-

 国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子
国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子

 二级病原微生物安全实验室
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业

谷禾健康
土壤作为地球上生物多样性最丰富的栖息地之一,不仅是植物生长的基础,也是众多微生物的”家园”。
然而,这片看似平静的地下世界,却隐藏着威胁人类健康的”隐形杀手”——人类细菌病原体。如单核细胞增生李斯特菌、铜绿假单胞菌、炭疽杆菌、沙门氏菌、大肠杆菌、结核分枝杆菌复合体和鼠疫杆菌等危险病原体,都可能在土壤中找到生存的港湾。
据统计,2019年全球约有770万例死亡与人类细菌病原体有关,这些数字背后,土壤作为病原体的自然来源和传播媒介,扮演着不容忽视的角色。
人类病原体通过多种途径进入土壤:动物和人类排泄物的渗透、农业实践中的灌溉和粪便应用、空气传播的病原体和孢子沉积等。一旦进入土壤,这些病原体可能成为常驻微生物组的一部分,并通过生食摄入、直接接触、吸入或昆虫及野生动物传播等多种途径感染人类。
本文基于最近一篇发表在《Cell Host & Microbe》上题为“Soil microbial diversity associates with lower prevalence of human bacterial pathogens across global soils”的研究,发现土壤微生物多样性与病原体流行率在全球呈负相关,并揭示病原体丰度与致病力及全球死亡率模式显著相关。在全球变化情景下,多数优势病原体比例可能上升。研究还绘制了全球土壤主要人类病原体分布图,阐明其生物地理与生态特征,为制定监测与风险管理策略、降低疫情暴发风险提供依据。
这意味着,在肠道菌群检测中发现的某些异常或致病菌,并不一定源于短期感染,而很可能是长期环境暴露累积的结果。因此,病原菌的控制不能仅依赖益生菌调节或抗生素干预,更重要的是减少其从环境进入人体的路径。
如果肠道检测报告中出现如沙门氏菌、致病性大肠杆菌、类鼻疽杆菌、克雷伯氏菌等,应重点考虑其环境来源。土壤是这些病原体的重要储存库,日常园艺、农作活动或儿童玩土等行为,都可能在无意识中增加暴露风险。特别是在潮湿或热带地区,这类病原体更易长期存活并富集。
农产品则是连接土壤与人体的重要媒介。叶菜类直接接触土壤,根茎类附着土壤较多,而水果也可能因灌溉水或土壤飞溅受到污染。如果种植过程中使用未经充分处理的粪肥,或灌溉水源受到污染,再加上洪水或暴雨等极端气候事件导致土壤扰动,都会显著增加农产品携带病原体的风险。此外,动物粪便通过土壤进入食物链,或宠物将病原体带入家庭环境,也构成重要的传播路径。
值得特别注意的是,病原体具有明显的地区性与季节性特征。不同地区的土壤病原谱差异显著,例如热带地区更易出现类鼻疽杆菌等环境致病菌,而干旱地区则更利于孢子类病原体的长期存在。同样,季节变化也会影响病原体丰度,雨季和洪水之后往往出现显著上升,高温条件促进细菌繁殖,这些都直接影响人群暴露风险。
▸ 土壤中的人类细菌病原体
一项覆盖59个国家、1602个土壤宏基因组的全球研究,为我们揭示了土壤中人类细菌病原体的分布模式。
研究中样本地点的全球分布
Xiong C,et al.Cell Host Microbe.2026
研究人员共鉴定出80个被归类为潜在土壤栖息人类病原体的细菌分类群,包括类鼻疽假单胞菌(Burkholderia pseudomallei)、结核分歧杆菌复合群(mycobacterium tuberculosis complex)、铜绿假单胞菌(Pseudomonas aeruginosa)、肠沙门氏菌(Salmonella enterica)、克雷伯氏菌(Klebsiella spp.)和志贺氏菌(Shigella spp.)等。
• 优势病原体主导模式
在这80种病原体中,25种被确定为优势病原体—它们不仅广泛存在(出现在≥80%的样本中),而且在病原体群落中表现出较高的相对丰度。
令人惊讶的是,这25种优势病原体仅占所有病原分类群的31.3%,却贡献了总病原体序列的90.8%。这种”少数主导”的模式与土壤中优势细菌和真菌的分布规律一致,反映了微生物群落结构的普遍特征。这些优势致病类群属于23种病原体,分布在19个属中,如伯克霍尔德氏菌属(Burkholderia)、分支杆菌属(Mycobacterium)、诺卡氏菌属(Nocardia)、假单胞菌属(Pseudomonas)、沙门氏菌属(Salmonella)和克雷伯氏菌属(Klebsiella)等。
▸ 高致病性优势病原体的健康威胁
在这23种显性病原体中,有11种被列为每年导致770万人死亡的33种人类细菌病原体之首,包括铜绿假单胞菌、肺炎克雷伯菌、肠道沙门氏菌、大肠杆菌、鲍曼不动杆菌、军团菌属、沙雷氏菌属和气单胞菌属等。
• 结核分枝杆菌每年导致大量感染且可在土壤中存活
其中,结核分枝杆菌尤为引人注目,其每年可导致约1060万人感染和160万人死亡。研究发现其成员能够感染各种家养和野生哺乳动物,并能在土壤中以毒力形式存活和增殖,使用变形虫和野生动物等替代宿主维持生存。
• 麻风分枝杆菌、淋病奈瑟菌也能在土壤中存在
值得注意的是,一些传统上不被认为是土壤传播的病原体(如结核分枝杆菌、麻风分枝杆菌和淋病奈瑟菌)也在土壤中被检测到。虽然它们从土壤到人类的传播途径尚不清楚,但最新研究发现蚊子可能是溃疡分枝杆菌(M. ulcerans)的新传播途径,这提示我们许多病原体的传播机制仍有待探索。
这些发现强调了重新关注病原体在土壤中的存活机制及其向人类传播途径的重要性,这对于有效监测和风险管理至关重要。
▸ 病原体的全球分布
通过对21个陆地生物群落的分析,研究发现了人类病原体在不同气候和植被类型下的分布模式。数据显示,主要人类病原体的相对丰度在热带和寒冷农田、温带湿地、温带农田以及热带和温带森林中最高。
进一步分析发现,农田、湿地和森林生态系统,特别是在热带和温带地区,表现出最高的优势病原体相对丰度。而在热带和干旱农田中,人类病原体的多样性最为丰富。
• 热带地区病原体多样性和患病率最高
分布模式与基于流行病学数据的临床研究结果一致——热带地区人类病原体的多样性和患病率最高。例如,东南亚(热带地区)报告了大多数结核病(由结核分枝杆菌引起)和麻风病(由麻风分枝杆菌引起)病例。
为探究环境因素对优势人类病原体全球分布的影响,研究人员通过随机森林和相关性分析发现:年均降水量(MAP)、年均温度(MAT)、降水季节性(PSEA)及土壤pH是最关键因素;此外,海拔、土壤质地、有机碳、土壤生物多样性(含微生物物种与功能基因多样性)及人口密度也具有重要作用。
全球土壤中主要人类细菌病原体的环境驱动因素
编辑
Xiong C,et al.Cell Host Microbe.2026
1
降雨与土壤病原体丰度正相关
• 降雨能够增加病原体在土壤中的分布
研究结果显示,降雨与23种主要病原体中的19种(约82.6%)相对丰度显著正相关。类鼻疽杆菌(B.pseudomallei)在潮湿土壤中生长最好,类鼻疽病例的增加与异常强降雨事件有关。
降雨增加可以提高土壤孔隙水含量,促进病原体的运动,增强其活性和流行性,从而促进其在土壤中的分布。
• 植物覆盖率和生产力也影响病原体丰度
植物覆盖率和生产力与病原体丰度同样呈正相关,这可能反映了植物驱动的土壤有机输入和水分稳定性增加,为某些病原体分类群的持久性提供了有利条件。
相比之下,降水季节性(23种病原体中的19种)、海拔(16种)和土壤pH值(14种)与这些分类群呈负相关。
全球不同生态系统中主要人类病原体的分布模式
编辑
Xiong C,et al.Cell Host Microbe.2026
2
土壤pH值与病原体呈负相关
土壤pH值与最主要的病原体呈负相关,表明酸性土壤往往含有更高的病原体丰度。
• 酸性土壤会破坏原有微生物结构,增加病原体增殖
这与最近的研究证据一致——土壤酸化会改变微生物群落结构,破坏关键的微生物功能,从而削弱微生物组限制病原体增殖的能力。
分析还揭示了降雨和土壤pH值之间存在显著的关系,这意味着气候变化下一些地区的降水增加可能会通过提供最佳水分条件和进一步酸化土壤来促进病原体的丰度。同样,不可持续的农业实践(如过度施肥)会加速土壤酸化,创造有利于人类病原体扩散的条件。
3
温度升高:病原体扩散的”催化剂”
温度与最主要的人类病原体(包括洋葱伯克霍尔德菌群(Burkholderia cepaciaComplex)、结核分枝杆菌复合体和炭疽杆菌)的相对丰度呈正相关,这表明全球变暖可能会进一步增加全球土壤中人类病原体的流行率。
这一发现说明气候变化可能会加剧人类致病性疾病,并表明在未来的气候情景下,降雨量和温度增加的地区可能成为人类细菌病原体的”热点”,进一步增加暴露、感染和死亡的风险。
除主要病原体外,研究还确定了其他8种病原体的环境驱动因素,包括柠檬酸杆菌属、志贺氏菌属、嗜肺军团菌、屎肠球菌、蜡样芽孢杆菌和问号钩端螺旋体等。这些病原体在降雨、温度和土壤特性方面与主要病原体具有相似的生态偏好。这些关于病原体当前分布和环境偏好的研究结果可以作为监测未来流行和爆发的基线数据,例如在筛查伯克霍尔德菌潜在爆发时,应优先考虑高降雨地区。
土壤微生物组多样性与病原体流行的关系是本研究的核心发现。研究基于宏基因组数据评估微生物物种多样性及功能多样性,并通过相关性与结构方程模型(SEM)分析其与人类病原体的关联;同时结合扩增子测序确定细菌和真菌多样性,并在全球200个土壤样本中加以验证。
▸ 微生物多样性与病原体丰度负相关
相关性分析显示,微生物物种多样性和功能多样性与大多数显性病原体的相对丰度呈显著负相关,包括类鼻疽杆菌、结核分枝杆菌复合物、炭疽杆菌、大肠杆菌和肠道沙门氏菌等。
• 土壤中细菌和真菌的多样性有助于减少病原体
在23种主要病原体中,15种和14种病原体分别与微生物种类和功能多样性呈显著负相关。研究还发现,与农业土壤相比,天然土壤中土壤微生物组多样性与主要人类病原体的相对丰度之间存在更强的负相关关系。
土壤微生物多样性决定主要人类病原体的全球分布
编辑
Xiong C,et al.Cell Host Microbe.2026
SEM分析进一步表明,土壤微生物多样性与优势人类病原体的相对丰度呈显著负相关。全球200个土壤样本亦证实:细菌和真菌多样性及细菌丰度均与主要病原体显著负相关。该结果拓展了以往土壤微观实验结论,表明提高微生物多样性可能抑制植物和人类病原体(如大肠杆菌O157:H7和肠道沙门菌)的增殖。
小编解释
较高的微生物多样性通过增加竞争者、耐压型和遗传策略类群,加剧竞争、扩大生态位占用并提升功能冗余,从而抑制病原体建立;这突显了保护土壤生物多样性对土壤、环境和人类健康的重要性。
为验证已鉴定的病原体与疾病毒力之间的关联,评估这些生物体的致病潜力,研究分析了病原体相对丰度与致病特征之间的关系。
• 土壤主要病原体丰度至少关联一种人类疾病特征
结果显示,23种主要病原体中有18种(包括类鼻疽杆菌、结核分枝杆菌、铜绿假单胞菌、鲍曼不动杆菌和大肠杆菌)以及一些其他重要病原体(如柠檬酸杆菌、嗜肺军团菌、伯氏考克斯氏体和蜡样芽孢杆菌)等,其相对丰度均与至少一种与人类疾病密切相关的关键致病特征显著正相关。
这些致病特征包括所有细菌病原体的毒力基因,根据其致病性分为不同的功能组,如毒素合酶、细胞毒性和毒力活性等。
人类细菌病原体与致病特征的关系
编辑
Xiong C,et al.Cell Host Microbe.2026
▸ 毒力因子与抗生素耐药基因
研究使用三个单独的数据库(PHI、VFDB和KEGG)进一步鉴定了与这些病原体相关的毒力因子,并使用DeepARG鉴定了抗生素耐药基因(ARG)。
• 土壤病原体相对丰度影响毒力因子与耐药性
分析表明,在全球范围内,显性病原体(类鼻疽杆菌、结核分枝杆菌、铜绿假单胞菌和鲍曼不动杆菌)的相对丰度与其相关毒力因子和抗生素耐药性之间存在很强的正相关关系。
这些发现进一步支持了识别人类病原体方法的稳健性,并证实了土壤中检测到的病原体可能会导致严重的疾病和死亡。
宏基因组揭示潜在病原体及其致病特征
为从全基因组层面验证宏基因组分箱识别显性病原体的可靠性,研究筛选出7个高质量(完整性≥90%、污染≤5%)MAG作为潜在病原体,其全球分布与定制数据库结果一致,且相对丰度与毒素合成和毒力活性等关键致病特征呈正相关。
• 宏基因组方法有助于识别土壤病原体
其中两个高质量MAG(T073.8和SRR8879131.4)与分枝杆菌的平均核苷酸同一性(ANI)达83%–84%,被认定为潜在重要病原体,并与温度和降雨量呈正相关。
这两者均携带与结核分枝杆菌复合群相关的五类致病特征(毒素合成、细胞毒性、毒力活性、器官损伤和宿主细胞死亡)、抗生素耐药基因(efpA,多重耐药)及外毒素基因(如磷脂酶C、VFC0235)。
结果表明,宏基因组学流程可用于从环境样本中识别潜在新兴病原体。
研究进一步评估了土壤人类病原体分布与全球相关死亡的关系。基于既往研究和世卫组织数据,汇总204个国家和地区的死亡率,并在国家尺度匹配到53个国家的数据。
• 土壤病原体分布与相关疾病死亡率相关
结果显示,麻风分枝杆菌(r=0.51,p=8.3×10^-5)和结核分枝杆菌复合物(r=0.47,p=0.0004)的相对丰度与病原体相关的全球死亡率(或感染病例)显著相关。
此外,细菌病原体相关死亡率与致病特征亦呈显著或接近显著相关,包括毒素合酶(r=0.45,p=0.0006)和毒力活性(r=0.25,p=0.07)。这些结果表明,土壤中人类病原体的流行可能与人类健康密切相关,提示土壤或为其天然宿主。
▸ 病原体的双向传播:土壤⇌人类
土壤与人类之间的病原体传播可能是双向的。一方面,土壤作为天然宿主,病原体可能通过各种途径感染人类;另一方面,人类病原体也可能通过空气、水道或农业实践(如施肥)等生物和非生物媒介从受感染的人群沉积或传播到自然和农业系统中。
• 土壤病原体多处于休眠状态,条件适宜时可经农产品等传播
传播到土壤中的病原体可能以低丰度或休眠孢子的形式存在,在土壤环境中找到替代宿主以生存和增殖;一旦满足传播条件,它们可能通过农产品、野生动物、病媒和城市扩张等各种传播途径重新进入人群,构成健康风险。
研究结果强调,在评估和管理与人类细菌病原体相关的风险时,需要明确考虑土壤因素。如果在地方和区域范围内得到证实,监测栖息在土壤中的人类病原体的流行情况可以为追踪全球疾病负担提供信息,简化流行病学监测,并有助于识别传染病热点。
研究局限性
疾病结果受到许多因素的影响,如人口密度、国家发展水平、社会经济状况和区域医疗质量等,在制定公共卫生政策时也应考虑这些因素。本研究所代表的国家数量有限可能会限制观察;随着数据库的改进,临床结果的预测能力也可能提高。
此外,一些病原体(如肠沙门氏菌和单核细胞增生李斯特菌)通过生食感染人类的途径已经确立,但其他病原体(如结核分枝杆菌)从土壤传播给人类的途径仍然难以捉摸。
基于环境变量(如气候和土壤因素)与人类病原体相对丰度的显著相关性,研究绘制了其在全球土壤中的当前及未来分布,模型可解释58%–88%的空间变异。
结果显示,总体显性病原体、特定病原体(如类鼻疽杆菌和结核分枝杆菌复合群)及关键致病功能(毒素合成、毒力活性和细胞毒性)在撒哈拉以南非洲、南美和东南亚等地区呈高丰度分布。
▸ 预测地图的验证及其应用价值
预测的结核病综合征全球地图显示了与世界卫生组织估计的全球结核病地图可比的模式——大多数结核病病例发生在东南亚、西太平洋和非洲。
• 病原菌分布与其致病人群分布相似
同样,预测的类鼻疽杆菌全球分布图与基于人类和动物感染病例的假鼻锤杆菌全球分布模式一致,类鼻疽在东南亚和南亚、澳大利亚北部、太平洋和印度洋岛国、撒哈拉以南非洲以及中南美洲的许多热带地区高度流行。这些结果为研究方法的稳健性提供了额外支持。
编辑
Xiong C,et al.Cell Host Microbe.2026
值得注意的是,在干旱地区(包括北非、西亚、澳大利亚北部和中部以及美国西部)发现了病原菌多药耐药诺卡菌的相对丰度最高。这与美国诺卡氏菌感染人数在西南部干燥温暖气候中最高的报告相吻合,表明不同病原体对特定气候条件的适应性。
▸ 气候变化下的病原体分布预测
在持续的气候变化下,降水模式和温度正在发生变化,这可能会对人类病原体的分布产生深远影响。
• 未来一些病原体和致病特征可能呈上升趋势
研究分析了病原体相对丰度从当前气候情景到预测的未来气候情景的潜在变化,结果表明在未来的气候情景下,一些关键病原体和致病特征的比例在全球大多数地区呈上升趋势。
例如,结核分枝杆菌复合物和致病群毒素合酶在撒哈拉以南非洲、南美洲、西欧、南亚和东南亚的某些地区呈现出明显的增长趋势。这与最近关于环境中人类病原体丰度总体增加的报告相一致。
这些地图提供了全球主要人类病原体的空间定量信息,填补了关于生活在土壤中的人类病原体的全球分布和环境偏好的关键知识空白。
然而,随着未来气候变化情景下土壤微生物组多样性的预测数据可用,以及更多宏基因组序列(特别是来自采样中代表性较低的地区/国家,如非洲国家)的积累,预测模型可以进一步改进。
总体而言,全球研究揭示了土壤中人类病原体分布的几个关键发现:主要栖息在土壤中的人类病原体的全球分布由气候条件和土壤生物多样性决定,在更潮湿的环境中观察到相对丰度更高。
土壤微生物多样性与这些病原体的流行率(即相对丰度)呈负相关,表明土壤生物多样性在最大限度地降低人类病原体爆发风险方面发挥着关键作用。
这些结果共同强调了土壤生物多样性是一个潜在的重要生态因素,它塑造了栖息在土壤中的人类细菌病原体的全球生物地理学。
▸ 研究的关键启示
这项研究确定了广泛分布在各种生态系统类型中的主要病原体,并揭示了这些病原体的相对丰度与传染病的致病特征和全球死亡率之间的正相关关系。此外,研究展示了主要病原体的环境偏好,并预测了未来全球变化可能如何影响其空间分布。这些发现对未来的研究重点和感染风险的管理具有重要意义:
1.病原体传播途径再评估:多种主要病原体(如类鼻疽杆菌、沙门氏菌和炭疽杆菌)已知可通过环境(如农产品、动物媒介和尘埃)传播,而结核分枝杆菌复合体等则传统上被认为仅经人际传播。研究表明这些病原体可在土壤中长期存活,未来应重点开展与风险管理相关的研究,如其在土壤中的存活与分布机制,以及潜在的土壤—人类传播新途径。
2.健康风险预警与管理:如果土壤中病原体的相对丰度与当地感染率之间具有稳定关系,就可以用于预测特定环境条件下的疾病风险,例如洪水或强降雨后类鼻疽的发生概率。通过持续监测土壤病原体分布与微生物多样性,并结合临床数据,有望建立更加精准和动态的疾病预警体系。
3.综合方法应对多重社会挑战:将气候、土壤与社会经济因素纳入同一分析框架,有助于提升人类病原体风险评估的系统性与前瞻性。相关研究已弥补了关键病原体在全球分布及其与生物多样性关系方面的认知空白,为预测不同气候情景下的人群健康风险提供了重要依据。
从健康管理角度看,对检测结果的理解也需要升级。肠道菌群检测不应只关注“缺乏哪些有益菌”,更应关注是否存在环境来源的病原体,以及这些菌是否反复出现。如果某类病原体持续存在,更可能提示持续暴露,而非单纯治疗不足。因此,定期监测趋势并结合生活环境变化进行分析,比单次检测更具意义。
在更宏观层面,未来公共卫生管理需要加强对土壤微生物的系统性监测,例如建立区域性的土壤微生物数据库与风险地图,对高风险区域进行预警,并在农业生产中规范粪肥处理和灌溉水管理。同时,在城市规划和灾害管理中,应将洪水后土壤病原风险纳入评估体系。
总体来看,对病原菌的控制应从“治疗导向”转向“源头管理”。环境中的土壤是一个关键但长期被忽视的源头,真正有效的健康策略应覆盖三个层面:上游减少环境暴露,中游控制食物污染,下游调节肠道生态。只有在这一整体框架下,才能更有效地降低感染风险并实现长期健康维护。
参考文献:
Xiong C, Delgado-Baquerizo M, Liang J, Wang J, Yan Z, Jensen SO, Gao M, Sáez-Sandino T, Guirado E, Muñoz-Rojas M, Román R, Maestre FT, Singh BK. Soil microbial diversity associates with lower prevalence of human bacterial pathogens across global soils. Cell Host Microbe. 2026 Mar 31:S1931-3128(26)00119-8.

