-

 国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子
国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子

 二级病原微生物安全实验室
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
谷禾健康
Hungatella属隶属于厚壁菌门梭菌目梭菌科,是一种具运动性、严格厌氧、能形成内生孢子的杆菌,代表种包括 Hungatella hathewayi 和 Hungatella effluvii。是人类肠道中常见的细菌之一,谷禾菌群数据库显示,约90%的人群肠道中可检出该细菌。
这类细菌的不同菌株对革兰氏染色表现存在变异,例如Hungatella effluvii染色呈革兰氏阳性,而Hungatella hathewayi染色则为革兰氏阴性。提示同属细菌之间在细胞壁结构和染色特性上并不完全一致,菌株间差异较大。
它是人类肠道“重要”共生菌之一,具有较强的糖类和多糖(特别是糖胺聚糖,GAG)降解能力,能产生乙酸、丙酸等短链脂肪酸,同时在牛磺酸和氧化三甲胺(TMAO)等代谢通路中扮演重要角色。还有研究发现其与某些肿瘤化疗药物的敏感性相关,但证据仍属初步,需进一步验证因果性。
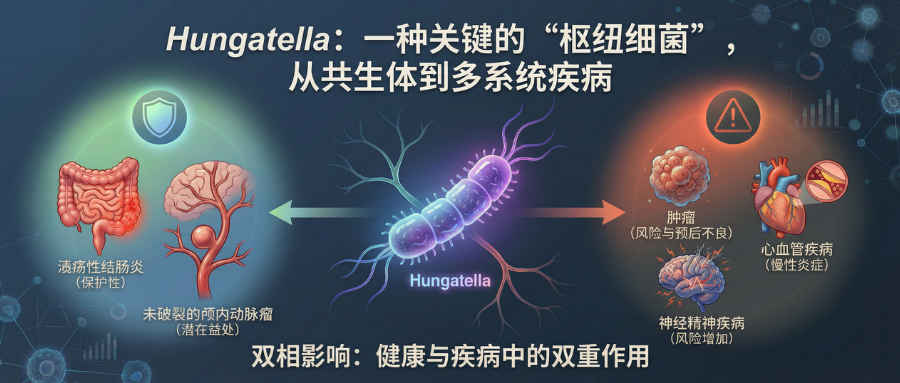
从目前的人群与动物研究来看,Hungatella在多种疾病中的丰度变化呈“双相”特点:一方面,在部分炎症性肠病(溃疡性结肠炎)和脑血管疾病(未破裂颅内动脉瘤)中表现出潜在保护作用;另一方面,在一些肿瘤、心血管及精神神经疾病中则与风险升高、预后不良或慢性炎症相关。
Hungatella是属于厚壁菌门、梭菌目、梭菌科下的一个属。Hungatella是人类肠道微生物群的成员之一,并具有重要的临床意义。
1
细胞形态
Hungatella属是一种能运动、严格厌氧、内生孢子(孢子形状为卵形至椭圆形,位于中央或近末端)的杆状细菌,长度为2-5μm,宽度为0.7-1.5μm。部分菌体还可产生粘稠胶囊。
固体琼脂培养基生长的Hungatella
编辑
doi: 10.3201/eid1011.040006.
▸ 革兰氏染色可变,取决于具体菌株
这类细菌的革兰氏染色结果因菌株而异,例如Hungatella effluvii染色呈革兰氏阳性,而Hungatella hathewayi染色则为革兰氏阴性。
一般而言,Hungatella菌落呈圆形,颜色为奶油色至灰白色,边缘稍不规则,凸起。通常,它们看起来有光泽且不透明。
注:37∘C 培养3天后,H.hathewayi 在哥伦比亚羊血琼脂培养基上形成灰白色、不透明、凸圆形、有光泽、边缘稍不规则的菌落。然而,在RCA培养基上,30°C 孵育24小时后,H.effluvii 形成奶油色、圆形、凸面和光滑的菌落,且边缘完整。
H.hathewayi经48小时培养后的菌落形态
编辑
doi: 10.3201/eid1011.040006.
2
营养和生长条件
▸ 生长环境与人体环境相近
菌株生长温度范围为10–45∘C(最佳温度为30–37∘C)、pH为5.0至8.5(最佳pH7.0)。生长可以发生在0–2.0%(w/v)NaCl 条件下,最佳条件是 0.5% NaCl,但在 3% (w/v) NaCl 条件下没有观察到生长。上述条件与人体内环境相近,说明 Hungatella可能适宜在人体肠道内定植与生长。厌氧培养的菌株通常需要24-72小时才能在RCA上形成菌落。
3
生态分布
Hungatella属分布广泛,构成正常人类微生物群的一部分,视为常见肠道成员之一。H.hathewayi(Clostridium hathewayi)最初从无胃肠道疾病的健康人粪便中分离并报道,随后也在术后血流感染患者中被检出。
编辑
另一种H.effluvii 菌株 DSM 24995T 是从用于处理益生菌制造业废发酵培养基的厌氧消化器获得的废水样品中分离出来的。
4
主要代谢与功能
▸ 糖类和多糖代谢
Hungatella属成员为糖分解菌,可发酵多种碳水化合物,包括苦杏仁苷、阿拉伯糖、纤维二糖、半乳糖、葡萄糖、乳糖、甘露糖、松三糖、蜜二糖、棉子糖、鼠李糖、核糖、水杨苷、山梨糖醇、淀粉、蔗糖和海藻糖,但不能将硝酸盐还原为亚硝酸盐。
葡萄糖发酵的主要终产物为乙酸盐和丙酸盐,并少量产生CO₂和H₂。
注:最新的功能组学研究表明,H.hathewayi在糖胺聚糖(GAG)代谢上具有非常突出的能力。
此外,Hungatella的过氧化氢酶、氧化酶和吲哚试验均为阴性,甲基红试验为阳性。其对七叶苷、淀粉和尿素的利用具有物种特异性,而所有已研究菌株均不水解明胶和酪蛋白。
▸ 与牛磺酸和氧化三甲胺含量相关
宏基因组关联研究及多个中国队列和相关动物模型的证据显示,Hungatella hathewayi 与血液中牛磺酸水平相关。但该关联尚不足以证明因果性,其在牛磺酸调控中的作用机制仍需进一步纵向研究和干预性试验加以证实。
而在另一项研究中,H.Hatheway产生氧化三甲胺(TMAO)的能力,与包括抑郁症在内的某些神经系统疾病有关。
▸ 可能会代谢5‑氟尿嘧啶
在结直肠癌队列中,H.hathewayi的肿瘤组织丰度与5-FU疗效下降及总体生存率下降呈相关关系。机制方面,研究提示可能通过CDX2/B-catenin通路的调控影响药物敏感性,但尚需因果性证据与纵向研究来证实。
5
耐药性
Hungatella属对青霉素、克林达霉素和莫西沙星的耐药性存在差异,但所有分离株均对美洛培南、β‑内酰胺/β‑内酰胺酶抑制剂及甲硝唑敏感。还发现利福昔明可抑制Hungatella hathewayi的生长。
▸ 抗生素体外敏感和实际临床药敏
•甲硝唑:迄今唯一在所有报道病例中均表现为敏感的药物。MIC 通常极低(如 0.032 μg/mL),无耐药报道,是治疗经验性厌氧覆盖首选。
•克林霉素:7/9例报道中Hungatella对其敏感。
•碳青霉烯类:约6例中表现为敏感,临床多作为重症败血症经验用药。
•β‑内酰胺/β‑内酰胺酶抑制剂:阿莫西林/克拉维酸、哌拉西林‑他唑巴坦在部分病例中敏感(4–5 例)。
•青霉素:多数早期H.hathewayi 菌株对其敏感,但不作为唯一经验药物。
•万古霉素:个别株报道为敏感,但Hungatella通常为厌氧 G+ 芽孢杆菌谱系,用万古霉素根除有限。
▸ 用药建议(基于有限病例):
•经验治疗严重感染:甲硝唑±第三/四代头孢或哌拉西林‑他唑巴坦或碳青霉烯。
•一旦确定为Hungatella:保持/调整为含甲硝唑方案通常可靠,再根据E‑test/MIC可逐步去除不必要的广谱药。
6
Hungatella属的重要物种
▸ Hungatella effluvii
细胞具运动性,杆状,长约2.0–4.0μm、宽1.0–1.5μm,革兰氏阳性。在pH6.0–8.0范围内可生长,最适pH为 7.0;适生温度为10–45℃,以30℃ 为最适。
菌株 UB-B.2ᵀ 的过氧化氢酶、氧化酶和吲哚试验为阴性,甲基红和脲酶试验为阳性;七叶苷、酪蛋白、淀粉和明胶水解阴性,不能还原硝酸盐,柠檬酸盐利用及 H₂S 产生均为阴性。
其对 L-吡咯烷酰芳香酰胺酶、脲酶和 β-吡喃半乳糖苷酶呈阳性,而对亮氨酸芳香酰胺酶、苯丙氨酸芳香酰胺酶、L-脯氨酸芳香酰胺酶、酪氨酸芳香酰胺酶、α-阿拉伯糖苷酶、β-甘露糖苷酶、七叶苷水解、β-D-岩藻糖苷酶、α-L-岩藻糖苷酶和碱性磷酸酶均为阴性。可利用半乳糖、蔗糖、葡萄糖、麦芽糖和甘露糖产酸。葡萄糖发酵的主要终产物为乙酸和丙酸。
DNA G+C 含量为 51.4 mol%。
遗传分析显示,Hungatella effluvii 与 Clostridium hathewayi(DSM 13479ᵀ)在 16S rRNA 基因水平的相似度为 97.84%,但全基因组亲缘性仅为 38.4%。在表型上,二者在七叶苷、淀粉和尿素水解等多项特征上存在显著差异。
▸ Hungatella hathewayi
细胞为严格厌氧、革兰氏阴性、具运动性的杆菌,宽0.7–1.0μm、长2.0–5.0μm,常成链排列,最长可达30个细胞。
生长pH范围为5.0–8.5,最适约7.0;温度范围15–45℃,最适约 37 ℃。菌株在含0–2.5%NaCl的培养基中生长最佳。
过氧化氢酶、氧化酶、吲哚、硝酸盐还原及 VP 试验均为阴性。葡萄糖发酵的主要终产物为乙酸、乙醇、H₂和CO₂。
能利用苦杏仁苷、阿拉伯糖、纤维二糖、果糖、半乳糖、葡萄糖、乳糖、麦芽糖、甘露糖、蜜二糖、松三糖、棉子糖、鼠李糖、核糖、水杨苷、山梨糖醇、淀粉、蔗糖、海藻糖和木糖产酸;不能利用麦芽三糖、糖原、肌醇、菊粉或甘露醇产酸。七叶苷和淀粉水解阳性,而酪蛋白、尿素、纤维素、明胶不被水解。
DNA G+C 含量为 50.7–50.9 mol%。
7
与人体及其他菌群的互作
▸ 与宿主的互作与潜在致病机制
(1)中性或有益方面
•作为肠道共生菌,可能通过产SCFA,维持结肠上皮能量与黏液层完整性;
•外泌GAG裂解酶,利用黏液层及 ECM 中的 GAG,为自身及共生菌提供碳源;在一定范围内维持黏膜更新与免疫稳态。
•牛磺酸产生:可能通过影响胆盐/抗氧化/血管平衡,对远端器官(如脑血管)产生系统性效应。
(2)有害或潜在有害方面
