-

 国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子
国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子

 二级病原微生物安全实验室
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
谷禾健康
在快节奏的现代生活中,我们面临着的食品安全挑战也在升级。全球化的食品供应链让美食跨越千山万水来到我们的餐桌,但也为病原菌的传播提供了更多机会。每一餐美食的背后,都可能潜藏着看不见的威胁。从夏日烧烤到家庭聚餐,食源性病原菌如同”隐形杀手”般悄然存在,威胁着我们的健康。
据世界卫生组织统计,全球每年有超过6亿人因食用被污染的食物而患病。在中国,食源性疾病同样影响着数百万人的健康。
有时候我们认为腹泻、呕吐只是”吃坏肚子”的小毛病,忍忍就过去了。然而这可能掩盖了严重的健康风险。食源性病原菌感染不仅会引起急性胃肠炎症状,还可能导致:
脱水和电解质紊乱:特别是对老人和儿童;
肠道菌群失调:破坏肠道微生态平衡;
免疫系统受损:降低机体抵抗力;
慢性健康问题:如肠易激综合征、炎症性肠病等后遗症。
产气荚膜梭菌、蜡样芽孢杆菌、肉毒杆菌、副溶血性弧菌、志贺氏菌等是最常见且危险的食源性病原菌。它们各具特色:有的耐高温,有的耐酸,有的产生致命毒素,有的传播力极强。
通过了解这些常见食源性病原菌的特性和传播规律,有助于我们识别和避免食用易受污染的食物,掌握正确的储存、烹饪和食用方式,同时监测肠道健康状况及时发现菌群异常变化。因为每个人的肠道菌群构成都是独特的,就像指纹一样。这意味着不同的人对同一种病原菌的易感性和抵抗力也不相同。有些人可能对某种病原菌特别敏感,而另一些人则相对耐受。
▶▷▶▷▶▷
▮ 产 气 荚 膜 梭 菌 ▮
产气荚膜梭菌(Clostridium perfringens)是一种革兰氏阳性杆状细菌,会引起食源性腹泻,是工业化国家常见的食源性致病菌之一,全年均有腹泻病例报道。它广泛存在于环境中,对人和动物均具致病性。
作为一种能够形成芽孢的厌氧细菌,产气荚膜梭菌在芽孢形成过程中会产生肠毒素(CPE),其A型到E型菌株与不同疾病相关。其中人类坏死性肠炎是一种致命的危及生命的疾病,会导致小肠溃疡。
该菌能在短期内快速繁殖、在相对较高的温度范围内生长、芽孢能抵抗恶劣环境条件、以及尽管是厌氧菌却具有耐氧能力,这些特性使得产气荚膜梭菌具有很强的环境适应性和致病潜力。
• 梭菌属中具有重要临床意义的致病菌种还有:
破伤风梭菌(Clostridium tetani)
肉毒梭菌(Clostridium botulinum)
巴氏梭菌(Clostridium baratii)
艰难梭菌(Clostridium difficile)
Clostridium argentinese
产气荚膜梭菌的常见栖息地是土壤、水、污泥、污水、食物(主要是生肉和冷冻肉)、受污染的设备等。产气荚膜杆菌也存在于人和动物的肠道中,尤其是老年人的粪便中。
产气荚膜梭菌需要多种氨基酸与维生素,最适生长温度为43–45°C。富含蛋白的肉类尤其适合其繁殖,特别是猪肉、牛肉和家禽产品;用酱料烹制的污染风险很高。
在加热和冷却过程中,如果食物在室温下没有足够的冷却温度,它会增殖并产生毒素。通常,学校食堂,医院,餐馆,监狱和集会是梭菌爆发的高风险场所。
产气荚膜梭菌在应激条件下孢子形成时产生肠毒素,已知可产生14种毒素与5类肠毒素(A–E型)。其还分泌4类细胞外毒素:α、β、ε、iota。
产气荚膜梭菌毒素和降解酶的作用
doi: 10.1080/21505594.2021.1886777.
A型:在人类中引起气性坏疽和腹泻相关的感染性腹泻,在动物中引起肠毒血症、坏死性肠炎和急性胃扩张。
C型:在人类中导致坏死性肠炎(空肠炎),在小猪、羔羊、小牛和鸟类中导致肠毒血症和坏死性肠炎。
B、D、E型:尚未证实致人类疾病;在动物可致肠炎与肠毒血症。
此外,该菌还能产生溶素O(θ毒素)与胶原酶(κ毒素),也需警惕。
与暴发有关的因素是大量准备食物、烹饪温度不够(食物的核心区域没有适当烹饪)、准备好的食物和食用期间受到污染。
集中爆发经常发生在提前准备大量食物的地方,如医院和学校食堂、监狱、聚会和研讨会。
案例调查显示,一道在室温下放置20小时且未再加热的羊肉菜是感染来源。产气荚膜杆菌还从羔羊肉、感染者粪便及尸检样本中被分离出。某校食堂因饮用制备后置于室温的受污染奶昔,导致77例腹泻。
产气荚膜梭菌只有在摄入含有约 10⁷-10⁹个营养细胞/克 的污染食物时才引起感染。
⑴第一阶段:胃肠道定植
-营养细胞抵抗胃酸
-在小肠内生长、增殖和产孢
⑵第二阶段:肠毒素作用机制
靶点结合:大量肠毒素与肠上皮细胞紧密连接处的 claudin蛋白 结合。
复合物形成:结合宿主膜蛋白,经构象变化形成 CPE复合物。
细胞损伤:
-细胞崩解和分离
-肠绒毛尖端损伤
-上皮细胞脱落
-绒毛缩短
膜损伤效应:
-膜上形成孔道
-电解质大量泄漏
-导致细胞溶解
-肠道吸收能力下降
-细胞骨架崩塌
⑶第三阶段:严重并发症
高浓度CPE (>10μg/ml):引起坏死(oncosis)
β-毒素作用:
-引起细胞毒性坏死性肠炎
-影响自主神经系统
-导致肠黏膜出血性坏死
-在真核细胞膜形成孔道
这种多重毒素协同作用使得产气荚膜梭菌食物中毒具有较强的致病性和复杂的临床表现。
主要症状多在食用重度污染食物后8–24小时出现。
常见症状包括腹部绞痛、水样腹泻、呕吐、恶心和发烧,大多数情况下通常在12至24小时内消退。致死率低于0.05%;但婴幼儿、老年人及慢性病患者风险较高;该菌可能侵入并导致严重溃疡,最终因急性脱水而死亡。
婴儿猝死综合征(SIDS)也是由产气荚膜梭菌与睡眠期间大脑的缺陷有关。
检测原理:产气荚膜梭菌食物中毒通过检测粪便样本中的肠毒素和营养细胞/芽孢数量来诊断。
⑴肠毒素检测
培养基法:熟肉培养基 → 巯基乙酸盐流体培养基(10ml) → Duncan-Strong产孢培养基
分子检测方法:通过PCR、多重PCR以及高通量测序手段检测样本中产气荚膜梭菌。例如谷禾健康依托高通量测序的tNGS技术可以特异性的识别粪便样本中的产气荚膜梭菌。
(来源:谷禾健康16S Plus版报告)
生物学方法
小鼠致死试验:评估毒素致死活性
Vero细胞毒性试验:检测细胞毒性效应
⑵免疫学检测
凝胶扩散法
对流免疫电泳
ELISA法
酶联免疫吸附试验进行CPE定量
注意事项:样本蛋白酶活性可降低IgG敏感性,需用1%血清白蛋白保护
⑶分子生物学方法
PCR技术:常规PCR、实时PCR
LAMP:环介导等温扩增
⑴治疗策略
一般治疗
抗生素使用原则
重症治疗
坏死性肠炎需要:
补液治疗
⑵预防控制措施
关键预防原则:烹调后立即食用
时间-温度控制
食品安全措施
▶▷▶▷▶▷
▮ 蜡 样 芽 孢 杆 菌 ▮
蜡样芽孢杆菌(Bacillus cereus)是一种致病性食源性微生物,广泛分布于自然界,如植物、土壤和昆虫及哺乳动物的胃肠道。
也常见于食品加工环境,其耐受性内生孢子可在恶劣条件下存活,污染原料与食品。
该菌可致两型食物中毒:腹泻型(在小肠产多种肠毒素)与呕吐型(摄入含耐热毒素的食物)。亦可引发机会性感染,如菌血症、败血症、肺炎、脑膜炎、胃炎、肝衰竭、肝坏死与脑水肿。
• 其他食源致病性芽孢杆菌:
B. subtilis(枯草芽孢杆菌):可致腹泻、呕吐;
B. licheniformis(地衣芽孢杆菌):常见食物中毒病因;
B. pumilus(短小芽孢杆菌):污染食品致胃肠炎。
• 其他特殊芽孢杆菌:
B. weihenstephanensis:强低温生长能力;
B. anthracis(炭疽芽孢杆菌):炭疽病原,极危;
B. mycoides(蕈状芽孢杆菌):形态特殊;
B. pseudomycoides(假蕈状芽孢杆菌):与蕈状近缘;
B. thuringiensis(苏云金芽孢杆菌):生物杀虫剂,偶可致病。
这些芽孢杆菌都具有形成芽孢的能力,使其在环境中具有很强的抵抗力和传播能力。
⑴基本特征
–革兰阳性菌
–产孢菌
–需氧至兼性厌氧
-有运动性
-杆状细菌
血培养的革兰氏染色蜡样芽孢杆菌
doi: 10.1128/CMR.00073-09.
