-

 国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子
国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子

 二级病原微生物安全实验室
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业

谷禾健康

2021年6月份,我们分享过一篇“全面认识——肺炎克雷伯菌(Klebsiella pneumoniae) ”的文章,当时也是发现该菌在肠道的人群检出率较高,基于想全面了解该菌,我们查阅整理了很多资料,包括统计了谷禾健康数据库中肺炎克雷伯菌的检出率等,向大家介绍了关于该菌的基本知识,人群分布,感染的症状,分类及来源,传播和易感因素,诊断方法以及治疗及预防。
本文我们要介绍的产气克雷伯菌(Klebsiella aerogenes),是肺炎克雷伯菌(Klebsiella pneumoniae) 的近源菌种,该菌是最近更名的,原来是叫产气肠杆菌。
革兰氏阴性菌的肠杆菌科(Enterobacterales)最初分为三个属:埃希氏菌属(Escherichia)、气杆菌属(Aerobacter)、克雷伯氏菌属(Klebsiella);其中气杆菌属包括A. aerogenes 和阴沟肠杆菌(A. cloacae)。
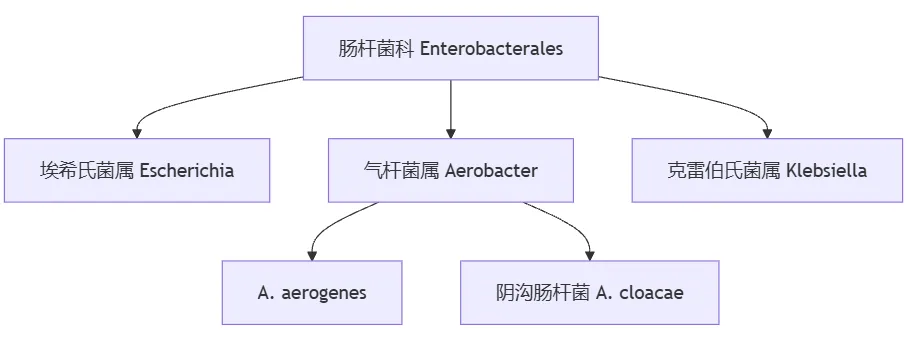
到 1960 年,Aerobacter 已被重新命名为肠杆菌属(Enterobacter)。最近,基于全基因组序列的比较细菌系统发育学表明,产气肠杆菌(E.aerogenes)与肺炎克雷伯菌(K.pneumoniae)的亲缘关系比与肠杆菌属的关系更密切。因此,以前称为产气肠杆菌的细菌更名为产气克雷伯菌(K.aerogenes)。
使用平均核苷酸同一性(ANI)将肠杆菌属的其余成员分为 22 个不同的系统发育组。这 22 个系统发育组统称为阴沟肠杆菌复合体(Ecc)。尽管细菌比较基因组学已经证明产气克雷伯菌和 Ecc 属于不同的系统发育组,但这些遗传差异的临床影响尚不清楚。
本文简单介绍和认识下产气克雷伯菌(K. aerogens) 的作为机会病原体的一些发现,其他着重分享一个有意思的研究结果,即肠道产气克雷伯菌产生高组胺,诱发IBS患者腹痛的证据,原因和干预建议。
K. aerogens
基本特征与临床意义
产气克雷伯菌(K.aerogens),属于肠杆菌科,是一种兼性革兰阴性厌氧菌。它广泛分布于环境中,存在于人体胃肠道中,也是医院常见的机会性病原体。当宿主免疫系统受损或肠粘膜受损时,可能导致呼吸系统、循环系统或泌尿生殖系统感染。
近年来,尽管关于大肠埃希菌和肺炎克雷伯菌的致病性和耐药性的报道越来越多,但关于产气克雷伯菌的报道很少。 既往关于该细菌的临床报告主要为呼吸道、胃肠道、泌尿道和血液感染。与其他肠杆菌科物种相比,产气克雷伯菌更容易导致患者感染性休克甚至死亡。
致病机制与毒力因子
与其他肠杆菌科病原体一样,克雷伯菌的毒力和耐药性很复杂,并受多种因素影响。产生的毒力因子和耐药基因可能因感染部位而异。
克雷伯菌产生的粘附素有助于其进入宿主细胞,细胞表面的荚膜多糖和脂多糖帮助细菌从吞噬作用中逃脱,而毒素或其他细胞外成分则引起粘膜损伤并通过循环传播。
耐药性与治疗策略
近年来,由于抗生素的过度使用,越来越多的克雷伯菌种出现了多药耐药性。不断有报道称,这种新兴病原体的爆发频率很高,其中一些与高死亡率有关,在世界各地的不同临床环境中,例如新生儿、老年和重症监护病房。
根据药敏试验结果表明,产气克雷伯菌分离株是对碳青霉烯类的非耐药细菌,对作为抗感染治疗的美罗培南特别敏感。此外,分离的产气克雷伯菌在亚胺培南中处于中间位置。然而,他们被发现对青霉素耐药,例如氨苄西林和阿莫西林。
一般克雷伯菌相关感染发展迅速,导致多器官衰竭甚至死亡,容易出现耐药菌株。因此,应尽早选择有效的抗生素,并确定剂量和疗程,以尽量减少副作用的发生。
本文主要的分享是基于基于目前发表于科学转化医学《Science Translational Medicine》上的一项有趣的研究,结合谷禾肠道菌群的检测实践检验综合分享。
先说该研究的主要结论:
1. 发现肠道细菌生产的组胺,可以诱发一部分 IBS 患者的腹痛
2. 确定了产气克雷伯氏菌是肠道组胺的主要产生者
组胺可以由许多细菌分泌,但到现在,导致肠道中组胺过量产生的特定微生物一直是个谜。
组胺是免疫细胞释放并参与局部免疫反应的化合物。它也是已知的疼痛介质。近年来,组胺与肠易激综合征(IBS)的关系越来越密切。人们还认为,“组胺不耐受”的各种症状特征可能是由于肠道中组胺分泌过多,压倒了身体分解和代谢组胺的能力。
在本文中,谷禾将带大家了解这些新发现并讨论其影响。首先,简单介绍一下背景知识,解释一下这项最新研究的动机。
一般低 FODMAP 饮食可减少 IBS 患者的尿液组胺。
低 FODMAP 饮食长期以来一直被认为能够帮助缓解 IBS 患者的症状。然而,这些碳水化合物加重肠道症状的确切机制尚不清楚。
2017 年,加拿大麦克马斯特大学的一组研究人员随机分配 IBS 患者食用低或高 FODMAP 饮食。饮食干预后,测量了尿液中的各种代谢物,发现,大约 1/3 的 IBS 患者在基线时尿组胺水平较高,而在低 FODMAP 饮食后,尿组胺水平降低了 8 倍。
饮食–微生物群相互作用导致肠道敏感性增加
基于他们最新研究的这些发现,研究人员将一名尿组胺含量高的 IBS 患者,一名尿组胺含量低的 IBS 患者,一名健康的对照受试者的肠道微生物群移植到三组独立的无菌小鼠中。
经过一段时间的定植期后,让微生物群在小鼠中扎根,它们进一步将每组小鼠一分为二,以接受低或高可发酵碳水化合物饮食。
结果发现只有用高尿组胺的 IBS 患者的微生物群定植并喂食高可发酵碳水化合物饮食的小鼠组增加了肠道敏感性。
产气克雷伯菌是IBS患者的主要组胺产生者
然后,研究人员回去更仔细地分析供体的粪便。尿组胺含量高的 IBS 患者的肠道微生物组产生组胺明显更多。他们着手确定哪些微生物可能是罪魁祸首。
研究人员总共分离了 164 种不同的肠道微生物,并测试了它们产生组胺的能力。
作者报告说:
在尿组胺水平高的 IBS 患者中,产气克雷伯菌是主要的组胺产生者。事实上, 产气菌产生的组胺比任何其他细菌分离物多 100 倍。
在 IBS 患者和低尿组胺患者中,屎肠球菌(Enterococcus faecium)和粪肠球菌(Enterococcus faecalis)是组胺的主要产生者。
但该论文没有包括测试的细菌及其产生组胺的能力的完整列表。例如,以前的研究强调Morganellamorganii、Raoultella planticola、 Raoultella ornithinolytica是肠道组胺的潜在生产者,但在结果或讨论中没有提到这种微生物。
在选择测试的微生物时,其他组胺生产者(也许是那些不太容易培养的)也完全有可能被遗漏。尽管如此,产气克雷伯菌(K.aerogenes) 在 IBS 患者的三个不同队列中含量很高,并且在低 FODMAP 饮食中减少。此外,用产气克雷伯菌定植的无菌小鼠产生的组胺是用另一种克雷伯氏菌定植的小鼠的 77 倍。
pH值很重要:为什么结肠酸度必不可少?
产气克雷伯菌是如何产生组胺的呢?查阅发现该菌使用一种称为组胺脱羧酶(HDC)的酶。HDC酶将组氨酸(饮食中的一种必需氨基酸)转化为组胺。(谷禾以前文章可以关注看下)
由于以前的研究发现这种酶的活性受 pH 值的影响,研究人员决定测试产气克雷伯菌产生的组胺是否取决于环境的酸度。
产气克雷伯菌在pH值为7.0时产生最多的组胺,而在pH值低于6.0或高于8.0时几乎没有组胺。
应该这是关键,因为结肠的 pH 值在 5.5 到 7.5 之间,具体取决于发酵水平。pH 值处于此范围的下限通常与更好的预防症状有关。换句话说,降低结肠的 pH 值(增加其酸度)可能是减少产气克雷伯菌产生肠道组胺的一种方法。
乳酸杆菌:一种防止克雷伯氏菌的方法
结肠 pH 值的主要决定因素之一是乳酸水平。当研究人员查看“人源化”小鼠的肠道样本时,他们发现从尿组胺含量高的 IBS 患者那里获得微生物群的小鼠的乳酸水平要低得多。与此同时,乳酸杆菌等产乳酸菌的水平较低。
为了进一步检查这种关系,研究人员培养了有或没有乳酸杆菌混合物的产气克雷伯菌。正如预期的那样,乳酸杆菌减少了组胺的产生,该实验室还在小鼠模型中证实了这些结果。
但本研究中使用的两种乳酸杆菌没有得到很好的表征。第一种是动物乳杆菌(L.animalis) 或鼠乳杆菌(L.murinus)的菌株,而第二种与嗜酸乳杆菌 (L.acidophilus) 和乳酸乳杆菌(L.lactis) 的关系更密切。
尽管如此,这表明一些乳酸杆菌可以调节产气克雷伯菌产生组胺,很可能是通过产生乳酸,从而降低pH值和组胺脱羧酶的活性。
但是要注意,许多乳酸杆菌对于有组胺不耐受或肥大细胞活化症状的人来说可能也是有问题的。稍后会详细介绍。
到目前为止,我们一直在谈论结肠中的克雷伯氏菌活动。但是小肠中的克雷伯氏菌呢?
小肠中的克雷伯氏菌
虽然一般小肠通常不会含有太多克雷伯氏菌,但至少有一项研究表明,克雷伯氏菌在有胃肠道症状的个体的十二指肠(小肠的上三分之一)中更为普遍,且它们可以破坏整体小肠微生物群结构。
初步证据表明,其中一些克雷伯氏菌可能起源于口腔。换句话说,唾液可能成为克雷伯氏菌持续传播到肠道的储存库,如果肠道环境允许,它们有机会将在那里蓬勃发展。
小肠部分的酸性也比结肠低得多。回肠(小肠最远端的部分)的 pH 值高达 7.4-7.8,这个水平可以支持产气克雷伯菌产生组胺。
也许小肠中产气克雷伯菌的机会性过度生长可能导致总体高水平的组胺。
细菌组胺将肥大细胞吸引到肠道
研究人员再次转向小鼠,以确定细菌组胺如何引起内脏疼痛。
他们发现细菌组胺通过组胺H4 受体发出信号,激活肥大细胞并将其吸引到结肠。这些肥大细胞贡献了更多的组胺和其他疼痛信号分子,触发肠道中的神经细胞并引起腹痛(详细可以看谷禾以前关于组胺的文章)。
因此,阻断 H4 受体可能代表了一种减轻高组胺患者症状的策略。这将防止肥大细胞募集到结肠和宿主产生额外的组胺。
如何检测细菌性组胺相关 IBS?
说到这,大家最感兴趣的应该如何知道你是否属于 IBS 患者的特定亚群呢?虽然需要更多的研究来准确定义此类 IBS 的特征,但指标可能包括:
有哪些干预措施?
以下我们整理了一些可能对患有这一特定 IBS 的人有帮助的潜在干预措施建议。(注意:此信息不应被视为医疗建议)。
低 FODMAP 饮食:低可发酵低聚糖、双糖、单糖和多元醇(FODMAP) 的饮食可以迅速降低肠道组胺水平,减少产气克雷伯菌的丰度,并暂时缓解腹痛。
恢复肠道酸度:较低的 pH 值被证明可以显著减少产气克雷伯菌产生的组胺。
以下可能对增加肠道酸度最有效,但组胺敏感个体可能无法耐受;仅在耐受性良好的情况下使用:
组胺敏感个体可能耐受性更好的其他干预措施:
1)肠道微生物群是组胺的重要来源:虽然传统上肥大细胞被认为是组胺的主要来源,但研究结果表明肠道微生物群是一个重要的来源。
2)产气克雷伯菌(Klebsiella aerogenes, K. aerogenes) 是 IBS 患者组胺升高的主要生产菌。产气克雷伯菌可以在肠道中募集和激活肥大细胞,导致腹痛,也可能参与以高组胺水平为特征的其他疾病,但需要更多的研究。
3)对于一部分组胺水平较高的肠易激综合征(IBS)患者来说,通过靶向治疗降低体内产气克雷伯菌的水平,可能会带来临床症状的改善。 产气菌或抑制组胺信号传导:将产气克雷伯菌或具有类似 HDC 活性的细菌鉴定为肠道中组胺的来源,可以指导饮食推荐、微生物群定向疗法或在患有慢性腹痛的 IBS 患者中使用H4 受体拮抗剂。
本文内容仅供学习和交流目的,不构成任何形式的医疗建议。
主要参考文献
De Palma, G. et al. Histamine production by the gut microbiota induces visceral hyperalgesia through histamine 4 receptor signaling in mice. Science Translational Medicine 14, eabj1895 (2022).
McIntosh, K. et al. FODMAPs alter symptoms and the metabolome of patients with IBS: a randomised controlled trial. Gut 66, 1241–1251 (2017).
Barcik, W. et al. Bacterial secretion of histamine within the gut influences immune responses within the lung. Allergy 74, 899–909 (2019).
Barcik, W. et al. Histamine-secreting microbes are increased in the gut of adult asthma patients. Journal of Allergy and Clinical Immunology 138, 1491-1494.e7 (2016).
Lucy et al. The gut bacteria that produce histamine in IBS – revealed. Lucymailing. 2022.
Barlow, J. T. et al. Quantitative sequencing clarifies the role of disruptor taxa, oral microbiota, and strict anaerobes in the human small-intestine microbiome. Microbiome 9, 214 (2021).
Atarashi, K. et al. Ectopic colonization of oral bacteria in the intestine drives TH1 cell induction and inflammation. Science 358, 359–365 (2017).
Koziolek, M. et al. Investigation of pH and Temperature Profiles in the GI Tract of Fasted Human Subjects Using the Intellicap(®) System. J Pharm Sci 104, 2855–2863 (2015).
Wesevich A, Sutton G, Ruffin F, Park LP, Fouts DE, Fowler VG Jr, Thaden JT. Newly Named Klebsiella aerogenes (formerly Enterobacter aerogenes) Is Associated with Poor Clinical Outcomes Relative to Other Enterobacter Species in Patients with Bloodstream Infection. J Clin Microbiol. 2020 Aug 24;58(9):e00582-20.
Gu, H., Cai, Q., Dai, X. et al. A case report of Klebsiella aerogenes-caused lumbar spine infection identified by metagenome next-generation sequencing. BMC Infect Dis 22, 616 (2022).
Morgado, S., Fonseca, É., Freitas, F. et al. In-depth analysis of Klebsiella aerogenes resistome, virulome and plasmidome worldwide. Sci Rep 14, 6538 (2024).
Niedzielin, K., Kordecki, H. & Birkenfeld, B. A controlled, double-blind, randomized study on the efficacy of Lactobacillus plantarum 299V in patients with irritable bowel syndrome. Eur J Gastroenterol Hepatol 13, 1143–1147 (2001).
Arellano, K. et al. Safety Evaluation and Whole-Genome Annotation of Lactobacillus plantarum Strains from Different Sources with Special Focus on Isolates from Green Tea. Probiotics Antimicrob Proteins 12, 1057–1070 (2020).
Shen, F. et al. Vinegar Treatment Prevents the Development of Murine Experimental Colitis via Inhibition of Inflammation and Apoptosis. J. Agric. Food Chem. 64, 1111–1121 (2016).
Wang, C. C. et al. Sodium butyrate enhances intestinal integrity, inhibits mast cell activation, inflammatory mediator production and JNK signaling pathway in weaned pigs. Innate Immun 24, 40–46 (2018).
Diakos, C. et al. n-Butyrate inhibits Jun NH(2)-terminal kinase activation and cytokine transcription in mast cells. Biochem Biophys Res Commun 349, 863–868 (2006).
Lewis, S. J. & Heaton, K. W. Increasing butyrate concentration in the distal colon by accelerating intestinal transit. Gut 41, 245–251 (1997).
Paulose, J. K., Wright, J. M., Patel, A. G. & Cassone, V. M. Human Gut Bacteria Are Sensitive to Melatonin and Express Endogenous Circadian Rhythmicity. PLOS ONE 11, e0146643 (2016).
Yang, C.-C. et al. Quercetin inhibits histamine-induced calcium influx in human keratinocyte via histamine H4 receptors. International Immunopharmacology 96, 107620 (2021).
谷禾健康
组胺是人体内的一种生物胺,最为人们所熟知的作用是作为过敏反应的介质,但也是神经系统、肠道、皮肤和免疫系统中重要的信号分子。
花粉之所以会引起人体过敏反应,并不是因为花粉本身,而是由于花粉颗粒携带了能引起过敏的抗原。接触过敏源后,体内的组织会释放组胺,作为一种神经递质,组胺会使毛细血管和微静脉的管壁通透性增加,导致局部组织水肿,造成皮肤发红、疹块;也会使气管平滑肌收缩,造成呼吸道狭窄,引起气喘、呼吸困难。
组胺主要由肥大细胞和嗜碱性粒细胞等免疫细胞产生和释放。一些细胞如树突状细胞或T细胞可以表达组氨酸脱羧酶,这是一种在刺激后合成组胺的酶。
尽管如此,一些食物中的微生物群和部分人体肠道微生物群也可以分泌组胺。表明组胺在肠道免疫调节中的潜在作用。事实上,最近的一项研究表明,组胺可以通过抑制肠道中白细胞介素18的产生来发挥抗炎作用。
组胺在免疫反应、神经传递和过敏反应中发挥着重要作用。在某些浓度范围内,组胺起着保护作用,对于维持健康状态至关重要。例如组胺可以刺激胃酸分泌,有助于消化;还会影响排便情况;组胺作为重要的神经递质,参与睡眠调节和体温调节。
然而,在较高浓度下,会导致包括组胺中毒和组胺不耐受或扩张血管,影响血压,导致免疫调节紊乱、胃肠道症状和神经系统疾病。组胺中毒是在食用被大量组胺污染的食物后发生的。如果个体由于基因突变而缺乏生物胺降解能力,或者正在服用单胺氧化酶抑制剂等抗抑郁药来减缓胺的降解,他们就会更容易出现组胺不耐受的症状。
组胺在体内与相应受体(H1、H2、H3、H4)结合而介导的各种症状。不同类型的组胺受体与该化合物之间的相互作用可能会引起多种影响,例如H1R主要介导过敏反应,而H2R具有抗炎作用,而H3R主要影响睡眠、认知障碍等精神健康。
肠道内的组胺对许多细胞过程的广泛影响会导致各种胃肠道疾病,包括食物过敏、肠易激综合征、炎症性肠病、结直肠癌等。谷禾将在本文带大家了解组胺在免疫及各种肠道疾病中的保护或致病作用。
组胺(C5H9N3)是一种重要的生物胺类神经递质,于20世纪初首次发现。从那时起,它的功能开始被逐渐发现并得到越来越详细的描述。
组胺存在于多种细胞类型中,是人体的关键信号分子,具有多种功能。例如作为神经递质或免疫反应调节剂。
拓展知识:组胺的发现
1903年,一位叫威廉·邓巴的医生证明了吸入花粉后的过敏反应并不是由花粉本身引起的,而是机体对花粉的反应引起的一种毒素的释放所造成的。
后来,亨利·戴尔于1910年在研究黑麦的毒性时,从麦角菌中提取出一种叫做组织胺的物质。他发现用组胺可以人工诱发荨麻疹。如果将组胺滴入气管,还会诱发哮喘发作。
★ 组胺的重要作用
介导过敏反应
组胺最被人熟知的是在过敏反应中的作用。例如在过敏性鼻炎、荨麻疹和血管神经性水肿等过敏性疾病中发挥重要的病理生理学作用。
当免疫系统对过敏原产生过度反应时,免疫细胞(如肥大细胞和嗜酸性粒细胞)会释放存储在细胞内的组胺。释放的组胺会结合到特定的受体上,引发一系列生理反应,例如瘙痒、肿胀、打喷嚏等。
扩张血管,影响血压
组胺对人心血管系统最突出的作用是扩张小血管。血管扩张使外周阻力降低,血压下降,并伴有潮红、头痛等症状。
组胺还增加毛细血管的通透性,使渗出增加,引起水肿,严重时甚至导致循环血量减少,可能引起休克。组胺对心脏的直接作用包括增强心肌收缩力、加快心率和减慢房室传导。
刺激胃酸分泌
首先,组胺可以刺激胃黏膜细胞中的H2受体,当组胺与H2受体结合时,会激活细胞内的信号转导通路,导致胃壁上的壁细胞释放胃酸。
其次,组胺还可以通过刺激嗜酸性细胞(胃窦细胞)来促进胃酸的分泌。嗜酸性细胞是胃黏膜中的一种细胞类型,它们包含大量的组胺。当胃黏膜受到刺激时,嗜酸性细胞会释放存储的组胺。释放的组胺通过与H2受体结合,刺激壁细胞分泌胃酸。
影响排便
一项研究中,发现细菌产生的组胺与小鼠结肠运动和粪便排出量增加有关,并且组胺受体拮抗剂治疗在很大程度上阻断了细菌组胺对结肠运动的影响。
收缩平滑肌
此外,组胺会诱导平滑肌细胞(包括支气管和肠道)收缩,哮喘患者对组胺比正常人敏感100~1000倍,组胺可引起支气管痉挛导致呼吸困难。组胺收缩胃肠平滑肌和子宫平滑肌还会引起痉挛性腹痛。
重要的神经递质
在中枢神经系统中,组胺作为一种神经递质,参与睡眠调节和体温调节等生理过程。组胺对感觉神经末梢有强烈的刺激作用,尤其对调节痛和痒的神经,该效应由H1受体所调节。
除此之外,有研究发现组胺还会影响伤口愈合、食欲和情绪等方面。
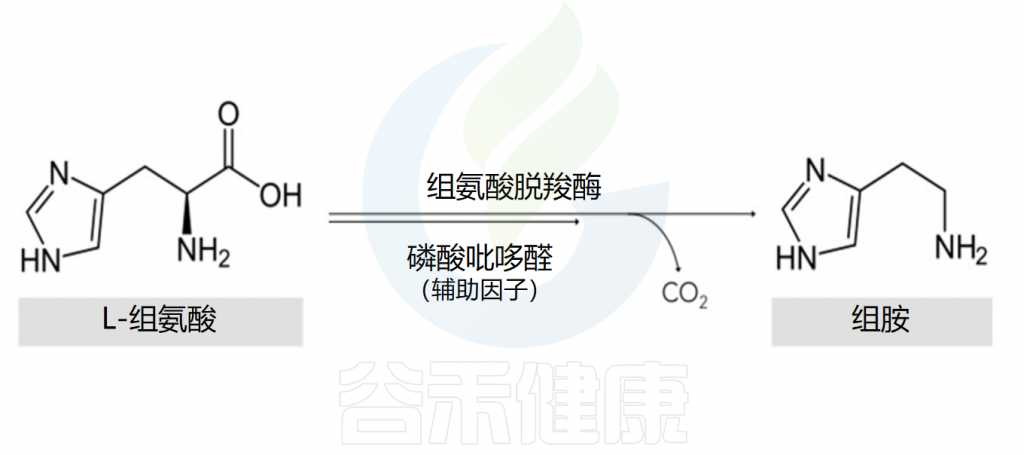
组氨酸是组胺的前体物质
组胺的主要细胞来源是肥大细胞和嗜碱性粒细胞。在细胞的高尔基体中,组胺是通过组氨酸脱羧酶将L-组氨酸氧化脱羧形成的,其辅助因子是吡哆醛(维生素B6)。
Comas-Basté O,et al.Biomolecules.2020
该反应的结果是组胺的形成,组胺随后与其他胺(如5-羟色胺)、蛋白酶、蛋白聚糖、细胞因子/趋化因子和血管生成因子一起储存在细胞质颗粒中,并在细胞致敏和脱颗粒后释放。
过敏时组胺会大量释放
肥大细胞的脱颗粒和组胺的释放主要是由于特异性抗原与FcRI受体结合以及对非免疫刺激(例如神经肽、补体系统的一部分、细胞因子、血小板活化因子)的反应。
IgE抗体是过敏性疾病期间肥大细胞脱颗粒的介质。IgE与其肥大细胞表面的高亲和力IgE受体的结合称为“致敏”。肥大细胞和嗜碱性粒细胞释放的组胺通过激活四种G蛋白偶联受体,即H1R、H2R、H3R(主要在大脑中表达)和H4R,发挥其生物活性。
细菌感染
某些细菌感染也可以引起组胺的释放。例如,胃幽门螺杆菌感染可以导致胃黏膜炎症,促使组胺释放,进而引发胃酸增多和溃疡等胃部疾病。
药物和化学物质
某些药物和化学物质也可以诱发组胺的释放。例如,非甾体抗炎药(如阿司匹林)和某些麻醉药物以及酒精可以引起组胺的释放,可能导致过敏样反应。
物理刺激
组胺释放也可由多种物理因素引起,如极端温度、创伤、摩擦等。这种情况下的组胺释放通常是局部性的,导致局部组织的充血和炎症反应。
运动和应激
剧烈运动、情绪激动和精神压力等也可以促使组胺的释放。这种情况下,组胺的释放通常是全身性的,可能导致血管扩张、心率增加和其他应激反应。
注:组胺也可以由其他细胞类型(例如胃肠嗜铬样细胞、组胺能神经元、树突状细胞、T淋巴细胞、血小板等)合成和释放。
食物和肠道微生物也是组胺的重要来源
一部分组胺通过食物进入人体或由肠道微生物产生。最受欢迎的富含组胺的食物是鱼和海鲜、发酵食品(如奶酪、葡萄酒、泡菜等)和一些蔬菜(如菠菜、茄子、番茄等)。
组氨酸主要在自溶或细菌过程中产生,因此高浓度的组胺主要存在于微生物发酵产物中。食品中生物胺形成的条件是游离氨基酸的可用性、脱羧酶阳性微生物的存在以及细菌生长和脱羧酶活性的条件。
影响细菌组氨酸脱羧酶活性的因素
细菌分泌脱羧酶受许多因素(例如,发酵碳水化合物、氧气或氯化物浓度的存在)调节。在酸性环境中,氨基酸脱羧酶活性的表达增加。
组氨酸脱羧酶活性较高的细菌种类有:
Morganella morganii,Eschericha coli, Hafnia alvei, Proteus vulgaris, Proteus milabilis, Enterobacter aerogenes, Raoultella planticola, Raoultella ornithinolytica, Citrobacter freundii, Pseudomonas fluorescens, Photobacterium damselae。其中一些细菌具有代谢组胺的能力。
我们将在下面的一个段落中展开讲述肠道中的组胺与微生物群。
在人体内,组胺降解主要有两种途径,涉及二胺氧化酶(DAO)或组胺-N-甲基转移酶(HNMT)。组胺降解酶缺失会导致组胺浓度过高甚至中毒,组胺不耐受患者就是组胺降解酶的酶活性降低或受到抑制,并且不足以灭活食物中的组胺并阻止其进入血液。
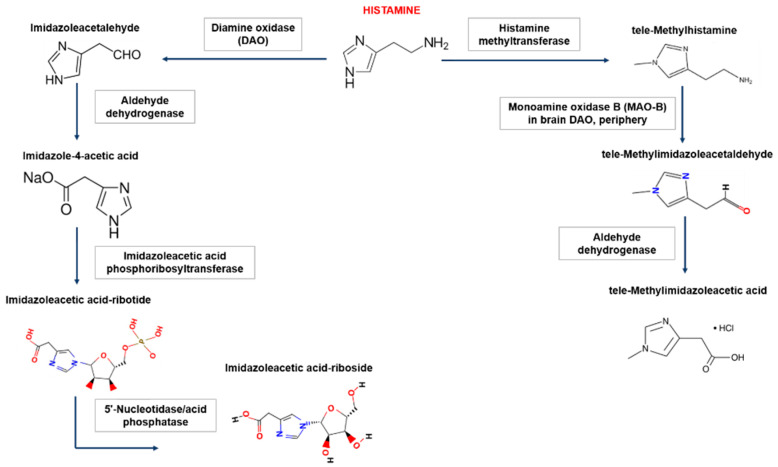
Shulpekova YO,et al.Nutrients.2021
组胺-N-甲基转移酶负责细胞内的组胺降解
组胺-N-甲基转移酶(HNMT)主要负责细胞内组胺的降解。HNMT在肾脏和肝脏以及脾脏、结肠、前列腺、卵巢、脊髓细胞、支气管和气管中的表达最高。一小部分组胺在HNMT的作用下转化为N-甲基组胺。
注:在原始形式中,约2%-3%的组胺被排泄。
二胺氧化酶负责细胞外的组胺降解
二胺氧化酶(DAO)是一种分泌蛋白,负责细胞外组胺的降解。DAO的最大活性记录在小肠、结肠、胎盘和肾脏中。DAO在细胞内囊泡中积累,然后进入细胞外空间,在那里降解组胺。绝大多数组胺通过DAO转化为咪唑乙酸。
保护身体免受组胺过高的侵害
二胺氧化酶执行“屏障功能”,从而限制组胺从肠道进入血液;组胺-N-甲基转移酶通过B族维生素和S-腺苷甲硫氨酸的参与使组胺甲基化,从而调节细胞内的组胺含量。
尽管二胺氧化酶和组胺-N-甲基转移酶这两种酶都存在于肠上皮中,但吸收组胺进入血流的主要障碍是 二胺氧化酶,组胺-N-甲基转移酶在此过程中仅发挥次要作用。在正常情况下,这种酶屏障足以防止组胺被吸收到血流中。
注:二胺氧化酶和组胺-N-甲基转移酶基因的多态性可能是组胺代谢个体差异的基础。在突触中,乙醛脱氢酶在组胺降解中发挥作用。
虽然健康人体内的组胺很快就会被二胺氧化酶降解,但即使在健康人体内,也可能会因摄入组胺含量高的食物(例如鲭鱼)导致血液中组胺浓度升高而出现严重症状——组胺中毒。
• 组胺中毒的症状
组胺中毒症状多种多样,可能包括口腔麻木、头痛、头晕、心悸、低血压、吞咽困难、脉搏微弱、荨麻疹、面部肿胀、潮红、呕吐、恶心和腹泻等。
组胺中毒的症状可能持续数小时或一天,但在极少数情况下,症状可能会持续数天。
• 组胺中毒的主要原因是食用鱼类和海鲜
不仅是鲭鱼,一些非鲭鱼(鲯鳅鱼、蓝鱼、沙丁鱼等)都含有大量组胺,此外发酵产品(奶酪、葡萄酒、罐头食品、腌制蔬菜和一些饮料)中组胺和血清素的含量也较高。
一项针对科学报告的分析显示,在98%的病例中,组胺中毒的原因是食用鱼类和海鲜,其余2%是由于食用奶酪等发酵食品。
鱼组织中的组胺是由多种细菌对游离组氨酸进行转化而产生的,这些细菌包括摩氏摩根菌(Morganellamorganii)、产气肠杆菌 (Enterobacter aerogenes), Raoultella planticola, Raoultella ornithinolytica和Photobacterium damselae。
鱼分解产生的其他物质会增强组胺的毒性并促进吸收
组胺中毒的发病机制不能仅用组胺的作用来解释。与口服等效剂量的纯组胺相比,鲭鱼毒性较高。还提出了其他机制,例如控制组胺的酶的抑制或增强、导致肥大细胞脱颗粒的物质的存在、其他组胺激动剂的存在。
鱼分解过程中产生的其他物质(尸胺和腐胺)会增强组胺的毒性,这些物质还会抑制肠道单胺氧化酶和二胺氧化酶,或从与粘膜粘蛋白的结合中释放组胺,促进其吸收。
组胺中毒后的干预及治疗
在生理值范围内。组胺中毒的病程通常是轻微的、自限性的、持续时间较短且不需要治疗。
对于严重的组胺中毒病例,需要进行治疗干预——支持性容量疗法和氧气疗法,并给予口服抗组胺药和支气管扩张剂。
组胺不耐受是由于组胺摄入量与机体清除能力不相称,导致组胺过度蓄积,并与相应的物质结合而出现症状的病理过程。
与组胺中毒不同,组胺中毒的严重程度与营养物质中组胺的含量成正比,组胺不耐受的发病机制与先天性或后天性缺乏中和组胺的酶有关。在健康患者中,肠上皮细胞具有由二胺氧化酶和组胺-N-甲基转移酶创建的酶屏障。该屏障可防止血液中外源性组胺的过度吸收。如果这些酶被抑制或减少,即使摄入少量的组胺也可能出现组胺不耐受的症状。
• 组胺不耐受的症状
组胺不耐受的神经系统症状包括头痛。除头痛外,其他重要症状包括弥漫性胃痛、绞痛、胀气和腹泻。
组胺不耐受的患者,在摄入组胺或酒精含量高的食物时或摄入后会出现流鼻涕、鼻塞等症状,极端情况下甚至会出现支气管哮喘发作、支气管收缩、咳嗽、喘息并伴有肺功能下降。
组胺不耐受患者还会出现皮肤瘙痒、发红、肿胀甚至荨麻疹。而患有组胺不耐受的女性会出现与月经周期相关的痛经和头痛。
• 影响组胺降解酶活性的因素
在胃肠道疾病(炎症性肠病、寄生虫感染、微生物菌群失调、代谢吸收不良)中,损伤的肠上皮细胞会减少二胺氧化酶的产生,或其他生物胺、酒精或药物也会抑制二胺氧化酶。
二胺氧化酶活性降低可见于慢性肾功能衰竭、病毒性肝炎、晚期肝硬化和慢性荨麻疹(一种典型的组胺相关疾病,对内源性组胺的耐受性降低)的患者。
二胺氧化酶降解能力下降还可能是由于缺乏其辅助因子、维生素B6、维生素C、铜和锌而导致的。
• 肠道微生物会影响组胺不耐受的发生
肠道微生物群也会影响组胺不耐受的发生。最近的研究表明,组胺不耐受可能是由肠道微生物群的改变引起的。肠道中大量的组胺分泌细菌可能会导致组胺不耐受。健康人体内双歧杆菌的数量较多。而在血清二胺氧化酶活性降低的人群中观察到变形菌属数量较多。
组胺不耐受人群存在肠道菌群失调
在组胺不耐受人群中观察到肠道菌群失调,与健康个体相比,与肠道健康相关的普氏菌科(Prevotellaceae)、瘤胃球菌(Ruminococcus)、粪杆菌(Faecalibacterium)和普拉梭菌(Faecablibacterium prausnitzii)的比例明显较低。组胺不耐受人群还具有显著更高丰度的组胺分泌细菌,包括葡萄球菌属和变形杆菌属、属于肠杆菌科的几个未识别属,以及产气荚膜梭菌和粪肠球菌。
需要注意的是,几项研究表明,即使在没有二胺氧化酶缺陷的个体中,大量的组胺产生细菌也会导致肠道内高水平组胺的积累、随后在血浆中的吸收以及不良反应的出现。
低组胺饮食
组胺不耐受疗法的基础是基于低组胺含量食物的消除饮食。一般来说,公认的饮食指南尚未制定;但是,建议避免食用富含组胺的食物如奶酪、油性鱼类和贝类,以及生发酵肉制品、腌菜、发酵豆制品、葡萄酒和啤酒、鸡蛋、巧克力和蘑菇。
避免食用易刺激组胺释放的蔬果
此外,应避免食用会刺激内源性组胺释放的蔬菜和水果(如菠菜、西红柿、柑橘类水果、草莓、茄子、鳄梨、木瓜、香蕉、猕猴桃、菠萝和李子)。


