-

 国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子
国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子

 二级病原微生物安全实验室
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业

谷禾健康
炎症性肠病(Inflammatory Bowel Disease, IBD)是一种复杂的、多因素驱动的慢性胃肠道炎症性疾病,研究显示其发病率在全球范围内持续上升。
研究表明,IBD的病因涉及遗传易感性、环境因素、肠道微生物动态变化以及宿主免疫调控的复杂相互作用。近年来,随着多组学技术和临床研究的深入,科学界对IBD在临床前阶段有了更深刻的认知框架。
前瞻性队列研究证实,基于高风险人群的纵向队列数据揭示,在临床确诊前5-10年,个体已呈现特征性的亚临床病理改变:肠道屏障功能渐进性损伤、微生物生态结构失衡、抗微生物抗体谱系异常,糖基化修饰的系统性改变等。比如,粪便钙卫蛋白等生物标志物的动态变化模式,为追踪疾病自然史提供了可量化的监测指标。
当这些亚临床病理改变突破代偿阈值,便表现为典型的IBD临床症状。
本文依据现有研究资料,系统性地阐述了IBD的定义、分类、流行病学特征、多重危险因素,重点剖析了临床前IBD的启动与发展阶段,涵盖肠道屏障功能、微生物生态、宿主免疫应答、蛋白质组学及糖基化修饰等多个层面的动态变化;在此基础上,我们还将探讨当前靶向菌群的干预策略与未来预防医学新范式,为IBD的早期识别与精准管理提供理论依据。
目 录
1. 炎症性肠病(IBD)概述
2. IBD的危险因素与决定因素
3. 临床前IBD:从启动到扩展
4. IBD综合发展模型方向
5. 治疗措施与预防机会
6. 结论与展望
IBD的定义与主要形式
炎症性肠病(IBD)是一组以胃肠道慢性、复发性炎症为特征的免疫介导的疾病。
其主要包括两种类型:
病变部位和病理特征
克罗恩病的炎症可累及从口腔到肛门的整个消化道,呈节段性、非对称性分布,炎症可穿透肠壁全层,导致狭窄、穿透(瘘管)等并发症。
溃疡性结肠炎的炎症通常局限于结肠黏膜和黏膜下层,自直肠开始呈连续性、弥漫性向近端蔓延。
尽管两者在病变部位和病理特征上有所区别,但它们共同的病理生理基础是宿主对肠道微生物群产生异常且持续的免疫反应,最终导致组织损伤。
肠道免疫系统在稳态和炎症状态下显著差异
在健康状态下(稳态),肠道上皮细胞形成完整的物理屏障,潘氏细胞(Paneth cell)分泌抗菌肽,杯状细胞(Goblet cell)分泌黏液,共同抵御微生物入侵。固有层中的免疫细胞,如调节性T细胞(Treg)和分泌IgA的浆细胞,共同维持对共生菌的免疫耐受(如下图 左边)。
编辑
doi.org/10.1038/s41590-025-02197-5
在IBD炎症状态下,这种平衡被打破。肠道屏障受损,潘氏细胞和杯状细胞功能下降,导致抗菌物质和黏液减少。
病原菌如克雷伯菌属(Klebsiella)增殖,共生有益菌菌(如普拉梭菌 F. prausnitzii)减少。大量炎症细胞(如TH1/TH2/TH17细胞、炎症性巨噬细胞)浸润肠道组织,分泌促炎细胞因子,并产生IgG抗体,导致慢性炎症和组织破坏(如上图右边)。
流行病学趋势
历史上,IBD主要被认为是西方工业化国家的疾病。然而,进入21世纪,IBD的全球发病率和患病率呈现出显著的上升趋势,尤其是在亚洲、南美和东欧等新兴工业化国家和地区,IBD已成为一个全球性的健康挑战。这种流行病学转变清晰地表明,除了遗传背景外,与工业化和城市化相关的生活方式、饮食结构和环境暴露等因素在IBD的发病中扮演着至关重要的角色。
针对不同种族和移民群体的研究也发现,IBD的临床表现、遗传易感性和治疗反应存在差异,这进一步凸显了基因-环境相互作用在IBD发病中的复杂性。
IBD的病因是多因素的,涉及遗传、环境、免疫和微生物等多个层面。这些因素相互交织,共同决定了个体患病的风险和疾病的表型。
遗 传 易 感 性
IBD具有明显的家族聚集性,遗传因素在其发病中起着基础性作用。单卵双生子研究显示,克罗恩病的一致性高达50%。通过全基因组关联研究(GWAS),目前已发现超过300个与IBD相关的易感基因位点。这些基因大多与免疫功能相关,特别是那些调节宿主与微生物相互作用的通路。例如:
值得注意的是,大多数遗传位点仅赋予个体一定概率的疾病风险,这也表明IBD是一种多基因疾病,其发病需要遗传与环境因素的共同作用。此外,不同种族人群的遗传背景差异也导致了IBD易感基因频率和效应的不同。
环 境 因 素
环境因素被认为是近年来IBD发病率快速上升的主要驱动力。这些因素在生命早期和成年期均可发挥作用,深刻影响肠道微生态和免疫系统的发育与功能。
✦ 饮食
西式饮食,即高脂肪、高糖、高加工食品和低纤维的饮食模式,被认为是IBD的重要风险因素。食品乳化剂(如羧甲基纤维素)会破坏肠道黏液屏障,改变微生物群组成,促进炎症。相反,富含纤维的饮食则有助于产生具有抗炎作用的短链脂肪酸(SCFAs)。
✦ 抗生素使用
尤其是在生命早期,抗生素的广泛使用会严重扰乱肠道微生物群的正常定植和发育,降低菌群多样性,增加IBD的长期风险。
✦ 感染
特定的肠道感染,如沙门氏菌、弯曲杆菌、诺如病毒,被发现与IBD的发病风险增加有关。这些感染可能作为“触发事件”,在遗传易感个体中打破免疫耐受。
✦ 吸烟
吸烟是影响IBD的明确环境因素之一。有趣的是,吸烟会增加克罗恩病的风险并加重其病情,但对溃疡性结肠炎却显示出一定的保护作用,其机制尚不完全清楚。
✦ 卫生假说
生命早期过度清洁的环境可能导致免疫系统未能充分接触微生物,从而发育不全,更易在日后对无害抗原产生过度反应。
✦ 其他因素
还包括空气污染、非甾体抗炎药(NSAIDs)的使用、社会经济地位以及心理压力等。
IBD的遗传和环境决定因素
编辑
doi.org/10.1038/s41590-025-02197-5
人群的免疫反应强度呈钟形分布。IBD可发生于免疫反应过弱(如NOD2缺陷导致对微生物抵抗力差)或过强(如IL-10调节缺陷导致对微生物耐受性差)的两个极端。
左下角的曼哈顿图代表遗传易感性,右下角的图标代表饮食、吸烟和微生物等环境因素。这些因素的相互作用决定了个体是否会滑向疾病的两端。
炎症反应的遗传与环境决定因素
个体的免疫反应强度存在连续的变异谱,从免疫缺陷到免疫超敏。IBD可以在这个谱系的两个极端发生。
编辑
◄ 免疫缺陷
一方面,某些遗传缺陷(如NOD2、ATG16L1功能丧失)会导致肠道先天免疫和屏障功能减弱,使宿主对微生物的抵抗力下降,易受机会性病原菌感染而引发炎症。
► 免疫超敏
另一方面,另一些遗传背景(如IL-10、IL-10R功能缺陷)则导致免疫调节功能受损,对正常的肠道共生菌产生过度的免疫反应,即免疫耐受丧失,从而引发炎症。
▲ 健康平衡
健康个体则处于两者之间的平衡状态,既能有效清除病原体,又能耐受共生菌。个体的遗传组成决定了其对特定环境暴露(如饮食、感染)的反应阈值和反应模式。
因此,IBD的发生是独特的遗传背景与特定环境因素相互作用(G×E)的结果,这也解释了为何暴露于相似环境中的人群只有少数遗传易感者会发病。
大量证据表明,IBD的临床诊断之前存在一个漫长的临床前阶段,在此期间,一系列病理生理变化已悄然发生。理解这一阶段是实现IBD早期预测和预防的关键。这一阶段可进一步细分为风险期、启动期和扩展期。
临床前IBD的分期模型
一个被广泛接受的模型将IBD的发展过程分为几个连续的阶段:
风险期
个体携带遗传易感基因,并可能在围产期、婴幼儿期及童年期暴露于某些环境风险因素(如抗生素、感染、母亲吸烟等)。这些因素共同启动了免疫系统的易感状态,但个体并无任何症状或病理改变。
临床前启动期
在某个未知的“触发事件”(如特定感染或饮食改变)后,疾病开始启动。这一阶段可持续多年(甚至在诊断前10年),其特征是出现亚临床的病理变化,如肠道通透性增加、微生物失调、以及针对微生物或自身抗原的低水平抗体应答。这些变化通常是无症状的。
临床前扩展期
在临床诊断前约2年,疾病进入扩展期。此时,病理变化加剧,全身性和肠道局部的炎症标志物(如CRP、粪便钙卫蛋白)显著升高,患者可能开始出现非特异性症状并开始出现要就诊的症状。这一阶段代表了从亚临床状态向临床显性疾病的过渡。
临床诊断期
当炎症累积到一定程度,导致明显的组织损伤和持续的临床症状(如腹痛、腹泻、便血)时,患者最终被诊断为IBD。
IBD临床前阶段的演变模型
编辑
doi:10.1038/s41575-023-00854-4.
IBD风险因素、发病途径和累积风险的时间演变
编辑
doi:10.1038/s41575-023-00854-4.
该图谱从生命阶段(怀孕至成年)的角度,展示了遗传风险和环境风险如何随时间累积。
在临床前阶段,微生物组扰动、屏障功能丧失、免疫功能障碍、代谢变化和糖基化改变等途径相互作用,共同推动IBD累积风险的增加,直至最终诊断。这也提示了不同生命阶段的预防干预窗口。
肠道屏障功能障碍与通透性改变
肠道屏障是维持肠道内环境稳态的第一道防线,其完整性在IBD发病中至关重要。
在临床前阶段,肠道屏障功能就已出现障碍。肠道屏障由物理屏障、化学屏障、生物屏障、免疫屏障构成。
☾ 物理屏障
由单层肠上皮细胞(IECs)及其间的紧密连接蛋白(如Claudin, Occludin)构成。
编辑
研究发现,在克罗恩病高风险亲属中,即使没有症状,其肠道通透性(以乳果糖/甘露醇比值LMR衡量)也显著增加,并且这种高通透性是未来发病的独立危险因素。
肌球蛋白轻链激酶(MLCK)的异常激活被认为是导致紧密连接开放、通透性增加的关键机制之一。
☾ 化学屏障
主要指覆盖在上皮表面的黏液层和由潘氏细胞分泌的抗菌肽(AMPs)。
编辑
黏液层由杯状细胞分泌的黏蛋白(主要是MUC2)构成,能有效隔离细菌与上皮细胞的直接接触。
在IBD中,黏液层变薄、不连续,其糖基化修饰也发生改变。同时,潘氏细胞功能受损,导致α-防御素等AMPs分泌减少,削弱了对微生物的控制能力。
☾ 生物屏障
编辑
肠道菌群中有益菌通过竞争营养、产生抑菌物质等方式抑制致病菌生长。另外通过产生短链脂肪酸,维持肠道酸性环境等。
☾ 免疫屏障
编辑
包括分泌型IgA(sIgA)和上皮内淋巴细胞(IELs)等。sIgA能中和并清除入侵的微生物,而在IBD中,针对特定病原菌的IgA反应可能不足或异常。
屏障功能的早期损害,使得肠腔内的微生物及其产物(如LPS)更容易易位至固有层,从而持续激活下方的免疫系统,启动炎症级联反应。
编辑
doi.org/10.1038/s41575-022-00604-y
肠道微生物群失调
肠道微生物失调是IBD的核心特征,既是疾病的结果,也可能是其驱动因素。在临床前阶段,微生物群的改变就已经出现。
✎ 菌群结构改变
IBD患者的肠道微生物多样性显著降低。
产短链脂肪酸菌,如普拉梭菌(Faecalibacterium prausnitzii)、罗氏菌属(Roseburia)等厚壁菌门细菌丰度减少。
而一些具有潜在致病性的需氧或兼性厌氧菌,如大肠杆菌(特别是黏附侵袭性大肠杆菌AIEC)、克雷伯菌属等变形杆菌门细菌,具核梭杆菌(Fusobacterium nucleatum)等则异常增殖。
✎ 真菌与病毒的改变
除了细菌,肠道真菌和病毒的失调也参与IBD。例如,白色念珠菌(Candida albicans)、马拉色菌(Malassezia)等真菌在IBD患者肠道中增多,某些高毒力菌株可通过分泌念珠菌溶血素(Candidalysin)等毒素加剧炎症。同时,噬菌体和某些肠道病毒的组成和丰度也发生改变。
编辑
doi.org/10.1038/s41579-025-01163-0
✎ 功能与代谢改变
微生物失调导致其功能和代谢产物的改变。丁酸等短链脂肪酸的产生减少,削弱了其对肠上皮细胞的营养支持和对免疫细胞的抗炎调节作用。相反,一些有害代谢物,如硫化氢、或诱导DNA损伤的吲哚胺类物质可能增多。胆汁酸的代谢也发生紊乱,影响了FXR、TGR5等核受体的信号,进而影响免疫调节。
这种失调的微生物群落,一方面无法提供维持肠道稳态所需的有益信号,另一方面其本身或其产物又成为持续刺激免疫系统的炎症来源。
编辑
doi.org/10.1038/s41579-025-01163-0
基因-微生物相互作用
特定的遗传背景与特定的微生物失调相结合,是驱动IBD发病的核心机制。研究表明,IBD相关的易感基因突变,往往导致宿主对特定微生物的反应异常。
NOD2与微生物
携带NOD2功能缺陷变异的个体,其肠道中脆弱拟杆菌(Bacteroides fragilis)等特定共生菌的调节能力下降,无法有效诱导Treg细胞,导致免疫耐受受损。
近期研究进一步揭示,粪肠球菌(Enterococcus faecium)分泌的DL-内肽酶SagA能够产生激活NOD2的信号分子MDP。在健康个体中,这一通路通过髓系细胞的NOD2激活,诱导IL-1β分泌,进而促进ILC3和CD4+ T细胞产生保护性的IL-22,促进组织修复。
