-

 国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子
国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子

 二级病原微生物安全实验室
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业

谷禾健康

胃肠道除了在食物消化和吸收中发挥作用外,还是一个主要的内分泌器官,产生多种生物活性肽和胺类物质,作为肠道激素。
大多数肠道激素来源于肠内分泌细胞,但来源于其他肠上皮细胞类型的生物活性肽也与代谢调节有关。
为什么说肠道激素重要呢?我们在日常生活中胃口好不好都可能与它相关,它能影响食欲和食物摄入,通过影响大脑中的食欲中枢,减轻食欲或刺激饱腹感。
其实,肠道激素在多种器官的多种代谢活跃组织中协调关键的生理过程,包括胰腺、肝脏、脂肪组织、肠道和中枢神经系统等。从局部控制营养吸收和调节肠道生长、肠道运动和液体分泌,到远距离调节胰岛素释放、脂肪细胞功能。
近日,来自英国剑桥大学威康Trust-MRC代谢科学研究实验室团队发表了一篇文章在《Nature Reviews Gastroenterology & Hepatology》期刊,主要讲述了参与代谢调节的肠道激素,讨论了它们在胃肠系统内外的作用。

肠道激素由多种肠内分泌细胞和非肠内分泌细胞产生,它们对不同刺激的反应不同,并分泌特定的激素组合。
肠内分泌细胞通过一系列感觉转运蛋白、离子通道和受体感知各种营养和非营养刺激,使其成为营养吸收和肠道激素反应之间的有效联系。
这些肠道激素信号传导过程可以作为治疗靶点;例如,GLP2受体激动剂用于治疗短肠综合征,并且GLP1受体激动器用于治疗2型糖尿病和肥胖。此外,激素类药物的组合可用于最大限度地提高疗效,同时最大限度地减少不良反应。
因此,深入了解肠道内分泌系统介导的复杂器官间串扰,有助于设计基于或靶向肠道激素及其受体的更有效药物,并将其治疗潜力扩展到肥胖和糖尿病之外的疾病。
肠内分泌细胞沿着胃肠上皮的长度广泛分布。肠道内的一些细胞会分泌一些激素来应对不同的刺激。
■ 肠内分泌细胞可分为两大类:
以前我们认为这些肠内分泌细胞只会分泌单一类型的激素,但现在我们知道,实际上它们通常会同时分泌多种不同的激素,所以它们之间的分泌有一些重叠的部分。
■ 肠道中不同部位的激素表达也会有所不同,如:
肠道特定区域的激素产生与激素的功能作用及其反应的感觉输入有关。
决定肠道不同区域肠内分泌细胞激素特征的因素在很大程度上仍然未知。来自不同肠段的肠道类器官似乎保留了与起源相关的激素特征,这表明肠道干细胞的效力有限。
■ 肠道细胞功能受限的机制与激素分泌的发育演化
然而,随着细胞的发育成熟,肠内分泌细胞的激素特征会发生变化,并沿着隐窝-绒毛轴分化。
举个例子,回肠L细胞在肠隐窝中共表达GLP1和肽YY(PYY),但当这些细胞到达绒毛时,前胰高血糖素转录下调,分泌素转录上调。一旦这些激素分泌出来,它们有两条路:
■ 激素在肠道中的分泌调节及其影响
肠道激素控制多种代谢活性组织的广泛生理过程,包括胰腺、肝脏、脂肪组织、肠道和中枢神经系统。这些激素大多具有不止一种生理作用,许多生理作用由不止一种激素发挥。如下表:
调节新陈代谢的主要肠道激素的特征

Bany Bakar R, et al., Nat Rev Gastroenterol Hepatol. 2023
■ 肠内分泌细胞如何与营养物质密切接触
这就需要我们先来了解一下,肠内分泌细胞的结构特点,大多数肠内分泌细胞是开放型细胞,也就是说它们有一个顶端表面,带有向肠腔延伸的微绒毛,以及一个基底外侧表面,负责在刺激时通过胞吐释放分泌颗粒。
因此,这些细胞与消化和吸收的营养物质直接接触,把它们推向产生肠道激素反应的理想位置,将营养吸收的变化传达给身体其他部位。
它怎么传达呢?