Kovacova-Hanuskova E,et al.Allergol Immunopathol.2015
补充二胺氧化酶
研究还发现补充二胺氧化酶可以改善组胺不耐受患者的生活质量。已开发出基于肠溶猪肾提取物的含二胺氧化酶食品添加剂。豆芽也被用作二胺氧化酶的来源,这种酶的活性是未发芽种子的250倍。
施用抗组胺药物
纠正组胺不耐受的药物方案基于使用组胺受体阻滞剂、肥大细胞膜稳定剂和其他具有抗组胺活性的药物。
在这方面,H1R拮抗剂是最有效的,特别是依巴斯汀。每天服用20毫克该药物,持续12周,46%的患者出现临床缓解。
一些生物活性物质有助于增强组胺降解
维生素C被用作纠正组胺不耐受的佐剂。每日剂量300-500毫克可增强组胺降解并抑制肥大细胞脱颗粒。
天然类黄酮(非瑟酮、山奈酚、槲皮素、芦丁和木犀草素)和活性生物碱小檗碱在体外抑制肥大细胞脱颗粒。这些都有助于减轻组胺不耐受。
虽然组胺含量过低的情况相对较少见,但也可能会对身体产生一些影响,包括:
• 伤口愈合减慢
组胺在人体中起着重要的调节作用,其能够帮助调节纤维细胞生长因子的生成,该成分可以加速局部肉芽的生长,帮助促进伤口的愈合。因此组胺含量较低时,伤口愈合能力也会减弱,速度会随之减慢。
• 消化不良
组胺可以通过刺激胃部,进而起到促进胃酸分泌的效果。若身体中没有组胺,则会导致胃酸减少,无法有效消化分解食物,进而引起消化不良的问题。
• 情绪不良
组胺还可以作为神经递质帮助调节中枢神经系统,而一旦没有组胺,则会影响中枢神经系统稳定性,引起睡眠异常、激素水平紊乱等情况,进而导致情绪不良。
• 血压升高
人体缺少组胺还会导致血压升高。因为组胺可以促进血管扩张,增加血管的通透性,所以具有降低血压的作用。若没有该成分,则血管会由于舒张能力减弱,因此出现血压升高的问题。
• 免疫力下降
组胺对免疫细胞,如吞噬细胞、T细胞、B细胞等,起到增强活性的功效,因此组胺还具有增强免疫功能的作用。若人体缺少该物质,则人体免疫力会下降,无法有效抵抗细菌、真菌等有害物质的侵入。
组胺对人体许多生理功能都可以起到积极作用,但是人体中的组胺也不能过量,否则容易导致过敏反应,出现头晕、皮肤瘙痒、胸闷等不适症状。
此种情况需及时就医,并且在医生的指导下服用抗组胺药物,例如盐酸西替利嗪片、氯雷他定片等药物,进行抗过敏治疗。
★ 组胺的免疫调节因受体的不同而存在区别
先天性和适应性免疫系统的细胞都可以被组胺调节。组胺的调节取决于它与组胺受体的四种亚型的结合,这些亚型按发现时间顺序命名——H1R-H4R,在多种细胞类型中差异表达,进而产生不同的作用。
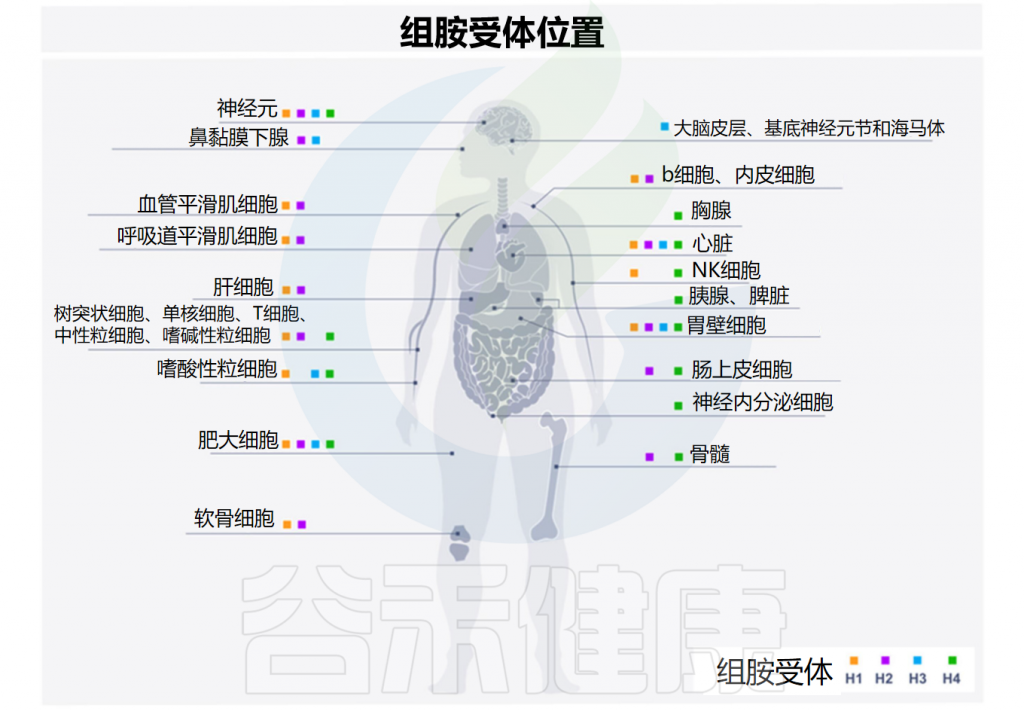
Smolinska S,et al.Metabolites.2022
1
H1R
H1R受体存在于几乎所有组织的内皮细胞和血管平滑肌细胞中(它们的密度在胃肠道中特别高),以及下丘脑、肾上腺髓质和免疫活性细胞(单核细胞、中性粒细胞、T细胞和B细胞)中。
• H1R受体介导的主要作用
H1R刺激的主要作用是炎症、全身血管舒张、血管通透性增加、支气管收缩、回肠收缩和昼夜节律周期调节。
外周H1R介导的影响包括鼻漏、支气管收缩、过敏反应、结膜炎和荨麻疹,而中枢相关的H1R影响包括食物和水摄入的调节、抽搐、注意力和睡眠调节。
• H1R是过敏反应的主要受体
IL-3、IL-4和组胺可上调H1R基因表达,而H1R激活导致许多与过敏性即时型超敏反应相关的特征,例如发红、瘙痒和肿胀。
例如小鼠模型中H1R的激活会诱导IFN(干扰素)产生增加,这与1型辅助性T细胞的增殖有关,并诱导促炎作用。
研究表明,瘙痒因子(例如神经生长因子、信号素 3A)的表达受组胺H1R的调节。在小鼠模型和特应性皮炎患者中,使用H1R拮抗剂可降低IL-31(白细胞介素-31)水平,这与瘙痒的发作有关。
2
H2R
与H1R类似,H2R的表达存在于多种组织和细胞中,包括脑、胃壁细胞、平滑肌细胞、T和B细胞、树突状细胞和心脏组织。H2R受体位于突触后,主要通过环腺苷单正膦(cAMP)传递信号并与Gαs偶联。
不同组胺受体下游的信号通路
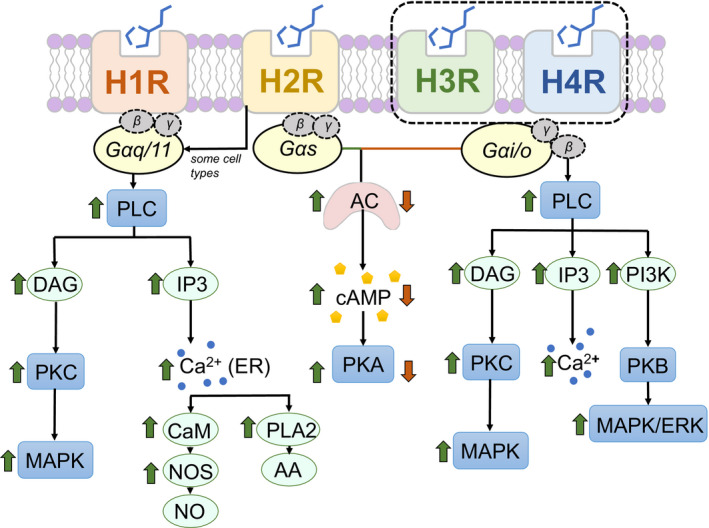
Sudarikova AV,et al.Physiol Rep.2021
H2R刺激的主要作用是外分泌(主要是盐酸)、心动过速、平滑肌细胞松弛、抗炎作用、抗体合成等免疫活动。
• H2R具有抗炎作用
在组胺与H2R结合期间,H2R刺激通过抑制单核细胞或巨噬细胞和肥大细胞产生IL-12、IFN-γ、TNF-α细胞因子,而IL-10分泌增加,从而引起抗炎作用。
• H2R影响对微生物的免疫反应
组胺(内源性和外源性)通过H2R显著改变对微生物的先天免疫反应。特别地,树突状细胞对微生物配体的反应被组胺以H2R依赖的方式显著改变。
H2R敲除的小鼠免疫系统紊乱以及胃缺陷(胃酸分泌减少),此外还观察到认知能力下降和伤害感受异常。
• 注意:不同组胺受体可能具有相反的作用!
H2R以与H1R不同的方式传输信号,H1R和H2R的激活对各种生物过程具有相反的影响。在T细胞介导的免疫反应中,H1R激活促进Th1极化,而H2R激活则抑制Th1极化。
在平滑肌收缩中也观察到H1R和H2R激活的相反作用。H1R和H2R拮抗剂分别抑制和加剧组胺引起的轻度哮喘患者的支气管痉挛。这些数据表明,组胺可能会产生相反的作用,具体取决于被激活的特定组胺受体。
3
H3R
• H3R主要存在于神经系统中
H3R受体在神经系统细胞中表达,特别是在大脑皮层,基底神经节神经元和海马体中。H3R位于含组胺神经元的突触前区域。它们的功能是调节组胺以及其他神经递质(如多巴胺、去甲肾上腺素、γ-氨基丁酸、乙酰胆碱和血清素)的合成和释放。
• H3R受体影响睡眠、认知障碍等精神健康
H3R受体表达和激活的变化在睡眠-觉醒周期障碍、注意力缺陷多动障碍、癫痫和认知障碍以及炎症的发展中发挥重要作用。
H3R缺陷小鼠表现出行为反应和运动的变化,以及伴有肥胖、食欲过盛以及瘦素和胰岛素水平升高的代谢综合征。
• H3R可能会促进炎症反应
H3R刺激可增加促炎活性以及免疫细胞呈递抗原的能力。H3R敲除也可导致神经炎症性疾病的严重程度和T细胞中IFN-诱导蛋白10的表达增加。
因此,组胺H3R拮抗剂的使用有可能用于预防或抑制炎症性疾病(例如呼吸系统疾病)的发展。
4
H4R
H4R受体是最近才发现的,它们的作用尚未完全了解。H4R主要存在于免疫细胞(嗜酸性粒细胞、嗜碱性粒细胞、肥大细胞、自然杀伤细胞、树突状细胞、单核细胞和T细胞)中,也存在于脾脏、胸腺、骨髓、胆管、胰腺、肠上皮细胞和神经内分泌细胞中。
• H4R会增强过敏和炎症反应
与其他类型的组胺受体相比,H4受体在中枢和周围神经系统中表达并不显著。H4R通过与蛋白质Gα/io相互作用介导,参与炎症和超敏反应的发展。
H4R介导的肥大细胞活化,促炎细胞因子和趋化因子IL-6、TNF-α、TGF-β1、RANTES、IL-8、MIP-1α和MCP-1均被表达。研究发现H4R激活显著增强结肠炎、放射性结肠炎、肠道缺血/再灌注损伤和过敏反应的炎症反应。
因此,选择性H4R阻滞剂的开发可能代表了治疗炎症性肠病的一种有前途的方法。
• H4R的激活还会导致瘙痒
H4R的激活已被证明会导致瘙痒。而H1R和H4R拮抗剂的联合治疗可能通过协同抑制瘙痒和皮肤炎症对慢性皮炎产生显著的治疗效果。
此外,H4R通过激活Th2细胞并产生 IL-31,可能引发过敏性皮炎的发生。H4R和H3R的激活增加了乙酰胆碱对肠道蠕动的影响。H4R受体还参与消化性溃疡的形成和癌变。
//小结
如前所述,组胺特异性受体的组织定位、功能和对组胺的亲和力不同。组胺调节广泛的代谢过程,具有促炎和抗炎作用,具体取决于受体亚型和受刺激细胞的类型。
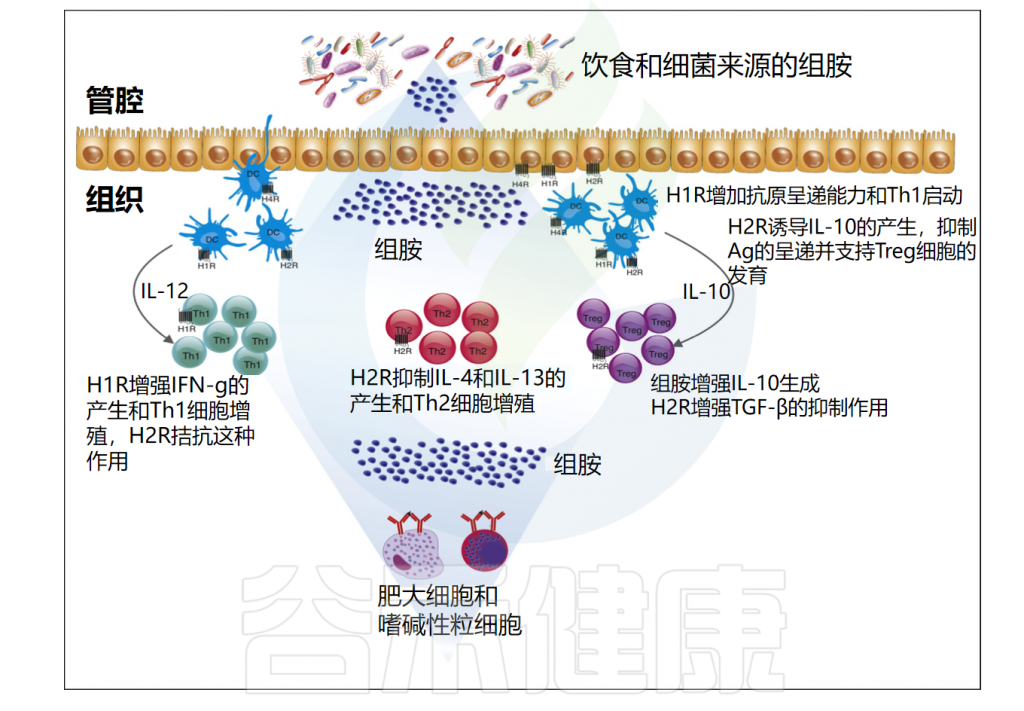
Barcik W,et al.Curr Opin Immunol.2017
组胺对免疫调节的各种作用似乎与其受体及其各自的细胞内信号的差异表达和调节有关。此外,这些受体对组胺的亲和力的差异在组胺和组胺受体药物配体的生物效应中发挥着关键作用。组胺受体在多种病理生理状况中发挥重要作用,是治疗过敏的有效靶点。
组胺因其在速发型超敏反应中的作用而广为人知。在胃肠道内,组胺以相对较高的浓度存在,特别是在炎症过程中。组胺水平的增加改变了宿主与微生物群的免疫相互作用,并导致体内平衡的破坏,导致许多难以应对的肠道疾病的发展。
★ 组胺的水平会影响肠道稳态
肠道中的组胺水平受到宿主过敏和炎症反应的影响,除了宿主微生物群的产生外,还会以某种方式改变降解或合成组胺的酶的活性及其饮食摄入量。
此外,在刺激产生组胺的免疫细胞时,可以增加内源性组胺的水平。所有这些都会影响肠道稳态,导致组胺积累,并影响特定疾病。组胺还可能对肠道寄生虫和细菌感染产生一定影响。
注:除H3R外,所有组胺受体都在人体肠道中表达。从量的角度来看,H4R表达的丰度明显低于H1R和H2R。
粘膜内的组胺
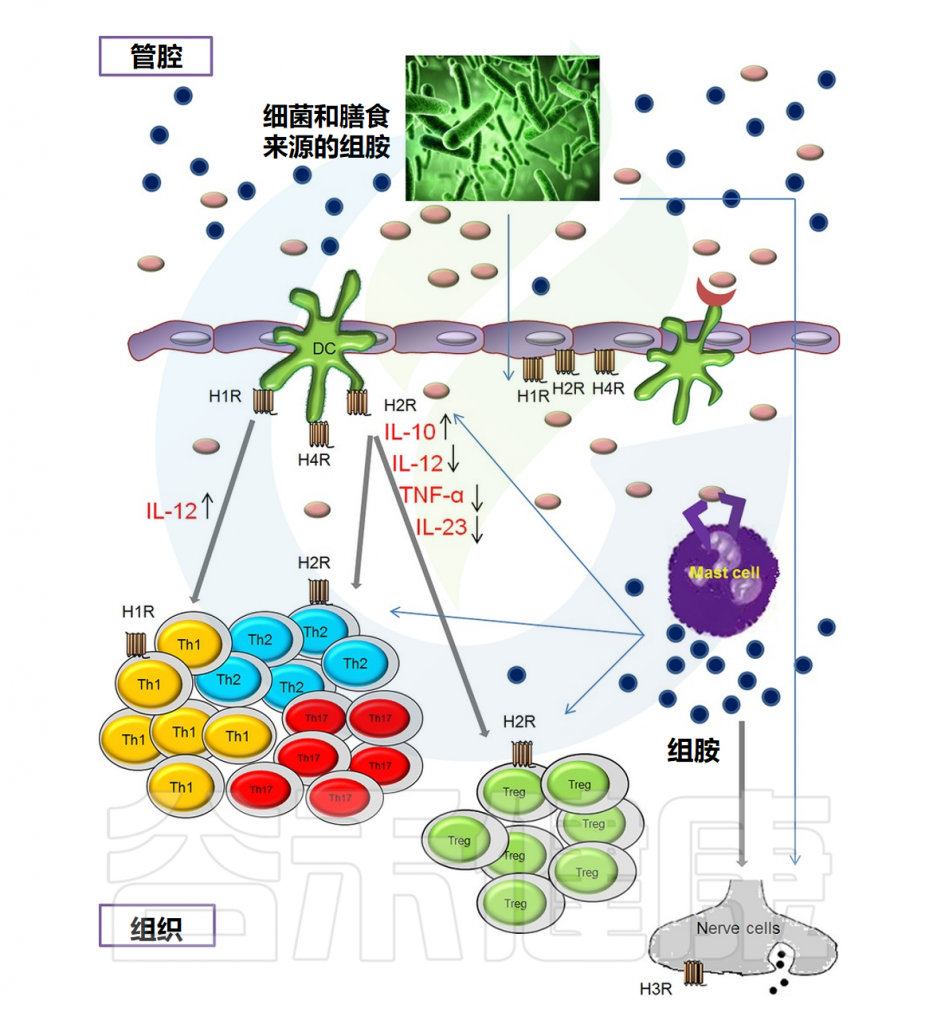
Smolinska S,et al.Allergy.2014
• 食物中的一些菌株会分泌组胺
除哺乳动物细胞外,还发现许多不同的细菌菌株能够在组氨酸通过组氨酸脱羧酶作用后分泌组胺。
最有力的证据就是那些能够在食品中分泌组胺的细菌。它们在奶酪、肉罐头、乳制品以及啤酒和葡萄酒发酵过程中产生组胺。鲭鱼中毒就是在储存或加工过程中处理不当,细菌代谢了鱼类中的组氨酸,导致组胺含量过高。
• 部分肠道微生物也会分泌组胺
迄今为止,对产生组胺细菌的研究主要集中在从食物样品中分离的菌株。肠道微生物群的组胺生成能力直到最近才被研究,数据仍然有限。
2021年发表的一项荟萃分析展示了人类肠道微生物组中具有假定组胺分泌能力的物种 ,其中许多属于广泛报道的组胺产生属,例如摩根菌属(Morganella),乳杆菌(Lactobacillus),葡萄球菌属(Staphylococcus), Photobacterium和巴氏梭菌(Clostridium)。
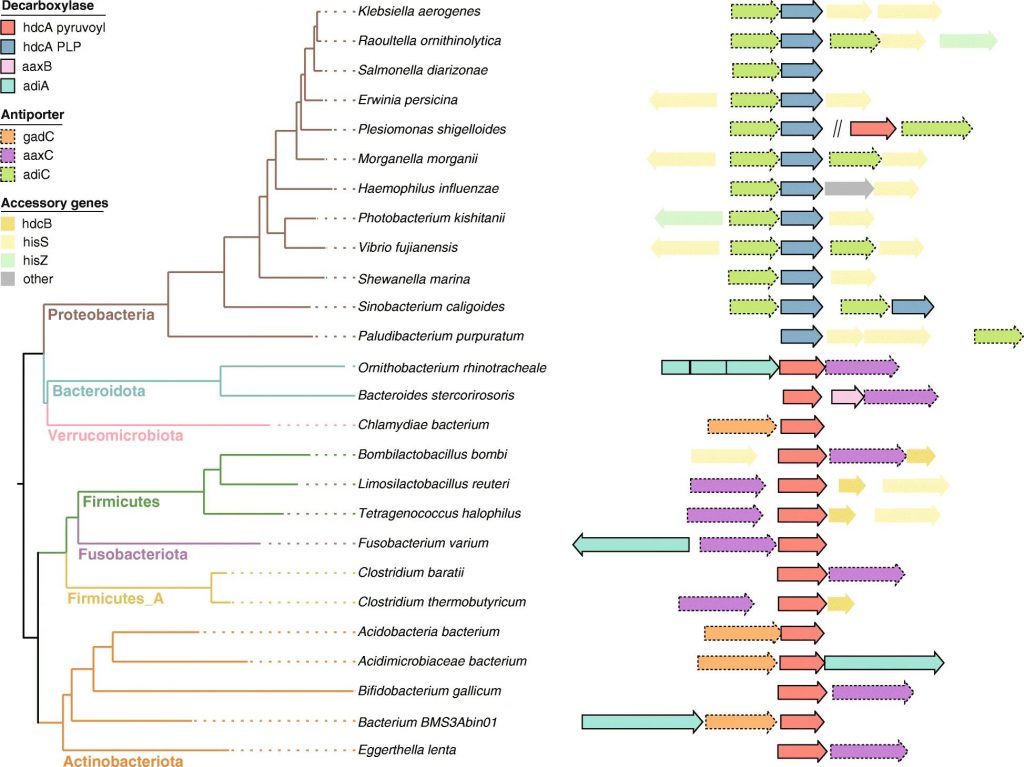
Mou Z,et al.BMC Genomics.2021
• 一些细菌具有组氨酸脱羧酶活性,也可能产生组胺
需要注意的是,一些具有组氨酸脱羧酶活性并与组胺中毒有关的细菌,可能也是潜在的组胺产生细菌。包括:Morganella morganii,
Eschericha coli,
Hafnia alvei,
Proteus vulgaris,
Proteus milabilis,
Enterobacter aerogenes,
Raoultella planticola,
Raoultella ornithinolytica,
Citrobacter freundii,
Pseudomonas fluorescens,
Photobacterium damselae。
证据表明细菌衍生的组胺具有多种影响,例如对宿主健康和食品安全。
• 抑制了促炎细胞因子的分泌
初步体外研究表明,组胺抑制人单核细胞衍生的树突状细胞中趋化因子和促炎细胞因子的分泌。
小鼠研究表明,给予分泌组胺的鼠李糖乳杆菌具有抗炎作用,各种白细胞介素和肿瘤坏死因子α的分泌减少。这种作用在缺乏组胺2受体的动物中消失,表明微生物群衍生的组胺可能具有免疫调节作用。
• 组胺的含量对于免疫效果至关重要
与鼠李糖乳杆菌相比,另一种乳杆菌能够分泌大约100倍多的组胺,服用这种乳杆菌会导致动物体重减轻和健康状况恶化。谷禾认为,微生物分泌的组胺量可能是决定其作用性质的关键。
最近的研究还表明,肠道中细菌释放的组胺不仅会引起宿主免疫系统的局部调节,而且还会在远处的粘膜部位(例如肺部)产生免疫学后果。例如成年哮喘患者肠道中分泌组胺的细菌丰度有所增加。
由于许多食物中含有组胺,一部分肠道微生物群也可以产生组胺,研究发现组胺在肠道疾病中可能发挥多种作用,下面讲述了一些与组胺相关的肠道疾病,组胺在其中发挥着有益或有害的作用。
