-

 国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子
国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子

 二级病原微生物安全实验室
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业

谷禾健康
阴道是一个复杂的动态环境,会随着女性生命阶段的变化而不断调整,对于青春期、育龄期(包括未怀孕、产前和产后)及更年期的健康至关重要。
阴道微生态具有独特的特征,如糖原和黏蛋白含量高、pH值低、细胞免疫活跃,以及影响复杂微生物群的激素信号波动。例如,雌激素影响阴道上皮的屏障功能和糖原可用性,而糖原分解在塑造阴道微生物群中发挥关键作用。同时,激素调节粘液分泌,增强对病原体的抵抗力,而黏蛋白又为阴道微生物提供附着基质和营养来源。
尽管传统上认为阴道微生物群主要由乳杆菌主导的五种典型群落状态(CST I–V)构成,但新兴研究突出了微生物群的多样性,包括共生微生物和潜在致病微生物,同时揭示了阴道微生物群的时空动态。最近提出的27种宏基因组CST(mgCST)为潜在群落功能提供了关键线索,使研究重点从“有哪些微生物”转向“它们在做什么”。
本文采用生态系统视角,探讨阴道黏膜、微生物群、宿主及环境因素之间的多维关系。重点分析激素如何影响阴道微生物群及环境变化,强调月经周期中微生物群落的动态特征,以及阴道环境变化对定植抵抗力的影响。不同的阴道生态失调状态与乳杆菌丰度及微生物多样性变化密切相关。
此外,我们还将研究阴道中微生物与免疫细胞的相互作用,探讨免疫系统如何调节局部环境。通过宿主与驻留微生物之间的反馈循环,提升对阴道健康与疾病的理解。这些复杂相互作用的理解对于推动女性阴道健康至关重要。
众所周知,生殖激素信号传导可以驱动生理和环境变化,从而在塑造阴道微生物组方面发挥关键作用。
女性一生中阴道微生物群组成的变化
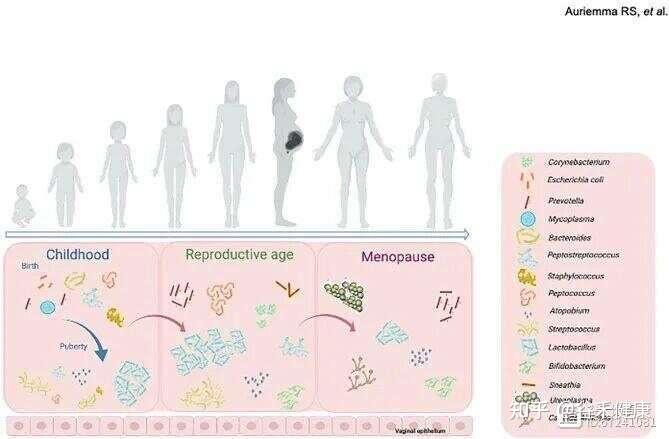
月经周期包括月经期、卵泡期(增殖期)、排卵期和黄体期,其特征是雌激素、孕激素、黄体生成素及促卵泡激素的波动,这些激素共同调控子宫内膜增厚及卵子释放。
阴道环境对月经周期及整个生殖生理过程中的荷尔蒙变化高度敏感,激素水平的波动会影响泌尿生殖系统对感染的抵抗力或易感性。宿主激素与微生物的相互作用构成了“微生物内分泌学”的研究基础,用以揭示激素对细菌生长与毒力的影响。本节将探讨激素分泌背景下阴道黏膜的变化,并分析其对营养供给、屏障功能及微生物群落结构的影响。
月经影响营养供应和阴道微生态稳定
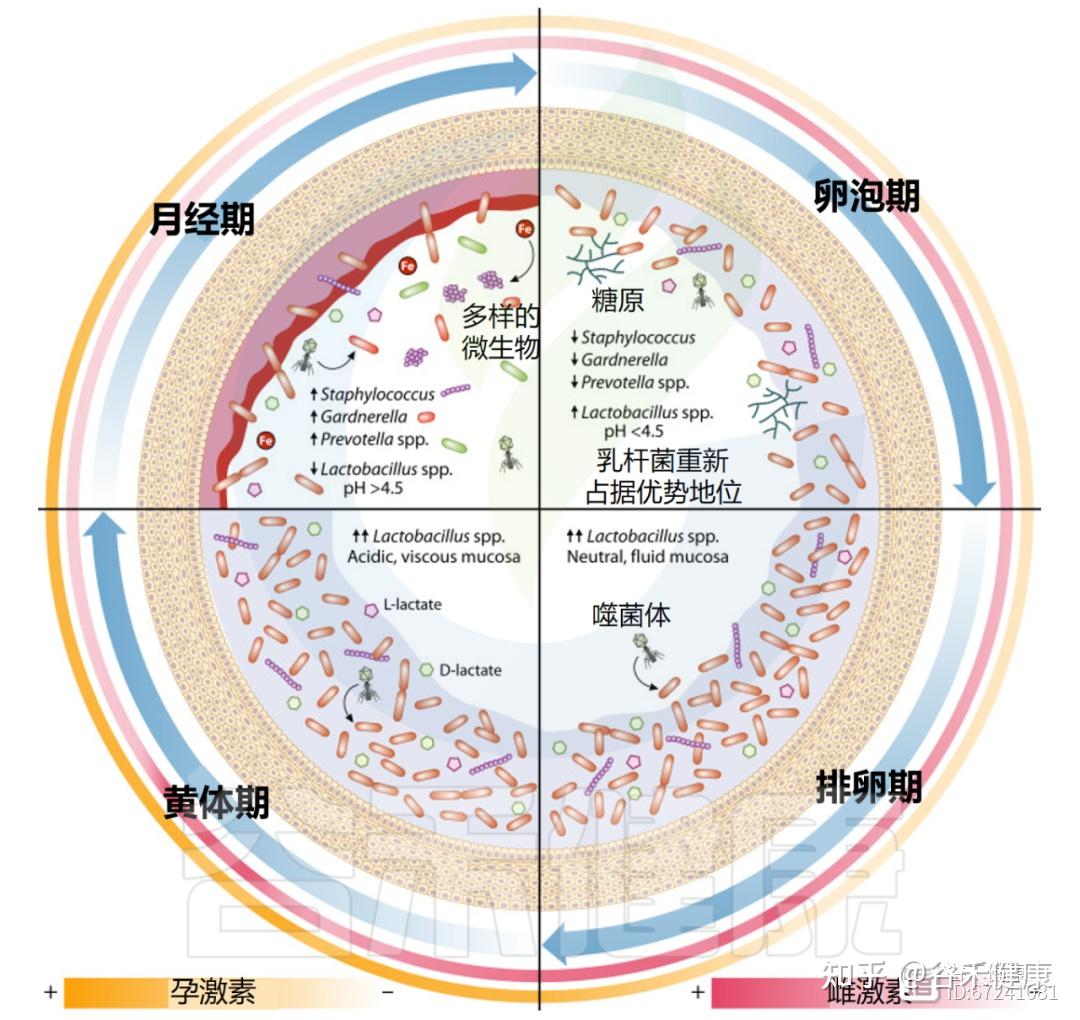
月经期间,阴道微生物群发生显著变化,细菌多样性和丰富度增加。虽然这些变化源于月经期的生理波动,但个体差异明显。
据此,研究提出了四种阴道群落动力学(VCD),用于描述月经周期中微生物群的时间变化。VCD基于“稳定共生”与“持续失调”的特征,其中超过80%的日常样本分别由卷曲乳杆菌(Lactobacillus crispatus)、詹氏乳杆菌(Lactobacillus jensenii)、惰性乳杆菌(Lactobacillus iners)或高多样性群落占主导。低于80%生态阈值的个体则归为“不稳定”群落亚组。
另有约12%个体形成“月经相关失调”亚组,其益生性群落在增殖期、排卵期和黄体期占主导,而月经期间出现可检测的生态失调。
◮ 月经期乳杆菌减少,微生物稳定性较差
其他研究亦观察到月经期乳杆菌减少、菌群多样性上升。在一项为期16周、涉及32名育龄女性的纵向研究中,月经被认为与最低的微生物群稳定性相关,部分个体出现由卷曲乳杆菌主导向惰性乳杆菌或链球菌主导转变的现象。
这些变化可能源于糖原沉积减少及经血流入的影响。血液中的血红素铁可促进L.iners生长,从而取代L.crispatus优势;与此同时,碱性血液流入使阴道pH升至7.2–7.4,进一步改变微生物生态。这种环境转变促使金黄色葡萄球菌、链球菌等机会致病菌的短暂定植,以及细菌性阴道病(BV)相关菌如加德纳菌、普雷沃氏菌、Fannyhessea vaginae、Sneathia amnii、微小脲原体、Veillonella montpellierensis与消化链球菌等的增长。
目前尚无证据表明微生物多样性的变化仅由单一因素——如铁、糖原或pH变化——直接驱动。更可能的是,激素波动、营养变化及血流剪切力共同作用,塑造了月经期阴道微生态的动态特征。
注:血流剪切力是指血液流动时对血管内皮细胞产生的摩擦力,是影响血管功能和结构的重要机械力。
月经激素信号影响阴道微生态
增殖期:影响上皮屏障与糖原可用性
月经期普雷沃氏菌和加德纳菌的增多可延续至增殖期初,此时雌激素水平仍较低;而在卵泡晚期至排卵前,雌激素达到峰值,与乳杆菌重新占优势相吻合。
◮ 雌激素影响阴道上皮屏障与糖原可用性
雌激素的作用超越月经周期,在青春期其分泌上升,促使阴道上皮成熟增厚,形成由富含糖原的多层细胞构成的屏障。顶层细胞周期性脱落,释放糖原至阴道腔内。卵泡晚期乳杆菌优势的恢复与糖原可用性增加密切相关,并在整个黄体期保持稳定。
雌激素和乳杆菌属
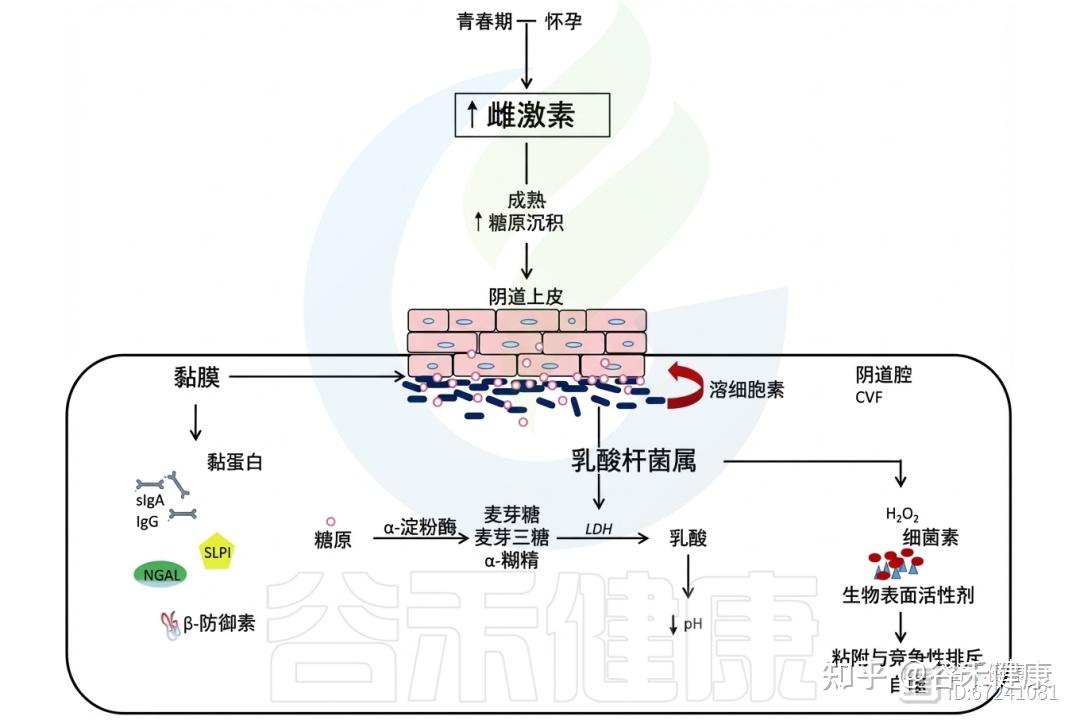
糖原是一种由α−1,4和α−1,6糖苷键组成的复杂分子,其在阴道中的浓度随月经周期波动且个体差异明显。有学者提出阴道乳杆菌可能将糖原作为碳源。然而,由于糖原结构复杂,微生物无法直接吸收,需经细胞外酶分解为糊精、麦芽糖、麦芽三糖等可利用的小分子糖,形成供微生物共享的“公共资源”。
阴道液中含有宿主和微生物来源的糖原水解酶,如α-淀粉酶和支链淀粉酶,分别裂解α−1,4和α−1,6糖苷键,其中宿主α-淀粉酶在阴道腔与宫颈内膜中含量丰富。
◮ α-淀粉酶有助于维持乳杆菌主导的环境
糖原的微生物利用最初被认为主要由保护性的乳杆菌完成,多项研究发现糖原水平、乳杆菌丰度及低阴道pH值之间存在正相关。但并非所有乳杆菌株都能利用糖原。
最新研究表明,各类阴道群落状态(CST)样本中均存在编码糖原降解酶的细菌基因,显示糖原可作为共生菌和病原菌在定植或感染期间的能量来源。研究发现,乳杆菌丰富的阴道样本中α-淀粉酶水平较高,而在乳杆菌减少或患有细菌性阴道病(BV)的个体中则显著降低,提示α-淀粉酶的产生有助于维持乳杆菌主导的环境。
相反,另一研究发现,当L.iners取代L.crispatus成为主要菌时,孕早期阴道分泌物中α-淀粉酶水平提高约5.4倍,可能反映宿主为恢复L. crispatus优势和稳定微生物群所作的代偿性反应。
◮ 糖原分解在塑造阴道微生物群中发挥关键作用
除乳杆菌外,阴道中的其他共生或致病微生物,如加德纳菌、无乳链球菌、白色念珠菌和阴道毛滴虫,也能表达糖原降解酶,这表明糖原分解对多种菌群的代谢至关重要。
值得注意的是,不同微生物的糖原酶在不同pH条件下活性最强:部分在低pH下最优,适应乳杆菌占优势的环境;而另一些则在pH 6–7的较中性环境中活跃。这意味着不同菌种可在月经周期或微生态失调等条件变化时,灵活利用糖原。
糖原降解酶的分泌还促进了“营养交叉喂养”,使无法直接分解糖原的菌种(如粪肠球菌)能利用分解产物生存。
尽管糖原可用性显然在塑造阴道微生物群结构与功能中发挥了关键作用,但其具体机制仍存在重要知识空白。进一步研究糖原供给与群落动态的因果关系,以及探明糖原是促进乳杆菌生长的驱动力,还是乳杆菌优势反过来维持糖原丰富的上皮屏障,将有助于全面理解糖原代谢对阴道健康的深远影响。
排卵期和黄体期:影响阴道粘膜屏障
排卵发生在增殖期雌激素激增之后,标志着黄体期的开始,其特点是黄体酮升高和雌激素的低次级峰值。
此阶段的激素变化显著影响阴道黏膜屏障的物理特性。该屏障由富含无机离子、防御素和免疫球蛋白的黏蛋白糖蛋白水凝胶组成,统称宫颈阴道黏液(CVM)。黏蛋白分为表面锚定型和分泌型,两者均高度O‑糖基化,形成带负电的大分子屏障。阴道及宫颈外上皮细胞主要表达MUC1和MUC16,而宫颈内皮与杯状细胞分泌MUC5B、MUC5AC及MUC6。
尽管MUC4在宫颈内高表达,近期蛋白质组学研究仅检测到MUC1和MUC16,未发现MUC4肽,其在CVM中的作用仍不明确。
阴道微生物组对女性性激素的反应
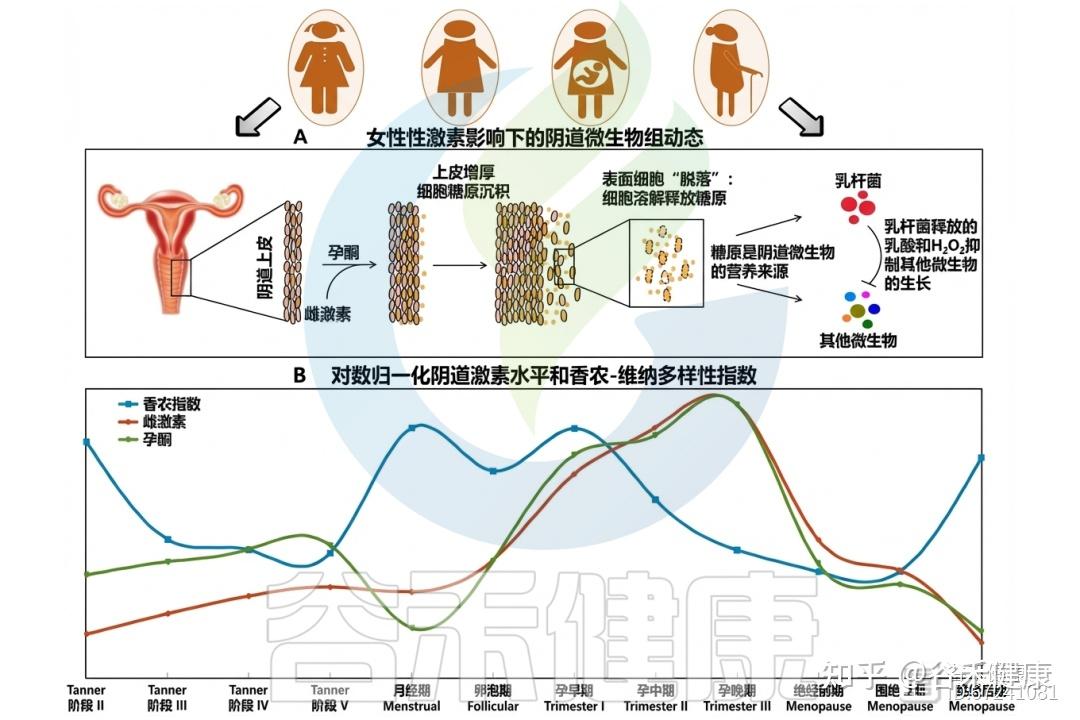
◮ 激素影响粘液的分泌量调节抵抗病原体能力
在排卵期,雌激素促使MUC5B分泌增加,使黏液稀薄、水样且pH更中性,有利于精子穿透。至黄体期,黄体酮上升使黏液减少,质地变黏稠、带负电,形成“黄体酮效应”,强化屏障功能,防御病原体入侵。CVM中含IgG,可抑制HIV‑1运动,其中以L.crispatus为主的群落较L. iners或失调型更具抗病毒力。
此外,噬菌体被发现可附着于黏膜表面,进一步阻止病原体定植。妊娠期,MUC5B与MUC5AC形成宫颈黏液栓,将富菌阴道环境与无菌子宫隔离,并富含抗菌肽(乳铁蛋白、溶菌酶、钙卫蛋白)、免疫球蛋白及具防御性的黏弹性结构。在小鼠模型中,缺失Muc5B会削弱屏障功能,增加感染与早产风险。
◮ 黏蛋白为阴道微生物提供附着基质和营养来源
黏液除了充当感染屏障外,还为阴道微生物提供附着基质和营养来源。无乳链球菌(S.agalactiae)和粪肠球菌(E.faecalis)通过表达可直接结合黏蛋白的菌毛结构,增强在阴道及肠道的定植能力。
同样,许多葡萄球菌和乳杆菌也能结合黏液,乳酸菌(LAB)中特有的黏蛋白结合结构域被认为参与黏液的附着或降解。感染后,黏蛋白及杯状细胞分泌通常上调,依赖NF‑κB信号通路,是维持黏膜屏障的先天免疫反应。
然而,部分微生物在长期适应过程中进化出可降解黏蛋白糖链、用于营养获取的糖苷酶。在阴道环境中,此类酶(如葡萄糖苷酶)的活性主要见于细菌性阴道病相关病原体,它们会破坏黏膜屏障,降低其黏度并增加感染风险。
◮ 细菌性阴道炎或菌群失调会影响酶活性
研究一致发现细菌性阴道炎女性中糖苷酶和唾液酸酶活性升高,而宏转录组分析显示CST IV群落患者的唾液酸酶表达最强。这种与细菌性阴道炎或菌群失调相关的酶活性增加,会导致N和O连接糖链耗竭,削弱黏膜屏障完整性并加剧炎症。
唾液酸酶主要由Gardnerella和Prevotella属产生,但近期研究发现其转录本在所有阴道CST类型中均较高,提示其作用可能超越病原入侵,参与群落代谢调控。支持这一观点的证据包括唾液酸酶促进微生物群共生与营养交互,如梭杆菌的唾液酸酶产物促进G. vaginalis生长与阴道定植。
类似的黏蛋白分解产物交叉喂养机制在肠道中更为常见,如Akkermansia muciniphila以黏液为主要营养源,分泌多种唾液酸酶和岩藻糖苷酶,助长与产丁酸梭菌的共生;双歧杆菌产生的唾液酸酶同样推动其与其他肠道双歧杆菌的共代谢。
阴道中,A.muciniphila与S.agalactiae的共栖可增强后者的持久性,代谢模型显示两者存在潜在的营养交换,但这种互作究竟源于免疫调节、物种交流或代谢互馈仍需进一步研究。
产后与绝经激素波动影响微生物群
◮ 产后和绝经后雌激素变化影响阴道微生物群结构
产后和绝经后阶段虽是关键的生理时期,却仍是生殖健康研究的薄弱环节。产后数天内雌激素水平急剧下降,常伴随出血、子宫收缩和疲劳等症状。更年期通常发生于45岁后,表现为雌激素分泌减少、月经不规律、阴道干燥及感染易感性增加。
这两个阶段雌激素下降均会削弱乳杆菌的优势。产后阴道微生物群常向多样化状态转变,L. iners、F. vaginae、G. vaginalis、Finegoldia magna和Prevotella spp. 等菌种显著富集,部分个体在分娩后一年的微生物组仍保持高多样性。同样,在绝经后,雌激素与黄体酮减少导致乳杆菌丰度下降,而无乳链球菌、表皮葡萄球菌及丙酮棒状杆菌等细菌性阴道病(BV)相关菌增多。
激素替代疗法是绝经后常用的治疗手段,可促使上皮成熟、提高碳水化合物供应以支持天然微生物群,并降低微生物多样性。然而,仍然需要更多研究扩大我们对这些人群阴道健康的理解。
阴道菌群CST分型最初源于对来自育龄女性进行16S rRNA测序的分析。结果确定了五种典型群落状态(CST I–V):CST I以L.crispatus为主,II、III和V分别以L. gasseri、L.iners和L.jensenii为主;CST IV则呈高度多样性,以Prevotella、Dialister、Fannyhessea、Gardnerella、Megasphaera、Peptoniphilus、Sneathia、Eggerthella、Aerococcus、Finegoldia和Mobiluncus等厌氧菌为特征。

