-

 CNAS L23010
CNAS L23010

 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业 二级病原微生物安全实验室
二级病原微生物安全实验室- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业 二级病原微生物安全实验室
近日,美国布兰代斯大学Piali Sengupta和Michael P. O’Donnell研究小组发现,肠道细菌产生的神经递质调节宿主的感觉行为(DOI:10.1038/s41586-020-2395-5)。相关论文于2020年6月17日在线发表在《自然》杂志上。该研究结果表明,肠道细菌产生的神经递质模拟了同源宿主分子的功能从而越过宿主调控感觉决定,这促进了宿主和微生物的适应性。


动物与包括微生物在内的多种生物体存在互惠共生关系。过去认为一些细菌产生的具有生物活性的神经递质可以调节其宿主的神经系统的活动和行为。然而这种“微生物-脑”的信号传输机制和它的生理相关性在很大程度上是未知的。在这篇文章中,研究人员通过使用秀丽隐杆线虫为模式生物,结果显示在肠道内定居共生菌普罗维登斯细菌产生的神经调节剂酪胺,绕过了宿主酪胺生物合成的要求并操纵了宿主的感觉行为。细菌产生的酪胺大概率被宿主酪胺β羟化酶转化为章鱼胺。反过来,章鱼胺靶向ASH伤害感受神经元上的OCTR-1章鱼胺受体以调节厌恶性嗅觉反应。研究人员确定了普罗维登斯细菌酪胺生物合成所需的基因,并表明这些基因对于调节宿主行为是必需的。而在普罗维登斯菌的定殖实验中发现,秀丽隐杆线虫在进行食物选择时,优先选择这类细菌,这种选择偏好性需要细菌产生的酪胺和宿主章鱼胺信号。结果表明,肠道细菌产生的神经递质模仿宿主同源分子的功能,以超越宿主对感觉决定的控制,从而促进宿主和微生物的适应性。
