-

 国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子
国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子

 二级病原微生物安全实验室
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
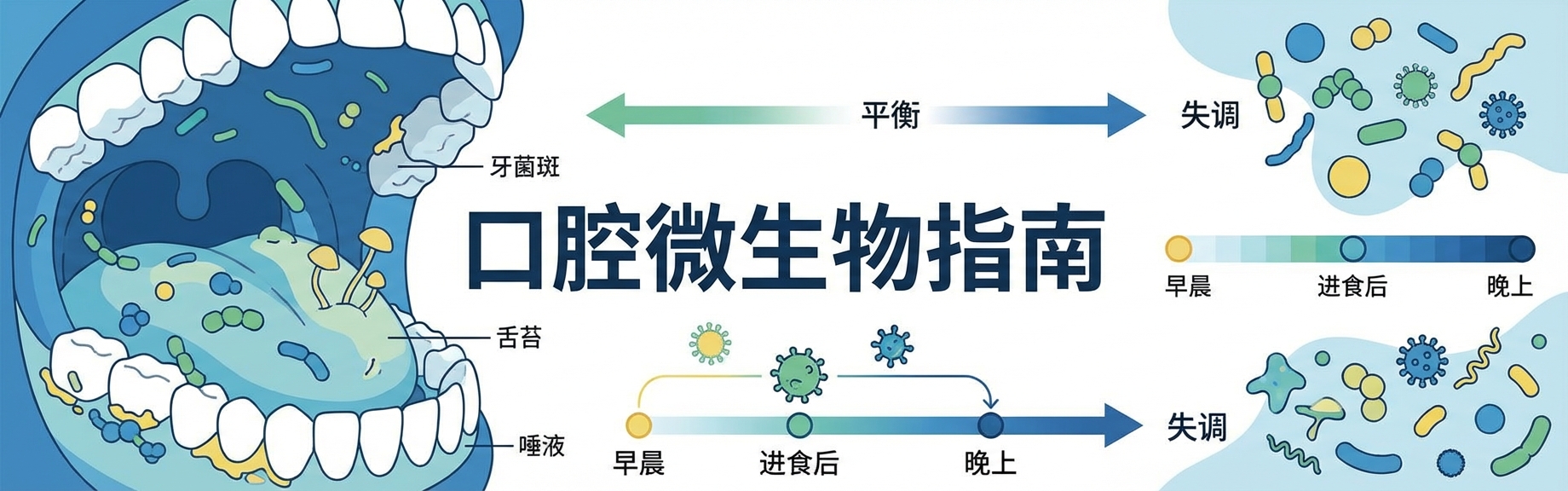
当我们谈论人体微生态时,最先想到的往往是肠道。但你是否知道,我们的口腔其实是人体内多样性仅次于肠道的第二大微生物栖息地?
近年来越来越多研究提示,口腔作为人体与外界环境交互的重要门户,栖息着高度复杂和动态的微生物群落,即口腔微生物组。这一微生态系统在维持宿主口腔乃至全身健康有着比想象中更深的联系。
现代人常见的龋病、口臭、牙周炎等问题,很多时候都不是某一种“坏菌”突然出现,而是原本受控的微生态关系发生倾斜。
想真正理解口腔菌群,关键不只是记住有哪些细菌,而是理解它如何定植、如何维持平衡,又如何在失调时影响口腔与全身。
本文将从口腔微生物组的组成、建立过程、共生稳态与失调机制出发,帮助大家建立一个更清晰的理解框架。
编者按:
本文是一篇基于近年综述与代表性研究整理的科普性综述,重点讨论口腔微生物组的构成、稳态与失调,以及其与部分全身疾病之间的研究联系。
需要特别说明的是,本文涉及癌症、认知衰退与痴呆、类风湿性关节炎、糖尿病等内容时,所引用的许多证据目前仍以观察性研究和相关性研究为主,不能简单理解为直接因果关系。不同主题的证据成熟度并不一致,相关机制也仍在持续研究中。
全文分上下两部分,本文为上篇;下篇将进一步讨论口腔微生物组与系统性疾病的研究进展、潜在机制、干预策略及临床意义。
从生态学视角来看,口腔微生物组是一个由细菌、古菌、真菌、病毒和原生动物等多种微生物组成的复杂生态系统。这些微生物以生物膜的形式存在牙面、舌背、黏膜、龈沟等不同部位,与宿主长期共处,并共同维持口腔环境的稳定。
换句话说,口腔微生物并不是偶然闯入的污染者,而是从出生起就开始定植的长期居民。
超700种菌:谁构成了口腔群落的核心骨架?
据估计,一般健康的人类口腔及上呼吸消化道中可发现超过700种细菌。
注:其中约58%已被正式命名,16%虽可培养但尚未命名,另有26%仅作为“不可培养的系统发育型”被识别。
在这些细菌中,厚壁菌门(Firmicutes)、拟杆菌门(Bacteroidetes)、变形菌门(Proteobacteria)、放线菌门(Actinobacteria)、梭杆菌门(Fusobacteria)和螺旋体门(Spirochaetes)是六个最主要的门类,占据了口腔细菌群落的绝大部分。
口腔微观:不同位置,住着不同的微生物
口腔独特的解剖结构,包括非脱落的硬组织表面(如牙齿、修复体)和持续脱落的软组织表面(如颊黏膜、舌、腭),以及流动的唾液,为不同微生物提供了多样化的生态位。这些生态位在物理化学条件(如pH、温度、氧化还原电位、营养可得性)上存在显著差异,从而赋予了口腔“空间异质性”,并塑造了具有高度位点特异性的微生物群。
例如:
口腔核心微生物组
尽管个体间差异巨大,但在健康人群中,不同口腔位点存在着相对稳定的“核心微生物组”,这些核心成员负责维持口腔的功能稳定性和生态平衡。例如,研究发现,在健康个体的多个口腔位点中,Streptococcus mitis、Gemella haemolysans、Granulicatella adiacens 几乎普遍存在。
除了细菌,口腔中还存在“核心真菌组”,其中念珠菌属(Candida)最为常见,其次是枝孢属(Cladosporium)、短梗霉属(Aureobasidium)等。
口腔病毒组(virome)则以噬菌体为主,它们通过调节细菌宿主的种群动态和基因交换,在塑造口腔微生物群落结构中发挥着关键作用。
古菌(archaeome)虽然丰度较低,但产甲烷菌如Methanobrevibacter oralis在牙周炎等病理状态下丰度会增加,提示其可能参与疾病过程。
综上,口腔菌群是一个多物种、多领域、高度结构化和功能化的微生态系统。它与宿主共同进化,形成了一个被称为“共生体”的超级有机体。理解这个微生态系统的组成、结构及其与宿主的相互作用,是揭示其在口腔及全身健康与疾病中作用的基础。
健康的口腔里,微生物群落是如何运转的?