谷禾健康
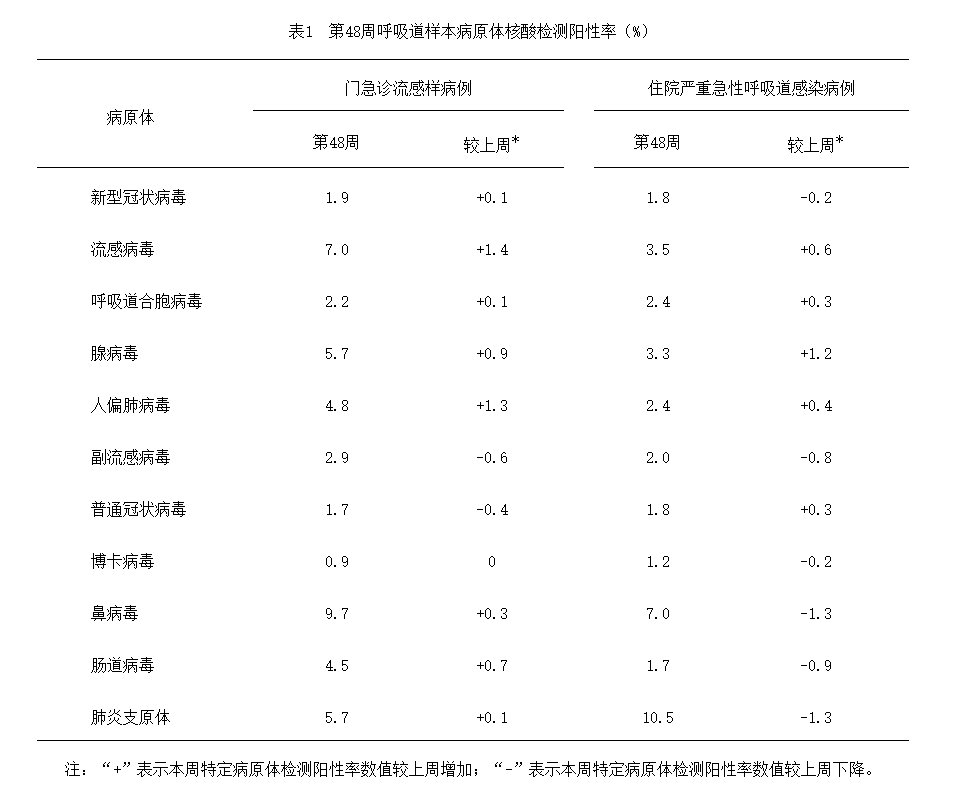
随着冬季到来,天气逐渐转冷,大部分呼吸道病原体在外界存活时间延长,同时人体的免疫力下降,呼吸道感染性疾病进入高发季节。
2024年第48周(11月25日—12月1日),对全国哨点医院(不含港澳台)采集的门急诊流感样病例和住院严重急性呼吸道感染病例的呼吸道样本,开展了多种呼吸道病原体检测,检测结果如下表所示:
呼吸道病原体检测阳性率(%)
来源:中国疾病预防控制中心
呼吸道主要病原体检测阳性率区域差异

来源:中国疾病预防控制中心
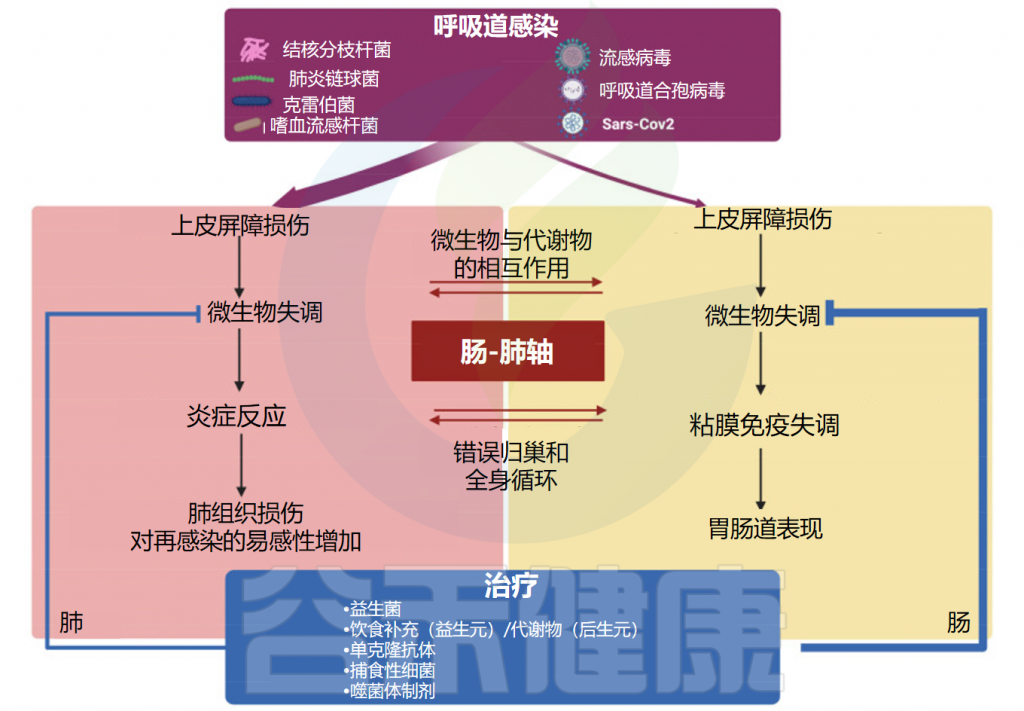
呼吸道感染是最常见的传染病类别,也是全球发病率和死亡率的主要原因之一。2019年冬天爆发新型冠状病毒(SARS-CoV-2)就对人类产生了深远且持续的影响,影响了健康、日常生活、经济和心理等多个方面。
呼吸道是一个复杂的器官系统,分为不同的区域;上呼吸道包括鼻腔、咽和喉,而下呼吸道则由传导气道(气管和支气管)、小气道(细支气管)和呼吸区(肺泡)组成。曾经被认为是无菌器官的健康肺现在被描述为拥有自己的特定微生物种群——肺微生物群。
肺微生物群被认为是不断吸入和消除的细菌的短暂定居点。这些共生细菌作用于免疫系统,诱导保护性反应并防止病原体入侵和定植。同时,它们通过生产抗菌物质直接抑制病原体的生长。共生菌与常驻上皮细胞和免疫细胞之间的持续互作支持肺稳态。
肺部微生物群被视为“肺部健康状况的镜子”:多项研究表明,在肺部疾病发生期间,肺部细菌组成和肺部环境会发生巨大变化。另一方面,肺与肠道进行连续的双向串扰,并且任一部位微生物群组成的改变都会导致远端疾病的发展和进展。
在本文中,我们将重点关注宿主-微生物群相互作用在健康和最常见的呼吸道细菌(结核分枝杆菌、肺炎链球菌、肺炎克雷伯菌和流感嗜血杆菌)和病毒(流感病毒、呼吸道合胞病毒和严重急性呼吸系统综合症冠状病毒)感染中的作用,描述肺微生物群在生理防御以及疾病环境中的变化。
最后,本文总结了基于微生物群的治疗方法在呼吸道感染中的作用,包括新型微生物基础疗法(益生菌和膳食补充剂)以及微生物靶向疗法(抗菌单克隆抗体和噬菌体),有助于重塑常驻微生物群落并恢复呼吸道健康。
doi: 10.3390/ijms25074051.
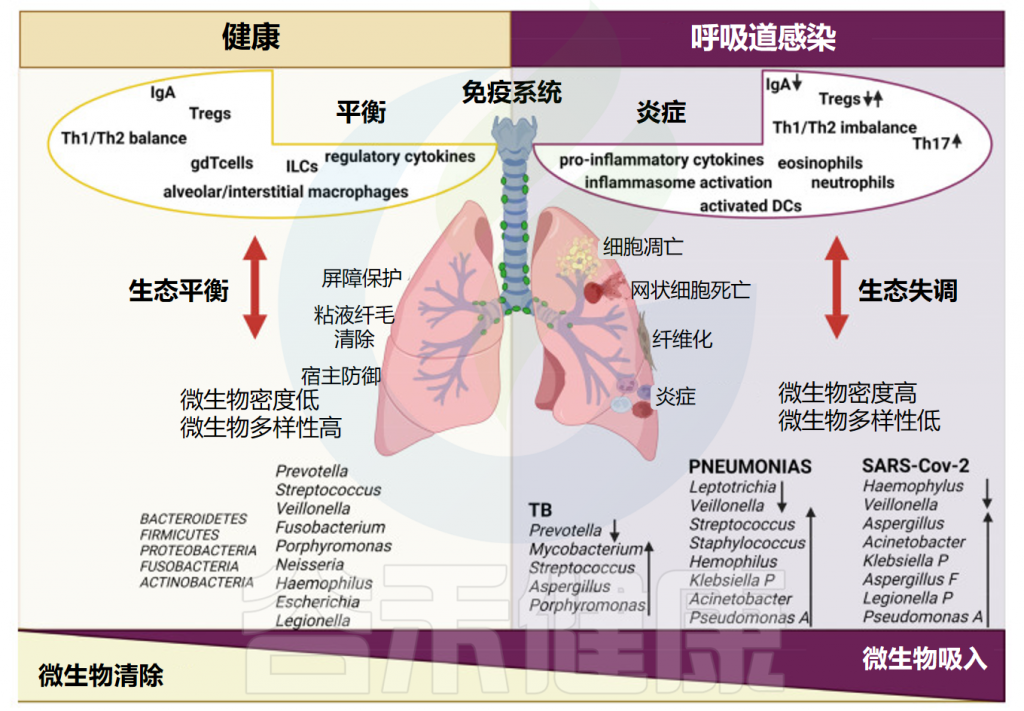
肺部微生物群是指存在于肺部的微生物群落,包括细菌、病毒、真菌和其他微生物。尽管肺部长期被认为是无菌的,但近年来的研究表明,健康的肺部也存在复杂的微生物群,其组成和功能对维持肺部健康和免疫平衡至关重要。
★ 肺部相比肠道微生物较少,但也存在
肺部环境通常不适合细菌群落的发育,导致与肠道环境相比,细菌复制率相对较低,生物量也较低(103-105VS1011-1012个细菌/克组织)。
健康个体的上呼吸道(URT)微生物群更复杂一些,其中口腔共生菌普遍存在。相比之下,下呼吸道(LRT)的微生物群生物量较低,其组成受上呼吸道微吸入的影响,同时因粘膜纤毛活动和先天免疫功能的作用,局部微生物复制受到限制。
大量研究表明,健康的肺部含有拟杆菌门(Bacteroidetes)和厚壁菌门(Firmicutes)的微生物群,其中普雷沃氏菌(Prevotella)、链球菌(Streptococcus)、奈瑟菌(Neisseria)、嗜血杆菌属(Haemophilus)、梭杆菌属(Fusobacterium)和Veilonella是最丰富的属。
16S rRNA 分析确定了健康URT和LRT之间微生物群的空间差异。从口腔到下肺不断发现与链球菌属的细菌重叠。然而,葡萄球菌通常栖息在上呼吸道,而普雷沃氏菌属和韦荣氏菌主要栖息在下呼吸道。
★ 呼吸道微生物定植在出生后就开始
呼吸道定植在出生后24小时内立即开始。在健康状态下,来自链球菌属(Streptococcus)、葡萄球菌属(Staphylococcus)、普雷沃氏菌属(Prevotella)、莫拉氏菌属(Moraxella)、嗜血杆菌属(Haemophilus)、乳杆菌属(Lactobacillus)、棒状杆菌属(Corynebacterium)和Dolosigranulum的物种代表了生命早期呼吸道的初始定植者,有助于平衡和动态的微生物群落。
从新生儿到成熟期,呼吸道中的细菌数量逐渐增加,多样性和功能能力受到分娩方式、母乳喂养、抗生素使用和烟雾暴露等因素的影响。动物模型研究显示,出生后前两周内细菌量明显增加,菌群从γ‐变形菌纲和厚壁菌门逐步转向拟杆菌门。
★ 微生物可以通过空气吸入和扩散进肺部
微生物迁移是通过吸入空气中的细菌、沿粘膜表面直接扩散发生的,后者被认为是健康受试者中占主导地位和普遍存在的途径。事实上,下呼吸道的细菌群落在很大程度上类似于口腔微生物群的组成。
与微生物群落具有高度抗性的高生物量粘膜(即口腔、肠道)的微生物相比,肺微生物组的动态性质可能是一个重要的独特特性。
★ 影响肺部微生物群的因素
影响肺微生物组成的因素主要有以下几个方面:
1.宿主因素
-年龄
-性别
-体重
-遗传因素
-健康状况(基础疾病)
2.环境因素
-空气质量
-吸烟
-职业暴露
-居住环境
3.生活方式因素
-饮食
-口腔卫生
-运动
-抗生素使用
4.其他因素
-季节变化
-医院环境暴露
-抗生素的使用
-疫苗接种
此外还有局部微生物竞争、宿主上皮细胞相互作用和免疫细胞活化等。术语“菌群失调”表示健康肺微生物组的三个决定因素(微生物迁移、微生物消除和繁殖率)平衡的变化,肺部菌群失调与许多不良生物学事件有关,并参与呼吸系统疾病的发生和发展。
★ 肺部微生物群的变化与疾病存在相关性
比较患病肺和健康肺发现,肺部微生物组成存在显著差异,疾病状况与细菌多样性的丧失有关,或者与一小群分类群的优势有关。微生物菌群失调是各种肺部疾病的特征,其中微生物多样性减少可能与疾病进展有关。
肺微生物组的生长条件在病理过程中发生了显著变化,导致适合受伤气道的疾病和患者的微生物群落。例如肺部富集的口腔厌氧菌群(如Prevotella和Veillonella)与炎症增强和免疫反应改变导致的感染易感性增加有关。
此外,肺微生物群成分的改变对某些个体造成更大的感染风险。肺微生物群的改变与多种肺部疾病的恶化有关,不同的细胞免疫反应与暴露于各种肺部微生物有关。例如在慢性肺部炎症中,来自病理性人支气管肺泡系统的假单胞菌(Pseudomonas)和乳杆菌(Lactobacillus)的富集与Th17型反应的增强相关。一些致病体还会诱导严重的不依赖性Toll样受体2的气道炎症和肺部免疫病理学。
呼吸道微生物组成影响人类和小鼠的炎症反应,调控肺γδ T细胞产生IL-17及肺泡巨噬细胞的分化,并影响抵抗呼吸道感染的能力。副流感嗜血杆菌通过TLR4激活促炎反应,阻碍皮质类固醇通路,诱导炎症性Th2途径,最终导致支气管反应。
健康和呼吸道感染期间的肺微生物群和免疫
doi: 10.3390/ijms25074051.
肺部感染包括病原体的获得、传播和侵入下呼吸道。
肺炎是肺实质的炎症。在病因学上,它被归类为社区获得性肺炎(先前健康的个体的感染)或医院获得性肺炎(住院个体在入院后48小时内的感染)。革兰氏阳性肺炎链球菌(Streptococcus pneumoniae)是大多数社区获得性肺炎病例的原因,也是2岁以下儿童、老年人和免疫功能低下个体患病的主要原因。
★ 不同人群易感的病原体有所差异
由流感嗜血杆菌和肺炎克雷伯菌引起的肺炎易在50岁以上的慢性阻塞性肺病或酗酒患者中传播,而由肺炎支原体和肺炎衣原体引起的肺炎则在儿童中更为广泛。由呼吸道合胞病毒(RSV)和腺病毒引起的病毒性肺炎在健康人群中很少见,而由流感病毒引起的肺炎仍然是老年人和基础疾病患者高死亡率的原因。
在成人中,肺炎链球菌(S.Pneumonia)、乳杆菌(Lactobacilli)和罗斯氏菌属(Rothia)的优势与肺炎有关。新生儿中肺炎链球菌、流感嗜血杆菌或莫拉菌的定植与患支气管炎的风险较高有关。在流感患者中还发现假单胞菌增多。
主要病原体检测阳性率年龄组差异