秀丽隐杆线虫(Caenorhabditis elegans,C. elegans)是一种无毒无害、可以独立生存的线虫。其个体小,成体仅1.5mm长,为雌雄同体(hermaphrodites),在20℃下平均生活史为3.5天,平均繁殖力为300-350个。自1965年起,科学家Sydney Brenner利用线虫作为分子生物学和发育生物学研究领域的模式生物。进入21世纪以来,已经有六位科学家利用秀丽隐杆线虫为实验材料解开了生命科学领域的重大秘密而获得了诺贝尔奖。2013年12月17日,据国外媒体报道,科学家认为,如果延长蠕虫生命的科学应用到人类身上,人类活到500岁将不只是梦想。秀丽隐杆线虫的两个基因通路,使通过遗传相互作用进行抗衰老治疗的成为可能。
背景
肠道定植细菌和宿主神经系统之间的化学沟通的调节途径大部分还是未知的。秀丽隐杆线虫是近年来出现的一种研究宿主-微生物化学沟通的强大实验系统。野外线虫的主要食物来源于其肠道内定殖的不同的致病菌和非致病菌菌群。接触致病菌会改变秀丽隐杆线虫的嗅觉行为,但肠道内的共生菌是否也能调节宿主行为还不清楚。
实验设计
线虫的制备:
所有的实验都使用的雌雄同体的秀丽隐杆线虫,所有应用于线虫的菌株都保持在在NGM平板上20℃条件下。分别以10
ng/μl注射SRV-11p::octR-1(PMOD110)和SRG-47p::ocTR-1(PMOD111)质粒。以20 ng/μl注射tbh-1p::tbh-1a::sL2::mCherry(PMOD115)质粒。以30 ng/μl注射UNC-122p::mCherry共注射标记,获得转基因菌株。接着将线虫培养在NGM平板上,以获得不同细菌型或基因型的线虫。
主要的实验:
长时程趋化实验和短时程趋化实验分别用于分析线虫对食物和环境选择的偏好性。响应1-辛醇和2-壬酮的SOS分析用于测定其对100%1-辛醇和100%2-壬酮的反应。肠道细菌细胞的量化、定量测定肠道菌落形成单位、荧光强度测量反应了细菌的变化趋势。通过细菌宏基因组测序得到的组装序列,使用普罗维登斯菌株JUb39 的TyrDC和adcA基因序列blastp比对至Nr数据库初始筛选,然后以粪肠球菌TyrDC和线虫tdc-1基因组序列作为tblastn搜索查询序列,最后使用从这些tblastn查询得到的氨基酸序列构建最终的系统发育树。Metaboseek MS分析软件分析高效液相色谱质谱法数据,并集成代谢网络分析。所有的统计分析都是在原始的、非归一化的数据上进行的。
