-

 CNAS L23010
CNAS L23010

 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业 二级病原微生物安全实验室
二级病原微生物安全实验室- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业 二级病原微生物安全实验室

谷禾健康
高脂饮食会诱发肥胖、胰岛素抵抗、葡萄糖耐受不良和脂肪变性,而脂肪性肝炎向纤维化和肝细胞癌(HCC)的进展主要与高膳食胆固醇有关。非酒精性脂肪性肝病(NAFLD)是一类代谢疾病,可从脂肪肝进展至脂肪性肝炎,进一步导致肝硬化甚至肝细胞癌(HCC),胆固醇被认为是疾病发展过程中主要的脂毒性分子。
近日,香港中文大学于君团队及其团队研究人员在《Gut》上发表了题为“Dietary cholesterol drives fatty liver- associated liver cancer by modulating gut microbiota and metabolites(膳食胆固醇通过调节肠道微生物群和代谢产物驱动脂肪肝相关的肝癌,DOI:10.1136/gutjnl-2019-319664)”的研究论文,阐述了膳食胆固醇在NAFLD–HCC发生中的作用及其相关的分子机制。这项研究证明了长时间的高胆固醇饮食可以引起小鼠自发性NAFLD-HCC(非酒精性脂肪肝肝细胞癌)。抑制胆固醇可以恢复肠道微生物群,完全阻止NAFLD-HCC的发展。

背景
非酒精性脂肪性肝病(NAFLD)是代谢综合征的肝脏表现,包括从单纯性脂肪变性到非酒精性脂肪性肝炎(NASH)的一系列肝脏病理。NASH可进展为肝硬化、终末期肝功能衰竭和肝细胞癌(HCC)。目前,非酒精性脂肪肝是世界范围内发病率和医疗负担的主要原因。脂肪毒性会导致NASH、纤维化/肝硬化,甚至HCC。