•当肠道屏障受损(阑尾炎、缺血、肿瘤、术后创面等),Hungatella 可由肠腔进入血流/腹腔,引起:菌血症、脓毒症、肝脓肿、胆囊炎、坏死性筋膜炎等;
•在合并其他菌(如 E. coli、Prevotella、Streptococcus constellatus)的多菌种感染中,则常与严重组织坏死、长期住院相关。
▸ 与其他肠道菌群的互作
•与Bacteroides的互补/竞争:
H.hathewayi拥有与B.thetaiotaomicron接近甚至更强的GAG降解能力,但使用的是完全不同的输运与调控网络(PTS/ABC vs SusC/D)。意味着在利用宿主黏液多糖这一生态位上,Hungatella可能与 Bacteroides形成功能冗余或在特定饮食/炎症条件下出现竞争/互补替代。
•向其他菌群“输出”寡糖与代谢中间体:
将大分子GAG裂解为寡糖/二糖,这些小分子自身能摄取利用,也可被缺乏初级外泌酶的共生菌进一步利用——这在微生态学上类似“初级降解者+次级利用者”的食物网结构。
1
影响炎症性肠病的风险
编辑
研究发现,Hungatella、Acidaminococcaceae及另外15个菌群是多种克罗恩病(CD)和溃疡性结肠炎(UC)亚型的保护因素,而Terrisporobacter、Anaerostipes及其他23个菌群与不同CD和UC亚型的风险增加相关。
▸ Hungatella有助于降低溃疡性结肠炎风险
分析显示,有9个肠道菌群对溃疡性结肠炎各亚型及肠外表现具有保护作用。其中,Lachnospiraceae 科 ND3007 组可降低溃疡性直肠乙状结肠炎风险;Ruminococcaceae UCG011和Hungatella属可降低溃疡性全结肠炎风险;Lachnospiraceae 科 ND3007 组和 Hungatella 属共同降低左侧溃疡性结肠炎风险。Prevotellaceae 科、Eubacterium fissicatena 属和 Ruminococcus gnavus 组则降低合并原发性硬化性胆管炎(PSC)的溃疡性结肠炎患者的风险。
▸ 与其他益生菌组合使用作为结肠炎的保护因子
Hungatella属和Lactobacillus属还可降低克罗恩病(CD)相关关节病变风险,而Barnesiella属则增加其发生风险。
在对比溃疡性直肠乙状结肠炎与左侧结肠炎时,Lachnospiraceae 科 ND3007 组在两者中均为保护因子,而 Hungatella 仅与左侧结肠炎相关,据此推测 Hungatella 可能参与降结肠炎症的起始。实验研究显示,克罗恩病术后吻合口处 Hungatella 丰度升高,提示其可能参与常见术后并发症的发生。
若肠道菌群在术后感染的发病机制中发挥促成作用,则针对肠道微生物群的靶向治疗或预防干预有望降低克罗恩病患者术后并发症风险。
2
丰度过高可能增加结直肠癌风险
编辑
近几十年来,早发结直肠癌(yCRC)发病率不断升高,但其肠道微生物特征仍知之甚少。既往研究多聚焦于老年发病型CRC(oCRC),尚不清楚老年患者的微生物特征能否外推至年轻患者。为此,有研究整合了迄今规模最大的 yCRC 肠道宏基因组数据,涵盖两个独立队列,发现CRC相关微生物组。
▸ H.hathewayi在结直肠癌患者中富集
分析显示,如Clostridium symbiosum、Peptostreptococcus stomatis、Parvimonas micra和Hungatella hathewayi在老年和年轻患者中均显著富集。并且在老年发病结直肠癌(oCRC)和早发结直肠癌(yCRC)中,观察到具核梭杆菌(Fusobacterium nucleatum)、脆弱拟杆菌(Bacteroides fragilis)和大肠杆菌(Escherichia coli)的菌株水平模式相似。
几乎所有与oCRC相关的宏基因组通路在年轻患者中均有方向一致的变化。值得注意的是,与对照组相比,oCRC和yCRC的相关毒力因子(fadA、bft)均为富集。此外,基于微生物组的分类模型在老年和年轻发病患者中对结肠癌状态的预测准确性相似,强调了不同年龄组微生物特征的一致性。
▸ H.hathewayi影响5‑氟尿嘧啶代谢进而干扰治疗效果
此外,刚才前文中也有讲到过的一项研究发现H.hathewayi在结直肠癌组织中的丰度显著增加,其高水平与整体生存率降低相关。
并且H.hathewayi处理明显减弱了5-氟尿嘧啶(5-FU)对结直肠癌(CRC)细胞系HCT116和HT29的增殖抑制和凋亡诱导作用,并增强了耐药细胞系 HCT116/5‑FU 和 HT29/5‑FU 对 5‑FU 的耐药性。机制上,H.hathewayi下调CDX2表达并促进 β‑catenin 在细胞核内的积累。CDX2 过表达可逆转H.hathewayi介导的细胞生长增强和对 HCT116/5‑FU、HT29/5‑FU 细胞凋亡的抑制,同时抑制 β‑catenin 的表达及其核内富集。
综上所述,结直肠癌组织中H.hathewayi高丰度不仅与较差生存预后相关,还可通过调控 CDX2/β‑catenin 信号通路影响结直肠癌细胞对5‑FU的耐药性。
3
未破裂动脉瘤患者中丰度较低
未破裂颅内动脉瘤(UIA)是一种危及生命的脑血管疾病。在两组中国UIA患者及接受人类供体粪便移植的对照个体和小鼠中进行了病例对照宏基因组全域关联研究。
注:非破裂颅内动脉瘤的全球患病率为3.2%,中国的社区调查表明中国成年人非破裂颅内动脉瘤患病率高达7%。颅内动脉瘤是一种极为凶险的脑血管疾病,一旦破裂其早期致死率为40-50%。
▸ 颅内动脉瘤患者中H.hathewayi减少,且与牛磺酸正相关
研究团队发现一群Clostridium属的物种包括C.bartlettii, C.nexile和C.bolteae在颅内动脉瘤组显著富集。然而,C.hathewayi(在2014年被更名为Hungatella hathewayi)却是唯一一个在颅内动脉瘤肠道中丰度降低的Clostridium属成员。
UIA患者的肠道微生物改变
编辑
doi: 10.1038/s41467-020-16990-3.
具体而言,在未破裂颅内动脉瘤(UIA)患者中Hungatella hathewayi 的丰度显著下降,并与人及小鼠体内的牛磺酸水平呈正相关。灌胃补充H.hathewayi可使小鼠血清牛磺酸水平恢复正常,并保护其免于颅内动脉瘤的形成与破裂;补充牛磺酸同样可逆转颅内动脉瘤的进展。上述结果提示,H.hathewayi相关的牛磺酸耗竭可能是UIAs发病机制中的关键因素。
此外,与单纯移植动脉瘤患者粪便的小鼠相比,补充H.hathewayi可显著降低颅内动脉瘤的总体发生率。Kaplan–Meier分析进一步证实,H.hathewayi 补充降低了动脉瘤破裂风险。同时,H.hathewayi 还能抑制脑血管炎症反应、细胞外基质重塑、MMP‑9 活性以及脑血管平滑肌细胞凋亡。
综上,H.hathewayi参与牛磺酸合成并提高血浆牛磺酸水平,从而降低颅内动脉瘤的发生率和破裂率。
4
在一些精神疾病中丰度较高
▸ 慢性精神分裂症患者中Hungatella丰度增加
编辑
研究共纳入115例处于阳性和紊乱症状缓解期的精神分裂症患者及119名对照者,评估了32项外周血标志物。
研究发现:在慢性精神分裂症患者中,Hungatella 的丰度显著增加,相比健康对照组其相对丰度更高。患者的肠道菌群多样性整体下降,而 Hungatella 等部分优势菌属呈富集状态。
差异丰度属与免疫炎症标志物、肠毒素、脂质谱成分和胰岛素抵抗水平的变化相关。此外,还观察到差异丰度属与认知障碍、阴性症状严重程度增加以及社交功能恶化存在若干相关性。甲烷短杆菌属(Methanobrevibacter)丰度与阴性症状、认知和社会功能水平的相关性似乎受白细胞介素-6和RANTES水平介导。
而Hungatella与注意力表现的相关性则受肠毒素水平介导。研究结果表明,精神分裂症患者肠道微生物群组成的改变与临床表现、肠道通透性、亚临床炎症、脂质谱改变以及葡萄糖稳态受损相关。亚临床炎症和肠道通透性受损可能介导了肠道微生物群改变与精神病理症状和认知障碍之间的关联。
▸ 产后抑郁的女性中H.hathewayi的含量较高
编辑
在另一项研究中,发现产后抑郁症状女性中 Hungatella hathewayi含量较高。在不同组学和横断面分析中观察到多种菌种丰度差异:中度且加重抑郁症状组中 Hungatella hathewayi丰度较高(FDR<0.25);高度且减轻抑郁症状组中 Bacteroides clarus 丰度较高(FDR<0.25)。
在妊娠晚期,与“低且稳定”组相比,“爱丁堡产后抑郁量表(EPDS)高且下降”组中 Bacteroides clarus、B. faecis 和 B. xylanisolvens 的相对丰度较高;而“EPDS 中且上升”组中 Hungatella hathewayi、Lachnospira SGB5076 和 Streptococcus thermophilus 的相对丰度较高。
既往研究也发现,H.hathewayi在非妊娠重度抑郁障碍人群中水平升高。其可能机制与产生三甲胺-N-氧化物(TMAO)的能力有关;TMAO 与包括抑郁在内的多种神经系统疾病有关,循环 TMAO 可激活促炎信号通路,从而诱发神经炎症。
5
冠状动脉病变中丰度升高
▸ 川崎病急性期合并冠状动脉病变儿童中富集
编辑
