-

 国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子
国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子

 二级病原微生物安全实验室
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
谷禾健康
阴道微生物组在女性阴道健康方面起着重要作用。使用高通量宏基因组和16S rRNA 测序,已在人类阴道中鉴定出250多种细菌。其中,乳杆菌是健康阴道中最常检测到的微生物,其中包括卷曲乳杆菌(Lactobacillus crispatus)、惰性乳杆菌(Lactobacillus iners)、詹氏乳杆菌(Lactobacillus jensenii)和格氏乳杆菌(Lactobacillus gasseri)。
乳杆菌被认为通过产生有机酸、过氧化氢 (H2O2)、细菌素和其他抗菌化合物来防止病原体入侵,从而对阴道生态有益。月经、怀孕、性行为、阴道冲洗和不受控制地使用抗生素等因素会迅速改变微生物群落。阴道生态系统的破坏的特征是乳杆菌物种的枯竭和非乳杆菌微生物的过度生长。
如果把阴道微生态比作一座城市,惰性乳杆菌(Lactobacillus iners)就像那位“随时上线的值班员”:它无处不在、反应灵敏,却不总是最强的守护者。与经典的“稳定派”L.crispatus不同,L.iners 更像过渡期的掌舵者——在月经、性行为、抗生素、妊娠激素等因素影响时占据主导地位,帮助群落从一种状态向另一种状态转变。
但这份灵活也带来代价:它产酸与抑菌能力较弱,黏附与免疫调控独具一格,因而既能伴随健康,也常与生态失调、细菌性阴道病、性传播感染、妊娠不良风险相伴。认识并理解这位“双刃剑”角色,或许是我们走向更精准的女性生殖健康管理、重塑“好菌优势”的关键一步。
惰性乳杆菌(Lactobacillus iners)是阴道微生物群中最普遍的成员,然而,其在复杂的阴道微环境中的作用尚不完全清楚。鉴于其在乳杆菌中具有独特属性,且对多种不良生殖结局的影响差异明显,了解其基本特征有助于更全面地认识其在女性健康中的作用。
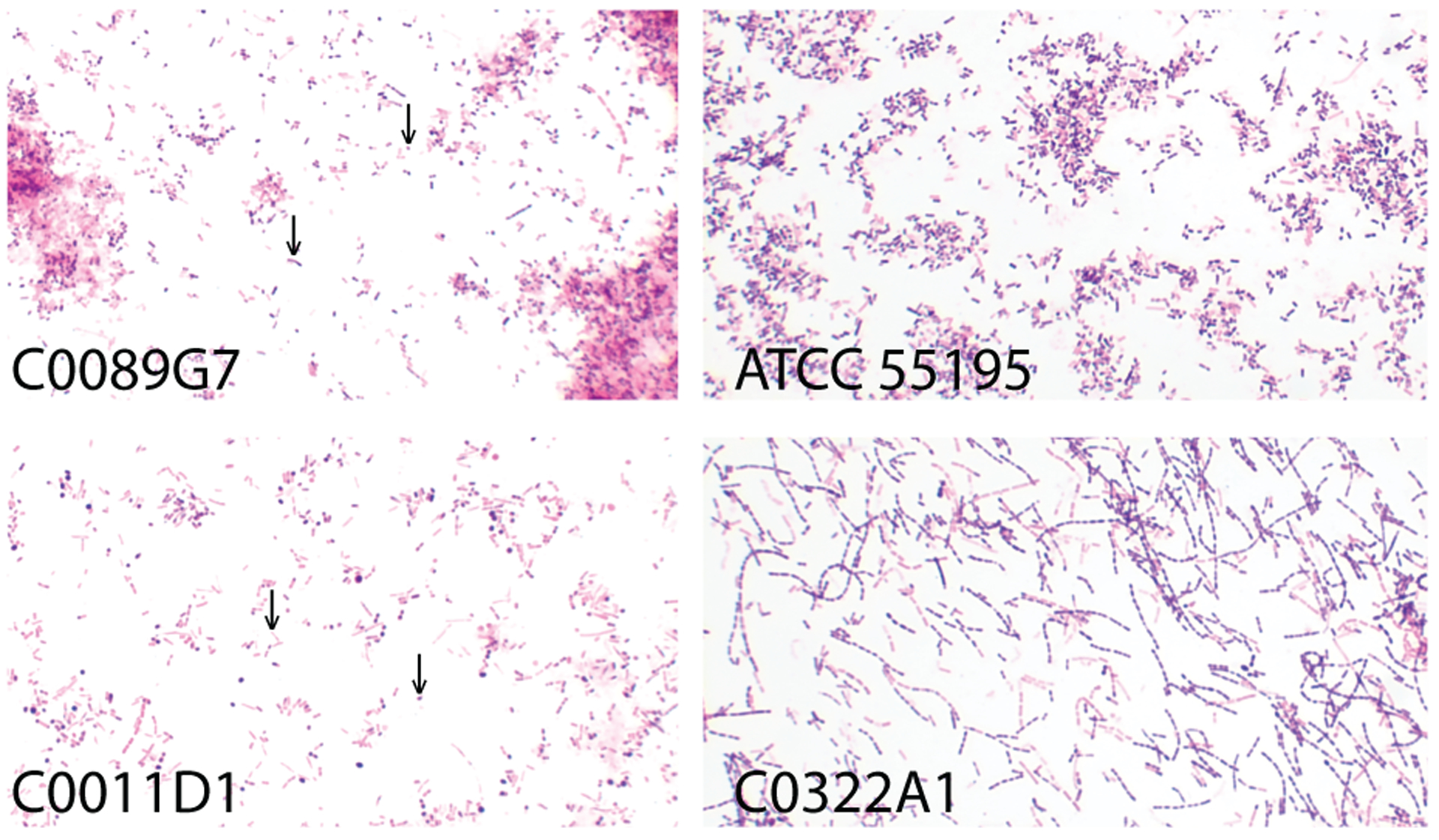
惰性乳杆菌(L.iners)于1999年才首次在阴道和泌尿道标本中被发现;因无法在常规MRS琼脂的标准条件下像其他乳杆菌那样生长而长期被忽视。
1
形态特征
• 革兰氏染色具有可变性
惰性乳杆菌(L.iners)起初被归为革兰氏阳性、杆状、非孢子形成的兼性厌氧菌,但多项研究表明其并非总呈典型的革兰氏阳性染色,形态亦常似球杆菌。一些报道其多呈革兰氏阴性、短小杆状,耐酸性弱(在pH3培养基中不可存活),这或致其早期在培养与显微观察中被忽视。
L. iners 出现“时而革兰氏阳性、时而革兰氏阴性(或变异性)”主要有以下原因:
–细胞壁肽聚糖层很薄:电镜研究显示其PG层显著薄于典型乳杆菌,导致在革兰染色中难以稳定保留结晶紫,呈现革兰阴性或“变异性”外观(可见杆菌或球菌样形态)。
-形态可塑性与菌株差异:同一菌株内及不同分离株之间均可见细胞形态与染色表现的变动,提示菌株层面的结构与功能差异会影响染色结果。
-环境与基因表达的影响:在不同阴道微环境(如细菌性阴道病样环境 vs. 乳杆菌占优势环境)中,L.iners 的基因表达(如调控细胞形态的rodZ等)会改变细胞形态与壁性质,从而影响革兰染色表现。
-方法学与判读偏差:Nugent评分依赖革兰染色形态计分,而L. iners 的“革兰阴性样”表型易被误判为加德纳菌等阴性形态,导致分类偏差,放大了其“阴性”出现的频率。
L. iners 培养分离株的革兰氏染色
Holm JB,et al.Curr Infect Dis Rep.2023
• 厌氧条件下生长能力显著强于其他乳杆菌
该菌在血琼脂上厌氧孵育 24 小时可形成小而光滑、圆形、半透明、无色素的菌落;在添加1–5%绵羊或人血的MRS琼脂上多数分离株亦可生长。
研究证实,L. iners 在厌氧条件下可在MRS琼脂上持续生长至少7天,显著长于其他乳杆菌。
2
基因组和功能
• 较小的基因组对环境波动适应能力较差
惰性乳杆菌(L.iners)的基因组很小,平均为1.28 Mbp,而L.crispatus为2.25 Mbp,这已经处于乳杆菌属的较低基因组大小范围内。相应地,L.crispatus泛基因组计数4300个基因,而L.iners的基因组为2300个基因,平均GC含量为 ~33.3%。
如此低的基因组大小强烈表明一种寄生性、宿主依赖性的生活方式。此外,人们认为较小的基因组预示着对环境波动的脆弱性更高。
• 碳水化合物和氨基酸代谢基因减少
基因组分析表明,L. iners碳水化合物和氨基酸代谢相关的基因数量严重减少,而它维持了主要核心代谢蛋白的保守基因和来自宿主或群落的必需化合物的膜转运基因。确定了三个潜在的核心基因(inerolysin、ZnuA 和 hsdR)与L.iners对阴道环境的特异性适应密切相关。
inerolysin是一种异常的成孔型胆固醇依赖溶素,在酸性环境中活性高,能在膜上形成孔洞,或为稳定获取宿主营养所必需;高亲和力锌摄取蛋白 ZnuA 维持金属离子稳态,可能介导对阴道上皮的强黏附;I 型限制性内切酶亚基 hsdR 被认为在细菌性阴道病期间参与抗噬菌体防御。
• 独特的肽聚糖层可能有助于吸收营养
此外,L. iners 携带完整的肽聚糖(PG)合成与水解酶基因。与其他乳杆菌物种相比,惰性乳杆菌细胞膜独特而薄的肽聚糖(PG)层可能更容易吸收营养或分泌蛋白质,从而提供必需的营养物质或应对阴道环境的快速变化。
3
产酸和过氧化氢能力较弱
• 相较其他乳杆菌抵抗病原体能力较弱
乳杆菌是阴道主要产酸者,能降低pH并抑制潜在致病菌。但不同物种的产酸能力存在差异:L.crispatus、L.gasseri、L.jensenii可由糖原发酵生成D-乳酸与L-乳酸;而L.iners缺乏D-乳酸脱氢酶,仅产L-乳酸,因此其L/D比值最高。
乳酸异构体对宿主免疫的影响不同;较高的L/D比值可上调 EMMPRIN 并激活 MMP-8,促进细胞外基质分解,利于细菌穿越宫颈并诱发上行感染。此外,D-乳酸对外源细菌的抑制强于L-乳酸,说明L.iners在防御病原入侵方面较弱。
• L.iners为主时阴道pH值偏高
研究发现,L.crispatus优势时阴道pH持续偏低;而 L.iners过度生长并不能维持低 pH,且在低 pH 环境中表现较弱。细菌性阴道病常伴随 pH 升高,此时多见加德纳菌与L.iners共存而少见其他乳杆菌。
L.iners 亦缺乏通过丙酮酸氧化产生 H2O2 的机制,而 H2O2 被认为有助于抑制厌氧菌定植。因此,当病原菌挑战阴道环境时,L.iners无法抵抗病原菌的过度生长和pH值的升高,而它可能会持续存在生态失调。
4
营养需求
L. iners基因组异常小、代谢受限,却有更复杂的营养需求,使其能适应多样阴道生态位。激素和其他因素的波动可能会影响阴道环境,导致粘液和糖原产生、pH 值和微生物种类发生变化,这可能为L.iners提供必需的营养。
• 主要对葡萄糖、麦芽糖进行糖酵解
基因组分析表明,除能从葡萄糖、麦芽糖、海藻糖和甘露糖产生酸外,这些菌株不能从L-阿拉伯糖、D-阿拉伯糖醇、环糊精、糖原、N-乙酰葡糖胺、乳糖、甘露醇、松三糖、蜜二糖、甲基β-D-吡喃葡萄糖苷、普鲁兰、棉子糖、核糖、鼠李糖、山梨糖醇、蔗糖、塔格糖、海藻糖或D-木糖产生酸。
• 编码inerolysin使其在失调环境下更具竞争优势
惰性乳杆菌(L.iners)对外源氨基酸的依赖性更高。并且L.iners已被证明是迄今为止已知的唯一编码一种形成孔的溶细胞毒素 inerolysin 的乳杆菌物种。
L.iners产生 inerolysin 的能力可能是影响其从阴道环境中获取营养能力的最重要因素之一。与在平衡的微生物环境中相比,超过10%的编码L.iners中 inerolysin 的基因在生态失调中的表达更高。该毒素可直接从宿主组织/细胞释放资源,使L.iners在营养匮乏和不利条件(如细菌性阴道病,其他乳酸杆菌难以定植)下具竞争优势。
注:尽管在乳杆菌中没有发现铁摄取系统,但在乳杆菌中检测到了能够催化亚铁离子并结合原卟啉IX形成血红素的铁螯合酶。
5
粘附能力
惰性乳杆菌(L.iners)对宿主细胞的粘附被认为通过阻断病原微生物在阴道上皮细胞的结合位点的机制在排除病原微生物方面发挥作用。
• 中性pH值下L.iners与人纤连蛋白的结合强于其他乳杆菌
尽管L.iners缺乏乳杆菌属的大部分主要粘附分子,但它仍然对阴道上皮细胞表现出很强的粘附能力。纤连蛋白是阴道上皮细胞外基质中的一种不溶性糖蛋白。L.iners基因组编码一种纤连蛋白结合蛋白,该蛋白含有金黄色葡萄球菌致病菌株共有的基序(纤连蛋白结合蛋白 A),从而介导金黄色葡萄球菌对宿主细胞的粘附和侵袭。研究证明,在更中性的 pH 值下,L.iners与人纤连蛋白的结合明显强于其他乳杆菌物种,这可能导致L.iners在阴道中持续存在,尽管存在病原体或用抗生素治疗。
• L.iners独特的粘附能力会影响其保护作用
一项体外研究报告说,L.iners可能会增加引起细菌性阴道病(BV)的G.vaginalis的粘附。研究还证明,L.iners产生溶孔素,这是一种常见于病原菌中的成孔蛋白,可以增强粘附能力。这些发现表明,L.iners独特的粘附功能降低了对健康阴道微生物组免受病原菌侵害的保护。
6
抗菌和免疫特性
L.iners是最常见且持久的阴道共生乳杆菌,能高度适应复杂且动态的阴道环境。环境波动时,其他乳酸杆菌或难以存活,而 L.iners 借助基因组层面的灵活调控——如特异性碳水化合物摄取、纤连蛋白结合、噬菌体防御与惰性溶血素合成——维持稳定丰度。其在多种条件下的生存优势使其在微生物组过渡期常成优势种。
• L.iners在不同环境下可能存在免疫保护也可能扰乱稳态
多项研究已证实 L.iners 具备多种免疫特性,其细胞因子调控、外源营养摄取和噬菌体防御等特性,有助于在不利条件(包括 BV)下生存。L.iners还能限制有害菌获取铁等关键营养,并通过激活上皮先天免疫抑制其生长;此外,通过阴道上皮细胞之间的特定分子相互作用观察到 L.iners 的抗炎作用。
以 L.iners 为主的阴道微生物组与上皮应激反应的诱导相关。研究发现,L. iners(而非 L. crispatus)显著上调人原代阴道上皮的模式识别受体通路并提升 TNF mRNA,显示其对宿主免疫的物种特异调节。有人提出,L. iners 在压力下可激活 TLR 信号、提高 HSP70、抑制自噬,从而扰乱上皮稳态并削弱对潜在病原的识别与应答。相反,多种参与上皮抗菌防御的分子(如 NGAL、钙卫蛋白、透明质酸)又更易被其诱导。
综上,L.iners 既能对抗非生理威胁、促进恢复至更健康状态,又表现促炎特性,且在不同环境下并不总是共生角色。
阴道微生物群在维持女性健康的阴道微环境方面起着重要作用,其组成受种族、年龄、妊娠、阴道疾病等多种因素的影响。作为全球育龄女性阴道微生物组中最常见的乳杆菌成员,Lactobacillus iners兼具“普遍性”与“复杂性”。
一方面,乳杆菌总体被视为女性阴道健康的关键守护者,通过乳酸与抗菌物质产生抑制病原体、维持低pH与生态稳态;另一方面,L.iners又与经典的有益乳杆菌(如L.crispatus、L.jensenii、L.gasseri)显著不同:其基因组较小、代谢能力相对受限、主要产生L-乳酸、能在高低pH及BV阳/阴性环境中存活,因而常被视为生态扰动后的“过渡型”定植者。
流行病学与多组学研究提示,L. iners主导的微生物群(CST III型)在多个人群中广泛存在,较缺乏乳杆菌的多样化菌群仍具一定保护性,但与L.crispatus主导状态相比,其对病原体(包括念珠菌)定植与部分不良结局(如阴道生态失调、性传播感染及妊娠不良)的防护可能次优;同时,行为与社会学因素(如无保护性行为、性伴数增加)可能与其主导状态相关。
阐明L.iners在阴道健康与疾病中的双刃剑角色,并识别可干预的致病特征与潜在生物标志物,至关重要且紧迫。
1
L.iners与阴道生态失调
证据强度:较高
与肠道菌群相比,健康个体阴道微生物环境的一个典型特征是其细菌多样性极低。健康绝经前妇女有五种主要的群落状态类型(CST),即L.crispatus 主导的 CST I、L. gasseri 主导的 CST II、L.iners 主导的 CST III 和 L. jensenii 主导的 CST V,而 CST IV 的特征是缺乏乳杆菌属。阴道生态失调以细菌多样性高和厌氧菌混合物为特征,经常与多种妇科疾病有关。
• L.iners在孕妇中占比均较高,但健康孕妇中更多
一项研究从中国广州中山大学附属第六医院招募了一组 95 名孕妇。该队列包括 34 名健康孕妇(健康)和 61 名患有至少一种疾病的孕妇。
参与者患有的疾病也被分为妊娠糖尿病(GDM)(n=21)、母体相关不良妊娠结局(MAPO, n=29)、胎儿相关不良妊娠结局 (FAPO, n=13)、阴道感染(n=6)、肝脏或肾脏异常(n=14)、贫血(n=8)、血栓形成倾向(n=4)和辅助受孕(n=11)。
中国孕晚期孕妇阴道微生物组组成及结构
Wang X,et al.NPJ Biofilms Microbiomes.2025
研究发现,在物种层面,健康孕妇的阴道微生物组呈现出独特的分布模式,其中L.iners主导的CST-III型占比高达50%,显著高于疾病组的31.15%,但在所有组中,L.iners所主导的分型占比均不低。
• L.iners为主的阴道菌群保护性不及L.crispatus
L.iners常见于健康女性、阴道生态失调(如 BV)患者,甚至抗菌治疗者。多项研究表明,L.crispatus与健康相关,而以L.iners为主的群落有时难以提供足够保护以预防失调。
L.iners的存在与较高水平的促炎因子有关,例如白细胞介素1α、白细胞介素18、巨噬细胞迁移抑制因子和肿瘤坏死因子α,它们负责激活阴道炎症反应。
• L.iners适应能力较强,BV患者丰度可能也不低
尽管L.iners的作用仍有争议,但其丰度相对稳定,不易被病原体替代。细菌性阴道炎时,L.iners常与潜在有害菌共存,而L.crispatus则很难做到。
其基因组支持对失调环境的适应,可能与代谢、细胞溶解及抗噬菌体防御基因随阴道环境变化而调控有关。其在多种条件下的强生存力提示其既是宿主防御的重要成员,也是可维持与恢复阴道微生物组的持久共生乳杆菌。
2
L.iners与细菌性阴道病
证据强度:较高
细菌性阴道病(BV)是育龄妇女中最常见的阴道炎类型。它的特点是乳杆菌属显著减少或消失,伴随着以厌氧菌和兼性细菌为主的更多样化的微生物群的出现,如加德纳菌属、普雷沃氏菌属和阴道曲霉属。
• L.iners是能在细菌性阴道病中存在的乳杆菌
L.iners通常是唯一可以在细菌性阴道病(BV)期间检测到的与BV相关细菌共存的阴道乳杆菌物种。凭借对基因组功能的动态调控,它能在剧变的阴道环境中持续存在;L.iners基因表达的增加可能导致琥珀酸和其他短链脂肪酸的产生以及 BV 环境中 pH 值的增加。
为了适应BV环境,L.iners可以增加inerolysin和粘蛋白的表达,促进甘油的产生和相关代谢酶的表达,从而保证其从外源获取营养物质。此外,噬菌体是导致 BV 期间乳杆菌种类突然减少的原因之一,而 L.iners 可以上调I型 RM 系统和 CRISPR 等防御系统,以及其特异性 hsdR 基因,以抵抗 BV 期间噬菌体的入侵。
最近的一项研究发现了三种由名为Lactobacillus paragasseri的人类肠道菌株产生的细菌素活性肽。这些细菌素对乳杆菌具有很强的选择性抑制活性,而L. crispatus、L. jensenii和L. gasseri仅受到轻微抑制,表明这些乳杆菌衍生的抑制剂可以与甲硝唑联合使用,以改善目前的BV治疗。
• L.iners有助于预测细菌性阴道病的状态
由于 L.iners 在 BV 中的共存与其他乳杆菌物种不同,因此L.iners的流行率可以作为微生物指标来预测BV的发作或中间状态。此外,L.iners对甲硝唑具有耐药性,并且被发现它是主要的乳杆菌物种,即使在用甲硝唑处理BV后也是如此。
与BV期间几乎缺失的保护性乳杆菌相比,L.iners竞争力更强,能在失衡微生物组中共存。有人认为它促成 BV 与非 BV 状态的过渡。值得注意的是,治疗后微生物组常仍以L.iners为主,而未转为L.crispatus优势。因此,L.iners持续存在或致长期阴道失调,尤见于反复治疗后。但仍需研究其究竟是转化生物标志物,还是 BV 的促成因素。
3
L.iners与早产
证据强度:一般,存在矛盾结果
在怀孕期间维持阴道微生物组中乳杆菌属的自然健康平衡尤为重要。早期的研究证实,怀孕期间阴道内的高雌二醇水平和随之而来的高糖原水平导致更强的阴道酸化,从而随着妊娠的进展促进乳杆菌物种的流行。
• L.iners的丰度在孕中期和晚期显著下降
许多研究表明,以惰性乳杆菌(L.iners)为主的阴道微生物组更有可能在怀孕期间转向生态失调。在之前的研究中,发现健康孕妇的L.iners丰度在妊娠中期和晚期显著下降,而 L.crispatus 的丰度在妊娠中期与孕早期相比有所增加。
此外,研究发现,阴道清洁度和白细胞酯酶活性阳性的升高与 L.iners 而非 L.crispatus 的丰度增加相关。
• L.iners的丰度与早产之间存在一定联系
越来越多证据表明,细菌性阴道病(BV)是不良妊娠结局的重要病因,尤以早产(PTB)为甚。以L.iners占优势的“中间微生物群”被视为 PTB 的潜在风险因素。
有研究人员认为,在怀孕早期健康女性的阴道涂片中检测到的L.iners可能与早产(PTB)有关。另有研究也证明了这一点,报告说,妊娠16周时以L.iners为主的阴道微生物组是短宫颈和早期 PTB(<34 周)的危险因素,而 L.crispatus 优势在种族更多样化的队列中对 PTB 具有保护作用。
来自不同国家的最新研究表明,L. iners 与早产患病率增加之间存在关联。L. iners也被证明是黑人孕妇中最丰富的乳杆菌物种。然而,大多数研究并未确定乳杆菌占主导地位的阴道微生物组与黑人孕妇的 PTB 之间存在显著关系。
相反,三项研究报告说,L.iners与降低 PTB 风险有关。因此,L. iners 与 PTB 风险之间的关联还存在争议(表1)。此外,有限的样本量、样本采集时间、PTB 定义的差异、种族和地理差异、菌株鉴定方法的差异,以及遗传异常或 PTB 病史等复杂的临床条件,都是影响结果的混杂因素。
L.iners和早产的研究主要结果
Zheng N,et al.Front Cell Infect Microbiol.2021
基于L.iners的特性及其在作为唯一乳杆菌时对病原体保护不足,一些研究者认为它并不直接致孕期感染。事实上,L.iners优势的阴道微生物组较不稳定,孕期易向与BV相关的CST-IV转变;其还可能通过调节局部炎症与影响宫颈完整性,削弱化学与机械黏膜屏障,从而增加早产风险。但仍需进一步研究阐明二者的潜在机制。
4
L.iners与生物膜形成
证据强度:一般
生物膜是紧密附着在表面上的细菌结构,已知它们比浮游细胞对宿主免疫反应和抗生素治疗更具抵抗力。阴道上皮的生物膜与感染密切相关;大量证据显示BV伴随致密的多菌生物膜,其中加德纳菌(G.vaginalis)为主导菌。一般认为,加德纳菌启动生物膜形成并促进其他 BV 相关菌附着,进而增厚生物膜;其生物膜还能阻挡抗生素渗透,保护其他菌株。BV 的高复发被广泛归因于此类生物膜既能保护细菌免受抗生素治疗,甚至作为病原体再生的宿主。
• L.iners对病原菌的粘附抑制不如其他乳杆菌
阴道本土乳杆菌被认为可以通过空间位阻或粘膜中的受体掩蔽来防止病原菌的定植。先前的研究使用乳杆菌益生菌方法试图清除多微生物生物膜,从根本上阻止细菌毒力并抑制人体阴道感染。
据报道,植物乳杆菌(Lactobacillus plantarum)可显著降低 HT-29 细胞系中大肠杆菌、鼠伤寒沙门氏菌、金黄色葡萄球菌和铜绿假单胞菌的粘附,这使其成为治疗 BV 的潜在抗生物膜剂。体外生长的阴道加德纳菌生物膜被罗伊氏乳杆菌RC-14取代,并在有限程度上被L.iners取代。
研究发现,L.crispatus大大降低了G.vaginalis菌株对宫颈上皮细胞的粘附,包括来自健康女性和患有BV的女性。有趣的是,L.iners显著降低了健康女性对G.vaginalis菌株的粘附,但却增强了致病性G.vaginalis的粘附,表明L.iners可能可以与BV相关的G.vaginalis共存,并可能有助于G.vaginalis主导的生物膜形成。
此外,众所周知,念珠菌属,主要是白色念珠菌,可以形成厚而坚韧的生物膜,这大大增加了治疗复发性外阴阴道念珠菌病期间对抗真菌药物的耐受性。有研究报道了各种乳杆菌在共培养时抑制白色念珠菌生物膜形成和生物膜相关基因表达的能力。鼠李糖乳杆菌可下调白色念珠菌生物膜相关基因表达。相反,与 L.iners 共培养导致生物膜相关基因表达(ALS3 和 ECE1)上调,表明 L.iners 的存在可能表明阴道生态失调的转变;因此,它不应用作白色念珠菌感染的益生菌干预措施。
进一步了解阴道共生乳杆菌与生物膜结构和功能之间的相互作用对于确定生物膜相关感染的新治疗方法极为重要。
5
L.iners与性传播感染
证据强度:低
先前的研究报告称,以L. crispatus为主的阴道微生物组与较低的性传播感染患病率有关,而细菌性阴道病与性传播感染风险升高有关,例如沙眼衣原体、人类免疫缺陷病毒(HIV)、淋病奈瑟菌、巨细胞病毒和单纯疱疹病毒2型感染。
• 以L.iners为主的阴道微生物群受病原体感染风险更高
有研究报告说,基线时以L.iners为主的阴道微生物组显著增加了一年后感染沙眼衣原体的风险。以乳杆菌为主的阴道微生物组中缺乏D-乳酸可能会通过调节宫颈完整性来增加 HIV 横贯宫颈阴道粘液的能力。
许多研究报告称,HPV阳性女性的阴道微生物多样性较高,乳杆菌种类丰度较低。与L.crispatus相比,以L.iners为主的阴道微生物组与高危 HPV 感染相关。可以推测,阴道生态失调可能会影响宿主对 HPV 感染的先天免疫力,导致发育不良/宫颈癌。
这些发现表明,L.iners可能表现出类似于BV的阴道微生物组组成的快速变化,并且可能是性传播感染下动态阴道环境的有价值的生物标志物。
6
L.iners与不孕症
证据强度:较低
阴道生态失调,包括pH值升高、菌群多样性增加、细菌性阴道病、外阴阴道念珠菌病和滴虫性阴道炎,被认为是不孕症的危险因素。还有研究报告说,L.iners的丰度与不孕率增加有关。
• L.iners丰度可能对怀孕率存在影响
近期有研究报道,以L.iners为主的阴道微生物组与输卵管性不孕和沙眼衣原体感染相关。作为过渡物种,L.iners可能在治疗或体外受精(IVF)所致的高雌激素环境下,促使异常与正常微生物组相互转换。
7
L.iners与月经周期
证据强度:一般
人类微生物组项目显示,包括阴道在内的各部位微生物群在同一受试者内的时间变化小于个体间差异。月经周期是扰动阴道微生物多样性的关键因素。L.crispatus通常在育龄妇女的阴道中占主导地位,而 L.iners 在月经周期中过度生长并取代L.crispatus。
• L.iners在月经期间明显增加,有助于判断月经时间点
最近的一项研究报告说,L. iners是卵泡期复发最频繁的微生物;L.iners和CST IV型在围排卵期占主导地位;在黄体期,最常见的类型是 CST IV。事实上,L.iners的丰度在月经期间显著增加,通常与G.vaginalis或Atopobium vaginae的增加同时;然而,在没有干预的情况下,它们随后在月经后减少。
由于阴道生态的动态变化是在同一个体月经周期的不同时间点表征的,因此相对于月经周期的采样时刻对于阴道微生态分析非常重要。
L.iners作为改善生殖健康的靶点
鉴于L. iners在阴道微生物组中的高检出率、其在多种阴道环境状态中的存在以及其功能可塑性(无论是通过遗传多样性、维持多菌株、差异基因表达还是两者兼有),特定的L. iners菌株、基因或产物可能成为治疗或预防性调节阴道微生物群的广泛适用靶点。
在关于阴道乳杆菌对半胱氨酸依赖性的报告中证明,L.iners缺乏L.crispatus、L.gasseri和L.jensenii中存在的外源性半胱氨酸摄取转运系统。相反,L.iners依赖于摄取外源性L-胱氨酸,然后在细胞内将其转化为半胱氨酸。在模拟细菌性阴道病(BV)样菌群的体外实验中,使用胱氨酸摄取抑制剂和甲硝唑处理促进了L.crispatus的生长,而单独使用甲硝唑处理则导致L.iners的扩增。因此,胱氨酸摄取抑制剂或其他特异性抑制L.iners的试剂未来可能作为细菌性阴道病治疗方案的补充,以调节微生物群组成,促进非L.crispatus乳杆菌属的扩增。
总的来说,卷曲乳杆菌(L.crispatus)与最佳阴道健康状况相关,部分原因在于其全球普遍存在以及与积极健康结果的相关性。然而,由于宿主生理或免疫学原因,卷曲乳杆菌的优势状态可能并非某些个体能够合理实现的状态。确实存在稳定的以无害乳杆菌(L.iners)为主的微生物群,这些微生物群可能具有特定的菌株水平特征,即宏基因组亚种,并且可能对某些个体而言是最优的。
一种新的干预措施可能是将与细菌性阴道病(BV)风险增加相关的不稳定乳杆菌微生物群调节为由对变异有抵抗力的无害乳杆菌菌株组成的稳定微生物群。
本文涵盖了L. iners的主要特性、健康影响及其在未来女性健康中的潜在作用:
一、L.iners的主要特性
普遍性与过渡性:是育龄女性阴道中最常见的乳杆菌之一,常出现在生态扰动后的过渡阶段,可在健康与失调状态(如BV)中共存。
基因组与代谢:基因组最小(约1.28 Mbp),代谢能力受限,依赖宿主营养;缺乏D-乳酸脱氢酶,仅产L-乳酸,产酸与过氧化氢能力弱于L.crispatus。
特异功能基因:携带inerolysin(成孔毒素)、ZnuA(高亲和锌摄取)和hsdR(I型限制性内切酶亚基)等,有利于在酸性、营养匮乏和噬菌体压力下生存。
免疫调控:可上调上皮PRR通路与炎症因子(如TNF),在不同环境下呈现抗炎与促炎双重效应。
二、对女性健康的影响
阴道生态:以L.iners为主(CST III型)的群落相较L.crispatus主导状态保护性较弱;在细菌性阴道病中常与厌氧菌共存,能耐受甲硝唑并在治疗后持续占优,或与生物膜形成、复发相关。
生物膜与病原体黏附:对G.vaginalis的黏附抑制弱,甚至增强致病株黏附;与白色念珠菌共培养可上调ALS3/ECE1,提示其不宜作为念珠菌感染的益生菌干预。
妊娠与早产:孕期以L.iners为主的群落较不稳定,易向BV相关CST-IV转变;可能通过局部炎症与宫颈完整性影响,增加早产风险(证据存在异质性)。
性传播感染(STI)与HPV:L. iners主导与较高的STI风险和高危HPV相关;多样性升高、乳酸杆菌减少的状态或削弱对HPV的先天免疫。
月经周期动态:卵泡期与围排卵期更易见L.iners增多,提示其对激素与环境波动高度敏感。
三、未来在女性健康中的作用与方向
风险分层与生物标志物:作为生态扰动与STI动态环境的指示菌,用于BV“中间状态”、治疗后复发风险及HPV/早产风险的预测与监测。
精准干预靶点:围绕其特征通路(inerolysin、金属离子摄取、CRISPR/限制修饰系统、黏附因子)设计干预,限制致病性而非简单清除。
益生策略优化:更倾向于重建L. crispatus优势的“功能性益生”与微生态移植;避免在念珠菌感染等场景使用L. iners相关干预。
个体化管理:结合月经周期、妊娠阶段、性行为模式与种族/地域差异,动态评估菌群状态,制定个体化预防与治疗方案。
总而言之,惰性乳杆菌(Lactobacillus iners)在波动环境中具生存与一定保护能力,但其较弱的产酸/抗菌功能与独特黏附、免疫调控特性,关联BV、STI、HPV风险升高以及妊娠不良与不孕。未来应将其作为风险标志与精准干预靶点,聚焦功能替代与群落重建,推动女性生殖健康的个体化管理与预防治疗升级。
主要参考文献
Wang X, Jiang Q, Tian X, Chen W, Mai J, Lin G, Huo Y, Zheng H, Yan D, Wang X, Li T, Gao Y, Mou X, Zhao W. Metagenomic analysis reveals the novel role of vaginal Lactobacillus iners in Chinese healthy pregnant women. NPJ Biofilms Microbiomes. 2025 May 30;11(1):92.
Zheng N, Guo R, Wang J, Zhou W, Ling Z. Contribution of Lactobacillus iners to Vaginal Health and Diseases: A Systematic Review. Front Cell Infect Microbiol. 2021 Nov 22;11:792787.
Holm JB, Carter KA, Ravel J, Brotman RM. Lactobacillus iners and genital health: molecular clues to an enigmatic vaginal species. Curr Infect Dis Rep. 2023 Apr;25(4):67-75.
Kim H, Kim T, Kang J, Kim Y, Kim H. Is Lactobacillus Gram-Positive? A Case Study of Lactobacillus iners. Microorganisms. 2020 Jun 29;8(7):969.
Novak J, Ravel J, Ma B, Ferreira CST, Tristão ADR, Silva MG, Marconi C. Characteristics associated with Lactobacillus iners-dominated vaginal microbiota. Sex Transm Infect. 2022 Aug;98(5):353-359.
Bautista C. T., Wurapa E., Sateren W. B., Morris S., Hollingsworth B., Sanchez J. L. (2016). Bacterial Vaginosis: A Synthesis of the Literature on Etiology, Prevalence, Risk Factors, and Relationship With Chlamydia and Gonorrhea Infections. Mil. Med. Res. 3, 4.
Alonzo Martinez M. C., Cazorla E., Canovas E., Martinez-Blanch J. F., Chenoll E., Climent E., et al. (2021). Study of the Vaginal Microbiota in Healthy Women of Reproductive Age. Microorganisms 9 (5):1069.
Borgdorff H., Armstrong S. D., Tytgat H. L., Xia D., Ndayisaba G. F., Wastling J. M., et al. (2016). Unique Insights in the Cervicovaginal Lactobacillus Iners and L. Crispatus Proteomes and Their Associations With Microbiota Dysbiosis. PloS One 11 (3), e0150767.
Campisciano G., Florian F., D’Eustacchio A., Stankovic D., Ricci G., De Seta F., et al. (2017). Subclinical Alteration of the Cervical-Vaginal Microbiome in Women With Idiopathic Infertility. J. Cell Physiol. 232 (7), 1681–1688.

谷禾健康
麦角硫因 (EGT) 是一种含硫的抗氧化剂,由某些微生物合成,大量存在于蘑菇、发酵食品和其他膳食产品中。人体无法自主合成,需要通过饮食摄取和积累。
麦角硫因(EGT)由哺乳动物细胞输入,可以在哺乳动物组织中积累到低毫摩尔浓度,在那里它与保护健康作用有关。 同样,许多宿主相关微生物输入EGT,EGT通过其抗氧化特性增强细胞内氧化还原稳态。某些细菌物种也编码降解 EGT 的酶。事实上,最近的研究发现 EGT 可以被人类粪便细菌的复杂群落代谢。 粪便群落的 EGT 代谢因人而异,表明微生物组组成的个体间差异可能影响 EGT 代谢。
许多与宿主相关的微生物都会吸收EGT,通过其抗氧化特性增强细胞内氧化还原稳态。某些细菌物种也编码降解 EGT 的酶。因此研究人员提出假设并进行验证:EGT是否会先被部分菌种代谢生成某种可用物质,再由后续菌种接力还原,从而获能(促进ATP合成与生长)?
