-

 国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子
国家认可委 CNAS L23010 认可项目:微生物宏基因组 | 16S rRNA扩增子

 二级病原微生物安全实验室
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业
国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业- 联系电话:+13336028502
- +400-161-1580
- service@guheinfo.com
二级病原微生物安全实验室 国家高新企业 | ISO9001认证 | 肠道健康精准检测高新技术研发中心 | 专精特新企业

谷禾健康
宠物 营养 健康
在全球范围内,猫犬早已不再只是传统意义上的“宠物”,而成为许多家庭中重要的“毛孩子”。这种“宠物人性化”的趋势,极大地推动了宠物主人对爱猫爱犬健康的关注,促使他们寻求超越传统温饱的、更精细化、科学化的喂养方案。
如今,延长宠物寿命、提升健康福祉已成为宠物主人与兽医共同的目标。从过去仅满足基本能量与营养需求的狗粮,到如今注重护理、饮食与功能性食品的综合方案,宠物饲养理念正日益进步。
与人类相似,犬猫的健康同样受到饮食与肠道微生物组的深刻影响。肠道微生物组作为宿主体内重要的代谢与免疫调节中心,其稳态或失衡直接决定整体健康水平。
最近,宠物饮食中越来越多地引入功能性食品,以促进肠道微生物组的良性调节。这些食品包括益生菌、益生元、合生元、后生元,以及多不饱和脂肪酸和植物营养素等必需营养成分。
功能性食品虽非药物,却体现了“食疗同源”的理念,通过日常摄入实现主动健康干预。对犬类而言,这类食品可视作“第四餐”,在常规日粮之外帮助优化生理机能、增强免疫力、延缓衰老,并辅助疾病管理。

本文结合多篇核心研究,探讨了功能性营养干预在调节犬猫肠道微生物组及其在营养代谢、免疫成熟、神经行为调控等方面的作用,同时分析了肠道生态失调作为多种疾病(包括胃肠道、皮肤、代谢及神经系统疾病)共同病理基础的分子机制。
在此基础上,系统总结了各类功能性营养成分的关键特性:益生菌的菌株特异性免疫调节;益生元对特定菌群的选择性促进及短链脂肪酸(SCFAs)的生成;合生元的协同增效作用;后生元的创新应用潜力;多不饱和脂肪酸(PUFAs)尤其是EPA和DHA在炎症调控中的核心角色;以及植物营养素的多靶点抗氧化与抗炎效应等。

狗和猫与人类相伴已经超过一万年,作为伴侣动物,它们在人类社会中扮演着非常重要的角色。最近的一份报告估计,全球大约已经有4.77亿只狗和3.77亿只猫被当作宠物饲养。
★ 宠物有助于人身心健康
除了作为人类的伙伴,宠物还有助于个人身心的整体健康。根据一些证据,养宠物有助于减压、缓解抑郁、降低血压、预防心脏病,并提升自尊与幸福感。此外,哮喘、过敏和肥胖等疾病的发生风险也可能随之降低。
并且随着社会经济的发展与人文关怀的提升,犬猫在家庭中的地位发生了根本转变——它们被视为家庭成员,其健康与福祉备受关注。
这种“宠物人性化”趋势推动了宠物健康管理理念的革新。现代兽医学已从单纯的疾病治疗转向以预防、维护健康和提升生命质量为核心的主动健康管理,目标在于实现“健康老龄化”和延长健康寿命。
★ 肠道微生物组:一个多功能“超级器官”
在这一过程中,功能性营养学作为一种非侵入性、可持续的干预手段,其科学与临床价值日益凸显。功能性食品或营养品,是指除提供基础营养外,经科学验证能改善特定生理功能或降低疾病风险的食品或活性成分。其作用机制虽多样,但越来越多的研究表明,关键靶点在于寄居于宿主肠道内的复杂微生物生态系统——肠道微生物组。
作为身体的“隐形器官”,肠道微生物组深刻影响犬猫的营养吸收、免疫防御、新陈代谢及情绪行为。其平衡是健康的基石,而失调则被认为是多种犬猫常见疾病的关键驱动因素。
从急性腹泻、慢性肠病,到特应性皮炎、肥胖、糖尿病、骨关节炎及认知功能障碍,几乎都与菌群失衡密切相关。因此,通过精准营养干预调节肠道微生物组、恢复生态稳定,已成为现代兽医营养学中最具潜力的研究与应用方向之一。
1
营养代谢中心:发酵、合成与转化
犬猫的消化系统无法完全分解复杂碳水化合物,尤其是膳食纤维。这一过程主要由结肠中的厌氧菌群完成,它们通过发酵将纤维转化为短链脂肪酸(SCFAs),为宿主提供关键代谢产物。
• 产生丁酸盐:结肠上皮的重要能量,抗炎物质
丁酸盐(Butyrate)主要由厚壁菌门中的Clostridium簇IV和XIVa(如Faecalibacterium、Roseburia)合成,是结肠上皮细胞超70%的能量来源。
它还可以抑制组蛋白去乙酰化酶(HDACs),发挥抗炎和抗肿瘤作用,并上调紧密连接蛋白(Occludin、Claudin-1、ZO-1)表达,强化肠道屏障。
• 产生丙酸盐:调节脂质合成,影响血糖
丙酸盐(Propionate)主要由拟杆菌门产生,被吸收至肝脏后可抑制糖异生、调节脂质合成,从而改善血糖稳态与胰岛素敏感性。
• 产生乙酸盐:含量最高的短链脂肪酸
乙酸盐(Acetate)是含量最高的SCFA,可由多种菌群生成,既为外周组织供能,也是产丁酸菌合成丁酸的重要前体。
• 代谢胆汁酸,合成一些必需维生素
此外,肠道菌群还负责代谢胆汁酸,将肝脏合成的初级胆汁酸转化为次级胆汁酸(如脱氧胆酸LCA和石胆酸DCA)。这些次级胆汁酸作为信号分子,通过激活法尼醇X受体(FXR)和G蛋白偶联胆汁酸受体1(TGR5),调控宿主的脂质、葡萄糖代谢及炎症反应。
同时,菌群还能合成犬猫必需的维生素K、多种B族维生素(生物素、叶酸、核黄素)及氨基酸等,维持营养与代谢平衡。
2
免疫系统的“首席教官”
肠道相关淋巴组织(GALT)是机体最大的免疫器官,而肠道菌群在免疫系统的发育、成熟和调控中发挥着关键作用。
• 诱导免疫耐受
在新生期,肠道菌群的定植是建立口服耐受的关键,帮助免疫系统对食物抗原和共生菌保持耐受,从而预防过敏和自身免疫疾病。
• 塑造黏膜免疫
菌群及其代谢产物(尤其是短链脂肪酸)可刺激上皮细胞和浆细胞分泌大量sIgA,后者能中和毒素并阻止病原体附着,是黏膜免疫的第一道防线。
• 调节T细胞分化
菌群组成深刻影响辅助性T细胞(Th)亚群平衡。分段丝状菌(SFB)促进促炎性Th17细胞分化,提高对胞外菌的防御力;而产丁酸梭菌则诱导调节性T细胞(Treg)分化,Treg通过分泌IL-10与TGF-β抑制过度免疫反应,维持免疫稳态。肠道菌群失衡常伴随Treg/Th17比例紊乱,导致免疫异常。
3
“肠-脑轴”的核心信使
肠道与大脑之间通过“肠-脑轴”形成复杂的双向通讯网络,而肠道菌群是其中的核心信号调节者。
• 神经递质的合成与调控
肠道菌群可直接生成多种神经递质,如GABA、去甲肾上腺素和多巴胺,或通过代谢膳食前体物质(如色氨酸)影响中枢血清素水平。菌群代谢产物(如丁酸)还能调节肠嗜铬细胞产生血清素,占全身总量的90%以上。
• 迷走神经通路
菌群代谢物可促使肠上皮细胞释放信号分子,这些信号经迷走神经传递至大脑,进而影响情绪与行为。
• 调节HPA轴
肠道菌群稳态对下丘脑-垂体-肾上腺(HPA)轴的发育与应激反应至关重要。无菌动物常表现出过度应激反应,而补充特定益生菌可恢复其正常。研究还发现,具有攻击性或恐惧行为的犬只,其肠道菌群结构与健康犬存在显著差异。
功能性食品在犬猫营养中具有重要意义,它不仅提供基础能量与必需营养素,还能通过特定的生理活性成分,支持机体健康、延缓衰老并预防疾病。
近年来,越来越多研究证实,富含抗氧化剂、ω-3脂肪酸、植物营养素、益生元及功能性蛋白的配方,可改善犬猫的免疫功能、皮肤与毛发状况、肠道健康及认知能力。
1
功能性食品与犬类健康
多项研究探讨了功能性食品在犬中的作用,相关结果见下表。


• 低聚果糖等益生元改善整体健康
例如,成年雄性比格犬摄入富含低聚果糖(FOS)的饲粮后,粪便中氨及梭菌浓度降低,而需氧菌含量上升,改善整体健康。