肠内分泌细胞的化学感受能力归因于它们表达一系列感觉转运蛋白、离子通道和受体,这些受体解释了它们对各种营养刺激的反应能力,包括葡萄糖、消化脂质、碳水化合物和蛋白质,以及非营养刺激,如胆汁酸、肠道激素、肠道神经递质,微生物产物和炎性胞质分裂。
■ GPCR——肠内分泌细胞通过其检测营养物质吸收
G蛋白偶联受体(GPCR)检测到广泛的营养和非营养刺激,肠内分泌细胞差异表达一系列GPCR,这些GPCR主要位于其基底外侧表面,能够检测出胆汁酸、游离脂肪酸(FFAs)和氨基酸在上皮层内的消化和吸收过程中的变化。
与肠道激素释放密切相关的GPCR包括:
各种脂质感应受体,包括:
相反,葡萄糖在通过顶端膜上的钠偶联葡萄糖转运蛋白SGLT1吸收时激活肠内分泌细胞。SGLT1对钠离子与葡萄糖的共转运直接触发质膜去极化,进而激活电压门控钙通道。由此产生的钙流入进一步使肠内分泌细胞质膜去极化,并启动分泌颗粒的释放(下图)。
调节肠内分泌细胞活性的营养诱导机制
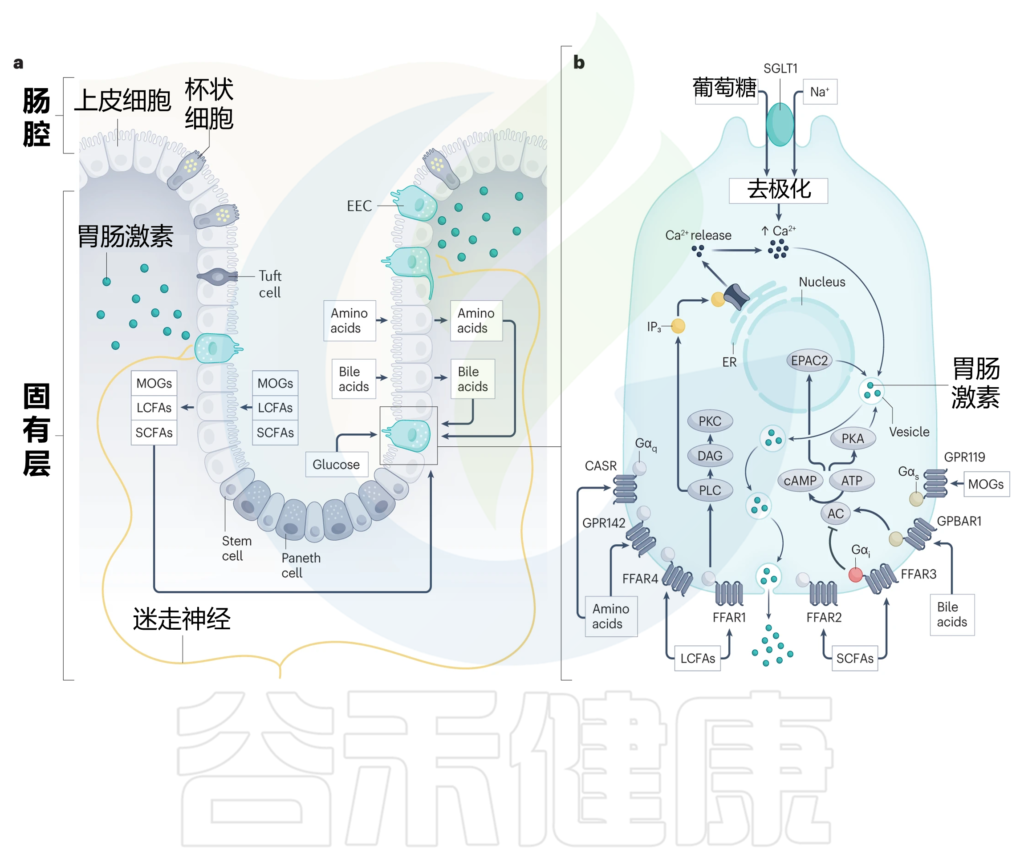
Bany Bakar R, et al., Nat Rev Gastroenterol Hepatol. 2023
肠道激素GDF15、FGF19(小鼠中的FGF15)、鸟苷素和尿鸟苷素不是由肠内分泌细胞产生的,而是由肠细胞和其他上皮谱系产生的。对这些激素生物合成和释放的途径了解相对较少。
肠内分泌细胞衍生的肽激素是由激素原转化酶的活性产生的,这些酶共同储存在囊泡中,囊泡与细胞膜融合以响应分泌刺激;而其他细胞类型中激素的产生则不同,它们并不经过传统的囊泡激素原转化酶和调节的囊泡分泌机制。
■ 肠道激素与二甲双胍疗效的关联
这些激素的生物合成与它们的mRNA水平有关,mRNA对特定刺激有反应。举例来说,FGF19(小鼠中的FGF15)的表达仅限于回肠末端,并通过核法尼糖样受体FXR被胆汁酸上调。在人体中,饭后胆汁释放后90-120分钟内FGF19的循环水平上升。在小鼠中,GDF15在多个器官中表达,但其在肠道中的表达受到细胞应激途径的增加的影响,就像在二甲双胍治疗过程中观察到的那样。实际上,治疗2型糖尿病患者的二甲双胍可能导致循环GDF15水平升高,从而产生该药物减肥效果。
■ Proguanylin:胃切除后水平升高?