在健康状态下,口腔微生物组与宿主之间维持着“共生”或“健康平衡”的和谐关系。
在这个状态下,多样化的微生物群落通过协同作用,共同执行多种有益功能。它们参与营养物质的初步消化与转化,例如,某些口腔细菌能够代谢膳食中的硝酸盐,产生具有血管舒张和抗菌作用的亚硝酸盐和一氧化氮,这不仅有助于调节血压,还能抑制致病菌的生长。
此外,共生微生物通过“定植抵抗”机制,占据口腔内的生态位,与外来病原体竞争营养和附着位点,从而构成了抵御感染的第一道生物防线。
口腔微生物组与宿主健康之间的复杂相互作用
Dame-Teixeira et al., 2025, 第1章
一旦打破生态平衡,问题悄然出现
这种微妙的平衡可能被多种因素打破,导致向“失调”状态的转变。
年龄、感染、环境因素、生活习惯(如高糖饮食、吸烟、饮酒)以及抗菌药物的不当使用,都会对口腔微生物组施加选择压力,破坏其多样性和稳定性。
例如,高糖饮食会促进产酸菌(如变形链球菌)的过度生长,导致口腔环境酸化,从而引发龋齿。同样,宿主的全身性疾病,如糖尿病、高血压、肥胖等代谢综合征,也会通过改变唾液成分、影响免疫应答等方式,加剧口腔微生物的失调。
近年来,随着高通量技术的发展,我们对口腔微生物组的认识正在迅速深化。自2010年以来,相关研究文献数量呈指数级增长(下图)。
1950年~2023年间,以“口腔微生物组”
为主题的已发表论文数量变化趋势
编辑
Dame-Teixeira et al., 2025, 第1章
图中显示,自2005年左右,尤其是2010年以后,相关研究呈现指数级增长。
这些研究从单纯的物种鉴定(它们是谁?)发展到功能分析(它们在做什么?),并整合宏基因组学、宏转录组学、宏蛋白质组学和代谢组学等多组学数据,逐步帮我们理解整个口腔微生态奠定了证据基础。
这些进展不仅揭示了口腔微生物组的复杂性,也为开发基于调节微生物组的个体化预防和治疗策略提供了新的契机。
人类口腔微生物组的建立是一个从生命最早期开始,贯穿一生的动态过程。它并非与生俱来,而是在出生后通过与环境的持续互动逐步获得和演替的。这一过程受到一系列内在(宿主相关)和外在(环境相关)因素的复杂调控,共同塑造了每个个体独特而又具有共性的口腔微生态系统。
人类从胎儿到老年的生命历程示意图
编辑
改编自 DOI: 10.1007/978-3-031-79146-8_2
生命早期的初始定植
出生后,喂养方式成为塑造口腔微生物组的另一个重要因素。婴儿吃进去的不仅是营养,也是微生态的“种子”与“养料”。
母乳:不仅是营养的来源,更是一个动态的生物系统,含有丰富的微生物、人乳寡糖(HMOs)、免疫球蛋白和抗菌蛋白。母乳喂养能够将母亲的特定菌株(如双歧杆菌属)传递给婴儿,为其肠道和口腔“播种”有益菌 。
HMOs作为益生元,能够选择性地促进有益菌(尤其是Bifidobacterium infantis)的生长,同时抑制病原体的黏附 。因此,与配方奶喂养的婴儿相比,母乳喂养婴儿的口腔微生物组通常具有更高的多样性,其中部分变化被认为更接近健康相关模式。
辅食:随着固体食物的引入,婴儿的口腔微生物组会经历又一次重大转变,多样性显著增加,以适应更复杂的碳水化合物和营养物质。
从婴儿期到成年的演替过程
随着个体的成长,口腔的物理环境也在不断变化,这为微生物的演替提供了新的生态位。其中,牙齿的萌出是一个重要阶段。
► 无牙期:以软组织表面为主
在无牙期,口腔内只有软组织黏膜表面,主要定植的是早期定植菌,例如:
► 约 6–10 个月:第一颗乳牙萌出
大约在6-10个月时,第一颗乳牙的萌出为微生物提供了独特的“非脱落表面”,使得能够形成更稳定、更复杂的生物膜(牙菌斑)。
随着乳牙列的完全建立,口腔微生物的多样性显著增加,例如下列菌群开始繁盛:
编辑
► 混合牙列期与恒牙期:厌氧菌定植条件增强
进入混合牙列期和恒牙期,口腔的解剖结构变得更加复杂,为更多厌氧菌的定植创造了条件。
牙龈沟的形成提供了一个低氧、富含蛋白质(龈沟液)的微环境,特别适合严格厌氧和蛋白水解菌(如牙周病相关菌群)的生长。
在这一过程中,微生物群落的组成从以革兰氏阳性兼性菌为主,逐渐向包含更多革兰氏阴性厌氧菌的成熟群落演替。
► 青春期:激素变化驱动的第二次转变
青春期是另一个重要的转变阶段。性激素(如雌二醇、孕酮和睾酮)水平的急剧上升,会直接或间接地影响口腔微环境。
这些激素可以被某些口腔细菌利用作为生长因子,同时也会改变牙龈组织的血管通透性和炎症反应,导致所谓的“青春期牙龈炎” 。
这期间,龈下微生物组中与牙龈炎相关的菌群丰度会增加,例如:
成人口腔内不同生态位及其优势细菌类群
编辑
DOI: 10.1007/978-3-031-79146-8_2
牙菌斑和舌部具有高微生物多样性,而口腔黏膜的多样性相对较低。
影响微生物组获得与建立的其他因素
除了上述生理阶段的变化,还有多种因素持续塑造着口腔微生物组的构成。
✦遗传与宿主防御:身体“筛选”哪些菌容易留下
宿主的遗传背景决定了其免疫系统的特征,如唾液中抗菌蛋白(溶菌酶、乳铁蛋白等)的含量和活性、主要组织相容性复合体(MHC/HLA)的类型等。这会影响哪些微生物更容易定植下来,并长期保留下来。
例如,唾液淀粉酶基因(AMY1)拷贝数较高的人,其唾液中普雷沃菌属和卟啉单胞菌属的水平也更高。