乳杆菌优势群落通常具保护作用,惟CST III(以L.iners为主)被认为是一种过渡状态,反映阴道微生物群在健康、亚健康及细菌性阴道病(BV)状态间的动态变化。
阴道微生物群落在不断变化
随着新技术的应用与研究视角的拓展,CST分类体系不断完善。VALENCIA工具通过样本相似性对群落进行分层,将原有五类CST扩展为多级结构,提升分辨率,涵盖高、低丰度的特征物种及复杂混合群落的细节。
另一项研究采用混合成员主题模型分析群落结构,结合纵向变化探讨妊娠影响并识别亚群落。基于孕期与非孕期的大规模阴道拭子样本,共确定九种亚群落,其中四种以乳杆菌(L. crispatus、L. jensenii、L. iners、L. gasseri)为主,其余五种为非乳杆菌群落,富含链球菌、Prevotella、Gardnerella、Corynebacterium、Fannyhessea vaginae和Finegoldia等。
◮ 阴道菌群分型有助于动态平衡与功能多样性
宏基因组学和宏转录组学的进展促进了物种水平的功能解析,更深入揭示了阴道微生物群的时空动态。最新提出的27种宏基因组CST(mgCST)为潜在群落功能提供了关键线索,使研究焦点从“有哪些微生物”转向“它们在做什么”。
尽管CST方法在特定时点揭示了群落结构特征,但其静态视角限制了对动态变化和多样性的理解。为此,提出基于CST并结合纵向采样的VCD系统,以反映月经周期中细菌与噬菌体群落的协同变化。研究发现,“持续稳态”多与CST I和V(>80%检出率)相关,而“持续失调”则常见于CST III和IV。
阴道微生物组与健康及疾病状态关系
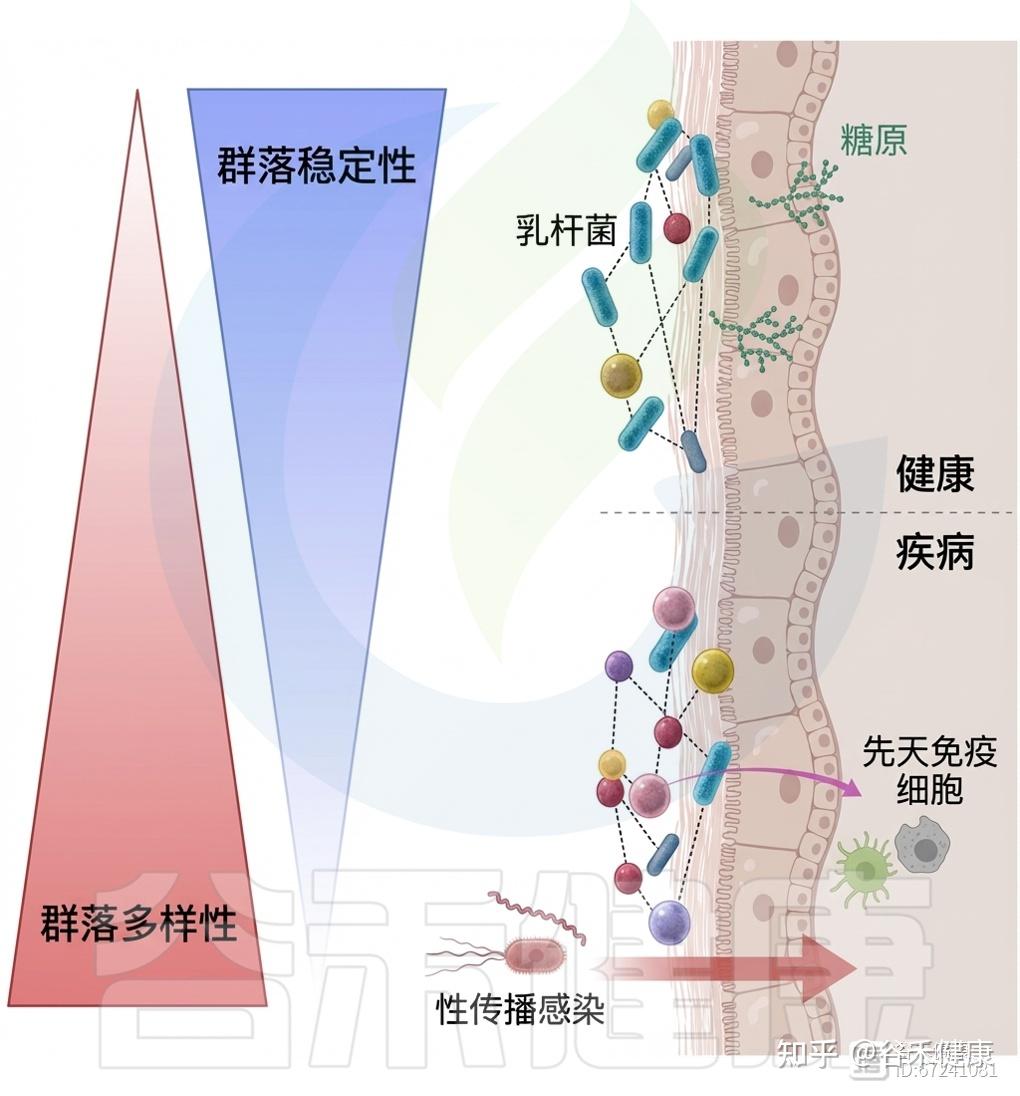
VCD分析进一步显示,不稳定或月经期失调状态下大肠杆菌丰度为稳态群落的两倍,且S. agalactiae、L. iners和U. parvum在这些状态中更为丰富。噬菌体鉴定显示,“稳态”个体的噬菌体丰度较其他类型高出约10倍,尽管群落多样性较低。这些研究为理解阴道微生态的动态平衡与功能多样性提供了新视角。
定植抗性:乳杆菌的保护作用
阴道低pH主要源于宿主与微生物共同将葡萄糖和糖原发酵为乳酸,从而使环境酸化。除上皮细胞产生的L-乳酸外,研究表明D-乳酸占阴道乳酸的大部分。
◮ 乳杆菌降低pH并增强黏膜屏障,形成保护环境
乳酸是乳酸菌(LAB)糖发酵的主要代谢产物,其中L. crispatus、L. gasseri和L. jensenii可产生高水平D-乳酸,而L. iners因缺乏D-乳酸脱氢酶而不具此能力。
微生物来源的乳酸通过降低pH并增强黏膜屏障,形成有利于乳杆菌定植、抑制其他细菌生长的保护性环境,是定殖抵抗的重要机制之一。然而,有研究发现,当pH升高时,乳酸对致病性厌氧菌不具显著抑制作用,提示乳酸产量与环境酸化对维持阴道定殖抵抗至关重要。
◮ 阴道pH变化可能是微生物群落失调的重要指标
阴道pH变化被视为细菌群落失调的重要指标之一,pH升高(>4.5)常与加德纳菌、普雷沃菌属和Sneathia等厌氧菌的增殖相关。能降解阴道带负电黏膜屏障的细菌可能削弱黏液层,从而导致pH升高。
此外,月经周期同样影响阴道pH,因糖原可用性、黏膜完整性及碱性经血的共同作用,短暂的pH升高可能促进潜在致病菌生长。通常,乳杆菌占优势时阴道pH < 4.5,但近期研究发现相当比例的乳杆菌型mgCST个体出现酸化减弱。
具体而言,L. crispatus的宏基因组亚型mgSs2与pH > 4.5相关,推测与第二种D-乳酸脱氢酶缺失有关。在包含1890个样本的研究中,16/27 mgCST以乳杆菌为主,但仅31%的样本pH < 4.5,69%的样本pH > 4.5,提示临床上存在不同程度的菌群失调。
部分乳杆菌可产生细菌素和过氧化氢,有助于定殖抵抗,但最新证据表明阴道中过氧化氢水平有限,其抗菌作用可能主要来自酸性环境或其他尚未明确的机制。
阴道生态系统在健康状态下维持着以乳杆菌为主的动态平衡,其低pH和代谢环境可有效抑制致病菌生长。然而,当这种平衡受到破坏时,便会出现不同类型的生态失调状态。常见的失调形式包括细菌性阴道炎(BV)、需氧性阴道炎(AV)、外阴阴道念珠菌症(VVC)、细胞溶解性阴道炎(CV)等。
这些状态常伴随乳杆菌减少、病原菌或真菌过度生长、阴道pH升高及黏膜防御功能减弱。阴道生态失调不仅影响局部微生物群结构和生理功能,还与性传播感染、高危妊娠及宫颈病变等一系列不良健康结局密切相关。因此,识别并理解不同类型的阴道生态失调及其微生态机制,对女性生殖健康的维护具有重要意义。
细菌性阴道炎(BV)
细菌性阴道炎(BV)是生育年龄女性中最常见的阴道炎类型,全球患病率为23%–29%。细菌性阴道炎与乳杆菌属数量偏低以及厌氧细菌过度繁殖有关,这些细菌来自阴道加德纳菌(Gardnerella)、普雷沃菌属(Prevotella)、Sneathia、动弯杆菌属(Mobiluncus)、Fannyhessea和Finegoldia等属。
携带细菌性阴道炎相关微生物的个体感染性传播感染(STI)和艾滋病病毒(HIV)、不良妊娠结局及宫颈发育不良的风险增加。虽然不建议孕期常规筛查细菌性阴道炎,但荟萃分析显示,阳性患者早产风险是正常孕妇的两倍,自然流产风险高出九倍。
◮ 短链脂肪酸在细菌性阴道炎中增加可能有害
细菌性阴道炎的特征是管腔内乳酸减少,短链脂肪酸(SCFAs)丰度增加,这些脂肪酸通常由微生物群中的厌氧成员通过发酵和氨基酸分解产生。肠道中的SCFAs已被证明具有抗炎作用;然而,近期研究表明,SCFA暴露(代表BV相关代谢产物)会诱导阴道上皮中促炎细胞因子的产生,并降低上皮屏障完整性。
研究还表明,肠道中的SCFA增加了罗伊氏乳杆菌(Lactobacillus reuteri)对原噬菌体元件的诱导。长链脂肪酸(LCFAs)在哺乳动物黏膜表面很常见,并已在从阴道腔采集的拭子样本中检测到。体外研究表明,LCFAs,特别是油酸,可以抑制L.iners及其他BV相关细菌的生长,并可能促进L.crispatus及其他有益乳杆菌的生长。
这些数据表明,长链脂肪酸和脂肪酸代谢可能作为阴道群落结构的潜在驱动因素发挥重要作用。
需氧性阴道炎(AV)
与细菌性阴道炎(BV)不同,阴道群落也可能被需氧性机会性病原体主导,这些病原体导致促炎标志物如白介素(IL)-6的增加,这种情况称为需氧性阴道炎(AV)。
◮ 大肠杆菌、无乳链球菌等可以成为需氧性阴道炎的病原体
需氧性阴道炎(AV)的患病率低于BV,仅占总人口的7%至12%。与Nugent评分用于BV诊断类似,AV诊断采用湿式相位对比显微镜评分系统。诊断标准依据乳杆菌的存在、炎症细胞的数量和上皮的形态。
AV最常见的病原体包括大肠杆菌(E.coli)、无乳链球菌(S.agalactiae)、粪肠球菌(E.faecalis)、肺炎克雷伯菌(Klebsiella pneumoniae)、凝血酶阴性葡萄球菌(如表皮葡萄球菌)和金黄色葡萄球菌。
外阴阴道念珠菌症(VVC)
念珠菌属,主要是白色念珠菌,是阴道微生物群中最常见的真菌之一,像许多细菌一样,可以作为共生菌在基础层面存活。念珠菌的过度生长可能引发外阴阴道念珠菌症(VVC),其症状包括阴道酸痛或不适、阴道瘙痒及异常分泌物。
◮ 阴道微生态失衡引起念珠菌过度生长
与阴道微生物群的其他成员一样,环境因素和群落动态会影响念珠菌的生长,其相互作用往往由乳杆菌及宿主因素驱动。例如,乳杆菌形成的生物膜有助于维持念珠菌的共生酵母状态,而非致病的菌丝形式。
尽管VVC及念珠菌对免疫系统的影响已有充分研究,但共生真菌在阴道微生物组中的具体作用仍需进一步探索。
细胞溶解性阴道炎(CV)
◮ 乳杆菌过度生长和乳酸水平过高也不健康
细胞溶解性阴道病(CV)是由乳杆菌过度生长和乳酸高于正常水平引起的,主要由卷曲乳杆菌(L.crispatus)引起。CV表现出类似的症状,经常被误诊为VVC,导致治疗不当,往往导致持续的症状。
细胞溶解性阴道病是一种有趣的病理学,表明群落平衡是生态平衡的一个更重要的因素,而不是纯粹的乳杆菌优势。
治疗阴道炎的方法
对有症状的细菌性阴道炎(BV)和好氧性阴道炎(AV)患者,推荐的治疗方法是使用处方抗生素。虽然抗生素通常是首选治疗,但滥用可能破坏阴道内细菌的自然平衡,影响致病菌和有益菌,增加对继发感染(如外阴阴道念珠菌症)的易感性。
◮ 通过微生物干预也有助于治疗阴道疾病
因此,已提出使用鼠李糖乳杆菌(Lactobacillus rhamnosus)或卷曲乳杆菌(L.crispatus)菌株的益生菌作为BV的替代治疗方案,以及选择性靶向BV相关微生物的重组噬菌体衍生内溶素。
对于细菌性阴道炎、好氧性阴道炎和细胞溶解性阴道炎是否应视为传染病,或其细菌群落状态是否是正常波动,仍存在不同意见。一些讨论建议对根据Amsel标准或Nugent评分被诊断为临床BV的患者,尤其是那些表现出可变或不一致症状的患者,进行更多关注,甚至对无症状但可检测到的细菌物种(如G. vaginalis)进行干预。
随着研究进展,应继续探讨这些问题,特别是在无症状个体的“生态失调或非最佳”微生物群落中引入抗生素时。
拓展:阴道环境中未充分研究的微生物
我们目前对阴道微生物群落的理解主要由优势物种定义。然而,在大型数据集中,稀有丰度物种的代表性通常较差,或被归为“其他”类别。关键物种生态的概念是阴道微生物组领域的一个新兴而未充分研究的方向,表明低丰度物种可能对微生物群落产生与其丰度不成比例的影响。
◮ 一些低丰度物种可能在阴道生态中起重要作用
近期的研究开始关注这些低丰度物种对阴道生态系统的重要性。例如,A. muciniphila促进无乳链球菌的定植并可能影响出生结果;奇异菌科(atopobiaceae)及Fannyhessea vaginalis在宫颈癌患者中常见,并与癌症免疫生物标志物增加相关;Sneathia与西班牙裔个体的宫颈癌和HPV发病率升高有关;Mobiluncus mulieris的细胞外囊泡刺激阴道和宫颈细胞中促炎细胞因子的产生;F.magna则被证明能激活中性粒细胞并触发中性粒细胞外陷阱(NET)释放等。
注:除了在性传播感染之外的阴道中发现的低丰度细菌种类外,栖息在这种环境中的病毒经常被忽视,尽管它们可能在阴道中发挥着重要作用。
◮ 噬菌体可能是阴道微生态的重要组成部分
研究评估阴道病毒组表明,只有4%-6%的总读数属于真核病毒,其余的94%-96%与噬菌体对齐。这些临床研究揭示了细菌与其在阴道中各自噬菌体之间的关联,表明噬菌体群落的组成是细菌性阴道炎(BV)的重要预测因子,并将特定噬菌体(如芽孢杆菌病毒Camphawk和Pony)与BV诊断联系起来。
噬菌体的分类与环境中宿主细菌的存在密切相关,已发现低噬菌体多样性与乳杆菌优势群落相关,而高噬菌体多样性则与非乳杆菌优势相关。
另一项研究指出,妊娠中期阴道中的噬菌体来自乳杆菌、链球菌、葡萄球菌和大肠杆菌,而非BV相关微生物,但特定噬菌体家族与CST之间并无联系。尽管噬菌体与阴道细菌群落之间存在某种关联,但尚不清楚噬菌体是否驱动了细菌群落的变化,或者特定噬菌体的存在是否仅是变化的伪影。
未来的研究,包括对阴道微生物组的纵向采样,将有助于阐明病毒与细菌群落之间的动态关系。
阴道具备一个强大的免疫细胞库,这些免疫细胞在维持阴道内环境的平衡和健康方面发挥着显著作用。这个多样化的细胞库不仅能够识别和应对潜在的威胁,还能够有效地影响炎症反应和免疫功能。
宿主免疫细胞与阴道炎症反应
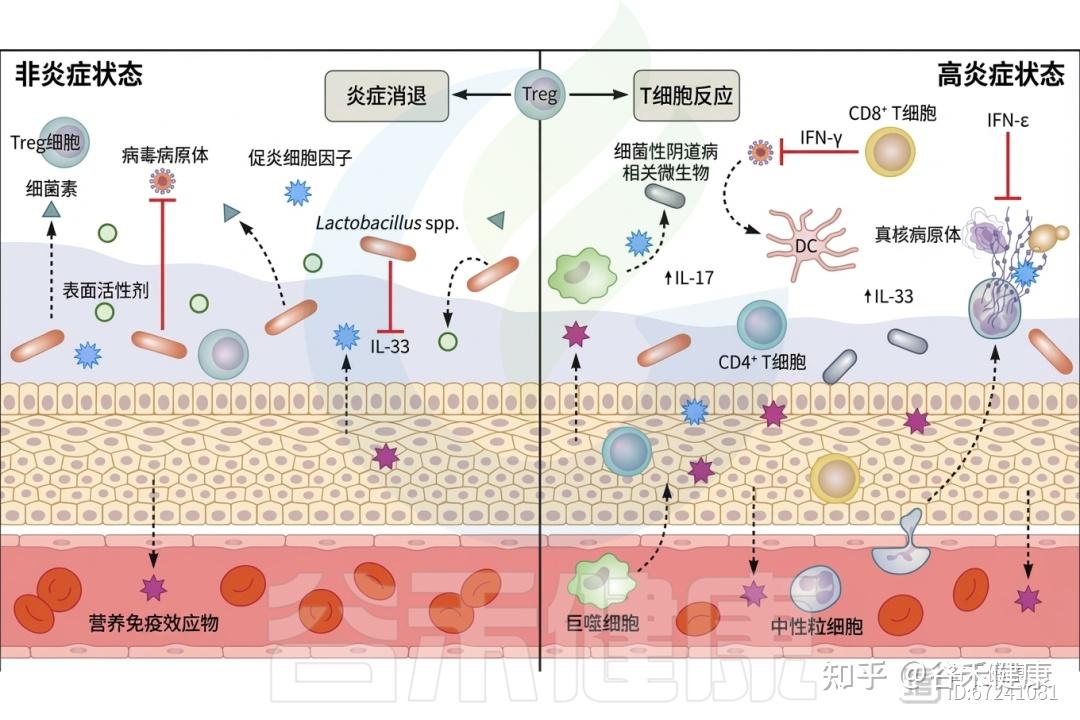
T细胞占比较高
其中CD8+ T细胞和CD4+ T细胞占阴道白细胞的50%。T细胞主要在性传播感染的研究中受到关注。CD8+组织驻留记忆淋巴细胞负责监测病原体入侵,并在识别后释放干扰素γ(IFN-γ),启动免疫细胞的招募。
◮ 调节性T细胞是阴道免疫稳态的关键参与者
调节性T细胞(Treg)是参与适当免疫反应启动的重要细胞,能够成功招募T细胞、自然杀伤细胞和树突状细胞(DC)。Treg在监测阴道炎症中发挥关键作用,激活后可抑制炎症细胞因子的产生、分泌抗炎性IL-10,并产生颗粒酶B,这是一种丝氨酸蛋白酶,有助于控制免疫相关的组织损伤和促进炎症消退。HIV阳性个体的Treg数量减少,伴随炎症粘膜(Tim)CD8+ T细胞的扩张,从而加剧阴道环境中的炎症。
此外,细菌性阴道炎(BV)阳性个体的Th17细胞中检测到了与BV阴性个体相比的T细胞功能变化和促炎标志物表达增加。这些慢性炎症状态会增加对性传播感染和BV等继发感染的易感性,以及感染HIV的风险。
其他免疫细胞
其他白细胞,如中性粒细胞、单核细胞、巨噬细胞和树突状细胞(DC),也在调节阴道免疫中发挥协同作用。
◮ 中性粒细胞具备多种杀菌机制
在阴道病原体入侵过程中,中性粒细胞通常是首批被招募的免疫细胞,具备多种杀菌机制,包括脱颗粒、吞噬作用和NETs的形成。NETs可以捕获病原体,其形成(NETosis)是针对病毒、真菌和寄生虫阴道感染的反应。
