-

 国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子
国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子

 二级病原微生物安全实验室
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业

谷禾健康

在我们日常的护肤和美容过程中,我们经常听到关于皮肤的各种话题,从保湿到抗衰老,从痘痘到过敏…
随着科学的不断进步和技术的发展,人们开始逐渐发现,皮肤上隐藏着一个神秘的世界——皮肤微生物群。它在维护我们的皮肤健康方面扮演着举足轻重的角色。
皮肤微生物群由各种细菌、真菌等微生物组成,它们聚集在毛囊、汗腺、皮脂腺等地方,形成一个庞大的生态系统。它们在皮肤表面形成了一道坚固的屏障,阻止了有害菌的入侵。除了提供保护作用外,皮肤微生物群还参与调节角质层的代谢,协助皮肤的水分平衡,并对免疫系统起到了重要的调节作用。
皮肤微生物群的平衡易受到许多因素的干扰。个人的生活方式(过度清洁)、饮食习惯(高糖高脂的饮食)等可能直接影响皮肤微生物的结构和组成,进而引发皮肤问题。外界环境中的污染物、紫外线辐射、气候变化等也会对皮肤微生物群产生影响,从而引发皮肤干燥、过敏、炎症等问题。肠道微生物群的失衡可能导致身体免疫系统的异常反应,进而影响皮肤的健康。
了解皮肤微生物群的特征及其与其他因素的相互关系,对于制定精确的治疗和护肤策略具有重要意义。

图源:Getty Images
本文我们来了解一下整个生命中皮肤微生物组,探讨皮肤微生物群的功能,包括保护屏障、免疫调节等,阐述了皮肤微生物与宿主的相互关系,微生物群在皮肤病中的影响,同时也介绍一些基于微生物群的保持皮肤健康的方法,以及皮肤微生物群在不同领域的应用前景和潜力。
-本文主要内容如下-
编辑
-正文-
皮肤是暴露于外界环境的重要器官,它可以调节体温、防止感染、保护内脏器官等。
皮肤表面是一个酸性、富含盐分、干燥、有氧的环境,而形成毛囊皮脂腺单位的内陷则相对厌氧,甚至富含脂质。
▼
皮肤是身体最大的器官。一个成年人的皮肤平均面积约为1.5-2.0平方米。皮肤除了作为外界与生物体之间的物理和化学屏障的功能外,还作为许多微生物的栖息地。通常,一个人的皮肤上有大约 1000 种细菌。
皮肤微生物组由多种微生物组成,包括细菌、真菌、病毒、螨虫等。
皮肤微生物群通过参与皮肤中发生的基本生理过程,对于维持皮肤屏障、抵御病原体入侵、增强免疫系统、分解天然产物等方面发挥着重要作用。
皮肤和微生物群的结构

编辑
上图可以看到,皮肤由两层组成,即真皮和表皮,具有不同的、专门的生态位或微环境。
▼
皮肤微生物成员和功能可能因皮肤的各种特殊生态位或微环境而异:
具有高密度的毛囊和皮脂腺,例如面部(额头、鼻翼、耳后)、胸部和背部。通常呈高酸性,其特点是细菌可以消耗脂质,需要或可以在厌氧条件下生存,例如:
Corynebacterium minutissimum(微小棒状杆菌)
Cutibacterium
肘部,膝盖,生殖器,肚脐,腹股沟等部位。温和的酸性环境,温度和湿度较高,导致体味的细菌喜欢在这样的环境生活,例如:
Corynebacterium (棒状杆菌)
Staphylococcus (葡萄球菌)
例如手掌等部位。生物量最低,但细菌多样性却最高。
最不稳定的是足部微生物群。足部皮肤上细菌的平均数量从足背表面的103CFU/cm2到第四趾裂处的107CFU/cm2不等。
脚跟底部的真菌居多, 例如:
Malassezia(马拉色菌属)
Aspergillus (曲霉属)
Cryptococcus (隐球菌属)
Rhodotorula (红酵母属)
Epicoccum (附球菌属)
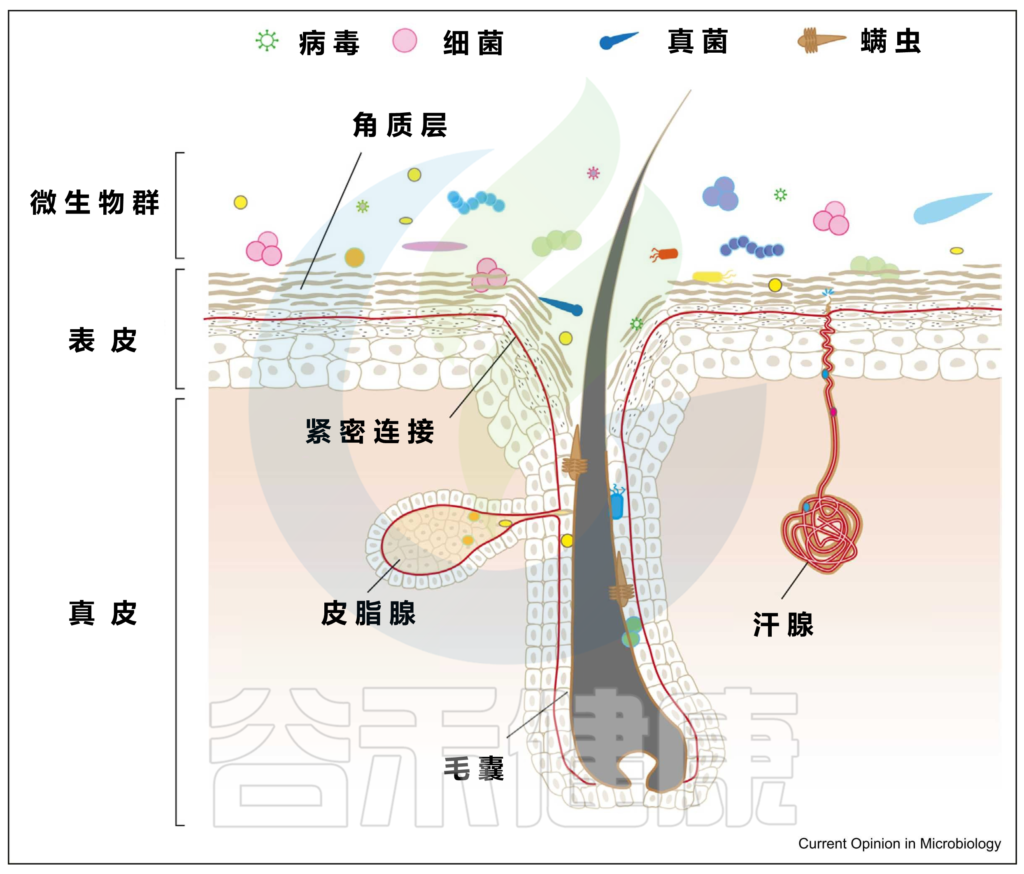
▼
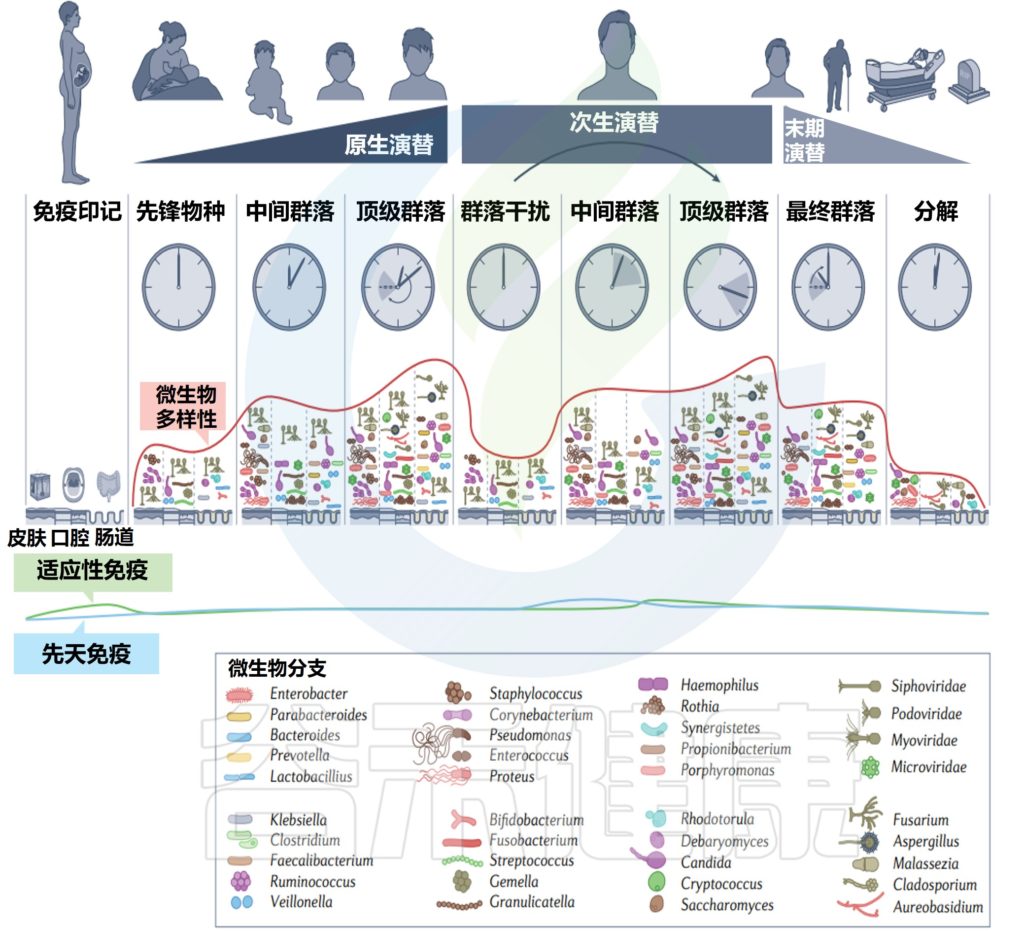
在一生中,随着个人皮肤免疫系统的成熟和激素驱动汗液和皮脂腺的发育,皮肤的生理机能会发生变化。这些变化与突出的皮肤微生物群的相对丰度的变化和整体微生物群落多样性的变化有关。
作为与环境的直接接触面,皮肤也不断地与我们周围的地方和人分享微生物。下图总结了人类一生中皮肤微生物组的变化,并强调了在与年龄相关的关键阶段皮肤微生物组的破坏会影响疾病发展的风险。
皮肤及其微生物组在整个生命周期中的动态平衡
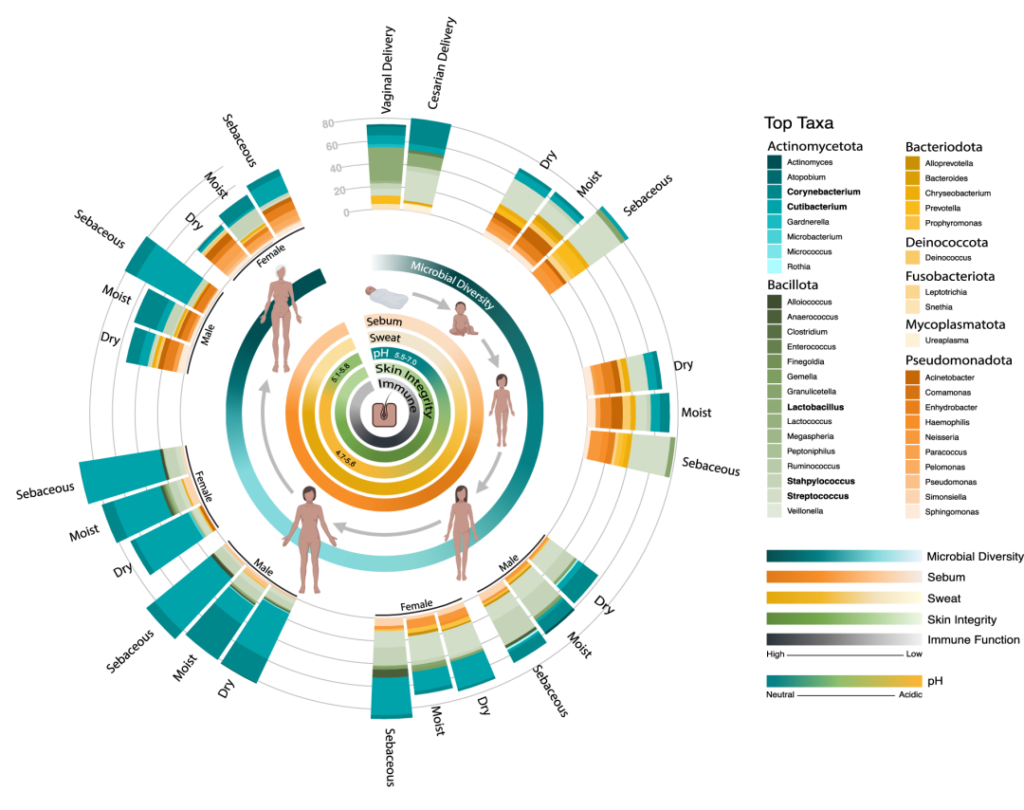
doi.org/10.1042/BST20220216
内圈代表相对微生物多样性、皮脂生成、汗液生成、表面pH值、皮肤完整性和终身免疫功能。微生物组16s测序数据显示了每组前 10 个微生物类群的平均相对丰度。
出生
皮肤微生物组在出生时就已开始定植,并受到多种因素的影响,如:分娩方式、母亲微生物群、抗生素治疗、卫生条件、营养缺乏、住房、动物/宠物接触和环境暴露等。
阴道分娩新生儿的皮肤微生物组以阴道相关菌群为主,主要是乳杆菌,普雷沃氏菌,白色念珠菌。
剖宫产新生儿的微生物群中含有母体皮肤相关微生物,包括葡萄球菌、链球菌、棒状杆菌,Cutibacterium等。
这些初始群落是短暂的,不过物种定殖的顺序和时间会影响菌株后面的相互作用。这些优先效应可以塑造未来的菌群结构,并对皮肤、微生物组和整体健康产生长期影响。
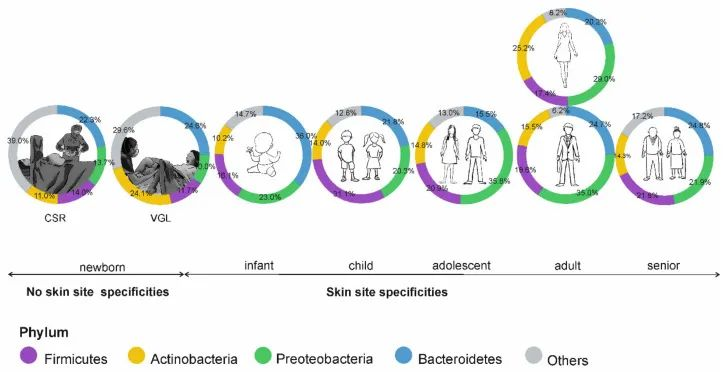
doi: 10.3390/microorganisms9030543
皮肤微生物组的年龄依赖性特异性;CSR剖宫产,VGL阴道分娩。
婴儿期和儿童期
婴儿期,最初接触微生物会促进免疫发育,并通过促进角质形成细胞的适当分化和表皮修复来加强皮肤屏障。
新生儿和婴儿皮肤含水量更高,pH值更高,皮脂生成受到抑制,表皮更新更快,抗菌性能更强。在3-6个月内,微生物分类群与皮肤代谢功能(如脂质生成和pH)之间的联系建立起来。
早期皮脂生成减少与棒状杆菌、Cutibacterium、马拉色菌丰度降低,葡萄球菌、链球菌增加以及以念珠菌为主的真菌生物群落有关。
随着儿童年龄的增长,皮肤进一步酸化并产生更多的皮脂脂质,这促使了酸敏感链球菌(acid-sensitive streptococci)的逐渐减少和整体群落多样性的增加。
在整个儿童时期,皮肤会继续携带来自照顾者的不同微生物群。然而随着年龄的增长,年龄较大的孩子具有更高的皮肤微生物多样性,以及更多来自农村或城市环境的微生物,母婴微生物组之间的相似性逐渐下降。
一旦这种平衡破坏,则可能与更大的炎症有关,并可能增加儿童患特应性皮炎和过敏的风险。
青春发育期
青春期标志着皮肤微生物群的下一个重大转变。驱动身体和性发育的激素也直接促进皮肤的结构和功能变化,如皮脂和顶泌汗液的产生,导致了随后微生物组成的变化。
横断面和纵向研究都表明,Tanner阶段的皮肤微生物组组成发生了明显变化。与V期的年轻人相比,I期的儿童链球菌、拟杆菌和假单胞菌的相对丰度更高,细菌和真菌的多样性也更高。
在年轻成人的皮肤微生物组中主要存在亲脂菌群,如棒状杆菌、痤疮角质杆菌和马拉色菌。与皮脂生成和痤疮相关。
青春期早期和晚期皮肤、微生物组和体味产生的差异
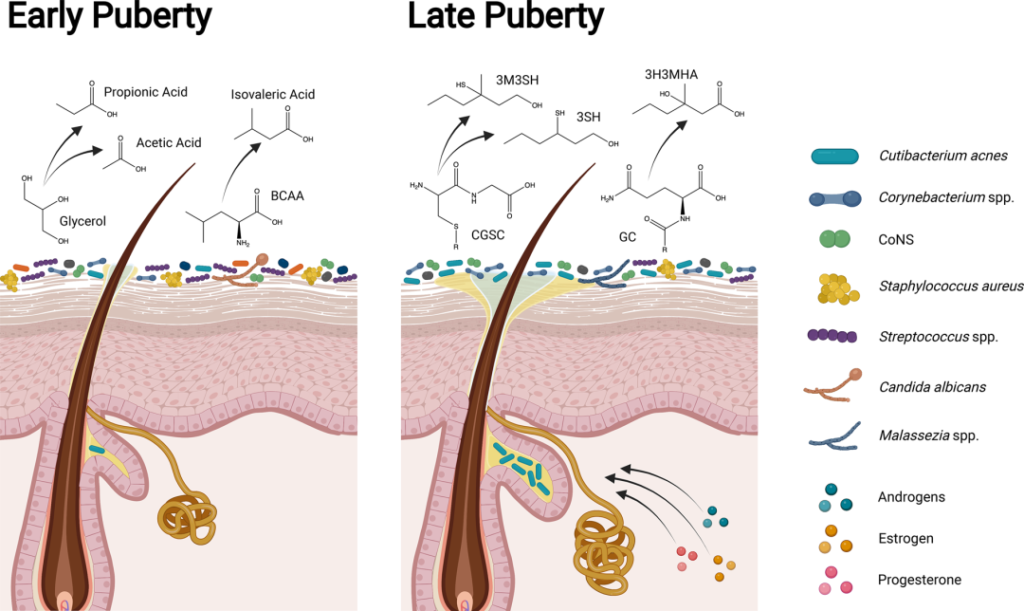
doi.org/10.1042/BST20220216
在儿童期和青春期早期(Tanner阶段I至II),皮肤微生物组高度多样化,体味与凝固酶阴性葡萄球菌属(如表皮葡萄球菌和人型葡萄球菌)产生挥发性脂肪酸(如丙酸、乙酸和异戊酸;酸味)和硫(臭鸡蛋味)有关。随着青春期的发展,类固醇激素促进皮脂腺和顶泌汗腺的发育,改变皮脂中的脂质类型,增强皮肤屏障。
在青春期后期(Tanner IV至V期),脂质生成增加和脂质含量改变与亲脂性类群主导的皮肤微生物组有关。虽然汗液和皮脂成分仍会分解为挥发性脂肪酸,但年轻人的体味与棒状杆菌属更为相关。皮脂和汗液成分代谢为硫烷基烷醇(如3-硫烷基己醇和3-甲基-3-磺基己醇;洋葱味)和挥发性有机化合物(如3-羟基-3-甲基己酸;类孜然味)。
成年期
成年皮肤微生物组在几年内是稳定的。微生物-微生物相互作用网络、持久的成人皮肤生理学和有弹性的皮肤免疫力维持了平衡的成人皮肤微生物群。
成年皮肤微生物群以角质杆菌、棒状杆菌、葡萄球菌、马拉色菌为主。
一旦成年后,成熟和持久的皮肤生理机能,会促进皮脂的产生、汗液成分和表面pH值的一致性,这些共同提供了稳定的身体部位微环境和营养库。免疫系统那时候也成熟了,这些内在特征使皮肤上的大部分微生物群能够在日常环境变化的情况下持续存在。
年龄增长
随着年龄的增长,皮肤会发生明显的变化,包括胶原蛋白合成下降、细胞外基质断裂和皮肤细胞再生减少,皮肤皱纹也就出现了。
随着皮肤屏障的变化,它可能会失去保持水分的能力,导致天然保湿因子(NFM)产生的补偿性增加。NMFs既能吸收水分,又能促进细菌增殖和粘附在皮肤上。随后,NMFs的增加与许多分类群的更丰富有关,如棒状杆菌、微球菌、链球菌、厌氧球菌,同时角质杆菌的减少。皮肤微生物多样性也广泛增加。
女性更年期后皮脂细胞面积和皮脂生成的减少,与角质杆菌的减少以及棒状杆菌、链球菌、不动杆菌和棒状杆菌丰度的增加有关。
在男性中,皮脂分泌下降的速度明显较慢,因此随着年龄的增长,它们保持着更丰富的角质杆菌。
随着年龄增长,免疫系统功能也会慢慢下降。老年人维持低度炎症状态,免疫防御受损和潜在致病菌(如β-溶血性链球菌)增加,皮肤感染的风险大幅增加,难以清除感染。
衰老会改变皮肤结构、功能和微生物定植
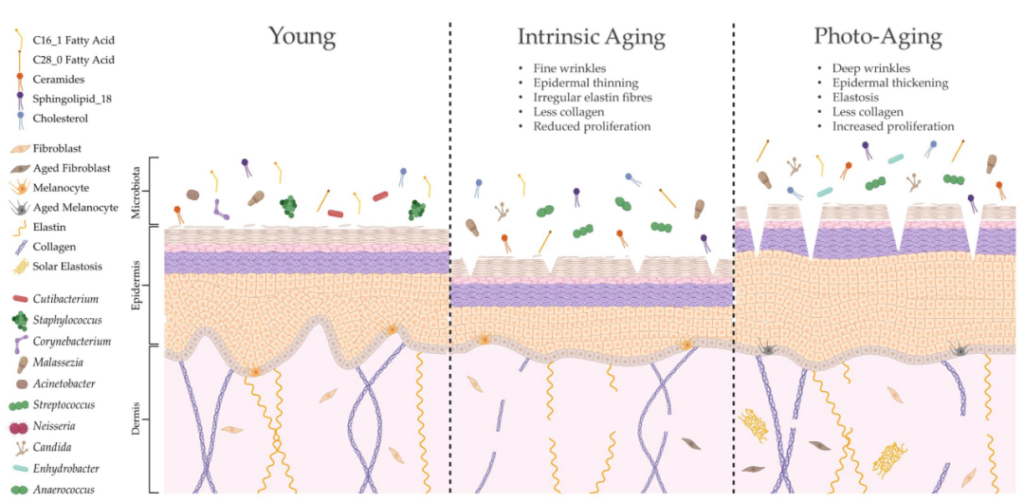
doi.org/10.3390/ijms24043950
内在衰老和光老化会导致皮肤结构和生理的不同变化,导致微生物组成的显著变化。这种改变的皮肤微生物组可能是由脂质成分的特定修饰形成的,这可能进一步导致与年龄相关的皮肤异常。
以上是皮肤微生物组在整个生命周期中的变化情况,那么皮肤微生物组是稳定的吗?它有可能受到哪些因素的影响?我们来看下一章节。
持续暴露于各种外在和内在因素会影响这个皮肤生态系统的平衡。
皮肤结构决定了皮肤微生物组的组成,个体特征取决于宿主的年龄、性别和健康状况等。个人生活方式和所处环境也会影响皮肤上微生物的数量和组成。微生物组的组成可能会随着宿主健康状况的恶化、衰老、甚至居住或职业的改变而改变。皮肤的物理和化学特性影响特定微生物群的优势、它们的比例以及它们之间的相互关系。
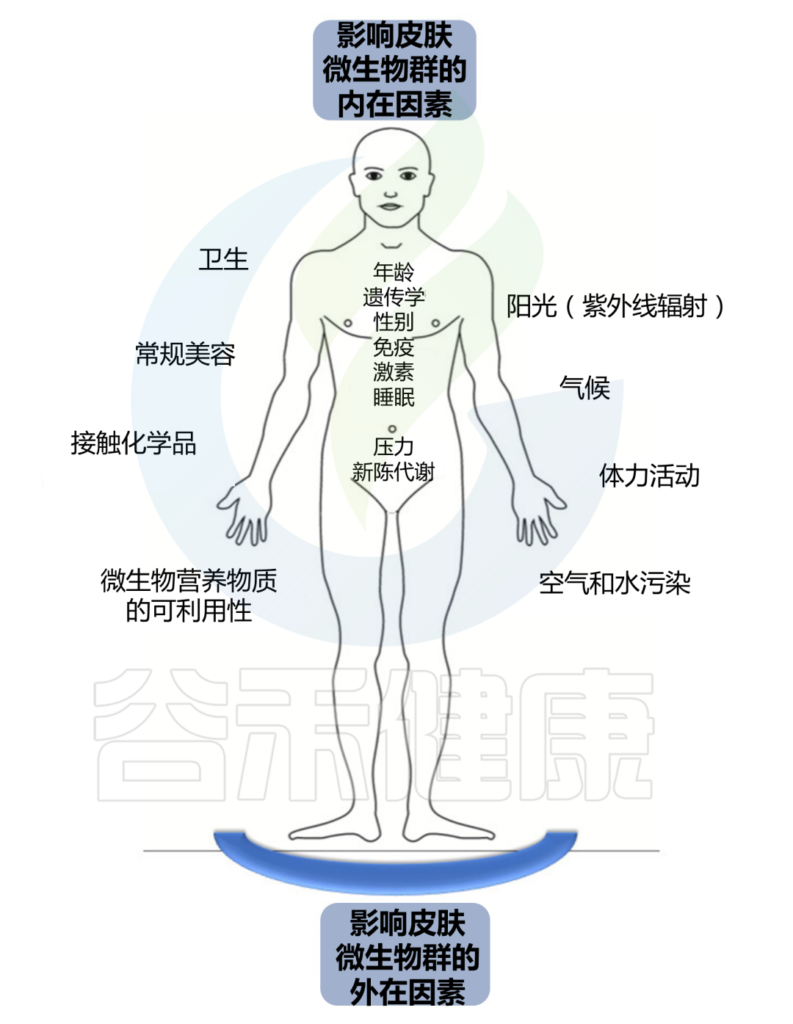
Skowron K, et al., Microorganisms. 2021
我们分为外在因素和内在因素两大块。
外 在 因 素
▼
紫外线辐射,对皮肤细胞有破坏和抗菌作用。大多数与年龄相关的皮肤病是由光老化引起的。皮肤光老化表现为:皱纹、局部色素沉着、毛细血管扩张、干燥和粗糙。这些与表皮和真皮中各种细胞和组织的病理生理变化有关。
皱纹作为光老化最明显的临床特征,主要是由于真皮成纤维细胞减少,以及胶原蛋白和弹性蛋白合成速度减慢但分解速度加快所致。皮肤光老化不仅影响美观,还会损害正常的皮肤屏障功能,增加皮肤炎症性疾病甚至恶性肿瘤的风险。
紫外线:破坏作用
皮肤强烈暴露于紫外线辐射可能会增加其感染的易感性,并加剧相关症状,例如单纯疱疹病毒。
紫外线辐射也可能影响皮肤微生物群的遗传变异,扰乱健康的微生物组结构。
皮肤暴露在紫外线下导致蓝藻菌数量总体增加,乳酸杆菌科和假单胞菌科数量减少。
紫外线:抗菌作用
阳光和紫外线也有效抑制了金黄色葡萄球菌和痤疮丙酸杆菌的生长。痤疮丙酸杆菌数量的减少与卟啉的产生减少有关。
微生物也可以抗紫外线辐射
皮肤微生物组对太阳辐射和紫外线辐射的抵抗力各不相同。一些细菌可以保护皮肤免受紫外线辐射的破坏。皮肤表面的蓝细菌和乳酸杆菌降低了色素沉着的强度和光老化相关损伤的发生。
共生马拉色菌对紫外线辐射表现出高度敏感性,尽管它们有能力合成类似紫外线过滤器的物质——pityriacitrin。
关于紫外线辐射对皮肤微生物群的影响详见谷禾之前的文章:
▼
化学空气污染物,包括 O3、颗粒物(PM 2.5:≤2.5 μm;PM 10:≤10 μm)、挥发性有机化合物和二氧化氮(NO2)等温室气体,是已知的外部暴露组的组成部分,增加过敏性疾病发生和恶化的风险。
空气污染物N2O干扰共生微生物,在对头葡萄球菌和结核棒状杆菌的负面影响大于对金黄色葡萄球菌的负面影响的情况下,有可能发生微生态失调。
烧烤烟雾中较多的成分——多环芳烃,在推动皮肤微生物群分化成不同类型中的作用
多环芳烃来源可以分为自然源和人为源:自然源指火山爆发、森林火灾等自然现象释放到环境介质中的;人为源则是由于人类生产生活活动中化石燃料(煤、油等)不充分燃烧造成的。
我们生活中例如室内外烧烤烟雾中存在较多,在烧烤的过程中,燃料的不完全燃烧或肉类食品脂肪的高温热解均可以产生大量多环芳烃类化合物。
一项研究揭示了多环芳烃暴露与皮肤微生物组分化成不同皮肤类型之间的关联。
皮肤微生物组分化为两种细胞类型(cutotype 1 和 cutotype2)。Cutotype 2与45岁以下受试者的皮肤干燥和色素沉着过度有关。多环芳烃暴露量高与皮肤干燥和cutotype 2有关,cutotype 2富含具有潜在生物降解功能的物种,相关网络结构完整性降低。
cutotype 1中精氨酸生物合成途径中的优势类群、关键功能基因和代谢产物之间的正相关性表明,来自细菌的精氨酸有助于合成聚丝蛋白衍生的天然保湿因子(NMFs),为皮肤提供水合作用,并可解释正常皮肤表型。
这项研究揭示了多环芳烃在推动皮肤微生物群分化成不同类型中的作用,这些类型在分类学和代谢功能上存在广泛的差异,并可能随后导致皮肤与微生物之间的相互作用变化,从而影响人体皮肤的健康。
也就是说:暴露于空气污染后皮肤微生物组组成的变化,可能导致皮肤干燥和炎症的恶化。
▼
全球变暖和极端天气事件等气候变化相关因素,会影响皮肤维持体内平衡的能力,在许多皮肤疾病的发病机制中发挥作用。
全球变暖可能破坏皮肤微生物组
温度和湿度的升高与皮肤上细菌的总体生长有关。
较高的气温和金黄色葡萄球菌的生长之间可能存在关联:
在一项以人群为基础的每月皮肤和软组织感染(SSTI)发病率研究中,SSTI 的时间变化与平均温度和比湿度显着相关。在美国 SSTI 的回顾性分析中(n = 616,375),在气温较高的南部地区,社区获得性耐甲氧西林金黄色葡萄球菌的感染率较高。
温度每升高1˚C,皮脂的产生就会增加10%,这反过来可能会增加微生物的生长,包括角质杆菌和马拉色菌。
极端天气可能引发皮肤病
气候变化导致极端天气事件发生的频率不断增加,包括热浪、干旱、野火、暴雨、洪水和飓风。
例如,洪水的最初影响阶段,经常有创伤与继发性伤口感染的相关风险,包括:嗜水气单胞菌( Aeromonas hydrophila)、创伤弧菌、副溶血性弧菌、Burkholderia pseudomallei等感染。
除了对皮肤病的直接影响外,极端天气事件的额外影响还包括冲突加剧、被迫迁移、心理健康恶化以及传染病的更大传播,所有这些都进一步增加了皮肤病的风险。
▼
农村和城市居民皮肤微生物组的差异,可能与不同程度地接触农业或畜牧业中的土壤、水和生物质中的微生物有关。即使皮肤与土壤和植物材料的短期接触,也会导致手部微生物组的变化以及酸杆菌Acidobacteria和拟杆菌的丰度增加。
在芬兰进行的一项研究结果表明,城市和乡村环境对 1-4 岁儿童的皮肤微生物群有显著影响。这种效应在青少年(14岁)中消失,这直接归因于该年龄段的户外活动时间有限。然而,在其他国家获得的研究结果并未证实这种趋势,表明其他因素(文化差异)也影响皮肤微生物组。
角质杆菌属在农村成年人的背部皮肤上更常见,而Trabulsiella属细菌在城市居民的手和前臂上更丰富。
农村环境的特点是微生物多样性很高
棒状杆菌和角质杆菌属数量的减少,以及假单胞菌和不动杆菌数量的增加,主要发生在与各种农场动物接触的农场工人身上。
封闭空间环境中的微生物有城市和工业区的特点
随着室内城市化的发展,与人类皮肤相关的真菌和细菌的相对丰度也在增加。此外,潜在致病真菌的数量也在增加,包括曲霉菌、马拉色菌、念珠菌等。
由于卫生习惯和西方生活方式,皮肤的细菌多样性降低。许多皮肤共生菌(如表皮葡萄球菌、乳酸杆菌、伯克霍尔德菌Burkholderis、痤疮梭菌)消失,取而代之的是葡萄球菌、棒状杆菌、角质杆菌(Cutibacterium)和微球菌Micrococcus。
▼
不同的动物物种含有独特的微生物群,与动物的持续接触会影响健康人皮肤细菌群落的组成和多样性。例如家养狗和家庭主人共享微生物群。菌群结构受季节的影响,但不受狗的性别、年龄、品种或皮毛类型的影响。
宠物肠道菌群与主人的肠道菌群也会产生关联,详见:
▼
皮肤与衣服的长时间接触也很重要,这会导致微生物的传播,并形成所谓的纺织品和挥发性微生物组。反过来,织物微生物组的组成会受到洗涤和干燥的影响。附着在纤维上的微生物可以利用污垢或皮脂化合物作为基质,并产生挥发性物质作为副产品,从而产生难闻的气味。
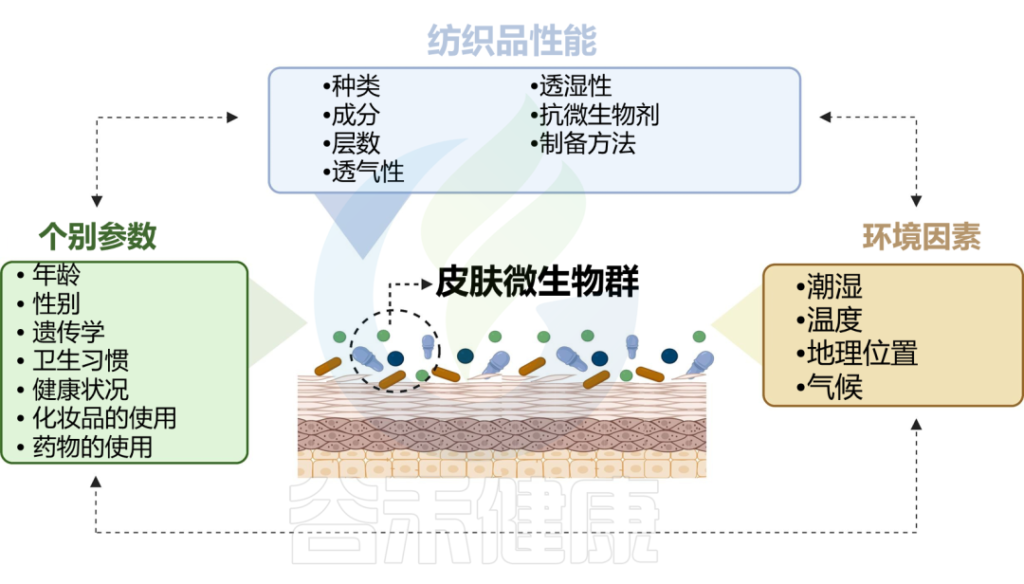
doi.org/10.1016/j.ejpb.2023.05.004
纺织纤维的性质可以直接影响微生物的附着、生长和定植
葡萄球菌属在几乎所有纺织纤维中显示出显着的固定性。Staphylococcus hominis对棉花的亲和力较高,在粘胶纤维和羊毛中不生长。
羊毛促进了许多菌群生长,包括表皮葡萄球菌、Enhydrobacter、角质杆菌、微球菌属。
聚酯为角质杆菌、Enhydrbecter、微球菌属提供了最大的生长环境。
棒状杆菌属无法在棉花、丙烯酸、羊毛、粘胶、尼龙、羊毛和聚酯上进行竞争,这解释了只有少量棒状杆菌属才能从破旧的衣服中分离出来。
合成纤维由于其疏水性和较差的吸附能力,通常抵抗微生物定植。
天然纤维更容易受到微生物定植的影响,因为它们具有高保湿性能,并且它们的聚合物键更容易被微生物酶获取。天然纤维可以以碳水化合物或蛋白质的形式为微生物提供营养和能量来源,支持微生物生长和定植。
与棉花相比,亚麻纺织品对金黄色葡萄球菌和表皮葡萄球菌表现出强烈的抑制作用,同时对角质形成细胞产生细胞毒性。
▼
化妆品旨在改善皮肤,减缓衰老过程。这些产品可能有助于皮肤微生物组的多样化,尤其是当定期或长期使用时。
化妆品中含有的活性成分可能有利于或抑制某些微生物的生长
N-乙酰氨基葡萄糖是刺激皮肤微生物群的化合物之一,它是透明质酸的前体,常见于护肤品中。
保湿产品可以降低皮肤水分流失的强度,并可以增加皮肤微生物群多样性(α多样性高是健康皮肤微生物群的标志),同时减少皮肤细胞剥落。它们的脂质化合物促进亲脂性细菌的生长,如葡萄球菌和角质杆菌。另一方面,皮肤水合水平的提高会降低皮脂含量,并可能减少角质杆菌数量。
化妆品成分的作用持续数周,个体的反应可能差异很大。不合适的化妆品或不合适的应用会减少皮肤微生物组的多样性,从而对其产生负面影响,导致生态失调。洗发水或面霜等化妆品也可能会导致感染,有时会导致严重的健康后果,尤其是在儿童或免疫力下降的人群中使用。
内 在 因 素
▼
皮肤表面呈微酸性(pH值5.6左右)且干燥,但温度比体内低。
表皮细胞自身脱落机制影响菌群组成
表皮外层不断释放角质化皮肤细胞,导致皮肤每四个星期自我更新一次。每小时有 500-3000 个细胞从1cm2的皮肤脱落,这意味着一个成年人每小时释放 600,000-100 万个或更多细胞。由于约 10% 的脱落细胞含有细菌,这种机制可能会显着影响微生物组的组成。
皮肤的厚度、表面褶皱的深度和位置,毛囊和腺体的密度都是影响宿主微生物群的关键因素。腺体释放的分泌物以不同的方式影响微生物,创造刺激或抑制微生物发育的条件。
皮脂腺:确保专性和兼性厌氧菌的最佳环境。这些腺体分泌的皮脂在皮肤上形成保湿、疏水的保护层,并且是微生物使用的脂质的来源。这些脂质水解产生的游离脂肪酸有利于细菌粘附到腺体表面并降低皮肤pH值,抑制金黄色葡萄球菌和化脓性链球菌等病原体的生长。
水分含量:潮湿的区域为许多微生物创造了有利的条件,如棒状杆菌属、葡萄球菌属等。相对干燥且温度波动较大的皮肤部位主要含有变形菌、拟杆菌、放线菌等。微生物的数量随着深层皮肤层中营养物质和水分含量的增加而增加。
▼
男性和女性微生物群之间物种组成的差异是由皮肤的性别特异性特性造成的,即皮肤厚度、毛发、汗液和皮脂腺的数量。女性多样性高于男性。更薄的皮肤、更低的 pH 值和更少的出汗量会导致更多的多样性。
对手部表面的微生物进行的一项研究表明,女性的物种多样性高于男性。在女性手上,肠杆菌和乳杆菌科的数量显著较高(300-400%),而在男性中,观察到更高浓度的角质杆菌和棒状杆菌。
关于不同年龄皮肤菌群构成不同,在前面第一章节已经详细阐述。
▼
在形成皮肤微生物组的遗传因素中,种族是次要的,但也有一定影响。最主要的是不同生活方式的差异。非洲和拉丁美洲男性头皮和腋下的Cutibacterium数量低于其他种族(高加索、非洲裔、东亚和南亚)。中国人皮肤微生物组与其他人群存在差异,比如Enhydrobacter在中国人的皮肤上较为常见。
▼
抑制细菌和减少炎症病变
口服米诺环素(用于治疗痤疮)降低了Cutibacterium、棒状杆菌、普雷沃氏菌、乳酸杆菌和卟啉单胞菌的丰度。
多西环素显著减少痤疮梭菌的数量(治疗6周后为1.96倍)。Snodgrassella alvi的数量也减少了(3.85倍)。另一方面,观察到Cutibacterium granulosum的数量显著增加(4.46倍)。
大环内酯类、四环素类和克林霉素用于治疗痤疮。用利美环素进行的脸颊皮肤治疗减少了角质杆菌的存在,并增加了链球菌、葡萄球菌、微球菌和棒状杆菌的数量。反过来,二甲胺四环素导致微生物组紊乱。
虽然氟喹诺酮类药物(培氟沙星)和大环内酯类药物(红霉素)显著减少了痤疮梭菌的数量,但只有纳氟沙星对凝固酶阴性葡萄球菌表现出抑制活性。
导致出现抗生素耐药性物种
例如痤疮梭菌和表皮葡萄球菌。大环内酯类药物的长期治疗痤疮,增加了痤疮梭菌分离株的数量,但对大环内酯的影响的敏感性降低。
据估计,红霉素和阿奇霉素耐药菌株的比例可能分别达到50%,甚至100%。从感染皮肤分离的G+细菌中,77.5%对青霉素耐药,28%对甲氧西林耐药。在所有测试的菌株中,31.9%对三种以上的抗生素不敏感。
儿童皮肤分析结果显示,36.4%的从皮肤表面分离的金黄色葡萄球菌菌株对甲氧西林有耐药性。此外,耐甲氧西林葡萄球菌(MRSA)是医院感染最常见的原因之一。
▼
肠道内表面和皮肤表面有一些有趣的相似之处:两者都被上皮细胞覆盖,上皮细胞维持着体内与外部环境之间的重要联系,充当第一道防线,在抵御外部病原体、调节免疫反应和抑制分解代谢物方面发挥着重要作用。
肠道和皮肤组织是宿主原核和真核共生微生物的两个主要生态位,因为它们的高细胞周转率决定了定植微生物组的低粘附和感染。
皮肤健康与肠道屏障的完整性有关。一些饮食代谢物可以直接吸收到皮肤中,其他通过肠道微生物代谢来做到这一点,这两者都可能有助于皮肤健康。
由于肠道通透性增加,肠道菌群或其代谢产物可能从肠道迁移到循环系统中并在皮肤中积聚,这可能会损害皮肤屏障并使其容易发炎。
肠道微生物群的变化还可能引发系统性炎症和异常免疫反应,从而破坏皮肤健康。
皮肤或肠道微生物群失调与免疫应答改变密切相关,与多种皮肤病相关,包括特应性皮炎、牛皮癣、寻常痤疮、甚至皮肤癌等,这在下一章节会详细讲述。
饮食强烈影响肠道微生物组的组成,影响代谢和免疫功能,间接影响皮肤健康。关于如何通过饮食调整在最后章节会讲到。
以上是影响皮肤微生物群的外在和内在因素,那么皮肤微生物群会如何影响人体健康呢?我们来看下一章节。
这里我们分为两个部分来阐述:
皮肤微生物群直接影响皮肤健康
我们知道,皮肤是由角质形成细胞的分层角质化上皮组成,这些上皮经历终末分化。这些物理结构通过增强屏障的化学和免疫学特征得到进一步强化。
皮肤微生物群影响皮肤屏障的各个方面,同时也直接与表面遇到的共生微生物和病原微生物相互作用。
皮肤微生物群介导多种屏障功能
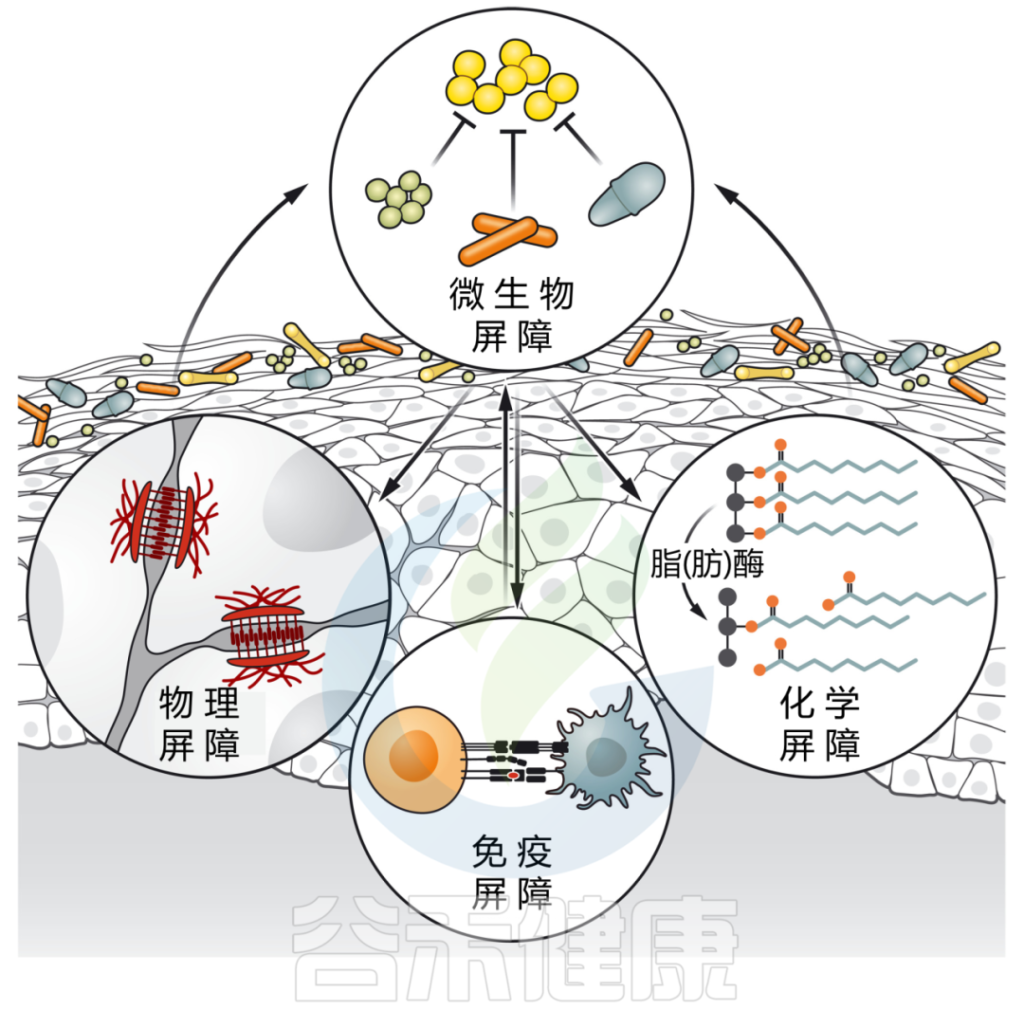
DOI: 10.1126/science.abo0693
微生物群强化皮肤屏障的多个方面:
皮肤微生物通过各种定殖抗性机制,包括资源排斥、直接抑制和/或干扰,形成对抗环境的第一道屏障。
皮肤微生物群也有助于物理皮肤屏障的分化和上皮化。微生物通过产生脂肪酶来增强皮肤的化学屏障,脂肪酶将皮脂甘油三酯消化为游离脂肪酸,从而增强皮肤的酸性,并限制瞬时和致病物种的定植。
最后,微生物刺激先天和适应性免疫防御,如抗菌肽的释放、新生儿耐受性的诱导和保护性免疫的发展。
接下来我们讨论微生物群到底如何与皮肤屏障的微生物、化学以及先天和适应性免疫成分相互作用。
▼
皮肤微生物群本身是抵御外来微生物和病原微生物入侵、定植和感染的屏障。
——直接竞争关系
皮肤微生物争夺资源,并进化出直接对抗对手的机制。
多种CoNS物种(凝固酶阴性葡萄球菌),如人葡萄球菌,产生具有独特化学性质的抗生素,并抑制皮肤病原体金黄色葡萄球菌。
其他物种,如头葡萄球菌,通过干扰金黄色葡萄球菌毒力所需的辅助基因调节因子(agr)群体感应途径来拮抗金黄色葡萄菌。
——拮抗机制与宿主抗菌反应协同作用
人型葡萄球菌和表皮葡萄球菌,可以产生共生衍生的AMPs,其发挥选择性抗菌活性,并与宿主衍生的AMPs协同作用,以抑制皮肤病原体的存活。
痤疮角质杆菌与产生硫肽抗生素角质霉素的特定菌株竞争,以维持其在人类毛皮脂腺单元中的生态位,从而限制金黄色葡萄球菌的定植。
皮肤微生物组内微生物之间的相互作用,可以驱动整体微生物群结构。
主要皮肤菌群产生的抑制其他微生物群,和/或潜在病原体的突出和最近鉴定的抗菌分子汇总在下表,分子作用机制也包括在内。
皮肤上关键的微生物与微生物相互作用

