-

 国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子
国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子

 二级病原微生物安全实验室
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业

谷禾健康

随着生活水平提升,我国养宠人士逐年增多。数据显示,我国城镇养宠人士达到6844万多人,且仍在不断增长。
其中养猫人群数占比59.5%,养狗人群数占比51.7%,养水族类占比8.3%,养爬行类宠物的占比为5.8%;啮齿类占比4.5%;鸟类占比3.6%。
养宠物,它们的健康绝对是重要的一方面。同时宠物的健康在一定程度上也会影响人们的健康。
胃肠道微生物在健康中起着重要作用。宠物的胃肠道 (GI) 微生物群也是与健康密不可分的代谢活跃器官。肠道微生物群有助于宿主新陈代谢、抵御病原体、影响免疫系统,并通过这些方式直接或间接影响宿主的大部分生理功能。
本文在这里主要介绍猫狗体内的肠道微生物,饮食与肠道微生物的关联以及肠道微生物的作用和对宿主健康的影响,并分析疾病中发生的变化。
在此基础上提出一些有助于恢复健康肠道微生物环境的建议,帮助宠物及主人拥有更好的健康。
本文主要从以下几个方面讲述
●猫狗体内的微生物
●肠道微生物的作用
●肠道菌群与宿主健康
●饮食对猫狗肠道微生物的影响
●宠物身上的微生物对主人的影响
●恢复猫狗肠道健康的方法
什么是微生物群?
微生物群是特定时间特定环境所有微生物有机体的总称,其组成包括非细胞结构的病毒(包括噬菌体)、原核生物中的真细菌和古细菌,以及真核细胞微生物。
微生物群对人类和宠物健康的广泛而深远的影响,胃肠微生物群现在被认为是具有独特代谢能力的器官。胃肠微生物群由位于消化道中的数万亿个细胞组成,从口腔开始,一直延伸到直肠。
微生物群的发育在出生前就开始了,微生物群会影响宿主健康的许多方面,包括生理学、解剖学、行为、繁殖和健康状况。
✦不同部位微生物群落不同
整个胃肠道的微生物群落不同,代表了每个肠段的微环境和生理功能,这已在狗和人类身上得到证实。
例如,对健康家犬的两项研究发现,小肠和大肠的微生物群之间存在显著差异,包括十二指肠中变形杆菌(Proteus)的比例高于结肠或直肠,以及大肠中毛螺菌属(Lachnospira)和瘤胃球菌属(Ruminococcus)的比例高于小肠。
健康狗不同部位的细菌群落组成
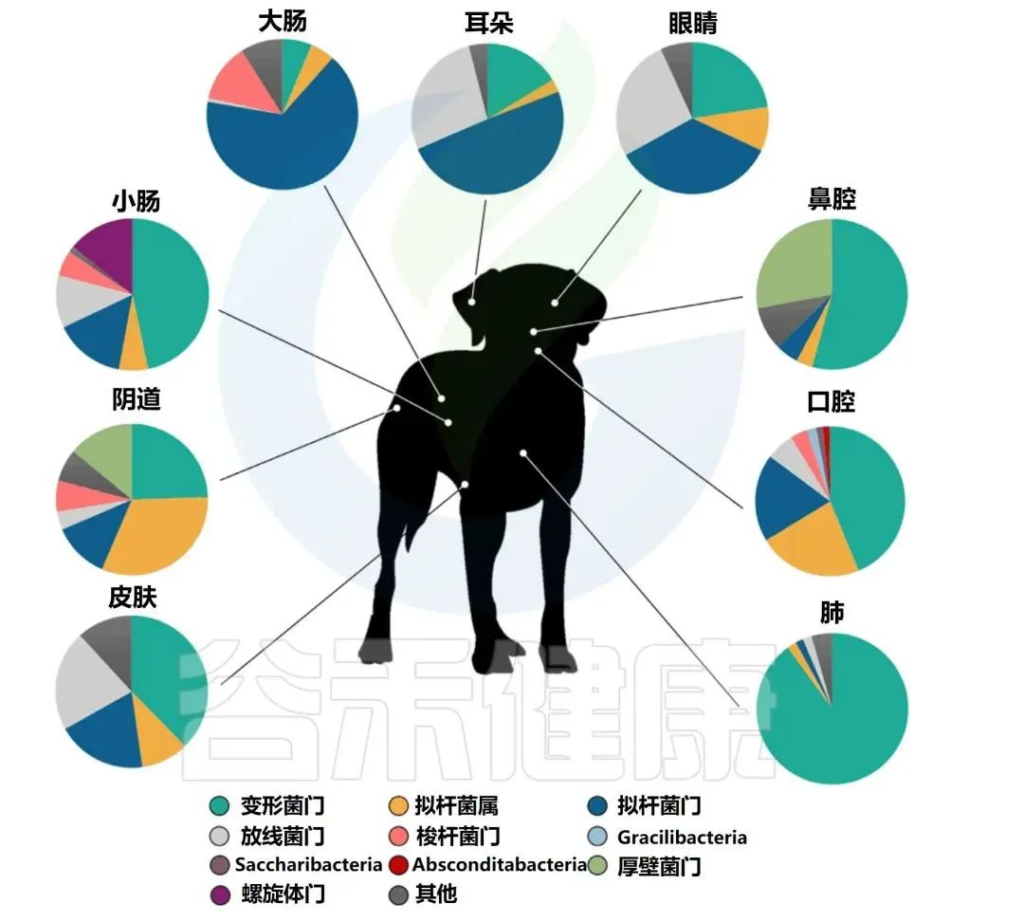
Pereira AM,et al.Top Companion Anim Med.2021
★ 主要肠道菌群
一般来说,猫和狗的肠道菌群组成相似。据报道,厚壁菌门(Firmicutes)、拟杆菌门(Bacteroides)、变形菌门(Proteobacteria)、梭杆菌门(Fusobacteria)和放线菌门(Actinobacteria)是猫和狗肠道中的主要微生物。
✦猫狗之间的细菌差异
然而,一些研究注意到猫与狗之间的肠道菌群还是存在某些差异。
• 猫比狗具有更高的α多样性
最近一项研究,基于粪便样本,比较46只猫和192只狗的肠道微生物组的差异,表明猫比狗表现出更高的α多样性。
与猫相比,在犬中增加的细菌门是肠球菌属、梭杆菌属、巨单胞菌属。
而在猫中的细菌门较多,包括另枝菌属(Alistipes)、双歧杆菌属(Bifidobacterium)、肉食杆菌属(Carnobacterium)、柯林斯菌属(Collinsella)、粪球菌属(Coprococcus)、脱硫弧菌属(Desulfovibrio)、普拉梭菌(Faecalibacterium)、颤螺菌属 (Oscillospira)、副乳杆菌属、消化球菌属、消化链球菌属、瘤胃球菌属和萨特氏菌(Sutterella).
在较早的分析中还观察到猫与狗之间的胃肠道微生物组更加多样化,然而,同一项研究也揭示了猫中大多数细菌丰度的个体间差异较小,而具有相同细菌属的猫数量较多。
✦猫狗之间的真菌差异
猫和狗之间的差异在真菌微生物组中也很明显,Nakaseomyces在狗中占主导地位,而酵母菌、曲霉属和青霉属在猫中含量更高。这种差异可能是由于微生物组对不同饮食的适应。
注:与猫的肉食性饮食相比,狗之间更大的个体间多样性可能是更多样化、杂食性饮食的产物。
✦ 狗和人的肠道菌群相似吗?
狗的微生物组比预期更接近人类。
欧洲分子生物学实验室 (EMBL) 研究人员发现,狗的微生物组的基因含量与人类肠道微生物组的相似性高于小鼠、猪的。
研究人员将小鼠、猪和狗微生物组中的基因内容映射到人类肠道基因目录中,发现分别有 20%、33% 和 63% 重叠。

注意
有证据表明,肠道中也有来自其他部位的微生物群。例如,已发现口腔微生物群的成员在患病的胃肠道下部定植。
由于品种、饮食、年龄、生活环境和不同研究分析方法等因素,同一物种的个体菌群比例也可能不同。然而,大多数研究只比较了少量动物,需要更大规模的队列研究来确认环境影响的存在并估计影响大小。
动物体内的肠道菌群数量庞大,类型复杂,与身体形成了一种“互惠互利”的共生关系。它们的生长依赖于身体提供的丰富营养和相对安全环境;同时它们在维护身体健康,促进正常发育等方面发挥着至关重要的作用。
正常菌群是能够在肠道中特定部位“安家落户”,即黏附、定植和繁殖,这个时候,菌群就能够在定植的部位表面形成一层“菌膜屏障”。
•维持微生态平衡状态
这层“菌膜屏障”就像是肠道表面的一层保护伞,对于流经消化道的外源性微生物(包括许多外源性病原体)形成了一种天然的隔绝,通过竞争、消化和分泌各种代谢产物和细菌素等,抵抗外源微生物定植和侵袭。
机体内的正常菌群通过这种拮抗作用,抑制并排斥机体不慎食入的病原菌在肠道的“安家落户”,维持体内微生态的平衡状态,使机体免于感染致病菌。
•提供能量
细菌代谢活动产生对细菌增殖和宿主都有用的能量和底物。主要活动是发酵酒精和不易消化的碳水化合物(淀粉、菊粉、纤维素、半纤维素、果胶和树胶)。
它发生在结肠中,它会导致产生气体和短链脂肪酸、乙酸盐、丙酸盐和丁酸盐,从而为肠上皮和其他组织提供能量。
•支持上皮细胞生长,利于物质吸收
短链脂肪酸合成导致肠道pH值降低,它支持肠上皮细胞的生长、增殖和分化。此外,当短链脂肪酸通过肠道吸收时,它们允许重新吸收Na+或K+离子。
微生物群的另一个重要作用是初级胆汁酸(胆酸和鹅去氧胆酸)和次级胆汁酸(脱氧胆酸和石胆酸)的转化,这对于肠道吸收膳食脂肪和脂溶性维生素至关重要。
共生菌对宿主免疫系统的诱导、塑造和功能具有重要作用,这反过来又对肠道结构的发育和共生菌病原体的鉴定具有重要意义。
微生物在局部和全身设置组织免疫基调的能力需要微生物的强直感应以及免疫系统先天和适应性成分之间的复杂反馈回路。肠黏膜是免疫系统与外部环境之间的界面。
•加快淋巴细胞、免疫球蛋白的增殖
几项研究的结果表明,无菌动物肠粘膜中的淋巴细胞密度较低,血清中的免疫球蛋白含量较低。这些动物接触共生微生物会导致肠道中淋巴细胞和免疫球蛋白以及血清中免疫球蛋白的快速增加。
1型和2型T辅助细胞的发育和调节受细菌的影响,细菌可以通过产生丁酸盐和乙酸盐来抑制NF-kB信号通路,并可以作用于与G蛋白偶联受体。
•诱导炎症介质产生和释放
免疫系统和微生物群之间存在动态关系。具有Toll 样受体的肠黏膜可诱导下降信号并引发炎症介质(细胞因子、吞噬细胞)的产生和释放。
•增强免疫细胞的杀伤力
此外,双歧杆菌(Bifidobacterium)能通过刺激免疫细胞产生重要的细胞因子白介素来促进动物机体内重要的免疫细胞——淋巴细胞的增殖、分化、成熟,增强免疫细胞的对病原体的杀伤力。
在健康条件下,宿主和微生物之间的串扰和交叉调节创造了细菌的稳态平衡,因此胃肠道保持健康并且没有潜在致病细菌的过度生长。
在那种情况下,存在一种共生状态。当这种体内平衡状态被破坏时,就会发生生态失调。在生态失调的情况下,细菌组成不平衡,肠道内的细菌代谢活动或细菌分布发生变化。
生态失调可以总结为以下三点:
减少细菌多样性;有益细菌的损失;病原体的过度生长。
✦肠道微生物失调可能与一些疾病有关
胃肠道微生物组直接或间接影响一系列生理和免疫功能。因此,胃肠道微生物组组成的破坏可能导致有害的后果,例如炎症性肠病、过敏、便秘、口腔疾病(即牙周病)、肥胖、糖尿病和肾脏疾病。
由于肠道微生物群影响胃肠道的环境栖息地,反之亦然,微生物相关的慢性肠病,特别是炎症性肠病的发病率和进展一直是研究的重点。
✦患有炎症性肠病的动物生物多样性降低
患有炎症性肠病的动物中更常见的生态失调的特点是微生物群落多样性普遍降低,尤其是产生短链脂肪酸减少。
•狗中大肠杆菌增加
与健康的狗相比,患有慢性肠病的狗的短链脂肪酸浓度和模式发生了改变,以及粪便微生物群的变化。
一项研究检查了来自患有炎症性肠病的狗的回肠和结肠粘膜微生物群样本,发现它们表现出附着在上皮细胞上或在这些采样部位侵入肠粘膜的肠杆菌科和大肠杆菌增加。
这些发现与最近的另一项研究一致,该研究还发现患有慢性肠病的狗结肠粘膜中的大肠杆菌增加,以及幽门螺杆菌和嗜黏蛋白阿克曼菌减少。
•猫中梭杆菌增多
对于猫,研究表明,与健康猫相比,有炎症性肠病的猫胃肠道微生物群发生了变化。一项研究还发现,患有胃肠道淋巴瘤的猫的回肠和结肠活检组织中梭杆菌属升高。然而,目前尚不清楚这种变化在胃肠道淋巴瘤发展中的任何作用。
有趣的是,最近有文献指出,与人类相比,嗜黏蛋白阿克曼菌在猫和狗的胃肠道中并不丰富,这表明这些细菌在动物粘液的微生物降解中作用还不明确。
✦改变食物可以有效减少肠病症状
两项内镜研究检查了饮食干预对诊断为炎症性的狗的影响。
这些研究发现,饮食的干预改变了位于十二指肠和结肠粘膜层的胃肠道微生物组,在几乎所有患有轻度疾病的狗中,仅改变食物就可以有效减少炎症性肠病的迹象。新型蛋白质和水解蛋白都是有效的,但水解蛋白在减少炎症性肠病症状方面稍微更有效。
此外,营养治疗对犬肠病微生物组的影响研究还分析了水解蛋白饮食对29只慢性肠病犬的微生物组和健康的影响。
参与这项研究的狗从目前的饮食改为治疗性水解蛋白饮食。在这种治疗性饮食2周后,69%的狗经历了慢性肠病的快速缓解,并持续了整个6周的研究。此外,慢性肠病的缓解与微生物群结构的改善和次级胆汁酸水平的增加有关。

•炎症性肠病改善的狗肠道微生物不同
另一项对患有炎症性肠病的狗进行的研究表明,炎症性肠病症状和体征消失的狗的微生物群与没有改善的狗不同。
那些对饮食有反应的狗的嗜胆菌属 (Bilophila)和伯克霍尔德菌(Burkholderia cepacia)占优势,拟杆菌丰富,而那些没有改善的狗的奈瑟菌科更丰富。伯克霍尔德菌尚未显示与狗的慢性肠病有关,拟杆菌可能具有保护性或毒性。
在患有食物敏感型慢性肠病的狗身上进行的一项研究发现,尽管微生物组成(β多样性)没有变化,但在喂食与合生元配对的水解蛋白食物时,细菌丰富度略有增加。
在动物中表现出胃肠道症状的食物过敏经常被误认为是炎症性肠病,因为症状重叠,最明显的是腹泻。
尽管在宠物和人类中开展的研究很少,但已发现在成年人中,花生和坚果过敏与较高的拟杆菌和梭菌、普雷沃氏菌和瘤胃球菌的减少有关。虽然坚果过敏可能与猫和狗不太相关,但这份报告表明微生物组的改变可能与过敏性疾病有关。
✦通过肠道微生物可以治疗过敏
由于过敏本质上是免疫疾病,并且肠道微生物组已被证明具有促炎和抗炎特性,这取决于定植生物体,因此针对胃肠道微生物组可能是治疗全身性过敏的一种合乎逻辑的方法,这已成为多项动物研究的主题。
✦益生菌治疗可以降低过敏反应
在狗中,发现添加益生菌会导致粪便中乙酸盐和丁酸盐浓度增加,以及改善细胞介导的对抗原攻击的免疫反应。然而,益生菌的免疫调节作用尚未在对食物反应性慢性肠病或腹泻的狗的研究中得到一致的证明。
经验证的犬特应性皮炎模型发现,早期接触益生菌(鼠李糖乳杆菌)可显著降低过敏原特异性免疫球蛋白lgE,并在生命的前6个月部分预防特应性皮炎。
这项研究的后续研究表明,在停用益生菌后,这些影响持续了3年。一项对确诊为特应性皮炎的狗进行的随机对照试验报告称,服用益生菌(清酒乳杆菌)2个月可显著降低所有接受益生菌的狗的特应性疾病严重程度。
不过应该指出的是,绝大多数评估使用益生菌治疗全身性过敏的研究都是在人类身上进行的。
猫和狗的便秘患病率没有得到很好的描述,很难量化宠物的排便频率。尽管如此,便秘可能是宠物中相对常见的临床问题,尤其是猫。

✦便秘与肠道微生物的关系还不确定
尚未确定便秘与胃肠道微生物组组成之间的明确联系,包括特定细菌群的丰度,以及肠道微生物群可能影响胃肠道感觉的确切机制和运动功能不清楚。尽管如此,证据表明存在联系,尽管主要来自人类研究,对动物的研究有限。
在一项针对20只猫的研究中,通过PCR分析的有限数量的细菌分类群在有和没有便秘的猫之间没有观察到显著差异。
对胃肠道微生物与便秘之间的机制关系的研究甚至更少,特定细菌类群对胃肠道运动的影响还远未得出结论。
•肠道细菌和产生的代谢物可能会影响肠道运动
一项体外研究表明,梭菌、乳杆菌、脱硫弧菌和甲基杆菌的丰度降低可能通过影响血清素受体来影响胃肠道运动,从而导致慢性便秘的发展。肠道细菌产生的短链脂肪酸,如乙酸盐和丁酸盐,也被认为是改变血清素可用性的一种手段,从而影响肠道的运动和分泌。
此外,某些胃肠道细菌菌株已被证明能从色氨酸中产生血清素。这不仅为治疗便秘开辟了一条新的途径,也为治疗受血清素水平影响的其他疾病开辟了新的途径。
✦纤维素可以调节猫的肠道微生物
对便秘可用的饮食疗法进行了回顾,主要关注纤维。纤维已被证明可以调节猫的肠道微生物群,尽管在对猫科动物便秘的纤维干预的有限研究中,没有进行微生物组分析。
✦益生菌可以改善肠道症状
一项针对便秘猫的干预试验评估了含有多种菌株的益生菌,包括:
Streptococcus thermophilus DSM32245;
Lactobacillus acidophilus DSM32241;
Lactobacillus plantarum DSM32244;
Lactobacillus casei DSM32243;
Lactobacillus helveticus DSM322422;
Lactobacillus brevis DSM27961;
Bifidobacterium lactis DSM32246;
B. lactis DSM32247
观察到猫慢性肠病指数显著下降,粪便评分和粘膜组织学有所改善。治疗后,链球菌和乳杆菌的数量显著增加,双歧杆菌和拟杆菌的数量有增加的趋势。不过这些猫的细菌分类群的上述变化对功能的影响尚未确定。
尽管关于猫和狗的数据有限,但一些综述总结了益生菌对人类便秘有益影响的研究。此外,最近的一项研究评估了益生菌对肠道蠕动的作用机制;第二项研究评估了益生菌对便秘患者自发排便的影响。
总体而言,这些研究支持使用益生菌治疗便秘、减少胃肠道转运时间、增加大便频率、改善大便稠度和改善胃肠道症状。
这些影响的潜在机制集中在肠腔环境、免疫系统、肠神经系统和中枢神经系统之间的相互作用,所有这些都高度相关并影响肠道运动。需要进一步研究益生菌对便秘猫和狗的微生物组的影响。
据报道76%的狗和68%的猫患有牙齿疾病,牙齿疾病是宠物中最常见的疾病。研究表明,临床上健康的狗和猫的口腔微生物群中始终有6种细菌门:放线菌门、拟杆菌门、厚壁菌门、梭杆菌门、变形菌门和螺旋体门。
已经评估了患有和不患有牙周病的猫和狗的口腔微生物群。总体而言,这些研究表明,牙科疾病与更多致病菌株替代与健康相关的菌株有关。
▸ 牙周疾病的猫
例如,在没有牙周病的猫中,研究表明最常见的采样门一直是拟杆菌门、厚壁菌门和变形菌门,而在患有牙周炎或牙龈炎的患者中,据报道致病菌的变异程度很高,但研究之间几乎没有相似之处。
▸ 牙周疾病的狗
另一方面,在没有牙周病的狗中,拟杆菌门、厚壁菌门和变形菌门占主要,尽管据报道放线菌在一些研究中占主导地位。
口腔疾病不同阶段微生物组成有所不同
对患有牙周病的狗的研究发现,根据口腔疾病的具体阶段,微生物组的组成会有所不同。
在一项对患有轻度牙周炎或牙龈炎的狗的研究中,毛螺菌科、梭菌目、消化链球菌科、消化球菌属和犬棒状杆菌属非常普遍。
另一项研究在患有牙周病晚期的狗中鉴定了血链球菌、消化链球菌属、大肠杆菌、奇异变形杆菌、韦荣氏球菌、金黄色葡萄球菌、唾液链球菌、放线菌属和粘性放线菌。牙周病已被证明与狗的全身性疾病有关。
✦饮食与牙周疾病
猫和狗的饮食之间的主要区别是食物是干的还是湿的。由于干粗磨食物的磨蚀性,通常喂食干粮以促进口腔健康。
尽管这些食物已被证明可以减少牙菌斑和牙龈炎,但尚未评估它们对微生物组的影响。

•喂养干粮的口腔微生物更多样化
一项针对只喂食干粮(高度精制、基于谷物的脱水口粮)或湿粮(罐装或新鲜肉类组合)食物的社区猫的研究表明,随着时间的推移,口腔微生物组存在差异,但无法确定是否这些差异影响了牙周病的风险。
然而,喂食干粮的猫的口腔微生物群更加多样化,富含与口腔健康和牙周病相关的细菌(牙龈卟啉单胞菌属和密螺旋体属的丰度更高)。
宠物肥胖是一个严重的问题。根据宠物肥胖预防协会最近的一份报告,60%的猫和56%的狗被归类为超重或肥胖。
肥胖还与多种其他疾病有关,例如糖尿病、骨关节炎、心血管疾病、皮肤病和寿命缩短。有几个因素会使动物易患肥胖,包括胃肠道微生物组、遗传、绝育、活动水平降低以及高脂肪和高能量饮食。
✦肥胖猫狗中的肠道微生物
一些报告发现,肥胖和瘦削的猫狗的胃肠道微生物组的组成不同。具体而言,与瘦狗相比,属于放线菌门和罗氏菌属的细菌在肥胖狗中的数量明显更多。
在猫中,与瘦猫相比,肥胖和超重猫的梭状芽孢杆菌、拟杆菌属和梭杆菌属的丰度较低,但属于肠杆菌科和梭菌簇IV组较高。
✦饮食对不同体重动物的肠道微生物影响不同
此外,多项研究发现,不同的饮食对肥胖动物和瘦动物的胃肠道微生物群有不同的影响。
例如,一项研究表明,与瘦狗相比,超重狗的微生物组对饮食干预更敏感。在4周的高蛋白/低碳水化合物饮食中,与瘦肉/正常狗相比,超重狗的微生物组成发生了显著的变化,这是由于乳酸杆菌、普氏杆菌、链球菌和Turicibacter的丰度变化较大所致。
✦肠道微生物与肥胖有关
肠道微生物组通过其对肠道的直接影响及其对远端器官的间接影响,与肥胖的发展有关。
胆汁酸利于脂质代谢
肠道微生物群已被证明会影响胆汁酸的代谢;细菌代谢产生的游离胆汁酸可以抑制细菌种群的生长,例如乳酸杆菌和双歧杆菌,它们被认为可以预防肥胖。
此外,肠道中的几种细菌(例如梭状芽孢杆菌、海莱蒙梭菌(Clostridium hylemonae)和Clostridium hiranonis)含有与胆汁酸代谢有关的基因。由于胆汁酸被认为有助于肠道激素分泌以及葡萄糖和脂质代谢,因此胃肠道微生物组对胆汁酸的影响也可能影响这些过程。
肠道微生物组破坏导致脂肪堆积
肠道黏膜屏障受到肠道微生物组的影响,这可能导致炎症增加,这是导致体重增加的已知因素。胃肠道微生物组的破坏已被证明会抑制脂蛋白脂肪酶,这会导致脂肪组织和肝脏、胰腺和心脏中的甘油三酯过度沉积。

✦肥胖的治疗
由于饮食、身体活动、遗传、新陈代谢和与体重增加相关的肠道微生物组之间的相互依赖关系,已经提出了许多饮食调整来减轻体重,包括消除高能量密度食物、改变饮食中的大量营养素组成(即高蛋白、高脂肪或高纤维饮食),并添加膳食甘油二酯和益生菌。
然而,目前针对猫和狗的体重管理指南并未考虑肠道微生物组。事实上,已经发现大约一半的肥胖猫和狗会在减肥计划后恢复体重,除非它们继续喂食专门配制的体重管理饮食而不是维持饮食,这表明可能需要持续喂养特定配方的饮食以维持体重减轻。
●狗减肥相关微生物变化
一些研究调查了与减肥相关的微生物群变化。
巨单胞菌和瘤胃球菌可能对体重减轻有负面影响
一项研究分析了18只食用限制性高蛋白/高纤维饮食的肥胖狗,18只狗中的8只也参加了锻炼计划。
两组的体重都有了相当的减轻,增加运动的组和单独饮食的组之间的微生物组没有差异。在这两组中,巨单胞菌的丰度都与体重下降呈负相关,而瘤胃球菌科的种群数量越少,体重下降越快(≥每周1%)。
此外,体重减轻较快的狗的乙酸和丙酸浓度降低。由于巨单胞菌和瘤胃球菌科产生这些酸,这些结果表明,产生这些短链脂肪酸的胃肠道微生物组可能对狗的体重减轻产生负面影响。
低脂肪饮食增加了肠道微生物多样性
一项类似的小型研究表明,17周的限制性低脂肪/高纤维饮食显著增加了6只肥胖比格犬的肠道微生物多样性。
在这项研究结束时,用这种饮食喂养的比格犬的Allobaculum数量也有所增加,梭状芽孢杆菌、乳酸杆菌和Dorea数量有所减少。
●猫的体重减轻与微生物群的关系暂不明确
然而,一项研究评估了中等蛋白质/高纤维饮食对肥胖猫体重和粪便微生物群的影响,结果与上述结果不一致。
八只成年雄性家猫进行了限制性饮食,以实现每周约1.5%体重的减重,持续18周。第18周时,平均体重比基线下降了近20%。尽管体重减轻与放线菌的比例更高和拟杆菌的比例更低相关,但微生物组多样性没有显著差异。
此外,一项类似的研究发现,与17只瘦猫相比,10周的标准减肥饮食对14只肥胖猫的粪便微生物群的影响很小。
因此,需要更多的研究来更好地了解猫的体重减轻与粪便微生物群之间的关系。

糖尿病在猫和狗中的患病率正在增加。虽然狗几乎只患有1型糖尿病,但猫更可能患有2型糖尿病。
各种模型系统中微生物组研究的进展表明,胃肠道微生物组在糖尿病等肠外疾病中发挥作用。改变的肠道微生物群组成与猫和狗的糖尿病的发展有关。
•1型糖尿病的狗肠道菌群失调
例如,最近的一项研究表明,患有自然发生的1型糖尿病的狗肠道菌群失调,粪便中浓度发生变化,表现出与患有2型糖尿病的人类相似的模式。
•糖尿病的猫肠道微生物多样性降低
此外,与同龄的健康猫相比,患有糖尿病的猫的肠道微生物多样性显著降低,产生丁酸盐的细菌也减少了。
✦药物通过微生物介导治疗糖尿病
用于治疗糖尿病的药物已被证明会影响胃肠道微生物组。在一项研究中,抗糖尿病药物二甲双胍通过显著提高拟杆菌门的丰度来改变胃肠道微生物组。
其他用于治疗糖尿病的药物(如阿卡波糖和二甲双胍)的功效也已被证明是通过微生物组介导的。基于微生物组的方法可能需要个性化,以有效管理与代谢相关的疾病,例如糖尿病,以解决微生物组组成和功能的个体间差异。
✦益生菌能改善糖尿病
•减少炎症反应
支持在糖尿病中使用益生菌的证据表明,益生菌可以减少炎症反应和氧化应激,增加胃肠道上皮细胞内粘附蛋白的表达,从而降低肠道通透性。已经提出这些机制导致胰岛素敏感性增加和自身免疫反应减少。
•降低发病率
一项针对256名女性的研究发现,妊娠早期的益生菌干预降低了妊娠糖尿病的发病率。含有双歧杆菌的益生菌也被证明可以改善糖尿病小鼠模型的葡萄糖耐量。
虽然不知道对猫和狗是否可行,但2型糖尿病或高血压患者严格遵循以高碳水化合物和高纤维植物为基础的素食饮食,其血红蛋白A1c降低,空腹和餐后血糖水平有所改善。这些影响还伴随着胃肠道微生物组中拟杆菌门和厚壁菌门比例的降低。
小结
由于大量营养素影响胃肠道微生物的组成,而微生物群的相关功能产物会影响宿主健康,因此通过营养来管理猫和狗的糖尿病是合乎逻辑的。
尽管目前数据有限,但正在进行几项研究以进一步研究益生元对代谢疾病的影响。总之,显然需要更好地了解不同的营养干预措施如何影响糖尿病中的胃肠道微生物组成和功能,这将为开发更有效的糖尿病营养干预措施提供信息。
慢性肾脏病 (CKD) 是猫和狗最常见的疾病之一,但迄今为止,关于这些物种的胃肠道微生物和肾脏疾病的研究很少。
✦慢性肾病的猫微生物丰富性和多样性降低
最近的一项研究表明,与健康猫相比,患有慢性肾脏病的猫的粪便微生物组的丰富性和多样性降低,这与先前对患有慢性肾脏病的人类肠道微生物的研究一致。
确定生态失调与肾脏疾病之间因果关系的研究正在进行中。潜在的微生物组成被认为会影响个体对慢性肾脏病的易感性,有证据支持在猫和狗中发现的这种联系。例如,据报道患有严重牙周病的动物患肾病的风险增加。
胃肠道转运时间慢、蛋白质同化受损和膳食纤维消耗减少等因素也与肾病患者的生物失调有关。
✦使用益生菌可能对减少肾结石有益
肾结石是猫和狗中常见的一种疾病,平均会使猫的寿命缩短约3年。动物的肾结石和膀胱结石大部分由草酸钙或磷酸铵镁(鸟粪石)组成,结石的形成是一个复杂的过程,涉及遗传和环境因素。
人类和动物的研究表明,草酸杆菌和乳酸杆菌通过草酸盐的降解防止结石形成。需要更多研究胃肠道微生物对猫和狗肾脏功能和肾脏疾病的影响,以及营养对这些微生物群的影响。
如前所述,猫和狗的肠道微生物群组成通常相似,但也有一些差异。肠道微生物组与宿主之间存在复杂的关系,这受到猫与狗的独特解剖学和生理学的影响。
关于猫和狗之间的消化和生理差异如何影响微生物组的证据是有限的。可以通过研究食物如何通过产生微生物后生元来影响每个物种的健康。
后生元——后生元是益生菌经加工处理后的益生菌代谢物成分统称,包括菌体与代谢产物。
食品、肠道微生物群和宿主之间的三方相互作用
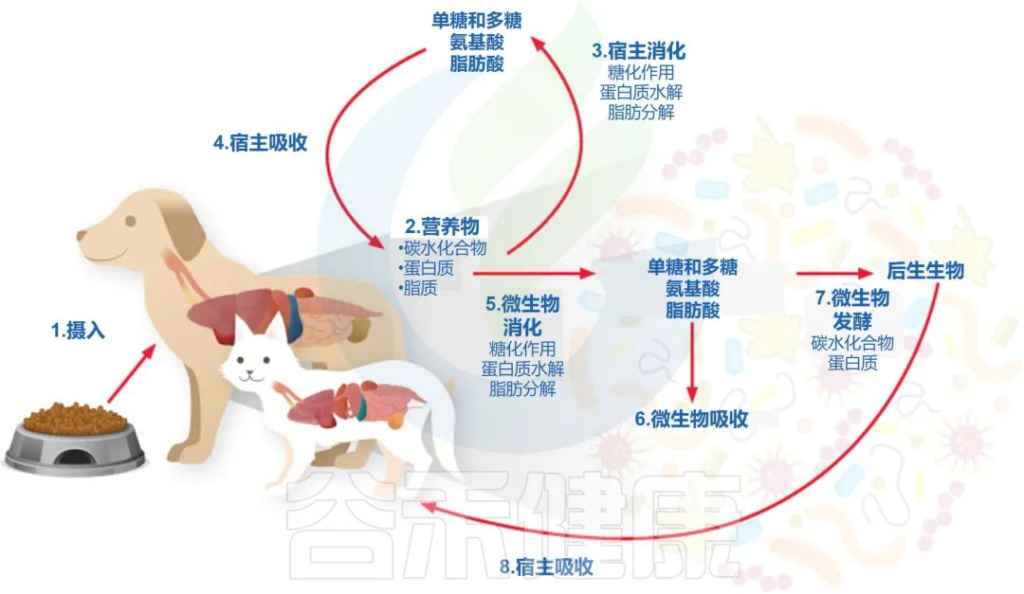
Wernimont SM,et al.Front Microbiol.2020
宠物食品、胃肠道微生物群和宿主之间的三方相互作用分8个阶段。
(1) 摄入:狗和猫以提供给它们的宠物食品的形式摄入碳水化合物、蛋白质和脂质等营养物质。
(2) 营养素:宠物食品中的营养素进入胃肠道,可供宿主和微生物组消化。
(3)宿主消化:宿主的消化包括糖解、蛋白水解和脂肪分解等过程,释放出单糖和双糖、氨基酸和脂肪酸。
(4)宿主吸收:通过宿主消化产生的单糖和双糖、氨基酸和脂肪酸可以被宿主细胞吸收利用。
(5) 微生物消化:未被宿主消化或吸收的营养物质可通过糖解、蛋白水解和脂肪分解被微生物组消化,释放出单糖和二糖、氨基酸和脂肪酸。
(6) 微生物吸收:微生物消化产生的单糖和双糖、氨基酸和脂肪酸可以被微生物组吸收和利用。
(7) 微生物发酵:超过宿主和微生物吸收能力的营养物质被绕过到下消化道,在那里它们可以进行微生物发酵以产生可以在胃肠道内局部影响宿主健康的后生元。
(8) 宿主吸收:微生物衍生的后生元也可以被宿主吸收,从而影响胃肠道以外部位的宿主健康。
当提供多种口味相似但营养素含量不同的食物,并允许通过在食物中自由选择喂养来自行选择常量营养素摄入量时,这导致在不同物种之间观察到显著不同的后生元。
例如,在28天的自由选择喂养期后,狗的循环血浆微生物代谢物均未发生变化,而猫的38种代谢物浓度中有16种发生了变化。
✦家猫的身体结构
家猫是一种专性食肉动物,因此从动物蛋白质的消耗中获取能量。猫科动物基因组高度保守区域的分析和饮食适应直系基因的评估揭示了淀粉和蔗糖代谢途径中基因家族的缺失,反映了它们的肉食性。
家猫特有的代谢和生理特性
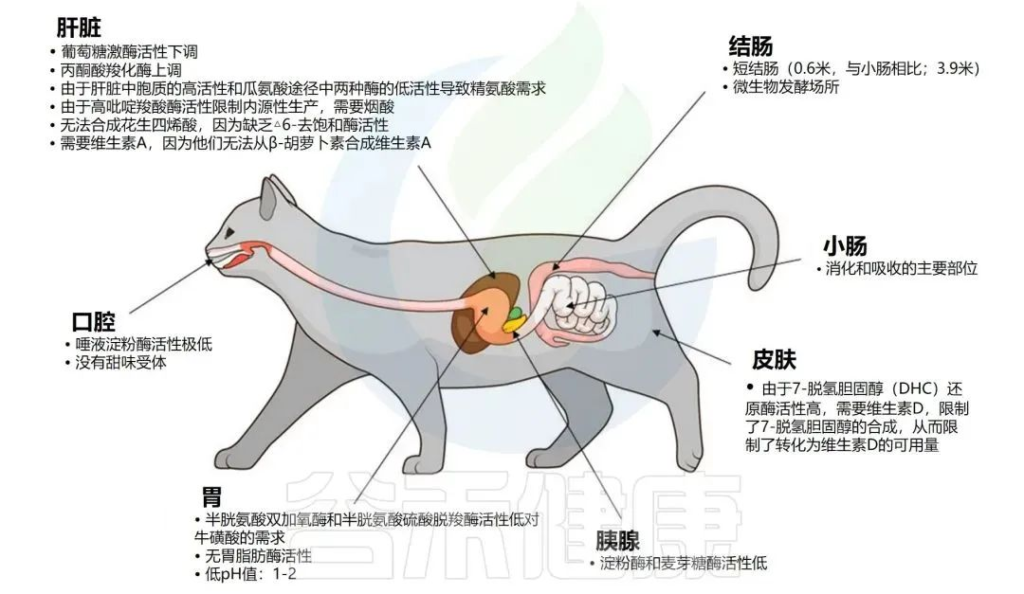
Butowski CF,et al.N Z Vet J.2022
✦家狗的身体结构
相比之下,狗被归类为兼性食肉动物,在营养上通常被视为杂食动物,因为它们消耗和利用植物和动物材料。通过驯化,狗保留了许多祖先特征,尽管已经观察到与人类文明一起适应碳水化合物含量较高的饮食。
家狗特有的代谢和生理特征
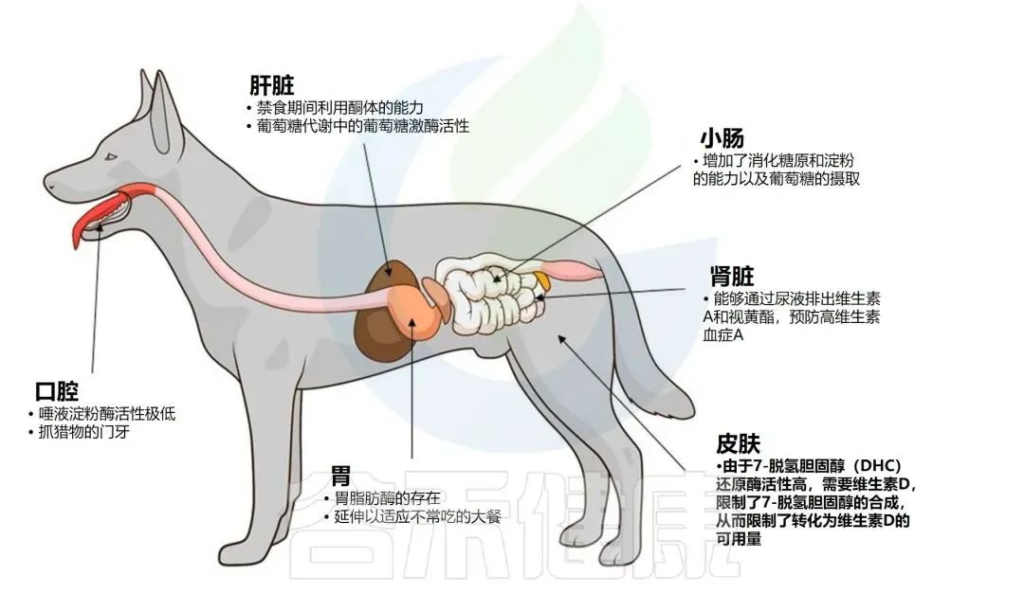
Butowski CF,et al.N Z Vet J.2022
✦不同的身体结构导致营养代谢不同
猫和狗之间的这些差异,无论是在首选的常量营养素摄入量还是在微生物组对选定的常量营养素摄入量的代谢反应方面,都为优化猫和狗的营养摄入带来了独特的挑战和益处。
▸
最近,低加工或生的高动物蛋白(肉类)饮食越来越受欢迎,更“天然”的宠物食品的需求,这些食品反映了动物的肉食性。这些宠物食品通常以生(新鲜或冷冻)、空气压缩或冻干形式出售。
这些高肉粮通常含有高粗蛋白,含有中高水平的粗脂肪,碳水化合物含量最低。从营养充足性的角度来看,生肉饮食可以制造完整和平衡的饮食。
✦狗吃生食的影响
一些研究评估了以肉类为基础的生食对健康狗肠道微生物的影响。与对照饮食相比,以肉类为基础的生食通常在常量营养素含量方面存在很大差异,蛋白质含量更高,碳水化合物和纤维含量更低。
不同饮食对狗粪便微生物组的影响

Pilla R,et al.Vet Clin North Am Small Anim Pract.2021
•厚壁菌和拟杆菌总体下降
喂食生食的狗的厚壁菌门和拟杆菌门总体下降。大多数细菌减少与膳食中碳水化合物产生短链脂肪酸相关,表明由于摄入量减少导致碳水化合物发酵减少。变形杆菌门、梭杆菌门和与蛋白质相关的属大量增加。
•微生物多样性增加
一项对6只狗喂食生食超过1年的研究发现,微生物的多样性增加 ,产气荚膜梭菌和梭杆菌的丰度增加,这两种细菌都可以从蛋白质中产生丁酸盐,表明对饮食的适应。
•梭状芽胞杆菌与蛋白质消化有关
尽管梭状芽胞杆菌科可能与胃肠道疾病有关,但有人认为,当给狗喂食富含蛋白质的饮食时,梭状芽胞杆菌的增加可能不会损害它们的健康,但与蛋白质消化有关。
此外,还发现梭状芽孢杆菌科与蛋白质消化率和更坚硬的粪便评分呈正相关,与粪便蛋白质含量负相关(即,梭状芽胞杆菌科越多,粪便中未消化的蛋白质越少)和粪便排出量越少。
在猫中,罐头饮食是一种额外的高蛋白替代品,通常单独喂养或与膨化饮食结合喂养。研究在猫身上使用潮湿的食物,它们的摄入会导致饮水量增加,减少自愿能量摄入量,并降低尿液比重,这可能对某些健康状况有益。
不同饮食对猫粪便微生物组的影响


Pilla R,et al.Vet Clin North Am Small Anim Pract.2021
研究评估了断奶到高蛋白/低碳水饮食小猫中的微生物,研究之间存在一定的一致性。
•高蛋白/低碳水饮食增加了物种多样性
有趣的是,高蛋白/低碳水饮食增加了物种多样性,在高蛋白/低碳水喂养的小猫中增加5个属是已知的丁酸盐生产者:梭状芽胞杆菌和真杆菌可以通过从碳水化合物产生丁酸盐,或通过从蛋白质产生丁酸盐,而Faecalibacterium、Ruminococcus和Blautia也是丁酸盐的生产者。
两种饮食方式的小猫之间的差异影响了代谢途径,包括与氨基酸生物合成和代谢相关的途径,表明蛋白质:碳水化合物的比例对微生物组功能有显著影响。然而,这种代谢物差异对整体健康的影响仍然未知。
小结
虽然饮食可以显著改变微生物的组成,并导致代谢途径和代谢物产生的变化,但这些变化通常比伴随疾病的变化要小得多。
在患病动物中,特别是那些患有胃肠道疾病(例如慢性肠病)的动物中,微生物组的多样性迅速降低,许多核心物种,如C.hiranonis、梭杆菌属和普拉梭菌减少。
养宠物益处也有很多,他可以让老人的心情更加舒畅,预防心绪管系统疾病,而且还在无形中增加运动量,起到强身健体的作用;也可能传播疾病。
与养狗相关的菌群:
养狗与室内灰尘中的细菌丰富度和多样性呈正相关。
最近关于宠物和人类微生物相互作用的研究表明,在家养狗会增加同居成年人之间的共同皮肤微生物群。
✦降低哮喘风险
研究表明,婴儿接触农场动物与降低日后发生哮喘的风险有关。最后,已证明围产期宠物暴露会影响肠道微生物群的组成差异,并保护儿童免受早期喘息。
许多科学家认为,婴儿和儿童需要接触无害的微生物,以“训练”他们正在发育的免疫系统来区分自身分子和外来分子,而不是过度反应并最终损害自身组织,与狗同住有助于这种免疫系统训练。
✦ 心血管疾病患者中,养宠物患者代谢综合征和肥胖风险小
一项研究分析了162 名冠心病患者的肠道微生物群,养宠物 ( n = 83) ,不养宠物 ( n = 79).
养宠物的患者患代谢综合征 (MetS) (OR = 0.462) 和肥胖症 (OR = 0.519) 的风险较低,养宠物的患者中发现了沙雷氏菌属Serratia和粪球菌属的优势,不养宠物的患者中瘤胃球菌属、肠杆菌科和Anaerotruncus的未知属占优势。
养宠物可能通过塑造肠道菌群成为预防代谢综合征的保护因素。
最近人们还发现了人畜共患的现象,并且可以在人与动物关系中的微生物交换中进行干预。
✦引起皮肤真菌病
皮肤真菌病(癣)由多种皮肤真菌引起,可感染人类和动物。感染源包括人类、家畜、宠物和野生动物(主要是兔子和啮齿动物)。
狗或猫可能表现出临床症状,由于主人经常接触宠物,会导致真菌病在宠物与家庭成员之间传染。
由于遗传因素、先前存在的皮肤病(如过敏性皮炎)或免疫功能受损状态,一些人更容易患上皮肤真菌病。
胃肠道微生物
胃肠道疾病是宠物与其主人比较常见的疾病。此外,沙门氏菌(Salmonella)和弯曲杆菌(Campylobacter)等特定细菌性肠道病原体是有充分记录的人畜共患病,对宠物与其主人都会带来健康影响。
注:感染沙门氏菌或弯曲杆菌会引起急性肠炎,主要症状有腹泻、发热、腹痛和血便,还伴有乏力、肌肉酸痛、视觉模糊、躁动不安和嗜睡。
狗还可能藏匿并脱落原生动物寄生虫蓝氏贾第虫的囊肿,或者携带有蛔虫。两者都是导致人类感染的常见潜在人畜共患疾病。
注意:感染蓝氏贾第虫会出现上腹部不适,有时伴有腹痛、腹鸣、胃肠胀气及慢性腹泻粪内带有粘液,量多并有臭味,重者可伴有脂肪。此外,尚可有厌食、低热、体重减轻、失眠、头痛及多汗等症状。但不发生贫血,亦无舌炎、口炎、手足抽搐等热带口炎。
其他微生物
✦巴尔通体病
巴尔通体病是一种媒介传播的人畜共患疾病,在世界范围内都有分布,可以感染人类和大量哺乳动物,包括小型伴侣动物(猫和狗)。
至少有十三种Bartonella已被确认为人类疾病的病原体。主要有B. clarridgeiae、B. elizabethae、B . henselae 、 B . koehlerae、B. quintana、B. rochalimae和B. vinsonii berkhoffii。
•家猫引起的巴尔通体病
猫是B. henselae的主要但不是唯一的宿主,B. henselae是人类猫抓病的病原体。
家猫也是B.clarridgeiae和B.koehlerae的主要宿主。人们认为受感染的猫很少会出现临床症状。然而,在受感染的猫中经常可以检测到慢性复发性菌血症,而复发性菌血症的潜在长期后果尚不清楚。
猫也可能感染B. bovis, 但家猫在这种巴尔通体的流行病学中的作用尚未明确。
•犬类引起的巴尔通体病
狗也可能是B. henselae的宿主,犬科动物被认为是B. vinsonii berkhoffii的主要宿主,可引起狗和人类的心内膜炎。
加利福尼亚州的土狼 等野生犬科动物和潜在的家犬已被描述为B. vinsonii berkhoffii的主要储存宿主,因为这些动物也会发生长期菌血症。
注:汉氏巴尔通体属最常感染宠物狗,普通狗也会被感染。
随着年龄或药物的影响,健康宠物的肠道微生物群动态平衡可能会被破坏,从而引发各种疾病。
了解宠物的肠道健康状况,最好的方法是进行肠道微生物检测,直观了解微生物的种类及数量。将得到的检测结果与同年龄、同品种的健康犬进行比较,可以比较全面地了解宠物的肠道微生物状况,来更好地进行对应的治疗。
在谷禾肠道菌群检测报告中,就有这样的特殊案例,来自宠物的肠道菌群。
报告显示,其样本菌群构成非常单一,实际测序深度很高,达到10万,但仅检出167种菌,绝大部分是大肠杆菌。
下面的表是根据人的菌群结果评估的,可以看到构成多样性非常低。
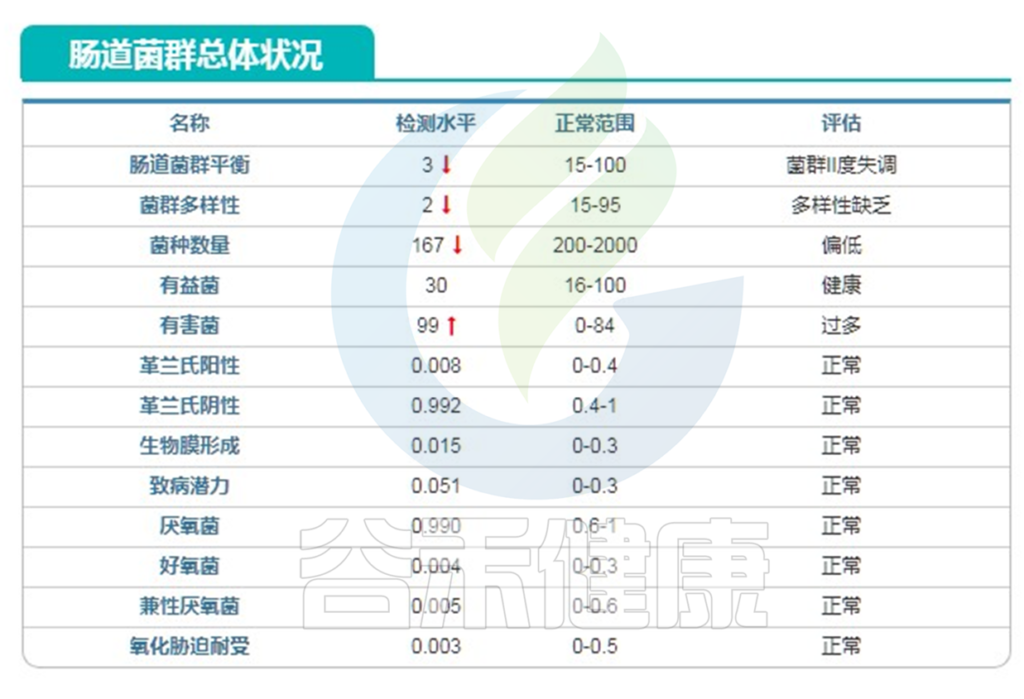
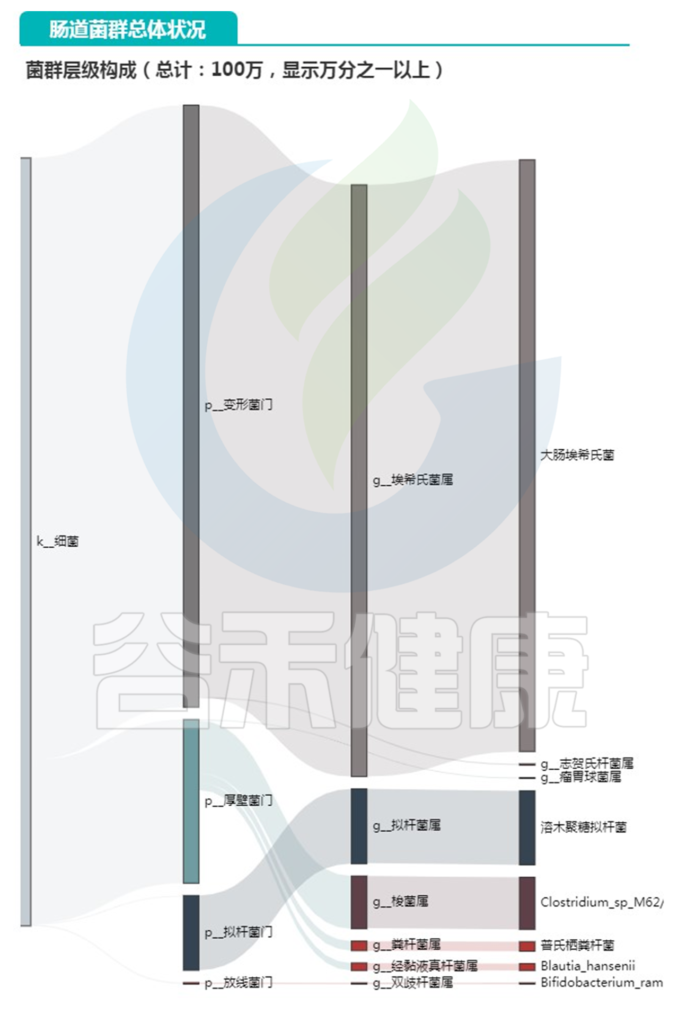
下面是主要菌门和属还有种的构成,种部分列出了注释有菌名称的丰度大于0.1%以上的菌。
下图是菌属构成表:
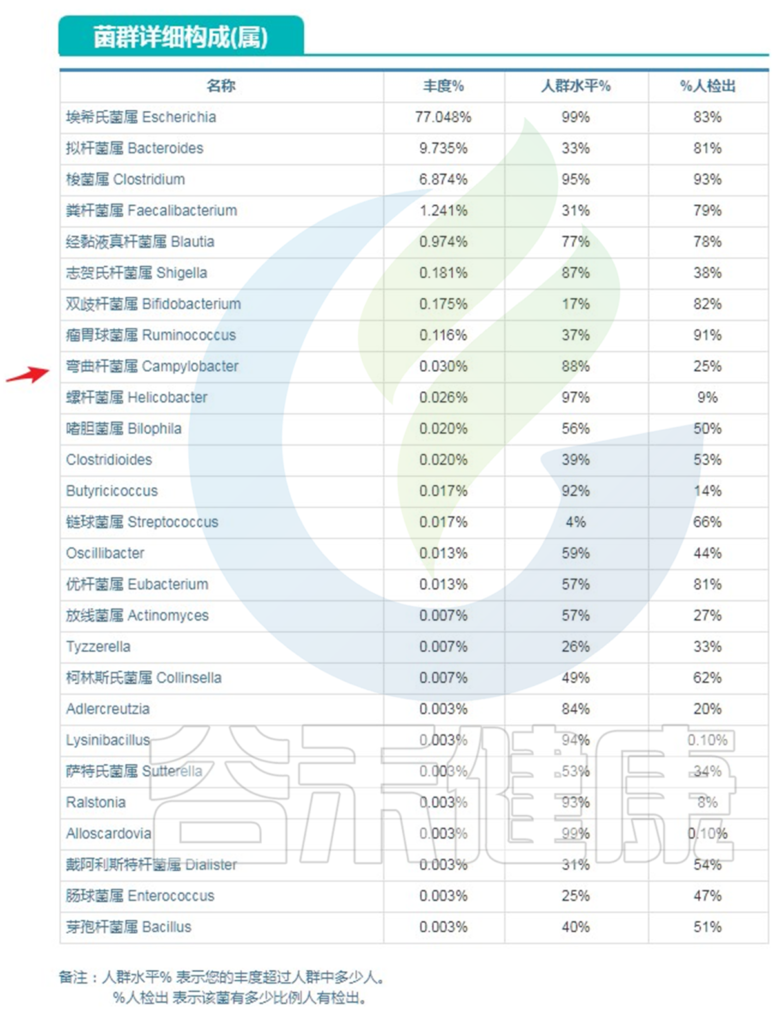
报告显示,菌属构成中弯曲杆菌占比较高。
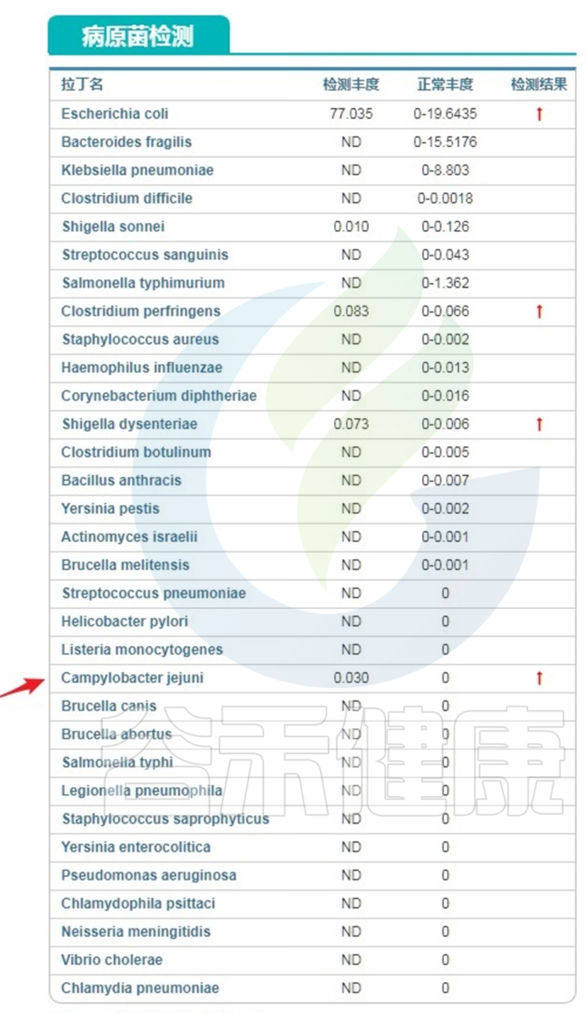
除此之外,其他的病原和机会致病菌主要检出了大肠杆菌,占比77%,属于严重超标,空肠弯曲杆菌,产气夹膜菌和痢疾志贺氏菌都有超标:
编辑
注:空肠弯曲菌(上图中红色箭头指向菌Campylobacter jejuni),有内毒素能侵袭小肠和大肠黏膜引起急性肠炎。
经过与送检者沟通后,我们了解的情况如下:
主人反映该宠物猫有腹泻,其有长期腹泻情况,因此送检了其本人和养的宠物猫的粪便样本做肠道菌群检测,了解菌群构成,找出腹泻原因,以便后续治疗。
了解猫狗的肠道微生物状况后,需要进行相应的治疗来帮它们恢复健康,谷禾在这里罗列了一些常见的治疗方式。
在疾病状态期间,所应用的疗法旨在改变微生物群落。
蛋白质、碳水化合物是大部分宠物必需的营养物质,是促进机体发生生长的重要营养素。宠物肠道中丰富的微生物菌群生长都离不开碳水化合物,前面也有提到一些饮食对疾病的影响作用。因此,合理控制宠物蛋白质、碳水化合物的摄入是维持肠道健康的关键。

