-

 国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子
国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子

 二级病原微生物安全实验室
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业

谷禾健康
健康的肠道菌群呈现出物种多样性高、结构稳定、功能良好等的特征,有益菌、中性菌和有害菌之间维持着动态平衡。然而,当这种平衡被打破,即发生菌群失衡,便可能成为多种急、慢性疾病的诱因或加剧因素。
目前谷禾积累了大量样本数据,通过对这些数据的深度分析,我们观察到多种多样的菌群状态,其中不乏一些表现出极端特征的案例。这些案例为我们理解菌群失衡的具体模式及其对健康的潜在影响提供了宝贵的窗口。
本文我们选取谷禾数据库中的一些检测报告,深入探讨几种典型的肠道菌群极端失衡状态,包括:菌群多样性严重低下、特定菌门/菌属的极端优势、益生菌缺乏、益生菌补充过量引发的新失衡等。
通过对这些案例的具体数据进行解读,我们希望能为相关研究人员和临床工作者及后端干预方提供更直观的参考,从而共同推动肠道微生态健康管理的发展。
编辑
( – 衡量微生态稳定的指标 – )
菌群多样性是评估肠道微生态健康状况的首要指标。它包含两个维度:一是物种丰富度,即菌种的数量;二是物种均匀度,即各种菌在群落中数量分布的均衡性。
一般情况下,一个高多样性的菌群生态系统更具弹性和稳定性,能够有效抵御外界干扰(如短期饮食改变、药物影响、病原体入侵),并维持正常的生理功能。相反,多样性低的菌群则显得脆弱,更容易在外界压力下崩溃,导致功能紊乱和疾病风险的增加。
案例一:
多样性荒漠化与病原菌疯长的恶性循环
这份报告显示,菌群多样性评分仅为3分(满分100),属非常低的水平。
编辑
<来源:谷禾健康肠道菌群检测数据库>
香农多样性指数仅为2.29(正常中位数约4,5左右)。
注:香农多样性(Shannon Diversity Index)是生态学中用于衡量生物群落多样性的一种常用指标。H′=−∑(pi⋅lnpi).
香浓多样性指数一个是反映了物种丰富度,一个是反映物种均匀度,综合考虑上述两者,数值越高越复杂。
这表明其肠道微生态环境极其单一,稳定性极差。这种荒漠化的肠道环境,为条件致病菌的过度繁殖提供了绝佳条件。
1)菌属构成分析
编辑
<来源:谷禾健康肠道菌群检测数据库>
可以看到,肠道内沙门氏菌属(Salmonella)的丰度非常高,达到了的68.365%,这其中主要是肠炎鼠伤寒沙门氏菌。
这是一种典型的致病菌,由于它的超高丰度,直接导致了变形菌门(Proteobacteria)的占比飙升至71.436%(正常人群中位数一般不超过10%,甚至低于5%)。
变形菌门被广泛认为是促炎菌门,其成员多为条件致病菌。沙门氏菌的肆虐,如同一场生态灾难,严重挤压了其他共生菌的生存空间,直接导致了有益菌(如双歧杆菌、乳杆菌)和大量核心功能菌属的缺失,这是其菌群多样性低下的直接原因。
编辑
<来源:谷禾健康肠道菌群检测数据库>
可以看到有益菌、核心菌属严重缺乏。
2)生理后果与炎症水平
这种极端失衡带来了严重的生理后果。报告显示,其肠道炎症水平高达95(重度炎症),肠道产气严重。这与沙门氏菌感染的典型症状(腹痛、消化不良、炎症反应)相吻合。
编辑
<来源:谷禾健康肠道菌群检测数据库>
同时,丁酸盐(1分,缺乏)、吲哚(2分,缺乏)等关键抗炎和维持肠道屏障功能的代谢物严重不足,进一步加剧了肠道损伤。
3)恶性循环的形成
这是一个典型的“病原菌疯长 → 多样性锐减 → 肠道功能崩溃 → 炎症加剧”的恶性循环。
此案例中我们看到了菌群多样性的重要性。当多样性降至冰点,肠道微生态的防御体系形同虚设,单一病原菌便可一家独大,主导整个群落,并引发严重的健康问题。
对于此类情况,干预策略不仅要考虑补充益生菌,更关键的是要针对性地清除优势病原菌,并创造条件帮助多样性恢复,当然这种较为复杂的情况是需要一个专业干预指导的过程。
上述是多样性很低,大量被病原菌占据的情况下,那么同样是多样性低的时候,有没有可能有一些相对“好”的菌群在竞争中脱颖而出呢?如果有的话,会发生什么呢?我们来看下一个案例。
案例二:
多样性低,大量普雷沃氏菌占据
接下来我们要看的这份报告是多样性低的另一种典型代表。其肠道菌群被普雷沃氏菌属(Prevotella)以61.025%的绝对优势所统治。
编辑
<来源:谷禾健康肠道菌群检测数据库>
普雷沃氏菌属通常与富含碳水化合物和植物纤维的饮食模式相关,常见于素食或以植物性饮食为主的人群。虽然在某些情况下,普雷沃氏菌属被认为是健康植物性饮食的标志,但当其丰度过高,形成单一优势时,问题便随之而来。
1)菌群多样性降低带来的影响
首先,普雷沃氏菌属的过度增殖同样挤压了其他核心菌属的生存空间,导致了菌群多样性的降低。
编辑
<来源:谷禾健康肠道菌群检测数据库>
报告显示,多个产丁酸的关键菌属如瘤胃球菌属(Ruminococcus)、罗氏菌属(Roseburia)、经黏液真杆菌属(Blautia)均处于缺乏状态。
丁酸是结肠上皮细胞的主要能量来源,具有强大的抗炎作用。这些菌的缺失意味着肠道抗炎能力的下降。
编辑
<来源:谷禾健康肠道菌群检测数据库>
2)促炎反应的风险
其次,高丰度的普雷沃氏菌属本身也与促炎反应相关。部分普雷沃氏菌菌株能够产生脂多糖(LPS),这是一种强烈的免疫刺激物,可引发炎症。