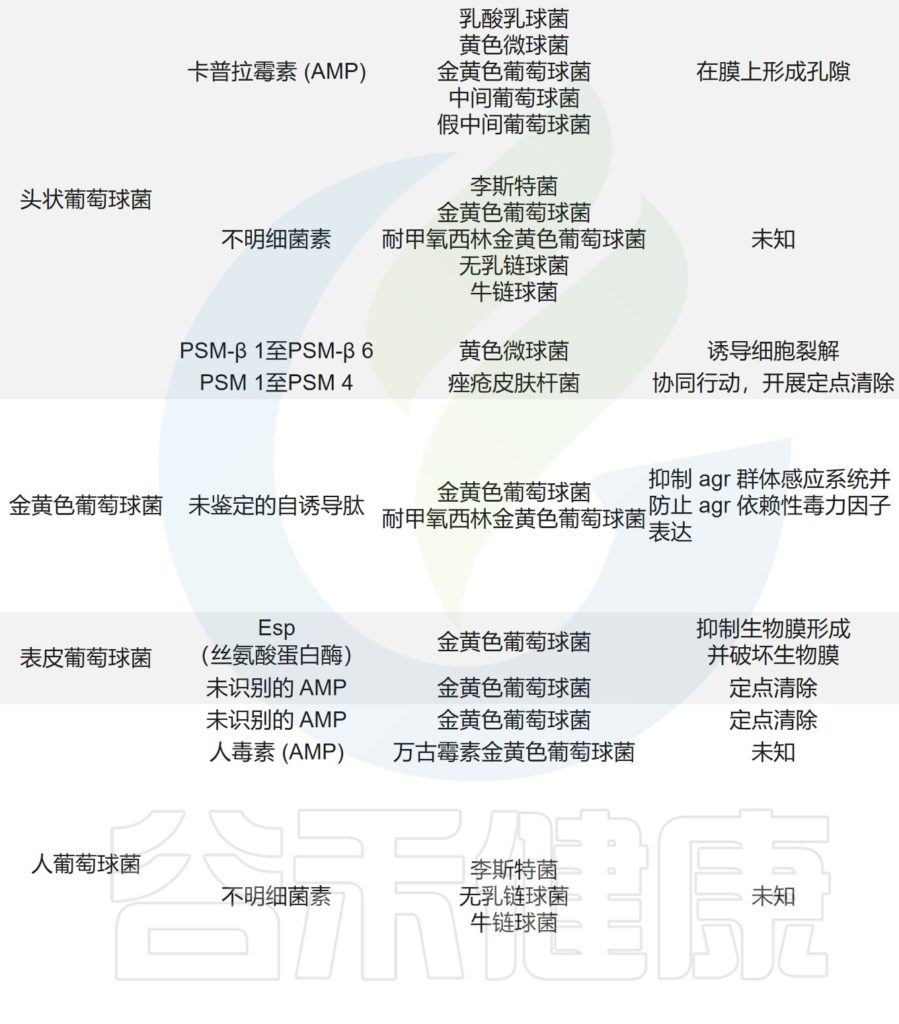
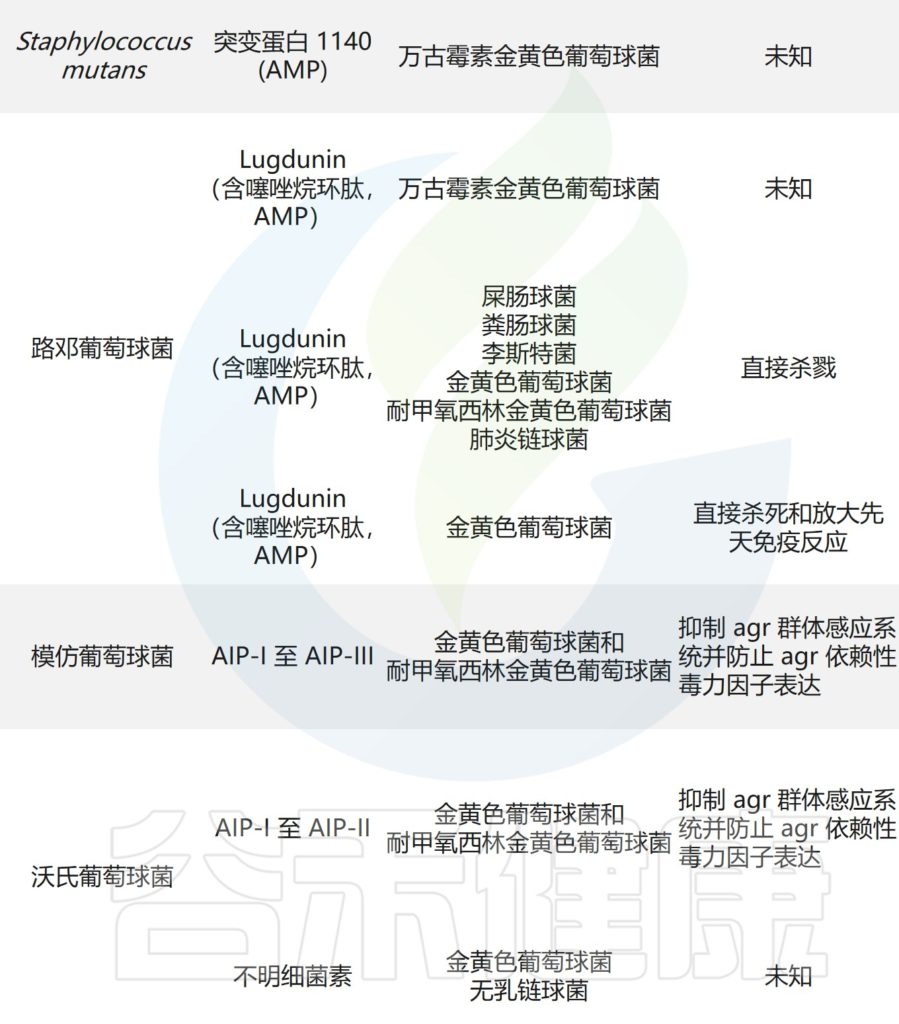
doi.org/10.1042/BST20220216
▼
角质细胞经历严格调控的终末分化程序,形成角质层,该过程由微生物群介导。微生物群通过角质形成细胞芳香烃受体(AHR)的信号传导促进分化和上皮完整性;还分泌鞘磷脂酶,将层状脂质加工成神经酰胺,神经酰胺是角质层的关键成分。
▼
酸性皮肤表面还产生了限制细菌定植的化学环境。痤疮角质杆菌和棒状杆菌都分泌脂肪酶,水解皮脂中甘油三酯中的游离脂肪酸。游离脂肪酸通过直接抑制细菌和刺激人β-防御素2(hBD-2)的表达,进一步增强皮肤免疫力。痤疮角质杆菌也直接与游离脂肪酸结合,这表明游离脂肪酸的存在促进了痤疮角质杆菌的定植。
▼
微生物可以刺激多种与先天免疫反应有关的反应,通常取决于代谢和炎症环境。例如,念珠菌的菌丝和酵母形式在皮肤中刺激不同的免疫反应。S. epidermidis在皮肤中引起的T细胞反应,需要菌体表面特定糖蛋白与宿主先天免疫细胞上的C型凝集素相互作用。
氧气的可用性也会影响皮肤表面宿主与微生物的相互作用。微氧耐性细菌痤疮角质杆菌生成短链脂肪酸,抑制组蛋白去乙酰化酶,后者可作为免疫系统的表观遗传调节因子,从而刺激炎症。
注:在皮肤中,短链脂肪酸具有促炎作用,这点和肠道中不同。SCFAs通过抑制HDAC8和HDAC9以及通过TLR信号通路刺激炎症。
皮肤微生物还通过刺激宿主产生抗菌肽和蛋白来增强皮肤免疫力,这些抗菌肽和蛋白起到天然抗生素的作用。
皮肤微生物群落还在创伤修复过程中协调先天免疫反应。在皮肤中的共生微生物群落会引发I型干扰素(IFN)反应。作为对微生物信号的反应,中性粒细胞会表达CXCL10,吸引活化浆细胞样树突状细胞(pDC)到损伤部位。pDC会产生I型干扰素,通过刺激成纤维细胞和巨噬细胞增长因子反应来加速创伤修复。
实际上,抗原呈递细胞向皮肤的募集是微生物群依赖性的。微生物通过需要IL-1R-MYD88信号传导的过程,在伤口修复和毛囊新生中增强皮肤再生。
▼
皮肤是各种适应性免疫细胞的家园,其中包括大量的常驻记忆T细胞,随时准备对各种环境刺激做出反应,包括致病微生物和共生微生物。
在婴儿早期,暴露于皮肤共生表皮葡萄球菌介导调节性T细胞(Tregs)流入皮肤。这种Treg迁移波与毛囊发育同时发生,需要毛囊角质形成细胞产生趋化因子。Tregs,以及皮肤中的许多其他免疫细胞亚群,最终位于毛囊附近,对在这个发育窗口期间检测到的微生物抗原具有特异性。
在一个平行的过程中,粘膜相关不变T细胞(MAIT)是在婴儿期在类似的时间限制的发育窗口中获得的。MAIT细胞在无菌小鼠中是不存在的,它们的发育需要维生素B2代谢产物,而这些代谢产物仅由细菌和真菌产生,而不是哺乳动物细胞。
在胸腺中,暴露于5-(2-oxopropylideneamino)-6-d-ribitylaminouracil(一种维生素B2的细菌代谢产物,从粘膜部位运输到胸腺),介导MAIT细胞扩增并靶向皮肤和粘膜部位。
微生物细胞表面分子也可以作为宿主的信号。大多数棒状杆菌的细胞膜中含有霉菌酸。棒状杆菌属霉菌酸在稳定状态下可以以IL-23依赖的方式促进γδT细胞的积累。然而,这种相互作用取决于环境,因为高脂肪饮食反而会促进皮肤炎症。因此,微生物暴露时存在的炎症环境影响皮肤内的免疫反应。
这些发现突出了微生物在皮肤免疫细胞的募集和刺激中发挥的关键作用。
以上是皮肤微生物群从物理、化学、免疫等多角度与皮肤之间的关联,如果说上述对局部组织微环境的相互作用,那么接下来我们从更系统的角度来看,皮肤微生物群通过与其他器官的交流,对全身健康产生的影响。
皮肤微生物群通过影响其他器官间接影响
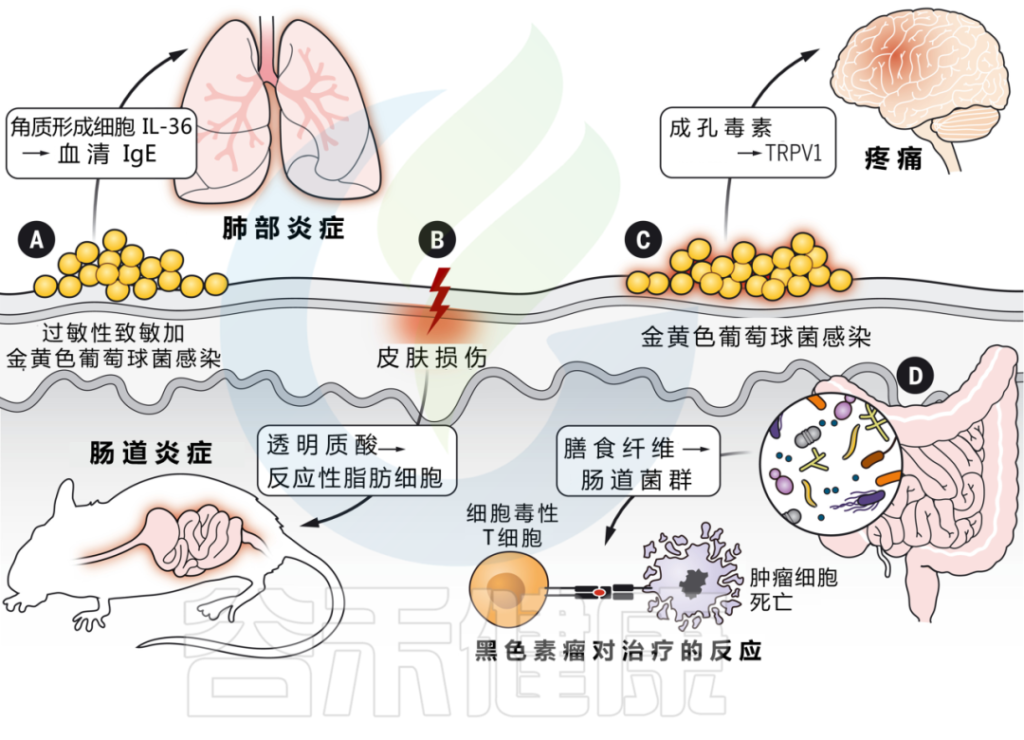
越来越多的证据表明,皮肤损伤和致敏会影响其他屏障部位,如肠道、肺部等。
▼
皮肤和肠道之间存在双向沟通。
为什么浅表皮肤损伤会引起肠道炎症?
浅表皮肤损伤会导致角质形成细胞全身释放 IL-33。IL-33 与 IL-25 协同作用,触发肠道内 ILC2 的激活,产生 IL-4。这反过来又刺激肠道中肥大细胞的扩张,在那里它们准备对食物过敏原做出反应并介导过敏反应。
在模拟炎症性肠病的小鼠模型中,皮肤受伤还会加剧肠道炎症。
皮肤和肠道之间的相互作用取决于损伤期间真皮中产生的透明质酸片段的产生,这些片段刺激肠道成纤维细胞,通过反应性脂肪生成的过程分化为促炎脂肪细胞。这些反应性脂肪细胞通过产生 AMP 和其他炎症介质来传播肠道炎症。
肠道微生物群变化也会影响皮肤炎症
在这两种情况下,肠道免疫网络的激活都会影响皮肤中炎症信号的振幅。
因此,肠道微生物组的改变可能会影响皮肤免疫力。
研究表明,饮食对肠道微生物组的影响,尤其是膳食纤维,对系统免疫有重要影响。皮肤先天免疫反应也与肠道有关,肠道中保护细菌性皮肤感染的AMPs的充分表达,取决于饮食中的维生素A。这些发现加强了我们对饮食在宿主免疫发展中重要性的分子理解。
▼
流行病学证据表明,许多患者经历了“特应性进军”,首先出现特异性皮炎,随后发展为过敏性鼻炎、食物过敏、哮喘。它们的先后出现意味着存在什么样的关联?
皮肤微生物群失调和金黄色葡萄球菌定植增加,与特应性皮炎的发作有关。
表皮暴露于金黄色葡萄球菌刺激角质形成细胞产生IL-36,从而提高血清IgE水平。
而缺乏IL-36受体的小鼠对金黄色葡萄球菌的反应不会产生升高的IgE,并且也可以免受过敏原特异性肺部炎症的影响。这些发现支持了皮肤暴露于微生物病原体作为全身炎症的起始。
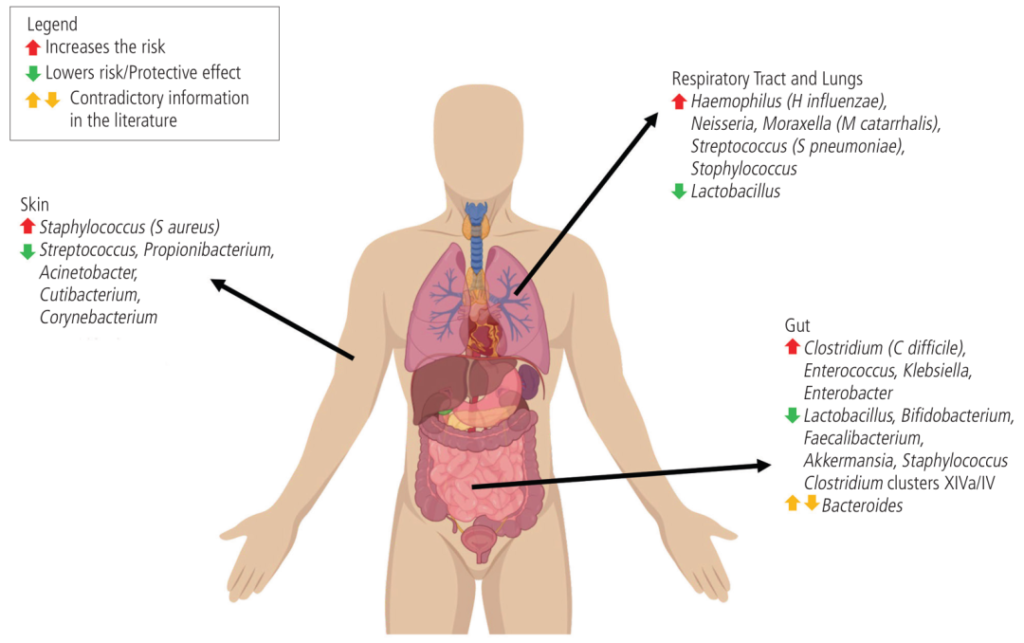
微生物组的变化与哮喘、过敏性鼻炎、特应性皮炎和食物过敏的风险有关
doi: 10.18176/jiaci.0852
在气道中,卡他莫拉克菌(Moraxella catarrhalis)、流感嗜血杆菌(Haemophilus influenzae)和肺炎链球菌水平较高与婴儿哮喘有关。
肠道艰难梭菌的比例高于双歧杆菌,这与更高的食物过敏率有关。
▼
神经免疫相互作用中的皮肤病原体
细菌可以直接激活皮肤中的感觉神经元,并通过产生造孔毒素引起疼痛。菌株水平的变化驱动着可变的反应,这取决于特定毒素和群体感应系统的存在。
关于群体感应,详见:
真菌(白色念珠菌)也可以直接激活皮肤中的感觉神经元。γδT细胞免疫需要刺激才能通过释放神经肽CGRP来控制皮肤念珠菌感染。
相反,引起坏死性筋膜炎的病原体化脓性链球菌,通过分泌链球菌溶血素S直接激活伤害感受器神经元,进而促进神经肽CGRP的释放并抑制化脓性链球菌的杀伤。在这种情况下,CGRP拮抗作用可防止坏死性感染。
皮肤与其他器官系统的交互作用是由微生物群介导的
DOI: 10.1126/science.abo0693
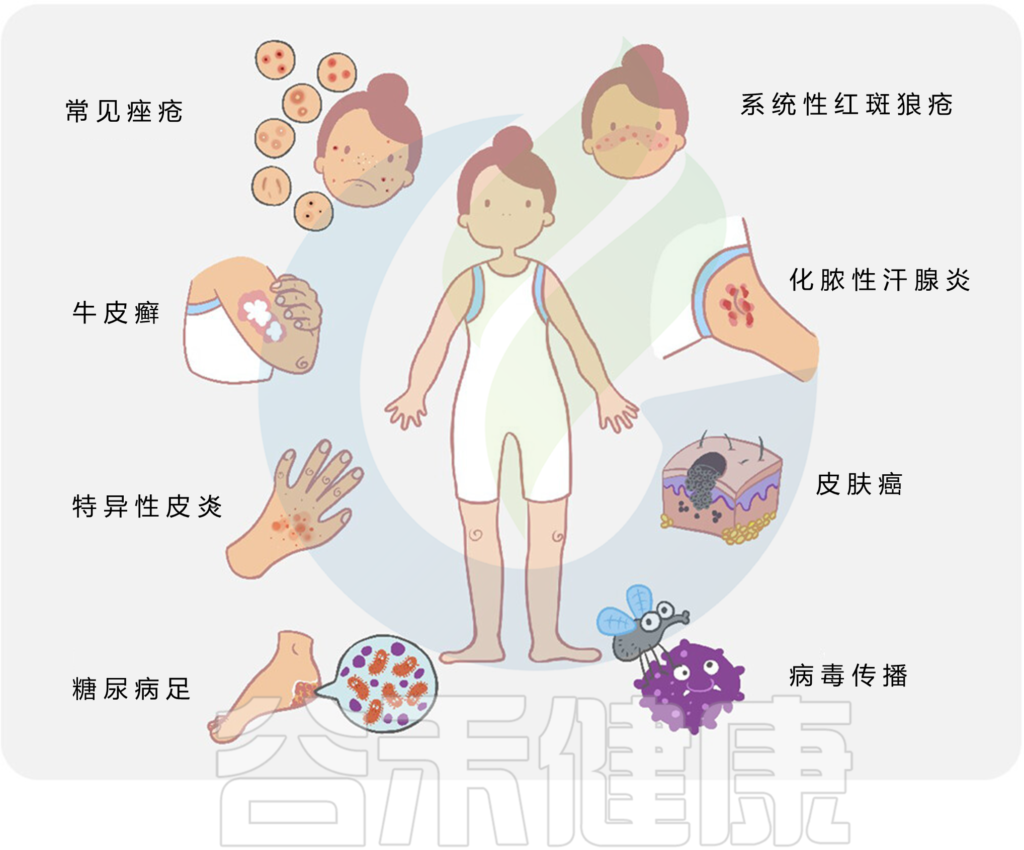
微生物与宿主相互作用和皮肤疾病
doi.org/10.1002/mlf2.12064
▼
痤疮患者,特别是那些症状严重的患者,表现出α多样性增加,四种革兰氏阴性细菌(即粪杆菌属、克雷伯氏菌属、臭杆菌属和拟杆菌属)的比例更高。
痤疮角质杆菌(C. acnes)的过度生长与痤疮发病机制有着长期的关联。宏基因组分析表明,痤疮患者中痤疮丙酸杆菌的菌株结构与健康个体不同,IV 型和 V 型菌株在受痤疮影响的皮肤中特别普遍。
痤疮丙酸杆菌通过多种不同方式参与痤疮发病机制的调节,它参与:
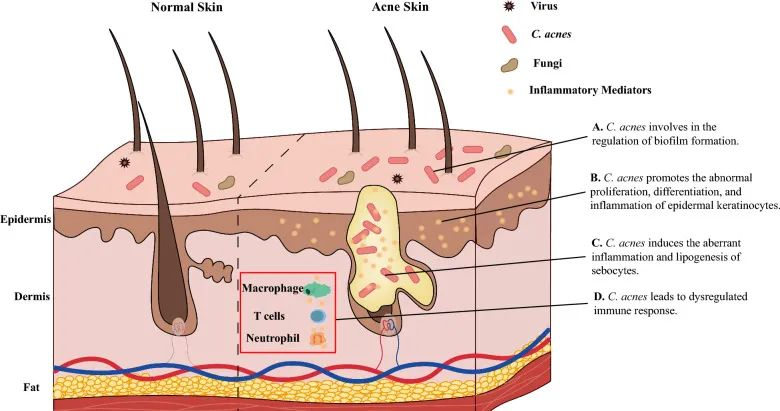
doi: 10.1186/s13578-023-01072-w
痤疮丙酸杆菌和表皮葡萄球菌在痤疮以及炎症后色素沉着过度中具有病理生理作用。
肠道微生物群在皮肤炎症和情绪之间起着中介作用
痤疮和胃肠道功能障碍之间的联系可能起源于大脑。支持这一假设的是压力引起的痤疮加重。实验动物和人类研究表明,压力会损害正常的肠道菌群,尤其是乳酸杆菌和双歧杆菌。心理应激源导致肠道微生物群产生神经递质(即乙酰胆碱、血清素、去甲肾上腺素),这些神经递质穿过肠粘膜进入血流,导致全身炎症。
痤疮中肠-脑-皮肤轴的拟议模型
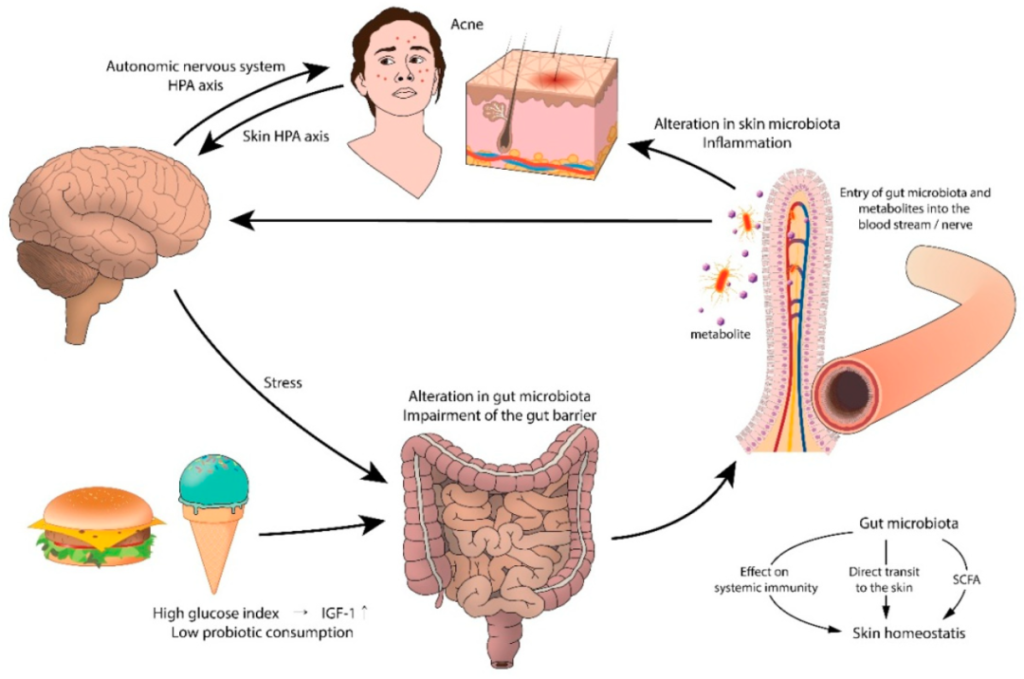
doi.org/10.3390/jcm8070987
西方饮食包括乳制品、精制碳水化合物、巧克力、饱和脂肪等,这些物质可能通过激活营养来源的代谢信号来加重痤疮。高脂肪饮食会降低肠道菌群水平,增加脂多糖的浓度,通过损害结肠上皮完整性和屏障功能、降低粘液层厚度和增加促炎细胞因子的分泌来引起全身炎症。
扩展阅读:
▼
皮肤干燥、斑块发痒和反复出现的湿疹是特异性皮炎的标志。
特异性皮炎引起的皮肤耀斑通常与更多的金黄色葡萄球菌丰度有关,金黄色葡萄球菌定殖的增加与CoNS数量的减少相关,CoNS本来会产生抗菌蛋白,它在特异性皮炎患者中数量少。
金黄色葡萄球菌在病变的真皮中更为普遍,这表明在剥皮过程中更容易接触到更深的皮肤层。
是什么引起金黄色葡萄球菌定植增加呢?
表皮葡萄球菌、痤疮杆菌和棒状杆菌属的丰度降低,它们通常对金黄色葡萄菌的入侵起作用。
特异性皮炎的菌群多样性低。共生细菌数量减少而导致的共生产生的AMPs的缺失,抵御病原体如金黄色葡萄球菌的能力下降,金黄色葡萄球菌定植增加。
与特异性皮炎相关的皮肤屏障缺陷会损害层状膜的完整性,改变皮肤的微生物群,并可能使金黄色葡萄球菌等有害细菌滋生。厌氧微生物的缺乏可能会降低关键的皮肤屏障活性,并促进潜在的感染。
金黄色葡萄球菌分泌毒力因子
金黄色葡萄球菌分泌几种毒力因子,包括纤连蛋白结合蛋白1(FBP1),α-和δ-溶血素,酚溶性调节素(psm)的蛋白家族等,所有这些毒素都会导致更高的炎症反应和更严重的症状。
皮肤稳态取决于复杂的宿主-微生物相互作用,包括金黄色葡萄球菌和特异性皮炎宿主细胞之间的相互作用,微生态失调会导致疾病的发展。
特异性皮炎的其他微生物群变化包括痤疮角质杆菌、棒状杆菌、Dermacoccus、微球菌、CoNS减少,链球菌和一些马拉色菌属增加。这些微生物变化似乎是暂时的,在特异性皮炎发作之前和期间,群落多样性丧失,金黄色葡萄球菌优势更大,在炎症消退后逐渐恢复到基线。
潜在益生菌治疗和预防AD的临床试验

doi.org/10.1016/j.phymed.2023.154824
▼
一般来说,皮肤破裂会导致炎症级联活动;然而,这种免疫反应在糖尿病皮肤中被破坏,也就是无法有效引起免疫反应。微生物组的改变可能会加剧疾病的严重程度。
糖尿病皮肤的菌群特征
糖尿病早期患者的皮肤细菌微生物群与健康人的非常相似。随着疾病的恶化,物种多样性和丰度发生动态变化。总的来说,糖尿病足的皮肤细菌微生物群的多样性低于健康足。因此,不太常见的微生物种类的变化,其中大多数只在健康的足部皮肤中发现,可以用来预测是否患有糖尿病。
糖尿病足皮肤中葡萄球菌的含量通常较低,金黄色葡萄球菌的比例较高。金黄色葡萄球菌的大量存在破坏皮肤微生物群平衡,可能会导致炎症变化,并增加皮肤感染的风险。
慢性溃疡相关菌群
铜绿假单胞菌和厌氧菌通常与深部慢性溃疡有关,但金黄色葡萄球菌通常与急性浅部溃疡有关。比较有慢性感染和没有慢性感染的糖尿病患者的微生物组的研究可以提供有关诊断标志物的信息,这些标志物可以用作发展为慢性损伤的可能性的指标。
▼
牛皮癣患者由于慢性炎症性皮肤病而出现中度至持续性皮肤斑块。许多遗传和环境变量之间的复杂组合导致皮肤过度活跃的炎症反应是病因。
牛皮癣皮肤菌群特征
牛皮癣患者皮肤样本在α、β多样性明显低于正常皮肤。下列菌群相对丰度和分类性能显著下降:
棒状杆菌在牛皮癣的发病机制中发挥重要作用
大量的研究表明,棒状杆菌属丰度上升,棒状杆菌有可能干扰干扰素信号系统,这可能导致皮肤微生物组的微生态失调。
乳制品和糖类的摄入是牛皮癣最常见的诱因之一,而肉类和鸡蛋则被列为次要的常见诱因之一。
关于牛皮癣与肠道菌群之间也存在很多相关性,此处不展开阐述,详见:
▼
皮肤黑色素瘤
黑色素瘤和正常皮肤样本之间的微生物组成和多样性存在显著差异。黑色素瘤样本中的梭杆菌和Trueperella水平较高。
棒状杆菌属与疾病严重程度相关,棒状杆菌水平与IL-17之间存在关系,IL-17可以通过增加IL-6和信号转导器和STAT-3来促进黑色素瘤细胞增殖。
相反,痤疮角质杆菌的细菌上清液增加了黑色素细胞的凋亡。
角质细胞皮肤癌
以皮肤微生物群为代表的生物屏障通过分泌抗微生物肽(AMP)(如组织蛋白酶LL-37和人β-防御素)来抑制病理生物和病原体入侵,从而与角质形成细胞和免疫细胞产生串扰。
研究人员认为金黄色葡萄球菌和鳞状细胞癌之间的联系不是偶然的,皮肤溃疡是有利于外源性搪塞/感染的致病过程的结果。金黄色葡萄球菌也可能参与鳞状细胞癌的发病机制,引起慢性局部炎症,涉及不同的致瘤阶段,包括促进生存、增殖、细胞转化、侵袭、血管生成、转移。
葡萄球菌毒素-α决定了参与炎症过程的局部细胞的分泌,进而导致活化B细胞的NF-Kβ的激活,从而增加不同细胞因子和趋化因子的表达,包括IL-1β、IL-6和IL-12。
其他因素如紫外线辐射(尤其是UVB)也是皮肤癌发生的主要危险因素之一。紫外线照射会改变皮肤微生物群,导致大量形成活性氧、细胞凋亡和炎症,与皮肤癌相关。
总的来说,许多常见的皮肤病,如痤疮、特异性皮炎、牛皮癣、皮肤癌等,都与皮肤微生物群的变化有关。
皮肤病中的关键微生物发现如下:

编辑
doi.org/10.1002/mlf2.12064
●
饮食对肠道微生物群的影响较大,皮肤和肠道微生物群是内在相关的,由宿主免疫系统介导。因此,肠道和皮肤可以通过饮食、微生物代谢产物、神经内分泌途径和中枢神经系统等途径相互作用,也就是说,饮食对皮肤也会产生较大影响。
饮食结构
西方饮食已被证明会破坏微生物组并导致皮肤病,从而对皮肤健康产生负面影响。相反,植物性饮食与更健康的皮肤有关。
以植物为基础的饮食是一种由多种蔬菜、水果、豆类、扁豆、豆类、坚果、种子、真菌和全谷物组成的饮食模式,并且限制或不摄入动物产品、加工食品或糖果。
这种饮食的饱和脂肪、反式脂肪和花生四烯酸含量较低,而抗氧化剂和 omega-3 脂肪酸含量较高,再加上其直接治疗作用,可减少炎症和皮肤症状。
植物性饮食与皮肤健康/疾病之间的关联
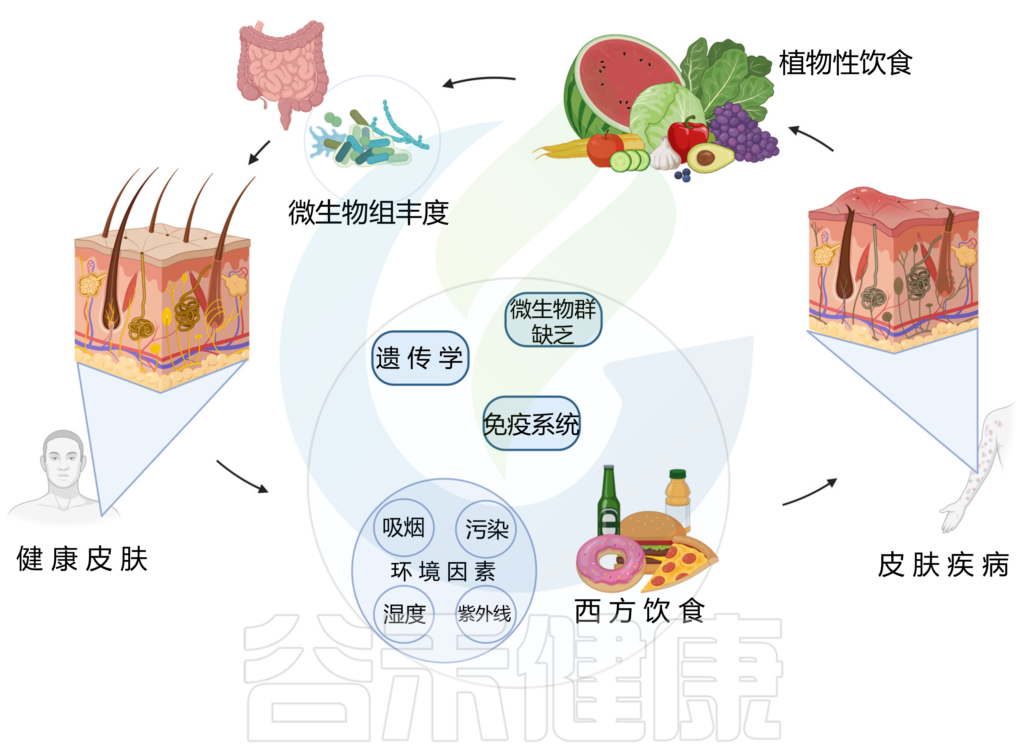
doi.org/10.3390/nu15132842
多项研究发现,植物性饮食对缓解牛皮癣、特异性皮炎、痤疮等皮肤问题有益。


doi.org/10.3390/nu15132842
食 物
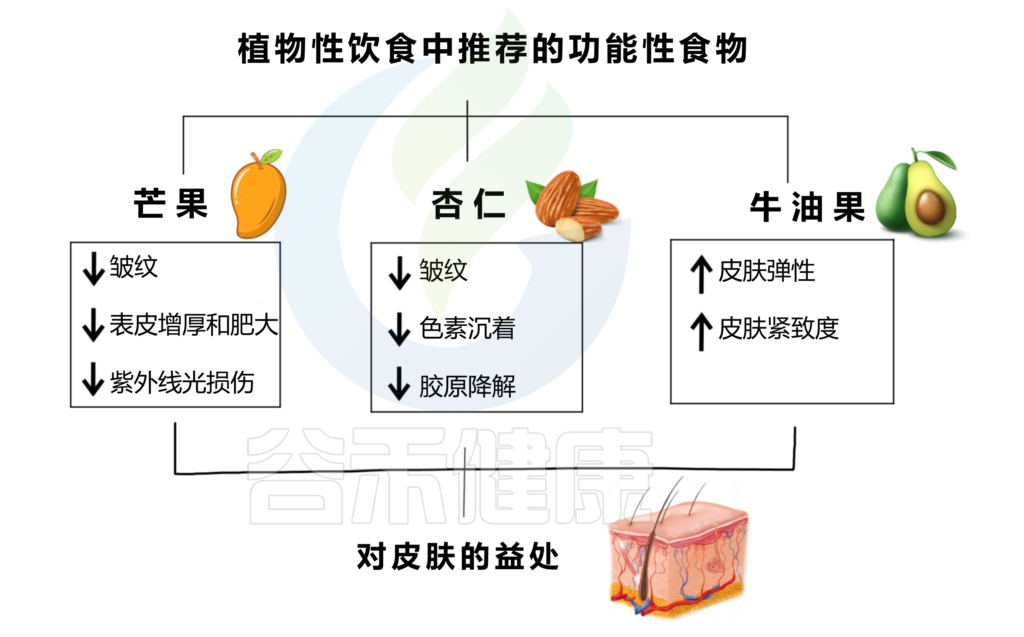
植物性功能性食品增强皮肤健康,减少皮肤老化迹象,并改善整体外观。下图是芒果、杏仁、牛油果及其对皮肤健康的积极影响。
编辑
doi.org/10.3390/nu15132842
芒果能够减少皱纹、表皮变薄和肥厚,防止 UVB 损伤。无论是果肉还是果汁,芒果酚酸的抗氧化特性和生物利用度都会得到保留,而果汁的呈现可能会增强其特性。从芒果干中提取的芒果提取物也可以减少UVB辐射引起的皱纹的形成。
杏仁富含α-生育酚(或维生素E)、脂肪酸、多酚,因此是一种具有抗氧化特性的食物,可以减少皱纹、色素沉着和胶原蛋白降解。
牛油果含有类胡萝卜素、单不饱和脂肪酸、酚类化合物,某些基因的表达,如胶原蛋白和弹性蛋白基因,在进行饮食调整后被诱导,因此可以促进皮肤弹性和紧致度的增加。
限制饮酒和甜食
酒精会使你的身体和皮肤脱水,这可能会使皮肤看起来更加干燥或有皱纹,许多含有酒精的混合饮料也富含糖,这都不利于皮肤健康。糖可能使胶原蛋白变硬,从而使皮肤老化,也可能带来炎症。
喝 水
对于每日饮水量较低的人(即那些本来就脱水的人)来说,增加饮水量对皮肤外观有积极影响,有助于维持皮肤水合水平。同时,尝试在食物中多加入黄瓜、芹菜、西葫芦、西瓜、草莓和花椰菜等,也可以适当补水。
●
适当的清洁和保湿可用于维持皮肤的生理pH值。据报道,早在1995年,与使用 pH8 的普通肥皂相比,使用酸性合成皂(pH 5.5-5.6)可显著减少非炎症和炎症病变。
从那时起,pH 值的变化通过皮肤屏障的完整性与痤疮的发病机制联系起来,建议使用 pH 值约为 5.5 的皮肤清洁剂。
为什么洗脸很重要?
脸每天面对风吹、紫外线、化妆/护肤品、屏幕等刺激,会积聚污垢、油脂和其他碎屑,如果不及时清除,可能会导致刺激和其他皮肤问题。
合适的洗脸方式
注意:
应该多久洗一次脸?
没有既定的指导方针,一般来说,最好每天洗两次脸。
如果皮肤干燥或敏感,可以在晚上用清洁去除污垢,然后在早上用温水冲洗脸。
即使当天不化妆或者不出门,污垢、油脂和其他不需要的碎屑仍然会在一天中积聚在皮肤上,因此最好在睡觉之前洗脸。
如果刚在健身房、参加高温瑜伽课或在户外徒步旅行,并且出汗较多,最好马上洗脸。
如果存在敏感问题或其他特殊的皮肤状况,请与医生沟通。
●
痤疮是一种慢性炎症性皮肤病,对于痤疮,护肤品有多种作用机制,包括:
1) 保护和改善皮肤屏障
2) 保护皮肤微生物组
3) 维持健康的皮肤 pH 值
4) 抵御紫外线伤害
保护皮肤屏障是皮肤化妆品改善痤疮管理的重要机制,临床上,屏障功能障碍表现为皮肤干燥、刺痛/烧灼/刺痛、紧绷、疼痛或刺激性皮炎等形式。这些被认为与经表皮失水 (TEWL) 有关,并且可以通过使用保湿剂至少部分缓解。
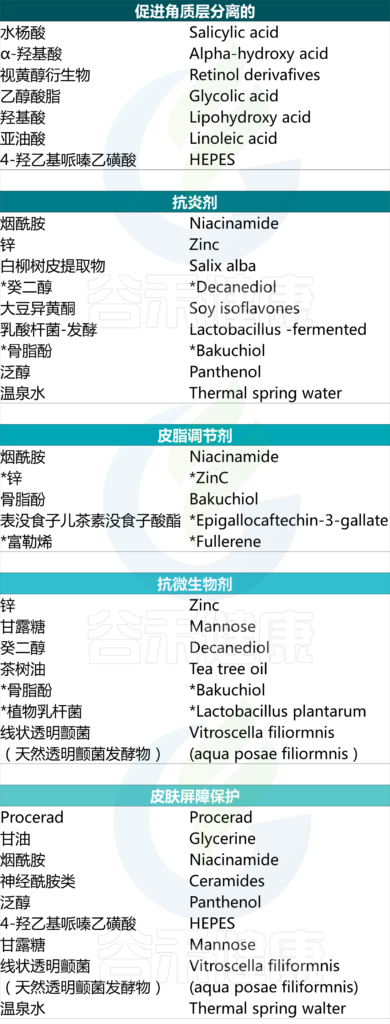
特别适用于痤疮的成分包括烟酰胺、视黄醇衍生物、水杨酸、神经酰胺、甘油、温泉水、泛醇等。
护肤品中可能存在的活性成分及其针对性作用
Kurokawa I, et al., Dermatol Ther (Heidelb). 2023
而皮肤微生物组的核心作用与表皮屏障功能一起,为优化护肤提供了强有力的支持。可以通过尝试恢复微生物组的多样性并通过下调先天免疫来抑制炎症。
总的来说,现有文献结果的总体趋势表明,护肤品可以改善整体皮肤健康,减少痤疮皮损,在处方治疗后维持痤疮清除,并且可能对减少表面皮肤油腻具有有益作用。
然而需要进一步研究才能更好地理解这一作用。在此情况下可能有益的成分包括但不限于:乙醇酸、LHA、亚油酸、烟酰胺、锌、吡罗克酮乙醇胺、procerad、Vitroscella filiformis.
注意:
痤疮的治疗管理需谨慎,一些基于类维生素A的治疗方案,可能会加剧皮肤干燥和刺激,这不仅可能导致屏障功能改变,而且还会增加深色皮肤患者继发性妊娠高血压综合征的可能性。
●
温泉水已被证明对膜流动性、皮肤屏障修复、抗自由基、抗氧化、抗炎和免疫调节特性以及增殖活性和衰老和保湿过程的调节有影响。
温泉水的水微生物群中的生物活性化合物可以改善特应性皮炎或红斑痤疮等皮肤病,并改善瘙痒和干燥症;还能增加对紫外线的防护,强化屏障功能,维持皮肤防御良好的稳态,修复受损皮肤,促进伤口愈合,改善皮肤状况,减少皮肤色素不均匀,防止皮肤老化。
未来,护肤品研发人员、水文学家、温泉中心之间的合作,将推动该行业更好地了解温泉水的水生生物群落对皮肤病的作用,并考虑将这种水生生物群落的衍生物纳入皮肤病配方(以发酵罐、裂解物、提取物等形式)。
●
某些益生菌菌株及其代谢物可能带来许多益处,如:改善皮肤屏障功能、减少炎症、改善易长粉刺或易湿疹的皮肤、抗皮肤光老化等。因此,近年来益生菌已成为护肤品中的流行成分。
益生菌分为口服和外涂。
口服益生菌
肠道微生物群的变化可能会引发全身炎症和异常免疫反应,从而破坏皮肤健康。口服益生菌直接作用于肠道微生物群,帮助恢复肠道微生物群的稳态,这在皮肤稳态中发挥着至关重要的作用。
益生菌在皮肤光老化中的作用
含有约氏乳杆菌和营养类胡萝卜素的膳食补充剂的摄入对紫外线暴露的长期和反复影响有益处,并且对光老化更有针对性。
含有长双歧杆菌和低聚半乳糖的膳食补充剂由于其抗炎和抗氧化特性,保护皮肤免受UVB诱导的光老化。它们也提高了血清中短链脂肪酸和乙酸盐的水平,可以增加和激活依赖于组蛋白乙酰化的皮肤固有Treg。
口服植物乳杆菌HY7714通过抑制JNK/AP-1信号通路的激活,降低了UVB损伤细胞中过量的MMP-13转录水平和MMP-2和MMP-9的活性。
口服清酒乳杆菌可以通过阻断MAPK信号通路来抑制AP-1的表达,以增加真皮成纤维细胞中的胶原蛋白并延缓皮肤光老化。
益生菌通过多种途径对抗皮肤光老化的作用
编辑
doi: 10.2147/CCID.S388954
局部益生菌(外涂)
局部益生菌于1912年首次被提出作为皮肤疾病的治疗方法。
在特定条件下,益生菌可以持续存在并成功定殖皮肤:
虽然益生菌对皮肤健康有一定益处,但益生菌使用的安全性可能存在一些限制,主要针对免疫系统较弱的人群,如婴儿、孕妇、老年人等。需要进一步的研究来证明益生菌作为皮肤病治疗和护肤品的功效,作用机制以及主要是局部使用益生菌的安全性。
●
过氧化苯甲酰 (BPO) 治疗可以调节痤疮患者的皮肤微生物群,治疗后细菌种类的数量和多样性均减少,接近健康组。也有研究人员认为,BPO治疗虽然降低了GAGS评分并降低了微生物多样性,但它也损害了痤疮的表皮屏障,这可以被认为是一种副作用。
抗生素
针对痤疮丙酸杆菌的抗生素一直是痤疮治疗的支柱。其中,大环内酯类、克林霉素和四环素类药物的处方最为广泛。
红霉素、罗红霉素、克拉霉素和阿奇霉素是大环内酯类药物。常用于痤疮的四环素类药物是多西环素、四环素和米诺环素。
异维甲酸是一种全反式维甲酸原药,是严重顽固性痤疮患者的最终选择,它抑制皮脂生成,它可以使痤疮患者的C.acnes/TLR-2介导的先天免疫反应正常化,也就是说,异维甲酸会间接影响皮肤微生物。
其他,比如抗生素治疗会降低皮肤伤口中的细菌密度并改变细菌组成,其次是RegIIIγ表达的降低,这可能有助于延迟伤口修复。
●
几项研究表明,互惠共生对维持微生物物种之间的新陈代谢很重要。我们不仅需要关注微生物组的转移,还需要关注潜在的交叉喂养和共同居住。
有研究表明,整个原始皮肤微生物组从一个皮肤部位移植到另一个部位。研究人员将能够在腋下产生气味的细菌转移到受试者的前臂,前臂上培养的双菌群样本产生强烈的气味,这表明引起气味的细菌可以从腋下传播到前臂。
这项研究表明,通过皮肤微生物组移植重塑人类气味,降低对传染病媒介的吸引力,从而阻断病毒传播,为传染病预防和控制提供了一条新的途径。
●
尽可能选择天然纤维材料,如棉、亚麻或丝绸等,这些材料通常具有良好的透气性和吸湿性,可以帮助皮肤保持干爽。同时,天然纤维材料也更加温和,减少与皮肤的摩擦和刺激。
避免合成纤维材料:尽量减少使用合成纤维材料,如尼龙或涤纶等。
保持衣物的清洁和卫生非常重要。定期清洗和消毒衣物,避免细菌、真菌和其他有害微生物的滋生。
●
吸烟会使皮肤最外层的微小血管变窄,从而减少血液流动并使皮肤更苍白。这也耗尽了皮肤对皮肤健康很重要的氧气和营养物质。吸烟还会损害胶原蛋白,也可能会增加患鳞状细胞皮肤癌的风险。
压力会通过肠-脑-皮肤轴影响皮肤健康,皮肤细胞的代谢和分泌作用可能发生变化,导致皮肤微生物群失去原本的平衡状态,皮肤更加敏感,并引发痤疮爆发和其他皮肤问题。
长期的压力会导致免疫系统的紊乱,使皮肤更容易受到各种外界因素的侵害,从而引发炎症、过敏等症状。
睡眠可以促进皮肤细胞的修复和再生,有助于提升肤色的均匀度。同时,在深度睡眠状态下,身体会产生更多的生长激素,促进皮肤中胶原蛋白和弹力纤维的生成,有助于减少皱纹的产生。因此,保证充足的睡眠对皮肤健康也相当重要。
适度的身体运动可以促进血液循环和新陈代谢,有助于清除毒素和废物,使皮肤更加健康。选择适合自己的运动方式,如散步、跑步、瑜伽等,坚持每周进行几次。
随着年龄的增长,我们的皮肤微生物群组成发生了很大变化且可预测。未来的研究将继续阐明动态皮肤微生物组在整个生命周期中的积极作用。
皮肤微生物群扮演着重要角色,它不仅是阻止致病菌进入皮肤的屏障,还可以通过调节免疫系统减少炎症等方式对皮肤疾病起到积极作用。肠道微生物组的研究为我们探索皮肤微生物群在其他生理系统中的作用提供了途径。确定哪些微生物及其代谢产物对维持人类健康和疾病至关重要。
了解皮肤微生物群和肠道微生物群之间的相互关系可以帮助我们更好地理解肠-皮肤轴的作用机制。
未来的研究需要更深入地了解皮肤微生物组的分子基础,包括微生物-微生物相互作用、微生物-宿主-微生物相互作用、环境因素-微生物相互作用以及不同细菌菌株之间的互动对宿主健康的影响,为调控皮肤微生物群的紊乱提供更专业的治疗方法。
微生物群研究的相关应用领域
护肤品可以改变皮肤上的分子和菌群多样性及微生物在皮肤上的动力学和结构。微生物群在护肤中的应用是一种先进、前沿的方法。
服装和皮肤之间的关系已成为探索纺织品如何通过调节皮肤微生物群,来治疗或缓解皮肤疾病的新领域。
考虑到生物活性纺织品的广泛应用,需要寻找创新技术和产品。对于抗菌纺织品,根据其对纺织材料和人体皮肤微生物群的主要影响,可以在下述生物医学研究方向中找到有价值的用途:
这些应用使得生物活性纺织品更加绿色环保、安全、高性能,能够提供更多的功能性和舒适性,满足人们对健康和舒适的需求。
通过利用皮肤微生物穿透皮肤屏障的能力,或许可以开发微生物活化免疫细胞来传递细胞因子、小分子化学物质或疫苗。
微生物在皮肤上引发免疫反应,并与皮肤免疫细胞进行相互作用,调节免疫应答和免疫耐受。这为开发新的免疫调节治疗策略提供了新的思路。
通过调节肠道微生物组来靶向皮肤健康是一种很有前景的替代疗法。对于一些慢性皮肤病患者来说,肠道微生物群的紊乱可能是病情加重或难以治愈的原因之一,而通过调整饮食、生活习惯、环境等因素,同时加入益生菌或其他补充剂等,调节肠道微生物群的平衡,增强皮肤对各种病原体的抵抗力;同时通过肠道菌群调节影响自身免疫反应,改善皮肤疾病的症状。
通过皮肤、肠道微生物群联合检测可以探索和评估微生物之间的相互关系,并在皮肤问题的中提供更全面有效、个性化的治疗方案。
主要参考文献:
Townsend EC, Kalan LR. The dynamic balance of the skin microbiome across the lifespan. Biochem Soc Trans. 2023 Feb 27;51(1):71-86.
Kurokawa I, Kobayashi M, Nomura Y, Abe M, Kerob D, Dreno B. The Role and Benefits of Dermocosmetics in Acne Management in Japan. Dermatol Ther (Heidelb). 2023 Jul;13(7):1423-1433.
Harris-Tryon TA, Grice EA. Microbiota and maintenance of skin barrier function. Science. 2022 May 27;376(6596):940-945.
Belzer A, Parker ER. Climate Change, Skin Health, and Dermatologic Disease: A Guide for the Dermatologist. Am J Clin Dermatol. 2023 Jul;24(4):577-593.
Patra V, Bordag N, Clement Y, Köfeler H, Nicolas JF, Vocanson M, Ayciriex S, Wolf P. Ultraviolet exposure regulates skin metabolome based on the microbiome. Sci Rep. 2023 May 3;13(1):7207.
Skowron K, Bauza-Kaszewska J, Kraszewska Z, Wiktorczyk-Kapischke N, Grudlewska-Buda K, Kwiecińska-Piróg J, Wałecka-Zacharska E, Radtke L, Gospodarek-Komkowska E. Human Skin Microbiome: Impact of Intrinsic and Extrinsic Factors on Skin Microbiota. Microorganisms. 2021 Mar 5;9(3):543.
Trompette A, Ubags ND. Skin barrier immunology from early life to adulthood. Mucosal Immunol. 2023 Apr;16(2):194-207.
Alashkar Alhamwe B, López JF, Zhernov Y, von Strandmann EP, Karaulov A, Kolahian S, Geßner R, Renz H. Impact of local human microbiota on the allergic diseases: Organ-organ interaction. Pediatr Allergy Immunol. 2023 Jun;34(6):e13976.
Xu H, Li H. Acne, the Skin Microbiome, and Antibiotic Treatment. Am J Clin Dermatol. 2019 Jun;20(3):335-344.
Zubeldia-Varela E, Barker-Tejeda TC, Obeso D, Villaseñor A, Barber D, Pérez-Gordo M. Microbiome and Allergy: New Insights and Perspectives. J Investig Allergol Clin Immunol. 2022 Oct;32(5):327-344.
De Almeida, C.V.; Antiga, E.; Lulli, M. Oral and Topical Probiotics and Postbiotics in Skincare and Dermatological Therapy: A Concise Review. Microorganisms 2023, 11, 1420.
Flores-Balderas, X.; Peña-Peña, M.; Rada, K.M.; Alvarez-Alvarez, Y.Q.; Guzmán-Martín, C.A.; Sánchez-Gloria, J.L.; Huang, F.; Ruiz-Ojeda, D.; Morán-Ramos, S.; Springall, R.; et al. Beneficial Effects of Plant-Based Diets on Skin Health and Inflammatory Skin Diseases. Nutrients 2023, 15, 2842.
Suellen Ferro de Oliveira C, Kekhasharú Tavaria F. The impact of bioactive textiles on human skin microbiota. Eur J Pharm Biopharm. 2023 Jul;188:66-77.
Santiago-Rodriguez, T.M.; Le François, B.; Macklaim, J.M.; Doukhanine, E.; Hollister, E.B. The Skin Microbiome: Current Techniques, Challenges, and Future Directions. Microorganisms 2023, 11, 1222.
Fernandes A, Rodrigues PM, Pintado M, Tavaria FK. A systematic review of natural products for skin applications: Targeting inflammation, wound healing, and photo-aging. Phytomedicine. 2023 Jul;115:154824.
Lee, H.-J.; Kim, M. Skin Barrier Function and the Microbiome. Int. J. Mol. Sci. 2022, 23, 13071.
Ito Y, Amagai M. Dissecting skin microbiota and microenvironment for the development of therapeutic strategies. Curr Opin Microbiol. 2023 Apr 3;74:102311.
Mourelle, M.L.; Gómez, C.P.; Legido, J.L. Hydrobiome of Thermal Waters: Potential Use in Dermocosmetics. Cosmetics 2023, 10, 94
Azzimonti, B.; Ballacchino, C.; Zanetta, P.; Cucci, M.A.; Monge, C.; Grattarola, M.; Dianzani, C.; Barrera, G.; Pizzimenti, S. Microbiota, Oxidative Stress, and Skin Cancer: An Unexpected Triangle. Antioxidants 2023, 12, 546
Leung MHY, Tong X, Shen Z, Du S, Bastien P, Appenzeller BMR, Betts RJ, Mezzache S, Bourokba N, Cavusoglu N, Aguilar L, Misra N, Clavaud C, Lee PKH. Skin microbiome differentiates into distinct cutotypes with unique metabolic functions upon exposure to polycyclic aromatic hydrocarbons. Microbiome. 2023 Jun 1;11(1):124.
Kengmo Tchoupa A, Kretschmer D, Schittek B, Peschel A. The epidermal lipid barrier in microbiome-skin interaction. Trends Microbiol. 2023 Jul;31(7):723-734.