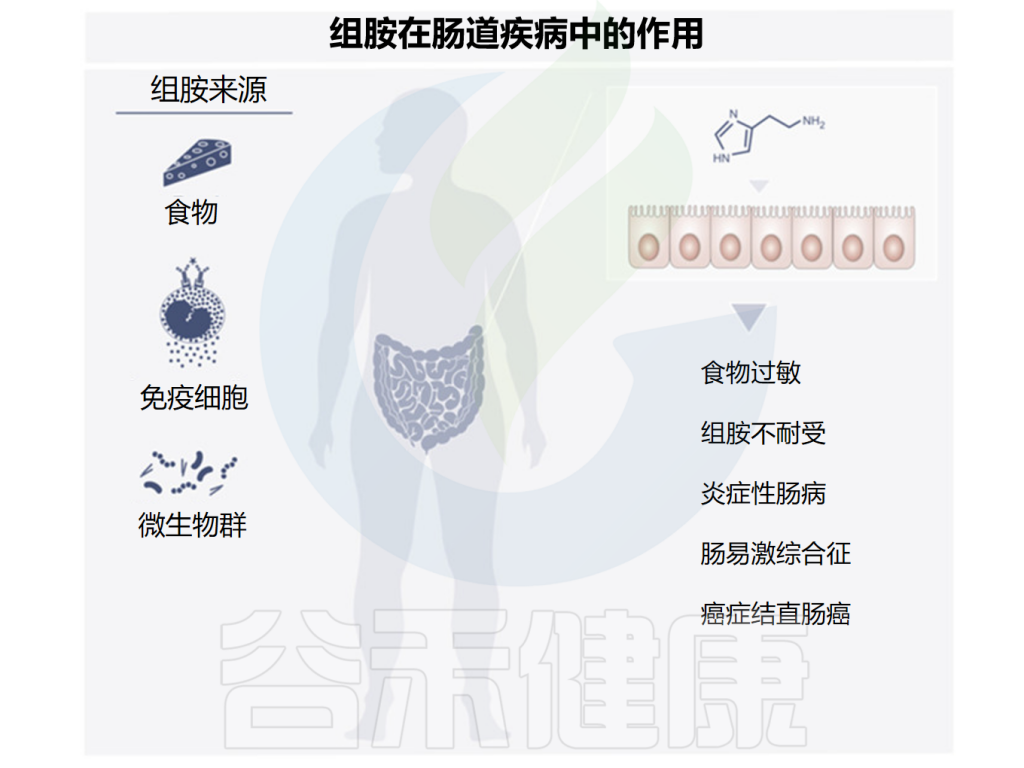
Smolinska S,et al.Metabolites.2022
消化道是接触大量不同分子的地方,这些分子中的一部分是潜在的过敏原,可能从而引起食物过敏。食物过敏可表现为轻度和重度症状,最严重的可能危及生命。
拓展:常见的食物过敏症状
食物过敏的症状表现在呼吸系统、消化系统、心血管系统和皮肤系统等。
皮肤反应:皮肤反应是最常见的食物过敏症状之一。它可以表现为皮疹、荨麻疹、瘙痒、红肿、水肿等。这些反应通常发生在口腔、面部、颈部和四肢等部位。
呼吸道症状:食物过敏还可以引起呼吸道症状,包括鼻塞、流涕、打喷嚏、喉咙痒、咳嗽、喉咙紧闭感和哮喘。严重的过敏反应可能导致呼吸困难和哮喘发作。
消化系统症状:食物过敏可以引起消化系统症状,包括腹痛、腹胀、恶心、呕吐和腹泻。这些症状还可能与肠道炎症反应、肠道收缩有关。
循环系统症状:在严重的食物过敏反应中,可能出现循环系统症状,如低血压、心悸、头晕和昏厥。这些症状是由于过敏反应导致血管扩张和血压下降。
全身症状:某些人可能出现全身性症状,如疲劳、焦虑、情绪波动和体重下降。这些症状可能与食物过敏引起的炎症反应和免疫系统的激活有关。
• 食物过敏时组胺分泌增加
IgE依赖性食物过敏是免疫系统紊乱的结果,使耐受性丧失。这导致将温和的食物抗原识别为病原体。组胺受体参与食物过敏的发展。研究表明,在人类中使用H2R拮抗剂会增加针对食物抗原的IgE的产生。在Th2细胞的刺激下,IgE与效应细胞(肥大细胞和嗜碱性粒细胞)上的Fcε受体结合。由于效应细胞的激活,组胺以及其他介质被释放。
在食物过敏受试者中,组胺分泌增加和肥大细胞数量增加已得到充分证明。嗜碱性粒细胞释放的组胺与皮肤点刺试验和食物激发呈正相关。与非过敏患者相比,食物过敏患者抗IgE介导的肥大细胞组胺释放增加。
此外,将食物过敏患者的活检与抗IgE(人类)抗体或过敏原一起孵育,会导致组胺释放增加9倍。
• 食物过敏的治疗
由于食物过敏的发生率不断增加,已经进行了大量研究以开发新的治疗和预防策略。严格避免食物过敏原是管理IgE介导的食物过敏的长期策略。
关于食物过敏发病机制的各个阶段也有许多研究,例如对Th2通路的影响、阻断IgE、抑制效应细胞和微生物治疗。长期免疫耐受应该是治疗食物过敏最理想的效果。
口服免疫疗法是食物过敏的发展治疗方法之一。它包括以每2-4周增加一次的剂量向患者施用过敏原,直到达到最大维持剂量。这个过程的结果是发展对食物的耐受性。该方法已用于对牛奶、鸡蛋、小麦、花生、坚果和贝类的食物过敏。
目前正在研究表皮和舌下免疫疗法。对牛奶和鸡蛋过敏的表皮免疫疗法也进行了临床试验。
炎症性肠病是特发性、慢性复发性肠道疾病。溃疡性结肠炎和克罗恩病是两种主要临床表现。
在克罗恩病中,炎症出现在弥漫性病变中,这些病变遍布消化道并深入穿透肠壁,可能影响所有层。相反,溃疡性结肠炎的炎性病变从直肠开始,向上但不超过结肠,并在黏膜浅表处保持。克罗恩病导致整个胃肠道的透壁性炎症,但其特征是模式不连续。与炎症为浅表的溃疡性结肠炎相比,溃疡主要局限于结肠粘膜。
• 炎症性肠病患者黏膜组胺水平升高
这两种疾病的发展都是遗传易感患者宿主免疫系统、肠道微生物群和环境因素之间复杂相互作用的结果。
炎症性肠病患者的黏膜组胺水平(而非血浆水平)升高。在患者尿液中发现与疾病活动度相关的N-甲基组胺水平升高。当受到抗原刺激时,源自活动性克罗恩病或溃疡性结肠炎切除结肠的肥大细胞能够释放比正常结肠更多的组胺。
• 炎症性肠病患者的组胺通路被破坏
克罗恩病和溃疡性结肠炎患者的组胺信号通路均被破坏,研究分析显示。炎症性肠病患者的组胺受体表达和功能活性发生改变。
在结肠炎模型中,H2R的阻断导致更严重的炎症性疾病。组胺主要抑制IFN-γ和TNF-α的分泌,这些细胞因子的基因表达与溃疡性结肠炎患者的H4R和H2R表达呈正相关。
发炎黏膜中组胺-N-甲基转移酶(HNMT)表达降低,二氨基氧化酶(DAO)多态性与炎症性肠病风险增加有关。使用H2R拮抗剂会增加克罗恩病患者的风险。
注:在小鼠结肠炎模型中,缺乏H2R或使用非莫替丁(H2R拮抗剂)抑制H2R的T细胞转移可加速体重减轻并增加疾病严重程度。
• 炎症性肠病的治疗
在许多情况下,获得缓解的唯一选择是根治性手术,切除发炎区域。潜在地,使用H1R和H4R拮抗剂同时使用H2R激动剂可能对炎症性肠病患者有益。
炎症性肠病患者还可接受抗炎药、类固醇、抗生素、氨基水杨酸盐治疗,或使用英夫利昔单抗(抗 TNF-α)进行生物疗法。
肠易激综合征是一种与腹部不适或疼痛有关的慢性疾病,其中吃的食物是更严重症状的触发因素。一些证据表明,肠易激综合征中肠道菌群发生了改变,这种改变会破坏黏膜的免疫应答。
• 内源性组胺与肠易激综合征的严重程度相关
研究显示,超过一半的患者因食用释放组胺的食物或富含生物胺的食物而出现胃肠道症状。内源性组胺的水平与肠易激综合征患者症状的严重程度绝对相关。活化的肥大细胞产生更多的组胺,这与肠易激综合征患者的腹痛相关。肠易激综合征患者的黏膜上清液的组胺含量高于健康受试者的上清液。
组胺水平更高,组胺受体表达也更高
肠易激综合征患者中组胺受体H1R和H2R的表达水平上调。我们推测:伴有组胺分泌或含组氨酸脱羧酶细菌的菌群失调可能与肠易激综合征的发生和加重有关。
给予特定微生物具有治疗作用,这也可能是微生物群变化导致疾病的原因。肠易激综合征没有特效治疗方法,减少炎症的药物正在使用中。
结直肠癌是第三常见癌症,也是癌症相关死亡的重要原因。与一般人群相比,炎症性肠病患者发生结直肠癌的终生风险会增加。
• 组胺可能具有潜在的抗肿瘤作用
肠道微生物群在结直肠癌中的作用已在谷禾之前的文章中详细描述过。而特定的肠道微生物代谢物可能与结直肠癌的病因有关。组氨酸脱羧酶缺乏已被证明通过CD11b+Gr-1+未成熟髓系细胞的积累促进炎症相关的结直肠癌,表明组胺具有潜在的抗肿瘤作用。
几种益生菌菌株,包括长双歧杆菌(Bifidobacterium longum)、嗜酸乳杆菌(Lactobacillus acidophilus NCFM)和鼠李糖乳杆菌GG,在结肠癌的小鼠模型中显示出有益作用。这些产生组胺的益生菌减少了结肠肿瘤的数量和大小。
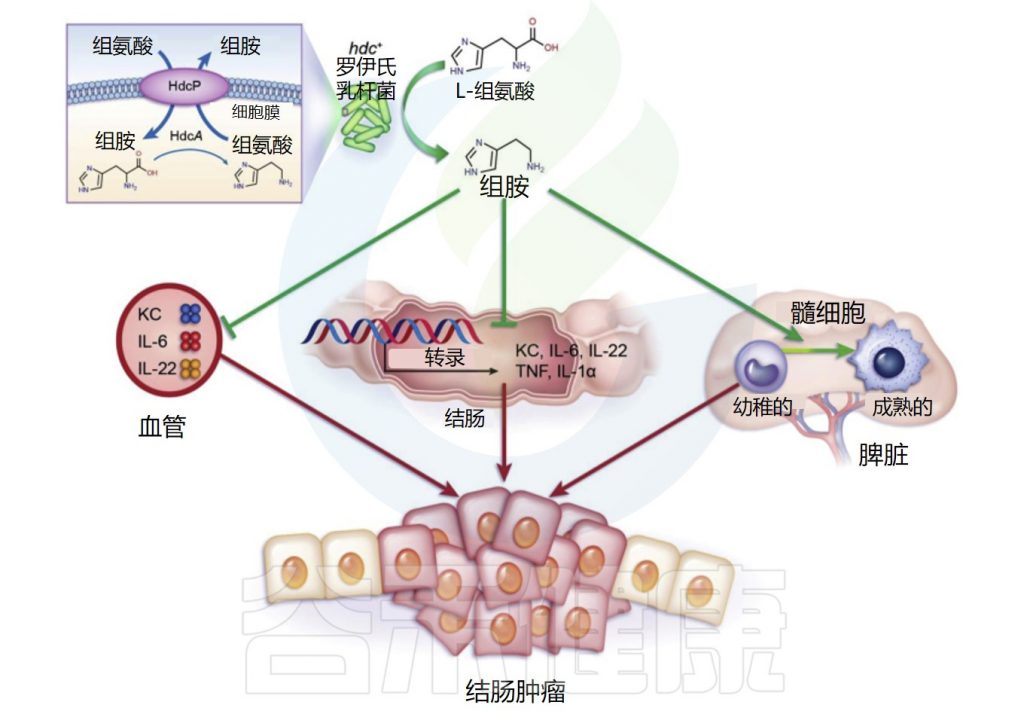
Gao C,et al.Am J Pathol.2017
罗伊氏乳杆菌(L. reuteri)抑制结肠粘膜中IL-22、IL-6、tnf-α和IL-1α 基因的表达,并减少血浆中促炎、癌症相关细胞因子、角质形成细胞趋化剂、IL-22和IL-6的数量。
此外,不能产生组胺的等基因组氨酸脱羧酶(HDC)缺陷罗伊氏乳杆菌突变体不能抑制致癌作用,这表明共代谢物组胺在抑制慢性肠道炎症和结直肠肿瘤发生中具有重要作用。
• 结直肠癌患者组胺含量和活性增加
在结直肠癌患者的结肠黏膜中,组氨酸脱羧酶活性和组胺含量均高于正常样本。在实验诱导的结直肠癌小鼠中,与野生型小鼠相比,组氨酸脱羧酶的缺失导致肿瘤发生增强,这也表明组胺具有抑制结直肠肿瘤发生的作用。
• 注意:过高的组胺含量可能对癌症不利!
需要注意的是,已发现肥大细胞在结肠癌中含量丰富,并促进小鼠化学诱导的结直肠癌的癌变,并且与人类结直肠癌患者的不良预后有关。与组胺通过H4R的促炎作用类似,H4R表达的缺失也导致小鼠化学诱导的致癌作用减少。
组胺作为一种重要信号分子,主要因其在诱导过敏症状中的作用而被识别,但它也参与非过敏性炎症反应。
组胺是肠道稳态的重要调节剂。通过增强分泌和蠕动,组胺促进潜在有毒物质的快速清除。这种生物胺对于肠道细菌的生存也至关重要。与健康个体相比,组胺不耐受患者的肠道菌群失衡或失调。
在某些浓度范围内,组胺起着保护作用,对于维持健康状态至关重要。然而,由于先天性或后天性缺乏组胺降解酶,以及肠道肥大细胞过度增生和过度活跃,使组胺浓度较高,会导致粘膜炎症性疾病,产生腹泻和腹痛。增强H2R表达或其细胞内信号,同时降低H1R和 H4R活性是提高粘膜免疫力的合理方法
组胺不耐受的现代治疗方法包括避免高组胺食物的摄入、影响组胺水平的药物(H1R阻滞剂、肥大细胞膜稳定剂和米氮平)和补充天然成分(维生素C、类黄酮、二氨基氧化酶强化补充剂和益生菌) 。
与此同时,使用具有释放组胺潜力的肠道微生物群提供了一种新的治疗视角。
主要参考文献
Smolinska S, Winiarska E, Globinska A, Jutel M. Histamine: A Mediator of Intestinal Disorders-A Review. Metabolites. 2022 Sep 23;12(10):895.
Sánchez-Pérez S, Comas-Basté O, Duelo A, Veciana-Nogués MT, Berlanga M, Latorre-Moratalla ML, Vidal-Carou MC. Intestinal Dysbiosis in Patients with Histamine Intolerance. Nutrients. 2022 Apr 23;14(9):1774.
Shulpekova YO, Nechaev VM, Popova IR, Deeva TA, Kopylov AT, Malsagova KA, Kaysheva AL, Ivashkin VT. Food Intolerance: The Role of Histamine. Nutrients. 2021 Sep 15;13(9):3207.
Krell T, Gavira JA, Velando F, Fernández M, Roca A, Monteagudo-Cascales E, Matilla MA. Histamine: A Bacterial Signal Molecule. Int J Mol Sci. 2021 Jun 12;22(12):6312.
Mou Z, Yang Y, Hall AB, Jiang X. The taxonomic distribution of histamine-secreting bacteria in the human gut microbiome. BMC Genomics. 2021 Sep 26;22(1):695.
Gao C, Ganesh BP, Shi Z, Shah RR, Fultz R, Major A, Venable S, Lugo M, Hoch K, Chen X, Haag A, Wang TC, Versalovic J. Gut Microbe-Mediated Suppression of Inflammation-Associated Colon Carcinogenesis by Luminal Histamine Production. Am J Pathol. 2017 Oct;187(10):2323-2336.
Kovacova-Hanuskova E, Buday T, Gavliakova S, Plevkova J. Histamine, histamine intoxication and intolerance. Allergol Immunopathol (Madr). 2015 Sep-Oct;43(5):498-506.
Barcik W, Wawrzyniak M, Akdis CA, O’Mahony L. Immune regulation by histamine and histamine-secreting bacteria. Curr Opin Immunol. 2017 Oct;48:108-113.
Smolinska S, Jutel M, Crameri R, O’Mahony L. Histamine and gut mucosal immune regulation. Allergy. 2014 Mar;69(3):273-81.

