-

 国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子
国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子

 二级病原微生物安全实验室
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
谷禾健康
突然全身奇痒,抓挠后出现红色或白色“疙瘩”,要命的是随着抓挠的进行,痒感丝毫未减,反而越抓越痒,这种情况有时会莫名其妙的出现,持续数小时甚至几天,此起彼伏,反反复复。
这就是我们常说的“荨麻疹”。荨麻疹又称风疹块,是一种免疫相关皮肤病,至少有20%的人一生中患过此病。可表现为皮肤瘙痒、血管性水肿或两者兼有。
根据荨麻疹的持续时间,可分为两种主要类型:急性荨麻疹(AU)和慢性荨麻疹(CU)。急性荨麻疹多为自限性疾病,持续时间少于6周,多与感染、药物或某些食物有关。慢性荨麻疹的特征是自发或可诱发,持续时间超过6周,包括慢性自发性荨麻疹(CSU)和慢性诱发性荨麻疹,病因大多时候不明确,它极大地影响了患者的生活质量,给他们带来了沉重的负担,本文重点围绕慢性荨麻疹展开。
慢性荨麻疹(CU)的发生发展被认为与免疫系统密切相关。相关研究表明,CU病变中有大量免疫细胞浸润,如肥大细胞、嗜酸性粒细胞和嗜碱性粒细胞。肥大细胞的激活是CU发生发展的关键因素,因为它会触发组胺的释放,从而导致疾病的特征性症状。
在慢性荨麻疹活动期,血液中的嗜酸性粒细胞和嗜碱性粒细胞被吸引到皮肤,与肥大细胞通过细胞因子等介质相互作用,影响IgE受体介导的肥大细胞脱颗粒。这些细胞间的相互作用对荨麻疹局部微环境的形成至关重要。
目前,慢性荨麻疹(CU)的主要治疗方法是第二代非镇静性H1抗组胺药(nsAH)。然而,目前的治疗对某些CU患者的疗效有限。近年来,几种创新的靶向治疗方案(如 dupilumab和Bruton酪氨酸激酶抑制剂)目前正在进行临床试验,用于治疗慢性荨麻疹。
令人惊喜的是,许多研究发现,肠道菌群失衡与某些皮肤病的发展有关。临床研究表明慢性荨麻疹患者的肠道菌群发生了改变,这表明肠道微生物在慢性荨麻疹的发展中发挥着作用。慢性荨麻疹患者中能够产生短链脂肪酸的拟杆菌、双歧杆菌等丰度显著下降,导致短链脂肪酸的免疫调节受损;免疫细胞出现功能障碍和表达异常;与此同时,肠杆菌等机会性病原体增多,脂多糖的促炎作用增强,使肥大细胞更易于激活,这些因素都可能在慢性荨麻疹的发生和发展中发挥重要作用。
在本文中,我们先带大家认识了解什么是荨麻疹及一些风险因素,慢性荨麻疹的一些发病机制,并通过查阅整理相关文献具体讲述了肠道微生物群与慢性荨麻疹之间存在怎样的互作,并探索了基于肠道菌群的潜在治疗策略。
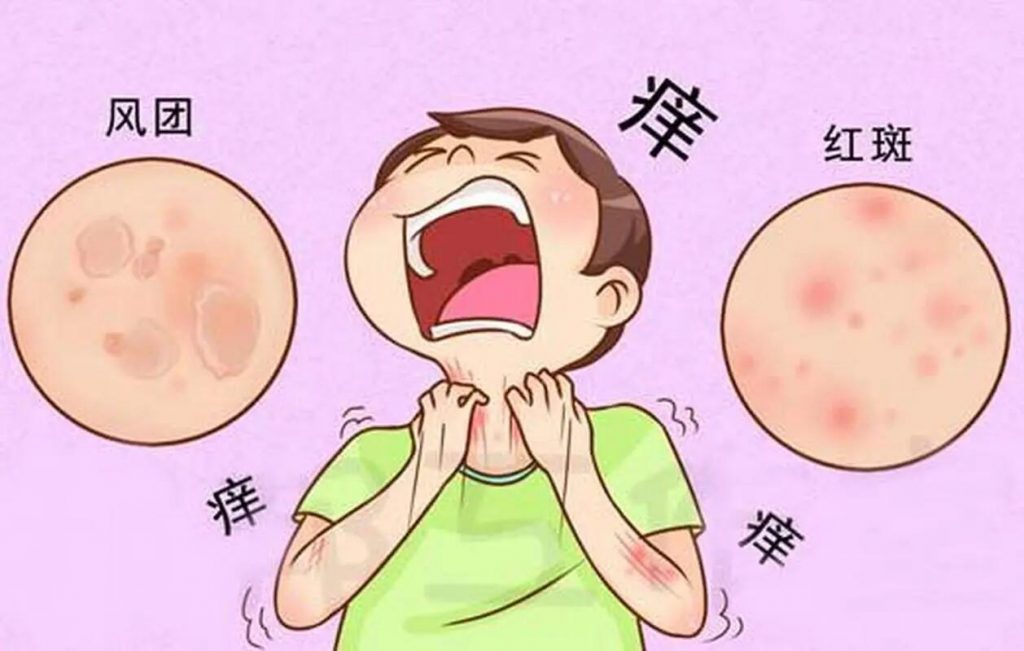
荨麻疹是一种炎症性皮肤病,以出现瘙痒性风团和血管性水肿(或两者兼有)为特征。
•风团是皮肤表面的红色或苍白肿胀,通常周围有红斑,可持续几分钟至24小时。风团通常很痒,但也会引起灼热感。
•血管性水肿是指皮肤或粘膜深层真皮层内的肿胀。肿胀通常呈皮肤色,但也可能呈红斑。血管性水肿可能无症状、发痒或引起灼痛。可能需要长达72小时才能消退。
2017年,全球荨麻疹发病率为1.6亿例新发病例。但每种荨麻疹亚型在不同人群中的患病率均不同,大约有20%的人一生中患过此病。
◮荨麻疹在不同人群中的发病率不同
急性荨麻疹(AU)在5岁以下儿童中的患病率最高,而慢性荨麻疹(CU)(尤其是慢性自发性荨麻疹)在30岁以上的女性中发病率最高。成年慢性自发性荨麻疹患者的年龄大于成年慢性诱导性荨麻疹患者(平均年龄约30-70岁vs约20-40岁),发病年龄也更晚(约30-50岁vs约20-35岁)。
◮女性的患病率高于男性
成人中,除胆碱能性荨麻疹(CholU)外,所有类型的荨麻疹在女性中的患病率均高于男性,而胆碱能性荨麻疹在男性成人和儿童中均更为突出;在较小的儿童中,女性患病率不存在或不太明显。
◮荨麻疹不会传染
一些研究但并非所有研究表明非白人患者中急性荨麻疹和慢性荨麻疹的患病率都较高。但荨麻疹不是感染性疾病,故不具备传染性。
所有类型的荨麻疹和急性荨麻疹的终生患病率分别为3–22%和6–19%。慢性荨麻疹的总体终生患病率为4.4% ,时点患病率(大多数研究中为1年患病率)从美国和欧洲的≤1.5%到墨西哥、韩国和中国的3–4%。
根据荨麻疹持续时间、频率和病因进行分类,可分为自发性荨麻疹(包括急性荨麻疹和慢性荨麻疹)、物理性荨麻疹、特殊类型的荨麻疹(例如接触性荨麻疹)以及与历史原因引起的荨麻疹相关的疾病(例如色素性荨麻疹)。
约80%的荨麻疹为自发性,10%为物理性荨麻疹,不到10%属于特殊类型。
◮急性荨麻疹
急性荨麻疹(AU)是指出现症状的持续时间少于六周的荨麻疹,急性发作,全身有瘙痒及大小不一的风团。约三分之二的自发性荨麻疹是急性的(过敏性或非过敏性)。
研究表明,患有过敏性疾病的人急性荨麻疹的发病率更高。50.2%的急性荨麻疹患者有花粉过敏、过敏性哮喘或特应性皮炎。
一项前瞻性研究,研究对象为800余名12~24个月大的特应性皮炎儿童,在未接受抗组胺药治疗的研究组中(n=396),18个月内16.2%的患者出现了急性荨麻疹。
◮慢性荨麻疹
慢性荨麻疹(CU)是指反复发作性的风团,伴或不伴血管性水肿,持续时间大于6周。慢性荨麻疹又可分为慢性自发性荨麻疹(CSU)和慢性诱导性荨麻疹(CIndUs)。其中,CSU约占CU的60%–80%。
慢性荨麻疹平均病程为3-5年,有些甚至可达20年以上,会导致瘙痒剧烈,睡眠障碍,继发心理社会问题,对生活质量影响较大。本文重点讲述的就是慢性自发性荨麻疹与肠道微生物群的联系以及可能提供的新型治疗方法。
除此之外,还有以下的荨麻疹类型:
◮物理性荨麻疹
物理性荨麻疹的共同特点是对不同的外部特定物理刺激(机械、冷、热、光)的荨麻疹反应,可分为以下几种。
皮肤性荨麻疹(人为荨麻疹):表现为用手搔抓或用钝器划过皮肤数分钟后沿划痕出现条状隆起,风团大多出现得很快,而且瘙痒剧烈,但一般约半小时后可自行消退。
注:皮肤性荨麻疹是物理性荨麻疹最常见的形式。
延迟压力性荨麻疹:与皮肤病性荨麻疹不同,这种类型的荨麻疹的典型病变是深部疼痛性肿胀,在暴露于垂直静态压力后4-8小时出现,并持续8-48小时。典型的受累部位是手掌和脚底,以及暴露于压力(例如坐在硬椅子上)时的臀部和背部。
压力性荨麻疹在男性中的发病率是女性的两倍,平均发病年龄为30岁。压力性荨麻疹可能会成为重体力劳动者的一种致残性疾病。
寒冷性荨麻疹:寒冷性荨麻疹是指接触寒冷后出现的荨麻疹反应。
热性荨麻疹:热性荨麻疹是一种罕见的物理性荨麻疹,由皮肤直接接触热物体或热空气引起。
日光性荨麻疹:日光性荨麻疹是由波长在280至760nm之间的光引起的。对于个别患者,诱发波长各不相同,但主要是紫外线引起的。
振动性血管性水肿:振动性血管性水肿是一种罕见的疾病,其中强烈的振动机械力(例如使用气动锤)会诱发血管性水肿。
◮特殊类型的荨麻疹
特殊类型的荨麻疹主要有以下几种:
胆碱能性荨麻疹:与物理性荨麻疹通过外部刺激引起症状不同,胆碱能性荨麻疹的病变是由于体温短暂升高引起的。
肾上腺素能性荨麻疹:肾上腺素能性荨麻疹是一种极为罕见的疾病,其特征是针尖大小的红色风团,带有白色光晕,与胆碱能性荨麻疹不同。风团是由压力引起的,而不是由运动或体温升高引起的。
这种疾病对β-肾上腺素能受体阻滞剂普萘洛尔的治疗有反应,它既可用于诊断目的,也可用于预防发作。
接触性荨麻疹:接触性荨麻疹是指化学物质与皮肤接触的部位出现风团。这种疾病可能严格局限于接触部位,如荨麻(荨麻疹或异株荨麻疹,荨麻疹由此得名),但可能会出现全身性症状,尤其是在IgE介导的过敏性接触性荨麻疹中。
水源性荨麻疹:水源性荨麻疹是一种独特的荨麻疹形式,需要与接触性荨麻疹相区别,因为水本身并不是致病因子,但会从角质层中释放出水溶性过敏原,然后扩散到真皮层后成为过敏原。
女性患此病的几率是男性的五倍,平均发病于青年时期。病变类似于胆碱能性荨麻疹,躯干上大多有针尖大小的风团。
◮早期症状
荨麻疹患者早期可在接触诱因,如过敏原或物理、化学刺激后,先有不同程度的皮肤瘙痒后出现风团,或以全身各部位风团(以躯干和四肢近端为主)为最初的表现。
注:风团是一种局限的隆起于皮肤、黏膜表面的中心性水肿性团块,是由于肥大细胞活化导致皮肤、黏膜小血管扩张及渗透性增加引起的。
◮典型症状
荨麻疹典型表现为突然自觉皮肤瘙痒,很快瘙痒部位出现大小不等的红色或苍白色风团,圆形、椭圆形或不规则形,可孤立分布或扩大融合成片,皮肤表面凹凸不平,类似于橘皮样外观,周围多伴有红晕。
数分钟至数小时内水肿减轻,风团变为红斑并逐渐消失,不留痕迹,单个风团的持续时间一般不超过24小时,但新风团可此起彼伏,不断发生。风团每天发作或间歇发作,持续时间≥6周即为慢性荨麻疹。
◮可能伴有的症状
▪ 病情严重者可伴有心慌、烦躁甚至血压降低等过敏性休克症状;
▪ 胃肠道黏膜受累时可出现恶心、呕吐、腹痛和腹泻等;
▪ 累及喉头、支气管时可出现胸闷、呼吸困难甚至窒息;慢性荨麻疹引起气管炎,气管炎常蔓延至喉部,引起喉炎。喉炎症状严重时甚至会导致窒息。
▪ 感染引起者可出现寒战、高热、甚至全身中毒症状。
▪ 心血管系统:患者出现发热、头痛、气短、四肢无力等症状,有时会突然消退,然后急剧加重。有些患者会得心肌炎。
急性荨麻疹(AU)多与食物药物过敏、感染等因素相关。已报告的AU风险因素包括人口密度高和个人及父母有过敏性疾病史。AU患病率较高或风险较高可能还与贫困和较低的社会经济地位有关。
对于多达95%的慢性荨麻疹患者来说,这种病症是“特发性的”,意味着没有明确的病因。但以下一些因素可能导致患慢性荨麻疹的风险上升。
1.感染
慢性自发性荨麻疹可能是感染引起,如幽门螺杆菌感染、病毒感染等。感染引起的炎症会刺激体内免疫系统,出现免疫反应,导致皮肤血管通透性增加,进而出现慢性自发性荨麻疹。
各种病毒(如病毒性上呼吸道病毒、肝炎病毒、柯萨奇病毒等)、细菌(如金黄色葡萄球菌所致的扁桃体炎、慢性中耳炎、幽门螺旋杆菌感染等)、真菌及寄生虫感染均可引起。
2.食物
如果本身属于过敏体质,在食用易过敏食物后,如海鲜、芒果后,可能会导致皮肤发生过敏反应,从而出现慢性自发性荨麻疹。
此外,腐败食物、某些食品添加剂如水杨酸盐、甲苯酸盐、亚硫酸盐等也可能引起荨麻疹。
3.药物
如果长期服用青霉素类、头孢类等药物,可能会导致身体产生过敏反应,从而诱发慢性自发性荨麻疹。
4.遗传因素
