-

 国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子
国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子

 二级病原微生物安全实验室
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业

谷禾健康
Akkermansia muciniphila(AKK菌)因其独特的黏蛋白降解能力和与宿主健康的复杂关系,目前已成为微生物学和医学研究的前沿热点。谷禾此前已对该菌进行过系统性介绍。
肠道重要菌属——Akkermansia Muciniphila,它如何保护肠道健康
本文将继续深入探讨AKK菌的研究前沿。
2004年,当这株微小的厌氧菌首次从人类粪便中被分离出来时,没人预料到它会凭借独特的黏蛋白降解能力(当然目前又陆续发现了几个黏蛋白降解菌),在此后近二十年间掀起一波又一波的研究热潮。
更引人关注的是AKK菌呈现出耐人寻味的双面性:它既是下一代益生菌的希望之星,与代谢健康、长寿紧密相连;又在某些特定情境下显现出促进疾病的潜在风险。这种复杂的功能属性,正是当前科研亟待厘清的关键议题。
三篇发表于顶级期刊的AKK菌研究为该领域带来了重要突破:
这三篇文献从不同维度构建了迄今为止最为全面和深入的AKK菌研究图景,涵盖了其基本生物学特性、复杂的基因组结构与菌株多样性、与宿主免疫系统的精密互作机制,以及在不同疾病模型中呈现的矛盾表型,系统解析了这些现象背后的科学逻辑。
因此,本文谷禾整合了这三篇文献的核心观点,以科研从业者的视角带领大家深入解构AKK菌的研究进展。我们希望与各位共同探讨一个关键问题:如何理性认知并合理应用AKK菌这个双面细菌,从而真正实现精准调控肠道健康的目标。
嗜黏蛋白阿克曼氏菌(Akkermansia muciniphila),简称为AKK菌,属于疣微菌门。这个名字是为了纪念荷兰微生物生态学家Antoon Akkermans博士,而“muciniphila”则源自拉丁语,意为偏爱黏蛋白。
注:Antoon Akkermans:荷兰瓦赫宁根大学(Wageningen University)微生物生态学家,对土壤和肠道微生物研究有重要贡献。
AKK菌的发现和命名具有里程碑意义
2004年,仅使用黏蛋白(mucin)作为唯一的碳源和氮源进行富集培养了该菌。AKK菌具备独特的酶系统,能够降解并利用黏蛋白作为其生存和生长的主要能量来源,这种自给自足的能力使其在竞争激烈的肠道环境中占据了稳定的一席之地。
其模式菌株为Akkermansia muciniphila MucT(亦写作Muc5),拥有多个菌株编号,如ATCC BAA-835、DSM 22959等,是目前研究中应用最广泛的菌株。
生态位
AKK菌在人体肠道中占据着一个非凡的生态位——肠道黏液层。这一层由宿主杯状细胞分泌的黏蛋白糖蛋白构成,是隔开肠道上皮细胞与肠腔内大量微生物的第一道物理和化学屏障。
从分布来看,AKK菌不仅存在于人类肠道,也广泛栖息于包括小鼠、牛、猪、兔等多种脊椎动物的胃肠道。
形态与基本特性
在显微镜下,AKK菌呈现为一种椭圆形、不运动、不产芽孢的革兰氏阴性严格厌氧菌。
AKK菌并非绝对厌氧
研究发现,在只有微量氧气的环境中,它不仅能存活,甚至可能生长得更好。这主要得益于其基因组编码的细胞色素 bd 氧化酶复合物等耐氧机制。
也正因为具备这种有限耐氧能力,AKK菌才能适应并定植于结肠黏液层——这里处在厌氧的肠腔与相对更富氧的上皮细胞之间,存在明显的氧梯度。
结构特征,驱动互作
AKK菌约2.66Mb的基因组,是其特殊生活方式的说明书。其中最引人注目的,就是黏蛋白利用基因簇(Mucin Utilization Loci, MULs)。
黏蛋白转运系统:把食物带回家再吃的策略
Grant等人的研究指出,这些基因编码了一套精密的黏蛋白转运系统,能将黏蛋白大分子吞入菌体内部一个叫黏蛋白体的特殊结构中进行降解。这种偏自私的代谢策略,有点像把食物带回家再吃,从而最大限度地减少了与肠道中其他微生物的营养竞争,进而巩固自身的生存优势。
编辑
此外,它的细胞表面也布满了武器。电镜观察到的菌毛样结构,特别是重要的外膜蛋白Amuc_1100,不仅参与黏附,更是与宿主免疫系统直接对话的关键分子。
其细胞壁的肽聚糖层含有非乙酰化的葡糖胺残基,这在革兰氏阴性菌中相当罕见,使其能被宿主的NOD1和NOD2受体识别,从而触发免疫应答。这些独特的结构,都为它与宿主之间复杂的相互作用埋下了伏笔。
生态分布:影响因素和趋势
AKK菌在人群中的分布呈现出鲜明的特征
根据人类肠道微生物组图谱对来自20个国家3268名健康人的数据分析,AKK菌在约40.3%的健康供体中被检测到,平均相对丰度为1.24%。这个数字看似不高,但其分布的倾向性却极具信息量。
AKK菌的分布图谱
编辑
Grant et al., Nature Microbiology
a) 在健康人群中,其丰度和患病率存在地理和性别差异。
b) 在不同疾病状态下,其丰度呈现显著变化,例如在IBD中减少,而在帕金森病中富集。
c) 其相对丰度随生命周期动态变化,在百岁老人中再次出现高峰。
d) 进化树揭示了AKK菌属内部复杂的系统发育关系。
从图1中我们可以清晰地看到:
-地域与生活方式
工业化人群中的丰度更高,这暗示着饮食或生活方式可能是其丰度的重要调节因素。
-年龄
AKK菌在婴儿期迅速定植,成年后逐渐下降,但在百岁老人中却意外地再次富集,被认为是健康长寿的潜在标志物。Luo等人的文章也系统总结了这一现象,指出AKK菌在人类中的年龄依赖性定植模式与在小鼠中的模式恰好相反,这提醒我们在选择动物模型时需格外谨慎。
-疾病状态
这是最能体现其双面性的一点。在炎症性肠病(IBD)患者中,AKK菌丰度显著降低(UC患者中为9.9%,克罗恩病患者中为14.3%);然而,在帕金森病(90.3%)、某些癌症(如黑色素瘤,65.9%)和动脉粥样硬化(83.3%)患者中,其丰度却异常增高。这种看似矛盾的分布模式,强烈暗示AKK菌的角色并非简单的好或坏,而是深度依赖于宿主的病理生理背景。
Tips:看到这些分布数据,首先想到的不是简单的因果关系,而是一个复杂的生态反馈回路。例如,在IBD中,AKK菌的减少可能是肠道炎症环境恶化、黏液层破坏的结果,而非原因。反之,在帕金森病中,其增多也可能是一种代偿性反应,或是神经退行性疾病引发的肠道环境改变所致。将AKK菌丰度作为疾病的生物标志物时,我们必须高度审慎,通过系统证据加以判别:该因素究竟是事件升级的关键驱动(促进因素),是对事态的响应性干预(缓解因素),还是与结果无显著因果关联的伴随出现(旁观变量)。
所以构建真实世界的特定疾病样本人群数据才能更有助于解析复杂的肠道微生态,类似的菌还有如活泼瘤胃球菌和普雷沃氏菌等相互矛盾的研究。
三篇文献都不约而同地强调了菌株水平多样性的重要性。
长久以来,大多数研究都围绕着模式菌株MucT(ATCC BAA-835)展开。然而,随着基因组学技术的发展,AKK菌家族的内部复杂性逐渐浮出水面。基于全基因组测序,研究人员已将AKK菌划分为至少四个系统发育群(phylogroups, AmI-AmIV)。
这些不同发育群的菌株虽然16S rRNA基因序列高度相似(>;99%),但其全基因组的平均核苷酸同一性(ANI)却可能低于95%,这已经达到了物种划分的界限。这意味着,我们过去所称的“A. muciniphila”很可能是一个包含多个物种或亚种的复合体。这种基因组上的差异,直接导致了功能上的多样性:
代谢能力差异
Loannou等人提到,AmII发育群的菌株拥有合成维生素B12的能力,而AmI发育群则不具备,这直接影响了它们的代谢产物谱。不同菌株对人类母乳寡糖(HMOs)的利用效率也存在显著差异。
对宿主影响的差异