为此,研究人员结合群落模型、分离菌株和共培养实验、代谢物时间序列实验、多队列宏基因组数据再分析等技术进行了研究和验证。完整重现了Clostridium symbiosum和Bacteroides xylanisolvens这两种细菌在厌氧环境下”EGT→TUA(thiourocanic acid)→还原产物(3-(2‑thione‑imidazol‑4‑yl)‑propionic acid)”的代谢接力过程。在对24份健康人粪便群落进行48小时培养的功能表型测试中,有18/24样本显示EGT代谢活性并产生TMA,且其中11/18检测到还原TUA产物;相应的宏基因组分析显示,具EGT代谢活性的样本中ergothionase基因显著富集。在四个独立结直肠癌队列的粪便宏基因组数据分析中,发现ergothionase基因在其中两队列中显著富集,在另外两队列中呈增加趋势。
这些发现揭示了饮食抗氧化分子在肠道微生物能量代谢与潜在疾病风险差异中的功能纽带,为未来通过调控微生物EGT代谢改善肠道健康提供了方向。
1
确定麦角硫因代谢菌和酶
已知麦角硫因酶(EGT trimethylammonia lyase)可将EGT裂解为TMA与TUA。Treponema denticola SP33 ergothionase (TdETL) 是已被表征的“参照”,用其氨基酸序列做同源搜索,在人肠道可培养菌株库里寻找麦角硫因(ergothionase)同源物,从而锁定了C. symbiosum CLOSYM_01531和C. symbiosum CLOSYM_03165。
通过LC-MS/LC-MS-MS对C. symbiosum+EGT-d9的培养上清与细胞组分进行非靶与靶向代谢组学分析,验证其将EGT裂解为TMA与TUA,并显示TUA主要分泌到胞外。异源表达验证实验(将CLOSYM_01531这个基因装进E. coli里表达),观察到EGT-d9在约6小时内被完全转化为TMA-d9与TUA。
2
小鼠粪便菌群
来自不同来源的雌性6周龄的C57BL/6小鼠(JAX、TAC、CR)做粪便群落48小时厌氧培养+EGT-d9,比较EGT代谢能力差异。对CR群落做非靶代谢组学,鉴定得到新的代谢物(m/z 173.0379),经标准品比对,确认为还原的TUA产物3-(2‑thione‑imidazol‑4‑yl)‑propionic acid
3
细菌共培养
通过对CR 和 TAC 粪便微生物组的16S rRNA 测序分析,发现CR群落中Bacteroides acidifaciens富集。研究人员选取与 B. acidifaciens 亲缘关系较近、且在人肠道常见并已知具备多糖与宿主营养代谢能力的代表物种 B. ovatus 与 B. xylanisolvens 作为候选,检验其是否具有将 TUA 还原为 3-(2thioneimidazol4yl)propionic acid 的活性。并测试还原过程对能量代谢与生长的影响。
单培养与共培养添加EGT-d9的C.symbiosum和B.xylanisolvens时,进行长时间跟踪。同时用表达CLOSYM_01531的工程E.coli替代C.symbiosum重复共培养过程。进一步证明接力关键在于EGT产生的胞外TUA与下游还原步骤的耦合。
4
健康人队列粪便菌群
对24份健康粪便样本做48小时厌氧培养+EGT-d9,查看EGT代谢能力和产物,发现有18/24个样本把EGTd9代谢掉了。在这18个样本中,对其中3个TUA还原能力强的样本做了时间跟踪,还原代谢轨迹。同时将其中23份宏基因组数据分为两组(有代谢活性 vs 无代谢活性),做配对分析,查看ergothionase基因富集情况。
5
结直肠癌队列宏基因组功能分析
基于EGT稳态与结直肠癌(CRC)的文献线索,汇总四个已发表CRC粪便宏基因组数据集,采用统一流程定量ergothionase基因丰度,然后进行组间差异分析。
▸ C.symbiosum可将EGT代谢为TMA与TUA,且TUA主要分泌到胞外
如上图所示,经EGT-d9处理的培养物的细胞沉淀物(C. symbiosum ATCC 14940)中检测到EGT-d9和TMA-d9,表明C. symbiosum ATCC 14940导入并代谢EGT。
在C. symbiosum ATCC 14940培养上清液中检测到一种单一的代谢物(m/z 171.0223,图C),对比未经EGT-d9处理的样品,m/z 171.0223显著上调(Log2(fold change) ≥ 2, p ≤ 0.05) ,经标准品对比(图D、E),确认为TUA。用或不用EGT-d9处理C. symbiosum培养物48小时后的EGT-d9、TMA-d9和TUA含靶向定量结果显示(图F),EGT‑d9下降与TMA‑d9、TUA上升在计量上匹配,符合EGT代谢为“两段”——TMA与TUA的表现。
将CLOSYM_01531异源表达于E. coli(图C),发现其可在约6小时内将EGT‑d9完全转为TMA‑d9与TUA(图D),空载对照(Ec_EV)无此活性(图G)。此外,C. symbiosum中还鉴定到第二个同源物CLOSYM_03165,异源表达同样具活性。
▸ 小鼠粪便菌群的代谢组学分析鉴定出“还原产物”
不同来源小鼠展现出群落差异,如图A,发现CR群落48小时内可将EGT‑d9完全代谢为TMA‑d9,TAC群落仅中等程度代谢并积累TUA,JAX群落则基本无代谢。
编辑
但在CR培养物上清液中并未检测到TUA,而是发现了一个新的代谢物(m/z 173.0379),且呈显著上调(log2(fold change) ≥ 3, p ≤ 0.05),经标准品比对,确认是3‑(2‑thione‑imidazol‑4‑yl)‑propionic acid,由TUA进行2e−/2H+加氢还原所得(图E)。CR群落可将外源TUA完全转为该还原产物,而JAX、TAC不能(图G)。
▸ TUA还原增强了B.xylanisolvens在厌氧条件下的ATP合成和生长
16S rRNA 分析CR和TAC样本的菌群,发现它们具有相似的菌群多样性(图A、B),且Bacteroides acidifaciens在CR群落中显著富集(图C)。研究人员又提出假设, B. ovatus和B. xylanisolvens或许可以将TUA还原为3‑(2‑thione‑imidazol‑4‑yl)‑propionic acid。
事实也的确如此,在添加了TUA的培养基中于厌氧条件下培养这两种菌48h,定量结果显示两种菌株都消耗了培养基中的TUA,并产生了等量的3‑(2‑thione‑imidazol‑4‑yl)‑propionic acid(下图A)。B. xylanisolvens约6小时内可将TUA完全转化,且产物主要在胞外(上图E、F)。在缓冲体系中,B. xylanisolvens因TUA或TUA+甲酸钠而ATP合成约提升4倍(下图B),并伴随还原产物累积(下图C);在厌氧环境中且缺乏其它电子受体(也就是能增强ATP合成的化合物)的最小培养基中,发现TUA显著提升其生长(下图D)。
▸ C.symbiosum和B.xylanisolvens互相利用EGT的代谢产物
鉴于C. symbiosum和B. xylanisolvens可以分别代谢EGT和TUA,研究人员先假设这两个菌种可以共同将EGT转化为3‑(2‑thione‑imidazol‑4‑yl)‑propionic acid,然后通过共培养实验进行验证。验证结果显示,C.symbiosum在单独培养时会将EGT‑d9转为TMA‑d9和TUA,不产生3‑(2‑thione‑imidazol‑4‑yl)‑propionic acid,这表明C. symbiosum不能还原TUA。B.xylanisolvens则对EGT‑d9无作为。两者共培养时,EGT‑d9消耗程度与C.symbiosum单独培养时相当,但TUA几乎不积累,反而是还原产物显著积累。(图E培养4天、图F培养7天)
代谢组火山图显示细胞沉淀和培养上清液中分别仅富集TMA‑d9与还原产物,符合跨物种接力还原产物的代谢模式“EGT→TUA→3‑(2‑thione‑imidazol‑4‑yl)‑propionic acid”(图I、L)。
用表达CLOSYM_01531的E.coli与B.xylanisolvens共培养,也再现了“EGT→TUA→3‑(2‑thione‑imidazol‑4‑yl)‑propionic acid”反应(图G)。
▸ 人群普遍性和个体差异
研究人员筛选了24名健康成人,获得其粪便样本。在添加EGT-d9培养基中共同培养48小时,观察EGT代谢活性差异。发现24个样本中,有18个样本在48小时内显著代谢EGT‑d9,且均产生TMA‑d9(图A-D),这表明麦角硫因介导的EGT代谢的一个特征是消除TMA。虽然在24个样本中都没有检测到TUA,但能够代谢EGT‑d9的18个样本里,有11个样本积累了还原产物(图D)。
对3个TUA还原能力强的样本做时间序列分析,发现6–12小时内TUA短暂出现并随EGT‑d9下降而上升,随后被转化为还原产物,这直接展示了跨物种接力还原产物的代谢模式。
▸ 肠道EGT稳态的改变可能与结直肠癌有关
编辑
研究人员量化了来自四个独立结直肠癌队列和健康对照队列的粪便宏基因组数据集中ergothionase基因的相对丰度,发现有两个CRC组存在ergothionase基因显著富集的现象,另两队列呈上升趋势,这提示EGT代谢能力可能是癌症相关肠道菌群的功能特征之一。
过去更多讲的是“单个细菌把EGT彻底分解”,现在发现EGT还能被“分工协作”重塑成能增强ATP合成的化合物。这项“分工协作“的主角分别是C. symbiosum和B. xylanisolvens,重点工作内容是,C.symbiosum将EGT代谢为TUA和TMA-d9,B.xylanisolvens将TUA还原为3-(2-thione-imidazol-4-yl)-propionic acid,同时提高了自己的ATP产量,促进了自己的生长。B. xylanisolvens作为一种益生菌,目前被认为与宿主健康密切相关。根据现有发现,通过添加TUA或在有“产TUA菌”背景下添加EGT,理论上有助于B. xylanisolvens的定植。
研究中虽然提示EGT→TUA轴与疾病生态有关,但当前缺乏动物与人体定植/功能结果的直接验证。
总而言之,如果在合适的群落背景下提升B.xylanisolvens的定植或TUA还原能力,理论上可以带来:更稳的肠道生态位占据、更高效的厌氧能量代谢、潜在更好的底物利用与有益代谢物输出,并可能通过改变供给-利用格局而影响与疾病相关的微生态失衡。
主要参考文献
Zhou Z, Jiang A, Jiang X, Hatzios SK. Metabolic cross-feeding of a dietary antioxidant enhances anaerobic energy metabolism by human gut bacteria. Cell Host Microbe. 2025 Aug 13;33(8):1321-1332.e9.
谷禾健康
白色念珠菌(Candida albicans),对于绝大多数健康人而言,它只是口腔、肠道和阴道黏膜上的一个安静成员,与我们和平共处,数十年而不引起任何症状,构成了人体复杂微生物生态的一部分。然而,一旦这种平衡被打破,无论是因为免疫力下降、抗生素滥用,还是其他环境因素的改变,白色念珠菌会迅速转变为一种危险的病原体,引发从浅表黏膜感染到致命性全身感染的多种疾病。
正是基于这种威胁的现实性和紧迫性,世界卫生组织已将其列为“重点优先关注对象”的人类真菌病原体,凸显了其巨大的临床挑战。
一般对白色念珠菌相关疾病的诊疗思维,在很大程度上遵循着“病原体-清除”的经典模式。然而,有些反复的治疗困境,如复发性外阴阴道念珠菌病的棘手性、抗真菌药物耐药性的攀升,以及与肠道念珠菌过度增殖相关的、复杂的亚健康状态,都迫使我们必须超越这种简化模型。
白色念珠菌的致病行为,并非由单一因素决定,而是宿主、真菌本身以及周围微生物菌群三者间复杂互动的最终结果。
本文将深入探讨白色念珠菌的形态可塑性;详细阐述其在不同人体生态位(口腔、胃肠道、阴道)中展现出的高度特异性的定植、感染机制与临床表现,如何在共生与致病之间维持微妙的平衡,揭示其在不同微环境下的生存策略,并阐述理解这对于我们理解机会性感染、开发新型治疗策略以及重新定义健康与疾病边界的深远意义。
同时,本文还将介绍白色念珠菌的现代检测方法,以及提供基于微生态调节、天然抗菌物质和生活方式改善的综合干预策略。
要理解白色念珠菌如何在人体内切换角色,我们首先需要认识这种微生物的基本特征。
从分类学角度来看,白色念珠菌属于真菌界、半知菌门、芽孢菌纲、隐球菌目、念珠菌属。
形态学特征:一种真菌的”三副面孔”
白色念珠菌能够在三种主要形态——酵母、假菌丝、菌丝之间可逆性转换。
正是这些独特的生物学特征,使得它能够在复杂多变的人体环境中既能稳定定植,又能快速响应环境变化。
白色念珠菌最显著的特征是其形态可塑性,能够根据环境条件在三种形态间自由转换:
酵母形态、假菌丝形态、真菌丝形态。
真菌丝是 “疾病活跃期” 的标志 —— 比如肠道真菌感染时,真菌丝会穿透肠壁,引发腹痛、腹泻等症状。真菌丝会强烈激活免疫系统:它的细胞壁成分(比如 β- 葡聚糖)会被宿主的模式识别受体识别,触发大量促炎细胞因子(IL-1β、TNF-α、IL-17)释放。
影响因素
温度、pH、营养等影响其形态转换
酵母与菌丝之间的转变是由通常与哺乳动物宿主相关的因素触发的,例如:至少 37℃ 的温度、血清的存在、N-乙酰葡萄糖胺(GlcNAc)或高水平的二氧化碳。
温度:最核心的信号之一。
环境温度(25℃)下,白色念珠菌以酵母形态为主;进入人体后(37℃),会显著促进向菌丝的转换 —— 这就是为什么它在体外温和,进入体内后可能变凶。
pH 值:
酸性环境(比如阴道 pH 3.8-4.5、胃酸 pH 1-3)有利于酵母形态;中性 / 弱碱性环境(比如肠道末端 pH 7.0-7.5、感染部位的炎症环境)会促进菌丝形成。
营养:
血清、葡萄糖等营养丰富时,白色念珠菌会加速生长并转向菌丝;营养匮乏时,更倾向于保持酵母形态 “休眠”。
利与弊
双刃剑的角色
虽然我们通常关注白色念珠菌的致病性,但它在体内的存在并非一无是处。在某些情况下,它甚至可能扮演有益的角色。
潜在益处
一些研究表明,白色念珠菌的定植可能有助于训练和调节我们的免疫系统。
例如,在小鼠模型中,预先定植白色念珠菌可以增强机体对某些细菌病原体(如艰难梭菌)的抵抗力,这可能是通过刺激IL-17等免疫信号通路实现的。
此外,它占据了特定的生态位,也可能在一定程度上阻止了其他更危险的病原体入侵,这是一种“生态位占领”的保护作用。
明确的风险
然而,其风险是显而易见且巨大的。一旦失控,它就是多种感染的罪魁祸首。最严重的情况是侵袭性念珠菌病,特别是念珠菌血症(血液感染),此时真菌进入血液并播散到全身各个器官,死亡率极高,可接近50%,尤其是在免疫力低下的重症患者中。这正是它被WHO列为重点关注对象的核心原因。
理解机会致病菌的新视角——共生毒力学派
为什么白色念珠菌这样一个与我们长期共生的微生物,会拥有如此强大的武器库?难道这些复杂的致病工具,仅仅是为了在宿主免疫力低下时趁火打劫吗?
一个名为“共生毒力学派”(Commensal Virulence School)的新理论,为我们提供了更深层次的解释。
这个理论认为,许多我们所知的“毒力因子”,其首要功能并非为了致病,而是在复杂的共生环境中为了生存和竞争。只有在特定条件下,这些因子才被二次利用,表现为致病性。
我们以菌丝形成为例来理解这个概念:
◖在微生物丰富的环境中(如健康的肠道)
这里充满了各种细菌竞争。白色念珠菌形成菌丝,并分泌念珠菌溶素,可能并非主要为了攻击宿主细胞,而是为了抑制周围的细菌,为自己抢夺生存空间和营养。在这种场景下,菌丝形成是一种竞争策略。
◗ 在微生物稀少的环境中(如抗生素处理后)
此时细菌竞争压力大大减小。如果白色念珠菌还大量形成菌丝,反而会过度刺激宿主免疫系统,招来杀身之祸。因此,在没有竞争压力时,保持温和的酵母形态反而更有利于生存。
这个理论解释了为什么在标准的小鼠实验中(肠道菌群被抗生素清除),酵母形态的白色念珠菌定植得更好;而在拥有完整菌群的小鼠模型中,能够形成菌丝的菌株反而更具优势。
带着共生毒力学派的崭新视角,我们重新审视了白色念珠菌的双重身份——它既是潜在的致病者,又是微生物社区中的平衡参与者。这种理解为评估其实际健康影响提供了基础,接下来让我们具体探讨白色念珠菌感染可能带来的各种风险,从常见浅表症状到严重系统性威胁。
– 常 见 症 状 –
白色念珠菌感染的常见症状包括多种表现形式,具体取决于感染部位和个体差异。
白色念珠菌过多可能相关的症状
消化问题
腹胀、胀气、腹泻、便秘等症状。这是因为念珠菌以糖和碳水化合物为食,在肠道过度生长导致的。
疲劳症状
身体疲劳:影响营养吸收,导致能量不足
精神疲劳:难以集中注意力,大脑感觉疲惫
脑雾:思维不清晰,难以清楚思考
异常渴望
对碳水化合物和甜食有强烈渴望。健康的人通常不会对甜食有这么大的吸引力。
其他症状
情绪波动等,因为念珠菌在全身扩散,影响不同系统。
注:
* 单个症状不能确诊念珠菌过度生长,多个症状同时出现,症状越多可能性越大。
* 以上是基于功能医学实践者和一些肠道专家临床发现,而非强科学证据。
除了上述体内过度生长可能引发的一般症状外,白色念珠菌感染往往会表现为更具体的临床症状。这些症状通常与感染部位相关,下面我们来详细了解一些临床上典型的感染类型及其表现。
白色念珠菌感染的表现与症状
∎ 浅表性及局部感染
此类感染通常发生在皮肤和黏膜,是临床上最常见的类型。
▸ 口腔念珠菌病
别名:鹅口疮。
常见人群:免疫力低下者,如新生儿、假牙老人、广谱抗生素/糖皮质激素治疗患者、HIV感染者。
典型症状:口腔、舌头、上颚或咽部黏膜出现白色/淡黄色凝乳状斑块,可刮除,下方黏膜红肿、糜烂或轻微出血。
其他形式 :萎缩性舌炎(舌乳头萎缩、舌面光滑发红);口角炎(嘴角红肿、皲裂)。
▸ 外阴阴道念珠菌病(VVC)
常见人群:育龄期女性(极为常见)。
主要症状:外阴/阴道剧烈瘙痒、烧灼感、性交痛、排尿困难。
典型分泌物:白色、稠厚、豆腐渣样/凝乳状。
检查表现:外阴红肿,阴道黏膜充血,附白色分泌物。
注意:部分女性为无症状携带者。
▸ 皮肤及甲感染
好发部位:温暖、潮湿皮肤褶皱,如乳房下、腹股沟、腋窝、臀部及指(趾)间。
皮损表现:边界清晰的湿润红斑,边缘可见卫星状丘疹或脓疱。
甲念珠菌病:甲板浑浊、增厚、变色;甲周红肿疼痛(甲沟炎)。
∎ 侵袭性及深部感染
当白色念珠菌突破皮肤黏膜屏障进入血液或深层组织时,可导致危及生命的严重感染。
▸ 食管念珠菌病
常见人群:免疫严重低下者(如晚期艾滋病、血液肿瘤化疗患者)。
症状:吞咽痛、吞咽困难、胸骨后痛。
检查:内镜见食管黏膜覆盖白色斑块/伪膜,去除后黏膜糜烂/溃疡。
▸ 胃肠道念珠菌病
受累部位:除食管外,胃、十二指肠及整个肠道。
表现:溃疡、炎症;可能与炎症性肠病(IBD)发病相关。
▸ 侵袭性念珠菌病与念珠菌血症
定义:念珠菌进入血流,播散全身器官(最严重形式)。
常见表现:念珠菌血症——持续高热(抗生素无效)、寒战、低血压、心动过速等脓毒症症状。
进展:多器官衰竭(如呼吸困难/ARDS、肾衰竭);播散至眼(眼内炎/失明)、心(心内膜炎)、CNS(脑膜炎/脑脓肿)、肝脾肾等,形成迁徙性脓肿。
人群:高危群体。
▸ 中枢神经系统念珠菌病
常见人群:新生儿 / 婴儿(尤需关注)。
表现:脑膜炎、脑脓肿或脑梗死;症状不典型,如间歇发热、嗜睡、喂养困难或抽搐。
浅表黏膜感染(口腔鹅口疮、外阴阴道念珠菌病)非常常见;而侵袭性念珠菌病虽相对少见,却与较高的病死率相关。
以上我们了解了白色念珠菌感染的广泛健康风险,从轻微的表面症状到致命的全身感染。接下来我们将深入探讨这种真菌是如何在人体特定部位安顿下来,以及它在不同微环境中采用的独特生存策略和致病机制。
– 口腔 ▪ 肠道 ▪ 阴道 –
白色念珠菌并非无处不在,它在我们体内有几个偏爱的“定居点”,主要是黏膜表面。这些地方环境各异,白色念珠菌却能凭借其超强的适应能力,在口腔、阴道和胃肠道这三个主要生态位中找到自己的生存之道。
口腔是白色念珠菌最早也是最重要的定植部位之一。在健康状态下,口腔中的定植是无症状的,即使真菌数量不少,也未必会引起炎症。这得益于口腔内复杂的平衡机制。
注:口腔也是个复杂的微生物社区,它是由 700 多种细菌和240 多种真菌物种以及古菌、阿米巴和鞭毛虫组成的复杂微生物组。
宿主的防御
唾液是第一道防线,它含有多种抗菌物质如:组蛋白、黏蛋白和分泌型免疫球蛋白A(sIgA),这些物质能聚集并清除真菌细胞。
真菌的策略
白色念珠菌也很聪明,它能利用自身的黏附蛋白(如Hwp1)与口腔上皮细胞紧密结合,抵抗唾液的冲刷。
口腔中的白色念珠菌定植是一个动态平衡的过程,受到宿主免疫、真菌自身特性和微生物群三方共同调控。
阴道是另一个白色念珠菌常见的定植部位,也是外阴阴道念珠菌病(VVC)的发源地。然而,在 20-30% 的女性中,阴道黏膜的定植通常是无症状的。这主要归功于阴道独特的微生态环境。
与口腔不同,健康的阴道微生物群相对简单,通常由乳杆菌占据绝对主导地位。这些乳杆菌通过多种方式构建起一道坚固的防线:
◆ 维持酸性环境:乳酸杆菌通过发酵产生大量乳酸,使阴道维持在pH 3.8-4.5的酸性环境。这种低pH值能有效抑制白色念珠菌从温和的酵母形态向侵袭性的菌丝形态转变。
◆产生抗菌物质:乳酸杆菌还会分泌多种武器,如细菌素、过氧化氢(H₂O₂)等,这些物质可以直接抑制或杀死白色念珠菌。
◆物理屏障与竞争:乳酸杆菌能黏附在阴道上皮细胞表面,形成一层生物屏障,直接与白色念珠菌竞争空间和营养,阻止其立足。
因此,当乳酸杆菌数量充足、功能正常时,白色念珠菌只能作为少数派潜伏下来。但当这道防线因抗生素使用、激素水平变化或过度清洁等因素被削弱时,白色念珠菌便会趁机大量繁殖,引发外阴阴道念珠菌病。
阴道菌群检测可以评估乳酸杆菌的占比和阴道微生态的健康状况。例如,在谷禾的阴道菌群检测报告中,可以看到乳酸杆菌比例极低,同时提示存在菌群失调,这正是白色念珠菌等机会性病原体易于滋生的环境。
如上图阴道菌群检测报告显示,乳杆菌比例较低,提示菌群失调。当乳杆菌减少时,阴道pH值上升,抵抗力下降,易引发感染,出现外阴瘙痒、灼热感、白带增多等症状。
肠道是系统性感染的源头,大多数健康人肠道都有白色念珠菌,但真菌只占微生物的0.1%左右(不过真菌细胞大,生物量不小)。它粘在肠上皮的黏液层上,用转录因子和蛋白酶帮忙降解黏蛋白。
肠道菌群通过“定植抗性”的机制,牢牢地控制着白色念珠菌的数量,防止其过度生长。
什么是定植抗性?
定植抗性,可以理解为健康肠道菌群天然具备的一种“排外”能力,它能有效阻止外来病原体的入侵和定植,同时也能压制体内常驻的条件致病菌(如白色念珠菌),不让它们过度生长、惹是生非。
这种抗性主要通过以下方式实现:
1
梭菌纲与丁酸盐的氧气消耗术
肠道,特别是结肠,是一个高度缺氧的环境。像Clostridium butyricum,Faecalibacterium Prausnitzii这类专性厌氧菌,它们在发酵膳食纤维时会产生短链脂肪酸,如丁酸盐。丁酸盐不仅是肠道细胞的能量来源,还能促进肠道上皮细胞消耗氧气,从而维持肠腔的缺氧状态。
Clostridia→丁酸盐→PPAR-γ→耗氧→缺氧
激活肠道上皮细胞核内的一个重要受体PPAR-γ,激活的PPAR-γ会促进上皮细胞进行脂肪酸的β-氧化,这是一个极其耗氧的过程。
上皮细胞大量消耗从肠道血管渗透过来的氧气,从而在紧邻肠腔的区域形成一个缺氧生态位。这个区域有效地阻止了氧气进入肠腔,维持了肠腔内部的严格厌氧状态。
这种缺氧环境非常不利于白色念珠菌,因为白色念珠菌需要氧气才能高效生长。一旦这个链条断裂(例如,抗生素杀死了Clostridia),氧气就会涌入肠腔,为白色念珠菌的疯狂增殖打开了门。
2
拟杆菌与抗菌肽的免疫联动
除了营造缺氧环境,肠道菌群还能直接刺激宿主免疫系统,生产武器来对抗白色念珠菌。在这方面,拟杆菌发挥着重要作用。
拟杆菌等共生菌能够刺激肠道中的潘氏细胞和上皮细胞,产生多种抗菌肽(AMPs),可以直接攻击微生物。其中,Cathelicidin这类抗菌肽(如人体的LL-37和小鼠的CRAMP)被证明对抑制白色念珠菌尤为关键。它们能够破坏真菌的细胞膜,导致其死亡。
3
营养争夺战
数以万亿计的肠道细菌与白色念珠菌之间存在着激烈的营养竞争。细菌消耗了大部分可利用的碳水化合物、氨基酸和微量元素,让白色念珠菌经常处于吃不饱的状态。
例如,研究发现,一些肠道菌群消耗掉山梨醇等糖醇类物质,而这类物质恰好是白色念珠菌偏爱的营养源。当菌群被破坏后,肠道中山梨醇水平升高,直接为白色念珠菌的生长添柴加火。
这场营养争夺战远比我们想象的更精细复杂。
在攻防战中,铁、锌等微量元素的争夺也至关重要。白色念珠菌有专门的系统来从宿主那里偷铁和锌,而我们的身体则会通过营养免疫策略,将这些元素隐藏起来,试图饿死病原体。
谷禾肠道菌群检测升级版的报告,既能能够通过分析菌群的代谢,间接评估体内微量元素如铁、锌等营养状况,又能检测白色念珠菌是否阳性,为我们理解宿主与微生物的互动提供了新的视角。
这里的白色念珠菌和铁、锌都处于正常。
假如当白色念珠菌异常增多,且铁或锌处于缺乏时,可能是由于白色念珠菌过度增殖,其专门的铁摄取系统高效工作,大量消耗了肠道中可用的铁,这是一种病原体与宿主争夺铁资源的表现。
这种组合可能提示:
潜在的肠道感染风险:铁缺乏可能削弱免疫力,可能伴随IL-17等炎症通路激活,关于IL-17的指标,在谷禾宏基因组检测报告中也有相关指示。
4
微生物间的外交博弈
肠道微生态就像一个复杂的社区,细菌与真菌之间的关系并不止敌对。
■ 与铜绿假单胞菌——铁之战
在某些环境下(如无粒小鼠模型),白色念珠菌会与细菌病原体铜绿假单胞菌(Pseudomonas aeruginosa)共存。铜绿假单胞菌会分泌物质抑制白色念珠菌的菌丝生长,而白色念珠菌则会抑制铜绿假单胞菌产生用于获取铁元素的铁载体,二者相互制衡。
■ 与沙门氏菌——资源博弈
在肠道炎症模型中,沙门氏菌(Salmonella)的定植会改变肠道环境,特别是通过竞争精氨酸,来抑制白色念珠菌的生长。
■ 与粪肠球菌——亦敌亦友
粪肠球菌(Enterococcus faecalis)与白色念珠菌的关系尤为复杂。一方面,粪肠球菌产生的细菌素EntV能够抑制白色念珠菌的菌丝形成和生物膜。但另一方面,在某些情况下,粪肠球菌又能与白色念珠菌协同作用,增强其在血液中的存活能力。
■ 与艰难梭菌——微妙平衡
一项研究发现艰难梭菌感染患者更易被白色念珠菌定植,而另一项研究则显示相反结果。
白色念珠菌可能通过两种方式保护机体免受艰难梭菌感染:①生态位抢占,阻止艰难梭菌定植;②刺激产生更多IL-17A,增强宿主抗感染能力。
虽然人类数据存在争议,但小鼠模型提供了令人信服的证据,证明白色念珠菌定植确实有助于预防艰难梭菌导致的死亡。
这个复杂的防御网络,就像一张无形的大网,将白色念珠菌牢牢地被限制在低水平的共生状态。
编辑
图:在稳态(左)下,肠道菌群(如梭菌和拟杆菌)抑制白色念珠菌生长。在失调(右)下,抗生素等因素破坏菌群,导致白色念珠菌过度增殖。
以上,我们从口腔、肠道和阴道这些主要定植部位了解了白色念珠菌如何作为正常共生菌维持生态平衡,却也潜藏着失控的风险。接下来我们来看一下白色念珠菌的感染途径。
– 感 染 途 径 –
白色念珠菌的感染途径多样,主要分为内源性感染和外源性感染两种方式。其中内源性感染是最主要的感染途径。
◭ 内源性感染途径
内源性感染指念珠菌来自患者自身定植部位。当机体免疫功能下降或微生态环境改变时,原本定植的念珠菌大量繁殖,突破屏障进入组织或血液,引起感染。
肠道感染
健康人肠道念珠菌定植率达20%-50%。正常情况下肠道菌群抑制其生长,但当微生态失衡时,如长期用抗生素或患慢性疾病等,念珠菌大量繁殖,产生毒力因子破坏肠道黏膜,引起感染,严重时可进入血液循环。
阴道感染
正常阴道内乳杆菌维持酸性环境抑制念珠菌。当微生态失衡,如抗生素使用、激素变化等,念珠菌大量繁殖引发感染。
* 肠道念珠菌也可通过肛门-阴道途径传播。
◮ 外源性感染途径
外源性感染指念珠菌来自体外,通过接触传播。虽然少见,但在特定条件下可发生。
直接接触
与感染患者皮肤黏膜接触,如孕妇患阴道念珠菌病时胎儿经产道分娩可能感染。
间接接触
通过污染物品传播,如毛巾、医疗器械等。公共场所或医院消毒不彻底易造成传播。
呼吸道传播
肺念珠菌病患者咳嗽排出含菌飞沫,健康人吸入可能感染,但传播效率低。
◴ 易感人群
念珠菌感染与免疫功能密切相关,易感人群包括如下:
以上,我们看到白色念珠菌的感染是从体内定植(如肠道或阴道微生态失衡)或外源接触(如污染器械)起步,特别在免疫缺陷、糖尿病或抗生素滥用等易感人群中更容易突破防线。然而,了解这些途径只是起点;接下来我们深入探讨白色念珠菌在人体内的生存策略与致病机制。
– 感 染 机 制 –
白色念珠菌:从黏附到破坏
白色念珠菌的攻击过程可以概括为“粘附-入侵-破坏”三部曲,每一步都有专门的工具。
1
黏附素(分子钩爪)
为了不在体液的冲刷下被清除,白色念珠菌首先要牢牢地抓住宿主细胞。它细胞壁上的多种粘附素蛋白(如ALS家族蛋白)就像强力胶,能特异性地结合到上皮细胞上。
2
水解酶(化学钻头)
一旦站稳脚跟,菌丝就会分泌多种水解酶,如分泌型天冬氨酸蛋白酶(SAPs)和磷脂酶。这些酶能降解组织中的蛋白质和细胞膜,为菌丝的物理穿透清扫道路。
3
念珠菌溶素(细胞打孔器)
念珠菌溶素(Candidalysin) 是白色念珠菌最直接、最暴力的生化武器。这种肽毒素由侵入性菌丝的顶端精准释放,它能在宿主细胞膜上打孔,导致细胞内容物泄漏,最终引发细胞死亡和剧烈的炎症反应。我们有时候感受到的灼热、疼痛等症状,很大程度上就是它在作祟。
4
生物膜(坚固堡垒)
白色念珠菌还能形成生物膜——一个由大量真菌细胞和它们分泌的胞外基质构成的三维结构。这个“堡垒”能帮助它们牢固地附着在粘膜或医疗设备(如导管)上,并且极大地增强对抗真菌药物和宿主免疫攻击的抵抗力,是导致慢性感染和复发的重要原因
宿主的防御:从物理屏障到免疫大军
面对真菌的入侵,我们的身体也绝非坐以待毙,而是启动了一套从宏观到微观的立体防御体系。
1
物理与化学屏障
完整的皮肤和粘膜是第一道防线。粘膜表面的粘液层、唾液和阴道的酸性环境都构成了重要的化学屏障。
2
先天免疫反应(快速反应部队)
➦上皮细胞
它们不仅是屏障,还是哨兵。上皮细胞能通过表面的模式识别受体(PRRs)识别出真菌,并释放出细胞因子和抗菌肽,拉响警报,招募免疫细胞。
➦吞噬细胞
中性粒细胞和巨噬细胞是免疫系统的主力军。它们被招募到感染部位后,会吞噬并杀死真菌。
➦NLRP3炎症小体
这是一个重要的信号平台。当念珠菌溶素损伤细胞膜时,会激活NLRP3炎症小体,进而产生大量的促炎因子,如IL-1β、IL-18,放大炎症信号,引起患有外阴阴道假丝酵母菌病(VVC)和复发性外阴阴道假丝酵母菌病的白带异常、疼痛、瘙痒等炎症症状。VVC患者NLRP3表达升高,基因多态性加剧炎症反应,在疾病发病中起关键作用。
因此,NLRP3炎症小体抑制剂成为治疗新策略,研究表明格列本脲等抑制剂可降低IL-1β表达,具有减轻VVC/RVVC症状和减少复发的治疗潜力。
3
适应性免疫反应(特种部队)
➦Th17细胞
这是对抗真菌感染中至关重要的T细胞亚群。它们产生的IL-17细胞因子,能够强力招募和激活中性粒细胞,是清除粘膜表面真菌的核心力量。然而,过度的IL-17反应也可能导致组织损伤,是念珠菌性阴道炎等疾病免疫病理损伤的一部分。
➦抗体
B细胞产生的抗体(如IgA)也能在粘膜表面发挥作用,它们可以包裹住真菌,阻止其黏附和入侵。
以上是肠道菌群构建的坚固防线以及宿主与真菌的攻防机制,那如此强大的防御体系,会不会被突破?是什么因素会影响它?下一章节我们继续来看。
– 风 险 因 素 –
药物的使用
▸ 抗生素
广谱抗生素是打破肠道和阴道微生态平衡的最常见、最主要的原因。抗生素在杀死有害细菌的同时,也会无差别攻击,大量杀伤我们体内的有益菌,尤其是对维持定植抗性至关重要的专性厌氧菌,如梭菌纲和拟杆菌。
▸ 化疗药物
用于治疗癌症的化疗药物(如5-氟尿嘧啶)不仅会抑制免疫系统,还会直接损伤肠道黏膜屏障,并对肠道菌群造成严重破坏,削弱对白色念珠菌的定植抵抗力,即使不使用抗生素也会产生这种效应。
减少的有益菌群 ⬇️
增加的风险菌群 ⬆️
化疗破坏具有保护作用的菌群,诱导肠道菌群衍生的短链脂肪酸消耗,进一步支持白色念珠菌定植。
癌症化疗患者即使未使用抗生素,也可能面临白色念珠菌过度生长的风险,需要进行预防性监测和干预。
▸ 非抗菌和非化疗药物
非抗菌、非化疗药物对白色念珠菌定植影响的研究证据不足,但初步证据显示其潜在重要性。
一项大型荟萃分析研究了来自 2000 多名患者(包括健康对照组、肠易激综合征患者或炎症性肠病患者)的宏基因组数据,评估了整体微生物组成、代谢通路和抗菌耐药基因的变化。
影响最大的非抗菌药物:
其中,质子泵抑制剂在微生物类群和代谢通路方面的关联变化数量最多。这项研究仅发现了相关性,而非因果关系,需要更多进一步的研究。
既然药物会破坏平衡,那有没有药物可以帮助重建平衡呢?