来源:中国疾病预防控制中心
★ 呼吸道病原体感染导致肺部疾病的发展
人体研究表明,不同菌株的早期定植与发生呼吸道感染的风险增加有关。因此,病原体感染和微生物相互作用都可能影响疾病的进程。从生态学上讲,感染的特点是微生物负荷增加和群落多样性减少,以及宿主炎症和组织损伤增加。上呼吸道微生物组的改变是导致肺炎的细菌感染发生的原因。
最近的一项研究描述了严重肺部感染中病毒复制的显著增加,以及轻度和重度疾病患者之间微生物相互作用的差异,尤其是与常见病原菌之间的关联。流感导致的高死亡风险主要归因于继发性细菌感染。病毒通过多种机制增加了宿主对细菌定植的脆弱性。
注:鼻内接种H1N1病毒以模拟人类流感的小鼠微生物组、肺转录组和代谢组显示出显著变化,这些变化在恢复期持续存在。这表明感染对微环境的稳态产生了长期影响,有利于潜在病原体。
结核病
结核分枝杆菌(Mtb)是导致肺结核(TB)疾病中肺部肺炎的原因。在小鼠和人类身上进行的研究表明,肺部微生物组在抵抗结核分枝杆菌感染方面发挥作用。
结核病患者肺部存在特殊的菌群富集
一些细菌菌株可能与结核病的发病、复发和治疗失败有关。例如假单胞菌的丰度与治疗失败的风险增加有关。与健康对照组相比,感染结核分枝杆菌的个体的微生物群多样性降低,并且经常表现出链球菌和假单胞菌的富集。
在结核病中,肺部和整个下呼吸道具有特殊的微生物特征:结核分枝杆菌、金黄色葡萄球菌和乳酸克鲁维酵母(Kluyveromyces lactis)在123名结核病患者的支气管肺泡灌洗液 (BALF)中高度富集,而副流感嗜血杆菌在未感染的肺部富集。
使用鼻咽拭子样本的研究表明,结核分枝杆菌感染显著改变了微生物群组成:与健康对照组相比,结核病患者的变形杆菌、γ-变形杆菌、假单胞菌和莫拉菌科增加,而芽孢杆菌和毛螺菌属减少。
肺泡微生物多样性降低可能由炎症环境引起
与健康对照组相比,结核病患者的肺泡微生物群多样性降低(链球菌和梭杆菌减少,结核分枝杆菌丰度增加)。这些变化可能是由炎症环境决定的,因为结核分枝杆菌可以释放毒力因子,从而抑制巨噬细胞的反应。
在另一项研究中,链球菌在结核病中显著增加,而结核病中的Th1反应可能是由奈瑟菌(Neisseria)和嗜血杆菌(Haemophilus)引发的。
新冠肺炎
2019年,一种新型冠状病毒严重急性呼吸系统综合症2(SARS-CoV-2)引起了新型冠状病毒病(COVID-19)大流行。疾病严重程度和死亡率因年龄及合并症而异,其中许多与肠道和肺部微生物改变有关。这表明菌群失调可以在一定程度上预测COVID-19的严重程度。
新冠病毒感染者肺部微生物群失调
一般来说,与健康或病情较轻的COVID-19个体相比,危重症COVID-19患者的肺部物种多样性降低,细菌负荷增加。不良的临床结果与支原体的下呼吸道富集有关。
一项研究描述了来自重症COVID-19患者的支气管肺泡灌洗液(BALF)样本,假单胞菌丰度显著更高,而来自COVID-19肺炎阴性患者的 BALF 的特征是流感嗜血杆菌和韦荣氏球菌等的富集。
据报道,20名已故 COVID-19 患者的肺组织微生物组以不动杆菌(Acinetobacter)为主,不动杆菌通常与引起肺炎的肺部感染有关。
在上呼吸道和下呼吸道中,宿主免疫系统对潜在的有害物质做出反应,并将它们与自身成分、外来非危险物质和有益共生微生物群区分开来。
◆ 气道上皮产生粘蛋白形成抵御病原体的屏障
气道上皮是与当地微生物群相互作用的场所,也是生物物理保护屏障。产生粘液的气道上皮分泌细胞是肺先天免疫系统的关键要素。一些数据表明,呼吸道微生物群调节塑造粘液产生的上皮屏障。具体来说,在小鼠模型中,证明了粘蛋白与肺部感染反应之间存在联系。
◆ 产生lgA阻止病原体的吸附
除粘蛋白外,气道上皮细胞还提供抗原特异性分泌型IgA屏障,能够保护气道表面。分泌型IgA的作用是阻止病原体的吸附,中和它们在粘膜上皮的毒性产物,介导感染上皮细胞中病毒的消除,并促进对病原体的杀伤。
多项研究证明,IgA在防御呼吸道感染中发挥重要作用:IgA缺陷小鼠对牛分枝杆菌卡介苗(BCG)鼻内感染的易感性增加。鼻内给药中和IgA后,接种流感病毒(IV)或呼肠孤病毒导致小鼠模型中肺部病毒滴度显著降低,肺炎严重程度减轻。值得注意的是,静脉注射抗原特异性聚合物IgA可通过血清中的IgA分泌保护小鼠免受流感感染。
◆ 分泌抗菌肽来对抗病原体
局部呼吸道还分泌称为抗菌肽(AMP)的保护性介质,包括溶菌酶、乳铁蛋白、脂质运载蛋白、过氧化物酶、氨肽酶、集合蛋白(表面活性剂蛋白A和表面活性剂蛋白D)、甘露聚糖结合凝集素(MBL)、cathelicidins和β-防御素。
值得注意的是,β-防御素和cathelicidins具有抗菌和免疫调节作用,并参与塑造微生物群组成。事实上,炎症反应的直接抗菌活性和免疫调节是由微生物群的成员触发的。
免疫细胞在呼吸道病原体感染中至关重要,是机体防御系统的核心。接下来,我们一起来了解一下不同免疫细胞与病原微生物感染之间的相互作用。
▸ 巨噬细胞
肺中存在不同类型的先天免疫细胞:肺泡巨噬细胞(AMs)是最具代表性的细胞,似乎是几种呼吸道感染发病机制的核心,包括结核分枝杆菌(MTB)、肺炎链球菌、鼻病毒、流感病毒(IV)和呼吸道合胞病毒(RSV)。
◆ 肺巨噬细胞失调影响呼吸道感染的死亡率和并发症
AMs启动白细胞募集,并使用几种病原体特异性机制直接消除病原体,例如分泌促炎细胞因子/趋化因子(IL-6、IL-8或CXCL10),启动I型IFN信号传导,增强模式识别受体的表达,以及抑制病毒基因组的核输出。
并发症发生率和死亡率的增加与肺泡巨噬细胞耗竭有关,无论是在实验过程中还是在自然过程中,呼吸道病毒感染都是相关的。此外还发现,重症COVID-19的一个一致特征是肺巨噬细胞失调。
▸ 树突状细胞
肺部的树突状细胞(DC)在防止呼吸道感染方面发挥作用,对病原体产生强大的适应性免疫反应。树突状细胞可以启动抗病毒CD8细胞毒性T细胞反应,导致病毒清除,还可以控制炎症反应的水平。
◆ 树突状细胞可作为继发性肺炎的生物标志物
炎症期间和之后的树突状细胞(DC)改变可用作继发性肺炎易感性的生物标志物,以及有希望的治疗靶点,以改善患者的预后。在结核分枝杆菌(MTB)感染中,DC在将先天免疫反应转变为适应性免疫方面发挥着多因素作用。值得注意的是,树突状细胞在防御SARS-CoV-2感染方面也发挥着关键作用。
▸ 黏膜相关不变T细胞
黏膜相关不变T(MAIT)细胞是非常规T细胞的一个亚群,对微生物感染进行免疫监视和免疫反应。
◆ 进行免疫监视和免疫反应
不同的研究表明,MAIT 细胞通过识别和杀死细菌感染的细胞(包括树突状细胞和肺上皮细胞)在结核分枝杆菌感染的免疫控制中发挥作用。
此外,MAIT细胞在人和小鼠中都需要细胞因子来响应结核分枝杆菌抗原,这可能是由于它们在感染部位募集。一项新的研究证实,急性SARS-CoV-2感染患者循环中的MAIT被激活,但频率降低,而它们在致命性COVID-19患者的肺部蓄积。
▸ 自然杀伤T细胞
不变自然杀伤T(iNKT)细胞在控制共生菌(包括机会性病原微生物群)中发挥作用,反过来,微生物群调节iNKT细胞。
在原发性呼吸道合胞病毒(RSV)感染期间,肺iNKT细胞的激活除了导致肺嗜酸性粒细胞增多和纤维化外,还导致抗病毒CD8 T淋巴细胞反应和病毒清除。
◆ 活动性结核病患者存在iNKT细胞缺乏
人类iNKT细胞缺乏可能是活动性/急性结核病发展的基础:与潜伏性结核病患者相比,活动性结核病患者的外周iNKT细胞较少,并且通过活动性结核病的治疗可以重建正常的iNKT细胞频率。据报道,重症 COVID-19 肺炎患者 iNKT 细胞减少,表明该亚群可能作为疾病严重程度的生物标志物发挥作用。
▸ γδT细胞
肺驻留γδT细胞是粘膜上皮屏障的主要T细胞成分,对维持肺稳态和影响多种肺部疾病的进展至关重要。γδT细胞是肺炎克雷伯菌感染中 IL-17A 的主要来源,也是宿主对急性铜绿假单胞菌肺部感染的早期免疫防御。
◆ γδT细胞对肺部感染进行早期免疫防御
在肺炎链球菌肺部感染期间,观察到活化的γδ T细胞数量显著增加。γδ T 细胞对冠状病毒感染的反应仍在研究中,之前关于SARS-CoV-2感染的报道显示对感染的靶标单核细胞系具有很强的溶细胞活性。
▸ 调节性T细胞
调节性T(Treg)细胞对于肺对空气传播的过敏原的免疫耐受以及减少对自身和非自身抗原的危险免疫反应至关重要。Treg耗竭通过抗原致敏在衣原体肺炎感染的发病机制中发挥作用。
◆ 调节性T细胞对肺炎链球菌肺炎具有保护作用
此外,Tregs通过与TGF-β通路相关的机制对肺炎链球菌引起的肺炎具有保护作用。其他研究使用呼吸道合胞病毒和甲型流感病毒的小鼠模型表明,Treg细胞的耗竭可能导致CD8+ T细胞亚群的迁移延迟。
最近的研究显示,COVID-19患者的Tregs数量显著减少,导致Treg/Th17比值失衡,这与呼吸衰竭风险相关。
此外,呼吸道微生物群的成员通过TLR反复暴露于病原体相关分子模式(PAMPs)和损伤相关分子模式(DAMPs),诱导树突状细胞和肺泡巨噬细胞的模式识别受体(PRR)耐受性。总之,呼吸道微生物群与气道上皮细胞和吞噬细胞形成正反馈回路,以实现免疫耐受并避免失控的炎症反应。
呼吸道中的宿主-微生物相互作用主要发生在粘膜部位。常驻微生物可以局部或全身引发免疫细胞(如上皮细胞、树突状细胞和中性粒细胞)的反应。这些物质能够进入循环并到达其他器官。
◆ 呼吸道微生物群与免疫和呼吸健康密切相关
呼吸道微生物组为宿主免疫系统提供重要信号,这些信号对免疫训练、器官发生和免疫耐受的维持至关重要。观察结果表明,在生命早期存在一个关键时期,正确的微生物群感知对免疫成熟和呼吸健康至关重要。
动物模型研究了微生物组在影响和管理宿主免疫系统中的作用,以及免疫系统在塑造微生物组中的作用。
例如,在小鼠模型中,拟杆菌门的成员相比流感嗜血杆菌减少了炎症、中性粒细胞募集和TLR2介导的细胞因子产生。
鼻内接种金黄色葡萄球菌导致单核细胞募集到肺部,并分化为抑制肺泡巨噬细胞的IV诱导炎症反应。
尽管肺炎链球菌被视为病原体,但它也是上呼吸道的一种共生菌。在小鼠鼻腔中同时定植肺炎链球菌和流感嗜血杆菌创造了炎症环境,导致C-X-C基序趋化因子配体2和中性粒细胞的高水平募集。协同反应依赖于肺炎链球菌产生的溶细胞毒素,表明其存在调节了对流感嗜血杆菌的免疫反应。遭受流感嗜血杆菌攻击的小鼠表现出严重的肺部疾病,显示出明显的中性粒细胞通路和高浓度的促炎细胞因子。
然而,如果小鼠在吸入共生普雷沃氏菌后进行预处理,炎症会显著减少,且不存在组织病理。腺病毒诱导的记忆肺泡巨噬细胞在小鼠急性鼻内感染中表现出主要组织相容性复合体II类(MHCII)表达增加,以及与宿主防御、趋化性、抗原呈递和糖酵解代谢相关的基因转录上调。
在呼吸道病原体引起的菌群失调期间,共生细菌受到干扰,致病菌可同时出现在肺和肠道中,因此可能导致两个部位的组织损伤。
肠道和肺之间的密切生理和病理联系主要依赖于宿主-微生物的串扰。事实上,肺和肠道细菌的成员可以通过血流成分和代谢物直接交换,从而促进这两个部位的健康或疾病。
肠-肺轴主要相互作用的示意图
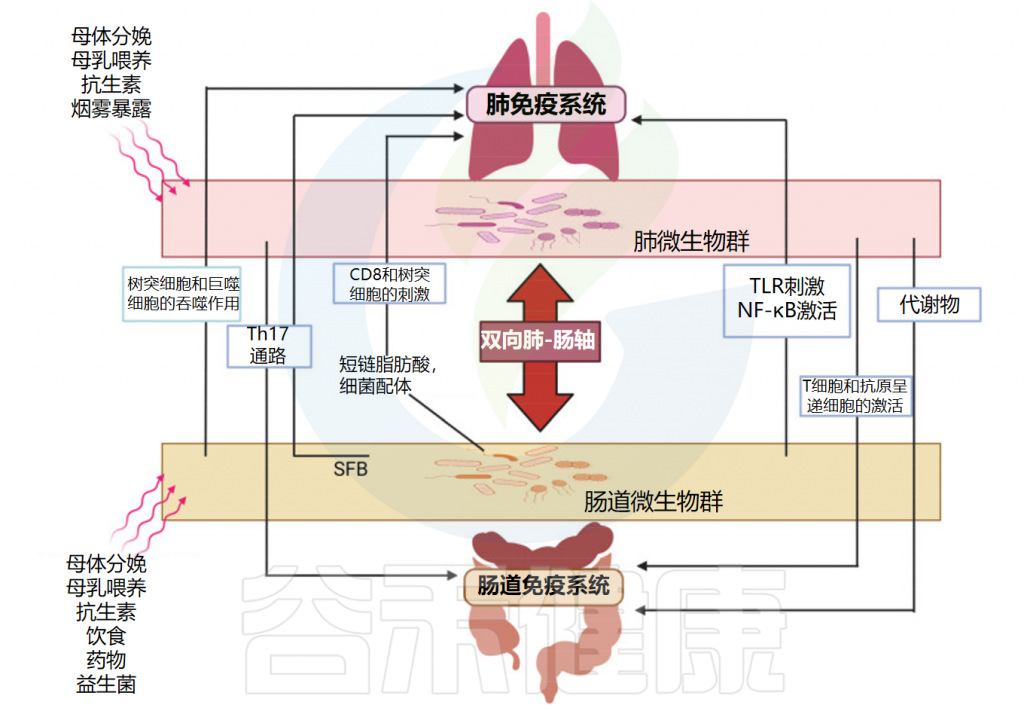
doi: 10.3390/ijms25074051.
呼吸道感染对肠道菌群的影响
研究发现,肺微生物群的改变调节肠道微生物群落,从而影响肠道信号传导。已知结核分枝杆菌感染会导致免疫系统失调,从而使肠道微生物组改变。
◆ 结核病患者肠道微生物群发生显著改变