•需要根据不同的宠物制定相应的饮食
不同年龄段的宠物对脂肪的吸收能力不同。老年犬猫由于肠道消化能力逐渐退化,不能较好地吸收脂肪。适量的功能性脂肪酸,如Omega-3脂肪酸可以增强宠物机体免疫力,预防炎症,减轻其胃肠道疾病的发生。
存在关于无谷物狗粮的争论。有些兽医认为不应该用任何谷物喂养宠物,有些人则说它可以适量喂养。常见的谷物包括大米、燕麦、玉米和小麦。
不建议吃的原因包括体重增加、胃肠道炎症增加、过敏等。如果狗不耐受谷物,可能会出现皮肤发痒、呕吐、胃部不适、脱毛、腹泻、感染和胀气。
换句话说,如果出现上述症状,可以尝试一下无谷物食物并监测症状是否有所改善。
骨汤含有多种有益营养素,但与肠道健康最相关的是胶原蛋白。胶原蛋白是一种结缔组织,有助于维持包括肠道和皮肤在内的身体器官的结构。
狗狗的肠道内部衬有一层叫肠粘膜的组织。粘膜的作用是防止食物分子和细菌等危险物质从肠道进入血液。
然而,粘膜可能会受损,导致肠道变得可渗透。换句话说,像细菌这样的物质可以通过。这就是所谓的“肠漏”。
通过给狗狗吃富含胶原蛋白的食物,比如骨头汤,可以加强粘膜中的结缔组织,从而减少肠漏程度。
宠物食品制造商在其产品中添加的一些成分会对狗的肠道菌群和整体健康产生负面影响。人工色素和抗氧化剂等食品添加剂减少肠道细菌的数量。
最好能阅读食物的配料表。
除了饮食之外,益生菌、益生元的施用也会影响和改变微生物群的组成。
益生菌和益生菌的使用在人类医学中广泛传播,以保持或恢复健康状况。
✦益生元治疗
▸ 益生元
根据2015年给出的第一个定义,它们是“一种不易消化的化合物,通过肠道微生物的代谢,调节肠道微生物群的组成或活性,从而赋予对宿主有益的生理影响”。
猫和狗接受益生元的治疗效果

Mondo E,et al.Open Vet J.2019
一些研究报告了在宠物饮食中添加益生元的好处。事实上,它们能调节肠道微生物群并保护动物免受肠道感染。
•益生元促进炎症性肠病缓解
几项研究评估了益生元治疗对诊断为炎症性肠病的狗的影响。这些分析的结果表明,多菌株益生元治疗促进了诊断为炎症性肠病的狗的临床缓解,并与其他一些益处相关,包括抗炎和抗增殖作用、增加紧密连接蛋白表达和上调多胺水平。
•益生元治疗改变肠病中微生物组成
关于微生物组的组成,用益生元治疗的炎症性肠病狗的粪杆菌属和乳酸杆菌显著增加。然而,与那些接受标准治疗的人相比,这些结果在研究之间并不一致。除了对炎症性肠病的有益作用外,多项双盲、安慰剂对照研究表明,益生元给药还可以减少猫和狗的腹泻。
✦狗使用益生元的疗效
有一些关于益生元在国内犬类饮食中的使用研究发现,在狗中,摄入1.5%的菊粉可以降低粪便的pH值并增加双歧杆菌的数量。
使用16S rRNA 测序表明,喂食相对少量膳食纤维的狗会改变肠道微生物群的结构,增加厚壁菌门的密度并减少梭杆菌的密度。另一项研究强调了低聚果糖的膳食补充剂如何诱导有益效果,例如双歧杆菌的生长,并且它提高了狗整个胃肠道中几种矿物质的消化率。
✦猫使用益生元的疗效
很少有关于猫科动物营养和益生元使用的研究。两项研究描述了猫饮食中典型的高蛋白饮食摄入主要诱导粪便梭状芽孢杆菌的生长。
发现在猫科动物饮食中添加低聚果糖会增加双歧杆菌浓度并减少大肠杆菌数量。同一项研究表明,在猫营养品中添加果胶提高了产气荚膜梭菌(Clostridium perfringens)和乳酸杆菌的浓度。
✦益生菌治疗
▸ 益生菌
数千年前,人类发现了发酵食品带来的好处。因此,益生菌的使用比益生元更古老。它们被定义为“当施用足够量时,可以为宿主带来健康益处的活微生物”。
无论是对人类还是宠物,益生菌对宿主健康都有许多有益的影响,尤其是在免疫系统的调节和压力状态下。益生菌在牲畜中的使用已广泛传播,而在宠物营养中的使用仍在发展中。
益生菌治疗效果的研究总结

Mondo E,et al.Open Vet J.2019
•益生菌促进了T细胞表达
对患有慢性肠病的狗进行了体外研究。这项研究证实了含有三种不同乳酸杆菌菌株的益生菌混合物对细胞因子表达的积极作用,主要是通过调节T细胞。
发现益生菌的施用促进了T细胞表达的增强。在这项研究中,为患有炎症性肠病的狗配制了一种类似于人类VSL#3的益生菌混合物。数据强调了T细胞靶点的增加、粪杆菌数量的减少以及生态失调的恢复。
从断奶到1岁,幼犬的饮食通常通过添加益生菌来补充。事实上,已经证明粪肠球菌在肠粘膜水平和全身水平都存在辅助作用。这些影响可能与断奶期间保护性免疫反应的改善有关。
当饮食改变不起作用时,患有慢性肠病的狗和猫会接受抗生素治疗。最常用的分子是甲硝唑和泰乐菌素。甲硝唑作用于细菌和原生动物。通常,它同时用于营养治疗和饮食改变,因此很难知道其真正的效果。
泰乐菌素用于治疗对泰乐菌素有反应的慢性腹泻,这通常会影响成年犬。许多研究强调了它的效率,但其作用机制仍然未知。
恩诺沙星是另一种用于肠病的抗生素。它是一种氟喹诺酮,可用于治疗肉芽肿性结肠炎。
●抗生素同时还会减少有益菌
需要注意的是,使用抗生素治疗某些疾病存在禁忌症。事实上,大量使用抗生素可能会减少有益菌的数量,促进潜在病原体数量的增加,并促进抗菌素耐药现象的发生。
一些研究人员表明,抗生素实际上会改变微生物组成,从而降低细菌多样性,因此它们会导致生态失调的发生。在过去的几年中,抗菌素耐药性的现象不断增加,并具有重要意义。人类与宠物的密切同居会导致抗菌素耐药性转移到对人类也有影响的细菌。
最近,已开发出一种新方法,该方法通过引入健康的细菌菌群来恢复结肠菌群。它被命名为粪便微生物群移植 (FMT)。
粪菌移植是一种医学非药物实验治疗,旨在恢复微生物多样性。它包括将粪便菌群从健康供体移植到受胃肠道疾病影响的患者中。
•粪菌移植后肠道生态失调得到改善
在人类医学中,该方法用于治疗艰难梭菌感染,有效率为87%–90%,并用于治疗炎症性肠病。
粪菌移植在兽医学中很有前景,但是应用的研究还很少。这种治疗方法已用于受胃肠道疾病影响的狗,如炎症性肠病和产气荚膜梭菌感染。
最近的一份病例报告描述了粪菌移植用于治疗一只患有难治性炎症性肠病的狗和一只患有无反应性慢性肠病的猫。在这两种情况下,粪菌移植后24小时内粪便稠度得到改善。
即使粪菌移植似乎是治疗生态失调的有效疗法,但仍需要进行更多的临床研究以更好地了解其机制和接受这些输注的患者的潜在风险。如今,只有有限的数据可用。
噬菌体是一种能够侵染细菌的病毒,是生物圈中最丰富的物种。
●噬菌体可以裂解消灭有害菌
噬菌体通过尾丝蛋白与宿主菌表面受体特异性结合侵染细菌使其裂解,精准消灭有害菌,不影响有益菌。30分钟内可杀灭99.9%的宿主菌,且不受细菌耐药性限制。面对细菌耐药、抗生素残留等问题,噬菌体作为天然的细菌杀手,是防治细菌性疾病的最佳选择。
调节肠道菌群
噬菌体一经发现,就被人类作为治疗细菌性疾病的利器之一。有研究表明,噬菌体可以裂解肠道中的有害菌例如沙门氏菌和大肠杆菌,降低了肠道有害菌数量,有益于肠道有益菌(如乳酸菌类)的繁殖和生长,起到了调节肠道菌群的作用。
建议
噬菌体能显著降低肠道内有害菌数量,提高肠道有益菌比例;并且有利于免疫器官的发育和体液免疫功能的提高。与抗生素相比,噬菌体在增强肠道活性,降低腹泻率方面,与抗生素差异不大,可以作为替代抗生素的候选产品之一。
研究表明,压力会对肠道微生物群的健康产生负面影响。当肠道微生物组受损时,它会导致压力增加,从而导致周期性模式。
压力会引起体内激素的变化,从而引起肠道微生物组的变化。特别是,压力会触发皮质醇激素的释放。皮质醇对身体有多种负面影响,包括血压升高和肠道健康受损。
狗的压力可以由多种因素引发,包括:
注意狗狗的压力迹象和症状。一些可能表示狗有压力的迹象包括:
可以为狗提供舒适的环境,以最大程度地减少压力。如果狗生病了,给它吃药,吃它最喜欢的食物,并给她一个舒适的地方睡觉。这会使狗平静下来,有助于减轻它的一些压力。
让狗狗进行体育锻炼
让你家狗狗参加各种体育锻炼,如散步、跑步、玩耍等。
研究表明,体育锻炼通过刺激体内内啡肽的释放来减轻压力。内啡肽是中枢神经系统释放的神经递质。它们的主要作用是减轻疼痛和增强快感。
身体也会在其他愉快的活动中释放内啡肽,体育活动对健康很重要。
定期对宠物进行身体检查
应该至少每年带狗狗去看一次兽医。兽医有专业知识来识别可能错过的狗的潜在健康问题。
让你的狗定期接受兽医检查可以降低疾病未被诊断的风险。如果不进行治疗,肠道疾病往往会加重,因为消化系统每天工作多次。
如果狗身体健康,兽医每年看一次就足够了。
如果你的狗患有髋关节发育不良或糖尿病等慢性疾病,应该多看兽医。
此外,随着肠道菌群检测技术的不断成熟,不久的将来通过宠物肠道菌群检测也能发现一些疾病风险,不用让宝贝宠物承受各类监测设备的心理困扰,简单便捷。
人体的研究表明肠道菌群影响并反映着身体的健康,同样肠道微生物群也对猫狗以及其他哺乳动物的健康至关重要。
对于猫狗的肠道菌群仍然有很多未知和亟待发掘的意义,有望成为一种非常有价值的检测和干预渠道。谷禾欢迎更多相关领域的合作伙伴共同推进和完善猫狗相关肠道菌群的研究和应用。
主要参考文献:
Pilla R, Suchodolski JS. The Gut Microbiome of Dogs and Cats, and the Influence of Diet. Vet Clin North Am Small Anim Pract. 2021 May;51(3):605-621. doi: 10.1016/j.cvsm.2021.01.002. Epub 2021 Feb 27. PMID: 33653538.
Wernimont SM, Radosevich J, Jackson MI, Ephraim E, Badri DV, MacLeay JM, Jewell DE, Suchodolski JS. The Effects of Nutrition on the Gastrointestinal Microbiome of Cats and Dogs: Impact on Health and Disease. Front Microbiol. 2020 Jun 25;11:1266. doi: 10.3389/fmicb.2020.01266. PMID: 32670224;PMCID: PMC7329990.
Mondo E, Marliani G, Accorsi PA, Cocchi M, Di Leone A. Role of gut microbiota in dog and cat’s health and diseases. Open Vet J. 2019 Oct;9(3):253-258. doi: 10.4314/ovj.v9i3.10. Epub 2019 Sep 1. PMID:31998619; PMCID: PMC6794400.
Pilla R, Suchodolski JS. The Role of the Canine Gut Microbiome and Metabolome in Health and Gastrointestinal Disease. Front Vet Sci. 2020 Jan 14;6:498. doi: 10.3389/fvets.2019.00498. PMID: 31993446; PMCID: PMC6971114.
Pereira AM, Clemente A. Dogs’ Microbiome From Tip to Toe. Top Companion Anim Med. 2021 Nov;45:100584. doi: 10.1016/j.tcam.2021.100584. Epub 2021 Sep 10. PMID: 34509665.
Butowski CF, Moon CD, Thomas DG, Young W, Bermingham EN. The effects of raw-meat diets on the gastrointestinal microbiota of the cat and dog: a review. N Z Vet J. 2022 Jan;70(1):1-9. doi: 10.1080/00480169.2021.1975586. Epub 2021 Sep 19. PMID: 34463606.

谷禾健康
肠道菌群 & 性激素
青春期是生命的一个关键阶段,与性成熟相关的生理变化有关,是一个受多种内分泌和遗传控制调控的复杂过程。
青春期发育可以在适当的时候,早熟或延迟。
青春期发育异常会给患者带来痛苦,也可能是潜在病理的征兆。

青春期是由激素驱动的,它会影响肠道微生物群。性别之间的肠道微生物群差异出现在青春期开始时,证实了微生物群和性激素之间的关系。在不同的研究中已经提出了肠道微生物群和性激素之间的双向相互作用。
本文介绍了性早熟发育,以及性早熟中肠道微生物群的变化,便于从儿科内分泌学的角度考虑性激素-肠道菌群轴的作用。
肠道菌群的改变可能发生在患有中枢性早熟的女孩身上的证据,为预测和预防性早熟提供了一个有趣的发现。
加深对性激素与微生物群变化作用之间联系的理解,可以通过提供儿科内分泌学的视角,在青春期疾病中实施微生物群靶向治疗。
在了解性早熟与肠道菌群的关系之前,我们先了解一下青春期发育的相关生理过程,以及关于性早熟的基本知识。
青春期特征是:
正常的青春期是由于下丘脑-垂体-性腺轴(HPG)的长期成熟活动所致。
下丘脑以脉动的方式将促性腺激素释放激素释放到垂体门静脉系统,在那里它刺激LH和FSH分泌(也是脉动的)。
注:
LH(促黄体生成素):主要刺激睾丸中的间质细胞和卵巢中的卵泡膜细胞分泌雄激素。
FSH(卵泡刺激素):主要刺激卵泡或生精小管形成雌激素、抑制素、卵子或精子。
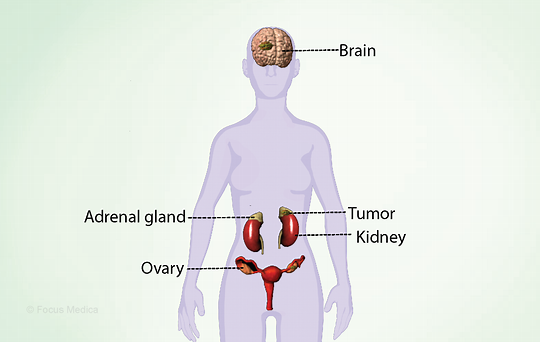
间质、管状和卵泡隔室通过旁分泌过程共同作用,产生雌激素,并调节性类固醇的产生和配子的发育。类固醇激素对促性腺激素释放激素和促性腺素分泌有内分泌负反馈作用。
FSH分泌被抑制素、孕酮和雌二醇的负反馈抑制。在成年女性受试者中,临界雌二醇浓度刺激LH激增,从而启动排卵。
HPG轴出生时短暂激活,青春期完全重新激活
出生时,由于缺乏抑制HPG轴的胎盘类固醇,这一轴会被激活,从而导致类固醇激素的产生增加,这是青春期继续进行的第一步。这种短暂的激活大约在出生后一周开始,几个月(大约6个月)后停止。
HPG轴在整个儿童期并不完全休眠,尤其是女性,其FSH浓度略高于男性。有时,也可以用超声波检查卵巢卵泡。在青春期,HPG轴经历完全重新激活。
最重要的GnRH抑制系统是γ-氨基丁酸能神经元(产生γ-氨基丁酸的神经元)和阿片受体;吻肽、神经激肽B和强啡肽A这三种存在于弓状核(ARC)中的神经肽被认为是影响GnRH释放的基本生成物,因为它们对男孩和女孩的青春期生理学有重要贡献。
厌食因子——瘦素:在青春期发挥重要作用
瘦素是一种主要由脂肪细胞产生的细胞因子,它作为一种厌食因子,通过抑制下丘脑神经肽Y(NPY),从而抑制食欲,在控制体重、食物摄入和能量平衡方面发挥重要作用。儿童时期必须达到正常的体重和组成,以避免青春期功能障碍。
除了瘦素与NPY的相互作用外,一些研究表明瘦素通过与KiSS-1基因直接相互作用而影响青春期和生殖功能。GnRH神经元缺乏瘦素受体,但KiSS-1神经元表达瘦素受体。瘦素直接刺激kisspeptin释放并介导GnRH的脉动释放。
青春期是一个复杂的过程,具有广泛的生理变化。调节青春期开始的机制涉及遗传、营养和环境相互作用。
胎儿营养异常以及内分泌系统可能导致发育改变,从而永久影响结构、生理和代谢。
生长关键时期激素和营养之间的相互作用对于代谢适应反应控制和青春期发育预期至关重要。
越来越多的证据表明,产前和产后早期是青春期开始规划的重要时期。各种研究表明,产前暴露于不利环境因素,如导致儿童出生SGA(小于胎龄)和/或IUGR(宫内生长受限)的因素,会影响青春期时机。儿童出生的SGA可能会经历几次青春期改变,如性早熟。
➡ 性早熟如何定义?
性早熟(PP)定义为女性青少年在8岁之前和男性在9岁之前出现变化:
性早熟患者表现出性和身体生长加快,同时出现生长突增。如果不治疗,骨骺生长加快可能会导致骨骺过早闭合,导致成年后身材矮小。
➡ 性早熟分类
根据潜在的生理病理过程,病理性性早熟分类如下:
• 中枢性性早熟
中枢性性早熟是最常见的性早熟形式,女孩多于男孩。一般异常位于大脑。
由于先天性或后天性中枢神经系统(CNS)损伤或单基因缺陷导致的HPG轴过早成熟,从而导致的中枢性性早熟(CPP)或促性腺激素依赖性性早熟(或真正的性早熟),也可能是特发性的。
• 外周性性早熟
一般异常不在大脑中,而是在睾丸、卵巢或肾上腺中。
外周性性早熟(PPP)或促性腺激素非依赖性性早熟(或假性早熟),由于遗传或肿瘤病因、分泌hCG的生殖细胞肿瘤(仅适用于男孩的人绒毛膜促性腺素)或外源性来源导致的性腺性激素或肾上腺激素分泌过多。
➡ 中枢性性早熟病例男女有别
据评估,每5000-10000名儿童中就有1名患有中枢性性早熟,女性的发病率是男性的10倍。
大多数女性中枢性性早熟病例似乎是特发性的,而男性中枢性性早熟患病率较高似乎通常是由病理性脑损伤引起的。特别是,下丘脑错构瘤是引起中枢性性早熟的最常见的脑损伤。
注:下丘脑错构瘤,是临床极为罕见的先天性脑组织发育异常性病变。
➡ 性早熟的临床检查评估
准确的个人和熟悉的病史、完整的体检、荷尔蒙和放射检查对性早熟诊断至关重要。
——青春期体征
临床检查应侧重于发育学数据、根据Marshall和Tanner分类对青春期体征的评估、过去6-12个月的生长模式,青春期体征(女孩的乳腺芽、男孩的睾丸体积、两者的阴毛)和其他青春期体征(痤疮、油性皮肤、勃起、男孩夜间排泄物、女孩阴道分泌物和月经出血)的进展率。
——基线LH水平
基线LH水平是诊断中枢性性早熟的有希望的生物标志物;基础晨LH值超过0.2 mUI/ml通常被认为是青春期的标志。在成瘾中,LH与FSH的比值高于0.6与中枢性性早熟有关。
——GnRH刺激试验
GnRH刺激试验仍然是鉴别中枢性性早熟的金标准,大于5 IU/L的LH截止峰值水平被广泛用于诊断中枢性性早熟。
——其他激素评估
其他激素评估应包括甲状腺测试、睾酮、雌二醇、17-羟基孕酮(17-OHP)、癌胚抗原(CEA)、癌抗原125(CA125)、甲胎蛋白和β-hCG,具体取决于患者的病史。
——骨龄
为了确定孩子的生物年龄,需要对非优势(左手)手和手腕进行骨龄X光检查。超过2.5标准差(SD)或超过2年的晚期骨龄更可能与病理性性早熟相关。
——盆腔超声
在女孩中,盆腔超声是评估卵巢早熟发育和排除卵巢囊肿或肿瘤的有用工具。
——脑部MRI检查
建议对患有中枢性性早熟的患者进行脑部MRI检查,以排除CNS损伤,应在男孩(<6岁)中进行常规检查。
性激素和肠道菌群之间的关系在最近的研究中得到了广泛的探讨,并且是一个不断扩大的研究领域,可能会为各种与性有关的疾病带来新的治疗选择。
20世纪80年代,最早的研究之一是在细胞水平上进行的,其中发现孕酮促进了拟杆菌和中间普氏杆菌的生长。
2012年一项研究在对91名女性的粪便样本进行测序时发现,无论健康状况如何,在怀孕期间,尤其是在孕晚期,当雌激素达到峰值时,肠道微生物组明显改变。
2015年,一项欧洲研究表明,健康男性的普氏拟杆菌丰度高于生育女性,而绝经后女性的微生物群与男性没有差异。
这两项研究都证明了雌激素和相关的雌性激素在调节肠道微生物组的组成方面是至关重要的。
因此,微生物群会影响雌激素水平,反过来,雌激素水平可能会受到微生物群组成和多样性的影响。
➤ 微生物群对雌激素的影响
微生物组能够通过表达B-葡萄糖醛酸酶(一种介导膳食和非膳食雌激素解偶联的酶)代谢雌激素。
未结合的雌激素可以通过作用于α和β雌激素受体而进入全身血流并具有代谢活性,这些受体在多种器官和组织中表达;雌激素活性不仅对生殖健康有影响,而且对心血管风险、代谢和骨稳态以及中枢神经系统也有影响。
微生物群多样性与绝经后妇女和男性的尿雌激素水平较高有关,而绝经前妇女的雌激素水平似乎不受微生物群组成的影响,这表明微生物群主要影响非卵巢雌激素水平。
►►
补充植物雌激素能够促进特定细菌的肠道定植,富含植物雌激素的饮食可能与亚洲绝经后妇女代谢综合征的风险降低有关。
➤ 反过来,性激素水平也可能影响微生物组的组成
雄激素过量,如多囊卵巢综合征患者,也可能导致生物失调和细菌多样性降低。多囊卵巢综合征(PCOS)是一种内分泌疾病,其特征是雄激素水平较高,雌激素水平较低,几项研究表明,多囊卵巢综合症患者的肠道失调与细菌多样性较低有关,导致丁酸生成减少,BMI升高,睾酮血清浓度升高。
此外,肠道微生物组通过产生有助于减少炎症反应的短链脂肪酸在决定胰岛素分泌方面发挥重要作用;菌群失调可能导致胰岛素抵抗和糖代谢改变,如多囊卵巢综合征;较高的胰岛素水平刺激卵巢产生雄激素,从而使多囊卵巢综合征的发病机制永久化。
➤ 微生物群及其代谢产物也可能影响肠道菌群定植
微生物群及其代谢产物也可能通过定殖阴道,以及一些作者认为的子宫内膜和胎盘,影响女性生育、怀孕、胚胎发育和分娩的各个阶段。微生物群的改变与促炎细胞因子的分泌和早产有关。剖宫产新生儿肠道菌群多样性较低,可能是因为他们没有通过阴道被母体肠道菌群定植。
➤ 性激素信号和脑-肠轴功能中的应激反应之间存在复杂的相互作用
我们知道,肠道和中枢神经系统之间的双向沟通途径,根据肠道细菌是大脑和内分泌系统之间的重要媒介。肠道菌群是调节脑-肠轴的中枢,肠道屏障短链脂肪酸,除了是能够调节肠道运动和伤口愈合的炎症调节剂外,还代表了微生物组和肠-脑轴之间的联系。
功能性胃肠疾病的发生率,如功能性消化不良和肠易激综合征,导致运动能力受损和/或敏感性改变,在女性中明显更高,这可能是因为性激素信号和脑-肠轴功能中的应激反应之间存在复杂的相互作用。
▸ 雌激素激活脑-肠轴的受体,间接干扰胃肠动力
具体而言,已观察到雌激素通过直接激活位于脑-肠轴的受体,并通过调节其他受体系统间接干扰胃肠动力和敏感性。在卵巢周期的卵泡期,当雌激素水平较高时,女性的胃肠动力降低。
此外,支持循环雌激素在胃排空延迟中起重要作用的假设,绝经前和绝经后女性接受激素替代疗法与未接受激素治疗的绝经后妇女相比,胃排空速度较慢,而绝经后的女性与同龄男性相类似。相比之下,睾酮或雄激素似乎对胃动力或胃超敏反应没有影响。
性激素和肠道菌群之间的相互关系
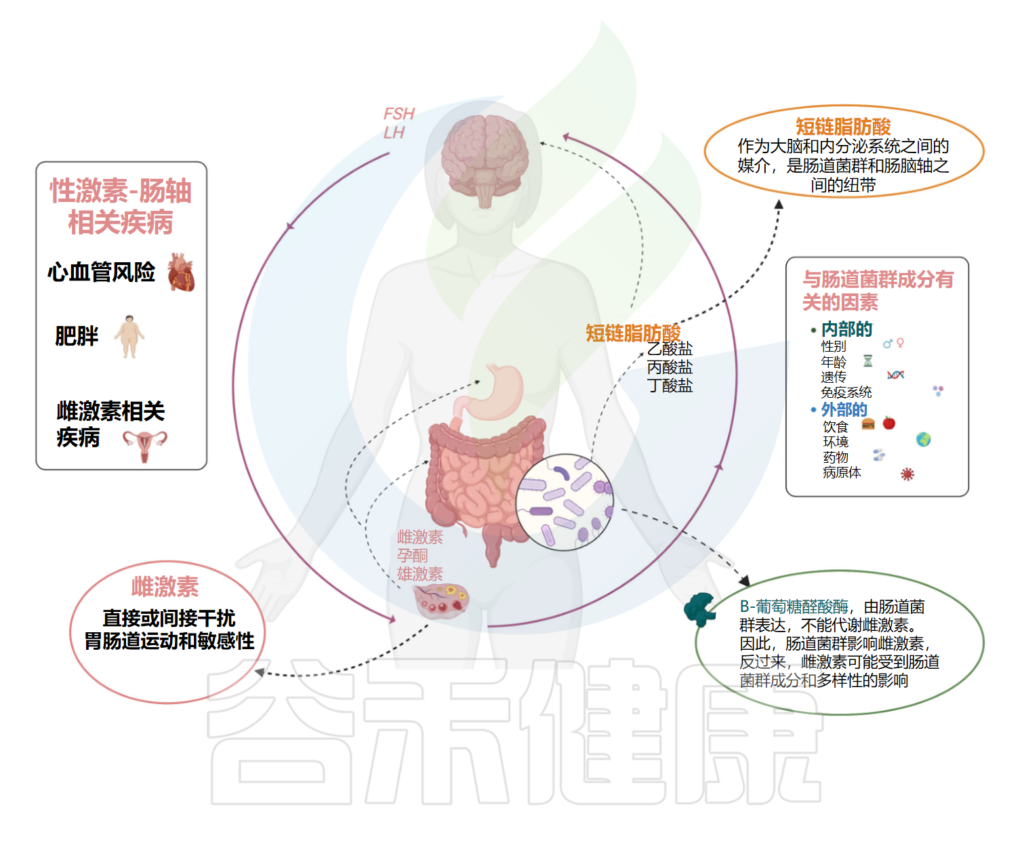
Calcaterra V, et al., Front Endocrinol (Lausanne). 2022
雌激素通过对核受体的作用,以及通过与膜G蛋白偶联的雌激素受体1 (ER 1)受体的激活,实现其快速、非基因组的作用机制。
▸ 雌激素干扰疼痛调节
雌激素受体在中枢神经系统和涉及内脏疼痛感知的通路中广泛表达,包括下丘脑、杏仁核和中脑,所有这些都已被证明向参与胃肠功能调节的迷走神经细胞发送广泛的投射。
具体而言,在外周内脏传入中,雌激素似乎通过改变离子通道的开放和受体表达的调节来调节伤害感受,以及激活胆酸速激肽-神经激肽1受体并诱导P物质的释放。
最后,除了干扰疼痛调节外,雌激素还参与中枢神经系统的内脏信息处理。
脑成像研究发现,与患有IBS的男性相比,患有IBS女性在对不良内脏刺激的反应中表现出更大的情绪回路激活,包括杏仁核和蓝斑。
众所周知,随着年龄的增长,微生物群的组成逐渐发生变化,需氧菌和兼性厌氧菌的数量普遍减少,专性厌氧菌种群数量增加。
传统上,普遍的观点是,在1-2岁之间,人类肠道菌群开始类似于成年人,主要由厚壁菌门、拟杆菌门、变形杆菌门和放线菌门的物种组成。然而,青少年和成人粪便微生物群在属水平上观察到许多差异。
非青春期和青春期菌群类别存在差异
一项研究评估青春期儿童的远端肠道菌群表明,双歧杆菌属和梭状芽孢杆菌属(已知定植于新生儿肠道并在2至18岁之间逐渐减少的物种)成员的丰度在青春期儿童中显著高于成年人。这些属的流行率以前在青春期前和青春期年龄组中没有被认识到。
2020年的一项横断面研究,通过16S rRNA测序分析了不同青春期阶段(5-15岁)的肠道菌群生物多样性。非青春期和青春期受试者之间的α或β多样性没有发现差异,但研究证明两组之间的细菌类群存在差异。
特别是,非青春期受试者的特征主要以下菌群:
另一方面,青春期组以β变形菌纲(Burkhollderiales)的流行率较高。
进一步分析血清性激素与菌群丰度之间的关系,结果表明:
睾酮水平与Adlercreutzia、Dorea、Clostridium和拟副杆菌属的丰度有关。作者假设这些细菌可能受到性激素的影响。
几项研究已经调查了肠道微生物与性类固醇激素之间的关系。Shin等人证明了肠道菌群特征与人类睾酮/雌激素状态之间的关系:
不动杆菌、多利亚菌、瘤胃球菌和巨单胞菌与睾酮水平显著相关,而Slackia菌和丁酸单胞菌则与雌二醇水平显著相关。
青春期两性之间的肠道菌群差异更加明显
据报道,肠道菌群似乎不受儿童性别的影响,但在青春期开始时就出现了差异。微生物门相关疾病也表现出性别偏见,支持肠道细菌与性别之间的关系可能是双重的。
最近的一项横断面调查发现,在青春期,两性之间的肠道菌群差异更加明显。
比较青少年和成年人的微生物群,发现双歧杆菌的数量,特别是在一些研究中,随着年龄的增长而减少,也报道了与拟杆菌门和厚壁菌门的年龄相关的联系。
Hollister等人比较了青春期和成人的肠道菌群组成,指出在青春期发育期间,女孩的肠道菌群逐渐发生变化,越来越像成年女性的肠道菌群,与她们的青春期发育程度成正比。
在雄性和雌性中,优势类群都是梭状芽孢杆菌和拟杆菌,表明微生物群成熟的要素是:
梭状芽胞菌和拟杆菌类菌群优势的变化
事实上,在青春期,梭状芽孢杆菌的相对丰度逐渐增加,拟杆菌的相对减少,逐渐类似于成人微生物群的组成。就门而言,随着青春期的进展,厚壁菌门的数量增加,而拟杆菌门的数量减少。
然而,尽管Hollister等人观察到男孩和女孩的肠道菌群变化,但由于男性青春期发育较晚,仅在女性青少年中获得了具有统计学意义的数据。
青春期,肠道菌群组成和预测的代谢谱存在性别显著差异
Yuan等人证实了这些数据,他们确定了不同青春期状态下两种性别的肠道菌群特征。对89名5-15岁的中国参与者进行了肠道菌群分析。参与者被分为青春期前和青春期组,男性(n=49)和女性(n=40)。
这项横断面研究表明,在青春期之前,肠道菌群组成和预测的代谢谱存在性别差异,在青春期变得更为显著。
实验结果表明,Dorea、Megamonas、Bilophila、Parabacteries、Phascolactobacterium属代表青春期受试者的微生物标记。
他们认为,性别依赖的肠道菌群多样性部分是由于性激素,部分是由于其他非激素影响因素。
“
以上是肠道菌群在生理性青春期发育中的作用的证据。而肠道菌群在病理性青春期中的作用越来越受到关注。
青春期到来的时间可能受到肠道菌群的影响,特别是某些梭菌属物种,包括调节宿主性激素水平的瘤胃球菌科、粪杆菌属和瘤胃球菌属。
肠道菌群影响雌激素代谢
Ruminococcus和Faecalibacterium的β-葡糖醛酸酶能够裂解雌酮和雌二醇,而拟杆菌物种只能代谢雌酮。
因此,尿液中雌激素-雌激素代谢物比率与瘤胃球菌的相对丰度呈正相关,与拟杆菌的相对丰量呈负相关。
根据这些数据,肠道菌群可能通过其雌激素代谢部分调节青春期的开始。
肠道菌群通过能够代谢雌激素的特定肠道微生物似乎能够调节青春期,但反过来也可能。性激素可以通过引导肠道菌群的成熟,直接影响特定菌群的生长。
肠道菌群代谢物影响内分泌系统
最近的研究发现,肠道菌群产生的代谢物可以影响人体内分泌系统,激活肠道神经系统。
一些关于微生物群功能的最佳研究强调了肠道微生物通过产生短链脂肪酸为宿主提供能量,包括丁酸和丙酸,这两种物质都参与胆汁盐代谢,并在脑肠轴中发挥重要作用。
已经证明,内源性受体游离脂肪酸受体(FFAR)2和FFAR3与短链脂肪酸相互作用,并在肠内分泌细胞中表达,肠内分泌细胞产生肽YY(一种厌食激素),从而参与调节宿主能量、食欲、脂肪组织储存和荷尔蒙平衡,影响青春期时间。
肥胖 & 青春期 & 肠道菌群失调
肥胖和青春期之间有着密切的联系;尤其是性早熟与体重指数(BMI)呈正相关。基于这一信息,并考虑到受性早熟影响的儿童往往肥胖,已假设肠道菌群可能参与性早熟的发病机制。
一项研究阐明了特发性中枢性性早熟(ICPP)患者(n=25)和健康女孩(n=23)之间肠道菌群的差异。
特发性中枢性性早熟肠道菌群与肥胖菌属相似
作者应用16S rDNA测序来比较两组之间的肠道菌群,观察到,特发性中枢性性早熟中鉴定的肠道属与肥胖相关的肠道属相似,特别是吉米格瘤胃杆菌属、示波杆菌属和XIVb梭状芽孢杆菌属。
考虑到微生物种类水平,患有特发性中枢性性早熟的女孩富含Rumicoccus bromii, Ruminococcus gnavus, Ruminococcus leptum,前两种是在肥胖人群中发现的;它们可以促进脂肪组织的能量吸收和增生,而据报道,Ruminococcus leptum会影响人体体重变化。
这些结果突出了肥胖、特发性中枢性性早熟和肠道菌群失调之间的关联。作者假设,肠道菌群失调会导致青春期前女孩发生类似于肥胖患者的过程,脂肪细胞的增殖和沉积会引发性早熟。
然而,肠道失调也可能导致下丘脑-垂体-性腺轴(HPGA)的早期激活。
不同的研究已经研究了肠道菌群在雌激素、FSH和LH分泌机制中的作用,但仍不清楚。
三种临床生物标志物(FSH、LH和胰岛素抵抗)与肠道菌群之间的关系
考虑到特发性中枢性性早熟女孩,作者证明了FSH与梭杆菌、LH与Gemmiger之间的正相关,LH与Romboutsia之间的负相关。此外,胰岛素抵抗与吉米格菌、瘤胃球菌、巨单胞菌和双歧杆菌呈正相关。
特发性中枢性性早熟:部分菌群高产短链脂肪酸
肠道菌群、激素分泌和肥胖之间的密切联系激发了对肠道菌群触发特发性中枢性性早熟机制的研究。
研究人员在工作中调查的特发性中枢性性早熟女孩的特征是与产短链脂肪酸相关的菌群:
短链脂肪酸诱导瘦素基因表达,激活HPGA轴
瘦素在女性青春期发病中的重要作用众所周知。瘦素是一种脂肪细胞代谢肽,参与其表达的基因与短链脂肪酸相关。
短链脂肪酸产生与特发性中枢性性早熟之间的关系可以通过高浓度短链脂肪酸诱导瘦素基因表达的机制来解释,瘦素基因激活下丘脑-垂体-性腺轴,从而导致青春期的开始。
Li等人招募了27名中枢性性早熟女孩、24名超重女孩和22名健康对照,以探讨肥胖与中枢性性早熟之间的关系。
肠道菌群代谢物激活HPGA轴
这项研究表明,中枢性性早熟患者表现出过多的Alistipes、Klebsiella、Sutterella,这通常出现在神经系统疾病患者中。这些微生物产生具有神经递质活性的代谢物(血清素和多巴胺),这些代谢物会引发早熟,激活下丘脑-垂体-性腺轴。
肠道菌群代谢物促进胰岛素抵抗
作者在中枢性性早熟和超重组中都发现了普雷沃氏菌;支链氨基酸的产生可促进胰岛素抵抗。这一机制可以解释中枢性性早熟患者肥胖的高发生率。
此外,在两组中,均观察到一氧化氮合成升高,这是一种重要的气体神经递质,可刺激促性腺激素释放激素的分泌并促进胰岛素抵抗。
这些情况,肠道菌群表达的改变,可以解释中枢性性早熟和肥胖之间的联系,如下图所示。
性早熟 – 肠道微生物组和肥胖:密切相关
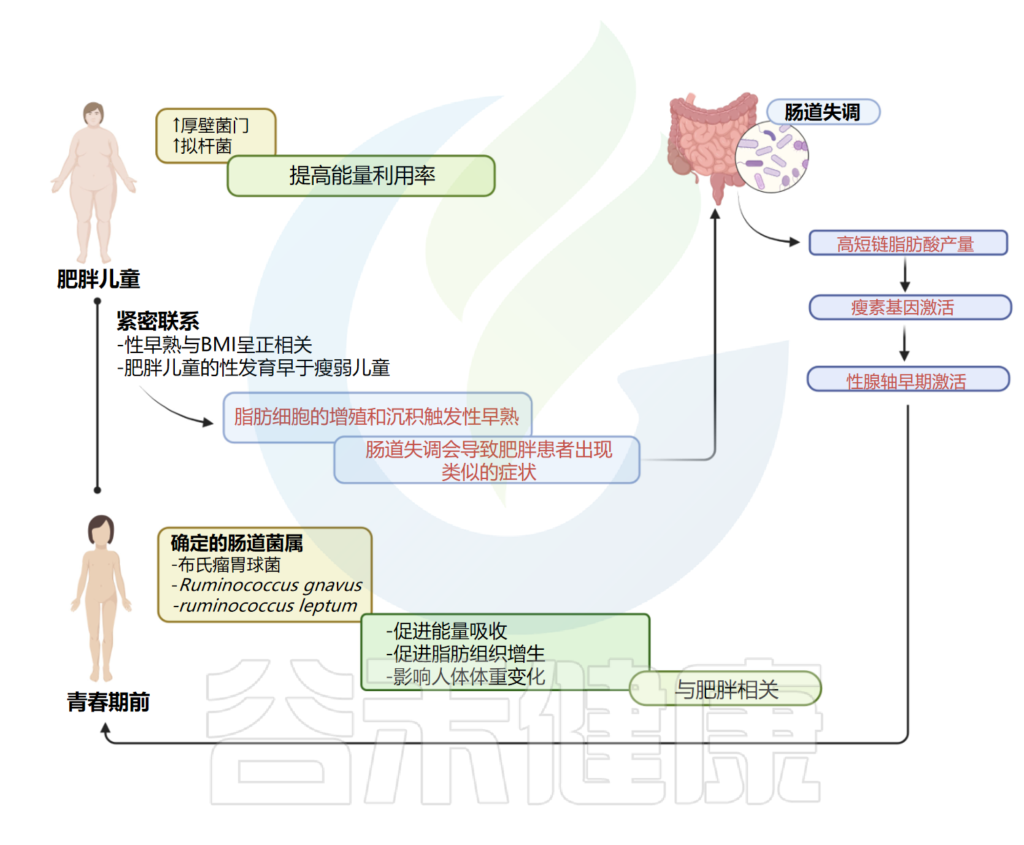
Calcaterra V, et al., Front Endocrinol (Lausanne). 2022
高脂饮食:过早激活HPG轴,诱导性早熟
大量营养素食物含量,如高脂肪摄入,可能会调节HPG轴的过早激活,诱导青春期的性早熟。
最近,在实验模型中,研究人员表明,高脂饮食对性早熟的影响受肠道微生物群和激素的相互作用调节。
断奶后的高脂饮食导致性早熟,下丘脑中的血清雌二醇、瘦素、脱氧胆酸和GnRH增加。特别是,GnRH与脱硫弧菌、Lachnoclostridium、GCA-900066575、链球菌、厌氧菌、双歧杆菌呈正相关,这表明这些细菌可能在促进性发育方面发挥作用。
此外,作者报道,“高脂饮食微生物群”移植促进了小鼠的性早熟,支持肠道菌群调节促进性早熟的局部和全身性类固醇水平。
性早熟治疗的最重要目标是保持成人身高并减少相关的心理社会压力。
促性腺激素释放激素激动剂,1个月或3个月储存制剂,是中枢性性早熟的标准护理。GnRH激动剂治疗被广泛认为是安全的。
最常见的不良反应包括局部皮肤反应和绝经后症状。需要定期验证青春期进展、生长速度和骨骼成熟。
PPP的治疗因发病机制而异,治疗的主要目的是消除性类固醇的内源性或外源性来源。
肾上腺和性腺肿瘤需要手术治疗。
其他生活中的注意事项如下,供参考。
生活方式
消除外源性雌激素(塑料、食品添加剂和杀虫剂)和其他内分泌干扰物(例如美容产品中的邻苯二甲酸盐)。薰衣草油和茶树油与乳突有关,尤其是男孩的男性乳房发育症。
营养
消除添加激素的乳制品、鸡蛋和肉类。考虑消除大豆、亚麻和芝麻,因为它们是植物雌激素,年轻组可能对这些非常敏感。
缺乏维生素 D 的 5-12 岁女孩面临青春期提前的风险是维生素 D 水平较高的女孩的两倍。
晒太阳是增加维生素 D 摄入量以预防和减缓青春期提前的一种方法。
其他食物补充包括:金枪鱼和鲑鱼等多脂鱼、橙汁、豆浆等。
肝脏解毒支持减少外源性激素作用。
食物
深色绿叶蔬菜、西兰花、甜菜、朝鲜蓟、姜黄、柠檬、蒲公英根等。
减少肾上腺激素输出的植物药:
刺五加 ;睡茄;红景天
父母的反应会影响孩子的应对能力
当孩子的家庭氛围不好,他们的病情无法得到家人的支持,会增加孩子的负面心理情绪和父母的养育压力。加强家庭成员之间的沟通至关重要。
一些导致性早熟的风险因素(例如性别和种族)无法避免。但是,有一些措施可以降低孩子出现性早熟的风险,包括:
不同的研究都提出了肠道菌群与性激素之间的双向相互作用。青春期期间,身体发育变化主要由激素驱动;因此,这一动态和过渡期是评估潜在激素对肠道菌群影响的机会。
不同青春期阶段的肠道菌群存在多样性,并且患有中枢性性早熟的女孩肠道菌群可能改变,这是预测和预防性早熟发育的一个有趣的发现。通过提供儿科内分泌学的观点,加深对性激素和微生物群变化作用之间的联系的理解,可以在青春期疾病中实施微生物群靶向治疗。
主要参考文献:
Calcaterra V, Rossi V, Massini G, Regalbuto C, Hruby C, Panelli S, Bandi C, Zuccotti G. Precocious puberty and microbiota: The role of the sex hormone-gut microbiome axis. Front Endocrinol (Lausanne). 2022 Oct 21;13:1000919. doi: 10.3389/fendo.2022.1000919. PMID: 36339428; PMCID: PMC9634744.
Li Y, Shen L, Huang C, Li X, Chen J, Li SC, Shen B. Altered nitric oxide induced by gut microbiota reveals the connection between central precocious puberty and obesity. Clin Transl Med. 2021 Feb;11(2):e299. doi: 10.1002/ctm2.299. PMID: 33634977; PMCID: PMC7842634.
Dong G, Zhang J, Yang Z, Feng X, Li J, Li D, Huang M, Li Y, Qiu M, Lu X, Liu P, Zeng Y, Xu X, Luo X, Dai W, Gong S. The Association of Gut Microbiota With Idiopathic Central Precocious Puberty in Girls. Front Endocrinol (Lausanne). 2020 Jan 22;10:941. doi: 10.3389/fendo.2019.00941. PMID: 32038493; PMCID: PMC6987398.

谷禾健康


什么是天然产物?
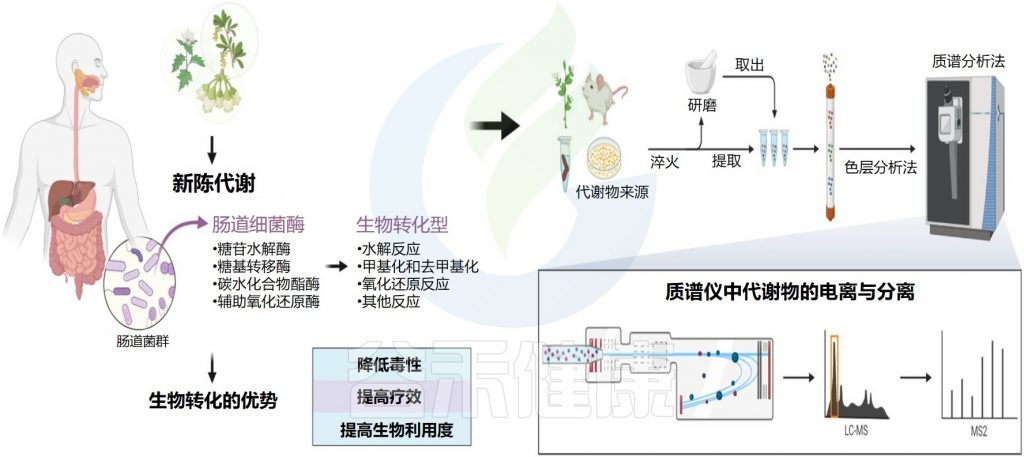
天然产物是指动物、植物、昆虫或微生物体内的组成成分或其代谢产物以及人和动物体内许多内源性的化学成分,是自然产生的小分子。值得注意的是,天然产物来源中,植物来源占85%以上。
谷禾在这里主要介绍植物来源的天然产物,包括营养素和药物。它们很容易与肠道微生物群相互作用,因为它们的成分复杂,在肠道中的停留时间长。通常,外源物质在小肠中的停留时间为1-6小时,在结肠中的停留时间为1-3天。特定的肠道微生物分解和转化天然产物,产生丰富的代谢物和功能化合物,具有宿主本身无法合成的生理活性。
天然产物在食品、药品、保健品领域均有运用。天然产物是极为重要的新药来源。近年来临床应用的药物,三分之二左右源自天然产物、天然产物衍生物以及天然的生物大分子,几乎涵盖了疾病治疗的各个方面。这在防治癌症及传染类疾病领域中的表现尤为明显。
天然产物在临床上得到了广泛的应用,其特点是多组分、多靶点、药效学物质多、作用机理复杂、生理活性多样。
对于口服天然产物,肠道菌群与临床疗效密切相关,但这种关系尚不明朗。肠道微生物在酶系统多样性引起的天然产物的转化和利用中起着重要作用。黄酮类化合物、生物碱、木酚素和酚类等有效成分不能通过人体消化酶直接代谢,但可以通过肠道微生物产生的酶转化,然后加以利用。因此,重点是通过肠道微生物群进行天然产物的代谢。
本文介绍了肠道微生物群及其对天然产物各组分生物转化的影响的研究,并强调了所涉及的常见菌群,反应类型,药理作用和研究方法。为临床疾病防治中的应用提供理论支持,以及日常的选择健康的药物提供一定的参考,并为基于肠道生物转化的天然产物研究提供新思路。
基于肠道微生物的天然产物的生物转化和代谢

Zhao Y,et al.Front Microbiol.2022
★人体肠道中的微生物
肠道微生物群由1000-1250种细菌组成,这些细菌以各种形式与人类相互作用,例如共生和寄生,这种相互作用通过微生物代谢物作为信号分子极大地影响人类健康。
肠道微生物构成了一个动态和多样化的微生态系统,这是抵抗致病细菌的天然屏障。肠道微生物具有丰富的酶系统,包括葡萄糖苷酶,还原酶,裂解酶,转移酶等,并大大扩展了人体内的代谢反应池。
✦肠道菌群影响口服给药的效果
口服给药是药物递送的首选途径,口服药物占市场最畅销药物的大部分。近年来,肠道微生物群对天然产物口服给药稳定性的影响受到广泛关注。
肠道具有丰富的细菌,有助于正常的消化功能,其中健康受试者中约98%的肠道微生物可分为四个门,厚壁菌门(Firmicutes),拟杆菌门(Bacteroidetes),变形菌门(Proteobacteria)和放线菌门(Actinobacteria)。
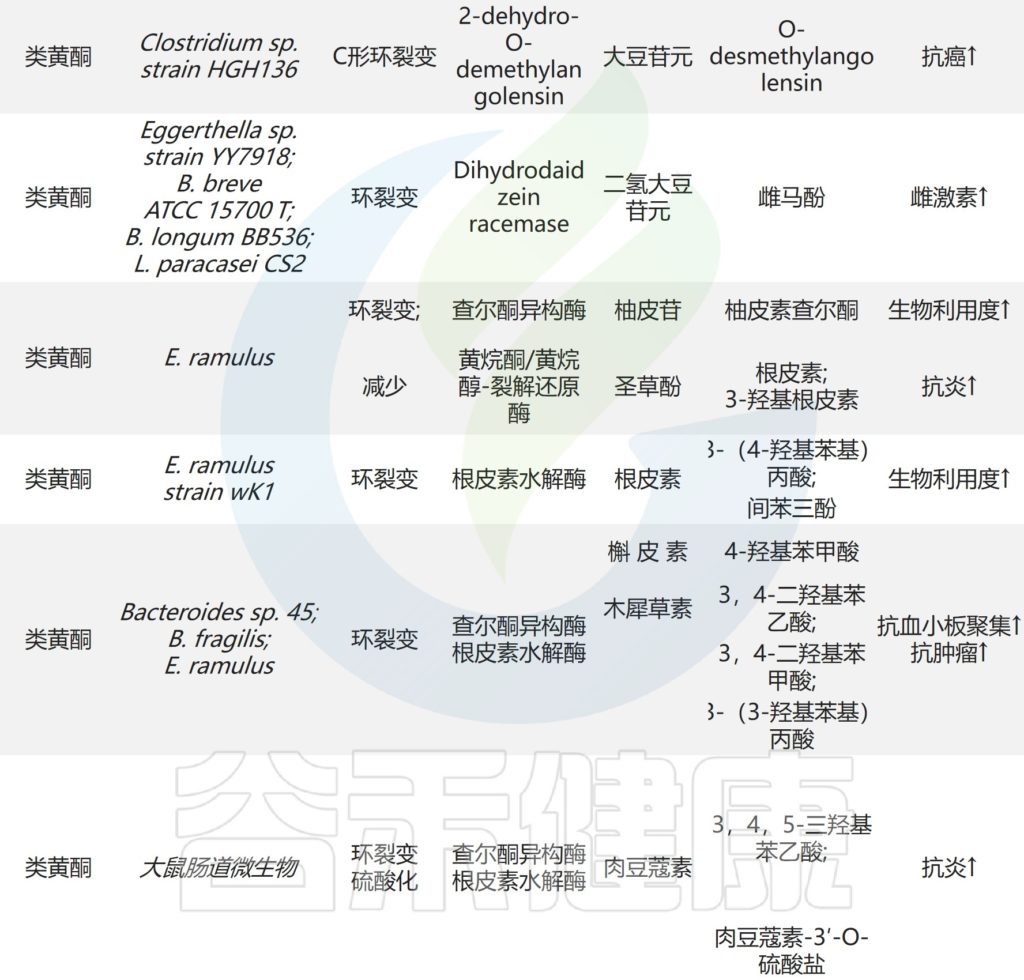
一些肠道微生物如大肠杆菌,双歧杆菌,真杆菌,乳酸杆菌,拟杆菌和链球菌参与天然产物的生物转化,其部分代谢物有利于肠道吸收并发挥显著的药理作用。
关键肠道微生物对天然产物的生物转化