有的。理解了丁酸盐-PPAR-γ通路的重要性后,科学家们正在探索新的治疗策略。例如,5-氨基水杨酸(5-ASA),一种常用于治疗炎症性肠病的药物,它本身就是一种PPAR-γ激动剂。理论上,使用这类药物可以直接激活肠道上皮细胞的耗氧能力,帮助恢复肠道缺氧环境,从而抑制白色念珠菌的生长。这为未来开发靶向微环境而非直接杀菌的新型抗真菌疗法提供了思路。
编辑
doi: 10.1016/j.mib.2020.05.006
饮食不健康
▸ 精制饮食的危害
现代西方饮食富含精制糖、饱和脂肪,但缺乏膳食纤维。这种饮食模式无法为产丁酸盐的菌如梭菌纲,提供足够的燃料。长期如此,会导致这类关键有益菌数量下降,削弱定植抗性。
研究表明,与喂食富含纤维的标准食物的小鼠相比,喂食精制饮食的小鼠更容易发生白色念珠菌的肠道过度生长和播散。
▸ 高糖饮食的双重打击
在高血糖或高糖饮食的情况下,白色念珠菌不仅获得了充足的能量来源,还会加剧炎症。在某些炎症环境下,高糖饮食会进一步促进白色念珠菌的致病性,形成恶性循环。
什么样的饮食能帮助控制白色念珠菌在正常范围,我们在后面09章节详细阐述。
免疫力下降
一个功能健全的免疫系统是控制白色念珠菌的一道防线。当这道防线因疾病或药物而出现问题时,即使微生物群没有发生大的变化,白色念珠菌也可能引发感染。
▸ 免疫抑制疾病
如HIV感染/艾滋病,会直接破坏免疫细胞(特别是CD4+ T细胞),导致机体对真菌的监控和清除能力严重下降。
▸ 药物性免疫抑制
癌症化疗、器官移植后使用的免疫抑制剂、长期大剂量使用糖皮质激素等,都会全面压制免疫系统的功能。
▸ 代谢性疾病
糖尿病是外阴阴道念珠菌病和皮肤念珠菌感染的重要风险因素。高血糖环境不仅为白色念珠菌提供了丰富的食物(葡萄糖),还会损害中性粒细胞等免疫细胞的功能,使其对抗真菌的能力大打折扣。
激素变化
激素波动,尤其是雌激素,会影响阴道环境。在怀孕、使用激素避孕或激素替代治疗期间,雌激素水平升高,会刺激阴道上皮中的糖原沉积,为 白色念珠菌的发展提供理想的基质。雌激素还会影响免疫反应,降低阴道黏膜的局部免疫防御,并产生有利于真菌持续存在的环境。
– 相 关 疾 病 –
糖尿病
高患病率与定植密度
多项研究证实,糖尿病患者口腔及皮肤的白色念珠菌定植率和感染率显著高于健康人群。
风险因素
高血糖环境为白色念珠菌的生长和毒力表达提供了有利条件。不良的血糖控制、免疫功能受损、长期糖尿病病程以及佩戴假牙等因素,均是促进糖尿病患者发生念珠菌感染的重要风险因素。
反复的念珠菌感染有时可能是未被诊断的糖尿病的早期信号之一 。在糖尿病患者中,真菌感染(尤其是足部感染)可能导致严重的并发症。
自闭症
自闭症儿童与粪便培养中白色念珠菌的重度生长之间存在显著关系。
白色念珠菌过度生长产生念珠菌溶血素、法呢醇等毒素,穿越血脑屏障直接损害神经系统。念珠菌产生的氨与肠道丙酸结合形成β-丙氨酸,干扰GABA神经递质系统,导致自闭症行为异常。
念珠菌激活NLRP3炎症小体,产生大量促炎因子,引起神经炎症。研究显示会导致海马和白质损伤,小胶质细胞和星形胶质细胞过度激活,影响认知功能和褪黑激素分泌。
念珠菌属可能增加肠道对血清素(5-羟色胺,5-HT)的产生,而大脑中的合成则较低(由于其前体色氨酸的消耗),导致自闭症儿童出现高血清素血症和行为异常。超过 25%的自闭症儿童存在全血血清素水平升高或高血清素血症。
自身免疫与炎症性疾病
白色念珠菌与宿主免疫系统存在微妙平衡,免疫失调可诱发自身免疫疾病。Th17/IL-17轴是抗真菌免疫的核心防线,但其过度激活也是银屑病、类风湿关节炎等自身免疫病的关键病理机制,形成双刃剑效应。
炎症性肠病(IBD)
在克罗恩病等IBD患者的肠道中,念珠菌属(包括白色念珠菌)的定植可能增加,普遍认为肠道真菌失调可能通过加剧肠道炎症,从而在IBD的病程中扮演了重要角色。
银屑病
银屑病是一种典型的IL-17介导的自身免疫性皮肤病。有理论认为,易感个体对皮肤或黏膜定植的白色念珠菌产生的过度免疫反应,可能是触发或加重银屑病皮损的因素之一。
一个极具说服力的旁证是,用于治疗银屑病的IL-17或IL-23抑制剂,其常见的副作用之一就是增加了患者发生粘膜念珠菌病的风险。
类风湿关节炎
关联证据间接复杂,RA患者存在针对念珠菌的Th17反应缺陷,但缺乏新的临床研究。
系统性红斑狼疮
有体外模型研究提示,白色念珠菌的成分(如β-葡聚糖)可能通过诱导中性粒细胞释放嗜中性粒细胞胞外诱捕网(NETs),从而加剧系统性红斑狼疮的疾病活动。
HIV/AIDS
在HIV感染者中,口腔咽部念珠菌病(鹅口疮)不仅是一种常见的机会性感染,更是HIV疾病进展、免疫功能严重受损的重要临床标志。此外,白色念珠菌在HIV患者肠道的定植,还可能加剧肠道菌群失调和全身性炎症状态,进一步促进疾病恶化。
白色念珠菌感染的检测对于疾病的诊断和治疗具有重要意义,目前,临床上常用的白色念珠菌感染检测方法主要包括直接镜检、培养鉴定、分子生物学检测等。
直接镜检
直接镜检是一种快速、简便的检测方法,通过显微镜观察标本中的酵母细胞和菌丝来初步判断感染。操作时将标本置于载玻片上,加生理盐水或10%氢氧化钾溶液,显微镜下可观察到圆形或椭圆形酵母细胞、出芽细胞、假菌丝。
该方法优点是快速、成本低,适合临床快速诊断;缺点是敏感性较低,早期或轻度感染可能出现假阴性,需结合其他方法综合判断。
培养鉴定
培养鉴定是诊断金标准。标本接种到SDA或显色培养基,37℃培养24-48小时,白色念珠菌形成白色奶油状菌落,显色培养基上呈翠绿色。通过形态学观察、生化反应试验(糖发酵、同化试验)或质谱技术进行鉴定。同时可进行药敏试验,采用纸片扩散法或微量稀释法检测抗真菌药物敏感性,为临床用药提供依据。
分子生物学检测
分子生物学检测包括PCR、实时荧光定量PCR(qPCR)、高通量测序等,敏感性高、特异性强、检测速度快,能快速准确检测标本中白色念珠菌核酸。
PCR技术
通过特异性引物扩增白色念珠菌基因组特定片段,琼脂糖凝胶电泳检测扩增产物。敏感性高,能检测少量病原菌,但易受污染影响导致假阳性。
实时荧光定量PCR
在PCR基础上加入荧光探针,实时检测荧光信号强度,定量检测核酸含量。敏感性高、特异性强、速度快、可定量,能检测病原菌数量,有助于判断感染严重程度和治疗效果。
高通量测序
一次性得到大量信息,为细菌、真菌等感染提供更深入、全面的分析。
谷禾肠道菌群检测升级版可精确识别到菌属甚至菌种水平,敏感性更高,病原微生物靶向检测涵盖细菌、真菌、病毒、寄生虫等,也包括白色念珠菌。同时也包括常见消化道病原体,耐药基因和毒力基因等,如幽门螺杆菌、大肠杆菌、艰难梭菌等。
谷禾宏基因组肠道菌群检测可检测到细菌、真菌、病毒、寄生虫等多种微生物,结合自主研发的多模态模型和机器学习算法,该技术可以建立微生物特征与宿主表型的关联,实现肠龄预测、菌群恢复力评估、定植能力分析、免疫指标评估等功能。
谷禾阴道菌群健康检测也可检测阴道微生态中的念珠菌,包括白色念珠菌,除此之外,还包括阴道菌群是否失调、乳杆菌分型,其他涵盖细菌性阴道病病原菌(如加德纳氏菌等)、需氧菌性阴道炎病原菌(如链球菌等)、生殖道感染病原菌(如沙眼衣原体等)等上百种相关病原菌。
药 物
如果确诊感染白色念珠菌后,一般以抗真菌药物治疗为主。
抗真菌药物类别及其作用机制
编辑
doi: 10.1007/s00253-025-13478-2
例如,制霉菌素和/或氟康唑最常用于恢复微生物群落的平衡或治疗自闭症儿童的肠道念珠菌过度生长。在连续70天、每天4次口服10万单位制霉菌素治疗后,自闭症儿童在儿童自闭症评定量表上的得分显著改善。
如果服用抗生素后出现念珠菌病怎么办?
一些抗生素的使用也可能导致肠道真菌包括白色念珠菌的过度生长,抗生素可以显著改变肠道代谢组和肠道菌群组成,导致有益细菌减少,从而为白色念珠菌提供更多生长空间,促进其定植、菌丝形成和潜在感染。
这在临床上常见于广谱抗生素使用后出现的念珠菌病,例如口腔或阴道念珠菌感染、甚至系统性感染。
那么是否需要停用抗生素?
不一定是立即停用。这取决于抗生素的使用原因。如果抗生素是为治疗细菌感染(如肺炎或尿路感染)而开的,且感染严重,贸然停用可能导致原发细菌感染恶化,甚至危及生命。在这种情况下,通常可能选择继续抗生素但同时启动抗真菌治疗来控制念珠菌。
如果抗生素不是必需的(例如预防性使用或非严重感染),往往建议停用或更换为窄谱抗生素,以减少对肠道菌群的破坏。这有助于恢复微生物平衡,降低念珠菌进一步生长的风险。
注:结合自身情况及遵医嘱。
也可以联合益生菌补充,且结合肠道菌群检测。在开始治疗前或过程中,进行肠道菌群检测,可以评估菌群多样性、特定细菌、真菌的丰度,例如,检测显示乳杆菌严重减少,可能优先推荐特定菌株补充,以针对性重建菌群。
益生菌 + 药物
一些研究表明,抗真菌药物与益生菌的联合治疗,在预防念珠菌属引起的肠道定植以及减少肠道症状方面显示出疗效。
对于患有念珠菌感染的婴儿,在接受抗真菌药物治疗(初始剂量为每天1 mg/kg,逐渐增加至每天最大剂量6 mg/kg,持续7天)和益生菌补充(如罗伊氏乳杆菌L. reuteri或副干酪乳杆菌L. paracasei)后,临床改善迅速。
新型抗真菌药物开发
通过改造现有药物结构,研制了新型三唑类药物(拉夫康唑、阿尔巴康唑、依沙康唑)和四唑类药物(奥替康唑),对念珠菌具有更强抗菌活性且细胞毒性更低。改性棘白菌素分子表现出更好的药代动力学特征和降低的耐药性风险。脂质体制剂(如两性霉素B脂质体)能有效降低药物毒性并增强药效。
抗菌肽技术
利用天然抗菌肽作为抗真菌药物的替代方案,这些短链、带正电荷分子能与真菌细胞膜相互作用,导致膜通透性增加。人α-防御素、乳铁蛋白、组蛋白-5、LL-37等抗菌肽已显示出对念珠菌的有效活性。
乳铁蛋白衍生肽(乳铁素)与两性霉素B联合使用可在低剂量下发挥协同抗真菌效果。
纳米技术应用
脂质体、固体脂质纳米颗粒、银纳米颗粒等纳米载体能精确递送抗真菌药物至感染部位,提高药物生物利用度并减少副作用。负载氟康唑的纳米系统对多种念珠菌表现出增强的抗菌活性。
这些创新方法为改善念珠菌感染治疗效果、减少对传统药物依赖提供了有希望的解决方案。
除了以上方式之外,鼓励保持良好卫生、控制血糖(如果有糖尿病)、避免滥用抗生素,并考虑饮食调整及其他天然植物提取物和补充剂等干预方式。
限 制 饮 食
减少高糖和高碳水化合物的摄入,避免零食,减少精制成分食品,前面我们知道这些食物可能促进念珠菌的生长或者说促进其致病性。
部分过敏人群需要避免特定食物:酒精饮料、面包,过敏性食物如牛奶、乳制品等。
在自闭症儿童中,碳水化合物,包括葡萄糖和甘露糖,对白色念珠菌的生长影响最大,增幅超过 1000%,此外,甘露醇、山梨醇、木糖、阿东醇、木糖醇也显著增加了白色念珠菌的生长,增幅为 100–200%。
编辑
doi: 10.1007/s00253-025-13478-2
同时,增加全食和有益食物的摄入,以支持身体的自然防御机制。
植物提取物
大蒜
新鲜捣碎的大蒜匀浆含有一种叫做大蒜素的化合物,具有抗真菌特性。大蒜影响白色念珠菌外表面的脂质组成,大蒜素通过抑制琥珀酸脱氢酶来抑制真菌生长。当大蒜提取物浓度增加时,抑制了白色念珠菌菌丝体产生并影响了SIR2基因表达。
定期食用大蒜可以帮助抑制白色念珠菌的生长,并促进肠道菌群的平衡。
肉桂
从肉桂中提取的精油能有效抑制白色念珠菌的多种关键致病能力,包括:阻止蛋白酶的产生、抑制菌丝管的形成、降低念珠菌对口腔上皮细胞的粘附能力。
此外,肉桂叶和树皮提取物抑制了 Fusarium graminearum、Fusarium proliferatum、 A. fumigatus、Trichophyton rubrum等。
肉桂树皮和叶子的精油都能通过破坏真菌膜结构而对耳念珠菌和白色念珠菌发挥抗真菌活性。
柠檬草
柠檬草油及其主要成分柠檬醛对白色念珠菌具有有效的抗真菌活性。它的作用方式包括:直接抑制悬浮的念珠菌,降低其生物膜中细胞的活力,以及作为气相剂改变真菌的细胞结构和表面形态。
椰子油
椰子油含有单月桂酸甘油酯,具有抗菌和抗病毒特性。口服单月桂酸处理可显著减少小鼠的口腔白色念珠菌感染和生物膜的形成。富含椰子油的饮食能减少小鼠中白色念珠菌的定植,并改变白色念珠菌细胞的代谢程序,降低其参与脂肪酸使用的基因表达活性。
姜
姜中的6-姜醇和6-姜酮可以抑制白色念珠菌的生物膜和菌丝形成。姜提取物能抑制白色念珠菌的生物膜形成。当姜提取物与氟康唑联合使用时,观察到协同效应,可改善耐药性念珠菌病的治疗效果。
海藻
多种海藻对白色念珠菌具有抗真菌活性。Ulva fasciata Delile海藻提取物能减少组织病理学变化和促炎细胞因子表达,通过减少炎症反应和增强细胞抗氧化防御发挥作用。
含有甾体、萜类化合物和单宁的海藻Gracilaria verrucosa提取物抑制生物膜形成。
海藻中的岩藻依聚糖(硫酸化多糖)对白色念珠菌具有强烈的抗真菌活性。
Fucus spiralis中的植物单宁能防止白色念珠菌的形态转变,降低其毒力和侵入宿主细胞的能力。
百里香
Thymus vulgaris是最重要的百里香品种,其精油含有百里香酚和百里香脑。百里香精油与氟康唑对浮游和生物膜形成的白色念珠菌和Candida tropicalis都显示出协同作用。
橄榄油
橄榄油主要含有油酸(占脂肪酸组成的55-83%),还含有亚油酸和亚麻酸。研究表明油酸对白色念珠菌表现出抗真菌活性。特级初榨橄榄油能改善肠道通透性,并减轻与炎症相关的组织病理学特征。
牛至油
牛至油以其抗菌特性而闻名,包括对白色念珠菌的抗真菌活性。服用牛至油补充剂或在烹饪中使用牛至油可以帮助减少白色念珠菌过度生长。
保哥果(Pau d’Arco)
保哥果是一种草本药物,源自南美洲树木的树皮。它具有抗真菌特性,可能有助于抑制白色念珠菌的生长。
茶树油
茶树油具有强大的抗真菌特性,可用于局部治疗念珠菌感染。稀释的茶树油可以涂抹在受影响的皮肤区域,或加入浴水中以获得舒缓效果。
几种植物提取物的油性制剂已被局部用于治疗外阴阴道念珠菌病,即柠檬香蜂草、大蒜、茴香、洋甘菊、姜、鼠尾草的油。
此外,如大蒜素、姜黄、莳萝油等分子显示出对阴道念珠菌病的有益效果。
绿茶、肉桂、大蒜、蜂胶、姜则显示出对口腔念珠菌病的抗念珠菌活性。
这些天然疗法可作为避免传统药物副作用或轻度感染人士的选择。
补 充 剂
维生素 D
维生素 D 是在皮肤暴露于阳光后合成的,或通过某些鱼类、蛋黄、乳制品和蘑菇等食物获得。
维生素D3具有显著的抗真菌作用。临床试验显示,每日补充维生素D3能降低烟曲霉诱导的IL-13反应,并通过抑制多种受体表达使细胞因子反应转向抗炎表型。维生素D3的脂溶性特点能破坏真菌细胞膜完整性发挥直接杀菌作用,同时影响白色念珠菌的代谢和生物合成。小鼠实验证实,维生素D3能降低各器官真菌负荷,减少炎症细胞浸润和炎症因子水平。
维生素 E
维生素E是存在于坚果、种子、蔬菜、油等食物中的脂溶性化合物,通过抑制NF-κB活性对白念珠菌发挥抗炎作用。研究显示,在两性霉素B中添加维生素C和E比单用两性霉素B效果更佳。
此外,当添加维生素 C 和 E 时,人类红细胞可免受两性霉素 B 引起的细胞毒性作用,表明这些维生素凭借其抗氧化特性,可对抗由两性霉素 B 诱导的自氧化作用。
Omega-3
Omega-3多不饱和脂肪酸含有α-亚麻酸(18:3 ω-3)、十八碳四烯酸(18:4 ω-3)、二十碳五烯酸(EPA;20:5 ω-3)、二十二碳五烯酸(22:5 ω-3)和二十二碳六烯酸(DHA;22:6 ω-3)。海洋哺乳动物的脂肪层和白鱼的肝脏含有 EPA 和 DHA 等长链脂肪酸。不同的研究表明,EPA 和 DHA 不仅有益于心脏、大脑和眼睛的健康,也有益于免疫系统。有证据表明,多不饱和脂肪酸及其酯类衍生物对多种口腔病原体有效,包括白色念珠菌。
硒
硒是通过植物进入食物链的微量元素,具有抗氧化、抗炎、抗菌作用。硒纳米颗粒能粘附并穿透白色念珠菌生物膜,通过替代硫元素破坏真菌细胞结构。研究显示,硒缺乏小鼠的肝脏和脾脏真菌负荷显著高于补硒小鼠,且硒缺乏会损害中性粒细胞的杀菌能力,表明硒在抗真菌感染中发挥重要作用。
锌
直接抗真菌作用:补锌通过破坏真菌生物膜结构(如锌载体触发抗真菌药物释放)及抑制毒力因子(干扰代谢酶活性),直接抑制白色念珠菌生长。
增强宿主免疫:锌元素提升巨噬细胞吞噬能力与T细胞活性,强化对真菌的免疫清除,同时减轻感染相关的炎症损伤(如降低TNF-α等因子)。
协同增效抗真菌药物:锌基载体(如V-ZIF)可靶向递送药物至生物膜深处,增强药物渗透性;并通过调控耐药基因,逆转白色念珠菌对唑类药物的耐药性。
注意:过量补锌可能被念珠菌利用促进生长,推荐成人每日8-11mg。与喹诺酮、四环素类同服会影响吸收,需间隔2–6小时。有明确感染时应以医生处方的抗真菌药为主,补锌以纠正缺乏为目的。
编辑
doi: 10.3390/microorganisms11061556
益 生 菌
最常见的有乳杆菌和双歧杆菌属,它们对恢复微生物群落的功效取决于失衡的原因。
乳杆菌
乳杆菌菌株(如L. acidophilus、L. jensenii、L. crispatus)和双歧杆菌属口服或阴道给药,帮助抑制生物膜形成。
乳杆菌还可能抑制白色念珠菌通过竞争营养物质以及产生乳酸和其他有机酸来降低 pH 值,从而抑制念珠菌的生长。
一项前瞻性研究对早产儿(249 例)进行了长达 12 个月的观察,这些儿童接受了 Lactobacillus reuteri(ATCC 55730;每日五滴;1 × 108 CFU)或 Lactobacillus rhamnosus(ATCC 53103;每日一粒;6 × 109 CFU)的治疗,并报告了在L. reuteri 给药后粪便中念珠菌的减少以及较少的胃肠道症状。
口服 Lactobacillus acidophilus(菌株 Rosell-11,5 × 109 CFU)治疗 2 个月(每天两次)减少了 22 名自闭症谱系障碍儿童的肠道念珠菌定植(45%的参与者处于“无糖饮食”状态),集中注意力和执行指令的能力也显著提高。
乳杆菌与乳铁蛋白组合、乳杆菌与甘露聚糖寡糖组合均显示协同效应,可改善阴道真菌感染症状并抑制白念珠菌粘附。
双歧杆菌
双歧杆菌可以代谢宿主饮食或粘膜中的不同类型碳水化合物,其发酵途径产生不同水平的乙酸和乳酸。接受乳杆菌和双歧杆菌菌株治疗的肥胖小鼠体重和胆固醇水平更低。它们真菌数量更少,肝脏形态得到恢复,肠道菌群得到有益调节。
益 生 元
发酵蔬菜
发酵蔬菜含有植物乳杆菌、短乳杆菌和Lactobacillus sakei 等。从泡菜中分离的三种植物乳杆菌菌株对白色念珠菌表现出抗真菌活性,通过真菌生长的显著减少得到证实。来自泡菜的乳杆菌产物通过增强小鼠的分泌型IgA水平来增强粘膜免疫力,并显示出抗白色念珠菌活性。
苹果醋(ACV)
苹果醋含有5%醋酸以及类黄酮和多酚。ACV能增强单核细胞对白色念珠菌的吞噬活性,并降低白色念珠菌的活力和生长。
酸奶
酸奶用保加利亚乳杆菌和嗜热链球菌发酵制成。每日摄入富含活L. acidophilus的酸奶可使复发性念珠菌阴道炎缓解。食用益生菌酸奶可减少女性真菌定植。
粪 菌 移 植
一项溃疡性结肠炎研究显示FMT后念珠菌减少。
一项开放标签试验显示,通过内镜每3个月进行FMT能显著改善自闭症行为症状(CARS评分)并将微生物群转变为健康状态。
MTT改良疗法(包括万古霉素预处理、肠道清洁、高剂量FMT和维持治疗)使胃肠道症状减少80%,核心自闭症症状持续改善长达2年。
生 活 方 式
编辑
doi: 10.3390/microorganisms11061556
戒烟
吸烟会减少唾液分泌,改变口腔菌群,并增加念珠菌在口腔中的定植。研究表明,吸烟者的口腔念珠菌感染风险增加了 7 倍。在活动性龋齿病变的吸烟者中,口腔念珠菌的存在可能性也更高。据信,由于中性粒细胞对真菌的活性降低,吸烟者白色念珠菌定植的患病率更高。
戒酒
过量饮酒是许多健康问题的主要风险因素。在小鼠中,长期摄入酒精会增加真菌菌群数量,并促进真菌β-葡聚糖向血液的转移,而使用抗真菌药物治疗则可减少肠道真菌负荷,降低β-葡聚糖转移,并改善乙醇引起的肝损伤。在随后的临床研究中,酒精相关性肝病患者的真菌多样性较低,且念珠菌属数量较多。此外,酒精性肝炎患者的血清抗中性粒细胞胞质抗体(ASCA)水平与死亡率增加相关,这表明治疗肠道真菌可能有益于酒精性肝炎患者,且ASCA可能有助于预测其预后。
运动
缺乏体力活动会导致多种健康问题并激活全身炎症标志物(如C反应蛋白、IL-6、TNFα和新蝶呤),而规律的体育锻炼则能改善生活质量、减少全身炎症并增强抗感染免疫反应。研究证据表明,适度运动能提高年轻男性中性粒细胞对白念珠菌的吞噬能力,同时体力训练还能显著增强巨噬细胞的自发移动性、趋化性、自发附着能力以及对白念珠菌的吞噬功能。
缓解压力
压力是一系列威胁身体稳态的事件,由外部或内部因素(应激源)引起。研究表明,暴露于社会应激源会改变结肠菌群稳定性,导致细菌易位和免疫调节,拟杆菌的相对丰度降低,梭菌的相对丰度增加。
一项实验研究表明,感染白色念珠菌并暴露于慢性多样化压力的大鼠肝脏和肾脏中的真菌负荷增加。此外,巨噬细胞产生的一氧化氮显著减少,表明慢性多样化压力损害了巨噬细胞对白色念珠菌的吞噬作用。
白色念珠菌作为一种典型的机会致病菌,充分体现了微生物与宿主关系的复杂性和动态性。从其独特的形态可塑性到精密的定植抗性机制,从共生毒力学派的理论创新到多元化的干预策略,我们对其认识正不断深化。
随着微生物组学、宏基因组测序和系统生物学技术的快速发展,未来的研究将更加精准地解析白色念珠菌与宿主微生态的互作机制。
肠道菌群检测作为一种技术手段,将成为评估个体微生态状态、预测感染风险和指导治疗决策的重要工具。通过高通量测序分析,我们能够实时监测菌群多样性变化、识别念珠菌过度生长的早期信号,并制定个性化的干预方案。精准微生态医学的兴起将使我们从单纯的“杀灭病原体”思维转向“重塑微生态平衡”的整体性策略。
注:本账号发表的内容仅是用于信息的分享,在采取任何预防、治疗措施之前,请先咨询临床医生。
主要参考文献:
Schille TB, Sprague JL, Naglik JR, Brunke S, Hube B. Commensalism and pathogenesis of Candida albicans at the mucosal interface. Nat Rev Microbiol. 2025 Aug;23(8):525-540.
Jawhara S. Healthy Diet and Lifestyle Improve the Gut Microbiota and Help Combat Fungal Infection. Microorganisms. 2023 Jun 11;11(6):1556.
Kumamoto CA, Gresnigt MS, Hube B. The gut, the bad and the harmless: Candida albicans as a commensal and opportunistic pathogen in the intestine. Curr Opin Microbiol. 2020 Aug;56:7-15.
Bays DJ, Savage HP. 0. Candida albicans gastrointestinal colonization resistance: a host-microbiome balancing act. Infect Immun 0:e00610-24.
Faustino M, Ferreira CMH, Pereira AM, Carvalho AP. Candida albicans: the current status regarding vaginal infections. Appl Microbiol Biotechnol. 2025 Apr 10;109(1):91.
Herman A, Herman AP. Could Candida Overgrowth Be Involved in the Pathophysiology of Autism? J Clin Med. 2022 Jan 15;11(2):442.
WANG Dongyue, LI Yunqing. The formation mechanism and prevention of candida albicans hyphae[J]. Journal Of Jining Medical University, 2020, 43(5): 356-361.
Sarika Parambath, Aiken Dao, Hannah Yejin Kim, Shukry Zawahir, et al., Candida albicans—A systematic review to inform the World Health Organization Fungal Priority Pathogens List, Medical Mycology, Volume 62, Issue 6, June 2024, myae045
Kumamoto CA, Gresnigt MS, Hube B. The gut, the bad and the harmless: Candida albicans as a commensal and opportunistic pathogen in the intestine. Curr Opin Microbiol. 2020 Aug;56:7-15.

谷禾健康
想象一下,你的肠道就像一个热闹的自助餐厅,而双歧杆菌这个家庭里的每个成员都是小顾客,每个小顾客对不同的碳水化合物有着独特的“口味偏好”,有些喜欢吃淀粉,有些偏爱果胶,还有些只喜欢母乳里的特殊糖类。
最近,科学家们通过分析来自全球不同地区的3083个人类肠道双歧杆菌非冗余基因组,构建了迄今最大规模的双歧杆菌代谢功能数据库-“菜单”。成功重建了68条单糖、二糖、寡糖和多糖的完整利用通路,揭示了这些肠道双歧杆菌的“饮食偏好”。
有趣的是,这些口味还会因为我们吃什么东西、住在哪里、年龄多大而变化。相信你也会好奇,这些住在你身体里的小顾客喜欢吃什么。
为了能快速识别小顾客的“口味偏好”,科学家们开发了Glycobif计算管道(https://github.com/Arzamasov/glycobif),用于预测双歧杆菌谱系的碳水化合物利用网络。
▸ 涵盖19种双歧杆菌的碳水化合物利用网络
首先构建双歧杆菌非冗余数据集。从公共数据库收集了相关基因组序列,根据自定义的标准做初步筛选,ANI≥99%聚类后,最终获得3083个非冗余基因组,涵盖19个双歧杆菌。
得到双歧杆菌非冗余数据集后,开始准备训练Glycobif预测模型,获得的3083个非冗余基因组里,有263个参考基因组,这些基因组被用作训练集。该模型会输出68条碳水化合物利用通路的存在与否。体外培养实验结果比较验证,Glycobif自动管道通路预测准确率94%,手动注释(manual)准确率95%。
转录组分析主要集中在Bifidobacterium catenulatum kashiwanohense(Bc.kashiwanohense)Bg42221_1E1菌株,确认基因表达模式,重点验证的通路有木葡聚糖(XGL)、母乳低聚糖(HMO)相关利用通路。
还对菌株进行了糖谱分析(Glycoprofiling),通过底物消耗模式进一步验证预测结果。
1
双歧杆菌成员之间的“口味差异”
★ 婴儿和成人双歧杆菌优势种及代谢偏好不同
对627个分离基因组+2456个宏基因组组装基因组(MAGs)的68条碳水化合物利用通路进行系统分析,图a展示的NMDS排序图,按物种和亚种聚类,分类学解释了91%的变异(“口味”差异),结果显示菌株水平变异性在不同类群之间存在差异(PERMANOVA R²=0.91, p=0.001),比如种之间显著分离的B.bifidum(婴儿肠道优势种)和B.adolescentis(成人肠道优势种)。
下图b显示不同物种(包括ANI>95%的亚种)之间在碳水化合物利用通路的数量(Predicted phenotypic richness)存在显著差异(GLM, p<2.2×10⁻¹⁶),例如Bl.nov可利用的通路总数明显低于其他亚种。
3083个双歧杆菌基因组中碳水化合物利用途径
Arzamasov AA,et al.Nat Microbiol.2025
图c展示了每个分类群(横轴)中能编码68种预测碳水化合物利用途径的基因组比例,颜色越深,表示在该分类群中有更高比例的基因组含有该碳水化合物利用途径;颜色越浅,则表示拥有该途径的基因组比例越低,可以直观地了解不同生物分类群中碳水化合物代谢能力的分布情况。
★ 几乎所有双歧杆菌都存在葡萄糖、半乳糖、果糖的代谢通路
发现核心通路(几乎所有成员都爱吃):在98%以上的基因组中存在的葡萄糖(Glc)、半乳糖(Gal)、果糖(Fru)、乳糖(Lac)和低聚半乳糖(GOS)利用通路,广泛保守通路(大多数成员都爱吃):在84%以上的基因组中存在的蔗糖(Scr)、麦芽糖(Mal)、低聚麦芽糖(MOS)、低聚异麦芽糖(IMO)、蜜二糖(Mel)、棉子糖家族低聚糖(RFO)和短链低聚果糖(scFOS)利用通路,除了B.bifidum。
长双歧杆菌婴儿亚种(Bifidobacterium longum subsp.Infantis)和长双歧杆菌长亚种(B. longum subsp. longum)是长双歧杆菌种内系统发育相近的亚种,但两者的碳水化合物利用通路存在明显差异,尤其是母乳低聚糖(HMO)利用能力,这与既往研究一致,Bl.infantis具有H1和FL1/2基因簇。这些基因簇使其能够利用多种HMOs,包括LNnT、2′FL、3FL、LDFT、LNFP I和SHMO(Sialylated HMO utilization),而Bl.longum则能力有限。
★ 不同菌株对母乳低聚糖的利用能力因独立进化而存在显著差异
将263个参考基因组基于68条代谢通路层次聚类后与核心基因构建的系统发育树做相关性分析,呈中等相关性(cophenetic correlation 0.58, permutation test, P < 0.001)。这些发现都表明功能特征并非完全由进化谱系决定,可能存在显著的水平基因转移或独立进化适应现象。
不同菌株对特定HMO的利用能力与其携带的基因簇高度相关。H1基因簇编码多个同源底物结合蛋白(SBPs)用于HMO转运。在研究范围内发现携带H1基因簇的菌株有Bl. Infantis ATCC 15697、Bl. suis Bg131.S11_17.F6和Bc. kashiwanohense Bg42221_1E1(下图a)。HPLC定量分析结果显示,这些菌株在24小时内消耗了72-86%的总HMOs。
双歧杆菌菌株中HMO利用能力的基因组和功能分析比较
Arzamasov AA,et al.Nat Microbiol.2025
★ 基因簇与母乳低聚糖的利用直接相关
PCA分析(上图d)清晰区分了携带不同基因簇(H1和FHMO)的菌株,这说明基因簇与HMO利用模式直接相关。
双歧杆菌菌株中HMO利用能力的基因组和功能分析比较
Arzamasov AA,et al.Nat Microbiol.2025
FHMO基因簇的存在决定了对复杂岩藻糖基化结构的利用能力,与H1基因簇相比,FHMO基因簇优先利用岩藻糖基化HMO,如图b、c,携带FHMO基因簇的菌株(Bl. suis Bg41121_2E1 和 Bc. kashiwanohense_A Bg42221_1D3)能够完全消耗2′FL,但对SHMO的利用能力有限。而缺乏任何已知的完整HMO利用基因簇的菌株(Bl. nov.和B. pseudocatenulatum)对HMO的利用能力最弱。
2
特别爱吃木葡聚糖的成员
★ B.kashiwanohense 特别擅长利用木葡聚糖
通过Glycobif计算管道预测到B.kashiwanohense等菌株编码完整的木葡聚糖利用基因簇(xgl)。体外培养结果验证B.kashiwanohense Bg42221_1E1在木葡聚糖培养基中很好生长(图d,OD600>0.5)。
转录组分析结果显示木葡聚糖相关的 xgl 基因表达量显著上调,如图e,log2FC均大于5,pvalue<0.01,补充材料Table21显示log2FC分布在6.65-9.14之间,相当于上调幅度约100-550倍。
Bc. kashiwanohense Bg42221_1E1 XGL代谢的
基因组和转录组学综合分析
Arzamasov AA,et al.Nat Microbiol.2025
3
环境影响饮食偏好
来自不同地理区域的30株双歧杆菌菌株中,仅在来自一个孟加拉国婴儿的两个分离菌株(Bc. kashiwanohense Bg42221_1E1 和 Bc. kashiwanohense_A Bg42221_1D3 )中发现了xgl基因簇。
★ 环境影响人体内双歧杆菌的种类和代谢能力
Bl.infantis是一种专门利用HMO的菌种,在非西方化人群中健康母乳喂养婴儿的肠道双歧杆菌中,其比例可高达90%。而西方化人群的婴儿往往缺乏Bl.infantis,取而代之的是利用HMO能力较低的菌种,如Bl. longum、Bifidobacterium breve和Bifidobacterium pseudocatenulatum。
代谢通路富集分析发现,3岁以下的孩子的肠道里的双歧杆菌主要“专攻”消化HMO,而3岁以上的孩子和成人的肠道里的双歧杆菌更擅长消化plant glycan(植物聚糖)。在3岁以下的孩子里也发现了地域差异,西方化群体的肠道双歧杆菌更擅长消化plant glycan(植物聚糖),非西化群体更擅长消化HMO,这说明即使在以母乳为主的3岁以下年龄段,不同地区的孩子肠道菌群已经开始体现出适应当地饮食环境的差异。
★ 同一菌种也能分化出不同的形态来适应饮食
发现同属于Bl.suis的两个分离菌株展现出两种不同的生态型,Bg131.S11_17.F6代表生态型1,Bg41121_2E1代表生态型2,生态型1专攻HMO相关,不能代谢阿拉伯糖和植物多糖,生态型2兼顾FHMO和含阿拉伯糖的寡糖和多糖,这种同一菌种内部存在的功能分化现象,就像一种互补策略,使得同一菌种能够适应婴儿不同的喂养阶段和饮食状态,体现了肠道微生物的精细化生态适应。
双歧杆菌的“个性”远比我们想象的更鲜明!不同种类甚至同一物种的不同菌株之间,对碳水化合物的利用能力差异巨大。而这种“饮食偏好”虽然与携带的基因相关,但并非固定,而是会随着宿主的生活环境、饮食习惯动态调整,形成独特的“功能优势”。
识别出B.longum物种内的独特分支(Bl.nov.),可以降解植物和真菌来源的α-葡聚糖,但缺乏乳糖-N-二糖(LNB)、半乳糖-N-二糖(GNB)、N-聚糖、HMO和T抗原(Tan)代谢通路。孟加拉国Bl.suis群菌株显示两种不同的生态型:第一种模拟Bl. infantis的HMO利用偏好,第二种优先利用FHMO,可代谢阿拉伯糖相关多糖。
在Bc.kashiwanohense菌株中发现了独特的xgl基因簇,该基因簇仅在110个B. catenulatum基因组中的3个中被识别。体外培养实验也验证了木葡聚糖利用能力,转录组分析显示xgl相关基因显著上调100-550倍。
研究证实不能仅基于物种名称预测菌株功能,名字一样,功能可能完全不一样,需要进行菌株特异性功能验证和完整基因簇的挖掘,才能精准指导益生菌开发,形成个性化产品。例如给婴儿补充益生菌,要选HMOs专家型,给成人或高纤维饮食人群吃,要选plant glycan专家型。
由于菌株存在的饮食偏好,还可以通过补充特定的益生元(如HMO)促进目标菌的生长。但也不能忽视生活背景、饮食习惯等环境因素影响,研究证实这些环境因素在塑造双歧杆菌功能多样性方面发挥着关键作用,因此,在设计个性化益生菌和益生元干预策略时,必须考虑宿主的地理背景、文化饮食习惯和生活方式等多重因素。
一句话总结:现在已经告别”一刀切”益生菌,想让你的益生菌“住得久、吃得好、干得棒”,先得弄清楚它们的口味,再决定请谁进门、请吃什么。
主要参考文献
Arzamasov AA, Rodionov DA, Hibberd MC, Guruge JL, Kent JE, Kazanov MD, Leyn SA, Elane ML, Sejane K, Furst A, Bode L, Barratt MJ, Gordon JI, Osterman AL. Integrative genomic reconstruction reveals heterogeneity in carbohydrate utilization across human gut bifidobacteria. Nat Microbiol. 2025 Jul 16.
