-

 国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子
国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子

 二级病原微生物安全实验室
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业

谷禾健康
“从来没吃过四环素类抗生素,为什么肠道宏基因组报告里会检出四环素耐药基因?”
很多人第一次看到这类结果时,都会有类似的困惑。“耐药风险”“耐药基因”“抗生素耐药相关通路”“ARGs”等这些是什么意思?
如果报告里还有“β-内酰胺类耐药基因”、“大环内酯类耐药基因”、“耐药风险中等”、“耐药相关基因丰度升高”,就更担心了。
这些担心可以理解,但从专业检测角度看,耐药基因检出并不等于感染,不等于临床耐药,也不等于一定吃过抗生素。它更多时候提示的是:在样本中的微生物群落里,存在某些与抗生素耐药相关的遗传信息。
这个信号有参考价值,但它还不是最终结论。它到底意味着什么,要继续看:
本文会用尽量通俗的语言,系统解释16S测序和宏基因组测序报告中的耐药风险、耐药基因、临床药敏之间到底是什么关系,以及拿到报告后应该如何看待。
报告里的“耐药风险”“耐药基因”,和临床上说的耐药,其实并不是一回事。首先我们来了解一下这些概念:
★ 耐药风险:是报告给出的提示
它提醒样本里有一些和耐药相关的信号,但还需要结合样本来源、病原体和临床情况判断。
★ 耐药基因:是DNA层面的线索
它说明样本中存在某些与抗生素耐受、降解、外排或修饰相关的遗传信息。但耐药基因存在,不代表一定表达,也不代表正在感染。
★ 表型耐药:是药敏试验看到的实际表现
也就是把活菌培养出来后,看它对某种抗生素是敏感、中介,还是耐药。临床常见的S、I、R和MIC,主要来自这个层面。
★ 临床耐药:是患者真实治疗中的结果
它不仅和细菌有关,还和感染部位、药物浓度、免疫状态、感染灶处理、是否混合感染等因素有关。
简单来说,这几个词的关系可以理解为:
-耐药风险,是提醒;
-耐药基因,是线索;
-表型耐药,是实验室证据;
-临床耐药,是最终综合判断(一般是临床医生下结论)。
编辑
前面我们了解了,耐药风险、耐药基因等大概是什么意思,这一章节,我们来看不同技术报告里的“耐药”有什么区别。
常见的检测包括:16S rRNA、宏基因组mNGS、靶向tNGS。这些技术的检测目标、覆盖范围、敏感性、成本和适用场景不同。
16S rRNA:耐药风险多为间接推断
16S rRNA测序常用于肠道菌群、口腔菌群、阴道菌群、皮肤菌群等微生态评估。
16S 看的是菌群结构,不是完整耐药基因
它主要是在讲:
需要强调的是:常规16S测序并不直接检测完整耐药基因。
因为16S测的是一个相对保守的标记基因,不是完整基因组。它可以帮助识别细菌组成和丰度,但不能像宏基因组那样直接读取大量功能基因。
编辑
<来源:谷禾健康肠道菌群检测数据库>
因此,16S报告里的“耐药风险”,多数情况下是基于样本中的细菌组成,再结合数据库中这些菌属或菌种常见携带耐药基因的背景,间接评估样本携带耐药基因的可能性和耐药风险高或者低。
换句话说,16S更适合告诉我们:菌群生态里有没有某些风险信号。
它不适合直接回答:这个抗生素到底能不能用。所以,如果16S报告写着“β-内酰胺类耐药风险升高”,更合理的理解是:样本菌群中存在某些与这类耐药背景相关的生态特征,需要结合具体情况看,而不是直接得出“头孢一定无效”。
适用场景
16S里的耐药风险,更适合用于菌群生态观察、健康人群菌群检测、长期趋势评估、阴道/肠道微生态研究、大规模队列或环境耐药组普查等场景,不适合作为临床感染用药的直接依据,只能提供病原菌耐药的风险提示。
编辑
宏基因组:可以发现耐药基因,但不等于药敏
宏基因组测序,也就是mNGS,通常是对样本中的总DNA进行测序。
全面探索病原体与耐药线索
它的优势是覆盖范围广,可以无偏倚地全面检测样本中所有微生物的耐药基因,理论上能发现所有已知耐药机制。
宏基因组可以把样本中的DNA片段与耐药基因数据库进行比对。如果某些序列与已知耐药基因高度相似,就可能被报告为相应耐药基因。
编辑
比如:
适用场景
相比16S,宏基因组对耐药基因的检测更直接,也更适合病因未明的复杂感染、免疫抑制患者感染、经验治疗失败、混合感染等需要全面探索的场景。
它像是在一个复杂样本里尽可能广泛地翻找线索,帮助临床发现罕见或意想不到的病原体和耐药机制。
宏基因组也有限制
它受整体测序深度影响,对于低丰度病原体中的特定耐药突变,敏感性往往不足,且数据量大、分析周期长、成本相对较高。
编辑
tNGS:聚焦重点靶标,敏感性更高
靶向高通量测序,也就是tNGS,是针对临床上最关注的高优先级病原菌和重要耐药基因,设计特异性引物或探针,只检测目标区域。
它不像宏基因组那样尽可能检测样本中所有微生物,而是更像带着清单去找重点目标。
这种设计有一个明显好处,测序深度集中在目标区域,所以敏感性显著提高,能够检测到低丰度病原体携带的耐药基因和耐药突变,结果也更容易解释,往往能直接对应到具体药物类别。
适用场景
非常适合已经怀疑特定病原体感染、需要快速获得精准耐药信息指导临床用药的场景,是平衡检测范围、敏感性和成本的务实选择。
编辑
总的来说,
这是很多人最困惑的地方。
“我明明没怎么吃抗生素,为什么报告里会有耐药基因?” 很多人以为耐药基因只有在医院、重症感染或长期抗生素使用者身上才会出现,那么耐药基因的出现,是不是已经很严重了?
先不用这样理解。
抗生素使用确实会促进耐药菌和耐药基因的富集,但它不是唯一原因。原因很简单:
耐药基因广泛存在于自然界和人体微生态中
抗生素并不是人类发明后才出现的。许多抗生素本来就来源于微生物,比如青霉素来自真菌,链霉素来自放线菌。微生物之间在自然界中长期竞争,产生抗菌物质,也进化出保护自己的机制。因此,耐药基因在土壤、水体、动物肠道、人类肠道中本来就可能存在。
人类大规模使用抗生素以后,确实加速了耐药基因的筛选和传播,但这不等于每一次耐药基因检出都能追溯到某个人近期吃过抗生素。
一个典型例子:四环素耐药基因
四环素类抗生素曾在畜禽、水产养殖中被广泛使用。长期选择压力下,动物肠道、粪便、养殖废水、养殖场土壤中,都可能富集四环素耐药菌或四环素耐药基因,比如常见的 tet类基因。
这些基因可能随着动物粪便进入土壤或水体,也可能通过养殖废水、农田粪肥、蔬菜表面、水产品和肉类加工处理等环节,进入更大的环境循环。人再通过食物、手部接触或环境接触,摄入少量相关菌或DNA片段。
编辑
所以,一个人的肠道宏基因组里检出 tet类四环素耐药基因,并不一定说明他近期吃过四环素。它也可能反映食物链、环境暴露或社区微生物交换。
日常接触,也可能留下低丰度信号
生活中类似的场景也不少。比如:
这些情况通常不意味着感染,也不意味着一定会耐药。但从检测角度看,它们可能让样本中出现一些低丰度的耐药基因信号。
质粒传播:细菌之间的移动U盘
还有一些耐药基因可以通过质粒等可移动遗传元件在细菌之间传播。
可以把质粒理解为细菌之间传递的小型U盘:一个细菌把抵抗某类抗生素的说明书传给另一个细菌,后者就可能获得新的耐药能力。
编辑
不过,检测到耐药基因,并不代表正在发生质粒转移,也不代表它已经转移到了致病菌身上。
报告通常只能提示存在相关基因或相关序列,是否具备转移能力、是否在病原菌中、是否表达,还需要更深入的分析或实验验证。
所以,没吃抗生素却检出耐药基因,并不奇怪,
耐药基因的来源可能很多,有些只是微生态里的背景信号,不能只用“有没有吃过抗生素”来解释。
但背景信号不等于没有意义。需要继续判断的是:这个基因只是背景信号,还是已经和某个明确病原体、某次感染、某类用药产生关系?
这就进入了下一个问题。
耐药基因不是一检出就自动决定能不能用药。它要影响用药判断,中间至少要看几层关系。
第一层:检出的是DNA,不一定是活菌
很多测序检测,尤其是宏基因组测序,看到的是样本里的DNA。DNA可能来自活菌,也可能来自死菌、破碎菌体,甚至是游离DNA片段。
比如感染治疗后,病原体可能已经被部分清除,但样本里仍然残留一些微生物DNA。这个时候,检测结果可能还能看到相关基因信号,但它不一定代表样本里还有大量活的耐药菌。
第二层:它是不是在真正的病原体身上?
这一步更关键。
同一个耐药基因,如果在普通共生菌身上,和在明确致病菌身上,意义完全不同。
比如,粪便样本里检出某个耐药基因,可能来自肠道里的普通共生菌;但如果在血液、脑脊液、肺泡灌洗液等感染相关样本中,同时检出明确病原体和关键耐药基因,临床意义就会明显增加。
问题在于,很多测序结果不一定能准确告诉你:
这个耐药基因到底属于哪个菌?
尤其是宏基因组短读长测序,样本DNA会被打碎成很多片段。报告可能识别出某个片段像耐药基因,也识别出样本里有哪些菌,但未必能百分之百确认这个基因就长在某个具体病原体身上。
第三层:基因存在,不代表正在发挥作用
耐药基因要真正产生耐药效果,通常需要被表达出来。
也就是说,DNA层面有这个基因,只说明细菌可能有这套说明书,但这套说明书有没有被读取,相关蛋白有没有产生,产生的量够不够影响药物作用,还要另说。
这也是为什么单纯看到一个耐药基因,不能直接等同于药敏试验里的“R”。
所以这里要分清,基因存在,是潜在能力;真正耐药,要看它有没有形成实际表现。
第四层:它和具体药物有没有明确对应关系?
有些耐药基因和药物类别的关系比较明确,比如某些碳青霉烯酶基因、mecA、vanA、vanB、mcr等,临床上会更重视。
但也有一些耐药基因属于较大的基因家族,或者只提示某一类耐药机制,并不能简单对应到某一个药一定无效。
比如,同样是β-内酰胺类耐药相关基因,不同基因、不同亚型、不同表达水平,对青霉素类、头孢类、碳青霉烯类药物的影响可能不同。
同样是四环素类耐药基因,不同tet基因对应的机制也可能不同,有的是外排泵,有的是核糖体保护蛋白。
所以,要去看它提示的是一个大类风险,还是足以影响具体药物选择的关键机制。
第五层:数据库和算法会影响检出的边界
耐药基因的识别,通常需要把测序片段和数据库进行比对。
但不同数据库收录的基因不同,命名方式不同,更新速度不同。不同实验室使用的分析流程、相似度阈值、覆盖度标准、报告规则也可能不同。
如果一个基因只匹配了很短一段,或者相似度不高,可信度就要谨慎。如果覆盖度高、相似度高、reads数也足够多,结果的可信度才会更强。
从背景信号,到用药线索,需要证据链
把这些层次串起来,就会发现:
所以,看到耐药基因后,不应该立刻下结论说这个药一定没用。更合适的理解是:报告提示样本里出现了一个耐药相关线索,接下来要判断它是否来自活菌、是否属于真正的病原体、是否是关键耐药机制等。
也就是说,从背景信号到影响用药,中间还差一整套证据链。下一步,真正需要看的就是:临床上到底怎么把这些证据串起来。
如果患者有明确感染,医生判断耐药性通常会综合病原体、样本来源、培养药敏、感染部位、患者状态和治疗反应。
首先看检出的病原体是否符合临床表现
判断病原体意义,要结合患者症状、感染部位、样本类型、采样质量、炎症指标、影像学表现、是否来自无菌部位、是否重复检出、菌量或序列丰度、既往病史和免疫状态。
其次,培养和药敏仍然重要
传统培养虽然有局限,比如生长慢、厌氧菌难培养、使用抗生素后阳性率下降,但它能获得活菌,是药敏试验的基础。
对于许多感染来说,培养阳性并结合药敏,仍然是指导抗感染治疗的核心依据。
药敏试验通常报告为S、I、R。
MIC是最低抑菌浓度,也就是抑制细菌可见生长所需的最低药物浓度。
但MIC也不是孤立指标。它要结合药物在人体内的药代动力学和药效学,也就是药物能否在感染部位达到足够浓度。
比如,有些药物在尿液中浓度很高,对尿路感染可能有效;但同样的药物在肺组织、脑脊液或脓肿内浓度不足,就未必适用。
第三,关键耐药基因需重视,但也要看归属
不同耐药基因的临床意义不同。有些基因具有较强提示意义,比如:
这些基因值得重视,但仍然要看它是否与对应病原体关联,是否有足够覆盖度和丰度,是否与临床培养药敏一致。
一个粪便样本里的低丰度tet基因,和一个血流感染样本中高可信度blaNDM基因,临床意义完全不同。
最后,还要看感染部位和药物可达性
临床用药不是只看敏感或耐药。
所以,耐药基因只是抗感染决策中的一个维度,不能单独决定治疗方案。
报告里的“高风险”,通常不是临床诊断等级,也不等于疾病严重程度。它更多是在说:根据这份样本和这套检测方法,报告发现了一些相对更值得注意的耐药相关信号。
那这个“高”,到底可能高在哪里?
可能是reads数比较高
报告会写 reads数。reads数可以简单理解为:有多少测序片段支持这个菌或这个基因的存在。
一般来说,reads数越多,说明检测到的证据越多;但它不能单独决定严重程度。因为reads数会受到很多因素影响,比如样本总DNA量、宿主DNA比例、测序深度、基因长度、样本保存质量等。
临床解读时,低reads结果尤其要谨慎。
编辑
<来源:谷禾健康肠道菌群检测数据库>
可能是占比更高
相对丰度表示某个菌或某个基因,在样本可识别微生物或基因中的占比。
它适合看结构变化,但不能直接等同于绝对数量。
比如,一个耐药基因的相对丰度升高,可能是它本身增加了;也可能是其他菌减少了,导致它的比例看起来变高。
所以,相对丰度高,说明它在这个样本里的占比更突出;但不等于体内一定有大量耐药菌。
在谷禾的检测报告中,会加入“在标本中浓度”、“信号强度”等指标,提供拷贝数、copies/mL、copies/g等。对判断病原负荷和变化趋势更有帮助。
编辑
<来源:谷禾健康肠道菌群检测数据库>
可能是比对结果更可靠
有些报告里还可能出现相似度、覆盖度、测序深度这类指标。
这些指标主要用来判断:测序片段和数据库里的目标基因,匹配得像不像、覆盖得全不全、证据够不够扎实。
如果一个耐药基因只匹配到很短一段,或者相似度不高,就要谨慎解读。
如果相似度高、覆盖度高、reads数也足够多,说明这个检出结果更可靠。
既然耐药基因不能直接等于感染,也不能直接等于药敏,那检出来到底有什么意义?
反复用药效果不好时,它能提醒尽早换思路
临床上有些患者,尤其是老年人、基础病多、反复感染的人,可能已经经历过好几轮抗生素治疗。
编辑
一开始是经验用药,效果不好换一种,还是不好,才进一步做培养、药敏,或者更深入的病原检测。
这个过程中,患者可能已经经历了较长时间的感染、反复发热、炎症指标不降,甚至住院时间延长。
如果前期就能看到一些耐药相关线索,至少可以提醒:这个人可能存在较高的耐药背景,常规经验用药未必合适,必要时应该尽早做培养和药敏,或者结合mNGS、tNGS等检测进一步判断。
某患者A,因反复腹泻、腹痛,使用抗生素后疗效不佳。
编辑
<来源:谷禾健康肠道菌群检测数据库>
谷禾宏基因组菌群检测报告提示,核心共生菌 Faecalibacterium prausnitzii 中检出 tet(W),提示四环素类耐药基因可能已进入稳定共生菌群。
肠道优势菌Bacteroides中检出 cepA、cblA。这类菌本就是肠道常驻菌,如果携带β-内酰胺类相关耐药基因,提示患者肠道厌氧菌群可能存在较稳定的β-内酰胺耐药背景;在后续腹腔、肠源性或混合感染风险评估中,经验性使用青霉素类、部分头孢类药物时就应更加谨慎,而不是简单继续轮换抗生素。
此外,Enterococcus 中检出 vanD,提示需关注糖肽类耐药风险;E. coli 中检出 arsC,提示菌群还可能存在环境金属抗性压力。该报告帮助医生从继续试药转向结合感染部位、病原学、药敏结果和菌群修复的综合管理。
在重症和疑难感染中,它能让判断更有方向
普通感染中,医生有时可以根据经验治疗,再根据疗效调整。
但在重症感染、免疫抑制患者感染、血流感染、重症肺炎、脑膜炎等场景里,试错成本会明显升高。
若能更早获得病原体和耐药相关信息,就可能帮助医生更快判断:哪些抗生素可能覆盖不足,哪些耐药机制需要警惕,是否需要联合用药,是否需要尽快补做培养和药敏,或者调整诊疗方向。
在健康管理中,它更偏向预防性参考
有些报告不是为了诊断一次急性感染,而是用于健康管理或微生态评估。
这类报告里的耐药风险,更多是预防为主。它可以提供一个背景信息:
比如老年人、长期住院者、肿瘤患者、反复尿路感染或呼吸道感染的人,如果报告提示耐药背景较高,就更不适合随意、反复地经验用药。
在感染控制中,它也可能有提示价值
有些耐药基因不仅影响个体治疗,也有传播风险。
比如某些碳青霉烯酶基因、万古霉素耐药相关基因、多黏菌素耐药基因,如果出现在医院环境、重症病房或高风险患者身上,就可能提示需要关注感染控制问题。
这时,耐药基因结果不仅是这个人用什么药,也可能是在提醒:这个耐药风险会不会在更大范围内传播。
可能帮助医院更早识别需要关注的人群或环境,配合隔离、手卫生、环境消毒、抗菌药物管理等措施,减少耐药菌传播。
对公共卫生耐药基因的监测
耐药基因库是公共卫生关注的重要内容。社区、医院、养殖业、污水系统中的耐药基因传播,可能增加未来耐药感染风险。因此,在群体水平监测耐药基因非常有意义。
但对个体来说,不能把公共卫生风险直接等同于个人疾病诊断。
所以,耐药基因检出的意义,不是把报告变成处方。更准确地说,它的价值在于:
误区一
× 检出耐药基因就是耐药感染
这是最常见误区。耐药基因是遗传信息,感染是临床疾病,二者不是一回事。
误区二
× 没检出耐药基因就一定不耐药
反过来也不能这样理解。可能数据库未收录相关机制,可能检测深度不足,可能耐药来自突变、表达调控、生物膜或其他非典型机制。
误区三
× 16S能准确判断用什么抗生素
常规16S rRNA测序主要看菌群组成,不是直接检测完整耐药基因。它报告里的耐药风险,很多时候是根据菌群结构和数据库背景做出的间接推断。
16S更适合做微生态评估和风险提示,不适合直接决定抗生素怎么用。
误区四
× 宏基因组能完全替代培养药敏
宏基因组很有价值,尤其在难培养、用药后、疑难感染中。但它不能完全替代培养和药敏,特别是在需要精准用药时。
误区五
× 耐药基因越多越需要治疗
健康人群或肠道菌群报告中,耐药基因多不代表需要治疗。滥用抗生素反而可能加重菌群紊乱和耐药选择压力。
对于没有感染证据的人来说,重点不是清除耐药基因,而是合理用药、减少不必要抗生素暴露、保护肠道微生态。
误区六
× 耐药基因说明生活不卫生
耐药基因广泛存在于环境和人体微生态中,不能简单归因于个人卫生问题。
误区七
× 耐药基因就是“超级细菌”
所谓“超级细菌”,通常指对多种抗生素耐药、治疗选择有限、可能导致严重感染的细菌。
耐药基因可能参与这种情况,但单独检出某个耐药基因,并不等于体内已经有了超级细菌。