在涉及双胞胎的研究中,遗传因素可以部分解释荨麻疹的易感性。几种基因的多态性(包括TNFRS11A 、TBXA2R和PLA2G4A)已被证明与荨麻疹易感性或多种非甾体抗炎药诱发的血管性水肿有关。
慢性荨麻疹(CU)的遗传易感性与IFNγ、IL-6、IL-17RA、IL-10、TGFβ、IL-6、肿瘤坏死因子 (TNF)、PTPN22、IL-1、IL-2和HLA I 类和 II 类等位基因的基因多态性有关。
例如,HLA-DR4与自身免疫性慢性自发性荨麻疹 (CSU)和其他自身免疫性疾病(例如类风湿性关节炎和1型糖尿病)密切相关。
5.呼吸道吸入物及皮肤接触物
常见呼吸道吸入物包括花粉、动物皮屑、粉尘、尘螨、真菌的孢子及一些挥发性化学品等,皮肤接触物如某些植物、动物毛发、昆虫叮螫、毒毛虫刺激等都可能会诱发荨麻疹。
6.物理刺激
如果皮肤受到冷热刺激、压力过大、摩擦、日光照射等物理刺激,可能会诱发荨麻疹,建议患者调整生活习惯,在生活中注意皮肤卫生,尽量使用温水清洗皮肤,避免使用刺激性的洗漱用品,以免加重荨麻疹症状。
7.自身原有疾病
研究发现,在诊断出自身免疫性疾病(包括类风湿性关节炎、系统性红斑狼疮、I型糖尿病和乳糜泻)之前和之后,分别约有80%和20%的患者被诊断为慢性自发性荨麻疹。
患有自身免疫性甲状腺疾病的患者,尤其是女性患者,患慢性自发性荨麻疹的风险要高得多。高达25%的慢性自发性荨麻疹(CSU)患者,尤其是自身免疫性荨麻疹标志物阳性的患者,有CSU家族史。
此外,患有消化性溃疡病和异常子宫出血的女性患者被证实具有更高的慢性荨麻疹 (CU)风险。
◮心理压力
心理压力是可能诱发荨麻疹的因素之一,如情绪波动、精神紧张、抑郁等可能会诱发荨麻疹。它通过影响免疫系统和神经内分泌系统,进而可能导致荨麻疹的发生或加重症状。
注:部分慢性荨麻疹患者可存在凝血功能和免疫功能异常。
急性荨麻疹一般发病数小时后会自行消退,而慢性荨麻疹的症状会持续六周以上,甚至可能持续数月至数年。由于反复发作,迁延不愈,慢性荨麻疹会严重影响患者及其家属的生活质量。
根据有无特定的诱发因素,慢性荨麻疹分为慢性自发性荨麻疹(CSU)和慢性诱导性荨麻疹(CIndU)。CSU约占慢性荨麻疹的2/3,因此,了解慢性自发性荨麻疹的发病机制就显得格外重要,有助于发现新的治疗靶点,为临床提供多样化的诊疗思路。
1
自身免疫在慢性自发性荨麻疹(CSU)中起关键作用,是CSU最主要的机制之一。肥大细胞(MC)通过I型自身免疫反应和IIb型自身免疫反应活化并脱颗粒,2种类型的CSU存在不同的临床特征。
慢性荨麻疹的表现和病理生理
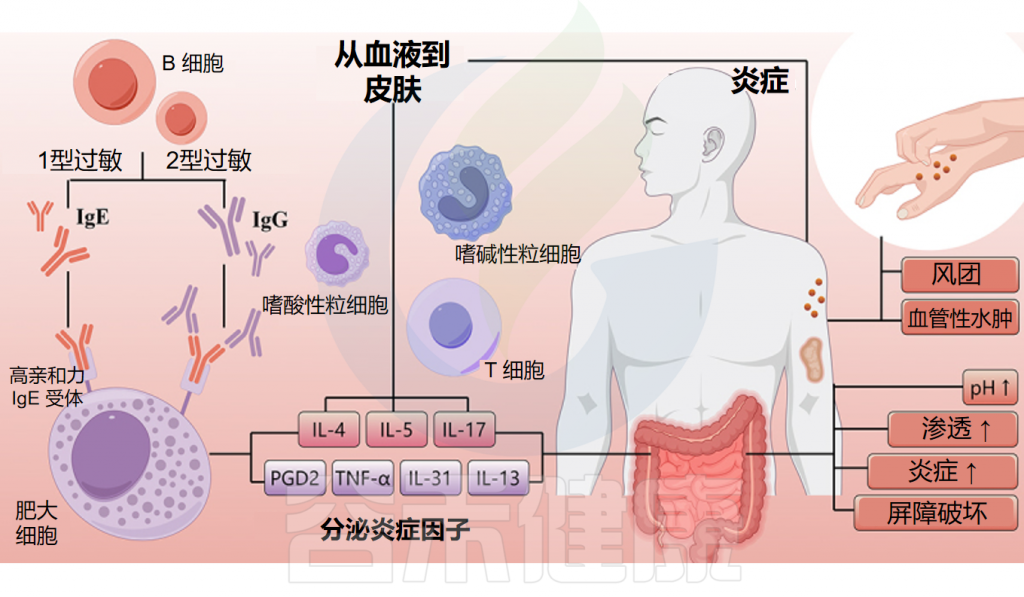
Cai R,et al.Gut Microbes.2024
◮自身过敏反应引发慢性自发性荨麻疹
I型自身免疫反应又称自身过敏反应,其特征是具有抗自身抗原的IgE抗体。IgE自身抗体通过其高亲和力受体抗IgE Fc受体I与相应的自身抗原交联导致肥大细胞的活化,激活下游的一系列生化反应,引发慢性自发性荨麻疹(CSU)。
目前已发现的针对自身抗原的IgE抗体包括抗甲状腺过氧化物酶(TPO)抗体、抗甲状腺球蛋白(TG)抗体、抗双链DNA(dsDNA)抗体、抗白细胞介素-24抗体、抗组织因子(TF)抗体等。
◮绝大部分CSU患者具有IIb自身免疫反应
IIb型自身免疫反应的特征是具有抗IgE的IgG自身抗体或抗FcεRI的IgG自身抗体。FcɛRI与补体C5a受体均在肥大细胞和嗜碱性粒细胞上表达,且被证实在IgG-抗FcɛRI介导的脱颗粒中发挥作用。

PURIST研究的结果表明,约88%的CSU患者至少具有3个IIb型自身免疫反应CSU诊断标准中的1个,不到10%的CSU患者3个标准全部符合,并且这些患者往往表现为更严重的病情、低水平的总IgE和高水平的TPO抗体。
IIb型自身免疫反应CSU的诊断标准:1)自体血清皮肤试验阳性;
2)体外嗜碱性粒细胞组胺释放试验或嗜碱性粒细胞活化标志物阳性;
3)针对抗IgE的IgG自身抗体或抗FcεRI的IgG自身抗体的免疫测定阳性。
◮患有自身免疫性疾病可能导致CSU风险增高
多种自身免疫性疾病可与慢性自发性荨麻疹(CSU)伴发,最常见的是桥本甲状腺炎(≥21%)和白癜风(2%)。CSU患者合并自身免疫性疾病与自身免疫性疾病家族史以及甲状腺功能减退/亢进的发生率较高有关,并且自身免疫性疾病与IIb型自身免疫反应CSU有关。
此外,全基因组关联研究结果显示CSU与自身免疫性疾病之间具有遗传重叠,CSU的易感遗传因素主要通过与自身免疫特征的关联来表现。
荨麻疹发病机制的关键途径
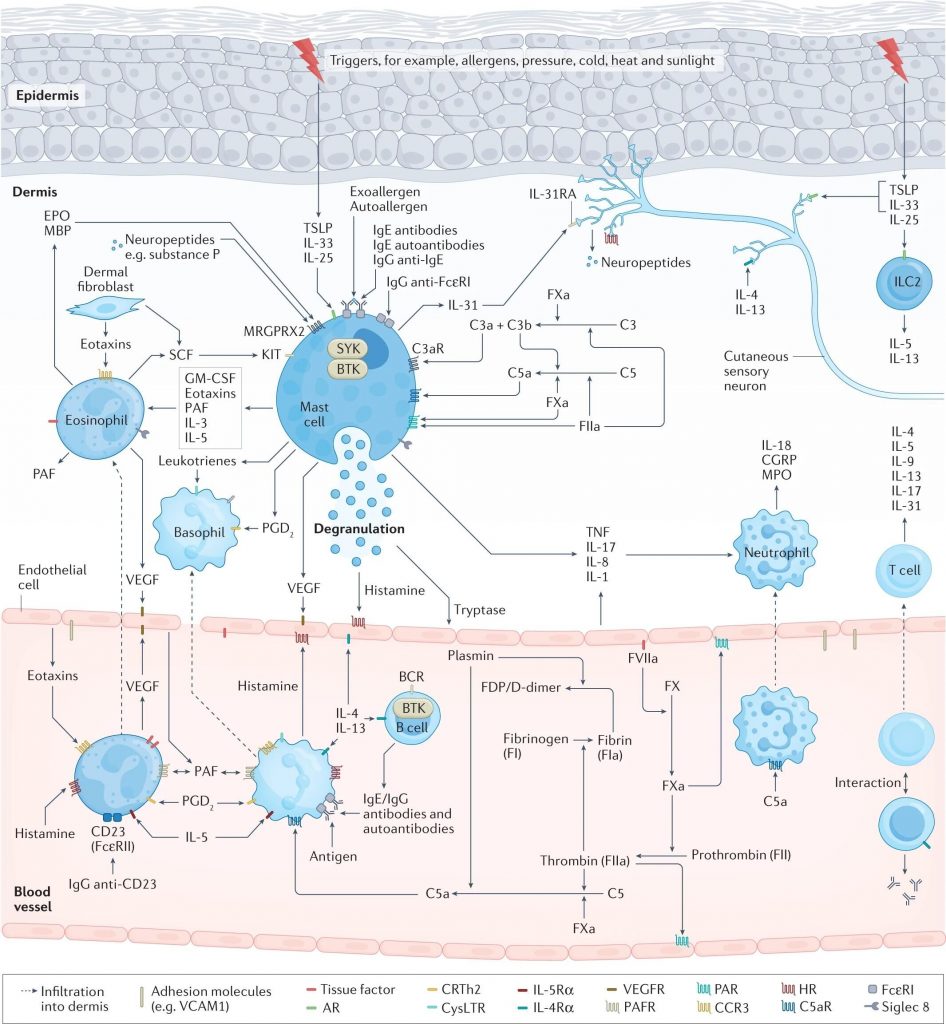
doi: 10.1038/s41572-022-00389-z.
肥大细胞活化和脱颗粒导致荨麻疹的症状和体征,这是由于组胺和其他介质的释放所致,这些介质激活皮肤感觉神经(瘙痒)、扩张皮肤血管(红斑)并诱导血浆外渗(水肿和其他免疫细胞的流入)。
在过敏性荨麻疹中,上皮细胞释放警报素、TSLP、IL-33和IL-25,皮肤驻留组2先天淋巴细胞(ILC2)活化,T 细胞(主要是辅助T细胞 2 (TH 2 细胞))极化并释放 TH2细胞因子,例如 IL-4、IL-5 和 IL-13,B 细胞产生过敏原特异性 IgE,随后过敏原交联肥大细胞表面的 IgE-FcεRI 复合物,从而触发肥大细胞活化。
慢性自发性荨麻疹(CSU)可能由于一系列复杂的多步骤相互关联的事件而出现,包括细胞浸润(主要是嗜酸性粒细胞、嗜碱性粒细胞、中性粒细胞和T细胞)、自身免疫(例如,IgE/IgG 组胺释放自身抗体)、神经源性炎症(通过皮肤瘙痒感觉神经介导的组胺依赖性和组胺非依赖性瘙痒信号通路)、补体级联的激活(例如,通过产生过敏毒素 C5a)和组织因子启动的凝血级联外在途径的激活。
2
◮CSU依赖于肥大细胞的信号激活
肥大细胞(MC)是慢性自发性荨麻疹(CSU)的关键效应细胞,CSU中风团和血管性水肿的发展依赖于肥大细胞激活信号和受体、信号通路、抑制性受体和介质。MC来自骨髓造血祖细胞,随后迁移到外周组织,并在干细胞因子和IL-3的影响下进行终末分化。
MC主要由针对自身变应原的IgE或FcεRI α链的自身抗体通过FcεRI激活,胸腺基质淋巴细胞生成素、血浆衍生外泌体、IL-33、IL-4、IL-13、IL-5、补体C5a和MAS相关G蛋白偶联受体-X2也可激活MC。
肥大细胞(MC)活化后释放的各种介质可导致感觉神经激活、血管扩张、血浆外渗以及参与荨麻疹的细胞募集,引起CSU的症状和体征(皮肤瘙痒和血管水肿)。
研究发现腺苷、动力相关蛋白1和琥珀酸受体1也可调节MC的活性,CSU患者的血浆腺苷水平升高,并且腺苷水平可预测重度CSU患者对非镇静H1抗组胺药是否有反应。
慢性荨麻疹机制模型
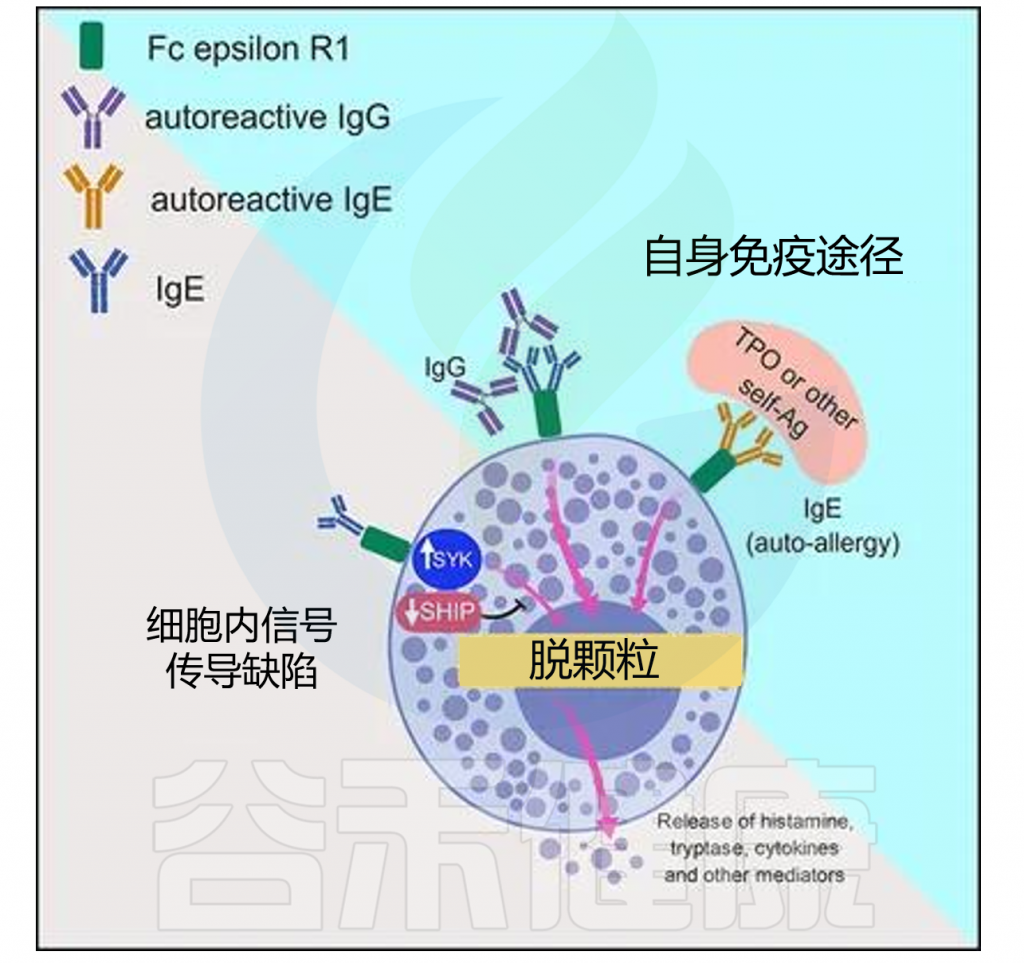
doi: 10.3389/fimmu.2019.00627.
◮嗜碱性粒细胞可以产生组胺,参与风团的诱导
嗜碱性粒细胞与肥大细胞一样,可以产生组胺、白三烯和细胞因子等,参与CSU风团的诱导。嗜碱性粒细胞还表达FcεRI,可被抗FcεRI自身抗体激活。在CSU患者中经常观察到外周血嗜碱性粒细胞减少,可能是嗜碱性粒细胞从循环迁移到组织的结果。
奥马珠单抗可能通过降低嗜碱性粒细胞上FcεRI和IgE的密度,使嗜碱性粒细胞从外周血迁移到组织的数量减少,外周血嗜碱性粒细胞增多,从而达到治疗慢性自发性荨麻疹(CSU)的作用。而血液循环中嗜碱性粒细胞数量降低则提示CSU患者对奥马珠单抗反应不佳。
◮嗜酸性粒细胞参与慢性荨麻疹的发病
嗜酸性粒细胞表达低亲和力的IgE受体FcεRII/CD23,可被IgG-抗FcεRII/CD23结合并激活。
嗜酸性粒细胞可能通过3种方式参与慢性荨麻疹(CU)发病:
1)产生干细胞因子,促进组织中肥大细胞的募集和局部成熟;
2)嗜酸性粒细胞蛋白主要碱性蛋白、嗜酸性粒细胞阳离子蛋白和嗜酸性粒细胞过氧化物酶可促进肥大细胞脱颗粒;
3)活化后表达抗组织因子(TF),TF是凝血级联反应中凝血酶的主要激活因子。此外,研究发现10%的CSU患者出现嗜酸性粒细胞减少症,且与IIb型自身免疫反应、疾病高活动度和治疗反应差有关。
◮T细胞在CSU中起调节作用
辅助性T(Th)1细胞/Th2和Th17相关细胞因子的水平在CSU中显著升高,并与疾病的活动度相关。
Th1反应通常与ASST阳性有关;Th2细胞产生IL-4、IL-5和IL-13等多种细胞因子,可刺激IgE的产生及诱导肥大细胞、嗜碱性粒细胞和嗜酸性粒细胞的激活。Th17通过分泌细胞因子IL-6促进肥大细胞的增殖,并诱导其趋化。
调节性T细胞(Tregs)靶向许多不同的免疫细胞亚群和组织,可防止炎症反应过度并支持组织修复和稳态。体内外证据表明Tregs通过OX40-OX40配体信号抑制肥大细胞脱颗粒。研究表明,血液循环中CD4 + CXCR5 +细胞比例、滤泡辅助性T细胞比例和Tfh/滤泡调节性T细胞的比值异常均与CSU严重程度呈正相关,提示这些细胞可能参与CSU发病的免疫机制。
◮免疫细胞相互作用共同促进CSU的发生和发展
慢性自发性荨麻疹的发病机制涉及包括免疫细胞在内的多种细胞(如血管内皮细胞、神经元)的共同参与。
先天免疫和适应性免疫在CSU中存在极其复杂的相互作用,肥大细胞是先天免疫和适应性免疫之间的重要纽带:活化的肥大细胞通过释放炎症介质和趋化因子[组胺、前列腺素D2、基质金属蛋白酶-9和趋化因子C-X-C模体配体1/2等]促进嗜酸性粒细胞、嗜碱性粒细胞、中性粒细胞和T细胞向皮肤迁移,来自肥大细胞的IL-4、TSLP是先天淋巴样细胞的共刺激细胞因子。
嗜碱性粒细胞和T细胞产生IL-4可影响肥大细胞的趋化;单核细胞通过趋化因子C-C模体配体2(CCL2)/单核细胞趋化蛋白-1(MCP-1)作用于肥大细胞和嗜碱性粒细胞,促进其激活;巨噬细胞可通过分泌IL-6促进MC增殖和成熟,并释放干扰素-λ1调节T细胞的发育。
B细胞产生的IgE/IgG抗体与肥大细胞和嗜碱性粒细胞的FcεRIα结合并导致一系列的活化反应,来自嗜碱性粒细胞的IL-4和IL-6可增强B细胞的存活和增殖;内皮细胞产生的SCF影响MC的发育和成熟,通过迁移抑制因子激活巨噬细胞,通过IL-6、IL-18、肿瘤坏死因子(TNF-α)和脂肪因子与中性粒细胞相互作用参与CSU的发病。
免疫细胞在CSU患者中的相互作用
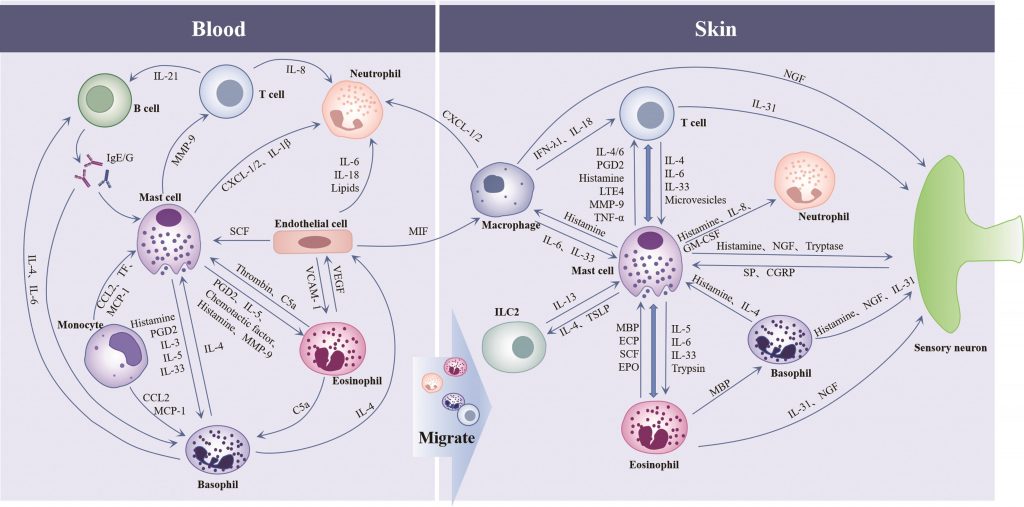
doi: 10.11817/j.issn.1672-7347.2023.230037.