谷禾健康

胃肠道除了在食物消化和吸收中发挥作用外,还是一个主要的内分泌器官,产生多种生物活性肽和胺类物质,作为肠道激素。
大多数肠道激素来源于肠内分泌细胞,但来源于其他肠上皮细胞类型的生物活性肽也与代谢调节有关。
为什么说肠道激素重要呢?我们在日常生活中胃口好不好都可能与它相关,它能影响食欲和食物摄入,通过影响大脑中的食欲中枢,减轻食欲或刺激饱腹感。
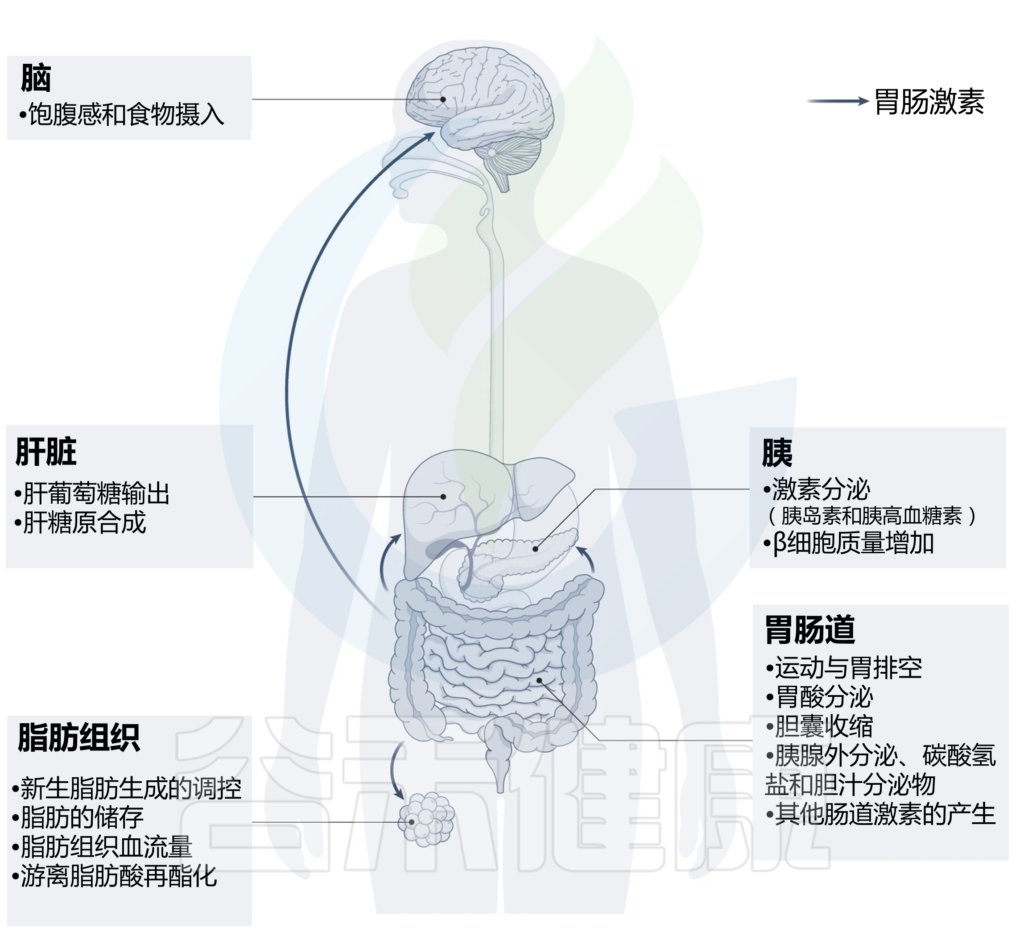
其实,肠道激素在多种器官的多种代谢活跃组织中协调关键的生理过程,包括胰腺、肝脏、脂肪组织、肠道和中枢神经系统等。从局部控制营养吸收和调节肠道生长、肠道运动和液体分泌,到远距离调节胰岛素释放、脂肪细胞功能。
近日,来自英国剑桥大学威康Trust-MRC代谢科学研究实验室团队发表了一篇文章在《Nature Reviews Gastroenterology & Hepatology》期刊,主要讲述了参与代谢调节的肠道激素,讨论了它们在胃肠系统内外的作用。

肠道激素由多种肠内分泌细胞和非肠内分泌细胞产生,它们对不同刺激的反应不同,并分泌特定的激素组合。
肠内分泌细胞通过一系列感觉转运蛋白、离子通道和受体感知各种营养和非营养刺激,使其成为营养吸收和肠道激素反应之间的有效联系。
这些肠道激素信号传导过程可以作为治疗靶点;例如,GLP2受体激动剂用于治疗短肠综合征,并且GLP1受体激动器用于治疗2型糖尿病和肥胖。此外,激素类药物的组合可用于最大限度地提高疗效,同时最大限度地减少不良反应。
因此,深入了解肠道内分泌系统介导的复杂器官间串扰,有助于设计基于或靶向肠道激素及其受体的更有效药物,并将其治疗潜力扩展到肥胖和糖尿病之外的疾病。
肠内分泌细胞沿着胃肠上皮的长度广泛分布。肠道内的一些细胞会分泌一些激素来应对不同的刺激。
■ 肠内分泌细胞可分为两大类:
以前我们认为这些肠内分泌细胞只会分泌单一类型的激素,但现在我们知道,实际上它们通常会同时分泌多种不同的激素,所以它们之间的分泌有一些重叠的部分。
■ 肠道中不同部位的激素表达也会有所不同,如:
肠道特定区域的激素产生与激素的功能作用及其反应的感觉输入有关。
决定肠道不同区域肠内分泌细胞激素特征的因素在很大程度上仍然未知。来自不同肠段的肠道类器官似乎保留了与起源相关的激素特征,这表明肠道干细胞的效力有限。
■ 肠道细胞功能受限的机制与激素分泌的发育演化
然而,随着细胞的发育成熟,肠内分泌细胞的激素特征会发生变化,并沿着隐窝-绒毛轴分化。
举个例子,回肠L细胞在肠隐窝中共表达GLP1和肽YY(PYY),但当这些细胞到达绒毛时,前胰高血糖素转录下调,分泌素转录上调。一旦这些激素分泌出来,它们有两条路:
■ 激素在肠道中的分泌调节及其影响
肠道激素控制多种代谢活性组织的广泛生理过程,包括胰腺、肝脏、脂肪组织、肠道和中枢神经系统。这些激素大多具有不止一种生理作用,许多生理作用由不止一种激素发挥。如下表:
调节新陈代谢的主要肠道激素的特征