然而,在IBD患者中,由于炎症环境中REG3等抗菌蛋白的过度产生,导致对REG3敏感的Efm被清除,从而使这一保护性通路中断。更重要的是,携带常见NOD2风险变异(如R702W)的个体,即使存在Efm,也无法有效启动这一保护性信号,导致炎症持续。这一发现为理解基因(NOD2变异)与微生物(Efm缺失或功能障碍)之间可能存在的相互作用机制提供了重要线索,提示它们在疾病发生与发展过程中潜在协同机制。
✨粪肠球菌
粪肠球菌在健康与IBD中的双重作用机制
编辑
doi.org/10.1016/j.chom.2023.08.002
健康状态下,粪肠球菌(E. faecium)通过其分泌的SagA激活髓系细胞的NOD2通路,最终诱导IL-22产生,促进组织再生和抑制炎症。
而在IBD中,一方面炎症环境产生的REG3蛋白会清除E. faecium;另一方面,NOD2的遗传变异会使该保护通路失效,导致炎症循环加剧。
ATG16L1与病毒
携带ATG16L1 T300A风险变异的小鼠,在感染鼠诺如病毒(MNV)后,其潘氏细胞会发生严重的细胞死亡和功能障碍,而野生型小鼠则不受影响。
这表明遗传易感性(ATG16L1变异)与环境触发(病毒感染)的结合,是导致特定细胞表型和病理变化的关键。
自身免疫与抗微生物体液应答
在IBD临床前阶段,体液免疫系统已经出现异常活化,表现为多种抗体水平的升高。这些抗体既可以靶向肠道微生物,也可以靶向宿主自身成分。
抗微生物抗体
在IBD诊断前数年,即可在患者血清中检测到针对肠道微生物成分的抗体。例如,抗酿酒酵母抗体(ASCA)是克罗恩病的经典标志物,其在诊断前5年甚至更早就已升高。
其他抗体还包括抗大肠杆菌外膜孔蛋白C(OmpC)抗体、抗鞭毛蛋白(CBir1, FlaX)抗体等。这些抗体的出现,反映了肠道屏障受损后,免疫系统与肠道微生物发生了异常的相互作用和免疫识别。
编辑
自身抗体
除了抗微生物抗体,一些自身抗体也在临床前阶段出现。例如,抗粒细胞-巨噬细胞集落刺激因子(GM-CSF)自身抗体在克罗恩病诊断前6年即可检出,并与更复杂的疾病表型相关。
部分研究提示,抗整合素αvβ6自身抗体在溃疡性结肠炎诊断前长达10年就已升高,并具有很高的预测价值。这些自身抗体的出现,标志着免疫系统打破了对自身组织的耐受,是疾病向自身免疫方向发展的重要证据。
蛋白质标志物的变化
利用高通量蛋白质组学技术,研究人员在IBD临床前患者的血清中鉴定出了一系列变化的蛋白质标志物。这些标志物反映了疾病早期潜在的病理过程。
◑ 炎症相关蛋白
在诊断前数年,全身性炎症标志物如C反应蛋白(CRP)和白细胞介素-6(IL-6)水平已轻度升高,提示存在潜在的炎症反应。
其他与炎症、免疫应答相关的蛋白,如肿瘤坏死因子受体(TNFR)、补体成分、脂多糖结合蛋白(LBP)等也发生改变。
◑ 趋化因子和细胞因子
趋化因子如CXCL9、CXCL11、CCL11在临床前阶段升高,这些分子与免疫细胞的招募和迁移有关,提示在临床症状出现前,免疫细胞的动态平衡可能已被打破。
◑ 组织重塑相关蛋白
基质金属蛋白酶(MMPs)如MMP10、MMP12等的水平变化,可能反映了疾病早期微小的组织损伤与修复过程。
这些蛋白质标志物的组合,有望构建预测模型,用于识别高风险人群,其预测准确性随着接近诊断时间点而提高。
糖基化修饰的改变
糖基化是蛋白质和脂质最常见和最复杂的翻译后修饰之一,形成的聚糖(Glycans)在细胞识别、信号转导、免疫调节中发挥关键作用。
在炎症性肠病(IBD)的发生和发展过程中,糖基化模式发生了显著变化。
N-聚糖和O-聚糖
聚糖主要分为N-连接聚糖(N-glycan)和O-连接聚糖(O-glycan)。
肠道不同部位的上皮细胞表达不同的O-聚糖核心结构,如结肠主要表达Core 3和Core 4。
常见的N-聚糖和O-聚糖结构
编辑
doi.org/10.1038/s41385-021-00466-8
该图展示了N-聚糖的核心与扩展形式(高甘露糖型、复杂型、混合型),以及O-聚糖的四种核心结构(Core 1-4)及其扩展形式。这些复杂的糖链结构是细胞功能的重要调节者。
炎症诱导的糖基化改变
在IBD的炎症黏膜中,一个普遍特征是出现不成熟或截短的聚糖结构。
例如,肿瘤相关的Thomsen-Friedenreich(TF)抗原(Galβ1-3GalNAc,即Core 1结构)在正常结肠中被掩盖,但在UC患者的炎症上皮中异常暴露。这种改变不仅影响细胞粘附,还可能通过与外源性凝集素(如花生凝集素PNA)结合,促进上皮细胞过度增殖,增加癌变风险。
编辑
关于凝集素可以详见我们之前的文章:
黏蛋白(Mucin)糖基化
MUC2是构成肠道黏液层的主要黏蛋白,其重度O-糖基化是维持黏液屏障功能的关键。
然而,在IBD中,MUC2的糖链变短,硫酸化水平降低,岩藻糖基化和唾液酸化模式也发生改变。这些改变削弱了黏液的保护功能,使其更容易被细菌降解,进而影响肠道的健康。
编辑
免疫细胞糖基化
免疫细胞的糖基化状态也影响其功能。例如,血清IgG的糖基化模式在IBD中发生改变,如半乳糖基化水平下降(agalactosylated IgG),这会改变其与Fc受体的亲和力,从而影响其促炎或抗炎功能。T细胞表面的聚糖分支(如β1-6分支)减少,会降低其激活阈值,使其更易被激活。
编辑
这些糖基化的改变,构成了IBD病理生理学中一个复杂但重要的层面,它们既是炎症的结果,也反过来驱动和维持炎症。
肠道稳态与炎症状态下的黏膜糖基化变化
编辑
doi.org/10.1038/s41385-021-00466-8
左图为稳态,具有完整的黏液层、正常的微生物群和成熟的细胞表面聚糖。
右图为炎症状态,黏液层降解,微生物失调,上皮细胞、免疫细胞和黏蛋白(Muc-2)的糖基化模式均发生显著改变,出现不成熟或异常的聚糖结构。
亚临床炎症
在IBD的临床前扩展期,一个关键特征是亚临床肠道炎症的出现。这可以通过非侵入性生物标志物——粪便钙卫蛋白(fecal calprotectin)的升高来检测。
钙卫蛋白是中性粒细胞释放的一种蛋白,其在粪便中的水平与肠道炎症程度密切相关。在克罗恩病高风险亲属中,基线粪便钙卫蛋白水平升高是未来发病的重要预测指标。
在这方面,谷禾肠道菌群检测报告(临床版)中基于肠道菌群特征预测了粪便钙卫蛋白这指标,若菌群检测提示有害菌过度增殖(如变形菌门)、有益菌减少(如双歧杆菌),且预测钙卫蛋白显著升高,常提示肠道可能存在活动性炎症(如 IBD、感染性肠炎)。
编辑
此外,一些高风险个体在接受胶囊内镜检查时,即使无任何症状,也可能发现小肠黏膜存在炎症性病变。这些亚临床炎症的存在,表明免疫系统的失调已经从分子和细胞水平进展到了组织层面,是疾病即将进入临床期的警报。
综合上述证据,可以构建一个多重组合的IBD发病模型。该模型强调,IBD并非由单一因素引起,而是遗传易感性与一系列环境“刺激”协同作用、累积效应的结果。
第一次刺激(遗传与早期环境)
个体出生时携带IBD易感基因(如NOD2、ATG16L1变异),这构成了发病的遗传基础。在生命早期,暴露于抗生素、不良饮食或特定感染等环境因素,导致肠道微生物群的早期定植异常和免疫系统的“错误编程”,使个体处于高风险状态。
第二次刺激(触发事件)
在随后的生命历程中,一次或多次环境经历,如特定的肠道感染(如诺如病毒)、饮食改变、药物使用或严重的心理压力,成为点燃炎症的“导火索”。这个触发事件在遗传易感个体中打破了原有的免疫稳态。
炎症的启动与自我维持
触发事件导致肠道屏障受损,微生物及其产物易位,激活了固有层的免疫细胞。在具有遗传缺陷的背景下(如NOD2信号通路受损),免疫系统无法进行有效的耐受或清除,而是产生持续的、过度的炎症反应。
例如,促炎细胞因子(TNF, IL-23)大量产生,而抗炎信号(IL-10, SCFAs)减弱。这种炎症环境本身又会进一步加剧微生物失调(如耗氧菌增殖)和屏障破坏(如上皮细胞死亡),形成一个“炎症-失调-屏障破坏”的恶性循环。
疾病的扩展与慢性化
随着时间的推移,这种自我维持的炎症循环导致免疫记忆的形成(如产生针对共生菌的致病性T细胞和IgG抗体)、组织重塑(如成纤维细胞活化导致纤维化)和淋巴结构破坏。当累积的损伤超过机体的代偿能力时,临床症状出现,疾病进入慢性、复发-缓解的病程。
在这个模型中,不同的基因-环境组合可能导致不同的疾病表型。例如,NOD2突变与特定细菌的相互作用可能主要导致小肠炎症,而ATG16L1突变与病毒感染的组合可能以潘氏细胞功能障碍为突出表现。这种异质性也解释了IBD患者在临床过程和治疗反应上的巨大差异(如下图)。
IBD的多重刺激疾病模型
编辑
doi.org/10.1038/s41590-025-02197-5
该图展示了从无疾病到临床前阶段再到晚期IBD的演变过程。遗传变异(如NOD2, ATG16L1)和环境因素(如感染、饮食)共同作用,在临床前阶段引发了微生物失调、免疫细胞活化(如IL-23/TNF产生)和早期上皮损伤。随着疾病进展,炎症加剧,大量免疫细胞浸润,肠道结构遭到严重破坏,形成晚期IBD。图中还标示了潜在的治疗干预靶点。
基于对IBD发病机制,特别是临床前阶段的深入理解,为开发新的治疗和预防策略可能提供了前所未有的机会。
当前治疗策略的挑战
目前的IBD治疗主要依赖于免疫抑制剂,包括糖皮质激素、免疫调节剂(如硫唑嘌呤)和生物制剂(如抗TNF、抗整合素、抗IL-12/23抗体)以及小分子药物(如JAK抑制剂)。这些药物通过抑制炎症反应来控制症状和促进黏膜愈合。但是这些治疗方法目前存在诸多局限:
–非特异性免疫抑制
大多数药物会全面抑制免疫系统,可能增加机会性感染和肿瘤的风险。
–原发性或继发性无应答
有相当一部分患者对初始治疗的疗效欠佳,或在治疗过程中疗效逐渐下降。
–症状缓解而非病因治疗
这些疗法主要通过控制炎症来改善临床症状,但对屏障功能缺陷、肠道菌群失调等根本病因的直接干预相对不足,停药后疾病容易复发。
–不良反应
长期用药可能引发多种药物相关的不良反应,需在临床实践中加强监测。
因此,开发更安全、更具针对性、能够实现无免疫抑制下长期缓解的疗法,是IBD领域的核心目标。
靶向肠道微生物的干预
鉴于微生物失调在IBD中的核心作用,靶向微生物的疗法成为研究热点。这些策略旨在恢复健康的肠道微生态,减少炎症刺激源。
靶向微生物组的干预策略
编辑
doi.org/10.1038/s41579-025-01163-0
微生物组分析可用于IBD的诊断前风险评估、诊断和预后预测。基于此,多种靶向微生物的干预措施正在被探索,包括饮食干预、益生菌、粪菌移植、限定细菌联盟、噬菌体疗法、真菌群调节以及更前沿的工程菌和酵母等。
🌈 饮食干预
特定饮食模式,如全肠内营养(Exclusive Enteral Nutrition, EEN),在儿童克罗恩病中显示出与激素相当的诱导缓解率,其机制可能与改变微生物组成和代谢、减轻肠道抗原负荷有关。
注:EEN 是一种液体单食谱疗法,要求患者在疗程期间完全排除所有固体食物和普通饮料,仅摄入特殊配方奶粉(或者说液体营养补充剂)作为唯一的营养来源。注意,它的蛋白质、脂肪、碳水化合物、维生素和矿物质均已科学配比,能够满足人体全部营养需求。切勿将其误解为普通的“清肠”或“断食”饮食。
—全肠内营养(研究最多的饮食干预措施之一)
全肠内营养(EEN),即在约8周内仅摄入营养完整的液体。EEN在儿童克罗恩病(CD)患者中显示出疗效,约80%的患者在这种饮食后进入缓解期。
目前尚不清楚EEN如何改善IBD症状,但有证据表明它会引起微生物组的变化,多项研究显示EEN饮食下微生物组的多样性降低。尽管微生物组多样性的减少通常与炎症状态相关,但在接受EEN的患者中,这种多样性的减少通常归因于EEN饮食成分的有限性。
对接受EEN饮食的儿童CD患者的粪便微生物组和代谢组分析发现,响应治疗的患者的两者均发生了变化:观察到与IBD发病机制相关的代谢物(包括微生物代谢物尸胺和三甲胺)减少,以及先前升高的氨基酸(丝氨酸、甘氨酸、丙氨酸)水平降低。
降低动物蛋白与动物脂肪摄入:研究表明,动物蛋白会促进肠道致病菌的生长,加重结肠炎。
个体化饮食:由于肠道菌群高度个体化,可通过肠道菌群检测制定更适合精准的饮食方案。
注意:虽然饮食干预是安全的辅助治疗手段,但对于活动期严重的 IBD 患者,饮食调整不建议替代药物治疗。应在专业医生或营养师的指导下进行,并定期监测体重、肠道菌群和营养状态。
编辑
注:现行美国胃肠病学会(AGA)指南指出,除非存在禁忌情况,否则所有IBD患者都可能从地中海饮食中受益。
—关注高发酵食品
一项临床试验比较了高纤维植物性饮食与高发酵食品饮食在健康成年人中的效果:吃发酵食品的个体具有更高的微生物多样性,同时伴随几种炎症标志物(包括IL-6、IL-10和IL-12b)的减少;而高纤维饮食组的微生物多样性保持稳定,微生物组编码的糖苷酶活性酶增加,特定短链脂肪酸亚群减少。
这些发现表明,特定的饮食干预(特别是增加发酵食品的摄入)可以改变微生物组的组成并减少肠道炎症。
这些例子强调了前临床模型在研究膳食添加剂的效果时的重要性,特别是在不同情况下的影响,以及在进行人体干预时考虑特定疾病状态的必要性。因此,这些方法应该被纳入临床研究中。
结合IBD的疼痛机制的营养饮食计划
为炎症性肠病患者制定饮食计划需要综合考虑疼痛机制、营养需求和个体差异。
-核心饮食模式
地中海饮食作为基础,采用高纤维、低饱和脂肪的饮食结构,富含坚果、油性鱼、水果、蔬菜和全谷物。
