-

 国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子
国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子

 二级病原微生物安全实验室
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业

谷禾健康

人体复杂的内部环境中栖息着种类繁多、功能各异的微生物群,这些微生物群落对人类的基本生理功能和各种疾病状态都产生着深刻而持续的影响。
人类胃肠道系统代表着宿主机体与宿主相关微生物群落之间最为重要和最大规模的生物接触界面。长期定植于这一特殊环境中的常驻细菌群体,经过漫长的进化历程,已经发展出高度专门化和精细调节的生物学机制,以实现对现有营养物质的最优化获取和高效利用,这些独特的适应性机制使得它们能够在激烈的生存竞争中胜过其他微生物,并在整个肠道的不同区域中成功建立起稳定而特定的微生物群落结构。
宿主与微生物群之间的众多复杂相互作用主要基于各种代谢物的交换和信号传递,因此无论是在生理稳态条件还是在各种疾病状态下,这些相互作用都会受到微生物代谢活动和环境中营养物质可用性的显著而持续的影响。
举例来说,结肠组织在正常生理稳态条件下维持着严格的厌氧环境特征,这种特殊的环境条件特别有利于那些主要通过底物水平磷酸化过程获取生存必需能量的专性厌氧微生物的成功定植和繁殖。
然而,当机体处于各种疾病条件下时,无论这些病理状态是由外来病原体感染引起,还是由非感染性自身免疫疾病如溃疡性结肠炎和克罗恩病等慢性炎症性疾病所导致,肠道上皮细胞的代谢模式都会发生显著变化,这些变化最终导致原本稀缺的氧气分子流入肠腔空间,从而为那些能够有效利用氧气进行有氧呼吸的兼性厌氧菌创造了有利的生长环境,促进了这类微生物的快速扩增和繁殖。
微生物对碳源和能量的高效获取能力始终是决定群落组成结构和功能表现的最主要驱动力量。肠道微生物群和各种肠道病原体经过长期的适应性进化,已经成功发展出多种多样的生物学机制来获得膳食来源的碳水化合物如糖醇类化合物和Amadori产物,以及来源于宿主机体的各种内源性代谢物如与粘蛋白分子密切相关的代谢产物。
注:Amadori产物是糖类与氨基酸或蛋白质发生非酶糖化反应(Maillard反应)的重要中间产物。
除此之外,微生物对各种关键微量营养素的成功获得,包括多种维生素化合物和重要的金属元素如铁、锰、铜、锌、钼、镍等,也在很大程度上控制和调节着微生物群落的整体结构组织。
本文将全面而深入地介绍肠道共生细菌和病原菌如何采用各种不同的生物学策略来获取和代谢肠道环境中丰富的宏量营养素,以实现高效的能量产生并促进其在宿主肠道中的成功定植和长期生存。
★ 不同细菌的能量代谢来源各异
环境细菌和人体相关细菌表现出惊人的代谢多样性。微生物生理学的一个关键决定因素是能量代谢,微生物根据其首选的碳源(有机物与CO2)、能源来源(阳光与化学反应)和氢供体来源(有机与H2O)可大致分类。
任何生物体要想存活,必须通过化学反应产生足够的吉布斯自由能来合成ATP并建立离子梯度。
注:吉布斯自由能可以简单理解为反应的”净可用能量”。
例如,硫杆菌属(Thiobacillus spp)等硫氧化细菌使用元素硫、硫化氢(H2S)或硫代硫酸盐作为电子供体,以氧为电子受体生成硫酸盐和其他氧化硫化合物;并通过电子传递链建立质子动力。
而大多数肠道共生菌降解有机化合物,从膳食和宿主来源(化学有机异养生物)中获得能量和代谢中间体。虽然肠道微生物营养策略多样,但人体结构严格限制了不同肠段栖息的细菌类型及其代谢模式。
▸ 肠道不同部位微生物群结构和代谢不同
人体已经进化出专门的机制,使特定微生物在肠道不同部位选择性定植,从而塑造微生物群落结构和代谢模式。
饮食和宿主因素决定了肠道微生物群的代谢

Muramatsu MK,et al.Cell Host Microbe.2024
★ 小肠以需氧和兼性厌氧菌为主,密度较低
小肠pH值从胃部酸性逐渐升至回肠末端的微酸性或中性,影响微生物群落多样性。十二指肠和空肠主要栖息耐酸需氧菌和兼性厌氧菌,如乳酸杆菌属(Lactobacillus)和链球菌属(Streptococcus)以及变形菌门(假单胞菌属)。
此外,肝胆系统释放的初级胆汁酸(BA)和牛磺酸或甘氨酸偶联的胆汁酸(CBA)以及潘氏细胞释放的抗菌肽抑制了小肠中许多细菌的生长。因此,小肠微生物密度远低于大肠,从近端至远端小肠的每克约103-7个细菌增加到结肠中的每克1011-12个细菌。
★ 大肠环境有利于专性厌氧菌定植
大肠塑造了与小肠截然不同的微生物群落结构和代谢模式。结肠的主要过滤器是缺氧环境,有利于专性厌氧菌定植。
结肠细胞大量消耗氧气进行β氧化,维持上皮缺氧状态,限制氧气从血管向肠腔扩散,促进厌氧厚壁菌门和拟杆菌门等专性厌氧菌定植。微生物产生的丁酸盐促进结肠细胞β氧化,形成宿主-微生物代谢反馈回路。由于氧气稀缺,兼性厌氧菌仅占次要地位。
大部分胆盐在回肠被吸收,剩余的初级胆盐由大肠微生物代谢。饮食营养在胃中分解后,单糖和氨基酸在小肠被吸收,而不可消化的碳水化合物(膳食纤维)和多元醇进入结肠,成为细菌的主要碳源和能源。
肠道微生物群的代谢构成了人体内最为活跃和复杂的生化反应工厂,这一庞大的微观生态系统依靠多元化的能量来源维持着自身的生存繁殖和功能发挥。肠道微生物主要是将宿主无法消化的复杂碳水化合物、膳食纤维、抗性淀粉以及内源性黏蛋白转化为可被利用的能量物质。
▸ 降解复杂多糖
由于大多数简单的营养物质在小肠中被吸收,因此大肠微生物群的许多成员主要碳和能量来源是膳食复杂多糖(纤维)。这类分子包括植物来源的聚糖,如纤维素、半纤维素、β-葡聚糖、菊粉和果胶,以及动物来源的糖原。
★ 拟杆菌等菌属具有强大的复合多糖降解能力
复杂多糖含有多样化的糖苷键,需要特异性糖苷水解酶降解。拟杆菌属等革兰氏阴性专性厌氧菌,如 Bacteroides spp. 和 Prevotella spp.,在复杂多糖的降解方面表现极强的能力。这些细菌拥有大量的碳水化合物活性酶(CAZymes),例如由不同多糖利用位点编码的碳水化合物结合蛋白、糖苷水解酶和多糖裂解酶。
拟杆菌淀粉利用系统(SUS)是多糖利用位点的经典例子:淀粉结合蛋白(SusD、SusE、SusF)识别并固定细菌表面的淀粉分子,糖苷水解酶(SusG)将其降解为寡糖,寡糖通过外膜蛋白SusC转运至周质,再被糖苷水解酶(SusA、SusB)进一步降解为单糖,最终转运至细胞质并发酵产生乙酸、琥珀酸和丙酸。拟杆菌编码众多SUS样系统和CAZymes,与膳食聚糖的多样性相匹配。
B.thetaiotaomicron中的淀粉利用系统(SUS)

Muramatsu MK,et al.Cell Host Microbe.2024
★ 毛螺菌和瘤胃球菌也能降解复杂多糖并产生短链脂肪酸
毛螺菌科(Lachnospiraceae)和瘤胃球菌科(Ruminococcaceae)等革兰氏阳性梭菌同样降解复合多糖并产生短链脂肪酸。
革兰氏阳性菌的降解机制涉及细胞外CAZymes和高亲和力转运蛋白(如ATP结合盒转运蛋白、主要促进子超家族转运蛋白和磷酸转移酶系统),底物特异性酶活性通常以基因簇形式编码。
注:虽然纤维素体降解植物细胞壁的能力在瘤胃微生物中常见,但在大多数人类肠道微生物中缺失,仅在农村和狩猎采集人群的微生物组中观察到此类活性。
▸ 代谢膳食糖醇和多元醇
多元醇是由糖类通过醛或酮基还原形成醇基而天然合成生成的糖醇。自然界中发现的主要多元醇包括赤藓糖醇、山梨糖醇、木糖醇、甘露醇和麦芽糖醇等天然形式,以及乳糖醇、异麦芽酮糖醇和氢化淀粉水解物等合成形式。