Zhao Y,et al.Front Microbiol.2022
▸
大肠杆菌(Escherichia coli),又叫大肠埃希氏菌,是一种革兰氏阴性,无孢子,兼性厌氧细菌,主要栖息在脊椎动物的肠道中。
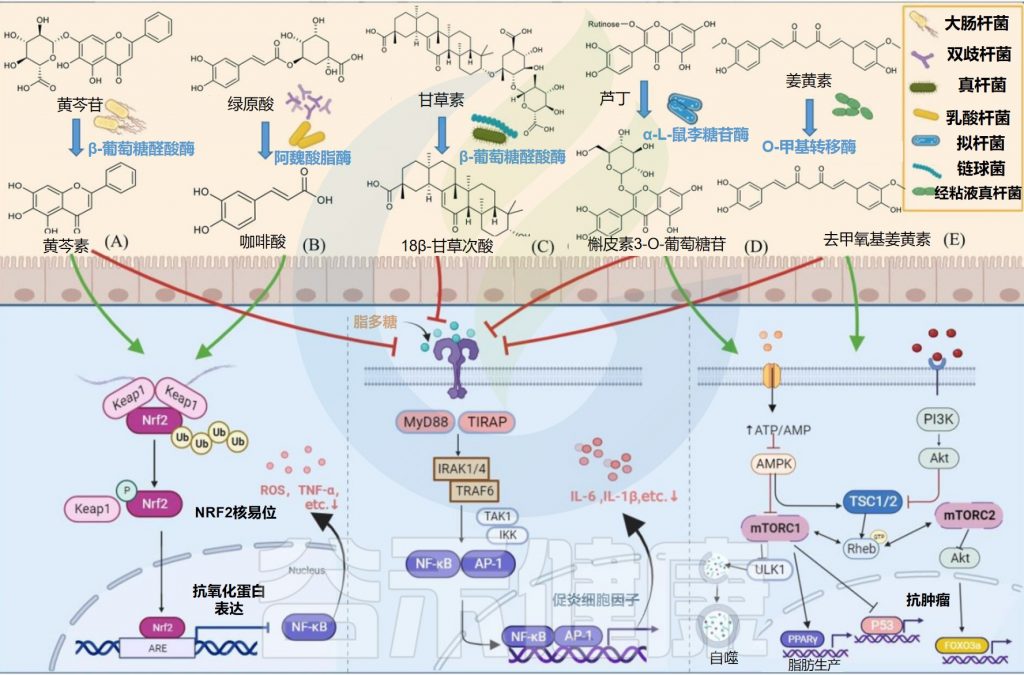
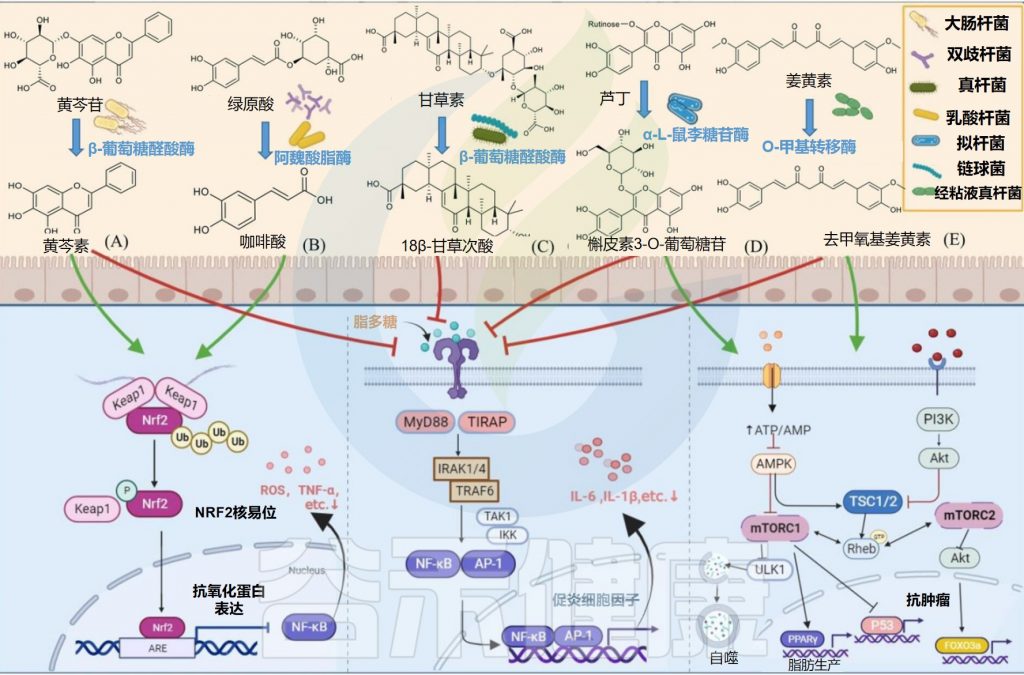
✦水解黄芩苷而具有抗炎抗氧化作用
部分大肠杆菌可以产生糖苷酶参与外源物质的转化,从而产生其有益作用。例如,E.coli HGU-3产生β-葡萄糖醛酸酶,水解黄芩苷中的糖苷键以产生黄芩素。
在相同剂量下,黄芩素比黄芩蒿素更有效地抑制组胺诱导的抓挠行为,并通过抑制Nrf2-ARE和NF-κB信号通路而具有抗炎和抗氧化作用。
✦在脂肪肝变性中有治疗益处
一些大肠杆菌菌株具有较高的姜黄素转化活性。E.coli DH10B的高表达NADPH依赖性姜黄素/二氢姜黄素还原酶(CurA)将姜黄素还原为二氢姜黄素(DHC)和四氢姜黄素(THC),其全基因组序列已经确定。
NADPH是一种辅酶,叫还原型辅酶Ⅱ,学名烟酰胺腺嘌呤二核苷酸,在很多生物体内的化学反应中起递氢体的作用,具有重要的意义。
二氢姜黄素和四氢姜黄素通过调节SREBP-1C和PPARα的mRNA和蛋白质表达水平来降低细胞中的甘油三酯水平,并以依赖性方式衰减肝脏脂肪生成;二氢姜黄素和四氢姜黄素在肝脂肪变性中比姜黄素具有新的治疗益处。
✦水解肉桂酸后有抗氧化和抗癌特性
E.coli Nu, E.coli MC和E.coli WC-1具有肉桂酰酯酶活性,可通过水解共轭羟基肉桂酸和游离羟基肉桂酸酯释放羟肉桂酸,在体外和体内均显示出抗氧化和抗癌特性。
目前,对大肠杆菌的遗传和生化特征的良好理解可能有助于在体外合成具有各种健康活性的天然产物衍生物。
▸
双歧杆菌(Bifidobacterium)是属于放线菌门的广泛而丰富的属,是人类肠道微生物群的首批定植者之一。人类肠道中最常见的双歧杆菌包括青春期双歧杆菌,B.angulatum, B.bifidum, B. breve, B.catenulatum, B.dentium, B.longum, B.pseudocatenulatum和B.pseudolongum,占成人微生物组的10%,它们与宿主健康有关。
✦预防急性肝损伤
某些种类的双歧杆菌可以通过表达阿魏酰酯酶产生酚酸。例如,动物双歧杆菌的阿魏酰酯酶可以将绿原酸(CHA)水解成咖啡酸(CAA)。咖啡酸可以通过增加Nrf2转录来预防对乙酰氨基酚诱导的小鼠急性肝损伤。
✦调节胆汁酸和降低胆固醇
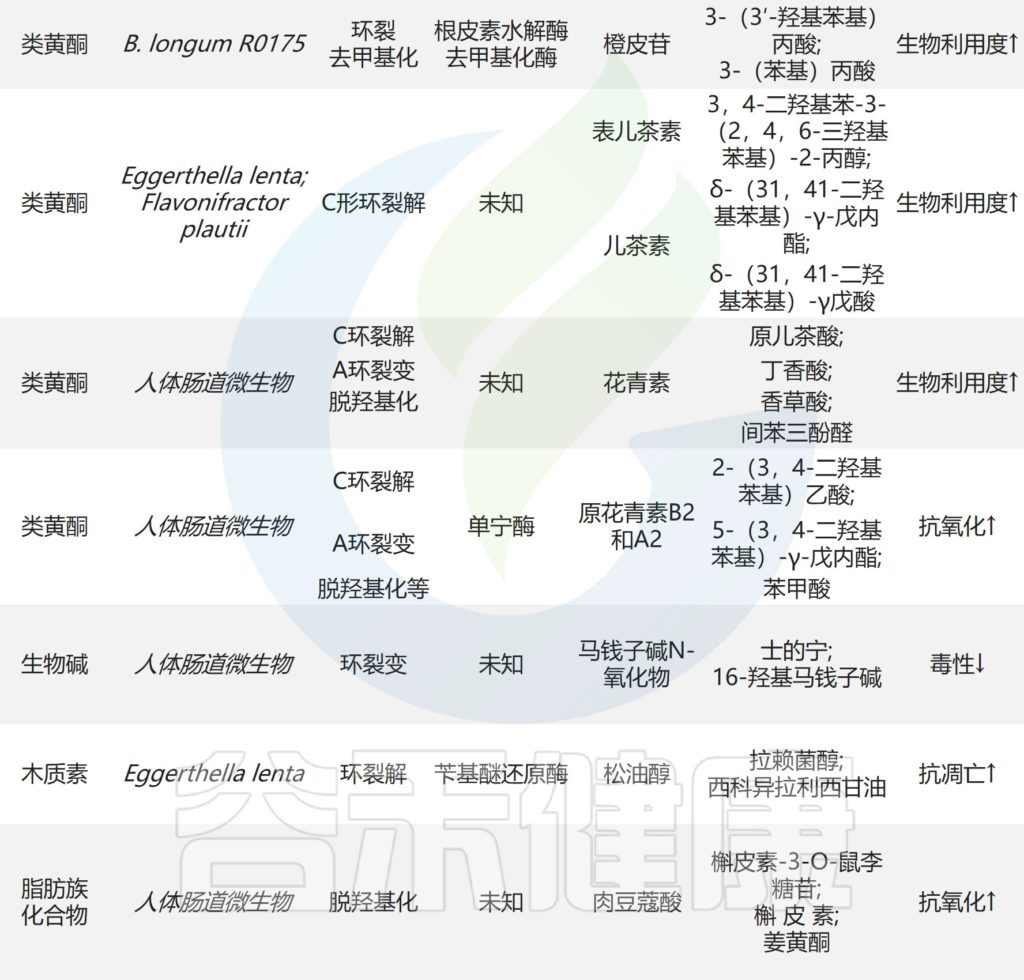
部分双歧杆菌的参与促进了黄烷酮,糖苷和皂苷在肠道中的代谢。B.longum R0175中的β-葡萄糖苷酶和去甲基化酶通过环裂解和脱甲基作用促进3-(3′-羟基苯基)丙酸和3-苯丙酸的产生。
B.longum SBT2928水解六种主要的人类和两种动物胆汁盐。因此,双歧杆菌可以调节胆汁酸代谢并降低体内胆固醇水平。
✦抗肿瘤和提高免疫
此外,B.breve ATCC 15700产生β-葡萄糖苷酶,在人参皂苷Rd的C-3和C-20位置裂解糖苷,生成脱糖基化的人参皂苷。
人参皂苷主要存在于人参和马铃薯药材中
人参皂甙的作用和功能是抑制肿瘤细胞的生长,诱导肿瘤细胞死亡,研究肿瘤的活性功能,肿瘤的异常逆行分化,提高人体免疫功能。
这些天然产物具有潜在的益处,代谢特征使双歧杆菌成为共生发展的主要候选者。
▸
真杆菌(Eubacterium)菌株的属是革兰氏阳性菌,它是人类肠道微生物群的核心属之一,并显示出在人类肠道的广泛定植。
一些真杆菌产生糖苷酶,还原酶等,并参与外源性物质的代谢。
✦抗炎抗氧化
E. ramulus是研究最广泛的类黄酮降解肠道细菌之一,它在人体肠道中很普遍。广泛存在于人体肠道中。来自E. ramulus的查尔酮异构酶和黄烷酮/黄烷醇裂解还原酶降解某些类黄酮产生查尔酮和二氢查尔酮。
二氢查尔酮及其代谢物具有抗炎和抗氧化作用,可以下调促炎细胞因子的分泌,并挽救脂多糖诱导的氧化磷酸化。
✦降解黄酮类化合物
研究了E.ramulus strain wK1对槲皮素和木犀草素的降解,发现静止细胞和酶制剂通过2,3位双键的还原和随后的环分裂将这些黄酮转化为3,4-二羟基苯基乙酸和3-(3,4-二羟基苯基)丙酸。
来自E.ramulus strain wK1的根皮素水解酶将邻近根皮素芳香A环的C-C键水解为3-(4-羟基苯基)丙酸和间苯三酚。
E.cellulosolvens ATCC 43171T可通过释放的葡萄糖部分的发酵促进类黄酮和葡萄糖苷的去糖基化。
注意:葡萄糖苷的去糖化仅由细菌酶催化。
✦预防气道过敏性炎症
Eubacterium L-8将甘草酸(GL)水解为18β-甘草次酸(18β-GA)。18β-甘草次酸通过抑制NF-κB磷酸化和增强Nrf2/HO-1通路来预防气道过敏性炎症。
NF-kB——核因子κB,参与细胞对外界刺激的响应,在细胞的炎症反应、免疫应答等过程中起到关键性作用
Nrf2/HO-1信号通路已成为细胞抗氧化应激的主要防御机制之一
这些代谢转化体现从真杆菌中获得的各种益处。然而,还需要进一步的体内研究,以最大限度地发挥真杆菌属的潜在益处。
▸
乳酸菌属(Lactobacillus)属于厚壁菌门,可以平衡微生物群落并保护胃肠粘膜。一些乳杆菌属富含代谢酶,如α-鼠李糖酶,单宁酶,没食子酸酯脱羧酶等,它们转化外源性物质。
✦抗炎、抗氧化作用
L. rhamnosus NCTC 10302具有β-葡糖苷酶和α-鼠李糖苷酶活性,通过水解,环裂变和脱氢将橙皮素7-O-葡萄糖苷和柚皮素-7-O-芸香糖苷转化为各自的糖苷元和3-(苯基)丙酸。
L. plantarum表达单宁酶水解没食子酸酯,具有短脂肪醇取代基的原儿茶酸酯和复杂的没食子单宁以产生没食子酸。没食子酸通过抑制MAPK/NF-κB途径并激活Akt/AMPK/ Nrf2途径,在脂多糖诱导的炎症和氧化应激中起保护作用。
MAPK是信号从细胞表面传导到细胞核内部的重要传递者。
Akt,蛋白激酶B,又称PKB或Rac,在细胞存活和凋亡中起重要作用。
✦预防代谢紊乱
观察到,没食子酸和邻苯三酚是由L.plantarum WCFS1中没食子甘氨酸代谢酶降解没食子甘肽而产生的。这项研究意味着益生元-益生菌相互作用在预防饮食诱导的代谢紊乱方面的潜在作用。
✦改善骨质疏松
具有大豆苷元还原酶活性的Lactobacillus sp. Niu-O16。二氢大豆苷元抑制NF-κB活化和MAPK磷酸化,从而改善骨质疏松症。
L.casei, L.plantarum和L.acidophilus影响白藜芦醇苷到白藜芦醇的去糖基化。这种转化对于提高白藜芦醇苷的生物利用度和生物活性非常重要。来自L.reuteri, L.helveticus和L.fermentum的阿魏酰酯酶水解绿原酸以释放咖啡酸。
这些发现为乳杆菌在促进健康的制药和食品应用中的作用开辟了新的视角。然而,潜在的转化机制值得进一步研究。
▸
拟杆菌属(Bacteroides)的成员是革兰氏阴性专性厌氧菌,占结肠中细菌总数的25%,在人类肠道细菌组中起着多种作用。在临床中经常检测到拟杆菌物种,例如脆弱拟杆菌,B.distasonis, B.ovatus和B.thetaiotaomicron。
✦抑制炎症反应
拟杆菌属拥有一系列水解酶,并通过转化外来物质参与与其微生物邻居的物种间关联。体外共孵育实验表明,某些拟杆菌物种参与类黄酮的生物转化。
Bacteroides sp. 45表达α-L-鼠李糖苷酶和β-芦丁糖苷酶,用于将芦丁水解成槲皮素3-O-葡萄糖苷,槲皮素和白花青素。
槲皮素3-O-葡萄糖苷比其他形式的槲皮素吸收得更好,并且可以通过抑制NF-κB和MAPK信号通路来抑制结肠炎小鼠的炎症反应。
Bacteroides sp. 54将槲皮苷代谢为羟基槲皮素和去甲基槲皮素。槲皮苷也被α-L-鼠李糖苷酶降解为槲皮素,并通过Bacteroides sp.45进行进一步的环裂解以产生3,4-二羟基苯甲酸。
β-葡萄糖醛酸酶由拟杆菌J-37表达,将甘草酸代谢为18β-甘草次酸。
天然产物在拟杆菌作用下进行生物转化,产生具有不同功能活性的代谢物。了解人体内天然产物的整个过程以评估其对人体健康的影响是很重要的。
▸
链球菌属(Streptococcus)是革兰氏阳性菌,呈球形或卵形,通常成链排列或成对排列,广泛存在于人类粪便和鼻咽中。
宏转录组学分析表明,磷酸转移酶系统主要由链球菌表达,这表明这些细菌是小肠中可用碳水化合物的主要利用者。
✦抗癌、抗过敏作用
Streptococcus LJ-22表达β-葡萄糖醛酸酶将甘草酸代谢为18β-甘草次酸-3-O-β-D-葡萄糖醛酸(GAMG)。GAMG对脂多糖诱导的RAW264.7细胞具有抗过敏活性。
甘草酸和甘草次酸均有一定的防癌和抗癌作用。甘草次酸可抑制原癌细胞的信息传递和基因表达.甘草酸对多种恶性肿瘤均有抑制作用。甘草次酸还具有抗病毒感染的作用,对致癌性的病毒如肝炎病毒,EB病毒及艾滋病毒的感染均有抑制作用。
此外,单宁酸被Streptococcus gallolyticus subsp.Gallolyticus(SGG)的单宁酶降解以产生邻苯三酚。SGG可能通过消除单宁酸对肿瘤细胞的毒性来促进结直肠癌的发展。因此,有必要进行进一步的体内研究,以确定消除这些单宁酸降解微生物是否可以支持结直肠癌的有效治疗。
✦降低血压,抗氧化
S.thermophilus GIM 1.321很高的β-葡萄糖苷酶生产能力,用于将果实花青素降解为绿原酸,咖啡酸和阿魏酸。
CAA和CHA(10/15毫克/千克/天)的施用可以降低血压并发挥抗氧化作用。
链球菌菌株可能是肠道中的共生、致病和机会性病原体,需要更多关于其对人类健康影响的信息。更好地了解链球菌如何代谢天然产物,可以调节肠道微生物组以提高治疗效果。
▸
经黏液真杆菌(Blautia)属物种严格无氧,不可移动,通常为球形或椭圆形,广泛存在于哺乳动物的肠道和粪便中。越来越多的证据表明,经黏液真杆菌的益生菌特性对天然产物的生物转化有影响。
✦影响生物转化
在类黄酮生物转化过程中,Blautia催化的反应包括去甲基化,去糖基化以及环裂解,其可能被相应的酶催化,例如O-糖苷酶和β-葡萄糖苷酶。
研究表明,Blautia sp. MRG-PMF1菌株分别将5,7-二甲氧基黄酮和5,7,4-三甲氧基黄酮转化为生物活性白杨素和芹菜素,对芳基甲基醚官能团具有水解能力。Blautia sp. MRG-PMF1还具有去糖基化活性,并且发现各种异黄酮,黄酮和黄酮被代谢成相应的糖苷元。
✦去甲基产物具有抗炎、抗癌、抗氧化能力
Blautia sp. MRG-PMF1在菌株进一步代谢具有雌激素作用的去甲基香菜素。该菌株还可以催化姜黄素以产生具有抗炎和抗癌特性的去甲氧基姜黄素。
此外,Blautia sp. AUH-JLD56能够单独生物转化牛蒡子苷或牛蒡子苷元,制成具有更好抗氧化能力的去甲基化产物。
最近,关于Blautia对草本植物和功能性食品的生物转化和代谢研究越来越多。探索Blautia的生物转化对于开发新的酶和生物活性代谢物具有重要意义。
复杂的微生物酶催化肠道中天然产物的代谢,产生有利于宿主利用/排泄的亲脂性和低分子量代谢物。与人类遗传学不同,肠道微生物组的特征是可改变的,使其成为优化治疗的潜在治疗靶点。
口服天然产品进入消化道后,首先会接触大量肠道微生物及其产生的活性酶。因此,天然产物的肠道生物转化可能发生在通过肝脏的第一次传递效应之前,也可以被运输到肝脏进行修饰/结合,然后排泄到肠道,与肠道微生物反应形成一系列代谢产物。
因此,与健康或疾病相关的特定菌株、特定代谢途径和特定酶的组合对于确定肠道微生物对宿主的影响非常重要。
✦水解提高生物活性和生物利用度
某些天然产物具有高分子量和低脂溶性,并且它们难以在肠道中被人体吸收并且具有低生物利用度。通过肠道微生物介导的水解,它们的物理性质发生了变化,它们的生物活性和生物利用度大大提高。
大多数糖苷具有低活性,被认为是“天然前体药物”。在与肠道微生物相互作用后,糖苷的糖基被去除,然后,糖苷部分被肠细胞吸收以发挥生理作用。水解反应是进一步转化所必需的,产物(例如糖)参与促进肠道微生物的生长和存活。
肠道微生物对天然产物的水解反应







Zhao Y,et al.Front Microbiol.2022
✦类黄酮
类黄酮是天然酚类化合物,大量存在于水果和蔬菜中。肠道微生物可能是黄酮类化合物(糖苷形式)功效的部分原因,由于存在水溶性糖成分,其生物利用度较低。
具有3羟基黄酮碱和平面环体系的黄烷醇构成了一类重要的黄酮类化合物。异鼠李素-3-O-新橙皮苷首先被Escherichia sp.23脱糖基化为异鼠李苷-3-O–葡糖苷,去糖基化成苷元异鼠李甙。
•肠道酶代谢比肠道微生物更快
肠道微生物和衍生酶共同控制通过体外测定确定的淫羊藿类黄酮的代谢。在目前的研究中,肠道酶代谢类黄酮的速度比肠道微生物更快。
来自Bacteroides thetaiotaomicron VPI-5482的α-L-鼠李糖苷酶可以水解淫羊藿定C的α-1,2糖苷键以产生芸香苷。
芸香苷,又叫芦丁。具有抗炎、抗氧化、抗过敏、抗病毒等功效。
•肠道酶在水解中起着重要作用
E. ramulus和Clostridium saccharogumia将花青素-氰苷3-葡萄糖苷转化为氰苷。人类肠道酶如β-葡糖苷酶在汉黄芩苷水解成汉黄苷素中起着关键作用。
茶氨酸菌素A是红茶中发现的一种生物活性儿茶素二聚体,通过人类粪便微生物群脱乙酰产生茶氨酸霉素C。
•空间位阻会限制酶降解
观察了黄烷醇的代谢差异,结果表明,在肠道微生物发酵过程中,空间位阻可能限制细菌酶降解部分黄烷醇环。
在肠道微生物的作用下,许多其他类黄酮也会发生水解反应。值得注意的是,考虑到类黄酮的结构差异,肠道微生物对类黄酮的降解程度差异很大,从而影响其生物可利用性。需要进一步研究肠道代谢在类黄酮生物利用度和吸收中的作用,以及可能的细菌类黄酮相互作用活性。
✦萜类化合物
萜类化合物是具有抗癌,抗炎和神经保护作用的最大一类天然产物。部分萜类化合物也可被肠道微生物水解。
萜类化合物广泛存在于自然界,是构成某些植物的香精、树脂、色素等的主要成分。如玫瑰油、桉叶油、松脂等都含有多种萜类化合物。另外,某些动物的激素、维生素等也属于萜类化合物。
肠道微生物也可以水解部分萜类化合物。京尼平苷在Eubacterium sp. A-44表达的β-葡萄糖苷酶的作用下产生京尼平。芍药苷在β-葡萄糖苷酶的作用下转化为PM-I,该酶由L. brevis和脆弱拟杆菌(B. fragilis)表达。通过与大鼠厌氧肠道微生物群孵育,芍药苷也被脱糖并脱苯基为小分子量的白花苷和酰基白花苷。
•肠道微生物中的酶促进水解作用
研究证明,几种具有酯酶的双歧杆菌物种可以在体外将白芍苷水解为苯甲酸。体外研究表明积雪草苷被糖苷键水解酶逐渐去糖基化,并产生相应的苷元。柴胡皂苷B1在β-葡萄糖苷酶和β-D-聚焦酶的作用下逐渐水解为原柴胡皂素和柴胡皂甙A,由Eubacterium sp. A-44表达。
除上述化合物外,萜类人参皂苷Rh2、阿迪普西洛苷I、罗汉果苷III和具栖冬青苷也可在肠道微生物的作用下发生水解反应。
肠道微生物在萜类化合物代谢中起着重要作用,其代谢产物对肠道微生物组和人体健康的影响有待进一步研究。
✦其他化合物
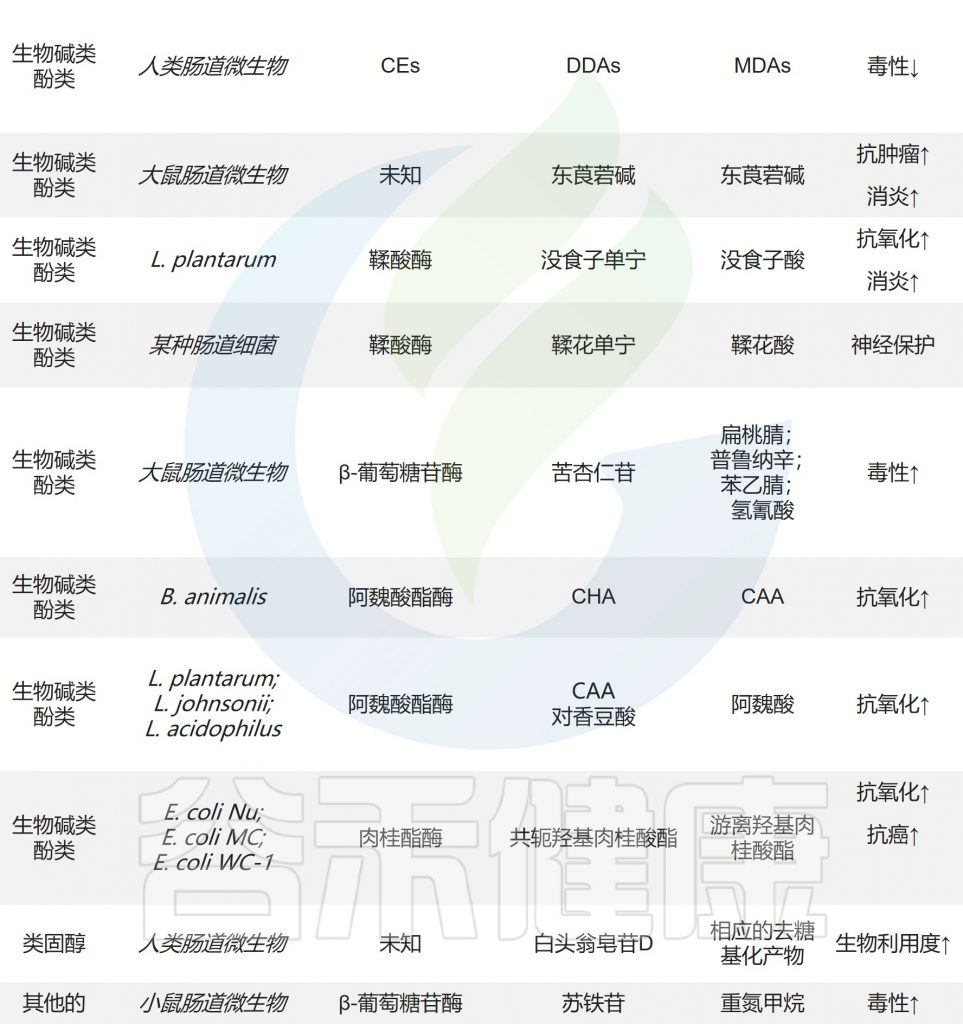
具有极低生物利用度的尿石素只有在来自Gordonibacter urolithinfaciens, Gordonibacter pamelaeae和Ellagibacter isourolithinifaciens的鞣酸酶作用下水解成鞣花酸和尿石等衍生物时才能发挥药理作用。
尿石素——存在于水果和坚果中的一类化合物鞣花单宁的天然代谢产物
从大黄中提取的蒽醌糖苷被肠道微生物水解为蒽醌苷元。Sennoside A是大黄提取物的主要成分,通过Bifidobacterium sp. strain SEN的β-葡萄糖苷酶代谢为大黄蒽酮。
大黄蒽酮具有泻下攻积、清热泻火、解毒、活血祛瘀、利胆退黄等功效。
在肠道微生物表达的羧酸酯酶(CE)的作用下,二酯二萜生物碱(DDAs,如乌头碱)水解C-8和C-14的酯键,产生毒性较小的单酯二萜生物碱。生物碱东莨菪碱、类固醇化合物白头翁皂苷D和半胱氨酸在肠道微生物的作用下也会发生水解反应。
小结
水解反应是肠道微生物代谢天然产物的重要步骤,是生物活性表达和进一步生物转化所必需的。
应重点关注参与该反应的特定微生物和酶,以充分了解天然产物的最终命运及其对人体健康的影响,并为个性化治疗提供依据。
肠道微生物可以通过亲核取代反应表达转移酶并在两种底物之间移动官能团。
肠道微生物将甲基添加到外源物质中需要化学活化的辅底物,如乙酰辅酶A、三磷酸腺苷或腺苷甲硫氨酸,而去甲基化需要可以进行亲核催化的辅因子。
甲基化修饰可以优化天然产物的生理活性,去甲基化可以释放极性基团以进一步结合和排出体外,并为肠道微生物的生长提供碳源。
下图显示了肠道微生物作用下天然产物的甲基化和去甲基化以及带来的影响。
肠道微生物对天然产物的甲基化和去甲基化



Zhao Y,et al.Front Microbiol.2022
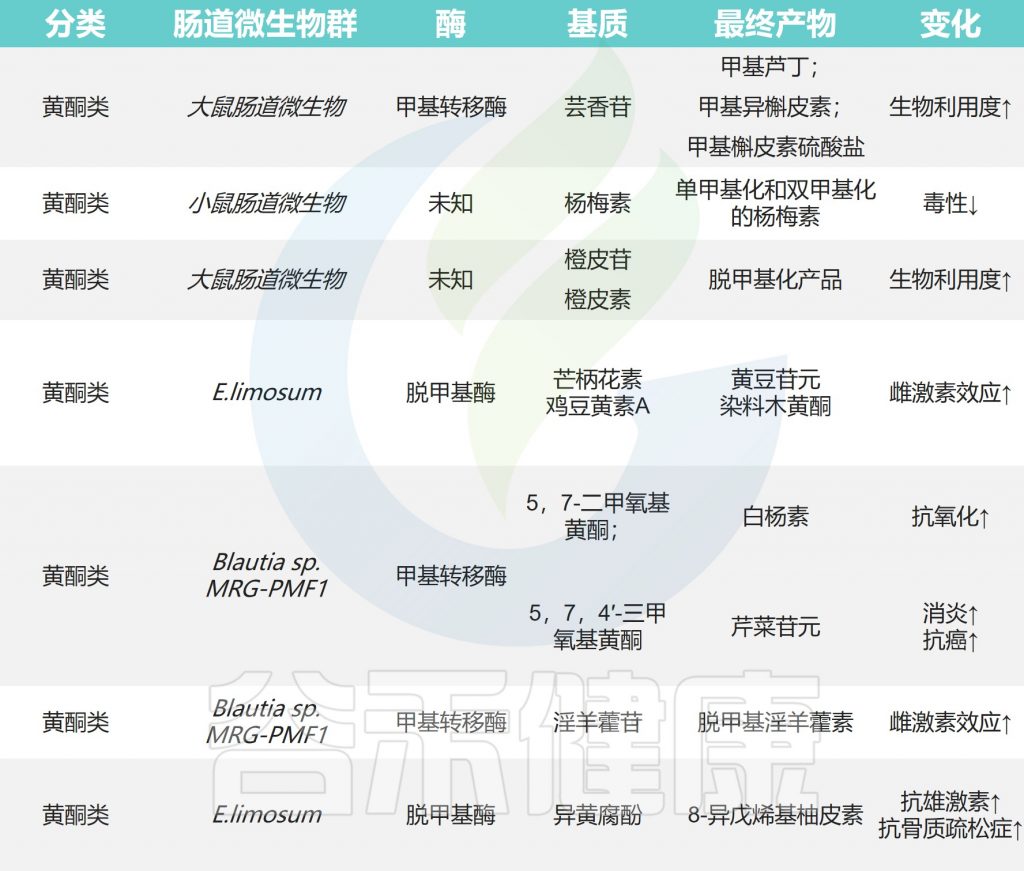
✦类黄酮甲基化后具有抗癌活性
类黄酮甲基化修饰可以在类黄酮结构中的C-2,C-3,C-4,C-5,C-6,C-7和C-8位置进行,甲基化类黄酮的生物利用度大大提高。
甲基化类黄酮具有显著的抗癌活性和对肝脏代谢的抵抗力。在大鼠口服芦丁后,在粪便样品中检测到许多甲基化代谢物,如甲基芦丁,甲基异槲皮素和甲基槲皮素硫酸盐。
作用过程
甲基化的异黄酮类化合物在E.limosum ATCC 8486的作用下进行去甲基化以产生大豆苷元和染料木黄酮。异氧杂环丁酚通过苌香产生去甲基化产物8-异辛基柚皮素。橙皮苷,5,7-二甲氧基黄酮,黄腐酚和5,7,4′-三甲氧基黄酮也可以在肠道微生物的作用下进行去甲基化反应。
✦生物碱具有抗癌性和抗病毒活性
生物碱是含氮化合物,由海洋和陆地生物生物合成,它们具有抗癌性和抗病毒活性。
绝大多数生物碱分布在高等植物,尤其是双子叶植物中,如毛茛科、罂粟科、防己科、茄科、夹竹桃科、芸香科、豆科、小檗科等。
在肠道微生物表达的酶的作用下,苦木中的主要生物碱成分准碳质酮被甲基化为准碳质碱丁基。异喹啉生物碱巴马汀通过体外厌氧培养产生去甲基产物,如哥伦比亚胺、药根碱、去甲基小檗碱和去甲基小蘖碱。通过离子阱电喷雾电离串联质谱法证明了肠道微生物对乌头碱的去甲基作用,并产生了去甲基乌头碱。
✦木酚素可以抗炎防心血管疾病
膳食木酚素是植物雌激素,主要存在于种子,坚果,豆类和蔬菜中。
木酚素在微生物作用下的一些转化
牛蒡苷可由Eubacterium sp. ARC-2菌株去甲基化为二羟基内酯和其他产物。从人类粪便细菌中分离出一种名为Lautia sp. AUH-JLD56的细菌,该物种可以有效地将牛蒡苷或牛蒡苷元转化为去甲基化代谢物3′-去甲基牛蒡苷(3′-DMAG)。
Secoisoolaricinol是亚麻籽中最常见的木脂素之一,可在Blautia producta、Gordonibacter和Lactonifactor longoviformis的存在下去甲基,形成肠内酯和肠二醇。
芝麻素通过甲基化、去甲基化和肠道微生物的其他反应代谢为哺乳动物木脂素内酯和肠二醇。罗汉松树脂酚和连翘脂素也可以去甲基化以产生内酯。水飞蓟宾A和B被人类粪便微生物群去甲基为去甲基水飞蓟宾A和脱甲基水飞蓟宾B。
✦其他化合物
多酚化合物姜黄素被Lautia sp. MRG-PMF1去甲基化,以产生代谢物去甲基姜黄素(DMC)和双去甲基姜黄素(bDMC)。二氢异铁酸的去甲基化产物,如二氢咖啡酸,也在粪便代谢物中获得。
研究发现,甲基化反应发生在甘草皂苷22β-乙酰氧基甘草酸糖链的内部和外部葡萄糖醛酸残基处,产生22β-乙氧基甘草酸甲酯。
丹参素,萜类化合物,二苯乙烯类化合物和类固醇化合物在肠道微生物的作用下经历甲基化和去甲基化。
小结
甲基化和去甲基化反应是肠道微生物代谢的重要途径。甲基化能引起染色质结构、稳定性及蛋白质相互作用方式的改变,从而控制基因表达。去甲基化则诱导了基因的重新活化和表达。
然而,介导这种反应的基因/酶尚未完全确定,还需要进一步的实验研究。
肠道微生物可以表达许多氧化还原酶并通过调节各种官能团转化天然化合物,如烯烃、羧酸衍生物、硝基、氧化物和a,b不饱和羧酸衍生物,这些官能团影响体内天然产物的活性。
下图显示了肠道微生物作用下天然产物的氧化和还原反应。
肠道微生物对天然产物的还原和氧化反应






Zhao Y,et al.Front Microbiol.2022
✦类黄酮
黄酮类化合物大豆苷元在Clostridium sp. strain HGH6和Lactobacillus. sp. Niu-O16的作用下还原为二氢大豆苷元和进一步的四氢大豆苷元。还原产物二氢染料木黄酮是由染料木黄酮在人类粪便细菌的作用下产生的。
通过分析,在厌氧条件下与槲皮素一起孵育,在大鼠肠液培养液中鉴定出脱氧代谢物山奈酚和双键氢化还原产物西叶素等化合物。
从Flavonifractor plautii ATCC 49531中发现了黄酮还原酶,该酶特异性催化黄酮/黄烷醇C环的C2-C3双结合的氢化作用,并在黄酮类化合物整个生物降解途径的初始步骤中起作用。
去甲黄腐酚,一种查尔酮化合物,被分枝杆菌还原为去甲-α,β-二氢黄腐酚
•药理作用
黄酮类化合物分布广泛,具有多种生物活性。
1.黄酮类化合物具有抗自由基和抗氧化作用;2.防治心血管疾病方面均具有良好的效果;3.抑菌抗免疫作用;4.对动物激素的调节作用。
✦生物碱
•小檗碱具有杀菌消炎的功效
由肠道微生物产生的硝酸还原酶催化生物碱中的醚和配位键反应。小檗碱(BBR)作为黄连的主要成分,可以通过肠道微生物表达的硝基还原酶还原为二氢小檗碱,这种还原产物具有很高的极性。
二氢小檗碱可以在肠道中被吸收,然后被氧化成原型小檗碱进入血液。二氢小檗碱在肠道中的吸收率是小檗碱的五倍。同时还发现,肠道微生物可以通过氧化将小檗碱转化为氧小檗碱。
小檗碱的功效和作用主要就是起到杀菌消炎的作用,抗菌谱比较广,主要用于治疗各种肠道感染和菌痢等。
•其他生物碱产物
血红素是小檗碱的一种新代谢产物,可能是一种值得探索的生物活性剂。黄连碱是一种天然的原小檗碱生物碱,具有与小檗碱相同的母体结构。口服黄连后,C-O键打开并断裂,随后发生还原反应,生成氢化小檗碱。阿南酰胺被小鼠和人类肠道微生物群还原为二氢阿南酰胺,生物利用度更高。
✦苯丙酸
•咖啡酸具有抑菌、抗病毒作用
咖啡酸(CAA)作为食品和饮料中的主要膳食多酚,在酯化后很容易进入结肠并与肠道微生物群发生反应。
苯丙酸通过C4双键还原和脱氢作用转化为3-羟基苯基丙酸,然后通过体外肠道微生物的β氧化迅速转化为3-苯基丙酸。苯丙酸也可以脱羟基化为间香豆酸或氢化为二氢咖啡酸。
咖啡酸片具有较普遍的抑菌效果,可用于抑菌、抗病毒。除此之外,还具有中枢兴奋、解毒凝血等作用。
丹参素是丹参的主要单体酚酸,通过肠道微生物群进行脱氢和脱氧,生成3-苯基-2-羟基丙酸、3-(3,4-二羟基苯基)-2-丙烯酸(咖啡酸)和3-(3,4-二羟基苯)丙酸。
✦其他化合物
甘草次酸通过盲肠中的Ruminococcus sp. po1-3的3β-羟基类固醇脱氢酶生成3-氧代甘草次甲酸。
•通便作用
番泻甙是一类天然蒽醌衍生物和二聚糖苷,首先被β-葡萄糖苷酶水解生成番泻甙-8-O-单糖苷,然后在体内被链球菌还原为具有通便作用的大黄蒽醌。
二萜类白藜芦醇被Slackia equolifaciens和Eggerthella lentaATCC 4305还原为二氢白藜芦醇。
此外,二酮姜黄,类固醇化合物地高辛和其他化合物马兜铃酸也可以在肠道微生物的存在下减少。肠道微生物黄酮还原酶和硝基还原酶具有特殊的催化选择性,填补了肠道微生物转化途径的关键空白。
然而,介导肠道微生物减少的特定基因和酶尚未完全确定。
肠道微生物对天然产物的其他反应
Zhao Y,et al.Front Microbiol.2022
如图中所示,肠道微生物也通过环分裂、硫化、芳构化和其他反应转化天然产物。
龙胆苦苷是一种天然的环烯醚萜苷,可通过肠道微生物β-葡萄糖苷酶水解为龙胆醛,然后通过N-杂环反应水解为含氮化合物。
京尼平丙酮醇的部分开环导致肠道微生物形成二醛。奎宁酸可以在肠道微生物存在下芳香化为马尿酸。
原花青素B2和A2与人类肠道微生物体外发酵,获得了主要的水解和环裂解代谢物,即苯甲酸、2-(3,4-二羟基苯基)乙酸和5-(3,4-二羟基苯)-γ-戊内酯。
口服木犀草素后,在大鼠粪便样品中检测到硫酸盐和氢还原代谢物。由肠道微生物促进的大豆苷元转化为雌马酚是另一个有趣的例子。
木犀草素,多以糖苷的形式存在于多种植物中,具有镇咳和祛痰作用。
雌马酚的作用:1.抗氧化,雌马酚属于多酚类物质,可作为氢/电子受体,清除自由基;2. 免疫调控,雌马酚的免疫保护作用可能是有其抗氧化作用所介导的,涂抹雌马酚对皮肤有免疫保护作用。
SesA是一种来自Sinomonas sp.no.22的芝麻素代谢酶,通过环裂解催化亚甲基从芝麻素或芝麻素单儿茶酚转移到四氢叶酸,产生芝麻素一儿茶酚或二儿茶酚和5,10-亚甲基四氢叶酸酯。
萜类化合物,类黄酮,苯酚,生物碱和脂肪酸都可以在肠道微生物的作用下进行生物转化反应,产生活性代谢物。
这些研究证明了各种肠道微生物组的巨大代谢潜力。天然产物的肠道微生物代谢及其在宿主健康中的作用应该是未来研究的重点。
小结
本节总结了来自单个反应的肠道微生物群介导的天然产物的生物转化。
但是,观察到一些限制。首先,考虑到肠道微生物的复杂性和肠道微生物酶的多样性,天然产物在肠道中经历复杂的转变。单个反应只能描述一定的新陈代谢过程。可以通过激活/抑制这一过程来优化治疗。
此外,考虑到肠道微生物含有各种潜在的多功能酶,可以预期肠道微生物会有更多的生物转化反应被天然产物所低估。
为了阐明肠道微生物代谢如何影响人类健康,研究人员应该将功能与基因和酶联系起来。
除了调节肠道微生物对天然产物的处置外,天然产物对肠道微生态学的调节作为潜在的功效机制也很重要。
越来越多的肠道微生物群研究逐渐揭示了大多数天然产品的高药理作用与低口服利用率之间的关系。大多数糖苷具有复杂的母体结构,很难被肠细胞吸收,从而限制了它们的组织特异性生物可及性。
这些化合物通过依赖于微生物/肠道微生物酶的降解反应转化为小分子代谢物/独特代谢物,从而对宿主产生广泛影响。肠道微生物还作用于膳食酚类物质,产生有助于宿主健康的功能代谢物。
✦增强抗肿瘤、抗炎、降脂作用
肠道微生物的生物转化促进了天然产物的治疗效果。人参皂苷对化合物K(CK)的典型代谢模型已被广泛报道,具有增强的抗肿瘤,抗炎和降脂作用。
人参皂苷的生物转化和代谢物化合物k的功效
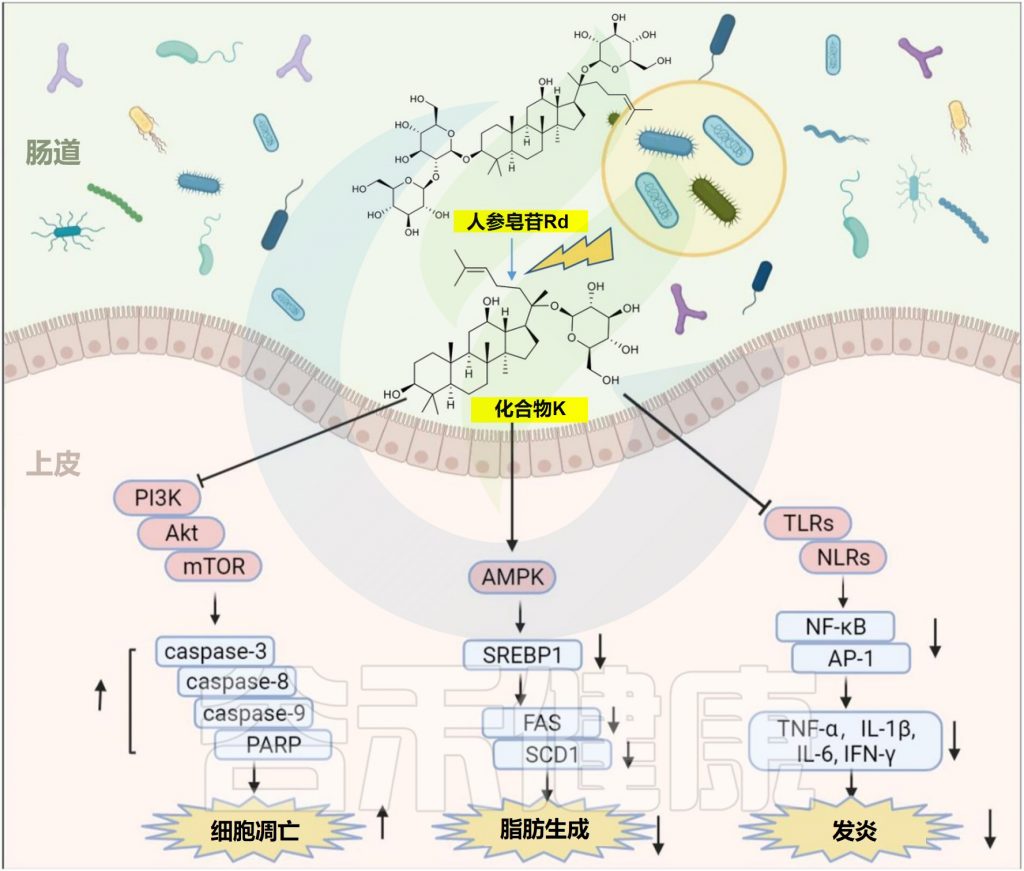