以肉类为主的犬粮中,单独或联合添加FOS和甘露寡糖(MOS),可提高回肠免疫球蛋白A水平,降低粪便中吲哚和酚含量。
测试了一种含多种成分的维持性饲粮(包括多种植物提取物及抗氧化剂)对12只成年犬的影响。6个月后,犬只氧化应激显著下降,活性氧代谢物减少,而视黄醇及血浆抗氧化力(BAP)升高,提示氧化平衡得以恢复,说明均衡饮食对维持犬的抗氧化状态至关重要。相反,摄入高血糖指数食物后可能导致氧化失衡。
注:进一步研究表明,在测试4种复杂碳水化合物(大麦、玉米、豌豆、大米)及葡萄糖后,豌豆的血糖指数最低(29%),大麦和大米较高(分别为51%和55%),支持它们被纳入平衡膳食。
• 富含抗氧化剂的食物有助于提升代谢和繁殖能力
有研究评估了一种富含抗氧化剂和功能性成分的配方(含水解蛋白、维生素、酵母提取物和植物提取物)对不同年龄雄犬繁殖力的影响。
实验持续4个月,结果显示,在45天内犬的游离甲状腺素和睾酮水平升高,代谢和生育能力改善,特别是2–7岁组的精子活力显著提高,表明富含抗氧化剂的饮食可增强生殖表现与内分泌功能。
• 含抗氧化剂与植物化合物改善认知衰老和行为障碍
此外,犬类模型还用于研究认知衰老、脑源性神经营养因子(BDNF)及饮食之间的关系。研究发现,给予含抗氧化剂与植物化合物(如鱼油、甜菜浆、木瓜、石榴、芦荟、葡萄及迷迭香等)的日粮,可显著提高脑源性神经营养因子(BDNF)水平,降低氧化应激,并改善神经内分泌平衡。
另一系列实验显示,富含功能性植物提取物及色氨酸的饮食能减轻犬的行为障碍,改善相关临床症状。
综上所述,功能性食品通过抗氧化、抗炎及神经保护等多重机制,不仅能优化犬的代谢与免疫状态,还可能成为支持认知健康和行为管理的有效营养干预手段。
• 功能性饮食还有助于改善狗的口臭、皮肤健康
口臭是一种常见于犬及人类的疾病,影响彼此关系。一项双盲、安慰剂对照交叉研究评估了专用功能性饮食对16只不同品种和年龄犬慢性口臭的改善作用。
注:该饮食含米饭、鱼粉、植物脂肪、鱼油、甜菜浆、矿物质、脱水酵母、低聚果糖、丝兰提取物、溶菌酶、生物类黄酮、胸腺提取物与黑肋骨等成分。
经气相色谱测定,犬只口腔中关键挥发性硫化物(甲基硫醇、硫化氢、二甲硫化物)在30天内显著下降,且停用饮食20天后仍维持改善,显示其持久疗效。
有研究进一步发现,一种含水解蛋白、植物提取物(包括茶树、百里香、大蒜、玫瑰果)及矿物质的功能性饮食,可显著缓解犬慢性外耳炎症状,减少药物使用需求。15只患犬接受该饮食与局部药物治疗(前7天使用抗炎滴耳剂OTOMAX,后83天仅靠饮食干预),90天后耳道阻塞、红斑、分泌物与异味均明显减轻,体现出其抗炎与抗氧化双重功效,为抗生素替代方案提供了潜在方向。
此外,研究了免疫调节饮食(IMMD)在犬利什曼病(CL)治疗中的辅助作用。与标准饮食(SD)组相比,IMMD组犬在3、6、12个月内均恢复了因CL降低的调节性T细胞水平,并在后期阶段呈现T辅助细胞数量正常化,表明该饮食可在药物治疗期间调节免疫反应。
另一项研究证实,IMMD还能有效减轻犬角膜结膜炎的典型症状,包括泪液减少、结膜炎症、角膜角化与色素沉积过多等。
综上所述,针对犬的功能性饮食还能在口腔健康、皮肤炎症及免疫调节等方面发挥作用,为临床营养干预提供了有益证据。
2
功能性食品与猫类健康
与犬相比,猫为严格食肉动物,具有独特的营养需求。多项研究已探讨功能性食品在猫营养与健康中的作用(见下表)。
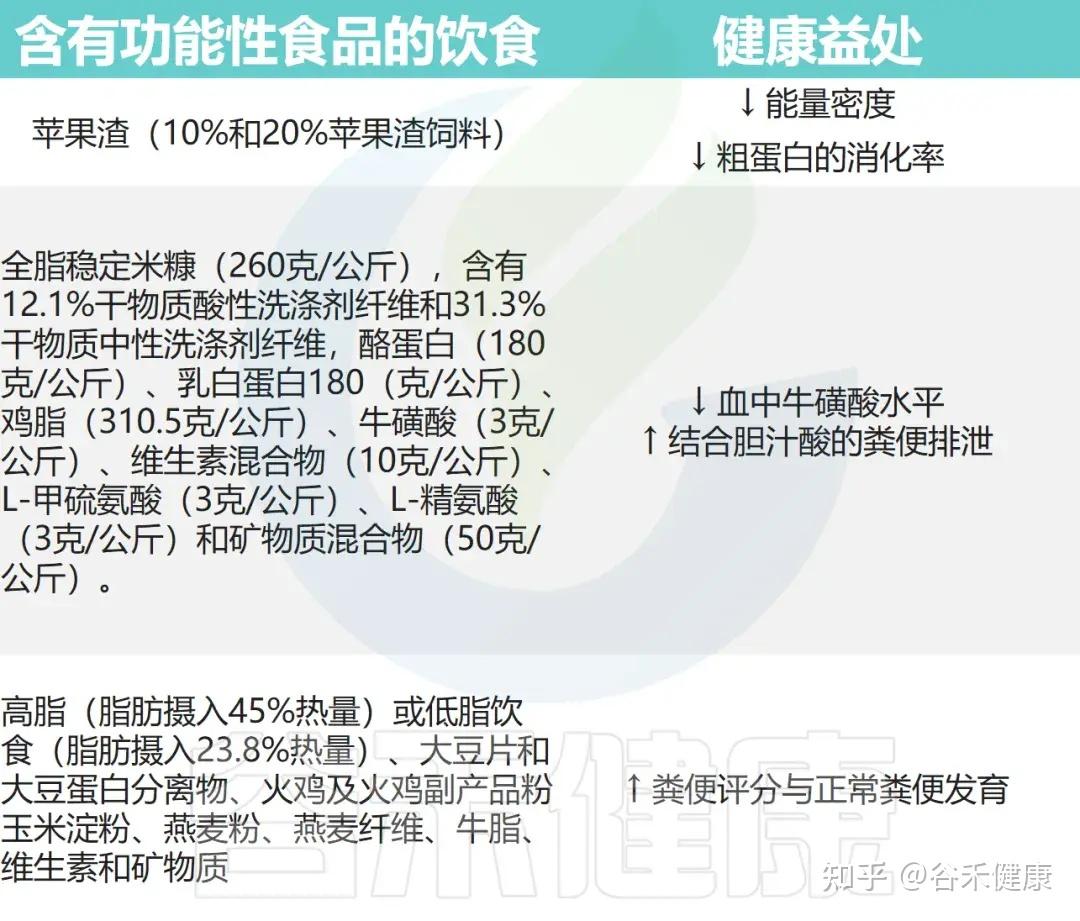
• 功能性食品改善了猫咪的腹泻
一项针对55只患慢性腹泻猫的随机、双盲、对照试验评估了高脂(23%)和低脂(10%)高消化性饮食的效果。该饮食由大豆片、大豆蛋白分离物、火鸡粉、玉米淀粉、燕麦粉、燕麦纤维、牛脂及维生素矿物质组成。结果显示,所有猫在第一周内粪便评分(FSa)均显著提高,3周后达到最佳改善,其中约三分之一的猫恢复正常排便。
• 含有纤维的饮食可能有助于猫咪减肥
苹果渣,即酿造苹果酒的剩余部分,可能是猫咪减肥饮食中的功能性食物。九只成年、绝育、肥胖的猫咪被喂食以肉类为主的饮食,并添加了不同程度的苹果渣。苹果渣分别用10%和20%,并未降低可口性,但确实降低了食物的能量密度。
因此,在肥胖猫的饮食中以有限的比例加入可入口纤维成分,是降低食物能量含量并维持生理食物摄入水平的好方法。
• 一些猫粮应适当提高牛磺酸含量
另一项研究显示,在纯化猫粮中添加26%全脂米糠后,猫的全血牛磺酸浓度显著低于含26%玉米淀粉的对照组。
这种降低与粪便中结合胆酸排泄增加及米糠中不可消化蛋白引发的后肠菌群变化有关,后者可导致牛磺酸降解。为避免缺乏,含米糠配方的猫粮应适当提高牛磺酸含量。
如前所述,调控肠道微生物组以维持或恢复生态平衡,是犬猫功能性食品的核心策略之一。根据作用机制与来源,目前应用最广的功能性成分主要包括:益生菌、益生元、合生元、后生元、多不饱和脂肪酸和植物营养素。它们各具优势,相互协同,共同守护宠物健康。
1
益生菌:直接补充“有益菌群
• 益生菌对狗的益处
研究表明,并非所有益生菌都适用于犬类。理想的菌株应具备宿主特异性,通常来源于犬只自身,以更好地适应其肠道环境。常见犬用益生菌包括:
乳杆菌属(Lactobacillus):
如嗜酸乳杆菌(L.acidophilus);
约氏乳杆菌(L.johnsonii);
发酵乳杆菌(L.fermentum);
鼠李糖乳杆菌(L.rhamnosus)。
双歧杆菌属(Bifidobacterium):如动物双歧杆菌(B.animalis)。
肠球菌属(Enterococcus):如屎肠球菌(E.faecium)的特定菌株(如SF68)。
注:但该属的使用需谨慎,因其部分菌株可能携带抗生素抗性基因。
多项研究表明,补充益生菌(如E. faecium SF68、B. animalis AHC7)可有效缩短犬急性腹泻的持续时间并改善粪便稠度。在慢性肠病(CE)中,它们可作为辅助疗法,帮助恢复菌群平衡、减轻炎症。
不同菌株的免疫作用各异。Lactobacillus casei 可诱导Th1型免疫反应(分泌IFN-γ),而Lactobacillus rhamnosus GG 更倾向促进Treg细胞分化。这种差异与细胞壁中不同的微生物相关分子模式(MAMPs)如肽聚糖、脂磷壁酸等有关。
多菌株组合制剂(如VSL#3)基于“菌种协同”理念。在犬炎症性肠病(IBD)研究中,使用VSL#3显著改善临床症状,降低肠道黏膜炎症评分,并增强Treg关键转录因子FoxP3的表达,证明其免疫调节作用。
• 益生菌对猫的益处
研究显示,健康成猫补充L.acidophilus DSM13241的膳食剂后,粪便中乳杆菌和嗜酸乳杆菌数量增加,而粪肠球菌和梭菌减少。
补充L.acidophilus CECT 4529能改善粪便质量。由S. boulardii和Pediococcus acidilactici组成的复合益生菌可提高粪便中丁酸和总短链脂肪酸含量,降低炎症标志物(髓过氧化物酶、钙卫蛋白)水平,并增强抗氧化酶活性(超氧化物歧化酶、谷胱甘肽)。
此外,口服地衣芽孢杆菌发酵产物可缓解腹泻,增加梭菌簇XIVa成员并减少慢性腹泻猫中C.perfringens数量。含八株乳酸菌的复合制剂可改善慢性便秘和特发性巨结肠猫的症状,并增加粪便中乳酸杆菌和链球菌。肠球菌E. faecium SF68在收容所猫中可显著降低腹泻率;另一粪源E. faecium菌株则改善肠道健康。
多种源自健康猫的菌株,如B.adolescentis、L.acidophilus、L.plantarum和L.rhamnosus,表现出对病原菌的益生活性和抗菌潜力。
• 益生菌的应用:
抗生素相关性腹泻:在犬只必须使用抗生素治疗时,同时或之后补充益生菌,有助于减少抗生素对肠道菌群的破坏,预防腹泻等副作用。
免疫调节:研究发现,给健康的犬只补充益生菌可以提高其免疫细胞(如白细胞)的吞噬活性,增强对病原体的防御能力。
压力引起的消化问题:对于寄养、旅行、更换环境等应激情况,提前补充益生菌有助于稳定肠道菌群,预防应激性腹泻。
!
