-

 国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子
国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子

 二级病原微生物安全实验室
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业

谷禾健康
土壤作为地球上生物多样性最丰富的栖息地之一,不仅是植物生长的基础,也是众多微生物的”家园”。
然而,这片看似平静的地下世界,却隐藏着威胁人类健康的”隐形杀手”——人类细菌病原体。如单核细胞增生李斯特菌、铜绿假单胞菌、炭疽杆菌、沙门氏菌、大肠杆菌、结核分枝杆菌复合体和鼠疫杆菌等危险病原体,都可能在土壤中找到生存的港湾。
据统计,2019年全球约有770万例死亡与人类细菌病原体有关,这些数字背后,土壤作为病原体的自然来源和传播媒介,扮演着不容忽视的角色。
人类病原体通过多种途径进入土壤:动物和人类排泄物的渗透、农业实践中的灌溉和粪便应用、空气传播的病原体和孢子沉积等。一旦进入土壤,这些病原体可能成为常驻微生物组的一部分,并通过生食摄入、直接接触、吸入或昆虫及野生动物传播等多种途径感染人类。
本文基于最近一篇发表在《Cell Host & Microbe》上题为“Soil microbial diversity associates with lower prevalence of human bacterial pathogens across global soils”的研究,发现土壤微生物多样性与病原体流行率在全球呈负相关,并揭示病原体丰度与致病力及全球死亡率模式显著相关。在全球变化情景下,多数优势病原体比例可能上升。研究还绘制了全球土壤主要人类病原体分布图,阐明其生物地理与生态特征,为制定监测与风险管理策略、降低疫情暴发风险提供依据。
这意味着,在肠道菌群检测中发现的某些异常或致病菌,并不一定源于短期感染,而很可能是长期环境暴露累积的结果。因此,病原菌的控制不能仅依赖益生菌调节或抗生素干预,更重要的是减少其从环境进入人体的路径。
如果肠道检测报告中出现如沙门氏菌、致病性大肠杆菌、类鼻疽杆菌、克雷伯氏菌等,应重点考虑其环境来源。土壤是这些病原体的重要储存库,日常园艺、农作活动或儿童玩土等行为,都可能在无意识中增加暴露风险。特别是在潮湿或热带地区,这类病原体更易长期存活并富集。
农产品则是连接土壤与人体的重要媒介。叶菜类直接接触土壤,根茎类附着土壤较多,而水果也可能因灌溉水或土壤飞溅受到污染。如果种植过程中使用未经充分处理的粪肥,或灌溉水源受到污染,再加上洪水或暴雨等极端气候事件导致土壤扰动,都会显著增加农产品携带病原体的风险。此外,动物粪便通过土壤进入食物链,或宠物将病原体带入家庭环境,也构成重要的传播路径。
值得特别注意的是,病原体具有明显的地区性与季节性特征。不同地区的土壤病原谱差异显著,例如热带地区更易出现类鼻疽杆菌等环境致病菌,而干旱地区则更利于孢子类病原体的长期存在。同样,季节变化也会影响病原体丰度,雨季和洪水之后往往出现显著上升,高温条件促进细菌繁殖,这些都直接影响人群暴露风险。
▸ 土壤中的人类细菌病原体
一项覆盖59个国家、1602个土壤宏基因组的全球研究,为我们揭示了土壤中人类细菌病原体的分布模式。
研究中样本地点的全球分布
Xiong C,et al.Cell Host Microbe.2026
研究人员共鉴定出80个被归类为潜在土壤栖息人类病原体的细菌分类群,包括类鼻疽假单胞菌(Burkholderia pseudomallei)、结核分歧杆菌复合群(mycobacterium tuberculosis complex)、铜绿假单胞菌(Pseudomonas aeruginosa)、肠沙门氏菌(Salmonella enterica)、克雷伯氏菌(Klebsiella spp.)和志贺氏菌(Shigella spp.)等。
• 优势病原体主导模式
在这80种病原体中,25种被确定为优势病原体—它们不仅广泛存在(出现在≥80%的样本中),而且在病原体群落中表现出较高的相对丰度。
令人惊讶的是,这25种优势病原体仅占所有病原分类群的31.3%,却贡献了总病原体序列的90.8%。这种”少数主导”的模式与土壤中优势细菌和真菌的分布规律一致,反映了微生物群落结构的普遍特征。这些优势致病类群属于23种病原体,分布在19个属中,如伯克霍尔德氏菌属(Burkholderia)、分支杆菌属(Mycobacterium)、诺卡氏菌属(Nocardia)、假单胞菌属(Pseudomonas)、沙门氏菌属(Salmonella)和克雷伯氏菌属(Klebsiella)等。
▸ 高致病性优势病原体的健康威胁
在这23种显性病原体中,有11种被列为每年导致770万人死亡的33种人类细菌病原体之首,包括铜绿假单胞菌、肺炎克雷伯菌、肠道沙门氏菌、大肠杆菌、鲍曼不动杆菌、军团菌属、沙雷氏菌属和气单胞菌属等。
• 结核分枝杆菌每年导致大量感染且可在土壤中存活
其中,结核分枝杆菌尤为引人注目,其每年可导致约1060万人感染和160万人死亡。研究发现其成员能够感染各种家养和野生哺乳动物,并能在土壤中以毒力形式存活和增殖,使用变形虫和野生动物等替代宿主维持生存。
• 麻风分枝杆菌、淋病奈瑟菌也能在土壤中存在
值得注意的是,一些传统上不被认为是土壤传播的病原体(如结核分枝杆菌、麻风分枝杆菌和淋病奈瑟菌)也在土壤中被检测到。虽然它们从土壤到人类的传播途径尚不清楚,但最新研究发现蚊子可能是溃疡分枝杆菌(M. ulcerans)的新传播途径,这提示我们许多病原体的传播机制仍有待探索。
这些发现强调了重新关注病原体在土壤中的存活机制及其向人类传播途径的重要性,这对于有效监测和风险管理至关重要。
▸ 病原体的全球分布
通过对21个陆地生物群落的分析,研究发现了人类病原体在不同气候和植被类型下的分布模式。数据显示,主要人类病原体的相对丰度在热带和寒冷农田、温带湿地、温带农田以及热带和温带森林中最高。
进一步分析发现,农田、湿地和森林生态系统,特别是在热带和温带地区,表现出最高的优势病原体相对丰度。而在热带和干旱农田中,人类病原体的多样性最为丰富。
• 热带地区病原体多样性和患病率最高
分布模式与基于流行病学数据的临床研究结果一致——热带地区人类病原体的多样性和患病率最高。例如,东南亚(热带地区)报告了大多数结核病(由结核分枝杆菌引起)和麻风病(由麻风分枝杆菌引起)病例。
为探究环境因素对优势人类病原体全球分布的影响,研究人员通过随机森林和相关性分析发现:年均降水量(MAP)、年均温度(MAT)、降水季节性(PSEA)及土壤pH是最关键因素;此外,海拔、土壤质地、有机碳、土壤生物多样性(含微生物物种与功能基因多样性)及人口密度也具有重要作用。
全球土壤中主要人类细菌病原体的环境驱动因素
编辑
Xiong C,et al.Cell Host Microbe.2026
1
降雨与土壤病原体丰度正相关
• 降雨能够增加病原体在土壤中的分布
研究结果显示,降雨与23种主要病原体中的19种(约82.6%)相对丰度显著正相关。类鼻疽杆菌(B.pseudomallei)在潮湿土壤中生长最好,类鼻疽病例的增加与异常强降雨事件有关。
降雨增加可以提高土壤孔隙水含量,促进病原体的运动,增强其活性和流行性,从而促进其在土壤中的分布。
• 植物覆盖率和生产力也影响病原体丰度