谷禾健康
恭喜!
杭州谷禾信息技术有限公司检测实验室
注册号:CNAS L23010
【新突破】
谷禾16S rRNA 扩增子检测项目正式通过中国合格评定国家认可委员会 CNAS 认可,纳入实验室认可范围。
【深扎根】
此前谷禾获认可的宏基因组项目(详情:杭州谷禾检测实验室获CNAS国家认可,微生物检测迈向更高台阶),顺利通过 CNAS 首次监督评审。
关于CNAS
CNAS 是“中国合格评定国家认可委员会”(China National Accreditation Service for Conformity Assessment)的英文缩写,是由国家认证认可监督管理委员会批准设立并授权的国家认可机构,统一负责对检测实验室、校准实验室、认证机构等合格评定机构进行认可。获得 CNAS 认可,表明该实验室具备按照相关国际和国家标准开展检测活动的技术能力和管理水平,其出具的检测报告具有权威性和公信力。
在我国合格评定体系中,CNAS 认可代表的是对机构能力的第三方评价。对于检测实验室而言,是围绕实验室的管理体系、技术能力、人员能力、设备设施、方法确认、质量控制、结果有效性、持续改进能力等方面进行系统评审。
CNAS 认可体系强调实验室活动的公正性、独立性和结果有效性。检测结果是否客观,检测过程是否受控,人员是否能够避免不当压力和利益干扰,都是实验室质量体系中的重要内容。
因此,实验室通过 CNAS 认可,意味着其在获认可范围内具备按照相应认可准则开展检测活动的能力,也意味着检测过程和结果具备更清晰的规范性、可追溯性和可信基础。
同时,CNAS 与国际认可互认机制相衔接,在相关互认范围内,有助于推动检测、校准、检验等合格评定结果在不同国家和地区之间获得更广泛的接受。
对高通量测序检测实验室来说,CNAS 认可不仅是一项资质成果,更是一种来自国家认可体系的能力确认,也是一份面向科研、产业与真实应用场景的可信承诺。
据目前公开认可信息及相关项目范围比对
谷禾获认可的16S rRNA 扩增子检测项目
为国内高通量测序检测类公司中
首个通过 CNAS 认可的同类项目
从宏基因组检测项目首次获得认可
到如今顺利通过监督评审;
从实验室基础质量体系建设
到16SrRNA扩增子检测项目纳入认可范围;
这是一份属于谷禾团队全体成员的喜报
更是一段长期秉持“工匠精神”
日常工作的阶段性回响
也意味着谷禾检测实验室在微生物组检测
和高通量测序检测能力建设的道路上
再次迈出了坚实且具有行业引领性的一步
16S rRNA 扩增子检测,是一条由多个高精细环节环环相扣组成的完整技术链路。通过对微生物特征基因片段进行富集、高通量测序和深度生物信息学分析,能够帮助洞察样本中微生物群落的物种组成、多度分布、多样性特征及结构演变。
然而,正因为这条技术链路横跨了分子生物学、微生态学与计算机信息学等多个学科,其全生命周期的复杂性与不确定性也呈指数级上升。
系统性鸿沟:标准化不足带来的数据可比性挑战
在当今蓬勃发展的微生物组研究领域,方法学标准化不足所导致的研究间可比性差,已成为横亘在科学探索与真实世界应用之间的一道系统性鸿沟,这不仅是对宝贵科研资源的巨大损耗,更是掣肘了微生态研究成果向产业化与临床端安全转化的核心通道。
这就要求实验室不仅在技术层面知道怎么做,更要在体系管理层面清晰、透明、有据可查。
比如说,数据质控阈值是如何经过数理统计学验证的,异常结果出现时的根本原因有没有相应的调查机制,涉及流程、软件版本或关键参数调整时,是否经过必要的技术确认等。
这些都构成了高通量测序检测项目的质量门槛。
谷禾的选择:以标准化支撑检测长期稳定
面对这一阻碍行业健康发展的底层痛点,谷禾健康团队深知,建立贯穿生命科学检测全周期的标准化操作规范(SOP)和严密质控体系,是让检测数据具备稳定性、可追溯性的关键,也是精准指导下游靶向干预、产品研发、功效评价,以及迈入临床级应用的关键基石。
为此,我们历经多年在底层技术上的持续深耕与无数次真实样本的反复打磨,系统性重构并确立了一套严苛、稳定的微生物高通量测序(涵盖16S rRNA及宏基因组)全流程检测技术规范。
正因如此,CNAS 认可对谷禾而言,不是一次单点项目申报,也不是把既有流程简单整理成文件,而是一次对检测能力、质量体系和长期稳定能力的系统性确认。
对于高通量测序实验室而言,认可体系的核心在于:将国际标准内化为员工工作习惯的能力,并在日复一日的枯燥运行中,始终保持检测能力的在册、受控、不走样。
高通量测序的底层逻辑是极其精密且连续的。为了确保发出的每一组测序数据的真实可靠,谷禾检测实验室严格围绕“公正、科学、守信、高效”的质量管理方针,从人员管理、设备量值、物料方法、环境监控、数据记录五大核心维度,构建了多维一体的立体防线,将标准彻底深化在底层秩序中。
在这套质量方针中,公正始终被放在最前面。对谷禾而言,公正意味着:
“ 在谷禾实验室体系,实验室人员,
从来不是简单完成标准操作的机器,
而是质量体系有血有肉的真正执行者。”
一份样本从接收到报告出具,中间会经过多个岗位。
有人负责核对信息,有人负责样本处理,有人负责核酸提取,有人负责文库构建,有人负责上机测序,有人负责数据分析…每个人手中的动作,可能只是流程中的一小步,但连在一起,就成为一份报告背后看不见的质量支撑。
CNAS 认可要求实验室关注人员能力、培训、监督、授权和持续评价。这些要求最终落到日常,各岗位都要知道:为什么这样做,这样做会影响什么?如果出现偏离,应该如何识别、记录、上报和处理等。
同样重要的,是检测人员在岗位职责内保持独立、客观的技术判断。
样本不会说话,数据也不会主动解释自己。守住每一道关口的,是实验室里那些反复核对编号的人,是每次操作前确认试剂状态的人,是发现异常后停下来追原因的人,是报告发出前逐项检查的人…
将CNAS要求落到谷禾检测的日常里,就是一套动态的人员能力验证模型。
质量管理不是把责任简单压给个人,而是让每个人在清晰的规则、充分的训练和持续的监督中,把事情做对、做稳、做久。
这种能力,不是某一次操作优秀,而是长期稳定;不是把标准挂在墙上,而是让标准融入每一次开盖、每一次移液、每一次复核、每一次签发…
这些看似重复的动作,最终都会落到客户所等待的那份结果里。也正因如此,我们愿意把看似细小的记录留得更清楚,把普通的日常做扎实,让质量体系真正落在每一天的工作里。
宁愿在发现疑点时多停一步,
也不愿把不确定带进样本流程。
高通量测序仪、PCR 仪、荧光定量 PCR 仪、生物安全柜、离心机、微量移液器、纯水仪……许许多多设备构成了一份肠道菌群检测报告背后的技术基础。
然而再精密的仪器,如果状态不可确认,也无法支撑可靠的结果。
一支微量移液器,可能只是实验台上最常见的工具,但它每一次吸取和转移,都关系到微量反应体系的稳定。
一台生物安全柜,不只是设备,更是样本保护、人员保护和污染控制的重要屏障。
设备会通过状态、记录和数据回应实验室的管理水平。因此,在谷禾实验室,设备会贯穿采购、验收、标识、使用、维护、核查、校准、异常处理和记录管理的全过程。
“一份看似普通的粪便样本,
背后都承载着一个人
对了解自身肠道微生态的期待。”
样本是检测的起点,也是客户信任进入实验室的第一站。
从采集、保存、运输,到实验室接收、编号、流转、处理和留存,每一步都需要被认真对待。
对每一份样本保持一致的接收、流转和处置要求,也是公正性在样本端的体现。无论样本来自何种项目、何种合作场景,进入实验室后,都应回到同一套受控流程中被认真对待。
样本要有清晰身份
试剂要有受控状态
耗材要适用于相应实验环节
关键物料要经过必要确认
外部提供的产品和服务也需要被纳入管理
成熟的实验室管理,是要把影响结果的因素尽可能管住。
与样本同等重要的,是各种高精密的试剂与反应耗材。高通量测序对试剂批次间的稳健性要求近乎苛刻。
磁珠吸附力的微量起伏、核酸提取试剂盒的微小原料波动,或者聚合酶的活性晃动,都会在敏感的测序仪中被指数级放大,进而干扰微生物群落的真实丰度。
为此,谷禾对所有生产投入品确立了严格的“状态受控”与“关键物料确认”原则。
“ 能说明的,用严谨的数据讲清楚。
不能过度推断的,我们严格守住边界。
把科学的不确定性放在它该在的位置。”
在菌群检测中,方法贯穿样本处理、核酸提取、扩增建库、测序、数据分析和报告审核等多个环节。每一个环节都可能影响最终结果。因此,方法要落实为“流程被理解、被执行、被记录、被复核、被持续改进”。
谷禾检测对方法的理解,是专业,也是克制。
专业,是因为检测方法必须经过充分确认。在检测项目建立和持续运行过程中,我们会围绕方法适用性、关键性能表现、操作边界和结果稳定性等方面开展系统评估,确保方法能够满足预期用途。
好的方法,不是让实验室显得复杂,而是让复杂工作变得稳定。
对于肠道菌群检测中涉及的分子生物学实验环节,谷禾检测关注的除了能不能检出之外,还有方法在不同样本状态、不同批次运行和不同实验条件下是否依然保持稳定。
哪些步骤必须严格一致,哪些条件不能随意改变,哪些异常信号需要暂停复核等等,都是由方法体系本身来约束。
在微生态检测领域,复杂性不仅来自实验台,也来自数据端。高通量测序产生的是海量数据,形成正式的报告之前,还需要经过数据质控、序列处理、分类注释、结果整合、专业审核等。
每一次涉及分析逻辑、版本状态或关键参数的调整,都需要经过相应评估和确认,确保升级带来的是能力提升,而不是结果波动。
CNAS评审专家们对谷禾检测实验室的实验方案、分析流程及关键参数控制,进行全方位、系统性的现场核查与技术确证。
专业的另一面,是克制,是边界。
报告表达必须基于检测数据本身,不能越过证据范围去制造确定性,也不能为了迎合期待而扩大解释。肠道菌群检测可以帮助客户了解肠道微生态的组成和特征,但它不应被包装成超出检测能力范围的承诺。
一次偶发异常结果背后的多轨复核
一处原始记录细节的补充确认
一项分析流程变更前后的评估
一份报告发出前的层层审查
…
它们隐身于幕后,
却真真切切地决定了
一份报告是否值得被托付信任
“ 好的实验环境,
让正确的事更容易发生,
让错误的事更不易发生。”
在微生物检测和基因扩增相关实验中,环境从来不是背景。环境本身,就是质量的一部分。
肠道菌群检测面对的是复杂微生物群落。样本之间、区域之间、人员操作之间,以及仪器状态、空间条件、输入物料等外部变量,都可能对结果稳定性产生影响。
对高通量测序而言,一些风险并不总是以明显故障的形式出现,它可能是设备运行状态中的细微漂移,也可能是空间流转中的潜在交叉干扰。
在空间管理层面,谷禾检测重视功能区域之间的边界感。
涉及核酸处理、扩增和测序相关的关键区域,需要通过合理分区、标识管理、人员与物料流转控制、清洁消毒和污染防控措施,尽可能降低交叉干扰风险。
该隔离的区域要隔离,该限制的进入要限制,该标识的状态要标识,该记录的条件要记录。环境条件不适合时,检测活动就不能被简单推进;存在潜在风险时,就需要先判断、先控制、先确认。
这些日复一日的环境控制,可以保护样本不被无关因素干扰,保护人员在安全条件下工作,也保护最终结果不被不必要的外部变量影响。
谷禾检测的环境,不仅是一间实验室的装修和空间,而是一种质量氛围:
环境会在每一份稳定的结果里,留下自己的痕迹。
应用十多年的底层技术迭代与平台进化
16S rRNA 扩增子检测,作为谷禾的高频基石技术,是谷禾在微生物组学领域扎根最深、应用场景最广、数据资产最雄厚的底层技术支撑。
在谷禾多年的肠道菌群检测、微生态研究服务和高通量测序的大规模实践中,16S rRNA 测序长期扮演着连接微生态样本与高维度生命科学大数据的关键纽带。
作为谷禾最早推向市场、经历了十余年应用与多轮标准化迭代的经典项目,它完整见证了海量复杂样本的自动化流转、实验反应体系的优化、生物信息学分析算法的迭代升级,数以万计科研与临床报告的交付等。
伴随着这一长周期的技术演进历程,谷禾在微生物组检测领域逐步构建起了严密的方法学验证体系,确立了标准作业程序(SOP)的底层逻辑。
一个高通量测序项目真正的成熟与强大,取决于它能否在大规模、跨批次、长周期的实际运行中,有效对抗各种高风险变量,始终保持极高的多中心稳定性和结果复现性。
纳入全球互认体系的底层逻辑升级
此次,16S rRNA 扩增子检测项目通过 CNAS 认可,是谷禾长期的技术沉淀与质量管理体系深度融合的必然结果,意味着谷禾持续运行的检测,全面正式跨入了全球多边互认的合规轨道。
16S rRNA 扩增子检测项目从企业经典向国际认可的跨越,是检测与质量的全面升级。在 CNAS 质量框架下,16S rRNA 项目的运行实现了多维一体的规范化:
经典,代表它经历过时间和样本的检验
认可,代表它进入了更规范、受控和可评价的质量框架
两者结合,构成了谷禾 16S rRNA 扩增子检测项目今天的底气
深度赋能全产业链伙伴
对于长期携手并进的科研学者、临床专家、生物医药企业以及大健康产业同仁而言,这一升级意味着:谷禾为全产业链合作伙伴的课题研究、队列分析、产品研发、商业化落地、功效评价、微生态应用,提供了坚实的数据底座。
-为科学研究、临床转化与大健康产业筑牢底座-
CNAS 认可是一项阶段性成果,也是一项持续性的要求。
从宏基因组检测项目获得认可,到顺利通过 CNAS 首次监督评审;从 16S rRNA 扩增子检测项目长期规模化运行,到正式纳入 CNAS 认可范围,谷禾走过的,是一条把技术做深、把体系做实、把数据做稳的长期主义之路。
这条路没有捷径。
– 它来自十多年的技术迭代,也来自每一天的样本、记录、复核和改进;
– 它来自实验平台的持续进化,也来自质量体系对每一个细节的长期约束;
– 它来自团队所有人对微生物组检测的专业投入,也来自谷禾对客户信任的持续回应
…
这一次,我们为团队感到骄傲;
同时也更清醒地知道,认可之后,标准只会更高,责任只会更重。
未来,谷禾将继续围绕微生物组检测和高通量测序技术,持续完善检测方法、质量控制、生物信息分析和数据管理流程,在每一次样本处理、每一轮数据分析、每一份报告审核中,保持对标准的敬畏和对结果的审慎。
因为我们深知样本的背后,连接着一个个科研问题、一项项转化探索、一次次产品验证,或者一个人对自身健康状态的认真关注。
样本和数据不会说话,但谷禾的选择,定义了数据的分量。
我们专注检测
坚持检测平台的中立性,不因商业压力或主观判断改变检测结果;
我们尊重数据
坚持科学严谨,不为了迎合期待而扩大解释;
我们重视体系
因为只有把标准融入日常,检测能力才能长期稳定地保持;
我们敬畏样本
坚持职业的同理心,因为每一份样本背后,都是一份真实的托付。
以此为新的起点,谷禾将坚定依托这套获得国家级认可的标准化检测体系,以扎实的技术积累和持续的体系建设,推动微生物组检测服务向更加规范、可靠、可持续的方向发展。
谷禾将用这份穿越周期的确定性,为国内外顶尖科研机构的多中心大队列研究、高分顶级临床文献的发表、精准个性化微生态干预方案的制定,乃至下游创新型产品开发,提供精准、可靠、经得起全球科学界与监管机构反复推敲的高质量底层数据支撑。
把看不见的标准
做进看得见的结果里
谷禾,与前沿科学同频
始终与全产业链伙伴同行
编辑

谷禾健康

在地球生态系统中,细菌、真菌、古菌、病毒以及真核生物共同构成了生命的基础,并在多个层面上发挥着重要的作用。人类和其他大型生物的反应往往可以直接观察到,但看不见的微生物的作用和影响也同样重要。
在微生物研究中,一直面临着一个棘手的难题:想要准确识别环境样本中的微生物,却常常遇到技术和成本障碍。传统上,研究人员需要分别测序16S和18S两种核糖体RNA,这不仅增加了研究成本,还可能因为使用”通用”引物导致鉴定结果产生偏差。
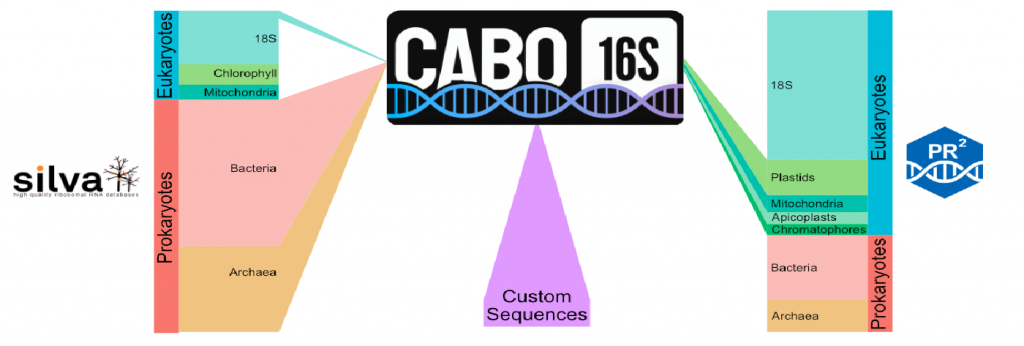
为解决这一问题,加州理工学院的研究团队在2024年10月创建了一个数据库——CABO-16S。这个数据库的独特之处在于将两个知名数据库(SILVA和PR2)中的微生物序列巧妙整合,让研究人员可以用更简单、更经济的方式获取微生物分类信息。
具体来说,CABO-16S将细菌、古细菌的序列与细胞器(如叶绿体)的16S rRNA序列结合,大大提高了微生物鉴定的准确性。特别是在海洋、湖泊和海草环境中,这个数据库在识别光合真核生物方面表现出色。
此外,CABO-16S还提供了一个灵活的框架,研究者可以添加专业的、尚未被其他数据库收录的微生物序列。本文以甲烷渗漏沉积物中的特定微生物为例,展示了如何通过添加精选序列来揭示微生物群落的细微差别。
这项创新不仅降低了研究成本,还为探索极端或独特环境中的微生物提供了新的可能性。
传统方法的不足
在过去20年中,小亚基(SSU)核糖体RNA(rRNA)的高通量测序用于研究陆地和海洋环境中的微生物生态学,并加深了对植物和动物微生物组的理解。通常16S rRNA基因用于原核生物鉴定,而优化的18S rRNA基因用于真核生物鉴定,ITS区域则被证明最适合真菌。
用单个PCR反应扩增原核和真核微生物是更为理想的,与使用单独引物进行16S和18S rRNA基因测序相比,可以将扩增子文库制备成本降低两到三倍。尽管一些“通用”引物 (515f/926r)可以扩增真核生物 18S rRNA 和 16S rRNA ,但同时准确分析来自 16S 和 18S rRNA 的真核生物和原核生物具有一定的难度。
◮ 真核生物被低估
首先,当试图扩增更广泛的目标群体时,引物与其模板之间的不匹配可能更常见,事实上,在模拟群落中,仅与反向引物的单个不匹配就会导致真核生物被低估3-8倍。
其次,18S序列通常比16S序列长160- 180bp,并且 PCR 和测序都偏向于较长的扩增子。天然样品可能含有较长的18S序列或较高比例的甲藻,这些甲藻往往存在错配,这表明在一些天然样品中,515f/926r引物扩增真核生物可能被严重低估。
◮ 不能完全可靠地表示相对分类丰度
最后,在大多数细菌中,16S rRNA基因拷贝数在1-15之间,在大多数古细菌门中平均只有1个拷贝,而在浮游植物中,18S基因拷贝数可以在1-12,000之间变化。尽管18S基因计数可能与生物量显著相关,但它们不能可靠地用于表示相对分类丰度。
CABO-16S的优势
◮ 同时分析原核和真核生物更全面、更精准
CABO-16S作为一个整合16S rRNA序列的创新数据库,为微生物生态学研究提供了更全面和精确的分类工具。CABO-16S将细菌、古细菌的序列与细胞器(如叶绿体)的16S rRNA序列结合,特别适合需要同时分析原核生物和真核生物(尤其是光合真核生物)的环境样本研究,尤其是海洋、湖泊和沿海生态系统。
◮ 可根据研究需求添加自定义序列
研究者可以根据自身研究需求,向数据库添加自定义序列,但需谨慎操作,避免引入分类学不一致性。对于特定环境或复杂微生物群落的研究,建议结合基因组系统发育信息来验证16S rRNA分类结果。
在具体使用时由于数据库仍在持续更新,研究者应关注SILVA和PR2数据库的最新版本,并及时更新CABO-16S数据库,以获得更准确的微生物分类信息。
◮ 获取SILVA数据
为构建 CABO-16S 数据库,下载最新版本的 Silva(138.2)序列 (SILVA_138.2_SSURef_NR99_tax_silva.fasta.gz) 以及映射的分类法(taxmap_slv_ssu_ref_nr_138.2.txt.gz)和质量值 (SILVA_138.2_SSURef_Nr99.quality.gz)。
去除了针尾值 < 50 或比对质量值 < 75 的所有序列。去掉鉴定为叶绿体、线粒体和真核生物的序列,随机选择100个真核生物作为外群重新添加。真核生物外群的分类学仅保留门水平。
对原核生物分类学进行清理,特别是在物种水平,以删除基于生物宿主、样本收集、不明确的细菌分组或属重复(即“Genus sp.”)的命名方案。为了便于直接比较,使用相同的方法构建了一个简化的 SILVA 数据库,但保留了鉴定为叶绿体的序列。
◮ PR2数据库获取叶绿体、染色质等序列
将质体、顶质体、叶绿体和染色质序列从PR 2数据库(v 5.0.0)添加到CABO-16S数据库中,用R包“pr 2database”(https://pr2database.github.io/pr2database/articles/pr2database.html)获取。
为了与SILVA分类中的7个分类等级相匹配,从PR2序列中删除了超群和亚群的等级。
◮ 将SILVA数据与PR2数据相结合
最后,将从甲烷渗漏 Sanger 测序(https://doi.org/10.6084/m9.figshare.27288090)中获得的定制的16S rRNA序列与来自 SILVA 和 PR2 的数据相结合,形成 CABO-16S 数据集的基础。
CABO-16S和简化的SILVA 138.2训练集是根据DECIPHER的建议和IDTAXA算法制作的。简而言之,在使用LearnTasa函数进行三次迭代训练之前,将过采样组被随机子集化为100个序列。
Kmer长度设置为8nt,以匹配RDP和QIIME2默认值。注意,使用全长16S rRNA参考序列用于训练;截断扩增子窗口的可能会略微提高准确性,但代价是可能产生模糊性。因此,我们呈现全长序列并从全长序列进行比较,并将是否截断的选择留给用户。
利用CABO-16S和SILVA-132.1对已发表的广泛来源16S rRNA序列进行分类比较,包括已知细菌分离株的模拟群落和环境样品。
对于所有比较的样品,使用古细菌/细菌引物(515 f/926 r)扩增16S rRNA基因的V4-V5区域,并在Illumina MiSeq平台上测序。在5个以上样本的环境数据集中,选择任意一组子样本进行分类比较。
对所有下载的原始序列进行了相同的处理,除了Needham和Fuhrman(2016)的数据,这些数据直接下载并使用了已经分析的OTU序列和观测矩阵。可重现的工作流程(https://github.com/emelissa3/CABO-16S commit 472d7fc)报告了用于从NCBI SRA上可用的原始FASTQ文件生成扩增子序列变体(ASVs)的全部细节和参数。
简言之,使用Cutadapt去除引物,然后修剪序列(240 f/200 r),合并12 bp重叠,去噪,并使用DADA2 进行比对。删除嵌合体,并通过IDTAXA的IdTaxa函数进行分类。
CABO-16S将常用的16S rRNA数据库统一整合,提供了一个用户可以轻松扩展的单一数据库,包含了数据库更新或未发表的序列。来自SILVA 138.2的389144个细菌和19213个古细菌16S rRNA序列用作CABO-16S数据库的初始框架,同时保留了来自SILVA的随机100个真核生物序列作为外群。
这些序列与来自PR 2数据库的细胞器16S rRNA基因的8540个16S rRNA序列相结合。最后,可以组合自定义序列,以最大限度地提高目标群落的分辨率;这里还添加了一组未发表的全长16S rRNA序列,这些序列是从甲烷渗漏沉积物的Sanger测序中获得的,以及一份精选的代表性seep-SRB1序列列表。
doi: https://doi.org/10.1101/2024.10.23.619938
以相当的准确度注释先前未分类的ASV
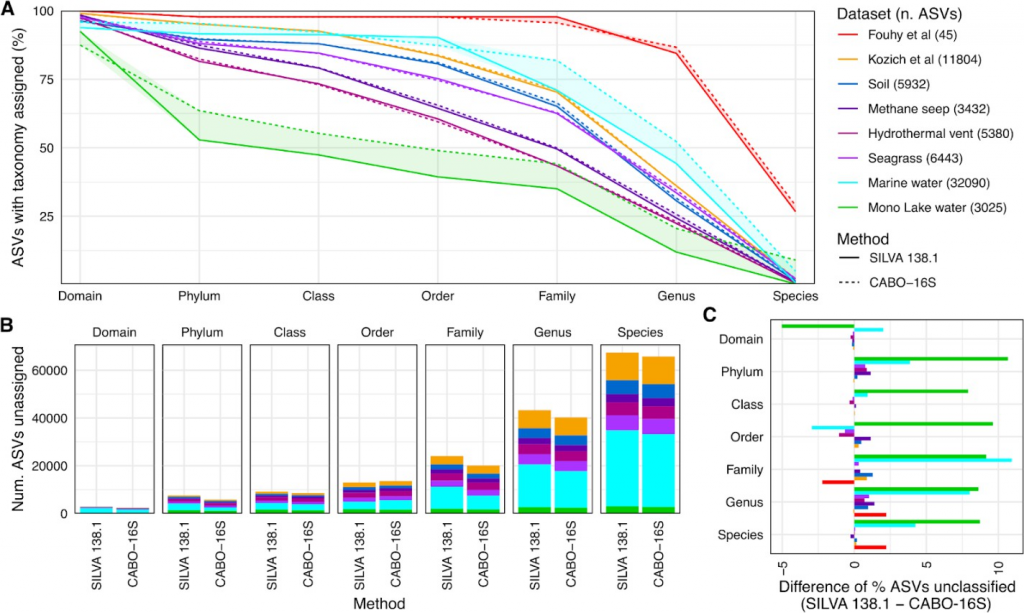
CABO-16S与SILVA 138.2比较了先前发表的代表不同系统的数据集,包括模拟群落和环境样本。该数据集包含基于模拟群落、哺乳动物肠道和住宅土壤、北方森林土壤、叶和周围沉积物、来自冷甲烷渗漏的深海沉积物、热液喷口沉积物、具有丰富浮游植物群落的海水以及最后来自以微藻Picocystis为主的封闭盆地湖泊的水的特征良好的基准集。合并数据集由64402个扩增子序列变体(ASVs)组成,单个数据集范围在45-32090个ASV之间。
CABO-16S的分类能力显著提高
在所有分类水平上接受分类分配的ASVs总数方面,CABO-16S数据库的表现优于未经修改的SILVA 138.2数据库。最大的差异是具有最多光合真核生物的数据集,如浅海草环境、海水和湖水。
例如,在海水数据集中,CABO-16S在门水平上比SILVA 138.2多分类了约10%的ASV。其他数据集差异不大,表明纳入PR2数据库的细胞器16S序列并没有对分类器继续准确预测细菌和古菌分类的能力产生有意义的影响。
唯一值得注意的例外是,SILVA 138.2中的海水数据集的分类率略高于CABO-16S,我们将其归因于PR2中的浮游植物目数比SILVA中的单一“叶绿体”标签增加。在这两个数据集中,绝大多数ASV都无法在物种水平上进行分类,尽管这可能部分是由于标记到物种水平的参考序列相对较少,特别是对于非人类环境特有的微生物。
doi: https://doi.org/10.1101/2024.10.23.619938
A)SILVA 138.2(虚线)和CABO-16S(实线)在给定分类水平(x轴)上按数据集(彩色线)分类的ASV百分比。随着ASV在较高级别(如域)缺乏注释,这些线也在不断减少,根据定义,ASV在较低级别(如物种)也缺乏注释。
B)A.缺乏分类的ASV的绝对数量。
C)两个数据库之间未分类的百分比差异。阳性百分比反映CABO-16S注释的ASVs比SILVA 138.2更多,反之亦然。
CABO-16S和SILVA都揭示了分类模糊性的两种类型之间的区别,这是妨碍分类注释的主要原因。模糊性是最常考虑的精度形式,其中一个序列位于两个或多个参考分类群之间,因此不能在选定的置信阈值(本研究中为40%)下分配给单个分类群。
IDTAXA和其他类似的分类器通过将序列分类到竞争参考分类群的最低共同水平来处理此类事件,有时会在冲突的分类等级中添加‘unclassified’未分类前缀。相反,一个序列可以被确信地分配给一个单一的分类单元,但是如果参考序列在给定的等级上缺乏注释,那么在该等级上仍然可能缺乏分类学。
这种情况会影响许多未培养的谱系,例如,候选门级辐射类群candidate phyla radiation(Patescibacter门)中的SR1 科在SILVA 138.2中没有属或种分类,所有121个序列仅在科水平上进行注释。
因此,缺乏属分类的SR1 ASV不是由于分类器的不确定性,而是由于分类学的不确定性。此外,一些谱系可能同时包含这两种不确定性来源,例如,在SILVA 138.2中,Desulfosarcinacae脱硫杆菌科下有53个序列标记为种水平、676个序列标记为属水平和345个序列标记为科水平。而脱硫杆菌科的ASV缺乏属水平注释可能是由于与仅标记到科级的一组序列有密切相似性(taxonomic ambiguity 分类学模糊性)或与不同属无法区分(classifier ambiguity 分类器模糊性)。
因此,我们依照IDTAXA惯例在分类器模糊的情况下加上“unclassified_”,以及在参考序列模糊的情况下在最低分类层级上添加“unspecied_”来区分两者。
CABO-16S能够分类出几乎占一半读数的真核叶绿体
在分类序列中,CABO-16S和SILVA在大部分数据集中都产生了相似的群落组成。事实上,由于CABO-16S古菌和细菌序列的非蓝藻部分与SILVA 138.2完全相同,因此预期会有这种一致性。然而,在含有光养真核生物的数据集中(例如海草、海洋和湖泊水柱数据集),CABO-16S数据库允许对真核叶绿体进行分类,这些叶绿体占某些样本读取量的近50%(Mono Lake数据集)。
迄今为止,海水数据集的浮游植物多样性是所有数据集中最大的;这种多样性中的大部分都可以用CABO-16S进行分类标记。在Mono Lake数据集中,通过与NCBI人工对比,剩余的未分类多样性可归因于浮游植物线粒体序列。虽然目前的PR2数据库包括大约1842个线粒体序列,但绝大多数(1782个 96.7%)属于Opisthokonta,只有22个序列属于泛植物界Archaeplastida(植物和许多藻类)。
虽然并非在所有真核细胞中都发现线粒体,但我们预计未来PR2的扩展将包括更多来自植物和藻类谱系的线粒体16S,这将改善这一问题。
CABO-16S与原始SILVA 138.2的每个数据集的组成
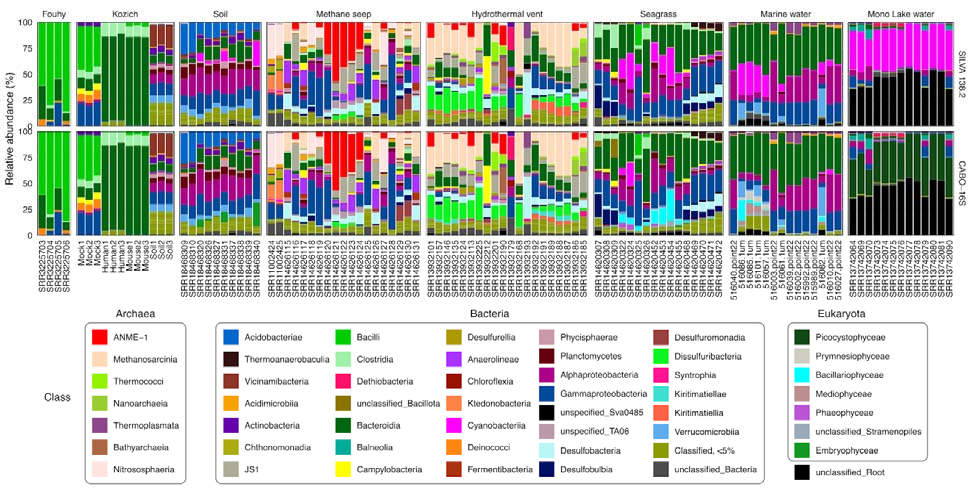
ASV被聚合到类级别(用不同颜色表示)。在样本中含量在5%以上的类别在图中展示。无法分配特定类别的ASV同样被汇总在最低注释级别。相对丰度<5%的剩余ASV被归为一类。
鉴定光合真核生物的精度更高
通过每个光养真核生物数据集对比不同ASV分类,以阐明数据库之间的差异点。对于富含真核浮游植物的沿海海水数据集,SILVA 138.2能够准确地对细菌群落进行分类,但是大部分reads(推测为真核生物)没有被分类到域水平或简单地注释为科水平的叶绿体。
扩增子序列变体序列得到进一步分类
而在CABO-16S数据集中,这些相同的质体ASV序列得到了进一步的分类。值得注意的是,序列的多样性并不总能对较低水平进行明确的分类注释,图中的分类注释显示在类或目水平上出现了许多歧义。
一些分类单元等级包括‘ _X ’后缀,这是PR2使用的中间占位符,类似于其他分类中使用的‘ Incertae Sedis ’。然而,通过质体分类获得的分类注释是有用的,因为主要的浮游植物类群(如硅藻、鞭毛藻、隐生植物等)是有区别的。
CABO-16S解析海洋数据集中的真核和细菌浮游植物
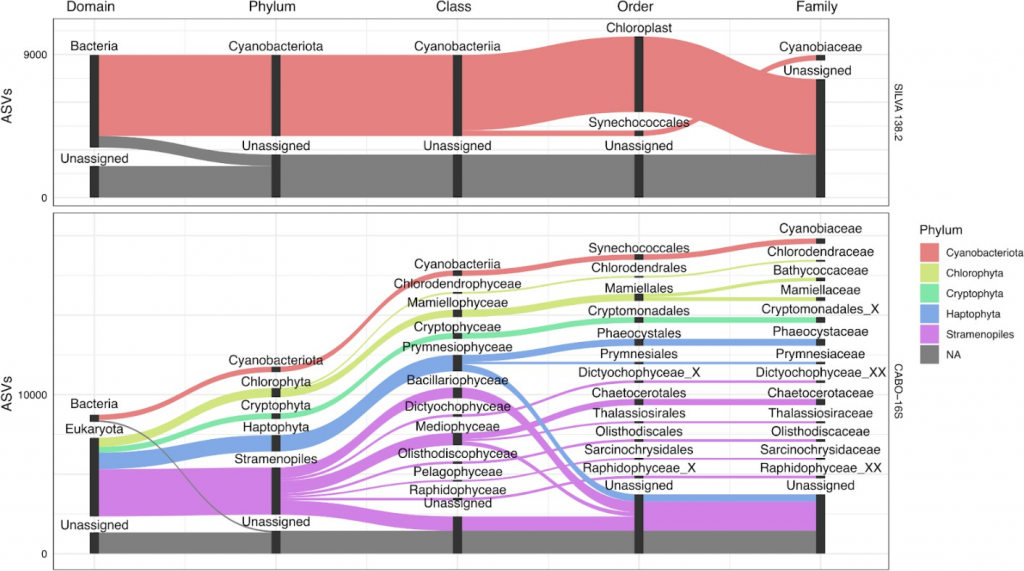
SILVA 138.2和CABO-16S的冲积图追踪了至少有100个ASV从域(最左侧)到科(最右侧)的ASV分类。对于每个层级,柱状图代表不同的分类群,每个柱状图的大小按比例反映了ASV的数量。条形图之间的空白是为了便于可视化。连接层级的流根据这些ASV的门级分类进行着色。仅显示了基于任一数据库的与蓝藻或质体序列相关的ASV。
自定义序列增加了多系进化枝的鉴定
向CABO-16S中添加自定义序列可以增加目前未纳入SILVA或PR2数据库中的物种的分类。增加了Sanger测序获得的甲烷渗漏沉积物序列和代表性SEEP-SRB1序列的精选列表。
SEEP-SRB 1是硫酸盐还原菌的多系分支,包括已知的ANME在甲烷厌氧氧化(AOM)过程中的共生性成员,如SEEP-SRB1a和SEEP-SRB1g,以及其他非共生性成员(SEEP-SRB1b、SEEP-SRB1c、SEEP-SRB1d、SEEP-SRB1e和SEEP-SRB1f)。
虽然目前在SILVA 138.2中被确定为属级进化枝,但这是这些生物的一个过于简化的分组。事实上,虽然一些成员,如SEEP-SRB1g和SEEP-SRB1c,被描述为物种水平的分支,但其他成员,如SEEP-SRB1a,更准确地描述为属水平的分支。
分辨率的提高揭示了不同SEEP-SRB 1亚组的不同分布
进一步使SEEP-SRB分类学复杂化的是SEEP-SRB1亚群之间不对称的系统发育距离——例如,基于基因组树,SEEP-SRB1g和SEEP-SRB1a可能位于不同的序列中。虽然用分类学分类纠正系统发育距离超出了本工作的范围,但这种历史命名惯例之间的冲突在环境微生物学中很常见并且难以解决。
然而,使用精确命名的组扩展数据库提供了一种避免这些差异的方法。因此,我们将SEEP-SRB亚群的代表性序列添加为SEEP-SRB1的“种”,但将SRB1g添加为“种”,这是SILVA 138.2中与SRB1g最相似的序列的分类名称。
将这些额外的SEEP-SRB序列纳入CABO-16S数据库,将部分环境SRB1组ASVs分解为各自的亚组。在甲烷渗漏和热液喷口数据集中,分辨率的提高揭示了不同SEEP-SRB 1亚组的不同分布。在喷口数据集中,只有一部分样本含有同营养型Seep-SRB1a和非同营养型Seep-SRB1d,这一区别无法用默认的SILVA 138.2数据库解决。与SILVA 138.2相比,CABO-16S如何将SILVA中注释为SEEP-SRB1、未分类的脱硫藻科或脱硫藻的ASVs分类时,分类分配的进一步差异变得清晰。
虽然CABO-16S对整个数据集中相对较小比例的ASV进行了不同的分类,但在特定环境中,如本研究中包括的沉积热液喷口,分类差异是显著的。有趣的是,一些SILVA 138.2分类为脱硫菌科的ASV在CABO-16S的较高级别上未分类(例如,unclassified_Desulfobacterales)或不同的分类。我们将这种差异归因于IDTAXA算法中的运行间差异,因为它对每次运行随机进行kmer的子采样,因此分类置信阈值边缘的一小部分ASV在运行之间会收到不同的分类。为了支持这一观察结果,将分析纳入归类为SEEP-SRB1、、Desulfosarcinaceae或LCP-80的ASVs,结果与一些归类为CABO-16S的asv在SILVA 138.2中未分类的结果大体一致。
CABO-16S通过添加特定序列,以更高的分辨率对SEEP-SRB1进行分类。