减少含添加剂、高盐、高糖和高饱和脂肪的食品,研究显示西方饮食模式通过增加氧化应激和免疫激活,加剧疼痛感知。
-个体化饮食调整(根据疼痛类型调整)
伤害性疼痛:活动期避免高纤维食物、坚果、种子和豆类;采用软食或流质饮食减少肠道机械刺激。
神经病理性疼痛:重点补充维生素B12、铜、锌等微量营养素,特别是术后患者。
中枢敏化性疼痛:增加富含抗氧化剂的食物,减少促炎食物,考虑肠-脑轴调节。
-关键营养干预
——维生素B12管理
高风险人群,比如回肠切除患者即使血清水平正常也应考虑补充。
缺乏者每周1000μg注射4周后每月维持,或口服1000-2000μg/天,改善神经性疼痛、疲劳,预防多维生素缺乏。
——维生素D
常规筛查和补充,改善骨健康和免疫调节。
——锌和铜
短肠综合征或长期腹泻患者特别需要关注
锌:慢性腹泻患者每日25-50mg,长期补充需同时补铜1-2mg
铜:减肥手术或短肠患者每日2-4mg,监测神经功能
——肉碱与能量支持
适用:慢性疲劳、神经性疼痛患者。
L-肉碱500-1000mg每日2-3次,可与辅酶Q10(100-200mg/天)联用。
食物质地优化
蒸煮、切碎、浸泡软化高纤维食物。
狭窄风险者控制食物颗粒<5mm,术后早期采用泥状食物
软化而非完全排除纤维,保留营养价值。
实施要点
基线全面评估→风险分层→个体化处方→多学科协作→4-8周随访调整(菌群动态监测)。
通过菌群数据指导精准营养干预,使其成为IBD综合治疗的核心组成部分。
🌈 益生菌
在炎症性肠病(IBD)中,益生菌的临床试验结果不一,这部分可能是由于现有益生菌在改变微生物组的组成和功能方面效果有限。
一种包含多种菌株的混合益生菌(如Lactobacillus spp.、Bifidobacterium spp.、Streptococcus spp.,即VSL#3),在诱导轻度至中度溃疡性结肠炎的缓解方面比安慰剂更有效,并且在预防复发性袋状结肠炎方面也显示出疗效。
E. coli Nissle 1917在溃疡性结肠炎患者中也表现出临床疗效,但相关研究较为有限。
新型益生菌菌株
近期研究发现,源自人乳的新型益生菌菌株短双歧杆菌SHMB 8001在DSS诱导的小鼠结肠炎模型中展现出显著治疗潜力。
该菌株通过多重机制发挥保护作用:增强肠道屏障功能(上调MUC2、occludin和claudin-1蛋白表达)、调节免疫炎症反应(上调IL-10并抑制IL-1β、IL-6和TNF-α)、提高粪便短链脂肪酸水平和结肠GPR43受体表达,以及重塑肠道微生物群(富集产短链脂肪酸菌属如双歧杆菌、乳酸杆菌)。
编辑
🌈 益生元
在炎症性肠病中研究最多的益生元包括低聚果糖和菊粉,其益处不一致。新研究有望通过引入合理设计的菌株群落,使其能够以条件依赖的方式定植或递送益生元,需要更多研究。
编辑
🌈 合生元
合生元通过益生菌与益生元的协同作用,在IBD治疗中展现出独特优势。临床前研究证实,多种合生元组合能有效减轻肠道炎症、调节免疫反应、增强屏障功能并改善菌群平衡。
编辑
未来需要更多研究来阐明作用机制、确定最佳组合方案,并探索其与常规疗法的协同效应。
🌈 后生元
后生元是指灭活微生物及其组分或代谢产物,能为宿主带来健康益处。这些成分通过调节免疫反应、抑制NF-κB通路、激活自噬等机制发挥抗炎作用,同时能重塑肠道菌群平衡。
编辑
相比活菌制剂,后生元具有稳定性高、安全性好、易于制剂开发等优势,为IBD治疗提供了新的微生物组靶向策略,但其临床应用仍需更多研究验证。
🌈 粪菌移植(FMT)
将健康捐赠者的粪便微生物群移植给患者,旨在重建健康的肠道生态。FMT在治疗复发性艰难梭菌感染中取得了巨大成功,但在IBD中的疗效尚不稳定,尤其是在克罗恩病中。其成功率受供体选择、移植方式以及受体宿主环境(如免疫状态)等多种因素影响。
在五项FMT的随机对照试验中,由于供体、给药方式和持续时间的不同,四项显示出统计学上的显著益处,缓解率在24%~53%之间。在FOCUS试验的长期随访中,35名在8周内通过每周FMT达到缓解的患者中,有34%在1年后仍保持缓解,这部分得益于自我启动的FMT(3名患者)或饮食改变(9名患者)。
间歇性低频FMT
在一项研究中,87%的患者能够通过每8周进行一次FMT维持临床缓解,持续达48周,尽管不到一半的患者实现了内镜缓解。
供体和受体特定因素
在一项针对UC的随机对照试验中,使用两个不同供体的反应率分别为10%和39%,这表明疗效存在差异。Roseburia inulinivorans和Eubacterium halli的富集,以及粪便短链脂肪酸的增加与缓解相关,而Escherichia spp.、Fusobacterium spp.和Candida spp.的增加则与缺乏反应相关。
FMT前Candida spp.丰度的增加与临床反应相关,而FMT后Candida spp.丰度的降低则表明疾病严重程度减轻。
在UC患者中,FMT前高丰度的白色念珠菌与FMT后细菌多样性增加和治疗成功率提高相关,表明真菌生态可能在细菌定植中发挥作用。
肠道菌群检测结合粪菌移植(FMT)可提高治疗炎症性肠病的疗效。通过持续干预和菌株动态管理,有助于用有益菌株替代疾病相关菌株,确保长期稳定性和定植。
🌈 限定细菌联盟与工程菌
基于对有益菌功能的理解,开发由特定功能的、明确的细菌菌株组成的活体生物药是更精准的策略。例如,包含产丁酸菌的菌群联盟或下一代益生菌。
关于下一代益生菌,详见谷禾文章:
更进一步,可以利用合成生物学技术,设计能够感知肠道炎症信号并原位分泌抗炎分子(如IL-10)或降解有害代谢物的“智能”工程菌。
🌈 噬菌体疗法
噬菌体(phages)是一种自我复制的病毒,能够感染细菌并利用其细胞机制进行复制。噬菌体针对多重耐药细菌的能力使其在炎症性肠病中的应用成为可能。
一种针对克雷伯氏肺炎菌的五种噬菌体组合在人体肠道IBD模型中有效抑制了肠道炎症,且健康志愿者食用后显示出良好的安全性和在下肠道的积累。
VE202是一种活性生物治疗产品,含有有益的梭状芽孢杆菌菌株,已在健康成年人中显示出安全性,为溃疡性结肠炎患者的潜在治疗试验铺平了道路(NCT05370885)。
一种包含10个菌株的18种厚壁菌门菌群(SER-301)在轻度至中度UC患者中测试,初步结果良好,但在2期试验中未能诱导临床缓解。
另一项涉及六种菌株的组合(MH002)的2a期随机对照试验显示,轻度至中度UC患者在使用该组合后,内镜Mayo评分改善了17%,并且粪便钙卫蛋白显著减少。
利用对细菌具有高度特异性的噬菌体,可以精准清除肠道中的特定病原菌(如AIEC、肺炎克雷伯菌),而不影响有益菌群,是一种极具潜力的微生物手术刀。
编辑
doi.org/10.1016/j.bpg.2025.102060
促进黏膜愈合的非免疫抑制疗法
与抑制炎症不同,直接促进肠道上皮修复和再生是另一条有吸引力的治疗路径,有望在不依赖免疫抑制的情况下实现黏膜愈合。
增强屏障功能
开发靶向紧密连接的药物,如MLCK抑制剂(如Divertin),可以直接增强上皮屏障的完整性,减少微生物易位。
靶向干细胞微环境
肠道干细胞(ISCs)是上皮再生的源泉。利用促进ISC增殖和分化的生长因子,如IL-22、GLP-2类似物(如替度格鲁肽),或通过移植间充质干细胞(MSCs)来改善干细胞微环境,是促进组织修复的潜在策略。
2024年2月23日,武田中国宣布,GLP-2(胰高血糖素样肽-2)类似物替度格鲁肽,正式获得NMPA批准,适用于治疗短肠综合征(SBS)成人和1岁及以上儿童患者。由此,替度格鲁肽由此成为中国首个治疗短肠综合征的GLP-2类似物。GLP-2的核心作用,则是调节胃肠道细胞的生长、增殖。
GLP-2当前唯一明确的适应症,是用于治疗短肠综合症,帮助患者吸收营养物质。短肠综合症主要病因是大规模小肠切除手术导致的肠道面积减少,从而引发营养吸收不良,造成其他综合症状。
目前,已有的治疗方式是通过肠外营养技术,为患者直接提高营养物质;或通过肠道修复增加对营养物质的吸收。
GLP-2药物恰好能够达成这一目标,其能够增加肠道上皮细胞,扩大接触面积,同时抑制肠胃蠕动,延长胃排空时间,从而增加肠道吸收。从临床数据来看,替度格鲁肽也的确给患者带来了新希望。
类器官移植
利用患者自身的干细胞在体外培养出“迷你肠道”(肠道类器官),再将其移植回受损的肠道黏膜,有望实现自体组织的再生和修复。这一技术虽然仍处于实验阶段,但展现了巨大的应用前景。
然而,促进再生也需警惕潜在风险,如过度增殖可能增加肿瘤形成的风险,以及再生与纤维化通路之间的重叠。因此,需要精准调控这些再生通路。
促进黏膜愈合的潜在治疗靶点
编辑
doi.org/10.1038/s41575-022-00604-y
黏膜愈合涉及多个层面,包括微生物群、饮食、肠道屏障、免疫细胞和干细胞微环境。图中展示了针对这些不同层面的潜在疗法,如通过饮食干预调节微生物,通过MLCK抑制剂增强屏障,通过IL-22或干细胞移植促进干细胞微环境的再生。
临床前干预与预防的展望
IBD漫长的临床前阶段为疾病预防提供了宝贵的时间窗口。未来的策略将聚焦于在高风险人群中进行早期筛查和干预。
风险分层
通过整合遗传风险评分、家族史、环境暴露史以及临床前生物标志物(如血清抗体、粪便钙卫蛋白、微生物组特征),可以识别出IBD的高风险个体。
早期干预
对于高风险人群,可以在临床前阶段采取干预措施。
例如,对于低风险个体,可以推荐改善饮食和生活方式;对于中高风险个体,可以考虑使用益生菌、益生元或靶向微生物的疗法。
对于即将进入临床期的极高风险个体,甚至可以考虑使用低剂量的免疫调节剂或靶向药物进行“截断治疗”,以阻止疾病的最终发生,类似于在1型糖尿病和类风湿关节炎领域已取得成功的预防性治疗。
实现这一目标需要更精准的生物标志物来动态监测疾病进程,以及更安全有效的早期干预手段。同时,必须关注并解决不同人群在疾病风险和医疗可及性上的差异。
炎症性肠病是一种由遗传与环境因素复杂相互作用驱动的慢性免疫介导疾病。其发病并非一蹴而就,而是经历了一个漫长的、多阶段的临床前演变过程。在这一过程中,肠道屏障功能的逐步丧失、微生物生态的持续失调、以及宿主免疫应答的异常活化和耐受丧失,共同构成了驱动疾病发展的核心病理生理学基础。基因-微生物相互作用,如NOD2通路的功能障碍,以及糖基化等翻译后修饰的改变,在这一过程中扮演了关键的调控角色。
未来的研究应致力于开发更灵敏、更特异的生物标志物组合,以实现对高风险人群的精准识别和疾病进程的动态监测。其中,肠道菌群检测作为评估微生物生态状态的重要手段,结合宿主免疫标志物和遗传风险评分,有望为临床前IBD的早期识别提供多维度的评估依据。如文中所述,特定菌群特征的变化可能先于临床症状出现,这为疾病风险分层和早期干预提供了潜在的时间窗口。
治疗上,应从单一的免疫抑制转向更加多元化和个体化的策略,包括靶向微生物组的生态疗法、直接促进黏膜修复的再生医学方法,以及在疾病极早期进行干预的截断治疗。科学界越来越关注在疾病自然史的早期阶段实施精准干预的可能性,通过整合多组学数据和动态生物标志物监测,有望在正确的时间对合适的个体实施优化的干预策略,从而改变IBD的疾病轨迹,推动临床实践从疾病治疗向风险预防的范式转变。
注:本账号内容仅供学习和交流,不构成任何形式的医疗建议。
主要参考文献:
Cadwell K, Loke P. Gene-environment interactions shape the host-microbial interface in inflammatory bowel disease. Nat Immunol. 2025 Jul;26(7):1023-1035.
Iliev ID, Ananthakrishnan AN, Guo CJ. Microbiota in inflammatory bowel disease: mechanisms of disease and therapeutic opportunities. Nat Rev Microbiol. 2025 Aug;23(8):509-524.
Deleu S, Sabino J. Cutting edge developments and novel targets in IBD: Microbiome in IBD. Best Pract Res Clin Gastroenterol. 2025 Sep;78:102060.
Vaghela, P., Dave, B., Dabhade, A. et al. Redrawing the gut map: evolving probiotic approaches to microbiota modulation in inflammatory bowel disease. Antonie van Leeuwenhoek 119, 51 (2026).
Villablanca EJ, Selin K, Hedin CRH. Mechanisms of mucosal healing: treating inflammatory bowel disease without immunosuppression? Nat Rev Gastroenterol Hepatol. 2022 Aug;19(8):493-507.