在DSS诱导的结肠炎小鼠模型中,研究人员发现不同的人源AKK菌株对肠道炎症的影响截然不同:一个菌株表现出保护作用,两个菌株没有效果,而第四个菌株甚至有加剧炎症的趋势。这为我们敲响了警钟:随意使用一种AKK菌株来治疗IBD,可能不仅无效,甚至有害。
抗生素抗性差异
模式菌株MucT携带多种抗生素抗性基因(如blaA, dfrA, sul, tetM, van)。但从健康人中分离的菌株,其抗性谱各不相同。考虑到AKK菌具有通过水平基因转移获取新基因的能力,将一个未经充分安全性评估的活菌株作为益生菌推向市场,存在传播抗生素抗性的潜在风险。
对于产业界而言,开发AKK菌产品时,可能要对菌株进行全面的功能和安全性评估(包括代谢能力、免疫调节特性、抗生素抗性谱、基因转移能力等)是不可或缺的关键步骤。
AKK菌能在竞争激烈的肠道环境中占据一席之地,并与宿主展开如此复杂的对话,其背后是一套精妙的分子机制。这不仅是它生存的智慧,也是我们理解其双面性的钥匙。
黏蛋白降解
AKK菌对黏蛋白的降解,远非简单的啃食。它拥有一整套工具箱——即种类繁多的碳水化合物活性酶(CAZymes)。这个过程极具策略性:
-外层突破
首先,通过表面的唾液酸酶和岩藻糖苷酶,切除黏蛋白聚糖链最外层的唾液酸和岩藻糖残基。
-核心瓦解
接着,动用半乳糖苷酶、己糖胺酶和硫酸酯酶等,逐步分解聚糖核心结构。
-内部消化
降解产物通过糖转运系统高效内化。AKK菌虽不具备拟杆菌式的典型PULs/MULs基因簇,但其膜转运蛋白能有效摄取降解后的单糖和寡糖,进行发酵,最终产生乙酸、丙酸等短链脂肪酸(SCFAs)。
注:
编辑
其中,丙酸的产生尤为重要。它不仅是其他肠道菌的能量来源,还能通过激活G蛋白偶联受体GPR41和GPR43,刺激肠道L细胞分泌胰高血糖素样肽-1(GLP-1),从而参与调节宿主的血糖稳态和食欲。这部分解释了AKK菌在代谢性疾病中的有益作用。
与免疫系统的多渠道对话
AKK菌与宿主免疫系统的互作,是一场多层次、多渠道的复杂对话。Grant等人的文章中这个信息量大的图,为我们生动地展示了这一过程。
AKK菌与宿主结肠黏膜的相互作用机制示意图
编辑
Grant et al., Nature Microbiology
有几个关键的互作途径:
编辑
图源:doi.org/10.1038/s41579-024-01106-1
AKK菌并非孤立地存在于肠道中,而是作为复杂微生物网络的一部分,与其他细菌、古菌甚至病毒发生着密切的相互作用。这些相互作用包括协同合作、竞争排斥和营养交换(交叉喂养),共同决定了肠道微生态的结构、功能以及对宿主健康的影响。
交叉喂养:生态系统中的资源分享者
尽管AKK菌被描述为自私的黏蛋白降解者,因为它倾向于将黏蛋白内化代谢,但这一过程实际上为其他微生物创造了丰富的资源。
为产丁酸菌提供底物
AKK菌降解黏蛋白释放的单糖(如岩藻糖)和产生的代谢产物(如乙酸盐、丙酸盐),可以被许多重要的产丁酸菌利用。
例如,在共培养实验中,AKK菌能支持产丁酸菌如Anaerostipes caccae、Anaerobutyricum hallii、Faecalibacterium prausnitzii、Roseburia等细菌的生长,并促进有益代谢物丁酸盐的产生。丁酸盐是结肠上皮细胞的主要能源,具有强大的抗炎和维持肠道屏障完整性的功能。
编辑
图源:doi.org/10.1038/s41579-024-01106-1
双向营养交换
AKK菌与其他细菌的相互作用可以是双向的。例如,它与A. hallii之间存在互惠共生关系:AKK菌为A. hallii提供黏蛋白降解产物和乙酸用于丁酸合成,而A. hallii产生的丁酸等代谢物有助于维持适宜AKK菌生长的肠道微环境。
某些研究还提示可能存在更直接的代谢物交换,如维生素或辅因子的共享,但具体机制仍在探索中。
协同作用:共同抵御疾病
A. muciniphila 可能需要其他微生物的存在来预防某些疾病。
1+1>2:微生物协同互作的抗病潜力
例如,在结直肠癌(CRC)小鼠模型 Apc 突变小鼠中,单独定植 A. muciniphila 或幽门螺杆菌导致肿瘤负担增加,而两者共定植则减少了肠道肿瘤数量。
A. muciniphila Muc 和狄氏副拟杆菌(Parabacteroides distasonis)表现出协同的抗结肠炎关系。
在癫痫 Kcna1⁻/⁻ 小鼠中,A. muciniphila Muc和 狄氏副拟杆菌的联合给药通过降低细菌交叉代谢产生的γ-谷氨酰转肽酶活性,在控制饮食条件下预防了癫痫发作。
这些发现强调了一个核心观点:微生物之间的相互作用网络,比单一菌株的存在与否更能决定疾病的结局。
编辑
图源:doi.org/10.1038/s41579-024-01106-1
不单单是AKK菌,而应评估菌群的整体效应
这种复杂的互作关系提示我们在应用层面需要更宏观的视野。
通过菌株脱除试验发现, A. muciniphila 的引入,与抗炎共生菌普拉梭菌(F. prausnitzii)丰度降低相关(在疾病诱导之前),对柠檬酸杆菌(C. rodentium)感染的易感性部分归因于这种细菌网络的变化。这意味着,即使 AKK 菌无法长期定植,它作为过客仍可能重塑肠道菌群的功能结构。
因此,未来评估益生菌应用时,不能仅盯着 AKK 菌本身,而应将其视为生态系统的一个扰动因子,充分重视个体间菌群基线的差异,深入评估其对肠道微生态结构和功能的长期重塑作用,以及宿主原有菌群对其行为的反向调控。
竞争与拮抗
在肠道有限的生态位中,竞争是不可避免的。
与黏液降解菌的竞争
在以MUC2为唯一碳源的体外共培养体系中,当AKK菌与其他的黏液降解菌如普通拟杆菌(Bacteroides vulgatus)、活泼瘤胃球菌(Ruminococcus gnavus)等一起培养时,AKK菌的生长会受到抑制,而其他细菌的生长则被促进。这表明在黏液降解这一功能上存在激烈的竞争。
对其他菌群的负向影响
在某些情况下,AKK菌的存在可能对其他有益菌产生负面影响。
例如,在一个定义的微生物群落中,AKK菌的存在与抗炎共生菌Faecalibacterium prausnitzii丰度的降低相关。但这种相关性并不一定代表因果关系——可能是:环境条件改变导致两者同时变化,或其他因素的间接影响特定疾病状态的反映(如炎症性肠病)。
与普雷沃氏菌的负相关
在人类肠型(enterotype)研究中,AKK菌通常在以Ruminococcus属为主的肠型中富集,而与以普雷沃氏菌为主的肠型呈负相关,这反映的是菌群组成的自然差异,不同肠型由遗传、饮食、地理位置等多因素决定,不是AKK菌排斥普雷沃氏菌,而是不同的生态位和代谢特征。
目前最核心、也最令人困惑的部分:AKK菌在不同疾病背景下的双面角色。三篇文献都花费了大量篇幅,尽力为我们揭示了这种环境依赖性。
AKK菌与结肠炎:保护、致病与宿主互作的复杂博弈
关于AKK菌与结肠炎的关系,研究结论充满了矛盾。
保护作用
在常规的DSS化学诱导结肠炎模型中,灌胃AKK菌或其蛋白Amuc_1100,通常能观察到保护效果。其机制包括上调紧密连接蛋白(如ZO-1、Occludin)以修复屏障、减轻内质网应激、促进Treg细胞应答等。Luo等人的综述也证实,活菌能够增加紧密连接蛋白的表达。
致病作用
然而,一旦宿主背景改变,情况就可能逆转。在遗传易感的Il10-/-小鼠(一种自发性结肠炎模型)中,单独定植AKK菌反而会加剧炎症,导致黏液层变薄和促炎细胞因子上调。这表明,在宿主免疫调节能力受损(如缺乏关键的抗炎因子IL-10)的情况下,AKK菌的黏蛋白降解活性可能弊大于利。
宿主蛋白的策反
Grant等人的研究揭示了一个更为精妙的机制。在溃疡性结肠炎患者中,一种名为Intelectin-1(ITLN1)的宿主蛋白会过度表达。这种蛋白会特异性地结合AKK菌,将其拉到更靠近上皮细胞的位置。这种亲密接触在炎症背景下,反而加剧了免疫反应和组织损伤。
感染:启动防御,还是放大风险?