编辑
Zhao Y,et al.Front Microbiol.2022
1 —降脂
在20μM时,化合物K通过激活AMPK减少人肝细胞癌细胞中的肝脂堆积
2–抗癌
在50μM时,化合物K通过上调半胱天冬酶-3,半胱天冬酶-8,半胱天冬酶-9和环磷酸腺苷依赖性蛋白激酶来抑制胶质母细胞瘤细胞的生长。
3–减弱炎症
化合物K通过自噬诱导和调节NF-κB、p38蛋白和JNK/MAPK信号来减轻巨噬细胞炎症和泡沫细胞形成。
✦姜黄素可以抑制炎症
姜黄素代谢物的生物利用度取决于微生物群依赖性。例如,去甲基姜黄素增加PPARγ表达,导致自噬和核转录因子κB抑制,随后抑制脂多糖诱导的炎症。
PPARγ——是一种过氧化物酶体增殖物激活受体
MAPK——即AMP依赖的蛋白激酶,是生物能量代谢调节的关键分子
去甲基姜黄素通过抑制炎症因子的分泌和MAPK和NF-κB途径的激活来减轻体内和体外的炎症反应。由于在其原型苯环结构中不存在甲基,去甲基姜黄素的化学稳定性增加,从而解释了姜黄素的强大有益作用。
姜黄素来源:姜黄素是从植物根茎中提取,主要来源于生姜,然后就是那些植物中,胡萝卜、番茄以及辣椒,咖喱,芥末中也含有一些姜黄素。
✦尿石素具有抗炎和神经保护等作用
值得注意的是,尿石素A(UA)是一种天然化合物,由肠道微生物从摄入的鞣花单宁和鞣花酸中产生,具有显著的抗炎和神经保护作用。
在1μM时,尿石素A足以降低聚(I:C)诱导的RAW264.7细胞中肿瘤坏死因子和MCP-1的产生,并使TLR3/TRIF信号失活。尿石素A提高高脂饮食小鼠的全身胰岛素敏感性并降低肝脏白细胞介素1β水平。
MCP-1,是一种由星形胶质细胞分泌的促进炎症的蛋白质。
Toll样受体3(TLR3)是TLR家族的成员,介导I型干扰素、促炎细胞因子和趋化因子的转录诱导
尿石素A通过降低皮层和海马体中白细胞介素6、白细胞介素1β和肿瘤坏死因子的水平来改善小鼠的认知障碍,并抑制神经炎症。
小结
这些研究强调了鉴定天然产物微生物代谢的重要性。此外,许多体外药理活性测量应与微生物代谢产物一起进行,微生物代谢产物实际上与体内的生化受体相互作用。
肠道微生物的组成,结构,功能和代谢物已成为天然产物发挥有益作用和降低毒性的潜在目标。
例如,肠道微生物可以通过羧基酯酶催化双酯型二萜生物碱的C-8和C-14的酯键水解,或催化C-8的酯交换以产生毒性较小的单酯二萜烯生物碱。
大肠杆菌的地高辛还原型菌株含有强心苷还原酶,它可以还原地高辛环上的α和β-不饱和内酯,并将其代谢为活性较低的二氢地高辛,从而抑制其可能的心脏毒性。
地高辛,主要用于心脏类疾病的治疗,对心脏的作用表现为正性肌力作用,减慢心率,抑制心脏传导。
然而,这种能力是有限的,并且50%的地高辛可以通过肠道微生物转化而失活。强心苷还原酶可能是地高辛失活的有效生物标志物,其表达可以被精氨酸抑制。
•饮食可能调节微生物代谢活动
饮食可以解释地高辛还原的个体间差异,并可能调节体内微生物代谢活动。相比之下,有毒化合物可以由肠道微生物产生。桦木素在来自肠道微生物的β-葡萄糖苷酶的作用下水解成致癌性重氮甲烷。
建议
应开发微生物肠道酶的小分子抑制剂,以在这个复杂的栖息地的特定转化中发挥调节作用。肠道微生物群转化的代谢物与前体物质之间的毒性差异值得进一步研究。
此外,过量的药物可能导致肠道微生物失衡和不良反应,不同剂量的天然产物对肠道微生物和新陈代谢的影响也需进一步研究。
考虑到肠道微生物可以增加宿主对药物或天然产物的复杂和可变反应,这一过程值得研究。
生物转化的方法
(1)肠液转化。转化产物的大规模制备可以通过肠液生物转化来实现;
(2)与宿主微生物群的样品一起孵育。原型药物和代谢物的类型和数量可以使用该方法检测。它具有准确表示个体的整个肠道微生物组的优点;
(3)代表性菌株的孵育。该方法具有高通量潜力,这对于大规模药物研究很有价值,并有助于有益代谢物的工业生产。
✦模型分析有利于认识微生物代谢
器官微生理系统,胃肠道类器官的各种预测/计算工具可能有助于提高我们对未来微生物代谢的理解。
此外,在动物模型中已经研究了天然产物代谢与肠道微生物之间的关系,其结果可用于研究代谢物的分布和形式。无菌/抗生素处理的动物与传统动物进行了比较,以证明肠道微生物在天然产物代谢中的关键作用。
这种方法的局限性在于人类和啮齿动物之间存在固有的胃肠道和微生物差异。在临床试验中从受试者收集的粪便进行详细微生物群和代谢物分析可以全面反映天然产物在体内的代谢过程,并用于解释个体差异。
✦增加测序技术的应用
增加测序技术的应用,以研究微生物的转录活性和代谢谱。通过使用单细胞法,可以表征肠道微生物的生理结构,以确定其代谢活性。
转录组测序技术(RNA-Seq)可以直接分析人体肠道中具有强代谢活性的微生物的基因表达谱。
单细胞方法,宏转录组学和宏基因组学的组合已被用于鉴定和表征肠道微生物群的活性亚群,并确定它们对天然产物的代谢反应。
肠道微生物群是编码各种代谢酶的基因库。大多数天然产物的生物活性和潜在健康益处的激活极度依赖肠道微生物作为底物加工工厂。
了解微生物如何独特地调整天然产物以及这些代谢物对宿主健康的影响。已经得出以下结论:
(1)肠道微生物可以改变天然产物;
(2)天然产物可以调节肠道微生物的组成和丰度;
(3)肠道微生物可以介导天然产物的多组分协同作用。
此外,肠道微生物的巨大可塑性和个体间差异是值得注意的。因此,需要提高对肠道微生物对天然产物代谢的生理,化学和微生物贡献的理解,以帮助解释天然产物反应的个体差异,并为个性化治疗提供支持。
确认生物转化中极其参与的象征性功能及其物质基础将有助于探索天然产物与肠道微生物群之间相互作用的治疗模式。
主要参考文献
Zhao Y, Zhong X, Yan J, Sun C, Zhao X, Wang X. Potential roles of gut microbes in biotransformation of natural products: An overview. Front Microbiol. 2022 Sep 29;13:956378. doi: 10.3389/fmicb.2022.956378. PMID: 36246222; PMCID: PMC9560768.
Abookleesh F. L., Al-Anzi B. S., Ullah A. (2022). Potential antiviral action of alkaloids. Molecules 27:903. doi: 10.3390/molecules27030903.
Agatonovic-Kustrin S., Kustrin E., Gegechkori V., Morton D. W. (2020). Anxiolytic terpenoids and aromatherapy for anxiety and depression. Adv. Exp. Med. Biol. 1260, 283–296. doi: 10.1007/978-3-030-42667-5_11.
Al-Ishaq R. K., Liskova A., Kubatka P., Büsselberg D. (2021). Enzymatic metabolism of flavonoids by gut microbiota and its impact on gastrointestinal cancer. Cancers 13:3934. doi: 10.3390/cancers13163934.
Augusti P. R., Conterato G. M. M., Denardin C. C., Prazeres I. D., Serra A. T., Bronze M. R., et al.. (2021). Bioactivity, bioavailability, and gut microbiota transformations of dietary phenolic compounds: implications for COVID-19. J. Nutr. Biochem. 97:108787. doi: 10.1016/j.jnutbio.2021.108787.
Berlinberg A. J., Brar A., Stahly A., Gerich M. E., Fennimore B. P., Scott F. I., et al.. (2022). A novel approach toward less invasive multiomics gut analyses: a pilot study. Microbiol. Spectr. 10:e0244621. doi: 10.1128/spectrum.02446-21.
Bess E. N., Bisanz J. E., Yarza F., Bustion A., Rich B. E., Li X., et al.. (2020). Genetic basis for the cooperative bioactivation of plant lignans by Eggerthella lenta and other human gut bacteria. Nat. Microbiol. 5, 56–66. doi: 10.1038/s41564-019-0596-1.
谷禾健康
健康是人生命活动的根本,而肠道健康更是其中的重要部分。随着现代生活方式改变,炎症性肠病发病率增加。
炎症性肠病是一种严重影响患者生活质量的疾病。在全球范围内影响数百万人,发病率高,给卫生保健系统带来了沉重负担。
●炎症性肠病的病因
炎症性肠病的病因尚不明确,可能与肠道免疫反应异常有关。但与其他炎症性疾病一样,炎症性肠病的病因是多因素的,涉及遗传和环境因素的参与。这些病因大多影响肠道菌群组成,说明微生物群在炎症性肠病发展中发挥重要作用。
肠道微生物群在维持宿主生理和免疫稳态方面发挥着关键作用。维持健康的肠道菌群对免疫平衡至关重要,多项研究证明,肠道菌群失调(菌群组成改变)与炎症性肠病有关。
●炎症性肠病引起的其他疾病
慢性结肠炎不及时治疗有可能引起肠穿孔、肠狭窄、大量便血、中毒性肠扩张甚至结肠癌等疾病,不过结肠癌的可能性是比较小的。
慢性结肠炎引起的并发症也是比较多的,有可能引起关节炎、肝功能障碍、皮肤病变等一系列并发症。
近年来,在健康和疾病中肠道微生物组成,以及通过使用益生菌调节肠道微生物群方面取得了实质性进展。通过益生菌或粪菌移植调节肠道菌群后,患者的炎症性肠病缓解,为肠道炎症疾病的治疗开辟了新的前景。
了解并关注肠道健康非常重要。本文带大家了解慢性肠道炎症的病因和机制,重点关注肠道失调在这一复杂疾病的免疫病理学中的作用。
同时还介绍了目前可用的肠道菌群导向治疗炎症性肠病的方法,并讨论了其对炎症性肠病长期缓解的作用。希望大家拥有更健康的肠道。
本文主要从以下几个方面讲述
●炎症性肠病的症状和病因
●炎症性肠病中的免疫
●炎症性肠病患者体内的菌群
●肠道微生物群失调与免疫失调
●慢性肠道炎症的治疗
炎症性肠病是肠道的一种慢性炎症状态,也可能影响其他器官,如果不治疗,会导致高发病率和死亡率。炎症性肠病可通过不同的临床表现发生,其主要亚型为克罗恩病和溃疡性结肠炎。
✦临床症状
克罗恩病和溃疡性结肠炎表现出明显的临床特征。
克罗恩病
在克罗恩病中,炎症病变是跨壁的(可能涉及肠壁的所有层),也不连续,正常的肠区与受影响的肠区交替出现。
虽然在大多数克罗恩病例中,病变发生在回肠或结肠的第一部分,但也可能位于胃肠道的其他区域(从口腔到肛门)。
溃疡性结肠炎
溃疡性结肠炎的炎症病变是连续的,通常累及直肠和近端结肠。与克罗恩病相反,溃疡性结肠炎只影响肠道上皮层。
两种炎症性肠病症状相似,包括腹泻、腹痛和直肠出血,通常伴有体重减轻、疲劳和发烧。
然而,在克罗恩病中,肉芽肿更常见于炎症区域,也可能出现脓肿,阻塞肠道狭窄和不同肠道区域之间以及皮肤和其他器官之间的瘘管(炎症连接)。
溃疡性结肠炎患者通常在诊断时表现为中度症状,但炎症病变的范围与疾病严重程度相关,而胰腺炎与更严重的疾病相关。
•肠道外的一些表现症状
肠外表现在克罗恩病和溃疡性结肠炎中都很常见,25%–40%的炎症性肠病患者都有肠外表现。
虽然这些表现可能出现在几乎所有的器官和系统中,但主要影响眼睛、皮肤、肝脏和关节。肺、胰腺和血管系统不太常见。
一些肠外表现与疾病活动有关,并通过治疗得到改善,如少关节炎和口腔溃疡。其他则是非依赖治疗的,如葡萄膜炎和强直性脊柱炎。
✦治疗方法
大多数溃疡性结肠炎患者只需要医疗,其中20%-30%需要手术治疗。克罗恩病通常是进行性的,约50%的患者在确诊后10年内需要手术治疗。
克罗恩病患者的死亡率是健康人群的1.3-1.5倍,不受影响区域的影响,主要由结直肠癌、低血容量、营养不良、贫血引起。
由于目前尚无完全治愈方法,炎症性肠病的治疗旨在改善症状和缓解疾病。
不同疾病类型和严重程度抗炎药的使用
溃疡性结肠炎的治疗
在活动期溃疡性结肠炎中,首先口服类固醇,然后是氨基水杨酸盐(ASA)。对于左侧结肠炎,通常使用外用制剂(栓剂或灌肠剂),而对于横向和上行结肠炎,口服制剂有时结合外用给药是有效的。
对于这种治疗无效的溃疡性结肠炎患者,考虑使用单克隆抗体,通常是英夫利西单抗(IFX)阻断炎症细胞因子肿瘤坏死因子α。为了保持病情缓解,ASA是首选药物。然而,对于ASA不能维持缓解的患者,也可以使用硫唑嘌呤(AZA)或IFX。
克罗恩病的治疗
对活动性克罗恩病的治疗可能遵循“自上而下”或“加速”策略。在自上而下的治疗中,治疗从高剂量免疫调节剂或生物制剂开始,剂量随症状改善而调整。
在强化治疗中,先用低剂量的低效药物,如果无效,则调整治疗以达到缓解,方法是使用未定义的更有效药物。
类固醇、氨基水杨酸盐和抗生素可以联合使用以达到缓解。如果出现肠梗阻、脓肿或瘘管等并发症,也可进行手术。
硫唑嘌呤通常用于维持治疗,有时与甲氨蝶呤联合使用。对这些方案无效的克罗恩病患者,需要进行生物治疗,单独使用英夫利西单抗或与之前的药物联合使用。
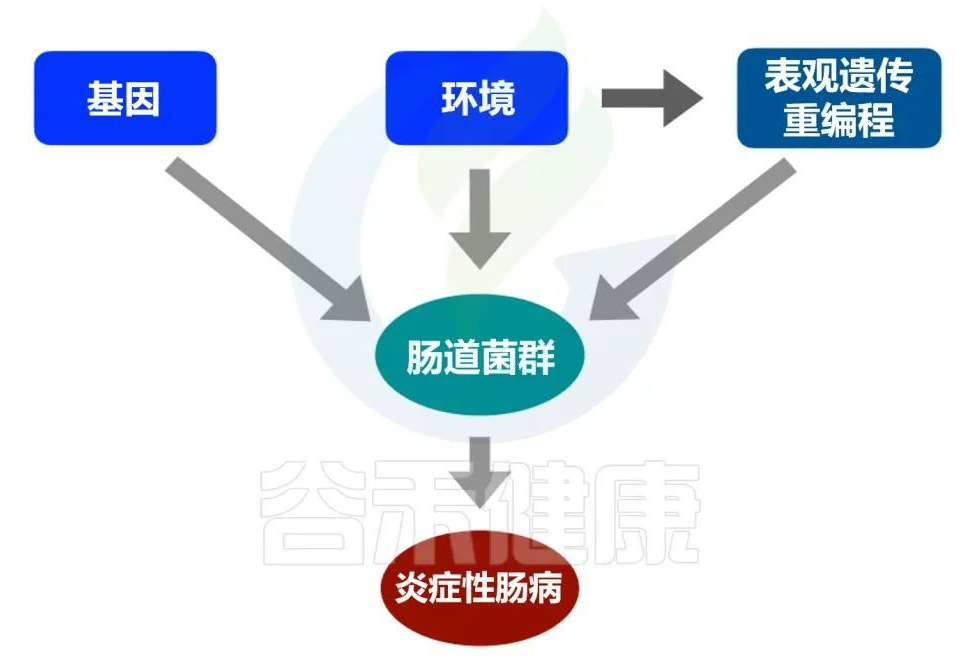
炎症性肠病被描述为一种多因素疾病,可能是由遗传物质和环境特征等不同易感因素造成的,这些因素反过来可能影响基因调控的表观遗传机制。
炎症性肠病的病因
编辑
Amelia S,et al.Elsevier.2022
基因和环境(在表观遗传调控的参与下)都可能影响微生物群的组成,进而可能触发炎症性肠病。
✦遗传因素影响炎症性肠病
遗传因素对炎症性肠病的重要性早已被提出。事实上,多达12%的患者有炎症性肠病家族史。在亲属中,炎症性肠病患病率明显高于普通人群,在兄弟姐妹中发现克罗恩病的易感性风险最高(30%-35%)。
注:全基因组关联研究有助于剖析炎症性肠病的遗传背景,迄今已确定240多个易感位点。克罗恩病和溃疡性结肠炎以及其他炎症性疾病共享大多数基因座。然而,除了非常早发病的炎症性肠病外,这些易感基因座仅占炎症性肠病估计遗传风险的一小部分(约20%)。
许多风险相关基因座参与了早期免疫应答期间宿主和微生物之间的相互作用,表明宿主-微生物相互作用和先天免疫在炎症性肠病的发展中起着核心作用。
影响炎症性肠病的基因
1996年确定的第一个与炎症性肠病(IBD)相关的遗传位点被指定为IBD-1。后来,在IBD-1位点发现了一个基因,该基因参与细胞内对细菌细胞壁成分胞壁二肽的传感。该基因被命名为“核苷酸结合寡聚化域-2”或“半胱氨酸天冬氨酸蛋白酶募集域15”(NOD-2/CARD-15)。
在NOD-2基因中发现了三种易感变异体,它们都导致核因子kB(NF-kB)激活功能丧失,这是一种参与促炎细胞因子表达的转录因子,对细胞内细菌的免疫发展很重要。尽管迄今为止,NOD-2风险变体在高加索人群中与回肠克罗恩病的相关性最强,但只有少数回肠疾病患者具有风险变体,而亚洲克罗恩病患者几乎没有这些变体。
影响先天免疫应答的其他遗传风险变体的例子包括ATG16L1(“自噬相关16样-1”)、LRRK2(“富含亮氨酸重复激酶-2”)和IRGM(“免疫相关GTPase M00”)基因,这些基因在自噬中起作用。
✦表观遗传(性状)与炎症性肠病有关
基因表达的表观遗传修饰也与炎症性肠病有关。影响基因表达的三个主要表观遗传过程是DNA甲基化、组蛋白修饰和非编码RNA。
DNA甲基化是指在胞嘧啶核苷酸中添加甲基(主要是CpG基序)。
其中,DNA甲基化和非编码RNA的作用在炎症性肠病中得到了最广泛的研究。这个过程影响基因的转录活性和表达水平。与健康对照受试者相比,炎症性肠病患者的基因甲基化程度增加。微RNA(miRNA)在转录和转录后水平介导RNA沉默和基因表达。
这些过程可能会干扰T细胞分化、Th17信号传导和自噬。与健康对照组相比,炎症性肠病患者以及活动性与非活动性疾病患者的miRNAs表达不同。
注意:在其他炎症性疾病中,如系统性红斑狼疮和类风湿性关节炎中发现了一些重叠的miRNA特征。环境因素强烈影响表观遗传调控,因此表明表观遗传是环境暴露和基因表达之间的联系。
✦环境因素
已确定炎症性肠病风险的不同环境决定因素,如吸烟、城市与农村生活、空气污染和饮食。
// 工业化与吸烟增加了风险
欧洲和北美的炎症性肠病发病率最高,但自21世纪初以来似乎趋于稳定。相反,在非洲、亚洲和南美洲的新兴工业化国家,随着这些国家进入更加西化的生活方式,炎症性肠病的发病率仍在增加。
吸烟似乎会增加炎症性肠病(尤其是克罗恩病)的风险,并与肠道通透性丧失有关。
// 饮食会影响炎症性肠病的风险
一些基于多中心欧洲前瞻性队列研究的研究试图将饮食习惯与炎症性肠病风险联系起来。
最近的一项文献荟萃分析也确定了炎症性肠病的其他环境风险因素:阑尾切除术或扁桃体切除术(CD)、软饮料消费(UC)、抗生素暴露、口服避孕药使用、维生素D缺乏和肠肝螺杆菌(enterohepatic Helicobacter)(IBD)。
// 微生物可能导致发病或炎症
环境因素也可能直接或间接影响肠道微生物群的组成。
长期以来,人们认为微生物在炎症性肠病发病中起着重要作用。并对炎症性肠病中特定致病生物的定植和肠道菌群组成进行了广泛研究。
禽分枝杆菌(Mycobacterium avium)的定植,副结核分枝杆菌(Mycobacterium tuberculosisycobactins)或带有大肠杆菌粘附侵袭性变体的副结核分枝杆菌似乎会促进易感宿主的慢性炎症,可能导致炎症性肠病的发展(尤其是克罗恩病)。
此外,肠道微生物群组成的改变(失调)可能使代谢失衡,导致炎症反应缺乏调节。
除环境因素外,宿主遗传也影响微生物群落组成。事实上,单卵双胞胎比双卵双胞胎显示出更相似的微生物群。炎症性肠病风险等位基因与肠道微生物群组成的关联研究表明,炎症性肠病的高遗传风险与罗氏菌属(Roseburia)的丰度较低相关,而肠杆菌科(Enterobacteriaceae)的丰度较高。
固有免疫是机体在种系发育和进化过程中形成的天然免疫防御功能,即出生后就已具备的非特异性防御功能,也称为非特异性免疫。
✦上皮细胞
上皮层和相关的抗菌机制对于限制肠道微生物群进入肠粘膜至关重要。当第一道防线受损时,就会发生微生物移位,并建立炎症机制。
覆盖肠上皮的粘液层在保持肠道微生物和上皮细胞之间的低接触方面起着重要作用。它由凝胶形成粘蛋白聚合而成,主要由特殊上皮细胞(即杯状细胞)分泌。
这些粘蛋白聚合物由于能够结合水而在肠腔中膨胀,形成肠粘液。小肠粘液由一层薄而不连续的粘液层组成,结肠粘液由两层密度不同的粘液组成:外层更疏松,细菌更易渗透,内层更坚固,通常无菌。
●粘蛋白缺少会增加结肠炎风险
小鼠粘蛋白2敲除被证明会发展为结肠炎并增加结直肠癌的风险。在这些小鼠中,发现许多细菌与上皮细胞直接接触。研究发现,克罗恩病患者的结肠中,黏蛋白表达减少,炎症区域MUC1表达减少,非炎症区域MUC3、MUC4和MUC5B表达降低。
黏蛋白(MUC)是一类主要由黏多糖组成的糖蛋白
最近,一项荟萃分析证实克罗恩病患者肠道中粘蛋白表达降低。黏液蛋白表达受损可能导致肠道微生物群与上皮衬里之间的接触增加,从而导致炎症。
•肠上皮细胞有助于维持稳态
肠道上皮细胞分泌抗菌肽(AMPs),在限制肠道病原体感染方面发挥重要作用,并有助于维持肠道内稳态,保持共生细菌和上皮细胞之间的低接触。
抗菌肽(AMPs)的例子包括再生蛋白、免疫球蛋白(Ig)A和防御素。在人类中,已经确定了十种防御素,分为两类:a-防御素(HD),主要由小肠的潘氏细胞、中性粒细胞和某些巨噬细胞群分泌;b-防御素(HBD),由上皮细胞分泌,以响应细胞因子或模式识别受体(PRR)的微生物识别。
克罗恩病中描述了防御素生产的失调。事实上,携带NOD2移码敏感变异体的回肠克罗恩病患者HD5和HD6的潘氏细胞表达降低。这可能导致微生物与上皮层接触增加,部分解释了克罗恩病敏感性增加的原因。相反,据报道,HBD2在结肠克罗恩病和溃疡性结肠炎的炎症区域表达增加,这可能是通过炎症刺激调节HBD2表达。
✦微生物分子的识别
通过上皮层里易位的微生物进入下层肠粘膜,被吞噬细胞(主要是单核细胞、巨噬细胞和树突状细胞)吞噬,并在吞噬小体中被进一步清除。这些细胞通过属于不同膜或细胞溶质蛋白家族的模式识别受体识别病原体相关分子模式(PAMP)。
PAMP是进化保守的微生物抗原
一旦识别,信号转导途径就会发生,导致转录因子的核移位和炎症细胞因子的产生。Toll样受体是一类广泛的模式识别受体,是肠道微生物群的关键免疫传感器,是通过PAMP识别的免疫系统的哨兵。它们在吞噬细胞的细胞质或内体膜以及肠上皮细胞和基质细胞中表达,激活后二聚并导致下游信号级联,导致NF-kB激活,推动参与树突状细胞成熟和T淋巴细胞增殖和激活的关键细胞因子的产生。
●炎症性肠病患者识别功能受到影响
炎症性肠病患者TLR信号似乎受到影响。与健康对照组相比,在活动期溃疡性结肠炎患者中,TLR2、TLR4、TLR5、TLR8和TLR9表达上调,而在静止期溃疡性结肠炎患者中,TLR5表达下调。
在结肠炎小鼠模型中,TLR2/6的抑制减缓了疾病进展,表明其与病理有关。据报道,在炎症性肠病患者的肠道中TLR6过度表达,可能会加剧T辅助因子Th1和Th17反应。
由于TLR3、TLR5和TLR7的多态性与更高的溃疡性结肠炎严重程度相关,其他TLR受体的失调似乎也会导致炎症性肠病。TLR下游信号传导导致细胞因子分泌,这可能对肠道内稳态有利或有害。
TLR诱导的细胞因子有助于肠道内稳态的例子包括IFN-a和b(后者诱导溃疡性结肠炎缓解、白细胞介素-15(通过调节性T细胞诱导控制肠道炎症和白细胞介素11(其刺激对溃疡性结肠炎有益)。
另一方面,TLR信号也可诱导促炎细胞因子,如IL-1b、IL-6、IL-8、TNF-a和IL-18(抑制杯状细胞成熟,促进溃疡性结肠炎病理。
✦炎性小体
——也称炎症小体,是由胞浆内模式识别受体(PRRs)参与组装的多蛋白复合物,是天然免疫系统的重要组成部分。
炎症小体激活异常也与炎症性肠病的发展有关。NLRP3炎性小体是细胞质模式识别受体的NLR家族成员,是肠内稳态的重要调节器。
NLRP3可在各种细胞类型中表达,如粒细胞、抗原呈递细胞(APC)、T和B淋巴细胞,其激活可在识别外源微生物抗原或内源性危险信号时触发。一旦触发,NLRP3介导炎症小体复合体的组装。
作用机制
在这个复合物中,caspase-1被激活,将白介素1β前体和白介素18前体分别裂解为分泌的IL-1b和IL-18。激活的半胱氨酸天冬氨酸蛋白酶1与半胱氨酸蛋白酶11一起裂解胃泌素D,使其发生聚合,从而导致细胞质膜孔的形成并导致细胞溶解,这是一种称为细胞凋亡的细胞死亡过程。
细胞溶解后,细胞内成分的释放诱导炎症细胞的募集,促进炎症。炎症激活分为两个步骤:在第一启动步骤中,NF-kB被激活,导致NLRP3和白介素1β前体合成的转录诱导;由微生物或危险信号触发的第二个激活步骤导致NLRP3炎性小体复合体的组装。
●炎症小体缺乏可能导致炎症恶化
NLRP3表达和炎症小体激活对维持肠道内稳态至关重要,因为据报道,NRLP3和炎症小体表达缺陷的小鼠对右旋糖酐硫酸钠(DSS)诱导的结肠炎的敏感性增强,病情恶化,死亡率更高。
然而,炎症小体激活的持续上调也可能导致白细胞介素1β和白细胞介素-18的过度生成,以及细胞凋亡的增加,从而导致炎症性肠病中观察到的炎症恶化。
事实上,60%的克罗恩病患者检测到NLRP3炎性体激活,并与长期溃疡性结肠炎疾病相关。据报道,克罗恩病患者中白细胞介素-18分泌增加,白细胞介素1β水平升高与炎症性肠病严重程度增加相关。此外,在携带TLR2风险等位基因的炎症性肠病患者中,共生肠道微生物诱导了细胞凋亡,使病情恶化。
注意
尽管有这些发现,NLRP3炎症组在炎症性肠病发病机制中的作用仍然存在争议,因为其他小鼠和人类研究也表明过度活跃的炎症组具有保护作用。
✦自噬
自噬是一种细胞更新过程,其中起源于内质网的小泡包围老化的细胞器和蛋白质,通过溶酶体融合促进其降解。
自噬机制影响其他细胞过程,如肽分泌和细胞内病原体降解。
●自噬损伤会使炎症失控
自噬相关克罗恩病易感性等位基因的存在导致自噬损伤。患有自噬相关风险变异体的克罗恩病患者的潘氏细胞表现出颗粒异常,可能影响其抗菌肽分泌途径,导致微生物增殖增加和炎症失控。自噬缺陷的巨噬细胞和树突状细胞清除细胞内病原体的能力较低,并可能通过炎症小体激活促进炎症。
✦固有淋巴细胞
固有淋巴细胞(ILC)是指不表达任何T、B或髓细胞标记物,但同时表达白细胞介素2和白细胞介素7受体的淋巴细胞。
它们在抵御微生物病原体方面起着核心作用,并且在组织内稳态方面很重要。固有淋巴细胞作用于先天免疫和适应性免疫之间的串扰,并对多种细胞类型分泌的细胞因子作出反应。
由于固有淋巴细胞作用于第一道防线,因此它们战略性地位于外部因素和传染源暴露程度较高的位置,如肠粘膜、皮肤和肺部。
固有淋巴细胞的三个主要亚组
ILC1细胞(Th1样细胞)表达转录因子T-bet,能够分泌IFN-g和TNF-a,并对抗细胞内病原体——这一亚群还包括NK(“自然杀伤”)细胞。
ILC2(Th2样细胞)表达GATA-3(“谷氨酰胺基tRNA转氨酶结合蛋白-3”)转录因子,分泌IL-4、IL-5、IL-9和IL-13,在抵御蠕虫方面很重要,也在过敏反应中起作用。
ILC3(Th17样细胞)表达转录因子RORgt,分泌IL-17、IL-22、GM-CSF(“粒细胞-巨噬细胞集落刺激因子”)和TNF-a,对细胞外病原体侵入粘膜部位具有重要作用。
然而,这些不同的表型保留了相当大的可塑性。粘膜ILC3细胞的主要功能是维持屏障完整性。
●固有淋巴细胞对调节稳态至关重要
如上所述,肠道粘膜巨噬细胞和树突状细胞对微生物的TLR感应导致下游释放促炎细胞因子,如白细胞介素1β和白细胞介素23。黏膜ILC3细胞通过分泌IL-22、GM-CSF和IL-17对这些介质作出反应。IL-22也由Th17和Th22细胞分泌,在体内稳态控制中发挥重要作用。
事实上,IL-22作用于上皮细胞,改善屏障完整性,从而防止细菌移位。该细胞因子还促进抗菌肽(如b-防御素)的产生和分泌,影响小鼠的微生物群组成,并增加宿主对病原微生物的耐药性。
除了产生白细胞介素22,ILC3细胞还能够通过直接接触和抗原递呈调节其他免疫细胞(如CD4+T细胞)的活性,因为这些细胞表达II类主要组织相容性复合物分子(MHC-II)。
克罗恩病中描述了ILC1和ILC3的肠道过度积累,在溃疡性结肠炎中观察到固有淋巴细胞活性增加。在小鼠中,由抗原呈递细胞分泌的白细胞介素1b和白细胞介素23诱导的ILC3刺激导致肠内累积的白细胞介素17和IFN-g分泌增加。缺乏ILC3细胞的小鼠未发生右旋糖酐硫酸钠诱导的结肠
这种免疫只针对一种病原体。它是人体经后天感染(病愈或无症状的感染)或人工预防接种(菌苗、疫苗、类毒素、免疫球蛋白等)而使机体获得的抵抗感染能力。
✦作用机制
巨噬细胞和树突状细胞消化吞噬微生物的抗原,并将其与MHC1类或2类分子结合,呈现给效应T细胞。
细胞膜标记物CD11b、CD11c和CD103的表达定义了树突状细胞的独立亚群,从而影响随后的T细胞反应。
T细胞是适应性免疫反应的关键参与者,根据周围组织环境的细胞因子特征,可以分化为效应器或调节亚型。抗原呈递细胞与T细胞相互作用的性质和调节促使Th细胞分化为不同的亚群,每个亚群都发挥着特定的作用:
Th1是产生IFN-g的细胞,对细胞内细菌和某些原生动物的免疫很重要;Th2是产生IL4、Il-5和Il-13的细胞,参与对蠕虫和寄生虫的免疫;Th17是IL-17A、IL-17F、IL-21和IL-22的产生者,参与中性粒细胞反应;Th1/Th17细胞同时产生IFNg和IL-17A,具有Th1和Th17细胞的特征。
虽然暴露于肿瘤生长因子-b(TGF-b)可促进调节性T细胞分化,但Th1细胞主要由IL-12和IFN-g诱导,而IL-4在IL-6存在的情况下可促进Th2细胞和TGF-b的分化,从而促进Th17细胞的分化。
✦炎症性肠病患者适应性免疫出现异常
在克罗恩病患者中,检测到巨噬细胞产生较高的白细胞介素12,以及白细胞介素12和白细胞介素18诱导的异常Th1免疫应答。此外,在炎症性肠病患者的肠道固有层中检测到Th17和Th1细胞群的丰度增加。
•促炎细胞因子增加
体外培养的克罗恩病和溃疡性结肠炎患者的活检均分泌了相当数量的IFN-g。这种细胞因子促进肠细胞凋亡和巨噬细胞活化,增加巨噬细胞产生肿瘤坏死因子,进一步加剧炎症。
炎症性肠病患者炎症粘膜的体外培养也报告了白细胞介素17A转录水平的升高和白细胞介素17A分泌的增加。白细胞介素17A诱导中性粒细胞向炎症部位募集,并介导炎症分子的上调,如诱导型一氧化氮合酶和白细胞介素1β。此外,白细胞介素17A诱导巨噬细胞产生促炎细胞因子。
注意
患有活动性疾病的炎症性肠病患者血液中的调节性T细胞减少,而肠粘膜中的调节性T细胞增加,功能正常。然而,肠固有层T细胞在炎症性肠病中对调节性T细胞功能无反应,这可以说明Th细胞缺乏调节。
胃肠道内微生物种群的建立是一个复杂的过程,涉及微生物和宿主的相互作用,最终形成稠密和稳定的种群。
✦肠道微生物与宿主互惠互利
肠道微生物群落或肠道内的微生物群落与宿主建立了共生关系,在健康环境中互惠互利。宿主提供了一个稳定且营养丰富的栖息地,而微生物群给宿主带来了重要的益处,例如未消化的饮食成分和肠粘膜产生的内源性粘液的发酵,以及短链脂肪酸、氨基酸和维生素的产生。
此外,微生物群通过防止外来生物入侵,例如通过产生代谢物和细菌素,抵抗病原体定植,并影响肠上皮和免疫系统的发育和内稳态。
肠道微生物群的组成受宿主因素的影响,如年龄、遗传、分泌产物(如胃酸和胆汁)、蠕动和肠道转运时间。另一方面,环境因素(如饮食)在生命周期内对宿主产生影响,不断调节微生物群落。
尽管肠道中的大多数微生物都存在于肠腔内,但与粘膜相关的微生物群对宿主来说非常重要,因为它对上皮和粘膜功能的影响比肠腔细菌更大,对炎症性肠病的发病机制的影响更大。
注意:由于采样限制,大多数人体肠道微生物群研究,包括炎症性肠病患者的研究,都分析粪便样本。尽管粪便样本可能不能准确反映整个胃肠道的微生物群落组成,但大多数微生物都是通过粪便途径离开的。因此,粪便微生物群组成的变化反映了胃肠相关效应。
健康成年人大肠粘膜微生物群的组成是一致的,但与同一个人的粪便微生物群不同。因此,对炎症性肠病患者粘膜相关微生物群的研究证实,远端胃肠道不同解剖部位的微生物组成没有显著差异。另一方面,比较炎症性肠病患者粪便微生物群相关粘膜的研究报告了不同的结果。
✦炎症性肠病患者肠道菌群多样性较低
与健康对照组相比,克罗恩病和溃疡性结肠炎患者的肠道菌群多样性较低。与健康个体相比,这种变化伴随着物种丰富度的降低(一种群落中物种总数的测量方法),在克罗恩病患者中尤为明显。
对大多数炎症性肠病患者的报告了厚壁菌门(Phylum Firmicutes)、拟杆菌门(Bacteroidetes)和变形杆菌门(Proteus)内特定细菌类群的丰度变化,随着被认为具有攻击性的菌群(如变形杆菌(Proteus)、梭杆菌属(Fusobacterium)和瘤胃球菌(Ruminococcus)的扩大,同时具有保护性的菌群(例如Faecalibacterium、罗氏菌属(Roseburia) 、毛螺菌科(Lachnospiraceae)和双歧杆菌属(Bifidobacterium spp.))的减少。
在厚壁菌门和梭状芽孢杆菌类中,属于Faecalibacterium、罗氏菌属(Roseburia) 、颤螺菌属(Oscillibacter)和粪球菌属(Coprococcus)的丁酸盐产生菌在炎症性肠病患者中普遍减少。
丁酸是一种短链脂肪酸,可被肠粘膜吸收,是结肠细胞的主要能量来源,提供高达70%的需求量。此外,丁酸具有抗炎作用,并积极调节肠道内稳态。
✦梭状芽孢杆菌缺乏可能引起肠道炎症
梭状芽孢杆菌(Clostridia)类成员的缺乏可能会使某些个体易于继发肠道炎症。相比之下,炎性多糖的产生菌——瘤胃球菌(Ruminococcus)(也属于梭状芽孢杆菌类)在炎症性肠病患者中富集。
✦肠道炎症患者拟杆菌、疣微菌等丰度减少
在拟杆菌门(Bacteroidaceae)中,克罗恩病和溃疡性结肠炎患者的拟杆菌丰度低于健康人。拟杆菌在活动性克罗恩病和溃疡性结肠炎患者中的减少比缓解期更明显。
在变性菌门变形菌门(Proteobacteria)内,肠道微生物群中的γ-变形菌纲(Gammaproteobacteria)、肠杆菌科( Enterobacteriaceae )成员的富集与炎症性肠病密切相关。
另一方面,在疣微菌门(Verrucomicrobia)内,炎症性肠病患者粪便样本中的嗜黏蛋白阿克曼菌(Akkermansia muciniphila)减少。在含有人类肠道微生物群的动物模型中,嗜黏蛋白阿克曼菌的存在降低了结肠组织学损伤和促炎介质的组织mRNA表达。
尽管一些研究表明炎症性肠病患者存在肠道微生物群失衡,但与克罗恩病和溃疡性结肠炎相关的失调似乎是特定疾病。
据报道,克罗恩病、溃疡性结肠炎和非炎症性肠病患者之间存在明显的微生物特征,无论炎症性肠病患者体内粘膜相关微生物群的稳定性如何。
炎症性肠病患者的粪便和粘膜真菌群均不平衡。与健康个体类似,炎症性肠病患者的真菌群主要由担子菌门(Basidiomycota)和子囊菌门(Ascomycota)两个门以及伞菌纲(Agaricomycetes)和酵母菌纲(Saccharomyces)两个类别组成。
✦克罗恩病中念珠菌占主导地位
主要差异在于较低的分类学水平。特别是对于克罗恩病, 念珠菌(Candidaspp)的丰度普遍增加,尽管研究中的优势种有所不同。
Candida tropicalis和Candida glabrata在克罗恩病患者中占主导地位,而 Leptosphaeria减少。
与健康受试者相比,克罗恩病中炎症和非炎症粘膜中的总真菌负荷显著增加。
Filobasidium uniguttulatum和Saccharomyces cerevisiae均与克罗恩病患者的非炎症粘膜相关,而Xylariales(子囊菌门)与炎症粘膜相关。
患有炎症性肠病的儿科患者有机会帮助研究疾病发病机制的生物学成分,尤其是在诊断过程中获得的样本。
–1 炎症性肠病中微生物群总体减少
一项研究分析了最近诊断为炎症性肠病的儿科患者粘膜相关菌群的主要细菌组成,报告了一些潜在有害细菌群的优势,或有益细菌种类的减少。
肠易激综合征患儿的粘膜与更多的需氧和兼性厌氧菌相关,而克罗恩病和溃疡性结肠炎中属于正常厌氧肠道微生物群的物种或组的总体减少,尤其是普通拟杆菌。
随后,通过细菌16S rRNA基因和真菌小亚单位核糖体区域的焦测序分析了治疗初期克罗恩病儿童的粘膜。虽然与对照组相比,罗氏菌属(Roseburia) 、Eubacterium和 Subdoligranulum、梭状芽孢杆菌类成员在克罗恩病中的丰度较低,但萨特氏菌(Sutterella)的丰度明显较高。
–2 特定细菌的丰富程度与疾病状态呈正相关
大型儿科新发克罗恩病队列研究,包括代表各种疾病表型的受试者,在开始治疗之前,调查了多个胃肠道位置的粪便和粘膜相关微生物群。
微生物群分析表明,粘膜相关生物并不局限于任何肠道位置,总体微生物组成主要通过样品类型和微生物多样性与对照组进行区分。疾病表型不足以区分患者。
在炎症性克罗恩病条件下,粘膜相关微生物群与物种多样性的总体下降和几个分类群丰度的变化密切相关。
E.coli、Pasteurellaceae、Veillonella parvula、Eikenella corredens和Fusobacteriaum nucleatum中特定细菌种类的丰富程度与治疗前新诊断的克罗恩病儿童的疾病状态呈正相关,该研究支持以上菌群为炎症性肠病进展驱动因素。
与克罗恩病呈负相关的菌群常见于:
长双歧杆菌(Bifidobacterium Longum) ↓↓↓
青春双歧杆菌(Bifidobacterium adolescentis) ↓↓↓
普拉梭菌(Faecalibacterium prausnitzii) ↓↓↓
罗氏菌属(Roseburia) ↓↓↓
直肠真杆菌(Eubacterium rectale) ↓↓↓
普通拟杆菌( Bacteroides vulgatus) ↓↓↓
Bacteroides caccae ↓↓↓
巴氏杆菌科(Pasteurellaceae)和奈瑟氏菌科(Neisseriaceae)在10岁以下患者中的丰度较高,但随着年龄的增长而减少。在诊断时收集的粪便中仅微弱地反映出新发的粘膜相关失调,这表明粘膜失调可能先于临床疾病,并独立于长期炎症发展。
–3 环境因素影响微生物的反应
一项关于儿童克罗恩病的前瞻性研究分析了患者在开始治疗之前和之后的粪便样本,首先使用抗生素,然后在8个月期间使用规定的配方食品或抗肿瘤坏死因子抗体。
在诊断时,肠道微生物群表现出特定细菌和真菌的变化,即埃希氏菌属(Escherichia)和韦荣氏球菌属(Veillonella)增加,阿克曼菌属( Akkermansia)、罗氏菌属(Roseburia) 、普雷沃氏菌(Prevotella)和真菌(包括念珠菌属)减少。
降低了肠道炎症反应中的失调。炎症、抗生素暴露和饮食独立影响肠道微生物群落的不同分类群。例如,真菌负荷随着疾病和抗生素的使用而增加,但随着饮食治疗而减少。
因此,虽然肠道内的失调在克罗恩病中很常见,但微生物的反应取决于环境因素。
✦开始治疗前的微生物群特征
在开始治疗之前,新诊断的溃疡性结肠炎儿科患者的粘膜微生物群也有特征。除了溃疡性结肠炎患者中疣微菌门(Verrucomicrobia)显著减少外,其他细菌种类的丰富程度与健康儿童相似。此外,在属水平上,溃疡性结肠炎患者中罗氏菌属(Roseburia)的相对丰度显著降低,而嗜血杆菌(Haemophilus)的相对丰量增加。
●疾病活动与部位影响肠菌群的组成和功能
肠菌群组成和功能的改变似乎与炎症性肠病患者的疾病活动、疾病行为和病变部位有关。尽管在研究之间或与疾病活动相关的研究中,微生物群组成的确切差异没有得到一致复制。
但已经观察到,在所有形式的克罗恩病中,下列菌群持续减少:
而下列菌群增加:
✦儿童患者的肠道菌群特征与成人患者相似
新发炎症性肠病儿童和成人常见的肠道菌群主要偏差
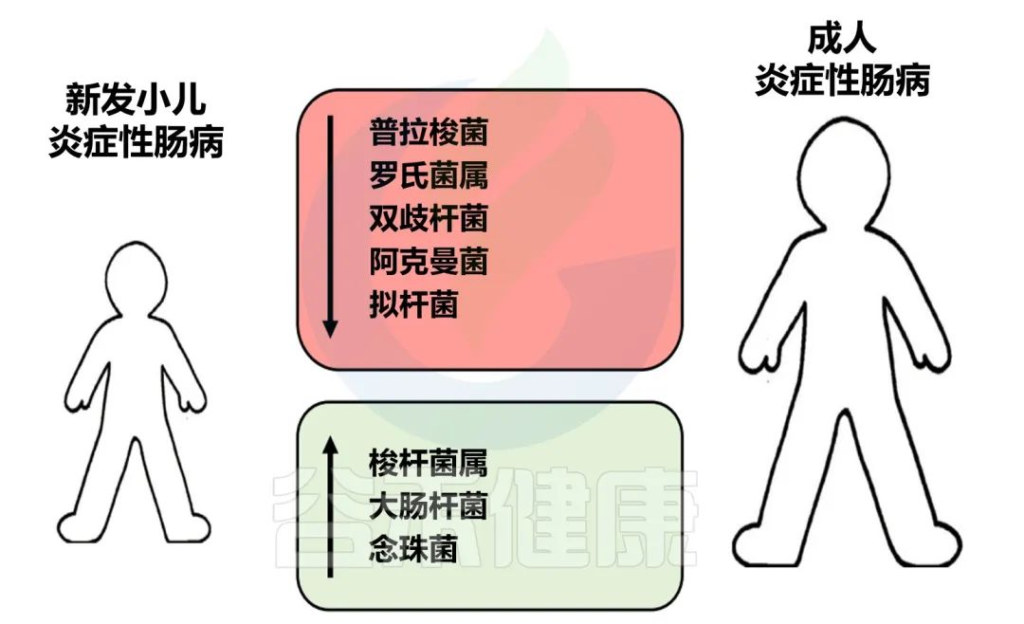
Amelia S,et al.Elsevier.2022
炎症性肠病患儿的肠道菌群特征与成人患者相似。因此,成人型炎症性肠病患者的生态失调可能在儿童期就已确立。
●肠道微生物失调反应炎症的严重程度
对儿科患者(包括治疗前新诊断的儿童)的研究得出结论,肠道生物失调反映了炎症的存在和严重程度。然而,虽然肠道微生物群的变化可能在炎症性肠病早期发生,并可能导致疾病的发生,但随着时间的推移,环境因素,包括炎症本身,可能通过改变肠道的代谢条件,进一步导致失调。
人类肠道微生物群失调是真正的致病因素还是仅仅是肠病炎症的后果,这个问题尚不清楚。但是肠道微生物被证明是炎症性肠病相关肠道炎症的重要因素。
在健康状态下,宿主对肠道微生物群的免疫反应仅局限于粘膜表面。据报道,炎症性肠病患者的粘膜相关微生物密度较高,在促进肠道炎症方面,被认为比粪便微生物群发挥更大的作用。
注意:比较炎症性肠病患者结肠粘膜炎症和非炎症区域的微生物群组成的研究试图阐明居民肠道微生物群对炎症免疫反应的可能影响,但结果相互矛盾。
一项研究发现,炎症性肠病患者体内粘膜炎症和非炎症区域的微生物群落组成没有显著差异。不过该项研究未根据炎症性肠病类型进行区分。
其他研究分析了克罗恩病和溃疡性结肠炎患者炎症和非炎症粘膜活检中的微生物群,发现同一个体内的微生物特征相似,但个体间差异很大。
✦克罗恩病患者中变形菌丰富
与溃疡性结肠炎患者或健康受试者相比,克罗恩病患者的变形菌门(Proteobacteria)显著增加。还报告说,在炎症克罗恩病组织中,埃希氏菌(Escherichia)和志贺氏菌(Shigella)更为丰富,而在非炎症克罗恩病组织里,Pseudomonas更为广泛。大肠杆菌和梭杆菌能够粘附粘膜,侵入肠上皮细胞,并可能加剧炎症。
拟杆菌和梭杆菌更常见
具核梭杆菌通过破坏上皮屏障和诱发异常炎症而加重结肠炎。相对于炎症性溃疡性结肠炎,拟杆菌门(Bacteroidetes)在克罗恩病炎症粘膜中更常见。克罗恩病粘膜活检中最常见的是普通拟杆菌(Bacteroides vulgatus),克罗恩病患者中脆弱拟杆菌(Bacteroides fragilis)的数量明显高于溃疡性结肠炎患者或健康受试者。脆弱拟杆菌与产生白细胞介素10的调节性T细胞的增加有关,这限制了促炎机制,从而有助于减少炎症。
除了拟杆菌外,梭杆菌门(Fusobacteria)在克罗恩病患者的炎症粘膜中比溃疡性结肠炎患者的炎症黏膜中更常见。相反,在发炎的溃疡性结肠炎粘膜中更频繁地检测到厚壁菌门(Phylum Firmicutes)和变形菌门(Proteobacteria)。与大肠杆菌类似,梭杆菌能够粘附粘膜,侵入肠上皮细胞,并可能加剧炎症。尤其是具核梭杆菌刺激结肠上皮细胞中肿瘤坏死因子的表达。
总的来说,这些研究结果表明,由于同一参与者的微生物群分布几乎没有变化,无论炎症状况或取样位置如何,因此粘膜相关微生物群没有发生明显的局部变化。
此外,根据引用的研究,粘膜微生物组成更依赖于个体间的变异,而不是疾病类型。
事实上,肠道微生物群组成的变化导致微生物代谢物的改变,这可能在炎症性肠病发病机制中起作用。
✦短链脂肪酸
短链脂肪酸是肠道微生物群通过发酵未消化的饮食成分和肠粘膜产生的内源性粘液而产生的。
短链脂肪酸影响肠上皮细胞生物能量学、微生物增殖、屏障和炎症功能。
短链脂肪酸影响受体激活与信号传导
短链脂肪酸被吸收到血液循环中,在那里它们可能与白细胞上的G蛋白偶联受体GPR43结合,阻碍其激活。GPR43信号传导抑制炎症反应,包括粘附分子和炎症介质的表达,以及白细胞趋化性。
缺乏GPR43的小鼠炎症反应加剧,外源性给予野生型小鼠短链脂肪酸对临床有益。短链脂肪酸还抑制NF-kB信号传导,在相对高浓度下,丁酸抑制I类组蛋白脱乙酰酶,后者是免疫和炎症的重要调节因子。
微生物群影响免疫细胞的激活
颤螺菌属 (Oscillospira)被认为是一种利用宿主衍生聚糖或从富含糖蛋白的饮食中获得的聚糖的丁酸生产菌。
在炎症性疾病中,尤其是克罗恩病中,颤螺菌的含量减少。微生物群似乎控制先天性和适应性免疫细胞激活的系统阈值。因此,持续接触常驻细菌发酵产物和代谢物对外周免疫系统的正确功能至关重要。
然而,不能排除常驻微生物在控制局部免疫中的直接作用,因为在其他研究中,粘膜炎症状态与局部粘膜相关微生物群的扰动有关,即同一炎症性肠病患者体内炎症粘膜和非炎症粘膜的微生物群组成差异。
✦色氨酸代谢物
对宿主的免疫反应至关重要的另一类代谢物是色氨酸代谢物。
色氨酸又称β-吲哚基丙氨酸,是人体的必须氨基酸之一。
缺乏色氨酸会导致结肠炎
使用小鼠进行的营养研究表明,饮食中缺乏色氨酸会导致结肠炎,并补充色氨酸以防止发炎。将失调的微生物组从喂食色氨酸缺乏饮食的小鼠转移到喂食正常饮食的无菌受体,足以引起结肠炎。
在小鼠中,色氨酸代谢菌株罗伊氏乳杆菌(Lactobacillus reuteri)可预防结肠炎。当它将色氨酸代谢成吲哚丙烯酸时,它是通过增强上皮屏障功能和减少炎症反应而起作用的芳烃受体的配体。
色氨酸影响上皮免疫
膳食色氨酸还可以通过增加调节微生物组组成并防止机会病原体感染的抗菌肽的产生来影响上皮免疫性。与炎症性肠病相反,在多发性硬化的鼠模型中,缺乏饮食色氨酸可防止中枢神经系统自身免疫,这是由于致脑病的T细胞反应受损和微生物组发生了深远的变化。
值得注意的是,饮食中色氨酸限制的保护作用在无菌小鼠中被取消,并且与色氨酸代谢物的原型传感器无关,这表明这些保护作用可以通过微生物组的代谢功能来介导。
✦结肠炎患者炎症部位细菌减少
研究表明,尽管溃疡性结肠炎和克罗恩病患者的成对活检样本之间的微生物群落组成具有高度的个体间变异性,但在每个疾病队列中,结肠发炎区域的细菌负荷通常低于非发炎区域。
克罗恩病和溃疡性结肠炎患者的炎症部位和非炎症部位所含的厚壁菌(以及相应的更多拟杆菌)均比非炎症性肠病对照样品少,但仅溃疡性结肠炎患者显著减少。
另一方面,在研究中包括的克罗恩病患者的大多数成对活检样本中检测到肠杆菌科成员,与溃疡性结肠炎和非炎症性肠病样本相比,肠杆菌属成员占克罗恩病总微生物群的比例增加了10倍。
肠杆菌科相对丰度的增加与NOD2风险等位基因计数呈正相关。NOD2基因内的多态性损害了这种细胞内受体作为细菌细胞壁成分的传感器在先天免疫中的作用,增加了克罗恩病的发病风险。
由于先天免疫受体(如NOD2和TLR4)的遗传多态性被认为是炎症性肠病发展的主要风险因素,宿主自身常驻微生物群的异常免疫反应被认为在促进炎症疾病进展方面起着关键作用。
✦白细胞介素放大炎症过程
与正常组织相比,在活动性炎症的溃疡性结肠炎组织中,厚壁菌门的数量较少,这主要是由于粪杆菌属(梭状芽孢杆菌目)的减少。事实上,白细胞介素22、白细胞介素17细胞(Th22细胞)和Th1细胞与厚壁菌门正相关。
白细胞介素17细胞渗入炎症性肠病患者的炎症肠道,在那里产生白细胞介素17A和其他细胞因子,触发并放大炎症过程。虽然Th17细胞因子在炎症性肠病发病机制中可能很重要,但Th17细胞也可能具有组织保护作用,主要取决于其通过产生白细胞介素22增强上皮屏障功能和反调节机制的能力。
✦结肠炎期间宿主-肠道微生物氧交换加强
事实上,众所周知,粪便细菌通过炎症细胞因子调节或刺激白细胞介素10的产生而表现出保护作用。相比之下,由于不动杆菌属的差异,蛋白杆菌门在炎症粘膜中更为丰富,这与Th22细胞和Th1细胞的数量呈负相关。
因此,在活动性炎症的溃疡性结肠炎患者的活检组织中,严格需氧不动杆菌属也扩大了,这表明结肠炎期间粘膜上皮的损伤可能会增加宿主-肠道微生物群的氧交换。
事实上,活动性炎症期间的生物失调状况可能与肠道内存在的径向氧梯度有关,这种氧梯度从较需氧粘膜界面延伸到大部分厌氧肠腔。宿主的氧合影响肠腔氧合,氧从宿主组织扩散到肠腔。在宿主组织恢复常氧后,腔氧浓度也降低,这表明氧气正被靠近粘膜界面的肠道微生物群消耗。粘膜较高的耐氧性和过氧化氢酶表达可能有利于肠道生态系统中的微生物竞争,因此,粘膜可能有利于耐气细菌群落的定植,尤其是来自蛋白质杆菌门的细菌。
与粪便微生物群相比,溃疡性结肠炎患者活检样本中富集的一些细菌类群在无胃肠道症状的研究对象的粘膜中也较高。
因此,在结肠炎活动性炎症期间扩张的细菌很可能在健康人的肠道菌群中正常存在,数量较少。例如,梭状芽孢杆菌和瘤胃球菌科在肠腔菌群中更为丰富,但与活动性结肠炎的样本相比,这些分类群在正常非炎症组织中也更为丰富。
✦炎症和肠道菌群失衡直接可能存在联系
内源性或外源性因素(如饮食、压力或感染)或它们的组合可能会引发亚临床肠道粘膜炎症,这取决于个体的遗传易感性,可能有利于特定促疾病肠道细菌的生长。
这些机会性微生物随后加剧了形态学和功能变化,导致病理后果,导致宿主的慢性炎症和临床症状。这些结果可能支持这样的假设,即炎症性肠病患者中观察到的整体生物失调在某种程度上可能是肠道环境紊乱的结果,而不是疾病的直接原因。
例如,作为炎症反应副产物生成的电子受体促进兼性厌氧菌(如肠杆菌科)的生长。因此,慢性炎症可能形成肠道微生物群,并进一步导致失调。
急性胃肠道粘膜感染的特点是与微生物群的显著变化相关的失调,以及具有增强的侵袭性和炎症特性的细菌的优势,这些特性可以直接加剧炎症和组织损伤,如γ-变形菌。
✦通过肠道菌群判断炎症状态
已有研究显示不同的肠道菌群可以通过免疫反应、影响肠道屏障以及通过胆汁酸转化等代谢途径影响免疫细胞相互作用,从而影响免疫调节。
借助这些方面的研究和临床数据,我们可以通过肠道菌群从几个方面来反映和了解免疫系统及自身的炎症状态。
病原菌的存在很大程度上会激发免疫系统的炎症,因此首先评估肠道菌群中是否存在异常的病原菌超标情况。
来自谷禾健康肠炎临床患者肠道菌群检测报告,结果显示血液链球菌(Streptococcus)及衣氏放线菌(Actinomyces israelii)超标,如下:
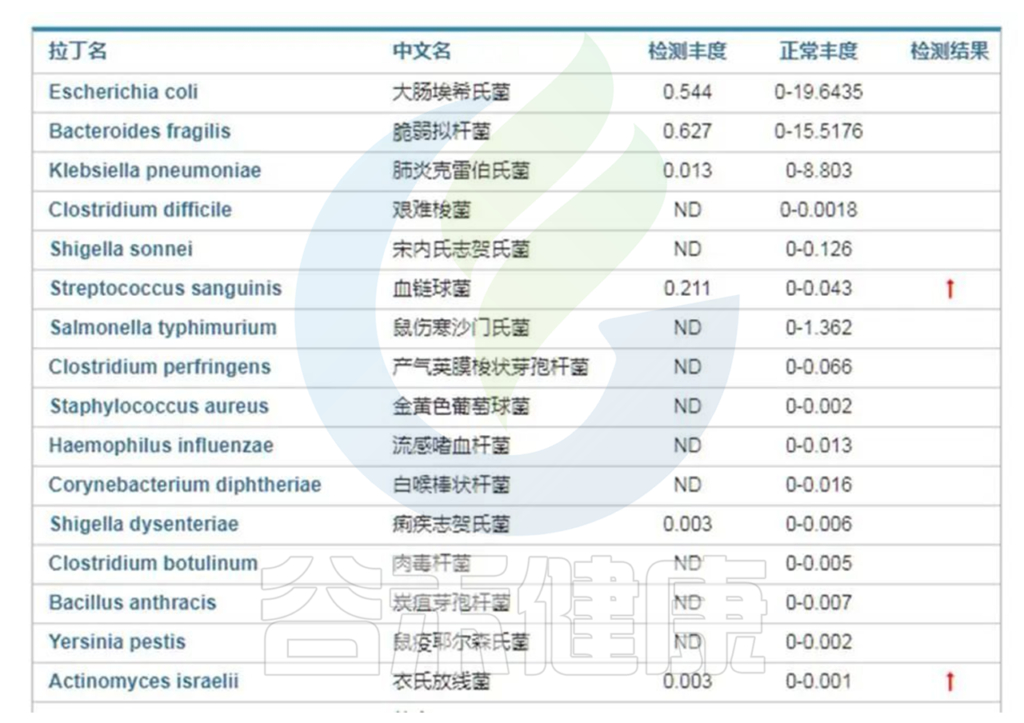
<来源:谷禾健康数据库>
类似的病原菌超标在没有严重导致感染症状的情况下也会诱发或刺激免疫系统炎症,如果免疫力低下就可能导致病原菌感染,如果持续存在超标也可能导致慢性炎症。
真菌微生物群在肠道炎症中的作用日益被认识。炎症性肠病患者和健康人之间的菌群组成不同,真菌菌群也不同,炎症粘膜中的真菌群与非炎症区域的真菌群是可以区分的。
✦炎症区域和非炎症区域真菌差异很大
白色念珠菌(C. albicans)和热带念珠菌(C.tropicalis)在炎症区域特别丰富,而在非炎症粘膜中则不存在。相比之下,S. cerevisiae和Saccharomyces castellii在炎症粘膜中较少出现。
光滑念珠菌(C.glabrata)也在克罗恩病粘膜中富集。光滑念珠菌是最重要的真菌机会性病原体之一,通过适应和免疫逃避策略促进炎症反应,这表明它可能在肠道炎症中起作用。
白色念珠菌也与炎症性肠病患者的粪便真菌有关。白色念珠菌可能是炎症性肠病中观察到的炎症过程的发起者,通过几种途径与酵母菌的主要成分和先天免疫反应的主要受体相结合。
此外,炭角菌目在炎症粘膜上更为丰富,而酿酒酵母在非炎症粘膜中更为丰富。炭角菌目的许多物种产生具有抗菌特性的化合物,可对抗人类病原体,如结核分枝杆菌、耶尔森菌、李斯特菌和沙门氏菌,以及具有细胞毒性和抗氧化活性的代谢物。因此,炭角菌目成员的存在可能反映了对炎症环境的适应。
✦真菌影响受体与免疫,导致炎症
真菌细胞壁成分与宿主免疫反应相关。与病原菌平行,真菌可以穿透被破坏的粘膜屏障,并通过其细胞壁成分激活TLR受体、Dectin-1(C型凝集素受体)、清道夫受体家族成员和固有层的补体系统。
这些受体的感应导致信号事件,依赖于白细胞介素17、白细胞介素22、CARD9(Caspase募集域家族成员-9)、ITAM(免疫受体酪氨酸基激活基序)、NFAT(活化T细胞的核因子)和NF-kB,导致严重的炎症表型。
由于文化传统、农业做法、社会经济地位和生活方式的改变,饮食本身在世界各地都在发生变化。炎症性肠病发病率的快速增加与工业化和暴露于环境因素,特别是饮食改变有关。营养和饮食模式影响免疫系统稳态,并可能通过不同的机制导致肠道炎症,包括肠道微生物群的调节。
饮食对肠道菌群组成有很大影响。目前关于饮食策略作为炎症性肠病主要治疗方法的证据越来越多。过去几年,针对炎症性肠病的潜在治疗效果,提出了几种饮食干预措施,即特定碳水化合物饮食、无麸质饮食、抗炎饮食。
然而,这些饮食要么不耐受,要么限制性很强,要么缺乏证据证明其对炎症性肠病的疗效。炎症性肠病的饮食干预研究最多的是独家肠内营养,这在儿科克罗恩病中常用,但饮食发挥其作用的确切机制尚不清楚。
独家肠内营养包括使用营养全面的液体饮食,而不是通常的固体和液体,通常长达8周,这是非常严格的,特别是在长期使用期间,可接受性有限。
✦抗炎饮食
你吃的食物在控制慢性炎症方面可能起到积极和消极的作用。
▸ 什么是抗炎饮食?
国际食品信息理事会基金会(International Food Information Council Foundation)营养传播副主任将其描述为一种饮食,重点关注高营养的食物,任何“营养密集”,“天然来源的维生素,矿物质和色素很多”的食物都是这种饮食的理想成分。
尤其是抗氧化剂, “它的主要参与者是水果,蔬菜,豆类,健康脂肪等食物,例如橄榄油和牛油果,包括鱼类,坚果和浆果等。
但是,抗炎饮食不仅是您的饮食,而且与食物的烹饪方式有关。应限制或避免高盐,饱和脂肪,糖和精制碳水化合物的食物。
在这里列举了一些抗炎食物和促炎食物,可以帮助更好地进行抗炎饮食。
•抗炎食物
很多种食物均具有抗炎特性,其中包括抗氧化剂和多酚含量高的食物。
1 浆果
浆果是小果实,富含纤维,维生素和矿物质
2 深海鱼
深海鱼是蛋白质和长链omega-3脂肪酸EPA和DHA的重要来源。
3 西兰花
西兰花是萝卜硫烷的最佳来源之一,萝卜硫烷是一种具有强大抗炎作用的抗氧化剂,可通过减少引起炎症的细胞因子和NF-kB的水平来对抗炎症。
4 牛油果
牛油果可能是少数值得冠以的超级食品之一,提供各种有益的化合物,可防止发炎并降低癌症风险。
5 辣椒
甜椒和辣椒中富含维生素C和抗氧化剂,具有强大的消炎作用。
6 蘑菇
蘑菇的热量非常低,并且富含硒,铜和所有B族维生素。它们还包含提供抗炎保护的酚和其他抗氧化剂。
7 葡萄
葡萄含有花青素,可减少炎症。此外,它们还可以降低多种疾病的风险,包括心脏病,糖尿病,肥胖症,阿尔茨海默氏病和眼疾。
8 姜黄
姜黄素是一种强大的抗炎营养素,因此受到了广泛的关注。姜黄可减轻与关节炎,糖尿病和其他疾病相关的炎症。
9 特级初榨橄榄油
特级初榨橄榄油是可以吃的最健康的脂肪之一。
它富含单不饱和脂肪,是地中海饮食中的主要食物,具有许多健康益处。
•促炎食物
在受伤或感染期间,身体会释放化学物质以帮助保护它并抵抗任何有害生物。但是,食用过多的促炎食物可能会导致慢性低度发炎。
1 糖和精制碳水化合物
摄入过多的糖和精制的碳水化合物与体内炎症增加以及胰岛素抵抗和体重增加有关。
2 人造反式脂肪
人造反式脂肪可能是您可以食用的最不健康的脂肪。它们是通过将氢添加到液态不饱和脂肪中而产生的,以使其具有更固态的脂肪的稳定性。
3 过量饮酒
虽然适度饮酒可带来一些健康益处。但是,较高的用量会导致严重的问题。大量饮酒可能会加剧炎症并导致“漏泄的肠道”,从而在整个身体内引发炎症。
4 植物油和种子油
概要一些研究表明,大量食用植物油中的omega-6脂肪酸含量可能会促进炎症。但是,证据不一致,需要更多的研究。
6 加工肉
加工肉富含AGEs等炎性化合物,其与结肠癌的强烈关联可能部分归因于炎症反应。食用加工肉会增加患心脏病,糖尿病,胃癌和结肠癌的风险。
✦FODMAP饮食
前不久提出了一种更有前景的直接针对肠道微生物群的营养方法,包括低可发酵低聚糖、双糖、单糖和(FODMAP)的饮食。
FODMAP是极易发酵但吸收不足的物质,分子量小,高渗效应导致肠道通透性增加。这些特性增强了肠道微生物群的发酵作用。
•FODMAP饮食的负面作用
富含FODMAP的饮食相关的症状是产气、腹痛、腹胀、抽筋、腹胀和腹泻。另一方面,近端结肠中FODMAP的快速发酵导致内腔中短链脂肪酸和乳酸的大量生成,进而影响粘膜屏障。
// 容易营养缺乏
尽管静止性炎症性肠病患者的胃肠道症状有潜在改善,但低FODMAP饮食与负面影响相关。由于饮食限制,营养缺乏的风险很高。
// 对肠道微生物有负面影响
另一方面,低FODMAP饮食意味着限制食品中天然存在的益生元,如果聚糖和低聚半乳糖,通过减少糖化细菌,特别是双歧杆菌,对肠道微生物群组成产生负面影响。
// 菌群数量减少
观察到,在静止期炎症性肠病患者中,低FODMAP饮食后,长双歧杆菌、青春期双歧杆菌和普氏双歧杆菌的数量减少。
由于双歧杆菌优先发酵果聚糖和低聚半乳糖,而普拉斯尼茨革兰阴性杆菌通过交叉喂养间接利用它们,因此,这种减少可能是由于到达肠道的可发酵底物数量的变化造成的。
在静止期克罗恩病患者中,低FODMAP饮食导致产生丁酸盐的梭状芽孢杆菌簇和嗜粘杆菌数量减少,这对粘膜相关微生物群的健康很重要,并且黏液溶解度瘤胃球菌扭矩的相对丰度增加,通常在克罗恩病患者中减少。
注意
长期服用低FODMAP饮食引起了一些担忧。除了评估饮食对炎症标记物或疾病活动的影响外,还需要对炎症性肠病患者低FODMAP饮食的充分性和安全性进行更多研究。
此外,关于低FODMAP饮食影响的少数研究样本量小,饮食应用的设计和时间不同,可能会影响结果的显著性。
营养干预可能在炎症性肠病症状管理和延长病情缓解方面发挥重要作用。然而,需要进一步的研究来更好地描述饮食、肠道微生物群和炎症性肠病之间的关系。
小结
饮食干预可能是挽救生命和降低医疗费用的可持续且具有成本效益的方式。但是,这些饮食干预措施的长期成功因人而异。成功与否取决于每个人的饮食建议是否可行,以及这些措施是否确实产生了预期的生理变化。
益生菌被定义为当摄入足够量时对宿主健康有益的活微生物。
益生菌作为操纵肠道微生物群内物种组成和代谢活动的一种手段,已被大量研究,以促进健康,预防或管理肠道疾病。
益生菌的作用可以是直接的,也可以是间接的,通过调节居民的微生物群或免疫系统。它们可以增强上皮屏障功能,增加对肠粘膜的粘附力,同时抑制病原体,促进对病原菌的竞争性排斥,产生抗微生物物质,并调节宿主的免疫系统。
例如,在感染或炎症状态下,益生菌可能会增加肠上皮细胞之间紧密连接的完整性,并防止肠细胞凋亡。此外,益生菌会产生短链脂肪酸和乳酸,降低肠上皮细胞的促炎反应,抑制潜在致病微生物的生长。
✦益生菌在炎症性肠病的作用
因此,使用益生菌调节肠道菌群,以对症和明确管理炎症性肠病具有巨大的治疗潜力。益生菌应从人类的肠道菌群中选择,不应对抗生素产生固有的耐药性。乳杆菌属(Lactobacillus)和双歧杆菌属(Bifidobacterium)的成员通常被认为是安全的,是功能性食品和补充剂中使用最多的益生菌,因为它们被证明对健康有益。
•益生菌对克罗恩病的作用暂不明确
使用益生菌治疗克罗恩病产生了相互矛盾的结果。一些研究已经成功地用益生菌治疗克罗恩病,如大肠杆菌(Escherichia coli)、鼠李糖乳杆菌( Lactobacillus rhamnosus)和布拉迪酵母菌(Saccharomyces cerevisiae boulardii)。
相反,也有报道称,鼠李糖乳杆菌未能维持克罗恩病的缓解。由于该研究领域缺乏精心设计的随机对照试验,目前关于益生菌诱导克罗恩病缓解的疗效和安全性的证据的确定性较低。
然而,最近公布的克罗恩病患者报告称,定期服用含有乳酸菌菌株组合(戊糖乳杆菌、短乳杆菌、植物乳杆菌、发酵乳杆菌、凯氏乳杆菌和林德纳乳杆菌)的产品导致其粪便中凯氏乳球菌丰度更高,症状的减轻和生活质量的提高。
L. kefiri具有很强的调节肠道微生物群组成的能力,导致与炎症反应和胃肠道疾病发病直接相关的几个细菌属显著减少。
✦益生元与合生元在炎症性肠病中的作用
益生元被定义为“通过选择性刺激结肠中一种或有限数量细菌的生长或活性,从而对宿主产生有益影响的非消化性食品成分,从而改善宿主的健康”。
益生元与短链脂肪酸(主要是醋酸盐、丙酸盐和丁酸盐)的生成增加有关,这可能有助于维持结肠内稳态。合生元是益生菌与益生元的结合,可能有利于活性克罗恩病患者,是治疗该疾病的潜在疗法。长期研究表明,服用合生元对活性克罗恩病有很好的疗效。
对最初接受氨基水杨酸盐和泼尼松龙治疗方案但未能缓解的患者进行合生疗法试验。合生元包括高剂量的益生菌短双歧杆菌、长双歧杆菌和干酪乳杆菌,以及作为益生元的木薯。
// 一定程度上改善克罗恩病症状
大多数患者的临床症状有所改善。车前草已被证明有助于腹泻患者,改善粪便粘稠度和黏度。其他随机对照试验评估了克罗恩病患者服用含有长双歧杆菌与商业益生元组合的合生元后,临床症状和组织学评分得到了有效改善,肠粘膜中的长双歧杆菌和其他种类的双歧杆菌定植增加。
此外,在3个月时,治疗组对粘膜白细胞介素18、干扰素(INF-g)和白细胞介素1b的影响不大,而TNF-a的表达在6个月时显著降低。粘膜中双歧杆菌物种的更高丰度可促进与参与调节促炎细胞因子途径的致病物种的竞争,从而刺激更具免疫调节性和耐受性的免疫反应。
// 缓解溃疡性结肠炎
大肠杆菌Nissle 1917是一种非致病性大肠杆菌菌株,可防止和对抗病原菌的定植。它减少结肠粘膜损伤,降低上皮通透性,有效维持溃疡性结肠炎缓解。
混合益生菌VSL#3由8株细菌组成,包括嗜酸乳杆菌、保加利亚乳杆菌、植物乳杆菌、干酪乳杆菌、嗜热链球菌、短双歧杆菌、婴儿双歧杆菌和长双歧杆菌。它下调宿主免疫反应,改善上皮屏障功能,增加粘液生成。
综合分析显示,VSL#3与安慰剂相比,在诱导缓解方面有显著益处的趋势,而与美沙拉秦相比,大肠杆菌Nissle 1917在维持缓解方面表现出非劣效性。
一种类似的合生元被证明在溃疡性结肠炎治疗中有用。肿瘤坏死因子、白细胞介素1a是驱动炎症和诱导防御素表达的炎症细胞因子,治疗后也显著降低。试验组的活检减少了上皮组织的炎症和再生。肿瘤坏死因子在活动性溃疡性结肠炎的发病机制中起着关键作用。因此,抑制炎症溃疡性结肠炎粘膜中肿瘤坏死因子的分泌是治疗疾病和预防复发的主要目标。
小结
虽然益生菌或合生元的使用对溃疡性结肠炎的治疗和维持有积极作用,但在克罗恩病中,只有合生元显示出明显的效果。
益生菌疗法可以通过与促进结肠中益生菌生长的益生元结合而潜在地得到改善。使用合生元的另一个优点是,益生元成分可以促进肠道内具有益生菌特性的本地生物体的生长。使用益生菌调节肠道细菌组成,并可能减少炎症或激活先天免疫,可用于治疗策略中,以恢复宿主肠道微生物群。
益生菌可能通过操纵微生物群来增加有益微生物的数量,减少可能有害的细菌,从而减少炎症触发因素,从而增加肠道生物多样性,改善炎症性肠病患者的症状。不过,不同微生物菌株和临床环境的益生菌效果不同。因此,对于大多数炎症性肠病患者来说,很难实现一般益生菌配方。
目前用于炎症性肠病的治疗策略会严重抑制免疫系统,从而导致严重感染和恶性肿瘤等风险。最近开发了一种基于微生物的免疫疗法,利用正常肠道微生物群的固有免疫调节特性,通过恢复免疫功能来治疗免疫相关疾病。
这种新的方法涉及位点特异性免疫调节剂,它是一种复杂的生物制剂,旨在以靶向、器官特异性的方式激活先天免疫应答。
▸ QBECO治疗
QBECO是一种来源于临床肠致病性大肠杆菌灭活分离物的试验性位点特异性免疫调节剂。
QBECO针对激活的巨噬细胞向胃肠道募集,大肠杆菌菌株通常会导致感染。免疫调节剂包含来自单个灭活微生物物种的成分,免疫系统将其识别为特定器官或组织中感染的常见原因,刺激新一波激活的固有免疫细胞的募集,并清除非生产性炎症的慢性来源。
因此,QBECO治疗不是抑制免疫功能,而是通过新免疫细胞的产生和动员,优化异常免疫反应,重建正常屏障功能和粘膜稳态,从而克服胃肠道免疫失调。
✦QBEOO减轻了炎症性肠病的病理
QBECO已经在诱导性结肠炎和炎症性肠病患者的实验模型中进行了测试。服用QBECO后,小鼠结肠炎的严重程度总体降低。该疗法减弱了肠道病理,改善了粘膜完整性,从而减少了胃肠粘膜的免疫细胞浸润,尤其是中性粒细胞的数量。
// 溃疡性结肠炎患者症状改善
在同一研究中,大多数接受QBECO治疗的UC患者在接受16周治疗后,其疾病在临床、内镜和组织学方面均有改善,粘膜完整性也有所改善。此外,治疗后1周内直肠出血明显减少。QBECO治疗还降低了结肠粘膜UC患者中中性粒细胞的数量,正如在诱导性结肠炎小鼠中观察到的那样。
// 克罗恩病患者症状也有改善
接受微生物QBECO治疗的克罗恩病患者也报告症状改善。在第一种临床方法中,十分之七的患者在至少三个月的治疗期间症状完全缓解。三名患者在停止药物治疗和QBECO治疗后持续出现临床病情缓解。此外,研究期间未报告严重不良事件。
最近对68名中重度克罗恩病患者进行的随机对照试验(RCT)评估了QBECO的安全性、疗效和耐受性。接受QBECO治疗的患者对治疗有良好的耐受性。
与安慰剂相比,到第8周时,疾病活动显著减少。之前接受肿瘤坏死因子抑制剂治疗的患者在第8周对QBECO反应的可能性较小,但随着治疗持续到第16周,症状有所改善,这表明可能需要更长的疗程才能在这些受试者中取得最佳结果。
此外,在随机对照试验期间QBECO治疗改善的克罗恩病患者产生IL-18,血清IFNg、IL-12p70和IL-17A水平较高,证实了对细菌刺激的有效免疫反应。
小结
利用基于微生物的免疫疗法,如QBECO,其功能是有效激活而不是抑制粘膜免疫功能,这是一种新的炎症性肠病治疗思路。然而,还需要进一步研究来评估这种新型免疫治疗方法在溃疡性结肠炎和克罗恩病治疗中的安全性和疗效。
用捐赠的粪便或纯化的粪便培养物进行粪便微生物群移植是目前治疗复发性艰难梭菌感染的常规方法。越来越多的人认识到微生物群失调在炎症性肠病发病机制中的作用,促使人们研究粪便移植作为一种潜在的炎症性肠病新疗法。
粪便移植改善炎症性肠病的证据进展迅速,前景看好。然而,关于这种治疗的疗效,有报道称结果相互矛盾。
注意:结果的多样性可能与不同的实验设计和方法学问题有关,包括供体选择、粪便类型、分娩方法、移植前准备、供体粪便处理方法、供体样本汇集以及给药频率和时间。
例如,粪便捐赠者样本的厌氧处理可保存可存活的厌氧菌用于移植,并可能影响临床结果。
最近的一项短期随机对照试验得出结论,使用经厌氧处理的供者样本进行粪便移植,与使用自体样本相比,8周后病情缓解的可能性更高,并且与本研究中观察到的治疗反应呈正相关的所有生物体都是厌氧菌(主要是专性厌氧菌)。Anaerofilum pentosovorans和粪拟杆菌物种的增加与治疗后疾病的改善密切相关。然而,从基线检查到第4周,微生物的数量发生了显著变化,在第8周之前保持稳定,但在12个月后发生了变化。
✦粪菌移植在溃疡性结肠炎中的作用
// 胃肠道症状改善,微生物群与健康者相似
一项前瞻性研究调查了溃疡性结肠炎患者粪便微生物群移植的临床疗效,结果表明,治疗后胃肠道症状改善,健康肠道微生物群重新繁殖。移植后,微生物群逐渐与健康献血者相似。克雷伯菌属和链球菌属的相对丰度下降,普氏菌属的相对丰度增加,表明这些属的变异可能是导致溃疡性结肠炎发病的重要因素。
// 微生物多样性显著增加
另一项研究表明,多次给药6周的多捐赠者密集剂量粪便微生物群移植是活动性溃疡性结肠炎患者的有效治疗方法。移植与微生物多样性显著增加相关,持续至治疗结束后8周。
此外,特定分类群与粪便微生物群移植结果相关。虽然梭状芽孢杆菌和Blautia属与治疗益处相关,但其他属(包括梭杆菌属和Sutterella属)的结果一直为阴性,这些属被认为与溃疡性结肠炎的发病机制有关。
粪便移植后病情缓解的溃疡性结肠炎患者粪便中的霍氏真杆菌(Eubacterium hallii)和inulivorans Roseburia富集,粪便样本中的短链脂肪酸水平升高。此外,未获得缓解的患者 Fusobacterium gonidiaformans、Sutterella wadsworthensis和埃希氏杆菌的富集。
建议
需要进一步研究来评估炎症性肠病患者粪便微生物群移植的长期缓解维持和安全性。
根据个体特征,使用各种特定肠道保护微生物菌株或其代谢物组合,可能比整个粪便微生物群移植更有效、更安全。
肠道微生物群失调似乎在炎症性肠病的发展中起着重要作用。发现可能导致慢性炎症的肠道微生物特征至关重要。
近年来,在鉴定不同微生物类群之间的健康平衡方面取得了重大进展。厚壁菌门和拟杆菌门中包含的某些微生物类群的丰富程度与健康肠道相关,有益或有害微生物之间的平衡强烈影响宿主健康。
影响宿主微生物群的遗传和环境因素需要进一步研究。为了开发个人定向治疗,还迫切需要更广泛地鉴定促进健康的微生物类群。
未来的另一个主要目标是选择个性化的益生菌或合生元疗法(或微生物特异性粪便移植)来治疗单个炎症性肠病患者,为实现长期缓解甚至完全治愈炎症性肠病做出巨大贡献。
主要参考文献:
Adamji, M., Day, A.S., 2019. An overview of the role of exclusive enteral nutrition for complicated Crohn’s disease. Int. Res. 17, 171–176.
Borisova, M.A., Achasova, K.M., Morozova, K.N., et al., 2020. Mucin-2 knockout is a model of intercellular junction defects, mitochondrial damage and ATP depletion in the intestinal epithelium. Sci. Rep. 10, 21135.
Buisson, A., Douadi, C., Ouchchane, L., et al., 2019. Macrophages inability to mediate adherent-invasive E. coli replication is linked to autophagy in Crohn’s disease patients. Cells 8, 1394.
Caldeira, L. de F., Borba, H.H., Tonin, F.S., et al., 2020. Fecal microbiota transplantation in inflammatory bowel disease patients: a systematic review and meta-analysis. PLoS One 15, e0238910.
Cox, S.R., Lindsay, J.O., Fromentin, S., et al., 2020. Effects of low FODMAP diet on symptoms, fecal microbiome, and markers of inflammation in patients with quiescent inflammatory bowel disease in a randomized trial. Gastroenterology 158, 176–188.e7.