Brazil JC, Parkos CA. Finding the sweet spot: glycosylation mediated regulation of intestinal inflammation. Mucosal Immunol. 2022 Feb;15(2):211-222.
Rudbaek JJ, Agrawal M, Torres J, Mehandru S, Colombel JF, Jess T. Deciphering the different phases of preclinical inflammatory bowel disease. Nat Rev Gastroenterol Hepatol. 2024 Feb;21(2):86-100.
Jang KK, Heaney T, London M, Ding Y, Putzel G, Yeung F, Ercelen D, Chen YH, Axelrad J, Gurunathan S, Zhou C, Podkowik M, Arguelles N, Srivastava A, Shopsin B, Torres VJ, Keestra-Gounder AM, Pironti A, Griffin ME, Hang HC, Cadwell K. Antimicrobial overproduction sustains intestinal inflammation by inhibiting Enterococcus colonization. Cell Host Microbe. 2023 Sep 13;31(9):1450-1468.e8.
Liu G, Wang X, Hao Y, Tu L, Mei Y, Zhang T, Wang Y. Bifidobacterium Breve SHMB 8001 Alleviates DSS-Induced Colitis in Mice Via Modulating the Intestinal Barrier and Gut Microbiota. J Food Sci. 2026 Feb;91(2):e70903.
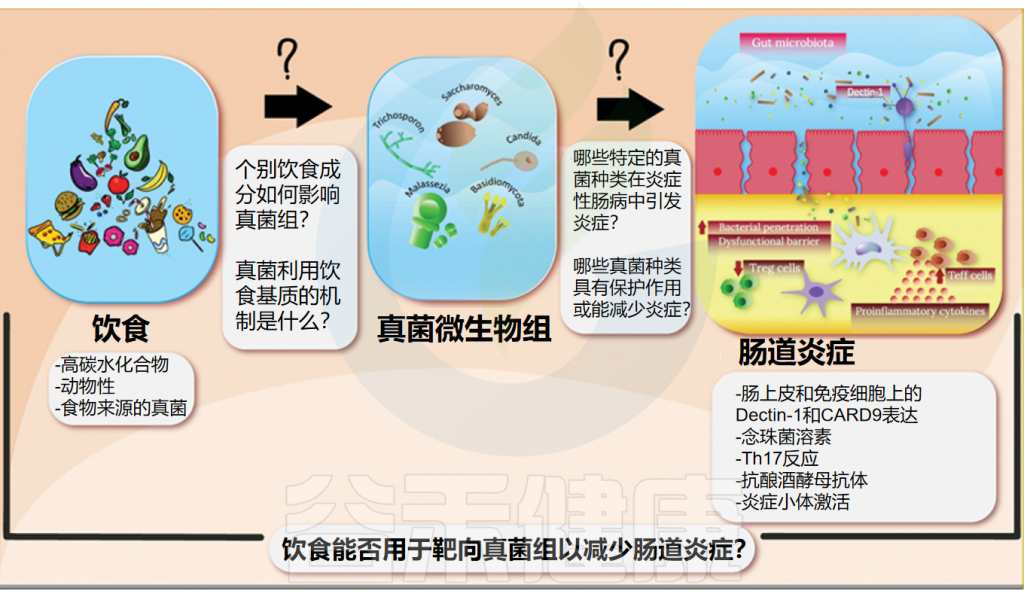
谷禾健康

有这样一些病人:经常性的腹痛、拉肚子、甚至便血、人也日渐消瘦,这很有可能是患有炎症性肠病。
炎症性肠病是一种慢性、非特异性的肠道炎症性疾病,因其顽固无法治愈且需要终身治疗的特点,又称为“绿色癌症”。目前全世界有超过1000万人患炎症性肠病。
在我国炎症性肠病的发病率也呈上升趋势,已经成为常见的肠道疾病,预计2025年患病人数将达到150万,且发病人群偏年轻化。如何预防和治疗炎症性肠病,成为了许多病友和大众关注的重点。
肠道微生物群是一个复杂而独特的实体,在宿主免疫、获取营养物质和强化肠道上皮方面发挥着重要的生理作用。与常见胃肠道疾病的预防、发病机制和进展有关。
虽然肠道微生物群主要由细菌主导,但先进的测序技术已经确定了共生真菌群落,统称为真菌群。大量研究和谷禾的检测数据发现,肠道炎症与微生物组成改变有关,称为肠道菌群失调。微生物群落改变与各种病理疾病有关,例如炎症性肠病(IBD),但它们作为炎症的原因或后果的作用仍是正在进行的研究主题。
而饮食在预防和管理各种疾病方面起着至关重要的作用,被认为是全身炎症的重要调节剂。近年来,随着饮食与肠道微生物关系的深入研究,我们逐渐认识到饮食不仅塑造了肠道细菌的组成,同样也在悄然影响着肠道真菌群落的结构和功能,包括真菌发酵中涉及的代谢途径和酶。
本文将探讨饮食如何调节肠道真菌群,并分析这一过程与炎症性肠病(IBD)之间的潜在联系,为未来饮食疗法在IBD治疗中的应用提供新的视角和研究方向。
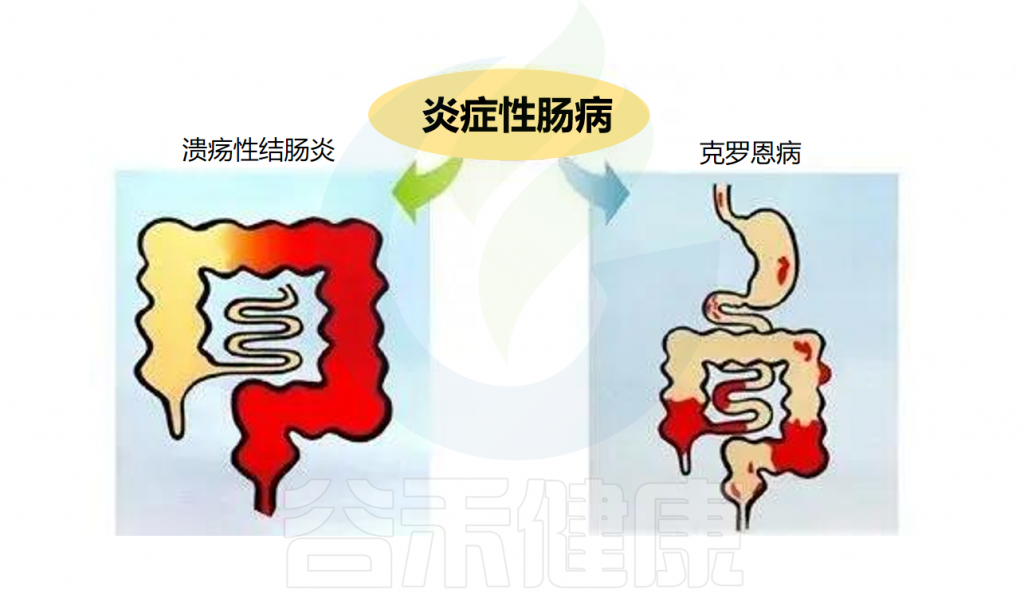
☆ 什么是炎症性肠病?
炎症性肠病(IBD)是一种慢性、非特异性的肠道炎症性疾病。包括溃疡性结肠炎(UC)和克罗恩病(CD)。克罗恩病涉及所有粘膜层,可在胃肠道的任何区域发现,并被细分为穿透性或狭窄性表型。溃疡性结肠炎累及直肠并向近端延伸,涉及粘膜和粘膜下层。
克罗恩病
-主要表现:腹泻、腹痛、大便带血、消瘦、乏力。
-可能还会出现:发烧 、关节痛、关节僵硬、眼睛红肿及疼痛、儿童生长或性发育延迟、皮肤红肿及发痛、口腔溃疡。
–并发症包括:瘘管、狭窄、脓肿、肠穿孔。
溃疡性结肠炎
主要表现:腹泻,多见黏液血便,常常伴发下腹部疼痛尤其以左下腹部疼痛多见。
可能还会出现:腹痛、腹胀、食欲不振、恶心、呕吐、发热、营养不良、关节炎、口腔溃疡。
并发症包括:下消化道大出血,肠穿孔、中毒性巨结肠及癌变。
☆ 炎症性肠病患病率有多高?
近几十年来,炎症性肠病(IBD)发病率逐年上升,目前已成为全球性疾病,具有显著的患者发病率和卫生保健负担。
在全球范围内,IBD的患病人数从1990年的332万例增加到2010年的490万例,增加了47%。据相关数据推测,预计到2025年,全球超过1000万人受到炎症性肠病的影响,我国炎症性肠病患者将达到150万,且以青壮年居多,发病高峰年龄为15至40岁。
注:在欧洲和北美,IBD已经逐步发展为常见疾病。比如在患病率最高的国家,UC和CD的患病率都已高达823/10万。在美国,目前大约有160万UC或CD患者,每年约7万人确诊为IBD。目前中国地区IBD发病率为1.96/10万-3.14/10万左右。
☆ 为什么会得炎症性肠病?
目前暂未明确炎症性肠病的发病机制,可能与多种因素有关:包括遗传因素、感染因素、过度反应的T细胞介导的免疫因素、环境触发因素、生活饮食因素和肠内微生物群因素。
炎症性肠病的日益流行引发了基础科学和临床研究的广泛关注,促使研究者们致力于开发有效的短期和长期治疗方案。这些研究的统一目标是显著减少患者的住院率,并同时提高患者的生活质量,以应对这一日益严重的健康问题。通过不断探索和创新,找到了一些有效的治疗方法,以改善患者的整体健康状况和生活体验。
▸ 药物治疗
抗炎药物短期效果好
炎症性肠病(IBD)的药物治疗是双管齐下的,旨在诱导或维持缓解。包括皮质类固醇和5-氨基水杨酸在内的抗炎药物可以通过多种方式给药,并且在及时诱导活动期病情缓解方面最为有效。
生物制剂可改善长期预后并提高疗效
相比之下,免疫调节剂和生物制剂起效较慢,但可以通过实现长期缓解维持来改变疾病轨迹。生物制剂是基于蛋白质的分子,可以阻断促炎细胞因子。
近年来,生物制剂已成为IBD治疗的基石,有证据表明,在IBD病程早期应用生物制剂可改善长期预后并提高疗效。此外,鉴于生物源化合物耗尽其开放性,生物疗法变得更具成本效益。
注:目前批准了四种不同类别的生物制剂:抗TNF制剂、抗整合素制剂、抗白介素12/23 IgG 1κ制剂和鞘氨醇-1-磷酸(S1P)抑制剂。
生物疗法的出现减少了炎症性肠病(IBD)相关的住院和腹部手术,然而,与IBD相关的总体发病率仍然很高。
▸ 饮食疗法
炎症性肠病(IBD)患者经常询问饮食策略来控制他们的胃肠道症状。饮食调节作为一种非药物疗法,具有显著的益处,其可以调节炎症和促进粘膜愈合并且基本没有副作用。
肠内营养
纯肠内营养(EEN)是一种通过消化道提供全面营养的方法,包括营养全面的流质饮食。在儿科克罗恩病患者中使用纯肠内营养(EEN)显示出与类固醇相似的功效,促进黏膜生长而无相关副作用。
部分肠内营养(PEN),允许一些食物摄入,比EEN耐受性更好,以克罗恩病排除饮食(CDED)为例。避免或减少动物/乳制品脂肪/蛋白质、小麦和食品添加剂的饮食相结合,允许逐渐重新引入水果、蔬菜和豆类以提高食物的灵活性。
特定碳水化合物饮食
一些针对成年IBD患者的饮食,如特定碳水化合物饮食(SCD),已经显示出减轻胃肠道症状的前景。SCD限制IBD患者难以吸收的复合碳水化合物,避免食用谷物、淀粉类蔬菜、乳制品、糖和甜味剂以及某些食品添加剂。
地中海饮食
地中海式饮食最近受到了关注,因为它限制较少,但却和SCD一样有效。这种饮食强调水果、蔬菜、坚果、鱼、全谷物、油性鱼和橄榄油,可以减少炎症,同时限制具有促炎症特性的动物脂肪/蛋白质和食品添加剂。
高膳食纤维并不总是有益
虽然膳食纤维通常被认为对健康有益,但一些炎症性肠病患者,尤其是在疾病活动期,对高纤维食物的耐受性降低。
在活动期疾病患者中证明,某些纤维,特别是低聚果糖(FOS),触发了促炎细胞因子IL-1β。而用来自非IBD或轻度感染患者的微生物群发酵FOS减少了IL-1β的分泌。但这种减少在大多数患有中度或重度IBD患者的微生物群中并不存在,这表明共生微生物群对FOS的发酵不足可能导致异常的免疫反应。
尽管饮食在炎症性肠病(IBD)发病机制中的作用已被认可,但我们对其对IBD进展的影响仍不够深入。目前尚未定义一种能够促进活动性疾病成年患者病情缓解的“IBD饮食”。需要加强对饮食对人类肠道影响的理解,特别是饮食与肠道微生物之间的关系及其在炎症中的作用。
最近的研究表明,肠道微生物群对不同纤维的反应存在相当大的个体间差异。菊粉和欧车前纤维的微生物群组成和功能差异很大,一些个体具有纤维抗性微生物群,而另一些个体具有纤维敏感性微生物群。这强调了个性化饮食建议的必要性,特别是在管理肠道健康和炎症方面。
人类肠道内居住着超过100万亿个微生物,包括细菌、古菌、病毒和真菌。这些微生物统称为肠道微生物群。肠道微生物群是一个复杂而独特的实体,与常见胃肠道疾病的预防、发病机制和进展有重要联系。
☆ 真菌虽然只占肠道微生物群的一小部分,但与多种疾病相关
然而,肠道中超过99%的微生物基因属于细菌,真菌只构成了常驻微生物群的一小部分,在不同的身体部位,包括皮肤、泌尿生殖系统、呼吸道和胃肠道,已经鉴定出390多种真菌。
尽管与细菌相比,真菌的多样性和丰度较低,但真菌的组成和功能与多种疾病有关,包括炎症性肠病、结直肠癌和代谢综合征。
☆ 肠道核心真菌群
然而,微生物组的研究主要集中在细菌上,而对真菌等其他重要的微生物组关注不够。真菌是单细胞或多细胞的异养真核生物,具有惊人的分解和获取营养物质的能力。
早期的研究集中在真菌对肠道健康的潜在危害上,随着人类微生物组项目的出现,真菌物种作为具有共生和互利关系的微生物组的必要组成部分获得了证明。
人体肠道内真菌的多样性和丰度较低,但个体之间存在相当大的可变性,然而,一些真菌物种在个体之间是共享的,这表明可能存在肠道核心真菌群落。
为了描述哪些真菌物种是本地的并有助于肠道健康,真菌群落被分为常驻和非常驻物种。常驻物种具有在缺氧的肠道环境中生长的内在能力,包括子囊菌门、担子菌门和接合菌门中的属,例如假丝酵母菌属(Candida)、马拉色菌属(Malassezia)和枝孢菌属(Cladosporium)。
相比之下,非常驻物种是通过非培养技术在肠道中分离出来的,常见的非常驻物种包括酵母属(Saccharomyces),曲霉属(Aspergillus),以及青霉菌属(Penicillium)等。非常驻物种通常是外部影响的产物,如短期饮食暴露或环境触发因素。这个亚群占分离真菌种类的三分之二。
念珠菌
常驻真菌中最常报道的是假丝酵母菌(Candida),假丝酵母菌又称念珠菌。包括白色念珠菌(C.albicans)、热带念珠菌(C.tropicalis)、近平滑念珠菌(C.parapsilosis),以及光滑念珠菌(C.glabrata)。
具有共生和致病双重特性
白色念珠菌是一种具有共生和致病双重特性,最广为人知的是作为肠道、口腔粘膜、阴道和血液的条件致病菌。它是每年数百万阴道和口腔感染病例的原因,当感染免疫低下的宿主时,导致显著的死亡率。
经历肠道诱导的转变更适合营养吸收
白色念珠菌当暴露于哺乳动物肠道时,会经历“胃肠诱导的转变”,这被认为部分是由于WOR1转录因子,导致更适合短链脂肪酸(SCFA)和N-乙酰氨基葡萄糖营养吸收的表型变化。