注意事项
不同益生菌菌株的免疫效应存在差异。选择时,应优先考虑针对犬或猫、有随机对照临床试验(RCT)支持并经同行评审的特定产品,同时关注其活菌数量(CFU)及保质期内的稳定性。
2
益生元:为有益菌提供“超级口粮”
益生元是“可被宿主肠道微生物选择性利用、并对宿主健康产生益处的底物”,通俗地说,就是肠道有益菌喜爱的“食物”,可促进其生长和繁殖。
• 常见的益生元
低聚果糖(FOS):存在于菊苣、洋葱、香蕉等植物中。
菊粉(Inulin):FOS的一种长链形式,主要来源是菊苣根。
低聚果糖(FOS)和菊粉主要被双歧杆菌高效利用,因此被称为“双歧因子”。研究表明,在犬猫日粮中添加1–2%的FOS或菊粉,可显著提高粪便中双歧杆菌丰度,同时降低粪便pH值和氨含量。
甘露寡糖(MOS):通常提取自酵母细胞壁。甘露寡糖除具益生元作用外,还能吸附并清除部分病原菌,特别适用于幼年动物及预防应激性腹泻。MOS能与带I型菌毛的革兰氏阴性菌(如沙门氏菌、大肠杆菌)结合,而不影响有益菌。
注:不同类型和剂量的益生元会影响短链脂肪酸(SCFAs)的产量和构成,例如抗性淀粉更易生成丁酸,而果胶则倾向于产生乙酸和丙酸。
• 益生元的应用:
改善粪便质量:补充低聚果糖、菊粉等益生元能显著改善犬的粪便稠度,减少腹泻和便秘。
调节菌群结构:研究一致表明,饲喂益生元能显著提高犬粪便中双歧杆菌和乳杆菌的数量,同时降低梭菌等潜在有害菌的比例。
增强免疫:通过增加短链脂肪酸的产量,益生元能间接调节免疫系统,减轻炎症。有研究显示,给怀孕母犬补充FOS,能提高其初乳中的免疫球蛋白水平,从而使幼犬获得更好的被动免疫。
代谢健康:在超重犬的研究中,补充益生元有助于改善葡萄糖代谢和胰岛素敏感性。
3
合生元、后生元:下一代微生态调节剂
合生元是益生菌与益生元的复合产品,旨在通过益生元为益生菌提供特定营养来源,提高其在肠道内的存活率与定植效率,从而实现协同增效。
后生元则代表微生态干预的进阶方向,将关注点从“活菌”转向其“功能分子”,包括灭活菌体(如热灭活的Lactobacillus acidophilus)、细胞组分(如细胞壁碎片、胞外多糖)及发酵代谢产物(如短链脂肪酸、有机酸、酶和细菌素)。
系统综述与荟萃分析显示,健康犬的短链脂肪酸(SCFA)水平普遍高于患病犬(如IBD个体),说明SCFAs可作为反映健康状态的功能性标志物。其他研究同样发现,慢性肠病犬的粪便SCFA含量下降。
而肽聚糖(PTDG)和磷壁酸(TEIA)是细胞壁的重要组成成分,具有显著的免疫调节作用。TEIA分为脂质型(LTEIA)和壁连型(WTEIA):前者通过糖鞘脂附着在细菌膜上,后者与肽聚糖链形成共价键。乳酸杆菌属的肽聚糖可抑制炎症性细胞因子的释放,其中来自L. rhamnosus的PTDG能增强感染致病链球菌小鼠的先天免疫反应。LTEIA还能通过诱导防御素和猫地霉素的产生,有效治疗皮肤感染并预防细菌及病毒感染。
• 合生元、后生元的应用:
合生元在应对复杂肠道疾病方面展现出显著潜力。研究表明,在犬慢性肠病中,含多种益生菌菌株与益生元(FOS)的合生元产品能迅速缓解腹泻、呕吐等症状,改善肠道组织的炎症评分,并通过上调紧密连接蛋白表达修复受损肠道屏障。
另一项针对顽固性慢性结肠炎犬的研究显示,富含MOS与FOS的高纤维处方粮结合多菌株益生菌混合物,可在约8.5天内显著缓解症状并恢复肠道菌群平衡。
后生元的优势在于其高安全性与稳定性,适合用于免疫低下或危重动物。临床上,可直接补充丁酸钠或丁酸钙以促进急性肠炎和炎症性肠病犬的肠道修复,或利用特定发酵产物实现靶向免疫调节。
4
多不饱和脂肪酸:抗炎与细胞结构的关键
多不饱和脂肪酸(PUFAs)是分子中含有多个双键的脂肪酸。其中,Omega-3(ω-3)和Omega-6(ω-6)两大系列是犬只自身无法合成,必须从食物中获取的必需脂肪酸。
• 主要类型与来源
①ω-6脂肪酸:
亚油酸(LA):广泛存在于玉米油、葵花籽油等植物油中,是维持皮肤和毛发健康的关键营养素。
花生四烯酸(AA):可由犬体内的亚油酸转化而来。
②ω-3脂肪酸:
α-亚麻酸(ALA):来源于亚麻籽油、奇亚籽等植物,但犬体内转化为长链ω-3的效率低。
二十碳五烯酸(EPA)与二十二碳六烯酸(DHA):主要来自鱼油(如三文鱼、沙丁鱼)、磷虾油及海藻,是发挥主要生理作用的活性ω-3。
• 作用机制
抗炎调节:ω-6与ω-3是类花生酸的前体——前者(AA)产生的分子多促炎,后者(EPA)则具抗炎或弱促炎作用。通过提高饮食中ω-3比例(降低ω-6:ω-3比值),可改变细胞膜脂肪酸构成,促进抗炎介质生成,减少过度炎症反应。
炎症消退机制:EPA和DHA还能生成消退素、保护素与脂氧素等“促炎症消退介质”,它们通过抑制中性粒细胞浸润、促进巨噬细胞清除凋亡细胞,加速组织从炎症向修复转变。
结构与功能作用:DHA是脑和视网膜的重要组成成分,对幼犬神经及视觉发育至关重要,也有助于维持老年犬的认知功能。
其他生理效应:可改善血脂、增强心血管功能并调节免疫反应。
• 多不饱和脂肪酸的应用:
骨关节炎(OA):富含EPA+DHA(干物质基础>2.5%或50–100 mg/kg/日鱼油补充)的饮食可显著减轻狗的关节疼痛和跛行,提升活动能力,机制与其抗炎作用相关。
皮肤健康与过敏:提高ω-3摄入可缓解特应性皮炎的炎症、瘙痒与红肿。
心血管健康:ω-3补充可改善犬的心肌功能,降低心律失常风险。
认知与发育:DHA促进幼犬大脑发育,并有助于延缓老年犬的认知衰退,改善学习与记忆。
肾脏健康:ω-3的抗炎特性可能有助于减缓慢性肾病进展。
5
植物营养素:来自大自然的“抗氧化卫士”
植物营养素是植物中具有生物活性的天然化合物,虽非传统维生素或矿物质,但具显著的抗氧化和抗炎作用。
• 主要类别与来源
①多酚类:
类黄酮:如槲皮素(苹果、洋葱)、儿茶素(绿茶)、花青素(蓝莓、蔓越莓)。
其他多酚:如白藜芦醇(葡萄皮)、姜黄素(姜黄)。
②类胡萝卜素:
类胡萝卜素是脂溶性色素,存在于红色、橙色和黄色的蔬菜和水果中。常见于犬类和人类膳食补充剂及食品中的类胡萝卜素有:α-和β-胡萝卜素、番茄红素、玉米黄素、隐黄素和叶黄素。
例如如β-胡萝卜素(胡萝卜)、番茄红素(西红柿)、叶黄素(菠菜)。
③植物甾醇
植物甾醇存在于油、蔬菜、水果、坚果、种子和豆类中。植物甾醇在降低高脂血症犬只的高血浆三酸甘油酯和胆固醇水平中起着重要作用。
这其中尤为重要的两个:
姜黄素(Curcumin)其抗炎机制不仅限于抑制NF-κB,还包括抑制COX-2、LOX、诱导型一氧化氮合酶(iNOS)等多种炎症相关酶的活性和表达。其主要限制在于口服生物利用度低。现代制剂通过将其与磷脂结合(形成植物体)、使用纳米颗粒或与胡椒碱(能抑制其在肝脏的快速代谢)联用,同时调节肠道菌群来提高其吸收率。
白藜芦醇(Resveratrol):作为一种SIRT1激动剂,它能模拟热量限制带来的部分健康效应,如改善线粒体功能、提高胰岛素敏感性、减轻氧化应激。在犬模型中,白藜芦醇显示出心脏保护作用。
注:抗氧化剂“鸡尾酒”疗法:在犬认知功能障碍(CCD)的研究中,单独使用某一种抗氧化剂(如维生素E)的效果往往有限。而将多种抗氧化剂(维生素E, C, L-肉碱, α-硫辛酸)与富含多种植物营养素(来自菠菜、番茄、葡萄、胡萝卜、柑橘)的饮食相结合,则显示出强大的协同效应。这可能是因为不同抗氧化剂在细胞的不同区室(水溶性/脂溶性)起作用,并能相互再生,形成一个抗氧化网络。
• 作用机制
抗氧化:机体在新陈代谢过程中会产生自由基,过多的自由基会攻击细胞,导致氧化损伤,这是衰老和多种慢性疾病的根源。植物营养素能直接中和自由基,或激活机体自身的抗氧化酶系统,从而保护细胞免受损伤。
抗炎:许多植物营养素能抑制关键的炎症通路(如NF-κB),减少促炎细胞因子的产生。
其他:抗癌、调节免疫、保护心血管等。
• 植物营养素的应用:
认知功能与抗衰老:一项针对老年比格犬的著名研究发现,饲喂富含多种抗氧化剂(包括来自菠菜、番茄、葡萄的多酚)的食物,并结合行为丰富化训练,能显著改善犬只的学习和记忆能力,延缓认知衰退。蓝莓和葡萄提取物也被证明能改善老年犬的认知表现。
骨关节炎:姜黄素、鳄梨豆非皂化物(ASU)等被研究用于辅助治疗犬OA,它们通过抗炎和抗氧化作用减轻关节疼痛和炎症。
癌症:一些植物营养素在体外实验中显示出抑制犬肿瘤细胞(如骨肉瘤细胞)生长的能力,如番茄红素、黄芩素等。但这方面的体内研究仍需加强。
心血管保护:葡萄籽提取物、槲皮素等在犬模型中显示出保护血管、抗血小板聚集的作用。
编者小结
以上五类功能性成分从不同的作用途径出发,以各自独特的机制在细胞、组织及系统层面协同发挥效应,共同促进并维持犬猫的整体健康与生理平衡,帮助其保持良好的生命状态和长期活力。
要将上述功能性成分有效地整合到临床实践中,需要充分考虑不同疾病的病理生理学特征,并依据现有的循证研究结果进行科学应用。同时,应结合个体差异和具体健康状况,选择合适的配方与干预方式,以确保营养策略在临床实施中的安全性与有效性。
1
慢性肠病(CE)/炎症性肠病(IBD)
饮食试验:采用水解蛋白或新型蛋白饮食进行6–8周的严格试验,是诊断和治疗食物反应性肠病(FRE)的金标准。
微生态干预:对于确诊的炎症性肠病(IBD)或抗生素反应性肠病(ARE),在饮食管理基础上应联合使用经验证的多菌株、高剂量益生菌或合生元,以恢复肠道菌群平衡并调节免疫反应,从而抑制异常炎症。
抗炎支持:在饲粮中补充富含EPA的鱼油(50–100 mg/kg/天)可有效辅助抗炎治疗。
纤维调整:依据病情(如结肠炎或小肠性腹泻)适当调整可溶性与不可溶性纤维比例,其中低聚果糖、菊粉等益生元纤维为首选。