从非乳杆菌主导的阴道微生物群个体采集的宫颈阴道拭子和灌洗液中发现,促炎细胞因子和中性粒细胞相关标志物丰度增加,而闭留蛋白和去雾化素-1表达减少,反映出上皮屏障的完整性降低。
在直接实验中,感染BV相关微生物M.mulieris和G.vaginalis的小鼠中,观察到中性粒细胞招募和激活增加。
◮ 阴道微生物群影响抗原呈递细胞的活化
抗原呈递细胞(APC),如巨噬细胞、树突状细胞(DCs)和朗格汉斯细胞(LCs),在调控阴道免疫应对病原体方面发挥着重要作用。
阴道微生物群落影响巨噬细胞的极化, 厌氧消化链球菌(Peptostreptococcus anaerobius)促进抗炎的M2极化;而在孕小鼠中,白色念珠菌则驱动胎盘巨噬细胞的促炎M1极化,可能增加与念珠菌病相关的不良妊娠结局风险。
树突状细胞(DCs)的成熟是通过模式识别受体的信号激活而实现的,暴露于细菌性阴道病(BV)相关细菌或从BV患者采集的宫颈阴道灌洗后,DC的激活明显高于健康个体。然而,DC的活化并不针对所有阴道微生物,暴露于L.crispatus并未能诱导活化。
DC对病毒感染的反应同样重要;黏膜下DC负责向CD4+ T细胞呈递抗原并诱导IFN-γ产生,从而促使针对单纯疱疹病毒2型(HSV-2)的保护性Th1反应。尽管DC对HSV-2感染无直接反应,与细菌相关的Prevotella timonensis却使LCs更易摄取HIV-1病毒颗粒。
◮ 阴道乳杆菌丰度低且多样性高的个体,促炎细胞因子水平更高
乳杆菌丰度较低、微生物多样性较高的个体促炎细胞因子丰度增加。尽管高阴道多样性与乳杆菌主导微生物群体的DC、单核细胞和巨噬细胞丰度无显著差异,但这些抗原呈递细胞(APC)群体的转录组却存在显著差异。
该信号的潜在来源可能与细胞包膜相关,因为转录组分析中的目标基因与脂多糖刺激APC反应密切相关。然而,许多与高多样性相关的生物并不产生脂多糖,因此该领域需要进一步研究。
免疫分泌因子
除了细胞免疫反应,抗菌肽、趋化因子和细胞因子等分泌因子也有助于增强宿主防御。
◮ 白细胞介素-33生成受阴道菌群影响
一个重要的分泌因子是由阴道上皮产生的警报素IL-33,其生成通常受到阴道菌群的抑制。然而,在生态失调时,上皮细胞分泌IL-33作为危险信号,抑制干扰素γ的产生,从而影响宿主对HSV-2(单纯疱疹病毒)的易感性。
◮ 干扰素ε与黄体酮受体的表达相关
另一种在阴道中特殊表达且关键于防止病原体入侵的分泌因子是干扰素ε(IFN-ε)。IFN-ε在阴道和宫颈内皮细胞中以组成型方式表达,但在子宫内膜中受到激素调节,其表达与黄体酮受体的表达相关,卵泡期IFN-ε水平较低,而黄体期水平较高。IFN-ε已被证明对衣原体感染及HIV、寨卡病毒和HSV-2的病毒感染具有保护作用。
◮ 细菌性阴道炎个体IL-17及抗菌肽水平不同
此外,在活动性性传播感染个体的宫颈阴道灌洗中观察到IL-17水平增加,且这一现象与Th17细胞数量无关。需要注意的是,IL-17可以由多种细胞类型产生,包括中性粒细胞、粘膜相关不变T细胞(MAIT)和巨噬细胞等。
在乳杆菌占主导的微生物群体与BV相关微生物相比,阳离子抗菌肽(CAMP)也存在差异表达。最近的阴道宏转录组荟萃分析显示,与非BV微生物组样本相比,从BV相关微生物组样本中涉及CAMP耐药性的基因谱有所增加。
◮ 不同微生物产生的表面活性剂抑制病原菌定植
在阴道中,微生物和宿主细胞均能分泌调节环境的表面活性剂,这影响微生物定植和宿主对感染的反应。宿主表面活性剂通过与真菌细胞的碳水化合物部分相互作用,促进其吞噬。
近期研究发现,L.crispatus产生的细菌表面活性剂可以减少念珠菌对上皮细胞的粘附,并抑制宿主细胞的沙眼衣原体感染。同样,L.gasseri产生的表面活性剂也能阻止耐甲氧西林金黄色葡萄球菌的生物膜形成,表明表面活性剂在阴道微生物群落的建立与形成中可能发挥直接作用。
小结
阴道微生物群与免疫细胞之间的相互作用在维持阴道健康中发挥着重要作用。阴道内的免疫细胞库,特别是T细胞,能够有效识别和应对病原体,同时调节炎症反应。CD8+ T细胞监测病原体,并释放干扰素γ以招募其他免疫细胞。
其他免疫细胞,包括中性粒细胞和抗原呈递细胞(APC),共同参与阴道免疫的调节。研究表明,低丰度乳杆菌且高多样性的微生物群体与促炎细胞因子水平升高相关,提示其在炎症反应中可能发挥关键角色。
此外,分泌因子如IL-33和干扰素ε等对于防止病原体入侵及调节免疫反应也至关重要。不同微生物产生的表面活性剂可有效抑制病原菌的定植,彰显了微生物群在维持阴道微环境中的显著影响。
总的来看,阴道微生物群和免疫细胞的动态平衡对于宿主健康至关重要,但还需进一步研究以深入理解其机制与临床意义。
营养免疫是宿主的一种防御机制,主要涉及抗菌肽,这些肽通过封存可用的营养金属离子,旨在抑制入侵病原体的生长。
营养免疫效应分子对阴道健康的影响
◮ 钙卫蛋白有效捕捉锌、铁等重要金属离子
例如,效应蛋白如S100家族蛋白、脂素和乳铁蛋白能够螯合金属并在宿主的防御机制中发挥重要作用。具体而言,钙卫蛋白是一种由S100A8和S100A9四异二聚体构成的蛋白质,能够有效捕捉锌、锰和铁等金属离子。
另一个例子是S100A7,这种蛋白质表现出对锌的高亲和力;而S100A12,即钙颗粒素C,则表现出对锌和铜的双重亲和力。此外,脂质运载白蛋(lipocalin),亦称lipocalin-2或中性粒细胞相关明胶酶脂素,以及乳铁蛋白,分别是结合铁载体和铁离子的有效铁螯合分子。
◮ 健康阴道中钙卫蛋白丰富,有助于抵抗病原体
值得注意的是,许多营养免疫效应分子由宿主的角质细胞、上皮细胞和先天免疫细胞表达,尤其是中性粒细胞,在炎症期间这些效应分子会大量存在。
在以惰性乳杆菌(L. iners)为主的群落中,研究发现lipocalin-2和钙卫蛋白的浓度明显增加,这指向其在维持阴道健康中的潜在作用。相对而言,有研究表明,与细菌性阴道炎患者相比,健康对照人群中的阴道脂肪钙蛋白更为丰富,这可能与细菌性阴道炎相关微生物的免疫抑制潜力有直接关系。在观察到的动态变化中,阴道乳铁蛋白水平最初在月经后立即上升,并且与阴道微生物多样性的增加相关联。
然而,近来的证据表明,在某些个体中,月经期间及之后的时间窗口与微生物多样性的增加有本质的联系,这可能成为连接这两者的重要线索。这些分子共同作用,可以限制生物利用性的养分金属,同时帮助减少病原微生物的入侵或扩散,包括白色念珠菌、淋病奈瑟菌,以及机会性病原体无乳链球菌(S.agalactiae)等。
效应分子还能影响先天免疫
除了在营养免疫中的重要作用外,S100家族的蛋白质还可以作为警报蛋白或损伤相关分子模式,起到激活和增强先天免疫反应的作用。
◮ S100蛋白影响免疫及炎症反应
例如,钙卫蛋白已被证明能够通过抑制Toll样受体4的活性促进炎症的发生,多种S100家族蛋白结合该受体进行高级糖基化终产物,进而激活转录因子NF-кB。这些数据表明,S100蛋白不仅能够作为金属螯合剂,还可能在功能上促进炎症的发生。
需要指出的是,炎症在阴道环境中是一个研究较少的重要领域,对阴道健康有着显著影响,但这方面的知识仍然存在巨大的空白。因此,未来的研究需要着重深入探讨一些关键问题,例如理解饮食对金属供应的影响、金属摄入对于宿主易感性或感染韧性的作用、在塑造微生物群落形成中的重要性,以及探究在复杂微生物群落中如何使营养金属的获取受到宿主的主动抑制。
阴道微生态是一个动态且错综复杂的生态系统,由微生物群落与宿主之间的相互作用塑造,这些相互作用会随着激素信号传导、屏障完整性和营养可用性的变化而波动。
阴道微生物群的早期研究基于相关数据,但现在的研究已开始深入探索群落互动及其变化的后果。关键研究领域包括:
(1)群落变化的驱动因素:宿主是否通过调节营养选择微生物群落,或微生物代谢导致环境变化?
(2)明确“阴道健康”的定义:健康是以微生物为主还是个性化的标准?应评估临床症状而非仅用Amsel和Nugent评分;
(3)群落优势的概念:微生物优势定义为>50%或>30%?不同群落的保护等级是否不同?
(4)低丰度物种的贡献:阴道并非乳杆菌的单一构成;低丰度微生物在其中起什么作用?
(5)研究真核微生物、古菌和噬菌体的贡献,因为这些在阴道中仍被忽视。
在这些研究领域的基础上,我们还需进一步探讨微生物群落与宿主之间的相互关系及其如何受到环境因素的影响。例如,激素波动可能改变阴道内的pH值,从而影响微生物的组成,进而影响免疫反应。
此外,研究需要关注细菌与宿主免疫系统之间的相互作用。微生物不仅在维持阴道生态平衡中发挥作用,还通过其代谢产物调节宿主的免疫状态。我们必须深入理解这些微生物代谢产物如短链脂肪酸、共生抗菌肽等如何影响免疫反应的强度与方向。
对于不同个体,微生物群的组成及其功能可能有所不同,这也提示我们必须考虑遗传因素、生活习惯和饮食在阴道微生物组发展中的作用。这些差异可能会导致对相同微生物群体的不同免疫响应或营养利用,进而影响女性的健康状况。
最后,随着对阴道微生物群的了解不断深化,跨学科的合作将变得愈发重要。微生物学、免疫学和营养学等领域的相互结合,能够为我们提供更全面的视角,帮助我们理解更复杂的生物系统。这种综合的研究方法将有助于填补当前知识空白,并推动我们在改善女性生殖健康方面取得新的进展。
主要参考文献
Landolt EF, da Conceição Mendonça J, Behler AE, Lumsdaine SW, Jafar T, Burcham LR. Exploring the vaginal ecosystem: insights into host-microbe interactions and microbial community dynamics. Infect Immun. 2025 Sep 9;93(9):e0049924.
Greenbaum S, Greenbaum G, Moran-Gilad J, Weintraub AY. Ecological dynamics of the vaginal microbiome in relation to health and disease. Am J Obstet Gynecol. 2019 Apr;220(4):324-335.
Amabebe E, Anumba DOC. The Vaginal Microenvironment: The Physiologic Role of Lactobacilli. Front Med (Lausanne). 2018 Jun 13;5:181.
Shen L, Zhang W, Yuan Y, Zhu W, Shang A. Vaginal microecological characteristics of women in different physiological and pathological period. Front Cell Infect Microbiol. 2022 Jul 22;12:959793.
Holm JB, France MT, Gajer P, Ma B, Brotman RM, Shardell M, Forney L, Ravel J. Integrating compositional and functional content to describe vaginal microbiomes in health and disease. Microbiome. 2023 Nov 30;11(1):259. doi: 10.1186/s40168-023-01692-x. Erratum in: Microbiome. 2024 Feb 6;12(1):21.
Ma B, Forney LJ, Ravel J. Vaginal microbiome: rethinking health and disease. Annu Rev Microbiol. 2012;66:371-89.
Torcia MG. Interplay among Vaginal Microbiome, Immune Response and Sexually Transmitted Viral Infections. Int J Mol Sci. 2019 Jan 11;20(2):266.
Kaur H, Merchant M, Haque MM, Mande SS. Crosstalk Between Female Gonadal Hormones and Vaginal Microbiota Across Various Phases of Women’s Gynecological Lifecycle. Front Microbiol. 2020 Mar 31;11:551.