在面对外来病原体入侵时,AKK菌的角色同样摇摆不定。
有益面:抗感染
AKK菌的某些组分或代谢物显示出抗感染的潜力。例如,其产生的三肽RKH能通过阻断TLR4信号通路,保护小鼠免于致死性脓毒症。Amuc_1100蛋白则能预防沙门氏菌引起的肝损伤。
有害面:缺膳食纤维就吃粘液层,病原体侵入
AKK菌的关键功能——降解黏蛋白,在特定条件下会为病原体开门。
在一项设计精巧的无菌小鼠实验中,研究者发现,缺乏膳食纤维的饮食会导致肠道中的AKK菌饥不择食,转而大量消耗黏液层。这层被削弱的物理屏障,使得致病菌Citrobacter rodentium能够轻易入侵,导致致命感染。
更具说服力的是,当研究者从这个菌群中移除AKK菌后,即使在无纤维饮食下,小鼠也能免于感染。这证明了AKK菌在这种情境下的内鬼角色。有趣的是,一旦恢复富含纤维的饮食,AKK菌的存在反而与较低的病原体载量相关,显示出保护作用。
编辑
Tips:同一种菌,两副面孔:有膳食纤维时,AKK菌是肠道卫士;没有纤维时,它却会啃食肠道黏膜。这个营养开关,让它在天使与魔鬼之间切换。这个发现具有极其重要的实践意义。告诉我们,补充AKK菌益生菌的同时,如果忽略了饮食管理(特别是保证充足的膳食纤维摄入),可能不仅无法获益,甚至可能损害肠道屏障,增加感染风险。这为精准营养与精准菌群干预的结合提供了强有力的理论依据。
癌症:抗肿瘤反应与促肿瘤微环境的动态博弈
在癌症领域,尤其是结直肠癌,AKK菌的形象同样复杂。
促癌风险
部分研究在结直肠癌患者的肿瘤组织和相应的小鼠模型中,都观察到了AKK菌的富集。有观点认为,它通过降解黏液屏障,可能为肿瘤的发生发展创造了条件。
抑癌潜力
然而,另一些研究则得出了相反的结论。AKK菌的EVs、乙酰转移酶Amuc_2172等组分,在小鼠模型中显示出抑制肿瘤生长的效果。
在免疫治疗中的助攻
AKK菌在癌症研究中最高光的时刻,莫过于其在免疫检查点抑制剂(如PD-1抗体)治疗中的作用。
多项研究一致发现,对PD-1治疗有反应的癌症患者(包括肺癌、肾癌等),其肠道中AKK菌的基线水平显著更高。更关键的是,在小鼠模型中,将无反应者粪菌移植给小鼠后,再补充AKK菌,能够重新恢复小鼠对PD-1治疗的敏感性。这表明AKK菌可能通过调节全身免疫状态,增强了抗肿瘤免疫应答,从而成为免疫治疗的增效剂。
代谢与神经系统疾病:更偏向有益的角色
相较于在炎症和感染中的摇摆不定,AKK菌在代谢性疾病中的有益作用,是目前证据最为一致、也最具转化潜力的领域。无论是动物模型还是初步的人体研究,补充活菌或巴氏杀菌的AKK菌,都被证明能够改善胰岛素敏感性、降低胆固醇、减少脂肪堆积。其机制与促进GLP-1分泌、调节脂肪酸代谢、减轻低度炎症等密切相关。
在神经系统疾病中,AKK菌的关联性再次变得复杂。它在帕金森病和多发性硬化症患者中常常富集,体外实验也显示它可能诱导α-突触核蛋白聚集或促炎反应。然而,也有研究将多发性硬化症患者中AKK菌的增多与较低的残疾程度联系起来,提示这可能是一种有益的代偿反应。这种矛盾性再次凸显了菌株差异和宿主背景的重要性。
面对AKK菌如此复杂,我们该如何从科学研究走向临床应用?有幸的是三篇文献为我们提供了一些思考和前瞻性的指导。
编辑
Grant et al., Nature Microbiology
影响AKK菌双重性的因素及潜在治疗策略。AKK菌的最终效应受到菌株差异、饮食、微生物互作和宿主状态的共同调节。理解这些变量,有助于我们选择最优的治疗应用方案,例如是使用活菌、灭活菌,还是分离的生物活性组分。
精准应用的策略选择
未来的AKK菌疗法,绝非一招鲜,吃遍天,而应是量体裁衣的精准策略。
活菌 vs. 灭活菌
巴氏杀菌的AKK菌是一个极具吸引力的选择。它在欧盟已被批准为新食品原料,安全性更高,避免了活菌定植和基因转移的风险。
多项研究证实,灭活菌依然保留了大部分有益的代谢调节功能,这可能归功于其热稳定的细胞壁成分(如Amuc_1100)。
整体 vs. 组分
直接使用分离的生物活性组分,是更为精准和安全的策略。例如,将Amuc_1100蛋白作为药物开发,或利用其EVs作为治疗载体。
Ioannou 等人的综述强调了 AKK 菌作为“酶学底盘”的巨大潜力。例如利用AKK菌的糖苷酶来改造血细胞表面的ABO血型抗原,展示了从基础研究到生物技术应用的转化潜力。
补充 vs. 内源调节
除了直接补充AKK菌,通过饮食干预(如补充富含多酚的食物、膳食纤维)来扶持宿主内源AKK菌的生长,也是一种温和而有效的方法。
关 键 挑 战
要把 AKK从研究热点真正推进到可精准应用的干预手段,还需要把菌群检测作为贯穿研发—临床—产业化全链条的基础设施,融入以下关键环节:
菌株筛选与鉴定
建立一个标准化的 AKK 菌株功能评价体系,涵盖其代谢谱、免疫调节能力、安全性等多个维度,是实现精准应用的前提。与此同时,需要配套规范化的菌群检测(如 16S/宏基因组与定量检测),用于界定不同菌株在不同微生态背景中的适配性与可重复性,避免同名不同效。
情境依赖性的机制阐明
需要更深入地研究,在不同宿主遗传背景、饮食模式和共存微生物群落的影响下,AKK 菌的功能会发生怎样的改变。这需要更复杂的动物模型(如人源化小鼠模型)和多组学技术的结合;其中,纵向菌群检测是识别“谁在场、谁在协同/拮抗、何时发生生态位迁移”的关键手段,也是将机制与真实个体差异对齐的必要条件。
安全性评估
必须对活菌制剂的抗生素抗性传播风险进行严格评估。开发不含抗性基因的工程菌株,或优先使用灭活菌及组分,是未来的方向。同时,应将菌群检测用于追踪干预后菌群结构扰动、耐药基因负荷变化以及潜在机会致病菌扩增等风险信号,实现从前期评估到使用中/使用后监测的闭环。
递送与生产工艺
作为严格厌氧菌,如何实现 AKK 菌的大规模、低成本培养,并开发出能保护其在通过胃肠道时保持活性的口服制剂,是产业化面临的技术瓶颈。
在这一过程中,菌群检测同样不可或缺:一方面用于生产端的质量控制(纯度、污染菌与批间一致性),另一方面用于应用端的效果评估与分层(基线菌群与 AKK 定植/丰度变化、关键功能菌群响应),从而把工艺参数—活性保持—体内生态学结果真正连接起来,提升可复制性与可监管性。
编辑
图源:doi.org/10.1038/s41579-024-01106-1
通过对这三篇力作的梳理,我们对Akkermansia muciniphila的认知,从一个模糊的有益菌形象,变得立体、丰满,也更加敬畏。它不再是一个简单的标签,而是一个充满动态和变数的生命体。
毒理学有一句名言:剂量决定毒性。在微生物学领域,我们或许可以引申为:情境决定属性。对于AKK菌而言,这个情境包含了我们吃下的每一口食物,我们基因中编码的每一个蛋白,以及我们肠道中与之共存的亿万菌群。
作为科研和从业工作者,我们的核心任务在于阐明这些情境依赖性效应的内在机制。未来研究需要超越简单的相关性分析,通过严谨的实验设计和先进的技术手段,明确AKK菌发挥特定生物学效应的必要条件、充分条件及其剂量-效应关系,解析宿主遗传背景、肠道微生态结构、代谢状态等因素对其功能的调控作用等。
探索之路,道阻且长,但充满希望。与各位同仁共勉。
主要参考文献