⑵生长条件
生长温度范围:8-55°C
最适温度:25-37°C
pH范围:4.9-9.3
耐盐浓度达7.5%
内生孢子耐高温、辐射、干燥和消毒剂
⑶污染源
-从原料到包装储存的全程食品加工污染
-生物技术设备和机器污染
-土壤栖息,易传播至蔬菜和农作物
⑷常见污染食品
-大米、小麦、面食、面粉
-乳制品、肉制品、香料
-婴儿食品、鱼类、汤类
-蔬菜和水果
⑸流行病学特点
-全球性分布
-年度持续发生
-与饮食文化相关的地区差异
⑹易感人群
老年人、低胃酸患者
⑺防控挑战
其孢子高耐受性与污染源多样性要求从源头到终端的全程管控,尤其要严格温度控制并预防交叉污染。
蜡样芽孢杆菌产生蛋白质毒素,即肉毒毒素和催吐毒素。
⑴肉毒毒素(腹泻毒素)
当摄入的营养细胞在小肠增殖时产生腹泻毒素。该毒素对蛋白酶敏感,如链霉蛋白酶、胃蛋白酶、胰蛋白酶和胰凝乳蛋白酶。
潜伏期通常为摄入后8–16小时,病程多持续24–48小时。症状以轻度水样腹泻和腹部痉挛为主;少数儿童或免疫低下者可出现血性腹泻与坏死性肠炎,进而致肝功能衰竭和脑水肿。
与肠道疾病相关的三种染色体编码肠毒素包括:
Hbl(溶血素BL):蜡样芽孢杆菌食物中毒的主要毒力因子,在小肠通过渗透溶解形成跨膜孔。
Nhe(非溶血性肠毒素):与 Hbl 同源,亦为三组分成孔毒素。
CytK(细胞毒素K):原型样β-桶成孔毒素,可致出血性腹泻与坏死性肠炎。
⑵呕吐毒素
呕吐型起病更急且更重,由小分子环状耐热肽引起,潜伏期为摄入受污染食物后约2–5小时。
其症状类似金黄色葡萄球菌食物中毒:恶心、呕吐、腹部痉挛,通常持续约24小时。催吐毒素作用机制尚不明,但高度耐热,可经受油炸、烘烤、煮沸和微波等烹调过程。
环境因素(温度、pH、大气成分、营养来源、食物稠度)亦影响毒素的产生。
⑴培养方法
实验室培养基如营养琼脂或血琼脂
⑵ELISA
ELISA技术在商业上被用于测定毒素,但它在评估蜡样芽孢杆菌的产毒活性方面不准确。它只检测一种毒素,溶血素BL或两种无毒蛋白质。
⑶反向被动乳胶凝集(RPLA)肠毒素试验
将样品煮沸以使其在生物学上稳定,从而产生阳性结果。该试验可识别溶血素B组分,但在存在高葡萄糖浓度的情况下,无法检测到毒素。
⑷PCR或高通量测序
通过检测蜡样芽孢杆菌DNA序列,来分析其丰度和毒力基因的存在。
蜡样芽孢杆菌食物中毒多为自限性,通常24–48小时内自行缓解。建议卧床休息并补充液体;重症时可在医生指导下使用抗生素,如克林霉素、万古霉素、庆大霉素或氯霉素。
蜡样芽孢杆菌耐热性强,常规烹调可杀灭营养细胞,但孢子仍可存活,故应在孢子萌发前切断污染。
食品应在储存前迅速冷却、食用前充分加热;芽孢杆菌属难以在高酸性环境存活。保持良好食品处理习惯与卫生。
▶▷▶▷▶▷
▮ 肉 毒 杆 菌 ▮
肉毒杆菌(Clostridium botulinum)是一种生长在缺氧环境下的细菌,在罐头食品及密封腌渍食物中具有极强的生存能力,是毒性最强的细菌之一。
肉毒杆菌中毒是由摄入肉毒毒素(BoNT)导致的致命性食源性麻痹性疾病,虽罕见但病死率高,微量毒素即可致命。
中毒来源包括食物中预先形成的BoNT或肠道内产生的BoNT。已知毒素分为A–G七型,可致人畜发病,其中A、B、E型与人类食源性肉毒中毒最相关。
肉毒杆菌为厌氧、产芽孢的革兰阳性菌,孢子广泛存在于土壤、污水、泥浆及湖海沉积物中,也见于陆生和水生动物肠道。
肉毒杆菌的孢子形成和发芽
doi: 10.1080/21505594.2023.2205251.
其孢子易污染食物并产生毒素。蜂蜜和糖浆曾被报道受其孢子污染,是婴儿肉毒中毒的主要来源。加工不当的罐头亦为重要来源,因孢子可耐受高温。其他高风险食物包括湖鱼、海鱼,以及烟熏、盐腌和发酵肉制品。
A型和B型毒素常见于土壤与生物肥料,故可能污染蔬果,尤其是加热不足的蔬菜制品。
症状通常在摄入后18–38小时出现,先累及胃肠道,常见腹痛、恶心、呕吐、腹泻,偶见便秘。随后出现神经系统表现:弛缓性麻痹、对称性下行无力、构音不清、吞咽困难、眩晕、视物模糊、口干及眼外肌无力。
婴儿肉毒中毒多见于1岁以下,因肠道菌群未成熟,毒素可在肠内产生。表现为哭声微弱嘶哑、上睑下垂、吮吸差、抬头困难与肌无力。
伤口型肉毒中毒为梭菌在伤口、脓肿等处定植,孢子发芽并产毒。其症状包括低血压、呼吸困难、眼肌麻痹、共济失调与瞳孔固定性散大,少见,通常在暴露后3–6天发病。
肉毒杆菌毒素曾在第二次世界大战期间被美国,日本军队用作生化武器。
1991年埃及因食用传统腌鱼 fesaikh 中的E型毒素发生大规模疫情,报告91例、死亡18例。
约90%的肉毒中毒病例见于美国,每年报告约9–10起暴发,常与家庭自制食品相关,尤以自制罐藏、瓶装及油浸食品为主。
口服或肠道内生成的肉毒毒素需先抵御胃酸,经肠道到达十二指肠与空肠。随后经内吞入血,分布至周围神经系统。
毒素进入神经元后作为锌依赖性蛋白酶,切割SNARE蛋白(可溶性N-乙基马来酰亚胺敏感因子附着受体)。其在神经肌肉接头终末结合并抑制乙酰胆碱释放,阻断神经传递,导致弛缓性麻痹。
肉毒毒素中毒的分子步骤
doi: 10.1038/srep23707.
⑴生物测定
小鼠生物测定是检测肉毒毒素最敏感、应用最广的方法:通过注射毒素评估其毒性,小鼠常在4小时内出现腹部颤动、蜂腰样腹部、四肢瘫
一个人表现出类似的肉毒中毒症状应立即置于重症监护与胃肠外营养供应。
如果怀疑有毒素,应在24小时内服用抗毒素,因为它能在到达神经肌肉接头之前中和血液中的所有游离毒素。
在伤口肉毒杆菌中毒的情况下,伤口应该用抗生素治疗,如青霉素。
以上都需要咨询医生确定。
食品应充分加热(≥121°C)并冷藏(≤4°C);加入柠檬酸等酸化剂可降低罐装食品中孢子形成的风险。
避免食用可疑的生或未充分加热食物,并避免其接触伤口。
▶▷▶▷▶▷
▮ 副 溶 血 性 弧 菌 ▮
副溶血性弧菌(Vibrio parahaemolyticus)是导致人类急性肠胃炎的主要病原体之一,通常发生在食用生的、未煮熟的和交叉污染的海鲜产品时。
该菌常见于热带海洋与沿海环境,可在滤食性贝类(牡蛎、蛤蜊、贻贝)肠道黏附并繁殖。除胃肠炎外,还可引起伤口与耳部感染,免疫低下者暴露海水时可发生败血症。
与之相关的人类致病弧菌还包括霍乱弧菌(Vibrio cholerae)与创伤弧菌(Vibrio vulnificus)。在日本及多国亚洲地区,副溶血性弧菌每年占食物中毒病例的约20%–30%。
其在全球范围内是胃肠炎的重要病因之一,亟需关注其流行情况、毒力因子及健康影响,并完善鉴定技术以降低风险。
–革兰氏阴性
-曲线杆状
-非孢子形成者的兼性厌氧菌
-轻度嗜盐,氧化酶阳性
-最佳温度30至35°C
-pH值范围为6.8至10.2。
副溶血性弧菌是海洋嗜盐菌,依靠单极鞭毛在水中游动,并可附着于鱼、蟹、虾、龙虾、浮游动物等水生动物表面。
当食用被副溶血性弧菌污染的生的和未煮熟的海鲜时,它会引起人类急性胃肠炎。当开放性伤口或伤口暴露于受污染的海水时,也会引起感染。
其他来源是副溶血性弧菌强毒株从海产品或设备到其他产品的交叉污染。
副溶血性弧菌按抗原性分为体细胞(O)和荚膜(K)抗原。其主要毒力因子包括粘附素、耐热直接溶血素(TDH)、TDH相关溶血素(TRH)及III型分泌系统(T3SS-1、T3SS-2)。
副溶血性弧菌的结构和毒力因子
doi: 10.3389/fmicb.2015.00437.