谷禾健康
在肠道菌群检测系列产品成熟之后,谷禾健康将深耕多年的微生物组学技术平台,延伸至关乎女性全生命周期健康的另一核心领域——阴道微生态。
还包括子宫颈沙眼衣原体、HPV、HSV、EB病毒、巨细胞病毒等。
谷禾阴道菌群检测报告引入科学前沿的菌群状态分型(CST)概念,将复杂的菌群构成归纳为几种易于理解的健康状态类型。
例如,以卷曲乳杆菌为主的CST-I型代表健康的稳定状态,而以加德纳菌等多种厌氧菌为主的CST-IV型则与细菌性阴道病高度相关。这为临床判断和干预效果评估提供了科学支持。
谷禾阴道菌群检测报告中包括阴道菌群总体评估、CST分型、致病菌表(细菌性阴道病,需氧菌性阴道炎,外阴念珠菌病等)、列出异常菌群及相关说明,菌群详细构成等(注:仅用于菌群科学研究和辅助参考,不直接用于临床诊断)。
…
…
…
…
阴道菌群检测让我们能够更全面地了解阴道微生物组的组成及其变化,以及它是如何随着时间的推移或对各种因素(如环境、激素变化、性活动和抗生素使用等)的反应而变化的。
注:本产品可辅助评估和筛查,不用作临床诊断。
05

