-

 国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子
国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子

 二级病原微生物安全实验室
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业

谷禾健康
肠道菌群 & 性激素
青春期是生命的一个关键阶段,与性成熟相关的生理变化有关,是一个受多种内分泌和遗传控制调控的复杂过程。
青春期发育可以在适当的时候,早熟或延迟。
青春期发育异常会给患者带来痛苦,也可能是潜在病理的征兆。

青春期是由激素驱动的,它会影响肠道微生物群。性别之间的肠道微生物群差异出现在青春期开始时,证实了微生物群和性激素之间的关系。在不同的研究中已经提出了肠道微生物群和性激素之间的双向相互作用。
本文介绍了性早熟发育,以及性早熟中肠道微生物群的变化,便于从儿科内分泌学的角度考虑性激素-肠道菌群轴的作用。
肠道菌群的改变可能发生在患有中枢性早熟的女孩身上的证据,为预测和预防性早熟提供了一个有趣的发现。
加深对性激素与微生物群变化作用之间联系的理解,可以通过提供儿科内分泌学的视角,在青春期疾病中实施微生物群靶向治疗。
在了解性早熟与肠道菌群的关系之前,我们先了解一下青春期发育的相关生理过程,以及关于性早熟的基本知识。
青春期特征是:
正常的青春期是由于下丘脑-垂体-性腺轴(HPG)的长期成熟活动所致。
下丘脑以脉动的方式将促性腺激素释放激素释放到垂体门静脉系统,在那里它刺激LH和FSH分泌(也是脉动的)。
注:
LH(促黄体生成素):主要刺激睾丸中的间质细胞和卵巢中的卵泡膜细胞分泌雄激素。
FSH(卵泡刺激素):主要刺激卵泡或生精小管形成雌激素、抑制素、卵子或精子。
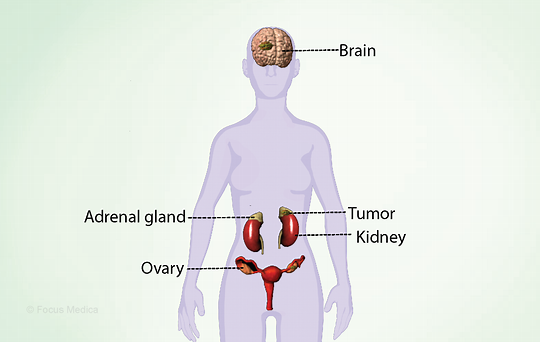
间质、管状和卵泡隔室通过旁分泌过程共同作用,产生雌激素,并调节性类固醇的产生和配子的发育。类固醇激素对促性腺激素释放激素和促性腺素分泌有内分泌负反馈作用。
FSH分泌被抑制素、孕酮和雌二醇的负反馈抑制。在成年女性受试者中,临界雌二醇浓度刺激LH激增,从而启动排卵。
HPG轴出生时短暂激活,青春期完全重新激活
出生时,由于缺乏抑制HPG轴的胎盘类固醇,这一轴会被激活,从而导致类固醇激素的产生增加,这是青春期继续进行的第一步。这种短暂的激活大约在出生后一周开始,几个月(大约6个月)后停止。
HPG轴在整个儿童期并不完全休眠,尤其是女性,其FSH浓度略高于男性。有时,也可以用超声波检查卵巢卵泡。在青春期,HPG轴经历完全重新激活。
最重要的GnRH抑制系统是γ-氨基丁酸能神经元(产生γ-氨基丁酸的神经元)和阿片受体;吻肽、神经激肽B和强啡肽A这三种存在于弓状核(ARC)中的神经肽被认为是影响GnRH释放的基本生成物,因为它们对男孩和女孩的青春期生理学有重要贡献。
厌食因子——瘦素:在青春期发挥重要作用
瘦素是一种主要由脂肪细胞产生的细胞因子,它作为一种厌食因子,通过抑制下丘脑神经肽Y(NPY),从而抑制食欲,在控制体重、食物摄入和能量平衡方面发挥重要作用。儿童时期必须达到正常的体重和组成,以避免青春期功能障碍。
除了瘦素与NPY的相互作用外,一些研究表明瘦素通过与KiSS-1基因直接相互作用而影响青春期和生殖功能。GnRH神经元缺乏瘦素受体,但KiSS-1神经元表达瘦素受体。瘦素直接刺激kisspeptin释放并介导GnRH的脉动释放。
青春期是一个复杂的过程,具有广泛的生理变化。调节青春期开始的机制涉及遗传、营养和环境相互作用。
胎儿营养异常以及内分泌系统可能导致发育改变,从而永久影响结构、生理和代谢。
生长关键时期激素和营养之间的相互作用对于代谢适应反应控制和青春期发育预期至关重要。
越来越多的证据表明,产前和产后早期是青春期开始规划的重要时期。各种研究表明,产前暴露于不利环境因素,如导致儿童出生SGA(小于胎龄)和/或IUGR(宫内生长受限)的因素,会影响青春期时机。儿童出生的SGA可能会经历几次青春期改变,如性早熟。
➡ 性早熟如何定义?
性早熟(PP)定义为女性青少年在8岁之前和男性在9岁之前出现变化:
性早熟患者表现出性和身体生长加快,同时出现生长突增。如果不治疗,骨骺生长加快可能会导致骨骺过早闭合,导致成年后身材矮小。
➡ 性早熟分类
根据潜在的生理病理过程,病理性性早熟分类如下:
• 中枢性性早熟
中枢性性早熟是最常见的性早熟形式,女孩多于男孩。一般异常位于大脑。
由于先天性或后天性中枢神经系统(CNS)损伤或单基因缺陷导致的HPG轴过早成熟,从而导致的中枢性性早熟(CPP)或促性腺激素依赖性性早熟(或真正的性早熟),也可能是特发性的。
• 外周性性早熟
一般异常不在大脑中,而是在睾丸、卵巢或肾上腺中。
外周性性早熟(PPP)或促性腺激素非依赖性性早熟(或假性早熟),由于遗传或肿瘤病因、分泌hCG的生殖细胞肿瘤(仅适用于男孩的人绒毛膜促性腺素)或外源性来源导致的性腺性激素或肾上腺激素分泌过多。
➡ 中枢性性早熟病例男女有别
据评估,每5000-10000名儿童中就有1名患有中枢性性早熟,女性的发病率是男性的10倍。
大多数女性中枢性性早熟病例似乎是特发性的,而男性中枢性性早熟患病率较高似乎通常是由病理性脑损伤引起的。特别是,下丘脑错构瘤是引起中枢性性早熟的最常见的脑损伤。
注:下丘脑错构瘤,是临床极为罕见的先天性脑组织发育异常性病变。
➡ 性早熟的临床检查评估
准确的个人和熟悉的病史、完整的体检、荷尔蒙和放射检查对性早熟诊断至关重要。
——青春期体征
临床检查应侧重于发育学数据、根据Marshall和Tanner分类对青春期体征的评估、过去6-12个月的生长模式,青春期体征(女孩的乳腺芽、男孩的睾丸体积、两者的阴毛)和其他青春期体征(痤疮、油性皮肤、勃起、男孩夜间排泄物、女孩阴道分泌物和月经出血)的进展率。