谷禾健康

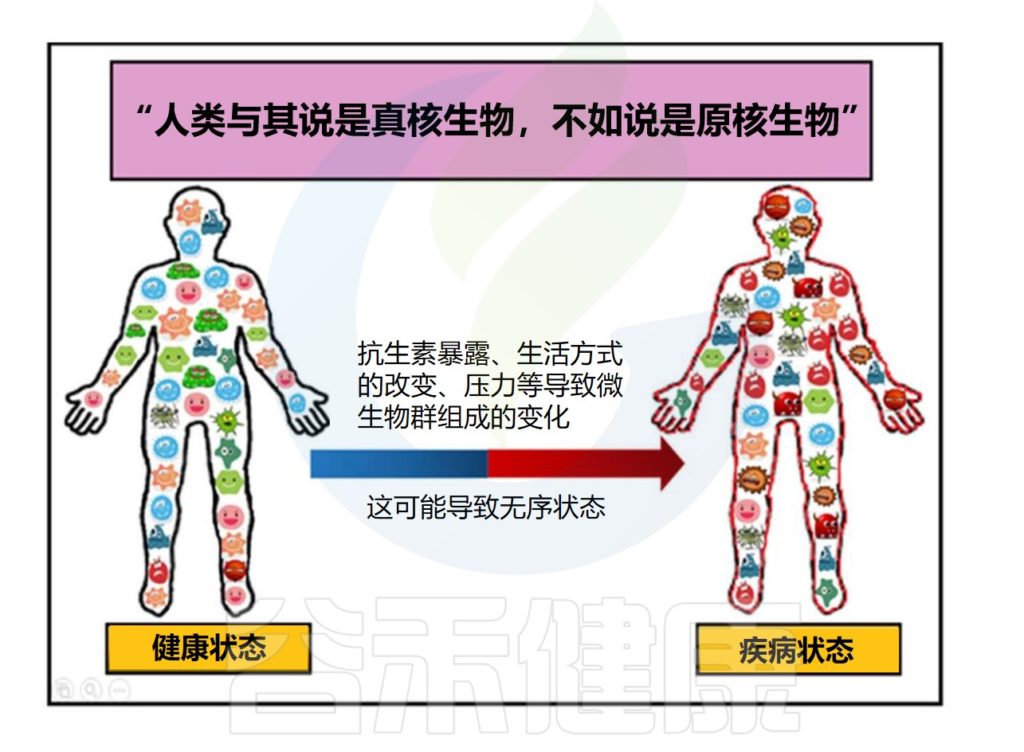
人类微生物组包含约 1000多 个常见物种,微生物细胞数量达 10-100 万亿个。微生物大多数遗传库分布在人类胃肠道内,与许多生理过程的发育和功能有内在联系,例如,肠道屏障完整性和稳态、营养、免疫和神经心理行为等。
人类肠道的菌群失调(即共生肠道微生物群的变化对宿主健康产生影响)通常与一系列疾病相关,其中许多疾病无法使用完善的医疗方案进行治疗。
在这种情况下,将健康捐赠者的粪便物质直接移植到受体的胃肠道中(称为粪便微生物群移植,简称粪菌移植,FMT)代表了一种操纵受体微生物群并赋予健康益处的治疗方法。许多国家食品和药物管理局最近的批准表明,粪菌移植和治疗性益生菌鸡尾酒疗法目前正在扩大研究和应用领域。
这种方法成功的关键是能够在患病患者的胃肠道内重建并将肠道微生物群恢复到理想的健康状态。据报道,粪菌移植在临床中治疗艰难梭菌感染成功率接近 90%。
最近发表了几项通过粪便微生物移植(FMT)来治疗炎症性肠病(IBD),特别是溃疡性结肠炎的随机临床试验,但是研究设计存在重大差异。这些包括给药剂量、给药途径和频率、安慰剂类型和评估终点的差异。总体结果看起来很有前景,但它们高度依赖于捐赠者和接受者因素。
近日,来自意大利、法国、英国、美国等 25 名IBD、免疫学和微生物学领域专家多次召开会议,通过对当前可用和/或已发布的数据进行深入评估来制定基于IBD 中与 FMT 相关证据的指南。所有成员对声明进行评估和投票,最终形成全体共识会议并生成拟议指南。
声明包括:
这份共识目的是为使用 FMT 评估、管理和潜在治疗 IBD 制定基于共识的声明和建议,以实现标准化实践。
今天谷禾与大家分享该共识或指南内容,并结合以往研究结果和相关经验阐述说明。我们也相信,在临床研究中明确考虑供体和受体的原有肠道微生物生态原则可以更好地指导FMT试验的设计并有助于提高 FMT 的疗效。
★
生态学是一门核心学科,研究集中构建生物群落的过程和机制,并调节其在空间和时间上的变化。
虽然说,生态学和微生物学在研究复杂群落的方向经常会有交叉部分,但两者在历史上都是作为独立的科学学科发展起来的,在概念、术语、综合和理论的使用上往往有所不同。
临床研究中的 FMT,个体之间粪便物质的移植符合社区合并的概念。这个概念指的是:
整个生物群落及其环境的大规模混合
虽然在宏观生物生态学(即植物和动物)中构成罕见事件,但群落合并在微生物系统中很常见,实际上,群落经常作为一个单元迁移。
这一原则与 FMT 治疗相一致,尽管该术语似乎尚未在临床研究中得到很好的确立。然而,合并后群落重新排列的结果(即植入)更常用于跟踪 FMT 功效。
基于来自供体、FMT前、FMT后接受者的粪便微生物组的微生物菌群监测,才能更好的确定与较高供体菌株植入相关的最重要的临床变量,通过不同的上消化道和下消化道引入的混合途径(即通过胶囊、肠镜检查、鼻胃管、鼻十二指肠管或上内窥镜检查和结肠镜检查),单独或结合进行 FMT 给药,会导致 FMT 后供体菌株植入率更高。
这一发现支持了繁殖压力(即引入的个体数量与独立引入的数量的组合)对于在新环境中成功引入非本地物种的重要性。
发生这种情况主要是因为:
i)初始微生物物种种群的大小可以直接影响其在非本地环境中持续存在的能力
ii)即使是短暂的入侵者也会对微生物组产生影响,有时会促进其在后续环境中的引入入侵尝试(例如,重复入侵尝试的重要性)
其次,在 FMT 给药前接受抗生素的患者中,可能供体菌株植入率更高。已知抗生素的使用会导致肠道微生物群失衡(即非生物紊乱)。这种干扰降低了受体胃肠道微生物群的生物抗性,有利于随后的供体群落植入。重要的是,这个个别研究结果,在推广这一原则时需要谨慎,因为 FMT 之前的抗生素剂量、类型和时间都会导致与 FMT 最终结果的差异。
第三,患有传染病的患者在接受单途径 FMT 给药后,微生物组植入量更大。这与以下观点一致:驻留微生物组的先前不稳定(即,在这种情况下,生物干扰)也会对肠道生物抵抗力产生负面影响。特别是,胃肠道中非本地物种(例如感染期间的致病菌)的存在可以促进其他非本地物种的后续入侵。
★
粪便微生物群移植(FMT)被定义为将健康捐赠者的粪便输注到受体的胃肠道中,以治疗与疾病相关的肠道菌群失调。
根据几项随机对照试验和荟萃分析的报告,FMT是治疗复发性艰难梭菌感染的一种既定且高效的治疗选择,最终制定了国际指南,以标准化其使用艰难梭菌感染的可行治疗方式。
继成功治疗艰难梭菌感染后,FMT 也在炎症性肠病 (IBD) 患者中进行了研究,首先是在非随机研究中,随后在随机对照试验中,尽管 FMT 方案和程序存在显着差异,但两者均显示出有希望的结果。
然而,采用 FMT 治疗 IBD 受到一些限制,包括招募捐赠者、准备粪便材料、确定最佳给药途径以及缺乏明确和既定的监管框架。
解决这些问题的潜在策略包括识别和使用可持续、可重复和标准化的方案,最终目标是改变肠道微生物组的组成。因此,建立最佳的FMT整体框架对于IBD的未来管理具有重要意义。
▼
共识过程是按照以下步骤制定的:
确定成员
确定了 25 名共识成员,他们在微生物学、免疫学、FMT 和 IBD 领域拥有公认的专业知识,并全部参加了专家小组。
注:德尔菲法是在20世纪40年代由O.赫尔姆和N.达尔克首创,经过T.J.戈尔登和兰德公司进一步发展而成的。
德尔菲法作为一种主观、定性的方法,不仅可以用于预测领域,而且可以广泛应用于各种评价指标体系的建立和具体指标的确定过程。
分工作小组
根据个人专业知识,每位成员被分配到四个工作组之一:
每个工作组提出了一份关键问题清单,并制定了与指定主题相关的声明。对于每个关键问题,最佳现有证据是通过对相关文献进行系统审查而获得的。声明以专家意见 (EO) 的形式发布。
详细陈述已上传至在线投票系统(http://scott.armstrong.delphi.stlouisintegration.com/delphi2/),并分发给专家组。
专家评分
每轮审查后,都会收集、处理并与专家小组分享专家的答复。
对于每项陈述,专家们被要求对他们的同意程度进行评分:
如果至少 80% 的受访者对每项陈述表示强烈同意或同意保留意见,则达成共识。
未通过此门槛的声明将在后续轮次投票中进行修改和再次评级,直至达成共识。小组专家于 2022 年 6 月 25 日齐聚罗马,对总体声明进行完善和最终批准。
最终达成共识
经过三轮投票,最终的累积声明达成共识。第一轮和第二轮后,分别有67%和79%的陈述通过了80%的同意门槛,而第三轮后100%的陈述达到了目标水平。
★
声明 A1
IBD 的确切病因目前尚不清楚;然而,其发病机制是多因素的,受遗传易感性、宿主粘膜免疫反应和环境(包括饮食和肠道微生物群)的影响。
评论:
IBD,如克罗恩病 (CD) 和溃疡性结肠炎 (UC),是一种慢性、复发性消化道炎症性疾病,是由于遗传易感个体肠道免疫系统和肠道微生物群之间失去稳态而导致的。由于对肠道微生物群的耐受性失调或将微生物与底层组织分开的上皮屏障破坏,导致不适当的粘膜免疫反应,可能会导致 IBD 的发展或持续。
声明 A2
肠道微生物群的组成、相对丰度、多样性和功能的改变(即菌群失调)促进IBD的发生和进展。
评论:
越来越多的证据表明,肠道微生物组组成的不平衡或“生态失调”是促进 IBD 发展影响最大的环境因素之一,因为改变的微生物群与宿主的相互作用可以触发和促进免疫与 IBD 相关的改变。大量证据表明,IBD 患者肠道微生物组存在特定的共同改变,这些改变与许多功能受损相关,包括短链脂肪酸代谢、氨基酸生物合成、氧化应激调节和毒素,可以单独或共同促进 IBD 的发展。
声明 A3
溃疡性结肠炎和克罗恩病患者的肠道微生物组特别缺乏普拉梭菌,这种细菌因其潜在的抗炎特性而被认可。
关于普拉梭菌详见:
肠道核心菌属——普拉梭菌(F. Prausnitzii),预防炎症的下一代益生菌
评论:
越来越多的临床和实验数据表明,共生微生物群在维持人类和实验性 IBD 的炎症过程中发挥着关键作用。
一些报告显示,IBD 期间微生物多样性下降,尤其是厚壁菌门和拟杆菌门。有趣的是,厚壁菌门的成员普拉梭菌显着减少,并且具有公认的抗炎活性。
相反,活动性 IBD 中的变形菌和放线菌通常升高,以及特定的大肠杆菌菌株也如此。在这种情况下,肠道微生物组的成分在炎症性肠病中发挥着至关重要的作用,这可能代表了一种与细菌加工相关的疾病。因此,肠道微生物组和宿主免疫反应之间存在功能失调的关系,从而引发和维持 IBD 的慢性炎症。
声明 A4
IBD 患者感染艰难梭菌的风险高于一般人群。
评论:
IBD 特异性危险因素,例如免疫抑制、炎症的严重程度和扩展以及 IBD 中观察到的肠道菌群失调,被认为是IBD 患者艰难梭菌感染高风险的主要原因。
IBD 患者并发艰难梭菌感染会导致住院时间延长以及结肠切除率和死亡率增加。鉴于这种情况可能会危及生命的并发症,建议在 IBD 发作期间和早期治疗干预期间筛查艰难梭菌。
艰难梭菌的选择较高毒力、抗生素耐药性和反复感染率增加的菌株使得IBD 中艰难梭菌感染的治疗更具挑战性。因此,建议采用个体化治疗方法来控制IBD 发作期间的艰难梭菌感染。
★
声明 B1
适合 IBD 实验性 FMT 的供体应接受血液和粪便检测,测试符合目前通过 FMT 治疗艰难梭菌感染的国家和国际指南,并且一般用于临床实践。
评论:
以 FMT 为目的的供体筛查的主要原则是避免潜在的传染病传播。用于筛查艰难梭菌感染 FMT 的指定血液和粪便参数已在溃疡性结肠炎患者的多项随机对照试验中被证明是安全的。应提供包含待测试的强制性国际参数的列表,而其他参数则应取决于地理区域(例如,热带地区)、患者的医疗状况或捐赠者的病史(例如,粪便钙卫蛋白增加史)。
除了血液和粪便检测外,还应通过多项调查问卷监测总体健康状况、饮食和心理状态,以避免任何潜在的非传染性不良事件。
声明 B2
粪便捐赠应该是自愿的,并且应告知捐赠者捐赠的潜在风险和/或好处。此外,每位患者必须提供书面知情同意书。
评论:
由于粪便采集程序是非侵入性的,并且可以在不受控制的环境中进行,因此不应允许捐赠者直接从粪便捐赠中受益,以避免样本欺诈。然而,根据国家规定,捐赠者可以获得时间和旅行费用的补偿。此外,捐赠者应了解捐赠的风险和益处,因为筛查过程可能会发现以前未知的疾病(例如艾滋病毒、结直肠癌)的诊断结果或其他疾病的易感性(例如与微生物群改变相关的疾病) 。此外,捐赠者应该知道他们可以随时撤回同意。
声明 B3
可以通过粪便库对供体进行管理,以供实验使用,符合适用于艰难梭菌感染的国家和国际指南和法规,并且一般用于临床实践。
评论:
粪便库能够以标准化方式处理粪便捐赠,这适合 IBD 的进一步临床和实验程序。在粪便库中,捐献者在给患者施用粪便之前经过严格筛选,FMT 制备是标准化的,与临床环境相比,制备 FMT 样本所需的成本和时间都可能减少。此外,粪便库拥有专业知识,因此总体上可能有助于优化 FMT 样本的质量。最后,粪便库可以为无法提供此项服务的 IBD 中心提供 FMT 样本。
声明 B4
捐赠者的粪便最好在粪便库现场或进行实验程序的地点收集,遵循国家和国际指南和法规。
评论:
捐赠者应收到有关如何收集粪便的明确说明,最好是在现场或粪便库收集。如果无法做到这一点,收集的粪便应储存在 4°C 下,并在收集后 6 小时内运送到临床地点或粪便库,由经过培训的人员进行处理。
声明 B5
根据实验方案,每个捐赠者都可以参与捐赠不同的 FMT 制剂。
评论:
供体粪便可用于制备不同的 FMT 制剂(例如,新鲜样品与冷冻样品,以及每个制剂的单个供体与多个供体)。到目前为止,建议使用冷冻 FMT 样品而不是新鲜制剂,主要是因为安全性。冷冻 FMT 样本可以被隔离,直到捐赠期结束时完成全面的捐赠者筛查。对于新鲜的 FMT,在完成完整的筛选过程之前对材料进行管理。因此,对于新鲜的 FMT,建议进行更定期的供体常规筛查和菌群评估。
声明 B6
应根据国家和国际准则和法规维护和存储捐助者信息登记册。
评论:
与 FMT 流程相关的捐赠者信息登记应由国家和国际医疗保健机构监管。来自捐赠者和接受者的信息应保存至少 10 年,或符合国家和国际法规。这些数据应提供给粪便库以获得长期安全数据。
声明 B7
患者不应直接使用粪便库来治疗 IBD。FMT 样本的提供应始终在治疗医疗保健提供者的指导下进行,并符合国家和国际指南和法规。
评论:
粪便库的使用应仅限于医疗保健提供者,因为 FMT 管理需要记录和适当的后续行动,以应对任何潜在的不良事件,而这些不良事件只能在医生的监督下安全地发生。因此,患者直接使用粪便库是不合适的。
声明 B8
FMT 的整个过程(从捐赠者筛选到粪便收集和 FMT 样本管理)应具有清晰的可追溯性。因此,应保留每个 FMT 样品的等分试样以进行测试,以防发生意外的不良事件。
评论:
FMT 过程的所有步骤都应进行登记,捐赠者样本的等分试样应保留并储存在 -80°C 下,以便在发生任何意外不良事件(例如病原体感染)时进行回溯。
据报道,FMT 后出现耐药大肠杆菌传播,这表明保留供体粪便以便在可能的不良事件发生后进行进一步分析的重要性,并保证在此类情况下及时干预。
声明 B9
关于供体 FMT 制备技术方面的共同协议,将有助于在全球范围内提供程序标准化和优化,从而促进结果的解释。
评论:
尽管研究设计不同,FMT 已显示出有希望的结果,尤其是在溃疡性结肠炎中。方案的变化包括不同的 FMT 输注途径(例如,鼻十二指肠、直肠、口服)、给药频率、对照安慰剂(例如,水、自体粪便材料)和终点测量。关于冷冻粪便材料制备、储存和给药量的剂量标准化将有助于未来 FMT 研究的解释和比较,包括结果。
声明 B10
需要进行研究,来确定与粪菌移植作为治疗炎症性肠病选择的临床疗效和整体治疗效果更好相关联的供体特征。
评论:
必须进行研究来确定供体标志物,以实现 FMT 在 IBD 中的最佳治疗效果和总体成功。因此,应该对微生物群、饮食模式(问卷)和其他方面(如药物使用、家族病史、心理状态和遗传背景)进行表征,以确定改善临床结果的潜在趋势。除了特定的供体标记外,还应该研究供体-受体的植入情况。
★
声明 C1
以前进行的随机对照试验一般规模较小,且方法学上存在异质性;因此,目前还不能得出明确的结论。
评论:
在成功用于治疗艰难梭菌感染后,FMT 也在溃疡性结肠炎患者中进行了研究,首先是在非随机研究中,然后在随机对照试验中,均取得了有希望的结果,尽管存在FMT 程序和测量结果存在显着差异。事实上,尽管这些研究和随后的荟萃分析强调供体 FMT 施用后令人满意的缓解率,已发表和/或可用的随机临床试验通常规模较小且方法学上异质,因为它们在粪便输注的时间、数量和途径、FMT 供体粪便与对照(假冒)的特征方面存在差异,使得结果难以从整体上解释结果,无法得出明确的结论。
声明 C2
对于患有IBD的轻度和重度复发或难治性难免性艰难梭菌感染,建议采用FMT作为治疗选择。
评论:
FMT 对于治疗无 IBD 患者的复发性艰难梭菌感染有效, 以及患有溃疡性结肠炎和克罗恩病的患者。然而,针对溃疡性结肠炎患者的研究报告称,单剂量 FMT 无法预防溃疡性结肠炎发作。没有足够的证据推荐 FMT 作为IBD首次艰难梭菌感染的治疗方法。
声明C3
FMT 可有效诱导轻度至中度溃疡性结肠炎的缓解;然而,没有足够的证据推荐 FMT 作为常规临床实践中溃疡性结肠炎的治疗方法,其使用通常应仅限于研究环境。
评论:
迄今为止,FMT 在诱导轻度至中度溃疡性结肠炎患者缓解方面显示出有希望的结果。然而,这些研究是在溃疡性结肠炎患者队列中进行的,样本量相对较小,而且研究设计之间存在差异,使得研究之间的比较难以协调。因此,没有足够的数据支持临床常规使用 FMT 来诱导溃疡性结肠炎患者缓解。但专家们一致认为,FMT可在特定情况下使用,应具体情况具体考虑,并与有关各方详细讨论。
声明C4
随机对照试验表明,在 FMT 后获得缓解的溃疡性结肠炎患者通常在 FMT 治疗后不会持续缓解超过 1 年。
评论:
溃疡性结肠炎中的 FMT 研究缺乏有关治疗效果和持久性的长期随访数据。因此,需要对治疗后的时间进行更持续的疾病监测随访。现有数据表明,复发是可能的,为了实现长期疗效,维持治疗可能是强制性的。然而,诱导治疗和维持治疗的输注次数和剂量均应进一步研究。
声明C5
反复输注和供体-受体移植可能对于 FMT 治疗溃疡性结肠炎的成功非常重要。
评论:
来自对溃疡性结肠炎进行 FMT 的现有随机对照试验的数据表明,重复输注很重要;然而,目前对于 FMT 成功所需的最少管理次数尚未达成共识。此外,FMT 在溃疡性结肠炎中的疗效似乎也依赖于受体,表明供体-受体移植至关重要。除了鉴定对 FMT 成功可能重要的供体标记外,还应研究受体标记及其与供体粪便物质/粪便抗原相互作用的重要性。
声明C6
FMT 后肠道微生物组组成多样性的增加,可能是 溃疡性结肠炎反应的标志。
评论:
除了临床和内镜缓解等临床结果外,还应评估微生物标志物,包括微生物组多样性的改变(即增加),并将其与未来研究中 FMT 的成功或失败相关联。这些标记物可用于预测患者的反应,这有可能实现精准医疗方法的个性化治疗。
声明C7
现有数据表明,FMT 对于轻度至中度溃疡性结肠炎诱导缓解的风险较低;然而,在使用 FMT 治疗 IBD 时已有严重不良事件的报道,包括疾病恶化。
评论:
已进行的随机对照试验中对患者安全性的评估显示出良好的结果,不良事件非常有限,其中大部分与给药方式有关。一般来说,鼻十二指肠给药比直肠给药灌肠产生的不良事件相对较多;因此,后者已被普遍认为是更安全的递送途径。然而,迄今为止,尚缺乏溃疡性结肠炎患者的长期 FMT 安全性数据。
事实上,在随机对照试验中,在有限数量的患者(<10%)中观察到了严重的不良事件。这些包括当通过上消化道给药进行 FMT 时误吸和疑似小肠穿孔。与 FMT 手术相关的最常见的严重不良事件是疾病恶化,需要住院治疗,在少数情况下需要结肠切除术。
在 Costello等人的试验中,观察到 43 名接受自体 FMT 的患者中有 2 名病情恶化,而 38 名接受捐赠者 FMT 的患者中有 1 名病情恶化。
在 Moayyedi 的试验中,38 名患者中有 2 名出现斑片状炎症和脓肿。其他显着的严重不良事件包括肺炎、艰难梭菌感染或其他形式的小肠结肠炎。
声明C8
随机对照试验尚未证明 FMT 和对照组在疾病恶化或轻微或严重不良事件引起的症状方面存在显着差异。
评论:
溃疡性结肠炎中的 FMT 试验对不良事件进行了评论;然而,由于对照组和治疗组之间没有发现显着差异,因此这些不良事件被认为是给药程序的结果,而不是加工后的供体粪便材料本身。
声明 C9
溃疡性结肠炎中 FMT 后,常见的不良事件是短暂的轻微胃肠道症状,如腹胀、腹泻和肠胃气胀。
评论:
FMT 试验中高达 83% 的患者观察到轻微不良事件,包括胃肠道症状,如短暂性腹泻、肠鸣、腹痛、腹胀和肠胃气胀。也有短暂发烧的报道。大多数不良事件在手术后几天内自然消失。
声明 C10
在临床实践中,没有足够的证据推荐 FMT 作为克罗恩病的治疗方法。迄今为止,其使用应仅限于研究环境。
评论:
克罗恩病中 FMT 的可用数据非常有限,主要包括病例报告和试点研究,而不是大型随机对照试验。这些研究显示不良事件,主要包括胃肠道症状,疾病发作报告为与 FMT 相关的严重不良事件。
Vermeire等人的一项初步研究显示,六名难治性 克罗恩病患者在 FMT 后第 8 周没有差异。
在评估 FMT 在维持克罗恩病缓解方面的效果时,Sokol等人报告称,与假手术组相比,FMT 组的急性发作发生率没有显着降低。
进一步的研究应包括优化该患者群体的缓解诱导和维持。大型随机对照试验必须推荐 FMT 作为克罗恩病患者的可行治疗方法。
声明 C11
在临床实践中,没有足够的证据推荐 FMT 作为贮袋炎(Pouchitis)的治疗方法。迄今为止,其使用应仅限于研究环境。
注:贮袋炎(Pouchitis)是保留肛门的大肠全切除术(或次全切除术)术后发生在患者回肠贮袋的非特异性炎症,是溃疡性结肠炎行回肠贮袋-肛管吻合术后最为常见的并发症。
本病病因不明。大约有1/3的患者在术后5年内可能出现这种难以解释的炎症,其中2/3的患者有复发的可能性。
评论:
据信,患有贮袋炎的患者也会出现菌群失调。目前,只有有限的研究使用 FMT 来治疗贮袋炎。然而,在已发表的文献和病例报告中,该手术大多被报道是安全的,但并不有效。
与溃疡性结肠炎中 FMT 的随机对照试验类似,所进行的研究也具有异质性,并评估了不同的结果测量。因此,需要进一步研究和优化方案来确定 FMT 在贮袋炎中的潜在用途。
声明 C12
关于 FMT 在克罗恩病和贮袋炎中的安全性数据不足。
评论:
基于缺乏大型随机对照试验和长期随访数据,目前无法得出FMT治疗克罗恩病和储袋炎的安全性结论。
声明 C13
需要进一步的研究来确定 FMT 在克罗恩病和贮袋炎中的有效性和安全性。
评论:
需要对克罗恩病和储袋炎的 FMT 进行进一步研究和优化,以评估疗效并生成(长期)安全性数据,以便将 FMT 引入临床环境。
★
展望D1
未来的研究需要确定精准的 FMT 供体和受体用于治疗 IBD 的最佳特征。
评论:
FMT 已被证明是一种有前景的溃疡性结肠炎治疗策略。然而,有效率似乎受到供体特异性、受体特异性和手术特异性特征的影响。此外,供体-患者移植获得了更多支持,有利于确定通用的“超级供体”。因此,需要进一步的研究来定义理想的患者和捐赠者特征及其最佳移植。
展望D2
有必要进行对照FMT试验,以优化IBD特定表型的疗效。
评论:
未来的研究在考虑 IBD 中 FMT 的结果时应考虑严格定义的患者表型。这些研究有可能确定 FMT 给药后与阳性反应或缺乏反应相关的特定表型。
展望D3
需要确定预测 IBD 中 FMT 反应的生物标志物。
评论:
除了确定供体和受体以及患者表型可能的最佳特征外,还需要确定免疫学和微生物生物标志物来预测对治疗的反应,以避免因 FMT 治疗无反应而造成时间、成本和不良事件的损失。
此类生物标志物可以通过 16S rRNA 扩增子测序、鸟枪法宏基因组测序、蛋白质组学和/或转录组分析来检测。通过识别此类生物标志物,有可能为 IBD 的 FMT 治疗设计更有针对性的策略,从而朝着更加精确、个性化的医疗方法发展。
展望D4
未来的研究需要确定基于 FMT 的 IBD 治疗的最佳配方和给药途径。
评论:
提高 FMT 功效的最佳剂量尚未确定。迄今为止,鼻十二指肠管输送以及通过结肠镜检查或直肠灌肠进行 FMT 输注已成为 IBD 研究最多的给药途径。
然而,最近的一项研究利用含有冻干粪便微生物群的口服胶囊作为其递送系统。这种给药途径侵入性较小,因此可能更受患者青睐。需要更多的随机对照试验来优化剂量和给药途径。
展望D5
需要进行研究来积累有关使用补充策略来提高 FMT 疗效的循证信息。
评论:
通过 FMT 调节肠道微生物组成的成功率可以通过补充策略来提高,例如捐赠者和接受者的支持性抗炎饮食, 通过优化FMT前的肠道准备,也可以通过抗生素预处理。如益生菌、益生元、合生元和后生元补充。
扩展阅读:
展望 D6
需要进行研究来评估 FMT 作为 IBD 的独立治疗方法或与当前可用的治疗方式相结合的作用。
评论:
除了评估补充策略(D5)之外,还需要进一步研究 FMT 与目前使用的 IBD 联合疗法(从皮质类固醇到生物制剂和 JAK 抑制剂)的组合。
这种针对免疫反应和肠道微生物组成的组合方法可能比单独的每种策略带来更高的缓解率。此外,应研究预防发病和/或术后复发,以及 FMT 对癌症治疗的影响,反之亦然。
★
本手稿的作者代表 IBD 各个方面的国际专家组,他们一致认为,在将 FMT 推广为公认的 IBD 治疗策略之前,有必要进行进一步的研究。人们普遍认为 FMT对于 IBD 患者是安全的,特别是对于溃疡性结肠炎患者。
文献中报道的大多数并发症主要与粪便输注的给药途径有关,而不是与感染传播有关。然而,为了避免不良事件的负担,应遵循针对艰难梭菌感染的 FMT 治疗现有的国际指南,来严格筛选捐赠者。
IBD 中的粪便微生物群移植 (FMT) 途径:从健康供体到恢复疾病患者的肠道微生物群
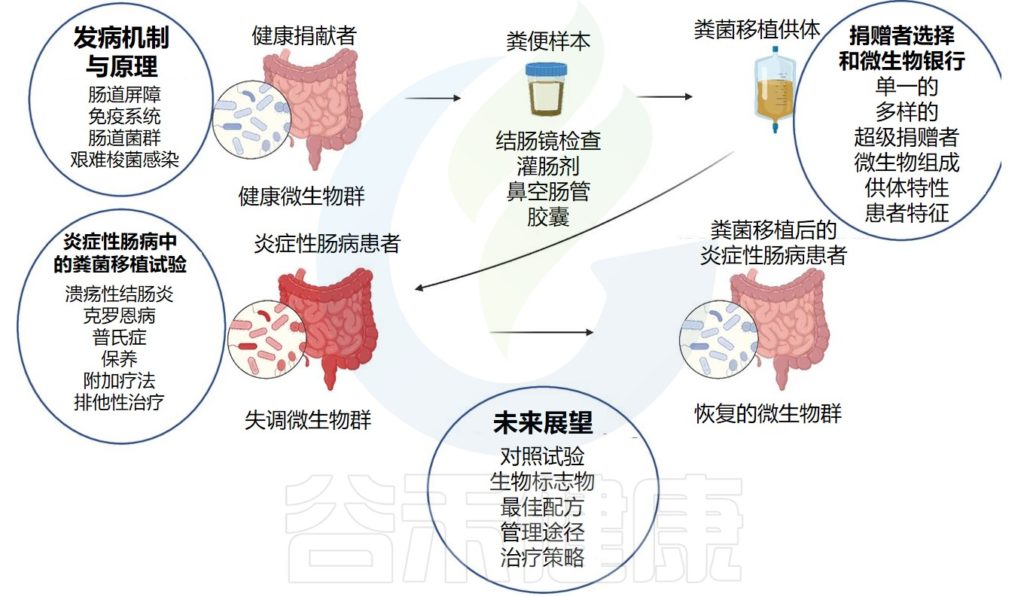
doi: 10.1136/gutjnl-2023-329948.
应投入粪便库以促进FMT研究,并应考虑通过实施永久捐赠者登记、通过微生物群特征和捐赠者健康状况筛选捐赠者粪便材料以及优化捐赠者粪便材料、粪便样本的储存,来治疗IBD的可能性。
粪便库应登记所有捐献者和患者数据,以便有效追踪并监测 FMT 施用后健康状况的变化(例如缓解/发作、心理状态)。
已经进行了几项 FMT 试点研究和随机对照试验来治疗 IBD,但使用的是异质研究设计。现有的结果,特别是在溃疡性结肠炎方面,是有希望的,但似乎依赖于供体和患者。然而,要将 FMT 纳入日常胃肠道实践中,还需要进一步研究来优化短期和长期成功率,并进一步评估安全性。
该方法应确定:最佳给药途径、剂量、频率、供体-受体植入、患者表型,以及 FMT 反应的免疫学和微生物组生物标志物的识别。
总而言之,这样将有助于 FMT 的标准化及其治疗溃疡性结肠炎的临床应用。对于克罗恩病和储袋炎,必须进行进一步的研究来评估其使用的(长期)安全性和有效性。尽管如此,这一系列研究可能会从优化其在溃疡性结肠炎中的使用所采取的措施中受益匪浅。
未来的工作包括对供体微生物群进行严格表征,以及调查 FMT 前后对 IBD 接受者的影响,这可用于最大限度地提高 FMT 功效并阐明作用机制。
通过研究供者和受者的支持性饮食、肠道准备、抗生素预处理、益生菌、益生元、合生元和生元后支持,以及与 IBD 治疗同时进行的联合治疗,进一步提高 FMT 的疗效至关重要。此外,识别与 FMT 成功预测相关的特定微生物菌株,可能会开发出明确的单菌株或多菌株益生菌。
参考文献:
Dini-Andreote F, Custer GF. Ecological principles of fecal microbiota transplantation. Trends Microbiol. 2023 Jun 8:S0966-842X(23)00162-2.
Lopetuso LR, Deleu S, Godny L, Petito V, Puca P, Facciotti F, Sokol H, Ianiro G, Masucci L, Abreu M, Dotan I, Costello SP, Hart A, Iqbal TH, Paramsothy S, Sanguinetti M, Danese S, Tilg H, Cominelli F, Pizarro TT, Armuzzi A, Cammarota G, Gasbarrini A, Vermeire S, Scaldaferri F. The first international Rome consensus conference on gut microbiota and faecal microbiota transplantation in inflammatory bowel disease. Gut. 2023 Jun 20:gutjnl-2023-329948.

谷禾健康

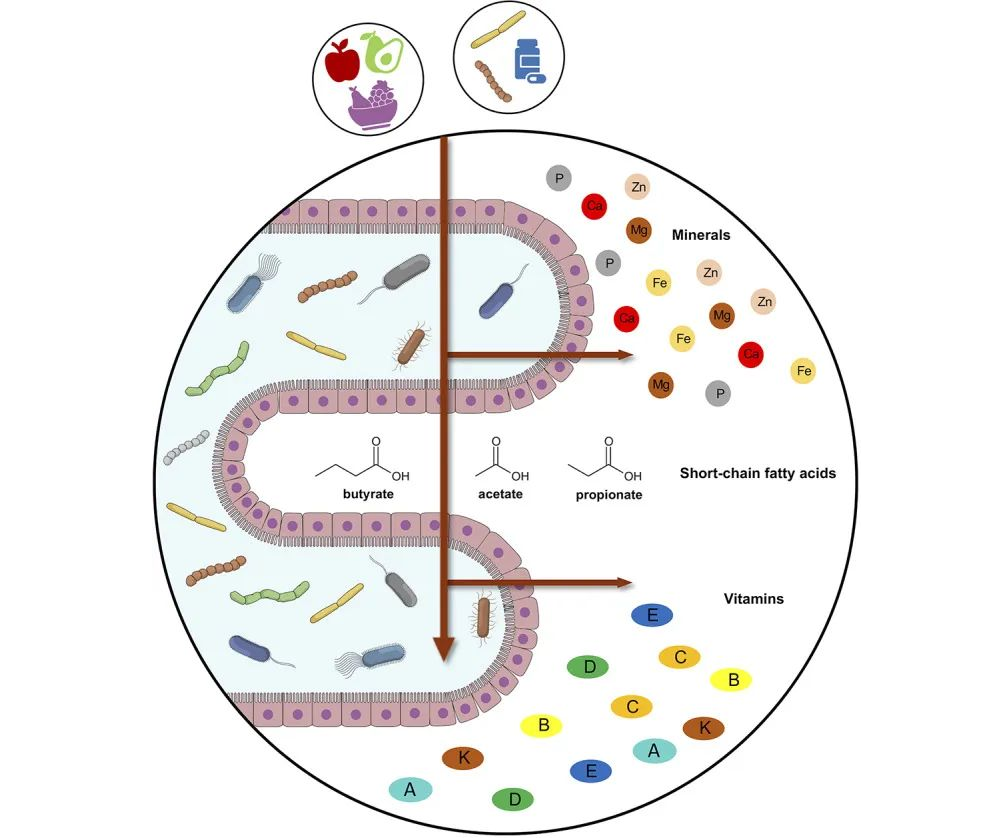
我们知道,人体的皮肤、口腔、肺部、肠道、阴道等都是微生物的栖息地,每个部位都有独特的微生物群组成。微生物群受到基因、饮食、环境和生活方式等多种因素的影响。
当然,人体微生物群的组成也会随着年龄的增长而发生变化。从婴儿期到老年阶段,微生物群的种类和数量都会发生变化,这些微生物与人体形成了错综复杂的共生关系,这对我们的健康和免疫系统功能产生重要影响。
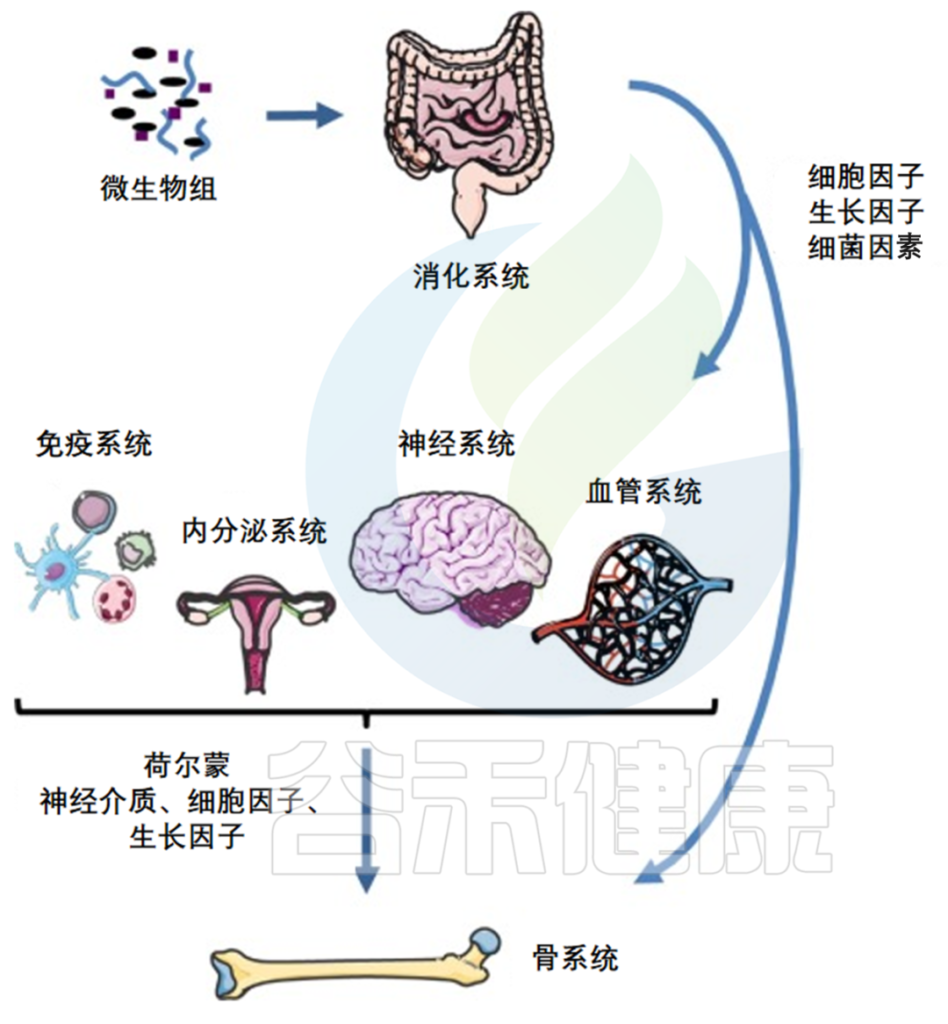
人体微生物群与发育、免疫、营养、神经、代谢稳态等方面有密切关联。反过来,宿主也会提供营养并促进健康和有弹性的微生物群的发展。
了解以上这些可以帮助我们更好地理解微生物与人类的共生关系,拓展对微生物多样性和生态系统的认知,为预防和治疗相关疾病提供新的思路。
本文讲述了不同年龄和不同部位的人体微生物群、影响微生物组成的各种因素、微生物与宿主的相互作用(包括对生理、疾病的影响及相关治疗中的作用)、及其对于健康管理和疾病治疗的价值。
目录
•人体不同部位的微生物群
•不同年龄下的微生物群
•影响微生物组成的因素
•微生物群对宿主生理的作用
•微生物群与疾病的关联
•微生物群与医学治疗
•结语
不同环境提供了不同的营养资源、温度、湿度、pH值等生态因素,这些因素会影响各种微生物的生存和繁殖。
多样化的全球微生物数据集
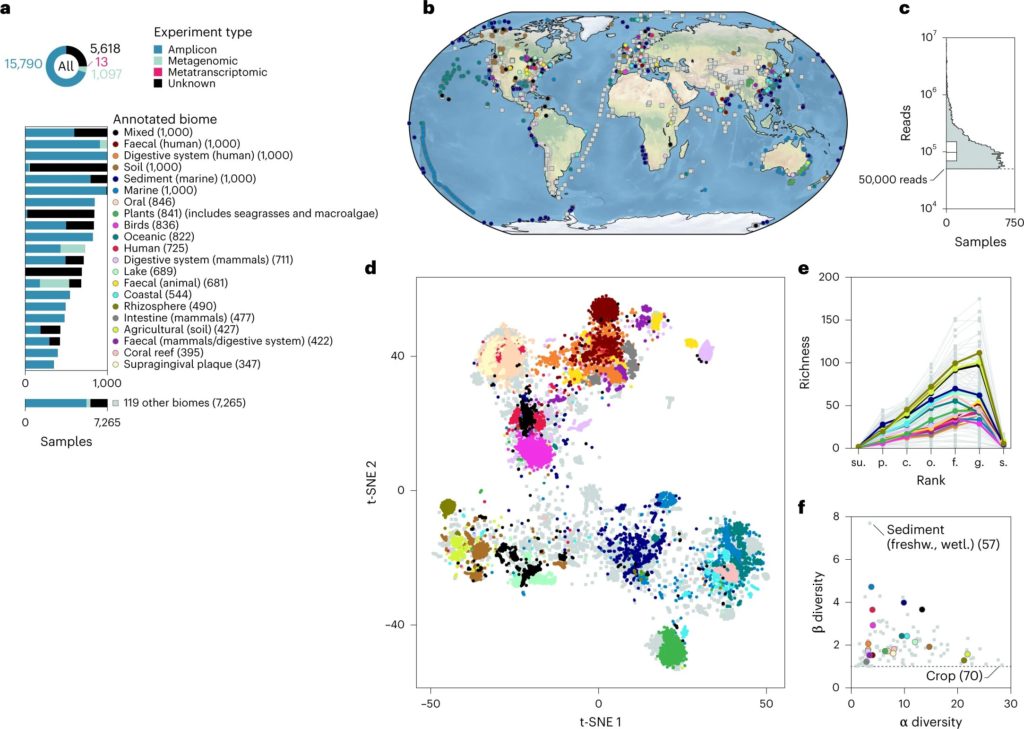
Hogeweg P,et al.Nat Ecol Evol.2023
a:样本来自截然不同的带注释的生物群落和研究设计;
b:样本的地理分布;
c:每个样本的分类注释读数总数(n = 22,518 个样本);
d:来自相似注释生物群落的样本根据 t-SNE 可视化中的分类概况(困惑度 = 500)聚集在一起;
e:类群丰富度因注释的生物群落和分类等级而异
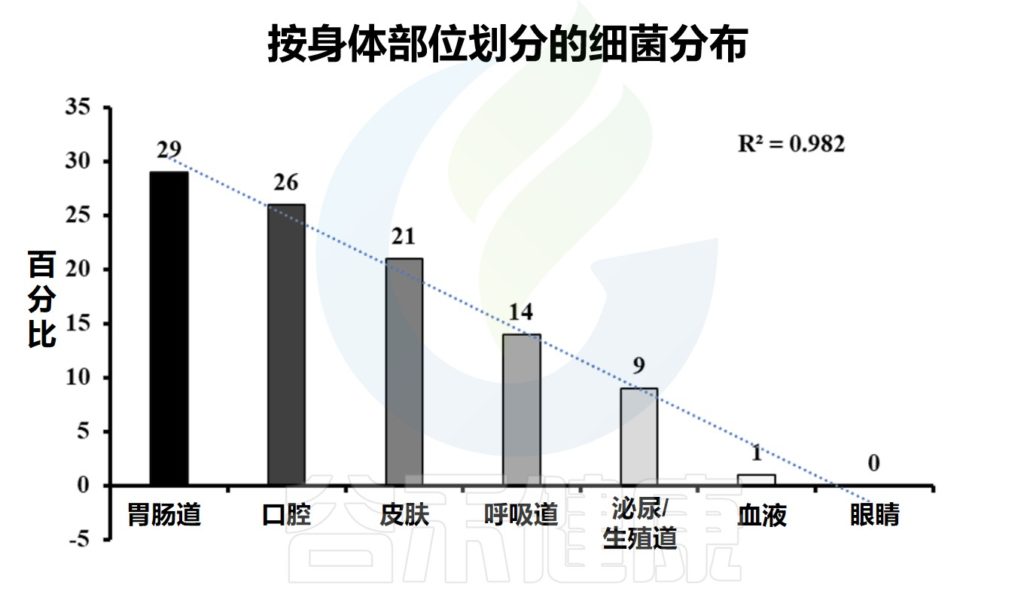
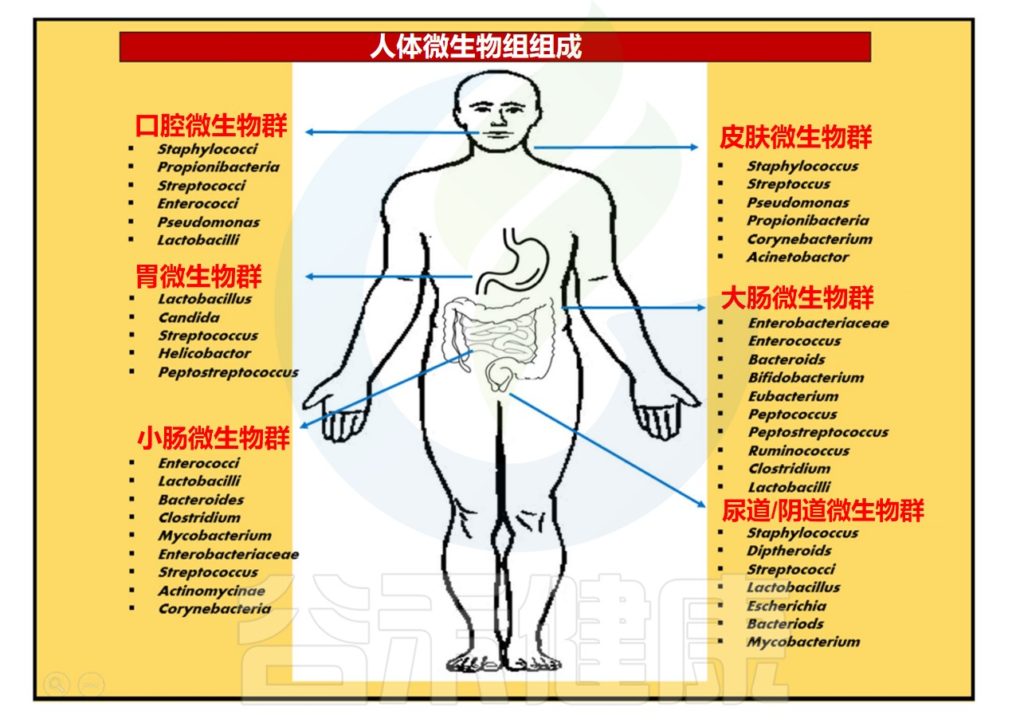
人体微生物群是指在人体内外生活的微生物群落,包括细菌、真菌、病毒等。这些微生物群落分布在人体的不同部位,如口腔、皮肤、肠道、生殖道、大脑等。人体的不同部位提供了不同的环境条件,适合不同类型的微生物生长和繁殖。
每个部位的微生物群落都有其独特的组成和功能,它们与人体之间存在着相互作用和影响。
注:肠道微生物是人体内最丰富、最多样化、功能最大的微生物群落
Ayariga JA,et al.Arch Microbiol.2022
Zaidi S,et al.Arch Microbiol.2023
在人体内,微生物群主要包括以下几个方面:
▼
肠道提供了温暖、潮湿、酸性和富含营养物质的环境,适合多种菌群的繁殖。此外,肠道还有大量的食物残渣和纤维素,为益生菌提供了生长的基质。
肠道中居住着高度多样化的微生物群落,其肠道内容物密度达到10^12个微生物/毫升,包含超过1000万个基因。
一般来说,肠道首先由兼性厌氧菌如肠球菌(Enterococci)和肠杆菌(Enterobacteria)定植,然后由专性厌氧菌定植。
肠道微生物群所拥有的基因编码了数千种微生物酶和代谢物。它们在消化、降解、消除有毒化合物、将难消化的复合糖聚合物转化为短链脂肪酸和维生素等多种功能中发挥着关键作用。
肠道菌群的分布
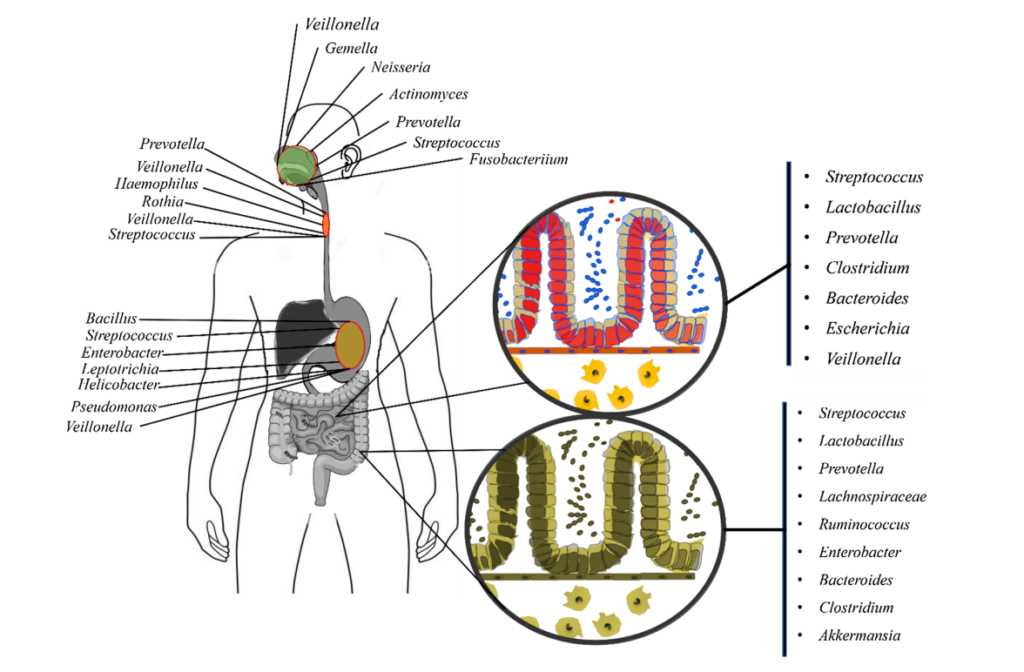
Ayariga JA,et al.Arch Microbiol.2022
▸ 空肠和回肠中主要的微生物群
通过分析空肠和回肠内容物,发现最丰富的群落是链球菌(Streptococci)、乳杆菌(Lactobacilli)、γ-变形杆菌、肠球菌(Enterococcus)和拟杆菌(Bacteroides)。
随着小肠远端向回肠推进,微生物群落变得更加复杂。回肠末端以梭菌科、毛螺菌科、消化链球菌科、瘤胃球菌科、肠杆菌科和拟杆菌科较丰富。
此外,十二指肠还含有与胃相似的菌属,包括肠杆菌科、链球菌科、韦荣氏球菌科和假单胞菌科。
▸ 结肠中主要的微生物群
结肠中栖息的微生物群丰富多样,主要包括放线菌门、拟杆菌门、厚壁菌门、变形菌门和疣微菌门。
与小肠相比,结肠黏液有更明确的层状组成。结肠黏液层具有物理清除细菌的内黏液层,并含有针对微生物群的免疫效应细胞。外层是松散的,为许多微生物提供了一个定植点。
嗜黏蛋白阿克曼菌(Akkermansia)、瘤胃球菌(Ruminococcus)和一些拟杆菌属是肠道黏液外层的居民。
★ 肠道核心微生物群
此外,从肠腔到粘膜存在氧梯度,并通过结肠向下移动,对结肠微生物组成产生影响。大部分细菌种类总是存在并形成稳定的核心微生物群。
这些核心微生物包括另枝菌属(Alistipes)、拟杆菌(Bacteroides)、经黏液真杆菌属(Blautia)、粪杆菌(Faecalibacterium)、瘤胃球菌属(Ruminococcus)、罗氏菌属(Roseburia) 、 普拉梭菌(Faecalibacterium prausnitzii)和颤螺菌属 (Oscillospira)。
▼
口腔是微生物群落多样性排名第二的地方,大约有700种不同亚群的细菌。
人体口腔包含牙龈、面颊、扁桃体、舌头、牙齿、软硬腭等多种微生物环境。口腔提供了温暖、潮湿和富含碳水化合物的环境,适合细菌的繁殖。此外,口腔还含有唾液,其中的酶可以帮助控制微生物的生长。
由于口腔内含有众多菌群,因此有自己的数据库——人类口腔微生物组数据库。健康人的唾液中含有Gemella、韦荣氏球菌属(Veillonella)、奈瑟菌属(Neisseria)、梭杆菌属(Fusobacterium)、链球菌属(Streptococcus)、普氏菌属(Prevotella)、Pseudomonas、放线菌属(Actinomyces) 等多个属,占总分类群的96%。
▼
早期的理论认为胃是一个无菌器官,不适合细菌生存,然而胃腔内幽门螺杆菌的发现打破了这一观点。
采用16S rRNA测序技术的研究进一步表明,胃内存在着链球菌(Streptococcus)、假单胞菌(Pseudomonas)、肠球菌、葡萄球菌(Staphylococcus)、以及变形菌门、放线菌门、厚壁菌门、拟杆菌门和梭杆菌门。
▼
呼吸道包括鼻腔、咽喉和肺部等部位。这些部位通常比较干燥,但仍然存在微生物的定居。
▸ 鼻腔中的微生物群
鼻腔是人体呼吸道的入口,也是微生物的第一个定居地。鼻腔内存在多种细菌,如葡萄球菌、链球菌等。这些细菌可以与宿主共生,帮助抵御潜在的病原体侵袭。
鼻腔内还有纤毛和黏液,可以帮助清除微生物
▸ 咽喉处的微生物群
咽喉是连接鼻腔和气管的部位,也是呼吸道的一部分。咽喉内存在多种细菌,包括厌氧菌和革兰氏阴性菌等。这些细菌参与了呼吸道的免疫调节和防御功能。
▸ 肺部的微生物群
正常情况下,肺部是相对无菌的环境。然而,在某些情况下,如免疫系统受损或存在呼吸道感染时,肺部可能会受到微生物的感染。常见的肺部微生物包括肺炎链球菌、流感病毒等。
▼
皮肤被认为是人体最大的器官。皮肤是一个动态的、复杂的生态系统,其中含有许多共生细菌。皮肤是人体最外层的保护屏障,同时也是微生物的栖息地。皮肤表面有油脂和汗液分泌物,提供了微生物生长所需的水分和营养物质。
注:研究表明,皮肤的生理特征,如温度、湿度、pH值、皮脂含量等,会影响和塑造微生物群。皮肤微生物群落的变异性和多样性还受到人口统计学、遗传学、区域环境波动等因素的影响,从而导致微生物群落结构的改变。因此,皮肤微生物群应该是独一无二的,因此可以作为“微生物指纹”。
最近的研究,在毛囊深处发现了大量细菌。棒状杆菌(Corynebacterium)和葡萄球菌(Staphylococcus)在特定的身体部位如脚底和腘窝繁殖良好。
✦湿润和干燥皮肤下的微生物群不同
大多数不同种类,以及不同相对丰度的厚壁菌门、变形菌门、拟杆菌门和放线菌门都被发现存在于干燥的皮肤中。
注:变形杆菌定植于深层的皮肤区域,可能参与控制宿主和环境之间的皮肤稳态。
棒状杆菌(Corynebacterium)是一种普遍存在于湿润和干燥皮肤的菌属,在表皮区比真皮区数量更丰富。
此外,Pelomonas spp是皮肤群落的核心共生生物之一。对金黄色葡萄球菌、表皮葡萄球菌、痤疮丙酸杆菌、马拉色菌等皮肤相关微生物进行分析,有助于阐明其复杂的分子机制及与皮肤的关联。
注:后三种被发现在湿润的皮肤占主导地位。
▼
阴道酸性较高,含有乳酸菌等益生菌。阴道微生物群被认为是预防许多泌尿生殖系统疾病的关键,可以防止病原体在阴道内定植。如艾滋病毒、细菌性阴道病和酵母菌感染。
然而,与月经周期相关的激素变化可以显著改变微生物群的组成,并在阴道微生物群的动态中发挥主导作用。
✦女性生殖道微生物群
测序表明,主要的乳杆菌属,如卷曲乳杆菌(L.crispatus)和惰性乳酸杆菌(L.iners)构成了一个“健康”的阴道微生物群。
这些物种产生乳酸、抑菌和杀菌分子,创造一个低pH的生态位,并通过竞争排斥提供保护。阴道微生物群的一部分在分娩时传递给婴儿,这反过来又推动了新生儿消化道中微生物群的初始定植。
✦男性生殖道微生物群
与女性生殖道和其他身体部位相比,男性生殖道微生物群的鉴定和研究一直较少。
男性下生殖道(即尿道和冠状沟)的微生物群主要由放线菌门、梭菌门、厚壁菌门、拟杆菌门和变形菌门组成,尽管受试者之间存在很大的差异。
注:与未行包皮环切术的个体相比,人工干预包皮环切术导致革兰氏阴性菌和厌氧菌的丰度减少。
▼
一系列突破性研究表明神经和神经胶质细胞内存在微生物。然而,生活在大脑中的共生细菌比肠道中的要少的多。
大脑常驻微生物的RNA测序显示,这些微生物属于肠道中常见的门,即厚壁菌门、变形菌门和拟杆菌门,它们可能影响情绪、行为或使个体易患神经系统疾病。
注:先前的研究表明,弓形虫可以侵入大脑,但不会引起明显的疾病。
大脑微生物群主要在黑质、海马体和前额叶皮层的星形胶质细胞内,这些发现有助于未来研究与神经精神疾病的关联。
▼
人类的血液通常被认为是无菌的,然而最近的研究表明,健康的人有一个血液微生物群。
✦血液中的微生物主要是其他部位转移而来
来自不同队列的测序数据描述了9770名健康人血液中的微生物。过滤污染物后,血液中有117种微生物,它们主要是来自胃(n=40)、口腔(n=32)和泌尿生殖系统(n=18)的共生菌,而不是医院血液培养中发现的病原体。
这些发现不支持人类血液固有稳定核心微生物群的观点。相反,它支持共生微生物从其他身体部位暂时和偶尔转移到循环中。
越来越多的证据表明,年龄与人类微生物群之间的关联很大,随着年龄的增长,人体内的微生物群落会发生变化。
人类相关微生物群从出生到死亡的变化
Martino C,et al.Nat Rev Microbiol.2022
美国一项研究集中测量了从儿童到老年人的粪便(a部分)、口腔(b部分)和皮肤(c部分)微生物群的细菌多样性,该项目包含21919个粪便、1920个口腔和998个皮肤微生物群样本。
α多样性,一种对样本中不同类型微生物数量的定量测量,通过Faith的系统发育多样性(PD)α多样性度量跨年龄测量。
UniFrac β多样性主坐标分析,一种用于比较微生物群落相似性的方法,其中空间上接近的点表示相似的样本,空间上远离的点表示不同的样本,按年龄着色。
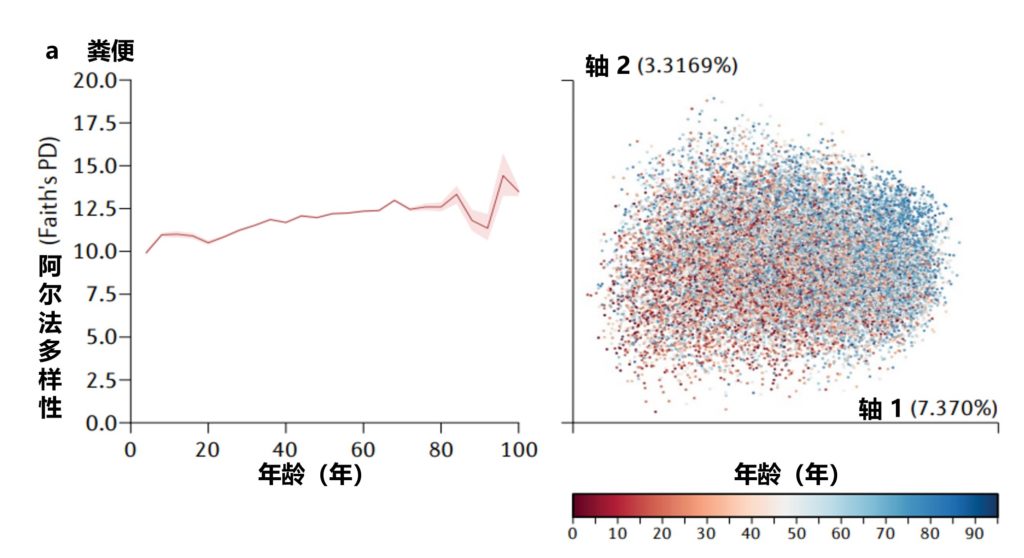
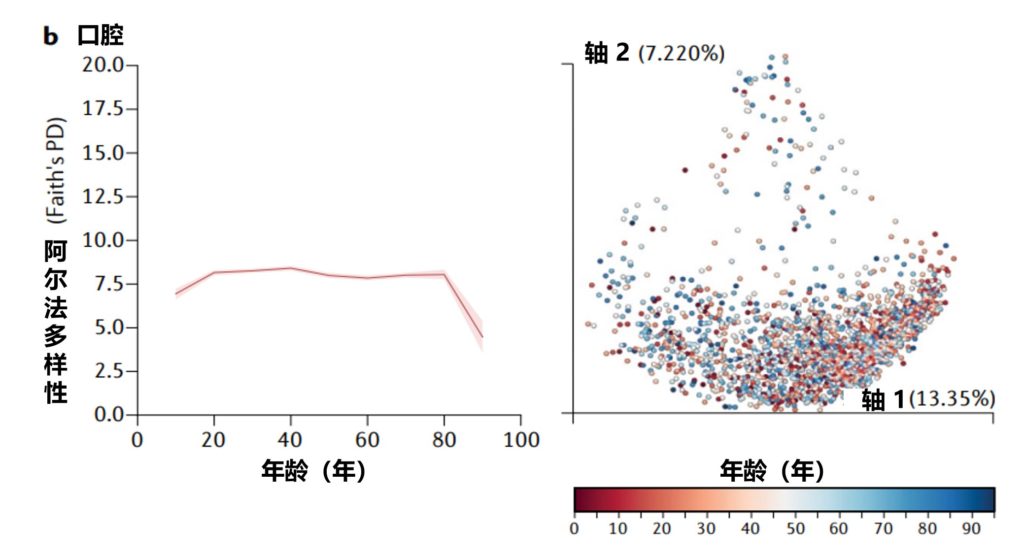
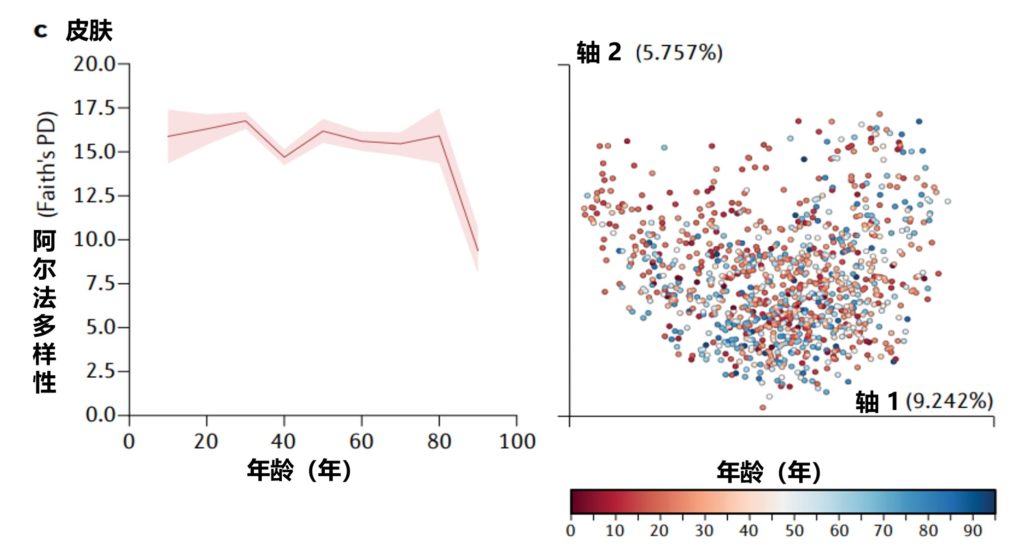
Martino C,et al.Nat Rev Microbiol.2022
▼
关于出生时获得的微生物群是否通过混合来源于阴道和粪便,或者阴道微生物群本身在出生时是否具有多能性,是否是微生物先驱的主要来源,存在一些争议。
无论确切的母体来源如何,这一阶段的特征是先锋细菌种类。包括下列菌群:
•Lactobacillus
•Enterobacter
•Escherichia
•Bacteroides
•Parabacteroides
•Prevotella
这些细菌定居在常规身体部位:肠道、口腔和皮肤。
许多先锋细菌是兼性厌氧菌,它们会消耗氧气,从而使专性厌氧菌能够在以后的环境中定居。先锋细菌进驻后,生命早期的微生物群逐渐开始形成。
双歧杆菌属(Bifidobacterium)在婴儿刚出生时占主导地位,直到在生命的第一年结束时,它们被双歧杆菌、梭状芽孢杆菌和拟杆菌的组合所取代。拟杆菌属的丰度增加,而双歧杆菌属等物种的丰度相对减少。
▼
健康成年人中的微生物群也会随着时间的推移而进化,不过功能和组成的进化以较稳定状态发生。饮食或是疾病会在一定时间内改变成年人的微生物群构成。
•不同季节饮食对微生物的影响
一个经过充分研究发生在几周到几年范围内变化的例子是饮食驱动的肠道微生物群的改变。
例如,坦桑尼亚哈扎部落在旱季食用富含肉类和块茎的饮食,但在雨季食用富含蜂蜜和浆果的饮食,拟杆菌等属中表现出较大的季节波动。
饮食对微生物群的影响也可能在人类健康中发挥作用,许多工作致力于了解特定的饮食成分和总体饮食模式如何影响微生物群及其对健康的影响。
•疾病在短期内改变微生物群
肠道中的许多疾病,如炎症性肠病,破坏了微生物群落,但没有达到新的稳定群落组成,而是在没有干预的情况下继续长期不稳定。
在皮肤上,特应性皮炎的特征是免疫介导的炎症引起的金黄色葡萄球菌大量繁殖和细菌多样性减少。在金黄色葡萄球菌大量繁殖期间观察到马拉色菌属的数量减少,反之亦然,真菌数量增加导致金黄色葡萄菌数量减少。
除了饮食和疾病,还有许多其他因素会在一定时间内影响成年人的微生物群,包括地理因素、压力、代谢情况等。
注:谷禾将在下一章节中具体讲述影响人体微生物群的因素
▼
衰老会影响细胞功能的各个方面,微生物群也不例外。随着年龄的增长,微生物群α多样性减少,β多样性增加。
•肠道
一般而言,老年人肠道中观察到的微生物群变化是年轻成年人中占优势和普遍的细菌属丰度减少,如双歧杆菌(Bifidobacteria), 拟杆菌(Bacteroides),乳杆菌(Lactobacillus), 抵御机会细菌爆发的能力降低。
•皮肤
在65岁及以上的人群中,genera Cutibacterium和Staphylococcus的皮肤细菌数量减少,同时观察到更多的Corynebacterium。
•口腔
在口腔部位,Rothia和Streptococcus spp.是核心口腔细菌群落,Porphyromonas,Treponema和Faecalibacterium spp.的数量持续减少。
由于微生物组的动态特性,它在空间和时间上会不断变化,还与个体的健康状况有关。
这些变化的程度和后果取决于性质、持续时间、扰动强度和微生物群的结构和稳定性。
已经发现许多因素影响微生物群的组成,如分娩方式,年龄、性别、地理位置、饮食、怀孕、昼夜节律、宿主遗传和社会经济地位,药物以及其他一些因素(益生元和益生菌补充剂,手术和非手术治疗)。
Zaidi S,et al.Arch Microbiol.2023
下面谷禾列举了其中一些对微生物群组成有重要影响的因素。
1
上一章节讲述了微生物群会随着年龄而变化。微生物群经历了一系列的发育阶段,它们的复杂性和丰富性提高,从新生儿期到断奶后的明显稳定。
这样的初级共生群落不断进化,变得更加多样化和稳定。一旦儿童达到3岁,其微生物群变得与成人的微生物群相似。
2
出生方式会决定了最初的微生物。在剖腹产和顺产婴儿之间有惊人的微生物差异。
√阴道分娩的婴儿微生物群和母亲更相似
在阴道分娩时,孩子接触到女性产道的微生物,导致母亲和孩子的微生物组成相似。这些婴儿的肠道中含有大量的乳杆菌(Lactobacillus)和普雷沃氏菌(Prevotella)。
√剖腹产分娩的婴儿微生物群主要来自母亲皮肤
相反,在剖腹产分娩的母亲和婴儿之间没有观察到大量微生物群重叠。通过剖腹产出生的新生儿从母亲的皮肤中获得细菌群,从而导致链球菌(Streptococcus)、棒状杆菌(Corynebacterium)和丙酸杆菌(Propionibacterium)较多。
剖腹产出生的婴儿在头六个月中乳杆菌的丰度较低,不像顺产的婴儿,在前六个月的时间里乳杆菌的百分比都在增加。
不过一旦儿童满3岁,乳杆菌检出率的这种差异就会消失。产后,尤其是拟杆菌和双歧杆菌在剖腹产出生的婴儿肠道内的定植也较晚,相反,他们的艰难梭菌水平增加。
注:剖腹产似乎是婴儿早期微生物群落破坏的原因之一。这种在剖腹产时定植的躁动扰乱了微生物与宿主的相互作用,这可能进一步表现为代谢紊乱的形式。在剖腹产分娩后的最初两年里,婴儿患特应性疾病的比例更高。
3
一旦婴儿出生,影响微生物群组成的最重要因素是婴儿饮食,要么是配方奶,要么是母乳。
√母乳喂养有助于诱导肠道微生物群成熟
饮食塑造了早期的微生物群,尤其是肠道中的微生物群。母乳中含有低聚糖,容易被乳酸菌和双歧杆菌(普遍存在于母乳喂养的婴儿肠道中)代谢,从而导致短链脂肪酸浓度上升。
这些短链脂肪酸进一步控制免疫系统过度表达免疫球蛋白G,并诱导新生儿肠道微生物群的成熟。
而在用配方奶粉喂养的婴儿中,常见的种类是肠球菌、肠杆菌、拟杆菌、梭菌和链球菌。
√母乳喂养下得婴儿免疫系统更完善
婴儿时期的微生物群定植似乎在整个儿童生长阶段的早期免疫发展中起着关键作用。因此,初始微生物群的组成是重要的,因为它可以防御可能由于免疫力低下而引起的多种疾病。
许多研究比较了母乳喂养和配方奶喂养的新生儿的肠道微生物群和粘膜免疫反应。观察到母乳喂养导致更稳定和更好的粘膜免疫反应。
相反,依赖配方奶粉的婴儿在以后的生活中发现免疫系统发育受损以及代谢不正常。在哺乳期间,影响母乳成分的生理和激素波动也可能影响微生物群的组成。
4
抗生素会扰乱微生物群结构。它们不仅对消化道上下段微生物的系统发育组成有不同的影响,而且对去除抗生素后微生物群落的恢复也有不同的影响。
√抗生素会减少微生物多样性
抗生素的使用导致肠道微生物多样性的减少,耐药物种的增加,宿主的应激反应和噬菌体基因的表达。
使用抗生素是一把双刃剑:它消除了病理微生物和有用微生物,最终导致生态失调。研究表明,一些抗生素如克林霉素、克拉霉素、甲硝唑和环丙沙星对微生物群结构的影响是长期的。
下面列举了使用一些抗生素后的微生物变化:
克林霉素可以持续2年而不恢复拟杆菌的多样性;同样,使用克拉霉素对抗幽门螺杆菌导致放线菌数量减少,然而环丙沙星已被提出导致鲁米诺球菌数量减少。
万古霉素是治疗艰难梭菌感染的最佳药物,但它也会引起肠道微生物群的改变,导致艰难梭菌感染的复发性感染,并诱导致病性大肠杆菌菌株的生长。
此外,万古霉素还会导致拟杆菌(Bacteroidetes)、Fuminococcus、普拉梭菌(Faecalibacterium)等肠道微生物群的减少,以及变形菌门(Proteobacteria)种类的增加。
特定抗生素对肠道菌群的影响和恢复时间取决于个体的生理状况。此外,围产期给孕妇服用抗生素也会影响新生儿的微生物群,因为其中一些抗生素可以穿过胎盘。
√抗生素的作用取决于身体部位
此外,抗生素的作用取决于身体部位。例如,与肠道相比,在抗生素治疗后,喉咙和唾液在更短的时间内恢复了最初的共生多样性。
抗生素还会干扰微生物组和免疫系统的相互作用,导致免疫紊乱,并增强宿主对病原体的易感性。抗生素的广泛使用推动了病原微生物耐药性的进化,导致耐药基因的流行增加。
5
膳食成分除了影响微生物组的功能外,还能调节其组成。
√不同饮食成分下的微生物群组成不同
饮食对于确定微生物群的形态、结构和多样性至关重要。素食饮食与健康、多样的微生物群有关,其特征是能够代谢不溶性碳水化合物的物种占优势,即瘤胃球菌(Ruminococcus)、罗氏菌属(Roseburia)和真杆菌(Eubacterium)。
而非素食饮食与厚壁菌门比例下降和拟杆菌门比例增加有关。随着肉类的摄入,微生物群代谢氨基酸,以短链脂肪酸的形式产生能量源,但也会形成产生不利影响的化合物。
在一项研究中,测定了150名健康的杂食性、素食性和纯素食性志愿者粪便中存在的微生物群的组成结构以及代谢组。研究表明,富含蔬菜的食物增加了纤维降解细菌的丰度,并导致粪便短链脂肪酸的产生。
对地中海饮食依从性降低的志愿者拥有较高百分比的有害微生物代谢产物,如酚类和吲哚衍生物,以及三甲胺N-氧化物。
这些例子表明,饮食调节微生物群的组成和功能,从而影响个体的代谢状态。
6
不同身体部位的微生物组成是不同的,那么当微生物群的位置从身体的一个部位交换到另一个部位时,优势微生物物种的生态或流行如何受到影响,这是一个有趣的研究。可见,器官相关微生物群既具有动态性,又具有可塑性。
器官特异性微生物群可以跨界到与身体其他部位相关的其他生态位,在此过程中,微生物承受与身体各器官相关的pH、温度、毒素、免疫细胞等变化。
然而,在给定的生态位上,微生物群的结构组成基本上没有受到干扰。
微生物群在不同器官内混合的机制在很大程度上是未知的。
√器官间的微生物联系可能有利于宿主平衡
从空间和生长的角度来看,跨生态位的微生物对特定器官的优势微生物群体构成了挑战,但不同微生物物种之间的竞争是有利于宿主的微妙平衡。宿主细胞的器官特异性微环境有利于与该器官相关的微生物群的优势种群,并防止微生物群生态中的无意干扰。
宿主和微生物群之间存在着复杂的相互作用。宿主提供了微生物群生存和繁殖的环境,而微生物群则对宿主的生理状态和代谢产生着重要的影响。
由于微生物群与其宿主之间存在高水平的串扰,因此对微生物与宿主之间相互作用的研究仍然具有挑战性,尽管如此,以代谢物为中心的研究已经认识到对宿主健康重要的各种微生物靶点。
▼
微生物群对人体食欲的影响是非常复杂的,因为不同类型的微生物群会产生不同的代谢产物,一些研究表明,肠道微生物群可以通过产生短链脂肪酸等代谢产物来影响人体的食欲和能量代谢。
短链脂肪酸是肠道微生物群代谢产物的一种,主要包括丙酸、丁酸和乙酸等。
▷短链脂肪酸影响神经系统进而影响食欲
关于短链脂肪酸在调节能量摄入和食欲中的作用已经有了详细的研究。研究表明,这些化合物也可以影响外周和中枢神经系统的活动。
不过目前尚不清楚是单一的短链脂肪酸驱动,还是这些化合物的组合被利用。
目前关于可发酵纤维在食欲调节中的作用的研究有限,但增加每日纤维摄入量在16-35克/天范围内可以帮助改善这种调节。
虽然短链脂肪酸在食欲调节中的确切作用机制尚不清楚,但已有研究表明,人类体内短链脂肪酸的存在可以触发短期食欲调节。
例如,人类结肠丙酸盐通过PYY和GLP-1介导的机制诱导短期食欲调节。
▼
短链脂肪酸对脂肪代谢的影响也是显著的。
▷调控脂质积累和瘦素分泌
研究表明丙酸盐可以防止脂肪和胆固醇生成,它可以通过抑制FARE 2信号的活性来阻止脂肪细胞中的脂质积累。
还有研究表明,乙酸盐可以刺激脂肪细胞中瘦素激素的分泌。这一关键信号调节食欲和能量平衡。其他研究表明,抑制脂肪分解可以减少游离脂肪酸从脂肪细胞向肝脏的转运。
在脂肪肝疾病中,已知来自脂肪细胞的脂肪积累贡献了肝脏中总脂肪酸的60%。直肠输注丙酸和乙酸显示血清脂肪酸水平降低40%。
因此,重要的是保持丙酸与乙酸的比例,以确保结肠乙酸对脂质储存的最佳贡献。
▼
几项临床研究指出,肠道中细菌过度增殖与骨矿物质密度(BMD)降低之间存在关联。小肠细菌过度生长综合征患者的骨矿物质密度值低,骨软化,其中一些患者具有高水平的促炎细胞因子TNF-α和IL-1,以及破骨细胞活化增加。
肠道微生物组和骨骼之间的联系
doi.org/10.1016/j.jbspin.2018.02.008
近年来,一些横断面的临床研究以及系统评价和荟萃分析均发现肠道菌群改变与调节骨量、骨髓生成,骨骼发育、骨代谢、骨质疏松、骨骼炎症、骨折风险以及骨癌有关。
▼
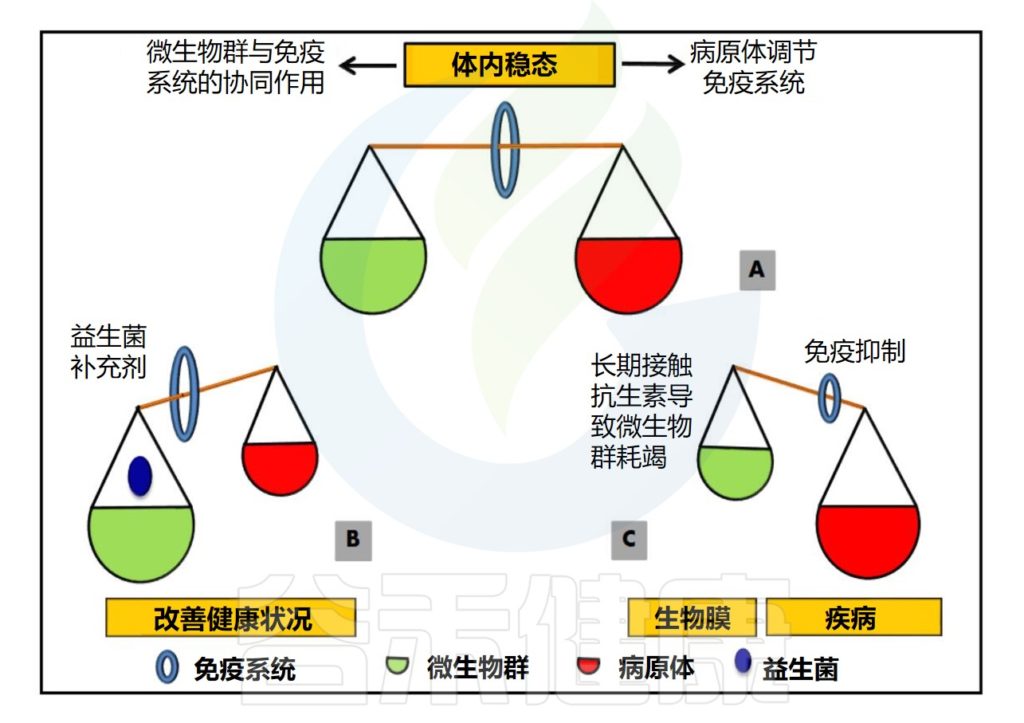
微生物群对宿主免疫系统的影响非常重要,可以通过多种机制来影响宿主的免疫系统。
▷直接影响免疫细胞功能
微生物群中的某些成分可以直接影响宿主免疫细胞的功能,如调节巨噬细胞和树突状细胞的活性,从而影响宿主的免疫反应。
▷调节免疫细胞分化和增殖
微生物群中的一些成分可以影响免疫细胞的分化和增殖,如调节T细胞的分化和功能,从而影响宿主的免疫反应。
▷影响肠道黏膜屏障
微生物群可以通过影响肠道黏膜屏障的完整性和功能来影响免疫系统。肠道黏膜屏障是宿主体内与外部环境之间的主要屏障,它可以防止有害物质和微生物进入宿主体内。微生物群通过增强肠道黏膜屏障的功能来促进免疫系统的正常功能。
总之,微生物群对宿主免疫系统的影响是非常重要的,它们可以影响宿主的免疫反应、调节免疫细胞的分化和增殖、以及影响肠道黏膜屏障的完整性和功能。
肠道菌群及其代谢产物对人体还有其他影响,谷禾罗列在下表中:


Ayariga JA,et al.Arch Microbiol.2022
微生物群不仅影响宿主的生理功能,研究发现病理状况也与微生物群的组成、功能和生长动态密切相关。
如肥胖、高血压、2型糖尿病、非酒精性脂肪肝以及胃肠道疾病、过敏、自闭症、神经退行性疾病甚至癌症等都被发现与微生物群生态失调相关。
Zaidi S,et al.Arch Microbiol.2023
▼
研究表明肠道微生物群的存在可以影响血压调节。
在高血压大鼠中,观察到肠道中的微生物多样性和丰富度显著下降。这种情况已知是由血管收缩和血管阻力引起的。
在无菌小鼠中观察到血管紧张素II对血压的影响,表明肠道微生物群在调节血压方面发挥作用。尽管肠道微生物群调节血压的机制尚不完全清楚,但人们认为这种情况可能是导致高血压发展的一个因素。
动物中的特定肠道微生物代谢物,如短链脂肪酸,可能是导致高血压的一个因素。
来自HELIUS队列研究表明,克雷伯氏菌属和链球菌属与血压呈正相关。已显示,Lactobacillus coryniformis可以改善血管功能和胰岛素敏感性。
注:乳杆菌(Lactobacillus)治疗不仅可以改善心血管疾病,还可以改善实验性自身免疫性疾病。
▼
一些研究表明,微生物群的失调可能与某些癌症的发生有关。
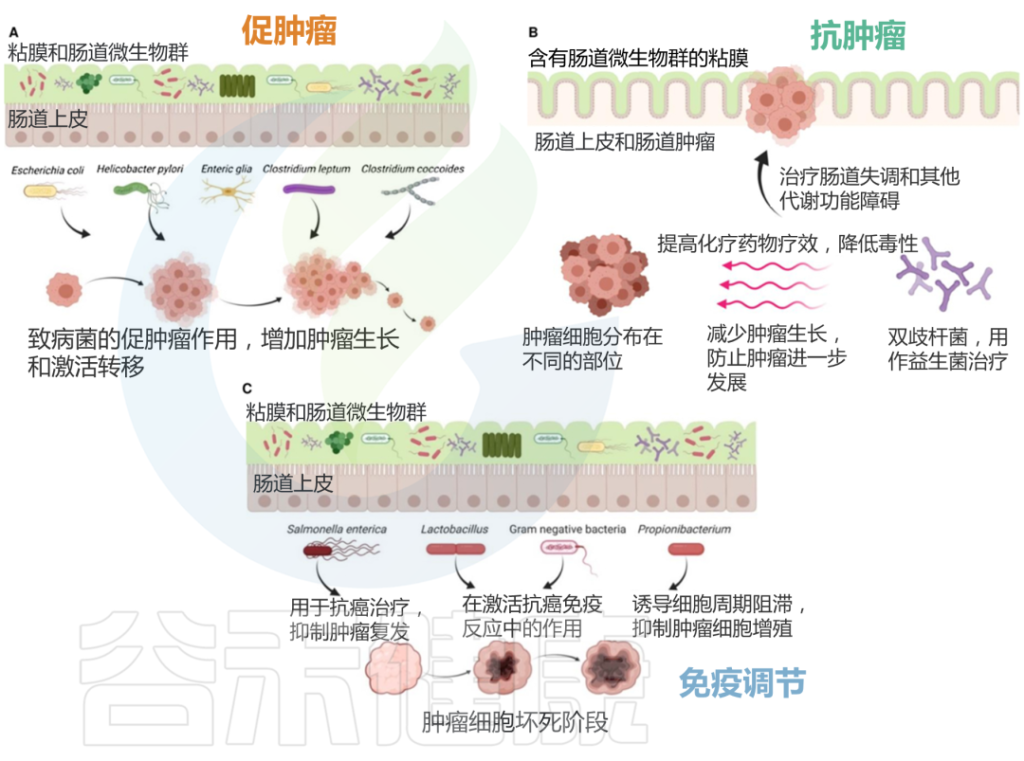
•结直肠癌
在与肠道微生物群相关的各种癌症中,对结直肠癌的研究最为广泛。
已经确定了肠道微生物群中的几种菌,这些细菌除了它们的致病性之外,还被假设对结肠直肠癌具有致癌作用,包括幽门螺杆菌、肝螺杆菌(Helicobacter hepaticus)、牛链球菌(Streptococcus bovis)、大肠杆菌、脆弱拟杆菌、败血梭菌(Clostridium septicum)、粪肠球菌、具核梭杆菌、厌氧消化球菌(Peptostreptococcus anaerobius)和牙龈卟啉单胞菌(Porphyromonas gingivalis),所有这些细菌都显示出潜在的致癌作用。
•肺癌
肺癌是常见的恶性肿瘤之一,迫切需要制定有效的肺癌治疗策略。研究表明,肠道和肺部微生物群之间通过淋巴和血液循环系统在双向轴上存在复杂的联系。
Martins D,et al.Pathobiology.2020
•乳腺癌
一项关于绝经后妇女的研究调查了乳腺癌与肠道代谢组学改变的相互关系。发现健康对照受试者和绝经后乳腺癌患者的肠道微生物组的组成和生物活性存在差异,其中绝经后乳腺癌患者的肠道宏基因组具有编码β-氧化、铁复合物转运系统和脂多糖生物合成的基因。
体外研究提供了支持肠道微生物群与乳腺癌转移进展之间联系的功能证据,其中微生物代谢物可以通过血液传播,影响乳腺癌细胞和免疫细胞的功能。
除此之外,分析唾液微生物组组成的变化有助于早期发现胰腺癌。另一种被称为产肠毒素脆弱拟杆菌的菌株与肠上皮细胞的致癌性有关。
▼
肠道微生物群的扰动可能导致炎症性肠病。同样,厚壁菌门相关细菌的不足和某些变形菌门数量的升高,可能导致粘膜免疫功能受损,这是引发慢性肠道炎症的主要原因,从而导致炎症性肠病的发生。
•肠易激综合征
肠易激综合征中厚壁菌门(Ruminococcus和Clostridium)数量增加,普拉梭菌和双歧杆菌种类数量减少。
•克罗恩病
此外,与克罗恩病患者或健康患者相比,回肠克罗恩病患者的普拉梭菌(Faecalibacterium Prausnitzii)数量明显减少,而大肠杆菌(Escherichia coli)数量过多。
▼
已经证明某些微生物可以通过感染特定的组织而引起疾病。
•细菌性相关组织感染
最早被人们了解的传染病是那些由制造毒素的细菌引起的传染病。白喉、肉毒杆菌和破伤风毒素分别与白喉棒状杆菌(Corynebacterium diphtheria)、肉毒梭菌(Clostridium botulinum)和破伤风梭菌(Clostridium tetani)引起的局部感染相关。
•细菌性腹泻和败血症
大肠杆菌、沙门氏菌、志贺氏菌、葡萄球菌和霍乱弧菌产生的肠毒素可导致由这些微生物引起的腹泻病。
革兰氏阴性菌脂多糖的脂质A部分具有强大的生物活性,可引起革兰氏阴性细菌性败血症的许多临床表现,包括发热、肌肉蛋白水解、血管内凝血失控和休克。
大多数致病的病原体都经过类似的途径,如呼吸道、胃肠道和生殖器官,这些途径被认为是通过与身体直接接触传播的;然而,其中一些微生物也可以通过与环境的间接接触获得,例如通过血液或水。
注:外科手术中的一些植入物也有可能引发细菌感染。
外科植入物引起的相关细菌感染
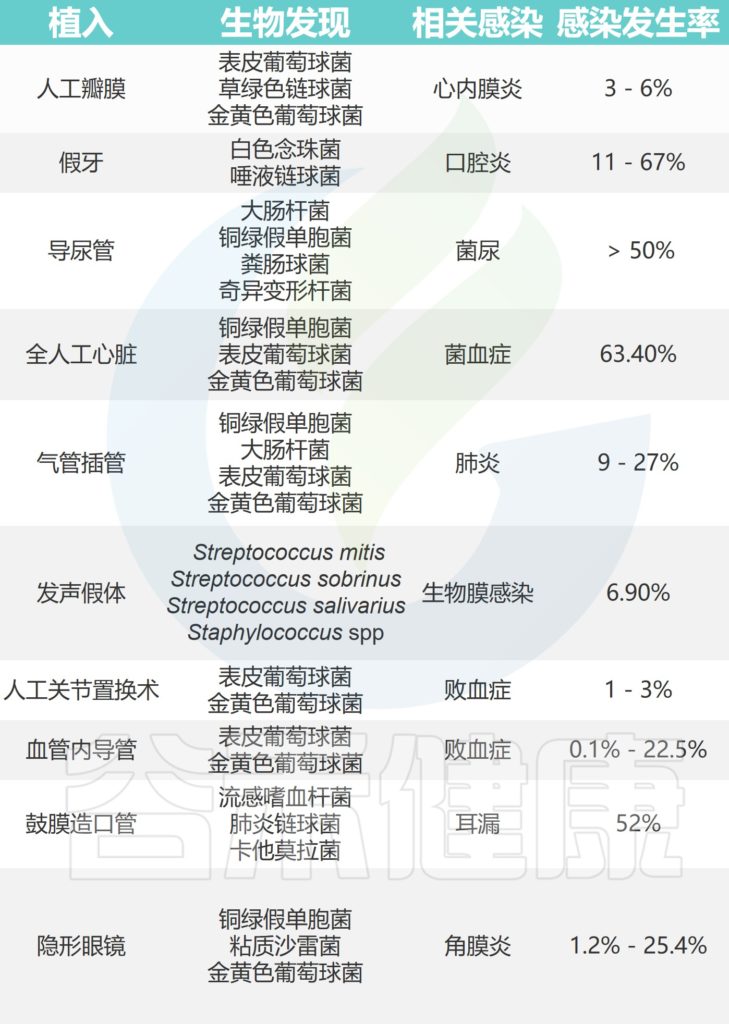
Zaidi S,et al.Arch Microbiol.2023
微生物群与其他一些疾病:
过敏
最近的研究表明Faecalibacterium、Bifidobacterium或Akkermansia的减少,加上Rhodotorula或念珠菌数量的增加,可能通过影响T细胞分化而使新生儿易过敏。
哮喘
同样,已发现罗氏菌属(Rothia)、毛螺菌属(Lachnospira)、韦荣氏球菌属(Veillonella)或普拉梭菌(Faecalibacterium )数量的减少会增加患哮喘的风险。
帕金森病
帕金森病最一致的发现是阿克曼菌(Akkermansia)的丰度增加。
其他组成特征包括双歧杆菌(Bifidobacterium)和乳杆菌(Lactobacillus)的丰度增加,丁酸生产菌(Roseburia)、(Faecalibacterium)和(Blautia)的丰度减少。
注意
微生物群评估有望在早期发现不同的疾病,如癌症、神经退行性疾病、代谢性疾病和自身免疫性疾病等。它采用非侵入性采样技术,同时降低了分析成本,从而使诊断过程变得可行。
对人类生理学、基因组学更好理解使我们的重点转向了针对患者的个性化/精准诊断和治疗。
个性化医疗的标志之一是对易感亚人群的特定疾病风险进行评估,从而可以对人群进行分层,提供更准确和更具成本效益的治疗。
微生物群在人类疾病和健康中的作用致使新的生物疗法发展,可以解决特定的疾病机制。微生物和免疫系统之间的相互作用是复杂的,它们的治疗可以提高患者的生活质量。
▼
每个人对医疗药物的反应也有很大的差异,可能与体内微生物群不同有关。不同微生物组在药物吸收、代谢、功效和毒性方面起着关键作用,并具有重大的健康影响。
地高辛
例如,地高辛是一种心脏糖苷类药物,专门用于治疗充血性心力衰竭。在微生物组研究进展之前,人们注意到一些患者能够化学还原地高辛,从而导致其失活,抗生素的使用导致血浆地高辛浓度增加两倍。
研究人员推断肠道细菌能够调节地高辛的代谢。最近的报告表明地高辛可被Eggerthella lenta灭活,而抗生素的摄入可使这种作用最小化,从而导致其在血浆中的浓度大幅增加。
对乙酰氨基酚
同样,对乙酰氨基酚,一种存在于许多镇痛药物中的化合物,在个体的临床作用中表现出明显的差异。这种个性化的反应最近被解释为与微生物组功能的差异有关。
他汀类药物
另一个微生物组驱动的个体化药物案例是他汀类药物和化疗药物,它们也被发现由于共生菌的作用而受到显著影响。
另一方面,药物可以通过微生物群转化为衍生物,这些衍生物可能具有非靶标效应。用于生产这些营养物质的底物的变化会影响肠道的代谢组学特征。这可能会对宿主产生不同的影响。
▼
根据上述数据,针对患者量身定制的微生物组操作似乎是多因素疾病更精确的微生物组特异性治疗的最佳替代方案。
如饮食干预、抗生素的应用、益生菌和益生元的使用以及粪便微生物群移植可用于调节微生物群以获得有利的反应。
微生物疗法的原理是通过调节宿主的微生物群,来促进身体健康和预防疾病。它可以包括多种形式,例如口服益生菌、粪菌移植、皮肤微生物移植等。
★ 肠道微生物群检测
要对人体的微生物群进行干预,首先要做的就是对体内的微生物群进行检测。肠道微生物群检测是一种通过分析肠道微生物群的组成和数量来评估患者的健康状况的方法。这种方法已经被证明对诊断肠道炎症、炎症性肠病等多项疾病有效。
口服益生菌改善健康
口服益生菌可以帮助恢复肠道菌群的平衡,从而改善肠道健康和免疫系统功能。
口服补充乳杆菌,可以提高耐受性,减轻儿童对牛奶的过敏,并通过过敏患者粪便中产生丁酸盐的细菌的活性恢复丁酸盐的最佳水平。
此外,酵母菌和乳酸菌可以将抗生素相关疾病的风险降低50%。益生菌持续治疗多种胃肠道和肠外疾病,如阴道感染、肠易激综合征、炎症性肠病和免疫增强。
食用合生元改善健康
此外,合生元是微生物群靶向治疗的另一种有效方法。包括在原始微生物群中引入新的微生物,补充足够的底物以促进新的所需微生物的生长。
例如,植物乳杆菌(益生菌)与低聚果糖(益生元)一起生长被发现可以减轻新生儿因败血症而死亡的数量。
麦角硫因是一种在蘑菇、豆类和谷物等食物中发现的著名抗氧化剂。研究证明麦角硫因被幽门螺杆菌利用来保护它免受宿主胃组织的侵害。
麦角硫因对人体有抗炎作用。更重要的是,麦角硫因的减少与心血管疾病、自身免疫性疾病和神经系统疾病的风险增加有关,这意味着肠道中的细菌会影响人类健康。
微生物移植改善健康
粪菌移植疗法的工作原理是通过引入健康的微生物群来替代患者肠道中的有害菌群,从而恢复肠道菌群的平衡。
这可以改善肠道健康和免疫系统功能,从而减少肠道炎症和其他与肠道微生物群失调相关的疾病的发生。
除了治疗肠道疾病外,粪菌移植疗法还可以用于治疗其他疾病,例如自身免疫性疾病、代谢性疾病和神经系统疾病等。皮肤微生物移植可以用于治疗某些皮肤疾病,例如顽固性湿疹和痤疮。
微生物群在我们身体的许多部位中存在着,如肠道、皮肤、口腔和生殖道等。这些微生物群与我们的身体密切相连,对我们的健康和疾病起着重要作用。
了解人体微生物群的组成、功能和影响因素,对于我们更好地了解自身健康以及疾病的预防和治疗具有重要意义。
主要参考文献
Zaidi S, Ali K, Khan AU. It’s all relative: analyzing microbiome compositions, its significance, pathogenesis and microbiota derived biofilms: Challenges and opportunities for disease intervention. Arch Microbiol. 2023 Jun 6;205(7):257.
Ayariga JA, Ibrahim I, Gildea L, Abugri J, Villafane R. Microbiota in a long survival discourse with the human host. Arch Microbiol. 2022 Nov 28;205(1):5.
Abenavoli L, Scarpellini E, Colica C, Boccuto L, Salehi B, SharifiRad J, Aiello V, Romano B, De Lorenzo A, Izzo AA, Capasso R (2019) Gut microbiota and obesity: a role for probiotics. Nutrients 11(11):2690.
Apparao Y, Phan CW, Kuppusamy UR, Sabaratnam V (2022) Ergothioneine and its prospects as an anti-ageing compound. Exp Gerontol 170:111982.
Aarnoutse R, Ziemons J, Penders J, Rensen SS, de Vos-Geelen J, Smidt ML (2019) The clinical link between human intestinal microbiota and systemic cancer therapy. Int J Mol Sci 20:4145.
Aggarwal N, Kitano S, Puah GR, Kittelmann S, Hwang IY, Chang MW (2022) Microbiome and human health: Current understanding, engineering, and enabling technologies. Chem Rev 123:31.
De Angelis M et al (2020) Diet influences the functions of the human intestinal microbiome. Sci Rep 10:1–15.

谷禾健康

维生素C是一种广泛存在于自然界中的水溶性维生素。维生素C在人体新陈代谢中具有多种重要功能,包括抗氧化、参与胶原蛋白合成、增强铁的吸收等。由于其众多生理益处,维生素C被广泛地应用于修复伤口、治疗感冒、癌症等多种疾病。
人体无法自行合成维生素C,需要从膳食来源(如水果、蔬菜)中获取。维生素C的代谢过程涉及多个酶和转运蛋白。肠道微生物群可能通过影响这些酶和蛋白的活性或表达,来影响维生素C的代谢吸收过程。
维生素C也可以直接调节肠道微生物群,或通过修复肠道屏障、改变氧化还原电位等方式间接对肠道微生物群的平衡起到调节作用。
个体之间的差异、饮食习惯和生活方式等因素各不相同,这些都可能对维生素C与肠道菌群的相互作用产生影响。
本文将从维生素C的结构、功能、吸收和代谢、与肠道菌群的关联等多角度,全面探讨维生素C的作用及其对人体健康的影响,同时介绍了一些维生素C的食物来源、人体需要的剂量、如何补充、注意事项等。
▼
本文主要内容:

编辑
维生素C,也称为抗坏血酸,是一种水溶性维生素,对人体健康非常重要。它在许多身体功能中起着关键作用,包括增强免疫力、抗氧化、胶原蛋白的合成等。
▼
维生素C的化学名称是L-抗坏血酸,它是一种有机化合物。无臭,味酸,易溶于水,微溶于乙醇,不溶于乙醚。
维生素C结构简单,化学式为C6H8O6:
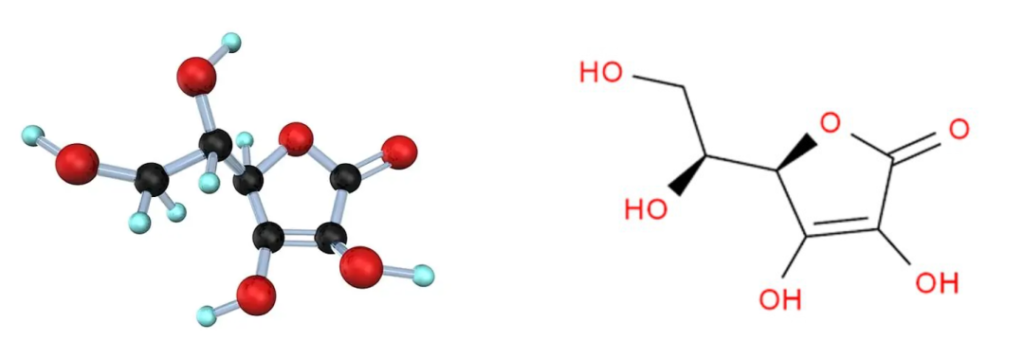
这种结构使维生素C具有抗氧化性质,能够捕捉自由基,并保护细胞免受氧化损伤。
注:体内的分子暴露于环境污染物、吸烟和慢性炎症等情况时,它们会变成自由基。自由基是不稳定的分子,会破坏细胞并导致疾病,维生素C可以通过中和自由基来减缓或预防某些健康问题。
由于其与葡萄糖的结构相似,维生素C可以在许多化学反应中取代葡萄糖,并且可以防止蛋白质的非酶糖基化。
▼
维生素C参与胶原蛋白、激素、肉碱的合成,促进铁离子的吸收,此外,它在免疫系统的功能和调节中发挥着重要作用,对维持内部环境的平衡和中枢神经系统的正常功能极为重要。
维生素C的大部分功能是由于其作为抗氧化剂和辅助因子的能力。由于人类缺乏L-gulono-1, 4-lactone氧化酶,无法自行合成维生素C,因此完全依赖于维生素C的饮食摄入。
▼
从食物中获取营养总是最好的。大约90%的日常需求来自蔬菜和水果,它们是这种维生素的极好来源,例如奇异果、橙子、芒果、草莓、红椒、青椒等。
一些维生素C含量较高的食物
注:单位“杯”是一个常见的非正式计量单位,美规和英规略有区别,大约是237毫升-250毫升左右,涉及到果蔬的份量时,一杯通常是指将果蔬切碎后填满一杯容器的量。
▼
维生素 C 对大脑健康非常重要。大脑在长期缺乏维生素 C 的情况下以牺牲其他组织为代价来保留维生素 C,并且可以维持比其他器官(例如肝脏和肾脏)高很多倍的浓度,如下图。
维生素C的分布在身体各器官之间差异很大
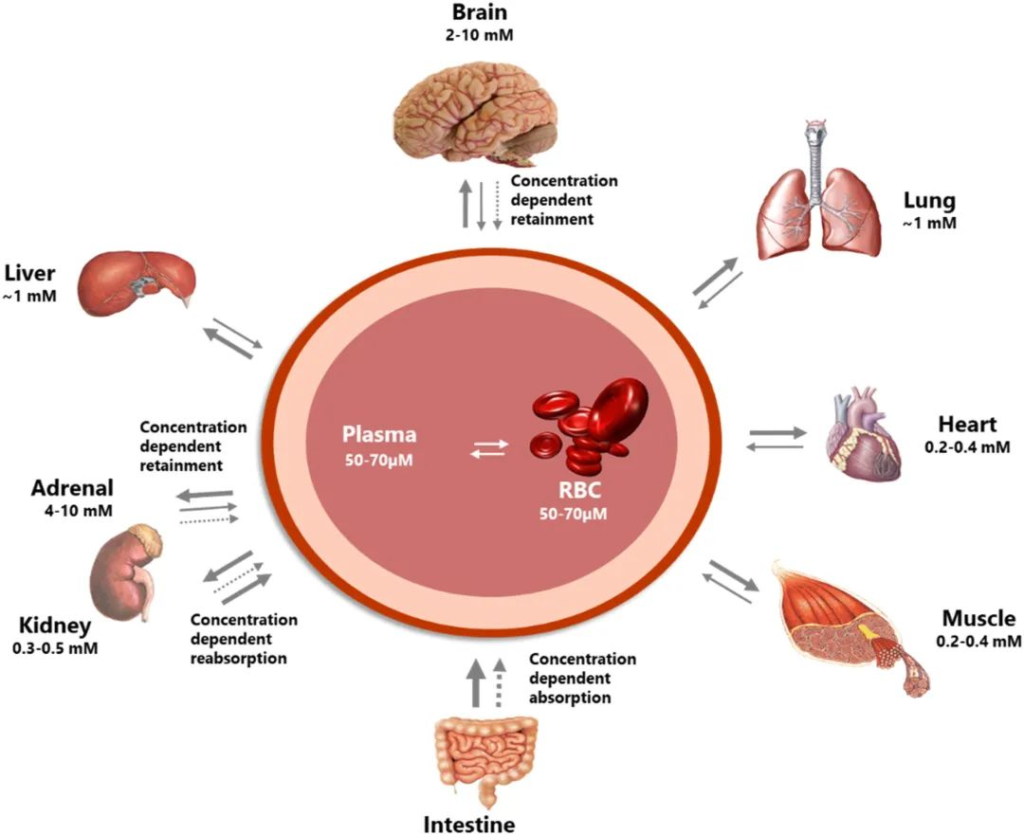
doi.org/10.1016/j.redox.2020.101532
维生素C供应不足时,首先保证大脑里的浓度,那么维生素C对于大脑而言有什么作用?
▼
维生素 C 对胶原蛋白的稳定作用对于形成整个身体的结缔组织框架至关重要;包括皮肤、骨骼、软骨、肌腱、韧带、血管等。
胶原蛋白生产的最后步骤取决于维生素 C,维生素 C 在前胶原脯氨酰和赖氨酰残基的羟基化中充当电子供体。
▼
维生素 C 可增强其他营养素的生物利用度,例如维生素 E 和非血红素铁,这可能会增强含维生素 C 的食物的健康效果。
维生素C经常添加到含铁的口服制剂中,以增加铁的吸收。
▼
维生素 C 是否可以预防或减轻包括普通感冒在内的感染的严重程度是一个有争议的话题。大多数证据都支持其好处。
▼
维生素C对正常骨骼发育至关重要。维生素C水平与骨骼健康之间存在正相关关系,如骨密度、骨折概率、骨转换标志物等。
▼
维生素 C 有助于维持健康的皮肤。
▼
肺部的维生素C水平是血液中的30倍。
维生素C在抵御氧化剂的同时也会被消耗,这表明即使是单剂量的维生素C,也能有效抵御肺部氧化应激的急性增加。
▼
根据现有证据,摄入足够的维生素 C 可能有助于保持健康的情绪。
焦虑
抑郁
▼
人类慢性低维生素 C 状态与神经退行性疾病有关。但是,尚未确定因果关系。
▼
维生素 C 可以通过抑制炎性细胞因子来减轻炎症。
▼
除上述主要的功能之外,在部分小型研究中提到的关于维生素C的功能如下:
助孕育:
助减肥:
降血压、防中风:
降血糖:
助排毒:
助抗癌:
牙周健康:
水溶性维生素在人体内储存较少,从肠道吸收后进入人体的多余的水溶性维生素大多从尿中排出。因此,摄入较多的水溶性维生素一般不会引起中毒现象,但是若摄入量过少,则会很快出现缺乏症状。
▼
刚缺乏的时候症状不明显,饮食中缺乏VC需要大约一个月的时间才会出现症状。
体内维生素C总含量低于300-400mg会出现明显症状。
维生素C的缺乏会出现什么症状?
由于维生素C功能的复杂性及其被不同还原剂的部分替代性,维生素C与坏血病症状的直接联系不容易确定。
如果发展为坏血病,典型症状是:
肌肉无力、牙龈肿胀和出血、牙齿脱落、瘀点出血、自发性瘀斑、贫血、愈合障碍、角化过度、虚弱、肌痛、关节痛和体重减轻(也可能因肿胀而出现矛盾的体重增加),而早期表现包括嗜睡、倦怠、易激惹,甚至呼吸困难等。
在生化上,维生素C血浆水平低于11μM被认为与坏血病的临床症状一致。
在专业医疗人员的指导下,补充维生素C可以轻松有效地逆转坏血病。许多症状可以在几周内轻松解决。富含维生素C的饮食将防止坏血病的发展。
什么人群更容易发生维生素C缺乏?
▼
NIH 认为,成人可耐受的上限是每天 2000 毫克,仅仅靠含有维生素 C 的食物几乎不可能达到这一上限,所以食物可以放心吃。服用补充剂则需注意剂量,可能存在过量的风险。
更高的剂量更有可能导致副作用。
维生素C过量可能会出现什么症状?
单次口服5-10克维生素C会产生短暂的渗透性腹泻和/或腹胀伴疼痛,不建议这样做。
随食物一起摄入可减少这些不良反应。
每天超过 2000 毫克的剂量可能会增加腹泻和肾结石的风险。如果有肾结石病史,每天摄入超过 1000 毫克可能会增加患结石的几率。
那么到底应该怎么补充?每日摄取多少维生素C 合适呢?
▼
科学界对维生素C的最佳剂量方案(摄入量和频率)最健康存在持续争论。
对于大多数健康人来说,通过食物可以获得足量的维生素 C。
维生素C摄入量标准在不同地区有所不同:
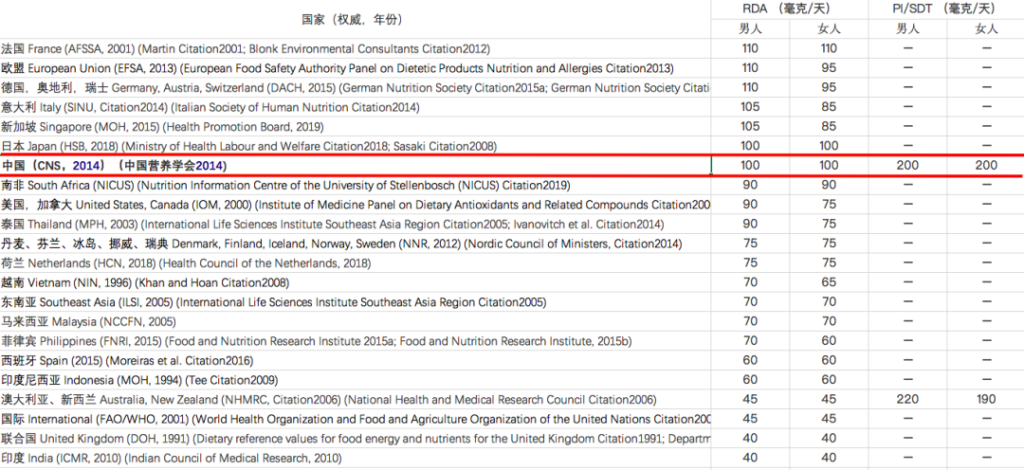
注:RDA – 推荐膳食摄入量,
PI – 建议摄入量,SDT – 建议膳食目标
在中国营养学会编著的《中国居民膳食指南》2022版中对维生素C的推荐摄入量:
中国居民膳食指南2022版
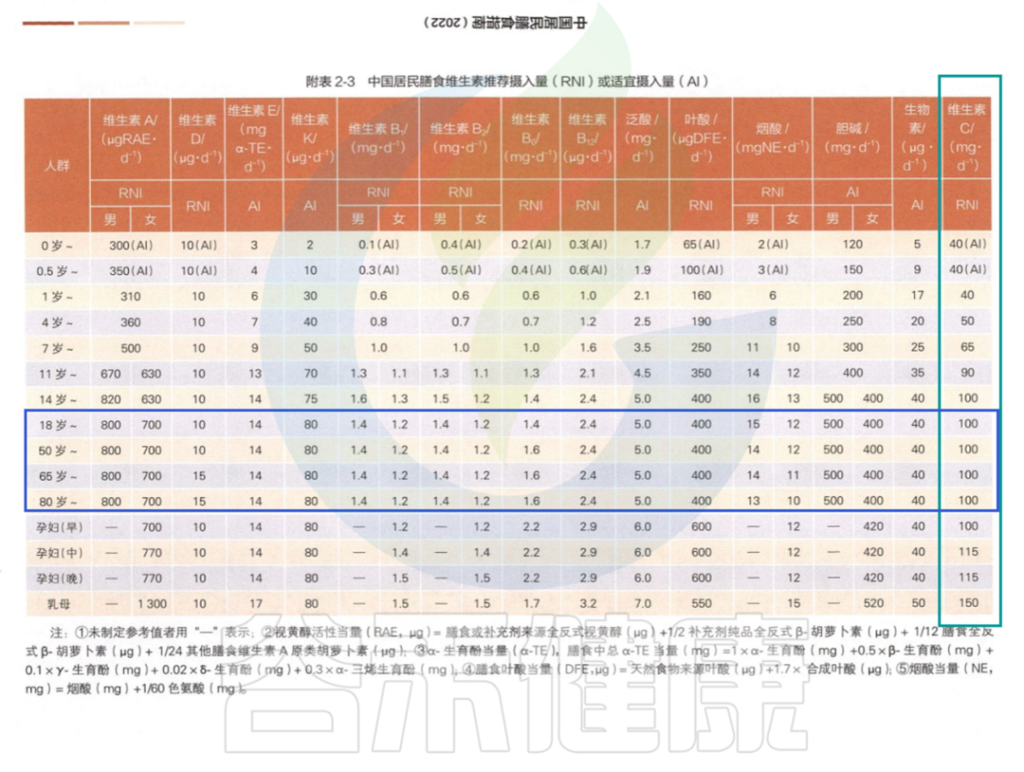
为什么不同地区的标准不一样呢?
这主要是由于RDA标准的基本前提从预防坏血病(~45 mg/d)至健康优化(~200 mg/d)。许多权威机构建议使用最低量的维生素C,但这可能无法满足不同亚群的健康需求。
例如,吸烟者和肥胖者比普通人群有更高的要求。一些国家的吸烟率继续上升,有证据表明,吸烟者每天至少摄入200 mg的维生素C。
随着全球肥胖率的增加,以及与肥胖相关的合并症,如代谢综合征、糖尿病和心血管疾病,需要更多地考虑适当的建议,以优化快速增加的亚健康人群中的维生素C状态。
以上是维生素C的摄入量标准,具体该如何补充,应该注意什么?详见下一章节。
▼
食物:如何才能最大程度地保留其维生素C?
——烹饪方式
长时间烹饪,特别是用大量水煮沸会导致维生素C浸出到水中,显著降低食物中的维生素C含量。
在少量水中蒸或煮,且持续时间较短,是保存维生素C的更温和的方法。
即使将所有外部因素消除到最低限度,也会发生损失,因为由于抗坏血酸氧化酶的存在,材料内部会发生氧化反应。因此,理想的加工方法是用最少量的水快速热灭活酶,然后快速冷却。
——长期保存条件
温度对储存稳定性也有很大影响。随着温度的升高,会出现更显著的损失。然而,在长期储存期间,即使维持短期储存期间仅发生少量损失的条件,维生素C的量也会显著减少。
损失主要是由于酶催化的氧化反应,其程度尤其取决于pH、材料完整性和温度。
总之,为了使水果和蔬菜的维生素C含量保持更长时间,最温和的方法是深度冷冻。
补充剂:一天中分几次服用
食物和许多补充剂中的维生素 C 是一种抗坏血酸的形式。
当肠道面临较低水平的抗坏血酸(即低于约 400 毫克)时,主动运输系统会吸收维生素 C(即,将营养物质通过肠道并进入血液,到身体需要的地方)。
一旦这些主动运输变得不堪重负,被动扩散就会接管吸收其余的维生素 C(这是一个相当低效的过程)。吸收并不像听起来那么容易,事实上抗坏血酸有吸收上限。
身体一次可以处理大约 300 – 400 毫克的纯抗坏血酸形式的维生素 C,更多量一下子难吸收。
所以如果能记得的话,一天中分几次服用比较合适。
▼
维生素C补充剂并非适合所有人。如果遇到以下任何情况,请首先与医生联系:
也不要认为维生素C服用越多越好,每天服用 1000 毫克或更多,实际上会使吸收率降低约 50%。
▼
维生素C可以增加某些药物的吸收,例如:
服用维生素C可以增加含铝药物(如磷酸盐粘合剂)对铝的吸收。这可能对有肾脏问题的人有害。抗酸剂中含铝:不要同时服用维生素C和抗酸剂。服用维生素C后至少等待两个小时,然后再服用抗酸剂。服用抗酸剂后等待四个小时服用维生素C。
维生素C可能会增加左旋甲状腺素的吸收。
补充维生素C会降低一些药物的疗效:
口服维生素C可能会降低这些抗病毒药物的作用。
当与维生素C一起服用时,烟酸和他汀类药物的影响可能会降低,这可能有益于高胆固醇的人。
高剂量的维生素C可能会降低人体对这种抗凝剂的反应。
其他还包括:
维生素C增加或减少药物副作用的风险
如果服用雌激素或基于雌激素的避孕药,维生素C可能会增加激素副作用的风险。这是因为维生素C可能会减缓雌激素离开身体的速度。
一些早期的研究认为, 维生素C可能有助于预防阿司匹林和非甾体抗炎药引起的胃部不适。
▼
维生素 C 和益生菌对肠道健康和免疫力有不同的好处,它们可以很好地互补,这意味着它们可以安全地一起服用。
一项针对学龄前儿童的双盲、随机、安慰剂对照初步研究中,发现益生菌与维生素 C 联合预防呼吸道感染的功效(URTI;33%,P =0.002)。
Lab4 益生菌和维生素 C 组合补充 6 个月的儿童显示,上呼吸道感染症状的发生率和持续时间有所减少,降低感染的严重程度。
注:Lab4 益生菌包含:嗜酸乳杆菌CUL21(NCIMB 30156)和CUL60(NCIMM 30157),双歧杆菌CUL20(NCIMB 30153)和动物双歧杆菌乳亚种CUL34(NCIMM 30172)。
注意:根据说明书剂量服用或遵医嘱。
以上并不是维生素C可能发生的相互作用的完整列表。在开始补充维生素C或调整摄入量之前,请与医生或药剂师沟通,让医生知道你正在服用的所有药物,包括处方药和非处方药、其他维生素或微量营养素、草药补充剂等。
▼
静脉注射和口服给药,这两者可能具有不同的药代动力学特征。
药理学模型显示,口服维生素C,即使是在非常大和频繁的剂量下,也只能适度地增加血浆浓度,从0.07 mM增加到最大0.22 mM。
而静脉注射剂量预计会导致血浆维生素C峰值水平比口服剂量高60倍以上,尿液浓度比口服剂量低140倍。
分子的实际生物利用度由许多因素控制,包括肠道和其他组织的吸收、肾脏的吸收和排泄以及其他患者特异性因素。
除了通过静脉给药和口服给药的浓度差异外,口服给药将维生素C直接输送到肠道微生物组,而不是通过血液;因此,它对肠道微生物的影响可能与动力学和浓度有关,这与影响血浆水平的动力学和浓度完全不同。
静脉注射维生素C常用于临床医疗环境中,用于治疗某些疾病或特殊情况下的高剂量补充,如感染、外伤、手术恢复等。专业医生会给予相应的建议。
口服给药适用于一般的日常维生素C补充,维持正常的维生素C水平。
有几种方法可以评估人体中的维生素C状态。这些包括测量血浆、尿液、组织、粪便中维生素 C 的浓度。
▼
检测血浆维生素C,血浆样品中维生素C的定量测定常见的有两种方法:酶法和色谱法。
酶促维生素C测定
有几种基于维生素C的酶促转化的商业试剂盒,产生可以用光光谱法检测的信号。通常,抗坏血酸氧化酶用于这种类型的测定。这些测定的常见方法是酶联免疫吸附测定(ELISA),它非常适合分批处理样品,但不太方便立即测定少数样品中的值。
根据欧洲外部质量评估计划(Instand EQAS)中报告的方法,基于酶的分析方法在医院中并不常规使用。如果临床对立即测定维生素C的需求增加,由于其直接的技术性质,这些基于酶的测定可用于护理点或集中平台。
色谱法测定维生素C
抗坏血酸和DHA的定量测量目前是通过高效液相色谱(HPLC)方法进行的。如果必须分析具有相似性质的多种化合物,或者如果存在许多可能干扰感兴趣化合物定量的物质,则HPLC方法是优越的。
将酸化样品注射到HPLC仪器中后,通过通过基于化合物的物理性质不同地保留化合物的柱来分离化合物。结果,在分离柱的末端,可以选择性地检测抗坏血酸和DHA,而不受其他化合物的干扰。
目前有两种方法可以检测分离后的抗坏血酸和DHA,一种是电化学检测,另一种是紫外线检测。这两种检测方法给出的结果相同,但由于相对技术简单,紫外线检测更广泛地用于日常检测。
其他检测技术,如荧光检测,需要在柱前对抗坏血酸和DHA进行化学改性,但使用较少。比色/荧光法可能会产生更高的DHA浓度,因为该方法缺乏特异性。
▼
可以查看近期体内维生素状况。
肠道菌群可以影响食物中的营养物质的吸收和利用。肠道菌群的失调可能会影响维生素C吸收,从而出现维生素C缺乏,引发一系列健康问题。因此,检测肠道菌群的状况,可以帮助我们更好地了解维生素C的吸收和利用情况。
与抽血检测不同,肠道菌群的评估更加反映一段时间 ( 一般2周左右 ) 的长期状态,如部分维生素无法在体内留存,需要每日补充,血液检测波动较大。
肠道菌群与维生素C的水平之间存在怎样的关联?
为什么肠道菌群检测报告可以了解维生素情况?
我们来看下一章节。
我们在日常生活会看到,同样吃食物,有些人的维生素吸收状况比较好,有些人就容易缺乏,这是为什么呢?
为什么大剂量补充对一些人的身体有益,而少数人因为过量出现了肾结石呢?
这其实都与肠道微生物群相关。
这里我们分为两个方面来讨论:
一个是肠道微生物群对维生素C的影响,
一个是维生素C的补充对肠道微生物群的影响。
维生素C在人体代谢过程中的吸收和利用,与肠道微生物群相关。了解肠道微生物群对维生素C的吸收和利用的影响,可以帮助我们更好地理解其与人体健康之间的关系。
▼
微量营养素使用各种特定的吸收途径和机制,既可以是被动的,也可以是主动的。
膳食维生素 C 很容易通过钠依赖性维生素 C 转运蛋白(SVCT1 和 SVCT2)在肠道中吸收,其他比如维生素A 、维生素D 的吸收通过小肠中的被动扩散发生。
肠道微生物群是人体肠道中的有效生物反应器,可将各种化合物转化为有益或有害的代谢物,因此对其生物利用度起着至关重要的作用。
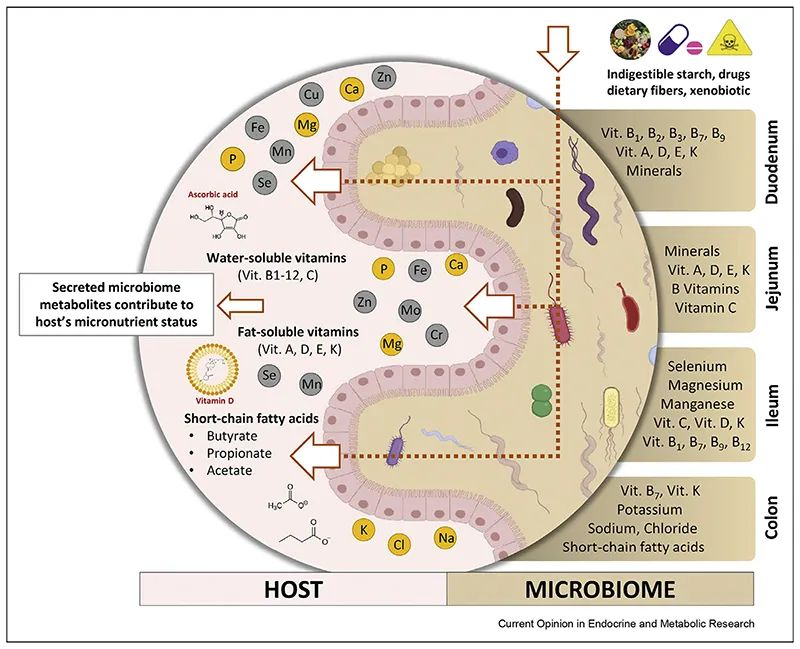
doi: 10.1016/j.coemr.2021.100285
胃肠道各部分理化特性的差异,以及位点特异性受体的存在,使得不同的维生素和矿物质能够沿胃肠道吸收。不同微生物在每个不同部分的定殖会影响当地环境,从而对微量营养素的生物利用度产生积极或消极的影响。
微生物可以干扰维生素C的吸收
微生物可以通过干预生物合成过程和调节吸收,来调节微量营养素的水平,包括维生素C。
来自革兰氏阴性菌的脂多糖降低SVCT-1的表达,进而降低SVCT-1-介导的维生素C的摄取。
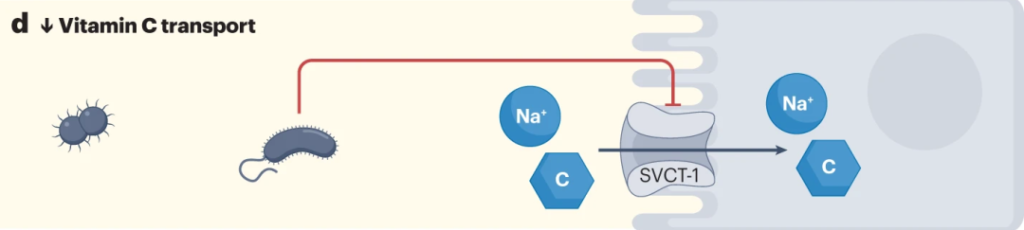
Chmiel JA, et al.,Nat Rev Urol. 2023
大肠杆菌可以通过释放脂多糖来减少宿主对抗坏血酸的摄取,从而增加NF-κB依赖性TNF的产生,其进而通过抑制SLC23A1和SLC23A2启动子(分别编码SVCT1和SVCT2),来降低钠依赖性维生素C转运蛋白SVCT1与SVCT2的表达。从而对维生素C的吸收率产生负面影响。
细菌可以与宿主竞争维生素C
大肠杆菌抗坏血酸转运蛋白对抗坏血酸的亲和力高于哺乳动物SVCT1,这表明细菌可以与宿主竞争维生素C。
因此,肠道菌群中大肠杆菌等革兰氏阴性菌占比较多的情况,可能不利于维生素C的吸收,这在谷禾肠道菌群检测报告中也是可以反映的。
▼
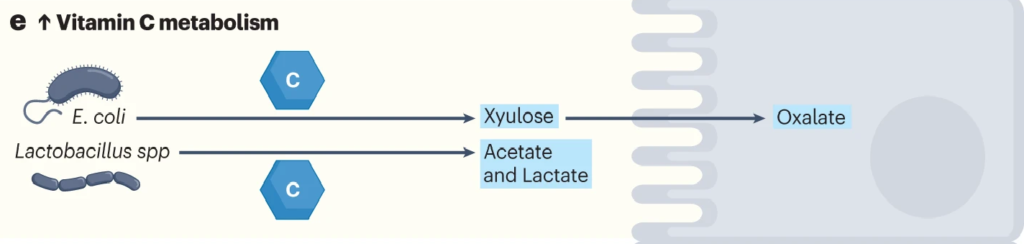
肠道细菌如大肠杆菌和乳酸杆菌代谢维生素C。
利用ula基因簇,大肠杆菌等细菌可以将抗坏血酸代谢为D-木酮糖,宿主细胞可以进一步加工木酮糖以产生草酸盐。
在肺炎链球菌、沙门氏菌、福氏志贺菌、粪肠球菌、肺炎克雷伯菌等病原菌中,也发现了相同的ula基因簇。
带ula的致病菌与草酸盐:携手制造结石
结石形成者(比如肾结石患者)微生物群更常见地富含携带ula基因簇的致病菌,并且这些患者在接受口服维生素C时,草酸盐增加的水平比非结石者增加得更多。高草酸盐水平增加了结石的风险。
因此,如果在肠道菌群检测报告中发现以上提到的致病菌占比较多,则有可能在代谢维生素C的时候产生的草酸盐过多,增加了结石的风险。
Chmiel JA, et al.,Nat Rev Urol. 2023
“
以上通过微生物群代谢维生素的机制,有助于我们更好地理解结石形成的机制。
既然有促进结石的细菌,自然也有降解的细菌:
一些乳酸杆菌可以将维生素C转化为乙酸盐和乳酸盐,这是一种无毒的代谢产物,通过生物能量途径增加微生物组的功能,并可能促进这些草酸降解细菌的定植。
Oxalobacter formigenes是一种革兰氏阴性厌氧细菌,可降解肠道草酸盐并促进原发性高草酸条件下肠道草酸盐的分泌。该菌在肠道定植可降低尿液或血浆中的草酸盐浓度。
Barone M, et al., Biofactors. 2022
我们看到生活中有人认为不能吃生菠菜,会得肾结石,但有些人每天吃生菠菜也没事…其实可能是因为人家的肠道菌群中致病菌较少,而降解草酸的菌群又在拼命干活…
因此,健康的肠道菌群结构非常重要,菌群在该干活的时候各司其职,井井有条,那么你的身体抵抗疾病的能力也随之提升。
以上是肠道菌群对维生素代谢吸收的影响,反过来,维生素C的补充也可以影响肠道菌群的组成。
二、
补充维生素C可以直接调节肠道微生物群,也可以通过改变氧化还原电位、修复肠道屏障,改善肠道条件,支持部分有益菌生长,防止有害菌泄漏到身体其他部位。
▼
维生素C直接调节肠道菌群
与安慰剂组相比,补充维生素C已被证明可以显着增加微生物生态系统的多样性,以及Collinsella的相对丰度和粪便水平的短链脂肪酸,特别是丁酸盐和丙酸盐。
在健康受试者中,每日高剂量维生素C补充(1000 mg/天):
下列菌群的相对丰度升高
下列菌群的相对丰度下降
一项观察性研究探讨了微量营养素维生素C对肠道微生物组组成和多样性的影响。结果表明,维生素C增加了肠道中双歧杆菌属的丰度,在科水平上,毛螺菌科和双歧杆菌科显著增加。
双歧杆菌属的成员是有益菌,例如增加ATP生成、调节免疫系统、黏膜屏障完整性、短链脂肪酸的产生,对健康有益,维生素C通过增加肠道有益菌促进健康。
研究发现,囊性纤维化患者维生素C摄入量的增加与厚壁菌门的丰度呈正相关,与拟杆菌门的丰度呈负相关。
▼
改变氧化还原电位
调节微生物组的机制类似于维生素B2:通过改变氧化还原电位,改善消化道中的厌氧/耐氧平衡,从而选择性地支持微生物生长,改善肠道条件。
与安慰剂相比,维生素C组粪便样本中的粪便pH值和氧化还原平衡降低。
▼
修复肠道屏障
我们知道,肠道屏障受损,细菌和有害物质有可能会穿过屏障进入血液循环,对人体产生各种负面影响,导致诸如过敏、炎症、自身免疫疾病等多种反应。
关于肠漏可以详见我们之前的文章:
而维生素 C 具有维持肠粘膜屏障完整性和修复粘膜屏障的作用。
这里介绍两种修复肠屏障机制。
——通过激活Notch 信号
Notch 信号影响细胞正常形态发生的多个过程,与许多人类疾病有关,包括IBD,因此被认为是癌症治疗的潜在靶点。Notch 信号通路的激活会改变紧密连接蛋白的表达并影响其分布的连续性,从而降低细胞屏障通透性。
豚鼠结肠组织相关基因检测表明,低剂量的维生素C可强烈激活Notch/Hes-1信号通路,对DSS诱导的结肠炎豚鼠的肠粘膜具有一定的保护作用。肠上皮受损时,Notch-1表达增加可促进上皮细胞增殖,有利于损伤部位的修复和重建。
——通过增加肠道胶原蛋白合成
增加维生素 C 摄入量的另一个潜在好处是肠道胶原蛋白合成增加,从而改善屏障功能。这一提出的机制与抗坏血酸的辅酶功能一致,即羟基化脯氨酸和赖氨酸以交联胶原蛋白。
例如,对具有吲哚美辛诱导的屏障功能障碍的 T84 人隐窝样上皮细胞系的研究表明,细菌通过跨细胞途径穿过上皮细胞,维生素 C 治疗可消除该途径。因此,肠道中抗坏血酸状态不佳可能会加剧屏障功能障碍,从而增加 LPS 衍生的革兰氏阴性菌的易位,从而加剧炎症。
维生素C修复肠道屏障后,肠道屏障可以正常发挥吸收、分解和转换营养物质等功能,同时可以帮助促进健康微生物群栖息和生长,从而促进整体健康。
以上我们了解到,维生素C可以通过多种方式影响肠道菌群,从而促进整体健康。那么当我们看到肠道菌群报告中菌群多样性较低,部分菌群失调,尤其是上面提到的毛螺菌科、肠球菌属、Collinsella、Gemmiger formicilis等,可以考虑通过补充维生素C来调节。
我们首先可以考虑通过食物补充,如卡姆果、针叶樱桃等维生素C含量很高的食物,或者一些常见的蔬菜水果例如:猕猴桃、番石榴(芭乐)、青椒、羽衣甘蓝等。也可以考虑通过维生素C补充剂调节。
人体维生素研究面临着许多挑战,维生素通常是通过食物摄入的,而食物中同时存在多种营养素,这使得难以研究单一维生素的作用;某些高剂量维生素的使用可能存在潜在的风险,进行研究时涉及到人体试验和干预,需要遵循伦理准则,并确保研究的安全性;不同人对维生素的需求和代谢能力存在个体差异,基因、环境和生活方式等因素都可能影响维生素的吸收、利用和代谢。
肠道菌群研究可以揭示不同个体之间菌群组成的差异,这有助于理解个体差异对维生素代谢和利用的影响。可以通过检测肠道菌群的组成和丰度,了解维生素的代谢情况,从而辅助评估维生素水平。
综合运用多种研究方法和技术,结合肠道菌群检测,可以考虑多个因素对维生素代谢和利用的综合影响,进一步理解维生素的作用机制,对人体健康具有重要意义。
注:本账号内容仅作交流参考,不作为诊断及医疗依据。
主要参考文献
Barone M, D’Amico F, Brigidi P, Turroni S. Gut microbiome-micronutrient interaction: The key to controlling the bioavailability of minerals and vitamins? Biofactors. 2022 Mar;48(2):307-314.
Chmiel JA, Stuivenberg GA, Al KF, Akouris PP, Razvi H, Burton JP, Bjazevic J. Vitamins as regulators of calcium-containing kidney stones – new perspectives on the role of the gut microbiome. Nat Rev Urol. 2023 May 9:1–23.
Pham VT, Dold S, Rehman A, Bird JK, Steinert RE. Vitamins, the gut microbiome and gastrointestinal health in humans. Nutr Res. 2021 Nov;95:35-53.
Li XY, Meng L, Shen L, Ji HF. Regulation of gut microbiota by vitamin C, vitamin E and β-carotene. Food Res Int. 2023 Jul;169:112749.
Yang Q, Liang Q, Balakrishnan B, Belobrajdic DP, Feng QJ, Zhang W. Role of Dietary Nutrients in the Modulation of Gut Microbiota: A Narrative Review. Nutrients. 2020 Jan 31;12(2):381.
Steinert RE, Lee YK, Sybesma W. Vitamins for the Gut Microbiome. Trends Mol Med. 2020 Feb;26(2):137-140.
Gomes-Neto JC, Round JL. Gut microbiota: a new way to take your vitamins. Nat Rev Gastroenterol Hepatol. 2018 Sep;15(9):521-522.
Rowland I, Gibson G, Heinken A, Scott K, Swann J, Thiele I, Tuohy K. Gut microbiota functions: metabolism of nutrients and other food components. Eur J Nutr. 2018 Feb;57(1):1-24. doi: 10.1007/s00394-017-1445-8. Epub 2017 Apr 9. PMID: 28393285; PMCID: PMC5847071.
Traber MG, Buettner GR, Bruno RS. The relationship between vitamin C status, the gut-liver axis, and metabolic syndrome. Redox Biol. 2019 Feb;21:101091.
Rozemeijer S, Spoelstra-de Man AME, Coenen S, Smit B, Elbers PWG, de Grooth HJ, Girbes ARJ, Oudemans-van Straaten HM. Estimating Vitamin C Status in Critically Ill Patients with a Novel Point-of-Care Oxidation-Reduction Potential Measurement. Nutrients. 2019 May 8;11(5):1031.
Otten AT, Bourgonje AR, Peters V, Alizadeh BZ, Dijkstra G, Harmsen HJM. Vitamin C Supplementation in Healthy Individuals Leads to Shifts of Bacterial Populations in the Gut-A Pilot Study. Antioxidants (Basel). 2021 Aug 12;10(8):1278.