植物覆盖率和生产力与病原体丰度同样呈正相关,这可能反映了植物驱动的土壤有机输入和水分稳定性增加,为某些病原体分类群的持久性提供了有利条件。
相比之下,降水季节性(23种病原体中的19种)、海拔(16种)和土壤pH值(14种)与这些分类群呈负相关。
全球不同生态系统中主要人类病原体的分布模式
编辑
Xiong C,et al.Cell Host Microbe.2026
2
土壤pH值与病原体呈负相关
土壤pH值与最主要的病原体呈负相关,表明酸性土壤往往含有更高的病原体丰度。
• 酸性土壤会破坏原有微生物结构,增加病原体增殖
这与最近的研究证据一致——土壤酸化会改变微生物群落结构,破坏关键的微生物功能,从而削弱微生物组限制病原体增殖的能力。
分析还揭示了降雨和土壤pH值之间存在显著的关系,这意味着气候变化下一些地区的降水增加可能会通过提供最佳水分条件和进一步酸化土壤来促进病原体的丰度。同样,不可持续的农业实践(如过度施肥)会加速土壤酸化,创造有利于人类病原体扩散的条件。
3
温度升高:病原体扩散的”催化剂”
温度与最主要的人类病原体(包括洋葱伯克霍尔德菌群(Burkholderia cepaciaComplex)、结核分枝杆菌复合体和炭疽杆菌)的相对丰度呈正相关,这表明全球变暖可能会进一步增加全球土壤中人类病原体的流行率。
这一发现说明气候变化可能会加剧人类致病性疾病,并表明在未来的气候情景下,降雨量和温度增加的地区可能成为人类细菌病原体的”热点”,进一步增加暴露、感染和死亡的风险。
除主要病原体外,研究还确定了其他8种病原体的环境驱动因素,包括柠檬酸杆菌属、志贺氏菌属、嗜肺军团菌、屎肠球菌、蜡样芽孢杆菌和问号钩端螺旋体等。这些病原体在降雨、温度和土壤特性方面与主要病原体具有相似的生态偏好。这些关于病原体当前分布和环境偏好的研究结果可以作为监测未来流行和爆发的基线数据,例如在筛查伯克霍尔德菌潜在爆发时,应优先考虑高降雨地区。
土壤微生物组多样性与病原体流行的关系是本研究的核心发现。研究基于宏基因组数据评估微生物物种多样性及功能多样性,并通过相关性与结构方程模型(SEM)分析其与人类病原体的关联;同时结合扩增子测序确定细菌和真菌多样性,并在全球200个土壤样本中加以验证。
▸ 微生物多样性与病原体丰度负相关
相关性分析显示,微生物物种多样性和功能多样性与大多数显性病原体的相对丰度呈显著负相关,包括类鼻疽杆菌、结核分枝杆菌复合物、炭疽杆菌、大肠杆菌和肠道沙门氏菌等。
• 土壤中细菌和真菌的多样性有助于减少病原体
在23种主要病原体中,15种和14种病原体分别与微生物种类和功能多样性呈显著负相关。研究还发现,与农业土壤相比,天然土壤中土壤微生物组多样性与主要人类病原体的相对丰度之间存在更强的负相关关系。
土壤微生物多样性决定主要人类病原体的全球分布
编辑
Xiong C,et al.Cell Host Microbe.2026
SEM分析进一步表明,土壤微生物多样性与优势人类病原体的相对丰度呈显著负相关。全球200个土壤样本亦证实:细菌和真菌多样性及细菌丰度均与主要病原体显著负相关。该结果拓展了以往土壤微观实验结论,表明提高微生物多样性可能抑制植物和人类病原体(如大肠杆菌O157:H7和肠道沙门菌)的增殖。
小编解释
较高的微生物多样性通过增加竞争者、耐压型和遗传策略类群,加剧竞争、扩大生态位占用并提升功能冗余,从而抑制病原体建立;这突显了保护土壤生物多样性对土壤、环境和人类健康的重要性。
为验证已鉴定的病原体与疾病毒力之间的关联,评估这些生物体的致病潜力,研究分析了病原体相对丰度与致病特征之间的关系。
• 土壤主要病原体丰度至少关联一种人类疾病特征
结果显示,23种主要病原体中有18种(包括类鼻疽杆菌、结核分枝杆菌、铜绿假单胞菌、鲍曼不动杆菌和大肠杆菌)以及一些其他重要病原体(如柠檬酸杆菌、嗜肺军团菌、伯氏考克斯氏体和蜡样芽孢杆菌)等,其相对丰度均与至少一种与人类疾病密切相关的关键致病特征显著正相关。
这些致病特征包括所有细菌病原体的毒力基因,根据其致病性分为不同的功能组,如毒素合酶、细胞毒性和毒力活性等。
人类细菌病原体与致病特征的关系
编辑
Xiong C,et al.Cell Host Microbe.2026
▸ 毒力因子与抗生素耐药基因
研究使用三个单独的数据库(PHI、VFDB和KEGG)进一步鉴定了与这些病原体相关的毒力因子,并使用DeepARG鉴定了抗生素耐药基因(ARG)。
• 土壤病原体相对丰度影响毒力因子与耐药性
分析表明,在全球范围内,显性病原体(类鼻疽杆菌、结核分枝杆菌、铜绿假单胞菌和鲍曼不动杆菌)的相对丰度与其相关毒力因子和抗生素耐药性之间存在很强的正相关关系。
这些发现进一步支持了识别人类病原体方法的稳健性,并证实了土壤中检测到的病原体可能会导致严重的疾病和死亡。
宏基因组揭示潜在病原体及其致病特征
为从全基因组层面验证宏基因组分箱识别显性病原体的可靠性,研究筛选出7个高质量(完整性≥90%、污染≤5%)MAG作为潜在病原体,其全球分布与定制数据库结果一致,且相对丰度与毒素合成和毒力活性等关键致病特征呈正相关。
• 宏基因组方法有助于识别土壤病原体
其中两个高质量MAG(T073.8和SRR8879131.4)与分枝杆菌的平均核苷酸同一性(ANI)达83%–84%,被认定为潜在重要病原体,并与温度和降雨量呈正相关。
这两者均携带与结核分枝杆菌复合群相关的五类致病特征(毒素合成、细胞毒性、毒力活性、器官损伤和宿主细胞死亡)、抗生素耐药基因(efpA,多重耐药)及外毒素基因(如磷脂酶C、VFC0235)。
结果表明,宏基因组学流程可用于从环境样本中识别潜在新兴病原体。
研究进一步评估了土壤人类病原体分布与全球相关死亡的关系。基于既往研究和世卫组织数据,汇总204个国家和地区的死亡率,并在国家尺度匹配到53个国家的数据。
• 土壤病原体分布与相关疾病死亡率相关
结果显示,麻风分枝杆菌(r=0.51,p=8.3×10^-5)和结核分枝杆菌复合物(r=0.47,p=0.0004)的相对丰度与病原体相关的全球死亡率(或感染病例)显著相关。
此外,细菌病原体相关死亡率与致病特征亦呈显著或接近显著相关,包括毒素合酶(r=0.45,p=0.0006)和毒力活性(r=0.25,p=0.07)。这些结果表明,土壤中人类病原体的流行可能与人类健康密切相关,提示土壤或为其天然宿主。
▸ 病原体的双向传播:土壤⇌人类
土壤与人类之间的病原体传播可能是双向的。一方面,土壤作为天然宿主,病原体可能通过各种途径感染人类;另一方面,人类病原体也可能通过空气、水道或农业实践(如施肥)等生物和非生物媒介从受感染的人群沉积或传播到自然和农业系统中。
• 土壤病原体多处于休眠状态,条件适宜时可经农产品等传播
传播到土壤中的病原体可能以低丰度或休眠孢子的形式存在,在土壤环境中找到替代宿主以生存和增殖;一旦满足传播条件,它们可能通过农产品、野生动物、病媒和城市扩张等各种传播途径重新进入人群,构成健康风险。
研究结果强调,在评估和管理与人类细菌病原体相关的风险时,需要明确考虑土壤因素。如果在地方和区域范围内得到证实,监测栖息在土壤中的人类病原体的流行情况可以为追踪全球疾病负担提供信息,简化流行病学监测,并有助于识别传染病热点。
研究局限性
疾病结果受到许多因素的影响,如人口密度、国家发展水平、社会经济状况和区域医疗质量等,在制定公共卫生政策时也应考虑这些因素。本研究所代表的国家数量有限可能会限制观察;随着数据库的改进,临床结果的预测能力也可能提高。
