-

 国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子
国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子

 二级病原微生物安全实验室
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业

谷禾健康
下一代益生菌
传统益生菌在维持肠道稳定性方面具有一定优势,但其作用效果受宿主遗传背景、菌株生物学特性以及个体肠道微生态环境等因素的制约,这也使得其在临床应用中呈现出多样化的效果,且在调控全身性疾病方面仍需进一步探索其潜在机制。
与传统益生菌不同,下一代益生菌(NGPs)采用基因工程和下一代测序技术来识别具有特定功能特性的细菌菌株。一些研究者认为,下一代益生菌可能在微生物治疗领域具有发展前景,为某些健康问题的管理提供新的思路。
下一代益生菌(NGPs)通过筛选具备特定功能的菌株,不仅在肠道内发挥作用,还通过其他途径影响全身的免疫系统、代谢过程和神经调控机制。相较于传统益生菌,NGPs在以下方面具有显著优势:
①多系统靶向性——不仅修复肠道屏障,还可通过肠脑轴、口腔、皮肤、呼吸道等通道调控局部与全身免疫反应;
②精准性与个体化潜力——结合基因组学与代谢组学,可实现更精准的宿主-微生物互动干预;
③作用机制的扩展性——不仅抑制病原体、抗炎,还能通过代谢产物和免疫调控网络实现更广泛的治疗效应。
当前,NGPs的主要种类包括但不限于:
嗜黏蛋白阿克曼菌(Akkermansia muciniphila)、普拉梭菌(Faecalibacterium prausnitzii)、丁酸梭菌(Clostridium butyricum)、Eubacterium hallii及若干经基因改造或筛选的菌株,它们在慢性炎症、胃肠道疾病、代谢综合征、皮肤健康、口腔健康、免疫相关疾病以及精神疾病等多种疾病的治疗与缓解中展现出潜在价值。
随着基因组学、代谢组学、表观遗传学及循证临床研究的迅速发展,新型益生菌有望成为个体化干预的新载体,将疾病管理从“治疗反应”转向“精准预测与预防”。
什么是益生菌?
“益生菌(probiotics)”一词源自希腊语/拉丁词根
oro(意为’为了’)与bios(意为’生命”),这主要体现了这些生物体在确保生命健康方面的作用。
益生菌是具有活性的微生物,主要是细菌和酵母,在给予足够剂量时具有健康益处, 益生菌已被证明能改善消化健康、免疫系统功能,甚至心理健康。
有效益生菌微生物的特征
doi: 10.1007/s11033-024-09398-5.
◮ 益生菌发挥效果的条件
同时益生菌要发挥效果,必须:
-忍受胃的酸性环境;
-活着抵达肠道;
-附着在肠道细胞上以发挥其益处;
-与有害细菌竞争;
-以及在肠道内定植到一定程度。
益生菌的发展历史
益生菌的概念源远流长,我们在这里简要梳理益生菌的起源及其随时间的演变。以下为历代发展要点的回顾。
◮ 古代起源
中东和中亚:两千多年来,发酵乳制品已被证明是一个值得关注的问题。中东和中亚被视为大约1万年前处于原始发酵状态的文明的摇篮。
亚洲:在古代中国和日本,也有纳豆和味噌等富含益生菌的食物,分别是发酵大豆和大豆酱。
◮ 19世纪及20世纪初:科学发现
益生菌一词直到19世纪末才进入科学界。随着微生物学的兴起,巴斯德揭示了发酵由微生物所驱动,从而解释了为何某些菌类存在是为了发酵。
1907年,梅契尼科夫被誉为“益生菌之父”,指出保加利亚农民的长寿与牛奶发酵有关,乳酸菌能阻止细菌损害宿主肠道,防止自毒与衰老。1908年,梅契尼科夫因在免疫学领域的研究荣获诺贝尔奖,进一步提升了公众对乳酸菌的认知。
◮ 20世纪中叶:益生菌产品的发展
在20世纪40–50年代,仿制食品成为益生菌引入的关键因素。自1935年起,日本率先开展商业益生菌饮料“养乐多”的生产;1939年,通过干酪乳杆菌(Lactobacillus casei)展示了益生菌在胃肠治疗中的潜力。
随后,研究扩展至其他菌株,甚至酵母菌也被探索,其中Saccharomyces boulardii已用于腹泻治疗。
◮ 现代科学中的益生菌
益生菌研究广泛开展,尤其关注其在医学上的潜在益处。自1970年代初起,益生菌的活性逐步增强,医学与微生物学的结合揭示了其积极影响的机制。此间,益生菌不再仅是传统发酵食品的成分,而是逐渐被视为可用于治疗特定健康问题的药物。
▸ 益生菌如何与微生物群相互作用
益生菌的作用机制错综复杂,涵盖众多对消化系统有益并增强免疫力的作用过程:
益生菌的作用机制
编辑
doi: 10.1007/s12602-025-10606-2.
1) 病原体的竞争排斥
益生菌通过竞争排斥来抑制病原微生物的生长:利用可用资源并争夺肠道受体位点,限制入侵生物的扩张。
此外,某些益生菌还能分泌抑制物(如细菌素),抑制致病菌增殖,从而提升整个细菌群落的健康水平。
2) 免疫反应的调节
益生菌还能通过诱导调节性T细胞来调控免疫反应,这些T细胞维持免疫平衡,避免对无害物产生过度反应。因此,益生菌在炎症并发症如炎症性肠病和自身免疫性过敏的管理中具有关键作用。
它们还支持健康的免疫反应,帮助抵抗感染、在必要时抑制过度炎症,并整体促进免疫健康。
3) 强化肠道屏障
益生菌的重要性在于提升黏膜质量与细胞间紧密连接的完整性,从而预防肠漏综合征。肠漏指肠壁的病理生理改变,导致有毒物质进入循环并引发炎症。Faecalibacterium prausnitzii和Akkermansia muciniphila这两种菌株被公认有助于维持肠道屏障功能,前者对黏膜覆盖也有保护作用。
4) 有益代谢产物的生成
益生菌能合成重要代谢产物,如短链脂肪酸(SCFAs)—丁酸、乙酸、丙酸等。证据显示SCFAs具有抗炎作用,促进肠道稳态、保护细胞完整性并缓解炎症,滋养结肠上皮,帮助维持健康的肠道黏膜。此外,SCFAs在调节与健康微生物群相关的免疫与代谢方面也发挥重要作用。
益生菌的作用机制
编辑
doi: 10.1007/s12602-025-10582-7.
5) 对肠-脑轴的影响
近期研究显示,益生菌通过肠脑轴影响心理状态的潜力。某些菌株能产生神经递质,如血清素与GABA,调节情绪与压力水平。
瑞士乳杆菌和长双歧杆菌已被证实可缓解焦虑、改善情绪,从而将健康微生物群与心理健康联系起来。
随着对人类微生物群落的深入了解,益生菌及其产品开发的研究日益广泛。借助现代微生物学工具,如PCR的16S rRNA检测、下一代测序(NGS)和生物信息学,我们能够更精准地识别和检测肠道细菌菌株。
最近,基因组测序与培养技术的进步促使从人类微生物组中分离出多种新型微生物,显示出良好的健康益处和益生菌潜力,因而有望发展成为下一代益生菌(NGPs)。
下一代益生菌(NGPs)的定义
下一代益生菌(NGPs)是指基于宏基因组学或16S rRNA测序等比较微生物组研究,遴选或工程改造的特定菌株。这些菌株在适当摄入量下,通过调节宿主肠道微生物群落结构或功能,对特定疾病状态的改善具有潜在的生物学效应和科学证据支持。与传统益生菌相比,NGPs针对性更强,而且有的已经累积了更多相关临床研究数据。
这类非传统益生菌包括嗜黏蛋白阿克曼菌(Akkermansia muciniphila)和普拉梭菌(Faecalibacterium prausnitzii)等,它们可改善肠道健康、降低炎症并增强肠道屏障。
下表总结了传统益生菌与下一代益生菌(NGPs)之间的主要区别。
▸ 对下一代益生菌(NGPs)的需求
益生菌的修复与预防作用已广泛被认可和应用,不仅有助于消化健康,也提升免疫与整体健康。传统定义多聚焦于含发酵菌株的产品,如乳杆菌和双歧杆菌,在大量应用中发挥作用。然而,随着人类微生物组的日益复杂与深入了解,经典益生菌未必能满足所有健康需求,因此出现了新型益生菌(NGPs),旨在更精准地解决更多健康问题。
下一代益生菌可能的治疗作用
编辑
doi: 10.1007/s11033-024-09398-5.
1) 针对复杂疾病
全球慢性疾病如肥胖、糖尿病、自身免疫性疾病和精神健康障碍的患病率持续上升,促使科学界逐步揭示微生物组在这些疾病中的作用。这为非处方的下一代益生菌(NGPs)提供了更有针对性的治疗潜力。
例如,Akkermansia muciniphila与健康个体的优良新陈代谢活性相关,并有助于维持黏膜屏障;而屏障的降解与代谢性疾病密切相关。Faecalibacterium prausnitzii 被视为抗炎菌株,突显了其在炎症性肠病(IBD)治疗中的潜力。
2) 个性化医疗与精准健康
下一代益生菌(NGPs)正与日益增长的个性化医疗需求融合:根据患者的基因、代谢或微生物组特征来治疗个体疾病。
与此同时,益生菌研究持续推进,致力于深入理解肠道微生物组,以实现精准、定制的益生菌处方。
3) 通过调节微生物群落提高疗效
下一代益生菌(NGPs)更接近于益生元,研究显示,一些NGPs能释放代谢产物,调控群落结构与活性,长期有助于健康。例如,它们可产生短链脂肪酸(SCFAs)如丁酸,具有显著的抗炎作用,对肠道环境至关重要。相比之下,传统益生菌尽管可提高SCFA水平,但由于功能特性,其贡献可能不及NGPs。
4) 心理健康与肠-脑轴
近期肠-脑轴研究显示,肠道微生物组的状态显著影响心理健康、情绪、应激反应与高级认知功能。传统益生菌在此领域的作用有限,新型益生菌则具更大治疗潜力。
例如,脆弱拟杆菌与植物乳杆菌可能影响脑功能。新型益生菌能合成多种功能性代谢产物,用于调节脑活动,治疗焦虑、抑郁及其他精神障碍等问题。
5) 肠道屏障完整性与免疫调节
对于慢性疾病与自身免疫过程,肠道病变引发的免疫过度激活会使有毒物质从肠道扩散入血。新型益生菌,如嗜粘蛋白阿克曼菌,通过增强肠道黏液层提升屏障防御,进而减轻全身炎症。
此外,NGPs 还可调控免疫细胞功能与促炎/抗炎反应的平衡,可能有助于治疗免疫相关疾病。
6) 在癌症治疗中的潜在应用
下一代益生菌(NGPs)在癌症管理领域的应用备受关注。研究表明,某些微生物组变体可增强免疫疗法的效果。通过使用特定菌株,可提升宿主免疫系统对肿瘤细胞的杀伤力,从而为癌症治疗中的益生菌辅助治疗提供新的可能性。
▸ 下一代益生菌的标准
与传统益生菌相比,下一代益生菌(NGPs)能更有效、更精确地解决复杂的健康问题。考虑到功能性、特异性和安全性,已提出了一些标准来界定新型益生菌。关键标准如下:
1) 菌株特异性和选择
下一代益生菌(NGPs)通常源自非传统菌株,相较于乳杆菌和双歧杆菌等传统益生菌,它们在特定健康领域具更大潜力。每种菌株的选择都以实现精确的健康目标为导向,如代谢、免疫或认知健康,以提升针对性治疗效果。
2) 作用理解
就NGPs而言,核心在于菌株对免疫反应的调控、益生代谢物的产生、肠道通透性的改变,以及与宿主微生物组的相互作用机制。多数NGP已被筛选或改造以产生代谢物(如SCFA、神经活性物质)并执行支持其健康益处的功能。
3) 临床疗效和针对性健康益处
应通过一致的前临床与临床研究证明NGPs在健康状况下的病情调节功效。在某些情境下,可能表现为提升胰岛素敏感性以改善代谢健康,或通过调控炎症来治疗自身免疫性疾病。这种功效使NGPs从一般肠道健康支持,扩展到糖尿病、炎症性肠病和精神疾病等特定疾病的治疗。
4) 生存能力和定植潜力
要让NGPs菌株成功定植,关键在于其能耐受胃肠道恶劣环境,并在可能的情况下实现内腔幼体的稳定定植,以确保长期的遗留效应。其在宿主微生物组中的建设性相互作用以及在胃酸环境中的存活能力,对疗效至关重要。
5) 安全性和最小不良反应
安全性备受关注,部分原因在于许多新型益生菌来自非传统来源,且人类使用的历史证据稀少。因此需进行严格的综合安全评估,包括毒性和长期安全性研究。应尽量减少并密切监测可能引发不良反应的机制,如免疫紊乱或对自然微生物群的破坏。
6) 个性化应用的潜力
随着个体微生物组的多样性,开发下一代益生菌(NGPs)时已将个性化纳入考量。这意味着选取或设计符合特定微生物组特征或个人健康状况的菌株。
个性化的NGP应用通过识别微生物失衡与健康风险,提供更定制化的干预,因此产生了在健康益处上更具针对性的NGP,成为个性化与精准医疗领域的宝贵资产。
小结
传统益生菌和新型益生菌之间的主要区别
编辑
总得来说,新一代益生菌(NGPs)相对于传统益生菌的科学、具体优势主要有以下几点:
•更广的靶向健康作用谱;
•更强的肠道微生态调控能力;
•更精准的作用机制与靶点;
•先进的制备与设计策略;
•改善药品监管与可控性前景
•增强个体化干预潜力;
•安全性与耐受性改进方向。
传统益生菌和新型益生菌的特征比较
编辑
doi: 10.1007/s12602-025-10582-7.
下一代益生菌(NGPs)中具有潜力的候选株正在逐步浮出水面,其中一些已在代谢调控、炎症抑制和跨系统健康影响方面显示出特别的潜力。如Akkermansia muciniphila、Faecalibacterium prausnitzii、Clostridium butyricum、Christensenella minuta等,初步研究显示它们在代谢调控、炎症抑制和肠-全身轴的潜在影响方面具有积极信号。
但这类证据尚处于早期阶段,且个体差异、长期安全性、给药策略与潜在不良反应尚需进一步验证。此外,NGP 需要经过严格的毒理学评估、长期随访与可重复的生产质量控制,才能避免潜在风险并确保可追溯性。因此,在扩展NGP候选池时,必须遵循安全优先、证据驱动、标准化与伦理法规合规的原则,强调并非所有物种都具备成为下一代益生菌的潜力,也不应过早推广到临床应用。
Akkermansia muciniphila
极具潜力
嗜黏蛋白阿克曼菌(Akkermansia muciniphila,Akk)是一种定居肠道黏膜的共生菌,作为下一代益生菌的有力候选。它在增强代谢与免疫反应方面发挥关键作用,并可能改善癌症治疗效果。
◮ 能利用黏蛋白,促进肠道屏障完整性
A. muciniphila能够利用肠道黏膜层的黏蛋白与糖蛋白作为唯一的氮碳来源,促进肠道屏障完整性并附着于黏膜,是一种有益的益生菌特征。
肠道黏膜中该菌含量下降会削弱屏障,增加毒素侵袭风险。它不仅参与葡萄糖、脂质和蛋白质代谢,还影响黏膜层的完整性与黏膜免疫反应,因此对肠道健康具有支持作用。
◮ 在肥胖、炎症性肠病、自闭症中都发挥作用
近年研究表明A.muciniphila在机体稳态与疾病中具有重要作用。其丰度与多种疾病相关,水平下降与肥胖、炎症性肠病、自闭症和2型糖尿病等相关。
它通过调节神经系统控制葡萄糖代谢,帮助抵御肥胖与糖尿病。研究还发现经巴氏杀菌的A.muciniphila可显著延长结肠长度、增强对肥胖与胰岛素抵抗的抵抗力,且炎症性肠病与代谢疾病患者常见其水平降低,提示潜在抗炎作用。此外,A.muciniphila还能增强某些抗癌治疗在动物模型中的效果。
总体而言,益生菌领域的最新证据支持A.muciniphila作为可行治疗靶点,其水平变化可作为疾病进展的潜在生物标志物。因此,A.muciniphila在癌症、代谢综合征、炎症性肠病及免疫相关疾病等领域具有广阔的临床潜力。
肠道重要菌属——Akkermansia Muciniphila,它如何保护肠道健康
Faecalibacterium prausnitzii
具有较高潜力
普拉梭菌(Faecalibacterium prausnitzii)是一种极具前景的下一代益生菌。
F.prausnitzii在肠道中的下降与微生物群失调及多种代谢疾病和慢性免疫介导疾病(如炎症性疾病和肥胖)相关,并常作为衡量年轻肥胖者是否患溃疡性结肠炎的生物标志物,这可能源于其促进黏蛋白与紧密连接蛋白合成、修复受损肠黏膜的能力。此外,它还能调控粘液分泌、肠道杯状细胞分化和糖基化,并维持黏液屏障完整性,因此被视为有益的益生菌,对肠道具有关键保护作用,其枯竭会削弱肠道免疫调控与抗炎能力。
◮ 重要的产丁酸菌、具有抗炎活性
F.prausnitzii被认定为肠道中最重要的丁酸生产者之一。丁酸是肠道上皮细胞的主要能量来源,调节肠道T细胞的活性,抑制病原体入侵,促进结肠癌细胞凋亡,预防肠道炎症,调节免疫系统,并帮助代谢综合征恢复。
此外,F. prausnitzii还会产生来自微生物抗炎分子(MAM)的肽,抑制宿主激活NF-κB通路。这些活性代谢物的优点包括抗炎活性、维持肠道屏障功能、肠道免疫稳态以及在结直肠癌细胞中诱导凋亡。
◮ 可作为癌症免疫治疗中的调节因子
目前,从健康人群中分离出的F. prausnitzii已被证明具有体外抗炎和免疫调节活性。此外,Faecalibacterium属候选物种已被研究为癌症免疫治疗中的调节因子。研究人员发现肠道中Faecalibacterium的浓度与癌症患者的存活率之间存在关联。
此外,科学家们发现,黑色素瘤转移期间,调控T细胞数量和促炎细胞因子IL-2、IL-8和IL-6的血液水平与肠道中Faecalibacterium的数量呈负相关。因此,F. prausnitzii被认为是黑色素瘤患者的关键治疗靶点和预后标志物。
肠道核心菌属——普拉梭菌(Faecalibacterium Prausnitzii),预防炎症的下一代益生菌
Clostridium butyricum
具有较高潜力
丁酸梭菌(Clostridium butyricum)是一种嗜氧、形成孢子的革兰氏阳性细菌,因其发酵非碳水化合物并高产丁酸而得名。
◮ 对肠道健康、免疫调节有益,甚至改善抑郁
丁酸梭菌,被认为对炎症性肠病有益,并起到免疫调节作用。它已被用于肠道健康支持、免疫调节和感染预防。丁酸梭菌还对肠上皮增殖和维持结肠健康至关重要。
有研究报道了丁酸梭菌的抗癌潜力,观察到其可显著抑制小鼠肠道肿瘤形成、减少肠道癌细胞增殖并诱导凋亡。此外,丁酸梭菌的益生菌株与抗抑郁药联合使用,可显著改善抑郁症状。
Christensenella minuta
具有较高潜力
Christensenella minuta已成为下一代益生菌(NGPs)候选者中的重要一员,显示出在代谢调控与炎症抑制方面的潜力。
◮ 改善能量平衡、降低炎症水平
C. minuta在肠道微环境中能够促进有益代谢产物如乙酸和丁酸的产生,改善能量平衡与脂质代谢,同时通过调节炎性介质和免疫途径,降低慢性炎症的水平。这些作用不仅局限于肠道,还通过肠-全身轴、代谢网络和免疫调控网络对全身健康产生潜在影响。
C.minuta可能通过以下途径发挥作用:增强肠道屏障功能、改变宿主基因表达与信号传导、促进短链脂肪酸等有益代谢产物的产生、以及调节脂肪组织与肝脏的炎症反应。
◮ 临床层面
研究表明,Christensenella minuta在2型糖尿病和肥胖等代谢紊乱以及炎症性肠病中的丰度显著下降。其相对丰度与低 BMI 指数相关的瘦表型呈正相关。
除此之外,支气管哮喘和过敏性疾病、肾结石、情感障碍、甲状腺癌、粘膜类天疱疮、多囊卵巢综合征和复发性口疮性口炎等疾病中C. minuta的丰度也较低。
总体而言,C. minuta 的证据虽仍处于初步阶段,但其在代谢健康与炎症控制方面展现的多维潜力使其成为有希望的下一代益生菌候选株。未来需要更多高质量的人体研究来确认其安全性、最优给药策略、长期效果及与其他干预的协同作用,以便将这一候选菌株尽快转化为可用于临床的干预工具。
Christensenella minuta——下一代益生菌候选者:改善代谢、减轻炎症
Eubacterium hallii
有潜力
Eubacterium hallii是另一种潜在的益生菌生物治疗制剂(NGP)。能利用多种碳源(包括糖和有机酸)并产生两种关键的短链脂肪酸(SCFAs):丙酸和丁酸。SCFAs对肠道健康至关重要,促进黏液生成、刺激肠上皮细胞增殖分化并维持上皮健康;SCFA不足会引发炎症。
注:据报道,E.hallii是新生儿肠道中丁酸的主要来源之一。
◮ 能与肠道其他细菌协作产生短链脂肪酸
某些E.hallii能发酵复杂碳水化合物,但也有依赖其他肠道微生物产物的情况。多项研究证实了这种交叉摄食机制在Eubacterium属SCFA生成中的重要性。
相关研究多在含复杂碳水化合物的培养基中将Eubacterium属与双歧杆菌共培养,能降解复杂碳水化合物的双歧杆菌株可产生1,2-丙二醇、乙酸酯与乳酸盐,随后被Eubacterium属吸收并转化为丁酸与丙酸。
这一交叉摄食模式明确揭示了肠道微生物与复杂碳水化合物的丁酸化协同及其在肠道生态系统中的作用。
◮ 可能改善肥胖和糖尿病
研究了E.hallii在肥胖和糖尿病中的作用。发现E.hallii能够代谢丁酸以激活G蛋白联结受体信号通路,改善GLP1和GLP2的产生,强化肠道屏障功能,同时不影响体重或食物摄入,并提升胰岛素敏感性和能量代谢率。因此,它在胰岛素敏感性方面可能安全且有效。
◮ 在体内对一些物质进行关键转化
最新研究表明,Eubacterium属在肠道中进行关键的代谢转化,对人体健康有积极影响。在这些效果中,有毒化合物解毒为更良性的形态似乎具有价值。
例如,观察到E.hallii将极为丰富的食品来源杂环芳香胺致癌物——2-氨基-1-甲基-6-苯基咪达唑(4,5-b)吡啶(PhIP)转化为生物学上无法获得的7-羟基-5-甲基-3-苯基-6,7,8,9-四氢吡啶(3′,2′:4,5 咪唑(1,2-α)嘧啶-5-m氯化物(PhIP-M1)。此外,PhIP在模拟远端和近端结肠微生物群存在下被E.hallii转化,导致其丰度分别增加了120倍和300倍。这表明它作为一种保护性治疗具有极佳的视角。
此外,在同一研究中,检测到了E.hallii的抗菌活性。作者证明,E.hallii能够将甘油分解为3-羟基丙烯醛(3-HPA),这种甘油以水溶液中的榘蛋白形式存在。
研究还发现Eubacterium属通过调节胆汁酸代谢谱,有助于肠道和肝脏健康。近年来,胆汁酸代谢或肠道微生物群的调节正被研究为肝细胞癌(HCC)和结直肠癌(CRC)的创新治疗策略。
Roseburia
有潜力
罗氏菌属(Roseburia)是一种革兰氏阳性、厌氧、弯曲杆状细菌。Roseburia物种能使用复杂多糖,产生短链脂肪酸(丙酸盐、丁酸盐、醋酸盐)。它对炎症、帕金森病和炎症性肠病(IBD)有益,并且被充分考虑为下一代益生菌(NGPs)。
◮ 降低动脉粥样硬化、改善肠道、肝脏健康
令人瞩目的是,最新研究显示,食用Roseburia和富含纤维的饮食的小鼠,其动脉粥样硬化的发生率降低了。这归因于高纤维饮食,介导了Roseburia丁酸的形成以减少动脉粥样硬化。
此外,最新研究显示 Roseburia intestinalis(细菌鞭毛的关键结构成分)在治疗酒精性脂肪肝和溃疡性结肠炎方面具有潜力。口服 Roseburia 肠源性鞭毛蛋白在酒精性脂肪肝模型中显著改善了肠道上皮完整性并抑制了肠道损伤风险。然而,关于 Roseburia 的临床证据仍然有限,需要进一步研究以明确其在多种人类疾病中的应用价值。
Bacteroides fragilis
有潜力,但同时存在安全隐患
脆弱拟杆菌(Bacteroides fragilis)是机会性病原体,然而,近期研究证实了非致毒性脆弱拟杆菌的益生菌特性。脆弱拟杆菌可以刺激宿主适应性免疫,抑制炎症,激活免疫系统的成熟,调节肠道微生物群,并通过多糖A(PsA)及该NGP的其他外膜囊泡维持肠道健康和稳态。
◮ 抑制其他致病微生物
先前研究显示,脆弱拟杆菌通过抑制其他致病微生物的生长与定居来抑制它们。在动物模型中,该菌能抑制艰难梭菌感染,且治疗后肠道微生物多样性提升、AKK菌丰度增加。
研究还表明,B.fragilis通过抑制凋亡、维持ZO-1和MUC-2完整性,从而提高肠道屏障功能,降低艰难梭菌的黏附性和定植性。因此,脆弱拟杆菌有助于维护肠道屏障的完整性。
其他研究还发现脆弱拟杆菌对沙门氏菌的易位具有竞争性抑制作用,机制包括抗菌蛋白 BSAP-1 的生成。BSAP-1 含有 MACPF 结构域,能裂解侵染宿主细胞的细菌。除了 BSAP-1 外,泛素样蛋白(BfUbb)及 VI 型分泌系统(T6SSs)等因素可能也在该竞争中发挥重要作用。
◮ 产生的荚膜多糖具有免疫调节作用
最新研究显示脆弱拟杆菌在结肠内代谢多种碳水化合物,生成八种荚膜多糖,其中多糖A(PSA)是一种独特的两性多糖,具免疫调节作用。
PSA被抗原呈递细胞内吞并处理后被T细胞识别,持续增强宿主免疫。PSA通过调节树突状细胞,促使初始T细胞分化为Treg,提升Foxp3和CD39表达,抑制IL-17并诱导IL-10产生,帮助治疗肠道炎症性疾病。
近期研究还报道脆弱拟杆菌通过增强免疫功能、抑制脂多糖信号、改善肠道微生物群活性并维持肠道屏障稳态来预防肠漏,对癌症患者有潜在益处。
SK08活细菌粉末是一种以脆弱拟杆菌为基础的活体生物药,已获中国食品药品监督管理局认可,目前处于临床试验阶段,属于治疗性生物药范畴。
Prevotella copri
有潜力
P. copri是拟杆菌门中的另一种新型益生菌,据报道可改善葡萄糖耐量和肝糖原水平。它被视为2型糖尿病、肥胖等代谢疾病的潜在靶点。
◮ 微生物组学基础
P. copri在健康人群和特定疾病患者(如类风湿关节炎、2型糖尿病)的肠道微生物群落中存在明显差异,已通过宏基因组学研究识别。
◮ 功能特性研究
•能产生短链脂肪酸(SCFAs),特别是丙酸;
•参与多糖代谢,具有特定的代谢功能;
•与肠屏障完整性和免疫调节相关;
◮ 临床相关性
•在类风湿关节炎(RA)中丰度降低,补充可能改善症状;
•与代谢相关疾病的关联有一定研究基础。
广泛存在于人群的双面使者——Prevotella copri与疾病和健康
Parabacteroides goldsteinii
有潜力
研究表明,P.goldsteinii也有望成为下一代益生菌。中山大学的程芳/陈红波教授团队探讨了肠道共生细菌P.goldsteinii衍生的外膜囊泡(OMVs)通过传递关键的抗炎分子(例如十五烷酸)来调节宿主的免疫系统。这些OMVs能够在炎症皮肤区域积聚,有效抑制IL-23/Th17轴,改善全身免疫稳态,从而显著缓解银屑病症状。
编辑
Su D,et al.J Control Release.2025
P.goldsteinii已被证实能够显著改善肠道和肺部炎症。强烈推荐用于肥胖改善。此外,P.goldsteinii也表现出显著的抗炎和胰岛素刺激特性。
Propionibacterium freudenreichii
有争议
费氏丙酸杆菌(Propionibacterium freudenreichii)主要用于乳制品行业,作为生产瑞士奶酪的食品级微生物。研究者正探索其特定的益生菌特性以实现新的健康益处。该菌已能产生丙酸,具有抗菌活性并可降低胆固醇。
另一项研究则显示,P.freudenreichii 对肠道屏障完整性与炎症状态的影响提示其在胃肠道健康方面的潜在应用。
工程菌株
具有较高潜力
工程菌株是尖端的下一代益生菌代表。经过改造的乳酸乳球菌和大肠杆菌等菌株通过产生外源蛋白或分子,可用于治疗结直肠癌、炎症性肠病甚至精神健康问题等疾病。
注:益生菌还通过CRISPR技术进行基因改造,这扩大了接种疫苗的治疗选择,并增强了免疫反应。
编辑
doi: 10.1007/s12602-025-10606-2.