在川崎病急性期合并冠状动脉病变(AKDCAL)的儿童中,LEfSe 分析显示Hungatella显著富集,而在无冠状动脉病变的 AKDNCAL 组未见此现象。
注:川崎病(KD)是一种急性全身性免疫血管炎,主要影响中小动脉,常见于儿科患者。
虽然总体α多样性在川崎病急性期合并冠状动脉病变与无冠状动脉病变川崎病间无显著差异,但Hungatella、Burkholderia-Caballeronia-Paraburkholderia和Clostridium_innocuum富集,被鉴定为与冠状动脉病变(CALs)相关的特征菌属。
6
其他疾病中的丰度变化
⑴丰度增加可能与婴儿湿疹风险升高相关
在一项香港队列研究中,肠道Hungatella hathewayi丰度随时间增加与1岁前发生湿疹的风险升高显著相关。
⑵有时可能引起菌血症或严重感染
Hungatella 在多数情况下为肠道共栖菌,但在特定易感人群或肠道屏障受损的情况下,也可能导致菌血症或其他严重感染。
H.effluvii和H.hathewayi偶见引起败血症、坏死性筋膜炎或阑尾炎相关菌血症等重症感染。并且比较基因组学提示Hungatella属内部遗传差异大,不同“种/亚种”在毒力和耐药基因上差异明显。
⑶伴便秘的帕金森病患者中增多
与健康对照相比,伴有便秘的帕金森病(PD)患者粪便中Hungatella显著增多,同时伴随Collinsella升高、Lachnospira和Fusicatenibacter降低,提示整体向“促炎型菌群”转变。
反映便秘严重程度的 Wexner 评分与 Hungatella 丰度呈正相关,即Hungatella越高,便秘越重。
在机制层面,目前证据支持其作为参与PD相关肠道菌群失调和便秘,可能通过影响 SCFA 平衡、肠道炎症与肠‑脑轴功能发挥作用,是帕金森病合并便秘中值得关注的潜在干预靶点。
⑷乳腺癌患者中Hungatella丰度增加
乳腺癌(BCa)患者微生物多样性减少,但从BCa患者采集的粪便样本中,三种特定微生物属——Acidaminococus、Tyzzerella和Hungatella的丰度得到了富集。
⑸新冠后遗症/康复患者中增加
在新冠后遗症或康复期人群中,有研究发现,其肠道中Hungatella的总体丰度持续处于升高状态。进一步的分析显示,这种Hungatella丰度的增加与多种炎症因子水平呈显著正相关关系,尤其是与 IL‑8、IL‑1β 等经典促炎细胞因子相关联,因此提示 Hungatella 的升高可能反映或参与了机体长期存在的慢性炎症过程,与新冠感染后的持续性炎症状态具有一定联系。
综合目前多学科证据,Hungatella 在疾病中的作用大致通过以下几点体现:
1.黏膜与多糖代谢:强糖胺聚糖降解能力一方面为自身及其他菌群提供碳源,支持黏膜更新和生态食物网;另一方面,若失衡或过度活跃,可能削弱肠黏液屏障,增加内毒素和炎症因子易位,驱动全身慢性炎症。
2.小分子代谢(SCFA、牛磺酸、TMAO 等):
-通过产乙酸/丙酸等 SCFA 参与能量代谢和局部抗炎;
-通过促进牛磺酸合成或维持其水平,对脑血管等远端器官产生保护作用(如预防颅内动脉瘤);
-通过产生氧化三甲胺(TMAO),参与动脉粥样硬化和神经精神疾病相关的促炎与代谢异常。
3.免疫与炎症信号:与多种炎性细胞因子(IL‑6、IL‑8、IL‑1β 等)水平相关,可能通过诱导肠炎和系统性低度炎症,连接肠道微生态与代谢综合征、心血管疾病及精神神经障碍。
4.表观遗传与肿瘤信号通路:在结直肠癌中,通过调控 DNA 甲基转移酶活性,影响抑癌基因甲基化,以及通过 CDX2/β‑catenin 通路干预细胞增殖和 5‑FU 化疗敏感性,表现出典型的“促肿瘤菌”特征。
5.肠‑脑/肠‑血管轴:在帕金森病、抑郁、精神分裂症、川崎病和颅内动脉瘤等疾病中的一系列关联,提示 Hungatella不仅是肠腔共栖者,更可能通过代谢物与炎症通路参与中枢神经和血管系统的病理过程。
总体而言,Hungatella 是一个功能高度复杂且菌株异质性很大的肠道菌属:
•在某些场景中(如未破裂颅内动脉瘤、部分溃疡性结肠炎亚型),它可能具有保护性作用;
•在另一些场景中(如结直肠癌、乳腺癌、部分心血管和精神神经疾病、婴儿湿疹、新冠后遗症),其丰度升高则更多与风险增加、预后不良或慢性炎症相关;
•而在免疫低下或屏障破坏条件下,还可作为机会致病菌引发严重感染。
现有研究多为关联性和部分机制探索,因物种鉴定精度、株间差异及干预研究仍有限,Hungatella 在各类疾病中的真正“因果角色”和可干预潜力仍需更多纵向、功能和临床试验加以澄清。
注:本账号内容仅供学习和交流,不构成任何形式的医疗建议。
主要参考文献
Rawat PS, Li Y, Zhang W, Meng X, Liu W. Hungatella hathewayi, an Efficient Glycosaminoglycan-Degrading Firmicutes from Human Gut and Its Chondroitin ABC Exolyase with High Activity and Broad Substrate Specificity. Appl Environ Microbiol. 2022 Nov 22;88(22):e0154622.
Yang Q, Tang W, Ren J, Li M, Zhao C. Unveiling the gut-heart potential connection: microbiota’s role in kawasaki disease and coronary artery lesions. Front Cell Infect Microbiol. 2025 May 29;15:1560083.
https://doi.org/10.1002/9781118960608.gbm01640Digital Object Identifier (DOI)
Li F, Yu C, Zhao Q, Wang Z, Wang Z, Chang Y, Xu Z, Han X, Li H, Liu Y, Hu S, Chang S, Tang T, Li Y. Exploring the intestinal ecosystem: from gut microbiota to associations with subtypes of inflammatory bowel disease. Front Cell Infect Microbiol. 2024 Jan 4;13:1304858.
Qin Y, Tong X, Mei WJ, Cheng Y, Zou Y, Han K, Yu J, Jie Z, Zhang T, Zhu S, Jin X, Wang J, Yang H, Xu X, Zhong H, Xiao L, Ding PR. Consistent signatures in the human gut microbiome of old- and young-onset colorectal cancer. Nat Commun. 2024 Apr 22;15(1):3396.
Huang Z, Wang C, Huang Q, Yan Z, Yin Z. Hungatella hathewayi impairs the sensitivity of colorectal cancer cells to 5-FU through decreasing CDX2 expression. Hum Cell. 2023 Nov;36(6):2055-2065.
Li H, Xu H, Li Y, Jiang Y, Hu Y, Liu T, Tian X, Zhao X, Zhu Y, Wang S, Zhang C, Ge J, Wang X, Wen H, Bai C, Sun Y, Song L, Zhang Y, Hui R, Cai J, Chen J. Alterations of gut microbiota contribute to the progression of unruptured intracranial aneurysms. Nat Commun. 2020 Jun 25;11(1):3218.
Hieta J, Benchraka C, Pärnänen K, Houttu N, Mokkala K, Lotankar M, Kataja EL, Lahti L, Laitinen K. Perinatal depressive and anxiety symptoms are associated with gut microbiota in pregnant women with overweight and obesity. Brain Behav Immun Health. 2025 Jun 19;47:101042.
Misiak B, Pawlak E, Rembacz K, Kotas M, Żebrowska-Różańska P, Kujawa D, Łaczmański Ł, Piotrowski P, Bielawski T, Samochowiec J, Samochowiec A, Karpiński P. Associations of gut microbiota alterations with clinical, metabolic, and immune-inflammatory characteristics of chronic schizophrenia. J Psychiatr Res. 2024 Mar;171:152-160.