Grant ET, Monzel E, Desai MS. Navigating the duality of Akkermansia muciniphila. Nat Microbiol. 2026 Jan;11(1):20-30.
Ioannou A, Berkhout MD, Geerlings SY, Belzer C. Akkermansia muciniphila: biology, microbial ecology, host interactions and therapeutic potential. Nat Rev Microbiol. 2025 Mar;23(3):162-177.
Luo Y, Lan C, Li H, Ouyang Q, Kong F, Wu A, Ren Z, Tian G, Cai J, Yu B, He J, Wright AG. Rational consideration of Akkermansia muciniphila targeting intestinal health: advantages and challenges. NPJ Biofilms Microbiomes. 2022 Oct 17;8(1):81.

谷禾健康


嗜粘蛋白-阿克曼氏菌(Akkermansia muciniphila,A.muciniphila),简称为 AKK,于 2004 年发现,是一种从人类粪便中分离出来的椭圆形革兰氏阴性细菌。
该细菌以荷兰微生物生态学家 Anton DL Akkermans 的名字命名,以表彰他对该领域的贡献。A. muciniphila 是人类肠道共生菌,并可以依靠肠粘液层的黏蛋白生存。
在过去十年中,越来越多的研究证明,在糖尿病、心血管疾病、疾病性肠病、神经疾病发现A. muciniphila丰度降低。最近的干预研究还证实如茶或富含多酚水果的饮食可增加其丰度,有助于改善糖尿病和肥胖个体的代谢功能。
总的来说,来自动物和人类研究的越来越多的证据表明A. muciniphila将成为下一代有临床应用前景的益生菌。尤其是它在预防和治疗糖尿病、肥胖症及癌症,这对未来的研究具有重要意义和发展。
A. muciniphila是一种厌氧、革兰氏阴性、无运动、没有内生孢子的卵圆形肠道细菌,其最适生长温度是37℃,最适生长pH为6.5,该菌的倍增时间大约是1.5 h。A. muciniphila属于疣微菌门,事实上,它是人类粪便样本中发现的属于这个特殊门的唯一物种。
值得注意的是,A. muciniphila具有一定的耐氧性,其暴露在空气中24 h,仍然有1%以上的存活率。此外,研究还发现低氧气浓度(纳摩尔)下可以显著促进A. muciniphila的生长。
A. muciniphila单独或者成对存在,很少成链生长,其代谢产物为乙酸盐、丙酸盐、1,2-丙二醇等。
在提供蛋白源的基础培养基中,加入N-乙酰氨基葡萄糖、N-乙酰半乳糖胺和葡萄糖作为能源,A. muciniphila可以生长,但是在果糖和纤维二糖等其他糖类作为能源的培养基中A. muciniphila不能生长,说明该菌的生长繁殖需要蛋白质。
A. muciniphila的特别有趣之处在于,与大多数其他有益细菌不同,它不完全依赖于我们的饮食来喂养。即使消化道中没有营养物质,它也会以粘膜层中的粘蛋白为食。
Akkermansia Muciniphila一般通过母体获得, 通常在儿童中大量存在,并随着年龄和疾病而减少。将其恢复到最佳水平可以带来许多好处。
肠道中低浓度的Akkermansia muciniphila可能表明黏液层较薄,从而导致肠道屏障功能减弱,此外还会增加细菌毒素的易位,炎症性肠病、哮喘、肥胖症和 II 型糖尿病患者的Akkermansia muciniphila浓度往往较低。
我们知道整个胃肠道 (GI) 的粘膜完整性对于维持人体健康至关重要。这其中,粘液屏障的完整性是胃肠道的第一道防线。肠道黏液层的周转包括黏液的合成、分泌和降解,这是一个需要调节和平衡的微妙过程,以确保黏液保持最佳的保护功能。
粘液由多种成分组成:水 (90%–95%)、电解质、脂质 (1%–2%)、蛋白质等。这种粘液是一种稀的、水性和粘弹性的分泌物,这要归功于被称为粘蛋白的特定粘液蛋白,它是粘液中存在的主要结构和功能成分,浓度为 1%–5%。
粘蛋白是一个大的、复杂的、糖基化的蛋白质家族,其特征是一个重要的元素,即“粘蛋白结构域”。它由一个蛋白质核心组成,该核心由含有氨基酸残基脯氨酸 (Pro)、苏氨酸 (Thr) 和丝氨酸 (Ser) 的序列组成,称为富含 PTS 的序列,通常串联重复,其中 Ser 和 Thr 广泛为 O-糖基化并赋予“瓶刷”状构象。
MUC2(黏糖蛋白2)的化学结构和肠道粘液的合成
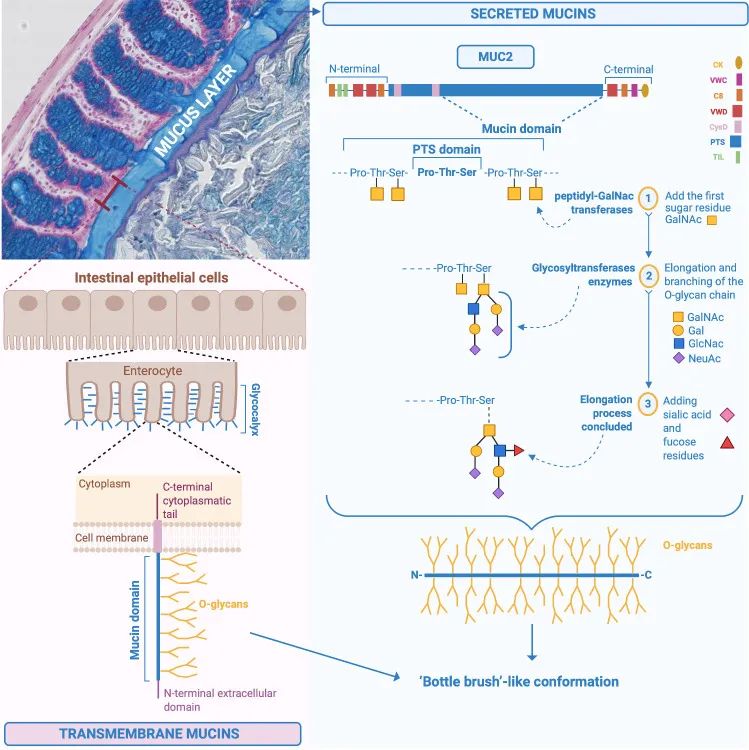

Paone P & Cani PD. Gut. 2020
肠道菌群在肠道中的分布梯度随其位置不同而变化;微生物密度从近端到远端肠道增加,每克肠道内容物的微生物细胞数量大约如下:
十二指肠中103 个
空肠中 104 个
回肠中 107个
结肠中1012 个
此外,微生物密度从上皮细胞向管腔增加,在后者中发现的细菌数量最多,与管腔相比,实际上很少有细菌物种能够很好地粘附和驻留在粘液层中。除了粘附这一重要功能外,肠道微生物群对调节肠道粘液层有很大贡献。
需要普及一个认知就是,当我们谈到构成微生物组的各种细菌物种时,有些菌似乎比其他细菌具有更显着的影响,比如肠道基石菌,核心菌,益生菌等,确保这些个体物种的最佳水平——不要太低,也不要太高——可以产生对健康深远的影响。这其中包括有益细菌的关键物种之一 Akkermansia Muciniphila。
谷禾肠道样本大数据库显示A. muciniphila缺乏或未检出情况在人群中很常见,尤其是那些有肠道问题的人。在健康个体中其约占肠道微生物群总数的0.5%–3%。
但是也有例外,在我们在最近的检测的一个56岁女士肠道内该菌的丰度占比超50%,菌群构成如下:


然而,好事太多也可能是坏事。它的名字——“muciniphila”的意思是粘液爱好者,指的是它独特的能力,降解黏蛋白和粘液层。过量的Akkermansia 将过度消耗粘液蛋白而存活下来,这是大多数其他细菌所缺乏的生存优势。在这种情况下,非粘液消耗物种的数量显着减少,导致物种多样性减少, Akkermansia 增殖异常,从而可能导致肠道屏障损伤,诱发肠道炎症、LPS 进入血液的增加、自身免疫性疾病有关,神经退行性疾病等。
该样本检测报告也显示菌群多样性低,肠炎和几项慢病注意风险:
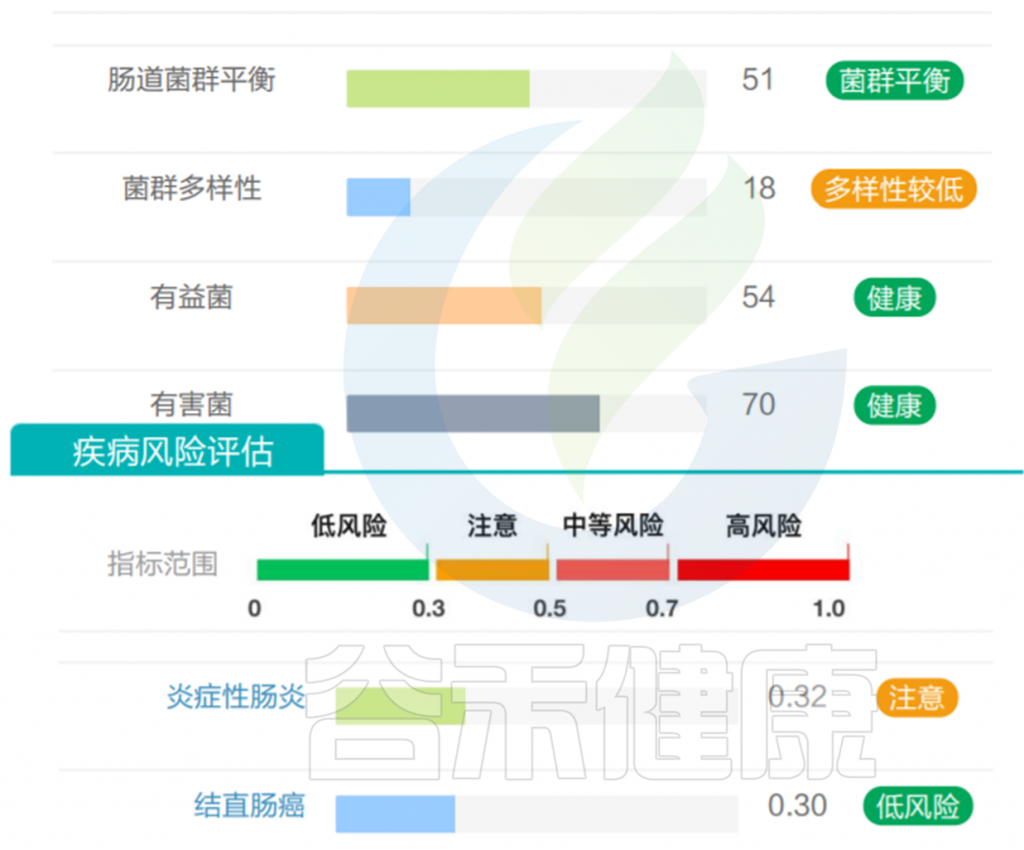

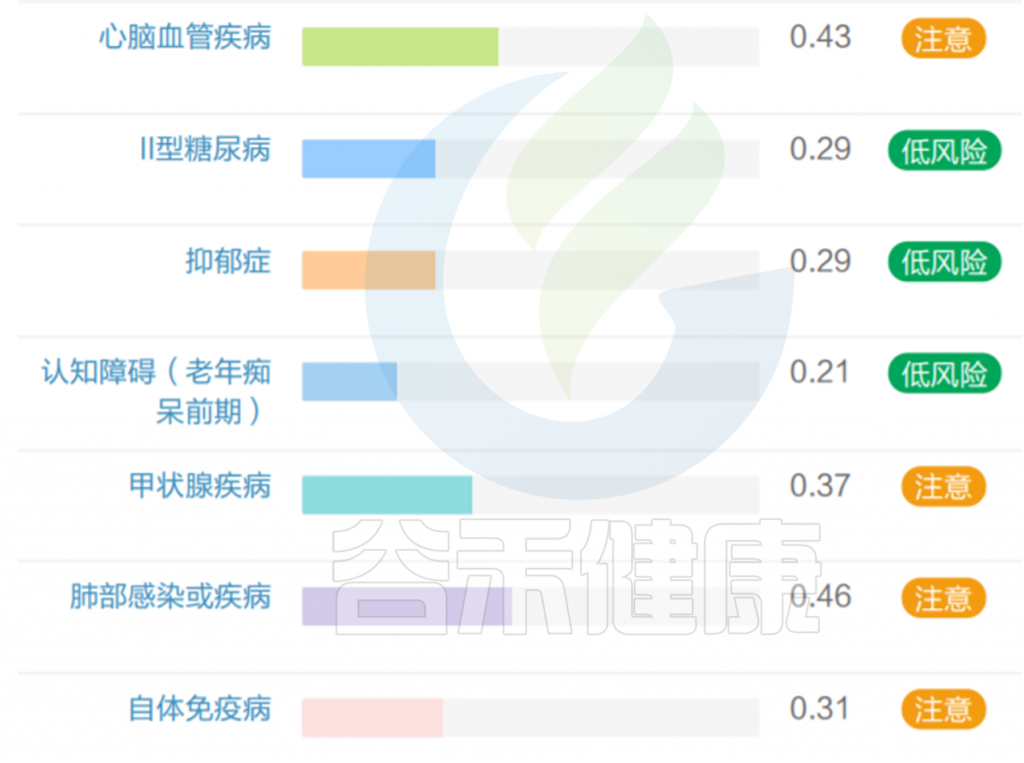

此外,在其它研究文献中也得出Akkermansia muciniphila丰度高的负面研究,如:
在中国人中,A. muciniphila菌的丰度与2型糖尿病呈正相关。
血红素在肠道中的过度增生,与A. muciniphila的粘膜溶解活性有关,在特定情况中,A. muciniphila参与促进代谢综合征。