——基线LH水平
基线LH水平是诊断中枢性性早熟的有希望的生物标志物;基础晨LH值超过0.2 mUI/ml通常被认为是青春期的标志。在成瘾中,LH与FSH的比值高于0.6与中枢性性早熟有关。
——GnRH刺激试验
GnRH刺激试验仍然是鉴别中枢性性早熟的金标准,大于5 IU/L的LH截止峰值水平被广泛用于诊断中枢性性早熟。
——其他激素评估
其他激素评估应包括甲状腺测试、睾酮、雌二醇、17-羟基孕酮(17-OHP)、癌胚抗原(CEA)、癌抗原125(CA125)、甲胎蛋白和β-hCG,具体取决于患者的病史。
——骨龄
为了确定孩子的生物年龄,需要对非优势(左手)手和手腕进行骨龄X光检查。超过2.5标准差(SD)或超过2年的晚期骨龄更可能与病理性性早熟相关。
——盆腔超声
在女孩中,盆腔超声是评估卵巢早熟发育和排除卵巢囊肿或肿瘤的有用工具。
——脑部MRI检查
建议对患有中枢性性早熟的患者进行脑部MRI检查,以排除CNS损伤,应在男孩(<6岁)中进行常规检查。
性激素和肠道菌群之间的关系在最近的研究中得到了广泛的探讨,并且是一个不断扩大的研究领域,可能会为各种与性有关的疾病带来新的治疗选择。
20世纪80年代,最早的研究之一是在细胞水平上进行的,其中发现孕酮促进了拟杆菌和中间普氏杆菌的生长。
2012年一项研究在对91名女性的粪便样本进行测序时发现,无论健康状况如何,在怀孕期间,尤其是在孕晚期,当雌激素达到峰值时,肠道微生物组明显改变。
2015年,一项欧洲研究表明,健康男性的普氏拟杆菌丰度高于生育女性,而绝经后女性的微生物群与男性没有差异。
这两项研究都证明了雌激素和相关的雌性激素在调节肠道微生物组的组成方面是至关重要的。
因此,微生物群会影响雌激素水平,反过来,雌激素水平可能会受到微生物群组成和多样性的影响。
➤ 微生物群对雌激素的影响
微生物组能够通过表达B-葡萄糖醛酸酶(一种介导膳食和非膳食雌激素解偶联的酶)代谢雌激素。
未结合的雌激素可以通过作用于α和β雌激素受体而进入全身血流并具有代谢活性,这些受体在多种器官和组织中表达;雌激素活性不仅对生殖健康有影响,而且对心血管风险、代谢和骨稳态以及中枢神经系统也有影响。
微生物群多样性与绝经后妇女和男性的尿雌激素水平较高有关,而绝经前妇女的雌激素水平似乎不受微生物群组成的影响,这表明微生物群主要影响非卵巢雌激素水平。
►►
补充植物雌激素能够促进特定细菌的肠道定植,富含植物雌激素的饮食可能与亚洲绝经后妇女代谢综合征的风险降低有关。
➤ 反过来,性激素水平也可能影响微生物组的组成
雄激素过量,如多囊卵巢综合征患者,也可能导致生物失调和细菌多样性降低。多囊卵巢综合征(PCOS)是一种内分泌疾病,其特征是雄激素水平较高,雌激素水平较低,几项研究表明,多囊卵巢综合症患者的肠道失调与细菌多样性较低有关,导致丁酸生成减少,BMI升高,睾酮血清浓度升高。
此外,肠道微生物组通过产生有助于减少炎症反应的短链脂肪酸在决定胰岛素分泌方面发挥重要作用;菌群失调可能导致胰岛素抵抗和糖代谢改变,如多囊卵巢综合征;较高的胰岛素水平刺激卵巢产生雄激素,从而使多囊卵巢综合征的发病机制永久化。
➤ 微生物群及其代谢产物也可能影响肠道菌群定植
微生物群及其代谢产物也可能通过定殖阴道,以及一些作者认为的子宫内膜和胎盘,影响女性生育、怀孕、胚胎发育和分娩的各个阶段。微生物群的改变与促炎细胞因子的分泌和早产有关。剖宫产新生儿肠道菌群多样性较低,可能是因为他们没有通过阴道被母体肠道菌群定植。
➤ 性激素信号和脑-肠轴功能中的应激反应之间存在复杂的相互作用
我们知道,肠道和中枢神经系统之间的双向沟通途径,根据肠道细菌是大脑和内分泌系统之间的重要媒介。肠道菌群是调节脑-肠轴的中枢,肠道屏障短链脂肪酸,除了是能够调节肠道运动和伤口愈合的炎症调节剂外,还代表了微生物组和肠-脑轴之间的联系。
功能性胃肠疾病的发生率,如功能性消化不良和肠易激综合征,导致运动能力受损和/或敏感性改变,在女性中明显更高,这可能是因为性激素信号和脑-肠轴功能中的应激反应之间存在复杂的相互作用。
▸ 雌激素激活脑-肠轴的受体,间接干扰胃肠动力
具体而言,已观察到雌激素通过直接激活位于脑-肠轴的受体,并通过调节其他受体系统间接干扰胃肠动力和敏感性。在卵巢周期的卵泡期,当雌激素水平较高时,女性的胃肠动力降低。
此外,支持循环雌激素在胃排空延迟中起重要作用的假设,绝经前和绝经后女性接受激素替代疗法与未接受激素治疗的绝经后妇女相比,胃排空速度较慢,而绝经后的女性与同龄男性相类似。相比之下,睾酮或雄激素似乎对胃动力或胃超敏反应没有影响。
性激素和肠道菌群之间的相互关系
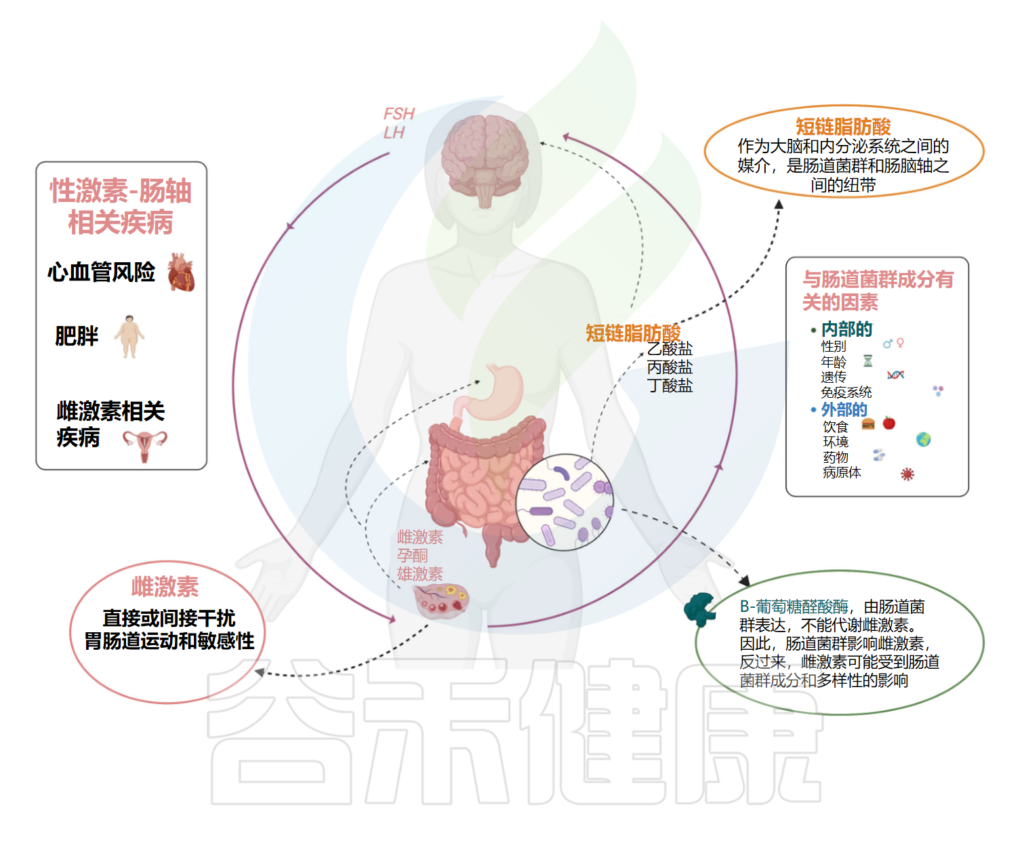
Calcaterra V, et al., Front Endocrinol (Lausanne). 2022
雌激素通过对核受体的作用,以及通过与膜G蛋白偶联的雌激素受体1 (ER 1)受体的激活,实现其快速、非基因组的作用机制。
▸ 雌激素干扰疼痛调节
雌激素受体在中枢神经系统和涉及内脏疼痛感知的通路中广泛表达,包括下丘脑、杏仁核和中脑,所有这些都已被证明向参与胃肠功能调节的迷走神经细胞发送广泛的投射。
具体而言,在外周内脏传入中,雌激素似乎通过改变离子通道的开放和受体表达的调节来调节伤害感受,以及激活胆酸速激肽-神经激肽1受体并诱导P物质的释放。
最后,除了干扰疼痛调节外,雌激素还参与中枢神经系统的内脏信息处理。
脑成像研究发现,与患有IBS的男性相比,患有IBS女性在对不良内脏刺激的反应中表现出更大的情绪回路激活,包括杏仁核和蓝斑。