3
◮神经元释放神经肽导致血管扩张、血浆外渗
在慢性自发性荨麻疹(CSU)中,神经元通过释放神经肽如P物质和降钙素基因相关肽(CGRP)等来调节肥大细胞的功能,可能导致血管扩张、血浆外渗。
◮免疫细胞释放炎症介质和细胞因子导致瘙痒
免疫细胞释放的炎症介质和细胞因子[如组胺、IL-31和神经生长因子(NGF)]等作用于神经元,可导致神经源性炎症和瘙痒症状。
MRGPRX2在皮肤肥大细胞(MC)上高度表达,可由神经肽如P物质激活,并导致MC释放组胺,进而引起CSU患者皮肤反应性的增高。另有研究发现CSU患者的焦虑与抑郁风险是对照组的6倍,CSU对焦虑或抑郁的影响与瘙痒和睡眠障碍有关,有效缓解瘙痒可以促进自然睡眠,从而改善CSU患者的情绪。
4
◮肠道菌群在CSU发病中发挥重要作用
近年来,肠道菌群与慢性自发性荨麻疹(CSU)之间的关联逐渐受到关注。研究发现CSU患者和健康对照组之间肠道微生物群的α多样性和β多样性存在差异。
通过16S核糖体RNA基因测序发现CSU患者由于有益菌(主要是产短链脂肪酸菌)的减少,导致短链脂肪酸的生成减少而有害代谢产物(脂多糖)的产生增加,从而通过促进Th2细胞的分化,IgE的产生激活肥大细胞,参与CSU的发病。
研究发现慢性荨麻疹(CU)患者血浆脂质代谢特征与健康对照组不同:CU患者血浆磷脂酰丝氨酸、磷脂酰乙醇胺和磷脂酰甘油水平显著升高,而磷脂酰胆碱水平显著下降。
肠道微生物与慢性荨麻疹之间的关联研究总结

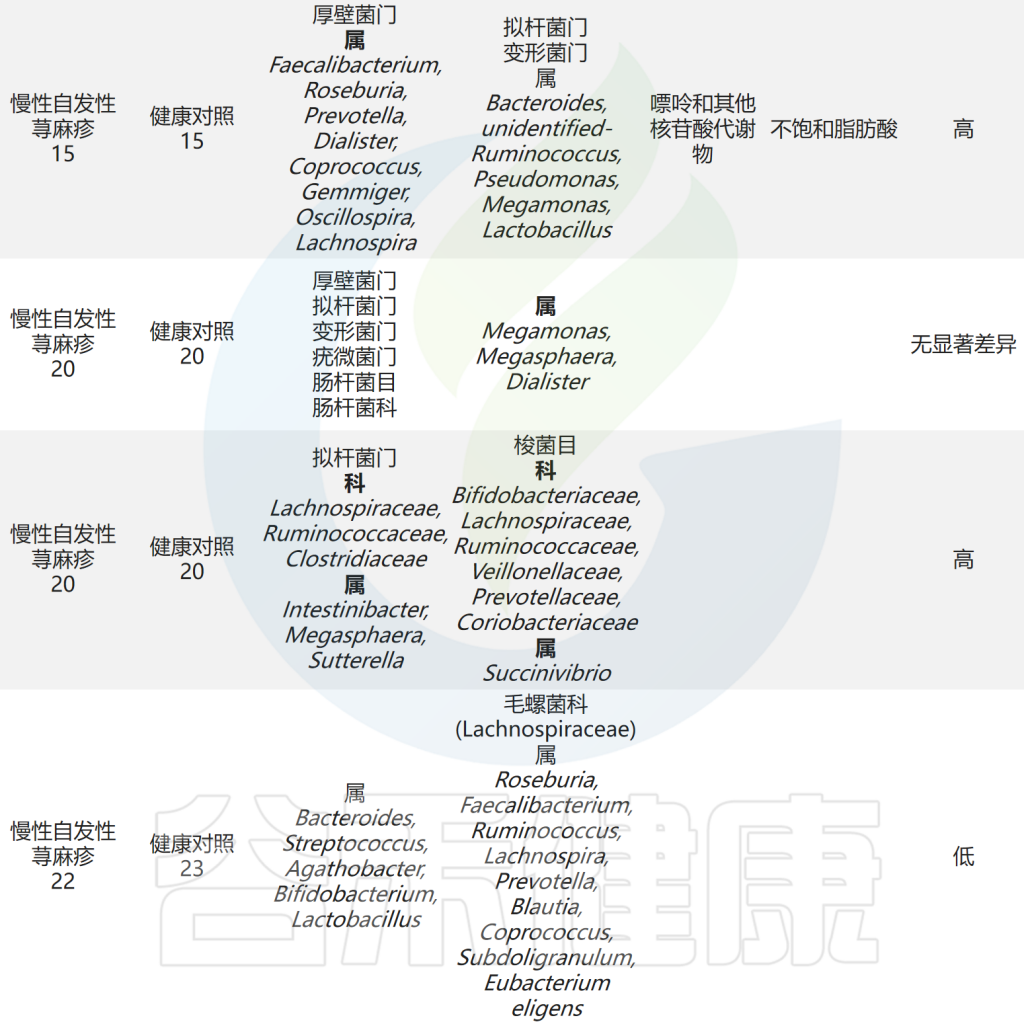
Cai R,et al.Gut Microbes.2024
以上研究表明CSU患者肠道微生物群组成和代谢物的紊乱以及它们之间的相互作用可能参与了慢性自发性荨麻疹(CSU)的发病。此外,研究人员发现毛螺菌科及其下属类群是抗组胺药治疗应答者和无应答者肠道微生物群的主要差异,应答者的毛螺菌科丰度高于无应答者,提示毛螺菌科是抗组胺药在CSU患者中的疗效预测标志。
我们将在下面的章节中具体讲述肠道微生物群在慢性自发性荨麻疹发病和治疗中的作用。
5
◮遗传因素在慢性自发性荨麻疹发病中占一定作用
部分慢性自发性荨麻疹(CSU)患者存在家族史,遗传因素在CSU的发病中可能存在潜在作用。研究通过飞行质谱法对ORAI钙释放激活钙调节因子1(ORAI1)多态性进行基因分型,发现ORAI1基因单核苷酸多态性(SNP)rs12320939和rs3741596与CSU易感性相关,ORAI1基因编码的Orai1参与肥大细胞脱颗粒、白三烯分泌、组胺释放以及TNF-α分泌。
补体C5a受体1(C5AR1)-1330T等位基因也与CSU易感性相关,可作为CSU的危险因素,Fcε受体Ia(FCER1A) rs2298805等位基因与CSU风险和总IgE血清浓度有关。
此外,研究发现经地氯雷他定治疗后出现镇静的患者中,组胺受体H1(HRH1) rs901865 G等位基因的频率明显高于rs901865 A等位基因,提示HRH1基因多态性可能与CSU患者地氯雷他定治疗后的镇静副作用有关。
◮环境因素参与CSU的发病
环境因素可能参与慢性自发性荨麻疹(CSU)的发病,细颗粒物(PM2.5)通过Gadd45b/促分裂原活化蛋白激酶4(MEKK4)/c-Jun氨基末端激酶(JNK)途径调节活性氧的产生,促进IgE介导的肥大细胞活化。
此外,人类广泛暴露于全氟和多氟烷基物质(PFAS)环境中,PFAS可能通过食物和饮用水被人体吸收,通过因果中介分析发现IL-4是全氟庚酸与CSU相关性的部分中介(14.8%),血清中的PFAS水平升高与CSU发病风险增加相关。因此,持续暴露在不利环境因素中可能会导致CSU的发病并加剧其病情。
◮非免疫学致病因素
非免疫学致病因素也参与慢性自发性荨麻疹(CSU)的发病。例如房屋尘螨可能穿过皮肤屏障,直接触发神经元释放P物质,导致肥大细胞脱颗粒;性激素可能通过遗传、微生物和环境因素的复杂网络导致女性的CSU患病率较男性高。
此外,研究发现凝血-补体系统可通过下游的肥大细胞或嗜碱性粒细胞激活轴参与CSU的发病。维生素D、金黄色葡萄球菌、热激蛋白和硫醇/二硫化物稳态等也被发现通过各种机制参与CSU的发生。
人们普遍认为慢性自发性荨麻疹(CSU)主要是一种免疫介导的炎症性疾病。I型和II型自身免疫,特别是针对自身过敏原的IgE抗体和针对IgE或其高亲和力受体(FcεRI)的IgG自身抗体的存在,被认为在CSU的发生和发展中发挥作用。
皮肤肥大细胞的活化、脱颗粒和介质释放对CSU的致病性至关重要。CSU的症状主要由组胺的释放引起,以及其他介质,包括前列腺素D2(PGD2)、肿瘤坏死因子(TNF)和各种白细胞介素(IL),例如 IL-4、IL-5、IL-13、IL-17 和 IL-31,它们会影响驻留的皮肤细胞和其他募集的靶细胞,如T细胞、嗜酸性粒细胞和嗜碱性粒细胞。
这些募集的细胞在肥大细胞、活化的内皮细胞、辅助T(Th)2细胞和其他细胞释放的趋化因子(例如 IL-5、补体3a、补体5a、TNF、IL-17 等)的作用下从血液迁移到皮肤中。
健康的肠道对整体健康至关重要,研究表明,肠道微生物群失衡可能导致皮肤病。菌群失衡通过炎症和免疫系统紊乱与牛皮癣、特应性皮炎和痤疮等皮肤病有关。慢性荨麻疹是一种免疫介导的皮肤病,也可能受到肠道菌群的影响。慢性自发性荨麻疹是慢性荨麻疹不同亚型相关研究的主要焦点。
最近的研究检查了慢性自发性荨麻疹(CSU)患者的肠道菌群,发现CSU患者和健康对照者(HC)之间的微生物组成存在显著差异。研究主要发现CSU患者的α多样性低于 HC。
注:肠道菌群的α多样性是指单个样本中的物种多样性,可以使用 Chao1 值、ACE 值和 Shannon 指数等指标来衡量。Beta多样性是对样本间多样性差异的比较。
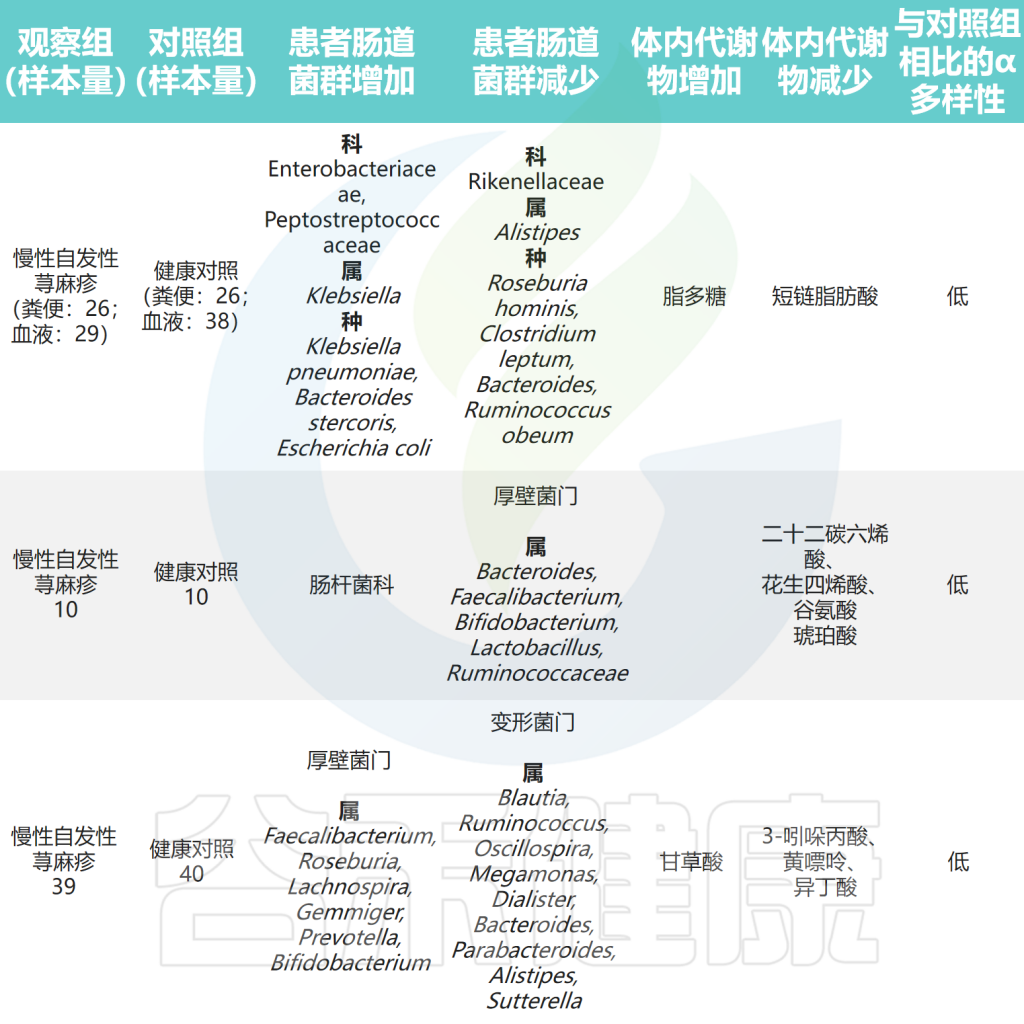
①患者中肠杆菌等机会性病原体水平升高
肠杆菌科(Enterobacteriaceae)是肠道菌群中促炎成员之一。初步研究探索了慢性荨麻疹患者肠道微生物组的变化。结果显示,肠杆菌科在慢性荨麻疹患者的粪便样本中更为普遍,还观察到慢性自发性荨麻疹(CSU)组的大肠杆菌(Escherichia coli)和克雷伯氏菌(Klebsiella spp.)的丰度较高。

最近的一项研究使用多组学分析分析了慢性自发性荨麻疹(CSU)患者肠道微生物组的组成,结果显示CSU患者的肠道菌群多样性较低,但肺炎克雷伯氏菌的水平较高。此外观察到Clostridium disporicum与低生活质量之间存在正相关性。
除上述结果外,还发现CSU组的乳酸杆菌属、Turicibacter属和Lachnobacterium属水平较高。采用孟德尔随机化分析探讨了肠道菌群与CSU之间的因果关系。该研究发现Verrucomicrobia门、Defluviitaleaceae科UCG011属和Coprococcus 属是荨麻疹的潜在危险因素。
②幽门螺杆菌感染人群更易患慢性荨麻疹
已知幽门螺杆菌(HP)定植于胃和十二指肠,可导致持续感染。荟萃分析显示,CU患者的HP感染患病率高于对照组,并且HP阴性患者比阳性患者更容易出现症状自发缓解。
另一项研究表明,根除幽门螺杆菌可使CU临床缓解的概率增加一倍。研究人员提出了一个假设,认为H. pylori的 kDa 混合蛋白质成分可能刺激人类肥大细胞脱颗粒。此外,一些细菌基因(如cagA、vacA和nap)可能编码特定蛋白质,触发增强促炎途径的免疫反应。
③拟杆菌、双歧杆菌有益菌水平下降
拟杆菌(Bacteroidetes)是肠道菌群的主要成分,是生产短链脂肪酸(SCFA)的物种之一,特别是乙酸和丙酸。研究发现CSU患者中拟杆菌的丰度低于对照组,这表明拟杆菌可能对CSU具有保护作用。
研究表明,拟杆菌主要通过产生荚膜多糖A和短链脂肪酸对人体免疫力产生调节作用。荚膜多糖A在维持和平衡免疫系统功能以及预防细菌和病毒感染方面发挥作用。乙酸盐和丙酸盐都是有效的抗炎剂,能够抑制中性粒细胞和巨噬细胞释放促炎细胞因子。
注:据报道CSU组厚壁菌门的数量也减少,厚壁菌门和拟杆菌门占肠道有益菌群的90% 。厚壁菌门可分解不溶性膳食纤维释放营养物质,促进肠道不同菌种增殖。
此外,研究发现健康人群中考拉杆菌属(Phascolarctobacterium)的含量较高。而患有慢性自发性荨麻疹和症状性皮肤划痕症(CSD)的患者,其Subdoligranulum和Ruminococcus bromii的含量明显较低。
对照组中的乳杆菌和双歧杆菌相对含量明显高于慢性荨麻疹患者,表明它们可能具有预防慢性荨麻疹的作用。这可能归因于它们诱导调节性T(Treg)细胞的能力。
此外慢性自发性荨麻疹患者肠道菌群中负责产生短链脂肪酸的毛螺菌科成员水平降低。在孟德尔随机化分析中,伯克氏菌目(Burkholderiales)和嗜木聚糖真杆菌属(Eubacterium xylanophilum)被发现具有预防荨麻疹的潜在作用。
如上所述,慢性自发性荨麻疹(CSU)患者的肠道微生物群发生了明显的变化。与对照组相比,CSU患者的肥大细胞更容易被激活,这可能与激活信号的阈值较低或抑制信号的暴露减少有关。
肠道微生物群的一些代谢物在其中也发挥着重要作用,脂多糖(LPS)和促炎细胞因子可使肥大细胞更易于激活,而短链脂肪酸(SCFA)可抑制肥大细胞激活。这些影响肥大细胞活化的因素与肠道菌群有关。
慢性荨麻疹中肠道菌群的改变及影响
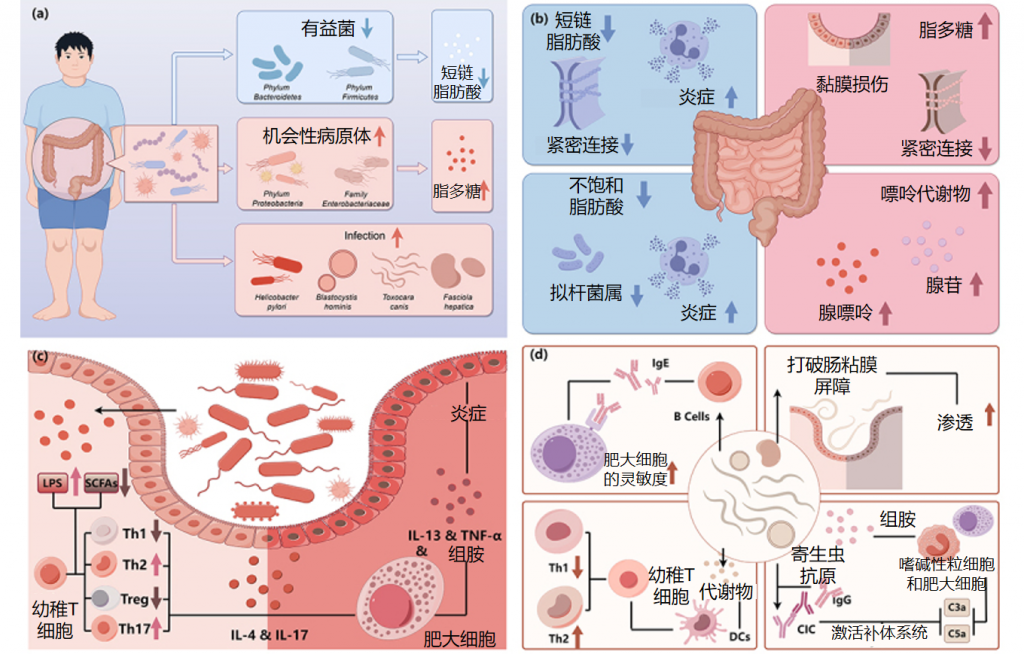
Cai R,et al.Gut Microbes.2024
①短链脂肪酸的免疫调节受损
短链脂肪酸通过增强前列环素E2和IL-10的产生,从而减轻炎症,同时促进Treg细胞的分化,抑制Th17细胞的活性,在免疫调节中发挥关键作用。通过动物实验发现,短链脂肪酸可以减轻肥大细胞驱动的皮肤炎症。
利用16S 测序和定量 PCR 识别了慢性荨麻疹患者的菌群。产生短链脂肪酸的有益细菌减少。短链脂肪酸(SCFA)不仅有助于维持免疫微环境,还能稳定肠粘膜上皮屏障。通过降低 pH 值来增强肠道菌群的多样性、维持上皮屏障的完整性以及调节粘液的分泌可以作为其他保护机制。发现慢性自发性荨麻疹患者的平均粪便pH值(7.17)高于对照组(6.7),这与短链脂肪酸的产生减少有关。
②短链脂肪酸减少进一步诱发免疫反应失常
慢性荨麻疹患者普遍存在细胞因子表达异常和调节性T细胞(Treg)功能障碍。Th2细胞释放细胞因子如 IL-4、IL-5 和 IL-13,这些细胞因子刺激IgE的产生并可激活肥大细胞、嗜碱性粒细胞和嗜酸性粒细胞。
此外,IL-4可增强 IgE 受体的表达。基于先前的证据提出一个假设,即短链脂肪酸的减少将进一步抑制 Treg 细胞的产生和功能,从而导致 Th1/Th2 比例失衡以及 IL-4 和 IgE 的产生增加。乳杆菌和双歧杆菌也可能通过 Treg 细胞的分化抑制炎症,从而发挥对慢性荨麻疹的保护作用。
与对照组相比,慢性荨麻疹患者的Th17细胞有所增加。CSU组中的Prevotella属比对照组组中的更丰富,并且在小鼠实验中显示出刺激 Th17 免疫反应并募集中性粒细胞以引发持续炎症。
③脂多糖促炎作用增强