更值得警惕的是,该案例中同时伴随着变形菌门中病原菌埃希氏菌属(25.553%)严重超标,影响肠道菌群平衡,同时诱发高炎症风险。
其II型糖尿病风险评估为0.83(高风险),这与菌群的长期失衡和促炎状态可能存在关联,因为慢性低度炎症是胰岛素抵抗和II型糖尿病的重要病理生理基础。
编辑
<来源:谷禾健康肠道菌群检测数据库>
3)生态平衡的重要性
此案例说明,即使是与健康饮食模式相关的菌属,其过度增殖也可能打破生态平衡,导致核心功能性菌群的缺失和炎症风险的升高。肠道健康的关键在于平衡而非某种超级细菌的绝对优势。
普雷沃氏菌属(Prevotella)和拟杆菌属(Bacteroides)的相对丰度则与长期饮食模式(植物性饮食vs.动物蛋白/脂肪饮食)紧密关联。当某一菌群异常增殖,打破了原有的平衡,就会导致一系列代谢和免疫问题。
这提示我们,饮食干预需要精细化,在保证膳食纤维摄入的同时,也需关注蛋白质、脂肪的均衡配比,避免长期单一的饮食结构导致菌群结构的极化。
#########################################
关于 多样性过低 的分析思路:
谷禾报告中,多样性分值<15 提示 “单一菌群”(菌群数量极少,功能单一);15-30 提示 “多样性较低”(需结合临床症状干预) 。
◆ 核心判断
若多样性低伴随核心菌属缺失、 有害菌比例升高,则提示菌群结构失衡,需重点干预。
◆ 原因分析
——外部因素
饮食单一化:长期高糖 / 高脂 / 精粮饮食(如仅吃精制米面),缺乏膳食纤维(<15g / 日),导致菌群可利用底物不足。
药物滥用:广谱抗生素(如头孢类)、化疗药物直接杀灭有益菌,破坏菌群平衡。
生活方式:久坐少动(肠道蠕动减慢,菌群代谢产物不足)、熬夜(皮质醇升高抑制厌氧菌生长)。
——内部因素
慢性疾病:溃疡性结肠炎、糖尿病、肥胖等疾病直接损伤肠道黏膜,或通过代谢紊乱抑制菌群增殖。
衰老:70 岁后肠道菌群多样性自然下降,核心菌丰度都会有所降低。
◆ 潜在危害与关联疾病
肠道功能异常
腹泻 / 便秘交替、腹胀、肠易激综合征(IBS)风险升高(核心菌减少导致肠道屏障功能下降)。
代谢与免疫风险
短链脂肪酸合成不足,增加肥胖、胰岛素抵抗、心血管疾病风险。
免疫细胞(如T细胞)活化不足,易引发反复感染(如呼吸道 / 肠道感染)。
疾病恶性循环
菌群失衡→消化吸收差→营养缺乏→免疫力进一步下降,形成 “菌群 – 疾病” 循环。
多样性过低的本质是菌群结构失衡,需结合数值、阈值、核心菌 / 有害菌比例及代谢功能综合判断,重点关注核心菌缺失和功能受损,而非单纯追求菌群数量。
#########################################
上述我们看到了多样性低引发的单一菌群大量占据的现象,这是不利的情况,那么,是否意味着多样性越高就越好呢?接下来,我们将通过一个数据库中的高多样性例子来探讨这个问题。
案例三:
多样性很高,是否是更健康的菌群结构?
通常情况下,我们认为多样性越高,肠道菌群结构就越趋向于稳定。然而,当多样性超出一定范围时,情况是否仍然如此呢?
这份报告,在数据库中属于多样性高的情况。
编辑
<来源:谷禾健康肠道菌群检测数据库>
从表面指标看,本案例的菌群呈现“高度多样”的特征:α多样性水平高。
一般成年人能测到的在500-1800种菌之间,平均香浓多样性指数就4-5的样子,这个多样性是显著超过平均值,这类结构通常意味着菌种丰富、生态位充足,理论上更具稳定性与抗干扰能力。
1)菌多但不够好的结构
然而,进一步结合平衡指数、有益菌、有害菌水平、核心菌属占比等指标,会发现这并不是一个典型的“稳态高多样性”,而更像一种“菌多但不够好”的结构。
我们来看一下菌群结构:
编辑
<来源:谷禾健康肠道菌群检测数据库>
结合整个报告来看,虽然多样性高但结构质量一般:核心/有益菌偏少 + 有害菌略偏多 + 还有几项机会型/胆汁相关菌偏高。可以说是一种看起来很热闹,但关键功能菌没站稳的生态结构。
2)核心菌属的缺乏
首先核心菌属占比水平 46(参考80–100):核心菌属不足是最核心的结构信号,报告也明确提示可能存在不常见菌增多或生态紊乱。
编辑
核心菌属累积占比不足,常被解读为常见稳定菌群缺乏,而由更多非核心菌或波动性菌群填充生态位,导致群落看似丰富,却可能更容易在饮食、压力、作息或抗生素等扰动下发生漂移。
3)有益菌与有害菌的失衡
有益菌水平 11(偏低,建议≥50) + 有害菌水平 52(略超参考上限50):说明功能性倾向不理想,“平衡指数29”也与此一致。
编辑
<来源:谷禾健康肠道菌群检测数据库>
在具体菌属层面:
关键产丁酸/抗炎相关核心属,粪杆菌属(Faecalibacterium)、罗氏菌属(Roseburia) 偏低的情况。这些菌常与丁酸盐生成、肠道屏障支持和抗炎稳态相关,缺乏时往往意味着肠道自我修复与抗炎能力的底盘不足。
4)机会致病菌的增多
机会型/胆汁相关菌脱硫弧菌属(Desulfovibrio)、嗜胆菌属(Bilophila) 偏高,且检出的沃氏嗜胆菌(Bilophila wadsworthia) 超过人群90%。这类菌群多常见于饮食结构(高脂/胆汁酸压力)、或近期扰动后的生态位变化。
综上,这是一个典型的多样性高≠更健康的案例:与案例一“病原菌极端优势导致崩盘”、案例二“单一优势菌占据生态位”不同,本案例更像“群落过于分散、关键功能菌不稳”的隐性失衡——需要把关注点从多不多转向核心功能菌是否到位、结构是否稳态。
#########################################