众所周知,随着年龄的增长,微生物群的组成逐渐发生变化,需氧菌和兼性厌氧菌的数量普遍减少,专性厌氧菌种群数量增加。
传统上,普遍的观点是,在1-2岁之间,人类肠道菌群开始类似于成年人,主要由厚壁菌门、拟杆菌门、变形杆菌门和放线菌门的物种组成。然而,青少年和成人粪便微生物群在属水平上观察到许多差异。
非青春期和青春期菌群类别存在差异
一项研究评估青春期儿童的远端肠道菌群表明,双歧杆菌属和梭状芽孢杆菌属(已知定植于新生儿肠道并在2至18岁之间逐渐减少的物种)成员的丰度在青春期儿童中显著高于成年人。这些属的流行率以前在青春期前和青春期年龄组中没有被认识到。
2020年的一项横断面研究,通过16S rRNA测序分析了不同青春期阶段(5-15岁)的肠道菌群生物多样性。非青春期和青春期受试者之间的α或β多样性没有发现差异,但研究证明两组之间的细菌类群存在差异。
特别是,非青春期受试者的特征主要以下菌群:
另一方面,青春期组以β变形菌纲(Burkhollderiales)的流行率较高。
进一步分析血清性激素与菌群丰度之间的关系,结果表明:
睾酮水平与Adlercreutzia、Dorea、Clostridium和拟副杆菌属的丰度有关。作者假设这些细菌可能受到性激素的影响。
几项研究已经调查了肠道微生物与性类固醇激素之间的关系。Shin等人证明了肠道菌群特征与人类睾酮/雌激素状态之间的关系:
不动杆菌、多利亚菌、瘤胃球菌和巨单胞菌与睾酮水平显著相关,而Slackia菌和丁酸单胞菌则与雌二醇水平显著相关。
青春期两性之间的肠道菌群差异更加明显
据报道,肠道菌群似乎不受儿童性别的影响,但在青春期开始时就出现了差异。微生物门相关疾病也表现出性别偏见,支持肠道细菌与性别之间的关系可能是双重的。
最近的一项横断面调查发现,在青春期,两性之间的肠道菌群差异更加明显。
比较青少年和成年人的微生物群,发现双歧杆菌的数量,特别是在一些研究中,随着年龄的增长而减少,也报道了与拟杆菌门和厚壁菌门的年龄相关的联系。
Hollister等人比较了青春期和成人的肠道菌群组成,指出在青春期发育期间,女孩的肠道菌群逐渐发生变化,越来越像成年女性的肠道菌群,与她们的青春期发育程度成正比。
在雄性和雌性中,优势类群都是梭状芽孢杆菌和拟杆菌,表明微生物群成熟的要素是:
梭状芽胞菌和拟杆菌类菌群优势的变化
事实上,在青春期,梭状芽孢杆菌的相对丰度逐渐增加,拟杆菌的相对减少,逐渐类似于成人微生物群的组成。就门而言,随着青春期的进展,厚壁菌门的数量增加,而拟杆菌门的数量减少。
然而,尽管Hollister等人观察到男孩和女孩的肠道菌群变化,但由于男性青春期发育较晚,仅在女性青少年中获得了具有统计学意义的数据。
青春期,肠道菌群组成和预测的代谢谱存在性别显著差异
Yuan等人证实了这些数据,他们确定了不同青春期状态下两种性别的肠道菌群特征。对89名5-15岁的中国参与者进行了肠道菌群分析。参与者被分为青春期前和青春期组,男性(n=49)和女性(n=40)。
这项横断面研究表明,在青春期之前,肠道菌群组成和预测的代谢谱存在性别差异,在青春期变得更为显著。
实验结果表明,Dorea、Megamonas、Bilophila、Parabacteries、Phascolactobacterium属代表青春期受试者的微生物标记。
他们认为,性别依赖的肠道菌群多样性部分是由于性激素,部分是由于其他非激素影响因素。
“
以上是肠道菌群在生理性青春期发育中的作用的证据。而肠道菌群在病理性青春期中的作用越来越受到关注。
青春期到来的时间可能受到肠道菌群的影响,特别是某些梭菌属物种,包括调节宿主性激素水平的瘤胃球菌科、粪杆菌属和瘤胃球菌属。
肠道菌群影响雌激素代谢
Ruminococcus和Faecalibacterium的β-葡糖醛酸酶能够裂解雌酮和雌二醇,而拟杆菌物种只能代谢雌酮。
因此,尿液中雌激素-雌激素代谢物比率与瘤胃球菌的相对丰度呈正相关,与拟杆菌的相对丰量呈负相关。
根据这些数据,肠道菌群可能通过其雌激素代谢部分调节青春期的开始。
肠道菌群通过能够代谢雌激素的特定肠道微生物似乎能够调节青春期,但反过来也可能。性激素可以通过引导肠道菌群的成熟,直接影响特定菌群的生长。
肠道菌群代谢物影响内分泌系统
最近的研究发现,肠道菌群产生的代谢物可以影响人体内分泌系统,激活肠道神经系统。
一些关于微生物群功能的最佳研究强调了肠道微生物通过产生短链脂肪酸为宿主提供能量,包括丁酸和丙酸,这两种物质都参与胆汁盐代谢,并在脑肠轴中发挥重要作用。
已经证明,内源性受体游离脂肪酸受体(FFAR)2和FFAR3与短链脂肪酸相互作用,并在肠内分泌细胞中表达,肠内分泌细胞产生肽YY(一种厌食激素),从而参与调节宿主能量、食欲、脂肪组织储存和荷尔蒙平衡,影响青春期时间。
肥胖 & 青春期 & 肠道菌群失调
肥胖和青春期之间有着密切的联系;尤其是性早熟与体重指数(BMI)呈正相关。基于这一信息,并考虑到受性早熟影响的儿童往往肥胖,已假设肠道菌群可能参与性早熟的发病机制。
一项研究阐明了特发性中枢性性早熟(ICPP)患者(n=25)和健康女孩(n=23)之间肠道菌群的差异。
特发性中枢性性早熟肠道菌群与肥胖菌属相似
作者应用16S rDNA测序来比较两组之间的肠道菌群,观察到,特发性中枢性性早熟中鉴定的肠道属与肥胖相关的肠道属相似,特别是吉米格瘤胃杆菌属、示波杆菌属和XIVb梭状芽孢杆菌属。
考虑到微生物种类水平,患有特发性中枢性性早熟的女孩富含Rumicoccus bromii, Ruminococcus gnavus, Ruminococcus leptum,前两种是在肥胖人群中发现的;它们可以促进脂肪组织的能量吸收和增生,而据报道,Ruminococcus leptum会影响人体体重变化。
这些结果突出了肥胖、特发性中枢性性早熟和肠道菌群失调之间的关联。作者假设,肠道菌群失调会导致青春期前女孩发生类似于肥胖患者的过程,脂肪细胞的增殖和沉积会引发性早熟。