Bany Bakar R, et al., Nat Rev Gastroenterol Hepatol. 2023
■ 肠内分泌细胞如何与营养物质密切接触
这就需要我们先来了解一下,肠内分泌细胞的结构特点,大多数肠内分泌细胞是开放型细胞,也就是说它们有一个顶端表面,带有向肠腔延伸的微绒毛,以及一个基底外侧表面,负责在刺激时通过胞吐释放分泌颗粒。
因此,这些细胞与消化和吸收的营养物质直接接触,把它们推向产生肠道激素反应的理想位置,将营养吸收的变化传达给身体其他部位。
它怎么传达呢?
肠内分泌细胞的化学感受能力归因于它们表达一系列感觉转运蛋白、离子通道和受体,这些受体解释了它们对各种营养刺激的反应能力,包括葡萄糖、消化脂质、碳水化合物和蛋白质,以及非营养刺激,如胆汁酸、肠道激素、肠道神经递质,微生物产物和炎性胞质分裂。
■ GPCR——肠内分泌细胞通过其检测营养物质吸收
G蛋白偶联受体(GPCR)检测到广泛的营养和非营养刺激,肠内分泌细胞差异表达一系列GPCR,这些GPCR主要位于其基底外侧表面,能够检测出胆汁酸、游离脂肪酸(FFAs)和氨基酸在上皮层内的消化和吸收过程中的变化。
与肠道激素释放密切相关的GPCR包括:
各种脂质感应受体,包括:
相反,葡萄糖在通过顶端膜上的钠偶联葡萄糖转运蛋白SGLT1吸收时激活肠内分泌细胞。SGLT1对钠离子与葡萄糖的共转运直接触发质膜去极化,进而激活电压门控钙通道。由此产生的钙流入进一步使肠内分泌细胞质膜去极化,并启动分泌颗粒的释放(下图)。
调节肠内分泌细胞活性的营养诱导机制
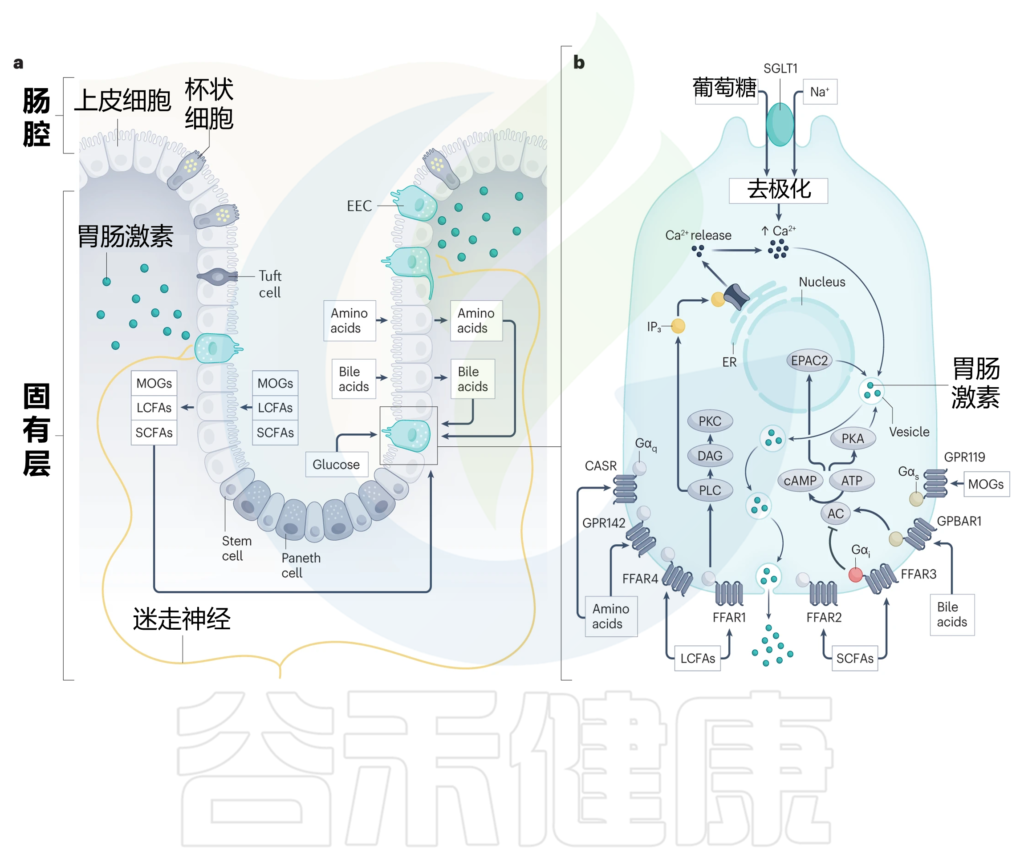
Bany Bakar R, et al., Nat Rev Gastroenterol Hepatol. 2023
肠道激素GDF15、FGF19(小鼠中的FGF15)、鸟苷素和尿鸟苷素不是由肠内分泌细胞产生的,而是由肠细胞和其他上皮谱系产生的。对这些激素生物合成和释放的途径了解相对较少。
肠内分泌细胞衍生的肽激素是由激素原转化酶的活性产生的,这些酶共同储存在囊泡中,囊泡与细胞膜融合以响应分泌刺激;而其他细胞类型中激素的产生则不同,它们并不经过传统的囊泡激素原转化酶和调节的囊泡分泌机制。
■ 肠道激素与二甲双胍疗效的关联
这些激素的生物合成与它们的mRNA水平有关,mRNA对特定刺激有反应。举例来说,FGF19(小鼠中的FGF15)的表达仅限于回肠末端,并通过核法尼糖样受体FXR被胆汁酸上调。在人体中,饭后胆汁释放后90-120分钟内FGF19的循环水平上升。在小鼠中,GDF15在多个器官中表达,但其在肠道中的表达受到细胞应激途径的增加的影响,就像在二甲双胍治疗过程中观察到的那样。实际上,治疗2型糖尿病患者的二甲双胍可能导致循环GDF15水平升高,从而产生该药物减肥效果。
■ Proguanylin:胃切除后水平升高?
Proguanylin在成熟肠细胞、一些杯状细胞和Paneth细胞中表达,在接受胃切除术或肠移植的患者中检测到的循环水平高于健康人。
Proguanylin释放到肠腔中,在那里被切割形成成熟的生物活性鸟苷肽。鸟苷由两个分子内二硫键稳定,这两个键对消化酶提供了一些保护,并从顶端方向靶向上皮膜鸟苷环化酶受体。Proguanylin和 prouroguanylin 也进入血液,但这些形式是如何分泌和激活的尚不清楚。
肠道激素控制许多胃肠道功能,这些功能涉及协调对食物摄入的反应,并确保摄入的食物被有效消化和吸收。
肠内分泌细胞检测管腔内容物的化学成分和局部营养吸收率,以及它们释放的激素控制过程,如胃酸分泌的调节、胃排空、胆囊收缩、胰腺酶和电解质的分泌、肠道运动和上皮屏障功能。
▸ 胃酸分泌
胃中特殊的壁细胞分泌酸有助于食物的消化和矿物质的吸收,并降低胃肠道感染的风险。胃酸的产生受到严格调节,以确保有效的食物消化,而不会损害胃和十二指肠粘膜。
胃窦G细胞分泌的胃泌素作用于肠嗜铬细胞上的胃泌素受体(称为CCKB或CCK2受体),刺激组胺的释放,组胺是一种强效的盐酸促分泌剂。胃窦和胃底D细胞的生长抑素对该系统进行微调,除了直接抑制壁细胞的酸释放外,生长抑素还能抑制胃泌素和组胺的释放。
其他几种肠道激素,包括胆囊收缩素(CCK)、氧调素、PYY和分泌素,通过不完全表征的机制抑制胃酸分泌,并募集涉及生长抑素的旁分泌回路。
▸ 胃排空
胃排空的速度决定了胃内容物输送到十二指肠的速率,并受到各种因素的调节,包括食物特性、肠道神经信号、肠道激素和血糖水平。胃排空率与食物消化率和营养吸收率密切相关,并受到调节,使得胃排空率仅反映小肠消化和吸收所接收营养的能力。
如果胃排空速率超过十二指肠吸收能力,营养物质会进入小肠并触发回肠制动,从而导致胃排空的反馈抑制,从而恢复平衡。主要位于回肠和结肠的肠道激素GLP1和PYY在回肠制动中起主要作用。回肠制动导致食物滞留在胃中,导致胃胀,从而激活发出饱腹信号的机械敏感迷走神经传入神经元。
▸ 胰腺和胆囊功能
一旦胃内容物进入小肠,其他激素就会被吸收以促进消化。由十二指肠 I 细胞产生的CCK刺激胰腺酶分泌,并通过刺激胆囊神经节中传出迷走神经神经递质的释放来促进胆囊排空。
分泌素在各种成熟的肠内分泌细胞中产生,刺激胰腺碱性电解质分泌,也与餐后热遗传学有关。鸟苷激活肠细胞中的鸟苷酸环化酶-cGMP信号,从而触发上皮氯化物和液体分泌。
这种途径被热稳定肠毒素劫持,是它们引发分泌性腹泻的能力的基础。鸟苷素-cGMP信号通路也与结直肠癌的抑制有关。
▸ 肠道运动
幽门以外胃肠道的运动由协调纵向蠕动波的肠神经系统和几种肠道激素之间的相互作用调节。
其中一种激素是胃动素,它存在于人类十二指肠中(尽管啮齿类动物不存在),并在胃中启动迁移运动复合体,特别是在餐间状态。胃动素受体激动剂,如低剂量红霉素,对胃轻瘫有治疗作用,一项研究表明,胃动素接收器的功能缺失变体在胃轻瘫患者中富集。
与调节小肠和大肠运动有关的其他激素包括:
粘膜血清素对肠道运动和液体分泌的重要性受到质疑,因为缺乏产生粘膜血清素所需的色氨酸羟化酶1的小鼠没有强烈的表型。然而,在一个模型中,产生血清素的肠嗜铬细胞被有条件地切除,这一发现强调了这些细胞确实在肠道转运控制中发挥作用。
▸ 肠道功能
除了控制肠道通道外,肠道激素还调节其他重要的肠道过程。例如:
GLP2刺激上皮生长,从而增加给定肠段的吸收能力。这种作用可能有助于GLP2治疗短肠综合征的疗效,
GLP1–GLP2双激动剂更有效。与血清素和神经降压素等其他肠道激素一样,GLP1和GLP2也会影响屏障功能并调节免疫系统。分泌血清素的肠嗜铬细胞与内脏疼痛有关,对肠道炎症、肝脏病理生理学和焦虑具有重要意义。
内 分 泌 胰 腺
我们知道饭后一段时间,人体内胰岛素的分泌量会增加,这其实也跟肠道激素有关。GLP1和GIP是两种主要的肠促胰岛素,占餐后胰岛素分泌的70%。
肠促胰岛素对葡萄糖刺激的胰岛素释放的影响可以通过以下事实来证明:口服葡萄糖比静脉注射葡萄糖(不刺激肠促胰岛素释放)能引起更大的胰岛素分泌——这种效应被称为肠促胰岛素效应。从生理上讲,肠促胰岛素调节对碳水化合物摄入的反应,并确保餐后健康的葡萄糖耐受性,无论摄入的葡萄糖负荷如何。
GLP1和GIP如何增加胰岛素分泌?
GLP1和GIP分别通过直接作用于GLP1受体和GIP受体来增加胰岛素分泌,这两种受体在胰腺β细胞中高度表达。
在β细胞中,GLP1和GIP受体主要募集Gαs并触发cAMP的升高。cAMP影响通过激活蛋白激酶A(PKA)和由cAMP-2激活的交换蛋白(EPAC2)调节胰岛素释放的几种下游途径。这两种效应物都调节离子通道活性以及参与葡萄糖依赖性胰岛素分泌的囊泡运输和融合。
GLP1和GIP具有额外的促胰岛素作用,但两者对肠促胰岛素作用的贡献有争议
GLP1和GIP对β细胞的作用不仅限于调控胰岛素分泌,还包括上调PDX1,促进胰岛素基因转录,增加β细胞质量并通过细胞周期和MAPK的激活促进新生。
基于这些作用,GLP1受体激动剂的长期治疗被认为是2型糖尿病的一种疾病改良治疗方法,可以通过增加或保留功能性β细胞质量来减轻β细胞功能障碍的进展。
注:迄今为止还没有证据表明GLP1受体促动剂调节人类的β细胞质量。
虽然说GLP1和GIP都刺激胰岛素分泌,但它们对胰腺α细胞分泌胰高血糖素的作用相反。
GIP刺激胰高血糖素分泌,而GLP1抑制分泌。
有证据表明,GLP1对胰岛素和胰高血糖素分泌的相反作用都有助于降低血浆葡萄糖。
GLP1通过激活GLP1受体介导的内在(非旁分泌)途径抑制α细胞,从而减少了胰高血糖素的分泌。
然而,其他研究结果表明,GLP1通过涉及胰岛内生长抑素的旁分泌机制间接抑制胰高血糖素的释放,并且GLP1和GIP的不同作用可以通过GIP对α细胞具有强大的直接刺激作用来解释,该作用掩盖了局部生长抑素的抑制作用。
肠促胰岛素的作用通常在2型糖尿病中受损,这种作用是糖尿病的后果而非原因。
在这种情况下,即使GIP的水平是超生理的,其作用也会减弱或缺失,而GLP1的促胰岛素作用相对保留。然而,GIP在2型糖尿病中保留刺激胰高血糖素分泌的能力,这会导致餐后高血糖。在各种研究中,GIP对胰腺β细胞的作用减弱归因于GIP受体表达减少、GIP受体下调或细胞内信号机制的受体后缺陷。
值得注意的是,2型糖尿病患者的血糖正常化提高了对GIP和GLP1的敏感性,这也许解释了为什么在双激动剂药物中在GL1中添加GIP对胰岛素分泌有额外的有益作用。
2型糖尿病是否与GLP1或GIP分泌缺陷有关尚不清楚。对2型糖尿病患者循环GIP水平的研究产生了矛盾的结果——高、低和不变的水平都有报道。相比之下,几项研究表明,2型糖尿病患者在混合餐后GLP1的分泌减少。
总的来说,2型糖尿病对肠道激素分泌的影响显然相对较小,这可能反映了这样一个事实,即:
肠内分泌细胞从隐窝干细胞中不断更新,在不利的代谢条件下,细胞损伤几乎没有时间积累。
然而,2型糖尿病对肠道运动和营养吸收的影响可能通过改变营养和胆汁酸刺激的局部递送而间接影响肠内分泌细胞的释放。
GLP1对胰岛素释放的影响比GIP大,GLP1已被开发为2型糖尿病的主要治疗靶点,并改变了2型糖尿病和肥胖患者血糖控制和体重减轻的临床治疗。
肝 脏
来自动物研究和临床试验的数据强调了GLP1和GLP1受体激动剂对肝脏代谢的多种有益作用。
GLP1受体是否在肝脏中表达有争议:
因此,在将GLP1对肝脏病理和功能的任何观察到的影响归因于肽对肝脏本身的直接影响之前,必须谨慎。
GLP1和GLP1受体激动剂对肝功能的间接作用
GLP1和GLP1受体激动剂对肝功能的间接作用是预期的,因为它们能够刺激胰岛素释放、降低胰高血糖素水平、改善葡萄糖稳态、减少胃肠道中乳糜微粒的产生以及减少热量摄入和体重。GLP1对肝脏影响的复杂性体现在它可以抑制肝脏葡萄糖产生并刺激肝脏葡萄糖摄取的发现上。20世纪90年代发表的研究将这些观察结果归因于胰岛素水平改变的间接影响,但后来的研究表明,胰岛素不是关键因素。
GLP1对肝脏脂质代谢有几种有益作用:
肝脏脂质代谢的这些改善可能至少部分归因于食物摄入的减少和随之而来的体重减轻。然而,一些研究结果表明,GLP1在调节肝脏脂质代谢方面具有额外的直接作用。
GLP1是否具有与体重减轻、胰岛素和胰高血糖素相关的肝脏作用之外的肝脏作用是一个有趣的问题。一项研究令人惊讶地表明,GLP19-36是GLP1的主要循环代谢产物,它独立于胰岛素的变化而抑制肝葡萄糖的产生,当GLP1受体被阻断时,这种作用仍然存在。这种作用的机制尚不清楚。一些证据表明,它涉及一种未鉴定的膜受体的激活,而其他研究结果表明,GLP19–36的降解产物(GLP128–36和GLP132–36)靶向PKA–β-连环蛋白–Wnt信号通路;这些降解产物可以在不涉及特定受体的情况下穿透质膜。需要进一步调查,以澄清和验证所涉及的机制。
无论GLP1对肝脏的有益作用是直接的还是间接的,GLP1受体激动剂正在被研究用于治疗非酒精性脂肪肝(NAFLD),这是肝脂代谢紊乱的最常见的慢性肝病之一。
GLP1受体激动剂:治疗非酒精性脂肪肝的希望
GLP1受体激动剂可以通过多种途径的组合来改善与非酒精性脂肪肝的发作和进展相关的基本代谢变化。
对非酒精性脂肪肝和非酒精性脂肪性肝炎(NASH)小鼠模型的研究表明,GLP1受体激动剂可降低肝脏脂肪含量,逆转脂肪变性,发挥抗炎作用,并改善纤维化的组织学严重程度。
此外,GLP1受体激动剂治疗非酒精性脂肪肝的临床试验(主要包括T2DM患者)产生了有希望的结果。几项研究表明,用GLP1受体激动剂治疗可降低肝脏脂肪含量,导致非酒精性脂肪性肝炎的组织学消退,并减缓纤维化的进展。然而,在这些临床研究中观察到体重和血红蛋白A1c水平显著降低,因此GLP1受体激动剂的作用是否独立于体重减轻和血糖控制的改善仍不清楚。
双激动剂疗法:研究非酒精性脂肪肝的新选择
双激动剂疗法,其中GLP1受体激动剂与直接抑制脂肪酸合成或靶向纤维化的第二分子结合,正在作为非酒精性脂肪肝的治疗选择进行研究;一个例子是GLP1–胰高血糖素组合肽。
其他肠内分泌激素(包括胃饥饿素和胃肠肽)对肝脏的影响尚不清楚。几项研究表明,GIP调节肝脏脂质含量,并可能参与非酒精性脂肪肝的发病机制。然而,GIP是否独立于其对胰岛素和脂肪组织的影响而直接影响肝脏中的脂质积聚(见“脂肪组织”)仍存在争议,因为GIP受体在肝脏中的表达仍未得到证实。几项动物研究表明,GIP信号传导的遗传或药理学破坏可以减少肝脏脂质积聚,但这些干预措施也可以防止饮食诱导的体重增加,因此研究结果不能证明GIP对肝脏有直接作用。
FGF19的多重代谢作用
与肠内分泌细胞衍生的肽不同,FGF19(小鼠中的FGF15)在回肠末端上皮中对胆汁酸吸收产生反应,是调节肝胆汁酸生物合成的重要抑制性反馈回路的基础。它还通过激活肝糖原合成和抑制糖异生等途径具有更广泛的代谢作用。
在不同器官中由肠道激素调节的过程
Bany Bakar R, et al., Nat Rev Gastroenterol Hepatol. 2023
脂 肪 组 织
脂肪细胞是哺乳动物的主要燃料储存室,它们的调节可以产生重要的代谢后果。在肠内分泌激素中,GIP与脂肪细胞功能的关系最为密切。
GIP与脂肪细胞功能:代谢调节的关键
几项研究表明,GIP促进白色脂肪组织(WAT)中的脂质储存,包括在大鼠中观察到,GIP通过增加脂肪组织葡萄糖摄取、从葡萄糖重新生成脂肪和将FFAs重新酯化为甘油三酯来支持脂肪组织中的脂质存储,以及在小鼠中敲除GIP受体可以预防饮食诱导的肥胖。
几项研究表明,脂肪组织中的GIP受体激活与胰岛素协同作用,增加脂质积累、葡萄糖摄取、新生脂肪生成和FFA再酯化。
脂肪细胞中的GIP受体类型:研究争议与需解决的问题
然而,这些发现需要在脂肪组织中哪些细胞类型表达GIP受体的争议中进行解释。单核转录组学表明,GIP受体不在成熟脂肪细胞中表达,而是在周细胞和间充质细胞中表达。考虑到许多脂肪细胞模型是由可以表达GIP受体的未成熟细胞在体外产生的,这些模型中的GIP受体表达是否能概括天然成熟脂肪细胞中的表达值得怀疑。
另一个必须考虑的因素是,GIP受体与Gαs偶联,Gαs下游cAMP水平升高通常与脂解而非脂肪生成有关,例如β3-肾上腺素能受体激活的脂解作用。
GIP的脂肪生成作用有益,改善脂质储存,减少异位脂肪积聚
有趣的是,GIP受体在小鼠和人类脂肪组织周细胞中表达的发现与GIP在体内显著增加脂肪组织血流量的证据一致,这一过程可以间接促进胰岛素依赖性作用,比如:清除甘油三酯、摄取葡萄糖。
无论潜在机制如何,GIP的脂肪生成作用都是有益的,因为它改善了白色脂肪组织中健康的长期脂质储存,减少了肝脏、心脏和胰腺等外周组织中的脂质溢出和异位脂肪积聚。
在2型糖尿病和肥胖的GLP1受体激动剂上添加GIP受体激动剂的代谢益处
早期的研究表明,GIP纯粹是致肥胖的,因此有利于GIP拮抗作用的翻译策略来实现减肥,而自从2018年以来发表的研究表明双重GIP受体-GLP1受体激动剂,替西帕肽在2型糖尿病患者中实现血糖控制和减轻体重方面比单独使用任一受体的激动剂更有效,该治疗改善了白色脂肪组织健康。替西帕肽治疗显著降低了肝脏脂肪含量以及内脏和皮下脂肪组织的体积,因此可能成为非酒精性脂肪肝或非酒精性脂肪性肝炎的重要治疗选择,尤其是在同时患有2型糖尿病或肥胖的患者中。
然而,对于是激动还是拮抗GIP受体,仍存在不清晰性;强有力的证据表明,任何一种方式都可以减轻肥胖和体重。
注:已经提出了几个假设来调和这些自相矛盾的观察结果,但还需要更多的研究来澄清这一领域。
尽管GIP在肠道白色脂肪组织轴中的确切作用机制仍有争议,但GIP在脂肪组织中的生理作用是明确的。在啮齿类动物中,棕色脂肪细胞组织(BAT)中GIP受体的表达已得到充分证实,给予GIP或替西帕肽可促进产热;替西帕肽也与支链氨基酸分解代谢增加有关。
然而,小鼠棕色脂肪细胞组织中GIP受体的条件性敲除并不能像全局敲除GIP受体那样防止饮食诱导的肥胖,这表明GIP对棕色脂肪细胞组织的影响在生理上并不是主要的。那主要的会是什么呢?
其他激素在肠道棕色脂肪细胞组织轴中发挥重要作用
例如,血清素抑制棕色脂肪细胞组织活性,而缺乏血清素产生所需的色氨酸羟化酶1的小鼠可以免受饮食诱导的肥胖。然而,鉴于血浆5-羟色胺是由血栓细胞缓冲的,肠道来源的5-羟色胺的内分泌作用值得怀疑,而且关于血浆5-羟色胺水平相对于用餐时间的日变化的报道一直不一致。相反,分泌素激活棕色脂肪细胞组织,这种激活与食欲的改变有关。
脑
肠-脑轴是胃肠道和大脑之间传递信息的渠道,包括神经、激素和营养信号;下丘脑核团和后脑是检测和整合这些外周信号的主要中心。肠-脑轴的紊乱已在多种疾病中被发现,包括炎症和功能性胃肠道疾病以及饮食障碍。
肠道激素在大脑的作用可以解释一些饮食失调和进食障碍的发生?
肠道向大脑发送关于饮食营养摄入状态的实时信号,肠道激素是肠道和大脑之间复杂相互作用的主要介质,这些相互作用是饱腹感(两餐之间不饿的感觉)和饱足感(一餐结束时吃得足够的感觉)的基础。许多肠道肽及其受体也在中枢神经系统中表达,并调节中枢回路,包括与食欲有关的回路。这种在中枢神经系统中的表达可能会使涉及肠道激素受体的药理学或全局遗传操作的研究的解释复杂化,因为其结果可能归因于对中枢神经系统回路的影响,而与肠道分泌的激素无关。
传入迷走神经神经元的细胞体位于结节神经节,并投射到孤束核(NTS),是介导肠道激素对食物摄入影响的主要途径。肠内分泌细胞通过局部升高的肠道激素水平以及可能的突触连接与传入迷走神经和内脏神经末梢进行交流。到达心室周围器官(例如,脊后和正中隆起区)和邻近细胞核(分别为下丘脑的NTS和弓状核)的循环肠道激素与这些信号汇合,以调节进食行为。几种肠道激素对大脑有着重要而独特的影响,下文将对此进行讨论。
CCK是在食欲调节中发挥作用的肠道激素。餐后释放CCK通过与CCK1受体结合激活传入迷走神经,向大脑提供反馈信号,从而减少进食。然而,根据小鼠研究,反馈信号的性质仍不清楚,因为动物可能因各种原因停止进食。在人类中,输注生理浓度的CCK可以抑制能量摄入并增加幽门收缩,而不会引起恶心;食物摄入减少被认为是幽门收缩的结果。在小鼠中,CCK通过传入迷走神经发出信号,激活中枢奖赏通路,并增加脂肪和糖的消耗。
CCK似乎会触发相对短暂的饱腹信号,PYY与餐后食物摄入的长期抑制有关。已知PYY的两种不同形式:PYY1-36和PYY3-36。PYY1–36通过二肽基肽酶IV转化为PYY3–36。两种形式的PYY都参与调节食物摄入,但它们表现出不同的特性和受体偏好。
GLP1和GLP1受体激动剂可以抑制食欲
GLP1受体激动剂主要通过作用于中枢神经系统内的GLP1受体来抑制食物摄入,但肠道来源的GLP1在大脑中是否达到足够高的水平以在生理条件下激活GLP1受体仍不清楚。
几项研究表明,迷走神经传入神经元的末端暴露于胃肠道中高水平的局部生物活性GLP1,介导内源性肠道衍生的GLP1的饱腹作用。
肠道来源GLP1控制食物摄入——不是主要的
在小鼠中,迷走神经神经元表达GLP1受体,而且响应机械刺激,外源性GLP1也可以增强由胃和肠道拉伸引起的饱腹感。然而,敲除肠道来源的GLP1不会影响小鼠的食物摄入或体重,这表明肠道来源的GLP1对控制食物摄入并不重要。
脑源性GLP1控制食物摄入——明显作用
虽然去除后脑GLP1表达也不会影响自由进食行为,但当化学遗传学激活时,它确实削弱了表达GLP1的神经元减少进食的能力,证明了脑源性GLP1在控制食物摄入方面的明显作用。
GLP1受体与氧调素的互动:探索胰岛素类似物的潜力
GLP1受体也被氧调素激活,氧调素是一种由肠道L细胞分泌的37个氨基酸的略微延伸的胰高血糖素类似物,在胰岛素受体处也具有活性。氧调节蛋白在体重管理和葡萄糖稳态方面具有有益作用,这一作用正开始通过使用GLP1受体和胰岛素受体双激动剂而得到利用;鉴于目前还没有发现特异性的尿囊素受体,这些作用被认为是由其对这两种受体的双重作用介导的。
减肥手术后,餐后GLP1、氧调节蛋白和PYY水平显著升高;循环水平足以对食物摄入和葡萄糖耐受性产生有益影响。然而,在小鼠中,Roux-en-Y胃旁路术的减肥作用在缺乏神经肽Y受体2型和GLP1受体的动物中仍然存在,这表明这些激素作用之外的机制也有助于减肥手术的益处。
神经降压素也与减肥手术的效果有关
神经降压素(Neurotensin)是一种13个氨基酸的肽,主要存在于大脑和胃肠道中。在啮齿类动物中,神经降压素直接进入大脑时会减少食物摄入,这种作用主要由NTSR1(神经降压素受体1)介导。
在一些研究中,Ntsr1敲除小鼠食欲增加,体重增加,对神经降压素的食欲抑制作用没有反应。
然而,在其他研究中,Ntsr1敲除降低了进食量并增加了活性,但对体重或组成没有影响。
脑源性神经降压素抑制食欲,外周的神经降压素对代谢的影响尚不清楚
一些研究表明,神经降压素的外周给药会导致啮齿动物的食物摄入量暂时减少。一些证据表明,这种影响可能归因于肠道中脂肪吸收的减少,但减少进食的确切机制尚不清楚。
另一种与减肥手术结果有关的激素是分泌素,因为它在手术后的餐后水平也会升高,尽管它在这些患者中的生理重要性尚不清楚。
来自非肠道内分泌细胞的肠道激素(GDF15、FGF19、uroguanylin)似乎也通过中枢途径调节食欲的作用。
最有力的证据是GDF15,它靶向后脑中有限的神经元群体,并导致由一系列毒性刺激(如化疗剂)诱导的恶心,这种恶心发生在癌症恶病质和妊娠剧吐中。
GDF15反应神经元表达GDF15受体,即GFRAL,并与脑干GLP1受体和CCK回路密切相关。有趣的是,GIP可以通过表达GIP受体的海马后区GABA能神经元抑制GDF15(和其他肠道激素,包括GLP1和PYY)引起的恶心。
Ghrelin:非凡食欲刺激者
胃饥饿素(Ghrelin)是唯一被广泛接受的刺激食物摄入的肠道激素。根据这种生物活性,血液中胃饥饿素的水平在用餐前升高,在进食后下降。
其生物学效应是通过生长激素促分泌受体(GHSR)介导的,GHSR存在于全身,包括胰腺、迷走神经传入神经元和大脑。几项研究表明,Ghrelin通过在下丘脑弓状核的中央和迷走神经传入神经的外周发挥其食欲产生活性。
然而,后一种机制是有争议的,一些研究表明,胃饥饿素的食欲产生作用不需要迷走神经传入信号。一些研究表明,胃动素也能促进食物摄入,但胃动素的生理学特征很差,因为它不存在于啮齿类动物中。
注:胃动素是一种与胃饥饿素有关的肽,由十二指肠中重叠的肠内分泌细胞群体产生。
INSL5:微弱但可能的食欲促进者
INSL5由远端结肠和直肠L细胞的一个子集分泌,对小鼠的研究表明它它具有食欲促进作用。它通过RXFP4(松弛素-胰岛素样家族肽受体4)发挥作用,该受体在大脑的关键进食中心表达。
INSL5的增食欲作用远低于胃饥饿素,这在一些研究中没有观察到,并且可能在体内被PYY所淹没,PYY是从相同的肠内分泌细胞平行分泌的。这些观察结果让人怀疑外周释放的INSL5是否在控制食物摄入方面具有重要的生理作用。然而,一项研究已经确定了腹内侧下丘脑中表达RXFP4的神经元在调节食物摄入和偏好方面的潜在作用。
胃肠道和身体其他活跃的组织之间有着双向的信息传递,而这个信息传递的改变与多种代谢性疾病有关。目前对于胃肠激素如何与这个复杂的调节网络相互作用的理解正在逐渐加深,但仍需要进一步的研究来完全揭开这个谜团。
肠道菌群的异常变化可能影响肠道激素的正常分泌,从而导致代谢相关疾病发生。因此,肠道菌群的稳定和平衡对于保持肠道激素的正常分泌和代谢健康非常重要。这些发现为我们深入了解肠道菌群和肠道激素的相互作用、开发相关治疗方法提供了新的方向。通过调节肠道菌群的结构和功能,我们可能能够干预肠道激素的分泌,从而实现对代谢相关疾病的治疗和预防。
通过开发创新、微创且耐受性良好的治疗方法,我们可以利用胃肠激素受体的转化潜能。GLP-1双重和三重激动剂疗法的临床试验已经证明了这种方法的价值。这个领域研究进展迅速,很可能会进一步细化现有的认识,解决关键的机制问题。期待揭示出胃肠激素控制的多个信号通路,可以成功地用于治疗2型糖尿病和肥胖,并且将这些基于胃肠激素的药物在其他代谢性疾病中推广应用。
主要参考文献:
Bany Bakar R, Reimann F, Gribble FM. The intestine as an endocrine organ and the role of gut hormones in metabolic regulation. Nat Rev Gastroenterol Hepatol. 2023 Aug 25.