在肝脏脂质种类中,胆固醇被认为是NASH发生过程中的主要脂毒分子。肝脏是调节全身胆固醇稳态的中心。膳食胆固醇对血浆和肝脏胆固醇稳态有重要影响。虽然膳食胆固醇在NASH进展中的作用已被论述,但长期胆固醇治疗在自发性和进行性NAFLD-HCC发展的作用和致病基础尚不清楚。一些研究表明,肠道菌群是一个有助于NAFLD发生和其发展为NAFLD-HCC的环境因素。微生物衍生的代谢物,如胆汁酸、短链脂肪酸和三甲胺和它们影响的信号通路可能有助于NAFLD的发展。特别是肠道菌群代谢产物3 -(4 -羟苯基)乳酸,对肝脏脂肪变性和肝纤维化具有共同的基因效应。
实验设计
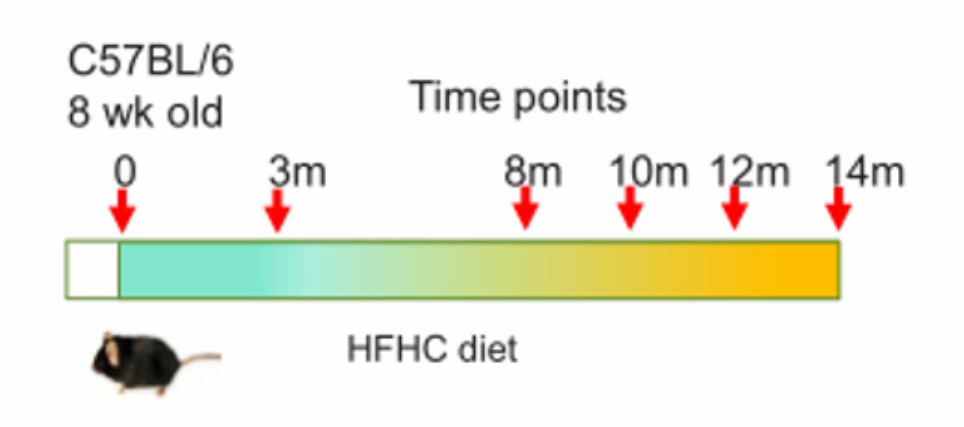
普通饲料(NC,18%脂肪,58%碳水化合物,24%蛋白质,0%胆固醇);高脂/低胆固醇饲料(HFLC,43.7%脂肪,36.6%碳水化合物,19.7%蛋白质,0.013%胆固醇);高脂/高胆固醇饲料(HFHC,43.7%脂肪,36.6%碳水化合物,19.7%蛋白质,0.203%胆固醇)
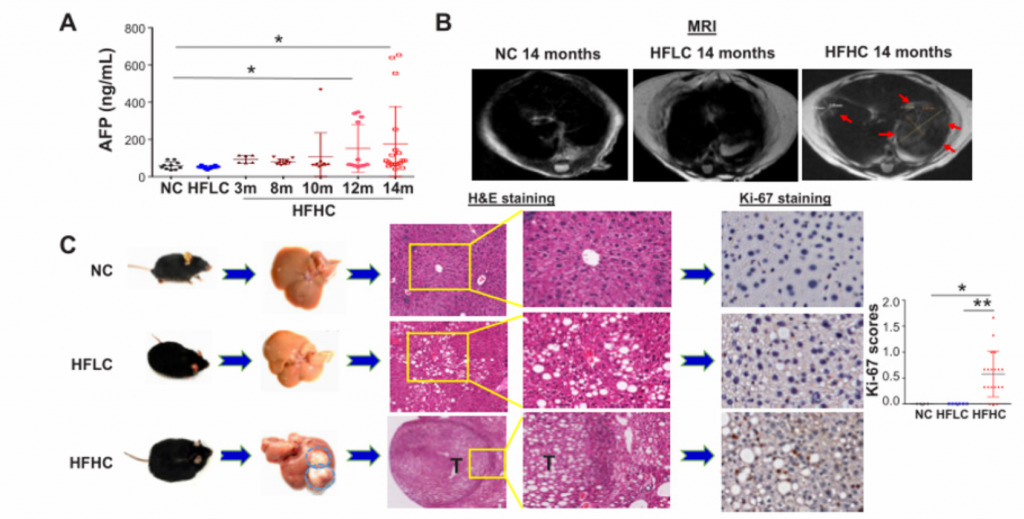

如图所示,以上饲料用于随机喂养8周龄雄性C57BL/6野生型同窝出生幼鼠14个月。阿托伐他汀(降胆固醇药)药物被用于喂食HFHC膳食7个月后的小鼠,持续7个月。然后分别在3、8、10和12个月龄处死。
粪移植实验:
取被喂养不同饲料的小鼠的1 g粪便样品用5ml磷酸盐缓冲生理盐水(PBS)混匀,然后将该混悬液2 0 0μL灌胃移植给无菌小鼠。于移植后8、10、14个月各组中随机抽取小鼠处死。
实验结束时,小鼠被禁食,取血清/组织。记录体重和内脏脂肪重量。肝脏被迅速切除并称重,评估表面结节的存在和大小。分离肝肿瘤,在液氮中快速冷冻,并将其保存在−80°C进行进一步实验。肠道菌群分析采用16SrRNA测序,血清代谢物分析采用液相色谱-质谱(LC-MS)代谢组学分析。