此外,一些病原体(如肠沙门氏菌和单核细胞增生李斯特菌)通过生食感染人类的途径已经确立,但其他病原体(如结核分枝杆菌)从土壤传播给人类的途径仍然难以捉摸。
基于环境变量(如气候和土壤因素)与人类病原体相对丰度的显著相关性,研究绘制了其在全球土壤中的当前及未来分布,模型可解释58%–88%的空间变异。
结果显示,总体显性病原体、特定病原体(如类鼻疽杆菌和结核分枝杆菌复合群)及关键致病功能(毒素合成、毒力活性和细胞毒性)在撒哈拉以南非洲、南美和东南亚等地区呈高丰度分布。
▸ 预测地图的验证及其应用价值
预测的结核病综合征全球地图显示了与世界卫生组织估计的全球结核病地图可比的模式——大多数结核病病例发生在东南亚、西太平洋和非洲。
• 病原菌分布与其致病人群分布相似
同样,预测的类鼻疽杆菌全球分布图与基于人类和动物感染病例的假鼻锤杆菌全球分布模式一致,类鼻疽在东南亚和南亚、澳大利亚北部、太平洋和印度洋岛国、撒哈拉以南非洲以及中南美洲的许多热带地区高度流行。这些结果为研究方法的稳健性提供了额外支持。
编辑
Xiong C,et al.Cell Host Microbe.2026
值得注意的是,在干旱地区(包括北非、西亚、澳大利亚北部和中部以及美国西部)发现了病原菌多药耐药诺卡菌的相对丰度最高。这与美国诺卡氏菌感染人数在西南部干燥温暖气候中最高的报告相吻合,表明不同病原体对特定气候条件的适应性。
▸ 气候变化下的病原体分布预测
在持续的气候变化下,降水模式和温度正在发生变化,这可能会对人类病原体的分布产生深远影响。
• 未来一些病原体和致病特征可能呈上升趋势
研究分析了病原体相对丰度从当前气候情景到预测的未来气候情景的潜在变化,结果表明在未来的气候情景下,一些关键病原体和致病特征的比例在全球大多数地区呈上升趋势。
例如,结核分枝杆菌复合物和致病群毒素合酶在撒哈拉以南非洲、南美洲、西欧、南亚和东南亚的某些地区呈现出明显的增长趋势。这与最近关于环境中人类病原体丰度总体增加的报告相一致。
这些地图提供了全球主要人类病原体的空间定量信息,填补了关于生活在土壤中的人类病原体的全球分布和环境偏好的关键知识空白。
然而,随着未来气候变化情景下土壤微生物组多样性的预测数据可用,以及更多宏基因组序列(特别是来自采样中代表性较低的地区/国家,如非洲国家)的积累,预测模型可以进一步改进。
总体而言,全球研究揭示了土壤中人类病原体分布的几个关键发现:主要栖息在土壤中的人类病原体的全球分布由气候条件和土壤生物多样性决定,在更潮湿的环境中观察到相对丰度更高。
土壤微生物多样性与这些病原体的流行率(即相对丰度)呈负相关,表明土壤生物多样性在最大限度地降低人类病原体爆发风险方面发挥着关键作用。
这些结果共同强调了土壤生物多样性是一个潜在的重要生态因素,它塑造了栖息在土壤中的人类细菌病原体的全球生物地理学。
▸ 研究的关键启示
这项研究确定了广泛分布在各种生态系统类型中的主要病原体,并揭示了这些病原体的相对丰度与传染病的致病特征和全球死亡率之间的正相关关系。此外,研究展示了主要病原体的环境偏好,并预测了未来全球变化可能如何影响其空间分布。这些发现对未来的研究重点和感染风险的管理具有重要意义:
1.病原体传播途径再评估:多种主要病原体(如类鼻疽杆菌、沙门氏菌和炭疽杆菌)已知可通过环境(如农产品、动物媒介和尘埃)传播,而结核分枝杆菌复合体等则传统上被认为仅经人际传播。研究表明这些病原体可在土壤中长期存活,未来应重点开展与风险管理相关的研究,如其在土壤中的存活与分布机制,以及潜在的土壤—人类传播新途径。
2.健康风险预警与管理:如果土壤中病原体的相对丰度与当地感染率之间具有稳定关系,就可以用于预测特定环境条件下的疾病风险,例如洪水或强降雨后类鼻疽的发生概率。通过持续监测土壤病原体分布与微生物多样性,并结合临床数据,有望建立更加精准和动态的疾病预警体系。
3.综合方法应对多重社会挑战:将气候、土壤与社会经济因素纳入同一分析框架,有助于提升人类病原体风险评估的系统性与前瞻性。相关研究已弥补了关键病原体在全球分布及其与生物多样性关系方面的认知空白,为预测不同气候情景下的人群健康风险提供了重要依据。
从健康管理角度看,对检测结果的理解也需要升级。肠道菌群检测不应只关注“缺乏哪些有益菌”,更应关注是否存在环境来源的病原体,以及这些菌是否反复出现。如果某类病原体持续存在,更可能提示持续暴露,而非单纯治疗不足。因此,定期监测趋势并结合生活环境变化进行分析,比单次检测更具意义。
在更宏观层面,未来公共卫生管理需要加强对土壤微生物的系统性监测,例如建立区域性的土壤微生物数据库与风险地图,对高风险区域进行预警,并在农业生产中规范粪肥处理和灌溉水管理。同时,在城市规划和灾害管理中,应将洪水后土壤病原风险纳入评估体系。
总体来看,对病原菌的控制应从“治疗导向”转向“源头管理”。环境中的土壤是一个关键但长期被忽视的源头,真正有效的健康策略应覆盖三个层面:上游减少环境暴露,中游控制食物污染,下游调节肠道生态。只有在这一整体框架下,才能更有效地降低感染风险并实现长期健康维护。
参考文献:
Xiong C, Delgado-Baquerizo M, Liang J, Wang J, Yan Z, Jensen SO, Gao M, Sáez-Sandino T, Guirado E, Muñoz-Rojas M, Román R, Maestre FT, Singh BK. Soil microbial diversity associates with lower prevalence of human bacterial pathogens across global soils. Cell Host Microbe. 2026 Mar 31:S1931-3128(26)00119-8.

谷禾健康
《植物性饮食》
植物性饮食能提供丰富的膳食纤维、多酚和其他生物活性化合物,适当和健康的植物性饮食能够促进有益肠道细菌的生长,增加菌群多样性和短链脂肪酸(SCFAs)的产生,有助于改善肠道屏障功能、减少炎症并降低慢性疾病风险。
然而,值得注意的是,植物性饮食也可能通过生食和未加工食品将环境微生物引入肠道,其中一些微生物可能是机会致病菌或携带抗生素抗性基因,这对健康构成潜在威胁。
此外,健康与不健康植物性饮食的区别也很关键。富含全谷物、水果、蔬菜和豆类的饮食与更有利的肠道菌群组成和代谢结果相关,而富含精制碳水化合物和加工食品的植物性饮食可能产生不利健康的效果。
本文一起了解下植物性饮食对肠道菌群的影响和塑造,尤其在多样性方面,以便更好地理解和实践植物性饮食,在享受植物性饮食带来的健康益处的同时,避免可能的健康风险。
植物性饮食主要由植物来源的食物组成。根据对动物产品的限制程度,可分为不同类型:
– 纯素饮食 (vegan)
仅包含植物性高纤维食物,如水果、蔬菜、谷物、豆类、坚果和种子,完全排除肉类、家禽、鱼类、乳制品、蛋类及所有动物副产品。
– 素食饮食 (vegetarian)
排除肉类和肉类衍生食品,但可能包含鸡蛋或乳制品等动物产品。
– 弹性素食 (flexitarian)
主要以植物性食物为主,但偶尔包含少量肉类或鱼类。
植物性饮食增加多样性和有益菌
研究表明,富含多种植物性食物的饮食始终与更高的肠道微生物多样性相关。总体而言,从植物中引入有益细菌,加上植物纤维,可能培养出更具弹性和功能多样性的肠道生态系统,最终可能导致人类更好的健康结果。
肠道微生物通过发酵促进消化