主要结果:
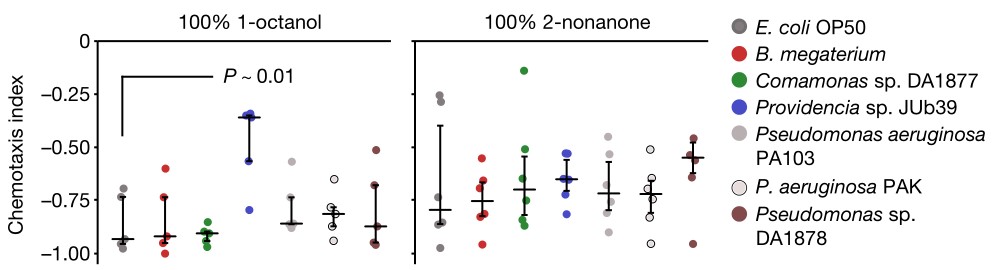
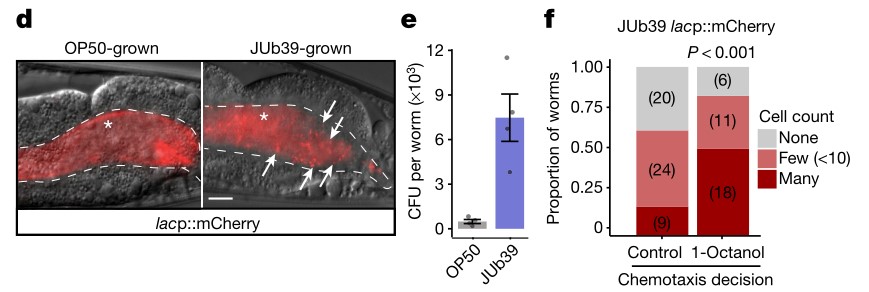
1肠道细菌改变宿主嗅觉行为。普罗维登斯菌定殖于秀丽隐杆线虫肠道内,定植程度与对辛醇的回避行为正相关。
注:以下的辛醇调节表示为对辛醇的回避行为减少的现象
a. 在长时程趋化实验分析中,秀丽隐杆线虫与这些指定的细菌菌株共培养,对一组有吸引力的挥发性气味表现出强烈的吸引力。纵坐标的chemotaxis index表示趋化指数,在辛醇(1-octanol)和壬酮(2-nonanone)的实验中,与在大肠杆菌OP50菌株共培养的线虫相比,与普罗维登斯JUb39菌株共培养导致线虫对辛醇的回避行为减少,但对壬酮的回避行为不减少。
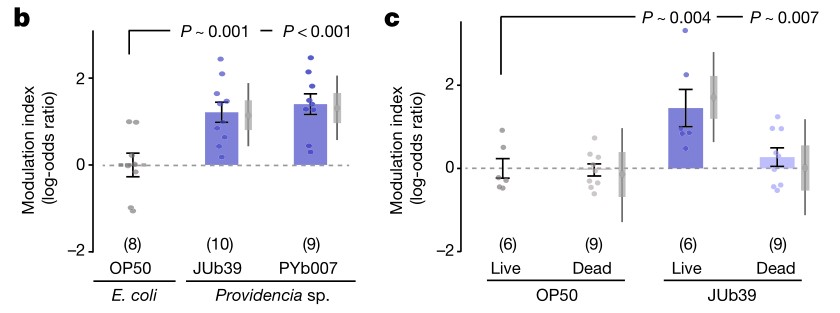
b.c. Modulation index表示线虫对100%辛醇的调节指数。Dead指用庆大霉素预处理过的细菌。PYb007指从堆肥中的线虫中分离出来的与普罗维登斯JUb39菌株有较远亲缘关系的 Providencia Rettgeri菌株。灰色虚线指在辛醇环境下生长的线虫与在不同环境下独立生长的线虫的对照。正值表示辛醇回避减少。使用庆大霉素预处理过JUb39菌株喂食线虫可以消除辛醇调节。而线虫必须摄取活的JUb39菌株才能诱导辛醇调节。
d.在成年线虫的后肠中使用红色荧光蛋白mCherry标记细菌。箭头指示完好的细胞。星号指示漫反射肠道荧光。虚线指示肠边界。喂食OP50的成虫的肠道只显示弥漫的肠道荧光而喂食JUb39菌株的成虫肠道内通常含有大量完整的杆状细胞,表达mCherry。JUb39细菌倾向于富集在后肠,此外被JUb39菌株定植的线虫没有表现出病原性感染的表型特征(如肛门肿胀),进一步证实JUb39菌株对线虫基本无致病性
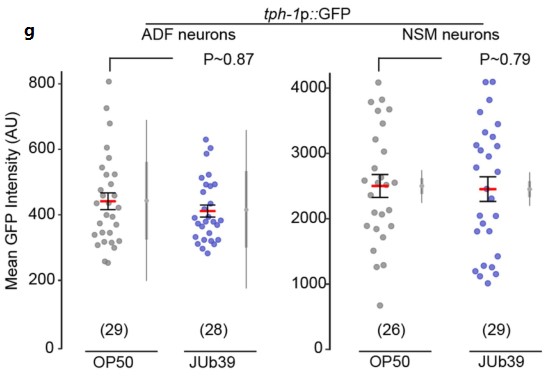