2
糖尿病与胰岛素敏感性
糖尿病(DM)在犬类中越来越常见,通常是1型糖尿病。自发性1型DM的犬只存在严重的胃肠道菌群失调和粪便中未结合胆汁酸水平的改变。此外,治疗DM的药物也会影响胃肠道微生物群。
个性化微生物治疗:益生菌在糖尿病治疗中具有潜力,可通过减少自身免疫反应、炎症及氧化应激来提高胰岛素敏感性,同时增强黏附蛋白表达以降低肠道通透性。例如,补充双歧杆菌已被证明可改善葡萄糖耐受性。由于个体间微生物组结构和功能差异显著,基于微生物组的糖尿病治疗需采取个体化策略以实现最佳效果。
饮食成分调整:部分植物活性物质具有调节胰岛素和血糖的潜力。例如,高脂饮食中以改性大豆蛋白代替牛肉可显著降低餐后胰岛素水平,可能与其碳水化合物和纤维特性相关。不同谷物、豆类及脂肪酸类型也会影响犬的胰岛素敏感性。
此外,多种植物可影响犬的糖代谢,如辣椒素、迷迭香、罗勒和绿茶提取物。研究表明,它们能降低血糖和胰岛素水平、增强胰岛素分泌,并通过多酚保护胰腺β细胞功能。
功能性食品开发流程
3
骨关节炎(OA)
骨关节炎的改善需要采用多模式的综合管理方法,通过多种干预手段协同作用以获得更好的治疗效果。
体重控制:是所有治疗方案中最重要的一环。减轻体重能直接降低关节负荷。
核心营养干预-高剂量EPA:应使用专门为关节健康设计的处方粮,或在现有日粮基础上添加足量的高品质鱼油,确保EPA+DHA的摄入量达到治疗水平。需告知主人,该干预起效较慢,需坚持8-12周。
辅助营养成分:可联合使用葡萄糖胺/软骨素和ASU作为软骨保护剂。高生物利用度的姜黄素、乳香提取物或新西兰绿唇贻贝提取物可作为额外的抗炎支持。
4
认知功能障碍(CCD)
核心:选择含有复合抗氧化剂(维生素E, C)和多种植物营养素(来自水果蔬菜提取物)的老年犬专用处方粮。
结构支持:DHA:确保日粮中DHA的含量充足。
能量支持:对于已有明显认知衰退症状的犬只,换用含有6-10% 中链脂肪酸(MCTs)的处方粮已被证明能显著改善其认知评分。
肠道支持:通过补充益生菌/益生元来维护老年犬的肠道健康,有助于减轻可能加剧神经炎症的“炎性衰老”。
针对肠道微生物组的功能性营养干预,将在未来犬猫的健康维护与疾病管理中扮演愈发关键的角色。从益生菌的菌株特异性免疫调节,到EPA对炎症通路的分子调控,再到抗氧化剂组合在延缓神经退行性病变中的协同作用,功能性营养已从一种辅助手段,发展成为现代循证兽医学中用于管理多种慢性疾病的、不可或缺的科学工具。
然而,我们必须清醒地认识到当前领域的挑战。最大的挑战来自于显著的个体差异性。不同品种、年龄、遗传背景和生活环境的动物,其基础微生物组构成和对营养干预的反应都可能截然不同。当前基于“群体平均值”的推荐方案,在个体应用中效果可能打折扣。
随着高通量测序成本的降低,多组学技术将使我们能够为每一只动物绘制出其独特的肠道微生态“指纹”。通过将这些个体化的多组学数据与临床表型、生活方式等信息相结合,并利用人工智能(AI)和机器学习算法进行深度分析,我们将能够:
实现疾病的早期预测与诊断:在临床症状出现前,通过识别特定的微生物组失调模式或代谢物谱异常,来预警疾病风险。
制定高度个性化的营养方案:根据动物特定的菌群缺陷(如缺乏某种产丁酸菌)或代谢异常,为其精准匹配最有效的益生菌菌株、益生元种类或后生元组合。
动态监测与方案优化:通过定期检测,评估干预效果,并对营养方案进行动态调整,形成一个“诊断-干预-监测-优化”的闭环管理。
综上所述,功能性营养学正引领着一场宠物健康管理的深刻变革——从“千篇一律”的传统模式迈向“量体裁衣”的精准时代。对临床工作者而言,这意味着需要将前沿的科学工具整合为个体化的诊疗方案。而对宠物主人来说,这更赋予了他们一种全新的能力:通过科学喂养,更主动、更深刻地参与到守护爱宠长期健康的责任中来。
主要参考文献:
https://www.sciencedirect.com/science/article/pii/S1756464623003444
Di Cerbo A, Morales-Medina JC, Palmieri B, Pezzuto F, Cocco R, Flores G, Iannitti T. Functional foods in pet nutrition: Focus on dogs and cats. Res Vet Sci. 2017 Jun;112:161-166.
Rindels JE, Loman BR. Gut microbiome – the key to our pets’ health and happiness? Anim Front. 2024 Jun 20;14(3):46-53.
Kates AE, Jarrett O, Skarlupka JH, Sethi A, Duster M, Watson L, Suen G, Poulsen K, Safdar N. Household Pet Ownership and the Microbial Diversity of the Human Gut Microbiota. Front Cell Infect Microbiol. 2020 Feb 28;10:73.
Yang, Q.; Wu, Z. Gut Probiotics and Health of Dogs and Cats: Benefits, Applications, and Underlying Mechanisms. Microorganisms 2023, 11, 2452. https://doi.org/10.3390/microorganisms11102452
Hernandez J, Rhimi S, Kriaa A, Mariaule V, Boudaya H, Drut A, Jablaoui A, Mkaouar H, Saidi A, Biourge V, Borgi MA, Rhimi M, Maguin E. Domestic Environment and Gut Microbiota: Lessons from Pet Dogs. Microorganisms. 2022 Apr 30;10(5):949.
Atuahene, D.; Mukarram, S.A.; Balouei, F.; Antwi, A. Gut Health Optimization in Canines and Felines: Exploring the Role of Probiotics and Nutraceuticals. Pets 2024, 1, 135-151.
Grześkowiak Ł, Endo A, Beasley S, Salminen S. Microbiota and probiotics in canine and feline welfare. Anaerobe. 2015 Aug;
Guo X, Wang Y, Zhu Z, Li L. The Role of Plant Extracts in Enhancing Nutrition and Health for Dogs and Cats: Safety, Benefits, and Applications. Vet Sci. 2024 Sep 12;11(9):426.
Caterino C, Aragosa F, Della Valle G, Costanza D, Lamagna F, Piscitelli A, Nieddu A, Fatone G. Clinical efficacy of Curcuvet and Boswellic acid combined with conventional nutraceutical product: An aid to canine osteoarthritis. PLoS One. 2021 May 28;16(5):e0252279.
Wernimont SM, Radosevich J, Jackson MI, Ephraim E, Badri DV, MacLeay JM, Jewell DE, Suchodolski JS. The Effects of Nutrition on the Gastrointestinal Microbiome of Cats and Dogs: Impact on Health and Disease. Front Microbiol. 2020 Jun 25;11:1266

谷禾健康
伴侣动物正越来越多地存在于人类生活中,提供陪伴、娱乐、工作和心理支持,人们逐渐倾将宠物视为家庭成员。
随着宠物经济的不断升温,宠物健康也同时受到广泛关注。肠道微生物组是一个重要的免疫和代谢器官,越来越多的研究表明,猫狗健康的奥秘很大程度上藏匿于它们肠道内由微生物组成的复杂生态系统,它就像体内的一个”微型器官”或”第二大脑”,深刻影响着消化、免疫、代谢甚至情绪和行为。
从胃肠炎到癫痫,从肥胖到心脏病,从皮肤过敏到肾病——越来越多的研究揭示,这些看似不相关的疾病,都与肠道微生态失衡有着千丝万缕的联系。
本文将带你解码肠道微生态对于维持宠物整体健康和预防疾病所起到的关键作用,并重点介绍了与肠道微生物组改变(生态失调)相关的特定疾病和综合性的探索,例如胃肠炎、癫痫、肾病、肥胖等疾病。此外,本文还分析了影响肠道微生物组的各种因素,如习惯性饮食、年龄、品种、抗生素使用等,并基于微生物组的菌落构成以及所产生的肠道代谢与不同疾病类型之间的内在关联,为您提供从”肠”计议的个性化健康干预新思路。让我们一起守护毛孩子的健康,从肠道微生物开始!