Alessandri G, van Sinderen D, Ventura M. The genus bifidobacterium: From genomics to functionality of an important component of the mammalian gut microbiota running title: Bifidobacterial adaptation to and interaction with the host. Comput Struct Biotechnol J. 2021 Mar 9;19:1472-1487.
Arboleya S, Watkins C, Stanton C, Ross RP. Gut Bifidobacteria Populations in Human Health and Aging. Front Microbiol. 2016 Aug 19;7:1204.
Stewart CJ, Ajami NJ, O’Brien JL, Hutchinson DS, Smith DP, Wong MC, Ross MC, Lloyd RE, Doddapaneni H, Metcalf GA, Muzny D, Gibbs RA, Vatanen T, Huttenhower C, Xavier RJ, Rewers M, Hagopian W, Toppari J, Ziegler AG, She JX, Akolkar B, Lernmark A, Hyoty H, Vehik K, Krischer JP, Petrosino JF. Temporal development of the gut microbiome in early childhood from the TEDDY study. Nature. 2018 Oct;562(7728):583-588.
Sakanaka M, Hansen ME, Gotoh A, Katoh T, Yoshida K, Odamaki T, Yachi H, Sugiyama Y, Kurihara S, Hirose J, Urashima T, Xiao JZ, Kitaoka M, Fukiya S, Yokota A, Lo Leggio L, Abou Hachem M, Katayama T. Evolutionary adaptation in fucosyllactose uptake systems supports bifidobacteria-infant symbiosis. Sci Adv. 2019 Aug 28;5(8):eaaw7696.
Arzamasov AA, Osterman AL. Milk glycan metabolism by intestinal bifidobacteria: insights from comparative genomics. Crit Rev Biochem Mol Biol. 2022 Oct-Dec;57(5-6):562-584.
Vatanen T, Ang QY, Siegwald L, Sarker SA, Le Roy CI, Duboux S, Delannoy-Bruno O, Ngom-Bru C, Boulangé CL, Stražar M, Avila-Pacheco J, Deik A, Pierce K, Bullock K, Dennis C, Sultana S, Sayed S, Rahman M, Ahmed T, Modesto M, Mattarelli P, Clish CB, Vlamakis H, Plichta DR, Sakwinska O, Xavier RJ. A distinct clade of Bifidobacterium longum in the gut of Bangladeshi children thrives during weaning. Cell. 2022 Nov 10;185(23):4280-4297.e12.

谷禾健康
内脏臭气杆菌(O.splanchnicus)是一种兼具共生与条件致病特性的细菌:它为严格厌氧的革兰氏阴性短杆菌,不运动、不产孢子。
O.splanchnicus能发酵多种单/二糖,如葡萄糖、果糖、半乳糖、阿拉伯糖、乳糖和甘露糖,还参与次级胆汁酸代谢;能合成多种关键代谢酶;可释放外膜/细胞外囊泡;对四环素、林可霉素、克林霉素、利福平、红霉素敏感,对氨基糖苷类和多粘菌素耐受,环丙沙星联合甲硝唑也可抑制其生长。
其丰度受宿主遗传与免疫状态、肠屏障、胆汁、饮食结构(红肉血红素、低碳高脂)、地域人群差异,以及抗生素、益生菌/益生元与植物活性成分等干预共同塑造。
O.splanchnicus作为一种重要的肠道微生物,与宿主的健康状况和各种疾病密切相关。需要注意的是,其健康效应具有环境依赖与双刃性:在健康稳定的微生态中多呈保护;在肿瘤等免疫抑制微环境或菌群失衡下,可能与不良结局相关。
O.splanchnicus通过短链脂肪酸与囊泡介导的免疫代谢调控,提升紧密连接与黏膜屏障、诱导IL-10与部分情境下的Th17反应,抑制NLRP3炎性小体,改善炎症性肠病,参与能量代谢与胰岛素敏感性,并可能以丁酸与苹果酸等代谢物影响结直肠癌的炎症与凋亡通路。
临床关联方面,O.splanchnicus的减少与1型糖尿病妊娠期炎症上升、炎症性肠病/克罗恩病、肠易激综合征、动脉粥样硬化、囊性纤维化、非酒精性脂肪性肝病(NAFLD)及儿童肥胖合并NAFLD等相关。
与此同时,在系统性红斑狼疮(SLE)、未破裂颅内动脉瘤、海绵状血管瘤、注意力缺陷多动障碍、急性冠状动脉综合征、慢性肾病以及代谢紊乱模型(glut1缺失)中呈富集。其在肠屏障受损、腹腔炎症时可成为机会致病菌,引发菌血症、假体关节感染、盆腔腹膜炎等。
本文全面总结了内脏臭气杆菌(O.splanchnicus)的研究历史、目前对其与健康和疾病相关性的认识、潜在作用机制以及未来研究和应用的前景等。
▸ 研究历史
内脏臭气杆菌(O.splanchnicus)是一种具有重要研究价值的细菌。它最初于1971年被分离出来,并于1975年被命名,他们从腹部脓肿样本中分离出来。由于其与拟杆菌属成员的许多共同特征,它最初被命名为Bacteroides splanchnicus。
与内脏臭气杆菌相关的研究时间表
Li J,et al.Microorganisms.2025
随着细菌分类学的不断进步,2008年,拟杆菌目Odoribacteraceae科中确定了一个新属Odoribacter。与脆弱拟杆菌相比,B.splanchnicus表现出许多独特的生化特征,其16S rRNA基因之间的同源相关性小于20%。因此,B.splanchnicus被重新归类为Odoribacter属中的一个新物种,并被正式命名为O.splanchnicus。
该属包括另外两个物种:从犬牙周炎中分离出的Odoribacter denticanis和从人类粪便中分离出的Odoribacter laneus。此外,还从盆腔腹膜炎患者的血液和腹膜脓液样本中分离出O.splanchnicus。
▸ 基本特征
内脏臭气杆菌(O.splanchnicus)是一种严格厌氧的革兰氏阴性杆菌,具有短杆状(0.7×1.0-5.0 um)。其形态多为单个或疏松聚集体,不运动、不产色素也不形成孢子;在37 ℃的厌氧环境下最适生长,通过透射电镜可见产生外膜囊泡。
▸ 代谢能力
该菌具有广泛代谢能力,以有机化合物为主要营养来源,参与次级胆汁酸的合成与代谢。它能发酵多种肠道常见糖类,如葡萄糖、果糖、半乳糖、阿拉伯糖、乳糖和甘露糖,是其重要碳源;但不能利用蔗糖、鼠李糖、海藻糖和水杨苷。它能产生大量的H2和H2S。当氨基酸是唯一可用的碳源时,该菌株专门使用赖氨酸来生产丁酸盐。
另一项研究表明,小鼠盲肠O.splanchnicus定植可显著提升乙酸、丙酸和丁酸水平,并经GPR43与GPR109a介导发挥作用。作为臭杆菌科成员,它还可能产生具抗菌活性的次级胆汁酸。
该菌合成多种关键代谢酶(磷酸酶、β-半乳糖苷酶、α-岩藻糖苷酶、N-乙酰-β-D-葡萄糖胺苷酶、谷氨酸脱羧酶),并在戊糖磷酸途径中具高活性(葡萄糖-6-磷酸脱氢酶、6-磷酸葡萄糖酸脱氢酶),有助于维持细胞氧化还原平衡与能量代谢。
▸ 抗生素敏感性
O.splanchnicus对多种抗生素表现出不同程度的敏感性。它对四环素类、林可霉素、克林霉素、利福平和红霉素高度敏感,但对氨基糖苷类和多粘菌素具有很强的耐药性。
这些不同的易感性可能归因于其细胞壁的独特结构或特定的抗性机制。此外,环丙沙星和甲硝唑类抗生素的联合治疗被证明可以抑制其生长。
内脏臭气杆菌(O.splanchnicus)的丰度受多重因素协同塑造,既包括宿主内在条件,也受到外部环境与干预的深刻影响。我们这里列举几个主要影响因素。
1
红肉中的血红素促进其生长
尽管内脏臭气杆菌(O.splanchnicus)的生长并不依赖血红素,但血红素的存在显著促进了其生长。血红素可能作为该菌株的重要生长因子或信号分子。值得注意的是,血红素是红肉中的关键成分,据报道,O.splanchnicus的丰度与红肉摄入量呈正相关,而与蔬菜摄入量呈负相关。
注:在饮食方面,采用低热量、低碳水化合物、高脂肪的饮食与这种细菌丰度的显著增加相关。
2
不同人群中差异较大
内脏臭气杆菌的富集在不同人群中呈现显著差异,受遗传背景、生活环境、饮食结构以及多种宿主相关与外部因素的共同影响。例如,在荷兰人的肠道中,该菌更易富集且相对丰度较高;而在南亚苏里南人中,其丰度与前者存在显著差异。值得注意的是,南亚苏里南人群2型糖尿病患病率较高,并且肠道中缺乏部分能够产生短链脂肪酸的有益菌群,其中可能包括内脏臭气杆菌,这种菌群构成差异或与其代谢特征和健康状况相关。
3
胆汁能促进其生长
此外,胆汁作为肠道消化液的关键组成部分,不仅能够支持并促进内脏臭气杆菌(O. splanchnicus)的生长,还对其生理适应具有积极影响。当培养条件中胆汁浓度提升至20%时,其生长环境得到明显优化,从而更有利于菌体的增殖与维持。
4
益生菌和益生元增加其丰度
在多种疾病相关模型中,益生菌和益生元可促进有益菌增殖、维持微生态平衡,并改善肠道健康。部分干预可显著提升O.splanchnicus丰度。例如,枯草芽孢杆菌发酵的黄芪(益生元/益生菌复合物)治疗高尿酸血症小鼠,既缓解症状,又显著增加该菌数量。
基于宏基因组分析证实,向ApcMin/+小鼠补充植物乳杆菌CCFM8661可提高O.splanchnicus丰度,并通过修复肠屏障缓解结直肠癌症状。
还有报道,在遗传性肥胖小鼠的标准饮食中添加紫檀芪可发挥抗肥胖作用、改善胰岛素敏感性、重塑微生物群,并显著增加O.splanchnicus丰度,且与脂肪量负相关。另有研究显示,单株乳酸菌L92预处理同样可上调该菌。然而,并非所有益生菌均能提升其丰度,提示该效应具有菌株或益生元特异性。
综上所述,O.splanchnicus的丰度变化反映了宿主—饮食—微生物三者的动态博弈,是评估肠道稳态与疾病风险的重要窗口。
宿主层面,遗传背景、免疫状态、胆汁分泌与肠道屏障完整性决定其定植与扩增潜力;生活方式与饮食结构(如红肉与蔬菜比例、能量与宏量营养素构成)通过底物供给与血红素、胆汁酸等信号分子间接调控其生长;同时,肠道微生态的群落互作、短链脂肪酸与次级胆汁酸代谢网络构成关键生态位约束。外源性干预如抗生素、益生菌/益生元与特定植物活性成分,亦可通过改变底物通量与免疫炎症微环境选择性富集或抑制该菌。
内脏臭气杆菌(O.splanchnicus)属隶属拟杆菌目,是人类肠道微生物群中常见的短链脂肪酸产生菌。其被认为是重要共生体,对宿主健康具有多重作用。
◮ 改善胃肠道炎症和肠道屏障功能
研究发现来源于内脏臭气杆菌(Odoribacter splanchnicus)的细胞外囊泡(Os-EVs)可显著缓解小鼠炎症性肠病(IBD),表现为减轻体重下降与腹泻出血、缩短的结肠长度恢复、组织学损伤减少。
机制上,细胞外囊泡(Os-EVs)通过提升紧密连接蛋白ZO-1与Occludin、减少上皮细胞凋亡,修复肠黏膜屏障,并抑制促炎细胞因子(IL-1β、IL-6、TNF-α)同时上调抗炎因子IL-10;关键在于其抑制NLRP3炎性小体(降低NLRP3、ASC及caspase-1裂解),且在NLRP3敲除小鼠中保护效应显著减弱,提示抑制NLRP3通路是核心机制,为益生菌来源囊泡干预IBD提供新思路。
◮ 诱导Th17细胞,影响免疫反应
内脏臭气杆菌(O.splanchnicus)在Tak1ΔM/ΔM小鼠中的高丰度及诱导肠道Th17细胞的能力表明其具有独立于特定免疫成分的功能相关性。Th17细胞在抵御感染和调控自身免疫中至关重要。通过诱导Th17,内脏臭气杆菌可使常规与无菌小鼠对化学诱导的结肠炎和结直肠癌产生抵抗力。
进一步的研究表明,除了Th17细胞诱导之外,内脏臭气杆菌(O.splanchnicus)主要刺激免疫细胞中 IL-10 的产生。体外研究表明,这种细菌释放外膜囊泡或产生特定的代谢产物,从而触发免疫细胞产生抗炎细胞因子IL-10,从而在肠道中发挥抗炎作用。
O.splanchnicus参与肠道健康的潜在机制
Li J,et al.Microorganisms.2025
◮ 通过代谢产生的丁酸、苹果酸影响结直肠癌
研究显示,O.splanchnicus通过代谢丁酸促进Foxp3/RORγt调节性T细胞增殖,并以淋巴细胞依赖方式诱导IL-10,从而抑制炎症。此外,其通过上调ZO-1与MUC2、促进杯状细胞生成并调节IL-10,强化肠道机械与黏膜屏障,缓解结直肠癌症状。
另一项研究发现,O.splanchnicus培养上清液中的代谢物苹果酸可诱导早期结肠癌细胞凋亡。然而,尽管O.splanchnicus总体有益,其在特定情境(如CRC患者的肠道微生物群)中的作用更为复杂:在非免疫型CRC患者中,高丰度与不良结局相关,包括血管与淋巴浸润增加及总生存期缩短。其他研究还报告,CRC患者的菌群丰富度与多样性下降,脆弱拟杆菌、O.splanchnicus等菌增多。
◮ O.splanchnicus对人体健康的影响存在两面性
O.splanchnicus的双重作用源于多重因素。在健康肠道中,免疫稳态良好,菌群与宿主免疫细胞保持平衡交互:它可诱导Th17细胞,有助于抗感染与维持稳态,并通过促进IL-10产生营造抗炎环境,避免过度免疫反应。
相较之下,在结直肠癌背景下,免疫抑制性的肿瘤微环境可能破坏其正常免疫调节功能。非免疫型CRC患者中该菌高丰度或反映免疫反应异常,可能与其他菌群与肿瘤细胞协同,促进肿瘤进展。宿主遗传与表观遗传差异会影响对该菌的免疫识别与应答,导致不同的疾病结局。
此外,肠道菌群的整体组成起着至关重要的作用。O.splanchnicus不是孤立的,在健康的肠道微生物群中,它可能与其他有益细菌相互作用以维持肠道稳态。然而,在CRC患者中常见的肠道菌群失调中,这些相互作用的平衡被破坏。其他促癌细菌的存在可能与O.splanchnicus协同作用,为肿瘤生长创造更有利的环境。
因此,更深入地了解内脏臭气杆菌(O.splanchnicus)在不同健康状态下的机制作用对于开发新的肠道疾病治疗策略至关重要。
高通量测序使人们在解析肠道微生物组及其与疾病关系时频繁识别到O.splanchnicus。其丰度在多种疾病中呈显著升降波动。在这里,我们总结了与内脏臭气杆菌(O.splanchnicus)的存在密切相关的疾病类型,无论是正相关还是负相关,旨在更深入地了解肠道微生物群在疾病机制中的作用。
O.splanchnicus丰度的变化与疾病关联
Li J,et al.Microorganisms.2025
最近的研究描述了肠道中内脏臭气杆菌(O.splanchnicus)相对丰度降低与各种病理状态之间的密切相关性,为微生物群与宿主健康之间错综复杂的相互作用提供了新的视角。
1
1型糖尿病中显著减少
具体来说,在一项关于怀孕期间患有1型糖尿病 (T1D) 的女性肠道微生物群的组成和功能变化的研究中,作者观察到显著的微生物群落重塑,这往往会加剧炎症反应。
◮ O.splanchnicus减少并且炎症反应加剧
这些表现为粪便钙卫蛋白和肠道脂肪酸结合蛋白水平显著升高,以及内脏臭气杆菌(O.splanchnicus)(FDR = 0.098)的丰度显著降低。这些改变可能会增加 T1D 女性发生妊娠相关并发症的风险。
2
多种肠道疾病中丰度降低
O.splanchnicus丰度下降与多种肠道疾病的病理状态密切相关,包括克罗恩病、炎症性肠病、结直肠癌和全结肠炎。
◮ 克罗恩病中O.splanchnicus的减少会影响抗炎反应
一项关于接受英夫利昔单抗治疗的克罗恩病儿科患者的微生物学变化的研究发现,与对照组相比,在接受治疗后O.splanchnicus和其他产生短链脂肪酸的细菌丰度显著且持续增加。
值得注意的是,O.splanchnicus可产生抗炎短链脂肪酸(如乙酸盐、丙酸盐和丁酸盐),对宿主具有抗炎作用。因此,O.splanchnicus丰度的减少可能会影响宿主的炎症反应,这导致克罗恩病和全结肠炎的恶化和慢性化。
◮ 肠易激综合征中O.splanchnicus丰度降低影响叶酸合成
在肠易激综合征患者中,O. splanchnicus的丰度与二氢叶酸的水平呈正相关,二氢叶酸是叶酸合成的关键中间体。肠易激综合征患者普遍存在的低叶酸水平表明O. splanchnicus在维持肠道代谢平衡和营养吸收方面具有潜在作用。
3
囊性纤维化、NAFLD等疾病中降低
◮ 囊性纤维化患者中显著降低
在其他几种疾病状态中也观察到O.splanchnicus丰度的减少。例如,囊性纤维化(CF)患者肠道微生物多样性显著降低的同时,该物种的相对丰度也大幅减少。进一步强调了其在维持健康的肠道微生物群平衡方面的关键作用。
◮ 非酒精性脂肪性肝病患者中丰度下降
同样,在非酒精性脂肪性肝病(NAFLD)患者中也观察到O.splanchnicus的丰度下降,这一变化与其肠道微生态失衡相一致。进一步地,在合并NAFLD的高体重儿童人群中,该菌的相对丰度呈现出更加显著的降低趋势,提示其减少可能与疾病状态的出现或加重相关。
◮ O.splanchnicus可能有助于抵抗艰难梭菌感染和移植物抗宿主病
一项多中心前瞻性队列研究对1500余名住院患者进行长期随访的结果显示,O.splanchnicus的存在与住院期间感染防控具有潜在关联。尤其当其与瘤胃球菌属(如Ruminococcus bromii)共同出现时,可能在医院环境中对抗艰难梭菌感染,表现出一定的保护作用。这一发现提示特定菌群组合在临床感染管理中的重要性与应用前景。
此外,在移植后植入阶段,O.splanchnicus的检出与急性移植物抗宿主病(aGVHD)风险降低相关。该相关性进一步表明,该菌可能在预防或缓解aGVHD方面发挥潜在作用,有助于改善移植相关并发症的发生率与患者预后。
这些发现表明,O.splanchnicus作为一种多功能肠道细菌,其丰度与各种疾病状态密切相关。它不仅影响肠道炎症和代谢活动,还可能调节全身炎症和代谢过程,从而对宿主的整体健康产生深远的影响。
某些细菌通过其代谢活动和与宿主的相互作用,对宿主健康产生积极影响,当存在于肠道中时,它们被归类为益生菌。
然而,在不同的疾病中,由于环境条件的变化、与其他微生物的竞争以及宿主免疫反应的变化,O.splanchnicus可能会引发感染、炎症和其他不良反应,从而对宿主健康产生不利影响。
1
系统性红斑狼疮患者中显著富集
使用宏基因组测序分析了117例未经治疗的系统性红斑狼疮(SLE)患者的粪便微生物群,其中52例还提供了治疗前和治疗后的样本,并将其与115例性别和年龄匹配的健康对照进行了比较。他们发现,O.splanchnicus在未经治疗的 SLE 患者的肠道中显着富集。
◮ 会产生一种类似自身抗体的肽加剧免疫异常
该细菌产生的肽与SLE患者体内特征性自身抗体Sm抗原的表位相似。这会导致免疫系统的误识别,并可能引发或加剧SLE患者的免疫异常。
2
动脉瘤和血管瘤患者中丰度较高
在病例对照研究中对未破裂颅内动脉瘤患者和对照组的肠道微生物群进行了宏基因组测序分析和比较。颅内动脉瘤患者肠道菌群结构的异质性显著降低,动脉瘤组O.splanchnicus明显富集。这表明这种细菌的存在与颅内动脉瘤的发病机制之间存在潜在联系。
◮ 可能作为海绵状血管瘤的生物标志物
海绵状血管瘤(CA)患者肠道中O.splanchnicus的丰度显著高于健康对照组。因此,肠道中这种细菌的丰度较高可能作为CA的生物标志物,其丰度的动态变化可能与CA的发病和进展密切相关。
3
多动障碍患者中丰度过高
对注意力缺陷多动障碍(ADHD)的研究也显示,受影响个体的肠道微生物群组成明显异常,特别是ADHD儿童中O.splanchnicus, Bacteroides caccae, Paraprevotella xylaniphila和Veillonella parvula的丰度明显高于年龄匹配的健康儿童。
◮ O.splanchnicus可能影响神经递质合成和多巴胺代谢
还有研究表明,多动症患者中O.splanchnicus的丰度显著升高,并可能影响神经递质合成、多巴胺代谢以及通过细胞因子释放调节炎症和神经发育。
4
急性冠状动脉综合征患者中增加
在急性冠状动脉综合征(ACS)患者中,肠道菌群内的O.splanchnicus丰度显著高于性别、年龄及临床特征匹配的对照人群。该差异提示O.splanchnicus的存在与ACS发生之间具有相关性,可能反映疾病相关的微生态失衡。
基于这一关联,O.splanchnicus有望作为ACS风险评估的生物学指标,为预防与管理提供新的方向。
5
慢性肾病、代谢疾病中富集
◮ 会产生吲哚和对甲酚等影响肾功能的物质
有研究分离并量化了慢性肾病患者产生尿毒症毒素前体的肠道细菌。他们发现,O.splanchnicus通过其特定的代谢机制,产生苯乙酰谷氨酰胺、吲哚和对甲酚,这些都是肾功能受损的重要标志物。这种细菌可能在肾功能下降或尿毒症毒素的积累中发挥重要作用。
◮ 脂肪堆积、葡萄糖耐量受损患者中丰度增加
在动物模型中,肠上皮缺乏glut1活性的条件敲除小鼠出现脂肪堆积、糖耐量受损及菌群紊乱,并伴随O.splanchnicus与Odoribacter laneus丰度升高,提示代谢疾病与肠道微生物群存在重要相互作用。
此外,O.splanchnicus的存在与功能性便秘以及共病抑郁和焦虑症状呈正相关,特别是与血清5-羟色胺水平相关。还观察到产生生长激素的垂体腺瘤患者肠道中O.splanchnicus显著富集。
6
肠道屏障受损时
在正常情况下,内脏臭气杆菌(O.splanchnicus)是正常的肠道菌群。然而,在特定条件下,如腹腔内炎症和肠道屏障受损,它可能作为机会性病原体并引起感染。
◮ 肠屏障受损时O.splanchnicus可引起感染
近期血培养病例报告将O.splanchnicus确认为急性阑尾炎继发菌血症的致病菌,提示其具有潜在致病性。通过分离厌氧革兰阴性杆菌并以16S rRNA测序确认,证实其为假体关节感染的病原体。另有多例人类感染报道,包括菌血症、免疫正常成人的坏疽性阑尾炎并菌血症及盆腔腹膜炎。在30例穿孔或坏疽性阑尾炎中,有12例在腹膜液、阑尾组织或脓肿中检测到O.splanchnicus,且常与其他需氧、厌氧菌共存。
尽管病例提示O. splanchnicus可致肠外感染(如菌血症),但其在腹腔感染中的确切作用与机制仍有待进一步阐明。
小编的话
总之,这些研究揭示了内脏臭气杆菌(O.splanchnicus)在各种疾病状态下的丰度变化与疾病的发生或进展之间存在显著关联。然而,需要强调的是,这种关联是由多种因素相互作用产生的,包括饮食、生活方式、遗传背景和其他环境影响。因此,我们不能将内脏臭气杆菌的存在单独归因于疾病发生的直接原因。
相反,这些发现强调了肠道菌群与宿主健康之间错综复杂的关系,同时也为未来的研究提供了方向,以进一步探索这些因素之间的相互作用机制。此类研究对全面理解发病机制、实现早期诊断并制定有效干预策略至关重要。
作为肠道微生物群成员,内脏臭气杆菌(O.splanchnicus)在多种疾病中的丰度、分布与功能活性显著变化,显示其作为生物标志物在诊断、预后评估及疗效监测中的重要潜力。
◮ 可能有助于诊断肝细胞癌
有研究使用宏基因组测序和液相色谱-质谱相结合,检查了68名肝细胞癌(HCC)患者、33名肝硬化(LC)患者和34名健康个体(NC)的肠道微生物群组成和血清代谢特征。
发现与非HCC患者相比,HCC患者的血清代谢物组成和肠道微生物群结构发生了显著变化。通过LEfSe和代谢途径富集分析鉴定与HCC相关的2个关键菌种(Odoribacter splanchnicus和Ruminococcus bicirculans)和5个关键代谢物(ouabain、牛磺去氧胆酸、糖去氧胆酸、茶碱和黄嘌呤)
◮ 肥胖的潜在生物标志物
此外,一项涵盖中国人群(n=2262)的研究比较了肥胖与正常体重者的肠道菌群差异,强调O.splanchnicus为与肥胖相关的三种潜在生物标志物之一。
◮ 肾癌和胰腺导管腺癌
还有发现O.splanchnicus某些特征或存在与肾癌(KC)风险升高相关,提示其为KC评估与诊断的危险因素与潜在标志物。通过功能与物种水平分析显示,胰腺导管腺癌(PDAC)患者中该菌丰度与健康对照及自身免疫性胰腺炎(AIP)患者显著不同,可用于区分PDAC与健康个体及AIP。
▷ 溃疡性结肠炎的标志物
基于生态与网络分析指出,溃疡性结肠炎(UC)患者中该菌的存在或丰度与疾病状态相关,并将其列为UC中具潜在诊断与治疗意义的四种“机会性病原体”之一,定位为生物标志物或治疗靶点。
总之,内脏臭气杆菌(O.splanchnicus)丰度的改变不仅是各种疾病状态下肠道微生物失衡的标志,而且也是一种重要的生物标志物,可能有助于调节宿主的健康状况和预防特定疾病。
作为重要的肠道微生物,O. splanchnicus因其在多种健康与疾病进程中的作用而备受关注。
◮ 百岁老人中O.splanchnicus丰度较高
一项涵盖32个长寿家系的16S rRNA与宏基因组研究显示,百岁老人的肠道菌群更具多样性,抗炎菌与潜在益生菌显著增多,尤其O. splanchnicus丰度更高。在日本百岁老人队列中以Spearman相关与FDR校正(q < 0.05)证实:其丰度随年龄增长而上升,提示肠道菌群组成可能影响寿命。
◮ 抗炎和改善代谢
肥胖者与正常体重者相比,肠道微生物多样性更低,短链脂肪酸重要产生者O. splanchnicus显著减少。鉴于短链脂肪酸在能量代谢、屏障强化与抗炎中的关键作用,恢复或提升O.splanchnicus丰度或为改善肥胖的新策略。
◮ 增强宿主抗肿瘤免疫
在肿瘤防治方面,O. splanchnicus有助于维持微生态平衡、降低癌症风险并提高疗效。研究发现,特异性药物或FMT可提升包括O. splanchnicus在内的菌群丰度,增强宿主抗肿瘤免疫(激活NK与T细胞),为“微生物群调节—增强抗肿瘤力”提供新路径。
相反,接受强化治疗的急性髓系白血病患者出现肠屏障受损与多样性下降,O. splanchnicus等特定菌群减少,强调治疗过程中维持菌群平衡的重要性。
◮ 有助于治疗慢性疼痛患者
此外,慢性疼痛患者的肠道微生物组发生了显著变化,其特征是α多样性较低,某些细菌种群相对丰度较高,包括O. splanchnicus。这些变化可能通过涉及肠脑轴的复杂机制影响疼痛感知和调节。
这些发现表明,肠道微生物群调节,特别是O.splanchnicus的丰度,可能有助于确定缓解慢性疼痛的新治疗靶点。
O.splanchnicus具有优异的抗炎特性和免疫调节作用。未来,以O.splanchnicus为重点的微生态制剂可能为食品、膳食补充剂和生物制药领域的消费和药用应用提供创新选择。
内脏臭气杆菌(O.splanchnicus)是人类肠道微生物群的重要成员,在维持肠道稳态和促进健康方面发挥重要作用。它在与炎症性肠病、非酒精性脂肪性肝病和系统性红斑狼疮等疾病相关的病理生理过程中发挥直接和间接的调节作用。然而,在特定的病理条件下,如腹腔内炎症加剧或肠道屏障功能受损,它可能作为机会性病原体,引发感染甚至败血症。
作为生物标志物,O.splanchnicus在肝细胞癌、肥胖、肾癌与溃疡性结肠炎中展现诊断、分层与疗效监测价值,部分情境下优于传统指标。应用前景上,特定益生菌药物与粪菌移植可提升其丰度并增强NK/T细胞抗肿瘤免疫;在慢性疼痛与长寿人群中与表型相关;其菌株与囊泡具抗炎与免疫调节活性,具备食品、营养与生物医药转化潜力。
未来,应以多组学与因果推断澄清其致益/致病边界与机制阈值,定义安全性与剂量-反应关系,解析与宿主遗传、饮食因子及菌群网络的互作规则,发展可控的菌/囊泡/代谢物精准干预与伴随诊断。在守护肠道稳态、代谢健康与免疫平衡的同时,审慎管理其机会致病风险,方能释放O.splanchnicus在健康促进与疾病防治中的最大价值。
主要参考文献
Li J, Xu J, Guo X, Xu H, Huang C, Nie Y, Zhou Y. Odoribacter splanchnicus-A Next-Generation Probiotic Candidate. Microorganisms. 2025 Apr 3;13(4):815.
Correction to: Odoribacter splanchnicus bacteremia secondary to acute appendicitis: a case report with review of literature. J Surg Case Rep. 2024 Jun 11;2024(6):rjae416.
Hiippala K, Barreto G, Burrello C, Diaz-Basabe A, Suutarinen M, Kainulainen V, Bowers JR, Lemmer D, Engelthaler DM, Eklund KK, Facciotti F, Satokari R. Novel Odoribacter splanchnicus Strain and Its Outer Membrane Vesicles Exert Immunoregulatory Effects in vitro. Front Microbiol. 2020 Nov 12;11:575455.
Lima SF, Gogokhia L, Viladomiu M, Chou L, Putzel G, Jin WB, Pires S, Guo CJ, Gerardin Y, Crawford CV, Jacob V, Scherl E, Brown SE, Hambor J, Longman RS. Transferable Immunoglobulin A-Coated Odoribacter splanchnicus in Responders to Fecal Microbiota Transplantation for Ulcerative Colitis Limits Colonic Inflammation. Gastroenterology. 2022 Jan;162(1):166-178.
Roth-Schulze AJ, Penno MAS, Ngui KM, Oakey H, Bandala-Sanchez E, Smith AD, Allnutt TR, Thomson RL, Vuillermin PJ, Craig ME, Rawlinson WD, Davis EA, Harris M, Soldatos G, Colman PG, Wentworth JM, Haynes A, Barry SC, Sinnott RO, Morahan G, Bediaga NG, Smyth GK, Papenfuss AT, Couper JJ, Harrison LC; ENDIA Study Group. Type 1 diabetes in pregnancy is associated with distinct changes in the composition and function of the gut microbiome. Microbiome. 2021 Aug 6;9(1):167.
Göker M, Gronow S, Zeytun A, Nolan M, Lucas S, Lapidus A, Hammon N, Deshpande S, Cheng JF, Pitluck S, Liolios K, Pagani I, Ivanova N, Mavromatis K, Ovchinikova G, Pati A, Tapia R, Han C, Goodwin L, Chen A, Palaniappan K, Land M, Hauser L, Jeffries CD, Brambilla EM, Rohde M, Detter JC, Woyke T, Bristow J, Markowitz V, Hugenholtz P, Eisen JA, Kyrpides NC, Klenk HP. Complete genome sequence of Odoribacter splanchnicus type strain (1651/6). Stand Genomic Sci. 2011 Apr 29;4(2):200-9.
Zhuang J, Zhuang Z, Chen B, Yang Y, Chen H, Guan G. Odoribacter splanchnicus-derived extracellular vesicles alleviate inflammatory bowel disease by modulating gastrointestinal inflammation and intestinal barrier function via the NLRP3 inflammasome suppression. Mol Med. 2025 Feb 11;31(1):56.
Morgan XC, Tickle TL, Sokol H, Gevers D, Devaney KL, Ward DV, Reyes JA, Shah SA, LeLeiko N, Snapper SB, Bousvaros A, Korzenik J, Sands BE, Xavier RJ, Huttenhower C. Dysfunction of the intestinal microbiome in inflammatory bowel disease and treatment. Genome Biol. 2012 Apr 16;13(9):R79.
Li H, Xu H, Li Y, Jiang Y, Hu Y, Liu T, Tian X, Zhao X, Zhu Y, Wang S, Zhang C, Ge J, Wang X, Wen H, Bai C, Sun Y, Song L, Zhang Y, Hui R, Cai J, Chen J. Alterations of gut microbiota contribute to the progression of unruptured intracranial aneurysms. Nat Commun. 2020 Jun 25;11(1):3218.

谷禾健康
清晨被肠绞痛惊醒,聚餐后腹胀如鼓,皮肤莫名冒出红疹…这些现代人熟悉的困扰,根源可能都指向同一个问题——肠道炎症。外卖高油盐饮食、熬夜加班压力、抗生素滥用,正让我们的肠道炎症悄悄出现。
肠道炎症不仅是消化系统问题,更是影响全身健康。肠道是人体最大的免疫器官(70%免疫细胞位于肠道),肠道炎症的严重性远不止于表面症状,当它持续存在时可能导致炎症性肠病、糖尿病、关节炎、抑郁焦虑等神经系统疾病、免疫相关疾病(如过敏)等非常多慢性疾病。研究表明,慢性肠道炎症患者的结直肠癌风险提高,自身免疫疾病患者的肠道通透性标志物连蛋白(Zonulin)水平显著高于健康人群。
正因为它如此重要,肠道炎症已经成为现代医学关注的重要问题。现代医学通过多维度指标评估肠道炎症水平,例如,直接生化标志物,肠道屏障功能等。在谷禾肠道菌群检测报告中,也有“肠道炎症”这项指标。
<来源:谷禾肠道菌群检测数据库>
本文深入解析肠道炎症的成因链条——从麸质敏感、工业种子油到慢性压力的触发机制;揭示其如何通过”肠-脑轴“、”肠-皮肤轴“等方式引发全身性疾病;并提供相关干预方案,涵盖屏障修复、菌群重建及创新生物制剂应用等。
▸ 炎症:是什么?如何作用?