Proguanylin在成熟肠细胞、一些杯状细胞和Paneth细胞中表达,在接受胃切除术或肠移植的患者中检测到的循环水平高于健康人。
Proguanylin释放到肠腔中,在那里被切割形成成熟的生物活性鸟苷肽。鸟苷由两个分子内二硫键稳定,这两个键对消化酶提供了一些保护,并从顶端方向靶向上皮膜鸟苷环化酶受体。Proguanylin和 prouroguanylin 也进入血液,但这些形式是如何分泌和激活的尚不清楚。
肠道激素控制许多胃肠道功能,这些功能涉及协调对食物摄入的反应,并确保摄入的食物被有效消化和吸收。
肠内分泌细胞检测管腔内容物的化学成分和局部营养吸收率,以及它们释放的激素控制过程,如胃酸分泌的调节、胃排空、胆囊收缩、胰腺酶和电解质的分泌、肠道运动和上皮屏障功能。
▸ 胃酸分泌
胃中特殊的壁细胞分泌酸有助于食物的消化和矿物质的吸收,并降低胃肠道感染的风险。胃酸的产生受到严格调节,以确保有效的食物消化,而不会损害胃和十二指肠粘膜。
胃窦G细胞分泌的胃泌素作用于肠嗜铬细胞上的胃泌素受体(称为CCKB或CCK2受体),刺激组胺的释放,组胺是一种强效的盐酸促分泌剂。胃窦和胃底D细胞的生长抑素对该系统进行微调,除了直接抑制壁细胞的酸释放外,生长抑素还能抑制胃泌素和组胺的释放。
其他几种肠道激素,包括胆囊收缩素(CCK)、氧调素、PYY和分泌素,通过不完全表征的机制抑制胃酸分泌,并募集涉及生长抑素的旁分泌回路。
▸ 胃排空
胃排空的速度决定了胃内容物输送到十二指肠的速率,并受到各种因素的调节,包括食物特性、肠道神经信号、肠道激素和血糖水平。胃排空率与食物消化率和营养吸收率密切相关,并受到调节,使得胃排空率仅反映小肠消化和吸收所接收营养的能力。
如果胃排空速率超过十二指肠吸收能力,营养物质会进入小肠并触发回肠制动,从而导致胃排空的反馈抑制,从而恢复平衡。主要位于回肠和结肠的肠道激素GLP1和PYY在回肠制动中起主要作用。回肠制动导致食物滞留在胃中,导致胃胀,从而激活发出饱腹信号的机械敏感迷走神经传入神经元。
▸ 胰腺和胆囊功能
一旦胃内容物进入小肠,其他激素就会被吸收以促进消化。由十二指肠 I 细胞产生的CCK刺激胰腺酶分泌,并通过刺激胆囊神经节中传出迷走神经神经递质的释放来促进胆囊排空。
分泌素在各种成熟的肠内分泌细胞中产生,刺激胰腺碱性电解质分泌,也与餐后热遗传学有关。鸟苷激活肠细胞中的鸟苷酸环化酶-cGMP信号,从而触发上皮氯化物和液体分泌。
这种途径被热稳定肠毒素劫持,是它们引发分泌性腹泻的能力的基础。鸟苷素-cGMP信号通路也与结直肠癌的抑制有关。
▸ 肠道运动
幽门以外胃肠道的运动由协调纵向蠕动波的肠神经系统和几种肠道激素之间的相互作用调节。
其中一种激素是胃动素,它存在于人类十二指肠中(尽管啮齿类动物不存在),并在胃中启动迁移运动复合体,特别是在餐间状态。胃动素受体激动剂,如低剂量红霉素,对胃轻瘫有治疗作用,一项研究表明,胃动素接收器的功能缺失变体在胃轻瘫患者中富集。
与调节小肠和大肠运动有关的其他激素包括:
粘膜血清素对肠道运动和液体分泌的重要性受到质疑,因为缺乏产生粘膜血清素所需的色氨酸羟化酶1的小鼠没有强烈的表型。然而,在一个模型中,产生血清素的肠嗜铬细胞被有条件地切除,这一发现强调了这些细胞确实在肠道转运控制中发挥作用。
▸ 肠道功能
除了控制肠道通道外,肠道激素还调节其他重要的肠道过程。例如:
GLP2刺激上皮生长,从而增加给定肠段的吸收能力。这种作用可能有助于GLP2治疗短肠综合征的疗效,
GLP1–GLP2双激动剂更有效。与血清素和神经降压素等其他肠道激素一样,GLP1和GLP2也会影响屏障功能并调节免疫系统。分泌血清素的肠嗜铬细胞与内脏疼痛有关,对肠道炎症、肝脏病理生理学和焦虑具有重要意义。
内 分 泌 胰 腺
我们知道饭后一段时间,人体内胰岛素的分泌量会增加,这其实也跟肠道激素有关。GLP1和GIP是两种主要的肠促胰岛素,占餐后胰岛素分泌的70%。
肠促胰岛素对葡萄糖刺激的胰岛素释放的影响可以通过以下事实来证明:口服葡萄糖比静脉注射葡萄糖(不刺激肠促胰岛素释放)能引起更大的胰岛素分泌——这种效应被称为肠促胰岛素效应。从生理上讲,肠促胰岛素调节对碳水化合物摄入的反应,并确保餐后健康的葡萄糖耐受性,无论摄入的葡萄糖负荷如何。
GLP1和GIP如何增加胰岛素分泌?