胃肠道(GI)拥有一个复杂的生态系统,由细菌、病毒、真菌和原生动物等各种微生物组成。其中细菌是主要成分,在狗和猫的粪便样本中占>98%的宏基因组测序读数。
16S rRNA基因测序是目前最常用评估猫狗肠道细菌的标准方法,能够深入分析微生物组成、健康与患病动物间的群落差异,以及对饮食或治疗干预的反应。
★ 厚壁菌门、变形菌门和拟杆菌门构成核心优势菌群
宠物肠道微生物群主要由严格或兼性厌氧细菌组成,尤其是在大肠中。狗和猫的粪便微生物主要由五个门组成:厚壁菌门(Firmicutes)、梭杆菌门(Fusobacteria)、拟杆菌门(Bacteroidetes)、变形杆菌门(Proteobacteria)和放线菌门(Actinobacteria)。其中厚壁菌门、变形菌门和拟杆菌门这三个占核心主导地位。
Garrigues Q,et al.Front Vet Sci.2022
★ 厚壁菌门下的重要菌属
在核心细菌群落中,许多主要分类群属于厚壁菌门。Clostridia纲为最丰富的分类群,主要包含三个Clostridium梭状芽孢杆菌群:IV(如Ruminococcaceae科、Faecalibacterium prausnitzii)、XI(如Peptostreptococcaceae科)和XIVa(如Lachnospiraceae科、Blautia spp.)。
除了Clostridia纲,厚壁菌门中其他常见纲包括Bacilli(主要由乳酸杆菌目组成,以链球菌属和乳杆菌属为主)和Erysipelotrichi(主要包括Turicibacter属、Catenibacterium属和Coprobacillus属)。
★ 拟杆菌门下的Bacteroides和Prevotella在狗中差异很大
拟杆菌门是猫狗粪便样本中另一个丰富的门,主要包括Prevotella、Bacteroides和Megamonas属,其中最丰富的Bacteroides和Prevotella在狗之间的丰度差异很大。
★ 梭杆菌属与健康状况紧密相关
在梭杆菌门中,梭杆菌属(Fusobacterium)与狗的健康状况有关,并且它被认为与人类结直肠癌和炎症性肠病相关。梭杆菌在肉食性动物肠道代谢中发挥关键作用,具有分解蛋白质和氨基酸产生支链挥发性脂肪酸的独特功能。在狗的胃肠道中,梭杆菌属(如F.perfoetens和F.mortiferum)大量存在,约占总相对丰度的20%。
★ 变形菌门与放线菌门大肠中数量相对较少
变形菌门(Proteobacteria)和放线菌门(Proteobacteria)也很常见。这些门通常是小肠的定植者,粪便样本中的数量较少。肠杆菌科(Enterobacteriaceae)的成员(例如,大肠杆菌Escherichia coli)是兼性厌氧菌,使得它们能够利用小肠中的氧气。在粪便样本中,它们的增加与许多疾病有关。放线菌门也与小肠有关,包括Corynebacteriaceae科(例如Corynebacterium spp.)和Coriobacteriaceae科(例如Collinsella spp.)。
肠道细菌能将饮食分子或药物转化为细菌衍生代谢物,因此肠道微生物群被视为重要的代谢器官。平衡的肠道微生物群通过调节免疫系统、抵御肠道病原体及提供维生素和营养物质,对宠物健康产生重要影响。
宠物肠道微生物群的组成受多种因素显著影响,包括饮食、年龄、品种、活动量、健康状况、益生菌和药物使用等,其中抗生素的影响尤为突出,这已被多种动物模型证实。
Shah H,et al.Microorganisms.2024
1
饮食
饮食已被认为是影响哺乳动物肠道微生物群生物多样性和功能特征的主要驱动因素之一。宠物饮食的类型与成分直接塑造肠道菌群结构,其营养物质(如膳食纤维、碳水化合物、蛋白质)作为微生物生长的底物,经菌群发酵后产生短链脂肪酸(SCFAs)、次级胆汁酸、氨基酸及脂溶性维生素等关键代谢物。
▸ 高纤维饮食促进狗肠道产短链脂肪酸菌群增殖
研究表明,饮食纤维含量是核心调控因素:高纤维饮食(28.1%)显著促进产短链脂肪酸(SCFAs)菌群(如Bacteroides、Prevotella、Faecalibacterium)增殖,而低脂饮食(8.6%纤维)则特异性富集Faecalibacterium。
注:SCFAs可通过抑制NF-κB通路等机制减轻肠道炎症。
饮食在狗的肠道微生物组中起至关重要的作用
Shah H,et al.Microorganisms.2024
▸ 高蛋白饮食会影响猫肠道微生物多样性
研究评估了断奶后使用高蛋白/低碳水化合物饮食 (HPLC)的小猫与断奶后使用中等蛋白质/中等碳水化合物 (MP/MC) 饮食的小猫的微生物组。
发现高蛋白/低碳水饮食增加了物种多样性,在高蛋白/低碳水喂养的小猫中增加5个属是已知的丁酸盐产生菌。
营养干预对猫狗肠道代谢的影响
Lyu Y,et al.Animals (Basel).2025
值得注意的是,菌群结构具有动态可塑性——犬类饲喂32周高纯度氨基酸与可消化淀粉饲料后,菌群组成发生显著偏移,但恢复基础饮食后即可回归初始状态。这种可逆性变化证实了饮食成分通过改变肠道底物微环境,直接驱动特定菌种(如产SCFA菌)的竞争性生长,进而重塑微生物组结构与代谢组功能。因此,精准设计膳食纤维与蛋白质比例,是调控宠物肠道微生态平衡的关键策略。
2
年龄
年龄是肠道微生物群落组成显著改变的重要决定因素,随着年龄增长微生物多样性明显下降。以炎症反应增强为特征的器官衰老现象导致肠道微生物组的组成和功能持续改变,从而影响机体的体力活动和药物摄入模式。
▸ 狗断奶后梭杆菌丰度增加
在犬类中,断奶后梭杆菌的存在更为普遍,6-10岁年龄组的具核梭杆菌(Fusobacterium perfoetens)丰度约为0.5-1岁年龄组的两倍(14.3%vs.7.2%)。虽然这种丰度的增加与狗的年龄呈正相关,但梭菌水平的增加与狗断奶后食用肉类食物之间也存在潜在关联。
▸ 猫肠道中双歧杆菌和乳杆菌丰度随年龄增长下降
同样地,在猫的研究中,第18至42周期间,四种最常见菌属的平均丰度发生了显著变化:双歧杆菌属(Bifidobacterium)、乳杆菌属(Lactobacillus)、普雷沃氏菌属(Prevotella)和拟杆菌属(Bacteroides)。
其中拟杆菌属和普雷沃氏菌属的丰度随年龄显著增加,而双歧杆菌属和乳杆菌属则显著下降。幼猫18周龄时,微生物组主要由乳杆菌属(35%)和双歧杆菌属(20%)构成。早期时间点(8-17周)乳杆菌属和双歧杆菌属丰度的升高可归因于哺乳/断奶期影响。至42周龄时,拟杆菌属占比达16%,普雷沃菌属占14%,巨球菌属(Megasphaera)占8.4%。巨球菌属的丰度随时间显著上升,从第18周的0.1%和第30周的0.2%增至第42周的8.4%。
3
品种
品种是微生物群个体间变异的主要来源。研究发现不同品种之间的微生物组成存在差异。
▸ 相同饮食下不同犬菌群丰度不同
例如,在对比马尔济斯犬、贵宾犬和迷你雪纳瑞三种犬种时发现,即使喂养相同的食物,在马尔济斯犬中,梭杆菌(Fusobacteria)数量丰富,而在贵宾犬中,厚壁菌(Firmicutes)和放线菌(Actinobacteria)数量丰富,且马尔济斯犬中厚壁菌门的丰度最低。
▸ 猫科和布偶猫存在肠道微生物差异
另一项研究通过16S rRNA基因测序技术证明了家猫与布偶猫肠道微生物组的品种差异:布偶猫组中肠球菌属(Enterococcus)、乳杆菌属(Lactobacillus)、链球菌属(Streptococcus)、罗氏菌属(Roseburia)和经黏液真杆菌属(Blautia)等有益微生物的丰度显著高于家猫组,这表明可针对特定品种设计益生菌制剂。
4
性别/绝育
大量研究表明,雄性和雌性之间的肠道菌群存在差异。这在动物和人类中都有观察到。例如,对非肥胖糖尿病(NOD)/ShiLtJ小鼠的研究发现,与雌性小鼠相比,某些细菌家族在雄性小鼠中更丰富,如Kineosporiaceae、Peptococcaceae、Porphyromonadaceae、 Veillonellaceae、 Lactobacillaceae、Peptostreptococcaceae、Bacteroidaceae、Cytophagaceae和Enterobacteriaceae。
而在人类中,双侧卵巢切除术与鲍氏梭菌(Clostridium bolteae)的较高存在相关。非卵巢系统性雌激素水平与粪便梭菌的丰度之间存在直接相关性,其中包括非梭状芽胞杆菌属和瘤胃球菌科的三个属。
▸ 绝育狗厚壁菌门相对丰度较高
在宠物研究中,发现厚壁菌门的相对丰度显示绝育雌性和阉割雄性之间的相似性,与正常雌性和雄性狗相比,这些群体的相对丰度明显更高。此外,雄性狗的厚壁菌门比例高于雌性狗。另一方面,与雌性相比,雄性狗的拟杆菌门较低,但高于阉割狗(绝育的雌性和阉割的雄性)。与绝育狗(绝育的雌性和绝育的雄性)相比,正常犬(雌性和雄性)的梭杆菌更高。
▸ 绝育后乳杆菌、双歧杆菌减少
还有一项研究发现,绝育后的雄性猫狗肠道中某些菌群数量会发生变化,如乳杆菌(Lactobacillus)、双歧杆菌(Bifidobacterium)等菌群数量减少,而肠球菌(Enterococcus)、大肠杆菌(Escherichia)等菌群数量增加。这些变化可能与代谢、免疫等方面有关。
此外,宠物的饮食、生活环境等因素也会对肠道微生物产生影响,因此在绝育后需要注意宠物的饮食和生活环境,以维护宠物的肠道健康。
5
运动
研究表明,体育活动与改善代谢健康之间存在密切联系。人类研究表明,体育活动会导致有益肠道微生物的增加,如梭菌。定期运动与丹毒丝菌科(Erysipelotrichaceae)含量较高之间存在正相关关系。