炎症是机体对有害刺激(如病原体、有毒化合物或受损细胞)的反应。它通过清除有害因子并启动修复过程发挥保护作用。
▸ 炎症的四个阶段
炎症过程包含四个不同阶段:
1.静息期,受损细胞释放介质启动炎症反应;
2.血管期,出现血管扩张和血浆外渗;
3.细胞期,中性粒细胞、单核细胞和巨噬细胞浸润,随后激活适应性免疫;
4.消退期,修复组织并清除碎片和炎症细胞。
因此,炎症本质上是一个具有明确起始、发展和结束阶段的生理过程。
慢性肠道炎症的各个阶段
Deraison C,et al.Annu Rev Pharmacol Toxicol.2025
急性炎症可能会发展成慢性炎症
然而,部分患者短期的炎症可能会发展为慢性炎症性疾病,炎症症状持续数月至数年。慢性炎症通常由于未能消除急性炎症的致病因子所致。这可能发生在病原体(如细菌、真菌、寄生虫)感染中,这些病原体能够抵抗宿主防御机制,或当刺激物持续存在且无法被内源性机制(如酶、炎症细胞吞噬作用)降解时。
自身免疫性疾病中也会出现慢性炎症,此时免疫系统错误地将自身蛋白质识别为外来抗原并引起组织破坏,如乳糜泻。正常情况下标志炎症过程结束的消退机制失调同样可导致慢性炎症。在肠道等黏膜表面,负责”处理”外来抗原和调节黏膜组织对微生物暴露反应的机制发生故障也可引起慢性炎症性疾病。这些故障机制包括屏障功能的各个层面——物理屏障(黏液)、细胞屏障(上皮细胞)和免疫屏障(常驻炎症细胞)。
▸ 与肠道炎症相关的疾病
急性回肠炎或结肠炎的发作通常与感染(病毒、细菌、真菌或寄生虫)相关。然而,在某些情况下,急性回肠炎或结肠炎可能是慢性炎症性疾病的最初表现,这些肠道慢性炎症性疾病包括炎症性肠病(IBD)、乳糜泻和肠易激综合征(IBS)。
▸ 炎症性肠病
炎症性肠病(IBD)是一种慢性肠道炎症性疾病,包括克罗恩病(CD)和溃疡性结肠炎(UC)两个主要亚型,通常表现为腹痛、腹泻和血便。
IBD的病程特点是急性炎症发作(活动期)与缓解期和复发期交替出现。在病因方面,普遍认为IBD是遗传易感性和环境因素共同作用的结果,两者协同导致肠道微生物群落改变和免疫反应失控。
▸ 乳糜泻
乳糜泻是一种遗传性免疫疾病。遗传易感个体摄入含麸质食物后,会发生小肠黏膜炎症和绒毛萎缩。小麦面筋蛋白和麦胶蛋白富含谷氨酰胺和脯氨酸,使其对蛋白酶降解具有抗性,在小肠内形成免疫原性大肽。然而,这些肽的存在并不足以引发疾病;患者还必须携带位于DQ位点的特定HLA II类基因。这种遗传易感性使肠道组织更易受到食物诱发的炎症因子影响。
▸ 肠易激综合征
肠易激综合征是一种伴有排便习惯改变的慢性腹痛综合征,尚未发现明确的结构异常或炎症细胞浸润。多项研究探讨了炎症征象与该综合征的关联性。部分研究显示抗炎治疗(如美沙拉嗪)未能缓解肠易激综合征症状,提示炎症可能并非症状成因。
然而,大多数肠易激综合征患者存在免疫细胞激活增强的证据。这些免疫细胞包括在患者结肠组织中发生脱颗粒的肥大细胞,以及在结肠活检中上调的多种促炎介质(组胺、多不饱和脂肪酸代谢物、蛋白酶)。因此,充分证据表明肠易激综合征可视为一种低度慢性肠道炎症性疾病,虽然炎症细胞浸润并非显著特征,但可能存在组织功能紊乱和黏膜稳态改变。
▸ 肠道炎症涉及的分子机制
肠道黏膜炎症状态的共同特征包括微生物群生物膜失调、上皮功能障碍、先天免疫细胞浸润和适应性免疫激活。
肠道炎症患者存在微生物群失调
虽然IBD、乳糜泻或IBS中的微生物群失调可能呈现不同的分类特征,难以识别共同的微生物分子机制,但所有炎症状态均表现出微生物物种多样性和丰富性的减少。
紧密连接分子降解和黏液产生减少
屏障功能改变是另一共同特征。肠道上皮细胞紧密连接分子(如Zonulin和occludin)的降解以及杯状细胞黏液产生减少均与肠道炎症相关。潘氏细胞和肠上皮细胞表达分泌的抗菌肽是肠道屏障功能的重要组成部分,在炎症时功能下降。
α-和β-防御素、溶菌酶以及elafin和分泌型白细胞蛋白酶抑制剂等抗菌肽蛋白酶抑制剂在炎症组织中表达下调。因此,促进紧密连接分子、黏液蛋白和抗菌肽的表达和功能有助于增强肠道屏障功能,被视为治疗炎症的有效方法。
微生物群失调和屏障功能改变共同促进肠道慢性炎症
微生物群失调与屏障功能改变导致微生物对组织的异常暴露增加,引起先天免疫细胞深度激活。活化的巨噬细胞吞噬微生物成分,通过Toll样受体9、磷酸二酯酶4和Janus激酶(JAK)等信号机制释放肿瘤坏死因子α(TNF-α)、白细胞介素(IL)-6、IL-12和IL-23,这些炎症介质和信号通路均被视为治疗慢性炎症的重要分子靶标。
组织损伤或侵袭激活后,肥大细胞、巨噬细胞和中性粒细胞共同释放储存在颗粒中的分子,包括炎症脂质介质(如前列腺素和白三烯)、活性氧(ROS)和蛋白酶,这些均构成炎症治疗的分子靶标。肠上皮细胞同样促进炎症性肠黏膜释放这些介质。
与慢性肠道炎症相关的机制
Deraison C,et al.Annu Rev Pharmacol Toxicol.2025
1
麸质——乳糜泻或麸质敏感人群的炎症
麸质蛋白是小麦、大麦、黑麦和燕麦的主要储能蛋白,位于谷物胚乳中,由麦谷蛋白和醇溶谷蛋白以1:1比例构成。
麸质会引起乳糜泻和麸质敏感患者炎症
麸质会导致乳糜泻和非乳糜泻麸质敏感性(NCGS)患者的炎症。乳糜泻是一种麸质自身免疫反应,表现为肠组织结构破坏、胃肠道不适及肠外症状(如皮疹和贫血)。
人群筛查显示乳糜泻为最常见的慢性胃肠道疾病之一,但因临床表现多样且模糊,绝大多数患者仍未被识别。虽然吸收不良和胃肠道症状较为人知,但多数患者实际表现为肠外症状,可累及神经系统、肝脏、皮肤、生殖和肌肉骨骼系统等全身器官,且可在不同年龄出现,更难确定诊断。
麸质会激活肠道肥大细胞并释放促炎分子
麸质与肠道CXCR3受体结合促进zonulin释放,引发易感个体肠道炎症。Zonulin分解上皮细胞间紧密连接蛋白,增加肠通透性和炎症。麸质还激活肠道肥大细胞释放组胺和类胰蛋白酶等促炎分子,使炎症持续存在。
需要注意的是,麸质并非对所有人都引起炎症。无乳糜泻或无非乳糜泻麸质敏感性(NCGS)的人通常能耐受饮食中的麸质。
注:多数晚期肠外表现可通过乳糜泻早期诊断预防。提高诊断率的方法是筛查高危人群,但无麸质饮食维持困难、社会限制且昂贵,可能降低生活质量,特别是症状轻微的患者。然而,证据表明即使无症状成年人也可能已有晚期组织学病变并从饮食治疗中获益,且诊断延迟会增加临床反应不良风险。
2
工业种子油
从大豆、玉米、油菜籽、棉籽和红花籽提取的高度加工工业籽油在标准饮食中占主要份额。这些油不到两个世纪前才引入人类饮食,与一些人体进化不匹配,会促进肠道炎症。
工业种子油烹饪会产生促炎产物
工业种子油富含ω-6脂肪酸,易受光和热损害。用其加工烹饪产生氧化炎症副产品,食用后引起肠道炎症。研究发现,富含工业种子油的饮食不仅会引起肠道炎症,还促进肠道病原体生长。
脂质饮食对肠道微生物群多样性的影响
doi: 10.3390/nu11020418.
脂质是哺乳动物正常发育和生存必需品,其化学性质细微差别影响生理和炎症反应。
不同的膳食脂质会影响肠道微生物群,可能促炎
研究显示饮食脂质对肠道微生物的影响:乳脂和玉米油饮食增加微生物多样性,橄榄油效果与低脂饮食相似;饮食脂质解释80.8%的微生物变异,形成三个不同聚类群;不同脂质类型特异性塑造肠道微生物群落。
总的来说,单不饱和脂肪酸消费与健康益处相关,包括降低消化系统癌症、2型糖尿病和炎症性肠病(IBD)发病率,橄榄油饮食可有效预防小鼠结肠炎。相比之下,虽然北美膳食指南推荐植物籽油中的ω-6多不饱和脂肪酸,但过量摄入是人类IBD风险因素,研究证实其加剧鼠结肠炎。
对于存在于乳制品和椰子油中的饱和脂肪酸,虽然数十年来被批评对健康不利,但欧洲前瞻性队列研究发现牛奶消费与IBD风险降低相关。然而,饲喂饱和脂肪酸通过与胆汁酸结合促进沃氏嗜胆菌(Bilophila wadsworthii)生长,增加IL-10-/-小鼠自发性结肠炎。但动物脂肪成分如丁酸具有抑制炎症、防止DSS-结肠炎和刺激结肠修复的作用。
3
非细胞碳水化合物
非细胞碳水化合物是指缺乏细胞壁的高碳水化合物密度食物,如面粉和糖,在饮食中大量存在并促进炎症性肠道微生物群。相比之下,细胞碳水化合物是含完整细胞的碳水化合物食物,如红薯和整个水果,通过促进抗炎肠道细菌生长来促进肠道健康。
doi: 10.2147/DMSO.S33473. Epub 2012 Jul 6.
现代食品中的无细胞致密碳水化合物从口腔开始产生炎症微生物群,最初引起牙周病。小肠暴露于口腔微生物群的脂多糖和其他病原体相关分子模式,并通过无细胞碳水化合物调节自身小细菌群的促炎性。膳食脂肪增强全身吸收后,炎症细菌化合物诱导瘦素抵抗和过度进食。
4
食品添加剂
超加工食品日益增多且含有大量食品添加剂。大多数添加剂缺乏长期安全性研究,而越来越多证据表明其对肠道炎症有害。
食品添加剂促进促炎细菌生长并增加肠道通透性
人造甜味剂三氯蔗糖刺激炎症肠道细菌的生长。海藻多糖角叉菜胶因其增稠和乳化特性被使用,但会加剧炎症性肠病。常见食品乳化剂羧甲基纤维素和聚山梨酯-80增加肠道通透性。增白剂二氧化钛在肠道引起炎症细胞因子反应。为防止肠道炎症,避免或减少这些普遍存在的食品添加剂应是首要任务。
5
剖腹产和配方奶喂养
剖腹产和奶粉喂养婴儿更易患肠道炎症
剖腹产深刻改变发育中婴儿肠道,用环境微生物而非母亲阴道有益微生物接种微生物组。剖腹产婴儿还通过胎盘循环暴露于预防术后感染的抗生素。这种早期抗生素暴露进一步扭曲婴儿肠道微生物群发育,使孩子易患肠道炎症和哮喘、肥胖等慢性炎症性疾病。
配方奶粉喂养对脆弱婴儿肠道同样可能有害。配方奶喂养的婴儿具有较高水平的促炎细菌类γ-变形菌门,增加肠道通透性和总细菌负荷。相反,母乳喂养增加婴儿肠道抗炎乳酸杆菌和双歧杆菌定植,降低肠道炎症和慢性炎症性疾病风险。
6
肠道感染
细菌、病毒、真菌和寄生虫等各类病原体引起的肠道感染会显著改变肠道微生物群的正常组成结构,从而创造出一个促进炎症反应的肠道内环境。
细菌和病原体感染会变相增加一些促炎细菌丰度
例如,甲型流感病毒感染过程中会通过多种机制减少具有保护作用的肠道有益细菌数量,这种保护性细菌的减少会瞬时性地增加机体对沙门氏菌属等致病菌的易感性,使患者更容易发生继发性细菌感染。
与此同时,幽门螺杆菌这种常见的胃部致病菌在感染过程中不仅会直接损害胃黏膜,还会通过复杂的相互作用机制增强普雷沃氏菌这种典型促炎细菌在肠道中的生长和繁殖能力。
食物中毒也会促进肠道炎症反应
此外,急性食物中毒事件除了造成胃肠道症状外,还可能通过刺激肠道免疫系统产生针对自身组织的炎症性自身抗体,这些异常的免疫反应不仅会引发持续性的肠道炎症状态,还可能进一步发展为肠易激综合征等功能性肠道疾病。
7
压力
人类的肠道对慢性压力非常敏感。糖皮质激素水平的长期升高驱动肠神经胶质细胞的炎症亚群的产生,通过集落刺激因子-1(CSF-1)促进单核细胞和TNF介导的炎症。此外,糖皮质激素通过TGF-β2引起肠神经元的转录不成熟、乙酰胆碱缺乏和运动障碍。
压力促使脂多糖进入血液导致炎症
慢性心理压力还会增加肠道通透性,并允许脂多糖(LPS),一种炎症细菌的副产物,进入血液循环;这会导致局部和全身炎症反应。应激还消耗保护性肠道粘膜层,并增加细菌粘附和渗透到肠上皮细胞中,引起肠道炎症。
不同居住条件下心理压力对肠道炎症的加剧作用
DOI: 10.1016/j.cell.2023.05.001
糖皮质激素治疗结肠炎的效果与心理压力相关
压力会导致炎症性疾病,而心理压力的典型全身介质——儿茶酚胺和糖皮质激素通常被认为具有抗炎作用。由于糖皮质激素受体激动剂泼尼松用于治疗IBD患者结肠炎,糖皮质激素信号传导诱导炎性肠神经胶质细胞和结肠炎恶化尤其令人困惑。
例如,长期压力或地塞米松治疗可能因负反馈抑制下丘脑-垂体-肾上腺轴而导致肾上腺皮质酮生产不足。然而,急性糖皮质激素受体抑制的有益作用与”肾上腺抑制”不符。或者,糖皮质激素抗炎和促炎作用间的平衡可能是暂时现象。
虽然急性糖皮质激素治疗改善DSS诱导的结肠炎,研究团队还发现持续升高的皮质酮水平有助于eGAPS的出现和产生TNF的单核细胞的积累。同样,临床观察已经证明,只有短期类固醇治疗才能在IBD中产生有益的结果。所以还是要强调了在炎症性疾病的临床管理中考虑患者心理健康的重要性,治疗效果可能因受影响个体的心理状态而异。
因此,心理状态的评估,结合减少压力,焦虑和抑郁的策略,可能是一个强大的和未充分利用的工具,以提高治疗的成功。
8
久坐不动的生活方式和过度训练
适量运动有助于促进抗炎细菌的生长
适度的运动可以促进有益的肠道细菌,包括产生抗炎性短链脂肪酸(SCFA)的细菌,如梭状芽孢杆菌属、罗氏菌属、毛螺菌属等,从而抑制肠道炎症。相反,久坐不动的生活方式与炎症分子向肠道的渗透增加有关。
然而,运动也并非越多越好。过度的体力活动会增加肠道通透性,对肠道微生物群产生负面影响。如果您患有慢性炎症性疾病或承受严重心理压力,应避免剧烈运动,专注于散步、瑜伽或游泳等温和活动。
9
昼夜节律紊乱和睡眠不足
昼夜节律是以约24小时为周期的内部生化过程,调节包括肠道健康和炎症在内的多项生理功能。当夜间蓝光照射和不规律睡眠等因素干扰昼夜节律时,会促进炎性肠道细菌生长,减少有益微生物,并增加肠道通透性和脂多糖向体循环的转运。
睡眠不足与昼夜节律紊乱促进炎性细菌增长
睡眠不足与昼夜节律紊乱密切相关,对肠道健康同样有害。仅两个晚上的部分睡眠剥夺就能改变肠道微生物群,增加促炎细菌种类,并加剧IBD的肠道炎症。因此,维持正常昼夜节律和充足睡眠对减少肠道炎症至关重要。
10
滥用抗生素
临床医生频繁开处抗生素会给患者带来虚假安全感,轻易就开抗生素已成为社会常态,但其对肠道健康的影响往往被低估。大量研究表明,抗生素可能对肠道微生物群产生长期有害影响,并诱导肠道炎症。
抗生素减少肠道菌群的多样性并导致肠道功能紊乱
抗生素会减少肠道共生细菌的多样性和丰度,使埃希氏菌和念珠菌等条件性致病菌过度生长,抗生素还增加了拟杆菌门/厚壁菌门的比率。抗生素对厌氧菌具有特异性,厌氧菌在人体肠道微生物群中占主导地位并发挥重要作用。克林霉素就是一个例子,这是一种相对广谱的抗生素,主要针对厌氧菌。克林霉素对肠道微生物群有很大的负面影响,表现为对病原体定植的抵抗力降低,导致艰难梭菌过度生长导致伪膜性结肠炎的高风险。克林霉素对肠道菌群的其他影响是胃炎和腹泻。正常肠道功能紊乱可导致腹胀和肠痛等症状。
抗生素可能导致长期的生态失调和肠道免疫下降
抗生素处理的微生物组释放岩藻糖和唾液酸等粘膜碳水化合物。这些物质水平的增加有利于机会性病原体鼠伤寒沙门氏菌和艰难梭菌在肠道内的扩张。肠出血性大肠埃希菌也被证明可以获取肠道微生物群从粘蛋白中释放的岩藻糖或唾液酸。
兵器抗生素导致的微生物的变化可以在停止使用抗生素后持续数月或数年。它们的不良影响在婴儿和幼儿中最为明显,他们正在经历肠道微生物群发育的关键时间窗口。
更令人担忧的是,耐药细菌在接触抗生素后会变得更具炎症性。抗生素还会增加肠道通透性,降低肠道天然免疫防御,从而间接促进肠道炎症。
11
非抗生素药物
除了抗生素会影响肠道微生态外,越来越多的证据表明普通常见药物也会对人体肠道健康产生影响。
胃肠道药物和抗抑郁药物对肠道微生态的影响尤为显著
质子泵抑制剂(PPI)用于治疗胃食管反流病和胃酸反流,通过减少胃酸产生来发挥作用。正常情况下,充足的胃酸能限制环境细菌进入肠道。PPI抑制胃酸产生后,更多细菌得以进入消化道并在小肠中增殖,导致细菌过度生长和肠道炎症,同时竞争资源减少产生抗炎短链脂肪酸的有益细菌生长。
选择性5-羟色胺再摄取抑制剂(SSRIs)除了引起便秘和食欲变化外,也会干扰正常的肠道微生物群结构。临床前研究表明氟西汀(百忧解)会抑制乳酸杆菌生长,同时显著增加Alistipes、各种Lachnospiraceae OTU、Lachnoclostridium和Anaerotruncus等菌属的丰度,从而加剧肠道菌群失调。
口服避孕药也会引发肠道炎症。事实上,它们的使用与克罗恩病(IBD的一种亚型)的风险增加有关。
12
环境毒素
工业界每年开发数千种新化学品,但现有环境化学品已造成的一系列健康危害。
环境毒素也会导致有害细菌增加并引发肠道炎症
肠道炎症是环境毒素暴露的最常见后果之一。双酚A(BPA)广泛存在于塑料水瓶、食品容器、儿童玩具和收银机收据中,通过增加有害肠道细菌和肠道通透性来促进肠道炎症。不含BPA产品中的替代增塑剂同样有害,甚至危害更大。
三氯生是洗手液和个人护理产品中使用的合成抗菌剂,也会加重肠道炎症。草甘膦作为农达除草剂的主要成分,能减少乳酸杆菌并促进耐除草剂大肠杆菌生长,表明其在肠道微生物组中发挥抗生素作用。
发炎的肠道会显著增加患多种慢性疾病的风险,包括炎症性肠病(IBD)和结肠癌等消化系统疾病,以及自闭症和阿尔茨海默病等神经系统疾病。肠道炎症作为一个关键的病理过程,与这些严重健康问题的发生和发展密切相关。
因此,积极阻止肠道炎症的发生和进展,对于有效预防这些慢性疾病的出现以及成功管理已有慢性疾病的症状和进程都具有至关重要的意义。通过控制肠道炎症,我们能够从根本上降低患病风险,改善整体健康状况。
1
过敏
促炎细胞因子会刺激诱导过敏性炎症
肠道炎症可能导致季节性过敏。有益和有害肠道微生物失衡会增加肠道促炎细胞因子的释放,这些细胞因子刺激肥大细胞、嗜碱性粒细胞和嗜酸性粒细胞活性,诱导过敏性炎症。生态失调相关的肠道炎症还与特应性皮炎和食物过敏密切相关。
相反,健康的肠道通过释放抗炎短链脂肪酸和调节性T细胞来抑制过敏性致敏,维持免疫平衡。
肠道微生物群与免疫系统之间的相互作用
doi: 10.3389/fimmu.2018.01584.
2
自身免疫性疾病
肠道屏障受损在免疫性疾病中发挥重要作用
肠道炎症是自身免疫性疾病发展的先导因素。1型糖尿病患者的循环脂多糖水平更高。Zonulin是肠道屏障受损时释放的蛋白质,自身免疫性疾病患者的Zonulin水平明显高于健康对照组,表明肠漏在自身免疫性疾病中发挥重要作用。
麸质敏感性和慢性应激是肠道炎症的两个主要触发因素,通常在自身免疫疾病发生之前出现。
3
关节炎
口腔和肠道微生物群发生炎症性改变时,会对关节炎的发生和进展产生重要影响。这些微生物群的改变主要表现为致龋细菌牙龈卟啉单胞菌的异常过度生长,同时伴随着有益细菌数量的显著减少,这种微生物失衡状态与关节炎病情的恶化和进展密切相关。
生态失调引起促炎细胞因子释放,导致关节和肌肉病理性改变
当肠道出现生态失调和肠道屏障功能受损导致肠道渗漏时,会在体内诱导广泛的全身性炎症反应。这种炎症反应会触发多种促炎细胞因子的大量释放,这些细胞因子进入循环系统后会直接作用于肌肉骨骼系统,最终导致关节、肌肉和骨骼组织的退化性改变。
有益的抗炎细菌和短链脂肪酸有助于减轻关节炎
然而,与此形成鲜明对比的是,当肠道中存在充足的抗炎细菌和短链脂肪酸时,这些有益成分已被科学研究证实能够有效减轻关节炎症状的严重程度,并且具有预防和阻止骨质流失的保护作用。
4
胃肠道疾病
炎症性肠病(IBD)
肠道炎症是炎症性肠病(IBD)的一个关键特征,包括溃疡性结肠炎和克罗恩病。一系列有害的肠道变化,包括生态失调和肠道通透性增加,会促进IBD的进展。
肠易激综合征(IBS)
科学研究已经深入阐明了肥大细胞在肠易激综合征(IBS)发病机制中发挥的关键作用。肥大细胞作为一类重要的免疫细胞,能够释放多种刺激性化合物,其中组胺是最主要的介质之一。这些刺激性化合物的释放会引起肠道组织的炎症反应,而持续存在的低度炎症状态在IBS的病理过程中扮演着至关重要的角色,直接影响着疾病的发生和发展进程。
在临床表现为腹泻为主的IBS或者混合型IBS的患者群体中,自身免疫机制同样可能在肠道炎症的产生和维持过程中发挥重要作用。特别是在这些特定类型的IBS病例中,患者既往经历的食物中毒事件可能会成为一个重要的触发因素。这些食物中毒事件会刺激机体的免疫系统产生异常的免疫反应,最终导致机体产生能够直接损害和攻击胃肠道组织的自身抗体,从而加重肠道的炎症状态和功能紊乱。
长期的肠道炎症会增加结直肠癌风险
结直肠癌是一种严重威胁人类健康的恶性肿瘤,其发病率和死亡率都相当高。大量的科学研究证据清楚地表明,预先存在的胃肠道炎症状态往往在结直肠癌的发生和发展过程中起到先导作用,也就是说肠道炎症通常会在结直肠癌实际形成之前就已经出现。
通过深入的研究分析,科学家们已经成功确定了多种能够引发结直肠癌发生的炎症触发因素。包括各种有害的肠道病原体感染,肠道内负责产生丁酸盐的有益细菌数量处于异常低下的水平,以及长期摄入高度加工和精制的典型西方饮食模式,这些因素都会通过不同的机制促进肠道炎症的发生,进而增加结直肠癌的患病风险。
肠道炎症还会引发憩室炎
憩室炎是一种肠壁刺激,由肠道炎症引发。其特征在于结肠中的生态失调和粪便钙卫蛋白升高,这是当胃肠道中存在活动性炎症时由白细胞释放的蛋白质。
5
心血管疾病
心血管疾病(CVD)是全球死亡的主要原因,药物和手术是传统医疗的基石。然而,越来越多的证据表明肠道和心脏健康之间存在重要联系,肠道靶向治疗可能是未来CVD的治疗方法。
生态失调会促进斑块形成和高血压
肠道内多种炎症过程影响心血管疾病的病程。生态失调通过释放脂多糖和肽聚糖等革兰氏阴性和阳性细菌的炎性细胞壁成分,促进动脉粥样硬化斑块形成和高血压,从而导致心血管疾病。菌群失调还会影响胆汁酸代谢,损害胆固醇和甘油三酯等血脂的排泄。
6
抑郁和焦虑
最新的科学研究表明,免疫系统在抑郁症和焦虑症的发生和发展过程中起着至关重要的作用。考虑到人类肠道系统拥有约70%的免疫细胞和免疫组织,集中了机体免疫系统的绝大部分功能,因此肠道内发生的炎症反应会对心理健康状态产生显著而深远的影响也就完全不足为奇了。
肠道炎症通过肠-脑轴诱导大脑产生神经炎症反应
肠-脑轴是连接肠道肠神经系统和中枢神经系统之间的一个复杂的双向信号传递网络系统,这个重要的生理网络在介导和调节肠道炎症状态与心理健康水平之间的密切关系方面发挥着关键作用。
当肠道内出现各种炎症刺激时,这些炎症信号会通过肠-脑轴这一重要通道向大脑传送相关信息,从而在大脑组织内诱导产生神经炎症反应,并且显著改变和影响各种神经递质的正常合成和分泌过程。这些深层次生化变化所带来的直接后果就是大脑神经活动模式发生异常改变,最终表现为抑郁症和焦虑症等精神心理疾病的临床症状。
抑郁症和焦虑症患者对抗肠道炎症的能力减弱
这种具有炎症特征的肠-脑轴信号传导异常主要是由肠道微生物群的生态失调状态和肠道屏障功能的严重障碍所触发和维持的,而这两种病理状态在患有抑郁症和焦虑症的患者群体中表现得极为常见和普遍。同时,患有抑郁症和焦虑症的人群还表现出对抗肠道炎症能力的明显降低和减弱,这种抗炎能力的下降主要是由于肠道内具有抗炎作用的短链脂肪酸水平显著降低所导致的。
7
神经系统疾病
肠道炎症可能是神经退行性疾病的“驱动器”
肠-脑轴在神经退行性疾病中发挥关键作用。肠道炎症被认为是帕金森病的”沉默驱动器”,可在症状出现前持续长达二十年。研究还发现,肠源性炎症反应促进淀粉样蛋白-β斑块沉积,导致阿尔茨海默病中的神经元变性和认知功能障碍。
自闭症、多动症患者菌群多样性减少,存在肠道炎症
神经发育障碍在全球患病率急剧上升;自闭症患病率已达到每59名儿童中有1名,截至2016年,11%的儿童和青少年被诊断为注意力缺陷/多动障碍(ADHD)。小儿急性发作性神经精神综合征(PANS)和链球菌感染相关的小儿自身免疫性神经精神障碍(PANDAS)是以抽搐和强迫症突然发作为特征的新兴神经发育障碍。
研究表明,生态失调和肠道炎症是这些疾病的共同致病因素。ADHD儿童表现出肠道微生物α多样性降低,即肠道细菌种类减少;而较高的多样性通常与更好的健康结果相关。自闭症谱系儿童和PANS/PANDAS儿童同样表现出生态失调。这些炎症变化改变了肠-脑信号传导,最终导致神经行为缺陷。
8
骨质疏松
肠道炎症会影响维生素D、钙的吸收
几条证据表明肠道炎症通过破坏骨生成和骨吸收细胞之间的平衡,导致骨量净损失,从而促进骨质疏松症。肠道炎症也会减少对关键的骨骼构建营养素的吸收,包括维生素D,钙和镁。
9
皮肤健康
肠道及其微生物通过肠-皮肤轴影响皮肤健康
我们已经写了许多关于肠道和皮肤健康之间联系的文章,由肠道-皮肤轴介导。就像肠-脑轴一样,肠-皮肤轴是一个连接肠道及其微生物与皮肤的信号分子网络。
肠道的炎症变化,包括生态失调和肠道通透性增加,与一系列皮肤病有关,包括痤疮、牛皮癣、红斑痤疮和湿疹。
10
代谢性疾病
肠道炎症和代谢失调之间存在双向加剧作用
代谢综合征、2型糖尿病(T2D)、肥胖症和肠道炎症之间形成了一个复杂而有害的恶性循环机制:血糖调节失调状态和体内过量脂肪组织的积累会显著促进肠道内炎症反应的发生和发展,而肠道炎症状态反过来又会进一步加剧和恶化机体的代谢功能障碍,从而形成一个持续恶化的病理生理过程。
肠道微生态失调会促进胰岛素抵抗和瘦素抵抗
肠道微生物生态系统的失调状态以及循环系统中脂多糖水平的升高共同诱导和促进胰岛素抵抗和瘦素抵抗的发生,而这两种激素抵抗现象正是代谢综合征、2型糖尿病和肥胖症发病机制中的关键病理特征和核心环节。
日常饮食中摄入的非纤维性碳水化合物以及环境中存在的各种致肥胖环境毒素暴露(包括双酚A和多种邻苯二甲酸盐化合物)都会对肠道微生物群落的组成和结构产生显著的改变和影响。这些肠道微生物群落的变化和改变会直接引起肠道组织内炎症反应的激活和维持,并且对代谢功能障碍疾病的发生、发展和进展过程产生重要的推动和影响作用。
肠道菌群受损时会驱动非酒精性脂肪性肝病
非酒精性脂肪性肝病(NAFLD)是一种特定的肝脏疾病状态,主要是指在很少饮酒或完全不饮酒的个体中,肝脏组织内出现多余脂肪物质的异常积聚和沉积现象,这种疾病状态与代谢综合征、2型糖尿病(T2D)以及肥胖症等代谢性疾病之间存在着非常密切和重要的关联性。
当肠道微生物群落结构和功能受到损害和破坏时,这种受损的肠道微生物生态系统似乎会成为驱动和促进NAFLD相关炎症反应发生和发展的重要因素和机制。与此相对应的是,各种有益的、针对肠道环境的靶向治疗方法和干预措施,例如通过补充益生菌等方式来调节和改善肠道微生物群落,能够有效地缓解和减轻肝脏组织内的炎症反应以及由此导致的肝脏功能障碍和损害。
肠道炎症作为多种慢性疾病的核心病理机制,其有效治疗需要采用多元化、个体化的综合干预策略。当前的治疗方法主要围绕恢复肠道微生物平衡、修复肠道屏障功能、调节免疫反应以及减少促炎因子等核心目标展开。
这些治疗手段包括但不限于营养干预、益生菌和益生元补充、抗炎药物应用、生活方式调整以及新兴的微生物移植疗法等。通过系统性地结合这些不同的治疗模式,能够从多个层面同时作用于肠道炎症的发病机制,从而实现更加全面和持久的治疗效果,最终改善患者的整体健康状况和生活质量。
1
药物治疗
目前用于治疗慢性肠道炎症的大多数药物要么针对在炎症反应早期阶段(即静息期和血管期)释放的炎症介质,要么针对炎症细胞的招募。
①氨基水杨酸类药物
氨基水杨酸类药物(包括柳氮磺吡啶和5-氨基水杨酸)通过作用于花生四烯酸代谢,抑制前列腺素和白三烯等脂质炎症介质的释放。然而,该类药物还具有其他作用机制,包括清除活性氧、抑制白细胞产生细胞因子、通过芳香烃受体依赖途径诱导调节性T细胞招募,以及与过氧化物酶体增殖物激活受体γ结合。
5-氨基水杨酸用于治疗溃疡性结肠炎,有助于维持病情缓解,但在克罗恩病患者中的应用仍存争议。一项研究显示,5-氨基水杨酸能有效减少乳糜泻患者组织器官型培养中的氧化爆发和炎症,但尚无减少组织炎症的报道。在肠易激综合征患者的研究中,5-氨基水杨酸治疗未能改善症状。
②皮质类固醇
20世纪50年代,皮质类固醇成为治疗IBD的首个药理方法,至今仍能使许多患者获得有效缓解,但其维持缓解的效果尚未得到证实。因此,皮质类固醇用于终止炎症发作和诱导缓解,但不推荐长期使用。
皮质类固醇对所有炎症相关途径的多效性作用解释了其疗效。然而,尽管努力开发更安全、耐受性更好的皮质类固醇,但其使用可能产生显著副作用,包括机会性感染、糖尿病、高血压、静脉血栓栓塞和骨质疏松症。此外,部分患者对皮质类固醇治疗存在抵抗性,必须考虑其他免疫调节方法。
③免疫调节剂
用于治疗肠道炎症的免疫调节剂包括硫唑嘌呤、甲氨蝶呤、钙调磷酸酶抑制剂和Janus激酶抑制剂,所有这些药物都旨在降低T淋巴细胞的活化或增殖,并且在减少炎症发作方面非常有效。然而,它们也引起许多不良反应,包括肝损伤、骨髓抑制和传染病。
④生物制剂
许多治疗肠道炎症(特别是IBD)的生物制剂针对细胞因子或整合素。由于TNF-α、IL-12和IL-23在IBD发病机制中的重要作用,抗TNF-α和抗IL-12/23抗体是治疗IBD的主要生物制剂。
抗TNF-α抗体可抑制炎症反应和组织损伤,用于治疗对皮质类固醇无反应的中重度UC或CD患者。通过剂量递增可实现抗TNF-α抗体的长期缓解。然而,高达40%的患者对TNF-α抑制剂无反应,且在开始治疗后仅1年就有20%-45%的患者出现继发性疗效丧失。因此,抗IL-12和抗IL-23在生物制剂初治和既往治疗患者中均有效,常作为二线治疗使用。
整合素通过与组织特异性细胞黏附分子结合介导白细胞归巢,在炎症细胞向炎症组织招募和滞留中起关键作用。抗整合素治疗通过阻断整合素与白细胞内皮细胞黏附分子或上皮细胞表达的E-钙黏蛋白结合发挥作用。抗整合素生物制剂针对黏附分子的α4或β7整合素亚单位。
所有生物制剂对应答者的缓解诱导效果均很好。然而,尽管具有选择性高、特异性强等优点,仍有相当一部分IBD患者出现原发性无应答、继发性应答丧失和治疗不耐受,因此需要其他治疗选择。
2
微生物群靶向治疗
微生物群失调既是炎症相关肠道病变的常见特征,也是导致长期炎症的早期事件。因此,针对微生物群的治疗是治疗肠道炎症的有前景选择。
微生物干预能够有效改善肠道炎症
基于微生物组的疗法包括饮食干预、益生元补充、单一或多菌株益生菌制剂、噬菌体疗法、粪便微生物群移植(FMT)和微生物组模拟物。多项证据表明针对微生物组的疗法对炎症性肠病、肠易激综合征以及一定程度上的乳糜泻有益。微生物组治疗的作用模式可能是多模态的,涉及直接影响生物膜组成和代谢、控制屏障功能以及影响黏膜免疫。
①微生物群相关修复
多项研究显示益生菌、益生元和后益生元在IBD和IBS患者中的潜在作用。它们安全且耐受性良好,尽管效果有时相当有限。在CD患者中,大多数研究表明益生菌无显著效果,而在UC患者或IBS患者中,对临床症状和缓解有益。
粪菌移植治疗IBD的潜力正被积极研究。在UC患者中,FMT在实现临床缓解方面优于安慰剂,一项关于FMT在IBD患者中疗效的荟萃分析显示,无论CD还是UC,临床缓解率均显著。
②干细胞移植
干细胞作为所有细胞类型的起源,包括上皮细胞,因此用于修复受损肠道组织并恢复功能具有理论依据。在重度难治性克罗恩病患者中,自体造血干细胞移植或同种异体骨髓间充质干细胞治疗均显示出积极效果,约半数患者实现临床缓解。
尽管大多数接受自体造血干细胞移植的患者在术后5年内复发,但80%复发者在再次移植后可重新达到缓解。因此,干细胞移植对CD患者的黏膜修复具有潜力,但其临床应用尚未获批。此外,干细胞移植还可利用自体肠道干细胞,经体外培养和检测后移植,该方法已在动物模型中成功应用,但在人类中的临床条件尚未确定。
③加强屏障功能
屏障功能障碍与肠道炎症密切相关,这种障碍在炎症性肠病、乳糜泻和肠易激综合征患者中均有发现。研究表明,即使炎症消退,屏障功能在之后数周内仍可能受损。清除急性炎症后,屏障功能的恢复是组织修复的关键,涉及黏膜微生物生物膜、黏液层、上皮单层、黏膜免疫系统和神经支配。因此,屏障功能可通过多种方式恢复。一些IBD治疗手段部分通过改善上皮屏障起效,如抗TNF-α治疗对克罗恩病有效。
肠道粘膜表面的解决和修复参与者及介质
Deraison C,et al.Annu Rev Pharmacol Toxicol.2025
临床上,恢复肠道屏障功能主要依赖饮食干预,包括免疫调节营养物、益生菌和微生物代谢产物。局部应用丁酸可改善难治性溃疡性结肠炎患者的屏障功能。新的理论认为,具有细胞功能失常遗传背景者更易发生自身免疫反应,此类细胞对炎症触发因素更为敏感。肠腔内容物常为炎症的主要触发因素,难以完全避免。因此,针对遗传性功能障碍的干预为新型治疗途径。
3
生活方式改善
①吃营养丰富的未经加工的饮食
营养丰富、未经加工的饮食提供了底物(即,可发酵纤维,多酚和其他营养素),你的肠道细菌需要茁壮成长,创造一个抗炎的肠道生态系统。特别关注益生元食物可以提高你的抗炎SCFA产生细菌的水平。避免非细胞碳水化合物,包括精制面粉产品和工业种子油。
对于患有严重肠道炎症的人,如IBD,自身免疫方案饮食可以提供显著的缓解。
②管理压力
慢性压力确实会损害肠道健康,因此压力管理对于减轻肠道炎症至关重要。冥想有助于调节压力反应,维持肠道屏障功能和炎症平衡。规律锻炼和良好睡眠同样有助于管理压力并支持肠道健康。
③控制合理使用药物
为减少肠道炎症,应谨慎使用抗生素和其他药物。目前对非抗生素药物对肠道健康影响的了解有限,但已有初步担忧。相比之下,抗生素对肠道的负面影响证据充分。如果您正在使用PPI,请谨慎使用,并了解如何通过饮食和生活方式调整来减少或停用抑酸药物。
④尽量限制接触毒素
环境毒素无处不在,但通过以下简单措施可以降低暴露并预防肠道炎症:
-购物时避免使用收据,减少接触。
-选择不锈钢或玻璃水瓶和食物容器,避免塑料制品。
–避免在家中和院子里使用杀虫剂。
这些措施有助于减少环境毒素的累积暴露,促进肠道和整体健康。
肠道掌控着全身的健康。从消化吸收到免疫防线,从情绪调节到慢性病预防,你的身体几乎所有系统,都绕不开它的“号令”。一旦肠道炎症悄然来袭,打破屏障和菌群的平衡,导致各种慢性疾病,比如炎症性肠病、结肠癌、糖尿病、哮喘,甚至影响大脑、皮肤和骨骼健康。
不健康饮食(精制碳水、工业油、食品添加剂),还有滥用抗生素、环境毒素、过度压力、久坐、睡眠紊乱、剖腹产及配方喂养、反复肠道感染、甚至是某些常见药物等都会影响肠道炎症。
总之,肠道健康是你和慢病拉开距离的“第一道防线”。面对日常的饮食、作息和环境时,不妨从肠道微生物组学的角度审视:这些是否有利于维持肠道菌群的多样性与稳态?