关于 “多样性是否过高” 的分析思路:
多样性过高的情况数据库中并不常见,且相对来说缺乏明确的关于过高的定义及统一的临床问题描述。但从菌群生态平衡的一般原理及现有知识推断,若假设存在 “多样性过高”(如菌种数量异常庞大、均匀性严重失衡),可能存在以下潜在问题:
1. 菌群结构稳定性下降
菌群多样性高通常伴随菌群间竞争激烈,若某类菌种因环境波动(如饮食、药物)出现适应性下降,可能导致整体结构趋于失衡,反而降低稳定性。
如同生态系统中物种过多但缺乏优势种,易因单一物种消失引发连锁反应,例如 “过度分散的菌群” 可能无法有效抵御外界致病菌入侵。
2. 有益菌丰度被稀释
若多样性过高伴随大量非优势菌种(如潜在致病菌、条件致病菌)过度繁殖,可能导致核心有益菌(如双歧杆菌、乳杆菌)的相对丰度被稀释,削弱其生理功能(如短链脂肪酸合成、免疫调节)。
3. 与特定疾病的潜在关联
现有研究多关注多样性低与疾病的关联(如炎症性肠病、代谢综合征),而多样性过高的临床数据较少。但部分罕见病或特殊生理状态下,可能出现菌群异常丰富伴随功能紊乱,例如:自身免疫病、慢性炎症等。
建议:若报告中多样性评分异常偏高,需关注是否存在检测误差或特殊生理状态(如长期摄入广谱益生菌导致菌群过度分散),并结合临床症状(如频繁腹泻、过敏)综合判断。
#########################################
多样性高并不天然等于更健康。当菌群看起来很丰富,但核心菌属占比不足、有益菌被稀释、机会型菌群趁势上升时,这种热闹但不稳的结构同样可能带来功能层面的隐患。
那么,较好的菌群结构应该长什么样?接下来,我们将通过案例四展示一份多样性相对较好的报告,看看多样性恰到好处时,菌群结构在平衡性、核心菌群与功能表现上会有哪些共同特征。
案例四:
多样性较好,核心菌群到位的相对稳态结构
在大多数情况下,它并不追求极端的高或低,而是呈现适度且稳定的多样性:核心菌属占比高、有益菌占优势、有害菌受控,整体更接近一种可持续的稳态。
1)多样性水平稳定
首先从整体生态稳定性来看,多样性水平稳定,说明菌群并未出现荒漠化的低多样性,也没有走向极端的异常发散,多样性在合理区间内往往意味着菌群具备更好的弹性,能更从容地应对饮食波动、作息变化等外界扰动。
编辑
编辑
<来源:谷禾健康肠道菌群检测数据库>
2)核心菌属充足:健康结构的标志
其次,本案例的菌群结构质量也较为理想。核心菌属85,达到报告建议的核心菌属累积占比最好超过80。核心菌属相对充足通常提示群落中以常见、稳定的共生菌为主体,生态位更稳,不容易被少数波动性菌群或机会型菌群抢位,因此也更接近健康人群的结构特征。
编辑
3)有益菌占优:功能性倾向良好
从功能倾向来看,本案例呈现出更符合健康菌群原则的比例:有益菌水平整体已处在建议线以上,既有足够的有益菌群支撑稳定性,也没有出现有害菌明显失控的信号。
最后看综合平衡指标,菌群平衡54。该值位于“中等”区间的偏高段,提示整体已较接近良好状态,但仍存在进一步优化空间。
编辑
<来源:谷禾健康肠道菌群检测数据库>
4)产丁酸菌正常:核心功能菌的支撑
具体菌属层面来看:
重要的核心菌属,包括产丁酸菌等,都处于正常范围(未标红)。
编辑
<来源:谷禾健康肠道菌群检测数据库>
拟杆菌属(Bacteroides)(处于人群79%):典型“基石菌/主导菌属”,在此报告里占比高且仍在报告正常范围内。一般代表复杂多糖降解与交叉喂养能力较强,是核心菌属占比的重要支撑点。
粪杆菌属(Faecalibacterium) (处于人群83%):关键产丁酸、抗炎相关核心菌,在参考范围内,属于加分项。
瘤胃球菌属(Ruminococcus)(人群80%):参与抗性淀粉/纤维素利用,处于正常范围。
总的来说,本案例展示了一个多样性良好且核心菌群充足的稳态结构,反映了肠道相对健康的特征。适度的多样性增强了菌群对外界变化的适应能力,而有益菌的优势地位则支持了整体稳定性。
案例五:
厚壁菌门过度占优势的结构
健康的肠道菌群中,各大菌门、菌属之间维持着相对稳定的比例。厚壁菌门(Firmicutes)和拟杆菌门(Bacteroidetes)是成年人肠道中最主要的两个菌门,它们的比例(F/B ratio)与能量代谢和体重密切相关。
接下来我们要看的是一个典型的门水平比例明显偏移的菌群失衡案例,其最突出的结构特征是——厚壁菌门显著过多,拟杆菌门被严重压缩。
1)门水平的异常信号:厚壁菌门一家独大
报告显示,该个体:F/B 比值 = 12.98(在谷禾数据库中,大多数人群参考范围:0.3–1.0)
这已经不属于轻微偏移,而是一个明确的门层面的生态极化结构。
编辑
<来源:谷禾健康肠道菌群检测数据库>
需要强调的是,厚壁菌门本身并非坏菌门,其中也包含多种产丁酸、支持肠道屏障的有益菌;但当其整体比例被放大到压倒性优势时,往往意味着特定饮食或代谢环境长期偏向,使菌群结构失去弹性。
2)结构层面的真实情况:不是厚壁菌多=结构好
如果继续看整体健康评估,会发现厚壁菌门水平的强大优势,并没有转化为结构质量优势:
编辑
<来源:谷禾健康肠道菌群检测数据库>
可以看到菌群平衡指数、菌群多样性、有益菌水平都偏低,有害菌较多,核心菌属严重缺乏。
这说明一个关键事实:
👉 厚壁菌门虽然数量巨大,但该个体并不是由高质量核心功能菌主导,而更像是由部分机会型或代谢偏向特定方向的菌反复扩增,形成的伪优势。
3)菌属层面:韦荣菌属异常升高
进一步看关键菌属,问题更加清晰:
厚壁菌优势并非由典型肠道核心功能菌主导,而是被高度富集的口腔 / 上呼吸道来源菌属明显拉高,其中以韦荣菌属(Veillonella)异常增多最为突出。
编辑
<来源:谷禾健康肠道菌群检测数据库>
韦荣菌属经典生态位在口腔、咽部、上呼吸道,常与牙菌斑、龋齿、口腔炎症、呼吸道感染高度相关,属于乳酸利用菌,常与链球菌属形成稳定的共生复合体。
该样本中并非单一菌属异常,而是出现了口腔—上呼吸道菌群组合特征:
链球菌属、嗜血杆菌属都偏高,这些菌群通常在口腔、鼻咽部常见的条件致病或共居菌,这些菌属理论上在肠道中正常情况下应是低丰度。
编辑
编辑
<来源:谷禾健康肠道菌群检测数据库>
它们的同步扩增,提示很可能存在:
换句话说,这是一个上消化道菌群下行 + 肠道生态位防御能力不足共同作用的结果。
4)为何这些上消化道菌群下行还能定植?核心菌、有益菌让位
多个产丁酸、抗炎核心菌属显著偏低或未检出:
编辑
<来源:谷禾健康肠道菌群检测数据库>
也就是说,厚壁菌门虽然多,但是真正负责肠道屏障修复、免疫调节的高价值成员并没有站稳生态位。
当这些核心菌群不足时,肠道就会变成一个空位过多、防御薄弱的生态系统,使原本不应成为优势菌的口腔 / 呼吸道菌有机会向下迁移并异常扩增。
5)功能与代谢结果:炎症与内毒素压力升高
这种结构性失衡,在功能指标中已经有所体现:
编辑
<来源:谷禾健康肠道菌群检测数据库>
当菌群结构过于偏移时,内毒素压力、慢性低度炎症和代谢负担往往同步上升。
###################################################
关于厚壁菌偏高属于哪种类型,判断思路:
厚壁菌门/拟杆菌门(F/B)比值是一个简单但粗糙的门水平线索,这在我们之前的文章有写过,详见:
当报告出现厚壁菌门明显占优时,可能需要追问:
这种偏高是短期饮食塑形造成的可逆偏移,还是已经与宿主代谢环境绑定,形成更稳定的结构性失衡? 实践中可按以下路径判断。
首先,先看结构是否还完整
我们上述展示的是一个相对极端的案例,若厚壁菌门只是偏高,但菌群多样性、平衡指数相对尚可,核心菌属并未明显减少,说明生态位仍然多元,系统并未被单一菌群占死。这种情况更常见于饮食诱导型厚壁菌偏高,即在长期高能量、高精制或膳食结构单一的环境中,厚壁菌因能量获取效率较高而被动扩增,但其优势具有一定可逆性。
进一步,拆解厚壁菌门内部的组成质量
厚壁菌门中既包含大量产丁酸、维护肠道屏障与免疫稳态的关键菌属,也可能包含机会型扩增、功能贡献有限甚至与炎症相关的成员。
若厚壁菌门偏高同时,经典产丁酸核心菌(如粪杆菌属、罗氏菌属、部分Blautia 等)仍有存在,提示结构功能尚未完全崩塌;
若厚壁菌门明显占优,但上述核心功能菌反而显著偏低或未检出,则说明这种数量优势并未转化为功能优势,更接近一种生态失衡型扩张。
还需要警惕另一种情况:
若厚壁菌门内部主要由韦荣菌属、链球菌属等非典型肠源菌占据(例如案例五报告中),而典型肠道核心菌普遍缺位,则这一厚壁菌优势更可能是一种来源错位导致的假性结构优势,而非真正的肠道功能性强化。
功能与代谢指标进行交叉验证
饮食诱导型厚壁菌偏高,往往仅伴随轻度产气、能量吸收效率升高等变化,炎症与内毒素状态未必明显异常。
而代谢型厚壁菌偏高,则更常见于炎症水平升高、胆汁酸代谢异常及肠道屏障薄弱等背景之下,提示菌群结构已与宿主的代谢环境形成相互强化的状态。
###################################################
案例六:
拟杆菌过多 ≠ 结构更健康
接下来我们来看一份非常典型的拟杆菌优势型的肠道菌群报告:
拟杆菌的丰度高达 85.613%,菌群整体倾向拟杆菌型。
编辑
<来源:谷禾健康肠道菌群检测数据库>
F/B 比值 0.08(显著低于谷禾数据库中的人群常见范围 0.3–1.0)
编辑
<来源:谷禾健康肠道菌群检测数据库>
仅从这个结果看,容易给人一种偏向植物性、有利代谢的直觉印象。但和厚壁菌偏高一样,拟杆菌偏高本身也只是结构线索,而非结论,同样需要进一步拆解其结构质量与功能背景。
1)整体生态:并非严重失衡型
菌群多样性自然是低的,有害菌也较多,但核心菌属处于正常水平。
编辑
<来源:谷禾健康肠道菌群检测数据库>
进一步看属水平,就会发现问题的关键并不在于有没有拟杆菌,而在于谁在过度主导生态位。
2)拟杆菌属极度集中,占据绝对生态位
其中多株典型拟杆菌(如 B. fragilis、B. vulgatus、B. uniformis 等)同时出现高丰度。
这意味着:碳源利用、蛋白/脂类代谢路径高度集中,其他功能型菌群(尤其是厚壁菌中的发酵—丁酸路径)参与度极低。
编辑
<来源:谷禾健康肠道菌群检测数据库>
图中可以看到,尤其是脆弱拟杆菌(Bacteroides fragilis)特别多,脆弱拟杆菌是人类肠道中的一种重要共生菌,具有复杂的双重角色。它分为两种亚型:不产肠毒素脆弱拟杆菌(NTBF)和产肠毒素脆弱拟杆菌(ETBF)。NTBF通常被认为是有益的,能够增强肠道屏障、调节免疫系统并抑制病原菌感染;而ETBF则通过分泌脆弱拟杆菌毒素(BFT)导致肠道上皮损伤,促进炎症和慢病发展,包括结肠炎、阿尔兹海默病、结直肠癌等。
3)厚壁菌门过低,丁酸通路成为结构短板
与拟杆菌高度集中相对应的是:
这类结构通常意味着:
功能指标很好地解释了这种结构的真实状态。
编辑
<来源:谷禾健康肠道菌群检测数据库>
这类组合非常符合拟杆菌垄断型结构的典型表现:
于是就形成了:
👉 虽不至于严重炎症,但长期胀气、消化负担偏重、肠腔压力较大 的状态。
4)拟杆菌过多,通常出于什么原因?