因为生活在结肠中的肠道微生物通过发酵胃和肠道酶无法降解的食物物质,对消化做出重要贡献。富含植物性食物的饮食能显著改善有益肠道菌群,直接影响人类健康。
最近的研究结果强调,植物性饮食富含益生元,如膳食纤维(包括抗性淀粉),它们作为有益肠道微生物的能量来源。
特定细菌发酵膳食纤维生成SCFAs
膳食纤维是一类植物性碳水化合物,包括非淀粉多糖、木质素和抗性淀粉,它们能逃脱人类上消化道内源性酶的消化。只有特定的肠道菌群,如F. prausnitzii、Roseburia、双歧杆菌和普雷沃氏菌,能在厌氧条件下发酵膳食纤维,产生SCFAs(如乙酸、丙酸和丁酸)作为主要代谢产物。
编辑
一项系统综述显示,纯素和素食饮食通过增加有益细菌同时减少有害细菌,在重塑肠道菌群方面发挥关键作用。在食用植物性饮食的个体中,观察到F. prausnitzii、Roseburia spp.、Coprococcus spp.和Ruminococcus spp.的水平增加,这些细菌都能产生SCFAs,促进更好的肠道健康、减少炎症和改善新陈代谢。
研究还发现普雷沃氏菌属的丰度增加,在某些情况下还有脆弱拟杆菌(Bacteroides fragilis),它们在纤维消化和免疫调节中起关键作用。但是要注意,不合理的植物性饮食也会使得普雷沃氏菌属或有些情况下的脆弱拟杆菌过度增殖,影响菌群平衡和进一步的消化免疫状况。
短链脂肪酸:肠道菌群的黄金代谢物
短链脂肪酸(SCFAs)已被广泛研究为关键的肠道菌群代谢物。人类由于缺乏必要的酶而无法发酵某些碳水化合物,而肠道菌群则能合成诸如丙酸-CoA转移酶和丙醛脱水酶等酶,将这些碳水化合物转化为SCFAs。
胃肠道中的主要SCFAs是乙酸、丙酸、丁酸(占总量的≥95%):
乙酸
通过Wood-Ljungdahl途径和乙酰-CoA途径合成,相关细菌包括乳酸杆菌属、双歧杆菌属、 Akkermansia muciniphila、拟杆菌属、普雷沃氏菌属、瘤胃球菌属、链球菌属等。
丙酸
由Phascolarctobacterium succinatutens、拟杆菌属、Dialister、Megasphaera elsdenii、韦荣球菌属、AKK、Coprococcus catus、Roseburia inulinivorans、Ruminococcus obeum、沙门氏菌属等通过琥珀酸、丙烯酸和丙二醇途径生成。
丁酸
主要由Faecalibacterium prausnitzii、Roseburia、Eubacterium rectale、Clostridium leptum、Eubacterium hallii、Coprococcus eutactus、Anaerostipes caccae、C. catus等通过丁酰-CoA:乙酸CoA转移酶和磷酸丁酰转移酶/丁酸激酶途径产生。
短链脂肪酸在维持肠道屏障完整性、调节免疫反应和支持代谢健康方面发挥着关键作用,因此与纯素和素食饮食相关的独特肠道菌群特征有助于产生抗炎作用和改善代谢结果。
并非所有植物性食物都同样健康
一些植物性食物可能增加健康风险。为解决这些问题,研究人员创建了:
另一项研究发现,较高的hPDI与Bacteroides cellulosilyticus和Eubacterium eligens的丰度增加相关,这两种细菌都有助于积极的代谢健康。
相反,较高的hPDI显示Ruminococcus torques和Ruminococcus gnavus的水平降低,这些细菌种类已知会引起炎症反应和不利的代谢结果。
高hPDI饮食促进有益微生物,降低代谢风险
高hPDI还与肠道微生物途径的富集相关,这些途径参与支链氨基酸(包括L-异亮氨酸和L-缬氨酸)的生物合成,以及丙酮酸发酵生成异丁醇,这些都与较低的代谢风险评分相关。相反,低hPDI与嘌呤碱基降解和脂质生物合成相关的途径相关,这与不太有利的代谢特征相关。
这些结果表明,坚持健康的植物性饮食能促进有益的肠道菌群和代谢健康,而不健康的植物性饮食可能产生相反的影响。
植物性饮食与肠道菌群的关系
一项大规模调查研究了杂食者、素食者和纯素者的肠道菌群特征,分析了来自5个独立全球队列的21,561人的数据。
研究表明,植物性饮食驱动肠道菌群组成,有利于参与纤维发酵和SCFAs合成的微生物物种。与杂食者相比,纯素饮食的人总体肠道微生物多样性较低,但有益物种(如毛螺菌科、Butyricicoccus spp.和Roseburia hominis)的较高,这些物种有助于减少炎症和增强肠道健康。
研究局限与未来方向
然而,该研究也存在局限性:参与者被分为杂食者、素食者或纯素者时,没有考虑饮食质量,例如全食物纯素与加工纯素的区别。尽管使用了健康植物性饮食指数(hPDI),但它没有考虑纤维摄入量的差异,而纤维摄入量可能对肠道菌群产生不同影响,并可能为积极的代谢健康提供机制见解。
大多数关于植物性饮食对肠道菌群影响的研究持续时间较短,从几天到13个月不等。虽然短期饮食干预可以快速改变微生物组成,但它们可能无法提供长期稳定性和更广泛健康影响的完整图景。未来需要更多长期研究来探索植物性饮食对肠道菌群的持久影响。
多酚:植物中的抗氧化战士
植物性饮食富含多种植物生物活性化合物,包括膳食纤维、多酚、类胡萝卜素、维生素和生物活性肽,这些化合物选择性地促进有益肠道细菌的生长和活性,从而有助于宿主健康。
据观察,富含叶菜类蔬菜、浆果、豆类、小扁豆和燕麦等植物性食物的饮食支持共生细菌的生长并提高SCFAs水平。
编辑
多酚是一类广泛而多样的天然化合物,大量存在于植物中,以其强大的抗氧化和抗炎特性为特征。它们可大致分为类黄酮和非类黄酮(酚酸和芪类)。
大多数膳食多酚在小肠中不被吸收,而是在结肠中进行微生物代谢,产生生物活性代谢物如尿石素,这些代谢物随后可被吸收到体内,并通过影响代谢、免疫甚至神经通路(通常通过复杂的肠-肝-脑轴)发挥显著的健康益处。
这些化合物似乎能增加人类肠道中益生菌如乳酸杆菌和双歧杆菌的丰度。据报道,来自浆果和蔓越莓等水果的多酚对梭状芽胞杆菌和大肠杆菌等致病菌具有抗菌活性,同时富集有益菌。
一些动物和人类研究表明,多酚可能将厚壁菌/拟杆菌比率转向更健康的平衡,这种平衡在肥胖和代谢紊乱中常常被破坏。通过增加SCFA水平和调节胆汁酸途径,多酚可以改善肠道屏障完整性,减少炎症,并支持代谢健康。
类胡萝卜素:未被充分探索的肠道调节剂
膳食纤维和多酚是研究最广泛的用于调节肠道菌群和促进肠道健康的生物活性化合物。相比之下,其他生物活性化合物如类胡萝卜素(β-胡萝卜素、番茄红素、虾青素、叶黄素、玉米黄质和岩藻黄质)在这方面的研究仍不充分。
类胡萝卜素具有抗炎和抗氧化特性,研究表明它们可能促进有益肠道细菌,增强肠道屏障功能,减少炎症。然而,大多数证据来自动物研究,剂量、干预持续时间各不相同,且对SCFAs产生的评估有限。
这突出了需要精心设计的人类干预研究,以更好地了解类胡萝卜素对肠道健康的影响。未来的研究应侧重于标准化的人体试验和综合多组学方法,以阐明特定类胡萝卜素和其他生物活性化合物影响肠道微生物多样性、屏障功能和宿主健康结果的机制。
不同植物性食物对肠道菌群的独特影响
不同的植物性食物含有独特的生物活性化合物组合,因此对肠道菌群有不同的影响:
-浆果类
富含花青素等多酚,促进双歧杆菌和乳酸杆菌生长。
-全谷物
含有丰富的膳食纤维和抗性淀粉,促进产丁酸菌如Faecalibacterium prausnitzii和Roseburia spp.