主要结果
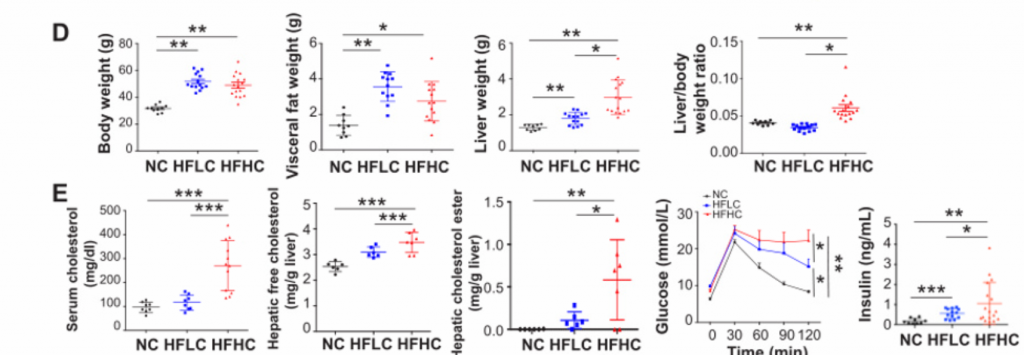
1. 膳食胆固醇可自发驱动NAFLD-HCC的发生。


A)[endif]喂养HFHC饲料的小鼠在第3、8、10、12、14个月检测的血清AFP水平以及喂养NC和HFLC饲料的小鼠在第14个月检测的血清AFP水平。在第10个月观察到AFP水平升高,在第12个月和14个月,与HFLC或NC饲料喂养的小鼠相比,HFHC饲料喂养的小鼠的AFP水平逐渐升高。
B) [endif]肝脏MRI影像。在第14个月时,喂养HFHC饲料的小鼠出现肝肿瘤,而其余两个没有
C) [endif]依次为小鼠肝脏的大体形态、镜下结构以及Ki67染色的免疫组化图像。HFHC小鼠肝脏切片组织学检查均为肝细胞癌,平均每只小鼠肝细胞癌细胞数为2.7±2.6,最大直径为4.1±5.0 mm。与HFLC喂养的小鼠相比,HFHC喂养的小鼠肝脏切片中Ki-67阳性细胞明显增多,表明HFHC喂养的小鼠细胞增殖增加。
D) [endif]依次为体重,内脏脂肪,肝脏重量,肝脏与体重的比率。随着肝癌的形成,喂养HFHC饲料的小鼠的这些指标的水平显著高于喂养NC饲料的小鼠。同时,喂养HFLC饲料的小鼠也表现出这些指标水平的上升。
E) [endif]依次为血清胆固醇、肝游离胆固醇、肝胆固醇酯含量、糖耐量试验及空腹胰岛素水平。与HFLC和NC相比,HFHC小鼠的这些指标的水平都有所上升。与NC相比,HFLC小鼠的葡萄糖耐量和空腹胰岛素也有所增加。
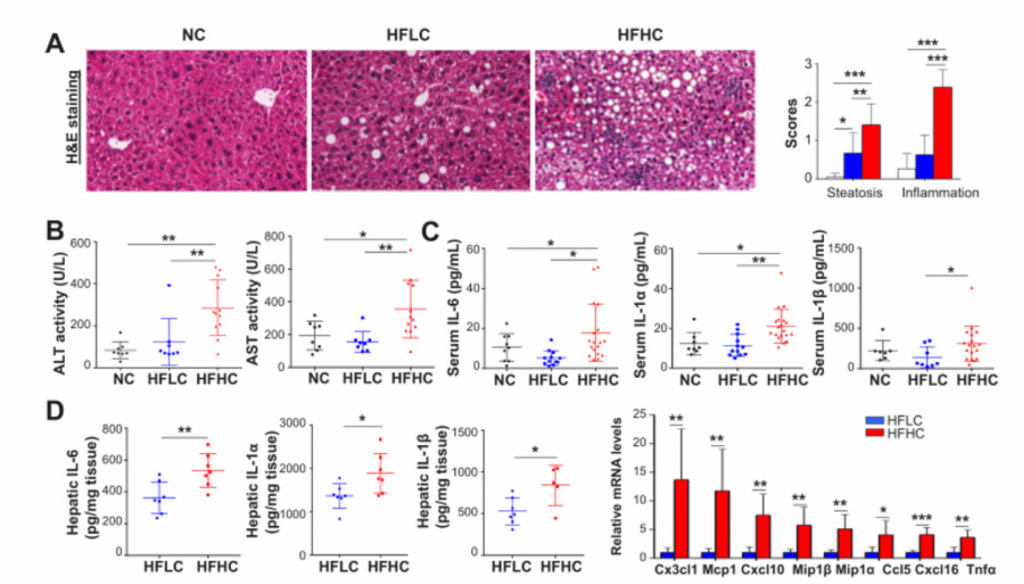
2. 高胆固醇膳食喂养的小鼠的非肝癌肝组织形成了NASH和纤维化。


A)HE染色后的肝脏切片组织检查。HFHC小鼠出现以脂肪变性和小叶炎症为特征的脂肪性肝炎,而在HFLC小鼠中只观察到脂肪变性。