A) SEEP-SRB1及其相关分类群在最低分类水平的相对丰度。对于甲烷渗漏和热液喷口数据集,左侧子面板显示SILVA 138.2分类,右侧子面板显示CABO-16S分类。y轴表示每个样本总读取次数的百分比。
B)冲积图显示了数据库中相同ASV序列的分类。每一列都是一个不同的等级(从科到种),用CABO-16S标记的物种等级为不同的颜色。流量高度反映asv的数量。
缩写:unc,未分类;unsp不明;sed,沉积物。请注意,我们区分了由于分类冲突(未分类)而缺乏注释与由于参考分类群的注释不完整(未指定)而缺乏注释,如上图所述。
添加自定义序列相关的挑战
虽然Seep-SRB1亚型的注释可以通过添加具有特定注释的已知序列来实现,但这些分支的分类学、层级命名系统和系统发育,进化史之间仍然存在差异。
其他小组可能需要比我们用于SEEP-SRB1的方法更复杂的方法。志贺氏菌属和埃希氏菌属是这种矛盾的象征,因为两者在进化上都有很深的重叠,但分类学继续使参考数据库层次结构复杂化。
几十年来,聚球藻等其他环境群体同样对纠正分类学和系统发育提出了挑战。对于此类群体,添加具有特定、系统发育正确层次结构的序列不太可能改善分类,因为解决歧义的LCA方法假设所有序列共享相同的层次结构。
因此,所有现有序列都需要按照所需的系统发育框架进行类似的重新分类,并需要额外的管理,以确保新的分类层次与序列相似性的兼容性。最终,纠正系统发育和分类学的可行性受到16S rRNA基因中嵌入的信号的限制,虽然基于基因组的系统发育和16S rRNA系统发育在很大程度上是一致的,但它们并不完全相同。
◮ 分类分配的错误或不一致
提高分辨率的另一个障碍是分类分配中的错误或不一致,即相似的序列具有冲突的名称;据估计,这类错误占SILVA序列的1.5-17%。
基于大多数序列被正确且一致地标记的假设,IDTAXA等方法结合了工具来识别和删除在训练期间与大多数相似命名的序列冲突的单个序列,并且也存在独立的工具。
然而,这种方法对于由许多序列代表的分类群效果最好,对于需要提高分辨率的环境谱系并不总是如此。分类器的分辨率和准确性也可以通过限制数据库只包括特定于采样栖息地的微生物来提高,正如许多动物微生物组已经成功做到的那样。
这种特定栖息地的训练集无疑是对特定系统进行集中研究的最佳方法。但是,要了解具有广泛分布的特定分类群(如SEEP-SRB)的环境背景,就需要使用诸如SILVA之类的通用数据库最大化分辨率的方法。
CABO-16S成功地将来自SILVA 138.2的细菌和原始16S rRNA序列以及来自PR2数据库的细胞器16S rRNA序列与自定义选择的序列结合起来。与SILVA 138.2相比,增加了扩增子序列变体(ASVs)的分类定位。
具体来说,通过添加PR2的质体序列,CABO-16S无需额外测序16S和18S引物,就能在海洋和湖泊水体中出色地鉴定光养真核生物。尽管一些16S序列,如来自植物和藻类的线粒体仍然很少,可能会影响特定环境的分类,但CABO-16S减少了未分配的光养生物的数量从而可以快速提取剩余的丰富序列。
CABO-16S也被构建为可以添加自定义序列。随着SEEP-SRB1多系分支序列的加入,我们看到热液喷口沉积物样本的分类分化增加。这有助于确定特定环境中综合征的可能性,并加深对AOM的社区的理解。尽管添加自定义序列必须谨慎进行,但考虑到SILVA中未指定序列的数量以及将多系分支限制在当前分类结构的难度,CABO-16S的这一功能使用户可以自由定制16S分类,并可能增加对特定环境的理解。
最后,CABO-16S提供了一个框架,可以随着SILVA和PR2数据库未来版本的发布而轻松更新。R脚本和工作区可在https://github.com/emelissa3/CABO-16S上获得。自定义序列和其他东西永久托管在Figshare (https://doi.org/10.6084/m9.figshare.27288090)。
参考文献:
CABO-16S – A Combined Archaea, Bacteria, Organelle 16S database for amplicon analysis of prokaryotes and eukaryotes in environmental samples.
Eryn M.Eitel, Daniel Utter, Stephanie Connon, Victoria J. Orphan, Ranjani MuralibioRxiv 2024.10.23.619938; doi: https://doi.org/10.1101/2024.10.23.619938

谷禾健康

当谈到肠道菌群研究时,16S测序是一种常用的方法,它在了解微生物组成和多样性方面非常重要且实用。
16S rRNA是细菌和古细菌中的一个高度保守的基因片段,同时具有一定的变异性。通过对16S rRNA基因进行测序,可以确定微生物群落中存在的不同菌属和菌种,并评估它们的相对丰度。
针对肠道菌群16s测序概括起来可以了解以下内容:
揭示微生物组成:通过16S rRNA测序,可以获得关于肠道菌群中存在的不同菌属和菌种的信息。这有助于了解菌群的组成,包括有益菌、有害菌和其他微生物的相对丰度。
探索微生物多样性:16S rRNA测序可以评估微生物群落的多样性水平。通过分析不同样本中的菌群组成和多样性指标,可以比较不同个体、不同健康状态或不同治疗干预对微生物多样性的影响。
鉴定潜在病原菌:通过16S rRNA测序,可以鉴定肠道中存在的潜在病原菌。这有助于了解潜在的疾病风险,并为相关疾病的预防和治疗提供指导。
监测治疗效果:在临床研究中,通过定期进行16S rRNA测序,可以监测治疗干预对肠道菌群的影响。这有助于评估治疗效果,并为个体化治疗提供依据。
指导个体化营养干预:通过16S rRNA测序,可以了解个体的肠道菌群状态,进而指导个体化的营养干预。根据菌群组成和功能,可以制定相应的饮食方案,促进肠道健康和营养吸收。
以上总结了16s对于了解肠道菌群的重要价值,今天本文主要分享和介绍16s测序相关的定义和术语,以及菌群命名的规则等。
正确的说法是16S rRNA(不是16s rRNA或16s rDNA)。
16S rRNA 代表16S核糖体核糖核酸 (rRNA),其中S(Svedberg) 是测量单位(沉降率)。该rRNA是原核核糖体小亚基(SSU)以及线粒体和叶绿体的重要组成部分。
下图显示了16S rRNA(简称16S)如何参与原核核糖体。
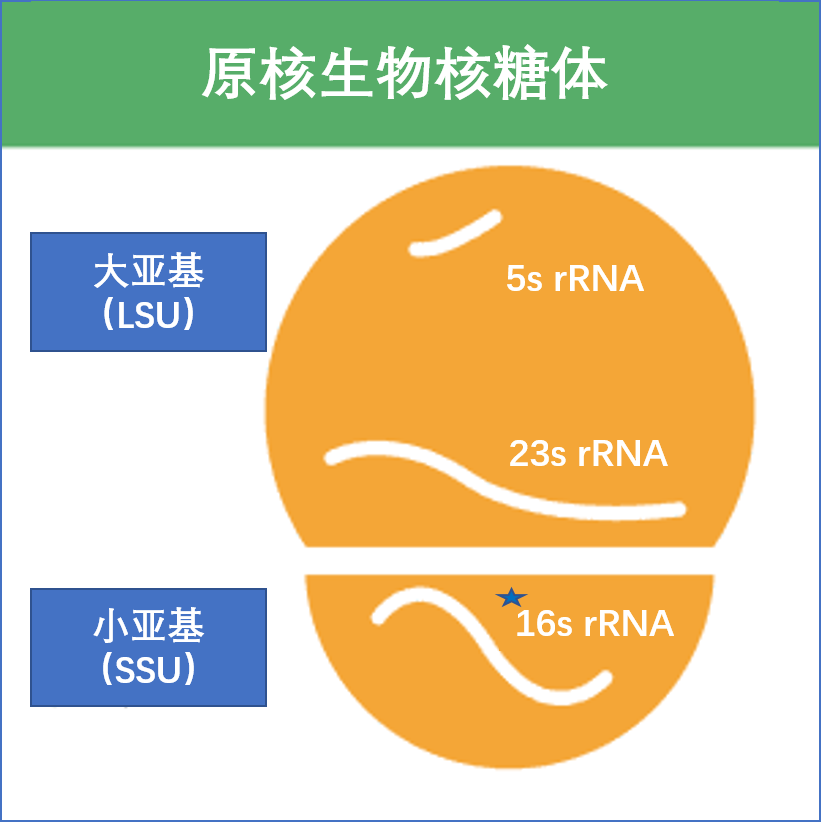
•16S rRNA在原核生物中发挥重要功能
16S rRNA是一种细菌和古细菌中常见的核糖体RNA分子。它在细胞中起着重要的功能,包括参与蛋白质合成的核糖体组装和识别启动子序列的功能。
由于16S rRNA在细菌和古细菌中具有高度保守的序列区域和变异的序列区域,因此它被广泛用于细菌分类和进化研究中。
相比之下,16S rDNA这个术语并不常用。rDNA是指编码核糖体RNA的DNA序列,因此16S rDNA可以理解为指代编码16S rRNA的DNA序列。
•科研中通常使用16S rRNA
在科研文献和学术讨论中,通常更常见地使用16S rRNA测序或16S ribosomal RNA测序。这种测序通常针对编码16S rRNA的DNA序列进行扩增测序,用来研究微生物的多样性和进化关系,特别是细菌和古细菌。
通过对16S rRNA基因进行测序,可以对微生物群落的组成和结构进行分析。这项技术在微生物学、生态学、医学和农业等领域有广泛的应用。
要用作DNA条形码,基因应具有以下特征:
•已知的细菌和古细菌都具有16S rRNA
它应该无处不在。否则,我们不能包括所有生物。已知细菌和古细菌的所有成员都具有16S基因。
它应该包含足够的系统发育信息。16S大约有1500bp长,不算太短也不算太长。
原核生物中发现的16S基因内的遗传变异足以用于广泛分类范围的系统发育分析。它成功地用于推断门之间的系统发育关系,同时也用于同一属的物种之间的比较。
•容易通过PCR扩增
它应该很容易通过PCR扩增。16S基因有多个保守区域可以作为引发位点。这成为基于NGS的短读长测序的一个显著优势。
经过多年的国内外研究和检测积累,我们拥有几乎所有已知细菌和古菌物种的16S序列数据库。通过在这些数据库中搜索16S序列,即使没有严格的分类学知识,也可以识别新分离的菌。
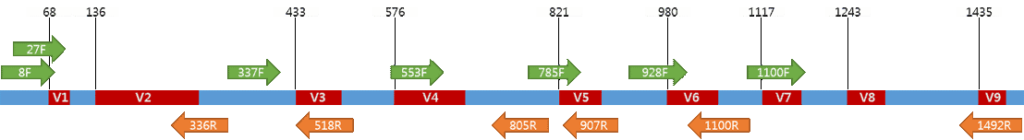
引物用于扩增
已知细菌16S之间的序列变异分布不均匀。在细菌中鉴定出9个高变区,分别命名为V1至V9。
从长度来看,全长16S长度为1.5kb左右,单菌落的16S全长sanger一代测序仍然是菌种鉴定的主要手段,纳米孔和Pacbio的三代测序可以高通量的获得全长序列,对于希望更高分辨率的分析菌种的研究有一定优势。
•三代测序建库成本较高,测序深度较低
三代的测序准确度目前逐渐改进,直接测序准确度可以在90%以上,纠错后可以提高到97~99%以上,已足够提供高精度的分类。三代目前主要问题在于建库成本相对较高,通过使用barcode可以降低部分但仍然偏高,此外普遍测序深度相对于二代测序要低许多。
•二代测序由于读长限制需要引物进行扩增
由于二代测序读长限制,无法使用高通量测序技术对16S rRNA整个基因全长进行测序,因此必须针对基因的某一片段设计引物进行扩增测序。
注:虽然有大量的文献研究不同片段的优缺点,但由于采用的样本类型、区域引物以及分析角度的不同,尚没有针对所有样本的最佳可变区片段的共识。
基于大量项目经验和文献调研, 目前最主要的可变区选择是V4区和V3V4区,V4区长度为256bp左右,加上两侧引物长度为290bp左右,使用双端2x250bp或2x150bp可以测通,此外如454、life、Illumina Hiseq 4000, Illumina Novaseq 6000的测序平台读长也可以主要涵盖该区段读长。
•16S V4 引物通用性是所有可变区中最高的
传统的认知中,普遍认为测序片段越长,测到物种数据就越多,故倾向选择16S V3V4。然而16S V4(515-806)引物通用性相对是所有可变区中最高的。且在大规模菌群调查研究中,如人体菌群研究HMP,地球微生物计划EMP,欧洲的FGFP、美国肠道计划AGP以及全球土壤菌群调查,都采用V4区作为检测区域。16S V4目前仍然是国际研究中使用最广泛和认可的检测区域。
细菌的命名法和命名受《国际原核生物命名法典》 (简称《原核生物法典》)的约束 ,但实际物种的分类则不然。例如,大肠杆菌(Escherichia coli)这个名称受《原核生物守则》的规范,但物种本身的特性和分类学分类不受规范。这意味着没有官方分类法,只有名称受到控制。
因此,“有效物种”、“正式物种”或“官方物种”等术语没有意义。只能验证名称,因此可以使用 “具有有效发布名称的物种”。
物种名称应由二项式系统的两部分组成:
(1)通用名称和(2)特定词缀。
如下示例:
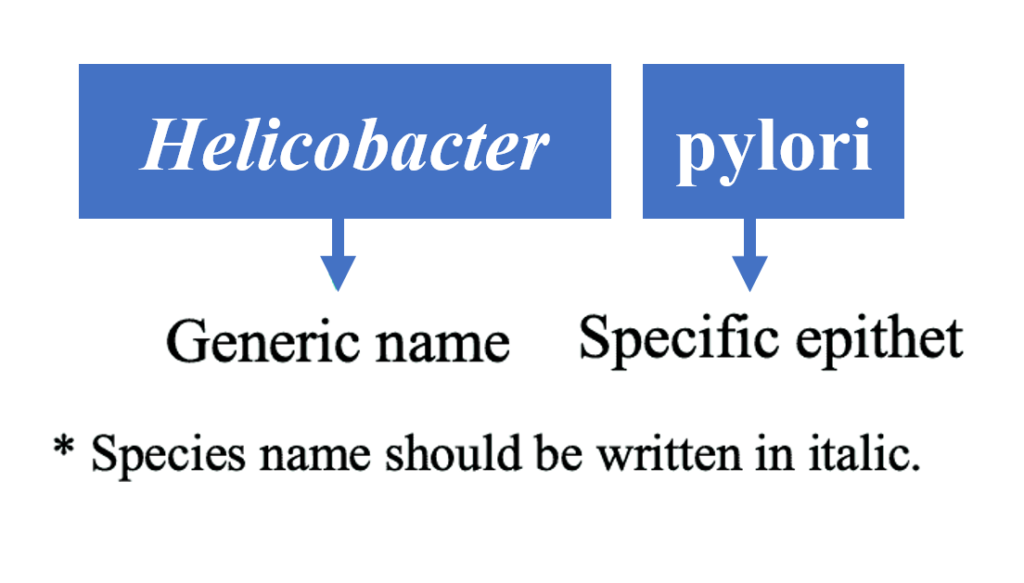
该组合代表“物种名称”,并且在命名系统中应该是唯一的。所有分类群的学名必须被视为拉丁文;物种等级以上的属群名称是单个词。
•生物分类按照不同等级
名称被组织成一个层次系统。
门(Phylum):门是细菌分类的最高级别之一,代表着细菌的大类别。细菌门包括了多个相关的纲和目。
纲(Class):纲是细菌分类的次级别,比门更具体。同一纲的细菌通常具有一些共同的特征和特性。
目(Order):目是细菌分类的更具体级别,比纲更详细。同一目的细菌通常具有更相似的形态、生理特征和生态习性。
科(Family):科是细菌分类的更具体级别,比目更详细。同一科的细菌通常具有更相似的基因组组成和代谢途径。
属(Genus):属是细菌分类的更具体级别,比科更详细。同一属的细菌通常具有相似的形态、生理特征和基因组组成。
种(Species):种是细菌分类的基本单位,代表着具有相似形态和生理特征的细菌群体。
亚种(Subspecies):亚种是种的更具体级别,用于进一步划分具有细微差异的细菌。
目前,门等级不受原核代码控制。提议将“门”纳入守则,但该提案尚未得到控制原核生物守则的机构国际原核生物系统学委员会(ICSP)的讨论和批准。
在细菌分类中,门、纲、目、科、属、种和亚种的命名规则和方法遵循国际细菌命名法。
以下是一些要注意的事项:
命名规则:细菌的命名应该是拉丁化的,以便在全球范围内使用和理解。通常使用斜体字体或以斜体书写。
门、纲、目、科、属、种和亚种的命名:这些分类级别的名称通常是基于拉丁词根、描述性词语、地名、科学家的名字或其他相关特征来命名的。例如,属名可以以科学家的姓氏命名,种名可以以形容词或名词来描述特定的特征。
•每个细菌分类级别的名称是唯一的
命名的唯一性:为了避免混淆,每个细菌分类级别的名称必须是唯一的。这意味着同一级别下的不同分类单元不能有相同的名称。
命名的正式发表:细菌分类的名称需要通过正式的科学出版物发表,以确保其被广泛认可和接受。这有助于维护分类的一致性和准确性。
命名的修订和更新:随着科学研究的进展和新的发现,细菌分类可能需要进行修订和更新。新的分类单元可能会被提出或已有的分类单元可能会被重新归类。这些修订和更新需要经过科学界的讨论和认可。
例如下面是假单胞菌菌名称和分类:
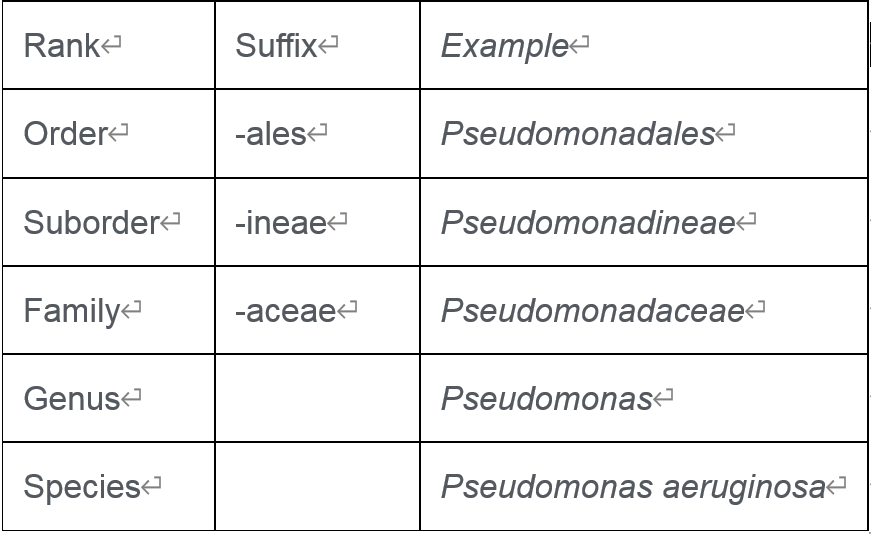
这些名称是由提出物种并将其分配给属的科学家给出的。例如, Thermacanus属已被提出,但并未将其归属于已知科。在NCBI分类学中,它属于芽孢杆菌目的暂定科“ Bacillales Family X. Incertae Sedis ” 。
EzBioCloud数据库将每个物种分配在完整的等级系统下,Thermicanus 被分配在暂定创建的家族“ Thermicanus_f ”下(请注意,这是一个无效名称,并且不是斜体)。当然,任何人都可以通过发表提案来提出Thermacanaceae家族的建议。
如前所述,名称由原核代码控制。要验证名称,该名称应包含在下述列表之一中。
▸ 批准的细菌名称列表
该论文由Skerman、McGowan 和 Sneath于1980年发表 ,标志着原核生物命名法的新开始。
此列表中的任何后续更改均由《国际系统与进化微生物学杂志》(IJSEM)(前身为《国际系统细菌学杂志》(IJSB))发布。
IJSEM中发布了两种类型的更新列表,即“通知列表”和“验证列表”。
▸ 通知列表
IJSEM是国际原核生物系统学委员会(ICSP)和国际微生物学会联盟(IUMS)细菌学和应用微生物学部的官方期刊。这意味着IJSEM是原核生物命名法的官方期刊。但是注意,没有官方的细菌分类法。
任何包含在IJSEM中发布的名称更改(新分类群或名称更改)的分类提案都将自动由IJSEM的列表编辑进行审核。如果论文符合《原核生物守则》的要求,则拟议的更改将被列入“通知列表”并被正式“验证”。
▸ 验证列表
原核代码还允许科学家有效验证在IJSEM以外的期刊上发表的名称。一旦名称被公布,就被称为“有效公布”。
任何关心科学的人都应该确保任何有效发布的名称都得到验证,方法是将出版物PDF发送给 IJSEM,并证明(对于物种和亚种)指定的类型菌株可以不受限制地从位于不同地区的至少两个公共培养物种保藏中心获得。
这可以由包括作者在内的任何人来完成。不这样做的带来的后果可能是有害的,因为该名称“有效发布”但从未“验证”。如果满足原核生物守则的所有要求(对于新物种和亚种,特别是规则27和30),IJSEM 的列表编辑将在验证列表中添加名称。
▸ 无效名称
不幸的是,在某些情况下名称无法验证。例如:
•一个名称在1980年之前被广泛使用,但在1980年没有被列入批准的细菌名称列表。
•一个名字在IJSEM之外发布,但从未被验证。
•没有人将PDF和进一步所需的文档发送给 IJSEM 进行验证。
•该出版物不符合准则的最低要求。
注:未有效公布的名称要用引号括起来,例如“Selenomonas Massiliensis”以区别于有效公布的名称。
•验证新物种的最低且唯一要求
根据原核生物守则, 验证新物种和亚种名称的最低且唯一的要求是:
•必须指定类型菌株。有时,描述新物种的论文不会提及哪个菌株是模式菌株。在这种情况下,名称无法验证。
•模式菌株应保藏于两个不同国家的两个培养物保藏中心。这确保了典型菌株仍然可用,例如当培养物保藏中心停止其活动或丢失菌株时。
•模式菌株必须通过培养物保藏中心可供任何人使用。专利菌株不能作为模式菌株。如果您想为某个菌株申请专利并限制其分布,则它不能作为物种或亚种的命名类型。
小结
以上总结起来如下:
确认命名规范:首先,确保您了解国际微生物学命名规范,如IJSEM是国际原核生物系统学委员会。这些规范包括命名原则、命名要求和规则,以确保新细菌名称的准确性和一致性。
检查已有命名:在确定您的新细菌名称之前,进行一次详细的文献调研,以确保该名称没有被其他研究人员使用或已经被正式命名。搜索相关数据库和文献,如国际细菌命名索引(International Journal of Systematic and Evolutionary Microbiology)和PubMed,以避免重复命名。
提交命名申请:一旦您确认您的新细菌名称符合命名规范并且尚未被使用,您可以准备提交命名申请。通常情况下,命名申请需要包括详细的描述、分类学特征、基因序列数据等相关信息。您可以向国际微生物学协会或相关的命名委员会(前面说的IJSEM)提交命名申请。
合作与审查:在提交命名申请之前,建议与其他研究人员、领域专家或命名委员会进行合作和讨论。这样确保命名申请符合规范。
审核和正式发布:一旦命名申请被提交,相关的命名委员会将对其进行审查。如果您的命名申请符合规范,经过审核后,新细菌名称将被正式发布,并被纳入相应的细菌命名索引和数据库中。
数据库很重要?
细菌名称的设计或命名是为了科学地组织,以提供基于系统发育的生物学见解;使用分层分类系统可以实现这一点。
•正确的名称对于解释微生物与其他数据的关联非常重要
例如,我们认为大肠杆菌与其他大肠杆菌物种比与芽孢杆菌物种有更多的表型特征。因此,使用正确的名称对于解释微生物组和其他类型的数据至关重要。
不幸的是,在当前的命名法中仍然有许多名称位于错误的位置。例如,Clostridium scindens因其在人类肠道菌群胆汁酸代谢中的重要作用而闻名,并且仍然保留着Clostridium的名称。然而,它甚至与梭状芽胞杆菌属并不接近 ,应归类为毛螺菌科的一个新属。
真正的梭菌属(Clostridium)属于梭菌科(Clostridiaceae)。例如Ruminococcus Sp(梭状芽胞杆菌)
梭状芽胞杆菌簇XIVa和IV 是已知在人类微生物组中发挥主要作用的细菌群。这些细菌簇在1994年才进入16S rRNA系统发育。
然而梭菌簇XIVa和IV 不代表正式命名法,也不表示单个分类群,例如属或科。自1994年以来,许多最初指定的属于梭状芽胞杆菌簇XIVa和IV的物种已被重新分类为新属。
•一些细菌存在错误的命名和分类
然而,这些簇中仍然存在错误分类的梭菌属物种,从而保留了旧名称。比如:Faecalibacterium prausnitzii(普拉梭菌),革兰氏阴性,对氧极度敏感,属于梭菌科,厚壁菌门。
该物种就是属于Clostridium clusterIV分组的Clostridium leptum group柔嫩梭菌类群,是该类群的最优势菌种。一般中文翻译柔嫩梭菌指的就是这个类群,其代表物种就是普氏栖粪杆菌,又名普拉梭菌。
以下是当前EzBioCloud数据库中对应于梭状芽胞杆菌簇XIVa和IV的分类群。
“Candidatus“是一个拉丁语词汇,通常用于微生物学中。它表示对一种微生物的命名,表示该微生物是一个新发现的候选物种,尚未被正式分类或命名。在中文中,”Candidatus“通常被翻译为”候选物种”或”候选菌种”。这个术语用于描述那些已经被发现但尚未被完全分类的微生物,通常是由于它们的特殊性质或难以培养。
总结来说,使用正确的细菌名称可以确保科学研究的准确性和一致性。细菌命名规范旨在为每个细菌物种提供唯一的、明确的名称,以避免混淆和误解。通过使用正确的名称,研究人员可以准确地标识和描述细菌物种,使得研究结果具有可比性,并促进科学交流和合作。
使用庞大的菌群的数据库,便于数据整合和共享。通过将新细菌与已知的细菌物种进行比对,可以确定其在分类学上的位置和相关信息。这有助于构建细菌分类树、建立物种间的关系,并为进一步的研究提供基础数据。
此外,正确的细菌命名和细菌样本数据库的比对可以促进研究进展和新的发现。通过对新细菌进行分类和命名,研究人员可以更好地理解细菌的多样性、生态学角色和潜在的应用价值。
人体肠道菌群健康检测需要依托本地化人群菌群样本检测的大数据库,区分菌群的组成和功能在个体间存在差异,并且与本地人体的生理状态、生化代谢、免疫炎症以及疾病风险等建立关联。通过建立大数据库,可以收集大量的菌群样本和相关数据,进行统计学分析和模式识别,从而揭示不同菌群组成与人体健康之间的关联。
总之,使用正确的细菌名称和拥有庞大的细菌样本数据库进行比对是确保科学研究准确性、促进数据整合和共享、推动研究进展和发现的关键步骤。
在某些情况下,即使一个物种是真实的并且具有科学重要性,也无法满足原核生物守则的基本要求。
根据正式命名法,这些物种的名字永远不会得到有效的公布。因此,分类学家创造了“Candidatus”一词来支持暂定命名。
对于Candidatus名称,没有纯培养物或分离株,因此没有典型菌株。由于Candidatus不属于正式命名法,因此不受原核法典管辖。
一个典型的例子是Candidatus Pelagibacter ubique该物种含有来自海洋的未培养或培养的细菌,可能是地球上最丰富的物种,因此它一定非常重要。
它从未得到验证的原因是该物种可以培养,但不能以培养物保藏所可用于长期储存的方式或规模进行培养。因此,无法满足该准则的最低要求。此外,特定的加词(ubique)格式错误。
并非所有Candidatus类群都得到了很好的表征,因为Candidatus的等级不受原核生物守则的规定。
•一般Candidatus的最低标准:
•提供全长高质量16S 序列,因为该基因是细菌和古细菌分类学的框架。
•未受污染且相当完整的基因组序列的可用性(通过域级核心基因的存在进行检查)。
•应提供最低限度的生态和代谢特征。
名称最好能代表该物种的典型特征。例如, Plasticicumulans这个名字的意思是“积累的塑料”。该属因积累生物塑料聚羟基脂肪酸酯而得名。
命名新分类单元的另一种流行方式是以人的名字命名。这个人应该有对科学做出贡献,尤其在微生物方面。著名的埃希氏菌(Escherich)和沙门氏菌(Salmon)是以微生物学家埃舍里奇(Theodor Escherich)和萨蒙(Salmon, D.E.)的名字命名的。
此外,以地理位置命名也很流行,但一般不鼓励。