谷禾健康

日常感觉疲劳,精力不好,稍微不注意就容易腹泻便秘,一不小心就感冒,更可怕的是,无论使尽各种方法依然减不下去的体重……
有以上状况的小伙伴注意,可能是慢性炎症在体内作怪。炎症是我们免疫系统的自然反应,也就是说身体和病原体斗争,试图自愈的过程。
根据时间和病理特征,炎症可以是急性和慢性的。
急性炎症来势汹汹,通常持续时间很短(几分钟到几天),包括淋巴细胞/嗜中性粒细胞和巨噬细胞迁移到炎症部位,刺激促炎细胞因子的释放【如:肿瘤坏死因子-α(TNF-α)、白细胞介素6 (IL-6)、高运动性蛋白B1(HMGB-1)】,以及细胞聚集,酶分解等。
NOD样受体(NLRs)(如NLRP3、NLRP1和NLRC4)的激活导致高度调节的蛋白复合物(称为炎症小体)的募集,其激活启动下游炎症细胞因子的产生,主要是白细胞介素1β(IL-1β)和白细胞介素18 (IL-18)对细胞应激的反应。
其他中介包括趋化因子、脂质介质、急性期蛋白如C-反应蛋白(CRP)、转录因子包括核因子κB(NF-κB)和主要免疫细胞类型。
然而,急性炎症如果不受控制,则可能发展为永久性疾病,导致组织损伤、血流动力学改变和器官衰竭。
慢性炎症就像温水煮青蛙,带来的损害缓慢,但是持久。事实上,慢性炎症与肥胖等非传染性疾病和相关的共病的发生有关。在这方面,肥胖导致脂肪细胞中的异常脂肪积累、免疫细胞浸润和促炎环境,从而破坏胰岛素信号级联诱导胰岛素抵抗。
炎症和氧化应激相互作用对于理解肥胖症的生理病理学至关重要,包括内质网功能受损、脂肪组织缺氧、线粒体改变和活性氧过度产生。
肠道微生物群与肥胖相关的低度炎症的发展有关,包括脂多糖易位和toll样受体4(TLR-4)结合,从而引发血液内毒素血症。
由此产生的未解决的免疫激活不仅影响局部组织,还影响全身生理学,即所谓的代谢性炎症。
本文讨论了不同营养因素对炎症的影响和最终调节,包括特定营养素(碳水化合物类型、蛋白质来源、结构脂肪酸、矿物质、微量元素)和生物活性化合物(多酚);饮食模式(即西方、地中海和北欧饮食);治疗性饮食(DASH饮食);常见烹饪原料(调味品和草药)等。
与人类炎症结果相关的营养因素

了解日常饮食中的营养物质对炎症的影响和调节,可以帮助我们在日常饮食中有意识地进行相应调整,从而更好地改善健康状况。
纵向和观察性研究表明,膳食维生素摄入量与炎症特征之间存在一些关联。
例如,维生素C和E或胡萝卜素的摄入与血清CRP浓度的概率成反比 > 美国成年人服用3 mg/L。
在横断面KORA研究中,剂量-反应分析显示,经常摄入超过78毫克维生素E/天的参与者的血清CRP水平比未接触任何额外维生素E来源的受试者低22%。
摄入含有维生素E和C以及B族复合维生素(B1、B2、B3、B5、B6、B9和B12)的膳食补充剂与女性血CRP水平降低相关。
饮食维生素K1(叶喹酮)摄入量变化的上三分位受试者(随访1年后)的IL-6和TNF-α血浆浓度比最低三分位组的受试者下降更大。
健康韩国成年人的膳食维生素B5摄入量与血清CRP浓度呈负相关。
消费 > 健康成人每天摄入310毫克的膳食胆碱(通常归入复合维生素B组),血液中CRP、IL-6和TNF-α浓度较低。
系统总结了探索维生素对炎症状态影响的临床试验结果(下表)。一些研究发现,补充维生素后有助于降低炎症。
表 临床试验:维生素和生物活性化合物抗炎作用
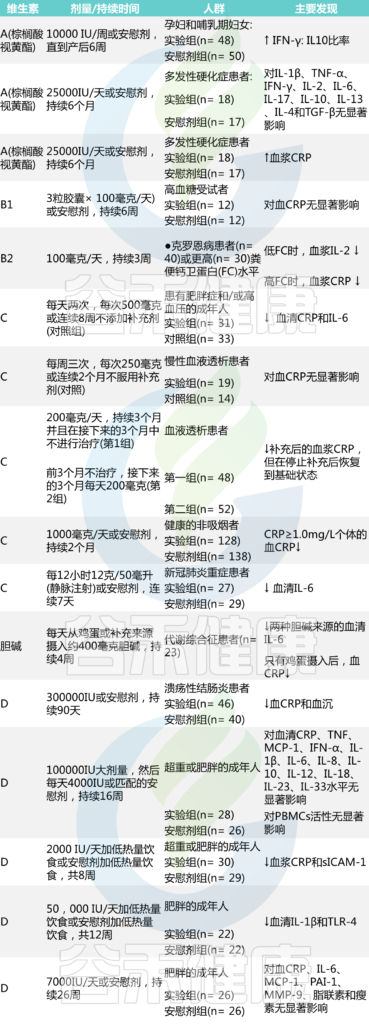

doi: 10.1007/s13679-022-00490-0
矿物质和微量元素对人体的结构、免疫和代谢功能至关重要。
高镁摄入与绝经后妇女体内潜在炎症标志物(CRP、sTNF-R2和IL-6)的血浆浓度降低有关。
在护士健康研究队列中,发现饮食来源的镁摄入量与血浆IL-6呈负相关。
一项嵌套病例对照研究报告,绝经后妇女的饮食锰与血清促炎细胞因子循环水平存在相反的关联。
据报道,淋巴细胞增殖和IL-2R表达的变化是健康男性轻度缺锌的早期标志。
膳食铜摄入量与成人血液CRP浓度直接相关。
反过来,肥胖相关炎症对肠道铁吸收的影响可能会加剧铁缺乏。
还显示了补充某些矿物质对人体抗炎作用的主要结果(下表)。
表 分析某些矿物质抗炎作用的临床试验



doi: 10.1007/s13679-022-00490-0
膳食碳水化合物对健康的影响取决于数量和质量特征。有趣的是,低碳水化合物饮食(总能量的20%)显著改善了糖尿病患者的亚临床炎症状态(血清IL-1Ra和IL-6水平较低)。
值得注意的是,坚持低碳水化合物饮食(占总能量的35%)可以降低肥胖女性的炎症标记物水平。
此外,低碳水化合物饮食的总体效果良好(≤ 30克/天)。此外,如其他地方报道的那样,与低脂饮食(总能量的24%)相比,极低碳水化合物饮食(占总能量的12%)可减少炎症反应。
血糖指数(GI)旨在根据对餐后血糖浓度的影响,从生理学上评估不同食物的碳水化合物质量。
有趣的是,高GI饮食(基于煮熟的意大利面,GI = 35)显著增加了瘦健康受试者单核细胞中NF-κB的激活率。
事实上,在糖尿病患者中,高GI饮食(GI > 70)诱导的负面代谢和炎症反应被低GI饮食(GI < 55)抵消。
此外,DIOGenes试验的结果表明,超重或肥胖受试者在减肥后,低GI碳水化合物(高GI碳水化合物的差异为15分)可以减少通过减肥饮食维持的低度炎症。
膳食纤维可能对健康有益,涉及一些免疫机制。因此,在糖尿病患者中,纤维摄入量等于或大于15 g/1000 kcal与血液CRP水平降低相关。
一项随机干预试验表明,从天然富含纤维的饮食或从补充剂中摄取纤维(30 g/天)可以显著降低瘦削正常血压参与者的循环CRP水平。
此外,中年成人膳食纤维摄入量(平均16.8克/天)和CRP血清浓度之间存在显著的负线性关系。
扩展阅读:肠道菌群与蛋白质代谢
膳食脂肪在生物体内引发许多基本功能;然而,过度消耗脂肪可能导致肥胖和相关的低度炎症过程。
事实上,临床证据表明,高脂饮食(即接近总能量的75%)会导致循环游离脂肪酸的过度生产和全身炎症。
一直以来,低脂饮食(占能量需求的25%)与糖尿病患者血浆IL-6水平降低相关。
越来越多的证据表明,膳食饱和脂肪酸(SFA)在肥胖和炎症之间起着重要的联系。
有趣的是,与摄入正常饱和脂肪的受试者相比,摄入超过10%能量作为饱和膳食脂肪的受试验者血清CRP水平升高(< 7%的热量摄入)。
同样,摄入膳食饱和脂肪酸(100毫升饱和脂肪含量为70%的乳脂)导致女性血浆CRP的脂质诱导升高,与肥胖状况无关。
单不饱和脂肪酸(MUFAs)被认为是一种健康的脂肪,油酸(OA)是日常营养中最常见的MUFA。
在这种情况下,一项针对日本人群的横断面流行病学研究报告,油酸摄入量(平均占总能量的6.94%)与血清CRP浓度之间存在显著的负相关。
对于不同剂量的MUFA治疗炎症特征的进一步对照试验是有保证的。
在过去的几年里,大量证据支持多不饱和脂肪酸(PUFAs)在预防心血管疾病和其他炎症性慢性疾病方面的有益作用。
在这种情况下,健康人n-3 PUFA二十碳五烯酸(EPA)和二十二碳六烯酸(DHA)的摄入量与血浆中可溶性TNF受体1和2的水平呈负相关。
此外,总膳食n-3 PUFA与女性CRP和IL-6的血水平呈负相关。
此外,一些临床试验评估了高PUFA饮食处方或通过补充PUFA对炎症结果的影响。例如,鱼油补充(38.2克/天EPA + 90天内的DHA)降低了高血压患者血液中促炎症标记物的水平。
健康的年轻人服用n-3 PUFA(2.5 g/天,2085 mg EPA和348 mg DHA)12周后,血清IL-6水平下降了14%。
在超重成人中,低(1.25 g/天)或高(2.5 g/天)剂量的n-3 PUFA补充4个月可以减少炎症反应(特别是血清IL-6和TNF-α浓度)。
反式脂肪酸(TFA)主要由植物油氢化或反刍动物衍生食品(包括乳制品和肉类)在工业上形成。
反式脂肪酸摄入量与女性血浆炎症生物标志物(包括CRP、VCAM-1、E-selectin)呈正相关。
在同一人群中,反式脂肪酸的摄入与可溶性TNF受体1和2的血浆水平呈正相关,主要是在体重指数较高的女性中。
男性服用TFA(占总脂肪的8%)后血清CRP浓度升高。
胆固醇过高可能会对健康产生有害影响,包括一些影响炎症状态的过程。
例如,在伊朗成年人中,血清CRP浓度的最高四分位数(5.9 mg/L)与饮食胆固醇的摄入量较高(189 mg/天)有关。
在大量具有代表性的中东人群中,发现饮食胆固醇和血浆CRP水平之间存在正相关。
膳食蛋白质的数量和质量是营养价值和身体/内分泌稳态的主要决定因素。
在Framingham心脏研究后代队列的参与者中,膳食蛋白质摄入量(尤其是植物来源的蛋白质)与血清炎症标记物(如IL-6和CRP)呈负相关。
此外,摄入高(总能量的30%)或低(总能量10%)蛋白质饮食会导致病态肥胖个体的血液CRP浓度降低。
关于蛋白质来源,以较高动物蛋白质摄入量(高脂肪和加工肉类水平)为特征的饮食与某些血液促炎标记物(如CRP、IL-6、TNF-a、IL-8、血清淀粉样蛋白a和糖蛋白乙酰化)呈正相关。
此外,RESMENA膳食研究的结果(30%的能量来自蛋白质)表明,动物和肉类蛋白质摄入量与炎症之间存在正相关,而蔬菜或鱼类来源的蛋白质对炎症状态没有显著影响。
扩展阅读:肠道菌群与蛋白质代谢
多酚是一大类生物活性分子,广泛存在于植物性食品中,具有强大的抗氧化和抗炎特性。在这种情况下,据报道,美国成年人的总黄酮摄入量与血清CRP浓度呈负相关。
在多种族队列中,黄烷酮消耗量与血液IL-6浓度呈负相关。同样,异黄酮摄入量较高(最高四分位数 = 1.61–78.8 mg/天)与健康绝经前妇女血浆CRP降低有关。
此外,在黄酮、黄烷酮和总黄酮摄入量较高的女性中,发现血清IL-8水平较低(五分位数最高 = 分别为264 ng/L、273 ng/L和276 ng/L)。
此外,台湾人群总黄酮摄入量和茶叶摄入量的增加与CRP水平呈负相关。
值得注意的是,许多随机临床试验已经测试了几种多酚的抗炎潜力,其结果总结如下:
表 分析某些多酚抗炎作用的临床试验

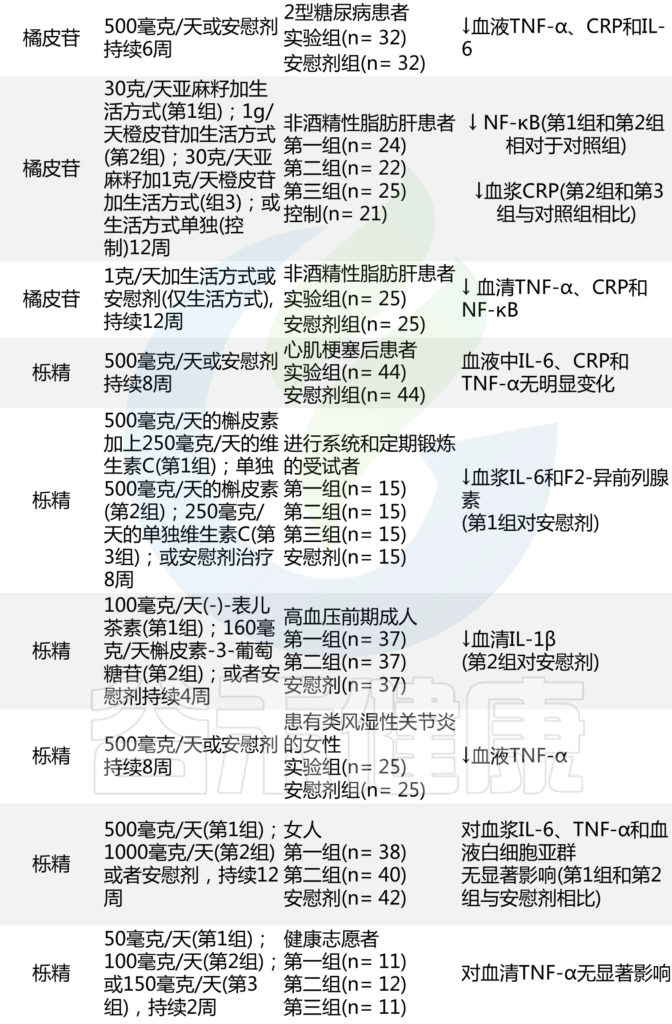

doi: 10.1007/s13679-022-00490-0
扩展阅读:
关于对炎症的影响,总摄入(中位数54 g/天)、未加工(中位数47 g/天)。
在多民族队列研究中,红肉和加工肉消费与血清CRP水平呈正相关。
在英国成年人中,食用加工肉与血清CRP水平增加有关(每天摄入50克以上,差异为38%)。
在调整后的模型中,在大量美国样本中,红肉消耗量与血液CRP显著相关。
在一项针对巴西人的横断面研究中,增加酸奶摄入量(中位数为10克/天)似乎会产生抗炎作用,而奶酪摄入量的增加(中位数10.7克/日)可能会加剧促炎状态。
在体重正常的青少年中,总乳制品和牛奶摄入量与血清IL-6浓度呈负相关。
ATTICA研究的结果显示,每周食用11-14份乳制品的人的CRP、IL-6和TNF-α血水平低于每周食用8份以下的人。
ATTICA研究结果显示,习惯性鱼类消费之间存在独立关联(> 每周食用300克鱼),并降低健康成年人的炎症标记物水平,包括CRP、IL-6、TNF-a、血清淀粉样蛋白a和白细胞计数降低。
在6年的随访中,食用鱼(约100克/周)可降低健康成人的内皮功能障碍和轻度炎症。
在表面上健康的日本人群中,高频率摄入鱼类与较低的外周血白细胞计数(慢性炎症标志物)相关。
事实上,随着每周摄入鱼的频率(0天、1-2天、3-4天或5-7天)的增加,全身炎症的标志显著降低(全身炎症的标志:中性粒细胞/淋巴细胞比率)。
近年来,食用昆虫被公认为具有抗炎和抗氧化特性的高价值食品。
例如,在健康成年人中,蟋蟀摄入量(25克/天)与通过微生物群调节减少全身炎症相关。
然而,需要对人类进行更多的研究来证实这些发现,以便推荐习惯性食用食用昆虫作为消炎疗法。
伊朗女性的水果和蔬菜摄入量与血清CRP水平呈负相关。
中国女性食用大量十字花科蔬菜(最高五分之一 > 140.6 g/天)显示循环中TNF-α、IL-1β和IL-6水平降低。
在一项随机交叉试验中,14天内食用十字花科蔬菜(14 g/kg体重)持续降低健康年轻人的循环IL-6.
在动脉粥样硬化的多种族研究中,经常食用坚果和种子(尤其是每周五次或五次以上)与较低水平的炎症标记物相关,包括IL-6和CRP。
在两个大的美国人队列中,与从未或几乎从未的频率类别的个体相比,每周坚果摄入五次或更多次的受试者的CRP和IL-6血浓度显著降低。
对随机对照试验的系统回顾和荟萃分析显示,摄入亚麻籽和相关营养衍生物系统地降低了肥胖受试者的循环CRP水平。使用同样的方法,急性高油酸花生摄入系统性地导致超重/肥胖男性餐后TNF-α浓度的下调。
研究表明,在健康饮食中添加杏仁(4周内每天56克)可以改善中国糖尿病患者的炎症和氧化应激。
一项随机试验还发现,在健康成年人中,食用杏仁(用杏仁替代对照饮食10–20%等量摄入4周)可以降低血清CRP水平。
事实上,在青少年和青年人中,杏仁喂养(每天56克,持续90天)后,血浆TNF-α和IL-6水平下降。
据报道,每天服用50毫升特级初榨橄榄油(EVOO),为期两周,可降低稳定型冠心病患者的血浆IL-6和CRP水平。
EVOO(50 mL)对正常血压的健康受试者具有急性餐后抗炎和抗氧化作用。
有趣的是,在糖尿病妇女中,谷类纤维的摄入与较低的CRP和TNF-R2血液水平呈负相关。
超重和肥胖受试者食用全麦小麦(8周内每天70克)后,血清TNF-α水平下降,血浆IL-10水平升高。
GRANDIOOS研究的结果表明,食用全麦小麦(每天98克,持续12周)可能会促进超重/肥胖和轻度高胆固醇血症患者的肝脏和炎症恢复力。
在中国中年女性中,食用大豆食品与炎症标志物(如IL-6、TNFα和可溶性TNF受体1和2)的循环水平呈负相关。
与习惯性饮食相比,一项为期6周的富含豆类的饮食营养试验(在所有干预阶段共摄入24包65克)显著降低了糖尿病患者的CRP浓度。
基于豆类的低热量饮食(每天160–235克,持续8周)持续降低超重/肥胖受试者的促炎状态并改善代谢特征。
在肥胖女性中,8周内补充绿茶提取物(450 mg/天)改善了氧化应激生物标记物,降低了IL-6循环水平。
3个月内饮用绿茶(379 mg/天)可降低肥胖、高血压患者的血清CRP和TNF-α浓度。
高咖啡消耗量(每天8杯)对习惯性咖啡饮用者的亚临床炎症产生了有益影响。
一直以来,在健康和糖尿病女性中,饮用咖啡与炎症标记物和内皮功能障碍呈负相关。
在年龄较大的非西班牙裔白人中,大量饮用咖啡(等于或超过2.5杯/天)的人全身炎症较低。
另一方面,来自ATTICA研究的分析报告称,中度至重度咖啡摄入后,炎症标记物(包括IL-6、TNF-α和CRP)增加(> 200毫升咖啡/天),强调剂量对结果的重要性。
补充蜂胶和限制热量饮食8周可以显著改善非酒精性脂肪性肝病患者的血糖稳态、肝纤维化评分和肝功能。
一项双盲安慰剂对照随机临床试验,44名非酒精性脂肪性肝病患者,用蜂胶和热量限制饮食(500千卡/天)干预,发现炎症因子降低,包括肿瘤坏死因子-α (TNF-α)、toll样受体-4 (TLR-4)和单核细胞趋化蛋白-1 (MCP-1)的血清水平以及肝酶和脂肪肝的严重程度显著降低。
现有证据表明,定期食用黑巧克力可能会减少炎症,尤其是对于每3天食用一份(20克)黑巧克力的消费者而言。
在一项随机平行临床试验中,与仅遵循一般生活方式指南的受试者相比,服用黑巧克力(8周内服用30克84%的黑巧克力)并保持健康生活方式的糖尿病患者的炎症标记物(CRP、TNF-α和IL-6)水平较低。
事实上,急性黑巧克力摄入(50克)通过增加IL-10的表达和减弱细胞内促炎性应激反应而引发抗炎症结果。
健康女性在摄入黑巧克力(一周内每天100克)后,CRP的血液水平较低,这在男性中没有发现。
在过去几十年里,几项调查已经确定了香料和草药在预防和治疗各种慢性病方面的有效作用。这些烹饪成分的多种健康特性归因于具有潜在抗炎特性的生物活性成分,如含硫分子、单宁、生物碱和酚类二萜。下表总结了探索香料对炎症状态影响的临床试验结果。
表 分析某些香料和烹饪成分抗炎作用的临床试验

doi: 10.1007/s13679-022-00490-0
益生菌、益生元和合生元是有益的微生物、底物(多糖和寡糖)或最终也可能缓解炎症症状的组合。
对于糖尿病患者,建议补充益生菌和合生元,通过持续降低循环中CRP和TNF-α的水平来减少炎症表现。
关于肠道疾病,最近有报道称,使用益生菌(基于乳酸杆菌和双歧杆菌)和合生元可以促进抗炎反应并平衡肠道内稳态。
短链脂肪酸(称为后生元的非活性细菌产物)的抗炎作用是通过抑制肠上皮细胞中的NF-κB通路、Treg细胞分化和促炎细胞因子阻断来介导的。例如,干酪乳杆菌DG和衍生后生物抑制肠易激综合征患者结肠粘膜中IL-8、IL-1α、IL-6和TLR-4的表达水平。
关于益生菌,益生元在此就不详述了,在前面的文章已有介绍,详见:
总的来说,植物性饮食已经证明可以改善肥胖相关的炎症状态。
值得注意的是,在北美,素食对血液CRP和IL-6水平的有益影响是由BMI介导的。此外,一项系统回顾和荟萃分析显示,素食饮食模式也降低了免疫生物标志物,如纤维蛋白原和白细胞总浓度。
对观察性和干预性试验的系统审查表明,北欧饮食(以北欧国家的主食为基础)对低度炎症缓解有积极影响。潜在机制包括代谢综合征患者的促炎症基因下调,尤其是TNFRSF1A和RELA。
南欧大西洋饮食(SEAD)是葡萄牙北部和西班牙加利西亚的传统饮食,其特点是摄入更多的鱼、牛奶、土豆、水果、蔬菜和橄榄油以及红酒。总体而言,SEAD依从性与炎症标记物(主要是CRP)的血浆浓度降低和心脏代谢风险降低相关。
就亚洲地区而言,健康的日本饮食模式(富含蘑菇、海藻、大豆制品和土豆、蔬菜、鱼类/贝类和水果)似乎可以发挥抗炎作用,改善当地消费者的心理健康。
一些中药已经显示出抑制促炎途径和控制炎症相关疾病。
墨西哥传统饮食(TMexD)已证明可以降低墨西哥裔女性的全身炎症和胰岛素抵抗风险。TMexD的特定食物包括玉米、豆类、辣椒、南瓜、番茄、仙人掌和洋葱,它们富含纤维、维生素、矿物质和辣椒素,具有潜在的抗炎和抗氧化特性。
在一项综合横断面研究中,旧石器时代饮食(基于蔬菜和水果、瘦肉、鱼类、坚果和钙来源的多样性消费)与人类较低水平的全身炎症和氧化应激相关。
在6周内,DASH饮食模式(以水果和蔬菜、低脂乳制品和复合碳水化合物的大量摄入为特征)降低了代谢综合征青少年的CRP循环水平。在女性成年人中,DASH饮食与伊朗人血清CRP水平较低有关,但与IL-17A浓度无关。定量评估显示,随访4周后,DASH饮食使CRP降低了13%。
PREDIMED试验的结果表明,地中海饮食(富含蔬菜和水果、纤维和维生素C和E)具有抗炎作用,因为它下调了动脉粥样硬化形成过程中涉及的细胞和循环炎症生物标记物。
在这个队列中,地中海饮食降低了血清CRP和IL-6水平,以及内皮和单核细胞粘附分子和促炎性趋化因子。此外,在随访1年后,Med饮食(包括EVOO和蔬菜)降低了心血管高危患者的血浆TNFR60浓度。长期(3年),PREDIMED试验通过与对照低脂饮食相比降低IL-1β、IL-6、IL-8和TNF-α水平,证实了Med饮食的抗炎作用。
总的来说,西式饮食(WTD)含有大量不健康的脂肪、精制谷物、糖和盐,会引发慢性代谢性炎症。在这方面,在护士健康研究I队列中,西式饮食与炎症和内皮功能障碍标志物呈正相关。此外,伊朗女性的西式饮食评分与CRP和IL-6促炎标记物呈正相关。
有趣的是,巴西妇女食用含有大量游离糖、总脂肪、膳食饱和脂肪酸、反式脂肪酸和钠的超加工食品(UPF)与血清CRP水平之间存在正相关。同样,超加工食品中上三分之一的巴西青少年(≥ 总能量的30%)与第1三分位青少年相比,循环IL-8浓度增加(≤ 15.9%的总能量)。
此外,瑞典类风湿关节炎患者的不良饮食质量(考虑到习惯性随意饮食,如糖果、蛋糕、软饮料和油炸土豆)与炎症增加有关,如血浆CRP和红细胞沉降率。
生物节律和营养分析(称为“时间营养”)的最新进展表明,一天中进食的时间可能会影响代谢稳态和免疫功能。
在这种情况下,在饮食质量较差的成年人中,不吃早餐与血清CRP浓度升高之间存在显著关联。在一项随机对照交叉试验中,不吃早餐会导致人类外周血单核细胞和单核细胞中NLRP3炎性体的更高活化。
间歇性禁食(IF),即个体连续或隔天禁食,改善了肥胖男性的全身炎症。然而,在超重或肥胖的女性中发现了间歇性禁食后,脂肪组织中巨噬细胞浸润(CD40+)和骨骼肌(CD163 +)的生物标志物短暂升高。
现有证据表明,限时饮食(TRE)是一种基于一天活动期总热量摄入巩固的替代时间营养方法,可能会调节多种代谢疾病风险因素,包括炎症。事实上,已经假设TRE作为定期营养计划的一部分,可能有助于减少炎症,并对免疫系统的某些组成部分产生保护作用。
有趣的是,代谢综合征患者在隔日禁食(ADF)后,血液CRP水平显著下降,ADF包括“禁食日”,热量摄入有限,而“喂食日”则是随意进食。此外,隔日禁食降低了健康非肥胖受试者血浆中sICAM-1(年龄相关炎症标记物)的水平。
此外,据报道,晚吃是指推迟用餐时间(通常是一天的主食或晚餐),可能会增加患心脏代谢疾病的风险。事实上,晚吃与腹部肥胖、炎症生物标志物(如IL-6和CRP)以及儿童的昼夜节律紊乱有关。
对精确变量(年龄、性别、身体表型、习惯性饮食摄入、体力活动水平和生活方式)以及个性化问题(遗传背景、表观遗传特征、微生物群组成、基因表达谱和代谢指纹)的综合分析可能有助于制定更个性化的治疗方案,以改善炎症的营养和药物管理。
例如,有证据表明,遗传变异可能通过与环境因素(如饮食)的相互作用,调节个体对与炎症相关的慢性和急性疾病的易感性,从而易诱发炎症状态。
表观遗传标记(包括DNA甲基化、miRNA表达和组蛋白修饰)在炎症基因转录中起着基础作用。
值得注意的是,基于微生物群的回归模型已经能够预测人类肥胖相关炎症状态,这可能是精确管理炎症性的有用工具。
具有促炎和抗炎作用的基因表达最终决定炎症的结果。
代谢组学是一种综合方法,可用于剖析炎症的局部和全身代谢后果,为炎症疾病的调节提供新的见解。
这些应用有助于阐明独特和特异的炎症代谢类型,扩大了我们对人类代谢复杂性和多样性的理解。
总的来说,这些新颖的科学见解正带来精确药物/营养战略,以预防和控制具有炎症背景的流行性慢性病。
营养物质对生命和健康至关重要,不仅有助于疾病预防、健康维护和疾病管理,而且可以抵御内源性和外源性有害因素,包括炎症/氧化应激或免疫系统功能障碍。
促炎
促炎营养因素包括大量食用富含简单碳水化合物、膳食饱和脂肪酸、TFA、胆固醇和动物蛋白的食物,以及习惯性不吃早餐和晚吃暴食。
抗炎
潜在的抗炎化合物包括MUFA、PUFA、抗氧化维生素和矿物质、生物活性分子(多酚)、特定食品(乳制品、全谷物、鱼类、油籽、水果和蔬菜、食用昆虫、豆类、绿茶和咖啡),烹饪香料(肉桂、姜、小茴香、大蒜和姜黄)和一些饮食习惯,包括间歇性禁食和限时进食。
由于研究之间存在不一致和差异,考虑到异质性的关键方面,包括人群类型(祖先)、最低和最高水平以及不利影响、烹饪方法、生理病理状态和干预时间,仍需在该领域进行进一步研究。
目前的证据有助于理解营养与代谢性炎症之间的关系,为慢病的控制和管理提供了新的见解和潜在目标。
主要参考文献:
Ramos-Lopez O, Martinez-Urbistondo D, Vargas-Nuñez JA, Martinez JA. The Role of Nutrition on Meta-inflammation: Insights and Potential Targets in Communicable and Chronic Disease Management. Curr Obes Rep. 2022 Oct 18. doi: 10.1007/s13679-022-00490-0. Epub ahead of print. PMID: 36258149.
Li C, Xu MM, Wang K, Adler AJ, Vella AT, Zhou B. Macrophage polarization and meta-inflammation. Transl Res. 2018 Jan;191:29-44. doi: 10.1016/j.trsl.2017.10.004. Epub 2017 Nov 3. PMID: 29154757; PMCID: PMC5776711.
Nikbaf-Shandiz M, Tutunchi H, Khoshbaten M, Nazari Bonab H, Ebrahimi-Mameghani M. Propolis supplementation in obese patients with non-alcoholic fatty liver disease: effects on glucose homeostasis, lipid profile, liver function, anthropometric indices and meta-inflammation. Food Funct. 2022 Oct 20. doi: 10.1039/d2fo01280d. Epub ahead of print. PMID: 36263703.
Rossi JF, Lu ZY, Massart C, Levon K. Dynamic Immune/Inflammation Precision Medicine: The Good and the Bad Inflammation in Infection and Cancer. Front Immunol. 2021 Feb 23;12:595722. doi: 10.3389/fimmu.2021.595722. PMID: 33708198; PMCID: PMC7940508.