g.在OP50或JUb39菌株环境下生长的年轻成虫的头部神经元tph-1p::GFP的荧光报告基因检测。发现tph-1p::GFP的表达没有明显变化。且a图中线虫在细菌含量较低的食物来源B. megaterium菌中的生长并没有改变辛醇调节。由此推断普罗维登斯菌对辛醇的调节不太可能是由于摄食状态的改变。
e.被喂食指定菌株生长的线虫的肠道细菌载量。点表示10条线虫的菌落形成单位中的细菌载量。在喂食JUb39菌株的线虫可以分离出大量的细菌菌落,但在喂食OP50菌株的线虫不能分离出大量的细菌菌落。这表明JUb39菌株很可能活在线虫肠道中。
f.在趋化试验中, 朝1-Octanol(100%辛醇)方向的线虫始终含有更多的肠道细菌。括号中的数字表示线虫的数量。
2. 发现了一种可能性即线虫产生的章鱼胺,而不是酪胺,其对于JUb39菌株介导的辛醇调节是必要的。细菌产生的酪胺在功能上补充了宿主衍生的酪胺在驱动感觉行为决定方面的损失。
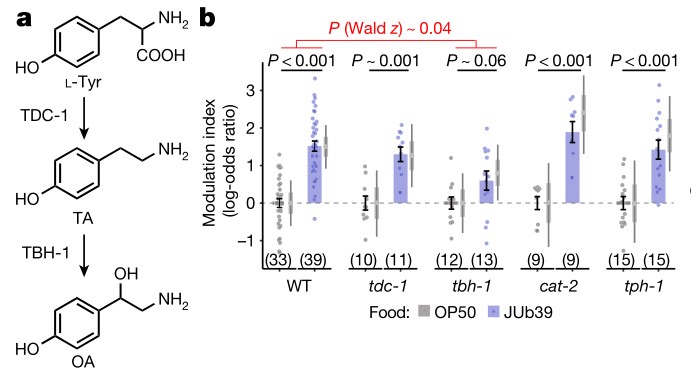
a.线虫体内酪胺(TA)和章鱼胺(OA)的生物合成。酪胺通过酪胺β-羟化酶(由TBH-1编码)转化为章鱼胺。
b.线虫对100%辛醇的调节指数。括号内的数字表示至少3个独立日内进行独立化验的次数。发现在JUb39菌株环境下生长的tdc-1和tbh-1突变体线虫都表现出辛醇调节,但tbh-1突变体线虫表现出的辛醇调节减弱。cat-2和tph-1的突变并不影响辛醇调节。WT表示野生型。
c.短程急性回避行为试验SOS分析。评估辛醇反应的单胺能调节。
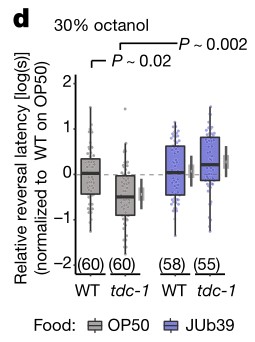
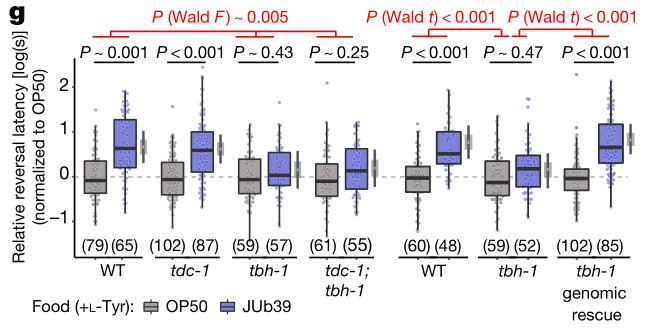
d.g.使用SOS分析进行了所有的辛醇行为实验。
d图中OP50菌株环境下生长的tdc-1突变线虫比野生型线虫对30%辛醇的反应更快,但在JUb39菌株环境下受到抑制。
g图中tbh-1的突变完全抑制了辛醇的调节,而tdc-1突变的线虫对辛醇的调节一如既往。此外tdc-1和tbh-1双突变线虫的辛醇回避行为与只有tbh-1突变的线虫相似。这些结果表明,缺乏宿主来源的章鱼胺是tbh-1突变线虫辛醇调节减少的原因。红色的P值指示指定基因型对JUb39菌效应大小的影响。