革兰氏阴性菌是肠道菌群的主要成分,其细胞壁上的脂多糖可通过 Toll 样受体4 刺激 IgE 诱导的肥大细胞脱颗粒和炎症介质的释放。在慢性自发性荨麻疹(CSU)患者的肠道中发现机会性病原体增加,而短链脂肪酸水平降低导致肠粘膜通透性增强,最终导致循环脂多糖水平升高。
在慢性荨麻疹患者的粪便中发现肠杆菌科细菌数量增加,肠杆菌科细菌是产生脂多糖的主要机会性病原体,可促进Th2细胞分化并产生IL-4。此外,高水平的肺炎克雷伯菌会增加受体小鼠的被动皮肤过敏性休克反应,并升高血液中的脂多糖水平。
④不饱和脂肪酸也对CSU的发病有影响
代谢组学分析表明,短链脂肪酸是主要代谢物,但不饱和脂肪酸(如花生四烯酸和二十二碳六烯酸)可能对慢性自发性荨麻疹(CSU)的发病也至关重要。这些脂肪酸能促进拟杆菌生长并具有抗炎作用。
研究进一步发现,不饱和脂肪酸花生四烯酸的氧化产物(如白三烯,参与免疫反应和炎症过程)与毛螺菌和吉米菌属(Gemmiger)与呈负相关,而毛螺菌的下降是慢性自发性荨麻疹患者肠道菌群特征。
寄生虫感染也被认为是导致慢性自发性荨麻疹(CSU)发病机制相关的肠道微生态异常的因素。多项研究已经探讨了体内寄生虫感染与CSU之间的关系,并进行了总结。
◮感染寄生虫的人群更易患CSU
与健康对照组相比,CSU患者更易诊断出原生动物感染,并且弓蛔虫血清阳性和尖线虫的致敏风险显著增加。此外,他们的片吸虫病和人芽囊原虫等位基因患病率也较高。在CSU儿童中,人芽囊原虫是最常见的寄生虫。

通过分析蛔虫科(Anisakis simplex和Toxocara canis)整个幼虫的粗提取物和分离成分,在荨麻疹与寄生虫感染之间的关联中, Anisakis似乎比Toxocara发挥更重要的作用,而原肌球蛋白是关键标志物。
◮蠕虫感染会导致肥大细胞和嗜碱性粒细胞激活
蠕虫感染可能通过涉及肥大细胞和嗜碱性粒细胞激活的各种途径影响慢性自发性荨麻疹(CSU)的发生和进展。寄生虫特异性IgE通过高亲和力IgE受体 (FceRI) 激活肥大细胞和嗜碱性粒细胞。诸如犬弓首蛔虫或肝片吸虫之类的蠕虫和诸如人芽囊原虫或蓝氏贾第鞭毛虫之类的原生动物能够诱导宿主产生针对其抗原的高水平特异性 IgE 抗体,这可导致宿主肥大细胞致敏和脱颗粒。
此外,蠕虫寄生会破坏人体的保护屏障,引发Th2免疫反应和组织修复。寄生虫本身会抑制Th1细胞的分化并促进Th2细胞的发育,而B细胞则通过产生 IgG1-3来辅助Th1反应,与寄生虫抗原形成免疫复合物(CIC)。该过程会激活补体系统并产生过敏毒素C5a和C3a来作用于肥大细胞,从而导致荨麻疹的发生。
慢性荨麻疹(CU)是一种常见且临床上具有挑战性的疾病。目前,已有一些标准化方案治疗慢性自发性荨麻疹(CSU)。其主要治疗方法是使用第二代非镇静性H1抗组胺药(nsAH)。
然而,这些治疗对CSU患者的有效性可能受到多种因素的影响,包括肠道菌群。一些患者表现出 nsAH 耐药性,nsAH 耐药性患者的肠道或全身炎症反应更为强烈。据推测,肠道菌群失调可能是 nsAH 耐药性的一个促成因素。
◮肠道菌群失调可能是一些患者使用nsAH疗效不佳的原因

对有无nsAH耐药性的慢性自发性荨麻疹(CSU)患者肠道菌群进行了比较研究,发现有 nsAH 耐药性的 CSU 患者肠道菌群中普雷沃氏菌(Prevotella)、巨单胞菌属(Megamonas)、大肠杆菌属(Escherichia)、琥珀酸弧菌属(Succinivibrio)、克雷伯菌属(Klebsiella)和Colidextribacter属的水平较高。相反,没有nsAH耐药性的CSU患者的Blautia属、Alistipes属和Anaerostipes属水平较低。
注:在另一项研究中,毛螺菌属被确定为nsAH特征的生物标志物。
◮幽门螺杆菌根除疗法有助于缓解CSU症状
荟萃分析显示,幽门螺杆菌(Hp)可能与CSU的发展和持续有关。研究发现,HP 根除疗法在缓解CSU症状方面具有显著效果。并且接受抗生素治疗以根除HP的CSU患者的缓解率明显更高,无论HP根除状态如何。
◮治疗会导致CSU患者的肠道微生物群发生改变
此外,目前的治疗也已被证明会导致CSU患者肠道微生物群发生改变。在接受奥马珠单抗治疗后,在菌类水平上观察到Alphaproteobacteria和Betaproteobacteria的相对丰度显著下降,在属水平上观察到Burkholderia、Rhodococcus和Sphingomonas 的相对丰度显著下降。这种减少可能是促成良好结果的一个因素。
而肠道菌群与慢性荨麻疹之间的相关性提供了一种有利的治疗途径。通过调节肠道微生态来治疗慢性荨麻疹是目前一个突出的研究重点。
针对肠道微生物群的慢性荨麻疹相关疗法
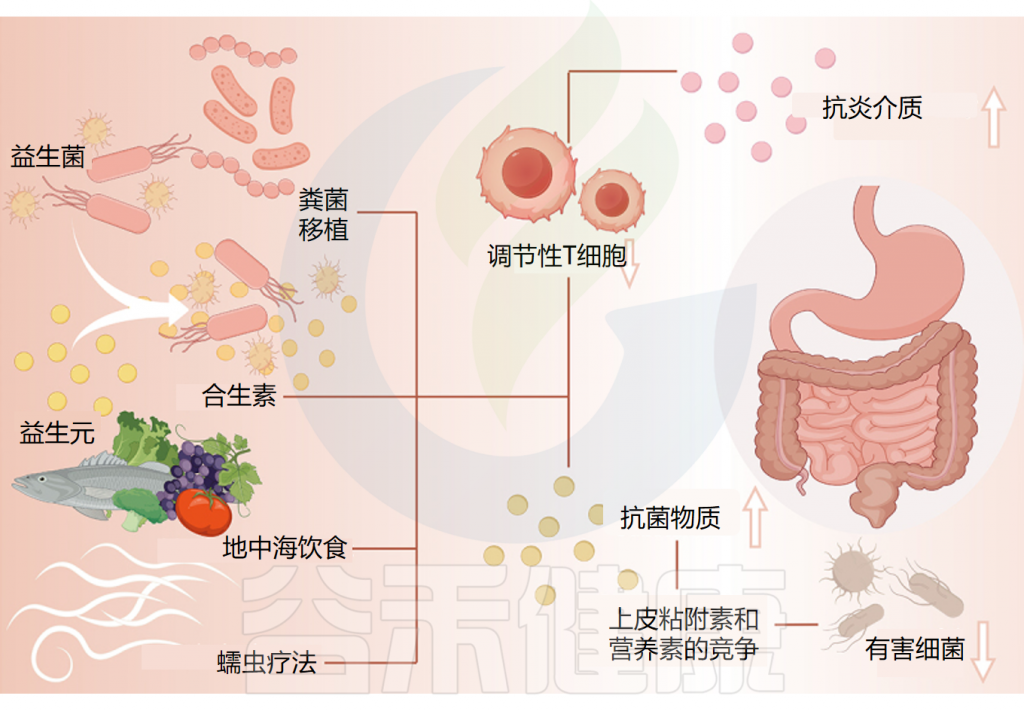
Cai R,et al.Gut Microbes.2024
可以通过几种机制来阐明益生菌对慢性荨麻疹的保护特性。
①减少炎症并促进抗炎环境
首先,有证据表明慢性荨麻疹(CU)患者的Treg细胞数量和功能均有所减少。研究表明,益生菌和相关化合物可以诱导 Treg 细胞的生成以对饮食抗原作出反应。并通过刺激抗炎介质的合成,这种诱导可以减少炎症并促进抗炎环境,从而作为针对CU的保护机制。
②分泌抗菌物质,改善肠道菌群失调
此外,益生菌通过分泌抗菌物质、抑制细菌毒素产生以及与病原体竞争上皮粘附和营养,维持肠道菌群平衡,改善肠道屏障功能,从而减少体内有害代谢物的产生。
由于慢性荨麻疹患者肠道菌群的改变,可能会导致肠道微生态失调,而服用益生菌可以改善肠道菌群的稳态。
③一些特定益生菌改善了慢性荨麻疹的严重程度
研究证据表明,某些肠道微生物群具有针对慢性荨麻疹(CU)的保护作用,有助于参与慢性荨麻疹的治疗和缓解。
研究了两种益生菌(唾液乳杆菌LS01和短双歧杆菌BR03)组合对难治性慢性自发性荨麻疹患者临床进展的影响。在益生菌干预期间,38名患者中,9名(23.7%)临床症状轻度改善;1名(2.6%)显著改善;1名(2.6%)荨麻疹症状完全消失。但27名患者 (71.1%) 的症状保持不变。
研究结果证实了这种益生菌组合在降低某些慢性自发性荨麻疹患者的症状评分方面的治疗效果,主要是那些尽管使用 H1 抗组胺药治疗但仍持续存在的患者,从而提高了他们的整体生活质量而没有副作用。
还进行了一项随机安慰剂对照研究,纳入了206名慢性荨麻疹儿童,两组儿童均使用了地氯雷他定干混悬剂。治疗组有104名儿童同时使用了益明佳®(六种菌的冻干混合物,浓度为每克5×109 CFU 活菌)。经过4周的监测,治疗组的风团大小和发作频率均显著减少,80.8%的患者具有明显效果(包括显著改善和完全缓解),而安慰剂组为 62.5%。
这些研究进一步证实了某些益生菌菌株组合对改善慢性荨麻疹症状有效,并为使用益生菌治疗慢性荨麻疹提供了支持。
益生元是指含有不可消化成分的补充剂或食物,它们不仅能选择性地促进有益的本土益生菌的生长或活性,还能增强免疫系统。
①服用益生元降低了有过敏风险婴儿的荨麻疹发病率

在一项前瞻性、双盲、安慰剂对照研究中,有过敏风险的足月婴儿在出生后前6个月内,42名婴儿被喂食含益生元(0.8g/100ml 半乳寡糖(scGOS)/低聚果糖(lcFOS))的低过敏性配方奶粉,50 名婴儿被喂食安慰剂(0.8 g/100 ml 麦芽糊精),随后完成了5年的随访。
研究证实,低聚糖益生元 (scGOS/lcFOS) 从生命早期开始,一直到5岁,对高风险婴儿的过敏性荨麻疹具有保护作用。scGOS/lcFOS组儿童的过敏性荨麻疹发病率(6%)低于安慰剂组儿童(38%)。
另一项对照试验中,父母有特应性皮炎病史的健康足月婴儿,以低过敏性配方奶粉喂养,并分为两组,干预组添加益生元(8g/升 scGOS/lcFOS),安慰剂组添加安慰剂(8g/升麦芽糊精)。6 个月干预期结束时,进行盲法随访,直至婴儿2岁。安慰剂组过敏性荨麻疹的累积发病率(10.3%)高于干预组(1.5%)。
综上所述,益生元单独应用已被证明能有效预防荨麻疹。还需在更大人群、更广泛年龄范围和多种益生元条件下进行长期研究,以评估其潜在预防作用。
合生元结合了益生菌和益生元的协同作用,兼具益生菌和益生元的特性,从而有益于宿主的健康。
①合生元的使用改善了患者生活质量,并减少了瘙痒等疾病症状
对42名慢性荨麻疹患者进行了为期8周的随机对照临床试验。通过计算机随机化,以1:1的分配比将患者分为对照组(仅抗组胺药)和干预组(抗组胺药+合生元)。干预组每天服用两次名为LactoCare的口服益生菌胶囊,其中含有大量有益细菌,包括鼠李糖乳杆菌、干酪乳杆菌、嗜酸乳杆菌、短双歧杆菌、保加利亚乳杆菌、长双歧杆菌、嗜热链球菌以及低聚果糖作为益生元。
对照组和干预组皮肤病生活质量指数改善率分别为44%和66%。同时接受联合治疗的患者可能会在瘙痒、荨麻疹数量和总评分方面有更显著的降低。
粪便微生物移植(FMT)在改变肠道微生物群方面非常有效,是一种可行的治疗方法。在FMT之后观察到的一系列疾病(炎症性肠病、糖尿病、癌症、肝硬化和肠脑疾病)的临床反应变化表明微生物群与宿主之间存在错综复杂的联系。
①粪菌移植改善皮肤炎症,减轻荨麻疹症状
研究表明,粪便微生物移植(FMT)在皮肤病学中具有治疗潜力。FMT抑制多种炎症细胞因子,迅速逆转皮肤表皮增厚。小鼠研究显示,FMT可提高肠道短链脂肪酸水平,恢复肠道微生物群至供体状态。此外,FMT降低IgE水平,平衡Th1/Th2反应,调控Treg细胞,减少肥大细胞、嗜酸性粒细胞和嗜碱性粒细胞,从而减轻特应性皮炎的过敏反应。
曾报道了一例病例,一名女性患者六年来反复出现阵发性胃部不适和荨麻疹,接受了四次经内镜肠内管结肠FMT。经过一年,患者症状明显改善,恢复正常社交和饮食。16S rRNA 测序分析显示,FMT 后肠道菌群发生显著变化,包括普雷沃氏菌增加,拟杆菌和粪杆菌减少,有助于维持肠道菌群稳态。
①地中海饮食缓解全身炎症和氧化应激
饮食是塑造肠道菌群、宿主代谢物和屏障免疫系统的关键因素。饮食变化会迅速影响肠道菌群的多样性,有利于特定细菌种群的增殖,同时影响肠道pH值、肠道通透性和细菌代谢物的产生,也可能引发炎症。
有研究表明,地中海饮食可能会影响肠道菌群,进而影响慢性荨麻疹。这种饮食的特点是混合了富含纤维的复杂碳水化合物、以抗动脉粥样硬化和抗炎作用而闻名的多不饱和脂肪酸,以及具有抗氧化特性的生物活性化合物,如黄酮类化合物、植物固醇、萜烯和多酚,是营养饮食的典范。
研究表明,地中海饮食的抗炎和抗氧化特性可能有助于缓解全身炎症和氧化应激,而这两者都是慢性自发性荨麻疹(CSU)的发病机制。此外,坚持地中海饮食与恢复肠道菌群平衡有关,拟杆菌门和特定有益梭菌群数量增加,而变形菌门和芽孢杆菌门数量减少。地中海饮食促进有益细菌及其代谢物的生长,同时缓解菌群失调并降低脂多糖水平。
注:慢性自发性荨麻疹(CSU)患者组的平均地中海饮食评分为5.40±1.88,而对照组的平均地中海饮食评分为6.30±1.39。CSU 患者7天内的荨麻疹活动评分与地中海饮食评分呈负相关,而荨麻疹控制测试评分呈正相关。
②利用蠕虫疗法减轻免疫反应
蠕虫疗法是一种利用寄生蠕虫(如线虫、吸虫和绦虫)来调节宿主免疫系统的治疗方法。其理论基础是寄生蠕虫在长期进化过程中,发展出一系列机制来逃避宿主的免疫攻击,从而在宿主体内生存。这些机制可以通过调节宿主的免疫反应,降低过度活跃的免疫反应,从而可能对某些免疫和炎症性疾病产生治疗效果。