谷禾健康

16S科研项目是一个完整的闭环,前期的课题项目设计方案、取样和重复实验设置决定了后期分析报告的数据完整性和项目类型。
想要拿到一手有利用价值的科研报告和项目数据,前期的实验方案设计和后续的分析都起着关键性的作用。
然而有时候拿到报告不知道如何去解读,这里为大家梳理一下16s科研项目的全过程,帮助大家更好的了解报告内容,快速获取关键信息。


实验方案设计就像一个总工程的设计图纸,决定了未来科研分析报告的类型走向,并且前期的分组设计的越详细,各种理化指标、生化指标、代谢物等信息准备越充分,后续报告的完整度越高。


明确项目课题类型
第一步要做的就是明确项目课题类型:
最常见的就是多分组之间差异分析比较:例如,要比较对照组、模型组、实验组,之间的差异结果。
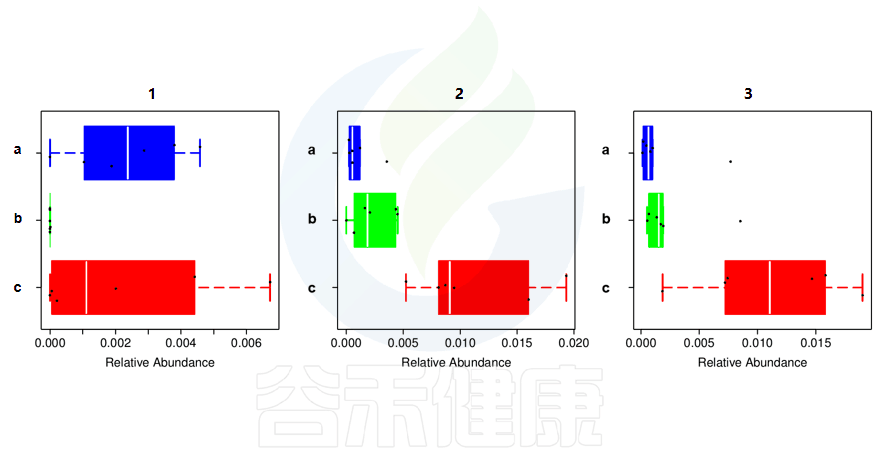

还有多分组中,任意两组之间比较:例如某实验设计了正常组、疾病组、用药组服用奥氮平、阿立哌唑、氨磺必利、利培酮,像比较不同的用药组和疾病组之间的菌群的差异结果,就用到了分组之间两两差异比较。
✦举个例子
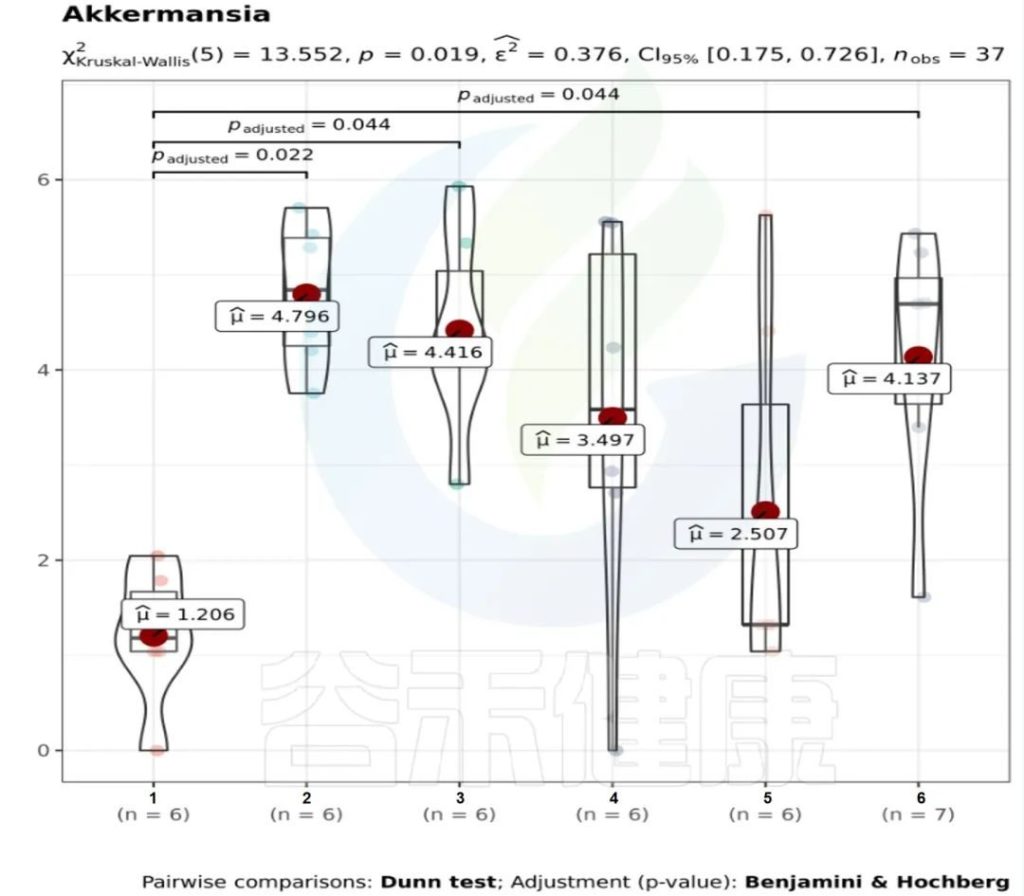
图中1组与3组、4组、6组 组间差异显著
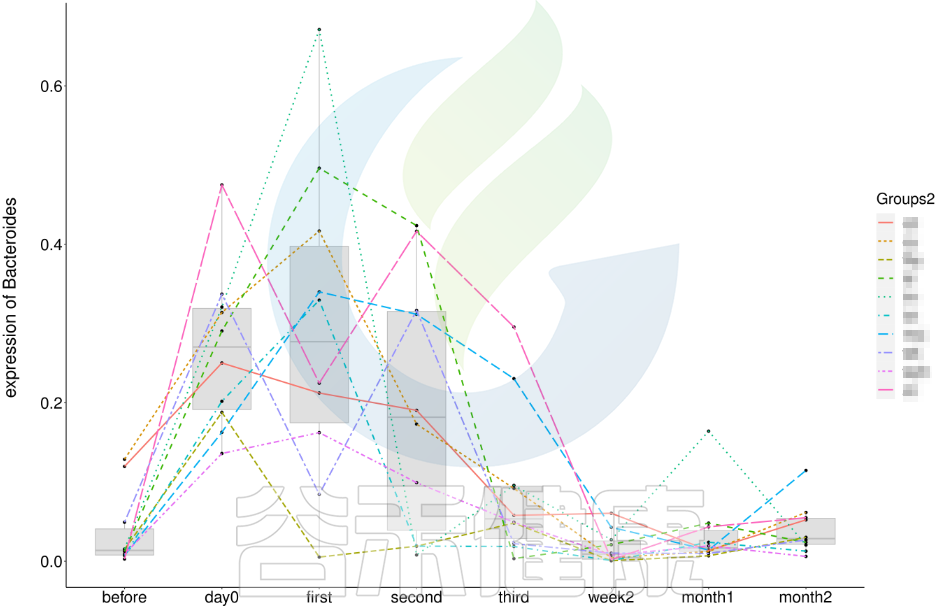
还有随时间的变化比较菌群之间的变化规律:例如在用药不同时间段包括3天,5天,2周,1个月,2个月,观察菌群的变化情况。如下图所示:
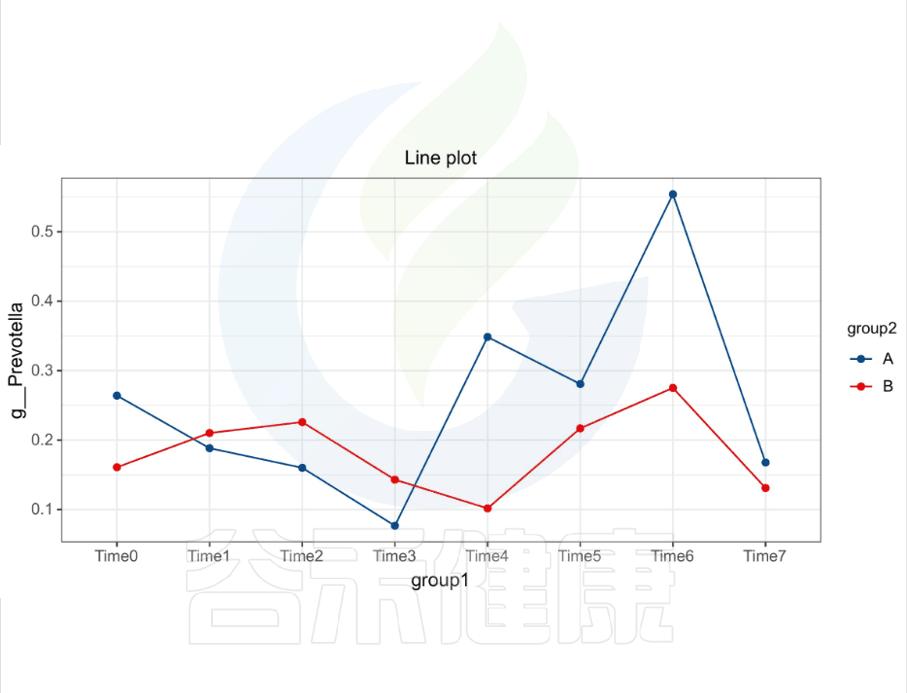
收集理化指标非常重要
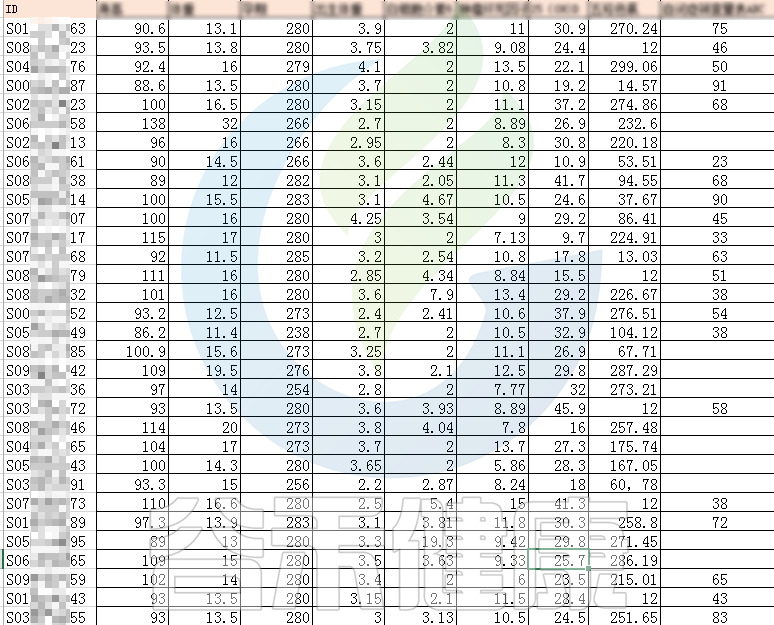
如果前期搜集好每个样本的相关理化指标,还可以计算这些指标与菌群之间是否具有相关性。
✦举个例子
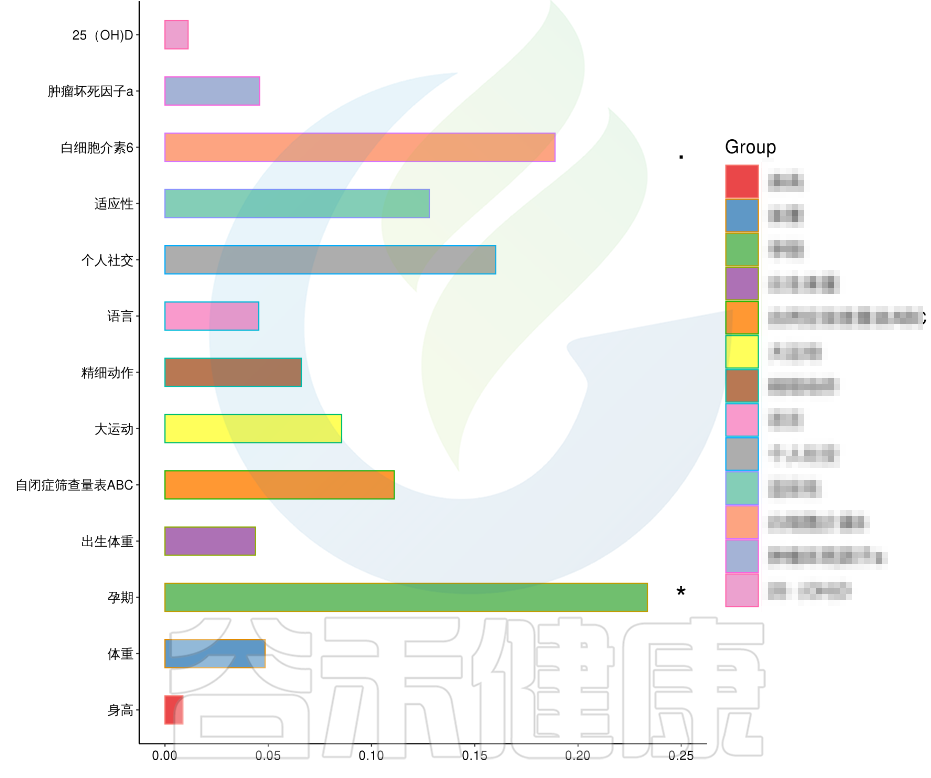
例如该项目比较自闭症儿童与正常儿童的菌群差异。客户在样本信息单里还详细搜集了母孕期的各种详细指标,例如孕期天数、出生体重、白细胞介素6、肿瘤坏死因子a、五羟色氨等数值型理化指标。
还搜集了是否顺产、是否妊娠高血压、是否孕期感染、是否妊娠糖尿病、是否先兆流产等因子型理化指标。其中0代表否,1代表是:
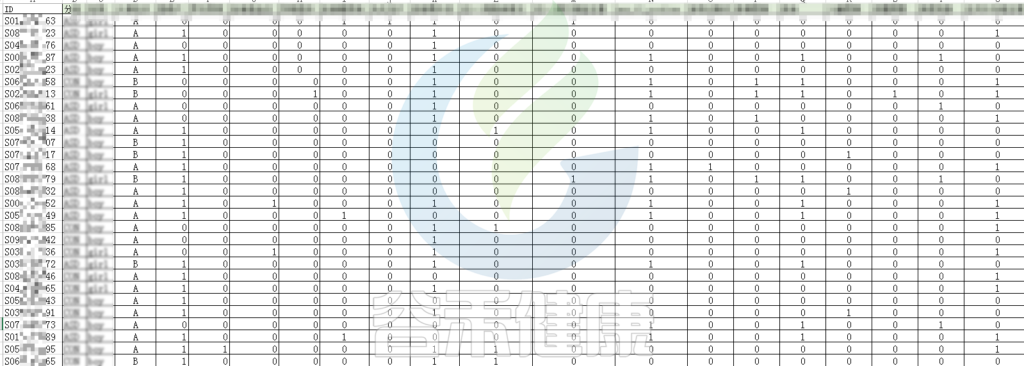
根据这些理化指标与菌群数据做相关性分析,从因子型的结果可以看出,自闭症(ASD)与正常儿童之间的分组与菌群之间相关性极显著**,其次是否有先兆流产的分组与菌群之间有显著相关性*,其他的包括是否喝牛奶、孕期是否感染、妊娠高血压都与菌群有相关性。
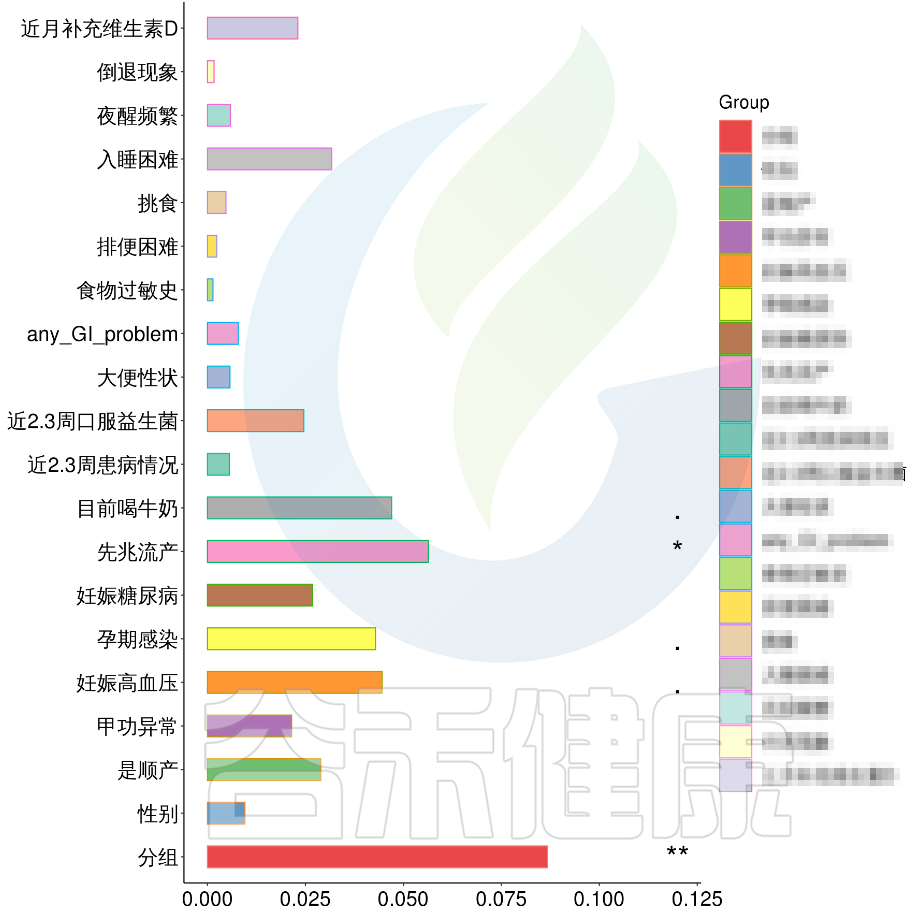
在数值型理化指标中,孕期的天数与菌群之间相关性显著*,其次是白细胞介素6与菌群之间有相关性。
小结
因此,前期搜集相关资料越详细充分,对分析报告的完整性也会有帮助,分析人员也会根据您的样本信息单提供的相关内容,做出个性化的分析和售后指导建议。
首先基于样本类型,最常见的环境样本来源是人体、动物、土壤、水体等。而人体中的肠道菌群样本是目前研究最广泛,可鉴定的物种也最为丰富,谷禾在肠道菌群与人体健康方面有深入研究,目前已完成超20万例临床肠道菌群样本检测,并构建了超过60万各类人群粪便样本数据库。
其他样本类型还包括人体/动物唾液样本、组织样本、尿液样本等。
▸ 粪便样本
目前粪便样本从采样到提取数据分析技术较为成熟、应用较为广泛,谷禾最早在15年就开发了针对粪便样本的取样管,也是最早致力于研发粪菌取样盒的公司,方便实验室、个人日常取样需求,实现了粪菌样本的常温运输。
谷禾取样管常温保存,取样也较为方便卫生,在家就可以轻松完成,相较于传统取样方法都有所升级。并且该取样管也有专利证书。该取样方法被大量客户采用并接纳,大大降低了采集粪便样本的难度,缩短了搜集样本的时间周期。
取样示意图