此外,它具有两类鞭毛用于运动,并可形成有助于在恶劣环境中生存的荚膜。感染多经粪-口途径,通过细菌表面粘附因子黏附宿主细胞。
TDH通过与红细胞膜结合并在其表面打孔致溶血;同时通过形成通道使细胞外Ca²⁺内流、细胞内Cl⁻分泌增加,导致渗透压升高、细胞形态与功能紊乱,最终细胞膨胀死亡。
TRH与TDH共同介导细胞毒性并诱发肠道感染;不耐热溶血素(TLH)主要用于种属鉴定,对红细胞溶血作用较弱。早期研究还显示,尿素酶是trh⁺菌株的重要辅助毒力因子,有助于在胃肠道定植与致病。
注:尿素酶指可以催化尿素,导致尿素分解成氨和二氧化碳的一种酶,在临床上明确幽门螺杆菌就是人体胃内能够产生大量尿素酶的细菌。
第三型分泌系统(T3SS 1)在组织细胞感染期间诱导自噬和细胞毒性。它连续引起自噬、细胞起泡、溶解最后导致死亡。
在我国东部沿海省份的部分暴发病例中,副溶血性弧菌感染相对较高。并且在亚洲、欧洲、非洲和美洲国家,类似病例的爆发已被频繁报道。
1950年,日本首次爆发副溶血性弧菌病,报告了272例急性胃肠炎,其中20人死亡。
1997年至1998年,美国报告了700多起食用受污染的生牡蛎的病例。
副溶血性弧菌食物中毒多发于夏季(6—10月),因温暖水域有利于其繁殖。
螃蟹、虾、龙虾、贝类、牡蛎、蛤蜊和金枪鱼是高风险的海产品,必须在食用前彻底煮熟。
副溶血性弧菌中毒典型的症状是水样腹泻、恶心、呕吐、腹部痉挛、发烧和发冷。进食受病原体污染的食物后,潜伏期约为12至24小时,并于5至7天内消退。
免疫功能低下的个体可能需要约10至15天的长时间才能恢复,因为疾病是自限性的,不需要药物治疗。
有些严重的患者可能会出现粘液或便血,血压下降,失去知觉,皮肤苍白、发紫,甚至死亡。此时应立即就医。
⑴菌株培养鉴定
使用含十二烷基硫酸钠(SDS)、烷基苯磺酸盐和胆汁盐的选择性增菌培养基可用于副溶血性弧菌的培养与分离。以碱性蛋白胨水为代表的增菌液具最佳pH和较高NaCl浓度,有利于弧菌生长。
⑵分子检测技术
副溶血性弧菌菌株的鉴定也可采用分子技术,如常规表型与生化试验、PCR及DNA检测。高通量宏基因组测序或靶向tNGS可用于分型与菌株区分。
该病多为自限性,但重症时需药物治疗。常用措施包括口服补液及应用四环素等抗生素;充足饮水以补充电解质、卧床休息亦有助于恢复。
副溶血性弧菌食物中毒的预防与控制:
–避免食用生或未熟透的贝类、牡蛎等海产品。
-处理海产品时注重个人卫生,生熟分开,防止交叉污染。
-皮肤有割伤或创口者应避免接触海水或微咸水,并尽量使用防水敷料。
▶▷▶▷▶▷
▮ 志 贺 氏 菌 ▮
志贺氏菌属(Shigella)是引起急性胃肠道感染的原因,这种感染会损害回肠和结肠上皮并引起细菌性痢疾。志贺氏菌感染多见于贫困、欠发达且缺乏卫生与医疗设施的国家和地区。
志贺氏菌病死亡率较高,尤以5岁以下营养不良儿童为甚。患者在腹泻期间因大量丢失电解质和血液而营养受损,可能致死。
• 志贺氏菌属分为四个血清群:
A群:痢疾志贺菌(S.dysenteriae)
B群:福氏志贺菌(S.flexneri)
C群:鲍氏志贺菌(S.boydii)
D群:宋内志贺菌(S.sonnei)
其中,S.dysenteriae 1型(志贺毒素产生菌)可引起严重的流行性痢疾,具有较高的致死率;而其他血清型通常引起相对较轻的痢疾症状。
–革兰阴性杆菌;无芽孢、无鞭毛(不运动)、无荚膜;兼性厌氧。
-最适生长温度为 37°C,生长范围约 10–40°C;可耐受一定盐度与酸性环境。
-低感染剂量:约 10–100 个菌体。
-对巴氏灭菌温度敏感(≥60°C 易被杀灭)。
-对紫外线和化学消毒剂敏感。
-能在不利的物理和化学条件下存活。
⑴病原菌储存宿主
–人类肠道是志贺菌的主要天然储存库;
-动物感染极为罕见,几乎不作为传染源。
⑵传播途径
–粪—口传播为主要途径
–人与人直接接触
–被污染的水和食物
-受污染的灌溉水可污染蔬果
⑶易感人群
–儿童和婴幼儿为高危人群
–免疫力低下者
–密切接触者(如家庭成员、托幼机构)
⑷流行特征
–发展中国家发病率较高
-与卫生设施不完善和个人卫生不良相关
-季节性:夏秋季高发
2003-2013年中国志贺氏菌的流行情况
doi: 10.3389/fcimb.2016.00045.
症状通常在摄入细菌后1–3天出现,视剂量可在12小时至7天内发病。
典型症状包括发热、食欲减退、腹痛、血性或水样腹泻、结肠炎、乏力与不适。大量水样便可致脱水,但较少见。伴随的厌食与营养不良需积极处理,是5岁以下儿童死亡的主要原因。少数患者可出现神经症状,如嗜睡、不自主运动和头痛。
这种疾病一般在5至7天内自行消退,但感染者可能会在很长一段时间内无症状地在粪便中排出细菌,并可能造成传播感染的威胁。
⑴侵入阶段
经口进入消化道
胃酸耐受:可抵抗低 pH
穿越小肠:因上皮更新快、液流迅速,难以定植
最终定植于大肠
⑵细胞侵袭机制
M细胞摄取:进入并被吞噬泡包裹
逃逸入胞:自吞噬泡逃出,进入细胞质
巨噬细胞:被吞噬后诱导其凋亡
⑶细胞内增殖与扩散
在大肠上皮内快速繁殖
经质膜包裹进行细胞间直接传播
继而感染邻近上皮细胞
⑴志贺毒素(主要由S.dysenteriae产生)
作用机制:经胞吞进入宿主细胞
毒性效应:抑制蛋白合成,致细胞死亡
临床表现:出血性腹泻
⑵脂多糖(LPS)
释放时机:细菌死亡破裂时释放
损伤效应:破坏肠道上皮组织
炎症反应:激发强烈的炎症应答
⑶炎症因子
IL-12释放:S. flexneri特别容易诱导IL-12产生
线粒体损伤:导致细胞坏死
组织破坏:引起肠道组织广泛炎症和溃疡
⑷病理损伤特点
组织破坏:主要由宿主免疫反应引起,而非病原菌直接作用
粘膜溃疡:免疫系统过度反应导致的继发损伤
炎症反应:局部组织出现严重炎症和水肿
⑴细菌培养方法
及时采样:出现痢疾症状后立即采集粪便标本
采集时机:感染初期阶段最佳,病原菌体外存活时间有限
样本处理:需要立即检测,避免病原菌死亡
⑵免疫学检测方法
酶免疫测定(EIA):成本低、操作简便。
⑶分子诊断技术
PCR/测序:基因检测快速、可靠,可区分不同志贺菌种。
⑷目标基因
ipaH基因:侵袭相关基因
virA基因:毒力基因
iral基因:调节基因
LPS基因:脂多糖合成基因
质粒DNA:毒力质粒
⑴抗菌治疗
首选:氟喹诺酮类(如环丙沙星)、大环内酯类(如阿奇霉素)
原则:尽早治疗,依据病情调整;监测耐药性
注意:具体用药请遵医嘱
⑵支持治疗
口服补液盐:轻中度脱水
静脉输液:重度脱水或电解质紊乱
营养支持:维持免疫功能
⑶环境与个人卫生
-完善卫生设施与排污系统
-安全处置粪便,防止污染
-保护水源,避免饮用水污染
-手卫生:勤洗手,使用肥皂或消毒剂
⑷食品安全
-生熟分开
-充分加热
-适当冷藏
-果蔬反复清洗
⑸重点人群
儿童与婴幼儿:加强护理与监测
食品从业人员:定期健康检查
集体生活人群:(学校、托幼机构等)
食源性病原菌虽“微小”,却以顽强的生存力、多样的毒力因子与隐匿的传播路径,对公众健康构成持续挑战。产气荚膜梭菌的耐受与产毒、蜡样芽孢杆菌的双型中毒与广泛污染、肉毒杆菌的致命神经毒素、以及志贺氏菌的低感染剂量与高传播力,共同勾勒出从农田到餐桌、从环境到个体的风险图谱。
守护肠道健康,关键在于两端同时用力:一端是系统性防控——完善卫生基础设施、保障饮水安全、规范食品加工与冷链管理、强化时间—温度控制与流程监测;另一端是个人与社区层面的良好习惯——勤洗手、生熟分开、充分加热、规范冷藏、创口防护与及时就医。面向重点人群(婴幼儿、老年人、免疫低下者及集体生活人群),还需更精准的健康教育与常态化监测。
科学认知让风险可见,规范行动让风险可控。以循证为底、以习惯为桥、以制度为盾,我们就能在快节奏与全球化的餐桌上,稳稳地守住每一个人的“肠道安全线”。
主要参考文献
Popoff, M. R.(2014). CLOSTRIDIUM | Detection of Enterotoxin of Clostridium perfringens. Encyclopedia of Food Microbiology, 474–480.