蠕虫疗法作为一种过敏免疫疗法,已被发现可治疗多种免疫和炎症疾病,如类风湿性关节炎和多发性硬化症。来自动物研究的有力证据表明,蠕虫不仅可以降低寄生虫特异性免疫反应,还可以调节自身的免疫力和过敏性炎症反应,改善代谢平衡。
蠕虫疗法有两种作用方式:(1)直接通过免疫系统影响;(2)通过改变肠道微生物群,从而对生物体产生影响。蠕虫通过操纵动物的自然免疫反应来逃避免疫识别和对抗。由于这种免疫调整,宿主体内先前存在的过敏或免疫相关疾病可能会缓解或完全消失。
但这种疗法目前主要在研究阶段,尚未被广泛接受或应用于临床实践。需要更多的研究来确定其安全性和有效性。
小结
通过使用益生菌、益生元、合生元、饮食调整和粪菌移植在改善肠道菌群中展现潜力,从而有助于治疗或改善慢性荨麻疹的症状,应在临床和科研中得到更多关注和应用。
尽管蠕虫疗法展示出了对慢性荨麻疹的治疗前景,但其过程尚不明确。临床试验样本量有限,难以评估安全性和有效性。此外,药物的临床应用障碍和生产成本高,限制了其快速临床应用。
肠道微生物群的组成和代谢物的改变对慢性荨麻疹的病理状态有显著影响。研究发现,慢性自发性荨麻疹(CSU)患者的肠道微生物组多样性减少,有益菌如拟杆菌、双歧杆菌丰度下降,而条件性致病菌肠杆菌等丰度上升,导致菌群失调。短链脂肪酸的生成减少,脂多糖的水平升高,引起肠道促炎和抗炎T细胞亚群失衡,转向促炎表型。这种促炎反应可能激活肥大细胞,使得免疫调节受损,加重慢性荨麻疹患者的症状。
通过针对肠道微生物群,有望开发新的治疗策略,如通过使用益生菌、益生元、合生元、饮食调整和粪菌移植来改善肠道菌群,有助于减少氧化应激,减轻炎症,从而治疗或改善慢性荨麻疹的症状。
然而目前关于肠道微生物群在慢性荨麻疹中的具体作用机制的研究仍相对不足。我们期待未来有更多的研究能够深入探讨和揭示肠道微生物群在慢性荨麻疹发病机制中的确切作用。这将为通过肠道微生物群治疗慢性荨麻疹提供新的视角和方法。我们相信这些基于肠道细菌的治疗策略在慢性荨麻疹的治疗和预防中具有巨大的应用潜力。
主要参考文献
Cai R, Zhou C, Tang R, Meng Y, Zeng J, Li Y, Wen X. Current insights on gut microbiome and chronic urticaria: progress in the pathogenesis and opportunities for novel therapeutic approaches. Gut Microbes. 2024 Jan-Dec;16(1):2382774.
Kolkhir, P., Giménez-Arnau, A.M., Kulthanan, K. et al. Urticaria. Nat Rev Dis Primers 8, 61 (2022).
Zuberbier T, Bernstein JA, Maurer M. Chronic spontaneous urticaria guidelines: What is new? J Allergy Clin Immunol. 2022 Dec;150(6):1249-1255. doi: 10.1016/j.jaci.2022.10.004. Erratum in: J Allergy Clin Immunol. 2023 Feb;151(2):580.
谷禾健康
组胺是人体内的一种生物胺,最为人们所熟知的作用是作为过敏反应的介质,但也是神经系统、肠道、皮肤和免疫系统中重要的信号分子。
花粉之所以会引起人体过敏反应,并不是因为花粉本身,而是由于花粉颗粒携带了能引起过敏的抗原。接触过敏源后,体内的组织会释放组胺,作为一种神经递质,组胺会使毛细血管和微静脉的管壁通透性增加,导致局部组织水肿,造成皮肤发红、疹块;也会使气管平滑肌收缩,造成呼吸道狭窄,引起气喘、呼吸困难。
组胺主要由肥大细胞和嗜碱性粒细胞等免疫细胞产生和释放。一些细胞如树突状细胞或T细胞可以表达组氨酸脱羧酶,这是一种在刺激后合成组胺的酶。
尽管如此,一些食物中的微生物群和部分人体肠道微生物群也可以分泌组胺。表明组胺在肠道免疫调节中的潜在作用。事实上,最近的一项研究表明,组胺可以通过抑制肠道中白细胞介素18的产生来发挥抗炎作用。
组胺在免疫反应、神经传递和过敏反应中发挥着重要作用。在某些浓度范围内,组胺起着保护作用,对于维持健康状态至关重要。例如组胺可以刺激胃酸分泌,有助于消化;还会影响排便情况;组胺作为重要的神经递质,参与睡眠调节和体温调节。
然而,在较高浓度下,会导致包括组胺中毒和组胺不耐受或扩张血管,影响血压,导致免疫调节紊乱、胃肠道症状和神经系统疾病。组胺中毒是在食用被大量组胺污染的食物后发生的。如果个体由于基因突变而缺乏生物胺降解能力,或者正在服用单胺氧化酶抑制剂等抗抑郁药来减缓胺的降解,他们就会更容易出现组胺不耐受的症状。
组胺在体内与相应受体(H1、H2、H3、H4)结合而介导的各种症状。不同类型的组胺受体与该化合物之间的相互作用可能会引起多种影响,例如H1R主要介导过敏反应,而H2R具有抗炎作用,而H3R主要影响睡眠、认知障碍等精神健康。
肠道内的组胺对许多细胞过程的广泛影响会导致各种胃肠道疾病,包括食物过敏、肠易激综合征、炎症性肠病、结直肠癌等。谷禾将在本文带大家了解组胺在免疫及各种肠道疾病中的保护或致病作用。
组胺(C5H9N3)是一种重要的生物胺类神经递质,于20世纪初首次发现。从那时起,它的功能开始被逐渐发现并得到越来越详细的描述。
组胺存在于多种细胞类型中,是人体的关键信号分子,具有多种功能。例如作为神经递质或免疫反应调节剂。
拓展知识:组胺的发现
1903年,一位叫威廉·邓巴的医生证明了吸入花粉后的过敏反应并不是由花粉本身引起的,而是机体对花粉的反应引起的一种毒素的释放所造成的。
后来,亨利·戴尔于1910年在研究黑麦的毒性时,从麦角菌中提取出一种叫做组织胺的物质。他发现用组胺可以人工诱发荨麻疹。如果将组胺滴入气管,还会诱发哮喘发作。
★ 组胺的重要作用
介导过敏反应
组胺最被人熟知的是在过敏反应中的作用。例如在过敏性鼻炎、荨麻疹和血管神经性水肿等过敏性疾病中发挥重要的病理生理学作用。
当免疫系统对过敏原产生过度反应时,免疫细胞(如肥大细胞和嗜酸性粒细胞)会释放存储在细胞内的组胺。释放的组胺会结合到特定的受体上,引发一系列生理反应,例如瘙痒、肿胀、打喷嚏等。
扩张血管,影响血压
组胺对人心血管系统最突出的作用是扩张小血管。血管扩张使外周阻力降低,血压下降,并伴有潮红、头痛等症状。
组胺还增加毛细血管的通透性,使渗出增加,引起水肿,严重时甚至导致循环血量减少,可能引起休克。组胺对心脏的直接作用包括增强心肌收缩力、加快心率和减慢房室传导。
刺激胃酸分泌
首先,组胺可以刺激胃黏膜细胞中的H2受体,当组胺与H2受体结合时,会激活细胞内的信号转导通路,导致胃壁上的壁细胞释放胃酸。
其次,组胺还可以通过刺激嗜酸性细胞(胃窦细胞)来促进胃酸的分泌。嗜酸性细胞是胃黏膜中的一种细胞类型,它们包含大量的组胺。当胃黏膜受到刺激时,嗜酸性细胞会释放存储的组胺。释放的组胺通过与H2受体结合,刺激壁细胞分泌胃酸。
影响排便
一项研究中,发现细菌产生的组胺与小鼠结肠运动和粪便排出量增加有关,并且组胺受体拮抗剂治疗在很大程度上阻断了细菌组胺对结肠运动的影响。
收缩平滑肌
此外,组胺会诱导平滑肌细胞(包括支气管和肠道)收缩,哮喘患者对组胺比正常人敏感100~1000倍,组胺可引起支气管痉挛导致呼吸困难。组胺收缩胃肠平滑肌和子宫平滑肌还会引起痉挛性腹痛。
重要的神经递质
在中枢神经系统中,组胺作为一种神经递质,参与睡眠调节和体温调节等生理过程。组胺对感觉神经末梢有强烈的刺激作用,尤其对调节痛和痒的神经,该效应由H1受体所调节。
除此之外,有研究发现组胺还会影响伤口愈合、食欲和情绪等方面。
组氨酸是组胺的前体物质
组胺的主要细胞来源是肥大细胞和嗜碱性粒细胞。在细胞的高尔基体中,组胺是通过组氨酸脱羧酶将L-组氨酸氧化脱羧形成的,其辅助因子是吡哆醛(维生素B6)。
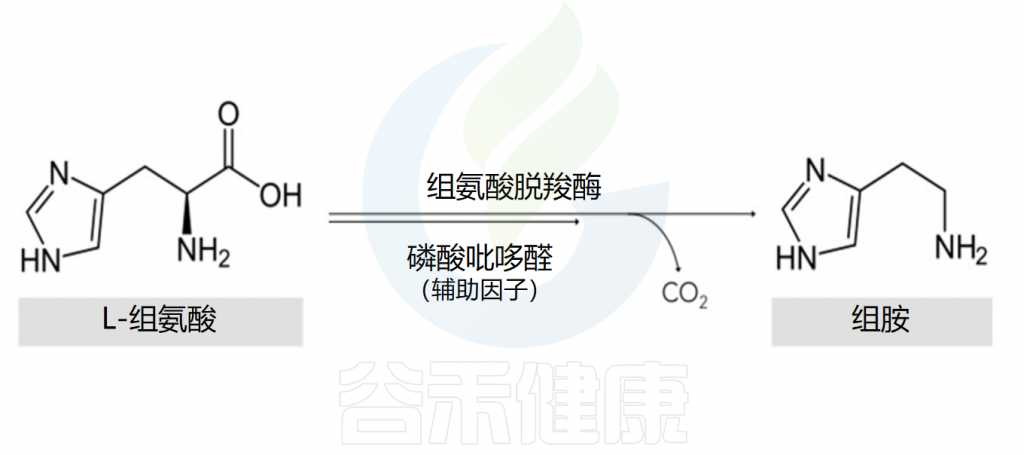
Comas-Basté O,et al.Biomolecules.2020
该反应的结果是组胺的形成,组胺随后与其他胺(如5-羟色胺)、蛋白酶、蛋白聚糖、细胞因子/趋化因子和血管生成因子一起储存在细胞质颗粒中,并在细胞致敏和脱颗粒后释放。
过敏时组胺会大量释放
肥大细胞的脱颗粒和组胺的释放主要是由于特异性抗原与FcRI受体结合以及对非免疫刺激(例如神经肽、补体系统的一部分、细胞因子、血小板活化因子)的反应。
IgE抗体是过敏性疾病期间肥大细胞脱颗粒的介质。IgE与其肥大细胞表面的高亲和力IgE受体的结合称为“致敏”。肥大细胞和嗜碱性粒细胞释放的组胺通过激活四种G蛋白偶联受体,即H1R、H2R、H3R(主要在大脑中表达)和H4R,发挥其生物活性。
细菌感染
某些细菌感染也可以引起组胺的释放。例如,胃幽门螺杆菌感染可以导致胃黏膜炎症,促使组胺释放,进而引发胃酸增多和溃疡等胃部疾病。
药物和化学物质
某些药物和化学物质也可以诱发组胺的释放。例如,非甾体抗炎药(如阿司匹林)和某些麻醉药物以及酒精可以引起组胺的释放,可能导致过敏样反应。
物理刺激
组胺释放也可由多种物理因素引起,如极端温度、创伤、摩擦等。这种情况下的组胺释放通常是局部性的,导致局部组织的充血和炎症反应。
运动和应激
剧烈运动、情绪激动和精神压力等也可以促使组胺的释放。这种情况下,组胺的释放通常是全身性的,可能导致血管扩张、心率增加和其他应激反应。
注:组胺也可以由其他细胞类型(例如胃肠嗜铬样细胞、组胺能神经元、树突状细胞、T淋巴细胞、血小板等)合成和释放。
食物和肠道微生物也是组胺的重要来源
一部分组胺通过食物进入人体或由肠道微生物产生。最受欢迎的富含组胺的食物是鱼和海鲜、发酵食品(如奶酪、葡萄酒、泡菜等)和一些蔬菜(如菠菜、茄子、番茄等)。
组氨酸主要在自溶或细菌过程中产生,因此高浓度的组胺主要存在于微生物发酵产物中。食品中生物胺形成的条件是游离氨基酸的可用性、脱羧酶阳性微生物的存在以及细菌生长和脱羧酶活性的条件。
影响细菌组氨酸脱羧酶活性的因素
细菌分泌脱羧酶受许多因素(例如,发酵碳水化合物、氧气或氯化物浓度的存在)调节。在酸性环境中,氨基酸脱羧酶活性的表达增加。
组氨酸脱羧酶活性较高的细菌种类有:
Morganella morganii,Eschericha coli, Hafnia alvei, Proteus vulgaris, Proteus milabilis, Enterobacter aerogenes, Raoultella planticola, Raoultella ornithinolytica, Citrobacter freundii, Pseudomonas fluorescens, Photobacterium damselae。其中一些细菌具有代谢组胺的能力。
我们将在下面的一个段落中展开讲述肠道中的组胺与微生物群。
在人体内,组胺降解主要有两种途径,涉及二胺氧化酶(DAO)或组胺-N-甲基转移酶(HNMT)。组胺降解酶缺失会导致组胺浓度过高甚至中毒,组胺不耐受患者就是组胺降解酶的酶活性降低或受到抑制,并且不足以灭活食物中的组胺并阻止其进入血液。