由于在胃肠道中吸收不完全,多元醇常用作低卡路里人造甜味剂。高脂肪饮食联合抗菌治疗会消耗氧敏感的多元醇降解梭菌,导致山梨糖醇不耐受。
★ 大肠杆菌能够利用甘露醇
甘露醇代谢在多种细菌中被广泛研究。在大肠杆菌中,甘露醇通过特异性磷酸烯醇式丙酮酸磷酸转移酶系统被吸收并磷酸化为甘露醇-1-磷酸,随后被甘露醇-1-磷酸脱氢酶转化为果糖-6-磷酸进入糖酵解途径。
另一途径是甘露醇以半乳糖基-甘露醇偶联物形式进入细胞,被β-半乳糖苷酶水解后,甘露醇通过甘露醇-2-脱氢酶转化为果糖,再被己糖激酶磷酸化。
★ 双歧杆菌代谢异麦芽酮糖醇并产生丁酸盐
除了甘露醇外,其他多元醇同样可被肠道微生物代谢。中等剂量的异麦芽酮糖醇和乳糖醇摄入后,肠道微生物群组成偏向双歧杆菌,为某些共生菌提供生长优势。体外实验显示双歧杆菌可代谢异麦芽酮糖醇,增加丁酸盐产生,这对维持结肠上皮细胞厌氧环境至关重要。
★ 沙门氏菌会与共生大肠杆菌竞争半乳糖醇
半乳糖醇天然存在于某些植物中,酵母也可产生。沙门氏菌通过gat操纵子利用半乳糖醇,包含磷酸转移酶系统gatABC将其转运并磷酸化,最终转化为磷酸二羟基丙酮和3-磷酸甘油醛。半乳糖醇是定植抗性的关键代谢物,沙门氏菌与共生大肠杆菌竞争这一有限资源。
▸ 粘蛋白:微生物的”应急储备”
肠道微生物群落还可分为管腔、黏液相关和上皮/隐窝相关群体。黏蛋白形成重要屏障,保护肠道上皮免受微生物侵害,其中MUC2是主要类型。小肠黏液层呈多孔状,含抗菌肽(如肠道α防御素)和凝集素(如RegIIIγ),限制微生物与上皮相互作用的同时允许营养吸收。
★ 食物缺乏时,某些细菌会消化黏蛋白
结肠黏液则形成细菌无法穿透的致密内层和松散外层,而嗜黏蛋白阿克曼菌(Akkermansia muciniphila)等可利用外层获取营养。在没有膳食纤维的情况下,肠道微生物开始消化粘蛋白,增加了患结肠炎的风险。
黏蛋白是富含脯氨酸、苏氨酸和丝氨酸的大糖蛋白家族。丝氨酸和苏氨酸残基作为N-乙酰半乳糖胺(GalNAc)的O-连接糖基化附着点,可被半乳糖和N-乙酰葡糖胺等单糖进一步修饰,形成长支链聚糖并用唾液酸、岩藻糖和硫酸盐残基末端修饰。
十二指肠、空肠和回肠的黏蛋白高度唾液酸化和硫酸化,结肠黏蛋白则含唾液酸化、硫酸化和岩藻糖化聚糖。近端结肠唾液酸化程度较高,远端结肠硫酸化程度较高。
类似膳食聚糖降解,黏蛋白的微生物消化需要多种降解菌(如嗜黏蛋白阿克曼菌及拟杆菌门、厚壁菌门成员)协同形成营养网络。降解过程由岩藻糖苷酶和唾液酸酶水解末端糖结构,硫酸酯酶去除硫酸盐基团引发。
★ 降解过程中可为其他细菌提供营养
去除这些末端结构可能会成为粘蛋白降解的潜在瓶颈,因为这些末端结构可以保护底层聚糖链免受糖基水解酶的降解,并且需要特定的酶。肠道微生物群的不同成员编码的酶可以去除这些结构并使其成为其他细菌的营养来源。
例如,扭链瘤胃球菌(Ruminococcus torques)很容易使用细胞外CAZymes消化肠粘蛋白上的聚糖。Bacteroides thetaiotaomicron通常更喜欢膳食聚糖而不是粘蛋白,并且在体外使用粘蛋白生长不佳。但R.torques产生的低聚糖的交叉喂养促进了B.thetaiotaomicron在粘蛋白上的生长。
同样,卵形拟杆菌(Bacteroides ovatus)和Roseburia intestinalis降解半纤维素成分β-甘露聚糖。而普拉梭菌(Faecalibacterium Prausnitzii)无法自行有效降解 β-甘露聚糖,但可以获得卵形拟杆菌和R.intestinalis释放的甘露寡糖。
这些例子强调了肠道营养网络的重要性以及定植于肠道的一些细菌之间的互惠关系。
唾液酸和岩藻糖降解—糖利用的例子
唾液酸和岩藻糖是复合多糖降解的常见中间体,为肠道微生物糖利用提供典型例子。
唾液酸的代谢
唾液酸(N-乙酰神经氨酸,Neu5Ac)是一种9碳单糖,通过α2-3/6键与半乳糖和GalNAc残基结合,覆盖胃肠道黏蛋白聚糖链末端。人乙状结肠MUC2分析显示,结肠中最常见表位是与α2-6 N-乙酰半乳糖胺醇结合的Neu5Ac,而小鼠中与GlcNAc结合的Neu5Ac更常见。
两物种的微生物群均编码特异性唾液酸酶,可识别特定糖苷键并去除末端Neu5Ac残基,启动糖苷水解酶的分解代谢。
部分细菌(如脆弱拟杆菌)可直接裂解和代谢唾液酸,而其他细菌仅能清除游离唾液酸分子。唾液酸转运(NanT)后,醛缩酶(NanA)将Neu5Ac代谢为丙酮酸和N-乙酰甘露糖胺(ManNAc)。ManNAc经ManNAc激酶(NanK)磷酸化,再由差向异构酶(NanE)转化为磷酸化N-乙酰葡糖胺(GlcNAc-6-P),最终代谢为果糖-6-磷酸进入糖酵解途径。
扭链瘤胃球菌(Ruminococcus torques)采用独特的唾液酸代谢策略获得竞争优势。其编码的分子内反式唾液酸酶在糖蛋白裂解时产生2,7-脱水-N-乙酰神经氨酸(2,7-anhydro-Neu5Ac)而非唾液酸,阻止其他利用唾液酸的细菌使用该底物。进入细菌细胞后,新型氧化还原酶(NanOx)将2,7-脱水-Neu5Ac转化为唾液酸。
肺炎链球菌和沙门氏菌等病原体中也检测到NanOx直系同源物和推定的2,7-脱水-Neu5Ac转运蛋白,表明该化合物可能是病原体体内定植的重要代谢物。
岩藻糖的代谢
L-岩藻糖是一种6碳脱氧糖,广泛存在于生命各分支。人和小鼠中,岩藻糖基转移酶Fut2负责胃肠道大部分岩藻糖基化,将岩藻糖附着在聚糖的基端α(1,6)和末端α(1,2)、(1,3)或(1,4)位。胃肠道中,岩藻糖通过肠道微生物分泌的α-L-岩藻糖苷酶从膳食或宿主黏蛋白聚糖中释放,随后被部分肠道微生物吸收代谢。
大肠杆菌的岩藻糖利用已被广泛研究,其编码岩藻糖利用操纵子(fucOAPIKR)。岩藻糖通过专用通透酶(FucP)导入细胞,经异构酶FucI转化为L-岩藻酮糖,再被激酶FucK磷酸化。岩藻酮糖1-磷酸被醛缩酶(FucA)裂解为乳醛和DHAP。
厌氧条件下,乳醛被氧化还原酶(FucO)还原为1,2-丙二醇;有氧条件下(如肠道炎症期间),乳醛转化为乳酸并氧化为丙酮酸进一步代谢。缺乏fucK和fucAO的大肠杆菌突变体虽能在小鼠肠道初步定植,但无法长期稳定定植,表明岩藻糖获取和利用能力是维持哺乳动物肠道环境的关键。多形拟杆菌和脆弱拟杆菌也利用岩藻糖在小肠中保持竞争优势。
有趣的是,共生肠道细菌可诱导小鼠肠道宿主Fut2表达,形成富含岩藻糖的生态位,既保护宿主聚糖又允许岩藻营养细菌扩增。无菌小鼠经常规小鼠粪便浆液处理可诱导小肠岩藻糖基化。广谱抗生素处理的常规小鼠杯状和柱状上皮细胞岩藻糖基化严重减少,表明共生菌在宿主Fut2表达和肠聚糖岩藻糖基化中发挥作用。
一些细菌的存在,如丝状细菌、多形拟杆菌和沙门氏菌等细菌可在无菌小鼠回肠诱导宿主岩藻糖基化。虽然肠道共生体诱导Fut2介导岩藻糖基化的信号分子仍不明确,但葡聚糖硫酸钠和霍乱毒素等促炎环境压力源可诱导岩藻糖基化。
除饮食偏好(高脂肪西餐vs富含纤维饮食)、食物类别(蔬菜vs肉类)和食品添加剂外,食物制备方法也影响微生物代谢。
▸ 食物制备方法会影响微生物代谢
法国科学家Maillard最早描述了一种导致烹饪中观察到的褐变效应的反应,称为美拉德反应。当单糖羰基与氨基酸氨基反应时产生Amadori产物,其聚合形成更复杂的类黑精。常见食品加工方法(加热、干燥)产生多种Amadori化合物,如果糖-谷氨酰胺(F-Gln)、果糖-苯丙氨酸(F-Phe)和果糖-天冬酰胺(F-Asn)。
注:美拉德反应(Maillard reaction)是食品工业中广泛存在的一种非酶褐变现象,由还原糖与氨基化合物(如氨基酸、蛋白质)在常温或加热条件下发生复杂反应,生成类黑精等棕色物质,并产生大量风味物质。类黑精是指含有半缩醛羟基的化合物(醛、还原糖)与含有氨基的化合物等经缩合、聚合反应生成的高分子量聚合物。