McLauchlin, J., & Grant, K. A. (n.d.). Clostridium botulinum and Clostridium perfringens. Foodborne Diseases, 41–78.
Miyamoto, K., & Nagahama, M. (2016). Clostridium: Food Poisoning by Clostridium perfringens. Encyclopedia of Food and Health, 149–154.
Chukwu, E. E., Nwaokorie, F. O., Coker, A. O., Avila-Campos, M. J., Solis, R. L., Llanco, L. A., & Ogunsola, F. T. (2016). Detection of toxigenic Clostridium perfringens and Clostridium botulinum from food sold in Lagos, Nigeria. Anaerobe, 42, 176–181.
Granum, P. E., & Lund, T. (2006). Bacillus cereus and its food poisoning toxins. FEMS Microbiology Letters, 157(2), 223–228.
Tewari, A., & Abdullah, S. (2014). Bacillus cereus food poisoning: international and Indian perspective. Journal of Food Science and Technology, 52(5), 2500–2511.
Parihar, H. S. (2014). Bacillus cereus. Encyclopedia of Toxicology, 353–354.
Batt, C. A. (2014). BACILLUS | Bacillus cereus. Encyclopedia of Food Microbiology, 124–128.
Carlin, F. (2016). Bacillus cereus and Other Bacillus sp. Causing Foodborne Poisonings, Detection of. Encyclopedia of Food and Health, 301–306.
Drobniewski, F. A. (1993). Bacillus cereus and related species. Clinical Microbiology Reviews, 6(4), 324–338.
Tewari, A., & Abdullah, S. (2014). Bacillus cereus food poisoning: international and Indian perspective. Journal of Food Science and Technology, 52(5), 2500–2511.
Marrollo, R. (2016). Bacillus cereus Food-Borne Disease. The Diverse Faces of Bacillus Cereus, 61–72.
Pavlik, B., Hruska, E., Van Cott, K. et al. Retargeting the Clostridium botulinum C2 toxin to the neuronal cytosol. Sci Rep 6, 23707 (2016).
Notermans, S. H. W., Stam, C. N., & Behar, A. E. (2014). CLOSTRIDIUM | Detection of Neurotoxins of Clostridium botulinum. Encyclopedia of Food Microbiology, 481–484.
Aureli, P. (2017). Botulism. International Encyclopedia of Public Health, 254–262.
McLauchlin, J., & Grant, K. A. (n.d.). Clostridium botulinum and Clostridium perfringens. Foodborne Diseases, 41–78.
Austin, J. W. (2016). Clostridium: Occurrence and Detection of Clostridium botulinum and Botulinum Neurotoxin. Encyclopedia of Food and Health, 155–159.
Lebeda, F. J., Dembek, Z. F., & Adler, M. (2018). Foodborne Botulism From a Systems Biology Perspective. Foodborne Diseases, 275–308.
Horowitz, B. Z. (2005). Botulinum Toxin. Critical Care Clinics, 21(4), 825–839.
Letchumanan, V., Chan, K.-G., & Lee, L.-H. (2014). Vibrio parahaemolyticus: a review on the pathogenesis, prevalence, and advance molecular identification techniques. Frontiers in Microbiology, 5.
Wang R, Zhong Y, Gu X, Yuan J, Saeed AF, Wang S. The pathogenesis, detection, and prevention of Vibrio parahaemolyticus. Front Microbiol. 2015 Mar 5;6:144.
Risk assessment tools for Vibrio parahaemolyticus and Vibrio vulnificus associated with seafood, Food and Agriculture Organization of the Unites Nations, ISSN 1726-5274.
Baker-Austin, C., Jenkins, C., Dadzie, J., Mestanza, O., Delgado, E., Powell, A., … Martinez-Urtaza, J. (2020). Genomic epidemiology of domestic and travel-associated Vibrio parahaemolyticus infections in the UK, 2008–2018. Food Control, 115, 107244.
Yaashikaa, P. R., Saravanan, A., & Kumar, P. S. (2016). Isolation and identification of Vibrio cholerae and Vibrio parahaemolyticus from prawn (Penaeus monodon) seafood: Preservation strategies. Microbial Pathogenesis, 99, 5–13.
Bolen, J. L., Zamiska, S. A., & Greenough, W. B. (1974). Clinical features in enteritis due to vibrio parahemolyticus. The American Journal of Medicine, 57(4), 638–641.
Mumy, K. L. (2014). Shigella. Encyclopedia of Toxicology, 254–255.
Ashkenazi, S. (2018). Shigella Species. Principles and Practice of Pediatric Infectious Diseases, 842–846.e2.