B)血清ALT和AST水平。HFHC小鼠ALT水平和AST水平显著高于HFLC和NC小鼠。
C) 细胞因子谱法测定喂养NC、HFLC和HFHC饲料的小鼠第14个月的血清IL-6、IL-1抑制因子和IL-1蛋白水平。这些指标的水平均有所上升。
D)在喂养NC、HFLC和HFHC饲料的小鼠第14个月时用ELISA法测定小鼠肝脏促炎细胞因子IL-6、IL-1α和IL-1β蛋白水平,RNA测序测定Cx3cl1、Mcp1、Cxcl10、Mip1β、Mip1α、CCL5、Cxcl16、TNFαmRNA水平。与NASH相关的促炎细胞因子Cx3cl1等在HFLC小鼠肝组织中显著上调。
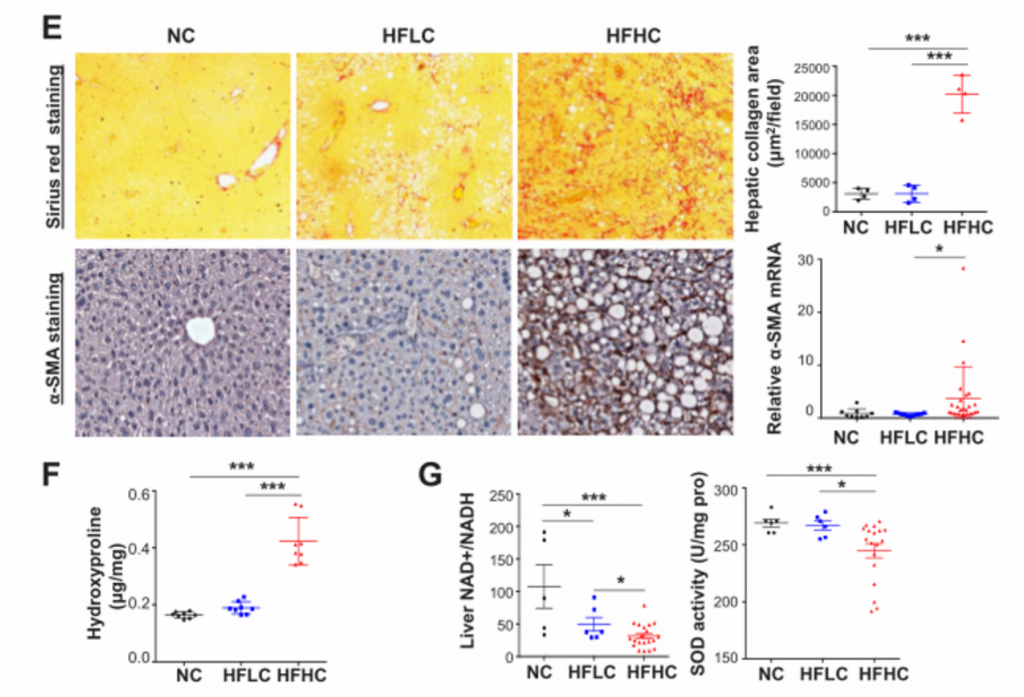
E) 从上至下依次为天狼星红染色和免疫组化染色。分别观察胶原沉积和α-SmA蛋白。RT-PCR法测定mRNA表达水平。HFHC小鼠的肝组织表现出严重的纤维化损伤,胶原分布面积明显增加。肝星状细胞活化表现为α-平滑肌肌动蛋白(α-SMA)的基因和蛋白水平升高。
F) 肝羟脯氨酸含量。其在HFHC小鼠中含量明显升高。
G) 肝脏的氧化应激检测。喂养HFHC饲料的小鼠的肝脏NAD+/NADH比值和SOD活性显著降低,提示膳食胆固醇可诱导肝脏的氧化应激。
3. 高胆固醇膳食引起小鼠肝脏依次发展为脂肪肝、脂肪性肝炎、纤维化和NAFLD-HCC。


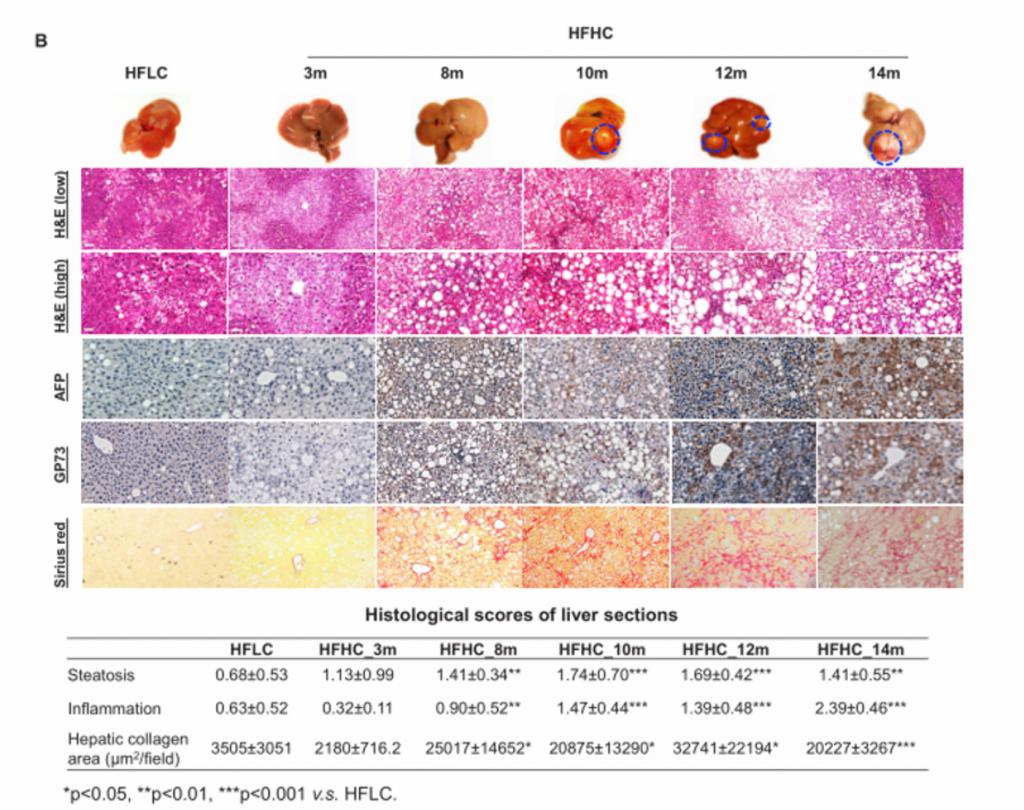
B)从上至下依次为HFLC喂养14个月和HFHC喂养3、8、10、12和14个月小鼠肝脏大体、肝脏肿瘤标志物AFP和GP73的H&E染色和免疫组化染色、天狼星红染色,脂肪变性和炎症的组织学评分以及天狼星红染色定量的肝胶原面积。HFHC小鼠肝脏组织学在3个月时表现为轻度炎症性脂肪变性,8个月时脂肪性肝炎伴纤维化,10、12、14个月时肝癌形成,而HFLC小鼠在3、8、10、14个月时仅表现为脂肪变性而没有进一步发展成肝癌。HFHC小鼠的肿瘤标志物免疫组化染色观察到阳性染色,HFLC小鼠没有。