谷禾健康

慢性子宫内膜炎(Chronic endometritis, CE)是一种慢性非特异性的子宫内膜炎症性疾病,常表现为无症状或非特异性临床症状,慢性子宫内膜炎经常被患者和妇科医生忽略。因此,寻找新的诊断标记对于改善慢性子宫内膜炎的预后至关重要。
针对上述问题,来自武汉大学人民医院生殖医学中心杨静团队的科研人员在《OBSTETRICS AND GYNECOLOGY》上发表了研究论文。
该研究针对慢性子宫内膜炎女性的阴道微生物特征进行深入研究,比较了98名接受子宫内膜活检进行不孕症常规临床检查的女性(49名诊断为CE的女性和49名非CE的女性)的阴道微生物组特征,并采用杭州谷禾核酸提取试剂盒及测序平台,使用16S rRNA基因扩增子测序分析阴道微生物组,对微生物标记物进行了鉴定。
该项研究确定了四个慢性子宫内膜炎的微生物标记物(肠杆菌,普雷沃氏菌,粪杆菌,Phascolarctobacterium),开发了针对慢性子宫内膜炎预测诊断分类器,并进一步探讨了阴道微生物组特征作为慢性子宫内膜炎诊断新工具的潜力。

英文题目:Vaginal microbiome dysbiosis as a novel noninvasive biomarker for detection of chronic endometritis in infertile women
中文题目:阴道微生物群失调作为检测不孕女性慢性子宫内膜炎的新型非侵入性生物标志物
期刊名:OBSTETRICS AND GYNECOLOGY
发表时间:2024年7月10日
▼ 什么是慢性子宫内膜炎?
慢性子宫内膜炎是发生在子宫内膜间质区的持续性细微炎症性疾病,其特征为子宫内膜间质区的浆细胞浸润,通常无症状或仅出现轻微的症状,如异常子宫出血、盆腔疼痛、白带增多等。
▼ 慢性子宫内膜炎有哪些不良后果?
大量研究表明,慢性子宫内膜炎与女性不孕症、反复流产、反复着床失败、子宫内粘连和辅助生殖技术的不良后果密切相关,此外慢性子宫内膜炎还是影响活产和足月分娩的客观因素之一。
▼ 慢性子宫内膜炎的发病率
慢性子宫内膜炎的发病率在不同人群中不尽相同:
其病理改变与内膜中微生物群的定性、定量改变有关。
近年来,人类微生物组学的研究表明,人体不同系统内微生物群与人类健康和疾病发病机制存在密切关系,刺激炎症并增加癌症等疾病发生的风险。作为人体主要的微生物组,阴道菌群对于女性生理和生殖健康起着重要作用。
◆阴道病原体感染是慢性子宫内膜炎的病因之一
Moreno等人通过微生物培养在慢性子宫内膜炎女性子宫内膜中发现了包括无乳链球菌和淋病奈瑟菌在内的细菌,并开发了一种诊断慢性子宫内膜炎的分子方法——逆转录聚合酶链反应试验,从这些病原体中识别DNA,用于慢性子宫内膜炎的诊断。
◆慢性子宫内膜炎:宫腔内致病菌增加
随着微生物组学技术的发展,传统观点认为宫腔内是无菌环境的观念也逐渐被推翻。16S和宏基因组测序等技术均证实宫腔内存在微生物的定植,健康女性以乳杆菌属占绝对优势,且宫腔内微生物与慢性子宫内膜炎的发病存在关联,慢性子宫内膜炎患者宫腔非乳酸杆菌为优势菌群且致病菌如加德纳菌、葡萄球菌、链球菌检出率增加。
在研究中也指出,诊断患有慢性子宫内膜炎的女性阴道微生物群生态失调。与子宫内膜冲洗样品相比,阴道分泌物可以作为诊断慢性子宫内膜炎的更无创和更经济有效的生物标志物。阴道宫颈黏液的免疫学指标已被认为是子宫内膜炎的潜在诊断生物标志物。然而,需要进一步评估阴道微生物组在慢性子宫内膜炎中的诊断潜力。
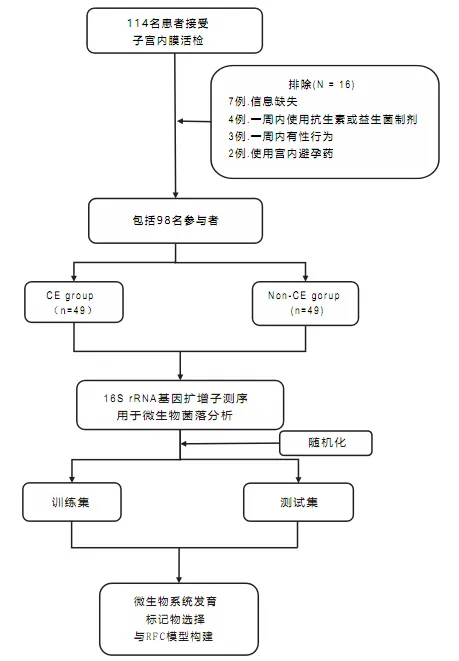
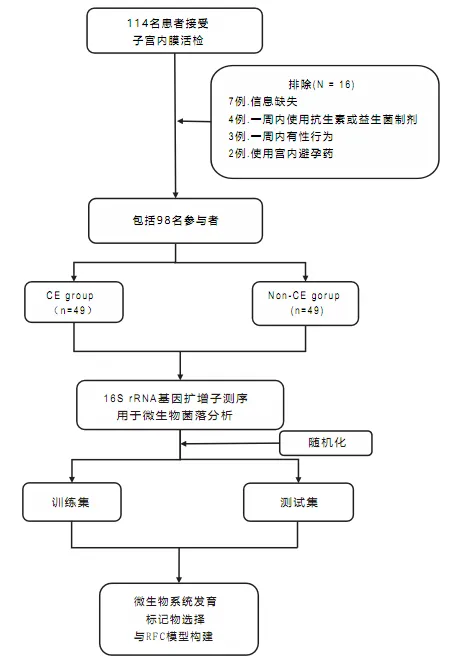
本研究于2023年6月至2023年11月在武汉大学人民医院生殖医学中心完成。研究集中于常规不孕症检查中接受子宫内膜活检的患者。最终共纳入49名患有慢性子宫内膜炎的女性,并与49名非慢性子宫内膜炎对照组相匹配。
慢性子宫内膜炎患者的选择标准如下:
排除标准为:
注:所有受试者均获得书面同意,并经武汉大学人民医院伦理委员会批准(批准通知号:WDRY2023- K090)。
该研究的设计和流程图如图所示:
编辑
在本研究中,所有样本均在子宫内膜活检前采集。用无菌生理盐水擦拭外阴。随后,使用无润滑剂的一次性无菌阴道窥镜完全暴露宫颈。使用两个无菌拭子从后穹窿收集阴道分泌物:一个使用Nugent评分法进行评估,另一个储存在−80°C下用于后续16S rRNA基因测序分析。
项目采用杭州谷禾核酸提取试剂盒(GHFDE100)提取阴道冲洗液样本的基因组DNA。用引物515F(5′- GTGCCAGCMGCCGCGGTAA- 3′)和806R(5′- GGACTACHVGGGTWTCTAAT- 3′)扩增16S rRNA的V4区。为了实现多重测序,将样品特异性对端6bp条形码集成到TrueSeq适配器中。纯化和定量后,整个DNA池在Illumina NovaSeq6000平台(Illumina)上按照协议进行测序。
使用Vsearch v2.22.1组装拼接双端序列,并以100%的相似性阈值将序列聚类为相同的扩增子序列变体(amplicon sequence variant, ASV)。利用QIIME2加权分类器获得ASV的分类信息。基于R包“VennDiagram”生成维恩图来说明分组间共有的和独有的ASV。
α-多样性是指群落内的物种多样性,以物种丰富度和归一化香农熵(均匀度)为其显著组成部分。物种丰富度采用Chao1指数进行量化,而Shannon多样性指数同时评估丰富度和均匀度。通过主坐标分析(PCoA)可视化两个组之间样本的微生物菌落结构。使用R的vegan包进行相似性分析(ANOSIM)评估微生物群落的差异。
此外,为了预测微生物的代谢功能,使用京都基因与基因组百科全书(KEGG)数据库与PICRUSt2 (Phylogenetic Investigation of Communities by Reconstruction of Unobserved States)进行代谢途径富集分析。组间的统计差异使用Kruskal-Wallis检验和Wilcoxon秩和检验进行评估,显著性水平定义为P < 0.05。
为了评估阴道微生物群组成对慢性子宫内膜炎(CE)的诊断潜力,应用随机森林算法,这是一种稳健的监督学习算法。使用R的pROC软件包生成的受试者工作特征(ROC)曲线评估模型的整体性能。曲线下的面积(AUC)量化了ROC曲线的判别能力。此外,还计算模型的特异性、敏感性和准确性。
连续变量的临床特征差异使用Student’s t检验进行评估,而分类变量则需要使用χ²检验或Fisher精确检验进行评估。统计分析使用SPSS版本27(IBM)进行,P < 0.05被认为具有统计学意义。
在98名参与者中,49名被诊断为CE,其标准是在30个随机选择的高倍视野中至少有一个视野中观察到超过5个MUM-1+/CD138+细胞。基线特征包括年龄、体重指数、抗穆勒激素水平和流产史,在两组之间显示了可比性(见表1)。
等级丰度曲线体现了物种丰富度和均匀度
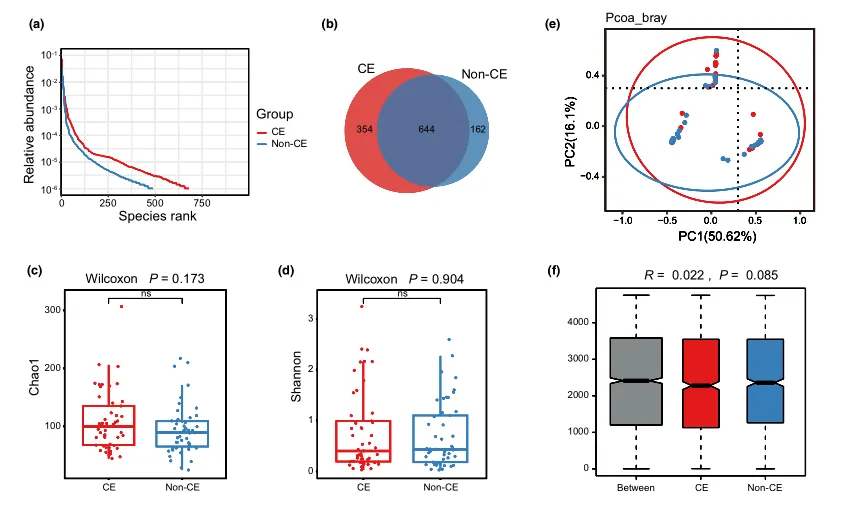
等级丰度曲线描述了两组微生物群落的物种丰富度和均匀度。曲线范围越宽表明物种丰度越高,曲线越光滑表明物种分布越均匀。
从曲线上看,CE组的物种丰富度和均匀度高于非CE组(图2a)。维恩图显示,两组共有1180个ASV中的664个,其中CE组特有的ASV为354个(图2b)。利用Chao1丰富度估计器和Shannon多样性指数量化的α-多样性结果显示,两组间差异不显著(Chao1的P = 0.1733, Shannon的P = 0.9043,图2c,d)。
为了说明样品之间的微生物组分布空间,进行了PCoA,如图2e所示。结果显示,两组样本的分布是对称的。
值得注意的是,ANOSIM结果显示组间差异大于组内差异,尽管没有达到统计学意义(R = 0.022, P = 0.085)(图2f)。
阴道微生物群的细菌多样性
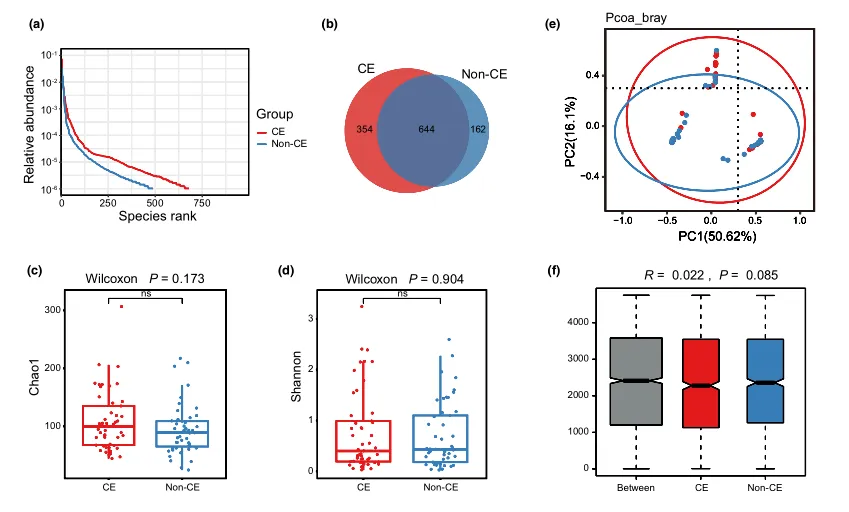
(a)用秩丰度曲线评价细菌的相对均匀度;
(b)显示组间重叠的维恩图;
(c)采用Chao1指数;
(d)Simpson指数估算口腔微生物多样性;
(e)使用Bray-Curtis通过主坐标分析计算β多样性;
(f)通过相似性分析计算了组间微生物群落的差异。
阴道微生物群的组成
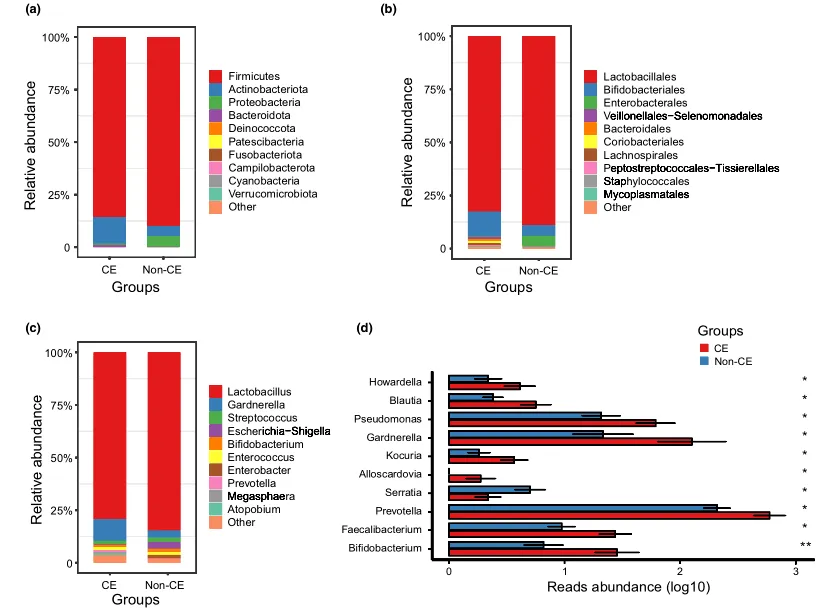
文中分析了与CE相关的阴道微生物组的分类组成和变化。门、科和属水平的平均细菌群落组成如图3a-c所示。值得注意的是,与非CE组相比,CE组有10个属的细菌显著富集,包括双歧杆菌、普雷沃氏菌和加德纳菌(均P < 0.05)(图3d)。
慢性子宫内膜炎患者阴道微生物群落的系统发育概况
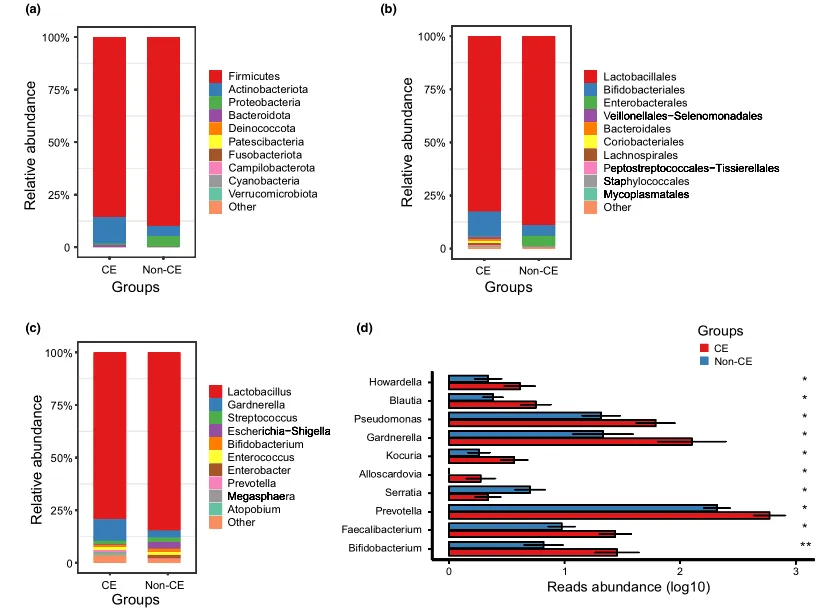
门(a)、科(b)、属(c)水平上细菌群落的平均组成。CE组与非CE组间差异有统计学意义(均 P < 0.05)。0.01 <矫正 P ≤ 0.05 ,标记为*。
代谢途径
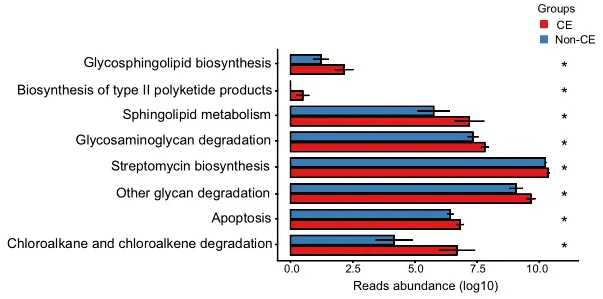
通过KEGG途径富集的功能分析揭示了CE组中几种上调的途径,如图4所示。具体而言,参与多糖生物合成和代谢的途径(包括鞘糖脂生物合成、糖胺聚糖降解和其他多糖降解过程)上调。
此外,萜类和聚酮类代谢、II型聚酮类生物合成、鞘脂质代谢、次生代谢物生物合成、链霉素生物合成、外源生物降解、氯烷烃和氯烯烃降解以及凋亡相关途径也上调。总共有8个KEGG模块在CE组显著富集,重点是代谢相关途径(均P < 0.05)。
KEGG代谢通路的丰度差异
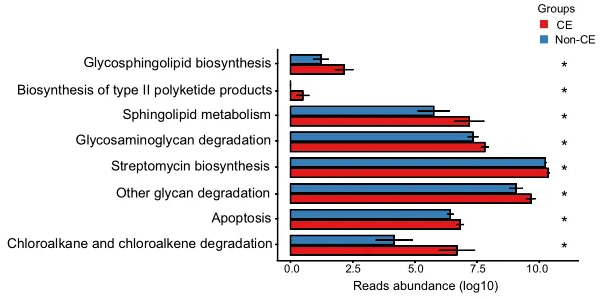
0.01< 校正P≤0.05,标记为*;0.001 <校正P≤0.01,标记为**。CE,慢性子宫内膜炎。
为测试阴道微生物组对慢性子宫内膜炎(CE)的诊断潜力,这里构建了一个随机森林分类器模型,用以区分CE样本和非CE样本。训练集用于模型训练,测试集用于评估模型性能。通过在发现阶段重复五次的10倍交叉验证,我们识别出四个最优标记 ASV 作为CE的标记集(图5a)。
这些选定ASV的对应细菌属包括:
随机森林模型在训练集队列上的AUC值为85.68%(95%置信区间[CI], 75.62%-95.73%),在测试集队列上的AUC值为83.26% (95% CI, 68.67%-97.85%)(图5b)。模型的特异性为82.76%,灵敏度为79.49%,准确率为80.88%。
这些发现表明,基于阴道微生物ASV标记的随机森林模型在区分CE和非CE队列方面表现出强大的诊断潜力。
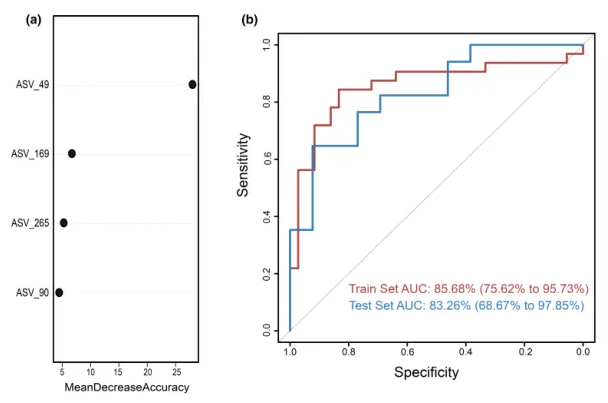
图5.通过随机森林模型鉴定用于临床诊断的微生物标志物。
(a)使用阴道样品中四种选定生物标志物的概况,来自随机森林模型的变量的平均降低准确度。
(b)接收器工作特征曲线基于组织样本的训练集和测试集。ASV,扩增子序列变体; AUC,曲线下面积。
◆阴道菌群的变化可引起宫腔的菌群失调
女性阴道中的微生物组是一个完整的连续体观点已被广泛接受。阴道细菌可以异位定植并改变上生殖道中的微生物组。局部微生物组的生态失调会导致微环境发生复杂的病理生理变化,最终扰乱与免疫反应、炎症和凋亡相关的各种过程,从而促进病理状况。
阴道和宫腔内的菌群变化有一定的同步性,阴道菌群的变化可引起宫腔内的菌群失调,从而影响女性生殖健康。
本研究旨在对有或没有慢性子宫内膜炎(CE)的女性阴道微生物组的群落结构和功能进行初步探索。此外,文章中还提出了基于阴道微生物ASV的新型非侵入性CE诊断标志物。
◆特定的菌群可作为新型非侵入性CE诊断标志物
与先前的研究结果一致,该研究中没有发现慢性子宫内膜炎组和非慢性子宫内膜炎组阴道微生物组之间的α-多样性或β-多样性有显著差异。
值得注意的是,微生物组的低组内变异情况表明其作为潜在诊断标记具有一定的稳健性。与非CE组相比,慢性子宫内膜炎组中几个低丰度属的富集程度不同,包括双歧杆菌,普雷沃氏菌和加德纳菌。
双歧杆菌
双歧杆菌以其保护作用而闻名,其高丰度在CE组中令人惊讶。双歧杆菌菌株作为益生菌用于肠道炎症、严重疾病和抑郁症患者的肠道微生物组调节已有很长的历史。然而,其他研究报告称,在一些不健康的情况下,包括子宫内膜异位症、高危人乳头瘤病毒和不孕症,阴道中双歧杆菌含量很高。这种不一致的潜在机制尚不清楚,需要进一步研究。
加德纳菌和普氏菌
加德纳菌和普氏菌是公认的阴道生态失调的指标。这些属的定植改变了免疫系统,并诱导了促炎反应。在宿主-微生物共培养模型中,加德纳菌与宫颈阴道屏障的破坏有关,同时促进小鼠中普雷沃菌属对上生殖道的侵入性感染。
总的来说,该研究结果表明,两组之间的阴道微生物组发生了实质性变化,突显了阴道冲洗液样本作为检测慢性子宫内膜炎的诊断工具的潜力。
◆阴道菌群的变化可引起宫腔的菌群失调
KEGG途径富集分析强调了关键代谢过程的上调,特别是CE组的聚糖生物合成和代谢。覆盖阴道上皮细胞的聚糖涂层是保护屏障的重要组成部分。乳杆菌与阴道上皮的粘附被认为是具有竞争性地抑制病原体,而上皮的糖胺聚糖在这些过程中起着至关重要的作用。
值得注意的是,糖胺聚糖的消耗与宫颈上皮细胞屏障功能的破坏有关,这可能会增加对局部细菌和上升感染的易感性。这表明CE患者的微生物组失调可能会对微生物侵入阴道的主要门户产生负面影响,并引发宫内炎症。
关于慢性子宫内膜炎(CE)的标准化定义和诊断指南仍存在争议,最被接受的诊断依赖于子宫内膜活检。基于选定的阴道微生物群构建了CE诊断模型。本文研究中,引入了一种新的随机森林模型,该模型可以根据ASV以更高的灵敏度和准确性将CE患者与使用阴道分泌物的健康个体区分开来。该研究结果表明,阴道微生物组的变化可以作为诊断CE的指标,为慢性子宫内膜炎提供了一种更容易获得和更方便患者的诊断选择。
然而,该研究也存在一定的局限性。首先,它主要侧重于检查有慢性子宫内膜炎和没有慢性子宫内膜炎的女性之间的成分差异,而不是研究与慢性炎症相关的免疫失调的潜在机制。其次,阴道微生物样本是在分泌中期收集的,这可能无法解释整个月经周期中微生物群组成的变化。第三,作为一项单中心调查,发现的普遍性可能有限。考虑到饮食和运动等因素,多中心方法可以提供更全面的解读。
最后,多种组学方法的整合(包括培养组学、元转录组学和代谢组学),可以提供对微生物群落功能的全面了解。这种更广泛的视角对于阐明阴道微生态系统内复杂的网络和阴道微环境的潜在变化至关重要。
阴道微生物生态失调是慢性子宫内膜炎的重要指标。该研究确定了与慢性子宫内膜炎发展相关的潜在候选细菌,并提出了潜在的机制。基于阴道微生物群的生物标志物有望成为检测慢性子宫内膜炎的非侵入性工具。需要进一步的研究来发现特定阴道微生物组在慢性子宫内膜炎中的功能,并确定具有诊断能力的强大阴道微生物标记物,以区分患有慢性子宫内膜炎的个体。

谷禾健康

慢性子宫内膜炎(Chronic endometritis, CE)是一种慢性非特异性的子宫内膜炎症性疾病,常表现为无症状或非特异性临床症状,慢性子宫内膜炎经常被患者和妇科医生忽略。因此,寻找新的诊断标记对于改善慢性子宫内膜炎的预后至关重要。
针对上述问题,来自武汉大学人民医院生殖医学中心杨静团队的科研人员在《OBSTETRICS AND GYNECOLOGY》上发表了研究论文。
该研究针对慢性子宫内膜炎女性的阴道微生物特征进行深入研究,比较了98名接受子宫内膜活检进行不孕症常规临床检查的女性(49名诊断为CE的女性和49名非CE的女性)的阴道微生物组特征,并采用杭州谷禾核酸提取试剂盒及测序平台,使用16S rRNA基因扩增子测序分析阴道微生物组,对微生物标记物进行了鉴定。
该项研究确定了四个慢性子宫内膜炎的微生物标记物(肠杆菌,普雷沃氏菌,粪杆菌,Phascolarctobacterium),开发了针对慢性子宫内膜炎预测诊断分类器,并进一步探讨了阴道微生物组特征作为慢性子宫内膜炎诊断新工具的潜力。