白色念珠菌的形态转变机制尚未完全了解,初步研究认为宿主上皮细胞在其中起调节作用。
马拉色菌
马拉色菌属主要以皮肤的定植者和病原体而闻名,也是肠道内第二普遍的真菌种类。其中,M.globosa、M.restricta、M.sympodalis和M.pachydermatis是肠道中最普遍的物种。
马拉色菌经常定植于富含皮脂的区域
几乎所有种类的马拉色菌属不合成脂肪酸,而是依赖外部来源的脂质,这解释了它们在人类皮肤富含皮脂的区域增殖。
在肠道内,脂质可从胆汁盐中获得,胆汁盐由胆汁酸通过肝细胞合成。人们对马拉色菌如何在人类肠道中定植仍然知之甚少,相互竞争的假设包括从人体皮肤间接暴露或由于婴儿期摄入母乳。
在炎症性肠病患者中更多,并可能导致黏膜炎
无论机制如何,马拉色菌都是真菌定植的主要参与者,最近的研究表明它与肠道共生有关。然而,在某些情况下,马拉色菌具有致病潜力,会诱发有害的、异常的免疫反应。
在皮肤上,它与脂溢性皮炎和花斑癣的发病机理有关。与肠道相比,马拉色菌在胰腺导管腺癌组织中的相对丰度更高,通过刺激模式识别受体甘露糖结合凝集素(MBL)与肿瘤发生有关。与健康对照者相比,在炎症性肠病患者中发现的马拉色菌相对比例更高,并且被认为会导致粘膜炎。
酵母菌
酵母菌被认为是人类肠道的非常驻真菌,通常是短暂的定植者和饮食习惯的结果。其中最主要的物种是酿酒酵母。它有助于面包发酵、啤酒酿造,可用作食品添加剂。它在肠道微生物组稳态中的作用经常被忽视。
有助于减轻胃肠道疾病引起的腹泻
最近的研究表明,酵母菌确实可以在肠道中茁壮成长,代谢被称为粘蛋白的高度糖基化蛋白质。布拉氏酵母菌(S.cerevisiae var. boulardii),为酿酒酵母变种,是一种众所周知的益生菌酵母物种,已知有助于解决各种胃肠道疾病,包括旅行者腹泻和抗生素相关性腹泻。研究表明克罗恩病(CD)患者的结肠通透性降低之后,使用布拉氏酵母菌有助于治疗CD。
胃肠道承载着一系列微生物,它们共同形成复杂而动态的生态系统。最近,人们开始关注细菌-真菌的相互作用,因为它们与疾病有关。
细菌-真菌存在互惠共生和竞争
细菌-真菌相互作用可能是互利的,然而,建立特定生态位或营养的竞争可能导致细菌和真菌的失衡。有各种各样的机制参与细菌-真菌的相互作用和拮抗作用。
一项研究表明,纤维发酵过程中细菌产生的短链脂肪酸本身具有抗真菌的特性。另一项研究发现细菌细胞壁合成过程中分泌的肽聚糖片段直接结合并灭活白色念珠菌,阻止其从出芽酵母转变为细长的菌丝。
稳定的肠道细菌群有助于抵抗病原真菌
一个稳定多样的肠道细菌群落调节其真菌对应物。在肠道炎症或长期使用抗生素期间,肠道中的细菌数量减少,使其容易受到侵袭性真菌物种的影响。这在一项涉及用万古霉素治疗的小鼠研究中得到强调,该研究显示在没有细菌种类的情况下存在播散性白色念珠菌感染。因此,病原真菌不仅受宿主免疫的调节,还受各种共生细菌的调节。
真菌和细菌相互作用以优化它们的微环境。肠道微生物产生一种称为生物膜的细胞外基质,以防止抗菌因子。体外研究表明,热带念珠菌(C.tropicalis)、大肠杆菌(E.coli)和粘质沙雷氏菌(Serratia marcescens)共生作用,形成比它们单独产生的能力要厚得多的生物膜。
扫描电子显微镜揭示了这三种生物体之间的密切相互作用,粘质沙雷氏菌使用菌毛将大肠杆菌和热带念珠菌连接在一起。
众所周知,炎症性肠病(IBD)患者体内的微生物群发生了改变,以往的研究主要集中在细菌的相对失衡上。IBD患者的细菌生物多样性降低,来自厚壁菌门的细菌相对减少,来自变形杆菌门的细菌相对增加。
炎症性肠病患者的真菌群落主要由子囊菌门和担子菌门的真菌组成,最常见的属包括酵母属、德巴利酵母(Debaryomyces)、青霉属、曲霉属,以及念珠菌属。
☆ 不同IBD表型真菌群落存在差异
炎症性肠病涉及疾病表型的异质性集合。疾病的表型和炎症的程度与独特的菌群特征有关。在克罗恩病(CD)中,与仅发生在结肠的疾病相比,霉菌种群因累及回肠的疾病而异。特别是,保留回肠的CD显示出较高的念珠菌和德巴利酵母菌,而曲霉菌属(Aspergillus)和毕赤酵母菌(Pichia)在回肠受累的CD中的丰度较高。
在疾病发作期间,与对照组相比,担子菌门与子囊菌门的比率增加。在物种水平上,处于活动发作期的克罗恩病患者显示出念珠菌属、Gibberella moniliformis、Alternaria brassicicola和新型隐球菌(Cryptococcus neoformans)的相对丰度更高。
当根据疾病程度分类时,菌群组成的变化在溃疡性结肠炎(UC)中最为突出。直肠炎患者的青霉菌相对丰度最高,随着疾病程度向近端进展呈负相关。与直肠炎患者相比,毕赤酵母菌(Pichia)在左侧结肠炎患者中的比例过高。
☆ 白色念珠菌增加,导致促炎细胞因子释放
随着与炎症性肠病发作相关的独特微生物组特征得到进一步验证,真菌致病因子在促炎状态中的作用逐渐被认可。
与健康对照相比,炎症性肠病(IBD)患者肠道中白色念珠菌的丰度增加。在菌丝形式中,白色念珠菌上调内皮转化酶1(ECE1)基因,导致ECE1蛋白的丰度更高。这种蛋白质经过蛋白水解加工以释放其活性形式,称为念珠菌溶血素。
念珠菌溶血素是一种31个氨基酸的溶细胞肽,以破坏上皮细胞质膜和刺激编码促炎细胞因子的转录因子而闻名。念珠菌溶血素还损害巨噬细胞的功能,对其抗真菌能力产生负面影响。与健康对照相比,一组克罗恩病患者表现出对念珠菌溶血素特异性的IgA介导的反应水平较低。这表明炎症性肠病患者的高免疫原性真菌病原体没有得到充分控制,并可能导致其促炎状态。
☆ 可作为炎症的替代标志物
作为炎症性肠病中真菌分析的副产品,某些物种已被确定为炎症的替代标志物。与同一克罗恩病(CD)患者体内的发炎组织相比,发现酿酒酵母在非发炎组织中的相对丰度更高。活跃发作的克罗恩病患者表现出更高水平的抗酿酒酵母抗体(ASCA)。
抗酿酒酵母抗体(ASCA)也被用于疾病预测,与对照组相比,在后来诊断为CD的患者中发现的ASCA 水平更高。此外,ASCA已被推断为疾病严重程度的预测指标,与ASCA IgG阴性的CD患者相比,ASCA IgG阳性患者的手术干预率更高。
对患有克罗恩病患者进行定期筛查抗酿酒酵母抗体(ASCA)尚未渗透到临床医学的常规实践中,但它作为一种辅助的、无创的疾病活动和严重程度的衡量方法继续显示出前景。
肠道真菌通过先天和适应性免疫系统影响宿主免疫,这是由真菌细胞壁成分和肠道免疫细胞的模式识别受体(PRR)之间的相互作用引发的。
真菌激活先天免疫和适应性免疫
真菌的吞噬作用导致各种免疫反应的激活,以努力控制真菌的定居。当这些调节机制功能障碍或缺失时,真菌定植不受抑制,导致肠道微生物结构紊乱和肠道炎症。
通过几个检查点,先天和适应性免疫系统可以区分共生真菌或致病真菌。鉴于白色念珠菌可作为共生因子或致病因子,通常用于免疫学分析,以帮助区分这些不同的免疫反应。蛋白质相关分子模式(PAMPs)白色念珠菌和其他真菌与肠道单核吞噬细胞(MNPs)上的toll样受体(TLRs)、C型凝集素受体(CLRs)和NOD样受体(NLRs)相互作用。
例如,巨噬细胞甘露糖受体1(MMR)、树突细胞特异性ICAM3抓取非整合素(DC-SIGN)和巨噬细胞诱导型C-型凝集素(MINCLE)识别某些真菌细胞壁上富含甘露糖的结构。常见的MNPs C型凝集素受体包括dectin-1和dectin-2。dectin-1与真菌β-葡聚糖结合后刺激构象变化,暴露其尾部的免疫受体基于酪氨酸的激活基序(ITAM),随后被SRC家族激酶磷酸化。这一过程使得磷酸化的ITAMs能够与脾酪氨酸激酶(Syk)的SH2结构域结合,激活含有胱天蛋白酶募集结构域的CARD9,并引导Th-17细胞的诱导,动员中性粒细胞进入肠道。
共生和致病形态分别通过不同的免疫途径影响肠道炎症
dectin-1与白色念珠菌或曲霉菌上的β-葡聚糖结合,促进生物活性IL-1β从非活性形式pro-IL-1β通过非经典caspase-8炎性小体激活。重要的是,caspase-8炎性小体的激活不依赖于真菌物质的内化。因此,dectin-1能够在识别外部真菌细胞壁成分后,通过激活IL-1β产生快速有效的反应。IL-1β是IL-1家族的一员,以诱导全身炎症而闻名,同时也能防御外来抗原。作为对真菌识别的反应,IL-1β刺激辅助性T细胞,动员中性粒细胞,并增强吞噬作用,以改善肠道中真菌病原体的清除。
当白色念珠菌处于共生酵母形态时,其负责炎症小体和IL-17通路的PAMPs被隐藏。当它转变为致病菌丝形式时,炎症小体通路和IL-17信号通路被激活。特别是,TLR会分泌细胞因子,如IL-1β、IL-23和IL-6,促进CD4+ T细胞激活进入Th17谱系。Th17细胞则分泌IL-17,招募巨噬细胞、中性粒细胞和抗菌肽,产生有效的抗真菌反应。因此,致病真菌上的PAMPs与宿主树突状细胞上的PRR之间的独特相互作用决定了炎症反应。
共生真菌通过训练免疫系统增加对病原体的抵抗
真菌群在宿主免疫中发挥重要作用。参与真菌识别的巨噬细胞和单核细胞在接触共生真菌(如酿酒酵母)后会获得训练,进一步接触细菌或真菌时,TNF-α和IL-6的产生会增加。
真菌共生菌直接参与稳定肠道稳态,白色念珠菌能够在宿主应激时脱落其致病菌株,保护宿主免受细菌和真菌病原体的侵害。
此外,真菌共生对结肠炎相关的致癌作用也有保护作用。与野生型小鼠相比,真菌耗尽的小鼠在施用氧化偶氮甲烷-葡聚糖硫酸钠(AOM/DSS)后结肠肿瘤发生率更高,而补充外源性IL-18则显著降低肿瘤发生率。因此,真菌共生通过炎症小体途径在预防结肠炎及其相关癌症中发挥重要作用。
鉴于肠道微生物组的广泛多样性及其对人类健康的独特、多模式影响,优化肠道微生物组以改善健康的机制已成为许多新兴研究的焦点。饮食不仅是胃肠道内许多微生物成分的载体,也是其主要营养来源。因此,饮食调节可以通过微生物群组成和功能的变化间接传播起到促炎或抗炎作用。
例如,以植物为基础的饮食与肠道细菌向更具抗炎性的转变有关,其特征是更高的普拉梭菌(Faecalibacterium prausnitzii),直肠真杆菌(E.rectale)和两形真杆菌(Eubacterium biforme),均为丁酸盐生产者。
注:丁酸盐对人体健康的重要性已经在谷禾之前的多篇文章中有过具体讲述。
相反,以肉类为基础的饮食与更易引发炎症的特征相关,这些特征包括活泼瘤胃球菌属(Ruminococcus gnavus)和柯林斯氏菌物种(Collinsella species)的增加。
然而迄今为止,大多数饮食微生物组研究关注点主要集中在细菌种群的组成和功能变化上。鉴于真菌群落在肠道健康中的重要性,有必要进一步研究饮食调节对这一重要群落的影响。这一更广阔的视角会增强我们对饮食如何针对微生物群的所有关键成员的理解,为它们对肠道健康和疾病的影响提供更全面的见解。
可消化的碳水化合物,如单糖、二糖和淀粉多糖,在被身体消化分解为能量并吸收。相比之下,大多数不易消化的碳水化合物或纤维与胃肠道微生物群相互作用,主要在结肠中进行发酵。这个发酵过程产生气体、短链脂肪酸和其他代谢物。
• 念珠菌与近期摄入的碳水化合物呈正相关
研究表明念珠菌与最近的碳水化合物摄入呈正相关。此外,发现高果糖、葡萄糖、麦芽糖和蔗糖的摄入增加了内皮细胞的粘附假丝酵母物种(白色念珠菌,热带念珠菌,以及克柔念珠菌)到上皮细胞。此外,减少饮食中的单糖和淀粉已被证明可以减少肠道中念珠菌的慢性过度生长。
• 高碳水化合物饮食的真菌变化
观察到,与健康的日本人相比,健康的印度成年人的粪便微生物群中含有更高比例的念珠菌和普雷沃氏菌,这可能是由于印度成年人对膳食植物多糖的摄入量较高。
高碳水化合物饮食增加了五个真菌属:
Pleurotus ↑↑↑
Kazachstania ↑↑↑
Auricularia ↑↑↑
Paraphaeosphaeria ↑↑↑
Ustilaginaceae ↑↑↑