谷禾健康

牛奶蛋白过敏(CMPA)是婴儿最常见的食物过敏类型之一。粪便病原菌培养显示产气荚膜梭菌阳性率超过30%,明显高于其他细菌。因此推测产气荚膜梭菌定植可能是婴儿牛奶蛋白过敏的发病因素之一。
一项真实世界的研究,杨敏团队从我国一家大型三甲医院(广州市妇女儿童医疗中心)招募患有腹泻、粘液便和/或血便的 0-6 个月婴儿的研究中,通过飞行质谱法证实了用于检测产气荚膜梭菌的粪便病原体培养物,并通过PCR鉴定了潜在的毒素基因。随访12个月后,记录牛奶蛋白过敏和食物过敏的诊断。通过Pearson相关分析评估相关性。
这项研究表明产气荚膜梭菌肠道定植在婴儿中很常见,与婴儿牛奶蛋白过敏和食物过敏的发生有关。
产气荚膜梭菌是一种厌氧、革兰氏阳性芽孢杆菌,是一种可引起胃肠道或皮肤和深层组织感染的细菌。产气荚膜梭菌的定植可能导致侵袭性胃肠道感染,产生肠毒素,可能破坏肠上皮细胞间的紧密连接,造成上皮损伤,引起肠道菌群失衡,导致宿主免疫反应异常。
本文我们来详细看一下这项研究过程及相关结果,结合产气荚膜梭菌的生物学特性、毒素等相关知识,深入探讨产气荚膜梭菌定植对婴幼儿过敏相关疾病及其他健康问题的影响,了解这些可以帮助我们更好地了解其在婴幼儿免疫系统发育中的作用,为临床上预防和治疗相关疾病提供新的思路和方法。
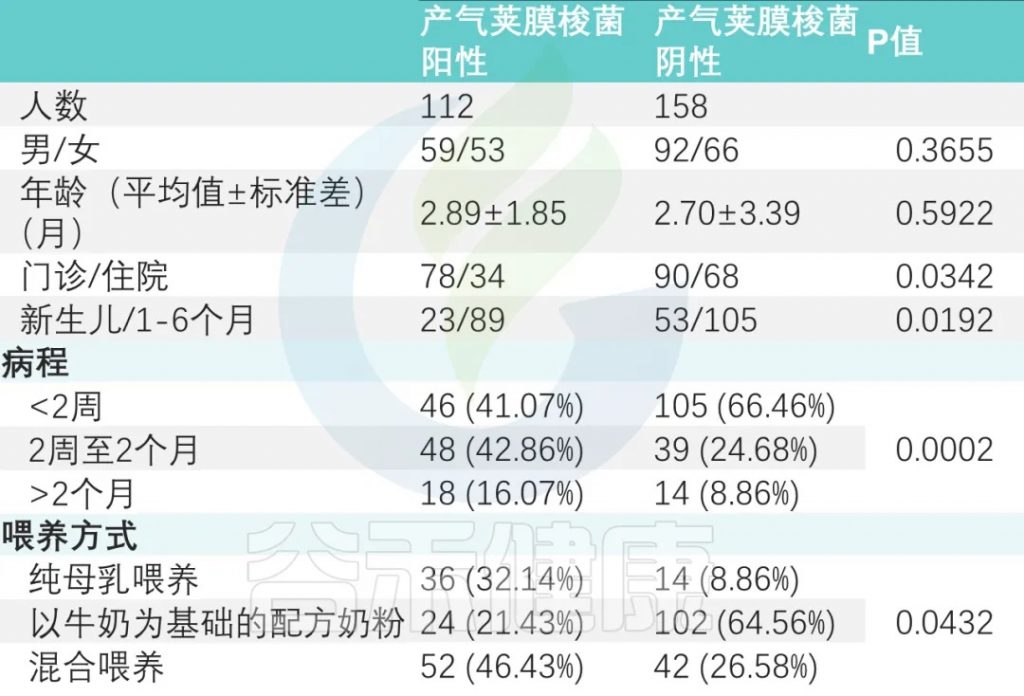
在这项广州妇女儿童医疗中心的研究中,共招募了358名0-6个月的婴儿。对这些婴儿粪便样本进行病原培养,其中128名产气荚膜梭菌呈阳性。
共有270名婴儿(44.07% 为女孩;平均年龄为 2.78 ± 2.84 个月)参加了为期 12 个月的跟踪调查。
注:
▪ 2020年1月1日至2021年12月31日期间因胃肠道症状住院或门诊的所有0-6个月大的婴儿
▪ 排除先天性代谢性疾病、免疫缺陷性疾病和恶性肿瘤的患者

doi: 10.1186/s13099-023-00572-x
专门针对食物过敏和牛奶蛋白过敏的临床诊断,其中包括湿疹、过敏性鼻炎以及食物蛋白诱导的过敏性直肠结肠炎等相关病症。食物过敏是根据症状、饮食排除和口服食物挑战进行临床诊断的。
病程差异
产气荚膜梭菌阴性组与产气荚膜梭菌阳性组显著差异(P < 0.001)。
喂养方式
纯母乳喂养组、母乳添加配方奶喂养组和纯配方奶喂养组产气荚膜梭菌定植阳性率分别为 32.14%、46.43%、21.43%(P<0.05)(下表)。
doi: 10.1186/s13099-023-00572-x
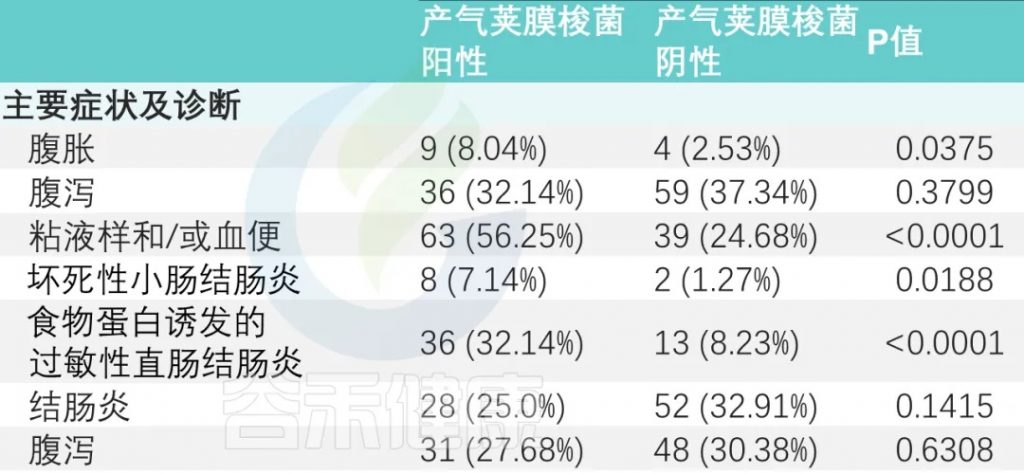
主要症状及诊断
主要症状是腹胀、腹泻和粘液便和/或血便。
有或没有产气荚膜梭菌定植的婴儿,腹胀和粘液样便和/或血便的发生率存在显著差异(P<0.05)。
与产气荚膜梭菌阴性组相比,阳性组NEC(坏死性小肠结肠炎)和FPIAP(食物蛋白诱发的过敏性直肠结肠炎)发生率显著升高(下表)。
抗生素
甲硝唑给药率:产气荚膜梭菌阳性组高于阴性组(33.9% vs. 10.76%)(P < 0.0001)。
血常规检查 (231名患者)
两组之间白细胞增多、血小板增多和中性粒细胞减少事件的数量存在显著差异。
粪便常规检查 (267次)
粪便潜血阳性率为37.45%(100/267),
粪便白细胞阳性率为8.2%(22/267)。
两组比较差异有显著性(P <0.001)(下表)。
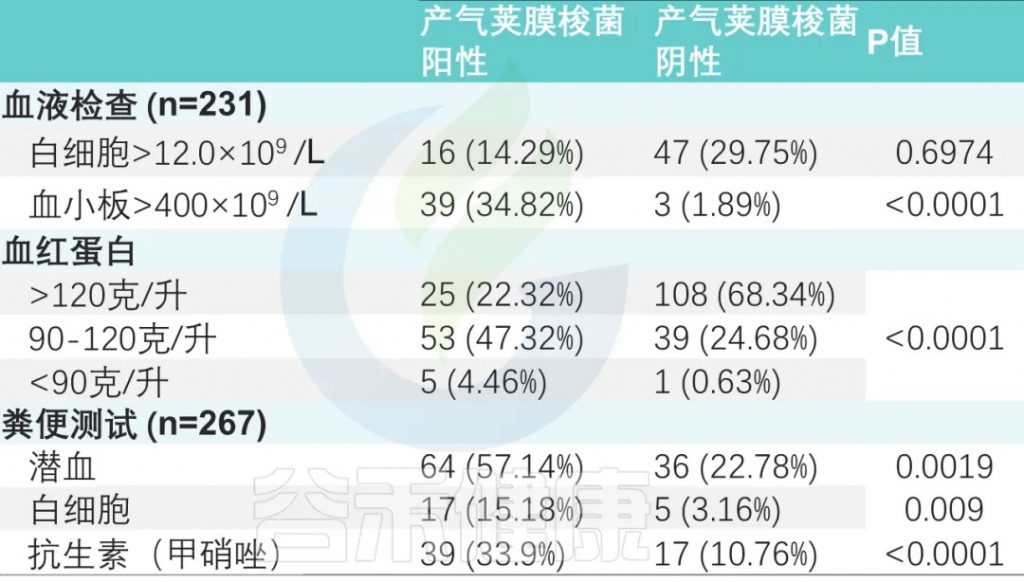
在 30 名患者不同时间点(0、2、4、12 和 24 周)的 85 份粪便样本中检测到了潜在的特异性毒素基因。
产气荚膜梭菌携带潜在特异性毒素基因的出现率如下:
没有携带其他毒素的分离株。
对83份样本进行基因型鉴定,其中:
17例患者中同时检测到A基因型和C基因型。
经过12个月的随访调查,根据症状、饮食排除和口服食物挑战,临床诊断如下:
24.44%的病例(66/270)食物过敏,其中18.5%的儿童(50/270)患有牛奶蛋白过敏。
在产气荚膜梭菌阳性组队列中:
这两个比例都显著高于产气荚膜梭菌阴性组 [分别为7.59%(12/158)和5.06%(8/158)] (P<0.0001)
(下图)。
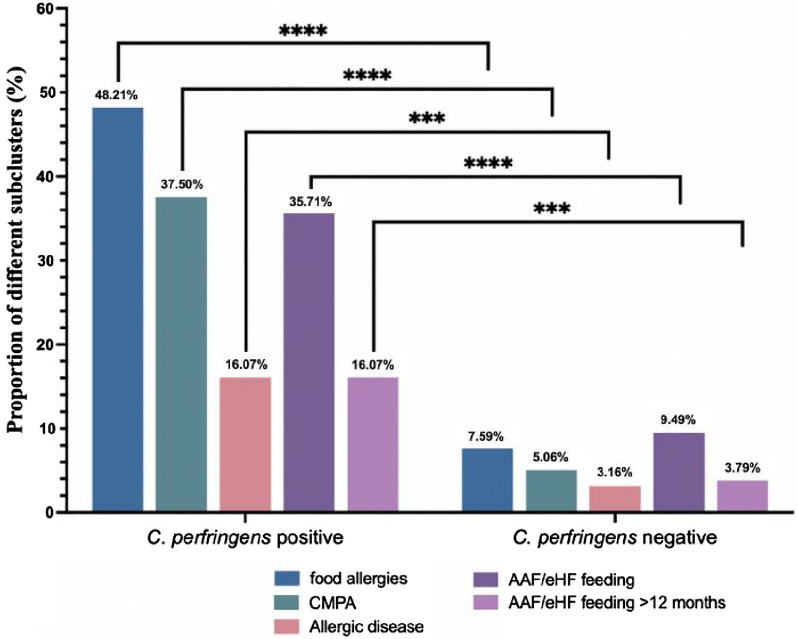
共有8.52%(23/270)的儿童被诊断为过敏性疾病,其中:
共有20.37%(55/270)的儿童喂养氨基酸配方奶粉(AAF)或深度水解配方奶粉(eHF),其中8.89%(24/270)喂养时间超过12个月。
产气荚膜梭菌阳性组氨基酸配方奶粉/深度水解配方奶粉利用率为35.71%,高于阴性组9.49%(P < 0.0001)。两组中喂养氨基酸配方奶粉和/或深度水解配方奶粉超过12个月的儿童人数分别为 16.07%和 3.79%(P = 0.0005)
为了探索产气荚膜梭菌阳性与食物过敏之间的相关性,进行了皮尔逊相关分析,生成了热图,显示食物过敏与产气荚膜梭菌阳性显著相关,粪便潜血、粪便中的白细胞、抗生素使用、血小板增加,血红蛋白减少(P <0.0001)。
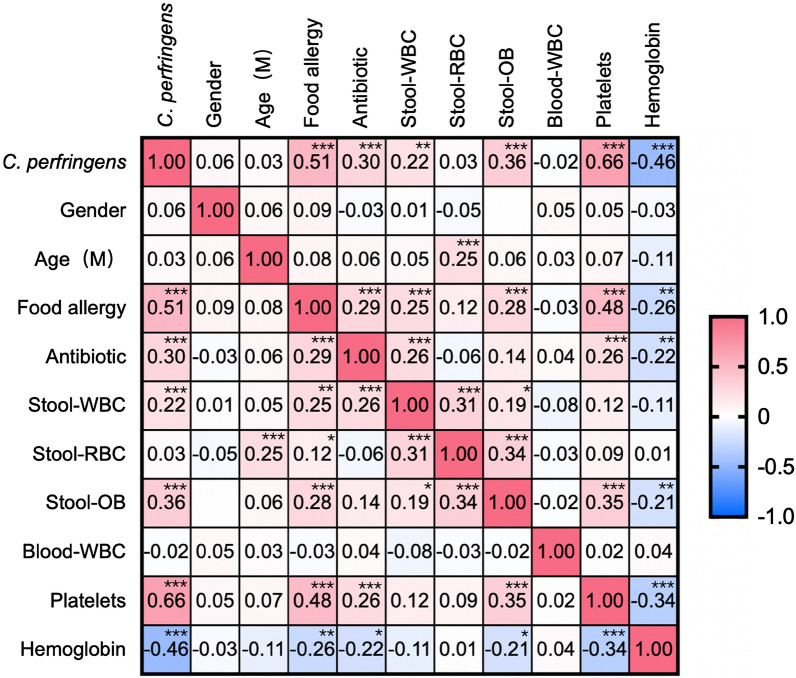
据报道,血小板与食物过敏反应有关,并且与过敏反应的严重程度相关。我们的研究显示食物过敏与血小板之间呈正相关(P<0.0001),这与报道的研究一致。婴儿食物过敏的常见症状包括腹泻和血便,常被误诊为急性或慢性肠炎而采用抗生素治疗。持续便血可能导致贫血,这可以解释本研究中食物过敏与抗生素使用(P<0.0001)和血红蛋白水平(P=0.0001)之间的相关性。