Elsayed S, Zhang K. Human infection caused by Clostridium hathewayi. Emerg Infect Dis. 2004 Nov;10(11):1950-2.

谷禾健康

在研究肠道菌群或复杂微生物样本构成时,“门”(Phylum)是细菌分类的高级分类单位之一。
细菌分类依次为门纲目科属种亚种,最大的分类层面是门,以前写过人群肠道菌群构成主要是以拟杆菌门和厚壁菌门为主,大部分人群占比超90%(个别的变形菌门或放线菌门占比也很高),作为两大“细菌王国”,这两大王国有着不同的生态和代谢属性,又有着相似的营养和生存需求,它们如同天平的两端的砝码,不同的人有着不同倾斜,即使同一个人不同时期不同身体状态下天平的倾斜都不一样。
拟杆菌门被比喻为肠道微生物群落的“多面手”,有超过7000种不同的革兰氏阴性菌组成,主要来自拟杆菌属、副拟杆菌属、普氏菌属和卟啉单胞菌属。它们的脂多糖和鞭毛蛋白与细胞受体相互作用,并通过细胞因子合成增强免疫反应。
拟杆菌门作为多糖降解联盟的成员,尤其是拟杆菌属和普雷沃氏菌属,具有多种可以利用多糖的聚糖和糖苷酶。它们有助于分解膳食纤维和淀粉释放能量,并且它们可能是丙酸盐的主要来源。
拟杆菌门一些细菌的功能还包括胆汁酸的解离。肠道微生物群,特别是肠道拟杆菌,一定程度上也具有使初级胆汁酸解离和脱水并将其转化为人结肠中次级胆汁酸的能力。它们也参与蛋白质分解过程中有毒产物的释放。比如氨,组胺,硫化氢等,这些高浓度的产物会对肠黏膜产生毒性。
该群体的成员具有一些可能有助于抑制炎症的活动,但它们也有可能促进炎症,其中一些已知是机会病原体。
厚壁菌门细菌大多为革兰氏阳性菌,通过短链脂肪酸合成在宿主的营养和代谢中发挥关键作用。
通过其代谢产物,厚壁菌门细菌与其他组织和器官间接连接并调节饥饿和饱腹感。厚壁菌门由大量功能更加多样化的核心细菌组成。从代谢的角度来看,共生梭菌簇 XIVa 和 IV 通过产生短链脂肪酸在宿主和肠道稳态中发挥着重要作用,使肠道通透性正常化,参与脑肠轴调节。许多厚壁菌门的能力与宿主的体重有关。
F/B比率升高或降低被视为生态失调,前者通常与肥胖,代谢障碍相关,这可能与从食物中提取热量增加、脂肪沉积和脂肪生成、胰岛素敏感性受损有关;而后者则与炎症性肠病,抑郁,阿尔茨海默病等相关,这可能与短链脂肪酸尤其丁酸的生成减少,蛋白代谢物组胺,脂多糖累积等相关诱导的免疫炎症反应相关。
目前许多研究工作都集中在识别特定的微生物特征,特别是与肥胖、2型糖尿病和炎症相关的微生物特征。
我们从谷禾肠道菌群检测数据库中随机抽取5万例样本,统计的人群厚壁菌门/拟杆菌门(F/B)比率如下:
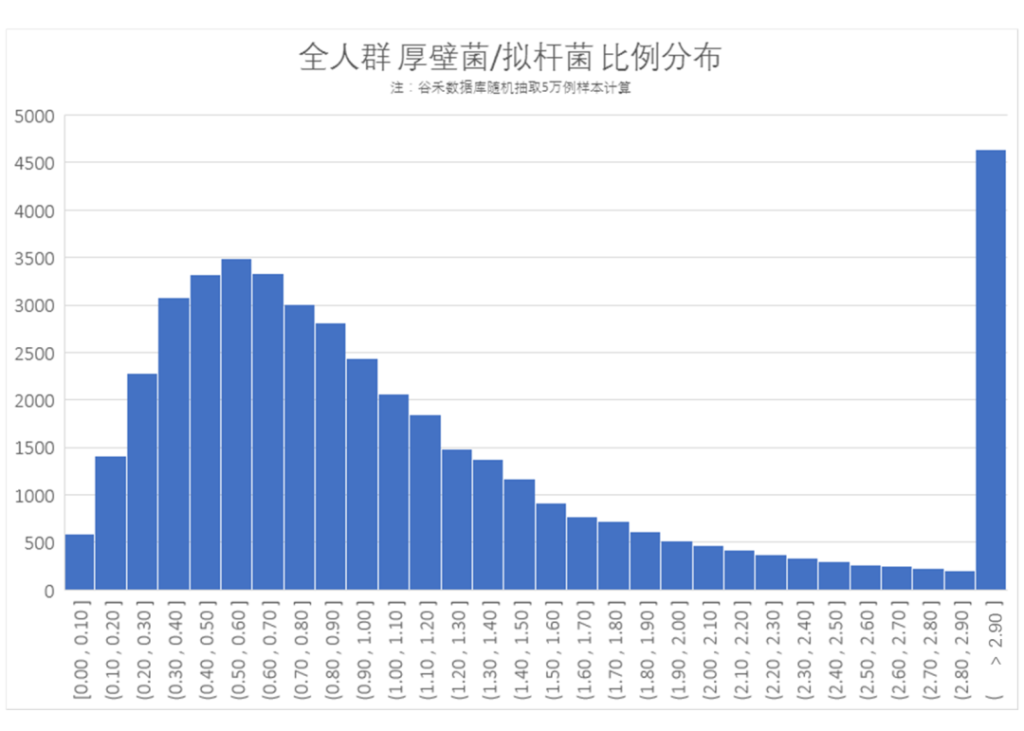
可以看出大部分人群的厚壁菌/拟杆菌比率在0.30-1.0之间(谷禾健康数据库随机抽样5万例样本),表明国人大部分人群肠道菌群的构成中拟杆菌占比多一点;同时抽样人群中1/10人厚壁菌/拟杆菌比率大于2.9,表明也有相当一部分人群厚壁菌门占比较高。
一般而言,从出生到成年,厚壁菌门/拟杆菌门的比例不断增加,并随着年龄的增长而进一步改变。该比率在婴儿、成人和老年人之间存在差异。它可能与生命不同阶段细菌谱的整体变化有关。
厚壁菌门/拟杆菌门比率是一个考虑高水平分类等级(即门)的指数,同一群体的受试者之间厚壁菌门和拟杆菌门的相对丰度差异很大。这可能是由于许多与生活方式相关的因素造成的,包括饮食、疾病状态、食品添加剂和污染物、抗生素消耗、体力活动等影响胃肠道微生物群组成的因素。
由于菌群是一个复杂的生态体系,门层面的简单划分只能提供一个最简单的线索,想要了解肠道菌群需要从最主要的核心菌属,常见的有害菌属以及大量功能菌属的构成来更加准确的分析,此外菌群的代谢通路也提供了不同角度的信息。
Wexler AG,et al.Nat Microbiol.2017
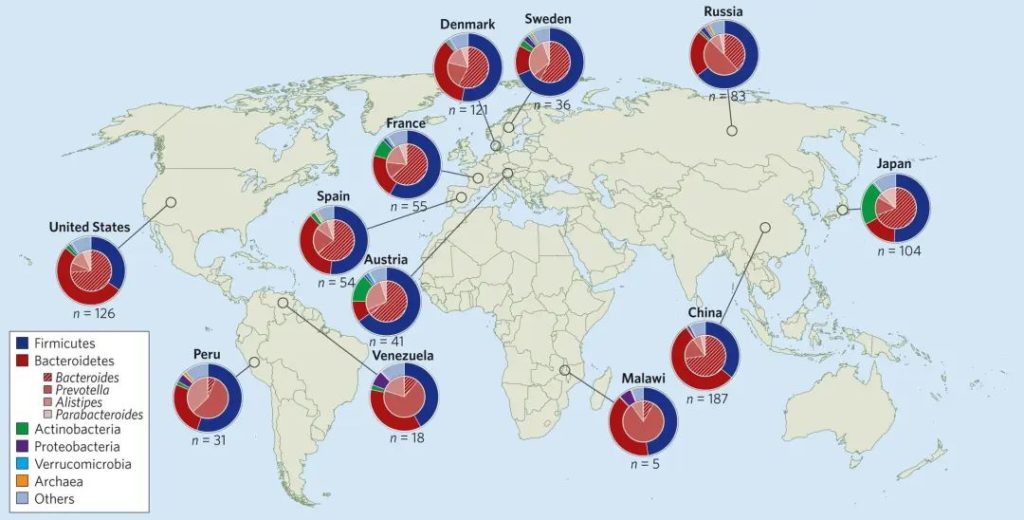
大多数健康成人微生物群仅由两个细菌门控制——革兰氏阳性厚壁菌门(许多属)和革兰氏阴性拟杆菌门(主要是拟杆菌属、另枝菌属、副拟杆菌属和普氏菌属)——它们共同构成了微生物中的大多数细菌分类群。
此外还有其他分类群包括变形菌门、放线菌门、梭杆菌门、疣微菌门、产甲烷古菌、真核生物(原生生物和真菌)和其他更短暂的定植者。
健康个体中发现的类群在家庭成员(例如父母及其子女)之间往往最相似,而在不同文化和地理空间(例如西方社会与非西方社会)之间则最不相似。
厚壁菌门(Phylum Firmicutes)是细菌的一个门,大多数具有革兰氏阳性细胞壁结构。然而,一些细菌,如巨球菌属、梳状菌属、月单胞菌属和嗜发酵菌属,具有多孔的假外膜,导致它们被染成革兰氏阴性。
科学家们曾经将厚壁菌门分类为包括所有革兰氏阳性细菌,但最近将它们定义为相关形式的核心组,称为低 G+C 组。与放线菌相反。它们有圆形细胞,称为球菌(单球菌),或杆状细胞(芽孢杆菌)。厚壁菌门缺乏脂多糖,而是向宿主呈现厚厚的肽聚糖层;研究人员已经发现了200多种不同类型的属于厚壁菌门的细菌。
★ 厚壁菌门中有许多有益的菌
厚壁菌门的成员包括许多有益的菌。比如乳酸菌是一种常见于酸奶和其他发酵乳制品中的益生菌,属于该门。这些微生物会产生醋酸盐(另一种促进健康的短链脂肪酸)以及乳酸和抗菌物质,以防止病原体定植。
又比如普氏栖粪杆菌(Faecalibacterium prausnitzii),又名:普拉梭菌,是人类肠道菌群中最重要的细菌之一,占健康人粪便样本中检测到的细菌总数的5-15%,是丁酸的重要生产者之一,具有抗炎作用,维持细菌酶的活性,保护消化系统免受肠道病原体的侵害。
★ 厚壁菌门中也存在一些致病菌
但是一些致病菌也属于该门。例如,产气荚膜梭菌是一种引起胃肠道感染的细菌。还有金黄色葡萄球菌,它是一些严重感染的常见原因。
丁酸盐是研究最深入的短链脂肪酸之一,是肠道上皮细胞营养的主要来源。丁酸盐的消耗与肠道屏障完整性受损有关。
注:研究支持,长寿、健康的老年人肠道内的丁酸细菌含量明显较高。
人类肠道包含一个非常密集的代谢活跃微生物群落。然而,只有有限数量的肠道细菌为丁酸生产者。人类肠道中大多数丁酸合成细菌分布于厚壁菌门。在厚壁菌门中,毛螺菌科、瘤胃球菌科、真杆菌科和梭状芽孢杆菌科是四个重要的丁酸合成科。
梭菌目的簇 IV、XIVa、XVI 和 I 包含大多数丁酸盐生产菌种。其中两个重要的物种,直肠真杆菌和普拉梭菌分别属于梭菌簇 XIVa 和梭菌簇 IV,占健康成人粪便样本中肠道菌群总数的12-14% 。此外,还有罗氏菌属,粪球菌属,瘤胃球菌属, Anaerostipes ,丁酸弧菌属和梭菌属。
分布在XIVa 簇中的Butyricicoccus pullicaecorum、Subdolicapsulu variabile、Anaaerotruncus colihominis和分布在 IV 簇中的Papillibacter cinnamivorans是其他产丁酸菌种。
据报道,属于韦荣氏球菌科、热厌氧杆菌科 III 的Megasphaera elsdenii和Caldocellum saccharolyticum也能合成丁酸盐。
最近,对人类样本的高通量宏基因组测序表明,Rawsonibactera saccharolyticus是瘤胃球菌科中的一个新物种,并且肠单胞菌AF211编码丁酰辅酶A:用于合成丁酸的乙酰辅酶A转移酶。
许多厚壁菌门细菌可以产生内生孢子,其耐干燥并且可以在极端条件下生存。孢子在细菌生长中起着重要的作用。细菌的孢子是一种休眠状态,可以在不利环境下存活,并在适宜条件下重新发芽成为活跃的细菌。这种休眠状态使得细菌能够在恶劣条件下存活并传播。
★ 内生孢子可以帮助细菌在恶劣条件下存活
内生孢子由细菌的DNA和部分细胞质组成,周围有一层非常坚韧的外层。它们可以在没有营养的情况下生存,并且能够抵抗紫外线、干燥、高温、极端冷冻和化学消毒剂。
厚壁菌门的代谢偏好
科学家发现,人类肠道微生物群虽然独特,但都可以按照几个分类划分下,称为肠型。第三种肠道型,被称为“谷物爱好者” ,通常以瘤胃球菌(厚壁菌门的成员)为主。
肠型往往可以追溯到长期的饮食习惯。顾名思义,具有“谷物爱好者”微生物组类型的人倾向于消耗更多全谷物中的抗性淀粉和膳食纤维。
✦ 高纤维饮食的人体内厚壁菌更丰富
高纤维饮食可以增加厚壁菌门的丰度并减少拟杆菌的丰度 ,从而增加肠道内短链脂肪酸的浓度。
而葡萄籽原花青素显著降低了肥胖大鼠肠道菌群中厚壁菌门的含量,提高了拟杆菌门的含量,显著降低了厚壁菌门与拟杆菌门比值。
✦厚壁菌偏向从膳食蛋白和循环尿素中获取氮
在厚壁菌门成员中,偏好尿素氮的属往往是菊粉的疯狂使用者,即使用菊粉和尿素合成自己的氨基酸。这包括一些脲酶阴性菌属,它们可能通过交叉喂养获得尿素氮。
此外,在厚壁菌中也看到了一些属更喜欢从膳食蛋白质中获得氮,而其他菌属更喜欢循环尿素。
静脉注射尿素以提高循环尿素浓度后,偏好尿素的厚壁菌以及阿克曼菌的丰度大幅增加。
膳食蛋白质和循环尿素是厚壁菌的主要氮原料,而分泌的宿主蛋白质为拟杆菌提供氮。
!
如果厚壁菌门过高意味着什么?