GLP1和GIP分别通过直接作用于GLP1受体和GIP受体来增加胰岛素分泌,这两种受体在胰腺β细胞中高度表达。
在β细胞中,GLP1和GIP受体主要募集Gαs并触发cAMP的升高。cAMP影响通过激活蛋白激酶A(PKA)和由cAMP-2激活的交换蛋白(EPAC2)调节胰岛素释放的几种下游途径。这两种效应物都调节离子通道活性以及参与葡萄糖依赖性胰岛素分泌的囊泡运输和融合。
GLP1和GIP具有额外的促胰岛素作用,但两者对肠促胰岛素作用的贡献有争议
GLP1和GIP对β细胞的作用不仅限于调控胰岛素分泌,还包括上调PDX1,促进胰岛素基因转录,增加β细胞质量并通过细胞周期和MAPK的激活促进新生。
基于这些作用,GLP1受体激动剂的长期治疗被认为是2型糖尿病的一种疾病改良治疗方法,可以通过增加或保留功能性β细胞质量来减轻β细胞功能障碍的进展。
注:迄今为止还没有证据表明GLP1受体促动剂调节人类的β细胞质量。
虽然说GLP1和GIP都刺激胰岛素分泌,但它们对胰腺α细胞分泌胰高血糖素的作用相反。
GIP刺激胰高血糖素分泌,而GLP1抑制分泌。
有证据表明,GLP1对胰岛素和胰高血糖素分泌的相反作用都有助于降低血浆葡萄糖。
GLP1通过激活GLP1受体介导的内在(非旁分泌)途径抑制α细胞,从而减少了胰高血糖素的分泌。
然而,其他研究结果表明,GLP1通过涉及胰岛内生长抑素的旁分泌机制间接抑制胰高血糖素的释放,并且GLP1和GIP的不同作用可以通过GIP对α细胞具有强大的直接刺激作用来解释,该作用掩盖了局部生长抑素的抑制作用。
肠促胰岛素的作用通常在2型糖尿病中受损,这种作用是糖尿病的后果而非原因。
在这种情况下,即使GIP的水平是超生理的,其作用也会减弱或缺失,而GLP1的促胰岛素作用相对保留。然而,GIP在2型糖尿病中保留刺激胰高血糖素分泌的能力,这会导致餐后高血糖。在各种研究中,GIP对胰腺β细胞的作用减弱归因于GIP受体表达减少、GIP受体下调或细胞内信号机制的受体后缺陷。
值得注意的是,2型糖尿病患者的血糖正常化提高了对GIP和GLP1的敏感性,这也许解释了为什么在双激动剂药物中在GL1中添加GIP对胰岛素分泌有额外的有益作用。
2型糖尿病是否与GLP1或GIP分泌缺陷有关尚不清楚。对2型糖尿病患者循环GIP水平的研究产生了矛盾的结果——高、低和不变的水平都有报道。相比之下,几项研究表明,2型糖尿病患者在混合餐后GLP1的分泌减少。
总的来说,2型糖尿病对肠道激素分泌的影响显然相对较小,这可能反映了这样一个事实,即:
肠内分泌细胞从隐窝干细胞中不断更新,在不利的代谢条件下,细胞损伤几乎没有时间积累。
然而,2型糖尿病对肠道运动和营养吸收的影响可能通过改变营养和胆汁酸刺激的局部递送而间接影响肠内分泌细胞的释放。
GLP1对胰岛素释放的影响比GIP大,GLP1已被开发为2型糖尿病的主要治疗靶点,并改变了2型糖尿病和肥胖患者血糖控制和体重减轻的临床治疗。
肝 脏
来自动物研究和临床试验的数据强调了GLP1和GLP1受体激动剂对肝脏代谢的多种有益作用。
GLP1受体是否在肝脏中表达有争议:
因此,在将GLP1对肝脏病理和功能的任何观察到的影响归因于肽对肝脏本身的直接影响之前,必须谨慎。
GLP1和GLP1受体激动剂对肝功能的间接作用
GLP1和GLP1受体激动剂对肝功能的间接作用是预期的,因为它们能够刺激胰岛素释放、降低胰高血糖素水平、改善葡萄糖稳态、减少胃肠道中乳糜微粒的产生以及减少热量摄入和体重。GLP1对肝脏影响的复杂性体现在它可以抑制肝脏葡萄糖产生并刺激肝脏葡萄糖摄取的发现上。20世纪90年代发表的研究将这些观察结果归因于胰岛素水平改变的间接影响,但后来的研究表明,胰岛素不是关键因素。