然而,肠道失调也可能导致下丘脑-垂体-性腺轴(HPGA)的早期激活。
不同的研究已经研究了肠道菌群在雌激素、FSH和LH分泌机制中的作用,但仍不清楚。
三种临床生物标志物(FSH、LH和胰岛素抵抗)与肠道菌群之间的关系
考虑到特发性中枢性性早熟女孩,作者证明了FSH与梭杆菌、LH与Gemmiger之间的正相关,LH与Romboutsia之间的负相关。此外,胰岛素抵抗与吉米格菌、瘤胃球菌、巨单胞菌和双歧杆菌呈正相关。
特发性中枢性性早熟:部分菌群高产短链脂肪酸
肠道菌群、激素分泌和肥胖之间的密切联系激发了对肠道菌群触发特发性中枢性性早熟机制的研究。
研究人员在工作中调查的特发性中枢性性早熟女孩的特征是与产短链脂肪酸相关的菌群:
短链脂肪酸诱导瘦素基因表达,激活HPGA轴
瘦素在女性青春期发病中的重要作用众所周知。瘦素是一种脂肪细胞代谢肽,参与其表达的基因与短链脂肪酸相关。
短链脂肪酸产生与特发性中枢性性早熟之间的关系可以通过高浓度短链脂肪酸诱导瘦素基因表达的机制来解释,瘦素基因激活下丘脑-垂体-性腺轴,从而导致青春期的开始。
Li等人招募了27名中枢性性早熟女孩、24名超重女孩和22名健康对照,以探讨肥胖与中枢性性早熟之间的关系。
肠道菌群代谢物激活HPGA轴
这项研究表明,中枢性性早熟患者表现出过多的Alistipes、Klebsiella、Sutterella,这通常出现在神经系统疾病患者中。这些微生物产生具有神经递质活性的代谢物(血清素和多巴胺),这些代谢物会引发早熟,激活下丘脑-垂体-性腺轴。
肠道菌群代谢物促进胰岛素抵抗
作者在中枢性性早熟和超重组中都发现了普雷沃氏菌;支链氨基酸的产生可促进胰岛素抵抗。这一机制可以解释中枢性性早熟患者肥胖的高发生率。
此外,在两组中,均观察到一氧化氮合成升高,这是一种重要的气体神经递质,可刺激促性腺激素释放激素的分泌并促进胰岛素抵抗。
这些情况,肠道菌群表达的改变,可以解释中枢性性早熟和肥胖之间的联系,如下图所示。
性早熟 – 肠道微生物组和肥胖:密切相关
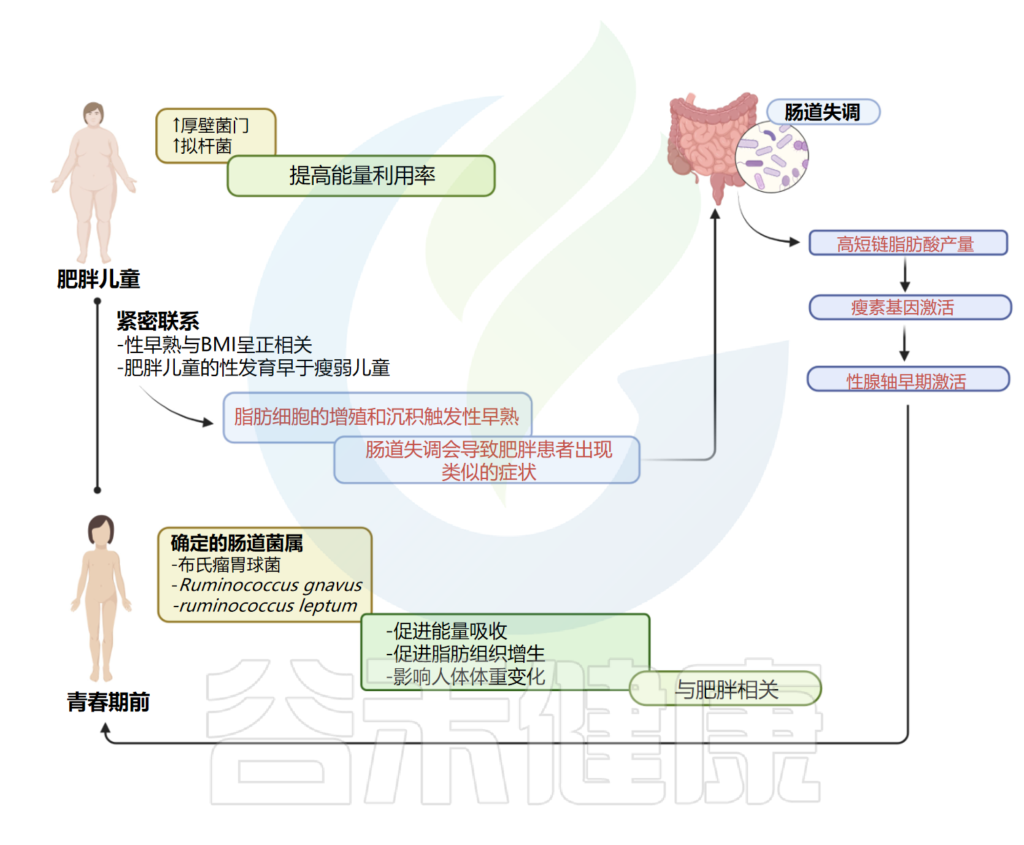
Calcaterra V, et al., Front Endocrinol (Lausanne). 2022
高脂饮食:过早激活HPG轴,诱导性早熟
大量营养素食物含量,如高脂肪摄入,可能会调节HPG轴的过早激活,诱导青春期的性早熟。
最近,在实验模型中,研究人员表明,高脂饮食对性早熟的影响受肠道微生物群和激素的相互作用调节。
断奶后的高脂饮食导致性早熟,下丘脑中的血清雌二醇、瘦素、脱氧胆酸和GnRH增加。特别是,GnRH与脱硫弧菌、Lachnoclostridium、GCA-900066575、链球菌、厌氧菌、双歧杆菌呈正相关,这表明这些细菌可能在促进性发育方面发挥作用。
此外,作者报道,“高脂饮食微生物群”移植促进了小鼠的性早熟,支持肠道菌群调节促进性早熟的局部和全身性类固醇水平。
性早熟治疗的最重要目标是保持成人身高并减少相关的心理社会压力。
促性腺激素释放激素激动剂,1个月或3个月储存制剂,是中枢性性早熟的标准护理。GnRH激动剂治疗被广泛认为是安全的。
最常见的不良反应包括局部皮肤反应和绝经后症状。需要定期验证青春期进展、生长速度和骨骼成熟。
PPP的治疗因发病机制而异,治疗的主要目的是消除性类固醇的内源性或外源性来源。
肾上腺和性腺肿瘤需要手术治疗。
其他生活中的注意事项如下,供参考。
生活方式
消除外源性雌激素(塑料、食品添加剂和杀虫剂)和其他内分泌干扰物(例如美容产品中的邻苯二甲酸盐)。薰衣草油和茶树油与乳突有关,尤其是男孩的男性乳房发育症。
营养
消除添加激素的乳制品、鸡蛋和肉类。考虑消除大豆、亚麻和芝麻,因为它们是植物雌激素,年轻组可能对这些非常敏感。
缺乏维生素 D 的 5-12 岁女孩面临青春期提前的风险是维生素 D 水平较高的女孩的两倍。
晒太阳是增加维生素 D 摄入量以预防和减缓青春期提前的一种方法。
其他食物补充包括:金枪鱼和鲑鱼等多脂鱼、橙汁、豆浆等。
肝脏解毒支持减少外源性激素作用。
食物
深色绿叶蔬菜、西兰花、甜菜、朝鲜蓟、姜黄、柠檬、蒲公英根等。
减少肾上腺激素输出的植物药:
刺五加 ;睡茄;红景天
父母的反应会影响孩子的应对能力
当孩子的家庭氛围不好,他们的病情无法得到家人的支持,会增加孩子的负面心理情绪和父母的养育压力。加强家庭成员之间的沟通至关重要。
一些导致性早熟的风险因素(例如性别和种族)无法避免。但是,有一些措施可以降低孩子出现性早熟的风险,包括:
不同的研究都提出了肠道菌群与性激素之间的双向相互作用。青春期期间,身体发育变化主要由激素驱动;因此,这一动态和过渡期是评估潜在激素对肠道菌群影响的机会。
不同青春期阶段的肠道菌群存在多样性,并且患有中枢性性早熟的女孩肠道菌群可能改变,这是预测和预防性早熟发育的一个有趣的发现。通过提供儿科内分泌学的观点,加深对性激素和微生物群变化作用之间的联系的理解,可以在青春期疾病中实施微生物群靶向治疗。
主要参考文献:
Calcaterra V, Rossi V, Massini G, Regalbuto C, Hruby C, Panelli S, Bandi C, Zuccotti G. Precocious puberty and microbiota: The role of the sex hormone-gut microbiome axis. Front Endocrinol (Lausanne). 2022 Oct 21;13:1000919. doi: 10.3389/fendo.2022.1000919. PMID: 36339428; PMCID: PMC9634744.