★ 肠道共生菌和病原菌均可利用美拉德反应产物
人肠道分离株肠单胞菌AF211(毛螺菌科成员)可代谢果糖赖氨酸(F-Lys)并产生丁酸盐、乳酸盐和氨。F-Lys通过ABC转运蛋白输入,被YhfQ磷酸化形成6-磷酸果糖-赖氨酸,再被6-磷酸果糖-赖氨酸脱糖酶(Yhfn)转化为6-磷酸葡萄糖(G6P)和赖氨酸。
肠道病原体沙门氏菌通过fra操纵子(fraRBDAE)利用F-Asn作为碳氮源。F-Asn首先被果葡天冬酰胺酶FraE转化为果糖-天冬氨酸(F-Asp),再由F-Asp转运蛋白FraA导入。随后F-Asp被FraD磷酸化形成F-Asp-6-磷酸,再被脱糖酶FraB代谢为G6P和天冬氨酸。
▸ 发酵是能源生产的主要模式
发酵作为厌氧环境中的关键生化过程,构成了肠道微生物能源生产的主要模式,这一代谢途径在维持微生物群落的生存和功能发挥方面起着至关重要的作用。
肠道微生物会使用大量的分子,特别是通过复合多糖降解释放的单糖以及氨基酸(Stickland 反应),进行发酵。
注:Stickland反应为专性厌氧细菌的梭菌属中常见的一种反应。
★ 发酵产生乳酸、短链脂肪酸等产物
发酵过程中需在细菌细胞内平衡氧化还原反应,避免还原当量(如NADH)积累。维持适当的NAD+/NADH比率对氧化还原稳态和NAD+依赖性酶功能至关重要。
最简单的发酵中,葡萄糖在Embden-Meyerhof-Parnas途径中转化为两个丙酮酸分子,净产生两个ATP和四个还原当量。丙酮酸代谢物用于恢复NAD+/NADH平衡,如将丙酮酸还原为乳酸(乳酸发酵)或将乙醛还原为乙醇(乙醇发酵)。肠道发酵的常见终产物包括乳酸、甲酸、琥珀酸及短链脂肪酸(乙酸、丙酸、丁酸)。
一些发酵途径广泛存在,如从丙酮酸产生乙酸的Pta-AckA途径。铁氧还蛋白氧化还原酶、丙酮酸甲酸裂解酶或丙酮酸脱氢酶将丙酮酸转化为乙酰辅酶A,再经磷酸乙酰转移酶转化为乙酰磷酸。乙酸激酶活性从乙酰磷酸和ADP生成ATP,同时产生乙酸。
发酵是肠道共生细菌产生细胞能量的主要方式
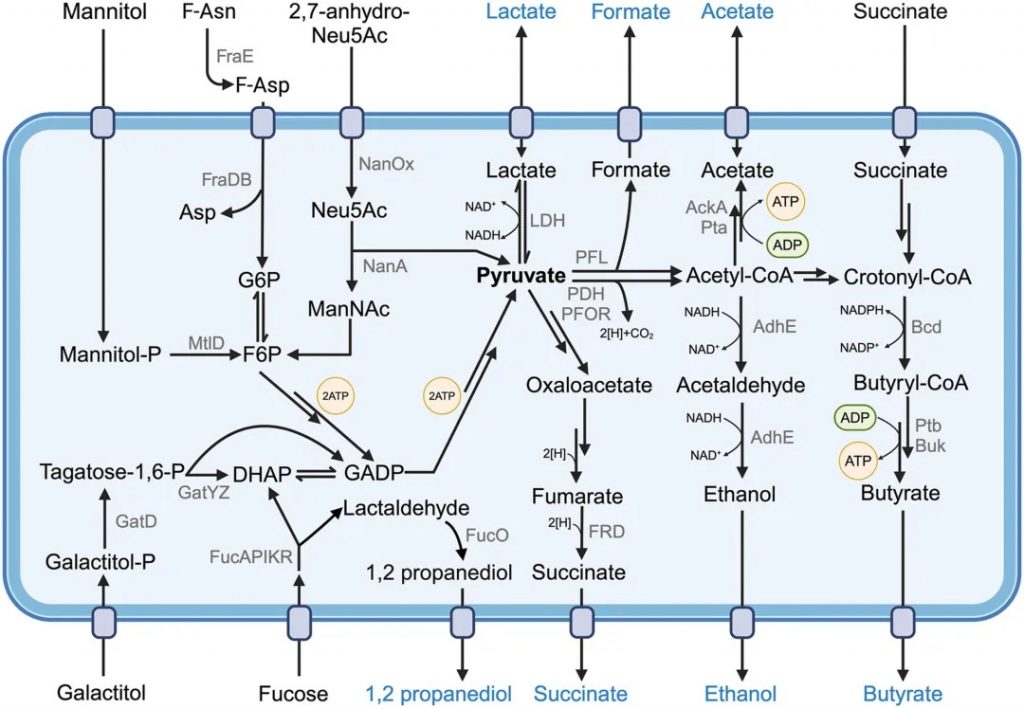
Muramatsu MK,et al.Cell Host Microbe.2024
★ 短链脂肪酸变化可反映肠道健康状态改变
丁酸主要由毛螺菌科和瘤胃球菌科成员通过聚糖直接降解或交叉发酵乳酸、乙酸、琥珀酸产生。两个乙酰辅酶A分子融合形成乙酰乙酰辅酶A,经数步转化为巴豆酰辅酶A和丁酰辅酶A,平衡NAD+/NADH池。类似Pta-AckA途径,丁酰辅酶A产生ATP并形成丁酸。另外,琥珀酸可在多步过程中还原为4-羟基丁酰辅酶A,再转化为巴豆酰辅酶A和丁酰辅酶A。戊二酸和赖氨酸也可转化为巴豆酰辅酶A。
短链脂肪酸的产生是关键代谢输出,指导结肠上皮进行β氧化,维持肠腔厌氧环境。该过程在肠道病原体感染、口服抗菌剂或高脂饮食时被破坏,导致肠道微生物群显著改变、丁酸产生减少和兼性厌氧菌扩增。
★ Stickland反应是肠道代谢物的重要来源
Stickland反应是某些肠道细菌产生能量的特殊发酵过程。在此反应中,成对氨基酸同时脱氨,一个被氧化,另一个被还原,产生ATP和NAD+。该反应主要见于梭菌属,已鉴定出特定的供体氨基酸(丙氨酸、缬氨酸、丝氨酸)和受体氨基酸(甘氨酸、脯氨酸、天冬氨酸)组合。氧化反应中供电子氨基酸转化为羧酸并产生ATP;还原反应中受电子氨基酸将NADH氧化为NAD+,产生乙酸、5-氨基戊酸、异己酸等代谢物。
编码甘氨酸还原酶途径基因的生物体可通过甘氨酸还原产生ATP、氨和乙酸。氨基酸氧化或还原可补充其他能量产生形式。丙氨酸和丝氨酸氧化脱氨产生的丙酮酸可用于TCA循环。氨基酸还原发酵通过再生NAD+等高能电子载体辅助糖酵解。编码和使用红杆菌氮酶(RNF)样复合物的生物体可将氨基酸代谢与质子驱动力产生结合以提高ATP产量。在厌氧菌具核梭杆菌(Fusobacterium nucleatum)中,RNF复合物的破坏导致ATP生成减少、氨基酸代谢减少和毒力受损。
肠道细菌中的Stickland发酵和硫代谢

Muramatsu MK,et al.Cell Host Microbe.2024
Stickland反应产生大量在小鼠肠道和人血中发现的代谢物。人肠道共生梭状芽孢杆菌(C.sporogenes)通过丝氨酸脱水酶氧化发酵丝氨酸和还原精氨酸产生ATP。由脯氨酸和芳香族氨基酸Stickland反应形成的还原代谢物(5-氨基戊酸、苯丙酸、3-(4-羟基苯基)丙酸、吲哚丙酸)也存在于人血中,表明还原Stickland反应衍生代谢物可离开肠道进入循环系统。这表明Stickland反应是哺乳动物肠道和远端部位代谢物的重要但被低估的来源。
★ 艰难梭菌的胶原蛋白降解与脯氨酸利用策略
许多依赖Stickland反应的细菌具有蛋白水解作用,可通过细胞外蛋白酶快速降解蛋白质。肠道病原体艰难梭菌导致宿主释放胶原蛋白应对毒素介导的炎症。胶原纤维主要由脯氨酸、甘氨酸和羟脯氨酸组成。艰难梭菌降解这些纤维,将反式-4-羟基-L-脯氨酸转化为L-脯氨酸,通过prd操纵子进行还原发酵产能。缺乏4-羟脯氨酸脱水酶HypD的突变菌株致病力受损。其他梭状芽胞杆菌如Paeniclostridium spp.在体外与艰难梭菌竞争脯氨酸,影响其发酵能力。
综上所述,这表明反式-4-羟基-L-脯氨酸的利用是艰难梭菌用于在体内获得脯氨酸的一种策略,并且脯氨酸发酵在艰难梭菌的体内存活中发挥作用。
呼吸作用过程中,电子通过电子传递链转移至外源电子受体,通过酶复合物质子泵活动或标量化学形成质子动力。
厌氧呼吸,特别是富马酸还原,在肠道细菌中常见。伯克氏菌科(Burkholderiaceae)、爱格氏菌属(Eggerthella)和丹毒丝菌科(Erysipelotrichaceae) 成员对膳食电子受体表现出种属特异性利用。
▸ 胆汁酸的利用
胆汁酸(BA)是消化系统重要组成部分。肝脏产生的主要胆汁酸包括胆酸(CA)和鹅去氧胆酸(CDCA),分泌前与牛磺酸或甘氨酸结合。人体内牛磺酸与甘氨酸结合胆汁酸的比例取决于饮食,牛磺酸主要来源于肉类、鱼类和贝类。
★ 胆汁酸有助于发酵过程中的能量转化
胆汁酸(BA)经细菌活动进一步修饰,包括转化和解偶联。在胆酸(CA)向脱氧胆酸和CDCA/熊去氧胆酸向石胆酸的多步转化中,7-羟基被去除,使梭菌科和爱格氏菌属(Eggerthella)能够将NADH转化为NADPH,有助于发酵过程中维持理想的NAD+/NADH比率。共生微生物从BA中解离牛磺酸,导致肠道游离牛磺酸浓度升高。胆盐水解酶通过催化类固醇部分C-24位置与BA氨基酸侧链间的酰胺键水解促进解偶联。
★ 肠道存在多种胆盐代谢细菌,包括致病菌
肠道微生物群含有许多编码不同底物特异性胆盐水解酶的细菌。厚壁菌门和放线菌门成员可降解大多数结合胆汁酸,而拟杆菌门菌株偏爱牛磺酸结合胆汁酸,双歧杆菌属和约氏乳杆菌(Lactobacillus johnsonii)乳杆菌等富含胆盐水解酶。结合胆汁酸的解离高度依赖于产胆盐水解酶微生物的位置:小鼠近端小肠中的乳酸杆菌开始解离,而人类中解离直到末端回肠和结肠才发生。
释放的牛磺酸可被不同肠道微生物利用。沃氏嗜胆菌(Bilophila wadsworthia)是肠道微生物群的亚硫酸盐还原致病菌,虽仅占正常肠道微生物群的0.01%,但与多种临床疾病相关。当IL-10缺陷小鼠饲喂低脂饮食并补充牛磺酸结合胆汁酸时,会导致肠道B.wadsworthia种群激增。B.wadsworthia利用异化亚硫酸盐还原酶复合物(Dsr)从有机磺酸盐释放的亚硫酸盐产生H2S。