GLP1对肝脏脂质代谢有几种有益作用:
肝脏脂质代谢的这些改善可能至少部分归因于食物摄入的减少和随之而来的体重减轻。然而,一些研究结果表明,GLP1在调节肝脏脂质代谢方面具有额外的直接作用。
GLP1是否具有与体重减轻、胰岛素和胰高血糖素相关的肝脏作用之外的肝脏作用是一个有趣的问题。一项研究令人惊讶地表明,GLP19-36是GLP1的主要循环代谢产物,它独立于胰岛素的变化而抑制肝葡萄糖的产生,当GLP1受体被阻断时,这种作用仍然存在。这种作用的机制尚不清楚。一些证据表明,它涉及一种未鉴定的膜受体的激活,而其他研究结果表明,GLP19–36的降解产物(GLP128–36和GLP132–36)靶向PKA–β-连环蛋白–Wnt信号通路;这些降解产物可以在不涉及特定受体的情况下穿透质膜。需要进一步调查,以澄清和验证所涉及的机制。
无论GLP1对肝脏的有益作用是直接的还是间接的,GLP1受体激动剂正在被研究用于治疗非酒精性脂肪肝(NAFLD),这是肝脂代谢紊乱的最常见的慢性肝病之一。
GLP1受体激动剂:治疗非酒精性脂肪肝的希望
GLP1受体激动剂可以通过多种途径的组合来改善与非酒精性脂肪肝的发作和进展相关的基本代谢变化。
对非酒精性脂肪肝和非酒精性脂肪性肝炎(NASH)小鼠模型的研究表明,GLP1受体激动剂可降低肝脏脂肪含量,逆转脂肪变性,发挥抗炎作用,并改善纤维化的组织学严重程度。
此外,GLP1受体激动剂治疗非酒精性脂肪肝的临床试验(主要包括T2DM患者)产生了有希望的结果。几项研究表明,用GLP1受体激动剂治疗可降低肝脏脂肪含量,导致非酒精性脂肪性肝炎的组织学消退,并减缓纤维化的进展。然而,在这些临床研究中观察到体重和血红蛋白A1c水平显著降低,因此GLP1受体激动剂的作用是否独立于体重减轻和血糖控制的改善仍不清楚。
双激动剂疗法:研究非酒精性脂肪肝的新选择
双激动剂疗法,其中GLP1受体激动剂与直接抑制脂肪酸合成或靶向纤维化的第二分子结合,正在作为非酒精性脂肪肝的治疗选择进行研究;一个例子是GLP1–胰高血糖素组合肽。
其他肠内分泌激素(包括胃饥饿素和胃肠肽)对肝脏的影响尚不清楚。几项研究表明,GIP调节肝脏脂质含量,并可能参与非酒精性脂肪肝的发病机制。然而,GIP是否独立于其对胰岛素和脂肪组织的影响而直接影响肝脏中的脂质积聚(见“脂肪组织”)仍存在争议,因为GIP受体在肝脏中的表达仍未得到证实。几项动物研究表明,GIP信号传导的遗传或药理学破坏可以减少肝脏脂质积聚,但这些干预措施也可以防止饮食诱导的体重增加,因此研究结果不能证明GIP对肝脏有直接作用。
FGF19的多重代谢作用
与肠内分泌细胞衍生的肽不同,FGF19(小鼠中的FGF15)在回肠末端上皮中对胆汁酸吸收产生反应,是调节肝胆汁酸生物合成的重要抑制性反馈回路的基础。它还通过激活肝糖原合成和抑制糖异生等途径具有更广泛的代谢作用。
在不同器官中由肠道激素调节的过程
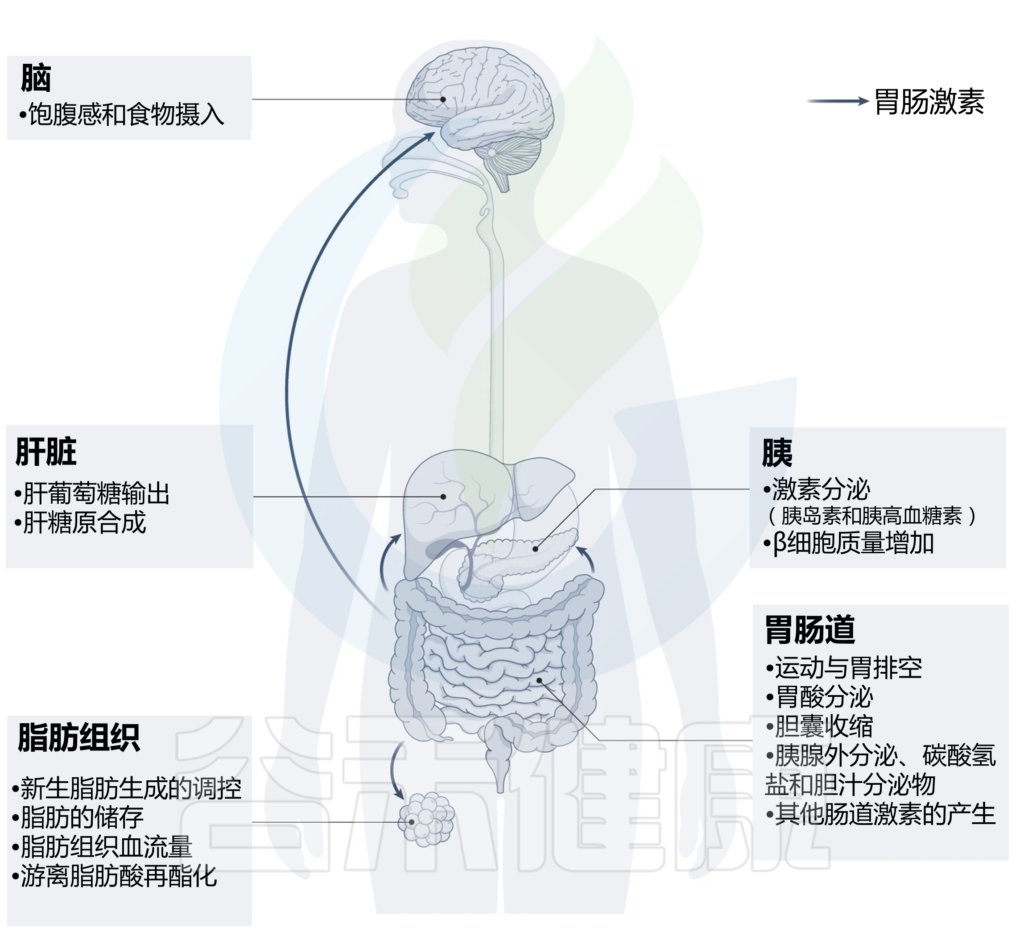
Bany Bakar R, et al., Nat Rev Gastroenterol Hepatol. 2023
脂 肪 组 织
脂肪细胞是哺乳动物的主要燃料储存室,它们的调节可以产生重要的代谢后果。在肠内分泌激素中,GIP与脂肪细胞功能的关系最为密切。
GIP与脂肪细胞功能:代谢调节的关键