✦社会与环境接触:微生物在人与人/动物之间流动
口腔微生物会通过接触持续传播。
婴儿早期主要受母亲影响,随着成长,兄弟姐妹、照料者、宠物和集体环境的作用也会逐渐增强。
编辑
✦药物使用:抗生素改变菌群起跑线
抗生素的使用,尤其是在生命早期,会对口腔微生物组的建立产生深远影响。
广谱抗生素会无差别地杀灭有益菌和有害菌(好坏一起杀),可能导致菌群失调,并为机会致病菌(如念珠菌)的过度生长创造条件。
✦口腔卫生与生活方式:直接改变牙菌斑与口腔环境
口腔卫生习惯(如刷牙、使用牙线)直接影响牙菌斑的量和组成。
不良的口腔卫生会导致菌斑堆积,促进厌氧菌和致病菌的生长。吸烟和饮酒等生活方式因素也会通过改变口腔的氧化还原电位、pH值和免疫反应,导致微生物组失调,增加牙周病和癌症的风险。
✦成年稳定与老年再变化:从平衡到更易失衡
进入成年期后,健康口腔微生物组通常进入相对稳定阶段。
然而,随着年龄的增长,生理性衰老(如唾液流速下降、免疫功能减退)以及慢性疾病和多重用药的累积效应,会再次改变这种平衡,可能导致机会性病原体(如肠杆菌科、葡萄球菌属)的丰度增加,从而增加老年人患口腔及全身性疾病的风险。
总的来说,口腔微生物组并非一成不变,而是在多种因素作用下不断调整。接下来,需要进一步理解它与宿主之间如何形成并维持这种共生平衡。
经过数千年的共同进化,人类与栖居其身的微生物群落形成了一种深刻的互利共生关系。种共生关系主要通过微生物稳态和宿主稳定性的相互支撑来实现。
微生物稳态:群落内部的协作与拮抗
在健康状态下,口腔微生物并不是各自为战,而是形成了一个相互作用网络,这个网络让菌群具备两种能力:
这种稳态主要靠两类关系共同维持:互利协作与相互拮抗。
↗互利关系:代谢协作与交叉喂养
口腔微生物的主要营养来源于宿主饮食、唾液和龈沟液中的大分子,如糖蛋白和蛋白质。
单个微生物菌种往往“吃不完整”,不具备完全降解这些复杂底物的能力,因此需要不同物种之间通过酶促互补进行代谢合作。这种现象被称为“交叉喂养”,即一个物种的代谢产物成为另一个物种的营养底物。
——典型例子:产酸菌 × 耗酸菌
编辑
这个协作带来两点好处:
整体上,这类代谢网络会提升群落效率,增强了其稳定性。
↘拮抗关系:竞争与抑制
口腔生态位很拥挤,共生菌为了守住地盘,也会用各种方式压制竞争者,尤其是潜在致病菌。这些拮抗作用是维持微生物平衡的关键。
——产生细菌素:定向“抑菌武器”
许多共生菌能够产生小分子的抗菌肽,即细菌素,它们可以特异性地杀灭或抑制亲缘关系较近的细菌。
例如,唾液链球菌(Streptococcus salivarius)产生的Salivaricin(一种细菌素)可以抑制引起咽喉炎的化脓性链球菌(S. pyogenes)。
从无龋个体中分离出的Streptococcus dentisani也能产生细菌素,有效抑制变形链球菌(S. mutans)和具核梭杆菌(F. nucleatum)等致病菌的生长。
——产生过氧化氢:用“氧化压力”压制对手
多种共生链球菌(如S. sanguinis, S. gordonii, S. oralis)能够通过丙酮酸氧化酶产生过氧化氢。
过氧化氢(H₂O₂)对许多无法有效应对氧化应激的厌氧致病菌(如P. gingivalis)和产酸菌(如S. mutans)具有强大的抗菌活性。这种机制在早期菌斑形成和应对糖冲击时尤为重要。
——产碱缓冲 pH:把酸中和掉
除了交叉喂养,一些细菌还能通过代谢氨基酸(如精氨酸)和尿素来产生氨(NH₃),从而中和酸性产物,提高局部pH值。
例如,精氨酸脱亚胺酶系统(ADS)在多种链球菌和放线菌中存在,它降解精氨酸产生氨,不仅能缓冲pH,其代谢过程本身还会消耗精氨酸,从而抑制依赖精氨酸的S. mutans的毒力表达。
——群体感应信号的干扰
微生物通过释放和感知被称为“自诱导物”的信号分子来进行种内和种间的通讯,这一过程称为群体感应(QS)。QS系统调控着生物膜形成、毒力因子产生等多种群体行为。
一些共生菌能够干扰致病菌的QS信号。例如,部分S. gordonii菌株可以抑制S. mutans的CSP信号肽,从而削弱其形成生物膜和产生细菌素的能力。
口腔健康条件下微生物间的相互作用机制
编辑
doi: 10.1007/978-3-031-79146-8_3
维持口腔稳态的关键:定植抵抗与免疫稳态
宿主与口腔微生物组之间的共生关系,不仅体现在微生物群落内部的自我调节,更体现在其与宿主免疫系统的协同作用上,共同维持宿主的稳定与健康。
◉ 定植抵抗:常驻菌先占位,外来菌难落脚
健康的常驻微生物群通过“占位”效应,物理性地占据了口腔黏膜和牙齿表面的附着位点,从而阻止了外来病原体的入侵和定植。这种现象被称为“定植抵抗”。
它主要通过两种方式实现:
这种由微生物组提供的天然屏障,是宿主抵御感染的第一道防线。
◉ 免疫稳态与耐受:不是不管,而是受控共存
口腔黏膜持续暴露于大量的微生物及其抗原之下,但通常不会引发剧烈的炎症反应,这表明宿主免疫系统已经进化出一种与共生微生物和平共处的机制,即免疫耐受。
这种耐受并非免疫系统的“无视”,而是一种主动的、精细调控的平衡状态:既控制微生物,又避免过度炎症伤害自己。
共生菌如何帮助维持这种平衡?