牛磺酸转运入细胞后,B.wadsworthia利用两个基因簇(ald-tpa-sarD和adhE-islA-islB)代谢牛磺酸。牛磺酸首先被Tpa和Ald转化为磺基乙醛,再被SarD转化为乙硫磺酸盐。异羟乙基磺酸进入细菌微区室后,IslAB形成甘油自由基酶,促进C-S键裂解,将其转化为乙醛和亚硫酸盐。乙醛被AdhE代谢成乙酰辅酶A,再通过Pta-AckA途径转化为乙酸,而亚硫酸盐被Dsr系统用作亚硫酸盐呼吸中的电子受体。
▸ 硫酸酯酶和硫酸盐的利用
多糖常被硫酸盐修饰,需释放硫酸盐基团才能接触糖基团。GlcNAc和半乳糖的硫酸化发生在不同羟基位置,特别是GlcNAc的6-羟基(6S-GlcNAc)和半乳糖的3-、4-或6-位置。利用O-糖核心结构中这些硫酸盐基团需要能识别并裂解特定糖苷键的碳水化合物硫酸酯酶。
★ 硫酸酯酶表达菌释放硫酸盐供硫酸盐还原菌利用
表达硫酸酯酶的细菌释放的硫酸盐通过交叉喂养被硫酸盐还原细菌(SRB)利用。SRB存在于约50%的人群中,进行异化硫酸盐还原,将硫酸盐还原为腺苷-5′-磷酸硫酸盐、亚硫酸盐,并进一步还原为H2S。与缺乏成熟硫酸酯酶的B.thetaiotaomicron 菌株相比,当与从肠道粘蛋白中释放硫酸盐的B.thetaiotaomicron菌株共定植时,其在体内的相对丰度增强。H2S具有剧毒,宿主将其解毒为硫代硫酸盐,后者可用作Desulfovibrio spp.的电子受体。
▸ 外部电子转移
寻找合适的电子供体和受体对是细菌产能的关键。部分细菌已开发利用外源性电子受体的策略。外部电子转移(EET)连接细胞质氧化还原反应,将电子转移至与细菌直接接触或远端的外部电子受体。该过程已在环境细菌中广泛描述,近期在哺乳动物微生物群和肠道病原体中也有报道。
★ 细菌具有直接电子转移和介导电子转移两类
革兰氏阴性菌的胞质膜、肽聚糖层和外膜对EET构成物理屏障。为克服这些障碍,电活性细菌进化出两类方法:直接电子转移和介导电子转移(MET)。直接电子转移主要见于环境生物体如Shewanella oneidensis,电子直接传输至与外膜接触的外部电子受体。
MET则需要电子穿梭介导电子从细菌转移至外部电子受体。铜绿假单胞菌释放苯嗪(含氮杂环化合物)将电子转移至分子氧,植物乳杆菌利用醌1,4-二羟基-2-萘甲酸还原铁。P.prausnitzii与上皮细胞相关,该专性厌氧菌可能使用黄素和硫醇作为细胞外电子穿梭来减少宿主组织释放的氧气。
★ 细菌获取外部电子的方式影响其环境适应性
在革兰氏阳性肠道病原体单核细胞增生李斯特菌中,基于黄素的EET(FLEET)途径将NADH脱氢酶(Ndh2)产生的电子穿梭至质膜中脂溶性去甲基甲萘醌衍生物,再转移至膜结合脂蛋白(PplA)上的黄素或黄素单核苷酸基团,最终传递给末端电子受体。
研究表明,单核细胞增生李斯特菌厌氧条件下代谢糖醇需要FLEET通路,ndh2突变体在小鼠肠道定植能力受损。FLEET通路基因的直系同源物在数百种厚壁菌门和人类病原体中均有发现。这些研究突出了电子受体的重要性,表明细菌已进化出复杂机制获取外部电子受体池,以增强其在特定环境中的生长能力。
▸ 肠道炎症时的能量代谢
肠道炎症会导致微生物群水平变化,变形菌门(尤其是兼性厌氧肠杆菌科)丰度增加。呼吸电子受体的释放是这些群落变化的关键驱动因素,重塑了肠道细菌的能量代谢。
★ 电子受体的释放影响肠道菌群的能量代谢
例如,沙门氏菌(Salmonella)诱导胃肠炎时,活性氧(ROS)将宿主产生的硫代硫酸盐氧化为连四硫酸盐。沙门氏菌编码连四硫酸盐利用基因簇(ttrRSBCA),实现厌氧连四硫酸盐呼吸。炎性ROS和活性氮(RNS)分解产生硝酸盐,为沙门氏菌提供高能电子受体。
肠道炎症改变了末端电子受体的可用性

Muramatsu MK,et al.Cell Host Microbe.2024
★ 炎症导致氧气泄漏促进需氧病原体增殖
此外,炎症相关的结肠细胞代谢变化使氧气泄漏至肠腔,支持肠道病原体生长。电子受体的可用性不仅实现高效的电子传输产能,还可利用发酵终产物(如琥珀酸、乳酸和1,2-丙二醇),将其完全氧化为二氧化碳。
类似的机制正在驱动非感染性结肠炎期间肠道微生物群的水平变化。硝酸盐和氧气呼吸有助于共生肠杆菌科细菌的繁殖,例如非感染性结肠炎小鼠模型中的大肠杆菌、肠杆菌属和克雷伯氏菌属。上皮细胞释放的活性氧被过氧化氢酶解毒,过氧化氢酶是一种产生分子氧的反应,支持大肠杆菌的呼吸。
此外,口服抗菌治疗期间产生丁酸盐的梭状芽胞杆菌的耗竭会改变结肠细胞代谢;氧气的流入导致肠杆菌科种群的扩大。
呼吸电子受体可用性增加引起微生物群水平变化表明,碳和氮并非细菌生长的限制因素,而能量代谢和从有限底物库产能的能力才是肠道微生物群组成和功能的关键决定因素。
本文介绍了共生菌和病原菌在肠道中获取营养素和产生能量的机制。细菌可氧化有机和无机化合物产生生长必需的能量。在不同微环境中的产能能力是群落结构的关键决定因素。
人体肠道包含多个动态生态位,肠道微生物群在受宿主营养摄入、栖息地过滤器和疾病炎症影响的环境中调节能量产生。
人类微生物组研究传统上侧重于识别与健康和疾病相关的关键细菌种类(如炎症性肠病、艰难梭菌感染、抗生素相关菌群失调等)。但宿主-微生物和微生物间相互作用及其代谢机制同样重要。研究显示,细菌有效利用特定营养物质的能力(无论独立利用还是通过细菌营养网络)是微生物群组成和功能的关键驱动因素。
对宿主健康而言,微生物执行的代谢功能非常重要。这在致病性菌群相关的生态失调中显而易见,共生微生物因面临新代谢环境和成员间相互作用挑战而异常增殖并致病。
发现疾病相关代谢途径对个性化医疗具有重要意义。未来,随着对肠道微生物代谢功能理解的不断深入,我们有望开发出更加精准的个性化医疗策略,包括恢复特定代谢缺陷的微生物疗法,以及深入理解宿主-微生物代谢相互作用的分子机制。通过调节肠道微环境、优化营养网络以及恢复关键代谢途径,我们将能够更有效地治疗与微生物群失调相关的疾病,为人类健康开辟新的治疗途径。
主要参考文献
Muramatsu MK, Winter SE. Nutrient acquisition strategies by gut microbes. Cell Host Microbe. 2024 Jun 12;32(6):863-874.
Yersin S, Vonaesch P. Small intestinal microbiota: from taxonomic composition to metabolism. Trends Microbiol. 2024 Oct;32(10):970-983.
Murdoch CC, Skaar EP. Nutritional immunity: the battle for nutrient metals at the host-pathogen interface. Nat Rev Microbiol. 2022 Nov;20(11):657-670.
Miller BM, Bäumler AJ. The Habitat Filters of Microbiota-Nourishing Immunity. Annu Rev Immunol. 2021 Apr 26;39:1-18.
Salzman NH, Hung K, Haribhai D, Chu H, Karlsson-Sjöberg J, Amir E, Teggatz P, Barman M, Hayward M, Eastwood D, Stoel M, Zhou Y, Sodergren E, Weinstock GM, Bevins CL, Williams CB, Bos NA. Enteric defensins are essential regulators of intestinal microbial ecology. Nat Immunol. 2010 Jan;11(1):76-83.