在一项研究中,比较了成年结核病患者与健康对照的肠道微生物组,发现厚壁菌门、变形菌门和疣微菌门减少,而放线菌门、拟杆菌门和梭杆菌门增加。
另一项研究分析了新发和复发性结核病患者,报道了拟杆菌门、普雷沃氏菌属和毛螺菌属的减少,而放线菌门和变形菌门的富集。
最后,在一组受影响的儿童中,观察到放线菌门和厚壁菌门的减少,而包括拟杆菌属、双歧杆菌属、Dorea属、粪杆菌属、瘤胃球菌属和普氏粪杆菌(F.prausnitzii)属、肠球菌属和普氏菌属的数量增加。
◆ 结核病患者的微生物代谢也变得不平衡
结核病患者可能导致微生物代谢物的产生不平衡,例如短链脂肪酸(SCFA),这可能会重置肺微生物组和通过“肠-肺轴”的免疫反应。这些发现也可能解释了结核分枝杆菌在胃肠道中的定植和肺结核患者发生肠结核的原因。普氏粪杆菌(F.prausnitzii)还被描述具有抗炎作用,可抵御胃肠道疾病。
很少有研究分析人类呼吸道病毒感染期间发生的肠道微生物群改变的性质。在流感期间,尽管肠道中明显不存在病毒,但一些患者仍会出现类似胃肠炎的症状。
◆ 流感病毒感染患者的肠道菌群也存在失衡
一项针对H7N9病毒感染患者的研究显示,拟杆菌门(包括经黏液真杆菌属(Blautia)、罗氏菌属(Roseburia)和瘤胃球菌(Ruminococcus))减少,但厚壁菌门和变形菌门以及属(包括埃希氏菌(Escherichia)、梭菌属(Clostridium))增加。
另一项在流感亚型H1N1患者中进行的研究报告了放线菌门和厚壁菌门的减少,以及包括Dorea、粪杆菌、瘤胃球菌、链球菌在内的属,放线菌科和微球菌科的富集。
在最近的一篇综述中,11项不同的研究报告了与健康对照相比,确诊或疑似呼吸道感染(RTI)患者的肠道微生物组改变。总之,患者的肠道微生物组改变始终是多样化的,包括一些菌的枯竭和富集。
◆ 新冠病毒感染可在肠道中检测到病毒RNA
与甲型流感病毒(IAV)和呼吸道合胞病毒(RSV)的情况相反,在SARS-CoV-2(新型冠状病毒)感染期间,即使病毒RNA不再存在于呼吸道中,也可以在肠道中检测到病毒RNA,从而指出消化道是病毒复制和活性的潜在部位。
◆ 产丁酸抗炎菌减少,机会性病原体增多
SARS-CoV-2感染患者的肠道菌群失调与COVID-19疾病进展和严重程度以及COVID-19后综合征相关。它的特点是双歧杆菌和粪杆菌等抗炎菌减少,丁酸盐生产者丰度降低,例如瘤胃球菌科和毛螺菌科的几个属。
相反,还存在炎症相关微生物群(包括链球菌和放线菌)的富集,以及机会性细菌病原体的过度生长。
最近的一篇论文也表明,SARS-CoV-2感染会导致小鼠肠道微生物组失调,以及潘氏细胞和杯状细胞以及屏障通透性标志物的改变。同样,从96名 COVID-19患者收集的微生物组样本显示,抗菌素耐药物种在内的机会性病原菌属大量繁殖,这种肠道菌群失调与肠道细菌的继发性血流感染有关。
◆ 通过促炎因子和食物摄入减少影响肠道菌群
肺部感染的细胞免疫反应会导致食欲不振,从而改变肠道微生物组和代谢。研究发现,消除CD8+细胞可以阻止食物摄入量的减少并逆转肠道微生物群的变化,这可能是通过分泌TNF-α实现的。实际上,在呼吸道合胞病毒感染期间,中和这种细胞因子可以减少体重减轻并缓解肠道微生物群的扰动。
总的来说,炎性细胞因子的释放和食物摄入量减少是急性病毒性呼吸道感染影响肠道微生物群的可能机制。另一种机制涉及浸润的CD4+ T细胞或全身 IFN释放,它改变了上皮细胞的代谢,导致营养物质的积累,肠腔的微生物为此竞争。这些发现可能在呼吸道病毒感染期间的肠道菌群失调和胃肠道疾病中发挥作用。
肠道微生物群对呼吸道感染的影响
◆ 炎症性肠病患者通常存在肺部受损
肠道和肺之间的重要联系主要体现在炎症性肠病(IBD)患者的肺部受累范围广泛且严重,从亚临床改变到明显的慢性炎症性肺病。气道受累的症状最常见于有长期IBD病史的患者,这与肠道发育不良或全身性不当免疫反应有关。
◆ 肠道菌群失调与呼吸道感染易感性增加相关
除了慢性疾病,肠道菌群失调还与呼吸道感染的易感性增加有关。临床观察强调了健康肠道微生物群在预防病毒呼吸道感染方面的重要性。在肾移植受者和同种异体造血干细胞移植(HSCT)患者中,产丁酸盐的肠道细菌减少与病毒呼吸道感染的风险和发生率增加相关。
在COVID-19(新冠病毒感染)患者中也发现了肠道菌群失调和肠道代谢产物的变化,这与炎症反应和疾病并发症相关。值得注意的是,将COVID-19患者的粪便移植到无菌小鼠体内会导致肺部炎症,并在多重耐药肺炎克雷伯菌感染期间导致更糟糕的结果,这表明微生物群可能直接导致疾病后遗症。
此外,肠道菌群还可以调节血管紧张素转换酶2(ACE2)受体的结肠表达。这些证据可能有助于解释肠道菌群失调患者(如老年人、免疫功能低下患者和有其他合并症的患者)疾病易感性和胃肠道症状的增加。
◆ 肠道微生物及其代谢物可以调节肺部免疫反应
在几种细菌和病毒呼吸道感染的实验模型中,肠道共生菌的缺失或通过抗生素耗竭导致微生物传播、炎症、器官损伤和死亡率增加。这些影响大多与肠道微生物群塑造全身免疫的能力有关。由肠道微生物及其代谢物(如SCFAs)触发的免疫细胞和细胞因子可以到达体循环,并调节肺部、健康和疾病中的免疫和炎症反应。
此外,据报道,炎症性肠病患者的肠道淋巴细胞缺乏组织特异性;这可以解释IBD患者肠外器官存在炎症的原因。菌群失调介导的炎症也会导致粪便钙卫蛋白、血浆C反应蛋白、IL-6和IL-8的循环水平升高,这可能导致肺部感染期间的并发症发生。
肠道菌群失调也可能通过减少营养吸收和能量可用性来影响肺部感染的结果,这反过来会干扰患者产生有效免疫反应的能力。
关于肠道微生物群如何影响呼吸道感染的机制,动物研究提供了更多见解。
在抗生素处理的小鼠中,腹膜巨噬细胞中IFN-γRI、MHC-I、CD86和CD40分子的表达在对病毒感染的早期反应中减弱,这表明肠道微生物群在病毒在宿主体内复制之前就向先天免疫反应发出信号。
Th1、IgA和巨噬细胞对呼吸道病毒感染的反应建立取决于肠道微生物。直肠TLR刺激为IL-1β和IL-18分泌提供信号,恢复了抗生素处理小鼠的肺CD4+和CD8+ T细胞对感染的反应。
最近报道了一种常见细菌种类,即分段丝状细菌(SFB)在肠道中的定植,重新编程了肺泡巨噬细胞,使其增殖、补体产生和吞噬作用增强,从而增强了对流感病毒、呼吸道合胞病毒和SARS-CoV-2的保护。
总体而言,这些发现证实了肠道微生物刺激在先天抗病毒免疫反应激活中的重要性。在感染模型中,抗生素消耗肠道微生物群增加了结核分枝杆菌的负担和传播。菌群失调特别降低了肺树突状细胞中先天受体和巨噬细胞诱导的C型凝集素的表达,导致对幼稚T细胞的刺激功能受损,从而减少了感染小鼠的效应细胞和记忆T细胞数量。
◆ 肺部重叠感染可能受肠道微生物群的影响
肺部重叠感染可能是肺部原发感染诱导的肠道微生物组成改变的进一步后果。对继发性细菌感染的易感性增加,特别是由肺炎链球菌和金黄色葡萄球菌诱导的细菌感染,经常发生在患有呼吸道病毒感染的儿童和老年人中,导致并发症和死亡。
流感期间的肠道菌群失调通过改变短链脂肪酸的产生导致肺炎链球菌重叠感染。在接受甲型流感病毒条件微生物群的小鼠中,乙酸盐的产生减少,改变了肺泡巨噬细胞的杀菌活性,降低了肺对继发性肺炎链球菌感染的防御能力,并促进了重叠感染小鼠的死亡。肠道疾病也可能导致重症 COVID-19 患者伴随或继发性细菌感染。
因此,局部肺部或肠道菌群失调,改变了微生物间相互作用的动力学以及微生物代谢,可能会增强潜在致病细菌物种的增殖。
揭示肺和肠道之间的复杂相互作用有助于更好地了解共生微生物群作为各种呼吸道传染病的治疗靶点。
使用益生菌(微生物)、有利于其生长的产品(如益生元)或微生物代谢物(如后生元)的给药可以通过与病原微生物的直接竞争、改善上皮屏障功能或免疫调节,在呼吸系统疾病期间提供宿主保护。
病毒感染下的治疗
◆ 使用益生菌能够减少流感风险和发生率
在一项系统评价中,评估了益生菌在大规模健康受试者中预防病毒性呼吸道感染(RTI)的疗效。乳杆菌(Lactobacillus)是最常用的益生菌,其次是双歧杆菌(Bifidobacterium)和乳球菌(Lactococcus)。
大多数研究表明,益生菌的使用与病毒RTI的风险和发生率降低相关。然而,在临床表现、病毒载量和免疫学结局的改善方面尚无共识。益生菌鼠李糖乳杆菌和短乳杆菌也与流感感染发生率的减少有关。
在新型冠状病毒(COVID-19)背景下,一项针对SARS-CoV-2诱发肺炎的ICU患者的回顾性研究显示,与单独使用标准治疗相比,使用乳杆菌、双歧杆菌和链球菌属的益生菌混合物治疗与死亡率降低相关。
◆ 益生菌增强了免疫反应,有助于提高生存率
几份报告描述了口服益生菌对病毒性呼吸道感染结果的影响,也提供了机制见解。在感染流感病毒或呼吸道合胞病毒之前服用益生菌,如乳杆菌、双歧杆菌、肠球菌或乳球菌,可缓解症状并提高生存率。肺部和鼻腔冲洗液中的病毒载量也有一定程度的降低。
从机制上讲,益生菌可以通过参与免疫细胞并诱导特定的细胞因子/趋化因子产生来引发针对病毒呼吸道感染的保护性反应,尽管其作用似乎是高度菌株特异性的。
研究表明,益生菌给药后,自然杀伤细胞活性增加,浸润性巨噬细胞和中性粒细胞减少,支气管肺泡灌洗液中的病毒特异性IgA/G滴度增加。粘膜乳杆菌抑制呼吸道合胞病毒复制并降低血炎细胞(如粒细胞和单核细胞)的比例。
用副流感病毒对用益生菌治疗的小鼠肺淋巴细胞进行离体刺激,导致IFN-α和IFN-β的高度表达。病毒感染期间IL-10的上调和IL-6的相应减少也由益生菌给药引发。除了这些保护作用外,经鼻给药鼠李糖乳杆菌GG和嗜酸乳杆菌L-92还增加了IL-1β和单核细胞趋化蛋白1细胞因子以及趋化因子eotaxin和M-CSF的水平。
注意:尽管益生菌具有令人满意的安全性,但它们的使用可能与体弱人群感染或发病率的较高风险有关。因此,人们对使用非活微生物越来越感兴趣。动物研究证明了热灭活益生菌在呼吸道感染中的有益作用,尽管它们的全球影响似乎仅次于活益生菌。
◆ 通过饮食补充益生元提高了抗病毒感染的能力
肠道微生物群可以通过产生短链脂肪酸来调节免疫反应。摄入微生物可利用的膳食纤维(益生元),促进特定共生微生物的多样性和活性的增加,导致对微生物代谢物产生不同影响,进而对宿主对感染的反应产生不同影响。
高纤维(可发酵菊粉)饮食通过两种互补机制传达对流感的保护。小鼠表现出增强的选择性激活巨噬细胞的骨髓生成,这些巨噬细胞促进CXCL1介导的中性粒细胞募集到气道的能力有限,从而导致感染期间组织免疫病理学受限。
同时,饮食衍生的短链脂肪酸刺激CD8+ T细胞抗病毒活性。这种作用是由丁酸盐通过游离脂肪酸受体(FFAR)介导的。同样,高纤维(可发酵果胶)饮食通过刺激肺上皮细胞中的I型IFN 反应来防止呼吸道感染,这种保护是由乙酸盐通过GPR43介导的。
此外,有研究报道,流感感染期间口服醋酸盐补充剂以FFAR2依赖性方式加强了肺部对继发性肺炎球菌感染的防御能力,并降低了重叠感染小鼠的致死结局。同样,鼻内醋酸盐增加了鼻病毒感染期间的干扰素依赖性反应,降低了肺病毒载量。
有趣的是,一项评估益生元(低聚半乳糖和聚葡萄糖)预防新生儿病毒性呼吸道感染功效的试验表明,与益生菌相比,益生元显示出更好的有益效果,这可能与对预先存在的有益细菌生长的直接刺激作用有关。
细菌感染下的治疗
通过调节微生物群对肺部细菌感染又有怎样的效果,让我们接着往下看。大多数研究分析了益生菌在预防和治疗ICU患者肺部感染中的疗效。
◆ 口服益生菌降低了细菌性肺部感染的概率
口服或口咽给予干酪乳杆菌和鼠李糖乳杆菌,可以减少铜绿假单胞菌或相关病原体对下呼吸道的定植和感染。一项研究观察到,在脓毒症患者中,使用短双歧杆菌、副干酪乳杆菌和低聚半乳糖的混合物后,呼吸机相关性肺炎(VAP)的发生率降低。
对感染肺炎链球菌的小鼠研究表明,口服不同的益生菌,如乳杆菌属和链球菌属的菌株,可增加对感染的抵抗力,降低肺细菌负荷,提高存活率。
◆ 鼠李糖乳杆菌等益生菌增强了肺部的免疫反应
鼠李糖乳杆菌的使用通过增加 Foxp3 + Treg 和减少促炎IL-6来增强抗炎反应。在气管内给予其他乳杆菌菌株后,在感染的小鼠中也观察到这种抗炎特征。
活的或灭活的益生菌长双歧杆菌 51A的给药通过增强肺泡巨噬细胞中活性氧的产生和减少促炎性TNF-α和IL-6来刺激肺炎克雷伯菌的肺清除。然而,只有活的益生菌诱导了IL-10水平的同时升高,主要是由醋酸盐介导的。
经鼻或口服接种能有效激活Nod2受体的细菌菌株(分别为脆弱乳杆菌、金黄色葡萄球菌和表皮葡萄球菌,或罗伊氏乳杆菌、粪肠球菌和解黄酮梭菌)可保护小鼠免受肺炎链球菌或肺炎克雷伯菌感染。
补充乳杆菌还可以恢复肺部树突状细胞介导的抗结核分枝杆菌免疫。在感染结核分枝杆菌的小鼠中,口服A.muciniphila或A.mucinihila介导的棕榈烯酸通过表观遗传抑制肿瘤坏死因子,强烈抑制了结核病感染。
呼吸道益生菌
虽然大多数用作益生菌的共生菌起源于肠道,但已经尝试使用呼吸道共生菌。特别是,在婴儿小鼠中鼻内施用Corynebacterium pseudodiphteriticum能够改善呼吸道合胞病毒原发感染和继发性肺炎链球菌重叠感染的特征,降低病原体负荷和肺损伤。据我们所知,迄今为止还没有人类研究评估过呼吸益生菌(即滴注或雾化到下呼吸道的活菌群)的潜力。
在动物模型中,细菌B.bacteriovorus和M.aeruginosavorus的鼻内给药大大降低了呼吸道肺炎克雷伯菌的负担。它们随后被宿主先天免疫机制消除,没有产生不良影响,这预示了它们可能用于治疗人类细菌性肺炎。
◆ 单克隆抗体特异性对抗细菌病原体
使用单克隆抗体(mAb)选择性消耗机会性细菌病原体,通过靶向和灭活细菌及其毒力因子或毒素,成为对抗传染病的一种有前景的方法。
这些抗体因其靶标特异性,不会对本身微生物群产生不良影响,且不太可能导致广泛耐药性。此外,多价mAb经过工程改造,能够发挥多种抗菌作用,包括灭活毒力因子、促进补体沉积和激活先天免疫。
Gremubamab(MEDI3902;AstraZeneca)是一种双特异性人IgG1 mAb,能选择性结合铜绿假单胞菌毒力因子,用于预防高危患者的医院肺炎。该单克隆抗体促进中性粒细胞清除细菌,并防止其附着在气道上皮细胞。Gremubamab的预防性和治疗性给药在急性铜绿假单胞菌肺炎中表现出高度保护作用。
单克隆IgM抗体帕诺巴单抗(AR-101,Aerumab;Aridis Pharmaceuticals)正在临床开发中,用于治疗医院获得性肺炎中的铜绿假单胞菌。帕诺巴单抗的给药减少了急性铜绿假单胞菌感染小鼠模型中的细菌负荷,并减轻了肺部炎症。
另外两种抗体已被检测用于预防和治疗金黄色葡萄球菌肺炎。舒拉妥珠单抗(MEDI4893;Aridis Pharmaceuticals)特异性结合并灭活金黄色葡萄球菌的成孔α-毒素,这是一种关键的毒力因子。在针对金黄色葡萄球菌定植的机械通气ICU患者的临床试验中,单克隆抗体显著缩短了部分患者的肺炎、住院时间和ICU住院时间。
◆ 噬菌体及其内溶素有助于治疗细菌感染
噬菌体疗法在治疗细菌病原体的呼吸道感染方面受到了临床关注,因为它能够在不影响宿主微生物群的情况下选择性地消除目标细菌,与抗生素相容性好,免疫原性低。
噬菌体制剂在治疗铜绿假单胞菌、肺炎克雷伯菌、鲍曼不动杆菌或大肠杆菌方面显示出很强的临床疗效,目前正在临床上评估用于治疗人类呼吸道感染。四种针对铜绿假单胞菌呼吸道感染的专性裂解噬菌体混合物被用于成功治疗肺炎和肺气肿患者。
噬菌体衍生的内溶素或溶酶在对抗细菌感染方面可能优于全噬菌体制剂,因为其细胞壁靶标的保守性使得耐药性难以产生。在鼻咽定植小鼠模型中,噬菌体内溶素的给药降低了肺炎链球菌的滴度,并有效保护小鼠免受致命性肺炎球菌肺炎或铜绿假单胞菌感染,显示出其在预防和治疗呼吸道细菌感染中的潜力。
呼吸道感染是最常见的病毒或细菌来源的传染病之一,造成严重社会和经济负担。研究发现呼吸道感染会导致肺部微生物群的多样性和组成发生变化,进而影响宿主与病原体的相互作用、炎症信号和免疫细胞因子的产生,最终影响疾病的进展和结果。
肺和肠道之间的重要性和复杂的串扰以及肠-肺轴与呼吸健康的紧密联系越来越受到认可,特别强调了肺-肠轴在调节炎症和免疫反应中的双向通信作用。然而,对涉及肠-肺轴的机制的理解,特别是在呼吸道感染的情况下才刚刚开始。需要更好地了解微生物组、呼吸道粘膜和潜在免疫途径之间的相互作用。
迄今为止,大多数用作益生菌的共生菌都来自胃肠道。由于肠道微生物群在影响免疫系统方面起关键作用,它会影响局部和全身(肺部)对病原体的反应。因此,改变肠道微生物组的微生物群靶向疗法(如益生菌、益生元和噬菌体疗法)已被证明对急性和慢性呼吸系统疾病都有益处。
然而,最近的研究表明,与肠道微生物群落相比,肺微生物群与肺免疫变化的相关性始终更密切。此外,肺共生菌的实验性调节直接和持续地改变了肺免疫反应,表明它们可能具有作为局部应用益生菌预防和管理呼吸道感染的潜力。
值得注意的是,致病细菌的抗生素耐药问题。下呼吸道感染是多重耐药感染导致死亡的主要原因,通常与金黄色葡萄球菌、肺炎克雷伯菌、肺炎链球菌、鲍曼不动杆菌和铜绿假单胞菌等重点病原体相关。同时,新变种的出现使呼吸道病毒感染愈加难以控制。
未来的研究应进一步揭示肺和肠道微生物群在健康和疾病状态下的具体机制,特别是它们如何通过代谢产物和免疫细胞影响呼吸道的免疫稳态。此外,需要更多的临床数据来验证这些微生物组干预措施在改善呼吸道感染预后方面的有效性。通过深入理解肺-肠轴的复杂相互作用,有望开发出新的预防和治疗策略,以减轻呼吸道感染的负担并改善患者的整体健康状况。
主要参考文献
Marrella V, Nicchiotti F, Cassani B. Microbiota and Immunity during Respiratory Infections: Lung and Gut Affair. Int J Mol Sci. 2024 Apr 5;25(7):4051. doi: 10.3390/ijms25074051.
Wypych T.P., Wickramasinghe L.C., Marsland B.J. The influence of the microbiome on respiratory health. Nat. Immunol. 2019;20:1279–1290.
Mathieu E., Escribano-Vazquez U., Descamps D., Cherbuy C., Langella P., Riffault S., Remot A., Thomas M. Paradigms of Lung Microbiota Functions in Health and Disease, Particularly, in Asthma. Front. Physiol. 2018;9:1168.
Sommariva M., Le Noci V., Bianchi F., Camelliti S., Balsari A., Tagliabue E., Sfondrini L. The lung microbiota: Role in maintaining pulmonary immune homeostasis and its implications in cancer development and therapy. Cell. Mol. Life Sci. 2020;77:2739–2749.
Natalini J.G., Singh S., Segal L.N. The dynamic lung microbiome in health and disease. Nat. Rev. Microbiol. 2023;21:222–235.
Wu B.G., Segal L.N. The Lung Microbiome and Its Role in Pneumonia. Clin. Chest Med. 2018;39:677–689.
Yagi K., Huffnagle G.B., Lukacs N.W., Asai N. The Lung Microbiome during Health and Disease. Int. J. Mol. Sci. 2021;22:10872.
Belizário J., Garay-Malpartida M., Faintuch J. Lung microbiome and origins of the respiratory diseases. Curr. Res. Immunol. 2023;4:100065.
谷禾健康
假单胞菌属(Pseudomonas)是最多样化和普遍存在的细菌属之一,其物种存在于沉积物、临床样本、植物(或植物根际)、患病动物、水、土壤、海洋、沙漠等,这反映在它们多变的代谢能力和广泛的适应环境的潜力上。
假单胞菌属是一类革兰氏阴性的好氧或微需氧的细菌,属于 Proteobacteria(变形菌门),代表种包括:
Pseudomonas aeruginosa(铜绿假单胞菌)
Pseudomonas putida (恶臭假单胞菌)
图源:CDC
假单胞菌属是机会性病原体,是全世界医院获得性感染的主要原因。它可以侵入多个器官系统,包括血液、肺、软组织(皮肤、肌肉和肌腱)、身体的其他部位。症状取决于感染部位。
与假单胞菌属感染相关包括免疫系统受损患者的感染和医疗器械相关感染等,如刚接受过手术的人和免疫功能低下的人,艾滋病或糖尿病患者。
假单胞菌属与人体健康相关,其中一些菌株可以引起感染和疾病,如铜绿假单胞菌(Pseudomonas aeruginosa),可以引起呼吸道、肠道、泌尿道和皮肤感染。
免疫抑制个体和住院患者的铜绿假单胞菌肠道携带率显着升高,因此感染和抗生素相关性腹泻的风险增加。
假单胞菌属与其他菌属如葡萄球菌、普雷沃氏菌属、不动杆菌属、芽孢杆菌属、肠杆菌属、肠球菌属等菌属有互作关系。
假单胞菌不被认为是肠道微生物组的典型成员,并且假单胞菌在肠道定植与发生肺部感染和死亡率的风险增加有关。
研究表明,假单胞菌与肠道屏障功能障碍和感染有关,可能导致败血症和多器官功能障碍综合征。此外,假单胞菌与炎症性肠病的发生也有关系,它在IBD患者的肠道中检测到丰度更高,而且它参与引起其他菌群的失衡会影响疾病的进展。
细菌有可能在人体内的不同部位之间转移,防止肠道定植或肠道到其它器官尤其肺部的传播,可能是预防重症患者假单胞菌感染的有效策略。
本文主要分享假单胞菌属(Pseudomonas)的生态,分类,致病因素以及与人体相关健康特性和可能的生活干预措施。
假单胞菌属(Pseudomonas),是革兰氏阴性、好氧菌属,于 1894 年被描述,是最多样化和普遍存在的细菌属之一。有些物种对人类、动物或植物具有致病性。
▼
假单胞菌属(Pseudomonas)属于假单胞菌科,包含200多个有效描述的物种。该属的成员表现出大量的代谢多样性,因此能够在广泛的生态位中定居。
它们易于体外培养,并且越来越多的假单胞菌菌株基因组序列可供使用,这使得该属成为科学研究的绝佳焦点。
Photograph by Kateryna Kon
研究最好的物种包括铜绿假单胞菌。直的或略微弯曲的棒状但非螺旋状,0.5–1.0 × 1.5–5.0 µm。由一根或数根极鞭毛运动;很少不动。在某些物种中,也可能形成短波长的侧鞭毛。有氧,具有严格的呼吸型代谢,以氧为末端电子受体;在某些情况下,硝酸盐可用作替代电子受体,从而允许厌氧生长。
大多数物种(不是全部)无法在酸性条件(pH 4.5 或更低)下生长。大多数物种不需要有机生长因子。氧化酶阳性或阴性。过氧化氢酶阳性。化学有机营养的。
▼
——分布广
假单胞菌属是变形菌门的一个庞大而多样的属。从北极苔原到热带雨林,从干旱的土壤到雨云,几乎在地球的每个角落都可以找到该属的成员。这种令人难以置信的环境适应性归功于假单胞菌非凡的新陈代谢多样性。
——野蛮生长
假单胞菌可以在 0 至 42 °C 的温度范围内生长,甚至可以在更极端的温度下存活。它们对营养的需求很少,可以利用多种碳源。尽管假单胞菌在有氧环境中生长最佳,但它们也可以利用氮进行厌氧呼吸。
它们可以作为自由生活的浮游细胞或作为生物膜群落的成员生活,并且具有将微生物信号和环境线索转化为特定生态位过程的非凡能力。
基因组分析表明,许多其他因素有助于假单胞菌属的多样性和适应性。除了赋予调节和代谢灵活性的常见基因的大量等位基因差异外,水平基因转移影响了致病性假单胞菌属的能力。
▼
第一个假单胞菌基因组测序于 2000 年,有 630 万个碱基对,是当时最大的细菌基因组测序。基因组包含大量参与分解代谢、运输、流出、运动和信号响应调节的基因。
事实上,铜绿假单胞菌基因组中超过 8% 的基因被认为参与了调控,这远远超过了在任何其他细菌基因组中观察到的百分比。很明显,假单胞菌成功的关键在于它可以表达其基因的可塑性,这是由多层监管复杂性提供的。