▸ 其他样本
土壤样本也相对较为容易提取出DNA,但需要注意的是土壤样本的菌群特征容易受植物腐殖质基因的影响和干扰,所以提取时要进行纯化。
而口腔、组织、尿液等样本,由于DNA含量较少,在实验阶段提取相对较为困难,所以提前准备样本时,尽量多取一些,并且可以多取几个重复,尽量避免扩增不出来的情况。
并且这些样还很容易受到环境样的污染,所以在实验阶段,可以取空白样本,和阳性样本ST做对照,数据分析时可以用来纯化样本,排除来自环境的干扰序列。
✦组间差异分析需重复取样
要做组间差异分析时,每组要重复取样,才能做组与组之间的统计检验。理论上,每个组至少3个样就满足基本的统计差异分析需求。所以在重复取样时,每个分组至少取3个样。取样时要保证每个分组内部的样本一致性,如果组内样本之间的个体差异性较大,则会影响后期组间差异结果分析。
✦举个例子
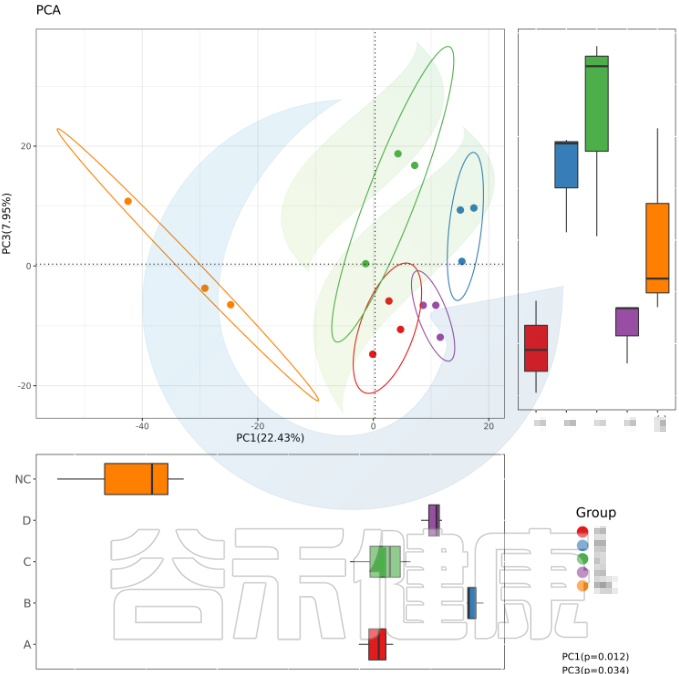
例如从该图可以看出,分组之间组间差异较大,并且组内的样本之间较为接近和相似。
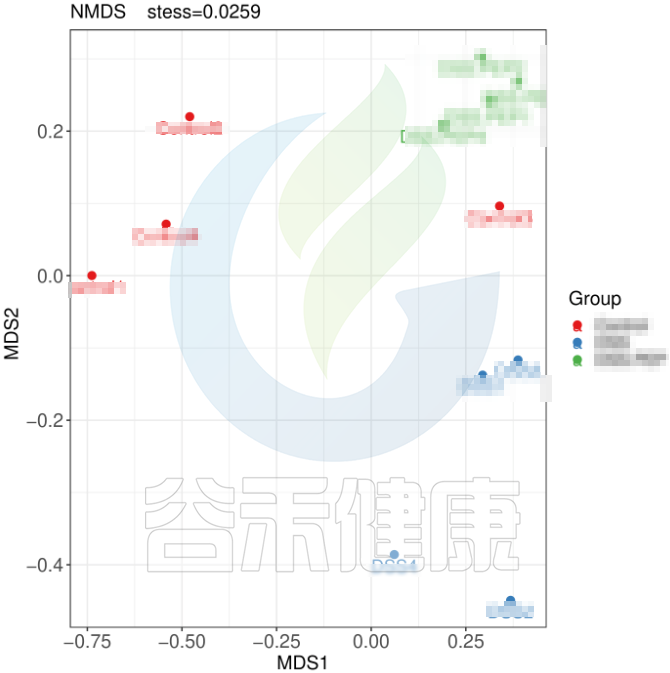
但从该图可以看出,Control组中Control3样本明显与组内的其他样本差异较大,与DSS组内的样本较为相近,这样就对后期组间差异分析的时候会产生影响,需要将该样本去除。
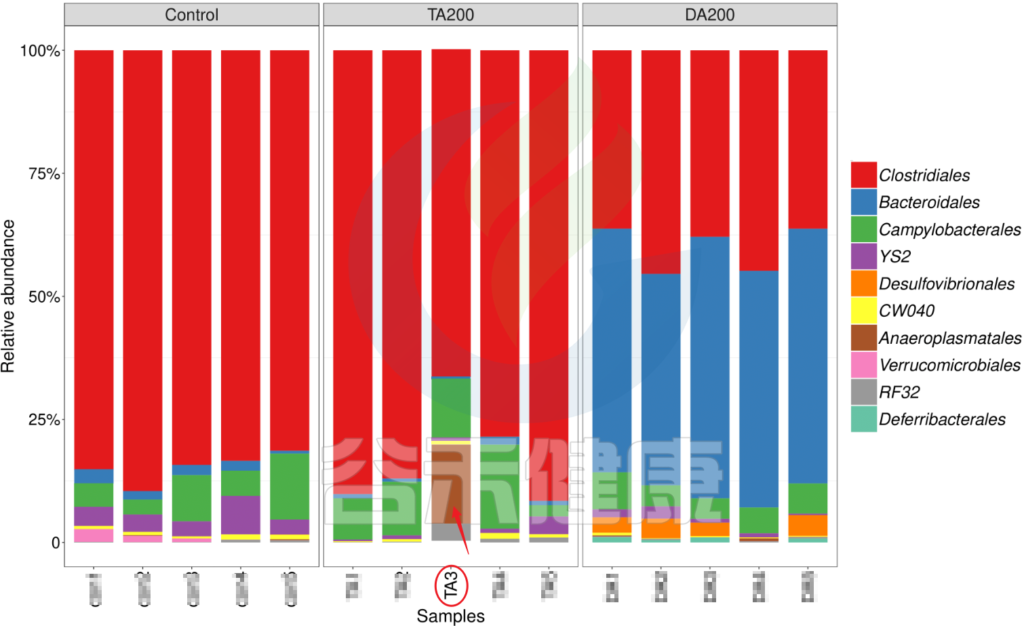
又例如在该图中,TA200组中的TA3样本的Anaeroplasmatales物种丰度含量非常高,该样本与组内的其他样本明显差异较大,该样本可能受到环境污染等其他因素干扰,这样就没有办法保证组内样本的均一性,也会影响分组之间的差异分析统计结果,再后期分析的时候建议把该样本去掉重新分析。
建议
为了便于后期数据整理修改,每个分组需要保留一定量的重复样本,假如每个分组只取了3次重复,假如其中有一个样本质量不好需要去除,该分组只剩2个样本,则不满足每组至少3个样的分组条件,整体就没有办法做组间差异分析统计。
所以这里建议每个分组至少取5个样做重复,一般6到10个样就能分析出比较完善的结果。具体分组和组内的重复取样数量视具体的实验设计方案而定。
在经费允许的情况下,建议多取一些重复。假设每组取50到100个重复或者以上,得到的分析结果就基本可以涵盖该分组情况所有的菌群构成情况,可以较为全面的研究分组之间的菌群构成差异情况。
当拿到16S科研分析报告以后,面对纷繁复杂,各式各样的图表分析结果犯了难,不知道如何从这么多的图表中入手,快速找到报告中需要的图表结果。
这里对16S科研分析结果抽丝剥茧,概括出报告中的主要几大内容板块。
•16S科研分析究竟是在做什么?
16S rDNA 是一种对特定环境样品中所有的细菌进行高通量测序,以研究环境样品中微生物群体的组成,解读微生物群体的多样性、丰富度及群体结构,探究微生物与环境或宿主之间的关系的技术。
主要是对原始数据进行拼接过滤得到的优化序列,降噪方法得到ASV,再对ASV进行物种注释,注释到门、纲、目、科、属、种各层次上的分类结果。
通过ASV表计算Alpha多样性,样本内的多样性指数,Beta多样性,样本间相似性的指标。
对ASV表进行功能预测,例如Picrust2功能预测分析、Bugbase菌群表型特征分析,FAPROTAX生态功能预测等。
得到的每个样的数据结果,根据客户提供的分组情况和理化指标,进一步做组间差异分析,以及和环境理化指标之间做关联分析,相关性分析,比较分组之间是否有差异,差异是否显著,来验证分组是否合理,和环境宿主之间是否有关联性。
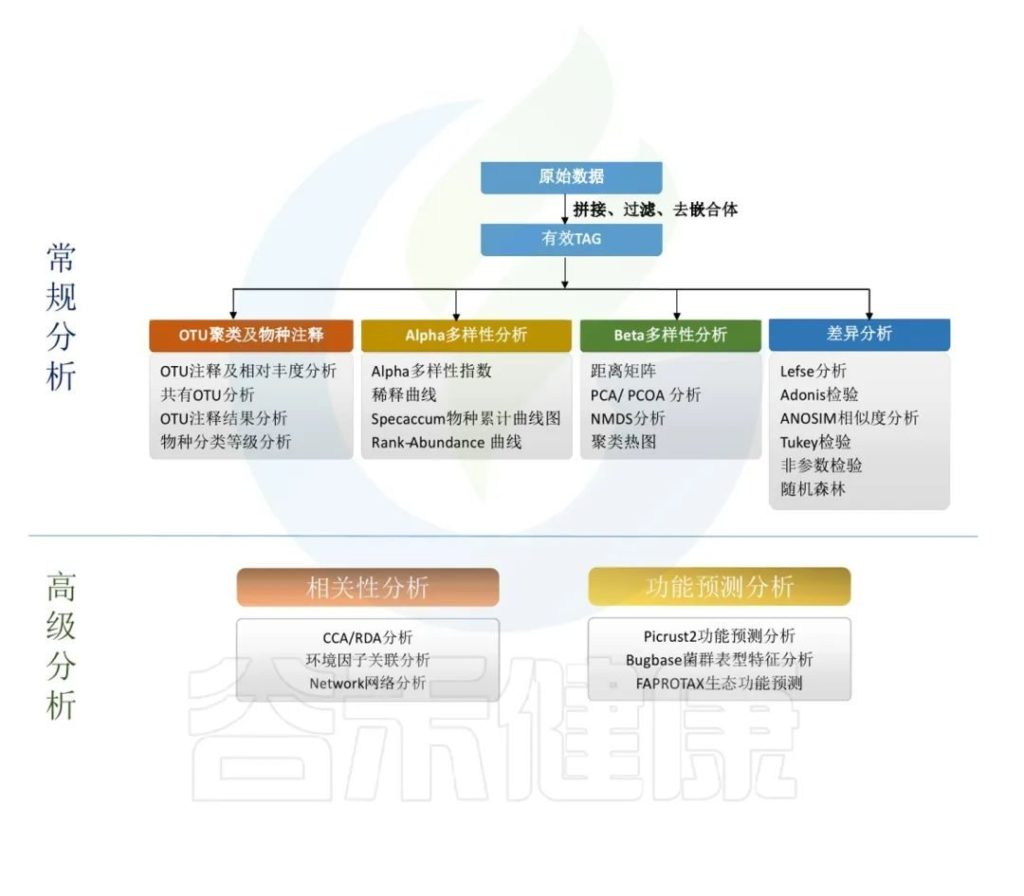
原始数据处理
Illumina NovaSeq测序平台测序得到的双端数据Raw PE,经过拼接和质控,根据一定的标准过滤掉低质量数据、接头或PCR错误,得到Raw Tags。再经过去重复序列,去singleton序列,过滤嵌合体,得到可用于后续分析的有效数据 Effective Tags。
OTU(ASV) 表生成
微生物多样性分析中最重要的就是OTU特征表,一切后续分析都围绕OTU表来进行。生成OTU除了传统的聚类的方法(一般按照97%的相似度进行聚类),现在最新用到的技术的是降噪的方法得到ASV。
简单来讲ASV就是在去除了错误序列之后,将Identity的标准设为100%进行聚类,常见的有DADA2、Deblur、Unoise三种降噪方法。项目里用到的是UNOISE2降噪方法获得ASV数据。
物种的分类与注释
采用QIIME2训练分类器方法对ASVs代表序列进行分类学注释,默认选用SILVA138数据库进行物种注释。并在各个分类水平上:domain(域),phylum(门),class(纲),order (目),family(科),genus(属),species(种)对每个样本的群落组成统计。
alpha多样性
Alpha多样性主要反映样本内多样性。对ASV表进行计算可以获得每个样本的simpson,ace,shannon,chao1以及goods_coverage等指数,alpha多样性指数用来来评估样本菌群物种的丰富度(richness)和多样性(diversity)
beta多样性
Beta多样性反映的是样本间多样性,Beta多样性是衡量个体间微生物组成相似性的一个指标。通过计算样本间距离可以获得β多样性矩阵,基于OTU的群落比较方法报告中给出了,欧式距离、bray curtis距离、Unweighted UniFrac距离和Weighted UniFrac距离等。
功能预测
得到群落的微生物组成之后,也可以对群落功能组成进行预测,常用的16S功能预测的相关软件有PICRUSt2、FAPROTAX、BugBase。
PICRUSt2用来预测功能,通常指的是基因家族,PICRUSt2支持基于多个基因家族数据库的预测,报告中包括了KEGG同源基因,KO直系同源物,EC酶分类编号,MetaCyc途径的丰度,CAZy碳水化合物活性酶数据库,GMM是肠道代谢模块和GBM是肠脑模块。
FAPROTAX是原核的微生物注释代谢或其他生态相关的功能(例如硝化,反硝化,发酵)的一个数据库和软件。FAPROTAX预测的功能主要集中在海洋、湖泊等环境样本微生物的功能,特别是硫、碳、氢、氮的循环功能。
BugBase能进行表型预测,其中表型类型包括革兰氏阳性(Gram Positive)、革兰氏阴性(Gram Negative)、生物膜形成(Biofilm Forming)、致病性(Pathogenic)、移动元件(Mobile Element Containing)、氧需求(Oxygen Utilizing,包括Aerobic、Anaerobic、facultatively anaerobic)及氧化胁迫耐受(Oxidative Stress Tolerant)等7类。
以上这些部分,我们通过数据处理分析,得到了每个样本相关的大量数据结果,包括每个样本的序列统计、ASVs表格、物种分类注释统计、alpha多样性指数、beta多样性指数、功能预测等。这些数据主要集中在报告里的这些内容:
▸ 科研分析报告结果文件夹
01_pick_otu/ 文件夹主要是对样本ASV表格统计
02_sequence_statistic/ 文件夹是对样本序列数据的统计
03_diversity-metrics / 文件夹是对样本的alpha多样性指数、beta多样性指数的统计
04_Taxonomic/ 文件夹是对物种分类注释的统计(门到种水平)
Picurst2/ 文件夹是Picrust2功能预测得到的每个样本的相关功能预测数据
Groups/ 文件夹下是对组间差异分析结果

红框是样本个体的相关数据统计,Group是分组比较
根据以上常规分析得到的相关数据进行作图,其路径也在对应文件夹下,可以打开 分析报告.html 有相关分析的图表和对应文件的详细介绍和路径说明。
★拿到样本后需要进行统计分析
当我们拿到这些样本大量的数据结果,之后关键的一步就是做对这些数据进行处理,做统计分析,比较分组之间的差异结果,找出菌群和环境之间的关联性等,对数据进一步做研究,找出课题方案对应的结果。
不同的数据用到的统计检验方法也不太一样,接下来我们对报告中的不同的分析结果对应的统计差异分析方法进行介绍说明。
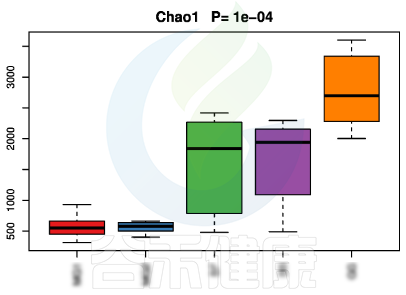
▸ alpha多样性
alpha多样性指数组间差异统计分析用到的检验方法是:单因素方差分析(如果只有两个分组,用Wilcoxon秩和检验,3个及以上的分组用Kruskal-Wallis 检验),图上方显示P值
▸ beta多样性
beta多样性指数的统计检验方法有ANOSIM相似性分析和Adonis多元方差分析,这两种都是基于距离矩阵的检验方法。
✦Anosim相似性分析
Anosim分析是一种非参数检验,用来检验组间的差异是否显著大于组内差异,从而判断分组是否有意义。
报告中给出了加权距离和非加权距离的Anosim结果图,图中给出了R值和P值。
R值用于比较不同组间是否存在差异,R-value 介于(-1,1)之间,R-value > 0,说明组间差异大于组内差异。R-value < 0,说明组间差异小于组内差异。R只是组间是否有差异的数值表示,并不提供显著性说明。
统计分析的可信度用 P-value 表示,P< 0.05 表示统计具有显著性。
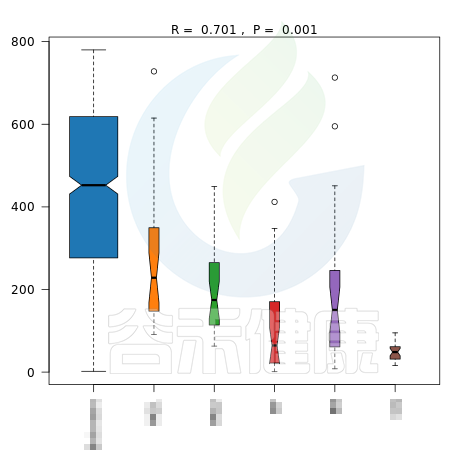
图中能看出R>0,说明组间差异大于组内差异,P<0.05 ,说明差异显著,证明该分组情况效果较好。
✦Adonis多元方差分析
Adonis多元方差分析,其实就是PERMANOVA,亦可称为非参数多元方差分析。
其原理是利用距离矩阵(比如基于Bray-Curtis距离、Euclidean距离)对总方差进行分解,分析不同分组因素对样品差异的解释度,并使用置换检验对其统计学意义进行显著性分析。
它与Anosim的用途相似,也能够给出不同分组因素对样品差异的解释度(R值)与分组显著性(P值)。
报告中PCoA bray距离、PCoA weighted_unifrac距离、PCoA unweighted_unifrac距离的图片右下角有给出PERMANOVA检验的P值和R值。
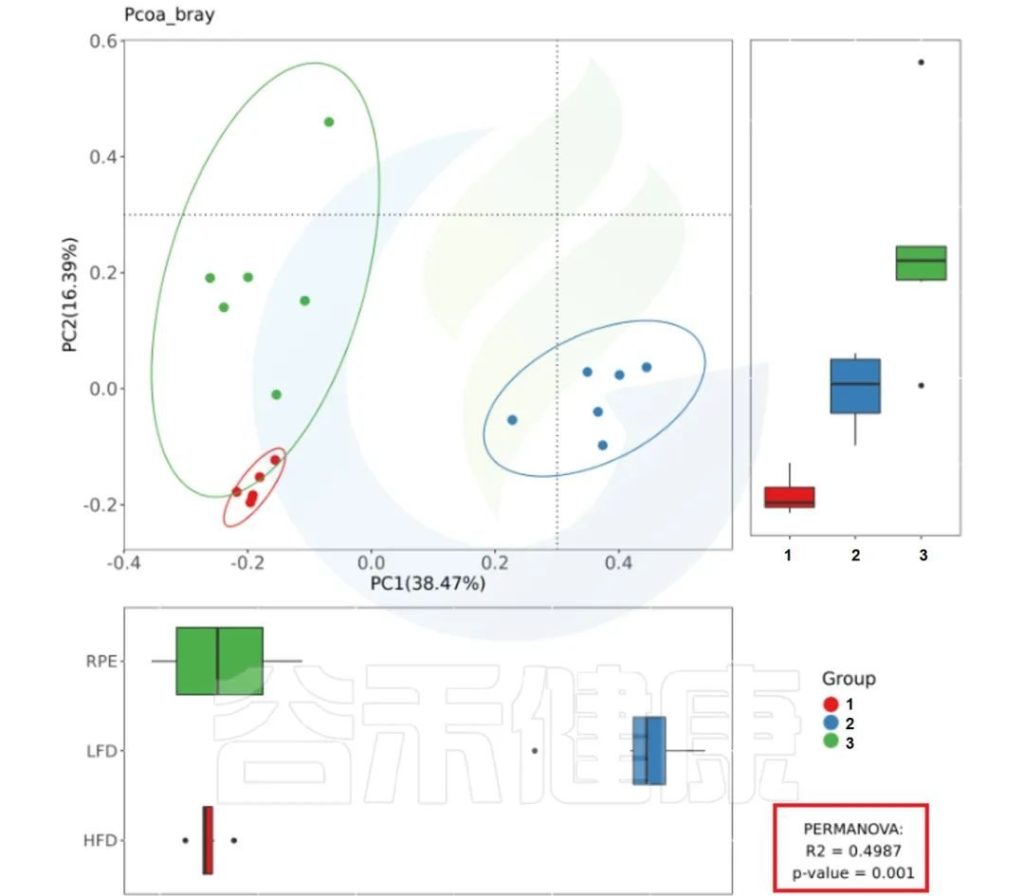
图中看出PCoa bray距离得到的检验P<0.05 组间差异显著,并且分组之间区分较为明显。
PCoa bray距离的PERMANOVA检验结果路径:
多组间检验结果:
Groups/betadiv/pcoa_bray_analysis/PERMANOVA.result_all.csv
两组间检验结果:
Groups/betadiv/pcoa_bray_analysis/ PERMANOVA_paired_result.csv