共生微生物的某些分子信号(MAMPs,可理解为“微生物特征信号”)会被宿主的模式识别受体识别(如Toll样受体,TLRs),持续地“教育”和“校准”宿主的免疫系统。在健康状态下,这种相互作用倾向于诱导一种“促耐受”的免疫环境,主要体现在:
——诱导调节性T细胞(Treg)
共生菌及其代谢产物(如短链脂肪酸)可以促进Treg细胞的分化。
Treg细胞通过分泌如IL-10、TGF-β等抗炎细胞因子,来抑制过度的炎症反应,防止对共生菌产生有害的免疫攻击。
——促进分泌型IgA(sIgA)产生
sIgA是黏膜免疫的主要抗体。它能够“包裹”微生物,阻止其黏附到上皮细胞表面并侵入组织,同时不激活强烈的炎症反应。
这种机制常被理解为温和的免疫排斥:既能控制微生物数量,又能避免组织损伤。
——强化上皮屏障
共生菌能够刺激上皮细胞产生黏蛋白和抗菌肽(AMPs),增强黏膜的物理和化学屏障功能,进一步限制微生物与宿主免疫细胞的直接接触。
口腔微生物群与宿主免疫系统的相互作用机制
编辑
doi: 10.1007/978-3-031-79146-8_3
口腔微生态具备定植抵抗、免疫耐受和屏障防御等多重稳定机制,但并不是建立起来就能一直维持下去,真正日复一日改写这套系统的,往往是那些看似普通的生活因素——尤其是饮食。下一章我们来具体了解。
人类饮食变迁与口腔微生物组的共同进化
🪨 旧石器时代
高纤维、低精制糖
→ 菌群多样性更高,龋病较少
🌾 新石器时代
谷物进入主食,碳水摄入增加
→ 产酸相关菌增多,龋病开始上升
🏭 工业革命至今
精制糖和加工食品大量增加
→ 产酸、耐酸菌占优势,微生态更易失衡
人类历史进程中饮食结构的变化与口腔微生物组的演变
编辑
doi: 10.1007/978-3-031-79146-8_4
宏量营养素与微量营养素调控口腔微生态
食物中的宏量营养素(碳水化合物、蛋白质、脂肪)和微量营养素(维生素、矿物质)通过多种途径影响口腔微生物组和宿主反应。
◯ 碳水化合物:一把双刃剑
碳水化合物是口腔微生物最主要的能量来源,但其类型和加工程度决定了其对口腔健康的影响。
——可发酵碳水(尤其精制糖/蔗糖):更易推动失调
可发酵碳水化合物,特别是蔗糖等精制糖,能被口腔中的糖酵解细菌(如变形链球菌、乳杆菌)迅速代谢,产生大量有机酸(主要是乳酸),导致牙菌斑pH值急剧下降。
频繁的酸攻击会破坏牙釉质的矿物质平衡,引发脱矿,最终形成龋洞。
——蔗糖:不仅产酸,还让菌斑黏更厚难清除
变形链球菌还能利用蔗糖合成不溶性的胞外多糖(EPS),这些黏性的葡聚糖构成了生物膜的骨架,不仅促进了细菌的黏附和聚集,还形成了一个扩散屏障,使酸性产物更难被唾液中和,从而加剧了局部的酸性微环境。
——复杂碳水/膳食纤维:整体更友好
复杂碳水化合物,如全谷物、蔬菜和豆类中富含的膳食纤维,对口腔健康则更为有益。这些纤维不易被口腔细菌直接发酵,因此不会引起pH的剧烈下降。
更重要的是,富含纤维的食物通常需要更多的咀嚼,这能刺激唾液分泌,增强唾液的冲刷和缓冲作用,有助于清除食物残渣和中和酸性物质。
编辑
◯ 脂肪和蛋白质:来源与平衡是关键
脂肪和蛋白质对口腔微生物组的影响相对复杂,且与其来源密切相关。
——脂肪:不同类型,炎症倾向不同
饱和脂肪和反式脂肪,常见于加工食品、动物脂肪和部分植物油中,被认为是促炎性的。
不饱和脂肪酸,特别是来自鱼油、坚果和橄榄油的ω-3多不饱和脂肪酸,具有抗炎特性。
——蛋白质:既是宿主营养成分,也可能被某些致病菌利用
蛋白质是构成唾液和龈沟液的重要成分,也是许多蛋白水解型牙周致病菌(如P. gingivalis)的主要营养来源。
在牙周炎状态下,富含蛋白质的龈沟液为这些病原体的生长提供了有利条件。
然而,饮食中的蛋白质来源同样重要。有研究指出,过量摄入动物蛋白可能与更高的炎症水平相关,而植物蛋白则可能更有益于健康。
编辑
doi: 10.1007/978-3-031-79146-8_4
◯ 微量营养素:免疫与修复的调节剂
维生素和矿物质等微量营养素虽然需求量小,但在维持口腔组织完整性和调节免疫功能方面发挥着不可或缺的作用。它们的缺乏会直接损害口腔健康。
——维生素C(抗坏血酸)
是胶原蛋白合成所必需的,对维持牙龈和牙周膜的结构完整性至关重要。
严重缺乏维生素C会导致坏血病,其典型症状之一就是牙龈出血和牙齿松动。
维生素C还是一种重要的抗氧化剂,有助于中和炎症过程中产生的活性氧(ROS),保护牙周组织免受氧化损伤。
——维生素D
在钙磷代谢和骨骼健康中起着核心作用。它能促进牙槽骨的矿化,并具有重要的免疫调节功能,可以抑制过度的炎症反应。
维生素D缺乏与牙周炎的严重程度增加和骨质疏松相关。
——B族维生素
参与多种细胞代谢过程,其缺乏可导致多种口腔黏膜疾病,如舌炎、口角炎和复发性阿弗他溃疡。
——抗氧化剂
抗氧化剂(维生素A、E、类胡萝卜素、多酚等)广泛存在于水果、蔬菜、茶和红酒中。这些物质能够清除自由基,减轻氧化应激,从而保护细胞免受损伤,对预防牙周炎和口腔癌具有积极意义。
综上所述,饮食不仅是口腔疾病(尤其是龋病和牙周炎)发生的重要风险因素,也是维持口腔微生态平衡和宿主健康的关键调控手段。一个均衡的、富含全食品、纤维、维生素和抗氧化剂,同时限制精制糖和饱和脂肪的饮食模式,是促进口腔共生、预防疾病的基石。
胃肠道是人体内最庞大、最复杂的微生物生态系统,从口腔延伸至肛门。传统上,口腔和肠道被视为两个相对独立的微生物栖息地,由胃酸、胆汁和物理距离等屏障隔开。然而,近年的研究揭示了两者之间存在着密切而动态的联系,这一联系被称为“口腔–肠道轴”(Oral-Gut Axis)。
通过该轴,口腔微生物及其代谢产物能够影响肠道微生态和宿主健康,反之亦然。理解口腔–肠道轴的机制,有助于我们从更整体的角度看待多种慢性炎症性疾病、代谢性疾病乃至癌症的相关研究线索。
口腔与肠道微生物组的生物地理学特征
——口腔和肠道的菌群,并不一样
在健康状态下,胃肠道沿线的微生物群落表现出显著的体位特异性。每一段消化道的环境都不一样,比如:
这些差异决定了什么样的细菌适合住在哪里。
比如,胃里的环境酸性很强,细菌数量相对较少;而到了结肠,细菌数量就会大幅增加,种类也更加丰富。
胃:约10³-10⁴/mL 结肠:10¹¹-10¹²/g
人体消化道不同部位的微生物群落分布示意图
编辑
doi: 10.1007/978-3-031-79146-8_5
——但它们之间并不是完全隔绝的
尽管存在显著的体位特异性,但在健康个体中,口腔和肠道之间仍存在一定程度的微生物重叠。研究发现,一些常见的口腔细菌,如:
也可能在肠道中被检测到,尽管丰度通常很低。这表明,口腔微生物向肠道的“转位”是一个生理现象。
问题不在于会不会进入,而在于它们进入后能不能存活、定植和扩张。
口腔微生物向肠道的转位机制
——口腔微生物主要通过两条途径到达肠道
编辑
——为什么健康人通常没事?