谷禾健康

Pantoea(泛菌属):是肠杆菌科内一个高度多样化和多变的菌属,也是肠道内神秘的“善恶兼备” 的细菌。
▸直到最近,人们对Pantoea还是一个模棱两可的认知。用研究人员的话来说:
“这些物种既是植物,动物,人类的机会病原体,又能生产在除草剂,重金属螯合剂,抗生素等方面发挥着非常关键的作用。”
▸该菌无论命名还是功能都很有的“故事性”
它最初被命名为:
Bacillus agglomerans
后来被称为:
Enterobacter agglomerans
1989 年将所有归类为类似的物种创建了一个名为Pantoea属(泛菌属)。
▸泛菌属物种在自然界中无处不在
已从污染物质、土壤、水、植物(作为附生植物或内生植物)、种子、水果(例如菠萝、橘子)以及人和动物的胃肠道、乳制品、血液和尿液等分离出来。
▸与皮肤、软组织、骨骼、关节感染有关
通常是在被污染的物体造成穿透性创伤,或摄入受污染的动植物后导致人类机会性感染,尤其是当免疫系统受损时。
泛菌属有 20 多个物种,其中90%以上的感染与2个物种有关:
▸成团泛菌(Pantoea agglomerans,P. agglomerans)是一种罕见的机会性人类病原菌
由植物材料的伤口感染或医院获得性感染引起,主要发生在免疫功能低下的个体中。 患者年龄从早产儿到老年人与复杂产科等人群。
P. agglomerans自发性菌血症还与胃食管反流病、活动性恶性肿瘤和终末期肾病患者有关。而另一方面,P. agglomerans被用于生产“Andrimid”,一种新型的、有前途的乙酰辅酶A羧化酶抑制剂,一种强效的广谱抗生素。
▸Pantoea dispersa参与机会性感染
同样,关于Pantoea dispersa(P. dispersa)也一样,越来越多证据显示该菌参与机会性感染,特别是在受损的老年患者以及婴儿中。印度中部首次报告了涉及该物种的新生儿败血症病例。相继的, 2014 年,有报告病例P. dispersa引起菌血症,再次被认为是机会主义入侵者。
▸其它的还有Pantoea ananatis (P. ananatis,菠萝多源菌)和Pantoea calida (P. calida)
这些是机会主义的多重耐药菌微生物,通常会导致医院感染。尼泊尔的一份最新报告表明,上面两种菌可能是两个儿科病例的感染原因,尤其是在术后期间。
▸Pantoea brenneri (P. brenneri)和Pantoea dispersa (P. dispersa )最近被鉴定为人类临床样本中的一种新物种,也被报道对人类具有致病性。
应该注意的是,根据报告只有大约 20% 的病例能够在培养中分离出泛菌属物种。虽然泛菌属菌血症可能是短暂的,但其引起的临床恶化的不容忽视。
▸早期诊断和正确使用抗生素可以预防潜在的并发症
值得注意的是,大多数报道的泛菌属(Pantoea)感染对广谱抗生素敏感。除了偶尔的严重临床过程外,大多数关于Pantoea 物种可以通过适当的抗生素和支持性管理进行治疗。
目前全世界范围报告了越来越多的泛菌属导致的感染。而且在我们肠道菌群检测实践中也发现许多健康状况不良的人群,尤其幼儿和老年患者中检出高丰度的Pantoea菌,这也是本次调查和讨论该菌的原因之一。
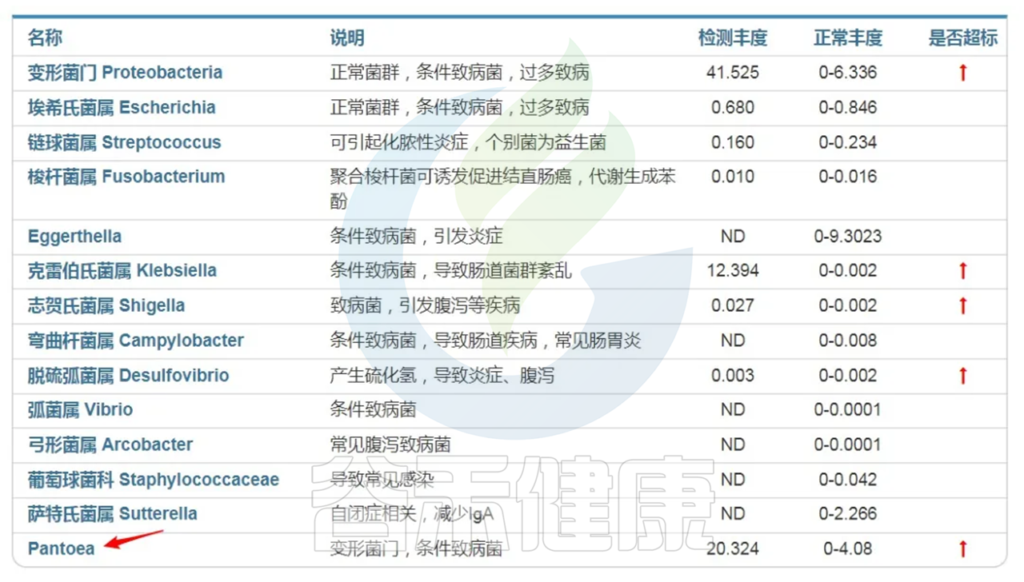
<来源:谷禾健康数据库>
该例报告是一个4个月幼儿做完直肠和乙状结肠切除手术一个月后的菌群检测,菌群结果显示有害菌较高,其中包括Pantoea,丰度高达20%显示超标,该检测结果在临床上有重要的用药管理价值。
泛菌属是在 1989年 提出的,泛菌属以前包括在肠杆菌属中。肠杆菌属经历了重大的分类修订。
随着该属的划定,新泛菌属物种的鉴定在过去几年中大大扩展。 Pantoea 与 Tatumella 和Erwinia 密切相关,这三者形成一个单系群,嵌套在其他肠杆菌属中,即埃希氏菌属、沙门氏菌属、柠檬酸杆菌属、肠杆菌属、克雷伯氏菌属。
这个单系群的基础谱系包含第二群,其中包含许多植物致病群,包括 Dickeya、Pectobacterium 和 Brenneria,以及内共生菌 Sodalis .
doi.org/10.1093/femsre/fuv027
泛菌属有 20 个物种,由 13 个不同的 DNA 杂交组组成。 其中P. agglomerans和P. dispersa是该属中最先发现的两个物种,这两个物种是与新生儿感染有关的最常见物种。
下图 Pantoea 菌株的邻接系统发育,基于由部分 rpoB、gyrB、atpD 和 16S rRNA 基因组成的连接数据集。
doi.org/10.1093/femsre/fuv027
与每个代表性类型菌株相邻的符号表示该物种组内的菌株从临床环境(红色圆圈)、作为附生植物或病原体的植物宿主(绿色正方形)或从自然环境(紫色三角形)中分离出来。
仅来自植物:
P. deleyi、P. anthophila、P. allii、
P. cypripedii、P. wallisi、
P. rodasii 、P. rwandensis;
仅来自临床:
P. conspicua、P. brenneri、
P. septica 、 P. eucrina;
仅来自室外环境:
P. gavinae
从自然环境或加工产品中鉴定出 P. calida、P. dispersa 和 P. gavinae 的分离株。此前,已经提出了其他几个物种,包括 P. citrea、P. punctata 和 P. terrea;然而,这些后来使用多位点序列分析 (MLSA) 方法重新分类为 Tatumella 属。
菌株分型和群体遗传学研究对于流行病学目的和识别具有重要表型的菌株(如对植物或人类的毒力)是必要的。例如,重要的是确定P. agglomerans菌株感染人类或引起植物特定疾病的能力是否不同。已有研究使用荧光扩增片段长度多态性 (fAFLP) 或脉冲场凝胶电泳对成团泛菌菌株进行了区分。
使用d-酒石酸盐可以将P. agglomerans与其他Pantoea物种区分开来,而myo-肌醇和内消旋酒石酸盐仅用于P. agglomerans和密切相关Pantoea物种。
目前,泛菌属内仍然存在相当大的分类多样性,其中一些欧文氏菌的分离株仍未分组。仍有许多工作要做,用以澄清该属的系统发育分类。
泛菌属基因组的测序有助于人们探索分离株在不同环境中茁壮成长的遗传因素。几种泛菌属分离株的基因组,如P. agglomerans、P. stewartii、P. vagans、P. ananatis,揭示了从 4.5 到 6.3 MB 的基因组大小和 52-55% 的 G+C 含量。
尽管迄今为止还没有对泛菌属的广泛系统比较或进化基因组分析,但在更具体的分析中发现的一些常见基因组特征包括酰基高丝氨酸内酯和其他群体感应基因, 植物生长促进基因, DNA 修复基因、致病因素以及 IV 型和 VI 型分泌系统。
最近使用测序的泛菌属和欧文氏菌基因组对 VI 型分泌系统 (T6SS) 进行了比较基因组分析,结果表明一种 T6SS 变体在来自不同环境的泛菌属分离物中普遍存在。特别是一个基因座,T6SS-1 基因座,包含两个高度保守的核心区域,它们与包含hcp和vgrG的可变区域交替,这些区域编码分泌的效应蛋白。
hcp和vgrG岛包含在Pantoea菌株的保守区域,并在抗菌、真菌细胞壁降解和动植物发病机制中与已知作用的基因同源。然而,这些结构域在来自不同环境的分离物中的存在表明可能存在遗传多样性和适应性的先天能力,这种适应性也反映在泛菌属的质粒中。这也是该物种在自然界中广泛存在的原因。
对至少 20 个分离株的比较基因组研究导致鉴定了大泛菌属质粒家族 (LPP-1),其范围从 280 到 789 kb,发现分布在代表 7 个不同物种的 20 个泛菌属分离株中,包括:
P. agglomerans, P. vagans, P. eucalyptii,
P. anthophila, P.stewartii, P . ananatis,
P. cypripedii
质粒编码位点与各种糖、碳水化合物、氨基酸和有机酸的代谢和运输,以及铁和氮的同化、抗生素和重金属抗性、宿主定植、发病机制和抗菌作用有关。这说明了该质粒家族的可塑性,这可能有助于不同物种群的成员之间保持能力多样性。
基因组比较虽然主要限于一小群分离株,但已经对赋予关键生存和宿主关联能力的毒力、抗性和代谢决定因素的性质产生了重要的见解。额外基因组的可用性将使更大的比较基因组分析成为可能,从而可以确定导致物种水平多样化的特定进化过程。此类研究还将允许评估个体泛菌属的致病潜力鉴于目前使用一些分离物作为生物防治、生物修复和治疗剂。
泛菌属物种在自然界中无处不在。已从污染物质、土壤、水、植物(作为附生植物或内生植物)、种子、水果(例如菠萝、橘子)以及人和动物的胃肠道、乳制品、血液和尿液等分离出来。
已从各种土壤和水环境中分离出泛菌属的分离株,并与不同的宿主相关联:
Alyssa M. W. et al,EMS Microbiology Reviews,2015
泛菌属( Pantoea )是肠杆菌科中一组呈黄色、杆状的革兰氏阴性菌。它们是兼性厌氧菌,氧化酶阴性。非包膜、非孢子形成的活动杆菌,具有周毛鞭毛,在营养琼脂中形成光滑、半透明和凸面的菌落。
Pantoea 可以利用 D-木糖、D-核糖、麦芽糖、D-半乳糖、D-甘露糖、D-果糖、海藻糖和 D-甘露醇作为能量代谢的碳源。
革兰氏染色上的成团泛菌(一种革兰氏阴性杆菌)
来源:doi.org/10.1155/2020/7890305
第一批的一些成员被认为是植物病原体,导致多种农业相关植物出现瘿、萎蔫、软腐和坏死,但从那时起,泛菌属菌株经常从许多水生和陆地环境中分离出来,以及与昆虫、动物和人类。
▸ 泛菌属会感染人类吗?可能带来什么样的后果?
人类通过摄入受感染的植物或刺突而接触。它也可能导致人类机会性感染,尤其是当免疫系统受损时,可能导致伤口、血液、肠道和尿路感染。
据报道,这种革兰氏阴性杆菌会导致新生儿 ICU 中的机会性血流感染。Pantoea 可作为社区获得性感染(包括职业接触)或医院获得性感染引起人类感染。
临床案例中报告了广泛的疾病已被归咎于泛菌属,包括脓毒性关节炎、骨髓炎、菌血症和败血症以及腹膜炎等,尽管其中大多数疾病的直接原因尚未得到证实。然而,几次新生儿暴发导致多起因败血症性休克和呼吸衰竭而死亡。
它们还可能引起过敏和过敏性肺炎。在儿童,尤其是新生儿中,感染好发于呼吸道,导致呼吸衰竭导致高死亡率。
脓毒性关节炎或滑膜炎是外源性成团 P. agglomerans感染的常见临床结果。
▸ 可能感染哪些部位和器官?
各种部位和器官都会受到泛菌的影响,导致:
伤口感染、滑膜炎、脓毒性关节炎、骨髓炎、血流感染、腹膜炎、胆石症、眼内炎、心内膜炎、泪囊炎、尿路感染、脑膜炎、脑脓肿、呼吸道感染。
值得注意的是,已从免疫功能正常和免疫功能低下的患者中鉴定出泛菌菌株,患者年龄从早产儿到老年人不等。P. septica、P. calida、P. dispersa、P. ananatis、P. agglomerans、P. eucalyptii菌株已常规从人体伤口、骨折、血液和其他液体、皮肤和表面拭子、粪便、囊肿和脓肿中分离出来,以及来自尿道、气管和口咽部的拭子。
▸ 泛菌属临床分离株实际上可能属于其他属/种
一些工作表明,这些和许多其他已被标记为泛菌属菌株的临床分离株实际上是被错误识别的。一项研究表明,临床和动物分离株报告为泛菌属的成员(最常见的是P.agglomerans)实际上属于其他泛菌属物种组,甚至其他属,如肠杆菌属。在临床分离株被证实为泛菌属的情况下,它们属于多个物种群,包括P. septica、P. calida、P. brenneri、P. eucalyptii、P.agglomerans.
为了试图了解泛菌属分离株的致病潜力,以及是否有证据表明谱系间宿主关联和/或宿主特化的演变,已经使用经过验证的临床和环境泛菌属分离株的集合进行了几项 MLSA 研究,以评估哪些临床和环境分离物聚集在一起。
▸ 临床分离株 & 环境分离株
临床和环境分离株的独立系统发育聚类(分为致病组和非致病组)被认为强烈支持谱系特异性宿主适应,正如其他肠道菌如大肠杆菌所见。在对泛菌进行的所有系统发育研究中,许多物种群的环境和临床分离株,包括P.agglomerans、P. ananatis、P. eucalyptii不会在每个物种群内形成独特的集群,而是混合在一起。
这种系统发育结构通常表明分离株具有未知的宿主关联能力,临床分离株可能具有定殖植物宿主的潜力,环境分离株可能具有定殖人类宿主潜力。
此外,P. agglomerans的分裂分解分析揭示了分离株之间的实质性重组,展示了在具有不同能力的个体分离株之间转移遗传决定因素的能力。
▸ 对植物和临床泛菌属分离株的毒力潜力评估
还使用功能宿主测定法对植物和临床泛菌属分离株的毒力潜力进行了评估。一项研究使用玉米、洋葱和果蝇的定量生长试验检测了来自环境和临床来源的多种泛菌的毒力潜力,结果表明临床分离株能够在两种植物宿主中生长,与环境分离株相当。临床分离株没有明显的生长或宿主定植模式,并且无法通过系统发育或分离来源预测宿主生长。
一项单独的研究评估了五种临床和五种植物相关的P. agglomerans在大豆和含胚鸡蛋中的菌株定量。临床菌株和植物菌株都能够在大豆植物上附生建立,并且在胚蛋中临床菌株和植物分离株之间的毒力没有差异,这表明所有P. agglomerans分离株可能具有相同的毒力潜力。
这两项研究表明,泛菌属分离株的宿主定植能力仍然不可预测,大多数分离株的植物或动物宿主范围/宿主关联能力未知。值得注意的是,Pantoea还被发现菌株与陆生和水生动物有关,包括鸟类、鱼类、无脊椎动物、熊和反刍动物,这可能暗示了涉及动物宿主的生活史。
在 1994 年 1 月 1 日至 2005 年 6 月 1 日期间在荷兰对 6,383 名患者进行的一项研究中,125 名患者 (2%) 报告了泛菌属定植,但没有聚集。
常见的定植部位包括气管、泌尿道和肠道。另一个病例系列指出,在 2005 年 1 月至 2006 年 12 月期间,科威特在 2005 年 1 月至 2006 年 12 月期间, 入住两个不同ICU 的 1,665 名新生儿中,有 5 名因泛菌属菌株导致院内血流感染。
在来自土耳其的一项单中心研究中,该研究评估了泛菌属的临床和微生物学特征从 2000 年到 2015 年,在儿科患者中,新生儿感染占 34.7%(23 个泛菌属分离株中有 8 个) 。
此外,临床医生更加关注从患者体内分离出的P .dispersa,目前,由它引起的人类感染临床报道的病例逐渐增多。
2003年报道一例71岁德国女性急性髓细胞白血病、多发性骨髓瘤并发呼吸道感染病例。该患者支气管肺泡灌洗液培养出P. dispersa。
2006年报道一例,两名成年患者出现关节感染,关节液培养显示P. dispersa生长。
2013年报道一例P. dispersa从 2 名患有早发性败血症的印度新生儿中发现。
2014年,日本报道一例中线相关性血流感染是由P. dispersa 引起的。
2019年报道一例从一名急性胆管炎患者身上分离出P. dispersa的病例。在这项研究中,它是第一个从中国肝细胞癌患者身上发现的P. dispersa
谷禾菌群检测案例:一例发育迟缓儿童的菌群检测中发现,其肠道菌群构成中泛菌属病理性滋生。
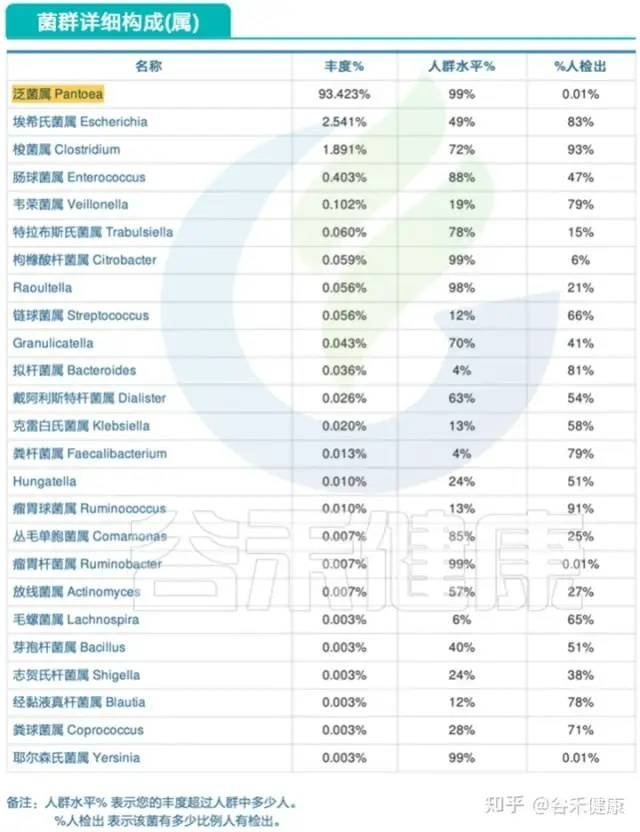
详见:真实案例 | 儿童发育迟缓肠道菌群检测的应用
目前从我们的检测实践中发现,在部分幼儿和中老年人的肠道菌群中发现泛菌属的高丰度水平,而且有的高丰度伴有菌群紊乱的迹象,希望有更深入临床研究关注人群肠道中泛菌属的健康特性。
儿 童
早产和相关的未成熟免疫系统导致相对免疫功能低下状态是新生儿泛菌属引起的医院获得性感染的主要危险因素。
截至 2020 年 7 月,在英国文献中报告的所有新生儿泛菌感染中,75%(40 人中的 30 人)发生在早产儿。这些婴儿中的大多数至少有一种合并症。迄今为止,文献中确定的合并症包括:
新生儿重症监护病房感染风险因素
新生儿重症监护病房中泛菌相关血流感染的暴发与肠外营养、静脉输液、婴儿配方奶粉、血液制品和麻醉剂的污染有关。追踪在新生儿重症监护病房中没有聚集的感染的起源是具有挑战性的。这些被定义为“散发”感染。尚无关于泛菌属垂直传播的确凿证据的记录。
儿童感染部位
在新生儿中,血液和脓液是最常见的分离泛菌属的样本。泛菌属菌血症的常见来源包括中心静脉导管和呼吸道。泛菌属菌血症的其他原因包括尿路感染、腹膜炎和皮肤感染。
在一项 6 年以上在儿童医院就诊的患者从正常无菌部位培养的 53 例成团泛菌感染的儿科病例。分离物包括来自血流的 23 个、来自脓肿的 14 个、来自关节/骨骼的 10 个、来自泌尿道的 4 个、来自腹膜和胸部的各 1 个。P. agglomerans 与植物材料的穿透性创伤和导管相关菌血症最相关。
在 2000 年至 2015 年在土耳其一家三级保健儿科医院进行的一项单中心回顾性研究中,从总共 15 个分离株中抽吸,最常见的Pantoea标本包括脓液(6 个标本,42.8%)、尿液(3 个标本,21.4%)、气管(3个样本,21.4%)和血液(3个样本,21.4%)。
成 人
感染进展:皮肤感染->深入骨骼->腹膜炎和脓毒症
皮肤感染作为伤口重复感染发生,或者当皮肤发生穿透性创伤导致慢性炎症反应时,有害物可能进入皮肤。感染可能深入骨骼,导致化脓性关节炎、脊椎间盘炎或胫骨骨炎,并可能进展为腹膜炎和脓毒症。
风险因素:植物材料伤口感染
P. agglomerans 的伤口感染通常是在用植物刺、木片或其他植物材料刺穿或撕裂皮肤,随后接种植物驻留细菌,主要是在从事农业和园艺工作或儿童玩耍时。在许多感染病例中,由于大多数菌血症感染具有多种微生物性质,因此很难辨别出这种细菌。
风险因素:接触被细菌污染的医疗设备或液体
除了导致上述结果的植物材料伤口感染外,接触被细菌污染的医疗设备或液体,特别是在免疫受损个体中,是导致菌血症爆发的主要感染原因。
工厂污染的带有静脉输液瓶的螺旋盖是 1970 年至 1971 年在美国婴儿和儿童流行的一个已证明的例子。在一项研究中,已发现该生物体作为粉状婴儿温和配方传播的机会性病原体的潜在候选者。
此外,成团泛菌菌血症与静脉输液污染、全胃肠外营养、麻醉剂和血液制品有关。在巴西的一家医院中,发现用于静脉水合的 0.9% NaCl 溶液连接的转移管是医院内 P.agglomerans 暴发的源头。
在成人中,医院获得性P. agglomerans感染已确定污染源的例子包括接受血液透析或血浆置换术的患者,这是由抗凝剂柠檬酸葡萄糖 46% 溶液的污染引起的输血后的败血症。
泛菌属还可以通过职业接触有机粉尘而发生。Pantoea agglomerans以大量定植棉花和棉花植物而闻名,它与棉花热有关,棉花热是一种在静脉内吸毒者中常见的良性发热综合征。
自发性或散发性菌血症:与胃食管反流病和抗酸剂有关,可能通过摄入蔬菜或水果引入
尽管血流感染通常与受污染的血管内产品和医疗设备相关,从而导致爆发,但P. agglomerans也可引起自发性或散发性菌血症。自发性菌血症与胃食管反流病 (GERD) 和服用抗酸剂密切相关。由于泛菌属物种普遍存在于植物上,因此有可能通过摄入蔬菜或水果引入,在胃食管黏膜病变(如 GERD)和/或没有保护性的情况下,导致疾病发生。
Pantoea引起的感染的临床结果包括感染骨骼、关节、滑膜引起化脓性关节炎、骨髓炎或滑膜炎,包括但不限于心内膜炎、眼内炎和皮肤感染。
新生儿泛菌感染最常见的表现是迟发性败血症。 除Habsah 等人报告的受污染的肠外营养导致新生儿重症监护室爆发之外,2005 年迄今仅有 2 例早发性脓毒症在新生儿人群中被报道,与绒毛膜羊膜炎和胎膜延长破裂有关。
患有泛菌属菌血症的新生儿最常出现肺部症状。迄今为止,文献中报道的泛菌感染的系统性临床特征包括在下表中。
Pantoea是新生儿重症监护室遇到的一种相对罕见但具有潜在危险的医院获得性感染,尤其是在有合并症的早产儿中。
▸ P. agglomerans 和 P. dispersa 是新生儿最常见的致病菌种
定植往往发生在呼吸道、泌尿生殖道和胃肠道。易位到血液中会产生侵袭性感染。对常规一线抗生素的敏感性因多药耐药率高而变化。
▸ P. dispersa具有引起不同系统感染的能力
例如呼吸系统、关节系统、血液系统和消化系统。患者通常会出现由相应感染病变引起的高烧等症状。此外,已知P .dispersa不仅会在免疫功能低下的患者中引起感染,还会在免疫功能正常的患者中引起感染。
▸ 自发性菌血症还与潜在疾病有关
自发性菌血症还与恶性肿瘤、糖尿病、慢性病毒性肝炎、脑血管意外、充血性心力衰竭、自身免疫或结缔组织疾病、慢性肺阻塞性疾病和终末期肾病等潜在疾病有关。
注:细菌内毒素导致细胞因子的产生,是感染的罪魁祸首。菌血症的体征和症状包括但不限于非特异性病理生理反应,例如胃肠道症状、发热、白细胞计数变化、贫血、血小板减少、弥散性血管内凝血、低血压和休克。
其他记录在案的儿科患者发生医源性暴发的例子包括但不限于由肠外营养液引起的呼吸衰竭败血症和用于静脉补液的受污染转移管引起的具有胃肠道症状的败血症。
▸泛菌属的血培养和分离是诊断血流感染的金标准
通过分子诊断测试,如基质相关激光解吸/电离飞行时间质谱 (MALDI-TOF MS)、16s RNA 基因测序、多位点序列分析 (MLSA),可以实现快速的物种水平鉴定和抗生素敏感性。
Pantoea感染是新生儿重症监护病房中相对罕见但具有潜在危险的机会性病原体,特别是在早产儿中,因此准确的物种水平鉴定和适当的定向治疗至关重要。
▸注意比较Pantoea菌的植物来源和临床菌株
使用多位点系统发育分析和荧光扩增片段长度多态性指纹识别来寻找区分基因型/表型标记。研究发现,大量来自培养物保藏中心的临床分离株在序列分析后被错误地命名为P. agglomerans。
在日常工作中,用常规方法难以准确识别Pantoea。采用传统方法,Pantoea的培养特性与肠杆菌科相似。菌落是兼性厌氧的。
▸通过部分16SrRNA或宏基因组测序鉴定Pantoea是一种有用的诊断工具
这些方法具有比传统表型方法更高的性能特征。16S rRNA 基因在细菌中普遍存在,这主要是由于它在细菌中的保守性,并且由于大约 1500 个碱基对 (bp) 基因座提供的系统发育信号。此外,16S rRNA 基因序列不限于一组已知细菌,新分离物可以与一组相关细菌建立连接。
成团泛菌已表现出对多种抗生素的耐药性,包括早代青霉素、早代头孢菌素、广谱头孢菌素和抗假单胞菌青霉素、氟喹诺酮类、氨基糖苷类、TMP-SMX和四环素。
关于有效的抗菌治疗,在成人患者自发性菌血症的队列研究中研究了抗菌药物敏感性。
100%的分离株对头孢噻肟、头孢他啶、哌拉西林-他唑巴坦、亚胺培南、环丙沙星、庆大霉素和阿米卡星敏感。61%对头孢唑林敏感,56%对氨苄青霉素敏感,33%对磷霉素敏感。
菌血症患者接受有效经验性抗生素治疗的临床治疗成功率高达100%。
据报道,使用多粘菌素 B 固定化纤维柱直接血液灌流 (PMH-DHP) 疗法从革兰氏阴性菌中去除脂多糖以及抗生素,可成功治疗由P. agglomerans感染引起的肺小细胞癌患者的败血症。
在包括儿科和新生儿在内的病例系列中,所有成团聚球菌分离株均显示出对阿米卡星、庆大霉素、美罗培南和甲氧苄啶-磺胺甲恶唑的抗菌敏感性,92.5% 的分离株对广谱头孢菌素和半合成青霉素敏感,62.3% 对超广谱头孢菌素,只有47.2%为氨苄青霉素。
众所周知,广谱抗生素的使用会增加医院环境中定植的风险,并增加出现抗生素耐药性的风险。胃肠道定植和后来的易位可能成为该生物体的储存库。
管理新生儿泛菌属血流感染的成功取决于及时识别和早期给予适当的抗生素治疗以及常规支持性护理。由于大多数报道的泛菌属。新生儿血流感染与中心静脉导管有关,应注意识别这种关联。
▸ 抗菌药物选择
根据临床案例治疗和研究报告建议新生儿科医生在选择抗菌药物治疗时应注意感染部位、婴儿胎龄及其合并症。对于发生感染而无中心静脉导管或其他合并症的足月或晚期早产儿,氨基糖苷类药物(庆大霉素或阿米卡星)联合氨苄青霉素是合适的一线治疗方法。
根据文献调查,这种微生物是导致极早产儿合并症(呼吸机依赖型支气管肺发育不良和中心静脉导管)迟发性败血症的病原体,有临床研究推荐碳青霉烯类作为肺炎或菌血症的一线治疗。对于局限于泌尿生殖道的单纯性感染,氨基糖苷类药物可作为一线治疗。
▸ 替代疗法
对于对碳青霉烯类耐药的泛菌属菌株,甲氧苄氨嘧啶/磺胺甲恶唑是一个不错的选择。新生儿的这种药物需要在整个治疗过程中定期监测血清胆红素水平和肝酶。
许多泛菌菌株显示出惊人的环境多功能性和适应性,并具有多种生物合成和生物降解能力,可用于农业、环境和临床环境中的潜在有用应用。
Pantoea agglomerans 是一种植物来源的 γ‐变形菌纲的菌,具有许多有益的特性,可用于预防和/或治疗人类和动物疾病、对抗植物病原体、促进植物生长和环境的生物修复。它生产多种抗生素(herbicolin、pantocins、microcin、agglomerins、andrimid 等),可用于对抗植物、动物和人类病原体或用于食品保存。
▸ 来自成团泛菌的 IP-PA1:愈合、镇痛、防止感染、过敏、癌症等
日本研究人员已经证明,由他们分离并描述为“来自成团泛菌的免疫增强剂1(IP-PA1)”的P. agglomerans 的低分子量脂多糖具有极广谱的愈合特性,主要是由于它能够通过巨噬细胞活化维持体内平衡。
最近,由P. agglomerans IG1 产生的免疫增强剂 IP-PA1 被证明可以增强小鼠和鸡的免疫相关功能,以对抗细菌和寄生虫感染,并增强免疫抑制的恢复。将 IP-PA1 施用于患有 B16 黑色素瘤(一种皮肤癌)的小鼠,存活期显着延长。
IP-PA1被证明可有效预防和治疗广泛的人类和动物疾病,如肿瘤、高脂血症、糖尿病、溃疡、各种传染病、特应性过敏和压力引起的免疫抑制;它还显示出很强的镇痛作用。重要的是,大多数这些效果可以通过安全口服 IP-PA1 来实现。
IP-PA1 也被研究用于巨噬细胞活化,以防止感染、过敏和癌症,以及逆转化疗引起的免疫抑制,这反映了Pantoea-衍生天然产物作为治疗剂。
▸ P.agglomerans——替代化学肥料的环保生物接种剂的理想候选者
据记载,Pantoea 菌株对土壤和水的各种化学污染物(包括石油烃和有毒金属)具有生物降解活性。P. agglomerans 通过生物膜的形成防止有害工业污染物渗透到土壤的更深处,并具有从废物中产生氢气的能力。因此,这种细菌似乎是一种有价值的生物修复剂,在某些情况下,它可以作为一种廉价的能源形式获得。
▸ P. ananatis 应用于食品冷冻,控制害虫,改良水稻
P. ananatis 的细胞外冰成核剂已经过测试并应用于食品的冷冻以获得所需的质地,以及食品的冷冻干燥。
P. ananatis 的冰成核菌株也显着降低桑葚幼虫的耐寒性,因此这些菌株具有作为害虫生物控制剂的潜力。
此外,瑞士研究人员对水稻进行基因改造以生产“黄米”的成就归功于P. ananatis。在这种情况下,来自P. ananatis(一种被鉴定为Erwinia uredovora的菌株)的八氢番茄红素去饱和酶被用于将 β-胡萝卜素生物合成途径引入水稻。
▸ 泛菌属——开发成生物防治产品,生物修复
一些泛菌属分离株产生抗微生物剂,并已开发成商业生物防治产品。例如 BlightBan C9-1 帮助控制苹果和梨树的火疫病,而其他具有生物修复潜力,能够在不产生有毒副产品的情况下降解除草剂。
Pantoea通过分解有毒物质来增强昆虫的适应性,甚至在限氮条件下促进植物根系的固氮。此外,这种细菌群在不同环境中竞争和生存的能力使其许多成员对生物防治和生物修复都特别有吸引力。
Pantoea具有独特的生物降解能力,包括降解除草剂和其他有毒化合物的代谢途径,为有用产品的开发和商业化提供了机会。
Pantoea 是肠杆菌科中一个高度多样化的群体,其成员分布在水生和陆地环境中,并与植物、昆虫、人类和动物有关。
Pantoea 目前被认为在临床环境中存在问题,然而,围绕单个菌株的宿主关联和致病能力仍然存在很多不确定性。
鉴于没有已知的病原体生物标志物可用,而且仍然需要确定那些能够实现特定生态位定殖的遗传决定因素,包括可能决定宿主定殖能力和宿主特异性的任何因素。确定这些遗传因素的性质仍然是一个有前途的研究方向,无疑将有助于揭示这种多用途、广泛利基细菌群的全部能力。
此外,一些分离株已被用作免疫增强剂。IP-PA1被证明可有效预防和治疗广泛的人类和动物疾病,用于开发治疗黑色素瘤、感染、过敏和逆转免疫抑制的支持药物。
因此,无论是环境还是临床都应保持对泛菌属的高度敏感和关注,有效的检测技术将帮助我们拓宽认知,推进对该菌内不同菌种与宿主的致病或共生关联以及感染生态位的确定。此外,不应拒绝对使用这些生物及其相关产品,但是应该注意在使用泛菌生物制剂时提供安全预防措施。
主要参考文献:
Dutkiewicz J, Mackiewicz B, Lemieszek MK, Golec M, Skórska C, Góra-Florek A, Milanowski J. Pantoea agglomerans: a mysterious bacterium of evil and good. Part II–Deleterious effects: Dust-borne endotoxins and allergens–focus on grain dust, other agricultural dusts and wood dust. Ann Agric Environ Med. 2016;23(1):6-29. doi: 10.5604/12321966.1196848. PMID: 27007514.
Dutkiewicz J, Mackiewicz B, Lemieszek MK, Golec M, Milanowski J. Pantoea agglomerans: a mysterious bacterium of evil and good. Part IV. Beneficial effects. Ann Agric Environ Med. 2016 Jun 2;23(2):206-22. doi: 10.5604/12321966.1203879. PMID: 27294621.
Dutkiewicz J, Mackiewicz B, Kinga Lemieszek M, Golec M, Milanowski J. Pantoea agglomerans: a mysterious bacterium of evil and good. Part III. Deleterious effects: infections of humans, animals and plants. Ann Agric Environ Med. 2016 Jun 2;23(2):197-205. doi: 10.5604/12321966.1203878. PMID: 27294620.
Cruz AT, Cazacu AC, Allen CH. Pantoea agglomerans, a plant pathogen causing human disease. J Clin Microbiol. 2007 Jun;45(6):1989-92. doi: 10.1128/JCM.00632-07. Epub 2007 Apr 18. PMID: 17442803; PMCID: PMC1933083.
Dutkiewicz J, Mackiewicz B, Lemieszek MK, Golec M, Skórska C, Góra-Florek A, Milanowski J. Pantoea agglomerans: a mysterious bacterium of evil and good. Part II–Deleterious effects: Dust-borne endotoxins and allergens–focus on grain dust, other agricultural dusts and wood dust. Ann Agric Environ Med. 2016;23(1):6-29. doi: 10.5604/12321966.1196848. PMID: 27007514.
Dutkiewicz J, Mackiewicz B, Lemieszek MK, Golec M, Milanowski J. Pantoea agglomerans: a mysterious bacterium of evil and good. Part IV. Beneficial effects. Ann Agric Environ Med. 2016 Jun 2;23(2):206-22. doi: 10.5604/12321966.1203879. PMID: 27294621.
Dutkiewicz J, Mackiewicz B, Kinga Lemieszek M, Golec M, Milanowski J. Pantoea agglomerans: a mysterious bacterium of evil and good. Part III. Deleterious effects: infections of humans, animals and plants. Ann Agric Environ Med. 2016 Jun 2;23(2):197-205. doi: 10.5604/12321966.1203878. PMID: 27294620.
Cruz AT, Cazacu AC, Allen CH. Pantoea agglomerans, a plant pathogen causing human disease. J Clin Microbiol. 2007 Jun;45(6):1989-92. doi: 10.1128/JCM.00632-07. Epub 2007 Apr 18. PMID: 17442803; PMCID: PMC1933083.
Mani S, Nair J. Pantoea Infections in the Neonatal Intensive Care Unit. Cureus. 2021 Feb 3;13(2):e13103. doi: 10.7759/cureus.13103. PMID: 33643749; PMCID: PMC7886163.
M, Golec M, Milanowski J. Pantoea agglomerans: a mysterious bacterium of evil and good. Part III. Deleterious effects: infections of humans, animals and plants. Ann Agric Environ Med. 2016 Jun 2;23(2):197-205. doi: 10.5604/12321966.1203878. PMID: 27294620.
Cruz AT, Cazacu AC, Allen CH. Pantoea agglomerans, a plant pathogen causing human disease. J Clin Microbiol. 2007 Jun;45(6):1989-92. doi: 10.1128/JCM.00632-07. Epub 2007 Apr 18. PMID: 17442803; PMCID: PMC1933083.
Coutinho TA, Venter SN. Pantoea ananatis: an unconventional plant pathogen. Mol Plant Pathol. 2009 May;10(3):325-35. doi: 10.1111/j.1364-3703.2009.00542.x. PMID: 19400836; PMCID: PMC6640510.
Walterson AM, Stavrinides J. Pantoea: insights into a highly versatile and diverse genus within the Enterobacteriaceae. FEMS Microbiol Rev. 2015 Nov;39(6):968-84. doi: 10.1093/femsre/fuv027. Epub 2015 Jun 24. PMID: 26109597.
Brady C, Cleenwerck I, Venter S, Vancanneyt M, Swings J, Coutinho T. Phylogeny and identification of Pantoea species associated with plants, humans and the natural environment based on multilocus sequence analysis (MLSA). Syst Appl Microbiol. 2008 Dec;31(6-8):447-60. doi: 10.1016/j.syapm.2008.09.004. Epub 2008 Nov 12. PMID: 19008066.
Tsubouchi H, Matsumoto N, Yanagi S, Arimura Y, Nakazato M. Successful treatment of sepsis due to Pantoea agglomerans by polymyxin B-immobilized fiber column direct hemoperfusion therapy in a small cell lung carcinoma patient. Respir Med Case Rep. 2016 Oct 6;19:155-158. doi: 10.1016/j.rmcr.2016.10.004. PMID: 27752466; PMCID: PMC5061076.
Luziatelli F, Ficca AG, Bonini P, Muleo R, Gatti L, Meneghini M, Tronati M, Melini F, Ruzzi M. A Genetic and Metabolomic Perspective on the Production of Indole-3-Acetic Acid by Pantoea agglomerans and Use of Their Metabolites as Biostimulants in Plant Nurseries. Front Microbiol. 2020 Jul 14;11:1475. doi: 10.3389/fmicb.2020.01475. PMID: 32765438; PMCID: PMC7381177.

谷禾健康


You are what you eat
国庆放假除了玩玩玩,当然少不了吃吃吃,把各种一直想吃的都提上日程,应该是对假期最起码的尊重了…
有时候我们喜欢吃的并不是食材本身,而是喜欢融入食材中的“味道”,不知从什么时候开始,我们对“重口味食物”越来越上瘾,而“重口味食物”往往意味着过甜、过咸、过辣……
我们知道饮食可以影响肠道菌群,在之前的文章里也多次提到,诸如食物种类,饮食方式,饮食习惯等对肠道菌群的影响。
之前写过关于饮食对肠道菌群的文章:
间歇性禁食 & 肠道菌群 & 心血管疾病
利用饮食精准干预肠道微生物群
饮食-肠道菌群对心血管疾病的相互作用
正视暴饮暴食、厌食症等饮食失调问题
深度解析 | 炎症,肠道菌群以及抗炎饮食
饮食习惯或将引领新的健康革命
深度解读 | 饮食、肠道菌群与健康
本文试着从不同饮食口味的角度「包括甜、咸、辣」,来了解一下糖、盐、辣椒等对肠道菌群的影响,对健康的潜在影响机制,以及相关食用小妙招。
如果说厨房里只能有一款调料,那一定是盐。盐,号称”百味之王”,同时也是一把”双刃剑”。身体缺不了它,食用不当也会给身体带来很多伤害,例如体重增加,高血压,慢性肾病等。

氯化钠,“盐”,是日常食物的重要组成部分,对身体的体内平衡起着至关重要的作用。
盐参与调节人体内水分的均衡分布,增强神经肌肉兴奋性,为使机体内酸碱平衡和血压正常功能,保证体液的正常循环,参与胃酸的形成,促使消化液的分泌,能增进食欲。
一般成人每天摄入3克食盐就可以维持基本生理需求。2022年颁布的《中国居民膳食指南》中也将食盐摄入量再次严格限制,每人每日食盐摄入量从原先的6克以内改为不高于5克。
过多的盐摄入量,可能给健康带来危害。
饮食中高盐含量会导致肠道免疫系统的变化。血液中含有过多的钠会导致体内免疫细胞产生较少的能量,还会影响机体正常代谢和细胞的渗透压等,引发心脑血管疾病和代谢性疾病,业已知道盐摄入过量是心脑血管疾病的主要风险因素之一。
doi.org/10.1016/j.biopha.2020.111156
◣ 高盐饮食和体重增加相关
或许会有这样的疑惑,盐含的热量极少,为什么会与体重增加有关?
这个问题不在于热量,确实盐一点热量都没有。然而摄入大量的盐会导致体重暂时增加,是因为它会导致身体保留水分。
盐摄入过多会增加口渴感。身体消耗的额外液体被用来稀释身体无法足够快速排出的多余钠。
小鼠研究表明,高盐饮食也可能使身体对瘦素产生抵抗力。
注:瘦素是一种激素,负责防止饥饿,让你感到充实和满足。
高盐饮食也可能间接导致体重增加,因为它们通常与大量的超加工食品有关。高度加工的食物导致摄入更多的热量并增加体重。
当热量相等时,高盐饮食不会比低盐饮食增加或减少更多的体脂。
含盐量较高的食物:零食、薯条、西式快餐、披萨、油炸食品、加工食品、调味品,甚至面包,也含有较高的热量。
因此,如果你的饮食中含有大量这些经过高度加工的食物,那么胃很可能会感觉不到满足感和饱腹感,最终导致摄入更多的热量并增加体重。
◣ 高盐饮食和夜尿症相关
如果你有夜尿症,钠摄入量可能是罪魁祸首。
在一项研究中,当那些夜间排尿频繁且饮食中摄入高钠的人减少了钠的摄入量时,他们报告说夜间上厕所的次数减少了,而良好的睡眠习惯提高了生活质量。
在用餐期间和白天大量喝水可以帮助排出体内的钠,导致夜间小便减少。
◣ 高盐饮食和心血管疾病的关系
说到高盐饮食,就会联想到高血压。
有些人可以吃含钠量很高的食物,但血压水平不会有任何有意义的变化。其他人吃同样的饮食可能会导致高血压,这是为什么呢?
可能与“盐敏感性”有关。“盐敏感性”的人,如果从低钠饮食转向高钠饮食,血压会增加。盐敏感性的潜在原因很多,从遗传到环境。
研究表明,对盐敏感的人比中度敏感的人患高血压的可能性至少高40%.
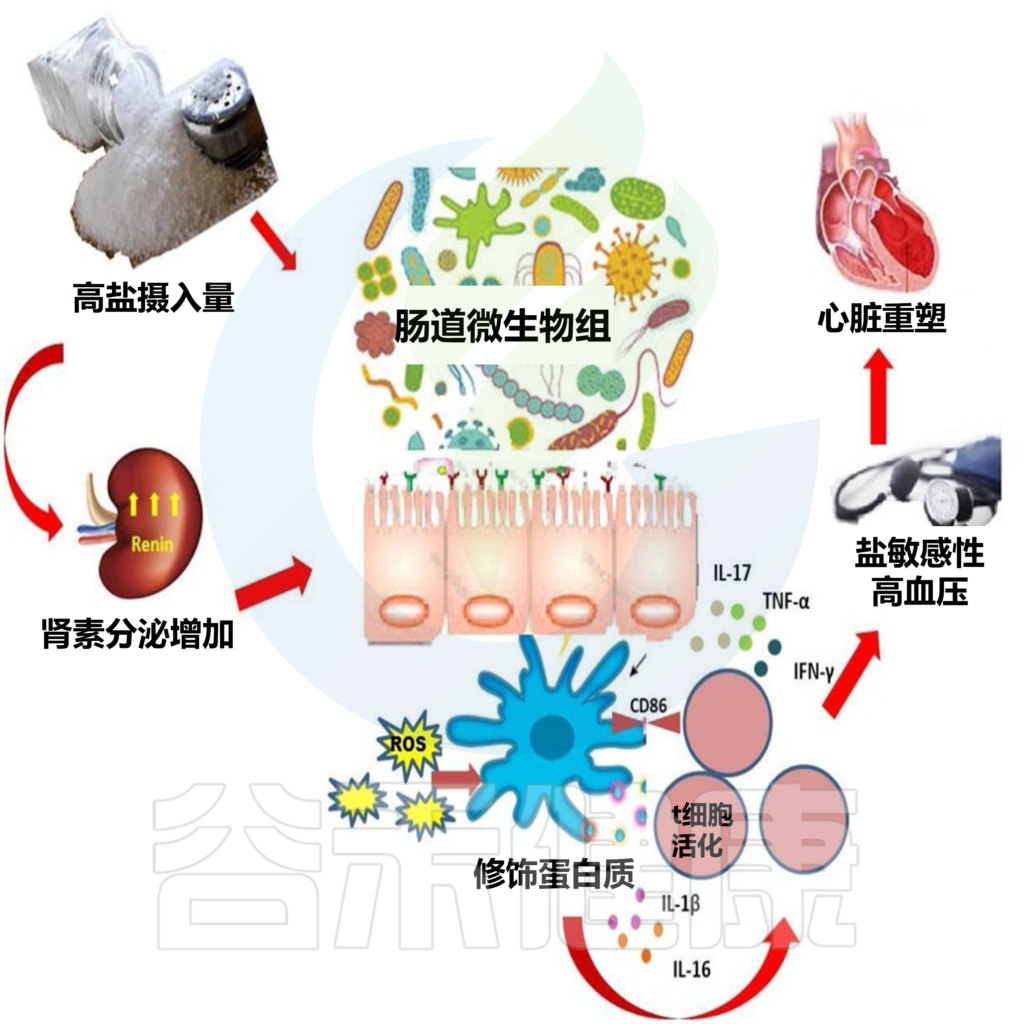
doi.org/10.1016/j.biopha.2020.111156
临床研究发现,较高的钠摄入量与心血管疾病和相关死亡有关。以下是重点研究:
研究人员测量了来自 32 个国家/地区的 10,000 多名成年人在 24 小时内排泄的钠量(这是盐摄入量的良好替代品)。平均每天接近 4,000 毫克钠。然而,范围很大,从巴西亚诺马莫人的每天 200 毫克到日本北部的 10,300 毫克。盐摄入量较高的人群平均血压较高,并且随着年龄的增长血压升高幅度较大。
两项预防高血压试验 (TOHP) 于 1987 年至 1995 年进行。他们测试了生活方式改变对血压的影响,例如减肥、压力管理、营养补充剂和摄入更少的钠。在每项研究中,在 18-36 个月内减少钠摄入后,血压略有下降。试验结束多年后,研究人员对参与者进行了调查,发现:
平均 10-15 年后,减钠组的 TOHP 参与者心脏病发作或中风的可能性降低 25%。需要手术打开或绕过胆固醇阻塞的冠状动脉,或死于心血管疾病的可能性降低。
参与者饮食中钾与钠的比例越高,患心血管疾病的机会就越低。这表明包括增加钾和降低钠的策略可能是对抗高血压的最有效方法。
◣ 高盐饮食与慢性肾病进展有关
对诊断为慢性肾病的患者进行的一项系统评价发现,每天摄入超过 4600 毫克的高钠盐与慢性肾病的进展有关。
与每天 2300 毫克的适度钠摄入量相比,每天摄入少于 2300 毫克的低钠盐没有显着影响。
一般来说,指南通常建议适度而不是低钠限制以防止慢性肾病的发展。
对于慢性肾病的整体管理,建议每日钠摄入量少于 4000 mg,对于伴有体液潴留或蛋白尿症状的慢性肾病,建议每日钠摄入量少于 3000 mg.
◣ 高盐饮食与骨质疏松症有关
身体通过排尿流失的钙量随着你摄入的盐量而增加。如果血液中钙供应不足,它会从骨骼中流失。因此,高钠饮食可能会产生额外的不良影响,即骨质疏松症。
一项对绝经后妇女的研究表明,两年内髋骨密度的下降与研究开始时24小时尿钠排泄有关,而且与骨质流失的联系与钙摄入量的联系一样强烈。
其他研究表明,减少盐的摄入量会导致钙的正平衡,这表明减少盐的摄入可以减缓随着年龄增长而发生的骨骼中钙的流失。
◣ 高盐饮食可能和胃癌相关
世界癌症研究基金会和美国癌症研究所的结论是,盐和咸的食物都是“胃癌的可能病因”。
高盐饮食通过肠道菌群影响健康
我们吃的东西和其中的盐,在某个阶段到达我们的肠道,是否会影响肠道微生物?
肠道菌群最有可能参与体内盐分的吸收,钠通过钠质子交换剂 3 (NHE3) 在结肠中被高度吸收。在小鼠 NHE-3 缺失时观察到肠道微生物环境的改变和血压的降低。
高盐消耗会影响蛋白质的消化并改变肠道菌群的多样性,菌群变化例如:棒状杆菌科的增加,乳酸杆菌的减少,详见下表:
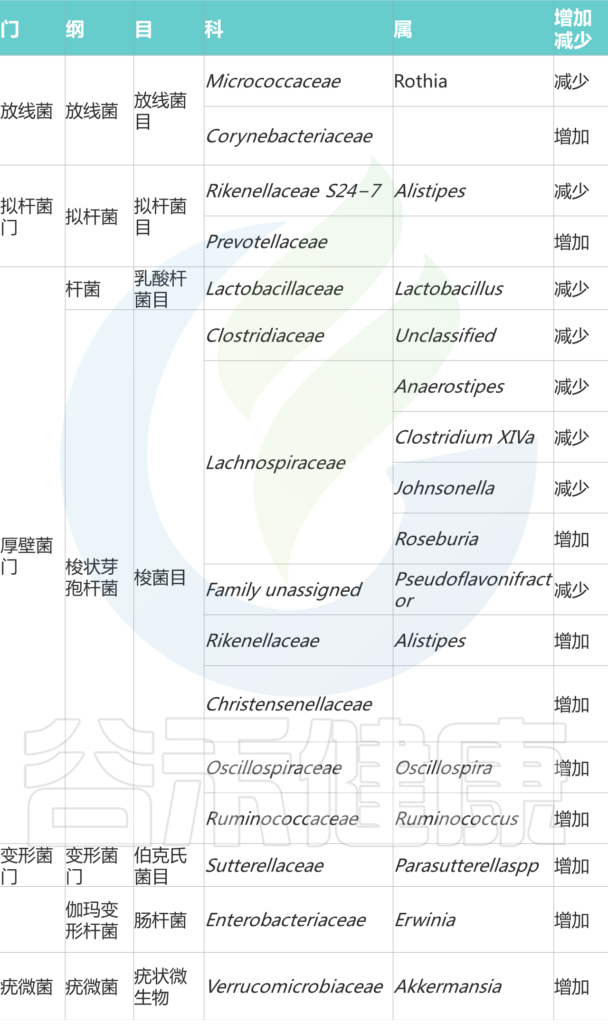
doi.org/10.1016/j.biopha.2020.111156
在大鼠模型中,食盐摄入降低了鼠乳杆菌的丰度,并增加了促炎性脾Th17细胞数量。
反过来想,作为一种益生菌疗法,每天服用鼠乳杆菌可显著减少治疗大鼠的Th17细胞并改善血压。
另一项研究发现,8周的高盐摄入显著改变了小鼠的肠道微生物组成。结果显示,拟杆菌和变形杆菌分别显著减少50.53%和2.96%,厚壁菌显著增加42.77%。
同时,在高盐饮食喂养的小鼠中,发现下肠中的短链脂肪酸水平显著降低,这归因于细菌发酵受到抑制。
饮食中钠的适度减少可以增加循环中的短链脂肪酸,从而增加肠道微生物群。短链脂肪酸水平的增加反过来降低血压并改善动脉顺应性。
以上是高盐饮食带来的危害,然而最近的研究表明高盐饮食也有可能带来某些益处。
▸ 高盐饮食:介导NK细胞和肠道微生物群之间的相互作用,诱导有效的肿瘤免疫
发表在《Science Advances》的一项研究报道了高盐饮食通过抑制PD-1表达,同时增强IFNγ和血清马尿酸水平,诱导自然杀伤(NK)细胞介导的肿瘤免疫。盐与次优剂量的抗PD1抗体联合使用可增强肿瘤免疫。
虽然高盐饮食诱导的肿瘤免疫随着肠道微生物群的减少而减弱,但高盐饮食小鼠的粪便微生物群移植恢复了与NK细胞功能相关的肿瘤免疫。
高盐饮食增加了双歧杆菌的数量,并导致肠道通透性增加,导致双歧杆菌在肿瘤内定位,从而增强NK细胞功能和肿瘤消退。瘤内注射双歧杆菌激活NK细胞,抑制肿瘤生长。
这些结果表明,高盐饮食通过潜在的平移作用调节肠道微生物组,诱导NK细胞依赖性肿瘤免疫。
▸ 高盐饮食:抵消高米饭饮食带来的肥胖影响
一项研究发现,高盐摄入可改善与大米饮食相关的代谢变化,包括粪便微生物群组成的变化。
研究将小鼠分为三组(n = 9),分别喂食正常饮食(ND)、高米饭饮食(HRD)、补充高盐(HRS)的高米饭饮食12周。
与正常饮食的小鼠相比,喂食高米饭饮食的小鼠厚壁菌门与拟杆菌门的比率(p < 0.01)和变形菌门与拟杆菌门的比率(p <0.001)显着增加。然而,高盐摄入减弱了这些影响,尽管变形杆菌的比例没有减少。
高盐摄入降低了高米饭饮食引起的体质量和白色脂肪组织重量的增加。另外,高盐饮食并没有逆转葡萄糖耐量和胰岛素抵抗的增加。
高盐饮食改变了高米饭诱导的微生物组成,高盐饮食调节高米饭饮食诱导的PPAR-γ和脂质代谢相关蛋白表达的增加。
注:PPAR-γ——过氧化物酶体增殖物激活受体-γ
此外,在白色脂肪组织中,高盐饮食可以逆转高米饭饮食诱导的脂联素的减少和PPAR-γ表达的增加。在体外,高氯化钠浓度也显著降低了3T3-L1细胞的分化和调节脂质代谢,而不会引起细胞毒性。
▸ 适度增加盐摄入量对中枢神经系统自身免疫病具有多方面和潜在的有益影响
研究人员用自发 EAE 小鼠模型评估了高盐饮食对启动中枢神经系统自身免疫所必需的早期致病事件的影响。
高盐饮食消耗增加了糖皮质激素皮质酮的循环血清水平。皮质酮增强了脑内皮细胞上紧密连接分子的表达,促进了血脑屏障 (BBB) 的收紧,从而控制了炎症性 T 细胞进入中枢神经系统。
注:需要进一步的研究来证实以上结果,以确保高盐摄入不会加剧高碳水化合物饮食引起的代谢紊乱。
以上高盐饮食带来的益处尚处于动物模型研究阶段,不宜擅自应用在个人健康。
调整烹调方法减少食盐摄入量应当注意烹饪时少放盐 , 控制烹调时和餐桌上的用盐,一家3口每日用盐不宜超过 250 克 , 也就是全家每日总共用一小汤勺(约8克 )。
大多数新鲜水果和蔬菜天然含钠量低。鲜肉比午餐肉、培根、热狗、香肠和火腿含钠量低。
习惯咸味的人, 为满足口感的需要,可在烹制菜肴肘放少许醋,提高菜肴的鲜香味,帮助自己适应少盐食物。
或者可以考虑有些时候用无盐香草和香料代替,例如迷迭香、百里香、大蒜粉,辣椒粉等。
还要注意减少酱菜 、 腌制食品以及其他咸食品的摄入量。
从燕麦粥到汤面到土豆,一切都可以以“速食”的形式获得。通常,这些速食食品比非速食食品含盐量高得多。例如,普通即食燕麦片含有近200毫克的盐。
例如,添加了盐的冷冻蔬菜可以在蒸或煮之前用漏勺冲洗一下。这个简单的步骤可以减少高达23%的盐量。
糖或甜味食物无处不在,很多人喜欢吃糖或者甜味食物,它在许多人心中占据重要的位置,甜食可能会唤起我们的舒适和美好的回忆。
然而需要注意的是,它让人上瘾的同时,可能会喂养有害菌,有害菌大量增殖的同时有益菌也在减少……
糖,正在悄悄地破坏着肠道菌群原有的平衡。

研究表明腹泻和某些糖的消化不良有关,患有乳糜泻、克罗恩病和慢性腹泻的人在肠道中产生异常高量的粘液,这阻碍了消化,阻止了这些淀粉和二糖的吸收。
糖摄入可能会扰乱微生物群的平衡,增加促炎性,降低调节上皮完整性和粘膜免疫的能力。
大量摄入添加糖和加工食品,包括典型的西方饮食,都会损害有益菌,使我们更容易受到有害物质的影响。
肠道内层很薄,没有肠道内有益菌产生的保护性粘膜层,一些东西会渗入血液中,这就会导致炎症。
也就是说,持续摄入过量糖可能会带来肠漏、慢性炎症增加和各种健康问题的风险。
身体无法分解和吸收的多余糖分将留在肠道中发酵。这种糖通过大肠的速度更慢,喂养了有害细菌和酵母,并导致气体的积累。这种气体会导致抽筋、痉挛和疼痛。
乳糖是牛奶中的糖分。当身体不能产生分解乳糖所需的酶时,就会发生乳糖不耐症,导致胀气、腹胀和其他消化不适。
同样,高果糖玉米糖浆会抑制消化,因为身体也不能分解它。果糖停留在肠道中,导致胃肠胀气和不适。
虽然吸收水分是大肠的主要工作,但糖可以将水分吸入大肠,或者至少阻止水分被正常吸收。这可能会导致腹胀或肠道沉重。
消化是由肝脏刺激的。果糖只能由肝脏处理,所以摄入的所有果糖都被一次性送到肝脏,使其超载并造成潜在的损害,进而损害消化。
果糖无法刺激胰岛素,而胰岛素又无法抑制“饥饿激素”。结果,饱腹荷尔蒙瘦素也没有被刺激,这导致吃得更多。
这就容易导致体重增加、腹部肥胖、胆固醇升高、血糖升高等各种代谢疾病。
而这些代谢疾病的发生离不开肠道菌群的运作,接下来我们来了解一下,糖是如何通过影响肠道菌群,从而引发代谢疾病的?
糖通过影响肠道菌群引发代谢疾病的机制
一项新的研究发表在8月29日在线出版的《细胞》杂志上,科学家们发现:
糖可能会破坏肠道菌群,从而耗尽关键的免疫细胞,导致肥胖等代谢综合征。
简单来说高糖饮食带来的影响如下:
高糖饮食
↓↓↓
刺激Faecalibaculum rodentium 的生长
↓↓↓
挤走了特定菌群SFB, SFB丢失
↓↓↓
改变了吸收膳食脂肪的方式
↓↓↓
出现“代谢综合征”的特征
注:SFB是小鼠肠道中的特定细菌,即分段丝状细菌。
我们来看下研究过程:
研究人员给小鼠喂食含蔗糖和麦芽糊精的高糖饮食,高糖饮食刺激了Faecalibaculum rodentium的生长,这种细菌基本上挤走了SFB。SFB的突然丢失在小鼠肠道内引发了连锁反应,最终改变了动物吸收膳食脂肪的方式。
吸收脂肪的改变导致小鼠肥胖,并出现“代谢综合征”的特征。代谢综合征如高血压、高血糖和胰岛素抵抗,共同增加了患心脏病、中风和2型糖尿病的风险。
也就是说:SFB在某种程度上可以防止代谢综合征和过度体重增加。那么,SFB是如何发挥作用的?
研究发现,随着小鼠逐渐失去SFB,它们的Th17细胞总数也下降,体重增加,出现胰岛素抵抗和葡萄糖不耐症等代谢综合征的症状。
这里不得不提一下,该过程中重要的细胞——Th17.
SFB会与免疫系统进行“对话”,促进产生一种特定类型的免疫细胞——Th17。
可以这么说,Th17细胞是一种保护小鼠免受代谢疾病影响的“盔甲”。
这些免疫细胞释放影响肠壁的蛋白质,防止多余脂肪被组织吸收进入血液。这些免疫细胞产生的分子可以减缓肠道对‘坏’脂质的吸收,并减少肠道炎症。也就是说,它们保持肠道健康,保护身体不吸收致病的脂质。
当喂食高脂肪、高糖饮食时,小鼠肠道中的Th17细胞发生了什么变化?
小鼠迅速发展出代谢疾病的几个特征(体重增加、葡萄糖不耐受),并显示出肠道Th17细胞的减少。
更具体地说,研究人员发现Th17水平的下降是由于饮食诱导的肠道微生物群的变化。已知促进Th17的细菌被其他种类的肠道细菌所取代。尤其是糖似乎增加了有害菌的数量,最终降低了Th17的水平。
也就是说,糖通过干扰微生物组分间接破坏了这种盔甲。
研究人员认为,只要动物保持高水平的诱导Th17的肠道细菌,高脂饮食就不会导致负面的代谢效应。
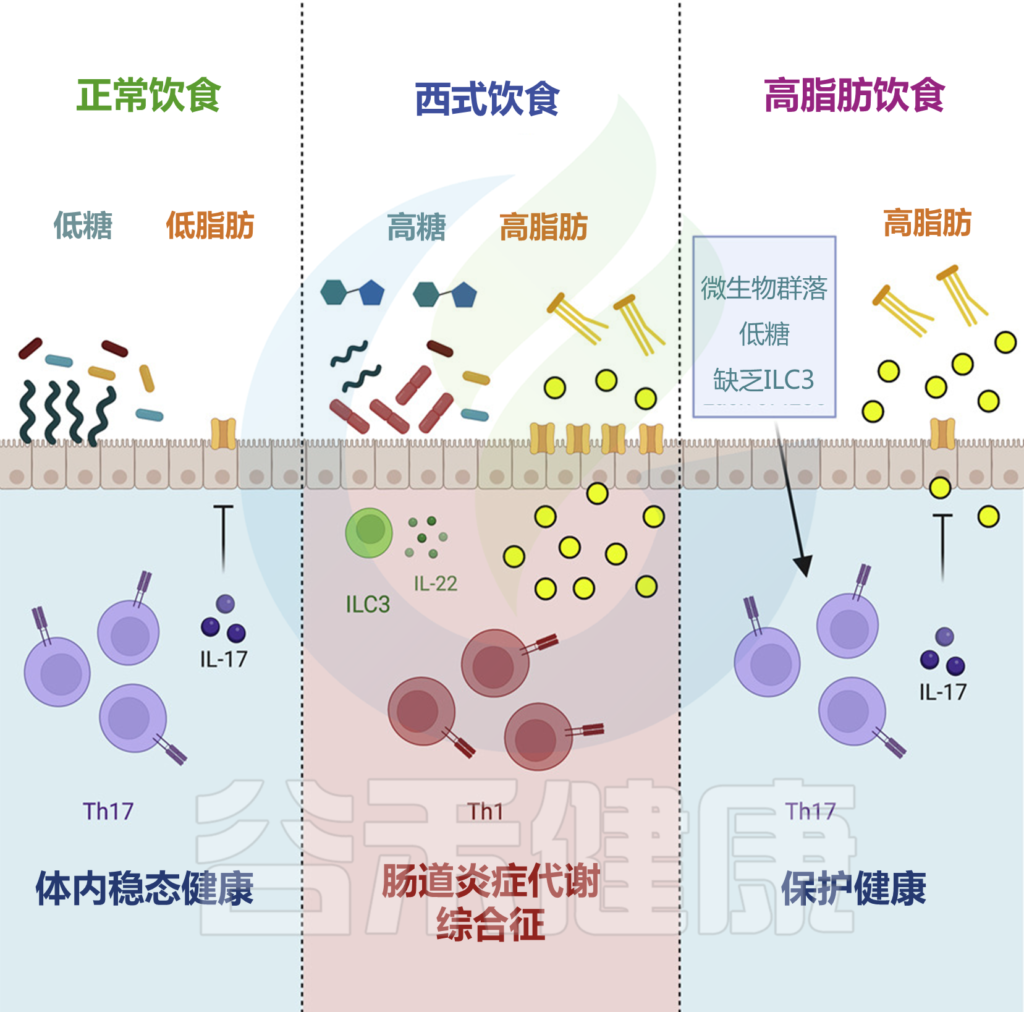
doi.org/10.1016/j.cell.2022.08.005
如果没有健康的肠道菌群,减糖也不能避免疾病
在另一个实验中,研究小组从一组小鼠中去除了SFB,然后喂它们无糖、高脂肪的饮食,发现这些小鼠尽管没有吃糖,但体重也增加了,并患有代谢疾病。那么,是什么导致体重增加呢?
本质上,如果没有健康的肠道菌群,小鼠就不会产生足够的Th17细胞,因此也就缺乏前面所说的“盔甲”。
研究小组发现可以通过两种方式提供这种盔甲:
这表明,如果小鼠的肠道中已经耗尽了SFB,那么减少糖的摄入并不能帮助小鼠避免代谢疾病。
如果把这一发现放到人类身上,那就意味着如果一个人的肠道微生物群已经被破坏,那么减少摄入糖不一定会有帮助。
我们可以想象,一些流行的饮食干预,如尽量减少糖,可能只对微生物群中有某些细菌种群的人有效。
研究人员说,可能需要额外的干预措施来恢复这些人的肠道细菌或Th17细胞。
当然也需要更多的研究来了解类似的机制是否在人体肠道中起作用。
通过以上研究,我们可以大致了解糖对肠道菌群的影响,及带来的危害,因此,一定程度上减少糖的摄入会带来好处。
重新控制饮食习惯,试着对糖说不
当对糖强烈的渴望袭来时,试着忽略它,深呼吸几次,然后问自己:

意识到微生物只是利用你来喂养它自己,是很好的第一步,也就是重新控制饮食习惯。
当你减少糖的摄入量时,这些以糖为食的微生物会进入饥饿模式,与此同时,你对糖的渴望会变得更加强烈,可能会“说服自己”去得到它。

编辑
识别糖的触发因素
有时候会在特定的情况下渴望吃糖。
想想上一次吃甜食是什么时候,是感到有压力吗?疲劳?抑郁?焦虑?压抑?紧张?……
可以把这些糖分触发因素想象成某个按钮。这个按钮一触碰就想吃甜食类食物,每个人的按钮可能不一样,有些人是焦虑,有些人是疲劳…
了解属于你自己的糖分触发因素,会帮助你在渴望到来时安然度过。有意识地关注你的渴望,是减少 对甜食不受控的一个强有力的方法。
喂养健康的肠道细菌
相对较为健康的做法是多吃真正的食物,并在饮食中加入更多的植物,来自植物的益生元可以喂养我们的微生物群,例如:
菊苣、朝鲜蓟、蒲公英嫩叶、芦笋、大蒜、洋葱、苹果、浆果类、香蕉、菠菜、羽衣甘蓝、胡萝卜、番茄、山药、黄瓜、芝麻菜等。
酸奶、泡菜、味噌、开菲尔、康普茶等。
尽可能避免加工和包装食品、含糖食物、酗酒等。
养成良好的生活方式
研究表明,如果你在吃饭时看电视或刷手机看各类小视频等,可能会吃得更快,嚼得更少,吃得更多。
吃饭时,试着把手机放在一边,关掉电视,有意识地彻底咀嚼每一口食物,并真正品尝味道,可能会吃的更少,消化更好。
看到一些新闻或对即将截止的工作任务感到压力,你会很难从膳食中吸收营养。因此,坐下来吃饭之前,试着深呼吸让自己平静下来。
其他还包括:适当运动、规律作息、保持充足的睡眠、适量沐浴阳光,保持良好的心情等方式。
扩展阅读:
菌群多样性是如何形成的,与健康的关系,如何改善?
20种有效改善肠道健康的科学方法
此外,减少糖的摄入并不意味着转向人工甜味剂。经常食用人工甜味剂可能会对微生物群产生负面影响,导致代谢紊乱和肥胖。
扩展阅读:
你的焦虑可能与食品添加剂有关,警惕食品添加剂引起的微生物群变化
现代人的嗜辣程度日益走高,很多人已经到了无辣不欢的境界。
「辣」准确地说并不是一种味觉,可以说是痛觉,是舌、口腔和鼻腔粘膜受到刺激产生的辛辣、刺痛、灼热的感觉。
「辣」让人一边痛不欲生,一边欲罢不能。
「辣」对身体的影响褒贬不一。有人说吃辣上火,腹泻,长痘痘;有人说吃辣抗氧化,助消化,降胆固醇…