北京协和团队发现Akkermansia muciniphila可能参与了系统性红斑狼疮的发生发展,致自身免疫亢进特征。
通过细菌16S rRNA测序表明其属于疣微菌门(Verrucomicrobia),与Verrucomicrobium spinosum有92%序列相似性,是现在唯一已知的人类肠道疣微菌门成员。其中文译名尚无统一,在文献中以阿克曼黏细菌、艾克曼黏细菌等名字出现。
为了揭示A. muciniphila的功能潜力,对其基因组进行了测序和注释。发现Akkermansia muciniphila在其相对较小的基因组中编码了大量黏蛋白降解酶,黏蛋白被其降解后其下游聚糖副产物可以交叉饲养其他肠道细菌。
根据DNA-DNA杂交(Digital DNA-DNA hybridiztion,dDDH)将23个Akkermansia属的菌株分为4种,分别为:
Akkermansia sp.CAG
Akkermansia sp.KLE
Akkermansia. muciniphila
Akkermansia. glycaniphila
但是发现它包含许多候选粘蛋白酶编码基因,但缺乏编码典型粘液结合域的基因。
种内存在多样性。在整个基因组中发现的大量噬菌体相关序列,表明病毒在该物种的进化中发挥了重要作用。此外,发掘了 37 个胃肠道宏基因组中,确定Akkermansia序列的存在和遗传多样性。在 37 个中,11 个包含 16S 核糖体 RNA 基因序列,与A. muciniphila 的相同 > 95% , 称为Akkermansia-like 微生物。
瓦赫宁根大学微生物实验室从蟒蛇体内分离培养出于A. muciniphila 基因序列具有94.4%相似度的新菌株,并且发现它们也具有相似的特性,将该微生物命名为Akkermansia glycaniphila。
Akkermansia muciniphila是一种肠道中的黏蛋白降解细菌。粘蛋白是肠上皮黏液层的重要组成部分。粘蛋白由杯状细胞产生肠上皮,是高度糖基化的分子,还包括丝氨酸、苏氨酸和半胱氨酸,它们在肠组织表面形成透明的粘液层,是大量肠道微生物栖息的主要部位。
A. muciniphila主要定植在胃肠道的外黏液层,以胃肠道的黏蛋白作为自身生长的碳和氮来源,其消耗黏蛋白与杯状细胞再生黏蛋白能够达到动态平衡,从而维持黏液层稳定。A.muciniphila以及其分泌物,如囊泡(Extracellular vesicles, EV)通过与结肠上皮细胞Toll受体(Toll-like receptor,TLR)结合及调节紧密连接蛋白的表达等来维持肠道稳态,从而改善高脂饮食诱导的肥胖和炎症性肠病等疾病。
此外,粘蛋白还覆盖在呼吸道、消化道和泌尿生殖道的细胞表面。许多癌症(胰腺癌、肺癌、乳腺癌、卵巢癌、结肠癌和其他组织)都会增加粘蛋白的产生。粘蛋白也在肺部疾病中过度表达,例如哮喘、支气管炎、慢性阻塞性肺病 (COPD) 或囊性纤维化。
Akkermansia muciniphila 高度适应从粘蛋白降解中生存——例如,它需要苏氨酸才能生长,并且具有多种适合利用粘蛋白寡糖中的特殊糖类的酶,如唾液酸酶和岩藻糖苷酶,甚至硫酸酯酶。表明宿主动物和AKK菌之间的共同进化进入了高级阶段,并暗示了这些疣状微生物对宿主的潜在功能意义。由于其向其他细菌物种提供粘蛋白降解产物,该物种也是肠道重要的共生菌属。
Akkermansia muciniphila 能够使用粘蛋白作为碳、氮和能量的唯一来源。尽管宿主肠道层中的许多黏液降解微生物可能具有致病性,但A. muciniphila不一样。Akkermansia muciniphila从粘蛋白发酵中产生乙酸盐、丙酸盐和乙醇,以调节宿主的生物学功能,包括宿主免疫反应和脂质代谢。由于A. muciniphila保留在外黏膜层并且不会渗透到内层,因此它的定植是有益的,因为细菌会促进外黏蛋白层的自然更新。
除了基因序列多样性外,这种细菌还表现出多种生物学功能,包括:
促进肠道屏障完整性;
调节免疫反应;
抑制炎症;
交叉喂养他微生物群;
降低某些心脏病风险,如胰岛素抵抗、总血胆固醇和脂肪组织储存;
防止体重增加。

DOI: (10.1111/jam.14911)
促进肠道屏障
肠道屏障是一个多层复杂的系统,它允许营养吸收,同时防止微生物及其产物的易位。肠道屏障的破坏导致管腔内容物进入血流,激活免疫反应并诱发炎症。
粘液覆盖外肠上皮细胞层,起到物理保护作用,防止微生物和有害化合物的渗透。除了降解粘蛋白外,A. muciniphila还被发现可以刺激粘蛋白的产生 。
A. muciniphila在防止人类肠道致病菌增殖中的作用
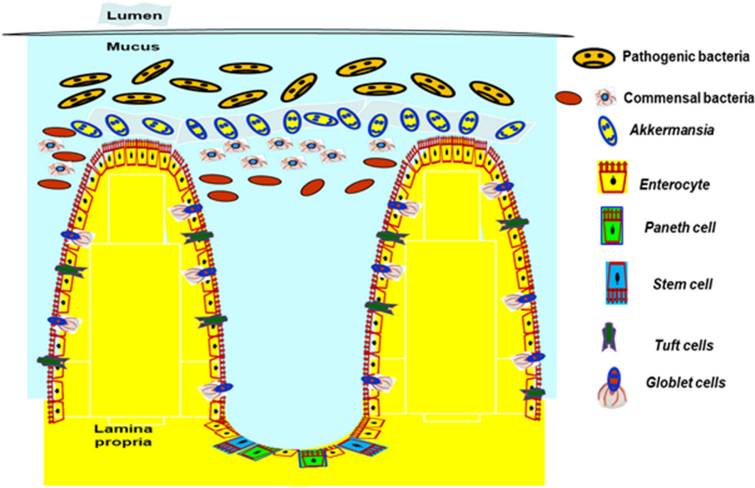
Kalia VC, et al., Indian J Microbiol. 2021
今年发表在Gut Microbes研究得出,虽然A. muciniphila 是一种粘蛋白降解细菌,但是通过促进分泌型 IEC 的分化,反而增加了粘液的产生。A. muciniphila处理以 Gpr41/43 依赖性方式增强了ISC 增殖,加速肠上皮再生,促进 IEC 发育并维持肠道稳态。
注释:哺乳动物肠上皮细胞(IEC)更新较快,每 3-5 天补充一次。所有类型的 IEC 都源自肠干细胞(ISC)
粘蛋白的周转率为 6-12 小时,内层的周转率约为 1 小时,据信其分泌受神经、激素和旁分泌作用的调节。
在动物模型中,A. muciniphila补充剂使结肠粘液层的厚度增加了大约 3 倍,明显高于由有益细菌植物乳杆菌引起的厚度增加。此外,在体外,发现A. muciniphila通过直接与肠细胞结合来改善肠细胞单层完整性 。
A. muciniphila如何促进粘液厚度尚不清楚。原因之一可能是A. muciniphila通过从降解的粘蛋白中制造短链脂肪酸来刺激粘液周转率,这是合成和分泌粘蛋白的宿主上皮的优选能源。事实上,A. muciniphila补充剂增加了小鼠产生粘蛋白的杯状细胞的数量。
抑制炎症
有证据表明A. muciniphila可以调节炎症。在加速衰老的小鼠模型中补充这种细菌可以减轻炎症 。其他研究也显示了A. muciniphila在不同小鼠模型中的抗炎特性,包括无菌模型、肝损伤模型和肥胖模型。
A. muciniphila可以减少由牙龈卟啉单胞菌引起的瘦或肥胖小鼠的炎症。
此外,A. muciniphila分泌的囊泡能够降低TLR4(Toll-like receptor 4, TLR4)的表达,从而调节NF-κB通路,减少促炎因子IL-6、IL-8的分泌。
调节免疫
A. muciniphila在小鼠体内平衡期间,在诱导肠道驻留 T 细胞方面发挥着宿主相关的作用。
在一项针对围产期 HIV 感染儿童和青少年的横断面研究中报告说,A. muciniphila丰度与升高的IL-6 和可溶性 CD14 相关 。
A. muciniphila降解黏蛋白,释放可以被周围非黏液降解细菌利用的短链脂肪酸(SCFAs)、寡糖等代谢物。SCFAs能抑制组蛋白脱乙酰基酶,从而调节NF-κB通路,减少炎症因子的表达。丁酸可以促进Treg细胞的外周活化,增加结肠中Treg细胞亚群的数量,抑制促炎免疫细胞CD4+T淋巴细胞、CD8+T淋巴细胞活性。