-豆类
含有棉子糖家族低聚糖,促进双歧杆菌增殖
-坚果和种子
富含不饱和脂肪酸、膳食纤维和多酚,增加 Akkermansia muciniphila等有益菌
-十字花科蔬菜
部分研究显示有调节拟杆菌丰度的潜力,含有硫代葡萄糖苷,经肠道菌群代谢为具有抗癌活性的异硫氰酸酯。
因此,多样化的植物性饮食能够提供更广泛的生物活性化合物,支持更丰富和多样化的肠道菌群,从而带来更多健康益处。
除了提供营养价值和塑造肠道菌群的生物活性次生代谢物外,植物性饮食还作为影响微生物多样性的来源。最近的研究首次提供了水果和蔬菜微生物群可能传播到人类肠道的证据。
在这项研究中发现的人类肠道与水果/蔬菜之间的共享微生物包括假单胞菌(Pseudomonas)、鞘氨醇单胞菌(Sphingomonas)、气单胞菌(Aeromicrobium)和泛菌(Pantoea)。
尽管这些细菌中的一些具有促进植物生长的特性,但它们可能在人类肠道中引起感染,特别是在免疫功能低下的个体中。
通过植物性饮食将土壤微生物群转移到人类肠道
编辑
图示显示微生物从土壤转移到植物,最后植物性饮食促进植物微生物群直接转移到人类肠道。
另外一项采用物种水平鉴定的跨国队列研究发现,食用植物性饮食的个体,特别是纯素者,肠道中通常与土壤和植物相关的微生物(包括Enterobacter hormaechei、Citrobacter freundii、Raoultella ornithinolytica和肺炎克雷伯菌)的较高。这是由于摄入更多的生的和最低限度加工的植物性食物,这些食物携带环境微生物。
编辑
在农业环境中,这些细菌通过溶解磷等营养物质、固氮、产生植物激素(包括吲哚-3-乙酸)以及作为植物病原体的生物防治剂,发挥着至关重要的作用。另一方面,它们在人类中可能是共生菌或机会致病菌,引起从尿路感染到败血症的广泛感染。这些细菌还携带抗菌素耐药基因,导致对抗生素的强烈耐药性。
下面总结了植物微生物在植物中的作用
和对人类的可能潜在风险(仅供参考)
编辑
我们食物中的微生物群落可能受到不同食品加工方法和更广泛的农业实践的影响。微生物从植物到人类的传播表明,植物及其土壤环境可以作为这些细菌的储存库。食用未加工或最低限度加工的植物性饮食可能是将共生菌或机会致病菌转移给人类的潜在载体。烹饪可能是使食物中天然存在或通过农业实践引入的抗菌化合物变性的途径。
小鼠模型研究:生食与熟食的微生物差异
一项对小鼠模型的研究表明,小鼠食用生的或熟的植物性饮食后,肠道菌群存在显著差异。根据饮食的不同,小鼠的肠道细菌负荷有所不同;食用生块茎的组比食用熟块茎的组细菌负荷更低。这种微生物负荷的下降与微生物细胞损伤增加和微生物活性降低有关。
人体研究
一项短期人体研究检查了在生食和熟食植物性饮食之间切换如何影响肠道菌群组成。结果表明,食用熟食后48小时内,肠道菌群发生了显著变化。食用生食和熟食的个体之间细菌类群丰度的差异突出了食品加工对肠道菌群结构和功能的影响。
农业实践通过减少整体多样性和选择性地支持化学/抗生素抗性细菌物种,改变土壤微生物组成。农业环境中的抗生素耐药性主要由肥料、废水等的应用驱动。
肥料作为抗生素耐药细菌的储存库
农业中广泛使用肥料是抗生素耐药细菌及其抗生素抗性基因的主要储存库。根际、内生圈和叶际内的微生物群落密切相关,形成一个统一的生态系统,对人类食物链至关重要。
抗生素耐药细菌在植物中的传播及其对人类的影响
抗生素耐药细菌可以侵入植物根部,进入茎和叶组织,或与内生菌交换抗性基因,促进植物内抗生素抗性的扩展。
人类通过食用可食用植物获得这些抗性基因。长期摄入这些抗性基因可能与人类致病性增加有关,因为这些基因可能通过水平基因转移被肠道的一些机会致病菌获得。然而,对与植物相关的抗生素耐药组及其向人类转移的理解仍处于初级阶段。
这些发现强调了土壤、植物和人类肠道中微生物群落的相互联系。因此,维持土壤健康对于健康的植物至关重要,而健康的植物又有助于平衡的肠道菌群。
多篇研究警告说,缺乏适当饮食规划的植物性饮食可能导致关键营养素(如维生素B12、维生素D、铁、钙、锌、二十碳五烯酸(EPA)和二十二碳六烯酸(DHA))的缺乏,可能损害肠道和整体健康。
考虑到遗传因素和微生物多样性的差异,即将进行的研究应阐明植物性饮食对个体肠道和代谢健康的影响。
营养建议
对于遵循植物性饮食的人,建议关注以下营养素的摄入:
-维生素B12
主要存在于动物性食物中,植物性饮食者应考虑强化食品或补充剂
-铁
植物性来源的铁(非血红素铁)吸收率较低,建议与富含维生素C的食物一起食用以提高吸收率
-钙
可从强化植物奶、豆腐(用钙盐制作)、绿叶蔬菜中获取
-Omega-3脂肪酸
可从亚麻籽、奇亚籽、核桃中获取ALA,以及其他富含DHA/EPA等食物
-锌
全谷物、豆类、坚果是主要来源,发酵食品可提高锌的生物利用率
编辑
✦ 如何开始并坚持健康的植物性饮食?
对于想要尝试植物性饮食的人,建议循序渐进:
-从餐盘开始
将餐盘的一半分配给蔬菜和水果,四分之一给全谷物,四分之一给植物蛋白来源
-增加多样性
尝试不同种类的植物性食物,每周至少尝试一种新的水果、蔬菜或全谷物
-注重全食物
优先选择完整的、未加工的食物,减少精制食品和添加糖
-合理搭配
结合不同植物蛋白来源(如豆类和谷物)以获得完整的氨基酸谱
-适当烹饪
虽然生食有其益处,但适当烹饪可以减少潜在病原体风险,同时提高某些营养素的生物利用率
✦ 平衡植物性饮食与食品安全
为了最大限度地发挥植物性饮食的益处同时减少风险:
-清洗生食
彻底清洗所有生水果和蔬菜,去除表面的土壤和微生物
-适当烹饪
对可能含有较多环境微生物的食物(如芽菜、生豆类)进行适当烹饪
-注意高风险人群
免疫功能低下者应格外小心生食,考虑咨询医生或营养师
-多样化来源
从不同来源购买农产品,减少单一来源的风险
-考虑有机选择
在可能的情况下,选择有机种植的农产品,减少农药暴露
编辑
✦ 个性化植物性饮食策略
每个人的肠道菌群都是独特的,因此对植物性饮食的反应也可能不同。建议:
通过这些策略,我们可以充分利用植物性饮食对肠道菌群和整体健康的益处,同时最大限度地减少潜在风险,踏上个性化的健康之旅。
主要参考文献
Anand R, Sahil R, Jain M, Maurya GK, Kharat AS. Plant-Based Diet as a Precursor to Human Gut Diversity. J Nutr. 2025 Nov 26:101251.
Fackelmann G, Manghi P, Carlino N, Heidrich V, et al., Gut microbiome signatures of vegan, vegetarian and omnivore diets and associated health outcomes across 21,561 individuals. Nat Microbiol. 2025 Jan;10(1):41-52.
Ramaboli MC, Ocvirk S, Khan Mirzaei M, et AL., Diet changes due to urbanization in South Africa are linked to microbiome and metabolome signatures of Westernization and colorectal cancer. Nat Commun. 2024 Apr 20;15(1):3379.