虽然口腔细菌确实可能到达肠道,但健康人的肠道并不会轻易让它们安家。
在健康状态下,肠道自身的防御机制——包括完整的黏膜屏障、强大的免疫系统(肠道相关淋巴组织,GALT)以及由常驻肠道菌群构成的“定植抵抗”
这些机制会共同作用,限制外来细菌在肠道中长期生存。因此,口腔菌在健康肠道中通常只是匆匆过客。
口腔–肠道轴失调与疾病
当宿主健康状况发生改变时,口腔–肠道轴的平衡可能被打破,导致口腔细菌在肠道中的“异位定植”,这与多种疾病的发生发展密切相关。
✦ 肠道疾病:口腔细菌可能火上浇油
在炎症性肠病(IBD,包括克罗恩病和溃疡性结肠炎)患者中,肠道黏膜屏障功能受损,通透性增加。
研究发现,IBD患者的肠道中,常常能检测到更多口腔来源的细菌,例如:
具核梭杆菌不仅在口腔中发挥作用,还可能通过口腔–肠道轴影响全身健康。
编辑
<来源:谷禾口腔菌群检测数据库>
动物实验也提供了进一步证据。
如果把患有牙周炎小鼠的口腔菌群,移植给无菌小鼠,后者会出现更明显的肠道炎症,尤其是与Th17细胞相关的免疫反应增强。而这种反应,正是IBD的重要发病机制之一。
提到口腔细菌与疾病的关系,具核梭杆菌(F. nucleatum)是一个非常典型的例子。
研究发现,具核梭杆菌在结直肠癌(CRC)组织中高度富集,并通过其黏附素FadA与肿瘤细胞的E-cadherin结合,进一步激活Wnt/β-catenin信号通路,促进肿瘤增殖。
简单说,就是它可能帮助肿瘤细胞长得更快、扩增得更多。这也说明,某些原本生活在口腔里的细菌,一旦进入异常环境,可能不只是旁观者。
✦ 肝脏疾病:口腔细菌也可能影响肝脏
在肝硬化和非酒精性脂肪性肝病(NAFLD)患者中,也观察到肠道中口腔细菌的富集。
这可能与肝功能受损导致胆汁酸代谢异常和肠道屏障功能下降有关。口腔细菌及其产物(如LPS)通过门静脉系统进入肝脏,可能加剧肝脏的炎症和纤维化进程。
✦ 系统性炎症性疾病:可能牵动全身免疫
在类风湿关节炎(RA)等自身免疫病中,口腔–肠道轴也可能扮演了重要角色。
牙周炎是类风湿关节炎的已知风险因素。口腔病原体(如P. gingivalis)不仅能在口腔局部诱导产生针对瓜氨酸化蛋白的抗体(ACPA),还可能转位至肠道,进一步扰乱肠道免疫系统,加剧全身性自身免疫反应。
口腔中的慢性炎症,可能通过肠道这个“中转站”,放大成全身性问题。
✦ 口腔+肠道菌群:可能提示病情轻重
研究通过对唾液和粪便样本的微生物组成进行分析将牙周炎患者分为不同的口腔-肠道谱型。
牙周炎患者的口腔-肠道微生物谱聚类分析
编辑
doi: 10.1007/978-3-031-79146-8_5
某些谱型与更严重的临床症状(如高比例深牙周袋、牙齿缺失)相关,提示口腔与肠道微生物的特定组合可能预示着疾病的严重程度。
临床意义与展望
口腔–肠道轴的发现,为我们从一个更宏观、更系统的角度理解健康与疾病提供了新的框架。它强调了口腔健康对于维持肠道微生态平衡和预防相关疾病的重要性。
临床上,牙周炎患者的肠道微生物组中,与牙周病相关的口腔类群数量明显更高,且某些口腔菌(如F. nucleatum)在粪便中的丰度与牙周病的严重程度相关。
这提示,粪便中的口腔微生物特征可能成为评估口腔健康状况和预测相关系统性疾病风险的非侵入性生物标志物。
近年来,口腔微生物组的一个特定代谢功能受到了科学界的广泛关注:
将膳食中的硝酸盐(NO₃⁻)
还原为亚硝酸盐(NO₂⁻)
这一过程不仅是口腔微生态系统内部复杂代谢网络的一部分,更通过一个被称为“肠-唾液循环”的途径,对人体的口腔及全身健康产生深远的有益影响。
口腔细菌不仅是潜在的致病者,更是维持宿主生理稳态的重要合作伙伴。
硝酸盐的来源、肠–唾液循环
硝酸盐是自然界中广泛存在的含氮化合物,我们日常饮食中的绿叶蔬菜,如菠菜、生菜、芝麻菜、甜菜根,是其最主要的来源。
当我们摄入这些富含硝酸盐的食物后,约25%的硝酸盐会被肠道吸收进入血液循环。随后,一个有趣的循环开始了:血液中的硝酸盐被唾液腺主动摄取并浓缩,其浓度可比血浆高出10倍以上,然后随唾液分泌回口腔。
编辑
在口腔中,这些来自饮食和唾液的硝酸盐,被特定的口腔细菌利用。这些细菌拥有效率极高的硝酸盐还原酶,能将硝酸盐(NO₃⁻)还原为亚硝酸盐(NO₂⁻)。随后,含有亚硝酸盐的唾液被吞咽,进入胃肠道。在胃的酸性环境中,亚硝酸盐可以被非酶促地转化为一氧化氮(NO)和其他活性氮物种(RNS)。
部分未被转化的亚硝酸盐被小肠吸收进入血液,成为全身NO的循环储备库。最终,血液中大部分的NO和亚硝酸盐会被重新氧化为硝酸盐,再次进入肠-唾液循环。这一完整的通路确保了口腔和全身持续获得由微生物介导产生的亚硝酸盐和NO。
硝酸盐-亚硝酸盐-一氧化氮通路在口腔和全身健康中的作用示意图
编辑
Washio and Takahashi, 2025
这个关键步骤,人体自己做不了→依赖菌群
这里有一个很重要的点:
哺乳动物细胞自身并不擅长把硝酸盐变成亚硝酸盐,因此,这一关键步骤完全依赖于口腔中的微生物。
研究表明,口腔中的亚硝酸盐产生能力存在显著的个体间差异,这主要取决于口腔微生物组中“亚硝酸盐产生菌”的数量和活性。
口腔中的亚硝酸盐产生菌
通过琼脂覆盖培养法等技术,研究人员从唾液、牙菌斑和舌苔中分离并鉴定出多种具有硝酸盐还原能力的细菌。这些细菌大多是口腔中的常见共生菌,而非罕见物种,这表明亚硝酸盐的产生是健康口腔微生态的一项普遍功能。主要的亚硝酸盐产生菌包括:
例如奈瑟菌属,是口腔微生物组中的一种重要细菌,在硝酸盐还原过程中发挥关键作用。通过谷禾口腔菌群检测报告,可以了解到其具体丰度及其在口腔微生物组中的表现,部分奈瑟菌属如下:
编辑
<来源:谷禾口腔菌群检测数据库>
——舌苔后部是“核心工作区”
这些菌主要定植在舌背后部的低氧区域,这里为硝酸盐还原酶的活性提供了理想环境。研究发现,舌苔的亚硝酸盐产生活性通常高于牙菌斑,凸显了舌在这一通路中的核心地位。
编辑
持续摄入富含硝酸盐的饮食,不仅能提供底物,还能作为一种益生元,选择性地增加口腔中硝酸盐还原菌(如奈瑟菌属和罗氏菌属)的丰度,从而正向增强整个系统的亚硝酸盐生成能力。
亚硝酸盐对口腔健康有什么好处?