La Rosa SL, Ostrowski MP, Vera-Ponce de León A, McKee LS, Larsbrink J, Eijsink VG, Lowe EC, Martens EC, Pope PB. Glycan processing in gut microbiomes. Curr Opin Microbiol. 2022 Jun;67:102143.
Eberl C, Weiss AS, Jochum LM, Durai Raj AC, Ring D, Hussain S, Herp S, Meng C, Kleigrewe K, Gigl M, Basic M, Stecher B. E. coli enhance colonization resistance against Salmonella Typhimurium by competing for galactitol, a context-dependent limiting carbon source. Cell Host Microbe. 2021 Nov 10;29(11):1680-1692.e7.
Schaus SR, Vasconcelos Periera G, Luis AS, Madlambayan E, Terrapon N, Ostrowski MP, Jin C, Hansson GC, Martens EC. Ruminococcus torques is a keystone degrader of intestinal mucin glycoprotein, releasing oligosaccharides used by Bacteroides thetaiotaomicron. bioRxiv [Preprint]. 2024 Jan 16:2024.01.15.575725.

谷禾健康

你可知道,为什么明明摄入了很多的蛋白质,仍然可能出现氨基酸缺乏的情况吗?谷禾今天就带你深入了解这一现象背后的各种奥秘和原因,帮助你更好地理解蛋白质的摄入与氨基酸利用关系。
造成这种情况的原因可以主要分为几个方面,我们将逐一详细讲述:
一、内源性氨基酸的产生不足。这些氨基酸是在体内合成的,主要通过蛋白质的分解与转化而来。然而,内源性氨基酸的生成受到多种因素的影响,比如身体的整体代谢状态、营养状况、肝脏的功能等。如果这些影响因素出现问题,就会导致体内内源性氨基酸的产生不足,从而影响整体的氨基酸平衡。特别值得注意的是,肠道菌群在这个过程中扮演着关键角色,它们不仅参与氨基酸的合成,还能影响宿主的代谢状态。如果肠道菌群失衡,可能会影响内源性氨基酸的产生效率。
二、外源性蛋白质的消化和吸收率不高。外源性蛋白质来自我们摄入的食物,经过消化过程转化为氨基酸。但这个过程的效率不总是很高,因素如食物的种类、加工方式、个人的消化功能等都会影响外源性蛋白质的消化和氨基酸的吸收。健康的肠道菌群能够提高蛋白质的消化效率,产生有益的代谢物,并维持肠道屏障功能,从而促进氨基酸的吸收,如果肠道菌群失衡,消化吸收不良,即使摄入了足够的蛋白质,身体也无法有效利用其中的氨基酸。
三、氨基酸的消耗过大,导致消耗量超过了摄入量。这就像一个游泳池,放水速度比注水速度快,那么即使持续地注入水,也很难增加池内的水量。而肠道菌群的代谢活动也会影响氨基酸的消耗速率,某些菌群可能会竞争利用宿主的氨基酸资源。
每个人对氨基酸的需求和消耗情况是不同的,这和个体的活动水平、身体状况以及生理需求都有很大关系,同时,每个人的肠道菌群组成都是独特的,这种个体差异会影响到氨基酸的代谢和利用效率。有些人可能会在日常生活中消耗掉大量氨基酸,导致最终的总量不足。因此,在摄入氨基酸时,需要综合考虑个体需求的差异,关注肠道健康状况,以确保氨基酸的充分吸收和利用,满足身体所需。
本文我们一起来了解一下,蛋白质在胃肠道内的消化过程,详细讨论影响氨基酸消化和吸收的关键因素,肠道菌群如何影响氨基酸的吸收,探讨根据不同人群的特点(如素食者、运动人群、孕妇等)来优化蛋白质的摄入策略,以及如何通过调节肠道菌群来提高氨基酸的利用效率,从而更好地把握蛋白质摄入与氨基酸利用的关系,制定有效的营养策略。
▸ 氨基酸的定义
氨基酸(amino acid)是含有氨基和羧基的一类有机化合物,是促进人体生长,维持身体正常代谢,影响生命活动的重要物质,也是合成机体所需抗体、激素、酶类的原料。
如果氨基酸代谢异常,可能导致生长发育、全身稳态受损,甚至死亡。总的来说,氨基酸作为生命体的最基本组成单位,在人体扮演着重要角色。
▸ 氨基酸与蛋白质的关系
蛋白质我们都知道,人体必需的营养素之一。它是有机大分子,是构成细胞的基本有机物,是生命活动的主要承担者。可以这么说,没有蛋白质就没有生命。
而氨基酸是蛋白质的基本组成单位。蛋白质和氨基酸之间的关系是:氨基酸通过脱水缩合形成肽键,进而连接形成肽链,肽链折叠盘旋形成具有空间结构的蛋白质。
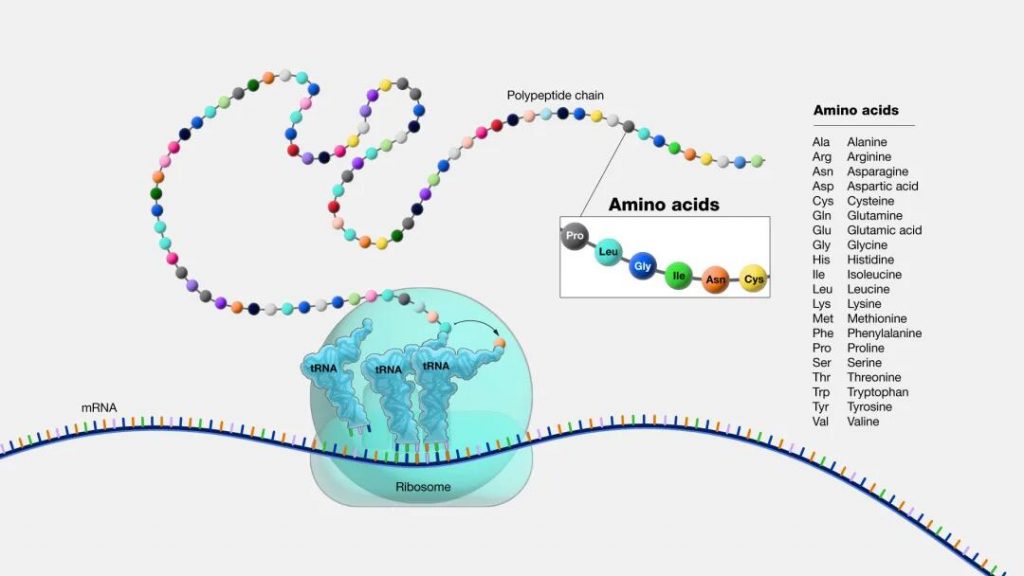
图源:National Human Genome Research Institute
氨基酸的种类、数目、排列方式、多肽链的盘曲、折叠方式及其空间结构的不同导致了蛋白质结构的多样性,其结构的多样性导致了功能的多样性。
通俗地讲:氨基酸是构成蛋白质的小分子“积木”。想象一下,蛋白质就像一串珠子项链,而每颗珠子就是一个氨基酸。它们通过不同的组合和排列形成各种蛋白质。
通常,我们体内的氨基酸以游离氨基酸总量计算,包括两部分,一是内源氨基酸,即体内组织蛋白降解产生或自身合成的非必需氨基酸;二是外源氨基酸,即食物蛋白质经消化吸收而来。两者共同分布于身体各处,参与代谢,总称为氨基酸代谢库。
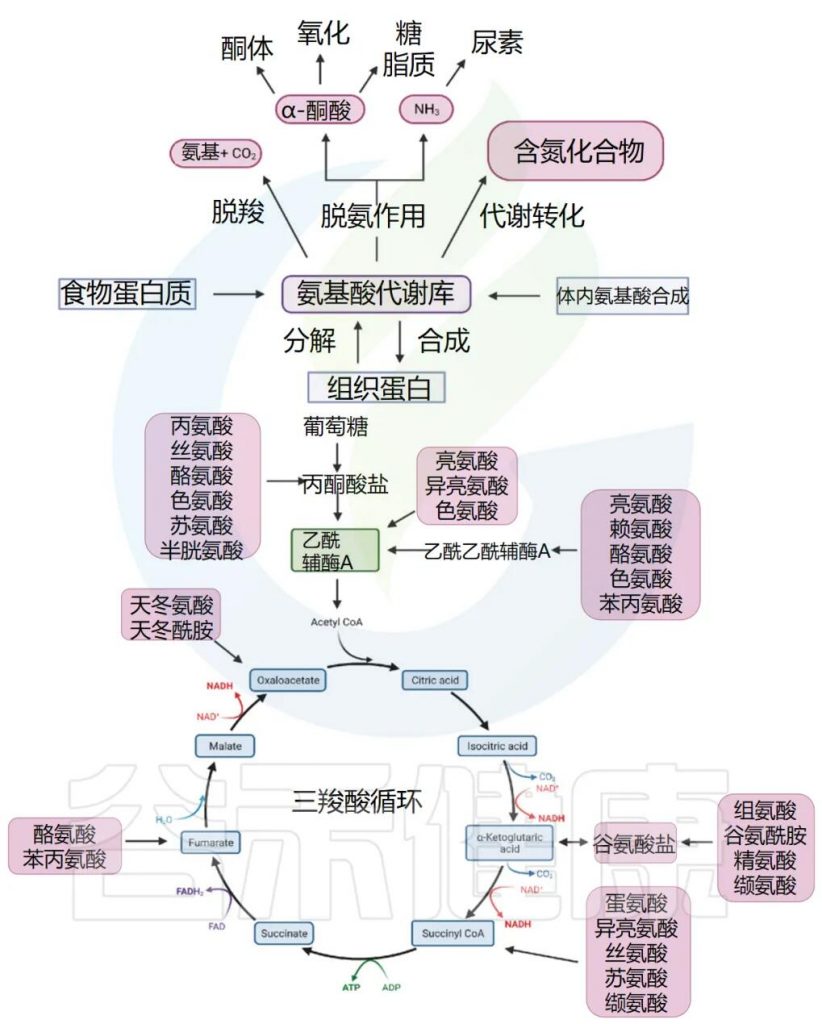
1
内源性氨基酸的合成
◆ 大多数非必需氨基酸可通过转氨基作用合成
转氨基作用指的是一种α-氨基酸的α-氨基转移到一种α-酮酸上的过程,可以看成是氨基酸的氨基与α-酮酸的酮基进行了交换。生成相应的α-氨基酸;而原来的α-氨基酸则转变为相应的α-酮酸。
例如,天冬氨酸是三羧酸循环中间产物草酰乙酸经转氨作用得到,谷氨酸是三羧酸循环中间产物α-酮戊二酸经转氨作用得到,丙氨酸是糖酵解生成的丙酮酸经转氨作用得到。
一般有以下几类:
1) 谷氨酸族:以a-酮戊二酸为起始物,可合成Glu,GIn,Pro,Arg(称为谷氨酸族);
2) 天冬氨酸族:以草酰乙酸为起始物,可合成Asp,Asn,Met,Thr,Lys,le(称为天冬氨酸族);
3)丙氨酸族:以丙酮酸为起始物,可合成Ala,Val,Leu(称为丙氨酸族);
4)丝氨酸族:以甘油酸-3-磷酸为起始物,可合成Ser,Gly,Cys(丝氨酸族);
5)芳香族氨基酸:以赤藓糖-4-磷酸、磷酸烯醇式丙酮酸为起始物,可合成芳香族氨基酸(Phe,Tyr,Trp);
6)组氨酸(His):由核糖-5-磷酸衍生,核糖-5-磷酸通过磷酸戊糖途径产生。
◆ 通过必需氨基酸转化
从上述合成的过程可以发现,氨基酸的种类发生了改变,数量并没有发生改变。那么,有没有不是通过转氨基途径合成氨基酸呢?答案肯定是有的,如下所示是半胱氨酸的合成示意图:
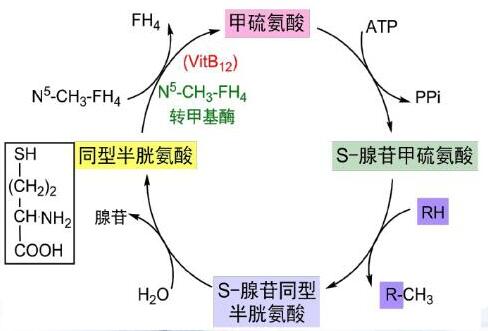
半胱氨酸在体内可以由甲硫氨酸转化而成,而这一过程必须有维生素B6参与。