这项真实世界研究结果首次表明,产气荚膜梭菌在有胃肠道症状的中国婴儿中定植率很高,并且与食物过敏和牛奶蛋白过敏的发生有关。
产气荚膜梭菌广泛存在于健康人和动物的胃肠道中。
本研究显示,有胃肠道症状的婴儿中有33.52%有产气荚膜梭菌定植,新生儿和1~6月龄婴儿产气荚膜梭菌阳性率分别为30.25%和45.88%。不同的定植率可能是由于不同的患者群体、分娩方式、喂养方法和胃肠道疾病状况所致。
产气荚膜梭菌和艰难梭菌是可能与婴儿胃肠道感染和过敏相关的致病性梭菌。
先前的研究表明,产气荚膜梭菌定植可能导致多种病理状况,从无症状感染到严重危及生命的败血性休克,例如气性坏疽、食物中毒、坏死性肠炎、抗生素相关性腹泻、菌血症、肠毒血症和严重血管内感染溶血。
研究重点是产气荚膜梭菌感染及其产生一系列致命毒素(七种主要毒素型 A-G)和酶(例如卵磷脂酶、纤维蛋白酶、透明质酸酶、胶原酶和 DNA 酶)的能力,这些酶有助于其侵袭性。
大多数已确定的腹泻相关病原体是病毒,但在近 80% 的报告病例中并未发现特定病原体。这些病例的病因可能包括产气荚膜梭菌胃肠道感染和饮食/环境因素。
目前还没有关于产气荚膜梭菌定植是否影响儿童食物过敏发生的报道。在112名确诊产气荚膜梭菌定植婴儿队列中,其中 48.21% 出现食物过敏,37.5% 出现牛奶蛋白过敏,这两种患病率均显著高于产气荚膜梭菌阴性组(7.59% 和 5.06%),并且显著高于一般儿科人群中报告的水平。
此外,相关分析显示,食物过敏与该研究队列中产气荚膜梭菌定植、抗生素使用、粪便潜血、粪便白细胞、血小板和血红蛋白水平显著相关。该研究结果清楚地表明,产气荚膜梭菌在出生后第一年定植可能会促进食物过敏和牛奶蛋白过敏的发生。
研究表明,产气荚膜梭菌肠毒素(CPE)在促进紧密连接分解和诱导上皮损伤方面发挥着关键作用,从而导致微生物群变化并触发宿主免疫反应。免疫细胞群的变化很大程度上是由环境和微生物抗原驱动的。
关于产气荚膜梭菌定植与食物过敏的研究很少,但结果并不一致。有报道,过敏婴儿的拟杆菌和/或克雷伯氏菌定植率较高,而产气荚膜梭菌/丁酸梭菌定植率较低。
总之,研究人员推测产气荚膜梭菌定植会因其肠毒素而引起侵袭性胃肠道感染,导致紧密连接分解、上皮损伤、微生物群变化、宿主免疫反应异常,并最终导致食物过敏。
需要进一步的研究来阐明产气荚膜梭菌定植的婴儿发生食物过敏的潜在免疫学和分子机制。
总的来说,产气荚膜梭菌定植为婴儿牛奶蛋白过敏和食物过敏提供了一个思路,应用肠道菌群检测可以帮助识别食物过敏的风险,从而采取相应的预防措施。
此外,对于产气荚膜梭菌定植的抵抗力、空间和特定营养物质的竞争或毒素功能的研究,也能为进一步探索肠道菌群与过敏关系及预防和治疗提供了新的视角。
通过深入研究肠道菌群的变化及其与过敏疾病的关联,这可能涉及有效的药理学抑制剂/化合物的合理设计,调节补充剂/益生菌等各种方式对肠道菌群加以改善,有望为未来开发更加个性化的过敏预防和治疗策略提供重要参考。
这里我们来系统地了解一下产气荚膜梭菌。
产气荚膜梭菌是一种革兰氏阳性,产毒厌氧细菌,有荚膜、非运动、无鞭毛,属于梭菌属,是人类和动物肠道固有的机会病原体。
芽胞形成能力强,可在不利环境下存活
可以形成在传播过程中至关重要的孢子。产气荚膜梭菌孢子对高温、氧气或低营养水平等应激环境具有极强的抵抗力。当遇到有利条件时,内生孢子迅速萌发,产气荚膜梭菌的世代时间最短,为 6.3 分钟。在缺氧环境下生长迅速,能快速繁衍和侵袭宿主组织。对抗生素和化学消毒剂具有一定耐受性。
这些特征有利于其在不同的环境生态位中生存,包括土壤、粪便、污水、食物以及人类和动物的肠道。
这种“生存能力”意味着产气荚膜梭菌与许多感染和疾病有关,包括食源性细菌性疾病等。
具有多种水解酶
可分解蛋白质、脂肪等营养物质,为自身生长提供营养。
产气荚膜梭菌产生大量胞外降解酶,例如蛋白酶(例如梭菌蛋白酶)、透明质酸酶(mu毒素)、胶原酶和糖苷内切酶。
内切-N-乙酰半乳糖酰胺酶 EngCP,在 A 型菌株引起的气性坏疽期间很重要,而两种锌金属蛋白酶则导致 G 型菌株引起的禽坏死性肠炎。
最广为人知的产气荚膜梭菌降解酶是神经氨酸酶,它从宿主细胞表面或粘液中的各种唾液酸糖结合物上产生游离的神经氨酸。产生三种神经氨酸酶,分别为NanJ、NanI、NanH。
产生多种细胞外毒素
已知它能分泌超过20 种毒素,每种毒素类型都与特定疾病相关
A型和C型毒素被认为会引起人类疾病。
A型毒素是导致大多数与产气荚膜梭菌相关的食物中毒和非食源性腹泻疾病的主要因素。根据CDC对食源性疾病暴发的流行病学监测数据,产气荚膜梭菌占5%的暴发事件,10%的疾病病例,4%的住院率。
男性的发病率略高(65%),大多数病例发生在20-49岁人群中。
C型毒素与1944年-1949年在德国战后出现的地方性坏死性肠炎,以及在巴布亚新几内亚高地出现的猪痢有关。严重营养不良会增加对 C 型感染的易感性。
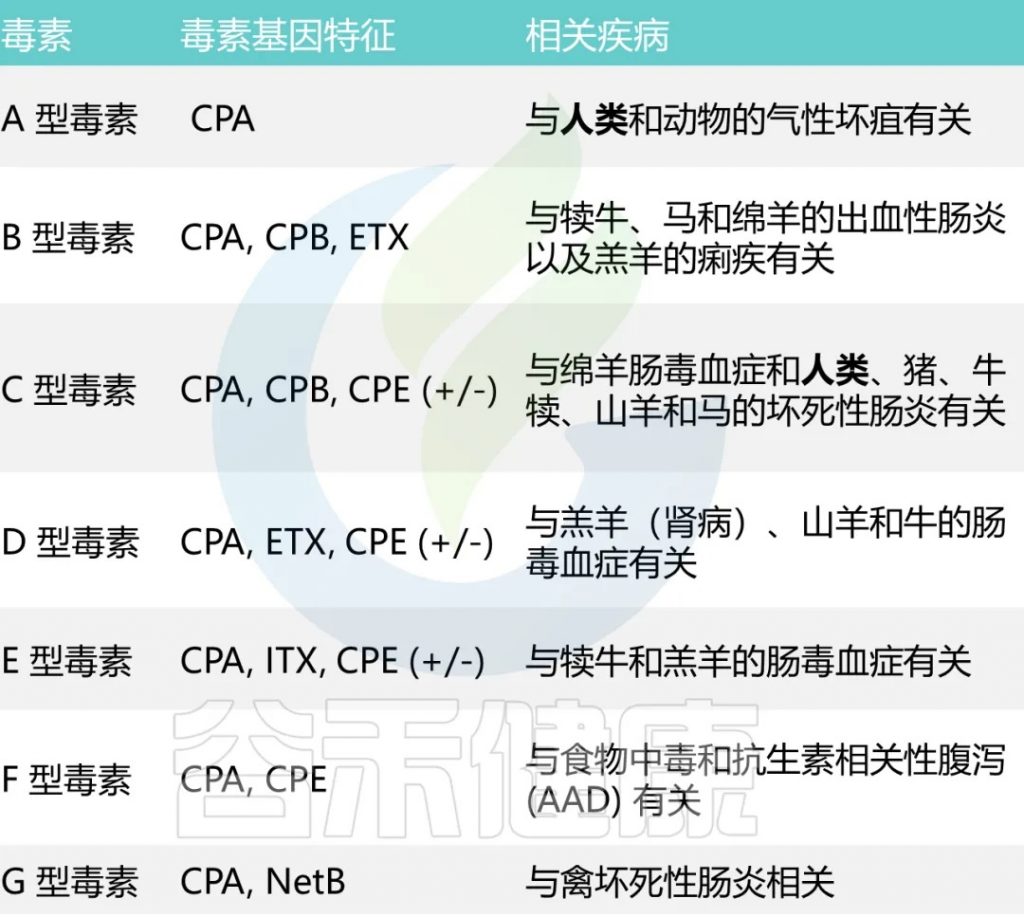
CPE基因,在孢子形成后细胞裂解时产生肠毒素CPE,可能会导致胃肠炎。
产气荚膜梭菌主要毒素的结构域示意图
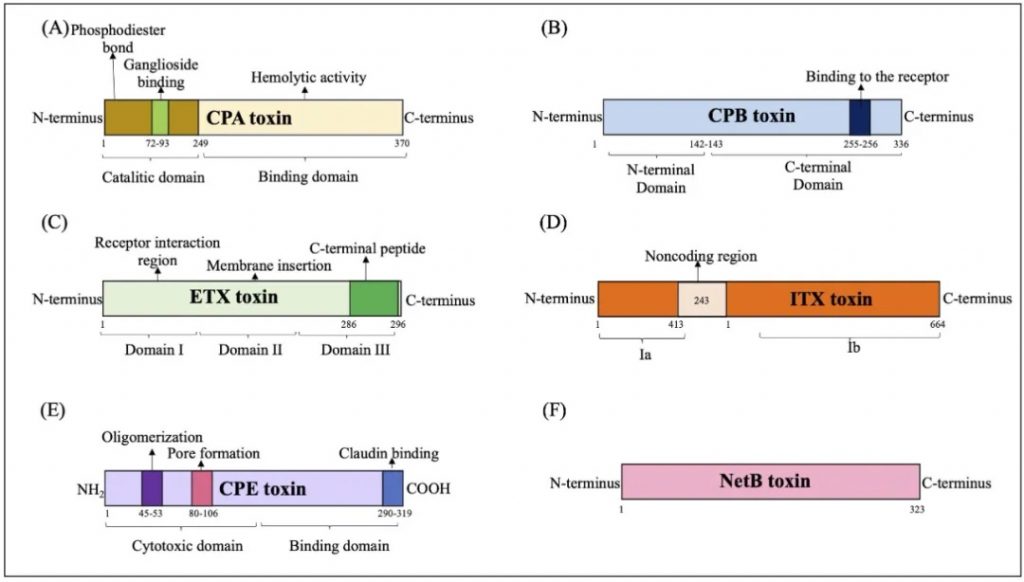
doi.org/10.1080/22221751.2024.2341968
(A) CPA 毒素:CPA毒素与GM1a相互作用,水解磷脂酰胆碱(PC)和鞘磷脂(SM),导致二酰甘油(DAG)和神经酰胺(CER)的形成,并激活原肌球蛋白激酶A受体(TrKA),并触发细胞内信号级联与IL-8释放。磷脂酰肌醇 3 (IP3) 的激活促进细胞质内钙 (Ca+) 进入。