可能的原因:
– 不良的饮食习惯
– 生态失调
– 消化不良
– 胃酸过少
– 代谢疾病
可能的治疗方法和注意事项:
– 平衡共生细菌
– 当厚壁菌门含量较高时,考虑主要使用双歧杆菌益生菌和布拉氏酵母菌
– 乳杆菌属和芽孢杆菌属(益生菌中发现)可以提高厚壁菌门的数量
– 优化饮食结构:低脂肪饮食可能有助于厚壁菌门水平正常化
滋养肠道内厚壁菌门的食物
纤维和淀粉,是这些细菌的能量来源。富含水果、蔬菜、豆类和全谷物的饮食也是理想的。
研究表明,地中海饮食与更丰富的厚壁菌门有关,包括普拉梭菌。另一方面,富含动物蛋白、脂肪和糖的西方饮食会对有益的这些细菌产生负面影响。
如果想增加肠道中厚壁菌门的丰度,那么应该多吃高纤维食物。以下是一些可以添加到饮食中的食物,以支持您的肠道微生物组并促进厚壁菌门和丁酸盐的产生:
-苹果
-蒜
-大麦
-菊苣根
-燕麦
-亚麻籽
-洋葱
拟杆菌是肠道生态系统中非常成功的竞争者,表现出相当大的营养灵活性以及对宿主和肠道环境施加的压力做出反应的能力。很难权衡肠道拟杆菌对宿主是否产生负面或正面影响。
拟杆菌是复杂碳水化合物的主要降解者,迄今为止调查的所有生态系统中都普遍发现该属,在土壤以及人类和动物肠道中尤其占主导地位。大多数已测序的拟杆菌基因组编码大量多糖降解酶(CAZymes),表明代谢重点是聚糖降解。
CAZymes 在碳水化合物活性酶数据库中分为类别和家族,其中降解酶存在于糖苷水解酶(GH)、多糖裂解酶(PL)、碳水化合物酯酶(CE)和辅助活性酶 (AA) 类别中。
拟杆菌还通过与免疫系统的相互作用,在激活T细胞介导的反应中发挥作用,从而有助于宿主的整体健康。
✦ 防止特定炎症反应
基于多项研究,发现脆弱拟杆菌通过产生称为两性离子多糖的多糖来激活CD4+ T细胞。CD4+ T细胞的产生会刺激白细胞介素10的释放,从而防止特定的炎症反应和脓肿形成。
肠道免疫保护取决于拟杆菌、白细胞介素36信号和巨噬细胞的发育。拟杆菌的这种作用需要其保守的共生定植因子的多糖利用位点。
✦ 抑制胃肠道中的病原体
与胃肠道中发现的其他一些细菌相比,研究表明拟杆菌门的一些成员对各种抗生素和胆汁等物质具有耐药性,这给了他们竞争优势。
除了这种竞争优势之外,这些细菌还被证明在调节胃肠道中其他正常菌群和病原体的数量方面发挥着重要作用。
拟杆菌可以积极改善肠道环境,使其更适合自身和其他微生物。
✦降低肠道内氧水平从而促进厌氧菌生长
例如,许多编码细胞色素bd氧化酶,据推测该酶可以降低细胞内氧水平,进而降低肠道氧水平,从而允许严格厌氧菌生长,否则这些厌氧菌会被氧气的存在杀死。
这种耐受和降低氧气水平的能力可能有助于拟杆菌传播到新宿主,这可能是它们在哺乳动物中如此广泛传播的主要原因。
✦改变肠道的营养状况
它们还可以通过促进宿主的生理变化来诱导某些食物来源(例如岩藻糖基化糖蛋白)的产生,或通过从糖蛋白中释放岩藻糖和唾液酸残基来改变肠道的营养状况,这些残基可以被其他微生物消耗,包括病原体。
通过比较缺乏自身微生物的无菌动物和与单个拟杆菌属菌株和突变体单相关的无菌动物(出生时无菌但后来被微生物定殖),已经鉴定了许多这些拟杆菌属依赖性的肠道环境变化。
✦拟杆菌拥有多糖利用位点有助于代谢
尽管人们可能会想象肠道中的生命充满了微生物的过多食物选择,但在结肠(细菌密度最高的地方),简单、容易获得的糖大部分已经被消耗或吸收。其中的残留物是复杂的长链多糖,它们不能自由吸收,也不能被人类酶促消化。对于许多细菌来说,这些复杂的多糖同样难以处理,并且无法跨膜运输。
然而,早期研究证实拟杆菌是熟练的聚糖降解剂,具有识别和代谢十多种植物和宿主来源的多糖的不同寻常的能力。拟杆菌属物种通过基因簇来实现这一点,这似乎是拟杆菌特有的,称为多糖利用位点(PUL)。
多糖利用位点可以确定拟杆菌可以占据哪些代谢生态位,甚至可以确定它们在肠道内的生物地理位置。多糖利用位点被如此分类是因为它们的susC和susD的同系物,以及其他成分(糖苷水解酶、多糖裂解酶、糖基转移酶、碳水化合物酯酶),它们对于分解多种植物或宿主来源的聚糖很重要。
肠道细菌的其他谱系,包括革兰氏阳性放线菌,降解和输入膳食多糖的途径远不那么复杂,这在很大程度上是由于缺乏外膜屏障。例如,短双歧杆菌使用多结构域、细胞表面锚定酶(而不是一系列单独的酶),可以结合并降解聚合物,例如淀粉、支链淀粉和糖原。
拓展
比较基因组分析已在每个拟杆菌属中鉴定出数十种不同的多糖利用位点。例如,Bacteroides thetaiotaomicron是第一个对其基因组进行测序的拟杆菌属成员,将其 6.26 Mbp 基因组的近五分之一专用于88个不同的多糖利用位点。
拟杆菌不是同时表达所有多糖利用位点以捕获周围环境中可能存在的任何多糖,而是通过大量SusR样调节因子、混合双组分系统和涉及胞质外 σ/抗的跨包膜信号通路来严格调节其表达。
也许是由于在跨外膜运输之前需要部分降解长多糖链,B. thetaiotaomicron和其他肠道拟杆菌已经成功地将适当的多糖利用位点转录与其相应多糖分解中间体的丰度(而不是丰度)耦合起来。
这使得肠道细菌能够快速适应营养可用性的变化。糖分解物质形成甲酸盐、乙酸盐、乳酸盐、丙酸盐和琥珀酸盐作为主要发酵产物。拟杆菌可能是结肠中丙酸盐形成的主要贡献者。
由于多糖利用位点激活是肠道环境的产物,富含植物源性多糖和糖蛋白的饮食将导致多糖利用位点暂时上调,从而降解这些膳食营养素。然而,当这些底物稀缺时,例如在禁食期间或某些低纤维饮食期间(如高糖、高脂肪的西方饮食),亲糖拟杆菌会改变其转录谱以消耗宿主衍生的聚糖,这些暂时的转换可能是导致某些细菌在婴儿期最初在肠道定殖并在我们一生中持续存在的主要因素。
根据最近使用动物模型的研究,拟杆菌属可以在相对较短的宿主婴儿期主要依靠宿主聚糖生存,但当在多个宿主世代的时间内被迫以低纤维饮食为生时,它们最终可能会灭绝。
有的菌会通过破坏中性粒细胞趋化性等机制帮助逃避免疫系统(例如,伤寒沙门氏菌)和逃避宿主补体系统(例如肺炎克雷伯菌)。
相比之下,拟杆菌使用一种称为相变的过程,以开/关方式控制其荚膜多糖生物合成位点的表达。例如,脆弱拟杆菌从八个生物合成位点产生不同的荚膜多糖,其表达受到可逆启动子区域和反式位点抑制剂的严格调控,限制多个位点的同时表达,从而使脆弱拟杆菌的同基因群体共同表现出广泛的多样性。当脆弱拟杆菌漏入通常无菌的腹膜腔时,可能会发生腹膜炎。
注:绝大多数人类腹膜炎病例都是由脆弱拟杆菌引起的。
目前,拟杆菌门中研究的最充分的,包括拟杆菌属和普氏菌属以及卟啉单胞菌属。
✦ 卟啉单胞菌属可导致急性坏死溃疡性牙龈炎
卟啉单胞菌属的成员作为共生体或机会性病原体生活在人类口腔中。牙龈卟啉单胞菌引起临床上侵袭性牙龈炎,可导致急性坏死溃疡性牙龈炎或扩展到口腔和面部组织,这种情况称为坏疽性口炎或口腔癌。
✦ 普雷沃氏菌也是一种条件性致病菌
普氏菌属是一个大属,包括50多个不同的物种。大多数可以从口腔和肠道中分离出来。普雷沃氏菌属存在于人类中,帮助分解蛋白质和碳水化合物食物。也可作为条件致病菌,引起牙周和牙齿、肠道炎症、类风湿性关节炎、细菌性阴道炎等问题。
大型基因组计划,包括美国国立卫生研究院(NIH)资助的人类微生物组项目(http://nihroadmap.nih.gov/hmp/)和欧盟资助的 MetaHIT 项目 (http://www.metahit.eu)对50多种人类来源的拟杆菌门的拟杆菌和普氏菌分离株进行了测序。
基因组草图可从 Genbank 公开获得,并揭示这些物种拥有多种糖苷酶,这些糖苷酶具有预测的对抗植物和宿主来源的多糖的活性,以及维生素和辅因子合成的许多途径。
拟杆菌门的基因组大小相对较大,例如B.thetaiotaomicron 为 6.26 Mb。迄今为止在拟杆菌属中发现的16S rRNA基因的最高拷贝数为7个(在普通拟杆菌中)。
✦ 拟杆菌的组成和代谢受到饮食调节
在全基因组水平上,拟杆菌属与副拟杆菌属共享超过1000个蛋白质家族的核心,但与卟啉单胞菌属和普雷沃菌属共享较少数量的核心家族蛋白质家族。它们的组成和代谢活动在很大程度上受饮食调节。
拟杆菌与高脂肪和蛋白质的摄入量有关,而普雷沃氏菌与富含植物的饮食(高水平的复合碳水化合物和水果和蔬菜的摄入量)有关。因此,拟杆菌属和普雷沃氏菌属物种在人体内具有其特定的栖息地,因为它们大多数是拮抗的。
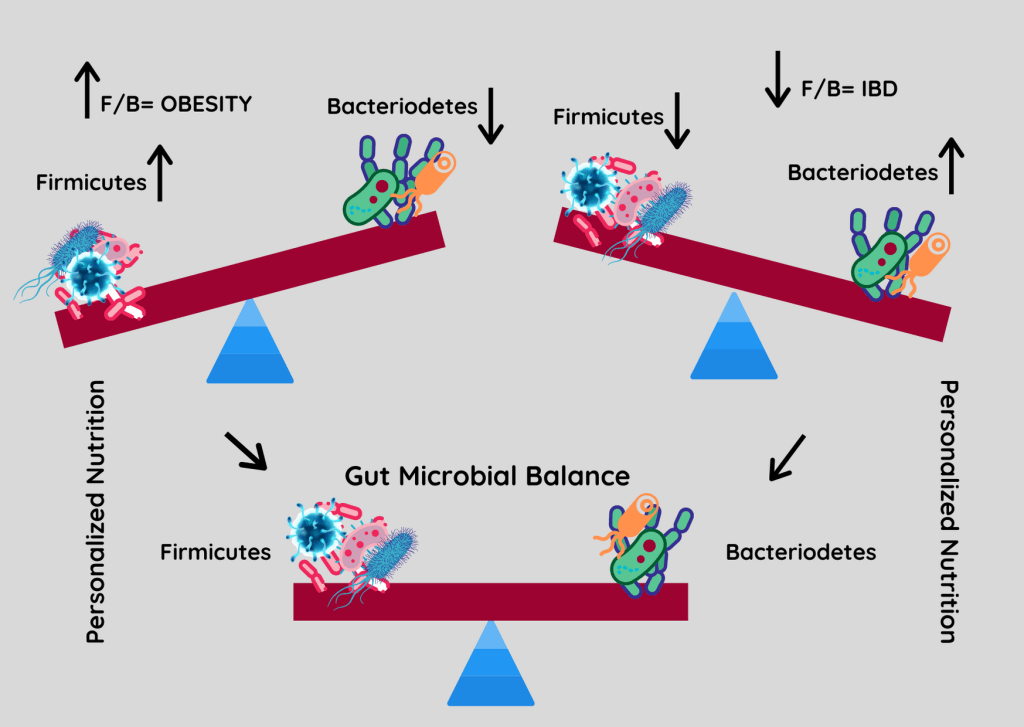
厚壁菌门与拟杆菌门之间的比率(F/B)与维持体内平衡有关,该比率的变化可能导致各种病理。