Li Y, Shen L, Huang C, Li X, Chen J, Li SC, Shen B. Altered nitric oxide induced by gut microbiota reveals the connection between central precocious puberty and obesity. Clin Transl Med. 2021 Feb;11(2):e299. doi: 10.1002/ctm2.299. PMID: 33634977; PMCID: PMC7842634.
Dong G, Zhang J, Yang Z, Feng X, Li J, Li D, Huang M, Li Y, Qiu M, Lu X, Liu P, Zeng Y, Xu X, Luo X, Dai W, Gong S. The Association of Gut Microbiota With Idiopathic Central Precocious Puberty in Girls. Front Endocrinol (Lausanne). 2020 Jan 22;10:941. doi: 10.3389/fendo.2019.00941. PMID: 32038493; PMCID: PMC6987398.
谷禾健康
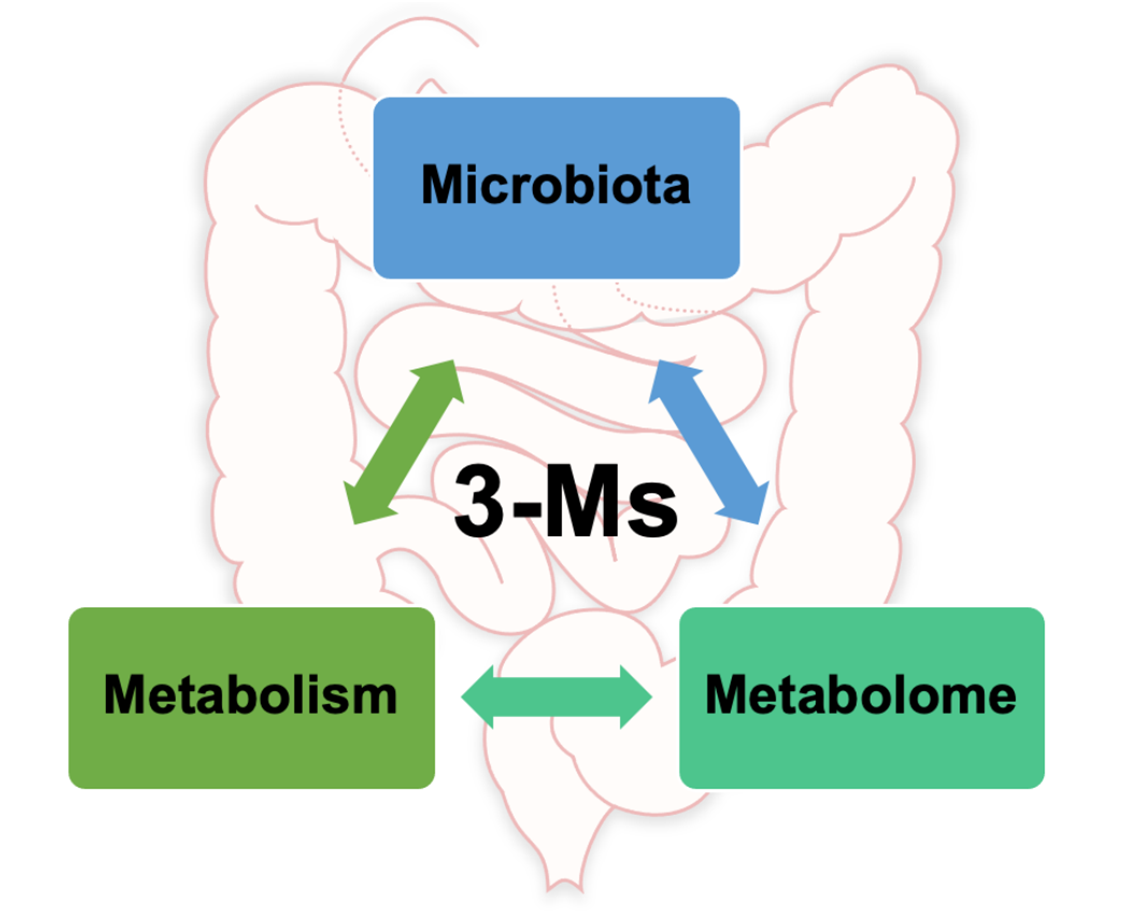
在过去的研究中,健康科学的突破表明人类肠道菌群可以受到环境因素的影响(饮食、生活方式等),而环境因素反过来又可以影响人类健康和疾病。
现在,更全新的概念肠道3M已经提出,即微生物组、代谢和代谢组(microbiota, metabolism, metabolome),对人类健康具有重要意义。
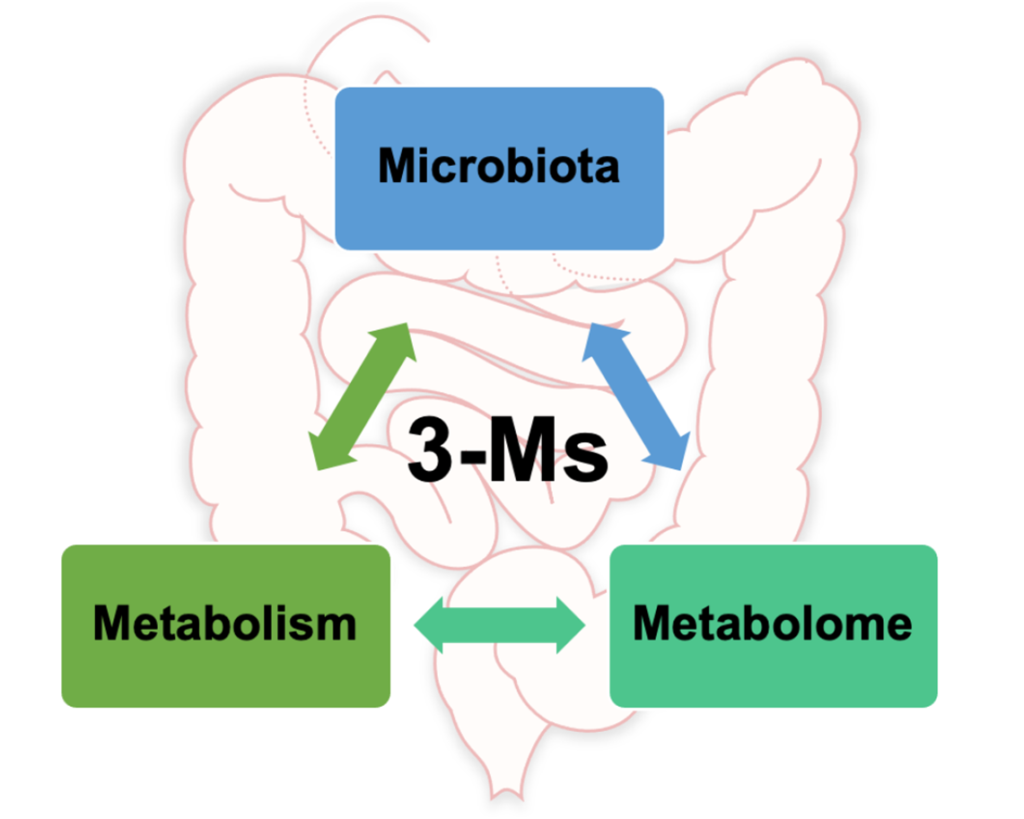
本文重点介绍3M相关的主要概念,提供了关于调节肠道健康的全面更新概述,有助于我们对肠道与人类健康和疾病之间的关系的基本理解。同时也定义了3M相互作用的挑战及讨论未来前景。
在阐述微生物群之前首先了解一下它的发展历史。
关于微生物群的历史
Dey P, et al., Free Radic Biol Med. 2021
人类微生物群
随着下一代高通量16S rRNA测序技术的出现,与人类健康和疾病相关的以微生物群为中心的研究有了巨大的增长。
数量
目前的估计表明,一个体重为70公斤的人平均含有39万亿细菌,而人体细胞则为30万亿。此外,在遗传水平上,在整个人类微生物群中基因数量超过1000万个基因,比人类基因(2万-2.5万)还要多很多,仅肠道微生物群基因就有330万。由于与生命各个方面相关的宿主-微生物相互作用的数量巨大,人类微生物群关联通常被称为“超有机体”。
分布
肠道细菌在肠道长度上的定植偏好是区域特异性的,可能是由缺氧肠腔内宿主的理化参数的变化(如pH值、营养有效性、pO2、胆汁酸、抗菌剂、粘蛋白)决定的。此外,在肠的特定区域内,管腔和绒毛隐窝之间的微生物种群多样性可能不同。比如说,厚壁菌和拟杆菌在盲肠中占优势,而隐窝主要由变形杆菌门和脱铁杆菌门定植。
“核心微生物群”
当然,由于与环境的不同相互作用,包括饮食模式的差异,文献中的微生物概况往往不一致。因此,人类微生物组项目的一个关键目标是确定一个“核心微生物组”,作为所有个体的普遍微生物分类学特征,该特征大部分保持不变。

谷禾健康
饮食的改变会改变人体的微生物群,对健康造成潜在的影响。根据以往的研究,饮食驱动的微生物群变化不仅可以导致失调,还可以塑造生活史特征,推动人类进化。考虑到环境、健康和进化是相互关联的,那么饮食驱动的微生物群干扰是否会产生超出其对人类健康直接影响的后果?
最近德国科学家指出,饮食结构的变化会导致微生物组宿主基因配置不匹配,从而影响人类的寿命和繁殖成功。 这些失配也可引起可遗传的全功能体(holobiont)应激反应,这促使全生命周期在变化的营养环境中重新建立平衡。
该研究介绍一个前瞻性的模型,其中的微生物组在现代人类适应环境变化中发挥作用。假设饮食变化重新配置了整合人类微生物群中表观遗传控制的平衡健康状态。这种重新配置是一把双刃剑。一方面,它可能通过调节完整生物的生命史特征(如寿命和繁殖)的表达而增加疾病的风险和严重性。另一方面,它有助于重新调整全功能体(holobiont)系统与新的营养环境,利用宿主基因和微生物组之间的不匹配产生的压力。
简单笼统的说,就是该机制框架将两个迄今联系不充分的辩论联系起来:全球气候变化对饮食的影响,以及微生物组,表观基因组和宿主之间的复杂关系。