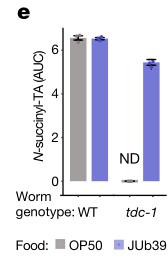
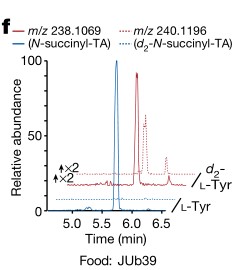
e.f. HPLC-MS进行代谢组成分析和N-琥珀酰酪氨酸的定量分析。喂食OP50菌株的tdc-1突变线虫缺乏N-琥珀酰酪氨酸。而5-羟色胺(N-acetyl serotonin)代谢不变。当喂食JUb39菌株时,这些酪胺衍生的代谢物被恢复。结果表示普罗维登斯细菌与线虫联合产生的酪胺,可以弥补tdc-1突变线虫内源性酪胺产生的不足。
3. 在真核生物和细菌中,生物胺通常由AADC产生。普罗维登斯细菌的多个AADCs产生的酪胺对于调节野生型线虫回避辛醇的行为是必要的,也是理由充分的。
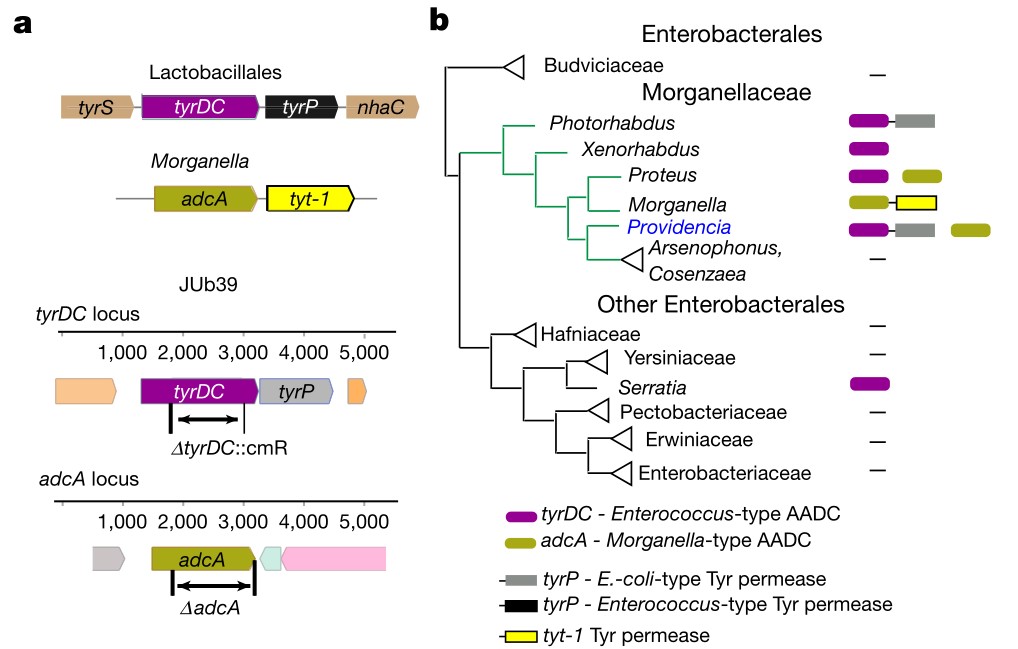

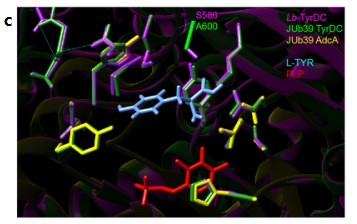
a.从上至下分别描述了乳杆菌的tryDC位点,Morganella菌的adcA位点,对JUb39菌株的tyrDC和adcA位点(分别为ΔtyrDC::cmr和ΔadcA)中设计了缺失。革兰阳性菌肠球菌和乳杆菌的酪胺生成主要是通过位于操纵子的两个基因tyrDC和tyrP介导的,它们分别编码AADC
tyrDC和酪氨酸通透酶。该操纵子可被l-Tyr诱导。全基因组测序证实JUb39菌株和PYb007菌株中均含有tyrDC和tyrP的同源操纵子。在Morganellaceae科的其它成员中也发现了tyrDC同源物。
b.系统发育分析。Morganellaceae科的菌株以前曾被报道在某些条件下产生酪胺,尽管没有可辨别的tyrDC同源物。但在摩根氏菌中鉴定了一个编码AADC的基因(这里命名为adcA),分别位于编码Tyt1家族酪氨酸渗透酶基因上游的操纵子中,与tyrDC肠球菌和人类GAD67有大约29%和27%的序列同源性。在普罗维登斯菌基因组中也存在adcA同源物(包括JUb39),但不与酪氨酸转运蛋白相邻。结论是,普罗维登斯菌编码至少两个可能产生酪胺的AADC,系统发育不一致提示,摩根科植物中的tyrDC和adcA基因可能都是通过水平基因转移而丢失或获得的。