Sem Photograph by David M. Phillips
自 2000 年以来,已测序的 1000 多个假单胞菌基因组中的绝大多数都是铜绿假单胞菌的临床菌株。
总的来说,铜绿假单胞菌基因组的主要部分(约 4000 个基因)在所有菌株中都是保守的,代表了“核心基因组”。多达另外 20% 的基因驻留在共同代表“附属基因组”的基因组岛上。正是这个附属基因组赋予了铜绿假单胞菌的可塑性,并包含许多参与代谢、毒力和抗生素抗性的基因。
由于已在测序分离株的附属区域鉴定出大约 10 000 个独特基因,因此估计铜绿假单胞菌泛基因组可以接近甚至超过 100 000 个基因,这意味着假单胞菌这一物种的遗传库将远远超过人类。
该属的分类学多年来一直存在争议,因为许多最初包含在假单胞菌属中的细菌类群已被重新归类为其他属或来自不同类别的变形杆菌的物种。
可以根据16S rRNA、gyrB、rpoB和rpoD基因的序列分为三个谱系和13个群。
临床常见菌种主要包括:
• 铜绿假单胞菌(Pseudomonas aeruginosa)
• 斯氏假单胞菌(Pseudomonas stutzeri)
• 恶臭假单胞菌(Pseudomonas putida)
• 荧光假单胞菌(Pseudomonas fluorescens)
• 门多萨假单胞菌(Pseudomonas mendocina)
• 产碱假单胞菌(Pseudomonas alcaligenes)
• 假产碱假单胞菌(Pseudomonas pseudoalcaligenes)
其它菌种见附录一
致病性菌株见附录二
▼
详见附录三
此外,铁是细菌必需且稀缺的营养素,也是重要的信号因子。铜绿假单胞菌已经开发出多种铁吸收系统来隔离足够的铁以维持其生存,在毒力因子的释放和生物膜的形成中具有重要的调节作用。
▼
假单胞菌适应进化的特性对人类既有益又有害。
——有益
一方面,我们利用假单胞菌的力量进行生物修复和生物防治。例如,荧光假单胞菌和P. protegens已被证明在害虫控制和作物保护方面特别成功,它们被认为可以战胜和/或对抗植物病原体。
假单胞菌的分解代谢能力也被用于杀虫剂、重金属和碳氢化合物(例如石油泄漏)以及许多其他污染物的生物降解和/或解毒。
——有害
另一方面,某些种类的假单胞菌对植物和动物具有致病性,导致极难根除的感染。例如,铜绿假单胞菌是世界范围内医院获得性感染最常见的原因之一,并在使用消毒剂和抗生素后存活下来。
它可以在许多解剖部位引起感染,包括皮肤、大脑、眼睛、耳朵、泌尿道、肺部等。免疫抑制的个体,尤其是那些有过度烧伤、囊性纤维化或中性粒细胞减少症的个体,尤其处于危险之中。(这在后一章节会详细介绍)
▼
一般非特异性症状(许多疾病中常见的症状)包括:
假单胞菌感染的症状取决于它们发生在身体的哪个部位。
假单胞菌(Pseudomonas)可引起以下器官感染:
耳朵(外耳炎或游泳者的耳朵):
疼痛、水肿(肿胀)、压痛和分泌物
皮肤:
假单胞菌通过压疮、烧伤和手术伤口或从血流进入菌血症患者的皮肤,还可能引起毛囊炎等。
具体可表现为:脓肿(感染肿块)、皮疹、脓液、皮肤溃疡、坏死组织(皮肤变黑,表明组织正在死亡)。
骨骼或关节:
受影响区域行动不便、关节或背部疼痛、肿胀、疼痛或发热。
这些通常是由于静脉内 (IV) 药物使用或感染的手术伤口或受伤造成的伤口细菌传播到皮肤、骨骼和关节的结果。
伤口:
伤口有分泌物或结痂(通常是黄绿色),可能有甜味或难闻的气味。
消化道:
阑尾炎(由于治疗导致白细胞低的人发生肠道感染)腹痛、腹胀和腹泻的症状。
肺:
假单胞菌可能会导致严重的肺炎,通常发生在住院、已经生病和免疫功能低下的人身上。它可引起囊性纤维化患者的慢性肺部感染(遗传病),支气管扩张(气道永久性扩张),或慢性阻塞性肺病。
具体可表现为:呼吸急促、胸痛、咳痰(伴有白色、黄色或绿色粘液)和充血。
泌尿道:
排尿疼痛、尿液难闻或有甜味、尿急、尿液浑浊。
因此,假单胞菌感染的特征在于广泛的症状。
▼
紧急情况下:呼吸急促、心率加快、脉搏微弱、意识模糊、发烧、发抖或剧烈疼痛是败血性休克的症状,是对血流感染的全身反应。
在极少数情况下,心脏瓣膜可能会受到感染,尤其是在静脉注射药物的人群中。
脏东西的细菌可以进入血液,会传播到心脏并感染心脏瓣膜,这种情况称为心内膜炎。
使用静脉注射药物的人出现新的杂音(心音异常)可能会怀疑与假单胞菌相关的心内膜炎。
▼
未经治疗、治疗不足或复发的假单胞菌感染会导致更广泛的感染,例如败血症,这可能是致命的。
更可能出现严重并发症的高危人群包括:
值得注意的是,假单胞菌属内的抗生素耐药性正在增长。这些细菌正在寻找新的方法来避免死于以往最强大有效的抗生素。
假单胞菌属与人体健康相关,其中一些菌株可以引起感染和疾病,如Pseudomonas aeruginosa 绿脓杆菌,又称铜绿假单胞菌,可以引起呼吸道、泌尿道、皮肤感染。虽然不是正常口腔菌群的成员,但在口腔疾病和牙齿矫正治疗中可能会富集。
▼
假单胞菌属不被认为是肠道微生物群落的典型成员,肠道定植假单胞菌属与发展肺部感染和死亡风险增加有关。
假单胞菌属的过度生长可能会导致肠道炎症和系统性炎症反应。
假单胞菌属感染会引起机体的Th17细胞反应和系统性炎症反应,因此,干预Th17细胞免疫反应可能是缓解肺炎的有效方法。此外,假单胞菌属也可以通过产生GABA来影响蛋白质代谢和细胞稳态,从而影响宿主的摄食行为。
▼
假单胞菌属与其他菌属如大肠杆菌、沙门氏菌等有关,与肠道微生物群的平衡和健康密切相关。
假单胞菌属与其他菌属如葡萄球菌属和普雷沃氏菌属等,共同存在于囊性纤维化患者的肺部感染中。
假单胞菌属与其他菌属如不动杆菌属(Acinetobacter)等一起被发现在冷鲜乳中,产生脂肪酶和蛋白酶,参与乳制品和肉类的变质。
假单胞菌属与其他菌属如Bacillus、Enterobacter、Enterococcus等一起参与了肠-肺轴的微生物和免疫调节,影响呼吸道疾病的发生和发展。
▼
与假单胞菌属相关的疾病包括:免疫系统受损患者的感染和医疗器械相关感染等。免疫抑制个体和住院患者的铜绿假单胞菌肠道携带率显著升高,因此感染和抗生素相关性腹泻的风险增加。
研究表明,假单胞菌属与肠道屏障功能障碍和感染有关,可能导致败血症和多器官功能障碍综合征。假单胞菌属与肺部疾病也有关联,如支气管扩张症。
肺部疾病
研究发现,肺部假单胞菌属感染会影响肠道菌群,而通过调节肠道菌群可以影响肺部疾病的治疗效果。
肠道定植通常先于肺部感染,并且相同菌株经常在肠道和肺部中发现,表明肠道充当了假单胞菌属的储存库,可以传播到肺部和其他感染部位。
肠道微生物群的免疫调节对呼吸道疾病的发生有影响,肠道共生菌通过屏障效应提供菌群抵抗力,从而保护肠道生态环境不被改变。
以肺部假单胞菌、肠道拟杆菌和肠道酵母菌为特征的高肠-肺相互作用集群,与加重的恶化,整体支气管扩张严重相关。
炎症性肠病(IBD)
假单胞菌属与炎症性肠病的发生也有关系。
研究表明,IBD患者肠道中假单胞菌属的种类和数量与非IBD患者存在显著差异。
其中,克罗恩患者中假单胞菌属的检出率明显高于非IBD患者。此外,假单胞菌属的多样性在活动性IBD患者中也明显降低。这些结果表明假单胞菌属可能与IBD的发病机制有关。具体而言,假单胞菌属可能通过影响肠道菌群的平衡和多样性,导致肠道免疫系统的异常反应,从而引发IBD的发生。
因此,调节肠道菌群可能是预防和治疗肠道疾病的一种新策略。
▼
铜绿假单胞菌(Pseudomonas aeruginosa)是一种普遍存在的革兰氏阴性细菌,存在于许多不同的环境中,包括不同的生命来源,如动物和人类。
铜绿假单胞菌能够以最低的营养需求生存并耐受各种身体条件,这使得该生物体能够在社区和医院环境中持续存在。
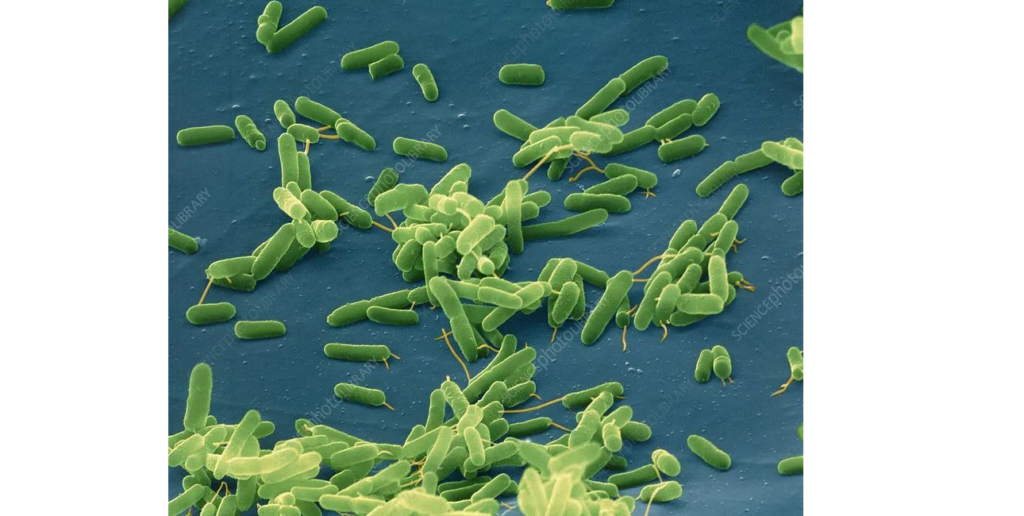
图源:EYE OF SCIENCE / SCIENCE PHOTO LIBRARY
引发感染
铜绿假单胞菌是一种机会性病原体,这意味着它利用宿主防御系统中的一些漏洞来引发感染。
铜绿假单胞菌是世界范围内医院感染的罪魁祸首,其医学和兽医重要性日益增加,并且主要在免疫系统受损的患者中引起感染。
这种细菌几乎从不感染未受损的组织,但如果组织防御以某种方式受损,它就会发起攻击。
铜绿假单胞菌可能引起尿路感染、呼吸系统感染、皮炎、软组织感染、菌血症、骨和关节感染、胃肠道感染和各种全身感染,特别是在严重烧伤患者和免疫抑制的癌症和艾滋病患者中。
在免疫缺陷的人群中,如严重烧伤患者、人工呼吸机使用者和支气管扩张症患者。
在囊性纤维化患者中,铜绿假单胞菌是慢性肺部感染的主要来源,导致患者的发病率和死亡率增加。
铜绿假单胞菌感染会导致肺部炎症和肺损伤,引起细胞浸润和肺泡水肿。在肺移植患者中,铜绿假单胞菌的分离率更高,但与非囊性纤维化肺移植患者相比,死亡率没有增加。铜绿假单胞菌感染会导致肺血管通透性增加,从而引起肺水肿。
耐药性
铜绿假单胞菌显示出对抗生素的高内在耐药性,并具有获得新耐药机制的非凡能力。由于临床治疗选择有限,多重耐药 (MDR) 铜绿假单胞菌分离株的流行率不断上升是一个全球性健康问题。
对来自西班牙两个不同地区的儿童粪便样本中铜绿假单胞菌的发生率、抗菌素耐药性表型、毒力基因型和遗传谱系表征数据显示:从1,443 个粪便样本中,有72份分离分离出来铜绿假单胞菌,并检测到低水平的抗菌素耐药性:头孢他啶 (8%)、头孢吡肟 (7%)、氨曲南 (7%)、庆大霉素 (3%)、环丙沙星 (1%) 和亚胺培南 (1%);对美罗培南、阿米卡星、妥布霉素、左氧氟沙星和粘菌素敏感,此外还发现了四种多重耐药菌株。
定植
人类粪便样本中的定植率范围为 2.6% 至 24 %。关于铜绿假单胞菌分离株在健康人粪便中定殖的数据很少。
其他研究表明,肠易激综合征患者的小肠和粪便样本中铜绿假单胞菌的定植率很高。迄今为止,关于铜绿假单胞菌分离株在健康人粪便中的定植及其抗菌素耐药性和毒力特征的数据相对较少。
尽管肠道微生物群作为抵御肠道病原体的屏障,但铜绿假单胞菌通过产生一系列令人印象深刻的毒力因子,克服了肠道微生物群和先天免疫系统介导的定植抵抗。
铜绿假单胞菌凝集素 LecA 和 LecB 以及细胞外附属物,如鞭毛和菌毛,在细菌附着到宿主上起着重要作用,并与粘附有关。
此外,铜绿假单胞菌携带属于双伙伴分泌(TPS)家族(也称为Vb型分泌系统,T5bSS)的大型蛋白质系统,TPS系统主要由分泌的TpsA效应蛋白及其TpsB伙伴转运蛋白组成。TpsA1 (CdiA2, 573 kDa) 和 TpsA2 (CdiA1, 361 kDa) 通过接触依赖性生长抑制系统 (CDI) 以及粘附和生物膜形成促进细菌竞争。
因此 TPS 系统效应子被指定为对革兰氏菌有益的主要毒力决定因素阴性病原体。
▼
铜绿假单胞菌等的特殊能力引起如此多样化感染的假单胞菌属是因为它们能够产生名副其实的毒力因子库,包括毒素、生物膜、蛋白酶和溶血素。
考虑到铜绿假单胞菌的医学重要性,假单胞菌领域的大部分研究工作都致力于了解影响这些毒力因子释放的调节、生物合成和环境因素。
此外,它们还可以获得赋予进一步抗菌素耐药性的移动遗传元件;因此,可以鉴定出泛耐药菌株。
▼
假单胞菌感知和适应环境的能力导致了一系列活动,例如许多酶和其他生物分子的分泌。虽然这些毒素、降解酶和抗菌剂可以将假单胞菌的环境转变为理想的微生物生态位,但它们可能对植物和动物宿主造成严重后果。例如,绿脓素是一种蓝/绿色色素毒素,它赋予铜绿假单胞菌培养物特有的颜色,并作为一种抗菌剂可以杀死竞争性微生物。然而,它也会破坏真核细胞过程,这会对人类细胞产生不利影响。
▼
过去几年的工作提供的证据表明,群体感应是一种通用的调节机制,它允许细菌以种群密度依赖的方式发起统一、协调的反应,以完成即使不是不可能,也很难完成的任务。
群体感应系统广泛存在于假单胞菌中,人类机会性病原体铜绿假单胞菌,属于研究最广泛的细胞间通讯系统。在这种生物体中,群体感应非常复杂,由两个相互关联的 N-酰基高丝氨酸内酯 (AHL) 依赖性调节回路组成,这些调节回路由非 AHL 相关信号分子和许多调节剂进一步调制,这些调节剂在转录过程中起作用和转录后水平。这种遗传复杂性可能是导致铜绿假单胞菌具有巨大环境多样性的关键因素之一。
过去几年的工作表明,群体感应对于一系列毒力因子的表达以及铜绿假单胞菌中生物膜的形成至关重要,因此代表了设计用于治疗铜绿假单胞菌的新型药物的有吸引力的目标感染。
此外,细胞间通讯能力也在许多额外的假单胞菌中得到证实。铜绿假单胞菌,因此可以用于设计新型药物来治疗铜绿假单胞菌感染。
长时间住院、创伤ICU住院、消毒干预等与肠道假单胞菌过度生长有关,而TAP暴露与抗菌剂干预则与肠道假单胞菌过度生长有负相关。
注:TAP的暴露是指肠道上皮细胞上的TAP暴露在肠道腔中,从而使肠道内的微生物可以通过TAP进入肠道上皮细胞内部。一些研究表明,TAP的暴露与肠道微生物群失调和某些疾病(如炎症性肠病)有关。而抗菌剂干预可以抑制肠道内细菌的生长,从而减少TAP的暴露,因此与假单胞菌肠道过度生长有负相关。
对于免疫功能低下的患者,应特别注意水的纯度,有些水中可能含有假单胞菌等细菌。
研究发现用海洋益生元岩藻依聚糖补充饮食可以通过抑制分泌性毒力因子 (TpsA/CdiA) 与粘蛋白的相互作用并促进有益种群的生长,从而介导铜绿假单胞菌从肠道菌群失调和去定植中的早期恢复。
芪归饮是一种用于治疗铜绿假单胞菌感染的中药。在门和属水平上显著抑制脱铁杆菌和粘孢子菌的过度积累。鉴定了11种潜在的代谢产物,这些代谢产物在铜绿假单胞菌感染中异常表达,服用芪归饮显著逆转。
此外,部分研究表明以下物质也可以抑制肠道内的假单胞菌属。
假单胞菌属相比于其它细菌引起人们广泛的研究兴趣,该属不仅包括人类病原体铜绿假单胞菌,还包括一系列与植物致病性、生物修复和环境微生物学相关的其他重要物种。
假单胞菌被广泛用作模型原型细菌,用于研究宿主-病原体相互作用、细胞-细胞通讯系统、进化生物学、基因调控和代谢网络、分泌系统、抗生素(耐药性)、生物修复、生物膜、细菌基因组学以及与微生物学和分子生物学广泛相关的许多其他主题。
假单胞菌属因其复杂的信号和调节系统,特别是细胞密度依赖性群体感应而被广泛研究。未来只有了解复杂调节回路中涉及的机制,才能确定新型抗菌方法的可能目标。铜绿假单胞菌中的锌、铁稳态是替代抗菌策略的一个有希望的目标。
由于其复杂性和适应性,它存在于无数的环境和临床生态位中,以及它在不同层面(种内、种间和与其宿主)相互作用的能力,假单胞菌属很可能在未来许多年内吸引更多研究人员。我们期待未来有更好的解决假单胞菌属,尤其人类病原体铜绿假单胞菌的抗菌策略。
附录一:
其他菌种
附录二:
致病菌菌株
附录三:
可能参与的代谢