谷禾健康

“铲屎官”们都希望自己的宠物有一个健康的身体。但是猫狗都不会说话,平时我们只能从它们的精神状态来判断它们是否健康,但这并不准确。去宠物医院又不太方便,很多猫咪和狗狗还会对抽血等检查有所抗拒。
肠道微生物检测在人类中的应用已经相对成熟,而猫咪和狗狗与人同为哺乳动物,身体结构具有一定的相似性,近年来关于猫狗等宠物肠道微生物的研究也越来越多。
宠物体内具有大量的微生物,包括细菌、真菌以及病毒,稳定的微生物生态平衡对健康成长意义重大。肠道微生物群有助于宿主新陈代谢、抵御病原体、影响免疫系统,并通过这些方式直接或间接影响宿主的行为、情绪等。
许多因素会影响微生物群的生态平衡,包括饮食、年龄、种族、是否绝育等。当体内微生物平衡被打破或者病原微生物入侵时,宠物的机体便遭受破坏,进而影响健康。

本文主要从以下四个方面讲述
Part 1: 猫狗体内的微生物群
Part 2: 影响猫狗微生物群的因素
Part 3: 宠物肠道微生物群影响健康
Part 4: 恢复宠物肠道菌群的措施
微生物群是微生物的复杂集合,包括细菌、病毒、真菌、古细菌和原生动物。猫狗体内的微生物群包括肠道微生物群和皮肤微生物群和口腔微生物群等。
宠物胃肠道微生物群对其健康的影响最为重要,也是数量最庞大的微生物群落。它们可以帮助宠物消化食物、合成维生素、维持肠道黏膜屏障等。
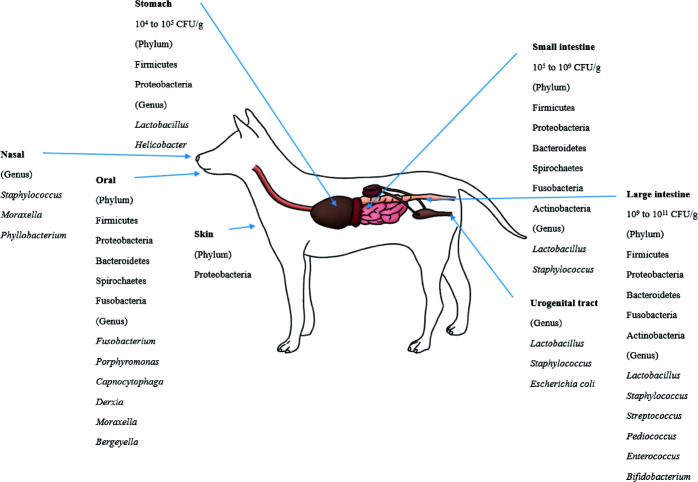
胃中的细菌计数在10^4和10^5 CFU/ml 之间;在十二指肠和空肠中,细菌计数通常较低 (10^5 CFU/ml),但在某些狗和猫中可达到10^9 CFU/mL。回肠含有越来越多的不同微生物群,大多数为10^7 CFU/mL;结肠中的细菌计数介于10^9和10^11 CFU/g。
狗狗身上的微生物群落
Lee D,et al.J Anim Sci Technol.2022
猫咪身上的微生物群落

Lee D,et al.J Anim Sci Technol.2022
狗肠道内的细菌
狗的核心细菌主要五个主要门组成:厚壁菌门(Firmicutes)、梭杆菌门(Fusobacteria)、拟杆菌门(Bacteroidetes)、变形杆菌门(Proteobacteria)和放线菌门(Actinobacteria)。
狗肠道菌群的一个特征是梭杆菌丰度高
狗与人类和小鼠具有相似的变形杆菌和放线菌的相对丰度,但它是三个物种中唯一具有高丰度梭杆菌的。使其成为狗肠道微生物群一个独有的特征。


Garrigues Q,et al.Front Vet Sci.2022
•厚壁菌
厚壁菌门是肠道微生物群中最丰富的三大细菌门之一,具有高度的物种多样性。它们的主要功能之一是在肠道中产生丁酸盐,丁酸盐被结肠细胞用作能量来源。
厚壁菌门的另一重要类别是杆菌,主要由乳杆菌属和链球菌属组成。乳酸杆菌产生乳酸和乙酸,能够刺激免疫功能并在抗原耐受性中发挥重要作用。
•拟杆菌
狗中第二个最主要的细菌是拟杆菌。
拟杆菌能够使用各种类型的底物进行发酵(其中包括蛋白质和各种碳水化合物),拟杆菌还可以使用聚糖与肠道组织相互作用,提供保护免受病原体侵害。
注:在患有炎症性肠病的狗中观察到该菌的丰度减少。
•梭杆菌
与人类不同,梭杆菌是成年犬肠道微生物群的三个主要细菌门之一,更具体地说是梭杆菌属,约占总相对丰度的20%。
虽然梭杆菌与人类的胃肠道疾病有关,但这种门在健康的狗中很常见。此外,由于梭杆菌在狗和猫体内的丰度高于人类,并且由于它们能够将蛋白质降解为氨基酸和肽,因此推测梭杆菌(Fusobacterium)是食肉动物肠道代谢的关键细菌。
•变形菌
变形菌门多种多样,包括一些机会性病原体,如大肠杆菌(Escherichia coli)、沙门氏菌(Salmonella)和弯曲杆菌(Campylobacter),它们对宿主的健康有潜在影响。
虽然变形菌的丰度增加确实与生态失调和炎症性疾病有关,但这些细菌也被证明在健康的狗中大量存在。
变形菌具有多种功能,包括蛋白质、碳水化合物和维生素的代谢,但与拟杆菌一样,它们的主要功能似乎是维持肠道厌氧环境以实现正常微生物组功能。
•放线菌
放线菌门是狗中数量最少的门,约占成年狗微生物群的4%。该门的相对丰度在幼犬中甚至更少,研究发现56日龄以下幼犬的粪便中放线菌的含量不到1%。
该门的一个重要属是双歧杆菌。在人类中,双歧杆菌是婴儿肠道的首批定植者之一,在宿主的全身和粘膜免疫以及母乳低聚糖降解中发挥关键作用。
注:虽然在1至7周大的幼犬中观察到了双歧杆菌,但在任何年龄较大的犬中均未检测到,这表明它是幼犬肠道的特定细菌。
狗其他部位的微生物群
•口腔微生物群
在口腔微生物中,拟杆菌门(60%)是最主要的,其次是变形菌门(20.8%)、厚壁菌门(11.4%)、梭杆菌门 (4.7%)和螺旋体门 (1.7%)。
在属水平上,口腔微生物群包括卟啉单胞菌(Porphyromonas)(39.2%)、 梭杆菌(4.5%)、二氧化碳嗜纤维菌(Capnocytophaga)(3.8%)、德克斯氏菌(Derxia)(3.7%)、莫拉菌(Moraxella)(3.3%) 和伯格菌(Bergeyella)(2.7%)。
•鼻腔微生物群
在健康狗的鼻腔微生物群中, 莫拉氏菌(Moraxella)是最丰富的物种,其次是叶杆菌属(Phyllobacterium)、葡萄球菌属(Staphylococcus)和心杆菌(Cardiobacteriaceae)。
•皮肤微生物群
在皮肤微生物群中,最主要的细菌是变形杆菌和草酸杆菌科(Oxalobacteraceae)。
•阴道微生物群
从狗的阴道中分离出最常见的细菌是乳杆菌、大肠杆菌和假中间葡萄球菌(Staphylococcus pseudointermedius)。
猫肠道内的细菌
关于健康猫肠道细菌的研究较少。家猫是专性食肉动物,依赖大量摄入动物组织来满足其营养需求。这导致其对低葡萄糖和高蛋白质代谢的适应。
与人类或其他哺乳动物相比,猫不太依赖肠道微生物群通过微生物发酵获取能量。尽管如此,稳定和平衡的微生物群对于维持肠道健康仍然至关重要。
猫胃肠道中厚壁菌门(68%)占主导地位,其次是变形菌门(14%)、拟杆菌门(10%),梭杆菌(5%)和放线菌(4%)。
猫胃肠道中的主要细菌群

Lyu Y,et al.Front Microbiol.2020
然而,这些细菌群的百分比通常因物种和个体具有差异。这些差异可能是由动物的生活环境或使用的不同实验方法引起的。
★ 猫和狗的肠道菌群差异
一项研究比较了喂养适合的饮食下狗和猫的粪便微生物群,发现猫的物种数量比狗多,暗示其多样性更高;然而,需要更多的研究来证实这一发现。
具体来说,变形菌在狗体内更丰富,而放线菌在猫中的相对丰度更高。
猫其他部位的微生物群
一项研究调查猫口腔微生物的水平。主要是变形菌门(75.2%),其次是拟杆菌门(9.3%)、厚壁菌门(6.7%)、螺旋体门(1.8%)、梭杆菌门(1.3%)和放线菌门(0.6%)。
猫狗身上的真菌
在真菌水平,在狗粪便中检测到子囊菌(Ascomycota)、担子菌(Basidiomycota)、球囊菌(Glomeromycota)和接合菌(Zygomycota) 。
而子囊菌门是唯一在猫肠道内检测到的真菌。
•一些真菌感染对猫狗有害
皮肤癣菌病是指皮肤和毛发的浅表真菌感染,常见病原体有:
•犬小孢子菌(影响猫、狗和人类皮肤上层的真菌)
•须毛癣菌、疣状毛癣菌和毛癣菌(影响30%的犬,尤其是梗类犬)
•石膏样小孢子菌(是一种与土壤有关的皮肤癣菌)
•马拉色菌是猫狗动物皮肤菌群中最普遍的真菌。M.pachydermatis被称为存在于皮肤微生物组中的酵母,但它也可以充当可引起皮炎的病原体。
•隐球菌是一种常见的猫狗真菌感染,主要通过呼吸道感染,可以引起呼吸道症状、眼部症状等。
•曲霉菌是一种常见的环境真菌,可以在猫狗身上生长,引起皮肤感染、呼吸道感染等。
需要注意的是,不同的真菌感染症状和治疗方法可能不同,如果发现宠物出现异常症状,应及时就医。
猫狗身上的病毒
家养的猫狗身上一般不存在病毒,或是之前通过疫苗产生免疫,已经存在抗体。猫狗身上的病毒对其基本都是有害的。
•狗狗容易感染的病毒
犬瘟热病毒、犬细小病毒、犬副流感病毒、犬腺病毒1型(传染性肝炎)、犬腺病毒2型(传染性喉气管炎)、犬冠状病毒、狂犬病毒。
一些病毒感染的临床表现
(1)犬瘟热病毒(狗瘟)
潜伏期3—9天,传播方式:直接接触,间接接触
双相热:体温先升高,再降低,然后再升高。 体温升高到39.5~41度,持续1~3天 ,然后下降到正常之后再升高,持续时间不定。
呼吸道症状:眼鼻流脓性粘液、喷嚏、咳嗽、严重时出现肺炎、以腹式呼吸为主。
消化道症状:食欲降低、呕吐、排水样便或粘便,严重时血便,病犬极度脱水,消瘦。
神经症状:抽搐、痉挛 。可能先从嘴角开始,表现为嘴角皮肤抽搐。也可能先从四肢开始抽搐,开始是一肢轻微抽搐,然后加重,频率增加,最后四肢或全身抽搐。
皮肤角化症:鼻部角化干裂、脚垫角化形成硬脚垫病。
眼睛损伤:临床上以角膜炎、结膜炎为特征 ,角膜变白,重者可以出现穿孔、角膜溃疡、失明。
(2)犬细小病毒
出血性肠炎型:接触感染潜伏期3~14天,平均5~7天。呕吐,粪便稀薄,呈喷射状或出现血便,味腥臭。
心肌炎型:40日龄左右的幼犬 ,无明显临床症状,突然衰竭死亡是幼犬患本病的唯一体症。
呼吸道传染病,主要感染幼犬, 发病急,传播快。出现咳嗽、流涕、发热
(4)犬腺病毒1型(传染性肝炎)
潜伏期2~5天。体温升高,随后降低。呕吐,腹痛腹泻。多数两周内死亡,致死率10~25%。
肝炎病变:蓝眼睛(角膜水肿和前葡萄膜的炎症)。肝脏增大,色淡;胆囊壁增厚,出血,呈黑红色。
潜伏期在7天内
呼吸道症状:持续性高热、先是阵发性干咳,后是湿咳并有痰液,浆液性至粘液性鼻漏、扁桃体炎、喉气管炎和肺炎。
潜伏期1~8天,感染途径是消化道。
胃肠道症状:反复呕吐 ,粪便由糊状、半糊状至水样,橙色或绿色,含黏液或血液 (血便),随着日龄的增长,死亡率降低。
•猫咪容易感染的病毒
猫瘟(猫泛白细胞减少症)、猫病毒性鼻气管炎、猫杯状病毒和猫冠状病毒。
一些病毒感染的临床表现
猫瘟(猫泛白细胞减少症 ):
潜伏期为2-9天 ,传染途径是接触带病毒的尿粪或经吸血昆虫及蚤类
发热为40℃左右,顽固性呕吐 ,腹泻后期带有血液,呈咖啡色 ,脱水、眼球下陷 ,循环血液中的白细胞减少 。
猫病毒性鼻气管炎:
潜伏期约2~6天
阵发性喷嚏和咳嗽 ,羞明、流泪、结膜炎,鼻腔分泌物增多 ,鼻液和泪液初期透明,后变为粘脓性。
急性病历通常持续10~14天。成年猫死亡率较低。
猫杯状病毒:
潜伏期约2—3天 ,传播方式:直接接触,间接接触。
浆液性和粘液性鼻漏。口腔溃疡是该病特有的症状,并且有时是唯一的症状。
注意
猫狗的真菌病和病毒通常只在宠物之间传播,但也有可能会对人类健康造成影响。
这些真菌和病毒可以通过接触患病宠物或宠物的环境而传播给人类,引起皮肤感染、呼吸道感染或是消化系统感染等疾病。
特别是对于免疫力较弱的人群,如老年人、儿童、孕妇等,感染的风险更高。
例如近日陕西西安,一名9岁女孩突然发高烧到40度,伴随寒颤四肢发凉,头痛精神差。
经检查确诊猫抓病,猫抓病是由汉氏巴尔通体感染引起的感染性疾病,主要宿主就是猫狗,部分孩子被抓咬也很容易感染,被抓伤后一定要及时处理。

因此,为了自己和家人的健康,我们需要注意以下几点:
1.定期带宠物进行检查,确保宠物健康;
2.及时给宠物接种疫苗,预防宠物感染病毒;
3.定期给宠物清洁并驱虫,避免宠物身上携带寄生虫;
4.在和宠物玩耍后洗手,避免细菌病毒通过手传播。
饮食、年龄、种族、生长环境、绝育、疾病状况和相关疗法在内的多种因素已被证明会影响猫狗的微生物组成。
Garrigues Q,et al.Front Vet Sci.2022
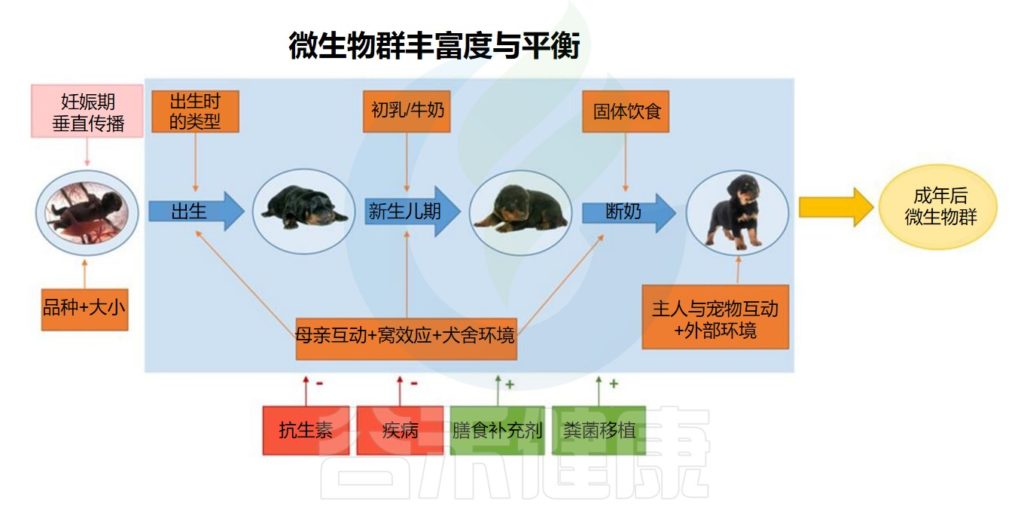
1
饮食
饮食已被认为是影响哺乳动物肠道微生物群生物多样性和功能特征的主要驱动因素之一。
家犬目前被认为是杂食动物,商业宠物食品的配方旨在提供均衡的营养摄入,同时补充高浓度的纤维和碳水化合物。
而结肠较短的家猫仍然被认为是专性食肉动物。尽管如此,家猫的商业饲料富含植物来源的成分。
✦常量营养素含量对狗微生物群组成影响显著
一项研究发现,与具有相似常量营养素含量的传统(动植物混合)膨化饮食相比,仅用植物来源的蛋白质制备的膨化饮食不会显著改变狗的微生物组。
另一项针对健康狗的研究测试了宏量营养素成分的重大变化,其中包括4种为减肥、肾脏疾病、低脂肪或抗过敏而配制的处方饮食。减肥饮食在常量营养素(更高的蛋白质和纤维)方面发生了最剧烈的变化,并导致微生物群组成发生了最大的变化。
✦高蛋白低碳水饮食更适合猫
研究评估了断奶后使用高蛋白/低碳水化合物饮食 (HPLC)的小猫与断奶后使用中等蛋白质/中等碳水化合物 (MP/MC) 饮食的小猫的微生物组。
发现高蛋白/低碳水饮食增加了物种多样性,在高蛋白/低碳水喂养的小猫中增加5个属是已知的丁酸盐生产者。
罐头饮食是一种额外的高蛋白替代品,通常单独喂养或与膨化饮食结合使用。研究表明,猫食用罐头会增加水的摄入量,从而减少自主能量摄入量并降低尿液比重,这可能对某些健康状况有益。
高蛋白饮食对猫肠道微生物组成的影响


Pilla R,et al.Vet Clin North Am Small Anim Pract.2021
✦以生肉为主的饮食
近年来,用生肉饮食(RMBD)代替更传统的商业干粮来喂养猫狗已经变得越来越流行。生肉饮食提供重要的健康益处,包括减少牙齿疾病和清新口气、缓解关节炎、增强免疫反应、更健康的皮肤和闪亮的皮毛。
然而生肉饮食也会改变宠物体内的微生物组成,增加接触人畜共患病原体的风险。
以生肉为主的饮食对猫狗肠道微生物的影响
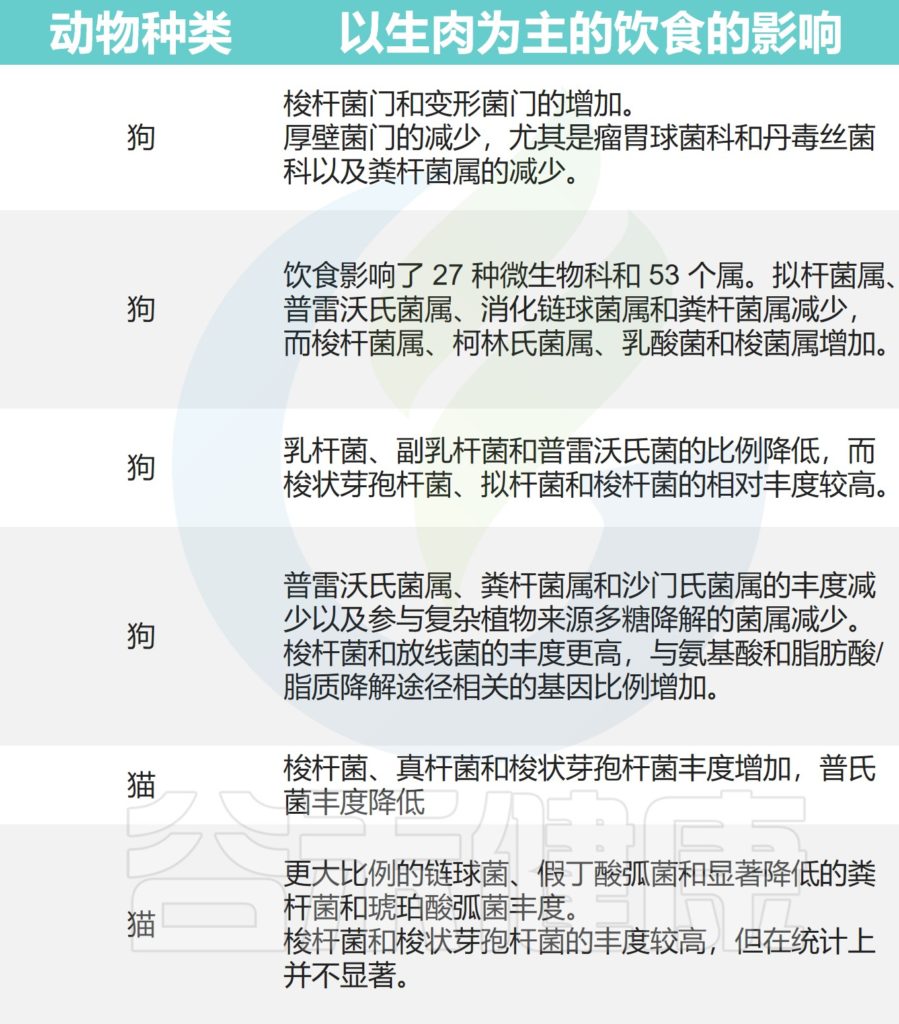
Alessandri G,et al.Microb Biotechnol.2020
总而言之,这些研究证据强调了不同饮食成分对猫狗肠道菌群的塑造作用。需要根据宠物具体的状况选择合适的饮食。
2
年龄
在影响肠道微生物群的众多因素中,年龄是对微生物组成影响最大的因素之一。肠道微生物群的发育在出生时就开始了,并且其组成会随着宿主生命的不同阶段而不断演变。
关于与年龄相关的差异,常见的发现是随着年龄的增加,肠道微生物组多样性减少,乳酸杆菌增多。在一些研究中,观察到梭杆菌随着年龄的增长而显著下降,这与在人类中的结果相反。
梭杆菌在人类百岁老人中的流行率(阳性样本的百分比)并不低于老年人和年轻人。
接下来以狗为例,具体表述一下不同生命阶段微生物群的差异。
幼犬从出生到成年菌群的变化
Garrigues Q,et al.Front Vet Sci.2022
✦产前暴露和出生时肠道定植
最初人们认为,哺乳动物的子宫内胎儿期是无菌的,在分娩后的最初几个小时内,通过接触母亲的阴道、皮肤和摄入母乳来接种微生物。
细菌可能在子宫内就从母体转播给胎儿
这种想法最近受到了挑战,因为分子技术的出现允许检测不同哺乳动物胎盘、子宫或羊水中的细菌,细菌可能在子宫内从母亲传播给胎儿。
通过分析胎粪和胎盘样本的微生物群组成,探索了狗胎儿宫内细菌定植的可能性:在出生后立即收集的86.5%的胎粪样本和57%的胎盘样本中检测到细菌。
在幼犬和人类中,分别来自厚壁菌门和变形菌门的葡萄球菌属、链球菌属和Neisseria zoodegmatis是从胎粪和胎盘中分离出的最常见的细菌。
葡萄球菌似乎是母犬子宫内膜微生物群中最常见的属之一,而链球菌更多地存在于母犬的阴道中,这证实了通过阴道分娩出生的幼犬胎粪微生物群部分类似于母犬阴道,并支持微生物的潜在经胎盘转移。
✦幼崽哺乳期乳杆菌丰度增加
出生后,新生小狗的胃肠道很快被微生物定植并且非常不稳定。在生命的前2天,微生物群大约60%的细菌群落的厚壁菌门为主。
注:这个时期微生物群的低微生物丰度和多样性促进了外部细菌的潜在定植。
虽然厚壁菌门在最初2日时主导肠道,但在出生后的头几周其相对丰度显著下降,梭状芽孢杆菌属从2日龄幼犬中鉴定出的总序列的10%开始下降, 在3周时下降到1%。
尽管厚壁菌门的相对丰度总体下降,但幼犬肠道中乳杆菌科的丰度却增加了100倍。结合这些细菌消化牛奶低聚糖和产生乳酸的能力,这表明幼犬肠道丰度的增加不仅与氧稳态有关,还与幼犬在整个新生儿期摄入的母乳有关。
从2天到21天,细菌丰富度显著增加。这些信息表明,幼犬胃肠道细菌的重要变化发生在生命的最初几周,甚至在幼犬开始吃固体食物之前。
✦断奶引起的微生物群变化
狗的断奶被描述为幼犬饮食从母乳逐渐过渡到固体饮食,通常发生在3周大左右,结束于8周左右,此时小狗与母亲分开并且没有无法再喝奶了。
断奶标志着幼犬肠道菌群建立和发展的重要一步,因为新型食物的到来促进了某些菌群的丰度和活动。
拟杆菌丰度增加
如前所述,拟杆菌从第2天的 <1% 丰度增加到第56天的39%, 并一直增加到成年。
拟杆菌是狗肠道中一种多糖降解菌,多糖对于断奶后的幼犬来说是必不可少的,因为它们的饮食开始主要由富含复合碳水化合物的宠物食品组成。
梭杆菌丰度增加
虽然不像拟杆菌那么重要,梭杆菌在断奶后也看到其相对丰度增加,与狗的年龄呈正相关。
梭杆菌可以发酵蛋白质和氨基酸以产生短链脂肪酸和支链挥发性脂肪酸。可以假设断奶后梭杆菌丰度的增加与肉类产品的摄入有关。
✦成年后肠道微生物群丰度趋于稳定
犬科动物和猫科动物的肠道微生物群在整个成年期保持稳定,到老年时会发生进一步变化。衰老通常与营养、生活方式和生理学的持续改变有关,伴随着免疫系统的衰退(免疫衰老),导致慢性低度炎症,并对肠道微生物群落产生影响。
3
品种
品种是微生物群个体间变异的主要来源。专门探索狗粪便微生物群差异的研究发现不同品种之间的α或β多样性没有差异,但微生物组成存在差异。
✦相同饮食和居住条件下不同犬菌群丰度不同
在马尔济斯犬中,梭杆菌(Fusobacteria)数量丰富,而在贵宾犬中,厚壁菌(Firmicutes)和放线菌(Actinobacteria)数量丰富。即使在相同条件下饲养并接受相同饮食时也是如此。
然而,由于在这些研究中观察到的大多数狗都超过1岁,该品种对成长中狗肠道细菌群的影响仍需要更多探索。
✦同一窝的幼崽菌群更相似
此外,与来自不同窝的幼崽相比,来自同一窝的小狗显示出更相似的双歧杆菌种群。这些发现支持这样一种观点,从母亲到她的后代的垂直传播在新生儿肠道微生物群的生物多样性和组成方面发挥着关键作用。
4
绝育
绝育对猫狗健康的影响是一个备受关注的话题,目前有一些研究表明,绝育可能会对宠物的肠道微生物组成产生影响。
一项研究发现,绝育后的雄性猫狗肠道中的某些菌群数量会发生变化,如乳杆菌(Lactobacillus)、双歧杆菌(Bifidobacterium)等菌群的数量会减少,而肠球菌(Enterococcus)、大肠杆菌(Escherichia)等菌群的数量会增加。这些变化可能与代谢、免疫等方面有关。
需要注意的是,目前绝育对宠物肠道微生物的影响还需要更多的研究来证实,而且不同的绝育方式(如手术绝育、化学绝育等)可能会对肠道微生物产生不同的影响。
此外,宠物的饮食、生活环境等因素也会对肠道微生物产生影响,因此在绝育后需要注意宠物的饮食和生活环境,以维护宠物的肠道健康。
5
健康状况及病原体感染
消化系统疾病如腹泻、便秘、胃肠炎等会对猫狗微生物群产生影响,破坏肠道微生物群的平衡,导致有益菌的减少,有害菌的增加。
免疫系统疾病如自身免疫性疾病、过敏等也会对猫狗微生物群产生影响,破坏肠道微生物群的平衡,导致免疫系统功能异常。
炎症性肠病期间肠道微生物群的紊乱
Hernandez J,et al.Microorganisms.2022
✦感染寄生虫改变肠道微生物群
许多肠道寄生虫被证明会引起宠物肠道微生物群的显著改变,其中贾第鞭毛虫是一种普遍存在的肠道寄生虫,可导致腹泻,其带来的改变最为明显。
贾第鞭毛虫与许多细菌群落的丰度呈正相关,例如普雷沃氏菌(Prevotella)和厌氧螺菌(Anaerobiospirillum succiniproducens)。这些细菌会导致肠道屏障粘液的脆弱化。这种脆弱化使肠贾第鞭毛虫更容易切割屏障并允许更多肠道病原体在肠道定植。
此外,在22周大的幼犬中,贾第鞭毛虫的高负荷也与约氏乳杆菌(Lactobacillus johnsonii)的减少相关。
这种细菌是幼犬特有的,并且由于免疫调节、病原体抑制和上皮细胞附着特性,可能在幼犬肠道健康的早期发育中发挥重要作用。
✦病毒感染影响肠道微生物群
犬细小病毒是影响狗的最常见病原体之一,会导致腹泻、出血性肠炎和幼犬死亡。在一项研究中,四只幼犬在6周大时自然感染了犬细小病毒,肠道微生物群发生严重改变,变形杆菌丰度增加,主要是肠杆菌科(Enterobacteriaceae),拟杆菌(Bacteroidetes)和梭杆菌(Fusobacteria)丰度减少。
有趣的是,犬细小病毒阳性幼犬的这些细菌变化并不是永久性的。在感染后2周(一旦从临床细小病毒病中恢复),微生物组成又恢复到与非犬细小病毒感染组相似的组成。
6
其他外源化学物质
宠物受益于先进的兽医护理,并接受与人类相同类别的药物治疗(如抗生素、抗炎药、疫苗等)。与此同时,其中的化学成分也会对宠物的肠道微生物群产生影响。
✦药物改变了宠物肠道菌群丰度
近年来,一些研究评估了某些异生素对人类和宠物狗肠道微生物群的影响。给健康狗服用奥美拉唑会导致厚壁菌门和梭杆菌门的比例升高,胃幽门螺杆菌数量减少,十二指肠总细菌数量增加。
双酚A的影响可以推断到人类,因此,宠物被认为是人类健康的生物哨兵。
✦食品内的化学物质对宠物肠道菌群也有影响
除了出于医学原因服用外源性药物外,人类和宠物狗每天都会接触到影响宿主和肠道微生物群的环境化学物质。研究证明了双酚A(一种广泛存在于食品罐头中的内分泌干扰化学物质)对宠物肠道细菌组成的影响。
在许多细菌属和物种中发现了干扰,包括拟杆菌属、均匀拟杆菌、瘤胃球菌属、罗氏菌属、巨单胞菌属、梭杆菌属、链型杆菌属(Catenibacterium)和普氏粪杆菌。
这里的健康我们分为两部分来讲,一个是影响宠物自身的健康,一个是与主人之间的健康关联。

1-影响宠物自身健康
微生物群可以影响宿主的许多方面,包括生理、行为、繁殖和健康。
胃肠道微生物群促进食物分解以及代谢物的产生,如短链脂肪酸、次级胆汁酸等。微生物群还将营养物质和代谢物释放到体内,影响免疫细胞和炎症反应等。
这里我们从宠物的肠道菌群参与消化吸收、影响肠道屏障、影响代谢、免疫、神经系统等多方面来了解肠道菌群具体如何影响宠物健康。
参与消化吸收
肠道微生物可以分解和代谢宠物食物中的一些难以消化的成分,如纤维素、淀粉等,从而帮助宠物消化和吸收营养物质。
例如,碳水化合物发酵导致短链脂肪酸的产生,短链脂肪酸可以为肠道上皮细胞的生长发育提供营养支持,促进肠道上皮的生长与分化。
蛋白质发酵产生短链脂肪酸、氨基酸和支链脂肪酸。
√ 影响微量元素的合成吸收
同时,肠道菌群还参与合成动物生长发育必需的氨基酸、维生素等,如B族维生素、维生素K、烟酸、泛酸等。
此外,肠道中微生物还能够能促进机体对钙、铁、镁、锌等多种离子的吸收,这些离子对于促进身体某些结构的生长与发育,如:骨骼、牙齿等,对体内氧的输送等有重要作用。
影响肠道屏障
胃肠道微生物群促进定植抵抗,通过竞争排斥提供针对潜在病原体的微生物屏障。这层“菌膜屏障”就像是一层保护伞,对于不慎食入的病原菌进行抑制并排斥,维持体内微生态的平衡状态。
宠物肠道中一些有益菌可以与肠道上皮细胞相互作用,促进上皮细胞的修复和再生,增强肠道屏障的完整性,减少有害物质的渗透。
影响免疫功能
√ 影响先天免疫和适应性免疫
例如,在健康成年猫中补充嗜酸乳杆菌(Lactobacillus acidophilus)可增加外周血粒细胞的吞噬能力并降低血浆中内毒素的浓度。
此外,双歧杆菌(Bifidobacterium)能通过刺激免疫细胞产生重要的细胞因子白介素来促进动物机体内重要的免疫细胞——淋巴细胞的增殖、分化、成熟,增强免疫细胞的对病原体的杀伤力。
√ 刺激宿主产生各种抗菌化合物
例如抗菌肽。微生物代谢物也可以诱导抗菌肽表达:例如,短链脂肪酸和石胆酸可通过不同途径诱导抗菌肽LL-37的表达,这在哺乳动物针对侵袭性细菌感染的先天免疫防御中起着关键作用。
肠道微生物可以与宠物的免疫系统相互作用,调节免疫系统的功能,从而帮助宠物应对病原体的侵袭。
影响代谢
肠道微生物可以产生一些有益的代谢产物,如短链脂肪酸等,这些代谢产物可以维护肠道黏膜屏障的完整性,抑制有害菌的生长,从而维护肠道健康。
肠道微生物组进行的代谢对宿主的影响


Ziese AL,et al.Vet Clin North Am Small Anim Pract.2021
√ 影响代谢相关疾病
•肥胖
肠道微生物群通过其对肠道的直接影响及其对远端器官的间接影响,与肥胖的发展有关。
肠道微生物群会影响胆汁酸的代谢;细菌代谢产生的游离胆汁酸可以抑制细菌种群的生长,例如乳酸杆菌和双歧杆菌,它们被认为可以预防肥胖。
•糖尿病
各种微生物研究表明,胃肠道微生物群在糖尿病等肠外疾病中发挥作用。改变的肠道微生物群组成与猫和狗的糖尿病的发展有关。
例如,最近的一项研究表明,患有1型糖尿病的狗肠道菌群失调,粪便中浓度发生变化。
此外,与同龄的健康猫相比,患有糖尿病的猫的肠道微生物多样性显著降低,产生丁酸盐的细菌也减少了。
•肾脏疾病
慢性肾脏病是猫和狗最常见的疾病之一,有研究发现肾脏病与肠道微生物群之间也存在关联。
与健康猫相比,患有慢性肾脏病的猫的粪便微生物组的丰富性和多样性降低,这与先前对患有慢性肾脏病的人类肠道微生物的研究一致。
影响神经系统
越来越多的证据表明肠道微生物群与身体的主要神经内分泌系统,即下丘脑-垂体-肾上腺(HPA)轴之间的密切相互作用,并将其称为“脑-肠微生物轴”。
√ 影响情绪和行为
该系统控制应对压力的各种身体过程,从而影响宠物的行为和情绪,如焦虑、抑郁等。有研究发现肠道微生物群的失衡会导致猫狗出现抑郁情绪。
√ 记忆力较弱的狗放线菌相对较多
在一项研究中,测量了宠物狗的肠道微生物群组成,并检查了它与年龄和记忆力之间的联系。
表现较差(即记忆力较弱)的狗的粪便样本中放线菌(Actinomycetes)相对较多。这一结果与人类阿尔茨海默病患者中某些放线菌的高丰度一致。
注:该研究的局限性包括犬数量相对较少、用于测试行为关联的样本量较小以及品种和喂养方式差异较大。
因此,宠物的肠道微生物群作为一项重要的健康指标,可以比较全面地了解宠物的肠道微生物组种类、丰度及整体健康状况,包括菌群失调、疾病风险的评估等;还可以在一定条件下判断宠物的情绪和行为。
这些数据可以帮助兽医或宠物主人更好地了解宠物的状况,并根据检测结果制定相关的治疗或饮食计划。
注:相比于普通的检查,肠道微生物检测只需提取少量粪便,不用带宠物去医院,更加方便与安全。
2-宠物菌群与主人健康的关联
研究发现宠物的肠道微生物群与其主人的肠道微生物群存在一定的相似性,因此,通过观察宠物的肠道微生物群情况,可以一定程度上反映出主人的肠道微生物状况及健康。
具体来说,以下是一些可能的方式:
√通过宠物的肠道微生物群反映主人的菌群
研究表明,宠物和主人的肠道微生物群中存在一些共生菌群,如双歧杆菌、乳酸菌等,因此可以通过宠物的肠道微生物群反映主人的肠道微生物群状况。
√通过宠物的微生物群反映主人的疾病风险
宠物与其主人共享家庭环境,因此暴露于相同的环境因素。一些研究表明,与没有疾病的狗相比,患有非传染性疾病的狗的主人更容易患上述疾病。据报道,与非糖尿病犬相比,患有糖尿病的犬的主人患2型糖尿病的风险增加。
宠物和主人的肠道微生物群健康状况与一些疾病的风险存在一定的关联,如肥胖、糖尿病、心血管疾病等,因此可以通过宠物的肠道微生物群反映主人的疾病风险。
√反映主人的生活方式
宠物和主人的生活方式也会对肠道微生物群产生影响,如饮食、运动、应激等,因此可以通过宠物的肠道微生物群反映主人的生活方式。
在谷禾肠道菌群检测报告中,就有这样的特殊案例:
报告显示,该宠物样本菌群构成非常单一,实际测序深度很高,达到10万,但仅检出167种菌,绝大部分是大肠杆菌。
下面的表是根据人的菌群结果评估的,可以看到构成多样性非常低。
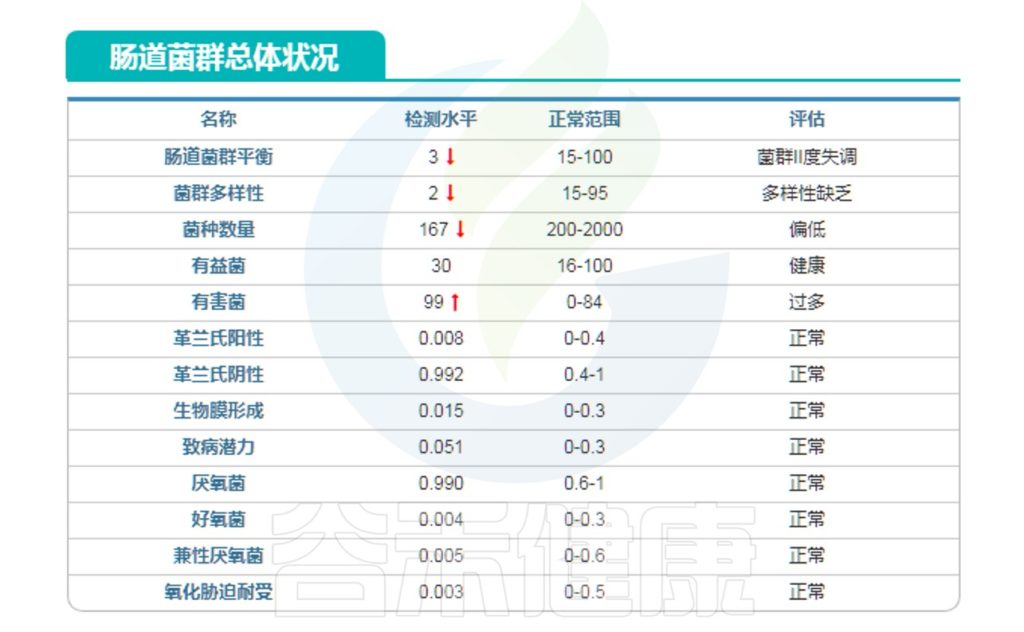
下面是主要菌门和属还有种的构成,种部分列出了注释有菌名称的丰度大于0.1%以上的菌。
下图是菌属构成表:
可以看到,菌属构成中弯曲杆菌占比较高。
除此之外,其他的病原和机会致病菌主要检出了大肠杆菌,占比77%,属于严重超标,空肠弯曲杆菌,产气夹膜菌和痢疾志贺氏菌都有超标:
注:空肠弯曲菌(上图中红色箭头指向菌Campylobacter jejuni),有内毒素能侵袭小肠和大肠黏膜引起急性肠炎。
经过沟通后,我们了解的情况如下:
主人反映该宠物猫有腹泻,其主人也有长期腹泻情况,因此送检了其本人和宠物猫的粪便样本做肠道菌群检测,了解菌群构成,找出腹泻原因,以便后续治疗。
因此,宠物的微生物群组成在一定程度上可以算是主人的健康哨兵。
随着年龄、生活方式或药物的影响,健康宠物的肠道微生物组平衡可能受到影响甚至会被破坏,从而引发各种疾病。我们应该采取何种方法来恢复其健康呢?
肠道微生物群可以通过饮食调整、抗菌剂、益生元和益生菌以及粪菌移植进行调节。这些治疗方法每一种都具有不同的优点和副作用。因此必须考虑潜在的疾病,因为没有针对所有类型生态失调的治疗方法。
饮食是影响宠物肠道微生物群的重要因素之一,不同的饮食组成会对宠物肠道微生物群的生态平衡产生不同的影响。
根据不同宠物制定相对应的饮食
蛋白质、碳水化合物和脂肪是大部分宠物必需的营养物质,是促进机体生长的重要营养素。合理控制这些营养素的摄入是维持肠道健康的关键。
不同年龄段的宠物对营养素的吸收能力不同。老年犬猫由于肠道消化功能逐渐退化,不能较好地吸收,需要根据不同的宠物状态制定相对应的饮食。
适量的功能性脂肪酸,如Omega-3脂肪酸可以增强宠物机体免疫力,预防炎症,减轻其胃肠道疾病的发生。
注意宠物粮成分
宠物食品制造商在其产品中添加的一些成分会对猫狗的肠道菌群和整体健康产生负面影响。
人工色素和抗氧化剂等食品添加剂减少肠道细菌的数量。在购买前最好能阅读食物的配料表,选择适合自家宠物当前阶段的粮食。
谷禾在此提出了两种饮食建议供大家参考
•“无谷物狗粮”
存在关于无谷物狗粮的争论。有些兽医认为不应该用任何谷物喂养宠物,有些人则说它可以适量喂养。
不建议吃的原因包括体重增加、胃肠道炎症增加、过敏等。如果狗不耐受谷物,可能会出现皮肤发痒、呕吐、胃部不适、脱毛、腹泻、感染和胀气。
换句话说,如果出现上述症状,可以尝试一下无谷物食物并监测症状是否有所改善。
•在饭菜中加入骨汤
骨汤含有多种有益营养素,但与肠道健康最相关的是胶原蛋白。胶原蛋白是一种结缔组织,有助于维持包括肠道和皮肤在内的身体器官的结构。
狗狗的肠道内部衬有一层叫肠粘膜的组织。粘膜的作用是防止食物分子和细菌等危险物质从肠道进入血液。
然而,粘膜可能会受损,导致肠道变得可渗透。换句话说,像细菌这样的物质可以通过。这就是所谓的“肠漏”。
通过给狗狗吃富含胶原蛋白的食物,比如骨头汤,可以加强粘膜中的结缔组织,从而减少肠漏程度。
研究发现压力会引起体内激素的变化,从而引起肠道微生物组的变化。特别是,压力会触发皮质醇激素的释放。皮质醇对身体有多种负面影响,包括血压升高和肠道健康受损。
保持良好的心情有利于宠物健康成长,以下是一些有助于宠物减轻压力的方法:
1.提供足够的运动和活动:适当的运动和活动可以帮助宠物消耗能量,缓解压力。主人可以带宠物出门散步、玩耍等。
2.提供安全感:宠物可能会因为分离焦虑或其他原因感到不安。主人可以给宠物提供一个安全的环境,例如提供一个温暖舒适的床铺、一个安静的角落等。
3.提供适当的社交环境:宠物需要与其他宠物和人类进行社交互动。主人可以带宠物参加社交活动,例如去公园或参加宠物聚会等。
4.适当的训练:训练可以帮助宠物建立自信和独立性,从而减少压力。
益生菌、益生元的施用也会影响和改变微生物群的组成。益生菌和益生元已经在人类中广泛运用,在宠物中也同样适用,现在许多宠物粮中已经添加了益生菌。
许多在宠物健康方面研究的益生菌属于乳杆菌属、双歧杆菌属和肠球菌属。
益生菌可以通过多种机制改变常驻微生物组,包括通过代谢相互作用刺激常驻细菌的生长,改变病原菌的丰度,或通过与宿主上皮细胞和上皮免疫系统的相互作用间接影响健康。
益生菌对狗的治疗作用

Mondo E,et al.Open Vet J.2019
一些研究报告了在宠物饮食中添加益生元的好处。事实上,它们能调节肠道微生物群并保护动物免受肠道感染。
猫和狗接受益生元的治疗效果

Mondo E,et al.Open Vet J.2019
注意
一些人建议益生菌中使用的细菌最好来自宿主同一物种的肠道;然而,对于目前市场上用于猫和狗的大多数益生菌而言,情况并非如此,并且没有研究将狗或猫来源的益生菌的功效与来自其他物种的市售菌株进行比较。
对市售兽医益生菌的审查揭示了质量问题,包括标签不准确和生存能力差; 其他人对安全性提出了担忧,还需要更多研究来了解在猫和狗中使用益生菌的效果。
当饮食调整不起作用时,患有慢性肠病的狗和猫会接受抗生素治疗。
常用的抗生素:
甲硝唑
甲硝唑对粪便微生物组和代谢组有显著影响,无论是单独使用还是与水解蛋白饮食结合使用。
施用甲硝唑后,微生物组丰富度降低。微生物组的变化伴随着粪便乳酸杆菌增加、粪便和血清中氧化应激标志物增加以及胆汁酸转化受损。
泰乐菌素
泰乐菌素用于治疗对泰乐菌素有反应的慢性腹泻,这通常会影响成年犬。许多研究强调了它的效率,但其作用机制仍然未知。
恩诺沙星
恩诺沙星是另一种用于治疗肠病的抗生素。它是一种氟喹诺酮,可用于治疗肉芽肿性结肠炎。
使用抗生素同时还会减少有益菌
需要注意的是,使用抗生素治疗某些疾病存在禁忌症。事实上,大量使用抗生素可能会减少有益菌的数量,促进潜在病原体数量的增加,并促进抗菌素耐药现象的发生。
在停用抗生素后,微生物的变化可能会持续数年,而一些细菌类群的数量可能永远不会恢复到初始状态。
一些研究人员表明,抗生素实际上会改变微生物组成,从而降低细菌多样性,因此它们会导致生态失调的发生。在过去的几年中,抗菌素耐药性的现象不断增加,并具有重要意义。人类与宠物的密切关系甚至会导致抗菌素耐药性细菌转移人体。
调节肠道微生物群的一种新方法是“粪便微生物群移植”。粪菌移植是指将来自供体的粪便物质溶液施用到受体中,主要是为了改变受体的微生物组成。
该手术可通过十二指肠镜检查、鼻胃管/鼻空肠管、结肠镜检查、灌肠或口服胶囊进行。
患有肠病的狗经粪菌移植后得到改善
报告证实一只患有难治性炎症性肠病的贵宾犬在粪菌移植后也产生了积极作用。这只狗通过灌肠接受了九次粪菌移植。6个月后,狗的临床炎症性肠病活动指数和粪便稠度得到改善。
另一项研究描述了一只患有慢性结肠炎和艰难梭菌感染的8个月大的法国斗牛犬。这只狗接受了单次口服粪菌移植。排便频率和粪便稠度在2至3天后显著改善,至少6个月未观察到复发。
对患有急性腹泻的犬只进行一次粪菌移植效果非常成功。使用粪菌移植代替抗生素可以预防负面后果,例如微生物多样性降低、特定细菌类群发生变化、丰度和代谢转变。
粪菌移植对宠物其他慢性疾病也具有治疗效果
粪菌移植在治疗患有慢性肠病或胰腺外分泌功能不全等慢性疾病的狗时也显示有效的结果。需要注意的是,在粪便移植几天后病情有所好转,但随后往往会复发。因此,在大多数情况下可能需要多次粪菌移植。
如今,在动物实践中,粪菌移植有可能改善与生态失调相关的急性和慢性疾病的健康状况。由于对其使用的研究较少,其标准化需要进一步研究。
噬菌体是一种能够侵染细菌的病毒,是生物圈中广泛分布,但是关于这方面的研究和应用还较少。
噬菌体可以裂解消灭有害菌
噬菌体通过尾丝蛋白与宿主菌表面受体特异性结合侵染细菌使其裂解,精准消灭有害菌,不影响有益菌。
30分钟内可杀灭99.9%的宿主菌,且不受细菌耐药性限制。
面对细菌耐药、抗生素残留等问题,噬菌体作为天然的细菌杀手,是防治细菌性疾病的最佳选择。
有研究表明,噬菌体可以裂解肠道中的有害菌例如沙门氏菌和大肠杆菌,降低了肠道有害菌数量,有益于肠道有益菌(如乳酸菌类)的繁殖和生长,起到了调节肠道菌群的作用。
噬菌体还有利于免疫器官的发育和体液免疫功能的提高。
研究宠物的微生物群不仅有益于其自身的健康,也有益于它们主人的健康,因为伴侣动物与人类具有相同的生活环境、相似的饮食模式和微生物群落。
对于猫狗的肠道菌群仍然有很多未知和亟待发掘的意义,有望成为一种非常有价值的检测和干预渠道。谷禾欢迎更多相关领域的合作伙伴共同推进和完善猫狗相关肠道菌群的研究和应用。
主要参考文献
Tian T, Zhou Y, Xu Y, Xu Y. Intestinal microbial 16S sequencing and LC-MS metabonomic analysis revealed differences between young and old cats. Heliyon. 2023 May 19;9(6):e16417.
Sinkko H, Lehtimäki J, Lohi H, Ruokolainen L, Hielm-Björkman A. Distinct healthy and atopic canine gut microbiota is influenced by diet and antibiotics. R Soc Open Sci. 2023 Apr 26;10(4):221104.
Garrigues Q, Apper E, Chastant S, Mila H. Gut microbiota development in the growing dog: A dynamic process influenced by maternal, environmental and host factors. Front Vet Sci. 2022 Sep 2;9:964649.
Deschamps C, Humbert D, Zentek J, Denis S, Priymenko N, Apper E, Blanquet-Diot S. From Chihuahua to Saint-Bernard: how did digestion and microbiota evolve with dog sizes. Int J Biol Sci. 2022 Aug 1;18(13):5086-5102.
Li Y, Ali I, Lei Z, Li Y, Yang M, Yang C, Li L. Effect of a Multistrain Probiotic on Feline Gut Health through the Fecal Microbiota and Its Metabolite SCFAs. Metabolites. 2023 Feb 3;13(2):228.
Pilla R, Suchodolski JS. The Gut Microbiome of Dogs and Cats, and the Influence of Diet. Vet Clin North Am Small Anim Pract. 2021 May;51(3):605-621.
Balouei F, Stefanon B, Sgorlon S, Sandri M. Factors Affecting Gut Microbiota of Puppies from Birth to Weaning. Animals (Basel). 2023 Feb 6;13(4):578.
Kubinyi E, Bel Rhali S, Sándor S, Szabó A, Felföldi T. Gut Microbiome Composition is Associated with Age and Memory Performance in Pet Dogs. Animals (Basel). 2020 Aug 24;10(9):1488.
You I, Kim MJ. Comparison of Gut Microbiota of 96 Healthy Dogs by Individual Traits: Breed, Age, and Body Condition Score. Animals (Basel). 2021 Aug 18;11(8):2432.