谷禾健康

在20世纪初,Elie Metchnikoff(著名生物学家,酸奶之父)发现了有益的肠道微生物,该微生物可使肠道健康正常化并延长寿命,后来被称为“益生菌”。益生菌是指“以适当的剂量给予宿主健康有益的活生物体” 。
益生菌如今越来越受欢迎,主要是因为它们对我们整体健康的重要性。益生菌主要存在于人体肠道内,通过维持肠道微生物平衡,在宿主体内发挥有益作用。
不久前发表于《自然•化学》的一项最新研究成果表明,“吃土”也能调节肠道微生物组和治疗肠炎。
究其原因是许多益生菌来源于土壤微生物 (SBO)。细菌在土壤中有着惊人的多样性和丰富性,它们在土壤生态系统中的作用方式与它们在肠道生态系统中的作用相似。
注:该研究中的“吃土”并不是直接食用土壤,而是“人造土”,通过人工合成的方法构建的土壤仿生材料。事实上,自然界土壤中的微生物与肠道菌群中的微生物并不完全相同,且土壤中可能存在有害物质,如重金属、农药等,因此,直接食用土壤可能会引起不必要的健康问题。
考虑土壤微生物的多样性和复杂性,为了让大家更好地了解来源于土壤微生物的益生菌,本文将介绍其中最常见的土壤益生菌及其作用。希望对未来益生菌的选择和恢复肠道微生态提供一定价值。
▼
土壤微生物一词涵盖了土壤中高度多样化的细菌(和其他生命形式),这些细菌在土壤中自然存在。在引入工业化和现代农业之前,我们每天都会定期接触这些细菌。近年来,一些土壤微生物已被分离并用作益生菌。
许多土壤微生物的一个关键特征是它们可以形成孢子。当条件不太有利时,土壤微生物会形成小孢子,这是细菌的一种休眠形式,具有坚硬的保护性外涂层。在这种形式下,细菌对热、酸和大多数抗生素具有高度抵抗力。
土壤微生物和肠道微生物组息息相关
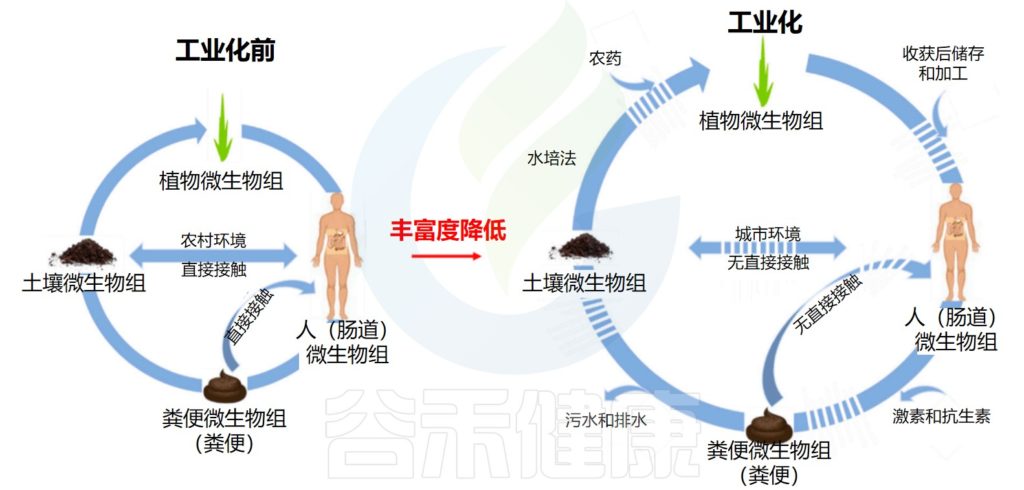
doi: 10.3390/microorganisms7090287
▼
• 通过触发抗体来帮助调节免疫系统;
• 对抗肠易激综合征,调节肠道功能;
• 减轻炎症;
• 治疗腹泻、腹痛和腹胀;
• 帮助消化和营养吸收;
• 防止感染;
• 支持情绪和心理健康;
• 支持平衡肠道微生物群;
• 促进新陈代谢,加速脂肪流失;
• 缓解过敏症状
▼
以下是益生菌中最常用的八种土壤微生物:
Bacillus coagulans(凝结芽孢杆菌)
Bacillus subtilis (枯草芽孢杆菌)
Bacillus clausii(克劳氏芽孢杆菌)
Bacillus indicus (印度芽孢杆菌)
Bacillus licheniformis(地衣芽孢杆菌)
Enterococcus faecium (屎肠球菌)
Enterococcus faecalis(粪肠球菌)
Clostridium butyricum(丁酸梭菌)
注:印度芽孢杆菌是Bacillus属的微生物,原产地为中国。
在下文中介绍了八种来源于土壤微生物的益生菌在人类临床试验中的作用,以确定哪些菌株有较好的治疗前景。
01
凝结芽孢杆菌 (Bacillus coagulans)
凝结芽孢杆菌(Bacillus coagulans)是一种产生L-乳酸的革兰氏阳性细菌。最适生长温度为45-50℃,最适pH为6.6-7.0。它表现出乳酸杆菌科的许多典型特征,但与大多数乳酸菌不同,凝固芽孢杆菌可以形成孢子。
图源:JBMbio
六种凝结芽孢杆菌菌株在人体临床试验中得到充分研究:
✦六种不同菌株显示能够调节肠道微生物群、改善肠易激综合征
GBI-30, 6086
Bacillus coagulans GBI-30, 6086 (“Ganeden BC30”):通过至少10项人类随机对照试验,这是研究最深入的土壤菌株之一。GBI-30、6086已被证明可以改善肠易激综合征 、免疫功能并且可能有益地调节肠道微生物群。
体外研究还表明,它可能有助于果糖和乳糖的消化。
LBSC (DSM 17654)
Bacillus coagulans LBSC (DSM 17654):两项随机对照试验表明该菌株对肠易激综合征患者和急性腹泻或腹部不适患者有益。
MTCC 5856
MTCC 5856(“LactoSpore”):两项随机对照试验表明,该菌株可显著减轻肠易激综合征患者的腹胀、腹痛和抑郁症状。
SANK 70258
Bacillus coagulans SANK 70258 (“Lacris-S”):
一项随机对照试验和一项开放研究发现,该菌株可改善健康成人的大便频率,减少大便次数,且无不良影响。
最近一项使用人体肠道模型系统的研究发现,该菌株抑制了肠杆菌科中的促炎细菌,并增加了健康个体中产丁酸盐的毛螺菌科的丰度。
注:自1966年以来,该菌株已在日本用作食品成分。
SNZ 1969
Bacillus coagulans SNZ 1969:一项安慰剂随机对照试验发现,这种菌株显著改善了结肠转运时间并减少了轻度间歇性便秘人群的肠道不适。它也被证明对细菌性阴道病有效。
另一项使用SNZ 1969以及克劳氏芽孢杆菌和枯草芽孢杆菌的随机对照试验发现,胃肠道不适的成年人的嗳气、腹胀、疼痛和总体症状有所减轻。
Unique IS-2
Unique IS-2(“ProDURA”):四项人类随机对照试验发现该菌株对儿童和成人的细菌性阴道病和肠易激综合征均有效。该菌株还被证明可以在体外代谢果糖。
所有这六种菌株都作为食品成分进行了安全评估,并获得了食品药品监督管理局的“公认安全”(GRAS) 状态。
▸ 小结
这些凝结芽孢杆菌菌株似乎是安全的、耐受性良好的,并且可能对患有细菌性阴道病或肠易激综合征的患者特别有益。
枯草芽孢杆菌(Bacillus subtilis)是一种形成孢子的革兰氏阳性细菌,被认为是人类肠道的正常居民。
枯草芽孢杆菌天然存在于纳豆中,纳豆是一种由发酵大豆制成的日本传统食品。
在引入抗生素之前,枯草芽孢杆菌培养物就已在世界范围内广泛用作免疫刺激剂,以帮助治疗泌尿道和胃肠道疾病。
B.subtilis可以产生一种降解草酸盐的酶,也可能产生少量维生素K2。

图源:eol.org
三种枯草芽孢杆菌菌株已证明对人类有治疗作用,并已通过作为食品成分的安全性测试:
✦三种不同菌株显示能够改善人体健康状况
DE111
Bacillus subtilis DE111:在训练期间对运动员进行的两项随机对照试验发现枯草芽孢杆菌DE111具有良好的耐受性。女运动员的体脂率下降幅度更大;男性运动员发现血液中的炎症标志物减少,对身体机能没有影响。
其他安慰剂随机对照试验发现枯草芽孢杆菌DE111改善了肠道不规律,增加了抗炎免疫细胞群的活性,降低了空腹血糖并降低了健康成人的总胆固醇。
该菌株被食品药品监督管理局公认为安全。
CU1
Bacillus subtilis CU1(“LifeinU”):一项随机对照试验发现,为期10天的B. subtilis CU1补充剂耐受性良好,并改善了老年人的免疫功能(粪便和唾液 中的免疫球蛋白)。
它没有表现出任何抗生素耐药性,并被证明在体外不存在产毒活性。
MB40
Bacillus subtilis MB40 (“OPTI-BIOME”) :
一项随机对照试验发现,该菌株在健康成人中具有良好的耐受性,可降低男性受试者的腹胀强度和胃肠道症状,并适度改善总体健康状况。
该菌株被食品药品监督管理局公认为安全 。
在亚洲对另外两种菌株进行了深入研究:
R0179
Bacillus subtilis R0179(“Medilac-S”):
该菌株存在于亚洲益生菌制剂Medilac-S中,该制剂还含有Enterococcus faecium R0026。几项安慰剂随机对照试验发现,两种菌株的制剂减少了肠易激综合征相关的腹痛,并改善了结肠镜检查的肠道。
最近一项针对临床试验的荟萃分析得出结论,Medilac-S也可有效诱导溃疡性结肠炎的缓解。
两项随机对照试验还发现,分离的枯草芽孢杆菌R0179具有良好的耐受性,并在通过人体胃肠道后存活下来。
TO-A
Bacillus subtilis TO-A (“BIO-THREE”) :
这种枯草芽孢杆菌菌株与E. faecalis T-110和C. butyricum TO-A相结合,作为益生菌在亚洲得到了相当深入的研究。在随机对照试验中,这三种菌株已被证明可以减少手术患者的术后感染并缩短急性腹泻患儿的住院时间。
▸ 小结
通常研究的枯草芽孢杆菌菌株似乎是安全的、耐受性良好的,并且没有不良反应的报道。一些菌株似乎对改善免疫功能和肠道规律性特别有益。
克劳氏芽孢杆菌(Bacillus clausii)是一种形成孢子的革兰氏阳性细菌,以其对金黄色葡萄球菌和艰难梭菌的抗菌活性而闻名。它被认为是一种共生微生物,已从健康个体的小肠和粪便中回收。