并降低了布氏白粉菌属(Blumeria)。
相反,低碳水化合物饮食导致了肠道真菌群落的显著变化,减少了:
Blumeria ↓↓↓
Agaricomycetes ↓↓↓
Malassezia ↓↓↓
Rhizopus ↓↓↓
Penicillium ↓↓↓
增加了Ustilaginaceae。
一些真菌群也可以产生短链脂肪酸,并呈正相关
多项研究表明念珠菌有助于将复杂的碳水化合物发酵成单糖,单糖是其他微生物的能量来源。对猪的研究发现,饮食中碳水化合物含量的差异与不同的真菌种群和短链脂肪酸(SCFA)产量有关。具体来说,低碳水化合物饮食的猪具有较低水平的乙酸盐、丁酸盐和总短链脂肪酸,一些真菌属与这些短链脂肪酸正相关,包括Tomentella,Metschnikowia和Loreleia。
人类研究中膳食碳水化合物调节对肠道真菌的影响


Buttar J,et al.Gut Microbes.2024
因此,膳食碳水化合物的摄入可能会影响肠道真菌种群及其代谢活动,未来的研究应侧重于阐明不同碳水化合物影响真菌群落的确切机制。
肠道微生物群高度参与蛋白水解发酵,产生短链脂肪酸、支链脂肪酸(BCFA)、气体和有害的腐败代谢物,如氨、胺和硫化氢。
• 真菌对氨基酸的高利用率提高其在肠道中的存活率
一项体外研究发现,氨基酸利用率与真菌生长呈正相关。氨基酸可以作为白色念珠菌的碳源,因为这种真菌物种产生蛋白酶并具有寡肽转运蛋白,使白色念珠菌能够通过代谢途径调节适应不同的环境。
在另一项研究中,发现酿酒酵母(S.cerevisiae)和无冠构巢曲霉(Aspergillus nidulans)使用转运蛋白,如氨基酸-多胺-有机阳离子(APC)超家族和主要促进剂超家族(MFS)来利用氨基酸,这可以提高真菌在肠道中的存活率。
• 高蛋白饮食与念珠菌丰度呈负相关,与酵母菌呈正相关
很少有研究评估富含蛋白质的饮食对人类的影响。研究了影响肠道真菌群落稳态的因素,发现乳制品消费是一种高蛋白食物来源,与念珠菌呈负相关,与酵母菌呈正相关。
膳食脂肪主要在小肠中被消化和吸收,并在小肠中代谢,为身体提供能量和各种脂肪酸。一项人类研究表明,脂肪会影响肠道真菌群落。具体来说,证明了念珠菌与总饱和脂肪消耗量呈负相关。
• 脂肪酸可能抑制真菌的生长
体外研究表明,不同真菌物种将脂肪酸和脂肪醇代谢为碳源和能源的效率差异很大。脂肪酸可以抑制真菌的生长。在专注于食品防腐的研究中,发现饱和脂肪酸、不饱和脂肪酸和氧化脂质具有抗真菌特性。
由于缺乏研究调查脂肪对人类肠道真菌群落的影响,这些饮食成分对肠道真菌群落的影响仍然不清楚。然而,在一项小鼠研究中,与喂食标准食物的小鼠相比,喂食高脂肪饮食的小鼠几种真菌类群有所增加,包括Alternaria, Saccharomyces, Septoriella和Tilletiopsis。
动物模型中饮食调节对肠道菌群的影响

Buttar J,et al.Gut Microbes.2024
虽然膳食脂肪对能量代谢至关重要,但它们对肠道真菌群落的影响还不太清楚。未来的研究还需要深入研究膳食脂肪在塑造肠道真菌群落中的作用。
已经发现某些饮食习惯和模式显著影响肠道真菌群落的组成。城市化和饮食习惯的变化会导致中国不同地区人群真菌组成的差异。

这项研究确定了食用酪乳和蓝莓茶与Tetrapissipora blattae、Sugiyamaella lignohabitans、Kazachstania naganishii等的存在之间的相关性。
此外,红茶和猪肉的消费与镰刀真菌属(Fusarium spp.)的四个物种的组成有关。
在另一项研究中,发现杏仁和开心果的摄入与念珠菌和青霉菌属呈负相关。此外,对动物性饮食和植物性饮食进行了比较分析,结果显示动物性饮食组中Neosartorya fischeri(一种源自青霉属的真菌)的丰度更高。
• 素食者体内易出现食源性真菌
研究表明,超过60%的素食者粪便样本中存在食源性真菌,如青霉菌和曲霉菌,以及潜在的致病真菌,如镰刀菌和马拉色菌,但这些真菌类群在食用西方饮食的个体中很少发现。小鼠研究进一步支持饮食调节对真菌群落特征的影响。与标准食物相比,喂食加工饮食的小鼠的总体真菌多样性降低。
• 酵母菌可能随饮食来源进入体内
另一个值得注意的方面是我们的食品中普遍存在常见的肠道真菌,例如酿酒酵母(S.cerevisiae)和汉斯德巴氏酵母菌(Debaryomyces Hansenii)。这些真菌常见于乳制品、面包、果皮(包括葡萄)、普洱茶和奶酪中,因此很难确定这些真菌物种是真的存在于肠道中,还是在自然界中更短暂。
研究发现,食用不含酿酒酵母的饮食导致酿酒酵母水平从基线时的86%大幅降低到无酿酒酵母饮食2天后ITS读数的 0.1%。这表明酿酒酵母的丰度高度依赖于这种真菌物种的饮食来源。
因此,探索特定真菌物种对疾病潜在影响的研究应考虑食物来源的真菌分类群的潜在短暂性。这种考虑很重要,因为它可能会影响这些物种在健康和疾病中的重要性。
• 利于真菌群落稳态和健康的饮食
最近的营养学研究利用Mycobiome 饮食来选择性地改变肠道中真菌的组成。Mycobiome 饮食的特点是消耗瘦肉或植物蛋白、单不饱和脂肪或多不饱和脂肪、富含纤维和抗性淀粉的食物,同时避免加工的高糖食物。
遵循Mycobiome饮食28天导致念珠菌的丰度下降了72.4%,尤其是白色念珠菌和热带念珠菌。此外,Mycobiome饮食增加了一些有益真菌物种的浓度,包括Pichia kluyveri和Galactomyces geotrichum。
这种饮食除了对肠道菌群的有益影响外,还使小肠细菌过度生长患者的细菌过度生长减少,并改善了所有参与者的疲劳、睡眠、腹胀、便秘和腹泻。
尽管研究表明,真菌群落随着饮食而变化,但我们对饮食对这些微生物功能潜在影响的了解仍然有限。据了解,细菌和真菌群落都产生代谢物,这些代谢物不仅形成它们自己的群体,而且对其他群落成员和各种宿主细胞群体施加影响。
作为对高纤维饮食的响应,肠道细菌上调功能途径和酶,使纤维发酵产生短链脂肪酸,特别是丁酸。
真菌作为真核生物,表现出与细菌等原核生物不同的代谢特征。真菌具有发酵植物细胞壁的代谢途径和酶,一些真菌种类在食品工业中用于乙醇生产,并通过发酵果胶、纤维素和木质素等纤维产生乳酸和气体。
真菌在降解纤维素纤维方面优于细菌。真菌产生更多的甲烷、乙酸盐和甲酸盐,而细菌产生更多的丁酸盐和丙酸盐。
重要的是要考虑代谢物在调节肠道细菌和真菌之间的作用。代谢物,如胆汁酸,可以影响微生物的相互作用,因为真菌有能力通过胆汁酸的转化对细菌的生存力产生不利影响。
!
小贴士
饮食对改变真菌群落组成的影响可能不直接归因于饮食成分对肠道真菌的影响。相反,这些影响可能是间接的,源于肠道细菌的调节,进而影响真菌的组成。
由于阐明饮食如何调节肠道真菌群落背后的机制的证据有限,有必要进行进一步的研究来辨别饮食对肠道真菌群落的直接和间接影响。
肠道微生物组是一个复杂而独特的实体,能对环境、饮食和疾病的变化做出反应。它充满了来自各个种群的微生物,通过复杂而动态的关系相互作用。虽然细菌是微生物组稳态中最重要的部分,但真菌在其中也起着不容忽视的作用,参与机会性病原体的调节和原本无法获得的营养物质的发酵。
菌群结构的改变与菌群失调有关,菌群失调是一种与包括炎症性肠病(IBD)在内的各种慢性疾病有关的发炎免疫状态。菌群概况因IBD表型和严重程度而异,菌群标志物有望作为疾病进展的指标。
还需研究饮食对非细菌微生物群的调节作用,以及参与真菌发酵和膳食成分利用的代谢途径和酶的机制。深入了解饮食如何影响肠道菌群的组成和功能至关重要。这些知识将帮助确定饮食疗法是否应该专门针对肠道真菌,从而可能减轻炎症并改善炎症性肠病患者的预后。
主要参考文献
Buttar J, Kon E, Lee A, Kaur G, Lunken G. Effect of diet on the gut mycobiome and potential implications in inflammatory bowel disease. Gut Microbes. 2024 Jan-Dec;16(1):2399360.
Rinninella E, Raoul P, Cintoni M, Franceschi F, Miggiano GAD, Gasbarrini A, Mele MC. What is the healthy gut microbiota composition? A changing ecosystem across age, environment, diet, and diseases. Microorganisms. 2019. Jan 10;7(1):14.
Liang G, Cobián-Güemes AG, Albenberg L, Bushman F. The gut virome in inflammatory bowel diseases. Curr Opin Virol. 2021. Dec;51:190–198.
Nacer S, Haddad F, Tahiri M, Hliwa W, Badre W, Choukri EM, Jai SR, Erguibi D, Boufettal R, Chehab F. P51 predictive factors of surgery in crohn’s disease. Gut. 2022.
Wang R, Li Z, Liu S, Zhang D. Global, regional and national burden of inflammatory bowel disease in 204 countries and territories from 1990 to 2019: a systematic analysis based on the global burden of disease study 2019. BMJ Open. 2023. Mar. 13(3):e065186.
Sigall-Boneh R, Levine A, Lomer M, Wierdsma N, Allan P, Fiorino G, Gatti S, Jonkers D, Kierkuś J, Katsanos KH, et al. Research gaps in diet and nutrition in inflammatory bowel disease. A topical review by D-ECCO working group [dietitians of ECCO]. J Crohns Colitis. 2017.
Raimondi S, Amaretti A, Gozzoli C, Simone M, Righini L, Candeliere F, Brun P, Ardizzoni A, Colombari B, Paulone S, et al. Longitudinal survey of fungi in the human gut: ITS profiling, phenotyping, and colonization. Front Microbiol. 2019. Jul 10;10:1575.
Dalle F, Wächtler B, L’Ollivier C, Holland G, Bannert N, Wilson D, Labruère C, Bonnin A, Hube B. Cellular interactions of Candida albicans with human oral epithelial cells and enterocytes. Cell Microbiol;2010;12(2):248–271. doi: 10.1111/j.1462-5822.2009.01394.x.