几项研究表明,GIP促进白色脂肪组织(WAT)中的脂质储存,包括在大鼠中观察到,GIP通过增加脂肪组织葡萄糖摄取、从葡萄糖重新生成脂肪和将FFAs重新酯化为甘油三酯来支持脂肪组织中的脂质存储,以及在小鼠中敲除GIP受体可以预防饮食诱导的肥胖。
几项研究表明,脂肪组织中的GIP受体激活与胰岛素协同作用,增加脂质积累、葡萄糖摄取、新生脂肪生成和FFA再酯化。
脂肪细胞中的GIP受体类型:研究争议与需解决的问题
然而,这些发现需要在脂肪组织中哪些细胞类型表达GIP受体的争议中进行解释。单核转录组学表明,GIP受体不在成熟脂肪细胞中表达,而是在周细胞和间充质细胞中表达。考虑到许多脂肪细胞模型是由可以表达GIP受体的未成熟细胞在体外产生的,这些模型中的GIP受体表达是否能概括天然成熟脂肪细胞中的表达值得怀疑。
另一个必须考虑的因素是,GIP受体与Gαs偶联,Gαs下游cAMP水平升高通常与脂解而非脂肪生成有关,例如β3-肾上腺素能受体激活的脂解作用。
GIP的脂肪生成作用有益,改善脂质储存,减少异位脂肪积聚
有趣的是,GIP受体在小鼠和人类脂肪组织周细胞中表达的发现与GIP在体内显著增加脂肪组织血流量的证据一致,这一过程可以间接促进胰岛素依赖性作用,比如:清除甘油三酯、摄取葡萄糖。
无论潜在机制如何,GIP的脂肪生成作用都是有益的,因为它改善了白色脂肪组织中健康的长期脂质储存,减少了肝脏、心脏和胰腺等外周组织中的脂质溢出和异位脂肪积聚。
在2型糖尿病和肥胖的GLP1受体激动剂上添加GIP受体激动剂的代谢益处
早期的研究表明,GIP纯粹是致肥胖的,因此有利于GIP拮抗作用的翻译策略来实现减肥,而自从2018年以来发表的研究表明双重GIP受体-GLP1受体激动剂,替西帕肽在2型糖尿病患者中实现血糖控制和减轻体重方面比单独使用任一受体的激动剂更有效,该治疗改善了白色脂肪组织健康。替西帕肽治疗显著降低了肝脏脂肪含量以及内脏和皮下脂肪组织的体积,因此可能成为非酒精性脂肪肝或非酒精性脂肪性肝炎的重要治疗选择,尤其是在同时患有2型糖尿病或肥胖的患者中。
然而,对于是激动还是拮抗GIP受体,仍存在不清晰性;强有力的证据表明,任何一种方式都可以减轻肥胖和体重。
注:已经提出了几个假设来调和这些自相矛盾的观察结果,但还需要更多的研究来澄清这一领域。
尽管GIP在肠道白色脂肪组织轴中的确切作用机制仍有争议,但GIP在脂肪组织中的生理作用是明确的。在啮齿类动物中,棕色脂肪细胞组织(BAT)中GIP受体的表达已得到充分证实,给予GIP或替西帕肽可促进产热;替西帕肽也与支链氨基酸分解代谢增加有关。
然而,小鼠棕色脂肪细胞组织中GIP受体的条件性敲除并不能像全局敲除GIP受体那样防止饮食诱导的肥胖,这表明GIP对棕色脂肪细胞组织的影响在生理上并不是主要的。那主要的会是什么呢?
其他激素在肠道棕色脂肪细胞组织轴中发挥重要作用
例如,血清素抑制棕色脂肪细胞组织活性,而缺乏血清素产生所需的色氨酸羟化酶1的小鼠可以免受饮食诱导的肥胖。然而,鉴于血浆5-羟色胺是由血栓细胞缓冲的,肠道来源的5-羟色胺的内分泌作用值得怀疑,而且关于血浆5-羟色胺水平相对于用餐时间的日变化的报道一直不一致。相反,分泌素激活棕色脂肪细胞组织,这种激活与食欲的改变有关。
脑
肠-脑轴是胃肠道和大脑之间传递信息的渠道,包括神经、激素和营养信号;下丘脑核团和后脑是检测和整合这些外周信号的主要中心。肠-脑轴的紊乱已在多种疾病中被发现,包括炎症和功能性胃肠道疾病以及饮食障碍。
肠道激素在大脑的作用可以解释一些饮食失调和进食障碍的发生?