谷禾健康


↑出现以上症状,可能是身体缺乏蛋白质的信号。
★ 蛋白质有多重要?
蛋白质不仅是构建人体组织的主要原料,而且对新陈代谢至关重要。膳食蛋白质及其代谢产物氨基酸是人类的必需营养素。
蛋白质提供能量,调节代谢;我们的皮肤、骨骼、毛发都有它的参与;当然免疫系统也少不了它。
★ 肠道菌群参与蛋白质代谢
肠道微生物群介导蛋白质代谢和宿主免疫反应之间的相互作用,在代谢过程中发挥重要作用。
小肠细菌能代谢部分氨基酸,进而影响宿主整体氨基酸的代谢。大肠拥有更丰富的菌群和更长的蠕动时间。
一方面,进入大肠的氮营养素会影响大肠菌群的代谢和群落结构;另一方面,大肠菌群也能广泛参与氮营养素的代谢与利用,生成许多代谢产物,进而影响人体健康。
因此,本文详细讨论蛋白质对人体的影响,肠道菌群与蛋白质的相互作用,肠道微生物群代谢产物的影响,文末有人体对蛋白质的需求量及相关食物补充说明。
我们先来看一下蛋白质的消化分解过程。
蛋白质的消化分解
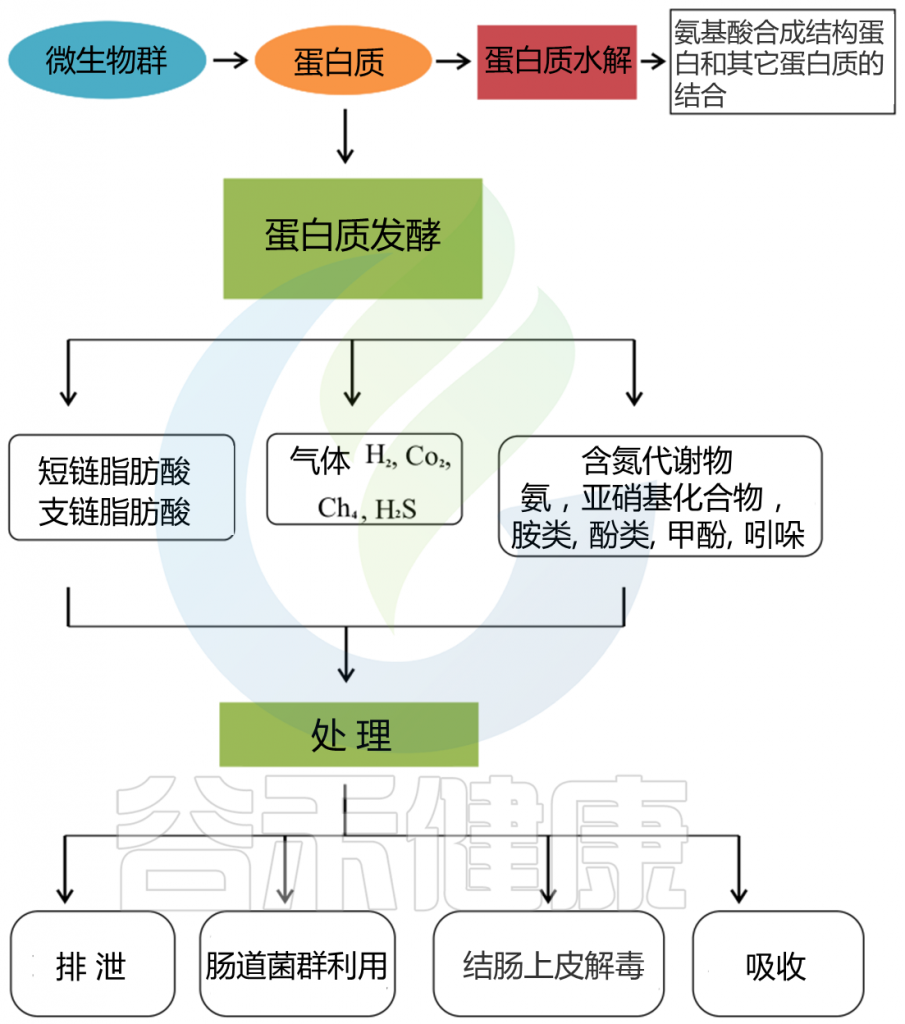
小肠上皮细胞消化膳食蛋白质,然后吸收氨基酸和肽是一个高效的过程。高可消化的蛋白质可能部分逃脱小肠的消化,大量的含氮物质可能从小肠转移到大肠。未消化的蛋白质和多肽通过肠道微生物和残留的胰腺蛋白酶进行蛋白水解,产生大量的微生物代谢物。
胃肠道中的蛋白质分解代谢物
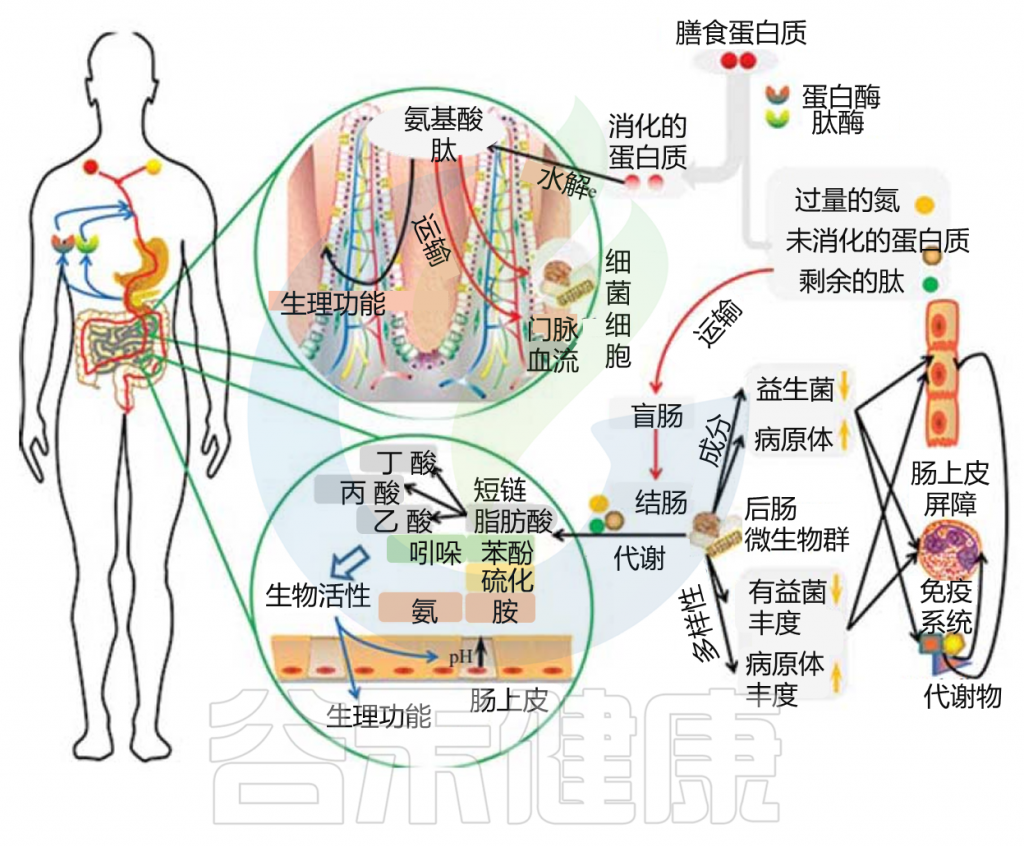

Zhao J, et al., Curr Protein Pept Sci. 2019
这些微生物代谢物中有些是中间产物,有些是最终产物。大量的最终产物主要是短链脂肪酸、氨、多胺、硫化氢、酚类和吲哚类化合物。
这些细菌代谢物中的一些可以被运输到结肠细胞,并根据其在腔内的浓度对这些上皮细胞产生有益或有害的影响。一些细菌代谢产物被输送到门脉血,对肝脏和周围器官组织产生各种生理作用(这些代谢物在健康中发挥的作用详见本文后面章节)。
肠道微生物群在膳食蛋白质分解代谢中作用
宿主饮食对肠道微生物群的分布和活性有很大影响。膳食蛋白质或氨基酸调节可作为调节发酵细菌的一种方法。
在研究不同动物蛋白质来源(牛肉、鸡肉和鱼类)对人类粪便微生物群的影响的研究中,与牛肉蛋白孵育2天后,产气荚膜梭菌的数量显著增加,双歧杆菌的数量显著减少。
微生物分泌的酶直接降解分解蛋白
肠道微生物群在促进氨基酸调节以及氨基酸消化和吸收过程中的分泌方面起着至关重要的作用。
蛋白质的转换主要发生在肠道中,结肠细菌降解内源性或外源性蛋白质的效率较高。
拟杆菌属(Bacteroides)、梭状芽胞杆菌属(Clostridium)、丙酸杆菌属(Propionibacterium)、梭杆菌属(Fusobacterium)、乳酸杆菌属(Lactobacillus)和链球菌属(Streptococcus)在蛋白质水解过程中发挥着重要作用。
粪便微生物群与结肠微生物群具有不同的蛋白水解活性。粪便微生物的活性仅在球状蛋白质的蛋白质水解方面较高。
发酵过程中,复杂蛋白质首先被各种细菌肽酶、蛋白酶和内肽酶切割,释放出游离氨基酸和短肽。氨基酸和短肽然后进行发酵。蛋白质发酵产生支链脂肪酸(2-甲基丁酸酯、异丁酸酯、异戊酸酯)、有机酸、气体(H2和CO2)以及微量酚、胺、吲哚和氨。
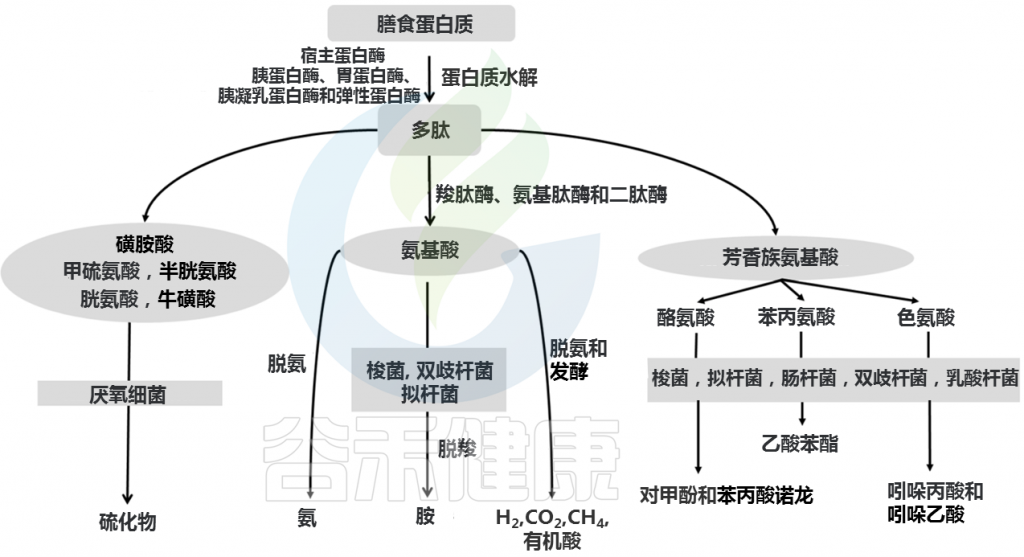

Yadav M, et al., Arch Microbiol. 2018
肠道微生物参与氨基酸代谢
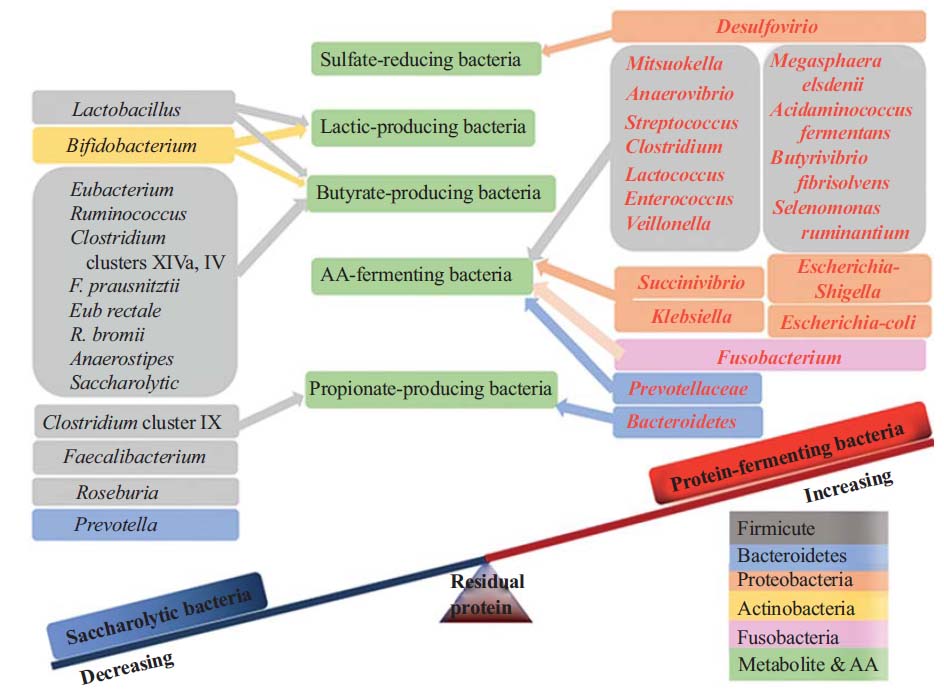
一些菌群可能在肠道中的氨基酸代谢中发挥重要作用,例如梭杆菌属、拟杆菌属和韦荣球菌,Megasphaera elsdenii, Selenomonas ruminantium.
肠道内的梭菌属细菌(赖氨酸或脯氨酸利用的基础细菌)是氨基酸发酵的关键驱动因素,而消化链球菌属细菌是谷氨酸或色氨酸利用的关键驱动因素。
其他例如瘤胃细菌,Selenomonas ruminantium, Megasphaera elsdenii, Prevotella ruminicola, Misuokella multiacidas, Butyrivibrio fibrisolvens, Streptococcus bovis等含有极为活跃的二肽基肽酶和二肽酶。
微生物从头合成氨基酸
肠道微生物群在产生氨基酸方面也起着关键作用,这包括从头生物合成。
例如,一些体外研究项目表明,瘤胃细菌种类,如Streptococcus bovis,Selenomonas ruminantium, Prevotella bryantii,在生理肽浓度存在的情况下参与氨基酸的从头合成。

Lin R, et al., Amino Acids. 2017
体内研究还表明,微生物衍生的赖氨酸(一种必需氨基酸)被吸收并并入宿主蛋白质。在无菌和常规化大鼠体内,将15NH4CL中的15N并入赖氨酸的比较表明,检测到的所有15N赖氨酸均来自微生物来源。在后续研究中,这些研究人员确定约75%的微生物15N标记的赖氨酸被小肠吸收。
此外,其他体内研究项目表明,微生物衍生的赖氨酸被吸收并整合到宿主蛋白质中。与动物研究相一致,六名健康男性的样本以15NH4Cl的形式口服15N与标记肠道微生物来源的微生物蛋白和苏氨酸有关,这些微生物来源存在于活体门静脉血流中。
对18岁以上的人样本进行了检查,这些人被提供了氮充足的饮食,并发现微生物衍生的赖氨酸和苏氨酸对游离血浆赖氨酸和苏氨酸库有显著贡献。
此外,Gill等人报告说,大肠中微生物群的富集是通过与必需氨基酸(EAA)生物合成相关的基因进行的,该生物合成基于人血浆池产生的前体。
蛋白质或氨基酸摄入对肠道微生物的影响
膳食蛋白质是均衡饮食的重要组成部分。人类无法合成多种氨基酸,因此这些氨基酸必须从食物中获取以维持健康。胃肠道微生物群组成和功能的差异可能是由于膳食蛋白质的变化。
进入结肠的蛋白质量和尿液中检测到的发酵代谢物量取决于蛋白质的消化率,蛋白质消化由蛋白质水平和来源的共同作用决定。
当进展到结肠的远端部分时,碳水化合物被消耗,pH值增加,蛋白质发酵变得更加有效。在消化过程中,蛋白质摄入量的增加总是与消化率的相对降低有关,通常会导致更多可发酵的大量营养物质进入结肠。
不同形式蛋白对肠道微生物群影响不同
蛋白质来源主要来自植物或动物来源,每种类型都具有独特的消化率和不同的降解模式,具体取决于所涉及的微生物。结肠中的细菌属参与蛋白质代谢,主要包括可能致病的拟杆菌属、大肠杆菌属和梭菌属。抑制这些潜在病原体通常与通过减少肠毒素和有害微生物代谢物的释放来恢复微生态系统平衡有关。
考虑到蛋白质来源在消化中的作用,动物来源的蛋白质,如乳品和动物蛋白的消化率普遍高于植物蛋白。然而,酪蛋白的热分解显著降低了蛋白质的消化率,增加了蛋白质的发酵程度。下面就详细来看蛋白质来源如何调节肠道微生物群。
植物蛋白
与动物蛋白相比,植物蛋白通常具有较低的蛋白质消化率,因为植物细胞壁不能消化。大豆和花生蛋白都在调节肠道有益细菌组成方面有积极的作用。
添加20%花生蛋白的饲粮改变了大鼠肠道菌群多样性,增加了双歧杆菌,降低了肠杆菌和产气荚膜梭菌的数量。增加的双歧杆菌有助于产生更多的微生物代谢物,包括乙酸和乳酸,导致肠道pH值降低,抑制有毒代谢物,如胺和苯并吡咯。
大豆已被广泛应用于人类和动物,它可以改变肠道微生物的组成,增加了埃希氏杆菌属和丙酸杆菌。系统发育分析显示,埃希氏杆菌属与志贺菌的同源性最高,两者都能在不损害肠道黏膜的情况下调节盐和水的代谢。但大豆中含有的抗营养因子会对生产性能产生负面影响,限制了其在非反刍动物中的应用。
植物蛋白被用于动物饲料行业,因为它的价格普遍低于动物蛋白,并且在食品安全方面具有一些优势。
动物蛋白
与植物蛋白相比,动物蛋白对于牲畜来说是高度易消化的。酪蛋白、脱脂奶粉和鱼粉通常用作猪的饲料,它们可以被消化成丰富的蛋白质底物并在到达大肠之前被吸收。这些动物蛋白的代谢特性有利于宿主健康,特别是,对环境应激导致仔猪断奶后腹泻的预防作用及一些植物蛋白中的抗营养因子。
酪蛋白可以被近端肠道中的宿主酶消化,从而减少大肠中细菌的降解。具体而言,酪蛋白可以增加乳杆菌和双歧杆菌,并减少粪便中葡萄球菌、大肠菌群和链球菌的数量。此外,酪蛋白可以调节直肠真杆菌和Marvinbryantia formatexigens的减少。
补充脱脂干牛奶可以增加总厌氧菌和需氧菌,而膳食鱼粉可以减少需氧菌,包括大肠菌群,并增加厌氧菌的数量,如乳酸杆菌属。
此外,动物蛋白的特点是 短链脂肪酸减少,肠道 pH 值和氨浓度增加。
显然,蛋白质来源对肠道微生物群的影响并不一致,需要更多的研究来评估不同蛋白质类型对蛋白质消化率和代谢以及肠道微生物群组成的影响。
蛋白质摄入量与肠道菌群
膳食蛋白质的浓度是影响蛋白质发酵和肠道微生物组成的主要因素。
更高的水平和未消化的蛋白质会导致致病微生物的增加,从而增加代谢疾病的风险。