例如,特定厚壁菌门或拟杆菌门物种丰度的增加分别导致肥胖和肠道炎症。尽管人们非常关注 F/B 比率,但应该记住,这一比率可能会受到其他门的增加的影响,但是其他门的菌群失调增加或减少并不一定会改变 F/B 比率。
Stojanov S,et al.Microorganisms.2020
肥胖是一种复杂的、多因素的疾病,其原因包括宿主遗传背景、体力活动减少和食物摄入过多等多种因素。在过去的几十年里,肠道微生物群被认为是有利于脂肪储存、体重增加和胰岛素抵抗的额外因素。
世界范围内的研究正在不断寻找其致病因素。厚壁菌门和拟杆菌门的改变首先在肥胖动物和受试者中被描述,这些动物和受试者表现出厚壁菌门丰度增加,而拟杆菌门却减少了。
✦ F/B 比率较高时相同食物会吸收更多能量
当F与B的比例较高或体重增加和肥胖状态时,这两个细菌门影响我们体重的原因是它们调节我们吸收的脂肪量。想象一下两个人都消耗2000卡路里,但F与B的比例不同。F/B 比率较高的一种会吸收更多的卡路里,因此会比另一种增加更多的体重。因此,重点应该是平衡F与B的比例,一方面防止肥胖,另一方面预防炎症性肠病的发生。
当受试者接受热量限制饮食一年后,他们的拟杆菌门丰度增加,厚壁菌门/拟杆菌门比例正常化,同时体重减轻。这些研究得到了对喂食高脂肪或高纤维饮食的动物的研究的支持,这些研究显示厚壁菌门和拟杆菌门的丰度分别较高。
基于这些结果以及从肥胖动物和人类身上获得的其他结果,有人提出厚壁菌门比拟杆菌更能有效地从食物中提取能量,从而促进有效吸收热量和随后的体重增加。
肥胖双胞胎的微生物组富含编码营养转运蛋白的基因,而瘦双胞胎的微生物组富含编码与碳水化合物代谢相关的酶的基因。
最近发表的一项系统综述显示,大多数研究都支持 F/B 比率增加与肥胖之间的关系。
对肥胖和非肥胖日本受试者的粪便样本进行的一项研究显示,他们的 F/B 比率存在显著差异。厚壁菌门的百分比为 37.0 ± 9.1%(非肥胖者)和 40.8 ± 15.0%(肥胖者),而拟杆菌门的百分比为 44.0 ± 9.8%(非肥胖者)和 37.0 ± 14.0%(肥胖者)。
在61名乌克兰成年人中也报告了类似的结果,其中 F/B 比率与体重指数(BMI)显著相关。F/B 比率≥ 1 的个体超重的可能性比F/B 比率<1 的个体高出23%。
同样,卡塔尔受试者(37名肥胖者和36名瘦者)表现出肠道微生物群的改变;肥胖和瘦受试者的 F/B 比率分别为 2.25 ± 1.83 和 1.76 ± 0.58。在哈萨克族和比利时学童中,肥胖组的F/B比值显著高于对照组。
✦ 肥胖人群与正常人群拟杆菌存在差异
然而,两个拟杆菌属的存在差异:与肥胖组相比,正常组中拟杆菌属的高出20%,而肥胖组中普雷沃氏菌属比正常组高出19%。拟杆菌属和普雷沃氏菌属都属于拟杆菌门。
同一群体的受试者之间厚壁菌门和拟杆菌门的相对丰度差异很大。这可能是由于许多与生活方式相关的因素造成的,包括饮食、体力活动、食品添加剂和污染物、抗生素消耗等影响胃肠道微生物群组成的因素。
这种高度变异性可能解释了在比较正常体重和肥胖受试者之间的微生物群时观察到的矛盾结果,使得很难将厚壁菌门/拟杆菌门比率与确定健康状况联系起来。
重要的是要观察肠道微生物种群结构的整体组成,将其作为肥胖和肥胖相关病理(例如GSD)的指标,而不仅仅是厚壁菌门/拟杆菌门的比率。然而,在我们看来,可以修改这个比率,例如引入表征拟杆菌门底下的拟杆菌属或普氏菌属与梭状芽胞杆菌 XIVa 簇菌的比例。
!
F/B 比改变导致肥胖的争议
然而,与这些结果相反,许多研究没有观察到该参数的任何改变,甚至报告肥胖动物和人类中厚壁菌门/拟杆菌门的比例降低。
事实上,在大多数研究中,肥胖患者比瘦受试者表现出更少的细菌多样性,这表明在科、属或种水平上存在其他组成变化,这可能比厚壁菌门/拟杆菌门比率更相关。
✦肥胖增加可能与长期暴露脂多糖有关
关于这一点,代谢性内毒素血症的假说提出,肥胖增加和全身炎症的发展可能是由于长期暴露于脂多糖所致,脂多糖是一种源自革兰氏阴性细菌的促炎分子,它会通过肠腔进入血液。
这一假设与肥胖症中所报道的拟杆菌门丰度下降的情况并不相符,因为该门是肠道微生物群中革兰氏阴性菌的主要群体。
这种差异可以通过这样的事实来解释:来自拟杆菌门细菌的脂多糖的内毒素活性被认为低于来自其他革兰氏阴性细菌(例如属于变形菌门的细菌)的内毒素活性。有趣的是,在肥胖受试者或动物中也观察到变形菌门的增加,并且在无菌小鼠中施用变形菌门成员肠杆菌会导致这些动物出现肥胖和胰岛素抵抗。
✦ F/B比率增加与短链脂肪酸产生无关
另一方面,厚壁菌门/拟杆菌门比例的增加与肥胖个体中观察到的短链脂肪酸产生无关。事实上,据报道,拟杆菌门主要产生乙酸和丙酸,而厚壁菌门则产生更多的丁酸。丁酸盐被认为是一种促进健康的分子,因为它具有增加胰岛素敏感性、发挥抗炎活性、调节能量代谢和增加瘦素基因表达的能力。
结肠中的丙酸盐刺激L-肠内分泌细胞释放GLP-1和 PYY,从而抑制食欲。它还可能到达门静脉循环,主要被肝脏捕获,参与肝糖异生并减少参与脂肪酸和胆固醇从头合成的酶的表达。
乙酸也被吸收并到达全身循环和周围器官,包括脂肪组织、肌肉和大脑。在肝脏中,与丙酸盐相反,它刺激肝脏合成脂质,导致血脂异常。在大脑中,它激活副交感神经系统,分别促进胰腺和胃粘膜分泌胰岛素和生长素释放肽。
这些事件会导致脂肪储存和食欲增加,从而导致肥胖。根据这些结果,醋酸盐通常被认为更容易导致肥胖。肥胖个体中厚壁菌门/拟杆菌门比例的增加意味着这些受试者中丁酸盐的产量较高,丙酸盐和乙酸盐的产量较低,这一发现与这些短链脂肪酸各自的抗肥胖和致肥胖作用部分矛盾。一种解释是,肥胖个体中产生丁酸的细菌减少,并逐渐被属于同一门的其他细菌取代,导致结肠腔中丁酸的产生量降低。例如,葡萄球菌属和罗伊氏乳杆菌的丰度增加(均来自厚壁菌门)
肥胖状态还与A. muciniphila (疣微菌门)丰度较低有关,这是一种粘蛋白降解细菌,有助于稳定肠道屏障功能、分泌抗菌肽和控制炎症。
✦生理及病理状况都会影响肠道微生物群
另一方面,性别、年龄、宿主遗传学的差异。男性和女性的肠道微生物群存在差异,例如男性中拟杆菌-普雷沃氏菌群的水平较高,而女性中厚壁菌门的比例较高。然而,个别研究也发现女性中拟杆菌门的丰度最高。
此外,存在病理状况(糖尿病、心血管疾病、炎症性肠病等)。例如,Oddi括约肌松弛与胆管结石有关,可能是由于肠内容物回流增强改变了微环境所致。
治疗(抗生素、二甲双胍等)通过富集Akkermansia muciniphila以及几种产生短链脂肪酸的微生物群(丁酸弧菌、两歧双歧杆菌等)来改变肠道微生物群组成。
总之,这些导致厚壁菌门/拟杆菌门比率并不是与肥胖相关的微生物群失调的有力标志。
我们建议与肥胖相关的肠道微生物组研究不应该研究肥胖本身的分类标志物,而应侧重于确定用于将患者分层为亚组的分类标志物。引入微生物组患者分层将通过直接操纵患者微生物组来制定个性化治疗决策,从而改善肥胖的管理。
炎症性肠病代表小肠和大肠炎症反应引起的肠道疾病,由两种疾病组成:克罗恩病(CD)和溃疡性结肠炎(UC)。
溃疡性结肠炎是由结肠和直肠(从大肠末端开始到肛门结束的腔室)的长期炎症引起的,而克罗恩病是由结肠或回肠(小肠的最后部分)或两者的长期炎症引起的。
✦ 炎症性肠病患者厚壁菌门丰度下降
肠道微生物群会导致这些疾病。具体来说,当厚壁菌门减少或拟杆菌门相对于厚壁菌门增加时(两种情况下肠道菌群失调),就会导致慢性炎症和消化系统疾病,如克罗恩病和溃疡性结肠炎。
一项研究检查了克罗恩病和溃疡性结肠炎患者以及健康对照活检组织中的微生物多样性。观察到厚壁菌门丰度下降;更准确地说,克罗恩病患者中梭状芽胞杆菌类细菌减少,但溃疡性结肠炎患者或健康个体中则没有减少。
相反,与溃疡性结肠炎患者和健康个体相比,克罗恩病患者的拟杆菌丰度显著增加(分别为74.97%、64.31%和67.41%)。
✦肠道微生物变化与炎症性肠病严重程度相关
肠道微生物群的变化也与克罗恩病和溃疡性结肠炎的疾病活动度和严重程度相关。例如,与患有非活动性疾病的患者相比,患有活动性疾病的溃疡性结肠炎患者中厚壁菌门的含量较低。同样,与患有非侵袭性疾病的克罗恩病患者相比,患有侵袭性疾病的克罗恩病患者的厚壁菌门丰度显著降低。
具有抗炎作用的普氏粪杆菌减少
普氏粪杆菌(F. prausnitzii)是一种厚壁菌门物种,在炎症性肠病患者中减少,其减少与克罗恩病术后六个月复发相关。