辣椒素的吸收部位:
辣椒素在胃和小肠上部被被动吸收的效率超过80%。
吃辣的健康益处
• 支持正常的血液循环,有利于心血管健康,以及降低患心脏病的风险。
• 促进新陈代谢和脂肪燃烧。一些研究表明,随着时间的推移,食用辛辣食物与体重增加较少之间存在联系,因为辛辣香料可能会增加卡路里燃烧。
• 增强消化健康,支持肠道微生物群中的有益菌生长。
• 提供抗氧化剂,包括抗氧化应激的酚类化合物。
• 减少炎症,包括胃肠道,以及影响关节和动脉的炎症。
• 通过表现出抗菌活性来支持免疫功能。
• 可能降低慢性病风险,包括高血压、高低密度脂蛋白胆固醇、二型糖尿病、心脏病和中风。
• 降低某些癌症的风险,包括食道癌和直肠癌等。
• 潜在地降低了过早死亡的风险。在一项研究中,与每周吃少于一次辛辣食物的成年人相比,那些每周吃六到七天辛辣食物的人受益于降低14%的风险总死亡率。
……
看到这里全是吃辣的益处,是不是想赶紧吃吃吃,先别急,这也要看辣的程度。
辣椒素(CAP)是辣椒的主要辛辣成分。
接下来我们来了解一下,不同剂量的辣椒素引起的肠道菌群变化,以及肠道菌群在辣椒素对抗疾病(肥胖,糖尿病等)中的作用。
一项研究评估了辣椒素对胃肠道健康的影响,并研究辣椒素是否调节短链脂肪酸和肠道微生物群的组成。
以40、60和80 mg/kg的剂量给小鼠施用辣椒素。
高剂量的辣椒素会损害胃肠道组织
对照组小鼠的结肠组织显示出丰富的杯状细胞和具有整齐绒毛的健康隐窝结构;
而辣椒素干预组小鼠显示出炎性细胞浸润,以及隐窝和杯状细胞的丧失。
与对照组中的小鼠相比,用60 mg/kg 辣椒素处理的小鼠显示出产生粘液的杯状细胞的损失;
在80 mg/kg 辣椒素处理的小鼠中,炎症细胞浸润明显。
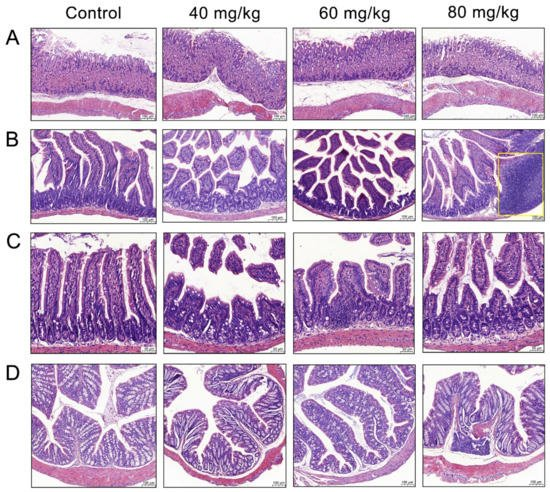
doi.org/10.3390/foods11050686
辣椒素引起的炎症反应,与剂量有关
辣椒素影响胃和回肠中的抗炎细胞因子水平,但不会导致严重的炎症损伤
炎症反应与胃肠道损伤密切相关。研究表明,辣椒素诱导小鼠胃肠道炎症的主要特征是炎症细胞因子水平升高,尤其是IL-10、IL-1β和TNF-α。
注:IL-10是一种关键的细胞因子,可以减少炎症介质的释放,并显示抗炎特性。
IL-1β和TNF-α是引起粘膜炎症和肠屏障损伤的重要促炎细胞因子。
高剂量辣椒素可能导致空肠和结肠的炎症损伤
结果表明,80 mg/kg 辣椒素干预组可导致所有胃肠组织中IL-10水平降低,空肠和结肠中IL-1β和TNF-α水平升高。
多样性下降
与对照组相比,40 mg/kg 辣椒素组的香浓指数显著下降(p<0.001),Chao1指数略有下降。同时,与对照组相比,60 mg/kg辣椒素组显著降低了Chao1指数。
使用加权UniFrac距离的PCoA测量β多样性,揭示了对照组和不同剂量辣椒素组之间的不同菌群。
在门的水平上,对照组小鼠中:
与对照组相比,40 mg/kg 辣椒素干预显著降低了拟杆菌的丰度至34.17%,但增加了厚壁菌的丰度(56.64%)(p<0.0001)。
80 mg/kg的辣椒素干预显著降低了拟杆菌的相对丰度,增加了放线菌和变形杆菌的丰度。
变形杆菌的存在是肠道微生物群稳态失衡的标志,与腹泻症状和炎症密切相关。
辣椒组的优势细菌标记物
对照组、40、60和 80mg/kg 辣椒素干预组中分别发现了优势细菌标记物。
差异最大的属包括丁酸杆菌属、乳杆菌属、粪杆菌属、科氏杆菌属_UCG_002、双歧杆菌属、Rikenellaceae_RC9_肠组、拟杆菌属、Alistites属和Dubosiella属。
高剂量增加了:双歧杆菌和粪杆菌的比例
对具有显著差异的选定属的相对丰度进行了分析,表明辣椒素处理以剂量依赖的方式增加了双歧杆菌和粪杆菌的比例,但仅在80 mg/kg 辣椒素干预组中显示出显著变化。
Faecalibacterium被认为是胃肠道疾病的生物指示剂,并与丁酸生成呈正相关。这可能是60和80 mg/kg 辣椒素组小鼠盲肠丁酸水平显著升高的原因。
在辣椒素干预组中,尤其是在40 mg/kg 辣椒素干预的组中,乳酸杆菌和Alistites的相对丰度显著降低。
与对照组相比,40 mg/kg 辣椒素干预显著提高了Dubosiella的比例,但减少了拟杆菌、丁酸单胞菌和Rikenellaceae_RC9_gut_group的丰度。
注:有研究曾报道,Dubosiella可以抑制小鼠的肥胖。
80 mg/kg 辣椒素干预后:Coriobacteriaceae_UCG_002的丰度增加。
注:Coriobacteriaceae_UCG_002可以通过产生必需氨基酸和发酵膳食蛋白而对宿主有利。
辣椒素的抗肥胖作用
辣椒素已被证明能够引起饱腹感,减少热量摄入,增加能量消耗,并增强脂肪氧化,这反过来可能导致体重减轻。
厚壁菌/拟杆菌 ↑↑↑↑
大量研究表明,在以辣椒素作为补充的高脂肪诱导小鼠模型中,厚壁菌/拟杆菌的比率会更高。
发现肠道菌群失调可减少拟杆菌,并增强在肥胖人类和动物肠道中观察到的分泌革兰氏阴性病原体的厚壁菌和脂多糖。
AKK菌 ↑↑↑
最近,还研究了辣椒素的抗肥胖作用,与肠道微生物群的变化、喂食高脂肪辣椒素小鼠中变形菌门的减少以及对宿主新陈代谢有益的粘液降解细菌Akkermansia muciniphila 的高丰度有关。
Faecalibacteria ↑↑↑
辣椒素对肠道和微生物群有消炎作用。辣椒素可以增加Faecalibacteria,从而有助于防止肥胖,调节血糖水平,防止肠道炎症。
产丁酸菌 ↑↑↑
此外,饮食中的辣椒素可以诱导产生丁酸盐的瘤胃球菌科和拉氏螺旋菌科的水平增加,但也可以刺激盲肠产丁酸细菌和丁酸盐水平的升高,以抑制结肠CB1受体,并减少LPS的生物合成。
注:丁酸盐的好处:保护肠道内壁,有助于肠漏的愈合,保护大脑和神经系统,它能增强免疫系统,更有效地对抗感染。
刺激微生物群减少饥饿激素——饥饿素
胃内产生的一种肽被称为“饥饿激素”.它是肠道在肠道微生物的帮助下产生的一种激素。它不仅能影响食欲,还能促进身体储存脂肪的能力。
辣椒素通过刺激微生物群向身体发送信号,告诉它减少饥饿素,从而帮助你控制饥饿。
综上所述,辣椒素是一种有效的抗肥胖化合物。补充后,它会激活肠道内的某些受体,称为TRPV1受体。一旦打开,这些受体向身体发送信号,告诉身体增加肠道菌群Akkermansia muciniphila。更高比例的Akk菌促进减肥和调节血糖水平,因此有助于控制糖尿病和肥胖症。
辣椒素抗糖尿病作用
大量体内和体外研究表明,辣椒素在改善葡萄糖代谢方面发挥着重要作用。早些时候已经证明,在糖尿病大鼠中,系统性辣椒素激活可导致辣椒素敏感细胞变性和葡萄糖诱发胰岛素分泌的长期变化。
补充辣椒素(100 mg/kg)的雄性肥胖Zucker大鼠的血浆CGRP水平升高,同时通过辣椒素诱导的感觉神经脱敏改善糖耐量。
新生辣椒素治疗SD大鼠(50 mg/kg)胰岛素介导的糖代谢增加,通过辣椒素诱导的含有神经肽的感觉神经增强体内胰岛素敏感性。
患有糖尿病的雄性Wistar大鼠以1 mg/kg·天的剂量长期服用辣椒素8周,表明辣椒素具有利尿作用,并增加了尿液表皮生长因子水平。含有高酚和辣椒素含量的红辣椒茎(9.7 mg/g,DW)具有较强的α-淀粉酶和α-葡萄糖苷酶抑制作用。
所有这些发现表明,辣椒素敏感结构一定参与了调节胰岛素分泌和血糖。
一项随机双盲临床试验表明,含有辣椒素的辣椒补充剂(5 mg/d辣椒素)定期改善妊娠期糖尿病妇女的餐后高血糖和高胰岛素血症以及空腹脂代谢紊乱。
膳食辣椒素通过对肠道微生物群的调节作用影响葡萄糖稳态和肥胖的拟议途径
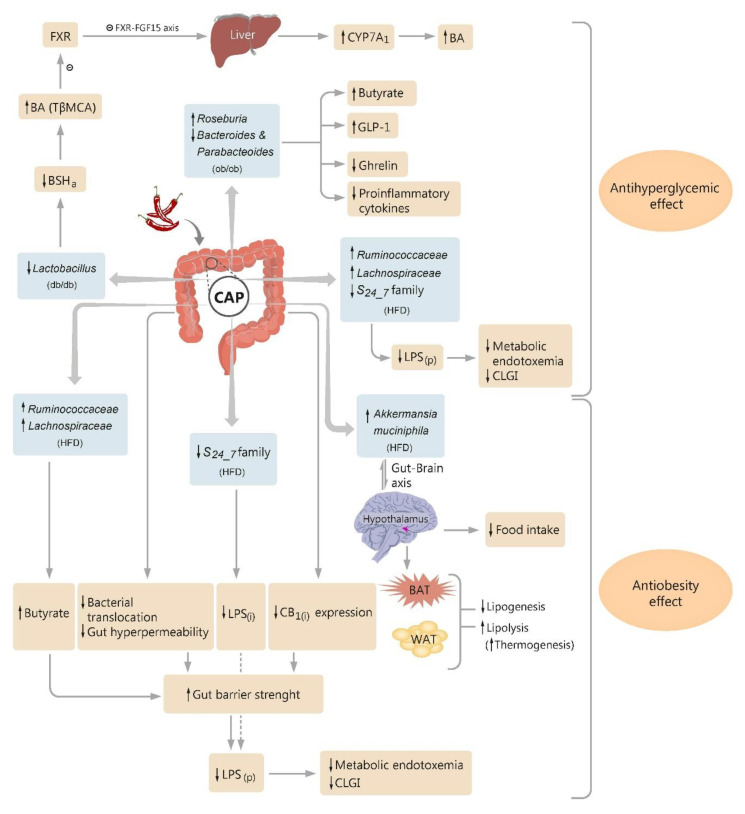
doi: 10.3390/molecules25235681
辣椒素可降低2型糖尿病小鼠体内乳酸杆菌的丰度(db/db),从而降低胆汁盐水解酶活性(BSHa),增加肠道中结合胆汁酸(BA)的水平,尤其是法尼样X受体(FXR)拮抗剂牛磺酸-β-鼠胆酸(TβMCA)。FXR信号发生改变,肠肝FXR-FGF15轴(FGF15成纤维细胞生长因子15)也受到抑制,导致胆固醇7α-羟化酶(CYP7A1)表达上调,肝BA合成增强。在肥胖糖尿病小鼠中,辣椒素增加Roseburia并抑制拟杆菌和副杆菌的丰度,随后粪便丁酸水平和血浆胰高血糖素样肽-1(GLP-1)增加,血浆总ghrelin和促炎细胞因子减少。
辣椒素通过调节肠-脑(下丘脑)轴,最终针对棕色脂肪组织、白色脂肪组织和小鼠食物摄入量,在高脂饮食喂养的小鼠中发挥抗肥胖作用。辣椒素减少了能够分泌LPS(i)(肠道细菌脂多糖)的革兰氏阴性病原体的数量,如S24_7科成员,并增加了高脂饮食小鼠中产丁酸菌的数量(例如,瘤胃球菌科和Lachnospiraceae),从而增加了粪便丁酸盐。辣椒素可减弱高脂饮食小鼠肠道通透性增加和细菌移位,并抑制肠道大麻素受体1型(CB1(i))的表达。
通过这些途径,辣椒素增加了这些肥胖小鼠的肠道屏障强度,同时减少了肠道菌群改变所产生的高水平LPS(i),从而降低了高水平血浆循环LPS.
TRPV1通道在感觉异常中起着核心作用,并在糖尿病动物模型中显示出高表达水平。
“恶性循环假说”指出,肥胖期间TRPV1对感觉神经的激活可能导致持续的神经肽物质P(SP)和CGRP释放,从而阻止胰岛素介导的葡萄糖摄取,最终导致代偿性高胰岛素血症(下图)。因为CGRP可以减少胰岛素分泌,而SP可以诱导胰岛素抵抗,它们会引发一个恶性循环,从而导致2型糖尿病发病。
而辣椒素作为一种TRPV1激动剂,可能会打破这种恶性循环,并有可能改善胰岛素分泌和胰岛素敏感性。
辣椒素抗肥胖、抗糖尿病和抗高血压的可能机制
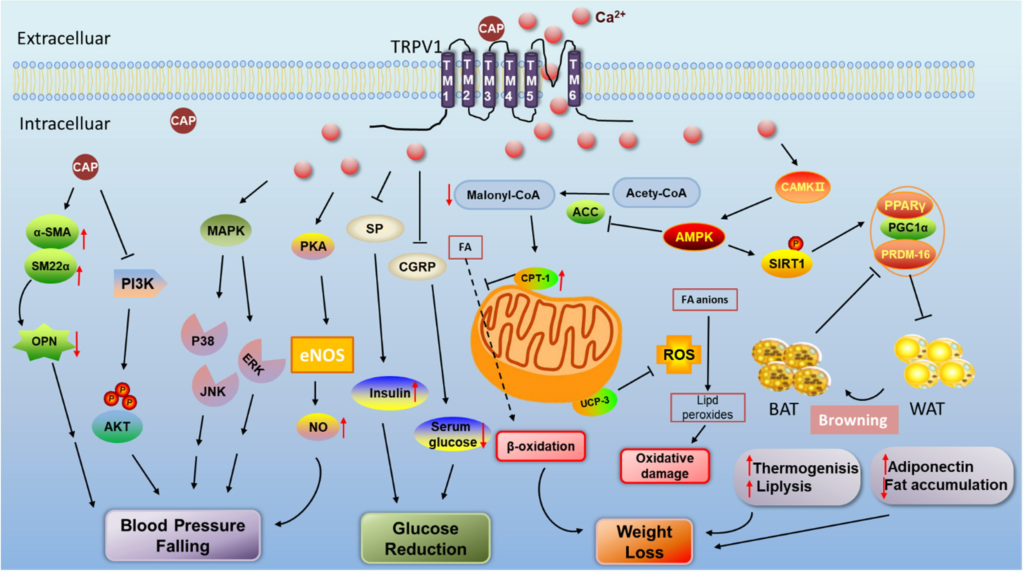
doi.org/10.1080/10408398.2021.1884840
抗肥胖机制:
通过磷酸化激活AMPK抑制ACC,抑制ACC降低丙二酰辅酶a浓度,导致CPT-1抑制解除,脂肪酸(FA)氧化增加,减少肌肉内脂质堆积。此外,辣椒素(CAP)没有增加HFD喂养的动物中UCP3的表达,但增加了正常肌肉细胞中的表达,因此影响产热。此外,CAP可能增加PRDM-16的表达,并促进其与PPARγ的相互作用,以及增加PGC-1α的表达来触发BAT分化和WAT褐变,从而增加产热和能量消耗来对抗肥胖。
抗糖尿病机制:
TRPV1在感觉神经上的激活可能导致神经肽P物质(SP)和降钙素基因相关肽(CGRP)的持续释放,从而阻断胰岛素介导的葡萄糖摄取。
抗高血压机制:
CAP诱导TRPV1激活Ca2+内流和PKA介导的内皮一氧化氮合酶(eNOS)磷酸化。此外,通过CAP激活TRPV1,通过增加α-平滑肌肌动蛋白(α-SMA)和SM22α的表达,减少骨桥蛋白(OPN)的表达,抑制PI3K/Akt信号通路,抑制高血压期间血管平滑肌细胞(VSMC)的表型转化,从而减轻颅内小动脉重塑。
辣椒素抗高血压作用
几项动物研究显示了辣椒素或辣椒摄入量与高血压之间的密切关系。
辣椒素对体内血压影响的关键研究综述

doi.org/10.1080/10408398.2021.1884840
一项包含9273名健康成年人的横断面研究表明,女性参与者中高频率的辛辣食物消费与高血压的低风险显著相关,但男性参与者中没有。
在1991年至2011年的中国健康与营养调查中,一项对13670名20-75岁成年人的队列研究表明,每1000人中不食用辣椒或每天食用1-20、20.1-50、> 50.1克辣椒的高血压发病率分别为30.5、33.4、31.9和24.0,表明辣椒食用量与高血压风险呈负相关。
辣椒素抑制高血压的机制可能涉及一些关键酶和不同的信号通路。
包括通过TRPV1激活、PKA活性激活和eNOS磷酸化增加以及p38/MAPK途径释放血管舒张神经肽。此外,抑制PI3K/AKT途径,刺激尿钠排泄和利尿,ACE抑制活性和L型钙2+平滑肌细胞中的通道抑制也涉及抗高血压机制。
因此,食用辣椒素可能是一种潜在的抗高血压干预手段。
辣椒素在炎症性肠病中的作用
克罗恩病患者的回肠粘膜碎片显示,Faecalibacterium prausnitzii的丰度较低,这与6个月后内镜下复发有关。众所周知,Faecalibacterium prausnitzii具有抗炎特性,在外周血单核细胞培养物和结肠炎动物模型中,分别减少促炎细胞因子的产生,并增加抗炎细胞因子IL-10的分泌。
扩展阅读:
肠道核心菌属——普拉梭菌(F. Prausnitzii),预防炎症的下一代益生菌
富含 CAP 的饮食可能对克罗恩病产生有益的影响,因为它们会增加厚壁菌门/拟杆菌门的比例和粪杆菌的丰度,从而将免疫平衡改变为对食物抗原和共生细菌更具耐受性的状态。
辣椒素抗癌症作用
体外和体内研究表明辣椒素对不同种类的癌症具有抗癌作用,如胃癌、结肠癌、前列腺癌、胰腺癌、肺癌、乳腺癌、膀胱癌等。
辣椒素潜在的抗癌机制可能与其对肿瘤细胞凋亡、自噬和转移的影响有关。
辣椒素通过诱导凋亡和抑制血管生成来抑制各种永生化或恶性细胞系的生长。
辣椒素可以通过调节其广泛的分子靶标来调节细胞增殖和凋亡,这些分子靶标包括转录因子、生长因子及其受体、细胞因子、酶和基因。
除了细胞凋亡之外,辣椒素在防止体液细胞转移方面也起着重要的作用。
体内研究评估辣椒素在各种癌症中的抗癌机制

doi.org/10.1080/10408398.2021.1884840
总之,辣椒素的抗癌机制如下图,包括细胞凋亡的激活、细胞生长停滞、细胞自噬以及血管生成和转移的抑制。
辣椒素可能的抗癌作用机制:
因此,辣椒素有可能成为一种预防和治疗癌症的新疗法。
辣椒素抗癌的关键机制
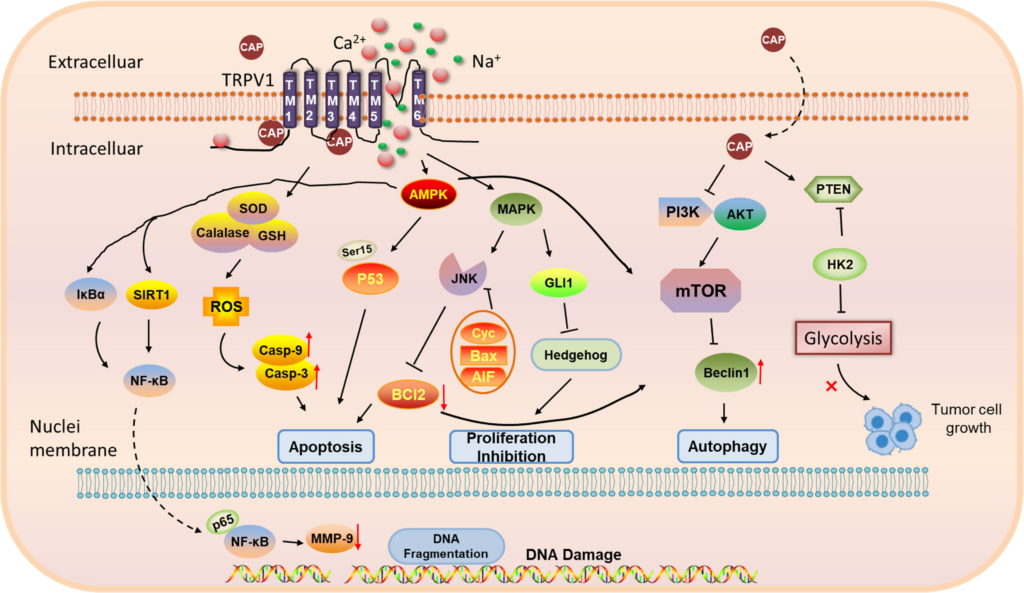
doi.org/10.1080/10408398.2021.1884840
CAP通过抑制Akt/mTOR途径抑制Akt磷酸化并诱导自噬。此外,CAP可以增加磷酸酶和张力蛋白同源物(PTEN)的表达,导致己糖激酶-2(hk2)表达的减少,从而抑制肿瘤细胞糖酵解。此外,TPPV1激活了Ca2+内流可以激活MAPK,进而阻断Hedgehog通路,抑制细胞增殖。此外,细胞内GSH水平的降低可能导致ROS的增加,进而激活线粒体死亡途径。CAP可能上调促凋亡基因,包括Cyc、AIF、Bax和裂解的caspase-3和-9,同时下调抗凋亡基因BCl2。P38和JNK MAPK通路以及AMPK/p53通路的激活也参与诱导细胞周期阻滞和凋亡。此外,CAP通过AMPK-SIRT1和AMPK-IκBα信号通路抑制NF-κB p65,从而抑制肿瘤细胞的迁移和侵袭,并引起基质金属蛋白酶-9 (MMP-9)的下调。
辣椒素的认知干预作用
在动物研究中,辣椒素在认知功能中的作用是有争议的。一些研究表明辣椒素具有神经毒性。
然而,辣椒素也被证明对认知功能或老年痴呆症有积极作用。例如,已经证明辣椒素对大鼠大脑中应激诱导的阿尔茨海默样变化具有预防作用。
具体而言,辣椒素可以部分减轻冷水应激诱导的大鼠空间记忆保留缺陷、LTP抑制、树突形态异常和突触相关蛋白丢失。
此外,辣椒素可以降低含辣椒素的高脂饮食喂养的SD大鼠患阿尔茨海默的风险。
关于食用辣椒与认知功能之间关系的流行病学研究有限。
目前,一项针对338名40岁以上参与者的调查表明,高水平的辣椒素饮食可能对中老年人的认知功能和AD血清Aβ水平产生有利影响,中国健康与营养调查(CHNS)在4582名中国成年人中收集了15年的数据,这些数据支持辣椒摄入量与认知功能之间存在正相关。
研究表明,与非消费者相比,那些累积平均辣椒摄入量超过50克/天的人,其总体认知功能的回归系数(和95%CI)为−1.13 (−1.71至0.54),自报记忆力差和自报记忆能力下降的比值比(和95%CI)分别为2.12(1.63–2.77)和1.56(1.23–1.97).
与食物一起摄入的辣椒素可以通过胃肠道中的非主动过程被迅速吸收。在被运输到门静脉,然后进入人和啮齿动物的全身后,约5%的未改变的辣椒素穿过血脑屏障并进入脑组织。
辣椒素受体TRPV1被证实可增加海马胰岛素信号通路,从而抑制GSK-3β,防止ad相关的tau蛋白过度磷酸化。
此外,自噬在β-淀粉样蛋白的生成和代谢中也起着重要作用,tau的组装及其功能障碍可能导致阿尔茨海默病的进展。
总之,辣椒素可以通过抑制tau蛋白过度磷酸化来减轻阿尔茨海默样神经病理改变和认知障碍,这表明它可能是一种有前途的阿尔茨海默治疗干预方法。
扩展阅读:
阿尔茨海默病de饮食-微生物-脑轴
辣椒素抗抑郁作用
膳食辣椒素可改善LPS诱导的抑郁样行为小鼠的抑郁样行为,如厌恶刺激、快感缺失和绝望等得到缓解。
辣椒素可以恢复抑郁症相关微生物群的异常变化。特别是在属水平上,辣椒素增加了某些关键微生物的相对丰度,如瘤胃球菌、普雷沃菌、 Allobaculum, Sutterella, Oscillospira.
相关分析显示,微生物群落组成的变化与抑郁行为改善、5-HT下降和TNF-α水平密切相关。
注:5-HT是一种代表性的单胺类神经递质,涉及调节几种生理活动和行为,包括与情绪和焦虑有关的活动和行为,并且低水平的5-HT已被证明与抑郁有关。
这些结果表明,膳食辣椒素可以调节肠道菌群的结构和数量,并在预防抑郁方面发挥重要作用。
▸ 健康的辛辣食物有哪些?
▸ 什么人群不适合吃辣?
虽然辛辣食物不会引起溃疡,但在部分人中也会引发腹痛。
一项研究特别强调,经常食用辛辣食物会引发一些消化不良患者的上消化道症状。对于肠易激综合征(IBS)患者,辛辣食物也会引发症状。
对于炎症性肠病(或IBD-克罗恩病或溃疡性结肠炎)患者,辛辣食物也会引发一些症状。
如果患有肛裂,可能会感觉到烧灼感。一项研究表明,辛辣食物会加重与肛裂相关的症状。
其他患胃酸倒流、胃灼热、腹泻、胃痛、怀孕期间的晨吐或恶心等人群,则需注意谨慎吃辣。
▸ 准备辣椒食物要注意什么?
准备辣椒时要戴手套,或者至少处理完后要彻底洗手。
保护眼睛和其他敏感区域。切辣椒时考虑戴眼镜。洗手前不要揉眼睛、鼻子或嘴巴等部位。
▸ 如何适应辛辣食物?
从微辣的食物开始,每周吃点辣的,舌头会慢慢习惯这种感觉,身体就像对酒精和咖啡因产生耐受性一样,慢慢也会对辣椒素产生耐受性。
▸ 一不小心吃太多辣如何缓解?
——牛奶或酸性饮料中和辣
辣的受不了的时候,可以喝点牛奶缓解,牛奶中的脂肪和蛋白质会中和食物中的香料,如果没有牛奶,可以喝冰水或酸性饮料。
普通的一杯水不会对解辣有帮助,因为水会将辣椒素扩散到嘴里,而柠檬水、橙汁或葡萄汁之类的酸性饮品都可以。
——不同质地的食物分散注意力
不同食物的质地可能会分散你对辣的注意力,吃口卷饼、饼干、面包丁之类的固体食物,它们本身其实并不能对抗辣椒素,但会给舌头一些不同的东西来关注。
——碳水化合物有助于吸收辣椒素
淀粉类碳水化合物会形成一道屏障,使辣椒素更难进入味蕾。
——吃饭的时候用嘴呼吸
每一口之间,慢慢呼气,把辣吹走,让嘴冷却下来。如果真的很痛,想象自己真的把辣椒片吹向空中,可能有助于减轻压力。
——提前吃点抗酸剂来防止胃不舒服
可以提前吃点抗酸剂或吃点小零食,比如三明治或土豆泥来填饱肚子,让辣椒素进入肠道后有所保留。注意不要过量使用抗酸剂,吃太多会扰乱胃产生酸的方式。
——记住熬过15分钟
辛辣食物的辣劲儿只需要15分钟就可以消散。如果感觉嘴巴着火一样,只要提醒自己这种感觉不会永远持续下去,不用害怕。
主要参考文献:
Naqvi S, Asar TO, Kumar V, Al-Abbasi FA, Alhayyani S, Kamal MA, Anwar F. A cross-talk between gut microbiome, salt and hypertension. Biomed Pharmacother. 2021 Feb;134:111156. doi: 10.1016/j.biopha.2020.111156. Epub 2021 Jan 2. PMID: 33401080.
Do MH, Lee HB, Oh MJ, Jhun H, Ha SK, Park HY. Consumption of salt leads to ameliorate symptoms of metabolic disorder and change of gut microbiota. Eur J Nutr. 2020 Dec;59(8):3779-3790. doi: 10.1007/s00394-020-02209-0. Epub 2020 Mar 3. PMID: 32125529.
Czesnikiewicz-Guzik M, Müller DN. Scientists on the Spot: Salt, the microbiome, and cardiovascular diseases. Cardiovasc Res. 2018 Aug 1;114(10):e72-e73. doi: 10.1093/cvr/cvy171. PMID: 30052920.
Na SY, Janakiraman M, Leliavski A, Krishnamoorthy G. High-salt diet suppresses autoimmune demyelination by regulating the blood-brain barrier permeability. Proc Natl Acad Sci U S A. 2021 Mar 23;118(12):e2025944118. doi: 10.1073/pnas.2025944118. PMID: 33723078; PMCID: PMC7999868.
National Academies of Sciences, Engineering, and Medicine; Health and Medicine Division; Food and Nutrition Board; Committee to Review the Dietary Reference Intakes for Sodium and Potassium. Dietary Reference Intakes for Sodium and Potassium. Oria M, Harrison M, Stallings VA, editors. Washington (DC): National Academies Press (US); 2019 Mar 5. PMID: 30844154.
Ma Y, He FJ, Sun Q, Yuan C, Kieneker LM, Curhan GC, MacGregor GA, Bakker SJL, Campbell NRC, Wang M, Rimm EB, Manson JE, Willett WC, Hofman A, Gansevoort RT, Cook NR, Hu FB. 24-Hour Urinary Sodium and Potassium Excretion and Cardiovascular Risk. N Engl J Med. 2022 Jan 20;386(3):252-263. doi: 10.1056/NEJMoa2109794. Epub 2021 Nov 13. PMID: 34767706; PMCID: PMC9153854.
Rizvi ZA, Dalal R, Sadhu S, Kumar Y, Kumar S, Gupta SK, Tripathy MR, Rathore DK, Awasthi A. High-salt diet mediates interplay between NK cells and gut microbiota to induce potent tumor immunity. Sci Adv. 2021 Sep 10;7(37):eabg5016. doi: 10.1126/sciadv.abg5016. Epub 2021 Sep 10. PMID: 34516769; PMCID: PMC8442882.
Kawano Y, Edwards M, Huang Y, Bilate AM, Araujo LP, Tanoue T, Atarashi K, Ladinsky MS, Reiner SL, Wang HH, Mucida D, Honda K, Ivanov II. Microbiota imbalance induced by dietary sugar disrupts immune-mediated protection from metabolic syndrome. Cell. 2022 Sep 15;185(19):3501-3519.e20. doi: 10.1016/j.cell.2022.08.005. Epub 2022 Aug 29. PMID: 36041436.
Xia J, Gu L, Guo Y, Feng H, Chen S, Jurat J, Fu W, Zhang D. Gut Microbiota Mediates the Preventive Effects of Dietary Capsaicin Against Depression-Like Behavior Induced by Lipopolysaccharide in Mice. Front Cell Infect Microbiol. 2021 Apr 27;11:627608. doi: 10.3389/fcimb.2021.627608. PMID: 33987106; PMCID: PMC8110911.
Wang F, Xue Y, Fu L, Wang Y, He M, Zhao L, Liao X. Extraction, purification, bioactivity and pharmacological effects of capsaicin: a review. Crit Rev Food Sci Nutr. 2022;62(19):5322-5348. doi: 10.1080/10408398.2021.1884840. Epub 2021 Feb 16. PMID: 33591238.
Xiang Q, Tang X, Cui S, Zhang Q, Liu X, Zhao J, Zhang H, Mao B, Chen W. Capsaicin, the Spicy Ingredient of Chili Peppers: Effects on Gastrointestinal Tract and Composition of Gut Microbiota at Various Dosages. Foods. 2022 Feb 25;11(5):686. doi: 10.3390/foods11050686. PMID: 35267319; PMCID: PMC8909049.

谷禾健康

今年夏天的高温相信大家都有所感受,连续数十日的高温让我们感到非常炎热,但随着逐渐入秋,天气也渐渐变冷,我们需要适当地增添一些衣物。
环境温度不仅与我们的生活相关,对人体的健康也有重要影响。最近的研究将环境温度的变化与代谢、肠道微生物以及抗癌免疫反应联系起来。
谷禾将这些研究进行了一些整理归纳,主要讲述环境温度引起的全身代谢变化如何影响抗肿瘤免疫反应。我们还描述了温度变化期间肠道微生物组和免疫代谢之间的相互作用,并涵盖了环境温度调节肿瘤进展的已知机制。
帮助人们认识环境温度对身体的影响,有助于更好地对自己的健康进行管理。甚至可能有助于发现代谢疾病和癌症的治疗新方法。
本文主要分为以下四部分进行讲述
● 环境温度与脂肪代谢
● 不同温度下的免疫反应
● 环境温度对肠道微生物群的影响
● 环境温度与癌症及免疫治疗
环境温度的差异会影响生物体的物理需求、代谢活动和肠道微生物群。
脂肪组织
脂肪组织是指由大量群集的脂肪细胞构成,聚集成团的脂肪细胞由薄层疏松结缔组织分隔成小叶。
葡萄糖和脂质的摄取是癌细胞的基本能量来源,脂肪组织在环境温度变化期间调节能量平衡方面起着重要作用。
脂肪组织包含几种不同类型的细胞,主要分为白色脂肪组织(WAT)和棕色脂肪组织(BAT)。
●白色脂肪组织
白色脂肪组织(WAT)由单泡脂肪细胞构成,通常主要负责以甘油三酯的形式储存脂质,是我们人体最大的能量储存库。
单泡脂肪细胞,细胞中央有一大脂滴,胞质呈薄层,位于细胞边缘,包绕脂滴。
●棕色脂肪组织
棕色脂肪组织(BAT)是指动物体内呈棕色的脂肪组织。其中的脂肪细胞体积较小,胞质中有多个较小的脂滴,并有较多的线粒体。细胞核呈圆形,位于细胞中央。
在寒冷环境中,棕色脂肪组织消耗较快,能产生大量热量、温暖流经其周围的血流,有利于御寒。
利用葡萄糖和脂肪通过解偶联氧化磷酸化(主要由解偶联蛋白-1(UCP-1)介导)产生热量,这一过程称为非颤抖产热。
注:大量的研究表明棕色脂肪组织不仅具有御寒功能,而且还会燃烧多余脂肪和糖分,产生热量,防止体内储存过多的脂肪。
低温下脂肪组织会褐变
在长时间的冷暴露后,棕色脂肪细胞也会出现在白色脂肪细胞中,这一过程通常称为WAT褐变。
由于拥有大量UCP-1阳性线粒体,这些棕色脂肪细胞的产热能力高于白色脂肪细胞。
解偶联蛋白-1(UCP-1)是唯一在棕色脂肪组织(BAT)中表达的解偶联蛋白质,UCP-1的主要功能是参与棕色脂肪组织的产热调节和能量代谢来维持机体的能量代谢平衡。
✦促进WAT褐变的因素
一些因素可以促进WAT褐变,从冷暴露、耐力锻炼和饮食习惯,到通过β3-肾上腺素能受体激活的交感神经刺激,以及微生物群的改变。
冷暴露诱导β3-肾上腺素能受体信号的激活可促进葡萄糖、脂肪酸和富含甘油三酯的脂蛋白从内部贮存器摄取到棕色脂肪组织,然后将其用作非颤抖产热的能量来源。
温度影响下的脂肪组织
热中性区
什么是热中性区?
热中性区是指在环境温度的某一范围内,内温动物耗氧量最低,并且不随环境温度而变化,是代谢的稳定区。小鼠的热中性区在有光照期时为29°C,在黑暗时为33°C。
内温动物——通过自身体内氧化代谢产热来调节体温的动物
因此,20-22°C(室温,RT)的房屋温度对小鼠来说是一个较温和的寒冷环境,通过能量资源的比例分配,以实现产热。
✦热中性区下代谢减弱
由于交感神经活性受到抑制,耗氧量和产热率降低,与室温饲养的小鼠相比,热中性导致脂肪组织“变白”。
在没有解偶联蛋白-1的情况下,热中性会导致小鼠肥胖表型,很可能是由于在此温度下缺乏较高的代谢。
✦棕色脂肪组织比白色脂肪组织代谢更强
与白色脂肪组织相比,即使在温暖温度下处于基础状态,棕色脂肪组织也能增加葡萄糖摄取、乳酸释放和代谢活性。
值得注意的是,经过热中性处理的白色脂肪细胞在冷暴露后可以重新获得产热能力,这一过程部分由不同温度条件下的转录和表观遗传调控介导。
活体细胞类型特异性分析表明,白色脂肪细胞经历了白化诱导的染色质变化,从而能够保护其表观遗传记忆免受先前的寒冷影响。
不过还需要进行进一步研究,以充分了解导致脂肪组织温度驱动变化的细胞机制,以及特定皮下脂肪与内脏脂肪库在肿瘤生长过程中发生褐变的偏好。
温度变化影响的其他器官
温度变化导致不同器官的代谢重编程
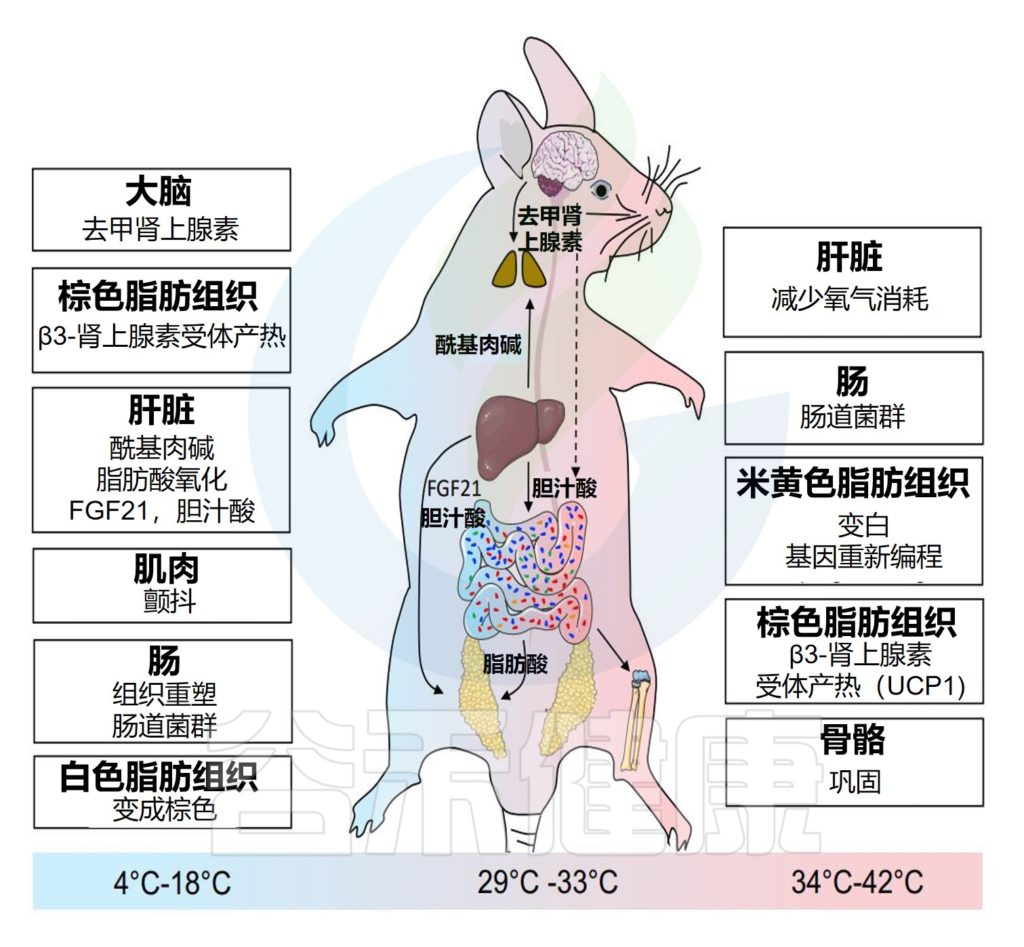
Wang H,et al.FEBS J.2022
生物体在不同环境温度下的生物物理需求,包括它们在不同组织和肠道微生物群中的代谢活动都不同。
▷寒冷环境
皮肤神经元感受到的冷刺激会激活交感神经系统,该系统负责棕色脂肪组织中去甲肾上腺素的局部生成。
交感神经系统(SNS)对整体生理功能具有调节意义: 在环境急剧变化时,交感神经系统可以动员机体许多器官的潜力以适应环境的变化。
去甲肾上腺素(NE),它既是一种神经递质,也是一种激素。
冷暴露也会引发肌肉颤抖,从而促进棕色脂肪组织产热。在长时间寒冷暴露期间,白色脂肪组织的褐变也通过各种机制参与生热,包括刺激肝脏FGF21和胆汁酸(BA)。肝脏酰基肉碱代谢的增加有助于肝脏和其他组织中的脂质氧化。
胆汁酸是胆汁的重要成分,在脂肪代谢中起着重要作用。
此外,寒冷诱导的食欲和代谢变化导致肠道微生物群落的厚壁菌/拟杆菌比率发生变化,厚壁菌的数量超过拟杆菌(从室温下的72.6%到低温下的35.2%),粪便和盲肠中几乎没有疣状杆菌门,影响到各种器官。
▷温暖环境
热暴露会导致微生物群发生相反的变化,通过增强多胺的产生,对骨重塑有显著的有益影响。肝脏热适应后,静息耗氧量降低。随着温度的升高,白色脂肪细胞可以经历表观基因重编程。
冷暴露(4oC-18oC);热中性温度(29°C-33°C);热暴露(≥34°C)。
建议
谷禾查阅到一项研究比较了温和与炎热环境下的运动脂肪的消耗率。结果显示,热环境显著降低了脂肪氧化率。
可以说,在相对适宜的温度下比高温下运动的减肥效率更好。对于那些试图在运动中增加脂肪氧化的人来说,不建议在高温下运动,同时也可以避免中暑。
免疫细胞监测并响应环境代谢线索以及各种内源性触发因素,导致其功能改变。人类和动物研究表明,不同的环境温度可以改变细胞和体液方面的免疫反应。
免疫系统和生物体的致热反应之间的相互作用可以在生命史理论的背景下进行观察,该理论认为生物程序之间资源的优先次序取决于环境。
在恶劣环境中,资源从增长和繁殖项目转移到维护项目。有趣的是,各种维护程序之间也存在资源竞争,其中对寒冷的代谢反应需要与其他耗能高的程序(如免疫反应)进行能量平衡。
不同温度下的免疫细胞
环境温度对小鼠和人类的代谢和免疫影响

Wang H,et al.FEBS J.2022
✦寒冷使活性降低,免疫反应受限
寒冷降低了单核细胞上的主要组织相容性复合物II类(MHCII)并使其活性降低,这反过来又抑制了自身免疫过程中致病性T细胞的启动。
这导致T细胞细胞因子表达减少,从而减轻神经炎症。这些数据表明,由于小鼠免疫系统的能量可用性降低,资源优先用于产热,导致免疫反应受限。
虽然这种竞争对自身免疫有明显的保护作用,但它也可以使寒冷期间对某些病毒感染的易感性增加,这值得进一步研究。
✦热中性环境下增强了免疫细胞
此外,热中性环境增强了免疫细胞在肿瘤微环境中的渗透。这与不断积累的证据一致,即反复寒冷暴露会抑制小鼠的免疫活动,而温暖会激发更大的抗病毒免疫反应。
有趣的是,处于热中性环境的小鼠在骨髓中积累LyG6+单核细胞,但在循环血液中减少,从而对动脉粥样硬化产生保护作用。
注意
虽然这些数据有力地支持了能量资源优先化会限制有利于增加产热的免疫反应的观点,但其作用还不能完全确定。
有证据表明,寒冷环境温度对免疫系统既有抑制作用,又有支持作用,这在一定程度上取决于寒冷暴露的时间长短。
几项研究表明,虽然短期冷刺激会降低人类淋巴增殖反应和Th1细胞因子的产生,但它也会引发炎症反应和免疫抑制特征基因。根据小鼠的数据,长期适应冷暴露会导致抗炎反应,这意味着冷适应期间免疫反应的变化可能具有普遍重要性。
免疫反应与脂肪组织的相互作用
免疫反应同时也会反过来调节瘦鼠和肥胖鼠的脂肪代谢。
抗炎信号的募集与脂肪中的白色脂肪生成密切相关。白色脂肪组织的低度炎症是肥胖的标志,与白色脂肪组织中促炎症(M1型)巨噬细胞的浸润和激活以及褐变能力的降低有关。
✦白色脂肪组织促使巨噬细胞极化
早期的报告表明,冷暴露后,白色脂肪组织被数量增加的嗜酸性粒细胞浸润,这可能会促使巨噬细胞极化,从促炎状态转变为抗炎状态。
白色脂肪组织驻留巨噬细胞可通过几种潜在机制发挥其褐变作用。M1型极化巨噬细胞通过结合整合素α4和血管细胞粘附分子1(VCAM-1)粘附到脂肪细胞,导致持续抑制。
✦巨噬细胞吸收并降解白色脂肪组织
一种称为交感神经元相关巨噬细胞的巨噬细胞亚群吸收并降解白色脂肪组织交感神经末梢网络释放的去甲肾上腺素。交感神经相关巨噬细胞在肥胖中增加,这些细胞摄取去甲肾上腺素的消融增加了褐变。
此外,在热中性状态下,小鼠棕色脂肪组织中巨噬细胞的浸润和相应的促炎细胞因子IFN-γ、TNF-α、IL-1β和IL-6增加。
不同温度下的免疫细胞数量
•冷暴露下免疫细胞数量变化暂不确定
在人类中,每日30–60分钟冷暴露(冷水游泳14°C-18°C)三周后,辅助T细胞(CD4+)和细胞毒性(CD8+)T细胞计数没有显著变化;然而,在同样的间歇性感冒六周后,T淋巴细胞增加。另一方面,短期(20–60分钟)冷暴露导致外周CD4+数量下降。
•室温下T细胞增殖受到抑制
与热中性温度相比,在室温条件下,髓源性抑制细胞(MDSC)通过上调β3-肾上腺素能受体,部分抑制T细胞增殖。
如上所述,冷暴露通过降低单核细胞MHCII的表达来减少T细胞启动。此外,据报道,急性热应激会增加自然杀伤细胞(NK细胞)的数量,;而慢性热应激抑制脾脏NK细胞的活性,并增加Th2与Th1的比值。
髓源性抑制细胞(MDSC)是一类未成熟的免疫细胞,有免疫抑制功能,可强力抑制T细胞
NK细胞是一种细胞毒性淋巴细胞,对先天免疫反应至关重要
✦全身热疗可能会增强免疫作用
小鼠和人类对高温(39°C-43°C)的免疫反应包括上调树突状细胞中的T细胞启动标记物,增强Toll样受体4(TLR4)+巨噬细胞,以及增强淋巴细胞向淋巴的转运。
这种免疫调节伴随着高温后热休克蛋白水平的增加及其与免疫细胞上热休克蛋白受体的相互作用。
热休克蛋白(HSP)是在从细菌到哺乳动物中广泛存在一类热应激蛋白质。当有机体暴露于高温的时候,就会由热激发合成此种蛋白,来保护有机体自身。
在人类癌症患者中,局部热疗不会改变细胞因子水平,然而,全身热疗会升高IL-1、IL-6或TNF-α,表明全身热疗可能对免疫治疗有益。
冷暴露下脂肪组织对免疫的影响
脂肪组织是一个高度代谢活性的器官,储存和释放脂质代谢产物。
•脂质总体成分变化
在对腹股沟白色脂肪组织进行的脂质组学/RNA序列组合分析中,短期(3天)冷暴露导致脂质成分的总体变化:特别是甘油磷脂和鞘脂的富集,以及产热机制、脂肪酸代谢、三酰甘油酯和甘油磷脂合成的转录组学变化。
•氨基酸大幅增加
然而,慢性(10天)冷暴露通过富集三羧酸循环中间产物,导致小鼠棕色脂肪组织和皮下白色脂肪组织中的线粒体葡萄糖氧化,这在内脏白色脂肪组织中未观察到。
短期冷暴露也会改变血浆氨基酸库,导致谷氨酰胺和支链氨基酸大幅增加,如谷氨酰胺含量、脯氨酸、色氨酸和苯丙氨酸,可以作为棕色脂肪组织产热的能源。
•有助于免疫细胞的激活
此外,研究还指出,其他代谢机制也参与其中,如脂肪酸、肌酸和钙的无效循环对冷适应产热的影响。脂肪组织中的生热诱导脂解可能有助于外周循环和重要代谢器官中免疫细胞的招募和激活。
在瘦个体中,脂肪组织相关巨噬细胞体积小,在脂肪细胞中稀疏分布,但在肥胖个体中,它们积累脂质并聚集。
其他潜在的代谢信号,如脂肪酸、氨基酸、缺氧和脂肪细胞应激,被认为有助于免疫细胞和脂肪细胞在不同温度下的相互作用;不过还需要更多的研究来更好地理解它们的重要性。
人体微生物群分布于身体的各个部位,包括呼吸道、皮肤以及肠胃。
肠道微生物
胃肠道微生物群落丰富多样,主要由放线菌、拟杆菌、厚壁菌、变形杆菌和疣状杆菌门的严格厌氧菌组成,肠道病毒和真菌种类繁多。
✦保护性免疫作用
肠道微生物群除了在食物消化和宿主生理调节中的作用外,越来越多的证据表明,它在增强癌症患者抗肿瘤免疫治疗中的保护性免疫作用。
环境因素对微生物的影响
一些环境因素,如营养素、盐和温度,影响微生物群的组成、定殖和代谢活动。
生命的组成部分天生对温度敏感。温度过高时,蛋白质变性,核酸失去碱基配对,质膜变得过于流动。温度太低时,一切都会变慢:酶工作效率低下,核酸形成不方便的二级结构,质膜变硬。
因此,微生物和其他有机体调整其细胞过程,使其在特定温度范围内生长,并在超出其最佳温度范围时作出响应。
✦环境温度会改变肠道微生物的功能
最近研究表明,环境温度变化会改变肠道微生物组的多种生物功能。反过来,肠道微生物群在多个层面上参与全身代谢。
一方面,适应低温的微生物群增加了营养素的吸收,从而影响从饮食和储存中获取能量,并调节免疫反应。另一方面,它通过调节棕色脂肪组织和白色脂肪组织褐变和产热来影响能量消耗。
✦肠道微生物对温度的适应能力如何?
•具有耐热性
大肠杆菌和其他肠杆菌科成员具有耐热性;该科中的许多物种在比宿主温度低和高的温度下都能较好地生存。
例如,耶尔森氏菌将在接近0°C的温度下继续生长,而实验室大肠杆菌菌株能从大约8°C生长到42°C,并很容易在高达48°C或更高的温度下生长。
•功能灵活
变形菌门被认为是功能灵活的,能对许多环境压力作出反应。此外,这一组的致病成员,如沙门氏菌, 耶尔森氏菌属, 假单胞菌和致病性大肠杆菌,明确响应宿主温度,将其作为上调毒力基因的环境线索。
这些温度响应基因在类似发烧的42℃温度下比在37℃温度下更容易上调,铜绿假单胞菌中的温度响应酶同样显示出在45°C的温度范围内提高效率。
艰难梭菌是另一种主要的人类肠道病原体,在37°C和41°C下在体外同样生长良好。总之,这些观察结果表明肠道病原体既能耐受也能利用宿主温度的变化。
不同温度下的肠道微生物
1
低温
一些实验研究了哺乳动物肠道微生物群对冷暴露和宿主体温过低的反应性。
•厚壁菌和短链脂肪酸增加
长期的寒冷暴露导致A.muciniphila几乎完全枯竭,同时增加了Lachnospiraceae、Clostridiaceae、Ruminococcaceae家族的丰度和短链脂肪酸的相对产量,使其不易受到高脂肪饮食诱导的肥胖的影响。
除了食物摄入量显著增加外,冷暴露还刺激棕色脂肪组织中的脂蛋白加工和胆固醇向胆汁酸的肝脏转化,这有助于微生物群重塑。
•新陈代谢良好
低温小鼠的新陈代谢健康状况也较好,不易受到高脂肪饮食诱导的肥胖的影响,这些特征可以通过将粪便微生物群移植到室温下保存的小鼠中来复制。
2
高温
•肠通透性增加
热应激(>40°C)可导致肠上皮损伤,导致细菌产生的脂多糖的肠通透性增加,从而引发局部和全身免疫反应,并与体重指数增加、胰岛素敏感性受损和脂肪褐变减少相关。
•厚壁菌和短链脂肪酸减少
与在哺乳动物冷应激期间观察到的厚壁菌和短链脂肪酸的丰度增加相反,多项研究表明,肠道厚壁菌随着热应激而减少,肠道微生物群的总体α多样性也随之减少。
令人惊讶的是,在多种宿主中都观察到了这种下降,包括放热动物和吸热动物。总之,这些数据表明,体温对肠壁厚菌的影响是一致的,这是由体温本身或宿主食欲或新陈代谢的保守变化引起的。
•影响随时间积累
此外,在反复的热应激循环后,沙鼠的微生物群落α多样性下降,表明热应激的后果可能会随着时间的推移而积累。
3
适宜温度
•肠道菌群组成发生变化
暴露在温和的温度(34°C)下也会引起肠道微生物群组成的变化。
以下的菌群数量会增加:
Turicibacter ↑↑↑
Akkermansia ↑↑↑
Parabacteroides ↑↑↑
以及下列菌属数量的减少:
Butyricococcus ↓↓↓
Peptococcaceae ↓↓↓
Ruminiclostridium ↓↓↓
•影响免疫反应和肠道稳态
在卵巢切除的老年雌性小鼠(绝经后骨质疏松症模型)中,温热条件下的微生物群移植均能逆转卵巢切除引起的胫骨转录组变化,并增加骨膜骨形成。
这种影响在一定程度上是由多胺的增加所介导的,多胺的产生可以影响骨重塑,但也可能具有免疫细胞功能。除了代谢调节的改变外,热应激诱导的热休克蛋白主要由肠道微生物群诱导,以响应细胞应激。
这些蛋白质作为肠道“看门人”,在免疫反应和肠道内稳态维持中具有若干关键功能,包括变性蛋白质的重折叠和从肠道中清除受损多肽。
免疫热应激和肠道微生物反应
✦体温升高有助于清除感染
为了应对感染和先天免疫刺激,大多数哺乳动物都会发烧:故意提高体温。发烧时,下丘脑通过触发棕色脂肪组织中的非颤动产热,对前列腺素水平升高(受刺激巨噬细胞释放的炎症脂质介质)作出反应,从而通过与低温时相同的基本机制产生代谢热。
即使是体温过高者也会改变他们的行为,在感染期间寻找更高的温度;事实上,感染后体温升高1.5至5°C在动物界非常普遍。核心体温升高的假定进化益处是通过直接抑制微生物生长或在更高温度下刺激免疫反应来限制病原微生物的活动。实验和观察证据都表明,发烧反应确实有助于更快地清除感染。
在抗生素开发之前,热疗甚至被用于治疗人类梅毒症状。
✦发烧会影响肠道微生物
尽管发烧对传染病有影响,但发烧对肠道微生物群的影响知之甚少。
一般来说,先天性炎症反应与肠道蛋白杆菌水平增加和α多样性降低有关,尤其是当炎症变为慢性时。然而,急性发热反应对哺乳动物微生物群的影响相对未知。
最近的一项研究发现,感染新型冠状病毒(SARS-Cov2)的患者的肠道微生物群出现发烧依赖性变化,包括细菌热休克蛋白的增加,这表明发烧温度确实会影响人体微生物群。
注意
虽然上述证据表明温度适应下的微生物群对宿主免疫调节有重要作用,但最近的研究强调,在物种层面上,人类和小鼠具有不同的肠道微生物群组成。在某些情况下,这些差异在家庭层面上也很明显。
例如,在人类中,拟杆菌门主要由拟杆菌科和瘤胃球菌科的厚壁菌门组成。相反,在小鼠中,拟杆菌主要由S24-7家族组成,而厚壁菌则由梭状芽孢杆菌目组成。
根据这些报告,可以设想不同的方法来克服使用小鼠微生物群作为人类替代物的挑战。例如,通过将人类肠道微生物群移植到无菌小鼠中来建立小鼠模型,以及发现小鼠物种和人类微生物群之间的功能同源物。
虽然在动物模型中的研究表明,微生物群在不同环境温度下调节免疫代谢方面起着关键作用,但人类肠道微生物群在这方面的重要性有待进一步研究。
自2008年以来,世界各地新发癌症病例的数量显著增加,但各国和世界各地不同类型癌症之间存在着相当大的异质性。
这些差异可归因于遗传差异、环境因素、寿命以及其他癌症诱因,包括社会行为、经济发展和医疗系统的进步。
环境温度与肿瘤存在联系
环境温度可能通过各种生理过程,如代谢和内分泌变化,以及免疫反应和肠道微生物群的变化,促进肿瘤的发生。
癌症和恶性细胞经历代谢改变,主要通过加速糖酵解代谢获得能量以促进其增殖、存活和迁移。然而,肿瘤微环境和宿主大环境中的葡萄糖缺乏也会导致癌细胞的代谢重编程,如脂质代谢的激活,从而改变肿瘤进展和耐药性。
•癌症中棕色脂肪组织活性较高
积累的证据表明棕色脂肪组织与癌症之间存在联系;然而,关于这是正相关还是负相关,有相互矛盾的证据。癌症患者在正电子发射断层扫描/计算机断层扫描中显示18F氟脱氧葡萄糖(18FFDG)摄取量较高,表明与健康对照组相比,棕色脂肪组织活性较高。
•温暖环境中免疫细胞渗透性更高
研究发现,由于免疫细胞在更温暖的环境中的渗透性更高,居住在室温中的小鼠的癌症生长速度比居住在热中性温度(大约30°C)下的小鼠更快、更具攻击性。
虽然对这些效应的分子解释有待进一步研究,但热疗是一种众所周知的癌症治疗替代策略,因为癌细胞在高达45°C的高温下死亡。然而,由于特殊的副作用(烧伤、水疱、腹泻和呕吐)和治疗效率的限制,这种方法在癌症患者中没有广泛应用。
•寒冷环境可能导致更高的癌症风险
相比之下,另一项研究报告,具有非典型强棕色脂肪组织活性的癌症患者预后更好。在动物研究中,在寒冷条件下饲养小鼠会导致脂肪组织褐变,以支持产热,并增加可能促进癌症的细胞因子分泌。
注意:小鼠在环境温度变化期间被广泛用于人类生物学建模,因为它们拥有与产热相关的一组相似的基因。
然而,人类和小鼠在生理体温调节方面存在差异,部分原因是不同的体型,以及生活温度的差异。人类通常倾向于在热中性区内进行活动,而大多数实验室啮齿动物居住在其热中性区以下,需要更多的能量来产生热量。
当试图将啮齿动物的数据转换为人类数据时,这可能会带来重大挑战,因为小鼠和人类的能量消耗差异反映在免疫系统的功能上。
因此,在试图将小鼠的临床前数据转化为人类的治疗药物时,需要仔细考虑环境温度作为可能有助于机体对抗肿瘤治疗反应的因素。
低环境温度有助于通过分泌谷氨酰胺的巨噬细胞促进肿瘤进展。在肝脏和脂肪组织中,慢性冷应激(>10天)触发转录激活物过氧化物酶体增殖物激活受体γ辅激活物和转录因子过氧化物酶体增殖物活化受体,这两种受体在调节肿瘤发生中都起着重要作用。
此外,冷适应导致的代谢活性增加与表观遗传改变有关,表观遗传变化可能导致更高的癌症风险。
与这些报告相反,最近的一项研究表明,冷暴露通过阻碍癌细胞中基于糖酵解的代谢来抑制各种类型的实体肿瘤的生长。具体来说,循环葡萄糖作为肿瘤生长、侵袭和转移的能量来源,对肿瘤糖酵解至关重要。因此,寒冷导致棕色脂肪组织中葡萄糖摄取量的增加,将使其不太适合肿瘤处理。
研究报告称,提供过量的葡萄糖,或删除UCP-1,可以消除冷诱导的肿瘤抑制。这项研究与上述研究之间不一致的原因值得进一步调查,可能部分取决于冷暴露的确切时间和强度、宿主微生物群以及饮食。
癌症的免疫治疗
✦不同温度下抗肿瘤免疫反应功效不同
与热中性的30℃相比,约22℃的标准室温下的慢性(轻度)冷应激会加速肿瘤生长;因此,抗肿瘤免疫反应的功效根据温度而显著不同。
在小鼠模型中,观察到肿瘤微环境和引流淋巴结中的抗肿瘤效应CD8+ T细胞显著增加,而调节性T细胞和免疫抑制细胞在30℃下均减少,这表明仅在22℃下饲养小鼠会导致抗肿瘤免疫反应的显著抑制。
我们还观察到,如果肿瘤在免疫缺陷小鼠中生长,这种作用就会消失,这暗示了适应性免疫反应的作用。
寒冷或温暖环境温度对肿瘤微环境的影响
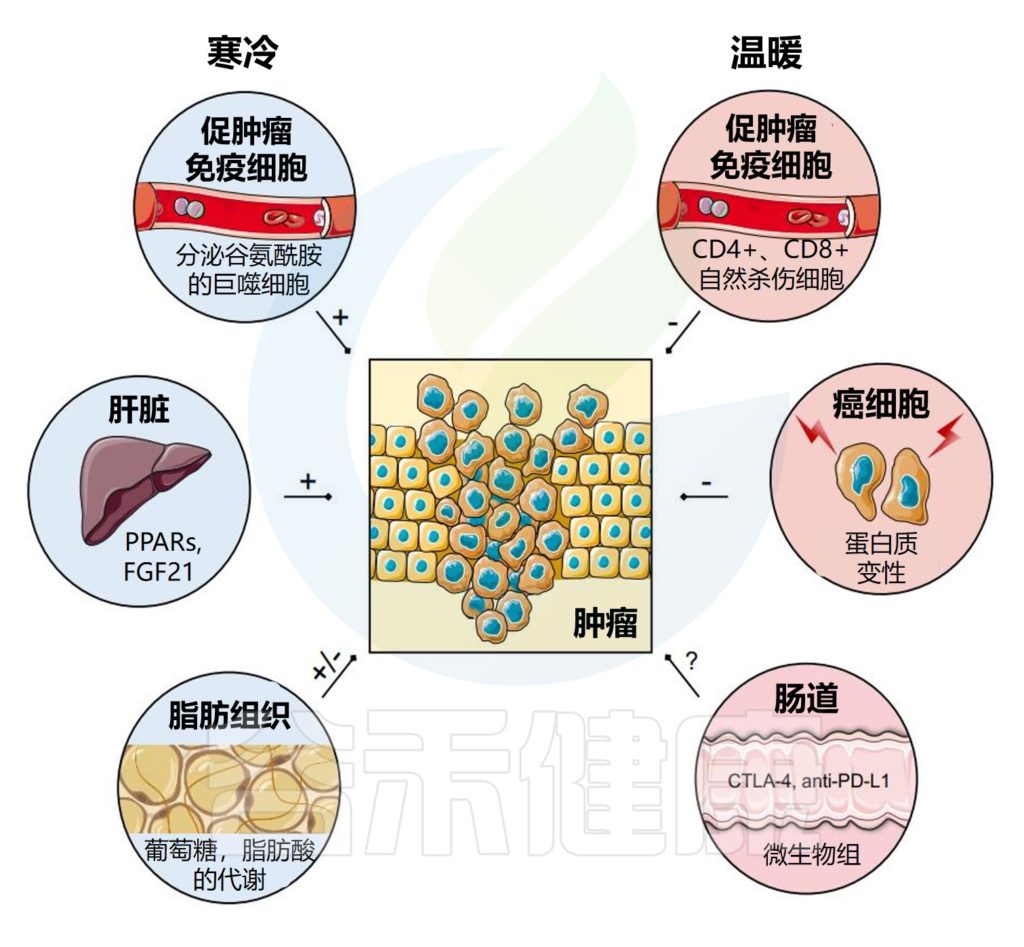
Wang H,et al.FEBS J.2022
温度诱导的免疫系统重编程可能阻碍或有利于抗肿瘤免疫治疗。
免疫细胞
冷暴露会增加分泌谷氨酰胺的巨噬细胞和促肿瘤细胞因子向肿瘤微环境的渗透,而温暖会促进激活的免疫细胞向肿瘤微环境渗透,如辅助性T细胞、细胞毒性T细胞和NK细胞。
肝脏
在肝脏和脂肪组织中,慢性寒冷诱导的激素和代谢重编程既可以触发肿瘤生长,如FGF21和脂肪酸代谢;或与肿瘤竞争营养物质,如葡萄糖。
然而,在热疗过程中,可以通过野生蛋白变性和激活细胞凋亡选择性地杀死癌细胞。
肠道菌群
此外,温度引起的肠道菌群变化可能有利于抗肿瘤免疫疗法的疗效,例如CDLA-4或抗PD-L1。
✦肾上腺素能应激的程度取决于温度
我们发现房屋温度对免疫疗法(检查点抑制剂抗PD-1)的效果有显著影响。乳腺肿瘤和黑色素瘤在22℃时几乎没有反应,但在30℃时有显著反应。
我们继续证明,当用β-肾上腺素能受体拮抗剂(β-阻断剂)处理小鼠(在22℃时)时,这种差异也消失了,证实了肾上腺素能应激的程度取决于室温。
这些问题对于解释研究肾上腺素能应激作用的实验结果和开发克服应激的策略以改善小鼠对免疫或细胞毒性治疗的反应具有重要意义。
小结
改进免疫监测对于癌症免疫治疗至关重要,因为癌症免疫治疗可以提高免疫介导的癌细胞清除率。肠道微生物群产生多种小分子和代谢物,在人体免疫反应和代谢健康方面发挥着不可或缺的作用。
越来越多的证据支持肠道微生物群在肿瘤生长中的作用,影响抗肿瘤免疫和各种免疫检查点抑制剂抗癌免疫效果的效率,包括环磷酰胺、CTLA-4阻断抗PD-L1功效。
微生物分子,如丁酸盐和戊酸盐,通过增加其mTOR活性和表观遗传重编程,增强细胞毒性T细胞和嵌合抗原受体T细胞的活化。因此,了解由于环境温度变化引起的肠道微生物群变化在多大程度上参与了癌症免疫代谢非常重要。
环境温度变化会影响抗肿瘤免疫反应。温度变化期间肠道微生物组和免疫代谢之间的相互作用也会改变。
虽然最近已经清楚肠道微生物群的改变会导致免疫系统的失调,但这种相互作用的确切性质仍有待确定。
通过脂肪组织和微生物群之间温度介导的相互作用调节免疫反应的机制进行了深入研究,这可能有助于阐明代谢疾病和癌症的治疗方法。
主要参考文献:
Wang H, Ülgen M, Trajkovski M. Importance of temperature on immuno-metabolic regulation and cancer progression. FEBS J. 2022 Sep 24. doi: 10.1111/febs.16632. Epub ahead of print. PMID: 36152006.
Hylander BL, Gordon CJ, Repasky EA. Manipulation of Ambient Housing Temperature To Study the Impact of Chronic Stress on Immunity and Cancer in Mice. J Immunol. 2019 Feb 1;202(3):631-636. doi: 10.4049/jimmunol.1800621. PMID: 30670578; PMCID: PMC6352311.
Chevalier C, Stojanović O, Colin DJ, Suarez-Zamorano N, Tarallo V, Veyrat-Durebex C, Rigo D, Fabbiano S, Stevanović A, Hagemann S, Montet X, Seimbille Y, Zamboni N, Hapfelmeier S, Trajkovski M. Gut Microbiota Orchestrates Energy Homeostasis during Cold. Cell. 2015 Dec 3;163(6):1360-74. doi: 10.1016/j.cell.2015.11.004. PMID: 26638070.
Huus KE, Ley RE. Blowing Hot and Cold: Body Temperature and the Microbiome. mSystems. 2021 Oct 26;6(5):e0070721. doi: 10.1128/mSystems.00707-21. Epub 2021 Sep 28. PMID: 34581596; PMCID: PMC8552956.
Li, X. Y., Wenes, M., Romero, P., Huang, S. C. C., Fendt, S. M. & Ho, P. C. (2019) Navigating metabolic pathways to enhance antitumour immunity and immunotherapy, Nat Rev Clin Oncol. 16, 425-441.