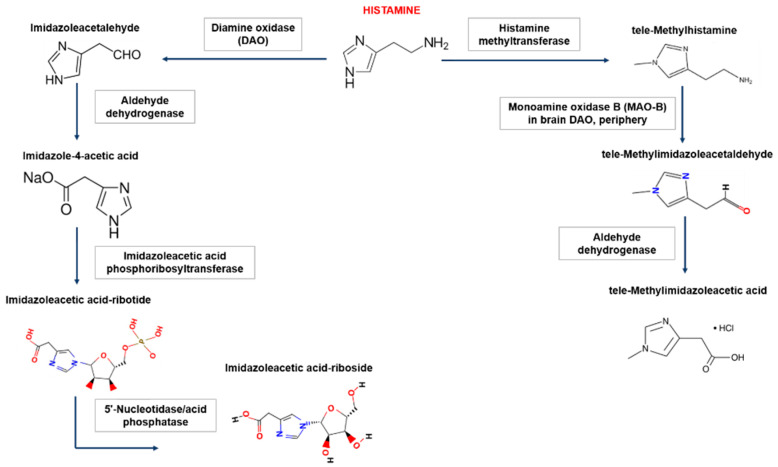
Shulpekova YO,et al.Nutrients.2021
组胺-N-甲基转移酶负责细胞内的组胺降解
组胺-N-甲基转移酶(HNMT)主要负责细胞内组胺的降解。HNMT在肾脏和肝脏以及脾脏、结肠、前列腺、卵巢、脊髓细胞、支气管和气管中的表达最高。一小部分组胺在HNMT的作用下转化为N-甲基组胺。
注:在原始形式中,约2%-3%的组胺被排泄。
二胺氧化酶负责细胞外的组胺降解
二胺氧化酶(DAO)是一种分泌蛋白,负责细胞外组胺的降解。DAO的最大活性记录在小肠、结肠、胎盘和肾脏中。DAO在细胞内囊泡中积累,然后进入细胞外空间,在那里降解组胺。绝大多数组胺通过DAO转化为咪唑乙酸。
保护身体免受组胺过高的侵害
二胺氧化酶执行“屏障功能”,从而限制组胺从肠道进入血液;组胺-N-甲基转移酶通过B族维生素和S-腺苷甲硫氨酸的参与使组胺甲基化,从而调节细胞内的组胺含量。
尽管二胺氧化酶和组胺-N-甲基转移酶这两种酶都存在于肠上皮中,但吸收组胺进入血流的主要障碍是 二胺氧化酶,组胺-N-甲基转移酶在此过程中仅发挥次要作用。在正常情况下,这种酶屏障足以防止组胺被吸收到血流中。
注:二胺氧化酶和组胺-N-甲基转移酶基因的多态性可能是组胺代谢个体差异的基础。在突触中,乙醛脱氢酶在组胺降解中发挥作用。
虽然健康人体内的组胺很快就会被二胺氧化酶降解,但即使在健康人体内,也可能会因摄入组胺含量高的食物(例如鲭鱼)导致血液中组胺浓度升高而出现严重症状——组胺中毒。
• 组胺中毒的症状
组胺中毒症状多种多样,可能包括口腔麻木、头痛、头晕、心悸、低血压、吞咽困难、脉搏微弱、荨麻疹、面部肿胀、潮红、呕吐、恶心和腹泻等。
组胺中毒的症状可能持续数小时或一天,但在极少数情况下,症状可能会持续数天。
• 组胺中毒的主要原因是食用鱼类和海鲜
不仅是鲭鱼,一些非鲭鱼(鲯鳅鱼、蓝鱼、沙丁鱼等)都含有大量组胺,此外发酵产品(奶酪、葡萄酒、罐头食品、腌制蔬菜和一些饮料)中组胺和血清素的含量也较高。
一项针对科学报告的分析显示,在98%的病例中,组胺中毒的原因是食用鱼类和海鲜,其余2%是由于食用奶酪等发酵食品。
鱼组织中的组胺是由多种细菌对游离组氨酸进行转化而产生的,这些细菌包括摩氏摩根菌(Morganellamorganii)、产气肠杆菌 (Enterobacter aerogenes), Raoultella planticola, Raoultella ornithinolytica和Photobacterium damselae。
鱼分解产生的其他物质会增强组胺的毒性并促进吸收
组胺中毒的发病机制不能仅用组胺的作用来解释。与口服等效剂量的纯组胺相比,鲭鱼毒性较高。还提出了其他机制,例如控制组胺的酶的抑制或增强、导致肥大细胞脱颗粒的物质的存在、其他组胺激动剂的存在。
鱼分解过程中产生的其他物质(尸胺和腐胺)会增强组胺的毒性,这些物质还会抑制肠道单胺氧化酶和二胺氧化酶,或从与粘膜粘蛋白的结合中释放组胺,促进其吸收。
组胺中毒后的干预及治疗
在生理值范围内。组胺中毒的病程通常是轻微的、自限性的、持续时间较短且不需要治疗。
对于严重的组胺中毒病例,需要进行治疗干预——支持性容量疗法和氧气疗法,并给予口服抗组胺药和支气管扩张剂。
组胺不耐受是由于组胺摄入量与机体清除能力不相称,导致组胺过度蓄积,并与相应的物质结合而出现症状的病理过程。
与组胺中毒不同,组胺中毒的严重程度与营养物质中组胺的含量成正比,组胺不耐受的发病机制与先天性或后天性缺乏中和组胺的酶有关。在健康患者中,肠上皮细胞具有由二胺氧化酶和组胺-N-甲基转移酶创建的酶屏障。该屏障可防止血液中外源性组胺的过度吸收。如果这些酶被抑制或减少,即使摄入少量的组胺也可能出现组胺不耐受的症状。
• 组胺不耐受的症状
组胺不耐受的神经系统症状包括头痛。除头痛外,其他重要症状包括弥漫性胃痛、绞痛、胀气和腹泻。
组胺不耐受的患者,在摄入组胺或酒精含量高的食物时或摄入后会出现流鼻涕、鼻塞等症状,极端情况下甚至会出现支气管哮喘发作、支气管收缩、咳嗽、喘息并伴有肺功能下降。
组胺不耐受患者还会出现皮肤瘙痒、发红、肿胀甚至荨麻疹。而患有组胺不耐受的女性会出现与月经周期相关的痛经和头痛。
• 影响组胺降解酶活性的因素
在胃肠道疾病(炎症性肠病、寄生虫感染、微生物菌群失调、代谢吸收不良)中,损伤的肠上皮细胞会减少二胺氧化酶的产生,或其他生物胺、酒精或药物也会抑制二胺氧化酶。
二胺氧化酶活性降低可见于慢性肾功能衰竭、病毒性肝炎、晚期肝硬化和慢性荨麻疹(一种典型的组胺相关疾病,对内源性组胺的耐受性降低)的患者。
二胺氧化酶降解能力下降还可能是由于缺乏其辅助因子、维生素B6、维生素C、铜和锌而导致的。
• 肠道微生物会影响组胺不耐受的发生
肠道微生物群也会影响组胺不耐受的发生。最近的研究表明,组胺不耐受可能是由肠道微生物群的改变引起的。肠道中大量的组胺分泌细菌可能会导致组胺不耐受。健康人体内双歧杆菌的数量较多。而在血清二胺氧化酶活性降低的人群中观察到变形菌属数量较多。
组胺不耐受人群存在肠道菌群失调
在组胺不耐受人群中观察到肠道菌群失调,与健康个体相比,与肠道健康相关的普氏菌科(Prevotellaceae)、瘤胃球菌(Ruminococcus)、粪杆菌(Faecalibacterium)和普拉梭菌(Faecablibacterium prausnitzii)的比例明显较低。组胺不耐受人群还具有显著更高丰度的组胺分泌细菌,包括葡萄球菌属和变形杆菌属、属于肠杆菌科的几个未识别属,以及产气荚膜梭菌和粪肠球菌。
需要注意的是,几项研究表明,即使在没有二胺氧化酶缺陷的个体中,大量的组胺产生细菌也会导致肠道内高水平组胺的积累、随后在血浆中的吸收以及不良反应的出现。
低组胺饮食
组胺不耐受疗法的基础是基于低组胺含量食物的消除饮食。一般来说,公认的饮食指南尚未制定;但是,建议避免食用富含组胺的食物如奶酪、油性鱼类和贝类,以及生发酵肉制品、腌菜、发酵豆制品、葡萄酒和啤酒、鸡蛋、巧克力和蘑菇。
避免食用易刺激组胺释放的蔬果
此外,应避免食用会刺激内源性组胺释放的蔬菜和水果(如菠菜、西红柿、柑橘类水果、草莓、茄子、鳄梨、木瓜、香蕉、猕猴桃、菠萝和李子)。


Kovacova-Hanuskova E,et al.Allergol Immunopathol.2015
补充二胺氧化酶
研究还发现补充二胺氧化酶可以改善组胺不耐受患者的生活质量。已开发出基于肠溶猪肾提取物的含二胺氧化酶食品添加剂。豆芽也被用作二胺氧化酶的来源,这种酶的活性是未发芽种子的250倍。
施用抗组胺药物
纠正组胺不耐受的药物方案基于使用组胺受体阻滞剂、肥大细胞膜稳定剂和其他具有抗组胺活性的药物。
在这方面,H1R拮抗剂是最有效的,特别是依巴斯汀。每天服用20毫克该药物,持续12周,46%的患者出现临床缓解。
一些生物活性物质有助于增强组胺降解
维生素C被用作纠正组胺不耐受的佐剂。每日剂量300-500毫克可增强组胺降解并抑制肥大细胞脱颗粒。
天然类黄酮(非瑟酮、山奈酚、槲皮素、芦丁和木犀草素)和活性生物碱小檗碱在体外抑制肥大细胞脱颗粒。这些都有助于减轻组胺不耐受。
虽然组胺含量过低的情况相对较少见,但也可能会对身体产生一些影响,包括:
• 伤口愈合减慢
组胺在人体中起着重要的调节作用,其能够帮助调节纤维细胞生长因子的生成,该成分可以加速局部肉芽的生长,帮助促进伤口的愈合。因此组胺含量较低时,伤口愈合能力也会减弱,速度会随之减慢。
• 消化不良
组胺可以通过刺激胃部,进而起到促进胃酸分泌的效果。若身体中没有组胺,则会导致胃酸减少,无法有效消化分解食物,进而引起消化不良的问题。
• 情绪不良
组胺还可以作为神经递质帮助调节中枢神经系统,而一旦没有组胺,则会影响中枢神经系统稳定性,引起睡眠异常、激素水平紊乱等情况,进而导致情绪不良。
• 血压升高
人体缺少组胺还会导致血压升高。因为组胺可以促进血管扩张,增加血管的通透性,所以具有降低血压的作用。若没有该成分,则血管会由于舒张能力减弱,因此出现血压升高的问题。
• 免疫力下降
组胺对免疫细胞,如吞噬细胞、T细胞、B细胞等,起到增强活性的功效,因此组胺还具有增强免疫功能的作用。若人体缺少该物质,则人体免疫力会下降,无法有效抵抗细菌、真菌等有害物质的侵入。
组胺对人体许多生理功能都可以起到积极作用,但是人体中的组胺也不能过量,否则容易导致过敏反应,出现头晕、皮肤瘙痒、胸闷等不适症状。
此种情况需及时就医,并且在医生的指导下服用抗组胺药物,例如盐酸西替利嗪片、氯雷他定片等药物,进行抗过敏治疗。
★ 组胺的免疫调节因受体的不同而存在区别
先天性和适应性免疫系统的细胞都可以被组胺调节。组胺的调节取决于它与组胺受体的四种亚型的结合,这些亚型按发现时间顺序命名——H1R-H4R,在多种细胞类型中差异表达,进而产生不同的作用。
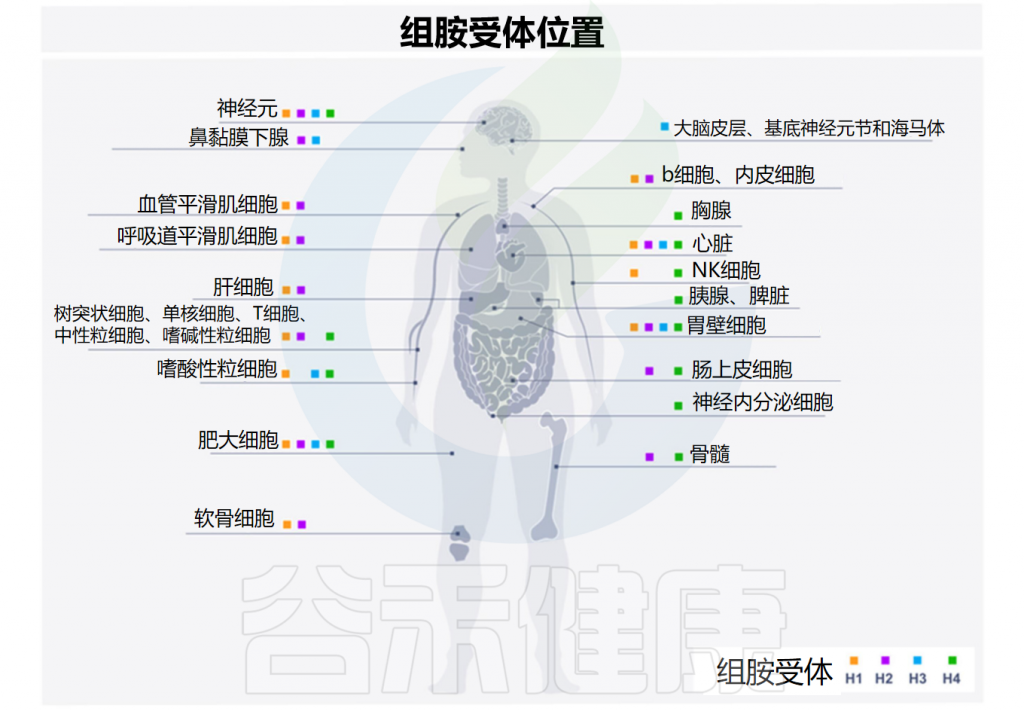
Smolinska S,et al.Metabolites.2022
1
H1R
H1R受体存在于几乎所有组织的内皮细胞和血管平滑肌细胞中(它们的密度在胃肠道中特别高),以及下丘脑、肾上腺髓质和免疫活性细胞(单核细胞、中性粒细胞、T细胞和B细胞)中。
• H1R受体介导的主要作用
H1R刺激的主要作用是炎症、全身血管舒张、血管通透性增加、支气管收缩、回肠收缩和昼夜节律周期调节。
外周H1R介导的影响包括鼻漏、支气管收缩、过敏反应、结膜炎和荨麻疹,而中枢相关的H1R影响包括食物和水摄入的调节、抽搐、注意力和睡眠调节。
• H1R是过敏反应的主要受体
IL-3、IL-4和组胺可上调H1R基因表达,而H1R激活导致许多与过敏性即时型超敏反应相关的特征,例如发红、瘙痒和肿胀。
例如小鼠模型中H1R的激活会诱导IFN(干扰素)产生增加,这与1型辅助性T细胞的增殖有关,并诱导促炎作用。
研究表明,瘙痒因子(例如神经生长因子、信号素 3A)的表达受组胺H1R的调节。在小鼠模型和特应性皮炎患者中,使用H1R拮抗剂可降低IL-31(白细胞介素-31)水平,这与瘙痒的发作有关。
2
H2R
与H1R类似,H2R的表达存在于多种组织和细胞中,包括脑、胃壁细胞、平滑肌细胞、T和B细胞、树突状细胞和心脏组织。H2R受体位于突触后,主要通过环腺苷单正膦(cAMP)传递信号并与Gαs偶联。
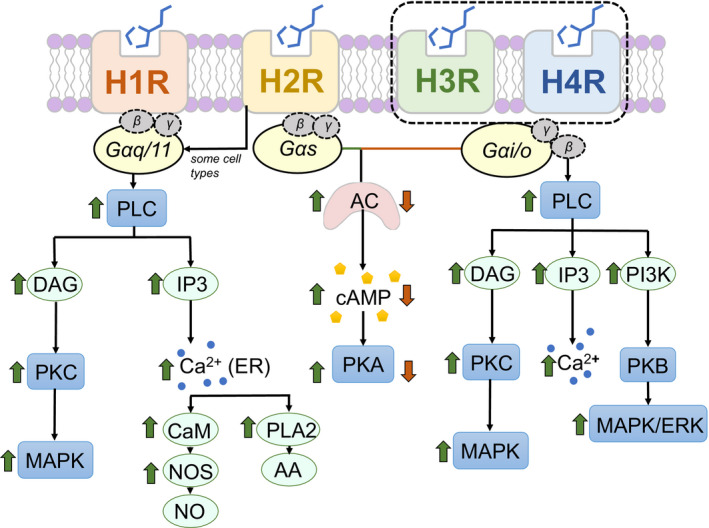
不同组胺受体下游的信号通路
Sudarikova AV,et al.Physiol Rep.2021
H2R刺激的主要作用是外分泌(主要是盐酸)、心动过速、平滑肌细胞松弛、抗炎作用、抗体合成等免疫活动。
• H2R具有抗炎作用
在组胺与H2R结合期间,H2R刺激通过抑制单核细胞或巨噬细胞和肥大细胞产生IL-12、IFN-γ、TNF-α细胞因子,而IL-10分泌增加,从而引起抗炎作用。
• H2R影响对微生物的免疫反应
组胺(内源性和外源性)通过H2R显著改变对微生物的先天免疫反应。特别地,树突状细胞对微生物配体的反应被组胺以H2R依赖的方式显著改变。
H2R敲除的小鼠免疫系统紊乱以及胃缺陷(胃酸分泌减少),此外还观察到认知能力下降和伤害感受异常。
• 注意:不同组胺受体可能具有相反的作用!