谷禾健康

大多数细菌对我们的生活是有帮助和必要的,某些细菌可以帮助消化,为身体提供能量,分解毒素,保护肠道,增强免疫力等,从而有益健康;也有一些细菌会给我们的健康带来一些危害。
然而有些菌并不是天生注定就是致病菌,也许正常状况下,它只是体内默默地存在着,但可能在某种特定条件下突然变身,彰显其致病威力。
事实上,许多严重的疾病是由健康个体的皮肤、粘膜或肠道中常见的细菌引起的。在这些情况下,致病菌根本不是专性病原体,而是遵循新的生态轨迹的共生体,通常会迁移到与宿主不再和谐相处的侵入性生态位。
细菌从共生到致病的转变在肺炎、脑膜炎、全身感染和医院获得性感染等疾病中发挥重要作用。当然,宿主环境扰动可能会提供感染机会。
因此,我们需要对致病菌的个性、不同环境下的状态有个基本的了解。
在谷禾肠道菌群健康检测报告中,我们会看到关于有益菌,有害菌,致病菌的检测:
有小伙伴有疑问,这里既显示有害菌又有致病菌,有害菌不是致病菌吗?
携带致病菌就一定会生病吗?
条件致病菌是如何引起感染的?
有害菌很多,该如何改善?
…
本文也将围绕着这些问题,展开一些讨论。
致病菌是引起疾病的细菌,也称为病原菌。当它进入身体时,就会破坏细胞或干扰身体的正常活动,人可能会患上轻微疾病或致命疾病。
✦ +
致病菌通过多种方式导致宿主生病。最明显的方法是在复制过程中直接破坏组织或细胞,通常是通过产生毒素,使病原体到达新组织或离开它复制的细胞。细菌毒素是已知的最致命的毒物之一,包括著名的例子,如破伤风、炭疽等。
然而,对宿主的损害通常是通过强烈的或有时是过度的免疫反应自行造成的,这种免疫反应会不加选择地杀死受感染和未感染的细胞并损害宿主组织。免疫系统过度反应的典型例子包括乙型肝炎中的肝硬化和肝癌。
一些病原体受益于宿主的免疫反应,可以在受感染的宿主内传播或增加它们向未感染宿主的传播。
流感主要通过它引起的打喷嚏和咳嗽产生的气溶胶传播。
霍乱弧菌在肠道粘膜中引发强烈的炎症反应,导致水样腹泻,并确保其在环境中的释放,从而感染更多的宿主。
✦ +
从概念上来讲,致病菌和病原菌意思差不多。需要注意的是,这里我们报告用的两个词分别是:
肠道致病菌,病原菌。
✦ +
正常情况下,肠道内是存在少量的病原菌,但是其丰度相对较低,丰度低于健康人群98%以下,不一定会导致疾病的发生。
这里我们可以了解一下关于“细菌感染”:
细菌感染:
病原菌侵入宿主体内并引起病理变化称为“感染”。
也就是说,如果报告中只是检出极少的病原菌,同时并没有症状,那么可能只是表面有病原菌摄入,需要注意饮食和生活卫生,这并不能称之为病原菌感染,因此不需要过于恐慌。
如果报告中病原菌检出已经超过98%人群,则代表可能存在感染的风险,需要结合相应症状和具体菌群丰度比例综合进行判断。
此外,如果出现多种病原菌或病原菌丰度水平很高,则需要引起注意。
✦ +
需要注意的是,谷禾肠道菌群检测报告中的疾病风险,是综合了多项指标判别的,并不只是考虑了致病菌。
慢病是每一种病单独构建模型,不一定和有害菌或菌群平衡指标直接有关。
下面我们来看一些致病菌,这些致病菌一旦感染可能会给人体带来较大危害。
✦ +
★ 炭疽杆菌 Bacillus anthracis
炭疽杆菌是革兰氏阳性、非运动、兼性厌氧、孢子形成和杆状细菌。
炭疽杆菌是一种人畜共患病的病原体,是一种专性病原体,因为细菌的繁殖周期只发生在合适的宿主中。
编辑
图源:tvmdl.tamu.edu . by Mallory Pfeifer
炭疽杆菌的发病机制主要由两种毒力因子引起:三方外毒素和聚γ-d-谷氨酸(γ-DPGA)。
人类可以通过四种不同的方式感染炭疽:吸入、摄入、通过皮肤和注射。
吸入性炭疽被认为是最致命的炭疽形式。如果不进行治疗,病死率接近95%,而立即干预可以将病死率降至50%。
胃肠道炭疽的发生是由于食用了来自感染炭疽的动物制备不当的肉类或肉制品。死亡率是可变的,通过适当的抗生素治疗可以达到≤40%.
皮肤炭疽通常通过皮肤接触受感染的动物或动物产品而发生。这种形式的炭疽病占全球人类病例的 95%。治疗后病死率<1%。潜伏期为 3-7 天。临床表现从轻度到重度不等。
抗生素疗法用于治疗炭疽感染。青霉素、强力霉素、环丙沙星。抗生素治疗必须在接触后立即开始,因为其有效性会随着毒血症的进展而降低。炭疽疫苗在治疗中也很重要。在确诊或疑似接触炭疽孢子的情况下,建议进行 60 天抗生素治疗。
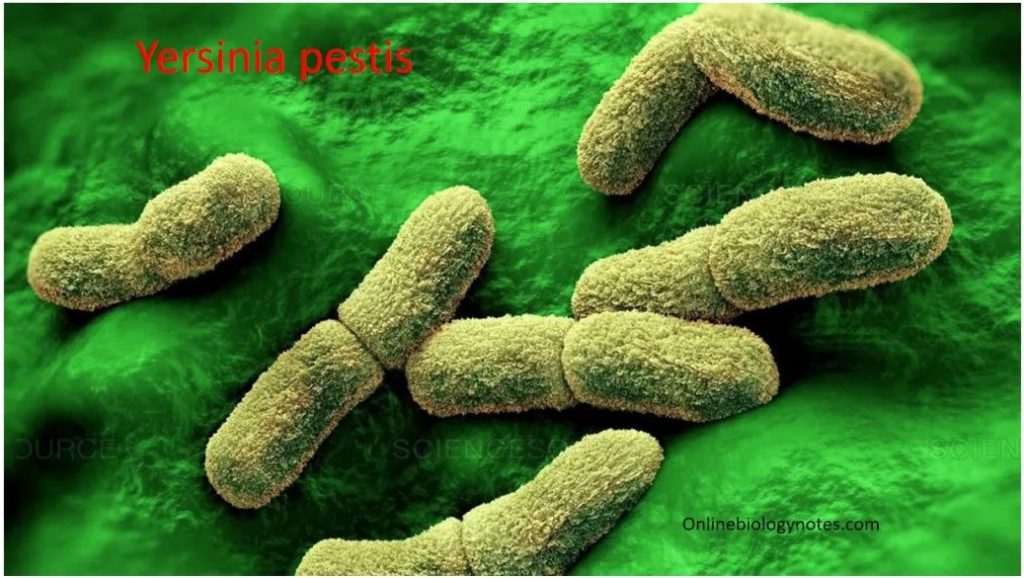
★ 鼠疫耶尔森菌 Yersinia pestis
鼠疫耶尔森菌是一种小型、非运动的革兰氏阴性细菌,属于肠杆菌科。鼠疫耶尔森氏菌是鼠疫的病原体,鼠疫是一种罕见但高度致命的人畜共患病。
图源:onlinebiologynotes
大多数人类感染是由于与受感染的动物接触或被受感染的跳蚤叮咬造成的。
鼠疫在全球超过 25 个国家流行。尽管有有效的抗生素治疗,流行地区的死亡率仍超过 10%,这主要是由于发病机制的快速发展。
鼠疫主要以三种形式发生:肺鼠疫、腺鼠疫和败血症鼠疫。
鼠疫最严重的表现,发展最快的是肺鼠疫,在没有治疗的情况下死亡率接近100%.
肺鼠疫通过呼吸道飞沫在人与人之间传播。经过 2-4 天的潜伏期后,疾病的各种症状包括发烧、头痛、恶心、不适、呕吐、咳痰带血、呼吸困难和胸痛。如果在症状出现后 24 小时内给予适当的抗生素治疗,死亡率可降低高达 50%。
腺泡形式是最常见的,腺鼠疫是鼠疫的主要形式,占病例的 80-95%。死亡率为 10–20%。腺鼠疫由受感染的跳蚤叮咬引起。腺鼠疫的特点是形成淋巴结(淋巴结肿大)。通常的潜伏期从 2 到 6 天不等,有时更长。
如果腺鼠疫没有在适当的时间得到诊断和治疗,它会通过血液传播细菌而发展成败血性鼠疫。这种形式的瘟疫也可能是由传染性跳蚤叮咬通过受损的皮肤或粘膜直接进入鼠疫耶尔森氏菌引起的。通常的潜伏期为 2-7 天,但这种类型的鼠疫甚至在临床表现出现之前就可能导致死亡。
败血性鼠疫的症状包括腹痛、皮肤和其他器官出血。皮肤和其他组织可能会坏死,尤其是鼻子、手指和脚趾。此外,可以观察到发烧、腹泻、呕吐和虚弱。败血症性鼠疫以高菌血症为特征,并伴有危险的内毒素血症。
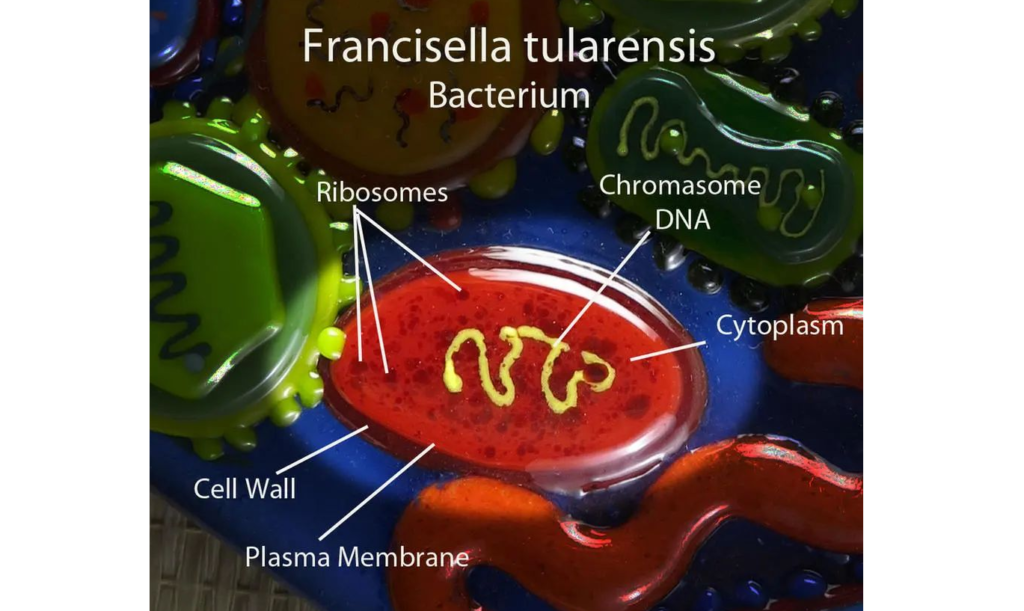
★ 土拉弗朗西斯菌 Francisella tularensis
土拉弗朗西斯菌是一种革兰氏阴性、非运动、非产孢球杆菌。它是一种小的细胞内病原体,具有高毒力和低感染剂量(1-10 个细胞)的特点。
细菌可以通过接触受污染的水进一步传播。吸入受感染的气溶胶,或直接接触受感染动物的组织和液体,也可用作细菌传播的途径。
细菌进入体内后,在局部繁殖,引起溃疡和坏死,然后侵入血液和淋巴管,扩散至肝、脾、肺、肾、浆膜、骨髓等淋巴结和器官,引起多发性凝固性坏死灶。
图源:DeviantArt
土拉弗朗西斯菌不仅容易在巨噬细胞中生长,而且还可以感染许多其他细胞类型,如上皮细胞、肝细胞、肌肉细胞和中性粒细胞。
有六种主要形式的土拉菌病,根据症状分类:肺病、腺病、溃疡腺病、口咽病、伤寒病和眼腺病。
该病的潜伏期通常为暴露后3-5天。
溃疡腺形式是最常见的,是节肢动物媒介叮咬或在与受感染动物接触期间通过皮肤获得感染的结果。
细菌通过磨损进入生物体后,可能会发生腺体形式的土拉菌病。
食用未煮熟、受感染的食物或受污染的水后,可能会形成口咽形式,之后细菌会感染咽部。患者常出现发热、咽痛、颈淋巴结肿大伴耳下淋巴结受累。
在疾病的严重形式中,可能会出现由于肠溃疡导致的胃肠道出血。
当细菌进入循环系统并从另一个感染部位扩散到肺部时,就会出现继发性肺炎性土拉菌病。症状包括高烧、干咳、胸痛和肺门淋巴结肿大;也可能出现肺部浸润或胸腔积液。
伤寒形式的主要症状是发烧、发冷和严重疲劳。然后出现呕吐、腹泻、谵妄和腹痛。临床表现还包括全身疲劳、败血症和死亡。伤寒形式是最难诊断的,因为它的一般症状没有明显的外部病变或区域淋巴结肿胀。
★ 肉毒杆菌 Clostridium botulinum
革兰氏阳性菌,厌氧菌,可运动,致病菌。
它会引起一种罕见但严重的疾病。产生一种特殊类型的外毒素,通过抑制神经肌肉连接的活动来影响神经系统。因此,肉毒杆菌毒素表现为一种神经毒素,它会阻止神经递质的释放。
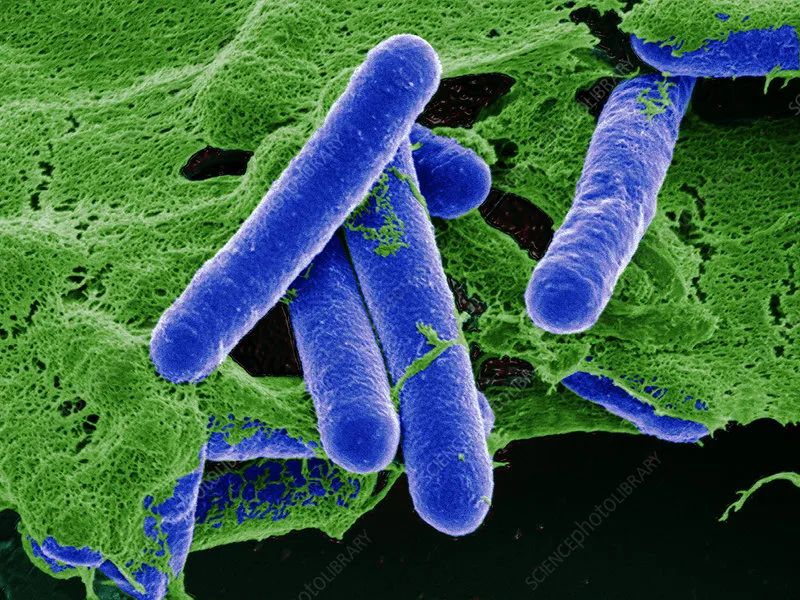
图源:Science Photo Library
肉毒中毒的症状通常始于控制眼睛,面部,嘴巴和喉咙的肌肉无力。
这种无力可能会蔓延到脖子,手臂,躯干和腿部。肉毒杆菌中毒还会削弱呼吸所涉及的肌肉,从而导致呼吸困难甚至死亡。
分为五种传播类型:
预防:
包括减少微生物污染水平,酸化,减少水分水平,以及尽可能破坏食物中所有肉毒杆菌孢子。
易感食物包括罐装芦笋、绿豆、油蒜、玉米、汤、熟橄榄、金枪鱼、香肠、午餐肉、发酵肉、沙拉酱和熏鱼。食用前,考虑将这些食物煮沸10分钟。
以上列举了一些常见的致病菌。致病菌与非致病菌并不是绝对的,是一个动态作用的过程。我们知道肠道中还有大量其他菌群,这些菌群在特定条件下,也可能转为致病菌,我们把这类细菌称为条件致病菌,接下来章节我们来详细了解一下条件致病菌的特点。
正常菌群与宿主、其他菌群之间,通过营养竞争、代谢产物的相互制约等因素,维持着良好的生存平衡。
然而,在一定条件下这种平衡关系被打破,一部分平时看起来正常的细菌就开始“作妖”,变成可以致病的细菌,这就是所谓的 “条件致病菌”。
条件致病菌包括引起肺炎、血流感染、脑膜炎和其他疾病的细菌。它们存在于环境中,可以通过皮肤上的伤口或吸入含有细菌的灰尘进入人体。
健康的免疫系统可以对抗许多病原体,但如果它受损,条件致病菌通常会引起感染。
与免疫系统受损相关的因素有:
遗传易感性;
癌症化疗;
给予免疫抑制药物以防止移植后排斥反应;
艾滋病感染;
严重营养不良;
长期抗生素治疗;
天生免疫系统较弱的婴儿和老年人;
导致白细胞生成减少的骨髓疾病;
怀孕;
正所谓 “橘生淮南则为橘,生于淮北则为枳”。某些细菌在肠道内是乖乖成长的,可一旦冲破束缚,例如在肠漏的情况下,细菌或其代谢物泄露到循环中,可能会变身“有害菌”,可以通过诱导慢性或急性炎症反应,导致疾病发生,包括损害肝脏和胰腺等重要器官,肝癌和胰腺癌可能与细菌易位有关。
如果没有健康的器官和器官系统来适当调节正常的身体功能,条件致病菌就有下手的机会,利用此环境推动疾病的发生。
手术通常涉及切口和伤口,这些切口和伤口为病原体进入身体创造了入口。这也为条件致病菌创造了有利条件。
长期使用会破坏体内正常的微生物群,杀死有益细菌,并导致条件致病菌增殖。
其实细菌(即便是致病菌),它们要生存下来并没有我们想象的容易,生活在复杂的群落中,多个物种和菌株存在相互竞争。
例如,在共生葡萄球菌中,分泌蛋白酶的表皮葡萄球菌菌株,会抑制金黄色葡萄球菌的生物膜形成和鼻腔定植。此外,生物膜破坏介导与其他物种的相互作用,并增加金黄色葡萄球菌对宿主免疫反应成分的易感性。可以看到宿主和共生细菌发出协同反应以排除其他物种。
然而你以为只有宿主和其他菌会带来威胁吗?不,另一种威胁来自与它们同居的同一物种的另一个谱系,同种内部也会互相竞争。这种种内争夺优势的斗争在金黄色葡萄球菌中得到了例证。
关于细菌之间如何交流详见:
金黄色葡萄球菌
Staphylococcus aureus
金黄色葡萄球菌是一种需氧的革兰氏阳性细菌,通常在健康人的鼻腔和皮肤上定植。
但当存在于假体关节和静脉输液管等内部装置中时,可能会导致感染,感染可能导致严重的败血症。
铜绿假单胞菌
Pseudomonas aeruginosa
铜绿假单胞菌是一种需氧、不发酵、高度运动的革兰氏阴性菌。
当感染宿主时,铜绿假单胞菌需要铁。因此铜绿假单胞菌合成了两种铁载体:pyochelin和pyoverdin。铜绿假单胞菌随后将这些嗜铁细胞秘密地存在于细胞的外部,与铁紧密结合并将铁带回细胞。铜绿假单胞菌还可以利用肠杆菌素中的铁来满足其铁需求。
其优先代谢是呼吸,通过将电子从葡萄糖(还原的底物)转移到氧(最终的电子受体)来获得能量。当处于厌氧状态时,铜绿假单胞菌使用硝酸盐作为末端电子受体。
形成生物膜,附着在金属,塑料,医疗植入材料和组织表面。
症状:
肺部感染:发烧和发冷、呼吸困难、胸痛、疲倦、咳嗽
尿路感染:强烈尿频冲动、小便疼痛、尿液中难闻的气味、尿液混浊或带血、骨盆区域疼痛
伤口感染:伤口发炎、漏液
耳部感染:耳痛、听力下降、外耳发红或肿胀、发热
也可能是囊性纤维化患者或机械呼吸机患者呼吸道感染的重要原因。
传播:
当暴露于受该菌污染的水或土壤中时,可以传播到医院患者。还可以在医院中通过被污染的手,设备或表面从一个人传播到另一个人。
治疗:
抗生素治疗
注:铜绿假单胞菌感染通常难以治疗,因为该细菌对许多抗生素具有抗性,并且具有形成生物膜的非凡能力。
易感人群:
免疫力低下人群
使用呼吸机患者、使用导管等装置、有手术或烧伤的伤口患者
预防:
患者和护理人员应保持双手清洁,避免感染。尤其是在护理伤口或触摸医疗设备之前和之后要彻底洗净双手。每天打扫房间。避免共享个人物品。
艰难梭菌
Clostridium difficile
革兰氏阳性,厌氧菌。经过氨基酸发酵,以产生ATP作为能量来源,并且还可以利用糖。
产生两种毒力因子:在70%的菌株中发现肠毒素(毒素A)和在所有菌株中发现的细胞毒素(毒素B)。毒素通过糖基化使Rho-gtpase失活,破坏肠上皮细胞的紧密连接,导致细胞旁通透性增加,从而导致体液分泌(腹泻)、粘膜损伤和炎症。
艰难梭菌存在于整个环境中,包括土壤,空气,水,人类和动物的粪便以及食品(例如加工肉)中。
疾病症状:
艰难梭菌通常影响住院的老年人。
常见症状有:严重腹泻、发热、排便频繁、胃部压痛或疼痛、食欲不振、恶心。
更严重的症状可能包括:
水样腹泻,每天10至15次;
脱水、心跳加快、体温升高、食欲不振、严重的腹部绞痛和疼痛、粪便中有脓液或血、体重下降、腹部肿胀、肾功能衰竭等。
传播:
通过粪-口途径在人与人之间转移。感染通常在医院发生。
该菌可以从粪便传播到食物,然后传播到其他物体表面,如果不洗手或者不正确洗手,就容易感染。
治疗:
抗生素(万古霉素,非达霉素)治疗。患有艰难梭菌感染的人容易脱水,可能需要住院治疗。但是,使用抗生素治疗可能会攻击体内有益细菌。
其他治疗感染的方法,包括:
脆弱拟杆菌
bacteroides fragilis
无芽孢,专性厌氧革兰氏阴性杆菌,人类结肠正常菌群的一部分。
BF毒素引起剧烈的炎症和“细胞间附着物的丧失”,从而引起典型的腹痛和腹泻。
某些脆弱类芽孢杆菌菌株无毒,甚至对其宿主生物有益。
疾病症状:
菌血症、阑尾炎、褥疮、化脓性关节炎、脑膜炎、喉咙发炎、上呼吸道感染、皮肤感染、心内膜炎、软组织感染、心包炎
该菌是正常肠道菌群成员,但是如果转移到周围组织中也会引起腹腔内感染。
这些感染包括产后子宫内膜炎,盆腔脓肿,会阴切开后软组织感染,微管卵巢脓肿和盆腔炎。
菌血症定义为血液中细菌的存在。脆弱拟杆菌是血液培养后最常见的厌氧菌。细菌在腹部,软组织和女性生殖道中的存在是最常见的菌血症来源。
该菌不是人体皮肤微生物菌群的一部分。如果这种细菌进入任何组织,则会引起皮肤感染,例如坏疽和坏死。糖尿病患者还容易受到该菌引起的皮肤感染的影响。
该菌还与引起心包炎,心内膜炎,脑膜炎,阑尾炎和咽喉发炎有关。
传播:
如果细菌被转移到任何其他人体组织,则可能导致该组织的疾病;可能由于外伤,割伤,烧伤,异物侵入或由于不当的外科手术做法造成感染。
治疗:
单一药物治疗或组合治疗
易感人群:
患有腹腔内感染的患者,糖尿病患者易感染
预防:
产气荚膜梭状芽胞杆菌
Clostridium perfringens
革兰氏阳性细菌,是嗜温菌,最适生长温度为37℃,产生内生孢子的非运动性菌。
通过无氧呼吸产生能量,使用硝酸盐作为其电子受体。可能导致许多胃肠道疾病,严重程度从轻微的肠毒血症到致命的气性坏疽。
还具有进行糖酵解和糖原代谢所需的所有酶,利用各种糖酵解酶将糖化合物分解为更简单的形式。
可以在人体肠道,污水和土壤中正常发现。
图源:Food Safety News
疾病症状:
食源性疾病(食物中毒)的最常见原因之一。
关于食源性疾病详见:正值夏季,警惕食源性疾病,常见的食物中毒的病原菌介绍
大多数感染了产气荚膜梭菌的人在食用受污染的食物后6-24小时内会出现腹泻和胃痉挛。这种疾病通常突然发作,持续不到24小时。
这种感染通常不会引起发烧或呕吐。
传播:
营养和食品卫生状况较差的地区人群;
糖尿病和动脉粥样硬化等患者;
产气荚膜梭菌感染也与多发性硬化症有关
危险因素:
常见感染来源包括肉类,禽类,肉汁和其他不安全温度下的食物。
爆发往往发生在医院,学校食堂,监狱和疗养院,以及带有餐饮的活动。最常发生在11月和12月。
治疗:
大多数人未经抗生素治疗即可恢复。
如持续腹泻应多喝水,多休息。
对于源自深层伤口的感染, 必须尽可能清洁该区域,并应使用抗生素。
预防:
将食物煮至安全温度;
食物煮熟后应保持在大于60度或低于4度
可以将热食直接放入冰箱,从冰箱拿出来后加热食用。
结核分枝杆菌
Mycobacterium tuberculosis
革兰氏阳性,专性需氧菌,无运动能力,无孢子形成,细胞内生长的细菌。
通常感染单核吞噬细胞。
在感染的潜伏期,结核分枝杆菌利用一系列效应蛋白将宿主免疫系统弄混,并使其生活方式驻留在肉芽肿中,肉芽肿是宿主为应对持续感染而建立的复杂和有组织的免疫细胞结构。肉芽肿中的结核分枝杆菌通常被限制在具有免疫能力的宿主中,但是当宿主免疫力受损时,它可能导致结核病复发。
常见的症状:
低烧、盗汗、疲劳、厌食(食欲不振)、体重下降。肺结核患者通常会产生咳嗽,并伴有低烧发冷,肌痛(疼痛)和出汗。
潜伏期没有症状。
引发疾病:
传播:
吸入飞沫(咳嗽或打喷嚏)
治疗:
抗生素治疗
预防:
肺炎链球菌
Streptococcus pneumoniae
柳叶刀形,革兰氏阳性,兼性厌氧菌,α-溶血性,条件致病菌。
该菌通过胞外酶系统获得大量的碳和氮,胞外酶系统允许多糖和己糖胺的代谢,并对宿主组织造成损害并使其定植。
荚膜多糖的组成和数量在毒力中起主要作用。产生最大量多糖的菌株可能是最强毒的。
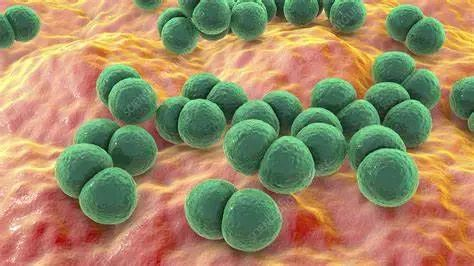
图源:Science Photo Library
疾病症状:
从无症状的咽部定植到粘膜疾病(中耳炎,鼻窦炎,肺炎)再到侵袭性疾病(通常在无菌部位的细菌;菌血症,脑膜炎,脓胸,心内膜炎,关节炎)
传播:
通过呼吸道飞沫直接进行人与人接触,上呼吸道携带细菌的人自动接种。
易感人群:
治疗:
抗生素;静脉注射疗法
预防:
无乳链球菌
Streptococcus agalactiae
革兰氏阳性双球菌,不耐酸,不形成孢子,不易动,兼性厌氧的条件致病菌。
通常称为B组链球菌(GBS),是四种Beta-溶血性链球菌之一,可导致血琼脂上细菌菌落周围宽阔清晰区域显示的血细胞完全破裂。
使用葡萄糖作为能源。该细菌能够通过氧化磷酸化合成ATP。
是一种异养菌,能够导入多种碳源。能够将不同的碳源发酵成多种副产物,如乳酸,乙酸盐,乙醇,甲酸盐或乙酰丙酮。
需要许多氨基酸才能生长,因为它不存在任何TCA循环来合成氨基酸。
毒力因子: 多糖胶囊, β溶血素毒素
属于生殖道的正常菌群。5-20%的女性阴道定植。
疾病症状:
尿路感染, 新生儿和幼儿败血症,脑膜炎
患有基础疾病的成年人:肺炎,心内膜炎,皮肤和软组织感染等。
传播:
GBS阴道或直肠定植的母亲所生的婴儿中,有1%至2%的婴儿发生早发感染。
通过孕妇在怀孕或分娩期间的生殖器官和/或肠道,以及来自其他新生儿或妇产医院的医院工作人员。
治疗:
抗生素治疗
易感人群:
预防:
流感嗜血杆菌
Haemophilus influenzae
革兰氏阴性杆菌。有6种囊化血清型(指定为a至f)具有不同的囊化多糖。
该菌在人的鼻子和喉咙中,通常不会造成伤害。但有时会移动到身体的其他部位并引起感染:
肺炎(肺部感染),菌血症,脑膜炎,喉咙肿胀,蜂窝织炎(皮肤感染);
引起儿童耳部感染和成人支气管炎;
较不常见的感染包括心内膜炎和骨髓炎。