英文题目:Vaginal microbiome dysbiosis as a novel noninvasive biomarker for detection of chronic endometritis in infertile women
中文题目:阴道微生物群失调作为检测不孕女性慢性子宫内膜炎的新型非侵入性生物标志物
期刊名:OBSTETRICS AND GYNECOLOGY
发表时间:2024年7月10日
▼ 什么是慢性子宫内膜炎?
慢性子宫内膜炎是发生在子宫内膜间质区的持续性细微炎症性疾病,其特征为子宫内膜间质区的浆细胞浸润,通常无症状或仅出现轻微的症状,如异常子宫出血、盆腔疼痛、白带增多等。
▼ 慢性子宫内膜炎有哪些不良后果?
大量研究表明,慢性子宫内膜炎与女性不孕症、反复流产、反复着床失败、子宫内粘连和辅助生殖技术的不良后果密切相关,此外慢性子宫内膜炎还是影响活产和足月分娩的客观因素之一。
▼ 慢性子宫内膜炎的发病率
慢性子宫内膜炎的发病率在不同人群中不尽相同:
其病理改变与内膜中微生物群的定性、定量改变有关。
近年来,人类微生物组学的研究表明,人体不同系统内微生物群与人类健康和疾病发病机制存在密切关系,刺激炎症并增加癌症等疾病发生的风险。作为人体主要的微生物组,阴道菌群对于女性生理和生殖健康起着重要作用。
◆阴道病原体感染是慢性子宫内膜炎的病因之一
Moreno等人通过微生物培养在慢性子宫内膜炎女性子宫内膜中发现了包括无乳链球菌和淋病奈瑟菌在内的细菌,并开发了一种诊断慢性子宫内膜炎的分子方法——逆转录聚合酶链反应试验,从这些病原体中识别DNA,用于慢性子宫内膜炎的诊断。
◆慢性子宫内膜炎:宫腔内致病菌增加
随着微生物组学技术的发展,传统观点认为宫腔内是无菌环境的观念也逐渐被推翻。16S和宏基因组测序等技术均证实宫腔内存在微生物的定植,健康女性以乳杆菌属占绝对优势,且宫腔内微生物与慢性子宫内膜炎的发病存在关联,慢性子宫内膜炎患者宫腔非乳酸杆菌为优势菌群且致病菌如加德纳菌、葡萄球菌、链球菌检出率增加。
在研究中也指出,诊断患有慢性子宫内膜炎的女性阴道微生物群生态失调。与子宫内膜冲洗样品相比,阴道分泌物可以作为诊断慢性子宫内膜炎的更无创和更经济有效的生物标志物。阴道宫颈黏液的免疫学指标已被认为是子宫内膜炎的潜在诊断生物标志物。然而,需要进一步评估阴道微生物组在慢性子宫内膜炎中的诊断潜力。
本研究于2023年6月至2023年11月在武汉大学人民医院生殖医学中心完成。研究集中于常规不孕症检查中接受子宫内膜活检的患者。最终共纳入49名患有慢性子宫内膜炎的女性,并与49名非慢性子宫内膜炎对照组相匹配。
慢性子宫内膜炎患者的选择标准如下:
排除标准为:
注:所有受试者均获得书面同意,并经武汉大学人民医院伦理委员会批准(批准通知号:WDRY2023- K090)。
该研究的设计和流程图如图所示:
编辑
在本研究中,所有样本均在子宫内膜活检前采集。用无菌生理盐水擦拭外阴。随后,使用无润滑剂的一次性无菌阴道窥镜完全暴露宫颈。使用两个无菌拭子从后穹窿收集阴道分泌物:一个使用Nugent评分法进行评估,另一个储存在−80°C下用于后续16S rRNA基因测序分析。
项目采用杭州谷禾核酸提取试剂盒(GHFDE100)提取阴道冲洗液样本的基因组DNA。用引物515F(5′- GTGCCAGCMGCCGCGGTAA- 3′)和806R(5′- GGACTACHVGGGTWTCTAAT- 3′)扩增16S rRNA的V4区。为了实现多重测序,将样品特异性对端6bp条形码集成到TrueSeq适配器中。纯化和定量后,整个DNA池在Illumina NovaSeq6000平台(Illumina)上按照协议进行测序。
使用Vsearch v2.22.1组装拼接双端序列,并以100%的相似性阈值将序列聚类为相同的扩增子序列变体(amplicon sequence variant, ASV)。利用QIIME2加权分类器获得ASV的分类信息。基于R包“VennDiagram”生成维恩图来说明分组间共有的和独有的ASV。
α-多样性是指群落内的物种多样性,以物种丰富度和归一化香农熵(均匀度)为其显著组成部分。物种丰富度采用Chao1指数进行量化,而Shannon多样性指数同时评估丰富度和均匀度。通过主坐标分析(PCoA)可视化两个组之间样本的微生物菌落结构。使用R的vegan包进行相似性分析(ANOSIM)评估微生物群落的差异。
此外,为了预测微生物的代谢功能,使用京都基因与基因组百科全书(KEGG)数据库与PICRUSt2 (Phylogenetic Investigation of Communities by Reconstruction of Unobserved States)进行代谢途径富集分析。组间的统计差异使用Kruskal-Wallis检验和Wilcoxon秩和检验进行评估,显著性水平定义为P < 0.05。
为了评估阴道微生物群组成对慢性子宫内膜炎(CE)的诊断潜力,应用随机森林算法,这是一种稳健的监督学习算法。使用R的pROC软件包生成的受试者工作特征(ROC)曲线评估模型的整体性能。曲线下的面积(AUC)量化了ROC曲线的判别能力。此外,还计算模型的特异性、敏感性和准确性。
连续变量的临床特征差异使用Student’s t检验进行评估,而分类变量则需要使用χ²检验或Fisher精确检验进行评估。统计分析使用SPSS版本27(IBM)进行,P < 0.05被认为具有统计学意义。
在98名参与者中,49名被诊断为CE,其标准是在30个随机选择的高倍视野中至少有一个视野中观察到超过5个MUM-1+/CD138+细胞。基线特征包括年龄、体重指数、抗穆勒激素水平和流产史,在两组之间显示了可比性(见表1)。
等级丰度曲线体现了物种丰富度和均匀度
等级丰度曲线描述了两组微生物群落的物种丰富度和均匀度。曲线范围越宽表明物种丰度越高,曲线越光滑表明物种分布越均匀。
从曲线上看,CE组的物种丰富度和均匀度高于非CE组(图2a)。维恩图显示,两组共有1180个ASV中的664个,其中CE组特有的ASV为354个(图2b)。利用Chao1丰富度估计器和Shannon多样性指数量化的α-多样性结果显示,两组间差异不显著(Chao1的P = 0.1733, Shannon的P = 0.9043,图2c,d)。
为了说明样品之间的微生物组分布空间,进行了PCoA,如图2e所示。结果显示,两组样本的分布是对称的。
值得注意的是,ANOSIM结果显示组间差异大于组内差异,尽管没有达到统计学意义(R = 0.022, P = 0.085)(图2f)。
阴道微生物群的细菌多样性
(a)用秩丰度曲线评价细菌的相对均匀度;
(b)显示组间重叠的维恩图;
(c)采用Chao1指数;
(d)Simpson指数估算口腔微生物多样性;
(e)使用Bray-Curtis通过主坐标分析计算β多样性;
(f)通过相似性分析计算了组间微生物群落的差异。
阴道微生物群的组成
文中分析了与CE相关的阴道微生物组的分类组成和变化。门、科和属水平的平均细菌群落组成如图3a-c所示。值得注意的是,与非CE组相比,CE组有10个属的细菌显著富集,包括双歧杆菌、普雷沃氏菌和加德纳菌(均P < 0.05)(图3d)。
慢性子宫内膜炎患者阴道微生物群落的系统发育概况
门(a)、科(b)、属(c)水平上细菌群落的平均组成。CE组与非CE组间差异有统计学意义(均 P < 0.05)。0.01 <矫正 P ≤ 0.05 ,标记为*。
代谢途径
通过KEGG途径富集的功能分析揭示了CE组中几种上调的途径,如图4所示。具体而言,参与多糖生物合成和代谢的途径(包括鞘糖脂生物合成、糖胺聚糖降解和其他多糖降解过程)上调。
此外,萜类和聚酮类代谢、II型聚酮类生物合成、鞘脂质代谢、次生代谢物生物合成、链霉素生物合成、外源生物降解、氯烷烃和氯烯烃降解以及凋亡相关途径也上调。总共有8个KEGG模块在CE组显著富集,重点是代谢相关途径(均P < 0.05)。
KEGG代谢通路的丰度差异
0.01< 校正P≤0.05,标记为*;0.001 <校正P≤0.01,标记为**。CE,慢性子宫内膜炎。
为测试阴道微生物组对慢性子宫内膜炎(CE)的诊断潜力,这里构建了一个随机森林分类器模型,用以区分CE样本和非CE样本。训练集用于模型训练,测试集用于评估模型性能。通过在发现阶段重复五次的10倍交叉验证,我们识别出四个最优标记 ASV 作为CE的标记集(图5a)。
这些选定ASV的对应细菌属包括:
随机森林模型在训练集队列上的AUC值为85.68%(95%置信区间[CI], 75.62%-95.73%),在测试集队列上的AUC值为83.26% (95% CI, 68.67%-97.85%)(图5b)。模型的特异性为82.76%,灵敏度为79.49%,准确率为80.88%。
这些发现表明,基于阴道微生物ASV标记的随机森林模型在区分CE和非CE队列方面表现出强大的诊断潜力。
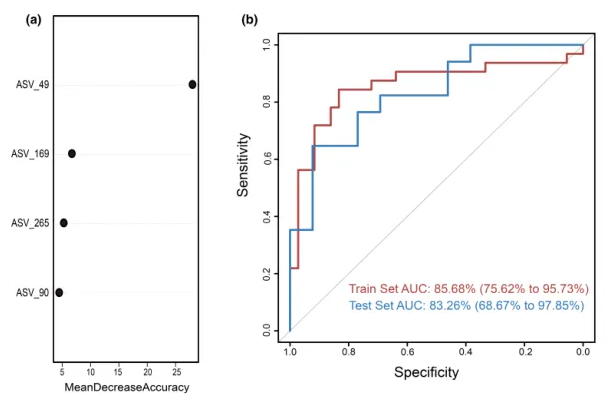
图5.通过随机森林模型鉴定用于临床诊断的微生物标志物。
(a)使用阴道样品中四种选定生物标志物的概况,来自随机森林模型的变量的平均降低准确度。
(b)接收器工作特征曲线基于组织样本的训练集和测试集。ASV,扩增子序列变体; AUC,曲线下面积。
◆阴道菌群的变化可引起宫腔的菌群失调
女性阴道中的微生物组是一个完整的连续体观点已被广泛接受。阴道细菌可以异位定植并改变上生殖道中的微生物组。局部微生物组的生态失调会导致微环境发生复杂的病理生理变化,最终扰乱与免疫反应、炎症和凋亡相关的各种过程,从而促进病理状况。
阴道和宫腔内的菌群变化有一定的同步性,阴道菌群的变化可引起宫腔内的菌群失调,从而影响女性生殖健康。
本研究旨在对有或没有慢性子宫内膜炎(CE)的女性阴道微生物组的群落结构和功能进行初步探索。此外,文章中还提出了基于阴道微生物ASV的新型非侵入性CE诊断标志物。
◆特定的菌群可作为新型非侵入性CE诊断标志物
与先前的研究结果一致,该研究中没有发现慢性子宫内膜炎组和非慢性子宫内膜炎组阴道微生物组之间的α-多样性或β-多样性有显著差异。
值得注意的是,微生物组的低组内变异情况表明其作为潜在诊断标记具有一定的稳健性。与非CE组相比,慢性子宫内膜炎组中几个低丰度属的富集程度不同,包括双歧杆菌,普雷沃氏菌和加德纳菌。
双歧杆菌
双歧杆菌以其保护作用而闻名,其高丰度在CE组中令人惊讶。双歧杆菌菌株作为益生菌用于肠道炎症、严重疾病和抑郁症患者的肠道微生物组调节已有很长的历史。然而,其他研究报告称,在一些不健康的情况下,包括子宫内膜异位症、高危人乳头瘤病毒和不孕症,阴道中双歧杆菌含量很高。这种不一致的潜在机制尚不清楚,需要进一步研究。
加德纳菌和普氏菌
加德纳菌和普氏菌是公认的阴道生态失调的指标。这些属的定植改变了免疫系统,并诱导了促炎反应。在宿主-微生物共培养模型中,加德纳菌与宫颈阴道屏障的破坏有关,同时促进小鼠中普雷沃菌属对上生殖道的侵入性感染。
总的来说,该研究结果表明,两组之间的阴道微生物组发生了实质性变化,突显了阴道冲洗液样本作为检测慢性子宫内膜炎的诊断工具的潜力。
◆阴道菌群的变化可引起宫腔的菌群失调
KEGG途径富集分析强调了关键代谢过程的上调,特别是CE组的聚糖生物合成和代谢。覆盖阴道上皮细胞的聚糖涂层是保护屏障的重要组成部分。乳杆菌与阴道上皮的粘附被认为是具有竞争性地抑制病原体,而上皮的糖胺聚糖在这些过程中起着至关重要的作用。
值得注意的是,糖胺聚糖的消耗与宫颈上皮细胞屏障功能的破坏有关,这可能会增加对局部细菌和上升感染的易感性。这表明CE患者的微生物组失调可能会对微生物侵入阴道的主要门户产生负面影响,并引发宫内炎症。
关于慢性子宫内膜炎(CE)的标准化定义和诊断指南仍存在争议,最被接受的诊断依赖于子宫内膜活检。基于选定的阴道微生物群构建了CE诊断模型。本文研究中,引入了一种新的随机森林模型,该模型可以根据ASV以更高的灵敏度和准确性将CE患者与使用阴道分泌物的健康个体区分开来。该研究结果表明,阴道微生物组的变化可以作为诊断CE的指标,为慢性子宫内膜炎提供了一种更容易获得和更方便患者的诊断选择。
然而,该研究也存在一定的局限性。首先,它主要侧重于检查有慢性子宫内膜炎和没有慢性子宫内膜炎的女性之间的成分差异,而不是研究与慢性炎症相关的免疫失调的潜在机制。其次,阴道微生物样本是在分泌中期收集的,这可能无法解释整个月经周期中微生物群组成的变化。第三,作为一项单中心调查,发现的普遍性可能有限。考虑到饮食和运动等因素,多中心方法可以提供更全面的解读。
最后,多种组学方法的整合(包括培养组学、元转录组学和代谢组学),可以提供对微生物群落功能的全面了解。这种更广泛的视角对于阐明阴道微生态系统内复杂的网络和阴道微环境的潜在变化至关重要。
阴道微生物生态失调是慢性子宫内膜炎的重要指标。该研究确定了与慢性子宫内膜炎发展相关的潜在候选细菌,并提出了潜在的机制。基于阴道微生物群的生物标志物有望成为检测慢性子宫内膜炎的非侵入性工具。需要进一步的研究来发现特定阴道微生物组在慢性子宫内膜炎中的功能,并确定具有诊断能力的强大阴道微生物标记物,以区分患有慢性子宫内膜炎的个体。