谷禾健康

嗜胆菌属
嗜胆菌属 (Bilophila)是变形菌门,脱硫弧菌科的一种厌氧、革兰氏阴性、耐胆汁、过氧化氢酶阳性杆菌。目前通过数据库发现在越来越多的人群,尤其男性,甚至低龄儿童中检出了高丰度的该菌。
该菌被认为是“喜欢动物脂肪喜欢胆汁”的微生物——在以动物为基础的饮食,尤其富含肉类和乳制品脂肪时,其肠道中Bilophila丰度会增加和积累。
目前为止,该属下研究最多和证据充分的菌种是沃氏嗜胆菌(Bilophila wadsworthii, B.wadsworthii),它是从穿孔和坏疽性阑尾炎患者的临床材料中回收的第三大最常见的厌氧菌。
Bilophila是机会致病菌,其丰度的增加的负面影响对肠道炎症的影响已经得到证实,B. wadsworthia与高脂肪饮食协同促进更高炎症反应、肠屏障功能障碍和胆汁酸代谢异常,导致更高的葡萄糖代谢异常和肝脂肪发生。
2021年《Cell》子刊最新研究发现生酮饮食会加剧间歇性缺氧引起的认知障碍,并认为这是由肠道菌群介导的。在生酮饮食和间歇性缺氧的双重条件下,Bilophila大量繁殖富集,进而对海马体功能造成损伤,增加认知障碍风险。
此外,证据显示Bilophila 在便秘,白赛病,重症感染,脑脓肿,帕金森,结直肠癌,卵巢癌患者中增加。
但是Bilophila属中的一些物种是有用的共生细菌,促进宿主新陈代谢并有利地塑造免疫反应。来自mSystems上发表的一项最新研究,发现Bilophila的基因组中存在遗传密码扩张现象,使三甲胺(TMA)甲基转移酶中的一个终止密码子可编码吡咯赖氨酸,从而导致嗜胆菌属可代谢TMA且不产生氧化三甲胺(TMAO)。该研究提示,嗜胆菌属可能通过“绕行”TMAO的生成,以降低动物性饮食诱导的心血管疾病风险,但是这一结论需要更多的样本支持和验证。
Bilophila 作为正常菌群存在于人类粪便中,偶尔也存在于唾液和阴道中。来自人类的分离物通常是β-内酰胺酶阳性,因此该菌对某些β-内酰胺抗生素具有抗药性。部分的菌株也对克林霉素有抗药性。
本文基于文献调查和谷禾数据库讨论和介绍肠道重要菌属——嗜胆菌属 (Bilophila)。
嗜胆菌属 (Bilophila)是变形菌门,脱硫弧菌科除了脱硫弧菌属(Desulfovibrio) 的第二类重要菌属。一种革兰氏阴性厌氧菌,包括 B. wadsworthia,可引起腹腔内和其他感染。
Bilophila 属下代表物种是:Bilophila wadsworthia。该菌最初从坏疽和穿孔性阑尾炎患者的感染中分离出来。后来在包括来自阴囊脓肿、下颌骨髓炎和腋窝化脓性汗腺炎的胸水、关节液、血液和脓液的临床组织样本中也逐渐发现。
Bilophila wadsworthia 是一种革兰氏阴性、专性厌氧、过氧化氢酶阳性、耐胆汁和解酶杆菌。单独或成对出现;偶尔观察到长丝。没有观察到孢子形成。菌落宽度约为 0.7 μm,长 1.0-10.0 μm,细胞呈多形性,细胞壁不规则,约 75% 的菌株为脲酶阳性。DNA G + C 含量约为59.2。
已鉴定菌种:
Bilophila wadsworthia
Bilophila sp. 4_1_30
基于核糖体 RNA 的系统发育研究表明 Bilophila与 脱硫弧菌科另外一个成员 脱硫弧菌属(Desulfovibrio)物种关系最密切。
基于数据库和文献Bilophila和其他肠道菌属的关系如下图:
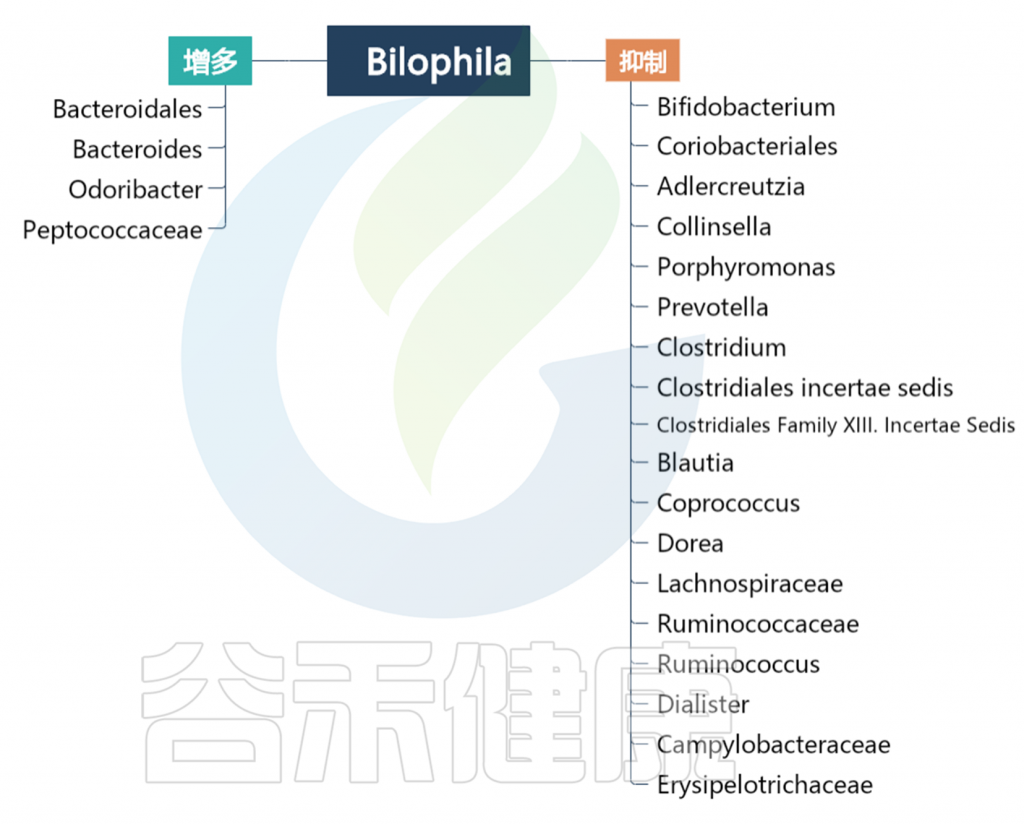

编辑
▸ 代谢牛磺酸,嗜好胆汁
该菌具有呼吸型新陈代谢,化学有机异养和非发酵。代谢蛋白底物,但不代谢碳水化合物。蛋白底物的主要产物是乙酸,含有少量至痕量的琥珀酸。能将硝酸盐还原为亚硝酸盐。
硫化物由含硫氨基酸和亚硫酸盐产生,有时由硫代硫酸盐产生,但Bilophila不会还原硫。过氧化氢酶强阳性。在甲酸盐存在的情况下,B. wadsworthia利用牛磺酸作为电子受体产生乙酸盐并将磺酸盐硫还原为硫化物。
Bilophila是机会致病菌,培养实验等得出硫,蛋白胨,L-牛磺酸,丙酮酸以及胆汁酸是B. wadsworthia扩张的基础。其代谢产物主要为硫化物,硫化氢,乙酸盐,琥珀酸等。
胆汁酸是胆固醇在肝脏分解以及肠肝循环中的一组代谢产物。在肝脏中,牛磺酸与甘氨酸一起用于结合胆汁酸以产生初级胆汁酸。胆汁酸经历肠肝循环,并被微生物群转化为次级胆汁酸。饱和动物源性脂肪先前已被证明可促进牛磺酸结合胆汁酸的产生。至少在高脂环境中,已经提出增加牛磺酸结合胆汁酸的产量是B. wadsworthia扩张的基础。
所有高脂肪喂养小鼠脂多糖(LPS)生物合成和牛磺酸代谢途径的活化显着提高,而涉及氨基酸、糖、淀粉和氮代谢的许多途径显着减少。
B. wadsworthia进一步加剧了高脂饮食情况下的胆汁酸失调,也表明该菌可能加强了高脂饮食诱导的代谢障碍和宿主功能障碍,特别是炎症和屏障功能障碍的一种机制。
▸ 喜欢动物脂肪和乳脂
小鼠和人类实验表明以动物脂肪和乳源性饱和脂肪为基础的饮食可以增加耐胆汁菌Bilophila wadsworthia.
研究人员称并没有刻意去衡量Bilophila的丰度,但当比较哪种细菌含量增加最快时,Bilophila排在首位。
不同的饮食可以快速且可重复地改变肠道微生物群的组成和功能。
《Nature》杂志的一项研究发现,当人们从素食转变为以肉类和奶酪为主的饮食结构上时,他们肠道里的细菌Bilophila几乎立即增加,但植物性为主的饮食结构可以降低该菌群的数量。一项人群研究实验,在清肠后 5 至 7 天收集素食者和杂食者的粪便样本。发现Bilophila和Lachnoclostridium与膳食动物蛋白呈正相关,与膳食植物蛋白来源呈负相关。
《Cell Metabolism》发表了一项研究,发现经常伴随红肉的饱和脂肪,还可能让人变肥胖,脂肪还会出现炎症。与之相反,以鱼肉为代表的白肉所包含的不饱和脂肪则健康很多。线性判别分析(LDA)表明,拟杆菌, Turicibacter和嗜胆菌(Bilophila)属细菌在猪油组小鼠肠道中显著增多,而在鱼油组小鼠中,主要是双歧杆菌和另一种菌 (Bifidobacterium,Adlercreutzia),乳酸菌(lactic acid)等增加。
当前西方化饮食(低纤维、高糖、高脂肪和高动物蛋白)饮食持续增加粪便中Bilophila丰度,所以在越来越多的人群水平中发现Bilophila的富集。研究表明Bilophila可以将亚硫酸盐还原为硫化氢(H2S气体),诱发炎症以及免疫和代谢障碍。这对那些尝试许多流行的“品牌”饮食(例如生酮、旧石器时代、食肉动物等)的个体具有临床意义。
▸ 产生硫化氢
硫代喹诺酮糖是葡萄糖的磺酸衍生物,在菠菜和生菜等所有绿色蔬菜中都能找到。专门的细菌配合利用磺基糖产生硫化氢(H2S)。
这种气体以臭鸡蛋味著称,也对人类健康有不同的影响:在低量的情况下,硫化氢可以对肠黏膜产生抗炎消炎作用。另一方面,肠道微生物产生的硫化氢增多,与慢性炎症性疾病和甚至癌症有关。
研究表明,与葡萄糖等喂养肠道内大量微生物不同,硫代喹诺酮糖刺激肠道微生物组中非常特定的关键微生物的生长。这些关键生物包括Eubacterium rectale物种的细菌,它是健康人常见的肠道微生物之一。E.rectale细菌通过发酵硫代喹诺酮糖,产生一种硫化合物,即二羟基丙烷磺酸盐,而这种硫化合物又是其他肠道细菌如Bilophila wadsworthia的能量来源。
Bilophila wadsworthia最终会通过一种新陈代谢途径从二羟基丙烷磺酸盐中产生硫化氢,这种途径也是最近才发现的。
以往认为主要是由于富含肉类或脂肪的饮食导致肠道中的硫酸盐和牛磺酸含量增加,已知它们是微生物的硫化氢来源。目前发现来自菠菜和藻类等绿色食物的硫代奎诺糖也有助于肠道内气体的产生,这让人感到意外。
病原菌,与肠道炎症有关
B. wadsworthia具有固有的促炎特性。然而,尚不清楚这种B. wadsworthia特征在多大程度上在宿主代谢障碍的发展中发挥作用。炎症是代谢综合征的一个重要特征;因此,尚不清楚与B. wadsworthia相关的代谢障碍是否仅继发于B. wadsworthia-驱动炎症。
为了解决这些问题,研究用广泛使用的免疫抑制剂环孢素抑制了喂食高脂饮食(HFD)的小鼠的炎症。
发现环孢素 (Ci) 有效地消除了高脂饮食喂养小鼠的炎症反应,无论它们是否含有低密度或高密度的B. wadsworthia,因此,可以清楚地推断出B. wadsworthia 的直接代谢作用。
高脂饮食 —— B. wadsworthia过度积累
为了更好地确定更高密度的B. wadsworthia是否会影响代谢功能,特意通过灌胃将B. wadsworthia给予小鼠。
结果强调高脂饮食不是B. wadsworthia在宿主肠道中茁壮成长所必需,但却是B. wadsworthia 的持续增长必不可少的。这个结果比较有意义,表明B. wadsworthia的过度积累离不开高脂饮食的喂养。
B. wadsworthia 诱发的全身炎症相关疾病
B. wadsworthia增加了高脂饮食诱导的代谢综合征,这是一种通常与低程度全身炎症相关的疾病。同时,B. wadsworthia已被证明会加剧遗传易感小鼠的肠道炎症并在野生型动物中诱发全身炎症。较高的炎症状态的特征是消瘦和体重减轻,而代谢综合征的特征是低度炎症和较高的体重指数。
因此,一个关键问题是:
B. wadworthia 如何影响两种相反的病理?
通过药理学抑制炎症,揭示了B. wadsworthia直接诱导对宿主代谢功能的负面影响的能力。
具体来说,不同的代谢障碍,即葡萄糖清除率降低和脂肪肝表型,受B. wadworthia的影响,它们并不完全依赖于其促炎特性。
尽管如此,B. wadworthia驱动的炎症仍然是一个重要因素,它进一步使平衡向更强的代谢功能障碍倾斜。因此,这可以解释为什么B. wadworthia能够在两种截然不同的环境中发挥病态作用。
B. wadsworthia通过恶化高脂饮食诱导的肠道炎症、抑制参与代谢稳态的途径、有利于增加 LPS 的产生和易位,以及减少微生物群的丁酸盐产生来作用于宿主和微生物群。这些与B. wadsworthia相关的改变中的大多数被鼠李糖乳杆菌CNCM I-3690 给药完全或部分逆转。
导致认知障碍
2021年cell最新研究,喂食生酮饮食并间歇性缺氧的小鼠的肠道微生物群中,嗜胆菌属(Bilophila)的细菌浓度急剧增加。他们还发现,沃氏嗜胆菌(Bilophila wadsworthia)沃氏嗜胆菌损害海马体,导致小鼠的认知能力下降。
减轻心血管疾病
目前导致心血管疾病(CVD)发生的心血管风险因素并没有减少,反而在增加。因此,想要防止这种疾病发生,仅靠控制传统风险因素是不够的。虽然许多二级预防患者的传统风险因素控制较好,但仍会出现新的心血管事件。
已证实肠道菌群导致胆汁酸功能性改变,决定了其与法尼酯X受体或G蛋白偶联胆汁酸受体(TGR5)等潜在受体的结合。
动物性饮食是三甲胺(TMA)的主要来源,肠道菌群代谢TMA产生的氧化三甲胺(TMAO)与心血管疾病风险的增加密切相关。
来自mSystems上发表的一项最新研究,发现嗜胆菌属(Bilophila)的基因组中存在遗传密码扩张现象,使TMA甲基转移酶中的一个终止密码子可编码吡咯赖氨酸,从而导致嗜胆菌属可代谢TMA且不产生TMAO。
该研究提示,嗜胆菌属可能通过“绕行”TMAO的生成,以降低动物性饮食诱导的心血管疾病风险。这个结果让临床看到了机会,后续这一机制希望能尽快完成临床实验验证。
扩展阅读:
饮食-肠道微生物群对心血管疾病的相互作用
结直肠癌
结直肠癌是最常见的恶性肿瘤之一,与年龄和生活习惯密切相关,饮食风险因素包括红肉、加工肉类以及酒精,另一个风险因素是炎症性肠病,包括克罗恩病和溃疡性结肠炎。
根据已有报道发现结直肠癌患者中生成硫化物的细菌增加:
美国黑人的结肠直肠癌发病率显著高于非西裔白人,研究假设:硫酸盐还原菌在结肠粘膜中的丰度可能是导致美国黑人结肠直肠癌发病风险较高的环境因素。
无论是结肠直肠癌患者还是健康人,美国黑人的结肠硫酸盐还原菌丰度均高于非西裔白人;沃氏嗜胆菌(Bilophila wadsworthia)特异性dsrA在美国黑人结肠直肠癌患者中高于健康美国黑人;美国黑人日常摄入的脂肪及蛋白质显著高于非西裔白人,多种饮食组分与硫酸盐还原菌的丰度较高相关。
扩展阅读:
结直肠癌防治新策略——微生物群
白塞病(BD)
贝塞特氏病(Behctet,白塞病,BD)是一是一种顽固的多系统性炎症性疾病,可导致不可逆转的失明。
微生物因子被认为是造成这种疾病的原因,但其潜在机制仍不清楚。来自活动性BD患者的粪便样品富含Bilophila spp,一种硫酸盐还原细菌(SRB)和一些机会病原体。活跃的BD患者的荚膜多糖转运系统,氧化还原过程,III型和IV型分泌系统也有所增加。
网络分析表明,富含BD的硫酸盐还原细菌和机会性病原体彼此呈正相关。动物实验表明,粪便微生物群与BD患者粪便的移植增加了包括IL-17和IFN-γ在内的炎性细胞因子的产生。
帕金森病进展
对比分析了帕金森患者与对照的菌群组成及代谢功能差异,发现帕金森相关的特定菌群组成变化与疾病严重程度相关,特定的菌群代谢功能也与疾病表型相关,例如:Hoehn-Yahr分级与Paraprevotella菌属呈负相关,与嗜胆菌属(Bilophila)呈正相关;
帕金森相关菌群变化与甲硫氨酸等9种菌群代谢产物的变化相关,菌群产生泛酸的能力与帕金森患者的特定非运动症状相关。
扩展阅读:
肠道微生物与帕金森以及相关影响因素
重症病人
许多重症医学科室住院患者有严重感染。这部分是由于危重患者经常接受各种各样的药物治疗,这些药物影响肠道菌群多样性,部分是由于患者病情不稳定,可导致缺氧病变、炎症、上皮完整性破坏、运动障碍、腔内pH值变化或肠道免疫功能受损。在一项多中心研究中,ICU患者的菌群显示与炎症密切相关的微生物丰度增加,如拟杆菌属、梭菌属和嗜Bilophila菌属。
脑脓肿:病例报告
研究报道了一例20 年慢性中耳炎病史的患者并发胆脂瘤和脑脓肿的病例。使用对比材料的 CT 扫描显示右小脑半球有三个脓肿腔。
胆脂瘤标本的培养物和脑脓肿的脓液中分离出了大量的 Bilophila wadsworthia、Bacteroides fragilis 和 Prevotella oris。不存在需氧菌。患者接受了开颅手术,最大的脓肿与胶囊一起被切除。抗菌治疗包括青霉素加甲硝唑和后来的增效素。治疗的结果是患者完全治愈并完全康复。这是在慢性中耳炎和脑脓肿中分离出 B. wadsworthia 的一个典型案例。
衰老相关
Cell 子刊一项研究分析了251位18-80岁个体十二指肠菌群多样性,发现多样性受年龄、伴随疾病和用药情况多因素影响,与衰老过程呈负相关;
十二指肠核心菌群随衰老进程变化,变形菌门相对丰度增加,大肠杆菌和克雷伯菌增加,拟杆菌进行性降低;该研究显示克雷伯氏菌属增加仅与药物服用相关,梭菌属和嗜胆菌属增加则与伴随疾病相关。
症状性手骨关节炎
我国湘雅医院2021年纳入1388名平均年龄为61.3岁的受试者分析研究症状性手骨关节炎患者的肠道菌群变化,发现粪便菌群的β-多样性(而非α-多样性)与手骨关节炎显著相关;手骨关节粪便菌群中有更高的嗜胆菌属(Bilophila)与脱硫弧菌属、更低的罗斯氏菌属。
功能分析显示,手骨关节炎患者的粪便菌群中,与氨基酸、碳水化合物及脂质代谢相关的通路发生显著变化。
“
降 低
鼠李糖乳杆菌CNCM I-3690 (Lr) 的每日口服管饲导致粪便 B. wadsworthia 丰度显着降低。同样,鼠李糖乳杆菌CNCM I-3690 能够进一步减少盲肠和小肠中的 B. wadsworthia 扩张。表明除了通过阻止其在体内扩张来抑制与 B. wadsworthia 相关的代谢功能障碍外,它还通过其调节胰岛素水平的固有能力来改善代谢功能。
菊苣根纤维(低聚果糖和菊粉)是益生元营养素,可滋养有益的肠道微生物,尤其是双歧杆菌。众多人体临床研究表明,这些益生元增加了肠道内的有益双歧杆菌,同时可以减少Bilophila菌。
大枣与巴豆霜合用可减缓巴豆霜的快速利尿作用。研究发现大枣合用高剂量巴豆霜后Bilophila水平降低。巴豆霜为大戟科植物巴豆Croton tiglium L.(CT)的干燥成熟果实经去皮制霜后制得的炮制品,具有峻下冷积、逐水退肿、豁痰利咽、蚀疮的功效。因其有大毒,临床应用受到一定限制。
酵母 β-葡聚糖 (Y-BG) 是一种以其免疫调节作用而闻名的膳食补充剂,在健康小鼠中进行了为期 4 周的膳食补充剂,发现 2% 的 Y-BG 颗粒物诱导强大的肠道微生物群落变化,包括粪便Bilophila丰度的显着减少。
水果和蔬菜富含纤维、维生素 B、维生素 C、β-胡萝卜素、钾和生物活性化合物,如番茄红素和白藜芦醇。食用水果可以在维持肠道菌群平衡和改善肠道生态方面发挥作用。健康成年人食用十字花科蔬菜(西兰花、花椰菜)显示出拟杆菌门与厚壁菌门的比例呈正变化,硫酸盐还原菌(SRB) ,包括嗜胆菌和脱硫弧菌丰度下降。
扩展阅读:
常见水果对肠道菌群、肠道蠕动和便秘的影响
在患有轻度便秘的健康成年人中,食用菊粉可能会导致厌氧菌、嗜胆菌和双歧杆菌的相对丰度发生变化。特别是,食用菊粉后Bilophila丰度的下降与较软的粪便和便秘特异性生活质量指标的有利变化有关。
扩展阅读:
排便困难?便秘反复?不要忽视肠道菌群
“
增 强
主要参考文献
Natividad JM, Lamas B, Pham HP, Michel ML, Rainteau D, Bridonneau C, da Costa G, van Hylckama Vlieg J, Sovran B, Chamignon C, Planchais J, Richard ML, Langella P, Veiga P, Sokol H. Bilophila wadsworthia aggravates high fat diet induced metabolic dysfunctions in mice. Nat Commun. 2018 Jul 18;9(1):2802. doi: 10.1038/s41467-018-05249-7. PMID: 30022049; PMCID: PMC6052103.
Kivenson V, Giovannoni SJ. An Expanded Genetic Code Enables Trimethylamine Metabolism in Human Gut Bacteria. mSystems. 2020 Oct 27;5(5):e00413-20. doi: 10.1128/mSystems.00413-20. PMID: 33109749; PMCID: PMC7593587.
Yazici C, Wolf PG, Kim H, Cross TL, Vermillion K, Carroll T, Augustus GJ, Mutlu E, Tussing-Humphreys L, Braunschweig C, Xicola RM, Jung B, Llor X, Ellis NA, Gaskins HR. Race-dependent association of sulfidogenic bacteria with colorectal cancer. Gut. 2017 Nov;66(11):1983-1994. doi: 10.1136/gutjnl-2016-313321. Epub 2017 Feb 2. PMID: 28153960; PMCID: PMC5575988.
Sun Y, Ma Y, Lin P, Tang YW, Yang L, Shen Y, Zhang R, Liu L, Cheng J, Shao J, Qi T, Tang Y, Cai R, Guan L, Luo B, Sun M, Li B, Pei Z, Lu H. Fecal bacterial microbiome diversity in chronic HIV-infected patients in China. Emerg Microbes Infect. 2016 Apr 6;5(4):e31. doi: 10.1038/emi.2016.25. PMID: 27048741; PMCID: PMC4855070.
Baron EJ. Bilophila wadsworthia: a unique Gram-negative anaerobic rod. Anaerobe. 1997 Apr-Jun;3(2-3):83-6. doi: 10.1006/anae.1997.0075. PMID: 16887567.
Baldini F, Hertel J, Sandt E, Thinnes CC, Neuberger-Castillo L, Pavelka L, Betsou F, Krüger R, Thiele I; NCER-PD Consortium. Parkinson’s disease-associated alterations of the gut microbiome predict disease-relevant changes in metabolic functions. BMC Biol. 2020 Jun 9;18(1):62. doi: 10.1186/s12915-020-00775-7. PMID: 32517799; PMCID: PMC7285525.
Wei J, Zhang C, Zhang Y, Zhang W, Doherty M, Yang T, Zhai G, Obotiba AD, Lyu H, Zeng C, Lei G. Association Between Gut Microbiota and Symptomatic Hand Osteoarthritis: Data From the Xiangya Osteoarthritis Study. Arthritis Rheumatol. 2021 Sep;73(9):1656-1662. doi: 10.1002/art.41729. Epub 2021 Aug 6. PMID: 33760399; PMCID: PMC8457181.
Leite G, Pimentel M, Barlow GM, Chang C, Hosseini A, Wang J, Parodi G, Sedighi R, Rezaie A, Mathur R. Age and the aging process significantly alter the small bowel microbiome. Cell Rep. 2021 Sep 28;36(13):109765. doi: 10.1016/j.celrep.2021.109765. PMID: 34592155.
Marina M, Ivanova K, Ficheva M, Fichev G. Bilophila wadsworthiain brain abscess: case report. Anaerobe. 1997 Apr-Jun;3(2-3):107-9. doi: 10.1006/anae.1997.0084. PMID: 16887572.
Chen Y, Zhang S, Zeng B, Zhao J, Yang M, Zhang M, Li Y, Ni Q, Wu D, Li Y. Transplant of microbiota from long-living people to mice reduces aging-related indices and transfers beneficial bacteria. Aging (Albany NY). 2020 Mar 16;12(6):4778-4793. doi: 10.18632/aging.102872. Epub 2020 Mar 16. PMID: 32176868; PMCID: PMC7138539.
Wu YT, Shen SJ, Liao KF, Huang CY. Dietary Plant and Animal Protein Sources Oppositely Modulate Fecal Bilophila and Lachnoclostridium in Vegetarians and Omnivores. Microbiol Spectr. 2022 Apr 27;10(2):e0204721. doi: 10.1128/spectrum.02047-21. Epub 2022 Mar 14. PMID: 35285706; PMCID: PMC9045121.
So SY, Wu Q, Leung KS, Kundi ZM, Savidge TC, El-Nezami H. Yeast β-glucan reduces obesity-associated Bilophila abundance and modulates bile acid metabolism in healthy and high-fat diet mouse models. Am J Physiol Gastrointest Liver Physiol. 2021 Dec 1;321(6):G639-G655. doi: 10.1152/ajpgi.00226.2021. Epub 2021 Oct 13. PMID: 34643089.
Wu YT, Shen SJ, Liao KF, Huang CY. Dietary Plant and Animal Protein Sources Oppositely Modulate Fecal Bilophila and Lachnoclostridium in Vegetarians and Omnivores. Microbiol Spectr. 2022 Apr 27;10(2):e0204721. doi: 10.1128/spectrum.02047-21. Epub 2022 Mar 14. PMID: 35285706; PMCID: PMC9045121.

谷禾健康

疫苗是公共卫生领域最伟大的成就之一,每年可预防数百万儿童疾病和死亡病例。然而,许多疫苗的功效在地理和社会经济不同地区的婴儿之间可能存在很大差异。
有研究发现,肠道微生物组组成的差异已成为解释免疫结果差异的主要因素之一。
在本篇文章中,谷禾总结了各位学者的研究。关于肠道微生物群对生命早期免疫的影响,重点关注微生物群中具有促进健康和免疫调节特性的两个重要成员:双歧杆菌和拟杆菌。
此外还罗列了一些具有免疫刺激特性的微生物产物、与宿主的相互作用以及它们对婴儿疫苗反应和功效的影响。
通过本文,可以让大家更好地了解肠道微生物对免疫接种的影响,让人们更有针对性地选择免疫接种,更好地保护我们的健康。
本文出现的专业名词
疫苗是指用各类病原微生物制作的用于预防接种的生物制品。其中用细菌或螺旋体制作的疫苗亦称为菌苗。疫苗分为活疫苗和死疫苗两种。
常用的疫苗:常用的活疫苗有卡介苗,脊髓灰质炎疫苗、麻疹疫苗、鼠疫菌苗等;常用的死疫苗有百日咳菌苗、伤寒菌苗、流脑菌苗、霍乱菌苗等。
免疫球蛋白(Ig)——指具有抗体活性或化学结构,与抗体分子相似的球蛋白
辅助T细胞(Th细胞)在免疫反应中扮演中间过程的角色:它可以增生扩散来激活其它类型的产生直接免疫反应的免疫细胞
调节性T细胞(Tregs)是维持机体免疫耐受的重要因素之一,通过主动调节的方式抑制存在于正常机体内潜在的自身反应性T细胞的活化与增殖,从而调节机体的免疫力
G蛋白偶联受体(GPCRs)是一大类膜蛋白受体的统称
Toll样受体(TLR)是参与非特异性免疫(天然免疫)的一类重要蛋白质分子,表达在巨噬细胞、树突状细胞和上皮细胞表面,可识别多种类型的病原体相关分子模式或损伤相关分子模式
人类胃肠道微生物群由细菌、病毒、古生菌和真菌组成的复杂群落,其组成沿胃肠道长度变化。
微生物定植在出生时开始,并在整个生命过程中不断变化,生命的前1000天是生态系统结构最脆弱和最不稳定的时期。
✦胃肠道微生物对健康至关重要
胃肠道微生物群有助于粘膜和全身免疫系统的成熟,抵抗病原体定植,消化膳食成分,并提供微量营养素。
胃肠道微生物群的结构和功能紊乱与各种疾病有关,包括代谢紊乱、神经退行性疾病,过敏,自身免疫性疾病,和癌症。
值得注意的是,影响微生物组的因素与影响疫苗免疫反应的因素相似,强调了免疫与胃肠道微生物群之间的相互关系。
在生命早期,影响胃肠道微生物群发育的其他因素是分娩方式,早产,营养(母乳与配方奶),早期使用抗生素或益生菌和卫生等。
✦ 分娩方式对早期微生物群的影响
在出生期间和出生后,婴儿会接触到促进免疫系统成熟的环境抗原和微生物,其性质取决于分娩方式。
●阴道分娩
阴道分娩的婴儿暴露于母体阴道和粪便微生物群中,导致微生物分布以埃希氏菌、乳酸杆菌、拟杆菌和双歧杆菌为主。
●剖宫产分娩
相比之下,剖宫产分娩的新生儿与母体皮肤和医院微生物的接触更多,并且经常被链球菌、葡萄球菌和肠球菌定植。
✦早期用抗生素对免疫系统发育有负面影响
早期使用抗生素治疗会对这种最初的定植和随后的免疫系统发育产生负面影响。这些早期生活障碍会在短期内导致感染风险增加。从长远来看,与免疫和代谢相关疾病(如特应性)的风险增加有关。
因此,在正确的时间由正确的微生物定殖对于有效建立免疫防御和稳态至关重要。
✦母乳喂养对肠道微生物群有益处
母乳喂养提供了抗菌肽、母体抗体和先天免疫因子,它们促进了对新生婴儿的被动保护,并提供了塑造婴儿微生物群的关键膳食成分。
人乳寡糖可增强双歧杆菌的定殖和持久性(占总微生物群落的80%),这在以配方奶喂养的婴儿中不太明显(5-30%)。
人乳寡糖(HMOs)是人类母乳中仅次于乳糖和脂肪的第三大固体成分,有助于预防人类细胞、组织和小鼠中的链球菌感染。
人乳寡糖的微生物代谢导致短链脂肪酸的产生,短链脂肪酸被与膜结合的特定G蛋白偶联受体识别,由免疫细胞在全身和胃肠道中表达,是关键用于发展免疫耐受性。母乳还直接通过母乳微生物组引入微生物,从而进一步播种婴儿肠道。
断奶和从母乳或以配方食品为基础的营养过渡到固体食物推动了胃肠道微生物群的重大变化,双歧杆菌种类减少,并引入了瘤胃球菌、阿克曼氏菌和普雷沃氏菌。
✦疫苗效果存在差异
许多许可疫苗的功效在人群之间存在差异,在某些情况下,转化为几乎没有效果。
许多内在因素导致疫苗反应的这种变异性,包括年龄、遗传(占受者之间变异的 20-40%)、贫血和性别。
此外,广泛的外部因素会极大地影响疫苗接种结果,包括疫苗成分和免疫方案,预先接触病原体和慢性炎症,暴露于母体抗体,营养状况和地理位置等。
✦肠道微生物有助于促进免疫
人们越来越认识到婴儿胃肠道微生物群在疫苗免疫中的作用。胃肠道微生物群已被证明可通过多种机制促进对疫苗的体液和细胞免疫反应的有效刺激。
肠道微生物群的免疫调节

Jordan A,et al.Lancet Microbe.2022
微生物群刺激对于产生免疫球蛋白 (IgA) 的B细胞的发育和成熟、通过促进IgA类别转换的记忆浆细胞以及生发中心的发育至关重要。此外,通过介导浆细胞样树突状细胞产生I型干扰素,微生物群增强了抗原特异性T细胞反应。
肠道微生物群和免疫系统之间的共生关系,以及遗传和环境影响,可以解释个体对疫苗的免疫反应的可变性。
抗生素引起的新生小鼠微生物组紊乱,以及使用免疫缺陷的无菌幼崽,导致对不同佐剂和减毒活疫苗的体液反应受损,其特征是Th1和Th17反应降低,IgG和IgM生产。
注:辅助性T细胞1(Th1)主要为对抗细胞内细菌及原虫的免疫反应。
辅助性T细胞17(Th17)是一种新发现的能够分泌白介素17的T细胞亚群。
IgG是免疫球蛋白G,是免疫球蛋白中的一种,来源于浆细胞,具有抗病毒、抗菌和免疫调节的功能。
IgM是免疫球蛋白M,由于B淋巴细胞在抗原如病毒或细菌等病原微生物等的刺激下转化为浆细胞,产生能与相应抗原发生特异性结合的抗体,而形成免疫球蛋白。
值得注意的是,在通过施用特定的鞭毛大肠杆菌菌株或粪便微生物群转移来恢复微生物群后,这种损害是可逆的。
某些细菌科、属和物种的丰度与人类对疫苗的免疫反应差异有关,无论是正面的还是负面的。
// 不同地区间的研究
对来自加纳、巴基斯坦、孟加拉国和荷兰的接种轮状病毒疫苗的有反应和无反应儿童的微生物谱比较研究表明,与同一队列的无应答者相比,来自不同低收入和中等收入国家的应答者的微生物群组成与荷兰婴儿微生物群更为相似(即梭菌群XI和变形菌的丰度更高,拟杆菌门的数量更低)。
小结
由于胃肠道微生物组在激活和抑制免疫反应以及随后对疫苗免疫的影响方面具有多因素作用,因此研究不同的微生物群调节干预措施以最大限度地提高疫苗效力。
益生元、益生菌和抗生素
益生元——指一些不被宿主消化吸收却能够选择性地促进体内有益菌的代谢和增殖,从而改善宿主健康的有机物质。
益生菌——通过定殖在人体内,改变宿主某一部位菌群组成的一类对宿主有益的活性微生物。
✦益生元和益生菌对疫苗免疫有积极作用
对小鼠接种流感和霍乱疫苗的研究报告表明,不同的益生元与疫苗接种后的全身免疫反应之间存在正相关,与双歧杆菌和乳酸杆菌的丰度增加以及短链脂肪酸的产生有关。
已经研究了它们对疫苗接种反应的影响。系统评价总结了26项使用益生菌对人类进行干预研究的结果,以提高17种不同疫苗的功效,其中一半的研究显示出积极的结果。
婴幼儿使用益生菌效果更好
与成人相比,在新生儿和幼儿(0-16 周龄)中使用益生菌乳酸杆菌和双歧杆菌菌株的试验显示成功率更高,接种流感、白喉、轮状病毒和脊髓灰质炎疫苗后对体液免疫的影响最大。
然而,不同研究(包括使用的细菌菌株)的设计缺乏一致性,因此难以得出可靠的结论。
✦抗生素对疫苗的影响暂不明确
抗生素——指由微生物(包括细菌、真菌、放线菌属)或高等动植物在生活过程中所产生的具有抗病原体或其他活性的一类次级代谢产物,能干扰其他生活细胞发育功能的化学物质。
人体研究调查了抗生素介导的微生物耗竭对流感、脊髓灰质炎、轮状病毒、破伤风和卡介苗免疫反应的影响。没有观察到疫苗免疫原性改善或显著降低,这与抗生素使用、微生物群紊乱和免疫介导疾病的增加有关。
总之,某些细菌种类与婴儿疫苗应答者的体液或细胞免疫呈正相关或直接诱导增强。双歧杆菌是增强疫苗接种反应的良好候选者,也是健康早期肠道微生物群的关键成员。
其他可以刺激先天和适应性免疫反应的核心肠道微生物群成员,如拟杆菌属,也正在成为疫苗研究中的新型微生物群疗法。
扩展阅读:抗生素对微生物组及对人体健康的影响
双歧杆菌和拟杆菌的免疫调节
✦ 双歧杆菌减少炎症
在小鼠肥胖模型中,给予假链状双歧杆菌通过恢复调节性T细胞 (Treg) 和B淋巴细胞的平衡状态来减少全身炎症,并降低促炎细胞因子白细胞介素IL-17A和肿瘤坏死因子TNF的浓度。
白细胞介素——指在白细胞或免疫细胞间相互作用的淋巴因子,它和血细胞生长因子同属细胞因子。两者相互协调,相互作用,共同完成造血和免疫调节功能。
肿瘤坏死因子——血清中出现一种能使多种肿瘤发生出血性坏死的物质。
类似的免疫稳态特性已归因于双歧杆菌菌株,其通过体外刺激树突状细胞,诱导Th17谱并增强Treg细胞从幼稚淋巴细胞的分化。
双歧双歧杆菌DSM 20082裂解物刺激外周T细胞可增加CD8+ T细胞的细胞毒活性,而对CD4+ T细胞活性没有任何影响。
尽管支持这些免疫调节特性的机制尚不清楚,但已经提出了几个目标。
包括膳食发酵产物(例如,在人乳寡糖和其他复合碳水化合物代谢后),导致产生短链脂肪酸(即乙酸盐)和其他代谢副产物,这些副产物直接与宿主免疫细胞受体相互作用并促进其他共生体的交叉喂养细菌,如拟杆菌属和大肠杆菌。
长双歧杆菌亚种产生免疫调节化合物和蛋白质,如细胞外蛇毒蛋白,它不可逆地灭活促炎蛋白酶。
双歧杆菌MIMBb75之前已经证明其表面有肽聚糖水解酶TgaA,它通过启动白细胞介素-2生成和单核细胞衍生的树突状细胞激活,促进调节性T细胞扩增。
●双歧杆菌增强了免疫抗病毒反应
在新生仔猪中,长双歧杆菌AH1206菌株增强了肠道IL-10的产生,而动物双歧杆菌亚种乳酸Bb12、婴儿双歧杆菌MCC12和短双歧杆菌MCC1274促进了免疫成熟和免疫稳态。
更重要的是,MCC12和MCC1274 菌株在接种轮状病毒疫苗后增强了B细胞和抗病毒反应,表明双歧杆菌的免疫改变特征对某些菌株具有特异性。
●放线菌和双歧杆菌对人体免疫反应起促进作用
在人类中,几项研究强调了高丰度放线菌门和某些双歧杆菌菌株与对不同疫苗的免疫反应增加之间的正相关关系。
孟加拉婴儿胃肠道微生物组中高浓度的婴儿长双歧杆菌亚种与CD8+ T 细胞和CD4+ T 细胞反应增加以及接种卡介苗、破伤风和乙型肝炎疫苗后IgG滴度升高相关。
接种卡介苗、破伤风和乙型肝炎疫苗后,IgG滴度更高,在口服脊髓灰质炎疫苗接种后,双歧杆菌丰度高的中国婴儿表现出脊髓灰质特异性IgA应答增加。在疫苗补充研究中,B.longum BB536显示通过诱导干扰素-γ分泌来增强婴儿的Th1反应。
干扰素-γ——免疫干扰素,是由有丝分裂原刺激T淋巴细胞产生的。干扰素是一种高效的抗病毒生物活性物质,又是一种具有广泛免疫调节作用的淋巴因子。
扩展阅读:如何调节肠道菌群?常见天然物质、益生菌、益生元的介绍
✦拟杆菌影响疫苗免疫反应
拟杆菌属是婴儿期肠道定植的主要属,并且在整个成年生活中占主导地位。某些物种和菌株具有重要的互惠作用,从产生抗菌分子到通过分解不同聚糖提供营养。
关于拟杆菌和疫苗的反应性,存在相互矛盾的结果:
在加纳进行的轮状病毒试验发现,疫苗应答与拟杆菌门丰度呈负相关,而对巴基斯坦婴儿的一项类似研究显示,疫苗应答者中拟杆菌门的浓度增加。
观察到尼加拉瓜婴儿的疫苗反应者和无反应者中不同拟杆菌菌株的丰度存在差异。
注:由于样本量小,这些发现在多次调整后没有统计学意义,强调需要进行更大规模的研究来探索这些关联。
考虑到拟杆菌在诱导稳态免疫启动中的突出作用,可以预期拟杆菌和疫苗反应之间存在联系。
Bacteroides thetaiotaomicron中独特的寡糖结构具有佐剂特性,以依赖CD4+ T 细胞的方式诱导由乙型肝炎病毒疫苗引发的乙型肝炎病毒抗原特异性抗体浓度升高,这表明这种寡糖可作为大肠杆菌中毒性更大的脂多糖的替代佐剂。
短链脂肪酸是由肠道微生物群的不同成员通过发酵膳食复合碳水化合物(包括母乳或益生元中的碳水化合物)产生的。
短链脂肪酸在其一元羧酸碳链中含有少于六个碳原子,其中醋酸盐 (C2)、丙酸盐 (C3) 和丁酸盐 (C4) 最为普遍。它们是水溶性的,可以直接被不同的细胞吸收、转运或相互作用,包括肠上皮细胞、交感神经元和免疫细胞。
✦ 短链脂肪酸的多种益处
它们提供许多有益的健康影响,包括肠细胞的能量来源、加强上皮屏障、改变代谢过程、抑制肠道病原体生长、离子吸收的介质,以及作为肠道和全身免疫调节途径中的信号分子。
双歧杆菌和拟杆菌产生的乙酸,以及拟杆菌产生的丙酸盐,将在下面更详细地讲述。
●作为主要能源
乙酸盐可以酶促转化为乙酰辅酶A,并被许多不同的微生物群成员用来生产丁酸盐,并作为三羧酸循环中的主要能源。
●激活调节免疫细胞,增强疫苗反应
T细胞中细胞内乙酰辅酶A的增加会激活mTOR,从而驱动Th1和Th17 T细胞的分化。乙酸盐还可以激活B细胞、T细胞亚群、中性粒细胞、巨噬细胞、树突细胞和肠上皮细胞上表达的G蛋白偶联受体43(GPR43),导致固有层调节性T细胞增殖,和调节自身抗体的产生和边缘区B细胞。
mTOR是一种丝氨酸/苏氨酸蛋白激酶,对激素、生长因子、营养物质,能量和压力信号做出反应,是细胞代谢,生长和存货的中心调节因子。
GPR43的激活影响中性粒细胞趋化性和脱粒,以及脂肪组织中巨噬细胞产生肿瘤坏死因子。此外,缺乏GPR43的树突细胞无法诱导B细胞的类别转换。
没有产生短链脂肪酸的胃肠道细菌的小鼠的浆细胞分化减少,并且在稳态和病原体特异性抗体反应方面存在缺陷。乙酸盐可通过在体外增强针对霍乱毒素的抗原特异性IgA和IgG的产生以及刺激浆细胞分化所必需的树突状细胞中的信号分子来增强疫苗反应。
●改变巨噬细胞浓度
丙酸盐诱导小鼠调节性T细胞的分化和增殖以及白细胞介素10的表达。丙酸盐激活GPR15和GPR43还通过降低组蛋白去乙酰化酶6和9的表达以及抑制 NF-κB信号传导来增加结肠调节性T细胞的浓度。
丙酸盐激活GPR41会改变骨髓造血功能,导致巨噬细胞和树突状细胞前体浓度升高,并扭曲Th2分化。
✦短链脂肪酸是流感疫苗的潜在佐剂
一项调查短链脂肪酸对甲型流感病毒感染影响的研究揭示了短链脂肪酸(包括丙酸盐)通过激活GPR43来限制感染严重程度和伴随的肺炎球菌二重感染的重要作用。
GPR43被认为是甲型流感病毒进入的辅助受体
这意味着短链脂肪酸与该受体的结合抑制了病毒的进入和复制,这表明流感疫苗具有潜在的佐剂特性。
扩展阅读:你吃的膳食纤维对你有帮助吗?
胞外多糖是一些特殊微生物在生长代谢过程中分泌到细胞壁外、易与菌体分离、分泌到环境中的水溶性多糖,属于微生物的次级代谢产物。
胞外多糖是单糖或寡糖簇,包括形成同多糖或杂多糖的葡萄糖、果糖、半乳糖、岩藻糖和鼠李糖。
✦改变代谢物环境浓度,提供保护
这些可以分泌到肠道环境中或与亲本细菌的细胞壁有关。胞外多糖的表达增强了对宿主细胞的粘附,提供了对消化和环境压力的保护,并促进了生物膜的形成和胃肠道中的长期定植。
来自不同双歧杆菌菌株的胞外多糖可以被其他微生物发酵,从而改变代谢物环境和短链脂肪酸浓度。胞外多糖可以通过巨噬细胞和树突状细胞表面表达的特定模式识别受体(如TLR1、TLR2或TLR6)被识别为微生物相关的分子模式。
✦促进抗炎反应
受体的激活,取决于胞外多糖的化学物理特性(例如,分子量或电荷),导致产生不同的促炎和抗炎细胞因子谱,并以菌株特异性方式分别分化幼稚T细胞。
脆弱拟杆菌的表面多糖A可激活巨噬细胞上的Toll样受体2,并诱导调节性T细胞的扩增和抗炎白细胞介素10的产生,从而在病毒感染期间促进强烈的抗炎反应。
TLRs是一种模式识别受体家族,在先天免疫反应中起着重要作用。
✦增强对病毒的抵抗力
此外,多糖A激活结肠树突状细胞的Toll样受体2和肿瘤坏死因子分泌,增强了对病毒感染的天然抵抗力。
来自长双歧杆菌BCRC14634的胞外多糖被证明可增强巨噬细胞产生白细胞介素,与同基因的胞外多糖阴性突变体相比,来自B.longum亚种longum 35624的胞外多糖显示通过抑制促炎细胞因子产生来抑制促炎性Th17细胞的扩增。
在某些情况下,胞外多糖的存在与免疫沉默效应和逃避适应性B细胞反应有关,如在短双歧杆菌UCC2003中观察到的。
此外,这种双歧杆菌菌株和胞外多糖的存在也与较低丰度的促炎IFN-γ、肿瘤坏死因子和IL-12相关。
研究表明,青春双歧杆菌IF1-03通过增加巨噬细胞分泌IL-10来增加抗炎免疫反应,增加调节性T细胞浓度,这需要通过细胞外信号调节激酶或丝裂原活化蛋白激酶和NF-κB途径激活Toll样受体2和信号转导。
值得注意的是,在青春芽孢杆菌IF1-11产生的胞外多糖的情况下观察到相反的效果,它模拟巨噬细胞分泌高浓度的促炎性白细胞介素6、白细胞介素-17A 和转化生长因子-β,以及少量的白细胞介素10,随后将T细胞偏向Th17细胞。
转化生长因子-β(TGF-β)是属于一组新近发现的调节细胞生长和分化的TGF-β超家族
小结
这些研究展现了依赖于菌株的胞外多糖的免疫调节能力。以前使用乳酸杆菌的工作衍生的胞外多糖表明它们可以作为新型疫苗佐剂。因此,在特定疫苗接种后需要进一步测试关注拟杆菌和双歧杆菌相关胞外多糖的作用。
细菌细胞外囊泡是球形的膜衍生结构,大小从10纳米到 400 纳米不等,其中包含来自母细胞膜和周质的各种成分。
✦功能和分布
根据其膜组成和结构进行区分,其含量受环境因素(如培养条件或营养胁迫)的影响。这种影响可能导致DNA、RNA、脂多糖、酶、肽聚糖、毒素、信号分子、代谢物和毒力因子的数量和质量差异。
细菌细胞外囊泡并不局限于胃肠道,并且已在血液中检测到,它们可以从那里轻松进入不同的组织,包括大脑。
一项研究详细概述了不同的细菌膜外囊泡、它们的特性、功能和潜在应用。由共生细菌产生的细菌膜外囊泡有助于复杂微生物群落成员之间的合作和共养相互作用,并作为胃肠道微生物群成员与宿主之间跨界串扰的中介。
✦促进免疫反应
细菌细胞外囊泡可以激活免疫细胞并促进针对囊泡本身和亲代细胞的免疫反应。
膜脂、蛋白质(包括与微生物相关的分子模式)、危险信号和胞外多糖可以结合并激活模式识别受体和toll样受体,触发上皮细胞以外的不同免疫反应。
来自脆弱拟杆菌的囊泡携带荚膜多糖A,它激活黏膜下树突状细胞上的Toll样受体2,细胞外囊泡随后以肌动蛋白依赖性方式内化,导致白细胞介素10浓度增加和T细胞极化偏斜向调节性T细胞分化。
发现表明源于拟杆菌的胞外囊泡以物种特异性的方式在人类粘膜和血液树突状细胞中诱导白细胞介素10依赖性免疫调节反应。
✦为疫苗提供了多种可能性
细菌细胞外囊泡的非复制性及其内在的佐剂性、热稳定性以及对低pH值和酶降解的抵抗力,为疫苗设计和交付提供了多种可能性。
它们可以在不注射的情况下直接给药至粘膜部位(例如胃肠道和呼吸道),从而降低给药成本并降低与肠外分娩相关的潜在不良反应的风险。
此外,含有免疫原性成分的细菌细胞外囊泡可以促进强烈的先天性和适应性免疫反应,并提供大量针对传染病的保护。
✦呈递抗原,降低毒性
来自病原菌的细菌细胞外囊泡已成功用于疫苗制剂,一些针对霍乱弧菌和B Neisseria meningitidis的细胞外囊泡疫苗已获得许可,其中B Neisseria meningitidis疫苗显示出针对淋球菌的潜在跨物种保护作用。
其他研究表明,由微生物群成员产生的细菌细胞外囊泡,包括生物工程细菌细胞外囊泡,已被用于传递病原体的抗原。
源于拟杆菌的、表达不同鼠疫耶尔森菌抗原的细菌细胞外囊泡在体内诱导了特异性和强免疫反应,包括血清IgG和粘膜IgA,它们能够清除鼠疫感染。
来自突变的非致病性大肠杆菌的细菌细胞外囊泡对抗原特异性T细胞反应具有佐剂特性,并降低了毒性。
人类肠道微生物群正在成为疫苗反应性的重要决定因素,其中双歧杆菌和拟杆菌能够影响免疫和个体疫苗免疫反应。
这两个属是发育中的健康婴儿肠道微生物群的重要组成部分,但极易受到早期生活干扰的影响,例如剖腹产、配方奶与母乳以及抗生素的使用。
专注于这些促进健康的菌群,并利用它们的免疫调节特性,可能会导致更安全的方法来增强婴儿免疫力和疫苗效力。使用整个细菌或其产物和代谢物来调节免疫反应的新策略是可能的,例如在癌症中观察到对免疫检查点抑制剂的反应增强。
未来展望
对来自健康婴儿微生物组的关键菌株及其副产物和代谢物有更深入的了解和表征,可以催生新一代安全、无针和经济的疫苗促进疗法。关键特征,如剂量、疫苗设计和给药时间,将是重要的考虑因素。
主要参考文献:
Jordan A, Carding SR, Hall LJ. The early-life gut microbiome and vaccine efficacy. Lancet Microbe. 2022 Sep 8:S2666-5247(22)00185-9. doi:
10.1016/S2666-5247(22)00185-9. Epub ahead of print. PMID: 36088916.
Praharaj I, John SM, Bandyopadhyay R, Kang G.
Probiotics, antibiotics and the immune responses to vaccines.Philos Trans R Soc Lond B Biol Sci. 2015; 37020140144.
Ciabattini A, Olivieri R, Lazzeri E, Medaglini D.
Role of the microbiota in the modulation of vaccine immune responses.Front Microbiol. 2019; 101305.
Hill DL, Carr EJ, Rutishauser T, et al.Immune system development varies according to age, location, and anemia in African children.Sci Transl Med. 2020; 12eaaw9522.
Parker EPK, Bronowski C, Sindhu KNC, et al.Impact of maternal antibodies and microbiota development on the immunogenicity of oral rotavirus vaccine in African, Indian, and European infants.Nat Commun. 2021; 127288.