必需氨基酸还可以通过苯丙氨酸羟化酶催化合成非必需氨基酸酪氨酸,如果缺少这种酶,转化过程很难进行,苯丙氨酸就会转化为苯丙酮酸,这就是苯丙酮尿症的原因。
2
影响内源性氨基酸合成的因素
▸ 营养因素
•必需氨基酸的充足供应
虽然非必需氨基酸可以在体内合成,但合成过程中往往需要以必需氨基酸为前体或底物。如果必需氨基酸缺乏,可能会影响非必需氨基酸的合成途径,导致非必需氨基酸合成减少。
例如,当饮食中缺乏蛋氨酸时,可能会影响半胱氨酸的合成,因为半胱氨酸可以由蛋氨酸转化而来。
•能量供应
非必需氨基酸的合成需要消耗能量,因此充足的能量供应对于非必需氨基酸的合成至关重要。如果能量摄入不足,身体会优先利用能量来维持生命活动的基本需求,从而可能减少非必需氨基酸的合成。
在饥饿或能量摄入不足的情况下,身体会分解蛋白质来提供能量,这会导致非必需氨基酸的合成减少。
•维生素和矿物质的供应
某些维生素和矿物质是参与非必需氨基酸合成的酶的辅助因子或激活剂。例如,维生素B6是参与氨基酸代谢的多种酶的辅酶,缺乏维生素B6可能会导致谷氨酸合成谷氨酰胺的过程受阻。锌、镁等矿物质也可能参与某些氨基酸合成酶的活性调节。
▸ 生理因素
•年龄和生长发育阶段
不同年龄和生长发育阶段的人对非必需氨基酸的合成能力可能不同。婴幼儿和青少年时期,身体生长发育迅速,对蛋白质和氨基酸的需求较大,非必需氨基酸的合成能力也相对较强。
随着年龄的增长,身体的代谢功能逐渐下降,非必需氨基酸的合成能力可能会减弱。因此老年人可能需要更多的优质蛋白质补充来满足身体对氨基酸的需求。
•健康状况和疾病状态
某些疾病可能会影响非必需氨基酸的合成。例如,肝脏疾病可能会影响氨基酸的代谢和转化,导致非必需氨基酸的合成减少。肾脏疾病可能会影响氨基酸的排泄和重吸收,从而影响体内氨基酸的平衡。
此外,一些慢性疾病如糖尿病、癌症等也可能影响非必需氨基酸的合成,因为这些疾病会导致身体的代谢紊乱和营养消耗增加。
•激素水平
激素可以调节身体的代谢过程,包括氨基酸的合成和分解。例如,生长激素可以促进蛋白质的合成和细胞的生长,从而可能增加非必需氨基酸的合成。胰岛素可以促进葡萄糖的摄取和利用,同时也可以促进氨基酸的摄取和蛋白质的合成,可能对非必需氨基酸的合成有一定的促进作用。
相反,一些应激激素如皮质醇可能会促进蛋白质的分解,从而可能减少非必需氨基酸的合成。
▸ 环境因素
•饮食结构和饮食习惯
长期的不良饮食结构和饮食习惯可能会影响非必需氨基酸的合成。例如,高糖、高脂肪、低蛋白的饮食可能会导致蛋白质摄入不足,从而影响非必需氨基酸的合成。过度饮酒、吸烟等不良习惯也可能对身体的代谢功能产生不良影响,从而影响非必需氨基酸的合成。
此外,一些特殊的饮食限制,如素食主义、低蛋白饮食等,需要更加关注非必需氨基酸的合成问题,以确保身体的正常代谢需求。
•运动和体力活动
适度的运动和体力活动可以促进身体的代谢功能,运动可以增加肌肉的蛋白质合成,从而可能增加对非必需氨基酸的需求和合成。
然而,过度的运动和体力活动可能会导致蛋白质分解增加,从而影响非必需氨基酸的合成。此外,运动过程中如果没有及时补充足够的营养物质,也会影响非必需氨基酸的合成。
•环境压力和应激
长期处于高压力、高应激的环境中,会影响身体的代谢功能,从而影响非必需氨基酸的合成。在应对环境压力和应激时,身体会优先利用能量和营养物质来维持生命活动的基本需求,从而可能减少非必需氨基酸的合成。
当这些因素受到影响时,会导致内源性氨基酸的产生不足。这种不足会进一步影响人体内氨基酸的整体水平,使得总氨基酸的水平较低。
还有一些氨基酸人体不能合成或合成速度远不能适应人体需要,必须通过食物中的蛋白质补充,也称为必需氨基酸。
1
蛋白质和多肽在肠道内的消化
以前,蛋白质消化和氨基酸吸收被认为是截然不同的独立过程;现在人们认识到,蛋白质分解成更小的单位,随后被肠细胞吸收,并将氨基酸释放到门静脉血液中,这是一个高度组织和紧密结合的过程。
消化涉及食物基质、自身物理过程和哺乳动物及细菌酶的化学过程之间的多种相互作用,所有这些都发生在胃肠道的复杂环境中。
•大分子蛋白质先被胃蛋白酶水解成多肽
摄入的食物在口腔中经过咀嚼发生物理分解,食物与唾液等液体混合,产生可被吞咽进入胃部的食物丸。胃蛋白酶在不同的裂解点水解蛋白质,形成更小的多肽,胃酸使蛋白质变性并部分展开,帮助胃蛋白酶更好地进入它们的键。
随后,剩余的蛋白胨和多肽被转移到小肠中,在小肠中胰腺产生和分泌多种蛋白酶和肽酶,如肠黏膜的胰蛋白酶、羧肽酶。上皮细胞的刷状缘膜结合酶包括许多氨基肽酶和一种羧肽酶,并作用于胰腺消化后留下的任何寡肽。
•多肽再被切割成各个氨基酸
胰腺蛋白酶将多肽切割成寡肽和氨基酸。胃蛋白酶、胰蛋白酶、凝乳胰蛋白酶和弹性酶都属于内肽酶,它们攻击某些内部键,产生大的多肽。
注:胃蛋白酶在芳香氨基酸如苯丙氨酸、色氨酸和酪氨酸旁切割消化键,而胰蛋白酶在碱性氨基酸精氨酸和赖氨酸旁切割键。
在这些内肽酶作用后剩余的寡肽受到外肽酶的攻击,如羧基肽酶A和B,它们每次从多肽或蛋白质的羧基或氨基端切割一个氨基酸。
羧基肽酶从肽链的羧基端切除氨基酸,羧基肽酶A优先释放缬氨酸、亮氨酸、异亮氨酸和丙氨酸,羧基肽酶B优先释放碱性氨基酸如精氨酸和赖氨酸。
2
氨基酸的吸收
蛋白质消化的产物,主要是游离氨基酸、二肽和三肽,通过几种运输机制从小肠腔吸收。
肽和氨基酸通过不同的转运体被小肠肠细胞吸收。空肠近端是肽和氨基酸吸收的主要部位,但小肠的其他部位也具有显著的运输能力。
短肽(二肽和三肽)通过肠细胞顶膜中的H+依赖性肽转运体(如PepT1)转运进入肠细胞,并在上皮细胞中进一步水解为氨基酸。
•不同的氨基酸有特定的转运系统
游离氨基酸(FAAs)被碱性、阳离子、中性氨基酸以及亮氨酸和甘氨酸转运体以梯度依赖的方式通过肠上皮细胞的刷状边界膜吸收。有许多刷状边缘(顶端膜)和基底侧膜转运系统,特定于特定的氨基酸或氨基酸群。
例如,浓缩氨基酸转运蛋白在顶膜上表达,用于吸收中性氨基酸(B0AT1,广义中性氨基酸转运蛋白 1,SLC6A19)、阳离子氨基酸(囊胚中性和阳离子氨基酸转运蛋白,SLC7A9)、甘氨酸和脯氨酸 (PAT1,质子氨基酸转运蛋白1,SLC36A1;SIT,系统亚氨基转运蛋白,SLC6A20) ,阴离子氨基酸(兴奋性氨基酸转运蛋白3(EAAT3),SLC1A1)和β-氨基酸(PAT1,SLC36A1,TauT,牛磺酸转运蛋白,SLC6A6)。
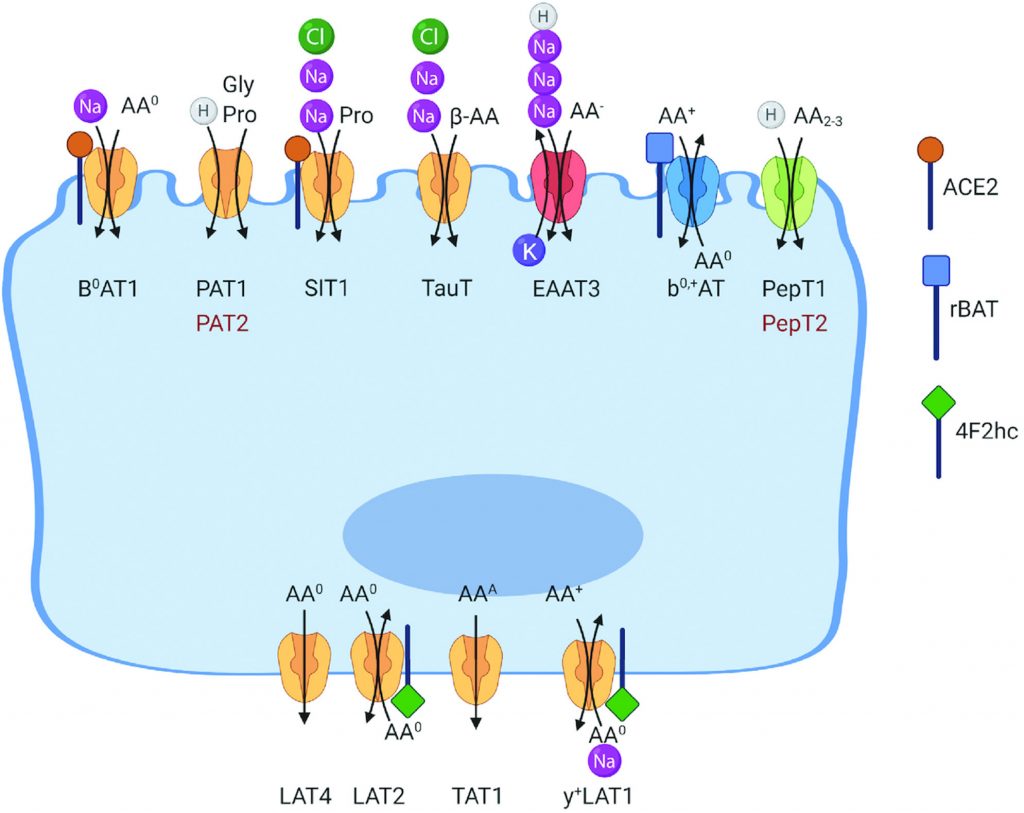
doi: 10.1093/jn/nxab342.
•小肠可以适应性地上调氨基酸吸收能力
人体从膳食蛋白质中获得氨基酸的净正流量是至关重要的,因此小肠具有适应性地上调其氨基酸吸收的能力。个体的膳食蛋白质摄入量和生理状态(影响氨基酸需求)都可能随着时间的推移而改变,氨基酸吸收机制可以在组织和细胞水平上进行相应的调整。例如转运蛋白上调和粘膜增生的共同作用可导致氨基酸吸收能力比禁食状态增加数倍。
继基底外侧膜转运到间质液后,氨基酸通过绒毛毛细血管进入门静脉。门静脉直接供应肝脏,在那里氨基酸可以进一步代谢或运输到其他器官和组织。
3
影响膳食氨基酸消化和吸收的因素
虽然膳食蛋白质通常在胃肠道内被消化,但它们并不一定被完全消化和吸收。此外,不同的氨基酸从蛋白质中释放出来,并在不同程度上被肠道吸收。因此,有必要了解膳食蛋白质和氨基酸的消化和吸收率。影响膳食氨基酸消化和吸收的因素主要有以下几个方面:
▸ 食物因素
•蛋白质的来源和性质
不同来源的蛋白质其氨基酸组成和结构不同,消化和吸收的难易程度也有所差异。例如,动物蛋白(如肉类、蛋类、奶类)通常含有较高比例的优质蛋白质,其氨基酸组成与人体需求较为接近,且消化率较高。