肠道向大脑发送关于饮食营养摄入状态的实时信号,肠道激素是肠道和大脑之间复杂相互作用的主要介质,这些相互作用是饱腹感(两餐之间不饿的感觉)和饱足感(一餐结束时吃得足够的感觉)的基础。许多肠道肽及其受体也在中枢神经系统中表达,并调节中枢回路,包括与食欲有关的回路。这种在中枢神经系统中的表达可能会使涉及肠道激素受体的药理学或全局遗传操作的研究的解释复杂化,因为其结果可能归因于对中枢神经系统回路的影响,而与肠道分泌的激素无关。
传入迷走神经神经元的细胞体位于结节神经节,并投射到孤束核(NTS),是介导肠道激素对食物摄入影响的主要途径。肠内分泌细胞通过局部升高的肠道激素水平以及可能的突触连接与传入迷走神经和内脏神经末梢进行交流。到达心室周围器官(例如,脊后和正中隆起区)和邻近细胞核(分别为下丘脑的NTS和弓状核)的循环肠道激素与这些信号汇合,以调节进食行为。几种肠道激素对大脑有着重要而独特的影响,下文将对此进行讨论。
CCK是在食欲调节中发挥作用的肠道激素。餐后释放CCK通过与CCK1受体结合激活传入迷走神经,向大脑提供反馈信号,从而减少进食。然而,根据小鼠研究,反馈信号的性质仍不清楚,因为动物可能因各种原因停止进食。在人类中,输注生理浓度的CCK可以抑制能量摄入并增加幽门收缩,而不会引起恶心;食物摄入减少被认为是幽门收缩的结果。在小鼠中,CCK通过传入迷走神经发出信号,激活中枢奖赏通路,并增加脂肪和糖的消耗。
CCK似乎会触发相对短暂的饱腹信号,PYY与餐后食物摄入的长期抑制有关。已知PYY的两种不同形式:PYY1-36和PYY3-36。PYY1–36通过二肽基肽酶IV转化为PYY3–36。两种形式的PYY都参与调节食物摄入,但它们表现出不同的特性和受体偏好。
GLP1和GLP1受体激动剂可以抑制食欲
GLP1受体激动剂主要通过作用于中枢神经系统内的GLP1受体来抑制食物摄入,但肠道来源的GLP1在大脑中是否达到足够高的水平以在生理条件下激活GLP1受体仍不清楚。
几项研究表明,迷走神经传入神经元的末端暴露于胃肠道中高水平的局部生物活性GLP1,介导内源性肠道衍生的GLP1的饱腹作用。
肠道来源GLP1控制食物摄入——不是主要的
在小鼠中,迷走神经神经元表达GLP1受体,而且响应机械刺激,外源性GLP1也可以增强由胃和肠道拉伸引起的饱腹感。然而,敲除肠道来源的GLP1不会影响小鼠的食物摄入或体重,这表明肠道来源的GLP1对控制食物摄入并不重要。
脑源性GLP1控制食物摄入——明显作用
虽然去除后脑GLP1表达也不会影响自由进食行为,但当化学遗传学激活时,它确实削弱了表达GLP1的神经元减少进食的能力,证明了脑源性GLP1在控制食物摄入方面的明显作用。
GLP1受体与氧调素的互动:探索胰岛素类似物的潜力
GLP1受体也被氧调素激活,氧调素是一种由肠道L细胞分泌的37个氨基酸的略微延伸的胰高血糖素类似物,在胰岛素受体处也具有活性。氧调节蛋白在体重管理和葡萄糖稳态方面具有有益作用,这一作用正开始通过使用GLP1受体和胰岛素受体双激动剂而得到利用;鉴于目前还没有发现特异性的尿囊素受体,这些作用被认为是由其对这两种受体的双重作用介导的。
减肥手术后,餐后GLP1、氧调节蛋白和PYY水平显著升高;循环水平足以对食物摄入和葡萄糖耐受性产生有益影响。然而,在小鼠中,Roux-en-Y胃旁路术的减肥作用在缺乏神经肽Y受体2型和GLP1受体的动物中仍然存在,这表明这些激素作用之外的机制也有助于减肥手术的益处。
神经降压素也与减肥手术的效果有关
神经降压素(Neurotensin)是一种13个氨基酸的肽,主要存在于大脑和胃肠道中。在啮齿类动物中,神经降压素直接进入大脑时会减少食物摄入,这种作用主要由NTSR1(神经降压素受体1)介导。
在一些研究中,Ntsr1敲除小鼠食欲增加,体重增加,对神经降压素的食欲抑制作用没有反应。
然而,在其他研究中,Ntsr1敲除降低了进食量并增加了活性,但对体重或组成没有影响。
脑源性神经降压素抑制食欲,外周的神经降压素对代谢的影响尚不清楚
一些研究表明,神经降压素的外周给药会导致啮齿动物的食物摄入量暂时减少。一些证据表明,这种影响可能归因于肠道中脂肪吸收的减少,但减少进食的确切机制尚不清楚。
另一种与减肥手术结果有关的激素是分泌素,因为它在手术后的餐后水平也会升高,尽管它在这些患者中的生理重要性尚不清楚。
来自非肠道内分泌细胞的肠道激素(GDF15、FGF19、uroguanylin)似乎也通过中枢途径调节食欲的作用。
最有力的证据是GDF15,它靶向后脑中有限的神经元群体,并导致由一系列毒性刺激(如化疗剂)诱导的恶心,这种恶心发生在癌症恶病质和妊娠剧吐中。
GDF15反应神经元表达GDF15受体,即GFRAL,并与脑干GLP1受体和CCK回路密切相关。有趣的是,GIP可以通过表达GIP受体的海马后区GABA能神经元抑制GDF15(和其他肠道激素,包括GLP1和PYY)引起的恶心。
Ghrelin:非凡食欲刺激者