Zhao J, et al., Curr Protein Pept Sci. 2019
未被小肠吸收的残留含氮化合物将被转移到远端肠道,并被该位置微生物代谢。微生物代谢物的数量和种类均受膳食蛋白质摄入量的影响。
有些微生物代谢物是有毒的,例如硫化氢、氨和吲哚化合物,并可能对宿主健康产生负面影响。有些代谢物是参与宿主各种生理过程的生物活性分子。
此外,由于肠道微生态系统的稳态破坏和有益菌的减少,高浓度的蛋白质补充会导致潜在病原体数量增加。膳食蛋白质改变的肠道微生物群,通过调节肠道屏障功能、肠道运动和免疫系统来影响宿主代谢。
结肠中过量蛋白质发酵影响肠道微生物群变化
未消化蛋白质的增加与蛋白质摄入水平高、蛋白质消化率相对降低和氨基酸组成不平衡有关。结肠中多余蛋白质的降解始于细菌蛋白酶和肽酶将蛋白质水解成较小的肽和氨基酸。这些含氮化合物会影响肠道微生物群,尤其是结肠。
-3.png)
稍低蛋白浓度有助于减少大肠杆菌,但蛋白浓度过低会增加潜在致病菌
相对低浓度的膳食蛋白质会减少致病菌增殖的底物量。例如,人类饮食中低浓度的蛋白质导致粘膜表面的大肠杆菌群落减少。在较低蛋白质条件下肠道微生物的变化会产生毒性较小的含氮细菌代谢物,例如多胺。
然而,当饮食中蛋白质的浓度过低而不能满足宿主的基本需要时,就会增加潜在病原体的丰度,减少有益菌的数量。
具体来看一项关于蛋白浓度变化对肠道菌群影响的研究。
断奶动物
蛋白水平适量:
当断奶动物饲喂100-200g/kg的膳食蛋白质时,粪便中需氧菌和厌氧菌的数量会增加,这些蛋白质水平会导致肠道中乳酸杆菌的增加以及大肠菌群和葡萄球菌的减少。
蛋白水平过高:
然而,当膳食蛋白质水平大于 200 g/kg 时,致病菌数量会增加,例如大肠菌群、链球菌和芽孢杆菌。
蛋白水平过低:
低浓度的膳食蛋白质会减少产丁酸菌(这些菌群是抗癌和其他肠道疾病的抗炎剂),包括乳酸杆菌、双歧杆菌、saccharolytic(糖酵解菌)。
成年动物
成年动物中,在蛋白质水平的微小变化下,肠道微生物群相对稳健。例如,当成年动物分别用 190 g/kg 和 150 g/kg 蛋白质喂养时,粪便样本中的细菌数量几乎没有差异。
在成年动物中,只有当蛋白质水平发生重大变化时,才能观察到微生物群的变化。
低蛋白饮食还与低浓度的氨、血浆尿素氮和回肠食糜中的短链脂肪酸含量有关。肠道中的氨不是来自宿主脲酶活性,而是来自蛋白水解和微生物活动。因此,回肠食糜中低浓度的氨可能表明蛋白质代谢降低。
蛋白质发酵产物对肠道微生物的影响
大部分膳食蛋白质在小肠中消化,但蛋白质摄入过多会导致结肠输入量增加。结肠中多余蛋白质的降解始于蛋白质被细菌蛋白酶和肽酶水解成更小的肽和AA,这些蛋白酶和肽酶在中性至碱性pH下更为活跃。这些残余蛋白质不仅提高了肠道pH值,而且还可供结肠微生物进一步代谢。
然而,碱性pH有利于病原体增殖,对肠道健康不利。此外,由于向肠道微生物群过量供应膳食蛋白质而导致的蛋白质发酵也被视为肠道疾病发展的一个因素。
虽然有报道称结肠蛋白输注过多会产生有害影响,仍有一些争议,但似乎较低量的蛋白到达远端肠道可能会降低腹泻的发生率,并抑制病原菌的增殖,特别是在应激情况下,如感染高压。
蛋白质的发酵通常与蛋白质发酵细菌以及潜在致病菌的生长有关。通过培养基分析,至少拟杆菌属和丙酸杆菌以及各种杆菌被鉴定为蛋白水解细菌,其中一些细菌的基因组中携带丝氨酸和其他蛋白酶的基因。
氨基酸为细菌提供代谢底物
饮食和内源性来源的蛋白质和氨基酸对于肠道微生物群的生长和存活是必要。
蛋白质水解后,释放的肽和氨基酸被宿主和胃肠道中的细菌用于各种代谢活动,虽然一些细菌缺乏使用细胞外氨基酸的特定转运体。
胃肠道中许多细菌的理想氮源是氨基酸和NH3,可能是因为它们缺乏必要的肽转运蛋白。然而,如瘤胃前置杆菌等细菌利用肽作为氮的唯一来源。这些肽最初被输送到瘤胃前置杆菌的细胞中,然后直接使用或水解成FAAs。通常,肽、氨基酸和NH3可用于细菌细胞成分的生产或通过各种途径分解代谢。这些过程可能对身体产生有利或不利的影响。
蛋白质摄入与肠道屏障
肠道是一个具有高度蛋白水解活性的部位,主要由微生物群介导。这些蛋白水解代谢物以及肠道微生物(可通过蛋白质水平改变)通过调节紧密连接蛋白和Zonulin在调节肠粘膜屏障中发挥重要作用。
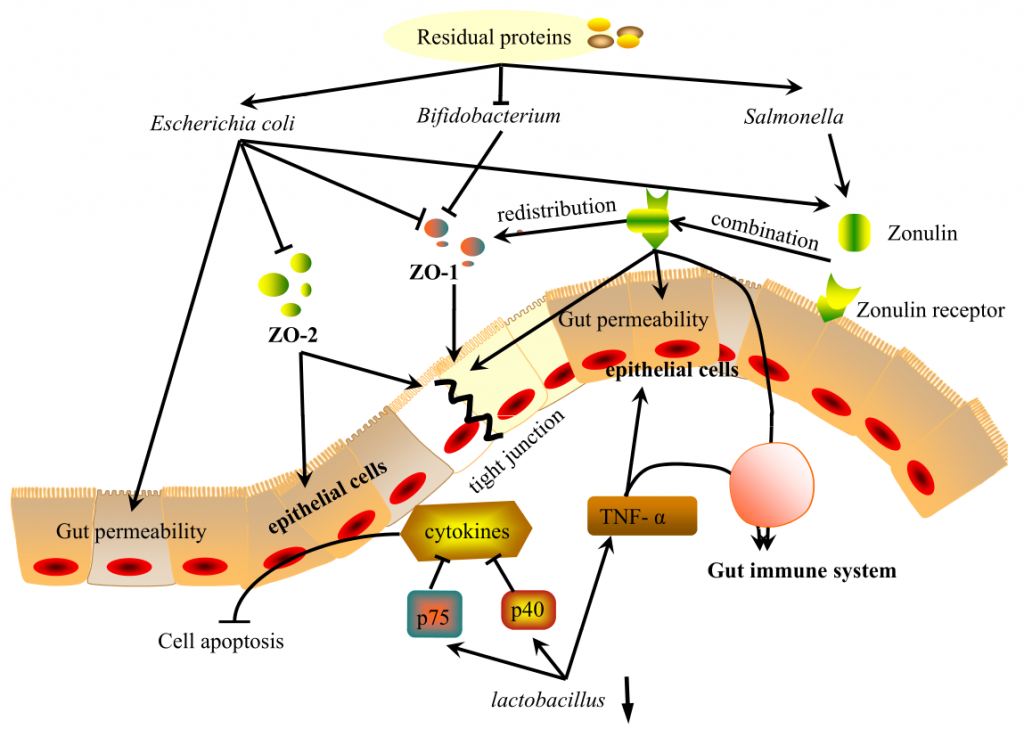

Ma N, et al., Curr Protein Pept Sci. 2017
肠道微生物通过调节紧密连接蛋白ZO-1和ZO-2的表达来影响肠道上皮屏障。研究表明,在高蛋白饮食(HPD)中增加双歧杆菌可有效缓解结肠上皮细胞膜屈曲,上调ZO-1表达,有利于紧密连接修复。
相反,低蛋白饮食(low-protein diet, LPD)或正常蛋白饲粮(normalprotein diet, NPD)通过降低肠上皮通透性,降低大肠杆菌水平,从而上调ZO-1的表达。抑制大肠杆菌生长也可上调ZO-2的表达,从而保护肠上皮细胞,修复上皮屏障。
连蛋白Zonulin是唯一已知的细胞间紧密连接的生理调节剂,可能是微生物群和紧密连接蛋白之间的一种可能的调节剂。
Zonulin可逆且快速地调节肠道通透性,其在遗传易感个体中的上调可能导致免疫介导的疾病。其释放将由细菌数量的变化触发。
在上皮细胞中结合其受体后,该复合物导致紧密连接的开放并增加肠道通透性。
研究人员选择了4种细菌菌株作为兴奋剂,包括3种大肠杆菌菌株和1种沙门氏菌菌株,它们都是病原体,HPD增加。它们都释放了Zonulin并改变了细胞旁通透性。同时,观察到ZO-1从紧密连接复合物重新分布到细胞单层。
此外,不同的细菌会导致不同水平的Zonulin分泌,这会在不同程度上改变肠道屏障功能,反映微生物和肠道之间的急性调节。
氨基酸不均衡会诱发肠道问题
膳食蛋白质中的氨基酸可通过调节肠道微生物群组成间接影响肠道形态。此前有研究报道,低赖氨酸玉米蛋白会导致肠道绒毛高度降低,隐窝深度加深,提示肠道黏膜的损伤是由于饲粮中氨基酸比例失衡所致。
小肠上皮细胞可将氨基酸和其他营养物质吸收到血液和器官中,营养不良对肠道微生物群的干扰和影响可能导致肠道形态受损。
断奶时观察到的腹泻可以通过低蛋白质水平和平衡氨基酸组成来缓解。然而,饮食中氨基酸的不平衡可能损害肠道形态,增加腹泻风险。因此,饮食中氨基酸失衡会抑制蛋白质吸收,导致肠上皮增生。
注意蛋白过敏或不耐受诱发的肠道问题
大豆中的粗蛋白质相对较高,约为40%,主要由球蛋白组成,球蛋白由伴大豆球蛋白和大豆球蛋白组成。这些球蛋白具有免疫原性和热稳定性。
大豆蛋白补充剂中抗原复合物的形成,如大豆球蛋白、β-伴大豆球蛋白和免疫球蛋白抗体,可能是刺激过敏反应和腹泻的原因。大部分膳食蛋白质在消化后降解为肽和氨基酸。
然而,还有一种可能性是,其他分子如一种抗营养因子,可以通过肠上皮细胞进入血液和淋巴。大豆中具有抗原活性的抗营养因子可刺激宿主的免疫应答。
免疫反应由具有IgE分泌的T淋巴细胞介导,其对肠道形态具有负面影响,导致绒毛损伤和隐窝细胞增殖。
此外,上皮细胞中消化酶活性的降低可能与吸收功能失调和肠道微生物群组成多样性减少有关。
由于其抗原性,膳食大豆蛋白可能有利于病原体增殖,这些抗原也可能是对粪便微生物群一致性产生不利影响的因素。
对大豆蛋白的超敏反应可能导致对病原体过度生长的更高易感性,主要包括蛋白质代谢细菌以及肠致病性大肠杆菌。
大豆蛋白和其他植物蛋白通常含有抗营养因子,包括可溶性非淀粉多糖含量升高,可能增加断奶仔猪腹泻的发生率。
在大豆蛋白质代谢过程中,会产生各种毒素,如组胺、尸碱和腐胺,这些毒素会破坏肠壁,增加肠上皮细胞的通透性。当病原体在管腔内定植时,可能发生肠道微生态失调。
大豆蛋白被认为是一种相对健康的蛋白质来源。大豆经加工处理后,抗原活性有所降低,抗营养因子显著降低。Pepsin-hydrolysate conglycinin (PTC)是一种治疗后从球蛋白中提取的小肽,具有降低高血压、抗氧化和免疫刺激等药理和生理活性。
伴大豆球蛋白是大豆贮藏蛋白的组分之一,经消化和酶处理后以PTC的形式发挥作用。PTC可以抑制病理性大肠杆菌的生长,无时间依赖性,并且在 大肠杆菌E. coli O138激发后也被激活以预防疾病。
伴胰球蛋白-胃蛋白酶水解物参与调节肠道微生物群落平衡和维持肠道完整性。当肠道微生态系统的稳态受到干扰时,生物活性肽可在预防与肠道疾病相关的疾病中发挥重要作用。PTC还通过产生胃蛋白酶介导的双歧杆菌生长刺激肽促进双歧杆菌的生长。然而,PTC对E. coli O138的抑制作用的潜在机制尚不清楚,需要进一步的研究来阐明。
酪蛋白与肠道微生物群
酪蛋白具有特殊的氨基酸组成,可以诱导肠道氨基酸转运蛋白基因的转录水平。肠道微生物群的组成和功能也受到氨基酸平衡改变的影响。
CAT1
阳离子氨基酸转运蛋白(CAT1)参与赖氨酸和精氨酸的转运。研究表明,对十二指肠和回肠中的CAT1没有显著影响。相反,与空肠中的膳食玉米醇溶蛋白相比,补充酪蛋白的CAT1表达水平更高。
因此,可以合理地得出结论,酪蛋白可促进赖氨酸和精氨酸的积累,这与补充酪蛋白的赖氨酸含量高于膳食玉米醇溶蛋白的报告一致。
肠道中赖氨酸和精氨酸的变化可能有助于酪蛋白对肠道微生物群的影响。CAT1的表达随着饮食中赖氨酸水平的增加而上调。
EAAC1
兴奋性氨基酸载体1(EAAC1)参与天冬氨酸和谷氨酸的转运,其作用类似于CAT1。EAAC1在十二指肠中的表达水平被膳食玉米醇溶蛋白下调,而在空肠和回肠中,补充酪蛋白的EAAC1表达水平高于玉米醇溶蛋白或大豆蛋白。
PePT1
此外,肽转运蛋白1(PePT1)的表达也受到不同蛋白质来源的调控。与植物蛋白相比,膳食酪蛋白在所有肠段都有更高的PepT1表达。
因此,具有平衡氨基酸组成的膳食酪蛋白可以促进氨基酸和肽转运蛋白的表达。这种表达可以加速功能性氨基酸在肠道环境中的运输。
赖氨酸、天冬氨酸和谷氨酸在管腔中的浓度受不同蛋白质来源的不同处理的影响。这些变化可以影响特定代谢物的衍生和转化途径,改变生理功能,改变肠道内的微生态平衡。
氨基酸还通过合成细菌蛋白质以及与肠道微生物群的相互作用对微生物组成起重要作用。
菌群发酵蛋白质主要副产物
短链脂肪酸
短链脂肪酸(SCFA)是细菌在大肠内代谢的最终产物,主要成分为乙酸、丁酸和丙酸。短链脂肪酸的基质主要来自膳食纤维和抗性淀粉。
然而,未消化的蛋白质也是短链脂肪酸产物的底物。膳食蛋白质在大肠中释放的几种氨基酸是短链脂肪酸合成的前体。肠道微生物可以从甘氨酸、丙氨酸、苏氨酸、谷氨酸、赖氨酸和天冬氨酸中产生乙酸盐。
谷氨酸和赖氨酸可以合成丁酸,丙氨酸和苏氨酸可以合成丙酸。
短链脂肪酸产物的数量和形态主要取决于营养物质的底物可得性、肠道菌群组成和肠道传递时间。短链脂肪酸是已知的氧化和使用作为燃料的结肠上皮。
此外,SCFA可以靶向游离脂肪酸受体(FFAR)配体转运到结肠肠细胞,FFAR代谢感知的激活在调节肠道脂肪沉积和营养物质吸收中发挥重要作用。
短链脂肪酸与肠道血流相互作用,促进5-羟色胺的释放,这可能受到肠道-脑内分泌轴的调节。这有助于增加肠道运动和离子转运,从而改变肠道菌群组成和免疫防御。
SCFA参与多种生理过程,在维持肠道完整性、葡萄糖稳态和食欲调节中发挥重要作用。除了为结肠细胞提供能量外,丁酸盐也是SLC5A8的底物。该蛋白编码基因抑制组蛋白去乙酰化酶的活性,而组蛋白去乙酰化酶是一种表观遗传调节因子,在多种肿瘤中诱导细胞凋亡。丁酸可以通过上调宿主防御肽的表达来刺激中性粒细胞群体,有利于破坏病原体,增强抗病能力。
氨
氨在大肠中以毫摩尔浓度存在。与远端结肠相比,近端结肠的特点是pH值低,碳水化合物含量高。因此,从升结肠到降结肠,氨浓度升高,这与远端结肠的蛋白质代谢率高于近端结肠是一致的。
大肠中的氨浓度主要是与氨基酸脱氨和尿素水解有关的微生物代谢物。肠道菌群可利用氨,氨可被上皮细胞吸收。尿素在肠道内的水解是通过细菌脲酶活性进行的。
虽然幽门螺杆菌的脲酶活性已经得到了充分的研究,但关于其他大肠微生物的脲酶活性的信息却很少。最近的一项研究报道,部分氨被谷氨酰胺合成酶的活性与l -谷氨酸缩合,使谷氨酰胺合成。这是一种可能的机制,以控制氨细胞内浓度在结肠细胞。
硫化氢 (H2S)
硫化氢(H2S)是蛋氨酸、半胱氨酸等含硫氨基酸发酵产生的一种微生物代谢物。H2S也来源于无机硫酸盐和亚硫酸盐添加剂的还原,以及肠道磺胺类物质的分解代谢。
肠道微生物能够通过脱硫酶从半胱氨酸碳链中获得能量。甲硫氨酸可转化为α-酮丁酸酯、氨和甲硫醇。这些代谢物归因于一个特定的分类组的细菌,包括大肠杆菌、肠沙门氏菌、梭菌和产气肠杆菌,通常在大肠中发现。
肠杆菌、肠球菌、肠链球菌、梭杆菌和真细菌能够发酵含硫氨基酸。
多胺
多胺是由精氨酸、鸟氨酸和蛋氨酸等氨基酸前体的结肠细胞产生的聚阳离子分子。从结肠癌中分离出来的结肠上皮具有高的多胺合成能力,这可能是由于肿瘤细胞持续有丝分裂需要高的多胺。
肠道微生物可从鸟氨酸、精氨酸、赖氨酸、酪氨酸和组氨酸等氨基酸前体产生腐胺、胍胺、尸胺、酪胺和组胺等多种多胺。
多胺参与细菌细胞的生长、增殖、分泌和运输活动。大肠生态系统中能够产生多胺的大肠细菌有多种,包括拟杆菌属(Bacteroides)、乳杆菌属(Lactobacillus)、Veillonella、双歧杆菌属(Bifidobacterium)和梭状芽胞杆菌属(Clostridium)。因此,肠道菌群组成是影响结肠多胺分布和分配的因素之一。
苯酚和吲哚
结肠中苯丙氨酸、酪氨酸和色氨酸等芳香族氨基酸可被特定的肠道微生物代谢为酚类和吲哚类化合物。
然而,芳香氨基酸的代谢与结肠微生物群的特定组分之间的相互作用还没有充分的研究。一些代谢产物如酚和吲哚,被怀疑为共致癌物和结肠癌促进剂。
已知发酵芳香氨基酸的厌氧菌包括大肠中的拟杆菌、乳酸菌、双歧杆菌、梭状芽胞杆菌和胃链球菌。与结肠中的其他氨基酸相比,芳香氨基酸在细菌中代谢缓慢。它们可以生产一系列的酚类和吲哚类化合物作为最终产品,如对甲酚、吲哚、酚和粪臭酚。
据报道,酪氨酸可产生苯酚和对甲酚,而苯乙酸和色氨酸在人体浆发酵过程中分解为吲哚乙酸盐和吲哚。
在一项研究中,远端结肠中的酚类化合物浓度增加,表明大肠远端区域的氨基酸代谢较高。吲哚和酚类代谢物的浓度取决于细菌产生率和结肠吸收率之间的平衡。酚类化合物似乎大部分被结肠内容物吸收。它们在从管腔转移到血液和肝脏的过程中部分代谢,最后通过尿液排出。
对无菌和常规小鼠血浆代谢物的对比分析表明,无菌小鼠的色氨酸和酪氨酸水平至少增加了1.5倍。涉及这些芳香族氨基酸的细菌代谢产物包括硫酸吲哚氧基、硫酸苯酯、硫酸对甲酚和苯丙酰甘氨酸,仅在常规小鼠中发现。
关于酚类和吲哚类化合物对结肠上皮细胞的影响知之甚少。体外实验表明,苯酚会降低屏障功能的完整性。需要注意的是,当苯酚浓度高于1.25 mM时,它会损害结肠上皮细胞。
如果蛋白质摄入过量,或者肠道里消化蛋白质的菌群构成不理想,那么蛋白质发酵发生的胺,硫化氢,苯酚等肠道毒素过量积累,会诱发肠道炎症,便秘,腹胀等问题,因此正确摄入适量优质蛋白对于维持肠道菌群和身体健康很重要。
蛋白质摄入要因人而异
实际上,我们摄入的一些蛋白质可以转化为葡萄糖,特别是当蛋白质的消化速度快且人体糖原储存量低时,有些蛋白质可以被微生物群发酵,特别是当蛋白质的消化速度慢时。
每个人对蛋白质的需求量各不相同,比如经常运动人群和久坐人群对蛋白质的需求量不一样,孕妇、老人、儿童、肥胖等群体都有各自不同的需求。当然,每个人的不同状态对蛋白质的需求量也都不同,比如爬山的一天,和在家刷剧的一天,也是不一样的。
因此,我们说最佳每日蛋白质摄入量取决于你的身高体重、健康状况、目标、身体活动水平。
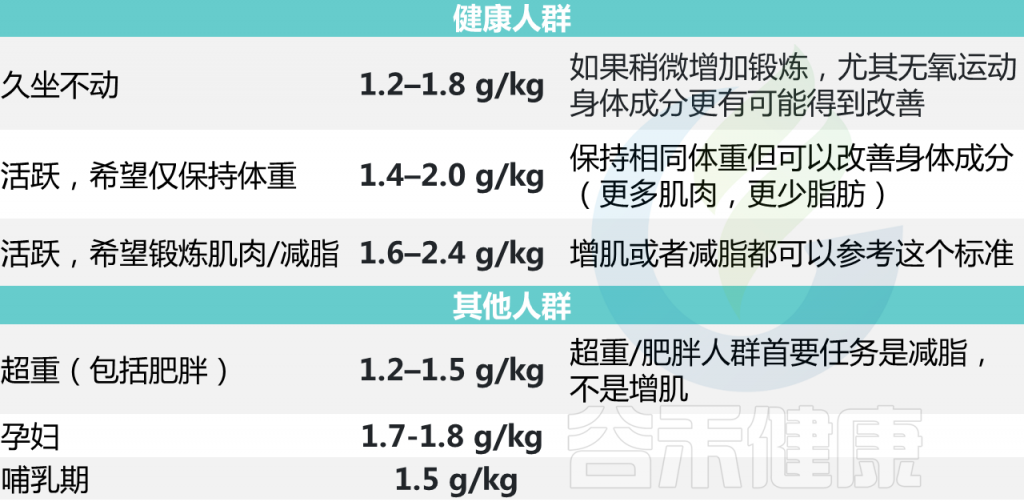