谷禾健康

人类微生物组研究正在从描述关联发展到了解整个微生态对人类的影响。虽然存在挑战,但在应用数据驱动的微生物组诊断和干预方面正在取得进展,这可能会在未来十年内带来精准医学的突破。
本文我们来探讨关于微生物组的研究进展及其对人类健康的影响及应用。
随着对微生物组的理解深入,发现巨大的潜力
过去 20 年,我们对微生物组在人类宿主生理学中作用的理解发生了重大转变。
开创性研究表明,与人类细胞和基因相比,原核生物为人类“全生物”贡献了相同数量的细胞和多倍的基因,同时影响哺乳动物从消化和新陈代谢过程到免疫甚至大脑功能。
微生物组的变化与越来越多的人类疾病有关,包括癌症。这些发现使人们普遍希望,积累的关于宿主-微生物组相互作用的知识,将迅速转化为人类疾病的治疗方法。
对微生物组的认识开始客观,同时意识到复杂性
随着该领域的稳步成熟,科学家们开始更好地了解整个微生物组。他们意识到这并不像他们最初想的那么简单。随着他们开始了解微生物之间的相互作用是多么复杂,以及将微生物组用于人类治疗有多么大的挑战,最初围绕它的兴奋和炒作开始平息。
面临挑战,需要综合多方面的努力
要克服这些挑战,许多研究者需要进行长期、数据驱动的研究。他们要综合各种看起来不相干的科学和医学学科,包括进化论、人类学和营养学等等,还要结合新技术的开发和优化,以及注意事项和混杂因素的识别和最小化,共同推动了这一探索。他们的目标是要弄清楚宿主和微生物组之间的互动关系和分子机制,以期找到因果关系。虽然充满了挑战,但只要大家协作一致,一定可以获得成果。这些成果将在未来十年内实现对可利用的治疗性微生物组目标进行更广泛和可重复的识别和调节。
微生物组研究中的挑战和新兴方法
doi: 10.1371/journal.pbio.3002053
本世纪第一个十年,研究微生物组结构表征、对生理功能的重要性,由此衍生到各个领域
事实上,本世纪微生物组研究的第一个十年,集中于证明肠道微生物组对生理功能的重要性,以及在各种生理和疾病背景下基于下一代测序的微生物组群落结构表征。
这带来了微生物组与各种健康结果之间的关联、相关性和预测的识别,报告规模和细节的不断提升。与此同时,该领域广泛描述了微生物特征响应多种因素的动态调节,例如饮食、宿主免疫 、基因组学和昼夜振荡。
▼
随着这些重大进展,人们也认识到了主要的技术和概念障碍,这些障碍对微生物组发现的解释、概括和转化到临床具有挑战性。
使微生物组相关观察结果的解释复杂化的技术挑战之一是:不同研究之间样本收集、处理和分析方法的广泛差异。
此外,在微生物组处理和分析级联的每个步骤中发生的污染和注释错误可能会引入伪影和偏差,这些伪影和偏差难以解决并使真实和虚假生物信号之间的区别复杂化。
解决方案:将真实信号与混杂因素分开
了解与这些步骤中的每一个相关的范围、限制和混杂因素,对于将真实信号与混杂因素分开至关重要,特别是在微生物丰度低或无微生物丰度的样本。更好地协调这些方法,加上多种技术和生物控制,可以更准确地解释可以在人群、地域、性别和种族之间推广和复制。
然而,如果不冒技术可变性和多样性减少的风险,微生物组处理和分析标准化就不能过度执行,而技术可变性和多样性构成了推动研究创新的引擎。
▼
大多数人类微生物组研究面临的一个概念挑战是,微生物群落结构中内在的和生理上重要的个体间变异性。
如何在大样本人群数据上去识别和消除这些差异,应该是目前微生物组特征分析面临的巨大分析挑战,因为它们代表了一种固有的、难以解决的“噪音”。
解决方案:基于深度人工智能的方法等创新方法
可以识别和利用不同的微生物组配置来探索微生物组对个体人类表型的贡献,范围从血糖反应到补充剂和食物携带相同疾病易感位点的个体之间的不同疾病表现。
总而言之,使用基于深度人工智能的方法等创新方法应对技术和生物变异性的挑战,将加深微生物组知识与精准医学的合理整合。
▼
一个同样艰巨的挑战涉及从微生物组-人类特征关联到因果关系证明的进展。事实上,在建议与临床结果相关、相互关联和预测的许多微生物群中,只有少数基因产物和代谢物可以证明对生理和疾病状态有因果影响,而我们预计一部分微生物特征是先发于疾病病理改变的特征,而有一部分特征是继发于以疾病状态为特征的环境变化。
越来越多的工具正在开发中,通过揭示因果关系和机制洞察力,将前者“驱动”微生物组改变与后者“乘客”改变区分开来。
解决方案:无菌或使用抗生素的动物模型或其他改进的技术,测试单个细菌菌株或群落的影响
一种经常使用的方法依赖于微生物群耗尽(通过抗生素治疗)或微生物群缺乏(无菌)小鼠模型,这些模型允许引入单个细菌菌株或群落以测试它们对宿主生理学的影响。
抗生素杀菌模型局限性:多因素混杂,难以预测
抗生素是现代医学的基石,因为它们能够在感染期间抑制病原体的生长。抗生素的广泛特征是它们的抑制机制是抑菌(生长停止)或杀菌(致死),并且它们的活性范围是宽的或窄的。大多数抗生素的作用主要在物种单一培养中进行了研究,尽管临床环境中的治疗不可避免地会对人类肠道上的多物种群落产生意想不到的后果。抗生素会对肠道细菌造成附带损害,并降低肠道微生物群的多样性,由此可能出现艰难梭菌感染倾向增加和癌症治疗后死亡率增加。
抗生素扰动期间的群落动态很难预测,因为环境可以通过种间相互作用、pH 调节和代谢转化改变抗生素的作用。使用多种治疗方案,例如可变剂量、顺序安排多种化合物和同时使用鸡尾酒治疗使预测进一步复杂化。
同时多种药物可产生协同或拮抗作用, 由此杀死的程度分别大于或小于单独药物的总和,但是他们对肠道菌群的扰动和影响研究还甚少,检测中面临这些用药信息准确跟踪记录也较困难。而且抗生素导致的多重耐药细菌的定植也会导致抗生素耐药基因转移到微生物群中,最终会导致对各种有效抗生素的耐药性增加,并增加致命感染的风险。
其他新技术:改进的体内模型、类器官和片上器官平台
随着更多具有人类代表性的实验平台的出现,野鼠体内的微生物群与圈养外的家鼠相似,正迅速受到关注。这些改进的体内模型得到了合成人类共生群落的发展、以前“无法培养”微生物在菌株水平分辨率下的改进培养和表征,以及通过非靶向代谢组学和蛋白质组学对生物活性微生物效应因子或代谢物的无偏见阐明的补充。
类器官和片上器官平台能够研究人体组织环境中的单个细菌、细菌群落及其生物活性产物。
注:
类器官(organoid)是指由人体组织或干细胞等体外培养出的三维结构,能够模拟某些人体器官的结构和功能。类器官的制备通常是通过将干细胞或组织细胞在特定的培养条件下进行分化和自组装,形成类似于人体器官的结构。类器官的制备可以为疾病研究和药物筛选提供更真实、更可靠的模型。例如,肝脏类器官可以用于测试药物代谢和毒性,肠道类器官可以用于研究肠道微生物和肠道疾病,而大脑类器官可以用于研究神经退行性疾病等。
片上器官平台(organ-on-a-chip platform)是一种仿生学技术,它是通过微流控技术、微加工技术和3D打印技术等手段,将人体组织和器官的细胞培养在微型芯片上,模拟人体器官的生理、病理过程,以实现高通量、高精度的药物筛选、毒性测试和疾病研究。这种技术可以模拟人体器官的微环境,如细胞间相互作用、细胞信号传递、细胞分化、细胞迁移、细胞凋亡等生理和病理过程,从而更好地研究疾病的发生和发展,以及评价药物的安全性和有效性。
凭借这些新技术获得的能力,该领域正在揭示基因组测序之外的功能读数,解码难以培养和低生物量微生物组的贡献,并评估重要但研究不足的非细菌共生王国及其错综复杂的相互作用组网络。
事实上,对共生和机会病毒(包括噬菌体)、真菌和寄生虫的潜在功能及其对细菌共生生态系统和人类宿主的影响的不断探索,构成了正在推进的研究领域,当然肠道细菌还是最主要的。对这些特征不明确的共生微生物的研究需要进一步开发各种工具,包括改进的计算参考数据集、分子开发工具和体内定植模型。
通过对微生物分子和功能见解的不断扩展,可能会在微生物组研究的下一个时代开发广泛的诊断、预后和治疗人类应用。
治疗性微生物组使用的一个有前景的例子:
粪便微生物组移植 (FMT)
除了治疗常见的艰难梭菌感染之外,显示出对肝病、炎症和免疫相关疾病的积极改善效果
促进微生物组研究领域的蓬勃发展
失败的试验和风险强调了对机制的了解的重要性
为了避免人体微生物研究领域中早期技术失误:
为了辅助人类疾病的诊断和治疗需要:
作为向前迈出的积极一步,微生物组研究能够做到以下:
早期检测
识别和利用微生物组粘膜,免疫和全身信号,以便通过离散的疾病特征进行早期检测和患者分层。
治疗层面
“一刀切”的传统观念正在发生改变,人们不再简单地认为所有人的微生物组都可以通过相同的方式进行调节,而是转向将微生物组视为“信号中心”的观点,也就是说更加注重研究和借鉴微生物群落的信号传递机制,以更好地治疗相关疾病。
微生物组被视为特定环境的中继或缓冲器,整合环境和内源性信号,影响携带健康或疾病易感遗传特征的个体的生理和疾病表现。
一个备受关注的话题是“益生菌干预法”。证据表明益生菌可能有助于治疗和预防传染病,改善过敏,降低血糖,联合免疫治疗提高治疗效果。益生菌治疗会改变腔内微生物群,这种改变进一步会显着影响全身代谢,包括胰岛素抵抗,炎症,代谢等。
将微生物组作为一个高度模块化的“指纹”,有助于开发合理的、针对具体情况的和数据驱动的干预措施,例如个性化营养、精准益生菌、代谢物补充(“后生元”疗法)和定向病原体抑制模式。
这些方法的验证和整合,加上对大量微生物群及其相关生物活性产品对人类宿主的影响的持续探索,将有助于实现该领域改变人类健康的巨大前景。
参考文献
Puschhof J, Elinav E. Human microbiome research: Growing pains and future promises. PLoS Biol. 2023 Mar 17;21(3):e3002053.

谷禾健康

我们知道,口服抗糖尿病药是治疗糖尿病的有效方式之一。然而,患者对抗糖尿病药的反应程度各不相同,例如,有些患者在长期使用阿卡波糖后会产生耐药性。
阿卡波糖通常在饭前口服。它抑制人α-葡萄糖苷酶达到降血糖作用,包括唾液和胰淀粉酶,从而减缓复杂碳水化合物的消化,并降低餐后血糖水平的升高。
大部分阿卡波糖剂量在肠道中仍未被吸收,导致与肠道微生物组的强烈相互作用。
近期发表在《自然新陈代谢》杂志上的一项研究揭示了阿卡波糖“诱导降解失活”的新机制:肠道菌群可以将阿卡波糖降解,从而破坏阿卡波糖的药效。

首先鉴定肠道细菌K. grimontii TD1,在阿卡波糖失活中起重要作用。
进一步在K. grimontii TD1 中鉴定出一种阿卡波糖首选葡萄糖苷酶 Apg ,它可以将阿卡波糖降解为小分子,并失去抑制剂功能。
由于阿卡波糖被肠道细菌降解,相对较大的人群可能存在阿卡波糖耐药性风险,这也是非抗生素耐药性的一个临床相关例子。
研究过程
研究人员招募了 393 名2型糖尿病患者,发现:
阿卡波糖的治疗效果存在显著的个体差异。
在接受α-葡萄糖苷酶抑制剂阿卡波糖治疗的2型糖尿病患者队列中,无应答者的粪便样本显示出明显高于有应答者的药物降解。
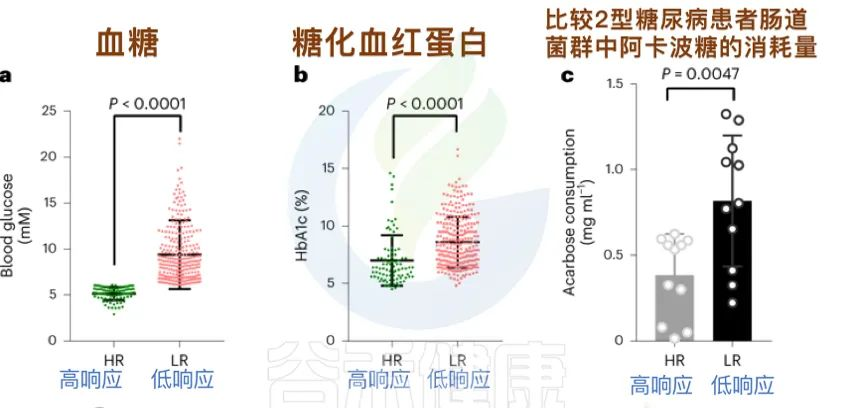
Tian J, et al., Nat Metab. 2023
这指向了微生物介导的药物降解途径。
为了分离阿卡波糖降解细菌种类,作者在逐步增加阿卡波糖浓度的条件下,对药物没反应的人的粪便样本进行培养,以富集细菌。
Tian J, et al., Nat Metab. 2023
这种方法导致分离出18种阿卡波糖降解剂,所有这些降解剂都属于Klebsiella grimontii。
Klebsiella grimontii 是兼性厌氧菌,属于变形菌门,肠杆菌科,产酸克雷伯菌复合体的一种,主要分布于小肠。
产酸克雷伯菌是人类肠道中的共生菌群,但也是一种机会致病菌,在近年来的临床分离的微生物样本中呈逐年上升的趋势。产酸克雷伯菌复合体可产生许多毒素,因其携带较多的毒力因子,并且具备携带和获得耐药基因的能力,故未来可能成为威胁人类健康的潜在病原菌,需要更多研究。
K. grimontii 这种兼性厌氧细菌主要定植于小肠,巧合的是,阿卡波糖主要靶向小肠上皮细胞中的多种淀粉酶和葡萄糖苷酶,这在空间上有利于其与K. grimontii TD1 的相互作用以及该菌株的代谢降解。
▶▷▶▷
事实上,在糖尿病小鼠模型中:
K. grimontii TD1菌株削弱了阿卡波糖的降血糖作用。
作者制作了一个T2DM小鼠模型。将小鼠分为四组,并用磷酸盐缓冲盐水、阿卡波糖、K. grimontii TD1或阿卡波糖和K. grimontii TD1处理。
阿卡波糖单独可以降低淀粉负荷后的血清葡萄糖水平;然而,这种作用在接受阿卡波糖和K. grimontii TD1的组中不太明显。这些发现表明,K. grimontii TD1降低了阿卡波糖的抗糖尿病作用。
由此,确定了降解阿卡波糖的细菌。
利用转录组学,研究人员在以淀粉为唯一碳源的最低培养基中培养了K. grimontii TD1,使细菌生长依赖于淀粉酶。通过添加阿卡波糖来抑制淀粉酶活性,迫使K. grimontii TD1上调有助于降解阿卡波糖的基因,这可以通过RNA测序进行量化。
同时,利用蛋白质组学,作者分离出细胞裂解物和具有高阿卡波糖降解活性的分离部分。这些组分中的蛋白质通过质谱法鉴定。
转录组学和蛋白质组学的发现都指向一个共同的候选者:一种新的葡萄糖苷酶,作者将其命名为阿卡波糖首选葡萄糖苷酶(简称Apg)。这是由K. grimontii TD1引起的阿卡波糖失活的关键。
在体外测定中或当在大肠杆菌中异源表达时,该酶能够将阿卡波糖水解为阿卡波糖葡萄糖和阿卡波糖。这两种产物在很大程度上都不能抑制猪α-淀粉酶。
很有意思的是,该酶在pH为7.5和温度为37°C的厌氧条件下表现最好,这些条件与人类肠道的生理学相似。

Tian J, et al., Nat Metab. 2023
那么这就引发了一个问题:
阿卡波糖如何成为细菌葡萄糖苷酶的底物,但又是人类葡萄糖苷酶(如α-淀粉酶)的抑制剂?
通过3D分子建模,作者发现葡萄糖苷酶Apg催化位点的残基与人类α-淀粉酶中的残基位置不同,这使得阿卡波糖能够水解裂解。
阿卡波糖的降解产物太小,无法有效地占据人类α-淀粉酶的配体结合口袋,导致酶的抑制作用降低。
这种独特的蛋白质结构不仅有助于理解这种酶的独特催化机制,而且为设计能够在体内阻断这种酶的抑制剂提供了潜在的靶点。
在上述2型糖尿病患者队列中,超过98%的阿卡波糖治疗患者携带具有Apg样酶的细菌,相比之下,约14.2%的健康个体携带Apg+细菌。
地理位置、性别、BMI等因素决定了其患病率和丰度的个体间差异,在分析中,Apg的丰度在不同人群之间有所不同,在中国人中最为普遍。
这些发现表明,肠道微生物群中的Apg和Apg样酶可能会影响许多患者的药物疗效和治疗成功率。
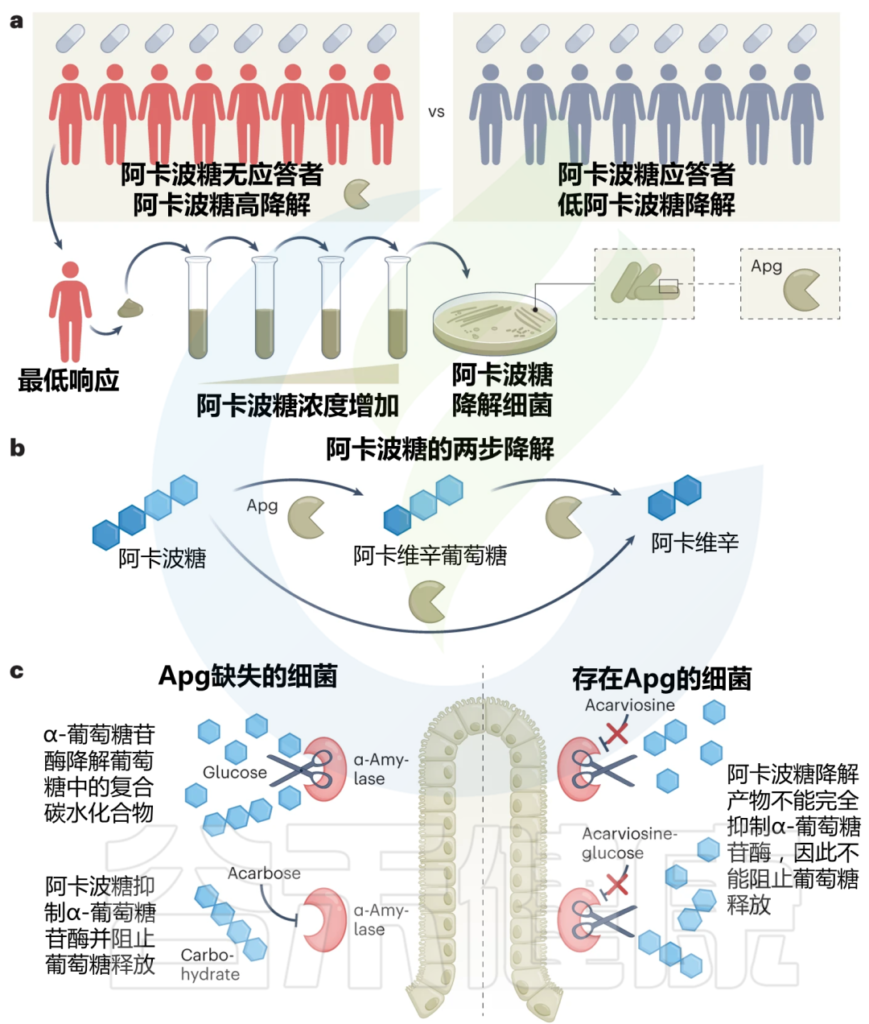
Brauny MM, et al., Nat Metab. 2023
▶ 01
这与已经描述的涉及阿卡波糖磷酸化的途径不同。阿卡波糖激酶在人类微生物组中广泛存在。这表明阿卡波糖抗性是一种常见的防御策略,在不同的肠道微生物中独立出现。为什么阿卡波糖抗性是肠道中的一个有益特征尚不清楚;也许是为了解毒相关的内源性化合物,或者成功地确保复合碳水化合物作为食物来源。
▶ 02
其他阿卡波糖降解葡萄糖苷酶以前也有描述,但从未与处理结果直接相关。由于碳水化合物代谢的基因经常过度表达,并且在肠道细菌的基因组中表现出相当大的差异,可以想象,很快就会发现更多这样的酶。
▶ 03
预计其他α-葡萄糖苷酶抑制剂以及其他基于碳水化合物的药物和食品添加剂,都可能会经历类似的过程。因此,这项研究可能只是冰山一角,很快可能会有更多这样的观测结果。
▶ 04
这种方法只需要关键细菌能够被培养。在目前的研究中,作者在纯培养中工作,但这种方法在细菌群落的培养中也应该是可行的。
总的来说,很大一部分2型糖尿病患者可能由于高水平的K. grimontii TD1而对阿卡波糖产生耐药性。对于阿卡波糖反应低的患者,可以考虑调节肠道微生态或靶向代谢酶的策略,以减少耐药性的发生。肠道微生物群的分析可用于辅助指导未来的治疗。
当然,K. grimontii TD1不太可能是肠道微生物群中唯一可以降解阿卡波糖的成员,可以对阿卡波糖降解肠道细菌以及相关代谢机制进行持续研究,从而更全面地了解微生物组与阿卡波糖的相互作用。
在一定条件下,肠道菌群可能利用一种主要机制,或几种共存方式来支持其生存,这需要更多的临床病例分析和深入研究来阐明其潜在机制,从而指导精准医疗。
微生物药物代谢很重要。更好地了解微生物组编码的药物降解途径,对于药物开发、个性化治疗及有效的治疗策略等方面具有巨大潜力。
本文提供了一种研究思路,微生物研究人员可以从中学习如何通过实验设计鉴定药物降解酶,这有助于开发个性化及有效的治疗策略。
药物开发相关人员可以考虑微生物代谢途径对药物吸收、代谢和治疗效果的影响,并在此基础上加强药物设计和开发过程中的微生物学考虑。
一些具体实验细节设计,详见原文:
Tian J, Li C, Dong Z, et al., Inactivation of the antidiabetic drug acarbose by human intestinal microbial-mediated degradation. Nat Metab. 2023 May;5(5):896-909.
谷禾健康
假单胞菌属(Pseudomonas)是最多样化和普遍存在的细菌属之一,其物种存在于沉积物、临床样本、植物(或植物根际)、患病动物、水、土壤、海洋、沙漠等,这反映在它们多变的代谢能力和广泛的适应环境的潜力上。
假单胞菌属是一类革兰氏阴性的好氧或微需氧的细菌,属于 Proteobacteria(变形菌门),代表种包括:
Pseudomonas aeruginosa(铜绿假单胞菌)
Pseudomonas putida (恶臭假单胞菌)
图源:CDC
假单胞菌属是机会性病原体,是全世界医院获得性感染的主要原因。它可以侵入多个器官系统,包括血液、肺、软组织(皮肤、肌肉和肌腱)、身体的其他部位。症状取决于感染部位。
与假单胞菌属感染相关包括免疫系统受损患者的感染和医疗器械相关感染等,如刚接受过手术的人和免疫功能低下的人,艾滋病或糖尿病患者。
假单胞菌属与人体健康相关,其中一些菌株可以引起感染和疾病,如铜绿假单胞菌(Pseudomonas aeruginosa),可以引起呼吸道、肠道、泌尿道和皮肤感染。
免疫抑制个体和住院患者的铜绿假单胞菌肠道携带率显着升高,因此感染和抗生素相关性腹泻的风险增加。
假单胞菌属与其他菌属如葡萄球菌、普雷沃氏菌属、不动杆菌属、芽孢杆菌属、肠杆菌属、肠球菌属等菌属有互作关系。
假单胞菌不被认为是肠道微生物组的典型成员,并且假单胞菌在肠道定植与发生肺部感染和死亡率的风险增加有关。
研究表明,假单胞菌与肠道屏障功能障碍和感染有关,可能导致败血症和多器官功能障碍综合征。此外,假单胞菌与炎症性肠病的发生也有关系,它在IBD患者的肠道中检测到丰度更高,而且它参与引起其他菌群的失衡会影响疾病的进展。
细菌有可能在人体内的不同部位之间转移,防止肠道定植或肠道到其它器官尤其肺部的传播,可能是预防重症患者假单胞菌感染的有效策略。
本文主要分享假单胞菌属(Pseudomonas)的生态,分类,致病因素以及与人体相关健康特性和可能的生活干预措施。
假单胞菌属(Pseudomonas),是革兰氏阴性、好氧菌属,于 1894 年被描述,是最多样化和普遍存在的细菌属之一。有些物种对人类、动物或植物具有致病性。
▼
假单胞菌属(Pseudomonas)属于假单胞菌科,包含200多个有效描述的物种。该属的成员表现出大量的代谢多样性,因此能够在广泛的生态位中定居。
它们易于体外培养,并且越来越多的假单胞菌菌株基因组序列可供使用,这使得该属成为科学研究的绝佳焦点。
Photograph by Kateryna Kon
研究最好的物种包括铜绿假单胞菌。直的或略微弯曲的棒状但非螺旋状,0.5–1.0 × 1.5–5.0 µm。由一根或数根极鞭毛运动;很少不动。在某些物种中,也可能形成短波长的侧鞭毛。有氧,具有严格的呼吸型代谢,以氧为末端电子受体;在某些情况下,硝酸盐可用作替代电子受体,从而允许厌氧生长。
大多数物种(不是全部)无法在酸性条件(pH 4.5 或更低)下生长。大多数物种不需要有机生长因子。氧化酶阳性或阴性。过氧化氢酶阳性。化学有机营养的。
▼
——分布广
假单胞菌属是变形菌门的一个庞大而多样的属。从北极苔原到热带雨林,从干旱的土壤到雨云,几乎在地球的每个角落都可以找到该属的成员。这种令人难以置信的环境适应性归功于假单胞菌非凡的新陈代谢多样性。
——野蛮生长
假单胞菌可以在 0 至 42 °C 的温度范围内生长,甚至可以在更极端的温度下存活。它们对营养的需求很少,可以利用多种碳源。尽管假单胞菌在有氧环境中生长最佳,但它们也可以利用氮进行厌氧呼吸。
它们可以作为自由生活的浮游细胞或作为生物膜群落的成员生活,并且具有将微生物信号和环境线索转化为特定生态位过程的非凡能力。
基因组分析表明,许多其他因素有助于假单胞菌属的多样性和适应性。除了赋予调节和代谢灵活性的常见基因的大量等位基因差异外,水平基因转移影响了致病性假单胞菌属的能力。
▼
第一个假单胞菌基因组测序于 2000 年,有 630 万个碱基对,是当时最大的细菌基因组测序。基因组包含大量参与分解代谢、运输、流出、运动和信号响应调节的基因。
事实上,铜绿假单胞菌基因组中超过 8% 的基因被认为参与了调控,这远远超过了在任何其他细菌基因组中观察到的百分比。很明显,假单胞菌成功的关键在于它可以表达其基因的可塑性,这是由多层监管复杂性提供的。

Sem Photograph by David M. Phillips
自 2000 年以来,已测序的 1000 多个假单胞菌基因组中的绝大多数都是铜绿假单胞菌的临床菌株。
总的来说,铜绿假单胞菌基因组的主要部分(约 4000 个基因)在所有菌株中都是保守的,代表了“核心基因组”。多达另外 20% 的基因驻留在共同代表“附属基因组”的基因组岛上。正是这个附属基因组赋予了铜绿假单胞菌的可塑性,并包含许多参与代谢、毒力和抗生素抗性的基因。
由于已在测序分离株的附属区域鉴定出大约 10 000 个独特基因,因此估计铜绿假单胞菌泛基因组可以接近甚至超过 100 000 个基因,这意味着假单胞菌这一物种的遗传库将远远超过人类。
该属的分类学多年来一直存在争议,因为许多最初包含在假单胞菌属中的细菌类群已被重新归类为其他属或来自不同类别的变形杆菌的物种。
可以根据16S rRNA、gyrB、rpoB和rpoD基因的序列分为三个谱系和13个群。
临床常见菌种主要包括:
• 铜绿假单胞菌(Pseudomonas aeruginosa)
• 斯氏假单胞菌(Pseudomonas stutzeri)
• 恶臭假单胞菌(Pseudomonas putida)
• 荧光假单胞菌(Pseudomonas fluorescens)
• 门多萨假单胞菌(Pseudomonas mendocina)
• 产碱假单胞菌(Pseudomonas alcaligenes)
• 假产碱假单胞菌(Pseudomonas pseudoalcaligenes)
其它菌种见附录一
致病性菌株见附录二
▼
详见附录三
此外,铁是细菌必需且稀缺的营养素,也是重要的信号因子。铜绿假单胞菌已经开发出多种铁吸收系统来隔离足够的铁以维持其生存,在毒力因子的释放和生物膜的形成中具有重要的调节作用。
▼
假单胞菌适应进化的特性对人类既有益又有害。
——有益
一方面,我们利用假单胞菌的力量进行生物修复和生物防治。例如,荧光假单胞菌和P. protegens已被证明在害虫控制和作物保护方面特别成功,它们被认为可以战胜和/或对抗植物病原体。
假单胞菌的分解代谢能力也被用于杀虫剂、重金属和碳氢化合物(例如石油泄漏)以及许多其他污染物的生物降解和/或解毒。
——有害
另一方面,某些种类的假单胞菌对植物和动物具有致病性,导致极难根除的感染。例如,铜绿假单胞菌是世界范围内医院获得性感染最常见的原因之一,并在使用消毒剂和抗生素后存活下来。
它可以在许多解剖部位引起感染,包括皮肤、大脑、眼睛、耳朵、泌尿道、肺部等。免疫抑制的个体,尤其是那些有过度烧伤、囊性纤维化或中性粒细胞减少症的个体,尤其处于危险之中。(这在后一章节会详细介绍)
▼
一般非特异性症状(许多疾病中常见的症状)包括:
假单胞菌感染的症状取决于它们发生在身体的哪个部位。
假单胞菌(Pseudomonas)可引起以下器官感染:
耳朵(外耳炎或游泳者的耳朵):
疼痛、水肿(肿胀)、压痛和分泌物
皮肤:
假单胞菌通过压疮、烧伤和手术伤口或从血流进入菌血症患者的皮肤,还可能引起毛囊炎等。
具体可表现为:脓肿(感染肿块)、皮疹、脓液、皮肤溃疡、坏死组织(皮肤变黑,表明组织正在死亡)。
骨骼或关节:
受影响区域行动不便、关节或背部疼痛、肿胀、疼痛或发热。
这些通常是由于静脉内 (IV) 药物使用或感染的手术伤口或受伤造成的伤口细菌传播到皮肤、骨骼和关节的结果。
伤口:
伤口有分泌物或结痂(通常是黄绿色),可能有甜味或难闻的气味。
消化道:
阑尾炎(由于治疗导致白细胞低的人发生肠道感染)腹痛、腹胀和腹泻的症状。
肺:
假单胞菌可能会导致严重的肺炎,通常发生在住院、已经生病和免疫功能低下的人身上。它可引起囊性纤维化患者的慢性肺部感染(遗传病),支气管扩张(气道永久性扩张),或慢性阻塞性肺病。
具体可表现为:呼吸急促、胸痛、咳痰(伴有白色、黄色或绿色粘液)和充血。
泌尿道:
排尿疼痛、尿液难闻或有甜味、尿急、尿液浑浊。
因此,假单胞菌感染的特征在于广泛的症状。
▼
紧急情况下:呼吸急促、心率加快、脉搏微弱、意识模糊、发烧、发抖或剧烈疼痛是败血性休克的症状,是对血流感染的全身反应。
在极少数情况下,心脏瓣膜可能会受到感染,尤其是在静脉注射药物的人群中。
脏东西的细菌可以进入血液,会传播到心脏并感染心脏瓣膜,这种情况称为心内膜炎。
使用静脉注射药物的人出现新的杂音(心音异常)可能会怀疑与假单胞菌相关的心内膜炎。
▼
未经治疗、治疗不足或复发的假单胞菌感染会导致更广泛的感染,例如败血症,这可能是致命的。
更可能出现严重并发症的高危人群包括:
值得注意的是,假单胞菌属内的抗生素耐药性正在增长。这些细菌正在寻找新的方法来避免死于以往最强大有效的抗生素。
假单胞菌属与人体健康相关,其中一些菌株可以引起感染和疾病,如Pseudomonas aeruginosa 绿脓杆菌,又称铜绿假单胞菌,可以引起呼吸道、泌尿道、皮肤感染。虽然不是正常口腔菌群的成员,但在口腔疾病和牙齿矫正治疗中可能会富集。
▼
假单胞菌属不被认为是肠道微生物群落的典型成员,肠道定植假单胞菌属与发展肺部感染和死亡风险增加有关。
假单胞菌属的过度生长可能会导致肠道炎症和系统性炎症反应。
假单胞菌属感染会引起机体的Th17细胞反应和系统性炎症反应,因此,干预Th17细胞免疫反应可能是缓解肺炎的有效方法。此外,假单胞菌属也可以通过产生GABA来影响蛋白质代谢和细胞稳态,从而影响宿主的摄食行为。
▼
假单胞菌属与其他菌属如大肠杆菌、沙门氏菌等有关,与肠道微生物群的平衡和健康密切相关。
假单胞菌属与其他菌属如葡萄球菌属和普雷沃氏菌属等,共同存在于囊性纤维化患者的肺部感染中。
假单胞菌属与其他菌属如不动杆菌属(Acinetobacter)等一起被发现在冷鲜乳中,产生脂肪酶和蛋白酶,参与乳制品和肉类的变质。
假单胞菌属与其他菌属如Bacillus、Enterobacter、Enterococcus等一起参与了肠-肺轴的微生物和免疫调节,影响呼吸道疾病的发生和发展。
▼
与假单胞菌属相关的疾病包括:免疫系统受损患者的感染和医疗器械相关感染等。免疫抑制个体和住院患者的铜绿假单胞菌肠道携带率显著升高,因此感染和抗生素相关性腹泻的风险增加。
研究表明,假单胞菌属与肠道屏障功能障碍和感染有关,可能导致败血症和多器官功能障碍综合征。假单胞菌属与肺部疾病也有关联,如支气管扩张症。
肺部疾病
研究发现,肺部假单胞菌属感染会影响肠道菌群,而通过调节肠道菌群可以影响肺部疾病的治疗效果。
肠道定植通常先于肺部感染,并且相同菌株经常在肠道和肺部中发现,表明肠道充当了假单胞菌属的储存库,可以传播到肺部和其他感染部位。
肠道微生物群的免疫调节对呼吸道疾病的发生有影响,肠道共生菌通过屏障效应提供菌群抵抗力,从而保护肠道生态环境不被改变。
以肺部假单胞菌、肠道拟杆菌和肠道酵母菌为特征的高肠-肺相互作用集群,与加重的恶化,整体支气管扩张严重相关。
炎症性肠病(IBD)
假单胞菌属与炎症性肠病的发生也有关系。
研究表明,IBD患者肠道中假单胞菌属的种类和数量与非IBD患者存在显著差异。
其中,克罗恩患者中假单胞菌属的检出率明显高于非IBD患者。此外,假单胞菌属的多样性在活动性IBD患者中也明显降低。这些结果表明假单胞菌属可能与IBD的发病机制有关。具体而言,假单胞菌属可能通过影响肠道菌群的平衡和多样性,导致肠道免疫系统的异常反应,从而引发IBD的发生。
因此,调节肠道菌群可能是预防和治疗肠道疾病的一种新策略。
▼
铜绿假单胞菌(Pseudomonas aeruginosa)是一种普遍存在的革兰氏阴性细菌,存在于许多不同的环境中,包括不同的生命来源,如动物和人类。
铜绿假单胞菌能够以最低的营养需求生存并耐受各种身体条件,这使得该生物体能够在社区和医院环境中持续存在。
图源:EYE OF SCIENCE / SCIENCE PHOTO LIBRARY
引发感染
铜绿假单胞菌是一种机会性病原体,这意味着它利用宿主防御系统中的一些漏洞来引发感染。
铜绿假单胞菌是世界范围内医院感染的罪魁祸首,其医学和兽医重要性日益增加,并且主要在免疫系统受损的患者中引起感染。
这种细菌几乎从不感染未受损的组织,但如果组织防御以某种方式受损,它就会发起攻击。
铜绿假单胞菌可能引起尿路感染、呼吸系统感染、皮炎、软组织感染、菌血症、骨和关节感染、胃肠道感染和各种全身感染,特别是在严重烧伤患者和免疫抑制的癌症和艾滋病患者中。
在免疫缺陷的人群中,如严重烧伤患者、人工呼吸机使用者和支气管扩张症患者。
在囊性纤维化患者中,铜绿假单胞菌是慢性肺部感染的主要来源,导致患者的发病率和死亡率增加。
铜绿假单胞菌感染会导致肺部炎症和肺损伤,引起细胞浸润和肺泡水肿。在肺移植患者中,铜绿假单胞菌的分离率更高,但与非囊性纤维化肺移植患者相比,死亡率没有增加。铜绿假单胞菌感染会导致肺血管通透性增加,从而引起肺水肿。
耐药性
铜绿假单胞菌显示出对抗生素的高内在耐药性,并具有获得新耐药机制的非凡能力。由于临床治疗选择有限,多重耐药 (MDR) 铜绿假单胞菌分离株的流行率不断上升是一个全球性健康问题。
对来自西班牙两个不同地区的儿童粪便样本中铜绿假单胞菌的发生率、抗菌素耐药性表型、毒力基因型和遗传谱系表征数据显示:从1,443 个粪便样本中,有72份分离分离出来铜绿假单胞菌,并检测到低水平的抗菌素耐药性:头孢他啶 (8%)、头孢吡肟 (7%)、氨曲南 (7%)、庆大霉素 (3%)、环丙沙星 (1%) 和亚胺培南 (1%);对美罗培南、阿米卡星、妥布霉素、左氧氟沙星和粘菌素敏感,此外还发现了四种多重耐药菌株。
定植
人类粪便样本中的定植率范围为 2.6% 至 24 %。关于铜绿假单胞菌分离株在健康人粪便中定殖的数据很少。
其他研究表明,肠易激综合征患者的小肠和粪便样本中铜绿假单胞菌的定植率很高。迄今为止,关于铜绿假单胞菌分离株在健康人粪便中的定植及其抗菌素耐药性和毒力特征的数据相对较少。
尽管肠道微生物群作为抵御肠道病原体的屏障,但铜绿假单胞菌通过产生一系列令人印象深刻的毒力因子,克服了肠道微生物群和先天免疫系统介导的定植抵抗。
铜绿假单胞菌凝集素 LecA 和 LecB 以及细胞外附属物,如鞭毛和菌毛,在细菌附着到宿主上起着重要作用,并与粘附有关。
此外,铜绿假单胞菌携带属于双伙伴分泌(TPS)家族(也称为Vb型分泌系统,T5bSS)的大型蛋白质系统,TPS系统主要由分泌的TpsA效应蛋白及其TpsB伙伴转运蛋白组成。TpsA1 (CdiA2, 573 kDa) 和 TpsA2 (CdiA1, 361 kDa) 通过接触依赖性生长抑制系统 (CDI) 以及粘附和生物膜形成促进细菌竞争。
因此 TPS 系统效应子被指定为对革兰氏菌有益的主要毒力决定因素阴性病原体。
▼
铜绿假单胞菌等的特殊能力引起如此多样化感染的假单胞菌属是因为它们能够产生名副其实的毒力因子库,包括毒素、生物膜、蛋白酶和溶血素。
考虑到铜绿假单胞菌的医学重要性,假单胞菌领域的大部分研究工作都致力于了解影响这些毒力因子释放的调节、生物合成和环境因素。
此外,它们还可以获得赋予进一步抗菌素耐药性的移动遗传元件;因此,可以鉴定出泛耐药菌株。
▼
假单胞菌感知和适应环境的能力导致了一系列活动,例如许多酶和其他生物分子的分泌。虽然这些毒素、降解酶和抗菌剂可以将假单胞菌的环境转变为理想的微生物生态位,但它们可能对植物和动物宿主造成严重后果。例如,绿脓素是一种蓝/绿色色素毒素,它赋予铜绿假单胞菌培养物特有的颜色,并作为一种抗菌剂可以杀死竞争性微生物。然而,它也会破坏真核细胞过程,这会对人类细胞产生不利影响。
▼
过去几年的工作提供的证据表明,群体感应是一种通用的调节机制,它允许细菌以种群密度依赖的方式发起统一、协调的反应,以完成即使不是不可能,也很难完成的任务。
群体感应系统广泛存在于假单胞菌中,人类机会性病原体铜绿假单胞菌,属于研究最广泛的细胞间通讯系统。在这种生物体中,群体感应非常复杂,由两个相互关联的 N-酰基高丝氨酸内酯 (AHL) 依赖性调节回路组成,这些调节回路由非 AHL 相关信号分子和许多调节剂进一步调制,这些调节剂在转录过程中起作用和转录后水平。这种遗传复杂性可能是导致铜绿假单胞菌具有巨大环境多样性的关键因素之一。
过去几年的工作表明,群体感应对于一系列毒力因子的表达以及铜绿假单胞菌中生物膜的形成至关重要,因此代表了设计用于治疗铜绿假单胞菌的新型药物的有吸引力的目标感染。
此外,细胞间通讯能力也在许多额外的假单胞菌中得到证实。铜绿假单胞菌,因此可以用于设计新型药物来治疗铜绿假单胞菌感染。
长时间住院、创伤ICU住院、消毒干预等与肠道假单胞菌过度生长有关,而TAP暴露与抗菌剂干预则与肠道假单胞菌过度生长有负相关。
注:TAP的暴露是指肠道上皮细胞上的TAP暴露在肠道腔中,从而使肠道内的微生物可以通过TAP进入肠道上皮细胞内部。一些研究表明,TAP的暴露与肠道微生物群失调和某些疾病(如炎症性肠病)有关。而抗菌剂干预可以抑制肠道内细菌的生长,从而减少TAP的暴露,因此与假单胞菌肠道过度生长有负相关。
对于免疫功能低下的患者,应特别注意水的纯度,有些水中可能含有假单胞菌等细菌。
研究发现用海洋益生元岩藻依聚糖补充饮食可以通过抑制分泌性毒力因子 (TpsA/CdiA) 与粘蛋白的相互作用并促进有益种群的生长,从而介导铜绿假单胞菌从肠道菌群失调和去定植中的早期恢复。
芪归饮是一种用于治疗铜绿假单胞菌感染的中药。在门和属水平上显著抑制脱铁杆菌和粘孢子菌的过度积累。鉴定了11种潜在的代谢产物,这些代谢产物在铜绿假单胞菌感染中异常表达,服用芪归饮显著逆转。
此外,部分研究表明以下物质也可以抑制肠道内的假单胞菌属。
假单胞菌属相比于其它细菌引起人们广泛的研究兴趣,该属不仅包括人类病原体铜绿假单胞菌,还包括一系列与植物致病性、生物修复和环境微生物学相关的其他重要物种。
假单胞菌被广泛用作模型原型细菌,用于研究宿主-病原体相互作用、细胞-细胞通讯系统、进化生物学、基因调控和代谢网络、分泌系统、抗生素(耐药性)、生物修复、生物膜、细菌基因组学以及与微生物学和分子生物学广泛相关的许多其他主题。
假单胞菌属因其复杂的信号和调节系统,特别是细胞密度依赖性群体感应而被广泛研究。未来只有了解复杂调节回路中涉及的机制,才能确定新型抗菌方法的可能目标。铜绿假单胞菌中的锌、铁稳态是替代抗菌策略的一个有希望的目标。
由于其复杂性和适应性,它存在于无数的环境和临床生态位中,以及它在不同层面(种内、种间和与其宿主)相互作用的能力,假单胞菌属很可能在未来许多年内吸引更多研究人员。我们期待未来有更好的解决假单胞菌属,尤其人类病原体铜绿假单胞菌的抗菌策略。
附录一:
其他菌种
附录二:
致病菌菌株
附录三:
可能参与的代谢

谷禾健康
花青素-肠道微生物群-健康轴
经常听到一些专家在介绍食品或保健品时说富含花青素,那么究竟什么是花青素?对我们的健康又有何益处?
花青素是在植物组织中发现的天然水溶性色素,水果、蔬菜、花卉中的呈色物质大部分与之有关,是类黄酮的一个亚类,迄今为止已鉴定出700多种结构不同的花青素。
花青素是一种重要的营养素,具有多种生物活性和保健作用,如抗氧化、抗炎、抗癌、降血压、降血糖、保护视力及心血管健康。
花青素在消化过程中还会影响肠道中的细菌代谢。花青素及其衍生物还通过调节肠道屏障功能和有益细菌的定植来促进肠道健康,从而更好地抵御病原体,改善营养代谢和整体免疫反应。
在本文中,谷禾简要介绍了不同类型的花青素,它们常见的饮食来源和通过胃肠道(包括胃、小肠和结肠)的吸收和代谢。
本文还总结了膳食花青素对健康的直接影响,以及肠道微生物群作为“媒介”时的影响。花青素-微生物的相互作用可能成为肥胖、2型糖尿病、心血管疾病、神经退行性疾病、炎症性肠病、癌症、慢性肾脏病和骨关节炎等疾病的治疗新途径。
目录
1.常见的花青素及膳食来源
2.膳食花青素在人体内的吸收和代谢
3.花青素对肠道微生物群的影响
4.花青素-微生物群对肥胖的改善作用
5.花青素-肠道菌群对糖尿病的影响
6.花青素改善心血管疾病
7.花青素降低神经退行性疾病风险
8.花青素对其他疾病的影响
9.日常补充膳食花青素的一些建议
花青素又称花色素,是自然界一类广泛存在于植物中的水溶性天然色素,是花色苷水解而得的有颜色的苷元。
水果、蔬菜、花卉中的主要呈色物质大部分与之有关。在植物细胞液泡不同的PH值条件下,花青素使花瓣呈现五彩缤纷的颜色。
▼
已知现有的花青素多达几百种,食物中重要的有6种,即天竺葵色素、矢车菊色素、飞燕草色素、芍药色素、牵牛花色素和锦葵色素。
花青素具有2-苯基苯并吡喃结构,具有完全不饱和的C环和3位的羟基。
植物性食物中主要花青素的结构
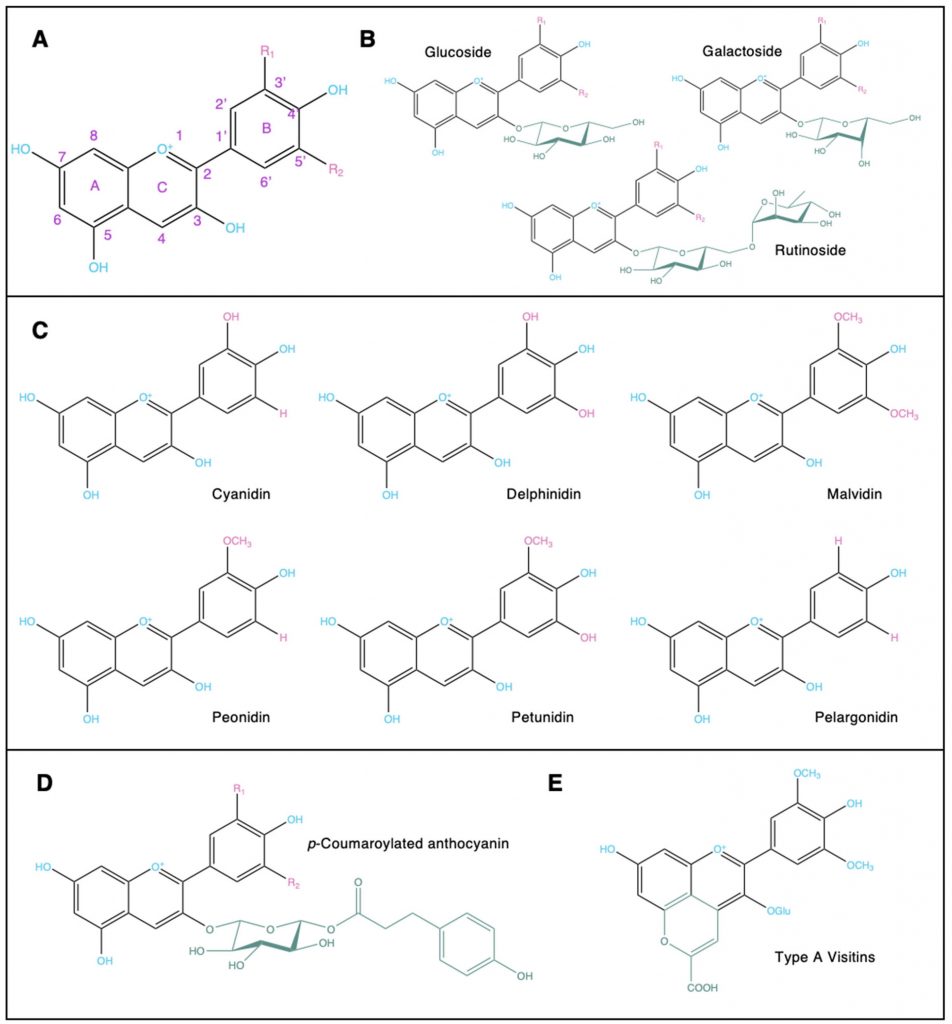

Liang A,et al.Crit Rev Food Sci Nutr.2023
(A)花青素中的2-苯基苯并吡喃骨架 (B)附着在糖基化花青素上的常见糖部分 (C)植物性食物中常见的花青素 (D)酰化花青素 (E)吡喃花青素
▼
植物性食品中花青素的浓度和组成高度依赖于植物品种、生长区域、植物部位、加工方法和测量方法。
花青素在各种红色、紫色和蓝色水果(如浆果、石榴、血橙和红梅)中含量丰富,这些水果代表了人类饮食中花青素的主要摄入。
常见水果中的花青素
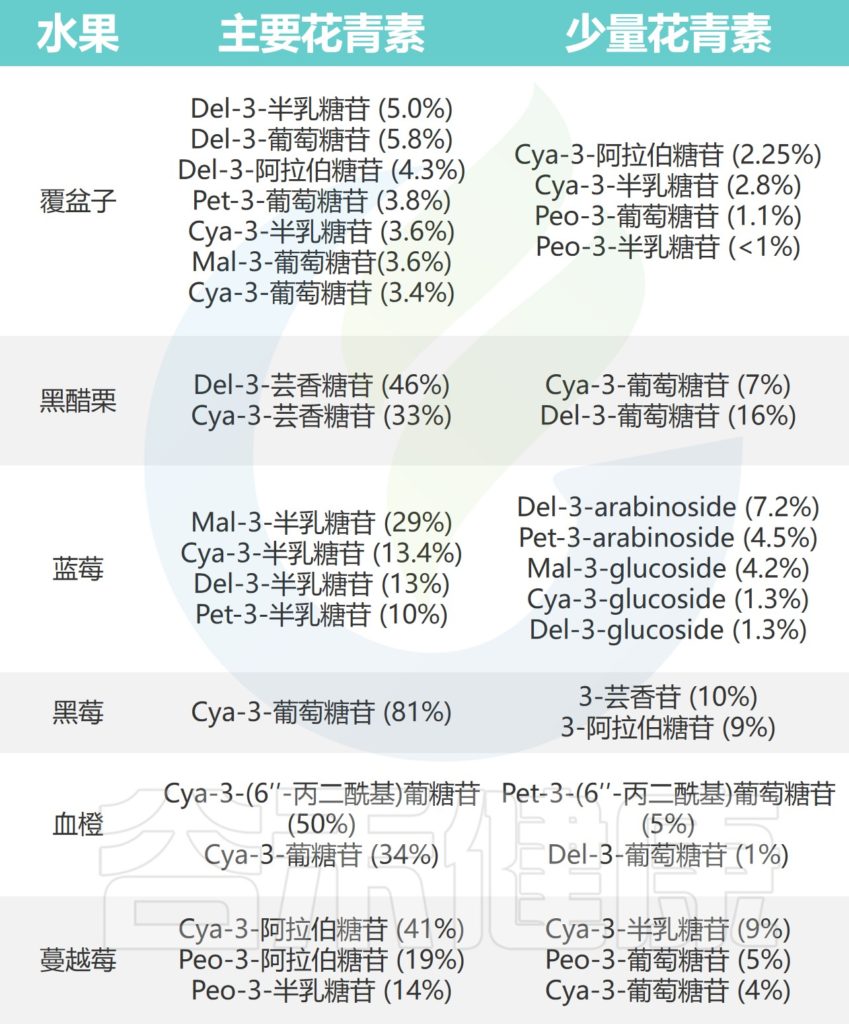
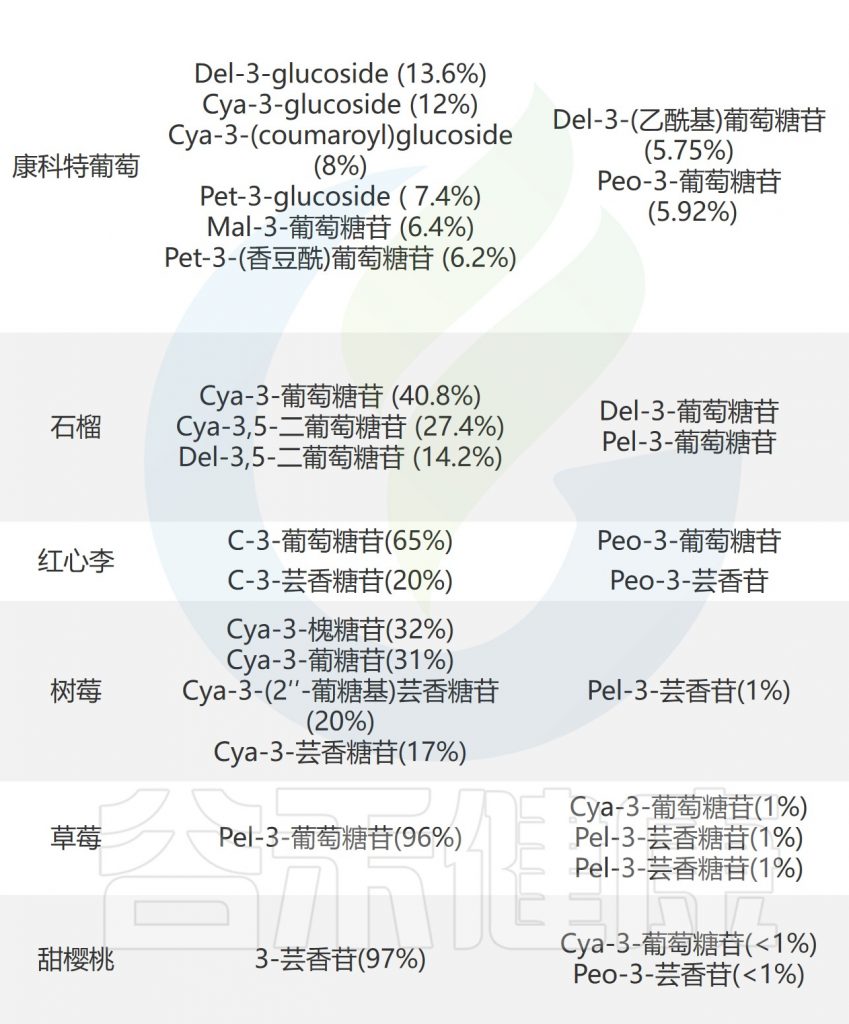