综合结构与功能特征,可能是以下背景之一或叠加:
此外也有其他非饮食因素的可能:
因此,这种时候,如果强行引入新菌、猛加益生菌或突然大幅调整底物结构,结果有可能不是很理想,拟杆菌可能会再次快速适应,继续占住生态位,新来者被挤走。
总的来说,需谨慎评估拟杆菌过高的情况,避免简单地将其丰度与健康状态等同。
益生菌补充是改善肠道健康的常用手段,能够有效促进消化、增强免疫力和维持微生态平衡。我们的数据库中也显示出两种极端情况的风险:一方面,益生菌的严重缺乏可能导致肠道菌群失衡,增加消化不良和感染的风险;另一方面,过量补充益生菌则可能引发肠道不适,甚至导致菌群的进一步失调。
案例七:
益生菌严重缺乏
接下来我们要看的这份报告最突出的特征,在于典型益生菌的系统性缺失。
从整体结构看,菌群尚未完全崩盘,但支持肠道屏障、免疫调节和代谢稳态的关键益生菌未能有效定植,是该案例的核心问题。
1)核心表现:双歧杆菌、乳杆菌几乎全部检不出
在益生菌模块中,几乎所有常见且功能明确的益生菌,比如双歧杆菌、乳杆菌等均显示为 ND.(ND为未检出)
编辑
<来源:谷禾健康肠道菌群检测数据库>
单个益生菌未检出其实在成年人中并不少见,但当整个双歧杆菌、乳杆菌体系几乎集体缺位时,已不太能用正常个体差异来解释,而更像是一个明确的益生菌底盘不足的结构。
2)指标层面的对应信号:有益菌偏低、核心菌属不足
这一菌属层面的缺失,也体现在综合指标上:
编辑
<来源:谷禾健康肠道菌群检测数据库>
这种组合特征十分典型,不是单一病原菌爆表,而是益生菌长期缺席后,被其他机会型或功能偏向不利的菌群逐步填补生态位,使结构看似还能运转,但稳定性和健康收益明显下降。
3)功能后果:屏障与代谢压力开始显现
在功能与代谢相关指标中,也能看到益生菌缺乏可能带来的连锁反应。
炎症信号偏高,如 LPS、肠道炎症水平超标;肠道屏障不足。
编辑
<来源:谷禾健康肠道菌群检测数据库>
蛋白腐败代谢相关产物(如对甲酚、苯酚)偏高。
编辑
<来源:谷禾健康肠道菌群检测数据库>
这些情况往往与双歧杆菌、乳杆菌不足导致的短链脂肪酸合成下降、屏障支持能力减弱有关。
总的来说,这类情况的干预重点,通常不在单纯追求多样性,而在于重建益生菌生态位,并为其提供长期可持续的生存环境。
如果说这个案例的问题在于——益生菌缺位,生态位长期空置,那么接下来这个案例呈现的,则是另一个方向的偏移——益生菌占位过强,反而挤压了其他核心菌群的生存空间。
案例八:
益生菌补充过犹不及
此案例的报告中,乳杆菌属丰度达到了19.076%,远超正常人群平均水平。
编辑
<来源:谷禾健康肠道菌群检测数据库>
这很有可能是长期或大剂量补充特定乳杆菌制剂的结果。虽然其有益菌高达57,看似健康,但细究之下却隐藏着新的失衡。
首先,单一菌种的极高丰度本身就是一种多样性失衡的表现。尽管其整体多样性评分为39(中等),但这种由外源补充菌株主导的伪多样性可能并不稳定。一旦停止补充,菌群结构可能迅速改变。
编辑
其次,功能上的失衡已经显现。报告显示其肠道产气严重(91,超标)。乳杆菌是主要的产乳酸菌,在发酵碳水化合物时会产生气体。当其数量过多时,过量的气体产生会导致腹胀、消化不良等症状,这与报告中的评估结果相符。
编辑
<来源:谷禾健康肠道菌群检测数据库>
此外,其对甲酚(p-Cresol)水平过高(95,过多)。对甲酚是蛋白质发酵产生的有害代谢物,被认为是尿毒症毒素之一,与慢性肾病、结直肠癌等风险相关。虽然乳杆菌不直接产生对甲酚,但其过度增殖可能改变了整个肠道环境的pH值和代谢网络,为其他能够产生对甲酚的菌群(如部分梭菌)创造了更有利的生长条件。
总的来说,益生菌并非多多益善。盲目、过量地补充单一或少数几种益生菌,可能会人为地制造出新的菌群失衡,导致产气过多、有害代谢物积累等问题。益生菌的补充应更具针对性,最好在检测评估的基础上,根据自身缺乏的菌种进行精准补充,并注意适时调整剂量,同时配合益生元和多样化饮食,以支持整个微生态系统的健康与平衡。
通过对以上极端案例的分析,我们可以清晰地看到,肠道菌群的健康远不止有益菌与有害菌的简单二元对立。它是一个关乎多样性、平衡性、功能性的复杂多维系统。无论是病原菌入侵导致的多样性崩溃,还是长期单一饮食或不当补充导致的比例失衡,亦或是关键功能菌群的缺失,最终都会导向肠道功能的紊乱和疾病风险的升高。
这些案例也为我们提供了重要的启示:
肠道菌群检测的意义在于评估微生态健康。菌群多样性是关键指标,关注物种丰富度和均匀度可及时识别失衡状态。此外,结合关键代谢物(如丁酸盐、短链脂肪酸)的检测,能够全面评估肠道健康。
在临床和机构干预中,针对性策略至关重要。干预不仅应补充益生菌,还需清除优势病原菌,并创造条件以恢复菌群多样性,避免一刀切的方法。饮食干预应精细化,关注膳食纤维、蛋白质和脂肪的均衡,以防止长期单一饮食导致的菌群极化。此外,干预后需进行长期监测与评估,以确保肠道菌群的稳定性和健康,及时调整干预措施,确保最佳效果。
在研究与发展方向上,深入探索微生物与宿主之间的相互作用机制,将有助于更好地利用这些微生物促进健康。
随着研究的深入,我们对肠道菌群的认识仍在不断拓展。未来,结合宏基因组学、代谢组学等多组学技术,我们将能更全面、更深入地描绘个体的肠道健康图景。作为行业的先行者,我们的使命是持续优化检测技术和模型,提升准确度。同时利用大样本数据库将这些复杂的生物信息转化为更加清晰、可执行的健康指导,帮助更多人通过管理好自己的内在肠道微生态,构筑起坚实的健康防线。
注:本文所引用的案例均为谷禾肠道菌群检测数据库中的真实检测数据,但为保护用户隐私,个人信息已做脱敏处理。所有分析和结论仅供参考和学术探讨,不构成医疗诊断或治疗建议。任何健康问题,请务必咨询专业医生。

谷禾健康

—变形菌门
—γ变形菌纲
—肠杆菌目
—肠杆菌科
—埃希氏菌属
埃希氏菌属 (Escherichia),是一种常见的细菌。其中最著名的种是大肠杆菌(Escherichia coli),大肠杆菌是一种厌氧菌,通常生活在人和动物的肠道中。在成人的横断面研究中,大肠杆菌是超过90%个体肠道微生物组的成员,是最早在新生儿出生时定植的细菌之一。
埃希氏菌属细菌是一类适应于肠道环境的细菌,其代谢能力丰富,既可以进行好氧代谢,又可以进行发酵代谢。
大肠杆菌在正常情况下对人体无害,但某些菌株也可能引起感染和疾病。例如,某些毒力菌株的大肠杆菌可引起食物中毒,导致胃肠道症状,如腹泻、呕吐和腹痛。此外,大肠杆菌也可能引起泌尿系统感染和其他感染性疾病。