这些假设的动态不必只与正在进行的气候变化相耦合,也不必只适用于人类全息生物。相反,微生物群-表观基因组-免疫轴中的类似动力学可能是对过去非人为气候变化的反应,并对我们和其他物种的进化轨迹作出了贡献。
气候变化可以通过多种途径影响人类健康。到目前为止,关于气候变化对人类健康的影响的公众关注和科学研究主要集中在气候灾难(例如洪水,热浪和流行病)产生的CO2排放和健康风险上。尽管仍然相对罕见,但随着气候变化,这些灾难预计会变得更加频繁和严重。
在这些事件中暴露于危险条件的风险很高,幸存者可能会遭受长期的健康危机(例如,运动功能丧失)的困扰。随着时间的推移,这些风险会积累。饮食就是其中一种途径。
气候变化和经济发展压力会影响许多影响人类饮食的因素,包括作物产量,耕作方式和饮食习惯。农作物的生产耕作以及烹饪方式可能在正在改变以适应人类的饮食习惯。 气候变化,对饮食有明显影响。 例如,对玻利维亚八个Aymara社区的案例研究发现,农民在介绍他们认为更坚固的农作物的同时,种植了他们认为更易受气候影响的作物,例如块茎旱金莲(一种不一样的土豆)。
饮食变化对人类健康和进化有何后果? 虽然气候变化与饮食之间的联系在文献中可能已经很好地确立,但饮食变化转化为健康后果并帮助产生新的表型性状的机制仍需要阐明。
饮食摄入可能通过影响寄主的表观遗传学和寄主的微生物组而影响健康。 此外,宿主与微生物组之间存在复杂的表观遗传关系,表观遗传现象可能会影响进化。
在此基础上,德国科学家Francesco Catania和Jan Baedke提出了一个模型,该模型考虑了表观遗传机制与人类微生物组之间的相互作用以及它们与宿主遗传学的串扰。 并且认为,将人类概念化为灵活的生态系统或整体生物可能有助于揭示与饮食有关的变化对人类健康的某些后果,并提供对人类后代的预测历史特征和适应能力。
在该模型中,气候变化引起的饮食变化会通过营养不良增加健康风险。 同时,这些过程促使微生物组重新配置,从而触发跨代对环境变化的适应性反应。 作为实现这些短期和长期影响的关键途径,我们确定了微生物组-表观基因组-免疫轴。
饮食、人类健康和表观遗传学饮食对人类健康有重大影响。不同的饮食和饮食模式可以调节个体间的变异性(例如,增加心血管疾病的风险)。饮食有助于出生体重,这是婴儿和儿童健康以及成人疾病易感性的主要预测因素。饮食也会影响寿命和繁殖。例如‘饮食限制’,在不饥饿的情况下减少食物摄入量可以上调应激反应和先天免疫的相关途径,下调不同物种间生长和繁殖的相关途径。
饮食也可以有长期的影响,代际后果。由于营养不良(即营养过剩或营养不足)导致免疫状态受损的父母的后代本身也经常受到免疫系统的干扰。
此外,为了应对父母饮食能量的增加,后代表现出较高的促炎性和抑制的抗炎性。然而,在某些条件下,饮食的改变也可能对后代的健康产生相反的积极影响。在一个例子中,一组瑞典北部人的孙子孙女经历了几代人的作物短缺器,他们的死亡率和心血管疾病的风险较低,与前几代人相比,这些研究中的一些暗示了饮食对人类健康影响的表观遗传机制。
近年来,微生物组群已成为环境因素(如营养模式)与人类健康之间的一个新中介。人类微生物群包括定植于人类皮肤、胎盘、子宫、精液、肺、唾液、口腔粘膜、结膜,尤其是胃肠道的所有细菌、古细菌、真菌、原生生物和病毒。如果我们将人类视为一个完整的生态系统及其微生物群。那么,微生物群提供了大量的生态系统服务。通过生态系统服务,刺激免疫系统并塑造其发育,有助于神经系统的发育和功能,微量营养素的合成,营养素的消化和吸收,以及能量调节。
这些生态系统服务的可靠性取决于宿主与其微生物组之间的有效串扰,而这种串扰受饮食模式的影响。因此,根据最近的建议,气候变化引起的饮食或生活方式的改变也可能影响宿主的健康状况。
事实上,人类健康状况的变化肠道微生物群(例如,由于在食品生产中越来越多地使用抗生素)与各种疾病的发生和发展有关,如自身免疫疾病,癌症,代谢紊乱和精神障碍,如抑郁症,通过所谓的“肠-脑轴”。然而,这种关系的分子基础以及表型与微生物组的组成和丰度之间联系的范围和强度仍然不清楚。
已经提出了微生物群建立、稳定性和跨代传播的多种进化和生态模型。人类宿主与其驻留微生物群之间的广泛串扰表明它们已经共同进化或可能已经共同适应,即人类微生物群已经与宿主共同进化,适应人类生态系统并发展互利。与此观点一致,在人类生态系统中只有一小部分已知细菌菌群存在 。整体全息图关联也可能反映了一种单方面的适应过程(例如,宿主已适应其微生物,但微生物未适应其宿主)。
但是如何实现微生物组与人类宿主之间的共同进化或单方面的适应或共同适应?