图源:ecdc.europa.eu
易感人群:
5岁以下和65岁以上感染风险增加,免疫力低下的(如艾滋病患者,癌症患者)感染风险增加。
传播方式:
咳嗽或打喷嚏;新生儿可以通过吸入羊水或与含有该菌的生殖道分泌物接触而感染。
治疗:
服用抗生素
预防:
婴儿接种Hib疫苗
衣氏放线菌
Actinomyces israelii
直径为1μm的革兰氏阳性杆菌,厌氧细菌,它是肠道正常菌群的一部分,条件致病菌。
浸润性,组织渗透/破裂;可引起慢性化脓性感染,放线菌病。
通过在各种手术(牙科,胃肠道),抽吸或病理性疾病(例如憩室炎)过程中破坏粘膜屏障来建立感染。
感染部位:
口腔,宫颈,面部疾病是最常见形式,有时感染可能发生在胸部(肺放线菌病),腹部,骨盆或身体其他部位。
当细菌进入人体时,它可以在软组织上形成脓肿。随着脓肿随着时间的流逝而扩大,它会穿透皮肤表面,引起皮肤溃疡。这些脓肿或肿块通常会影响头和颈部,并且会引起肌肉痉挛,阻止下巴正常运动。
其他常见症状包括:
发烧,体重减轻,咳嗽,胸痛和窦腔过度引流。症状可能发展缓慢,但是早期治疗是迅速康复的关键。
易感人群:
治疗:
抗生素;如果与宫内节育器有关,则须卸下
预防:
良好的口腔卫生和定期看牙医可能有助于预防某种形式的放线菌病。
嗜肺军团菌
Legionella pneumophila
具有一定铁含量的细胞内多形革兰氏阴性细菌,条件致病菌。
普遍存在于水环境中,例如饮用水系统、温泉、冷却水,可引起人类感染军团菌肺炎。
通过抽吸或直接吸入到达肺部后,会附着在呼吸道粘膜上。

图源:apotheekteirlinck.be
症状:
高烧(可能到40℃或更高),发冷,咳嗽,肌肉酸痛和头痛。
传播:
可以在人类制造的水系统(例如空调)中繁殖。大多数人吸入含有军团菌细菌的微小水滴时会被感染。这可能来自淋浴,水龙头或漩涡浴池中的喷雾,或者来自大型建筑物中通风系统中的水。
如何避免致病菌感染?
尽快恢复免疫力,减少致病菌感染的发生率。
个人应通过煮熟鸡蛋和肉类来避免传染源,饮用巴氏杀菌乳制品,避免感染个人和医院的潜在感染源,避免与粪便、灰尘或农场动物接触,这些是主要的感染源。
不要饮用未经处理的水,例如直接来自湖泊或河流的水。避免在国外饮用自来水。使用瓶装水或滤水器。
不要共用针头、注射器或其他药物注射设备。
有害菌和肠道内的其他共生菌共同构成菌群微生态,也是大部分人群肠道内常见的菌群。
有害菌是相对而言的。正常肠道菌群也包含许多有害菌,但有害菌比例或个别菌属丰度超标可能预示着肠道菌群的健康状况受到破坏。
有害菌过多会影响肠道内环境,如pH值,含氧量以及肠道内毒素等,可能会导致出现一些机会感染和机会致病菌入侵,进而诱发炎症和疾病。
谷禾肠道菌群检测报告中的有害菌包含了原发致病菌和条件致病菌,以及属内主要菌种为致病菌的属。
为便于统计,我们在计算的时候统一按照属层级进行计算比例。
报告中的有害菌包括了以下的菌属:韦荣氏球菌属、葡萄球菌科、变形菌属、弓形菌属、弯曲菌属、螺杆菌属、厌氧螺菌属以及弧菌属等。
具体每个菌相关介绍详见:
全面认识——肺炎克雷伯菌 (Klebsiella pneumoniae)
慢病是每一种病单独构建模型,不一定和有害菌或菌群平衡指标直接有关。
肠道菌群中的有害菌过多,也就是说肠道菌群趋向于失衡,这会给身体带来诸多麻烦。
当肠道内的有害菌增多时,起初可能会有些症状出现,例如:
随着时间的推移,如果没有采取相应的干预措施,有害菌变得过多,则可能会对身体方方面面产生影响:
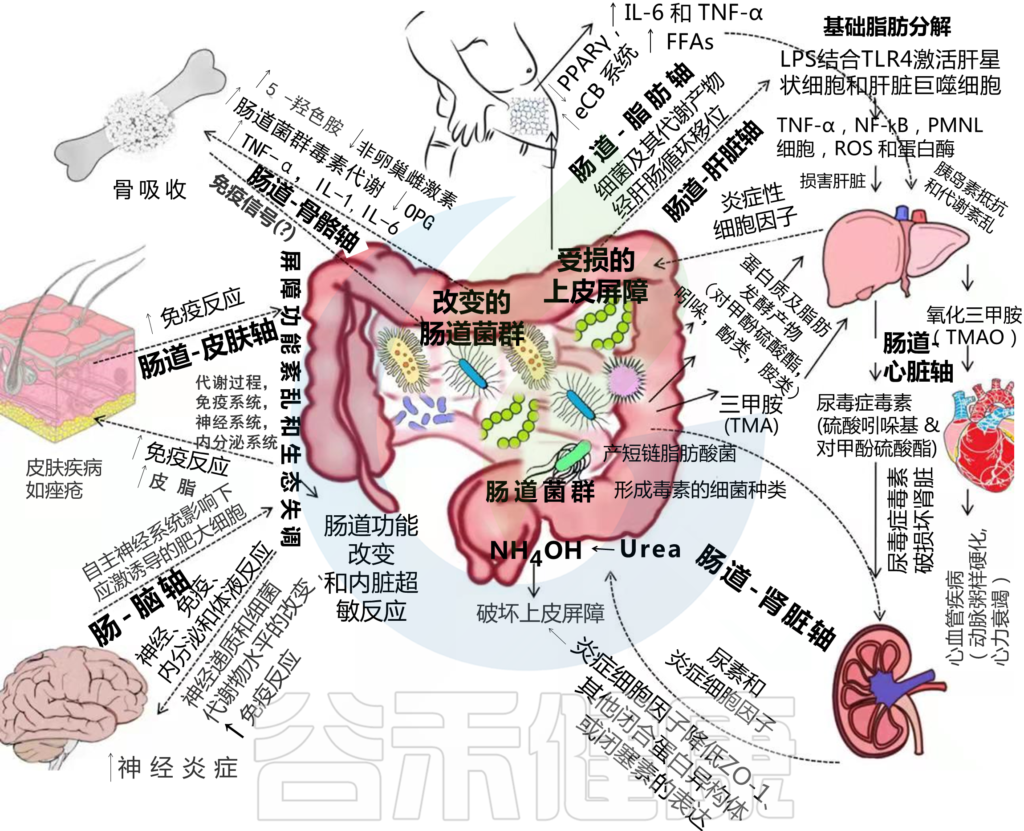
特殊类型的细菌会产生一种化学物质,肝脏会将其转化为三甲胺- n -氧化物(TMAO)。
TMAO产生的增加会导致血管中胆固醇的积聚,从而可能导致心脏病。
详见:
TMAO产生的增加也与慢性肾脏疾病有关。也会导致肾结石的发展。
与健康对照组相比,终末期肾病患者体内的TMAO浓度可高出20倍。
对终末期肾病患者的类似研究表明,从普雷沃氏菌向拟杆菌转变,产丁酸菌减少。