小结
无论来自天然还是合成来源,这些新一代菌株都代表益生菌的未来前沿。特定有益代谢物如丁酸、抗菌肽和免疫调节化合物可作为复杂疾病的治疗干预,提供传统益生菌难以实现的疾病特异性与个性化策略。因此,研究将聚焦新一代益生菌的关键点,推动医学在预防与治疗上的新前景。
下一代益生菌(NGP)在治疗领域的前景日益广阔,标志着其不再局限于传统的肠道健康维护,而是在疾病预防、诊断与治疗中发挥更为精准和多样的作用。通过基因改造或天然/合成菌株的多样化开发,NGP 能在人体内定向表达有益代谢物、抗微生物肽以及免疫调节因子,为难治性炎症、代谢疾病、免疫相关疾病等提供新的干预路径。
临床应用将持续扩展,覆盖免疫、神经、泌尿生殖、心脏、代谢、呼吸、皮肤、口腔及体重管理等领域。下面是当前新型益生菌已涉足的方向:
下一代益生菌在各种健康疾病中的应用
编辑
doi: 10.1007/s12602-025-10606-2.
胃肠道健康
下一代益生菌可能聚焦于已被证实对特定胃肠道问题有益的单一菌株,而非传统益生菌组合的多菌株混合,从而实现更个性化、针对性的治疗。
随着DNA测序技术的发展,下一代测序能够对肠道微生物群进行更为细致的分析,帮助理解胃肠道疾病患者的微生物组结构,并识别潜在的治疗重点菌群或菌株。
•炎症性肠病(IBD):包括Akkermansia muciniphila和Faecalibacterium prausnitzii,两者都与减轻肠道炎症和促进粘膜修复有关。抗炎分解代谢物(如SCFA)的合成有助于调节与IBD相关的炎症,包括克罗恩病和溃疡性结肠炎。
•肠易激综合征(IBS):某些益生菌可以缓解腹胀、疼痛和排便异常等症状。其中一些包括婴儿双歧杆菌和丁酸梭菌(Clostridium butyricum),它们可以对抗炎症并提高肠道内的健康运动能力,在这种情况下,它们在治疗IBS相关疾病方面非常有用。
•抗生素相关性腹泻:非致病性细菌大肠杆菌Nissle 1917通过引入有益细菌来缓解抗生素相关性腹泻和其他胃肠道问题,从而恢复肠道健康。
代谢健康
代谢综合征(MS)是一组特征性疾病,包含胰岛素抵抗、血脂异常、肝肾功能异常、高血压、血糖稳态紊乱等。如今,益生菌正被视为潜在的代谢疾病生物治疗药物,作为具有健康促进作用的功能性食品成分,并具备对抗特定疾病的潜力。
小鼠研究显示,A. muciniphila 与体重、脂肪量变化相关,新型益生菌菌株能够有效调节葡萄糖、脂质和体重的平衡。其改变必需代谢物谱,包括短链脂肪酸、维生素、 多种脂肪酸、氨基酸及胆汁酸代谢物等。
•肥胖和体重管理:肠道菌群的一些调节剂会影响促进脂肪储存、脂肪燃烧和饱腹感的因素。如前所述,使用Akkermansia muciniphila已被证明可以改善胰岛素反应,缓解脂肪堆积,从而治疗肥胖。
•2型糖尿病:新型益生菌可能有助于血糖控制,因为它们会产生SCFA,SCFA已被证明可以降低血糖水平并增强胰岛素敏感性。两种有益的微生物,Akkermansia muciniphila和鼠李糖乳杆菌,抑制胰岛素抵抗,已被推荐用于控制2型糖尿病。
免疫系统调节
健康的微生物群通过迷走神经直接或间接调节免疫系统。由于肠道微态的稳定性,益生菌可调节免疫环境,有助于自身免疫疾病的治疗。
•自身免疫调节:如脆弱拟杆菌通过产生多糖A调节促炎与抗炎反应,从而为治疗类风湿性关节炎和多发性硬化症等自身免疫疾病提供干预,目标是调节免疫与减轻自身免疫。
•过敏与哮喘:部分新型益生菌能调节免疫耐受,降低过敏反应。鼠李糖乳杆菌在给药后表现出抗过敏、抗哮喘特性,透过增强调节性T细胞并降低过度免疫反应来发挥作用。
•感染:部分新型益生菌具抗感染作用,如丁酸梭菌具有抗菌特性,能够增强对感染的免疫防御,但需在宿主体补偿机制恢复后方能实现。
心理健康
•抑郁与焦虑:仍有被称为“心理益生菌”(psychobiotics)的新型益生菌(NGP)与情绪和认知改善相关”。瑞士乳杆菌和长双歧杆菌已显示抗焦虑与抗抑郁效应,可能通过产生血清素和GABA等神经递质、以及降低全身炎症来实现。
•压力管理:与GBA(脑-肠-轴)相互作用的NGP 能通过维持微生物群平衡、调节 HPA 轴来减轻生理压力对身心的影响,关联情绪与压力相关的精神状态。
新型益生菌的治疗益处
编辑
doi: 10.1007/s12602-025-10582-7.
皮肤健康
健康的皮肤微生物群通过调节免疫细胞与炎症维持体内平衡。湿度、pH、温度、脂质、营养、运动、药物、手术以及身心压力等变量共同影响皮肤微生物组的日常波动。
表皮屏障缺陷与免疫失调与特应性皮炎(AD)密切相关,且具有遗传易感性和环境影响的个体更易发病,体现了 AD 的复杂病理生理。最新研究显示局部治疗可改善 AD 患者的微生物群,帮助清除有害菌、促进有益菌生长,从而平衡微生物群。
•湿疹与皮炎:益生菌已逐步用于炎症性皮肤病的治疗。鼠李糖乳杆菌与长双歧杆菌能改善屏障功能、降低炎症,可能对湿疹与皮炎患者有益。
•痤疮与牛皮癣:部分新型益生菌具抑炎特性、可抑制免疫活动,有助于治疗痤疮及银屑病等疾病;有助于控炎、保护屏障功能,对皮肤健康有益,减少炎症发作。
口腔健康
根据既有文献,口腔内约有700多种细菌构成口腔微生物群。免疫抑制、激素治疗及不良饮食等因素可引发牙釉质侵蚀与牙周疾病。
益生菌作为一种新兴且具成本效益的微生物疗法,能抑制口腔病原体并降低粘附,延长假体装置的使用寿命。
•牙龈疾病与龋齿:最新口腔研究显示,某些益生菌通过重新定植口腔菌群来预防口腔疾病,抑制有害生物。例如,唾液链球菌与罗伊氏乳杆菌通过抗炎、维持口腔菌群平衡及抑制致病生物膜,对牙龈疾病与龋齿具有积极作用。
变形链球菌在龋齿形成中扮演重要角色,能产生胞外多糖并在酸性条件下存活。研究了鼠李糖乳杆菌GG与发酵乳杆菌KU200060对变形链球菌KCTC 5316的最小抑菌浓度(MIC),抗菌效果分别为25%与12.5%。唾液链球菌DB-B5为从健康成年女性龈上牙菌斑分离的菌株,具备产生新型细菌素等益生菌特性。
•口臭:部分益生菌能降低挥发性硫化合物,这是口臭的主要成因。通过平衡口腔微生物群,可维持口腔健康环境,减少口臭发生。
呼吸健康
•上呼吸道感染:鼠李糖乳杆菌GG及其他新型益生菌在预防肺部感染方面已显示有效。它们可通过调节免疫反应来增强上皮黏膜对呼吸道病原体的防御,具备免疫抑制潜力。这些益生菌已用于预防普通感冒与流感等疾病。
•哮喘管理:一些NGP可通过降低全身炎症、促进免疫耐受来缓解哮喘,并帮助调节在寒冷天气下的异常免疫反应,通常用于慢性呼吸道疾病的治疗。
肝脏健康
•非酒精性脂肪肝(NAFLD):新型益生菌(NGPs)通过调整肠道脂质代谢相关微生物群,促进肝脏健康,抑制脂肪积聚。像Akkermansia muciniphila等菌株能减少肝脏氧化应激与炎症的证据正在积累,但仍需更多临床试验验证。
•肝脏排毒与健康维护:某些菌株通过改善肠道环境、降低进入肝脏的微生物内毒素,提升肝脏排毒能力,从而增强肝功能并降低全身炎症。
癌症预防与治疗
•癌症预防:特异性NGPs可通过产生抗氧化与抗炎应激因子来抑制癌症诱导效应,具有抗癌潜力。研究显示,干酪乳杆菌通过调节肠道菌群和免疫监测,减少结直肠癌风险。
•癌症治疗支持:NGPs可减轻化疗相关副作用(如黏膜炎)并支持免疫功能。据报道,益生菌通过增强免疫反应与改善肠道健康,抑制肿瘤细胞增殖,或作为标准肿瘤治疗的辅助手段。
新型益生菌(NGPs)已从单纯的肠道守卫者发展为多系统、多靶点的免疫与代谢调控工具。基于对肠道微生物组及其代谢产物的深入理解,NGPs显示出在慢性炎症、代谢疾病、免疫性疾病、皮肤与神经系统疾病等领域的广阔应用前景。它们不仅能够改善局部屏障功能与炎症状态,还可能通过肠脑轴、肠-皮肤轴等通路实现对远端器官的调控,从而带来更全面、个体化的健康管理方案。
尽管前景光明,NGPs的临床转化仍面临若干挑战:菌株的稳定性与可控性、个体微生环境差异、长期安全性评估、以及标准化的制备、给药剂量与疗效评估体系尚需完善。
未来发展的关键方向包括:
总之,NGPs具备成为下一代个体化健康管理核心元素的潜力。通过持续的基础研究与循证临床探索,未来的益生菌干预有望实现更高的治疗精准度、更广的适应证和更持久的健康收益。
主要参考文献
Gupta MK, Srivastava R. Gut Microbiome Interventions: From Dysbiosis to Next-Generation Probiotics (NGPs) for Disease Management. Probiotics Antimicrob Proteins. 2025 Aug;17(4):2629-2652.
Vijayaganapathi A, Mohanasrinivasan V. A Review of Next-Generation Probiotics-As a Gateway to Biotherapeutics. Probiotics Antimicrob Proteins. 2025 Aug;17(4):1985-1997.
Al-Fakhrany OM, Elekhnawy E. Next-generation probiotics: the upcoming biotherapeutics. Mol Biol Rep. 2024 Apr 15;51(1):505.
Maftei NM, Raileanu CR, Balta AA, Ambrose L, Boev M, Marin DB, Lisa EL. The Potential Impact of Probiotics on Human Health: An Update on Their Health-Promoting Properties. Microorganisms. 2024 Jan 23;12(2):234.
Santacroce L, Charitos IA, Bottalico L. A successful history: probiotics and their potential as antimicrobials. Expert Rev Anti Infect Ther. 2019 Aug;17(8):635-645.
Meng N, Liu Q, Dong Q, Gu J, Yang Y. Effects of probiotics on preventing caries in preschool children: a systematic review and meta-analysis. J Clin Pediatr Dent. 2023 Mar;47(2):85-100.
Evivie SE, Huo GC, Igene JO, Bian X. Some current applications, limitations and future perspectives of lactic acid bacteria as probiotics. Food Nutr Res. 2017 May 3;61(1):1318034.
Haghshenas B, Kiani A, Mansoori S, Mohammadi-Noori E, Nami Y. Probiotic properties and antimicrobial evaluation of silymarin-enriched Lactobacillus bacteria isolated from traditional curd. Sci Rep. 2023 Jul 5;13(1):10916.
Silva DR, Sardi JdCO, de Souza Pitangui N, Roque SM, da Silva ACB, Rosalen PL (2020) Probiotics as an alternative antimicrobial therapy: current reality and future directions. J Funct Foods 73:104080.
Mejía-Caballero A, Salas-Villagrán VA, Jiménez-Serna A, Farrés A (2021) Challenges in the production and use of probiotics as therapeuticals in cancer treatment or prevention. J Ind Microbiol Biotechnol 48(9–10):kuab052.