Shah H,et al.Microorganisms.2024
▸ 运动与饮食一起影响了狗肠道菌群
几项临床试验研究了运动和饮食干预对犬肠道菌群的影响。在利用商业高蛋白、低脂肪和高纤维干粮的减肥计划中,运动会影响肠道微生物群的组成。在与运动相关的活动中,葡萄糖胺的添加会影响雪橇犬微生物组的组成。食用葡萄糖胺的雪橇犬的乳杆菌科(Lactobacillaceae)和Anaerovoracaceae减少。在补充葡萄糖胺的狗中,Sellimonas、Eubacterium brachy和Parvibacter的丰度降低,尤其是在运动后。
6
健康状况
▸ 消化系统疾病破坏肠道微生物群平衡
消化系统疾病如腹泻、便秘、胃肠炎等会对猫狗微生物群产生影响,破坏肠道微生物群的平衡,导致有益菌的减少,有害菌的增加。
▸ 感染寄生虫改变肠道微生物群
许多肠道寄生虫被证明会引起宠物肠道微生物群的显著改变,其中贾第鞭毛虫是一种普遍存在的肠道寄生虫,可导致腹泻,其带来的改变最为明显。
贾第鞭毛虫与许多细菌群落的丰度呈正相关,例如普雷沃氏菌(Prevotella)和厌氧螺菌(Anaerobiospirillum succiniproducens)。这些细菌会导致肠道屏障粘液的脆弱化。这种脆弱化使肠贾第鞭毛虫更容易切割屏障并允许更多肠道病原体在肠道定植。
此外,在22周大的幼犬中,贾第鞭毛虫的高负荷也与约氏乳杆菌(Lactobacillus johnsonii)的减少相关。
▸ 病毒感染降低拟杆菌丰度
犬细小病毒是影响狗的最常见病原体之一,会导致腹泻、出血性肠炎和幼犬死亡。在一项研究中,四只幼犬在6周大时自然感染了犬细小病毒,肠道微生物群发生严重改变,变形杆菌丰度增加,主要是肠杆菌科(Enterobacteriaceae),拟杆菌(Bacteroidetes)和梭杆菌(Fusobacteria)丰度减少。
7
抗生素等外源药物
抗生素通过作用于细菌保守靶点(如核糖体、细胞壁)发挥广谱抗菌作用,但其非选择性抑制会显著扰动宠物肠道微生态平衡。
▸ 抗生素会改变猫狗肠道菌群组成与代谢能力
研究表明,抗生素暴露可导致犬猫肠道菌群结构与功能发生多维度改变:菌群组成方面,常见有益菌如产短链脂肪酸(SCFA)的Faecalibacterium、Blautia和Roseburia丰度显著降低,而潜在致病菌(如Enterobacteriaceae、Enterococcus spp.及Escherichia spp.)增殖。
代谢功能方面,碳水化合物发酵能力下降导致SCFA产量减少,胆汁酸代谢紊乱表现为初级胆汁酸(胆酸、鹅去氧胆酸)累积与次级胆汁酸(脱氧胆酸、石胆酸)耗竭。
▸ 使用抗生素可能导致长期菌群失衡
值得注意的是,甲硝唑、泰乐菌素及阿莫西林-克拉维酸等常用抗生素还可降低粪便维生素与抗氧化物质浓度,同时升高氧化应激标志物(如核糖酸)。这些扰动具有长期持续性——猫停用克林霉素2年后仍存在菌群(如Prevotellaceae减少)与胆汁酸代谢异常,幼猫抗生素暴露后菌群失衡可持续至成年期。菌群失调进一步增加耐药菌定植风险(如艰难梭菌),并可能通过破坏肠屏障功能诱发慢性肠病。
值得注意的是,益生菌干预(如猫源Enterococcus hirae)或特定酶制剂(如核糖酶)可能缓解部分不良反应,提示在临床抗感染治疗中需综合评估菌群修复策略。
与人类和其他动物类似,狗和猫肠道代谢库的主要贡献者是蛋白质、碳水化合物和脂肪。这些常量营养素在消化过程中分解,产生各种代谢物,包括气体(例如CO2、CH4)、有机酸(例如乳酸、丙酮酸)、生物胺(例如组胺、血清素)、短链脂肪酸(例如丁酸盐、乙酸盐)、支链脂肪酸(BCFAs,例如异丁酸酯、异戊酸酯)和支链氨基酸(例如异亮氨酸、缬氨酸)。
猫和狗肠道中的典型代谢物
Lyu Y,et al.Animals (Basel).2025
这些代谢物在维持宿主生理过程中起着关键作用。肠道还包括来自细菌细胞壁成分、多胺、挥发性有机化合物、植物次生代谢物和膳食胆碱的各种代谢物。这些代谢物具有高度动态性,经历吸收、分布或从体内排泄等各种过程。这种动态性质强调了肠道代谢的复杂性及其对整体健康的深远影响。通过检测猫狗肠道中的主要代谢物及微量元素,也可以一定程度上反映其健康状况。
一般来说,从产量和健康影响的角度来看,犬科和猫科动物结肠中的主要代谢物主要是短链脂肪酸、次级胆汁酸以及支链脂肪酸和吲哚衍生物,它们分别来自碳水化合物、脂肪和蛋白质的发酵。
1
短链脂肪酸
短链脂肪酸(SCFA)是结肠内特定厌氧菌(例如罗氏菌属、拟杆菌属)发酵膳食纤维和抗性淀粉产生的主要细菌代谢产物,这些短链脂肪酸对人类和伴侣动物都表现出许多健康益处,包括抗炎、免疫调节、体重管理、糖尿病管理和癌症预防,以及对心脏和肝脏等器官的保护作用。
▸ 猫和狗可通过蛋白质发酵产生短链脂肪酸
此外,SCFA可以降低肠道pH值,防止氨等有害代谢物的吸收。值得注意的是,与大多数杂食性哺乳动物通过膳食纤维和抗性淀粉发酵产生短链脂肪酸不同,狗和猫作为食肉动物,其结肠中具有更丰富的蛋白质发酵细菌(如产气荚膜梭菌),能够产生短链脂肪酸。虽然蛋白质对SCFA产生的直接贡献不如纤维重要,但它可以通过改变肠道微生物组组成、转运时间和特定氨基酸的利用来间接影响该过程。
乙酸盐、丙酸盐和丁酸盐是三种主要的短链脂肪酸,因其多样化的生物活性而被广泛研究。肌组织可以利用乙酸盐作为能量来源。丙酸盐被肝脏从门静脉血吸收并转化为琥珀酰辅酶A,琥珀酰辅酶A参与能量产生、柠檬酸循环、酮代谢和血红素合成。丁酸盐被结肠氧化,是结肠细胞的首选能量底物。
▸ 营养和疾病状况会影响体内短链脂肪酸含量
在各种饮食干预和补充策略的犬猫营养研究中,SCFA的产生经常被报道。在最近的许多狗和猫研究中,来自多种食品成分的植物来源的低聚糖(例如,果糖、半乳糖、甘露糖和木糖低聚糖(FOS、GOS、MOS、XOS))和各种类型的可发酵碳水化合物(例如,果胶、菊粉和抗性淀粉)一致表明SCFA的产生显著增加。
具体而言,研究表明,当成年犬的饮食中添加0.5% FOS和GOS、0.15% MOS、1%菊粉和1%抗性淀粉时,丁酸盐、乙酸盐或戊酸盐的水平升高。在猫中,当动物喂食0.4% XOS时,粪便丁酸盐和丙酸盐浓度与乳酸菌种类丰度的增加呈正相关。
疾病状态的发生和发展通常也会影响SCFA的产生。例如,几项研究表明,患有炎症性肠病(IBD)的犬科动物和猫科动物经常表现出肠道菌群失调,导致产生SCFA的细菌(例如双歧杆菌、Roseburia)下调,随后导致特定SCFA的合成减少。
2
次级胆汁酸
次级胆汁酸,如胆酸和鹅去氧胆酸,是在肝脏中产生并通过胆道器官释放到消化系统中的脂质分子。肠道微生物群将这些宿主来源的胆汁酸转化为次级胆汁酸,这些胆汁酸构成了粪便中发现的大部分胆汁酸,是狗和猫的主要肠道代谢产物。
胆汁酸的一个基本作用是促进脂肪的消化和吸收。除了消化功能外,胆汁酸还充当信号分子,影响葡萄糖和脂质代谢、胰岛素敏感性、炎症反应、免疫功能和甲状腺调节的能量消耗。
▸ 次级胆汁酸可以反映营养状况
主要由碎牛肉(75%)组成的高蛋白、高脂肪、低纤维饮食(46.7%蛋白质、33.1%脂肪、3.4%纤维、干物质)已被证明可以提高狗的粪便脱氧胆酸水平。
与饲喂湿粮(46%蛋白质、27%脂肪、4.7%纤维)的猫相比,喂食干粮(33%蛋白质、7%脂肪、9.3%纤维、干物质)的猫的粪便总胆汁酸减少了3倍。这一发现意味着高脂肪和低纤维饮食可以诱导健康猫的粪便总胆汁酸增加。
▸ 食物中蛋白质和纤维含量影响猫胆汁酸水平
最近的一项研究评估了猫的23种粪便胆汁酸,结果显示,将7.9%高消化抗性淀粉饮食(37.6%蛋白质、18.9%脂肪、3.2%纤维)与0.5%低消化抗性淀粉饮食(38.0%蛋白质、19.0%脂肪、3.1%纤维)进行比较时,胆汁酸谱发生了显著变化。具体来说,12种胆汁酸表现出显著增加,而3种表现出减少。在次级胆汁酸胆酸盐和牛磺胆酸盐硫酸盐以及次级胆汁酸12-脱氢胆酸盐和3-脱氢胆酸盐中观察到最明显的变化。因此,尽管宏量营养素含量相似,但食品加工技术及配方对微生物(次级)胆汁酸代谢有显著影响。
在一项交叉研究中,制定了一种老年干粮(31.1%蛋白质、16.8%脂肪、1.3%纤维)含有特定成分(即燕麦粒、西兰花和番茄渣),可防止与年龄相关的认知能力下降,并与商业干粮(32.7%蛋白质、19.1%脂肪、1.4%纤维)进行了比较。老年干粮与粪便次生胆汁盐7-α-羟基-胆甾烯酮、脱氢石胆酸盐、脱氧胆酸盐、异熊去氧胆酸盐、石胆酸盐和熊去氧胆酸盐的显著减少以及粪便初级胆汁盐胆酸盐的增加有关。
一般来说,饮食类型和营养成分会影响犬科动物和猫科动物的肠道微生物群,从而进而改变粪便胆汁酸库。具体而言,在患有慢性肠病和胰腺外分泌功能不全的狗中观察到粪便石胆酸、脱氧胆酸和总次级胆汁酸浓度较低,粪便胆汁酸代谢失调和熊去氧胆酸降低与猫慢性肾病中的新型微生物特征相关。需要进一步的研究来阐明这些改变与饮食因素以及肠道微生物群之间的关系。
3
支链脂肪酸和吲哚代谢物
肠道微生物组可以改变氨基酸的代谢,产生支链脂肪酸(BCFA)和吲哚衍生物等代谢物。这些化合物影响各种生理过程,包括免疫功能和炎症。
▸ 支链脂肪酸影响肠道微生物组及健康
肠道中BCFA的存在会显著影响其环境,从而影响肠道微生物组的组成和活性。此外,这些脂肪酸充当信号分子,与肠道内壁和免疫细胞进行交流,最终帮助维持肠道屏障的完整性,调节免疫反应和炎症,并与认知功能和心理健康有关。
▸ 膳食蛋白质和纤维含量会影响支链脂肪酸浓度
然而,由于狗和猫的肉食性,与人类和其他杂食性哺乳动物相比,它们具有更高的膳食蛋白质需求量和较低的结肠蛋白发酵水平。因此,BCFA的浓度远小于短链脂肪酸(即占SCFA的5%-10%)。狗和猫中最重要和最丰富的BCFA是异丁酸盐和异戊酸盐,它们分别由缬氨酸和亮氨酸产生。BCFA的产生与饮食中的粗蛋白含量和氨基酸浓度直接相关。