免疫相关的皮肤病——牛皮癣
银屑病(俗称牛皮癣)是一种免疫介导的慢性炎症性皮肤病。研究表明银屑病和肠道炎症之间存在关联。一项研究评估了来自巴西的21名银屑病患者与24名健康对照者的肠道微生物群组成和多样性。与对照组相比,银屑病患者的 Akkermansia muciniphila 数量有所减少。
扩展阅读:牛皮癣看似皮肤病,实则关系到肠道
交叉喂养
除了A. muciniphila外,粘液层中还常见其他微生物。产生丁酸盐的细菌不具有降解粘液的能力,而是利用被粘蛋白降解的物种(如A. muciniphila)降解的碳和氮,这些微生物包括细菌如Faecalibacterium prausnitzii,罗斯氏菌等其产生抗炎短链脂肪酸(SCFA)。
产生的乙酸能够促进粪厌氧棒状菌(Anaerostipes caccae),霍氏真杆菌(Eubacterium hallii)。
报道共培养A. muciniphila与非粘液降解丁酸盐产生细菌F. prausnitzii、A. caccae和Eubacteriumhallii导致同养生长和丁酸盐的产生。因此,A. muciniphila不仅自身在保护肠道上皮方面发挥重要作用,而且还支持抗炎肠道微生物群。
A. muciniphila和其他细菌之间的相互作用
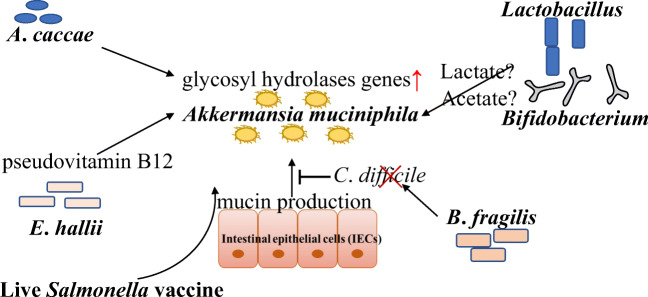
Hagi T, et al., Appl Microbiol Biotechnol. 2021
有些细菌不能降解粘蛋白,因此非粘蛋白降解细菌受益于粘蛋白降解细菌。
A. muciniphila产生来自粘液和短链脂肪酸的糖,如乙酸盐和丙酸盐。非粘液降解细菌,如Anaerostipes caccae, Eubacterium hallii and Faecalibacterium prausnitzii ,利用A. muciniphila从粘液中降解的糖进行生长。
A. muciniphila的粘蛋白降解支持了Anaerostipes caccae的生长和丁酸生产。Anaerostipes caccae诱导AKK中粘蛋白降解基因的表达增加,核糖体基因的表达减少。
防止体重增加
普遍发现A. muciniphila在瘦个体中更为丰富。在超重个体中,细菌的丰度较低。
在小鼠中,与肥胖小鼠的丰度相比,瘦动物粪便中A. muciniphila丰度增加之间的相关性表明了这种意义,并且观察到在数周内每天向高脂肪饮食诱导的肥胖小鼠喂食Akkermansia显然可以逆转这种诱导的肥胖以及改善上皮完整性和相关效应。
也有一些数据表明,A. muciniphila干预可能成为人类受试者肥胖改善方案的基础。但是强调只有活的Akkermansia菌才能在小鼠中产生这种效果,但是发现高温预处理杀菌的细胞甚至纯化的Akkermansia蛋白,与活细胞治疗的效果相比,Akkermansia的热杀死细胞似乎对高脂肪饮食肥胖小鼠具有相同甚至增强的有益效果。
此外,A. muciniphila通过分解粘蛋白产生醋酸盐这样的 SCFAs,对我们的体重产生积极影响。因为乙酸盐可以阻止我们感到饥饿,从而防止体重增加。
A.muciniphila治疗不会显著改变饮食诱导的肥胖小鼠的肠道微生物组成,但它可以逆转HFD诱导的代谢紊乱,包括脂肪质量增加、代谢内毒素血症、脂肪组织炎症和胰岛素抵抗,这表明它可能用于预防或治疗肥胖和相关的代谢紊乱。
人类肠道微生物群中丰富的A.muciniphila可预防疾病
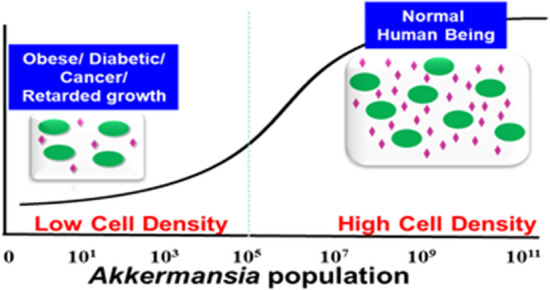
Kalia VC, et al., Indian J Microbiol. 2021
改善代谢
A. muciniphila补充剂能够恢复肥胖和 2 型糖尿病小鼠的粘液厚度,其中高脂肪饮食治疗破坏了肠道粘液;该治疗还导致血清脂多糖 (LPS) 显着减少,这是一种代谢性内毒素血症,并改善了代谢状况。
对肥胖小鼠施用Akkermansia还通过减少循环中的炎性脂多糖和减轻胰岛素抵抗而导致观察到的“代谢性内毒素血症”减少作为小鼠肥胖的心脏代谢并发症,如动脉硬化。至少来自小鼠模型的结论Akkermansia效应得到了许多不同研究的支持。
A. muciniphila及其衍生产物对宿主代谢的影响
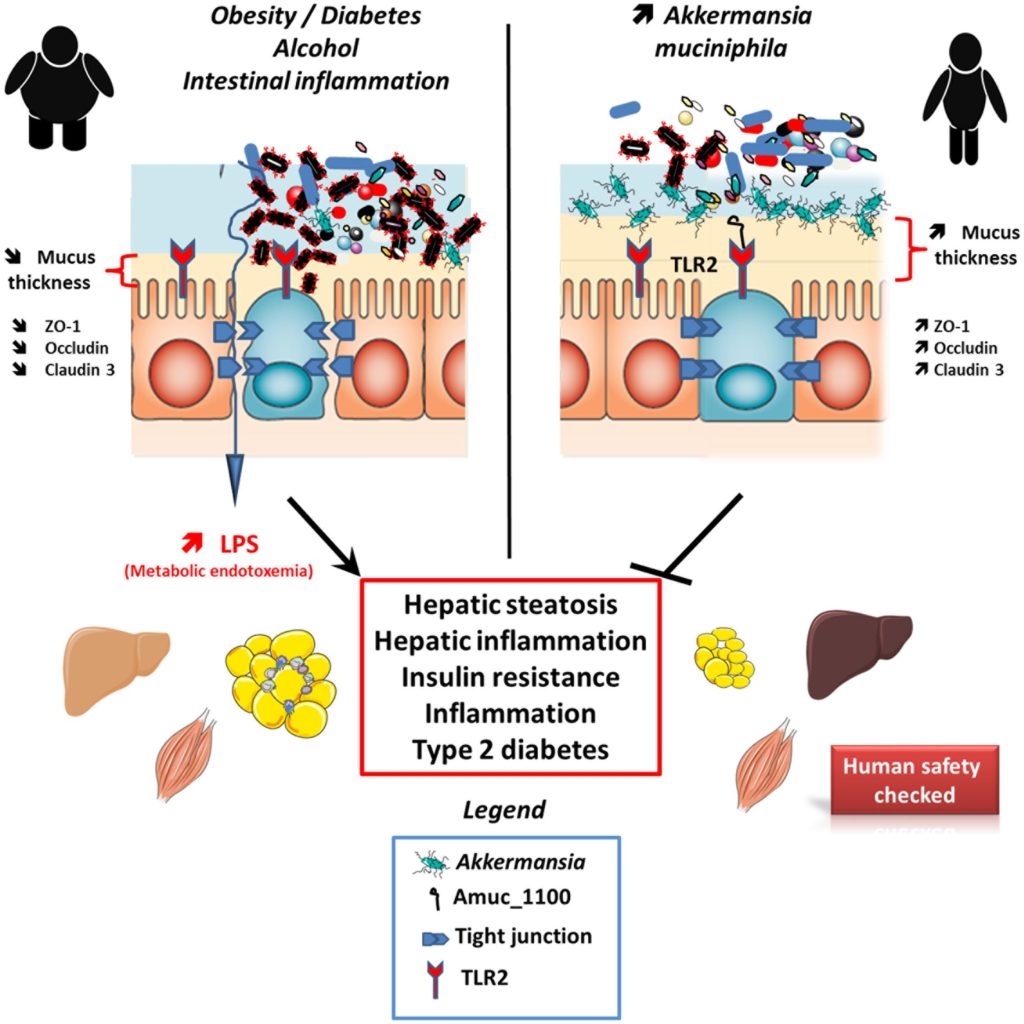
Cani PD,et al.,Front Microbiol. 2017
非酒精性脂肪性肝
与野生型小鼠相比,在NAFLD小鼠中观察到A. muciniphila丰度显著降低,而导致A.muciniphila丰度增加的干预措施改善了代谢参数。使用益生元(如低聚果糖)可以恢复A.muciniphila的丰度,并改善相关疾病。
酒精性肝病
酒精性肝病(ALD)由脂肪变性、纤维化和肝硬化发展为急性酒精性脂肪性肝炎,死亡率高,但治疗方法非常有限。结果发现,内毒素血症水平升高的酒精性肝病患者肝损伤程度增加。乙醇消耗引起的肠道紧密连接的破坏导致炎症信号通过病原体相关的分子模式,如脂多糖。
研究ALD患者队列时,发现与健康受试者相比,ALD患者粪便A. muciniphila数量减少。给乙醇喂养的小鼠口服粘液粘菌A. muciniphila可恢复该细菌的消耗,减少肠漏,增加Claudin-3和Occludin的紧密连接表达。因此,炎症和肝损伤促进细胞因子白介素1β (IL-1β)和肿瘤坏死因子α (TNF-α)明显减少,导致MPO +中性粒细胞浸润,改善肝损伤。
扩展阅读:深度解析 | 肠道菌群与慢性肝病,肝癌
改善血糖和胰岛素抵抗
A. muciniphila在糖耐量正常的人群中比糖尿病前期或 II 型糖尿病人群中的数量更多。
事实上,这些细菌已被证明可以增强称为二甲双胍的抗糖尿病药物的作用。在喂食高脂肪饮食的小鼠中,二甲双胍治疗增加了A. muciniphila的数量 并改善了它们的血糖水平。
同样,喂食高脂肪饮食的小鼠口服A. muciniphila但不服用二甲双胍,对葡萄糖的耐受性也有所提高。因此,这种细菌通过改变肠道微生物群,可能有助于在未来找到控制 2 型糖尿病的新方法。
扩展阅读:2型糖尿病如何做到可防可控?肠道菌群发挥重要作用
癌症免疫治疗
Akkermansia muciniphila也可以积极影响宿主对癌症免疫疗法的反应。研究发现,抗生素的消耗与对免疫治疗 PD-1 阻断剂的反应较差有关。进一步研究确定较低水平的Akkermansia muciniphila确实对反应率有直接影响,给抗生素治疗的小鼠口服细菌补充剂,发现它恢复了它们对免疫疗法的反应。根据这项研究,有人建议维持健康的肠道菌群,主要包括Akkermansia muciniphila,对于提高癌症免疫治疗有效性很重要。
此外,Akkermansia muciniphila通过刺激 DCs 产生 IL-12 并促进 CCR9 + CXCR3 + CD4 +的积累来增强 ICB 功效 上皮肿瘤和淋巴结中的 T 细胞。