谷禾健康

你敢相信吗?从粪便中提取出具有强烈粪臭味的物质,当用酒精稀释上千倍后,脱胎换骨变成了一种香味。这就是一种吲哚衍生物——3-甲基吲哚(又名粪臭素)
吲哚,是所有花香类原精的关键成分,这种物质在低剂量1-3%浓度时,具有放松的作用,协同苯基酯,能强效抗痉挛,增添感官愉悦和魅力。然而,如果简单将吲哚等同于香水,未免太少看它的功效。
在谷禾特定肠道菌群检测报告中炎症部分或毒素部分有吲哚这个指标。
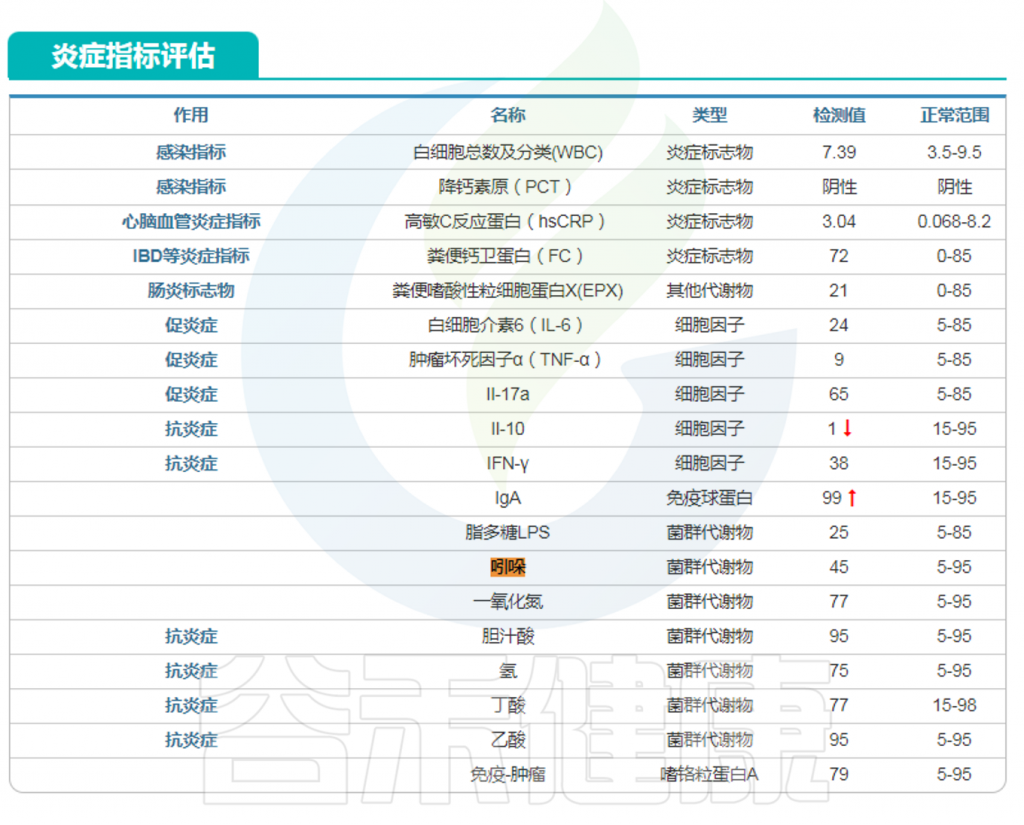
<来源:谷禾肠道菌群检测数据库>

<来源:谷禾肠道菌群检测数据库>
很多人可能对与菌群和健康相关的血清素,色氨酸,脂多糖,组胺等指标比较熟悉,但是对吲哚还是比较陌生,实际上吲哚与肠道微生物群和人类健康和生活关系密切,例如,吲哚衍生物可以影响胃肠道的蠕动,可能会影响便秘或腹泻等状况;吲哚及其衍生物已被证明可以加强肠道屏障功能,有助于防止病原体和有害物质进入血液。
事实上,由吲哚衍生而来的各种化学物质,对人体神经调节和抗病原微生物,有极为积极的作用。在人体中存在着许多这样神奇的吲哚类物质。内源性吲哚及其衍生物是源自肠道微生物群的色氨酸代谢物,具有一系列生物活性。
最近的研究表明,吲哚有助于维持人体肠道的生物屏障,主要通过激活AhR和PXR受体来发挥抗炎活性,从而影响免疫系统的功能,显著改善肠道健康(炎症性肠病、出血性结肠炎、癌症结直肠癌),并进一步促进人类健康(糖尿病、中枢系统炎症和血管调节)。
作为细胞间信号分子,吲哚还控制细菌生理学的各个方面,例如产吲哚细菌的孢子形成、质粒稳定性、耐药性、生物膜形成和毒力。相比之下,许多不产生吲哚的细菌、植物和动物产生多种加氧酶,可能会干扰吲哚信号传导。
吲哚还是身体神经系统与内分泌系统正常与否的风向标。所谓的体臭,其实就是人体内吲哚含量增高的表现。吲哚在细菌生理学、生态平衡以及人体健康中发挥着重要作用。
然而,吲哚的毒性影响也不容忽视。过量的吲哚会对情绪和行为产生不利影响,硫酸吲哚酚是一种吲哚衍生物,具有肾毒性和心血管毒性。羟吲哚还可能降低神经元兴奋性。
吲哚与肠道微生物群之间的相互作用以及吲哚对人类健康的影响是一把双刃剑。本文主要讲述了内源性吲哚的生物合成、吲哚对肠道功能及微生物群的影响,为吲哚与疾病的相关性提供了科学依据。
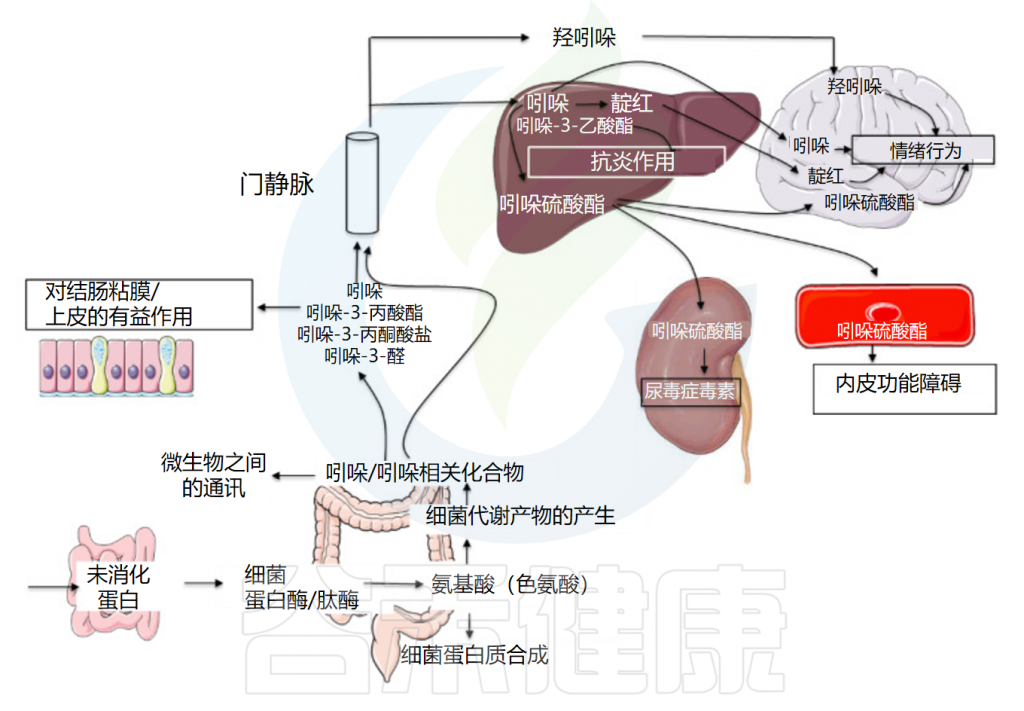
Tennoune N,et al., Microorganisms.2022
吲哚(Indole)是一种有机化合物,是吡咯与苯并联的化合物,又称苯并吡咯。吲哚及其同系物和衍生物广泛存在于自然界,主要存在于天然花油,如茉莉花、苦橙花、水仙花、香罗兰等。
而内源性吲哚及其衍生物是来源于肠道菌群的色氨酸代谢物,具有一系列生物活性。内源性吲哚类物质包括吲哚本身以及吲哚的代谢产物,如吲哚-3-甲酸、吲哚-3-丙酸、硫酸吲哚酚等。
吲哚相关化合物的结构
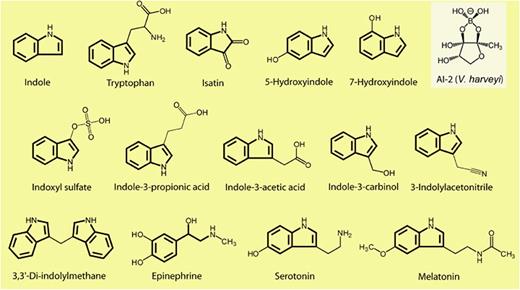
Lee JH, Lee J.FEMS Microbiol Rev.2010
肠道菌群产生吲哚类物质
细菌蛋白酶和肽酶从可用的膳食中释放氨基酸,包括色氨酸。而色氨酸是吲哚的主要前体。大部分色氨酸在小肠中被吸收,约4%~6%的色氨酸在结肠中被肠道菌群分解代谢为吲哚。
色氨酸作为一种必需氨基酸,主要从富含蛋白质的食物中获取,是唯一含有吲哚的氨基酸。
肠道中的色氨酸通过三个主要途径进行代谢:1) 犬尿氨酸途径(摄入的色氨酸的95%);2)细菌色氨酸代谢(4-6%);
3) 血清素途径 (1-2%)
✦ 肠道菌群主要通过色氨酸酶将色氨酸转化为吲哚
肠道细菌主要通过色氨酸酶(TnaA)的作用将色氨酸转化为吲哚;色氨酸酶TnaA可逆地将色氨酸降解为吲哚、丙酮酸和氨。
值得注意的是,在哺乳动物中,吲哚完全来源于细菌代谢活动,因为宿主细胞不具有产生这种化合物的代谢能力。
色氨酸也可以通过肠道微生物群直接或间接转化为几种吲哚相关化合物,包括吲哚-3-丙酮酸、吲哚-3-乳酸、吲哚-3-丙酸、吲哚-3-乙酰胺、吲哚-3-丙烯酸、吲哚乙醛、吲哚-3-乙酸、3-甲基吲哚(粪臭素)等。
✦ 内源性吲哚产生的机制
大肠杆菌(E. coli)是研究最广泛的吲哚合成生物体,它可以使用多种机制(抑制、转录衰减和反馈抑制)来调节色氨酸代谢中的色氨酸操纵子(Trp operon)和tna操纵子的表达。
在色氨酸缺乏的条件下, trp操纵子的表达升高,而由TnaC(24个氨基酸前导肽,以前称为TnaL)、TnaA(色氨酸酶)和TnaB(通透酶)组成的tna操纵子的表达受到抑制,因为转录终止因子(Rho)依赖性终止发生在tna操纵子中。
结果,当色氨酸水平较低时,TnaA和TnaB的表达以及吲哚的产生受到抑制。在富含色氨酸的条件下,转录终止因子依赖性转录终止被消除,因此吲哚产量增加。
因此,细胞外色氨酸和其他氨基酸直接影响吲哚的产生。
// 拓展:渗透酶通过转运也会影响吲哚水平
此外,三种渗透酶(Mtr、TnaB和AroP)在不同环境的色氨酸转运中发挥不同的作用,因此可以直接影响吲哚的水平。
Mtr通透酶主要负责转运吲哚,而TnaB通透酶对于色氨酸的摄取至关重要。尽管色氨酸的生产成本高昂,但细胞仍然利用色氨酸途径大量生产和分泌吲哚。
由色氨酸产生的吲哚可以通过细胞膜蛋白进行转运。例如,大肠杆菌和霍乱弧菌在丰富的培养基中可以分泌高达0.6 mM的吲哚。
在大肠杆菌中,外排蛋白AcrEF部分负责吲哚的输出,因为AcrEF突变体的吲哚排泄低于其野生型菌株。Mtr通透酶主要负责输入吲哚,因为吲哚不被mtr突变体吸收。
然而,最近有人提出,吲哚由于其疏水性,可能可以直接扩散通过细胞膜。因此,有必要清楚地了解吲哚的进出口情况。
大肠杆菌中吲哚生物合成和吲哚信号传导
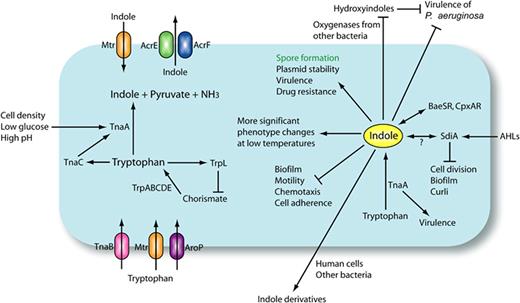
Lee JH, Lee J.FEMS Microbiol Rev.2010
人体粪便中吲哚的浓度较高,通常范围为0.25毫摩尔浓度~1.1毫摩尔浓度,但可能由于人群之间肠道微生物群组成的差异以及膳食蛋白质消耗水平不同而存在差异区别。
注:由于吲哚似乎是通过结肠上皮吸收的,结肠中吲哚的浓度可能高于粪便中记录的浓度,远端大肠的浓度高于近端部分的浓度。
尽管大多数生物体都含有色氨酸代谢途径,但只有能编码tnaA的细菌才能合成吲哚。表达色氨酸酶(tnaA)的细菌,例如大肠杆菌(E. coli)、梭菌属(Clostridium spp.)、拟杆菌属(Bacteroides spp.)和普通变形杆菌(Proteus Vulgaris),可催化色氨酸直接转化为吲哚。
✦ 梭状芽孢杆菌代谢产生吲哚-3-丙酸
已经证明吲哚-3-丙酸是由梭状芽孢杆菌(Clostridium sporogenes)产生的。粪臭素(3-甲基吲哚)是由梭菌属和拟杆菌属产生的。
此外,产孢梭菌(C. sporogene)可以将色氨酸转化为吲哚-3-丙酮酸(IPyA),并在吲哚-3-丙酮酸脱羧酶的催化下产生吲哚-3-乙醛(IAAld)。然后,IAAld 被厚壁菌门、变形菌门、拟杆菌门和放线菌门的成员脱羧,产生吲哚乙酸 (IAA)。
✦ 一些病原菌也可以产生吲哚
色氨酸代谢很复杂,许多菌株参与吲哚衍生物的制造。总的来说,数据支持迄今为止有至少85种细菌已被证明可以产生吲哚。
这些细菌包括许多病原菌,例如创伤弧菌、流感嗜血杆菌、多杀巴氏杆菌(pasteurella multocida)、四种志贺氏菌菌株、植物克雷伯菌(Klebsiella planticola)和普通变形杆菌。值得注意的是,一些革兰氏阳性菌株,包括B.alvei和粪肠球菌(E.faecalis)也产生吲哚。
✦ 一些拥有tnaA基因的细菌却无法产生吲哚
有趣的是,一些细菌,如杀鲑气单胞菌(Aeromonas salmonicida)、宋氏志贺氏菌(Shigella sonnei)、克氏耶尔森氏菌(Yersinia kristensenii)和Seudovibrio spp.,已经失去了合成吲哚的能力,尽管这些菌株的染色体上有一个tnaA基因同源物。
例如,与正常志贺氏菌菌株不同,一些在tna操纵子中具有点突变、插入或缺失的志贺氏菌不产生吲哚,这可能是由于某些适应性优势。虽然只是推测,但一些个体可能通过利用当地细菌群体的信号来避免产生吲哚的成本。
其他可以产生吲哚的细菌:
Alkaliphilus metalliredigens
Bacillus alvei
Clostridium novyi
Clostridium limosum
Clostridium tetani
Corynebacterium acnes
Desulfitobacterium hafniense
Nocardioidessp.
Oribacterium sinus
Propionibacterium acnes
Enterococcus faecalis
Symbiobacterium thermophilum
Aeromonas hydrophila
Aeromonas liquefaciens
Aeromonas punctata
Bacteroides thetaiotaomicron
Bacteroides sp.
Brachyspira hyodysenteriae
Burkholderiasp.
Chromobacterium violaceum
Chryseobacterium gleum
Citrobactersp.
Citrobacter intermedius
Citrobacter koseri
Desulfovibrio vulgaris
Edwardsiella tarda
Enterobacter liquefaciens
Escherichia coli
Escherichia fergusonii
Flavobacteria bacterium
Flavobacteriumsp.
Fusobacterium nucleatum
Fusobacterium polymorphum
Haemophilus influenzae
Haemophilus somnus
Haloarcula marismortui
Halogeometricum borinquense
Hyphomonas neptunium
Klebsiella ornithinolytica
Klebsiella planticola
Micrococcus aerogenes
Morganella morganii
Paracolobactrum coliforme
Pasturella multocida
Pasturella pneumotropica
Photobacterium harveyi
Porphyromonas asaccharolytica
Porphyromonas endodontalis
Porphyromonas gingivalis
Prevotella intermedia
Proteus inconstans
Proteus vulgaris
Providencia alcalifaciens
Providencia rettgeri
Providencia rustigianii
Providencia stuartii
Rhizobium leguminosarumbv.trifolii
Saccharomonospora viridis
Shigella boydii
Shigella dysenteriae
Shigella flexneri
Sphaerophorus varius
Stigmatella aurantiaca
Treponema denticola
Vibrio alginolyticus
Vibrio cholerae
Vibrio fluvialis
Vibrio harveyi
Vibrio hollisae
Vibrio mediterranei
Vibrio nigripulchritudo
Vibrio orientalis
Vibrio parahaemolyticus
Vibrio splendidus
Vibrio tapetis
Vibrio tubiashii
Yersinia frederiksenii
Yersinia intermedia
吲哚类物质的生成可能受到环境因素的严重影响,例如膳食成分、肠道微生物群、碳源、温度、pH值以及生理状态。
★ 肠道微生物群和饮食对吲哚生成有重要影响
吲哚类物质是肠道微生物群产生的膳食色氨酸的代谢产物。因此,肠道微生物组成以及饮食模式的变化会影响其产生。
研究发现吲哚-3-丙酸(IPA)与肠道微生物群的α多样性呈正相关,肠道微生物组的组成可以解释大约20%的IPA循环水平的变化。
高纤维和富含多酚的饮食可以提高吲哚含量
多项研究发现,吲哚-3-丙酸的循环水平与纤维摄入量相关,这可能是通过肠道微生物群的变化来解释的。
此外,富含多酚的饮食会导致肾功能正常受试者的血清IPA显著增加,但肾功能受损的受试者不会显著增加。与此相一致的是,饲喂菊粉60天的猪的盲肠和结肠中微生物组组成的β多样性显著增加,同时IPA含量也显著增加。
地中海饮食会导致吲哚-3-丙酸水平升高
不同的饮食结构也会改变吲哚的水平。例如,地中海饮食仅治疗4天后就导致吲哚-3-丙酸水平升高,而快餐(即薯条和汉堡)则引起相反的结果。
此外,正常饮食喂养的同窝小鼠的血浆IPA水平显著高于生酮饮食喂养的小鼠。一项针对117名超重成年人的研究发现,摄入炸肉会降低肠道微生物群落的丰富度,并导致IPA浓度下降。
发酵食品可能会降低吲哚含量
通过对14名健康男性进行随机交叉研究设计,探讨了发酵乳制品对人类血清代谢组的影响。与牛奶组相比,酸奶摄入组餐后血中吲哚-3-丙酸和吲哚乙酸浓度较低。
同样,C反应蛋白水平轻度升高的健康超重男性在接受抗炎膳食混合物后,饮用500mL餐后奶昔(由300mL蛋奶冻、150mL奶油干酪和50mL鲜奶油组成)。受试者在后期表现出吲哚-3-丙酸血浆浓度降低。
总之,我们推测不同的饮食结构会改变肠道微生物的组成,从而导致吲哚类物质产生的个体差异。
• 细胞群体密度影响吲哚分泌
细胞外吲哚浓度取决于细胞群体密度,其中大肠杆菌和霍乱弧菌在指数早期开始产生吲哚。浓度在固定相中达到最高水平(在富集培养基中高达0.6mM吲哚),并在固定相期间稳定维持。
• 碳源影响吲哚产量
其次,有报道称葡萄糖抑制吲哚生物合成。TnaA的分解代谢抑制得到证实,因为tnaA基因的转录在碳限制期间受到抑制。
此外,tnaAB操纵子被大肠杆菌中的分解代谢调节蛋白环AMP复合物激活。因此,当大肠杆菌数量较多且碳源减少时,会产生相对大量的吲哚。
• 温度和pH影响吲哚生物合成
温度和pH也是影响大肠杆菌吲哚生物合成的重要环境因素。温度从30°C转变到43°C会诱导大肠杆菌中tnaAB基因的表达,但大肠杆菌在44.5°C时失去吲哚生物合成的能力。
注:与37°C相比,吲哚信号传导在较低温度(30°C)下对大肠杆菌基因表达、生物膜形成和抗生素耐药性的控制作用更为显著。
此外,低pH值会抑制大肠杆菌中吲哚的产生,而TnaA是pH9.0时最容易诱导的蛋白质之一。
• 生理状态影响吲哚生成
例如年龄、性别以及个体的生理状态都可能影响内源性吲哚的产生。睡眠模式、压力水平和运动习惯等生活方式因素都可能对内源性吲哚的产生产生影响。
此外,特定的健康状况和疾病,如肠道炎症、肝脏疾病等,可能会改变内源性吲哚的生成或代谢。
• 药物的使用也会影响吲哚生成
一些药物可能影响吲哚类物质的生成。例如,一些抗生素和非甾体抗炎药物被发现可以改变肠道微生物群的组成和功能,从而影响吲哚类物质的生成。
此外,一些药物可能直接干扰色氨酸代谢途径,从而影响吲哚类物质的生成。
小结
因此,细胞群密度、碳源、温度和pH等环境条件直接会影响吲哚的浓度;而饮食、药物的使用以及生活方式可能通过影响肠道微生物群或其他器官间接影响吲哚的生成。
肠道屏障的功能与肠道健康密切相关,在健康中发挥着至关重要的作用。肠道屏障将宿主与肠腔中的微生物隔离,并限制微生物和分子从肠腔的运动。
一旦肠道屏障受损,肠道通透性增加,细菌及其产物(如内毒素)将跨膜并激活单核巨噬细胞系统,促进大量炎症因子的产生,如白细胞介素6和肿瘤坏死因子-α(TNF-α),导致慢性微炎症状态。
吲哚对于控制肠道屏障功效至关重要,包括调节炎症和免疫反应、影响上皮通透性、粘液产生和紧密连接(TJ)形成。
▼
✦ 吲哚减轻非甾体抗炎药引起的肠道炎症
研究了使用吲哚作为非甾体抗炎药肠病的治疗方法。吲哚降低粪便钙卫蛋白浓度以及吲哚美辛诱导的脾和肠系膜淋巴结中中性粒细胞的浸润。
吲哚与吲哚美辛联合使用可减少非甾体抗炎诱导的粘膜转录组变化。粪便钙卫蛋白是肠道炎症的中性粒细胞特异性生物标志物。而中性粒细胞被认为是非甾体抗炎肠病发病的关键因素,可通过释放多种炎症介质引起炎症和组织损伤。
这就说明吲哚能够减轻吲哚美辛引起的小鼠肠道炎症,对非甾体抗炎药肠病具有潜在的免疫调节作用。
此外还发现吲哚减少促炎细胞因子IL-8的产生和趋化因子TNF-α激活的核因子κB(NF-κB)的表达,并诱导抗炎细胞因子IL-10的分泌。
研究表明,共生菌可以限制鼠伤寒沙门氏菌诱导的NF-κB激活,减轻小鼠炎症。因此,吲哚可能是益生菌减少肠道炎症的信号。
✦ 细菌产生的吲哚衍生物增强免疫屏障
吲哚乙酸
卵形拟杆菌(Bacteroides ovatus)产生的吲哚乙酸(IAA)增加了CD103+ /CD11c+免疫群体的丰度,而 CD103+ /CD11c+免疫细胞对于维持肠道免疫稳态和诱导耐受性免疫反应至关重要。
此外,吲哚乙酸与树突状细胞上的芳基氢受体结合并驱动IL-22的产生。在炎症性肠病个体中,IL-22 刺激上皮再生并减少炎症,并且在肠道炎症的调节中发挥关键作用。
注:除卵形拟杆菌外,其他拟杆菌产生的吲哚也可以激活免疫细胞上的AhR,从而产生IL-22。
在脂多糖存在的情况下,肠上皮细胞中表达的Reg4通过增加乳酸菌及其代谢物吲哚乙酸的比例来维持免疫稳态,从而促进肠组织中IL-35 +B调节细胞的产生和积累。这些结果表明吲哚乙酸可以调节IL-35 的产生,并影响肠道免疫屏障。
吲哚-3-乳酸
罗伊氏乳杆菌(Lactobacillus reuteri)产生吲哚-3-乳酸(ILA),ILA是Trp的吲哚衍生物,可激活CD4+ T细胞中的AhR并下调转录因子ThpoK,使CD4 + T细胞分化为具有免疫调节功能的CD4+ CD8αα +双阳性上皮内T淋巴细胞(DPIELs)。通过这些机制,吲哚-3-乳酸可以促进肠道屏障功能并减少炎症。
生活在母乳喂养婴儿胃肠道中的婴儿长双歧杆菌亚种也可产生吲哚-3-乳酸,从而显著减弱TNF-α和脂多糖诱导的肠上皮细胞中促炎细胞因子IL-8的增加。吲哚-3-乳酸还增加AhR靶基因CYP1A1和NRF2靶向基因谷胱甘肽还原酶2、超氧化物歧化酶2和NAD(P) H脱氢酶的 mRNA 表达,这可能是母乳喂养婴儿肠道炎症的重要调节剂。
此外,另一项研究表明,婴儿双歧杆菌分泌的吲哚-3-乳酸对未成熟肠道具有抗炎作用。这些数据为生产吲哚-3-乳酸益生菌的生产和饮食建议提供了重要的见解。
吲哚-3-丙酸
吲哚-3-丙酸(IPA)下调肠上皮细胞介导的炎症细胞因子TNF-α,同时上调配体蛋白编码mRNA,从而调节肠道屏障功能,缓解肠道炎症。
吲哚-3-丙酸还通过激活AhR显著诱导肠上皮细胞上的IL-10受体配体结合亚基(IL-10R1)来调节肠道免疫稳态。
IL-10R1是抗炎细胞因子IL-10的受体,IL-10通过IL-10R1发送抗炎信号,抑制多种细胞(包括肠上皮细胞)过度释放促炎介质。这进一步说明了吲哚-3-丙酸对肠道免疫屏障的保护作用。
吲哚-3-醛
乳杆菌(Lactobacillus)产生的吲哚-3-醛(IAld)有助于AhR依赖性IL22转录,IL-22产生者抑制炎症并保护粘膜表面的免疫生理学。吲哚-3-醛可以替代益生菌,在感染或化学损伤期间保护和维持粘膜完整性。
吲哚-3-醛可用作菌群处理和肠道菌群失调期间的支持疗法。乳酸菌还通过IAld诱导的AhR刺激LPL分泌IL-22,从而激活pSTAT3通路并抑制IL-1β和TNF-α分泌,以保护粘膜免疫屏障.
吲哚对肠道免疫的影响
Ye X,et al.Front Immunol.2022
服用非甾体抗炎药时,肠道微生物组以革兰氏阴性菌为主,这种转变会导致肠道损伤。吲哚和吲哚美辛的共同给药可以维持甚至增加厚壁菌门的重要成员,特别是对肠道稳态至关重要的C. cluster XIVa和C. cluster IV。似乎它可以防止拟杆菌的任何增加,这种变化可以减少肠粘膜损伤。
吲哚可通过增殖MUC基因的表达来增加HCT-8细胞对去甲肾上腺素介导的肠出血性大肠杆菌(EHEC)沉降的抵抗力。
吲哚-3-醛(IAld)激活AhR以产生IL-22,IL-22调节粘膜反应,使混合微生物群落存活并抑制白色念珠菌(Candida albicans)定植
✦ 减轻克雷伯菌属的细胞毒性
吲哚调节多种肠道细菌的毒力因子。吲哚增强替利霉素向替利伐林的转化。两者均由克雷伯菌属(Klebsiella spp.)产生,不同之处在于替利伐林是一种细胞毒性降低的吲哚类似物,并且替利伐林与上调孕酮X受体(PXR)反应性解毒基因结合并抑制微管蛋白定向毒性。因此,吲哚以多种方式减轻克雷伯菌属的细胞毒性。
✦ 调节霍乱弧菌和大肠杆菌的毒力基因表达
肠道益生菌代谢产生的吲哚在肠腔内浓度较高,减少了肠道大肠杆菌和啮齿类柠檬酸杆菌(Citrobacter rodentium)致病基因的表达。
吲哚还用作ToxR激动剂,调节肠道内霍乱弧菌(Vibrio cholerae)的毒力基因表达和生物膜产生。吲哚、吲哚甲酸和吲哚乙酸还可下调致病性大肠杆菌菌株的毒力和志贺氏毒素的产生。
此外,吲哚和7-羟基吲哚显著改变铜绿假单胞菌(Pseudomonas aeruginosa)的毒力基因表达,从而降低毒力因子并降低集群运动性。
铜绿假单胞菌用7-羟基吲哚治疗后,胃肠道中有更好的清除率。因此,吲哚可能有助于作为针对肠出血性大肠杆菌等病原体的治疗剂。
✦ 增强肠道机械屏障
吲哚通过诱导许多涉及内皮细胞的基因的表达,包括紧密连接、粘附分子连接(AJ)、肌动蛋白细胞骨架和黏蛋白形成,在体外增强内皮细胞的屏障功能。
将人肠上皮细胞系HCT-8暴露于吲哚以测量其基因表达的变化。结果显示,高剂量的吲哚增强了肠上皮细胞HCT-8中与黏蛋白产生、粘膜屏障增强和上皮间抵抗相关的分子谱。因此,吲哚加强了机械屏障。
✦ 增强肠道粘液屏障
吲哚-3-丙酸通过增加MUC2和MUC4以及杯状细胞分泌产物(TFF3和RELMβ)来增强粘液屏障功能。
吲哚-3-丙酸还促进IL-10释放,IL-10的信号转导增强粘液屏障功能并维持上皮细胞的稳态。
细菌中的吲哚-3-丙烯酸(IA)可增强IL-10的产生和MUC基因的表达,而IL-10对于维持杯状细胞中MUC的产生至关重要。因此,吲哚-3-丙烯酸通过 IL-10间接影响粘液屏障。
吲哚通过AhR或PXR调节肠道屏障
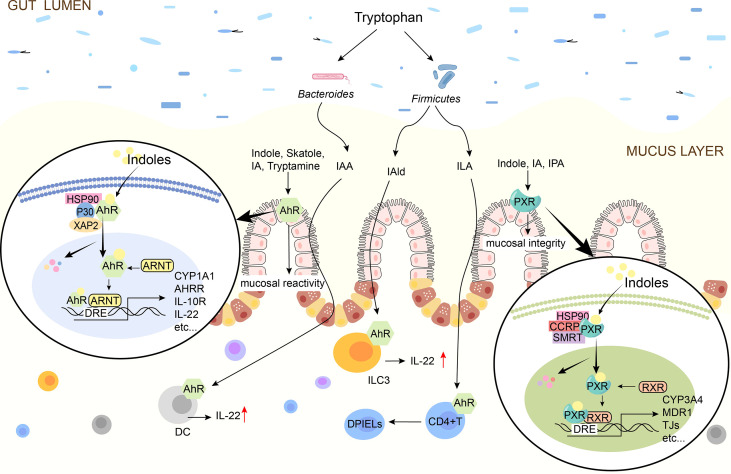
Ye X,et al.Front Immunol.2022
研究发现吲哚在细菌中发挥多种生物学作用,包括孢子形成、耐药性、毒力因子、质粒稳定性和生物膜形成等。
吲哚可以作为细菌间的信号分子
事实上,吲哚可以作为一种细菌信号,参与同一物种内以及不同物种之间的通讯。它在细菌生态系统中充当细胞间和细胞内信号。
注:吲哚还诱导了S. aurantiaca的孢子形成。
通过遗传筛选,在大肠杆菌中研究了吲哚作为细胞外信号分子的可能。结果,四个基因(astD、cysK、gabT和tnaB)被证明在稳定期通过自身产生的细胞外信号的积累而被激活。
在后续研究中,吲哚被证实是astD、gabT和tnaB激活所需的细胞外信号分子。
吲哚作为细胞间信号分子存在哪些争议?