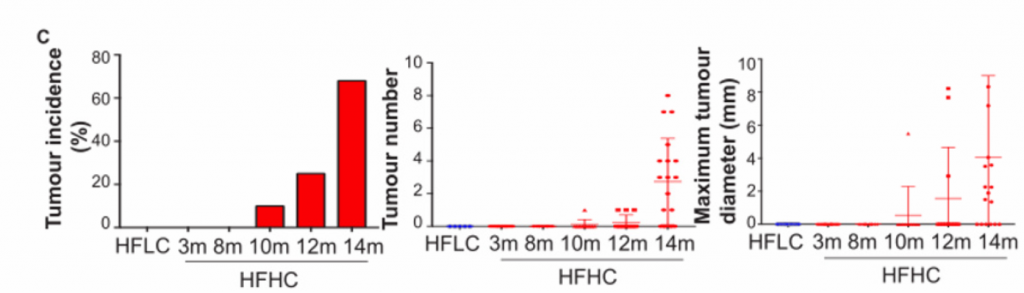
C)在HFHC膳食控制下的不同时间点的小鼠的最大肝肿瘤的肿瘤发病率、肿瘤数量和最大肿瘤直径。
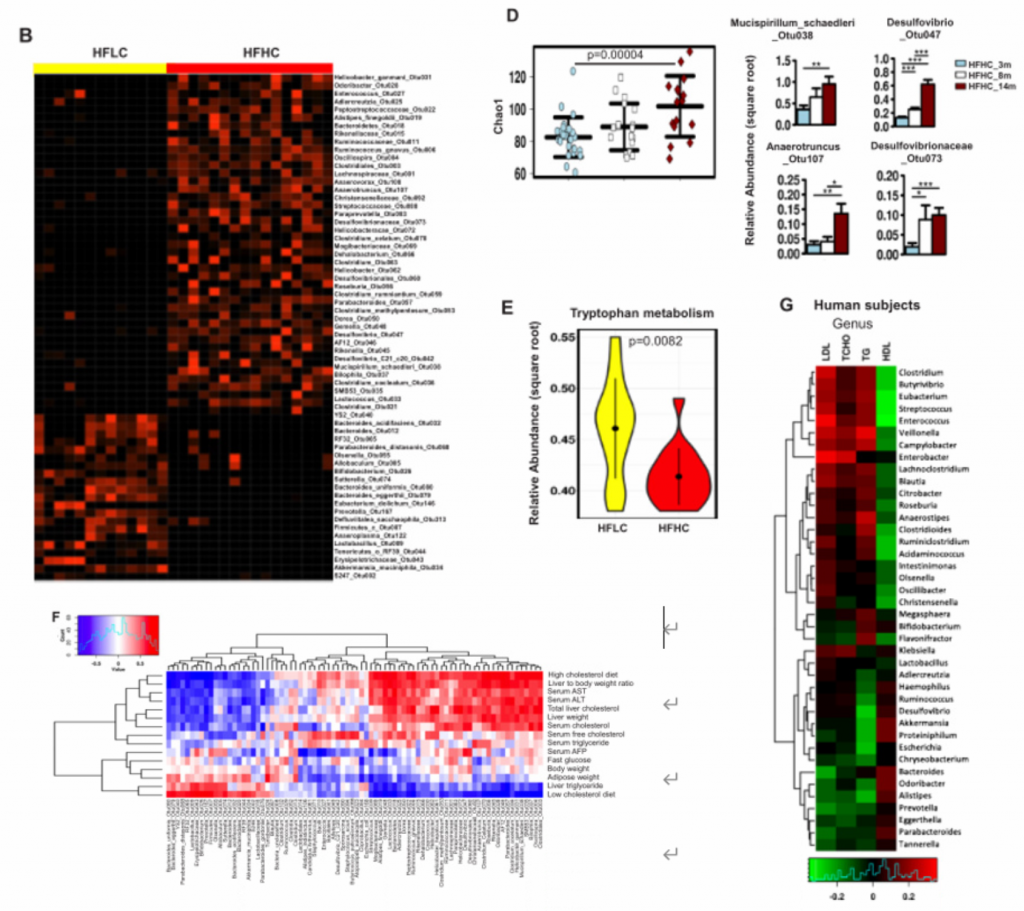
4. NAFLD-HCC的肠道菌群失调与胆固醇水平相关。






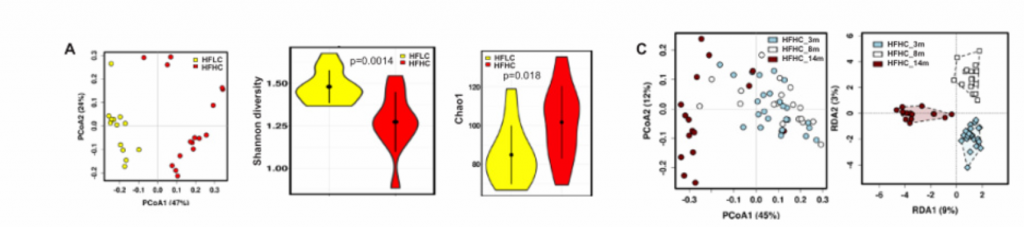
A)14月龄的HFLC和HFHC小鼠之间的肠道微生物的PCoA分析、香浓多样性和丰富度分析,PCoA分析中P<0.001. HFHC小鼠的微生物多样性更低,丰富度更高。
B)14月龄的HFLC和HFHC小鼠的粪便细菌热图。
C)HFHC膳食控制3、8和14个月后的肠道微生物的PCoA和冗余分析。发现HFHC_3m、HFHC_8m和HFHC_14m的小鼠的肠道微生物群落明显聚类,这表明肠道微生物组成随NAFLD-HCC进展阶段的变化而变化。
D)在HFHC饲养3、8 ~ 14个月,用Chao1指数测定微生物群落丰富度,发现其逐渐升高。随着NAFLD-HCC进展,细菌丰富度依次增加。
E)喂养HFLC和HFHC的小鼠肠道菌群的色氨酸代谢能力。HFHC小鼠肠道菌群的色氨酸代谢能力降低。综上,高胆固醇膳食导致肠道菌群失调和微生物色氨酸代谢受损。
F)细菌丰度与小鼠表型的相关性。
G)在59例高胆固醇血症患者和39例健康人的研究中,人血清总胆固醇、甘油三酯、低密度脂蛋白胆固醇和高密度脂蛋白胆固醇(横轴)与细菌(纵轴)基因组测序的相关性。双歧杆菌和类杆菌与血清总胆固醇、低密度脂蛋白胆固醇呈负相关,与高密度脂蛋白胆固醇呈正相关。
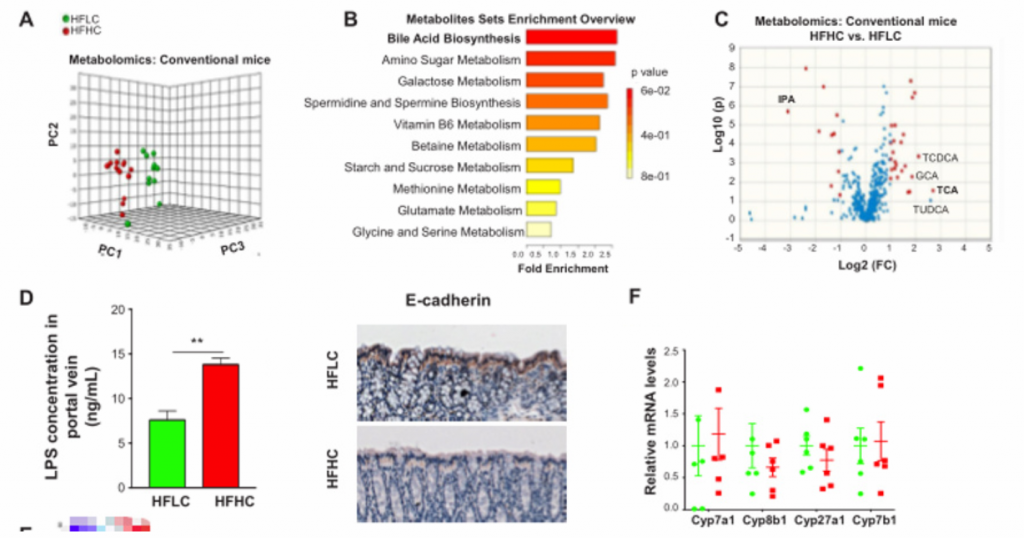