c. 以乳杆菌tyrDC的晶体结构为指导进行蛋白质模拟,结果表明JUb39 tyrDC与乳杆菌tyrDC具有大多数已知的催化位点。JUb39 tyrDC在A600处含有一个替换(乳杆菌tyrDC中的S586),能增强酪氨酸乳酸杆菌对酪氨酸的特异性催化活性。由此推测JUb39 tyrDC可能由酪氨酸产生酪胺。
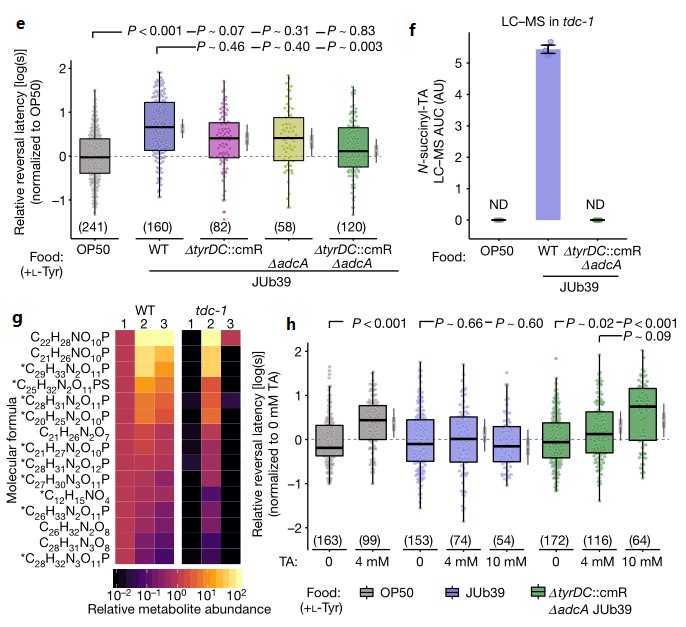
e.在不同基因型细菌环境下生长的野生型线虫在NGM+0.5%l-Tyr培养基中生长期间对100%辛醇回避行为的SOS测定。h图为在此基础上补充了指定浓度的酪胺。
在单个细菌突变体的培养中,辛醇调节减弱,而野生型线虫在双敲除ΔtyrDC::cmrΔadca基因JUb39菌株环境下生长则取消了辛醇调节。
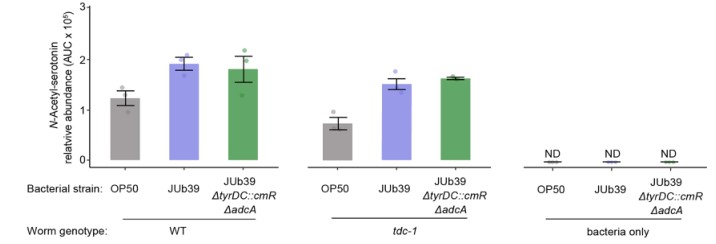
g.f. N-琥珀酰酪胺的含量测定(f);(g)检测到的其它酪胺衍生物的平均丰度,1指示OP50,2指示JUb59,3指示ΔtyrDC::CmrΔADCA JUb39。与野生型JUb39相比,比较代谢组学分析表明,在双敲除ΔtyrDC::cmrΔadca基因JUb39菌株环境下生长的tdc-1突变线虫不产生N-琥珀酰酪胺或其他酪胺衍生代谢物。
h.在双敲除ΔtyrDC::cmrΔadca基因JUb39菌株环境下生长的野生型线虫在补充酪胺后辛醇调节得以恢复。此外,虽然JUb39菌株环境下生长的野生型线虫不会因为补充酪胺而增加对辛醇的回避行为,但补充酪胺足以诱导在OP50菌株环境下生长的野生型线虫的辛醇调节。
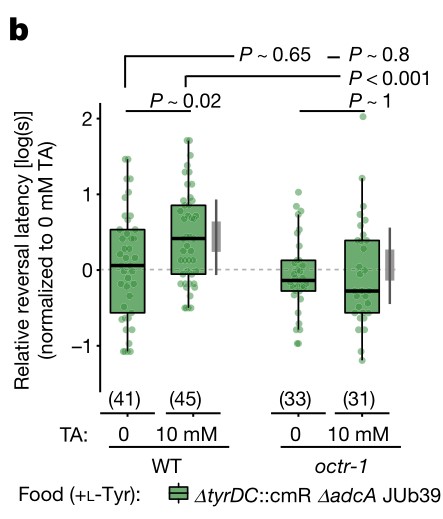
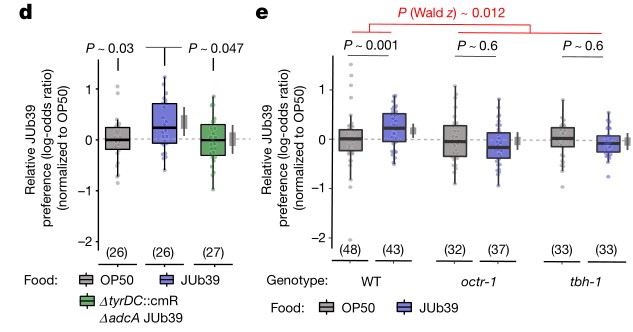