只有两种菌株组合已在人类身上进行过研究:
✦两种不同菌株组合显示对过敏、肠易激综合征有益
Bacillus clausii OC、NR、SIN和T
自1958年以来,这四种菌株已被广泛研究并用作药物制剂Enterogermina(美菌纳)。这些菌株已被证明具有酸和胆汁抗性,并且在人类单次口服给药后可在粪便中恢复4-12天。
这些菌株可能对那些有过敏症、减少鼻塞和抗组胺药需求的人特别有益。
它也被证明可以减少幽门螺杆菌抗生素治疗期间的副作用。
一项针对40名诊断为“小肠细菌过度生长”患者的研究发现,服用美菌纳一个月可使 47%的患者的葡萄糖呼气试验结果恢复正常。
值得注意的是,这个比率与许多用于小肠细菌过度生长治疗的抗生素所见的正常化率相当。只有一名患者报告了便秘的副作用。
尽管美菌纳有许多记录在案的益处和总体安全性,但仍有零星的克劳氏芽孢杆菌败血症病例报告。大多数报告都是在老年人、重病患者或免疫功能低下的个体中使用益生菌。
Bacillus clausii UBBC-07
在一项针对印度急性腹泻儿童的安慰剂对照随机试验中,该菌株被证明可以改善粪便稠度并缩短腹泻持续时间。该菌株经过了毒理学研究,被确定可供人类安全食用。
▸ 小结
克劳氏芽孢杆菌已在许多随机、安慰剂对照试验中证明了疗效,并且可能对过敏或肠易激综合症患者特别有益。
脓毒症的个别病例报告表明,老年人、重病或免疫功能低下的人以及最近服用广谱抗生素的人可能禁忌使用它。
印度芽孢杆菌(Bacillus indicus)是一种形成孢子的细菌,因其能够有效刺激免疫系统并产生高水平的类胡萝卜素、维生素和喹啉而受到吹捧。
✦暂未有毒性或致病性的发现
体外和动物研究未能发现B.indicus HU36的任何潜在毒性或致病性,该菌株是益生菌中最常用的菌株,并且没有关于人类感染的报道。
▸ 小结
关于人类印度芽孢杆菌的科学研究,目前还缺乏人类证据。
地衣芽孢杆菌(Bacillus licheniformis)是一种革兰氏阳性、产芽孢的细菌,通常用于工业酶生产。
在所有基于土壤使用的益生菌中,地衣芽孢杆菌可能是最具争议的。
✦可能具有一定毒性
根据一些报道,地衣芽孢杆菌可能是一种机会性病原体,可导致免疫功能低下的宿主感染。
然而,其他报告得出的结论是,地衣芽孢杆菌的毒力非常低。地衣芽孢杆菌存在于韩国传统消费的许多发酵食品中,并且已从健康人类志愿者的胃肠道中分离出来,因此至少某些菌株似乎是共生的。
虽然已经出现了与大量该物种的分离相关的急性自限性胃肠炎病例, 对肠上皮细胞的毒性或直接影响尚未得到证实。很难确定这些报告病例中的物种(数量相当有限)是否积极参与了感染,或与一种不明病原体一起分离。
只有四项关于分离的地衣芽孢杆菌的人体研究,没有一项提供菌株信息,并且只有一项是随机和安慰剂对照的。
▸ 小结
虽然含有地衣芽孢杆菌的传统发酵食品几乎是安全的,但目前没有足够的证据支持大剂量补充地衣芽孢杆菌。最好谨慎行事,避免使用含有该物种的益生菌。
✦发酵碳水化合物
Enterococcus faecium是革兰氏阳性菌。虽然它不形成孢子,但它可以耐受多种环境条件。E. faecium是人类肠道的正常居民,发酵碳水化合物并产生乳酸作为副产品。
粪肠球菌的一些菌株可以在传统发酵食品中找到。
by Dennis Kunkel Microscopy
✦可能致病
然而,粪肠球菌也可能致病,引起新生儿脑膜炎和心内膜炎等疾病。这种细菌的许多菌株已经产生了抗生素抗性和毒力因子,使其能够聚集并形成生物膜。
在美国,80-90%的医疗器械相关感染可归因于耐抗生素粪肠球菌。因此,人们担心将其用作益生菌。
已经研究了许多不同的菌株,但只有两种菌株值得强调,它们已经在多项人体试验中进行了研究:
M-74
Enterococcus faecium M-74:该菌株有许多与之相关的“随机”临床试验,但每一个菌株都服用益生菌和50微克硒。
许多益生菌指出这些研究表明粪肠球菌可以降低胆固醇,但缺乏硒对照组意味着硒单独提供治疗效果是完全合理的。
R0026
Enterococcus faecium R0026 (“Medilac-S”):
该菌株存在于亚洲益生菌制剂Medilac-S中,该制剂还含有枯草芽孢杆菌R0179,并在国内得到了很好的研究。
如前面所述,多项随机对照试验发现,双菌株制剂可减少肠易激综合征相关的腹痛,并改善结肠镜检查的肠道准备,还可以有效诱导溃疡性结肠炎的缓解。
▸ 小结
除了Medilac-S中的菌株外,很少有精心设计的随机对照试验证明了粪肠球菌的功效。
鉴于其可能获得多重抗生素耐药性并引起感染。谨慎使用高剂量的分离粪肠球菌。
Enterococcus faecalis是一种革兰氏阳性菌。与Enterococcus faecium类似,它不能形成孢子,但对恶劣的环境条件相当耐受。
✦在肠道免疫中起重要作用
粪肠球菌是人类肠道的正常居民,存在于大约90-95%的人中。这种细菌通常是生命早期定植于人类胃肠道的首批微生物之一,在肠道免疫系统的发育中发挥着重要作用。
T-110
Enterococcus faecalis T-110:这种粪肠球菌菌株与枯草芽孢杆菌TO-A和丁酸梭菌TO-A结合使用作为益生菌(BIO-THREE)。这三种菌株一起可以减少手术患者的术后感染并缩短急性腹泻患儿的住院时间。
YM0831
Enterococcus faecalis YM0831:一项交叉研究发现,对健康人类受试者单次施用该菌株可显着改善血糖对蔗糖耐量试验的反应。
▸ 小结
除了BIO-THREE中的菌株外,很少有精心设计的随机对照试验证明了Enterococcus faecalis作为益生菌的功效,还需要更多的研究。
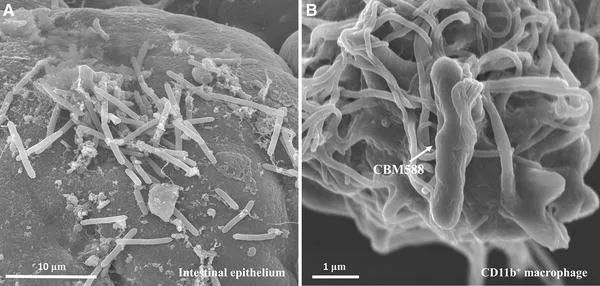
✦丁酸梭菌是肠道重要的菌属
丁酸梭菌(Clostridium butyricum)是人类胃肠道的天然居民。它是肠道中发酵膳食纤维并产生有益的短链脂肪酸丁酸盐的众多细菌之一。丁酸盐作为结肠上皮细胞的能量来源,有助于维持肠道屏障,具有抗炎作用,并可预防结肠癌。
doi.org/10.1007/s00535-015-1084-x
三种菌株已在人体中得到充分研究:
✦三种不同菌株显示能够调节肠道稳态、减轻肠炎
CBM 588
Clostridium butyricum CBM 588:这种丁酸梭菌菌株早在1963年就用作益生菌,用于一种名为Miyairisan的药物制剂中。它已经过安全性评估,不携带任何编码任何已知毒素或毒力因子的基因。
两项随机对照试验发现,该菌株可减少接受幽门螺杆菌根除治疗的患者的腹部症状和腹泻发生率的副作用。
另一项针对溃疡性结肠炎患者的安慰剂对照随机对照试验发现,接受MIYAIRI 588治疗的受试者中,患上肠结肠炎的人数较少。
尽管其影响在统计学上并不显著,而安慰剂组则服用乳糖,这会加重许多人的结肠炎。
TO-A
Clostridium butyricum TO-A:这种丁酸梭菌菌株与粪肠球菌T-110和枯草芽孢杆菌TO-A结合使用,作为益生菌在亚洲得到了广泛使用和深入研究。
A Tai Ning
该菌株包含在一种名为阿泰宁的产品中,最近在一项大规模、多中心、的随机对照试验中显示,该菌株对以腹泻为主的肠易激综合征具有显著益处。
该研究包括200名患者。4周后,与安慰剂组相比,益生菌组在整体肠易激综合征症状、大便频率和生活质量方面有显著改善。
几项研究发现丁酸梭菌降低了早产儿败血症和腹泻的发生率,并改善了抗原特异性免疫治疗。在动物研究中,丁酸梭菌已被证明可以改善肠道稳态并减轻结肠炎。
▸ 小结
丁酸梭菌是肠道重要的菌属,在随机试验中似乎是安全且耐受性良好的,并且帮助增加丁酸盐水平的同时,极少引起毒性。
➣土壤基益生菌的安全性仍存在争议,值得进一步关注
支持者声称它们是益生菌,可以使肠道功能正常化、帮助消化、有益地刺激免疫系统并帮助重新播种肠道微生物群。他们还称赞土壤微生物具有抵抗胃酸的能力并且不需要冷藏。
反对者认为,由于它们形成孢子的性质,它们会迅速增殖,与我们常驻的肠道微生物竞争,在某些情况下,它们甚至可能致病。由于它们的孢子形成能力和对大多数抗生素的天然抗性,意外的过度生长将很难治疗。
在随机、安慰剂对照的人体临床试验中,许多单独的土壤微生物菌株已被证明是有益的,几乎没有报告不良反应。
然而,其他菌株的临床证据有限或没有临床证据,可能会导致免疫系统受损的人感染。我们需要进行更细致的讨论——考虑每个特定物种、菌株和配方,而不是广泛地给土壤微生物贴上好或坏的标签。
基于土壤的微生物得到了很好的研究,并且已经在随机临床试验中显示出对一些症状有效,但仍需更多研究来确定其安全性和有效性。建议选择使用高质量和经过临床测试的菌株,在服用益生菌制剂前最好咨询医生的建议。