谷禾健康

Faecalibacterium prausnitzii(F. prausnitzii),普氏栖粪杆菌,又名:普拉梭菌,是人类肠道菌群中最重要的细菌之一,占健康人粪便样本中检测到的细菌总数的 5-15%,是丁酸的重要生产者之一,具有抗炎作用,维持细菌酶的活性,保护消化系统免受肠道病原体的侵害。
已经证实,患有慢性便秘、乳糜泻、肠易激综合征和炎症性肠病(包括克罗恩病和溃疡性结肠炎)的个体中这种微生物的数量减少。此外,在 2 型糖尿病、结直肠癌和银屑病患者中观察到较低水平的F. prausnitzii。
F. prausnitzii 数量的变化可能是人类肠道菌群失调的迹象,科学家们将其视为下一代益生菌的候选者。
Faecalibacterium prausnitzii,革兰氏阴性,对氧极度敏感,是柔嫩梭菌类群的优势菌,属于梭菌科,厚壁菌门。该物种属于Clostridium cluster IV分组的Clostridium leptum group柔嫩梭菌类群,是该类群的最优势菌种,占64%左右。一般中文翻译柔嫩梭菌指的就是这个类群,其代表物种就是普氏栖粪杆菌,又名普拉梭菌。
普拉梭菌的扫描电子显微镜图像
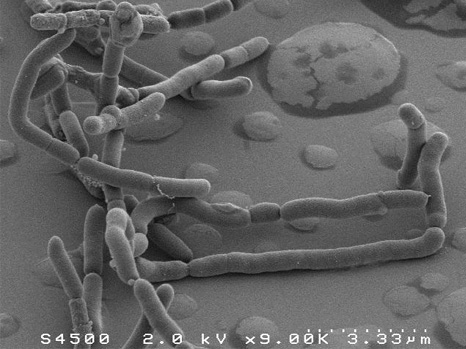
Plateforme MIMA 2, T. Meylheuc
直到 2000 年代中期,16S rRNA 文库的高通量测序和粪便 DNA 的宏基因组分析显示F. prausnitzii是人类肠道中最丰富的细菌之一,人们才完全意识到该物种对人类健康的重要性细菌总数的 5–15%。
在婴儿早期,Faecalibacterium prausnitzii 的数量非常低,并在原定殖细菌建立后增加。同时,在克罗恩病 (CD)和溃疡性结肠炎 (UC) 等各种炎症性肠病 (IBD) 以及结直肠癌 (CRC) 和 2 型糖尿病中观察到 F. prausnitzii 水平降低。
F. prausnitzii 生长繁殖肠道理化调节
F. prausnitzii 生长的最佳 pH 值范围在 5.7 和 6.7 之间,这是在结肠中发现的 pH 值范围。尽管在 5-5.7 的 pH 值范围内菌株之间的耐受性存在差异,但在 3.5 和 4.5 的 pH 值之间没有观察到生长。这表明 pH 值影响F. prausnitzii 沿肠道的分布。
该菌在健康受试者和肠道疾病患者的十二指肠(pH 范围 5.7-6.4)和回肠末端中也被检测到。据报道,溃疡性结肠炎和克罗恩病患者经常出现酸性大便,因此仍有待证明肠道局部 pH 值是否会调节F.prausnitzii 在肠道疾病(如 IBD)患者中的丰度和组成。
F. prausnitzii 对胆汁盐生理浓度的轻微增加也高度敏感,这为克罗恩病患者表现出的F. prausnitzii丰度减少提供了一个合理的解释,因为这些患者的胆红素浓度增加,尤其是那些有回肠疾病受累和接受过肠切除术的患者。
此外,据报道,分离株之间的耐受性存在差异,尤其是胆盐浓度为 0.1% (wt/vol) 时,表明胆汁盐浓度的变化可能决定F. prausnitzii亚型组成的变化。由于克罗恩病患者的胆盐成分也发生改变,需要进一步研究以确定F. prausnitzii是否对某些类型的胆盐成分具有更高的敏感性,并确定不同的胆汁盐谱是否会改变 F. prausnitzii 亚型组成。
F. prausnitzii对氧极为敏感,但它能够通过在黄素和半胱氨酸或谷胱甘肽存在的情况下使用细胞外电子转移来承受肠粘膜中发现的低水平氧气 。最近,已经证明,当与这些抗氧化剂和菊粉作为冷冻保护剂一起配制时,菌株 A2-165 可以在环境空气中保持活力 24 小时。因为氧气梯度在定义结肠中微生物的空间组织方面具有重要作用,确定F. prausnitzii亚型之间的氧耐受性是否存在差异,以及它是否与粘膜的发炎状态相关,将会很有趣。
F. prausnitzii的必需营养素的可用性和有效性可能会影响其在肠道中的分布。
F. prausnitzii的全局营养需求已经通过人工策划的代谢重建进行了调查,该重建确定了几种氨基酸和维生素作为生长因子。体外实验表明,F. prausnitzii可以通过代谢交叉喂养直接或间接地代谢纤维。乙酸盐消耗是F. prausnitzii生产丁酸盐的主要驱动力。当这两个物种在低聚果糖上一起生长时,青春双歧杆菌释放的醋酸盐的消耗已被证明会刺激F. prausnitzii。F. prausnitzii可以代谢来自宿主和饮食来源的底物,菌株之间存在一些差异。
F. prausnitzii分离株可以使用简单碳水化合物生长良好,但菌株之间在发酵更复杂碳水化合物的能力方面存在一些差异,例如那些来自宿主或饮食的碳水化合物,尽管大多数F. prausnitzii菌株都能够发酵菊粉,但研究结果表明,其中只有两种菌株可以在该基质上生长良好。这支持观察到该益生元在营养干预中对该物种的刺激,并表明只有F. prausnitzii种群的一些成员受到菊粉的选择性刺激。
该物种的菌株利用肠腔中发现的其他多糖的能力有限,例如阿拉伯半乳聚糖、木聚糖和可溶性淀粉。大多数分离株可以在苹果果胶上生长,并且能够使用一些果胶衍生物。体外研究表明,在生理条件下,F. prausnitzii可以在某些类型的果胶发酵中发挥关键作用,并且可以与其他肠道细菌成功竞争这种底物。
最近一项基于普拉梭菌 A2-165菌株功能代谢图的研究预测其无法合成氨基酸丙氨酸、半胱氨酸、蛋氨酸、丝氨酸和色氨酸。通过对其他F. prausnitzii菌株基因组的进一步分析,已经观察到维生素和辅助因子(例如:生物素、叶酸、烟酸、泛酸、吡哆醇和硫胺素)的营养缺陷型,并且菌株之间似乎存在与核黄素生产有关的一些差异,这可能是由于菌株间差异。
相比之下,该物种已被预测为钴胺素生产者。有证据表明,一些 IBD 患者易患钴胺素缺乏症,但尚未确定这种情况的原因。由于缺乏一致的临床数据表明 IBD 患者易患这种缺陷,因此确定它是否与肠道中钴胺素生产者的消耗有关会很有价值。
F. prausnitzii与肠道微生物群其他成员的关系
F. prausnitzii在肠道中与C. coccoides组和 Bacteroidetes 的几个成员共同发生。有人提出F. prausnitzii可能依赖其他物种(如拟杆菌)进行交叉饲养。在共培养实验中,观察到F. prausnitzii 的发酵活动过程中,多形拟杆菌(B. thetaiotaomicron)正在发酵果胶。表明后者产生的乙酸盐促进了F. prausnitzii 的生长。此外,B. thetaiotaomicron对果胶的初始发酵可以释放果胶衍生物,然后可以被F. prausnitzii使用。
最近对大鼠模型的研究表明,F.prausnitzii 需要预先存在后,B.thetaiotaomicron才能在肠道定植。已经反复观察到无法维持F. prausnitzii单相关动物模型 并且还描述了一种小鼠模型,其中F. prausnitzii植入胃肠道需要用大肠杆菌预先制备,表明F. prausnitzii和其他菌群存在密切关系。在 IBD 患者中发现了这两种物种之间的相关性,根据疾病位置观察到正相关或负相关。这表明一个种群对另一个种群的影响,尽管还不能排除宿主因素的影响。
F. prausnitzii 的分类学和系统发育
已经确定Faecalibacterium属与厚壁菌门、梭菌类和瘤胃球菌科中的梭菌群 IV的成员有关。目前,F. prausnitzii是唯一成功分离的粪杆菌属物种。
最近对分离株的系统发育表征确定该物种包括两个系统发育群,它们具有 97% 的 16S rRNA 基因序列相似性。其中分类群 I 的减少是肠道炎症的更敏感标志物。迄今为止,大部分可用的基因组和生理数据是使用分类群 II 菌株收集的。
F. prausnitzii基因组之间的平均核苷酸同一性水平非常低,而基因组的可塑性却很高。两个基因组可以根据功能基因互补的差异进行分离,尽管这种划分与基于保守基因系统发育的分离不完全一致,突出了水平基因转移在塑造F. prausnitzii 中的重要性基因组。两个基因组之间的差异主要在于与碳水化合物分解代谢相关的基因(例如分类群 I中预测的唾液酸酶基因)和氨基酸的互补,以及防御机制。
值得注意的是,最近对健康和患病肠道样本中物种多样性和丰度的研究表明,存在其他F. prausnitzii系统发育型,并且不能排除Faecalibacterium属内其他物种的存在。这些已经通过分析粪便样本中总体细菌群落的分子方法估计,代表大约 2% 的粪杆菌序列,并使用物种特异性引物。有趣的是,罕见的系统发育型主要是从患有肠道疾病的受试者身上发现的。
产生丁酸,维持肠道稳态
Faecalibacterium prausnitzii 一直被报道为肠道中发现的主要丁酸盐生产者之一。丁酸盐在肠道生理学和宿主健康中起着至关重要的作用。丁酸盐结肠细胞的主要能量来源之一,能够保持肠道内壁的完整性,防止病原体通过肠道进入人体。它刺激绒毛的生长 (肠道屏障上的微观挤压)并促进粘蛋白的产生,粘蛋白是一种覆盖肠道内壁的保护性凝胶。此外,丁酸盐通过增加紧密连接蛋白的合成和抗菌肽的产生来参与结肠屏障的增强。
肠道屏障依赖于 T 连接蛋白来防止肠道内容物泄漏到血液中。丁酸盐通过激活 AMP 活化蛋白激酶 (AMPK) 来促进紧密结合的 T 连接蛋白。
丁酸盐可以通过调节肠腔 pH 值来调节肠道微生物群,这对产生短链脂肪酸的细菌有益,丁酸盐可以保持上皮缺氧并限制硝酸盐呼吸依赖性细菌的过度生长以维持肠道稳态。
丁酸盐是由某些细菌在乳酸发酵过程中产生的。然而,乳酸也是硫酸盐还原菌的首选能源,硫酸盐还原菌会产生硫化物并抑制结肠细胞对丁酸的氧化。IBD 中产生丁酸盐的菌的消耗可能会削弱已经脆弱的肠上皮细胞,导致共生或低致病性细菌的入侵,随后激活免疫活性细胞。
产生丁酸,抗炎抗肿瘤
丁酸盐可以通过抑制 NF-κB 转录因子激活、上调 PPARγ和抑制干扰素γ来减轻肠黏膜炎症。
此外,F. prausnitzii已被证明可以产生许多具有抗炎特性的物质,包括一种 15 kDa 的蛋白质“微生物抗炎分子”(MAM),它能够抑制肠上皮中的 NF-κB 通路细胞和预防小鼠 IBD 模型中的结肠炎 。而且,丁酸盐可能通过抑制组蛋白脱乙酰酶活性来抑制炎症反应,导致组蛋白过度乙酰化和抑制 NF-κB 活性。
Faecalibacterium prausnitzii 上清液已被证明可以抑制促炎性 IL-8 免疫蛋白的产生。
F. prausnitzii 还通过激活产生 IL-10(一种抗炎蛋白)的 T 细胞来帮助对抗炎症。
F. prausnitzii 的抗炎机制
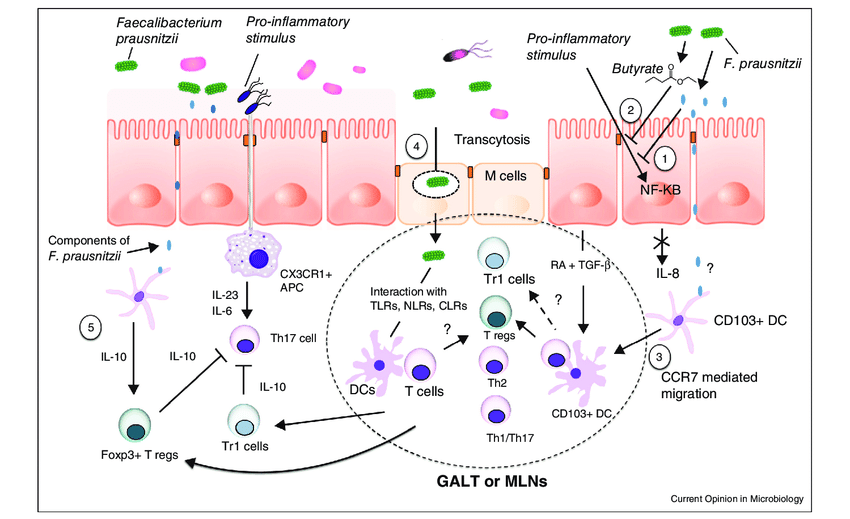
额外的抗炎特性归因于该菌还通过其诱导耐受性细胞因子谱的能力(促炎细胞因子如 IL-12 和 IFN-γ 的分泌非常低,而抗炎细胞因子 IL-10 的分泌增加)。这些抗炎作用部分与能够阻断 NF-κB 激活、IL-8 产生和调节性 T 细胞产生上调的分泌代谢物有关。
这些微生物可以产生莽草酸,一种常见于植物中的酸,能够防止脂多糖 (LPS) 引起的炎症。LPS 是一些细菌外膜上的毒素,可以激活免疫系统并引起炎症。F. prausnitzii 还可以制造水杨酸,这有助于防止细菌形成生物膜,这是一些引起感染的微生物的常见特征。
体外研究表明,丁酸盐还具有抗肿瘤作用,包括减少肠上皮细胞中肿瘤坏死因子 (TNF) 的分泌,并诱导肿瘤细胞的分化和凋亡,从而抑制肿瘤生长。
F. prausnitzii 及其代谢物对小鼠结肠炎发挥保护作用,改善了肠道菌群失调,增加了细菌多样性和产生短链脂肪酸菌的丰度,降低了血清 TNF-α 和Proteinbacteria,酸杆菌门和拟杆菌。这些发现将为F. prausnitzii 的抗炎抗肿瘤作用提供进一步的证据,其具有治疗 IBD 的潜力。

下一代益生菌特性
大多数商业益生菌不包括有益健康的优势肠道微生物菌株,这就是为什么这些益生菌菌株不会在人类肠道中定殖并且它们的作用仅在很短的时间内持续存在的原因。如今,人们对使用共生细菌作为潜在的益生菌剂越来越感兴趣。
使用厌氧细菌和 EOS(氧敏感) 细菌是成功分离可能影响肠道稳态的代表性菌株的先决条件。Faecalibacterium prausnitzii从健康志愿者分离株:F. prausnitzii作为新一代益生菌在使用中向前迈进了一步。
但是下一代共生益生菌必须满足与传统益生菌相同的标准。它们应该:
(i) 被隔离和充分表征
(ii) 达到安全要求,例如可接受的抗生素抗性或缺乏裂解和粘附能力
(iii) 作为益生菌考虑,在被隔离之前对宿主表现出有益的影响
关于F. prausnitzii,虽然对其安全性知之甚少,但该物种作为下一代益生菌具有明显的潜力。到目前为止,所有F. prausnitzii分离株具有一些特征,例如:
(i) 缺乏上皮细胞粘附、质粒、抗微生物和溶血活性
(ii) 存在 DNAse 活性。部分的F. prausnitzii菌株产生的 D-乳酸微弱,不会对宿主产生代谢有害影响,无法降解黏蛋白。
仅使用参考菌株F. prausnitzii A2-165在体外和体内分析了其有益的抗炎作用和生物膜形成菌株 HTF-F。由于益生菌特性通常是菌株特异性的,需要单独研究来评估其他F. prausnitzii分离菌株的抗炎特性。
人体微生物组中 F. prausnitzii 的丰度受其健康状况和结肠环境的影响。这一点尤其重要,因为在患有消化系统疾病的患者中发现了低于平均水平的F. prausnitzii。
克罗恩病和溃疡性结肠炎
肠道微生物群可能通过两种机制驱动致病性,即“促炎”菌群的扩张或微生物群保护性化合物的限制。
健康肠道和IBD患者中 F. prausnitzii
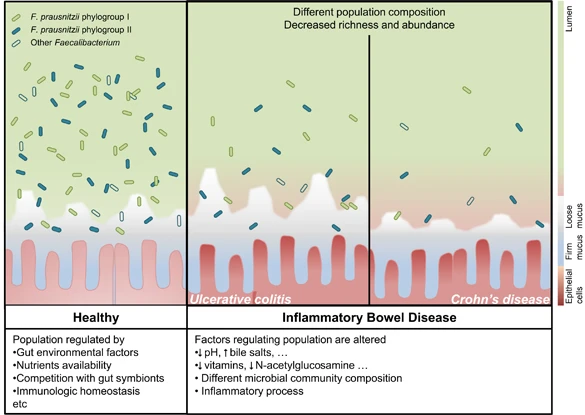
在IBD患者中,肠道环境的改变可能会影响F. prausnitzii 的种群组成和负荷。这些差异可以通过监测来鉴别IBD亚型。
在过去的十年的研究已发现,相当数量的降低的F. prausnitzii水平与某些形式的IBD相关。虽然F. prausnitzii水平与溃疡性结肠炎活动和隐窝炎之间的联系是有争议的,在克罗恩病中F. prausnitzii枯竭,尤其是在疾病发作和克罗恩病的回肠部位。
◥ 治疗后F. prausnitzii恢复
在粪便和活检样本中使用的各种证明方法表明,对克罗恩病患者有效的各种抗炎和抗菌治疗,包括大剂量皮质醇、英夫利昔单抗、干扰素-α2b 和利福昔明,都能够恢复正常水平的F. prausnitzii。
因此,有人提出F. prausnitzii的消耗不是克罗恩病的致病事件,而是产生过量活性氧 (ROS) 的粘膜炎症的结果。这导致粘膜相关和管腔 EOS 培养物显着减少,包括F. prausnitzii。
如何解释Faecalibacterium prausnitzii在溃疡性结肠炎和克罗恩病患者中减少?