(B) CPB 毒素:CPB 与血小板内皮细胞粘附分子 1 (PECAM-1) 结合,随后释放三磷酸腺苷 (ATP) 并形成允许离子交换进出细胞的孔。
(C) ETX 毒素:ETX 毒素与蛋白质“髓磷脂和淋巴细胞”(MAL) 相互作用,形成活性孔,诱导离子跨细胞膜运输和交换。
(D) ITX 毒素:Ib与脂解刺激脂蛋白受体(LSR)受体的结合介导其进入宿主细胞,通过内吞作用促进Ia进入通道的形成,随后肌动蛋白丝解聚,产生形态变化和改变细胞通透性。
(E) CPE 毒素:CPE毒素与紧密连接蛋白受体结合,通过离子交换和渗透失衡促进细胞表面孔的形成。
(F) NetB 毒素:通过形成允许 Na+、Cl–y 和 Ca2+等离子进入的七聚体亲水孔来识别细胞膜中的无胆固醇区域。
有研究培养从 333 名婴儿的粪便样本中分离出的产气荚膜梭菌,其中29.4% 的婴儿在出院前已被产气荚膜梭菌定植。
三个因素与携带概率呈负相关:
产气荚膜梭菌在母乳中生长不良,说明母乳喂养在早产新生儿护理中的重要性。
长期使用抗生素或持续气道正压通气(CPAP)氧气治疗会在早产儿肠道中为产气荚膜梭菌创造一个不利的环境,从而降低携带的可能性。
产气荚膜梭菌分离物在营养丰富的培养基或母乳中生长,可以作为单一培养物,也可以与婴儿双歧杆菌共培养,婴儿双歧杆菌被选为代表典型的竞争性肠道物种。
食源性疾病
产气荚膜梭菌是食源性疾病(食物中毒)的常见原因,产生肠毒素导致急性胃肠炎。食源性产气荚膜梭菌会导致:腹泻、腹部绞痛、胀气和腹胀。
通常,人们在摄入细菌及其孢子后,当微生物在肠道中形成孢子时,就会形成毒素,通常在 6-24 小时内就会出现症状。腹泻突然发生,但通常症状会在 24-48 小时内消失。
非食源性胃肠道感染
这种类型的感染通常归因于抗生素的使用。但有些人即使不服用抗生素,也会因产气荚膜梭菌而出现偶发性胃肠道症状。免疫系统较弱的人患此类疾病的风险最大。
非食源性产气 荚膜梭菌胃肠道感染导致:
这种类型的产气荚膜梭菌感染通常发生在最近服用过抗生素的60 岁以上成年人中。症状与食源性梭菌感染相似,但持续三天到几周不等。
这种类型的梭菌感染可导致严重脱水(体内水分流失)。然而,人们通常会康复。
一项研究表明,与健康个体 [5%] 相比,IBD 成年患者[19-27%] 产气荚膜梭菌的患病率更高,这表明与 IBD 存在潜在关联。
产气荚膜梭菌具有大量HMO消化所需的糖苷水解酶,表明它具有婴儿肠道机会性定植的可能性,从而导致临床疾病。
皮肤和深层组织感染
产气荚膜梭菌与严重外伤后可能发生的严重皮肤和深层组织感染(气性坏疽)有关。
从受污染的伤口(例如复合性骨折、产后子宫)开始,感染会在 1-3 天内扩散。
皮肤或深层组织产气荚膜梭菌感染(气性坏疽)会导致:
有时,感染仅导致厌氧性筋膜炎或蜂窝织炎。
其他感染
如乳腺炎、子宫内膜炎等。
促进定植
定植能力,即体内持续和增殖/存活的能力,是一些产气荚膜梭菌感染(例如F型菌株非食源性人类胃肠道疾病)的重要特征,产气荚膜梭菌定植通常涉及生长/生存和粘附所需的营养获取。例如,F型非食源性胃肠道疾病即使在腹泻时也能持续长达数周,这表明肠道粘附和营养获取对于这些 F型菌株在腹泻时稳定定植于肠道是必需的。
肠道粘附
为了在胃肠道中定殖,产气荚膜梭菌使用多种分子策略,包括产生唾液酸酶。虽然产气荚膜梭菌产生三种唾液酸酶,但 NanI 是由持续存在于肠道中的 F 型非食源性胃肠道疾病菌株产生的。这种相关性表明 NanI 对产气荚膜梭菌慢性肠道疾病有重要贡献。
NanI 的蛋白酶激活可能进一步促进产气荚膜梭菌菌株引起的定植。
增加营养物质的获取
多项体外研究支持 NanI 对与肠道感染相关的产气荚膜梭菌菌株的生长和存活的贡献。例如,NanI在宿主粘蛋白或培养的 Caco-2 细胞存在的情况下支持F 型非食源性疾病菌株 F4969 的体外生长和存活。这种生长促进涉及 NanI 生成的唾液酸的产生和利用,然后唾液酸可以被产气荚膜梭菌代谢。
可能涉及几种潜在的机制:
NanI 可能通过修饰肠细胞表面来促进产气荚膜梭菌粘附,从而允许暴露用于结合的未知受体,和/或减少该表面上唾液酸的负电荷。
大批量烹制并在不安全温度下保存的食物,通常会导致产气荚膜梭菌食物中毒的爆发。通常与产气荚膜梭菌食物中毒有关的特定食物包括:
产气荚膜梭菌食物中毒的爆发往往发生在为大量人群提供服务并且将食物保持在适当温度可能很困难的环境中,例如医院、学校食堂、监狱、疗养院和提供餐饮的大型活动。
组织感染
食物中毒
将食物煮至安全的内部温度杀死细菌。使用食物温度计进行检查,尤其是整只家禽和大块烤肉。
如果煮熟的食物不尽快食用,请将其保持在60 ℃或更高温度或4℃或更低温度。
烹饪食物或将其从保持安全内部温度的器具中取出后 2 小时内,将剩菜冷藏在4℃或更低的温度下。如果食物暴露在32℃以上的温度下(例如炎热的汽车或野餐),请在 1 小时内冷藏。
热食可以直接放入冰箱。
将大锅食物(例如汤和炖菜)和大块肉类(例如烤肉)分成小份,以帮助其在冰箱中快速冷却。
食用前将剩菜重新加热至74℃或更高温度。
主要参考文献
Huang KY, Liang BS, Zhang XY, Chen H, Ma N, Lan JL, Li DY, Zhou ZW, Yang M. Molecular characterization of Clostridium perfringens isolates from a tertiary children’s hospital in Guangzhou, China, establishing an association between bacterial colonization and food allergies in infants. Gut Pathog. 2023 Oct 8;15(1):47.