吲哚是否是细胞间信号分子一直存在争议。已经提出了对群体感应信号分子要求的许多标准,如下所示:
1.假定的信号必须在特定阶段产生,吲哚主要在固定相中产生。
2.假定的信号必须在细胞外积累并被特定受体识别。吲哚的化学性质是众所周知的,并且在大多数吲哚调节情况下,化学互补被证明,其中吲哚在稳定期积累并且是已知的细胞外信号。该受体由AcrEF输出并由Mtr输入。
3.假定的信号必须累积并产生一致的响应,吲哚已被证明可以控制孢子和生物膜的形成。
4.重要的是,假定的信号必须引起超出信号代谢或解毒所需的生理变化的反应。吲哚已被证明可以控制毒力、生物膜和质粒稳定性,这些与吲哚无关代谢。
因此,根据这些标准,吲哚有可能成为群体感应分子。
由于上述四个标准仅适合规范的群体感应信号,例如AHL,Monds&O’Toole又添加了两个更普遍适用且具有实用价值的标准:
1.表型变化所需的生理相关信号浓度对细胞无毒。大肠杆菌和霍乱弧菌产生高达0.6mM的吲哚,对细胞无毒,并在该生理浓度下控制许多表型。
2.信号网络在社区层面是自适应的。尽管由于进化实验的困难而难以证明,但我们假设细胞间信号吲哚可能对微生物群落有益,尽管吲哚的生产对个体来说成本高昂。
例如,吲哚在产生吲哚的细菌大肠杆菌和霍乱弧菌中增加了质粒稳定性、耐药性。相反,在不产生吲哚的细菌中,吲哚通过干扰群体感应系统,降低了真菌的细胞生长和铜绿假单胞菌的毒力。因此,我们推测产生吲哚的细菌可能利用吲哚来对抗其他细菌和真核生物。
吲哚和tnaA基因也影响病原菌的毒力。色氨酸酶活性与致病性大肠杆菌有关,因为色氨酸酶活性是LEE1启动子完全激活所必需的。
吲哚会增加肠出血性大肠杆菌中毒力相关的EspA和EspB蛋白(LEE4基因产物)的分泌以及附着和消除病变的形成。
在霍乱弧菌中,吲哚和tnaA基因可能通过诱导毒力相关的分泌蛋白来增加对盘基网柄菌(Dictyostelium discoideum)的抗性。
此外,在流感嗜血杆菌分离株中,大多数血清型 (94-100%)呈吲哚阳性,而无害分离株中只有70-75%呈吲哚阳性。结果表明,吲哚的产生是必要的,但不足以对该菌株产生毒力。
值得注意的是,吲哚通过抑制毒素的产生来减轻克雷伯菌属(Klebsiella spp)的细胞毒性,还通过降低细胞运动性和聚集性来减少李斯特菌(L. monocytogenes)的相关毒力。
吲哚还通过改变基因表达来减少铜绿假单胞菌(不产生吲哚的细菌)中毒力因子的产生。
吲哚降低毒力基因的表达
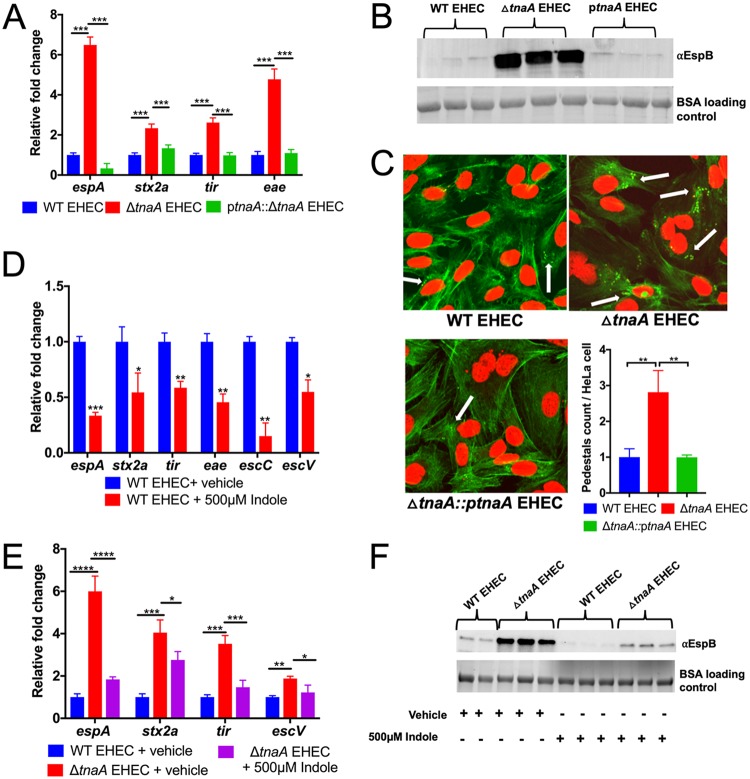
Kumar A,et al.mBio.2019
研究表明,吲哚通过在大肠杆菌中诱导输出基因(mdtEF和acrD)来增加耐药性,其中吲哚通过双组分信号转导系统(BaeSR和CpxAR)发挥作用。
这些双组分信号系统可以用作吲哚传感器。该结果证实了另一项研究,其中吲哚通过BaeSR和CpxAR诱导spy(球形体蛋白Y)基因的表达。
此外,有人认为GadX(AraC型转录因子)、Hfq(sRNA功能的全局调节因子)和RpoS(应激和固定相西格玛S)对吲哚诱导的mdtEF表达是必需的。因此,吲哚可能与多种调节因子相互作用。
增强大肠杆菌的质粒稳定性
吲哚增强大肠杆菌中的质粒稳定性。研究表明,来自大肠杆菌质粒ColE1的小非编码RNA与TnaA结合,有助于防止质粒丢失,而吲哚则延迟细胞分裂。
大肠杆菌中的吲哚还通过抑制gadABCEX、hdeABD和ymgB等耐酸基因来降低耐酸性。大肠杆菌可能会在弱碱性肠道菌群中存在吲哚的情况下关闭耐酸性基因,因为在通过酸性胃存活后不再需要耐酸性蛋白。
降低了运动性
此外,吲哚是一种化学排斥剂,会降低运动性,可能是由于细胞分裂干扰,而肾上腺素和去甲肾上腺素是一种化学引诱剂,会增加大肠杆菌O157: H7的运动性。
此外,吲哚降低细胞对上皮细胞的粘附,而肾上腺素和去甲肾上腺素增加细胞粘附。由于细菌粘附和定植到上皮细胞对于感染很重要,因此假设这也会对细菌毒力产生不同的影响。
吲哚对其他微生物的影响如下所示:

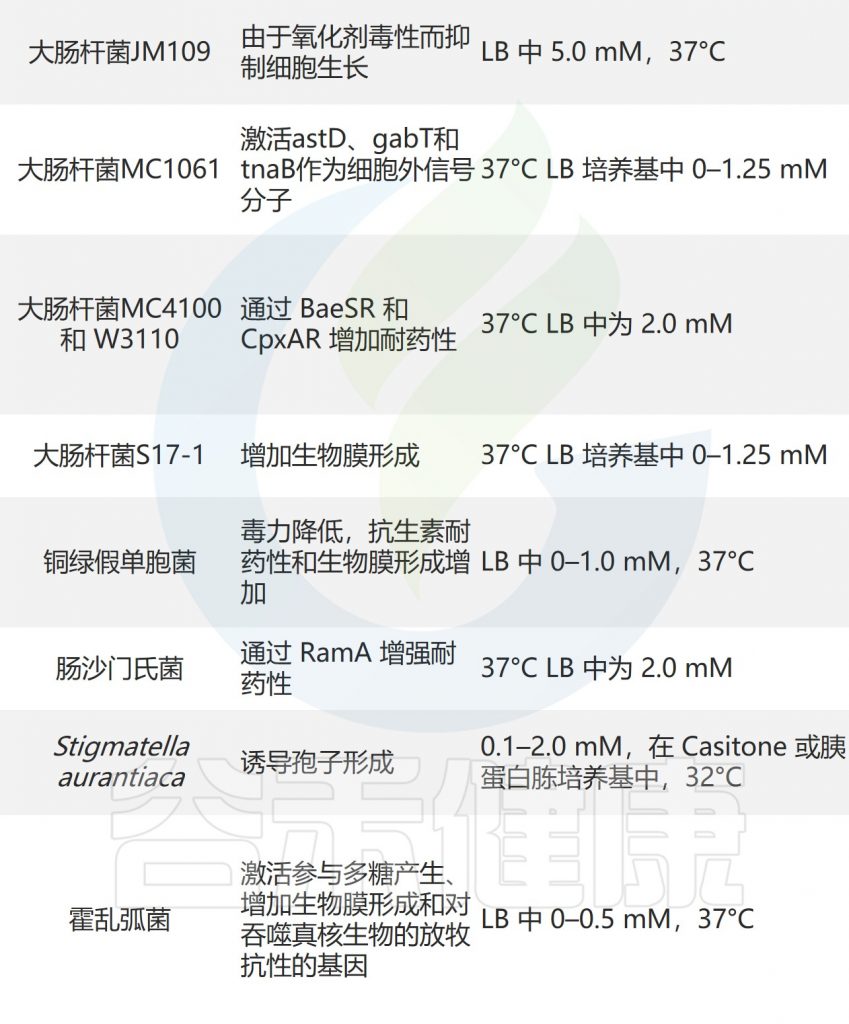
Lee JH, Lee J.FEMS Microbiol Rev.2010
吲哚类物质被肠上皮细胞吸收并扩散到血液中,从而循环到全身并影响各个系统,例如肠道及相关疾病的调节。
谷禾在这里总结了它们对炎症性肠病、出血性结肠炎、结直肠癌、糖尿病、中枢神经系统炎症和血管调节的影响。
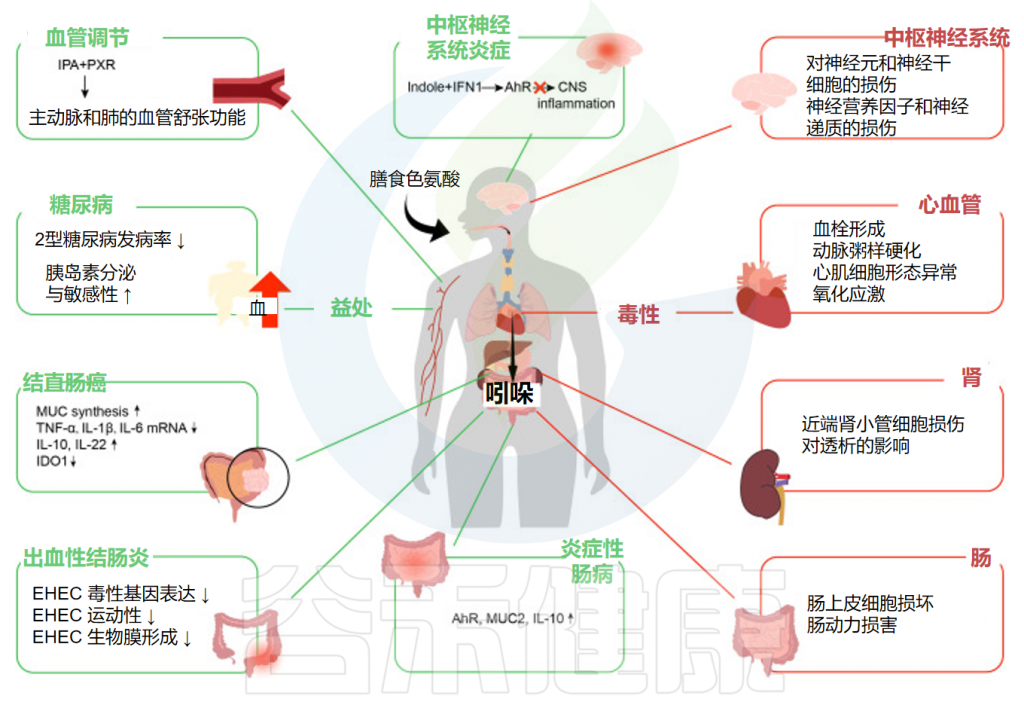
Ye X,et al.Front Immunol.2022
炎症性肠病是克罗恩病、溃疡性结肠炎等一组疾病的总称,是临床上重要的慢性胃肠道炎症性疾病。大量研究表明,吲哚对缓解炎症性肠病症状有一定的作用。
✦ 吲哚类物质增加了抗炎细胞因子
炎症性肠病与粘液层有关,炎症性肠病患者的内部粘液层较薄,MUC2糖基化较低。结果表明,吲哚-3-丙烯酸甲酯(IA)处理小鼠结肠球体可显著增加AhR激活和MUC2基因表达。
即使在脂多糖刺激的共培养物存在下,IA仍能保持其对MUC2基因表达的影响并增加IL-10的产生。杯状细胞产生MUC依赖于IL-10。因此,细菌产生的 IA 有可能增加IL-10的产生和MUC基因的表达,这可能对炎症性肠病患者有利。
吲哚对于肠出血性大肠杆菌的发病机制至关重要。出血性结肠炎是一种出血性腹泻,可导致溶血性尿毒症综合征,由人类病原体大肠杆菌O157:H7引起。
✦ 吲哚减少了肠出血性大肠杆菌的定值
根据表面相关EHEC的DNA微阵列研究,吲哚降低了EHEC中与表面定植和致病性有关的基因表达。
吲哚可以被加氧酶氧化,产生新的种间生物膜信号,从而以多种方式影响相同的表型(生物膜产生)。大肠杆菌K-12和肠道内其他共生菌产生的吲哚可能会限制大肠杆菌K-12生物膜的产生,并通过改变SdiA的表达来降低非致病性大肠杆菌的运动性,从而影响EHEC趋化性和粘附力。粘附测定证实,吲哚可在体外减少EHEC与上皮细胞的附着。
总之,吲哚通过改变肠出血性大肠杆菌毒性基因的表达、减少其运动性和生物膜形成以及减少粘附来减轻肠出血性大肠杆菌感染并改善出血性结肠炎。
根据越来越多的数据,肠道吲哚,特别是色氨酸代谢物,似乎在结直肠癌中发挥着重要作用。吲哚刺激AhR,它可以直接作用于肠道干细胞,维持MUC合成并改善肠道屏障功能。
然而,在炎症相关结直肠癌发生模型中,阻断吲哚-AhR信号通路会显著增加TNF-α、IL-1β和IL-6 mRNA水平。
此外,吲哚衍生物色胺会抑制吲哚胺2,3-双加氧酶(IDO1),该酶参与肿瘤免疫耐受。这些发现表明肠道菌群产生的吲哚可能会减缓结直肠癌的进展。
与健康个体相比,结直肠癌患者的吲哚/色氨酸比率较低,犬尿氨酸(kyn)/色氨酸比率较高。在结直肠癌患者中,犬尿氨酸和IDO1的表达增加,吲哚的产生减少。
吲哚产生的下降削弱了对结肠癌的抑制作用。总体而言,有证据表明微生物色氨酸-吲哚代谢途径的改变在结直肠癌的发病机制中发挥着重要作用。
较高的吲哚-3-丙酸血清浓度可降低患2型糖尿病的可能性,并增强胰岛素分泌和敏感性。
研究发现,与喂食对照饮食的大鼠相比,喂食吲哚-3-丙酸饮食的大鼠的空腹血糖水平显著降低。根据其他研究,吲哚可以调节小鼠结肠成纤维细胞释放胰高血糖素样肽-1(GLP-1)。
✦ 吲哚调节胰高血糖素样肽-1的释放
吲哚在短时间内增加GLP-1的释放,在较长时间内减少其分泌。这些作用结果的产生是因为吲哚可以影响L细胞的两个关键化学途径。
在某种程度上,吲哚可以抑制电压门控K+通道,增加L细胞诱导的动作电位持续时间,并导致Ca2+进入大量增加,从而快速促进GLP-1的分泌。吲哚抑制了NADH脱氢酶,随着时间的推移减缓ATP的生成并降低GLP-1的释放。
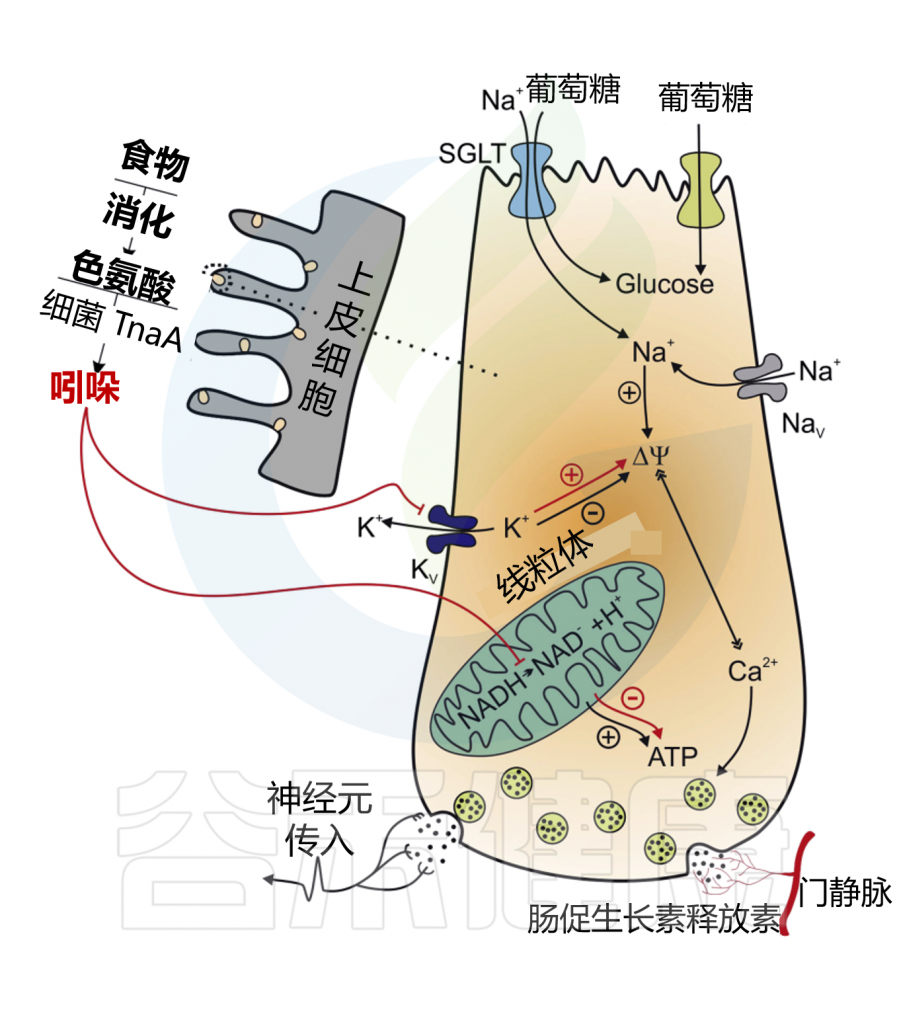
Chimerel C,et al.Cell Rep.2014
肠道内的吲哚可能还会影响食欲,同时,吲哚-3-丙酸具有强大的抵抗氧化应激的能力,这表明这种代谢物可以保护β细胞免受代谢和氧化应激相关的损伤以及淀粉样蛋白积累。因此,肠道微生物群代谢产生的吲哚-3-丙酸对2型糖尿病具有保护作用。
体外实验发现,吲哚丙酸也是嗜肺军团菌(Legionella pneumophila)生长的强效抑制剂,且抑菌效果随着吲哚丙酸浓度和暴露时间的增加而增强。因此,吲哚丙酸可以抑制细菌引起的肺部炎症。
HIV阳性患者通常更容易感染结核病,这是死亡率增加的重要因素,两种疾病的共同感染可能会因抗逆转录病毒药物和抗结核药物之间的相互作用而使治疗复杂化。
值得注意的是,吲哚丙酸可能是此类治疗中潜在的药物成分。有研究发现,接受抗逆转录病毒治疗的HIV患者体内吲哚丙酸含量显著下降,这是区分HIV感染者与健康人的重要标志物。
这种作用的机制尚不清楚,但部分原因是肠道屏障受损导致脂多糖和脂多糖结合蛋白易位,从而加剧全身炎症反应。此外,接受抗逆转录病毒治疗的HIV患者也容易出现非酒精性脂肪性肝病/非酒精性脂肪性肝炎,进一步说明了吲哚丙酸在各种疾病中的多种治疗潜力。
相比之下,吲哚丙酸是一种小分子代谢产物,具有良好的药代动力学特性,很容易被宿主吸收,充分发挥治疗作用。
未来,吲哚丙酸有望通过其作为抗结核病和抗逆转录病毒药物的补充的潜力来改善某些疾病的现有治疗选择。
吲哚丙酸可以触发孕烷X受体(PXR),一种生物量激活的核受体,存在于包括血管内皮在内的各种组织中,以调节内皮功能。
吲哚丙酸通过PXR调节主动脉和肺动脉导管中激动剂诱导的内皮依赖性松弛。这种调节主要是由于内皮一氧化氮合酶产生的一氧化氮的改变,而内皮一氧化氮合酶受到吲哚丙酸介导的PXR激活的抑制。
然而,破坏肠道微生物群并降低吲哚丙酸丰度的抗生素治疗会通过改变内皮PXR途径来改变吲哚丙酸的血管舒张作用。
而微生物代谢的吲哚丙酸补充可以提高全身吲哚丙酸水平并刺激PXR表达,从而逆转抗生素治疗引起的主动脉和肺动脉中激动剂诱导的内皮依赖性血管舒张的增强。
注:吲哚与血管的相互作用可能会显著改变传统传染病或结肠手术相关的抗生素治疗,导致微生物群落紊乱。
尽管吲哚对于改善肠道甚至全身疾病至关重要,但它们中的一些仍然会产生负面影响,例如硫酸吲哚酚(IS)、吲哚基-β-d-葡萄糖苷和吲哚乙酸。
硫酸吲哚酚是最重要的肾毒性代谢产物之一,其肾毒性已在临床研究中得到广泛证实。此外,一些吲哚类物质还会对胃肠系统、神经系统和心血管系统产生毒性。
越来越多的证据表明吲哚和吲哚化合物对大脑代谢、生理和宿主行为有影响。在拥有产生吲哚的大肠杆菌的大鼠中,吲哚的慢性过量产生已被证明会增强焦虑样行为和抑郁。
另一项研究还发现,给予常规大鼠肠道内吲哚可以激活一个称为迷走神经背侧复合体的大脑核。通过比较与非吲哚产生大肠杆菌菌株或吲哚产生大肠菌菌株单体相关的小鼠,发现肠道微生物群的长期高吲哚产生增加了慢性压力对整体情绪行为不利影响的敏感性。
✦ 可能影响儿童自闭症的发病率
在人类中,一项研究发现,患有自闭症谱系障碍的儿童在大便样本中吲哚水平较健康儿童低,而3-甲基吲哚水平升高。 这两种吲哚与患者中更常见的梭菌属细菌有关。
肠道微生物代谢活性产生的血浆吲哚升高与肝性脑病有关,这是由肝功能障碍引起的神经精神障碍,以及个体意识、行为和个性的变化。
此外,NutriNet-Santé观察性前瞻性研究的结果表明,尿液中吲哚和吲哚化合物浓度与反复发作的抑郁症状呈正相关。这种相关性提出了这样一个假设:肠道微生物过度产生这些化合物可能在人类情绪障碍的出现中起到作用。
硫酸吲哚酚由肠道菌群和宿主共同代谢。肠道菌群用酶分解食物中的色氨酸生成吲哚,然后由门静脉携带至肝脏,在细胞色素P450酶和磺基转移酶的作用下转化为硫酸吲哚酚。
硫酸吲哚酚通过上调IRF1表达、抑制动力相关蛋白1表达并干扰线粒体自噬通量来诱导肠上皮细胞(IEC)损伤。硫酸吲哚酚通过以浓度依赖的方式增加活性氧的释放,在IEC-6细胞中引起氧化应激。
硫酸吲哚酚还抑制NRF2激活,降低抗氧化防御细胞系统并抑制血红素加氧酶-1、NAD(P)H 脱氢酶和超氧化物歧化酶的表达。
在IEC-6细胞中,硫酸吲哚酚处理可显著增加TNF-α的释放、环氧合酶-2和诱导型一氧化氮合酶的产生以及硝基酪氨酸的合成,表明肠上皮细胞是硫酸吲哚酚诱导的肠道炎症的靶点。
与硫酸吲哚酚一起培养的结肠显示出收缩性降低,这表明该毒素可能对结肠平滑肌细胞产生有害影响并导致肠道蠕动受损。
微生物群代谢物硫酸吲哚酚在中枢神经系统疾病的发生中也具有致病作用。众所周知,慢性肾病和心血管疾病常常是由硫酸吲哚酚引起的。此外,肾功能不全会导致尿毒症毒素在大脑中积聚,导致中枢神经系统功能异常。
✦ 损伤神经元和神经干细胞
其次,硫酸吲哚酚可损伤神经元和神经干细胞,损害神经营养因子和神经递质,诱发氧化应激和神经炎症。例如,通过作用于中枢神经系统胶质细胞,硫酸吲哚酚促进神经炎症并表现出促炎作用。
小鼠表现出情绪障碍和神经元退化的行为证据,如焦虑、抑郁和认知功能障碍。这些相应的器质性病变伴随着这些行为变化。这也表明硫酸吲哚酚和中枢神经系统疾病之间似乎存在病理联系。
吲哚对神经和神经精神疾病的影响