◥ pH值
健康的肠道 pH 值 (5.7–6.7) 为 F. prausnitzii 的生长提供了最佳条件,而高酸性环境会抑制 Faecalibacterium prausnitzii 的生长。尤其是,溃疡性结肠炎和克罗恩病患者的大便呈酸性,这可能有助于解释为什么他们的微生物群中缺乏 F. prausnitzii .
◥ 胆汁
胆汁及其成分水平略微升高也会影响 F. prausnitzii 丰度。胆汁是肝脏分泌的一种物质,可在消化过程中分解脂肪。胆汁盐已被证明可以抑制 Faecalibacterium prausnitzii。
研究人员假设这种对胆汁的敏感性可以解释为什么这些微生物在克罗恩病患者中较少。患有这种炎症性疾病的人往往有较高水平的胆红素,胆红素是一种赋予胆汁颜色的色素,特别是如果他们已经切除了部分肠道。
溃疡性结肠炎中, F. prausnitzii有缺陷的肠道定植,这在缓解期短、既往有频繁复发史和结肠炎大范围扩展的患者中更为严重。此外,复发后,F. prausnitzii在维持缓解时逐渐恢复,1年后达到接近参考种群的值。
基于F. prausnitzii菌株特性的治疗探索
科学研究人员提出假设,即F. prausnitzii 的高定植可能会阻止溃疡性结肠炎中粘膜炎症过程的重新激活。这一假设应在未来的研究和增加肠道F. prausnitzii 的策略中得到证实需要在患有广泛疾病的患者中对人群进行验证。
F. prausnitzii菌株还可以利用N-乙酰氨基葡萄糖,一种在肠粘膜中发现的糖蛋白的成分。有趣的是,用这种化合物进行治疗可能会改善克罗恩病,因为它可以作为炎症、受损肠道软组织的愈合因子。因此,鉴于F. prausnitzii能够发酵这种碳水化合物,探索在接受这种治疗的克罗恩病患者中恢复这种有益肠道细菌的效果将是有意义的。
镇痛特性
慢性腹痛是肠易激综合征或炎症性肠病等肠道疾病患者的常见症状之一。研究人员表明,在炎症性肠病的患者中,肠道中的Faecalibacterium prausnitzii细菌的丰度降低。
这些研究人员已经证明,由于压力动物模型(产生绞痛起源的内脏超敏反应),F. prausnitzii丰度的减少与内脏超敏反应的出现有关。对于人类,这种超敏反应会导致肠道不适或腹痛。
科学家已经证明,通过给这些动物服用F. prausnitzii,它们恢复了正常的绞痛敏感性。
结直肠癌
丁酸盐的抗炎功能包括抗癌特性。这种短链脂肪酸是一种组蛋白脱乙酰酶 (HDAC) 抑制剂:它抑制癌细胞的活性和生命周期。这促使研究人员调查 Faecalibacterium prausnitzii 在结直肠癌中的作用。
结直肠癌患者产丁酸盐的细菌较少,包括Faecalibacterium prausnitzii。有人提出,由于丁酸盐产量较低,肠道内壁细胞可能更容易受到损伤,这可能会增加患癌症的风险。
扩展阅读:结直肠癌防治新策略——微生物群
乳腺癌
F.prausnitzii 通过抑制IL-6/STAT3通路抑制乳腺癌细胞的生长。菌群代谢物与菌群结合(如Faecalibacterium与磷胆碱结合)可能是乳腺癌的一种新的检测方法。
糖尿病
糖尿病是一种代谢性疾病,其中由于胰岛素功能受损,身体失去控制血糖的能力。升高的循环葡萄糖会损害血管并增加患心脏病的风险。
许多针对人类的研究已经确定了肠道微生物组的失衡(生态失调),这会削弱肠道内壁,并使不良分子进入体内,从而破坏其他器官。科学家们认为,生态失调先于并促进了糖尿病的发展。
糖尿病患者的厚壁菌门水平较低,包括 Faecalibacterium prausnitzii 和其他可强化肠道内壁的产丁酸盐细菌。
研究表明,脂多糖是在某些革兰氏阴性细菌上发现的促炎分子,当肠道屏障薄弱时,它会进入血液并损害胰腺中的B 细胞功能,产生胰岛素的细胞。
过敏反应
Faecalibacterium prausnitzii通过调节肠道微生物群和短链脂肪酸的产生来缓解屋尘螨引起的过敏性哮喘。
补充F. prausnitzii可阻断嗜酸性粒细胞、中性粒细胞、淋巴细胞和巨噬细胞的流入,减轻病理变化。此外,活的和死的F. prausnitzii给药都降低了白细胞介素 IL-4、IL-5、IL-13 和免疫球蛋白 G1 的水平,提高了调节性 T 细胞 (Treg) 的比例,改善了微生物生态失调并增强了短链脂肪酸生产。
Faecalibacterium prausnitzii部分通过肠道微生物群调节和短链脂肪酸产生发挥抗哮喘作用,表明其作为预防过敏性哮喘的益生菌剂的潜力。
肝病
研究人员研究了口服F. prausnitzii治疗对高脂肪喂养小鼠的影响。与高脂肪对照小鼠相比,F. prausnitzii处理的小鼠肝脏脂肪含量、天冬氨酸氨基转移酶和丙氨酸氨基转移酶较低,肝脏中脂肪酸氧化和脂联素信号传导增加。肝脏脂质组学分析显示,几种甘油三酯、磷脂和胆固醇酯的含量减少。内脏脂肪组织中的脂联素表达增加,皮下和内脏脂肪组织对胰岛素更敏感。
且F. prausnitzii治疗增加了肌肉质量,这可能与增强线粒体呼吸、改变肠道微生物群组成和改善肠道完整性有关。总的来说,F. prausnitzii治疗可改善肝脏健康,减少小鼠脂肪组织炎症,需要进一步研究以发现其治疗潜力。
其它
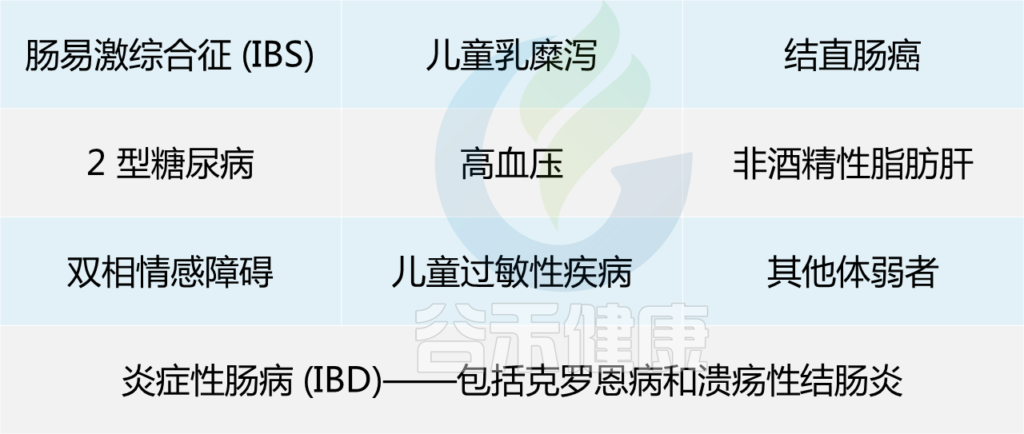
肠道菌群中F. prausnitzii的消耗与微生物失调有关,同时伴随着广泛的代谢和/或免疫介导的慢性疾病,包括银屑病、高血压、心脏和肾脏疾病。
在以下患者中已显示低水平的F. prausnitzii:
综上所述,F. prausnitzii 对人体肠道和健康非常重要,以至于它被称为肠道中的“关键物种”。现在问题来了:如何增加肠道中这种细菌的数量?
能否添加到酸奶中补充?
不能。
添加到酸奶中或作为补充剂的典型细菌在暴露于空气(氧气)时能够存活。然而,F. prausnitzii是“氧敏感的”,它们在暴露于空气后几分钟内就会死亡。研究人员将这种有益细菌视为“未来的益生菌”,目前正在进行研究以找出它可以轻松储存并暴露在空气中几个小时而不会死亡的方法。所以目前没有办法服用益生菌F. prausnitzii补充剂。
除此之外还能做什么呢?
饮食。
高动物肉、高动物脂肪、高糖、高度加工食品和低纤维饮食(典型的西化饮食)会降低F. prausnitzii 的数量,而高纤维、低肉的饮食会增加 F. prausnitzii 的数量。
——高纤维
我们可以做的第一个提高 F. prausnitzii 的数量的办法, 就是增加饮食中的纤维。增加膳食纤维会增加丁酸盐,丁酸盐与结肠健康有关,具有抗炎和抗癌作用。
高纤维是包括:全谷物、蔬菜、水果、坚果、种子和豆类。此外,还要主要吃多样化的植物性饮食,也就是说大量的植物性食物。多样性似乎很重要——不同类型的纤维喂养不同的细菌。
——益生元
菊粉型果聚糖和阿拉伯木聚糖等益生元可增加F. prausnitzii的数量。

一项研究表明,猕猴桃胶囊的摄入增加了功能性便秘患者的Faecalibacterium prausnitzii 丰度。
虽然F. prausnitzii可能是肠道中一种重要的有益细菌,但是它与其他有益细菌的关系,它们是交叉喂养的。研究表明青春双歧杆菌是利用菊粉主要物种并刺激Faecalibacterium prausnitzii的增加。
主要参考文献:
Heinken, A., Khan, M. T., Paglia, G., Rodionov, D. A., Harmsen, H. J. M., & Thiele, I. (2014). Functional Metabolic Map of Faecalibacterium prausnitzii, a Beneficial Human Gut Microbe. Journal of Bacteriology, 196(18), 3289–3302. doi:10.1128/jb.01780-14
Rivière, A., Selak, M., Lantin, D., Leroy, F., & De Vuyst, L. (2016). Bifidobacteria and Butyrate-Producing Colon Bacteria: Importance and Strategies for Their Stimulation in the Human Gut. Frontiers in Microbiology, 7. doi:10.3389/fmicb.2016.00979
Lopez-Siles, M., Duncan, S. H., Garcia-Gil, L. J., & Martinez-Medina, M. (2017). Faecalibacterium prausnitzii: from microbiology to diagnostics and prognostics. The ISME Journal, 11(4), 841–852. doi:10.1038/ismej.2016.176
Moreno-Indias, I., Sánchez-Alcoholado, L., Pérez-Martínez, P., Andrés-Lacueva, C., Cardona, F., Tinahones, F., & Queipo-Ortuño, M. I. (2016). Red wine polyphenols modulate fecal microbiota and reduce markers of the metabolic syndrome in obese patients. Food & Function, 7(4), 1775–1787. doi:10.1039/c5fo00886g
Richardson, D. P., Ansell, J., & Drummond, L. N. (2018). The nutritional and health attributes of kiwifruit: a review. European Journal of Nutrition. doi:10.1007/s00394-018-1627-z
Blatchford, P., Stoklosinski, H., Eady, S., Wallace, A., Butts, C., Gearry, R., … Ansell, J. (2017). Consumption of kiwifruit capsules increases Faecalibacterium prausnitzii abundance in functionally constipated individuals: a randomized controlled human trial. Journal of Nutritional Science, 6. doi:10.1017/jns.2017.52
Lopez-Siles M, Duncan SH, Garcia-Gil LJ, Martinez-Medina M. Faecalibacterium prausnitzii: from microbiology to diagnostics and prognostics. ISME J. 2017 Apr;11(4):841-852. doi: 10.1038/ismej.2016.176. Epub 2017 Jan 3. PMID: 28045459; PMCID: PMC5364359.
Ramirez-Farias C, Slezak K, Fuller Z, Duncan A, Holtrop G, Louis P. Effect of inulin on the human gut microbiota: stimulation of Bifidobacterium adolescentis and Faecalibacterium prausnitzii. Br J Nutr. 2009 Feb;101(4):541-50. doi: 10.1017/S0007114508019880. Epub 2008 Jul 1. PMID: 18590586.
Varela E, Manichanh C, Gallart M, Torrejón A, Borruel N, Casellas F, Guarner F, Antolin M. Colonisation by Faecalibacterium prausnitzii and maintenance of clinical remission in patients with ulcerative colitis. Aliment Pharmacol Ther. 2013 Jul;38(2):151-61. doi: 10.1111/apt.12365. Epub 2013 Jun 3. PMID: 23725320.
Munukka, E., Rintala, A., Toivonen, R. et al. Faecalibacterium prausnitzii treatment improves hepatic health and reduces adipose tissue inflammation in high-fat fed mice. ISME J 11, 1667–1679 (2017). doi.org/10.1038/ismej.2017.24