每个人对益生菌的反应都不同。反应将取决于服用的益生菌、剂量、肠道中已有的微生物以及肠道和免疫系统的健康状况。
可以通过微生物测序技术来检测使用益生菌前后的肠道菌群组成及相关健康状况。再选择针对自身特定健康状况的益生菌菌株。
同时也期待更多关于土壤微生物的单个菌株和配方在不同情况下影响肠道微生物群和肠道环境的研究。
注:本账号内容仅作交流参考,不作为诊断及医疗依据。
主要参考文献:
Mu Y, Cong Y. Bacillus coagulans and its applications in medicine. Benef Microbes. 2019 Jul 10;10(6):679-688. doi: 10.3920/BM2019.0016. Epub 2019 Jun 17. PMID: 31203635.
Jäger R, Purpura M, Farmer S, Cash HA, Keller D. Probiotic Bacillus coagulans GBI-30, 6086 Improves Protein Absorption and Utilization. Probiotics Antimicrob Proteins. 2018 Dec;10(4):611-615. doi: 10.1007/s12602-017-9354-y. PMID: 29196920; PMCID: PMC6208742.
Acosta-Rodríguez-Bueno CP, Abreu Y Abreu AT, Guarner F, Guno MJV, Pehlivanoğlu E, Perez M 3rd. Bacillus clausii for Gastrointestinal Disorders: A Narrative Literature Review. Adv Ther. 2022 Nov;39(11):4854-4874. doi: 10.1007/s12325-022-02285-0. Epub 2022 Aug 26. PMID: 36018495; PMCID: PMC9525334.
Ianiro G, Rizzatti G, Plomer M, Lopetuso L, Scaldaferri F, Franceschi F, Cammarota G, Gasbarrini A. Bacillus clausii for the Treatment of Acute Diarrhea in Children: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Nutrients. 2018 Aug 12;10(8):1074. doi: 10.3390/nu10081074. PMID: 30103531; PMCID: PMC6116021.
Kovács ÁT. Bacillus subtilis. Trends Microbiol. 2019 Aug;27(8):724-725. doi: 10.1016/j.tim.2019.03.008. Epub 2019 Apr 15. PMID: 31000489.
Marzorati M, Van den Abbeele P, Bubeck S, Bayne T, Krishnan K, Young A. Treatment with a spore-based probiotic containing five strains of Bacillus induced changes in the metabolic activity and community composition of the gut microbiota in a SHIME® model of the human gastrointestinal system. Food Res Int. 2021 Nov;149:110676. doi: 10.1016/j.foodres.2021.110676. Epub 2021 Aug 30. PMID: 34600678.
Kawarizadeh A, Pourmontaseri M, Farzaneh M, Hosseinzadeh S, Ghaemi M, Tabatabaei M, Pourmontaseri Z, Pirnia MM. Interleukin-8 gene expression and apoptosis induced by Salmonella Typhimurium in the presence of Bacillus probiotics in the epithelial cell. J Appl Microbiol. 2021 Jul;131(1):449-459. doi: 10.1111/jam.14898. Epub 2020 Dec 7. PMID: 33058340.
Muras A, Romero M, Mayer C, Otero A. Biotechnological applications of Bacillus licheniformis. Crit Rev Biotechnol. 2021 Jun;41(4):609-627. doi: 10.1080/07388551.2021.1873239. Epub 2021 Feb 16. PMID: 33593221.
Gudiña EJ, Teixeira JA. Bacillus licheniformis: The unexplored alternative for the anaerobic production of lipopeptide biosurfactants? Biotechnol Adv. 2022 Nov;60:108013. doi: 10.1016/j.biotechadv.2022.108013. Epub 2022 Jun 22. PMID: 35752271.
Gök ŞM, Türk Dağı H, Kara F, Arslan U, Fındık D. Klinik Örneklerden İzole Edilen Enterococcus faecium ve Enterococcus faecalis İzolatlarının Antibiyotik Direnci ve Virülans Faktörlerinin Araştırılması [Investigation of Antibiotic Resistance and Virulence Factors of Enterococcus faecium and Enterococcus faecalis Strains Isolated from Clinical Samples]. Mikrobiyol Bul. 2020 Jan;54(1):26-39. Turkish. doi: 10.5578/mb.68810. PMID: 32050876.
Stoeva MK, Garcia-So J, Justice N, Myers J, Tyagi S, Nemchek M, McMurdie PJ, Kolterman O, Eid J. Butyrate-producing human gut symbiont, Clostridium butyricum, and its role in health and disease. Gut Microbes. 2021 Jan-Dec;13(1):1-28. doi: 10.1080/19490976.2021.1907272. PMID: 33874858; PMCID: PMC8078720.
Chen D, Jin D, Huang S, Wu J, Xu M, Liu T, Dong W, Liu X, Wang S, Zhong W, Liu Y, Jiang R, Piao M, Wang B, Cao H. Clostridium butyricum, a butyrate-producing probiotic, inhibits intestinal tumor development through modulating Wnt signaling and gut microbiota. Cancer Lett. 2020 Jan 28;469:456-467. doi: 10.1016/j.canlet.2019.11.019. Epub 2019 Nov 14. PMID: 31734354.