蛋白质的消化是一个有效的过程,从鸡蛋中粗蛋白的97%消化率到谷物中的≥70%不等。
蛋白质的结构也会影响消化。例如,胶原蛋白等结构紧密的蛋白质较难被消化酶分解,而球状蛋白等结构较为松散的蛋白质则相对容易消化。研究表明,与酪蛋白相比,乳清蛋白的消化和吸收速度更快。
•食物的加工和烹饪方式
适当的加工和烹饪可以提高蛋白质的消化率。例如,加热可以使蛋白质变性,使其结构变得松散,更容易被消化酶作用。此外,研磨、破碎等加工方式可以增加蛋白质与消化酶的接触面积,促进消化。
然而,过度加工和烹饪可能会破坏蛋白质的结构和营养成分,降低其消化率。例如,高温油炸、长时间炖煮等可能会导致蛋白质发生美拉德反应,产生难以消化的化合物,同时也会破坏一些必需氨基酸。当被吸收时,通常不被利用,而是被分解并随尿液排出。
•抗营养因子
抗营养因子是一类能够干扰人体对营养素的吸收和利用的物质。食品中的胰蛋白酶抑制剂、单宁和植酸盐等化合物会显著降低蛋白质和氨基酸的消化率。此外,还可能会与蛋白质结合,形成难以消化的复合物,影响蛋白质的消化和吸收。
▸ 消化系统因素
•消化酶的分泌和活性
胃液中的胃蛋白酶、胰液中的胰蛋白酶、糜蛋白酶等消化酶在蛋白质的消化过程中起着关键作用。消化酶的分泌量和活性受到多种因素的影响,如年龄、健康状况、饮食等。老年人的消化酶分泌量可能会减少,导致蛋白质消化能力下降。某些疾病(如胰腺疾病、胃肠道疾病)也会影响消化酶的分泌和活性。
此外,消化酶的活性还受到pH值、温度等环境因素的影响。例如,胃蛋白酶在酸性环境下活性较高,而胰蛋白酶等则在碱性环境下活性较高。
•胃肠道的生理状态
胃肠道的蠕动、分泌、吸收等生理功能对氨基酸的消化和吸收至关重要。胃肠道蠕动过慢会导致食物在胃肠道内停留时间过长,影响消化和吸收;而蠕动过快则可能使食物来不及充分消化就被排出体外。
胃肠道的分泌功能也会影响消化。例如,胃液、胰液、胆汁等消化液的分泌不足会影响蛋白质的消化。此外,胃肠道的吸收面积和功能也会影响氨基酸的吸收。例如,小肠黏膜的损伤会导致吸收面积减少或吸收功能障碍。
▸ 个体因素
•年龄
婴幼儿的消化系统尚未发育完全,消化酶的分泌和活性较低,胃肠道的吸收能力也较弱。随着年龄的增长,消化系统的功能逐渐成熟,但老年人的消化和吸收功能又会逐渐下降。
不同年龄段的人对蛋白质的需求和消化吸收能力不同,因此在饮食中应根据年龄特点选择合适的蛋白质来源和摄入量。
•健康状况
一些疾病会影响蛋白质的消化和吸收。例如,胃肠道疾病(如胃炎、胃溃疡、肠炎等)、肝脏疾病、肾脏疾病等都可能会影响消化酶的分泌、胃肠道的蠕动和吸收功能。此外,一些慢性疾病(如糖尿病、心血管疾病等)也可能通过影响代谢和营养状态间接影响蛋白质的消化和吸收。
营养不良、消瘦、肥胖等也可能影响蛋白质的消化和吸收。营养不良可能导致消化酶的合成减少、胃肠道黏膜萎缩等,从而影响消化和吸收功能;而肥胖可能会引起胃肠道激素分泌异常、肠道菌群失调,也会影响蛋白质的消化和吸收。
•遗传因素
个体的遗传差异可能会影响蛋白质的消化和吸收。例如,某些人可能存在特定的基因突变,导致消化酶的活性或结构异常,从而影响蛋白质的消化。此外,遗传因素还可能影响胃肠道的生理功能、肠道菌群的组成等,进而影响蛋白质的消化和吸收。
▸ 其他因素
•饮食中的其他成分
饮食中过多的膳食纤维、脂肪等成分可能会影响蛋白质的消化和吸收。膳食纤维可以促进肠道蠕动,但过多的膳食纤维可能会吸附蛋白质和消化酶,影响蛋白质的消化。脂肪可以延缓胃排空,使蛋白质在胃中停留时间延长,有利于胃蛋白酶的作用,但过多的脂肪可能会引起消化不良。
•药物和补充剂
某些药物会影响蛋白质的消化和吸收。例如,抗生素可能会破坏肠道菌群,影响消化酶的合成和活性;抗酸药会改变胃肠道的pH值,影响消化酶的活性。一些补充剂,如蛋白粉、氨基酸补充剂等,如果使用不当也可能会影响蛋白质的消化和吸收。
在使用药物和补充剂时,应注意其对蛋白质消化和吸收的影响,并在医生或营养师的指导下合理使用。
•肠道菌群
肠道微生物群在氨基酸的调节、消化和吸收过程中发挥着至关重要的作用,它们通过多种方式来影响氨基酸的利用效率。在下一章节中,我们将详细探讨肠道微生物群如何在氨基酸的消化、吸收及利用中发挥作用,并分析其对整体健康的影响。
氨基酸对各种生物活性化合物的合成至关重要,这些化合物在信号通路和代谢中发挥关键的调节作用。肠道微生物群在促进氨基酸调节以及氨基酸消化和吸收过程中起着不可缺少的作用。
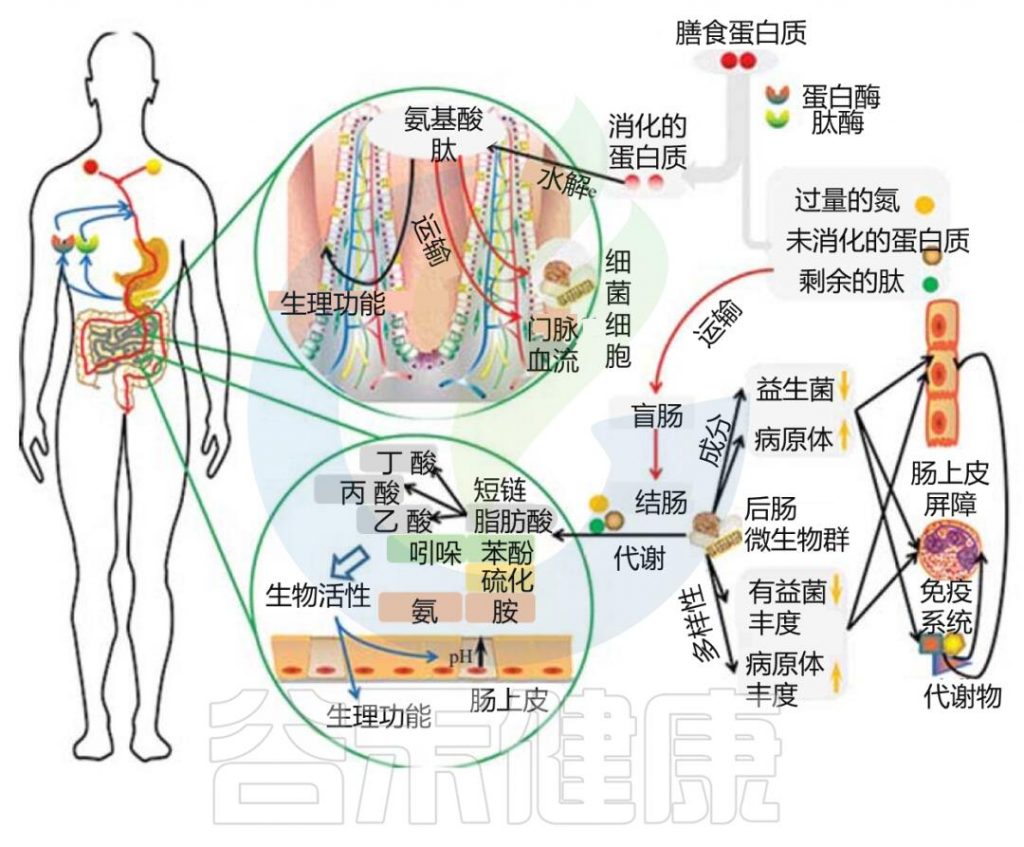
doi: 10.2174/1389203719666180514145437.
1
参与氨基酸代谢和吸收
过去对肠道微生物群和氨基酸相互作用的研究表明,许多肠道细菌参与了胃肠道中蛋白质的消化和氨基酸的吸收。
•一些肠道细菌能够产生肽酶,用于降解蛋白质
蛋白质的转换主要发生在肠道中,结肠细菌降解内源性或外源性蛋白质的效率较高。拟杆菌属(Bacteroides)、梭状芽胞杆菌属(Clostridium)、丙酸杆菌属(Propionibacterium)、梭杆菌属(Fusobacterium)、乳酸杆菌属(Lactobacillus)和链球菌属(Streptococcus)在蛋白质水解过程中发挥着重要作用。
其中一些细菌可以直接代谢氨基酸,并具有分泌各种蛋白酶和肽酶的能力。发酵过程中,复杂蛋白质首先被各种细菌肽酶、蛋白酶和内肽酶切割,释放出游离氨基酸和短肽。然后对氨基酸和短肽进行发酵。产生支链脂肪酸(2-甲基丁酸酯、异丁酸酯、异戊酸酯)、有机酸、气体(H2和CO2)以及微量酚、胺、吲哚和氨。
肠道内的梭菌属细菌(赖氨酸或脯氨酸利用的基础细菌)是氨基酸发酵的关键驱动因素,而消化链球菌属细菌是谷氨酸或色氨酸利用的关键驱动因素。

Yadav M,et al.Arch Microbiol.2018
其他例如瘤胃细菌,Selenomonas ruminantium, Megasphaera elsdenii, Prevotella ruminicola, Misuokella multiacidas, Butyrivibrio fibrisolvens, Streptococcus bovis等含有极为活跃的二肽基肽酶和二肽酶。
•代谢氨基酸的基因广泛分布于人类肠道细菌

研究了来自不同系统发育群体的肠道细菌中相关代谢酶及其同源物的分布情况。对代表10个门的380种肠道细菌的基因组做了blast比对。
结果表明,大多数这些酶广泛分布在肠道细菌中,但有些酶表现出不同的分布模式。例如,L-天冬酰胺酶在拟杆菌门(Bacteroidetes)和假单胞菌门(Proteobacteria)中普遍存在,而芳香族氨基酸转氨酶和精氨酸脱亚胺酶主要存在于厚壁菌门(Firmicutes)和放线菌门(Actinomycetota)中。
支链氨基酸转氨酶仅在拟杆菌门和厚壁菌门中观察到,而前消旋酶主要在厚壁菌门中普遍存在。此外,大多数拟杆菌表现出快速消耗天冬酰胺(Asn)的能力,而L-脯氨酸降解菌仅在属于厚壁菌门的毛螺菌科和梭菌科中发现。
这些结果表明,氨基酸代谢基因广泛分布在人类肠道微生物中,并编码消化代谢蛋白质氨基酸的关键酶。
2
从头合成氨基酸
肠道菌群除了参与蛋白质的降解外,还可以合成氨基酸,包括从头合成。
体外研究表明,瘤胃中的细菌种类可以在不同的肽浓度下合成氨基酸。包括Streptococcus bovis,Selenomonas ruminantium, Prevotella bryantii。
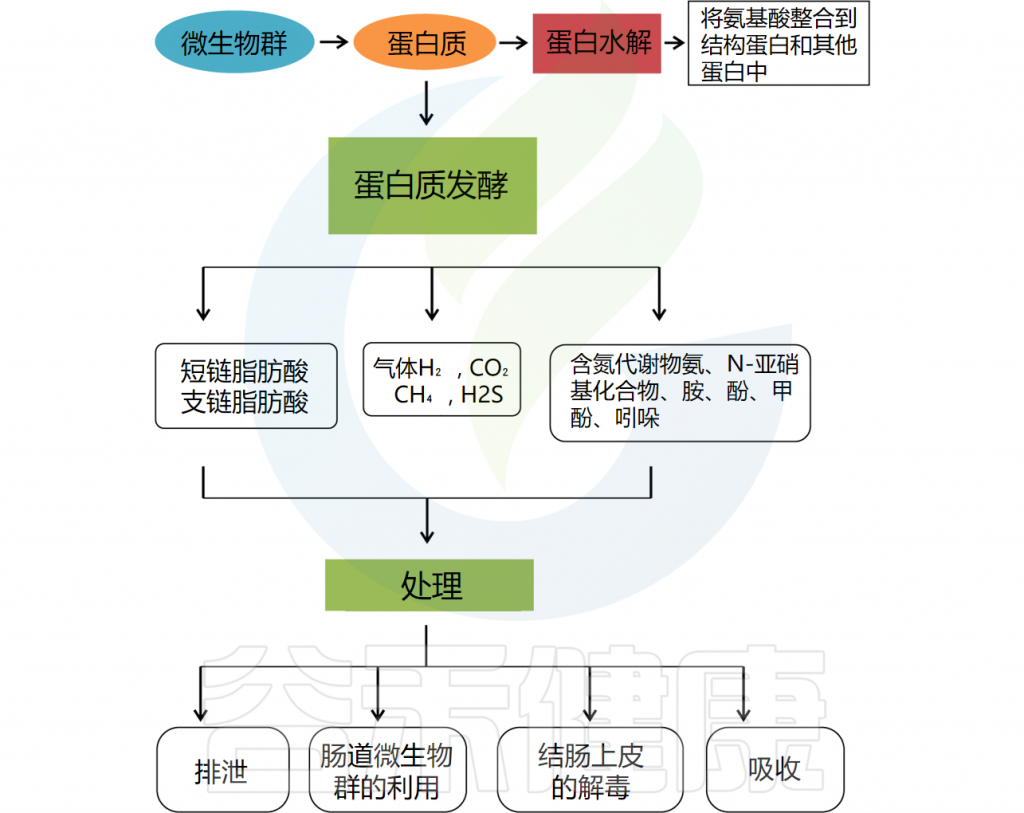
doi: 10.2174/1389203721666200212095503.