不同分类水平下的检验方法
在很多分析报告当中,例如在不同疾病的肠道菌群分组中,本身样本个体之间肠道菌群的物种多样性,丰富度差异并不大,alpha多样性组间差异并不显著,beta多样性分组间区分不是很明显,这样就需要进一步找出分组之间的差异物种或者差异功能来进行分析。
对于不同分类水平的物种和功能预测结果用到以下几种检验方法:
Tukey检验
Tukey主要应用于3组或以上的多重比较,适合于各组例数相等的每两两分组之间比较。
Tukey检验的一个重要的优点是非常简单,而且所需实验样本相对较少。
其检验结果的可信度达到95%的置信水平时,最少的情况下只需6个样本进行验证(改善前3个样本、改善后3个样本)。
•举个例子
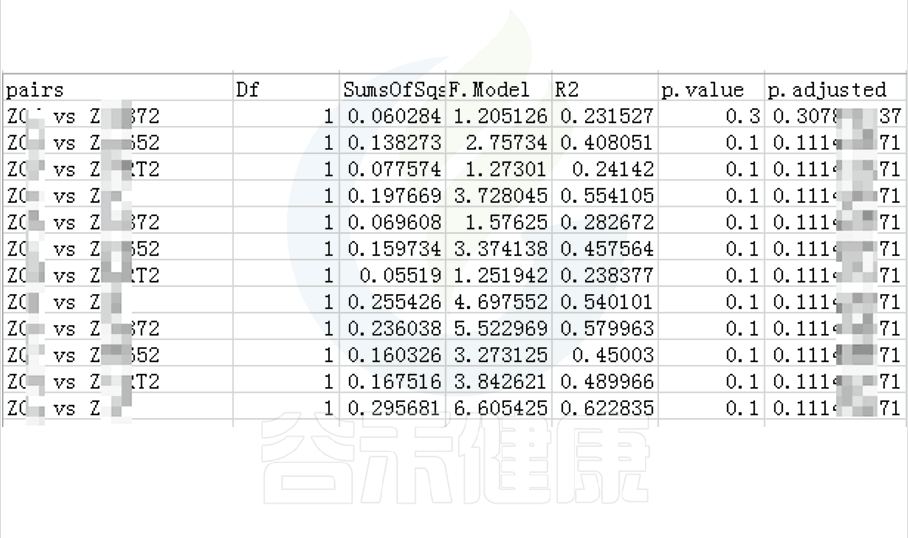
图中的字母代表显著性差异的字母表示法,只要含有相同的字母,就表明两组之间没有显著性差异。
例如a和ab含有相同字母“a”,表示两组之间没有显著性差异。ab中的“b”表示这一组和其他含有字母b的组(比如bc)没有显著性差异,但是a和bc就有显著性差异了。
图中只展示Tukey检验差异显著的物种或功能,如果数量较多,则只展示前10个。
路径:Groups/diff_analysis/TukeyHSD/
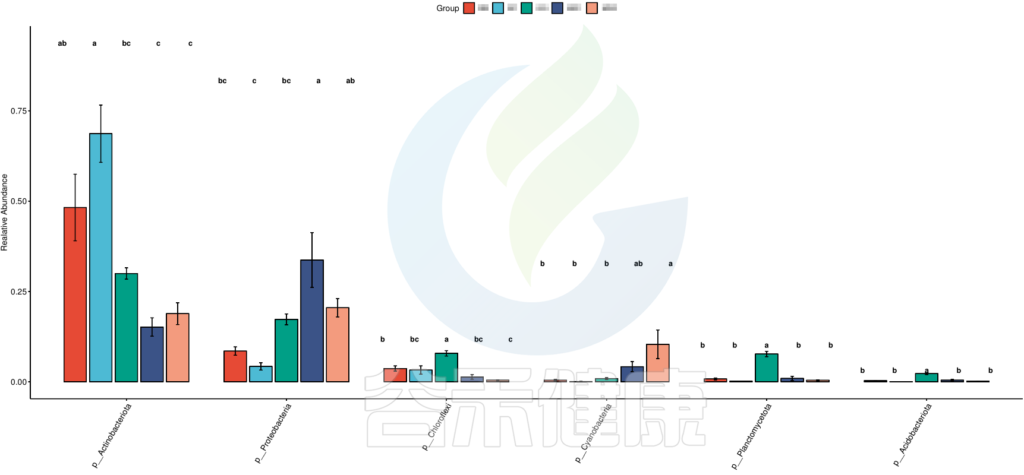
图中显示的都是Tukey检验组间差异显著的物种,依次按照丰度从高到底排列,如果差异结果较大,则显示前10个物种。例如在该图中,Tukey检验结果,门水平物种Actinobacteriota在BB与MG1组、BB与MG2、BF与GG组、BF与MG1组、BF与MG2组,这些分组之间组间差异显著。
组间差异箱型图
组间差异箱型图用到的检验方法是通过单因素方差检验(只有两个分组,用的是Wilcoxon秩和检验,3个及以上的分组用的是Kruskal-Wallis 检验),Var检验和one-way相结合,筛选出组间差异性物种。
路径:Groups/diff_analysis/TaxaMarkers
图中每一个箱型图代表一个组间差异显著的物种
图中显示的都是统计方法得到的差异显著的物种,图中能看出这3个物种分组之间差异显著。
命名格式是,例如:Cen_Nitrosopumilus 指的是,当前分类水平(属水平)的名字 g__Nitrosopu 加上一级分类水平(科水平)的名字 f__Cenarchaeaceae 的前 3 个字母简写Cen,如果当前水平没有注释到名字则以全称的名字表示。
统计结果表:Groups/diff_analysis/TaxaMarkers/ xxx.Groups.sig.meanTests.csv
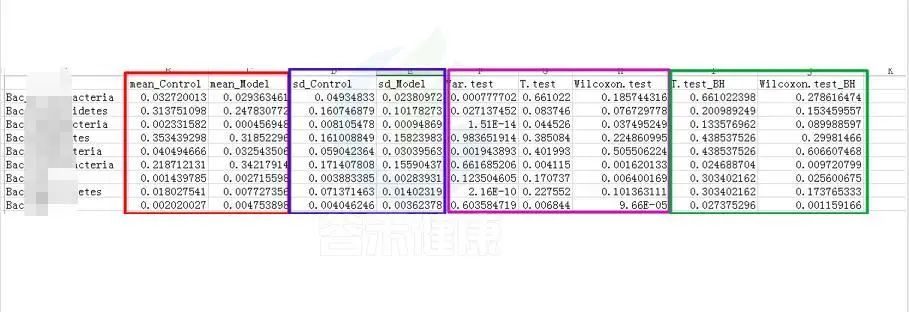
例如这是一个表格的截图
红框 mean_ 是分组组间的平均值
蓝框 sd_ 代表组间的标准差
粉色 .test 代表不同统计检验结果的P-value P值,这里有var检验 T 检验 Wilcoxon检验(或Kruskal-Wallis 检验)
绿色 _BH 例如Wilcoxon.test_BH代表Wilcoxon.test检验BH矫正的Q-value,Q值
UnivarTest检验(单因素方差分析)
单因素方差分析是指如果只有两个分组,用Wilcoxon秩和检验,3个及以上的分组用Kruskal-Wallis 检验。
路径:Groups/diff_analysis/UnivarTestXXX
Groups\diff_analysis\UnivarTestKEGG\figure 文件夹下有做成柱状图、箱型图和单个物种之间的图,其中有横着排列和竖着排列的,有用原始值计算的,还有对原始值取log后进行统计的。图中只展示Univar 检验组间差异显著的物种/功能。
统计结果表:Groups/diff_analysis/UnivarTestXXX/ UnivarTest_sign.txt
•举个例子
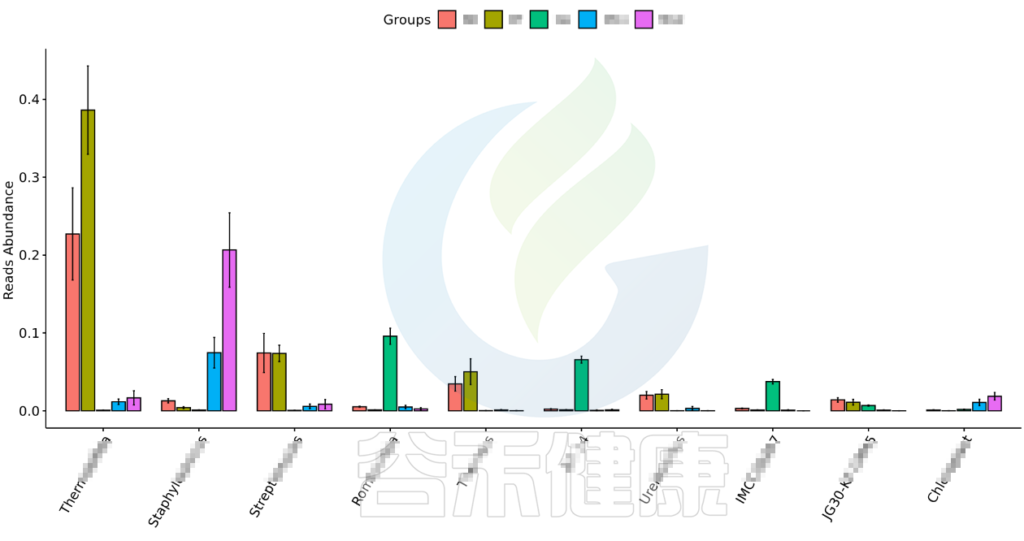
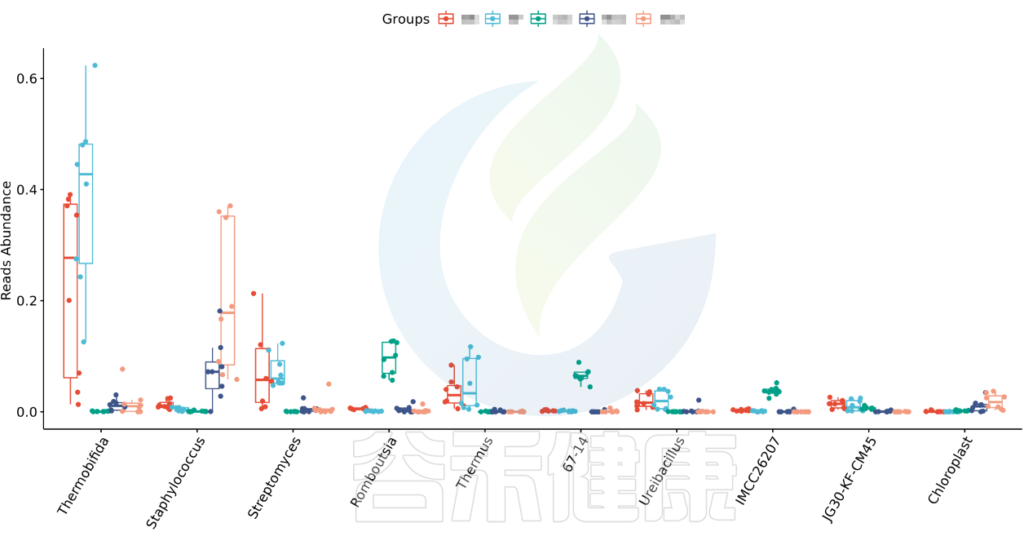
图中显示的是该统计检验差异显著的物种的柱状图或箱型图,按照丰度从高到低排列,如果差异物种/功能较大,则只显示前10个。例如该图中Therobifida、Staphylococcus、Streptomyces等物种用Kruskal-Wallis 检验得到的组间显著差异物种。
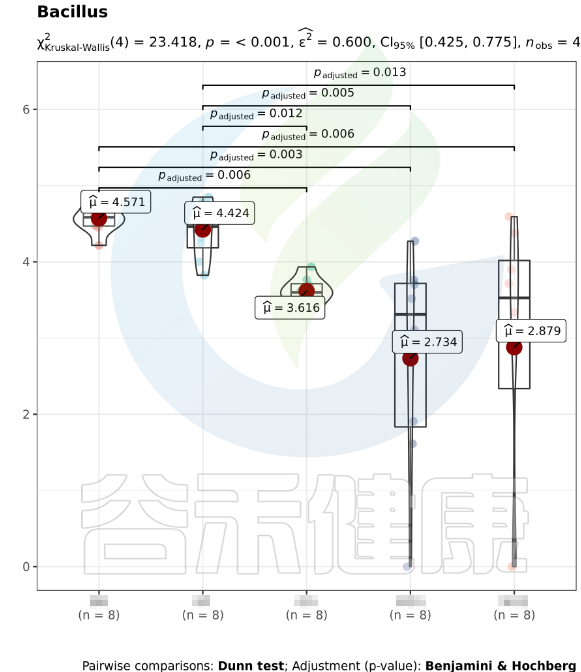
该图展示了Bacillus物种Kruskal-Wallis 检验差异结果,所有分组中P<0,001 多组间差异显著,两组间BB与GG、BB与MG1、BB与MG2、BF与GG、BF与MG1、BF与MG2,组间差异显著。
LEfse分析
LEfse分析即LDA Effect Size分析,是一种用于发现和解释高维度数据生物标识(基因、通路和分类单元等)的分析工具,可以进行两个或多个分组的比较,它强调统计意义和生物相关性,能够在组与组之间寻找具有统计学差异的生物标识(Biomarker)。
LEfSe用到的统计分析方法是将线性判别分析与非参数的Kruskal-Wallis以及Wilcoxon秩和检验相结合。
LEfse分析结果中一般会出现两个图,一张表( LDA值分布柱状图、进化分支图以及特征表)。
LDA值分布柱状图
这个条形图主要为我们展示了LDA score大于预设值的显著差异物种,即具有统计学差异的Biomaker,默认值为2.0(看横坐标,只有LDA值的绝对值大于2才会显示在图中);柱状图的颜色代表各自的分组,长短代表的是LDA score,即不同组间显著差异物种的影响程度。
路径:
Group/Lefse_Analysis/out_formant.cladogram.png
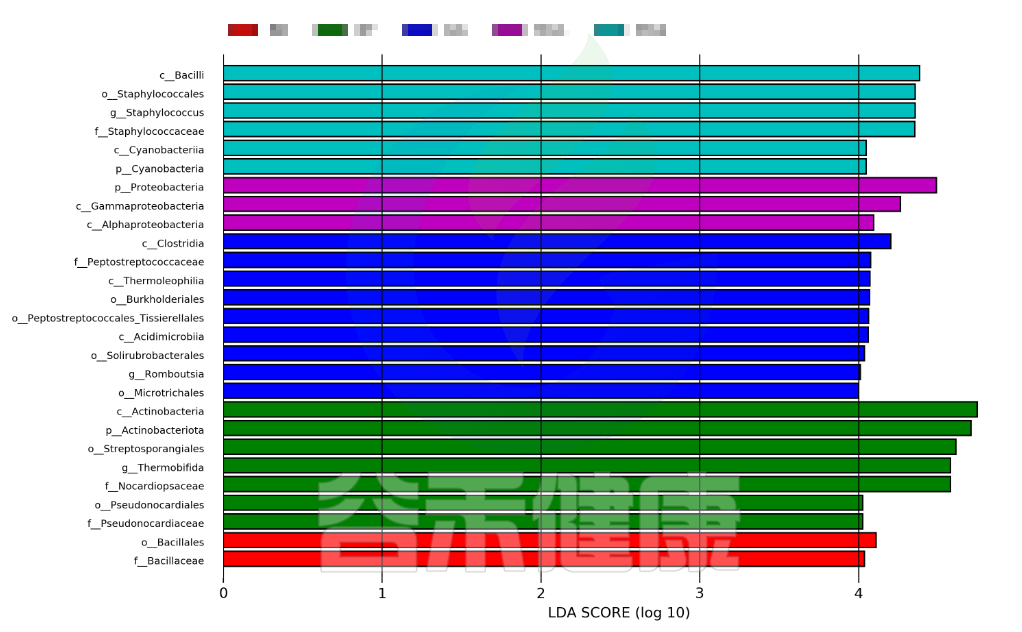
图中展示了不同分组特有的Lefse组间差异标记物,例如BB组的标记物是目水平的Bacillales和科水平的Bacillaceae,不同的分组标记物也不同,图中如果只展示了部分分组,则代表只有部分分组通过Lefse分析筛选出组间差异标记物。
进化分支图
小圆圈: 图中由内至外辐射的圆圈代表了由门至属的分类级别(最里面的那个黄圈圈是界)。不同分类级别上的每一个小圆圈代表该水平下的一个分类,小圆圈的直径大小代表了相对丰度的大小。
颜色: 无显著差异的物种统一着色为黄色,差异显著的物种Biomarker跟随组别进行着色,红色节点表示在红色组别中起到重要作用的微生物类群,蓝色节点表示在蓝色组别中起到重要作用的微生物类群。
未能在图中显示的Biomarker对应的物种名会展示在右侧,字母编号与图中对应(为了美观,右侧默认只显示门到科的差异物种)。
路径:Group/Lefse_Analysis/out_formant.png
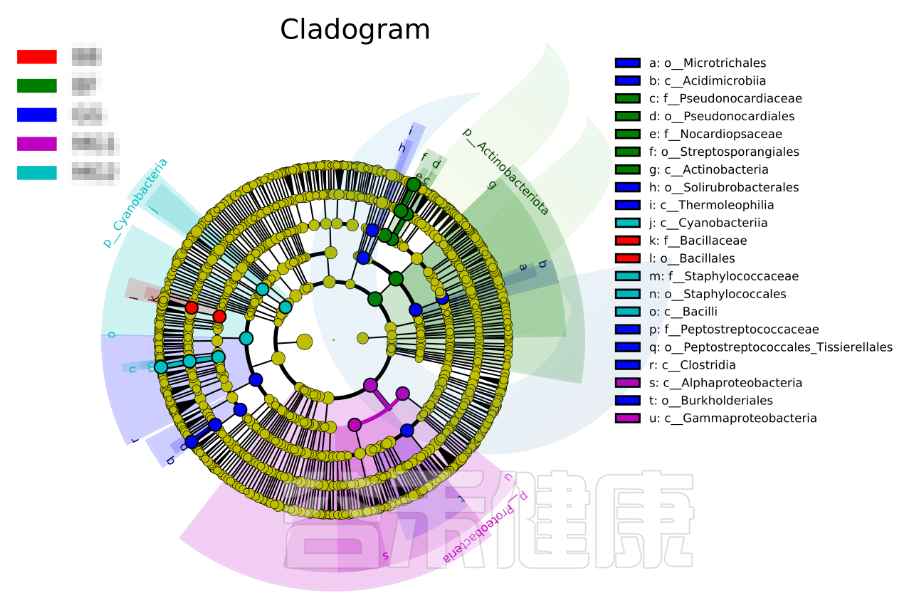
图中右侧展示了分支图中的字母对应的物种信息,例如a 代表GG组的标记物目水平的Microtrichales ,b代表GG组的标记物刚水平的Acidimicrobiia。在分支图的最外层显示的是各分组门水平物种的标记物,例如BF组的是Actinobacteriota、MG1组的是Proteobacteria、
MG2组的是Cyanobacteria
特征表
路径:Group/Lefse_Analysis/out_formant.res.csv
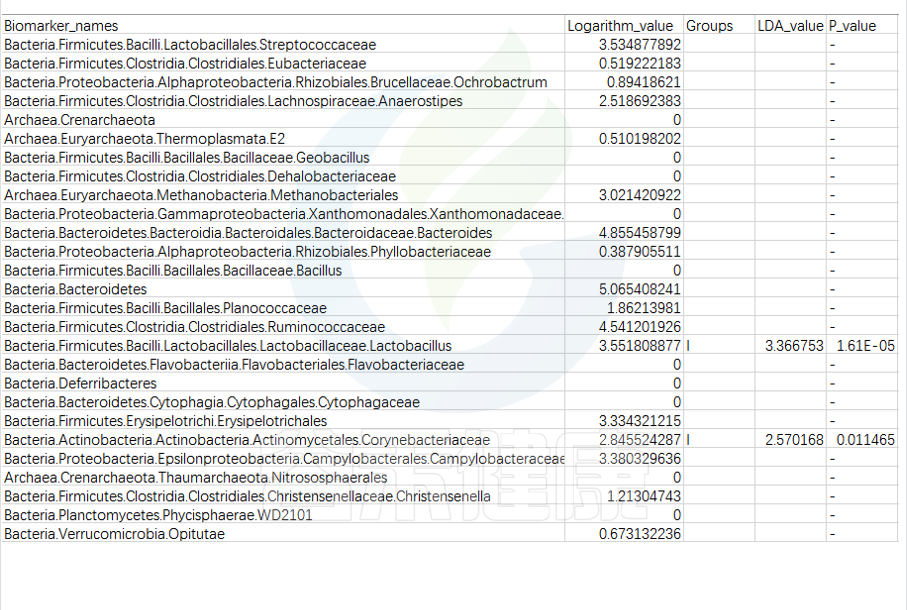
第一列是样本中从门到属水平所有分类单位的列表
Lefse会逐一判断这些分类单位的在分组之间是否具有统计学显著性差异。
第二列:各组分丰度平均值中最大值的log10,如果平均丰度小于10的按照10来计算;如果该分类单位未体现出显著组间差异,则后三列为空。
对于具有统计学差异的分类单位:
第三列:差异基因或物种富集平均丰度最高的分组组名;
第四列:LDA差异分析的对数得分值;
第五列:Kruskal-Wallis秩和检验的p值,若不是Biomarker用“-”表示。
默认LDA>2,P<0.05
通常根据第4列的LDA差异分析对数得分值和第五列的P值,可以描述组间具有显著差异的分类单位统计学效力强弱。
metagenomeSeq
metagenomeSeq是用R开发的一个包,metagenomeSeq的基本思想,用normalization实现分类注释时的biases处理,同时用零膨胀高斯分布(zero-flated Gaussian distribution)处理了测序深度所带来的影响,在此基础上,利用线性模型找到存在的差异所在。
路径:Groups/diff_analysis/ metagenomeRXXX
metagenomeSeq 差异显著物种/功能 热图
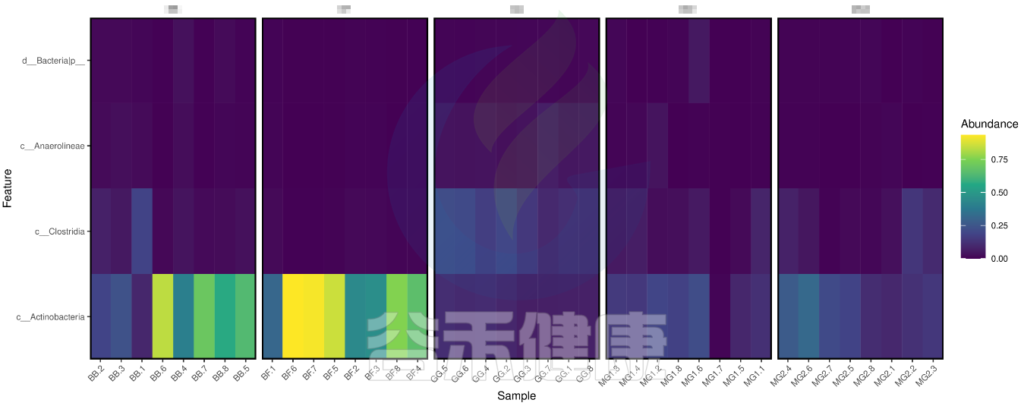
图中颜色越深相关性越小,颜色越接近黄色相关性越大,从图中能看出Actinobacteria物种与BB组和BF组相关性较大。
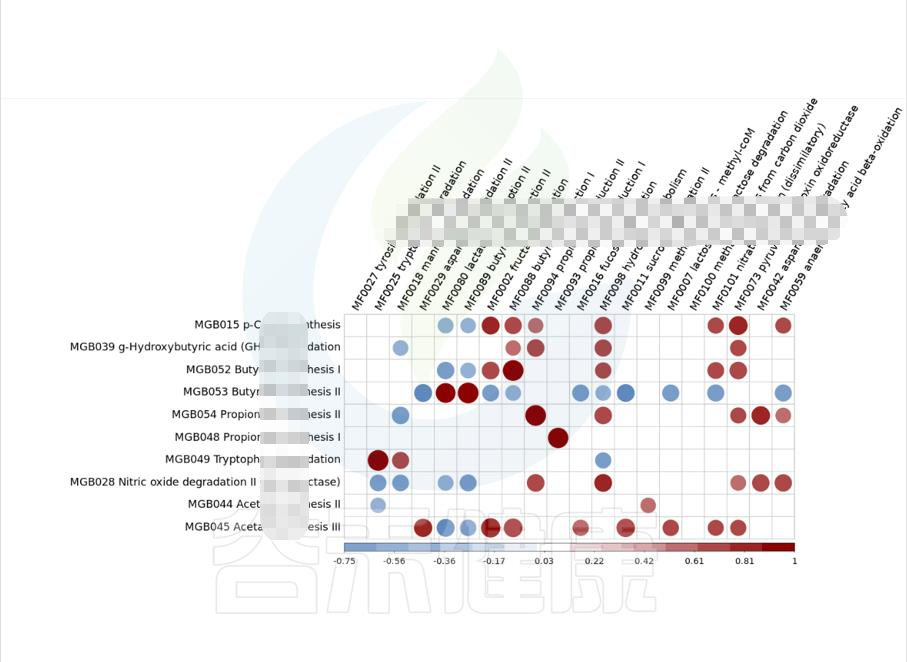
metagenomeSeq差异菌属于功能代谢关联分析
图中红色代表正相关,蓝色代表负相关,颜色越深,圆圈越大,相关性也越大,例如图中能看出MGB049余MF0025 之间成正相关,且相关性较大。
随机森林模型
一种非线性分类器,随机森林属于集成类型的机器学习算法,挖掘变量之间复杂的非线性的相互依赖关系。通过随机森林重要性点图,可以找出分组间差异的关键物种/功能。
反映了分类器中对分类效果起主要作用的特征,按重要性从大到小排列。
Error rate:表示使用下方的特征进行随机森林方法预测分类的错误率,数值越高表示基于特征分类准确度不高,可能分组之间特征不明显。分值越低证明分组效果比较好。
•举个例子
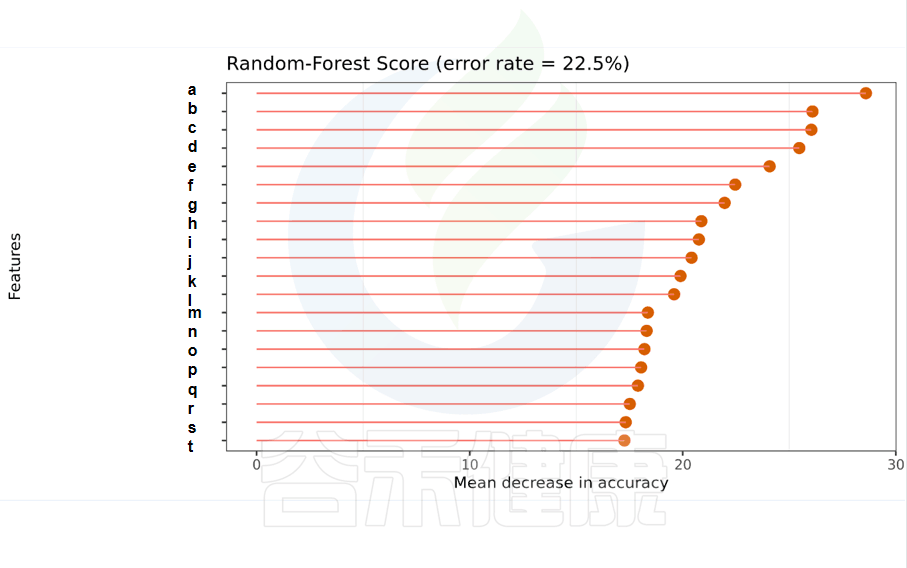
图中按照随机森林模型效果筛选出的对分组效果有重要性作用的物种,按照重要性从高到低进行排列,例如图中最终要的是a,依次往下是b、c等。错误率较小,表明该分组效果较好。
ROC曲线
ROC曲线分析是一种常用的统计学分析方法,在医学研究中主要用于评价诊断试验的效能。在16S测序报告中,我们通过绘制ROC曲线,并计算ROC曲线下面积(AUC),来确定分组对于菌群是否有诊断价值。
ROC曲线图是反映敏感性与特异性之间关系的曲线。ROC曲线下的面积值在1.0和0.5之间。在 AUC>0.5的情况下,AUC越接近于1,说明诊断效果越好。
AUC在0.5~0.7时有较低准确性,AUC在0.7~0.9时有一定准确性,AUC在0.9以上时有较高准确性。AUC=0.5时,说明诊断方法完全不起作用,无诊断价值。AUC<0.5不符合真实情况,在实际中极少出现。
•举个例子
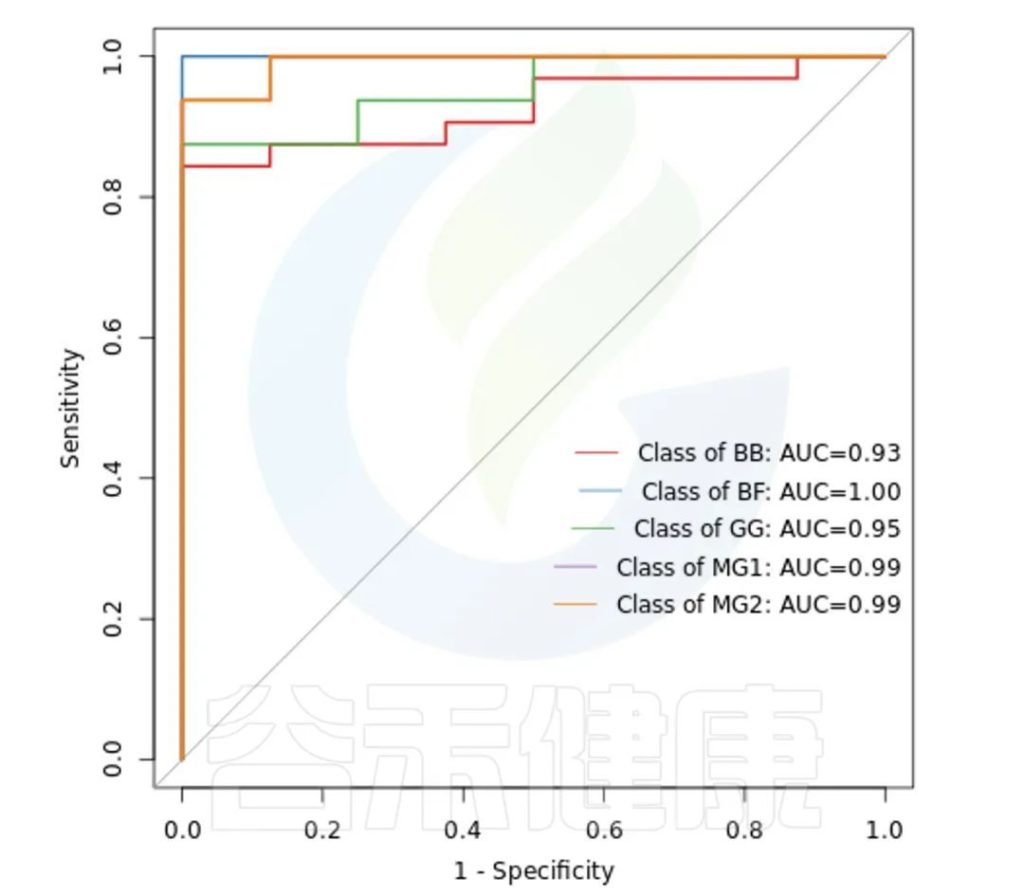
从图中能看出各分组的AUC都大于大于0.9,各分组的分组效果较好,BF组AUC等于1,该分组效果最好,可能样本之间较为相近,并且跟其他分组组间差异也比较大。
以上是组间统计差异的方法介绍,其他的还包括关联分析。
例如客户提供了每个样的相关理化指标数据,想计算这些指标与均属之间有什么相关性,就可以做一下分析。
关联性分析
✦相关性热图
图中X轴代表属水平物种,Y轴代表代谢指标,红色代表正相关,蓝色代表负相关,**代表相关极显著P<0.01,* 代表相关性显著P<0.05相关性具有统计学意义。
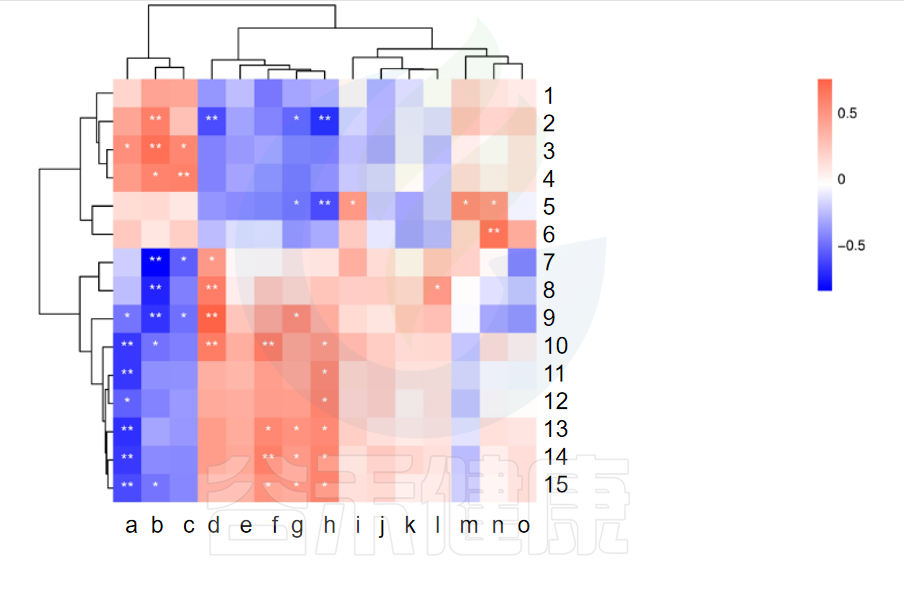
例如从该图中能看出6与n物种成正相关,并且相关性极显著**,7与b物种成负相关,并且相关性极显著**。
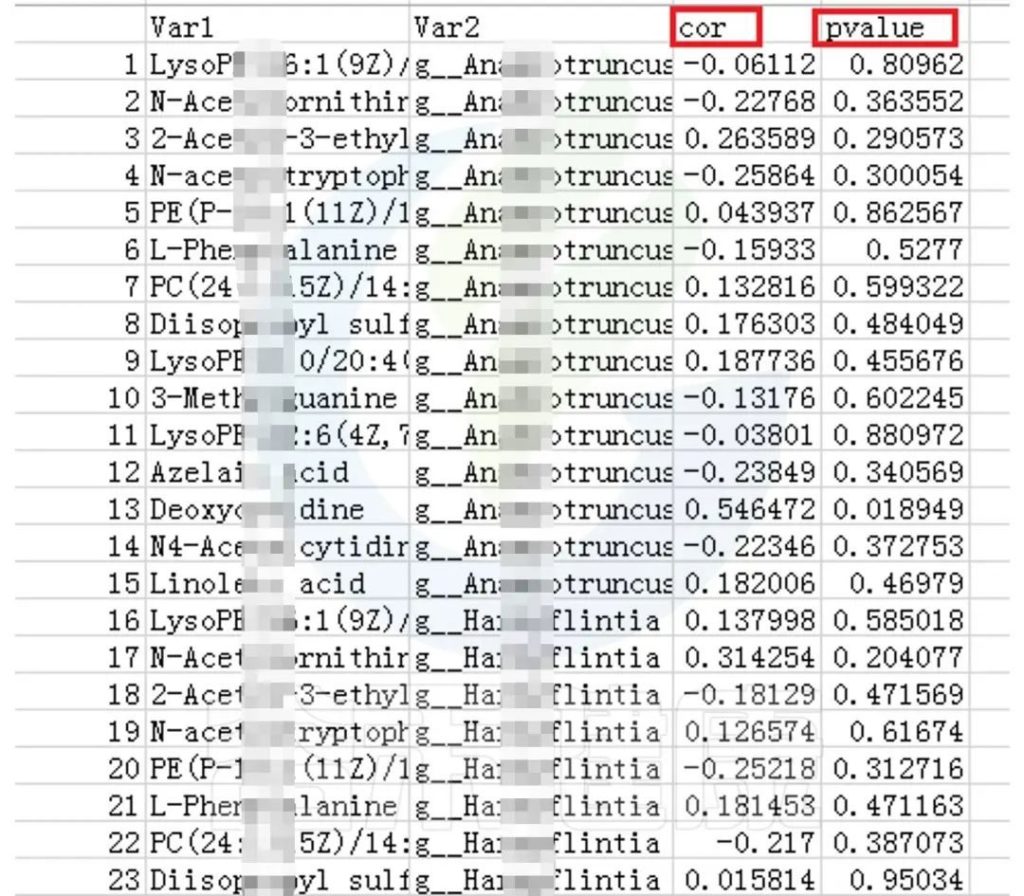
可以得到表格:任意菌属和代谢的相关性的值和P值
✦CCA图
可以分析样本、菌群、理化指标之间的关联关系。图中使用点代表不同的样本,从原点发出的箭头代表不同的环境因子。
箭头的长度越长,表示环境因子的影响越大;夹角越小,代表相关性越高。样本点与箭头距离越近,该环境因子对样本的作用越强。
图像中坐标轴标签中的数值,代表了坐标轴所代表的环境因子组合对物种群落变化的解释比例。
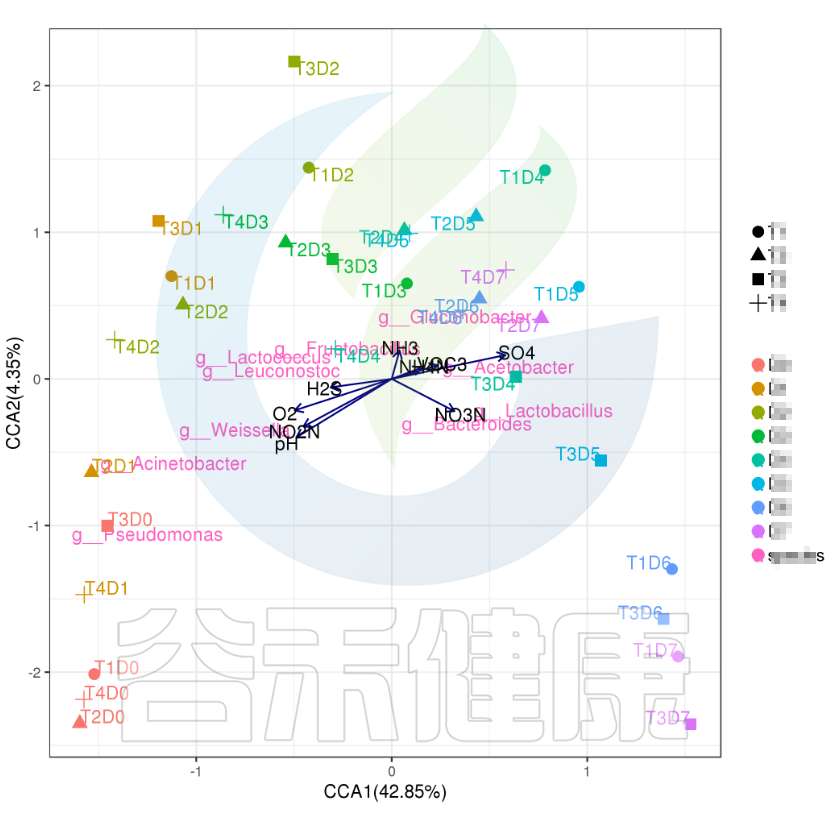
例如从图中能看出pH 、NO2N、02与 Acinetobacter、Weissella等物种成正相关,与T3D0、T1D0、T4D0等D0组的样本成正相关。
✦RDA 冗余分析