从丰富多样、天然未加工的饮食入手,善用益生菌和益生元,科学运动,重视深度睡眠,减少压力,限用药物,远离环境毒素,肠道就会回馈你健康的身体和愉悦的心情。而现代医学的营养干预、生物制剂、微生态移植和干细胞等创新治疗,也在不断拓宽肠道炎症的管理边界。
肠道炎症,正走向现代医学的中心,未来,肠道炎症指标将如同血压、血糖一样成为常规监测项目,在疾病症状出现前数年就能识别风险并及时干预。谷禾肠道菌群检测为这一转变提供了强有力的技术支撑,通过分析肠道微生物组成和功能,基于大样本量数据库及最新的算法模型,评估肠道炎症水平。健康管理也正从症状出现后治疗向风险早期识别的根本转变。通过持续的肠道健康监测和精准干预,我们能够预防很多慢性疾病。
面对肠道炎症的复杂病理机制,未来的医疗模式将迎来跨学科融合,整合营养学、微生物学和免疫学等多学科知识,基于个体化检测结果制定多维度干预策略,为患者提供系统化解决方案。
主要参考文献
Pelaseyed T, Bergström JH, Gustafsson JK, Ermund A, Birchenough GM, Schütte A, van der Post S, Svensson F, Rodríguez-Piñeiro AM, Nyström EE, Wising C, Johansson ME, Hansson GC. The mucus and mucins of the goblet cells and enterocytes provide the first defense line of the gastrointestinal tract and interact with the immune system. Immunol Rev. 2014 Jul;260(1):8-20.
Ohno H. Intestinal M cells. J Biochem. 2016 Feb;159(2):151-60.
Gassler N. Paneth cells in intestinal physiology and pathophysiology. World J Gastrointest Pathophysiol. 2017 Nov 15;8(4):150-160.
Bäckhed F, Ley RE, Sonnenburg JL, Peterson DA, Gordon JI. Host-bacterial mutualism in the human intestine. Science. 2005 Mar 25;307(5717):1915-20.
Gill SR, Pop M, Deboy RT, Eckburg PB, Turnbaugh PJ, Samuel BS, Gordon JI, Relman DA, Fraser-Liggett CM, Nelson KE. Metagenomic analysis of the human distal gut microbiome. Science. 2006 Jun 2;312(5778):1355-9.
Qin J, Li R, Raes J, Arumugam M, Burgdorf KS, Manichanh C, Nielsen T, Pons N, Levenez F, Yamada T, Mende DR, Li J, Xu J, Li S, Li D, Cao J, Wang B, Liang H, Zheng H, Xie Y, Tap J, Lepage P, Bertalan M, Batto JM, Hansen T, Le Paslier D, Linneberg A, Nielsen HB, Pelletier E, Renault P, Sicheritz-Ponten T, Turner K, Zhu H, Yu C, Li S, Jian M, Zhou Y, Li Y, Zhang X, Li S, Qin N, Yang H, Wang J, Brunak S, Doré J, Guarner F, Kristiansen K, Pedersen O, Parkhill J, Weissenbach J; MetaHIT Consortium; Bork P, Ehrlich SD, Wang J. A human gut microbial gene catalogue established by metagenomic sequencing. Nature. 2010 Mar 4;464(7285):59-65.
Laurikka P, Nurminen S, Kivelä L, Kurppa K. Extraintestinal Manifestations of Celiac Disease: Early Detection for Better Long-Term Outcomes. Nutrients. 2018 Aug 3;10(8):1015.
Sturgeon C, Fasano A. Zonulin, a regulator of epithelial and endothelial barrier functions, and its involvement in chronic inflammatory diseases. Tissue Barriers. 2016 Oct 21;4(4):e1251384.
Frossi B, De Carli M, Calabrò A. Coeliac Disease and Mast Cells. Int J Mol Sci. 2019 Jul 11;20(14):3400.
Losurdo G, Piscitelli D, Pezzuto F, Fortarezza F, Covelli C, Marra A, Iannone A, Amoruso A, Principi M, Ierardi E, Di Leo A. T Helper Lymphocyte and Mast Cell Immunohistochemical Pattern in Nonceliac Gluten Sensitivity. Gastroenterol Res Pract. 2017;2017:5023680.
Abulizi N, Quin C, Brown K, Chan YK, Gill SK, Gibson DL. Gut Mucosal Proteins and Bacteriome Are Shaped by the Saturation Index of Dietary Lipids. Nutrients. 2019 Feb 16;11(2):418.
Spreadbury I. Comparison with ancestral diets suggests dense acellular carbohydrates promote an inflammatory microbiota, and may be the primary dietary cause of leptin resistance and obesity. Diabetes Metab Syndr Obes. 2012;5:175-89.
Martínez Steele E, Baraldi LG, Louzada ML, Moubarac JC, Mozaffarian D, Monteiro CA. Ultra-processed foods and added sugars in the US diet: evidence from a nationally representative cross-sectional study. BMJ Open. 2016 Mar 9;6(3):e009892.
Nickerson KP, McDonald C. Crohn’s disease-associated adherent-invasive Escherichia coli adhesion is enhanced by exposure to the ubiquitous dietary polysaccharide maltodextrin. PLoS One. 2012;7(12):e52132.
Rodriguez-Palacios A, Harding A, Menghini P, Himmelman C, Retuerto M, Nickerson KP, Lam M, Croniger CM, McLean MH, Durum SK, Pizarro TT, Ghannoum MA, Ilic S, McDonald C, Cominelli F. The Artificial Sweetener Splenda Promotes Gut Proteobacteria, Dysbiosis, and Myeloperoxidase Reactivity in Crohn’s Disease-Like Ileitis. Inflamm Bowel Dis. 2018 Apr 23;24(5):1005-1020.
Braniste V, Jouault A, Gaultier E, Polizzi A, Buisson-Brenac C, Leveque M, Martin PG, Theodorou V, Fioramonti J, Houdeau E. Impact of oral bisphenol A at reference doses on intestinal barrier function and sex differences after perinatal exposure in rats. Proc Natl Acad Sci U S A. 2010
JPascal M, Perez-Gordo M, Caballero T, Escribese MM, Lopez Longo MN, Luengo O, Manso L, Matheu V, Seoane E, Zamorano M, Labrador M, Mayorga C. Microbiome and Allergic Diseases. Front Immunol. 2018 Jul 17;9:1584. an 5;107(1):448-53.
Mao, Q., Manservisi, F., Panzacchi, S. et al. The Ramazzini Institute 13-week pilot study on glyphosate and Roundup administered at human-equivalent dose to Sprague Dawley rats: effects on the microbiome. Environ Health 17, 50 (2018).
Han H, Li Y, Fang J, Liu G, Yin J, Li T, Yin Y. Gut Microbiota and Type 1 Diabetes. Int J Mol Sci. 2018 Mar 27;19(4):995.
Chen, J., Chia, N., Kalari, K. et al. Multiple sclerosis patients have a distinct gut microbiota compared to healthy controls. Sci Rep 6, 28484 (2016).
谷禾健康
近年来,肠道屏障通透性异常增加(intestinal hyperpermeability)的临床诊断呈上升趋势,其病理生理机制涉及多个层面的复杂相互作用。
肠道屏障就像一堵城墙,一旦“城门大开”,各种“异类分子”如病原信号(PAMPs)、脂多糖(LPS)、未消化的蛋白碎片就会趁机溜进固有层和血液,掀起一场免疫风暴。
肠道固有层宛如“安检总站”,这里的树突状细胞和巨噬细胞负责利用“雷达”——比如Toll样受体——扫描入侵分子,然后拉响警报,释放大量炎症信号分子(TNF-α、IL-1β、IL-6)和趋化因子,引发局部炎症反应。
被激活的免疫细胞及其炎症介质通过门静脉循环和淋巴系统进入全身循环,触发远端器官的炎症反应,这种”肠-器官轴”的病理连接可能是类风湿关节炎、强直性脊柱炎、系统性红斑狼疮等自身免疫性疾病以及过敏性疾病、神经退行性疾病的潜在发病机制。
同时,肠道屏障功能受损导致肠道微环境稳态失调,有益共生菌群数量下降而潜在致病菌过度增殖,形成肠道菌群失调,进一步破坏肠道屏障功能并释放更多内毒素,加剧炎症反应,形成病理性正反馈循环。
造成肠道屏障告急的“罪魁祸首”很多:慢性压力、滥用抗生素、非甾体消炎药、高脂快餐、食品添加剂、各种感染和遗传易感性都可能“推波助澜”。理解这套复杂机制,有助于我们探索科学高效的“修复之路”。
本文介绍了目前肠道屏障受损的成因、健康影响和尝试干预改善过度渗透的肠屏障的一些方法。研究表明,预防肠道屏障受损以及在屏障受损时开发治疗方法至关重要。
肠上皮细胞是外界环境和体内环境之间最大的接触部位。胃肠道上皮屏障的功能是防止外来抗原和微生物进入,同时允许吸收必需的营养物质、水和电解质。
肠道屏障由物理屏障、生化屏障、免疫屏障和微生物屏障组成。
肠道屏障
Macura B,et al.Clin Exp Med.2024
▸ 物理屏障
物理(上皮)屏障是整个肠道屏障的重要组成部分。它由单层特化细胞组成:肠上皮细胞、杯状细胞(产生粘蛋白)、潘氏细胞(产生抗菌肽和蛋白质)、肠内分泌细胞、M细胞和肠干细胞。
这些细胞每3-5天更新一次。上皮细胞具有多种功能,并且紧密相连。
▸ 生化屏障
生化屏障由粘液构成,覆盖并保护上皮细胞免受病原体和有害物质侵害。
粘液是一种富含水分(90%–95%)、电解质、脂质(1%–2%)和蛋白质的稀薄分泌物,其中粘蛋白(1%–5%)为其主要结构和功能成分。
doi: 10.1007/164_2016_107.
上图展示了小肠和大肠粘液的生成与分布,包括内外粘液层的结构,并标示了杯状细胞从粘液产生到分泌和扩散的全过程(步骤1至7)。
粘液分泌是一个复杂的过程:杯状细胞在从隐窝底部迁移时用Muc2填充其分泌囊泡,并含有其他成分,例如IgG结合蛋白(FCGBP)的Fc片段、氯通道附件1(CLCA1)、酶原颗粒蛋白16(ZG16)和前梯度同源物2(AGR2)。这些囊泡与顶膜融合后,通过胞吐作用释放内容物,实现粘液分泌。
最后,粘蛋白可以分为两种不同的类型:跨膜粘蛋白和形成凝胶的粘蛋白。
▸ 免疫屏障
免疫屏障与肠道中淋巴组织的存在有关,称为肠道相关淋巴组织(GALT)。GALT系统分布于肠道粘膜及粘膜下层紧邻上皮细胞处,包括上皮内淋巴细胞、派伊尔集合淋巴结和淋巴细胞簇。
GALT系统含有抗原呈递细胞、T细胞、B细胞、浆细胞、巨噬细胞、肥大细胞和粒细胞,并且分泌型IgA(sIgA)主要在肠道中合成。
▸ 肠道屏障的功能-紧密连接调节通透性
大多数膳食蛋白质通过肠上皮细胞进行内吞作用。溶酶体降解导致蛋白质分解成更小的肽,从而避免激活免疫系统。液体和溶质在细胞之间运输。这种转运由紧密连接(TJ)调节。
紧密连接构建蛋白通过特殊的封闭小带蛋白(ZO)与肌动蛋白结合,肌动蛋白是细胞骨架的一部分。 这些蛋白质是调节紧密连接功能的分子的靶元件,因此负责调节肠屏障的通透性。
肠道屏障在外源性或内源性因素的作用下会受到损害,从而导致其正常功能被破坏。
▸ 饮食因素
不良饮食模式
高脂肪、高糖、高盐饮食
过度加工食品和快餐
缺乏膳食纤维的精制食品
过量饮酒
特定食物成分
人工添加剂、防腐剂
过量的乳化剂
某些人工甜味剂
过敏原食物(因人而异)
▸ 药物因素
抗生素:破坏肠道菌群平衡
非甾体抗炎药(NSAIDs):直接损伤肠粘膜
质子泵抑制剂:长期使用影响胃酸分泌
化疗药物:对肠道上皮细胞有毒性作用
▸ 生活方式因素
压力相关
慢性心理压力
睡眠不足或质量差
过度疲劳
运动相关
久坐不动的生活方式
过度剧烈运动(可能增加肠通透性)
▸ 环境因素
空气污染和化学毒素暴露
重金属污染
农药残留
吸烟
▸ 疾病和感染
肠道感染(细菌、病毒、寄生虫)
炎症性肠病
自身免疫疾病
糖尿病等代谢性疾病
▸ 年龄因素
衰老过程中肠道屏障功能自然下降
婴幼儿肠道屏障发育不完善
▸ 空气、水、食物污染等破坏了肠上皮屏障
近几十年来,城市化与全球化带来了巨大的环境变化,污染和气候变化,工业和家庭中常用的化学化合物,生态系统变化,不健康的饮食以及兴奋剂,主要是酒精,烟草和电子烟,可能会破坏皮肤和粘膜表面的上皮屏障。
空气、水、食物污染、微塑料、纳米颗粒、家用化学品和烟草烟雾是常见的上皮屏障破坏因素。这些因素复杂交互,但对生物体的影响取决于其组合及个体易感性。
▸ 微塑料和纳米颗粒会减少肠道粘液产生
近年来,食品和饮用水中的化学污染显著增加,即使是常用的食品添加剂、杀虫剂和饲料添加剂也在其中。最近,关于微塑料和纳米颗粒污染食品的报道越来越多。这些物质可以很容易地穿透组织并与细胞结构相互作用。
微塑料会改变蛋白质结构,与细胞膜脂质相互作用,诱导炎症基因转录,增加促炎细胞因子,导致内质网和线粒体功能障碍,甚至引发细胞因氧化应激而死亡。
纳米颗粒也能引起类似的蛋白结构变化和细胞膜相互作用。纳米塑料已被证明可以诱导炎症基因的转录,增加促炎细胞因子的水平,并改变某些蛋白质的表达。微塑料在胃肠道内吸收率高,可在体内外环境中积累。动物实验证实,聚苯乙烯微塑料会损伤肠道屏障并减少粘液分泌。
下表总结了被认为对肠道屏障状态具有有害影响的主要因素:微塑料,纳米颗粒,洗涤剂和乳化剂,肠道微生物群状态和饮食。
doi: 10.1007/s10238-024-01496-9
暴露于环境因素可以直接削弱肠道上皮屏障的完整性并改变微生物组结构。上皮屏障渗漏和肠道菌群失调常同时出现,促进炎症发生,部分原因是由于免疫细胞紧邻上皮层。
上皮屏障受损时,免疫系统被激活,引发局部炎症,炎症可通过免疫细胞及其分泌的细胞因子扩散至其他系统和器官。因此,多种疾病的发展与上皮屏障渗漏相关。肠上皮受损后,机会性病原体易定植,菌群多样性下降。微生物可移位至更深组织,导致微炎症。此外,化合物暴露还能诱导免疫系统表观遗传变化,促进某些疾病的发生。
▸ 生态失调→上皮损伤→慢性炎症
大量证据表明,微生物组对肠道上皮健康至关重要。正常微生物群调节紧密连接蛋白表达、血管生成、通透性和免疫功能。上皮屏障渗漏时,细菌和病原体穿透上皮,引发炎症并参与多种疾病。过敏性疾病的上升可能与菌群失调和多样性下降有关。
微生物群受到饮食、药物(特别是抗生素、精神药物、质子泵抑制剂)及遗传等影响。与人体共生的非致病性菌群有助于免疫调节,有利于预防慢性疾病并维持免疫平衡。
健康的微生物群通过调控屏障通透性、紧密连接和局部免疫,维持上皮屏障功能。缺乏免疫调节因子,如短链脂肪酸,可导致免疫系统过度激活、上皮炎症和疾病。这种反应主要由TH2细胞、ILC2和嗜酸性粒细胞介导,肥大细胞和巨噬细胞等也参与其中。而菌群失调、上皮损伤和慢性炎症会形成恶性循环,阻碍上皮再生。
同时损伤上皮中的炎症持续激活免疫细胞,可导致炎症扩散至其他器官。
微生物群变化、肠道屏障受损与疾病相互依存,但其因果关系仍有争议。尚不清楚是菌群失调导致屏障功能障碍和疾病,还是疾病先引发炎症和屏障损伤,进而加重细菌移位和器官损害。
因此,肠漏究竟是疾病的起因还是结果?此外,有研究表明,呼吸道或肠上皮屏障破坏以及其他屏障(如血脑屏障、血管内皮屏障)减弱,可能会促进代谢及自身免疫性疾病的发生。
▸ 胃食管反流病患者食管壁存在渗漏
胃食管反流病(GERD)最常见的症状是烧心和反酸,影响全球超过10%的成年人。胃镜检查显示,只有约30%的患者存在明显糜烂,其余70%(非糜烂性反流病,NERD)虽无可见损伤,但仍有症状。
GERD就像胃酸不断冲刷食管墙壁,表面虽完好但可能已“渗漏”,问题可能在于连接细胞的“水泥”。研究发现,GERD患者食管中即使无表面破损,其“水泥”成分也发生了变化:部分蛋白(如E-钙粘蛋白)减少,而其他蛋白(如CLDN-1和桥粒蛋白)则增加。
▸ 功能性消化不良存在十二指肠黏膜屏障渗漏
功能性消化不良(FD)是一种常见的胃肠道疾病,表现为早饱、腹胀、上腹痛或灼热,且常规检查无法发现明确病因。
最新研究发现,FD的核心问题在于十二指肠黏膜屏障渗漏,不仅离子,大分子也能穿透。该屏障损伤与肥大细胞和嗜酸性粒细胞增多,以及ZO-1和钙粘蛋白等紧密连接蛋白减少有关,说明低度炎症和屏障受损是FD的主要致病机制。
▸ 肠易激综合征与“肠漏”有关
肠易激综合征(IBS)是一种常见的功能性胃肠疾病,典型表现为反复腹痛和排便习惯改变(如腹泻、便秘或交替)。
尽管具体病因不明,但大量证据表明,其核心机制与“肠漏”有关。研究发现,几乎所有类型的IBS患者都存在肠道屏障通透性增加的问题。这具体表现为:肠道细胞间的“水泥”(即紧密连接蛋白ZO-1等)被破坏或减少,同时存在低度的黏膜炎症(如肥大细胞活化)。
这种屏障功能的损害,使得肠道内容物更容易“渗漏”过去,激活免疫系统,最终导致了IBS特有的腹痛和排便紊乱等一系列症状。
▸ 高血压患者肠道屏障功能障碍患病率较高
近年来,越来越多的研究关注肠道微生物群与高血压之间的关系。大量证据显示,肠道菌群的生态失调不仅可能促进高血压的发生和发展,还可能加重病情。
与此同时,高血压本身也会对肠道微生物群的组成产生影响,进一步加剧生态失调。研究表明,维持适当和健康的微生物群状态有助于血压的正常化,改善心血管系统的健康,而理想的血压水平同样有助于微生物群落的维持与恢复。
已有结果显示,高血压患者较非高血压患者更易出现肠屏障功能障碍,肠屏障的损伤被认为可能与高血压的发病机制密切相关。这一发现为进一步研究血压调控与肠道健康之间的复杂联系提供了新的思路,也提示改善肠道屏障功能或许有助于高血压的防治。
▸ 炎症性肠病患者即便无症状也会出现肠道通透性增加
炎症性肠病(IBD)患者的肠道微生物群特征主要表现为促炎细菌数量的明显增加,同时多种细胞因子的表达也发生了显著变化。这些变化会导致机体免疫反应失衡,加剧炎症进程。
值得注意的是,在患者尚未出现明显的临床症状之前,尽管处于无症状期,肠道的通透性实际上已经发生了异常改变。这表明肠道屏障功能的损伤可能早于症状表现,为疾病的早期诊断和干预提供了理论依据。
▸ 肠道屏障和菌群稳定影响肝脏健康
研究数据显示,肠上皮细胞和血管屏障的损伤在非酒精性脂肪肝的发展过程中起着关键和必要的作用。肠上皮屏障功能的破坏,使得肠道内容物更容易进入血液循环,进而促进肝脏炎症和脂肪堆积。
此外,肠道菌群的失调以及肠道菌群结构的破坏与这一过程密切相关。肠道菌群异常会导致有害菌数量增加,正常菌群比例下降,其产生的代谢产物也会对肠道细胞产生毒性作用,影响细胞功能和屏障完整性。
这些机制不仅涉及非酒精性脂肪肝的发生与发展,在酒精性肝病中同样存在。酒精摄入可加重肠上皮与血管屏障的损伤,进一步削弱肠道屏障功能,从而成为酒精性肝病患者肠上皮功能障碍的主要原因之一。由此可见,肠道屏障及菌群的稳定性对于肝脏健康至关重要。
▸ 免疫性疾病患者存在菌群失调和屏障渗漏
类风湿性关节炎患者表现为血清zonulin水平升高、肠道菌群失调和屏障渗漏,强直性脊柱炎患者可见亚临床肠道炎症。潘氏细胞对肠道菌群失调的激活可能是疾病早期症状的原因,并证实了脊椎-关节轴的存在。
▸ 帕金森、自闭症患者存在胃肠道症状
在帕金森病中,患者在诊断前数年就表现出胃肠道症状。α-突触核蛋白是帕金森病的典型蛋白质,在肠道中合成,然后通过迷走神经运送到中枢神经系统。相比之下,自闭症谱系障碍儿童会出现肠道症状,如便秘、腹痛和腹泻。对肠道微生物群的研究表明,微生物群多样性与神经系统疾病、拟杆菌减少以及肠粘膜中紧密连接蛋白表达减少之间存在负相关关系。
▸ 糖尿病患者的肠道通透性发生变化
研究显示,肠道通透性变化早于1型糖尿病的临床症状出现,并且与紧密连接蛋白和生态失调的改变,尤其是丁酸盐产生菌减少密切相关。2型糖尿病患者的肠道菌群失调加重了屏障损伤,并促进了炎症状态,导致胰岛素抵抗和β细胞功能障碍。
此外,2型糖尿病患者的高血糖有助于持续的促炎状态。肥胖则与慢性低度炎症密切相关,主要表现为脂肪组织中促炎巨噬细胞的活性增加。代谢紊乱在肥胖人群中也更为常见。
doi: 10.1007/s10238-024-01496-9
在间接或直接诊断试验中,可以评价肠屏障通透性水平。
▸ 口服物质测试
间接检测方法包括口服试剂后测定其在血液或尿液中的浓度,最常用的是糖。乳果糖/甘露醇(L/M)试验是评估小肠通透性的常用方法,通过测量尿中两者的排泄量进行判断。乳果糖为大分子,仅在细胞间连接受损时吸收,甘露醇为小分子能自由通过肠屏障。L/M试验非侵入性且灵敏度高,此外还可使用其他单一或多种糖类的检测方法。
▸ 测量内源物质水平
此外,还可以测量血液中内源性物质的水平。这样的生物标志物包括例如连蛋白、脂肪酸结合蛋白(FABP)、瓜氨酸、胰高血糖素样肽(GLP)-2、脂多糖、脂多糖结合蛋白(LBP)或粪便α1抗胰蛋白酶(AAT)。然而,这些测试不像糖测试那样敏感。
▸ 新型影像技术
新的成像技术,特别是共聚焦激光显微内镜,允许在体内评价静脉注射荧光素作为造影剂后肠屏障的完整性。共聚焦激光显微内镜目前用于诊断胃肠道癌症,以及肠易激综合征或乳糜泻。放大1000倍可以检测肠上皮内的病理变化。
▸ 肠道菌群检测
肠道菌群检测可以提供非常重要的间接线索和风险评估。肠道菌群与肠道屏障健康密切相关。菌群测序可以发现一些可能导致或指示肠道屏障通透性增加的风险因素,例如:嗜黏蛋白菌,AKK菌以及扭链瘤胃球菌等菌对的丰度,以及产丁酸菌减少,特定有害菌增多:某些细菌(如部分革兰氏阴性菌)会产生脂多糖(LPS),这是一种强烈的炎症触发物,会破坏肠道屏障。菌群多样性降低:通常一个多样性低、不稳定的菌群生态系统,其维护屏障功能的能力也会下降。
总之,肠道菌群检测可以揭示与肠道屏障功能受损相关的菌群失衡状态,提供有力的间接证据和风险预警。
功能性胃肠道疾病(FGID)在临床上发病率高、症状慢性且治疗效果有限,已成为重大公共卫生问题。多种病理机制被提出,其中屏障通透性受损是常见特征。局部免疫激活与屏障损伤的先后关系及其在症状发生中的作用仍需进一步研究。
目前的研究表明,治疗这些疾病的一个很有前景的策略,就是直接针对这道受损的屏障进行修复。
这主要包括两大类治疗方法:
一是直接使用药物来加固和修复肠道细胞间的连接,恢复屏障的完整性;
二是通过靶向调节局部的免疫反应,减轻炎症,从而间接保护和修复肠道屏障。
对屏障功能的直接影响
①谷氨酰胺可修复部分人群的肠道通透性
谷氨酰胺是一种非必需氨基酸,是肠细胞和结肠细胞的重要能量来源。谷氨酰胺在接受腔内禁食的Caco-2细胞中保持蛋白质合成和对离子和4 kDa葡聚糖分子的细胞旁渗透性。它还可降低大分子渗透性(40 kDa),其剥夺可促进TNF-α诱导的细菌易位,而不影响TEER。这些和其他体外研究表明,谷氨酰胺补充通过干扰泄漏途径恢复了对离子和小分子增加的细胞旁渗透性。
然而,在人身上的研究结果却不那么一致,有时甚至相互矛盾。这可能是因为补充谷氨酰胺的时机很关键——最好是在肠道屏障还没受损或者刚刚受损时就介入。
不过,最近一项针对腹泻型肠易激综合征(IBS-D)患者的研究带来了好消息:口服谷氨酰胺显著改善了患者的症状,并成功修复了他们受损的肠道通透性。这表明,对于特定人群,谷氨酰胺可能是一种修复“肠漏”的有效治疗选择。
②MLCK抑制剂有助于减轻肠漏
如前所述,通过肠上皮细胞的泄漏途径和细菌移位由肌球蛋白轻链激酶(MLCK)控制。因此,MLCK抑制剂可能是治疗或预防屏障功能改变的潜在药物。
MLCK的物质是控制肠道屏障“阀门”的关键开关。当它被激活时,肠道细胞间的连接就会松开,导致“肠漏”和细菌穿透。因此,开发能抑制MLCK的药物(即MLCK抑制剂)就成了一种非常有前景的治疗思路。
目前,这类药物在动物实验中已有效预防压力或毒素引起的肠道通透性增加及内脏高敏。如果能在人类中安全应用,MLCK抑制剂有望成为治疗“肠漏”相关胃肠疾病的新选择。
③Zonulin肽抑制剂可能修复通透性并预防肠炎
研究发现,Zonulin是一种调节肠道通透性的关键物质。当肠道受麸质或特定细菌等刺激时,Zonulin释放,可打开细胞间紧密连接,增加肠道通透性,导致“肠漏”。
基于此,科学家开发了Zonulin肽抑制剂(如醋酸拉唑肽),能有效阻断Zonulin作用,“锁上”肠道屏障。动物实验显示该药物可修复通透性并预防肠炎,但其在人类中的疗效尚需验证。
临床试验中,虽然乳糜泻患者用药后自觉症状改善,但肠道通透性并无明显下降。因此,Zonulin抑制剂是否能真正用于胃肠道疾病治疗,还需进一步研究确证。
对免疫激活的影响
研究发现,FGID中的炎症主要与肥大细胞、嗜酸性粒细胞和T细胞的轻度活化有关。因此,目前的治疗尝试大多集中在稳定“肥大细胞”上,因为它在肠易激综合征(IBS)等疾病中扮演了核心角色。
①药物治疗
一些药物已经显示出潜力:
-5-氨基水杨酸:可以减少炎症细胞的浸润。
-酮替芬和色甘酸二钠:作为肥大细胞稳定剂,它们在改善IBS和功能性消化不良(FD)的症状方面取得了良好效果,但其具体如何修复肠道屏障的机制还有待阐明。
此外,一些靶向神经调节和抑制肥大细胞的新型药物(如CRF拮抗剂、大麻素类物质)在动物实验中也表现不错,但它们在人体中的实际效果还需要进一步的临床研究来验证。
②缓解肠道炎症的药物
生物治疗主要包括促炎细胞因子抑制剂和整合素拮抗剂:抗TNF治疗、抗IL-12/23治疗、抗整合素治疗。生物疗法,尤其是抗TNF疗法是有效的治疗方法,但对TNF抑制剂的原发性无效或部分患者的继发性无效需要寻找新的治疗方案。
重要的是,炎症环境可能会对上皮干细胞的功能产生负面影响,因此可能会阻碍健康上皮的重建。 肠漏的治疗应该开始尝试确定肠屏障损伤的原因,然后消除这个因素。在肠道屏障恢复之前,使用缓解肠道炎症的药物似乎是合理的。
③恢复肠道微生物群
益生菌是活的微生物,对宿主的健康有有益的影响。益生菌管理减少肠渗漏,其中,影响肠道免疫调节和抗炎作用(例如,sIgA产生的增加)、抗炎作用、加强上皮屏障(例如,粘蛋白和短链脂肪酸(SCFA)的合成增加,以及限制病原微生物生长的细菌素(例如β防御素)的产生。
益生菌还可以增加构成紧密连接的蛋白质的合成。对肠道屏障状态具有保护作用的益生菌主要有鼠李糖乳杆菌GG、乳双歧杆菌BB-12、嗜酸乳杆菌、植物乳杆菌、双歧杆菌、大肠杆菌等。
恢复正常肠道微生物群的另一种方法是粪便微生物群移植(FMT)。该疗法涉及将从健康供体获得的胃肠道微生物群转移到受体的胃肠道。实施前需准确评估患者和供体的肠道及菌群状况,避免盲目移植导致免疫反应和屏障损伤。
④低FODMAP饮食并增加多酚摄入
FODMAP饮食以短链碳水化合物和多元醇为主,这些成分吸收不良且易发酵,因渗透性强会增加肠腔含水量。FODMAP对肠道微生物群和屏障功能有益,但IBS或IBD患者采用低FODMAP饮食更能减少气体和肠腔扩张,减轻症状。
研究显示,维生素A和D可多途径改善肠道屏障,充足摄入能提升微生物多样性、强化紧密连接、调节免疫反应。
膳食纤维通过发酵生成短链脂肪酸(如丁酸盐),有助于加强粘蛋白和紧密连接,调节免疫,防止屏障通透性增加。
多酚类物质有助于增强紧密连接和黏液分泌,提高抗氧化力,富含多酚的饮食有助于降低肠道屏障障碍风险。
部分草药如普洱茶、芙蓉、蒲公英、姜黄等具备调节微生物群、强化屏障和抗炎作用,但其机制和疗效尚不完全明确,需注意提取纯度。
蘑菇富含维生素D、多酚和多糖,是益生元来源,可通过调节儿茶酚胺及炎症反应影响肠道微生物群。
⑤适当体力活动可降低肠道屏障障碍风险
众所周知,体育活动或锻炼可以增加肠道微生物群的多样性,增强SCFA的产生并刺激抗炎机制,主要是有氧运动,如跑步,骑自行车,健身运动,可以降低肠道屏障功能障碍的风险。
研究结果表明,体力活动以依赖于运动强度的方式调节肠道屏障通透性水平。定期、中等强度的体育锻炼对肠道上皮状态和肠道屏障完整性有积极影响。运动增加了肠道微生物组的多样性,有助于维持肠道屏障的完整性,还显示出抗炎和抗氧化作用。适度的体力活动可能是治疗肠漏的一个有利因素。
但是,高强度的体力活动,典型的竞争性或极限运动,与胃肠道疾病和肠道通透性增加的症状的高发病率相关。这是身体的应激源,由于体内血液的重新分配,体温过高和脱水,导致肠细胞缺氧。
近几十年来,过敏性和自身免疫性疾病的发病率显著增加,例如哮喘、特应性皮炎、过敏性鼻炎、慢性鼻窦炎、食物过敏、乳糜泻和炎症性肠病。全身性和代谢性疾病,如糖尿病、肥胖症、多发性硬化症、类风湿性关节炎、红斑狼疮、强直性脊柱炎以及阿尔茨海默病、帕金森病、慢性抑郁症和自闭症谱系障碍也成为日益严重的健康问题。
现代生活中的食品添加剂、抗生素滥用、环境污染物、慢性压力和过度依赖加工食品等因素正是导致这一代人屏障功能普遍衰退的隐形杀手。面对这一挑战,当前的科学建议首先聚焦于预防屏障损伤,通过建立标准化检测体系、开发肠道友好型产品替代有害化学物质、推广个性化营养干预等手段来实现。
食品工业也将从”填饱肚子”向”滋养健康“转型、医疗健康从”治疗疾病”向”维护屏障”转变、营养保健从”营养补充”向”功能修复“升级等多个行业带来了前所未有的发展机遇。
通过将传统医疗手段与肠道屏障修复、微生物群调节相结合,不仅能显著提高疾病治疗效果,更重要的是实现从“治病”到”防病”的医疗模式转变。
主要参考文献:
Macura B, Kiecka A, Szczepanik M. Intestinal permeability disturbances: causes, diseases and therapy. Clin Exp Med. 2024 Sep 28;24(1):232.
Farré R, Vicario M. Abnormal Barrier Function in Gastrointestinal Disorders. Handb Exp Pharmacol. 2017;239:193-217.
Brandl C, Bucci L, Schett G, Zaiss MM. Crossing the barriers: Revisiting the gut feeling in rheumatoid arthritis. Eur J Immunol. 2021;51(4):798–810.
Ramakrishna BS. Role of the gut microbiota in human nutrition and metabolism. J Gastroenterol Hepatol. 2013;28(Suppl 4):9–17.
Bäumler AJ, Sperandio V. Interactions between the microbiota and pathogenic bacteria in the gut. Nature. 2016;535(7610):85–93.
Ostaff MJ, Stange EF, Wehkamp J. Antimicrobial peptides and gut microbiota in homeostasis and pathology. EMBO Mol Med. 2013;5(10):1465–83.
Hansson GC. Role of mucus layers in gut infection and inflammation. Curr Opin Microbiol. 2012;15(1):57–62.
Gerbe F, Legraverend C, Jay P. The intestinal epithelium tuft cells: specification and function. Cell Mol Life Sci. 2012;69(17):2907–17.
Anderson JM, Van Itallie CM. Physiology and function of the tight junction. Cold Spring Harb Perspect Biol. 2009;1(2):a002584.
Drąg J, Goździalska A, Knapik-Czajka M, Matuła A, Jaśkiewicz J. Nieszczelność jelit w chorobach autoimmunologicznych. Państwo i Społeczeństwo. 2017;17(4):133–46.
Camilleri M. Leaky gut: mechanisms, measurement and clinical implications in humans. Gut. 2019;68(8):1516–26.
Wood Heickman LK, DeBoer MD, Fasano A. Zonulin as a potential putative biomarker of risk for shared type 1 diabetes and celiac disease autoimmunity. Diabetes Metab Res Rev. 2020;36(5):e3309.
Chelakkot C, Ghim J, Ryu SH. Mechanisms regulating intestinal barrier integrity and its pathological implications. Exp Mol Med. 2018;50(8):1–9.
Wright SL, Kelly FJ. Plastic and human health: a micro issue? Environ Sci Technol. 2017;51(12):6634–47.
Yee M-L, Hii L-W, Looi CK, et al. Impact of microplastics and nanoplastics on human health. Nanomaterials.2021;11(2):496.
Holloczki O, Gehrke S. Can nanoplastics alter cell membranes? ChemPhysChem. 2020;21(1):9–12.
Holloczki O, Gehrke S. Nanoplastics can change the secondary structure of proteins. Sci Rep. 2019;9(1):16013.
Xu M, Halimu G, Zhang Q, Song Y, Fu X, Li Y, Li Y, Zhang H. Internalization and toxicity: a preliminary study of effects of nanoplastic particles on human lung epithelial cell. Sci Total Environ. 2019;694:133794.
Jin Y, Lu L, Tu W, Luo T, Fu Z. Impacts of polystyrene microplastic on the gut barrier, microbiota and metabolism of mice. Sci Total Environ. 2019;649:308–17.