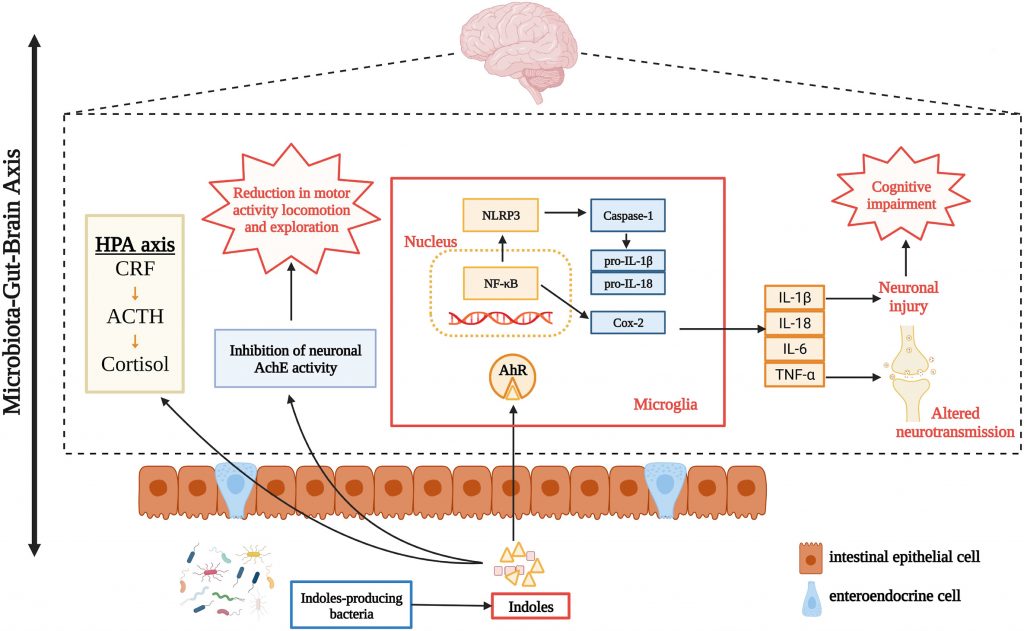
编辑
Zhou Y,et al.Neuropharmacology.2023
硫酸吲哚酚对大脑产生有益或有害的影响取决于含量:
通过比较患有抑郁症的志愿者和健康参与者,鉴定出了22种尿液代谢物,并且两组受试者之间的丰度存在差异。患有严重抑郁症的个体尿液中硫酸吲哚酚浓度低于健康人。在志愿者中,血清硫酸吲哚酚浓度与精神焦虑和相关的基于功能磁共振成像的神经特征有关。
在实验性自身免疫性脑脊髓炎的小鼠模型中,每日腹腔注射硫酸吲哚酚(10mg/kg)可调节星形胶质细胞活性,并通过芳基烃受体对中枢神经系统发挥抗炎作用。相反,当大鼠口服较高剂量(100和200mg/kg)的硫酸吲哚酚时,观察到空间记忆受损以及运动和探索活动减少。
另一项研究发现,单次腹腔注射更大剂量(800mg/kg)的硫酸吲哚酚会引起与神经元坏死相一致的大脑组织学变化。硫酸吲哚酚在15-60微摩尔范围内使用时,会诱导原代星形胶质细胞产生自由基氧,并导致海马神经元细胞死亡。在10微摩尔浓度下,硫酸吲哚酚通过氧化应激诱导人星形胶质细胞凋亡。
在慢性肾病(CKD)患者中,硫酸吲哚酚与心血管健康和全因死亡率相关。硫酸吲哚酚是血液透析患者发生心血管疾病的重要因素。在血液透析中,发现游离形式硫酸吲哚酚水平与成纤维细胞生长因子23呈正相关,与C-C基序趋化因子15、补体成分C1q受体、串珠素(Perlecan)、博来霉素水解酶、分化簇166抗原和信号淋巴细胞活化分子家族成员5呈负相关。
这些蛋白质在血管修复和内皮生长中发挥着至关重要的作用。硫酸吲哚酚还可以通过增加血小板过度活跃、提高血浆促凝血水平和产生促凝血颗粒来导致血栓形成和动脉粥样硬化。硫酸吲哚酚通过增加炎症、心脏纤维化、心肌细胞增殖来影响心脏组织。
并且强有力的证据表明,硫酸吲哚酚在肾功能不全的情况下积累时是有害的。硫酸吲哚酚会损害近端肾小管细胞并诱导炎症和纤维化发展。肾脏通过肾小管分泌实现硫酸吲哚酚的高清除率,而硫酸吲哚酚与血浆蛋白结合超过90%,受到蛋白结合的限制,并且血液透析患者的血浆水平相对较高,这也表明与肾脏疾病密切相关。
啮齿动物口服新霉素(一种广谱抗生素)会降低脑内羟吲哚含量,因此表明羟吲哚至少部分源自肠道微生物群代谢活动。
有趣的是,最近在人类粪便样本中发现了羟吲哚,从而证实肠道微生物是这种代谢物的来源。在人类粪便中存在的细菌代谢物中,羟吲哚被发现是主要的芳基烃受体激活剂之一。
大鼠海马切片的体外实验表明,羟吲哚可能与电压门控钠通道相互作用,增加产生动作电位的阈值,从而大大降低神经元的兴奋性。
大胆推测
这些结果说明羟吲哚可能与已知的神经抑制化合物具有一些共同特征,从而能够降低神经元的兴奋性
吲哚及其衍生物可以影响肠道健康甚至是全身其他器官的健康,而过高或过低的水平都可能带来不利影响,导致一些疾病的发生。
调节人体内吲哚水平的机制主要有3种:首先,它可以通过调节与吲哚产生相关的肠道微生物群的丰度来影响吲哚的水平。
先前的研究发现,特定的肠道微生物群成员在色氨酸转化为吲哚的过程中发挥着不可或缺的作用,例如产孢梭菌(Clostridium sporogenes),它已被证明可以通过氧化和还原途径产生吲哚乙酸和吲哚-3-丙酸。在大肠杆菌中也发现了吲哚活性转运蛋白,在乳杆菌中也发现了色氨酸酶的表达,并有助于将色氨酸转化为吲哚。
其次,饮食调整可以增加或减少色氨酸的摄入量,从而间接影响体内的吲哚水平。
最后,直接作用于AHR受体激动剂的药物可能会发挥与吲哚类似的作用。
益生菌可以通过多种机制与体内微生物群相互作用,对宿主健康产生有益影响。
临床试验表明,益生菌可以抵消肠道微生物失衡造成的负面影响,并可导致吲哚产生属及其衍生物的富集,如乳杆菌(Lactobacillus)。在另一项临床试验中,具有抗炎作用的双歧杆菌属在益生菌治疗后的丰度也显著增加。
益生菌还有可能成为神经和精神疾病的治疗辅助药物。在一项针对抑郁症和海马体功能变化的研究中,接受益生菌治疗的小鼠表现出肠道微生物生态失调的显著缓解,以及较低的肠道炎症介质如IL-1β、TNF-α和iNOS的表达水平。此外,情感行为也被检测到与肠道微生物群组成变化相关的显著改善。
益生元是宿主细菌可以选择性利用并提供健康益处的底物。常见的益生元包括低分子量碳水化合物、多酚和多不饱和脂肪酸。
在实验室和临床试验中,益生元(包括菊粉和低聚半乳糖)已被证明可以显著提高产生吲哚的益生菌(例如双歧杆菌和乳酸杆菌)的丰度。在对肠道微生物失衡进行一定纠正后,吲哚作为具有调节作用的配体,对炎症反应具有抑制作用。
合生元是益生菌和益生元的混合物。在大鼠实验中,合生元治疗逆转了肠道微生物特定菌群丰度的变化。
经过合生元(乳杆菌、双歧杆菌、链球菌和菊粉的组合)治疗的患有慢性肾病的大鼠中,肠道中产生吲哚的产气荚膜梭菌的丰度降低至正常水平。此外,合生元治疗可通过改变肠道微生物丰度来有效调节粪便中吲哚及其衍生物的水平。
合生元治疗由于其对吲哚水平的间接影响,可能成为许多神经系统和神经精神疾病的辅助治疗。最近一项针对阿尔茨海默病患者的临床实验表明,在使用合生元后,神经炎症有所减轻。
值得注意的是,合生元干预的改善效果优于单独的益生元或益生菌干预,这表明复合干预的组合更有可能成为临床治疗的实际应用。
粪便微生物移植(FMT)在先前的几项研究中已被证明对缓解胃肠道微生物失调具有不可否认的作用。通过FMT将产生吲哚的益生菌(乳酸杆菌、双歧杆菌等)移植到患者的肠道中,可以改善肠道微生物群。据观察,肠道中产生吲哚的微生物群的丰度增加,吲哚及其衍生物的水平受到间接影响。
吲哚是色氨酸代谢途径上的关键配体,可以调节和抑制神经和精神疾病的炎症。粪菌移植已被证明在各种疾病和临床治疗中对改善患者的肠道生态起着至关重要的作用,它可以通过吲哚在中枢神经系统疾病治疗中间接发挥作用,并被认为是一种潜在的辅助疗法。
肠道微生物通过色氨酸调节吲哚水平。色氨酸是人体必需氨基酸之一,需要完全依赖饮食摄入。
以下是一些富含色氨酸的常见食物:
蛋白质食物:肉类(牛肉、猪肉、鸡肉、火鸡)、鱼类(鳕鱼、鲑鱼、金枪鱼)、贝类(蛤蜊、扇贝)、蛋(鸡蛋、鸭蛋)、豆类(大豆、黑豆、红豆)、奶制品(牛奶、乳制品)等。
蔬菜和水果:菠菜、花椰菜、蘑菇、芝麻、松子、香蕉、菠萝、樱桃、草莓、蓝莓等。
谷物和豆类:燕麦、大麦、小麦、黑米、糙米等。
坚果和种子:核桃、杏仁、腰果、葵花子、南瓜子、亚麻籽等。
研究表明,0.4%膳食色氨酸可显著减少衰老小鼠的神经退行性病变和神经炎症。在此过程中,补充色氨酸可以通过改变肠道微生物组成并增加阿克曼氏菌等有益细菌的丰度来增强肠粘膜屏障功能。肠道菌群产生的吲哚类物质(如吲哚乙酸、吲哚-3-丙酸)显著升高,可以激活AHR,从而抑制神经炎症。
通过血清靶向代谢组学分析和16s rRNA测序对全身色氨酸代谢和肠道微生物组进行了分析,进一步证实了微生物群-肠-脑轴在调节色氨酸饮食产生吲哚的潜在作用。
虽然色氨酸饮食是一种可能的治疗策略,但过多的色氨酸摄入也被证明具有有害作用,应注意适量摄入。
此外,据报道,间歇性禁食还可能通过调节肠道微生物组成来影响血浆吲哚,从而对神经和神经精神疾病的预后产生有益影响。饮食调节可能是改善吲哚含量的一种简单但有效的方法。
最近的综合研究表明,吲哚通过作用于靶细胞来调节炎症反应和细胞生理。近年来,开发吲哚类作用靶点的药物一直是一个重要的研究热点。
吲哚及其衍生物充当AHR的配体,并通过AHR间接调节星形胶质细胞和小胶质细胞的炎症。因此,直接作用于AHR的激动剂也可以作为内源性AHR配体(如吲哚)的替代品,并参与色氨酸AHR介导的炎症抑制途径。
体外合成的AHR配体2-(1′H-吲哚-3′-羰基)-噻唑-4-羧酸甲酯在一项临床研究中被证明具有抑制自身免疫性脑脊髓炎进展的能力。对AHR具有激动作用的药物有可能取代色氨酸代谢产物,并作为外源性AHR配体激活自限制途径和修复神经和神经精神疾病。
除了靶向AHR,影响吲哚作用途径中其他靶点的药物也可能具有治疗作用,这需要在未来进行更多的研究。
由于吲哚是肠道细菌常见的代谢产物,其重要的生物学作用不容忽视。吲哚与肠道菌群的稳态和肠道健康直接相关,吲哚还可以间接影响其他系统和人体的整体健康。
吲哚对人类健康具有有益影响,通过激活免疫细胞释放抗炎因子如IL-22来调节肠道屏障并帮助维持肠道稳态,抑制包括肠出血性大肠杆菌在内的病原菌的定植,以及增加MUC表达以增强粘液屏障功能。
此外,吲哚在调节肠道微生态方面也起着关键作用。它们抑制有害菌株,并以影响基因表达的方式改变肠道致病菌的毒力,这有助于缓解出血性结肠炎等疾病。同时,吲哚及其衍生物在激活AhR和PXR介导的抗炎途径中至关重要。例如吲哚-3-丙酸通过PXR介导肠细胞TNF-α的下调,并上调编码生长抑素的mRNA,以调节炎症中的肠道通透性和肠道屏障功能。这意味着吲哚及其衍生物有前景的治疗途径。
尽管如此,吲哚也是一把双刃剑,少数衍生物也存在一定的缺点。例如,吲哚肝脏代谢产生的硫酸吲哚酚在高浓度时具有肾毒性和心血管毒性,可通过促进氧化应激、炎症等病理变化而导致多系统功能障碍。
吲哚在肠道稳态和人类健康中发挥着至关重要的作用。许多肠道细菌被鉴定为将色氨酸代谢为吲哚。尽管如此,仍可能存在未鉴定的细菌,因此未来需要使用代谢组学和宏基因组学来进一步表征未知的吲哚产生细菌及其相关代谢途径。
不同浓度的吲哚发挥着不同的生理功能。健康成年人的粪便吲哚浓度是已知的,而人类肠道和血液中许多吲哚衍生物的浓度仍然未知。缺乏在不同环境中测定吲哚及其衍生物浓度的定量方法,未来需要开发简单快速的测定方法。
未来仍需进一步研究吲哚与人类健康之间的关系。需要研究多种吲哚组合对宿主生理学的影响,并将这些发现与临床环境相结合,以开发相关疾病的新疗法。
主要参考文献
Zhou Y, Chen Y, He H, Peng M, Zeng M, Sun H. The role of the indoles in microbiota-gut-brain axis and potential therapeutic targets: A focus on human neurological and neuropsychiatric diseases. Neuropharmacology. 2023 Nov 15;239:109690.
Tennoune N, Andriamihaja M, Blachier F. Production of Indole and Indole-Related Compounds by the Intestinal Microbiota and Consequences for the Host: The Good, the Bad, and the Ugly. Microorganisms. 2022 Apr 28;10(5):930.
Ye X, Li H, Anjum K, Zhong X, Miao S, Zheng G, Liu W, Li L. Dual Role of Indoles Derived From Intestinal Microbiota on Human Health. Front Immunol. 2022 Jun 17;13:903526.
Lee JH, Lee J. Indole as an intercellular signal in microbial communities. FEMS Microbiol Rev. 2010 Jul;34(4):426-44.
Jiang H, Chen C, Gao J. Extensive Summary of the Important Roles of Indole Propionic Acid, a Gut Microbial Metabolite in Host Health and Disease. Nutrients. 2022 Dec 28;15(1):151.
Kumar A, Sperandio V. Indole Signaling at the Host-Microbiota-Pathogen Interface. mBio. 2019 Jun 4;10(3):e01031-19.
Chimerel C, Emery E, Summers DK, Keyser U, Gribble FM, Reimann F. Bacterial metabolite indole modulates incretin secretion from intestinal enteroendocrine L cells. Cell Rep. 2014 Nov 20;9(4):1202-8.
Lu Y, Yang W, Qi Z, Gao R, Tong J, Gao T, Zhang Y, Sun A, Zhang S, Ge J. Gut microbe-derived metabolite indole-3-carboxaldehyde alleviates atherosclerosis. Signal Transduct Target Ther. 2023 Oct 4;8(1):378.