口腔微生物产生的亚硝酸盐及其下游产物一氧化氮,对维持口腔健康具有多重益处,尤其是在预防两大主要口腔疾病——龋齿和牙周炎方面。
◉ 预防龋齿
亚硝酸盐的防龋机制是多方面的:
——直接抗菌作用
亚硝酸盐在酸性条件下可质子化形成亚硝酸(HNO₂),后者能穿透细菌细胞膜,干扰其能量代谢和关键酶的功能。这对主要的致龋菌——变形链球菌(S. mutans)具有显著的抑制作用。
——抑制产酸
糖吃进去后,细菌产酸的能力可能会被部分压住。
龋齿的一个关键过程,就是细菌分解糖分后产酸,使口腔环境变酸,进而腐蚀牙齿。亚硝酸盐能够抑制牙菌斑微生物的糖酵解过程,从而减少有机酸的产生,减缓菌斑pH值的下降。研究表明,摄入富含硝酸盐的蔬菜汁可以显著缓冲食糖后的口腔酸性环境。
龋齿的发生与口腔内多种细菌的活动密切相关,通过监测口腔内的龋齿相关菌群丰度及是否超标,可以更好地掌握口腔微生物组的动态变化。
编辑
<来源:谷禾口腔菌群检测数据库>
——帮助菌群恢复平衡
亚硝酸盐对不同细菌的敏感性不同,它能抑制产酸/耐酸菌的生长,同时可能对硝酸盐还原菌影响较小或无影响。这种选择性压力有助于将失调的、致龋的微生物组向更健康的、共生的方向逆转。
——与乳酸代谢的生态协同
韦荣菌属等细菌在利用乳酸的同时进行硝酸盐还原。这一过程既消耗了致龋的乳酸,又产生了具有保护作用的亚硝酸盐,形成了一个精妙的负反馈调节环路,有助于维持口腔pH稳态。
◉ 预防牙周炎
亚硝酸盐和一氧化氮(NO)对牙周健康同样有益。
首先,它们对多种主要的牙周致病菌,如牙龈卟啉单胞菌(P. gingivalis)、中间普雷沃菌(P. intermedia)、具核梭杆菌(F. nucleatum),也具有抗菌活性。
其次,NO作为一种重要的信号分子,具有强大的血管舒张和抗炎作用。通过改善牙龈组织的微循环血流,NO有助于输送营养和免疫细胞,清除代谢废物,从而促进牙龈炎症的消退和组织修复。临床研究已表明,摄入富含硝酸盐的莴苣汁可以减轻慢性牙龈炎患者的牙龈炎症症状。
亚硝酸盐的影响,不止停留在口腔
通过肠-唾液循环,口腔微生物组的硝酸盐代谢功能超越了口腔局部,对全身健康,特别是心血管系统,产生了系统性的积极影响。
最核心的作用是调节血压
由口腔产生的亚硝酸盐进入循环系统后,在缺氧组织中被有效还原为NO。NO是内皮细胞产生的一种关键的血管舒张因子,能够松弛血管平滑肌,降低外周血管阻力,从而降低血压。
大量临床试验证实,补充膳食硝酸盐(如饮用甜菜根汁)能够显著降低高血压患者的血压。
编辑
一个很关键的证据:抗菌漱口水打断这条通路
重要的是,当使用抗菌漱口水清除口腔中的硝酸盐还原菌后,这种降压效应会明显减弱甚至消失,这证明了口腔微生物组在这一过程中的核心地位。因此,一个功能活跃的口腔硝酸盐还原菌群,可以作为内源性NO生成途径(依赖内皮NO合酶)的一个重要补充,尤其是在衰老或疾病状态下内皮功能受损时。
也就是说,口腔微生物组的功能状态,可能真的会影响全身血压调节。
其他潜在好处也在被关注
NO还具有改善运动表现、保护胃黏膜、增强认知功能等多种潜在益处。
例如,补充硝酸盐可以降低运动时的耗氧量,提高肌肉收缩效率,从而提升耐力运动表现。
那硝酸盐会不会有风险?
很多人会担心一个问题:硝酸盐和亚硝酸盐会不会形成致癌物,比如亚硝胺?
这个担忧曾经非常常见。
然而,目前的主流科学观点认为,对于通过蔬菜摄入的硝酸盐,这种风险极低。因为蔬菜中富含维生素C和多酚等抗氧化剂,能够有效抑制亚硝胺的形成。因此,从健康角度看,摄入富含硝酸盐的蔬菜所带来的益处远大于其潜在风险。
总之,口腔微生物组的硝酸盐代谢功能是宿主-微生物共生关系的一个典范。它不仅通过局部抗菌和pH调节作用维护口腔健康,还通过肠-唾液循环系统性地贡献于心血管健康。
这也带来了一个很有意义的启发:
将硝酸盐视为一种重要的益生元,将硝酸盐还原菌视为潜在的益生菌,为通过饮食和微生态干预来预防和管理慢性疾病提供了新的思路。
唾液是口腔环境的命脉,它不仅仅是水的简单混合物,而是一种复杂的生物流体,富含多种有机和无机成分。
在维持口腔微生物组与宿主之间的共生关系中,唾液扮演着不可或缺的守护者角色。它通过机械清洁、免疫防御、缓冲调节和组织保护等多重机制,主动塑造和调控着口腔微生态的平衡,是抵御口腔及全身感染的第一道关键防线。
唾液的物理与机械功能
冲洗与清洁
我们每天都在不断分泌和吞咽唾液,伴随着吞咽动作,唾液能够有效地将食物残渣、脱落的上皮细胞以及未牢固附着的微生物从口腔中清除,这一过程被称为“唾液清除”。
据估计,一个健康成年人每天仅通过吞咽唾液,
就能清除掉大约 800亿(8×10¹⁰)个细菌!