普氏粪杆菌在体内和体外均表现出抗炎作用。在患有诱导性结肠炎的小鼠中,活普氏粪杆菌(或其上清液)降低了结肠炎的严重程度并纠正了生态失调。此外,普氏粪杆菌对血液单核细胞的体外刺激导致IL-10分泌增加以及IL-12和干扰素-γ(IFN-γ)分泌减少。
拟杆菌在炎症性肠病中增加
相比之下,拟杆菌在炎症性肠病中通常会增加,并与其进展和发展相关。对炎症性肠病患者和健康个体肠道炎症和非炎症区域的粘膜活检显示,炎症性肠病样本中厚壁菌门减少,拟杆菌门丰度增加,而肠杆菌科仅在克罗恩病患者中增加。活检还显示,与非发炎区域相比,发炎区域的细菌失调程度更高。
另一项对克罗恩病和溃疡性结肠炎患者进行粘膜活检和粪便菌群构成的研究证明了肠道微生物群和 F/B 比率的改变。克罗恩病和溃疡性结肠炎患者的F/B比值均较对照组显著降低。
此外,种、属和科的组成也发生了变化。与克罗恩病患者和对照组相比,溃疡性结肠炎患者中拟杆菌和乳酸菌的丰度较高;与对照组相比,溃疡性结肠炎患者中大肠杆菌的丰度也有所增加。相反,克罗恩病患者中球状梭菌减少。
✦ 不同拟杆菌种类与不同炎症性肠病程度相关
不同的拟杆菌与不同程度的炎症性肠病相关。在经过抗生素预处理且存在转化生长因子(TGF)βRII和IL-10R2信号传导缺陷的小鼠系中,在用普通拟杆菌和多形拟杆菌灌胃后观察到严重的溃疡性疾病。
与显示 F/B 比率对肥胖影响不大的研究类似,并不是在所有炎症性肠病病例中都观察到 F/B 比率降低。炎症性肠病患者粪便微生物分析显示,拟杆菌门、厚壁菌门和疣微菌门的丰度下降,而变形菌门、放线菌门和梭杆菌门的丰度增加。在48名克罗恩病患者中,粪便样本中的拟杆菌细菌丰度不到健康个体的10%,梭菌亚群 XIVab(厚壁菌门)也显著降低。
然而,大多数研究表明,拟杆菌因内毒素而表现出促炎特性,并影响细胞因子的产生,从而导致炎症性肠病。此外,厚壁菌门细菌具有抗炎作用,可以缓解炎症性肠病的进展。
2型糖尿病(T2DM)是一种代谢性疾病,其主要原因是与肥胖相关的胰岛素抵抗。然而,其他一些因素,如精神压力、感染和遗传倾向也可能导致糖尿病。
肥胖和糖尿病的特征都是慢性低度炎症状态,并伴有多种炎症介质(例如肿瘤坏死因子和白细胞介素)的异常表达和产生。
最近的研究基于大规模16S rRNA 基因测序、定量实时PCR (qPCR) 和荧光原位杂交 (FISH),显示了肠道微生物群组成与糖尿病等代谢疾病之间的关系。
有人提出,肠道微生物群引导增加肠道对单糖的摄取,并指示宿主增加肝脏产生与胰岛素抵抗相关的甘油三酯。
✦ 2型糖尿病患者中产丁酸厚壁菌门丰度降低
权威研究显示主要的丁酸盐合成菌厚壁菌门在2型糖尿病患者中显著减少。多项人体研究支持这样一个事实:与健康受试者相比,2型糖尿病受试者中合成丁酸的厚壁菌门丰度显著降低。
对这些患者的粪便微生物群分析显示,与正常耐糖受试者相比,新诊断或长期患有糖尿病的受试者中瘤胃球菌科和毛螺菌科等科的丰度有所下降。
还报告了罗氏菌属(Roseburia)和普拉梭菌(Faecalibacter prausnitzii)的丰度在健康对照中比2型或1型糖尿病患者更高。
对粪便样本进行深入的宏基因组学和宏蛋白质组学分析证实,在患有糖尿病前期 (Pre-DM) 或初治2型糖尿病 (TN-T2D) 的个体中,产生丁酸盐的菌含量显著降低。与正常耐糖个体相比, 患有糖尿病前期个体中的普拉梭菌的水平低于正常耐糖个体和初治2型糖尿病个体。
肠道微生物组可以反映整个身体的健康状况。厚壁菌门和拟杆菌门是结肠的主要门,可以影响与肥胖相关的疾病,这些疾病也是乳腺癌的危险因素。
✦ 乳腺癌患者F/B比值比健康人群低很多
因此,研究对乳腺癌患者和健康对照者的血清中提取细菌细胞外囊泡,使用微生物组测序分析厚壁菌门和拟杆菌门。分析乳腺癌的预后因素和血清学检测结果与 F/B 比值的相关性。发现乳腺癌患者的 F/B 比值比健康对照者低三倍。
此外,乳腺癌的危险因素,如空腹血糖,被发现与F/B比值有关。F/B比值可以作为乳腺癌的危险因素,并可能作为解释影响乳腺癌发生的潜在机制的线索之一。
健康对照者和乳腺癌患者中 F/B 的比率
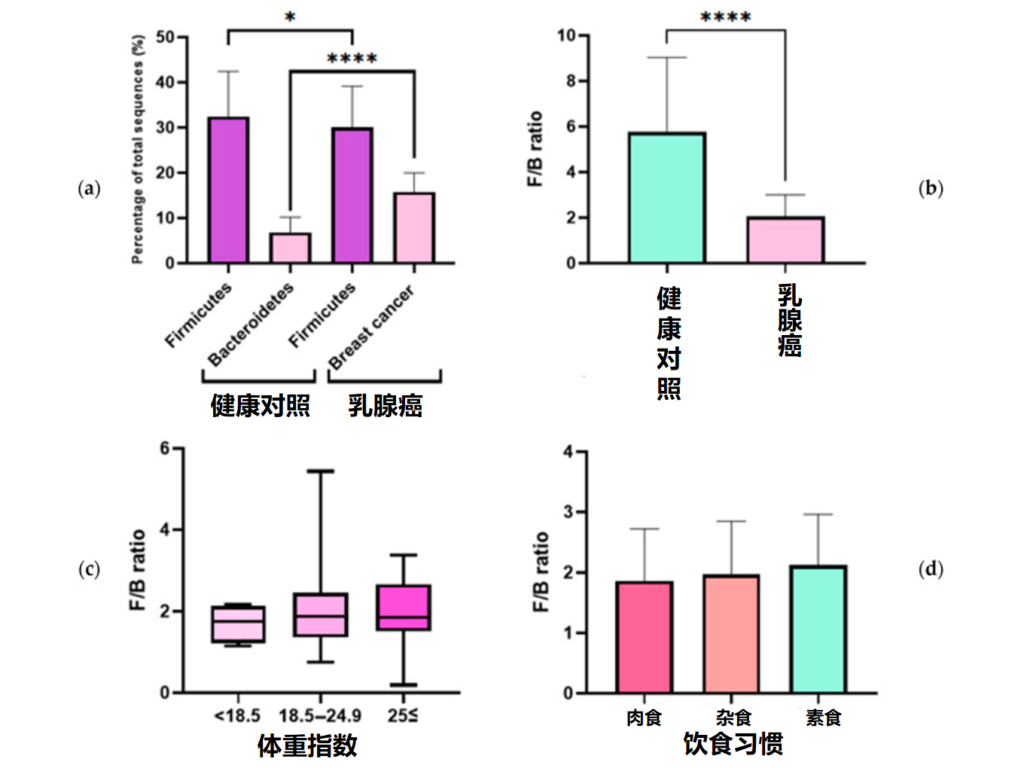
An J,et al.J Clin Med.2023
(a)健康对照组和乳腺癌患者中厚壁菌门和拟杆菌门占总微生物群的比例;(b)健康对照者和乳腺癌患者的F/B比率;(c)F/B比与体重指数之间的关系;(d)F/B比与饮食习惯的关系(*p≤0.05,**** p≤0.0001)。
调查研究发现中国饮食中的动物脂肪和蛋白质含量高于印度饮食中的全谷物和植物性素食。而印度参与者微生物组中拟杆菌的比例几乎是中国参与者的四倍,分别为16.39%和4.27%(p=0.001)。
✦ 动物产品食用较少与拟杆菌数量较多有关
据推测,印度人体内拟杆菌门的丰度较高是由于他们对动物产品的消耗较少;表明饮食中动物产品含量较低与拟杆菌数量较多有关。
对欧洲儿童和布基纳法索儿童的微生物群的比较也显示了由于饮食习惯不同而导致的微生物群组成的差异,欧洲儿童的饮食富含动物蛋白、糖、淀粉和脂肪,纤维含量低。以素食为主的饮食主要由碳水化合物、纤维和非动物蛋白组成。欧洲儿童微生物组富含厚壁菌门和变形菌门,而放线菌门和拟杆菌门在非洲儿童中较多。
✦ 营养不良儿童的肠道微生物多样性较低
研究发现,与同一地区中高收入家庭的健康儿童相比,孟加拉国贫困社会经济地位家庭的营养不良儿童的肠道微生物群多样性较低,其特点是拟杆菌门的相对丰度较低,变形菌门占主导地位。
注:拟杆菌门的存在率较低众所周知,它们可以消化复杂的膳食材料,从而提高从各种食物中提取能量,而潜在致病性变形菌的较高含量可能有助于解释营养不良儿童的健康状况不佳。
✦ 增加纤维摄入量可以增加厚壁菌门的数量
一项研究将西方饮食的美国儿童与食用大米、面包和扁豆等植物性饮食的孟加拉国儿童进行了比较。在食用西方饮食的美国儿童中,厚壁菌门/拟杆菌门的比例减少了三倍。
另一项研究要求参与者增加纤维摄入量并避免西方减肥食品。研究表明厚壁菌门:拟杆菌门比率增加了。
通过分析三名健康受试者6-12个月的肠道宏蛋白质组,进一步证实了这些结果。在这项研究中,参与碳水化合物运输和代谢的蛋白质占检测到的蛋白质的10%以上,构成了所有测试对象中发现的核心宏蛋白质组的一部分。
糖酵解途径尤其引人注目,并鉴定出几种相关酶。在为每个门的每个COG功能类别分配光谱命中后,很明显,厚壁菌门和放线菌门负责活跃的碳水化合物代谢,而拟杆菌门则显示出更多的混合功能。
这些例子反映了将某些门与特定饮食广泛联系起来主要挑战是需要考虑涵盖整个微生物组的微生物的状态和相互作用动态。
✦ 当F/B比率增加时
即更多厚壁菌门,拟杆菌较少。或两者兼而有之,厚壁菌门增长更快。可能导致以下疾病:
-肥胖(BMI较高)
-高血压中的肠道菌群失调
-自闭症谱系障碍
-慢性疲劳综合症
-肠易激综合症
-干燥综合症
-类风湿性关节炎
✦当F/B比率下降时
即厚壁菌门较少,更多拟杆菌。或两者兼而有之,厚壁菌门减少得更快。可能导致以下疾病:
-系统性红斑狼疮
-乳糜泻
-克罗恩病
-老龄化
我们审查评估了饮食干预如何影响微生物组成的相关文章。总体而言得出以下建议供参考:
✦ 增加植物蛋白摄入,避免摄入过量动物蛋白
总体而言,蛋白质摄入量,尤其植物蛋白的摄入与肠道拟杆菌门,细菌多样性呈正相关。乳清和豌豆蛋白可以增加双歧杆菌和乳酸菌的数量,而乳清还可以减少脆弱拟杆菌和产气荚膜梭菌的数量,这两种细菌都是致病性的。
此外,豌豆蛋白被发现可以增加短链脂肪酸水平,短链脂肪酸被认为具有抗炎作用,并在粘膜完整性中发挥作用。然而,一项研究发现,高动物蛋白/低碳水化合物饮食会导致罗氏菌属和直肠真杆菌(这两种有益细菌)减少。动物蛋白可能含有大量脂肪,而脂肪也会影响肠道微生物群。
✦ 尽量摄入不饱和脂肪
当研究人员观察脂肪时,发现并不是所有的脂肪都是平等产生的。在一项研究中,小鼠被喂食猪油或鱼的脂肪。喂猪油的小鼠中拟杆菌属和嗜胆菌属(Bilophila)增加,而双歧杆菌和Adlercreutzia、乳酸菌(乳杆菌和链球菌)和疣微菌属(Akkermansia muciniphila)在鱼油喂养的小鼠中增加。
猪油喂养的小鼠还出现白色脂肪组织炎症和胰岛素敏感性受损。喂食鱼油的小鼠总体表现更好,研究人员得出结论,肠道微生物群促进了高饱和脂肪饮食的整体炎症反应。
✦ 高纤维的饮食可以减少炎症
碳水化合物,包括可消化的(淀粉和糖)和不可消化的(纤维),可能是研究最多的常量营养素,因为它们具有改变肠道微生物组的能力。