H2R以与H1R不同的方式传输信号,H1R和H2R的激活对各种生物过程具有相反的影响。在T细胞介导的免疫反应中,H1R激活促进Th1极化,而H2R激活则抑制Th1极化。
在平滑肌收缩中也观察到H1R和H2R激活的相反作用。H1R和H2R拮抗剂分别抑制和加剧组胺引起的轻度哮喘患者的支气管痉挛。这些数据表明,组胺可能会产生相反的作用,具体取决于被激活的特定组胺受体。
3
H3R
• H3R主要存在于神经系统中
H3R受体在神经系统细胞中表达,特别是在大脑皮层,基底神经节神经元和海马体中。H3R位于含组胺神经元的突触前区域。它们的功能是调节组胺以及其他神经递质(如多巴胺、去甲肾上腺素、γ-氨基丁酸、乙酰胆碱和血清素)的合成和释放。
• H3R受体影响睡眠、认知障碍等精神健康
H3R受体表达和激活的变化在睡眠-觉醒周期障碍、注意力缺陷多动障碍、癫痫和认知障碍以及炎症的发展中发挥重要作用。
H3R缺陷小鼠表现出行为反应和运动的变化,以及伴有肥胖、食欲过盛以及瘦素和胰岛素水平升高的代谢综合征。
• H3R可能会促进炎症反应
H3R刺激可增加促炎活性以及免疫细胞呈递抗原的能力。H3R敲除也可导致神经炎症性疾病的严重程度和T细胞中IFN-诱导蛋白10的表达增加。
因此,组胺H3R拮抗剂的使用有可能用于预防或抑制炎症性疾病(例如呼吸系统疾病)的发展。
4
H4R
H4R受体是最近才发现的,它们的作用尚未完全了解。H4R主要存在于免疫细胞(嗜酸性粒细胞、嗜碱性粒细胞、肥大细胞、自然杀伤细胞、树突状细胞、单核细胞和T细胞)中,也存在于脾脏、胸腺、骨髓、胆管、胰腺、肠上皮细胞和神经内分泌细胞中。
• H4R会增强过敏和炎症反应
与其他类型的组胺受体相比,H4受体在中枢和周围神经系统中表达并不显著。H4R通过与蛋白质Gα/io相互作用介导,参与炎症和超敏反应的发展。
H4R介导的肥大细胞活化,促炎细胞因子和趋化因子IL-6、TNF-α、TGF-β1、RANTES、IL-8、MIP-1α和MCP-1均被表达。研究发现H4R激活显著增强结肠炎、放射性结肠炎、肠道缺血/再灌注损伤和过敏反应的炎症反应。
因此,选择性H4R阻滞剂的开发可能代表了治疗炎症性肠病的一种有前途的方法。
• H4R的激活还会导致瘙痒
H4R的激活已被证明会导致瘙痒。而H1R和H4R拮抗剂的联合治疗可能通过协同抑制瘙痒和皮肤炎症对慢性皮炎产生显著的治疗效果。
此外,H4R通过激活Th2细胞并产生 IL-31,可能引发过敏性皮炎的发生。H4R和H3R的激活增加了乙酰胆碱对肠道蠕动的影响。H4R受体还参与消化性溃疡的形成和癌变。
//小结
如前所述,组胺特异性受体的组织定位、功能和对组胺的亲和力不同。组胺调节广泛的代谢过程,具有促炎和抗炎作用,具体取决于受体亚型和受刺激细胞的类型。
Barcik W,et al.Curr Opin Immunol.2017
组胺对免疫调节的各种作用似乎与其受体及其各自的细胞内信号的差异表达和调节有关。此外,这些受体对组胺的亲和力的差异在组胺和组胺受体药物配体的生物效应中发挥着关键作用。组胺受体在多种病理生理状况中发挥重要作用,是治疗过敏的有效靶点。
组胺因其在速发型超敏反应中的作用而广为人知。在胃肠道内,组胺以相对较高的浓度存在,特别是在炎症过程中。组胺水平的增加改变了宿主与微生物群的免疫相互作用,并导致体内平衡的破坏,导致许多难以应对的肠道疾病的发展。
★ 组胺的水平会影响肠道稳态
肠道中的组胺水平受到宿主过敏和炎症反应的影响,除了宿主微生物群的产生外,还会以某种方式改变降解或合成组胺的酶的活性及其饮食摄入量。
此外,在刺激产生组胺的免疫细胞时,可以增加内源性组胺的水平。所有这些都会影响肠道稳态,导致组胺积累,并影响特定疾病。组胺还可能对肠道寄生虫和细菌感染产生一定影响。
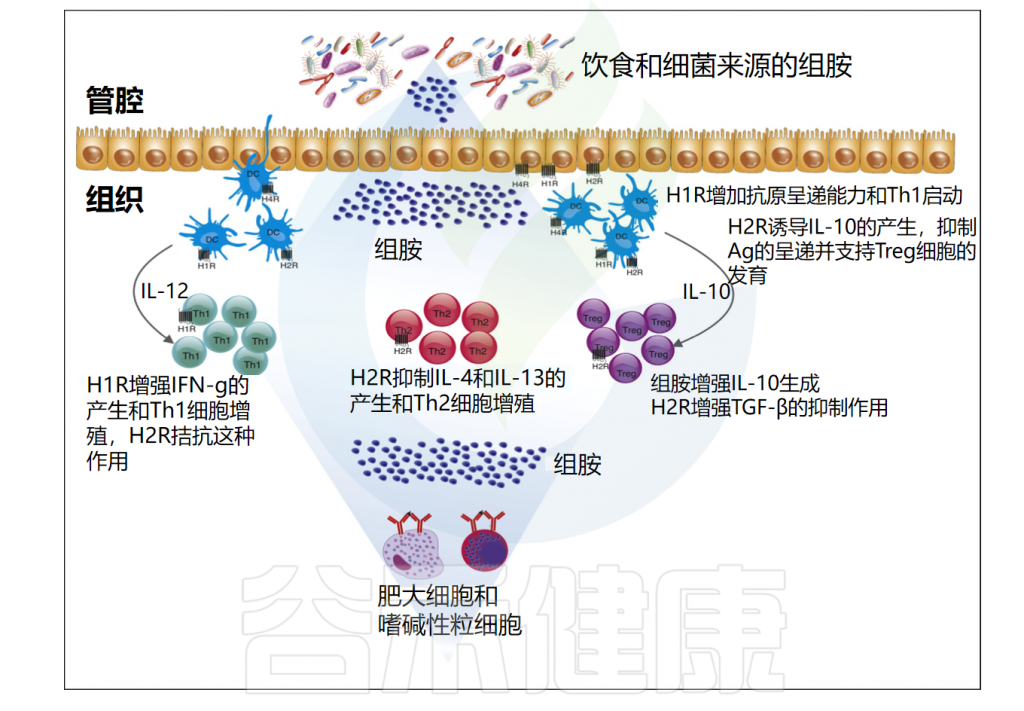
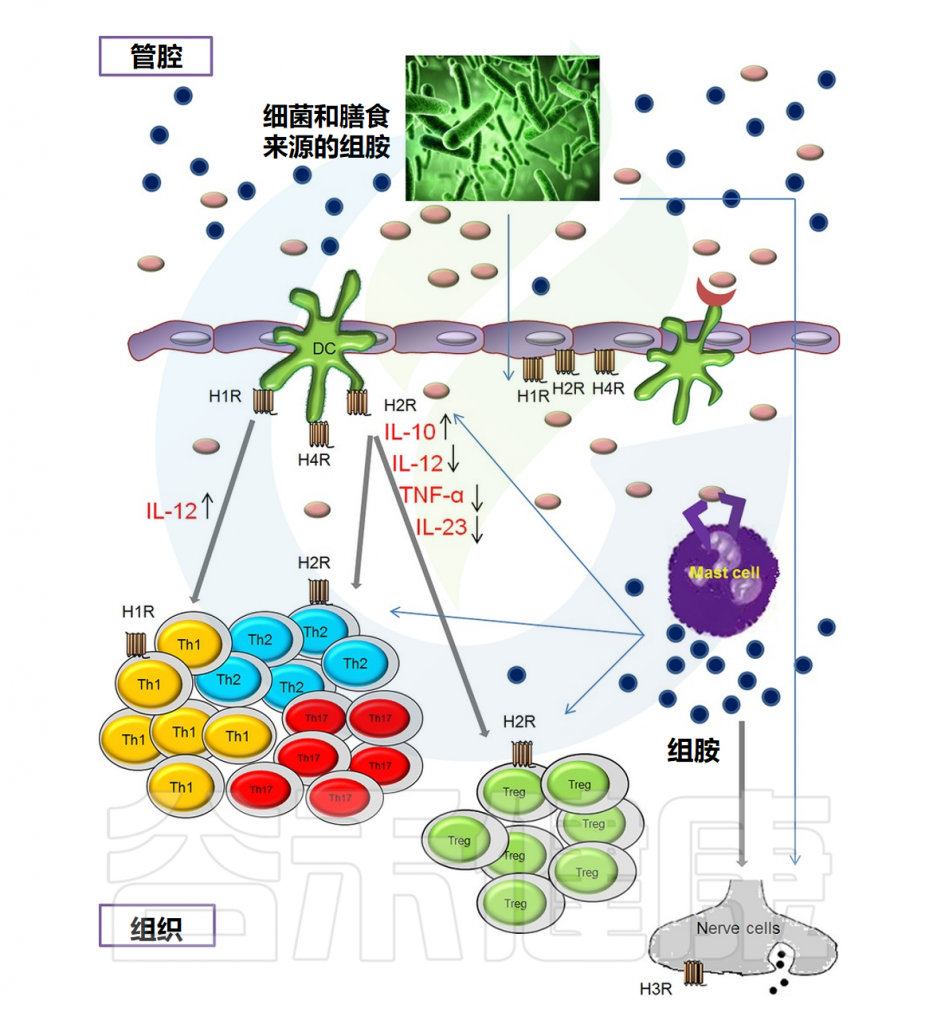
注:除H3R外,所有组胺受体都在人体肠道中表达。从量的角度来看,H4R表达的丰度明显低于H1R和H2R。
粘膜内的组胺
Smolinska S,et al.Allergy.2014
• 食物中的一些菌株会分泌组胺
除哺乳动物细胞外,还发现许多不同的细菌菌株能够在组氨酸通过组氨酸脱羧酶作用后分泌组胺。
最有力的证据就是那些能够在食品中分泌组胺的细菌。它们在奶酪、肉罐头、乳制品以及啤酒和葡萄酒发酵过程中产生组胺。鲭鱼中毒就是在储存或加工过程中处理不当,细菌代谢了鱼类中的组氨酸,导致组胺含量过高。
• 部分肠道微生物也会分泌组胺
迄今为止,对产生组胺细菌的研究主要集中在从食物样品中分离的菌株。肠道微生物群的组胺生成能力直到最近才被研究,数据仍然有限。
2021年发表的一项荟萃分析展示了人类肠道微生物组中具有假定组胺分泌能力的物种 ,其中许多属于广泛报道的组胺产生属,例如摩根菌属(Morganella),乳杆菌(Lactobacillus),葡萄球菌属(Staphylococcus), Photobacterium和巴氏梭菌(Clostridium)。
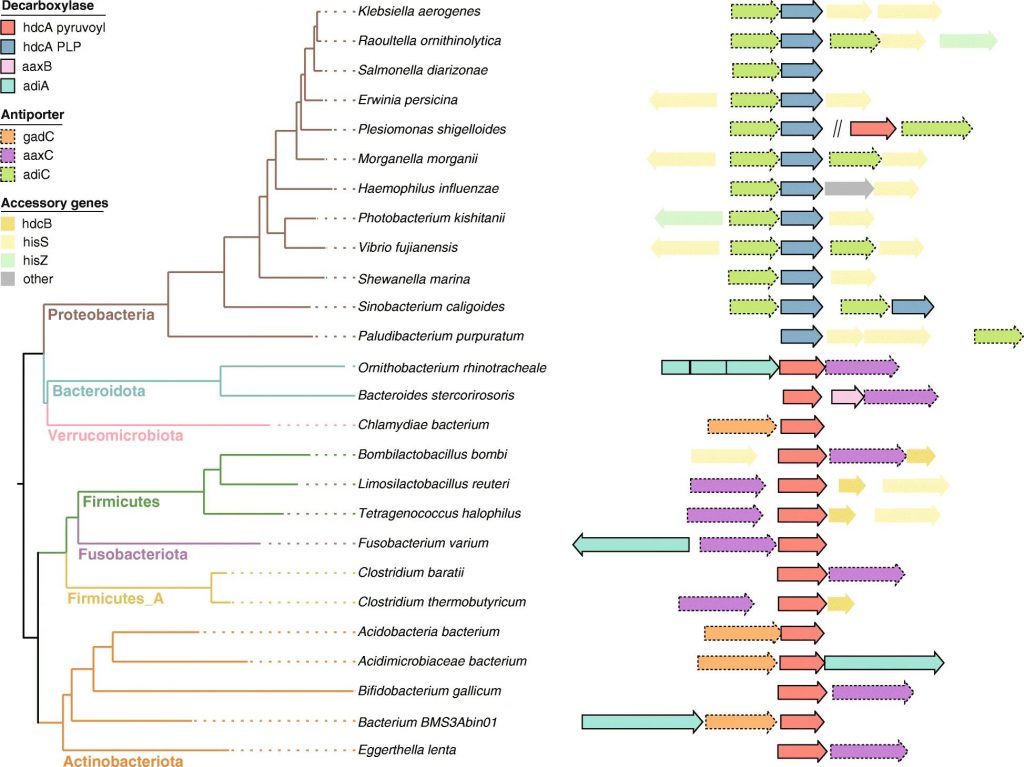
Mou Z,et al.BMC Genomics.2021
• 一些细菌具有组氨酸脱羧酶活性,也可能产生组胺
需要注意的是,一些具有组氨酸脱羧酶活性并与组胺中毒有关的细菌,可能也是潜在的组胺产生细菌。包括:Morganella morganii,
Eschericha coli,
Hafnia alvei,
Proteus vulgaris,
Proteus milabilis,
Enterobacter aerogenes,
Raoultella planticola,
Raoultella ornithinolytica,
Citrobacter freundii,
Pseudomonas fluorescens,
Photobacterium damselae。
证据表明细菌衍生的组胺具有多种影响,例如对宿主健康和食品安全。
• 抑制了促炎细胞因子的分泌
初步体外研究表明,组胺抑制人单核细胞衍生的树突状细胞中趋化因子和促炎细胞因子的分泌。
小鼠研究表明,给予分泌组胺的鼠李糖乳杆菌具有抗炎作用,各种白细胞介素和肿瘤坏死因子α的分泌减少。这种作用在缺乏组胺2受体的动物中消失,表明微生物群衍生的组胺可能具有免疫调节作用。
• 组胺的含量对于免疫效果至关重要
与鼠李糖乳杆菌相比,另一种乳杆菌能够分泌大约100倍多的组胺,服用这种乳杆菌会导致动物体重减轻和健康状况恶化。谷禾认为,微生物分泌的组胺量可能是决定其作用性质的关键。
最近的研究还表明,肠道中细菌释放的组胺不仅会引起宿主免疫系统的局部调节,而且还会在远处的粘膜部位(例如肺部)产生免疫学后果。例如成年哮喘患者肠道中分泌组胺的细菌丰度有所增加。
由于许多食物中含有组胺,一部分肠道微生物群也可以产生组胺,研究发现组胺在肠道疾病中可能发挥多种作用,下面讲述了一些与组胺相关的肠道疾病,组胺在其中发挥着有益或有害的作用。
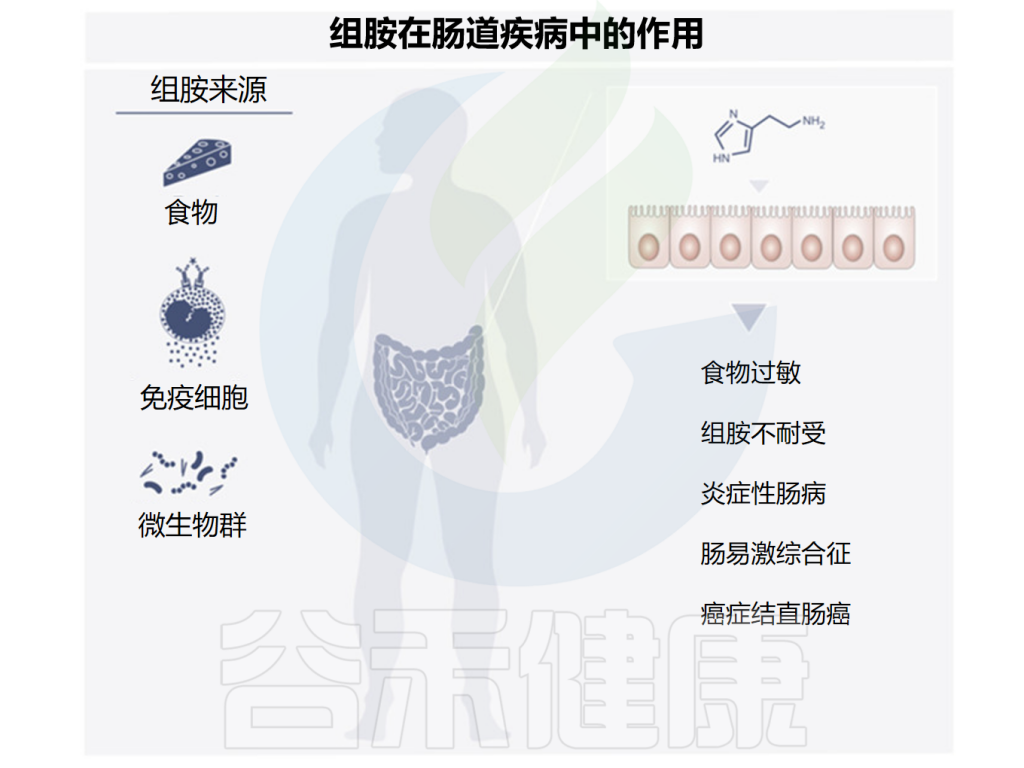
Smolinska S,et al.Metabolites.2022
消化道是接触大量不同分子的地方,这些分子中的一部分是潜在的过敏原,可能从而引起食物过敏。食物过敏可表现为轻度和重度症状,最严重的可能危及生命。
拓展:常见的食物过敏症状
食物过敏的症状表现在呼吸系统、消化系统、心血管系统和皮肤系统等。
皮肤反应:皮肤反应是最常见的食物过敏症状之一。它可以表现为皮疹、荨麻疹、瘙痒、红肿、水肿等。这些反应通常发生在口腔、面部、颈部和四肢等部位。
呼吸道症状:食物过敏还可以引起呼吸道症状,包括鼻塞、流涕、打喷嚏、喉咙痒、咳嗽、喉咙紧闭感和哮喘。严重的过敏反应可能导致呼吸困难和哮喘发作。
消化系统症状:食物过敏可以引起消化系统症状,包括腹痛、腹胀、恶心、呕吐和腹泻。这些症状还可能与肠道炎症反应、肠道收缩有关。
循环系统症状:在严重的食物过敏反应中,可能出现循环系统症状,如低血压、心悸、头晕和昏厥。这些症状是由于过敏反应导致血管扩张和血压下降。
全身症状:某些人可能出现全身性症状,如疲劳、焦虑、情绪波动和体重下降。这些症状可能与食物过敏引起的炎症反应和免疫系统的激活有关。
• 食物过敏时组胺分泌增加
IgE依赖性食物过敏是免疫系统紊乱的结果,使耐受性丧失。这导致将温和的食物抗原识别为病原体。组胺受体参与食物过敏的发展。研究表明,在人类中使用H2R拮抗剂会增加针对食物抗原的IgE的产生。在Th2细胞的刺激下,IgE与效应细胞(肥大细胞和嗜碱性粒细胞)上的Fcε受体结合。由于效应细胞的激活,组胺以及其他介质被释放。
在食物过敏受试者中,组胺分泌增加和肥大细胞数量增加已得到充分证明。嗜碱性粒细胞释放的组胺与皮肤点刺试验和食物激发呈正相关。与非过敏患者相比,食物过敏患者抗IgE介导的肥大细胞组胺释放增加。
此外,将食物过敏患者的活检与抗IgE(人类)抗体或过敏原一起孵育,会导致组胺释放增加9倍。
• 食物过敏的治疗
由于食物过敏的发生率不断增加,已经进行了大量研究以开发新的治疗和预防策略。严格避免食物过敏原是管理IgE介导的食物过敏的长期策略。
关于食物过敏发病机制的各个阶段也有许多研究,例如对Th2通路的影响、阻断IgE、抑制效应细胞和微生物治疗。长期免疫耐受应该是治疗食物过敏最理想的效果。