谷禾健康

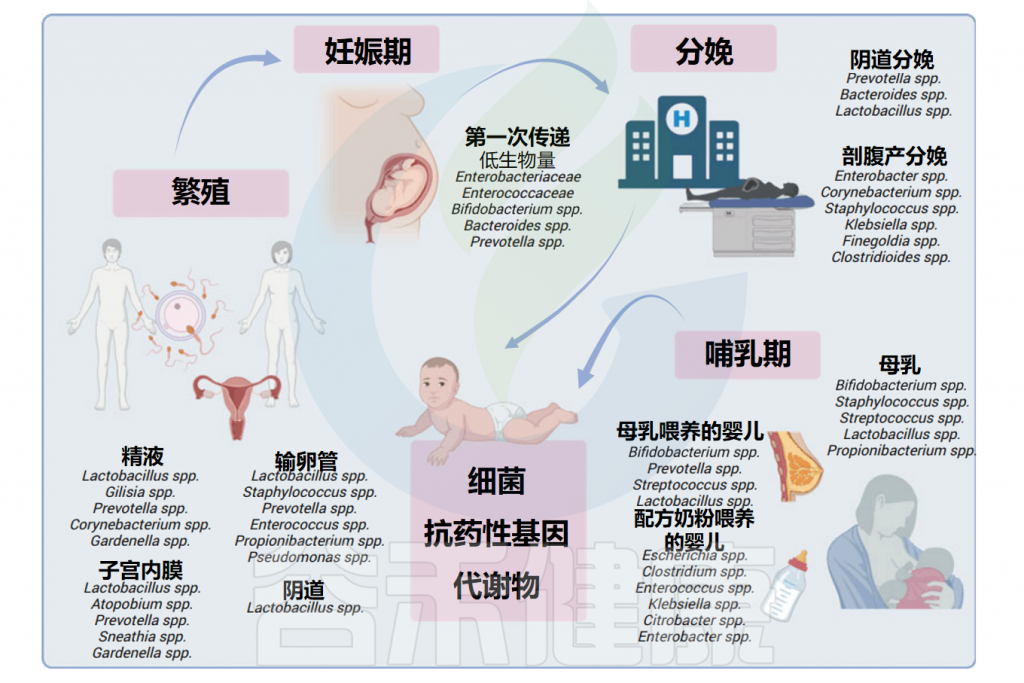
众多的文献和谷禾之前写过的文章都阐述了微生物群与人类健康密切相关。这种联系甚至可能在还未出生的时候就已经开始形成。微生物群的存在和活动从母体怀孕前到幼年的过程中,持续地参与和调节着人体的各种生理功能,从而在每一个生命阶段都扮演着至关重要的角色。
在受孕之前,男性和女性生殖道中的微生物就具有关键作用,它们可以在受孕期间产生影响。例如女性阴道、子宫颈内膜、子宫、输卵管和卵巢中的微生物群,其特征是生物量低,并且与生殖和子宫健康相关。精液微生物群可能会干扰受孕和早产。据报道,怀孕期间典型的精液细菌(如嗜酸乳杆菌)的存在率较高与早产有关,而另一种常见的精液细菌——卷曲乳杆菌(Lactobacillus crispatus)被确定为具有保护作用。
在怀孕期间,母体微生物的变化与所经历的生理和激素变化同步。阴道微生物多样性降低,乳杆菌成员增加,潜在地加强了它们的免疫保护功能,而较低水平的乳杆菌与早产风险增加相关。妊娠三个月后肠道中促炎的微生物更丰富,产短链脂肪酸的微生物减少。同时妊娠期口腔微生物群的特征是活菌数量增加,同时致病菌水平较高。需要注意的是,口腔生态失调与妊娠期糖尿病、先兆子痫和胎儿紊乱的风险增加相关。
此外,母婴间的垂直传播与婴儿健康相关。一些有益菌种(双歧杆菌、乳杆菌)可以从母亲传播到婴儿体内并定植。但与此同时,一些抗生素抗性基因也会随着微生物的转移,从而使新生儿对一些药物具有耐药性,这将不利于一些新生儿疾病的治疗。
因此了解从受孕到生命最初几年期间关键的微生物群非常重要。本文总结了有关人类生殖微生物群的现有证据,以及生命早期的微生物集群,重点是对婴儿发育和健康结果的潜在影响。
研究还发现:包括特定益生菌和其他益生菌混合物在内的一些饮食策略可能会成为潜在的工具,调节母婴微生物群,并在整个生命周期中实现健康结果。
★ 生命早期的微生物群对代谢和免疫非常重要
生命的前1000天(即从怀孕到出生后的前2年)对于先锋微生物在人体内的定植以及免疫系统的发育和成熟都是至关重要的。因此,这一时期被认为是一个“关键时期”,在此期间,任何事件都将对代谢、免疫和微生物行为产生关键影响,从而影响人类健康。
而对逐步形成的新生儿微生物群的任何改变和破坏都有可能在短期和长期增加个体患病的风险和倾向。
▸ 阴道微生物群
母体微生物群是新生儿最相关的产前和产后微生物来源。
阴道微生物群是一个复杂的生态系统,大约由200多种细菌组成,最主要的阴道种群是乳杆菌(Lactobacillus),在阴道中已检测出超过20种乳杆菌,占细菌群落的近70%。以及其他不太丰富的细菌,如普雷沃氏菌属、链球菌属、拟杆菌属和韦荣氏球菌属。
✦ 阴道微生物群主要分为五种类型
阴道微生物群主要分为五种类型,它们在组成和丰度方面都不同,每个类型都会以一种细菌为主导:
卷曲乳杆菌(CSTI),加氏乳杆菌(CSTII),惰性乳杆菌(CSTIII),多元组(CSTIV表现出较低的乳杆菌属丰度)和詹氏乳杆菌(CSTV)。
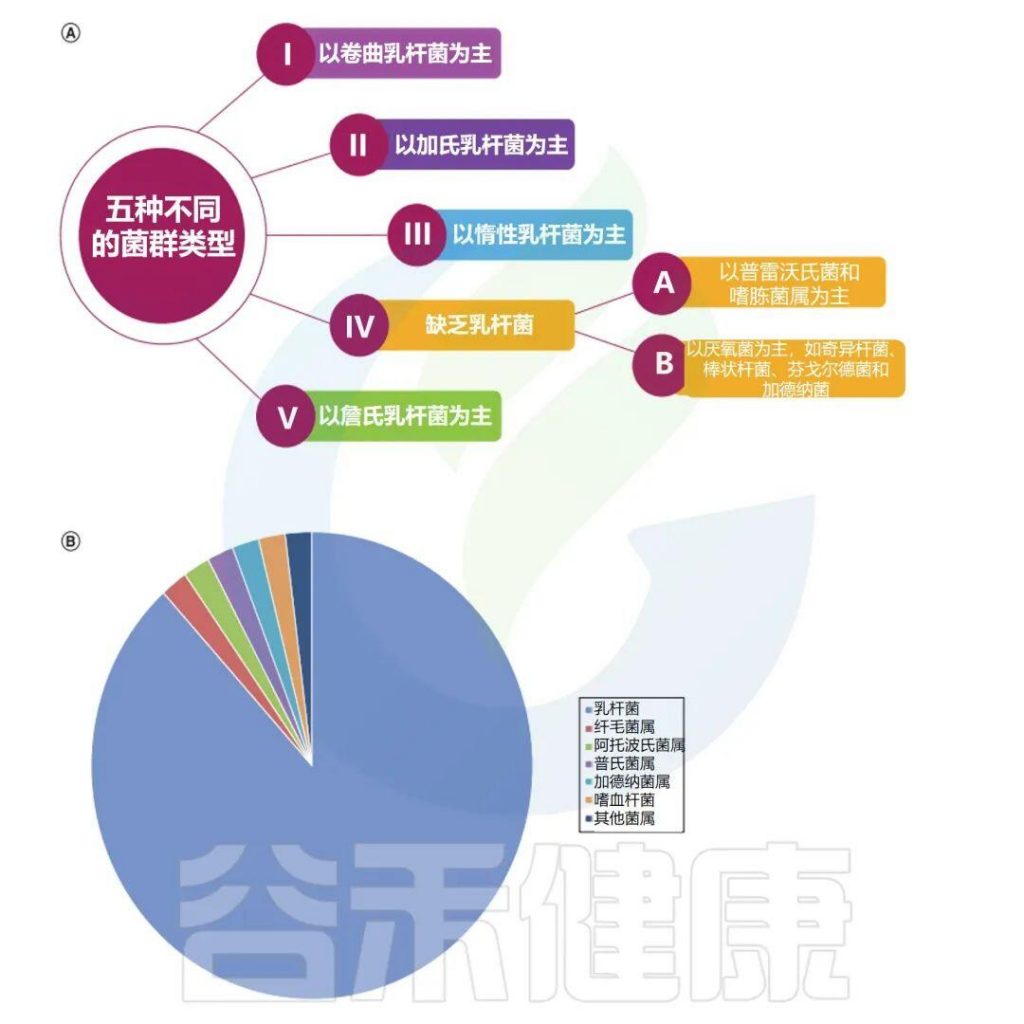
阴道微生物群会因地理位置、种族和卫生条件而异。例如,在美国的非裔美国人和西班牙裔妇女中CSTIV的丰度较高,这些成员在维持低pH值和代谢物分泌中起重要作用,以防止病原体在阴道中定植。
✦ 阴道微生物群会随激素水平、分娩、母乳喂养而变化
越来越多的研究证据表明阴道微生物群是阴道健康的重要生物标志物。然而,阴道微生物群并非一成不变。事实上,阴道微生物群显示出时间动态变化,有些群落发生变化,有些则相对稳定,这取决于核心菌群。

最近一项涉及比利时3345名妇女(18-98岁)的研究显示,阴道微生物群与激素变化(雌激素水平和避孕药的使用)正相关。
相反,阴道微生物群与分娩和母乳喂养等因素呈负相关。此外,同一项研究表明,富含阴道加德纳菌(Gardnerella)、普雷沃氏菌属(Prevotella)和拟杆菌(Bacteroides)的阴道微生物群与更年期、月经卫生和避孕药具使用有关。
▸ 子宫、输卵管的微生物群对生殖健康也有影响
女性生殖系统含有对女性健康有影响的细菌,这些细菌也存在于阴道之外。先前的几项研究表明,女性上生殖道(UFRT)中存在微生物群,包括子宫颈内膜、子宫、输卵管和卵巢,其特征是生物量低,并且与生殖和子宫健康相关。
✦ 子宫内膜也以乳杆菌为主,但与阴道菌群有所不同
子宫内膜微生物群的特征是大量乳杆菌(Lactobacillus),其次是加德纳菌(Gardnerella), 普雷沃氏菌属、Atopobium和Sneathia,这也在阴道中发现。然而,似乎子宫内膜细菌群与阴道中的细菌群有些不同,这表明这两种微生物群是相关的,但并不相同。
✦ 输卵管中的菌群
此外,输卵管中也有一些细菌定居,如乳杆菌属、葡萄球菌属、肠球菌属、普雷沃氏菌属和丙酸杆菌,与假单胞菌属。
▸ 男性生殖道的微生物群
之前很少有人研究男性生殖道中的微生物群。最近的研究显示,精液中存在微生物群,其中芽孢杆菌门(Bacillota)是最主要的门,其次包括拟杆菌门、假单胞菌门和放线菌门,其中最常见的属有乳杆菌属、普雷沃氏菌属、Gillisia、棒状杆菌属和加德纳菌。
✦ 男性的生殖道微生物群对生育健康有一定影响
阴道加德纳菌在阴道微生物群中的优势可能与男性炎症有关,这或与阴道定植/感染有关。有人认为,男性和女性生殖微生物群之间存在联系,影响生育、生殖健康和妊娠。
事实上,之前的大多数关于精液微生物组研究都集中在不孕不育问题上,突出了微生物特征和多样性的差异。据报道,男性微生物群受到包括环境在内的不同因素的影响。此外,精液微生物群可能会干扰受孕和早产。据报道,怀孕期间典型的精液细菌(如嗜酸乳杆菌)的存在率较高与早产有关,而另一种常见的精液细菌——卷曲乳杆菌(Lactobacillus crispatus)的优势已被确定为具有保护作用。
总之,女性和男性的生殖微生物组与生殖健康密切相关。研究显示,微生物可作为生物标志物,有助于开发诊断和治疗不孕症的新工具。
生殖过程发生在“微生物环境”下,这些微生物及其代谢物需得到更多关注,因为它们与孕前阶段密切相关。
在怀孕期间,妇女的生理和代谢有所变化,以便为胎儿提供最佳子宫内环境,促进其健康发育。妊娠期间的内分泌、免疫和代谢变化,会营造一种促炎状态,反映在阴道、肠道和口腔的微生物群变化中。
研究微生物标志物与妊娠结局之间的关系,有助于预防和调整与妊娠并发症相关的不利微生物变化。
▸ 阴道微生物群
据报道,怀孕后阴道微生物多样性降低,而乳杆菌成员增加,潜在地加强了它们的保护功能。
✦ 较低水平的乳杆菌与早产风险增加相关
就个别物种而言,卷曲乳杆菌与来自足月妊娠的样本相关。进一步证明,病史包括反复尿路感染的妇女表现出早产风险增加。在足月分娩和早产分娩之间,已经观察到阴道微生物群在组成、稳定性和多样性方面的差异。
加特纳菌(Gardnerella)、Atopobium和Uraplasma spp.的存在,以及较低水平的乳杆菌成员或较高水平的白色念珠菌的存在,已被发现与早产风险有关。
因此,在妊娠期间检测阴道微生物组内的异常可以用作预测早产可能性的微生物生物标志物,特别是在妊娠前三个月检查阴道微生物组。

最近的一项研究报告称,无抗生素的阴道微生物群移植(VMT)和供体移植解决了复发性流产后的阴道生态失调问题。这些研究表明,VMT是一种潜在的治疗具有不良生育和妊娠结局的严重阴道生态失调的方法,然而,还需要进一步的研究。
▸ 肠道微生物群
虽然在妊娠的前三个月期间,肠道微生物群的组成保持稳定,并且类似于孕前微生物群,但是从妊娠的前三个月结束起,其组成发生了根本的变化。
✦ 妊娠三个月后促炎的微生物更丰富,产短链脂肪酸的微生物减少
这些变化反映了更具促炎性的特征,包括妊娠晚期假单胞菌门(肠杆菌科成员)和放线菌门(主要是双歧杆菌成员)的增加,以及短链脂肪酸(SCFA)生产者的减少。
肠道微生物群与多种妊娠表型有关,包括体重增加、轻度炎症和胰岛素抵抗。
✦ 怀孕期间的饮食、营养、抗生素的使用会影响后代的微生物
已经有研究广泛证明,怀孕期间暴露于抗生素会改变母亲的微生物生态系统,从而改变后代的微生物生态系统。怀孕期间使用抗生素会增加阴道内葡萄球菌物种,此外,肠道以外的其他区域的潜在微生物转移可能与过敏和肥胖的风险增加有关。
此外,微生物群在怀孕期间受到母亲的饮食和营养状况的影响,也对其后代的微生物群组成产生影响。人体研究表明,妊娠期女性肠道微生物群的变化易受孕前母体最大体重指数(身体质量指数)的影响,也易受妊娠期体重增加的影响。较低的双歧杆菌属物种在超重和肥胖的母亲以及在怀孕期间体重增加过多的母亲中观察到。
另一项研究报告了怀孕期间体重状况的类似变化和较低水平的拟杆菌物种,以及更丰富的葡萄球菌和大肠杆菌物种在超重孕妇中被发现。
母体肠道微生物群的这种变化可能与通过阴道分娩出生的新生儿的肠道定植过程的差异有关。
▸ 口腔微生物群
怀孕还会引起口腔微生物群的变化。比较孕妇和非孕妇口腔中几种物种的丰度时,已经报道了显著的差异。
✦ 怀孕时口腔中的微生物数量增加,致病菌水平也较高
妊娠期口腔微生物群的特征是活菌数量增加,同时致病菌水平较高,例如牙龈卟啉单胞菌(Porphyromonas gingivalis),Aggregatibacter actinomycetemcomitans和假丝酵母(Candida genus)。
✦ 口腔生态失调与妊娠期并发症存在一定关联
越来越多的证据表明口腔感染和与妊娠并发症(包括早产)之间的联系。还有证据表明,妊娠期口腔生态失调与妊娠期糖尿病、先兆子痫和胎儿紊乱的风险增加相关。
这些数据表明,口腔微生物群在孕产妇和新生儿结局中也起着关键作用,包括妊娠并发症、早产风险和早期胎儿/新生儿微生物定植。
越来越多的证据表明了细菌从母亲到孩子的垂直传播。垂直传播是一个显著的过程,其中细菌菌株在母亲和新生儿之间交换。
研究人员探索了各种母体菌群来源,包括粪便、母乳、阴道微生物群和皮肤,以揭示这些关键细菌菌株的来源。影响婴儿肠道定植的关键因素包括出生时的胎龄、分娩方式、产前暴露、遗传、抗生素的使用和是否纯母乳喂养。
▸ 特定的垂直传播——例如双歧杆菌
双歧杆菌属(Bifidobacterium)是从母亲传播到婴儿肠道的最常见的微生物,主要传播源是母乳和粪便,出生时的胎龄、出生地、分娩方式和抗生素使用等变量可能会影响双歧杆菌的状态。
✦ 双歧杆菌等菌属从出生时自母亲传播到婴儿体内并定植
已知胃肠道微生物的早期正常定殖有助于免疫系统的发育,并有助于通过调节环境来维持体内平衡。