研究观察到,当狗和猫补充膳食纤维(例如低聚果糖、菊粉)时,BCFA浓度,特别是异丁酸盐浓度降低。这表明结肠环境和微生物组功能已从蛋白质发酵转变为纤维发酵。同样,胃肠道疾病可促使BCFA生成量下降,诊断为慢性肠病的猫科动物的异丁酸盐和异戊酸盐水平降低就证明了这一点,这一结局可能与潜在的微生物群失调有关。
▸ 不同剂量的吲哚衍生物对狗健康影响不同
另一组重要的氨基酸代谢物包括吲哚衍生物。这些化合物来源于氨基酸色氨酸,起信号分子的作用,对免疫调节(如T细胞反应)、抗炎过程和维持肠道内的屏障功能至关重要。
尽管如此,各种吲哚衍生物对健康的影响并不一致,并且明显依赖于剂量。某些化合物(如硫酸吲哚)在肠道内过度积累可导致不良反应,如炎症性肠病和肾功能障碍。与BCFA类似,吲哚衍生物(例如吲哚、吲哚-2-酮、吲哚-丙烯酸酯、吲哚-3-乙酸酯)随着蛋白质摄入量的增加而增加,而在健康狗中,纤维补充剂会减少或趋于减少,但饮食与吲哚衍生物之间的相互作用相当复杂。
在最近的一项研究中,比较了不同蛋白质水平(19.99%、25.34%、45.77%)对狗肠道健康的影响,吲哚和吲哚-2-酮的粪便水平随着蛋白质消耗的增加而显著增加,而吲哚-乳酸、吲哚-乙酸、吲哚-丙酸和2-氧吲哚-3-乙酸酯的水平随着蛋白质消耗水平的增加而显著降低。色氨酸是吲哚的前体氨基酸,随着蛋白质消耗的减少而增加。
4
维生素
维生素与肠道菌群的互作是猫狗肠道微生态健康的重要组成部分,是评估消化系统功能的关键指标。通过分析维生素的合成能力和代谢能力,可以早期识别微生态失衡状态,预防营养吸收障碍。
▸ 维生素A影响视觉、皮肤健康
维生素A是宠物体内重要的营养成分,参与视觉、皮肤健康、免疫系统功能以及细胞生长和分化的调节。它有助于维持视⼒,特别是在夜间视力方面,⽀持正常的生长和发育,促进皮肤和被毛的健康。
▸ 维生素D影响钙的吸收及骨骼健康
维生素D主要影响钙和磷的代谢,帮助维持骨骼和牙齿的健康。它促进钙的吸收和利用,对猫狗的生长、发育以及整体骨骼健康至关重要。
▸ B族维生素能影响大部分生理过程
B族维生素参与猫狗几乎所有的生理过程,这些水溶性维生素无法在体内大量储存,需要持续补充。B族维生素缺乏会导致神经系统异常、皮肤病变、贫血、消化不良和免疫功能下降等多种问题。
▸ 维生素C影响免疫及皮肤弹性
维生素C是一种强效的抗氧化剂,对宠物的健康至关重要。它有助于增强免疫系统,促进伤口愈合,支持皮肤和被毛的健康,同时还参与胶原蛋白的合成,保持关节健康和皮肤弹性。
对狗和猫的研究表明,肠道微生物组和功能的变化不仅决定着胃肠道疾病,而且还与其他器官系统的疾病密切相关,例如慢性肾病、肠胃炎、神经系统疾病和肥胖等。虽然在许多疾病中仍需要探索确切的潜在机制,但一些微生物途径现在已得到公认,会影响健康和疾病,其中一些可以使用不同的方法直接评估。
1
急性和慢性胃肠炎
急性胃肠炎(AGE)发作时(尤其细菌/病毒性感染),犬猫肠道菌群呈现多样性骤降与致病菌暴发性增殖。
▸ 急性胃肠炎时肠道菌群多样性会骤降
一项针对健康犬、急性非出血性腹泻犬、急性出血性腹泻犬以及特发性炎症性肠病(IBD)犬的16S rRNA基因测序研究显示,患有急性腹泻的犬,尤其是急性出血性腹泻犬,其菌群中的细菌类群发生了最显著的变化。与健康犬相比,急性出血性腹泻犬的大肠杆菌、Sutterella和产气荚膜梭菌含量显著增加,同时伴随产短链脂肪酸(SCFA)的核心菌群(如Faecalibacterium、Blautia)减少。
这种失衡直接削弱SCFA合成能力,导致肠屏障修复障碍与炎症因子(如IL-8、TNF-α)释放增加。值得注意的是,抗生素治疗虽抑制病原菌,但进一步降低菌群多样性,延长恢复周期。
患有胃肠道疾病的狗和猫的微生物变化
doi: 10.3748/wjg.v20.i44.16489.
▸ 肠道炎症宠物体内变形菌门细菌增加,拟杆菌属减少
慢性炎性肠病(CE)是一种多因素疾病,肠道免疫和环境因素(饮食、微生物群)之间的相互作用引发并驱动慢性肠道炎症,导致受影响狗呕吐、腹泻、食欲改变或体重减轻。
患有CE的犬表现出粪便细菌丰富度和多样性的显著降低。炎症性肠病中常见的微生物组变化是厚壁菌门和拟杆菌门内的细菌群减少,而变形菌门内的细菌群增加,此外,Lachnospiraceae和Clostridium亚群多样性减少与IBD有关。
不同的微生物学研究表明,肠道变形菌门的增加和梭菌目的减少与肠道炎症有关。持续胃肠道症状的犬受试者中梭菌属、梭杆菌属和拟杆菌属丰度降低,同时肠杆菌科丰度升高,肠杆菌科的成员被认为是致病的,因为它们有能力在胃肠道内引起先天免疫反应。
Shah H,et al.Microorganisms.2024
对患病犬和健康对照组的微生物种群进行比较分析发现,前者的双歧杆菌、乳酸杆菌、链球菌和潜在致病性大肠杆菌水平均有所升高。在胃肠道中,驻留细菌可以将各种化合物释放到周围的肠道环境中,包括脂多糖、淀粉样蛋白和其他免疫原性混合物。免疫激活,无论是通过脂多糖分泌还是与抗炎微生物相比促炎微生物的增加,都会形成肠道菌群失调,进一步导致肠道炎症。
2
神经系统疾病
脑肠轴(GBA)是一个由内分泌、免疫和神经元介质组成的网络,在肠道和大脑之间形成错综复杂的相互作用。肠道微生物群、中枢神经系统(CNS)和肠道神经系统(ENS)共同调节GBA。胃肠道中产生的某些神经活性物质可以穿过血脑屏障(BBB)和肠道粘膜屏障进入中枢神经系统。
▸ 肠道菌群可能通过神经递质影响犬类癫痫
癫痫是犬类最常见的神经系统疾病之一。肠道微生物群(GM)衍生的短链脂肪酸(SCFAs)可能会影响癫痫发作的可能性,通过调节兴奋性/抑制性神经递质、神经炎症、氧化应激以及心理社会压力来实现。
肠道菌群紊乱可加剧癫痫严重程度,其核心机制为菌源性炎症激活免疫应答,通过肠神经系统(ENS)影响中枢神经功能。肠道微生物群组成的变化与狗的神经系统疾病(如不明原因的脑膜脑脊髓炎以及攻击性和恐惧症等行为问题)有关。
代谢物在大脑的兴奋/抑制平衡和癫痫发生的作用
Shah H,et al.Microorganisms.2024
▸ 攻击性行为犬巨单胞菌属和Catenibacterium升高
攻击性行为犬的粪便中Catenibacterium与Megamonas丰度异常升高,该菌群同时参与初级胆汁酸代谢失调,可能通过腹痛-应激通路诱发神经兴奋。
老年犬的梭杆菌门丰度随年龄增长下降,而放线菌门减少与认知功能提升正相关。致病菌如大肠杆菌和假单胞菌可合成γ-氨基丁酸(GABA)等神经活性物质,其透过血脑屏障后干扰神经递质平衡——GABA作为关键抑制性神经递质,其水平下降将直接降低神经元兴奋阈值,促进癫痫发作。
生酮饮食(KD)可通过重塑菌群结构(如提升阿克曼菌、粪便拟杆菌丰度),显著降低发作频率。而犬类研究显示KD能同步调节代谢组(如降低促瘤因子)与菌群(减少丹毒丝菌目、乳杆菌目,增加双歧杆菌目等)。
3
肾病
猫和狗的慢性肾病带来了重大的临床挑战,新兴研究强调了肠肾轴在其发病机制和管理中的关键作用。
肠-肾轴包含肠道微生物组、肠道屏障、微生物代谢物和肾脏生理学之间相互作用的复杂网络。该轴的破坏,特别是以肠道菌群失调和微生物代谢失调为特征,是导致慢性肾病(CKD)发病机制的重要因素。
▸ 肠道菌群失调导致毒素积累会加剧肾功能障碍
肠道微生物群组成和功能的改变会引起相关微生物代谢失调,导致尿毒症毒素积累和氨基酸、胆汁酸和脂肪酸谱的破坏。这些紊乱反过来会加剧肾功能障碍和全身炎症。
▸ 慢性肾病也会反向加剧尿毒素生成菌增殖
慢性肾病(CKD)通过多重途径驱动肠道菌群失调:肾功能下降导致含氮废物(如尿素、肌酐)肠腔蓄积,促进变形菌门中尿毒素生成菌(如大肠杆菌、假单胞菌)增殖,同时抑制产短链脂肪酸(SCFAs)的厚壁菌门生长;饮食限制(如低磷、低蛋白处方粮)往往会减少膳食纤维摄入,剥夺益生菌代谢底物,加剧菌群多样性下降;药物副作用(如磷结合剂、抗生素)进一步破坏肠黏膜屏障。
这种失调形成恶性循环:菌群紊乱减少SCFAs(尤其丁酸)合成,削弱其抗炎与维护肠屏障功能,导致内毒素易位激活全身炎症,加速肾纤维化;而尿毒素蓄积又进一步改变肠道微环境,促进条件致病菌定植。
▸ 通过菌群有望改善慢性肾病
而干预菌群-肾脏轴有望改善CKD进程:临床研究证实,补充特定益生菌组合(如含双歧杆菌的制剂)可降低CKD犬猫血清尿素氮,其机制涉及抑制变形菌门过度增殖、恢复SCFAs水平及减轻尿毒症毒素积累;直接补充丁酸盐或高纤维饮食促进产丁酸菌(如Faecalibacterium)生长,改善肾小球硬化评分;此外,粪菌移植(FMT)在CKD犬模型中显示可部分逆转菌群失调,延缓eGFR下降速率。这些证据凸显了以肠道菌群为靶点的营养干预在宠物CKD管理中的临床应用潜力。
4
过敏/皮肤病
对肠道微生物的深入研究发现,肠道菌群与皮肤之间存在内在关联,被称之为“肠-皮肤轴”(Gut-Skin Axis)。肠道菌群及其所产生的代谢物通过免疫调节影响皮肤的屏障功能,诱发皮肤炎症。
▸ 慢性皮炎犬艰难梭菌、丹毒丝菌等促炎菌增多
研究表明,慢性皮炎(如犬特应性皮炎cAD)与肠道之间存在关联。cAD犬的肠道菌群α多样性(香农指数、均匀度)显著低于健康犬,且梭杆菌属、巨单胞菌属等有益菌丰度下降,而艰难梭菌、丹毒丝菌等促炎菌增殖;这种失衡通过”肠-皮肤轴”激活全身免疫反应,表现为Th2型细胞因子升高和血清IgE浓度增加,肠道菌群失调加剧皮肤瘙痒和损伤。
cAD犬肠道代谢功能异常涉及氨基酸生物合成的代谢途径异常富集,而短链脂肪酸(SCFAs)合成通路受抑制,削弱其抗炎和屏障修复作用。