胃饥饿素(Ghrelin)是唯一被广泛接受的刺激食物摄入的肠道激素。根据这种生物活性,血液中胃饥饿素的水平在用餐前升高,在进食后下降。
其生物学效应是通过生长激素促分泌受体(GHSR)介导的,GHSR存在于全身,包括胰腺、迷走神经传入神经元和大脑。几项研究表明,Ghrelin通过在下丘脑弓状核的中央和迷走神经传入神经的外周发挥其食欲产生活性。
然而,后一种机制是有争议的,一些研究表明,胃饥饿素的食欲产生作用不需要迷走神经传入信号。一些研究表明,胃动素也能促进食物摄入,但胃动素的生理学特征很差,因为它不存在于啮齿类动物中。
注:胃动素是一种与胃饥饿素有关的肽,由十二指肠中重叠的肠内分泌细胞群体产生。
INSL5:微弱但可能的食欲促进者
INSL5由远端结肠和直肠L细胞的一个子集分泌,对小鼠的研究表明它它具有食欲促进作用。它通过RXFP4(松弛素-胰岛素样家族肽受体4)发挥作用,该受体在大脑的关键进食中心表达。
INSL5的增食欲作用远低于胃饥饿素,这在一些研究中没有观察到,并且可能在体内被PYY所淹没,PYY是从相同的肠内分泌细胞平行分泌的。这些观察结果让人怀疑外周释放的INSL5是否在控制食物摄入方面具有重要的生理作用。然而,一项研究已经确定了腹内侧下丘脑中表达RXFP4的神经元在调节食物摄入和偏好方面的潜在作用。
胃肠道和身体其他活跃的组织之间有着双向的信息传递,而这个信息传递的改变与多种代谢性疾病有关。目前对于胃肠激素如何与这个复杂的调节网络相互作用的理解正在逐渐加深,但仍需要进一步的研究来完全揭开这个谜团。
肠道菌群的异常变化可能影响肠道激素的正常分泌,从而导致代谢相关疾病发生。因此,肠道菌群的稳定和平衡对于保持肠道激素的正常分泌和代谢健康非常重要。这些发现为我们深入了解肠道菌群和肠道激素的相互作用、开发相关治疗方法提供了新的方向。通过调节肠道菌群的结构和功能,我们可能能够干预肠道激素的分泌,从而实现对代谢相关疾病的治疗和预防。
通过开发创新、微创且耐受性良好的治疗方法,我们可以利用胃肠激素受体的转化潜能。GLP-1双重和三重激动剂疗法的临床试验已经证明了这种方法的价值。这个领域研究进展迅速,很可能会进一步细化现有的认识,解决关键的机制问题。期待揭示出胃肠激素控制的多个信号通路,可以成功地用于治疗2型糖尿病和肥胖,并且将这些基于胃肠激素的药物在其他代谢性疾病中推广应用。
主要参考文献:
Bany Bakar R, Reimann F, Gribble FM. The intestine as an endocrine organ and the role of gut hormones in metabolic regulation. Nat Rev Gastroenterol Hepatol. 2023 Aug 25.

谷禾健康
热门综艺《向往的生活》第四季中,嘉宾岳云鹏的“吃了就困”,“吃了睡,睡了吃”…意外抢镜。
他说自己,吃完饭就犯困 ↓↓↓

对于 “饭后就困” 这件事,网友们表示太真实,自己也一样 ↓↓↓

生活中,我们会发现很多人吃完就会感到疲倦,想睡觉,那么,
为什么吃完饭会想睡觉?
所有人都是这样的吗?
有没有可能通过一些方法改善?
……
饭后感到疲倦的程度可能会因人而异,因为它可能取决于许多因素,包括年龄、健康状况、食物的数量和类型、一天中就餐时间等等。
本文将归纳整理“餐后疲劳”的一些原因,从而帮助预防和改善餐后疲劳。
关于人们饭后感到困倦的原因,科学家们有许多假设。
根据一项实验假设,困倦的原因之一与下丘脑有关。这个假设主要在动物身上进行了测试。
科学家认为几个下丘脑区域,例如视交叉上核 (SCN)、外侧下丘脑 (LH) 和下丘脑腹内侧核 (VMH),与睡眠、清醒和食物摄入的调节有关。
睡眠和进食之间有很强的双向互动。
我们知道,身体需要能量来运作和生存。人体从食物中获取能量,食物通过消化过程分解并转化为燃料或葡萄糖,然后大量营养素为身体提供能量。这个消化代谢过程触发体内的各种反应。
以下列举的每一种原因都可能与餐后疲倦有关,虽然每个原因都是不同的,但餐后疲劳可以由这些因素的任意组合引发。
01 糖和精制碳水化合物
单糖和精制碳水化合物会迅速分解成葡萄糖,这可能会引发更突然和明显的疲劳。
研究表明,高血糖会抑制控制清醒的食欲素。食欲素在下丘脑中最为活跃。
这里要提到orexin/hypocretin(食欲素/下丘脑分泌素)。
食欲素是一种调节清醒和食欲的神经递质。大脑在下丘脑中包含大约 10,000 到 20,000 个神经元,但这些神经元的轴突延伸到整个大脑和脊髓,那里也有食欲素的受体。
它有什么作用?
科学表明,大脑食欲素神经元刺激清醒、警觉、进食、寻求奖励和健康的葡萄糖平衡。食欲素被认为是迄今为止最重要的唤醒刺激剂。
因此,将白面包等精制碳水化合物换成高纤维(低升糖指数)碳水化合物对整体健康更好。
02 炎症和食物敏感性
研究人员认为,有些人饭后感到疲倦的另一个可能原因与炎症有关。TNF和IL-1b等炎性细胞因子似乎会抑制促进清醒的食欲素。