除了大肠杆菌,埃希氏菌属还包括其他一些菌株,如致病性的Escherichia albertii 和 Escherichia fergusonii。这些菌株在临床上相对较少见,但仍然可能引起感染和疾病。
本文我们来了解大肠杆菌的特性,代谢途径,和其他菌群相互作用等,可以更好地从肠道菌群的复杂性中窥探其在人体健康中的重要角色,并为预防和治疗相关疾病提供指导。
埃希氏菌属(Escherichia)是革兰氏阴性细菌中的一类,其中最为知名的是大肠杆菌(Escherichia coli)。
以下是埃希氏菌属的一些特点:
埃希氏菌属细菌主要存在于人类和动物的肠道中,是正常肠道菌群的一部分。
– 适宜温度:埃希氏菌属细菌一般生长在温度范围为20°C-45°C之间,最适生长温度约为37°C。
– 厌氧/需氧:大多数肠杆菌属细菌为好氧生物,但也有一些菌株可以在缺氧条件下生长。
– 好氧代谢:埃希氏菌属细菌能够利用氧气进行呼吸代谢,产生能量。
– 发酵代谢:埃希氏菌属细菌也可以进行发酵代谢,利用糖类等有机物产生能量。
埃希氏菌属目前包括:
序列比较表明,E. fergusonii 和 E. albertii 与大肠杆菌密切相关,而其余三个物种在进化上可能更为遥远。
志贺氏菌菌株在系统发育上与某些大肠杆菌菌株的关系比某些大肠杆菌菌株之间的关系更为密切。
– 大肠埃希氏菌属,又叫大肠杆菌(Escherichia coli):是埃希氏菌属中最为重要的菌株之一,广泛存在于人类和动物的肠道中。大肠杆菌具有多种毒力因子,包括肠毒素和肠出血性大肠杆菌毒素。
– 弗氏埃希氏菌(Escherichia fergusonii):是埃希氏菌属的另一种菌株,也存在于人类和动物的肠道中,但其致病性较低。
肠道感染的传播途径通常是粪口传播,可以通过人传人、直接接触动物或其环境,或者通过食用受污染的食物或水。
内源性感染途径也是可能的(例如细菌从肠道转移到血液),导致肠外疾病,并且更常见于免疫功能低下的人群,或患有肝硬化或接受化疗等基础疾病的人。
最重要的肠道病原体是肠沙门氏菌、大肠杆菌的某些菌株、志贺氏菌和小肠结肠炎耶尔森氏菌。在急性腹泻期间从粪便标本中分离出的肠杆菌科细菌,可能反映了粪便菌群的剧烈变化,而不是症状的原因。
代表菌大肠杆菌
大肠杆菌(Escherichia coli)是一种常见的革兰阴性细菌,存在于人类和动物的肠道中。大肠杆菌的大多数菌株对人类无害,但也有一些菌株具有致病性,可以引起不同程度的疾病。以下是一些主要的致病毒株和它们的具体信息:
大肠杆菌O157:H7
– 特点:产生肠出血性大肠杆菌毒素(Shiga毒素),引起严重的肠道感染。
– 病症:可导致急性胃肠炎、腹泻、腹痛、呕吐和血便,严重时可引起溶血性尿毒症综合征(HUS)。
– 传播途径:食物污染(尤其是未煮熟的牛肉)、接触感染(粪口传播)等。
大肠杆菌O26、O45、O103、O111、O121和O145(非O157致病毒株)
– 特点:产生肠出血性大肠杆菌毒素,与O157:H7类似,但毒力较弱。
– 病症:引起类似O157致病毒株的症状,包括腹泻、腹痛、呕吐和血便,也可引发HUS。
– 传播途径:主要通过食物污染(尤其是未煮熟的肉类、生奶和生菜)传播。
大肠杆菌O104
– 特点:产生肠出血性大肠杆菌毒素,与O157:H7类似。
– 病症:引起急性胃肠炎、腹泻、腹痛、呕吐和血便,也可导致HUS。
– 传播途径:主要通过食物污染(尤其是未煮熟的食品和生鲜蔬菜)传播。
大肠杆菌O111
– 特点:产生肠毒素,但不产生肠出血性大肠杆菌毒素。
– 病症:引起急性胃肠炎、腹泻、腹痛和呕吐。
– 传播途径:主要通过食物污染(尤其是未煮熟的肉类和生菜)传播。
需要注意的是,以上只是一些常见的致病毒株,大肠杆菌还有其他致病毒株,具体病症和传播途径可能有所不同。

总体而言,埃希氏菌属细菌是一类适应于肠道环境的细菌,其代谢能力丰富,既可以进行好氧代谢,又可以进行发酵代谢。大肠杆菌作为埃希氏菌属中最重要的代表菌株之一,具有多种致病因子,可能引起胃肠道感染和其他相关疾病。
埃希氏菌属与其他菌群共同维持肠道的稳态平衡,参与食物消化和营养吸收过程。它可以与厌氧菌共生,分解单糖、二糖和复杂多糖,为肠道提供能量和营养物质。
此外,埃希氏菌属还参与清除氧气,维持肠道的厌氧环境。埃希氏菌属在婴儿肠道中占据主导地位,可能是因为其具有上述功能,能够适应婴儿肠道的特殊环境。
然而,某些埃希氏菌属和梭状芽孢杆菌属的菌株与严重的胃肠道疾病(如炎症性肠病和腹泻)有关,这为肠道菌群成分与宿主免疫系统之间的相互作用提供了新的见解。
肠杆菌其外膜上存在一种叫做脂多糖(LPS)的分子。
LPS由三个部分组成:脂A、核心多糖和O抗原。
埃希氏菌属通过合成LPS来保护自身免受外界环境的伤害。LPS能够与宿主的免疫系统相互作用,引发炎症反应。
在健康的情况下,埃希氏菌属产生的LPS与宿主的免疫系统保持平衡,不会引发过度的炎症反应。然而,在某些情况下,如肠道菌群失调或感染,埃希氏菌属产生的LPS可能会超过宿主免疫系统的调节能力,导致炎症反应过度,对肠道和整体健康产生负面影响。
短链脂肪酸包括乙酸、丙酸和丁酸,它们是肠道内的重要能量来源,并对肠道菌群和健康产生影响。
虽然埃希氏菌通常不是短链脂肪酸的主要产生者,但它的存在和代谢活动可以对肠道微生物群落的整体代谢产生影响,并间接影响短链脂肪酸的生成。
埃希氏菌属的影响不仅局限于肠道,还可以通过肠-脑轴(MGB)对大脑功能产生影响。研究发现,埃希氏菌属的异常增加与焦虑、抑郁等精神疾病的发生有关。
大肠杆菌可以产生多种神经递质,如去甲肾上腺素、多巴胺、5-羟色胺和乙酰胆碱。这些神经递质可以通过影响神经元活动和调节情绪、压力反应等方式,对神经系统功能产生影响。
其次,大肠杆菌可以通过调节肠道微生物组的组成,影响神经递质的合成和释放。例如,它可以改变肠道微生物组中产生神经递质的菌株的丰度,从而影响神经递质的水平和功能。
此外,大肠杆菌还可以通过调节肠道微生物组与神经系统之间的相互作用,影响神经递质的信号传导和神经系统的功能。
增强:
抑制:
产生肠毒素
某些埃希氏菌株能够产生肠毒素,包括肠毒素A、肠毒素B等。这些毒素能够破坏肠道黏膜细胞,导致腹泻和肠道炎症,引起食物中毒或感染性腹泻。
破坏肠道屏障完整性