口服免疫疗法是食物过敏的发展治疗方法之一。它包括以每2-4周增加一次的剂量向患者施用过敏原,直到达到最大维持剂量。这个过程的结果是发展对食物的耐受性。该方法已用于对牛奶、鸡蛋、小麦、花生、坚果和贝类的食物过敏。
目前正在研究表皮和舌下免疫疗法。对牛奶和鸡蛋过敏的表皮免疫疗法也进行了临床试验。
炎症性肠病是特发性、慢性复发性肠道疾病。溃疡性结肠炎和克罗恩病是两种主要临床表现。
在克罗恩病中,炎症出现在弥漫性病变中,这些病变遍布消化道并深入穿透肠壁,可能影响所有层。相反,溃疡性结肠炎的炎性病变从直肠开始,向上但不超过结肠,并在黏膜浅表处保持。克罗恩病导致整个胃肠道的透壁性炎症,但其特征是模式不连续。与炎症为浅表的溃疡性结肠炎相比,溃疡主要局限于结肠粘膜。
• 炎症性肠病患者黏膜组胺水平升高
这两种疾病的发展都是遗传易感患者宿主免疫系统、肠道微生物群和环境因素之间复杂相互作用的结果。
炎症性肠病患者的黏膜组胺水平(而非血浆水平)升高。在患者尿液中发现与疾病活动度相关的N-甲基组胺水平升高。当受到抗原刺激时,源自活动性克罗恩病或溃疡性结肠炎切除结肠的肥大细胞能够释放比正常结肠更多的组胺。
• 炎症性肠病患者的组胺通路被破坏
克罗恩病和溃疡性结肠炎患者的组胺信号通路均被破坏,研究分析显示。炎症性肠病患者的组胺受体表达和功能活性发生改变。
在结肠炎模型中,H2R的阻断导致更严重的炎症性疾病。组胺主要抑制IFN-γ和TNF-α的分泌,这些细胞因子的基因表达与溃疡性结肠炎患者的H4R和H2R表达呈正相关。
发炎黏膜中组胺-N-甲基转移酶(HNMT)表达降低,二氨基氧化酶(DAO)多态性与炎症性肠病风险增加有关。使用H2R拮抗剂会增加克罗恩病患者的风险。
注:在小鼠结肠炎模型中,缺乏H2R或使用非莫替丁(H2R拮抗剂)抑制H2R的T细胞转移可加速体重减轻并增加疾病严重程度。
• 炎症性肠病的治疗
在许多情况下,获得缓解的唯一选择是根治性手术,切除发炎区域。潜在地,使用H1R和H4R拮抗剂同时使用H2R激动剂可能对炎症性肠病患者有益。
炎症性肠病患者还可接受抗炎药、类固醇、抗生素、氨基水杨酸盐治疗,或使用英夫利昔单抗(抗 TNF-α)进行生物疗法。
肠易激综合征是一种与腹部不适或疼痛有关的慢性疾病,其中吃的食物是更严重症状的触发因素。一些证据表明,肠易激综合征中肠道菌群发生了改变,这种改变会破坏黏膜的免疫应答。
• 内源性组胺与肠易激综合征的严重程度相关
研究显示,超过一半的患者因食用释放组胺的食物或富含生物胺的食物而出现胃肠道症状。内源性组胺的水平与肠易激综合征患者症状的严重程度绝对相关。活化的肥大细胞产生更多的组胺,这与肠易激综合征患者的腹痛相关。肠易激综合征患者的黏膜上清液的组胺含量高于健康受试者的上清液。
组胺水平更高,组胺受体表达也更高
肠易激综合征患者中组胺受体H1R和H2R的表达水平上调。我们推测:伴有组胺分泌或含组氨酸脱羧酶细菌的菌群失调可能与肠易激综合征的发生和加重有关。
给予特定微生物具有治疗作用,这也可能是微生物群变化导致疾病的原因。肠易激综合征没有特效治疗方法,减少炎症的药物正在使用中。
结直肠癌是第三常见癌症,也是癌症相关死亡的重要原因。与一般人群相比,炎症性肠病患者发生结直肠癌的终生风险会增加。
• 组胺可能具有潜在的抗肿瘤作用
肠道微生物群在结直肠癌中的作用已在谷禾之前的文章中详细描述过。而特定的肠道微生物代谢物可能与结直肠癌的病因有关。组氨酸脱羧酶缺乏已被证明通过CD11b+Gr-1+未成熟髓系细胞的积累促进炎症相关的结直肠癌,表明组胺具有潜在的抗肿瘤作用。
几种益生菌菌株,包括长双歧杆菌(Bifidobacterium longum)、嗜酸乳杆菌(Lactobacillus acidophilus NCFM)和鼠李糖乳杆菌GG,在结肠癌的小鼠模型中显示出有益作用。这些产生组胺的益生菌减少了结肠肿瘤的数量和大小。
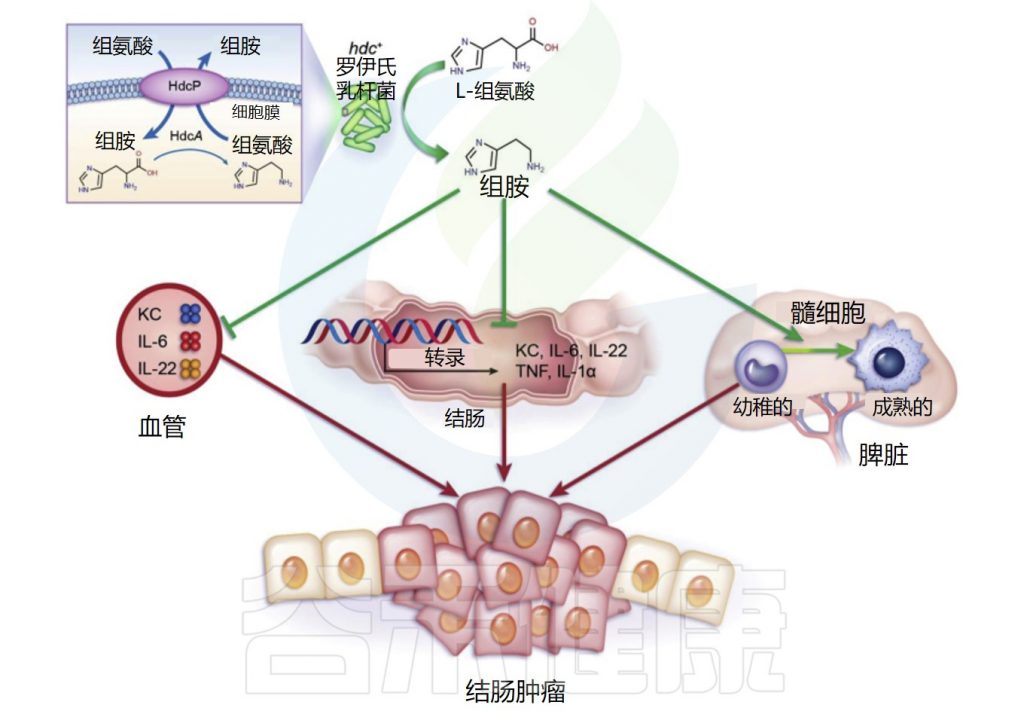
Gao C,et al.Am J Pathol.2017
罗伊氏乳杆菌(L. reuteri)抑制结肠粘膜中IL-22、IL-6、tnf-α和IL-1α 基因的表达,并减少血浆中促炎、癌症相关细胞因子、角质形成细胞趋化剂、IL-22和IL-6的数量。
此外,不能产生组胺的等基因组氨酸脱羧酶(HDC)缺陷罗伊氏乳杆菌突变体不能抑制致癌作用,这表明共代谢物组胺在抑制慢性肠道炎症和结直肠肿瘤发生中具有重要作用。
• 结直肠癌患者组胺含量和活性增加
在结直肠癌患者的结肠黏膜中,组氨酸脱羧酶活性和组胺含量均高于正常样本。在实验诱导的结直肠癌小鼠中,与野生型小鼠相比,组氨酸脱羧酶的缺失导致肿瘤发生增强,这也表明组胺具有抑制结直肠肿瘤发生的作用。
• 注意:过高的组胺含量可能对癌症不利!
需要注意的是,已发现肥大细胞在结肠癌中含量丰富,并促进小鼠化学诱导的结直肠癌的癌变,并且与人类结直肠癌患者的不良预后有关。与组胺通过H4R的促炎作用类似,H4R表达的缺失也导致小鼠化学诱导的致癌作用减少。
组胺作为一种重要信号分子,主要因其在诱导过敏症状中的作用而被识别,但它也参与非过敏性炎症反应。
组胺是肠道稳态的重要调节剂。通过增强分泌和蠕动,组胺促进潜在有毒物质的快速清除。这种生物胺对于肠道细菌的生存也至关重要。与健康个体相比,组胺不耐受患者的肠道菌群失衡或失调。
在某些浓度范围内,组胺起着保护作用,对于维持健康状态至关重要。然而,由于先天性或后天性缺乏组胺降解酶,以及肠道肥大细胞过度增生和过度活跃,使组胺浓度较高,会导致粘膜炎症性疾病,产生腹泻和腹痛。增强H2R表达或其细胞内信号,同时降低H1R和 H4R活性是提高粘膜免疫力的合理方法
组胺不耐受的现代治疗方法包括避免高组胺食物的摄入、影响组胺水平的药物(H1R阻滞剂、肥大细胞膜稳定剂和米氮平)和补充天然成分(维生素C、类黄酮、二氨基氧化酶强化补充剂和益生菌) 。
与此同时,使用具有释放组胺潜力的肠道微生物群提供了一种新的治疗视角。
主要参考文献
Smolinska S, Winiarska E, Globinska A, Jutel M. Histamine: A Mediator of Intestinal Disorders-A Review. Metabolites. 2022 Sep 23;12(10):895.
Sánchez-Pérez S, Comas-Basté O, Duelo A, Veciana-Nogués MT, Berlanga M, Latorre-Moratalla ML, Vidal-Carou MC. Intestinal Dysbiosis in Patients with Histamine Intolerance. Nutrients. 2022 Apr 23;14(9):1774.
Shulpekova YO, Nechaev VM, Popova IR, Deeva TA, Kopylov AT, Malsagova KA, Kaysheva AL, Ivashkin VT. Food Intolerance: The Role of Histamine. Nutrients. 2021 Sep 15;13(9):3207.
Krell T, Gavira JA, Velando F, Fernández M, Roca A, Monteagudo-Cascales E, Matilla MA. Histamine: A Bacterial Signal Molecule. Int J Mol Sci. 2021 Jun 12;22(12):6312.
Mou Z, Yang Y, Hall AB, Jiang X. The taxonomic distribution of histamine-secreting bacteria in the human gut microbiome. BMC Genomics. 2021 Sep 26;22(1):695.
Gao C, Ganesh BP, Shi Z, Shah RR, Fultz R, Major A, Venable S, Lugo M, Hoch K, Chen X, Haag A, Wang TC, Versalovic J. Gut Microbe-Mediated Suppression of Inflammation-Associated Colon Carcinogenesis by Luminal Histamine Production. Am J Pathol. 2017 Oct;187(10):2323-2336.
Kovacova-Hanuskova E, Buday T, Gavliakova S, Plevkova J. Histamine, histamine intoxication and intolerance. Allergol Immunopathol (Madr). 2015 Sep-Oct;43(5):498-506.
Barcik W, Wawrzyniak M, Akdis CA, O’Mahony L. Immune regulation by histamine and histamine-secreting bacteria. Curr Opin Immunol. 2017 Oct;48:108-113.
Smolinska S, Jutel M, Crameri R, O’Mahony L. Histamine and gut mucosal immune regulation. Allergy. 2014 Mar;69(3):273-81.