有害细菌将纤维转化为脂肪酸。身体可能会把它们沉积在肝脏中,如果不及时治疗,可能导致代谢综合征的发展。
肝硬化患者中韦荣球菌属和链球菌增多。
肝细胞癌与肠道大肠杆菌过度生长有关,患者微生物群多样性增加,与产丁酸菌属(如Alistipes)减少有关,而致病性产脂多糖菌(如克雷伯氏菌)增加。
研究表明,肠道菌群在宿主的代谢和疾病状态中起着重要的作用。特别是2型糖尿病,其病因复杂,包括肥胖、慢性低度炎症,受肠道微生物群和微生物代谢产物的调节。
在2型糖尿病患者普遍具有相对高丰度的特定属:
Blautia、Coprococcus、Sporobacter、Abiotrophia、Peptostreptococcus、Parasutterella、Collinsella
2型糖尿病患者肠道菌群详见:
谷禾数据库统计发现,在有害菌属的丰度水平分布上,肥胖人群要高于对照人群。进一步对具体菌属进行分析,发现肺炎克雷伯氏菌的丰度水平肥胖人群更高。
肥胖患者肠道菌群详见:
肠道有许多直接与大脑沟通的神经末梢,称为肠脑轴。肠道炎症和菌群失调与心理健康不良有关。过多的有害肠道细菌会导致:
详见:
不良的肠道细菌会影响你的整体健康。它会增加消化问题的风险,这些症状通常伴有体重减轻和腹痛。比如:
肠道上皮、免疫系统和共生细菌之间的串扰是启动全身炎症反应的关键。有益菌和有害菌的失衡,抗炎和促炎细胞因子之间的失衡,包括白细胞介素(IL)-1β、肿瘤坏死因子、干扰素(IFN)-γ、白细胞介素-6、白细胞介素-12和白细胞介素-17,在参与RA发病机制的炎症过程中起着核心作用。
为了支持肠道菌群成分变化在类风湿性关节炎发病和进展中起重要作用的假说,已经提出了肠道菌群与关节炎相关的几种机制。
这些包括调节宿主免疫系统(触发T细胞分化)、通过作用Toll样受体(TLR)或NOD样受体(NLR)激活抗原呈递细胞(APC)、通过酶促作用促进肽的瓜氨酸化、抗原模拟和增加肠粘膜通透性。关于对APCs TLRs表达的影响,这可能导致Th17/Treg细胞比率失衡,这种局部免疫反应可能导致系统性自身免疫。
有害菌过多会导致肠道通透性和肠道屏障破坏。肠黏膜屏障功能障碍可能导致血清脂多糖 (LPS) 水平升高,从而导致代谢性内毒素血症。早期研究表明,LPS 可促进体内股骨的骨质流失和体外破骨细胞的。
与骨骼疾病相关的肠道菌群变化如下:

编辑
详见:
鉴于具有高度稳定性的平衡肠道微生物群与宿主的免疫系统具有共生相互作用,能够抑制有害菌增长。然而不稳定的状态例如肠道紊乱,慢性疾病,由遗传易感性、化学物质或肠道病原体感染引起的肠道炎症会导致有害菌增加。
其他包括饮食、生活方式、环境等因素也会影响肠道微生物群的分类和功能组成。例如,西方饮食、高糖饮食,饮食结构过于单一,加工食品过多摄入等不健康的饮食方式,睡眠不足,作息不规律,不运动等不良生活习惯,压力过大,服用药物等因素都会导致有害菌增多。
有害菌和肠道内的其他共生菌共同构成菌群微生态,如果有害菌过多,通常我们可以通过服用益生菌或益生元的方式首先增加有益菌的比例,相应的有害菌比例就会降低。
双歧杆菌和乳酸杆菌有助于发酵碳水化合物,同时会产短链脂肪酸,有助于维持良好的消化系统。
乳酸杆菌
乳酸杆菌菌株产生乳糖酶。它有助于分解乳制品中的乳糖,有助于维持肠道的酸度水平,对于吸收关键矿物质至关重要。
双歧杆菌
保护肠壁;维持肠道的酸度;限制产生硝酸盐菌的生长;生产 B 族维生素和维生素 K 等。
益生菌抑制其他菌群的生长:
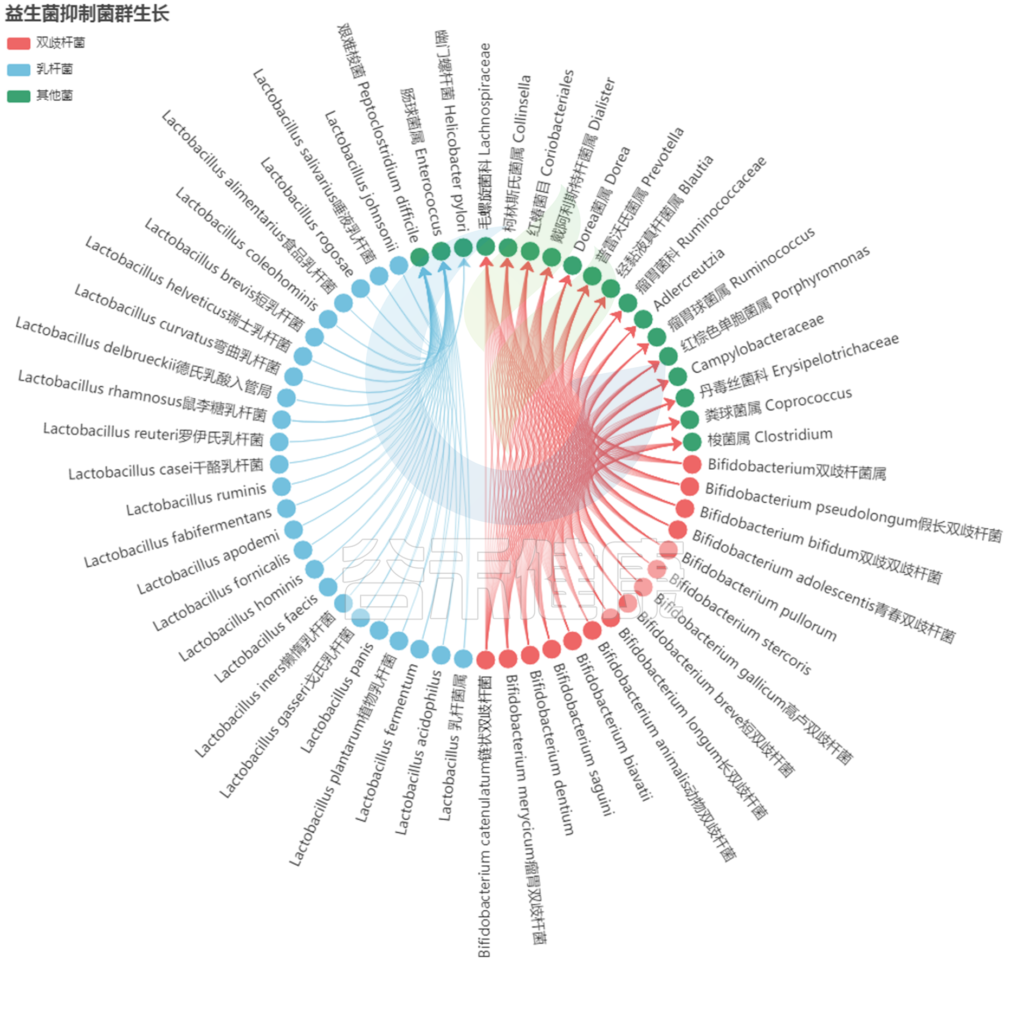
编辑
<来自谷禾健康数据库>
除了直接服用益生菌这种方式之外,有些食物中也富含益生菌,如:
乳制品:酸奶、牛奶、开菲尔等
发酵食品:泡菜、酸菜、味噌汤、豆豉等
益生元是一种可溶于水的可溶性纤维,可以作为益生菌的“食物”。
最广泛认可的益生元包括低聚果糖 (FOS)、菊粉和低聚半乳糖 (GOS) 等。
此外还包括抗性淀粉、果胶寡糖 (POS)、多酚等。
含益生元的食物包括:菊芋、青香蕉或青香蕉粉、大麦和燕麦、魔芋根、菊苣根、牛蒡根、亚麻籽、海藻、苹果、土豆等。
关于益生菌,益生元的补充详见:
想要持久的改善菌群结构降低有害菌水平就需要改善生活方式,适当增加抗性淀粉等膳食纤维并规律饮食和睡眠,增加运动等。
下面介绍一些日常生活中可以自行调整的饮食及生活方式。
尝试食用多种食物,避免每天食物一样
饮食多样性更有利于菌群维持健康平衡。体内的微生物群就好比一群挑剔的孩子,每个孩子都会去吃自己喜欢的食物。当你吃下各种食物时,就相当于喂食了各种微生物。
如果可以的话,一个星期的饮食中可以摄入 40 种及以上不同类型的天然食物,尽可能地提高肠道多样性。
减少西方饮食
西方饮食(其特点是大量摄入脂肪、蛋白质、糖、盐和加工食品),可能增加有害菌,与伴随的导致自身免疫疾病发展的微生物变化之间的联系越来越明显。
尝试地中海饮食
其他可以参考地中海饮食结构摄入,适量食用纤维,多吃各种颜色的水果和蔬菜。将红肉的摄入限制在每月两三次,可以将其视为一种“奢侈品”,每周食用两到三次家禽。它的饱和脂肪和胆固醇比红肉少得多,选择健康的脂肪,每周可以食用两次三文鱼等,做菜选择橄榄油,适量食用坚果(杏仁、巴西坚果、榛子、松子、开心果和核桃都是非常健康的坚果类型),它们是单不饱和脂肪的重要来源。
尝试抗炎饮食
很多种食物均具有抗炎特性,其中包括抗氧化剂和多酚含量高的食物。肠道抗炎饮食推荐的食物包括:浆果类,西兰花,牛油果,辣椒,姜黄,洋葱,大蒜等。
详见:
避免膳食纤维过多或过少
膳食纤维细菌发酵的产物(短链脂肪酸、乙醇和乳酸)过多会破坏细菌。纤维也会让人“上瘾”,其发展方式:随着发酵破坏细菌,需要越来越多的纤维来形成粪便。
但是膳食纤维摄入不足或突然停止所有纤维摄入,也会发生菌群失衡,导致便秘或其他肠道问题,从而导致有害菌增加。
减少摄入加工和包装食品
食用的加工食品越多,饮食越无菌,加工食品会减少我们体内有益细菌的数量,相应的有害菌会逐渐增多,破坏肠道菌群原有的平衡。
具体来说,防腐剂,比如聚山梨酯80和羧甲基纤维素(CMPF),它们是许多加工食品中常见的乳化剂,直接改变了肠道微生物群的组成。
食品添加剂对菌群的影响详见:
你的焦虑可能与食品添加剂有关,警惕食品添加剂引起的微生物群变化
尽量避免含糖饮食
对于我们大多数人来说,糖在我们的饮食中太普遍了。
在现代饮食中,糖无处不在,而且形式多种多样。我们大多数人现在都知道,过度消费“游离糖”的精制糖并不健康。糖也是造成菌群失调的主要因素。然而在忙碌的现代生活中,很难不过度摄入糖分。
高糖食物的常见罪魁祸首包括碳酸饮料、能量饮料、糖果、饼干、甜点、蛋糕、果汁和谷物等,它们都会导致菌群失调。
还有一些食物,含有的糖分更加隐蔽。这些包括:即食食品、腌泡汁、酸辣酱和泡菜、一些酒精饮料、调味酱、白面包、白米饭和土豆。
糖对菌群的影响详见:
吃八分饱
少吃多餐,每三到四个小时吃一顿小餐或吃零食,以补充精力。
间歇性禁食
间歇性禁食是一种越来越流行的健康实践,研究发现间歇性禁食导致肠道菌群结构改变,进食时机和频率可以一定程度上改善生活方式和心血管代谢,防止2型糖尿病和心血管疾病的发生。
关于间歇性禁食详见:
维生素维持肠道菌群稳态和减少肠道炎症以预防癌症的机制;肠道菌群帮助吸收营养,并参与维生素代谢。几项观察表明,微生物群失调和维生素缺乏是相互关联的。
例如:
补充维生素 C 可减少肠杆菌科细菌的数量,增加乳酸杆菌的丰度,抑制有害菌的生长,促进有益菌的增加。
维生素 D 的缺乏会增加拟杆菌门、变形杆菌门和螺杆菌科的丰度。
维生素E对变形菌有抑制作用,而维生素E(和纤维)的摄入量较低与Sutterella水平较高相关。

详见维生素的文章:
当肠道渗漏也就是屏障受损时,大量的有害菌及微生物代谢毒素、食物中的有毒物质逃离肠道,涌入血液循环,这可能会产生炎症并导致组织损伤,器官从感染到炎症再到功能缺失,甚至是到癌症。
关于什么情况会导致肠漏,肠漏带来的危害详见:
即使是很小的压力也会触发体内激素和化合物的释放。压力会慢慢积累,如果你是一个压力大的人,经常烦躁,愤怒,那么会对微生物群产生负面影响,扰乱 HPA 轴。不过这是双向的,肠道中的某些细菌菌株也会影响体内神经递质的方式。
适当给自己减压,可以帮助菌群恢复平衡。减压的方式包括:
农村环境和微生物群与过敏患病率的降低有关。通过暴露于农村室内灰尘来调节肠道微生物群可以改善过敏预防。
城市儿童和小鼠暴露在城市灰尘提取物中,肠道菌群向拟杆菌类的变化是明显的。相比之下,农村儿童和接触农村粉尘提取物的小鼠肠道菌群分别富含普雷沃氏菌属和梭状芽孢杆菌属。
环境对菌群的文章详见:
环境污染物通过肠脑轴影响心理健康,精神益生菌或将发挥重要作用
睡眠质量与肠道菌群组成之间存在双向关系。

编辑
拟杆菌门和厚壁菌门的丰度与睡眠质量呈正相关,而毛螺菌科(Lachnospiraceae)、棒状杆菌(Corynebacterium)、Blautia等几种菌与睡眠质量测量值呈负相关。
睡眠不足或者其他因素如受伤、食物摄入、压力、昼夜节律和运动等,可致肠屏障损伤和细菌移位,增加感染易感性,激活HPA轴从而影响菌群。
详见睡眠对肠道菌群的影响的文章:
抗生素
正常情况下,强大的免疫系统会追捕并消灭病原体,但在系统较弱的情况下,人体的免疫防御系统无法控制病原菌的生长。
服用抗生素会杀死体内的许多好细菌和坏细菌。在瑞典的一项临床试验中,研究人员发现,在服用抗生素仅一周后,一些参与者在一整年后就破坏了微生物组。
抗生素使用对肠道菌群变化的不同影响
Yang L, et al., AMB Express. 2021
我们之前写过抗生素对菌群的影响文章,详见:
其他包括非甾体抗炎药、质子泵抑制剂 (PPI)等也会影响肠道菌群的组成。
限制饮酒量
酒精诱导的胃肠道菌群组成和代谢功能的变化可能有助于建立酒精诱导的氧化应激、肠道对菌群产物的高通透性和随后发展的酒精性肝病和其他疾病之间建立明确联系。如果体内酒精过多,肝酶可能没有足够的能力对其进行处理。过量的酒精会在身体的其余部分循环,产生负面影响。
坚持适量运动
运动锻炼与肠道微生物群组成之间可能存在密切关联。经常中等强度的耐力运动对肠道微生物产生最有益的影响,促进健康和抗炎细菌增加;长期运动的人菌群多样性更高。
运动改善菌群多样性,增加菌群种类,有益于提高菌群稳定性,降低有害菌的相对比例。
运动对菌群的影响详见:
肠道微生物组如何影响运动能力,所谓的“精英肠道微生物组”真的存在吗?
在我们的检测实践中,以上的这些干预调节方式可以有效的定向改善特定有害菌和致病菌的超标,并最终带来整体健康状况的改善。
主要参考文献:
Janik E, Ceremuga M, Niemcewicz M, Bijak M. Dangerous Pathogens as a Potential Problem for Public Health. Medicina (Kaunas). 2020 Nov 6;56(11):591. doi: 10.3390/medicina56110591. PMID: 33172013; PMCID: PMC7694656.
Sheppard SK. Strain wars and the evolution of opportunistic pathogens. Curr Opin Microbiol. 2022 Jun;67:102138. doi: 10.1016/j.mib.2022.01.009. Epub 2022 Feb 12. PMID: 35168173.
Balloux F, van Dorp L. Q&A: What are pathogens, and what have they done to and for us? BMC Biol. 2017 Oct 19;15(1):91. doi: 10.1186/s12915-017-0433-z. PMID: 29052511; PMCID: PMC5648414.
Kinnula H, Mappes J, Sundberg LR. Coinfection outcome in an opportunistic pathogen depends on the inter-strain interactions. BMC Evol Biol. 2017 Mar 14;17(1):77. doi: 10.1186/s12862-017-0922-2. PMID: 28288561; PMCID: PMC5348763.
Engen PA, Green SJ, Voigt RM, Forsyth CB, Keshavarzian A. The Gastrointestinal Microbiome: Alcohol Effects on the Composition of Intestinal Microbiota. Alcohol Res. 2015;37(2):223-36. PMID: 26695747; PMCID: PMC4590619.
Kouzu K, Tsujimoto H, Kishi Y, Ueno H, Shinomiya N. Bacterial Translocation in Gastrointestinal Cancers and Cancer Treatment. Biomedicines. 2022 Feb 4;10(2):380. doi: 10.3390/biomedicines10020380. PMID: 35203589; PMCID: PMC8962358.