谷禾健康
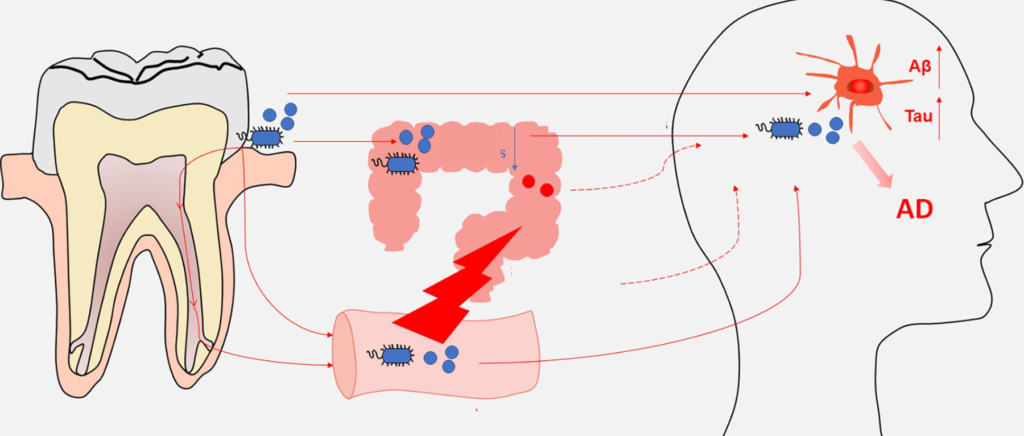
在个体中,每个微生物栖息地都表现出独特的微生物种群模式。迄今为止,关于微生物组相关疾病的研究主要集中在器官特异性微生物组上。然而,器官间的微生物网络正逐渐成为生理功能和病理过程中的重要调节因子和治疗机会。
在正常情况下,人体维持动态平衡,各个身体部位的微生物可以通过直接相互作用或间接作用于系统循环中的炎性物质、细胞因子和代谢物来相互影响。在这其中,口腔和肠道是人体中两个最为重要的微生物栖息地,它们在微生物组相关疾病中起着关键的作用。
近年来,随着网络的发展和生活水平的提高,大众对心理健康产生了极大关注,同时精神类疾病也面临巨大挑战,患病率高、致残率高、治疗率低。许多患者无法获得及时有效的救治,导致病情日益严重。药物治疗只能暂时缓解症状,容易出现复发,甚至形成对药物的依赖,带来更加严重的后果。因此,对于人体微生物与精神健康之间的研究越来越多,尤其是以肠道菌群为研究靶点,探索和验证以肠道菌群为靶点治疗精神疾病的潜力。
此外,大量高质量的研究证据以及实践检测发现,精神疾病往往与功能性胃肠病共存,并可能相互影响和形成负向循环,其共同的病理生理基础是“菌-肠-脑轴”的异常互动。例如,精神疾病患者通常会出现胃肠道症状,特别是腹疼痛、饱胀、恶心、反酸、消化不良和腹泻等,以至于他们会在消化内科或外科咨询就诊。反过来,功能性胃肠疾病患者则伴随着睡眠障碍、社交障碍、焦虑和抑郁等症状,他们将前往心理科寻求帮助。同时,研究也发现患有精神类疾病的人除了胃肠道症状外,常常还伴有口腔问题,如牙周病或口臭,而患有牙周炎或牙周病的人更容易出现认知或精神障碍。
目前,在功能性胃肠病、口腔疾病与精神疾病的临床诊疗中存在一些问题,医患双方对于其临床表现的认知和重视程度不高,对病理生理机制的理解不够全面,对干预方式的整体观念和综合视角不足,导致目前该类疾病的疗效不理想。急需让精神疾病患者和医务工作者了解这些相关联系,并在治疗过程中予以关注。
在讨论和传播关于脑肠轴与精神疾病相关的知识和科普的基础上,本文主要拓展了对于口-肠轴或口-肠-脑轴在精神健康障碍中的重要性的理解,尤其是探讨了基于口肠微生物群的联合治疗神经精神疾病的解决方案和预防措施等。
口腔是继肠道之后微生物组的第二大定植区。此外,它还是呼吸道和消化道的门户。口腔微生物群的改变可能导致或预示各种口腔和全身疾病。
▼
食物的消化很少发生在口腔中。然而,通过咀嚼或咀嚼的过程,食物才能很好的通过上消化道运输到胃和小肠。咀嚼是食物经历的第一个机械过程。咀嚼时下颌的运动是由咀嚼肌(咬肌、颞肌、翼内肌、翼外肌和颊肌)。包围和支撑牙齿的牙周膜的敏感性,而不是咀嚼肌肉的力量,决定了咬合的力量。
理论上来说,咀嚼对于充分消化来说并不是必需的,但是咀嚼对于吞咽和运输又有很大的影响,从这个意义上说咀嚼确实有助于消化,因为它可以将食物切成小颗粒并将其与唾液腺分泌的唾液混合。唾液润滑和润湿干燥的食物,而咀嚼则使唾液分布在整个食物团中。舌头抵住硬腭和脸颊的运动有助于形成圆形的食物团。
嘴唇和脸颊
嘴唇是围绕嘴的两个肉质褶皱,外部由皮肤组成,内部有粘膜。粘膜含粘液分泌腺,它们与唾液一起确保言语和咀嚼的充分润滑。
脸颊,即嘴的两侧,与嘴唇连续,具有相似的结构。脸颊的皮下组织(皮肤下的组织)有一个明显的脂肪垫;这种垫对于婴儿来说特别大,被称为吸吮垫。在每个脸颊的内表面上,与第二上臼齿相对的地方有一个轻微的隆起,标志着腮腺管的开口,从位于耳朵前面的腮腺唾液腺引出。在这个腺体的后面有四到五个分泌粘液的腺体,其导管在最后一颗臼齿的对面打开。
口腔顶部
口腔顶部呈凹形,由硬质和软质形成上颚。硬腭由两块腭骨的水平部分和上颌骨或上颌的腭部分形成。硬腭覆盖着厚厚的、略显苍白的粘膜,该粘膜与牙龈连续,并通过坚固的纤维组织与上颌骨和腭骨结合。软腭与前面的硬腭是连续的。其后面与覆盖鼻腔底部的粘膜连续。软腭由坚固、薄的纤维片、腭腱膜、舌腭肌和咽腭肌组成。
口腔底
只有当舌头抬起时才能看到口底。中线是一个突出的、升高的粘膜皱襞(舌系带)将每个嘴唇与牙龈结合在一起,其两侧都有一个轻微的褶皱,称为“舌系带”舌下乳头,下颌下唾液腺导管从中开口。从每个舌下乳头向外和向后延伸的是一个脊(舌下皱襞),标志着舌下(舌下)唾液腺的上边缘,该腺体的大部分导管都通向此处。
牙龈
牙龈由粘膜组成,粘膜通过厚纤维组织与颌骨周围的膜连接。牙龈膜上升,在每颗牙齿的牙冠基部(暴露部分)周围形成一个颈圈。牙龈组织富含血管,接收来自牙龈的分支肺泡动脉;这些血管由于与牙槽或牙槽的关系而被称为牙槽,也供应牙齿和上颌和下颌的松质骨,牙齿位于其中。
牙齿
牙齿是口腔中坚硬的白色结构。不同脊椎动物的牙齿通常用于咀嚼,有时是专门化的牙齿。咀嚼对于食肉动物来说并不像对于草食动物那么重要。人类是杂食动物(吃植物和动物组织),其牙齿在功能和结构上属于食肉动物和食草动物的牙齿所达到的极端专业化之间。
每颗牙齿有牙冠和压根。它们具有不同的功能。口腔的不同部位以及不同动物的牙冠和牙根的形状各不相同。下颌一侧的牙齿本质上是另一侧牙齿的镜像。上牙与下牙不同,但又互补。
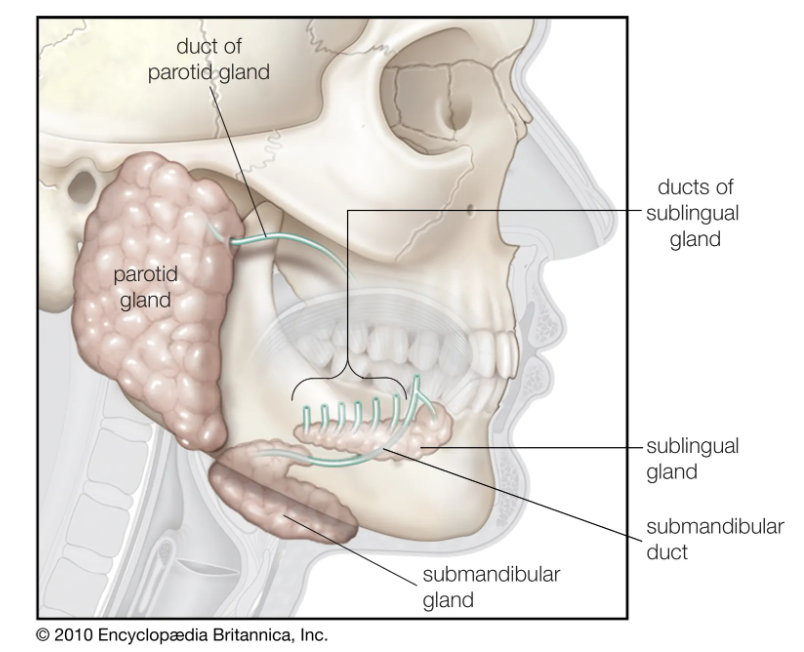
舌头
舌头是位于口底的肌肉器官,是一个极其灵活的结构,是言语、咀嚼和吞咽等运动功能的重要辅助器官,也是口腔微生物聚集的地方。它与脸颊一起,能够引导和保持食物在上牙和下牙之间,直到咀嚼完成。舌头的运动有助于在口腔内产生负压,从而使婴儿能够吸吮。作为一种外周感觉器官,舌头尤其重要,它含有一组特殊的上皮细胞,称为味蕾,将刺激从口腔传送到中枢神经系统。此外,舌头的腺体会产生一些吞咽所需的唾液。
舌头由大量交织的横纹肌组成,其中散布着脂肪。覆盖舌头的粘膜在不同区域有所不同。舌头通过其外在肌肉附着在下颌、舌骨(下颌和喉部之间的U 形骨)、颅骨、软腭和咽部。它与口腔底部和口腔相连会厌(作为喉盖的一块软骨板)由粘膜褶皱形成。
唾液腺
食物被品尝并与几组腺体分泌的唾液混合。除了许多分泌唾液的微小腺体外,还有三对主要的唾液腺:腮腺、颌下腺和舌下腺。唾液腺是最大的一对,位于面部侧面、每只耳朵的下方和前方。腮腺被包裹在鞘中,当发炎时,如腮腺炎,腮腺可以限制肿胀的程度。下颌下腺呈圆形,位于下颌骨内侧附近,位于胸骨乳突肌(下颌突出的肌肉)前面。这舌下腺直接位于覆盖舌头下方口腔底部的粘膜下方。
唾液腺中含有很多分泌细胞,分泌细胞在圆形囊中呈簇状排列,称为腺泡,附着于导管的自由分支系统。分泌细胞可以是浆液型或粘液型。后一种类型分泌粘蛋白,粘蛋白的主要成分粘液; 前者是含有淀粉酶的 水状液体。
腮腺的分泌细胞为浆液型;颌下腺,有浆液性和粘液性两种类型,浆液性细胞与粘液性细胞的数量为四比一。舌下腺的腺泡主要由粘液细胞组成。
唾液腺由自主神经系统的两个部分(交感神经和副交感神经)控制。这副交感神经 供应调节腺泡细胞的分泌并导致血管扩张。受规管的职能交感神经包括腺泡细胞的分泌、血管的收缩以及可能的肌上皮细胞的收缩。
正常情况下,无论口腔中是否有食物,唾液的分泌都是恒定的。24小时内分泌的唾液量通常为1-1.5升。当有东西接触牙龈、舌头或口腔内壁的某些区域时,或者咀嚼时发生时,唾液分泌量增加。刺激物质不一定是食物——嘴里的干沙,甚至在空嘴时移动下巴和舌头都会增加唾液流量。这种对口腔粘膜的直接刺激与唾液分泌增加的结合被称为无条件唾液反射。当一个人了解到特定的视觉、声音、气味或其他刺激通常与食物相关时,仅该刺激就足以刺激唾液流量增加。这种反应被称为条件性唾液反射。
唾液
唾液溶解一些咀嚼的食物并充当润滑剂,促进后续食物顺利通过消化道。唾液还含有一种称为淀粉酶(ptyalin)的淀粉消化酶,它可以启动酶水解过程;它将淀粉(一种含有许多连续链结合的糖分子的多糖)分解成双糖麦芽糖分子。
许多食肉动物,例如狗和猫,其唾液中没有淀粉酶;因此,他们的天然饮食中含有很少的淀粉。物质必须处于溶液中才能刺激味蕾;唾液的成分各不相同,但其主要成分是水、与血浆中常见的无机离子,以及许多有机成分,包括唾液蛋白质、游离氨基酸以及溶菌酶和淀粉酶。尽管唾液呈弱酸性,但其中所含的碳酸氢盐和磷酸盐可充当缓冲剂,并在正常条件下保持唾液的pH 值或氢离子浓度相对恒定。
唾液中碳酸氢盐、氯化物、钾和钠的浓度与其流速直接相关。碳酸氢盐浓度与血液中二氧化碳分压之间也存在直接关系。血液中氯化物的浓度从低流速时的 5 毫摩尔/升到高流速时的 70 毫摩尔/升不等。类似情况下的钠浓度从每升 5 毫摩尔到每升 100 毫摩尔不等。血液中钾的浓度通常高于血浆中的浓度,每升高达 20 毫摩尔,这就是唾液在快速流动时产生尖锐金属味的原因。
唾液的持续流动使口腔和牙齿保持湿润,并且相对清除食物残渣、脱落的上皮细胞和异物颗粒。通过去除可用作细菌培养基的物质,唾液抑制细菌的生长。唾液具有保护功能,因为溶菌酶具有溶解或溶解某些细菌的能力。因此唾液的分泌与口腔菌群构成和变化紧密。
唾液的分泌还提供了一种机制,使某些有机和无机物质可以从体内排出,包括汞、铅、碘化钾、溴化物、吗啡、乙醇以及某些抗生素,如青霉素、链霉素和金霉素。虽然唾液不是生命所必需的,但唾液的缺乏会导致许多不便,包括口腔粘膜干燥、细菌过度生长导致口腔卫生不良、味觉大大减弱以及言语困难。
▼
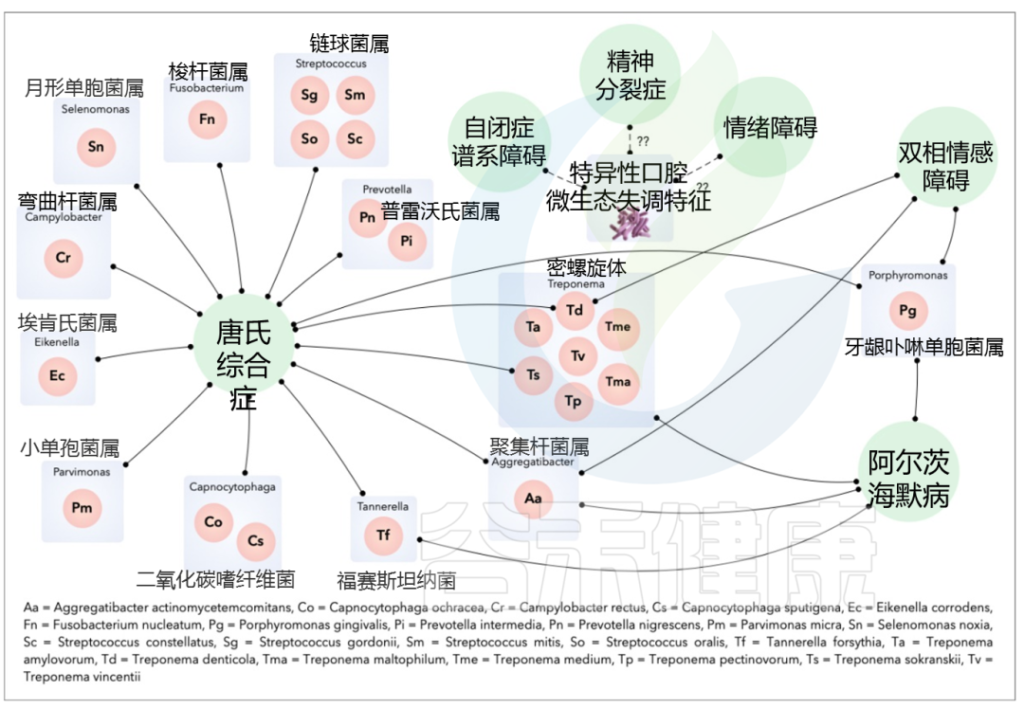
与牙周炎有关:
如普氏菌属、卟啉单胞菌属、密螺旋体属、聚集杆菌属、梭杆菌属等。
与龋齿有关:
如变形链球菌 、乳酸菌、奈瑟菌属等。
与精神障碍潜在相关的主要口腔细菌种类如下:
doi.org/10.3390/microorganisms9071450
口腔和肠道之间存在密切的解剖和生理联系。口腔和肠道都是消化系统的一部分。由于胃酸的pH值较低,恶劣的化学环境使得许多口腔细菌难以在肠道内定殖。然而,疾病、药物、衰老等各种情况都会促进口腔细菌在肠道中的定植。
▼
研究人员已经提出了细菌从口腔转移到肠道的两种可能途径:
胃肠道途径
人类每天要吞下大约1.5升唾液,这些唾液富含口腔细菌。来自牙周炎患者和健康对照者的唾液微生物可以在小鼠肠道中存活至少 24 小时,暗示口腔菌群影响肠道菌群的重要途径。然而,胃酸和碱性胆汁对口腔微生物群在肠道中的定位构成了很大的瓶颈,关于口腔微生物群是否可以通过肠内途径定植肠道存在激烈争论。最近的一项研究表明,没有证据表明口腔细菌在健康成人的远端肠道中定植。相反,另外的研究得出的结论是,至少三分之一的口腔微生物群可以解决健康成年人的肠道问题,肠癌和类风湿性关节炎患者比健康人有更多的口腔到肠道微生物传播。胃炎、炎症性肠病、结直肠癌等肠道疾病,使易位的口腔细菌在肠道内定植和扩张。
由上所述,唾液含有粘液(由水、脂质和蛋白质如粘蛋白组成),可以保护微生物群免受胃酸影响,从而在胃肠道中存活。口腔管饲与牙周炎相关的唾液可加重小鼠糖尿病模型、结肠炎、精神症状、老年痴呆症和骨质疏松症。据估计,患有严重牙周炎的患者每天会吞咽大约大量的牙龈卟啉单胞菌 (P. gingivalis),如果进入肠道定制会改变肠道菌群 。
然而,由于胃肠道的屏障功能和胃的酸性,摄入的口腔细菌很少到达并定植在健康的肠道中。但是这两道屏障也可能受损,以下三种情况下,口腔菌群可以趁机进入肠道。
方式一:肠道微生物群破坏
我们知道,肠道微生物群作为一个重要的屏障,可以阻止摄入的口腔细菌在肠道中定植,一旦健康肠道微生物群被破坏,则会导致口腔细菌在肠道的定植增加。例如,抗生素万古霉素用于治疗细菌感染,扰乱肠道微生物组成,从而为口腔细菌在肠道中定植和扩张提供便利。
方式二:胃功能不全
由于长期使用质子泵抑制剂而导致胃功能不全的患者,其口腔细菌如嗜血杆菌属、链球菌属、韦荣氏球菌属的肠道定植显著增加。
胃炎和胃手术也可能导致摄入的口腔细菌暴露于胃液的减少,研究表明,患有胃炎或接受过胃手术的人的肠道微生物组成发生了改变,特定口腔微生物组分类群的相对丰度在肠道中显著增加,如链球菌属、韦荣氏球菌属、肠杆菌科等。
方式三:某些菌耐酸
某些口腔细菌,如牙龈卟啉单胞菌,具有耐酸性,因此可以通过胃屏障进入肠道。
血液途径
当一个人患上牙周病时,牙周袋表皮的撕裂、日常的口腔卫生习惯(激烈刷牙/用牙线)、侵入性的牙齿手术等情况,都可能导致口腔细菌传播至全身循环(菌血症)。
此外,日常牙科活动(如用力咀嚼、刷牙)和牙科手术(如洗牙和牙根平整、牙齿矫正、拔牙)造成的口腔机械损伤可能会使口腔细菌扩散到体循环中。此外,牙周炎导致牙周袋血管化和牙龈溃疡,使牙周病原体很容易进入血液。血液途径可能是口腔梭杆菌到达结肠肿瘤的首选途径,而不是肠道途径。
研究表明,牙周炎引发口腔细菌传播至肝脏和脾脏。此外,口腔细菌具有入侵免疫细胞(如巨噬细胞和树突状细胞)并在其内部存活的能力,这就好像口腔细菌能够利用宿主免疫细胞作为特洛伊木马,从口腔传播到肠道粘膜。
以上是口腔菌群进入肠道的几种方式。研究发现,绝大多数口腔细菌可转移到肠道。
免疫细胞迁移路线
一些口腔细菌可以在树突状细胞和巨噬细胞等免疫细胞内存活,表明口腔细菌可能劫持宿主免疫细胞作为“木马”从口腔粘膜传播到肠道粘膜。此外,来自口腔淋巴结引流的免疫细胞可以迁移到其他淋巴组织,包括但不限于肠道。口腔致病菌反应性 T 辅助细胞 17(Th17) 可以迁移到发炎的肠道。在肠道中,口腔来源的 Th17 细胞可被易位的口腔致病菌激活并导致结肠炎的发展。
以上我们了解到口腔-肠道传播是影响肠道微生物组成的重要过程,因此可能通过改变肠道微生物群间接影响中枢神经系统功能,我们在下面展开阐述。
▼
口腔-肠道轴在正反馈回路中具有引发全身炎症的倾向。失调的口腔微生物群不仅直接引发炎症,还可以改变肠道微生物群的组成、功能和微生物代谢产物,从而导致促炎级联反应,进一步加剧口腔炎症。
口腔微生物群也有可能通过代谢产物和细菌素的释放影响肠道微生物群。在自然环境中,细菌产生具有抗菌活性的细菌素和肽,以便与其他细菌争夺营养物质。
唾液微生物组影响肠道微生物群
将重度牙周炎患者的唾液灌胃移植到小鼠体内可改变肠道菌群,肠道菌群的β多样性与对照组有显着差异,卟啉单胞菌和梭杆菌增多,阿克曼菌减少,表明唾液微生物群可以通过肠道途径改变肠道微生物群。
通过灌胃将牙周炎患者的唾液移植到患有结肠炎的小鼠体内可以加速结肠炎,并改变与炎症性肠病相关的微生物群,如Blautia、幽门螺杆菌和瘤胃球菌。
牙龈卟啉单胞菌影响肠道菌群
牙龈卟啉单胞菌是牙周炎最重要的致病菌之一。它也是研究最多的影响肠道菌群的口腔致病菌。研究牙龈卟啉单胞菌对肠道菌群的影响,首先要明确以下问题:
下面将从这三个方面进行讨论。
牙龈卟啉单胞菌定植于小鼠口腔并影响口腔微生物群
一般认为牙龈卟啉单胞菌可以在小鼠口腔内定植。接种P. gingivalis 7 天后,小鼠口腔内可检测到P. gingivalis 。在小鼠牙龈停止外用牙龈卟啉菌4周和8周后口腔内仍可检测到牙龈卟啉单胞菌DNA,提示牙龈卟啉单胞菌可在小鼠牙龈内定植增殖。
在小鼠口腔中接种牙龈卟啉单胞菌会导致可培养的共生细菌总量增加,并改变口腔微生物群的质量组成,增加口腔微生物多样性并允许潜在的机会性物种的定植。
牙龈卟啉单胞菌可以在肠道定植吗?
由于胃液和胆汁的恶劣环境,牙龈卟啉单胞菌能否在肠道定植尚不确定。为了模拟体外胃环境,将牙龈卟啉单胞菌暴露于人工胃液 (AGJ) 中。在 pH 5 下暴露于 AGJ 2 小时后,只有 1% 的牙龈卟啉单胞菌浮游细胞存活,这相当于饭后立即的 pH 值。牙龈卟啉单胞菌生物膜的形成显着提高了存活率。几乎 100% 的细胞在 pH 5 下存活。因为牙龈卟啉单胞菌在口腔内与多种细菌形成复杂的生物膜,所以这个体外实验的结果不能推导到体内。
有的研究人员试图通过使用无菌小鼠来解决这个问题。他们通过在无菌小鼠口中擦拭人类唾液,开发了人类口腔微生物群相关小鼠模型(HOMA),还通过灌胃无菌人类粪便悬浮液开发了人类微生物群相关小鼠模型(HMA)老鼠。然后,他们cohoused HOMA 和HMA 模型。与 HMA 模型相比,同居模型显示小肠中的卟啉单胞菌增加,Turicibacter减少这个结果表明卟啉单胞菌在与肠道微生物群竞争小肠定植方面发挥了关键作用。
牙龈卟啉单胞菌影响肠道菌群
大量研究表明,牙龈卟啉单胞菌可以改变肠道菌群组成, 持续时间范围为P. gingivalis口服灌胃后 2 天一次到 10 周的重复应用P. gingivalis。
大多数研究使用口服管饲,而有些人将P. gingivalis应用于口腔,一项研究使用静脉注射。三项研究报告α多样性没有显著变化,并且有两个报告称 α多样性降低。五项研究表明β多样性存在显著差异,一项研究表明β多样性没有差异。
P. gingivalis 属于拟杆菌门,在门水平上,一些研究显示拟杆菌的比例增加,而有些人则相反。厚壁菌门是肠道微生物群中的另一个主要门,一项研究显示厚壁菌门的丰度增加,以及其他显示厚壁菌门减少的研究。
有趣的是,牙龈卟啉单胞菌在野生型小鼠 (WT) 和链脲佐菌素诱导的小鼠 (STZ) 中诱导了一些肠道微生物群的相反变化。乳酸杆菌的丰度在 WT 小鼠中减少,但在 STZ 小鼠中增加。Turicibacter的丰度在 WT 小鼠中增加,但在 STZ 小鼠中减少。这表明高血糖可能会影响细菌生长并改变小鼠肠道菌群的组成。
具核梭杆菌影响肠道菌群
据报道,具核梭杆菌( F. nucleatum ) 参与牙周病和根尖病变的发展,会影响肠道微生物群。具核梭杆菌感染牙髓诱发大鼠磨牙根尖周炎后, 2周时可在肠道中检测到具核梭菌,并改变肠道菌群,证实大肠感染。
伴放线放线杆菌影响肠道菌群
伴放线放线杆菌Aggregatibacter actinomycetemcomitans (A. actinomycetemcomitans )经常在严重牙周炎中检测到,并与局部侵袭性牙周炎有关。在管理A之后。给小鼠灌胃 6 周伴随放线菌,肠道中的Turicibacter属显着减少。该属与丁酸的生产有关,丁酸盐的减少与胰岛素抵抗有关。
肠道微生物群影响口腔微生物群
由于口腔与肠道在物理上相连,一些研究人员报告说,肠道微生物群的变化也会影响口腔微生物群。发现不同类型的肠道微生物群与不同类型的口腔微生物群相关。经过长期富含脂肪的饮食,小鼠的肠道微生物群概况可分为三种类型:糖尿病抵抗型、中间型和糖尿病敏感型。只有糖尿病敏感小鼠的牙周微生物群显示出丰富的普氏菌属和坦纳氏菌属,它们是主要的牙周病原体,表明肠道微生物群和口腔微生物群存在相互作用。同样,糖尿病导致口腔微生物群变得更具致病性。高血糖发作后,口腔微生物群中肠杆菌科、气球菌、肠球菌和葡萄球菌的水平升高,这些菌群通常与牙周炎有关。
在了解口腔微生物群是如何影响精神障碍类疾病之前,我们先来看一下,口-肠轴是如何影响大脑的。
这部分内容我们分成几个部分来详细阐述:
doi.org/10.1016/j.jdsr.2022.11.003
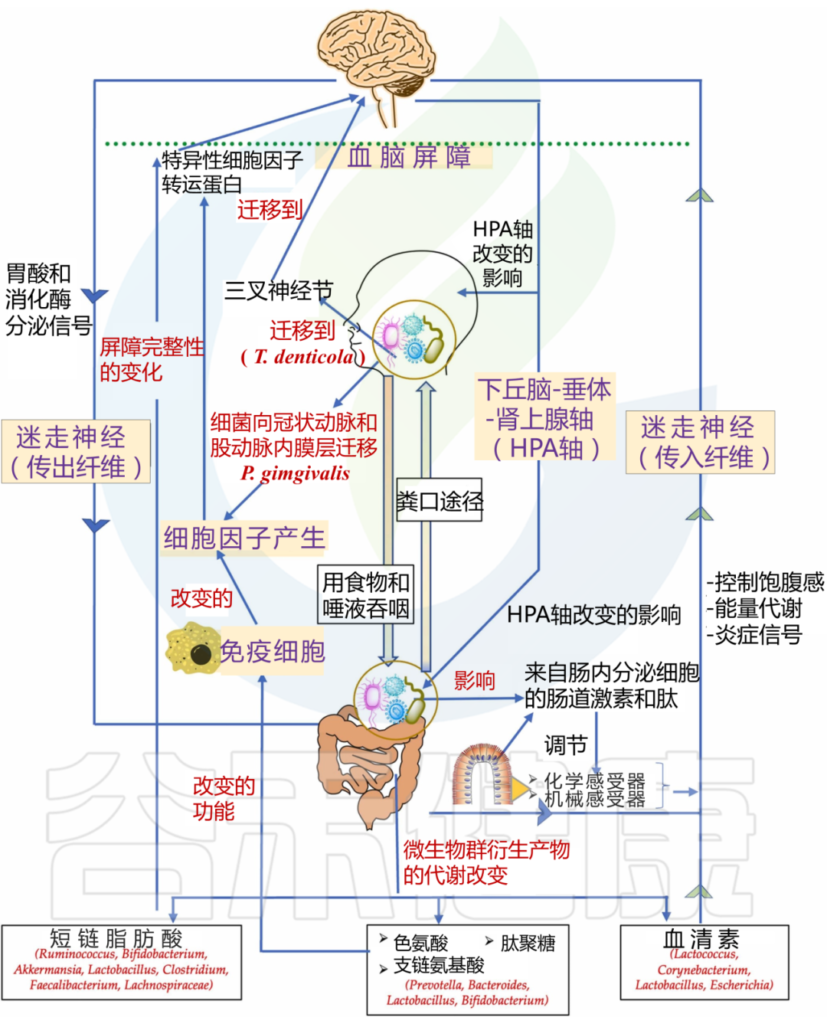
口腔微生物群如何影响大脑?