4. 普罗维登斯细菌的酪胺靶向感觉神经元。线虫头部两栖器官中的双侧ASH伤害性神经元表达多种酪胺和章鱼胺受体,这些单胺对辛醇的调节需要一部分受体。普罗维登斯细菌酪胺的产生和宿主章鱼胺的信号都是发生食物偏好所必需的。
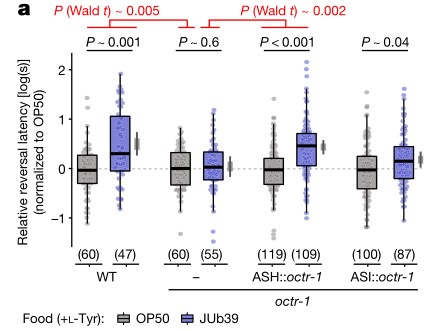
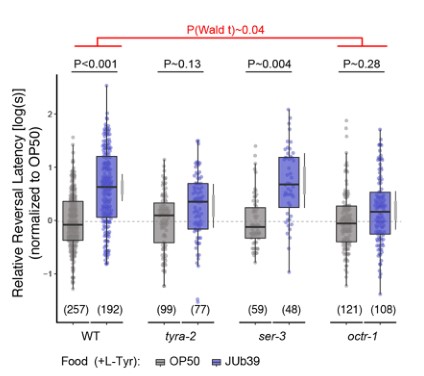
a. SOS测定。在ASH神经元表达的章鱼胺受体编码基因中,octr-1而不是ser-3的突变在不改变肠道定植的程度下取消了JUb39菌株介导的辛醇调节。但在ASH神经元表达的octr-1互补DNA完全恢复了辛醇调节。
b. SOS测定。当线虫在添加了酪胺的ΔtyrDC::cmR ΔadcA JUb39菌株环境中生长时,octr-1突变线虫也会缺乏辛醇调节。
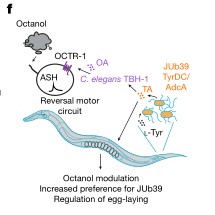
c. JUb39菌株和其他普罗维登斯菌株会产生酒精异戊醇,这种酒精在浓缩时对线虫是有害的。线虫在食物选择实验中可能会优先选择对这类酒精产物的回避行为的减少的这些细菌。
d.e. 在指定菌株环境下生长的野生型和突变型线虫对测试菌株JUb39的相对偏好指数,事实上,在JUb39菌株环境下生长的线虫更喜欢JUb39菌株,而在OP50菌株环境下生长的线虫在短期食物选择试验中只显示出对JUb39菌株的轻微偏好,在octr-1和tbh-1突变的线虫中也是如此。
f. JUb39菌株在线虫肠道内产生的酪胺(TA)通过宿主章鱼胺(OA)和octr-1章鱼胺受体发挥作用,以减少ASH神经元介导的对JUb39菌株产生的回避行为,并改变摄食选择。
结论:
研究人员认为,综上所述,肠道普罗维登斯细菌产生的神经递质酪胺可以推翻宿主依赖的酪胺合成,从而调节线虫的多条单胺能途径,改变线虫的行为和生理。研究人员推测,普罗维登斯菌一旦定植于秀丽隐杆线虫肠道内的,就会促进更多的消耗,从而导致细菌稳定的联系和传播。由于普罗维登斯菌是线虫的主要食物来源,这种联系可能是互惠互利的。
这些结果描述了一种途径,即由天然共生细菌产生的神经递质通过补充或补偿关键的宿主生物合成酶的活性来指导宿主的感觉行为决定,从而改变宿主和微生物的适应性。