唾液流速是关键
高流速的唾液能更快地稀释和冲走食糖后产生的酸,以及减少细菌在牙面的停留时间,从而降低龋病的风险。相反,唾液分泌减少(口干症)的患者,其口腔细菌载量通常更高,患龋齿、牙周病和口腔念珠菌病的风险也显著增加。
润滑口腔,帮助吞咽和说话
除了冲洗功能,唾液还是口腔的重要润滑剂。
唾液中的黏蛋白等糖蛋白赋予了唾液黏弹性,使其能够在口腔软硬组织表面形成一层润滑膜。这层膜有几个非常实际的作用:
唾液作为口腔“守护者”的多重功能
编辑
DOI: 10.1007/978-3-031-79146-8_7
唾液的化学防御:不让口腔变酸
缓冲pH调节
细菌导致蛀牙的一个核心武器是:产酸。而唾液的一个重要任务,就是把口腔的酸碱度(pH值)维持在中性范围(大约 6.5 – 7.5 之间)。
唾液里自带了一套碳酸氢盐缓冲系统。当我们吃了酸性食物,或者细菌开始大量产酸时,这套系统就会迅速出动,把酸中和掉。
有意思的是,当你咀嚼食物时,唾液流速会加快,里面的碳酸氢盐浓度也会跟着升高,中和酸的能力就变得更强。
此外,一些口腔细菌也能通过分解唾液中的尿素产生氨,辅助提高pH值,共同维持口腔环境的稳定。
唾液和口腔微生物之间并不是简单的“防守与进攻”关系,很多时候更像是一种动态协作。
免疫防御:非特异性与特异性免疫
唾液并不是机械地“冲走细菌”,它本身还含有大量具有抗菌和免疫作用的分子,它们协同作用,构成一个强大的防御网络。
◖溶菌酶:直接破坏细菌细胞壁
溶菌酶能够水解革兰氏阳性菌细胞壁的肽聚糖,导致细菌裂解死亡。它还具有非酶依赖的抗革兰氏阴性菌活性,并能促进细菌聚集,便于清除。
◖乳铁蛋白:不给细菌“吃铁”
乳铁蛋白是一种铁结合蛋白,通过螯合游离的铁离子,剥夺了细菌生长所必需的铁元素,从而发挥抑菌作用(营养免疫)。此外,它还具有不依赖铁的直接杀菌和抗病毒活性。
◖乳过氧化物酶系统:抑制细菌产酸和代谢
该系统利用细菌代谢产生的过氧化氢(H₂O₂)和唾液中的硫氰酸盐(SCN⁻)作为底物,生成具有强氧化性的次硫氰酸(OSCN⁻),后者能抑制多种细菌的糖酵解酶,从而阻断其产酸和生长。
简单来说,就是让细菌“没法高效产酸、没法顺利生长”。
◖组蛋白、抗菌肽:广谱抗菌抗真菌
组蛋白是一类富含组氨酸的阳离子肽,具有广谱的抗真菌(特别是抗白色念珠菌)和抗菌活性。它们通过破坏微生物细胞膜的完整性或干扰其内部代谢来发挥作用。
防御素和 Cathelicidin (LL-37) 这些是小分子的阳离子抗菌肽,主要由上皮细胞和中性粒细胞分泌。它们能像分子飞镖一样插入微生物的细胞膜,形成孔道,导致细胞内容物泄漏而死亡。
◖sIgA:温和但精准的免疫控制者
分泌型免疫球蛋白A (sIgA)是唾液中最主要的抗体,也是黏膜适应性免疫的核心。
它的特别之处在于:不是通过强烈炎症来“消灭一切”,而是通过更温和的方式控制微生物。
sIgA能够特异性地结合入侵的微生物和毒素,通过免疫排斥机制阻止它们黏附到宿主细胞表面,并促进它们以聚集体的形式被唾液清除。sIgA在不引发强烈炎症反应的情况下控制微生物,是维持黏膜稳态的关键。
唾液中的非特异性与特异性免疫防御
编辑
DOI: 10.1007/978-3-031-79146-8_7
唾液会给口腔表面加上一层保护膜
唾液中的蛋白质和糖蛋白(如黏蛋白、富含脯氨酸蛋白、软蛋白等)会选择性地吸附在牙齿和口腔黏膜表面,形成一层厚度仅为几百纳米的无细胞有机薄膜,分别称为获得性釉质膜和黏膜膜。
这层唾液膜具有双重功能:
它作为一道物理屏障,保护牙釉质免受酸的直接侵蚀,减缓脱矿过程。同时,膜中的钙结合蛋白(如软蛋白)有助于将钙和磷酸盐离子富集在牙齿表面,促进再矿化(相当于帮助牙齿进行微小损伤的自我修复)。对于软组织,黏膜膜则提供了润滑和保湿,防止干燥和机械损伤。
这层膜还有一个非常有意思的生态学功能:
它会影响微生物最初如何在口腔表面定植。
不同的唾液蛋白会选择性地结合不同的细菌,相当于为特定微生物提供落脚点。这意味着,唾液膜并不是单纯被动地让细菌附着,而是在一定程度上选择性地决定谁先成为第一批定植者。
例如,α-淀粉酶可以结合某些链球菌,而富含脯氨酸蛋白则为另一些细菌提供受体。
而最早附着的那批细菌,往往会影响后续整个牙菌斑生物膜的形成顺序和组成。
总的来说,唾液是维持口腔微生态平衡的重要保障。可一旦唾液分泌减少或功能受损,原本稳定的菌群关系就可能被打破,口腔也会由健康走向失衡。接下来,我们就看看口腔微生态失调是如何发生的。
在健康状态下,口腔微生物组与宿主维持着一种动态的共生平衡。然而,当这种平衡被宿主、环境或微生物自身的因素打破时,就会导致失调。
失调并非简单地指有害细菌的增加,而是一个更复杂的生态学概念,指微生物群落的组成、结构和/或功能发生有害的改变,从而增加了宿主患病的风险。
口腔微生态失调是龋病、牙周炎等主要口腔疾病的核心驱动力,并与多种系统性疾病密切相关。
失调的生态学假说
理解口腔失调的关键在于“生态学假说”(Ecological Plaque Hypothesis)。该假说由Philip Marsh提出,这个假说认为,口腔疾病并不是单纯由菌太多引起,也不只是由某一种坏菌单独造成,而是局部环境发生改变后,整个微生物群发生了致病的重组。
简单来说,它强调三件事:
1
疾病的本质是生态失衡
在健康的口腔里,一些潜在致病菌其实本来就可能存在,只是数量很低,而且被共生菌群和宿主防御系统共同压制着。
问题不在于它们有没有,而在于潜在致病菌会不会在合适的环境下突然占上风。
2
环境变化是失调的起点
当口腔局部环境出现明显变化时,比如:
原有的微生物平衡就会被打破。某些原本不占优势的微生物,会因此获得更适合自己的生存条件。
1
菌群演替会推动疾病不断加重
一旦新的环境开始偏向某些致病相关微生物,它们就会逐渐增殖,并进一步改变周围环境。
这种改变又会继续压制共生菌、扶持致病菌,形成一个自我强化的恶性循环,最终推动疾病发生和进展。
龋病和牙周炎,就是两个典型例子。
——在龋病中:高糖让口腔变酸
如果长期频繁摄入糖分,口腔中的细菌就会不断把糖发酵成酸。于是,口腔环境反复酸化。
这种酸性环境抑制了对酸敏感的共生菌(如S. sanguinis),同时促进了耐酸和产酸菌(如变形链球菌和乳杆菌)的生长。这些菌的增殖又会产生更多的酸,使失调状态加剧,最终导致牙齿脱矿,龋病发生。
编辑
——在牙周炎中:缺氧和炎症改变生态位
牙周炎的过程则有所不同。
当菌斑长期堆积,牙龈沟内环境会逐渐变得更缺氧,同时炎症反应会带来更多组织分解产物和龈沟液。
这就为一些厌氧、蛋白水解型细菌创造了有利条件,比如牙龈卟啉单胞菌等。
这些细菌的毒力因子又会进一步刺激和放大宿主炎症反应,造成组织破坏、牙周袋加深,进而为更多厌氧菌提供栖息空间。
于是,牙周炎也进入了“环境改变—菌群演替—炎症加重”的循环。
牙周炎的病理过程不仅涉及到常见的细菌,还包括一些厌氧菌和蛋白水解型细菌的增殖等。这些细菌的丰度和比例变化能综合辅助评估牙周炎的风险。
编辑
<来源:谷禾口腔菌群检测数据库>
口腔微生态失调有哪些表现?