根据美国癌症研究中心新研究显示Akkermansia muciniphila提升免疫检查点抑制剂的效果,还能产生大量调节T细胞反应的肌苷。食用高纤维食物可以有效地调节肿瘤微环境中的单核巨噬细胞的组成,而食用得越多,Akkermansia muciniphila的肠道菌数量就会更多。
该菌还能产生大量环二腺苷酸,即使是无菌小鼠,仅仅植入Akkermansia muciniphila,它们也能有更强的抗癌反应。未来,或许一种通过食物和环二腺苷酸联合的促进抗癌疗法就在眼前。
神经退行性疾病
肌萎缩性侧索硬化症(amyotrophic lateral sclerosis, ALS)是一种复杂的神经退行性疾病,患者通常会出现运动神经元的过早死亡,导致诊断后的平均生存时间不足5年。然而,针对肌萎缩性侧索硬化症的药物所做的巨大努力却收效甚微。
急性ALS小鼠模型经抗生素治疗后显示,微生物群失调在渐冻症的进展和恶化中起着重要作用。
通过挑选11株与ALS相关的微生物菌株,包括A.muciniphila和单接种到经抗生素治疗的ALS小鼠中,只有A.muciniphila与运动神经元改善相关。
其他疾病,例如,帕金森病与对照组的肠道微生物组组成显著不同, Akkermansia 丰度增加。
年龄
年龄和地理环境等因素影响人体A. muciniphila的数量。从婴幼儿时期开始A. muciniphila在人类胃肠道定植,并在一年后迅速接近成年人的水平。随着年龄增长A. muciniphila在人体内的数量逐渐降低。
不同的是,另一研究发现大于60岁人群A. muciniphila的数量升高。此外,另外研究发现在百岁长寿老人中A. muciniphila数量显著增加,提示A. muciniphila水平的升高可能有助于延缓机体的衰老。
对两种类型的早衰综合征小鼠模型,外用该菌A. muciniphila可显著提高小鼠的健康寿命。
地理位置
另外,不同地理位置A. muciniphila在人群中定植也存在差别。如通过细菌16S rRNA分析发现,智利健康受试者粪便中的微生物菌群和巴布亚新几内亚受试者有明显不同;但与美国和阿根廷受试者接近;A. muciniphila在智利受试者的丰富度最高。
中国南方的男性和女性定植率没有差异,但是中国南方人群的A. muciniphila定植率低于欧洲人群。暨南大学理工学院在广州地区人群的A. muciniphila定植率为89.31%,平均丰度为5.825 lg(CFU/mL)。肠道消化模拟不同时间点的RT PCR结果显示,A. muciniphila在模拟人类肠道微生物菌群环境中具备生存能力;对Bifidobacterium属和Bacteroides属具有抑制效果,对Lactobacillus属具有促进作用。
2021年在Nature发表的文章中,研究人员将古肠道样本数据和一组来自于已测序的工业化及非工业化生活方式下现代人群的粪便样本数据进行对比。比起非工业化样本和古粪便,包括Akkermansia muciniphila(可降解人体粘液)在内的BloSSUM类群在工业化样本中更丰富。
工业化生活下微生物群落中的粘液降解菌和基因更普遍,这可能是由西方饮食导致的。
药物
药物是影响肠道A.muciniphila丰度的重要因素之一。首先,抗菌药物作用后可以直接影响肠道A.muciniphila的数量。
在小鼠出生后早期用万古霉素进行干预可以有效预防1型糖尿病,显著降低小鼠血糖值以及减轻胰岛炎症;而对8周龄小鼠未见明显预防效果。万古霉素作用于幼儿时期的小鼠使A. muciniphila成为胃肠道优势菌群,该研究提示A. muciniphila可能是推迟1型糖尿病的关键菌群。
对60例6~11个月的婴儿使用阿奇霉素治疗3 d后,婴儿A. muciniphila数量显著减少。此外Dubourg等的研究中,2例成人患者经过抗生素(强力霉素、哌拉西林、羟氯喹、亚胺培南等)治疗后,A. muciniphila数量显著增加,在肠道菌群的比例均大于40%。
有趣的是,在另外一项小鼠模型中也发现类似现象,给小鼠灌胃四联抗生素(氨苄青霉素,万古霉素,新霉素和甲硝唑)可显著逆转免疫抑制剂导致的A. muciniphila数量减少。这些研究表明抗菌药物等药物使用会影响肠道内A. muciniphila数量的变化,其对人体代谢产生的影响需进一步研究。
最新的研究发现,代谢综合征相关治疗药物对A.muciniphila丰度的影响可能参与其作用机制。如二甲双胍通常用作治疗代谢综合征(如肥胖和2型糖尿病)的一线用药。
对14例服用二甲双胍的糖尿病患者进行肠道菌群分析,与未服用二甲双胍的糖尿病患者以及健康受试者相比,A. muciniphila显著增加。在体外实验中,收集二甲双胍治疗组和对照组小鼠的粪便,发现二甲双胍培养组A. muciniphila的比例显著增加。
在另外一项研究使用另外一种代谢病治疗药物利拉鲁肽(Liraglutide)。对高脂饮食诱导的肥胖小鼠进行皮下注射给药,连续15 d给药后处死小鼠,收集肠道内容物进行菌群分析,发现治疗小鼠肠道中A. muciniphila显著增加。此外,抗肿瘤药物也可以影响肠道A.muciniphila的丰度。如在胰腺导管腺癌(Pancreatic ductal adenocarcinoma, PDAC)异种移植的小鼠模型中发现,吉西他滨(Gemcitabine)治疗后使A. muciniphila在肠道中的数量显著增加(5%~33%)。
饮食方式
饮食对肠道A.muciniphila影响最显著。
与正常饮食组相比,高蛋白饮食显著降低大鼠肠道A. muciniphila。
Akkermansia 的数量随着短期(3 周)低FODMAPs饮食而减少。这不足为奇,因为 FODMAP 基本上与益生元纤维相同。
Akkermansia在生酮饮食中显着增加,并且与 Parabacteroides一起,可能是减少酮症癫痫发作的原因。
小鼠实验表明,A.muciniphila数量受到高脂饮食的强烈影响,高脂饮食三周后A.muciniphila丰度开始下降,并且A.muciniphila丰度与体脂呈负相关。
与高脂高糖饮食组相比,小鼠食用热带水果8周后,可以显著增加A. muciniphila在胃肠道中的丰度;同时干预后可以降低高脂高蔗糖饮食喂养小鼠的内毒素血症,改善其胰岛素敏感性。
饱和脂肪含量高的饮食与较高的炎症发生率有关。而富含多不饱和脂肪酸的饮食,如亚麻籽和鱼油中的那些,已被证明可以抵抗炎症,并且与更瘦的人有关。
这些膳食脂肪也会影响肠道中A. muciniphila的水平。

在一项研究中,老鼠被喂食由不同脂肪组成的不同饮食。一组喂猪油,另一组喂鱼油十一周,结果很有趣。
在鱼油喂养组中,A. muciniphila的丰度增加,乳酸杆菌的丰度也增加。
然后将两组的粪便材料移植到新的小鼠组中,在接受鱼油喂养小鼠粪便移植的小鼠中,它们的A. muciniphila水平增加,炎症水平降低。然而,接受猪油喂养小鼠移植的小鼠的炎症水平很高。
因此,摄入的膳食脂肪类型也会极大地影响肠道中A. muciniphila的数量。
我们已经知道多酚可以减少炎症并有助于预防许多疾病,因此高多酚食物可以增加Akkermansia.