谷禾健康
当”健康中国2030″战略深入人心,当消费者从”治病”思维转向”防病”理念,整个医疗健康行业正在迎来前所未有的发展契机。
随着精准医疗时代的到来,用户需求正呈现出前所未有的细分化和专业化特征:
– 临床端:医生迫切需要具备更高灵敏度和特异性的诊断早筛等工具,以实现疾病的早期识别和精准分层;
– 机构端:健康管理机构正在寻求具有独特技术壁垒和差异化优势的检测产品,以构建竞争差异化;
– 消费端:用户不再满足于标准化的方案,而是渴望针对个体症状或后端干预的精准指导检测。
在这样的市场需求下,谷禾作为菌群检测行业先行者和深耕者,经过不断的技术测试和研发,完成了产品的全面升级和多形态布局。
从最初的16SrRNA测序技术科研应用起步,
到如今构建起涵盖
肠道菌群检测专业版
16S+tNGS靶向检测
宏基因组精准健康检测
针对特定人群的检测报告的完整产品矩阵,
再到技术平台向 女性阴道微生态
宠物肠道健康 等新兴领域的拓展,
谷禾以”场景导向、需求先行“的产品策略,
持续拓宽微生态检测技术的应用疆域。
本文将带您深入了解这一产品生态背后的技术逻辑与应用思考,每一个产品的诞生,都见证着微生态检测技术从”标准化”走向”个性化”、从”通用型”迈向”专业化”的发展轨迹。
01
肠道菌群检测专业版(16S rRNA测序),作为谷禾健康最早推向市场、历经十余年打磨的经典产品,凭借其成本效益、高效性与成熟度,为大规模人群的健康筛查、慢病风险评估、营养干预以及诸多科研项目基线建立,提供了坚实、可靠且极具价值的数据基础。
16S检测专注于细菌和古菌的16S rRNA基因,能够快速勾勒出肠道菌群的整体结构,包括多样性、核心菌属构成、有益菌与有害菌比例等关键菌群相关指标,评估肠道菌群失衡风险,评估健康风险、营养代谢及免疫情绪等多维度健康指标,提供个性化健康管理建议。
基于这一庞大数据资源,我们能通过菌群信息判别菌群的平衡或失调状态,进而判别个体的菌群与健康风险及与饮食、生活方式等关联。
数据的力量
在样本量达到一定量级时,
就会发生质的飞跃
📊 以结直肠癌模型为例:
<来源:谷禾健康肠道菌群检测数据库>
这也就是大样本量赋予16S检测
从 “观察” 到 “预见” 的核心能力
主要包括:健康总分评估、慢病风险预警、
肠道屏障及代谢物、神经递质分析、
个性化营养评估等内容。
健康总分评估
报告提供综合性的健康评分,直观反映客户肠道微生态的整体状态。
菌群整体分析
整体评估肠道菌群平衡、菌群多样性、有益菌、有害菌等指标,还包括核心菌群的丰度,这些核心菌群的减少往往与免疫力下降、肠漏、炎症等问题直接相关。
慢病风险预警
依托海量数据库和先进算法模型,16S检测能够评估与消化系统疾病、代谢类疾病(如肥胖、2型糖尿病)、心血管疾病、肝病甚至部分精神心理问题(自闭症、抑郁症等)等多种疾病相关风险。这为健康管理提供了强有力的早期干预工具,将健康管理从治疗推向预防。
肠道屏障及代谢物、神经递质分析
个性化营养评估
报告能分析菌群对不同营养物质(如膳食纤维、蛋白质、脂肪、维生素、微量元素)的代谢能力,为用户提供饮食建议。
无论是院内还是院外的健康管理,“先检后干预”的科学思路是健康管理的基础理念。通过全面系统的健康检测,准确评估个体健康状况,制定针对性的干预措施。谷禾的肠道菌群检测报告,针对菌群异常、营养代谢失衡以及疾病风险等问题,都会提供相应的干预建议。
市场应用场景
适用人群
专业应用
健康管理升级
多维度健康评估+个性化干预方案,提升服务深度
临床辅助
基于大样本数据库疾病预测模型,助力辅助诊断,辅助用药
渠道共赢
周期短(3-5天)+全流程可控+资质保障,实现快速业务整合
科研赋能
海量200万数据库+研究成果,支持学术前沿探索
谷禾肠道菌群检测专业版
凭借其10多年的发展和应用
已成为理解肠道微生态的成熟且普及的工具
这为许多常规健康管理和慢病预防场景
是谷禾所有产品中不可或缺的基石
通过多种技术分析方法的持续迭代
特别是依托我们的国家发明专利技术
谷禾实现了重要的技术突破
我们能够将16S的物种识别精度大幅提升
在成本可控的前提下
实现了更高的检测精度
⚠️ 技术局限性
但是我们同样坦诚其技术局限性和边界,由于16S针对细菌或古菌的保守基因的扩增,它没有覆盖病毒、真菌、寄生虫等,16S技术还是很难更进一步精确到“菌株”水平,也难以精确到毒力/耐药基因的判别。
🔄 技术进化:从局限到突破
正是基于对16S技术边界的清醒认知,特别是对于个别需要判断毒力基因或者幽门螺杆菌、艰难梭菌、致病性大肠杆菌等病原体以及其特定毒株分型的临床需求,考虑到宏基因组检测的高成本现状,谷禾历经两年研发,成功开发了粪便样本的靶向消化道测序技术(16S+tNGS),为精准病原体检测提供了更加经济高效的解决方案。
02
谷禾16S+tNGS技术结合了超多重PCR和高通量测序的优势,旨在提供比传统16S rRNA测序和宏基因组测序更优、更全面的病原体及耐药基因检测方案。
传统16S + 病原体精准分型 = 全新升级
它在保留16S报告的基础上
以接近16S的成本和周期
用靶向测序技术在原16S的基础上
增加了125 种消化道病原体的检测
还包括耐药基因和毒力基因等
如幽门螺杆菌、大肠杆菌、艰难梭菌分型
弥补了传统16S无法检测
非细菌/古菌病原体的不足
这是一款突破传统16S检测瓶颈而生的产品
编辑
值得一提的是,谷禾在tNGS技术的研发道路上并非一帆风顺。这项看似成熟的技术,在不同应用场景下却面临着截然不同的挑战难度。
tNGS对血液和上呼吸道样本检测较简单,因其主要含病原体。但在消化道特别是粪便样本检测时,技术难度大幅增加。
“两年磨一剑,突破粪菌检测技术壁垒”
这两年谷禾团队需要解决一系列前所未有的技术难题:
这正是消化道微生态检测的技术壁垒所在。与呼吸道样本不同,肠道环境的复杂性要求我们必须在技术层面实现更精准的信号识别与干扰排除。
经过无数次的实验优化和迭代
我们最终突破了这一技术瓶颈
为大家带来真正可靠的
消化道病原体检测解决方案
谷禾16S+tNGS产品特点
弥补16S检测技术边界
以接近16S的成本
实现靶向病原体精准检测
谷禾16S+tNGS报告内容涵盖所有16S报告的内容,即包括健康总分评估、慢病风险预警、肠道屏障及代谢物、神经递质分析、个性化营养评估等。此外还包括常见消化道病原体,例如:
细菌病原体
…
病毒
真菌、寄生虫、其他病原体
…
毒力基因
…
耐药基因
…
也包括相关病原微生物的解释
…
例如,通过检测幽门螺杆菌毒力基因组合,可判断是否需立即治疗,避免对弱毒株患者的不必要抗生素使用。强毒株感染会损伤胃黏膜,增加胃炎和溃疡风险,早期预警能在胃黏膜不可逆损伤前提供治疗窗口期。
注: 由于该技术是检测粪便中的幽门螺杆菌,当浓度低于检测下限(50 copies/mL)时,可能出现假阴性结果。因此,对于临床症状明显但检测结果为阴性的患者,建议结合其他检测方法。
✎ 谷禾16S+tNGS精准检测
一次检测即可实现对消化道病原体的全面筛查和耐药基因的精准识别,提供科学依据,最大化治疗效果,最小化治疗风险。
03
宏基因组检测项目以环境中所有微生物基因组为研究对象,通过对样本中的全基因组DNA进行高通量测序,能够多维度全面解析肠道微生物组。
基于持续积累的20万+肠道宏基因组数据库,我们可以系统性地挖掘微生物群的功能基因谱,并预测其代谢潜力。
结合自主研发的多模态模型和机器学习算法,该技术可以建立微生物特征与宿主表型的关联,实现肠龄预测、菌群恢复力评估、定植能力分析等功能。这些分析结果有助于为个性化益生菌干预、精准营养调控及FMT供体筛选等应用场景提供分子水平的参考依据。
宏基因组流程——从随机打断到精准重构
优 点
缺 点
不适合大规模筛查
宏基因组更适合宿主含量不高的样本
谷禾以往在科研领域深耕十余年,积累了丰富的科研项目经验和数据分析能力,几年前,谷禾成功实现了从科研到大健康应用的技术转化,推出了谷禾宏基因组精准检测。
对于一些特定需要深度检测的应用场景,如健康管理机构的差异化服务需求,或者临床应用需求,宏基因组精准健康检测提供了另一种专业的技术选择。
宏基因组数据库的物种涵盖范围和菌株构成,直接影响着宏基因组物种鉴定分类的准确性和分类精度。
针对宏基因组数据库不完善的问题,谷禾整合了最新的NCBI refseq数据库,涵盖细菌、病毒、真菌和寄生虫,结合自研多元统计模型和机器学习算法,极大提升了物种鉴定和功能注释的准确性。
物种精准鉴定
分辨率更加精细,可达“种”和“菌株”水平,并对复杂的多菌种感染进行精细化解构。
肠道功能评估
肠道基础功能:包括蛋白质发酵能⼒、消化吸收效率、肠道产气情况、肠道屏障完整性、肠道炎症状态等,在菌群整体评估指标中也增加了包括菌群恢复力、革兰氏阴性菌、好氧菌等指标。
功能基因分析
– 全面评估菌群的基因功能潜力
如次生代谢产物合成通路、维生素合成能力、碳水化合物利用能力等,为个性化营养和精准干预提供科学证据。
耐药基因
– 分析耐药基因
不局限于已知靶点,可鉴定出各种已知和新型耐药基因,全面评估耐药基因的种类和数量,例如,在人体肠道宏基因组中发现了大量β-内酰胺酶等耐药基因。
– 追踪耐药基因的传播途径
通过比较不同环境(如土壤、水体、动物和人体)中耐药基因的分布情况,可推测耐药基因的来源和传播途径。
– 辅助指导耐药风险评估和防控策略
宏基因组学评估环境和宿主中耐药基因的分布特点,识别高风险区域和人群,为制定针对性监测和干预措施提供依据。
毒力基因
宏基因组检测技术在病原微生物毒力基因研究中具有独特优势,可在基因组水平系统分析其毒力基因组成及调控网络,加深对致病机理的理解。
免疫炎症分析
肠道菌群通过调控免疫平衡维持健康,⽽炎症标志物则作为评估机体炎症程度的关键指标。
宏基因组测序对测序深度要求较高,当数据量不足时,一些低丰度的真菌、寄生虫等病原体可能覆盖不到;然而,若要获得足够的测序深度来确保全面覆盖,则会显著增加测序成本,同时对分析能力和计算资源提出更高要求。
因此,宏基因组检测看似”简单粗暴”,只要更多数据量,实则真正的挑战在于,如何在成本与深度之间找到最优平衡。
谷禾持续迭代升级自有数据库
整合最新的NCBI refseq数据库
并结合十余年积累的临床样本数据
让相对较小的测序量
也能获得高精度的物种鉴定结果
同时,谷禾致力于深入挖掘
数据背后的生物学意义
通过专业团队的生物信息学分析
从宏基因组数据中构建
炎症状态、消化功能异常等关键健康指标
通过机器学习算法将海量基因信息
转化为实用的健康评估结果
通过这种”数据挖掘+算法迭代“
尽可能为大家控制成本的同时提供
媲美高深度测序的检测精度
极力追求技术创新与商业价值的完美结合
宏基因组报告中的解读更详细,还整理了一些评估指标,检测指标的总结等。
其他谷禾肠道菌群检测专业版的内容,宏基因组报告里面也都涵盖了,包括慢病风险、菌群代谢物及神经递质代谢、个性化营养等板块。
…
…
个性化饮食板块也在谷禾16S版本的基础上进行了迭代升级。
…
宏基因组检测并不常用于常规检测,其高昂的成本和复杂的数据分析决定了它更适用于关键时刻。
特定菌群感染的判别
对于一些复杂的多菌种感染,宏基因组能够更精细化鉴定感染菌群的构成,为临床辅助诊疗提供依据。
真菌与病毒感染的深度判别
相比传统培养,宏基因组学诊断真菌感染的敏感性和特异性更高,适用于一些真菌感染疾病。也可能鉴定出可疑的新病原体,为后续的病原学研究、药物和疫苗开发奠定基础。
一图看懂以上谷禾三大产品线
04
在肠道菌群检测系列产品成熟之后,谷禾健康将深耕多年的微生物组学技术平台,延伸至关乎女性全生命周期健康的另一核心领域——阴道微生态。
还包括子宫颈沙眼衣原体、HPV、HSV、EB病毒、巨细胞病毒等。
谷禾阴道菌群检测报告引入科学前沿的菌群状态分型(CST)概念,将复杂的菌群构成归纳为几种易于理解的健康状态类型。
例如,以卷曲乳杆菌为主的CST-I型代表健康的稳定状态,而以加德纳菌等多种厌氧菌为主的CST-IV型则与细菌性阴道病高度相关。这为临床判断和干预效果评估提供了科学支持。
谷禾阴道菌群检测报告中包括阴道菌群总体评估、CST分型、致病菌表(细菌性阴道病,需氧菌性阴道炎,外阴念珠菌病等)、列出异常菌群及相关说明,菌群详细构成等。
…
…
…
…
阴道菌群检测让我们能够更全面地了解阴道微生物组的组成及其变化,以及它是如何随着时间的推移或对各种因素(如环境、激素变化、性活动和抗生素使用等)的反应而变化的。
注:本产品可辅助评估和筛查,不用作临床诊断。
05
随着“它经济”的蓬勃发展和“科学养宠”理念的深入人心,宠物已成为家庭的重要成员。然而,面对“毛孩子”们无法言说的病痛,如反复腹泻、顽固皮肤病、食欲不振、呕吐等,传统兽医诊断往往面临挑战。
从宠物医院的实际经营来看,慢性疾病正成为他们面临的核心挑战。慢性肾病、老年痴呆、精神类疾病等病症不仅治疗费用昂贵,而且现有手段往往无法覆盖,特别是小型诊所更是心有余而力不足。
在与许多B端合作伙伴的深度交流中我们发现
宠物腹泻,肾病以及其他疾病等正在增加
后期医疗费用高昂让宠物主人无能为力
异常行为严重影响生活质量和主人养宠体验
情感难舍却不得不放弃…
因此,迫切需要一种更加
科学、经济、精准的健康管理方式
既能降低医疗成本
又能提供个性化的健康方案
还能避免过度医疗
这就需要我们从根本上
重新思考宠物健康管理的方法论
从”治疗导向“转向”预防导向“
✲✲✲✲✲✲✲✲✲✲✲✲✲✲✲✲✲✲✲✲✲✲✲✲✲✲✲✲✲✲
谷禾凭借在人类健康领域积累的深厚微生物组学经验,战略性延伸至动物医学领域,推出宠物菌群精准检测服务。
我们致力于解码宠物肠道微生态的奥秘,为兽医临床、宠物营养和家庭养护提供科学依据,开启宠物健康管理的精准化新时代。
从人类微生态到宠物微生态,不是简单的复制,而是技术能力的升维应用。人类肠道微生态的复杂性研究为我们提供了强大的算法基础和数据分析能力,以及多年来在宠物菌群科研中的持续投入,这些经验在宠物领域的应用中展现出了独特的技术优势。
研究表明,宠物肠道菌群不仅影响消化吸收功能,更与免疫调节、神经系统、皮肤健康等多个生理系统密切相关,成为宠物整体健康状况的重要晴雨表。
谷禾正有序推进构建涵盖不同品种、年龄、健康状况犬猫的肠道菌群数据库,结合最新的机器学习算法,实现对宠物肠道微生态健康状况和营养进行精准评估。
宠物菌群报告展示采用更温馨活泼的配色,通过可视化图表和情感化设计,让复杂数据更直观,帮助主人轻松了解爱宠健康。
谷禾宠物菌群检测报告中包括菌群评估(整体指标)、肠道基础功能评估(屏障功能、炎症水平、代谢状态)、菌群代谢物评估(短链脂肪酸等)、炎症免疫评估(促炎、抗炎等指标)、营养饮食评估(维生素、微量元素)等。
…
…
症状相关菌群分析,包括腹泻、呕吐、过敏等。
…
菌群代谢物评估(短链脂肪酸等)。
…
…
从多维度全面评估宠物肠道健康状况,为宠物主人提供科学的健康管理依据和个性化调理建议。
自2012年成立以来,谷禾健康始终扎根于肠道微生态领域。我们不仅是国家高新技术企业和专精特新企业,更是通过中国合格评定国家认可委员会(CNAS)认可评审,成为CNAS认可的微生物检测实验室。同时,谷禾也拥有几十项国家发明专利以及在国际顶级期刊发表的研究成果,已经服务和合作150多家顶级医院与机构,积累了超过200万的样本数据库,这既是我们的底气,也是您成功的保障。
作为菌群检测行业的先行者和深耕者,谷禾始终坚守科学严谨的初心,在技术研发、质量管控、数据安全等各个环节持续深化建设。
从样本储存运输的标准化流程,
到阳性对照、阴性对照的严格设置;
从仪器校准溯源的精准把控,
到人员素质培训的持续提升,
从数据安全保障的多重防护,
到人机料法环的全方位管控,
每一处细节都是我们
以科研匠心在守护谷禾检测命脉。
正是出于对科学研究价值的深度认同,谷禾设立了『人体肠道菌群开放基金』,从心梗脑梗风险监测到儿童自闭症干预,从肿瘤免疫治疗到妊娠期健康管理,通过阶段性的递进式合作模式,我们已成功孵化近百个前沿研究项目,并在国际权威期刊
《Gut》、《Advanced Science》、《Clin Transl Oncol》等发表突破性成果。
已开展申请项目
未来,谷禾将持续投入研发力量,在青少年抑郁症、代谢综合征、心脑血管、老年阿尔茨海默病、过敏相关免疫疾病以及特定肿瘤(如胰腺癌、肝癌)等前沿领域继续深耕,进行模型的深度开发与优化。
同时,谷禾正积极推进临床营养检测评估中心建设,通过与首科等权威机构合作,深入开展儿童精神发育、老年营养监测等特定人群研究,未来加入社区筛查项目,长期追踪社区人群的菌群状况,通过对稳定人群的菌群纵向研究更加深度挖掘菌群与健康之间的关联性,为精准医疗和个体化营养干预提供更加坚实的科学依据。
从科研到应用的全链条能力建设,让我们能够更好地赋能下游产业。通过携手更多科研院所、医疗机构和行业伙伴,谷禾将持续探索菌群检测技术在医疗大健康生态系统中的创新应用,与业界伙伴携手共进,推动行业健康发展。
欢迎有相关方向的人群或者
科研、临床、干预机构咨询合作
谷禾健康
为什么在劳累一天后,我们的大脑会疯狂渴望一块巧克力蛋糕,而不是一盘清爽的西兰花?为什么明知奶茶不健康,那种甜腻的口感却能带来无与伦比的慰藉?
过去,我们总把这归结为“压力大”、“嘴馋”或是“意志力薄弱”。但科学的聚光灯正照向一个我们长期忽略的角落——我们的肠道。
你可能不知道,你的每一次口腹之欲,背后都可能是一场发生在肠道里的“生物战争”。你体内的微生物菌群,这些数量比我们自身细胞还多的微小生命体,它们有着自己的“生存法则”和“饮食偏好”。
你吃的每一口食物,都在“投票”:你选择高纤维的蔬菜,就是在为你体内的“有益菌”投票,它们会回报你稳定的情绪和清晰的饱腹感信号。而你选择高度加工的甜食,则是在为那些渴望糖分的“捣蛋菌”投票,它们会分泌出让你更加渴求垃圾食品的化学物质。
这不仅仅是关于消化:这些菌群产生的代谢产物,如短链脂肪酸、胆汁酸甚至神经递质,能像信使一样,顺着“肠-脑轴”这条信息高速公路直达大脑,直接影响你的心情、味觉乃至成瘾行为。
高度加工食品的特点是脂肪和糖含量升高、纤维含量降低,同时含有大量盐和食品添加剂,维生素等微量营养素减少。而饮食中膳食底物(纤维、糖、脂肪和蛋白质等)比例的改变会扰乱微生物组-肠道-大脑轴。
越来越多的证据表明,微生物群在调节饮食相关行为及代谢紊乱的行为合并症方面发挥至关重要作用。了解肠道微生物群影响宿主食欲和新陈代谢的机制,将有助于更好地理解肥胖等饮食失调和代谢失调情况,从而开发新的生物治疗策略。
本文将为你揭示,现代工业化饮食是如何通过扰乱我们体内的“微生态平衡”,来悄无声息地影响我们的行为的。理解了这场“肠脑对话”,你将找到一把全新的钥匙,去解锁困扰已久的饮食难题,并最终学会如何“喂养”出一个更健康、更快乐的自己。
现代化进程推动了人类生活方式的变革,饮食行为作为基本生存需求也随之发生复杂多样的演变。工业革命的食品加工技术、信息时代的个性化营养、全球化的饮食文化融合、城市化的快节奏生活、科技进步的精准营养指导,以及社交媒体对饮食观念的重塑,这些现代化力量共同催生了当代社会中数量庞大、类型繁多的饮食行为模式。
这些饮食行为呈现出鲜明的双重性特征:既有科学化营养管理、个性化健康需求满足等积极趋势,也存在高糖高脂成瘾性消费、不规律进食习惯、过度依赖加工食品等不健康倾向。深入理解这些饮食行为的成因、特征及健康影响,对构建科学合理可持续的现代饮食体系具有重要意义。
Part 01
不健康的饮食行为
以下列举了一些在当代社会中普遍存在的不健康饮食行为模式,这些行为可能正在悄悄影响着我们的身体健康,快来看看自己是否存在这些问题。
高糖高脂类
高脂肪高糖饮食:现代食品的发展使得高糖高脂食品变得极其便利和诱人。这类饮食方式不仅包括传统的高脂肪高糖饮食如奶茶、甜品、油炸食品和快餐组合,还延伸出了更加隐蔽的形式。
精制糖依赖:已成为现代人普遍面临的健康威胁。从早晨的含糖咖啡开始,到下午的甜点小憩,再到深夜的糖分慰藉,精制糖以各种形式渗透到日常生活的每一个角落。含糖饮料、糖果、精制糕点的过量摄入不仅导致血糖波动,还可能引发类似药物依赖的生理和心理反应。
反式脂肪摄入:人造奶油、酥饼、部分快餐食品中含有的反式脂肪酸,以其独特的口感和保存特性被广泛使用,但其对心血管健康的危害已被科学研究充分证实。
高盐高钠类
重口味饮食:现代人对味觉刺激的追求催生了重口味饮食的盛行。这种饮食方式表现为过度依赖调味料、腌制食品、咸菜等高钠食物,以及对麻辣、酸辣等强烈味觉刺激的偏好。
加工食品依赖:火腿肠、方便面、罐头等便利食品为了延长保质期和增强口感,往往含有超出日常所需的钠含量。这些”隐形盐”的大量摄入与高血压、心血管疾病的发生密切相关。
不规律进食类
现代生活节奏的加快导致了饮食时间的严重失调。这种失调不仅体现在进食时间的不规律,更表现为进食量和频率的极端化。
暴饮暴食:在工作压力、社交焦虑、情感困扰的驱动下,许多人选择通过短时间内大量进食来获得心理慰藉,这种压力性进食行为已成为现代都市生活的普遍现象。
过度节食:在追求”完美身材”的社会压力下,极端限制热量摄入、长期营养不均衡的节食行为不仅损害身体健康,还可能导致进食障碍等严重心理问题。
夜宵文化:深夜大量进食高热量食物不仅干扰正常的生物钟,还会增加消化系统负担,影响睡眠质量。
跳餐习惯:不吃早餐的现象在年轻人中非常普遍。这种习惯不仅影响一天的能量供应和代谢稳定,还可能导致午餐和晚餐的过度补偿性进食。
营养结构单一化饮食
碳水化合物过量:表现为主要以米面等精制谷物为主食,缺乏粗粮、杂粮的摄入。这种饮食模式不仅导致B族维生素和纤维素摄入不足,还可能引发血糖波动和胰岛素抵抗。
蛋白质缺乏:在一些盲目追求素食的人群中尤为明显。长期素食但营养搭配不当,特别是缺乏对植物蛋白互补性的理解,可能导致必需氨基酸摄入不足。
维生素矿物质不足:主要源于新鲜蔬果摄入不足。现代人对加工食品、外卖食品的依赖,使得天然维生素和矿物质的摄入量远低于推荐标准。
Part 02
较健康的饮食行为
与上述不健康的饮食行为形成鲜明对比的是,在现代社会中同时也涌现出了一些相对更加科学合理、对身体健康更为有益的健康饮食方式和饮食理念。
均衡营养类
地中海饮食:地中海饮食作为被广泛认可的健康饮食模式,以橄榄油、鱼类、坚果、全谷物、蔬果为主要构成,其对心血管健康的保护作用已被大量科学研究证实。
DASH饮食:通过低钠高钾的营养配比,强调蔬果、全谷物、瘦肉蛋白的摄入,不仅有助于控制血压,还能改善整体代谢健康。
彩虹饮食:鼓励人们每天摄入多种颜色的蔬果,通过颜色的多样性确保植物化学物质和营养素的全面摄入。
全食物饮食:强调选择未加工或轻加工的天然食物,避免过度加工食品中的添加剂和人工成分,回归食物的天然营养价值。
一些饮食对肠道微生物群和免疫的影响
doi: 10.3390/nu11102393.
植物性饮食类
素食主义:现代素食主义已经从单纯的不摄入动物产品,发展为注重营养搭配、科学规划的健康生活方式。
弹性素食:以植物性食物为主,适量摄入动物产品的饮食模式,既满足了营养需求,又减少了环境负担。
轻食主义:以低热量、低脂肪、高纤维的食物为主,追求营养均衡而非饱腹感。典型食物包括沙拉、轻汤、蒸煮食品等,强调”吃得少而精”。
功能性饮食类
抗炎饮食:通过富含抗氧化剂的食物,如浆果、绿叶蔬菜、深海鱼类等,帮助人体对抗慢性炎症,预防相关疾病。
益生菌饮食:强调发酵食品、酸奶、泡菜等有益菌群的摄入,促进肠道健康,这与现代医学对肠道微生物群重要性的认识高度一致。
低升糖指数饮食:通过选择血糖反应较低的食物,帮助维持血糖稳定,对糖尿病防治具有重要意义。
▸ 膳食成分影响微生物代谢和饮食行为
√ 膳食成分和加工程度会影响微生物和代谢
膳食成分组成而非热量负荷本身是微生物组结构和功能的主要驱动因素。人类队列和动物模型研究一致表明,不同的营养素特征会引起特定的微生物和代谢反应。
对照喂养研究显示,营养素比例相同但纤维含量和加工程度不同的等热量饮食会产生截然不同的微生物特征、发酵活性和宿主能量利用效率。工业化饮食减少了可发酵化合物的可用性,而这些化合物对维持多样化和有弹性的微生物群落至关重要。
由于肠道微生物组影响免疫、代谢、神经传递和内分泌等多种宿主过程,饮食的重大变化可能改变肠道微生物组的代谢输出,对宿主生理和行为产生重要影响。
√ 肠道微生物可以通过肠脑轴反向影响饮食行为
肠-脑轴对摄入的营养物质(纤维、糖、脂肪和蛋白质)进行差异感知,并将有关营养、能量和饱腹感的信息传递到大脑,以协调摄食行为。这些行为包括饥饿时觅食和饱腹时停止摄入。该系统通过专门的感觉途径整合来自胃肠道的化学和物理影响,从而维持能量稳态。
肠道微生物组作为对饮食敏感且高度可塑的微生物群落,在结肠中密度最高,通过将膳食底物代谢成生物活性化合物在宿主营养中发挥关键作用。由于肠道微生物组对饮食的反应和代谢能力远超人体自身,它能够显著影响肠脑信号传导和摄食行为。然而,这种饮食敏感性可能是双刃剑:在进化的低能量环境中发挥优势的机制,在现代工业化产生的高能量密集且易消化食物环境中可能带来危害。
饮食驱动的微生物组-肠-脑轴调节摄食行为
Liow YJ,et al.Trends Endocrinol Metab.2025
√ 现代饮食的大量加工会导致饮食行为改变
现代饮食改变了纤维、糖、脂肪和蛋白质的比例,并通过大量加工提高小肠营养物质消化率。这些变化增加了工业化饮食破坏微生物介导营养传感网络的可能性,导致饮食行为改变和能量失衡。这些饮食-微生物组相互作用可能为工业化生活方式相关的流行病(包括饮食失调和代谢疾病)提供了现有模型中基本未解释的隐藏信号层。
▸ 膳食常量营养素的化学感觉编码
√ 葡萄糖信号传感
葡萄糖传感发生在门静脉肠系膜系统内,代谢信号通过迷走神经和脊髓传入神经编码。门静脉葡萄糖代谢的迷走神经元通过结节神经节将信号传递至孤立道核,并通过调节纹状体多巴胺释放增强糖偏好。平行的门静脉-脊髓轴确保强大的营养信号编码,凸显了糖检测在摄食调节中的重要性。
然而,葡萄糖缺乏检测涉及非迷走神经机制。孤立道核后区是感知葡萄糖缺乏的中心枢纽,迷走神经切断术不能消除葡萄糖缺乏引起的摄食反应,而损伤该回路则能消除此反应,表明葡萄糖缺乏检测可能依赖中枢糖感受器或脊髓传入神经。此外,葡萄糖通过非迷走神经途径抑制刺鼠相关肽(AgRP)神经元,支持其他内感受机制的参与。
注:刺鼠基因相关蛋白——一种与食欲控制密切相关的神经肽。
√ 脂质信号传感
相比之下,脂质传感显示出对迷走神经传入神经的严格依赖。迷走神经切断术消除脂质摄食反应,表明脂质代谢信息必须通过肠-脑信号传导而非中枢直接检测传达。十二指肠脂肪需要迷走神经输入抑制AgRP神经元,强化了迷走神经信号的重要性。关键介质是油酰乙醇酰胺,这是近端肠道合成的脂质衍生分子,与PPARα受体结合并调节纹状体多巴胺信号,可能通过投射至孤立道核后区的迷走神经传入发挥作用。
长期高脂肪饮食会破坏油酰乙醇酰胺信号,削弱肠脑通讯和脂肪衍生的多巴胺释放,这种适应不良反应可能导致代偿性暴食。糖和脂肪信号参与重叠但不同的回路,同时出现时会放大多巴胺释放,促进超出热量需求的过度消耗。
√ 蛋白质信号传感
蛋白质衍生的信号参与迷走神经和非迷走神经途径,具体取决于蛋白质形式和生理环境。蛋白胨等水解蛋白激活肠道化学感受器并触发胆囊收缩素(CCK)释放,刺激迷走神经传入并激活食欲调节的肠脑信号。
迷走神经切断术或CCK1受体阻断可消除此效应,证实迷走神经依赖性。相反,完整蛋白质通过非迷走神经反馈调节摄食,可能通过血浆氨基酸传感或肝脏传入神经实现。肝门传感器以剂量依赖方式检测循环氨基酸,在氨基酸缺乏时迷走神经传感器敏感性增加约100倍,放大代偿驱动。这种肝脏中心机制能根据营养可用性动态调节蛋白质摄入。
▸ 胃胀的机械感觉编码
机械感觉提供独立于营养成分的肠脑信号传导层,传达胃扩张和进食量信息。表达催产素受体的迷走神经感觉神经元作为机械传感器,对胃和十二指肠牵拉做出反应,将机械信号传递至孤立道核。
√ 机械敏感传入神经影响饱腹感和能量平衡
这些机械敏感传入神经影响饱腹感和能量平衡控制回路,激活时抑制食物摄入并诱导昏睡样状态,表现为能量消耗减少和下丘脑-垂体-肾上腺轴活动增加。除迷走神经通路外,脊髓传入神经也传递机械感觉信号,背根神经节介导的通路将GLP-1介导的胃松弛信号传递至下丘脑外侧。
然而,机械感觉通路并非孤立运作,而是与肠内分泌细胞的化学感觉输入相互作用。产生5-羟色胺的肠嗜铬细胞既对管腔刺激物反应,也促进肠道蠕动并加速肠道转运以助毒素清除。此外,胰高血糖素样肽-1(GLP-1)和CCK释放细胞可减缓胃排空,可能与迷走神经机械传感器协同延长饱腹感。
√ 营养信号和机械信号共同调节进食行为
营养信号和机械信号间的相互作用精细调节进食行为,促进对不同膳食成分的适应性反应。
通过整合来自营养成分的化学感觉输入和编码胃扩张的机械感觉信号,肠脑轴传入分支确保摄食行为根据摄入食物的代谢和物理特性动态调节。脑干迷走神经和脊髓传入通路的汇聚实现对食欲和能量平衡的精细调节。肠内分泌细胞作为综合枢纽,将管腔营养信号转化为激素和神经输出,塑造肠道蠕动和摄食行为,进一步完善能量稳态的内感受控制。
饮食对肠道微生物组组成和功能的影响远超遗传因素。比较植物性与动物性饮食、地中海与西方饮食、非洲农村高纤维与欧洲城市饮食以及熟食与生食的研究中,均可观察到饮食对肠道微生物组的显著影响。
√ 肠道微生物组有助于评估生活方式的健康性
肠道微生物组对饮食的敏感性使其成为评估现代生活方式健康影响的重要标志和靶点。特别是向工业化饮食的转变可能对与饮食行为相关的肠道微生物组产生显著的组成和功能变化。
与传统饮食相比,工业化饮食的特征是纤维含量急剧减少,糖、脂肪和蛋白质含量增加。肠道微生物组代谢部分抵抗小肠消化的营养物质,产生多种生物活性小分子代谢物,使工业化饮食与其他饮食产生区别。在动物模型和人类研究中,饮食成为肠道微生物生态学的核心决定因素,不仅重塑群落组成,还重塑介导微生物-宿主相互作用的基因组、转录组和代谢组学程序。
√ 微生物代谢影响食欲和摄食行为
肠道微生物组在接触膳食底物时产生的代谢物包括短链脂肪酸(SCFA)、氨基酸代谢物、次级胆汁酸和神经递质,其产生取决于管腔中常量营养素的可用性。这些代谢物改变局部肠道环境、粪便和血浆代谢组特征以及全身能量调节,并对宿主生理学产生其他影响。
重要的是,这些代谢物与外周和中枢神经元相互作用,影响宿主进食行为。下图表概述了膳食底物的肠道微生物代谢如何产生影响食欲调节和摄食行为神经回路的生物活性代谢物。接下来我们将介绍微生物与各主要膳食成分的相互作用。
微生物代谢膳食底物及对宿主进食行为的影响
Liow YJ,et al.Trends Endocrinol Metab.2025
1
膳食纤维
工业化饮食的特征是纤维含量相对较低,精制碳水化合物含量较高。这种膳食底物模式促进了支持炎症和奖励敏感代谢途径的微生物环境,而抑制了促进饱腹感和代谢平衡的途径。
√ 能否发酵会影响宿主的饮食选择
膳食纤维对肠道微生物组的塑造程度取决于其可发酵性:可发酵纤维(如菊粉)更强烈地调节微生物组成和活性,而不可发酵纤维(如纤维素)代谢最少,对宿主能量贡献很小。
纤维素虽能诱导肠扩张,但代谢惰性相对较高,虽然它可激活迷走神经信号,但与脂肪和糖等代谢活性营养素相比作用较弱。有趣的是,微生物发酵能力可能影响宿主食物偏好:最近研究发现,小鼠更偏好其常驻微生物无法发酵的膳食纤维,而非可发酵纤维。因此,发酵性不仅控制微生物和代谢结果,还控制对膳食纤维的行为反应。
富含纤维的食物会诱导肠道蠕动、胃排空和肠内分泌信号传导的急性变化。在短期内,纤维消耗通过胃胀气和延迟营养吸收来增加饱腹感,从而减少膳食量。在数天至数周内,持续摄入纤维会重新配置微生物组组成和代谢信号通路,增强肠内分泌激素分泌,并导致食欲调节和能量稳态的持续改变。长期以来,高纤维饮食与较低的能量摄入和体重调节有关。
√ 膳食纤维代谢产生的短链脂肪酸是肠脑通讯的重要信号分子
膳食纤维发酵是短链脂肪酸(主要是丁酸盐、乙酸盐和丙酸盐)的主要来源。短链脂肪酸已成为肠脑通讯的重要调节剂。丁酸盐通过激活迷走神经传入神经元、降低孤立束核和背侧迷走神经复合体活性以及抑制下丘脑神经肽Y(NPY)表达的食欲神经元来抑制食物摄入。这种食欲抑制作用需要迷走神经完整性。
乙酸盐和丙酸盐腹腔注射也具有显著厌食作用。在禁食小鼠中,全身注射短链脂肪酸导致食物摄入量剂量依赖性减少,丁酸盐效果最强,其次是丙酸盐和乙酸盐。这种短链脂肪酸诱导的食欲抑制由迷走神经传入神经元介导,迷走神经传入神经元表达游离脂肪酸受体(FFAR2、FFAR3),为微生物代谢物提供了独立于经典肠内分泌信号通路调节宿主摄食行为的途径。
√ 短链脂肪酸影响肠内分泌激素进而影响饮食行为
短链脂肪酸不仅直接作用于迷走神经传入神经,还刺激肠内分泌细胞通过激活肠内分泌L细胞上的FFAR2释放激素,调节摄食行为和能量稳态。其中,CCK、GLP-1和PYY是肠脑信号传导和饱腹感的关键调节因子。
短链脂肪酸及其对宿主食欲和新陈代谢的影响
doi: 10.3945/jn.116.240481.