口腔微生物组的失调可以体现在多个层面,这些改变共同构成了疾病的微生物学基础。
◎ 物种组成改变
这是失调最直观的表现。
在龋病中,可见变形链球菌、乳杆菌和部分放线菌的丰度增加。
在牙周炎中,则表现为“红色复合体”(P. gingivalis, T. forsythia, T. denticola)和“橙色复合体”等厌氧菌群的富集,而与健康相关的链球菌、放线菌等共生菌比例下降。
◎ 多样性变化
失调对微生物多样性的影响因疾病而异。
在龋病中,由于强烈的酸性选择压力,微生物多样性通常会降低,少数耐酸菌占据主导地位。
在牙周炎中,由于牙周袋的形成创造了新的生态位,并且炎症反应提供了更丰富的营养,微生物多样性反而会增加。
◎ 功能代谢改变
失调不仅是物种的替换,更是整个群落功能的转变。
在龋病相关的生物膜中,糖酵解和产酸相关的代谢通路被上调。
在牙周炎相关的微生物组中,则表现为蛋白水解、氨基酸发酵、毒力因子(如LPS、牙龈蛋白酶)合成以及炎症诱导相关通路的富集。
◎ 生物膜空间结构改变
失调也会改变生物膜的物理结构。
例如,在致龋生物膜中,大量的胞外多糖(EPS)形成了一个致密的、酸性的微环境。
在牙周炎中,生物膜结构变得更加疏松,但更具侵袭性,细菌更容易从生物膜中脱落并侵入宿主组织。
这说明,疾病并不只是菌种名单变了,连它们在口腔里如何组织起来也变了。
是什么在推动口腔失调?
多种因素可以作为触发或加剧口腔失调的“生态压力”。
► 宿主因素
饮食:如前所述,高糖饮食是龋病失调的主要驱动力。频繁吃糖,比偶尔一次大量摄糖更容易反复制造酸性环境。
免疫状态:宿主免疫功能低下或免疫反应失调(如过度炎症)会无法有效控制致病菌,从而导致失调。例如,糖尿病患者的高血糖状态和受损的中性粒细胞功能,使其更容易发生严重的牙周炎。
唾液功能:唾液流速降低和缓冲能力下降会削弱口腔的自洁和中和酸的能力,为产酸菌的生长创造有利条件。
激素水平:青春期、妊娠期和更年期的激素波动会改变口腔环境,影响特定微生物的生长,可能暂时性地导致失调。
► 环境与行为因素
口腔卫生不良:不良的口腔卫生导致菌斑大量积聚,是牙龈炎和牙周炎失调的始动因素。
如果清洁不到位,微生物生物膜就会越来越成熟,生态环境也会越来越偏向致病状态。
吸烟与饮酒:吸烟会造成口腔局部缺氧,抑制有益的需氧/兼性厌氧菌,促进厌氧病原体的生长。酒精则可能直接影响微生物代谢并损害黏膜屏障。
药物使用:广谱抗生素会破坏正常的微生物群落,为机会性感染(如念珠菌病)打开大门。其他药物(如抗高血压药、抗抑郁药)可能通过引起口干,而间接导致失调。
►关键病原体:不是数量最多,却可能最会“带节奏”
“关键病原体假说”是生态学假说的延伸,它强调某些低丰度的微生物能够通过不成比例的巨大影响力来操纵宿主反应和微生物群落,从而引发失调。
牙龈卟啉单胞菌(P. gingivalis)是典型的关键病原体。
它虽然在菌斑中数量不多,但能通过其独特的毒力因子(如牙龈蛋白酶)破坏宿主的补体系统和中性粒细胞功能,造成局部免疫抑制。这种“被操纵”的炎症环境既能保护牙龈卟啉单胞菌自身免于被清除,又能为整个厌氧菌群落的生长提供营养(源自组织破坏和炎症渗出),最终导致一个协同致病的失调群落的形成。
它不只是自己受益,还把整个微生态都往致病方向带偏了。
总之,口腔微生态的失调是一个由多因素驱动的、复杂的生态学过程。它不仅仅是“坏细菌”战胜“好细菌”的简单故事,而是整个微生态系统在内外压力下失去稳态、功能转向致病方向的系统性失败。
未完待续,下期我们将继续讨论:
注:本账号内容仅供学习和交流,不构成任何形式的医疗建议。
主要参考文献
Dame-Teixeira N, Do T, Deng D. The Oral Microbiome and Us. Adv Exp Med Biol. 2025;1472:3-9.
Azevedo MJ, Kaan AM, Costa CFFA, Sampaio-Maia B, Zaura E. Acquisition of the Oral Microbiome. Adv Exp Med Biol. 2025;1472:13-29.
de Barros Santos HS, Pagnussatti MEL, Arthur RA. Symbiosis Between the Oral Microbiome and the Human Host: Microbial Homeostasis and Stability of the Host. Adv Exp Med Biol. 2025;1472:31-51.
Santonocito S, Polizzi A, Isola G. The Impact of Diet and Nutrition on the Oral Microbiome. Adv Exp Med Biol. 2025;1472:53-69.
Colombo APV, Lourenço TGB, de Oliveira AM, da Costa ALA. Link Between Oral and Gut Microbiomes: The Oral-Gut Axis. Adv Exp Med Biol. 2025;1472:71-87.
Washio, J., Takahashi, N. (2025). Nitrite Production from Nitrate in the Oral Microbiome and Its Contribution to Oral and Systemic Health. In: Dame-Teixeira, N., Deng, D., Do, T. (eds) Oral Microbiome. Advances in Experimental Medicine and Biology, vol 1472. Springer, Cham.
Heller D, Nery GB, Bachi ALL, Al-Hashimi I. Positive Role of Saliva in the Oral Microbiome. Adv Exp Med Biol. 2025;1472:103-118.