5. 膳食中的胆固醇通过诱导代谢产物的改变而促进NASH-HCC的进展。胆固醇通过调节宿主血清代谢物,以及至少部分通过增加TCA和降低IPA,促进了NASH-HCC的进展。


A) 主成分排序分析表明,膳食胆固醇含量对血清代谢产物有显著影响
B)HFHC小鼠的具有差异的富集代谢物的通路分析。胆汁酸合成是关键。
C) [endif]血清代谢组学火山图,指示了HFLC和HFHC小鼠的代谢产物的异常值。初级胆汁酸包括牛磺胆酸(TCA)、TUDCA、甘胆酸(GCA)和TCDCA,这些也是HFHC小鼠中异常上调的代谢物。IPA是微生物色氨酸代谢的产物,也是HFHC小鼠中异常下调的代谢物。
D) [endif]HFLC和HFHC饲料喂养3个月的小鼠门静脉LPS浓度和14个月的小鼠结肠组织E- cadherin表达。两者相比,HFHC小鼠门静脉血清脂多糖(LPS)浓度升高,结肠E-钙粘蛋白丢失。提示膳食胆固醇可损害肠道屏障功能
E) [endif]微生物与代谢物的相关性分析。红色呈正相关,蓝色呈负相关。
F) [endif]HFLC和HFHC饲料喂养小鼠肝脏组织中Cyp7a1、Cyp8b1、Cyp27a1和Cyp7b1 mRNA水平。高胆固醇膳食不能改变肝脏中细胞色素P450 (Cyp)7a1、Cyp8b1、Cyp27a1、Cyp7b1等胆汁酸合成酶的mRNA表达。
G) [endif]油红O染色显示,TCA加重胆固醇诱导的人LO2细胞甘油三酯的积累,IPA抑制胆固醇诱导的肝癌细胞、HKCI-2和HKCI-10细胞甘油三酯的积累。
H) [endif]IPA抑制NASH-HCC细胞系的细胞增殖。
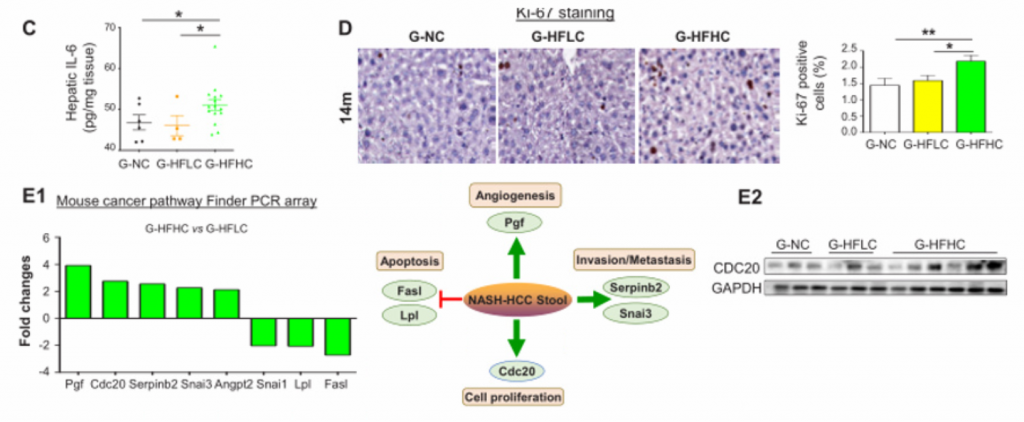
6. 膳食胆固醇调节的肠道菌群通过诱导代谢产物的改变,促进NAFLD和肝细胞的增殖,从而促进胆固醇诱导的NAFLD-HCC的形成。