此外,体内研究还表明,微生物衍生的赖氨酸(一种必需氨基酸)被吸收并并入宿主蛋白质。在无菌和常规化大鼠体内,将15NH4CL中的15N并入赖氨酸的比较表明,检测到的所有15N赖氨酸均来自微生物来源。在后续研究中,这些研究人员确定约75%的微生物15N标记的赖氨酸被小肠吸收。
对18岁以上的人样本进行了检查,这些人被提供了氮充足的饮食,并发现微生物衍生的赖氨酸和苏氨酸对游离血浆赖氨酸和苏氨酸库有显著贡献。此外,有报告说,大肠中微生物群的富集是通过与必需氨基酸(EAA)生物合成相关的基因进行的,该生物合成基于人血浆池产生的前体。
3
对氨基酸吸收的影响
肠道微生物群还可以通过以下几种方式影响氨基酸的吸收。
▸ 调节肠道环境影响氨基酸吸收
•改变肠道pH值
肠道菌群的代谢活动可以改变肠道的pH值。不同的氨基酸在不同的pH值下具有不同的离子化状态,从而影响其吸收。例如,在酸性环境下,一些氨基酸更容易以阳离子形式存在,而在碱性环境下则更容易以阴离子形式存在。
某些肠道菌群如乳酸菌可以产生乳酸,降低肠道pH值,使一些氨基酸更容易被吸收。而另一些细菌如大肠杆菌等则可以产生碱性物质,升高肠道pH值,可能会影响某些氨基酸的吸收。
•影响肠道通透性
肠道菌群可以通过调节肠道上皮细胞之间的紧密连接来影响肠道通透性。正常情况下,肠道上皮细胞之间的紧密连接可以阻止大分子物质和有害物质进入体内,但也会限制一些氨基酸的吸收。
一些有益的肠道菌群如双歧杆菌可以增强肠道上皮细胞之间的紧密连接,维持肠道屏障功能,防止有害物质进入体内,同时也可以促进一些氨基酸的选择性吸收。而一些有害的致病菌则可能破坏肠道上皮细胞之间的紧密连接,增加肠道通透性,导致氨基酸吸收紊乱和肠道炎症。
▸ 与宿主相互作用影响氨基酸吸收
•竞争吸收位点
肠道菌群和宿主细胞可能会竞争氨基酸的吸收位点。一些肠道细菌可以表达与宿主细胞相似的氨基酸转运蛋白,从而与宿主竞争氨基酸的吸收。
例如,某些大肠杆菌可以表达与人体小肠上皮细胞相似的赖氨酸转运蛋白,从而竞争赖氨酸的吸收。这种竞争会导致宿主明明摄入了较多的蛋白质,但实际对氨基酸的吸收量却不多。
•调节宿主基因表达
肠道菌群可以通过产生代谢产物、分泌信号分子等方式调节宿主基因的表达,从而影响氨基酸的吸收。
例如,一些短链脂肪酸如丁酸可以作为信号分子,调节肠道上皮细胞中氨基酸转运蛋白的表达。此外,肠道菌群还可以通过调节肠道免疫系统、内分泌系统等间接影响氨基酸的吸收。
4
补充益生菌增加了植物蛋白的吸收

该实验研究了益生菌补充剂对植物蛋白(如豌豆蛋白)中氨基酸吸收的影响。
研究发现,副干酪乳杆菌LP-DG®(每天50亿 CFU)给药显著增加了摄入豌豆蛋白后蛋氨酸(+20.0%)、组氨酸(+40.4%)、缬氨酸(+21.5%)、亮氨酸(+23.3%)、异亮氨酸(+26.0%)、酪氨酸(+16.0%)、总支链氨基酸(+22.8%)和总必需氨基酸(+16.0%)的含量。
而副干酪乳杆菌 LPC-S01(每天50亿 CFU)给药显著增加了蛋氨酸(+16.3%)、组氨酸(+49.2%)、缬氨酸(+24.7%)、亮氨酸(+25.2%)、异亮氨酸(+26.1%)、酪氨酸(+11.6%)、总支链氨基酸(+26.8%)和总必需氨基酸(+15.6%)最大浓度。
这项研究表明,通过补充益生菌可能是一种重要的营养策略,可以改善餐后血液氨基酸的变化,并克服植物蛋白的组成缺陷。益生菌可以通过优化肠道菌群结构,提高植物蛋白的消化吸收率。
氨基酸在人体营养和生理活动中至关重要,人们对蛋白质的需求实际上是对氨基酸的需求。我们每天从食物中摄取蛋白质,正是为了满足这些生理活动所必需的氨基酸。那么氨基酸被人体吸收后主要有哪些去处呢?让我们继续来看。
1
氨基酸吸收后的利用
人体的游离氨基酸主要有三条去路:
一是合成人体所需的多肽和蛋白质,多肽和蛋白质是机体生命活动的执行者。蛋白质合成过程较复杂,需根据特定基因上携带的遗传信息,经复制、转录、翻译等一系列过程,以各种氨基酸为原料装配成人体蛋白质。
二是经脱氨、脱羧等分解代谢途经生成氨、α-酮酸和胺类、CO2。其中,生成的α-酮酸可进一步代谢,经氨基化作用生成非必需氨基酸供机体利用。或氧化供能(当非蛋白质能量来源不足时,氨基酸可能被用于能量代谢,以满足能量需求)。氨基酸脱氨作用产生的游离氨,在正常情况下,主要在肝脏中合成尿素,随尿排出体外,少部分氨在肾脏中以铵盐形式由尿排出。
三是转变成其它含氮化合物,比如经一系列酶催化转变成嘌呤、嘧啶、肌酸等(赖氨酸合成肉碱,精氨酸合成肌酐,蛋氨酸作为甲基供体)。
!
注意
一些氨基酸代谢产生出具有特殊重要生理功能的小分子化合物,比如:谷氨酸脱氨分解生成γ-氨基丁酸(GABA),GABA是抑制性神经递质,可抑制中枢神经;组氨酸脱羧分解产生组胺,组胺是一种强烈的血管扩张剂,引起平滑肌收缩,并增加毛细血管通透性;色氨酸脱羧分解产生5-羟色胺(又叫血清素),也是一种抑制性神经递质,直接影响神经传导。
•不是所有氨基酸都能被吸收利用
人体内的氨基酸除了被利用外,也会有部分无意流失。每天摄入大量的蛋白质进入胃肠道;虽然其中一些物质被消化和吸收,但相当数量的氨基酸在胃肠道中分解代谢或被纳入微生物蛋白质,这些蛋白质与一些未消化的蛋白质一起在粪便中从体内流失。
2
氨基酸的重吸收
生物体氨基酸稳态的一个重要因素是发生在肾皮层的肾小球滤过/肾小管重吸收机制。
每天都会不可避免地流失一些氨基酸,如尿素、肌酐和氨。当蛋白质成分增加超过必需替代或氨基酸用于糖异生时,尿素的量会增加。
•肾小管重吸收受损会导致氨基酸的流失增加
肾小管重吸收是氨基酸循环的一部分。它由于肠道中发现的几乎相同的一组转运蛋白介导,当肾小管重吸收受损时,可能导致氨基酸流失增加,即使摄入了较多的蛋白质,氨基酸的总含量仍缺乏。
3
不同人群氨基酸需求不同
多年来,人们进行了大量的工作来确定人类对膳食氨基酸的需求。粮农组织、世卫组织和联合国建议,每公斤体重0.83克优质蛋白质是每日膳食蛋白质的“安全”摄入量,即体重70公斤的参考男性每天摄入58克蛋白质,体重57公斤的参考女性每天摄入47克蛋白质。
但越来越多的研究和谷禾数据表明,由于个体消化吸收差异以及不同食物和饮食习惯的代谢影响,每个人对蛋白质的需求量各不相同。
•不同人群对氨基酸的需求和利用都不同
每个人的不同状态对蛋白质的需求量也都不同:在成长中的儿童中,需要氨基酸来维持体内蛋白质和生成新的组织,而在成人中,需要氨基酸来补充体内丢失的氨基酸。对于孕妇和哺乳期妇女来说,还需要额外的氨基酸来促进胎儿的生长和产奶量。健身人群则需要大量的蛋白质来合成肌肉。
因此,成人所需的膳食氨基酸不是固定值,每日最佳摄入量取决于你的身高体重、健康状况、目标、身体活动水平。
•消化率较低的人群需要摄入更多的蛋白质
素食主义者对蛋白质的需求较高,因为植物蛋白在生物利用度上通常不如动物蛋白。植物蛋白消化率为60-80%,而动物蛋白消化率超过90%。
此外一些植物含有抑制蛋白质消化和吸收的抗营养因子,如胰蛋白酶抑制剂、凝集素、植酸盐和单宁、硫苷,会导致素食主义者的蛋白质消化和吸收率偏低。而消化率较低的人群需要摄入更多的蛋白质,以确保满足身体的营养需求。
•注意摄入蛋白的氨基酸构成
另外,一些蛋白质也含有限制性氨基酸,限制性氨基酸是指食物所含必需氨基酸的量与人体所需的蛋白质必需氨基酸的量相比,比值偏低的氨基酸。
一种食物里特别缺少某一种氨基酸,即使其它的氨基酸含量很高,因为这个氨基酸导致它的各个氨基酸比例不平衡。人体对这种食物的吸收依然不理想。多样性饮食结合不同的膳食蛋白可以帮助弥补它们各自的不足。
比如说,赖氨酸是最常见的限制性氨基酸,特别是在谷类中,如小麦和水稻。坚果往往也含有赖氨酸作为限制性氨基酸。另一方面,豆类含有足够的赖氨酸,可以弥补,但缺乏硫氨基酸,如蛋氨酸和半胱氨酸。
通过整合微生物组学、宏基因组学、代谢组学和蛋白质组学数据,我们可以构建更完整的个体营养代谢图谱,有望建立更精确的营养-微生物-代谢互作网络模型,为精准营养干预提供依据。
益生菌在提高氨基酸吸收率方面的应用潜力。新型益生菌的筛选和开发将更加注重其对氨基酸代谢的调节作用,有望开发出更有针对性地提高氨基酸的生物利用度的微生态制剂。
随着精准医疗的发展,个性化的氨基酸需求评估和补充策略将成为研究热点,这需要我们更深入地了解个体差异对氨基酸代谢的影响。基于机器学习的预测模型将帮助我们更准确地评估个体的氨基酸需求,这需要综合考虑肠道菌群特征、代谢指标和临床表型等多维度数据。通过这些数据的综合分析,我们可以构建出更加个性化的营养干预方案,优化个体健康和疾病预防。
主要参考文献
Li TT, Chen X, Huo D, Arifuzzaman M, Qiao S, Jin WB, Shi H, Li XV; JRI Live Cell Bank Consortium; Iliev ID, Artis D, Guo CJ. Microbiota metabolism of intestinal amino acids impacts host nutrient homeostasis and physiology. Cell Host Microbe. 2024 May 8;32(5):661-675.e10.
Jäger R, Zaragoza J, Purpura M, Iametti S, Marengo M, Tinsley GM, Anzalone AJ, Oliver JM, Fiore W, Biffi A, Urbina S, Taylor L. Probiotic Administration Increases Amino Acid Absorption from Plant Protein: a Placebo-Controlled, Randomized, Double-Blind, Multicenter, Crossover Study. Probiotics Antimicrob Proteins. 2020 Dec;12(4):1330-1339.
Neis EP, Dejong CH, Rensen SS. The role of microbial amino acid metabolism in host metabolism. Nutrients. 2015 Apr 16;7(4):2930-46.
Hou Y, Yao K, Yin Y, Wu G. Endogenous Synthesis of Amino Acids Limits Growth, Lactation, and Reproduction in Animals. Adv Nutr. 2016 Mar 15;7(2):331-42.
Gorissen SHM, Trommelen J, Kouw IWK, Holwerda AM, Pennings B, Groen BBL, Wall BT, Churchward-Venne TA, Horstman AMH, Koopman R, Burd NA, Fuchs CJ, Dirks ML, Res PT, Senden JMG, Steijns JMJM, de Groot LCPGM, Verdijk LB, van Loon LJC. Protein Type, Protein Dose, and Age Modulate Dietary Protein Digestion and Phenylalanine Absorption Kinetics and Plasma Phenylalanine Availability in Humans. J Nutr. 2020 Aug 1;150(8):2041-2050.
Bröer S, Gauthier-Coles G. Amino Acid Homeostasis in Mammalian Cells with a Focus on Amino Acid Transport. J Nutr. 2022 Jan 11;152(1):16-28.
Mariotti F, Gardner CD. Dietary Protein and Amino Acids in Vegetarian Diets-A Review. Nutrients. 2019 Nov 4;11(11):2661.
Abdallah A, Elemba E, Zhong Q, Sun Z. Gastrointestinal Interaction between Dietary Amino Acids and Gut Microbiota: With Special Emphasis on Host Nutrition. Curr Protein Pept Sci. 2020;21(8):785-798.
Fan P, Li L, Rezaei A, Eslamfam S, Che D, Ma X. Metabolites of Dietary Protein and Peptides by Intestinal Microbes and their Impacts on Gut. Curr Protein Pept Sci. 2015;16(7):646-54.