人类喂食大量葡萄糖、果糖和蔗糖,双歧杆菌水平增加,拟杆菌减少。相反,当喂食糖精(一种人工甜味剂)时,拟杆菌丰度增加,罗伊氏乳杆菌减少,微生物发生与天然糖完全相反的变化。
纤维被认为是一种不易消化的膳食成分,有时被称为益生元,因为它通过刺激有益肠道细菌的生长而对宿主有益。
来源包括大豆、菊粉、未精制的小麦和大麦、生燕麦和不可消化的低聚糖,例如果聚糖、聚葡萄糖、低聚果糖 (FOS)、低聚半乳糖 (GOS)、低聚木糖 (XOS) 和阿拉伯低聚糖 (AOS)。
低纤维饮食与较低的细菌丰度有关,而富含这些不易消化的碳水化合物的饮食会增加双歧杆菌和乳酸杆菌。较高的纤维摄入量导致促炎细胞因子IL-6和胰岛素抵抗减少。
✦ 发酵食品可以增加双歧杆菌和乳杆菌水平
除了常量营养素和纤维对肠道细菌的影响外,含有乳酸菌的发酵食品(例如发酵乳制品)也被发现可以增加双歧杆菌和乳酸杆菌的水平。
✦ 多酚具有抗氧化活性能够减少一些致病菌
最后但并非最不重要的是多酚,包括儿茶素、黄酮醇、黄酮、花青素、原花青素和酚酸(存在于水果、种子、蔬菜、茶、可可制品和葡萄酒等食品中),被发现可以增加拟杆菌,减少金黄色葡萄球菌、鼠伤寒沙门氏菌和梭菌等致病菌。
拟杆菌门的成员被假设可以降低细胞内的氧水平,从而有利于厌氧菌的生长,从而促进肠道平衡的维持,并且它们被认为是关键的聚糖降解细菌比厚壁菌门等其他类群更能代谢多酚。
多酚可以改变肠道环境,它们可以被肠道细菌分解代谢,将它们转化为多种化合物比原始化合物具有更强的抗氧化活性。
厚壁菌门/拟杆菌门(F/B比率)作为一个指标在很多关于肠道菌群的研究中被提及和表征,从疾病发生进展,到药物饮食干预,涉及肠道菌群的研究,首先会描述细菌门水平的变化,而人类或动物肠道细菌约90%都属于厚壁菌门与拟杆菌门,而它们的比率也被认为是肠道菌群动态平衡的标志。
拟杆菌门主要为革兰氏阴性菌,能激活T细胞介导的免疫反应,降解多糖保护肠道和其他病原菌,厚壁菌门主要为革兰氏阳性菌,尤其产丁酸的菌,但是更容易吸收能量。
但是肠道菌群受到抗生素,药物,年龄以及分析注释数据库等的影响,此外随着肠道菌群研究队列的扩大,以及对菌群构成与不同疾病状态和药物饮食等因素的相关性研究的深入,F/B比率作为一个相对粗放的指标已经无法准确反映复杂的肠道菌群状况。我们需要更加深入和全面的通过菌属甚至菌种的构成变化来解释和分析肠道菌群特征,并进而更准确的解释肠道菌群和健康及生活方式的关联。
主要参考文献
Stojanov S, Berlec A, Štrukelj B. The Influence of Probiotics on the Firmicutes/Bacteroidetes Ratio in the Treatment of Obesity and Inflammatory Bowel disease. Microorganisms. 2020 Nov 1;8(11):1715.
Mariat D, Firmesse O, Levenez F, Guimarăes V, Sokol H, Doré J, Corthier G, Furet JP. The Firmicutes/Bacteroidetes ratio of the human microbiota changes with age. BMC Microbiol. 2009 Jun 9;9:123.
Gui X, Yang Z, Li MD. Effect of Cigarette Smoke on Gut Microbiota: State of Knowledge. Front Physiol. 2021 Jun 17;12:673341.
Grigor’eva IN. Gallstone Disease, Obesity and the Firmicutes/Bacteroidetes Ratio as a Possible Biomarker of Gut Dysbiosis. J Pers Med. 2020 Dec 25;11(1):13.
Vaiserman A, Romanenko M, Piven L, Moseiko V, Lushchak O, Kryzhanovska N, Guryanov V, Koliada A. Differences in the gut Firmicutes to Bacteroidetes ratio across age groups in healthy Ukrainian population. BMC Microbiol. 2020 Jul 22;20(1):221.
Indiani CMDSP, Rizzardi KF, Castelo PM, Ferraz LFC, Darrieux M, Parisotto TM. Childhood Obesity and Firmicutes/Bacteroidetes Ratio in the Gut Microbiota: A Systematic Review. Child Obes. 2018 Nov/Dec;14(8):501-509.
Hidalgo-Cantabrana C, Delgado S, Ruiz L, Ruas-Madiedo P, Sánchez B, Margolles A. Bifidobacteria and Their Health-Promoting Effects. Microbiol Spectr. 2017 Jun;5(3).
An J, Kwon H, Kim YJ. The Firmicutes/Bacteroidetes Ratio as a Risk Factor of Breast Cancer. J Clin Med. 2023 Mar 13;12(6):2216.

谷禾健康
一般在肠道菌群健康检测报告中,会有主要菌属构成比例,类似下图:
图源:谷禾肠道菌群健康检测报告我们可以看到图中列举了拟杆菌门,厚壁菌门…等。很多小伙伴拿到报告之后,对于这些细菌门表示困惑。
这些细菌门都代表了什么意思?
某个细菌门比例高了会怎么样?
某个细菌门比例高,是好是坏?
是不是与某些疾病相关?
…….
实际上,这几大类门的丰度和构成比例变化与人类健康和疾病发生息息相关。
两个主要门类,拟杆菌门和厚壁菌门,构成人类肠道细菌的大部分。七岁以后,拟杆菌和厚壁菌之间的比例相对稳定,而它们比例紊乱可能会导致肥胖和糖尿病等代谢综合征。
本文,我们就给大家详细介绍一下这其中最常见的几大细菌门:
拟杆菌门、厚壁菌门、变形菌门、放线菌门。
为什么选这四大细菌门?
依据自然属性分类,人类肠道菌群已经鉴定出细菌的几十个门,包括:拟杆菌门、厚壁菌门、变形菌门、放线菌门、疣微球菌门、梭杆菌门、蓝藻菌门、螺旋体门等。 但是其中98%的肠道菌可以归属为前四类,拟杆菌门、厚壁菌门、变形菌门和放线菌门,大多数人这4个门的菌占据肠道细菌数量的99%。 了解这四大细菌门,可以说你对大部分人体内肠道菌群的有了一定的了解。
▲ 拟杆菌 :革兰氏阴性菌
不形成孢子 厌氧菌 杆菌 ▲
它们具有外膜、肽聚糖层和细胞质膜。它们无氧呼吸的主要副产物是乙酸、异戊酸和琥珀酸。
包括碳水化合物的发酵、含氮物质的利用以及胆汁酸和其他类固醇的生物转化。大多数肠道细菌是糖酵解的,这意味着它们通过碳水化合物分子的水解获得碳和能量。
据估计,只有约 2% 的单糖通过上消化道到达结肠。因此,单糖可能不是拟杆菌的主要能量来源。然而,来自植物纤维的多糖,如纤维素、木聚糖、阿拉伯半乳聚糖和果胶,以及植物淀粉,如直链淀粉和支链淀粉,在结肠中更为普遍。
多糖还被证明可以诱导来自拟杆菌的多种葡糖苷酶活性,包括负责海带多糖降解的 β-1,3-葡糖苷酶活性以及多种 α、β-1,4 和 α、β–1,6 木糖苷酶和葡糖苷酶活性。大部分拟杆菌成员蛋白质组包括水解这些多糖的蛋白质。
表现出相当大的营养灵活性和对宿主和肠道环境施加的压力做出反应的能力。很难说肠道拟杆菌是否对宿主产生负面或正面影响。拟杆菌能够通过预防可能定植和感染肠道的潜在病原体感染来使宿主受益。由但是,在短链脂肪酸存在下,弱酸性 pH 值会抑制生长。诸如脆弱拟杆菌之类的致病物种已经开发出逃避宿主免疫系统的策略。
作为多糖降解联盟的成员,它们有助于从膳食纤维和淀粉中释放能量,它们很可能是丙酸盐的主要来源;然而,它们也参与了蛋白质分解过程中有毒产物的释放,这个群体的成员有一些可能有助于抑制炎症的活动,但他们也有可能促进炎症,有些是已知的机会性病原体。