素食主义者对蛋白质的需求较高,因为植物蛋白在生物利用度上通常不如动物蛋白。植物蛋白消化率为 60-80%,而动物蛋白消化率超过90%.
注意植物蛋白的较难消化吸收
因为植物含有抑制蛋白质消化和吸收的抗营养物质,如胰蛋白酶抑制剂、植酸盐和单宁。虽然烧熟后确实降低了抗营养物质的浓度,但并不能完全消除它们。
另外,植物性蛋白质也含有限制性氨基酸,这些氨基酸是必须氨基酸但数量太少,以至于不够蛋白质的合成所需。
注:限制性氨基酸是指食物所含必需氨基酸的量与动物所需的蛋白质必需氨基酸的量相比,比值偏低的氨基酸。比如说一种食物里特别缺少某一种氨基酸,即使其它的氨基酸含量很高,因为这个氨基酸导致它的各个氨基酸比例不平衡。人体对这种食物的吸收依然不理想,这种氨基酸就叫限制性氨基酸。
结合不同的植物性蛋白质可以帮助弥补它们各自的不足。
摄入蛋白注意其氨基酸构成
比如说,赖氨酸是最常见的限制性氨基酸,特别是在谷类中,如小麦和水稻。坚果往往也含有赖氨酸作为限制性氨基酸。另一方面,豆类含有足够的赖氨酸,可以弥补,但缺乏硫氨基酸,如蛋氨酸和半胱氨酸。
蛋白质能否被人体吸收利用,还应该考虑蛋白质的氨基酸组成。
因此,了解一下评估蛋白质的另一个标准:氨基酸的组成。
氨基酸组成
蛋白质由氨基酸组成,身体可以合成其中一些氨基酸,而另一些则不能。
需要但不能合成、需要从食物中摄取的九种氨基酸被称为必需氨基酸(EAA)。
氨基酸:成人的需求和不同食物的含量
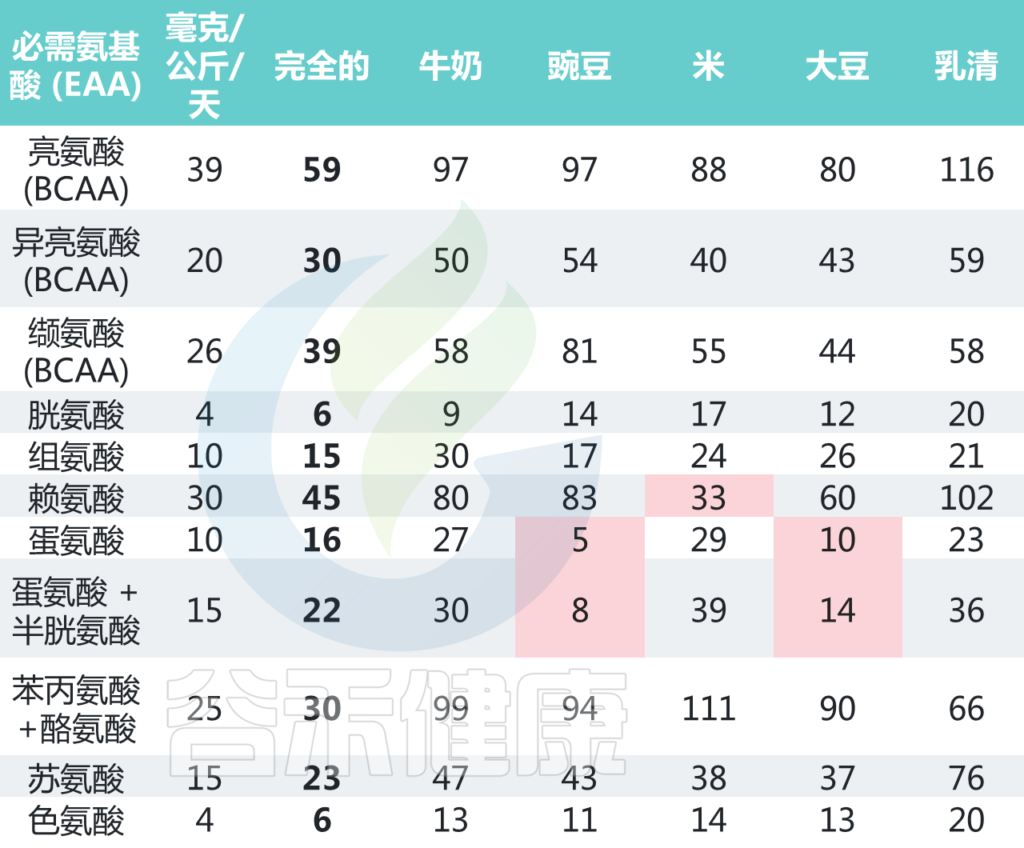
World Health Organization. Protein and Amino Acid Requirements in Human Nutrition
人群范围蛋白质摄入量如何?
使用谷禾检测人群,基于肠道菌群分析营养膳食摄入构成,图中给出了蛋白质占总能量摄入的占比,正常比例在10~20%范围。可以看出0~3岁蛋白质占比较高,3~15岁蛋白质占比较低,80~100岁比例又进一步下降。
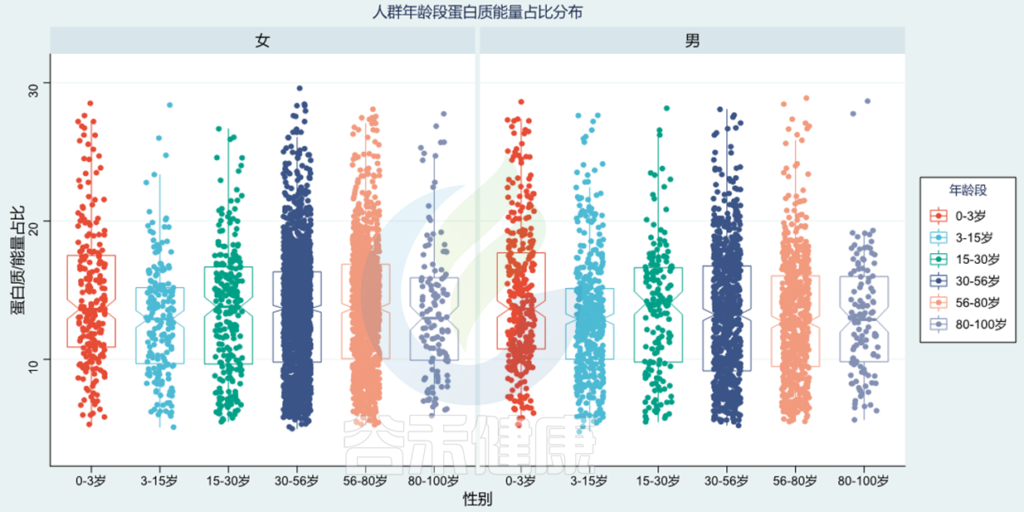
<谷禾健康肠道菌群检测数据库>
结 语
蛋白质代谢与肠道菌群密切相关。膳食蛋白质在小肠中被蛋白酶和众多肽酶代谢,膳食蛋白质释放的氨基酸可用于肠道微生物合成蛋白质。这反过来又促进了饮食、微生物群和宿主之间的氮循环和利用。
不同的细菌有不同的代谢能力,依靠我们摄入的营养物质繁殖生存,其中拟杆菌属(Bacteroides)、梭状芽胞杆菌属(Clostridium)、丙酸杆菌属(Propionibacterium)、梭杆菌属(Fusobacterium)、乳酸杆菌属(Lactobacillus)和链球菌属(Streptococcus)在蛋白质水解过程中发挥着重要作用。通过肠道菌群检测评估的蛋白质摄入水平显示不同人蛋白质摄入及营养占比差异很大,过高或者过低均伴随肠道菌群问题,包括过低导致潜在病原体数量增加,有益菌减少,同时导致部分氨基酸缺乏,过高又导致例如大肠菌群、链球菌和芽孢杆菌增殖,蛋白质发酵产生的副产物如,腐胺,苯酚,硫化氢等增诱发便秘,腹痛等健康问题。
除了蛋白质的摄入量,不同的蛋白形式,如植物蛋白,乳蛋白、动物蛋白以及蛋白质与其他营养摄入的相对比例构成,如膳食纤维、抗性淀粉、脂肪、微量元素等的比例都会影响肠道菌群进而反过来影响其消化降解,改变氨基酸的合成和代谢,最终影响宿主营养代谢健康。
总之,我们提倡膳食平衡,没有特殊情况不建议盲目补充蛋白粉等,建议从膳食结构调整保证营养均衡,从而使各种营养都能达到生长发育和身体健康的生理需要量,这同时要关注肠道菌群状况,因为他们是相辅相生。
常见食物中氨基酸含量见附录。
附录
供人类食用的植物性食品中总氨基酸的含量
(肽结合氨基酸加上游离氨基酸) mg/g
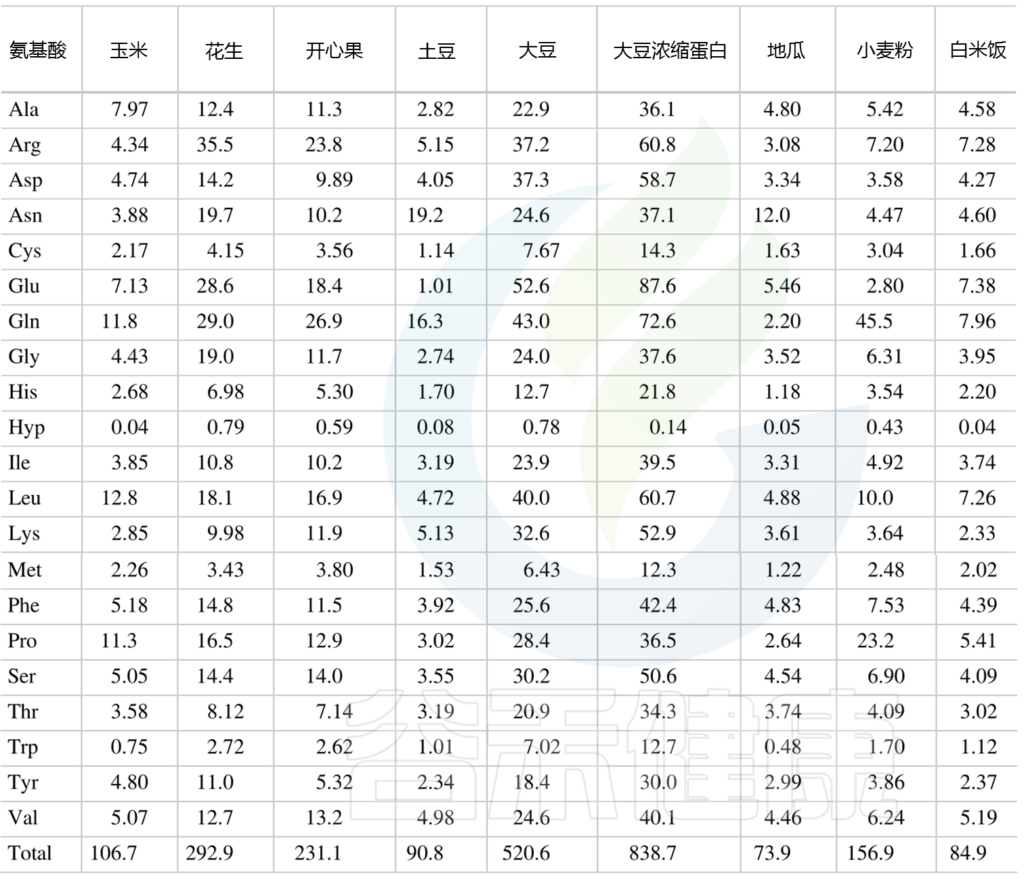
Amino Acids in Nutrition and Health, 2021
人类动物性食品中总氨基酸的含量
(肽结合氨基酸加上游离氨基酸)mg/g
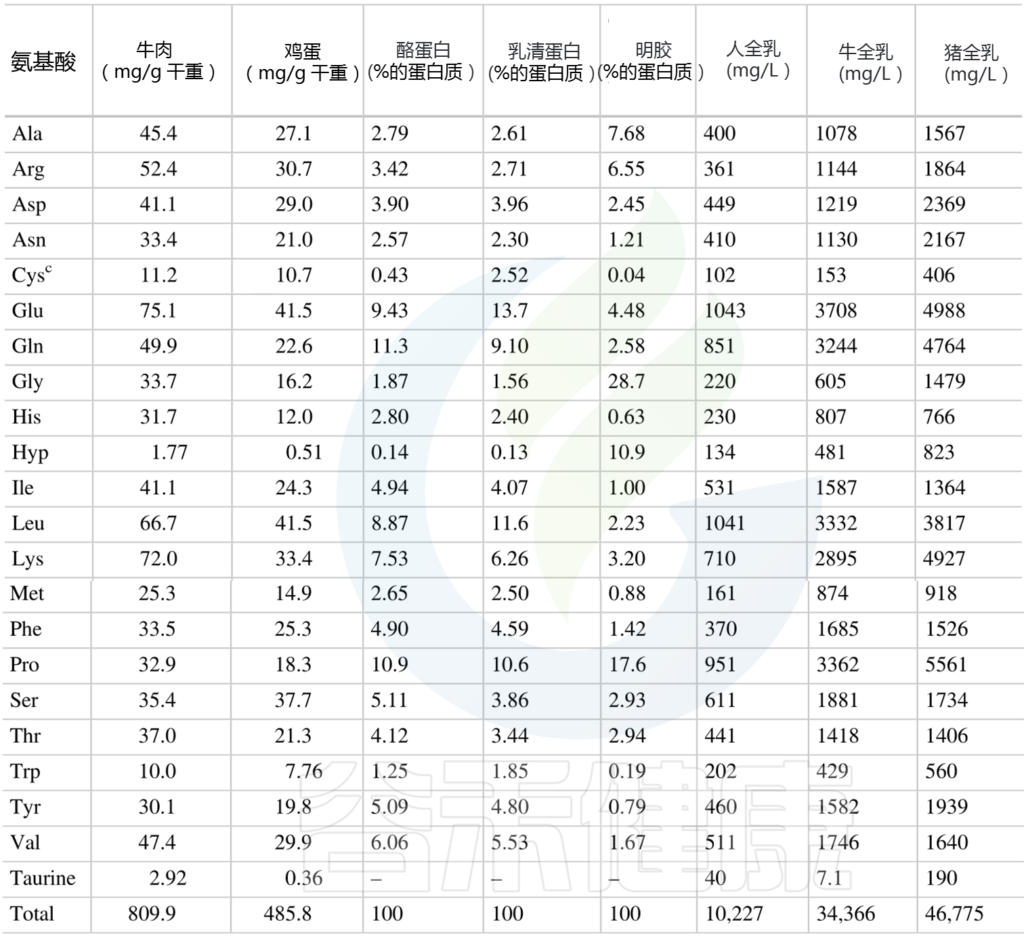
Amino Acids in Nutrition and Health, 2021
主要参考文献:
向上滑动阅览
Abdallah A, Elemba E, Zhong Q, Sun Z. Gastrointestinal Interaction between Dietary Amino Acids and Gut Microbiota: With Special Emphasis on Host Nutrition. Curr Protein Pept Sci. 2020;21(8):785-798.
doi: 10.2174/1389203721666200212095503. PMID: 32048965.
Ma N, Tian Y, Wu Y, Ma X. Contributions of the Interaction Between Dietary Protein and Gut Microbiota to Intestinal Health. Curr Protein Pept Sci. 2017;18(8):795-808.
Yadav M, Verma MK, Chauhan NS. A review of metabolic potential of human gut microbiome in human nutrition. Arch Microbiol. 2018 Mar;200(2):203-217. doi: 10.1007/s00203-017-1459-x. Epub 2017 Nov 29. PMID: 29188341.
Zhao J, Zhang X, Liu H, Brown MA, Qiao S. Dietary Protein and Gut Microbiota Composition and Function. Curr Protein Pept Sci. 2019;20(2):145-154.
Lin R, Liu W, Piao M, Zhu H. A review of the relationship between the gut microbiota and amino acid metabolism. Amino Acids. 2017 Dec;49(12):2083-2090. doi: 10.1007/s00726-017-2493-3. Epub 2017 Sep 20. PMID: 28932911.

谷禾健康

有没有发现我们吃的食品正在变得越来越鲜艳,让人有食欲,“低脂”、“无糖”等字眼出现的频率越来越高,然而这其中必然会用到各类食品添加剂,如防腐剂,甜味剂,乳化剂,着色剂,香料等等。
可以看到孩子们喝的牛奶都是纯白无瑕的,事实上天然牛奶往往没有那么白;孩子们吃的五颜六色的糖果,糕点等也会结合一些人工色素,因此儿童比成人更容易接触到食用色素。

食品添加剂会诱发菌群失调,通过微生物群肠脑轴导致肠道疾病,代谢性疾病等各类问题的发生。这个过程是如何发生的?各类添加剂有什么不同的作用?为什么儿童学习、记忆受到影响?为什么与情绪也有关系?…
本文为大家解答相关疑惑。
首先,我们先来看看:食品添加剂都有哪些种类,它们起到什么样的作用,以及如何查看食品中是否存在添加剂。
工业界使用合成色素使其产品更有吸引力,使用防腐剂有助于保持其功能,适当pH值等特性,因此在大多数情况下,食品添加剂是不可或缺的存在。常见的食品添加剂及其功能用途见下表。


IFIC & FDA

谷禾健康
乳酸菌属益生菌是使用最广泛的益生菌之一。罗伊氏乳杆菌( L. reuteri ) 是一种经过充分研究的益生菌,可以在大量哺乳动物中定殖。
罗伊氏乳杆菌是一种革兰氏阳性杆状细菌,已在各种食物中发现,尤其是肉类和奶制品。在人类中,罗伊氏乳杆菌存在于不同的身体部位,包括胃肠道、泌尿道、皮肤和母乳。罗伊氏乳杆菌的丰度因个体而异。
可能的优势
● 可能促进皮肤光泽和头发浓密
● 可能有助于对抗感染
● 可能减少炎症并加强免疫
● 可能会改善肠道健康(减少 IBS 和 IBD 症状)
● 可能降低胆固醇
可能的副作用
● 可能对组胺不耐受的人不利
● 可能导致某些人体重增加
● 缺乏大规模临床研究
● 长期补充的安全性未知
罗伊氏乳杆菌(Lactobacillus reuteri)名字来自德国微生物学家 Gerhard Reuter,他在1960 年代在人类肠道和粪便样本中发现了它。1960 年代被发现时,罗伊氏乳杆菌自然存在于 30-40% 的人体内,现在大约降至10-20%。科学研究者将这种下降与生活方式的改变联系起来。我们不像以前那样吃发酵食品,如酸菜,而是使用防腐剂,这会杀死食物和体内的细菌。
罗伊氏乳杆菌在发酵过程中能够产生葡聚糖和果聚糖。其中一种葡聚糖,α-1,4/1,6 葡聚糖,分子量为 40 MDa,支化度约为 16%,似乎是一种饱腹感诱导剂,对胰岛素和血糖水平有良好的影响在人类。葡聚糖不会在胃和空肠中降解,而是在结肠中完全降解。由于其慢淀粉特性,这种葡聚糖可能是烘焙应用中一种促进健康的成分。
不同菌株的Lactobacillus reuteri已被证明具有不同的生理作用。例如,Lactobacillus reuteri DSMZ 17648用于治疗幽门螺杆菌(H. pylori),而Lactobacillus reuteriNCIMB 30242 用于治疗高胆固醇。
但是,罗伊氏乳杆菌补充剂尚未获得批准用于医疗用途,而且有的益处和副作用缺乏可靠的临床研究。法规制定了补充剂的制造标准,但不保证它们是安全或有效的。
大多数关于Lactobacillus reuteri的研究是在动物或细胞中进行的。临床研究很少,而且大多数是低质量的、小规模的或可能存在偏见的。此外,使用的确切菌株因不同研究而异。
尽管罗伊氏乳杆菌在人类中是正常的,但并不是每个人的胃肠道中都有它。口服补充剂可以增加和补充胃肠道罗伊氏乳杆菌,然而它不一定会长期留在那里。
同样,罗伊氏乳杆菌的良好来源是乳制品和肉类,素食者和大多数素食者都避免食用这些食品,因此补充剂很重要。在母乳喂养时服用罗伊氏乳杆菌补充剂的女性更有可能将这些有益细菌转移给婴儿。
为消化和吸收而建,胃肠系统的某些部位已发展为对微生物定植不利。这方面的例子可以在由小肠上部胃酸和胆汁盐引起的低 pH 条件下看到。因此,在胃肠道定植的第一步就是在这样的环境中生存。幸运的是,罗伊氏乳杆菌对低 pH 值和胆汁盐具有抵抗力。这种抗性被认为至少部分取决于其形成生物膜的能力。