患有皮炎的犬皮肤微环境改变(如pH升高、经皮水分流失增加),导致菌群多样性下降,致病菌如伪中间葡萄球菌和棒状杆菌过度定植,进一步破坏皮肤物理屏障并诱发慢性炎症。
5
肥胖
宠物肥胖已成为日益严重的健康问题,这与久坐的生活方式、高热量饮食、遗传倾向及绝育等多种因素相关,而肠道菌群的重要性尤为关键,还与犬的心脏病、糖尿病和骨科疾病存在关联。
▸ 肥胖和超重犬菌群多样性降低
肥胖和超重犬普遍存在粪便菌群多样性降低,肥胖犬体内放线菌门(Actinobacteria)和罗斯氏菌属(Roseburia)的数量更多,且放线菌纲(Actinobacteria)的存在更为明显。
瘦犬的肠道菌群主要由厚壁菌门(Firmicutes)的微生物组成,而肥胖犬的肠道菌群则主要由变形菌门(Proteobacteria)的细菌构成,其中革兰氏阴性菌通过产生较高水平的肠道脂多糖(LPS),诱发肥胖犬出现慢性炎症。
另一方面,超重犬的丹毒丝菌纲(Erysipelotrichi)丰度更高,这主要是由于真杆菌属(Eubacterium spp.)的差异所致,与肥胖犬相比,超重犬的双歧杆菌目(Bifidobacteriales)丰度显著降低,气单胞菌目(Aeromonadales)丰度略有增加,且普雷沃氏菌属(Prevotella copri)和梭菌属(Clostridium)的检出率更高。
▸ 巨单胞菌属和瘤胃球菌与减重呈负相关
此前研究还发现,与正常犬相比,超重犬的梭杆菌门,尤其是产气荚膜梭菌的数量更多。正常体重犬的瘤胃球菌科相对丰度也高于肥胖犬。巨单胞菌属的丰度与减重速率呈负相关,与减重较慢的犬相比,快速减重的犬粪便中瘤胃球菌科的数量明显减少,丙酸和乙酸水平也较低,这表明肥胖犬的粪便中存在能产生丙酸和乙酸的细菌,且由于这些细菌能通过产生短链脂肪酸(SCFAs)从饮食中获取更多能量,可能使其对减重的反应较差。
干预研究表明,补充低聚果糖(FOS)可提升菌群多样性,促进双歧杆菌增殖并增加抗炎性丁酸合成,为肥胖管理提供新策略。
6
心脏健康
狗经常表现出充血性心力衰竭(CHF),其特征是肺水肿、胸腔积液或腹水引起的腹胀导致的急性呼吸窘迫。即使心脏病得到很好的控制,患有充血性心力衰竭的狗,尤其是那些患有右侧充血性心力衰竭(RCHF)的狗,也经常因为严重的恶病质和进行性食欲不振而被处死。
▸ 肠道菌群失调指数和狗二尖瓣病之间存在联系
越来越多的研究表明,肠道微生物群及其代谢物在心血管疾病的发展和病程中发挥作用。患有充血性心力衰竭的狗增加了大肠杆菌和未表征的肠杆菌科物种。在另一项研究中,观察到肠道微生物组与狗粘液瘤性二尖瓣病(MMVD)之间存在潜在关系。肠道菌群失调指数与MMVD的严重程度成正比,并且与肠道中关键的胆汁酸转化器Clostridium hiranonis呈负相关。
次级胆汁酸是肠道微生物发酵初级胆汁酸产生的重要副产物。次级胆汁酸可能会促进有益细菌的生长,但会抑制有害物种。例如,次级胆汁酸,如DCA,可促进梭杆菌的生长,而LCA可促进粪杆菌的生长。相反,DCA阻碍了大肠杆菌的生长。这些发现表明,肠道微生物群、肠道微生物群产生的代谢物与狗MMVD的病理生理学进展之间存在潜在的相互作用。
▸ 三甲胺可能是肠道菌群影响心血管疾病的关键物质
而另一方面,短链脂肪酸有助于调节血压。大量的胆碱、肉碱和磷脂酰胆碱被一些肠道细菌分解成前体三甲胺(TMA)。TMA的下游信号传导和代谢导致氧化三甲胺(FMO)的产生,FMO被认为是治疗心血管疾病的标志物。
肠道微生物群通过代谢影响心血管健康
Shah H,et al.Microorganisms.2024
在急性心肌梗死动物模型中,肠道微生物组,特别是Tissierella soehngenia属、Synergistetes门、Spirochaetes门、Lachnospiraceae家族和Syntrophomonadaceae家族,表现出更大的趋势。然而,对心血管系统的研究尚未探索,需要考虑更多基于机制的研究来发现与心力衰竭相关的风险。
随着年龄、生活方式或药物的影响,健康宠物的肠道微生物组平衡可能受到影响甚至会被破坏,从而引发各种疾病。我们应该采取何种方法来恢复其健康呢?
肠道微生物群可以通过饮食调整、益生菌及益生元以及粪菌移植进行调节。这些治疗方法每一种都具有不同的优点和副作用。因此必须考虑潜在的疾病,因为没有针对所有类型生态失调的治疗方法。
1
饮食调整
饮食是影响宠物肠道微生物群的重要因素之一,不同的饮食组成会对宠物肠道微生物群的生态平衡产生不同的影响。
▸ 根据不同年龄和营养状况挑选合适的食物
蛋白质、碳水化合物和脂肪是大部分宠物必需的营养物质,是促进机体生长的重要营养素。合理控制这些营养素的摄入是维持健康的关键。
不同年龄段的宠物对营养素的吸收能力不同。老年犬猫由于肠道消化功能逐渐退化,不能较好地吸收,需要根据不同的宠物状态制定相对应的饮食。
2
益生菌
▸ 益生菌促进了整体的微生物平衡病减少了炎症
益生菌的使用通常被认为对维持和促进宠物的胃肠道健康很有价值。宠物益生菌产品的普及突显了宠物主人对益生菌补充剂对动物健康的潜在益处的兴趣和意识不断增强。此外,它在狗和猫等伴侣动物中的使用改善了肠道微生物群组成,促进了整体微生物平衡,从而增强了免疫反应,减少了肠道炎症,增加了肠道屏障功能,并防止肠道病原体定植。
对猫狗施用益生菌干预的效果
Shah H,et al.Microorganisms.2024
3
粪菌移植
粪菌移植(FMT)是一种新兴的治疗方法,通过将健康供体的粪便转移到受者体内,为患病个体提供健康的微生物群。
▸ 粪菌移植在改善菌群失调方面效果最好
研究表明,在人类以及患有感染性和慢性胃肠道疾病的犬中,粪菌移植在改善菌群失调方面比其他调节微生物组的治疗方法更有效。与接受甲硝唑治疗的腹泻和炎症性肠病(IBD)患犬相比,接受粪菌移植灌肠治疗的腹泻患犬和仅接受皮质类固醇治疗的炎症性肠病患犬,其粪便菌群与健康犬的粪便菌群更相似,且胆酸和初级胆汁酸的比例有所降低。对于因产气荚膜梭菌A毒素引起的腹泻患犬,抗生素治疗无效,但通过灌肠接受粪菌移植后,腹泻成功得到解决。
在一项随机临床试验中,对于犬细小病毒(CPV)引起的急性出血性腹泻且存活的幼犬,使用粪菌移植可加快临床恢复速度,缩短住院时间。此外,患有慢性肠炎(CE)的患犬在接受单次粪菌移植灌肠治疗后,其犬炎症性肠病活动指数(CIBDAI)评分显著降低。
随着肠道微生物组研究的深入,我们越来越认识到这个”隐形器官”对宠物健康的深远影响。肠道微生物组与猫狗的胃肠炎、癫痫、肾病、肥胖、心脏病等多种疾病密切相关。
饮食结构、年龄变化、品种差异、抗生素使用等因素都会深刻影响菌群平衡,进而影响宠物的整体健康状况。当菌群失衡时,有益菌如双歧杆菌、乳杆菌减少,而致病菌如大肠杆菌、肠球菌增加,这种变化不仅影响肠道功能,还会通过肠-脑轴、肠-肾轴、肠-皮肤轴等途径影响神经系统、泌尿系统和皮肤健康。
值得欣慰的是,肠道菌群具有可调节性。通过科学的饮食调整、合理的益生菌补充、甚至粪菌移植等手段,我们可以有效地重建和维护宠物的肠道微生态平衡。未来,随着个性化营养学的发展,我们期待能够基于宠物的品种、年龄、健康状况和菌群特征,制定更加精准的健康管理方案。让我们携手守护毛孩子的健康,从肠道微生物开始,为它们创造更加美好的生活!
主要参考文献
Shah H, Trivedi M, Gurjar T, Sahoo DK, Jergens AE, Yadav VK, Patel A, Pandya P. Decoding the Gut Microbiome in Companion Animals: Impacts and Innovations. Microorganisms. 2024 Sep 4;12(9):1831.
Lyu Y, Pu J, Deng B, Wu C. Gut Metabolome in Companion Animal Nutrition-Linking Diets to Health. Animals (Basel). 2025 Feb 24;15(5):651.
Bartochowski P, Gayrard N, Bornes S, Druart C, Argilés A, Cordaillat-Simmons M, Duranton F. Gut-Kidney Axis Investigations in Animal Models of Chronic Kidney Disease. Toxins (Basel). 2022 Sep 7;14(9):626.
Pilla R, Suchodolski JS. The Role of the Canine Gut Microbiome and Metabolome in Health and Gastrointestinal Disease. Front Vet Sci. 2020 Jan 14;6:498.
Song H, Mun SH, Han DW, Kang JH, An JU, Hwang CY, Cho S. Probiotics ameliorate atopic dermatitis by modulating the dysbiosis of the gut microbiota in dogs. BMC Microbiol. 2025 Apr 22;25(1):228.
Honneffer JB, Minamoto Y, Suchodolski JS. Microbiota alterations in acute and chronic gastrointestinal inflammation of cats and dogs. World J Gastroenterol. 2014 Nov 28;20(44):16489-97.