这个问题对于理解饮食对人类健康和进化的影响至关重要。共进化意味着微生物组必须世代遗传和/或忠实获得。但是这是怎么做到的呢?
微生物群的构型在生命的早期就受到很大影响。在成年人中,微生物群保持一定程度的灵活性,这种灵活性可能在一定程度上受到环境条件和因素的调节,如宿主的遗传背景、行为和生活方式、性别和年龄。因此,微生物组分的跨代复发至少在一定程度上依赖于生命早期展开的生物过程。
在人类中,母体环境似乎有助于婴儿肠道的细菌定植。然而,这种母体贡献是部分的,因此不足以保证类似于遗传信息的可靠传播。稳定的(环境遗传的)文化和行为模式可以弥补宿主-微生物群关系的跨代稳定性不足。 例如,稳定的喂养方式,分娩方式和卫生习惯中的文化模式会导致婴儿期相似的微生物连续定居。 与其他动物一样,社会互动和网络可能会打开微生物传播的渠道。
最后,宿主遗传学也可能在微生物组(尤其是肠道)的获取,维持和稳定中发挥重要作用。 基因决定的免疫特性,连同微生物细胞之间的竞争,可能有助于调节和维持给定生态位空间中微生物种群的适当组成和水平。总之,垂直和水平传播以及宿主基因有助于微生物组的跨代稳定性。
宿主表观基因组对微生物定殖的贡献与将表观遗传变化(如DNA甲基化、组蛋白修饰、非编码RNA的调控)与微生物群定殖和功能性联系起来的研究一致。这也与研究改变微生物群表观遗传修饰的报告一致与疾病的发生有关。
宿主表观基因组既可以形成,也可以由获得的微生物群形成。获得的微生物群可以影响宿主的食欲、摄食行为和食物选择,进而影响基因调节和调节免疫反应。
因此,建议微生物群不仅可以通过各种模式和渠道稳定地传播和获取。 它也可以重塑宿主表观基因组。 这种表观遗传重塑会影响后代的发育,从而有效地将当前的环境变化(如饮食变化)与宿主的发育,代谢和免疫过程的未来变化联系起来。
表观和微生物组重塑的这种相互过程可能会在各个世代之间展开,直到整个全功能系统达到平衡状态(宿主-微生物组匹配)为止。

全功能生物的微生物群-表观基因组-免疫轴。(1) 气候变化改变了人类的饮食。(2) 与气候变化相关的饮食变化会影响微生物组(例如,影响微生物多样性、成分或代谢物)、宿主的表观基因组(例如,通过摄入改变基因表达的化合物)和宿主免疫系统(例如,营养不足可延迟对病原体的免疫反应)。(3) 在人体内,微生物群、宿主表观基因组和宿主免疫系统之间的相互作用是广泛而动态的。宿主DNA和环境会影响这种串扰(例如,表观遗传因素标志着微生物定殖的宿主生态位,这一过程也由免疫系统决定)。饮食介导的效应可以使微生物组、表观基因组和免疫系统之间的关系失去平衡,并在世代内和世代间对全息生物特性产生影响。(4) 同时,人类全息生物通过潜在的下游反馈积极地改变其环境(例如,通过文化生态位构建)
扩展上述命题,假设微生物物种的优势和复发及其在人类中的功能部分反映了可以跨代传递生态信息的保守的表观遗传调控过程。这些表观遗传过程可直接(例如,通过生态位特异性基因调控)和/或间接(例如,通过免疫系统的可塑性选择性)指导宿主的微生物定殖。
进一步假设“微生物组表观基因组免疫轴”对与气候变化相关的饮食、饮食习惯和压力相关的营养变化敏感。充分的饮食改变改变了宿主的表观基因组,进而改变了宿主的微生物组。同时,它们影响微生物的定殖过程,并通过这一过程影响宿主的表观基因组。这些动态变化在宿主的基因组成和非基因成分之间产生了不同程度的不匹配,从而产生了两个关键效应和权衡。
首先,全功能体内的生态系统服务可能被破坏,有害表型可能出现。这与通常提出的异常肠道菌群组成与肥胖症、炎症性肠病、和其他疾病之间的联系一致。然而,改变常驻或定植微生物群,如抗生素使用、饮食变化和人为影响也可能产生非疾病表型。
先前的研究表明,一方面,完整生物环境(即导致微生物组分和代谢能力失衡的环境)与生命早期先天免疫系统活性降低以及生命后期适应性免疫系统(AIS)的过度刺激有关。AIS过度刺激可能会增强免疫失火,从而促进自身免疫疾病的发生。
另一方面,AIS过度刺激可能会增强全功能体生物的防御机制,这种机制通常会随着年龄的增长而恶化,从而延长生物的寿命。AIS过度刺激导致的增强的免疫失火预计会降低妊娠至足月的几率。例如,自身免疫性疾病系统性红斑狼疮增加了妊娠丢失、临产前分娩和胎盘功能不全的风险。因此,在增加疾病的风险,饮食相关的改变也可能延长寿命,降低人类的生育成功率。
其次,宿主的基因组成与其微生物组之间的不匹配可能会产生生物内部的压力,这可能会对生物的组成部分施加不同程度的压力。这种压力类似于在宿主-病原体相互作用中或在本地物种和入侵物种之间的生物入侵过程中出现的压力。例如,宿主-微生物组错配可能会触发不期望的免疫反应,从而削弱宿主,同时在本地(即共同进化的)微生物组分受到破坏后,为定植创造生态位空间。因此,这种不匹配代表了获得适应性变化的第一步。
要理解这一进化过程,首先必须注意的是,寄主的基因组成和微生物群之间的不匹配可以由后代遗传。更具体地说,完整生物的表观基因组可能通过表观遗传修饰的直接传递或表观遗传标记的从头诱导从母亲传给胎儿。这些反复出现的表观遗传模式有利于后代完整生物中亲本微生物组群的重新配置。这意味着患有自身免疫性疾病的个体的后代可能通过非基因遗传表现出相同的特征。与此相一致的是,具有适度遗传力和共同环境风险因素的自身免疫性疾病通常会跨代复发。该模型的一个含义是,人类父母和后代寿命之间的正相关部分反映了跨代表观遗传,与先前的观察和其他物种的发现一致。
与上一代相比,根据宿主基因(其中一半遗传自母亲,另一半遗传自父亲)和环境(可能相对恒定或世代不同)之间的不匹配程度,后代完整生物反复出现的生理或免疫反应可能减弱或加剧。
在一个基本不变的环境中,表型性状的代际转换可能是有方向的,并趋向于局部最优。或者,当环境不断变化时,代际转换可能是波动和不规则的。最简单的情况是,代际环境基本不变,遗传变异性很小,全生命体内的压力有助于恢复生命史特征之间的失调前关系(即,生殖成功率的增加以寿命为代价)。
更一般地说,我们假设,人口的平均适合度有一种自然趋势,即在健康平衡中断时反弹(下图)。类似的动态也与“辅助基因流”有关,通过这种做法,合适的基因型被重新定位,以帮助当地人口跟上气候变化的步伐。这种非当地基因型的重新定位可能会导致繁殖抑制,从而降低人口相对于亲本人口的平均适应度。然而,这种健康状况的下降通常是暂时的。当地人口在几代之内恢复,可能获得比引进移民基因型之前更高的平均适应值。