例如从图中能看出pH与Helicobacer物种成正相关,相关性较大,pH与NC组有一定的相关性。
✦Envfit分析
回归拟合分析结果:
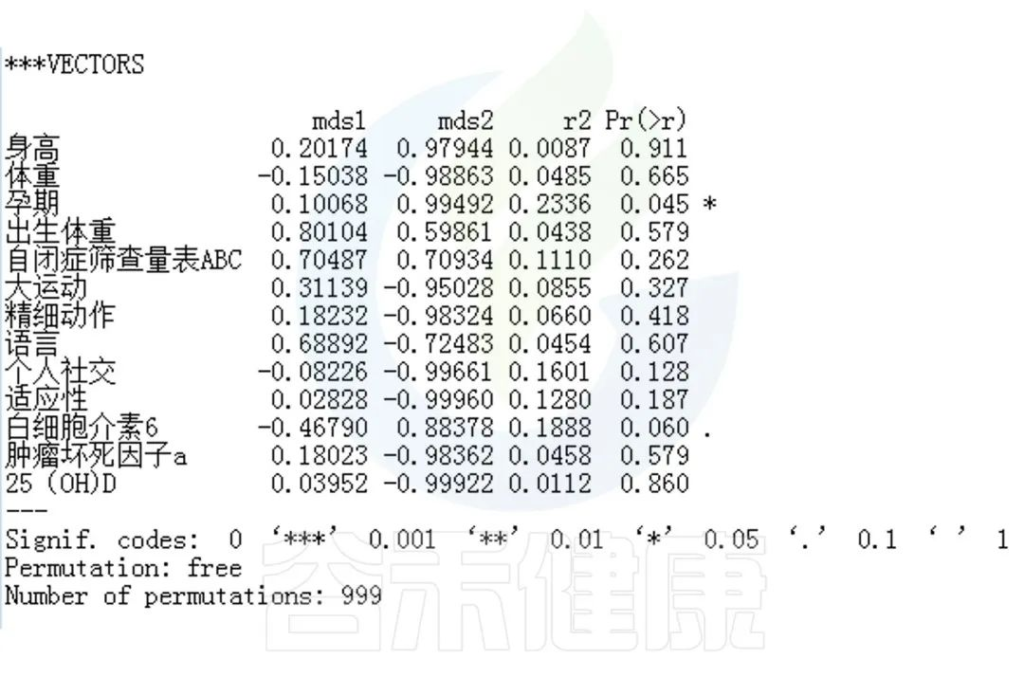
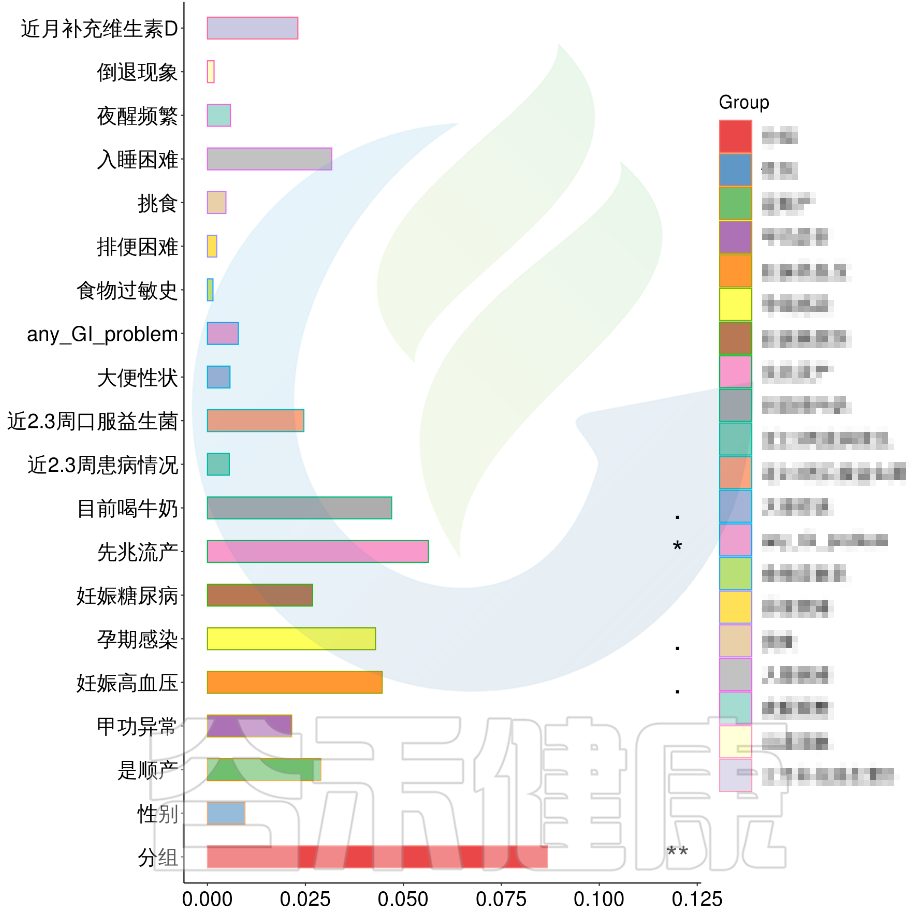
图中能看出ASD与正常儿童之间的分组与菌群之间相关性极显著**,其次是否有先兆流产的分组与菌群之间有显著相关性*,其他的包括是否喝牛奶、孕期是否感染、妊娠高血压都与菌群有相关性。
环境因子与功能/物种的相关性线形图P<0.05显著,图中红色点代表正相关,绿色点代表负相关,灰色相关性不显著。
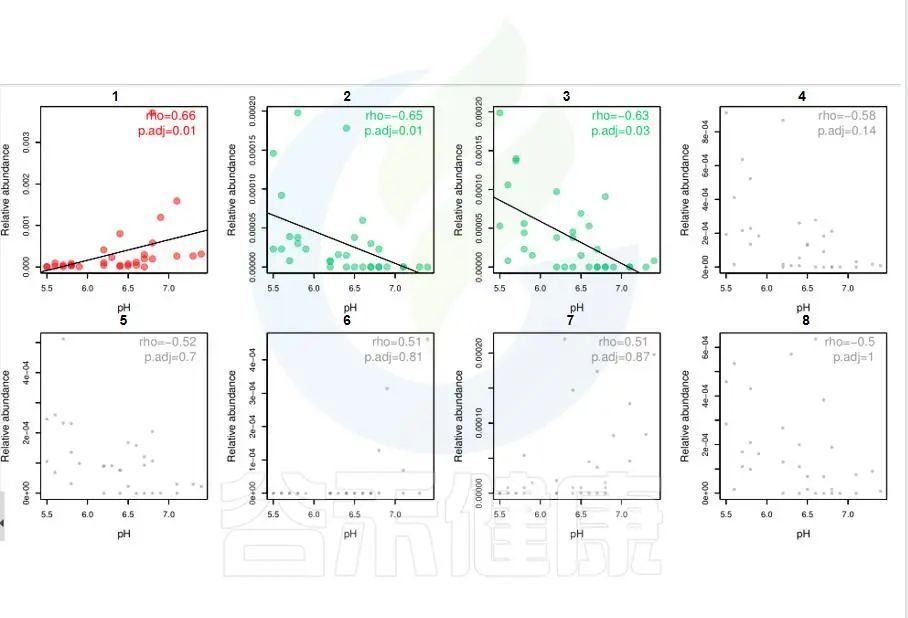
图中能看出pH 与Candidatus Rhabdochlamydia 之间成正相关,且相关性显著,pH 与Sinorhizobium、Euzebya 之间成负相关,切相关性显著。
✦Network网络分析
还可以做菌属之间的网络分析关联图,共发生网络图为研究复杂微生物环境的群落结构和功能提供了新的视角。
由于不同环境下微生物的共发生关系截然不同,通过物种共发生网络图,可以直观看出不同环境因素对微生物适应性的影响,以及某个环境下占互作主导地位的优势物种、互作紧密的物种群,这些优势物种以及物种群往往对维持该环境的微生物群落结构和功能稳定发挥着独特以及重要的作用。
•举个例子
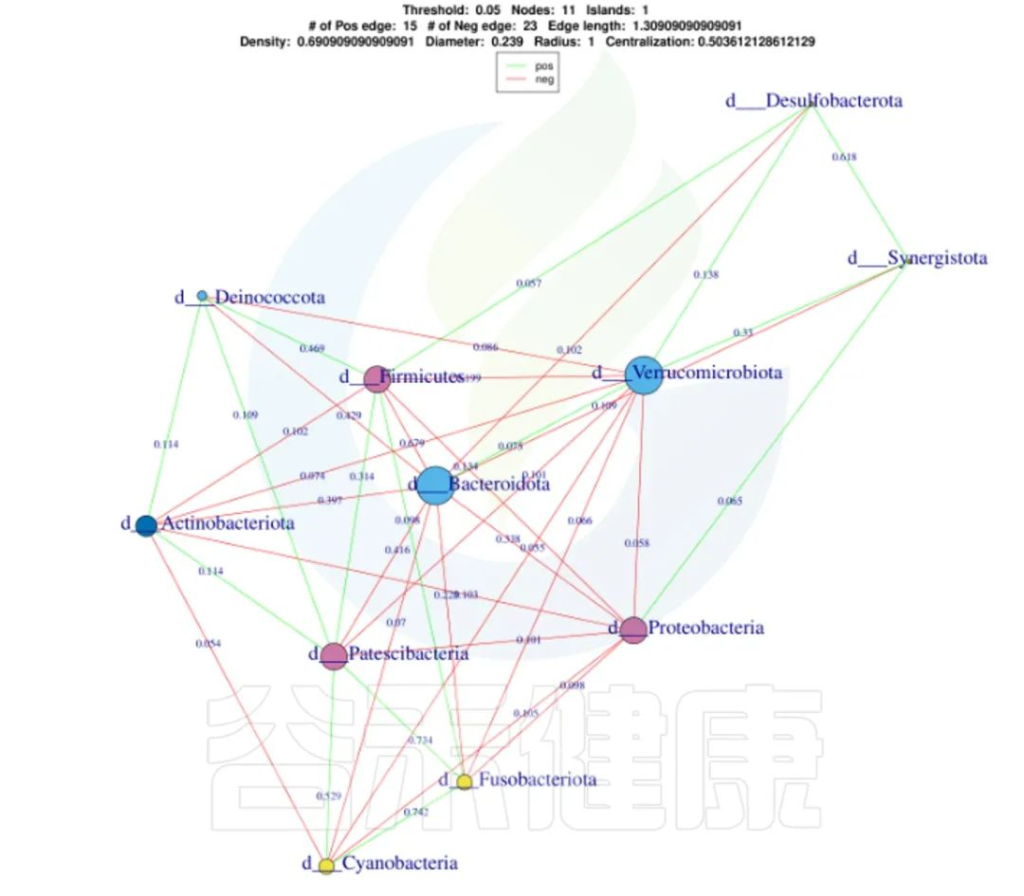
图中展示了相关性的物种,例如Bacteroidota、Actinobacteriota、Proteobacteria 这些物种与其他物种相关较大,图中这些物种与其他物种连线较多,字体比较大也代表相关性较强,例如Actinobacteriota与Deinococcota连线是绿色的代表这两个物种是负相关。
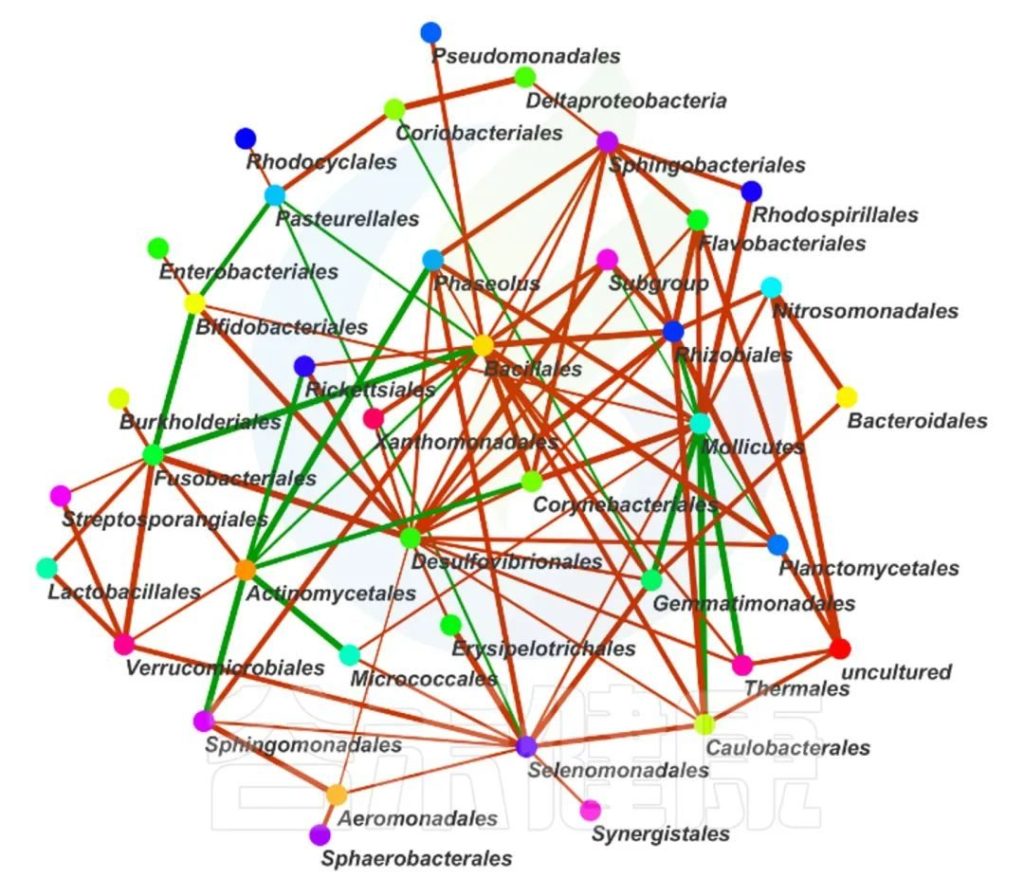
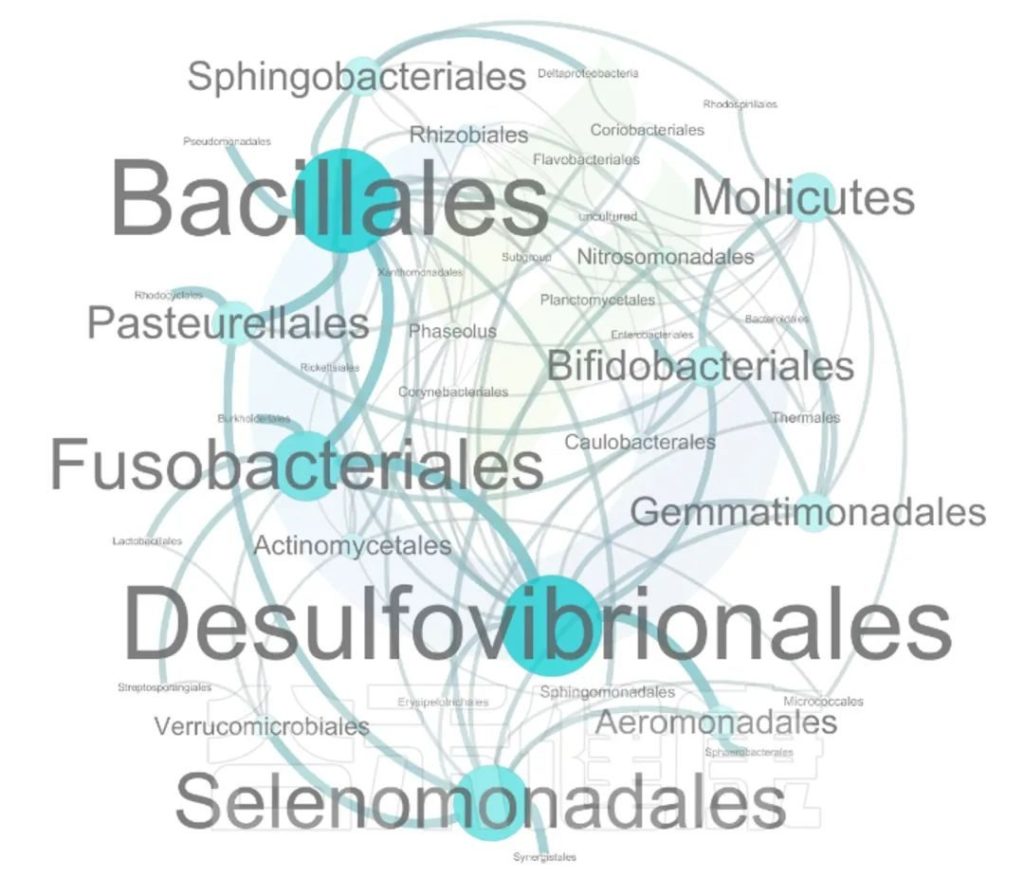
这两个图类似的物种相关性的图,用同一个数据做出来的,图中能看出Bacillales、Desulfovibrionales、Selenomonadales与其他物种相关性较强。
报告中已经基本都涵盖了16S科研数据分析所需要的图表、差异统计,以及相关性分析结果。如果在几种不同类型的统计方法对比之下有略微的差异结果,选取其中一组差异结果即可。
报告里涵盖了大部分16S所需要的图片,不过也有个别个性化的图需要单独用到软件去做,可以单独完成个性化图表生成。
随着16s分析报告的不断升级,报告中的图表以及相应的解读也会越来越精细完善,谷禾也将尽可能为大家的科研之路带来更多便利。


