗体绕过了毒性、耐药性发展和免疫系统早期清除的并发症,这在抗生素的情况下是可见的。使用单克隆抗体治疗细菌感染是近几年兴起的,在此之前,单克隆抗体主要用于治疗癌症、自身免疫性疾病或病毒感染。
在兔子身上进行实验研究表明,使用单克隆抗体bezlotoxumab对抗炭疽保护性抗原是成功的。奥比妥昔单抗的使用提高了接受致死剂量的炭疽芽孢杆菌孢子兔子的存活率;在一项 2655 名参与者的临床试验中,使用bezlotoxumab(抗艰难梭菌毒素的单克隆抗体) 治疗艰难梭菌感染可降低感染的复发率。进一步研究发现,从患者的 B 细胞中产生了一种单克隆抗体,用于对抗小鼠结核分枝杆菌的感染。
单克隆抗体比生产抗生素虽然成本更高,但有很许多好处 —— 相比较细菌素、噬菌体及其内溶素、单克隆抗体的主要优点之一是可以具有高度的靶标特异性,因此对微生物群的附带损害最小。对于这方面的研究,更多的是将用于改变医学上的使用。
◆ 益生菌被广泛的应用于此方面,如今已被证明其可以增加有益微生物的丰度,稳定微生物群落,从而减轻抗生素的影响。
益生菌是通过促进抗菌肽的产生、从而产生细菌素,通过争夺肠道黏膜上的营养物质和受体来抑制非共生菌的生长、增强肠道屏障功能和调节免疫力来发挥作用的,但使用益生菌可能不能够让肠道微生物群完全恢复如初。
◆ FMT更有利于恢复肠道中的微生物平衡。FMT已被广泛用于医疗性的重平衡艰难梭菌感染患者的微生物群,恢复其微生物群和代谢活性。
FMT还可以在抗生素治疗前利用患者自身的供体粪便,称为自体FMT。由于如疗效、成本和适用性等多种因素,使选择FMT成为了一个极具发展性的选择,但具体的使用还需要详细的研究来进行优化,并对其进一步进行肠道疾病以外的其他应用可能性的研发。
人体还可以通过合理的饮食调节和健康的作息安排来控制自身的微生物群,从长远来看,自身的调控会保持机体持续的健康,且对预防炎症疾病起到很好的作用。
以上对于肠道菌群在宿主代谢和免疫功能(如免疫发育、定植抗性、细胞信号传导) 中的重要性;抗生素在单个细菌细胞水平上的影响,其使用和滥用会导致耐药性的发生以及抗生素治疗的替代方法进行了说明。
在先进的组学技术的帮助下,宿主和微生物群之间复杂的相互作用现在变得清晰起来。抗生素会破坏微生物平衡,从而破坏细菌群落内相互关系,以及影响其与宿主之间的关联,并因此产生的耐药菌使临床治疗变得困难。
由于宿主和微生物群之间的这种复杂联系,目前对抗生素需要遵循严谨的使用规范,而;另一方面,对抗生素替代品的研究以及应用是重点,要同时满足限制替代品所附带的损害。研究出可以维持和改善宿主健康并在使用时尽量减少抗生素影响的治疗方法。
主要参考文献:
Abrams, G. D., & Bishop, J. E. (1967). Effect of the normal microbial flora on gastrointestinal motility. Proceedings of the Society for Experimental Biology and Medicine, 126(1), 301– 304.
Adedeji, W. A. (2016). The treasure called antibiotics. Annals of Ibadan Postgraduate Medicine, 14(2), 56– 57.
Adriaenssens, N., Coenen, S., Versporten, A., Muller, A., Minalu, G., Faes, C., Vankerckhoven, V., Aerts, M., Hens, N., Molenberghs, G., & Goossens, H., ESAC Project Group. (2011). European surveillance of antimicrobial consumption (ESAC): Outpatient antibiotic use in Europe (1997-2009). Journal of Antimicrobial Chemotherapy, 66(Suppl. 6), 3– 12.
Akhavan, B. J., Khanna, N. R., & Vijhani, P. (2020). Amoxicillin, StatPearls [Internet]. StatPearls Publishing.
Allegretti, J. R., Mullish, B. H., Kelly, C., & Fischer, M. (2019). The evolution of the use of faecal microbiota transplantation and emerging therapeutic indications. The Lancet, 394(10196), 420– 431.
Aagaard, K., Ma, J., Antony, K. M., Ganu, R., Petrosino, J., & Versalovic, J. (2014). The placenta harbors a unique microbiome. Science Translational Medicine, 6(237), 237ra65.
Blaser, M. (2011). Stop the killing of beneficial bacteria. Nature, 476(7361), 393– 394.
Brestoff, J., & Artis, D. (2013). Population genetic tools to dissect innate immunity in humans. Nature Immunology, 14(7), 676– 684.
Brugman, S., Klatter, F. A., Visser, J. T., Wildeboer-Veloo, A. C., Harmsen, H. J., Rozing, J., & Bos, N. A. (2006). Antibiotic treatment partially protects against type 1 diabetes in the bio-breeding diabetes-prone rat. Is the gut flora involved in the development of type 1 diabetes? Diabetologia, 49(9), 2105– 2108.
Bui, T., & Preuss, C. V. (2021). Cephalosporins, StatPearls [Internet]. StatPearls Publishing.
Cahenzli, J., Köller, Y., Wyss, M., Geuking, M. B., & McCoy, K. D. (2013). Intestinal microbial diversity during early-life colonization shapes long-term IgE levels. Cell Host and Microbe, 14(5), 559– 570.
Cao, F., Wang, X., Wang, L., Li, Z., Che, J., Wang, L., Li, X., Cao, Z., Zhang, J., Jin, L., & Xu, Y. (2015). Evaluation of the efficacy of a bacteriophage in the treatment of pneumonia induced by multidrug resistance Klebsiella pneumoniae in mice. BioMed Research International, 2015, 2015– 2019.
Cao, L. T., Wu, J. Q., Xie, F., Hu, S. H., & Mo, Y. (2007). Efficacy of nisin in treatment of clinical mastitis in lactating dairy cows. Journal of Dairy Science, 90(8), 3980– 3985.
Cash, H. L., Whitham, C. V., Behrendt, C. L., & Hooper, L. V. (2006). ‘Symbiotic bacteria direct expression of an intestinal bactericidal lectin’. Science, 313(5790), 1126– 1130.
Chaves, B. J., & Tadi, P. (2021). Gentamicin, StatPearls [Internet]. StatPearls Publishing.
Centre for Disease Control (CDC). (2017). Pediatric treatment recommendations.
Aloisio, I., Quagliariello, A., De Fanti, S., Luiselli, D., De Filippo, C., Albanese, D., Corvaglia, L. T., Faldella, G., & Di Gioia, D. (2016). Evaluation of the effects of intrapartum antibiotic prophylaxis on newborn intestinal microbiota using a sequencing approach targeted to multi hypervariable 16S rDNA regions. Applied Microbiology and Biotechnology, 100(12), 5537– 5546.
Arboleya, S., Binetti, A., Salazar, N., Fernández, N., Solís, G., Hernández-Barranco, A., Margolles, A., los Reyes-Gavilán, C. G., & Gueimonde, M. (2012). Establishment and development of intestinal microbiota in preterm neonates. FEMS Microbiology Ecology, 79(3), 763– 772.