CCK通过迷走神经元影响摄食行为,迷走神经切开术消除其厌食作用。微生物来源的丙酸盐通过直接上调肠内分泌细胞中的Cck mRNA和肽水平刺激回肠和盲肠中CCK表达。CCK塑造营养特异性偏好,包括脂肪偏好发展。阻断CCK信号传导消除对糖、脂肪和氨基酸的迷走神经反应,表明CCK对营养驱动的食欲行为至关重要。膳食纤维摄入促进微生物发酵并促进CCK释放,通过微生物活动将膳食纤维与肠脑信号传导联系起来。
GLP-1是食欲和血糖控制的关键调节因子,通过表达GLP-1受体的迷走神经传入神经发挥作用。这些神经元密集支配胃肠道并投射到孤立道核,介导GLP-1的厌食和代谢作用。内源性GLP-1还通过替代的肠交感神经回路发挥作用。GLP-1和PYY都受微生物来源短链脂肪酸调节,后者通过结肠L细胞上的FFAR2急性刺激其分泌。长期暴露时,短链脂肪酸还通过FFAR2/PAX4依赖性分化扩大L细胞群来增强PYY输出。PYY3-36通过与迷走神经传入神经上的Y2受体结合发挥厌食作用。
√ 特定微生物代谢膳食纤维改变宿主的食物偏好
尽管微生物短链脂肪酸和肠内分泌激素相互作用影响饱腹感和能量稳态,但膳食纤维的微生物代谢也可能通过替代途径影响宿主食物偏好。菊粉补充剂研究表明其将食物偏好转向脂肪而非糖,这种变化与特定微生物特征相关。受控研究显示,用特定拟杆菌定植的小鼠选择性发酵果聚糖,使宿主偏好从可发酵果聚糖转向不可发酵果聚糖,当微生物代谢中断时这种效应消失。
人体研究提供初步证据表明肠道微生物对纤维的发酵影响食物相关线索的神经处理。超重成人补充菊粉型益生元纤维改变大脑对高热量食物刺激的激活模式,减少奖励评估相关脑区激活。尽管这些神经变化未直接转化为食物摄入或偏好改变,但提示微生物代谢物可能影响大脑对营养丰富食物的评估。
综上所述,低纤维的工业化饮食减少短链脂肪酸的微生物产生,通过迷走神经传入神经减弱饱腹感信号传导并减少CCK、GLP-1和PYY等饱腹感激素分泌,从而破坏肠脑通讯。因此,膳食纤维缺失可能导致饱腹感下降,促进过度摄入并损害能量平衡。
膳食底物的微生物代谢控制宿主的进食行为
Liow YJ,et al.Trends Endocrinol Metab.2025
2
膳食糖
膳食糖在现代工业化饮食体系中占据了相当大的比例,现代食品中,有相当多的食品类别都含有大量的糖分成分。
因为糖在小肠中易被消化,大大减少了到达结肠肠道微生物群的糖量。然而,在糖摄入过量的情况下——如某些工业化饮食所预期的,未吸收的糖会溢出到达结肠,引发微生物组、免疫和神经生理学变化的级联反应,间接影响宿主摄食行为。这些机制包括短链脂肪酸产生改变、免疫-微生物组相互作用和迷走神经重塑。
√ 过量的糖到达结肠可能会抑制拟杆菌等纤维降解菌
葡萄糖通常几乎完全在小肠中被吸收,而果糖代谢变化更大。当果糖到达结肠肠道微生物群落时,通过抑制编码关键定植因子的roc基因来抑制关键的纤维降解细菌,如多形拟杆菌(Bacteroides thetaiotaomicron)。
这种转变破坏微生物群落稳定性,可能导致短链脂肪酸产生减少。鉴于短链脂肪酸作为代谢信号分子,通过迷走神经通路和神经递质调节参与肠脑轴,其可用性降低会扰乱摄食行为和代谢稳态。
√ 膳食糖影响辅助性T17细胞进而影响饮食行为
与直接促进微生物代谢产生生物活性代谢物的膳食纤维不同,膳食糖对微生物组与免疫相互作用产生更间接影响。其中一种机制是通过消耗辅助性T17(Th17)细胞,这种免疫细胞群对肠道稳态至关重要。高糖饮食有利于适应糖的微生物扩张,而牺牲Th17诱导的共生体,导致肠黏膜中Th17细胞丢失。
Th17细胞破坏减少IL-17产生,这种细胞因子直接作用于下丘脑神经元,通过增加POMC表达和激活厌食信号通路抑制食物摄入,从而通过抑制这种免疫来源的饱腹感信号改变摄食行为。同时,Th17细胞通过IL-17调节肠道脂质吸收,并下调CD36(关键脂质转运蛋白)。这种免疫调节功能丧失增加脂质吸收,表明微生物组介导的糖摄入代谢后果与免疫抑制和微生物破坏交织。
这些发现共同强调了膳食糖塑造饮食行为的独特但未被充分认识的途径——不是通过作为微生物底物,而是通过以驱动代谢和神经生理学效应的方式重新编程宿主-微生物相互作用。
尽管膳食糖与微生物组结构变化、免疫调节和迷走神经重塑相关,但证明这些变化如何转化为摄食行为改变的直接实验证据仍然有限。然而,这一途径对现代饮食具有重要意义,因为长期摄入糖可能通过持续的微生物组-肠-脑改变强化失调的摄食行为。
3
膳食蛋白质
蛋白质杠杆理论假设生物体优先考虑蛋白质摄入而非总能量摄入,当膳食蛋白质被稀释时,会导致脂肪和碳水化合物的代偿性过度消耗。这种现象在包括人类在内的各物种中均可观察到,强调蛋白质食欲是摄食行为的重要驱动因素。尽管研究主要集中在宿主机制上,如氨基酸感觉检测、激素反馈回路和控制蛋白质食欲的中枢神经通路,但肠道微生物组在塑造蛋白质驱动摄食行为中的作用在很大程度上仍未探索。
√ 微生物氨基酸合成可抑制宿主代偿性摄食
越来越多证据表明肠脑信号传导在蛋白质稳态中起核心作用,新发现暗示微生物对这一调节网络的贡献。肠道微生物通过调节氨基酸可用性缓冲宿主对抗膳食蛋白质稀缺:用产生必需氨基酸的细菌Acetobacter pomorum定植的果蝇表现出减少的蛋白质寻找行为,表明微生物氨基酸合成可抑制宿主代偿性摄食。
相比之下,乳酸杆菌缺乏必需支链氨基酸(亮氨酸、异亮氨酸、缬氨酸)的生物合成途径,在必需氨基酸剥夺时不会抑制蛋白质食欲,这与该食欲调节作用需要必需氨基酸微生物生物合成的假设一致。这表明微生物对代偿性摄食反应的抑制取决于特定的生物合成能力。
√ 不同饮食下的微生物群落会影响宿主的摄食偏好
微生物组驱动的与必需氨基酸相互作用也可能影响更广泛的常量营养素偏好。在草食性供体肠道微生物中定植的小鼠表现出血浆色氨酸升高和对蛋白质的更大偏好,而在杂食性或肉食性供体肠道微生物中定植的小鼠消耗更多碳水化合物。其他一些研究已将犬尿氨酸途径与微生物组驱动的色氨酸代谢对摄食行为的影响联系起来。
色氨酸向犬尿酸的微生物代谢通过涉及迷走神经传入和中枢食欲通路的肠脑回路调节摄食行为。慢性压力和节食史改变肠道微生物组成并降低管腔犬尿酸水平。这种消耗抑制迷走神经末梢,通过孤束核和室旁核过度激活导致食物摄入量增加。通过补充益生菌或直接给药犬尿酸恢复其水平,使摄食行为正常化,证实犬尿氨酸途径是强迫性暴饮暴食的微生物代谢调节剂。
√ 肠脑轴信号和微生物代谢共同调节蛋白质摄入
蛋白质稳态依赖于肠道来源神经肽,这些神经肽参与肠道和中枢回路调整摄食行为以响应氨基酸可用性,肠道微生物组通过促进营养平衡进一步塑造这一过程。微生物合成必需氨基酸时,其生物合成抑制宿主因蛋白质稀缺而产生的代偿性摄食。这些机制共同证明宿主驱动的肠脑信号传导和微生物代谢都调节蛋白质摄入。
√ 不同蛋白质对肠道微生物群的影响存在差异
蛋白质对肠道微生物群组成的影响因类型而异。食用动物蛋白(特别是红肉和奶制品中的蛋白质)可能导致耐胆汁厌氧菌(如拟杆菌门、Alistipes和嗜胆菌门)丰度增加。这些肠道微生物群改变诱导氧化三甲胺(TMAO)增加。
TMAO是一种具有促动脉粥样硬化潜力的化合物,在心血管疾病中发挥重要作用。
另一项研究表明,硫酸盐还原菌从膳食中的无机硫和硫酸化氨基酸(即蛋氨酸、半胱氨酸、胱氨酸和牛磺酸)中产生硫化氢,可能增加炎症性肠病风险。此外,动物蛋白发酵降低了双歧杆菌丰度和短链脂肪酸产生,可能进一步增加炎症性肠病风险。
而食用植物蛋白(如豌豆蛋白)会增加肠道共生双歧杆菌和乳酸杆菌,并减少致病性脆弱拟杆菌和产气荚膜梭菌。在小鼠模型中,西式饮食3周后补充大豆浓缩蛋白导致双歧杆菌科、梭状芽胞杆菌科和递叶杆菌科丰度显著增加,拟杆菌门水平降低。植物蛋白的发酵与双歧杆菌和乳酸杆菌丰度增加相关,从而刺激短链脂肪酸产生。
√ 优化饮食中纤维和蛋白质的比例可能有助于改善代谢和微生物群
膳食纤维和蛋白质对肠道微生物组组成和微生物衍生代谢物的影响有据可查,突显它们对宿主饮食行为的重要性。尽管不可消化碳水化合物是微生物介导短链脂肪酸产生的主要底物,但结肠中未消化的膳食蛋白质和氨基酸也可作为短链脂肪酸合成前体。
纤维和蛋白质衍生代谢物间的相互作用可能抵消与西方饮食相关的一些不良代谢影响,西方饮食通常纤维含量低,脂肪和蛋白质含量高。饮食的代谢和微生物结果不仅取决于单个常量营养素存在,还取决于它们的相对比例和相互作用。
将可发酵纤维与优质蛋白质来源共同给药,可通过增加微生物对可发酵底物暴露来减少直接能量吸收并调节肠道微生物组组成。同时,降低高脂肪饮食中蛋白质与纤维比例可改善血糖控制和血脂状况,同时有利于产生短链脂肪酸的细菌类群扩张。
确定优化纤维与蛋白质比例是否可将微生物代谢转向增加短链脂肪酸产生和减少蛋白水解副产物,可能揭示抵消与纤维缺乏西方饮食相关饮食失调和代谢功能障碍的策略。
4
膳食脂肪
膳食脂肪通过热量负荷或感觉奖励以外的机制影响宿主摄食行为,新证据表明肠道来源代谢信号与脂肪偏好和饱腹感调节有关。工业化饮食的高脂肪和低纤维含量迅速使肠道微生物群落转向胆汁耐受生物,同时减少纤维发酵物种,导致包括胆汁酸代谢改变在内的功能变化。
√ 膳食脂肪改变微生物胆汁酸代谢影响脂肪感应
胆汁酸是微生物组衍生的关键代谢产物,将饮食与宿主代谢和摄食神经控制相结合。胆汁酸传统上因其在脂质乳化和吸收中的作用而受到认可,现在被认为是参与肠道激素通路、迷走神经传入神经元和中枢食欲回路的生物活性信号分子。
饮食、肠道微生物组组成、胆汁酸组成和摄食行为间的相互关系表明存在动态调节系统,其中膳食脂肪改变微生物胆汁酸代谢和胆汁酸组成,进而调节脂肪感应、饱腹感和食物摄入量。
高脂肪饮食通过重塑肠道微生物组和胆汁酸库破坏这种调节平衡,对肠脑信号传导产生下游影响。这些转变部分由微生物解偶联和初级胆汁酸脱羟基化为二级形式驱动,改变肠道和肝脏中的受体信号传导。例如,暴露于西方饮食的小鼠表现出胆汁酸组成变化,降低参与代谢调节的关键受体激活,包括法尼醇X受体和G蛋白偶联受体5,导致肠道激素分泌受损和代谢功能障碍。在人类中,不受控制的进食行为与扭链瘤胃球菌(Ruminococcus torques)和双歧杆菌属(Bifidobacterium spp)富集以及参与胆汁酸代谢和神经递质产生的基因表达减少有关。
高脂肪饮食对肠道微生物群和粘液屏障的影响
doi: 10.3390/nu11102393.
√ 胆汁酸可以充当肠道信号影响饱腹感和摄食行为
胆汁酸组成对摄食行为的影响是动态的,因为胆汁酸还影响膳食脂肪的感知和代谢方式。由关键胆汁酸产生酶CYP8B1破坏引起的小鼠12α-羟基化胆汁酸丢失会改变肠道脂质感应。这种变化增强GPR119介导的肠道激素分泌,从而增加饱腹感并减少高脂肪饮食消耗。
虽然胆汁酸主要通过肠道激素信号传导和迷走神经激活调节脂肪消化和饱腹感,但内源性大麻素通过直接参与奖励途径加强脂肪偏好来扩展这种调节功能。内源性大麻素系统通过CB1受体介导的抑制和兴奋性输入微调多巴胺和血清素信号传导,抑制奖励相关区域的多巴胺释放,同时以特定区域方式调节血清素传递——这些机制共同塑造包括摄食和强化学习在内的动机行为。
√ 肠道微生物群通过调节内源性大麻素水平诱导脂肪摄入
饮食引起的肠道微生物组组成变化通过调节生物活性内源性大麻素的合成和降解影响这些过程,导致回肠和血浆内源性大麻素水平平行变化。因此,限制和间歇性暴露于高脂肪饮食会改变大脑内源性大麻素信号传导并诱导暴饮暴食样行为。
内源性大麻素信号传导对膳食脂肪摄入的选择性反应。在受控喂养模型中,肠道内安非他命和2-花生四烯酰甘油水平对脂肪有反应但对碳水化合物或蛋白质无反应,迅速升高。直接给药这些内源性大麻素足以诱导脂肪偏好,迷走神经切断术消除了它们在空肠中脂肪诱导的积累和行为反应,加强了肠脑轴作为脂肪驱动奖励信号传导渠道的作用。
一个关键问题是肠道微生物组本身是否合成影响宿主食物摄入的生物活性脂质介质。肠道细菌可将膳食中的ω-3多不饱和脂肪酸转化为内源性大麻素样分子,表明微生物代谢可能至少扩展了宿主脂质信号传导的范围。
5
盐
世界卫生组织推荐盐的最大摄入量为5g/天。高盐饮食是高血压、肾损伤和心血管疾病发展的主要危险因素之一。高盐饮食还通过直接损害胃粘膜导致上皮增生,与胃癌风险增加相关。此外,考虑到幽门螺杆菌CagA阳性菌株是胃癌风险的标志物,当暴露于高盐饮食时,胃幽门螺杆菌通过诱导CagA表达来改变其毒力。
√ 高盐饮食会减少乳杆菌等有益菌并促进炎症
几项小鼠模型研究表明,高盐饮食与乳酸菌属、Oscillibacter、Pseudoflavonifractor、Clostridium XIVa、Johnsonella和Rothia的丰度降低相关,同时Parasutterella、Erwinia、Christensenellaceae、Corynebacteriaceae和瘤胃球菌属的丰度增加。
特别是,与高盐饮食相关的乳酸菌属减少会增加Th17细胞,这是许多炎症性疾病的促炎标志。进一步研究表明,高盐饮食诱导的小鼠结肠炎恶化与乳酸菌属减少有关,导致保护性短链脂肪酸产生改变,这些变化改变了肠道免疫稳态,增加了对炎症损伤的易感性。高盐饮食可能导致肠道菌群组成改变,厚壁菌门/拟杆菌门比率增加,从而改变与肠道通透性和免疫稳态变化相关的短链脂肪酸产生。
6
微量营养素
微量营养素在塑造肠道微生物群中起着重要作用,而肠道微生物群又是介导其保护性健康影响的有效参与者。
√ 许多拟杆菌和梭菌都能合成B族维生素
肠道微生物群可以合成多种维生素,包括硫胺素(VB1)、核黄素(VB2)、烟酸(VB3)、生物素(VB7)、泛酸(VB5)、叶酸(VB9)等B族维生素以及维生素K。拟杆菌门、梭菌门和变形菌门的细菌许多具备核黄素和生物素生物合成途径,大部分梭菌门都能产生维生素B12,而拟杆菌门是B族维生素的主要产生者。
√ 维生素会影响肠道菌群组成并可能对免疫和炎症性肠病有益
另一方面,研究表明维生素D等维生素会影响肠道微生物群组成。维生素D通过调节肠道微生物组和增加有益细菌菌株丰度,对炎症性肠病患者产生积极影响。在3-6个月大婴儿的研究中,维生素D与Lachnobacterium增加和乳球菌减少相关,这些变化可能对免疫系统调节和哮喘/过敏性疾病发病率产生长期影响。
其他抗氧化维生素,如负责许多水果和蔬菜黄色、橙色和红色的类胡萝卜素,也可能影响肠道微生物群组成。最近一项人类植物疗法研究确定叶黄素是黑醋栗提取物粉末的成分,并证明叶黄素显著促进双歧杆菌和乳酸杆菌生长,同时减少拟杆菌属和梭状芽胞杆菌属等其他细菌种群。
另一方面,血清类胡萝卜素水平可能受肠道微生物群组成影响。研究肠道宏基因组改变与动脉粥样硬化的关联,提出健康对照血清中高水平β-胡萝卜素可能由于肠道微生物群产生这种抗氧化剂。该研究表明β-胡萝卜素的抗炎作用由肠道微生物群介导。
7
食品添加剂
随着加工食品发展,食品添加剂(如非营养性甜味剂和乳化剂)数量飙升。人造甜味剂几乎被添加到所有加工食品中,通常有助于提高稳定性和保质期,并改善味道和质地。
√ 人造甜味剂可能改变肠道菌群并诱导不良反应
食用人造甜味剂会改变肠道菌群,并在宿主中诱导微生物群介导的不良反应(如葡萄糖耐受不良)。无热量人工甜味剂改变了微生物代谢途径,并将这些变化与宿主对代谢疾病的易感性联系起来。
几项研究证实,人工甜味剂摄入增加了拟杆菌属和一些梭菌属丰度,降低了梭状芽胞杆菌属、双歧杆菌属和乳酸菌属丰度,同时空腹血糖水平升高和胰岛素反应受损。
√ 膳食乳化剂可能促进全身炎症和菌群失调
除人造甜味剂外,膳食乳化剂(如卵磷脂、脂肪酸的甘油单酯和甘油二酯)可在体外增加细菌跨上皮细胞易位,促进全身炎症,改变微生物群定位和组成。乳化剂摄入降低了肠道微生物多样性和拟杆菌门丰度,增加了阿克曼菌(Akkermansia muciniphila)等粘液降解细菌的丰度。这些微生物群改变易导致菌群失调和慢性肠道炎症,促进结肠炎和代谢综合征发生。
感知和品尝营养物质的能力在维持营养平衡、食用有益食物以及识别变质食物方面起着至关重要的作用。动物模型已证明,特定信号机制会影响不同口味(甜、苦、咸、酸和鲜)的反应性。因此,肠道微生物对这些信号传导机制的调节可影响宿主摄取特定营养物质,从而增加它们的食物底物和存活率。
√ 饮食失调会导致对应的味觉反应改变
患有饮食失调的个体具有与其表型相对应的味觉反应性改变。事实上,肥胖与对甜味和脂肪味的反应性降低有关,导致人们转向偏好这些味道浓度较高的食物。此外,神经性厌食症患者的味觉受损,这已被证明会随着体重增加而改善。
舌头上的味觉受体细胞检测传递味觉的信号,随后通过孤立束传递到丘脑,向其他大脑区域发出信号。这些味觉受体细胞还表达厌食激素如GLP-1、PYY和胆囊收缩素,表明存在通过胃肠道激素分泌导致食欲下降的外周信号通路。
注:PYY敲除小鼠对脂肪和苦味化合物的行为反应降低。在这些小鼠中重建唾液PYY浓度会改善它们对脂肪的反应,但不会改善苦味反应。此外,在味觉反应性增加的个体中检测到较高浓度的循环TNF-α、胰岛素样生长因子1和瘦素,表明这些特定循环激素可能在味觉感知中发挥作用。
▸ 肠道微生物影响味觉
很少有研究调查肠道微生物群在宿主营养和味觉感应中的作用。最近的研究表明,使用选择性培育的西方低糖精和高糖精消耗大鼠模型,天生更易摄入糖精的大鼠具有与不易摄入糖精的大鼠不同的肠道微生物群组成。
√ 肠道微生物群通过影响味觉受体影响饮食偏好
此外,对无菌小鼠的研究表明,肠道微生物群能够影响味觉受体,从而影响味觉。例如,据报道小鼠中肠道甜味受体增加,蔗糖偏好增加。此外,小鼠对舌FA转位酶的口服偏好和表达增加,但FA受体的肠道表达降低,表明脂肪信号转导减少。
关于肠道微生物群对营养和味觉感应以及味觉功能影响的知识表明,这些影响可通过免疫系统调节分化味觉受体细胞的持续供应来实现。这些细胞在味觉化合物检测中必不可少,并通过味蕾细胞直接或间接传递后续信号,从而产生口腔味觉感知。
√ 肠道微生物可能通过调节免疫功能影响味觉
分化味觉受体细胞的持续供应对正常味觉功能至关重要,而这种供应的中断可能对味觉信号传导有害。免疫系统激活导致舌头上味觉受体和味蕾细胞的细胞更新和寿命缩短,这可能通过哺乳动物toll样受体以及I型和II型IFN受体介导,因为它们位于味觉细胞中。几项研究进一步指出免疫系统功能与味觉相关。例如,IL-10敲除小鼠的味蕾和味觉受体细胞数量减少,并且对脂多糖诱导的炎症反应增加。此外,TNF敲除小鼠对各种苦味化合物的反应降低,但对其他口味的反应也降低。
细菌衍生毒素脂多糖会导致舌头炎症反应,同时味觉细胞寿命缩短和味觉偏好降低。此外,小鼠长期口服脂多糖会导致甜味受体表达降低和对蔗糖的反应降低。
值得注意的是,脂多糖给药还会导致包括快感缺乏和焦虑成分的疾病行为,因此在研究体内行为时可能是混杂因素。然而,脂多糖给药降低了舌味上皮中c-Fos的表达,表明存在局部抑制的细胞活性。此外,将脂多糖皮下注射到舌腹表面会导致白细胞募集,并通过主要感觉传入神经鼓索抑制钠诱导的味觉信号传导。
综合来看,这些结果表明肠道微生物群可通过免疫系统调节影响宿主的营养和味觉信号传导,从而影响味觉功能,而脂多糖是其中的潜在介质。
▸ 肠道微生物影响其他饮食相关行为
除了味觉功能外,其他与饮食相关的行为很可能受到肠道微生物群及其代谢物的影响。尽管尚未广泛研究与肠道微生物群的直接联系,但嗅觉功能、与食物相关的认知加工、与食物相关的冲动和强迫性以及食物摄入的社会方面很可能容易受到微生物组调节。
√ 肠道微生物可能在食物成瘾中起关键作用
最近有假设肠道微生物群在成瘾中起关键作用,这可能与有争议的食物成瘾概念特别相关。肠道微生物群已经与小鼠的可卡因使用和人类的酒精依赖有关。同样地,大麻的主要精神活性成分Δ9-四氢大麻酚的给药缓解了高脂肪饮食诱导的与肠道微生物群改变相关的脂肪量和体重增加。
关于肠道微生物群在食物成瘾中的作用,特别是其他与饮食相关的行为,仍有很多未知数,因此需要进行更多研究,以更全面地了解肠道微生物群在饮食相关行为中的作用。
√ 微生物可能影响饮食紊乱导致焦虑和抑郁
饮食和代谢紊乱通常与焦虑和抑郁等其他行为合并症相关,因此这些行为可能存在很强的双向相互作用。影响这些食物相关行为的微生物或微生物衍生代谢物也有可能会改变宿主的食欲和新陈代谢。
微生物群-肠-脑轴的影响在焦虑和抑郁等行为中也越来越受到认可。因此,关于焦虑、压力、抑郁、社交和其他行为的发现对于全面了解饮食和代谢紊乱至关重要。
最近的一项研究表明,住院时神经性厌食症患者的肠道微生物群组成和多样性与抑郁、焦虑和饮食失调精神病理学水平相关。此外,许多与微生物群-肠-脑轴相关的食物相关行为发现通常通过肠道微生物群与宿主代谢和食欲相互作用的途径介导,包括免疫系统调节、迷走神经信号传导和神经化学物质产生。
▸ 工业化饮食塑造的不良饮食行为
工业化社会中饮食失调的患病率不断上升,反映出饮食行为与调节饱腹感的内部信号脱节。超加工食品作为工业化饮食的标志,可以通过增强奖励敏感性和鼓励超越生理饱腹感信号的摄入来发挥作用。
√ 超加工食品导致微生物多样性减少和氧化应激
微生物代谢被视为连接工业化饮食与适应不良饮食行为的潜在因素。工业化饮食通过提供高度加工的饮食底物,改变肠腔中的资源及其对宿主健康的广泛影响,从而塑造肠道微生物组及其代谢输出。
肠道微生物代谢物的下游变化既是饮食行为改变的潜在驱动因素,也是其强化因素,特别是通过对奖励途径和炎症过程的影响。由饮食和生活方式塑造的工业化微生物组表现出微生物多样性减少和氧化应激相关途径丰富,反映了生态脆弱和促炎状态。
通过这些级联效应,工业化饮食与微生物组-肠-脑轴之间的相互作用有可能形成一个生化反馈循环,从而促进食物成瘾和其他病态饮食行为。
√ 当代高糖低纤维的饮食改变了微生物组成及其代谢物
尽管在工业化和非工业化环境中人类饮食差异很大,但工业化饮食通常含有较少的纤维、较多的糖、蛋白质和脂肪,以及较高的总体热量。常量营养素平衡的这些变化预计将改变与宿主信号通路相互作用的微生物组衍生代谢物。
例如,膳食纤维减少会减少短链脂肪酸的微生物产生,而短链脂肪酸是饱腹感和代谢调节的关键信号。因此,短链脂肪酸信号传导受损可能会削弱控制饱腹感的肠脑通路,从而促进暴饮暴食和热量过剩。
膳食糖的增加驱动了适应糖的微生物丰度增加,牺牲了纤维发酵微生物,并通过短链脂肪酸的丢失直接损害了食欲调节。升高的膳食蛋白质驱动微生物代谢产生神经活性氨基酸代谢物,如犬尿酸和酪胺,这些代谢物能够破坏宿主营养感知并加强寻求蛋白质的摄食行为。
同样,高膳食脂肪摄入量会增加参与胆汁酸代谢的胆汁耐受生物的丰度,这些变化有可能激活奖励途径并促进针对脂肪食物的强迫性饮食行为。
随着全球肥胖症和代谢性疾病的流行,传统治疗方法面临疗效有限和副作用明显的挑战。近年来,肠道微生物群作为人体”第二基因组”的重要作用日益受到关注,在调节宿主代谢和食欲控制方面展现出巨大治疗潜力。
研究表明,肠道微生物群不仅参与食物消化和营养吸收,更通过产生短链脂肪酸、胆汁酸代谢产物和神经递质前体等生物活性代谢物,直接影响宿主的能量平衡、糖脂代谢和摄食行为。这些发现使肠道微生物群及其代谢物成为代谢和食欲控制领域极具前景的治疗靶点。
肠道微生物代谢物影响宿主食欲和新陈代谢
doi: 10.3945/jn.116.240481.
√ 益生元能增加饱腹感并降低餐后血糖水平
益生菌和益生元已被证明是显著改善超重和肥胖个体新陈代谢的有效策略。啮齿动物研究表明,补充益生菌可以促进饥饿的恢复,特定细菌菌株会影响体重或超重肥胖相关症状。
宿主难以消化的膳食纤维等益生元会增加饱腹感并影响餐后葡萄糖和胰岛素浓度。据报道,它们还与能量摄入、体重、胰岛素分泌、循环脂质和炎症标志物的减少相关。
√ 肠道菌群代谢物影响宿主代谢及食欲和摄入
微生物代谢物通过多种途径影响宿主代谢,包括与胃肠道和外周组织的直接相互作用。这些相互作用包括:通过表观遗传机制影响宿主基因表达、影响肠道神经系统并直接诱导迷走神经信号传导、改变胆汁酸信号传导,以及影响整合宿主能量状态的中枢食欲通路。
肠道微生物代谢物调节宿主食欲和新陈代谢控制。摄入的营养物质被肠道微生物利用,导致微生物群组成和功能改变。这些肠道微生物影响胆汁酸代谢,并产生各种代谢物,包括短链脂肪酸、神经活性物质、小蛋白序列和毒素。
部分肠道微生物代谢物通过与肠道中肠内分泌L细胞或迷走神经上的受体直接相互作用,或通过肠上皮转位到外周循环中发挥作用。胆汁酸或短链脂肪酸刺激L细胞导致厌食激素PYY和GLP-1的释放,并可增加胰岛素、瘦素和生长素释放肽等外周激素浓度。这些激素介导的信号转导受免疫球蛋白影响,而免疫球蛋白又可受到刺激或与肠道微生物衍生的小蛋白序列(如ClpB)相互作用。这种蛋白质序列以及肠道微生物衍生毒素(如LPS)的影响受肠道屏障完整性高度影响。
脂多糖还能够诱发免疫反应,可能导致味觉受体细胞分化减少和味觉检测降低。这些通路累积影响食欲和新陈代谢,受下丘脑中促阿片黑皮质素和可卡因苯丙胺调节转录物以及含有神经肽Y和刺豚鼠调节肽的神经元高度调控,最终影响进食行为、饮食相关行为(如认知和冲动)以及与食欲和代谢障碍相关的行为合并症功能失调(如焦虑和抑郁)。
▸ 作用机制
①微生物群-肠-脑轴的核心作用
肠道微生物群通过微生物群-肠-脑轴影响宿主代谢和食欲控制,主要通过以下途径:
-神经途径:通过迷走神经和肠神经系统直接信号传导;
-激素途径:调节GLP-1、PYY、瘦素、胰岛素等代谢激素分泌;
-免疫途径:调节慢性低度炎症和免疫反应;
-表观遗传机制:通过表观遗传修饰影响宿主基因表达。
②关键代谢物及其功能
–短链脂肪酸(SCFA)是最重要的微生物代谢物:
乙酸盐、丙酸盐、丁酸盐由纤维发酵产生,刺激肠内分泌L细胞释放厌食激素(PYY、GLP-1),可穿越血脑屏障直接调节下丘脑食欲中枢,激活迷走神经传入信号,抑制食物摄入。
-胆汁酸代谢产物:调节脂质代谢和能量平衡。
-GABA(γ-氨基丁酸)作为肠道微生物产生的多种神经活性物质之一,与以下物质共同调节宿主代谢和食欲:
组胺:影响食欲和代谢;
乙酰胆碱:调节胃肠道功能;
色胺:参与情绪和食欲调节;
儿茶酚胺(去甲肾上腺素、多巴胺):调节奖励机制和食欲;
血清素:调节情绪、饱腹感和胃肠道蠕动。
-小蛋白序列:如ClpB,影响激素信号传导。
√ 工业化饮食通过破坏微生物群-肠-脑轴造成了以下负面影响:
促进菌群失调:有益菌减少,有害菌增加;
损害饱腹感信号:影响GLP-1和PYY分泌;
增强奖励反应:激活大脑奖励回路,促进过度进食;
诱发慢性炎症:破坏肠道屏障,引起全身炎症;
形成恶性循环:代谢物谱改变强化不良饮食行为。
▸ 应对措施
针对现代高度加工食品对人体健康和肠道微生物群带来的各种负面影响,目前研究和临床实践中主要采用两大类应对策略:一类是通过益生菌、益生元等方式进行的直接微生物调节方法,另一类是通过调整膳食结构和营养成分的饮食干预策略。
√ 直接微生物调节方法
调节肠道菌群的主要方法有四种:
1)施用活的有益细菌菌株(益生菌);
2)施用宿主难消化的膳食纤维,这些纤维经细菌发酵后刺激特定细菌生长(益生元);
3)施用靶向抗生素;
4)粪便微生物群移植。
益生菌治疗:给予特定有益菌株(如Akkermansia muciniphila、Lactobacillus等);
分泌菌株特异性代谢物,改善代谢内毒素血症;
需要持续补充以维持效果。
益生元补充:提供宿主难消化的膳食纤维(如菊粉、低聚糖)选择性促进有益菌生长;
增加SCFA产生,改善饱腹感信号。
靶向抗生素:选择性清除有害菌群;
需要精准识别病原菌;
避免影响有益菌群。
粪菌移植:从健康供体移植微生物群;
适用于严重菌群失调病例;
需要严格的安全性评估。
√ 饮食干预策略
“去工业化”饮食模式
高纤维摄入:每日至少25-35g膳食纤维;
最小化加工:选择全谷物、新鲜蔬果;
低能量密度:减少高糖高脂食品;
植物为主:增加植物性食物比例。
增加发酵食品摄入(酸奶、发酵蔬菜等);
多样化膳食纤维来源(不同类型蔬果、谷物);
控制精制糖和加工食品摄入;
规律饮食,避免暴饮暴食。
想象一下,未来的减肥和健康管理将是什么样子?不再是千篇一律的节食食谱,也不是痛苦的意志力考验。你的医生或营养师可能会给你开一份特殊的“处方”:一份为你量身定制的“菌群优化”饮食方案。
·早餐:可能不再是简单的牛奶鸡蛋,而是一杯富含特定益生元的营养奶昔,专门用来唤醒你肠道中负责传递饱腹感信号的菌群。
·午餐:你的外卖软件可能会根据你上周的菌群报告,为你优先推荐富含短链脂肪酸前体的菜品。
·零食:你甚至可以享用专门设计的“益生菌软糖”,它不仅能解馋,还能精准地为你的肠道“盟友”补充能量。
通过精准调控肠道这个“第二大脑”,我们不仅能从根源上解决肥胖、糖尿病等代谢难题,更能彻底重塑我们与食物的关系——让吃饭回归健康与愉悦的本质。
现代饮食“高糖高脂低纤维”的组合拳,不仅喂饱了我们,更喂养出了一支会“胁迫”大脑、破坏代谢平衡的“叛逆”菌群。喂养你的“好菌”:别再只喂饱你自己,更要喂饱你体内的有益菌。多吃富含膳食纤维的食物,比如燕麦、豆类、全谷物和五颜六色的蔬菜水果。它们是益生菌最爱的“口粮”,能帮你产生更多有益的代谢物,向大脑发送“我饱了”的信号。
好消息是,我们已经找到了扭转战局的钥匙。未来的医学,将不再仅仅关注疾病本身,而是会深入到肠道以及肠道的亿万微生物,这意味着,我们有望从根本上告别“管不住嘴”的困扰,让身体自然而然地渴望健康食物。这一前沿领域不仅为攻克肥胖和代谢综合征带来了曙光,更将引领一场全民性的饮食观念升级:善待你的肠道菌群,就是善待你自己。
主要参考文献
Liow YJ, Sarkar A, Carmody RN. Industrialized diets modulate host eating behavior via the microbiome-gut-brain axis. Trends Endocrinol Metab. 2025 Jul 17:S1043-2760(25)00125-0.
Samulėnaitė S, García-Blanco A, Mayneris-Perxachs J, Domingo-Rodríguez L, Cabana-Domínguez J, Fernàndez-Castillo N, Gago-García E, Pineda-Cirera L, Burokas A, Espinosa-Carrasco J, Arboleya S, Latorre J, Stanton C, Hosomi K, Kunisawa J, Cormand B, Fernández-Real JM, Maldonado R, Martín-García E. Gut microbiota signatures of vulnerability to food addiction in mice and humans. Gut. 2024 Oct 7;73(11):1799-1815.
van de Wouw M, Schellekens H, Dinan TG, Cryan JF. Microbiota-Gut-Brain Axis: Modulator of Host Metabolism and Appetite. J Nutr. 2017 May;147(5):727-745.
Rinninella E, Cintoni M, Raoul P, Lopetuso LR, Scaldaferri F, Pulcini G, Miggiano GAD, Gasbarrini A, Mele MC. Food Components and Dietary Habits: Keys for a Healthy Gut Microbiota Composition. Nutrients. 2019 Oct 7;11(10):2393.
Medawar E, Haange SB, Rolle-Kampczyk U, Engelmann B, Dietrich A, Thieleking R, Wiegank C, Fries C, Horstmann A, Villringer A, von Bergen M, Fenske W, Veronica Witte A. Gut microbiota link dietary fiber intake and short-chain fatty acid metabolism with eating behavior. Transl Psychiatry. 2021 Oct 1;11(1):500.
McDougle M, de Araujo A, Singh A, Yang M, Braga I, Paille V, Mendez-Hernandez R, Vergara M, Woodie LN, Gour A, Sharma A, Urs N, Warren B, de Lartigue G. Separate gut-brain circuits for fat and sugar reinforcement combine to promote overeating. Cell Metab. 2024 Feb 6;36(2):393-407.e7.
Li M, Tan HE, Lu Z, Tsang KS, Chung AJ, Zuker CS. Gut-brain circuits for fat preference. Nature. 2022 Oct;610(7933):722-730.
Tan HE, Sisti AC, Jin H, Vignovich M, Villavicencio M, Tsang KS, Goffer Y, Zuker CS. The gut-brain axis mediates sugar preference. Nature. 2020 Apr;580(7804):511-516.