Yao PY, Annamaraju P. Clostridium perfringens Infection. [Updated 2023 Aug 8]. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2024 Jan-.
McDonald AG, Lisacek F. Simulated digestions of free oligosaccharides and mucin-type O-glycans reveal a potential role for Clostridium perfringens. Sci Rep. 2024 Jan 18;14(1):1649.
Low KE, Smith SP, Abbott DW, Boraston AB. The glycoconjugate-degrading enzymes of Clostridium perfringens: Tailored catalysts for breaching the intestinal mucus barrier. Glycobiology. 2021 Jun 29;31(6):681-690.
Mehdizadeh Gohari I, A Navarro M, Li J, Shrestha A, Uzal F, A McClane B. Pathogenicity and virulence of Clostridium perfringens. Virulence. 2021 Dec;12(1):723-753.
Camargo A, Rámirez JD, Kiu R, Hall LJ, Muñoz M. Unveiling the pathogenic mechanisms of Clostridium perfringens toxins and virulence factors. Emerg Microbes Infect. 2024 Apr 9:2341968.
Arjomand Fard N, Wine E. Clostridium perfringens: A Potential Pathobiont in Inflammatory Bowel Disease. J Crohns Colitis. 2024 Feb 16:jjae019.
Kiu R, Shaw AG, Sim K, Acuna-Gonzalez A, et al., Particular genomic and virulence traits associated with preterm infant-derived toxigenic Clostridium perfringens strains. Nat Microbiol. 2023 Jun;8(6):1160-1175.

谷禾健康
民以食为天,食以安为先。
随着夏季来临,我们最近检测的报告中发现以上食源性致病菌的检出率和超标率也在逐步上升。
尽管许多人可能没有意识到,近年来,食源性疾病的发病率逐年升高,食源性病原体正在引起大量疾病,对人类健康和经济产生重大影响,已成为全球范围内的重大公共卫生问题。
食源性疾病是一个全球性问题,影响所有年龄和背景的人,尤其是 5 岁以下的人和的65岁老人以及生活条件差的人。我国细菌性食源性疾病每年发病人数可达9411.7万人次,根据美国疾病控制与预防中心 (CDC) 的估计,美国每年也有4800 万例疾病、128000 例住院治疗和 3000 例死亡是由食源性细菌引起的。澳大利亚,每年约有五分之一的人患有这些疾病,住院人数超过 30,000 人。
致病菌广泛存在于各种食品中,食源性致病菌是食源性疾病的首要病因,由其引起的食源性疾病长期以来一直是食品安全的主要威胁,尤其在发展中国家,形势更严峻。它们会导致 200 多种疾病——从腹泻到癌症。在我国,每年因副溶血性弧菌导致急性腹泻665.5万人,导致急性胃肠炎病例估计为728.1万人。
最常见的致病细菌包括:蜡样芽胞杆菌,空肠弯曲杆菌,肉毒梭菌,产气荚膜梭菌,坂崎肠杆菌,埃希氏菌,李斯特菌,沙门氏菌属,志贺氏菌属,Staphylococccus黄色葡萄球菌,弧菌属等。
定 义
食物中毒:指食用了被有毒有害物质污染的食品或者食用了含有有毒有害物质的食品后出现的急性,亚急性以及其它食源性疾病。
目前一些-国家和国际组织以及很少使用食物中毒的概念,经常使用的是“食源性疾病”,食物中毒仅是食源性疾病的一部分,是一大类最常见最典型的食源性疾病
食源性疾病:世界卫生组织认为,凡是通过摄食进入人体的致病因素,是人体患感染性的或中毒性的疾病,都称之为食源性疾病。
我国《食品安全法》定义食源性疾病:指食品中致病因素进入人体引起的感染性,中毒性疾病以及其它疾病。
食物中毒的症状
细菌性
食物中毒的症状因人而异,有些人的病情会比其他人更严重。但是,一些常见的症状是:
不太常见的症状也包括:
保持水分和休息通常是治疗食物中毒的唯一方法,因为身体可以自然地清除污染物。但是,在某些情况下,需用抗生素来帮助身体抵抗感染,特别是在单核细胞增生李斯特菌中毒的情况下以及当易感受伤害的人受到影响时。
病毒性
与细菌不同,病毒不会在食物上生长和繁殖。相反,病毒细胞存在于食物中,当被食用时,它们开始接管人体细胞并感染它们。
常见的食源性疾病病毒类型是:
诺如病毒是导致大多数与食品污染有关的疾病的罪魁祸首 。