谷禾健康
迄今为止,已经有了许多对呼吸道微生物组通过16S rRNA高通量测序的研究。这其中所有基于扩增子的研究的共同之处就是PCR的应用:
一是扩增待测序的目标标记基因,
二是为多样本的混合测序添加必要的索引序列。
这些步骤可以通过一步PCR或两步PCR完成,但没有研究说明两步PCR方案相关的实验室处理步骤是否会使样品比一步PCR方案更容易受到来自实验室的细菌DNA污染的影响。
本文
试图确定对16S rRNA V3V4与V4基因区域的一步或两步PCR的建库方案对上呼吸道和下呼吸道微生物组的影响
对收集的样本进行了三个设置下的lllumina MiSeq测序
设置1(两步PCR,V3V4区域)
设置2(两步PCR,V4区域)
设置3(一步PCR,V4区域)
分别对这三个设置产生的测序数据进行分析
结论
PCR步骤数量的差异会影响对呼吸道微生物群落的物种组成分析,且对上呼吸道(高细菌载量)的影响小于下呼吸道(低细菌载量),这表明PCR设置的偏差与样本生物量有关。
通过三个实验,即对模拟群落样品HM-783D、NCS样品、呼吸道样品采用三种PCR方案进行建库后的测序结果分析,研究这三种建库方案对其菌群描述的影响。
模拟群落样品HM-783D,来自20种不同细菌物种(17个属)的基因组DNA。
阴性对照样本NCS
呼吸道样品,从Bergen COPD微生物组研究中选择了23名研究对象,其中9名健康,4名患哮喘,10名患COPD(慢阻肺)。上呼吸道样本以漱口水(OW)为代表,下呼吸道样本以标本刷(PSB)和支气管肺泡灌洗液(PBAL)为代表。
三种PCR方案:
细菌DNA提取后通过三种不同的建库设置进行MiSeq测序,分别为
Setup1(两步PCR,V3V4区域);
Setup2(两步PCR,V4区域);
Setup3(一步PCR,V4区域)
下图完整的展示了三个PCR设置下的使用呼吸道样本的生物信息学过滤步骤:
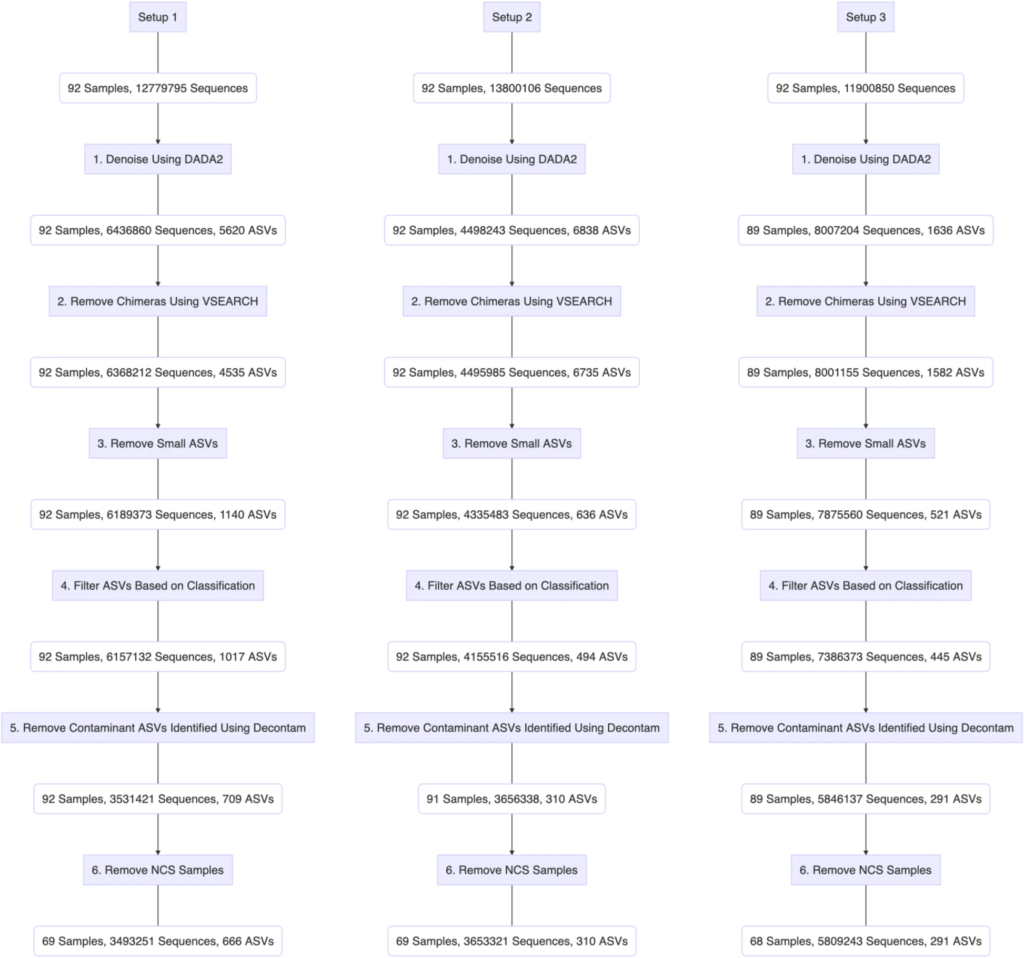
最终:
设置1:得到了666个ASVs
设置2:得到了310个ASVs
设置3:得到了291个ASVs
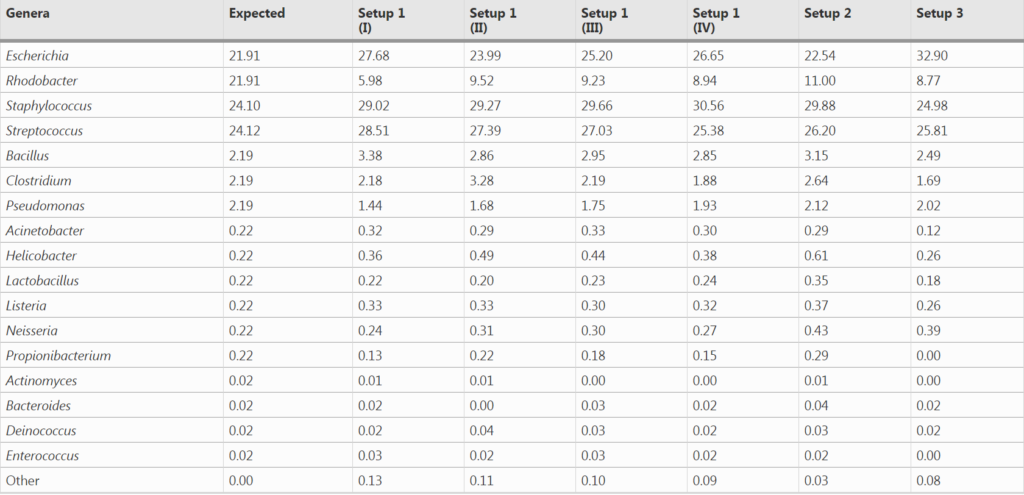
1. 对模拟群落样品HM-783D的分析
在设置1中进行了四次测序,设置2和设置3分别进行了一次。
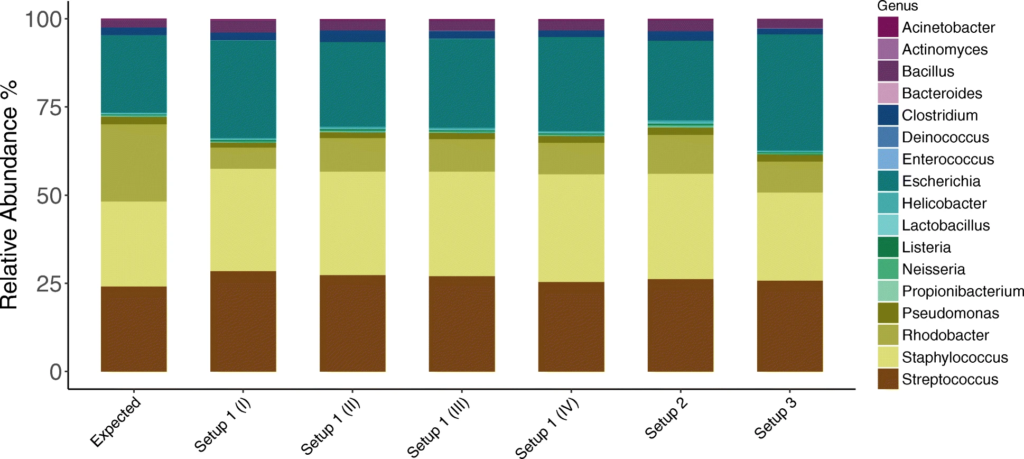
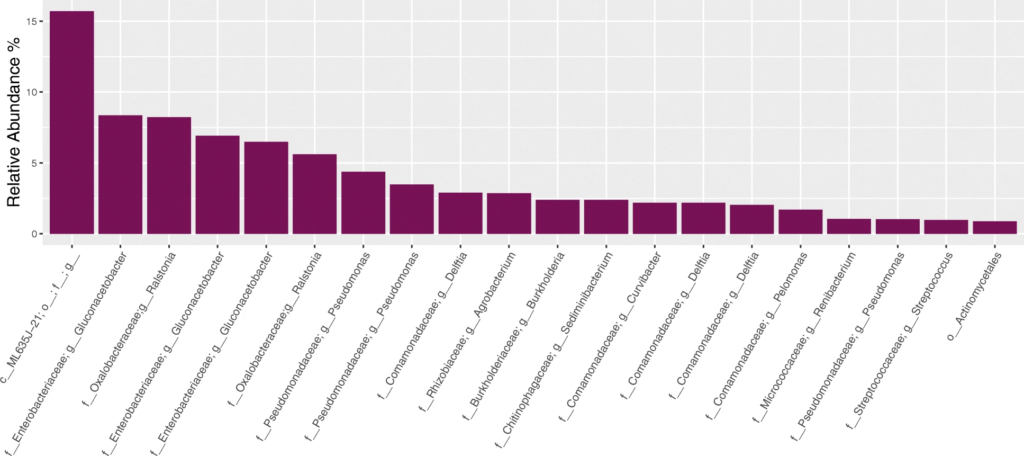
与预期丰度(Expected)相比,柱状图中观察到三种PCR设置下的各菌属的相对丰度与预期丰度相差不大,表中数据显示,三种PCR设置都在恢复高丰度物种方面具有最高的效率,但设置3在回收低丰度物种时的效率最低。
2. 对阴性对照样品的分析
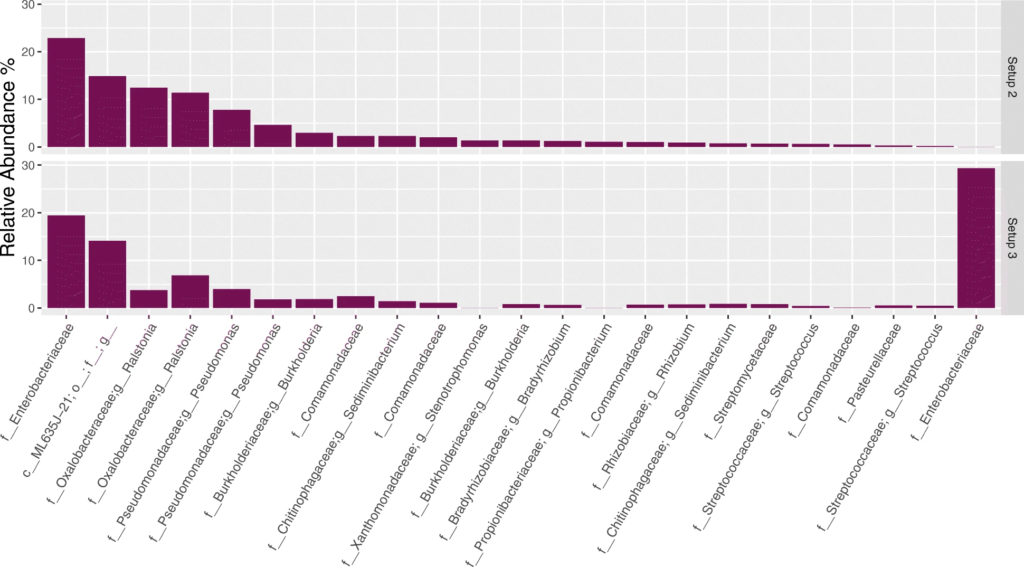
从上至下分别为设置123测序后,在NCS样品中观察到的20种最丰富的ASV。通过R包Decontam去除污染物,在设置23之间差异最大的是属于肠杆菌科的ASV,与后续的对水样品进行设置23下的测序分析结果相比较,发现大肠杆菌ASV就是在建库步骤中使用设置3的试剂时引入的污染物。
3. 对采集的呼吸道样品的分析
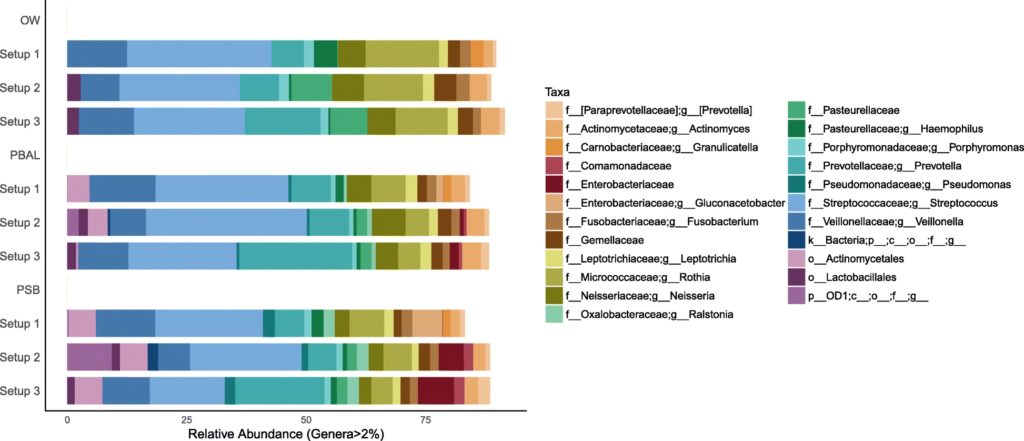
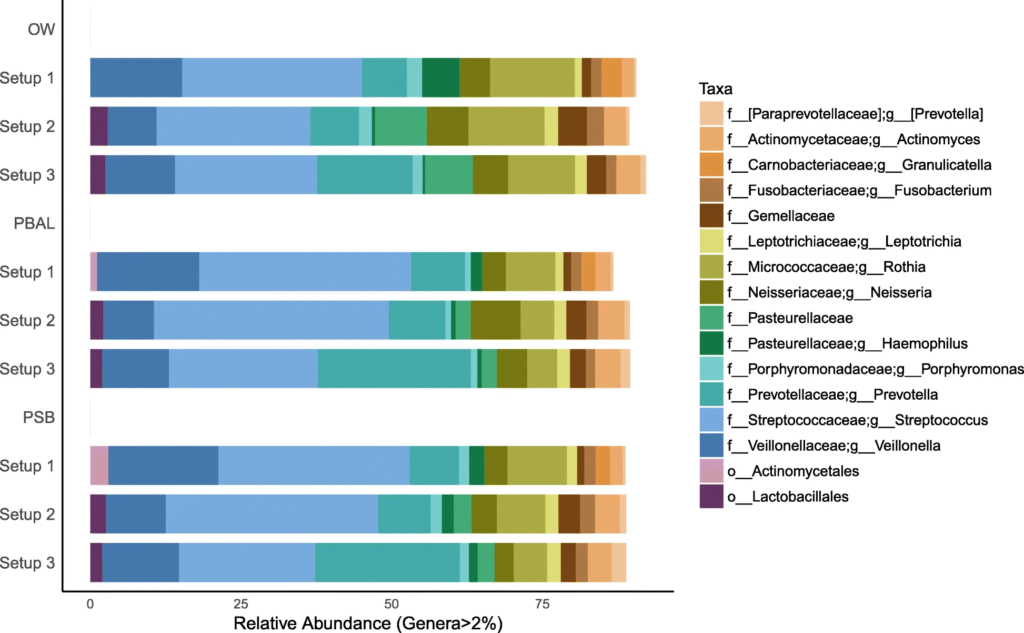
在去除污染物前后,代表为呼吸道菌群的链球菌,普雷伏氏菌,Veillonella和Rothia属的相对丰度变化不大,而去除污染物后,预测作为污染物代表的数量较少的物种被滤出。基于主坐标分析,发现高细菌载量的OW样品聚集在一起,低细菌载量PBAL,PSB一句设置23分离开。
去除污染物之前的三种类型样品的三种PCR设置下的物种分类
去除污染物后的
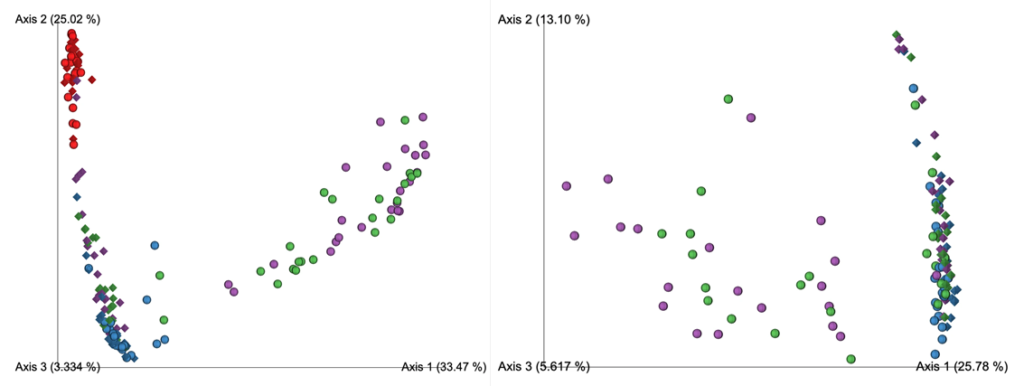
从左至右分别为去除污染物前后的未加权UniFrac距离的主坐标分析
OW:蓝色; PBAL:绿色;PSB:紫色;NCS:红色。
设置2(球形),3(菱形)
文章作者给出的结论是文库制备和测序方法的选择会对呼吸道微生物组的分析产生影响,且对上呼吸道的影响小于下呼吸道。靶向扩增子区域的差异(16S rRNA基因V3 V4与V4)并未表现出对细菌群落描述的重大影响。对于整篇研究存在的主要的局限性在于仅研究了DNA提取后的PCR步骤,污染或影响也可能来自于更前期的处理。
编者按
在使用测序技术进行的微生物研究中,测序偏差和污染物是一直存在的问题,也因此诞生了许多工具和计算方法用于尽可能的消除或降低这方面的影响。这篇研究也提醒了我们,在呼吸道微生物组的研究中,要注意上呼吸道与下呼吸道的菌群差异或相似可能不仅仅来源于样本自身,还可能掺杂着PCR方法选择上的影响。
参考文献:
Drengenes C, Eagan TML, Haaland I, Wiker HG, Nielsen R. Exploring protocol bias in airway microbiome studies: one versus two PCR steps and 16S rRNA gene region V3 V4 versus V4. BMC Genomics. 2021 Jan 4;22(1):3.
相关阅读:

谷禾健康


在现代测序技术的帮助下,微生物组研究的范围被扩大,通过16S rRNA测序或鸟枪法宏基因组测序可以生成大量的微生物组数据。而微生物群落研究中的一个重要问题是对这些微生物的归类,模拟和分析人类微生物群。
通常使用16S rRNA技术量化微生物群落的组成,但量化后的数据是偏斜的,带有过多的0。目前还缺乏对复杂的微生物群落测序数据的标准化的聚类分析方法。
近日,加拿大多伦多大学研究人员在《Microorganisms》上发表的一篇研究,针对上述问题构建了一个参数化的混合模型用于计算聚类分析的距离度量,模型根据观察到的OTU计数和估计的混合权重产生sample-specific的分布。这个方法可以准确的估计真实的0比例,从而构建一个精确的beta多样性度量。
大量的模拟研究表明,与一些被广泛使用的距离度量方法相比,当存在较大比例的0时,该方法取得了较好的聚类效果。
该研究人员提出了一种具有特定beta多样性度量的聚类算法,该算法可以解决稀疏计数数据遇到的有无偏差问题且能有效的度量样本距离,达到分层的目的。
微生物群落研究中的一个重要问题是对这些微生物的归类,它们是否能被划分为亚群。如果有,有多少组亚群,如何解释这个亚群。例如,这种分类是否区分了治疗方法、疾病或遗传类型。
为了回答这些问题,需要测量两个微生物群落之间的相似性。beta多样性是为了适应不同的目的而提出的,在评估群落之间的差异时提供不同的结果。对于微生物组成,beta多样性根据测量丰度来衡量群落之间的距离,丰度可以是观察到的计数,也可以是相对丰度,这些丰度是根据不同或距离度量计算出来的,以量化样本之间的相似性。
现如今,已经有许多非参数统计方法来量化距离度量。例如Euclidean和Manhattan距离是最常用的。其它beta多样性指标,例如Bray-Curtis距离、Jensen-Shannon距离、Jaccard指数、UniFrac距离(未加权的、加权的和广义的)也经常用于微生物组研究。
除了距离度量之外,还引入了用于生态关联推理的稀疏逆协方差估计(SPICE-EASI)的图形网络模型。然而这些方法都会有一定的局限性,例如SPIEC-EASI方法依赖于单一的方差-协方差矩阵,由于微生物群落结构复杂,可能无法完全恢复底层OTU网络。
于是,研究人员开发了一种创新的聚类方法,以混合模型而不是beta多样性度量作为距离度量,并将聚类算法应用于微生物群落数据来表征亚群。该算法还包括根据选择的内部指标选择最优聚类数,并将结果在几种距离度量和不同评估方法之间进行比较。通过全面的模拟研究和一个真实的帕金森病肠道微生物群数据集对该算法的性能进行了评估。
1. 构建混合模型
混合模型是一种概率模型,用于表示在无监督学习中经常使用的总体内的子群体。该模型关注单个OTU在种群中的分布,可以解决样本间的稀疏性问题。它参数化地模拟了计数的潜在分布,包括低计数OTU和极高计数。对于个体样本之间的成对距离,在L2范数距离中使用公式化的混合概率。
2. 距离度量
在确定混合模型分布后,使用概率分布通过样本之间的两两距离计算距离度量。为了进行比较,考虑了基于L2范数的三种距离度量(L2-PDF、L2CDF、L2-DCDF、L2-CCDF)。
除此之外还选择了一些其他的距离度量进行比较,即Manhattan距离和Euclidean距离,以及微生物组分析中特有的三个距离度量:Bray-Curtis距离、加权UniFrac距离和广义UniFrac距离。本研究不考虑未加权的UniFrac距离,因为它不包含类群丰度信息。
3. 聚类分析验证指数
这些指数用于衡量集群在集群内部和集群之间的可分离性表现很好。验证指标可以分为内部指标和外部评估,许多内部验证指标被用来选择最优聚类数。外部评估分数是在假设标签在建模阶段没有使用时,通过直接将划分结果与之前的标签进行比较来计算的。
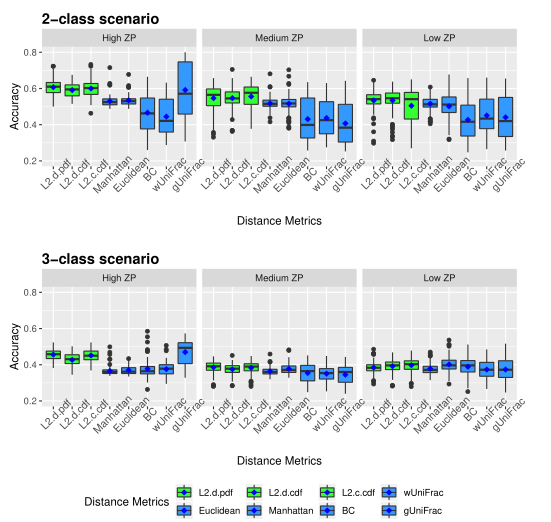
4. 用于聚类的分区算法(PAM)

使用混合分布的聚类过程的详细步骤:
为了测试该方法在聚类中的表现如何,研究人员推导了其准确性和Jaccard指数。
准确性是指聚类结果与真实的聚类指数的接近程度。它被定义为正确聚集的受试者所占的比例。
Jaccard指数衡量聚类结果与原始聚类标签之间的相似性,原始聚类标签定义为正确分类的主题数量(预测集与真实集的交集 )与两组总样本量(两集的并集)之比。
研究人员用类标签模拟数据来模拟OTU计数及其复杂的结构。研究人员考虑两个有两个子类和三个子类的场景,每个子类包含200个样本,总样本量分别为400和600。所有的结果被重复了100次。
下图展示了模拟数据的准确性。评估了三种不同的0的比例(ZP)情况,左中右分别为高ZP、中等ZP、低ZP。
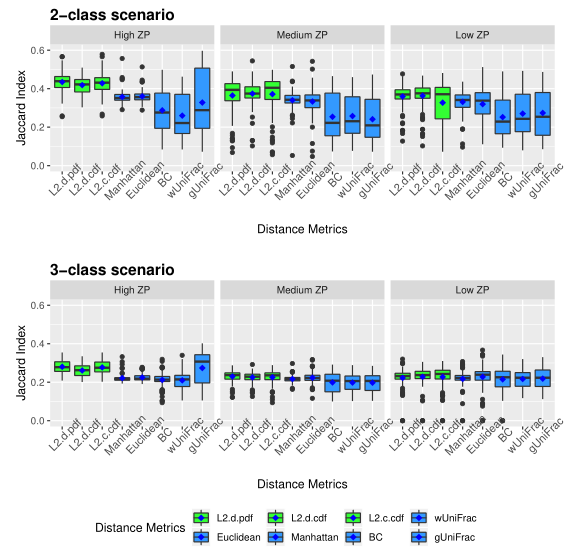
下图展示了模拟数据的Jaccard指数。同上图一样评估了三种不同的0的比例。
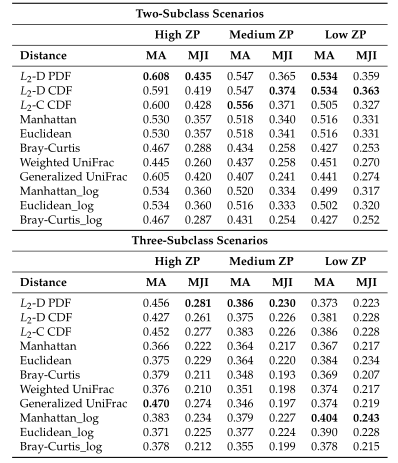
以上两图显示了具有不同ZP和子类数量的不同场景下模拟数据集的聚类结果。通过准确率和Jaccard指数对基于距离的算法性能进行了评估。填充颜色为绿色的箱形图为研究人员建议使用的距离度量。所有的距离都是根据相对丰度计算的。
Table1平均准确率(MA)和平均Jaccard指数(MJI)估计。粗体表示每个方案下的最佳情况。Log表示对输入的模拟数据进行了对数变换。
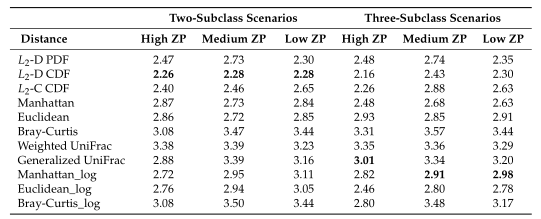
Table2所有模拟场景的平均集群数。根据Dunn内部指数计算出每次重复的最优聚类数。
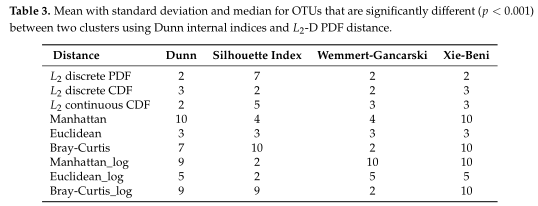
Table1 的结果是通过对每个场景中的100个重复结果求平均值计算得出的。观察得到在聚类算法中实现的距离度量(即绿色标识的箱形图)的准确率和Jaccard指数都优于其他距离度量,特别是在数据集中包含大量0时(高ZP)。在MA和MJI方面,L2范数也显示了其优势,在基于100次重复计算的L2范数在两个子类场景下的;平均准确率约为0.6秒,在三个子类场景下的平均准确率为0.45。而广义Unifrac距离(gUniFrac)在准确性估计中有很大的波动变化。
数据集为197名PD患者和130名对照的粪便样本的16S rRNA测序数据。首先过滤掉在80%的OTU中相对丰度都为0的OTU,因此,此次分析中共使用280个OTUs。将基于相对丰度计算的L2范数与其他三个距离度量进行了比较,包括对数变换和不变换的比较。
如Table3所示,距离度量采用各种内部验证指标(列名)进行灵敏度分析。对于Dunn和Xie-Beni指数,l2范数倾向于将数据聚类为两到三个亚群,而在有和没有对数变换的情况下,除了未变换的欧氏距离外,Manhattan、Euclidean和Bray-Curtis更倾向于聚类更多的亚群。(设置了最多聚类数目为10)
接着选择L2-D PDF范数作为进一步分析的例子。
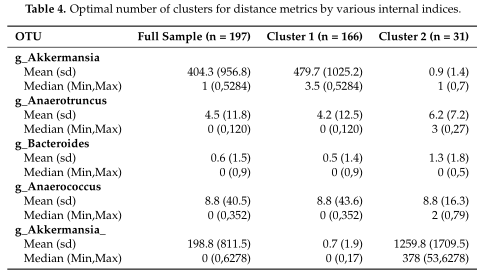
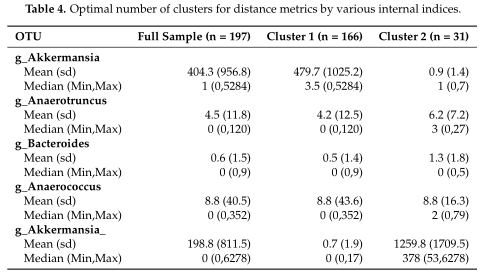
结果在Table4中展示,对数据集中的两个集群之间的OTUs进行了探索,得到了聚类之间差异最大的前5个OTU。
研究认为该聚类算法在高ZP和中等ZP的情况下表现最好,因此,当数据中出现大量的0时,建议使用。并且,在PAM框架下,文章中列出的那些距离度量都可以用作聚类的输入。
如模拟研究中显示的那样,在各种场景下,由混合模型计算的成对距离比其他距离度量表现的更好。但是与所有聚类方法一样,都有一个局限性,就是很难为每个新数据选择合适的内部指标,因此很难获得最优和最稳健的集群数。
此外,对于L2范数距离,在聚类中无法进行变量选择。但不可否认,该聚类算法结合了微生物测序数据的特殊距离,所介绍的聚类算法除了目前使用的方法之外,还可以被看作是分析微生物数据的一个很好的辅助工具。
研究人员指出,下一步会基于Dirichlet-Polyomial等模型进行分区,与文章中的方法进行比较,并努力将该方法扩展到其他微生物群落和疾病相关的数据上。
【参考文献】
Yang D, Xu W. Clustering on Human Microbiome Sequencing Data: A Distance-Based Unsupervised Learning Model. Microorganisms. 2020 Oct 20;8(10):E1612.