多酚是强大的抗氧化剂,可以对抗体内的自由基(与多种疾病有关的化合物,包括糖尿病、心脏病和癌症)。
有许多不同类型的多酚。其中一些不能很好地被身体吸收留在肠道中,这对 Akkermansia来说是个好消息,它们吞噬了多酚并因此增强其力量,从而达到双赢。
此外,丁酸钠和菊粉、北极浆果提取物、绿茶提取物和异麦芽寡糖、膳食多酚等这些益生元也能增加Akkermansia丰度。
含有高水平多酚和鱼油的食物是提高A. muciniphila菌最好的食物。
高多酚食品
多酚使食物一般颜色丰富多彩,尝起来略带苦味或酸味。泡了很长时间的茶尝起来很苦:浸泡时间越长,茶多酚含量越高。浓橄榄油的青草味或红酒的味道也是很好的例子。

在自然界中,苦味可以抵御想要攻击植物的昆虫。有超过8000种多酚。一些例子是红酒中的白藜芦醇、辣椒中的辣椒素、百里香中的百里酚、肉桂中的肉桂酸、迷迭香中的迷迭香酸。
食物中多酚的含量在很大程度上取决于其生长的土壤、成熟度以及种植、运输和制备方式,因此以下列表仅供参考。
高多酚食品:
蔓越莓、石榴、山莓、黑莓、蓝莓、草莓、樱桃、李子、亚麻种子、黑巧克力、栗子、红茶、绿茶、苹果汁、苹果、黑麦面包、核桃、榛子、红酒。

其他常见的高多酚食物包括葡萄、橄榄、菠菜、李子和桃子。一般来说,颜色越深越好,所以选择绿色的黑橄榄和葡萄。
油性鱼
Akkermansia 喜欢鱼油,但目前还不清楚它们喜欢鱼油的特定成分——例如脂肪酸——或者它们喜欢所有的部分。所以目前,最好吃鱼。
尽管鱼油现在是有争议的,一些科学家认为它的好处被夸大了。研究人员发现,食用整条鲑鱼的一组人的 DHA(一种脂肪酸)水平是食用鱼油的一组人的 9 倍。食用鱼类的同时既增加鱼油,同时增加优质蛋白质摄入,均有助于A. muciniphila。
试着每周吃两到三份鱼:
沙丁鱼、鲭鱼、鲱鱼、三文鱼、鳟鱼、鳀鱼、旗鱼、金枪鱼

补充膳食纤维,选择有机产品。
含纤维的食物:
生菊苣根、绿色蒲公英 、麦麸 、芦笋 、大蒜、香蕉 、洋葱 、韭菜,也存在于燕麦片、红酒、蜂蜜、枫糖浆、豆类和其他食物中。
避免高脂肪饮食和酒精
微生物多样性是我们肠道健康的主要目标。实现这一目标的最佳方法是均衡饮食,吃能让肠道菌群快乐的食物,从而你的感觉也会良好。
难点
A. muciniphila与机体健康联系紧密,有潜力成为新型益生菌。但目前A. muciniphila是专性厌氧菌且难以培养,成为新型益生菌,还有以下几点重要问题亟待解决:
首先,需研发高效快速培养A. muciniphila的新技术。目前实验室或小规模已经成功在实验室中获得活性A. muciniphila,并且能够稳定,但是这只是实验室规模生产,大规模的工业生产仍是需要解决的问题。比如培养基中存在蛋白质来源问题,是否会对人体可能有过敏反应,工业化生产时需研发合理的工业化生产培养基。
其次,需确定安全有效的治疗人群。目前A. Muciniphila 与改善肝功能、降低葡萄糖毒性、减轻氧化应激和抑制炎症有强关联,但是,在炎症性肠病、动脉粥样硬化等其他疾病中的干预,不同研究中存在不同的结果,对于其作为药物面对的安全问题,其应用仍待商榷。
最后,需要优化A. muciniphila的补充或给药方式。在体外模拟胃肠道的释放实验中,发现包封的 A. muciniphila活性比直接分散在模拟胃肠道的系统中活性更高,并且能够定向在肠道释放细菌,显著增加A. muciniphila对胃的抵抗力,对于实现在肠道的定向给药还需要更多的实验进行优化。
最后,尽管已经有研究证明有活性的A. muciniphila或者是巴氏消毒的A. muciniphila对人体是安全的,但目前A. muciniphila的临床研究较少,在临床上应用还需要更多的实验验证。
主要参考文献
Hasani A, Ebrahimzadeh S, Hemmati F, Khabbaz A, Hasani A, Gholizadeh P. The role of Akkermansia muciniphila in obesity, diabetes and atherosclerosis. J Med Microbiol. 2021 Oct;70(10). doi: 10.1099/jmm.0.001435. PMID: 34623232.
Zhang T, Ji X, Lu G, Zhang F. The potential of Akkermansia muciniphila in inflammatory bowel disease. Appl Microbiol Biotechnol. 2021 Aug;105(14-15):5785-5794. doi: 10.1007/s00253-021-11453-1. Epub 2021 Jul 27. PMID: 34312713.
Druart C, Plovier H, Van Hul M, Brient A, Phipps KR, de Vos WM, Cani PD. Toxicological safety evaluation of pasteurized Akkermansia muciniphila. J Appl Toxicol. 2021 Feb;41(2):276-290. doi: 10.1002/jat.4044. Epub 2020 Jul 28. PMID: 32725676; PMCID: PMC7818173.
Kalia VC, Gong C, Shanmugam R, Lin H, Zhang L, Lee JK. The Emerging Biotherapeutic Agent: Akkermansia. Indian J Microbiol. 2021 Dec 16:1-10. doi: 10.1007/s12088-021-00993-9. Epub ahead of print. PMID: 34931096; PMCID: PMC8674859.
Yu Y, Lu J, Sun L, Lyu X, Chang XY, Mi X, Hu MG, Wu C, Chen X. Akkermansia muciniphila: A potential novel mechanism of nuciferine to improve hyperlipidemia. Biomed Pharmacother. 2021 Jan;133:111014. doi: 10.1016/j.biopha.2020.111014. Epub 2020 Nov 26. PMID: 33246225.
Cheng D, Xie MZ. A review of a potential and promising probiotic candidate-Akkermansia muciniphila. J Appl Microbiol. 2021 Jun;130(6):1813-1822. doi: 10.1111/jam.14911. Epub 2020 Nov 15. PMID: 33113228.
Hagi T, Belzer C. The interaction of Akkermansia muciniphila with host-derived substances, bacteria and diets. Appl Microbiol Biotechnol. 2021;105(12):4833-4841. doi:10.1007/s00253-021-11362-3
Anhê, F. F et al. (2016). Triggering Akkermansia with dietary polyphenols: A new weapon to combat the metabolic syndrome?. Gut microbes, 7(2), 146–153.
Schade L, Mesa D, Faria AR, Santamaria JR, Xavier CA, Ribeiro D, Hajar FN, Azevedo VF. The gut microbiota profile in psoriasis: a Brazilian case-control study. Lett Appl Microbiol. 2021 Dec 12. doi: 10.1111/lam.13630. Epub ahead of print. PMID: 34897759.
Belzer, C et al. (2017). Microbial Metabolic Networks at the Mucus Layer Lead to Diet-Independent Butyrate and Vitamin B12 Production By Intestinal Symbionts. American Society for Microbiology: 8(5).
Caesar, R et al. (2015). Crosstalk Between Gut Microbiota and Dietary Lipids Aggravates WAT Inflammation Through TLR Signalling. Cell Metab: 22(4), pp 658-668.
Dao, M, C et al. (2015). Akkermansia muciniphila and Improved Metabolic Health During a Dietary Intervention in Obesity: Relationship with Gut Microbiome Richness and Ecology. Gut.
de Vos, W, M. (2017). Microbe Profile: Akkermansia muciniphila: A Conserved Intestinal Symbiont that Acts as The Gatekeeper of our Mucosa. Microbiology: 163(5).
Derrien, M et al. (2017). Akkermansia muciniphila and its Role in Regulating Host Functions. Microbial Pathogenesis: 106, pp 171-181.
Everard, A et al. (2013). Cross-Talk Between Akkermansia muciniphila and intestinal Epithelium Controls Diet-Induced Obesity. PNAS: 110(22), pp 9066-9071.
Geerlings, S, Y et al. (2018). Akkermansia muciniphila in the Human Gastrointestinal Tract: When, Where, and How? Microorganisms: 6(3).
Krishnan, A. (2016). Akkermansia muciniphila: What You Need to Know About this Bacterium. uBiome.
Naito, Y et al. (2018). A Next-Generation Beneficial Microbe: Akkermansia muciniphila. J. Clin. Biochem. Nutr: 63(1), pp 33-35.
Ottman, N et al. (2017). Pili-Like Proteins of Akkermansia muciniphila Modulate Host Immune Responses and Gut Barrier Function. PLOS One.
Pierre, J, F et al (2015). Cranberry Proanthocynaidins Improve the Gut Mucous Layer Morpohology and Function in Mice Receiving Elemental Enteral Nutrition. JPEN J Parenter Enteral Nutr: 37(3), pp 401-409.
Shin, N, R et al. (2013). An Increase in the Akkermansia spp. Population Induced by Metformin Treatment Improves Glucose Homeostasis in Diet-Induced Obese Mice. Gut: 63, pp 706-707.
Zhang, T et al. (2019). Akkermansia muciniphila is a promising probiotic. Microbial Biotechnology.
Ouyang J, Lin J, Isnard S, et al. The Bacterium Akkermansia muciniphila: A Sentinel for Gut Permeability and Its Relevance to HIV-Related Inflammation. Front Immunol. 2020;11:645. Published 2020 Apr 9. doi:10.3389/fimmu.2020.00645
Zhou K. Strategies to promote abundance of Akkermansia muciniphila, an emerging probiotics in the gut, evidence from dietary intervention studies. J Funct Foods. 2017;33:194-201. doi:10.1016/j.jff.2017.03.045
Naito, Y., Uchiyama, K. & Takagi, T. A next-generation beneficial microbe: Akkermansia muciniphila. J Clin Biochem Nutr 63, 33–35 (2018).
Derrien, M., Vaughan, E. E., Plugge, C. M. & de Vos, W. M. Akkermansia muciniphila gen. nov., sp. nov., a human intestinal mucin-degrading bacterium. Int. J. Syst. Evol. Microbiol. 54, 1469–1476 (2004).
Hansson, G. C. Role of mucus layers in gut infection and inflammation. Curr. Opin. Microbiol. 15, 57–62 (2012).
van der Lugt, B. et al. Akkermansia muciniphila ameliorates the age-related decline in colonic mucus thickness and attenuates immune activation in accelerated aging Ercc1-/Δ7 mice. Immun. Ageing 16, 6 (2019).
Lukovac, S. et al. Differential modulation by Akkermansia muciniphila and Faecalibacterium prausnitzii of host peripheral lipid metabolism and histone acetylation in mouse gut organoids. MBio 5, (2014).
Dao, M. C. et al. Akkermansia muciniphila and improved metabolic health during a dietary intervention in obesity: relationship with gut microbiome richness and ecology. Gut 65, 426–436 (2016).
Depommier, C. et al. Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: a proof-of-concept exploratory study. Nat. Med. 25, 1096–1103 (2019).
Png, C. W. et al. Mucolytic bacteria with increased prevalence in IBD mucosa augment in vitro utilization of mucin by other bacteria. Am. J. Gastroenterol. 105, 2420–2428 (2010).
Anhê, F. F. et al. A polyphenol-rich cranberry extract protects from diet-induced obesity, insulin resistance and intestinal inflammation in association with increased Akkermansia spp. population in the gut microbiota of mice. Gut 64, 872–883 (2015).
Pierre, J. F. et al. Cranberry proanthocyanidins improve the gut mucous layer morphology and function in mice receiving elemental enteral nutrition. JPEN J Parenter Enteral Nutr 37, 401–409 (2013).
Vinson, J. A. & Cai, Y. Nuts, especially walnuts, have both antioxidant quantity and efficacy and exhibit significant potential health benefits. Food Funct 3, 134–140 (2012).
Mao, B. et al. Effects of different doses of fructooligosaccharides (FOS) on the composition of mice fecal microbiota, especially the bifidobacterium composition. Nutrients 10, (2018).