Liang A,et al.Crit Rev Food Sci Nutr.2023
•花青素在不同酸碱度下颜色不同
花青素在水果中根据pH条件,它会显示红色(pH<3)、紫色(pH=7-8)及蓝色(pH>11)。
例如飞燕草色素通常存在于花朵和葡萄中,它们提供蓝紫色,而天竺葵色素为许多浆果和水果提供橙色和红色,包括树莓、草莓和石榴。
•酰化花青素具有更强的耐热性和耐光性
酰化花青素是通过糖基和有机酸的乙酰化酯键得到的,进一步扩大了食品中花青素的多样性。常见的酰化酸包括脂肪酸(乙酸、丙二酸、苹果酸、草酸和琥珀酸)和环状酸(对香豆酸、咖啡酸、芥子酸、没食子酸和阿魏酸)。
在葡萄中鉴定出酰化花青素,在黑胡萝卜、红薯和红甘蓝中发现了肉桂酸酰化的花青素。在红萝卜中发现了多达12种酰化花青素。
酰化花青素在加工过程中表现出更强的耐热性和耐光性,因此被广泛用作食品工业中的着色剂。
•吡喃花青素具有更强的抗氧化能力
吡喃花青素是另一组具有额外吡喃环的花青素,主要在红酒的酒精发酵和陈酿过程中产生。少量吡喃花青素天然存在于血橙、草莓和黑加仑等水果中。
吡喃花青素的主要类别包括A型和B型视黄素、甲基吡喃花青素、羟基苯基吡喃花青素和黄烷基吡喃花青素。
相较于单体花青素,吡喃花青素,在更大范围的pH条件下保持其颜色,这是因为额外的吡喃环可以防止亲核攻击和无色甲醇碱的形成。
吡喃花青素在二氧化硫漂白和热降解过程中也具有更大的稳定性,并且相对于单体花青素具有更强的抗氧化能力。
由于这些原因,吡喃花青素作为颜料和生物活性剂广泛用于食品加工。
膳食花青素是指可以通过饮食摄入的一类花青素,广泛存在于深红色、紫色或蓝色蔬菜水果中,如葡萄、蓝莓、紫甘蓝、紫薯、黑枸杞等。
膳食花青素的代谢和治疗活性取决于其生物可及性(可从食物基质中获得的量)和生物利用度(在人体肠道中的吸收)。这受到许多因素的影响,包括化学结构、与食物基质的相互作用、其他膳食化合物的存在以及个体遗传学和生理学差异。
★ 不同结构的花青素消化率存在差异
例如,具有双糖部分的糖基化花青素相对于单葡萄糖苷和苷元更耐消化。酰化花青素的消化率低于非酰化化合物,主要是因为极性降低和酰基的阻碍。吡喃花青素和聚合物花青素在刺激消化过程中更稳定。
花青素及其衍生物在人体内的消化吸收发生在胃、小肠和大肠中。
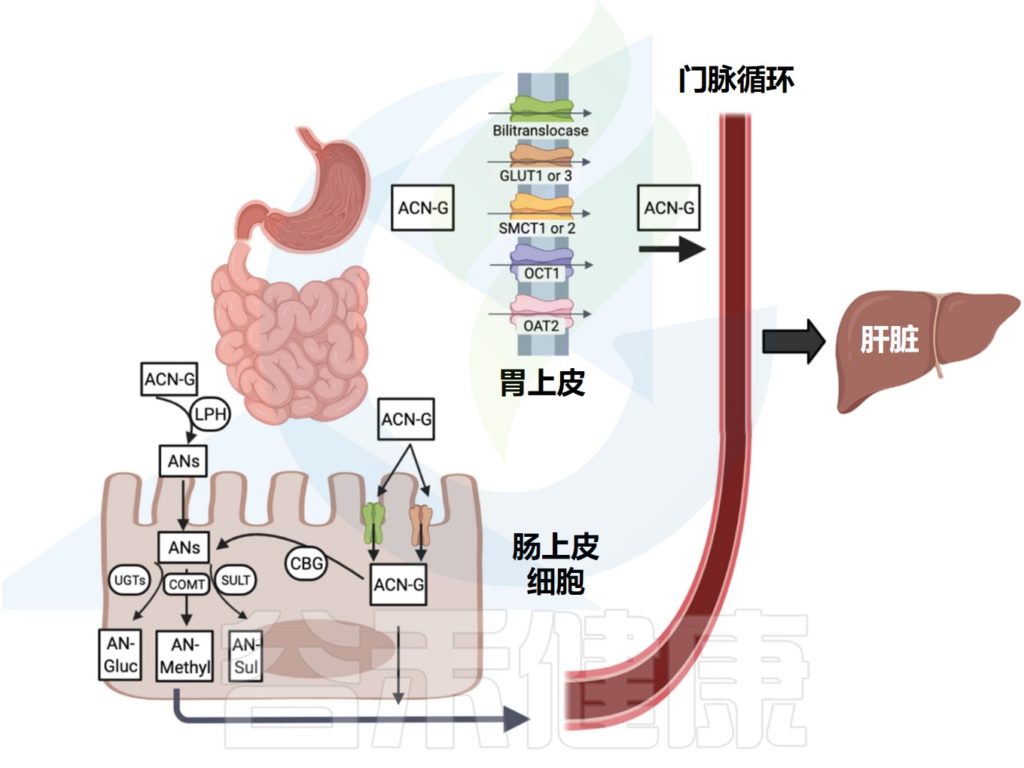
Liang A,et al.Crit Rev Food Sci Nutr.2023
▼
食用食物后第一个吸收花青素的部位是胃,在胃中,一小部分花青素穿过胃细胞屏障(大部分以完整的糖基化形式),并在血浆中检测到。
在胃内,花青素保持相对稳定,在大鼠胃肠道研究期间,从流出物中可回收总摄入花青素的75-79%,在体外研究期间可回收75-88%。
✦胃吸收效率受膳食成分影响
胃吸收效率也受到膳食成分的影响,在高葡萄糖 (>40mM)存在的情况下,花青素通过胃细胞屏障的摄取速率显著降低。
✦参与胃中花青素代谢的蛋白
胃胆转位酶是一种膜蛋白载体,可将完整的花青素转运穿过胃屏障。花青素的糖苷部分优先满足双转位酶提供的结构规格,因此无需去糖基化即可被吸收。
后来,葡萄糖转运蛋白GLUT1和GLUT3也参与了胃对花青素的摄取。花青素中的A环和C环以及葡萄糖部分在与人类GLUT1和GLUT3载体的跨膜结合中起重要作用。
其他参与花青素在胃中吸收的转运蛋白包括单羧酸转运蛋白(SMCT1和SMCT2)和有机阳离子/阴离子转运蛋白(OCT1和OAT2)。
胃以前被认为是花青素吸收而非代谢的部位,直到最近在胃上皮细胞中发现结合酶活性才证实了胃的代谢功能。
此外,红酒中花青素在体外消化(使用胃MKN-28和肠道Caco-2细胞)过程中代谢物malvidin-3-O-葡糖苷酸的存在提供了花青素在胃内代谢的证据。
▼
空肠(小肠中部)是花青素吸收的主要部位。花青素在小肠内的吸收通过膜转运蛋白促进的被动扩散或主动转运发生。
✦花青素的主动转运吸收
与胃吸收类似,完整的糖基化花青素被肠上皮细胞内的转位酶和葡萄糖转运蛋白(SGLT1和GLUT2)吸收。
SGLT1转运蛋白是一种在肠细胞顶端表面发现的钠-葡萄糖共转运蛋白1,而GLUT2转运蛋白是主要在上皮细胞基底外侧膜上发现的促进载体。这两种转运蛋白都与花青素的葡萄糖部分相互作用,并负责基于葡萄糖的花青素的肠道吸收。
除了己糖转运(糖、葡萄糖、果糖等简单碳水化合物的转运系统)外,最近发现有机阴离子转运多肽转运体(OATP)也参与了大鼠空肠上皮对完整β型酰化花青素的吸收,但其吸收率很低,仅为0.2%~2.2%。
✦花青素的被动扩散吸收
另一方面,当花青素被乳糖酶-根皮苷水解酶、β-葡萄糖苷酶、α-鼠李糖苷酶和β-葡萄糖醛酸酶等水解为其苷元形式时,花青素通过肠上皮的被动扩散吸收。
✦花青素的转运速率
最近使用Caco-2细胞(人类结直肠腺癌细胞)作为人类肠道屏障功能的模型,可以评估花青素的运输,并表明运输效率取决于附着在花青素骨架上的苷元或糖的类型,以及花青素是否为聚合物。
Caco-2细胞模型还估计,达到系统循环的花青素比例约为总膳食摄入量的1%。
✦肠道内花青素的代谢
在肠道内吸收的花青素经历第一阶段代谢,包括氧化、还原和水解,但它们发生的频率较低,在多酚代谢中也不太重要。
第二阶段的生物转化更加密集和快速,它们首先发生在小肠,然后发生在肝脏和肾脏。一旦被肠细胞吸收,花青素及其分解代谢物就会发生结合反应。
包括分别由UDP葡萄糖醛酸基转移酶(UGTs)、硫转移酶(SULT)和邻苯二酚-O甲基转移酶(COMT)催化的葡萄糖醛酸化、硫酸化和甲基化。
这些生物转化产生葡萄糖醛酸、硫酸盐和甲基化代谢产物,这些代谢产物由活性ATP结合盒蛋白转运,因为它们过于亲水,无法被动扩散到细胞中。
活性ATP结合盒蛋白——一个膜内在蛋白质超家族。将ATP水解释出的能量提供给各种分子进行穿膜转运。
花青素进入血液后,通过门静脉分布到肝组织,进行进一步的偶联反应,增加其在水中的溶解度,并最终促进其在尿液中的排泄。
▼
膳食中花青素的低生物利用度以及粪便和尿液样本中花青素的低回收率表明,花青素在结肠中会被降解。
例如,食用覆盆子后,总膳食花青素的40%从回肠液中回收。这些来自小肠的未被吸收的花青素通过细菌或化学降解进行生物转化,潜在地提高了它们的生物利用度,并允许通过结肠上皮壁吸收。
花青素的微生物降解需要多种细菌,如拟杆菌、Enterococcus casseliflavus、优/真杆菌属(Eubacterium)和梭状芽胞杆菌(Clostridium)等,它们产生各种酶,如β-葡萄糖苷酶、α-半乳糖苷酶和α-鼠李糖苷酶,用于微生物分解代谢。
✦原儿茶酸是最丰富的代谢物
由于这些相互作用,通过肠道微生物降解产生了各种代谢产物。代谢物原儿茶酸占花青素摄入量的73%,是花青素消耗后最丰富的人体代谢物。
桑椹的矢车菊素-3-O-葡萄糖苷和矢车菊苷-3-O-芸香糖苷的体外粪便发酵产生了原儿茶酸、对香豆酸和间苯三酚醛,而飞燕草素-3-O-芸香糖苷的发酵产生了没食子酸、丁香酸和间苯三酚醛。
✦其他花青素的代谢产物
桑椹的矢车菊素-3-O-葡萄糖苷和矢车菊苷-3-O-芸香糖苷的体外粪便发酵产生了原儿茶酸、对香豆酸和间苯三酚醛,而飞燕草素-3-O-芸香糖苷的发酵产生了没食子酸、丁香酸和间苯三酚醛。
在与结肠细菌孵育24小时后,Malvidin-3-O-葡萄糖苷会完全代谢为丁香酸。
如体内人体研究和体外研究所示,天竺葵素-3-O-葡糖苷被代谢为4-羟基苯甲酸。
六种主要花青素及其结肠代谢物


Liang A,et al.Crit Rev Food Sci Nutr.2023
★ 花青素的主要代谢物对人体健康有益
原儿茶酸
原儿茶酸是一种水溶性酚酸成分,并且是很多中药中的活性物质,其不仅具有抗血小板凝集、降低心肌耗氧量、提高心肌耐氧能力、减慢心率、抑菌、镇痛等药理活性,还具有抗氧化、抗肿瘤和神经保护作用。
没食子酸
没食子酸具有抗炎、抗突变、抗氧化、抗自由基等多种生物学活性;同时没食子酸具有抗肿瘤作用,可以抑制肥大细胞瘤的转移, 从而延长生存期;对肝脏具有保护作用。
丁香酸
丁香酸可以抑制炎症反应,减轻炎症症状;同时丁香酸具有抗菌作用,可以抑制多种细菌和真菌的生长;丁香酸还能够降血糖、降血脂。
✦肠道微生物在花青素的去酰化起重要作用
此外,人体肠道微生物组在花青素的去酰化过程中发挥重要作用,释放出具有更高生物利用度的酚酸。
花青素衍生的代谢物可能比母体花青素表现出更强的稳定性和生物活性,并可能调节人体肠道微生物组成。
肠道微生物群受年龄、体重、饮食模式、抗生素治疗和宿主遗传的影响,并且个体之间(以及个体内部)存在差异。这种变化可能会影响膳食花青素的生物利用度和代谢。
花青素及其结肠代谢物通过其生物活性直接改善健康,并通过重塑肠道微生物组的组成和操纵短链脂肪酸的产生间接改善健康。
在宿主的一生中,肠道微生物组的组成随着时间的推移而变化,以响应内部和外部刺激。因此,肠道微生物组成的动态平衡和多样性是正常肠道功能所必需的,包括维生素和短链脂肪酸的产生、脂质代谢、免疫防御和肠-脑轴相互作用等。
▼
花青素及其代谢物目前被认为是改变肠道微生物群的调节剂,主要是通过促进有益菌的增殖和抑制有害菌的生长,许多不同模型的研究都观察到这一点。
✦促进了有益菌的增殖
研究发现黑米中分离的矢车菊素-3-葡萄糖苷在体外发酵条件下对青春双歧杆菌(B. adolescentis)、婴儿双歧杆菌(B. infantis)、双歧双歧杆菌(B.bifidum)和嗜酸乳酸杆菌(L.acidophilus)的增殖具有促进作用。
培养48小时后还观察到生长培养基pH值的降低,这归因于由酚酸和短链脂肪酸组成的有机酸的产生。双歧杆菌和乳杆菌的数量增加。它们通过竞争底物和粘附位点以及降低结肠腔内pH值来对病原体产生抗菌作用。
研究发现,降低结肠内pH值可抑制肠杆菌科(Enterobacteriaceae)和梭状芽胞杆菌(Clostridia)等pH敏感有害菌群的过度增殖,并通过抑制前致癌物的形成来降低结直肠癌的风险。
✦抑制有害菌的生长
与上述发现一致,紫甘薯中提取的花青素增加了最知名的益生菌之一双歧杆菌(Bifidobacterium)和乳杆菌(Lactobacillus)的种群数量,但减少了溶组织梭状芽胞杆菌(C. histolyticum)和拟杆菌/普雷沃氏菌的种群数量。
溶组织梭状芽胞杆菌是与感染和炎症性肠病相关的最常见致病性物种之一。
▼
虽然许多研究表明花青素及其代谢产物促进有益菌群生长,抑制有害菌群生长,但对结肠细菌的促进作用存在差异。
对微生物群的不同影响可能是由于源自花青素的代谢物的差异。
花青素(单独和混合物)和没食子酸的孵育被证明可以增加双歧杆菌和乳酸杆菌的数量。而混合花青素和没食子酸显著诱导奇异菌属(Atopobium)的生长,并减少了潜在致病性溶组织梭菌。
没食子酸似乎是本研究中的关键代谢物,可抑制溶组织梭菌增殖。
在其他研究中,研究了没食子酸对梭菌、大肠杆菌和金黄色葡萄球菌等有害微生物的抗菌活性。
花青素对肠道菌群的调节作用



Liang A,et al.Crit Rev Food Sci Nutr.2023
▼
花青素的发酵有助于短链脂肪酸的产生,主要是乳酸、乙酸、丙酸和丁酸。短链脂肪酸通过调节细菌毒力基因表达和降低结肠中管腔pH值来抑制肠道病原体如沙门氏菌(Salmonella spp.)、大肠杆菌(E. coli)和李斯特菌(Listeria spp.)的入侵。
此外,短链脂肪酸在肠道生理事件中发挥着重要作用,如获取能量、调节pH、参与肠脑轴和保护肠道屏障功能。
花青素与肠道菌群作用的研究证据
花青素与肠道微生物的相互作用已经通过动物模型得到了广泛的研究。用黑甜樱桃对小鼠进行饮食干预发现,除了双歧杆菌和乳杆菌外,樱桃花青素的补充还促进了结肠中阿克曼氏菌(Akkermansia)的增殖。
阿克曼氏菌的丰富度被认为可以刺激粘液分泌,改善肠道屏障功能,从而降低炎症风险和免疫介导的疾病,如乳糜泻。
促进Coriobacteriaceae有助于健康
对小鼠进行为期六周的多种浆果饮食干预,结果发现放线菌(Actinobacteria)数量有所改善,这与补充蓝莓的研究结果一致。
放线菌门下的一些科,如双歧杆菌和Coriobacteriaceae,已被证明对健康有益。特别是,有人认为,在高脂肪饮食存在的情况下, Coriobacteriaceae在降低肥胖和肝脏相关疾病的风险方面发挥作用。
双歧杆菌和乳酸杆菌的丰度增加
与体内试验的结果一致,在参加为期六周的25克蓝莓能量饮料的健康男性中发现双歧杆菌和乳酸杆菌的丰度增加。
花青素、其代谢物和肠道微生物群间的相互作用
Liang A,et al.Crit Rev Food Sci Nutr.2023
小结
体外、动物和人体研究一致认为,花青素及其代谢物能够调节肠道微生物群。花青素会促进益生菌的生长,特别是双歧杆菌、乳杆菌和阿克曼氏菌,它们通过产生短链脂肪酸或通过竞争底物和定植位点来对抗有害和致病性菌株,从而有利于宿主健康。
然而,花青素对肠道微生物群的调节也可能依赖于其他相关的化学物质,如葡萄糖,因为与那些喂食高葡萄糖对照饮食的大鼠相比,在蓝莓补充后,在肠道中发现了较低水平的乳酸杆菌。
此外,不同的动物模型和志愿者群体在微生物组成方面存在较大差异,这是这些研究的另一个挑战。
因此,未来的临床研究重点应放在不同年龄和生理阶段的男性和女性不同亚类人群上,这可能会更深入地了解花青素在微生物群调节中的作用及其对特定人群的健康益处。
前面讲述了花青素对肠道菌群的调节作用,下面谷禾来具体讲讲花青素和肠道微生物群相互作用下对一些疾病的影响
肥胖和超重被定义为对健康构成威胁的脂肪组织的过度积累。脂肪组织中的代谢紊乱,如炎症、脂肪因子释放失调和脂肪生成增加,是肥胖和其他肥胖相关疾病的一些常见潜在特征。
有证据表明,与肥胖相关的疾病可能部分由肠道生态失调引起,包括微生物组成及其代谢物的变化。
最近,花青素与肠道微生物的相互作用被证明具有改善肥胖的作用。主要通过调节脂肪生成和脂肪分解,炎症反应的介导以及通过肠脑通讯控制食欲。
1
肠道微生物组影响脂质代谢,肠道菌群失调与血脂异常和肥胖等有关。
最近的一项体内研究显示,对高脂喂养的小鼠补充了200毫克/公斤体重的紫甘薯花青素(花青素和芍药苷酰基糖苷),改善了高脂饮食引起的体重增加和肝脏总脂肪积累。
√调节AMPK通路减少脂肪积累
他们认为,花青素抑制脂肪堆积的作用需要激活小鼠肝脏细胞AMPK通路。此外,在摄入花青素后,AMPK途径下游靶点乙酰辅酶a羧化酶、脂肪酸合成酶、甾醇调节元件结合蛋白也被下调,以减少小鼠肝细胞中的脂肪积累。
已知AMPK通路的上调在抑制脂肪生成中发挥重要作用,例如抑制脂蛋白脂肪酶 (LPL),这是一种可受肠道微生物群调节的酶。肠道菌群通过影响抑制脂蛋白脂肪酶,以相反的方式促进脂肪细胞中甘油三酯的储存。而活化的AMPK及其下游信号通路至少部分抵消了肠道微生物群促进的能量储存。
√操纵法尼醇X受体信号减轻代谢受损
此外,已知肠道菌群产生的次生胆汁酸通过胆汁酸受体法尼醇X受体(FXR)改变肝脏脂质代谢。胆汁酸-结肠微生物群相互作用的破坏可能导致肥胖易感性。
花青素被证明可以操纵肠道微生物群/法尼醇X受体信号,从而减轻高脂肪诱导的肝脏脂肪积累和脂质代谢受损。
黑米花青素(0.48g/kg)喂养12周使血清乳酸脱氢酶(LDH)、丙氨酸转氨酶(ALT)和天冬氨酸转氨酶(AST)含量正常化,这些指标是肝脏脂肪过度积累影响肝损伤的指标。
这些发现表明,膳食花青素可能通过改变肠道菌群来减轻高脂饮食引起的脂质代谢紊乱。
2
肠道微生物群参与调节炎症,在肥胖患者的脂肪组织中通常观察到系统性的低级别炎症并发症。
√经常摄入花青素的肥胖患者炎症标志物含量下降
在一项研究中,代谢综合征患者每天食用400克新鲜越桔,持续12周后,血清脂多糖、IL-6、IL-12和高敏C反应蛋白(hsCPR)浓度下降。
hsCPR是一种与肥胖相关的炎症标志物。
另一项动物研究显示,花青素的干预通过抑制21天大的小鼠白色脂肪组织中TLR4介导的NF-κB信号通路,降低了高脂肪诱导的促炎标志物IL-6和肿瘤坏死因子的表达。
此外,高脂肪喂养诱导下的粪便双歧杆菌减少,在妊娠期和哺乳期用花青素处理后恢复。
3
肠道微生物在调节涉及能量稳态的神经激素活动中发挥作用。由微生物群产生的花青素和短链脂肪酸会影响促食欲激素和厌食激素的释放。
√刺激肠促胰岛素激素分泌减少食物摄入量
飞燕草素3-芸香糖苷被证明可刺激释放胰高血糖素样肽-1,这是一种肠促胰岛素激素,可促进胰岛素分泌和饱腹感,从而减少食物摄入量。
√减轻肥胖引起的神经激素紊乱
肥胖会通过表观遗传修饰相关基因表达导致参与能量平衡的神经激素活动紊乱。食用花青素可能会恢复肥胖引起的这些不利影响。
在肥胖大鼠中,连续8周食用额外的樱桃和桑葚花青素可以减少体重增加,降低血清瘦素水平,增加脂联素。
与先前的研究一致,肥胖患者在食用富含花青素的水果6周后,发现瘦素受体表达增加,这表明补充花青素后瘦素敏感性有所改善。
此外,高脂饮食诱导的肥胖大鼠大脑皮层和海马体中的γ-氨基丁酸(GABA)水平下降。研究报道,与对照组相比,40天摄入黑大豆花青素减少了成年大鼠的体重增加。高剂量花青素可降低促氧神经肽Y (NPY)的表达,提高下丘脑GABA受体B (GABAB1R)的表达。
GABA是中枢神经系统中的一种抑制性神经递质,通过控制神经元的兴奋性、可塑性和额叶皮质的同步性,在摄食反应中起关键作用。
综上所述,花青素似乎通过多种方式与肠道微生物群相互作用,包括调节脂质代谢、炎症反应和参与能量平衡的神经激素活动,从而具有改善肥胖的作用。
注:关于健康和肥胖个体肠道中花青素代谢的差异,人们的理解有限。因此,未来的研究重点应放在花青素结肠代谢物及其对健康和肥胖受试者的健康影响上。
2型糖尿病是一种以高血糖为特征的代谢性疾病,由胰岛素抵抗逐渐发展而来。
许多研究已经报道了花青素及其代谢物的抗糖尿病潜力,这可能归因于以下单一或同时的作用,包括增强胰岛素敏感性,改善β细胞功能和延迟餐后血糖反应。
1
花青素可以通过自身或其微生物代谢物改善胰岛素敏感性。
脂肪组织能够分泌脂联素等脂肪细胞因子来调节葡萄糖稳态。
√花青素可以诱导脂肪细胞摄取葡萄糖
花青素及其微生物代谢物原儿茶酸已被证明可以通过上调葡萄糖转运蛋白4来增加人类脂肪细胞去除血浆葡萄糖的能力。
PPARγ的敲除消除了花青素的胰岛素增敏特性,这表明PPARγ可能在增加花青素诱导的脂肪细胞葡萄糖摄取中发挥作用。
PPARγ是一种参与葡萄糖处理和葡萄糖转运蛋白调节的核转录因子
√花青素还可以介导骨骼肌中的葡萄糖摄取
除了脂肪细胞外,骨骼肌中的葡萄糖摄取也可由花青素介导。
糖尿病小鼠口服紫玉米花青素提取物8周后,骨骼肌对葡萄糖的摄取增加,因为在肌肉细胞中发现葡萄糖转运蛋白4表达增加,并伴随AMPK通路的激活。增加AMPK活性是刺激葡萄糖转运蛋白4从细胞内囊泡转移到细胞膜的重要途径,从而促进葡萄糖流入细胞。
此外,在摄入原儿茶酸后大鼠肝脏中发现胰岛素受体底物1/磷脂酰肌醇3-激酶/蛋白激酶B-2 (IRS1/PI3K/AKT2) 信号通路的活性增加,表明肝脏胰岛素敏感性得到改善。
证据表明,花青素及其降解产物改善了脂肪细胞和肌肉细胞的胰岛素敏感性。
2
花青素还可以通过消除高血糖引起的氧化应激来保护胰腺β细胞的功能。
√改善高糖影响下的胰岛β细胞损伤
发现富含花青素的提取物可减轻高糖诱导的β细胞损伤,保护其分泌功能,并导致血清胰岛素水平升高。
一项体外研究(对与桑椹花青素孵育的高糖培养的β细胞)发现,花青素能够通过抑制β细胞中的凋亡相关蛋白,包括丝裂原活化蛋白激酶、细胞色素c和Bcl-2家族蛋白,消除高糖诱导的细胞内活性氧水平升高,并将β细胞活力恢复到84%。
除了完整的花青素外,丁香酸是花青素的主要微生物代谢产物之一,口服30天可改善四氧嘧啶诱导的糖尿病大鼠的胰岛素释放功能异常、血糖和糖蛋白水平升高。
丁香酸的抗糖尿病作用可能与保持β细胞的活力和功能有关。
3
在人体试验中也观察到了膳食花青素和微生物代谢物在调节餐前和餐后血糖中的作用,从而降低了患2型糖尿病的风险。
众所周知,血糖水平是动态的,全天都在变化。空腹和餐后血糖水平是诊断糖尿病的两个重要指标。空腹血糖值代表身体在一夜禁食后如何管理血浆葡萄糖,而餐后血糖则代表餐后的血糖反应。
√多摄入花青素表现出延迟的血糖反应
与未补充花青素的人相比,额外摄入150克混合浆果泥的参与者表现出延迟的血糖反应。
研究人员解释:餐后血糖降低至少部分是由于花青素对α-葡萄糖苷酶的抑制作用,α-葡萄糖苷酶是一种释放α-葡萄糖的酶。
该研究结果支持以下机制:花青素通过抑制胰腺α-淀粉酶和α-葡萄糖苷酶的活性,以及干扰葡萄糖转运蛋白(SGLT1和GLUT2),因为这些转运蛋白存在竞争关系,从而降低餐后血糖。
√降低了空腹血糖浓度
血清马尿酸是一种花青素的结肠代谢产物,在成年人食用越橘8周(200克/天)后显著增加。8周越桔 干预也降低了空腹血糖浓度。
然而,没有观察到血浆马尿酸与餐后血糖浓度之间存在任何相关性。
花青素可能通过抑制α-淀粉酶和α-葡萄糖苷酶活性以及与葡萄糖载体相互作用来调节餐后血糖,而花青素代谢产物马尿酸则由于微生物降解物的延迟出现而与改善空腹血糖控制有关。
小结
花青素及其代谢物的抗糖尿病作用已经在体外和体内研究中得到阐明。主要通过调节葡萄糖转运蛋白表达和易位来改善胰岛素抵抗,对胰岛β细胞及其分泌功能的保护作用,以及调节空腹和餐后血糖水平。
因此,由富含花青素的食物或补充剂组成的饮食干预可能是更好地控制患有2型糖尿病高风险个体和糖尿病患者血糖的替代途径。
心血管疾病是一类影响心脏和血管的疾病,最常见的形式包括动脉粥样硬化、冠心病、心律失常、心力衰竭和中风。
流行病学研究表明,高花青素摄入量与心血管疾病风险降低有关。花青素及其代谢物发挥保护作用,主要归因于抑制血小板过度活跃,缓解内皮功能障碍,改善脂质谱和逆转胆固醇转运。
1
花青素和原儿茶酸已被证明稳定血小板活性,从而防止血栓形成。
√摄入花青素抑制了血小板的过度聚集
一项研究发现,100μM和500μM没食子酸可以减少刺激物(二磷酸腺苷和血栓素A2激动剂U46619)诱导的血小板聚集、血小板-白细胞聚集和P-选择素表达。
P-选择素(一种参与细胞表面粘附的糖蛋白)的表达减少,表明血小板稳定性提高,血栓形成风险降低。
没食子酸抗血小板聚集的潜在机制是通过降低细胞内钙水平和下调蛋白激酶c-α(PKCα)/p38丝裂原活化蛋白激酶(p38MAPK)和Akt/糖原合成酶激酶3β (GSK3β)通路的活性。
另一项临床试验也支持了这种抗血栓作用,该试验发现,在健康志愿者中,200mL富含花青素和芸香苷的李子汁干预28天,可有效抑制人工刺激的血小板聚集,并降低P-选择素的表达。
此外,补充花青素28天后还观察到氧化应激生物标志物丙二醛水平降低,这表明血小板的抗机能亢进至少部分归因于花青素及其代谢物的抗氧化能力。
2
花青素及其代谢物具有血管内皮保护作用,内皮功能障碍被认为是动脉粥样硬化发展的早期指标。内皮功能受损可破坏血管收缩和血管舒张的止血作用,引发一系列生理事件,如促炎因子分泌、白细胞粘附等,从而引发动脉粥样硬化。
√减缓动脉粥样硬化
研究了花青素代谢物原儿茶酸对载脂蛋白e缺乏小鼠的内皮保护作用。原儿茶酸(0.03g/kg)喂养20周小鼠,动脉粥样硬化形成延迟,血管细胞粘附分子1 (VCAM-1)和细胞间粘附分子1(ICAM-1)表达显著降低。
这些粘附分子的减少可以减少单核细胞对血管内皮的附着,从而减缓动脉粥样硬化的早期发展。
√改善血管舒张情况
蓝莓花青素的血管保护特性已在临床试验中得到证实。健康志愿者在摄入蓝莓花青素(766-1791毫克)1至2小时和6小时后,内皮依赖性血管舒张得到改善。
3
许多研究也报道了富含花青素的饮食对血脂的有益影响。
√直接补充花青素缓解高脂血症
在高脂饮食诱导的肥胖小鼠中,每天补充150 mg/kg黑加仑花青素12周后,高脂血症得到缓解,相应的血清总胆固醇、甘油三酯和低密度脂蛋白水平降低。
另一项研究报告,50mg/kg原儿茶酸干预10周后,降低了先前由脂肪和果糖丰富的饮食引起的高脂血症雄性大鼠的血清甘油三酯、总胆固醇和低密度脂蛋白水平。
三项新指标,即血浆动脉粥样硬化指数(AIP)、动脉粥样硬化指数(AI)和冠状动脉危险指数(CRI),在50mg/kg原儿茶酸治疗后分别显著降低33%、52%和36%。
√通过调节菌群降低血脂
此外,花青素干预后观察到嗜黏蛋白阿克曼菌(Akkermansia muciniphila)和狄氏副拟杆菌(Parabacteroides distasonis)的富集,其与循环甘油三酯和低密度脂蛋白水平呈负相关。已知阿克曼菌可以积极调节肠道屏障完整性,防止脂多糖易位诱导炎症。
低密度脂蛋白(LDL)及其氧化修饰(oxLDL)通过刺激泡沫细胞的形成与动脉粥样硬化的发病机制有关。
最近发现狄氏副拟杆菌(Parabacteroides distasonis)通过产生琥珀酸和次生胆汁酸,如石胆酸和熊去氧胆酸,具有降血脂作用。
综上所述,花青素及其代谢物可能通过改善血脂来预防动脉粥样硬化和冠状动脉疾病。
小结
综上所述,花青素及其代谢物通过维持血小板稳定、保护血管内皮功能、改善血脂等发挥心血管保护作用。
然而,关于花青素心血管保护作用的人类和动物研究之间存在差距。因为动物中使用的浓度可能无法在人体中达到生理水平。
此外,临床研究的实验持续时间通常在4至12周之间,这可能无法捕捉到长期补充的影响。因此,未来可能需要进行长达一年的长期饮食干预研究。
帕金森病、阿尔茨海默病和肌萎缩侧索硬化症等神经退行性疾病是发生在神经系统中的疾病,导致日常活动协调困难、认知障碍和记忆丧失。
这些疾病的病因可能多种多样,但它们有一系列共同的潜在因素。这些因素包括但不限于长时间的氧化应激、中枢神经系统炎症、神经元钙信号失调和兴奋性毒性(即神经毒性)。
有证据表明,一些花青素代谢物,如原儿茶酸、香草酸、没食子酸和其他简单的酚酸,可以通过血脑屏障进入中枢神经系统,这表明这些物质可能直接在发生神经系统疾病的中枢神经系统中发挥作用。
1
花青素肠道代谢物与防止神经毒性和维持Ca2+体内平衡有关。
√没食子酸可以减轻谷氨酸诱导的神经毒性
谷氨酸是一种兴奋性神经递质,有助于大脑正常功能,而谷氨酸水平异常升高可引起谷氨酸兴奋性毒性。谷氨酸兴奋性毒性被认为是神经退行性疾病的主要原因之一。
没食子酸是能够穿透血脑屏障的代谢物之一。报道,25和50μg/ml 没食子酸预处理2小时可导致大鼠皮层神经元细胞中N-乙酰天冬氨酸(NAA)水平和微管相关蛋白2(MAP-2)表达增加,表明没食子酸处理减轻了谷氨酸诱导的大鼠神经元神经毒性。
在缺乏没食子酸的细胞中观察到的NAA和MAP-2的下调被认为是神经元损伤和丧失的直接指标。
此外,在没食子酸处理的细胞中也观察到钙结合蛋白(一种能够缓冲Ca2+内流的钙结合蛋白)的上调,表明没食子酸在保护神经元免受谷氨酸引发的钙失衡方面的作用。
√天竺葵素代谢物减轻神经兴奋性毒性
观察到4-羟基苯甲酸(HBA)是一种天竺葵素的代谢物,可在体外保护大鼠小脑颗粒神经元免受谷氨酸兴奋性毒性。当仅用谷氨酸处理的大鼠小脑颗粒神经元导致近50%的细胞损失时,4-羟基苯甲酸处理保留了细胞并且只有13%的细胞死亡得到证实,揭示了4-羟基苯甲酸在谷氨酸兴奋性毒性缓解中的作用。
注:原儿茶酸和4-羟基苯甲酸似乎发挥互补作用,表明它们的共同治疗可能比单一代谢物更有效。然而,本研究并未提供原儿茶酸和4-羟基苯甲酸的联合治疗,因此其最佳效果的混合比例值得进一步研究。
2
花青素及其代谢物的神经保护作用与它们在防止蛋白质聚集和促进自噬中的潜在作用有关。
蛋白质聚集被认为刺激神经元变性,与许多神经退行性疾病有关。例如,由聚集的淀粉样蛋白(Aβ)形成的斑块和由tau蛋白形成的神经原纤维缠结是阿尔茨海默病的标志。导致路易体形成的α-突触核蛋白(αS)的异常组装与帕金森病有关。
√原儿茶酸防止蛋白质异常聚集
48小时原儿茶酸处理(10至100 μM)对Aβ和αS原纤维聚集的抑制率分别为30–79%和70–80%。原儿茶酸治疗6天(100μM)也使预先形成的Aβ原纤维和αS原纤维不稳定65%和80%。
这些结果表明,原儿茶酸不仅能抑制Aβ和αS纤颤,还能逆转预先形成的纤颤,它们共同调节蛋白质相互作用,防止蛋白质异常聚集导致的细胞死亡
√香草酸的干预有助于治疗阿尔兹海默病
香草酸是黑芝麻色素释放的主要代谢物,它显示出对β-淀粉样蛋白的抑制活性。在用6μM香草酸处理24小时后发现一半(50%)的抑制,在50μM香草酸孵育时达到90%的抑制。
此外,较高浓度的香草酸(60μM)可使β-分泌酶活性降低60%。β-分泌酶是一种在神经元中催化淀粉样前体蛋白合成淀粉样肽的第一步酶,促进蛋白质聚集。抑制β-分泌酶活性被认为是治疗阿尔茨海默病的另一种方法。
3
花青素微生物降解产物的有益作用已通过调节神经元细胞凋亡信号通路被发现。诱导细胞死亡是神经退行性疾病的最终病理活动。
√花青素代谢物调控细胞凋亡信号
原儿茶酸在最近的动物研究中显示出抗凋亡能力。原儿茶酸通过上调大脑皮层中参与抗氧化活性的基因,如Sod2、Cat、Gpx1和Gsr,来阻止镉诱导的细胞凋亡。
原儿茶酸处理可以减轻镉引起的促炎生物标志物肿瘤坏死因子和IL-1β的表达增加,这表明原儿茶酸通过介导促凋亡蛋白和抗凋亡蛋白,增强抗氧化防御系统和减少炎症细胞因子的产生来保护皮质组织免受镉损伤。
总之,这些发现表明花青素代谢物不仅可以减轻导致神经元死亡的各种诱导剂,还可以调节凋亡信号,从而在多个层面上降低神经退行性病变的风险。
√花青素介导的肠道菌群变化减少了神经退行性疾病的风险
除了花青素及其代谢物有助于神经保护作用外,花青素介导的肠道微生物群变化间接减少了神经退行性疾病的发生。
研究表明,由于脂多糖诱导的神经炎症反应、血脑屏障渗漏增加和β-淀粉样蛋白错误折叠的发生率,肠道生态失调可能增加神经退行性疾病的易感性。
在高脂喂养大鼠中补充黑莓花青素(25mg/kg)17天可以抵消高脂饮食引起的部分生态失调,例如孢杆菌属(sporobacter)数量增加和瘤胃球菌(Rumminococcus)数量减少。
花青素相关的肠道菌群调节通过犬尿氨酸途径改变色氨酸代谢,从而与抗神经炎症活性相关。这种改变被证明可以增加犬尿氨酸的产生,犬尿酸是一种抗兴奋毒性分子,可以减轻神经退行性病变。
尽管花青素介导的色氨酸代谢对神经保护的证据有限,但肠道微生物群对色氨酸代谢的操纵很可能会影响肠-脑轴。
建议
花青素及其微生物代谢产物可能是治疗神经退行性疾病的有效分子。然而,由于大多数研究结果都是基于体外或动物模型,因此在未来的研究中需要进行大量的临床前和临床研究。
由于衰老是神经退行性变的主要危险因素,因此有必要以老年人为重点补充花青素,以探索其在认知改善中的作用和肠道微生物群。
▼
•花青素抑制了炎性细胞因子的产生
花青素可以通过调节微生物群来预防肠道疾病。在葡聚糖硫酸钠(DSS)诱导的结肠炎小鼠中,用枸杞中的花青素提取物(200 mg/kg/天)喂养持续至少一周,恢复了紧密连接蛋白的表达,抑制炎症细胞因子(如TNF-α,IL-6)的产生。
同样,食用富含花青素的马铃薯持续9周喂养结肠炎小鼠,缓解了结肠长度的缩短,降低了髓过氧化物酶(炎性粘膜标志物)水平和炎性细胞因子。
•微生物群是花青素调节炎症性肠病的介质
然而,这种作用在抗生素治疗的小鼠身上消失了,强烈表明微生物群作为花青素对炎症性肠病介质的作用。
▼
摄入花青素可通过多种潜在机制降低患非酒精性脂肪性肝病的风险,例如其固有的抗炎作用、脂质代谢和葡萄糖代谢的管理以及改变微生物群特性。
•改变了脂肪肝小鼠体内的微生物群
其他研究也证明了花青素改变微生物群的作用,但花青素来源的多样性可能导致不同的微生物种群发生改变。在高脂饮食诱导的非酒精性脂肪肝变性小鼠中,摄入桑树水提取物(0.2-0.6%)可减少促炎的Turicibacteres,并提高拟杆菌(Bacteroidales)与梭菌(Clostridiales)的比例。
总的来说,花青素的摄入可能会改变肠道微生物群的组成,从而改善肠道屏障功能和能量消耗,降低肥胖、炎症、胰岛素抵抗,并最终降低非酒精性脂肪性肝病及其代谢并发症的风险。
▼
先前的动物研究已经证明了花青素通过微生物群调节途径抑制肿瘤生长和癌症进展的潜力。
•花青素治疗后肿瘤计数显著降低
在结直肠癌小鼠模型中,用覆盆子花青素治疗可显著削弱结肠癌的发生,每只动物的肿瘤计数减少约90%,肿瘤发病率降低(从100%降至72%左右)。
肠道生态失调可能与结直肠癌的增加有关,而花青素似乎可以恢复这种平衡,并通过促进乳酸菌的生长、抑制肠球菌和脱硫弧菌来调节微生物群。
此外,口服越桔花青素可能通过微生物群加速免疫检查点抑制剂(即程序性死亡配体1,可以限制T免疫细胞反应激活的蛋白质)的肿瘤抑制特性。
研究人员提出,补充花青素可改善肠道物种多样性,诱导梭状芽孢杆菌、约氏乳杆菌和毛螺菌科的丰度增加。
▼
花青素有可能降低慢性肾脏疾病的风险。慢性肾脏疾病的特点是肠道生态失调和由此产生的尿毒症毒素。
在一项研究中,花青素显示出通过AMPK磷酸化减轻培养细胞中糖尿病肾病的能力,并在腺嘌呤诱导的慢性肾病小鼠模型中减轻症状。
▼
花青素和富含花青素的食品的抗炎作用也与减缓骨关节炎和骨修复的进展有关,主要是通过抑制MAPK信号通路和NF-kB信号通路。
认识了花青素对人体健康的重要性,那么我们在日常生活中应该如何去补充呢,谷禾在这里列出了一些建议。
水果:存在于葡萄、蓝莓、桑葚、血橙、草莓、樱桃、红心火龙果、山楂等水果中,与水果色泽有一定的关系,在外观上有所体现,水果是补充花青素的较好选择;
蔬菜:如紫甘蓝、紫薯、紫玉米、紫土豆、黑枸杞、茄子、紫苏、甜菜、萝卜、紫洋葱等蔬菜中,也含有较多的花青素;
谷物:大麦、高粱、黑米,以及豆类如绿豆、红豆、黑豆、黄豆、赤小豆等,其中也含有较多的花青素。
花青素易受热、光、氧等因素的影响而降解,因此建议选择新鲜的食物食用,以保证花青素的含量。
花青素在高温下易被破坏,因此建议采用轻微加热的烹饪方式,如蒸、煮、炖等,以尽可能保留花青素的含量。
花青素与其他营养素的搭配可以增强其吸收和利用效果。例如,与维生素C搭配可以增强花青素的抗氧化作用。
如果日常饮食中花青素的摄入量不足,可以考虑适当补充花青素保健品。但是,需要注意选择正规品牌和适量摄入。
虽然花青素对人体健康具有多种保护作用,但过量摄入也可能对健康造成负面影响。
因此,建议适量摄入花青素,不要过度依赖花青素保健品等补充剂。
总之,通过日常饮食摄入富含花青素的食物,可以有效补充花青素,发挥其保健作用。同时,需要注意花青素的摄入量和搭配方式,以充分发挥其保健作用。
总的来说,花青素的健康益处已被广泛研究,它们被认为可以有效减轻或预防一些疾病。
通过与肠道微生物相互作用了解花青素及其健康益处是很重要的。肠道菌群在花青素降解过程中发挥重要作用,并介导多种生理活动。
花青素也会改变肠道微生物组成,维持微生态平衡,操纵短链脂肪酸的产生、脂肪代谢、肠道上皮功能、与中枢神经系统的联系和免疫反应。
因此,花青素的潜在益处可能间接归因于其对肠道微生物群的调节作用及其随之而来的生理影响。由于这些原因,研究花青素和肠道微生物群的相互作用在慢性疾病预防和治疗中是很重要的。
主要参考文献
Liang A, Leonard W, Beasley JT, Fang Z, Zhang P, Ranadheera CS. Anthocyanins-gut microbiota-health axis: A review. Crit Rev Food Sci Nutr. 2023 Mar 17:1-26.
Akbari, G. 2020. Molecular mechanisms underlying gallic acid effects against cardiovascular diseases: An update review. Avicenna Journal of Phytomedicine 10 (1):11–23.
Agustí, A., M. P. García-Pardo, I. López-Almela, I. Campillo, M. Maes, M. Romaní-Pérez, and Y. Sanz. 2018. Interplay between the gut-brain axis, obesity and cognitive function. Frontiers in Neuroscience 12:155.
Cortés‐Martín, A., C. E. Iglesias‐Aguirre, A. Meoro, M. V. Selma, and J. C. Espín. 2021. Pharmacological therapy determines the gut microbiota modulation by a pomegranate extract nutraceutical in metabolic syndrome: A randomized clinical trial. Molecular Nutrition & Food Research 65 (6):2001048. 2001048.
Drago, L. 2019. Probiotics and colon cancer. Microorganisms 7 (3):66.
Foley, M. H., S. O’Flaherty, R. Barrangou, and C. M. Theriot. 2019. Bile salt hydrolases: Gatekeepers of bile acid metabolism and host-microbiome crosstalk in the gastrointestinal tract. PLOS Pathogens 15 (3):e1007581.
Hampel, H., R. Vassar, B. De Strooper, J. Hardy, M. Willem, N. Singh, … A. De Vos. 2020. The β-secretase BACE1 in Alzheimer’s disease. Biological Psychiatry 89 (8):745–756.

谷禾健康
我们的身体中有很多不同的器官,组织,腺体等会产生许多信号分子来精确控制和影响身体的反应和活动,这些信号分子包括激素、神经递质、生长因子、细胞因子等。它们可以促进或抑制细胞的生长和分化,调节细胞间的相互作用和通讯,影响身体的代谢和能量平衡,参与免疫系统的细胞和分子相互作用,调节身体的内环境和外环境的适应性等。
其中,激素是一类重要的生物活性物质,也可以说是信号分子,一般由特定的内分泌细胞,腺体或器官所产生,包括性激素、脂质激素、蛋白质激素、胰岛素、甲状腺激素、肾上腺素和去甲肾上腺素等。
这其中性激素是一类重要的激素,主要由生殖腺(如卵巢、睾丸)和肾上腺产生,包括雌激素、孕激素和雄激素等。它们在生殖系统中发挥重要作用,对女性月经周期、妊娠、分娩、哺乳等过程起调节作用,对男性性发育、精液生成、性欲等方面也具有重要影响。此外,性激素还与骨密度、心血管健康、认知能力等方面有关。
临床上性激素六项是生殖内分泌门诊常用的检查。性激素六项检查主要用于评估男女生殖内分泌功能,对月经失调、性功能障碍、不孕不育、生殖系统肿瘤、性染色体病等的鉴别和诊断及内分泌治疗的效果检测都具有重要的临床意义和参考价值。
近年来,越来越多的科学证据表明,肠道菌群与性激素(宿主的内源激素或外源补充激素)存在直接或间接的相互作用。性激素对于塑造肠道微生物群组成具有一定影响,与此同时,肠道微生物群本身也会影响性激素水平。
目前的研究主要基于横断面研究,基于纵向的检测肠道菌群发育以及激素水平变化的研究队列还较少,目前无法区分是性激素变化在前还是菌群变化在前,但是肠道菌群的失调和性激素的异常水平都与多种疾病的发生和发展有关,如肥胖症、炎症性肠病、乳腺增生、甲状腺疾病,妇科疾病、自身免疫疾病等。因此,保持肠道菌群和激素水平的平衡,对于维持人体的健康十分重要。
本文旨在阐述激素与肠道微生物群的相互作用,主要着重讲述性激素影响下的人体一些疾病和菌群的差异。
这些发现可能会在未来用于开发治疗激素紊乱相关疾病或与性别和激素活动相关的自身免疫性疾病、甲状腺疾病、代谢疾病甚至是压力等情绪状态的新疗法。
目录/contents
Part1:肠道微生物与激素的关联
Part2:肠道微生物通过激素对宿主的影响
Part3:性别差异下的肠道微生物群
Part4:肠道菌群与性激素的相互作用
Part5:性激素影响下的相关疾病
Part6:结语
肠道微生物群以多种方式影响动物和人类的行为,甚至可能影响人类的情绪状态和疾病状态。最近揭示了菌群的一个关键作用:调节激素分泌。
微生物和激素之间的相互作用会影响宿主的新陈代谢、免疫力和行为。这种相互作用是双向的,因为微生物群已被证明既受宿主激素影响又影响宿主激素。
肠道微生物群通过激素影响宿主情绪、免疫及代谢等
微生物群对宿主激素水平的影响可能是直接的,即微生物群产生激素,也可能是间接的,即微生物可能调节肾上腺皮质(控制焦虑和压力反应)的功能,或调节炎症和免疫反应。
Neuman H,et al.FEMS Microbiol Rev.2015
激素水平调节微生物群的生长、毒性
多种宿主因素(如饮食、运动、情绪、健康状况、压力和性别)会导致激素水平发生变化,进而对微生物群产生多种影响(包括生长、毒力和抵抗力)。
Neuman H,et al.FEMS Microbiol Rev.2015
文献中已记载激素和微生物群之间的相互作用