A)将喂养NC、HFLC和HFHC饲料的小鼠(14个月)的粪便移植到NC条件下的无菌小鼠(G- NC、G-
HFLC和G- HFHC)中。图中为G- NC组、G- HFLC组和GHFHC组灌胃无菌小鼠肝脏的大体形态和组织学检查。G- HFHC小鼠在14月龄时出现一个肝结节,而在8月龄和10月龄时未出现肝结节。组织学检查证实该结节发育不全,有小球状增生,细胞增殖增强。
B) [endif]8、10、14月龄时,G-NC、G-
HFLC和G-HFHC组无菌受体小鼠的肝脏甘油三酯含量、脂质过氧化反应和肝组织学改变。FMT后8、10、14个月,G-HFHC小鼠肝脏脂质积累明显增加,并伴有肝组织损害
C) [endif]G-NC、G- HFLC、G-HFHC组小鼠14月龄时的肝脏IL-6蛋白水平。
D) [endif]肝切片Ki-67染色。可见在14月龄时,G-HFHC小鼠中肝细胞增殖增加。但在FMT后8个月和10个月时肝细胞增殖并未见增加。
E) [endif]肿瘤通路检测。图中直方图向上表示基因表达上调,向下为下调。免疫印迹法验证CDC20(细胞增殖)表达上调。
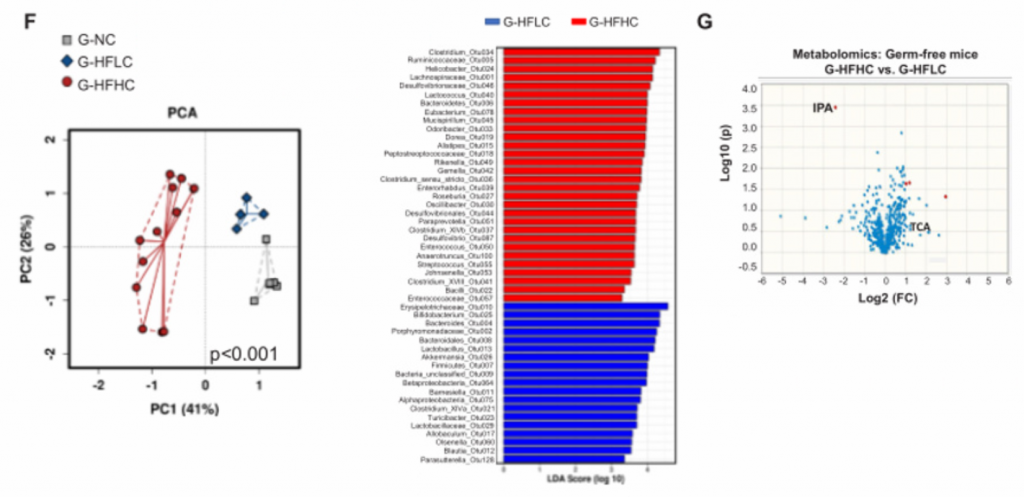
F) [endif]G-NC、G- HFLC、G-HFHC小鼠肠道微生物群的PCA分析和LDA直方图。在不同的膳食控制下,肠道菌群的差异很大。
G) [endif]G- HFLC和G- HFHC小鼠血清代谢组学分析。G- HFHC小鼠的IPA较G-HFLC小鼠降低,与喂养HFHC饲料的常规小鼠一致。
7. 降胆固醇治疗完全抑制了高脂血症小鼠NAFLD-HCC的形成。



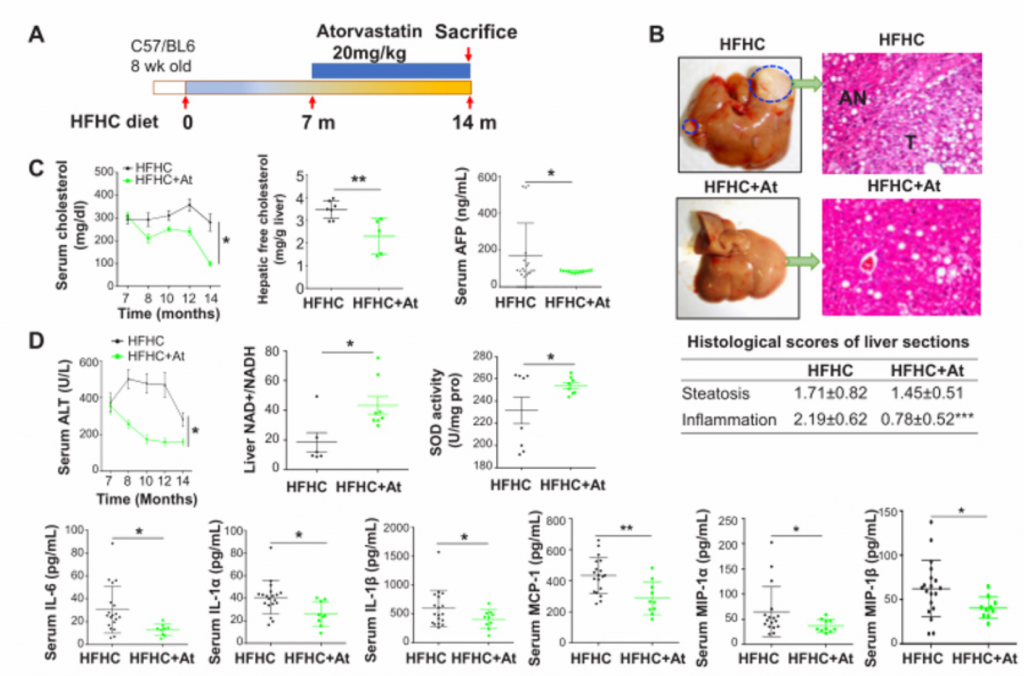
A)阿托伐他汀治疗喂养HFHC饲料的C57BL/6小鼠的示意图。给小鼠喂食HFHC 饲料7个月,并继续喂食HFHC饲料和阿托伐他汀7个月。(HFHCAt)