1
血液途径(细菌或其产物破坏血脑屏障)
口腔微生物群的改变可能导致炎症反应的增加,破坏血脑屏障,使得炎症介质和细菌代谢产物进入大脑,从而影响神经元的功能和存活。
如牙龈卟啉单胞菌通过血流迁移到冠状动脉和股动脉的内膜层。这种迁移可能会在体内产生急性炎症状态,导致炎性细胞因子的产生,这些细胞因子可以通过特定的细胞因子转运蛋白,穿过血脑屏障进入大脑。
2
神经途径
迷走神经和三叉神经复合体是口腔与大脑之间的重要神经连接。口腔微生物群的变化可能通过这些神经连接与中枢神经系统进行交流,影响大脑的功能和健康。
在阿尔茨海默病患者的三叉神经节和海马中发现了一种口腔细菌,即齿状密螺旋体(Treponema denticola)。这一发现在另一项针对小鼠的临床前研究中得到了支持,其中口腔T. denticola感染诱导了海马中淀粉样蛋白-β的产生。目前尚不清楚这些细菌是如何迁移到海马体的,人们认为口腔细菌可能是通过三叉神经途径到达大脑的。
牙周炎会诱发全身炎症,促炎细胞因子可以激活表达 TNF-α 和 IL-1 受体的内皮细胞,进而向紧邻脑内皮细胞的血管周围巨噬细胞发出信号。这些血管周围巨噬细胞随后与小胶质细胞通讯,从而导致小胶质细胞激活和随后的神经炎症。
牙周细菌细胞外囊泡,如外泌体,是免疫系统的有力刺激物,增加炎症负担。外泌体存在于大多数体液中,包括唾液,一项研究发现唾液中 CD9/CD81 外泌体水平降低与牙周病的发病机制有关。
因此,牙周细菌具有多种机制,可以将包括牙周炎在内的周围炎症转化为神经炎症,从而影响中枢神经系统的功能和行为。
牙周血管系统的渗透性增加导致脂多糖“泄漏”。脂多糖可激活下丘脑-垂体-肾上腺轴,从而增加应激激素和/或神经递质。这会影响肠道生理、栖息地、微生物组组成和细菌基因表达。
肠道微生物群的改变可能导致全身炎症,这不仅影响中枢神经系统,而且加剧其他炎症病理。
口腔微生物群影响和促进神经精神疾病结果的机制
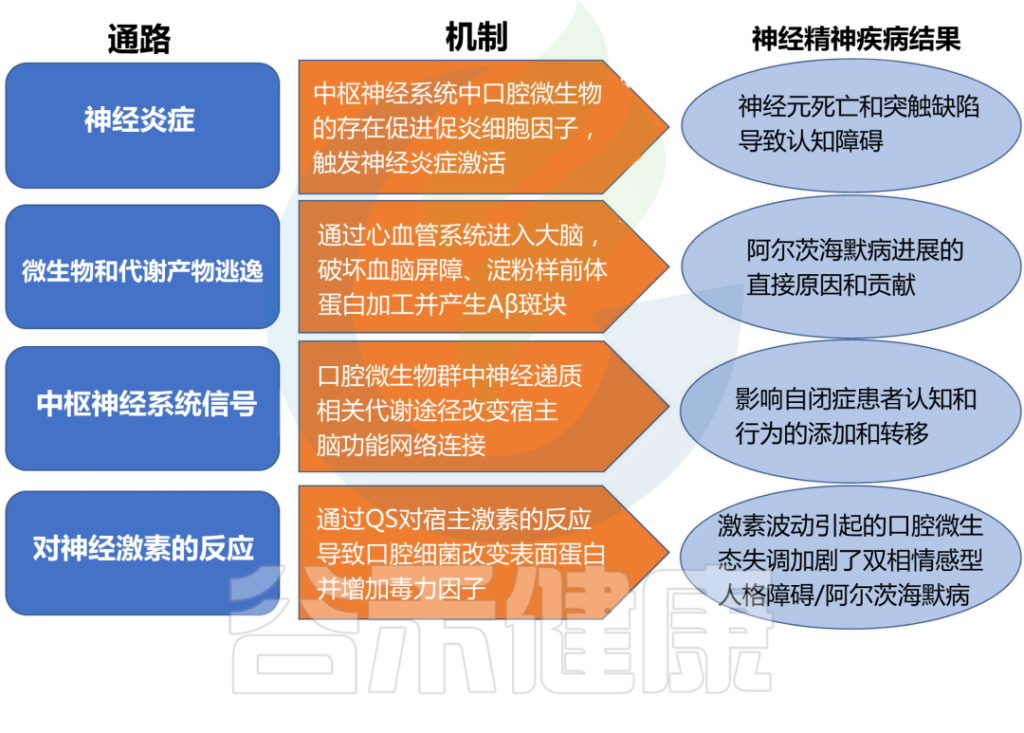
doi.org/10.3389/fpsyt.2022.810008
以上是口腔微生物群影响神经精神疾病的一些机制,口腔微生物群还可以通过肠道间接和大脑产生交流。
肠道微生物群如何影响大脑?
肠道作为“第二大脑”影响情绪和行为,大脑和肠道直接通过迷走神经和脊髓中的自主神经系统进行沟通。细菌通过迷走神经和肠神经系统传入神经元的刺激,在大脑和胃肠道微生物群之间建立直接的神经联系。
肠-脑轴的相互作用对我们的情绪和行为产生影响。肠道微生物的失衡和肠道疾病与焦虑、抑郁等精神疾病的发生和发展密切相关。
微生物群与大脑之间通过肠脑轴的分子通讯途径
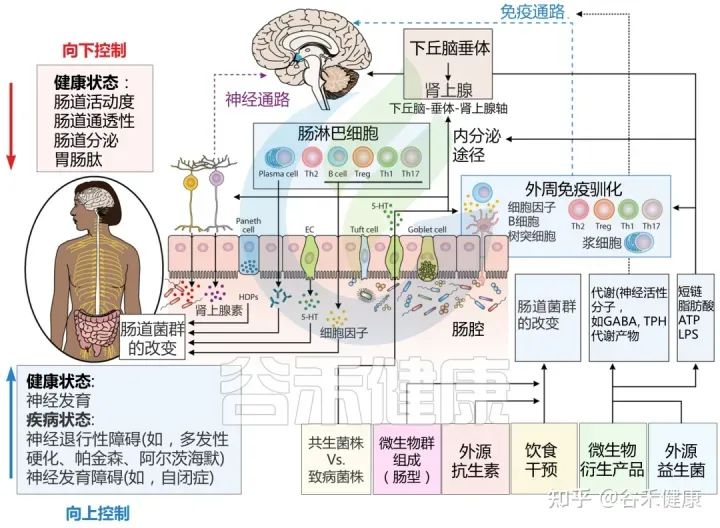
Sorboni SG, et al.,Clin Microbiol Rev. 2022
肠-脑轴的相互作用是一个复杂的系统,涉及到肠道微生物群、肠道黏膜屏障、免疫系统、神经递质和代谢产物等多种机制的相互作用和影响。这些在我们之前的很多文章已经详细阐述,此处就不展开,详见:
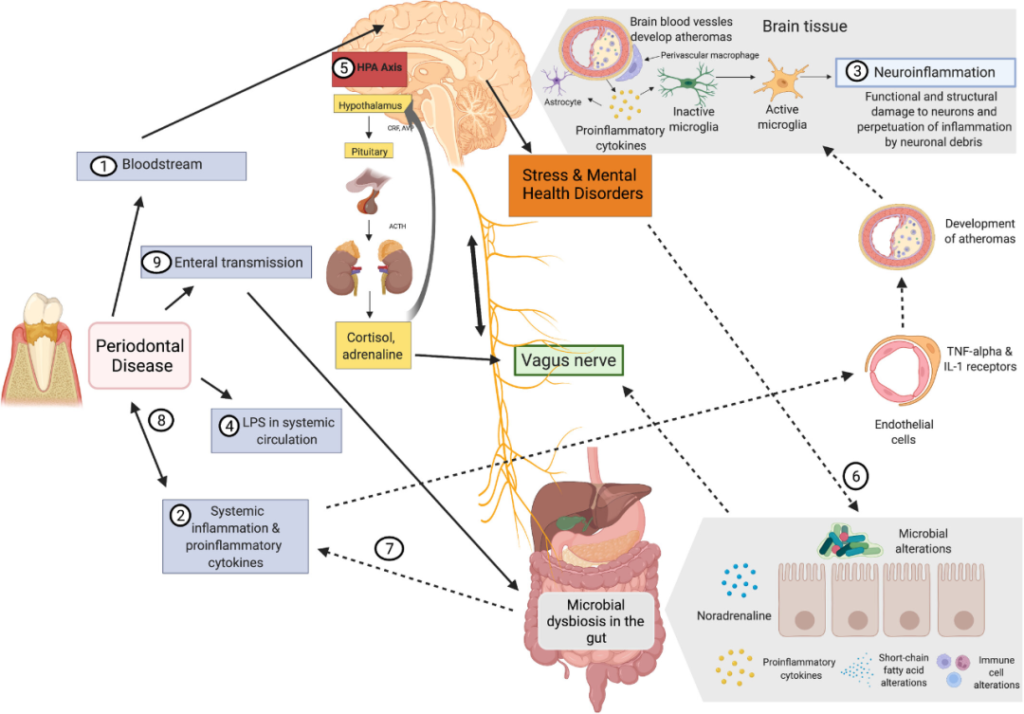
口腔和肠道微生物群之间的相互交流如何影响大脑?
doi.org/10.1111/prd.12452
1) 牙周病原体可以通过血液和受损的血脑屏障直接到达大脑。
2) 促炎细胞因子激活内皮细胞可以间接影响中枢神经系统。
3) TNF-α和IL-1内皮受体的表达激活小胶质细胞,导致炎症。
4) 牙周血管系统通透性的增加导致脂多糖的“渗漏”。
5) 脂多糖可以激活下丘脑-垂体-肾上腺轴,从而增加应激激素和/或神经递质。
6) 这会影响肠道生理、栖息地、微生物组组成和细菌基因表达。
7) 肠道微生物组的变化可能导致进一步的全身炎症,从而增强对中枢神经系统的影响。
8) 此外,它可能通过增加炎症负担来影响牙周病。
9) 口腔细菌通过唾液传播到肠道也可能影响肠道微生物组的组成和功能。
10)激素途径,例如,神经肽Y (NPY)、糖皮质激素、胰高血糖素样肽-1 (GLP-1)、食欲素-A、瘦素,口肠菌群的变化导致这些激素稳态或受体响应,进而影响精神系统。
以上,我们可以看到,口-肠轴对于大脑,也就是心理健康来说非常重要。这也就说明了口腔健康和心理健康之间存在着很大关联。
值得注意的是,口腔-肠道微生物组轴改善了胃肠道系统的发病机理和预后的预测。荟萃分析表明,口腔微生物组的变化与胃肠道癌的风险有关,包括CRC,PDAC和HCC,这可能是早期发现的潜在指标。已经验证了PDAC特定的口腔微生物模式作为PDAC生物标志物。两种口腔细菌物种长奈瑟菌(Neisseria elongata)和 轻型链球菌(Streptococcus mitis),同时富集可以将PDAC患者与健康受试者区分开来。
龋齿、严重牙周病、牙齿脱落是主要口腔疾病。口腔健康是一个全球被低估的健康问题。
口腔健康问题带来的经济负担
2017年,全球每年所有口腔疾病的负担达到了1830万,较1990年增加了19.9%;而全球经济负担为5440亿美元,其中1870亿美元是由生产力损失造成的。
间接成本,如生产力损失,可能归因于口腔健康对社交退缩和孤立、疼痛和咀嚼功能减弱、自尊心、缺乏口腔健康意识,对口腔医生的不信任所产生的影响。
口腔健康带来的健康问题与心理障碍并存
口腔健康对一般健康有影响,有证据表明口腔健康与冠心病、呼吸系统疾病、中风和糖尿病密切相关。这些疾病在患有心理障碍的人群中常见并存。
口腔健康问题和心理问题之间关联
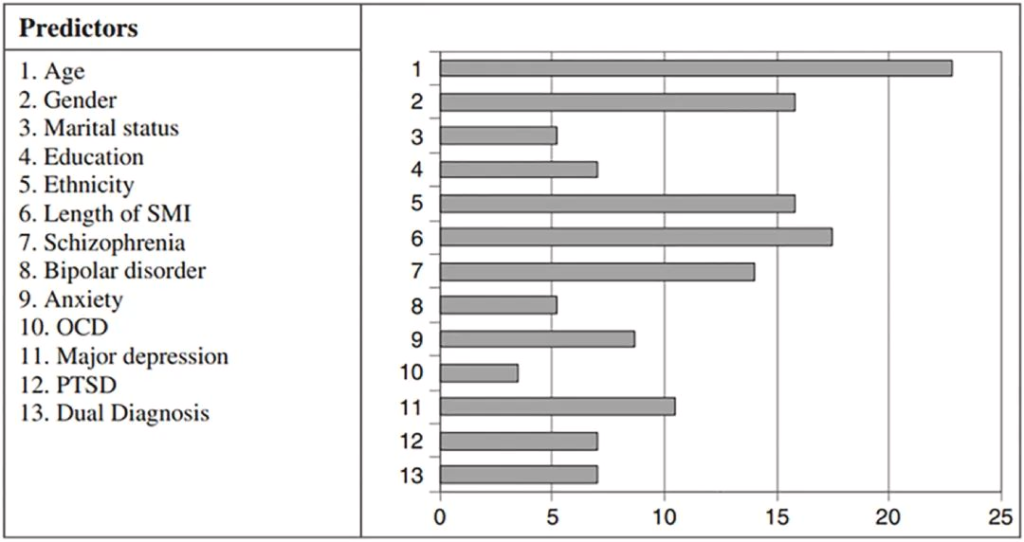
据报道,与普通人群相比,严重的精神障碍导致缺牙的风险高2.8倍。口腔健康不佳的预测因素和决定因素涉及心理障碍和生活方式因素,说明了心理和口腔健康中影响因素的复杂相互作用。
口腔健康状况不佳的预测因素
doi.org/10.1016/j.jobcr.2023.06.003
口腔健康状况不佳的决定因素
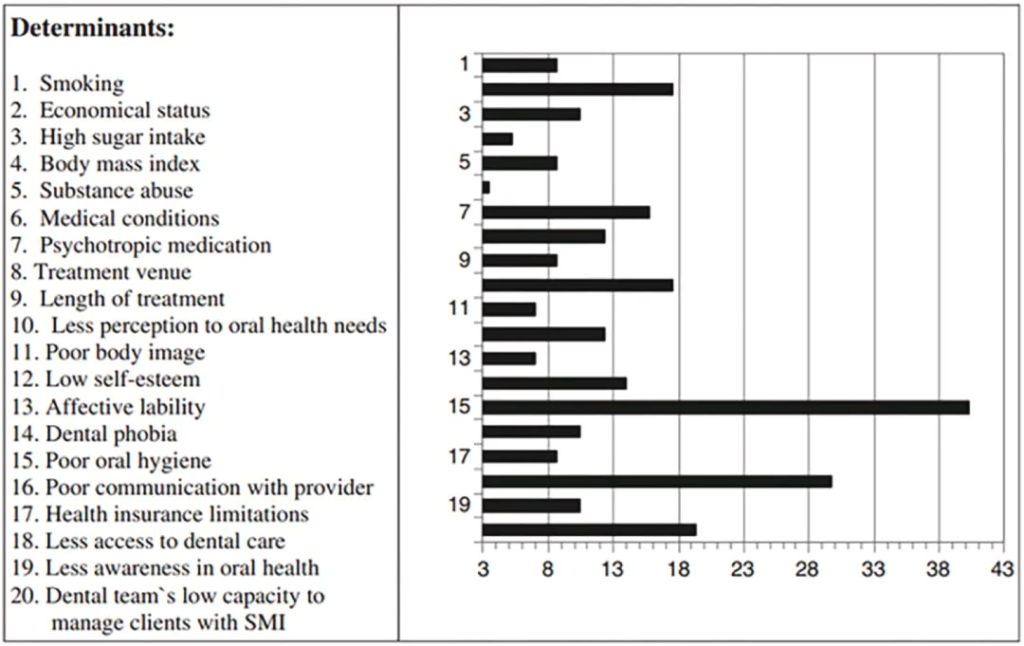
doi.org/10.1016/j.jobcr.2023.06.003
诊断口腔疾病时发现异常,应考虑是否存在精神障碍
常见的情况,如牙齿表面物质的损失,可能归因于许多基于其表现的精神障碍。
以上我们了解了口腔健康与心理健康之间的关联,接下来进一步深入探讨精神障碍与牙科疾病之间的关系。研究表明,精神障碍患者更容易出现牙科问题,而牙科疾病也可能与精神健康问题相关联。
▼
精神障碍涉及一个人的行为或心理模式;基于 DSM-5 中的标准。
最常见的疾病是抑郁症和焦虑症,影响世界人口的约 3.8%。最近,全球青少年抑郁和焦虑的患病率估计为 25-31%。心理健康状况不佳是全球日益严重的负担。这不仅仅涉及药物和住院等直接成本,还包括因失业或迅速退休而导致生产损失的收入损失。
常见的精神障碍包括抑郁症、焦虑症、躁郁症、精神分裂症、痴呆症、酒精和药物滥用障碍等。
常见的心理健康障碍及其常见症状

doi.org/10.1016/j.jobcr.2023.06.003
▼
牙周炎常见且危害较多
牙周炎是一种慢性疾病,影响牙齿结构周围的组织,伴有炎症障碍、退化,最终导致牙齿脱落。
除了牙齿脱落和随后的咀嚼功能障碍,牙周炎还影响整体健康。在全球范围内,牙周炎是最常见的疾病之一,有20-50%的人口受到影响。随着人口老龄化和越来越多的老年人选择保留天然牙齿,牙周炎的患病率预计将增加。
牙周炎是一种慢性炎症性疾病
牙周炎会使宿主全身长期暴露于促炎细胞因子和急性期蛋白中,中度至重度牙周炎患者的全身C反应蛋白水平升高。评估牙周炎全身炎症影响的研究表明,前列腺素E2、IL-1β、IL-6和TNF-α水平升高,牙周炎产生的炎症介质可以通过系统和神经途径延伸到大脑。牙周袋为大量牙周细菌进入系统循环和神经组织提供了独特的机会。
精神障碍及其与口腔疾病,特别是牙周炎的关系,在研究界越来越受到关注。其中一些关系被认为是双向的,这为未来的治疗、诊断和预防措施开辟了道路。
阿尔茨海默病
牙周炎和阿尔茨海默病之间存在显著关联
许多研究支持这种联系,并提出了几种解释,细菌移位导致全身炎症的机制似乎是合理的。
牙龈卟啉单胞菌(牙周炎的主要病原体之一)的DNA以及针对几种牙周炎相关细菌的抗体支持了这一点。
口腔微生物组通过血管、炎症/免疫、神经毒性和葡萄糖代谢途径,在已建立的生活方式因素和阿尔茨海默病风险之间是一个合理的因果中介:
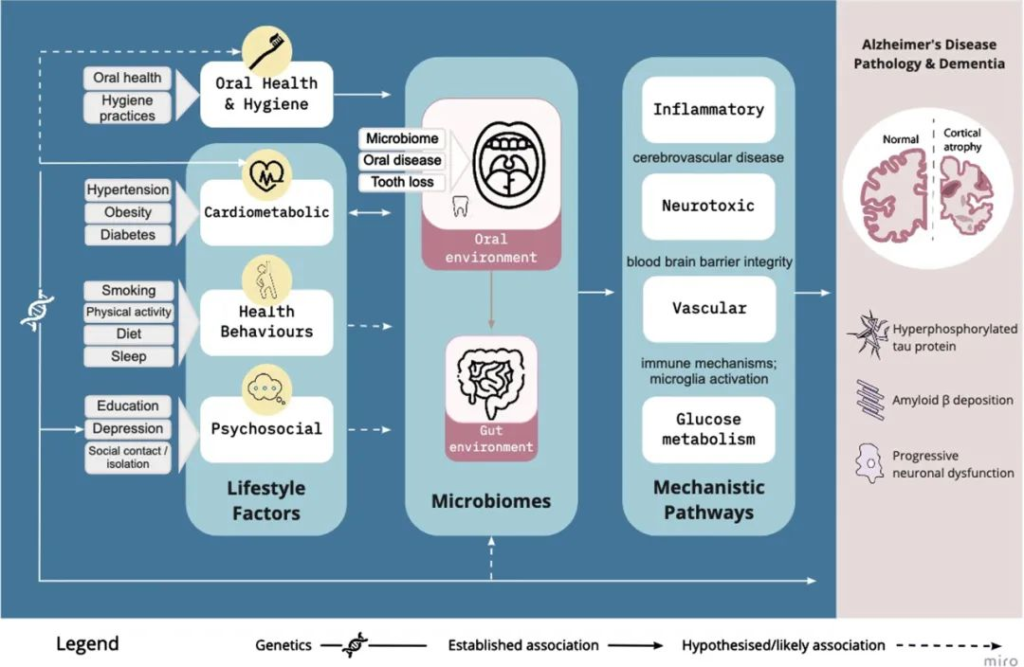
Loughman A, et al.,J Alzheimers Dis. 2023
牙周炎的严重程度与阿尔茨海默病之间存在关联
对这一证据的进一步支持归功于动物研究,其中小鼠受试者被给予活的牙龈卟啉单胞菌或其内毒素脂多糖。给药导致动物海马中学习和记忆功能明显下降,淀粉样蛋白-β斑块(一种与阿尔茨海默病相关的典型组织学发现)减少。
这些发现激发了抑制牙龈卟啉单胞菌蛋白酶的银杏蛋白酶抑制剂的开发。抑制剂导致斑块形成、细菌体积减少,并对海马细胞产生保护作用。
注:一个例子是用于治疗阿尔茨海默病的银杏蛋白酶抑制剂COR388,目前正处于2/3期临床试验中。
总之,文献表明,牙周炎是痴呆症的一个可改变的风险因素(特别是阿尔茨海默病),因此可以作为治疗和预防措施的目标。
抑郁症
细菌可能在牙周炎和抑郁症之间的关系中发挥作用。
抑郁症患者的口腔微生物组存在显著差异
一项16s高通量测序研究比较了符合 DSM-IV 抑郁症标准的年轻人 (n = 40) 和匹配对照 (n = 43) 的唾液微生物组的结构和组成,健康受试者和抑郁受试者之间的分类群在丰度上不同,奈瑟菌属(Neisseria)和变黑普雷沃氏菌(Prevotella nigrescens)的水平升高。
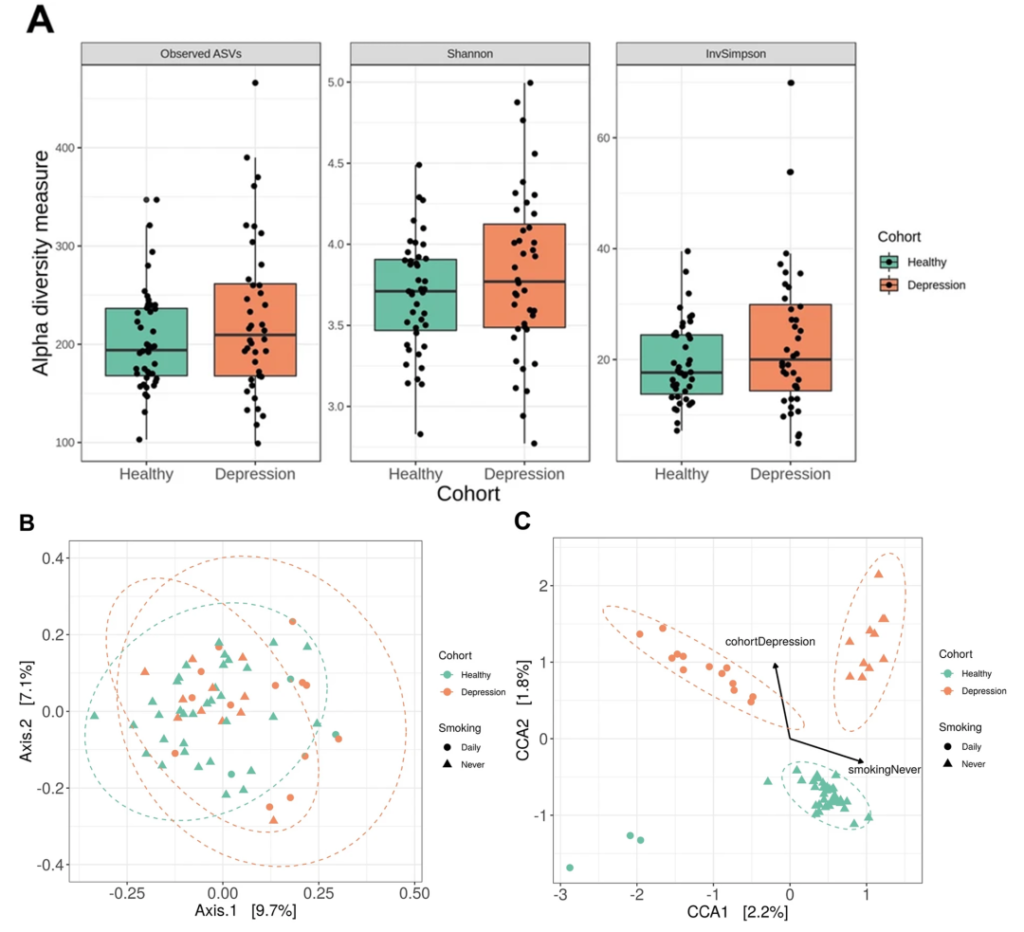
Wingfield B, et al.,Sci Rep. 2021
动物研究支持这样一种观点,即:
口服脂多糖或活的牙周炎相关病原体会导致包括大脑在内的系统炎症标志物升高和抑郁样行为。
有人提出了一种遗传关系。一项研究调查了串扰基因和神经肽在这两种疾病中的作用。
神经肽肾上腺髓质素、胰岛素样生长因子2、强啡肽原和抵抗素在牙周炎和抑郁症中相互表达,也在识别抑郁症中发挥作用。
牙周炎和抑郁症共病
抑郁症受到心理和社会因素的影响。牙周炎症状,如口臭、口腔卫生不良、缺牙、牙龈退缩,可能会出现社会孤立、羞耻、自尊下降等,产生负面影响,从而导致抑郁。
种植体周围炎也可能是抑郁症的危险因素
当牙齿丢失时,可能会被种植牙替代,然而,种植牙也可能发展为种植体周围炎,类似于牙周炎。最终,植入物可能会丢失。从牙周炎到种植体周围炎,可以推断出几种影响和疾病关系。牙周炎和种植体周围炎可能是抑郁症的可改变的危险因素,如果是这样,简单的牙周干预和口腔卫生指导可以预防或帮助治疗抑郁症。
双相情感障碍(躁郁症)
慢性炎症是双相情感障碍的一个因素。
2001年-2012年对双相情感障碍与牙周炎之间的关系进行了研究:
这些研究支持了双相情感障碍和牙周炎之间的可能关系,值得进一步研究。
帕金森病
一些研究报告了帕金森氏症患者牙周炎患病率的升高。
帕金森病会导致运动障碍和认知障碍,这是由于大脑黑质中产生多巴胺的神经元的神经元细胞死亡所致。
手抖和僵硬是常见的症状,这就很难保持足够的日常口腔卫生。这种疾病本身可以说是牙周炎的一个危险因素,然而,流行病学证据支持牙周炎会增加患帕金森病的风险。
与完全没有接受治疗或连续5年没有接受治疗的患者相比,在连续5年接受牙周治疗的患者中观察到了对帕金森病的保护作用。作者推测:
牙周炎相关病原体进入大脑,引发和维持的炎症最终会导致帕金森症。
进一步研究其机制关系,以及牙周治疗对已确定的帕金森氏症的影响,可能是未来的方向。
精神分裂症
关于牙周炎和精神分裂症之间关系的支持性文献很少。
一些研究报告称,精神分裂症患者患牙周炎的风险较高,服用抗精神病药物的患者患牙周炎风险更高。
血管紧张素转换酶基因的D等位基因是对抗精神分裂症和牙周炎的保护因子,并可能被证明是一种生物学联系。
口咽微生物组、唾液微生物组和牙周炎的作用被认为可能与精神分裂症有关,牙周炎强化了炎症在精神分裂症病理生理学中的作用。
口腔微生物群与精神分裂症之间的联系
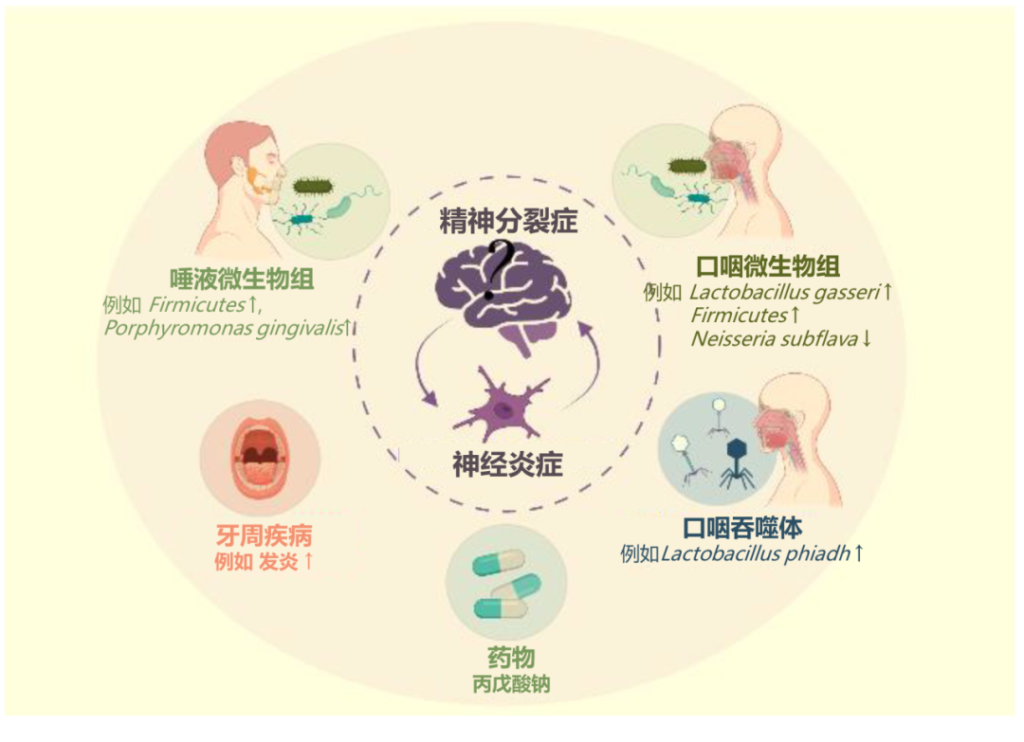
doi.org/10.3390/ijms23020846
唾液是一种对多种疾病具有诊断意义的体液,也可能用于精神分裂症的诊断,目前的证据仍然有限,还需更多研究。
精神分裂症患者唾液牙龈卟啉单胞菌较多
一项横截面研究发现,精神分裂症患者唾液中牙龈卟啉单胞菌的数量显著更高。此外,牙龈卟啉单胞菌细胞的数量与精神分裂症的精神病理学严重程度呈正相关。牙龈卟啉单胞菌可能导致神经炎症状态。
患有精神病发作的精神分裂症患者血清中炎症细胞因子的浓度升高,包括IL-12、干扰素 γ、肿瘤坏死因子α 和 C反应蛋白。低度慢性炎症状态可能会导致免疫系统异常,从而使精神分裂症患者易患全身性疾病。
精神分裂症患者口腔菌群在肠道定植
口腔常驻细菌口腔乳杆菌(Lactobacillus oris)、唾液链球菌(Streptococcus salivarius)可能在精神分裂症患者的肠道中定殖,导致口腔常驻细菌显著富集。
也有宏基因组研究发现,与精神分裂症相关的肠道细菌之间的共现相关性,大多数物种起源于口腔,口腔常驻细菌可能以协同的方式在精神分裂症患者的肠道定植。
与口腔疾病相关的精神障碍的管理
精神健康障碍患者应接受全面的口腔和牙周健康信息、卫生指导、教育和定期随访,以提高患者的意识、习惯。
多学科干预可以进一步改善依从性、牙齿恐惧、口腔健康和习惯,并有助于更积极的预后。
牙科医生应接受有关精神障碍的教育,以便更好地管理、沟通和识别这些患者,并与其他卫生专业人员合作。牙科应与现有的精神康复和预防计划相结合,实现一个全面、方便的多学科计划,将人体内部的整体关联考虑在内。
“胃肠精神病学”涉及两大系统,涉及的病种、症状均较多,而且各个病种间可能交互出现,因此症状也可能隐匿、多变,这给诊断和治疗带来巨大的困难,因此需要包括消化系统内外科、精神心理、营养及影像科的多学科讨论。
医生应该了解当前的研究状况,在治疗过程中应该详细了解患者的饮食习惯、生活习惯和肠道健康状况对患者的发病原因进行综合评价,并就基于肠道菌群的干预措施向患者提准确而明智的建议,将改善饮食、生活方式等改善肠道健康的方式纳入患者的治疗方案之中。以下给大家一些在选用常见的菌群改善或营养饮食方面的简单建议。
肠道菌群作为治疗精神疾病的潜在靶点,一直是近年来精神病学研究的热点。肠道细菌通过影响神经、免疫和内分泌,在肠道和大脑的交流中发挥关键作用。
微生物-肠-脑轴为精神病学的研究和治疗提供了一种新的范式。一项大规模的人群研究证实,许多精神疾病患者,特别是精神分裂症、双相情感障碍和重度抑郁症等重度精神疾病患者,比一般人群摄入的致肥胖营养物质和炎症性饮食更高。自闭症患儿也大多存在严重的偏食和挑食等不良饮食习惯。
虽然关于肠道菌群在精神类疾病发生中的具体作用还有待发现,但是营养和肠道健康领域已经成为精神疾病整体治疗中的一个重要组成部分。如在自闭症儿童中,存在严重的食物过敏现象,限制麸质饮食和酪蛋白饮食有助于自闭症症状的改善;诸多精神类疾病也存在偏食症状,如偏食促发自闭症的发生发展。IBS患者可能有与饮食成分消化不良相关的腹胀,如可发酵的低聚糖、双糖或单糖和多元醇饮食有关。因此,建议精神障碍人群的精准个性化的营养饮食。