大肠杆菌可以通过多种机制破坏肠道屏障的完整性,包括改变肠上皮细胞之间的连接、破坏肠上皮细胞与肠道黏膜之间的黏附、改变肠道黏膜的结构等。这些破坏作用导致肠道屏障的通透性增加,使得细菌、毒素和其他有害物质能够穿过肠道屏障进入血液循环系统,引发炎症反应和其他肠道相关疾病。
激活肠道黏膜免疫
大肠杆菌感染会激活肠道黏膜免疫系统,导致炎症反应和免疫细胞的活化。这些炎症因子和免疫细胞的活化会进一步破坏肠道屏障的完整性,加剧肠道屏障功能的损伤。
引发肠道菌群失调
大肠杆菌感染会导致肠道菌群的失调,即有益菌的数量减少,有害菌的数量增加。这种菌群失调会进一步破坏肠道屏障的完整性,增加肠道屏障的通透性。同时,肠道菌群失调还会影响肠道黏膜免疫系统的平衡,导致炎症反应的增加。
在一些重症患者中,肠道菌群失调常常表现为埃希氏菌属的过度生长,同时伴随着其他有害菌的增加和有益菌的减少。这种失衡的菌群状态与多种疾病的发生和发展密切相关,包括肠道感染、炎症性肠病、代谢性疾病等。
▼
埃希氏菌属与炎症性肠病(IBD)之间存在着密切的关联。多项研究表明,在IBD患者的肠道中,埃希氏菌属的数量明显增加。
埃希氏菌属属于肠道菌群中的一种细菌,它可以在肠道黏膜上富集并与宿主相互作用。埃希氏菌属的增加可能与肠道黏膜的损伤和炎症反应有关。埃希氏菌属可以通过黏附和侵入肠道上皮细胞,导致炎症反应的加剧。它可以释放一些毒素和代谢产物,损伤宿主细胞膜和上皮屏障,进一步加剧肠道黏膜的损伤。
埃希氏菌属还可以释放细胞毒性扩展底物谱β-内酰胺酶(ESBL),这种酶可以降解β-内酰胺类抗生素,导致抗生素耐药性的产生。抗生素耐药性的增加可能使得治疗IBD的抗生素疗效降低,从而加剧炎症反应。
▼
大肠癌与大肠杆菌(Escherichia coli)的关联机制是多方面的。
首先,一些致病性的大肠杆菌菌株,如粘附侵袭性大肠杆菌(AIEC)和肠致病性大肠杆菌(EPEC),分泌的环调素染色质蛋白(colibactin)和效应蛋白EspF与大肠癌的发展和进展有关。
其次,大肠杆菌感染破坏了肠道微生物的微环境,从而诱导大肠癌的发生。
此外,大肠杆菌感染还与慢性炎症相关,而慢性炎症是大肠癌的一个潜在风险因素。大肠杆菌还可以通过多种策略促进结肠癌的生长和发展。
肠道微生物组的失调也被认为是大肠癌发生的因素之一。一些研究还发现,大肠癌患者的肠道微生物组成与正常人有所不同,大肠杆菌等菌株的丰度增加与大肠癌的发生相关。
▼
在摄入被致病性产志贺毒素的大肠杆菌(STEC)污染的食物或水后,Stxs可能通过M细胞摄取和胞吞作用或细胞旁转运穿过肠上皮屏障。一旦进入粘膜下层,毒素就会激活先天免疫细胞,如中性粒细胞或单核细胞,它们作为“载体”细胞在血液中输送Stxs,还可能通过局部产生促炎细胞因子进一步加剧组织损伤。
最终,毒素被转移到肾小球内皮细胞和肾小管上皮细胞,这些细胞富含毒素受体Gb3。对主要靶器官肾脏的损伤会导致腹泻相关溶血性尿毒症综合征。
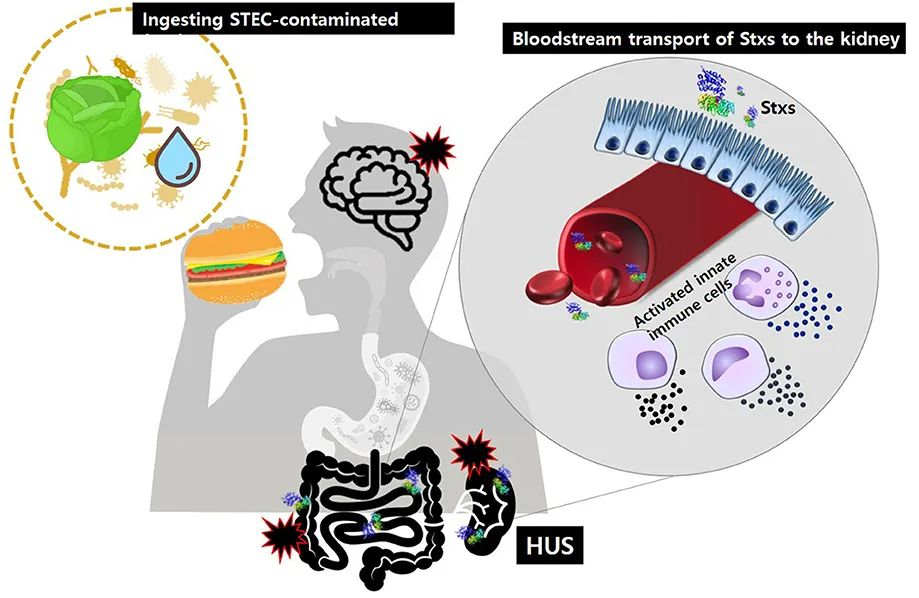
doi.org/10.3389/fcimb.2020.00273
▼
一些研究发现,在动脉粥样硬化患者中,肠杆菌属的丰度明显增加,而其他一些有益菌群的丰度明显降低。肠杆菌属的增加可能与动脉粥样硬化的发生和发展有关。
埃希氏菌属与糖尿病之间存在一定的关联。糖尿病患者往往存在高血糖状态,这为埃希氏菌属等细菌提供了良好的生长环境。研究发现,糖尿病患者的肠道菌群组成与健康人存在差异,其中埃希氏菌属的相对丰度较高。
埃希氏菌属可以产生多种促炎因子,如脂多糖和肽聚糖,从而引发宿主免疫反应,导致不同程度的肠道炎症。埃希氏菌属还可以破坏肠道上皮的完整性,导致低度炎症和自身免疫反应的发生,增加了1型糖尿病的风险。
研究还发现,糖尿病患者肠道菌群的改变与炎症、代谢紊乱等病理过程密切相关。
▼
二甲双胍的使用会导致埃希氏菌属的丰度增加
多项研究发现,二甲双胍治疗后,埃希氏菌属的数量明显增加。这种变化在2型糖尿病患者和健康人群中均有观察到。
埃希氏菌属的增加与二甲双胍的副作用有关
一些研究发现,埃希氏菌属的增加与二甲双胍治疗的胃肠道副作用有关。这可能是由于埃希氏菌属的存在导致了肠道内毒素的合成增加,从而引起胃肠道不适的症状。
埃希氏菌属的增加可能与肠道菌群的失衡有关
研究表明,二甲双胍的使用会导致肠道菌群的改变,包括埃希氏菌属的增加和其他有益菌的减少。这种菌群失衡可能与二甲双胍的疗效和副作用有关。
埃希氏菌属的增加可能与肠道炎症和代谢紊乱有关
一些研究发现,埃希氏菌属的增加与肠道炎症和代谢紊乱相关。这可能是由于埃希氏菌属的存在导致了肠道内炎症因子的释放增加,从而影响了机体的代谢功能。
▼
埃希氏菌属与非酒精性脂肪肝病(NAFLD)之间存在着密切的关联。多项研究发现,在非酒精性脂肪肝病患者中,埃希氏菌属的丰度明显增加。
埃希氏菌属可以产生乙醇,乙醇会破坏肠道屏障功能,导致肠道通透性增加。这可能是埃希氏菌属与非酒精性脂肪肝病发展相关的关键机制之一。
另外,埃希氏菌属的增加也与肝脏炎症和肝纤维化的发生相关。研究发现,在NAFLD患者中,埃希氏菌属的数量与肝纤维化的严重程度呈正相关。埃希氏菌属可能通过产生内毒素和诱导炎症反应,促进肝脏纤维化的发展。
肠道菌群失调和肠道屏障功能受损是NAFLD发展的重要因素。埃希氏菌属的增加可能导致肠道菌群失衡,进一步破坏肠道屏障功能,使内毒素进入血液循环,引发炎症反应,从而促进NAFLD的发展。
▼
埃希氏菌属与多动症存在相关性。研究发现,多动症患者的肠道菌群中有害菌如肠杆菌、大肠埃希氏菌和梭状芽孢杆菌的丰度显著增加,而有益菌如双歧杆菌、瘤胃球菌丰度显著降低。这些结果表明,某些埃希氏菌属可能与多动症的发展和症状有关。
罗伊氏乳杆菌DSM 17938已被证明具有有效的抗菌活性,调节免疫应答、减少肠道炎症。
植物乳杆菌CCFM1143对产肠毒素大肠杆菌引起的腹泻具有最明显的保护作用,通过调节炎症细胞因子、再平衡肠道菌群和调节短链脂肪酸生成的方式。
鼠李糖乳杆菌SHA113对耐药大肠杆菌QBQ009感染显示出有效的治疗效果。
短双歧杆菌FHNFQ23M3和双歧双歧杆菌FSDJN7O5可缓解产肠毒素大肠杆菌引起的腹泻症状。
荷花种子荚中的寡聚原花青素(LSPC)和益生菌(鼠李糖乳杆菌LGG和双歧杆菌Bb-12)对肠毒素性大肠杆菌(ETEC)感染的腹泻小鼠的保护作用。
阿拉伯木聚糖、菊粉和罗伊氏乳杆菌 1063 抑制粘膜中粘液的粘附侵袭性大肠杆菌。
主要参考文献:
Braz VS, Melchior K, Moreira CG. Escherichia coli as a Multifaceted Pathogenic and Versatile Bacterium. Front Cell Infect Microbiol. 2020 Dec 21;10:548492. doi: 10.3389/fcimb.2020.548492. PMID: 33409157; PMCID: PMC7779793.
Mirsepasi-Lauridsen HC, Vallance BA, Krogfelt KA, Petersen AM. Escherichia coli Pathobionts Associated with Inflammatory Bowel Disease. Clin Microbiol Rev. 2019 Jan 30;32(2):e00060-18. doi: 10.1128/CMR.00060-18. PMID: 30700431; PMCID: PMC6431131.
Sun J, Marais JP, Khoo C, LaPlante K, Vejborg RM, Givskov M, Tolker-Nielsen T, Seeram NP, Rowley DC. Cranberry (Vaccinium macrocarpon) oligosaccharides decrease biofilm formation by uropathogenic Escherichia coli. J Funct Foods. 2015 Aug;17:235-242. doi: 10.1016/j.jff.2015.05.016. PMID: 26613004; PMCID: PMC4657873.
Kim JS, Lee MS, Kim JH. Recent Updates on Outbreaks of Shiga Toxin-Producing Escherichia coli and Its Potential Reservoirs. Front Cell Infect Microbiol. 2020 Jun 4;10:273. doi: 10.3389/fcimb.2020.00273. PMID: 32582571; PMCID: PMC7287036.
Zhang S, Abbas M, Rehman MU, Wang M, Jia R, Chen S, Liu M, Zhu D, Zhao X, Gao Q, Tian B, Cheng A. Updates on the global dissemination of colistin-resistant Escherichia coli: An emerging threat to public health. Sci Total Environ. 2021 Dec 10;799:149280. doi: 10.1016/j.scitotenv.2021.149280. Epub 2021 Jul 27. PMID: 34364270.
Denamur E, Clermont O, Bonacorsi S, Gordon D. The population genetics of pathogenic Escherichia coli. Nat Rev Microbiol. 2021 Jan;19(1):37-54. doi: 10.1038/s41579-020-0416-x. Epub 2020 Aug 21. PMID: 32826992.
Leekitcharoenphon P, Johansson MHK, Munk P, Malorny B, Skarżyńska M, Wadepohl K, Moyano G, Hesp A, Veldman KT, Bossers A; EFFORT Consortium; Zając M, Wasyl D, Sanders P, Gonzalez-Zorn B, Brouwer MSM, Wagenaar JA, Heederik DJJ, Mevius D, Aarestrup FM. Genomic evolution of antimicrobial resistance in Escherichia coli. Sci Rep. 2021 Jul 23;11(1):15108. doi: 10.1038/s41598-021-93970-7. PMID: 34301966; PMCID: PMC8302606.
Khorsand B, Asadzadeh Aghdaei H, Nazemalhosseini-Mojarad E, Nadalian B, Nadalian B, Houri H. Overrepresentation of Enterobacteriaceae and Escherichia coli is the major gut microbiome signature in Crohn’s disease and ulcerative colitis; a comprehensive metagenomic analysis of IBDMDB datasets. Front Cell Infect Microbiol. 2022 Oct 4;12:1015890. doi: 10.3389/fcimb.2022.1015890. PMID: 36268225; PMCID: PMC9577114.
Wang J, Li R, Zhang M, Gu C, Wang H, Feng J, Bao L, Wu Y, Chen S, Zhang X. Influence of Huangqin Decoction on the immune function and fecal microbiome of chicks after experimental infection with Escherichia coli O78. Sci Rep. 2022 Oct 5;12(1):16632. doi: 10.1038/s41598-022-20709-3. Erratum in: Sci Rep. 2022 Nov 18;12(1):19901. PMID: 36198724; PMCID: PMC9534884.