整体人的健康和适应饮食变化的适应能力。 饮食变化会触发宿主微生物组失配,从而扰乱整个节肢动物(例如,在宿主体内触发不良的免疫反应),同时促进其适应性反应(例如,通过为新型微生物定殖创造适当的空间)。 箭头描绘了由于气候变化引起的饮食转变而引发的人类全生命周期的进化轨迹。 在第一阶段(红色箭头)描述了与饮食变化相关的整体全息内应力。 在这里,整体人面临着疾病的易感性,例如过敏性和自身免疫性疾病,这些疾病可能是表观遗传的。 在这种不稳定的健康状态下,整体类人承受着促进获得微生物,遗传和非遗传变异的压力,最终使整体类人达到了新的平衡(绿色箭头)
影响表型状态遗传的表观遗传机制有助于在新环境中建立最佳适应度。除此之外,还可能发生重新建立宿主-微生物和谐关系的基因变化。免疫、生殖和寿命相关的宿主基因是上述许多可塑性的基础。因此,这些基因的变化有助于重建生态失调前的宿主-微生物平衡。事实上,这些基因在人类适应中起着重要作用,是最常见的候选选择目标之一。
遗传变化也可能在全功能体生物的微生物成分中积累。应激诱导的突变提高了微生物的突变率,加速了有益基因变体的出现。此外,微生物组对应激(暴露于有毒物质)的快速适应和抗性细菌的选择可能使宿主的后代产生更高的毒性耐受性。这可能通过不同的微生物传播途径遗传。最后,微生物组对新饮食的适应可能解释了人类食用海藻的原因,]奶牛的草食进化,吸血蝙蝠的嗜血性。例如,有人认为,应对吸血蝙蝠新饮食所需的形态、免疫和生理适应的很大一部分并不是由于宿主的基因组适应,而是由普通蝙蝠功能核心微生物群中基因的正选择驱动的。因此,由于微生物组的适应性变化,全功能体生物可能进化出适应营养挑战的解决方案。宿主基因组的适应性变化可能伴随也可能不伴随微生物组的重组。
总而言之,与饮食相关的生物体内应激可能会促进宿主和微生物群的遗传和非遗传变化,这些变化可能会传染给下一代。虽然这些变化可能会增加疾病易感性,但也可能有助于重新调整宿主和微生物群的利益。宿主与其微生物群之间重新建立的和谐串扰(如果实现)表明了全息生物在新环境中的最佳综合性能。
除了对全基因组方法的扩展之外,该模型还提供了“健康”或“有利”微生物组的操作定义,即微生物联合体,不仅与宿主的生活方式、社会文化和环境环境相协调,而且在很大程度上,以及它的遗传背景。这些联盟不需要由固定物种组成。由于相当多的代谢冗余,具有相同功能的基因分布在许多细菌物种中。这使得一个“健康”的肠道微生物组群能够以多种不同的方式组合起来,并允许在宿主世代间丢失或重新发现微生物类群。这一定义有助于实施“精确”肠道微生物组群调节的最佳治疗策略。
微生物组通常被认为是许多慢性疾病(如自身免疫疾病)治疗干预的有效靶点,前提是改变的微生物组与某种疾病有因果联系,目前减轻与局部疾病相关的健康挑战的医疗干预形式有可能减轻或抑制生物体内的压力。
在该模型支持下,意味着医疗干预也延迟了潜在适应性变异的出现。基于医学干预放松了对疾病相关基因变异的自然选择这一观点,先前也得出了质量上类似的结论。最后,如果模型是正确的,目前未能解释表观遗传微生物背景和代际影响阻碍了我们对许多疾病的理解。通常与癌症等风险增加相关的特定基因变异,只有在与相关(微)环境因素相结合的情况下才可能是重要的。
到目前为止,很少有生物学家认为发育中有机体的微生物群是有偏见和快速适应性变异的潜在来源,尤其是在人类中。
该研究提出的大视野,跨时空观点需要理解全功能生物体既是一个免疫个体,也是一个进化个体。
为了研究这两者之间的复杂纠缠。首先需要掌握人类微生物变异的限度和范围,了解在环境变化面前(肠道)微生物群落的重组是如何广泛和迅速的。
第二个挑战是了解人类群体之间和内部肠道微生物变异性的差异是否会导致对气候变化相关营养挑战的不同健康和进化反应。
最后,为患者量身定制的医疗应用程序的开发不应将人类个体视为与气候变化相关的健康干预措施的唯一目标。相反,全功能生物体的集体应被视为潜在患者。它复杂的内部关系、串扰和权衡需要成为人们关注的焦点。
相关阅读:
参考文献:
Tomova, A., Bukovsky, I., Rembert, E., Yonas, W., Alwarith, J., Barnard, N. D., & Kahleova, H. (2019). The effects of vegetarian and vegan diets on gut microbiota. Frontiers Nutrition 6, 47
Meldrum, G., Mijatović, D., Rojas, W., Flores, J., Pinto, M., Mamani, G., Condori, E., Hilaquita, D., Gruberg, H., & Padulosi, S. (2018). Climate change and crop diversity: Farmers’ perceptions and adaptation on the Bolivian Altiplano. Environment, Development and Sustainability, 20(2), 703– 730.
Lynch, J. B., & Hsiao, E. Y. (2019). Microbiomes as sources of emergent host phenotypes. Science, 365(6460), 1405– 1409.
Catania Francesco,Baedke Jan,Fábregas-Tejeda Alejandro et al. Global climate change, diet, and the complex relationship between human host and microbiome: Towards an integrated picture.[J] .Bioessays, 2021.
Deutsch, C. A., Tewksbury, J. J., Tigchelaar, M., Battisti, D. S., Merrill, S. C., Huey, R. B., & Naylor, R. L. (2018). Increase in crop losses to insect pests in a warming climate. Science, 361(6405), 916– 919.