益生菌无论是在精神性疾病或者胃肠功能性疾病都表现良好的治疗效果,而且存在双向调节作用。近年来,关于肠-脑轴的研究则提供了更加充分的理论支持。因此,肠-脑-菌群轴被认为是能为精神病患者提供创新疗法的基础,其重点是在临床层面上系统性鉴别出精神益生菌(psychobiotics)。在临床前研究中发现,精神益生菌对行为、肠道通透性、神经活性有益处和减少促炎性及应激反应。
在啮齿动物中的实验表明,精神病益生菌可通过迷走神经、脊髓、神经内分泌系统起到抗抑郁及抗焦虑的作用。精神病益生菌在IBS患者中被广泛研究,他的作用包括缓解抑郁症状及慢性疲劳综合征等,可能与益生菌的抗炎及减少下丘脑-垂体-肾上腺轴活性相关。
虽然领域还处于起步阶段,存在着许多挑战,但是鉴于目前已有的初步证据,在精神类疾病的治疗中,补充益生菌或加入一些营养配方是合理且具有极大潜力的。
临床报道益生元对于慢性便秘、腹泻、IBS及焦虑、抑郁、自闭症等具有良好的疗效。益生元是被宿主微生物选择性利用以促进健康的物质,发挥抵抗病原体、调节免疫、增加矿物质吸收、改善肠道功能、影响代谢和饱腹感等作用。
益生元与益生菌还存在协同和互养的作用,如果聚糖、低聚半乳糖、抗性淀粉、维生素、植物多酚、海藻等益生元可通过不同机制维持益生菌在肠道中的活性,包括增强益生菌对氧气及活性氧的抗性、增强其对胃酸和胆汁酸的抗性。益生元可通过多种机制增强益生菌在肠道菌群中的益生功能,包括发酵产生的短链脂肪酸促进益生菌的定殖、与致病菌竞争性结合宿主上皮细胞上的受体。
无论是基础研究还是临床研究均表明,肠道菌群移植在治疗胃肠功能的同时,对肠道外疾病如精神神经系统包括焦虑和抑郁症、自闭症、帕金森病、阿尔兹海默症等均有良好的疗效。
上海市第十人民医院在《Lancet Gastroenterol Hepatol 》杂志上发表成果显示,8547例菌群移植治疗肠道功能合并自闭症、焦虑和抑郁等疾病的成果,均表现出良好的临床疗效和安全性。此外,在对IBS的5年长期随访中发现,菌群移植在改善胃肠功能障碍的同时,对精神心理也有很好的疗效。
菌群移植通过纠正肠道菌群的失衡,增加菌群的多样性,促进有益菌的定殖等,在改善胃肠道功能的同时,肠道内环境的改善,肠道菌群及其代谢产物可以通过自主神经系统、免疫系统调节中枢的活性,从而改善精神行为的异常症状。近年来,人源化的无菌动物模型的构建,更进一步证明肠道菌群移植对精神神经系统的干预作用,即将自闭症、焦虑或者抑郁症患者的粪便移植至无菌小鼠内,无菌小鼠产生临床类似的精神症状,而通过健康小鼠共喂养或者将健康人群的粪便移植至模型小鼠,其症状可以得到明显的改善。
无锡第二人民医院柳老师团队前期的研究结果显示,对菌群移植可以显著改善睡眠障碍合并抑郁症的患者,菌群移植治疗后,睡眠和抑郁症同时得以明显的改善。因此,菌群移植在功能性胃肠病合并精神症状的机体中,具有双向调节作用,为该类患者提供了整体治疗思路。
对于复杂胃肠疾病,心理医师的介入是非常关键的,需要心理医师,参与筛查、评估、诊断、治疗和随访。
同时患者家庭,亲属和朋友对“胃肠精神共病”需要有充分的认知和参与,认知越高参与越积极,治疗效果越好。如父母因素在儿童慢性便秘的病理生理和预后中有重要作用,神经质和存在抑郁症状的父母及父母的养育态度和培养行为与便秘严重程度相关;在这种情况下,基于家庭的认知干预也许有好处。
▼
抗菌药物是我们对抗病原体感染的第一道防线,致力于消除特定物种(即噬菌体治疗),或整个微生物群(即抗生素治疗)。
去除与神经精神疾病相关的微生物,可能是未来解决神经精神疾病症状或严重程度的新方法。口服抗生素已被证明对口腔微生物群影响很小,在口腔中局部或直接施用抗生素可能更有效。
▼
一项关于饮食与牙周炎之间关系的10000名NHANES参与者的横断面研究发现,富含水果、蔬菜、沙拉、水、茶的饮食模式,一定程度限制摄入可发酵碳水化合物、脂肪酸、蛋白质和高糖饮料的摄入,患牙周病的程度较低。
▼
益生菌作用的主要益生菌机制包括:
体外和体内证实了乳酸杆菌和双歧杆菌对调节与精神障碍相关的口腔微生物群(动物双歧杆菌、副干酪乳杆菌、嗜酸乳杆菌、鼠李糖乳杆菌、德氏乳杆菌)的作用。细菌竞争排除了一些病原体而不破坏生物膜结构(具核梭杆菌、牙龈卟啉单胞菌等)。
乳酸杆菌和双歧杆菌属可以帮助控制口腔中致龋链球菌的生长。
▼
使用各种益生元化合物(硝酸盐、β-甲基-d-半乳糖苷、N-乙酰基-d-甘露糖胺等)对口腔微生物组进行营养刺激,可能会诱导牙齿生物膜的组成和有益口腔细菌的生长,减少致病菌(P.gingivalis、A. actinomycetemcomitans、F.nucleanum)。
虽然益生菌的使用可能作为精神障碍患者的补充治疗手段,但有必要注意口腔微生物组稳态的多因素特征。
有研究发现,海藻提取物、n-3 PUFA、海参提取物和海洋细菌代谢物等海洋生物活性成分具有抑制口腔致病菌、消除炎症和抗肿瘤的作用。这一发现为通过使用这种生物活性成分(例如以口香糖或无糖片剂的形式包装)来预防和稳定精神障碍开辟了有趣的研究前景。
▼
来自健康捐赠者的微生物群被移植或播种到接受者体内。口腔微生物移植(OMT)可能能够作为预防龋齿的第一道防线或用于治疗牙周病,但这尚未在人体中进行过测试。未来应探索 OMT 在缓解神经精神疾病等全身性疾病症状方面的应用。
▼
牙膏和漱口水等口腔卫生产品通过限制某些物种的生长来管理口腔微生物群落,并且可能是促进所需口腔微生物定殖的一种方法。
例如,当前的牙膏采用具有抗菌特性的化学物质(例如氟化物)配制,除了促进牙釉质健康之外,氟化物已被证明可以降低总体微生物负荷和多样性。许多漱口水含有酒精来杀死微生物。
虽然这些是日常卫生习惯中用于预防和减少口腔疾病的工具,但它们也可能是帮助调节与神经精神疾病相关的微生物的工具。新的研究正在检查其他化合物,它们可以在不破坏微生物群共生平衡的情况下保持口腔卫生。
▼
在研究口腔疾病的预防措施和干预措施时,需要考虑的一个重要组成部分是微生物群产生的生物膜。
生物膜是口腔微生物在牙齿坚硬表面形成的细胞外基质。这些生物膜附着在牙齿表面,将微生物群包裹在分泌聚合物的保护层中,使微生物能够抵抗环境变化。
微生物群还能够通过基因表达模式改变生物膜表型以响应变化。正因为如此,口腔微生物生物膜能够抵抗去除和抗生素或用于抗菌治疗。因此,生物膜可能在移植成功或抗菌产品功效中发挥重要作用,并可能在神经精神疾病的发展和治疗反应中发挥潜在作用。
▼
口腔微生物群可以通过改变生活方式或饮食来调节,或者可能通过改变环境暴露来调节。
通过饮食和水源将环境微生物引入口腔是优先的,尽管这方面的研究有限。环境暴露可能在我们如何考虑调节口腔微生物群以解决系统健康问题方面发挥作用。
例如,城市规划通过引入自然“绿地”来增加土壤微生物的多样性,可能有助于增加对有益微生物的接触,因为接触这些环境微生物可能在神经精神疾病的治疗中发挥关键作用。
具体而言,暴露于土壤细菌分枝杆菌已被证明对宿主具有抗焦虑作用,因为宿主的免疫反应释放抗炎细胞因子,对减少身体和大脑中的炎症具有积极效果,这在焦虑和抑郁中是一个重要因素。
然而,通过口腔微生物群促进的机制尚未确定。尽管如此,实施提供健康和环境暴露的社会政策(例如,要求儿童在上学期间每天安排一点时间接触这些空间),可以进一步确保人们能够受益于环境微生物多样的地方。
口腔健康与精神障碍之间的相互作用可以从微生物群-口肠-脑轴的几个角度进行解释,包括微生物群失调、细菌迁移和神经炎症等。目前对于微生物组在口腔-肠道-脑轴中的作用已经有了一定的认识,这是一个重要的研究方向,从病理生理学到调节肠道微生物组对精神疾病的临床影响。进一步了解微生物组与精神疾病的发展和预后之间的紧密联系,还需要探究地理、种族、饮食模式、过往医疗状况、口腔护理以及胃肠道手术等因素如何改变微生物组的情况。
口肠微生物组还可被视为精神疾病潜在治疗的目标之一。例如,通过改变饮食习惯、使用益生菌或抗生素等手段来调节口肠微生物组,可能对精神疾病的治疗产生积极的影响。谷禾正在整合口腔和肠道微生物组数据,在提高检出率等方面获得了更全面的视角。
总的来说,人体微生物群检测有望为精神疾病的早期辅助诊断、辅助治疗和预防提供新的方法和策略。在一些细分疾病领域中仍需要进一步的研究和临床实践来推动其在临床上的应用。
相关阅读:
环境污染物通过肠脑轴影响心理健康,精神益生菌或将发挥重要作用
主要参考文献:
Skallevold HE, Rokaya N, Wongsirichat N, Rokaya D. Importance of oral health in mental health disorders: An updated review. J Oral Biol Craniofac Res. 2023 Sep-Oct;13(5):544-552. doi: 10.1016/j.jobcr.2023.06.003. Epub 2023 Jun 19. PMID: 37396968; PMCID: PMC10314291.
Wingfield B, Lapsley C, McDowell A, Miliotis G, McLafferty M, O’Neill SM, Coleman S, McGinnity TM, Bjourson AJ, Murray EK. Variations in the oral microbiome are associated with depression in young adults. Sci Rep. 2021 Jul 22;11(1):15009. doi: 10.1038/s41598-021-94498-6. PMID: 34294835; PMCID: PMC8298414.
Loughman A, Adler CJ, Macpherson H. Unlocking Modifiable Risk Factors for Alzheimer’s Disease: Does the Oral Microbiome Hold Some of the Keys? J Alzheimers Dis. 2023;92(4):1111-1129. doi: 10.3233/JAD-220760. PMID: 36872775; PMCID: PMC10200234.
Ball J, Darby I. Mental health and periodontal and peri-implant diseases. Periodontol 2000. 2022 Oct;90(1):106-124. doi: 10.1111/prd.12452. Epub 2022 Aug 1. PMID: 35913583; PMCID: PMC9804456.
Maitre Y, Mahalli R, Micheneau P, Delpierre A, Guerin M, Amador G, Denis F. Pre and Probiotics Involved in the Modulation of Oral Bacterial Species: New Therapeutic Leads in Mental Disorders? Microorganisms. 2021 Jul 6;9(7):1450. doi: 10.3390/microorganisms9071450. PMID: 34361886; PMCID: PMC8306040.
Maitre Y, Micheneau P, Delpierre A, Mahalli R, Guerin M, Amador G, Denis F. Did the Brain and Oral Microbiota Talk to Each Other? A Review of the Literature. J Clin Med. 2020 Nov 28;9(12):3876. doi: 10.3390/jcm9123876. PMID: 33260581; PMCID: PMC7760025.
Scassellati C, Marizzoni M, Cattane N, Lopizzo N, Mombelli E, Riva MA, Cattaneo A. The Complex Molecular Picture of Gut and Oral Microbiota-Brain-Depression System: What We Know and What We Need to Know. Front Psychiatry. 2021 Nov 2;12:722335. doi: 10.3389/fpsyt.2021.722335. PMID: 34819883; PMCID: PMC8607517.
Bowland GB, Weyrich LS. The Oral-Microbiome-Brain Axis and Neuropsychiatric Disorders: An Anthropological Perspective. Front Psychiatry. 2022 Mar 30;13:810008. doi: 10.3389/fpsyt.2022.810008. PMID: 35432038; PMCID: PMC9005879.
Martínez M, Postolache TT, García-Bueno B, Leza JC, Figuero E, Lowry CA, Malan-Müller S. The Role of the Oral Microbiota Related to Periodontal Diseases in Anxiety, Mood and Trauma- and Stress-Related Disorders. Front Psychiatry. 2022 Jan 27;12:814177. doi: 10.3389/fpsyt.2021.814177. PMID: 35153869; PMCID: PMC8833739.
Paudel D, Uehara O, Giri S, Yoshida K, Morikawa T, Kitagawa T, Matsuoka H, Miura H, Toyofuku A, Kuramitsu Y, Ohta T, Kobayashi M, Abiko Y. Effect of psychological stress on the oral-gut microbiota and the potential oral-gut-brain axis. Jpn Dent Sci Rev. 2022 Nov;58:365-375. doi: 10.1016/j.jdsr.2022.11.003. Epub 2022 Nov 17. PMID: 36425317; PMCID: PMC9678961.
Tan HE. The microbiota-gut-brain axis in stress and depression. Front Neurosci. 2023 Apr 14;17:1151478. doi: 10.3389/fnins.2023.1151478. PMID: 37123352; PMCID: PMC10140437.
Martin S, Foulon A, El Hage W, Dufour-Rainfray D, Denis F. Is There a Link between Oropharyngeal Microbiome and Schizophrenia? A Narrative Review. Int J Mol Sci. 2022 Jan 13;23(2):846. doi: 10.3390/ijms23020846. PMID: 35055031; PMCID: PMC8775665.