一项纵向研究表明长双歧杆菌(B.longum)和B. Brief的菌株似乎从出生到4个月一直存在于婴儿的微生物生态系统中。
通过宏基因组测序,我们深入了解了母体肠道到婴儿肠道的传播,外部因素的影响——地点、出生模式和抗生素的使用,每个因素都在形成婴儿微生物群中发挥作用。
双歧杆菌属物种表现出持久性和适应性。与母乳中发现的生物活性化合物(如母乳低聚糖)一起,在支持婴儿肠道微生物群和免疫系统的成熟方面发挥着至关重要的作用。
▸ 抗生素抗性基因转移
双歧杆菌等有益菌可以由母亲传递给婴儿并定植,与此同时,一些带有抗生素抗性基因的微生物也在母体到婴儿之间转移。
抗生素抗性微生物(ARMs)分布广泛,在人、动物之间传播,并通过动物性食物传播。它们携带抗生素抗性基因(ARG),整个细菌群体中的ARG组被称为抗性组。
✦ 耐药病原体可能从母体转移至新生儿体内
生命早期,尤其是围产期和产后,是婴儿肠道微生物群和免疫力发育的关键时期。此外,婴儿的免疫系统尚未成熟,因此他们特别容易获得和携带耐药病原体。这导致每年估计有214000名新生儿因抗生素抗性病原体引起的脓毒性感染而死亡。
婴儿肠道中抗生素耐药微生物菌株的主要来源尚不确定;虽然母体微生物群是主要来源,但其他个体和环境也是相关的,并可能导致婴儿耐药性。微生物在出生、哺乳和密切接触期间从母亲传播给后代。
✦ 母乳喂养、剖腹产等因素影响抗性基因的转移
各种围产期因素,如剖腹产、抗生素暴露、早产和母乳喂养会影响微生物从母亲到婴儿的垂直传播。一些研究阐明了抗生素抗性基因(ARG)在新生儿出生后不久就存在,表明ARMs的最初定殖发生在出生后不久,可能来自与母亲或医院环境的接触。一些研究表明,母亲向婴儿传播耐药细菌,包括各种抗生素耐药菌株,如大肠杆菌, 链球菌等。
众所周知,母乳喂养对婴儿健康有益,为婴儿提供营养和细菌。然而,对于发育中的婴儿来说,母乳似乎是耐药细菌和精氨酸的来源之一。在人乳中发现了耐药菌株。宏基因组分析揭示了母乳中高水平的精氨酸和遗传元素,类似于婴儿对母乳的抵抗力。这表明母亲通过抗性细菌的转移在发展婴儿的肠道抗性中起作用。
需要更多的研究来了解耐药细菌在母体与婴儿间的垂直传播,以理解其对婴儿最初肠道定植的影响。这种更深入的理解将有助于开发控制抗生素耐药基因在母婴之间传播的策略。
在出生时,除了来自环境的微生物之外,新生儿还会遇到来自阴道和母体肠道的数量巨大、种类繁多的微生物。
新生儿中的微生物群定植是一个与健康密切相关的敏感、动态过程。早期多样的微生物建立和特定短链脂肪酸的产生对免疫系统的成熟至关重要。
▸ 最早的新生儿微生物群
✦ 母体与婴儿之间的微生物接触最早可能始于子宫收缩时
不同的研究报告了通过培养依赖性和独立性方法确定的第一次粪便中存在细菌,即胎粪。一些研究人员认为,微生物接触可能最早始于子宫收缩和胎膜破裂期间,从而解释了胎粪中细菌的存在。
首个微生物群由变形菌门(Pseudomonadota)和芽孢杆菌门(Bacillota)的物种组成,如肠杆菌科(Enterobacteriaceae)和肠球菌科(Enterococcaceae)。
此外,还发现了其他菌属,包括双歧杆菌属、拟杆菌属和普雷沃氏菌属。与7天新生儿微生物群相比,首次通过的胎粪微生物群显示出更高的微生物多样性和丰富性,这表明新生儿出生时大量接触微生物,但只有少数能够在肠道中定植并持续存在,以便在出生后的第一年逐步进化。
▸ 影响新生儿微生物群的因素
肠道微生物群会随时间进化,变得更加多样。因此,新生儿定植是一个脆弱的、动态的和逐步的过程,它会受到许多母婴因素以及环境因素的影响。
其中,最相关的因素包括:孕龄、分娩方式(阴道分娩与剖腹产)、地点(医院与家庭)、喂养方式(母乳、混合膳食或婴儿配方奶粉)以及产妇健康状况、遗传、药物、接触异生物质或物理化学制剂的环境、兄弟姐妹、宠物以及人与人之间的水平微生物传播等。
✦ 抗生素的使用

分娩期抗生素预防可能会改变出生后第一周的肠道微生物群,包括放线菌的减少以及假单胞菌属和杆菌属比例的稳步增加。
另一项研究报道,分娩期间施用的抗生素也可以改变新生儿的口腔微生物群,从而导致在抗生素治疗后假单胞菌门的丰度更高。这可能会对儿童发育产生重要影响。
此外,据观察,在妊娠中期和晚期接受抗生素治疗的儿童患肥胖症的风险增加了84%,这可能是由于肠道微生物的变化引起的。
✦ 早产影响婴儿的肠道微生物群平衡
此外,有几项研究报道早产婴儿的微生物群是不平衡的。例如,已有研究表明,与足月婴儿相比,早产胎粪微生物组导致无菌小鼠粪便移植后免疫功能改变、生长受限和激素水平变化。
然而,早产儿有其他合并症,因为他们大多数是通过剖腹产出生的,在出生时和住院期间暴露于抗生素;他们也用母乳喂养,并在新生儿重症监护室呆了更长时间,这些因素可能都会影响早产婴儿的肠道微生物群。
✦ 母亲的健康状况也会影响婴儿的微生物群
也有越来越多的证据表明,子宫内炎症导致围产期并发症增加。婴儿健康和微生物群的母婴传播之间的关联可能受到诸如母亲糖尿病等因素的驱动、特应性疾病、饮食,身体质量指数和围产期抗生素使用。
研究人员观察到患有妊娠期糖尿病(GDM)前期的母亲所生婴儿的胎粪中含有丰富的细菌,这些细菌通常在成年糖尿病患者中观察到。母亲的高脂肪饮食影响婴儿微生物群,表现为拟杆菌属显著减少。
此外,婴儿微生物群影响免疫系统发育的分子机制尚未完全了解,尽管有越来越多的证据表明肠道微生物群在儿童期和成年期的疾病和健康中具有免疫调节活性。
✦ 顺产的婴儿微生物群与母亲肠道微生物更相似
先前已经证明,阴道出生的新生儿具有类似于阴道和肠道母体微生物群的微生物群,富含乳杆菌和普雷沃氏菌,尽管也存在其他细菌,如肠杆菌科,包括埃希氏菌属或克雷伯菌属。
而通过剖腹产出生的新生儿呈现出与口腔、皮肤和环境微生物群相似的独特微生物群。母体肠道似乎也是早期定植细菌的重要来源,因为在经阴道分娩的新生儿中,已发现72%的肠道细菌来自母体肠道,相比之下,经剖腹产出生的受试者中只有41%来自母体肠道,已显示双歧杆菌属在新生儿肠道定植过程中作为先锋细菌的成员,剖腹产出生的婴儿表现出拟杆菌属的延迟定植和多样性降低。
✦ 母乳喂养有助于母亲的微生物群在婴儿中定植
出生后,母乳喂养既支持了微生物的肠道定殖,又促进了免疫系统的成熟。据报道,母乳喂养和配方奶粉喂养的新生儿微生物群之间存在差异。
此外,母乳喂养的停止,而不是补充和固体食物的引入,对婴儿微生物群产生了关键影响。传统上,母乳被认为是无菌的。尽管如此,证据表明,母乳样本中存在活微生物,包括葡萄球菌和链球菌,是最丰富的微生物群,其次是乳酸菌、双歧杆菌和某些假单胞菌(Pseudomonas和Acinetobacter)。

最近的一项研究表明,婴儿肠道中27.7%的细菌来自母乳,10.3%来自乳晕皮肤,从而突显了母乳喂养对婴儿肠道微生物群的潜在影响。
此外,母乳和婴儿肠道之间共享的微生物和病毒种类也有被发现。
饮食是塑造我们肠道微生物群的一个关键因素。因此,建议进行饮食和营养咨询以及营养补充,以便在孕前和妊娠阶段达到适当的营养状态。最近有报道称,怀孕期间以及随后的母乳喂养期间,母亲的饮食对母亲和婴儿的微生物群都有重要影响。
★ 地中海饮食有助于改善母婴的健康水平
母亲的蛋白和纤维摄入量会影响母乳成分,如微生物群、脂质、母乳低聚糖(HMO)和microRNA表达谱。地中海饮食(MD)被证明在怀孕期间能够改善母婴健康结果,包括生长受限、胎儿和新生儿大脑发育、儿童神经发育。
妊娠期地中海饮食,主要是水果、蔬菜和豆类的摄入量增加,红肉的摄入量减少,与Acidaminoacaeae家族水平以及短链脂肪酸生产者的丰度较高有关。
植物蛋白、纤维和多酚的摄入量增加与瘤胃球菌属(Ruminococcus)、Christenselacae科、Dehalobacterium和真/优杆菌属(Eubacterium)以及短链脂肪酸生产菌的存在增加有关。
✦ 饮食不仅改善肠道微生物,对阴道、口腔微生物也有影响
大多数关于饮食-怀孕-微生物群的可用数据都集中在肠道微生物群上,但据报道,增加水果、维生素D、纤维和酸奶的摄入量与女性阴道卷曲乳杆菌(L.Crispatus)有关;纤维摄入与较高的异质性相关。
有限的证据表明饮食对阴道微生物群和妊娠结局的影响。在这种情况下,仍需确定哪些食物、食物成分、营养素和其他膳食化合物会影响围产期妇女的微生物群(肠道、口腔和阴道),以及对母婴结局的潜在影响。
对围产期饮食干预的研究兴趣日益增加,以及在怀孕期间使用益生菌、益生元和共生体来调节微生物群和促进“有利的微生物垂直传播”。有一些证据表明,在怀孕和哺乳期间服用益生菌可以起到保护作用,而益生元和其他益生菌的证据较少。
值得注意的是,脆弱和敏感人群需要特殊的安全性考虑,包括安全性和与妊娠、哺乳和婴儿期相关的不良反应的潜在报告。
▸ 益生菌对生育能力的影响
补充乳杆菌有利于重建阴道菌群平衡
阴道感染会影响乳杆菌的平衡,导致可能影响生育能力的细菌的生长。因此,补充乳杆菌应有助于重建平衡。
一项涉及健康女性的体外受精研究显示,在阴道定植卷曲乳杆菌(L.Crispatus)后,受孕结果很有希望。
补充益生菌有助于提高精子质量
在不孕症和辅助生殖医学的病例中,已报道了精液微生物群组成的差异,使用益生菌可能在精液特征和质量参数方面具有有益的效果。研究已经显示出Lacticaseibacillus rhamnosus CECT8361和B.longum CECT7347在弱精子症男性中,补充为期六周可以改善精子质量参数和活力,并降低DNA片段和活性氧(ROS)水平。
另一方面,有报道称,共生菌(Flortec, Lacticaseibacillus paracasei B21060 和阿拉伯半乳聚糖、低聚果糖和l-谷氨酰胺)对少弱畸形精子症患者精液质量和数量具有一定的改善作用。
此外,系统综述和荟萃分析表明,益生菌有利于精子活力。然而,关于益生菌对接受辅助生殖的女性的功效,存在相互矛盾的证据。这表明需要更多临床研究来证明益生菌对生育能力的影响。
▸妊娠期使用益生菌降低疾病风险
孕期服用益生菌为减少母婴健康风险提供了方案。现有研究主要集中在早产风险、减少生殖器感染、妊娠期糖尿病(GDM)和先兆子痫前期等方面,这些往往被视为次要健康结果。
益生菌有助于改善孕妇生殖器感染
孕妇经常患有复发性尿路感染(UTIs)、外阴阴道念珠菌病(VVC)和细菌性阴道病(BV)。这些问题与不利的母婴妊娠结果有关,包括早产风险和胎儿生长受限。益生菌已被提出作为治疗BV和UTI的预防性替代品,通过维持或恢复阴道微生物群或防止病原体在尿道定植。
先前的研究表明,益生菌组合,即嗜酸乳杆菌、乳双歧杆菌(B.lactis)和长双歧杆菌(B.longum),对孕妇B组链球菌(GBS)定植具有潜在的治疗作用。

此外,最近的研究表明益生菌菌株在治疗妊娠期GBS感染方面的有效性,其中一项临床研究报告称,补充鼠李糖乳杆菌GR-1和罗伊氏乳杆菌RC-14被证明对治疗GBS感染有效。
最近一项涉及36名GBS阳性女性的研究报告称阴道共生菌显著增加,对阴道GBS有影响。而一项对18项研究的系统综述和荟萃分析表明,抗生素和益生菌显著降低了细菌性阴道病(BV)的复发率,提高了治愈/缓解率。其他报告益生菌对阴道炎影响的研究也显示了类似的结果。
✦ 降低先兆子痫的风险

最近一项涉及70149名孕妇的研究发现,益生菌(嗜酸乳杆菌[LA-5],乳双歧杆菌[Bb12]还有鼠李糖乳酸杆菌GG [LGG])显著降低先兆子痫的风险。
这项研究还强调了“时机”(即何时进行益生菌干预)的相关性,因为在怀孕前或怀孕早期食用益生菌没有观察到任何影响。孕晚期(而非孕前或孕早期)摄入益生菌牛奶与降低先兆子痫风险显著相关(调整后的 OR:0.80(95% CI 0.68 至 0.94)p 值:0.007)。孕早期(而非孕前或孕晚期)摄入益生菌与降低早产风险显著相关(调整后的 OR:0.79(0.64 至 0.97)p 值:0.03)。
然而另一项系统综述报告称,与安慰剂相比,益生菌会导致肥胖受试者的先兆子痫风险增加。肥胖与先兆子痫的风险较高有关。因此,需要更多的研究来确定弱势群体中与益生菌相关的风险。
预防妊娠期糖尿病,改善胰岛素敏感性
鼠李糖乳杆菌(L.rhamnosus)GG和乳双歧杆菌(B.lactis)BB12与饮食相结合已被证明可以预防妊娠期糖尿病(GDM),降低血糖浓度和改善胰岛素敏感性。
另一种益生菌菌株,鼠李糖乳杆菌(L.rhamnosus) HN001菌株能够降低GDM的患病率,特别是在中老年妇女和那些以前患有妊娠期糖尿病的妇女中。
最近的一项系统综述包括30多项在怀孕期间进行的研究,表明益生菌干预对改善血糖控制、降低低密度脂蛋白胆固醇(VDL)和甘油三酯以及减少炎症标记物的潜在作用。
益生菌补充剂可能会降低妊娠期糖尿病(GDM)的发病率,并有助于控制健康孕妇和超重和肥胖妇女的血糖参数。
最近的荟萃分析显示,益生菌将GDM发病率降低了33%,当使用多种菌株益生菌组合时,效果更好。一项系统综述和荟萃分析还报告称,摄入酸奶(活微生物)可以降低GDM的风险,并降低空腹血糖。
减少产后压力和抑郁
越来越多的数据显示益生菌对抑郁、焦虑和情绪的潜在用途。一项研究(系统综述和荟萃分析)显示,益生菌在减少孕妇和哺乳期妇女的焦虑和抑郁方面具有积极作用。
其他研究报告了益生菌和对照组在抑郁评分方面没有差异,然而,在总体精神健康评分方面,报告了焦虑的降低。
▸ 出生后使用益生菌的有益作用
文献中有数据和研究表明特定益生菌的有益作用,例如布拉氏酵母菌,鼠李糖乳酸杆菌GG(LGG),罗伊氏乳杆菌和动物乳双歧杆菌(Bb12)。
婴儿早期使用益生菌的主要目标是:
1)预防腹泻和感染;
2)预防过敏;
3)预防坏死性小肠结肠炎和早产儿迟发性败血症、预防婴儿绞痛。
此外,一项系统综述强调了灭活益生菌的潜在益处,灭活益生菌在生命的早期和敏感期以及敏感人群是一种安全的替代品,如早产儿和儿科人群。
产后使用益生菌的另一个例子与患有乳腺炎的女性有关,包括感染性乳腺炎和葡萄球菌乳腺炎。
在探索微生物群与人类健康之间的复杂关系时,我们发现,从受孕到生命最初几年,微生物群在每一个生命阶段都扮演着不可或缺的角色。母体的微生物环境不仅影响着怀孕过程中的生理变化,还直接关系到婴儿的健康和发育。通过了解这些微生物的作用,我们可以更好地认识到早期干预的重要性,尤其是通过饮食和生活方式的调整来优化母婴微生物群。
随着科学研究的不断深入,我们有望开发出更有效的策略,以促进母婴健康,预防潜在的健康问题。未来的研究将继续揭示微生物群的奥秘,帮助我们在整个生命周期中实现更好的健康结果。
主要参考文献
Samarra A, Flores E, Bernabeu M, Cabrera-Rubio R, Bäuerl C, Selma-Royo M, Collado MC. Shaping Microbiota During the First 1000 Days of Life. Adv Exp Med Biol. 2024;1449:1-28.
Arzamasov AA, Nakajima A, Sakanaka M, Ojima MN, Katayama T, Rodionov DA, Osterman AL (2022) Human milk oligosaccharide utilization in intestinal Bifidobacteria is governed by global transcriptional regulator NagR. mSystems 7(5):e00343–e00322.
Calvo-Lerma J, Selma-Royo M, Hervas D, Yang B, Intonen L, González S, Martínez-Costa C, Linderborg KM, Collado MC (2022) Breast milk lipidome is associated with maternal diet and infants’ growth. Front Nutr 9:854786.
Canha-Gouveia A, Pérez-Prieto I, Rodríguez CM, Escamez T, Leonés-Baños I, Salas-Espejo E, Prieto-Sánchez MT, Sánchez-Ferrer ML, Coy P, Altmäe S (2023) The female upper reproductive tract harbors endogenous microbial profiles. Front Endocrinol 14:1096050.
Mikami K, Kimura M, Takahashi H (2012) Influence of maternal Bifidobacteria on the development of gut Bifidobacteria in infants. Pharmaceuticals 5(6):629–642.
Garcia-Mantrana I, Selma-Royo M, Gonzalez S, Parra-Llorca A, Martinez-Costa C, Collado MC (2020) Distinct maternal microbiota clusters are associated with diet during pregnancy: impact on neonatal microbiota and infant growth during the first 18 months of life. Gut Microbes 11(4):962–978.
García-Mantrana I, Bertua B, Martínez-Costa C, Collado MC (2016) Perinatal nutrition: how to take care of the gut microbiota? Clin Nutr Exp 6:3–16.
Li W, Tapiainen T, Brinkac L, Lorenzi HA, Moncera K, Tejesvi MV, Salo J, Nelson KE (2021) Vertical transmission of gut microbiome and antimicrobial resistance genes in infants exposed to antibiotics at birth. J Infect Dis 224(7):1236–1246.