Neuman H,et al.FEMS Microbiol Rev.2015
▸ 菌群通过影响神经激素前体调节肠道运动、情绪、食欲等
神经内分泌细胞响应神经元输入而分泌神经激素。尽管它们被分泌到血液中以发挥全身作用,但它们也可以充当神经递质。微生物群对行为的调节(例如小鼠的焦虑)被认为是通过神经激素前体(例如血清素、多巴胺)发生的。
最近,肠道细菌被证明可以产生和响应神经激素,例如血清素、多巴胺和去甲肾上腺素。
这些机制对于研究病原体的研究人员来说很有趣,因为它们可能会影响病原体对宿主防御反应的敏感性。例如,为了响应宿主肾上腺素,沙门氏菌会下调其对宿主抗菌肽的抵抗力并诱导关键的金属转运系统,从而影响细胞中的氧化应激平衡。
// 血清素
血清素,也称为 5-羟色胺 (5-HT),是大脑中的主要神经递质之一。然而,超过90%的哺乳动物宿主血清素都存在于肠道中。
肠道血清素的分泌受饮食影响,调节肠道运动、情绪、食欲、睡眠和认知功能。这种双重作用表明血清素可能将肠道(包括其微生物群)与宿主行为联系起来。
血清素与肠易激综合征和克罗恩病有关
大脑血清素可以通过血清素转运蛋白穿过血脑屏障进入血液,这表明肠-脑轴中存在另一个联系。血清素与肠易激综合征和克罗恩病等胃肠道疾病有关,这些疾病也与微生物群的差异有关。
// 多巴胺
多巴胺由芽孢杆菌(Bacillus)和沙雷氏菌(Serratia)等细菌产生 。
无菌小鼠中的游离管腔多巴胺水平显著低于常规小鼠,并且在接种表达β-葡萄糖醛酸酶的细菌后再次升高。
多巴胺不足与帕金森病有关
这些结果表明,在以多巴胺形成不足为特征的帕金森病等疾病中,肠道细菌与多巴胺水平之间可能存在相关性。
//γ-氨基丁酸
γ-氨基丁酸 (GABA) 是哺乳动物中枢神经系统中的主要抑制性神经递质,它也由微生物群产生,并可能影响宿主行为。这很有趣,因为中枢γ-氨基丁酸受体表达的改变与焦虑和抑郁的发病机制有关。
γ-氨基丁酸影响焦虑和抑郁
已经研究了由乳杆菌产生的γ-氨基丁酸,以试图大规模发酵安全的γ-氨基丁酸。因此,向小鼠施用鼠李糖乳杆菌会改变不同中枢神经系统区域中γ-氨基丁酸受体的表达,从而减少与焦虑和抑郁相关的行为。
▸ 菌群通过调节应激激素降低宿主焦虑行为
微生物群可以通过改变应激激素水平来帮助我们保持冷静和平衡。无菌小鼠具有升高的应激激素皮质酮和促肾上腺皮质激素 (ACTH) 血浆水平以响应轻度压力,增加与焦虑和压力相关的行为。
促肾上腺皮质激素通过进一步产生皮质类固醇在下丘脑-垂体-肾上腺轴中发挥重要作用。因此,两种特定物种,瑞士乳杆菌(Lactobacillus helveticus)和长双歧杆菌,降低了大鼠和健康人类的应激激素皮质醇水平和焦虑样行为。
此外,与对照组相比,长期接受益生菌鼠李糖乳杆菌治疗的小鼠在强迫游泳测试中皮质酮水平较低,抑郁行为也较少。
▸ 肠道菌群影响甲状腺激素的转化影响代谢
肠道菌群产生的代谢物对甲状腺和内分泌功能也是必不可少的。它影响硒、铁、锌和碘的吸收,这些都是甲状腺激素产生所需的重要矿物质。它制造短链脂肪酸(SCFAs)与甲状腺激素协同工作,以改善肠道屏障功能和加强细胞间紧密连接。
肠道菌群失衡是影响甲状腺激素转化的主要障碍。事实上,20%的T4(甲状腺素)在肠道中转化为活性的T3(三碘甲状腺原氨酸)形式,肠道菌群不健康,肠道内存在过多的病原体,肠道中甲状腺激素的转化也会减少。
此外当肠道屏障功能减弱时,毒素和未完全消化的食物残渣等颗粒会逃逸,导致肠胃不适和全身炎症。炎症会在组织水平损害甲状腺激素的转化。受体受损,身体也不能将T4激素转化为活性的T3形式。
▸ 细菌通过信息素影响宿主行为
信息素是在性识别、吸引力和交配行为以及攻击行为和支配行为中发挥重要作用的激素。
信息素也称为外激素,一种分泌到一个人体外并影响他人行为的化学物质。
共生细菌和信息素之间的这种联系也发生在哺乳动物身上。两种鬣狗的气味腺中有不同的细菌群落,这与气味分泌物中不同的挥发性脂肪酸特征有关。作者推测,共生细菌产生的代谢产物提供了物种特有的气味。
这进一步支持了细菌可以产生影响宿主行为的气味特征的观点。细菌也可以在配偶选择中发挥作用:雌性小鼠不会被感染沙门氏菌的雄性小鼠的尿液所吸引。
化学信号和嗅觉刺激也在人类行为中发挥作用,未来的研究将帮助我们了解细菌产生的气味是否会影响我们自身的相互作用,甚至可能影响进化。
▸ 肠道菌群代谢性激素影响健康
肠道菌群也影响雌激素,它们通过特定的酶对雌激素进行循环利用并产生雌激素代谢物。当肠道菌群失衡或不健康时,这些过程就会被破坏,从而导致健康问题。
例如,Prevotella intermedius吸收雌二醇和黄体酮,从而促进其生长。雌激素受体ER-β表达的变化也会影响肠道微生物群的组成。
雌二醇在卵巢中合成,是雌激素中含量最高的,活性最强的激素,经肝脏代谢后,通过胆道排泄到胃肠道,部分雌二醇被胃肠道重新吸收进入血液。肠道菌群可以通过改变雌二醇的活性和非活性状态,影响血清中类固醇激素的水平。
武汉大学人民医院转化医学研究所的李艳教授团队在cell metabolism上发表研究论文,发现绝经期前抑郁症患者肠道细菌Klebsiella aerogenes(产气克雷伯氏菌)的3β-羟基类固醇脱氢酶(3β-HSD)通过降解雌二醇诱导绝经期前女性出现抑郁症。
肠道微生物群作为人体最大的菌群库,动物和人类研究发现,肠道微生物群存在性别相关差异。
影响微生物群的重要因素(例如环境或激素)取决于不同的生命周期,并且在男性和女性中会有所不同。
Kim YS,et al.World J Mens Health.2020
需要注意的是,性激素对肠道微生物具有显著的影响,青春期、怀孕、更年期、多囊卵巢综合征和性腺切除术等各种因素会导致性激素水平(睾酮和雌二醇)发生变化,进而导致肠道菌群组成发生变化。
肠道微生物群与性激素之间的相互作用
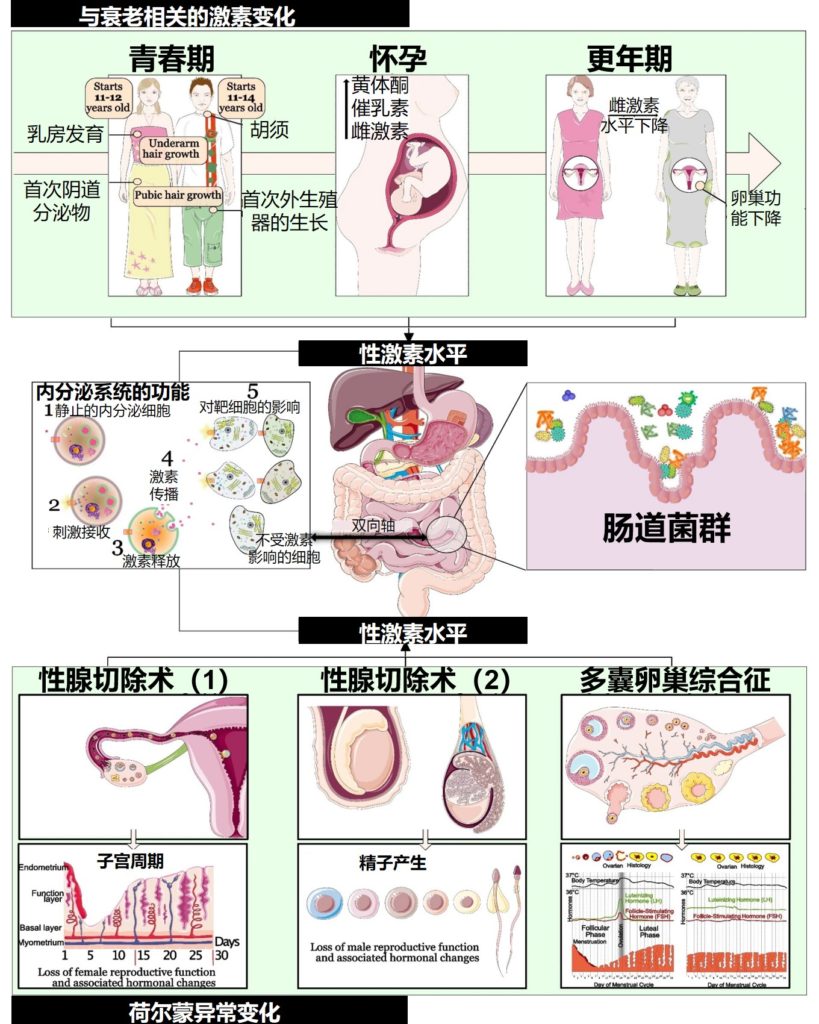
Santos-Marcos JA,et al.Biol Sex Differ.2023
▷婴儿时期肠道菌群组成与性别相关的差异
婴儿期的特点是参与人乳低聚糖代谢的细菌种类占主导地位(据估计,婴儿细菌微生物群的25-30%来自母乳)并且肠道微生物群多样性的差异减少。
特别是,这个年龄段的特点是被属于双歧杆菌(Bifidobacterium)和链球菌(Streptococcus)的几种物种定殖,例如长双歧杆菌和嗜热链球菌。
6个月至2岁之间的年龄段的特点是将固体食物引入婴儿的饮食中,并伴随着肠道微生物群结构和功能多样性的快速增加。这是婴儿生长的关键时期,肠道微生物群开始接近成人特征。
最近对人类婴儿进行的几项研究发现了他们出生后第一年不同时间点肠道微生物群落的潜在性别依赖差异。
男婴α多样性较低
例如,对出生后前30天新生儿粪便样本的分析显示,与女性相比,男婴的α多样性较低,梭状芽孢杆菌的丰度也较高,而肠杆菌的丰度较低。
男婴双歧杆菌更丰富
在新生儿肠道中最早和最丰富的细菌定植者中,已知双歧杆菌可为宿主肠道提供健康益处。改变在从出生后一天到六个月大,在男性和女性阴道分娩的婴儿中观察到双歧杆菌丰度。发现与女孩相比,男孩的双歧杆菌水平更高。
▷儿童时期肠道菌群组成与性别相关的差异
在出生后的前12个月,婴儿的肠道因饮食中引入固体食物而发生变化,随之而来的是微生物群落的变化。
特别是,在引入固体食物后,婴儿大肠中厌氧菌种群的初始定植在数量和组成上开始与成人相似。几项研究表明,在没有饮食变化或抗生素治疗等外部压力因素的情况下,与儿童相比,成年期的肠道微生物群更加稳定和有弹性。
这表明尽管出现了部分趋同,但仍存在差异,因此分析这一生命阶段与性别差异的关系很有价值。
男孩双歧杆菌与乳酸杆菌之间的正相关性比女孩更强
对来自哥伦比亚两个不同地点的277名1至5岁儿童(154名女孩和123名男孩)的评估(研究临床腹泻的存在)表明,粪便微生物群受到多种宿主因素的影响,包括年龄、健康状况、地点和性别。
有趣的是,与女孩相比,健康男孩的双歧杆菌和乳酸杆菌之间存在显著更强的正相关性,而在受腹泻影响的儿童中未观察到显着相关性。
▷青春期与性别相关的肠道菌群差异最明显!
青春期是发展的关键时期,性激素,如雌激素和睾酮,在这个年龄段人群的肠道微生物群中发挥重要作用。
在性激素的影响下异性间微生物群差距变大
一项针对青春期肠道微生物群组成的人类研究表明,与同性双胞胎相比,青少年(13-17岁)异性双胞胎的粪便微生物群差异更大。
最近的一项研究得出结论,与青春期受试者(5-15岁)相比,非青春期的α和β多样性没有差异。随着青春期的开始,梭状芽孢杆菌(Clostridium)的水平降低,而变形杆菌(Proteus)的丰度增加。
在青春期受试者中,发现Adlercreutzia、Ruminococcus、Dorea、Clostridium和Parabacteroides与睾酮水平有关。
考虑到人类这一生命阶段的可用数据有限——必须使用啮齿动物模型的研究作为替代。在小鼠身上进行的实验表明,肠道微生物群的性别差异出现在与性激素水平相关的青春期开始时。
动物实验中青春期的菌群性别差异
特别是,仅在10至13周龄的非肥胖糖尿病青春期后小鼠中检测到α多样性的差异,与雄性相比,雌性的水平显著更高。
此外,雄性小鼠表现出更高丰度的卟啉单胞菌科(porphyromonas)、韦荣球菌科、消化球菌科、乳杆菌科和肠杆菌科与女性相比。而当雄性小鼠被阉割后,这些差异就消失了。
▷成年后肠道菌群组成与性别相关的差异
肠道微生物组在生命的最初几年大部分时间都不稳定,但在成年期变得更加稳定,之后细菌丰富度和组成可能伴随着特定疾病会发生变化。
在整个生命周期中,随着免疫和神经系统的成熟,肠道微生物群的伴随发育是性别二态性的,导致成年男性和女性的不同微生物群落以及免疫和神经炎症通路。
注:年龄、环境(饮食和身体活动)和地理位置在内的多种因素似乎在这些性别依赖性肠道菌群差异中发挥了作用。
分析来自结肠癌研究对照组的82名受试者得出结论,性别与肠道微生物组的整体组成显著相关。然而,这项研究包括的参与者数量相对较少,年龄范围相当广泛(30-83 岁),这可能混淆了研究的结果。
年轻女性肠道微生物α多样性比同年龄男性更高
在包括美国、英国在内的四个地理区域以及来自哥伦比亚和中国的两个队列中评估了年龄在20至69岁之间的三个大型队列中年龄、性别和肠道微生物群α多样性之间的关系。
有趣的是,与同年龄段的男性相比,年轻的成年女性(20-45 岁)表现出更高的α多样性。此外,性别和α多样性之间的关联在年轻人中比在中年人中更为明显,而当参与者的平均年龄为60岁时,男女之间的α多样性没有差异。这可以解释为更年期女性的雌激素水平下降。而更年期之后男性与女性的激素水平差异变小,肠道微生物群的相似程度更接近。
总之,对来自不同国家的男性和女性进行的人口研究发现人体肠道微生物群组成存在性别相关差异。
这意味着,性激素的动态变化会影响肠道共生,而青春期时激素的变化最大,此时男性与女性的肠道微生物群之间的差异非常明显。
注:由于该研究所覆盖的样本量还较小,年龄之间的跨度比较大,仅作为研究参考,还不能用于诊断标准。
整个生命周期中男性和女性肠道微生物群组成的差异
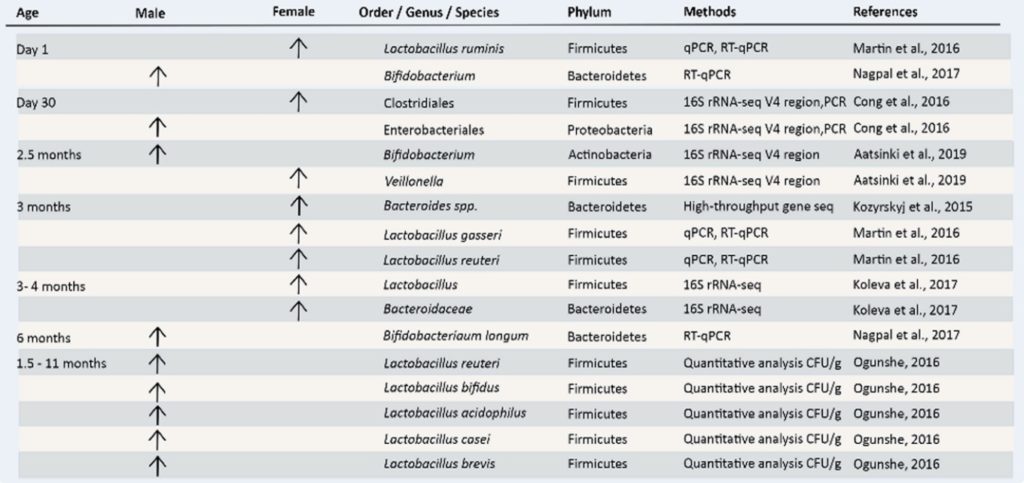
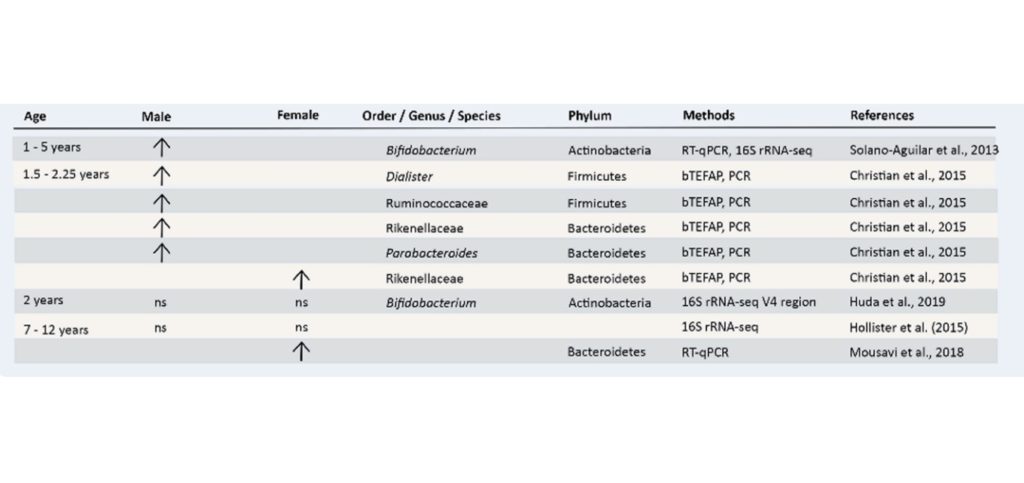
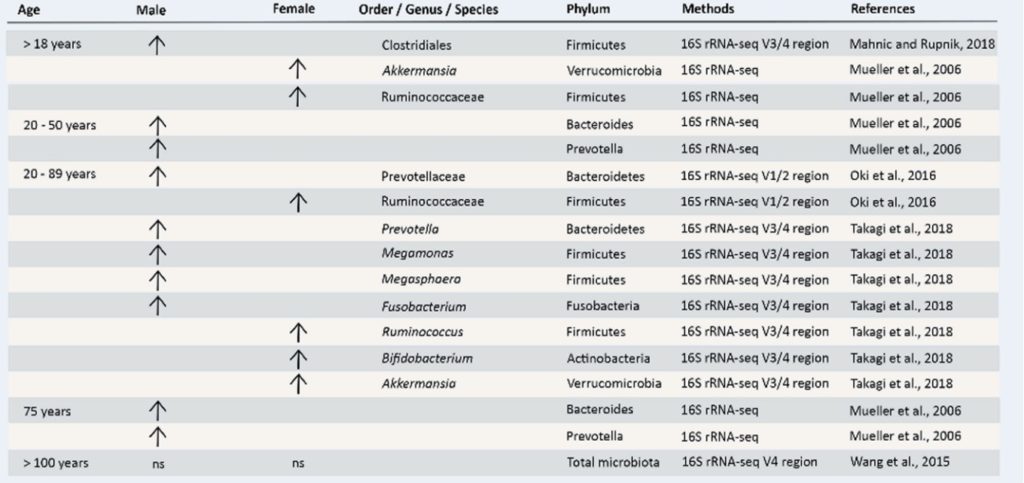
Valeri F,et al.Front Neuroendocrinol.2021
不仅肠道微生物群受性激素的影响,而且肠道微生物群本身也会影响激素水平。
▷ 微生物代谢雌激素进而影响多个器官
生殖内分泌功能涉及由复杂反馈机制控制的多种激素。卵巢、肾上腺和脂肪组织会产生雌激素。体内产生的或作为食物摄入的雌激素可以被肠道微生物代谢。
产生的代谢物再次影响宿主。性激素通过类固醇受体(包括雌激素受体β)直接调节细菌的新陈代谢。
具有雌激素代谢能力的肠道微生物群基因库
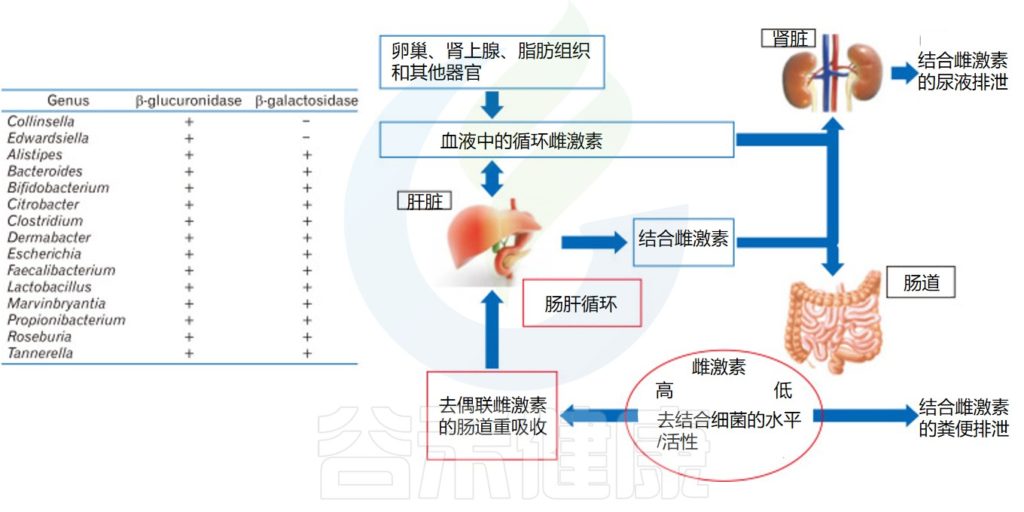
Yoon K,et al.J Neurogastroenterol Motil.2021
同时,具有β-葡萄糖醛酸酶活性的肠道微生物群解离胆汁中排出的结合循环雌激素。去结合使雌激素对系统的重吸收过程成为可能。去结合的雌激素通过雌激素受体循环并影响多个器官,不仅影响生殖器官,还影响骨骼系统、心血管系统和中枢神经系统。
雌激素循环途径
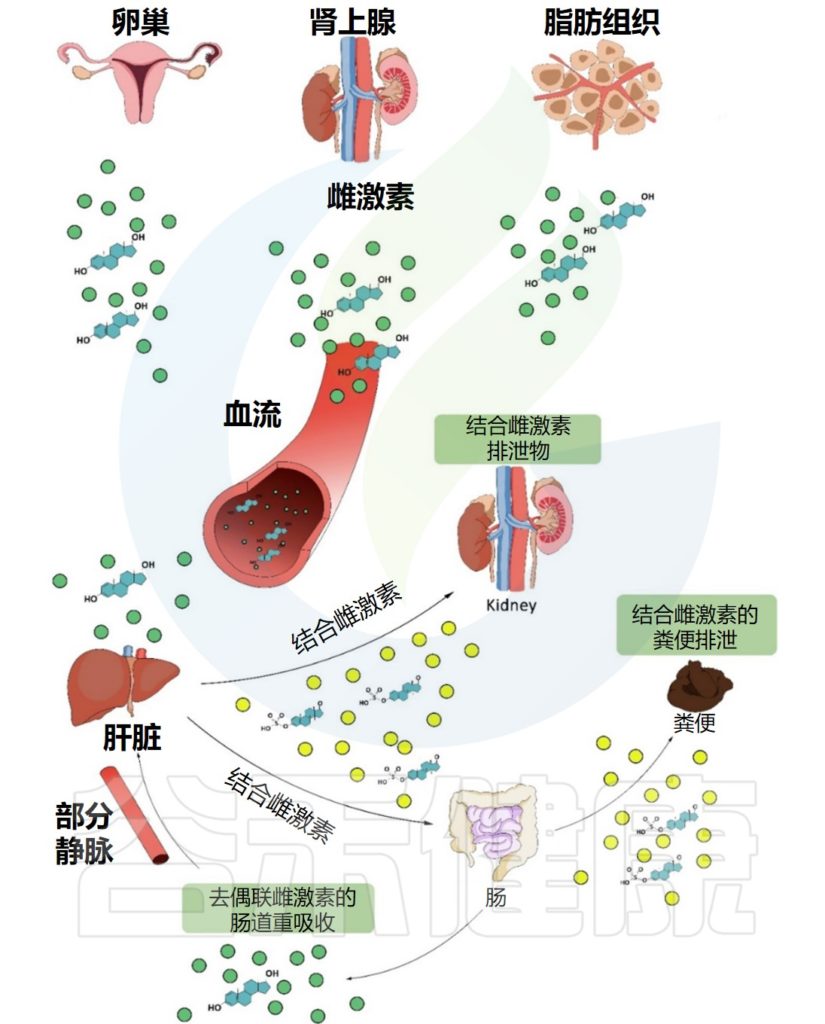
Valeri F,et al.Front Neuroendocrinol.2021
雌激素在几个组织中局部产生(主要是卵巢,但也有肾上腺和脂肪组织)。它们在血液中循环,到达多个靶器官,如肝脏和肾脏。一旦到达肝脏,它们就会通过葡萄糖醛酸化或磺化反应进行结合。
此外,结合雌激素被转化为水溶性分子,随后可以通过胆汁、尿液和粪便排出体外。某些部分的结合雌激素可以在肠道中被肠道微生物分解,并通过门静脉被重新吸收到肝脏中。
雌激素对发病机制具有影响
通常,雌激素与核受体结合,引起构象变化。通过肠道屏障通透性和免疫系统调节,中枢和外周改变与肠易激综合征 (IBS) 相关联。流行病学研究表明女性在肠易激综合征中比例更大,暗示性激素对其发病机制的影响。
雌激素受体作用的分子途径
Yoon K,et al.J Neurogastroenterol Motil.2021
▷ 微生物群影响雄激素的代谢
据报道,在雄激素代谢中,肠道微生物群是一个重要的调节因子。在小鼠的小肠内容物中,葡萄糖醛酸化睾酮和双氢睾酮含量很高。远端肠道具有高水平的游离双氢睾酮。
在年轻成年男性的粪便中观察到明显高水平的未结合双氢睾酮。在无菌小鼠中,葡萄糖醛酸化睾酮和双氢睾酮较高;然而,在远端肠道中发现游离水平非常低。这意味着肠道微生物群会影响双氢睾酮和睾酮的肠道代谢。
▷ 性激素高的人拥有更多样化的肠道微生物
血清睾酮升高的男性和雌二醇水平升高的女性分别拥有更多样化的肠道微生物群,许多细菌属与睾酮(Acinetobacter, Dorea, Ruminococcus和Megamonas)和雌二醇(Slackia和Butyricimonas)水平相关。
高雄激素的女性微生物群明显改变
在这方面,患有多囊卵巢综合征(高雄激素)的女性在妊娠早期到晚期的变化中,表现出明显的微生物群改变,变形菌(Proteobacteria)和放线菌(Actinobacteria)总体增加,丰富度降低。
肠道微生物群组成的性别差异在青春期增加,随着青春期的进展,女孩的肠道微生物群变得与成年人更相似。这些结果可能还表明肠道微生物群可能会影响青春期的时间,可能是通过调节宿主性激素水平。
雌激素水平与肠道微生物多样性密切相关
在男性和绝经后女性中,尿液中的雌激素水平与肠道微生物群丰富度和α多样性密切相关,而绝经前女性雌激素水平在月经周期中收集时变化很大,并未显示出这种关联。
▷ 微生物群可以预测粪菌移植后睾酮水平
最近报道,绝经后妇女的肠道菌群与男性比绝经前妇女更相似,绝经后妇女与同龄男性之间实际上没有显著差异。
这项研究还表明,性腺类固醇与微生物群差异之间存在关联,绝经前妇女的类固醇生物合成和降解途径丰富,并且与血浆睾酮水平显著相关。
此外,微生物群可以预测人类粪便转移后人类和(经抗生素处理的)雄性小鼠的循环睾酮水平。
综上所述,这些结果表明,肠道微生物群组成的差异与性别以及不同激素状态有关。
既然知道了肠道微生物群与性激素之间存在相互作用,那么它们具体是通过什么机制影响的呢?
性激素和肠道微生物组之间相互作用机制
He S,et al.Front Microbiol.2021
1
微生物群通过改变胆汁酸影响性激素水平
最近有人提出,肠道微生物群的部分性别差异可能取决于胆汁酸,因为男性的胆汁酸库比女性大。在肝脏中由胆固醇合成后,它们被肠道微生物群代谢为次级胆汁酸,从而改变微生物群的结构并导致各种病理。
因此,肠道微生物群通过依赖于法尼醇X受体(FXR)的机制调节回肠中成纤维细胞生长因子15(FGF15)和肝脏中胆固醇7α-羟化酶(CYP7A1)的表达,从而调节胆汁酸的二次代谢,并抑制胆汁酸在肝脏中的合成。
FGF15抑制肝脏中CYP7A1的表达,CYP7A1是一种催化和调节胆汁酸合成第一步的酶。
胆汁酸同时也能影响微生物群的结构和丰度
已经观察到胆汁酸的减少导致细菌增殖,并且法尼醇X受体抑制细菌过度生长。
几项研究证实了胆汁酸、性激素和肠道菌群组成之间的关系。通过这种方式,给大鼠服用胆汁酸引起的微生物群变化类似于高脂肪饮食引起的变化,以牺牲拟杆菌属为代价增加厚壁菌门的水平。
此外,粪便微生物群(来自瘦供体)的移植产生了与瘦供体相似的肠道微生物群和胆汁酸库的变化,而小鼠的性腺切除术也改变了胆汁酸模式。
由于睾酮是由胆汁酸衍生的,并且如上所述,胆汁酸水平会被微生物群改变,因此微生物群可能间接影响睾酮水平的说法是成立的。
2
微生物群通过酶促激活性激素
共生微生物群落可以通过其酶的活性影响性激素水平。通过这种方式,术语“strobolome”被创造出来,定义为肠道微生物群中能够从其无活性葡萄糖醛酸苷中激活雌激素的一组基因,这主要归功于β-葡萄糖醛酸苷酶,它将雌激素解离为活性形式。这些活性雌激素进入血液并作用于雌激素受体α(ERα) 和雌激素受体β(ERβ)。
同样,最近的一项研究得出结论,肠道微生物群参与双氢睾酮 (DHT) 和睾酮的代谢和肠道去葡萄糖化,从而导致最高水平的雄激素。
肠道微生物群在性别差异中的另一种可能作用机制可以在羟基类固醇脱氢酶 (HSD) 中找到,这些酶参与类固醇激素的代谢并控制类固醇与其核受体的结合,使它们充当激活剂或抑制剂。
✦ 肠道微生物还通过植物雌激素影响代谢
除了三种主要形式的雌激素(胆固醇衍生的类固醇激素)、雌二醇(E2,在绝经前的非孕妇中占主导地位)、雌酮(E1,在绝经后占主导地位)和雌三醇(E3,在怀孕期间占主导地位),还有一种称为植物雌激素的植物化合物,其结构和功能与雌激素相似。
植物雌激素是天然存在于植物中的化合物,植物雌激素包括异黄酮,如染料木黄酮和黄豆苷原,它们主要存在于大豆中,并在被肠道微生物群代谢后被激活。从这个意义上说,肠道微生物群允许从黄豆苷元中获得O-Desmethylangolensin (ODMA) 和雌马酚,这两种物质都具有雌激素活性。
与雌激素类似,植物雌激素通过影响细胞信号传导引起生理效应,因为它们可以通过激活或抑制雌激素受体α或雌激素受体β来诱导或抑制雌激素作用,并且还可能引发表观遗传效应和细胞内信号级联反应。
植物雌激素可以调节内分泌系统改善病症
与此相关的是,几项人类研究表明,植物雌激素可以通过调节内分泌系统来改善各种病症,包括更年期症状,并且可以逆转代谢性内毒素血症的症状。
在这方面,植物雌激素代谢物雌马酚通过促进雌激素的尿液排泄和改变女性的血液水平,与降低女性激素相关疾病的风险有关,而不产生O-Desmethylangolensin与肥胖有关。
植物雌激素在饮食中被消耗,因为它们出现在水果、蔬菜、豆类和一些谷物中。事实上,饮食成分对肠道微生物群生态系统具有急性影响。通过促进更多样化和更稳定的微生物系统的发展,以植物为基础的饮食似乎对人类健康更有益。
!
药物和益生菌治疗的性别差异
已有一些关于药物治疗和微生物群性别差异的报道。
▷男女服用同一种抗炎药后微生物变化有差异
一项人体研究调查了23名健康男性和女性在非甾体抗炎药诱导的肠道屏障功能和微生物组方面的差异。使用 16S 核糖体 RNA 测序确定了十二指肠和粪便微生物群组成。健康女性的肠道通透性较低,十二指肠和粪便微生物多样性高于健康男性。
两性服用吲哚美辛后肠道通透性增加。然而,只有雌性在摄入吲哚美辛后粪便微生物多样性下降,包括普氏菌丰度增加。十二指肠微生物群组成没有表现出性别特异性变化。
▷益生菌给药存在性别差异
如今,益生菌因多种原因而流行,并且有人认为其效果可能因性别而异。
一项研究报告说,将5种乳杆菌菌株的益生菌混合物给予易患狼疮的小鼠可改善肾功能,并在雌性和阉割的雄性小鼠中显示出抗炎作用,但在性腺完整的雄性小鼠中则没有,这表明益生菌给药存在性别差异。
另一项研究报告了金枪鱼油和海藻油混合物对肠道微生物群的抗衰老作用存在性别差异。雄性小鼠在特定混合油比例下表现出比雌性小鼠更好的抗衰老效果。
肠道菌群失调引发的疾病在两性之间表现不同。研究最多的疾病主要是代谢性疾病。研究发现,性激素等多种因素影响这些疾病,它们密切相关,相互作用。
▸ 全球肥胖发病率越来越高
肥胖症的体重指数(BMI)为30kg/m2或更高,在全球各国成人和儿童中的流行率都在增加。这种病理学是复杂的遗传、社会经济和文化相互作用的结果,会导致严重的健康、经济和社会问题。
科学研究表明,一些代谢紊乱的发生与身体脂肪的分布有关,而且这种分布表现出性别二态性。脂肪倾向于堆积在男性的躯干和腹部周围,而女性的脂肪一般在臀部和大腿周围。
•腹部肥胖
腹部肥胖,尤其是内脏肥胖,会导致血糖和甘油三酯升高、高密度脂蛋白胆固醇降低和低密度脂蛋白 (LDL) 颗粒的增加,以及炎症标志物的增加 。
•臀部肥胖
相反,臀部脂肪与保护性脂质和葡萄糖水平以及降低代谢风险有关,似乎通过长期脂肪酸储存和有益的脂肪因子水平(与瘦素和脂联素水平呈正相关,与炎性细胞因子水平负相关)发挥其保护作用。
肥胖中的微生物多样性和性激素
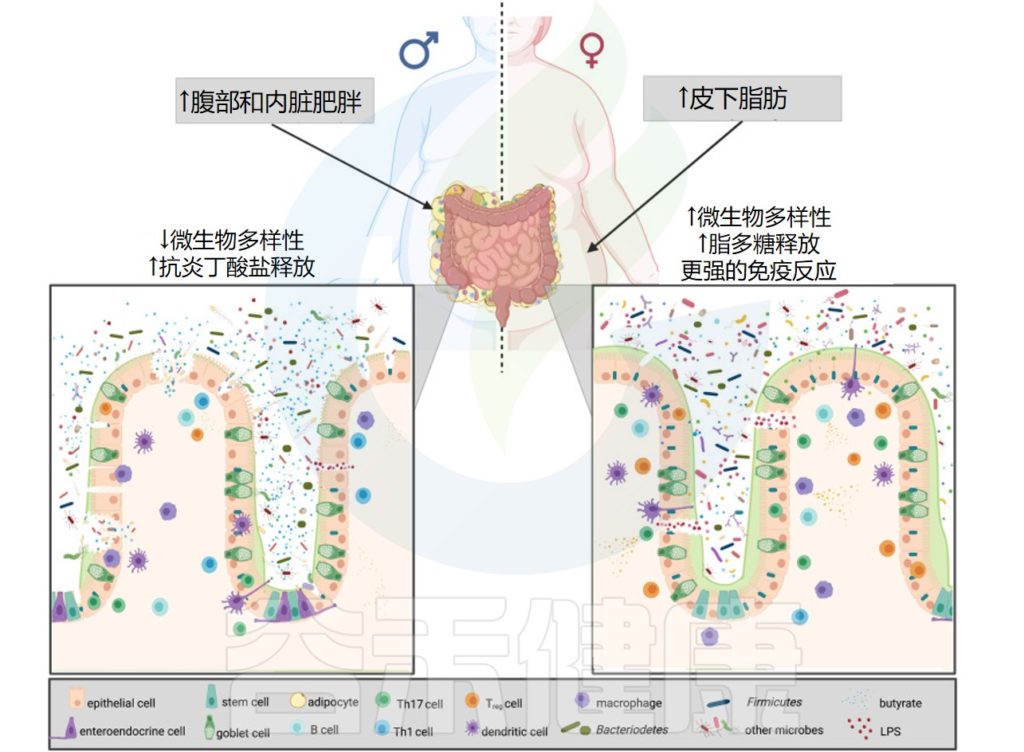
Brettle H,et al.Front Immunol.2022
在肥胖症中,拟杆菌门决定了疾病的严重程度。肥胖男性的物种丰富度较低,并且发现睾酮与厚壁菌门增加有关,因此会释放更多的抗炎丁酸盐。
另一方面,肥胖女性尽管微生物多样性更高,但雌二醇和拟杆菌增加,导致脂多糖释放更多,从而引发更强的免疫反应。
脂肪组织分布、性激素和肥胖的代谢紊乱
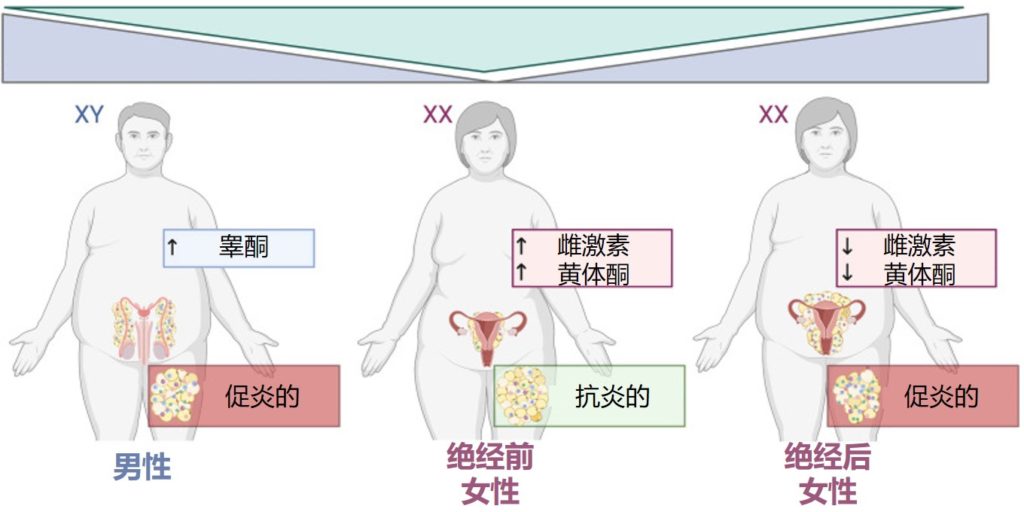
Brettle H,et al.Front Immunol.2022
与绝经前女性相比,男性和绝经后女性心血管风险增加,腹部/内脏肥胖和胰岛素皮下脂肪分布减少。男性和绝经后女性体内的脂肪组织比绝经前女性的脂肪组织更具促炎性。
√性激素调节身体脂肪分布
大量证据支持性类固醇调节身体脂肪分布的观点。在这方面,青春期激素的变化与不同性别之间的体重增加有关,这是由于男孩的瘦体重增加和女孩的脂肪量增加,以及男性和女性的脂肪分布。
此外,几项研究表明,一些基因参与了在体脂分布中观察到的性别二态性,以及性类固醇激素在这些基因调节中的潜在作用。
睾酮影响不同部位的脂肪堆积
在男性中,睾酮抑制腹内区域甘油三酯的摄取,并似乎促进它们在皮下区域的积累,同时导致儿茶酚胺刺激的皮下脂肪分解减少,而不是内脏脂肪分解。
这些过程似乎受到雄激素受体 (AR) 基因的影响,因为在雄激素受体敲除小鼠模型中,雄激素受体的缺失通过减少脂肪分解导致肥胖增加,尤其是晚期肥胖。
此外,caveolin-1蛋白(CAV1)似乎在脂肪堆积中起着重要作用,并且它受雌激素(雌二醇)和雄激素(双氢睾酮)的不同调节。
√性激素影响脂肪细胞功能
在细胞水平上,已经观察到性激素(雄激素和雌激素)对白色脂肪组织中脂肪细胞功能影响的差异,涉及脂肪细胞分化、脂肪分解、脂肪生成、胰岛素敏感性和脂肪因子产生/分泌等关键方面。
在这种情况下,睾酮和双氢睾酮调节小鼠间充质干细胞的分化,以雄激素依赖性方式分别促进和抑制它们向肌细胞和脂肪细胞的分化。
双氢睾酮增加脂肪分解
同样,在对人体细胞进行的体外研究中,双氢睾酮以雄激素受体依赖性方式抑制人间充质干细胞和人前脂肪细胞的脂肪形成分化,增加脂肪分解并减少脂质积累。
睾酮阻止脂肪细胞肥大和脂肪细胞生成
在阉割小鼠(雄性性腺功能减退症模型)中,脂肪量通过脂肪细胞肥大和脂肪生成增加,而当这些小鼠接受激素替代疗法时,睾酮阻止内脏和皮下脂肪量的扩张。
此外,致肥胖脂肪生成也通过抑制雄激素受体活性而升高。这项研究还显示了脂肪分布的不同调节,睾酮衍生的雌二醇和双氢睾酮分别阻止了内脏和皮下脂肪的增加。
√睾酮浓度较高利于减少内脏脂肪
睾酮浓度与向心性肥胖呈负相关,并且已发现睾酮治疗可减少具有雄激素缺乏症状和低血清睾酮水平的男性的内脏脂肪。
睾酮含量下降后脂肪更多的积累
此外,随着年龄的增长,睾丸激素的下降伴随着肥胖的增加,腹部脂肪的优先积累和内脏脂肪组织的更多积累。
还有报道称,内脏脂肪组织与生物可利用和游离睾酮呈负相关,皮下脂肪组织与性激素结合球蛋白 (SHBG) 呈负相关。
√雌二醇水平较低的女性易患向心性肥胖
在女性中,向心性肥胖与睾酮水平升高和雌二醇降低相关。更年期的荷尔蒙变化导致脂肪重新分布,与总脂肪和年龄无关,然而,一些研究表明,绝经后上半身脂肪的分布可能是由于衰老而不是绝经本身。
向心性肥胖——指患者体内脂肪沉积是以心脏、腹部为中心发展的一种肥胖类型。
最近发现,绝经前妇女的身体或躯干脂肪量与较低的总雌二醇和较高的游离雌二醇浓度有关,绝经前和绝经后妇女的总睾酮和计算游离睾酮浓度较高,性激素结合球蛋白和胰岛素样生长因子-I(IGF-I)浓度较低。
▼
代谢综合征是一种以腹部肥胖、胰岛素抵抗、高血压和高血脂为特征的病理状况,已遍布全球并导致2型糖尿病、冠心病和中风等疾病患病率上升。
性激素对肥胖、代谢综合征和2型糖尿病的影响

Santos-Marcos JA,et al.Biol Sex Differ.2023
√代谢综合征患者中男女的菌群差异
在代谢综合征患者中也观察到特定菌属的丰度差异。男性中韦荣氏球菌属(Veillonella)、Methanobrevibacter、Acidaminococcus、梭状芽孢杆菌(Clostridium)、罗氏菌属(Roseburia)和Faecalibacterium的丰度较高,而女性中嗜胆菌属(Bilophila)、瘤胃球菌属(Ruminococcus)和拟杆菌属(Bacteroides)的丰度更高。
√睾酮水平会影响代谢综合征
在睾酮对代谢综合征的影响中观察到的性别二态性似乎是雄激素受体依赖性的,并且已经提出了几种机制来解释睾酮水平与代谢综合征之间的关联。
男性中睾酮水平高不易患代谢综合征
在男性中,有证据表明睾酮与内脏肥胖、胰岛素抵抗和代谢综合症的发展呈负相关。沿着这些思路,在男性和啮齿动物中都报道了睾酮的雄激素受体介导的抗肥胖作用。
过量的睾酮可能导致女性胰岛素分泌受损
在女性中,据报道睾酮水平升高与胰岛素抵抗和葡萄糖耐受不良相关,因为它会降低全身葡萄糖的摄取。
关于睾酮对胰腺的作用,一项小鼠研究表明,雄激素受体调节男性胰岛β细胞生理,因此该受体的缺乏会降低葡萄糖刺激的胰岛素分泌并导致葡萄糖耐受不良。相反,有人提出,过量的睾酮可能通过雄激素受体依赖性机制导致女性胰岛β细胞功能障碍,胰岛素分泌受损。
√雌激素缺乏的女性代谢综合征患病率较高
雌激素水平似乎也影响代谢综合症的患病率。因此,大鼠卵巢切除术引起的雌激素耗竭导致大多数代谢综合征成分(脂质、葡萄糖、高密度脂蛋白和低密度脂蛋白)恶化,而在50岁以下的女性中,即经历更年期,其患病率增加。
卵巢切除的女性血糖和血压升高
此外,据报道,在接受过子宫切除术(通常伴随双侧卵巢切除术以预防后续卵巢癌)的女性中,血糖水平和高血压升高。
绝经导致性激素结合球蛋白水平下降,至少部分是由于雌激素减少,而睾酮水平在绝经期间没有改变 。从这个意义上说,绝经可以被认为是代谢综合征及其独立于年龄的所有独立组成部分的预测因素(风险因素)。
此外,已经描述了性激素结合球蛋白和代谢综合征之间的负相关,尤其是在绝经后妇女中。
▼
糖尿病包括一组疾病,根据其发展机制进行区分,这些疾病会降低调节血流中葡萄糖水平的能力,并导致长期高血糖症。
由于自身免疫和代谢过程,糖尿病有两种主要形式,即胰岛素依赖型糖尿病(1型糖尿病,T1D)和非胰岛素依赖型糖尿病(2型糖尿病,T2D)。
2型糖尿病的特征是胰岛B细胞产生的胰岛素不足和肝脏葡萄糖代谢受损,以及胰岛素抵抗,导致组织对胰岛素的反应性降低。
√糖尿病前期的症状表现出性别差异
空腹血糖受损 (IFG) 和葡萄糖耐量受损 (IGT) 作为2型糖尿病的前期阶段发生,表现出性别二态性,葡萄糖耐量受损在女性中更常见,而空腹血糖受损在男性中更常见。
有人提出,性激素可能是造成这种二态性的原因。事实上,更年期的雌激素治疗会降低空腹血糖并恶化葡萄糖耐量。
√男性的2型糖尿病发病率高于女性
此外,已证实男性的2型糖尿病发病率高于女性,这进一步支持了性激素参与这种病理学的发展。更年期2型糖尿病的风险增加,而更年期的激素治疗可能会延迟2型糖尿病的发作。
√睾酮水平影响男性2型糖尿病患病率
患有2型糖尿病的男性总睾酮和游离睾酮水平较低。与此相关的是,低水平的睾酮和性激素结合球蛋白与男性胰岛素抵抗和随后的2型糖尿病的发展有关。
低睾酮水平男性死亡率增加
此外,高水平性激素结合球蛋白和低水平睾酮的结合与2型糖尿病男性死亡率增加有关。其他研究表明,在患有2型糖尿病的男性中,低睾酮水平本身与死亡率增加相关,而睾酮替代疗法可能会提高这些男性的生存率。
同样,据报道,经过2年的睾酮治疗后,患有2型糖尿病的男性比例有所降低。此外,已发现前列腺癌的雄激素剥夺疗法会增加患糖尿病的风险。
患有2型糖尿病的男性睾酮水平往往较低
与上述一致,患有2型糖尿病的男性往往睾酮水平较低,而且他们中的大多数患有性腺功能减退症。事实上,许多研究已经证实,患有性腺功能减退症和低睾酮水平的肥胖2型糖尿病患者在接受睾酮替代疗法 (TRT) 后表现出改善的胰岛素抵抗和血糖控制。
√雌二醇水平高的男性患2型糖尿病的风险增加
关于女性荷尔蒙,雌二醇水平高的男性患2型糖尿病的风险增加,这种高雌二醇浓度与低性激素结合球蛋白浓度一起对男性患2型糖尿病的风险产生不利影响。
√女性睾酮水平较高可能易患2型糖尿病
与男性相反,女性的高睾酮水平与胰岛素抵抗和2型糖尿病相关。然而一项研究表明,中国女性性激素结合球蛋白值升高与患2型糖尿病的可能性降低相关,但雌二醇和睾酮水平与该族群中的2型糖尿病无关。
注:这些关于睾酮与2型糖尿病发病率之间关系的相互矛盾的结果可能是由于睾酮的测量,一些作者使用总睾酮而其他人使用游离睾酮,并且根据最近的一项研究,不同研究的分析方法可能不同。
此外,当性激素结合球蛋白浓度低于30nmol/L时,游离雄激素指数(FAI)不是游离睾酮的可靠指标,这将导致性激素结合球蛋白水平较低的女性可能出现研究错误。
因此,在女性中,总睾酮与2型糖尿病之间没有关联,尽管游离睾酮水平较高与2型糖尿病风险增加有关。
√雌二醇水平异常会增加2型糖尿病发病率
关于雌二醇,据报道患有2型糖尿病的绝经后女性的雌二醇水平高于健康女性。然而,基于早期初潮或绝经以及子宫切除术和卵巢切除术实践的大量证据表明,非生理性雌二醇水平(高于或低于正常值)可能是导致2型糖尿病发病率增加的原因。
月经初潮提前会增加2型糖尿病的风险
在这方面,月经初潮提前似乎会增加患2型糖尿病的风险。然而,一些研究表明,初潮早导致的2型糖尿病风险的部分原因可能是肥胖增加,因为早期月经初潮已被证明也与成年期体重指数的增加有关。
提前绝经和卵巢功能不全导致2型糖尿病风险增加
另一方面,提前绝经或卵巢功能不全会导致患2型糖尿病的风险增加。在接受双侧卵巢切除术的绝经后妇女中也观察到了类似的结果。
切除卵巢的女性2型糖尿病风险增加
最后,伴有双侧输卵管卵巢切除术(BSO)的子宫切除术显示2型糖尿病的风险高于子宫切除术本身。然而,其他研究表明子宫切除术与2型糖尿病风险增加有关,而双侧输卵管卵巢切除术本身或与子宫切除术一起并没有增加2型糖尿病的风险。
研究表明表明接受子宫切除术或卵巢切除术的女性患2型糖尿病的风险增加,但并未区分这两种情况是单独发生还是同时发生。
另一项研究表明,相对于完整女性,双侧卵巢切除的子宫切除女性的总睾酮和生物可利用睾酮水平较低,而保留卵巢的子宫切除女性的睾酮水平居中。
与此相关,绝经后妇女(子宫完整和子宫切除)的雌激素和孕激素激素疗法降低了糖尿病的发病率。
▼
肠易激综合征是一组持续或间歇发作,以腹痛、腹胀、排便习惯或大便性状改变为临床表现,胃肠道结构和生化异常的肠道功能紊乱性疾病。
√女性患肠易激综合征的比例更高
肠易激综合征 (IBS) 是一种具有性别差异的代表性疾病,通常在女性中的发生频率是男性的两倍。特别是,到三级中心就诊的患有严重症状的肠易激综合征患者的女性比例更高。
感染性结肠炎后肠易激综合征的风险增加,约10%的肠易激综合征患者在感染性结肠炎后开始出现症状。有趣的是,感染后肠易激综合征的发病率在女性中较高,这表明性别间肠道微生物群的差异可能在肠易激综合征的发病机制中发挥重要作用。
√肠易激综合征的症状存在性别差异
女性更有可能出现腹痛和便秘
肠易激综合征症状存在性别差异:具体而言,IBS-C在女性中占主导地位,而IBS-D在男性中占主导地位。
便秘型肠易激综合征(IBS-C)是肠易激综合征的一种亚型:至少25%的排便为硬粪或干球粪,且松散(糊状)粪或水样粪<25%。
腹泻型肠易激综合征(IBS-D):至少25%的排便为松散(糊状)粪或水样粪,且硬粪或干球粪<25%。
在对22项关于肠易激综合征症状性别差异的研究进行的系统回顾和荟萃分析中,发现女性比男性更可能出现腹痛和便秘相关症状。便秘型肠易激综合征在患有肠易激综合征的女性中的合并患病率为40%,而在患有肠易激综合征的男性中为21%。
男性出现腹泻症状更多
相比之下,患有肠易激综合征的男性比女性更有可能报告腹泻相关症状。腹泻型肠易激综合征(IBS-D)在患有肠易激综合征的女性中的合并患病率为 31%,而在男性中为50%。
√女性月经期间肠易激综合征症状加重
女性在月经期间报告更频繁和更严重的肠易激综合征症状,例如稀便、腹胀和腹痛加重,这些症状的增加可能与月经周期的变化有关。
在冰岛进行的一项基于人群的邮政研究表明,患有痛经的女性比没有痛经的女性更容易出现肠易激综合征症状。此外,更年期似乎与肠易激综合征症状的恶化有关。也就是说,女性在绝经后报告的腹痛比绝经前更严重。
此外,纤维肌痛、慢性疲劳综合征、慢性盆腔痛和偏头痛等经常与肠易激综合征重叠的慢性疼痛疾病的患病率在肠易激综合征女性中更高,这表明她们的症状与激素之间存在关联。
▼
甲状腺是重要的内分泌腺。甲状腺的主要功能是分泌含碘的甲状腺激素三碘甲状腺原氨酸(T3)和甲状腺素(T4)以及肽类激素降钙素,通过影响新陈代谢和组织发育来影响心血管和生殖疾病。
肠道微生物群在甲状腺疾病中起着至关重要的作用,包括桥本氏甲状腺炎 (HT) 和格雷夫斯氏病 (GD)。桥本氏甲状腺炎和格雷夫斯氏病分别是甲状腺功能减退症和甲状腺功能亢进症的主要原因。
√女性甲状腺疾病发病率高于男性
甲状腺疾病性别差异特别的显著,它的高发人群多为中年的女性,男性的患病率相对来说比较低,而之所以会这样是因为女性的内分泌活跃程度很高,没有一定的稳定性。
在平时压力大和情绪发生变化的情况下,会导致自身的免疫调节出现异常,致使甲状腺激素分泌过多,从而就会容易造成甲状腺疾病的发生。
√胆汁酸的组成可以反映甲状腺功能
胆汁酸是肠道微生物群的重要代谢产物,在甲状腺疾病中起着至关重要的作用。甲亢和甲减患者的血清胆汁酸谱不同,胆汁酸的组成可以反映甲状腺功能。甲状腺功能减退患者中最主要的胆汁酸是次级胆汁酸脱氧胆酸,而鹅去氧胆酸是甲状腺功能亢进患者中最主要的胆汁酸。
胆汁酸的组成和水平参与甲状腺激素的分泌,促甲状腺激素水平与亚临床甲减患者血清总胆汁酸呈负相关。
√短链脂肪酸与甲状腺激素相互作用影响女性情绪健康
短链脂肪酸是肠道菌群代谢的主要产物之一,短链脂肪酸与甲状腺激素三碘甲状腺原氨酸相互作用,并通过调节肠细胞基因转录来影响激素的分泌。例如,粪便短链脂肪酸酯可通过影响高脂血症和甲状腺疾病来影响女性的情绪健康。
▼
卵巢癌症的病因尚不清楚,可能与环境、生殖、行为和遗传因素有关。在这些因素中,雌激素水平的失调和雌激素的活性被认为是重要因素。
肠道微生物群可能通过影响雌激素水平参与卵巢的发育。发现17β-雌二醇治疗会改变卵巢癌小鼠模型的病理生理学,导致肿瘤提前发作、缩短总生存时间,并具有典型的乳头组织学特征。
√雌激素促进细胞粘附和迁移从而影响卵巢癌的病症
利用不同的卵巢癌症细胞系进行了一项体外研究,以确认雌激素受体在卵巢癌症病因中的需求。
他们的结果表明,雌激素治疗后,雌激素受体阳性细胞系的粘附和迁移能力增强,而雌激素受体阴性细胞系的粘附和移动能力没有显著变化。
这些结果表明,雌激素的作用与雌激素受体的表达直接相关,雌激素促进细胞粘附和迁移,从而影响卵巢癌症的转移和定植。
√微生物通过影响活性雌激素含量影响卵巢癌
如上所述,具有 β-葡萄糖醛酸酶活性的细菌可以介导雌激素的去结合,从而影响循环中活性雌激素的数量。结合雌激素水平和活性在卵巢癌病因学中所起的作用,我们可以推断肠道微生物组可能通过影响活性雌激素的量来促进雌激素驱动的疾病(如卵巢癌)的发展。
▼
绝经后骨质疏松症 (PMOP) 是一种由雌激素缺乏引起的骨质疏松症,会导致绝经后妇女骨折频率增加。
√雌激素缺乏易引起骨质疏松
目前的研究表明,肠道微生物群与骨重塑以及骨代谢疾病之间存在潜在的密切关系。
研究人员发现,在无菌小鼠中,性类固醇缺乏不能诱导破骨细胞因子表达增加、骨吸收激活,这表明肠道微生物组在性类固醇缺乏导致的骨质疏松中至关重要。
他们进一步证明,用益生菌鼠李糖乳杆菌或市售益生菌补充剂每周两次治疗性类固醇缺乏小鼠可以避免骨质疏松。这种情况的发生可能是由于肠道通透性的降低、肠道和骨髓炎症的抑制。
相比之下,补充非益生菌大肠杆菌菌株或突变鼠李糖乳杆菌并没有显示出对骨丢失的保护作用。
上述结果表明,肠道微生物群失调可能导致肠道通透性增加,并触发重要炎症通路的激活,从而导致性类固醇缺乏小鼠的骨质流失。
这些结果将雌激素缺乏引起的骨质疏松症与肠道微生物多样性、肠道通透性和炎症联系起来。
▼
√高雄激素可能导致多囊卵巢综合征
多囊卵巢综合征是育龄妇女常见的内分泌疾病,其病因可能与高雄激素血症、胰岛素抵抗和神经内分泌功能障碍有关。
一些研究关注多囊卵巢综合征患者的肠道微生物组,这些研究发现肠道微生物组与多囊卵巢综合征之间存在某种关联。然而,与多囊卵巢综合征相关的微生物的确切机制尚未确定。
√多囊卵巢综合征患者肠道微生物多样性较低
一些研究人员发现,患有多囊卵巢综合征的患者肠道微生物组的多样性明显低于健康对照组。在多囊卵巢综合征患者的肠道微生物组中,普通拟杆菌(Bacteroides vulgatus)显著升高,脱氧胆酸和熊去氧胆酸水平降低。
通过比较多囊卵巢综合征女性的粪便微生物组、肠上皮通透性和炎症状态,一些研究提出了一个假设,即肠上皮高通透性导致的毒血症与炎症、胰岛素抵抗和高雄激素血症有关。不过还需要更深入的研究来阐明内毒素血症的具体机制。
√调节肠道微生物有助于改善多囊卵巢综合征
菊粉和二甲双胍对多囊卵巢综合征的缓解作用与抗炎和肠道微生物群的调节有关,这可能有助于多囊卵巢综合症的潜在临床治疗。
环丙孕酮Diane-35(雌激素和孕激素)和益生菌可以帮助重建肠道微生物群的多样性,减少肠道菌群紊乱可以改善多囊卵巢综合征样大鼠的生殖功能。
还需要进一步的研究来确定改善肠道微生物组是否可以作为多囊卵巢综合征的有效治疗方法。
小结
根据性激素和肠道微生物组之间的相互作用,性激素相关疾病的可能治疗策略的显示在下图:
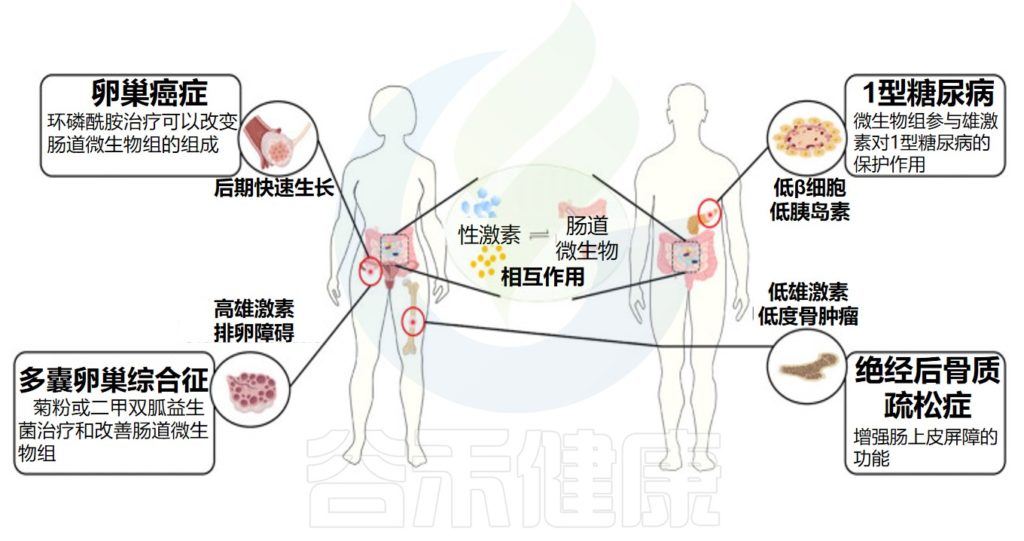
He S,et al.Front Microbiol.2021
在对性激素相关疾病治疗的过程中发现了肠道微生物群的变化,未来通过对肠道微生物的检测或许有助于判断性激素相关疾病的治疗效果。
性激素,特别是雌激素和睾酮,与肠道微生物群的结构和组成有关,性激素与肠道微生物群的相互作用对疾病的发展和性别差异具有重要影响。
肠道微生物群的特定变化可能有助于缓解激素紊乱相关的疾病,因此肠道微生物群被认为是一个重要治疗靶点。
在制定治疗策略时,确定导致疾病发病率中性别二态性的机制具有特殊的重要性,因为肠道微生物群的组成取决于与性激素的相互作用以及其他因素。治疗与这些疾病相关的肠道微生物群失调的疗法可能具有性别特异性效应。
总之,肠道微生物群与激素相互作用在疾病中的影响已经成为了一个备受关注的研究领域。肠道微生物群在激素变化及其带来的健康风险的重要作用及其检测的意义也被越来越多的人所认识。未来,随着相关研究的不断深入,肠道微生物群与激素相互作用的机制和应用价值也将会进一步得到探索和发展。
主要参考文献
Santos-Marcos JA, Mora-Ortiz M, Tena-Sempere M, Lopez-Miranda J, Camargo A. Interaction between gut microbiota and sex hormones and their relation to sexual dimorphism in metabolic diseases. Biol Sex Differ. 2023 Feb 7;14(1):4.
Li D, Sun T, Tong Y, Le J, Yao Q, Tao J, Liu H, Jiao W, Mei Y, Chen J, Liu Z, Wang G, Li Y. Gut-microbiome-expressed 3β-hydroxysteroid dehydrogenase degrades estradiol and is linked to depression in premenopausal females. Cell Metab. 2023 Apr 4;35(4):685-694.e5.
Brettle H, Tran V, Drummond GR, Franks AE, Petrovski S, Vinh A, Jelinic M. Sex hormones, intestinal inflammation, and the gut microbiome: Major influencers of the sexual dimorphisms in obesity. Front Immunol. 2022 Sep 27;13:971048.
Kautzky-Willer A, Harreiter J, Pacini G. Sex and Gender Differences in Risk, Pathophysiology and Complications of Type 2 Diabetes Mellitus. Endocr Rev. 2016 Jun;37(3):278-316.
Saklayen MG. The global epidemic of the metabolic syndrome. Curr Hypertens Rep. 2018;20(2):12.
Paschou SA, Papanas N. Type 2 diabetes mellitus and menopausal hormone therapy: an update. Diabetes Ther. 2019;10(6):2313–2320.
Valeri F, Endres K. How biological sex of the host shapes its gut microbiota. Front Neuroendocrinol. 2021 Apr;61:100912.
Kim YS, Unno T, Kim BY, Park MS. Sex Differences in Gut Microbiota. World J Mens Health. 2020 Jan;38(1):48-60.
Yoon K, Kim N. Roles of Sex Hormones and Gender in the Gut Microbiota. J Neurogastroenterol Motil. 2021 Jul 30;27(3):314-325.
He S, Li H, Yu Z, Zhang F, Liang S, Liu H, Chen H, Lü M. The Gut Microbiome and Sex Hormone-Related Diseases. Front Microbiol. 2021 Sep 28;12:711137.