B)HFHC小鼠的肝组织HE染色及肝脏大体形态,分别计算了其脂肪变性和炎症的组织学评分。在实验结束时(14个月),阿托伐他汀完全抑制了HFHC饮食诱导的NAFLD-HCC的形成,并改善了NASH的严重程度。
C)从左至右分别为血清胆固醇水平、肝脏游离胆固醇水平、血清甲胎蛋白水平,显著降低。
D) 丙氨酸氨基转移酶(ALT)、血清促炎细胞因子(IL-6、IL-1α、IL-1β、单核细胞趋化蛋白-1、巨噬细胞炎性蛋白-1α和巨噬细胞趋化蛋白-1β)显著降低。
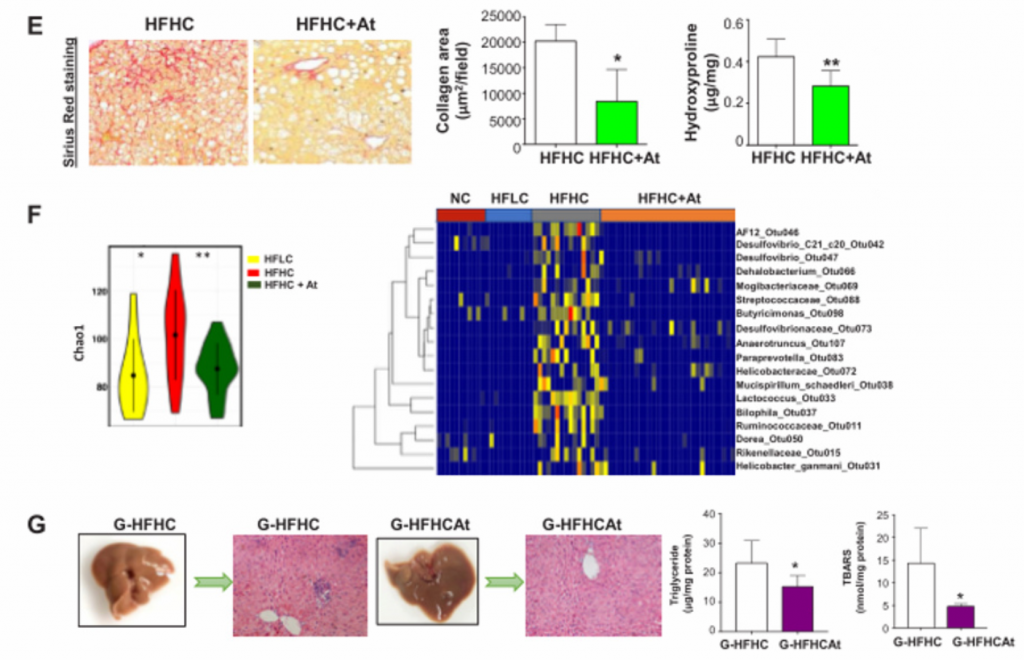
E)阿托伐他汀通过显著减少肝脏胶原沉积和羟脯氨酸含量来改善肝纤维化
F)用NC、HFLC和HFHC饲料喂养的小鼠,在阿托伐他汀治疗或不治疗的情况下,粪便中细菌的Chao1指数和热图。经阿托伐他汀治疗后,HFHC喂养的小鼠肠道菌群丰富度显著恢复。
G)被灌服HFHCAt(阿托伐他汀治疗的HFHC喂养小鼠)粪便的无菌小鼠的肝脏大体形态、组织学检查、甘油三酯和脂质过氧化含量。与G-HFHC组相比,G-HFHCAt组小鼠肝脏组织学有所改善,肝脏甘油三酯和脂质过氧化水平降低。
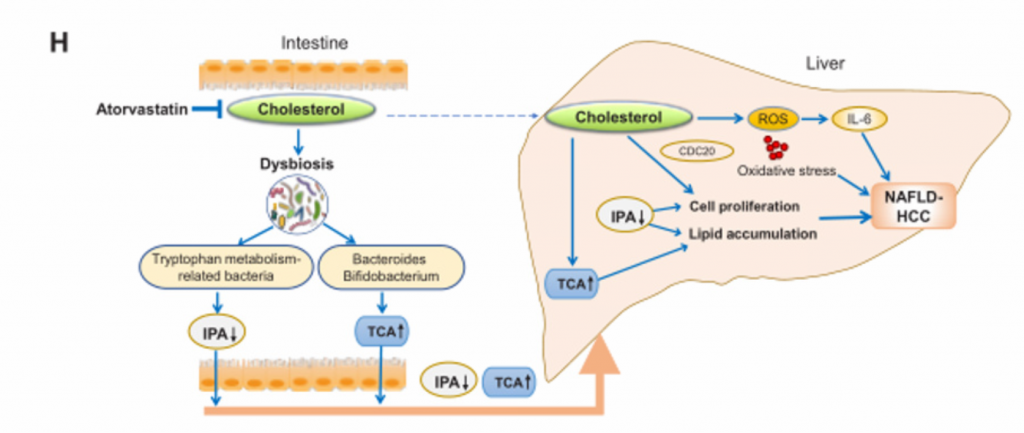
H)胆固醇诱导的NAFLD-HCC发生机制示意图。胆固醇破坏胆汁酸代谢和微生物色氨酸代谢,导致血清TCA增强,IPA降低,从而促进NAFLD-HCC的发展。
结论
长时间的高胆固醇饮食通过调节肠道微生物群,诱导雄性小鼠NAFLD-HCC自发和进行性发展。胆固醇破坏胆汁酸代谢和微生物色氨酸代谢,导致血清TCA增强,IPA降低,从而促进NAFLD-HCC的发展。降胆固醇治疗完全抑制了膳食胆固醇诱导的NAFLD-HCC的形成。这项研究强调,抑制胆固醇和调控肠道菌群及其相关代谢产物可能是预防NAFLD-HCC的有效策略。